o conceito de espécie_uma abordagem pratica
TRANSCRIPT

OS CONCEITOS DE ESPÉCIE: UMA ABORDAGEM PRÁTICA
Edio-Ernst KischlatInstituto de Geociências
Universidade Federal do Rio Grande do [email protected]
RESUMO
O entendimento da Biodiversidade pos-sibilita a avaliação da riqueza genética de de-terminada área e serve de base para propos-tas conservacionistas. Esta Biodiversidade éconseqüência direta da aplicação de um con-ceito de espécie. Pode-se enumerar seis dife-rentes tipos básicos de conceito existentes naliteratura: (1) o Conceito Tipológico, (2) o Con-ceito Biológico, (3) o Conceito Isolacionista,(4) o Conceito Evolutivo, (5) o Conceito Auta-pomórfico e (6) o Conceito Filogenético. Cadaum deles é reflexo de um determinado momen-to histórico-filosófico, e todos possuem limita-ções quanto a sua abrangência e aplicação.Alguns são eminentemente ontológicos, outros,bastante epistemológicos. A aplicação destesconceitos resulta em diferentes avaliações dabiodiversidade, tanto presente quanto pretérita.Assim, procurou-se diferenciar cada conceito,mostrando seu universo de aplicação e discu-tindo suas limitações quanto a cada universo.
PALAVRAS-CHAVE: Biodiversidade, Conceito deespécie, Conceito Tipológico, Conceito Biológi-co, Conceito Isolacionista, Conceito Evolutivo,Conceito Autapomórfico, Conceito Filogenético.
INTRODUÇÃO
A experiência adquirida em atividadesdocentes junto ao curso de graduação emCiências Biológicas mostra o interesse dos alu-nos em saber quantas espécies existem em
determinado universo, tratando �espécies�como algo concreto e natural, onde o pesqui-sador apenas teria o trabalho de enumerar di-versos nomes em uma listagem, representati-va da biodiversidade no espaço geográfico e/ou na coluna geológica. O reconhecimento deque espécies fazem parte da natureza é muitoantigo, possuindo raízes bíblicas (vide Gêne-sis, 1:11-27; 7:2-3,8-9,14-16; 8:1,17,19).Pode-se mesmo admitir uma grande contribui-ção destas passagens bíblicas na construçãodo Conceito Biológico de espécie, extensiva-mente ministrado no Ensino Médio.
Com a inserção da �Sistemática Filoge-nética� (Cladismo) no currículo de graduação,onde um novo raciocínio é proposto, a escolaevolutiva gradista clássica (a denominada �Teo-ria Sintética�) é contrariada, e novos conceitosapenas adicionam mais problemas no entendi-mento da biodiversidade espaço-temporal.
A Biodiversidade é muito discutida nosdias de hoje. É o seu entendimento que possi-bilita a avaliação da riqueza genética de deter-minada área, servindo de base, principalmen-te, para propostas conservacionistas, represen-tadas por uma legislação pertinente. Basica-mente, o estudo da biodiversidade inclui os se-guintes objetivos (Wiley e Mayden, 2000c):(1) inventário; (2) conservação e gerenciamen-to; e (3) educação do público quanto a sua im-portância, assim como a importância da Siste-mática e Taxonomia, que são áreas da Biologiaque se detêm no estudo da espécie. Assim, aespécie é a unidade atômica da biodiversida-de e seu conceito é crucial (Wiley e Maiden,2000c). Entretanto, as diferenças de aborda-gem, objetivando a conceituação da espécie e
Métodos de Estudo em Biologia 11
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia12
a aplicação destes conceitos, interferem na ava-liação da biodiversidade.
O presente artigo é, em grande parte,um resumo simplificado do livro de Wheeler eMeier (2000) Species concepts and phyloge-netic theory - a debate e tem como objetivo adivulgação, em português, das idéias ali conti-das visando o público universitário discente.Cerca de cinco conceitos (Biológico, Isolacio-nista, Evolutivo, Autapomórfico e Filogenético)são extensivamente defendidos por autoresque, na maioria, são os próprios proponentes.Análises e críticas são feitas pelos demais, for-mando um debate rico em argumentos e dis-cussões. Em cada conceito, as principais críti-cas são enumeradas e, na presença de falsea-mento, são contra-argumentadas. É claro queo citado livro é bem mais amplo em discus-sões. Alguns conceitos alternativos de espéciejá propostos (e.g., Conceitos Recognitivo, Eco-lógico, Coesivo, Nominalista) são descartadosou sinonimizados. Caso o leitor queira se apro-fundar no assunto, sua leitura é fundamental.Por outro lado, vários termos cladísticos bási-cos são utilizados no decorrer do texto, muitosdeles discutidos e definidos por Amorim (1997),que, por sua vez, deve ser consultado adnauseum pelo público que se inicia na Siste-mática Filogenética.
CONCEITO, CATEGORIA ENOMENCLATURA
A grande maioria dos biólogos não sedetém em questões sistemáticas. Seus proble-mas se direcionam aos demais campos da Bio-logia onde determinada espécie está incluídaapenas como instrumento de estudo. Suas ne-cessidades então restringem-se a uma deno-minação formal em que um ou mais indivíduos(=espécimens) utilizados são reconhecidoscomo formando uma unidade �natural� atravésde um argumento fenético ou molecular. Estu-dos etológicos e sinecológicos, por exemplo,
não se detém em conceitos de espécies. É ne-cessário apenas conhecer a denominação for-mal utilizada para determinados táxons (tantozoológicos quanto botânicos), geralmente se-guindo a opinião de outros autores.
Isto seria uma tarefa simples se, ao seavaliar a biodiversidade de determinado local,não se encontrasse grupos de espécimens quese desviam do registrado na literatura, ou mes-mo, diferentes grupos de espécimens denomi-nados sob um mesmo nome. No primeiro caso,existe um novo fênon (e.g., morfologia) nãodescrito, em que não existe um nome propos-to para denominá-lo. Tem-se assim uma espé-cie nova. No segundo caso, um dos fênons foisubavaliado e incluído em outro, sendo poten-cialmente também uma espécie nova.
Quando o problema passa a ser o ques-tionamento de ser ou não uma espécie nova, otrabalho torna-se sistemático. Até onde um fê-non aparentemente distinto mereceria o reco-nhecimento de ser uma espécie independen-te das demais e qual a melhor forma de sedenominá-la?
Conceito de EspécieO uso do termo �espécie� para objetos
inanimados (e.g., espécies minerais1 ) e anima-dos foi de uso corrente até o século XIX. Na-turalistas se ocupavam fazendo um inventáriodas espécies na natureza, e o método utiliza-do para a sua discriminação era o reconheci-mento das diferenças, baseando-se em carac-terísticas organolépticas. No que se refere àsespécies animadas, a morfologia tinha impor-tância fundamental, e classes de organismospoderiam ser descritas e delimitadas em rela-ção às demais.
Um conceito de espécie representa aforma de descrição e delimitação destas clas-ses de organismos segundo um critério defini-do. A aplicação de tal conceito resulta no reco-nhecimento de uma entidade concreta identifi-cável, consistindo de um conjunto de organis-mos (zoológicos ou botânicos) que seguem um
1 A duodécima edição do Systema naturæ de Caroli a Linné é composta de três tomos: o primeiro, em dois volumes, écomposto pelo Regnum animale (1766); o segundo é composto pelo Regnum vegetabile (1767); e no terceiro tomo -Regnum lapideum (1768) - são descritas as “espécies minerais”. Estas obras podem ser encontradas na página daGallica (http://gallica.bnf.fr/)
Métodos de Estudo em Biologia12
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 13
padrão qualquer. Estas entidades identificáveisrecebem o nome de �espécie� e comportam umtáxon (táxon-espécie) que pode receber umnome para a sua representação. O pardal e abatata são táxons-espécie, assim denominadossegundo critérios morfológicos, refletidos e es-tabelecidos há muito tempo no senso comumhumano, pois são reconhecidos universalmen-te como tais, independentemente do idioma edo momento histórico de cada nação ocidental.
Um conceito de espécie deve contem-plar os seguintes objetivos (Wheeler e Platnick,2000b): (1) reconhecer os tipos de organis-mos que se auto-perpetuam no planeta, as-sim como a quantidade destes tipos, tanto nopassado como no presente; (2) identificar oresultado dos diversos processos evolutivos;(3) descobrir os agrupamentos mínimos de or-ganismos (espécies), assim como sua históriacomum que possa ser resgatada; (4) determi-nar estas unidades mínimas (espécies), emconcordância com um sistema de classifica-ção que proporcione previsão na distribuiçãodos atributos destas espécies e comunicaçãoentre as pessoas.
CategoriaEm seu Systema naturæ, Linnæus (1758)
utilizou o nome �espécie� para a categoria demais baixa ordem (categoria-espécie) em seusistema de classificação. Posteriormente, acategoria-subespécie foi inserida neste sistemade classificação. Comumente, seu uso refere-seao reconhecimento nomenclatural de popula-ções geograficamente disjuntas (alopatria), quemostram determinada variação na comparaçãocom a população �principal�, sendo esta varia-ção insuficiente para o reconhecimento de umaespécie plena. Este procedimento traz embuti-do a idéia de pouca importância da subespéciegeográfica quando comparada com a subespé-cie �principal� (a que possui o nome �subespe-cífico� igual ao da espécie). Deve-se entenderque esta população �principal� não representa,necessariamente, a população maior ou de dis-tribuição mais ampla, mas simplesmente a po-pulação que foi descrita primeiro, em termos his-tóricos formais (que pode ser, eventualmente, ade distribuição mais limitada). Se determinada�subespécie� mereceria ou não ser reconheci-
da como uma espécie plena, é uma discussãobaseada na opinião pessoal de cada autor, e aliteratura está cheia de casos de contenda.
O uso da categoria-subespécie implicano reconhecimento de trinômios formais latinose representaria uma subunidade da espécie. Sea subespécie é reconhecida como tal, é porqueela possui características diagnósticas, pois a au-sência destas negaria sua existência. Mas é fatoque, dentro da literatura taxonômica, coexistembinômios formais (aqueles representativos so-mente de espécies, sem o reconhecimento desubespécies) e trinômios formais (aqueles deuma espécie com, no mínimo, duas subespé-cies). A contagem da biodiversidade, então, de-veria ser feita pela soma da quantidade de tri-nômios com a de binômios (excetuando-se osbinômios embutidos nos trinômios). Isto implicana utilização de duas unidades de contagem, oque é ilógico. Se a espécie é a unidade da bio-diversidade, ela não pode ser subdividida, a nãoser que seja reconhecida como comportandomais de uma espécie (cf. Wiley e Maiden, 2000c).Visando um nivelamento de categoria, ou se tra-tam todos os táxons-espécie sob a forma de tri-nômios, o que seria uma solução redundantepara os binômios, ou se eleva cada subespéciepara a categoria de espécie, com o uso dacategoria-superespécie para a reunião destasespécies. Embora seja um artifício utilizado paranivelar os táxons, isso acaba também sendoredundante no reconhecimento de superespé-cies monotípicas. Mesmo assim, o reconheci-mento de uma superespécie com várias espé-cies, ou de uma espécie com várias subespé-cies, representa apenas visões distintas de ummesmo problema: populações alopátricas comvariação morfológica.
Por outro lado, o uso das categorias deordem mais alta (gênero, família, ordem, clas-se) tem sido questionado quanto a sua utilida-de (vide Queiroz e Gauthier, 1992; Kischlat eSchultz, 2002). O problema não está em se re-conhecer táxons de ordem mais alta que a es-pécie (i.e., táxons mais inclusivos), mas, sim, emse relacionar categorias hierárquicas a estestáxons. Categorias hierárquicas trazem valoresrelativos embutidos que permitiriam, ao leitordesavisado, assumir certa importância, valori-zar morfologias e fazer comparações de todosos tipos (e.g., a �família� X-idæ de coleópteros
Métodos de Estudo em Biologia 13
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia14
com a �família� Y-idæ de aves). Comparações,em termos de quantidade de �famílias�, já fo-ram muito utilizadas tanto para a comparaçãoda biodiversidade entre distintas regiões quantopara avaliação de extinções periódicas no re-gistro fossilífero (vide Gaston, 1991; Benton,1989, 1995). Estudos desta natureza apresen-tam problemas na quantificação dos dados esuas conclusões estão sujeitas a reavaliações(vide Lane e Benton, 2003).
Outro problema no uso de categoriasreside no fato de que grupos reconhecidoscomo de ordem mais alta (e.g., a �classe� Aves),conforme sejam avaliados filogeneticamente,mostram que estão incluídos dentro de outrosde igual hierarquia (e.g., a �classe� Aves estáaninhada na �classe� Reptilia) ou mesmo de hi-erarquia inferior (a �classe� Aves está aninha-da na �ordem� Dinosauria) (cf. Romer, 1956).Discussões sobre as contrariedades do uso decategorias Linneanas podem ser encontradasdesde a década de 70 (vide Griffiths, 1976;Queiroz e Gauthier, 1994).
NomenclaturaÉ o método de denominação dos táxons
reconhecidos segundo um conceito. Linnæus(1758) utilizou a nomenclatura binomial latina,composta do nome da categoria-gênero e donome da categoria-espécie para a denomina-ção de táxons-espécie2 . O pardal3 recebeu onome �Passer domesticus� e a batata �Solanumtuberosum�. Estes nomes representam, for-malmente, em latim, os nomes �pardal� e �ba-tata�, que, por sua vez, são nomes informaisem português vernacular. �Pardal� e �Passerdomesticus�, assim como �batata� e �Solanumtuberosum�, são nomes representativos detáxons-espécie, definidos sob critérios morfo-lógicos. A formação de binômios formais se-gue regras claras encontradas nos diversoscódigos de nomenclatura4 e dá-se o nome�espécie-nominativa� ao binômio formalmenteproposto sob estas regras, seja representativode um táxon-espécie, seja subordinado (=si-
nonimizado) a um outro binômio formal, repre-sentativo do mesmo táxon-espécie mas comprecedência histórica. Por sua vez, a formaçãode nomes informais não possui regras e variano tempo e espaço geográfico segundo as cul-turas locais. Cantino et al. (1999) fazem umarevisão das propostas nomenclaturais paratáxons-espécie, muitas delas partindo do mo-delo Linneano.
O táxon-espécie é assumido como oagrupamento de organismos de mais baixa or-dem, potencialmente merecedor de um binô-mio formal latino, representando a unidade nacontagem da biodiversidade. O problema, en-tão, passa a ser o reconhecimento de agrupa-mentos de organismos que mereceriam estestatus nomenclatural.
Em relação a táxons mais inclusivos (i.e.,de mais alta ordem), existem propostas nomen-claturais que seguem, basicamente, dois tiposde definição: (1) definição por base-nodal, e(2) definição por base-estemática. A discussãosobre métodos nomenclaturais de definição detáxons mais inclusivos foge do presente objeti-vo, mas pode ser encontrada em Kischlat eSchultz (2002) e nas referências ali citadas.
OS CONCEITOS DE ESPÉCIEPROPOSTOS
O Conceito Tipológico LinneanoMayr (2000a) observa que desde a An-
tiguidade, de Platão e Aristóteles até Linnæus(1758) e autores do início do século XIX, a pa-lavra �espécie� exprimia a idéia de classes deobjetos, cujos membros compartilhavam cer-tas propriedades em comum, que as diferenci-avam de outras classes. Estas classes eramconstantes, não mudavam com o tempo, e qual-quer desvio na definição original representavaapenas acidentes que eram manifestações im-perfeitas de sua essência.
2 A categoria-espécie e a categoria-gênero são uninomiais, já o táxon-espécie é binomial.3 Seguiu-se os exemplos fornecidos por Mayr (2000a).4 International Code of Nomenclature of Bacteria (Sneath, 1992), International Code of Botanical Nomenclature (Greu-ter, et al., 2000), e International Code of Zoological Nomenclature (ICZN, 1999).
Métodos de Estudo em Biologia14
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 15
O Conceito Tipológico aborda a espé-cie como uma entidade que se diferencia deoutras espécies por características diagnós-ticas constantes. Assim, a espécie represen-tava uma entidade natural, imutável morfologi-camente, de origem Divinamente criada. Utili-zar uma amostra de determinada espécie, atra-vés de um ou mais exemplares, seria suficien-te para a sua representação. Esta representa-ção amostral continua nos dias de hoje, previs-ta pelos códigos de nomenclatura (séries-tipo).
Entretanto, a mutabilidade morfológicafoi colocada em questão. Darwin (1859) intro-duziu o conceito de evolução e a espécie pas-sou a ser mutável no tempo. Por outro lado,pode-se encontrar distintos fenótipos na natu-reza, causados tanto pelo dimorfismo sexualcomo também pela ontogenia, sazonalidade oumesmo variação genética geral (Mayr, 2000a).O resultado é que membros de uma mesmapopulação podem ser mais diferentes entre sido que com membros de outra população re-conhecidamente comportando uma outra es-pécie. Mayr (2000a) ainda enfatiza a existên-cia de espécies crípticas (sibling species), ex-tremamente semelhantes e virtualmente indis-tingüiveis, cujos indivíduos, quando coexistin-do, não intercruzam, mantendo seu genoma.
Assim pode-se caracterizar o ConceitoTipológico como: (1) baseado em atributos mor-fológicos, logo, espécies crípticas são passíveisde serem ignoradas; (2) independente do cri-tério reprodutivo (assexual ou sexual); (3) des-conhecedor do processo evolutivo, assim comodo processo de formação de espécies; e(4) desconhecedor da variabilidade morfológi-ca, tanto geográfica quanto temporal, induzin-do a se considerar múltiplos táxons-espécie,onde, potencialmente, existe apenas um.
O Conceito Biológico MayrianoMayr (2000a) define espécie biológica
como5 �grupos de populações naturais in-tercruzantes que são reprodutivamente iso-ladas de outros grupos�. O autor enfatiza quenão interessa o grau de diferenciação mor-
fológica, mas sim as relações genéticas. Es-pécies não podem ser definidas como clas-ses naturais, mas representam entidades con-cretas de indivíduos que podem ser descritase delimitadas. Cada espécie biológica é umagrupamento de genótipos em harmonia equalquer intercruzamento levaria ao desequi-líbrio. O status de espécie é uma propriedadede populações, não de indivíduos; e uma po-pulação não perde seu status de espécie sóporque um indivíduo a qual pertence cometeum �erro� e hibridiza. Uma população isoladageograficamente não intercruza com outra po-pulação, mas poderia ter a tendência de in-tercruzar, se o isolamento externo terminar.Este conceito enfatiza a proteção de um poolgênico harmonioso, sendo estritamente bio-lógico e possuindo significado apenas onde opool gênico de uma espécie entra em conta-to com o pool gênico de outra, numa certalocalidade e em um determinado tempo (re-presentando uma situação não-dimensional).Devido ao fato de que táxons-espécie possu-em uma extensão no tempo e no espaço, ostatus de populações não-contíguas deve serdeterminado por inferência. O Conceito Bio-lógico não é aplicável a organismos assexu-ados (agamotáxons), pois eles formam clonese não populações. Estes organismos mantémseu genótipo de geração a geração sem in-tercruzamento, e não precisam de mecanis-mos de proteção da integridade e harmoniado genótipo. O autor enfatiza que seu con-ceito é não-dimensional, e aceita certa arbi-trariedade no reconhecimento de populaçõesalopátricas, defendendo o uso dacategoria-subespécie. Quanto a aplicação emfósseis, a separação entre espécies tambémseria arbitrária em linhagens fósseis com mor-fologia contínua. Por outro lado, a incomple-teza do registro fossilífero proporcionaria suadelimitação prática, embora, de certa forma,igualmente arbitrária.
Pode-se concluir que o Conceito Bioló-gico é iminentemente ontológico. O reconhe-cimento de táxons-espécie é subjetivo e o usoda categoria-subespécie indicaria níveis de es-
5 “I define biological species as groups of interbreeding natural populations that are reproductively isolated from othersuch groups” (Mayr, 2000a:17).
Métodos de Estudo em Biologia 15
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia16
peciação incompleta ao lidar-se com variaçãomorfológica em populações alopátricas ou alo-crônicas. As críticas feitas a este conceito com-preendem (Willmann e Meier, 2000; Mishler eTheriot, 2000b; Wheeler e Platnick, 2000b;Wiley e Mayden, 2000b):
(1) O conceito é incompatível com es-pécies de reprodução assexuada, pois qualquerindivíduo seria uma espécie em separado, jáque são reprodutivamente isolados de outrosindivíduos. Mayr (2000c) reconhece que o con-ceito não foi proposto com vista à aplicaçãoem agamoespécies (i.e., espécies com repro-dução assexuada).
(2) Qualquer conceito deve ser compa-tível com a evolução e deve incluir critérios paradelimitação da espécie multi-dimesionalmente(alopatria e alocronia). A observação da pre-sença ou ausência de intercruzamento entreindivíduos de populações distintas (próximasou longínquas) é um argumento pouco práticoou mesmo impossível. Da mesma forma, é im-possível sua aplicação no reconhecimento deformas extintas (fósseis ou não). Mayr (2000c)reconhece que o status de populações alopá-tricas e alocrônicas pode ser determinado ape-nas por inferência indireta ou por critérios sub-jetivos (e.g., categoria-subespécie). A espécie,realmente, é multidimensional, mas a situaçãonão-dimensional é necessária para se determi-nar as suas reais propriedades biológicas.
(3) Sendo a delimitação da espécie re-flexo de uma situação não-dimensional (sim-patria e sincronicidade), esta delimitação esta-ria diretamente relacionada a um instante tem-poral inicial.
(4) O intercruzamento e a coesão (fluxogênico) de uma população representariam umacaracterística herdada (plesiomorfia). Assim,não representam critérios de distinção de es-pécies ou populações. Somente o isolamentoreprodutivo representaria argumento válidopara o reconhecimento, como espécie, de umconjunto de populações.
(5) O reconhecimento da espécie comoum mecanismo de proteção do equilíbrio e
harmonia do genótipo é por demais determi-nístico e implica em assumir a espécie comoum estágio final de preservação destes genóti-pos quanto a misturas posteriores. Em realida-de, o processo de especiação, na grande mai-oria dos casos, é uma conseqüência incidentaldo tempo de separação de duas populaçõesque acabam por adquirir caracteristicas distin-tas e incompatíveis reprodutivamente, carac-terísticas estas resultantes de diferentes forçasseletivas. Mayr (2000c) responde que a sele-ção natural age, sim, sobre o fenótipo, e aque-les que se desviam muito do ideal tornam-sevulneráveis ou são eliminados. Contudo, nãose entende porque algumas populações pare-cem possuir morfologia uniforme, enquantooutras são bastante variáveis. Mas como a dife-rença morfológica não é o critério utilizado nadistinção da espécie, tais diferenças na variabili-dade são irrelevantes na aplicação do conceito,embora sejam a causa de dificuldades conside-ráveis na discriminação de táxons-espécie.
(6) O conceito de espécie deve permitira avaliação da biodiversidade, permitindo mes-mo a comparação de agrupamentos distante-mente relacionados. A utilização da cate-goria-subespécie implica em reconhecer umanatureza politípica para a espécie em questão,reduzindo, arbitrariamente, o número de uni-dades da biodiversidade.
O Conceito Isolacionista HennigianoHennig (1950) preocupou-se em delimi-
tar a espécie no tempo. Meier e Willmann(2000a) definem6 espécie como �populaçõesnaturais, ou grupos de populações naturais,reprodutivamente isoladas; as espécies se ori-ginam pela dissolução de uma espécie este-mática7 em um evento de especiação, e dei-xam de existir, tanto por extinção quanto poruma nova especiação�. É a existência de umadescontinuidade reprodutiva específica quedefine a identidade da espécie e previne a tro-ca de informação genética entre as espé-cies-irmãs. Um evento especiativo é definidopela formação desta descontinuidade. Assim,
6 “Species are reproductively isolated natural populations or group of natural populations. They originate via the dissolution ofthe stem species in a speciation event and cease to exist either through extinction or speciation” (Meier e Willmann, 2000a:30).7 Espécie-ancestral.
Métodos de Estudo em Biologia16
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 17
�espécie� e �especiação� são conceitos relacio-nais. Uma espécie só existe na correlação comsua espécie-irmã, e na formação de uma des-continuidade reprodutiva específica entre elas.Se a espécie é definida com relação a uma des-continuidade reprodutiva específica, ela deixade existir durante um novo evento especiativo,quando uma nova descontinuidade é formada.O isolamento reprodutivo representa o �pontode não retorno� no processo evolutivo, sepa-rando grupos naturais de populações. Estas uni-dades reprodutivamente isoladas são táxonsapropriados a uma análise filogenética, poispossuem uma relação hierárquica.
O Conceito Isolacionista não é aplicávela agamotáxons já que cada organismo é, emsi, isolado reprodutivamente. Por outro lado, eleexpande o conceito biológico, especificandoos limites da espécie no tempo. É um conceitoque não se limita a argumentos fenéticos (e.g.,morfologia) mas enfatiza o isolamento repro-dutivo entre duas espécies-irmãs. Este isola-mento reprodutivo pode ser de natureza bio-lógica ou geográfica. Caracteres fenéticos ape-nas evidenciam os limites da espécie, mas nãoa sua existência. Em outras palavras, se idênti-cos indivíduos são retirados de duas popu-lações distintas (i.e., as duas populações sãoindistintas feneticamente), a conclusão deque as duas populações representam umamesma espécie é apenas uma hipótese.
Este processo pode ser melhor visuali-zado pelo modelo alopátrico de separação ge-ográfica8 de uma população ancestral em, pelomenos, duas subpopulações através da forma-ção de uma barreira geográfica (e.g., aberturade mares, levantamento de cadeia montanho-sa, formação de um rio, etc.) que pode, eventu-almente, ser datada. As relações tocogenéticas9
(Figura 1) entre os indivíduos destas duas subpo-pulações cessam, e cada subpopulação segueo seu caminho, diferenciando-se (Figura 2). Qual-quer simpatria entre espécies-irmãs representa
um evento posterior, conseqüência da retiradada barreira geográfica e dispersão. Hoje em dia,existem populações-irmãs em diversos estágiosde isolamento reprodutivo, algumas completa-mente, outras apenas com isolamento incipien-te, e outras ainda com isolamento inexistente.Estes estágios podem estar acompanhados dediferenciações fenéticas (e.g., morfológicas, eto-lógicas, etc.) ou não, independentemente de seutempo de isolamento. Assim pode-se assumirdois casos extremos: (1) populações fenetica-mente distintas e com isolamento reprodutivoincipiente ou inexistente; e (2) populações to-talmente isoladas reprodutivamente mas feneti-camente indistingüíveis. Na retirada da barreirageográfica e simpatria secundária, o caso(1) resultaria em hibridização cada vez maior,com apenas uma espécie comportando polimor-fismo fenético; no caso (2) tem-se a coexistên-cia de espécies crípticas (sibling species).
Figura 1. Processo de clivagem de uma espécie mos-trando as relações tocogenéticas entre os indivíduos.Sexos distintos estão representados por círculos chei-os ou não. Modificado de Hennig (1966).
8 Hennig (1966:51) observa que separação espacial (spatial separation) seria um termo mais acurado, podendo seraplicado também a parasitas que se tornam isolados ao colonizar um novo hospedeiro.9 Relações tocogenéticas (tokogenetic relationships) são as relações genéticas entre individuos, como numa árvoregenealógica, em que estes são descendência de dois outros de sexos opostos (Hennig, 1966:29-30). Este fenômenotambém é denominado reticulação (reticulation). Por outro lado, relações filogenéticas são as relações entre duaslinhagens independentes de organismos (Mishler e Theriot, 2000a).
Métodos de Estudo em Biologia 17
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia18
Figura 2. Divergência morfológica entre duas espéci-es-filhas em relação ao tempo, comparando-se com aespécie-estemática. Modificado de Meier e Willmann(2000a).
Concluindo, em relação ao ConceitoBiológico, o Conceito Isolacionista é igualmen-te ontológico, mas incorpora os seguintes fa-tos: (1) assume o processo evolutivo, explican-do o porquê e o modo de formação de espéci-es multidimensionalmente (i.e., alopatria e alo-cronia); (2) reconhece o monofiletismo10 de li-nhagens, inclusive de táxons supraespecíficos(i.e., mais inclusivos). As críticas feitas a esteconceito compreendem (Mayr, 2000b;Mishler e Theriot, 2000b; Wheeler e Platnick,2000b; Wiley e Mayden, 2000b):
(1) O conceito Isolacionista é, essencial-mente, uma versão do Conceito Biológico mo-dificado para acomodar uma perspectiva pa-leontológica, permanecendo subjetiva a avali-ação de populações parcialmente isoladasquanto ao seu status de espécie. Assim comoo Conceito Biológico, inclui o potencial repro-dutivo como critério principal, embora enfatizeseu isolamento. Este argumento é de difícil apli-cação, quer na natureza, devido a alopatria,quer em laboratório, pois a ausência ou pre-
sença de intercruzamento faz-se sob condiçõesartificiais. Temporalmente o isolamento repro-dutivo é impossível de ser testado. Meier eWillmann (2000b) observam que o isolamentoreprodutivo não é aplicável na seqüência tem-poral de gerações, mas experimentos de labo-ratório, entre organismos de duas populaçõesdistintas, podem provar a incompatibilidade, oque seria uma boa evidência da existência deduas espécies, assim como qualquer parapa-tria ou simpatria.
(2) A simples separação geográfica en-tre organismos implica em considerá-los espé-cies distintas. Em termos práticos, utilizam-se,sim, os atributos (caracteres) das populaçõespara o reconhecimento de espécies. Meier eWillmann (2000b) observam que se deve re-conhecer a diferença entre isolamento repro-dutivo e separação geográfica. Mecanismos deisolamento são intrínsecos aos organismos en-volvidos, enquanto que obstáculos geofísicosnão representam estes mecanismos: são ape-nas causadores de separação geográfica. Se-paração não necessariamente resulta em iso-lamento11 e qualquer fluxo gênico entre duaspopulações indica uma co-especificidade.
(3) A incompatibilidade perante a repro-dução assexuada, defendida no conceito, nãoseria real. A clonagem não é o único meio dereprodução de microorganismos. Existem pro-cessos tanto sexuais como assexuais que pro-movem a tocogenia, e a reprodução sexuadaseria apenas um destes processos tocogenéti-cos. Mesmo assim, a reprodução sexuada tro-ca apenas parte do genoma. Por outro lado, aseleção natural não trabalha apenas com a va-riabilidade oriunda da tocogenia, pois esta podeser mantida sem recombinação gênica. Damesma forma, semelhanças genéticas, morfo-lógicas e/ou ecológicas também podem ser
10 O termo “holofiletismo” foi proposto em substituição ao termo “monofiletismo” (Ashlock, 1971; Dubois, 1986). Oargumento utilizado é o de que grupos parafiléticos são também monofiléticos. Por outro lado, Mayr (2000b:96) criticaW. Hennig em ter proposto um novo conceito para o termo “monofiletismo”, proposto por Simpson (1945:17) sob basesgradistas, defendendo que o nome correto seria “holofiletismo”. Em outra publicação (Kischlat e Schultz, 2002) real-mente utilizei o termo “holofiletismo” mas, já que o presente artigo transcreve e traduz passagens de outros autores, otermo “monofiletismo” será aqui utilizado indiscriminadamente. Por sua vez, Amorim (1997:268) observa que o termo“monofilia” (assim como os demais relacionados) está semanticamente incorreto.11 Embora Willmann e Meier (2000:173) enfatizem esta diferença, um pouco antes, estes autores (Meier e Willmann,2000a:39) afirmaram que a separação geográfica possui um significado comparável ao do isolamento reprodutivo (“Inparticular, geographic separation then gains the same significance as reproductive isolation.”).
Métodos de Estudo em Biologia18
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 19
mantidas sem reticulação. Meier e Willmann(2000b) observam que não existiria qualquerconceito de espécie satisfatório para organis-mos que, primitivamente, não possuem meio-se (agamotáxons primários), pois tal conceitoesconderia as diferenças e criaria a ilusão deque existem unidades comparáveis entre as-sexuados (e.g., bactérias) e sexuados (e.g., in-setos). Por outro lado, o conceito pode ser apli-cável em agamotáxons secundários (e.g., dia-tomáceas) com reprodução sexual ocasional.
(4) O processo de especiação peripátri-ca12 representa o isolamento de uma peque-na população periférica. A utilização do Con-ceito Isolacionista implica em reconhecer tan-to a população periférica quanto a populaçãoprincipal como populações-irmãs, e mesmotratá-las como novas espécies após a separa-ção geográfica. Como qualquer novidade evo-lutiva levará menos tempo para se fixar na po-pulação periférica, a população principal con-tinuaria a ser praticamente a mesma quandocomparada à população ancestral, e seria umcontra-senso chamá-la de população-irmã (oumesmo denominá-la como uma espécie nova).Ela representaria, sim, uma população-materna.Na aplicação do conceito, não existiria uma de-limitação clara do término da espécie-ancestral.Meier e Willmann (2000b) argumentam que oConceito Isolacionista não é baseado em fê-nons; o uso destes serve apenas como reco-nhecimento e delimitação de espécies. Peripa-tria e dicopatria13 seriam casos extremos e arealidade encontra-se nos intermediários.
(5) Rejeição de qualquer reconhecimen-to de espécies baseado em anagênese (espe-ciação filética), tanto viventes quanto fósseis.Meier e Willmann (2000b) reconhecem a ana-gênese como mudanças consideráveis quepodem ocorrer dentro da espécie, e não como
argumento para o reconhecimento de distin-tas espécies em seqüência temporal (especia-ção filética).
(6) Impossibilidade de reconhecimen-to das espécies envolvidas no momento daseparação geográfica, quando não existemmudanças aparentes na extinção da espé-cie-ancestral e na origem das espécies-filhas.Meier e Willmann (2000b) observam que, sea delimitação de espécies for baseada em si-milaridade genética, qual seria a quantidadenecessária de mudança para se decidir sedeterminada linhagem constitui, ou não, amesma espécie, antes e depois do evento es-peciativo? Duas populações ou duas geraçõesde uma mesma população nunca são geneti-camente iguais e as freqüências gênicas vari-am continuamente, sendo impossível desig-nar limites não-arbitrários. Por outro lado, li-nhagens podem ser objetivamente delimita-das por eventos isolacionistas.
(7) A assunção de que a espé-cie-ancestral não sobrevive a eventos especia-tivos seria, em realidade, uma convenção me-todológica, pois o reconhecimento da transi-ção ancestral-descendente é problemático. De-vido a esta convenção, ao se analisar um uni-verso de espécies, estas devem ser tratadasapenas como termos14 , independentementedo fato de representarem, ou não, espé-cies-ancestrais. O padrão filogenético resultantecomporta o parentesco relativo entre os ter-mos e qualquer idéia de real ancestralidade éespeculativa, já que espera-se que o ancestralnão exiba qualquer apomorfia não comparti-lhada pelos descendentes.
(8) O reconhecimento relacional da es-pécie é problemático. Dentro de um contextopaleontológico, com um grande número de es-pécies extintas e em grande parte desconhe-
12 Denominação de Mayr (2000b:94), correspondente ao isolamento periférico, e ao segundo modo de especiaçãoalopátrica de Wiley (1981:48; “periferal isolates”). Compreende a divisão da população ancestral em duaspopulacões-filhas com grande desproporção no número de indivíduos. Assim, existe uma população-filha com grandeamplitude de distribuição geográfica e grande número de indivíduos, e outra, restrita geograficamente e com pequenonúmero de indivíduos.13 Denominação de Mayr (2000b:94), correspondente a vicariância ou especiação geográfica, e ao primeiro modo deespeciação alopátrica de Wiley (1981:43). Compreende a divisão da população ancestral em duas populacões-filhas,subiguais na amplitude geográfica e no número de indivíduos.14 A literatura anglófona utiliza “terminal” para os táxons terminais de um cladograma. Amorim (1997) utiliza a palavra“termo” em português.
Métodos de Estudo em Biologia 19
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia20
cidas. O reconhecimento de espécies por com-paração com suas espécies-irmãs seria ou ina-plicável ou apenas aplicável ao expandir, e des-figurar, o conceito de espécie-irmã. Meier eWillmann (2000b) observam que, ao se utilizargrupos mais distantemente relacionados como�espécies-irmãs�, o isolamento reprodutivo se-ria, até mesmo, mais suportado.
O Conceito Evolutivo WileyanoWiley e Mayden (2000a) reconhecem
que espécies evolutivas são indivíduos lógicos,com origens, existência e término. Represen-tam as maiores entidades tocogenéticas quefuncionam no sistema filogenético em analo-gia às entidades filogenéticas - os clados. Or-ganismos multicelulares são compostos porpartes (células), ligados pela mitose e manifes-tados através da ontogenia; espécies são com-postas de partes (organismos individuais), liga-das pela reprodução e manifestadas através datocogenia; clados são compostos de partes(espécies individuais), ligados pela especiaçãoe manifestados através da filogenia. Assim,ontogenia, tocogenia e filogenia não represen-tam apenas processos, mas resultados de pro-cessos. Espécies são o resultado de proces-sos históricos que são descobertos durante ocurso de uma investigação. Espécies assexua-das (agamoespécies) seriam semelhantes a in-divíduos multicelulares, sendo compostas porvetores clonais descendentes de um únicoancestral. Assim, como as células de organis-mos multicelulares, elas possuem relações mi-tóticas (e por vezes meióticas) que são idênti-cas nas suas relações de descendência.
Estes autores definem15 espécie evoluti-va como �uma entidade composta de orga-nismos que mantêm sua identidade peranteoutras iguais entidades pelo tempo e espaçoe possui seus próprios destino evolutivo e ten-dências históricas independentes�. Eles obser-
vam que muitas espécies são compostas porpopulações (partes) que trocam gens apenasocasionalmente, podendo ser temporariamen-te alopátricas o suficiente para incluir dezenasde gerações. Populações geográficas poderiamaté mesmo ser avaliadas cladisticamente,dividindo-as em diferentes espécies, já que re-presentam diferentes linhagens. No reconheci-mento de populações (linhagens) independen-tes, estas terão de possuir tendências indepen-dentes (i.e., variação e evolução distinta da en-contrada na população-irmã). Logo, é esperadaa descoberta de diferenças entre estas linha-gens, representativas de marcos de independên-cia, que podem ser trabalhados numa análisecladística de caracteres, servindo como verifi-cação de que este processo ocorreu na nature-za. Porém, se após algum tempo, os processostocogenéticos voltarem a atuar (simpatria/para-patria), as partes tendem a deixar de possuir umaidentidade, e uma análise cladística produziriapadrões sinapomórficos confusos e politomias.Contudo, mesmo na ausência de tocogenia (reti-culação) entre linhagens, a elucidação da histó-ria filogenética pode ser difícil, pois nem todas asevidências residem sob a forma de apomorfias:existem também sob a forma de reprodução, flu-xo gênico e posicionamento geográfico.
Os autores (cf. Wiley, 1981) também re-conhecem a sobrevivência da espécie-ancestralem alguns casos de especiação alopátrica comdesproporção entre o número de indivíduosdas subpopulações envolvidas (e.g., peripatriae microalopatria16 ). Nestes casos, as cadênci-as anagenéticas (de diferenciação) de cada umadas duas linhagens envolvidas serão distintas.A população maior possuirá uma anagênesemais lenta e qualquer novidade evolutiva de-morará mais tempo para se distribuir homoge-neamente. Nestes casos, de um modo geral,as espécies ancestrais são difíceis de seremencontradas, embora não necessariamente im-possíveis. É fato que a espécie-ancestral não
15 “An evolutionary species is an entity composed of organisms that maintains its identity from other such entitiesthrough time and over space and that has its own independent evolutionary fate and historical tendencies” (Wiley eMayden, 2000a:73).16 Correspondente ao terceiro modo de especiação alopátrica de Wiley (1981:51). Compreende o isolamento de umapequena parte da população ancestral no interior da área de distribuição original.
Métodos de Estudo em Biologia20
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 21
pode possuir autapomorfias, pois qualquer apo-morfia é compartilhada com as espécies-filhas(logo, representam sinapomorfias entre a espé-cie-ancestral e as espécies-filhas). Além da es-peciação alopátrica, este conceito tambémaceita a especiação parapátrica17 (cf. Wiley,1981), onde duas linhagens dão origem a umaterceira através da hibridização.
Wiley e Mayden (2000c) reconhecemainda que a anagênese, que é o mecanismoprimário da fixação do caracter, não está dire-tamente ligada à cladogênese; e a cladogê-nese pode não estar convenientemente acom-panhada pela anagênese (Figuras 4 e 5). As-sim, o evento especiativo isolacionista (clado-gênese) e a evidência fornecida pela transfor-mação de caracteres (anagênese) são proces-sos distintos.
Contrariando todos os demais conceitos,estes autores (cf. Wiley, 1981:54-58) reconhe-cem a hibridização18 e a segregação dehabitat19 como eventos especiativos. A hibridiza-ção, como mecanismo de especiação, tem algu-mas implicações e precondições: (1) reconheci-mento polifilético da nova espécie; (2) necessá-ria simpatria, ou parapatria, entre as reconheci-das espécies ancestrais; (3) necessário isolamentoreprodutivo entre o híbrido e as espécies ances-trais (pois o fluxo gênico deve ser evitado); e (4)quantidade mínima de espécimens sincrônicospermitindo o encontro entre os sexos, reconhe-cimento mútuo, coesão reprodutiva e fluxo gêni-co somente entre os híbridos, fixando a novacombinação de caracteres. Por outro lado, a se-gregação de habitat (especiação ecológica) re-presenta numa tendência de exploração ambi-ental que apenas certos indivíduos possuiriam.A fixação desta tendência só seria possível com
um isolamento tocogenético (evitando fluxo gê-nico) em um modelo microalopátrico.
Concluindo, em relação aos ConceitosBiológico e Isolacionista, o Conceito Evolutivoé igualmente ontológico, mas incorpora os se-guintes fatos: (1) independe do critério repro-dutivo; (2) espécies-ancestrais podem ser re-conhecidas em casos especiais, o que implicana dissociação da cladogênese perante a ana-gênese; (3) reconhece o monofiletismo comouma situação ideal, mas não necessariamentereal, no que se refere ao reconhecimento detáxons-espécie; (4) aceita uma eventual situa-ção parafilética e polifilética para a espécie,sendo a hibridização entre duas linhagens umaforma de especiação; (5) reconhece a análisecladística como ferramenta para avaliação davariação fenética; (6) reconhece o sucesso daanálise cladística de caracteres como verifica-ção de independência de linhagens (especia-ção) e seu fracasso representa tanto falseamen-to no reconhecimento da independência de li-nhagens, como também dificuldade no reco-nhecimento da realidade destas; (7) reconhe-ce a biodiversidade de um modo muito maisamplo, independente do critério reprodutivo eda distribuição geográfica. As críticas feitas aeste conceito compreendem (Mayr, 2000b;Willmann e Meier, 2000; Mishler e Theriot,2000b; Wheeler e Platnick, 2000b):
(1) A definição é vaga para se determi-nar o status de espécie de uma população iso-lada já que todas �mantém sua identidade pe-rante outras iguais entidades pelo tempo e es-paço�. Por outro lado, é impossível determinarse uma população �possui seus próprios desti-no evolutivo e tendências históricas indepen-dentes�, mesmo em formas fósseis.
17 “Allo-parapatric speciation” e “parapatric speciation” (cf. Wiley, 1981:54). Tais modos de especiação também podemser interpretados como especiação alopátrica onde a barreira geográfica foi retirada posteriormente, e aspopulações-irmãs voltaram a possuir tocogenia e hibridização. Wiley (1981) reconhece ainda especiações estasipátri-cas (“stasipatric speciation”; p. 55) e simpátricas (“sympatric speciation”; p. 56). A especiação estasipátrica é conside-rada um caso especial de microalopatria (Wiley, 1981:55). Por outro lado, a especiação simpátrica, como exposto porWiley (1981:56-57), comporta quatro casos: especiação por hibridização, especiação por apomixia (relacionada aoaparecimento de agamotáxons secundários), microalopatria e especiação ecológica.18 “Speciation by hybridization” sendo o primeiro modo de especiação simpátrica de Wiley (1981:56), Por outro lado,Wiley e Maiden (2000c:207) comentam apenas a poliploidia como mecanismo de isolamento reprodutivo essencialpara a especiação simpátrica.19 “Ecological sympatric speciation” sendo o quarto modo de especiação simpátrica de Wiley (1981:57).
Métodos de Estudo em Biologia 21
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia22
(2) Seria apenas uma expansão do Con-ceito Isolacionista, com aplicação independentedo processo reprodutivo e reconhecimento dasobrevivência da espécie-ancestral em umevento especiativo.
(3) Defende a análise cladística apenascomo um método de descoberta da espécie,mas não de suporte da sua existência, o queimplica em assumir que a realidade está alémdo método. Wiley e Mayden (2000c) obser-vam que grupos monofiléticos e espécies nãoexistem porque possuem apomorfias: apenassão descobertos pela existência destas apomor-fias. Assim, espera-se que alguns grupos mo-nofiléticos e algumas espécies não possam serdescobertos através da utilização de ferramen-tas analíticas hoje disponíveis, pois transcen-dem seus limites.
(4) Embora este conceito tenha sido pro-posto para trabalhar com a dimensão tempo-ral, a definição da espécie fica prejudicada pelaausência de parâmetros, pois não reconheceo evento especiativo Isolacionista e a evidên-cia fornecida pela transformação de caracte-res. Desta forma, a aplicação do conceito fun-ciona para tudo e não pode ser falseada. Wileye Mayden (2000c) reconhecem que não exis-te uma abordagem epistemológica simples paradescobrir uma espécie evolutiva, mas sim vári-as, utilizando-se a biogeografia, morfologia, eto-logia reprodutiva e coesão reprodutiva (ou au-sência desta coesão).
O Conceito Autapomórfico Mishler-Theriotiano
O Conceito Autapomórfico reconheceque as relações tocogenéticas entre os indiví-duos de duas linhagens, conseqüentes de umevento de especiação, se perduram de tal for-ma que, quando chegam a zero, este limiteencontra-se bem após o aparecimento de di-ferenciações entre estas linhagens (Mishler eTheriot, 2000a). Estes autores reconhecem
também que a habilidade de intercruzamento,potencial ou real, dos indivíduos não é um cri-tério filogenético válido, mas representa, sim,uma plesiomorfia, uma característica ancestralque se perdura. O argumento básico para oagrupamento de organismos em um sistemacladístico é a sinapomorfia, a característica com-partilhada pelos indivíduos de cada linhagem,que representa uma evidência empírica da es-pécie. Relações tocogenéticas podem ocorrerpor toda a hierarquia da vida e não seria umproblema específico da espécie, mas sim umadificuldade a mais no seu reconhecimento. Poroutro lado, não existe uma distinção clara en-tre populações com reprodução assexuada esexuada na maioria dos organismos. Bactériastrocam material genético por vários meios; di-atomáceas, cladóceros e rotíferos geralmenteproduzem clones, mas a reprodução sexuadapode ocorrer como resposta a mudanças am-bientais.
Mishler e Theriot (2000a) definem20 es-pécie como �o táxon menos inclusivo reco-nhecido em uma classificação filogenéticaformal; assim como todos os níveis hierár-quicos de táxons em tal classificação, orga-nismos são agrupados em espécies devidoa evidência de monofiletismo; táxons sãoconsiderados como espécies em vez de ou-tro nível mais alto porque são os menoresgrupos monofiléticos merecedores de reco-nhecimento formal, tanto por causa do mon-tante de suporte para seu monofiletismoquanto de sua importância em processos bi-ológicos que operam na linhagem em ques-tão�. O agrupamento de organismos, seguin-do o critério de monofiletismo, é o menos arbi-trário, embora ainda envolva decisões sobrehomologia e qualidade de suporte. A razão prin-cipal para fornecer uma classificação é a cons-trução de um guia de nomes para os gruposmonofiléticos que se deseja discutir ou regis-trar dados. É necessário denominar-se linha-gens distintivas como parte de um processo
20 “A species is the least inclusive taxon recognized in a formal phylogenetic classification. As with all hierarchical levelsof taxa in such a classification, organisms are grouped into species because of evidence of monophyly. Taxa are rankedas species rather than at some higher level because they are the smallest monophyletic groups deemed worthy offormal recognition, because of the amount of support for their monophyly and/or because of their importance in biologi-cal processes operating on the lineage in question” (Mishler e Theriot, 2000a:44-5).
Métodos de Estudo em Biologia22
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 23
de inventário, conservação e uso da biodiver-sidade; mas nem sempre existe a necessidadede denominação de todos os grupos monofilé-ticos descobertos, pois alguns podem ser(1) triviais em termos evolutivos (i.e., de curtaduração temporal ou caracterizados por apo-morfias pouco relevantes e neutras), (2) ser críp-ticos (i.e., marcados apenas por apomorfias mo-leculares ou químicas, de uso prático questio-nável), ou, mesmo, (3) pouco suportados (su-jeitos a revisão conforme novos táxons e ca-racteres sejam descobertos). Assim, reconhe-cimento como táxon-espécie para uma deter-minada linhagem monofilética nunca é auto-mática, mas sempre subjetiva.
O procedimento de análise, segundoMishler e Theriot (2000a), reconhece três fa-ses: (1) inicial, onde �Unidades TaxonômicasOperacionais� (UTO�s)21 são reconhecidas,tabulando-se caracteres (e seus estados) de-tectados em uma matriz de dados; (2) tradu-ção destes dados em um cladograma segun-do o método cladístico (i.e., as UTO�s devemser homogêneas em relação aos caracteres,ser conseqüência de processos de divergên-cia filogenética - e não de tocogenia - e os ca-racteres devem ser conspícuos, hereditários eindependentes); (3) baseando-se nas avaliaçõesdo suporte relativo de cada clado, propõe-seuma classificação, que por sua vez fornece abase para estudos evolutivos. Os táxons for-mais (incluindo táxons-espécie) são denomina-dos com base no suporte monofilético de cadalinhagem assim como pela sua utilidade no de-senvolvimento e discussão de processos.A presença de eventuais politomias em um cla-dograma seria evidência de intensa tocogeniaentre as UTO�s, sendo, então, falseada a hipó-tese inicial de que estas representam distintaslinhagens filogenéticas.
Após a análise de um complexo deUTO�s, a presença de autapomorfias é discri-minatória para o reconhecimento de espécies.
Contudo, pode-se reconhecer um grupo basalde UTO�s que apenas mostram plesiomorfias.Este agrupamento representa meramente umagregado de linhagens de natureza parafiléti-ca22 (Figura 3). Como nem sempre todas aslinhagens expressam características apomórfi-cas, não se pode identificá-las pela análise decaracteres. Na aplicação deste conceito, nãose pode, e nem se deve, reconhecer agrupa-mentos suportados apenas por plesiomorfias(i.e., metaespécies).
Figura 3. Na aplicação do Conceito Autapomórfico,após a análise filogenética, pode-se reconhecer umgrupo basal de UTO’s em politomia, que apenas mos-tram plesiomorfias. Este agrupamento representa umagregado de linhagens de natureza parafilética ecompõe uma metaespécie. Modificado de Willmanne Meier (2000).
Pode-se, assim, concluir que o Concei-to Autapomórfico de espécie está intimamen-te ligado ao método cladístico de análise (epis-temologia), sendo a espécie um subproduto(uma conclusão) da análise de uma matriz dedados através de um algoritmo. Caso exista umanova hipótese quanto às UTO�s, assim comonovas UTO�s e caracteres detectados e inseri-dos na análise, um novo resultado pode apare-cer. Em relação aos Conceitos Biológico, Isola-cionista e Evolutivo, o Conceito Autapomórfico
21 “Operational taxonomic units (OTUs)” (Mishler e Theriot, 2000a:48). Uma UTO pode compreender vários espéci-mens (ou, eventualmente, apenas um) representativos, por hipótese, da variabilidade morfológica de um termo. Nocaso destes espécimens representarem diferentes semaforontes e ambos os sexos, monta-se, também por hipótese,uma hipodigmia representativa da ontogenia e dimorfismo sexual do táxon-espécie a ser incluído na análise.22 Willman e Meier (2000b:113) utilizaram o nome “metaespécie” (metaspecies) para este agrupamento basal.
Métodos de Estudo em Biologia 23
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia24
é epistemológico, incorporando os seguintesfatos: (1) sendo baseado em caracteres fenéti-cos, pode ser aplicado a organismos indepen-dentemente do critério reprodutivo; (2) é práti-co, pois reconhece grupos monofiléticos se-gundo caracteres apomórficos discriminadosem uma análise objetiva; (3) reconhece a sub-jetividade de categorias Linneanas e a hierar-quização de grupos monofiléticos mais inclusi-vos; (4) assume que a tocogenia entre linha-gens é uma fonte conhecida de erros, que podeser desprezada se for mínima; (5) reconhece ahibridização de duas linhagens como uma ple-siomorfia, e não como uma forma de especia-ção, não sendo mesmo um critério filogenéti-co válido; (6) a falta de resolução em um cla-dograma (politomias) representa evidência detocogenia entre as UTO�s; (7) a espécie passaa ser uma conclusão sobre um universo anali-sado, e seu reconhecimento como tal (assimcomo sua denominação formal), segue argu-mentos práticos (e subjetivos). Críticas feitas aeste conceito compreendem (Mayr, 2000b;Willmann e Meier, 2000; Wheeler e Platnick,2000b; Wiley e Mayden, 2000b):
(1) A definição apresenta problemas:(a) �o táxon menos inclusivo reconhecido emuma classificação filogenética formal� depen-de da resolução do cladograma, conseqüen-cia da escolha dos caracteres, que, por sua vez,depende de critérios e interpretações pesso-ais; (b) as relações entre organismos são toco-genéticas, e não filogenéticas, sendo difícil agru-par organismos �em espécies devido a evidên-cia de monofiletismo�; (c) o reconhecimento decategorias Linneanas não é só subjetivo, massupérfluo e sem sentido dentro da sistemáticafilogenética (�táxons são considerados comoespécies em vez de outro nível mais alto�);(d) é totalmente subjetivo reconhecer quais são�os menores grupos monofiléticos merecedo-res de reconhecimento formal�. Mishler e The-riot (2000c) reconhecem que o ideal é se li-vrar definitivamente das categorias Linneanas,mas assumem que o sistema Linneano de clas-sificação ainda é previsto pelos diversos códi-gos de nomenclatura.
(2) Seria um conceito puramente ope-racional para se delimitar táxons-espécie, sen-do um retorno ao Conceito Tipológico. Um con-
ceito de espécie deve ser independente dométodo utilizado para reconstruir as relaçõesfilogenéticas já que o método cladístico de aná-lise necessita reconhecer as espécies antes daprópria análise. Assim, basicamente, dois pro-blemas existem: (a) certos termos podem nãoestar discriminados por apomorfias, ou porqueelas não existam, ou porque não tenham sidodetectadas pelo autor; (b) o reconhecimentode autapomorfias é conseqüência da polariza-ção dos estados dos caracteres segundo umgrupo-externo que é escolhido por hipótese,seja ele ad hoc ou conseqüência de uma aná-lise anterior. Se a espécie é uma conclusão daanálise em um universo finito (de UTO�s e decaracteres), uma nova análise sob um universomaior pode mostrar diferentes resultados. Sen-do o reconhecimento da espécie dependenteda análise filogenética, sua existência pode serquestionada, pois seria apenas uma constru-ção artificial na mente das pessoas. Mishler eTheriot (2000c) explicam que classificação éuma conveniência humana, superposta na hie-rarquia de grupos monofiléticos reais. Por essarazão, a classificação é conseqüência da re-construção filogenética. Por outro lado, o ele-mento básico da sistemática filogenética nãoé a espécie, mas o espécimen semaforonte.Por sua vez, o espécimen é portador não docaractere, mas de um estado que, por com-paração com outros espécimens semaforon-tes, é hipotetizado como formando uma sé-rie transformacional evolutiva de um carac-tere. Uma análise é imprescindível para se li-gar o espécimen semaforonte a grupos onto-genéticos, tocogenéticos, e, finalmente, filoge-néticos monofiléticos. Táxons, incluindo espé-cies, nunca são reconhecíveis antes de qual-quer análise.
(3) A espécie ancestral de outras duasnão possui, necessariamente, autapomorfias.Mesmo que o conceito negue a existência deuma espécie-ancestral, é fato que ela, um dia,existiu, e seus membros devem ser referidos aela. Por outro lado, o reconhecimento de umanova categoria alternativa de espécie (�meta-espécie�) não representa uma solução satisfa-tória para o reconhecimento de um agrupamen-to de linhagens basais sem autapomorfias, comrelações filogenéticas conflitantes ou ignoradas.
Métodos de Estudo em Biologia24
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 25
Mishler e Theriot (2000c) reconhecem quebinômios formais são construções humanasaplicadas como representativas de unidadesmonofiléticas, que são entidades reais. Enquan-to que espécies monofiléticas são diagnosticá-veis por apomorfias, alguns binômios formaisespecíficos apenas refletem unidadesnão-monofiléticas (i.e., metafiléticas23 ). Aespécie-ancestral seria um caso especial e de-veria apenas ser reconhecida numa classifica-ção utilizando uma convenção especial (i.e., me-taespécie), já que ela equivale à totalidade desuas espécies-filhas (cf. Hennig, 1966:72).
O Conceito Filogenético Wheeler-
Platnickiano
Wheeler e Platnick (2000a) reconhecemque os Conceitos Biológico, Isolacionista e Evo-lutivo possuem uma grande dependência so-
bre assunções originadas de processos evolu-tivos (ontologia). Por outro lado, existe a ne-cessidade de que um conceito se aplique igual-mente a todos os processos de especiaçãoconcebíveis, a fim de que se possa medir tantoa biodiversidade quanto os próprios mecanis-mos causadores de especiação. É ainda impor-tante que o conceito de espécie funcione comouma hipótese aberta ao teste crítico. O Con-ceito Filogenético se baseia na distribuição decaracteres observáveis e a diferença fundamen-tal em relação ao Conceito Autapomórfico serefere ao reconhecimento, tanto de caracteresautapomórficos, quanto plesiomórficos, na iden-tificação da espécie.
Estes autores definem24 espécie como�o menor agregado de populações ou li-nhagens, independente do tipo de repro-dução, diagnosticáveis por uma combina-ção única de estados de caracteres�. A es-
Figura 4. Aplicando o Conceito Filogenético, a origem de uma espécie está correlacionadacom a transformação de caracteres. Durante certo tempo, dois ou mais estados podem coe-xistir, marcando uma fase de variabilidade (polimorfia), onde se reconhecem apenas traços.A extinção desta polimorfia resulta numa distribuição constante de estados e a formação deuma nova espécie. O conceito é aplicável tanto à anagênese (A), quanto à cladogênese comsobrevivência da espécie ancestral (B) ou com formação de duas espécies-filhas e extinçãoda espécie ancestral (C). Modificado de Wheeler e Platnick (2000a).
23 Metafiletismo é o estado de um grupo que se apresenta sob dúvidas, não se sabendo se comporta um grupomonofilético ou parafilético (Mishler e Theriot, 2000c:183). Não confundir com Merofiletismo que é a denominação geralde um grupo que não compreende todas as espécies de uma mesma linhagem, reunindo os estados parafilético epolifilético (Kischlat e Schultz, 2002).24 “We define species as the smallest aggregation of (sexual) populations or (asexual) lineages diagnosable by aunique combination of character states” (Wheeler e Platnick, 2000a:58).
Métodos de Estudo em Biologia 25
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005
Métodos de Estudo em Biologia26
pécie filogenética, assim, é a unidade básicada nomenclatura científica formal, da classi-ficação Linneana e da evolução orgânica(=táxon-espécie). Espécies são simples hipó-teses. Sua aceitação ou rejeição está relacio-nada à evidência disponível e aos padrõesrevelados pelo seu estudo. Uma hipótesebem estabelecida pode ser rejeitada no futu-ro na presença de novas evidências.
Assim como o Conceito Autapomórficode Mishler e Theriot (2000a), a distinção inicialentre espécies ocorre anteriormente a, e obje-tivando, uma análise filogenética. Não é neces-sária a certeza da polaridade dos caracteresdetectados pois qualquer polaridade inferidarepresenta apenas uma hipótese. A distinçãode espécies é feita pela distribuição de esta-dos, e não apenas pelos estados apomórficos,já que são dependentes do grupo-externo hi-potetizado como tal.
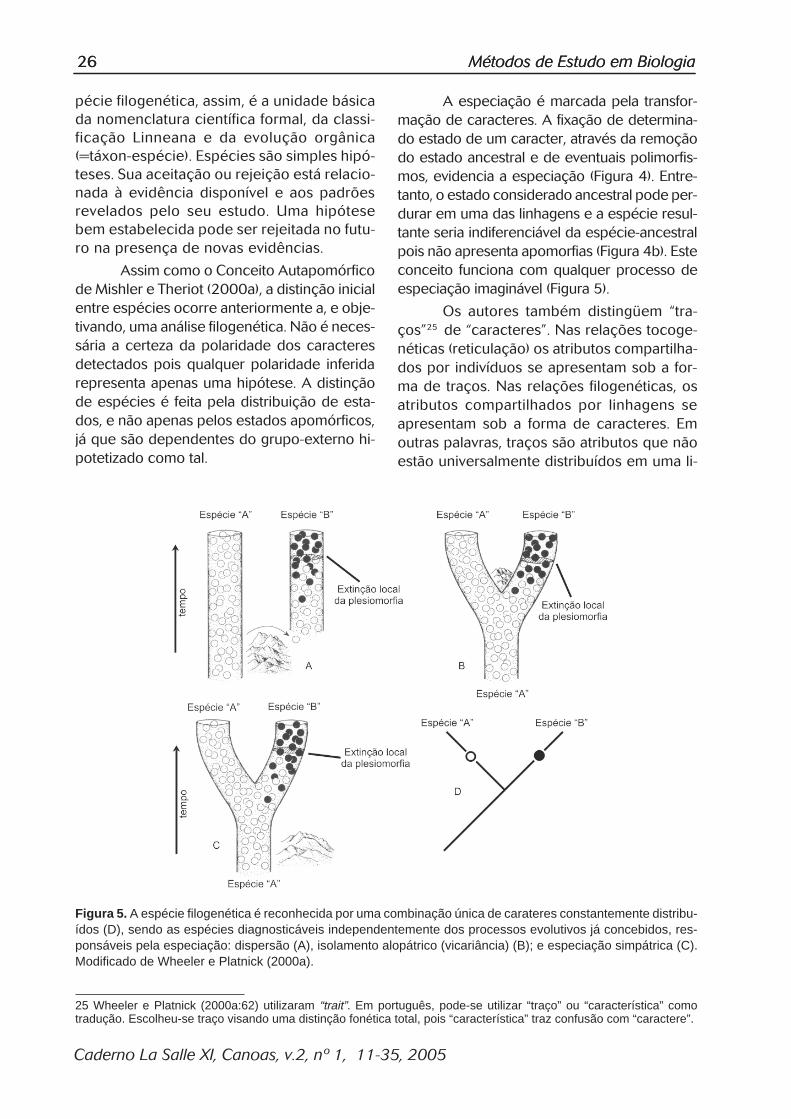
A especiação é marcada pela transfor-mação de caracteres. A fixação de determina-do estado de um caracter, através da remoçãodo estado ancestral e de eventuais polimorfis-mos, evidencia a especiação (Figura 4). Entre-tanto, o estado considerado ancestral pode per-durar em uma das linhagens e a espécie resul-tante seria indiferenciável da espécie-ancestralpois não apresenta apomorfias (Figura 4b). Esteconceito funciona com qualquer processo deespeciação imaginável (Figura 5).
Os autores também distingüem �tra-ços�25 de �caracteres�. Nas relações tocoge-néticas (reticulação) os atributos compartilha-dos por indivíduos se apresentam sob a for-ma de traços. Nas relações filogenéticas, osatributos compartilhados por linhagens seapresentam sob a forma de caracteres. Emoutras palavras, traços são atributos que nãoestão universalmente distribuídos em uma li-
Figura 5. A espécie filogenética é reconhecida por uma combinação única de carateres constantemente distribu-ídos (D), sendo as espécies diagnosticáveis independentemente dos processos evolutivos já concebidos, res-ponsáveis pela especiação: dispersão (A), isolamento alopátrico (vicariância) (B); e especiação simpátrica (C).Modificado de Wheeler e Platnick (2000a).
25 Wheeler e Platnick (2000a:62) utilizaram “trait”. Em português, pode-se utilizar “traço” ou “característica” comotradução. Escolheu-se traço visando uma distinção fonética total, pois “característica” traz confusão com “caractere”.
Métodos de Estudo em Biologia26
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 27
nhagem terminal (espécie), e caracteres sãoos atributos distribuídos por todos os mem-bros desta linhagem.
Pode-se concluir que, assim como oConceito Autapomórfico, o Conceito Filogené-tico é igualmente epistemológico. Em relaçãoaos Conceitos Biológico, Isolacionista, Evoluti-vo e Autapomórfico, incorpora os seguintes fa-tos: (1) procura se desprender de qualquer pro-cesso evolutivo, visando uma avaliação impar-cial do resultado destes processos; (2) reco-nhece a especiação como marcada pela trans-formação do caracter, e seu momento corres-ponde à morte do último indivíduo da popula-ção que seja portador de polimorfismo ou ple-siomorfia; (3) na medida em que reconhece aeventual sobrevivência da espécie-ancestral,aceita também uma natureza parafilética paraa espécie. Críticas feitas a este conceito com-preendem (Mayr, 2000b; Willmann e Meier,2000; Mishler e Theriot, 2000b; Wiley eMayden, 2000b):
(1) Seria outro conceito que retorna aoConceito Tipológico, segundo o qual a espé-cie é alguma coisa diferente de outra coisa,não havendo qualquer referência a um signi-ficado biológico. Representa apenas umaconstrução arbitrária da mente humana. Ado-tar a menor unidade adequada a uma análisecladística é por demais reducionista, pois le-varia a um aumento excessivo no reconheci-mento do número de espécies em todos osgrupos com isolamento e variação geográfi-ca. Platnick e Wheeler (2000) observam queo Conceito Filogenético proporciona base parase investigar o parentesco filogenético entreas unidades que ele discrimina, sem impor li-mites arbitrários a processos evolutivos que,conceitualmente, poderiam dar origem a es-tas unidades. Espécies filogenéticas, assimcomo espécies biológicas, são construções damente humana que representam apenas hi-póteses. Os táxons reconhecidos são, emmuitos casos, os mesmos reconhecidos peloConceito Biológico, mas, em outros, o Con-ceito Filogenético é mais amplo e preciso, poisnão esconde biodiversidade. É um conceitoque pode ser aplicado e testado nomundo-real. Qualquer conceito que esteja dis-
sociado de evidências observáveis, deve, sim,ser rejeitado em favor daquele que está aber-to a testes através de observação empírica.
(2) O Conceito Filogenético defendeuma independência perante processos evolu-tivos. Na construção do caracter é necessáriaa avaliação da similaridade (e as conseqüenteshipóteses homológicas). Esta avaliação é umprocesso sujeito a interpretações pessoais, poisapenas os estados de um caractere podem serobjetivamente observáveis. A relação transfor-macional evolutiva entre estes estados, visan-do à construção do caracter, comporta, sim,assunções de parentesco para a construção deuma hipótese, que, por sua vez, é difícil de sertestada. Platnick e Wheeler (2000) observamque minimizar assunções evolutivas não signi-fica ser anti-evolutivo. Este conceito reconhe-ce a existência da evolução como fundamen-tal, havendo uma história de descendência commodificações, refletida pelo significado dos pa-drões hierárquicos nos cladogramas. Existemainda assunções teóricas implícitas relaciona-das aos conceitos de caracteres e traços, po-pulações e polimorfismo ancestral. Caracteressão hipóteses e não apenas observações fe-néticas. Sua utilização e distribuição determi-nará se são úteis ou não como delimitadoresde espécies em análises cladísticas.
(3) Ao se considerar que o isolamentoreprodutivo é irrelevante para o reconhecimen-to de espécies, as espécies reconhecidas den-tro deste conceito seriam unidades potencial-mente tocogenéticas, inclusive no que se refe-re ao reconhecimento de diferentes espécieshumanas (e.g., pigmeus, aborígenes, etc.).Platnick e Wheeler (2000) observam que duaspopulações, temporariamente alopátricas, po-dem ter se transformado o suficiente para se-rem reconhecidas como espécies através datransformação de seus caracteres. Contudo, aausência futura da barreira geográfica pode per-mitir a introgressão (tocogenia) destas popula-ções, estabelecendo-se um polimorfismo. As-sim, o Conceito Filogenético pode reconhecerestas duas populações como espécies distin-tas em um primeiro momento, mas, em estági-os posteriores, na presença de simpatria e po-limorfismo, este Conceito hipotetizaria apenasa presença de uma mesma espécie. No caso
Métodos de Estudo em Biologia 27
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia28
do homem, anteriormente à miscigenação in-tercontinental dos últimos séculos, a distribui-ção de caracteres poderia sugerir a presençade mais de uma espécie humana. Mas a intro-gressão das várias populações humanas alo-pátricas sugere que já somos, ou logo sere-mos, uma espécie cosmopolita polimórfica.
(4) Se binômios supostamente represen-tam espécies, e espécies supostamente são oresultado de vários processos denominadoscomo especiativos, existiria alguma relaçãoentre a aplicação de binômios e o reconheci-mento de processos naturais, tais como a cla-dogênese. A adoção do Conceito Filogenéticoacaba por dissociar binômios de cladogênese,já que o evento especiativo está ligado à fixa-ção do caractere, e não à ramificação de umalinhagem. Esta dissociação implica: (a) no re-conhecimento da anagênese (Figura 4a) comoprocesso especiativo sempre que ocorrer a fi-xação de um estado de caractere, o que des-trói a distinção entre tocogenia e filogenia;(b) na destruição da base para o reconhecimen-to de que táxons-irmãos possuem o mesmotempo de origem (Figura 4c), já que a extinçãoda plesiomorfia em ambas linhagens e a fixa-ção dos estados apomórficos, podem ocorrerem tempos distintos; (c) na destruição da rela-ção entre a biogeografia de vicariância e a eco-logia histórica, já que não existe a necessáriacorrelação entre a nomenclatura e a história cla-dogenética (Figura 5b,d). Platnick e Wheeler(2000) afirmam: (a) Não existe certeza na de-terminação do mecanismo específico de espe-ciação de cada caso e um mesmo conceitodeve ser compatível a todos os métodos con-cebíveis e suas combinações. Na prática, a ana-gênese pode ser abordada da mesma formaque a cladogênese, onde a cada modificaçãodo caracter corresponderia a um evento cla-dogenético desconhecido. (b) Não existe for-ma de se saber que, quando a alopatria acon-tece, será imediatamente acompanhada datransformação do caracter. Embora possa serhipoteticamente possível chegar a estas con-clusões numa abordagem retroativa, não exis-tem evidências inequívocas de suporte. Damesma forma, não existem formas de se pre-ver em que se transformarão, no futuro, popu-lações hoje isoladas, a não ser que caracteres
já tenham sido transformados. Táxons-irmãosdevem ser assumidos como possuindo a mes-ma idade, que é o momento em que a trans-formação do caractere se acha completa, e nãoo tempo da cladogênese inicial (o isolamentoalopátrico geográfico). Por outro lado, a veloci-dade de transformação do caractere pode serdistinta (e.g., taquitelia, braditelia), o que repre-senta um problema anagenético.
(5) O reconhecimento tanto de espéci-es monofiléticas (Figuras 4b, 5b: �espécie B�)quanto de espécies parafiléticas (Figuras 4b,5b: �espécie A�) implica na dissociação do sta-tus ontológico de grupos monofiléticos (que sãoentidades reais para as quais existem ou nãoevidências epistemológicas de sua existência,i.e., apomorfias) do status ontológico detáxons-espécie (que são construções humanas,mesmo que documentadas pelas mesmas evi-dências epistemológicas). Platnick e Wheeler(2000) comentam que espécies são simpleshipóteses, e sua aceitação ou rejeição estãorelacionadas às evidências disponíveis e aospadrões revelados pelo seu estudo. Neste con-texto, não seria surpresa que uma hipótese hojeaceita fosse, no futuro, rejeitada na presençade novas evidências.
A Espécie e a BiodiversidadeAlgumas áreas da Biologia (e.g., Neonto-
logia, Ecologia, Etologia, Genética) trabalhamapenas com as espécies encontradas no atualmomento geográfico da Terra. Outras áreas (e.g.,Paleontologia) se detém, principalmente, no re-conhecimento de espécies no tempo geológi-co. Logicamente, cada profissional aborda seusproblemas com um conceito de espécie queseja compatível com seu universo de estudo.O ideal é que o conceito seja único e forneçasubsídios para uma abordagem tanto geográfi-ca atual como geográfica temporal. Ao neontó-logo não cabem questionamentos de espéciesancestrais, mas cabe o reconhecimento de hie-rarquização monofilética de linhagens que for-neça subsídios para uma classificação. Ao ecó-logo é necessário um entendimento da espéciequanto à presença de tocogenia e hibridizaçãoentre linhagens distintas em alopatria ou para-patria, além de simpatria entre espécies crípti-
Métodos de Estudo em Biologia28
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 29
cas. Mesmo ao geneticista humano é importan-te o reconhecimento de apomorfias e plesio-morfias na comparação de diferentes linhagensde DNA mitocondrial (mtDNA) e do cromosso-mo sexual masculino (y-DNA) (vide Cann et al.,1987; Vigilant et al., 1991; Ayala, 1995;Hammer, 1995; Ovchinnikov et al., 2000;Templeton, 2002).
Por outro lado, ao paleontólogo, apenascaracteres morfológicos estão disponíveis e,assim mesmo, às vezes, de maneira bem res-trita. O reconhecimento, tanto de dimorfismosexual, quanto de semaforontes, representaapenas hipóteses baseadas em uma quantida-de relativamente grande de espécimens e nacomparação com padrão ontogenético detáxons atuais. O conceito de espécie temporal(paleoespécie, cronoespécie) possui um com-ponente morfológico muito grande, sendo mui-to importante saber como foi feita a análise.A amostragem morfológica presente pode nãoser representativa da variabilidade morfológi-ca passada, e as paleoespécies denominadasrepresentam apenas uma tentativa de entendi-mento de um táxon-espécie, uma hipótese su-jeita a revisões. Conforme novos espécimenssejam encontrados, esta variabilidade pode sermelhor conhecida. Assim, podem ser individu-alizados outros táxons-espécie, que, da mes-ma forma, representam apenas hipóteses.
A pergunta que deve ser feita quandose tem por objetivo o reconhecimento da pa-leobiodiversidade não é a de quantasespécies existiam aqui no passado, mas quan-tas espécies do passado podem ser discerni-das dentro do registro disponível no presente.Dois espécimens (eventualmente completos ebem preservados) indiferenciáveis morfologi-camente, poderiam representar diferentestáxons-espécie se uma viagem no tempo fos-se possível. Entretanto, esta resolução pode nãoestar acessível utilizando-se as evidências dis-poníveis. Pode-se ter que utilizar um único bi-nômio formal latino englobando ambos os es-pécimens, considerando-os como representan-do apenas um táxon-espécie até que evidênci-as mostrem o contrário. Por outro lado, a pre-sença de morfologias relativamente distintasresulta em um reconhecimento de váriostáxons-espécie, representados por uma profu-
são de nomes formais latinos. Mas o posteriorentendimento de uma variabilidade morfológi-ca conseqüente de distintos estágios ontoge-néticos (semaforontes) e dimorfismo sexual,pode levar a uma reavaliação destes putativostáxons-espécie para apenas um. O que se deveenfatizar é que binômios formais latinos de pa-leoespécies estão sujeitos a revisões confor-me o conhecimento avança. Qualquer listagemde paleoespécies de determinado local ou dedeterminada linhagem monofilética mais inclu-siva é apenas uma avaliação do realmente re-gistrado.
A utilidade em se reconhecer espéciescomo unidades evolutivas (táxons-espécie)merecedoras de um nome formal tem váriasimplicações. É fato que existe atualmente umevento antropogênico que culmina em extin-ções, geralmente relacionado com a modifica-ção de ambientes ou caça/captura/pesca pre-datórias. Existe a preocupação da sociedadeem se delimitar áreas de preservação ambien-tal e de proteção a espécies endêmicas. Restaao governo avaliar as propostas e determinar,no que se refere a legislação, se tal área ne-cessita ser, ou não, preservada, ou se tal espé-cie necessita ser, ou não, protegida, visando-sea preservação do patrimônio genético nacio-nal. Estas ações governamentais se baseiam,quase que exclusivamente, no reconhecimen-to quantitativo ou qualitativo de espécies. Emtermos práticos, mais precisamente, no reco-nhecimento, pela comunidade científica, dosbinômios latinos formais encontrados em de-terminada área.
Daugherty et al. (1990) observam queo reconhecimento formal de apenas uma es-pécie de tuatara (Sphenodon puncatus) na NovaZelândia quase levou à extinção uma outra es-pécie relacionada (S. guntheri) encontrada emapenas uma pequena ilha deste país. O não re-conhecimento desta última espécie pela litera-tura conservacionista e pela legislação neoze-landeza, levou a não consideração de medidaslegais para a sua preservação (o binômio latinoformal estava incluído dentro da espécie me-lhor conhecida e de distribuição relativamentemais ampla). Por outro lado, dois outros táxonsrelacionados (S.p. reischeki e S. diversum, nanomenclatura dos autores) estão atualmente
Métodos de Estudo em Biologia 29
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia30
extintos porque foram incluídos em S. puncta-tus, ficando sem uma legislação visando a pro-teção de suas áreas de ocorrência.
O padrão alopátrico de distribuição en-tre os táxons supracitados e o uso do ConceitoBiológico de espécie (não-dimensional por de-finição) acabaram por considerar a variaçãomorfológica ou como insuficiente para a distin-ção específica, ou como representativa apenasde uma �subespécie�, sendo ambos negligen-ciados. Este problema alcança magnitude bemmaior quando não se possui conhecimento for-mal sobre os táxons-espécie de determinadolocal. Mesmo que os binômios formais propos-tos para determinada área representem ape-nas espécies-nominativas, passíveis de seremsubordinadas a um outro nome formal com pre-cedência histórica, já servem de base para umainvestigação preliminar, já que alguém, em cer-to momento histórico, acreditou que represen-tassem algo distinto o suficiente para receberum nome à parte.
Um argumento utilizado para se propora preservação de determinada área como deimportância ambiental é a quantidade relativade espécies (biodiversidade). Pode-se, ainda, re-conhecer que a qualidade desta biodiversidadelocal pode ser mensurada em termos relativos:(1) com o reconhecimento de determinadotáxon endêmico como representando uma uni-dade evolutiva independente e indivisível (=es-pécie), e não como uma �variação de menor im-portância� de um táxon de distribuição maisampla (�subespécie�), cujo desaparecimentoimplica em perda irreversível do patrimônio ge-nético nacional; (2) com o reconhecimento dadiversidade filogenética da biodiversidade emquestão (Wheeler e Platnick, 2000a). Este últi-mo argumento pode ser melhor examinado aose comparar duas áreas com o mesmo númerode espécies listadas (biodiversidade). A primei-ra seria composta por espécies filogeneticamen-te próximas, a segunda apresentaria igual nú-mero de espécies, mas estas representariam li-nhagens filogenéticas mais distantes entre si. Aárea com maior biodiversidade filogenética (i.e.,maior número de linhagens monofiléticas inde-
pendentes) é a segunda, e a sua conservaçãopassa a ser prioritária em relação à primeira, poispreservaria um patrimônio genético maior.
O conflito entre ConceitosPode-se perceber que duas correntes
de pensamento se antagonizam no reconheci-mento da espécie dentro de um cenário evolu-tivo (Cracraft, 2000; Mishler e Theriot, 2000b):(1) assunção da realidade26 da espécie (ontolo-gia), procurando-se argumentos teóricos desuporte para a formação destas espécies (Con-ceitos Biológico, Isolacionista e Evolutivo);(2) uso de métodos pelo qual se conclui a es-pécie (epistemologia), avaliando-se atributosempíricos (caracteres) e dando pouca ênfase,a priori, aos processos especiativos (ConceitosAutapomórfico e Filogenético). Este antagonis-mo filosófico não é incomum na ciência.Pode-se tanto concluir que indivíduos são se-melhantes porque pertencem à mesma es-pécie (ontologia), quanto que indivíduos per-tencem a mesma espécie porque são seme-lhantes (epistemologia). Ou se acredita que algorealmente existe, e se procura um entendimen-to, ou apenas se hipotetiza a existência destealgo, utilizando métodos para a sua comprova-ção ou falseamento. É fato que uma ontologiaque não possui base epistemológica lógica noestudo da espécie leva a um sistema quenão difere muito do criacionismo (Wheeler ePlatinick, 2000c).
Todos os conceitos procuram a defini-ção de uma unidade básica para a avaliação dabiodiversidade, entretanto, a abrangência qua-litativa e quantitativa desta biodiversidade va-ria. Os Conceitos Biológico e Isolacionista nãoreconhecem espécies com reprodução asse-xuada; já os Conceitos Evolutivo, Autapomórfi-co e Filogenético defendem um reconhecimen-to muito mais amplo da biodiversidade, inde-pendente do critério reprodutivo e da distribui-ção geográfica. Isso multiplica o número deespécies, pois a especiação é feita por alopa-tria e a reprodução assexuada faz parte de umgrande segmento de organismos. O Conceito
26 “Realidade” é a existência de algo independente da vontade de seu descobridor (Mishler e Theriot, 2000b:121).
Métodos de Estudo em Biologia30
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 31
Biológico, em especial, subestima a verdadei-ra biodiversidade, negligenciando populaçõesgeograficamente isoladas, denominando-ascomo representativas apenas de �subespé-cies�, passíveis de serem substituídas por es-pécimens de outras áreas caso sua populaçãoseja crítica (Wiley e Mayden, 2000c).
A importância da Sistemática Filogené-tica é a relação entre os eventos cladogenéti-cos (a história seqüencial dos clados) e a bio-geografia (história da biosfera). Os ConceitosIsolacionista e Autapomórfico se aplicam bemnesta correlação, ao reconhecerem o monofi-letismo de táxons-espécie terminais. Contudo,o problema passa a ser o reconhecimento for-mal das espécies-ancestrais: a ontologia Isola-cionista o suporta, mas a epistemologia Auta-pomórfica o rejeita. O Conceito Filogenético,propondo poucas assunções sobre a evoluçãoe sendo mais operacional, procura suportarepistemologicamente a espécie-ancestral, masacaba por questionar a correlação entre a cla-dogênese e a biogeografia de vicariância.
Pode-se resumir que, quando se traba-lha com organismos sexuados e presentes (evivos) numa determinada área, o Conceito Bio-lógico funciona, já que está intimamente liga-do à percepção empírica (senso comum). Quan-do esta área passa a ser grande o suficientepara comportar populações alopátricas, o Con-ceito Isolacionista passa a ser o mais eficiente,pois ele pode correlacionar eventos geográfi-cos relativamente recentes com a alopatria.Todas as neoespécies reconhecidas pelo Con-ceito Biológico também são reconhecidas peloConceito Isolacionista, assim como outras tan-tas tratadas como �subespécies�. O ConceitoIsolacionista refuta o intercruzamento, base doConceito Biológico, e propõe, como idéia prin-cipal, o isolamento reprodutivo e a vicariância.A extensão temporal de determinada neoes-pécie também pode ser avaliada se, e somen-te se, assumir anagênese pós-cladogênese emambas espécies-filhas. Na medida em que umaespécie-filha possua uma cadência anagenéti-ca excessivamente lenta (braditelia), ela passaa ser indiferenciável da espécie-ancestral. Aespécie-ancestral (=espécie estemática) pos-sui um componente bastante teórico, e seu re-
conhecimento no registro fossilífero fica con-dicionado à simpatria desta em relação àsespécies-filhas, mesmo assim com todas as di-ficuldades supracitadas no reconhecimento defósseis. Espécies extintas historicamente, des-de que alopátricas, também podem ser reco-nhecidas.
Tanto o Conceito Biológico quanto o Iso-lacionista, com componentes ontológicos mui-to fortes não funcionam com agamotáxons, e aaplicação direta em fósseis ou é impossível, ouapenas é especulativa, pois um fóssil nuncapossui todas as características disponíveis, sóencontradas em um organismo vivo.
O Conceito Evolutivo passa a utilizar umaespistemologia investigativa, mas assume queo método está aquém da teoria, e a aplicaçãodo conceito não vai muito além da aplicaçãodo Conceito Isolacionista, pois apenas passa areconhecer agamotáxons e espécies parafilé-ticas como táxons-espécie. O Conceito Evolu-tivo inicia o questionamento entre a correla-ção direta (i.e., sincronismo) entre o evento cla-dogenético e a anagênese, que são a base doConceito Isolacionista, mas se perde por umexcesso de ontologia, com múltiplos proces-sos de especiação e pouca detecção prática,que apenas repete os Conceitos Biológico eIsolacionista. Sua aplicação em fósseis tambémnão avança significantemente, ainda possuin-do um componente subjetivo na determinaçãode paleoespécies.
Os Conceitos Autapomórfico e Filoge-nético caminham juntos, com um componenteepistemológico forte, reconhecendo que diva-gações ontológicas nada ajudam a elucidar oproblema do reconhecimento da espécie. Sãoaplicáveis a todos os organismos reconhecidosou não pelos conceitos anteriores (agamo-táxons, alopatria, paleoespécies), mas são muitomais objetivos e acurados quanto à discrimina-ção dos táxons. Nestes conceitos, a espécierepresenta apenas uma hipótese, que pode sersustentada ou rejeitada no futuro através doacúmulo de informações, e não uma realidadenatural, que existe independente das informa-ções disponíveis. A diferenciação entre am-bas pode ser resumida pela qualificação napolarização dos estados dos caracteres. O
Métodos de Estudo em Biologia 31
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia32
Conceito Autapomórfico reconhece apenas autilidade de apomorfias e grupos monofiléticospara o reconhecimento de táxons-espécie; jáo Conceito Filogenético utiliza tanto a apomor-fia quanto a plesiomorfia, reconhecendotáxons-espécie tanto monofiléticos quanto pa-rafiléticos. O Conceito Autapomórfico é maisrestrito quanto à aplicação em fósseis, poisestes podem não mostrar apomorfias, sendoreconhecidos apenas como metatáxons. Já oConceito Filogenético reconhece paleoespéci-es independentemente de autapomorfias (es-pécies parafiléticas), assim como a independên-cia de eventos cladogenéticos e anagenéticos.O reconhecimento de espécies parafiléticasimplica numa correlação menos direta (i.e., di-acronismo) entre o binômio formal latino e abiogeografia de vicariância. Por outro lado, aindependência entre a cladogênese e a ana-gênese pode ser minimizada quando avaliadadentro do tempo geológico.
Pode-se assim concluir que o conceitode espécie evoluiu sobre cinco marcos bási-cos na comparação de organismos: (1) reco-nhecimento de variabilidade ontogenética (se-maforontes e dimorfismo sexual); (2) reconhe-cimento de variabilidade fenotípicanão-dimensional (simpátrica e sincrônica atu-al); (3) reconhecimento de variabilidade alopá-trica; (4) reconhecimento do tipo de reprodu-ção (sexuada ou assexuada); (5) reconhecimen-to de variabilidade fenotípica temporal conca-tenada (evolução). Se o problema do leitor foro reconhecimento de espécies sexuadas notempo presente e em uma área limitada, o Con-ceito Biológico é suficiente. Se a área for gran-de o suficiente para se reconhecerempopulações-irmãs alopátricas com variabilida-de fenética inter-populacional maior que a va-riabilidade fenética intra-populacional, o leitordeve recorrer ao Conceito Isolacionista. Se oproblema passa a ser o reconhecimento tam-bém de táxons-espécie assexuados (agamoes-pécies), o presente leitor deve recorrer ao Con-ceito Evolutivo. E se o problema passar a ser oreconhecimento também de táxons fósseis, oleitor deve recorrer ao Conceito Autapomórfi-co, no caso do táxon em questão mostrar au-tapomorfias. Caso contrário, o leitor deve re-correr ao Conceito Filogenético.
Entretanto, pode-se observar que todosos conceitos apresentam deficiências, quer porconseqüência de um excesso de teoria e as-sunções (que podem representar apenas cren-ças), quer por excesso de confiança quanto aum método. Contudo, a ausência de teoria e ade métodos resultam em crenças sem funda-mentos, representadas pelo Conceito Tipológi-co utilizado por Linnæus (1758). É fato que, atu-almente, a Sistemática Filogenética é o melhormétodo de análise disponível, mas ela não re-presenta a solução de todos os problemas. Muitopelo contrário, acaba revelando incongruênciasentre o que se quer fazer e o que se conseguefazer. Portanto, aos interessados sobre o assun-to, é importante o conhecimento sobre as defi-ciências que hoje pairam sobre o entendimentoda espécie e a quantificação da biodiversidade.
AGRADECIMENTOS
Meus agradecimentos às organizadorasdeste volume, Lílian de Lemos Timm e CristinaVargas Cademartori, que me deram a oportu-nidade de divulgar pontos de vista conflitantesno que diz respeito ao conceito de espécie.
REFERÊNCIAS BIBLIOGRÁFICAS
AMORIM, D. de S. Elementos básicos de Sis-temática Filogenética. Segunda edição. Ribei-rão Preto: Holos Editora, Sociedade Brasileirade Entomologia, 1997. xviii+276 p.
ASHLOCK, P.D. Monophyly and associatedterms. Systematic Zoology, Washington, v. 20,n. 1, p. 63-69, 1971.
AYALA, F.J. The Myth of Eve: molecular biologyand human origins. Science, Washington, v.270, n. 5244, p.1930-1936, 1995.
BENTON, M.J. Mass extinctions among tetra-pods and the quality of the fossil record. Philo-sophical Transactions of the Royal Society(Series B: Biological Sciences), London, v. 325,p. 369-386, 1989.
Métodos de Estudo em Biologia32
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 33
BENTON, M.J. Diversification and extinction inthe history of life. Science, Washington, v. 268,p. 52-58, 1995.
CANN, R.L.; STONEKING, M.; WILSON, A.C.Mitochondrial DNA and human evolution.Nature, London, v. 325, n. 6099, p. 31-36, 1987.
CANTINO, P.D.; BRYANT, H.N.; QUEIROZ, K. de;DONOGHUE, M.J.; ERIKSON, T.; HILLIS, D.M.;LEE, M.S.Y. Species names in phylogenetic no-menclature. Systematic Biology, Washington,v. 48, n. 4, p. 790-807, 1999.
CRACRAFT, J. Species concepts in theoreticaland applied Biology: a systematic debate withconsequences. In: WHEELER, Q.D.; MEIER, R.(Ed.). Species concepts and phylogenetictheory: a debate. 2000. p. 3-14.
DARWIN, C. On the origin of species bymeans of natural selection. London: JohnMurray, 1859.
DAUGHERTY, C.H.; CREE, A.; HAY, J.M.;THOMPSON, M.B. Neglected taxonomy andcontinuing extinctions of tuatara (Sphenodon).Nature, London, v. 347, n. 6286, p. 177-179,1990.
DUBOIS, A. A propos de l�emploi controversedu terme �monophyletique�. Nouvelles propo-sitions. Bulletin Mensuel de la SociétéLinnéenne, Lyon, v. 55, n. 7, p. 248-254, 1986.
GASTON, K.J. The magnitude of global insectspecies richness. Conservation Biology,Malden, v. 5, p. 283-296, 1991.
GREUTER, W.; MCNEILL, J.; BARRIE, F.R.; BUR-DET, H.M.; DEMOULIN, V.; FILGUEIRAS, T.S.;NICOLSON, D.H.; SILVA, P.C.; SKOG, J.E.;TREHANE, P.; TURLAND, N.J.; HAWKSWORTH,D.L. International Code of Botanical Nomen-clature (Saint Louis Code) adopted by the XVIInternational Botanical Congress, St Louis,Missouri, July-August 1999. Regnum Vege-tabile. Koeltz Scientific Books, Königstein. 2000.
GRIFFITHS, G.C.D. The future of Linnaeannomenclature. Systematic Zoology, Washington,v. 25, n. 2, p.168-173, 1976.
HAMMER, M.F. A recent common ancestry forhuman Y chromosomes. Nature, London, v.378, n. 6555, p. 376-378, 1995.
HENNIG, W. Grundzüge einer Theorie derphylogenetischen Systematik. Berlin: DeutscherZentralverlag, 1950.
HENNIG, W. Phylogenetic systematics. Urbana:University of Illinois Press, 1966. xvi+263 p.,69 figs.
I.C.Z.N. (International Comission on ZoologicalNomenclature). International code of zoologi-cal nomenclature. Fourth edition. InternationalTrust for Zoological Nomenclature. London:Natural History Museum, 1999. xxviii+306 p.
KISCHLAT, E.E.; SCHULTZ, C.L. Introdução àclassificação dos organismos: consideraçõessobre sistemática filogenética. In: HOLZ, M.;ROS, L.E. de (Ed.). Paleontologia do Rio Grandedo Sul. 2ª Edição, Centro de Investigação doGonduana, Univesidade Federal do Rio Gran-de do Sul, 2002. p. 11-24.
LANE, A.; BENTON, M.J. Taxonomic level as adeterminant of the shape of the Phanerozoicmarine biodiversity curve. The American Natu-ralist, Chicago, v. 162, n. 265-276, 2003.
LINNAEUS, C. Systema naturæ per regna trianaturæ, secundum classes, ordines, genera,species, cum characteribus, differentiis, synony-mis, locis. Tomus I. Editio decima, reformata.Holmiæ, Laurentii Salvii. 1758. ii+824 p.
LINNÉ, C. Systema naturæ per regna tria na-turæ, secundum classes, ordines, genera,species, cum characteribus, differentiis,synonymis, locis. Tomi I-III. Editio duodecima,reformata. Holmiæ, Laurentii Salvii. 1766-1768.
Métodos de Estudo em Biologia 33
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia34
MAYR, E. The Biological Species Concept. In:WHEELER, Q.D; MEIER, R. (Ed.). Speciesconcepts and phylogenetic theory: a debate.2000a. p. 17-29.
MAYR, E. A critique from the Biological SpeciesConcept perspective: what is a species, andwhat is not? In: WHEELER, Q.D; MEIER, R. (Ed.).Species concepts and phylogenetic theory: adebate. 2000b. p. 93-100.
MAYR, E. A defense of the Biological SpeciesConcept. In: WHEELER, Q.D; MEIER, R. (Ed.).Species concepts and phylogenetic theory: adebate. 2000c. p. 161-166.
MEIER, R.; WILLMANN, R. The HennigianSpecies Concept. In: WHEELER, Q.D; MEIER, R.(Ed.). Species concepts and phylogenetictheory: a debate. 2000a. p. 30-43.
MEIER, R.; WILLMANN, R. A defense of the Hen-nigian Species Concept. In: WHEELER, Q.D;MEIER, R. (Ed.). Species concepts and phyloge-netic theory: a debate. 2000b. p. 167-178.
MISHLER, B.D.; THERIOT, E.C. The PhylogeneticSpecies Concept (sensu Mishler and Theriot):Monophyly, Apomorphy, and PhylogeneticSpecies Concepts. In: WHEELER, Q.D; MEIER,R. (Ed.). Species concepts and phylogenetictheory: a debate. 2000a. p. 44-54.
MISHLER, B.D.; THERIOT, E.C. A critique from theMishler and Theriot Phylogenetic Species Con-cept perspective: Monophyly, Apomorphy, andPhylogenetic Species Concepts. In: WHEELER,Q.D; MEIER, R. (Ed.). Species concepts and phylo-genetic theory: a debate. 2000b. p. 119-132.
MISHLER, B.D.; THERIOT, E.C. A defense of thePhylogenetic Species Concept (sensu Mishlerand Theriot): Monophyly, Apomorphy, andPhylogenetic Species Concepts. In: WHEELER,Q.D; MEIER, R. (Ed.). Species concepts andphylogenetic theory: a debate. 2000c. p.179-184.
OVCHINNIKOV, I.V.; GÖTHERSTRÖM, A.;ROMANOVA, G.P.; KHARITONOV, V.M; LIDÉN,K.; GOODWIN, W. Molecular analysis ofNeanderthal DNA from the northern Caucasus.Nature, London, v. 404, p. 490-493, 2000.
PLATNICK, N.I.; WHEELER, Q.D. A defense ofthe Phylogenetic Species Concept (sensuWheeler and Platnick). In: WHEELER, Q.D;MEIER, R. (Ed.). Species concepts and phyloge-netic theory: a debate. 2000. p. 185-197.
QUEIROZ, K. de; GAUTHIER, J.A. Phylogenetictaxonomy. Annual Review of Ecology and Sys-tematics, Palo Alto, v. 23, p. 449-480, 1992.
QUEIROZ, K. de; GAUTHIER, J.A. Toward aphylogenetic system of biological nomenclatu-re. Trends in Ecology & Evolution, Amsterdam,v. 9, n. 1, p. 27-31, 1994.
ROMER, A.S. Osteology of reptiles. Chicago,London: University of Chicago Press, 1956.xxi+772 p. 248 fig.
SIMPSON, G.G. The principles of classificationand a classification of mammals. Bulletin of theAmerican Museum of Natural History, NewYork, v. 85, p. 1-350, 1945.
SNEATH, P.H.A. (Ed.). International Code ofNomenclature of Bacteria. Washington. 1992.
TEMPLETON, A.R. Out of Africa again and again.Nature, London, v. 416, p. 45-51, 2002.
VIGILANT, L.; STONEKING, M.; HARPENDING,H.; HAWKES, K.; WILSON, A.C. African popu-lations and the evolution of human mitochon-drial DNA. Science, Washington, v. 253, n.5027, p. 1503-1507, 1991.
WHEELER, Q.D; MEIER, R. (Ed.). Species con-cepts and phylogenetic theory: a debate. NewYork: Columbia University Press, 2000.Xiv+230 p.
Métodos de Estudo em Biologia34
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Métodos de Estudo em Biologia 35
WHEELER, Q.D; PLATNICK, N.I. The Phyloge-netic Species Concept (sensu Wheeler andPlatnick). In: WHEELER, Q.D; MEIER, R. (Ed.).Species concepts and phylogenetic theory:a debate. 2000a. p. 55-69.
WHEELER, Q.D.; PLATNICK, N.I. A critique fromthe Wheeler and Platnick Phylogenetic SpeciesConcept perspective: problems with alternati-ve concepts of species. In: WHEELER, Q.D;MEIER, R. (Ed.). Species concepts and phylo-genetic theory: a debate. 2000b. p. 133-145.
WILEY, E.O. Phylogenetics: The theory andpractice of phylogenetic systematics. NewYork, Chichester, Brisbane, Toronto, Singapo-re: John Wiley & Sons, 1981. xvi+440 p.
WILEY, E.O.; MAYDEN, R.L. The EvolutionarySpecies Concept. In: WHEELER, Q.D; MEIER,R. (Ed.). Species concepts and phylogenetictheory: a debate. 2000a. p. 70-89.
WILEY, E.O.; MAYDEN, R.L. A critique from theEvolutionary Species Concept perspective. In:WHEELER, Q.D; MEIER, R. (Ed.). Species con-cepts and phylogenetic theory: a debate.2000b. p. 146-158.
WILEY, E.O.; MAYDEN, R.L. A defense of theEvolutionary Species Concept. In: WHEELER,Q.D; MEIER, R. (Ed.). Species concepts andphylogenetic theory: a debate. 2000c. p.198-208
WILLMANN, R.; MEIER, R. A critique from theHennigian Species Concept perspective. In:WHEELER, Q.D; MEIER, R. (Ed.). Species con-cepts and phylogenetic theory: a debate.2000. p. 101-118.
Métodos de Estudo em Biologia 35
Caderno La Salle XI, Canoas, v.2, nº 1, 11-35, 2005

Cadernos La Salle
Métodos de Estudo Métodos de Estudo Métodos de Estudo Métodos de Estudo em em em em
BiologiaBiologiaBiologiaBiologiaLílian de L. TimmCristina Vargas Cademartori(Organizadoras)

Cadernos La Salle

Métodos de Estudoem Biologia
Centro Universitário La Salle
Canoas, 2005

Trabalhos submetidos em janeiro de 2004 e aceitos em junho de 2004.
Organizadoras: Lílian de L. TimmCristina Vargas Cademartori
Secretária: Aline Guerini Moura
Diagramação: Fábio Teixeira
Impressão:
CENTRO UNIVERSITÁRIO LA SALLE
Reitor – Nelso Antonio Borgidgnon, fsc
Vice-Reitor – Marcos Antonio Corbellini, fsc
Pró-Reitor Acadêmico – Marcos AntonioCorbellini, fsc
Pró-Reitor Administrativo – Renato Silva
Pró-Reitor Comunitário – Adelar Hengemuhle
Coordenadora de Extensão – Vera LúciaRamirez
Coordenadora de Pesquisa e Pós-Graduação –Ana Maria Machado Toaldo
Coordenador do Conselho Editorial – MarcosAntonio Corbellini, fsc
CONSELHO EDITORIAL
Alexandre Bandeira Monteiro e SilvaAna Maria Machado ToaldoCristina Vargas CademartoriElsa Gonçalves AvanciniGilberto Ferreira da SilvaMárcia Angelita TiburiMárcos Antonio Corbellini – CoordenadorRejane Silva PennaRoger Pierre Fabris HoefelStela Maris Herrmann
COMISSÃO CIENTÍFICA
Prof. Dr. Airton da Silva NegrineProf. Dr. Alexandre Bandeira Monteiro e SilvaProfª Drª Ana Maria Machado ToaldoProfª Drª Andrea RapoportProf. Dr. Bruno Godofredo GlaabProf. Dr. Cicero Galeno LopesProfª Drª Cláudia do Nascimento WyrvalkiProf. Dr. Cláudio José de Holanda CavalcantiProf. Dr. Claus Dieter StobäusProfª Drª Cristina Vargas CademartoriProfª Drª Delia CroviProf. Dr. Delmar BizaniProf. Dr. Edson Prestes e Silva JúniorProfª Drª Elaine Azambuja de LimaProfª Drª Elsa Gonçalves AvanciniProf. Dr. Gilberto Ferreira da SilvaProf. Dr. Henrique Carlos de Oliveira de Castro
CONSELHO CIENTÍFICO
Dr. Cláudio José de Holanda CavalcantiDra. Cristina Vargas CademartoriDr. Edio-Ernst KischlatDra. Maria TsaoDr. Sérgio Augusto de Loreto BordignonDra. Susi Missel PachecoDra. Trícia Kommers Albuquerque
EXPEDIENTE
FICHA CATALOGRÁFICA

APRESENTAÇÃO
A organização deste número da revista Cadernos La Salle expressa a tentativa dereunir, numa única publicação, métodos de estudo de uso corrente nas várias áreas dasCiências Biológicas. O primeiro Caderno do Curso de Biologia do UNILASALLE, com otema Métodos de Estudo em Biologia, surpreendeu por ter se constituído num trabalhointerinstitucional, agregando contribuições de profissionais da FEPAM, FZB-RS, PUCRS,UFRGS, ULBRA, UNISC, UNISINOS, UNIVATES e UNOCHAPECÓ, além daquelas igual-mente enriquecedoras produções de nosso corpo docente.
Esta proposta nasceu de um ímpeto, uma necessidade de compartilhar com nos-sos alunos as metodologias que levam à produção de conhecimento biológico. No de-correr do processo, compreendemos que o resultado deste esforço poderia atingir umpúblico mais amplo, abrangendo também recém-graduados, técnicos, alunos de pós-graduação ou mesmo profissionais de outras áreas que desejem obter um referencialmetodológico inicial. Com a crescente especialização e fragmentação do universo bioló-gico em inúmeras áreas e subáreas, que se justifica, pelo menos em parte, em funçãodas especificidades de seus objetos de estudo, as várias técnicas e métodos utilizadostornam-se cada vez mais de domínio restrito de seus usuários. Freqüentemente disper-sas em literatura especializada e, muitas vezes, com linguagem inacessível, tais metodo-logias tornam-se de difícil obtenção. Entendemos que esta publicação contribui parasolucionar tal problema, cumprindo também seu papel na divulgação científica. Consi-deramos que �comunicar ciência� também deve ser compromisso assumido por aquelesque fazem ciência. E parece que fomos bem sucedidas nessa primeira tentativa: são 23artigos a descreverem os métodos e suas aplicações nas áreas de Paleontologia, Zoolo-gia de Invertebrados, Zoologia de Vertebrados, Morfologia, Bioquímica e Ciências Ambi-entais. O primeiro artigo, todavia, embora não trate de metodologias específicas, abordauma questão fundamental e polêmica em biologia, o conceito de espécie, cuja compre-ensão afeta qualquer área do conhecimento biológico.
Ofertamos, então, à comunidade acadêmica e ao público em geral este primeiroCaderno de Biologia, produto da dedicação de colegas e amigos que prontamente acei-taram compor conosco esta coletânea de métodos e técnicas. Agradecemos a todos porsuas valorosas contribuições e também ao UNILASALLE, pelo apoio incondicional a estapublicação.
Profa. Cristina Cademartori e Profa. Lílian Timm Organizadoras

SUMÁRIO
1. CONCEITO DE ESPÉCIE
1.1. Os Conceitos de Espécie: Uma Abordagem Prática � Edio Ernst Kischlat ...........11
2. MÉTODOS DE ESTUDO EM PALEONTOLOGIA
2.1. Considerações sobre as Áreas de Estudo em Paleontologia, Métodos deColeta e Preparação de Vertebrados Fósseis � Lílian de L. Timm eEdio Ernst Kischlat ....................................................................................................................39
2.2. Métodos para o Trabalho com Microfósseis e Formas Atuais - Itamar IvoLeipnitz; José Luiz Lorenz Silva; Beatriz Leipnitz; Eduardo da Silva Aguiar;Carolina Jardim Leão e Luciana Giovanoni ...................................................................... 49
2.3. Metodologias Utilizadas em Conodontes � Ana Karina Scomazzon e ValescaBrasil Lemos ...............................................................................................................................59
2.4. Tafonomia: Processos de Fossilização e sua Relação com a Paleoecologia� Cristina Bertoni Machado ................................................................................................... 65
2.5. Confecção de Moldes e Réplicas de Vertebrados Fósseis � Cristina Vega-Diase Lisandro Ferreira Nunes ..................................................................................................... 73
2.6. Considerações sobre as Técnicas Utilizadas na Confecção de LâminasDelgadas de Vertebrados Fósseis � Lílian de L. Timm ................................................ 79
2.7. A Aplicação de Técnicas de Análise Multivariada à Taxonomia dePaleovertebrados: uma Alternativa à Visão Tipológica em Paleontologia� Marcos Machado ..................................................................................................................87
2.8. Métodos de Estudos em Palinologia do Quaternário e de Plantas Atuais� Soraia Girardi Bauermann e Paulo César Pereira das Neves ................................. 99
2.9. Folhas das Angiospermas: Taxonomia, Preservação e sua Aplicação naReconstituição das Floras e dos Climas do Passado � Tânia Lindner Dutrae Daiana Rockenbach Boardman ..................................................................................... 109
3. MÉTODOS DE ESTUDO EM ZOOLOGIA DE INVERTEBRADOS
3.1. Métodos para Estudos Biológicos em Poríferos � Jairo Luis Cândido ................. 123
3.2. Protocolos de Campo e Laboratório para a Caracterização das PlanáriasTerrestres Neotropicais (EUPLATYHELMINTHES: TRICLADIDA) �Fernando Carbayo ................................................................................................................. 131
3.3. Linhas de Pesquisa e Métodos para Estudos com Moluscos Terrestres� Ingrid Heydrich ................................................................................................................... 145
4. MÉTODOS DE ESTUDO EM ZOOLOGIA DE VERTEBRADOS
4.1. Métodos de Estudos em Peixes � Karin Martha Grosser eFernando Gertum Becker .................................................................................................... 161
4.2. Estudos com Avifauna: Métodos Gerais de Levantamentos de Espécies� Carla Suertegaray Fontana e André de Mendonça-Lima ...................................... 173

4.3. Introdução aos Métodos de Estudo dos Pequenos Mamíferos �Cristina Vargas Cademartori .............................................................................................. 183
4.4. Técnicas de Campo Empregadas no Estudo de Quirópteros �Susi Missel Pacheco.............................................................................................................. 193
4.5. Ecologia e Comportamento de Primatas: Métodos de Estudo de Campo� Vanessa Barbisan Fortes e Júlio César Bicca-Marques ......................................... 207
4.6. A Utilização de Armadilhas Fotográficas para o Estudo de Mamíferos deMédio e Grande Porte � Rosane Vera Marques e Fábio Dias Mazim .................. 219
5. MÉTODOS DE ESTUDO EM MORFOLOGIA E BIOQUÍMICA
5.1. Técnicas Rotineiras de Preparação e Análise de Lâminas Histológicas �Lílian de L. Timm .................................................................................................................... 231
5.2. Metodologias de Uso Corrente em Microbiologia � Francisco Fernandode Castilho Koller ................................................................................................................... 241
5.3. Coleta, Conservação, Seleção e Cultivo de Fungos Basidiomicetos Produtoresde Fenol Oxidases � Giovani André Piva ....................................................................... 251
5.4. Identificação Humana através de Marcadores Moleculares �Trícia Kommers Albuquerque ............................................................................................ 265
6. MÉTODOS DE ESTUDO EM CIÊNCIAS AMBIENTAIS
6.1. Utilização de Técnicas de Sensoriamento Remoto e Sistemas de InformaçãoGeográfica (Sig) na Biologia � Eduardo Périco, Gisele Cemin eClaudete Rempel ................................................................................................................... 271

CONCEITO DE ESPÉCIE
