métodos de avaliação de alimentos para aves - weby · a água influencia nos valores nutritivos...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS–GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
Métodos de avaliação de alimentos para aves
Bruno Duarte Alves Fortes
Orientador:
Prof. Dr. Marcos Barcellos Café
GOIÂNIA
2011

i
BRUNO DUARTE ALVES FORTES
Métodos de avaliação de alimentos para aves
Seminário apresentado junto à Disciplina
Seminários Aplicados do Programa de Pós-
Graduação em Ciência Animal da Escola de
Veterinária e Zootecnia da Universidade
Federal de Goiás.
Nível: Doutorado
Área de Concentração:
Produção Animal
Linha de Pesquisa: Metabolismo
nutricional, alimentação e forragicultura
na produção animal
Orientador:
Prof. Dr. Marcos Barcellos Café - UFG
Comitê de Orientação:
Prof. Dr. José Henrique Stringhini - UFG
Profª. Drª. Nadja Susana Mogyca Leandro - UFG
GOIÂNIA
2011

ii
SUMÁRIO
1 INTRODUÇÃO .....................................................................................................1
2 REVISÃO DE LITERATURA ................................................................................3
2.1 Composição química dos alimentos ................................................................. 3
2.2 Métodos químicos de análises dos alimentos .................................................. 4
2.2.1 Método de Weende ou sistema de análise proximal ..................................... 4
2.2.2 Matéria seca (MS) e teor de umidade (U) ..................................................... 5
2.2.3 Matéria mineral (MM) .................................................................................... 6
2.2.4 Proteína bruta (PB) ....................................................................................... 7
2.2.5 Extrato etéreo (EE) ........................................................................................ 8
2.2.6 Fibra Bruta (FB) ............................................................................................. 9
2.2.7 Extrativos não nitrogenados (ENN) ............................................................. 13
2.2.8 Método de Van Soest .................................................................................. 14
2.2.9 Fibra em detergente neutro (FDN) .............................................................. 15
2.2.10 Fibra em detergente ácido (FDA) .............................................................. 16
3 Metodologias de avaliação energética dos alimentos ....................................... 17
4 Metodologias de avaliação aminoacídica dos alimentos ................................... 23
4.1 Digestibilidade aparente e digestibilidade verdadeira dos aminoácidos ......... 23
4.2 Disponibilidade e digestibilidade dos aminoácidos ......................................... 24
4.3 Métodos para determinar digestibilidade dos aminoácidos com aves ............ 26
4.3.1 Método da alimentação forçada com galos ................................................. 26
4.3.2 Método da coleta ileal em aves ................................................................... 27
4.3.3 Método da coleta total de excreta com aves ............................................... 28
5 Tendências para análises dos alimentos .......................................................... 28
5.1 Novas tendências na avaliação de fósforo digestível em aves ...................... 28
5.2 Novas tendências na avaliação do nível energético ....................................... 30

iii
6 CONSIDERAÇÕES FINAIS ............................................................................... 33
7 REFERÊNCIAS .................................................................................................. 34

iv
LISTA DE TABELAS
TABELA 1- Diferenças percentuais dos teores de energia digestível para grãos e raízes, quando comparados em seus estados naturais e de matéria seca.. .......... 6
TABELA 2- Fatores de conversão do nitrogênio para proteína de diversos alimentos.. .............................................................................................................. 8
TABELA 3- Digestibilidade da fração fibra bruta de diversos alimentos para aves ............................................................................................................................. 11
TABELA 4- Conteúdo de carboidratos de alguns ingredientes de origem vegetal (%) ........................................................................................................................ 14
TABELA 5- Valores obtidos de energia metabolizável aparente (kcal/kg MS), corrigida para o balanço de nitrogênio (EMAn) e energia metabolizável verdadeira, corrigida para o balanço de nitrogênio (EMVn) pelo método de coleta total e pela técnica de alimentação forçada com galos .......................................................... 21
TABELA 6- Valores de energia metabolizável aparente corrigida (kcal/kg MS) das sojas, determinadas em cada idade das aves ...................................................... 22
TABELA 7- Coeficientes de digestibilidade do fósforo em alimentos para aves (matéria natural, %) .............................................................................................. 30

v
LISTA DE FIGURAS
FIGURA 1- Esquema analítico de Weende ............................................................ 4
FIGURA 2- Comparação entre os métodos de Weende e Soest ......................... 16
FIGURA 3- Locais onde são avaliados o aproveitamento dos aminoácidos nos animais monogástricos ......................................................................................... 25

1
1 INTRODUÇÃO
O uso rotineiro de novas tecnologias na alimentação de aves está
diretamente relacionado com a disponibilidade de dados de exigência dos animais
e também com os valores nutritivos dos ingredientes utilizados nas formulações
para as diversas fases de criação.
Os programas alimentares utilizados na avicultura têm como finalidade
conciliar a nutrição e a fisiologia nos diversos estágios de desenvolvimento das
aves. A capacidade digestiva das aves é influenciada pela idade, nota-se que
ocorre aumento no aproveitamento dos nutrientes presentes nas dietas com o
avanço da idade, em função do desenvolvimento do trato gastro intestinal desses
animais.
Para atender adequadamente às exigências nutricionais dos animais e
para que possam expressar o máximo do seu potencial, é imprescindível que se
formulem rações eficientes. Por isso, é necessário conhecer com maior precisão a
composição química juntamente com os valores energéticos dos alimentos
utilizados nas dietas avícolas.
Com o decorrer dos anos os nutricionistas vêm utilizando várias
ferramentas, como programas computacionais e tabelas de exigências
nutricionais, com o objetivo de conciliar a fisiologia dos animais com a
composição bromatológica dos alimentos utilizados nas rações.
O desenvolvimento da nutrição está associado ao conhecimento do
valor nutricional dos ingredientes que são utilizados nas rações avícolas, sendo o
valor nutricional de um alimento diretamente relacionado à sua composição
química e energética, entre outros fatores.
Em decorrência de inúmeros fatores que interferem na concentração de
nutrientes dos ingredientes, como a fertilidade de solo, o clima, a cultivar, o
armazenamento, a amostragem, os tipos de processamento e as substâncias
antinutricionais, faz-se necessário realizar análises bromatológicas ou físico-
químicas, que têm como principal objetivo a obtenção da composição química dos
alimentos, ou seja, a determinação de suas frações nutritivas.
Devido às grandes variações na capacidade de utilização dos
nutrientes pelos animais, em função das diferenças anatômicas e fisiológicas
existentes nos tratos digestório das várias espécies a avaliação da digestibilidade

2
dos alimentos é um parâmetro de grande importância, pois para determinar se um
alimento é eficientemente utilizado pelo animal, se faz necessário descobrir o seu
coeficiente de digestibilidade, servindo como base de cálculo para as exigências
das várias espécies.
De uma forma geral, pode-se concluir que os processos de avaliação
dos alimentos tradicionalmente empregados na avicultura são representados
pelos valores obtidos pelas análises bromatológicas, teores de energia e de
aminoácidos, sejam eles totais ou digestíveis.
Nesse contexto, objetivou-se com a presente revisão, discutir e abordar
as principais características dos métodos tradicionais de análises dos alimentos
assim como as novas técnicas de avaliação dos nutrientes presentes nos
ingredientes utilizados nas dietas avícolas.

3
2 REVISÃO DE LITERATURA
2.1 Composição química dos alimentos
Os ingredientes que compõem as rações avícolas representam a
porção mais onerosa da produção e o adequado fornecimento dos nutrientes é
essencial para o máximo crescimento e deposição de tecido muscular pelos
animais. A formulação das rações de aves possui o intuito de atender as
exigências nutricionais, para isso, se torna necessário o conhecimento da
composição química dos alimentos, bem como suas limitações nutricionais.
SANTOS et al. (2005), destacaram que o conhecimento dos dados de
composição química, dos valores de digestibilidade e da disponibilidade de
nutrientes são extremamente importantes no balanceamento de dietas.
Dentre os diversos problemas enfrentados pelos nutricionistas destaca-
se a variação na composição química de um mesmo tipo de alimento. Porém essa
variação é normal, principalmente em se tratando de alimentos de diferentes
condições de cultivo, solo, regiões, clima e cultivares.
O ideal ao se elaborar uma ração seria avaliar a composição dos
ingredientes disponíveis, porém isso demanda trabalho, tempo e custo. Com isso,
a utilização de tabelas de composição química de alimentos tem sido uma
alternativa (CALDERANO, 2008).
Tanto nas indústrias como nas instituições de pesquisa, a formulação
de rações já baseou-se em informações de composição de alimentos e de
exigências nutricionais estabelecidas no exterior, principalmente na Europa e
Estados Unidos (ROSTAGNO et al., 2005). Porém, essas tabelas, sob alguns
aspectos, deixavam a desejar quanto a sua aplicabilidade em condições
brasileiras, o que incentivou pesquisadores a elaborarem suas próprias tabelas
(ROSTAGNO et al., 1983; EMBRAPA, 1991).
Com isso vários trabalhos (BRUMANO et al., 2006; CAFÉ et al., 2000;
CARVALHO et al., 2004), têm sido desenvolvidos gerando dados que são
utilizados para atualizar e incluir novos alimentos às tabelas já existentes, como a
4
tabela atualizada de ROSTAGNO et al. (2011), possibilitando assim melhor
utilização dos alimentos na formulação de dietas.
2.2 Métodos químicos de análises dos alimentos
2.2.1 Método de Weende ou sistema de análise proximal
O método de Weende, também chamado Sistema de Análise Proximal,
foi criado por Henneberg em 1860, na Estação Experimental de Weende,
Alemanha. Segundo Henneberg, o alimento é composto de diversos nutrientes
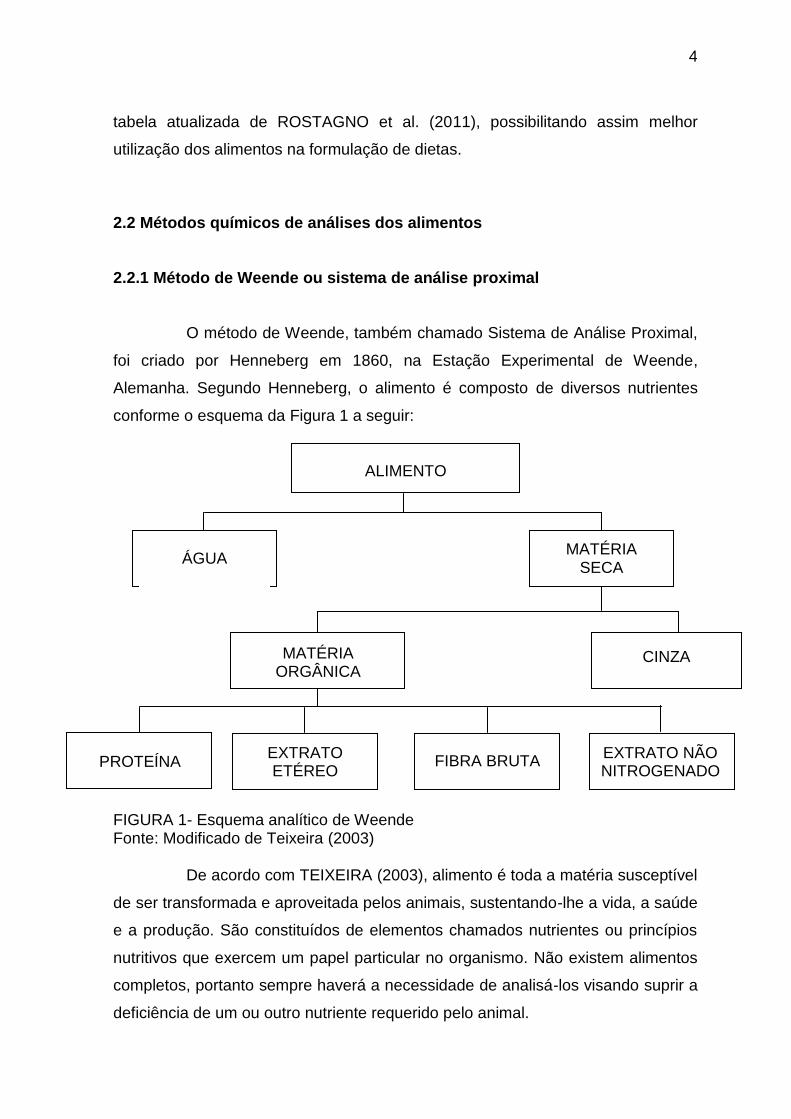
conforme o esquema da Figura 1 a seguir:
FIGURA 1- Esquema analítico de Weende Fonte: Modificado de Teixeira (2003)
De acordo com TEIXEIRA (2003), alimento é toda a matéria susceptível
de ser transformada e aproveitada pelos animais, sustentando-lhe a vida, a saúde
e a produção. São constituídos de elementos chamados nutrientes ou princípios
nutritivos que exercem um papel particular no organismo. Não existem alimentos
completos, portanto sempre haverá a necessidade de analisá-los visando suprir a
deficiência de um ou outro nutriente requerido pelo animal.
ALIMENTO
FIBRA BRUTA
CINZA
PROTEÍNA
MATÉRIA ORGÂNICA
EXTRATO ETÉREO
EXTRATO NÃO NITROGENADO
MATÉRIA SECA
ÁGUA

5
As análises laboratoriais visam separar os componentes dos alimentos
em frações de digestibilidade e metabolização previsíveis, com um custo analítico
baixo e através de métodos rápidos. Devem ser realizadas para fornecer um valor
nutricional aproximado da dieta utilizada, que é a mistura de todos os ingredientes
oferecidos a um animal.
O método de Weende consiste em fracionar o alimento em matéria
seca (MS), proteína bruta (PB), extrato etéreo (EE), fibra bruta (FB), extrativos
não nitrogenados (ENN) e cinzas ou matéria mineral (MM). Estes nutrientes
compõem as análises clássicas ou comumente feitas visando obter as
informações sobre um alimento qualquer.
Entretanto, o sistema de análise proximal ou método de Weende ao
longo do tempo apresentou-se insatisfatório, por não reconhecer as diversas
frações dos carboidratos, com características de solubilidade e degradação
distintas e não promover o fracionamento da fibra. No grupo dos extratos não-
nitrogenados encontram-se frações de naturezas diversas, como amido,
hemicelulose, pectina, lignina solúvel em álcali e os carboidratos solúveis em
água (SILVA & QUEIROZ, 2009).
2.2.2 Matéria seca (MS) e teor de umidade (U)
A obtenção do teor de matéria seca e de umidade é o primeiro passo
na determinação bromatológica de qualquer alimento, além de ser de grande
importância pois a preservação dos alimentos pode estar condicionada ao teor de
umidade do material; além disso, quando se compara o valor nutritivo de dois ou
mais alimentos, é necessário levar em consideração os respectivos teores de
matéria seca (SILVA & QUEIROZ, 2009).
A concentração de matéria seca dos alimentos é determinada por
secagem da amostra em estufa com circulação forçada de ar à temperatura de
55°C por 16 a 24 horas (pré-secagem), a 135°C por duas horas, 100°C por 24
horas ou a 105°C por 16 horas (secagem definitiva). Ressalta-se que a pré-
secagem é realizada apenas para alimentos com menos de 85% de matéria seca.
A água influencia nos valores nutritivos dos alimentos, ou seja, muitas
vezes as diferenças de valores energéticos entre dois alimentos estão em função
6
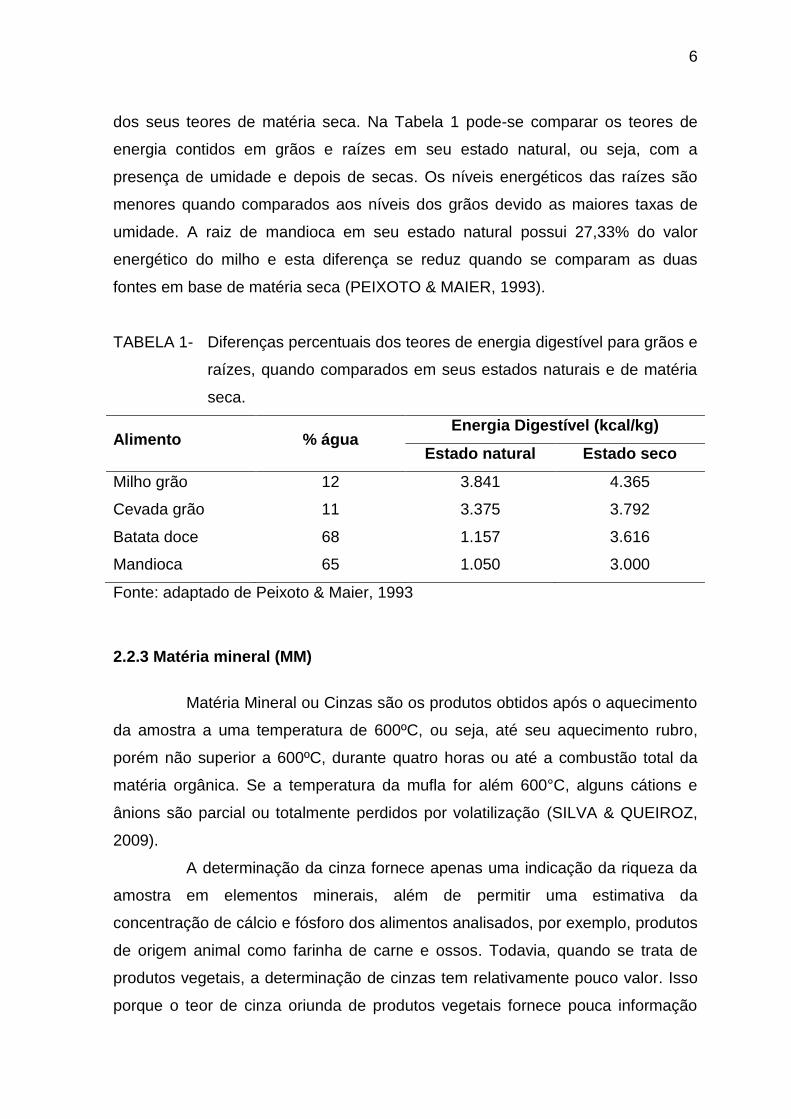
dos seus teores de matéria seca. Na Tabela 1 pode-se comparar os teores de
energia contidos em grãos e raízes em seu estado natural, ou seja, com a
presença de umidade e depois de secas. Os níveis energéticos das raízes são
menores quando comparados aos níveis dos grãos devido as maiores taxas de
umidade. A raiz de mandioca em seu estado natural possui 27,33% do valor
energético do milho e esta diferença se reduz quando se comparam as duas
fontes em base de matéria seca (PEIXOTO & MAIER, 1993).
TABELA 1- Diferenças percentuais dos teores de energia digestível para grãos e
raízes, quando comparados em seus estados naturais e de matéria
seca.
Alimento % água Energia Digestível (kcal/kg)
Estado natural Estado seco
Milho grão 12 3.841 4.365
Cevada grão 11 3.375 3.792
Batata doce 68 1.157 3.616
Mandioca 65 1.050 3.000
Fonte: adaptado de Peixoto & Maier, 1993
2.2.3 Matéria mineral (MM)
Matéria Mineral ou Cinzas são os produtos obtidos após o aquecimento
da amostra a uma temperatura de 600ºC, ou seja, até seu aquecimento rubro,
porém não superior a 600ºC, durante quatro horas ou até a combustão total da
matéria orgânica. Se a temperatura da mufla for além 600°C, alguns cátions e
ânions são parcial ou totalmente perdidos por volatilização (SILVA & QUEIROZ,
2009).
A determinação da cinza fornece apenas uma indicação da riqueza da
amostra em elementos minerais, além de permitir uma estimativa da
concentração de cálcio e fósforo dos alimentos analisados, por exemplo, produtos
de origem animal como farinha de carne e ossos. Todavia, quando se trata de
produtos vegetais, a determinação de cinzas tem relativamente pouco valor. Isso
porque o teor de cinza oriunda de produtos vegetais fornece pouca informação

7
sobre sua composição, uma vez que seus componentes, em minerais, são muito
variáveis (SILVA & QUEIROZ, 2009).
De acordo com TEIXEIRA (2003), a análise de minerais, tanto os
macros como os microminerais, atualmente é realizada com grande precisão pela
técnica de absorção atômica. Os macrominerais são expressos % dos
ingredientes e os microminerais na base de mg/kg de alimento ou ppm. As
análises mais comuns são para determinação de cálcio e fósforo.
2.2.4 Proteína bruta (PB)
As proteínas são nutrientes nitrogenados presentes em todas as células
vivas; portanto, são essenciais à vida de todo animal. Todos os animais
necessitam ingerir quantidades adequadas de proteína diariamente, além disso,
para aves e suínos, a quantidade é tão importante quanto a qualidade.
O termo proteína bruta (PB) envolve um grande grupo de substâncias
com estruturas semelhantes, porém com funções fisiológicas diferentes. As
proteínas dos alimentos vegetais possuem em torno de 16% de nitrogênio (N).
Baseado nesta concentração foi desenvolvido o método de determinação da
proteína nos alimentos, através da avaliação do conteúdo de N total da amostra.
Então, PB significa o nitrogênio total contido em um material analisado,
multiplicado pelo fator 6,25 (100 ÷ 16 = 6,25), segundo o método de Kjeldahl.
O método de Kjeldahl foi desenvolvido em 1883 pelo químico
dinamarquês, Johan Kjeldahl e é até hoje o método-padrão de determinação do
valor de nitrogenado dos alimentos. Este método consiste basicamente em três
passos: 1) digestão da amostra em ácido sulfúrico com um catalisador, que
resulta em conversão do nitrogênio em sulfato de amônia; 2) destilação da
amônia em uma solução receptora; e 3) quantificação da amônia por titulação por
volumetria com uma solução padrão. Este método determina o nitrogênio contido
na matéria orgânica, incluindo o nitrogênio protéico propriamente dito e outros
compostos nitrogenados não-protéicos, como aminas, amidas, lecitinas, nitrilas e
aminoácidos (SILVA & QUEIROZ, 2009).
No entanto, o teor de nitrogênio presente no alimento pode variar com o
tipo de proteína (Tabela 2). Verifica-se que existem diferenças na porcentagem de

8
nitrogênio nas diversas fontes de proteína, porém, estes valores não são
comumente utilizados. O fator 6,25 é aceito pela comunidade científica que
trabalha em nutrição animal (BERTECHINI, 2006).
Além do método Kjeldahl existem outras metodologias para
determinação de nitrogênio total como o método descoberto por Dumas (1831),
que consiste em aquecer a amostra a 700-800ºC, e depois medir o volume
nitrogênio gasoso. O método de Dumas apresenta vantagens em relação ao
método Kjeldahl, pois analisa as amostras (de até quatro gramas) em apenas
quatros minutos, sem utilizar reagentes corrosivos, quentes ou ácidos.
TABELA 2- Fatores de conversão do nitrogênio para proteína de diversos
alimentos.
Alimentos Fator
Arroz 5,95
Aveia, trigo 5,83
Algodão, girassol 5,30
Farinha de sangue 5,80
Leite 6,38
Milho 6,25
Soja 5,71
Sorgo 6,25
Fonte: adaptado de Bertechini, 2006
2.2.5 Extrato etéreo (EE)
O extrato etéreo é definido como todas as substâncias extraídas pelo
éter (ou clorofórmio): triglicerídeos, ácidos graxos livres, colesterol, lecitina,
clorofila, álcoois voláteis, resinas e pigmentos. Os lipídeos pertencem a um grupo
de substâncias que são extraídas por solventes orgânicos e são comumente
chamados de extrato etéreo. Além de reserva energética, os lipídeos
desempenham outras funções, por exemplo, estrutural (componente das
membranas celulares) e hormonal (BETERCHINI, 2006).

9
As gorduras ou lipídios são substâncias insolúveis em água, porém
solúvel em éter, clorofórmio, benzeno e outros solventes orgânicos chamados de
extratores. Gorduras, óleos, pigmentos e outras substâncias gordurosas solúveis
contidas em uma amostra seca são dissolvidos através da extração com o éter, o
qual é evaporado desta solução gordurosa.
O éter usado no processo é aquecido até tornar-se volátil e ao
condensar-se, circula sobre a amostra em análise, arrastando toda a fração
gordurosa e demais substâncias solúveis em éter. Este é recuperado em outro
recipiente, enquanto a gordura extraída é calculada por diferença de pesagem
(SILVA & QUEIROZ, 2009).
Considera-se que um grama de gordura produz 9,35 kcal de energia
bruta, quando medida em bomba calorimétrica, o que corresponde à
aproximadamente, 9 kcal de energia metabolizável (EM). Os alimentos com maior
teor de gordura têm valores energéticos mais altos, pelo fato dos lipídeos
fornecerem 2,25 vezes mais energia do que os carboidratos (SILVA & QUEIROZ,
2009).
O teor de gordura nos alimentos também influencia no armazenamento
dos produtos, uma vez que a gordura dos alimentos constitui uma fração instável
e é passível de rancificação.
2.2.6 Fibra Bruta (FB)
De acordo com SILVA & QUEIROZ (2009), o termo fibra bruta (FB) é a
parte dos carboidratos resistentes ao tratamento sucessivo com ácido e base
diluídos, representando a grande parte da fração fibrosa dos alimentos. TEIXEIRA
(2003), cita que a FB é composta principalmente de celulose com pequenas
quantidades de lignina e hemicelulose, sendo a celulose a maior fração.
Existem vários métodos para a determinação da fibra e da qualidade
dos alimentos, porém é necessário avaliar as limitações do método a ser
empregado em sua determinação. O método de Weende determina a FB através
do uso de uma amostra seca e desengordurada a uma digestão ácida com ácido
sulfúrico diluído e uma digestão alcalina com hidróxido de sódio diluído, com isso
tenta-se reproduzir o que usualmente ocorre no estômago e intestino dos animais.

10
Nesta fase ocorrem à remoção de proteínas, açúcares e amido,
deixando como resíduos: celulose e outros componentes polissacarídeos, além
da matéria mineral. Na etapa seguinte incinera-se o resíduo restando à matéria
mineral. A diferença entre as duas etapas constitui-se no que convencionalmente
se chama de fibra bruta (SILVA & QUEIROZ, 2009).
Outros métodos surgiram para melhorar as definições das frações
obtidas pelo método de Weende e entre elas a fibra bruta foi fracionada por Van
Soest e Wine (1967) em fibra detergente neutro (FDN), fibra detergente ácido
(FDA) e lignina.
Segundo WARPECHOWSKI (1996), tem sido adotado duas definições
de fibra da dieta: definição química e fisiológica. A definição química considera a
soma de polissacarídeos não amiláceos insolúveis (celulose, hemicelulose),
polissacarídeos não amiláceos solúveis (arabinoxilanos, β-glucanos e pectinas) e
lignina. A definição fisiológica considera todos os componentes da dieta
resistentes a degradação por enzimas endógenas de mamíferos (extensível as
aves), considerando esta uma definição mais ampla, pois inclui os
oligossacarídeos de reserva e qualquer componente resistentes à digestão
enzimática endógena (amido resistente, proteína da parede celular, etc.).
O principal problema quando se determina FB é a quantidade variável
de lignina que ocorre nos alimentos, a qual não é digestível, e que é removida
pela solução alcalina (básica) durante esta determinação. Esta lignina removida
juntamente com a hemicelulose vai fazer parte da fração extrato não nitrogenado
(ENN), que tem digestibilidade maior que a fibra bruta. No entanto, em vários
casos, a digestibilidade do ENN é inferior à da FB face à grande contaminação
com lignina, principalmente (SILVA & QUEIROZ, 2009).
A fibra da dieta exerce vários efeitos metabólicos e fisiológicos no
organismo animal, sendo diferenciados conforme as frações que a constituem,
solúvel ou insolúvel. Esses efeitos podem ser decorrentes de alterações em
funções fisiológicas, como a taxa de excreção endógena e a passagem do
alimento pelo trato gastrintestinal, alterações no bolo alimentar e digesta, tais
como a capacidade de hidratação, o volume, o pH e a fermentabilidade ou ainda,
por alterações nas populações e na atividade da microbiota intestinal (VAN
SOEST, 1994).

11
Segundo BERTECHINI (2006), as aves possuem baixa capacidade de
digerir materiais fibrosos devido à reduzida microflora existente no trato digestório,
sendo que as dietas devem ser concentradas. Estima-se que aves adultas são
capazes de digerir até 25% da fibra da ração, principalmente no ceco (Tabela 3).
TABELA 3- Digestibilidade da fração fibra bruta de diversos alimentos para aves.
Alimento % Fibra Bruta Digestibilidade (%)
Aveia 11,3 7
Cevada 5,5 11
Milho 1,9 13
Trigo 2,5 9
Fonte: adaptado de Bertechini, 2006
Segundo HETLAND et al. (2004), a fibra na dieta pode ser considerada
um componente diluidor da energia metabolizável, além de possuir efeitos
antinutricionais, dependendo de sua solubilidade, relacionados com a diminuição
do aproveitamento de nutrientes presentes nas dietas. Outra característica
relacionada ao alto nível de fibra nas rações de frangos de corte é o aumento da
umidade da cama provocada pela maior produção de excretas pelas aves.
As fibras podem ser classificadas de acordo com sua estrutura e
solubilidade em água. Quanto à estrutura, grande parte das fibras pertence ao
grupo de polissacarídeos, podendo ser carboidrato estrutural, que incluem os
constituintes da parede celular, ou carboidratos não estruturais, que incluem os
carboidratos presentes no conteúdo celular. Com base na sua solubilidade em
água, as fibras podem ser classificadas em solúveis e insolúveis.
Na fração insolúvel da fibra encontram-se a lignina, os polissacarídeos
não amiláceos (PNA) celulose e hemiceluloses insolúveis, os taninos, as cutinas e
outros compostos minoritários, enquanto que na fração solúvel da fibra são
encontradas as hemiceluloses solúveis e as substâncias pécticas (VAN SOEST et
al., 1991).
Geralmente a fibra da dieta é considerada como parte dos
componentes remanescentes da extração com solução de detergente neutro,

12
conhecida como fibra em detergente neutro (FDN), de acordo com o método
descrito por VAN SOEST & WINE (1967).
O aumento nos teores de fibra insolúvel na dieta pode provocar
diminuição no tempo de passagem da digesta pelo trato gastrintestinal, podendo
ser decorrente da estimulação física da fibra insolúvel sobre as paredes do trato
gastrintestinal (TGI), que tende a aumentar a motilidade e a taxa de passagem. O
aumento dos teores desta fração provoca também diluição da energia da dieta,
levando a um aumento compensatório no consumo para que atinja os níveis
energéticos exigidos para o crescimento, desenvolvimento e produção
(WARPECHOWSKI, 1996).
De acordo com LARBIER & LECLERQ (1994), o aumento da fibra
insolúvel na dieta aumenta linearmente a excreção endógena de nitrogênio e a
massa bacteriana na excreta, deduzindo-se que o consumo de fibra insolúvel
pode causar aumento na quantidade de substratos endógenos e exógenos,
disponíveis à fermentação bacteriana na região do ceco.
O teor de fibra solúvel na dieta está associado a uma maior viscosidade
da dieta, o que contribui para um trânsito mais lento da digesta no TGI e com
efeitos negativos sobre o desempenho animal (BEDFORD & CLASSEN 1992).
Esse aumento da viscosidade dificulta a ação de enzimas e sais biliares no bolo
alimentar, reduzindo a digestão e absorção dos nutrientes.
A disponibilidade dos nutrientes nos alimentos é freqüentemente
limitada pela presença de fatores antinutricionais. De acordo com THORPE &
BEAL (2001), trata-se de fatores com efeitos depressivos sobre a digestão e
utilização de proteínas, carboidratos, minerais e vitaminas. Estes fatores, por
exemplo, podem diminuir ou aumentar a exigência de vitaminas para o animal ou
mesmo estimular o sistema imune e causar danos por reação de
hipersensibilidade.
A maioria das dietas de frangos de corte, no Brasil, são constituídas de
alimentos de origem vegetal, entre eles os mais utilizados são o milho e o farelo
de soja. Contudo, esses alimentos apresentam constituintes que são indigeríveis
pelas aves, entre eles, os polissacarídeos não amiláceos (PNAs) e o ácido fítico
que são de grande importância.

13
LIMA et al. (2007) afirmam que o termo polissacarídeos não amiláceos
(PNAs) vem sendo freqüentemente utilizado para se referir à fibra bruta. Os
componentes da fibra dos grãos são basicamente PNA’s, no qual fazem parte da
estrutura da parede celular e não podem ser digeridos pelas aves, devido à
natureza de suas ligações, sendo resistentes à hidrólise no trato digestivo
(CONTE et al., 2002).
A classificação de carboidratos em estruturais e não estruturais refere-
se à função desempenhada nas plantas. Os carboidratos estruturais são
encontrados na parede celular dos vegetais e fornecem o suporte físico
necessário para o crescimento das plantas. A parede celular é composta de
pectina, celulose, hemicelulose, lignina, complexos fenólicos e proteína
(MERTENS, 1996).
Os carboidratos não estruturais estão localizados no conteúdo celular e
são encontrados em maior concentração nas sementes, folhas e hastes.
Representam as reservas de energia usadas para reprodução, crescimento e
sobrevivência durante períodos de estresse (MERTENS, 1996).
2.2.7 Extrativos não nitrogenados (ENN)
Segundo TEIXEIRA (2003), cerca de 75% da energia provém dos
carboidratos, sendo que, para os herbívoros, 25% são extrativos não
nitrogenados e o restante fibra bruta. Os extrativos não nitrogenados da dieta são
constituídos pelos açucares e amido dos alimentos. A divisão dos carboidratos do
alimento em fibra bruta e extratos não nitrogenados, pelas análises comuns
realizadas em laboratórios, é bastante empírica.
Dependendo do tipo de alimento, uma parte da lignina e da celulose
podem estar presentes nos extrativos não nitrogenados. A lignina é de natureza
puramente estrutural e, se presente nos extrativos não nitrogenados, diminui seu
potencial energético.
Os ENN são obtidos por meio de cálculos e são constituídos pelos
açucares, amidos, dextrinas, hemiceluloses, e variavelmente lignina. Essa fração
do esquema de Weende corresponde predominantemente aos carboidratos mais

14
digestíveis, ou como se diz comumente, “carboidratos solúveis” (PEIXOTO &
MAIER, 1993).
Na maioria dos vegetais, com exceção das sementes oleaginosas, o
carboidrato é em geral o principal componente (Tabela 4). Na análise proximal
dos alimentos, a fração carboidrato é representada por açúcares solúveis
(extrativo não nitrogenado) e fibra bruta que representa os carboidratos
estruturais (BERTECHINI, 2006).
TABELA 4- Conteúdo de carboidratos de alguns ingredientes de origem
vegetal (%).
Fontes ENN FDA FDN
Farelo de soja 45% 31,50 8,16 13,86
Farelo de trigo 53,50 13,85 40,59
Milho 72,08 3,54 11,75
Soja integral extrusada 24,00 11,40 15,70
Sorgo BT* 71,05 5,90 10,03
* Baixo tanino Fonte: adaptado de Bertechini, 2006
Para obtenção dos valores dos ENN é realizado o seguinte cálculo:
ENN = 100 - (% umidade + % PB + % FB + % EE + % MM)
A determinação do ENN, por seu cálculo, se ressente de todas as
imprecisões pessoais e imperfeições inerentes às outras determinações
laboratoriais, ou seja, acumula todos os erros que por ventura foram cometidos
(SILVA & QUEIROZ, 2009).
2.2.8 Método de Van Soest
O método de VAN SOEST (1967) divide os componentes da amostra
analisada em conteúdo celular, que compreende as frações solúveis em
detergente neutro, conforme preconiza o método e se tornou rotina freqüente nos
laboratórios de análises de alimentos (BERCHIELLI et al., 2001).

15
2.2.9 Fibra em detergente neutro (FDN)
O método para determinação do teor de fibra em detergente neutro dos
alimentos foi desenvolvido no início da década de 60, com o trabalho publicado
por GOERING & VAN SOEST (1970). Desde então, várias modificações ao longo
do tempo foram realizadas (VAN SOEST et al., 1991). Os reagentes usados para
análise de FDN (VAN SOEST & WINE, 1967) não dissolvem as frações
indigestíveis ou lentamente digestível dos alimentos, sugerindo que esse método
mede com mais acurácia as características nutricionais associadas à fibra.
A parede celular é chamada de fibra em detergente neutro (FDN) e
inclui proteínas insolúveis, hemicelulose e lignocelulose que engloba,
principalmente, as frações de lignina e celulose. Sob o aspecto nutricional, o
método de Van Soest separa melhor os diversos componentes das frações
fibrosas dos alimentos.
JERACI & VAN SOEST (1990) consideram a FDN uma medida
eficiente da fibra insolúvel da dieta que melhor representa a fração do alimento de
digestão lenta ou indigestível, e que esta função poderia ser uma medida
importante para caracterização de dietas para aves. Porém, o maior
inconveniente do método é a solubilização das substâncias pécticas, pectinas e β-
glucanas, que são substâncias freqüentemente presentes na parede celular
vegetal.
No sistema detergente a amostra é exposta primeiramente ao
detergente neutro (pH 7). Após a exposição ao detergente neutro, realiza-se uma
filtragem que separa o conteúdo celular, solúvel, da parede celular ou fibra em
detergente neutro. O conteúdo celular contém amido, proteínas, lipídeos e outros
compostos com alta digestibilidade. Portanto, a fibra em detergente neutro é
composta por: hemicelulose, celulose e lignina.
A fibra em detergente neutro é uma medida do conteúdo total de fibra
insolúvel do alimento e constitui o parâmetro mais utilizado para o balanceamento
de dietas uma vez que interfere na qualidade da mesma.

16
2.2.10 Fibra em detergente ácido (FDA)
A determinação da fibra em detergente ácido (FDA) foi desenvolvida
para evitar a solubilização lignina que ocorre no método da fibra bruta. Esse
método não utiliza álcali para isolar a fibra, propondo detergente ácido específico
a fim de solubilizar o conteúdo celular e as hemiceluloses, obtendo um conteúdo
insolúvel em detergente ácido, denominado fibra em detergente ácido.
Desta forma, a FDA isola, principalmente, a celulose e a lignina com
contaminação de pectina, cinzas e compostos nitrogenados, principalmente os
produzidos pela reação de Maillard, sendo estes resíduos a porção menos
digerível pelos microrganismos presentes no trato digestório de alguns animais.
A determinação de FDA é importante quando se deseja avaliar a
digestibilidade, pois o teor de FDA nos alimentos possui alta correlação positiva
com a digestibilidade da matéria seca.
Na Figura 2 podem ser observadas as principais diferenças entre
determinação de fibra bruta pelo método de Weende e de FDN e FDA pelo
método de Van Soest.
Método de Weende Fração química Método de Van Soest
Matéria mineral (1)* Matéria mineral solúvel
Conteúdo celular (Solúveis
em detergente neutro)
Extrato etéreo Lipídeos, pigmentos, etc.
Proteína bruta Proteínas, aminoácidos
Extrato não
nitrogenado
Açucares, amido e pectina
Hemicelulose
Parede
celular FDN
Álcali solúvel
Lignina FDA
Fibra bruta
Álcali
insolúvel
Celulose
Matéria mineral (2)* Cinza insolúvel (Sílica)
* Matéria mineral total do método de Weende = MM (1) + MM (2) FIGURA 2- Comparação entre os métodos de Weende e Soest. Fonte: Branco (2006)

17
3 Metodologias de avaliação energética dos alimentos
O conhecimento do valor energético dos alimentos é de fundamental
importância nutricional e econômica, para a formulação de rações que resultem
em ótimo desempenho dos animais. A energia liberada da oxidação dos
alimentos, assim como a oriunda do metabolismo energético como calor
produzido, é expressa em caloria (cal) o joule (J). Uma caloria é definida como a
quantidade de calor necessária para elevar um grama de água de 14,5 °C a 15,5
°C, um joule equivale a 0,239 cal, ou seja, uma caloria é igual a 4,18 joules
(SAKOMURA & ROSTAGNO, 2007).
Todos os alimentos possuem sua fração energética, geralmente
representada pelo teor de lipídios e carboidratos presentes, sendo esta
denominada de energia bruta dos alimentos. Praticamente todo o metabolismo de
energia corpóreo gera o que se denomina de incremento calórico, sendo este o
principal responsável pela homeostase da temperatura corpórea (LARBIER &
LECLERQ, 1994).
A energia bruta (EB) é o produto da oxidação total da matéria orgânica
de uma ração ou de um alimento medida em bomba calorimétrica (SAKOMURA &
ROSTAGNO, 2007). Os carboidratos fornecem 3,7 kcal/g (glicose) e 4,2 kcal/g
(amido); as proteínas 5,6 kcal/g e as gorduras 9,4 kcal/g de EB, respectivamente
(NRC, 1998).
A energia digestível (ED) representa a energia do alimento que é
absorvida após o processo de digestão nos animais. É determinada pela
diferença entre a EB do alimento consumido e a energia bruta das fezes. Para
aves, essa forma de energia não é usualmente utilizada em virtude da dificuldade
de separar as fezes da urina.
A energia metabolizável (EM) é a forma mais utilizada para formulação
de rações avícolas, sendo obtida pela diferença entre EB do alimento e a EB das
excretas (fezes e urina) e dos gases oriundos da digestão (insignificantes em
aves). Considerando que a energia perdida na forma de gases nos monogástricos
é muito baixa, tem sido desprezada nos cálculos da energia metabolizável
(SAKOMURA & ROSTAGNO, 2007).

18
Para as aves, a EM pode ser determinada e expressa como energia
metabolizável aparente (EMA), energia metabolizável verdadeira (EMV), energia
metabolizável aparente corrigida para o balanço de nitrogênio (EMAn) ou energia
metabolizável verdadeira corrigida para o balanço de nitrogênio (EMVn).
O método mais utilizado para a determinação da EMA é o tradicional de
coleta total de excretas, descrito por SIBBALD & SLINGER (1963), que considera
a quantidade de energia consumida subtraída da quantidade de energia
excretada pelas aves. Porém, existem outras metodologias para determinar a
energia dos alimentos em aves como: metodologia da alimentação precisa
proposta inicialmente por SIBBALD (1976), uso de indicadores com o intuito de
relacionar substâncias indigestíveis presentes nos alimentos e nas excretas
proposto por KOBT & LUCKEY (1972), equações de predições (método indireto) e
o sistema NIRS (near infrared spectroscopy), sendo que este fornece apenas
dados de EB para os ingredientes analisados.
O sistema NIRS para determinação dos componentes químicos dos
alimentos tem sido usado como rotina nos laboratórios nos últimos anos. O
princípio do NIRS foi desenvolvido por Karl Norris no início da década de 70. O
sistema tornou-se uma técnica de laboratório ideal por ser rápida, não necessitar
de reagentes químicos e não produzir resíduos. Além disso, não há necessidade
de preparar as amostras e vários nutrientes podem ser analisados ao mesmo
tempo.
A técnica é uma integração de espectroscopia de luz, estatística e
ciência da computação. Modelos matemáticos são construídos para relacionar a
composição dos grupos químicos ativos à absorção de energia na região do
espectro do infravermelho próximo (700-2500nm). Nessa região, são medidas
vibrações de átomos de hidrogênio ligados ao nitrogênio, oxigênio e carbono
(SAKOMURA & ROSTAGNO, 2007).
No cálculo da EMA considera que toda energia das fezes e urina é
derivada do alimento. Entretanto, a energia fecal é proveniente de resíduos do
alimento não digerido e da energia metabólica oriunda da bile, escamações das
células da parede intestinal e suco digestivo (SIBBALD & WOLYNETZ, 1984),
assim como a energia da urina compreende a energia de origem alimentar que
não foi utilizada, energia endógena de subprodutos nitrogenados dos tecidos, e a

19
metabólica de subprodutos nitrogenados do metabolismo protéico (SIBBALD,
1987).
A EMA pode ser corrigida segundo a quantidade de nitrogênio
excretado, objetivando comparar animais em estados fisiológicos distintos, esta
se denomina de EMAn. Sabe-se que o balanço de nitrogênio (BN) é obtido
quantificando o teor de nitrogênio ingerido versus o excretado, este resultado é
multiplicado por uma constante 8,22 kcal/g de nitrogênio retido pelo animal. O
fator de 8,22 é utilizado para aves pois representa a energia equivalente do ácido
úrico, quando este é completamente oxidado (NRC, 1981).
HILL & ANDERSON (1958) relataram que essa correção é usada para
contabilizar os efeitos variáveis de crescimento e de deposição de proteína
corporal entre as aves. Segundo LOPEZ & LEESON (2007) o valor de correção é
acrescentado à energia da excreta por cada grama de N retido, pois se o
nitrogênio não tivesse sido retido, teria sido excretado como ácido úrico.
Para NUNES (2003) é necessário corrigir os valores estimados de
energia pelo BN, pois durante o ensaio de metabolismo é impossível assegurar
que todas as aves apresentem a mesma taxa de crescimento. De acordo com
NERY (2005) o nitrogênio retido como tecido, se catabolizado, contribuirá para as
perdas de energia urinária endógena, portanto, variações na retenção de
nitrogênio contribuirão para variações nos valores de EMA.
A seguir tem-se as fórmulas para o cálculo da EMA e EMAn:
EMA = Ebing - Efec – Euri
MS ingerida
Onde:
EMA = Energia Metabolizável Aparente;
Ebing = Energia bruta ingerida;
Eexc = Energia fecal;
Euri = Energia urinária;
MS = Matéria seca ingerida.

20
EMAn = EMA - (BN x 8,22)
Onde:
EMAn = Energia metabolizável aparente corrigida pelo balanço de
nitrogênio;
EMA = Energia metabolizável aparente;
BN = Balanço de nitrogênio.
A EMV é obtida pela diferença entre a EB do alimento consumido e a
energia bruta da excreta (fezes e urina), corrigida pelas perdas de energia fecal
metabólica e urinária endógena.
A seguir tem-se as fórmulas para o cálculo da EMV:
EMV = Ebing - (Ebexc + EFm + EUe)
Onde:
EMV = Energia metabolizável verdadeira;
Ebing = Energia bruta ingerida;
Ebexc = Energia bruta excretada;
Efm = Energia fecal metabólica;
EUe = Energia urinária endógena.
Da mesma forma a EMV pode ser corrigida pelo BN gerando a EMVn,
tendo como base o cálculo demonstrado para a EMAn.
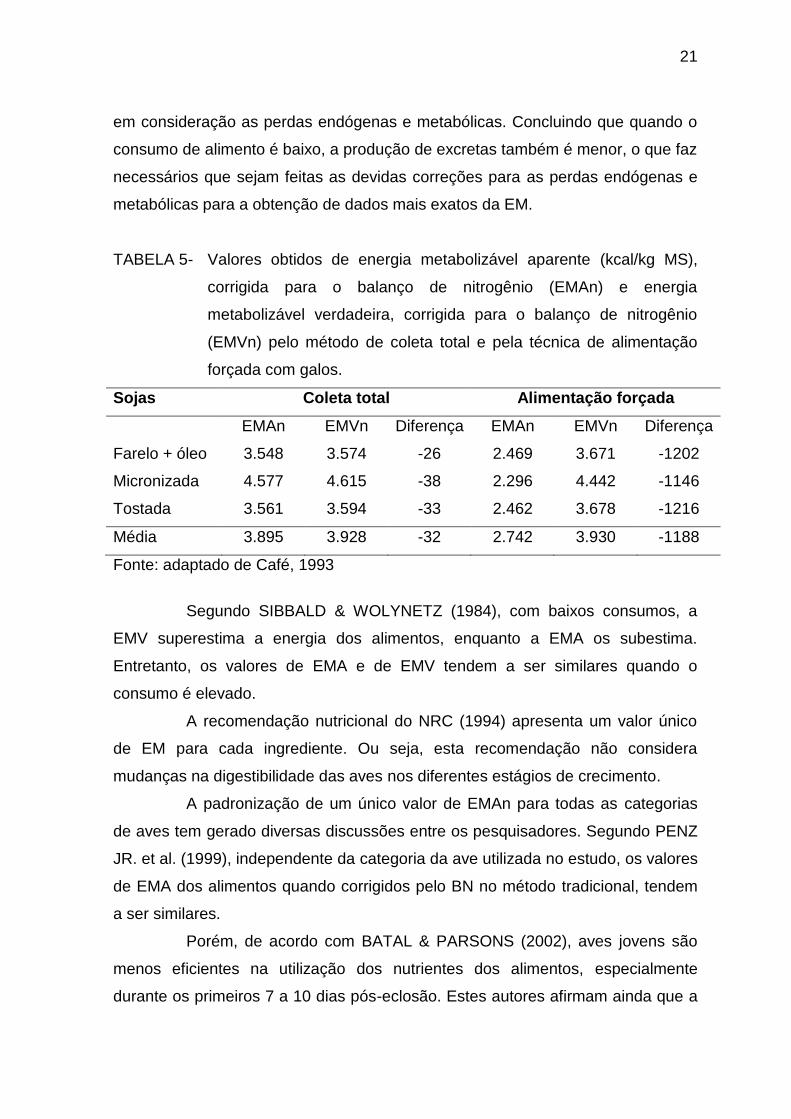
CAFÉ (1993) comparou os sistemas de EMAn e EMVn (Tabela 5)
através de ensaios realizados com galos pelo método de coleta total e pela
técnica da alimentação forçada ou precisa (SIBBALD, 1976). No método de coleta
total, no qual o consumo das dietas experimentais é ad libitum, verificou-se que as
diferenças entre os valores de EMAn e EMVn das sojas foram pequenas, o que
indica que neste método a correção das perdas endógenas e metabólicas são
pequenas ou inexpressivas (Tabela 5).
Porém, quando o autor comparou os valores de EMAn e EMVn obtidos
pela técnica de alimentação forçada, com o consumo fixo de 30 g, as diferenças
foram maiores. Os menores valores de EMAn foram explicados por não se levar
21
em consideração as perdas endógenas e metabólicas. Concluindo que quando o
consumo de alimento é baixo, a produção de excretas também é menor, o que faz
necessários que sejam feitas as devidas correções para as perdas endógenas e
metabólicas para a obtenção de dados mais exatos da EM.
TABELA 5- Valores obtidos de energia metabolizável aparente (kcal/kg MS),
corrigida para o balanço de nitrogênio (EMAn) e energia
metabolizável verdadeira, corrigida para o balanço de nitrogênio
(EMVn) pelo método de coleta total e pela técnica de alimentação
forçada com galos.
Sojas Coleta total Alimentação forçada
EMAn EMVn Diferença EMAn EMVn Diferença
Farelo + óleo 3.548 3.574 -26 2.469 3.671 -1202
Micronizada 4.577 4.615 -38 2.296 4.442 -1146
Tostada 3.561 3.594 -33 2.462 3.678 -1216
Média 3.895 3.928 -32 2.742 3.930 -1188
Fonte: adaptado de Café, 1993
Segundo SIBBALD & WOLYNETZ (1984), com baixos consumos, a
EMV superestima a energia dos alimentos, enquanto a EMA os subestima.
Entretanto, os valores de EMA e de EMV tendem a ser similares quando o
consumo é elevado.
A recomendação nutricional do NRC (1994) apresenta um valor único
de EM para cada ingrediente. Ou seja, esta recomendação não considera
mudanças na digestibilidade das aves nos diferentes estágios de crecimento.
A padronização de um único valor de EMAn para todas as categorias
de aves tem gerado diversas discussões entre os pesquisadores. Segundo PENZ
JR. et al. (1999), independente da categoria da ave utilizada no estudo, os valores
de EMA dos alimentos quando corrigidos pelo BN no método tradicional, tendem
a ser similares.
Porém, de acordo com BATAL & PARSONS (2002), aves jovens são
menos eficientes na utilização dos nutrientes dos alimentos, especialmente
durante os primeiros 7 a 10 dias pós-eclosão. Estes autores afirmam ainda que a
22
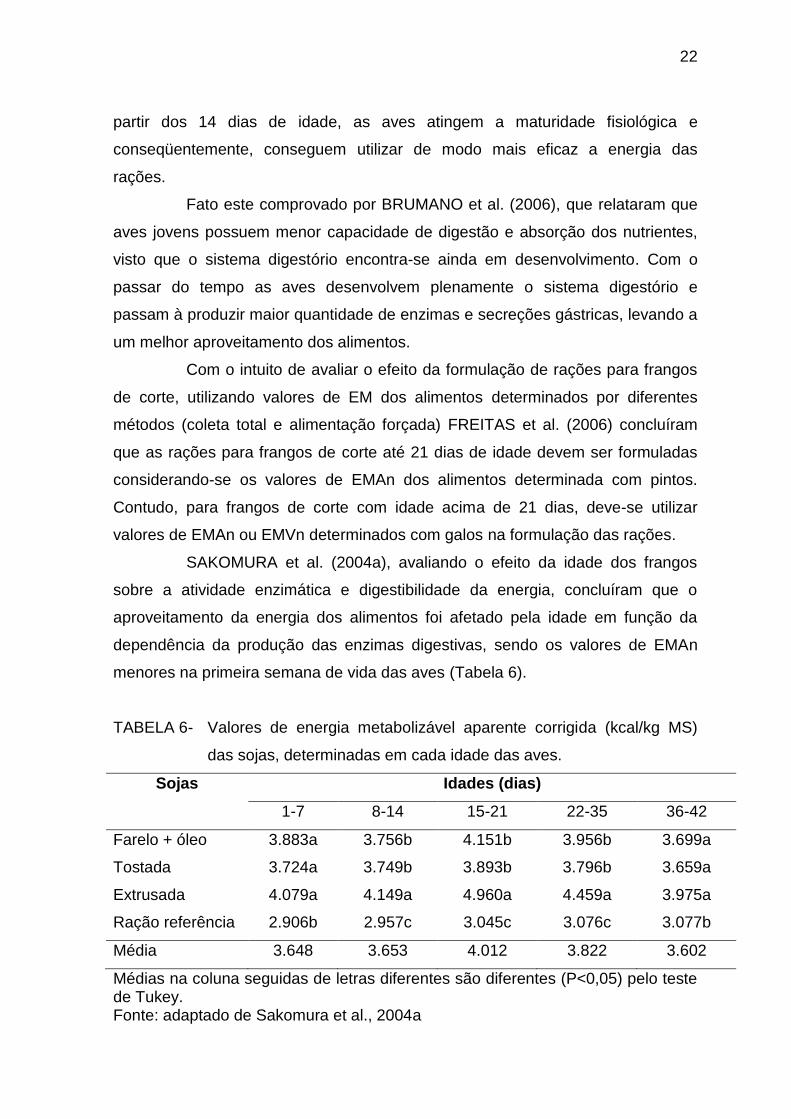
partir dos 14 dias de idade, as aves atingem a maturidade fisiológica e
conseqüentemente, conseguem utilizar de modo mais eficaz a energia das
rações.
Fato este comprovado por BRUMANO et al. (2006), que relataram que
aves jovens possuem menor capacidade de digestão e absorção dos nutrientes,
visto que o sistema digestório encontra-se ainda em desenvolvimento. Com o
passar do tempo as aves desenvolvem plenamente o sistema digestório e
passam à produzir maior quantidade de enzimas e secreções gástricas, levando a
um melhor aproveitamento dos alimentos.
Com o intuito de avaliar o efeito da formulação de rações para frangos
de corte, utilizando valores de EM dos alimentos determinados por diferentes
métodos (coleta total e alimentação forçada) FREITAS et al. (2006) concluíram
que as rações para frangos de corte até 21 dias de idade devem ser formuladas
considerando-se os valores de EMAn dos alimentos determinada com pintos.
Contudo, para frangos de corte com idade acima de 21 dias, deve-se utilizar
valores de EMAn ou EMVn determinados com galos na formulação das rações.
SAKOMURA et al. (2004a), avaliando o efeito da idade dos frangos
sobre a atividade enzimática e digestibilidade da energia, concluíram que o
aproveitamento da energia dos alimentos foi afetado pela idade em função da
dependência da produção das enzimas digestivas, sendo os valores de EMAn
menores na primeira semana de vida das aves (Tabela 6).
TABELA 6- Valores de energia metabolizável aparente corrigida (kcal/kg MS)
das sojas, determinadas em cada idade das aves.
Sojas Idades (dias)
1-7 8-14 15-21 22-35 36-42
Farelo + óleo 3.883a 3.756b 4.151b 3.956b 3.699a
Tostada 3.724a 3.749b 3.893b 3.796b 3.659a
Extrusada 4.079a 4.149a 4.960a 4.459a 3.975a
Ração referência 2.906b 2.957c 3.045c 3.076c 3.077b
Média 3.648 3.653 4.012 3.822 3.602
Médias na coluna seguidas de letras diferentes são diferentes (P<0,05) pelo teste de Tukey. Fonte: adaptado de Sakomura et al., 2004a

23
4 Metodologias de avaliação aminoacídica dos alimentos
O conhecimento preciso da composição de aminoácidos dos alimentos
é um dos principais fatores para o êxito da formulação de dietas balanceadas. Os
aminoácidos essenciais estão entre os nutrientes que mais impactam o
desempenho animal. Por isso, é de fundamental importância o conhecimento da
composição aminoacídica dos alimentos, bem como do seu aproveitamento pelos
animais (SAKOMURA & ROSTAGNO, 2007).
Sabe-se que as quantidades de aminoácidos digestíveis nos alimentos
são substancialmente menores que a quantidade total e, associado a isto, o
desenvolvimento de ensaios de digestibilidade mais rápidos, bem como o
aperfeiçoamento daqueles já existentes, tornou possível a condução de maior
número de pesquisas sobre este assunto (PARSONS, 1996).
4.1 Digestibilidade aparente e digestibilidade verdadeira dos aminoácidos
A digestibilidade aparente é definida como sendo a diferença entre a
quantidade de aminoácido consumida e quantidade destes nas fezes ou digesta
ileal. Já a digestibilidade verdadeira é determinada pela diferença entre a
quantidade de aminoácido consumida e nas fezes ou digesta ileal, sendo
consideradas as perdas endógenas dos aminoácidos que são subtraídas da
quantidade total de aminoácidos presentes nas fezes ou digesta ileal
(SAKOMURA & ROSTAGNO, 2007).
A avaliação da digestibilidade pode ser feita por duas metodologias,
pela coleta total (excreta) e pela coleta ileal (digesta). Na coleta total, a excreta é
influenciada pela microbiota cecal que modifica o perfil aminoacídico, pela perda
endógena e metabólica. A dificuldade em separar urina de fezes sem intervenção
cirúrgica torna a determinação da digestibilidade dos aminoácidos mais complexa
e com poucas certezas de quanto à digestão cecal contribui para a nutrição
(LEESON & SUMMERS, 2001).
As perdas endógenas podem ser determinadas utilizando-se animais
em jejum (SIBBALD, 1976) ou fornecendo dietas livres de nitrogênio
(ROSTAGNO et al., 1973, PAPADOPOULOS et al., 1986). Entretanto, PARSONS

24
et al. (1982) ressalta que a produção de aminoácidos, nas aves em jejum, pode
ser inferior a das aves alimentadas e essa variação pode levar a erros nos valores
de digestibilidade, principalmente quando o alimento possui baixa concentração
de aminoácidos.
PERTILLA et al. (2002), avaliando a digestibilidade de alimentos
protéicos (farelo de soja, colza e farinha de carnes e ossos) e a formulação com
base em aminoácidos totais ou digestíveis, observaram que frangos de corte
alimentados com dietas formuladas com base na lisina digestível apresentaram
melhor ganho de peso e composição de carcaça que as aves alimentadas com
aminoácidos totais.
4.2 Disponibilidade e digestibilidade dos aminoácidos
A disponibilidade dos aminoácidos inclui os processos de digestão,
absorção e metabolismo ou utilização dos mesmos. De acordo com SAKOMURA
& ROSTAGNO (2007), a disponibilidade de aminoácidos é definida como sendo a
quantidade de aminoácidos absorvidos e utilizados pelos animais. Pode ser
determinada em ensaios de crescimento, ou seja, sua utilização é avaliada pelo
crescimento dos animais.
Já a digestibilidade é determinada pela diferença entre a quantidade de
aminoácidos consumida e a excretada nas fezes. Em aves, determina-se os
aminoácidos metabolizáveis, pois os aminoácidos da urina estão incluídos no
cálculo. A digestibilidade é determinada com base no local em que é realizada a
coleta de material, ou seja, pode ser pelo método da coleta de excretas total ou
coleta ileal, sendo esta última mais precisa do que a coleta total, visto que o
conteúdo da dieta não sofre contaminação dos aminoácidos sintetizados pelas
bactérias do ceco, conforme Figura 3.
A disponibilidade e a digestibilidade dos aminoácidos podem ser
determinadas de diversas maneiras, por meio de métodos de avaliação in vitro e
in vivo. No método in vitro, a digestão biológica é simulada e os aminoácidos são
avaliados após a ação de enzimas proteolíticas.

25
1- Local para avaliar a disponibilidade dos aminoácidos
2- Local de medida da digestibilidade ileal dos aminoácidos
3- Local para medir a digestibilidade fecal dos aminoácidos
4- Local de avaliação da digestibilidade na excreta
FIGURA 3- Locais onde são avaliados o aproveitamento dos aminoácidos nos animais monogástricos Fonte: adaptado de Sakomura & Rostagno, 2007
A maior desvantagem deste método ocorre, pois não é possível
reproduzir as condições complexas e dinâmicas que ocorrem no trato digestório
do animal. Porém, os ensaios in vivo ou biológicos são considerados mais
precisos para avaliar a digestibilidade de aminoácidos dos alimentos, dentre eles
se destacam: o método da alimentação forçada com galos inteiros ou
cecectomizados, a alimentação ad libitum com galos ou pintos intactos para
determinar a digestibilidade ileal e a coleta total de excretas com pintos
(BRUMANO et al., 2006).
Proteína (AA) do alimento
Aminoácidos não absorvidos
Aminoácidos absorvidos
Íleo (2) Síntese de proteínas
Catabolismo
Urina Crescimento (1)
Fezes (3)
Parte distal do intestino
Excretas (4) (fezes e urina)
Digestão

26
4.3 Métodos para determinar digestibilidade dos aminoácidos com aves
A determinação da digestibilidade dos aminoácidos é realizada,
estimando-se a fração de aminoácidos da ração que é degradada durante a
passagem pelo intestino das aves. As técnicas mais utilizadas com aves têm sido
uma adaptação daquela descrita por SIBBALD (1976), para determinação da
EMV, e o método de coleta ileal com o uso de indicadores.
4.3.1 Método da alimentação forçada com galos
Este método tem sido bastante utilizado pelos pesquisadores para
determinar a digestibilidade dos aminoácidos por ser uma técnica precisa,
recomenda-se que a utilização de galos cecectomomizados a partir de 20
semanas de idade, de linhagens de corte ou postura conforme a técnica descrita
por PUPA et al. (1998).
Este ensaio tem a duração de 96 horas, sendo que nas primeiras 48
horas os galos são submetidos a um jejum para a limpeza do trato digestório e
nas 48 horas subseqüentes eles são forçados a ingerir 30 gramas do alimento
teste, normalmente fracionado em duas vezes (15g + 15g). A ingestão forçada é
feita com o auxílio de uma sonda esofágica de dimensões e características
específicas descritas por SIBBALD (1987), sendo introduzido diretamente no
inglúvio das aves.
Após a alimentação, é realizada a coleta de excretas durante 48 horas
em intervalos de 12 horas. Recomenda-se a utilização de bolsas plásticas (sacos
plásticos) que são presas à região pélvica dos galos para a coleta de excretas.
No entanto, alguns problemas podem ocorrer quando são utilizados os
ensaios in vivo, pois o período experimental é longo e oneroso e vários fatores
podem interferir no resultado, sendo um deles a produção dos chamados
aminoácidos exógenos, produzidos pela microflora localizada nos cecos (PUPA et
al., 1998).
A grande vantagem da utilização de animais cecectomizados é a
garantia do resultado da avaliação da digestibilidade dos aminoácidos não ser

27
influenciada pela fermentação das bactérias do ceco, conseqüentemente,
impedindo a degradação microbiana de aminoácidos endógenos no ceco.
4.3.2 Método da coleta ileal em aves
Este método tem sido o mais utilizado para determinar a digestibilidade
dos aminoácidos com aves em crescimento. Embora este método seja mais
preciso que o de coleta total de excretas para determinar a digestibilidade de
aminoácidos, apresenta algumas desvantagens como: ser mais trabalhoso que o
método de coleta total de excretas, exigir uma maior quantidade de aves por
unidade experimental (principalmente nas fases iniciais de criação) para que se
tenha uma quantidade considerável de amostra para realização das análises
laboratoriais (SAKOMURA & ROSTAGNO, 2007).
Este método utiliza-se um indicador indigestível, o Cr2O3 ou cinza
insolúvel em ácido (CIA), que é adicionado às dietas experimentais em
quantidade de 1 a 2% da dieta. As aves são submetidas a um período de
adaptação às dietas experimentais, em torno de sete dias. Ao final deste período,
as aves são abatidas para retirar o conteúdo presente no segmento do íleo
terminando a quatro centímetros da junção íleo-cecal. Após as coletas indica-se o
armazenamento em freezer a -70 °C e posteriormente a secagem por liofilização.
Após a liofilização as amostras são analisadas para quantificação dos teores de
aminoácidos e indicador nas dietas e digestas, com estes dados são calculadas a
digestibilidade dos aminoácidos.
A digestibilidade ileal verdadeira refere-se à quantidade de aminoácidos
presente na proteína/aminoácido dietético que foram absorvidos no intestino
delgado, considerando a redução dos aminoácidos endógenos presentes na
digesta ileal. São considerados aminoácidos endógenos as enzimas pancreáticas,
a mucina, a proteína bacteriana e aminoácidos componentes do epitélio celular. A
correção endógena é coerente, principalmente quando se faz uso de formulações
com inclusão de vários tipos de alimentos (BRITO, 2007).

28
4.3.3 Método da coleta total de excreta com aves
Este método foi descrito por SIBBALD & SLINGER (1963), baseado nos
princípios de HILL & ANDERSON (1958) e POTTER & MATTERSON (1960). O
método da coleta total baseia-se no princípio de mensurar o total de alimento
consumido e o total de excretas produzidas durante um certo período de tempo.
Vários critérios têm sido utilizados para definir o início e término das
coletas. O estabelecimento do mesmo horário para iniciar e terminar as coletas
baseia-se no fato de que parte das excretas que estavam no trato digestivo, no
início, são compensadas pelas perdas no final da coleta. Outra maneira é o uso
de marcador, por exemplo 1% de óxido férrico nas rações no primeiro e no último
dia de coleta para marcar o início e o término do período de coleta.
O método de coleta total de excreta em aves tem sido criticado em
virtude dos efeitos das bactérias do trato gastrointestinal final na excreção dos
aminoácidos (SAKOMURA & ROSTAGNO, 2007). De acordo com PARSONS et
al. (1982) cerca de 25% dos aminoácidos eliminados nas excretas das aves é de
origem microbiana. Com intuído de reduzir os efeitos dessas bactérias na parte
distal do intestino das aves vários procedimentos têm sido utilizados como, por
exemplo, o método da coleta ileal.
5 Tendências para análises dos alimentos
5.1 Novas tendências na avaliação de fósforo digestível em aves
As exigências de minerais, particularmente cálcio e fósforo são de
fundamental importância para o crescimento e desenvolvimento dos animais.
Quando as exigências nutricionais não são atendidas se observa redução no
desempenho zootécnico, piora na eficiência de utilização de alimentos,
mineralização óssea inadequada, discondroplasia tibial e raquitismo (NRC, 1994).
Têm-se verificado que as recomendações práticas de fósforo para máximo
desempenho são inferiores àquelas necessárias para máxima mineralização
óssea (CRENSHAW et al., 2001).

29
Os valores de exigência de fósforo para os animais podem ser
encontrados em diferentes tabelas (NRC, 1994; ROSTAGNO et al., 2011). Estas
referências, no entanto, consideram valores de fósforo total ou de
biodisponibilidade do fósforo nos alimentos, sendo baseados em trabalhos de
pesquisa realizados em sua maioria na década de 90, os quais não representam
necessariamente as linhagens genéticas, as condições tecnológicas e o nível de
conhecimentos atuais, o que levanta questionamento sobre a acurácia destas
informações (KNOWLTON et al., 2004).
Com o constante melhoramento e seleção genética dos animais e dos
alimentos, além da necessidade de se reduzir custos com suplementação de
fósforo e também a excreção deste mineral no ambiente, existe a necessidade de
reavaliação contínua da eficiência de utilização dos nutrientes e das exigências
nutricionais para obtenção do máximo desempenho nos sistemas de produção
intensiva (GOMES et al., 2004; ROSTAGNO et al., 2005)
Considerando que os coeficientes de digestibilidade verdadeira do
fósforo nos alimentos podem ser utilizados como parâmetros para determinar a
utilização do fósforo pelas aves (DILGER & ADEOLA, 2006; PETEY et al., 2006),
estes valores poderiam ser utilizados na determinação das exigências deste
mineral e assim manter atualizadas as tabelas de exigências de nutrientes nas
diferentes fases de produção.
A determinação dos valores de fósforo endógeno excretado pelas aves
é importante para tornar mais precisos os valores de digestibilidade do fósforo dos
alimentos. Diferentes técnicas para avaliar a excreção endógena de fósforo têm
sido empregadas, utilizando análises de regressão e dietas purificadas ou semi-
purificadas, com grande variação nos resultados (SAKOMURA & ROSTAGNO,
2007).
JONGBLOED et al. (1992) citaram que a digestibilidade é uma
metodologia prática e rápida para determinar a digestibilidade do fósforo nos
alimentos, através da condução de ensaios em gaiolas metabólicas, uma vez que,
para avaliar a disponibilidade deste mineral utilizam-se ensaios de crescimento
que demandam maior tempo e número de animais, além do sacrifício destes para
avaliar a deposição do mineral nos tecidos (ossos).

30
Na Tabela 7 são mostrados os coeficientes de digestibilidade do
fósforo, de alguns alimentos, determinados com aves.
TABELA 7- Coeficientes de digestibilidade do fósforo em alimentos para aves
(matéria natural, %).
Alimentos Conteúdo de P total
Coeficiente de digestibilidade
aparente – Aves
Milho 0,24 30
Farelo de Soja 0,59 42
Farelo de Trigo 0,94 37
Fosfato Bicálcico 18,50 90
Fonte: adaptado de Rostagno et al., 2007
5.2 Novas tendências na avaliação do nível energético
Tradicionalmente utiliza-se os princípios da energia metabolizável
aparente (EMA) energia metabolizável aparente corrigida pelo balanço de
nitrogênio (EMAn), energia metabolizável verdadeira (EMV) e energia
metabolizável verdadeira corrigida pelo balanço de nitrogênio (EMVn) na
avaliação dos valores energéticos dos alimentos. Porém novos conceitos
começam a ser formulados para aprimorar ou facilitar a determinação destes
valores. Um bom exemplo é a teoria da energia líquida (EL), que tem como
principal objetivo melhorar ou aprimorar a forma desta determinação energética.
A energia líquida é definida como a energia metabolizável (EM) menos
a perda de energia causada pelo incremento calórico (IC), que é o calor produzido
durante ingestão, digestão, metabolismo e excreção dos alimentos e nutrientes. A
energia que resulta destes processos é a energia verdadeiramente disponível
para mantença (ELm - energia líquida para mantença) e produção (ELp - energia
líquida para produção: crescimento e produção de ovos).
A principal diferença entre os sistemas de energia digestível (ED) ou
energia metabolizável (EM) e energia líquida (EL) é que os dois primeiros
expressam o potencial energético, enquanto o último expressa a energia útil e

31
inclui a eficiência com a qual cada nutriente pode ser utilizado (VASCONCELOS,
2009).
A relação entre a EL/EM é a eficiência de utilização da EM para
produzir EL. Os sistemas de energia líquida comumente utilizados baseiam-se
nos conteúdos de nutrientes digestíveis dos alimentos e dietas. Esses conteúdos
são utilizados para predizer o total de EL nas dietas completas através de
equações de regressão derivadas de resultados de experimentos de
digestibilidade e de respirometria (VASCONCELOS, 2009).
As dietas utilizadas nesses experimentos são formuladas de maneira
que se tenham grandes variações nas concentrações dos nutrientes; variações
maiores que aquelas que existem nas dietas comumente utilizadas. Isso é uma
forma de se certificar que as concentrações de nutrientes presentes nas dietas
práticas se encaixam nas concentrações das dietas teste, evitando problemas
advindos da extrapolação de concentrações maiores ou menores que aquelas
presentes nas dietas experimentais (MOENH et al. 2005).
A utilização do conceito de energia líquida consiste em uma nova
abordagem na formulação das dietas para aves, permitindo que os nutricionistas
descontem as perdas nas excretas sólidas, líquidas e gasosas, ou na forma de
calor. Apesar das vantagens deste sistema de energia ser amplamente
conhecidas na espécie suína, sobretudo nos países europeus, poucos trabalhos
foram conduzidos com aves.
De acordo com MOEHN et al. (2005), a energia líquida expressa a
energia que as aves realmente utilizam e determina a eficiência com que os
nutrientes são empregados. Por exemplo, as proteínas são substâncias que são
submetidas à síntese e ao catabolismo. Durante estes processos, uma
determinada quantidade de aminoácidos é utilizada e outra é perdida.
Para que estas transformações ocorram existe um consumo de energia.
Estes processos fazem com que a eficiência real de utilização da proteína para
deposição de tecido seja de apenas 54%. Já o amido e os lipídios são utilizados
com 74% e 90% de eficiência, respectivamente. As fibras, quando digeridas no
intestino grosso, são transformadas em ácidos graxos voláteis e eles têm um
índice de eficiência de transformação em tecido adiposo de 50%. Este valor é
inferior àqueles dos açúcares digeridos e absorvidos no intestino delgado.

32
Para determinação dos valores de energia líquida de dietas e
alimentos, a correta determinação da energia de mantença é extremamente
importante. A energia metabolizável para mantença (EMm) de frangos de corte foi
determinado em 141kcal/kg0,75 por SAKOMURA et al. (2004b) por meio da técnica
de abate comparativo com aves recebendo dieta de 3200kcal/kg. Porém, LONGO
et al. (2006), também através de abate comparativo encontraram o valores de
116,2Kcal/kg de EMm para aves criadas em temperatura de 23°C. Esses autores
encontraram efeito quadrático da temperatura sobre a energia e mantença. Já
NIETO et al. (1995) relatam valores que variam de 111 a 143Kcal/kg0,75 como
exigência de mantença de frangos de corte.
Assim, no sistema de energia líquida, o custo da energia
(reais/tonelada, dividido pela energia) é diferenciado, de modo que o custo/kcal
aumenta para alimentos ricos em proteínas, e é menor para aqueles ricos em
gordura, quando comparado com os sistemas que utilizam energia digestível ou
energia metabolizável.
Os menores valores de energia atribuídos à proteína e à fibra e os
maiores valores atribuídos à gordura e ao amido afetam a formulação das dietas,
levando à limitação da utilização de ingredientes caros, ricos em proteína. Neste
caso, menos proteína é empregada nas fórmulas, reduzindo seus custos e
diminuindo a excreção de nitrogênio através dos dejetos.
NOBLET et al. (2001) avaliaram a produção de calor em suínos
alimentados com dietas com diferentes níveis de PB e gordura. Verificou-se
redução na produção de calor nas dietas de menor conteúdo protéico ou quando
gordura era adicionada. Isso indica que a eficiência de utilização da EM para
energia retida depende da composição da dieta. Os autores concluíram que a
utilização da energia é melhorada quando os níveis de PB são reduzidos e
adicionado gordura à dieta; indicando a superioridade do sistema de EL em
relação à EM e ED.
Poucos são os trabalhos envolvendo este tema na literatura, visto a
difícil mensuração deste incremento calórico. Porém a sua determinação, desde
que correta, poderia trazer benefícios à nutrição como a utilização da quantidade
estritamente necessária requerida pela ave para a manutenção e crescimento
corporal.

33
6 CONSIDERAÇÕES FINAIS
Conhecer a composição química do alimento antes de formular a ração
é imprescindível para obtenção de resultados esperados. A utilização dos
diversos métodos de avaliação de alimentos é uma ferramenta de grande
aplicabilidade para estimativas da composição e disponibilidade das diferentes
frações dos alimentos, porém a correta interpretação dos resultados obtidos com
estas avaliações é determinante e exige conhecimento das diferentes
metodologias.
Os diferentes métodos proporcionam diferenças nas avaliações de
energia e aminoácidos para aves. Portanto, as variações devem ser consideradas
para uma melhor definição da metodologia a ser usada em função dos objetivos
da pesquisa. A produção constante de dados mais precisos de composição
nutricional dos alimentos é de fundamental importância para elaborar rações que
proporcionem melhores custos e produtividade.
Visto isso fica claro que o passo determinante na produção avícola a
médio e longo prazo é o aperfeiçoamento nas análises da composição nutricional
dos alimentos, bem como das exigências dos animais, sendo este um grandioso
desafio para os próximos anos.

34
7 REFERÊNCIAS 1. BATAL, A. B.; PARSONS, C. M. Effects of age on nutrient digestibility in
chicks fed different diets. Poultry Science, v.81, p.400-407, 2002.
2. BEDFORD, M. R.; CLASSEN, H. L. An in vitro assay for prediction of broiler
intestinal viscosity and growth when fed rye-based diets in the presence of
exogenous enzymes. Poultry Science, v.72, n.1, p.137-143, 1992.
3. BERCHIELLI, T. T.; SADER, A. P. O.; TONANI, F. L.; PAZIANI, S. F.;
ANDRADE, P. Avaliação da determinação da fibra em detergente neutro e da
fibra em detergente ácido pelo sistema ANKOM. Revista Brasileira de
Zootecnia, v.30, n.5, p.1572-1578, 2001.
4. BERTECHINI, A. G. Nutrição de Monogástricos. Lavras: Editora UFLA,
2006. 301 p.
5. BRANCO, A. F. Confinamento de bovinos, 1.ed., Santo Anastácio: Potensal
nutrição e saúde animal, 2006. 122 p. [Apostila]
6. BRITO, C. O. Avaliação de dietas formuladas com aminoácidos totais e
digestíveis e estimativas do crescimento e da deposição de nutrientes em
frangos de corte. 2007. 155 f. Tese (Doutorado em Zootecnia) – Departamento
de Zootecnia, Universidade Federal de Viçosa, Viçosa.
7. BRUMANO, G.; GOMES, P. C.; ALBINO, L. F. T.; ROSTAGNO, H. S.;
GENEROSO, R. A. R.; SCHMIDT, M. Composição química e valores de energia
metabolizável de alimentos protéicos determinados com frangos de corte em
diferentes idades. Revista Brasileira de Zootecnia, v.35, n.6, p.2297-2302,
2006.
8. CAFÉ, M. B. Estudo do valor nutricional da soja integral processada para
aves. 1993. 97 f. Dissertação (Mestrado em Zootecnia) – Faculdade de Ciências
Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal.
9. CAFÉ, M. B.; SAKOMURA, N. K.; JUNQUEIRA, O. M.; CARVALHO M. R. B.;
DEL BIANCHI, M. Determinação do valor nutricional das sojas integrais
processadas para aves. Revista Brasileira de Ciência Avícola, v.2, n.1, p. 67-
74, 2000.
10. CALDERANO, A. A. Valores de composição química e de energia de
alimentos de origem vegetal determinados com aves de diferentes idades.

35
2008. 50 f. Dissertação (Mestrado em Zootecnia) – Departamento de Zootecnia,
Universidade Federal de Viçosa, Viçosa.
11. CARVALHO, D. C. O.; ALBINO, L. F. T.; ROSTAGNO, H. S.; OLIVEIRA, J. E.;
JÚNIOR, J. G. V.; TOLEDO, R. S.; COSTA, C. H. R.; PINHEIRO, S. R. F.;
SOUZA, R. M. Composição química e energética de amostras de milho
submetidas a diferentes temperaturas de secagem e períodos de
armazenamento. Revista Brasileira de Zootecnia, v.33, n.2, p.358-364, 2004.
12. CONTE, A. J.; TEIXEIRA, A. S.; BERTECHINI, A. G.; FIALHO, E. T.; MUNIZ,
J. A. Efeito da fitase e xilanase sobre a energia metabolizável do farelo de arroz
integral em frangos de corte. Ciência e Agrotecnologia, Lavras. v.26, n.6,
p.1289-1296, 2002.
13. CRENSHAW, T. D. Calcium, phosphorus, vitamin D, and vitamin K in swine
nutrition. In: Lewis, A. J.; Southern, L. L. Swine Nutrition. 2.ed., Washington:
CRC Press, 2001. p.187-212.
14. DILGER, R. N.; ADEOLA, O. Estimation of true phosphorus digestibility and
endogenous phosphorus loss in growing chicks fed conventional and low-phytate
soybean meals. Poultry Science, v.85, p.661-668. 2006.
15. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro Nacional
de Pesquisa de Suínos e Aves (CNPSA) – EMBRAPA-CNPSA. Tabelas de
composição química e valores energéticos de alimentos para suínos e aves.
3.ed. Concórdia -SC, EMBRAPA-CNPSA, 1991, 97p. (Documento 19).
16. FREITAS, E. R.; SAKOMURA, N. K.; EZEQUIEL, J. M. B.; NEME, R.;
MENDONÇA, M. O. Energia metabolizável de alimentos na formulação de ração
para frangos de corte. Pesquisa Agropecuaria Brasileira, v.41, n.1, p.107-115,
2006.
17. GOERING, H. K.; VAN SOEST, P. J. Forage Fiber Analyses (Apparatus,
reagents, procedures, and some appli-cations). USDA - ARS Agric. Handbook.
Washington: US Govt. Printying Office, 1970. 20 p.
18. GOMES, P. C.; RUNHO, R. C.; D’AGOSTINI, P.; ROSTAGNO, H. S.;
ALBINO, L. F. T.; LOPES, P. S. Exigência de fósforo disponível para frangos de
corte machos e fêmeas de 22 a 42 e de 43 a 53 dias de idade. Revista Brasileira
de Zootecnia, v.33, p.1734-1746, 2004 (Supl. 1).

36
19. HETLAND, H.; CHOCT, M.; SVIHUS, B. Role of insoluble non-starch
polysaccharides in poultry nutrition. World’s Poultry Science Journal, v.60,
p.415-422, 2004.
20. HILL, F. W.; ANDERSON, D. L. Comparison of metabolizable energy and
productive energy determinations whit growing chicks. Journal of Nutrition, v.64,
n.3, p.587-604, 1958.
21. JERACI, J. L.; VAN SOEST, P. J. Improved methods for analysis and
biological characterization of fiber. Advances in Experimental Medicine and
Biology, v.270, p.245-263, 1990.
22. JONGBLOED, A. W.; MROZ, Z.; KEMME, P. A. The effect of suplementary
Aspergilus niger phytase in diets for pigs on concentration and apparent
digestibility of dry matter, total phosphorus, and phytic acid in different sections of
the alimentary tract. Journal of Animal Science, v.70, p.1159-1168, 1992.
23. KNOWLTON, K. F.; RADCLIFFE, J. S.; NOVAK, C. L.; EMMERSON, D. A.
Animal management to reduce phosphorus losses to the environment. Journal of
Animal Science, v.82, p.E173-E195. 2004 (Supplement).
24. KOBT, A. R.; LUCKEY, T. D. Markers in nutrition. Nutrition abstracts and
reviews, v.42, p.813-845, 1972.
25. LARBIER, M.; LECLERQ, B. Nutrition and feeding of poultry.
Loughborough: Nottingham University Press, 1994. 305p.
26. LESSON, S.; SUMMERS, J. D. Nutrition of the chicken. 4.ed. Ontario:
Guelph, 2001. 591 p.
27. LIMA, M. R.; SILVA, J. H. V.; ARAÚJO, J. A.; LIMA, C. B.; OLIVEIRA, E. R. A.
Enzimas exógenas na alimentação de aves. Acta Veterinaria Brasilica, v.1, n.4,
p.99-110, 2007.
28. LONGO, F. A.; SAKOMURA, N. K.; RABELLO, C. B.; FIGUEIREDO, A. N.;
FERNANDES, J. B. K. Exigências energéticas para mantença e para o
crescimento de frangos de corte. Revista Brasileira de Zootecnia, v.35, n.1,
p.119-125, 2006.
29. LOPEZ, G.; LEESON, S. Relevance of nitrogen correction for assessment of
metabolizable energy with broilers to forty-nine days of age. Poultry Science,
v.86, p.1696-1704, 2007.

37
30. MERTENS, D. R. Using fiber and carbohydrate analyses to formulate dairy
rations. In: Informational Conference with Dairy and Forages Industries, 1996.
Proceedings..., US Dairy Forage Resource Center, 1996. 81-91p.
31. MOEHN, S.; ATAKORA, J.; BALL, R. O. Using net energy for diet formulation:
Potencial for the canadian pig industry. Advances in Pork Production, v.16. p.
119-129, 2005.
32. NATIONAL RESEARCH COUNCIL – NRC. Nutritional energetics of
domestic animals and glossary of energy terms. 2.ed. Washington: National
Academy Press, 1981. 54 p.
33. National Research Council. Nutrient Requirements of Poultry. 9ed.
Washington: National Academy Press, 1994. 155 p.
34. NERY, L. R. Valores de energia metabolizável e de aminoácidos
digestíveis de alguns alimentos para aves. 2005. 100 f. Dissertação (Mestrado
em Zootecnia) – Departamento de Zootecnia, Universidade Federal de Viçosa,
Viçosa.
35. NIETO, R.; PRIETO, C.; FERNÁNDEZ-FÍGARES, I.; AGUILERA, J. F. Effect
of dietary protein quality on energy metabolism in growing chicken. British
Journal of Nutrition, v.74. p.163-172, 1995.
36. NOBLET, J.; LE BELLEGO, L.; VAN MILGEN, J.; DUBOIS, S. Effects of
reduced dietary protein level and fat addition on heat production and nitrogen and
energy balance in growing pigs. Animal Research, v.50. p.227-238, 2001.
37. NUNES, R. V. Digestibilidade de nutrientes e valores energéticos de
alguns alimentos para aves. 2003. 113 f. Tese (Doutorado em Zootecnia) –
Departamento de Zootecnia Universidade Federal de Viçosa, Viçosa.
38. PAPADOPOULOS, M. C.; EL BOUSHY, A. R.; ROODBEEN, A. E.;
KETELAARS, E. H. Effects of processing time and moisture content on amino acid
composition and nitrogen characteristics of feather meal. Animal Feed Science
and Technology, v.14, n.3-4, p.279-290, 1986.
39. PARSONS, C. M. Digestible amino acids for poultry and swine. Animal Feed
Science and Technology, v.59, n.1-3, p.147-153, 1996.
40. PARSONS, C. M.; POTTER, L. M.; BROWN, J. R. Effects of dietary protein
and intestinal microflora and excretion of amino acids in poultry. Poultry Science,
v.61, p.939-946, 1982.

38
41. PEIXOTO, R. R.; MAIER, J. C. Nutrição e alimentação animal. 2.ed.
Pelotas: UCPel, 1993. 169 p.
42. PENZ JR., A. M.; KESSLER, A. M.; BRUGALLI, I. Novos conceitos de energia
para aves. In: SIMPÓSIO INTERNACIONAL SOBRE NUTRIÇÃO DE AVES,
Campinas. Anais ... Campinas: FACTA, 1999, p.1-24.
43. PERTTILA, S.; VALAJA, J.; PARTANEN, K.; JALAVA, T.; VENALAINEN, E.
Apparent ileal digestibility of amino acids in protein feedstuffs and diet formulation
based on total vs digestible lysine for poultry. Animal Feed Science and
Technology, v.98, n.3-4, p.203-218, 2002.
44. PETTEY, L. A.; CROMWELL, G. L.; LINDEMANN, M. D. Estimation of
endogenous phosphorus loss in growing and finishing pigs fed semi-purified diets.
Journal of Animal Science, v. 84, p. 618-626, 2006.
45. POTTER, L. M.; MATTERSON, L. D. Metabolizable energy of feed ingredients
for the growing chick. Poultry Science, v.39, p.781-782,1960.
46. PUPA, J. M. R.; LEAO, M. I.; CARVALHO, A. U.; POMPERMAYER, L. G.;
ROSTAGNO, H. S. Cecectomia em galos sob anestesia local e incisão abdominal.
Arquivos Brasileiros de Medicina Veterinaria e Zootecnia, v.50, n.5, p.531-
535, 1998.
47. ROSTAGNO, H. S.; ALBINO, L. F. T.; DONZELE J. L.; GOMES, P. C.;
OLIVEIRA, R. F.; LOPES, D. C.; FERREIRA, A. S.; BARRETO, L. S. T.;
EUCLIDES, R. F. Tabelas brasileiras para aves e suínos: composição de
alimentos e exigências nutricionais. 3.ed. Viçosa: Universidade Federal de
Viçosa, 2011. 252p.
48. ROSTAGNO, H. S.; ALBINO, L. F. T.; DONZELE, J. L.; GOMES, P. C.;
OLIVEIRA, R. F.; LOPES, D. C.; FERREIRA, A. S.; BARRETO, S. L. T. Tabelas
Brasileiras para Aves e Suínos. Composição de Alimentos e Exigências
Nutricionais. 2.ed. Viçosa: Universidade Federal de Viçosa, 2005. 186. p.
49. ROSTAGNO, H. S.; BÜNZEN, S.; SAKOMURA, N. K.; ALBINO, L. F. T.
Avanços metodológicos na avaliação de alimentos e de exigências nutricionais
para aves e suínos Revista Brasileira de Zootecnia, v.36, suplemento especial,
p.295-304, 2007.

39
50. ROSTAGNO, H. S.; ROGLER, J. C.; FEATHERSTON, W. R. Studies on the
nutritional values of sorghum grains with varying tannin content for chickens. 2.
Amino acids digestibility studies. Poultry Science, v.52, p.772-776, 1973.
51. ROSTAGNO, H. S.; SILVA, D. J.; COSTA, P. M. A. Composição de
alimentos e exigências nutricionais de aves e suínos (Tabelas brasileiras).
Viçosa: UFV. 1983. 59p.
52. SAKOMURA, N. K.; BIANCHI, M. D.; PIZAURO JR., J. M.; CAFÉ, M. B.;
FREITAS, E. R. Efeito da idade dos frangos de corte na atividade enzimática e
digestibilidade dos nutrientes do farelo de soja e soja integral. Revista Brasileira
de Zootecnia, v.33, n.4, p.924-935, 2004a.
53. SAKOMURA, N. K.; LONGO, F. A.; RABELLO, C. B. V.; WATANABE, K.;
PELÍCIA, K.; FREITAS, E. R. Efeito do nível de energia metabolizável da ração no
desempenho e metabolismo energético de frangos de corte. Revista Brasileira
de Zootecnia, v.33, n.6, p.1758-1767, 2004b.
54. SAKOMURA, N. K.; ROSTAGNO, H. S. Métodos de pesquisa em nutrição
de monogástricos. Jaboticabal: Funep, 2007. 283 p.
55. SANTOS, Z. A. S.; FREITAS, R. T. F.; FIALHO, E. T.; RODRIGUES, P. B.;
LIMA, J. A. F.; CARELLOS, D. C.; BRANCO, P. A. C.; CANTARELLI, V. S. Valor
nutricional de alimentos para suínos determinados na Universidade Federal de
Lavras. Ciência e Agrotecnologia, Lavras, v.29, n.1, p.232-237, 2005.
56. SIBBALD, I. R. A bioassay for true metabolizable energy in feedingstuffs.
Poultry Science, v.55, p.303-308, 1976.
57. SIBBALD, I. R. Measurement of bioavailable energy in poultry feeding stuffs: A
review. Canadian Journal of Animal Science, v. 62, p.983-1048, 1987.
58. SIBBALD, I. R.; SLINGER, S. J. A biological assay for metabolizable energy in
poultry feed ingredients together with findings which demonstrate some of the
problems associated with evaluation of fats. Poultry Science, v.42, n.1, p.13-25,
1963.
59. SIBBALD, I. R.; WOLYNETZ, I. R. Relationships between apparent and true
metabolizable energy and the effects of a nitrogen correction. Poultry Science, v.
63, p. 1386-1399, 1984.
60. SILVA, D. J.; QUEIROZ, A. C. Análise de alimentos (métodos químicos e
biológicos). 3.ed. Viçosa: Universidade Federal de Viçosa, 2009. 235 p.

40
61. TEIXEIRA, A. S. Alimentos e alimentação dos animais. Vol. I. 5.ed. Lavras:
UFLA/Faepe, 2003. 241 p.
62. THORPE, J.; BEAL, J. D. Vegetable protein meals and the effects of enzymes.
In: BEDFORD, M. R.; PARTRIDGE, G. G. Enzymes in farm nutrition. London:
Cab International, 2001. p.125-143.
63. VAN SOEST, P. J. Nutritional Ecology of the Ruminant, 2.ed. Ithaca:
Cornell University Press, 1994. 476 p.
64. VAN SOEST, P. J.; ROBERTSON, J. B.; LEWIS, B. A. Methods for dietary
fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal
nutrition. Journal of Dairy Science, v.74, p.3583-3597, 1991.
65. VAN SOEST, P. J.; WINE, R. H. Use of detergents in the analysis of fibrous
feeds. IV. Determination of plant cell-wall constituents. Journal of the
Association of Official Analytical Chemists, v.50, p.50-5, 1967.
66. VASCONCELOS, C. H. F. REDUÇÃO DO TEOR DE PROTEÍNA BRUTA EM
DIETAS SUPLEMENTADAS OU NÃO COM L-GLICINA PARA FRANGOS DE
CORTE. 2009. 134 f. Tese (Doutorado em Zootecnia) – Escola de Veterinária,
Universidade Federal de Minas Gerais, Belo Horizonte.
67. WARPECHOWSKI, M. B. Efeito da fibra insolúvel da dieta sobre a
passagem no trato gastrintestinal de aves intactas, cecectomizadas e
fistuladas no íleo terminal. 1996. 125 f. Dissertação (Mestrado em Zootecnia) –
Faculdade de Agronomia, Universidade Federal do Rio Grande do Sul, Porto
Alegre.