prostaglandinas y función pineal : su participación en la ... · la glándula pineal. relaciones...
TRANSCRIPT

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis de Posgrado
Prostaglandinas y función pineal :Prostaglandinas y función pineal :Su participación en la síntesis ySu participación en la síntesis y
mecanismo de acción de la hormonamecanismo de acción de la hormonapineal melatoninapineal melatonina
Ritta, Mónica Nora
1981
Tesis presentada para obtener el grado de Doctor en CienciasQuímicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Ritta, Mónica Nora. (1981). Prostaglandinas y función pineal : Su participación en la síntesis ymecanismo de acción de la hormona pineal melatonina. Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_1675_Ritta.pdfCita tipo Chicago:
Ritta, Mónica Nora. "Prostaglandinas y función pineal : Su participación en la síntesis ymecanismo de acción de la hormona pineal melatonina". Tesis de Doctor. Facultad de CienciasExactas y Naturales. Universidad de Buenos Aires. 1981.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_1675_Ritta.pdf

UNIVERSIDAD DE BUENOS AIRES
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
Prostaglandinas y función Eineal.
Su participgpión en la síntesis y mgggpismode
acción de la hormona Eineal melatonina.
Autor: Lic. Mónica Nora Ritta.
Director de tesis: Dr. Daniel P. Carolnali.
Lugar de trabajo: Centro de Estudios Farmacológicoo y de Principios
Naturales. CONICET.
TESIS PRESENTADA PARA DPTAR AL TITULO DE
DOCTOR EN CIENCIAS QUIMICAS
-.1981.-1.-)\

AGRADECIMIENTOS
A1Dr. Daniel P. Cardineli, por haberme brindado constante
mente su apoyo, su estimulo y sus consejos.
Al Dr. Carlos P. Lentoe, quien hizo posible el inicio de mi
teree en el campode le investigación.
A la Lic. AneMaris Fuentes, por su generosa asistencia en
le preparación de este texto.
A mis compañeros de laboratorio, que me brindaron directa o
indirectamente su apoyo y colaboración haciendo posible este trek
bajo.
Al Centro de Estudios Farmacolñgicos y de Principios Natura
les (CEFAPRIN),por permitir que se llevara e cebo este investiga
ción.
Al Consejo Nacional de Investigaciones Cientificas y Técnicas
(CONICET)por lee becas que me otorgara para investigar sobre este
teme.
A todos aquéllos que de alguns manera contribuyeron pere que
este tesis pudiera concretarse.

A mis padres

ADP
AQEo
ARN
ATP
BSA
lb
G.6.
Ci
CoA
CUHÏ
Corticoaterons
DOPA
Dopanins (DA)
EDTA
ES
Eotradiol
fmol
FSH
GABA
GCS
ABREVIAÏURAS
Adrenocorticotrofins
Adenosina difosfato
Actividad especifica
Adenosins monofosfsto cíclico
Ácido ribonucleico
Adenosinatrifosfsto
Albúnina ds suero bovino
Grados centígrados
Carbono lb
concentración efectivo 50
Curie
Cosnzila A
Catecol-O-nstiltrsnaferasa
11(5,21-dihidroxi-h-pregneno-3,20-dions3.h-dihidroxifenilalanina
3,k-dihidroxifenilatilaninsAcidoetilendismino tetrsscitico
Error standard
1.3.5 (lO)-satrstrisno-3,17(¿ -diolFemtomol
Hormonafolículo estimulante
Ácido gamma-aninooutirico
Ganglio cervical superior
Hora
Tritio

hCG
HHT
NIGHT
HHH
HPETE
Indanctacina
LHRH
H
MAO
¡Ci
uCi
MDA
Helatonina
"9
ug
min
Gonndotrofina coridhica humana
Ácido hidroxieicosatetraenoico
¡Elda 12-hidrox1-5,B,lD-heptadecadienoico
Hidroxindol-U-metiItranoferaaa
Hipotfilamo nadiobaaal
¡tido hidroxiperoxieicoaatetraenoico
(N-p- [clorobenzoill -2-met11-5-netox1ndol-Jacético
Constante de disociacián
Constante de inhibicio’n
Hreba-Ringer-Bicarbonato
Hermanaluteinizante
Hormona liberadora de LH
Molar
Monoaninooxidaaa
Hilicurie
Microcurie
Ácido malondialdehfdo
N-acetil-S-netoxitriptanina
Hiligramo
Microgramo
Minuto
HililitroHilimal
Normal
Nanomolar

Norepinefrina (NE)
pg
PGís)
PHS
PRL
RPM
SAM
s.c.
Serotonins (S-HT)
SNAT
SNC
TCA
ÏCI
Testosterona
TRIS
TSH
Tx
3,h-oihidroxifeniletsnolsmins
Picogrsmo
Proatsglandine (e)
Suero de yegua preñsos
Prolactine
Revoluciones por minuto
S-adenosilnetionins
Subcutánea
S-hidroxitriptsmins
Serotonins-N-scetiltrsnsferese
Sistens nervioso central
Acidotricloroacético
tensión contráctil isonátrics
17 p -h1drox1-5 o(-snorostsn-3-ons
Tris (hidrosinetil)-sm1nonetsno
TirotrofinaTronboxsno

INDICE DE CONTENIDO
I - INTRODUCCION
1.1.- Ls Glinduls Pineal
1.2 - Participación de las Prostaglandinss en los MecanismosNeursles
11.3
II.“
- Estudio de sitios de union especificos para pro!
1.1.1 - Conceptos generales y aspectos morfo
lógicos
1.1.2 - Biosintssis y metabolismo de ls mela
tonins
1.1.3 - Control neural de ls ectividsd pinesl
1.1.“ - Efectos biológicos de la melatonins
Adrenér icos Centrales Periféricos
1.2.1 - Biosintesis y metabolismo de las pros
taglsndinas
1.2.2 - La unidh neuroefectora sdrenérgica
1.2.3 - Función de las prostaglandinas en los
mecanismos sdrenérgicos
Objetivos
¡1 - MATERIALESv Merooqg
Animales y Tejidos
Produccion de prostaglandinas in vitro por glag
dula pineal y HHHbovino y HHBde rata
taglsndinas en la glándula pineal bovina
Determinación de sitios de union especificos ps
rs AMPcen la glshdula pinesl de rats
¡,71
9
17
23
36
k2
k7
52
53
55
57
61

1105 ‘
IIOG
1107
IIaB ’
11.9 "
11.10
11.11
III.l
Determinacidh-ds actividades enzimáticas
11.5.1 - Catecol-D-metiltrsnsferaaa
11.5.2 - Honoaminooxidass
11.5.3 - Serotonina-N-acetiltransferssa
11.5.“ - Hidroxindol-O-metiltranaferasa
Dsterminaciones por análisis de saturación de
melstonins, PGE y AMPC2
11.6.1 - Helatonina
11.6.2 - Prostaglandina E2
II0603‘Determinación de la liberación de NEpor glán
dula pineal ds rata in vitro
Determinaciónde la actividad contríctil del
útero
Determinacioñ de ls agregación plaquetaris
Reactivos utilizados
Estadistica
III - RESULTADOS
Participación de las Prostaglandinss en el con
trol noradrenérgico de la gldhdula pineal
111.1.1 - Produccidh de prostaglandinas por la
glanduls pinsal bovina in vitro
111.1.2 - Caracterizacidh de sitios de union e!
pacíficos para prostaglandinas en ls
glándula pineal bovina
62
62
63
6h
66
67
70
71
73
7k
76
77
7B
81

III.2
III.1.3
111.1.“
III.1.5
III.1.6
III.1.7
Efectos de la indometacina sobre la
sintesis de melatonina y metabolismo
monoaminérgicode la glándula pineal
de rata
Efecto de laa prostaglandinas sobre
la actividad de SNAÏen la glándula
pineal de rata in vitro
Efecto de PGE2sobre la acumulacián
de AMPcy su union a sitios recepto
res intracelularea en ls glafiduls pineal de rata in vitro
Efecto de PGEZ, NE y bloqueantes de
le sintesis de prostaglandinas sobre
ls liberación de nelatonina por la
glándula pineal de rata in vitro
Efecto de PGE2sobre la liberación
de NE-JHpor la glándula pineel de
rate in vitro
Participación de las prostaglandinas en el me
canismo de acción de la hormona pineal melsto
nina
III.2.l - Inhibición por neletonine de le pro
ducción de prostaglandinas en HMSog
vino y de rata
III.2.2 - Inhibición por melstonins de efectos
vinculados s ls liberación de prostg
glandinas en útero de rata y plaque
tas humanas
le?
92
98
101
105
106
110
111
116

IV - DISCUSION
V - CONCLUSIONES
VI - REFERENCIAS

Fig. 2:
Fig. 3:
Fig. b:
W: Corte sagital de la región epitalámics en
los mamíferos. Relaciones anatómicas da la
la glándula pineal.
Relaciones celulares en la glándula pineal
de mamíferos.
Inervación simpática de la glándula pineal
de los mamíferos.
Vía neuronal y secuencia metabólica de cqg
trol de la sintesis de melatonina por la
luz ambiental.
: Bioaintesis y metabolismo de la melatonina.
: via neural para el control de la función oinesl.
: Vía metabólica del ácido araquidónico.
: Vías metabólicas de síntesis y metabolismo
de NE.
: Representación esquemática de la uniún neugg
efectora simpática.
: Efecto de la NE sobre la producción de PGE y
PGFpor glándula pineal bovina in vitro.
la?
lO
12
20
“3
“6
79

Fig. 11:
Fig. 12:
Fig. 13:
F19. 1h:
Fig. 15:
Fig. 16:
Fig. 17:
Fig. 18:
Fig. 19:
Asociación de PGF -3H y PGE -3H al sobreng2d. 2
dante de 900 x g de glándulas pineales bovi
naa en función del tiempo de incubación.
Efecto del CsH sobre la unión especifica de
PGs-JH a preparaciones crudas de membranas
pineales.
Union de PGEz-JH a membranas de glándula pinesl bovina comofunción de concentraciones
crecientes de PGEz-JHen el medio.
Unión de PGF2u_-3H a membranas de glándula oi
neal bovina comofunción de concentraciones
crecientes de PGF2“_-3Hen el medio.
Efecto del tratamiento con indometacina sobre
1a actividad de SNATy contenido de mslstoni
na pineal durante el periodo de oscuridad.
Efecto de las PGa sobre la actividad de SNAÏ
de glándula pineal de rata in vitro.
Efecto de las PGs sobre la actividad de SNAÏ
de glándula pineal de rata in vitro.
Efecto de las PGa sobre la acumulación de
AMP:in vitro.
Efecto de PGE2sobre la asociación ds AHPC
-3H a la quinasa de proteinas de glándula
pineal de rata in vitro (sitios de unión ds
aocupadoa).
la?
83
8k
85
86
97
99
100
102
103

Fig. 20:
Fig. 21:
Fig. 22:
Fig. 23:
Fig. 2k:
Fig. 25:
Efecto oe PGE sobre la ssociscióh de AMPC2
-3H s la quinsss de proteínas de gláhdula
pinesl de rsts in vitro (sitios de unión tgtales).
Efecto de ls melstonins sobre ls Hiberscioh
inducida por NE de PGE2 por HMBoe rsts ¿gVitro.
Tension isométrios desarrollada por útero de
rata castrads en función del tiempo de expo
sición s distintas concentraciones de melstgnina in vitro.
Fórmulas estructurales de la melstonins y de
ls indometscins.
Control neural de la actividad pinesl y meo!
nismos celulares que psrticipsn en ls sínte
sis de melstonins.
Posibles sitios de accion de ls melstonins
en neuronas del SNC.
10k
115
118
126
135
lhl

Tabla :
Ïabla :
Tabla :
Tabla :
Tabla :
Tabla :
Tabla :
Tabla :
INDICE DE TABLAS
[131
Cambios endócrinos producidos por la admi
nistración de melatonina en mamíferos. 29
Otros efectos de la melatonina en mamífe
ros. 3h
Efecto de 1a melatonina sobre 1a produccion
y liberación de PGs por explantos de glánqg
1a pineal bovina in vitro. BU
Efectos de la preincubscioh con tripsina, e!
posición a alta temperatura o incubación en
ausencia de indometacina sobre la unión de
PGEz-JHy Para“ -3H a sitios específicos englándula pineal bovina. 87
Distribucidh aubcelular de los sitios de unión
para PGa-JH en glándula pineal bovina. BB
BESOy H1 de una variedad de PGs en los estu
dios de competencia por los sitios de unión
para PGEz-JH (A) y PGF2°¿-3H (B) en membranas
de glándula pineal bovina. 90
Efecto de 1a indometacina sobre las sctividae
des de MADtipo A y tipo B en glándula pineal
de rata. 9“
Efectos de la indometacina sobre las actividg
des de CUHTy HIÜMTen glándula pineal de rata. 95

Tabla 9:
Tabla 10:
Tabla 11:
Tabla 12:
Tabla 13:
Tabla 1h:
Tabla 15:
Efectos de la indometacina sobre las activi
dades de MAO tipo A y tipo B y CUMTen HHB
de rata.
Efecto de la PGE sobre la liberación de me2
latonina al medio de incubación de glándulas
pinceles de rata.
Efecto de 1a NE, en presencia o ausencia de
indometacina, ácido acetilsalicilico o ácido
mefenánico sobre la üiberación de melatonina
al medio de incubacián de glándulas pineales
de rata.
Efecto de la PGE2sobre la liberacion de NE
-3H por glándula pineal de rata in vitro.
Efecto de la melatonina sobre la producción
y liberacion de PGs por explantos de HMBog
vino in vitro.
Efectos de concentraciones submicromolares
de melatonina sobre la liberacion espontá
nea de PGE2 por HMBde rata in vitro.
Inhibición por melatonina de la agregación
de plaquetas inducida por ADP.
96
106
107
109
113
116
119

I. INTRODUCCION

1.1 - LA GLANDULA PINEAL
I.l.l ConceEtosgenerales y asEectos morfológicos
Si se acepta 1a ubicuidad de acción como sinónimo de importan
cia y se consideran comoexistentes las interacciones descriptas de
la glándula pineal con otros órganos y sistemas, la pineal es, sin
duda, una de las glándulas endóbrinas de mayor influencia Fisiológi
ca. La primera descripción de este órgano se remonta al siglo III A.C.
y es adjudicada a Herófilo de Alejandría. Este consideró'a la glándu
la comouna válvula reguladora del flujo en los ventrículos cerebra
les. En el siglo II de nuestra era, Galeno de Párgamola describió
detalladamente y la llamó Hornarium (del griego, piña). Para Galeno
la glándula pineal constituía el sostén de un grupo de venas cerebra
les (las llamadas posteriormente "venas de Galeno").
En 1682, el anatomista inglés ThomasGibson la denominó glán
dula pineal (de "penís", por su situación sobre y entre los tubércu
los cuadrigéminos inferiores o "testes"). Para Descartes (1596-16502
en la glándula pineal tendria su expresión principal el alma. Esta
idea influyó en la vinculación alegada de la glándula pineal con en
fermedades mentales, que ha persistido hasta nuestros dias.
En poco más de una década (l95h-1965) el desarrollo de la in
vestigación pineal transformó en axiomática su participación en la
regulación neuroendócrina. Hitay y Altschule recopilaron en su li
orn "The Pineal Gland" (195h) la información en conjunto que sugerí:
el papel de la glándula en la regulación de la función gonsdal, en

la respuesta de los pigmentos de la piel de anfibios a la luz y en
la conducta animal.
La hormonapineal melatonina (S-metoxi-N-acetiltriptamina)
fue aislada e identificada en extractos de glándulas pineales bovi
nas por Lerner y col. (1959), comoconsecuencia de la purificación
del factor pineal aclarador de 1a piel de los anfibios. La melatoni
na es el prototipo de una familia de compuestos biológicamente acti
vos, los metoxindoles, producidos por la glándula pineal de todas
las especies de vertebrados conocidos, incluido el hombre. Su descu
brimiento y las investigaciones subsecuentes que generó proveyeron
los primeros elementos bioquímicos para el análisis experimental de
1a función pineal. La melatonina fue la primera sustancia que repro
duJo los efectos biológicos de los extractos pineales y que revirtió
las secuelas endócrinas de la pinealectomia.
Un año después del descubrimiento de Lerner y col., Axelrod y
Meissbach (1960) describieron la via biosintética de la melatonina
y confirmaron que la pineal de mamíferos y vertebrados inferiores
contiene todos los componentesnecesarios para dicha sintesis.
Estudios bioquímicos y anatómicos llevados a cabo en varios
laboratorios, demostraron un vinculo entre las variaciones de la
luz ambiental y 1a sintesis de melatonina. En 1965, Hoffmany Reiter
observaron que los fotoperiodos cortos inducian cambios gonadales
en el hamster, los que eran suprimidos por la pinealeotomia. En es
te mismo año Murtmany Axelrod (1965) introdujeron el término
"transductor neuroendócrino" para definir a 1a glándula pineal (es
decir comoa un órgano capaz de convertir una señal neural en res
puesta endócrina). El cúmulo de información obtenido durante la ú;

tima década indica que la glándula pineal es un "regulador de re
guladores" a nivel Jerárquíco superior de otros procesos de con
trol neuroendócrino (Reiter, 1980; Cardinali, 1981).
Embriológicamente, la glándula pinesl proviene de una evagi
nación del techo del diencéfalo y está ubicada entre las comisuraa
habenular y posterior; en el hombretal evaginación se evidencia
ya en el 20 mes de vida intrauterina (Fig. l). La glándula pineal
ae caracteriza por una gran variabilidad anatómica a través de la
escala zoológica (Ralph y col., 1979). En peces, anfibios y repti
les lacértidos contiene células fotorreceptoras semejantes a las
retinianas; éstas se observan también en los órganos parapinealea
presentes en algunos reptiles. anuros y lacértidos. Por el contra
rio, la pineal de los mamíferos ea típicamente endócrina y carece
de fotorreceptores desarrollados, observándose rudimentos de és
tos en serpientes y aves. Comoórgano circunventricular ls pineal
está constituida por células con funciones neurosecretorias, los
Einealocitos que, si bien no pueden considerarse estrictamente cg
lulas nerviosas, derivan del neuroepitelio embrionario.
Los restantes representantes del grupo de organos circuhveg
triculares son: la eminenciamedia. la neurohipófisis, el área
postrema y los órganos subfornical, subcomisural y vasculoso de la
lámina terminal.
Los pinealocitos representan el 85%de la masa pineal; el
resto está constituido por células con función de sostén (glis)
y por terminales nerviosos. Los pinealocitos emiten prolongacio
nes que contacten en Forma de pies terminales con los espacios
perivascularea o,en menorproporción,con el receso pineal o su

Fig. 1: Corte aagital de la región opitnlánica en lo. acuíferos.
Relacionan anatómical do la glándula pincel. (modifica
do de Quay. 197k).

prapinaal (Martenberg, 1968; Gregorek y col., 1977; Mataushima y
Reiter, 1977, 1978) (Fig. 2). A través de estos últimos pueden
alcanzar la cavidad ventricular. Noestá aún aclarado el comparti
miento, vascular o ventricular, hacia el cual la pineal secreta
sus productos.
El flujo sanguíneo de la glándula pineal es uno de los más
elevados del organismo, lo que de por sí niega 1a naturaleza ves
tigial que se le atribuyó durante años. Esta idea tuvo origen en
la calcificsción de 1a pineal humana, observada desde el desarro4
lla intrauterino. Los depósitos calcáreos no afectan, sin embargo,
1a función pineal. existiendo producción normal de melatonina en
seras humanosque presentan cslcificación pineal avanzada (Hurt
man y Cardinali, 197h).
Unode los principales problemas no resueltos por los anato
mistas es la identificación del sitio intracelular de producción y
almacenamiento de las hormonas pineales (Pévet, 1979). Se acepta
en forma general que los pinealocitos contienen una sustancia prg
bablemente glucoproteica, sintetizada en el retículo endoplásmico
granular y transportada y concentrada en los sáculos del retículo
de Golgi. La formación y liberación de estas vesículas muestra un
ritmo circadiano y depende de 1a integridad del sistema simpático
(Pévet, 1979). Si bien se ha propuesto que tales vesículas son el
sitio de almacenamientodel o de los principios endócrinos pinea
las, no existe aún prueba experimental a1 respecto. La posibilidad
de que la vesícula secretaria contenga tanto 1a sustancia proteica
comoun derivado indblico (melatonina o serotonina) constituye 1a
extensión a1 pinealocito del concepto de célula APUDy psraneurona
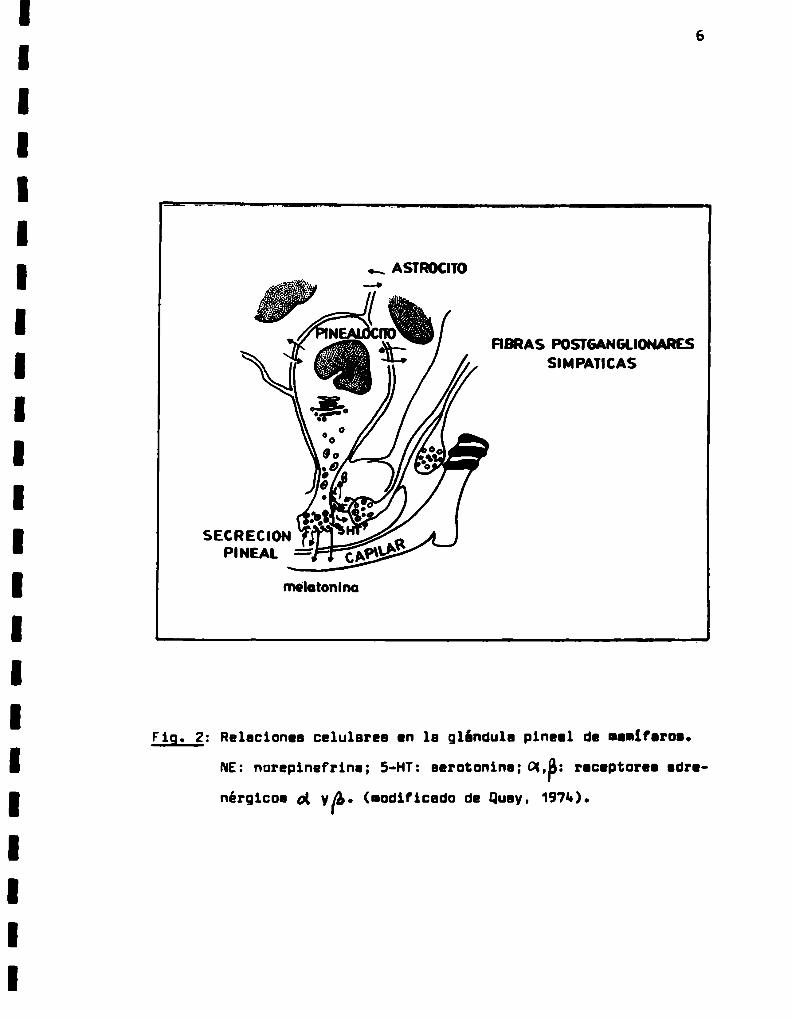
FIBRAS POSÏGANGUONARESSl MPAÏICAS
SECRECIONPINEAL -'
mlotonlm
F19. 2: Relaciones celulares en la glándula pincel de ¡enlfcrom
NE:norepinefrina; S-HT:serotonina: DLP: nclptorn adu
nérglcoa al yp. (modificada de Quay, 1974.).

(Fujita y Hogayashi, 1979). En efecto, existen evidencias autorra
diográficas que apoyanuna localización preferencial de la seroto
nina en la matriz de las vesículas secretorias de las células ps
renquimatosas pineales (Pévet, 1979).
La inervación pineal de los mamíferos sigue dos disposicio
nes anatómicas fundamentales. En la forma más simple, fibras sim
páticas postganglionares originadas en el ganglio cervical superior
alcanzan a la glándula por dos vias. Un grupo (nO l de la Fig. 3)
sigue la distribución de los vasos sanguíneos y termina en el espg
cio perivascular, en las cercanias de los procesos de los pineslo
citos; otro grupo penetrs.en la glándula por el polo caudal forman
do los nervios canarios (nO 2 de la Fig. 3). Estos terminales tie
nen una distribución parenquimstosa y perivascular. Dicho tipo de
inervación, que se describirá en detalle en ls sección 1.1.3 es la
forma más común en mamíferos y existe en los roedores.
La segunda disposición de la inervsción es la encontrada en
los primates y lsgomorfos (conejo). Consiste en una combinación
de 1a inervación simpática anteriormente mencionada y una inerva
ción parasimpática constituida por Fibras preganglionares que na
cen en el núcleo salival superior y siguen el curso de los nervios
petrosos superiores y del nervio facial. Algunas de dichas fibras
hacen sinapsis en pequeños ganglios ubicados en los nervios, mien
tras que otras terminan en neuronas ubicadas dentro de la glándula
pineal. La distribución de las fibras postganglionares parasimpá
ticas es semejante a las simpáticas, terminando la mayoria en los
espacios pericapilares. Los estudios sobre el pspel de la inerva
cifin en la regulación de la función glandular discutidos en la seg

NERVIOScommos
GLANDULA aPINEAL 7‘ '’ 1
l‘ u¡4' x '
7/ ///\¿‘ COMISURAHABENULAR// “Cá/ ‘e/ “ COMISURAPOSTERIOR
Fig. 3: Inervecibn simpática de le glándula pineel de los neniferoe.
Lee fibras provenientes del ganglio cervical superior
llegen e le glándula pineel por le pared de los veooe
sanguíneos (no 1) o e través de los nervios conerioe
(no 2). (modificado de Quay, 1971.).

ción 1.1.3, han sido realizados en Formacasi exclusiva en especies
que, comola rata y el hamster, cuentan con el primer tipo de dis
tribución.
1.1.2 Biosintesis y metabolismo de la melatonina
La biosintesis del derivado indólico melatonina comienza en
los pinealocitos con la captación del triptofano de la circulación
(Cardinali y Murtman, 197%; Minnemann y Murtman, 1976; Murtman y "qe
küüitl. 1977) (Fig. k y 5). El aumentodiurno del triptofano pincel
precede a1 aumento del eminoácido libre o total sérico, hecho que
sugiere que la glándula lo capta activamente (Sugden, 1979). Una
vez en el pínealocito, parte del triptofano es utilizado para la
síntesis proteica, mientras una fracción mayores metabolizada a
través de la via de los indoles. La ruta metabólica principal de
la serotonina es la oxidación del ciclo pirrólico a quinurenemina
por la enzima 2,3-dioxigenasa (Fujiuara y col., 1978)(Fig. k).
El triptofano es convertido en 5-hidroxitriptofano por acción
de la enzima triptofano hidroxilasa (Sitaram y Lees, 1978). En Cog
traste con otras regiones del cerebro, la tríptofano hidroxilasa
pineal está saturada por su sustrato (Sugden, 1979).
La desoarboxilación del S-hidroxitriptofano a serotonina es
catalizada por una descarboxilasa de relative especificidad, que
participa también en la biosintesis de catecolaminas. La serotoni
na es el indol presente en mayor concentración en el tejido pine
al y su contenido y velocidad de recambio ("turnover") es mayor
en la glándula que en otras áreas del sistema nervioso central
(SNC)(Quay, l97h). Uno de los Factores que determinan estas altas

10
Fino .0: Via neuronal y secuencia IItlhÓliCl de control de la ¡ln
teoia de aclatonina por la luz anhicntal.
TP: triptofano: HTP:S-hidroxitriptofano: S-Hï: otrotoninl;
HIAA:ácido 5-h1drox1ndolncét1co; HTDH:S-hidrouitriptofol;
NAS:N-acetllaerotoniná; HTOH:S-nctnxitriptofol: HIAfl: iq;
do S-netoxindolacótico; HIOMT:hidroxindol-O-netiltranafegg
aa; MAO:nonoaninooxidaea.

J=----------m
11
concentraciones de serotonina en la pineal es la gran actividad de
la triptofano hidroxilasa glandular (Sitarnm y Lees, 1978). Alredg
dor del 30%del total de lu serotonina almacenada en la pineal de
rata está localizada en los terminales nerviosos simpáticos; sin
embargo, sólo los pinealocitos son capaces de producir serotonina,
1a que luego es captada por los terminales (Jaim-Etcheverry y
Zieher, 1968). La serotonina pineal es el precursor de compuestos
biológicamente activos como1a melatonina y el 5-metoxitriptofol.
La serotonina liberada al espacio vascular puede, asimismo, tener
efectos propios (Quay, l97h).
La disminución del contenido de serotonina que se observa en
la fase de oscuridad es cansecuencia de cuatro vias metabólicas
independientes: l) desaminación oxidativa por la monoamino-oxidasa
(MAD),2) N-metilación, 3) oxidación por 2,3-dioxigenasa, h) libe
ración al espacio extracelular (Fig. h). La disponibilidad de la
serotonina para cualquiera de estas rutas depende del equilibrio
"serotonina granular :2 serotonina libre". Sólo la segunda está
sujeta a la metabolización.
La MADconvierte a la serotonina en 5-hidroxindol acetalde
hído, un intermediario inestable que es oxidado a ácido 5-hidroxig
dol acético,o reducido a S-hidroxitriptofol por una alcohol deshi
drogenasa (Fig. h); Comose verá más adelante, se han descripto en
el SNDdos isoenzimas MAD,denominadas A y B por su especificidad
de sustrato. En la glándula pineal coexisten también ambas isoenzi
mas (Neff y Yang, 197h). La enzima tipo A es dependiente de la i
nervación simpática intacta, ya que su actividad desaparece des
pués de la gangliectomia cervical superior, mientras que la MAO

J----------
12
ELNILLA_EMEAL
© .- mom-coo" "° GQ-M-COCH "° (cum-m.U ——.H H Hmmormo 5-».an W
gm n © (www-00% warm-com.-' (caen-wwe"; . .._ W©U fx H H
"° H .47 \ MEMTOMNA kms-mmuroum \, mDEWETLNIN c (a‘ -MFaxH
[amm] [animan] k a)! warm-N-“ ¡"Llr quA " flPaW &MEnmnuMnflunna
l 1 c o -( -NH-cocn
(ns) (w (en) tw.) m‘@fii a“! s N un“5%, A e s-ncroxmnenmQEEA ‘ ,
cnwumnw/‘\ammmusmramu PEMWKEMIHMEA
F19. 5: Bloeinteeie y meteholleno de le ¡eletonlne.
Le glándula pineel cerece de elete-ee enzlnáticoe de degre
declbn pere le ¡eletonlne y elnecene eólo une frecclón ¡1:2
na de le hormoneque elntetlze. El metabolismo central y p;
rlférlco (higado) difieren entre ei. comoee dlecute en le
sección 1.1.2; MIAA:ácido S-metoxindolecétlco.

13
tipo Ü permanece inalterada (Neff y Yang, 197h; Vacas y Cardinali,
19798)oEStd observación ha sido interpretada comolocalización neg
ronal y extraneuronal de la MADA y Ü respectivamente.
La conversión de serotonina en melatonina involucra dos pa
sos enzimáticos (Fig. A y 5). El primero es la N-acetilación por
una N-acetiltransferasa (serotonino-N-acetiltransferasa. SNAT)cg
lativamente inespecifica para dar N-acetilserotonina (Klein, 1978).
El grupo acetilo proviene del cofactor acetilcoenzima A (acetil
CoA)cuya concentración es controlada por une activa acetil-CDA
hidroxilasa (Namboodiri y cnl., 1979). Luego un grupo metilo es
transferido de la S-adenosil-metionina (SAM)al grupo 5-hidroxi
de la N-acetilserotonina, obteniéndose así melatonina; esta reac
ción es catalizaoa por la hidroxindol-Ü-metiltransferasa (HIÜMT)
(Axelrod y Meissbach, 1960). Los niveles elevados del cofactor S
adenosil-metionina se deben a las altas concentraciones de la en
zima L-metionína-S-adenosiltransferasa pineal, que cataliza, a
partir de ATPy L-metionina, la formación de S-adenosil-metionina
(Guchhait y Grau, 1978). Ademásde la N-scetil-serotonina. la
HIÜMTpuede metilar al ácido 5-hídroxindolacético, S-hidroxitrip
tofol y 5-hidroxitriptofano (Fig. A). Unode los productos forma
dos, 5-metoxitriptofol,es considerado comouna hormonapineal pre
sente en la circulación (Mullen y col., 1979).
La sintesis de la melatonina en los mamíferos no está res
tringida a la glándula pineal. La descripción inicial de posibles
fuentes extrapineales de melatonina, basada en el aislamiento de
HIUMTen la retina y glándula harderiana de roedores (Bardinali y
Murtman, 1972) y en la conversión in vitro de serotonina en mela
tonins por retina (Cardinali y Rosner, 1971) ha sido verificada

1h
inmunohistoquimicamente (Bubenik y col.. 1978). Luego de la pinea
lectomia se detectan niveles residuales de melatonina circulante en
rata (Üzaki y Lynch, 1976) y oveja (Hennauay y col.. 1977). Al menos
en una de los tejidos citados más arriba. 1a retina, la melatonina
puede tener una función regulatorio sobre la migración del pigmento
en las células pigmentarias retinianas (Cardinali, 197“; Pang y Yew.
1979). Las dos enzimas retinianas que catslizan la conversión de
serotonina en melatonina son sensibles a la luz.
La melatonina parece ser secretada por ls glándula pineal median
te simple difusión y no se ha descripto un mecanismoactivo de se
crecidh. El consenso general es que la melatonina presente en la pi
neal es un reflejo directo de la síntesis de la hormona (Minnemany
Murtman, 1976; Murtman y Moskouitz, 1977; Reiter, 1980). Durante los
ultimos 5 años se han dedicado importantes esfuerzos sl desarrollo
de un radioinmunoanálisis específico para la determinación de melstg
nina en tejidos y fluidos biológicos (Murtmany Moskouitz, 1977; A
rendt, 1978). La aplicación de esta técnica Junto a la cromatografía
gaseosa asociada a espectrometria de masa (Wilson, 1976) posibilitó
la descripción de un ritmo diario en la secreción de melatonina en
todos los vertebrados estudiados hasta el momentoactual (unas no
especies). Se estableció asi que 1a concentración de melatonina en
plasma, liquido cefalorraquídeo y orina es máximadurante el perío
do de oscuridad, sea 1a especie estudiada de actividad diurna o
nocturna (Üzaki y Lynch, 1976; Kennauay y col.. 1977; Murtman y
Moskouitz, 1977; Arendt, 1978; Rollag y col.. 1978; Mullen y col..
1979; Mithyachumnarnkul y Hnigge, 1980).
En el hombre, 1a concentración máxima de melatonina plasmá

15
tica es prepuberul (Silman y col., 1979). tn los adultos existen
también ritmos circanuales de la melatnnina circulante (Arendt,
1978).
Uomoya se ha mencionado, está en discusión el compartimieg
to primario (vascular o liquido cefalorraquideo) al cual 1a pi
neal de mamíferos secreta la melatonina (Fig. 5). En la rata, du
rante el periodo de oscuridad hay un incremento precoz de la con
centracíón de melatonina circulante, mientras que los niveles de
1a hormona en el liquido cefalorraquideo aumentan más tarde
(Uithyachumnarnkui y Knigge, 1980). Evidencias anatómicas en di
Ferentes especies tienden también a considerar comominoritaria
la posible via de secreción hacia las cavidades ventriculares ce
rebrales (Gregorek y col., 1977).
Alrededor del EDP7Ü%de la melatonina presente en el plasma
está asociada a albúmina (Cardinali y col., 1972). Por lo tanto,
sólo el hÜ%correspondiente a la hormona libre es capaz de llegar
al liquido cefalorraquideo atravesando la barrera hematoencefáli
ca. Sin embargo, datos recientes indican que 1a melatonina asocyg
da a la albúmina puede también ser transportada hacia el cerebro
(Pardridge y Mietus, 1980). El primer paso en el transporte de la
hormonahacia el liquido cefaiorraquideo seria su acumulación En
el plexo coroideo (Trentíni y col., 1979; Smulders y Uright, 1980).
La degradación metabólica de la melatonina ha sido estudiada
luego de la administración sistémica o intracerebral de la hormo
na radiactiva. La inyeoción intravenosa resultó en una rápida de
saparición de la melatonina radiactiva del torrente sanguíneo

16
(Hopín y col., 1961; Murtmany col., 196h; Roppert y col., 1979;
Cardinalí, 1981).
El hígado es el principal sitio de inactivación. El metoxig
dol se hídroxlla a un compuestocarente de actividad biológica, la
6-hidroximelatonina, el que se conjuga luego con ácido glucurónico
o sulfúrico, forma excretada por orina y heces (Cardinali, l97h;
Happers y Pévet, 1979)(Fig. 5). Alrededor de un 70-80% de la melg
tonina urinaria corresponde a sulfatos y el 5%a glucurónido. La
melatonína remanente permanece inalterada (< 1%), como ácido S-mg
toxindolacético (0.5%) o comoun metabolito no indólico (15%),1
dentificado comoN-acetil-S-metoxiquinurenamina (Hirata y col.,
l97h). El ácido 5-metoxindnlacético es producido periféricamente
en el hígado por desacetílación de la melatonina (Rogeuski y col.,
1979).
Fue en la rata donde se realizaron por primera vez los esta
dios cinétícos del metabolismo cerebral de la melatonina (Carding
li y col.. 19739.0espués de una única inyección intracisternal,
la melatonina desaparece rápidamente del cerebro, detectándose ag
lo un 5%de la dosis administrada 5 min después. Este dato sugie
re un rápido intercambio de la hormonadel líquido cefalorraqui
deo con la del compartimiento vascular.
La concentración de melatonina en el hipotálemo es h a 5 VE
ces mayor que en el resto del cerebro, tanto después de la inyec
ción intraventricular comointracisternal del compuestoradiacti
vo (Anton-Tay y Murtman, 1969; Cardinali y col., 1973 ¿L El metano
118m0de la melatonina en el cerebro es activo: 1 h después de la
inyección intracisternal, sólo un 30-h0%del compuesto original

17
está presente en el cerebro (Sardinali y cnl., 1973Q.Estudios 113
vados a cabo tanto in vitro comoin vivo indican que la melatonina
cerebral es oxidada a N-aceti1-5-metoxiquínurenamina, Formándose
comometabolito intermedio N-Formil-5-metoxiquínurenamina (Hirata
y col., 197h)(Fig. 5).
1.1.3 Control neural de la actividad gineal
Las vias neurales a través de las cuales el cerebro controla
la Función pineal se estudiaron en relación a los mecanismosque
median los efectos de la luz ambiental sobre 1a actividad de la
glándula (Cardinali y Murtman, 1975; Minneman y Murtman, 1976;
Murtmany Hoskowitz, 1977; Reiter, 1980; Cardinali, 1981). Si bien
en algunos reptiles y aves la glándula pineal (y probablemente o
tras zonas del SNC)responde directamente a 1a información lumí
nica que penetra a través de 1a calota craneana, en los mamíferos
la Fotorrecepción ha quedado restringida exclusivamente a los ojos.
Ya que la actividad pineal en los mamíferos está aún bajo dependen
cía del Fotoperíodo ambiental, Fue necesario que se desarrollara
una conexión humoral o neural entre la retina y la glinduln, rea
ponsable de la transmisión de la información ffitica. De las dos pg
sibilidades teóricas (humoral o neural), en los mamíferos se desa
rrollo la segunda. Ya hemosanalizado en la sección 1.1.1 algunas
de las características morfológicas de la neurona final de dicha
via. Comprendeaxonas simpáticos periféricos originados en el gas
glio cervical superior que arriban a la pineal por la pared de susarterias irrigantes y forman los nervios conarioa del polo caudal
de la misma (Happers, 1960, 1965) (Fig. 3). Comohemos visto, en

18
la pineal las Fibras terminan primariamente en los espacios peri
vasculares (Martemberg, 1968; Matsuahima y Reiter, 1977) y en las
cercanias de los elementos del parénquima (Matsushima y Reiter,
1978) (Fig. 2).
A través de estes terminales neurales, la luz inhibe la
producción de melatonina. Ls exposición de los animales a la luz
disminuye la actividad neural en los terminales, mientras que lo
opuesto se observa durante la fase de oscuridad (Klein. 1978).
Cualquier maniobra experimental que cause una activación importan
te del sistema nervioso simpático puede eliminar el efecto inhioi
torio de la luz y asi estimular la sintesis de melatonina. siempre
que tal estimulo se aplique en el momentoapropiado del dis. El
ciclo diario de luz y oscuridad, al cual 1a mayoria de los anima
les están expuestos, genera un ritmo paralelo en las funciones me
tabólicas pineales relacionadas con la sintesis de melatonina. E2
tas comprenden: el "turnuver" de norepinefrina (NE) en las termi
nales nerviosss (Brownstein y Axelrod, 197h), receptores q-(Va
cas, 1980) y /b -adrenérgicos (Romeroy col., 1975), actividad eléctrica pineal (Semmy Vollrath, 1979), niveles de adenilciclaaa
y AMPc(Dleshansky y Neff, 1978; Zstz, 1976). fosfodisstersso (Ols
shansky y NeFF, 1976), triptofsno hidroxilasa (Sitaram y Lees.
1978), HIÜMT(Murtman y Moskowitz, 1977), SNAT(Klein, 1978). serg
tonina (Quay, l97h) y contenido de melatonina (Reiter, 1980). asi
también como la concentración de la hormona pineal en plasma y
liquido cefalorraquideo (Üzaki y Lynch, 1976; Arendt y col., 1977;
Uurtman y Moskouitz. 1977; Arendt._197&¿_Rollaq_y_gnl¿4_jfllfl;
Vaughan v 001o. 1978 l: Uilson. 1978; Hallan y coi.. 1979; Rsppsrt

19
y col, 1979; Uithyschunnornkul y “nigga. 1960).
Los globos oculares deben estar presentes para que la pineal
responda a la iluminación ambiental, lo que indica que el fotorre
ceptor reside en el ojo. Sin embargo, los elementos fotorreceptores
clásicos de la retina parecen tener poca importancia; por ejemplo,
la pineal de rata en la cual los bastones han sido destruidos por
exposicion continua a 1a luz, es capaz de responder normalmente a
cambios en el fotoperiodo (Reiter y Klein, 1971). Si las retinas
son lesionados quirúrgicamente, pero el resto del ojo se mantiene
intacto, la función pineal no se inhibe por la luz (Reiter y Jong
son, 197h).
La via neural que vincula la retina con la glándula pineal
está resumida en 1a Fig. 6. Los impulsos neureles que proceden de
1a retina con destino a la glándula pineal llegan al cerebro a
través de axones de las células gonglionares retinianas que term;
nan en el núcleo supraquiasmático del hipotálamo (haz retino-hiog
talámico)(n0 1 de la Fig. 6). Esta proyección directa desde la r2
tina hasta el núcleo suprauuiasmático parece ser un componentegg
neral del sistema de lu visión de los mamíferos (Moore, 1973; Rel
ter, 1980). Las vías neurales eferentes del núcleo Supraquiasmá
tico, que se proyectan a través del hipotálamo,han sido caracte
rizadas topográficamente por estudios de lesiones estereutáxicas.
Tales lesiones indican que los haces de Fibras descendentes o as
cendentes en el área lateral del nipotálomo son críticos para la
funcion normal de lo pineal (Moore y cnl., 1968; Reiter, 1972;
Reiter, 1980). Lo información neural es transferida desde el nú
cleo supraquiasmático hacia el segmento toráclco de la médula eg

20
tracto retina -h¡potalámico
luzI
o'o IJ l
a
CD cinc hipotolámicn. núcleo --\ lateralsupraquiasmático ' .'
,-‘ ®I \1 ’ vi /.(céldas delasta‘“1' -1 internadohtorol)
glándula ganglioPmeal cervicgl
supenpr
Fig. 6: Via neural para el control de la función pineal.
Los números designan las distintas etapas de la via nou
ral descripto: en al texto (pag. 19).

21
pinal por una vía que se proyecta caudolmente a las áreas periven
tricular y tuberal del hipotálamo (nD 2 de la Fig. 6)(Szentagothai
y col., 1968; Suanson y Couan, 1975). Los oxnnes del área ventrotg
beral se proyectan al hipotálamo lateral (nO 3 de la Fig. 6) y fi
nalmente neuronas de esta localización proyectan sus axones a tra
vés del tallo cerebral directamente sobre células del asta intermg
diolateral de la médulaespinal torácica (neuronas simpáticas pre
ganglionares)(n0 h de la Fig. 6)(Ssper y col.,1976). Esta secuencia
de neuronas tiene comoúltima estación neuronal a1 ganglio cervical
superior (nO 5 de la Fig. 6)(Moore, 1978). Trabajos recientes su
gieren que la porción central de esta via neural es susceptible a
la manipulación Farmacológica ya que la infusión de drogas online;gicas en la zona del núcleo supraquiasmático o la inyección de a
gentes d -adrenérgicos en los ventrículos cerebrales modifican la
sintesis de melatonina en la pineal de rata (Alphs y col., 1980;
Zatz, 1980).
La glándula pineal de rata constituye un excelente modelo pg
ra estudiar el control neural de las respuestas celulares. La esti
mulación P-adrenérgica producida por la liberación de NEde los
terminales neurales pineales.desencadena una secuencia de fenómenos
que conducen al incremento de la sintesis de hormonas pineales.
Tal cascada metabólica comprende: cambios en el potencial de mem
brana (Sakai y Marks, 1972), incremento en actividad de adenilci
class (Davis, 1978; Weiss y Crayton, 197D) y niveles de N4Pc (De
guchi, 1973), Fosforilacibn de histcnas (Zatz, 1978). aumento en
la sintesis de ARN(Cardinali y col., 1973b;Zatz, 1978) y protei
nas (wurtman y col., 1969) y activación de las enzimas que sinteti

22
zan melatonina, SNAT(Klein y Meller, 197D) y HIUMT(Cardinali
y col., 1973 oEstos cambios bioquímicos explican el aumento de
melatonina en liquido cefalorraguideo y plasma que sigue a la e!
posición de los animales a la oscuridad (Hedlund y col., 1977).
En el presente trabajo de tesis se estudia la participación de
las prostaglandinas (PGs) en dicha secuencia metabólica. Comoag
tecedente de dichos estudios debe citarse la comunicaciónpreli
minar de Szabo y Friedhoff (1976) en la que la inyección de indo
metacina, comúnmenteutilizada para inhibir la sintesis de PGs,
bloques la estimulación de SNATdurante la oscuridad.
La inducción de SNATque ocurre tempranamente durante la fig
se de oscuridad se puede reproducir en cultivos de pineal por el
agregado de dibutiril-AMPC o agonistas /5-adrenérgicos. Durantela transición de la oscuridad a la luz, el "turnover" de NEdis
minuye y hay una caida marcada de la actividad de SNAT. Aún no se
conocen los mecanismos involucrados en tal disminución. Las evi
dencias disponibles demuestran que la reducción de la actividad
enzimática después de una exposición fugaz a la luz o luego de la
administración del antagonista fl-adrenérgico propranolol es muyrápida (5 min). Por el contrario, la vida media de la SNAT,deter
minada usando un inhibidor de la sintesis proteica, comola ciclg
heximida, es larga (alrededor de 60 min). Una posible explicación de
esta discrepancia fue dada en cultivos celulares de pinealocitos
donde se comprobó que la rápida reducción que sigue a la exposi
ción a la luz depende de la disminución abrupta de los niveles de
AMPC(Klein, 1978). Por consiguiente, el AMPCregula los niveles
de SNATno sólo actuando a nivel de la transcripción y transduc

23
ción sino en el mantenimiento de la actividad enzimática durante
el período de oscuridad.
Numerosasevidencias experimentales indican que la glándula
pineal puede ser super o subsensible a la estimulación por NE
(Minneman y Murtman, 1976; Hlein, 1978; Dleshansky y Neff, 1978;
Zatz, 1978). Tal respuesta está determinada por el grado y dura
ción de 1a exposición previa al transmisor. La incubación de la
glándula pineal con agonistas /5-adrenérgicos resulta en una rá
pida pérdida de la capacidad pineal para responder a la estimulg
ción adrenérgica subsecuente con aumento de la actividad SNAT.
Cuanto mayor es la interacción del agonista con el receptor, ya
sea por exposición prolongada in vivo o in vitro, menos sensible
se torna el sistema a una posterior estimulación. Los mecanismos
responsables del control de estos cambios en sensibilidad involg
cran tanto al sistema de la adenilciclasa comoa los pasos bioqui
micos posteriores a la sintesis de AMPC,de tal manera que la e
fectividad del AMPcgenerado es relativamente mayor en glándulas
supersensibles que subsensibles.(Minneman y Murtman, 1976; Zatz,
1977; Klein, 1978; Uleshansky y NeFF, 1975; .Zatz, 1978).
I.1.h Efectos biológicos de la melatonina
La melatonina secretada por la glándula pineal participa en
numerosos mecanismosneurales y neuroendócrinos, entre ellos el
control de las funciones gonadal, adrenal y tiroides, el sueño y
varios ritmos biológicos. La pineal constituye un componenteeseg
cial del sistema neuroendócrino, regulador de ritmos circanuales
en la actividad endócrina, especialmente el Ce la reproducción.
Si hamsters, de ambos sexos, son privados de luz ya sea por ex

2h
tirpación ocular o exponiéndolos a fotoperiodos de menos de 12.5 h
de luz por dia, éstos presentan regresión gonadal en 8-10 semanas
(Hoffman y Reiter, 1965; Berndtson y Desjardina, 197h; Temarkin y
col., 1976 a, 1976 b; Reiter, 1980). En hamsters pinealectomizados
la regresión gonadal asociada al fotoperiodo no se observa, indie
cando que los efectos de 1a oscuridad sobre el aparato neuroendó
crino son mediados por la pineal (Hoffman y Reiser, 1965; Blask y
col., 1979; Turek, 1979; Reiter, 1980).
Ratas pinealectomizadas sometidas a lZ-lh h de luz por dia
no presentan, salvo excepción, cambios endócrinos permanentes. E2
tas observaciones sugieren que, en ia rata. 1a ausencia de la
glándula pineal es compensadapor el desarrollo de un nuevo estado
de equilibrio entre las señales que modulan1a función hipotalámi
ca. Según Reiter (1977, 1978, 1979, 1980) la rata, un animal de
hábitos nocturnos en su hábitat natural.estó, en las condiciones
de luz del laboratorio (12-lh h de luz) sujeta a fotoperiodos que
determinan una depresión artificial de 1a actividad pineal. En
ciertas condiciones experimentales, comola desnutrición, anosmia
o el tratamiento neonatal con esteroides, el hipotálamo de la ra
ta es más sensible a la melatonina endógena o a la administrada en
forma exógena (Rsiter, 1977, 1978, 1979, 1980; Cardinali, 1981).
En las Tablas l y 2 se resumen los efectos de la melatonina
sobre las distintas funciones fisiológicas en mamíferos(Cardinali,
1981). En el hamster, hurón y, probablemente, la rata, la melatoni
na es capaz de contrarrestar los efectos de la pinealectomia si es
inyectada en el momentoapropiado del dia; por ejemplo el hamster
es insensible a la hormonaexógena durante la primera etapa del

25
periodo de luz (Tamarkin y col., 1976 a, 1976 b; Goldman y col.,
1979). Murtmany col. (1963) llevaron a cabo los primeros estudios
sobre la participación de la melatonína en la fisiología gonadal
de los mamíferos, demostrando que en ratas hembra- inyectadas con
melatonína existe hipotrofia e hipofuncionalidad ovárica. A partir
de este hallazgo, numerososestudios en diversas situaciones neurg
endócrinas revelaron que el tratamiento con melatonina atrasa la
apertura vaginal.e inhibe (si es inyectada durante el proestro) la
liberación de LHy la ovulación. Este y otros efectos de la mela
tonina sobre la actividad neuroendócrina están enumerados en la
Tabla 1 (pág. 29).
Aúnno han sido aclarados definitivamente el sitio o los
mecanismos celulares que median la acción neuroendócrins de la me
latonina. Se acepta que el cerebro constituye el principal sitio
de acción, sin excluirse sitios periféricos comolas glándulas eg
dócrinas y tejidos efectores de la hormona. Una serie de funciones
metabólicas centrales son modificadas por el tratamiento con mela
tonina (Tablas 1 y 2). Estas son: la sintesis de proteinas (Cardi
nali y col., 197ha) contenido de serotonina y ácido gammaaminobu
tirico (GABA)(Anton-Tay, 1971), la liberación y recaptacifin si
naptosomal de neurotransmisores, en especial de serotonina (Car
dinali, 1975; Cardinali y col., 1975'aila liberación y contenido
hipotalámico de LHRH(Hao y Ueisz, 1977; Leonardelli y col., 1978).
Tales observaciones bioquímicas se han acompañadode estudios ul
traestructurales que indican alteración por el tratamiento con me
latonina en la eminencia media y glándula pínual (Cardinali y
Freire, 1975; Freire y Cardinali, 1975; El-Domeiri y Das Grupta,

26
1976; Benson y Krasovich, 1977).
Si se inyecta melatonina-JH en la circulación general, la
hormona penetra en todos los tejidos, inclusive en el SNC(Murtman
y col., 19Gh). Cuando el compuesto marcado se administra en el li
quido cefalorraquideo por inyección intraventricular o intrscister
nal 1a melatonina se concentra en el hipotálamo y mesencéfalo (Ea;
dinali y col., 19735}Por cromatografía gaseosa asociado a espec
troscopia de masa (Hoslou y Green, 1973) e inmunohistoquimica (Bu
benik y col., 1976) se ha identificado melatonina en el hipotálamo
de la rata.
La descripción de mecanismosde captación saturables para
melatonina en el cerebro de la rata (Cardinsli y col., 1979o)'p1a_r¿
teo la posible existencia de receptores neurales para 1a hormona.
Recientemente se han descripto tales sitios de unión especificos
para melatonina en preparaciones de membranade cerebro, particu
larmente en el hipotálamo mediobasal (HMB)(Csrdinali y col.,
1978; Cardinali y col., 19795}El "binding" de melatonina resultó
dependiente del entorno iónico, ya que el agregado de H+ y Na+ ig
hibió dicha asociacion mientras que el Ca++la incremento. Los en
sayos con compuestos análogos de la melatonina indicaron que el
grupo 5-metoxi es necesario para que la unión sea significativa,
excepto en el caso de estar presente una amina primaria (Cardina
li y col., 1979aJ.La unión de melatonina a diversas zonas del ce
rebro es compatible con los efectos farmacológicos del indol pi
neal sobre el SNC, resumidos en las Tablas 1 y 2. Una de estas r5
giones es la glándula pineal donde también se detectan receptores
especificos para la hormona(Vacas y Csrdinali, 1980).

27
Si bien para la descripción de un receptor las propiedades
de los sitios de unión (especificidad, alto afinidad, baja capaci
dad, etc.) son un requisito sine gus non, es importante además la
caracterización ds una actividad biológica asociada a los mismos
en la mismapreparación celular. En el caso de la melatonina. ls
consideración de los sitios de unión comoreceptores se ha basado
en estudios correlativos con 1a respuesta nsuroendócrina a la ho;
mona. Vacas y Cardinali (1979t1demostraron que el cerebro de hamg
ter y rata la concentración de los sitios de unión varia durante
el dia en forma concomitante con la sensibilidad nsuroendócrina a
la hormona. De acuerdo con los datos neuroendbcrinos (Ïabla l) que
indican que 1a melatonina es activa en horas ds la tarde, el núme
ro total de sitios de unión de melatonina en cerebros de rata y
hamster fue, a las 2000 h, 3k-56% mayor que s las D700 h, sin que
la afinidad se modificase. Ya que se estimaron los sitios totales,
ocupados y desocupados, estos resultados indican que hay una mayor
disponibilidad de sitios receptores para la melatonina durante el
periodo de sensibilidad neuroendócrina a la hormona.
Un importante Factor regulatorio de la concentración de los
sitios es probablemente 1a secreción endógena de melatonina. La
exposición a altas concentraciones de melatonína durante la fase
de oscuridad determina ls pérdida de sensibilidad del sistema neu
roendbcrino a la hormona endógena por desensibilización ("down
regulation") de los sitios receptores, mientras que durante el pg
riodo de luz el número de sitios aumenta (ya que los niveles de
la hormonason bajos) hasta restaurarse la sensibilidad hacia el
Final de este periodo (Vacas y Cardinali, 1979hífleitor, 1980.).

26
Eataa diferenciaa en a1 núnaro de racaptoraa deaaparacan anta
cualquier alteracidh en el ritno diario da aecracidh de aalatoni
na, aaf comolo nacen laa raapueataa nauroandácrinaa (Vacaa y Car
dinali,1979 b).
En el preaante trabajo da taaia aa aatudia 1a participación
de laa PGa en el mecaniano da acción de 1a naiatonina. Tal hipotg
aie fue elaborada a partir da la obaarvacidn qua 1a lalatonina (N
aceti1-5-oetoxitriptanina) y la indonatacina (N-pd[clorobanzoiiJ-2-neti1-5-netoxindol-3-acetico) praaantan aiailitud aatructurai.
tienen propiedadea antig‘nicaa an común(Lavina y Ricabarg. 1975)
y aueatran daatacabia coincidanoia an aua afactoa nauroandócrinoa
(Tabla I, McCanny col.. 1976). Eapacificalanta aa trató da aluci
dar ai la aelatonina afecta 1a aintaaia de PGaan al HHHan laa
concentracionea afactivaa para aaturar aua aitioa racaptoraa caraorales.

29
Tabla l
Cambiosendócrinos producidos Eor la administración de melato
1.
nina en mamíferos.
Función Gonadal
Hamster
Inyectads en la última parte del periodo de luz o de oscuridad produce regresión gonadal y disminución de ls liberación de LH, PRL y FSH en machos (Tsmsrkin y col., 1976 b;Goldmany col., 1979). En hembras altera el ciclo e inhibelas descargas de LHy FSHen la tarde del proestro (Iamarkin y col., 1976 e; Andre y Parrish, 1979).
Implantads s.c. previene la regresión gonsdsl debida a exposición a fotoperiodos cortos (Hoffman, i979; Reiter,1980).
Pubertad retrasada en mechas (Rissman, 1980).
Rata
Apertura vaginal retardads (Collu y col., 1971). Inhibiciónde ls ovulación inducida por suero de yegua preñsds (PMS)en ratas inmadurss (Longnecker y Gallo, 1971).
Disminución del peso testicular (Mas y col., 1979), de laespermatogónesis (Honig y Regs, 1978) y de la testosteronaplasmática (Hinson y Peet, 1971). Inhibición de la esterqideogenesis testicular in vitro (Ellis, 1972) V de los efestos androgénicos de la testosterona (Alonso y col., 1978).
Disminución del peso oVárico y uterino (murtman y col.,1963).
Inhibe 1a respuesta hipofisaria de LH y FSHa LHRHin vitro en ratas prepuberales (Martin y col., 1977). No afeg

30
tu la respuesta a LHRHen ratos adultos (Hoguílevsky ycol., 1976; Martin y nnl., 1977).
Inyectada al Final de la Fase de luz aumenta la regresióngonadal en ratas desnutridas o anósmicas (Ülask y Nodelman,1980; Blask y col., 1980; Reiter y col., 1980).
Inyectada simultáneamente con testosterona inhibe la andrggenización de ratas hembrarecién nacidas (Reiter y col.,1975).Una inyección aguda o tratamientos cortos aumentan la secreción de PRL (Kamberi, 1973; Lu y Meites, 1973; Kramer yBen-ÓavidÍ—Ï975}Vaughan y’Eb1., 1978 b; Cardinali V cbiïï“’1979 b). La administración diaria en forma crónica a ratasano’amicaa inhibe la liberación de PRL (Blask y Nudelman,1980; Reiter y col., 1980).
Inhibe la liberación de LHy la ovulación en ratas con ciclos normales (Ying y Greep, 1973). Aumenta el contenidohipotalámico de LHRH(Leonardelli y col., 1978). Efectodependiente de la dosis sobre la liberación de LHen ratashembra castradas inyectadas con esteroides (Moguilevsky ycol., 1979). Disminuye la FSH plasmática en machos (Mas ycol., 1979).
Revierte los efectos de la pinealectomia sobre la liberación de LHy PRLdurante la lactancia tardía. Acorta elperiodo de gestación (Nir y Hirschmann, 1979).
Induce la liberación de LHen ratas con ciclos anovulatorios por exposición permanente a la luz (Mess y col., 1979).
Hurón
Regresión gonadal (Thorpe y Herbert, 1976).
Ütros mamíferos
Bloqueo del incremento postcastración de LHplasmática en

31
la oveja (Roche y col., 1970); no modifica los niveles dePRL(Driver y col., 1979).
Inhibición de 1a liberacion de PRLdespués oe 1a inyecciónintrayugular en bovinos (Padmanabhany col., 1979).
Nomodifica la liberación de PRLpor células hipofisariasbovinas in vitro (Padmanaohany col., 1979).
Inhibe la respuesta de LH a LHRHen perros (Yamashita ycol., 1978).
Inhibe la estimulacion de la sintesis de esteroides en Foliculos ováricos por hCG(coneja) (Young, 1979).
Disminuye (Nordlund y Lerner, 1977) o no modifica (Fideleffy col., 1976; Meinberg y col., 1980) la liberación de LHenel hombre.
Retrasa el comienzo o elimina 1a secreción luteal de progegterona en monos (Fiske y Mc Donald, 1975).
Involución gonadal en meriones (Vaughan y col., 1976), raton (Lynch y Epstein, 1976), foca (Rust y Meyer, 1969) yliebre (Hüderling y col., 1979).
Otras Funciones hipotélamo-hipofisarias
Rata
Disminuye1a sintesis de proteinas hipotalámicas (Cardinali y col., 197háLRevierte el incremento inducido por lapinealectomia del contenido hipotalámico de tubulina. Produce cambios ultraestructurales en eminencia media compatibles con neurosecreción y transporte axonal (Cardinaliy Freire, 1975).
Aumenta el contenido hipotalámico de serotonina y GABA(Anton-Toy y col., 1968; Anton-Tey, 197h). Restaura el ritmo oe 1a serotonina hipotalámica en hurones pinealectomiqg

32
dos (Yates y Herbert, 1979). Deprime la actividad de laMADhipotalámica (Urry y Ellis, 1975). Deprime la captación de serotonina, NE, dopamina y glutamato por sinaptosgmas hipotalámicos (Cardinali, 1975; Cardinali y col., 1975a). Aumenta1a liberación de hormonas hipofísiotropas (Hollander y col., 1977; Hao y Ueisz, 1977) o no la afecta(Brammer, 1979).
Revierte el efecto estimulatorio del 5-hioroxitriptofanosobre la liberación de hormona de crecimiento (Smythe y
Lazarus, 1973).
En el hombre, la premedicación con melatonina reduce 1arespuesta de 1a hormonade crecimiento a L-triptofano(Houlo y Lammintausta, 1979) o a la hipoglucemia insulinica(Smythe y Lazarus, 197h).Contrarresta los cambios'inducioos por 1a pinealectomia enla captación de eatradiol (Cardinali y col., 197bb;PedrozaGarcia y col., 197h). Modifica (Frehn y col., 197h) o noafecta (Mas y col., 1979) la Soi-reductasa hipotalámica.Aumenta1a liberación de vasopresina por la neurohipófisis(Lemay y col., 1979).
Función Tiroidea
Hamster
Deprime los niveles de tiroxina plasmática y el indice detíroxina libre. Nose modifica la triyodotironina (Vriendy col., 1979).
Rata
La inyeccion intraventricular de melatonina deprime la liberación de TSH(Relkin, 1978). En hipófisis in vitro noafecta la liberación de TSH(Brammery col., 1979). Dismi
nuye el contenido de AMPctiroideo y la liberación de tiroxina (Nir y col., 1978).

33
Función Adrenal
Rata
Disminuye el peso adrenal (Reiter y col., 1975). Disminuyela corticosterona plasmática (Motta y col., 1971). Inhibela producción de corticosterona in vitro (Dgle y Hitay,1978); estimula la via de la 5 A-Teouctasa (Dgle y Kitay,1978). Contrarresta la hipertensión en ratas pinealectomizadas (Holmes y Sudgen, 1975).
Páncreas
Rata
Bloques la liberación de insulina in vitro (Bailey y col.,197h). En el mono aumenta la glucemia (Burns, 1973).
Pineal
Contrarresta los cambios inducidos por la luz de los lípidos de pineal (Ebels y Prop, 1965). Modifica los ritmosdiarios de serotonina pineal (Fiske y Huppert, 1968). Modificacion ultraeetructural de los pinealocitas (rata: Freire
y Csrdinali, 1975; ratón: Benson y Hrasovich, 1977; hamster:El-Domeíri y Das Gupta, 1976) Aumenta el contenido pinealde tubulina (Freire y Cardinali, 1975).
Aumentala liberación de arginina vasotocina pineal (Pavely Godstein, 1979). Inhibe la captación oe tiroxina porslioes oe pineales bovinss in vitro (Csoy y Dillman, 1971).

__
3k
Tabla 2
Otros efectos de la melatonína en mamíferos
Sedación, inducción del sueño; cambios electroencefolográfiicos (rate, gato, mono, hombre) (Anton-Tay, 197h; Sampson,1975; Brailousky, 1976; Hüderlíng y col., 1979). Deprime laactividad motriz en ratas ciegas y anfismicas (Sackman y Reiter, 1977). Abolición de las crisis epileptiformes inducidaspor estimulación sensorial en el gato (Farriello y Bubenik,1976). La aplicación del anticuerpo anti-melatonina en lasuperficie cerebral desencadenaconvulsiones (rata) (Farrigllo y col., 1977).
Aumentode 1a incidencia y duración de la hibernación en a;dillas (Palmer y Riedsel, 1976). Respuesta homeostática aumentada ante cambios en 1a temperatura corporal (Hagelsteiny Folk, 1979). Cambios en tejido adiposo pardo en ratón(Lynch y Epstein, 1976) y rata (Hagelstein y Folk, 1979).Aumenta la hipotermia inducida por D-anfetamina en rata (VEhuda y Frommer, 1977). Depríme la sintesis de ácidos grasosy oxidación de la glucosa en grasa perirrenal de conejo(Murthy y Modesto, 1975).
Depríme la actividad del músculo liso (duodeno de rata,Quaste y Rahamimoff, 1965).
Inhibe la melanogénesis en Foliculo piloso de hamster ¿Evitro (Logan y weatherhead, 1980).
Deprime el contenido del Factor de crecimiento nervioso englándula submaxilar de ratón (Pérez-Polo y col., 1978).
Prolonga el tiempo de sobrevida en ratas hembra pinealectgmizadas portadoras de sarcoma de Yoshida (Lapin, 1979). Fogterga la aparición del sarcoma inducido por metilcolantrenoen ratón; deprime el crecimiento de tumores mamarios inducidos por dimetil-bencil-antraceno en rata (Aubert y col.,

IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII‘IIIIIIIIIIIIIIII¡iiIIIIII
35
1980). Deprime el crecimiento tumoral en hamsters inyectaUos con células neoplásioas de melanoma (El-Domeiri y DasGrupta, 1976). Actividad antimitótica in vitro sólo en a1gunas pruebas para drogas ontimitóticas (Banerjee y col.,1972; Banerjee y Margulie, 1973; Fitzgerald y Ueal, 1976;PoFFenbarger y Fuller, 1976). No compite con 1a colchioinapor los sitios de unión en la tubulina (Cardinali y Freire,1975; Poffenbarger y Fuller, 1976). Inhibición de procesosdependientes de proteínas contráctiles comoel transporteaxonal (Cardinali y Freire, 1975; Prevedello y col.. 1979)y el movimiento de gránulos pigmentarios (Malauísta, 1973;Pang y Yeu, 1979).

36
I.2 - PARTICIPACION DE LAS PRÜSTQQLANDINAS EN LDS MECANISMOS NEU
RALES ADRENERGICÜS CENTRALES Y PERIFERICÜS.
1.2.1 Biosintesis y metabolismode las prostaglandinas
Las prostaglandinas (PGs) constituyen un grupo de compuestos
derivados de ácidos grasos de amplia distribución en gran número
de tejidos. Fueron descriptas a principios de la década del 3Dy
son hoy una de las familias de compuestos biológicamente activos
más estudiadas. La actividad contróctil del útero humanoinducida
por semen de la misma especie descripta por Hurzrob y Lieb en 1930
fue la primera demostración de un efecto biológico asociado a las
PGs. Años más tarde, Von Euler (193h, 1935) y Guldblatt (1933, 1935)
describieron con más detalle tal acción farmacológica del plasma
seminal humanosobre el músculo liso. VonEuler también describió
los efectos producidos por el Fluído seminal de mono, oveja y cabra
y por extractos de vesículas seminales ovinas, adjudicóndolos a un
material liposoluble al que denominó"prostaglandina".
Recién en 1957 Fue posible aislar en forma cristalina dos
PGs, actualmente conocidas como PGE1 y PGF1°< (Bergstrüm y Sjüvall,
1957). Por ultramicroanálisis y espectroscopia de masa se determi
naron sus Fórmulas empíricas y la estructura completa se elucidó
por espectroscopia de masa asociada con cromatografía gaseosa
(Bergstrïm y col., 1963).
Las PGsson sintetizadas a partir de ácidos grasos insatu
rados (Sprecher, 1977). De acuerdo con el ácido graso precursor

37
se biosintetizan las PGsmono,bi y trienoicas, según los distig
tos grados de insaturación (Lands, 1979):
- la serie l, con una doble ligadura en la cadena lateral, deriva
del ácido graso esencial dihomo-gamma-linolénico(ácido B,ll,lh
eicosatrienoico );
la serie 2, con dos dobles ligaduras en la cadena lateral, deri
va del ácido graso esencial araquidónico (ácido 5,8,ll,lh-eico
satetraenoico);
- la serie 3, con tres dobles ligaduras, se Formaa partir del é
cido graso no esencial 5.8.11,lh,l7-eicosapentenoico.
El ácido araquidónico, precursor de las PGs dienoicaa, es
el más ampliamente distribuido en los fosfolipidos de membranay
en el SNCy proviene de 1a dieta y de la desaturación y elongación
de 1a cadena lateral del ácido linoleico (Fig. 7).
La molécula de PGs está formada por un ciclopentano y dos qa
denas laterales (Fig. 7). De acuerdo con los sustituyentes del ani
llo, las PGs se clasifican en seis grupos: A, B, C, D, E y F, de
las cuales las más estudiadas en sus efectos biológicos son la E
y F.
La biosintesis de PGsha sido estudiada utilizando precursg
res radiactivos, identificándose los metabolitos cromatográficameg
te. La producción endógena ha sido evaluada por ensayo biológico,
cromatografía en fase gaseosa, espectrometria de masa y radioinmu
noanálisis. Mediante estos métodos se determinó que la mayoria de
los tejidos animales producen PGs, con una única excepción consti
tuida por los glóbulos rojos maduros (Harris y Ramuell, 1979).
Las células de mamíferos liberan PGs en respuesta a estimu
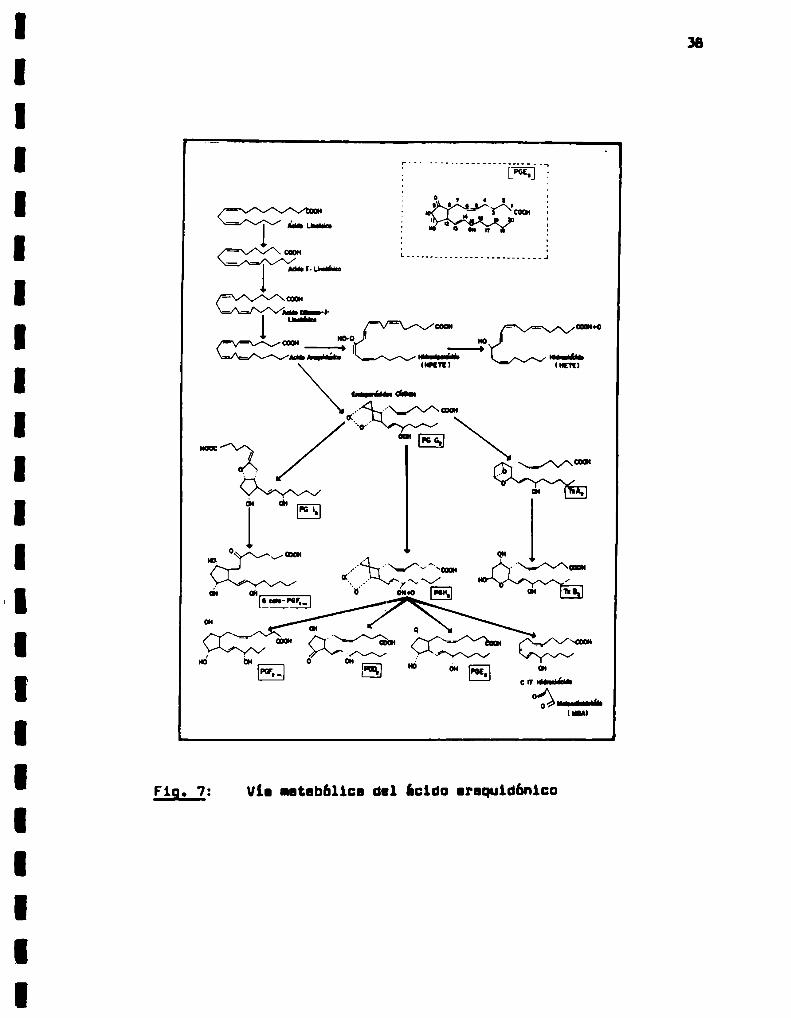
L. . . . . . . . . . . . . . . . . . . . . _. . . .
cuan-¡6hfl...“
F19. 7: Vio ntebbltca dal ¡cido araquidónico

39
los Fisiológicos, farmacológicos y patológicos y, dado que las PGs
no son almacenadas en las células, su biosintesis precede en forma
inmediata a su liberación. Por lo tanto, la determinación de pro
ductos en el medio de incubación es un indicador sensible de su
biosintesis.
La Formano esterificada de los ácidos grasos poli-insaturs
dos constituya el sustrato para la sintesis de PGs (Bergstrbm y
col., 196h; Moncaday Vane, 1978). Los ácidos grasos se hallan en
la membranacelular comoésteres del colesterol, mono. di y tri
glicéridos y lípidos fosfatldicos (Ramwelly col., 1977) y deben
liberarse por acción de la fosfolipass A2. Estalipsss hidrolizsza selectivamente el enlace del carbono 2 de los fosfoglicéridos.
Por ejemplo, en 1a corteza suprarrenal y glándula tiroides el soi
do sraquidbnico es liberado de fosfolipidos por un mecanismode
pEndiente de Ca++ estimulado por ACTH(Laychnck y col., 1977) y
tirotrofina (Haye y Jacquemin, 1977). Másadelante se discutirá la
liberación de PGs en respuesta a estimulación nerviosa en el SNC
y periférico.
Unavez liberado el ácido araquidbnico no esterificado es
metabolizado por dos tipos de enzimas: lipoxigenasa y complejo PG
sintetasa (Bergstrbm y col.. 196i; Dnrp y col.. 195“; Lands, 1979).
La lipoxigenasa cataliza la peroxidacibn del ácido araquidbnico en
carbono 5 dando origen a hidroperóxidos inestables (hidroxiperoxi
eicosatetraenoíco, HPETE),que se convierten en hidroxiácidos es
tables (hidroxieicosatetraenoico, HETE).Esta via metabólica para
lela a la formación de PGs tiene comoproductos a los recientemen
te descriptos leucotríenos (Borgeat y Samuelsson, 1979). Yamamoto

50
y col. (1977) demostraron que el sistema PG-sintetasa es de locali
zación microsomal y consta de dos fracciones: l) cicloxigenasa.
que cataliza la conversión de ácido araquidónico en el endoperóxi
do PGE2y éste último en PGH2(Fig. 7) v 2) endoperoxidasa, que
cataliza la conversión de PGH2en los-productos estables PGEZ.
PG PGDZ,un hidroxiácido de 17 carbonos (lZ-hidroxi-5,8,10F2u.'heptadecadienoico, HHT)y el ácido malondialdehido (MDA)(Fig. 7).
A pesar de ser PGG2el primer producto formado a partir del
ácido araquidónico y oxigeno molecular, es su derivado lS-hidrogi
lado (PGHZ)el producto acumulado en mayor proporción por acción
de la cicloxigenasa. Esta reducción es catalizada por una peroxi
dasa. El endoperóxido PGH2,sustrato de dos isomerasas (ll-cata yendoperóxidosintetasa), una reductaaa y dos sintetasas, da ori
gen a las distintas PGade la serie 2 (Fig. 7).
En 1971, Vanedescribió la acción inhibitoria sobre la sin
tesis de PGsde drogas antiinflamatorias no esteroides comoaspi
rina e índometacina. Actualmente, se considera que los efectos ag
tipiréticos, analgésicos y antiinflamatorios de estas sustancias
están mediados por la inhibición de la sintesis de PGs a nivel de
la cicloxigenasa. Ademásde los conocidos efectos terapéuticos,
las drogas tipo aspirina tienen en el hombrey otras especies e
fectos colaterales comoel daño renal e inhibición de la segunda
fase de agregación plaquetaria (Moncaday Vane, 1979). Estos efeg
tos han sido adjudicados a una inhibición de la cicloxigenasa. Q2
do que la indometacina previene la sintesislde todas las PGs Gong
cidas, esta droga ha sido utilizada ampliamentepara estudiar la
participación de las PGsen fenómenosfisiológicos y patológicos.

hi
Debe destacarse, sin embargo, que ls droga tiene otros efectos 52
bre distintas actividades enzimáticas tales comofosfolipasa, fos
fodiesterasa y quinasa de proteinas dependiente de AMPC(Flower,
197h; Hantor y Hampton, 1978), a los que es necesario considerar
en la interpretación de los resultados experimentales.
E1 endoperóxido ciclico PGH2es también sustrato de otra eg
zima distinta de la cicloxigenssa, que ha sido aislada de plaque
tas humanas y equinss (Moncaday col., 1976), la tromboxano-sintg
tasa (Fig. 7). Los productos de esta reacción son compuestos hate
rociclicos que incluyen s uno de los átomos de oxigeno, mientras
que el segundo forms la unión éter entre los carbonos 9 y ll. El
intermediario inestable es el TxA2y el compuestoestable el Tsz.Por acción de otra enzima, la prostsciclins-sintetasa, los
endoperóxidos se convierten en P612 o prostaciclina (Fig. 7). Enresumen, los endoperóxidos pueden seguir tres caminos: PGs esta
bles, tromboxanosy prostaciclinas. En ls4circulsción general las
PGs son de vida media muycorta y pierden su actividad biológica
como consecuencia de una oxidación seguida por reducción o por una
hidrólisis espontánea a nivel pulmonar. Tal degradación es mucho
más lenta en los tejidos, particularmente in vitro, representando
las PGs acumuladas en el medio de incubación un indicador confiable
de su sintesis y liberación.

‘2
1.2.2 La unión nooroofoctoro ooronirgioo
tanto en ootructoroo porifóricoo (oiotooo nervioso outónooo
oiopitico) cono on ol SNCext-ton vino nouronoloo. cofiniooo colo
ooronirglooo, quo olploon colo nourotronooiooroo o lo. cotocoloq¿
noo. Loo noorotronooiooroo cotocolo-inirgicoo oo localizacion coa
trol o portfirico Ion ointotizoooo según 1o vio tonto-notado on
lo F19. a. Lo ointoolo oo loo anti-oo ocurro on ol cuerpo neuro
nal y por un orooooo oo tronoporto donoolnooo oxoploolitico lol
onziooo migran o lo largo del ¡son boot. llegar o 1o. tor-¡noia
nooroloo (Koptn y Silborotoin. 1972; Ocho y Uorth. 1976),
Lo tiroolno oo ooptodo oo 1o circulacion por un loconiolo
oo transporta activo y oo conoontro on ol SIC. En ¡oo nouronoo
odronírgicoo oo hidroxllooo por 1o tirooino hidroullooo poro fo;
ont dihlorouifonllolonino (BOPA)(Lovitt y col.. 1965; Hoyor.
1980). Eoto onzioo oo oopoclfico poro 1o L-tirooino.oo hallo oo
jo control rogolotorio inhioitorio por ol producto final IE y q!
tolizo ol pooo lioitonto tonto on 1o ointooio control cono port
fñrico do NE(Lovitt y col.. 1965; Costo y Hoff. 1920; Mayor.
1960). El oogundo polo bio-intotico. 1o doocorhonilooibn oo L-QQ
PA o dopnlno eo cut-111o por lo doocorboxuooo oo olinoóoiooo o
rolbticoo con poco oopooificidod do ouotroto y coooz oo concor
boxllor toooiín o otroo olinoiclooo lovfigiroo (Mayor. 1960). Por
últioo, lo oopoolno oo conviorto on NEdontfo do lao voolcoloo
do loo torninoloo norooronirgicoo. por noción do lo om-ino- f5
hidroxilooo o oopnino-fi-Intoooo asociado o dicha voolcoloo.
Su oopooificidoo por ol oootroto oo boJo y puedo oxido: coolqoior
fonilotllooino o ou corroopondlonto fonilotonoloolno (Coylo y

53
¡BIOSINÏESIS DE NOREPINEFRINA
AROMAnco
T 7 HTU'IÏIQÉÉÜSA T Fl' L'AM'NOAC'DO
C-fi-COOH——o Ho ?_9_cw.|W' 0 ¡Fer loslnbdeHo H NH? (pterfdina reducida) Ho H "Hz (pifidoxcl
L-IIROSINA DOPA
+¡I ¡la DOPAMINA og! tll
N0©<|z-c|:—nnz fi-OXIDIASAl Ï:]@J|Z—C-NH2_’. 0 Cu ’ l
H0 H H (cgcorbato) H H
DOPAMINA L-NOREPINEFRINA
MEÏABOLISMO DE NOREPINEFRINA
W 9"Ho©g_c'_NH2 M. HO©C-COOHl
H0 H H H0 H
NOREPINEFRINA M111. mmonoxmmoeuco
lcom(ImIg com
CH30©€ -<ï—NHzH0 H H OH
MAO CH;O©<%_COOH\NORMEÏANEFRINA \ H0 H\
\ AC. 3-MEÏOXl-lo-HIDROXIMANIIIJCO\\\\
i l l
CH30© c- '—OHHo h H
3 -MEÏOXI-k-HIDROXIFENILGUCOL
‘ OHH
C
F19. B: Vias metabólica. de cinta-10 y "unan-no de E.

kh
Axelrod, 1972; Roeanberg y Lovenberg. 1960).
Una vez aintetizeda. le NEao almacena en lee veeiculaa ai
napticea donde forma un complejo aatable con el ATPy protainea
eepecificee. generico-ente lleeedae croeoqreninaa. Eetea veelculea
ae hallan praaentea an gran nünero en loa ter-inelee nervioaoe y
previenen le difueión de NEel eapecio einaptico y au eeteholiza
ción por la enziea degradative intraneuronel MAO(Fig. 9) (Anton
ïey y Hurt-an, 1971; Euler, 1972; Trendelanburg. 1979).
Heate el preaente no ha eioo totaleente eaclarecioo al Iaca
nieeo por el cual loa iepulaoa naurelee producen le liberacibn de
NE. A1deepolerizaree la lelbrane por le llegada de potencial de
accion propagado e lo largo del exao. eueente le per-aabilidao del
tereinel e loa ionea, entre elloe el Ce7*. El increeento del Ca’*
intracelular aa el pri-er paeo en la aecuencie que lleve e le li
beracion del neurotrenanieor (Kirpeker y Hieu. 1967; Uaetfall,
1977; Zinder y Phllerd, 1980).
Aparenteeente laa cetecolaninee eon liberadae de loa tar-ing
lea einpiticoe periféricoa y centrelee por un proceeo conocido co
eo exocitoeia (Zinder y Pollard, 1980). Eate reno-eno coneiate an
la fueibn de lee veeiculea con la eeeorane celular y le deacerge
de au contenido el eapecio extracelular.
La NEliberada tiene 5 caeinoe poaiblea a aeguir (Pio. 9):
l) interactuar con eua receptoree eepecificoe poeteinepticoa
(Ahlquiat, l9k6; Hinneean y col.. 1979).
2) aer receptade por el terminal eiepitico (receptecibn ne!
ronel o uptake I) y almacenada en lee veeiculaa (Ivereen.
1967; Írendalenburg, 1979).

“5
3) ser convertida en normetanefrina por acción de 1a CDMTsn
tes de pasar a la circulación (captación extraneuronal o
uptake II) (Iversen, 1967; Guldberg y Marsden, 1975).
h sufrir dessminación oxidativa por acción de la MAD,de loV
calización pre o postsináptica.
5 en el caso de los terminales periféricos o en zonas fuera deV
la barrera hematoencefálica pasar a la circulación como
NElibre. En el caso de zonas dentro de 1a barrera hemato
encefálics no hay difusión a 1a circulación.
En el cerebro de mamíferos se distinguen dos tipos de MAOcon
diferente afinidad por sustratos y que son inhibidas selectivamente
por distintas drogas. La MADtipo A utiliza como sustratos a la NE
y serotonina y es inhibida por la clorgilina. La MAOtipo B degrada
preferentemente a 1a feniletilamina y el deprenil es su inhibidor
selectivo ( Johnstone, 1968). Otras aminas comodopamina, triptsmina,
histamina y tiramina son buenos sustratos para ambas isoenzimas.
Por su localización en la cara externa de las mitocondrias, la MAD
parece regular 1a concentración de catecolaminas libres en el cito
plasma, tanto de las que acceden por captación comolas sintetizadaa
en el terminal y qmno están protegidos en las vesículas de almacena
miento. Comoya se ha mencionado en la sección 1.1,2, la desnervación
pinesl por gangliectomia cervical superior determina la reducción o
pérdida de la actividad de la MAOen los terminales ( MADA) mientras
que la MAOB permanece inalterada ( Snyder y col., 1965; Goridis y
NefF, 1972; Vacas y Cardinali, 1979 a; Vacas, 1980).
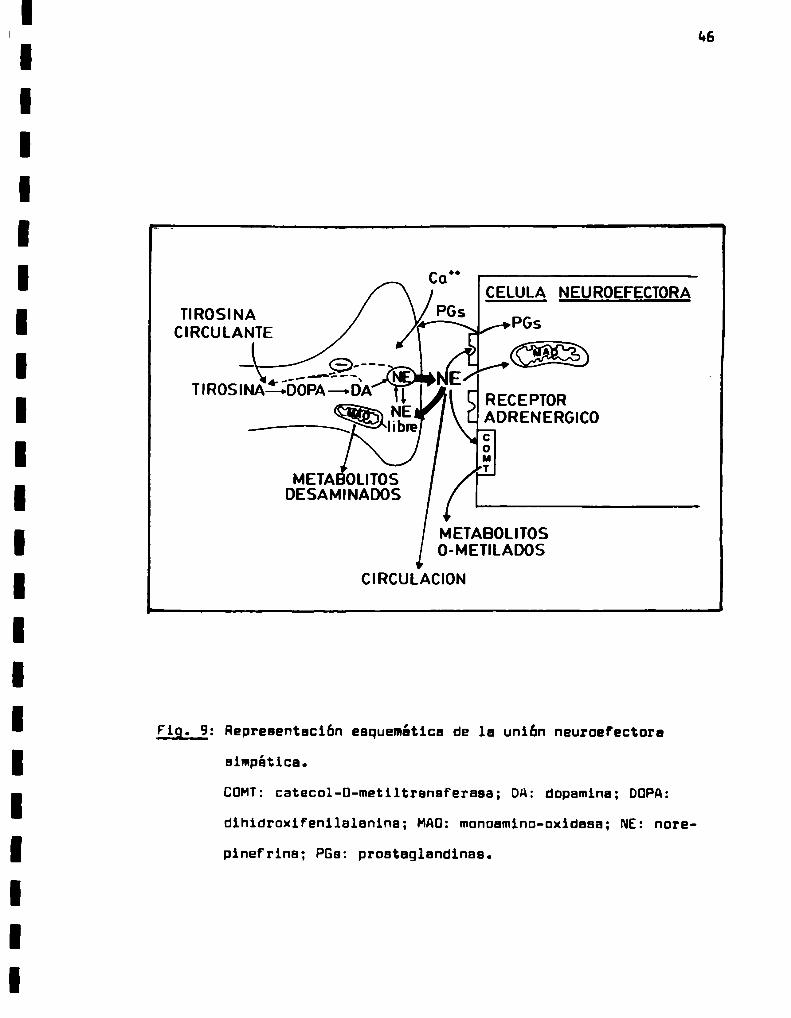
k6
c060CELULA NEUROEFECTORA
ÏIROSINACIRCULANTE /* P65/@
Ï'ROS'NA l RECEPTORADRENERGICO
META OLITOSDESAMINADOS
METABOLITOSO-METILADOS
CIRCULACION
Fig. 9: Representación eaquemática de 1a unión neuroefectora
simpática.
CDMT:catecol-D-metiltranaferaaa; DA: dopamina; DÜPA:
dihidroxifenilalanina; MAD:monoamino-axidaaa; NE: nore
pinafrina; PGa: prostaglandinas .

k7
1.2.} Función de las Prostaglandinas en los Hecanisaos Rdrenergicoe
Conoaecaniaso adicional a la retroalimentación que los neurg
transmisores ejercen sobre su propia sintesis y liberación. otras
sustancias liberadas por las neuronas adrenergicaa, o poetsinápticg
mente por la acción del neurotransmisor, actúan COIOaodulsdorss de
trsnseiaibn neural (Heatfsll, 1977). Nueerosss evidencias apoyen la
participación de las PGecolo reguladores locales an ls unión neurg
efectors adrenérgics (Fig. 9). Estas actúan de dos asnersa: l) audi
ficando la liberacibn del neurotrananisor (Bergetrda y col., 1973;
Uestfall, 1977); 2) cono eedisdorea de los efectos postsinápticos
del neurotransmisor (Hoffer y col., 1969; Partington y col.. 1960).
En el presente trabajo de tesis ee estudie ls influencia de las
PGs pineslea comonoduladores locales sobre la unión neureefectora
adrenbrgics en los pineslocitoe. Conoantecedente de las presentes
estudios nos referirenoa en pri-er lugar sl efecto que las PGeti;
nen sobre la liberación de NEpor los terainalea siepaticos en di
versos tejidos perifericoa.
En tejidos con inervacibn aiapética ha sido deaoatrade la
acción inhibitoria preaináptica da las PGe, en particular las de
la serie E. La reducción del eflujo efectivo de NEde los tejidos
eatiaulsdos eléctricsaente se interpretó en una priaers instancia
comodebida al efecto estieulatorio sobre la degradación eetabbli
cs da la cetecoleains. Sin encargo. estudios posteriores en core
zbn de cobsyo, no deeostraron alteraciones en las actividades de
HROo CÜHÏpor tratamiento con PGE2 (Hedqvist, 1975). Se deduJa
entonces que el susento del netsbolisao ds NEpor PGE2se debia
a una aayor disponibilidad de la catecolaains s las enziaae degrg

k8
dantes, probablemente por inhibición de ls recaptsción neuronal o
uptake I (Lands, 1979). Tal interpretacibn se vió avalada por los
datos experimentales derivados de varios sistemas (por ejemplo, bg
zo de gato). cuyos depósitos endógenos de NEfueron marcados con
NE-3H(Hedqvist. 1975).
Comoya se ha analizado. la liberación de NEpor impulsos ne;
viosos ea un proceso dependiente de Ca“; incrementando la concen
tración de este catión se revierte la inhibición por PGE2de la liberacibn de NEque sigue a ls estimulación nerviosa en bazo de ga
to y conducto deferente de cobayo (Hedqvist, 197“). Por otro lado,
la liberacion de NEpor tiramina, proceso independiente del CaN y
debido al desplazamiento de ls NEde las vesiculas adrenérgicaa
(Baldeasarini y Fischer. 1978), no es afectada por PGE2en modelos
comoel bazo de gato (Nestfall y Brsted. 197“) o el corazón de co
bayo (Hedqvtst, 1970 a). Estos resultados sugieren que le PGE2inhibe ls liberación de NEmodificando la disponibilidad de Ca“,
probablemente ocluyendo los canales de la membranaaxónica (Hedqvist,
1970 b) (Fig. 9).
El sitio de sintesis de las PGs endógenaa que participan en
ls regulación local de la liberación del neurotransmisor adrenér
gico no está bien establecido. Pueden ser liberadas directamente
por los nervios adrenérgicos comoresultado de ls liberación del
neurotransmisor; en ese caso la NEactuaria sobre la preainapaia
liberando a las PGs o tal liberación seria simultánea con la de
NE. Otra alternativa es que la NEactúe sobre la célula neuroefeg
tors postsináptica y produzca PGs comoconsecuencia de la interes
ción del neurotransmisor con su receptor. Los resultados experimeg

k9
tales obtenidos hasta el momentotienden a avalar este último meca
nismo.
Unaserie de evidencias indican que la postsinapsia es el sitio
de sintesis de laa PGs: a) la liberación de PGs pueden darse en res
puesta a diversos estimuloa que no sean nerviosos. Estos incluyen
neurohormonas, estimulación mecánica asociada a la contracción y
aún hipoxia (Meatfall, 1977), b) 1a administración de epinefrina eg
timula la liberación de PGa en bazo desnervsdo (Gilmore y col.. 1968).
Por otro lado, algunos datos sugieren que las PGapueden asr liberadas
también presinápticanente por laa neuronas adrenérgicaae a) as aisla
ron PGa de tejido nervioso puro comolos troncos del vago y esplínico
(Hedovist, 1976): b) propranolol y fenoxibenzamina an concentraciones
suficientes comopara bloquear la respuesta mecánica a la NE(respusg
ta postaináptica). no inhiben la liberación de PGainducida por la cg
tecolamina (Uestfall. 1977).
En relación al segundo de los mecanismosposibles por loa cuales
laa PGs actúan en la unión neuroefectora, es decir, comomediadores
postsinápticas del efecto de la NE, un ejemplo ea ofrecido por loa
órganos del aparato genital. El agregado simultáneo de PGs de las ag
rias E y F al medio de incubación con epinefrina o NE, determina una
potenciación del efecto de ambosagentes adrenérgicoa sobre el útero
aislado de rata (Clegg, 1966: Borda y col.. 1981). En el caso concrg
to de la sensibilidad a agonistas fi -adrenérgicos, se observó que
PG PGE1y PGE2no sólo son capaces de restaurar la respuestaFz-c'a metoxaminaen úteroa pretratados con indometacina (o sea tejidos
que tienen bloqueada la sintesis de PGs), sino que también está po
tenciada la acción estimulatoria de agentes oc-adrenérgicos reapeg

to a la observada en útero de animales ovariectomizsdos que no fue
ron pretrstsdos con indometacins (Borda y col.. 1981). Aparentemeg
te y de acuerdo con los datos disponibles hasta el momento, tanto
las PGs comolas cetecolsminas ejercen su acción sobre el útero a
través de un efecto primario a nivel de la adenilciclaes, con el
consecuente aumento del contenido tisular de AHPC(Vesin y Harbon,197“).
Estos dos mecanismos por los cuales las PGs afecten 1a unión
neuroefectora adrenérgica periférica (liberación del neurotransmi
sor o mediación de su efecto postsinaptico) están muchomenos estg
diados s nivel del SNC. Tsl hecho se debe a la marcada complejidad
estructural del SNCy a 1a imposibilidad de eliminar en forma efes
tiva y única los componentespre o postsinapticos. En los últimos
años se demostró le participación de las PGsen el control fiaiolé
gico del SNC, pero permanecen osouros los mecanismos de acción a
través de los cuales ejercen su efecto. Se detectaron PGsen diver
sas áreas del cerebro de mamíferos, especialmente en homogenstos
de zones ricas en terminales nerviosos (Hstsoks y col.. 1967) y su
liberación por corteza cerebral (Bradley y col., 1969); Ramuell y
Shaw, 1966). cerebelo (Coceani y Wolfe, 1965) y médula espinal (ng
ceani y col.. 1971). Enzimasparticipantes en ls sintesis de PGs
han sido también detectadas histoquimicamenteaen las células de
Purkinje del cerebelo (Ueetfell. 1977).
Avanzino y col. (1966) fueron los primeros en observar 1a ag
ción directa de las PGs sobre neuronas centrales y. años mas tarde.
Hoffer y col. (1969) demostraron que PGE1y PBE2aplicadas directa
mente en células de Purkinje sntsgonizsban la reduccifin de la velg

51
cidad de descarga inducida por NE pero no por AHPC(Folco y col..
1976). PGE1y PGEZ, pero no PGqu . estimulan la sintesis de AHPC,
usando comomarcador isotbpico adenoaina-3H, en alicea de corteza
cerebral, efecto potenciado por ls teofilina. inhibidor de la fos
fodieaterasa (Partington y col., 1980). Estos autorea también demos
trsron una disminución en la acumulación de AMPcsintetizado en reg
puesta a la NEen‘aliceg de hipotálamo y corteza cerebral de rata
incubadoa en presencia del inhibidor de la sintesis de PGa, indone
tacina.
En conjunto, los resultados discutidos sugieren que s semejan
za de lo que ocurre en la unión neuroefectora autonómica periférica,
las PGaparecen ejercer una función significativa en la regulación
de la transmisión sinñptica en el SNC.Desde el punto de vista nau
roendócrino, seta relación es particularmente relevante en el hipo
tálamo donde laa neuronas liberadoraa de LHRHdel HHHestan contro
ladaa por NEa través de la liberación de PGade la serie E (Ojeda
y col., 1979b). En el presente estudio se analizan laa interrela
ciones entre la estimulación por NEde la liberación de PGE2en
HMBy diversas concentraciones de melatonina en el medio de incudg
ción.

52
1.3 - DBJEÏIVDS
El presente trabajo de tesis examinediversos sspectcs
de 1a relscibn existente entre las PGay la función pinaal.
Son dos los objetivos principales de los experimentos pre
sentados s continuación:
1.- Estudiar ls participación de las PGaen 1a secuencia da
fenómenos metabólicos que vincula a la NE liberada de
los terminales neuralea pineales con la sintesis de 1a
hormonapineal nelstonina.
2.- Analizar le posible participación de las PGaan el ¡aca
nismo de acción de 1a hormona pineal melatonina en Give;
sas tejidos efectores, particularmente en el hipctálanu
mediobsssl (HHH).

]],'MATER| A LES Y METODOS
_|

11.1 - nniaaloa y tadiooa
Sa utilizaron rataa Hiatar adultaa (100-250 o). aantaoioaa an
un rbúiaan da luz diaria da lb h (0700 a 2100 h) an alhianta oliaa
tizado (22 1 2 C) y aliaantadaa con aliaanto bolonoaaoo 'Purino' y
agua ad libitua.I
Laa glánduloa pinaalaa y aoranalaa yloa HHHbovinoa aa obtuvig
ron an un aatadaro da lo zona dantro oa loa 20-30 ain daapuia dal
aaorificio oal oniaal. Para loa axoariaantoa da 'binoing' aa congo
loron an hialo aaco haato al aoaanto oa uaarloa (noraalaanta dentro
da loa 2“ h). Para loa axpariaantoa da libaracion do PGaloa taji
doa aa tranaportaron a1 laboratorio ao buffar ¡rabo-Ringar-oicarog
noto (HRS). pH 7.6.
Lo aonora hulana fua obtanida da donantaa qua no hablan ingari
do aapirino duroota la aaaana antarior a la vaoopontura. El ploaaa
rico an plaquataa oa obtuvo por oantrifugaoibn oa la aangra duranta
10 ain a 200xg y aa oJuatb la dilución a 250000 PIIQUOtII/II}.
Sa raalizó bajo aneataaia atiraa.da la rota una aaaana antaa
dal sacrificio (a fin da oaagorarao la doganoracibnda loa tar-ing
laa naurolao una vaz quo han aido oaparadoa ool ouarpo ganglionar).
Sa axpuaiaron laa glbnoulaa aalivaraa Iadianta una inciailn vantral
an al ouallo y aa laa ratraJo con al obJato da axtariorizar loa
afioculoa auprahioidaoa. Cada ganglio oarvioal auparior fua idanti
Picado an la bifuroaoiún oarotldaa antro aua ralaa intarna y anto;
no y aaboa ganglioa. daraoho a izquiardo. fuaron totalaanta raaaqg

5h
Ovaricctalia biietarai
La ovaricctalia bil-total un practicó 20 dias anto. del ¡anti
ficio (castración crónica) bajo anestesia atérna. Se realizó una
incisión longitudinal indio-interni. Il ubicó c1 ovario y ¡1 cxtgg
no utnrino proxi-nl a bota. Se iigó con hilo du cirugia a una. 5 II
de dicho extremo y le cortó ci cuerno. extraylndo nai el ovario y
oviducto y renpotnndo el útero odyaccntc.

55
II.2 - Producción de groataglandinaa in vitro Eor glándula Etneal
y HMBbovino y HMBde rata
11.2.1 - Glándula plnealgy HHBbovinos
La liberacion de PGa por glándula pineal v HHBbovinoa fue-a!
tuoiada in vitro. Loa tejidoa ae incubaron durante l n en buffer
HRB,pH 7.“, an preaancia de 0.1! de ¡cido aacórbico y diatintaa
concentracionea de NE(1-1000 uH) o lalatonina (0.01-1 un). Laa ig
cuoacionaa aa realizaron a 37 C bajo atabafera da carhógano (99! 02
5%002) y ae interruapieron por separación y congelación inaadiatadel aedio da incubación. Dicho aeoio fue acidificado a pH 3 con 01H
l N y extraido doa vacaa con 1 volulen de acetato da etilo, el que
aa evaporó bajo vacio en un evaporaoor rotatorio a k0 C. El reaiouo
ae raeuapandib an 10 al de buffer foafato de potaaio pHa. acidifi
candoaa la faae acuoaa coabinada a pH 3 con ClH l N. Loa bcidoa
graaoa fueron luego axtractadoa en l volumende cloroforao, rapi
tiendoae la operación doa vecaa aaa; La faaa clorofbraica fue eva
porada y el raaiduo ae raouapanoib en cloroforIo-aetanol (k:l) v
fue aplicado en al origen de una placa’de cromatografía recubierta
con ailica gel G. La cromatografía aa deaarrollb en un aiateaa de
aolventea cloroforao-aetanol-úcido acético-agua (90:9:l:0.65). co
rriindoaa paralela-ante placao con PGE2y PGFzg comoatandarda.La poaibibn de laa FBa de referencia fue viaualizada luego de ro
ciar laa placaa aacaa con una aolucibn de acido foafonolihdico al
15! en etanol y eosatiindolaa a 120 C durante 15 nin; loa Rf para
P662 y PGde fueron 0.61 y 0.4.2 raapectivanenta. Laa zonaa dalaa placaa con axtractoa del ¡odio da incubacibn correapondientaa
a loa Rf de loa atandarda fueron aluidaa con cloroforao-aatanol

56
(kzl) y iiavadaa a aaquadad bajo vacio. La actividad PGda aataa
fraccionaa fua aatiaada por al procadiaianto da Vana (196k). Loa
axtractda aa raauapandiaron an 0.2 ni da HRBy aa anaayarcn an ci
raa de aatúlagc y colon da rata auparfundidoa con KRBqua contanla
una aazcla da antagoniataa para laa aiguiantaa auatanciaa activaa:
acatiicclina, aaratonina. catacolaninaa a hiatanina. ïaahiin aa a
gragfi inddaatacina (1 ug/ai) a fin da pravanir ia producción andb
gana da PGa. Laa concentracidnaa fueron ¡adidaa con un tranaductdr
iaoabtricc y ragiatradaa an un poligrafa. La actividad PGfua ax.
praaada an aquivaiantaa da PGE2o PGF2¡_ cono pol-g dc tajido aacd/
h da incubación. Laa auaatraa axpariaantaiaa y aua cdntroiaa fuardn
analizadoa aianpra an 1a aiana praparacibn. La racuparaciún luago
da 1a axtracción y croaatcgrafia fua hD-SÜI, con un coaficianta da
variación intaranaayp da IIHOI dal 10%.
Eata procadiaianto biológico para 1a datarninacibn da PGafua
utilizado an laa atapaa iniciaiaa dai praaanta trabajo. antaa da
contaraa can a1 attado radioinnunalbgico. Alboa procediaiantoa,
biológico y radicinaunolbgico. aa uaan actuaiaanta para ai doana
de Püa (Vana, 1965; Haciauf y col.. 1975).
11.2.2 - HHHda rata
Sa incubaron an forma individual HHHda rata (3.5 - “.5 ng
paao húmedo) an 500 ul da buffer HRS, pH 7.“. qua contania giuccaa
(1 ag/Il) y 0.06 un da acido aacórbicd, a 37 C y bajo atnbafara da
carhbgand (DJada y col., i979b).Loa taJidoa fuaron praincubadoa dg
tanta 15 ain, al cabo da ica cuaiaa aa raaapiazb a1 nadia por und
fraacd qua contania naiatdnina (ifl'k- lo'ah). Daapubada 30 ain da
incubación, aa tdaaron alícuotaa (100 ul) dal India para avalulr 18
concentración de PGamediante radioinmunoanálisia ( ver sección II.
6 .2)¡

57
11.3 - Eetudio de eitioe de union ee ecificoe ere roete
en glindule Eineel bovino
Le preeencie de eitioe de unión eepecificoe pere PGeen le
glindule pineel bovino fue inveetigede en el eooreneoente de 900 e
g eegún el procedieiento deecripto por Hiebell y Uyngerden (1975)
pere el nioletrio de neeeter. Lee pineelee congeledee fueron eeccig
nedee en trozoe pequefloe y honogeneizedoe en buffer foefeto 0.02 H,
pH 7.“. en preeenoie de eeceroee 0.25 H. CeCl2 2 IH y lO ug/Il de
indoeetecine (buffer BI). El honogeneto ee oentrifugb e 900 x g og
rento lS ein e O C y elicuotee del eohreneoente fueron inouhedee
en triplicado con P6 oercede. Lee incuoecionee ee lleveron e cebo3en tuhoe de polieetireno pero eviter le edeorcibn de lee PGe- H el
vidrio. Se utilizeron loe eiguientee redioieótopoe: PGEZ-JH(A.E.
150Ci/I-ol) y Por“ -’H (me. 120cue-ol), cuyopurue redioquinice fue verificede por croeetogrefie en cepe delgeoe en el elote
Ie doecripto en le eección 11.2.1. Le reeccibn tipice de eeocieción
('binding') ee llevó e cebo en un volumentotel de 210 ul contenieg
do 200 ul de eobrenedente (3-9 eg de proteinee). PBEZ-JH(0.2 uni.
-’H (0.2 uCi, 1.7 x 10'12 no1) dieueltee9
1.5 x 10'12 lol) o Por“en 5 ul de etenol-BI (1:7, vol/vol). lee PGno Iercede (l x lO
lol en 5 ul de etanol-BI. 1:50 vol/vol) o 5 ul de BI. Le concepto!
ción proteice fue deterlinede por el ¡6todo de Loury y col. (1951).
En cede experieento ee incluyó une eerie perelele de Iezclee de o!
eccibn ein tejido. Aeenoe que ee coneiooe lo contrario. el tie-po
de inouoecibn fue de l hore e 37 C.
Le eeperecibn de lee PGelibree y unidee ee reelizb por edeq;
ción en carbon-gelatina (Hinhell y Uyngerden, 1975). A cede luce

tra aa agregó 0.5 nl da una auapenaibn da carbón (2.5! de carbón
Norit A y 0.1% de gelatina an borrar BI) v luago dc lO ainutoa aa
cantrifugb a 3000 x g durante 15 nin a 0 C, trae lo cual aa traa
vaab el aohranadanta a vialoa da cantallao. El 3Hfue cuantifica
do por eapactroaatria da cantollao liquido, pravio agragado da ag
lución de centellao toloeno-fóaforo (k g da 2,5-difaniloxazol y
50 ag da p-hio-2(5-faniloxazolil)-oanceno por litro da toluano)
con 30%(v/v) da Iritbn x-lflfl.
La corrección de 'quencning' fua raalizada automatica-anto
por ladio de un atandard axtarno; an aataa condicional la aficion
cia varió entre 50 y 605. La difarancia antro la PG-JHunida an ag
aoncia y an proaancia de PGno aarcada fue conaidcrada una aatila
ción de lo PGunida aapacificaaenta. La unión aapacifica conatitu
yó un Sh-BZ! da la total.
En una oario de cxpariaantoa praliainorao aa catableciaron
las condicionaa óptiaaa da adaorcibn.
La aeporacibn da PE libra dc unida ae realizo, an otra aorie
da experiaantoa. por filtrado rápido en papal de fibra da vidrio
Hhataon BFBbajo vacio parcial (Bennet y Snyder. 1975). Luago da
lavadoa con un total da lO al da buffer BI a k C. lo radiactividad
ronanonta an al filtro fua datarainada por eapoctrolatria da cant;
llao liquido. Loa raaultadoa obtanidoa utilizando cono natodo dc
separación la filtración rápida an papel da fibra da vidrio fueron
aaancialaante igualan a loa de adsorción en carbón-galatina.
Laa conatantca da equilibrio da la union dc alta afinidad da
PGaa coaponantaa pinaalaa foaron datarainadaa por incubacibn an
triplicado de aohranadantaa da 900 x g con diatintaa concantra

59
ciones de PGEz-JHo PGqu-BH. Luego de le treneforsscibn de losdstoe (Scstchsrd. l9k9) ls pendiente e intersecciones fueron cal
culados por análisis de regresión. En otro experisento se detersi
nsron las afinidades relativas de diversos análogos para los si
tios de alta afinidad de PGFZ“ o PGEZ.Por lo senos lo concentraciones del análogo fueron utilizadas sn csds caso.
Los valores de radiactividad unida especifica-ente se grafi
caron en papel log-probitoa y se calcularon las concentraciones
de los agentes que producen una inhibición del 50! ds ls unión de
' concentración efectiva 50). Las pendientes e inter50'
secciones se calcularon por analisis de regresión. Las constantes
PG-JH (ce
de disociación en el equilibrio de los ligsndos cospetidoree no
ssrcados se calcularon a partir de la fbrluls: H1 - GESD/l +
(JH-PG)/Kd, donde Hd es la constante de disociación de las PG-JH
calculadas a partir del grafico de Scstcnsrd.
Ls naturaleza de la rsdisctividad unida a proteinas se veri
ficb luego de aer extraido tres veces con 5 nl de acetato de eti
lo scidificado. Los extractos se llevaron a sequedad bajo N2y losresiduos se reaaspendieron en cloroforno-netsnol (kzl. vol/vol) y
crosstografiaron en capa delgada de silics gel G, usando coso elu
yente el sistens cloroforsoznetanol:ácido acéticozagus (90:9:1:
0.65). Másdel 96%de la radiactividad corrió paralela e loe stan
dards de PGs.
Conel objeto de estudiar la distribución intracelular de los
sitios de unión para PGaen la glándula pinesl. los nosogenstos
fueron procesadoa en una centrífuga refrigerada a fin de obtener
las siguientes fracciones: l) nuclear cruda (900 x g. lO sin);

2) nitocondrisl cruda (27000 x g, 10 sin): 3) microsomal cruda
(105000 x g, 60 sin) y h) fracción oitosbllca (105000 x g, sobrs
nsdsnts). Allcuntss ds las suspensiones de estas frsccionss ss {g
cubsron con lss PG-JHcono ys hs sido descripto y se detsrsinb ls
concentración prstsics por el sótano de Lasry y col. (1951). con
ssroslbúsins bovina cano standard.

61
Il.“ - Deterainacibn de los sitios de unibn aspecificoa para AHF
en la glándula Einaal da rata
Las glándulas pinceles ae preincubaron durante 10 ein en buf
far HRBa 37 C, cono ya ha sido descripto. Luego de este preincuog
ción, las glándulas fueron incubadas durante 15 ein an el mis-o
buffer en presencia de PGE2 56 nH. La deterainación da la unión
del nuclebtido ciclico a la quinssa de proteinas ae realizo según
la tficnica deacripta por Dufau y col. (1977).
Los estudios de unión de AHPGae llevaron a cabo incubando s
licuotas de 100 ul (Sk-51 ug de proteinas) del aohrenadsnte de 900
x g de honogenatos de pinealaa preparados en buffer fosfato de po
taaio 80 IM, pH 6.6, conteniendo acetato de magnesio lO IH. 2-ner
csptoetanol l MHy 1-astil-3-isohutil-xantina l IH, en presencia
de AHPc-3H(A.E. hu Ci/a-ol) en concentraciones que vsriaron entre
0.1 nH y 0.1 uH. La incubación se afectub a 6 C, durante 2 horas
para deterainar los sitios disponibles, o durante 16 horas para
la evaluación de sitios totales. Finalizada ls incubación. sa agrg
garon 2 nl del buffer Iris-61H y se filtro a través de papel Unat
nan GFB;
La rsdiactividad reasnenta sn los filtros se determinó por e!
pactronatria de centellao liquido, previo agregado de 2.5 al da eg
lución de cantelleo. La unión inespecifica determinada para cada
concentración del nuclebtido ciclico-JH en presencia de S uHde
MPc no marcado fue restada del valor de unión total para obtenerasi la unión especifica que fue. en todos los casos, esyor de un
80%. Los rssultadoa fueron referidos a Ig de proteinas que aa dc
tsrninaron según el metodo de Loury y col. (1951). Los datos fue
ron analizados según el método de Scstchard (19h9).

----------
62
11.5 - Datersinación ds actividades anti-áticos
11.5.1 Catecol-O-metiltransferasa
La catacol-U-Ietiltranafersas (CUHT).sal colo ls nonoalino
oxidsaa (una). son las snziaaa responsables del nstsóolisao ds do
pamina y NE(Fig. 8). La COHT,anzias citoaólica, cataliza la tran;
farsncia del grupo ¡stilo de 1a S-adanoailletionina (SAM).en pre
sencia de cationss divalentss cono cofactorea. a uno de los grupos
fanólicoa de las catscolsninaa para for-ar el derivado natoxifsnol
y S 4 iït ‘nk-inn. La actividad de CDHTfue determinada ss1diante la estimación de la transferencia ds grupos setilo- l'l':de la
SAM-¡“Cal ácido 3,k-dihidroxiósnzoico (Jarrot, 197%).
Los tejidos (glándula pinasl s ¡HBda rata) se honogsnsizaron
sn 250 ul de sodio hipotónico ds fosfato de sodio 10 en, pH 8.2.
Se incuóaron alicuotas ds 70 ul durante l h a 37 C con ácido dini
droxibenzoico 1 un, san-“c 2.7 x 10‘5n (me. ¡.7 ¡zi/no1) sn pre
sencia de "9012 5 en y en un Isdio buffer de fosfato da potasio80 IH, pH 6:0. La reacción as detuvo aadiante el agregado de 50 ul
de ClH 2 N. El producto nstilado ss extrajo con 500 ul de acetato
de stilo y se lsvó con 100 ul ds ClH 0.3 N. Finalmente aa evaluó
la radiactividad en una alícuota de 250 ul de fase orgánica por e;
pectroletria de csntallso liquido. previo agregado de solución ds
cantsllso. Los resultados se expresaron en peolsa de producto/h
ng ds proteina. Las proteinas ss evaluaron ssgón sl lótodo de Lau
ry y col. (1951).
11.5.2 Monosnino-oxidosa
La nonoanino-oxidssa (MAO)(Fig. a). snzisa nitocondrisl. das!

63
sino un ssplio sspsctro ds ssinss que posssn sl csroono-d-Istilsno
vscino s ls ssins no sustituids. dsndo ss! sl sldshido corrsspon
diente. Este conpussto ss reduce rápidsssnts s slcohol o ss oxids
s ácido. En expsrissntos in vitro, slrsdsdor dsl 80%dsl producto
ss hslls coso ácido, lo qus sugisrs que sos producto forssdo por ls
MADss oxidsdo s ácido por ls sldshido dsshidroqsnsss. Dsds ls dis
tribucion sitocondrisl ds ssts snziss. snilogs s ls ds ls MAO,ss
sugirió nus sabes sstsrisn scoplsdss.
Los glándulss pinsslss o HHHisdividuslss ss hosoqsnsilsron sn
160 ul ds buffsr fosfato ds sodio 10 un, pH6.2: ss incuosron sli
cuotss ds 50 ui dursnts 30 sin s 37 C sn prsssncis ds 0.15 IH ds
oioxslsto ds ssrotonins-uc (0.1 Ci/i ul) o 0.1 ¡ñ de f'snilstilssi
ns-th (0.05 Ci/5 ul), sustrsto ds ls HflDtipo n y MAOtipo B rss
psctivslsnts. Luego ds dstsnsr ls rsscccibn con 50 ul ds ClH 2 N.
los productos dssssinsdos ss sxtrstron con 500 ul ds scststo ds
stilo y ss lsvsron con 100 ul ds ClH 0.3 N según sl sitodo dsscrig
to por Jsrrot (197“). Los productos forsodos se svslusron por sdÉ
psctrosetris ds csntsllso liquido sn 250 ul ds ls fsss orgbnics.
previo sgrsgsdo ds 2.5 s1 de ls solucion ds csntellso.
Los rssultsdos ss sxprsssron coso psolss ds producto forssdo/
h/mg de proteínasn. Estss últisss ss svslusron ds'scusrdoucoo sl
sstodo dsscripto por Loury y col. (1951).
11.5.3 Serotonina-N-acetiltronsfersso
La ssrotonins-N-scstiltrsnsfsrsss (SNAÏ). ss uns ds lss dos
snzisss involucradossn ls siotssis ds sslstonins pinssl s psrtir
de ssrotonins (Fig. 6). Su sctividsd ss svslús por ls trsnsfsrsn
cis dsl grupo scstilo-1“C ds ls scstil-Con ssrcsds (A.E. k5 Ci/

6k
col) a ls triptasina para foraar la N-acetil-triptasins-lhc. pro
ducto extraido por partición de solventes según el aétodo descrig
to por Deguchi y Axslrod (1972). Les pineslaa se honogeneizsron en
forsa individual en 100 ul de buffer fosfato de potasio 36 sn, pH
6.5. que contenia 0.1 uso] de triptaains. Luego de una incubación
de lO cin a 37 C en presencia de 3.“ naolea de acetil-CoA-1“C, la
reacción sa detuvo por el agregado de 500 ul ds buffer borsto 0.5 H
pH lO. y el producto formadopor la transferencia enzinstics del
grupo ll'IZ-ecetilo a la triptasins se extraJo con 6 al de tolueno:
alcohol isoalilico (100:l; vol:vol). Se lavo dos veces con 500 ul
de buffer borsto y una alícuota de 5 al de la fase organica se cua!
tificb por espactronatris de centelleo liquido. luego del agregado
de lO nl de solución de centelleo.
Loa resultados aa expreaaron coso paolee de producto foraado/
h/Ig de proteins o por glándula. En los exparisantoe en que los r5
sultados se expresaron por Ig de proteina. estas fueron evaluadas
según el letodo deacripto por Loury y col. (1951).
11.5.“ Hidroxindol-Ü-metiltranaferaaa
La hidroxindol-D-aetiltransfersaa (HIUHÏ). es la enzima res
ponsable de la traneferencis del grupo metilo de la S-adenosilne
tionins s la N-ecetllserotonina, dandocono producto la aelatonina
(Fig. 6).
La actividad HIUHÏfue determinada por el procediliento des
cripto por axelrod y Ueisabach (1960). Glñndulaa pinesles indiai
duales sa hologeneizaron en buffer fosfato de sodio 50 an, pH 7.9
y slicuotas ds lDDul ee incubaron sn presencia de l ¡H de N-scs

65
tilserotonins y 30 uH de SAM-lhc. Luego de 30 nin de incubación e
37 C, le reacción se detuvo mediante el agregado de buffer borsto
0.5 H, pH 10. Ls melotonins-1“C formado, se extrajo con 8 el de
cloroforno. el que se lsvb dos veces con 1 nl de buffer horsto. Se
toleron 6 Il de ls rose orgánico y se llevaron e sequedad en estu
fe (60 C). El residuo se resuspendió en 500 ul de etanol, ogregfin
dose 10 nl de mezcle de centslleo y cusntificándose el producto r!
diectivo fornsdo por espectrometrle de centelleo liquido, previo
agregado de 10 el de solución de centslleo.
Los resultados se expresaron comopmoles de producto formsdo/
mgde proteins/h.

66
11.6 - Deterninaciones Eor análisis de saturación de melatonina.
Ez.ch11.6.1 - Melstonina
La concentración de melstonina en la glándula pineal o en el
medio de incubación de pinealea in vitro fue determinada por radig
inmunoanáliais, utilizando el antisuero desarrollado por inyección
a conejos del inmunogenoindonetacina-slbómina sérica humana(Levi
ne y Riceberg, 1975) (donación del Dr. Levine. Brandeis University.
HA, EE UU).El tejido se homogeneizó en 1 ml de ClH 0.1 N y se ex
trajo conusmI de cloroformo, el que fue lavado con 2 ml de NsHCD3
2 i y 2 ml de agua tridestilsda. llevándose s sequedad sto vacío.
En el caao de los medios de incubación de glándulas pineales. alf
cuotaa de 0.5 ml fueron extraídas con 6 nl de cloroformo, realizán
dose los mismoslavados sucesivos que para los extractos pineales.
El residuo fue resuependido en 0.5 ml de cloroformo y se lo sometió
a una particion cloroformo-n-heptano-agus, luego de agregar l ml de
buffer Iris-81H, pH 7.“ y 5 ml de n-heptano; en estas condiciones
la melstonina pasa a la fase acuosa. Para el rsdioinmunoanalisis se
utilizaron alícuotas de dicha fase acuosa que se incubaron con el
antisuero y de acuerdo con el método desarrollado por Levine y Ri
ceberg (1975). En un volumen final de 0.3 ml se incubsron durante
1 h a 37 C, volúmenes iguales de nelatonins fría (en el caso de ls
curva standard que abarcí un rango de JOB-10000pg/ml) o muestra,
antiauero y melatonina-BH.
Luego de detener la reacción colocando los tubos a 0 C duran
te 10 min, ae separó el complejo inmune formado mediante el agrega

67
do de 1 m1 de (NH“)250“ (sa), incubación de B h a h C y posterior
centrifugaciofi a 2500 RPM.E1 precipitado obtenido oe resuapendio
en 0.2 mi de NaUH0.1 N y la radiactividad se valoró por espectro
metria de centelleo liquido, previo agregado de solucion de centelleo.
La sensibilidad del ensayo fue de 30 pg/nuestra; los coefi
cientes de variación intra e inter ensayo para nuestras de 100 pg
fueron 3 y 5%respectivamente.
11.6.2 Prostaglandina E2E1 radioinmunoanalisis se llevó a'cabo con el antiauero del
Instituto Pasteur de Paris.
En un volumen final de 0.3 m1 se incubaron durante 1 h a tea
peratura ambiente y 20-2“ h a 0 C, volumenes iguales de PGE2(en
el caso de 1a curva standard, cuyo rango abarcá 30-1000 pg/nl).
muestra, antisuero y PGEz-BH(aproximadamente 12 pg/tubo). Luego
de esta incubaoián se separó la PGE2unida de la libre mediante elagregado de una suspensión de carbón-dextrán (250 mg de carbón, 25
mg de Dextrén en 100 ml de buffer fosfato di-monopotaaico 0.1 H,pH
7.a, 0.1% de gelatina), incubación a h C por 10 min y posterior cen
trifugacián (1500xg) a 1a misma temperatura, durante 10 min.
El sobrenadente se cuantificó por espectronetris de centalioo
liquido, previo agregado de solucion de centelleo.
La sensibilidad del ensayo es de 30 pg PGEZ/tUDDF108 00871‘cientes intra e interensayo para muestras de 100 pg fueron 3 y 7%
respectivamente.

¡1.6.3 AMP
Se incubaron an forma individual glándulaa pinaalaa da rata
durante lO ain a 37 C an 200 ul da buffer NRB,pH 7.5, qua conta
nla 122 ¡1 NaCl. k.9 dl KCl, 10.3 ¡4 NaZHPD“.1.2 ¡4 KHZPO‘. 1.2 ¡1
HgSO“,0.5 ¡H CaClz, 11.8 an glucosa. 1.1 ¡a acido aacbrhico,
0.13'IH EDÏAy 0.01 añ da inda-etacina. Luego da aata praincuhacibn
laa glándulas fuaron incubadaa duranta 15 ain an al aiaao nadia qua
contanla laa difarantaa PGaanwconcantracionaa qua variaron antra
l nHy l un. Al finalizar la incubación laa pinaalaa aa naaaganai
zaron an ácido tricloroacético (Ica) Si; a1 hanoganato aa cantrifig
96 a 1000 x g por 20 ain a 0 C y el anhranadanta aa lavb k vacaa
con 2 nl de diatilitar con al fin da alininar al ICA. El AHF:fue
detarainado por triplicado an la faaa acudaa libra da ÍCAda acuq;
do can la técnica da Braun y cal. (1971).
La quinaaa da protainaa fue aialada da glándulaa adranalaa ag
vinaa. Luagoda la aaparación da la aédula. laa cortazaa adranalaa
fuaron hanngenaizadaa an un hoaoganaizador tipo Polytron (Brickaann)
a O C, con 1.5 volúnanaa da buffer Tria-ClH. SD¡1, pH 7.40, conta
niendo aacaraaa 0.25 H; HC} 25 ¡H y HgCl2 5 IN. Eata honoganato aa
cantrifugb a 2000 x g duranta 5 ain y al aobranadanta a 5000 x g
durante 15 ain. El auhranadanta da aata últiaa cantrifugacibn aa
fraccionó an alicuotaa da 0.5 al, qua aa alaacanaron a -20 C. Eata
praparación fue daacnngalada y diluida con al buffer da anaayo
(Tria-ClH SCIan. pH 7.“. contaniando taofilina 8 ¡4 y Z-narcapta
atannl 6 an, an al aguanto de cada datarainacibn. Eata huffar aa
utilizó an todua loa paaoa pnatarioraa.

69
Cada tubo contania 50 ui de 1a dilución apropiada de la anzi
aa (1:8). 100 ui da una cantidad conocida de AHFc(entra 0.039 y
20 panico) o de hondganatd da taJido y 50 ul de AHPc-JH(aproxiaa
danente 3000 cpu). La cantidad de anzina ara tal que unia aproxi
aadaaanta a1 ha! dal AHPCaarcada.
La incubación aa realizó a k C durante 90 ainutoa. E1 ¡HPc
libra aa aaparb por al agragado da 200 ui de una auapanaión de
carbón-68a (Sigma, Fracción V) (500 Ig de carbfinzioo Ig da 85A,
para 10 al da huffar da anaayo). La radiactividad aa datcrlinó
por aapactrd-atria da cantaliao liquido. pravio agregada da ani!ción da cantailao.

7D
11.7 - Determinación de la liberación de NE-JHpor glándula Eineal_
de rata in vitro.
Se utilizó el procedimiento descripto por Pelayo y col. (1977).
consistente en la incubación de glándula pineal de rata in vitro con
NE-JHy la estimulación de su liberación por 80 un de H‘. Se incuba-"
ron cuatro pineales por grupo en un cilindro de poliestireno cerra
do en su parte inferior con una-malla de acero inoxidable. El buffer
de incubación consistió en una solución de Tyrode con ls siguiente
composición (mn): NaCI! 137; KCl: 2.68; CaC12: 1.78; MgClZ: 1.0k;
NaZHPO“:0.k2; NaHCDJ: 11.9; glucosa: 5.5; NazEDÏA: 0.069; ócido ss
córbico: 0.061. La temperatura se mantuvo a 37C.burbquóndose la so
lución con csrbogéno. Los depósitos intraneuronalaa de NE-JHse aar
caron por incubación de las glándulas en presencia de 0.5 un de E NE
3H (5 Ci/mmol) durante 3Gmin.Lss pineales fueron incubadss durante
50 min más en nuevas fracciones de buffer sin el radioisótopo. y
cuando el flujo espontáneo de NE-JHalcanzó el equilibrio. sa eatinu¿
ló la liberación de NEpor exposición a 80 mMde K‘. El intervalo sn
tre la primera estimulación por H‘ (Sl) y ls segunda (52) fue de 35
min. Se agregó PGE2 (10 a 100 nM) o vehiculo al medio de incubación
20 min antes de 52. Los resultados se expresaron como"liberaciónfraccional', o sea el S de la radiactividad tisular liberada luego
del estimuloks1 y SáL Asimismose estableció la relación 82/51 como
un indice de la inhibición de la liberación producida pcr PBEZ.

71
11.8 - Deterninacifin de la actividad contraotil del útero
Loa eatudioa ae realizaron in vitro con cuarnoa uterinoa de r!
taa virgenea ovariectolizadaa, ya que en experilentoe piloto fiataa
reeultaron laa aenaiblea al tratamiento con aelatonina que laa nul
tiparaa. Cada cuerno diaecado ae abrio longitudinal-ente aiguiando
la linea del aeaonetrio. Finalnente ae cortaron trozoa traneverea
lea de útero de aproximada-ente 3-“ al de longitud. El tejido
ee fijó a un aoporte tiaular por el extrano inferior. y el extra-o
auperior ae conectó a un tranaductor capaz de tranaforaar la activi
dad necánica del tejido en una aeflal albctrica. La aalida de eate
transductor ae conectó a un amplificador acoplado a un inscriptor.
de lodo de registrar en forma inladiata y continua la actividad q!
canica del tejido.
Cono ¡odio de incubación ae utilizó una eolución buffer KRB.
pH 7.a, que contania ll MHda glucoaa como auatrato netabblico. U
na vez montado el tejido ae aplicó una teneión oaaal de 500 ng. lo!
go de lo cual aeregiatró au actividad contráctil por un periodo de
60 nin. conaiderfindoae loa 10 prileroa cono control interno da di
cha actividad y loa 50 últiloa colo periodo experilental. La nora
de experiaentación fue analizada en periodos de lO nin y laa varia
cionea obeervadaa ae expreaaron cono S de cambio reapecto de loa
10 prineroa nin control, que fueron conaideradoa cono el lOOI de
actividad. En laa aeriaa de exparieentoa en laa que aa eatudib el
efecto de la aelatonina. el letoxindol estuvo preaente en el le
dio de incubación deade el comienzo de la hora de axparinentacibn.
Para valorar la actividad contráctil del útero de rata ovariegtomizada de 20 díaa ae tuvo en cuenta la tensión contractil ieoméF

72
trica (TCI): medida desde el nivel de tensidh de reposo hasta el
punto de mayortensión desarrollada (tensión pico); se consideré
1a TCI promedio de laa contracciones registradas en 10 min.

73
II-9-Daterlinación de la agregaciónElaguetaria
Se preincubaron 600 ul de plasma rico en plaguctac durante 30
nin en presencia de distintas concentraciones de nelatonina (50
100-200 uH). La actividad de 1a hormona pinaal sobre la agregación '
plaquataria ae evaluó en un agragónatro y ca utilizó cala agente
agregante ADP(concentración final 5 uH).
La agregación inducida por 5 un de ADPen auaencie de Delete
nina fue conaiderada COIOal ¡óxiao valor de agregación (100! de
trancuitancia).

7k
11.10 - REACTIVÜS UTILIZADOS
Sigma Chemical 60.:
- acetilaalicilico, acidoN-acetilaerotonina
adenoaina difoafato (ADP)
adenoaina 3',5'-monofoafato ciclico (AHPC)ascórbico, acido
2,5-aifeniloxazol
3,h-dihidroxibenzoico, ácido
p-bia-2(S-feniloxazolil)-benceno
indometacina
melatcnina
i-metil-J-iaobutil-xantina
L-nareplnefrina, bitlrtrata
aeroalbómina bovina (fraccion V)
aílica gel G
tacfilina
Pharmacia Fine Chemicals Cc.:
- Dextrán T 70
Amend Drug and Chemical Co.:
- Carbón Norit A, neutro.
Laa prostaglandinas (PGEZ, PGEI, PGFZO‘, lS-ceto-PGF2m , P612) fue
ron cedidaa por el Dr. John Pikee, UpJohn Co., Halanazoo, Michigan,
EEUU.

75
El acido nefanámico fue cedido por el Dr. Casareto, Parke Davis.
Lao sales y solventes utilizados fueron de grado analítico (Carlo Er
ba, Hollinckrodt, Merck).
NewEngland Nuclear, "399., EEUU:
acetil-coenzima A, (1-acet11-1“C)-, A.E.: “5 Ci/mmol
N-acet11-5-metox1triptamina, (2-am1noet11-JH)-. A.E.: 31.5 Ci/nnol
S-adenoail-L-netionina, (metil-lhc), A.E.: Q7.UCI/mmol
odenoaina 3',5' fosfato cíclico, sal de amonio [3H(Gá] . A.E.:
“2.5 Ci/mmol
lB-rennetnamina. (Lun-1%), HCl, A.E.: 1.7 mCi/molS-hidroxitriptamina, (2-1“c), bioxaloto, A.E.: h?.2 m81/mmo1
L-norepinefrina, 7-3H(N) , A.E.: S Ci/mmol
Proataglanoina E2 [s.s,e,11,12,1u,15-3H(Nïj , A.E.: 150 Ci/mol
Proataglandina F2K[5,6.8.9.11,12,1h,15-3H(N)] , A.E.: 120 Ci/mol

76
II..11- ESÍADISTICA
Para los anililil estadístico. no utilizaron lo. tonta de
Student y du Dunnnt un el culo de dos nuestros y el tout d. Seno
ffb (1959) o tant de ïukcy (19h9) para cunparacinncn múltiplco.

IIIRESU LTADOS

71
III.1 - PARTICIPACION DE LAS PRDSTABLANDINAS EN EL CÜNTRÜL NURADRE
NERGICÜ DE LA ELANDULA PINEAL
Comose ha analizado en 1a Introducción (Sección 1.2.2).
las PGs desempeñan un papel significativo en 1a modulación de
1a liberación y efecto postsináptico del neurotransmisor en
diversas uniones neuroefectoras adrenérgicas (Fig. 9). Const¿
tuyendo la glándula pineal un modelo por excelencia de tales
uniones neuroefectoras (Vacas, 1980), se decidió examinar el
papel de las PGs en ese sistema. Ya hemos mencionado como an
tecedente del presente estudio la nota preliminar de Szabo y
FriedhofF (1976) describiendo el efecto de 1a indometacina ag
bre la inducción de SNATen ratas durante el período de oscu
ridad.

78
III.1.1 - Producción de Erostaglandinas Eor la glándula Eineal bovina "in vitro"
El objetivo de la primera serie de experimentos Fue determi
nar si la glándula pineal in vitro producía PGsy si esta produc
ción se modificaba por la exposición al neurotransmisor NE.
En la Fig. lÜ se muestra la liberación de PGEy PGF por ex
plantos de glándulas pineales bovinas. La liberación espontánea o
inducida por NEde FEF resultó aproximadamente el doble que la de
PGE. E1 agregado de 10'“ - 10-6 M de NE al medio de incubación prg
dujo un incremento significativo an la liberación de ambas PGs.
En los experimentos en los que se idcubaron las glándulas pi
neales bovinas en presencia de melatonina (l mM)se observó una
disminucifin del 52% y 88% de la liberación de PGFZC!y PGE2rea
pectivamente (Tabla 3), resultados que evidencian el efecto inhi
bítorio del metoxindol pinaal sobre la producción de PGs.

79
a)
á A su
a: Í 2°°°r PGF I PGEm o '
e 8 l. F- :_l en ¡e l
09:3 Í4 '5‘ lZ 6- :22's 1000- I t . ’.4 c. I ___...l E 'o l4 \ ' lEB5 í lSÉ ¿a? (5) = 'a o- Á _J i A
NOREPlNEFRlNA - l 10 100 ' - l 10 ¡00(uM)
Fig. 10: Efecto de le NE entre le producción da PGEy PGFpor clan
dule pincel bovina in vitro.
Glendulea pinaelae bovinea fueron incubedee an praaancia
k - 10' H) durante 1 h a 37 C. Loao euaencie de NE (10'
reeultedce ea axpraaen an pg/Ig da tejido aaco.Hedia 3
1 es (n).
’p < 0.01 en releción el control (teat de Dunnat).

Teble 3: Efecto de le ecletonine sobre le producción y libereclbn
de PGepor explentoe de glándula pincel bovine ln vitro'
H3de P662
Control 526.1 t 15.6 (-5) 24.3.6 t 4.9.3 (5)
nelatonm. 10"» 250.1.t 63.2 (5)" 30.9 t 19.2 (5)"
’Glendulee p1nee1ee bovlnee fueron incubedee durente 1 h e 37 C
en preeencie o eueencie de eeletonine (10-3"). Loe reeultedoe
ee expreeen en ¡Ig/eg tejido eece Media Ï ES, n).
"p 4 0.05 en relación el control (tee: de Student).

Bl
III.l.2 - Caracterización de sitiog_gg unión EEEEpíFicospara Eros
taglandinas en 1a glándula pinggl bovina.
El objetivo de 1a segunda serie de experimentos Fue 1a evalua
ción de 1a posible existencia de sitios receptores para PGs en glág
nula pineal bovina. Se utilizaron para ello sobrenadantes de 900 x
g de homogenatos pineales en un buffer que contenía indometacina
(a fin de inhibir la producción endógena de PGs, la que aumenta ma;
cadamente comoconsecuencia del Fraccionamiento subcelular) (Himball
y Uyngarden, 1975).
En la Fig. 11 se resume el experimento de tiempo para la aso
ciación especifica de PGFde y PGE2a sus sitios de unión en glán
dula pineal bovina. A 37 C el "plateau" fue alcanzado en 30 min pa
ra ambas PES. La unión aumentó aproximadamente a1 doble luego de
incrementar la concentración de Ca4+ de D hasta 2 mM(Fig. 12).
Con el objeto de obtener información sobre la naturaleza de
los sitios de unión se preincubaron las muestras con tripsina. La
enzima proteolitica afectó la unión de PGE2-3Hy PGqu-BH, dismi
nuyéndola a 16-17%respecto de los controles. Asimismo, el calentg
miento de los sobrenadantes a ebullición, previo a 1a reacción,
destruyó los sitios de unión para ambas PGs. En el caso de preparar
los sobrenadantes con buffer que no contenía indometacina, el'"bing
ing" de las PGs-3Hdisminuyó hasta prácticamente anularse (Tabla h).
La posible metabolización de PGE2-3Hy Por2m -3H luego de su
asociación a los sitios receptores fue examinadapor medio del sig
tema cromatbgráfico descripto en Materiales y Métodos (Sección II.
l). En todos los casos, más del 98%de 1a radiactividad co-croma
tografió con los compuestos puros de referencia, lo que indica

82
que los sitios estudiados no corresponden a un sistema enzimática
de degradación de las PGs.
En la stla 5 se resumen los resultados de los experimentos
de localización subcelular de los sitios pineales de alta afinidad
para PGs. La unión Fue máiima en la fracción cruda mitocondrial,
donde precipitan también vesículas y restos de membranas.
La saturación de los sitios de unión para PGE2y PGFaxen glág
dula pineal Fue examinada en los experimentos resumidos en las Fi
guras 13 y 1h. El análisis de Scatchard (19h9) reveló una única
población de sitios de unión para PGEz-BHcon una Hd = 1.2 nM y
una concentración de sitios de 1-2 Fmoles/mgde proteina (Fig. 13).
En forma semejante. los experimentos de saturación utilizando PGqu
3H revelaron una única población de sitios de unión con Hd = 1.7 nM
y concentración de sitios de 2 Fmoles/mgde proteina (Fig. 1h)
Con el objeto de establecer la especificidad de la unión ob
servada, se incubsron cantidades constantes de PGEZ-SHy PGFZu-3H
con distintas concentraciones (como minimo h) de PGEl, PGE2y
PGF2c*no radiactivas. La concentración efectiva capaz de disminuir
al 50%la unión (BESO) de las PES-3Hy la constante de disociación
para los distintos inhibidores (Hi) se calcularon de acuerdo con el
procedimiento ya descripto en Materiales y Métodos (Sección 11.3).
PGEl y PGE2 desplazaron en Forma igualmente efectiva a PGEz-BHde
sus sitios de unión, pero Fueron 2-3 órdenes de magnitud menos po
tentes en sus efectos sobre la unión de PGF2°(-3Ha sus sitios reg
pectivos. Por su parte, PGF2C¡Fue unos 3 órdenes de magnitud me
nos eficaz que las PGs de la serie E para desplazar a la PGEz-JH
de sus sitios de unión (Tabla 6). Estos resultados indican que los
sitios de unión de PGE(PGE1y PGEé) son una entidad distinta e lo.d.

F19. 11:
83
6- ——0PGF2°¿<<"‘no!iso‘rnos
U
CL“5,2
l 41o ao 60
TlEMPO(min.)
asociación un PGF -3H y PGEz-JH a1 cobren-dante de 900 x g2*dc glándulas pincalco bovina! un función dal tic-po dc incu
bac16n o
La unión especifica de Pcs-JH o. dctnrninb comoII dolcrlbe
en Materiales y Métodos luego de incubar alícuota. de 200 ul
del sobrenadantc de 900 x g de pincel bovina en prenancia de
Pao-JH a 37 C durante los periodo. de tic-po indicado. un 1a
figura.
Cada punta es la Medi. de k determinaciones.

8h
PGEz PGon<DDUI ñ
8 I JÜ! I
’o I
(femtornoles/mgprot.)
1|
PG-‘HUNIDAESPECIFICA
SD(D I
O 0.5 2.0 O 0.5 2.0
Concentración de Co" en el medio (mM)
Fig. 12:3Efecto del CaH eobra 1a unión eapecifica de PGe- H e
preparacionea crudaa de neoprenaa pinealea.
Alicuotaa de laa preparecionea de aeabranea aa incuberon
cono ae deacribe en Heteriaiea y "¡todoo en auaencia y
en praaencia de 0.5 o 2 ¡H de Ce**. Loa reaultaooa ae a!
preaen colo Media 3 ES (n a k en cada grupo). Loa datoa
fueron analizaooa por análiaia de varianza aeguido por
el teat de Dunnatpara conparacionaa aúltiplea.
La variación intarenaayo fue lenor que el 10%.

85
PGEz-3HUNIDAESPECIFICA
RECEPTOR PARA PGE EN GLANDULA PINEAL BOVINA
(femtomoles)
'Ï‘Ov
” 2—J\D
(DD p
PGE2 -3H UNIDA(no42 M)
l l
5 10 15
[PGEz - 3H] (x10'9M)
F19. 13: Unión de PGE2-3Ha membranas de glándula pineal bovina3
como función de concentraciones crecientea de PGEZ-H
Cada punto ea la Media de cuadruplicadoa de muestra. A
1a derecha ae muestra el gráfico de Scatchard.

86
RECEPTOR PARA PGFz o< EN GLANDULA
zoí PINEAL BOVINA
g, 15 u.6'UJn.un
“J 1o
9‘zD:c7 5‘ o¡f
LE ° ¡o zo ao
g 3H-PGF2-cUNIDA(¡num
o 1 l I 1s ¡o 15 zo
[PGP2,L-3H1 (xlO'gM)
F19. 1h: ¿“73H a membranas de glándula pineel bovine.encomo función de concentraciones crecientes de PGFZaTJHen
Unión de PGF
el medio.
Cada punto es 1a Media de cuadriplicadoe de muestra. A le
derecha ee muestra el gráfico de Scatcherd.

87
Tabla k: Efectoa de 1a preincubación con tripaina. axpoaición a
alta temperatura o incubación an ausencia da inca-¡taci
3 -3H a citioa capacin y PGFZ‘na sobre la unión de P652
ficos en glándula pineal bovina’.
s DEtuumcmu
PGEz-JH Pilian -’H
Iripaina a} i 10 al. 1 9
Calor (100 c. 10 nin) ae t 10 99 1 12
sm indometacina 95 1 9 100
'Laa nueatraa fueron preincuhadaa a 100 C durant: 10 nin a con
10 ng de tripaina a 37 C durante 15 nin. Loa controlan fueron pc!
incubadoa a 37 C an auaancia de enzima protaolitica. Loc dataa aa
prelantan cono Media 3 ES (n - k).

'(auïfigdazuaïnñïaatuaagutzuog)
6TDZ'D; LS'ISD’D; 99’Erescata
GZOí‘O; av°zLD'D1DG'D3311365KDOOSDIIfiOC'D; Sï'íOS’O; IS’H43113651OOOLZ
IIUI'O; SG'O{Z'D; DE’Z¡ÜIIIU5l006
_*B.HE39dLCD’D; BZ’DLD'O; 59'0IDBOQIS
{íSI’D; hfi’I0050’0; ZS'OZittid5XUDUSOTavvt'o; 6L’Icatre-o; zc°zzattád6xnunca
BIBO’O; ¿L‘DZO'D; 6I'DiittadÜ!006
tngo:rapgopa-quantro:Ennat/catan;saidGI/aato-¿Hí-Z3fld
VOIJIOBdSBVGINnHï-Sd
°.autnnq
Inautd¡Inpuqtñuau
{-agd¡saduqtunapnotaynnotap¡otntaaqnaugtnnthaota
:5¡1091

B9
Tabla 5 (continuación)
’Loa homogenatoa de glándula pineal bovina ae fraccionaron por
centrifugación diferencial y. en laa diatintaa fracciones, se
midió la unión eapecifica de PGa-JHsegún ae describe en Hate
rialaa y Hétodoa. Alicuotaa da 200 ul de laa diatintaa fraccio3nea ae incubaron durante l n a 37 C con 10'12H de PG- H en pra
aencia o auaencia de l nH de PGa no ¡arcadaa.
Loa reaultadoa ae expreaan como Media Ï ES (n-k en cada grupo)
Loa datoa ae analizaron por analiaia de varianza aeguido por
teat de Tukey(l9k9) para conparacionea núltiplea.
"Diferancia aignificativa con reapacto al precipitado de 900 x g
(p 40.05) y el precipitado da 27000x g (p 40.01).
'°'Diferencia significativa con reapecto al precipitado de 900 x g
y el precipitado de 105000 x g (p < 0.01).

9D
ïabla 6: C650 y H1 de una variedad de PGa en loa eatudioa de con
petancia por loa altioa de unión para PGEz-JH(A) y-3H (B) en aenhranaa de glándula pineal bovina‘.PGFZ“
(a) Paez-3ta
PG 0550 (nH) K1 (nM)
El 5 0.65
E2 2 0.26
ru mou 130
(a) PGF -3H2.4
pl; c250 (nn) H1(nH)
t:1 125 22.5
E2 mua 180
F2“ 1.2 0.22
(continúe en la siguiente página).

91
Tabla 6 (continuación)
’Les preparaciones de ¡entrena se incubaron durante 1 h a 37 C con
cantidades constantes de PGEz-JHy PGF2.‘-3Hcon distintas conceg
traciones (colo ninino k) de P651, P582 y PGFZ.<no marcadas. Losdatos de unión especifica obtenidos se graficaron en papel log
probitos para determinar la concentración de 1a PGque cause uns
inhibición del 50%de la unión náxias de PG-JH (BESO). Las pen
dientes e intersecciones se calcularon por análisis de regresión.
La K1, constante de disociación del agente inhibidor de la unión
de PG-3H, fue calculada a pertir de la fórmula: H1 = ceso/1 +
+ (PG-3H)/Hd). donde Hd es la constante de diaociacibn para 1a
PG-JHrespectiva.

92
III.1.3 - Efectos de la indometncínï sobre la sintesis de me!atoni—
naAv el metabolismo moneaminérgíco de la glánduia pineal
de rata
El efecto estimulatorio de ln NEsobre la liberación de PGs
pineales descripto en lu Sección 111.1.1 planteó La pasiuilidad
que éstas actúen en 1a secuencia de Fenómenosdesencadenados por
la liberación del neurotrunSmisur de los tvrminales pineales y
que conducen a la estimulación pineal.
E1 objetivo de esta serie de experimentos fd« ‘a! r Tí
1a inhibición de la sintesis de PGsig_vivo afecta distintos paré
tros de 1a Función pineal. Comolos efectos de las PGs nueden ser
pre o postsinápticos, se utilizaron comoindicadores diversas ac
tividades enzimáticas, representativas de uno u otro sector, en a
nimales inyectados con indometacina, un conocido inhibidor de la
sintesis de PGs(ver Sección 1.2.1).
En una primera serie de experimentos los animales Fueron in
yectados con indometacina (5 mg/kg) o vehículo a las 1800 h del
dia previo y a las 0900 h del dia del experimento y Fueron sacri
ficados 1 h más tarde.
El tratamiento con la droga resultó en un incremento del 55%
de la actividad de MAGtipo A (presinóptica) y una disminución del
37%de la actividad de MADtipo B (postsináptica) (Tabla 7).
En la Tabla 8 se resumen los cambios en las actividades de
HIÜMTy CUMTdespués de la inyección de indumetacino. La actividad
de CÜMTno se afectó mientras que la de HIÜMTresultó disminuida
en un 79%. A titulo de comparación se resumen en la Tabla 9 los
cambios de dichas enzimas en el HMBde los mismos animales. Sólo

93
la actividad de HAUtipo A re5ultñ modificada por el tratuñieHLU
con indometacina, observándose un incremento del 66%(p'(0.01).
Las actividades de la MADtipo B y de la CENTno se afectaron por
el tratamiento.
Con el fin de examinar el efecto de la indnnrzaclno sobre la
activación de la sintesis de melatonina pineal durante la Fase de
oscuridad del ciclo diario, los animales recibieron una única do;
sis de la droga (5 mg/kg) o el vehiculo l h antes de Finalizar el
periodo de luz y Fueron sacrificados en la cuarta hora de oscuri
dad (ÜlÜÜ h), momento en el cual 1a actividad SNATy el contenido
de melatonina alcanzan los valores máíimos (Wilkinson y col., 1977).
La inyección de indometacina produjo una disminución signifi
cativa del 60%de la actividad de SNATinducida por la oscuridad
y redujo en un 27%el contenido de melatonina pineal (p< 0.05)
(Fig. 15).

9k
Teble 7: Efecto de le indonetecine eobre iae ectividedee de MAO
tipo A y tipo B en glánduie pineel de rete'.
nao
tipo A tipo B
VEHICULO 24.50 i 281 (a) 12621 1 1600 (6)
momcmcmn 3791 t 364.(7)" 7932 I 1573 (7)"
’Grupoe de retee recibieron indonetecine (5 ng/kg e.c.) e lee
1800 h del die enterior y e lee 0900 n del die del eecrificio y
fueron eecrificedee 1 h nee terde. Loe reeultedoe ee expresen en
peoiee/mg prot./n. Media Ï ES (n).
"p 4 0.02 en reiecibn ei inyectado con vehiculo (teet de Student)

95
stls 8: Efectos de 1a indonetacina sobre las actividades de CUHÏ
y HIOHÏen glándula pinssl de rsts’.
con Hum
VEHICULO 76:. 1 165 (a) 127 i 67 (6)
IuomEïncmA 915 1 21.9 (4.) 27 i 13 (5)"
’Lss rstss fueron tratadas comoen s1 experimento de ls ïshls 7.
Los resultados ss sxprsssn en pnolss/ng prot./h, Media 3 ES (n).
’°p«< 0.01 sn relación s1 inyectado con vehiculo (test de Student).

Tabla9:EfectosdelaindometacinasobrelasactividadesdeMAGtipoAytipo8yCDMT
enHHHderata'.
MAOAHAOBEDIT
VEHICULO17611.1Ï 1117.6(B)24.700.“t 1192.0(0)3051.0t ¡403.9(7) INDCHEÏRCINA29521.3t 277k.5(BW2kk59.81 1653.1(B)lo2k9.loÏ 36h.?(8)
'LnratasfuerontratadascanoenelexperimentodclaTabla13.Losresultadosseex
presancanomoles/Igprot./h,MediaÍ ES(n).
"p40.01enrelacióna1inyectadoconvehiculo(testdeStudent).
96

97
Ü vc"mmm(sm/Kg)
¿oooí 4000l:
t 2- 3000- g 30002 a‘-— a
E É;;7 E .s2 \
3' zooo- g 2000- ——2 % E
¡ooo- ' //, moo(1) :(0)/ (a)7 /
Contenido de Melatonina Pinool Actividad de SaroíoniM- N-AcomTransforma
Fig. 15: Efecto del tratamiento con indoaetaclna aobre la actividad
da SNATy contanido da aalatontna pinaal durante al pario
do da oscuridad.
Laa rataa fueron inyectadaa con indanatacina (5 ag/kg) a
laa 2000 h y aacrificadaa a laa 0200 h dal dia alguianta.
Loa raaultadoa aa expresan comoMedia Ï ES (n).
p < 0.05 en relación a1 inyactado con vehiculo (taat da
Student).

9B
III.l.b - Efecto de las Erostnglandinas sobre la actividad de
SNATen la glándula pineal de rata "in vitro”
El objetivo de esta serie de experimentos fue la caracteri
zación del efecto de las PGssobre la sintesis de melatonina ¿g
vitro. Se utilizó para ello el parámetro mássensible, la antivi
dad SNAT, la que se inhibió por indometacina en un 80%, mientras
que el contenido de melatonina cambió sólo en un 27% (Fig. 15).
En la Fig. 16 se resumen los efectos de las distintas PGs sE
bre la actividad de SNAT.Sólo PGE?la incrementó a todas las con
centraciones estudiadas, presentando una curva en campana con maxi
mo en 0.1 uM. PGEl sólo tuvo efecto a la máxima dosis usada (1 uM).
Con el objeto de determinar si el efecto estímulatorio de
PGE2sobre la actividad de SNATrequiere la integridad presinápti
ca, se incubaron glándulas pineales de ratas sometidas a gangliec
tomia cervical superior 7 dias antes, en preSEncia de distintas
concentraciones de PGEZ.A todas las dosis estudiadas (0.01 - 1 uM)
el efecto estimulatorio de PGE2Fue significativamente mayor en las
pineales desnervadas que en las glándulas de animales controles
(Fig. 17).

F19. 16:
99
:‘c
á 150''<a:uuu.ZÏ'<o:¡:5’- í:u1..f¿.5100z E¿t EZ cn
C2,2
É 3.tfi 3uJ E 50"C31:o v PGE|á2 96a.}l- ——9|5 Cotoz
2 Mil-¿Joe1 1/ l l 1 1
o c -9 -a --7 -6
[PROSTAGLANDINA].Iog M
Efecto ds los PGs sobre lo actividad de SNATdo glán
dula plneal ds rsto ln vitro.
Las glándulas pinceles ss lncubsron durant. 6 h sn ns
dlo IC 199 sn presencia ds los distintos FGs. Ls seti
vldad SNAÏse detsrllnó comoss dssorlho sn Hstorlslss
y Métodos. Los rssultsdos se expresan cono Hedls 3 ES
(n = 7 en csds grupo experimental).
’p 4 0.05 en relsclbn sl grupo control (C) (test de
Dunnet).

IÜÜ
4 200 *
É T
E la)
" * III \\\:! ¡r \E l’ \\g I \\\
á A Il \\
g É ¡00 ’ ¡II I \\ *ig ,’ ‘lE53? / ccsme ,13 ¿e x Ïo g 50r- lse IWe ¡5’2 I"g 174M
n ?l ¡l 1 n A 1" c " -9 -3 -7 - 6
[PROSTAGLANDINA Ez] Log M
Fig. 17: Efecto de las PGs sobre 1a actividad de SNRTde glándula
pineel de rata in vitro.
Lea glándulas pineales. obtenidas de animales gsnglieoto
mizados o sometidos a operación simulado 7 dias antea del
experimento, se incubsron durante 6 h en medio TC 199 en
presencia de P652 comose describe en Materiales y Méto
doa . Loa resultados se expresan como Media Í ES (n a 7
en csda grupo experimental).
'p < 0.01 en relación el grupo control (C) (test de Student

101
III.1.5 - Efecto de PGEasobre la acumulación de AMP y su uniáh a
sitios receptores intracelularea en la glándula Einealde rata "in vitro".
Cómoya Fue discutido en lo Introducción (Sección 1.1.3) 1a
inducción de la SNATestá mediada por la activación oc una adeuii
ciclasa; en respuesta a la estimulación {b-adrenérgico se pruduce
un incrementó del contenido de AMPÚpinoal (ÜJeshansky y JuFF,
1978; Zatz, 1978). Ya que la PGE2produjo activación de la SNAT
y ante la posibilidad de que dicho Fenómeno fuese medi du pc: HMPC
se estudió el efecto de la miamasobre el contenido glandular del
nucleótido cíclico y su asociación a 1a suhunidad reguladora dc la
quinasa de proteinas.
En la Fig. 13 se resumen lds efectos de las distintas PGs so
bre la acumulación de AMPcglandular. Sólo PGEZincrementó signifii
cativamente el ccntenidd del nucleótidu cicliLo en un rango de con
centracicnes que varió entre 0.01 y l um. PGE: (0.1 uM) determinó
una disminución significativa de los niveles (e AHPC.
La incubación de glándulas pineales con r.05 uMda PGE2dote;
minó una disminución del 37%en los sitios de unión desocupados de
1a quinasa de proteínas, sin modificar la afinidad de esta unión
(Fig. 19). Las constantes de disociación (HU = 3A-SBnM) son del
orden de las descriptas para diversos tejidos (Sanbdrn y col.,
1973; Talmadge y col., 1975; Marcus y 001., 1979).
Bajo condiciones de incubación similares, el número total de
sitios de unión para AMPcno fue alterado por lo presencia de PGEZ.
Sin embargo, la concentración de sitios en ambos grupos se redujo
a un tercio después de 16 h de incubación a 5 C (Fig. 20).

F19. 18:
102
*
30'- .a‘iE2
* *
4% 20- Pr:on<1tu cz ‘2 15cmF2“a: a
E, É PGlzo oÉ É PGE!< 8
10¡k
o 1 ’IIÍ 1 l Ll
-9 - 8 - 7 - 6
Í PROSTAGLANDINA] loq M
C
Efecto do las PG: sobre 1a oculuiocibn de AMPCpincelin vitro.
Las glándulas pinceles no incubaron duranto 15 nin en
presoncio o: lo. distintos PGCy lo concontrocibn do
AMPcse determinó por al procedimiento de unión o la
quinooo de proteinas doocripto on Notariales y H6todoo.
Loa resultado. oe oxprolon como Medio 1 ES (n a 7 poro
codo grupo experimental).
’p <.D.USen relación oi grupo control (oniiioil do [g
rionzo. tout de Dunnot).
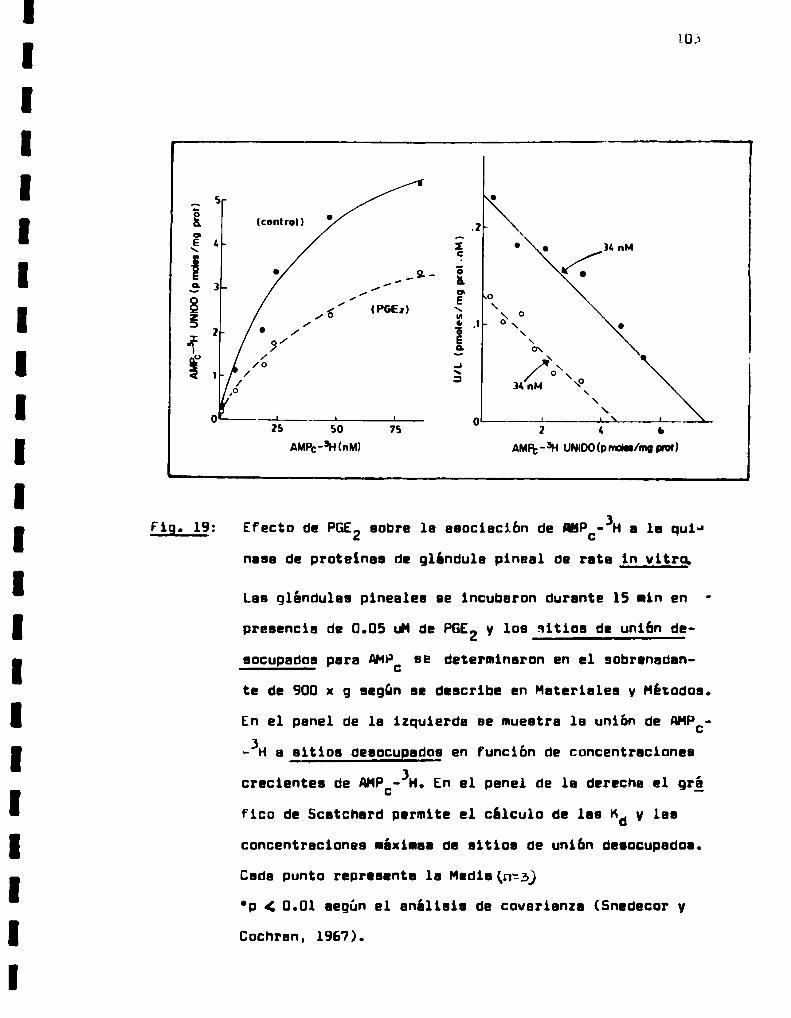
[Uh
A s
i (control)o A.
í ‘ E_ 2 - 7'ea a8 (PGEI) É2 vIa g .'f ’ á
g" 3‘ 3
\o l n 1 I \ 1
25 so 75 " 2 4 o
AMPc-’H(nM) AMPc-SHum00(pmua/mg pm)
Fig. 19: Efecto de PGE2sobre la asociación de flBPc-JH a la quie
naaa de proteinas de glándula pinesl de rata in vitro.
Las glándulas pinesles se incubsron durante 15 nin en
presencia de 0.05 uH de P662 y los sitios de unión de
socupados para AHPCse determinaron en el sobrsnadan
te de 900 x g según se describe en Materiales y Métodos.
En el panel de la izquierda se muestra ls unión de AHPC3
- H a sitios desocugados en función de concentraciones
crecientes de AHPc-JH.En el psnel de ls derecha el gré
fico de Scatcnard permite el cEICulo de las Ku y lasconcentraciones máximasda sitios de unión desocupados.
Cada punto representa 1a Madiaang)
’p <.D.Dl según el analisis de covarianza (Snadacor y
Cochrsn, 1967).

10h
ñ .mm \'éa g í.\ ‘6g ae Lo Ée \
0|
s %' Eo 3 //*\
s a MnM \x \o s3 \
‘\\\
l l l __ , __ -7, _._.,_o L0 so ¡zo o ; 2 '—'
MPC-SH (nM) AMPCJH UNIDO(pmoles/mgpm)
Fig. 20: Efecto de PGE2sobre la asociación de AHPc-JHa la quine
ss de proteinas de glándula pineal de rate in vitro.
Las glándulas pinceles se incuboron durante 15 sin en pr!
sencia de 0.05 uH de PGE y loa sitios de union totales2
para AMPcse determinaron en el sobrenadsnte de 900 x g
aegún ae describe en Materiales y Metodos. En el panel
de ls izquierdo se muestra le union de AHPc-JHa sitios
totales en función de concentraciones crecientes de AMP:-3H. En el panel de le derecha sl gráfico de Scetcnard
permite el cálculo de las Kd y las concentraciones de sitios de unión totales . Cada punto representa la Media
(n:39

105
111.1.6 - Efecto de 1a PGEZ,NEy blogueantea de 1a einteaia de
Eroatagiandinaa aobre 1a liberación de nelatonina por
1a glándula Eineal de la rata in vitro.
La aiguiente aerie de expariaentoa fue realizada con el chi;
to de caracterizar: a) el efecto de 1a PGE2aoore 1a liberación de
neiatonina, producto final de 1a aecuencia eatudiada en lua eaccig
nea ¡11.1.3 a 111.1.5: o) la interferencia. por tree agentee con
actividad inhioitoria de 1a ainteaie de PGe. en el eatinulo adre
nérgico de 1a liberación de neletonina in vitro.
En la Tabla 10 ae reaunen loa reaultadoa de 1a curva doaia
reapueata para dietintea concentracionea de PGEZ.Dicha PG incre-c
aantó en forma significativa 1a concentración de aelatonina libe
rada a1 ledio de incubación a partir de concentracionea 10' H.
El agregado de NE (10' H) a1 nedio de incubación de glándu
iaa pinaaiea auaentó en aproximadamente20 vecee 1a liberación de
aelatonina (table 11). Eate efecto fue innioido en foraa aignificg
“H de indowetacina. ácido acatiiaalitiva, aunqueparcial, por 10'
ciiico o acido nefenánico. A estas concentraciones los 3 agentes
inhiben en Forma total la sintesis de PGs (Flower, 197b).

ïabla 10:
106
Efecto de la PGE2sobre la liberación de nelatonina almedio de incubación de glándulas pinealea de rnta’.
pg de neiatonino liberados/pl
naci/6 h de incubación
CONTROL 2592 t 7m (6)“
-9 +Pal-:2 10 H 7372 — 923 (7)
iO'BH 5359 Í 501 (10)
10’7n 4.959 1 312 (5)
10's" unas t ¡.31 (7)
'Glándulna pinceles de rata fueron incubadac durante 6 h a 37 C un
medio TC 199 en presencia de PGE2 (10
ce expresan cono Media Í ES (n).
-6 - 10-9"). Los resultados
"p < 0.01. difiere en forma significativa de inn grupos con PGEZ.
análisis de varianza (test de Scheffé).

Table ll:
107
Efecto de lo NE, en presencia o ausencia de indonetscine.
ácido ocetilselicilico o ácido nefenánico sobre ls libe
ración de nelatonins s1 medio oe incubación de glándulas
pinesles de rsts’.
pg de nelstonins liberados!
pinsal/G h de incubscibn’
coumm 1229 t 331
nc 10's" 253w t nos"
NE10'5n, indonetocins 10"" 11136 3 953°"
m: món, ácido ocetiloolicilico 10'“n 88321 556'”
NE10-5", ácido nefenánico 10-“H lSkkO t 1290...
’Glándulss pineslee de rate se incubsron durante 6 h s 37 C en
medio IC 199 en presencia de las distintas drogas. Los resulto
dos se expresen cono Media Ï ES (n a 5 psrs csds grupo).
"Difiere en forma significativa de los demásgrupos (análisis de
varianza. test de Scheffé. p.< 0.01).
”’p <.D.Dl. difiere en forme significativa del grupo control, eng
lisis de vsrisnzs (lost de Scneffé).

108
III.1.7 - Efecto de la PGí1 sobre le liberación de NE-JHEor_glán«
gyla pineal de rata in vitru
El objetivo de esta serie de experimentos fue examinar el efec
to de PGE2en concentraciones Fisiológicas sobre la liberación deNE-JHpor la glándula piueal. Existe concluyente oemUeración que
luego de una inyección ¿n xixg de NE-JH(Cardinali y col., 1975 c)
o de la incubación in vitro con el compues;n marcado inelavo y co1.,
1977). el neurotransmiaor es captado por los terminales adrenérgicoa.
Ante un estimulo eléctrico o luego de la exposición d? la glándula a
una solución de ClH BOmH(que despolariza a los terminales). el neu
rotransmisor se libera por un proceso semejante al fisiológico.
Los resultados de este experimento se resumen en la Taola 12.
La PGE2(10'7M) results en inhibición de la liberación de NE. La in
oubación con cantidades menores oe PGE2(lÜ'BM) no alteró la libera
ción de neurotransmisor producida por BDmMoe H‘ en el medio.

109
Tabla 12: Efecto de la PGE2sobre la liberación de NE-JH por glándu
la píneal de rata in vítro’.
Liberación Fraccionai (x 10-3)
s. sn .1 ¿ 52/5l
Control 20.9 Í 2.a 13.3 Í 2.2 0.61. i 0.09
PGE2 10’8M 21.1. i 3.0 13.2 Í 3.o 0.51 - 0.10
9135210'7M 20.9 i ¿.9 5.9 i 1.3" 0.33 Í 0.05"
’Lna depósitos de NEen los terminales simpáticos fueron marcadas
por incubación con 0.5 uMde NE-JH durante 30 min. Luego de pro
ducirse el equilibrio en el eflujo espontáneo de radiactividad.
la liberación de neurotransmisor se estimuló por exposición duran
1 min a BÜ mMde H+. S1 corresponde a la primera exposición y 52
a 1a segunda, 35 min más tarde. La PGE2 o vehículo se agregó al
medio de incubación durante los 20 min previos a 52. Los resulta
dos se expresan como Media Ï ES de E determinaciones.
"p 4 0.05 en relación al control.

III.
llÜ
2 - PARTICIPACION DE LAS PRUSTAULHNDIHAS EN EL MECANISMO DE
ACCION DE LA HURMDNA PINEÑL HELHTuHINÁ
Los experimentos a analizar a continuación se originaron
a partir de la hipótesis que la melatonina y la indometocina
no sólo comparten similitudes estructurales e inmunogénicas
(Levine y Riceoerg, 1975), sino también un mecanismo bioquí
mico, la inhibición de la síntesis de PGs. Tal mecanismome
diaría los efectos neuroendócrinos de ambas sustancias de ng
table semejanza. Esta hipótesis está avalado por los observa
ciones resumidas en la Tabla 3.
El agregado de la hormona pineal melntonina inhibió, co
mola indometacína, ln sintesis de Püc en la glándula pinoal.
Dichos resultados sugieren la presencia de una retro-alimen
tación de la vía biosintética de lo melntonina a través de la
síntesis de PGs.

111
III.2.1 - Inhibición por melatonina de la producción de prostaglan
dinas en HHBbovino 1723 rata "in vitro".
Con el fin de verificar la presunta participación de las PGs
en el mecanismo de acción de la melatonina se incubaron HMBbovino
y de rata en presencia de distintas concentraciones de la hormona
pineal.
Se llevaron a cabo tres series de experimentos. En 1a Tabla
13 se resumen los efectos del agregado de melstonina al buffer de
incubación sobre la producción de PGE y PGF por el HMBbovino. avg
luada por ensayo biológico. Luego del agregado de l_mMde malatoni
na se detectó un marcadoefecto inhibitorio del efiujo de ambosti
pos de PGa. En presencia de 0.1 mMy 0.01 mMno se observó tal e
fecto inhibitorio (Tabla 13).
La concentración a la cual 1a meiatonina es activa en el HHB
bovino es varios órdenes de magnitud mayor que 1a Hd de los sitios
receptores detectados en 1a mismapreparación (Cardinali y col..
1979 a). Este hecho planteó 1a cuestionaoilidad del significado big
lógico de tal hallazgo. Decidimosentonces utilizar una preparación
experimental de HMBde rata, descripta por Ojeda y col. (1979 b),
como sensible para detectar cambios en el efluJo de PGE2producido
en respuesta a distintas catecolaminas. Asimismo, en esos experimeg
tos ae evaluó la concentración de PGEpor un radioinmunoanálisia eg
pacífico.
En 1a Tabla 1h se resumen los efectos de concentraciones suomi
cromolares de melatonina sobre 1a liberación espontánea de PGE2por
HMBde rata. La melstonina (0.01 - 100 laH) disminuyó un h7-6hï dichaliberación.

112
En otra serie de experimentos se estudió el efecto de distig
tas concentraciones de melatonina sobre la liberación de PGE2indg
cida por NEen HMBde rata (Fig. 21). Se utilizaron dos concentra
ciones de NE (1 y IDOuM) y distintas concentraciones (0.1 - 1000 uH)
de melatonina. Comoya Fuese demostrado por Ojeda y col. (1979 a),
el agregado de l uMde NE no afectó la liberación de PGE2 por el HHB;
sin embargo, en presencia de esa concentración de neurotransmisor. la
melatonina fue menos efectiva para disminuir la generación de PGEpor
HMB(compárese con 1a Tabla 1h).
Cuando las incubaciones ae llevaron a caoo en presencia de 100 uH
de NE, 1a liberación de PGE2aumentó al doble y 1a melatonina revirtiátal efecto estimulatorio. La dosis efectiva mínimade melstonina fue
0.1 - 1 uM, según los experimentos.
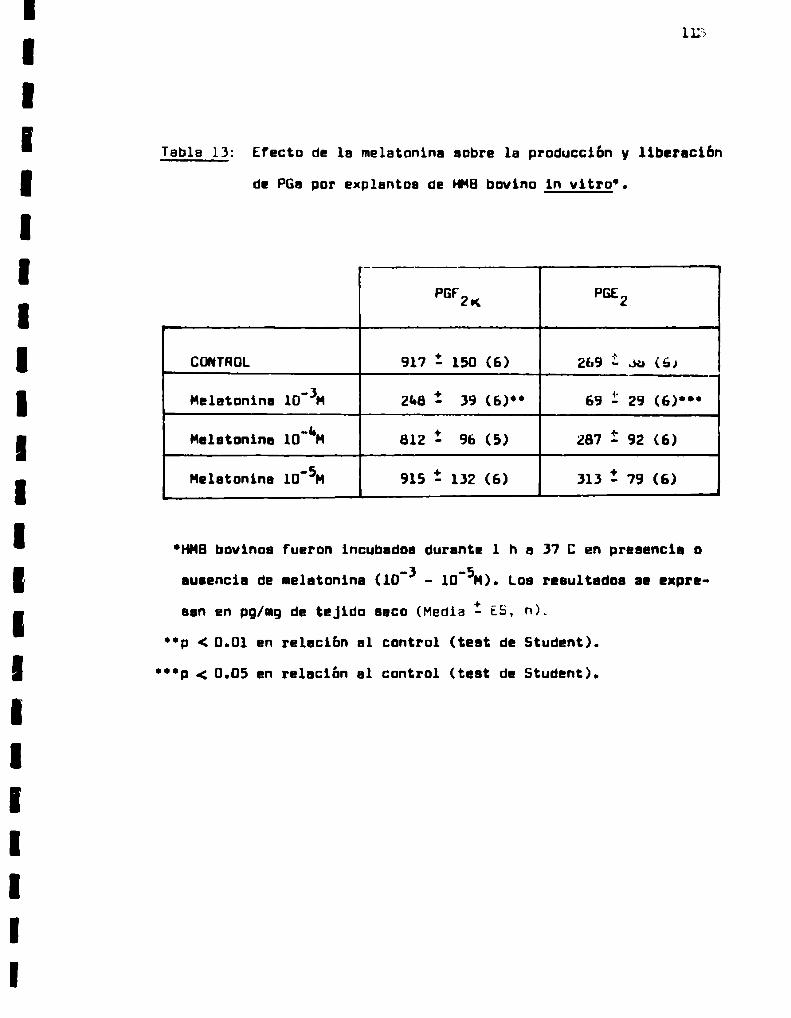
IES
Tabla 13: Efecto de la melatanina sobre la producción y liberación
da PGa por explantoa de HHBbovino in vitro’.
PGFZ" 9362
common. 917 1 150 (5) 259 i ¿a (5)
Halatonina 10"" 21.3i 39 (5)" 59 Í 29 (6)'“
Helatonina 1o“"n 312 i 96 (5) 257 t 92 <6)
Helatonina 10’5n 915 1 132 (e) 313 1 79 (6)
'HHBbovinos fueron incubadoa durante 1 n a 37 C en preaancia o
ausencia de aelatonina (10'3 - 10-5"). Loa raaultadoa aa axpra
aan en pg/ng de tejido aaco (Media Ï ES, n)»
"p < 0.01 en relación a1 control (teat de Student).
”’D < 0.05 en relación al control (teat da Student).
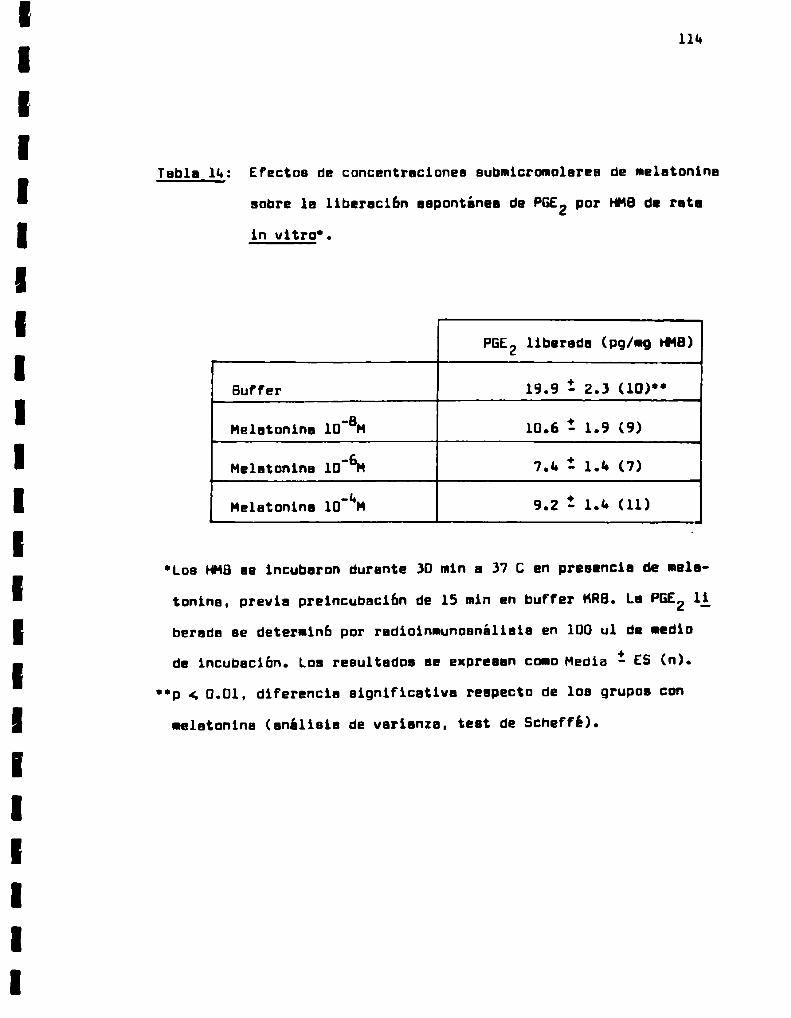
11h
Tabla 1h: Efectos de concentraciones aubnicromnlarea de nelatonina_aobre ia liberación eapontanea de PGE2por HMBda ratain vitro’.
PGE2 liberada (pg/ag HHH)
Buffer 19.9 1 2.3 (10)"
Helatonina lO'BH 10.6 1 1.9 (9)
Halatonina io‘E‘H 7.4. t 1.1. (7)
Helatonina 10'“M 9.2 1 1.a (11)
'Loa HHBae incubaron durante 30 min a 37 C en preaencia de mela
tonina. previa preincubación de 15 min en buffer HRB. La PGE2li
berada ae determinó por radiainmunoanáiiaia en 100 ul de ledio
da incubación. Loa reaultadoa ae expraaan cano Media 1 ES (n).
"p 4 0.01. diferencia significativa reapecta de loa grupca con
aelatonina (análiaia de varianza. teat de Scheffé).

,¿aga
- ¡WM vo‘n ¡o‘u‘ _ _ .
- - no"M ¡I‘M 10"" ¡0"MJ
115
I
1.o i
8 I .:5. I ‘\\\\‘- - Veh : \\\ - V0“g NE10-M I m NE|0"Mx. ' \\\
I :»\\a | \ '\\1:: \¿¿ \e ' \‘O I \ x N:9 I \\*\‘L I Em l<9 la ¡o la l
l
lV
PConcentración de Melatonina en el Medio
F19. 21: Efecto de la melatonina sobre la liberación inducida por
NE de PGE2 por HHBde rata in vitro.3
Se agregó melatonina (10' - 1D'BM)al medio de incubación
de los HMBde rata que contenía 10-6M de NE (Exp. 1) o
“M de NE (Exp. 2). Los resultados ae expresan como10'
Media I EE (n = 5 en cada grupo experimental).
Exg. 1 (panel izquierdo): las diferencias entre el grupo
tratado con NEy aquellos incubados con NEmás melatonina
son significativas. (p < 0.05. análisis de varianza, test
de Dunnet).
E¿E¿_g(panel derecho): las diferencias entre el grupo tra
tado con NEy los restantes fue significativa (p < 0.01. a
nálisis de varianza. test de Dunnet).

116
III.2.2 - Inhibición por la melatonina de efectos vinculados a la
liberacion de prostaglandinas en útero de rata y Elaguetas humanas.
Con el objeto de evaluar la hipótesis que la melatonina inhibe
1a sintesis de PGsen varios tejidos, se utilizaron otras dos prepg
raciones, útero de rata y plaquetas humanas, en las cuales ciertos
efectos biolóbicos estan mediados por las PGs. La vinculación de la
motilidad uterina oon la síntesis de PGs ha sido Fehacientemente eg
tablecida en los últimos 10 años. El útero de diversas especies sin
tetiza y contiene PGsde distintos tipos (Pickles y col., 1965; Ha
rim y Devlin. 1967; Csapo y col., 1973; Goldberg y Ramuell, 1975).
El aumento de 1a liberacion de PGs es paralelo al desarrollo de la
motilidad espontánea del útero (Vane y Williams, 1973; Harney y col..
197h) y sugiere la participación de las PGs endógenaa en los mecanig
mos que desencadenan tal motilidad (Williams y col., 197k).
Asimismolos inhibidores de 1a sintesis de PGs bloquean simulté
neamente la liberacidh y 1a motilidad uterina, fenómenos que son re
vertidos por el agregado de PGE2(Gimeno y col.. 1979).
De acuerdo con estos conceptos, el utero constituye un excelen
te sistema para examinar cambios biológicos vinculados a 1a libera
ción de PGs en presencia de melatonina.
Concentraciones de melatonina superiores a 10-7" deprimieron
la tensión isométrica desarrollada por el útero de ratas castradaa
en función del tiempo de exposición al metoxindol (Fig. 22).
Ütro sistema utilizado para examinar el efecto de la melatoni
na sobre mecanismos biológicos mediados por PGs fue el de las pla

117
quetas. En 1973, Smith y col., demostraron que el ácido araquidáni
co inducfa la agregación plaquetaria y que tal efecto era reverti
do por aspirina o indometacina (Silver y col., 1973), Estos efectos
de los inhibidores de la sintesis de PGssugieren la participación
de algún intermediario de la cascada del ácida arasuiddhico en el
proceso de agregación plaquetaria. De los compuestos cuyas fórmulas
se muestran en la Fig. 7, los que inducen agregación plaquetaria
son: P862 y PGH2 (Hamberg y col., l97h), PGE2 y PGFZG‘ (Salzman y
col.. 1976), tromboxano A2 (Smith y col.. 1977).
En 1977, Salzman y col. estudiaron el mecanismo de a9r893615“
en plasma humanorico en plaquetas y comprobaron que tal proceso
era inducido en ese sistema por PGGZ.
De acuerdo con estos datos, el plasma humanorico en plaquetas
constituye otro modelo óptimo para el estudio de los efectos de ls
melatonina sobre mecanismosdependientes de la sintesis de PGs.
En la Tabla 15 se resume el efecto de la melatonina sobre la
agregación de plaquetas humanas inducida pnr ADP. La hormona pine
al produjo una inhibición significativa de tal agregación en conceg
traciones de lfl-kM, sugiriendo que el metoxindol inhibe la sintesis
de PGs y compuestos relacionados en esta preparación.

F19. 22:
118
30
í o-o control20 o-..o MMWODIDMAMAmm ¡PM
¡0* n----aMm IWHM
\\0A\._ \\ \ \\\ \\\
n X‘\ \ *\ \ x‘l \ \' " \ \ ‘\
‘x \\ \\ \ \\ \ x\ \
Ï\decambm)
("l. l8 l 1/,
*‘—f7U
//”l
[pq
TCI
a: xx * (5)
-60- '\
BG J | l l lIO zo 30 4o 50
Ïiempo (min)
Tensión isonétrica desarrollada por útero de rsts castra
ds en función del tiempo de exposición s distintas concqg
traciones de melstonina in vitro.
Los resultados se expresan comoMedia Ï ES de los i ds
cambio respecto de los valores iniciales de TCI.
’p < 0.05, diferencia significativa respecto del control

e
i
IIIIIIIIIIIIsusIIIslsIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIli
119
Ïsbls 15: Inhibición por melatdnina de la agregación de plaqustss
hunsnaa inducida por ADP’.
Concentración de nelatonina í de Transnitancia
---- 100
0.05 un 100
0.1 un 73.5 "3“.9.’
0.2 ¡H u9.2 1 3.9-.
’El plasma rico en plaquetas obtenido según ae deacribe en Materia
les y Métodos, se preincubó durante 30 nin con melatonina antes
del agregado de 5 uH de ADP. Los resultados se expresan cano Me
dia Ï ES (n = k).
"p 4 0.05, diferencia significativa respecta del control (test de
Student).

IYDISCUSION

120
En la glándula pinesl, comoen toda estructura inervada por el
sistema simpático, las PGspueden participar en la secuencia de fe
nómenos metabólicos desencadenads por el neurotransmisor. Comose ha
analizado en ls Introducción (sección 1.2.2), existe abundante evi
dencia experimental que indica que las PGs son liberadas en la vecig
dad de la unión neuroefectora autonómica tanto en forma espontánea
comoen respuesta a estimulos diversos y que estos compuestos afectan
presinápticamente la liberación del neurotransmisor y postsináptics
mente sus efectos biológica (Nestfall. 1977). Los experimentos des
ariptos en la sección III.1.1 fueron diseñados para responder a lss
siguientes preguntas:
- ¿Se liberan PGs luego de la incubación de glándulas pineales con
NE7.
¿Existen sitios especificos de unión para las PGs en le glándula
pinea1?.
¿Modifica la inhibición de la sintesis-de PBs indicadores pre- y
postsinépticos de la actividad pinea17.
¿Tienen las PGsefectos directos sobre los pineslocitoa in vitro?
¿Afectan los inhibidores de la sintesis de PGsel efecto estimu
latorio de la NEsobre la producción de melatonina in vitro?
¿Modifica la PGE la liberación de NEpor los terminales adrenér2
gicos pineales7.
Los resultados obtenidos demuestran que la NElibera PGs de
las series E y F de la glándula pineai en forma parcialmente depen
diente de la dosis y en concentraciones consideradas comofisioló

121
gicas. La liberación de PGapor NEdescripts por primera vez en es
te trabajo, ha sido previamente observada en otras regiones inerva
das simpáticamente, por ej., riñón y bazo perfundidos de gato (Gil
more y col., 1966; Dunhamy Zimmermsn, 1970; Ferreira y col., 1973);
corazón perfundido de conejo (Junatad y uennmalm, 1973) y prepara
ción de nervio frénico-diafragma (Ramuell y col.. 1965) y de grsse
epididimsria de rata (Shaw y Ramuell, 1968).
6Men adelante) son lasLas concentraciones de NEutilizadas (10'
mismas que aumentan la sintesis de melstonina in vitro (Klein. 1978)
y por lo tanto sugieren que el efecto observado es fisiológico.
Estos resultados satisfacen, por lo tanto. un primer criterio
importante para la demostración de ls participación de lss PGs en
la secuencia de efectos metabólicos desencadansds por la interac
ción de la NEcon receptores {b-adrenérgicos de los pineslocitos.
La siguiente serie de experimentos tuvo comoobjetivo determi
nar la presencia de sitios de unión especificos para PGsen la
glándula pineal bovina. En 1972. Huenl y Homesofrecieron ls prime
ra evidencia experimental sobre un receptor para PGs en tejidos ds
mamíferos, demostrando ls unión especifica de PGEl-JH en uns fracciónparticulada obtenida de homogenatosde hepatocitos de rata. Poste
riormente ae observa la existencia de receptores para PGsen los
siguientes tejidos: útero de hamster (Uskeling y col., 1973). mem
branas de higado de rata (Smigel y Fleisoner. 197k). cuerpos lú
teos bovinos (Roo, 1973: Powell y col., 1975), ovinos (Powell y
col., 197“ a) y humanos(Powell y col.. l97h b). lenduls tiroides
bovina (Moore y Wolfe. 197“) y ovioucto de conejo (Uskeling y

122
Spilmsn, 1973).
En la glándula pineel bovina la concentración de sitios recep
tores fue máximaen la fracción precipitada a 27000 x g. correspon
diente a membranas y mitocondriss y aumentó mediante el agregado de
0afl (lo que también apoya 1a naturaleza membranoaade los sitios
de unión). Tales componentes capaces de unir a las PGs con alta afi
nidad y especificidad fueron sensibles a la preincubación con trip
sina. indicando que ensu constitución participa una importante frac
ción proteica.
Los resultados obtenidos en ausencia del agregado de indometa
cina a1 buffer de incubación indica que las PGs endógenas liberadas
durante la homogeneización ocupan en forma efectiva los sitios de
unión disponibles en membranasde glándula pineal. Esto resulta en
una depresión del "binding'l de la PG-JHcorrespondiente.
El análisis de saturación de los sitios pineales indicó una
población homogénea de sitios de unión para PGFZO‘y PGE2 con Hd s3
mejante-(l-Z nH) y dentro del orden de los detectados en otros te
jidos. Estos sitios receptores fueron especificos para las serias
E y F, por lo que, aparentemente, constituyen entidades separadas.
El 'binding" de PGEz-JHfue dependiente de la presencia de un gru
po cetónico en carbono-9 y no se modificó en ausencia de la doble
ligadura 5,6-cis; es decir. debe hablarse estrictamente de un re
ceptor para PGE. Sin embargo. existe en el SNCsuficiente informa
ción comopara sostener que son las PGs dienoicas, producidas a
partir del ácido araquidbnico, las que se sintetizan en foros casi
exclusiva (Mestfall, 1977); por lo tanto, el presunto ligando ando

123
geno de los sitios especificos pinesles detectados es probablemente
ls PGEZ.En el caso del "binding" de PGP“ -3" en glándula pinesl.éste requirió un hidroxilo en la posición q_ del carbono 9 y por lo
tanto 1a actividad de las PGE1y PGE2fue despreciable.Anslizados en conjunto. los resultados de la sección ¡11.1.2
indican que existen receptores celulares para las PGsen los pines
locitos y constituyen la primera demostración de sitios especificos
de unión para las PGs en región alguns del SNC.
Comose analizó en ls Introducción (sección 1.2.2), las PGs
ejercen efectos pre y postsinópticos sobre ls unión neuroefectors
autonómica. Estos efectos son en general inhibitorios sobre la presi
nspsis y estimulstorios sobre la postsinapsis. La siguiente serie de
experimentos tuvo comoobjetivo obtener información sobre tales fe
nómenos en 1a glándula pineal.
En los experimentos in vivo se recurrió a ls inyección de un
inhibidor de la sintesis de PGs, la indemetacina. dado que los e
fectos de las PGs inyectadas por vis sistémica son efineros (unos
pocos minutos) (Blackwell y col., 1975 s; Blackwell y col., 1975 b)
y. por lo tanto, pasibles de no detección en situaciones fisiológi
cas que requieren de la persistencia de la PGasociada s su sitio
receptor.
Desde el descubrimiento, en 1971, que la indometacins inhibe
la sintesis de PGs (Ferreira y col.. 1971), se ha acumuladoabun
dante información experimental sobre la inhibición de ls PG-sinte
tsss por drogas antiinflamatorios no esteroides (por 23., indone
tacins, ácido mefenómico). El prototipo de este grupo es ls indo

12h
metscina y ha sido ampliamente usada para elucidar el papel de las
PGaen diversos procesos Fisiológicos y patológicos (Flower. l97k;
Vane, 1978). En los presentes experimentos in vivo se administró
indometacina a ratas en dosis que producen una inhibición completa
de 1a sintesis de PGa (Vane, 1976; Flower. 1976). Tal tratamiento
fue realizado en dos situaciones experimentales: a) durante el dia,
para evaluar los cambios en enzimas vinculadas a la degradación de
la NE, como la MAOy CGMT;b) previamente al comienzo del periodo
de oscuridad, momentoen que se desencadenan los fenómenos metabó
licos que conducen s un aumento de 20 a 30 veces en la biosintesis
de melatonina pinesl.
Luego de la inyección de indometacina se observaron cambios
en parámetros pineales presinópticos (aumento en la actividad da
MAOtipo A) y postsinápticos (como la depresión de las actividades
de SNAT, HIDMTy MAOtipo B y la concentración de melatonina glsn
dular). Estos efectos pueden depender de una acción pinesl de la
droga o de su actividad sobre cualquiera de las etapas de la vis
neuronal descripta en las Fig. k y 6. Asimismo. la indometscina pg
dria, teóricamente, afectar la liberación de ciertas hormonas,sien
do estas últimas las responsables de los cambios observados.
Suponiendo que el sitio primario de acción de 1a indometscins
sea pineal, el efecto observado podria resultar de una acción sobre
la pre o postsinapais, o sobre ambossectores. Por ejemplo. 1a in
hibición por indometacina de la sintesis de melatonina sn el pines
locito puede ser consecuencia exclusiva de una liberación disminui
da de NE por aumento de 1a MADtipo A intraneuronal o alternativa
mente el resultado de la supresión de una señal modulatoria trans

125
aináptica (por ej. PGs) originada en la postsinspais comoconsecueg
cia de la interacción de la NEcon su receptor.
Varios son los mecanismosbioquimicoa que axplicsrlan los efes
toa de la indometacina sobre la glándula pineal. En primer lugar,
ai se considera que los efectos de este droga están limitados a ls
inhibición de la sintesis de laa PGs. los resultados daacriptos en
la sección 111.1.) sugieren que le prs y/o postainapsis son posi
bles sitios de acción de las PGaliberadas en la glándula pineal
por acción del neurotransmisor. Sin embargo, y comose ha señalado
en la Introducción (pág bl), la indometacins en concentraciones
submicromolsres modifica diversas enzimas, tales comofoafolipssa.
fosfodieatersas y quinsss de proteinas dependiente de ¡HPC(Flower.197“; Mentor y Hampton, 1978). Por lo tanto, un segundo mecanismo
de acción posible de la indometscina consiste en efectos de la mig
me sobre sistemas enzimáticoa no vinculados a las PGs.
Una tercera posibilidad es que la indometaoins. comoconse
cuencia de su similitud estructural con la melstonina (Fig. 23),
interfiera en la unión de la mismaa sus sitios receptores especi
ficos en la glándula pinesl y modifique el mecanismode retroali
mentación sugerido por los resultados da este trabajo y los resu
nidos en la Tabla I (Freire y Cardinsli, 1975).
Resultados previos (Vacas y Cardinali. dstoa no publicados).
no apoyan tal posibilidad. ya que la indometscina no fue un compe
tidor afectivo de la asociación de melstonina a sitios de unión en
membranaspinesles.
Un hecho que merece ser destacado es que a pesar de habarse
usado una dosis de indometocins que inhibe completamente a la PG

126
CH3o N—C CH3W“xN/ MELAÏONINA
H
CH30(DAMN \ CH INDOMETACINA
QA“Cl
Fig. 23: Fórmulas estructurales de la melatonina y la inaometacina

127
sintetaaa (Vane, 1976; Flower, 1978). 1a sintesis de melatonina
sólo resultó parcialmente afectada. Esta observación sugiere que
otros mecanismosalternativos, en los que no participan las PGa,
median la activación inducida por NEde la sintesis de melatonina
pineal, hecho corroborado an experimentos posteriores da esta Te
aia (sección III.l.6) en los que concentraciones aupramáximaade
indometacina, ácido acetilsalicilico o ácido mefenñmicosólo inhi
ben parcialmente el efecto astimuiatorio de NEsobre lalibaración
de melatonina.
La demostración definitiva del efecto de las PGaen la glán
dula pineal implica necesariamente el examende tales fenómenos ¿g
gitgg y en las concentraciones efectivas para la saturación ds los
sitios receptores previamente descriptos. Tal fue el objetivo ds
loa experimentos cuyos resultados se resumen en la sección 111.1.5.
Se utilizó comopreparación experimental el cultivo organotipico
de glándula pineal de rata. el que ha sido empleado en éste y o
tros Laboratorios con objetivos tan diversos comola alucidacibn
de la sintesis de aelatonina (Klein, 1978). el estudio de la poteg
cialidad fotorreceptora glandular (Cardinali y col., 1973a) o la rs
gulación de la translocacibn da receptores estrofilicos del compa;
timiento citoplasmatico al nuclear (Cardinali. 1977). En los pra
sentes experimentos la incubación con PGE2aumentó la actividad de
SNAÏde la glándula pineal de rata in vitro a partir de concentra
ciones l nH. Tal efecto presentó una curva en campana con rango e
fectivo entre lll].9 y lO-GHy máximoen 10'7H (Fig. 16). Elia obaqg
vación sobre ls actividad enzimática ae vio corroborado por expsrl
aentoa adicionales en los que el producto final melatonina fue de

128
terminado en el medio de incubación (Tabla 10). También en este ca
so PGE (ll).g - 10-6") modificó el parámetro estudiado; sin embar2
go, no se observó el efecto en campanadetectable en la actividad
SNAT.
Por lo tanto, nuestros resultados demuestran en conjunto un
efecto de PGE2sobre la sintesis de melstonins pinesl en la concen
tración del Ku de los receptores, compatible con los resultados ob
tenidos luego de inyectar indometscina in vivo o de agregar indoma
tscins, acido acetilsalicilico o ácido mefenámicoin vitro.
Un hecho de interés en el presente trabajo es la carencia re
lativa de efectos de la PGE1sobre la SNATen relación a la PGEZ.
Comolo indican los resultados de ls sección 111.1.2 , ambas PGs
se asocian a un sitio de unión comúncon igual afinidad aparente.
Sin embargo, desde el punto de vista del efecto biológico, la pr;
sencis de la doble ligadura 5,6-cis otorga una actividad 103 veces
mayor a la PGEZ.Esta diferencia puede depender de distintos factores, entre ellos una distinta velocidad de inactivsción por las en
zimas degradantes in vivo.
Comoya se na señalado, el efecto de PGE2sobre la actividad de
SNATfue significativamente mayor en glándulas pineales desnervadss
por gangliectomis cervical superior. Esta observación indica que sl g
facto eatimulatorio de las PGssobre ls unión neuroefectora pinesl no
requiere un componentepresináptico y es por lo tanto predominante-eg
te postsináptico. Ls potenciación del efecto de la PGE2por la gangliectomia cervical superior a todas las dosis estudiadas demuestra
que el desarrollo de supsrsensibilidsd que sigue a la desnervsción
no está restringido exclusivamente a los mecanismosmediados por rs

129
ceptores fi-adrenergicos.
Nuestros resultados están en concordancia con experieentos pr;
vioa en que se utilizó toxina colérica para activar a ls adenilciq
clssa mediante un mecanismoindependiente del receptor ¡b-adrenfig
gico (Zatz. 1977). En dichos experimentos la toxina colérics pradg
Jo un incremento mayor de la actividad de SNATen glándulas pinOl*
lea en las cuales se habia desarrollado supersensibilidad.
Es asimismo de interés comparar la aupereeneibilidad a PGE2
observada en nuestros experimentos con la descripta para el recep
tor P.-adrenérgico por Deguchi y Axelrod (1973). Estos autores ogservaron que glándulas pineales intactas cultivadas en presencia de
NEpresentaron el increeento maximoen actividad de SNATs una con
centraciñn de ¡o un, mientras que en las glándulas pinealee deeneg
vadas tal maximose detectó a una concentración unas 200 veces in
ferior (20 n"). A diferencia de lo observado en la Fig. 17 , o de
lo descripto para la toxina colerica (Zatz. 1977). la eáxiea acti
vidad SNATinducida por NEen pinesles intactas o desnervades fue
la misma. Tales observaciones indican la singularidad de la via es
tabólica de activación de la SNATen que participen las PGe. en
comparación a la mediada por el sieteea receptor (a-adrenergico1g
denilciclasa. Una prueba adicional de que ambas vias pueden ser
independientes lo constituye la inhibición parcial del efecto de
NEsobre la liberación de melatonins en presencia de concentracio
nes supranaxinas (para inhibir la sintesis de PGa) de indonetacina.
ácido acetilsalicllinn_n_ñcido nefenánico.(Tablall).
Si bien, comolos resultados en animales gangliectomizados lo
demuestran, el efecto estimulatorio de la PGE2sobre la sintesis da

130
melstonina pineal as ejercido a nivel postsináptico, ls P662 actúe
también a nivel presináptico. Tal efecto, inhibitorio de la liberg
ción de NE, está demostrado por los experimentos de la Tabla 12,
donde lÜO nfi de PGE deprimió en un “9% la liberación de NE induci2
de por un estimulo despolsrizsnte de H’. De preaominar este efecto
preeináptico, el resultado final seria una inhibición de la sinte
sis de melstonina, ya que tal sintesis es estimulsda por el neuro
transmisor. Sin embargo. lD'BH de PGEZ. una concentración que ya
ejerce el efecto estimulatorio de la producción de melatonina, ca
rsee de actividad sobre la liberación de NEpor los terminales s
drenérgicos. Esta observación está en concordancia con los result!
dos obtenidos in vivo luego de inyectar indometscina y avala la Cog
cluaión que de los dos efectos contrapuestos de las PGs en la unión
neuroefectora pinesl (inhibición preaináptica de la liberación de
NE;mediación postsináptica del efecto estimuletorio del neurotrang
misor) éste último es el que predomina.
Comofue analizado en la Introducción (sección 1.2.2) aún no
se han aclarado los mecanismosmoleculares a través de los cuales
las PGsactúan en el SNC.Tales agentes participan aparentemente
en la regulación del sistema adenilciclasa-AMPC, ya ses directameg
te s través de un receptor vinculado a la activación de la enzima
(Wolfe y Cocesni, 1979) o indirectamente modificando le liberación
del neurotransmisor (Uestfall, 1977).
En nuestra preparación de glándula pineal de rate in vitro la
PGE determinó un incremento del contenido pinesl de AMPcs partir2
de una concentración de 10 nM. Va que existe un vinculo demostrado

131
entre el sistema adenilciclasa-AMPC y 1a inducción de SNAT(Klein
y Meller, 197D; Ueiss y Crayton, 1970; Deguchi, 1973; Davis, 1978)
es posible que los resultados obtenidos luego del agregado de PGE2
a1 medio (aumento de la acumulación de AMPcy de actividad SUATde
glándula pineal in vitro) estén causalmente relacionados. Comoen
el caso de la actividad SNAT,también en el efecto sobre al AMPC
se observó una disociación entre la actividad de la PGE2y de ls
PGEI.
En 1968, Walsh y col. demostraron que el mecanismo a través
del cual el AMPCregula el metabolismo glucolitico en el músculo
liso incluye la activación de Quinasas de proteinas dependientes
de AHPC.En197Ü¡Huoy Greengard demostraron la presencia de dichas
quinasas en prácticamente todos los tejidos de mamíferos. El AHPC,una vez sintetizado. puede ser hidrolizado por una fosfodiestersss
a adenosina-5'-monofosfato o bien activar a la quinasa que foafori
la proteinas o enzimas afectadas a diversas funciones celulares.
Estas quinasas que han sido aisladas de distintos tejidos, son te
trámeros constituidos por dos subunidades regulatorias y dos subu
nidades cataliticas (Rubin y Rosen. 1975; Hrebs y Beavo, 1979). En
una célula no estimulads ambas subunidades forman un complejo ines
tivo. A través de un proceso de autofosforilación (Gresngsrd, 1978).
la subunidad catalitica fosforila a 1a regulatoria mediante una re
acción intramoleculsr. A1 aumentar los niveles de AMPcintracelular
éste se une a la aubunidad regulatoria, lo que disocia sl complejo
subunidsdcatalitica-subunidad regulatoria y 1a subunidsdcstaliti
ca. ahora libre, es capaz de catalizar la fosforilacibn de protei

132
nas. Con estos antecedentes se decidió explorar 1a unión del AMPc
a sus sitios receptores intracelulares (subunidad reguladora de is
quinass de proteinas) luego de la incubación con PGEZ.Hasta el momentono existisn datos sobre lss caracteristicas cinéticas de ls
asociación de AMPca sitios intracelulares en la glándula pinsal.
E1 análisis de ls unión del AMPca fracción sobrenadsnte de
900 x g en glándula pinesl de rata reveló la existencia de uns uni
ca población de sitios con una constante de afinidad (Hd a 36-58 nH)del orden de la descripts para diversos tejidos (Sanborn y col.,
1973; ïalmadge y col., 1975; Marcus y col., 1979). Es de destacar
que publicaciones recientes indican que la quinasa de proteinas pr;
sents dos poblaciones distintas de sitios de unión para AHPC,siendo
detectsble sólo una de ellas por el ensayo de filtración utilizadoen el presente trabajo (Dgreid y Dñskeiand, 1980). Luego de la in
cubación con 0.05 un de PGEZ,el número de sitios libres disminuyóen un 37%sin modificarse el númerode sitios totales. Es decir.
el AMPcsintetizado por acción de la PGE2in vitro se une s continua
ción a sus sitios receptores intracelulsres para desencadenar los
efectos fisiológicos que culminan con el aumentode ls sintesis de
melatonins. A pesar de la estimulación cercana al máximode la sin
tesis de AMPCproducida por 0.05 uMde PGEZ, persistib una fracción
residual elevada de sitios libres en la subunidad regulatoria de la
quinass de proteinas. Unaexplicación posible para este hallazgo es
que una fracción importante de los sitios de unión para AMPcperma
neció en compartimientos celulares no accesibles sl AMPcrecién fo;
mado. Otra posibilidad es que sólo una fracción de las células pines

133
les ses sensible a las PGs. Por último. tal "binding" residual po
dria también deberse al intercambio del AMPCendógeno unido con
AHPc-3Hdurante el periodo de incubación. Una observación semejan
te a la del presente trabajo Fue publicada por Marcus y col. (1979).
quienes detectaron 20-50%de sitios libres para AHPCen células óseas en cultivo luego de estimulación supramóximade la adenilciclasa
por PGE1u hormona paratiroidea.
En conjunto los resultados discutidos hasta aqui y presentados
en la sección III.1,indican que las PGsparticipan en la‘seouancia
de fenómenos desencadenadoa por la interacción de la NEcon recep
tores P-adrenérgicos en la glándula pineal. La NElibera PGEy PGF
de la glándula pineal. las que se unen a receptores celulares espa
cificos ubicados en los pinealocitos y probablemente en los termina
les neurales. La PGE2estimula, en las concentraciones efectivas pa
ra saturar sus sitios de unión, la sintesis de AMPC,la actividad deSNATy la liberación de melatonina observándose conoomitantemente un
aumento en la ocupación de sitios receptores para AMPcen la subuni
dad reguladora de la quinasa de proteinas. A una concentración 100
veces mayor que el umbral de la estimulación de la sintesis de mela
toninaIIa PGE2ejerce un efecto inhibitorio sobre la liberación deNE; quizás esta actividad inhibitoria constituya una señal negativa
de control que frena el sistema de producción de PGE2inducidd por
NE, una vez alcanzada cierta concentración limite de PGEzenla brecha sinóptica.
El papel estimulatorio de las PGsen la sintesis de melatoni
na pineal se ve avalado in vivo por la inhibición qee sigue a la
administrac ón de indcmetacina, un bloqueante de la sintesis de PGs,

13k
e in vitro por la inhibición del efecto estimulstorio de la NEso
bre la liberación de melatonina luego de la incubación en presencia
de indometacins, ácida acetilsalicilico o ácido mefenámico.
La Fig. 2h es una reelaborsción de la Fig. h y resume los meca
nismos celulares que participan en la sintesis de melatonina a la
luz de los hallazgos del presente trabajo de tesis.

135
__7_7“JP/"D-PGÜN;_Ü _4___<fi‘¡).__ Lu'n'ho; "‘¿‘Jw“pr»»m—T—55KEEÏÍÍÏthu
'\ ,
TSHT
Fig. 2h: Control neural de actividad pineal y mecanismoscelularea
que participan en la sintesis de melatonina

136
En la segunda parte de este trabajo de tesis se evaluó otro
aspecto del vinculo entre PGsy función pinesl: su participación
en el mecanismo de acción de la hormona pineal melatonina.
Comose ha analizado en la Introducción (sección 1.1.“). des
conocemos aún el mecanismo intimo de acción de la melstonins. Dicha
hormonaactúa a nivel de sitios receptores del SNC(Csrdinsli y
col., 1978, 1979a) en particular en el hipotálsmo y desencadena una
serie de fenómenos que conducen a la modificación funcional del eje
hipotólano-hipofisario (ïabla l).
En la Fig. 23 puede apreciarse la similitud estructural entre
la melatonina y el inhibidor de la sintesis de PGs. indometacins.
Comoya se ha expresado, tal similitud se manifiesta en la activi
dad inmunogénica cruzada cuando estas moléculas se acoplsn s la al
búsins. Asi. por ejemplo, el antisuero utilizado para el radioinqg
noanálisis de meletonins en el presente trabajo ha sido desarro
llado mediante la inyección del complejo indometacins-slbúmins (bg
vine y Riceberg, 1975).
Otra indicación de la posible similitud en mecanismosde sc
ción de la melatonins e indometacina la constituye la sorprendente
semejanza que presentan sus efectos neuroendócrinos. Ambasdrogas
inhiben ls liberación de LHinducida por estrógenos (Iamarkin y
col., 1976 a; McCann y col., 1976; André y Parrish, 1979). la li
bersción poatcastración de LH(Roche y col., 1970; McCsnn y col.,
1976) y el ritmo pulaótil de liberación de LHen ratas csstradas
(Mc Cenn y col., 1976; Moguilevsky y col., 1979).
Dos son las posibles interpretaciones que pueden ofrecerse pg

137
ra esta semejanza en los efectos neuroendbcrinos de ambas sustan
cias:
a) la nelatonins y la indometacins inhiben en forma seneJante la
sintesis de PGsa nivel del eje hipotúlano-nipofiasrio;
b) ls indonetscins ea un sgonists efectivo de la nelstonina a ni
vel de los sitios receptores para ls horeons en el HHB.Esta ul
tina posibilidad fue descartado por resultados previos de este La
boratorio ya que la competencia de indonetacina por el 'oinding'
de melatonina a sus sitios receptores cerebrales fue despreciable
(Csrdinali y col.. 1979a),
Decidimos entonces someter a verificación la primera de laahipótesis, determinando en distintas preparaciones de HHBy tejidos
periféricos el efecto de la neletonina sobre la producción y fenúqgnos vinculados e las PGs.
En le glándula pinesl y en el HHBbovinos. la nelatonina inhi
be la liberación de PGs E y F. En el HHHde rata. tal efecto se oo
servó e concentraciones suonicronolarea de la horeona. en el orden
del Hd de los sitios receptores para melatonins. Tal efecto enel HHBde rata fue observado tanto para la liberación espontánea de
PGE2 como pere la inducida por NE.
En el HHBel papel de las PGs en los necaniseos que regulan la
liberación de gonadotrofinas hs sido ampliamente documentado(Laorie
y col.. 1976; McCsnn y col., 1976; Ojeda y col.. 1979 B).Las obser
veciones iniciales fueron realizadas por Armstrongy Grinuicn
(1972):Ürczyk y Behrman(1972) y Tsafriri y col. (1972) quienes
demostraron que el bloqueo de la ovulación por 1a administración

138
de indometscina, ers revertido por el tratamiento con LHo PGs. Asi
mismo, Behrnan y col. (1972) demostraron que el LHRHtambién rever
tia el bloqueo de la ovulación por la administración de indonstsci
na en el dis del proestro. La observacion que le inyección intravsg
tricular (pero no intrahipofiaaria) de PGE2determina un ausento enlos niveles circulantes de LHen rstaa ovarlectomizadss. sugirió
que el principal sitio de accion de las PGspara estimular la libe
ración de LHera el SNC(Hsrns y col.. 197“). Esta hipbteaia se co!
probó en animales en diversas situaciones hormonales (ratas intac
tas caatradas de ambossexos, ratas hesora prepúberes y ratas cas
tradaa tratadas con estrógenos) en los que también se demostró lis
beracibn de LHluego de la inyección intraventricular de P662 (013da y col., 19796).
En experimentos in vitro Ojeda y col. (l979b)conprobsron que
dopamina y NE estimulan la liberación de LHRHpor HHH.ïal efecto
no se observó en el caso de agregar indonetacina al sedio de incu
bación. mientras que el agregado de P662 produjo le liberación de
LHRH.Este efecto estimulatorio de P682 sobre la liberacion de LHRH
es aparentemente ejercido sobre las neuronas productoras de LHRH
y no sobre las sferenciaa cstecolaninergicas,ya que distintos blo
queantes adrenérgicos no modificaron la liberación de LHinducida
por le inyección intraventriculsr de PGs (McCann y col.. 1976:
Ojeda y McCann, 1976). Comoen nuestros experimentos le nslatonina
inhibib en bajas concentraciones la liberacion de PGE2producidapor NEen explantos de HHH.y en vista de los resultados discutidos
precedentemente sobre el papel de PGE2cono intermediario en el ne

139
canismo por el cual NE libera LHRHen el HMS. se propone como hipo
teais que el conocido efecto antigonadal central de la melatonins
(Tabla l) depende de una inhibición de ls sintesis de PGs a nivel
hipotalámico. Datos previos de este Laboratorio (Cardinali y col..
1975 a) indican que ls melatonina aumenta la liberación de varios
neurotransmisores. entre ellos NE, en terminales simpáticos aisla
dos de hipotálamo de rata. Este efecto puede depender también de g
na inhibición de la sintesis de PGEZ,ya que como se ha analizado
en detalle en 1a Introducción (sección 1.2.3). y se demuestra en los
experimentos de la Tabla 12), las PGs de la serie E constituyen uns
señal trsnsináptica negativa para ls liberación de NEa nivel de q;
varsas sinapsis adrenérgicas. Por lo tanto, las consecuencias de la
inhibición de la sintesis de PGE2a nivel del HHHproducida por la
melatonins son complejas e implicarian tanto un aumento en la libe
ración de NEcomoel bloqueo del efecto de este transmisor a nivel
de 1a neurona productora de LHRH(Fig. 9 y 25).
Otra zona del SNCdonde se evaluó el posible efecto de la nel!
tonina sobre la sintesis de PGsfue la glándula pineal. Una caractg
ristica peculiar de la glándula pineal es su sensibilidad a la admi
nistración exógena de melatonins. La inyección de esta hormonaefes
ta la concentración de lípidos pinesles (Ebels y Prop, 1965). modi
fica los niveles de serotonina pineal (Fiske y Huppert. 1968) y prg
duce cambiosultrsestructurales en la glándula pinesl de varias ea
pecies (Freire y Cardinsli. 1975; El-Domeiri y Das Gupta, 1976; Seg
son y Hrasovich, 1977).
Recientemente se ha descripto en este Laboratorio la existen
cia de sitios receptores para la melatonins en la glándula pineal

lhÜ
bovina (Vacas y Cardinali. 1960). Los resultados de la sección III.
1.1 indican que la melatonina afecta. en altas concentraciones
(l mM).la sintesis de PGs en pineales bovinos. Si bien estas con
centraciones son farmacológicas, debe destacarse que es de esperar
en el sitio de producción una concentración de hormona mucho mayor
que en los tejidos efectores centrales y periféricos. En conjunto
los resultados previos y las presentes observaciones sugieren que
la melatonina puede controlar a través de un mecanismode retroali
mentación su propia sintesis en 1a glándula pineal. A semejanza de
lo que ocurre en el aparato neuroendócrino, 1a melatonina inhibió
también dos efectos vinculados a la sintesis de PGs en tejidos pe
riféricos. comola motilidad uterina y la agregación plaquetaria.
En conjunto, los resultados de la sección III.2 avalan la hi
póteais que la inhibición de 1a sintesis de PGs es el mecanismocg
mún que media los efectos de 1a indometacina y melatonina en diveg
sas preparaciones celulares. Esta observación es particularmente rs
levante en el SNC,sitio primario de acción de la melatonina para
modificar diversos fenómenosneurales y neuroendócrinos (Tablas l y
2). En concentraciones compatibles con el Hd de los sitios receptores especificos para melatonina descriptos en el SNC(Cardinali y
col., 1978, 1979 a; Vacas y Cardinali, 1979). 1a melatonina inhibe
la sintesis de PGEZ.A través de los efectos pre y postsinápticos
de esta inhibición pueden explicarse varias de las secuelas neuroeg
dócranas que siguen a la inyección de la hormonapineal (Fig. 25).
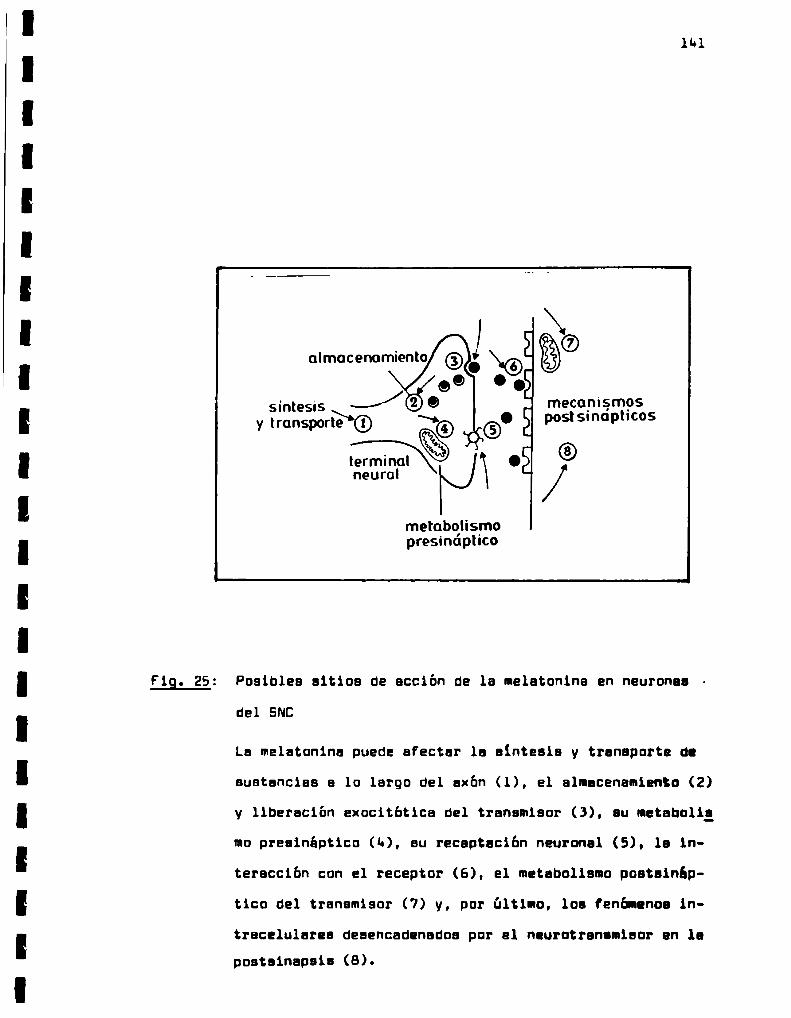
lbl
¿(asíntesis s—//
ytransporm N
ternúnal<ïÏÉ3 C! C)’Ñ\5/Ci
mecanismoso E postsmophcos
neural
metabolismopresinoptlco
Fig. 25: Posibles sitios de acción de la melstonina en neuronas
del SNC
La melatonins puede afectar 1a sintesis y transporta de
sustancias a lo largo del sxón (1), el almacenamiento (2)
y liberación exocitótica del transmisor (3), su nataholig
mopreaináptico (k), su recaptación neuronal (5). 1a in
teracción con el receptor (6). el metabolismopost-inap
tico del transmisor (7) y. por último, los fenómenosin
tracelulares desencadenadoapor el neurotransnisor en lapostsinspsis (B).

YCONCLUSIONES

lh2
Las PGa participan en la secuencia de fenómenos metabólicos que
vincula a la NEliberada de los terminales neurales con la sin
tesis de la hormonapineal melatonina porque:
NE libera PGEy PGFde explantos pineales bovinos.
existen sitios receptores para PGEy PGFZQ‘en fracciones li
bres de núcleos de honogenatos pinesles bovinos.
el bloqueo de la sintesis de PGs in vivo por inyección de in
dometacina inhibe la activación de la sintesis de melatonins
durante la fase de oscuridad del ciclo diario sn la rata.
el bloqueo de la sintesis de PGs in vitro por incubaciones
con concentraciones supramáximasde indometacina, ácido acetil
sslicilico o ácido mefenámicoinhibe la estimulación por NEde
la liberación de melatonina pineal.
el agregado de PGE2al medio de incubación y en concentraciones
Fisiolbgicas aumenta la sintesis de AMPC,ls ocupación de ls
subunidad reguladora de 1a quinass de proteinas dependiente de
AMP la actividad de SNATy la liberación de melatonina por lsc.glándula pinesl.
el agregado de PGE2en concentración 100 veces mayor que el um
bral para estimular la sintesis de melatonins produce inhibi
ción de ls liberación de NEde los terminales adrenérgicos pi
neales.
Las PGs participan en el mecanismo de acción de 1a hormona pineal
melatonina en tadidos efectores porque:

1h}
la melatonina, en cancentraciones farmacológicas, inhibe la
liberación espontánea de PGEy PGF en HMBy glándula pineal
bovinaa.
la meletonina inhibe, en concentraciones fisiológicas, 1a
liberación espontánea de PGE2en HMBde rata.
le melatonine inhibe. en concentraciones fisiológicas, le
liberación oe PGE2inducida por NE en el HHBde rata.
1a melatonina inhibe loa efectos fisiológicos atribuidos a
lee PGeen dos preparaciones utilizadas para evaluar la cin
teeia de PGa(motilidad uterina y agregación plaquetaria).
«9% WW

EIREFERENCIASI
I'l'l"llllllllllllllll
,.

1k“
Ahlquiat R.P. (l9h8). A study of the adrenotropic receptora.
Am.J. Physiol., 153: 586.
Alonso R.. Prieto L., Hernández C. y Has H. (1978). Antiandrogenic
effects of pinesl gland and nelstonin in csstrated and intact pre
pubsrsl Isle rsts. J. Endocr.. 22: 77.
Alpha L., Heller A. y Lovenberg U. (1980). Adrenergic regulation of
the reduction in acetyl coenzyneA: srylamina-N-acetyl-transfersse
activity ln the rat pineal. J. Neurochen.,23: 83.
AndréJ.S. y Parrish J. (1979). Inhibition of estrous ciclicity in
golden hamsters by melatonin administration on the day of prosstrus.
J. Exe. 2001.. 207: 61.
Anton-TayF. (1971). Pineal brain relationships. En: The Pinesl
Glsnd. G.E.U. Holstenholme, J. Knight (ed), Churchill. Londres,
p. 213.
Anton-TsyF. (197“). Helatonin: effects on brain function. Adv.
Biochen. Psychthsrnscol.. ll: 315.
Anton-Toy F.. Chou CS, Anton S. y Uurtmsn R.J. (1968). Brain sarn
tonin concentration: elevation following intrsperitoneal adminis
tration of melatonin. Science. 162: 277.
Anton-Tay F. y MurtmanR.J. (1969). Regional uptske of 3H-Ilelstonin
from blood or cerebrospinsl fluid by rat brain. Nature, 221: #7“.
Arendt J. (1978). Helatonin ssssys in body fluids. J. Neursl ïrsnsn..
suEEl. ¿2: 265.
Arendt 3.. Ustterberg L., Hayden T. Sizonenko P.C. y Psunier L.
(1977). Radioilnunoasssy of selatonin: hulan serum and csrsbrospir

l
le
nsl fluid. J. Neursl Trenam., sugl. 12: 265.
Armstrong D. y Grinuich D. (1972). Blocksde of spontsneous snd LH
induced ovulstion in rata by indonethscin. an inhibitor of prosta
glsndin biosynthesis. Prostsglandina, 1: 21.
Aubsrt Ch., anisud P. y Lecslvez J. (1960). Effect of pineslscto
my snd nelatonin on mammarytumor growth in Sprague-Dsuley rsts
under different cooitions of lighting. J. Neural Trsnsn.. 31: 121.
Avsnzino G. L., Bradley P. y Molstencroft J. (1966). Actions of
prostaglandins E1 , E2 y F2u on brain sten neuronas. Br. J. Phsrls9-0-1."a :
Axelrod J. y Weissbsch H. (1960). Enzylstic D-nethylstion of N
scetylserotonin to seletonin. Science. 131: 1312.
Bailey 0., Atkins T. y Hstty A. (197“). Melotonin inhihitio of
insulin secretion in the rst ano mouse. HornoneRes.. 2: 21.
ledesssrine.R. y Fischer J. (1976). Trsce anines snd sltsrns
tive nsurotransmitters in the central nervoue syatsl. Biochen.
Phenscol., gl: 621.
Bsnerjee S., Hen V.. Uinston H., Kelleher J. y Margulis L. (1972).
Helstonin: inhibitor of microtubule-bssed oral morphogsnssis in
Stentor-coeruleus. J. Protozol., 12: 108.
Bsnsrjse S..y Hsrgulis L. (1973). Hitotic srrest by sslstonin.
Exa. Cell. Rss., 2Q: 31h.
Behrnan H., Orcyk G. y Grsep R. (1972). Effect of synthstic gonsqg
tropin-releasing hormone(Sn-RH) on ovulation blocksoe by sspirin

1h6
end indonethscin. Proataglsndina, i: 2h5.
Bennet J.P. y Snyder S.H. (1975). Stereospecific binding of D
lisergic acid diethylamide (LBD)to brain membranas:relation
ships to serotonin receptors. Brain Res., 23: 507.
Benson B. y Hrssovich H. (1977). Circadian rhythm in the nu-oers
of grsnulated vesicles in mousepineaiocytes. Anat. Rec.. 167:
536.
Bergstrun 5.. Danielason H., Hlenoerg D. y Samuelsson 8. (l96k).
The enzimstic conversion of essential fatty acids into prostagleg
dins. J. Biol. Chen.. 239: PC “006.
Bsrgatrdm 5., Ryhage R., Samuelsson B. y SJbvall J. (1963). The
structure of prostaglandin El. F1“ end Flp . J. Biol. Chen" 238:3555.
Bergstrün S. y SJUvsli J. (1957). The isolation of prostaglandins.
Acta Chem. Scand., ll3 1086.
Bergatrüm 5., Farnabo L. y Fuxe H. (1973). Effect of prostaglandin
E2 on central and peripheral catecholamine neurones. Eur. J. Phar
macol., g¿: 362.
Berndtson U.E. y Desjsrdina C. (197“). Circulating LHy FSHlevels
and testicular function in hamsters during light deprivstion and
subsequent photoperiodic stinulation. Endocrinology, 22: 195.
Blackwell G.J., Flower R.J., Person H. y VaneJ. (1975 o). Factors
influencing the turnover of PG. Brit. J. Pharnscol.. 22: “67.
Blackwell G.J., Flower R.J. y Vane J.R. (1975 a). Rapid reduction
of PG-lS-hydroxygensseactivity in rat tiasues after treat-ent

1k?
Uith protein synthesis inhihitors. Brit. J. Pharmacol., 22: 233.
Blask D.E. y NodelmsnJ.L. (1980). An interaction hetueen the pi
neal gland and olfatory deprivation in poteotiating the effects of
nelstonin on the gonads, accessory sex organo, and prolactin in
male rate. J. Neurosci. Res., 2: 129.
Blask D.E., Nodelnan J.L., Leaden C.A., Richardson B.A. (1980).
Influence of exogenously sdninistered nelatonin on the reproductive
system snd prolactin levels in underfed male rats. Biol. ReErod..
g: 507.
Borda E., Agostini H., Sterin-Borda L.. Gimeno H. y Gimeno A.
(1961). Inhibitory effects of somecstecholsnines on contractions
of uterine strips isolated from estrous and spayed rats. Influence
of endogenous end exogenous prostaglandina on the action of Ietho
xemine. Eurog. J. Pharmacol.. gg: 55.
Borgeat P. y Sanoelsson B. (1979). Metsboliam of srachidonic acid
in polynorphonuclear leukocytes. Structural analysis of novel hy
droxylated conpounds. J. Biol. Chen.. 25k: 7665.
Bradley P.B., Samuels G.H. and ShawJ.E. (1969). Correlation of
prostsglsndins release from the cerebral cortex of cats with the
electrocorticogrsn, following stinulation of the reticulsr foros
tion. Brit. J. Phsrmacol., 22: lSl.\\- Brailousky S. (1976). Effects of melatonin on the photosensitive
epilepsy of the baboon, PaEio gagi . ElsctroenceEhsl. Clin. Neuro
Ehysiol., El: 31h.
- Branner G.L., Horley J.E.. Geller E., Vurviler A. y Hershan J.H.

le
(1979). Hypothalamus-pituitary-thyroid axis interactions with
pineal glend in the rat. Amer. J. Physiol., 236: E hifi.
Brown D.A., Albano J.. Ekins R.. Sgherzi A. y Tempion M. (1971).
A simple and sensitive saturstion asssy method for the measurement
of adenosine-B',5'-cyclic monophosphate. Biochem. J.. 121: 561.
Brownstein M.J. y Axelrod J. (197“). Pineel gleno: a Zh-hour rhythn
in norepinephrine turnover. Science, ¿93: 16}.
Bubenik G.A., BrownG.H. y Grota L.J. (1976). Differential lacalizg
tion of N-acetylsted inoolealkylamines in DNSand the Harderisn
gland using immunohistology. Brain Res.. 118: #17.
Bubenik G.A., Purtill R.A., Braun G.M. y Grotts L.J. (1978). Hele
tonin in the retina and the Harderien gland. Ontogeny, diurnal ve
ristions end melatonin treatment. Eng. Eye Res.. gg: 323.
Burns J.H. (1973). Serum sodium and potessium and blood glucose
levels in cynsmolgusmonkeysafter administration of melatonin. g.
Physiol., 232: 8k P.
Cody P. y Dillman R.D. (1971). Influence of cetechol and indole
smines upon pineal uptake of thyroxine. Neuroendocrinologx. Q: 226.
Cerdinali D.P. (197“). Heletonin and the sndocrine role of the pi
neel organ. Curr. T09. Exa. Endocr.. g: 107.
Cerdinsli D.P. (1975). Changes in hypothaiemic neurotransmitter og
take following pineslectomy, superior cervical gsnglionectomy or
meletonin administration to reta. Neuroendocrinology, ¿2: 91.
Csrdinali D.P. (1977). Nuclear receptor-estrogen complex in the
pinesl glsnd. Modulstion by sympathetic nerves. Neuroendocrinology.

_¿._. 333.
Csrdinali D.P. (1981). Helatonin. A mammalianpineal hormone. Endo
crine Reg. (en prensa).
Csrdinali D.P., Fsigón M.R.. Scachi P. y Hoguilevsky J. (1979 b).
Failure of melatonin to increase plasma prolactin levels in ova
riectomized rsts subjected to superior cervical ganglionectomyor
pinealectomy. J. Endocrinol.. gg: 315.
Cardinsli D.P. y Freire, F. (1975). Helatonin effects on orain.
Interaction uith microtubule protein, inhioition of fast axoplas
mic flow and induction of crystaloid and tubular formation in the
hypothalamus. Mol. Cell. Endocrinol., g: 317.
Cardinsli D.P., Freire F3, Nagle C.A. y Rosner J.M. (1975 b).
Effects of environmental lighting, superior cervical ganglionectg
myand adrenergic drugs on microtubule proteins levels of the rat
hypothalamus. Neuroenoocrinologv. ¿2: kh.
Cardinali D.P.. HyyppáH.T. y MurtmanR.J. (1973 a). Fate of intra
cisternally injecteo melatonin in the rat brain. Neuroendocrinology.
lg: 30.
Cardinali D.P., Lynch H.J. y MurtmanR.J. (1972). Binding of mela
tonin to humanano rat plasma proteins. Endocrinology. El: 1213.
Csrdinali D.P., Nagle C.A., Denari J.H., Bedes G.D.P.. Rosner J.H.
(1973 b). Lsck of effects of light on the rat pinesl in organ
culture. Gen. Come.Endocr., gi: 573.
Cardinali D.P.. Nagle C.A., Freire F. y Rosner J.M. (1975 s).
Effects of melatonin on neurotransmitter uptake ano release by

150
syneptoaone-rich homogenateeof the rat hypothalemua. Neuroendocri
"0109!, lg: 72.
Cardinali D.P., Negle C.A. y Rusner J.M. (197“ b). Effects of pines
lectony, superior cervical aympethectomyand meletonin administre
tion on the uptake of eatradiol by the rat neurohypophyeie. Acta
Phxaiol. Letinoamer., gg: 69.
Cardineli D.P.. Negle C.A. y Roaner J.H. (197%e). Incorporation of
3H-leucine into proteina in the hypothelemus and the anterior hypu
phyaia. Effects of pineelectomy, superior cervical ganglionectomy
and continuous expoeure to light end darkness. Neuroendocrinology,
lg: 7k.
Cardinali D.P., Nagle C., GómezE. y Rosner 3.". (1975 c). Norepi
nephrine turnaver in the pineel glend. Acceleration by estrediol
and teatoeterone. Life Sci.. ¿Ez 1717.
Cardineli D.P. y Roaner J.M. (1971). Hetebolism of eerotonin by
the rat retina ín vitro. J. Neurochem.,¿9: 1769.
Cardineli D.P.. Vacas M.I. y Estevez Bayer E. (1978). High effinity
binding of meletonin in hovine medial basal hypothelauue. IRCSMed.
523., É: 357.
Cerdinali D.P., Vacas M.l. y Estevez Boyer E. (1979 a). Specific
binding of melatOnin in bovine brain. Endocrinology. 103: “37.
Cerdineli D.P. y UurtmanR.J. (1972). Hydraxyindale-D-nethyl
transfereee in rat pineel, retina and Haraerian glend. Endocrino:
nclogy. 2;: 2k7.
Cardinali D.P. y NurtmanR.J. (1975). Control of meletonin eynthe
sie in the pineel organ. En: Frontiere in Piqsgl Phyeiology. ".0.

151
Altechule (ed). ïhe MITPresa, Cambridge, Mesa, o. 12.
Clegg P.C. (1966). The effect of prostaglandine on the response of
isolated smooth muscle preparation of aympathomimeticeubstancee.
En: Endogenoue aubetancee effecting the myometrium. V. Picklee y
R. Fitzpetrick (edo). p. 122.
Coceeni F., Puglisi L. and Lavere B. (1971). Prosteglanoine eno
neuronal activity in epinel coro end cuneate nucleua. Am.N.V.
Aced. Sci.. 180: 289.
Coceeni F. ano Molfe L.S. (1965). Proetaglendins ir\brain end the
release of prostaglandin-iike compounoefrom the cat cerebellor
cortex. Can. J. Phxeiol.l Pharmecoi., 22: aus.
Collu R., Fraechini F. y Martini L. (1971). The effect of pineel
methoxyinoolee on rat vaginal opening time. J. Endocrinols, gg:
679.
Coste E. y Neff N.H. (1970). Estimetion of turnover rates to atu
dy the metabolic regulation of the eteaoy state level of neuronal
monoeminee. En: Handbook of Neurochemietry. A. Lejthe (eo). Plenum
Press, NewYork, p. k5.
Boyle J.ï. y Axelroo J. (1972). Dopemine-¡B-hydroxyleoe in the
rat brain: developmentalcherecterietic. J. Neuroghernecoi., ¿2:
“#9.
Ceepo A.. Csepo E., Fey E.. Henze M. y Seleu G. (1973). ïhe delay
of epontaneous labor by neproxen in the rat model. Proeteglendine.
g: 327.
Devie G.A. (1978). ïhe response of edenoeine-J‘.5'—monphoephete

to noreplnephrlne ln the hypothalsmus ano plneal orqan of female
rsts ln proestrus or diestrus. Endocrinologx, 103: leB.
Deguchl T. (1973). Role of the fñ-adrenergic receptor in the ele
votlon of edenoslne cycllc 3',5'-monophosphste and lnouctlon ofserotonin-N-acetyltransferase in the rat plneal gland. Mol. Phar
gggg¿.. 2: 18h.
Deguchl T. y Axelrod J. (1972). Induction end superlnduction of
serotonln-N-acetyltrsnsferase by drugs end denervstion of the ret
pinesl. Proc. Not. Acsd. Sci. USA,El: 660.
Dorp D.A. Van, Beerthuls R.H., Nugteren D.H. end Vonkenen H. (196“).
The blosynthesls of prostaglandins. Blochtm. Bloghxs. Acta. 22: 20h.
Driver P.H.. Forbes J.H. y Scenes G.6. (1979). Hornones. feeding
and temperature in the sheep following cerebroventriculsr injecw
tions of neurotransmltters ano carbschol. J. Physlol.. 290: 399.
Dufau M.. Tsuruhara T., Horner H., Podestá E. y Catt J. (1977).
Intermediate role of adenosine-B'.5'-cycle monophosphsteand prg
tein klnsse during gonadotropin-inducedsteroioogenesis in testi
cular lnterotltlal cells. Proc. Nat. Aced. Sci. USA.23: 3hl9.
DunhamE.w. y Zimmerman 8.5. (1970). Release of prostaglandin
llke material from dog kidney during nerve stimulstlon. Am.J.
Physiol.. 19: 1279.
Ebels I. y Prop N. (1965). A study of the effect of melstonin on
the gonads. the oestrous cycle ano the pineal organ of the rat.
Acta Endocr.. 32: 567.
- El-Domeirl A.A. y Des Gupta T.H. (1976). The influence of plneal

153
ablation and administration of melatonin on growth and spread of
hamster melanoma. J. Surg. Dncol., Q: 197.
Ellis L.C. (1972). Inhioition of rat testicular androgen synthesis
in vitro by melatonin ano serotonin. Endocrinologv. gg: 17.
Euler U.S. Von (193k). Zur kenntnis der pharnakologischen uirkungen
von nativsekreten und extrackten münnlicher accesaorischer
geschlechtsdrüsen. Arch. EXE. Pathol. Pharmakol.. 175: 7B.
Euler U.5. Von (1935). A depresaor substance in the vesicular
glano. J. Phyaiol.. gg: 21 P.
Euler U.S. Von(1972). Synthesia, uptake ano storage of cstechoi
aminea in adrenergic nerves. The effects of oruga. En: Cstechol
amines. Handb. Exp. Pharn. H. Blaschko y E. Muscholl (ada). p. 186.
Farriello R.G., Buhenik G.A., BrownG.M. y Grota L.J. (1977). Epi
leptogenic action of intraventricularly injected antimelstonin sn
tibody. Neurologv, gl: 567.
Ferreira 5., MoncadaS. y Vane J. (1971). Indomethacin and aspirin
abolish proataglandin release from the spleen. Nature New8101..
31: 237.
Ferreira 5., MoncadaS. y VaneJ. (1973). Someeffects of inhibi
ting sndogenous prostaglandin formation on the responses of the
cat apleen. Brit. J. Pharnacol.. 31: kB.
Fideleff H.. Aparicio N.J., Guitelman A., Debeljuk L.. Mancini A.
y Cramer C. (1976). Effect of nelatonin on the basal and etimolo
ted gonadotropin levels in normal men ano postaenopausal uonen.
J. Clin. Endocr. Metab.. gg: lUlk.

IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII
le
Fiake 0.". y Huppert L.C. (1968). Helatonin action on pineal
varias with photoperiod. Science. 162: 279.
Fiake U.H. y MacDonald G.8. (1975). Eatrogen secretion and ita
relation to corpus luteum.fornation in monkeys(Hacaca fascicula
ria) given melatonin. Fertil. Steril.. gg: 609.
Fitzgerald T.J. y Veal A. (1976). Melatonin antagonizea colchicine
induced mitotic arrast. Exgerientia. 2g: 372.
Flower R. (l97k). Drugs which inhibit proataglandin hioayntheaia.
Pharmacol. Rev., gg: 33.
Flower R. (1978). Staroidal anti-inflannatory drugs aa inhibitors
of phoapholipaae A2. En: Adv. in Proataglandin and Thronooxane Res.
R. Paoletti y 8. Sanuelsson (eds). AcademicPresa, N.V..‘2. p. 105.
Folco 6.. Longiave 0., Berti F., Funagalli R. y Psoletti R. (1976).
Prostaglandin E2 and central cyclic nucleotidea. Adv. in Proataglandins and ThromboxaneRes. R. Paoletti y B. Sanuelason (ada). Acade
mic Press, N.V.. l, p. 305.
Frehn J.L., Urry R.L. y Ellis L.C. (197“). Effect of melatonin and
short photoperiod on ¿Sk-reductase activity in the liver and hypo
thalamua of the hamster and the rat. J. Endocr.. gg: 507.
Freire F. y Cardinali D.P. (1975). Effects of melatonin treatment
and environmental lighting on the ultraeatructural appearance turg
over and microtubule protein content of the rat pinaal gland. g.
Neural Iransm.. 21: 237.
Fujita M. y Kogayashi T. (1979): Current views on the paraneurone

155
concept. Trends Neuroaci., g: 27.
Fujiuara H., Shibata H., Uatanaoe V., Nukiua T., Hirata F.. Hizu
no N. y Hayaishi D. (1976). Indoleamine 2,3-dioxigenase formation
of L-kynurenine from L-tryptophsn in cultured rabbitt pineal gland.
J. Biol. Chem., ggg: 6081.
Gilmore N., VaneJ.R. y Uyllie J.H. (1968). Proatsglandin release
oy the spleen. Nature. 218: 1135.
GimenoM.F.. Sterin-Speziale N.. Landa A.. Bonacoass A. y Gimeno
A. (1979). Is the apontaneousmotility of isolated rat uterus
controlled oy Proataglsndin E7. Prostsglandlna, ¿1: 673.
Goldberg V.J. y Ramuell P.U. (1975). Role of proatsglandins in re
production. Physiol. Rev.. 22: 325.
Goldblatt H.w. (1933). A depressor substance in seminal fluid. g.
Soc. Chen. Ind.. gg: 1056.
Goldblstt H.U. (1935). Properties of humanseminal plasma. J. Ph!
siol.. gg: zoe.
Goldman8., Hall V., Hollister C., Roychoudhury P., Tsmsrkin L. y
Uestrom w. (1979). Effects of melatonln on the reproductive system
in intact and pinealectomized male hamsters maintained under vs
rious photoperiods. Endocrinologx, 10h: 82.
Goridia C. y Neff N.H. (1972). Evidence for specific monoamine
oxidases in humanaympsthetio nerve and pineal gland. Proc. Soc.
Eng. Biol. Med., lao: 573.
Greengard P. (1978). En: Cyclic Nucleotidesl Ehosghorilated Er
teina and neuronal function. ¿z k1.

156
- Gregorek J.C., Seibel H.R. y Reiter R.J. (1977). The pineal complex
and lts relationships to other splthalamlc structures. Acta Anat..
29: “25.
Guchhait R.B. y Grau J.E. (1978). Blosyntheais of S-adenosyl-L-me
thlonlne ln the rat plneal gland. J. Neurochen.. gl: 921.
Goldberg H. y Marsden M. (1975)} Catechol-0-methy1transferaae: Phar
macologlcal aspects and Physlological role. Pharmacol. Rev., gl: 135.
Hsgelstein H.A. y Folk G.E. (1979). Effects of photoperlod. cold
accllmation and melatunin on the white rat. Come.Biochem. Phlslol..
62 C: 225.
Hamberg M., Svensaon J. y Samuelsson B. (1975). Thromboxsnea: A neu
group of blologlcally active compoundaderived from prostaglandin
endoperoxldes. Proc. Nat. Acsd. Sci.l USA,gg: 299k-2998.
Hshberg M.. Svensson 3., Maksbsysshi I. y Samuelsaon 8. (197“).
Iaolation and structure of tuo prostaglandin endoperoxides that
cause plstelet aggregatlon. Proc. Nat. Acad. Sci. USA,ll: 3k5.
Harms P.. Ojeda S. y McCsnn S. (197h). Prostaglsndin induced re
lesae of pltultary gonsdotroplns: central nervous system and pitui
tsry sites of action. Endocrinologx, 22: lb59.
Harney P.J., Sneddon J.M. y Ulllisms H.I. (197“). The influence of
ovarlsn hormonesupon the motillty and proataglsndln production of
the pregnant rat uterus ln vitro. J. Endocr., gg: 3h}.
Harris R. y Rsmuell P. (1979). Cellular mechanisms of prostaglandln
action. Ann. Rev. Physlol.. 3;: 653.
Haye B. y Jacouemln C. (1977). Incorporatlon of [1“c] arschidonste

[57
in pig thyroid lipioa and prostaglandins. Biochim. Bioghxs. Acta.“87: 231.
Hedluno L.. Lischko H.M., Rollsg H.D. y Nisuender G.0. (1977). He
latonin daily cycle in plasma and cerebroaoinal fluid of calves.
Science. 195: 686.
Hedoviat P. (197Da). Studies on the effect of proataglandins E1
and E2 on the sympsthetic neuromuscular transmission in some ani
mal tissues. Acta Phxaiol. Scand., suEEI. 3h5: l.
Heoqvist P. (197D b). Antagonism by calcium of the inhibitory ac
tion of prostaglandin E2 on aympsthetic neurotransmiasion in the
cat spleen. Acta Physiol. Scand., gg: 269.
Hedqviat P. (197“). Interaction between proatsglandina and calcium
ions on noradrenaline release from the stimulsted guinea pig vas
deferena. Acta Phyaiol. Scand.. 29: 153.
Hedqvist P. (1976). Effects of proatsglandina on autonomic neuro
transmisaion. En: Prostaglandins: Physiologicall Pharmacological
and Pathological AaEects. S.M.M.Karim (ed). Baltimore University.
Park Press. p. 37.
Hirata F., Hsyaiahi 0., FokuysmaT. y Senoh S. (197“). In vitro
and in vivo formation of tuo neu metabolitea of melatonin. g.
Biol. Chem., 2k9: 1311.
Hoffer 8.3., Diggins G.R. ano BloomF.E. (1969). Proataglandina
E1 and E2 antagonize norapinephrine effects on cerebellar Purkinjecells: microelectrophoretic atuoyt-Science. 166: 1618.
HoffmannH. (1979). Photoperiod, pinesl melatonin and reproduction

158
in hamsters. Prog. Brain Res.. gg: 397.
HoffmanR.A. y Reiter R.J. (1965). Pineel glsnd: influence on go
neds of male hamsters. Science, ¿29: 1609.
Hollander C.S., Pressd R., Richardson 5.. Hirooks Y. y Suzuki S.
(1977). Melstonin moduletes hormonal release from orgsn cultures
of rat hypothslamus. J. Neurel Trsnsm.l sugl. 13: 369.
Holmes 5.o. y Sugden D. (1975). The effect of meletonin on pines
lectomy-induced hypertension in the rat. Brit. J. Phermscol., 29:360 P.
Iversen L. (1967). The ugtske end storage of noredrensline in
sxmgathetic nerve . CambridgeUniversity Press, Cambridge.
Join-Etcheverry G. y Zieher L.M. (1968). Cytochemistry of 5-hydroel
tryptamine et the electron microscope level II. Localization in the
sutonomic nerves of the rst pineel glsnd. Z. Zellforsch. gg: 393.
Jarrot B. (197k). Methods for enelyzing monosmineoxidsse snd cste
chol-O-methyltransfereee in nervous tissues. En: Research Methods
in Neurochemistry, vol. 2 (N. Marks y R. Rodnight, eds.). Plenum
Press, N.V., p. 377.
Johnston J.P. (1968). Someobservations upon a new inhibitor of
monoamineoxidsse in brain tissue. Biochem. Pharmac., ¿1: 1265.
Junstad M. y uennmalmA. (1973). Prostaglandin mediated inhibi
tion of norodreneline release at different nerve impulse frecuen
cies. Acta Physiol. Scand., QE: san.
Hamberi J.A. (1973). The role of brain monoamines and pineal indo

159
lee in eecretion of gonadotrophina and gonadutrophin-releasing
Factors. Prog. Brain Ree., 22: 261.
Kantor H. y Hampton M. (1978). Indomethacin in submicromolar con
centrations inhibita cAHP-dependentprotein kinaae. Nature. 276:
ahi.
Hao L.U.L. y Meiaz J. (1977). Release of gonadotrophin-releaeing
hormone (Gn-RH)from isolated. perifuaeo medial basal hypothalalue
by melatonin. Endocrinology, 100: 1723.
Happers J.A. (1960). The development, topographical relations and
innervation of the epiphyaia cerebri in the albino rat. Zetechrift
für zellforechung und mikroekogic anatomia, gg: 163.
Happera J.A. (1965). Survey of the innervation of the epiphyaia
cerebri and the acceaory pineal organ of vertebratea. En: Structu
re ano function of the egighyaia cerebri. J.A. Kappers y J.P.
Schade (eds.). Elsevier, Amsterdam,p. 87.
Kappers J.A. y Pévet P. (1979). The Eineal gland of vertebratee
including man, Elsevier. Amsterdam, p. 6k.
Karim S.H.M. y Devlin J. (1967). Prostaglandin content of amniotic
fluid during pregnancy ano labour. J. Obstet. Gynaecol. Brit.
Commonu., 22: 230.
Hataoka K., RammellP.w. y Jeeeup 5. (1967). Proetaglandina: loca
lization in aubcelullar particlee of rat cerebral corten. Science.
157: 187.
Hennauay D.J., Frith R.G., Phillipou 5., Matthews G.0. y Seamerk
R.F. (1977). A specific radioimmunoaasayfor melatonin in biologi

160
cal tissue and its validstion by gas chromatogrsphy-maasspectru
metry. Endocrinology, ¿glz 119.
Himbsll F.A. y UyngsrdenL.J. (1975). Prostaglendin specific bin
ding in hamster myometrisl low speed supernatant. Prostsglsndins,
2: bl}.
Hinson G.A. y Peet F. (1971). The influences of illuminstion, mela
tonin and pineslectomy on testicular function in the rat. Life Sci..
¿9: 259.
Hirpekar S. y Misu V. (1967). Release of noradrenaline by splenic
nerve stimulstion and its dependence an calcium. J.Ph!siol.. 199:
219.
Klein D.C. (1978). The pineal glsnd: s model of neuroendocrine
regulation. En: TheHigothalsmu. S.Reichelin, R.J.ledessarini,J.B.;
Martin (eds.). Raven Press, N.V., p. 303.
Klein D.C. y Ueller J.L. (1970). Meletonin synthesis: sdenasine-J'.5'
monophosphateend narepinephrine stimulste N-scetyltranafersse. Science.
199: 978.
Konig A. y Rega R. (1976). Effect of melatonin on the spermatogene
sis of unilaterally orchidectomized puberal rsts. J. Neursl Tranam..
suEl. 13: 373.
Kitsy y Atschule (195“). The Pineal Blend. Cambridge, Mass.: Harvard
University Press.
Hopin J.J., Pare C.M.B., Axelrod J. y Weissbsch H. (1961). The fate
of melstonin in animals. J. Biol. Chem., 236: 3072.
Hopin J.J. y Silberstein S.D. (1972). Axonsof synpsthstic neurona:

161
transport of enzymes in vivo and properties of axonai spronts
in vitro. Pharmacol. Rev., gg: 2h5.
Hoslow S.H. y Green A.R. (1973). Analysis uf pineal ano brain
indolealkyiamines by gas chromatography-mass spectrometry.
Adv. Biochem. Psychogharmaco ., Z: 33.
Houlu M. y Lammintausta R. (1979). Effect of melatonin on L
tryptophan and apomorphine-stimulateo growth hormone secretion
in man. J. Clin. qugpr. Met., 22: 7D.
Kramer J. y Ben-David M. (1978). Prolactin supression by (-)
9-tetrahydrocannabinol (THC): involvement of serotoninergic ano
dopaminergic pathways. Endocrinology. 103: #52.
ñrebs E. y Bravo J. (1979). Phosphorylation-dephosphorylation of
enzymes. Ann. Rev. Biochem.. gg: 923.
Hüderling 1.. Trocchi M.. Dellantonio M.. Spagnesi M. y Fraschini
F. (1979). Investigations on seasonal rhythms ano the effect of
melatonin in the alpine hare (LeEus timious timidus L.). Prog.
Brain Res.. 2g: b17.
Huehl F.A. Jr. y HumesJ.L. (1972). Direct evidence for a prosta
glandin receptor ano its application to prostaglandin measurements.
Proc. Nati. Acad. Sci.I USA, gg: “BD.
Hurzrok R. y Lieb C.C. (1930). Biochemical studies of human semen.
II - The action of semen on the humanuterus. Proc. Soc. Exa. Biol.
Med., gg: 266.
Labrie F.. Pelletier 6., Borgeat P., Drouin 3.. Ferland L. y Belen

162
ger A. (1975). Modeof action of hypothalamic regulatory hormones
in the edenohypophysis. En: Frontiers of Neuroenaocrinology. L.
Martini, U.F. Ganong (ads). Raven Press, NewYork, p. 163.
Lands U.(1979). The bioaynthesis end metaboliem of prostaglandine.
Ann. Rev. Phyeiol.. gi: 633.
Lapin V, (1979). Pineel influence on tumor. Prog. Brain Ree., gg:
523.
Leychock 5.8., Franeon R.C., Ueglícki U.B. y Rubin R.P. (1977).
Identification and partial cheracterization of phospholipeaee ln
isolated aorenocortical cells. The effect of eynacthen (cortico
tropin-(I-26)-tetracosapeutide) and calcium ians. Biochem.3.,
16h: 753.
LemeyA., Brouiellette A., Denizeau F. y Lavnie M. (1979). Melafig
nin-stimulated and serotonin-stimulated release of vasopresin from
rat neurohypophyeie in vitro. M01. Cell. Endocr., ¿3: 157.
Leonardelli J.. TramuG. y HermanaE. (1978). Melatonine et cellules
a gonadoliberine (LHRH)de l'hypothalamus du rat. Comet. Rendue Soc.
8101., 172: hBl.
Lerner A.B.. Case J.D. y Heinzelmen R.U. (1959). Structure of mela
tonín. J. Am. Chem. Soc., gi: GÜBk.
Levine L. y Riceberg L.J. (1975). Radioimmunoassay For melatonin.
Res. Comm.Chem. Pathol. Pharmacol., lg: 693.
Levitt M.. Spector 5.. Sjoerdsman A. y Udenfriend S. (1955). Eluci
detion of the rate-limiting step in nurepinephrine bioeyntheale in
the perfused guinea pig heart. J. Pharmacol. Exp. Ther., Ika: 1.
l"

163
Logan A. y Weatherhead B. (1980). Post-tyroeinaee inhioition of
melanogeneaieby melatonin in hair folliclea in vitro. J. Invest.
Dermat.. lg: #7.
Longnecker D.E. y Gallo D.G. (1971). The inhibition of PHSG-indu
ced ovulation in immature rata by meiatonin. Proc. Soc. Eng. Biol.
Med., 137: 623.
Loury Ü.H.. Roaeorough N.J., Farr A.L. y Randall R.J. (1951). Pro
tein measurementwith the Foiin phenol reagent. J. Biol. Chem.. 193:
265.
Lu H.H. y Míetes J. (1973). Effects of aerotonin precursora and me
latonin on aerumprolactin release in rate. Endocrinologx, 22: 152.
Lynch G.R. y Epstein A.L. (1976). Melatonin induced changes in go
nada. pelage and thermogenic characters in the uhite-footed mouse.
Peromucue leucogu . Coma. Biochem. Physio ., 53 C: 67.
Maclouf J.. Andriew J. y Dvey F. (1975). Validity of PGEradioimmung
assay by using PGEantisera with differential binding parameters.
FEBSLett.. gg: 273.
Malewieta S.E. (1973). The effects of colchicine and of cytochalaein
B on the hormone-induced movement of melanin granulea in frog dermal
melanocytes. En: Endocrinology. R. Sou, F. Ebling, I. Henderson
(2da.). Amsterdam, p. 286.
Marcus R., Avereen G. y Üvner F. (1979). Fluctuation of adenoeine
3',5'-monphosphate-binding site occupancy ae an index of hormone
dependent adenoeine-B'.5'-monophoaphate formation in bone cells.
Endocrinology. 10h: 7hh.

165
- Martin J.E., Engel J.N. y Klein D.C. (1977). Inhibition of the
in vitro pituitary response to luteinizing hormone-releaaingho;
moneby melatonin, serotonin and S-hyoroxytryptamine. Endocrino
logy, IDO: 675.
Mas M., Massa R., Hontagna A., Negri-Cesi P. y Martini L. (1979).
Role of the pineal glsnd in the control of gonadotropins and an
orogen-reduoing enzymeain the rat. Prog. Brain Res.. gg: 367.
MetsushimaS. y Reiter R.J. (1977). Fine structural features of
edrehergic nerve fibers ano endings in the pineal glano of the
rata, ground squirrel and Chinchilla. Am.J. Anat., Ika: “63.
MataushimaS. y Reiter R.J. (1978). Electron microscopio observa
tion on neuron like cells in the ground squirrel pineal gland.
J. Neural Transm., gg: 223.
MayerS. (1980). Drugs acting st synaptic and neuroeffector func
tional sites. En: The Pharmacological Besis of Therageutic .
Goodman Gilman, Goodman y Gilman (eds.). p. 62.
McCann S.M., Ojeda S.R.. Herms P.G.. uheaton J.E.. Sundberg.D.H.
y Fewcett C.P. (1976). Centrol of adenohypophyseal hormone secre
tion by prostaglandins. En: Suocellular Mechanismein Reeroductive
Neuroenooorinology. F. Naftolin, H.S. Ryan, J. Davies (eos.). El
sevier, Amsterdam, p. h07.
Mess 8., Trentini G.P., Ruzsaa C. y de Gaetani C.F. (1979). Some
endocrine effiects of the pineel gland and meletonin with special
reference to reproduction. Prog. Brain Res., 2g: 329.

m_._-—¡II.IIIIII.IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII
165
MinnemanK.P., Hegsïrand L. y Molinoff P.B. (1979). Simultaneous
determination of beta-l and beta-2 sdrenergic receptora in tissuea
containing both receptor subtypes. Mol. Pharmacol., ¿2: 286.
MinnemanH.P. y Uurtman R.J. (1976). The Pharmacology of the pine
al gland. Ann. Rev. Pharm. Toxicol., ¿9: 33.
Moguilevsky J., Fsigon M.R., Scacchi P. y Cardinali D.P. (1979).
Effect of melatonin and superior cervical ganglionectomy on lutei
nizing hormonerelease induced by eatrsdiol-progesterona in castrg
ted rata. Neuroendoorinology, gg: 163.
Moguilevsky J., Scacchi P. y Deisa R.P. y Siaeles V.Ü. (1976).
Effect of melatonin on the luteinizing hormonerelease induced by
clomiphene and luteinizing hormone-releasing hormone. Proc. Soc.
Biol. Med.. 151: 663.
Moncada5.. Needleman P.. Hunting S. y Vane J. (1976). Prostaglag
din endoperoxide and thromboxane generating systems and their se
lective inhibition. Prostaglandins, lg: 323.
Moncada S. y Vane J. (1978). Pharmacology and endogenous roles of
proataglsndinsendoperoxidea, thromboxaneA2 and prostscyclin.
Pharm. Rev.. gg: 293.
MoncadaS. y Vane J. (1979). Modeof action of aapirin-like drugs.
En: deaoces in Internal Medicine. H. Stollerman (ed). Vear Book
Medical Publishers, Inc.. p. 1.
Moore R.Y. (1973). Retinonypothalsmic projection in mammals: A
comparative atudy. Brain Res., 32: #03.

J
166
Moore R.T. (1976). Neural control of pineel function in mammals
and birds. J. Neural Transm., suEI. 13: #7.
Moore R.V., Heller A., Bhatnager R.H., Murtman R.J. y Axelrou J.
(1968). Central control of the pinesl gland: visual pathuey.
Arch. Neurology, ¿9: 208.
Moore M. y Uolff J. (1973). Binding of prostaglandin E1 to oeef
thyroid membranes. J. Biol. Chem.. ZhB: 5705.
Motta M., Schieffini 0.. Five F. y Martini L. (1971). Pineel prig
ciples and the control of sdrenocorticotrophin secretion. En: ïhe
Pineal Glenn. J. Knight (ed). Uolstenholme G.E.w., Churchill. Log
ares, p. 279.
Mullen P.E.. Leone R;M., Hooper J., Smith I.. Silmen R.E.. Finnie
M., Carter E. y Linsell C. (1979). Pineel S-methoxytryptophol in
men. Psychoneuroendocrinology. g: 117.
Murphy G.6. y Modesto R. (1975). Effects of melatonin on 1“LI-glu
cose metebolism in rabbit edipose tissue. Experientis, 2¿: 383.
Namboodiri M.A.A., Nemboodiri M.A., Meller J.L. y Klein D.C.
(1979). A comparative study of pineel scetyl-CoA hidroxylsse end
N-ecetyltrensferese activities. J. Neurochem.,2g: 31.
Neff N.H. y Yang H. (197“). Another look at the monoamino oxideses
end the monoamineoxidese inhibitor drugs. Life Sci.. ¿3: 2061.
Nir I. y Hirschmenn N. (1979). A possible role of the pincel gland
in pregnancy and fertility. Prog. Brein Res.. 2g: #21.
Nir 1., Hirschmenn N., Puder M. y Petrank J.S. (1978). Changes in

167
rodent thyroid hormones and cyclio AMPFollowing treatment with
pinesl indolic compounds.Arch. Int. Physiol. Biochem.. gg: 353.
Nordlund J.J. y Lerner A.B. (1977). The effects of oral melato
nin on skin color and on the release of pituitary hormonas. g;
Clin. Endocr. Hetsb.. gg: 768.
Üchs S. y North R.M. (1976). Axoplasmic transport in normal and
pathological systems. En: Physiology ano Pathobiology of Axoqg.
Maxman, S.G. (ed). Raven Press. NewYork, p. 251.
flgreid D. y Dúkeland S. (1980). Protein kinase II has two dis
tinct binding sites for cyclic AMPonly one of which is detec
table by the conventional membrane-filtrstion method. FEBSLett..
121: 3h0.
Ügle T.F. y Hitsy J.J. (1978). In vitro effects of melatonin and
serotonin on adrenal steroidogenesis. Proc. Soc. Exg. Biol. Med..
157: 103.
Ojeda S.R. y McCann S.H. (1978). The possible role of prosta
glandins in the control of pituitary hormonesecretion. En:
Curr. Stud. Hypothal. Funct. (H. Lederis, m.L. Veale, eos.).
Harger, Basel. vol. I: 106.
Ojeda S.R.. Naor Z. y Negro-Vilar A. (1979 a). The role of pros
taglandins in the control of 90nadotropin and prolactin secretion.
Prostsglsndina and Medicine, 2: 2h9.
Ojeda S.R.. Negro-Vilar A. y McCann S. (1979 b). Release of pro!
taglandin Es (PGE)by hypothalamic tissue: evidence for their in

IIIIIIIII.IIIIIIIIIIIIIIIIIIIllthllIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII
168
volvement in catecholamine-induced LHRHrelease. Endocrinology,
10h: 617.
- Dleshanaky M.A. y Neff N.H. (1978). Studies on the control of
pineal indole synthesis: cyclic nucleotides, adenylate cyclase
and phosphodiesterase. J. Neural Transm., sugl. 13: 97.
- Dzaki Y. y Lynch H.J. (1976). Presence of melatonin in plasma
and urine of pinealectomized rats. Enducrinology, 22: Gal.
- Psdmanabhan V.. Convey E.H. y Tucker H.A. (1979). Pineal com-'
poundsalter prolactin release from bovine pituitary cells.
Proc. Soc. Exg. Biol. Med., 160: 3b0.
- Palmer D. y Riedaael M. (1976). Responses of whole animal and
isolated hearts of groundsquirrels, Citellus lateralis to melg
tonin. Come. Biochem. Physiol., 53 C: 69.
- Pang S.F. y VewD.T. (1979). Pigment aggregation by melatonin in
the retinal pigment epithelium and choroid of guinea pigs, Cavia
Eorcellus. Exgerientia, 22: 231.
- Pardridge w.M. y Hietus L.J. (1980). Transport of albumin-bound
melatonin through the blood-brain barrier. J. Neurocnem.,gg:
1761.
- Partington C., Edwards M. y Caly J. (1980). Regulation of cyclic
AMPformation in brain tissue by 0(-adrenergic receptors: Reqisi
site intermediacy of proataglandins of the E series. Proc. Nat.
Acad. Sci.l USA,21: 302“.
- Pavel S. y Goldstein R. (1979). Further evidence that Melatonin

IIIIIIIIIIIIIIIIIIIIIIIIIIIIlliilllIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII
169
represente the releasing hormonefor plneal vasotocln. J. Lndocr.
gg: 1.
Pedroza García E., Cardinali D.P., Laboroa N.P., Bienere U.G., Na
gle C.A. y Rosner J.M. (1975). Binoing of estradíol to macromolecu
les ln the rat neurohypophysis. Effects of time of the day, pines
lectomy or melatonin administration. figuroendocrinology, ¿5: 17k.
Pelayo F.. Dubocovich M. y Longer S. (1977). Regulation of noraorg
naline release in the rat pineal through a negative feedback mechg
nism mediated by presynaptic q-adrenoceptors. Eur. J. Pharmscol.,
22: 317.
Perez-Polo J.R., Hall H., VaughanM.H. y Reiter R.J. (1978). Effect
of pinealectomy, blinoing. castration ano melatonin on nerve growth
factor levels in the eubmaxillary glands of mlce. Neuroscl. Lett.,
lg: 83.
Pevet P. (1979). Secretary processes ln the mammalianpinealocyte.
The plneal glano of vertebrates including man. Prog. Brain Res.,
gg: lh9.
Picklee V.R., Hall w.J., Best F.A. y Smith G.N. (1965). Prostaglsg
dins ln endometrium and menstrual fluio from normal and dysmeno
rrhoeic subjects. J. Obstet. Gynaecol. Brit. Commoqg.,lg: 185.
Poffenbsrger M. y Fuller G.M. (1976). Is melatonin a microtubule
inhibitor? EXE.Cell. Res., 103: 135.
Powell U., Hemmarstrüm 8.. Samuelsson B. y Sjüberg B. (197“ D).
Prostaglsndin F2<x receptor in humancorgora lutea. Lancet: 1120

170
Prevedello M.R., Ritta M.N. y Cardinsli D.P. (1979). Fast sxonal
transport in rst sciatic nerve. Inhibition by pineal índoles.
Neurosci. Lett., ¿2: 29.
Quests M.R. y Rahamimoff R. (1965). Effect of melatonin on spontg
neous contractions and response to 5-hyoroxytryptem1ne of rst leg
lsted duodenum.Br. J. Pharmscol., gi: b55.
Quay U.B. (197h). Pineal Chemistry (Thomás.<5prigfie1d).
Ralph C.L.. Firth B.T. y Scott Turner J. (1979). The role of the
pinesl body tn ectotherm regulation. Am.Zool.. ¿2: 273.
Rsmuell P.U., Lsovey E.M. y Sintetos A. (1977). Regulation of the
sraohidonic acid cascade. Biol. ReErod., ¿É:,7D.
Rammsll P.w. y Shaw J.E. (1966). Spontaneous and evoked release of
prostaglandins from cerebral cortex of snesthetized cats. Ann. J.
Physiol.. 211: 125.
RamwellP.U., ShawJ.E. y Kucharski J. (1965). Prostsglsndin; re
lease Fromthe rat phrenic nerve-oiaphrsgm preparation. Science.
1h9: 139D.
Rao Ch.V. (1973). Receptors for prostaglandins and gonsdotrophins
in the cell membranasof bovine corpus luteum. Prostaglsndins, g;
567.
Reiter R.J. (1972). Surgical procedures involving the pineal gisnd
which preven goneosl degeneration in adult male hamsters. Annsles
d'Endocrinologie, 33: 571.
Reiter R.J. (1977). En: The Pinesl - 1977. Eden Press, Montreal.

171
Reiter R.J. (1978). En: The Pineal - 1978. Lden Press, Montreal.
Reiter R.J. (1979). En: The Pineal - 1979. Eden Press, Montreal.
Reiter R.J. (1980). The pineal and its hormones in the control of
reproduction in mammals.Endocrine Rev., 1: 109.
Reiter R.J. y Johnson L.V. (197“). Depressant action of the pineal
gland on pituitary luteinizing hormoneand prolactin in male hams
ters. Horm. Res., g: 311.
Reiter R.J. y Klein D.C. (1971). Dhservations on the pineal gland.
the Harderian glands, the retina, and the reproductive organs of
adult female rata exposed to continuous light. J. Endocr.. El: 117.
Reiter R.J., Petterborg L.J., Trakulrungsi C. y Trakulrungai U.
(1980). Surgical removal of the olfatory nulos increases sensitiqi
ty of the reproductive system of female rats to the inhibitory
effects of late afternoon melatonin injections. J. EXE.2001., gig:“7.
Reiter R.J., Vaughan M.K., Vaughan G.M., Sorrentino S. y Donofrio
R.J.(1975). The pineal gland as an organ of internal secretion.
En: Frontiers of Pigggl Physiolqu¿ M.D. Altschule (ed). The HIT
Press, Cambridge, Haas.. p. 5h.
Relkin R. (1978). Use of melatonin and synthetic TRHto determine
site of pineal inhibition of TSHsecretion. Neuroendocrinology,
gg: 310.
Reppert S.M.,\Perlow M.J.. Tamarkin L. y Klein D.C. (1979). A
diurnal melatonin rhythm in primate cereorospinal fluid. Endocrino
logy, 10k: 295.

172
RissmanE.F. (1980). Prepubertal sensitivity to melatonin in male
hamsters. Biol. Regrod., gg: 277.
Roche J.F., Foster D.L., MarshF.J. (197D). Effect of castration
snd infusion of meiatonin on levels of luteinizing hormonein sera
and pituitaries of eues. Endocrinology, gl: 1205.
Rogauski M.A., Roth R.H. y Aghsjsnian G.H. (1979). Melstonin: descg
tylation to S-methoxytryptamineby liver but not brain sryl aoylani
dase. J. Neurochem., gg: 1219.
Rollag M.D., Morgan R.J. y Niswender G.0. (1978). Route of meiato
nin secretion in sheep. Endocrinology, 102: i.
RomeroJ.A., Zstz M., Hebabian J.U. y Axelrod J. (1975). Circsdian
cycles in binding of 3H-s).prenolol to p-adrenergic receptor sitesin rat pineai. Nature, 258: #35.
Rosenberg R.C. y Lovenberg w. (1980). Dopamine p-hidroxylase. En:
Essays in Neurochemistr! and Phsrmscology. M.B. Youdim, m. Loven
berg, D. Sherman y J. Lagusda (eos), p. 163.
Rubin G.5. y Rosen Ü.M. (1975). Protein phosphorviation. Ann. Rev.
Biochem.. gg: 831.
Rust C.C. y Meyer R.K. (1969). Hair color, molt ano testis size in
male short-tailed uessels treated with melatonin. Science, ¿92: 921.
SsckmsnJ.U. y Reiter R.J. (1977). Hyperactivity in the blind-anos
mic female rat: role of the pinesl glsnd. Physiol Behav., ¿9: 321.
Sakai H.H. y Marks B.H. (1972). Aorenergic effects on pinesl cell
membrsnepotential. Life Sci., ¿iz 285.

173
SalzmanE.U. (1977). Interrelation of prostaglanoin endoperoxioe
(PGGZ)ano cyclio AMPin human blood plateleta. Biochim. Biophxa.
Acta, b99: ha.
Salzman E.w., Lindon N.J. y Roovien R. (1976). Cyclic AMPin human
blood platelets: Relation to platelet prostaglanoin ayntheeis inqg
ced by centrifugation or surface contact. J. Cyclic Nucleotide Rea..
g: 25.
SampaonP.H. (1975). Behaviour ano pineal functioning. En: Fron
tiers of Pinggl Phyaiolqgl. M.D. Altschule (ed). The HIT Presa.
Cambridge, Mase.. p. 20k.
Sanborn 8., Bhalla R. y HorenmanS. (1973). The endometrial adenoqi
ne cyclic 3',5'-monophoaphate-oependent protein kinaee. Distribution
ano properties of islet-cell protein phosphokinase. Biochem.J.,
129: 551.
Saper C.B., Loeuy A.D., Suanaon L.U. y Couan U.M. (1976). Direct hy
pothalamo-autonomic connections. Brain Res., 117: 305.
Scatchard G. (19h9). The attraction of proteins for small moleculas
and ions. Ann. N.Y. Acad. Sci., gi: 660.
Scheffé H. (1979). The analysis of variance. Milley. N.Y.
SemmP. y Vollrath L. (1979). Electrophyaiology of the guinea-pig
pineal organ: sympathetically influenced celLa responoing differen
tly to light and darkness. Neurosci. Lett., ¿gz 93.
Shaw J.E. y Ramwell P.w. (1966). Release of prostaglanoin from rat
Epididymal Fat pad on nervous ano hormonal stimulation. J. Biol.
Chem., 2h3: 1h98.

17k
Silmen R.E.. Leone R.Mt. Hooper R.J.L. y Preece M.A. (1979). He
latonln, the pineal gland and humanpuberty. Nature, 282: 301.
Silver M.J., Smith J.B.. Ingerman C. y Hoceis J.J. (1973). Ara
chidonlc acid-induced humanplatelet aggregatlon and prostaglan
oin formation. Prootaglandine, i: 863.
Sltaram B.R. y Lees G.J. (1978). Diurnal rhythma and turnover of
tryptophan hydroxylase in the pineal glano of the rat. J. Neuro
chem., El: 1021.
Smigel M. y Fleicher J. (197h). Ceracterization ano locolization
of PGE1
Bioghys. Acta, 332: 358.
receptora in the plasma membraneof rat liver. Blochlm.
Smith J.B.. Ingerman C.M. y Silver M.J. (1977). Effects of arachi
donio acid and someof its metebolltee on platelets. En: Proeta
glandins ln Hematology. M.J. Silver, J.B. Smith y J.J. Hocoie (edo).
p. 277.
Smuldera A.P. y Urlght E.M. (1960). Role of choroid plexus ln
transport of melatonin between blood eno brain. Braln Ree.. 191:
555.
Smythe G.A. y Lazarua L. (1973). Growth hormone regulation by me
latonin and aerotonin. Nature, 2h“: 230.
Smythe G.A. y Lazerus L. (197k). Supreeeion of human growth horno
ne aecretion by melatonln and cyroheptadine. J. Clin. Invest.. 23:
116.
Snedecor 5.o. y Bochren w.G. (1967). Statistical Methods. Iowa
State University Preee.

175
Snyder S.H., Fischer S. y Axeirod J. (1965). Evidence for the
presence of monoamineoxidase in sympathetic nerve endings. Bio
chem. Pharmacol., li: 363.
Sprecher H. (1977). Biosynthesis of polyunsaturated fatty acids
and its regulation. En: Polyunsaturated Patty Acid . w. humany
R. Holman (eds). Chigago, 111.. p. 1.
Sudgen D. (1978). Circadian changes in rat pineal tryptophan con
tent: lack of correlation with aerum tryptophan. J. Neurochem.,
33: 811.
Swanson R. y Friedhoff A. (1976). Uork in Progress. Decrease of
serotonin-N-acetyltransferase activity in rat pineal organs after
treatment with prostaglandin aynthesis inhibitor indomethacin.
Prostaglandins. ¿iz 503.
Szentagothai J., Flerko 8., Mess B. y Halasz B. (1976). Hygothaia
mic Control of the Anterior Pituitary. Akademiai Hiado, Budapest.
Talmadge H.. Bechtel E., Salonkangaa A., Huber P.. Jungmann R. y
Eppenberger U. (1975). Characterization of calf-ovary adenosine
3',5'-monophosphate-dependentprotein kinases and adenosine-J',5'
monophosphate binding proteins. Eur. J. Biochem.. gg: 621.
Tamarkin L., Hutchinson 3.5. y Goldman8.0. (1976 a). Regulation
of serum gonadotropins oy photoperiod ano testicular hormone in
the Syrian hamster. Endocrinology, 2g: 1528.
Tamarkin L., Mastrom w.H., Hamill A.J. y Goldman 8.0. (1976 D).
Effects of melatonin on the reproductive systems of male ano fema

176
le hamsters: a diurnal rhythm in sensitivity to melatonin. Endo
crinology. 22: 153“.
Thorpe P.A. y Herbert J. (1976). Studies on the duration of the
breeding season and photorefractoriness in female ferrets pinealeg
tomized or treated with melatonin. J. Endocr., lg: 255.
Trendelenburg U. (1979). The extraneural uptake of catecholamines:
is it an experimental oddity or a physiologicel mechanism?Trends
in Pharmacol. Sci., l: b.
Trentini G.P., Mess 8.. de Gaetani C.F. y Ruzsas C. (1979). Pineal
brain relationship. Prog. Brain Res., 2g: 3hl.
Tsafiri A., Lindner H., an U. y Lamprecht S. (1972). Physiologicsl
role of prostaglandins in the induction of ovulation. Prostaglandins,
g: 1.
Turek F.w. (1979). Role of the pineal gland in photnpericdic-induced
changes in hypothalamic-pituitary sensitivity to testosterona feed
back in castrated male hamsters. Enducrinology, 10k: 636.
Turek F.w., Desjardins C., MenakerH. (1976). Differential effects
of melatonin on the testea of photoperiodic and nonphotoperiodic
rodents. Biol. Regrod.. ¿2: 9h.
Tukey J.w. (19h9). Comparing individual means in the analysis of
variance. Biometrics. 2: 69.
Urry R.L. y Ellis L.C. (1975). Monoamine-oxidaseactivity of the
hypothalamusand pituitary: alteratians after.pinealectomy, changes
in photoperiod or additiona of melatonin in vitro. Eerriential 31:
B91.

177
Vacas M.I. (1980). -a Unidad Glándula Pinesl-Ganglig_üerviosl Su«
Eerior comoun Modelode Integración figyroendócrina..ïesis Docto
ral. Facultad de Ciencias Exactas y Naturales. UBA.
Vacas M.I. y Cardinali D.P. (1979 s). Effects of castration and
reproductive hormonason pineal serotonin metabolism in rata.
Neuroendocrinology, gg: 187.
Vacas M.I. y Cardinali D.P. (1979 b). Diurnal changes in melatonin
Dinding sites of hamster and rat brain. Correlation with neuroendo
crine responsiveness to melatonin. Neurosci. Lett., ¿2: 259.
Vacas H.I. y Cardinsli D.P. (1980). Binding sites for melatonin in
bovine pineal gland. HormoneRes., ¿2: 121.
Vane J.R. (19GB). The use of isolated organ For detecting active
substances in circulating blood. Br. J. Pharmsc¿'Chemoter., gg: 360.
VaneJ.R. (1971). Inhibition of prostaglandin synthesis as a mecha
nism of action of aspirin—like drugs. Nature. 231: 232.
Vane J. (1976). The mode of action of aspirin and similar compounds.n
J. Allergy Clin. Immunnl., 2Q: 691.
VaneJ.u. y Uilliams H.I. (1973). The contrioution of prostaglandin
production to contractions of the isolated rat uterus. Brit. J.
Pharmacols, EQ: 629.
Vaughan M.K., Little J.C., Johnson L.Y., Blask D.E.. Vaughan G.M.
y Reiter R.J. (1978 D). Effects of melatonin and natural and synthg
tic analogues of arginine vasotocin on plasma prolactin levels in
adult male rata. HormoneRes., g: 236.

178
Vaughan G.M., Hc Donald A.. Jordon R.M., Allen J.P.. Bohmfalk
G.L.. Abou-SamraM. y Story J.L. (1978 a). Melatcnin concentra
tion in humanblood and cerebrospinal fluid: relationship to
stress. J. Clin. Endocr. Metsb., 22: 220.
Vaughan M.H., Vaughan G.M., Blaak D.E. y Reiter R.J. (1976).
Influence of melatonin, constant light or blinding on reproduc
tive system of gerbils (Merionesungulatus). Eerrientia, 2g: 13hl.
Vesin M.F. y Harbou S. (197“). The effects of epinephrine, prosta
glandins and their antagonists on adenosine cyclic-J’,5'-monophos
phate concentrations and motility of the rat uterus. Mol. Pharma
col., lg: #57.
Vriend J., Reiter R.J. y Anderson G.R. (1979). Effects of the pi
neal and melatonin on thyroid activity of male golden hamsters.
Gen. ComE. Endocr., gg: 189.
Makeling A., Kirton H.T. y Uyngarden L. (1973). Prostaglandin re
ceptora in the hamster uterua during the estrous cycle. Prosta
glandins, i: l.
Makeling A. y Spilmsn C. (1973). Prostaglandin specific binding
in the rabbit oviduct. Prostsglandins, k: #05.
ualsh D.A., Perkins J.P. y Hrebs E.G. (1966). An adenoaine-B‘,5'
monophosphate-dependentprotein kinase Fromraobit skeletal muscle.
J. Biol. Chem.. 2h}: 3763.
Uartenberg H. (1968). The mammalianpineal orgsn: Electron micros
copio studies on the fine structure of the pineslocytes, glial cells
and on the perivasCUlsr compartment. Zeitscnrift fur zellforachung

179
und Mikroskopic Anatomie, gg: 7h.
Ueinberg U., Meítzman E.D., Fukushima D.H., Janoel G.F. y Rosen
feld R.S. (1980). Melatonin does not supress the pituitary lutei
nizing hormoneresponse to lutainizing hormone-releasihg hormone
in men. J. Clin. Endocr. Met., g¿: 161.
Ueiss B. y Crayton J. (1970). Gonadal hormonas as regulators of
pineal adenyl-cyclase activity. EndocrinologL, 91: 527.
Ueatfall T.C. y Brated M. (197“). Specificitt of blockade of the
nicotine-inouced release of 3H-norepinephrine Fromadrenergic neu
rons of the goinea-pig heart by various pnarnacological agente.
J. Pharmacol. Exp. Ther., ¿22: 659.
Wilkinson M., Arendt J.. Bradkte J. y Ziegler O. (1977). Determina
tion of a dark-induced increase of pineal N-ncetyltransferase acti
vity and simulataneous radioimmunoassay of mulatonin in pineal, se
rumand pituitary tissue of the male rat. J. Endocr., 2g: 2h}.
Uílliama H.I., Sneddon J.M. y Harvey P.J. (197b). Prostaglandin
production by the pregnant rat uterus in vitro ano its relevance
to partirution. J.EPharmacol., gg: 207.
Wilson B.w. (1978). The application of mass apectrometry to the
study of the pineal glano. J. Naural Transm.. sugl. 13: 279.
Uithyachumnarnkul B. y Hnigge H.M. (1980). Malatonin concentration
in cerebrospinal fluid. peripheral plasma ano plasma of the con
fluena sinuun of the rat. Neuroendocrinology. gg: 382.
molfe L. y Coceani F. (1979). The role of prostaglandins in the
central nervous system. Ann. Rev. Physiol., il: 669.

UurtmanR.J. (1973). Biogenic amines and endacrine function. In
troduction: neuroenoocrine transducer ano mo1oam1nes.Fed. Proc..
¿2_: 1769.
MurtmsnR.J. y Axelrod J. (1965). The Pineal Glsnd. Scientific
American. 213: 50.
MurtmanR.J., Axelrod J. y Chu E.U. (1963). Melatonin, a pineal
substance: effect on the rat ovary. Science, IBI: 277.
Murtman R.J., Axelrod J. y Potter L.T. (196%). The uptake of 3H
melatonin in endocrine ano nervous tissues 31d the effects of
constant light exposure. J. Pharmacol. Exp. 'her., 1h}: 31k.
MurtmanR.J. y Cardinali D.P. (197b). The Pineal Organ. En: Text
hook of Endocrinolqgl. R. Williams (ed). w.a Saunders, Filadelfia,
p. 832.
Murtman R.J. y Moskouitz M.A. (1977). The Píneal Urgen. NewEngland
J. Med.. 296: 1329 E 1383.
MurtmanR.J.. Shein H,M.. Axelrod J. y Larin F. (1969). Incorpora
tion of 1L'El-tryptophaninto 1“fl-protein by cultured rat pineals:
stimulation by L-norepinephrine. Proc. Nat. Acad. Sci. USA,gg:
7h9.
Yamamoto 8., Ügino N., Ühki S. y Yoshimoto T. (1977). En: Enzxmo
logicel studigg on prostaglandinvbiosynthesis. Biochemicasaspects
of PG and thromboxanes. Norman Hharasch y Joseph Fried (eos). Aca
demic Press Inc., p. 1.
Vamashíta H.. Mieno M.. Shimizu T. y Yamashita E. (1978). Inhbition
by melatonin of the pituitary response to luteinizing hormonerelea

181
sing hormone in vivo. J. Endocr.. 29: #87.
Vates C.A. y Herbert J. (1979). The effects of different photope
riods on circadian 5-HTrhythms in regional brain areas and their
modulation by pinealectomy, melatonin and oestradiol. Brain Res.,
176: 311.
Yehuda S. y Frommer R. (1977). Circadian rhythms in melatonin e
ffects on D-amphetamine-induced hypthermia among rats kept at k C.
Preliminary results. J. Neural Transm.l suEl. 13: 607.
Ying S.V. y Greep R.Ü. (1973). Inhibition of ovulation by melato
nin in the cyclic rat. Endocrinology, 2g: 333.
Young, Lai E.U. (1979). In vitro effects of melatonin on hCGsting
iation of steroid accumulationby rabbit ovarian follicles. J. Ste
roid Biochem., lg: 71h.
Zatz M. (1977). Effects of cholera toxin on Supersensitivú and sub
sensitive rat pineal glanda: regulation of sensitivity in multiple
sites. Life Sci., gl: 1267.
Zatz M. (1978). Sensitivity and cyclic nucleotides in the rat pine
al gland. En: The Pineal Gland. I. Nir, R.J. Reiter, R.J. Nurtman
(eds). Springer Verlag, Viena, p. 97.
Zatz M. (1980). The pineal gland: shedding light on the internal
clock. Trends Pharm. Sci., l: 230.
Zinder Ü. y Pollard H. (1980). The chromaffin granule: recent stu
dies leading to a functional model for exocytosis. En: Neurochemis
try and Neurogharmacology, g: 125.