instituto politcnico nacional rivera.pdfalgunos protistas y la mayoría de las bacterias. la pared...
TRANSCRIPT



Instituto Politécnico Nacional CENTRO INTERDISCIPLINARIO DE INVESTIGACIÓN
PARA EL DESARROLLO INTEGRAL REGIONAL
DEPARTAMENTO AGROPECUARIO
Detección y Caracterización Molecular de Xanth das
T E S I S
PARA OBTENER EL GRADO DE:
MAESTRIA EN RECURSOS NATURALES Y MEDIO AMBIENTE
PRESENTA: FATIMA DEL ROSARIO RIVERA SOTO
Guasave, Sinaloa, México
UNIDAD SINALOA
omonas campestris pv. vesicatoria Asociacon Mancha Bacteriana de Cultivos de Tomate en
Sinaloa
2007

El presente trabajo se realizó en el Departamento Agropecuario en el Laboratorio de Biología Molecular del Centro Interdisciplinario de Investigación para el desarrollo Integral Regional CIIDIR-IPN Unidad Sinaloa del Instituto Politécnico Nacional (IPN) bajo la dirección de la Dra. Norma Elena Leyva López. El autor agradece al Instituto Politécnico Nacional por el apoyo en Infraestructura y económico brindado a través de su sistema de becas, así como al CIIDIR-IPN Unidad Sinaloa por el apoyo durante la realización de este proyecto.

i
CONTENIDO
Número de página CONTENIDO------------------------------------------------------------------------ i
ÍNDICE DE FIGURAS-------------------------------------------------------------- v ÍNDICE DE CUADROS----------------------------------------------------------- vii
GLOSARIO-------------------------------------------------------------------------- viii
ABREVIATURAS------------------------------------------------------------------ xii
RESUMEN--------------------------------------------------------------------------- xiv
ABSTRACT------------------------------------------------------------------------- xv
I. INTRODUCCIÓN----------------------------------------------------------------- 1
II. REVISION DE LITERATURA------------------------------------------------ 3
A. Bacterias-------------------------------------------------------------- 3
1. Morfología y estructura bacteriana --------------------- 3
2. Bacterias fitopatógenas----------------------------------- 5
3. Bacterias que causan enfermedad en el tomate---- 5
4. Xanthomonas campestris pv. vesicatoria
4.1. Origen ----------------------------------------------- 7
4.2. Clasificación actual de Xanthomonas
campestris pv. vesicatoria ------------------- 8
4.3. Morfología específica de Xanthomonas
campestris pv. vesicatoria.--------------------- 9
4.4. Sintomatología de la mancha bacteriana -- 9
4.5. Estructura y organización del genoma ------ 11
4.6. Importancia del gen hrp ------------------------- 12
4.7. Importancia del cultivo de tomate en Sinaloa-- 13
4.8. Importancia de la mancha bacteriana en
tomate ------------------------------------------------ 14
4.9. Ciclo de la enfermedad ------------------------- 15
4.10. Control de la enfermedad --------------------- 15
B. Caracterización fisiológica de Xanthomonas campestris
pv. vesicatoria -------------------------------------------------------- 16 1. Caracterización a nivel género---------------------------- 16
1.1. Fluorescencia ------------------------------------- 17

ii
1.2. Tinción Gram ------------------------------------ 18
1.3. Catalasa ------------------------------------------- 20
1.4. Oxidasa-------------------------------------------- 20
1.5. Crecimiento en YDC---------------------------- 21
2. Caracterización a nivel especie ------------------------ 21
2.1. Crecimiento en YS ----------------------------- 22
2.2. Hidrólisis de almidón -------------------------- 22
2.3. Licuefacción de gelatina ---------------------- 23
2.4. Hidrolasa de arginina -------------------------- 23
2.5. Fuentes de carbono --------------------------- 24
3. Caracterización a nivel patovar------------------------ 24
3.1. Crecimiento en Agar Tween y SX----------- 24
4. Prueba de patogenicidad -------------------------------- 25
C. Caracterización genotípica de Xanthomonas spp.--------- 26
1. Detección de Xanthomonas campestris pv.
vesicatoria por la PCR---------------------------------- 29
III. HIPÓTESIS ----------------------------------------------------------------- 29
IV. OBJETIVOS ---------------------------------------------------------------- 29
A. Objetivo general ---------------------------------------------------- 29
B. Objetivos específicos -------------------------------------------- 29
V. MATERIALES Y MÉTODOS ------------------------------------------- 30
A. Material vegetal --------------------------------------------------- 30
B. Metodología -------------------------------------------------------- 31
1. Pruebas simples para la identificación de
Xanthomonas -------------------------------------------- 31
1.1. Aislamiento y condiciones de
crecimiento para Xanthomonas------------ 31
1.2. Tinción Gram ---------------------------------- 31
1.3. Prueba de fluorescencia ------------------- 32
C. Caracterización molecular-------------------------------------- 32
1. Extracción de DNA de cepa bacteriana- 32
2. Electroforesis ----------------------------------- 33
3. Determinación de la concentración de DNA 34
4. Detección del patógeno por PCR -------------------- 34

iii
5. Purificación de bandas --------------------------------- 35
6. Ligación de productos de PCR ---------------------- 36
7. Transformación de células competentes 36
8. Extracción de DNA plasmídico ----------------------- 37
9. Análisis de restricción de los productos amplificados
por PCR ------------------------------------------------------- 37
10. Secuenciación, análisis y comparación
de secuencias-------------------------------------------- 38
D. Caracterización fisiológica ------------------------------------- 39
1. Caracterización fisiológica del género
Xanthomonas -------------------------------------------- 39
1.1. Catalasa --------------------------------------- 39
1.2. Oxidasa ---------------------------------------- 40
1.3. Crecimiento en YDC ------------------------ 40
2. Caracterización a nivel especie---------------------- 40
2.1. Crecimiento en YS -------------------------- 40
2.2. Hidrólisis de almidón ----------------------- 41
2.3. Licuefacción de gelatina ------------------- 41
2.4. Dihidrolasa de arginina --------------------- 42
2.5. Fuentes de carbono ------------------------- 42
3. Caracterización a nivel patovar ---------------------- 43
3.1. Crecimiento en medio Tween ---- 43
3.2. Crecimiento en medio SX -------- 43
4. Pruebas de patogenicidad -------------------- 43
VI. RESULTADOS Y DISCUSIÓN --------------------------------------- 45
A. Sintomatología --------------------------------------------------- 45
B. Pruebas simples de identificación de Xanthomonas
spp. ------------------------------------------------------------------ 46
1. Tinción Gram ---------------------------------------------- 47
2. Fluorescencia --------------------------------------------- 48
C. Caracterización molecular ------------------------------------- 48
1. Extracción de DNA ------------------------------ 48
2. Detección del patógeno por PCR ----------- 49
3. Identificación de Xanthomonas spp. por

iv
RFLP-PCR ---------------------------------------- 51
5. Análisis de secuencias ------------------------ 52
6. Análisis de restricciones ----------------------- 56
7. Análisis filogenético ---------------------------- 59
D. Caracterización fisiológica ------------------------------------- 61
1. Caracterización a nivel genero ----------------------- 61
1.1. Catalasa ---------------------------------------- 61
1.2. Oxidasa ----------------------------------------- 61
1.3. Crecimiento en YDC ------------------------- 61
2. Caracterización a nivel especie ---------------------- 62
2.1. Crecimiento en YS --------------------------- 62
2.2. Hidrólisis de almidón ------------------------ 63
2.3. Licuefacción de gelatina -------------------- 64
2.4. Hidrolasa de arginina ------------------------ 65
2.5. Fuentes de carbono ------------------------- 65
3. Caracterización a nivel patovar--------------------- 67
3.1. Crecimiento en medio Tween y SX ------ 67
D. Pruebas de patogenicidad ------------------------------------- 69
VII. CONCLUSIONES ------------------------------------------------------- 70
IX. BIBLIOGRAFÍA ----------------------------------------------------------- 71

v
ÍNDICE DE FIGURAS 1. Estructura bacteriana mostrando los principales organelos -- 4 2. Algunos síntomas causados por bacterias en el cultivo del tomate ----------------------------------------------------------------------- 7 3. Micrográfica de Xanthomonas campestris ------------------------ 9 4. Síntomas causados por Xanthomonas campestris pv. vesicatoria en cultivos de tomate------------------------------------- 10 5. Estructura de una bacteria Gram positiva------------------------ 18 6. Estructura de una bacteria Gram negativa------------------------ 19 7. Localización de los municipios de Sinaloa donde se realizó la colecta de muestras de tomate con síntomas de mancha bacteriana ------------------------------------------------------------------- 30 8. Restricción de fragmentos de 420 pb del gen hrp de
Xanthomonas spp. con la enzima HaeIII con los primer RST65/69----------------------------------------------------------------- 39
9. Síntomas de mancha bacteriana observados en cultivos de
tomate en Sinaloa ------------------------------------------------------ 45 10. Síntomas observados en cultivos de tomate--------------------- 46 11. Aislados bacteriológicos obtenidos de tejido foliar
sintomático de planas de tomate con mancha bacteriana --- 47 12. Fotografía con microscopio de la bacteria Gram negativa y
forma bacilar de aislados bacteriológicos de plantas de tomate --------------------------------------------------------------------- 47
13. Prueba de fluorescencia en medio B de King, con
exposición a luz ultravioleta (260 nm) ---------------------------- 48 14. DNA de aislados bacteriológicos de tejido de tomate--------- 49 15. Productos amplificados por PCR del gen hrp de
Xanthomonas spp. con los primers RST65/RST69------------ 50 16. Análisis de restricción de los fragmentos amplificados por PCR con los primers RST65/RST69----------------------------- 51

vi
17. Análisis de los sitios de restricción de las secuencias del gen hrp de Xanthomonas campestris pv vesicatoria--------- 58
18. Árbol filogenético construido mediante el método de alineamiento (CLUSTAR Method) de secuencias del gen hrp amplificado con el par de primers RST65/RST69 de Xanthomonas campestris pv vesicatoria de la cepa 75-3 reportada en el banco de genes---------------------------------- 60 19. Colonias de consistencia mucoide, de color amarillo, desarrolla en YDC, correspondiente a la cepa del Estado de Sinaloa a las 48 hr de incubación -------------------------------------------------------- 62 20 Crecimiento de un aislado del Estado de Sinaloa de
Xanthomonas campestris pv. vesicatoria después de 48 hr de incubación en medio YS a 35°C-------------------------------- 63
21. Pruebas de hidrólisis de almidón ----------------------------------- 64 22. Prueba de licuefacción de gelatina a aislados de tomate---- 67 23. Prueba de arginina en medio Thornley´s 2A -------------------- 64 24. Producción de ácido a partir de fructuosa como fuente de carbono--------------------------------------------------------------------- 66 25. Producción de ácido a partir de glucosa como fuente de carbono -------------------------------------------------------------------- 66 26. Producción de ácido a partir de celobiosa como fuentes de carbono -------------------------------------------------------------------- 67 27. Crecimiento en medio selectivo para patovares
“vesicatoria” de tomate------------------------------------------------ 67 28. Pruebas de patogenicidad en cultivo de tomate---------------- 69

vii
ÍNDICE DE CUADROS
1 Clasificación de bacterias fitopatógenas-------------------------- 6
2 Características usadas para diferenciar géneros de bacterias
patógenas en platnas capaces de crecen en medio estándar--------------------------------------------------------------------- 17
3 Pruebas bioquímicas para la diferenciación de Xanthomonas
spp..--------------------------------------------------------------------------- 22 4 Crecimiento de algunos patovares de Xanthomons
campestris pv. vesicatoria en medios semiselctivos------------- 24 5 Pruebas rápidas de patogenicidad--------------------------------- 27 6 Secuencias de los oligonucleótidos usados en la PCR para
detectar aislados de Xanthomonas campestris pv. vesicatoria------------------------------------------------------------------- 34
7 Secuencias de Xanthomonas campestris pv vesicatoria
obtenidas de los cultivos de tomate de los diferentes municipios de Sinaloa--------------------------------------------------- 54
8 Porcentaje de similitud entre secuencias del gen hrp de
Xanthomonas campestris pv vesicatoria clasificadas en el grupo A (X. axonopodis pv. vesicatoria)---------------------------- 55
9 Características culturales, morfológicas y fisiológicas de los aislamientos de la bacteria en estudio --------------------------------- 68

viii
GLOSARIO
Ácidos nucleicos. Moléculas formadas por macropolímeros de nucleótidos, o
polinucleótidos. Esta presente con todas las células y constituye la base
material de la herencia que se transmite de una a otra generación. Existen dos
tipos, el ácido desoxirribonucleico (DNA) y el ácido ribonucleico (RNA).
Ácido Ribonucléico. Ácido nucleico formado por nucleótidos en los que el
azúcar es ribosa y las bases nitrogenadas son adenina, uracilo, citosina y
guanina. Actúa como intermediario y complemento de las instrucciones
genéticas codificadas en el DNA. Existen varios tipos de ARN, relacionados
con la síntesis de proteínas; RNA mensajero (RNAm), RNA ribosómico (RNAr)
y el RNA de transferencia (RNAt).
ADNt. Ácido desoxirribonucleico de transferencia.
Ampicilina. Antibiótico derivado de la penicilina que interfiere con la síntesis de
la pared celular, impidiendo el crecimiento bacteriano. El gen asociado con la
resistencia a la ampicilina es muy usado en ingeniería genética como marcador
de selección.
Clonación molecular: Inserción de un segmento de DNA ajeno, de una,
dentro de un vector que se replica en un huésped específico.
Clones. Grupo de células o de organismos de idéntica constitución genética
entre si y con el antepasado común del que se producen por división binaria o
por reproducción sexual.
DNA. Ácido desoxirribonucleico de doble cadena formado por nucleótidos en el
que el azúcar es desoxirribosa, y las bases nitrogenadas son adenina, timina,
citosina y guanina. El DNA codifica la información para la reproducción y
funcionamiento de las células y para la replicación de la propia molécula de
DNA.

ix
DNA Ligasa (Polinucleotido ligasa). Es una enzima que une covalentemente
a los extremos 3´ con extremos 5´ de cadenas polinucleotídicas, para formar
una cadena continua. Participa en replicación y reparación de DNA.
Electroforesis. Técnica utilizada para la separación de moléculas (Proteínas o
ácidos nucleicos) a través de un campo eléctrico de acuerdo a su tamaño
molecular y carga eléctrica.
Enfermedad. Cualquier mal funcionamiento de las células y tejido del
hospedante, que resulta de la irritación continua por un agente patogénico o
factor ambiental y que lleva al desarrollo del síntoma.
Enzimas de restricción. Endonucleasas que reconocen una secuencia entre
4-8 pb en DNAs. El sitio de reconocimiento se llama sitio de restricción, y la
enzima rompe un enlace fosfodiester en la hebra de arriba y otro enlace
fosfodiester en la hebra complementaria. Son las tijeras de la ingeniería
genética que abrieron las puertas a la manipulación genética.
Fitopatógeno. Microorganismos que producen enfermedades en las plantas.
Inóculo: Patógeno o partes de él que hacen contacto con el hospedero.
In vitro. Literalmente significa “en el vidrio”. Se refiere a las acciones o
condiciones en el tubo de ensayo del laboratorio, investigado y manipulado fuera del organismo vivo.
Kilobase (Kb). Unidad empleada para medir la longitud de los fragmentos de
DNA, constituidos por una serie de bases. 1kb = 1000 bases.
Nucleótido. Monómero de los ácidos nucleicos integrado por la combinación
de una base nitrogenada (purina o pirimidina), un azúcar (ribosa o
desoxirribosa) y un grupo fosfato. Se obtiene como producto de la hidrólisis de
ácidos nucleicos por acción de nucleasas.

x
Oligonucleótidos. Secuencia lineal de nucleótidos (hasta 20).
Organismo. Entidad biológica capaz de producirse o de transferir material
genético, incluyéndose dentro de este concepto a las entidades
microbiológicas, sean o no celulares. Casi todo organismo está formado por
células, que pueden agruparse en órganos, y estos a su vez en sistemas, cada
uno de los cuales realizan funciones específicas.
Pared celular. Capa externa y rígida de las células de las plantas superiores,
algunos protistas y la mayoría de las bacterias. La pared celular vegetal está
constituida principalmente de celulosa, aunque también se presenta
emicelulosa, pectinas y puede tener lignina.
Pares de bases. Dos bases nitrogenadas (timina y adenina o citosina y
guanina) que se unen por puentes de hidrógeno en la molécula de DNA.
Patógeno. Microorganismo que tiene la propiedad de producir enfermedad en
los seres humanos, animales o plantas.
Patovar. En las bacterias, subespecie o grupo de razas que sólo puede infectar
a plantas de un cierto género o especie.
Plásmido. Fragmento circular de DNA bicatenario que contiene unos cuantos
genes y se encuentran en el interior de ciertas bacterias. Actúan y se replican
de forma independiente al DNA bacteriano y pueden pasar de unas bacterias a
otras. Se utilizan como vectores en manipulación genética.
Primers (Iniciador). Cadena corta de polinucleótido preexistente a la cual
pueden agregársele nuevos desoxirribonucleótidos por la acción de la enzima
DNA polimerasa.
Procariote. Célula u organismos carentes de una membrana definida, núcleo
estructural discreto con un solo cromosoma así como de otros compartimentos

xi
celulares. Las bacterias son procariotes.
(PCR) Reacción en Cadena de la Polimerasa. Técnica de análisis del
genoma mediante la amplificación limitada de porciones específicas del DNA,
aunque sean minúsculas. Es un método revolucionario de amplificación
exponencial del DNA por la intervención de una enzima termoestable.
Secuencia conservada. Una secuencia de bases de una molécula de DNA (ó
una secuencia de aminoácidos en una proteína) que ha permanecido
esencialmente sin cambios a través de la evolución.
Secuenciación. Determinación del orden de nucleótidos (secuencias de
bases) en una molécula de DNA o RNA, o el orden de aminoácidos en una
proteína.
Sustrato. Molécula que sufre transformaciones durante una reacción
catalizada por una enzima.
Tejido. Grupo de células similares organizadas en una unidad estructural y
funcional.
Transformación bacteriana. Uno de los procesos naturales de transferencia
de material genético de una bacteria a otra. Experimentalmente consiste en
introducir un fragmento de DNA en una bacteria para provocar en ella una
recombinación genética.
Transformación. Modificación permanente y heredable de una célula como
resultado de la incorporación de un DNA foráneo.
Vector. Portador que transfiere un gen de un huésped a otro. Sistema que
permite la transferencia, la expresión y la replicación de un DNA extraño en
células huésped para una posterior clonación o transgénesis. Se trata de una
molécula de DNA (plasmido bacteriano, microsoma artificial de levadura o de
bacteria) o de un virus defectuoso.

xii
ABREVIATURAS
A = Adenina
AN= Agar Nutritivo µg = Microgramo µl= Microlitro
µm = Micrometro
Blast = Basic Local Alingment Search Tool C = citosina dNTPs = Desoxiribonucleótidos (Desoxiribonucleótidos trisfosfato)
G = Guanina
hr = Hora
IPTG = (isopropil- β D- tiogalactopiranosida) KCl = Cloruro de potasio LB = Medio de cultivo Luria- Bertani M = Molar
mg = Miligramo
MgCl2 = Cloruro de magnesio
min = Minuto
ml = Mililitro mM = milimolar
N = Normal
NaCl = Cloruro de sodio
NaOH = Hidróxido de sodio
ng = Nanogramo nm = Nanómetro
°C = Grados centígrados pb = Pares de bases
PCR = Polymerase Chain Reaction (Reacción en caena de la polimerasa)
pmol = Picomol
RH= Reacción de hipersensibilidad
RFLP = Restricción fragments Length Polimorphism (Polimorfismos de longitud de fragmentos de restricción)

xiii
RNA = Ácido ribonucleico RNasa = Ribonucleasa
rpm = Revoluciones por minuto rRNA = Ácido ribonucleico ribosómico seg = segundos T = Timina
Taq polimerasa = DNA polimerasa, obtenido de la bacteria Thermus aquaticus
TE = Tris- EDTA
Tris = Tris [hidroximetil] aminometano
Tris- HCl = Tris [hidroximetil] aminometano – ácido clorhídrico tRNA = RNA de transferencia U = Unidad enzimática
X-Gal = (5-bromo-4cloro-3i-ndoli-β-D galactopiranosa)
YDC = Extracto de levadura dextrosa
YS = Caldo de levadura

xiv
RESUMEN
El tomate es la hortaliza más difundida en todo el mundo y la de mayor valor económico. En México se obtuvo una producción de 2, 148,130 ton en el 2005. En Sinaloa en el ciclo otoño-invierno 2005-2006 se obtuvo una producción de 537,177 ton con un rendimiento de 32.735 ton/ha. Las enfermedades constituyen un factor limitante en la producción de tomate en muchas partes del mundo. La mancha bacteriana es una enfermedad del tomate que causa considerables pérdidas en la producción, es causada por Xanthomonas campestris pv. vesicatoria y es clasificada dentro de cuatro especies, Xanthomonas euvesicatoria (Grupo A), X. vesicatoria (Grupo B), X. perforans (Grupo C) y X. gardneri (Grupo D). El objetivo del presente trabajo es detectar y caracterizar molecularmente aislados de Xanthomonas campestris pv. vesicatoria asociadas con cultivos de tomate de Sinaloa. Para el desarrollo de este trabajo de tesis se realizaron muestreos en los municipios de Guasave, Ahome, Mocorito, Sinaloa de Leyva, Salvador Alvarado, El Fuerte, Culiacán y Elota, obteniéndose un total de 174 muestras, de las cuales 15 aislados fueron detectados por PCR y de acuerdo al análisis de RFLP-PCR y secuenciación son miembros del grupo A (Xanthomonas euvesicatoria) de Xanthomonas campestris pv. vesicatoria. La caracterización fisiológica y bioquímica confirma que estos 15 aislados pertenecen a Xanthomonas campestris pv. vesicatoria. Mediante la prueba de patogenicidad se demostró que los 15 asilados fueron patógenos a plantas de tomate.

xv
ABSTRACT Tomato is one of the most important vegetable crops around the world and one
of the greater economic value. In México a production of 2, 148.130 ton was
obtained in the 2005. In Sinaloa in 2005-2006 a production of 537,177 ton with
a yield of 32,735 ton/ha was obtained. Many diseases and disorders can affect
tomatoes during the growing season. The bacterial spot is one of the most
common foliar diseases of the tomato in Mexico. This disease is caused by the
bacterium Xanthomonas campestris pv. vesicatoria and recently it was
classified in four different species, Xanthomonas euvesicatoria (Group A), X.
vesicatoria (Group B), X. perforans (Group C) and X. gardneri (Group D). The
objective of the present work was to detect and to molecular characterization of
Xanthomonas campestris pv. vesicatoria associated to tomato crops at Sinaloa.
174 Samples were collected in the counties of Guasave, Ahome, Mocorito,
Sinaloa de Leyva, Salvador Alvarado, El Fuerte, Culiacán and Elota. A total of
15 isolates yielded the expected product in PCR assays and according to the
RFLP-PCR analysis and sequencing of the PCR products allowed their
classification as members of group A (Xanthomonas euvesicatoria) of
Xanthomonas campestris pv. vesicatoria. The physiological and biochemical
characterization confirms that these 15 isolated belong to Xanthomonas
campestris pv. vesicatoria. By means of the pathogenicity assays was
demonstrated that the 15 isolates were pathogenic to tomato plants.

Rivera Soto Fatima del Rosario
I. Introducción. Dentro de la agricultura mexicana, la actividad hortícola es una de las más
dinámicas y con mayor capacidad exportadora. En el 2004, México obtuvo una
producción de alrededor de 2.1 millones de toneladas de tomate, 1.6 de papa, 1.8
de chile y 1.2 millones de toneladas de tomatillo (SAGARPA, 2004). Entre los
principales productos hortícola sobresale el cultivo del tomate (Licopersicum
esculentum Mill). Tradicionalmente la producción de esta hortaliza en el Estado de
Sinaloa ha sido un componente importante de la producción nacional y su
comportamiento ha sido muy similar. Por ello para este estado es un cultivo
económicamente importante y cubre una superficie de 26,530 has (SAGARPA, 2004). La enfermedad mancha bacteriana ocasionada por la bacteria
Xanthomonas campestris pv. vesicatoria, es una de las enfermedades más
importantes en los cultivos de tomate y chile en el Estado de Sinaloa, donde se
localiza con una distribución general y en forma epidémica, ocasionando pérididas
en la producción (Cruz et al., 1998). Al cultivo del tomate lo afectan diversos agentes patógenos, pero la
bacteria Xanthomonas campestris pv. vesicatoria está considerada como una de
las más peligrosa (Gitaitis, et al., 1992). La mancha bacteriana se caracteriza por
presentar lesiones necróticas en hojas, tallos y frutos. En verano y climas cálidos,
la mancha bacteriana podría causar severas defoliaciones de plantas ocasionando
pérdidas en la producción (Pohronezny et al., 1986). El uso de técnicas moleculares han contribuidao a una rápida y eficiente
detección de patógenos las cuales estan enfocadas en la manipulación del DNA.
El gen hrp que determina la hipersensibilidad y patogenicidad es necesario
para las bacterias patógenas, en la planta induce síntomas en hospederos
susceptibles y reacción de hipersensibilidad en hospederos resistentes o no
hospederos (Willis et al., 1990), y puede ser encontrado en algunas bacterias
fitopatogénas tales como Erwinia amylovora (Beer et al., 1991), Pseudomona
solanacearum (Boucher et al., 1987), P. syringae pv. phaseolicola (Lindgren et
al., 1986), y Xanthomonas campestris pv. vesicatoria (Bonas, 1991). En una
región altamente conservada del genoma de bacterias fitopatógenas es más fácil
1

Rivera Soto Fatima del Rosario
la selección de DNA específicos para la detección e identificación de un gran
número de especies o patovares de Xanthomonas.
Dada la importancia el cultivo del tomate en Sinaloa y el riesgo que
representa la presencia de cepas patogénicas de esta bacteria en cultivos de
tomate, se requiere de métodos de detección e identificación eficientes que
permitan detectar a tiempo la enfermedad. La identificación y clasificación de
bacterias se basó historicamente en las características fenotípicas, lo que ha sido
de base para diferenciarlas; sin embargo, las características moleculares son
esenciales para diferenciar las distintas especies bacterianas. La biología
molecular es una herramienta de gran impacto ante la problemática que
representa el poder caracterizar esta bacteria por métodos convencionales. Por lo
anterior, el objetivo del presente estudio es detectar y caracterizar diferentes
cepas de la bacteria Xanthomonas campestris pv. vesicatoria teniendo como
principal herramienta las técnicas moleculares.
2

Rivera Soto Fatima del Rosario
II. REVISION DE LITERATURA. A. Bacterias.
Dos clases de procariotes causan enfermedades en las plantas: las
bacterias y los mollicutes (Agrios, 2004). Las bacterias son células pequeñas
(0.5-3.0 µ), que carecen de una membrana alrededor de un núcleo; no poseen
organelos claramente definidos y delimitados por membranas, tales como
mitocondrias, cloroplastos, aparato de Golgi y lisosomas. En el medio ambiente
global, los procariotas son absolutamente esenciales en los ciclos de los
elementos, como Carbono, Oxígeno, Nitrógeno, Azufre (Montealegre, 2002). Las
bacterias se definen como seres generalmente unicelulares, sus dimensiones
medias ascilan entre 0.5 y 1 µm. Se conocen alrededor de 1600 especies de
bacterias (Goto, 1990). La inmensa mayoría de bacterias son organismos
estrictamente saprófitos y como tales benefician al hombre, ya que ayudan a
descomponer cantidades enormes de materia orgánica, que produce anualmente,
en forma de productos de desecho o que son resultado de la muerte de las
plantas y los animales. Varias especies producen enfermedades en el hombre,
entre ellas la tuberculosis, la neumonía y la fiebre tifoidea, y otras que causan
enfermedades tanto en el hombre como en los animales donde resaltan por su
importancia la brucelosis y el ántrax (Agrios, 1986). En la actualidad se reconocen
alrededor de 60 especies de bacterias causantes de enfermedades en plantas,
que incluyen aproximadamente 300 subespecies y patovares (Blancard, 1990).
1. Morfología.
La mayoría de las bacterias fotopatógenas tienen forma de bastón. Los
bastones son, más o menos cortos y cilíndricos y, en los cultivos jóvenes, tienen
una longitud que va de 0.6 a 3.5 µm. Las paredes celuares de la mayoría de las
especies de bacterias están cubiertas por un material vizcoso y gomoso que
puede ser delgado (“capa mucilaginosa”) o denso, formando una masa
relativamente amplia en torno a la célula (“cápsula”). La mayoría de las bacterias
poseen delicados flagelos en forma de filamentos que a menudo son
3

Rivera Soto Fatima del Rosario
considerablemente más largos que las células que los formaron. En algunas
especies bacterianas, cada especie presenta un solo flagelo (Figura 1), mientras
que otras poseen un ramillete de ellos en uno de sus extremos y todavía otras
poseen un flagelo perítrico, es decir, distribuidos sobre toda su superficie. Las
celulas bacterianas presentan paredes celulares delgadas, relativamente firmes y
un poco rígidas que al parecer son bastanes distintas a la menbrana citoplásmica
interna pero que en ocasiones parece intergradas y fusionarse con la capa
mucilaginosa externa o la cápsula. La pared celular in cluye el contenido de la
célula y permite la entrada al flujo de nutrientes y la salida de desechos, enzimas
digestivas y otros productos emitidos por la célula bacteriana. La membrana
citoplásmica determina el grado de permeabilidad selectiva de las distintas
sustancias que fluyen hacia el interior y desde el exterior de la célula; por el
citoplasma, que es una mezcla compleja de proteínas, lípidos, carbohidratos,
muchos otros compuestos orgánicos y por el material nuclear, que consiste en un
gran cromosoma circular de DNA que constituye el rpincipal armazón de material
genético de las bacterias y aparece como un corpúsculo esférico, elipsoidal o en
forma de pesas localizado dentro del citoplasma. Con frecuencia, las bacterias
tambien tienen copias individuales o multiples de otros cromosomas circulares
más pequeños denominados plásmidos (Agrios, 2004).
Figura 1. Estructura bacteriana mostrando los principales organelos.
http://www.monografias.com/trabajos/bacterias/bacterias.html)
4

Rivera Soto Fatima del Rosario
2. Bacterias fitopatógenas.
Las bacterias fitopatógenas se conocen desde 1882; son, con mucho, el
grupo más grande de procariotes fitopatógenos que mejor se conocen (Agrios, 2004). Este grupo de patógenos constituye el segundo en importancia, después
de los hongos, si tenemos en cuenta el número y gravedad de las enfermedades
que produce (Goto, 1990). La clasificación de las bacterias, al igual que la de
otros organismos, nunca es estática y continuamente se proponen cambios.
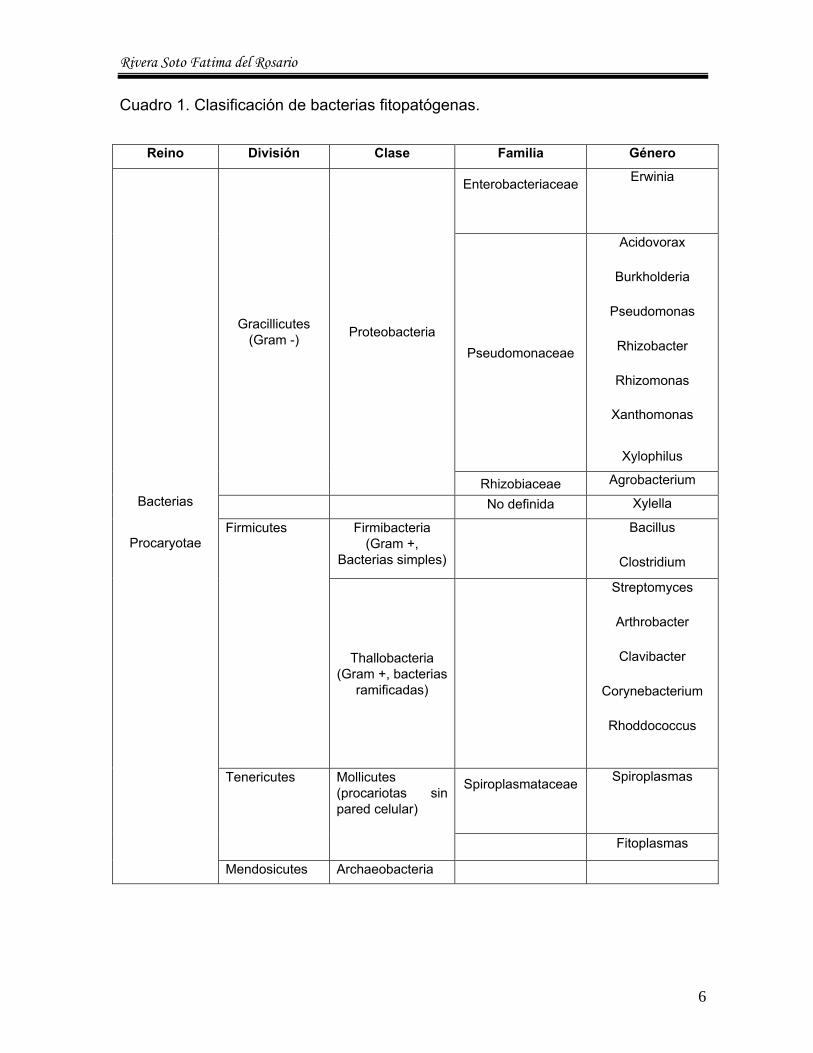
Actualmente, la clasificación de las bacterias se puede plantear, según se indica
en el Cuadro 1.
3. Bacterias que causan enfermedad en el tomate.
Existe una gran variedad de enfermedades causadas por bacterias que
dañan tallo, fruto y hojas en el cultivo de tomate; entre ellas se encuentran, la
mancha bacteriana (Xanthomonas campestris pv. vesicatoria), cancro bacteriano
(Clavibacter michiganensis subsp. michiganensis), la peca bacteriana
(Pseudomonas syringae pv. tomato), tizón foliar bacteriano (Pseudomona syringae
pv. syringae), la pudrición blanda (Erwinia carotovora subs. carotovora), la
marchités bacteriana del tomate (Ralstonia solanacearum) y la necrosis medular
(Pseudomonas corrugata) (Agrios, 2004) (Figura 2).
5
Rivera Soto Fatima del Rosario
Cuadro 1. Clasificación de bacterias fitopatógenas.
Reino División Clase Familia Género
Enterobacteriaceae
Erwinia
Pseudomonaceae
Acidovorax
Burkholderia
Pseudomonas
Rhizobacter
Rhizomonas
Xanthomonas
Xylophilus
Gracillicutes (Gram -) Proteobacteria
Rhizobiaceae Agrobacterium
No definida Xylella
Firmibacteria (Gram +,
Bacterias simples)
Bacillus
Clostridium
Firmicutes
Thallobacteria (Gram +, bacterias
ramificadas)
Streptomyces
Arthrobacter
Clavibacter
Corynebacterium
Rhoddococcus
Spiroplasmataceae
Spiroplasmas
Tenericutes Mollicutes (procariotas sin pared celular)
Fitoplasmas
Bacterias
Procaryotae
Mendosicutes Archaeobacteria
6

Rivera Soto Fatima del Rosario
A
A B C
FD
D
E
Figura 2. Algunos síntomas causados por bacterias en el cultivo del tomate. (A) Mancha
bacteriana, (B) Peca bacteriana, (C) Marchités y cancro bacteriano, (D) Tizón foliar
bacteriano (E) Pudrición blanda y (F) Marchités bacteriana del tomate.
Figura 2. Algunos síntomas causados por bacterias en el cultivo del tomate. (A) Mancha
bacteriana, (B) Peca bacteriana, (C) Marchités y cancro bacteriano, (D) Tizón foliar
bacteriano (E) Pudrición blanda y (F) Marchités bacteriana del tomate.
4. Xanthomonas campestris pv. vesicatoria. 4. Xanthomonas campestris pv. vesicatoria.
4.1. Origen. 4.1. Origen. La mancha bacteriana del tomate fue observada primeramente en
Sudáfrica y descrita por Doidge en 1920 (Jones et al., 1998). La mancha bacteriana del tomate fue observada primeramente en
Sudáfrica y descrita por Doidge en 1920 (Jones et al., 1998). Los agentes causales de los síntomas cancro bacteriano y mancha
bacteriana fueron identificados en 1921 como, Bacterium vesicatorium n. sp., por
Doidge y como Bacterium exitiosum n. sp., por Gardner & kendrick (Bradbury, 1984). Ambos estudios cloncluyerón que se trataba de una única especie
bacteriana que era capaz de infectar principalmente plantas de tomate y chile y
fue denominada por Doidge Bacterium vesicatorium. Posteriormente el agente
causal se denominó Pseudomonas vesicatoria en 1925 y Phytomonas vesicatoia
en 1930 y finalmente en 1939 recibió el nombre de Xanthomonas vesicatoria (Jones et al., 1998). El agente causal de la mancha bacteriana en plantas de
Los agentes causales de los síntomas cancro bacteriano y mancha
bacteriana fueron identificados en 1921 como, Bacterium vesicatorium n. sp., por
Doidge y como Bacterium exitiosum n. sp., por Gardner & kendrick (Bradbury, 1984). Ambos estudios cloncluyerón que se trataba de una única especie
bacteriana que era capaz de infectar principalmente plantas de tomate y chile y
fue denominada por Doidge Bacterium vesicatorium. Posteriormente el agente
causal se denominó Pseudomonas vesicatoria en 1925 y Phytomonas vesicatoia
en 1930 y finalmente en 1939 recibió el nombre de Xanthomonas vesicatoria (Jones et al., 1998). El agente causal de la mancha bacteriana en plantas de
7

Rivera Soto Fatima del Rosario
tomate y chile fue reclasificada en 1978 como un pathovar de la espécie X.
campestris, Xanthomonas campestris pv. vesicatoria (Sahin, 1997). Inicialmente, se creía que todos los aislados de Xanthomonas campestris
pv. vesicatoria, podían infectar plantas de tomate y chile, pero algunos aislados
son patógenos al tomate, otros solo a chile y otros a ambas especies (Canteros et
al., 1991). Se han reclasificado varias especies de Xanthomonas y entre ellas
subdividido Xanthomonas campestris pv. vesicatoria, que se dividió en dos
especies, Xanthomonas axonopodis pv. vesicatoria y Xanthomonas vesicatoria,
aunque esta propuesta ha sido controversial (Schaad et al., 2000; Young et al., 2001).
El patógeno es bastante variable en sus características fenotípicas y
genéticas siendo posible la distinción de hasta cuatro grupos (“A”, “B”, “C” y “D”),
en virtud de sus diferencias (Vauterin et al., 1995). X. gardneri también esta asociada a la mancha bacteriana de tomate y
chile (Jones et al., 1998). En 1957, esta bacteria fue identificada por primera vez
en Yugoslavia como P. gardneri (Sutic, 1957). La especie no fue considerada
sinónimo de X. vesicatoria, ya que mostraba diferencias en pruebas fisiológicas y
de patogenicidad. Por pruebas de hibridización de DNA, verificó que X. gardneri
estaba más relacionada con especies como X. campestris pv. carotae, X.
campestris pv. pelargonii y X. campestris pv. taraxaci y que Xanthomonas
campestris pv. vesicatoria representa otro grupo. A pesar de que ha sido validada
como especie, estudios recientes de aislados de Xanthomonas spp. patógenos a
hospederos de tomate y chile confirman que Xanthomonas gardneri (=grupo “D”),
es una entidad taxonómica distinta de Xanthomonas vesicatoria (=grupo “B”) y
Xanthomonas axonopodis pv. vesicatoria (=grupos “A” y “C”) (Jones et al., 2000).
4.2. Clasificación actual de Xanthomonas spp. De acuerdo con la actual clasificación filogenética de las bacterias, basadas
principalmente en comparaciones de secuencias nucleotídicas, principalmente de
rRNA 16S, el género Xanthomonas pertenece al phylum Proteobacteria, clase
8

Rivera Soto Fatima del Rosario
“Gammaproteobacteria” (Clase III), orden “Xanthomonadales” (Orden II) y familia
“Xanthomonadaceae” (Garrity & Holt., 2000). Estudios recientes ha propuesto la siguiente clasificación: Xanthomonas
euvesicatoria (Grupo A), X. vesicatoria (Grupo B), X. perforans (Grupo C) y X.
gardneri (Grupo D) (Jones et al., 2004).
4.3. Morfología específica de Xanthomonas spp. Xanthomonas campestris pv. vesicatoria es una bacteria baciliforme, con
dimensiones entre 0,7-1,0 x 2,0-2,4 µm, Gram-negativa, aerobica estricta, móvil y
con un flagelo polar (Jones, 1997). Se caracteriza por presentar colonias amarillas
en medio agar nutritivo (AN), mucoides y/o de producir pigmentos
(xanthomonadinas) (Stall, 1993). La mayoría de ellas crecen muy lentas en
medios de cultivo (Agrios, 2004) (Figura 3).
Figura 3. A. Micrografía de Xanthomonas campestris. B. Colonias de Xanthomonas
campestris pv. vesicatoria aisladas en AN (agar nutritivo).
4.4. Sintomatología de la mancha bacteriana en tomate. Los síntomas de la mancha bacteriana ocurren en toda la parte aérea de la
planta y se manifiestan en cualquier etapa fenológica del cultivo (Gitaitis et al., 1992). Se caracteriza por presentar lesiones necróticas en hojas, tallos y frutos.
9

Rivera Soto Fatima del Rosario
En verano y climas cálidos la mancha bacteriana causa severas defoliaciones de
plantas ocasionando pérdidas en la producción (Pohronezny et al., 1986). En
hojas los primeros síntomas se presentan como ampollas irregulares, con bordes
definidos de un color amarillo o verde claro a marrón oscuro o necrótico (Goode & Passer, 1980). En los frutos verdes aparecen pequeñas manchas acuosas
que sobresalen ligeramente, tienen halos blancos verduscos y se extienden hasta
alcanzar un diámetro aproximado de 3 a 6 mm (Agrios, 2004). Poco después los
halos desaparecen y las manchas se van tornando de color marrón y de textura
áspera (Jones, 1997). Estas lesiones tienden a ser hundidas en el centro y
elevadas por los márgenes (Gonder & Passer, 1980). Una enfermedad conocida
como peca bacteriana del tomate se parece a la mancha bacteriana pero es
ocasionado por la bacteria Pseudomonas syringae pv. tomato. Las lesiones que
causa en hojas, tallos y el fruto son similares (pero más pequeñas) que las que
causa Xanthomonas campestris pv. vesicatoria (Agrios, 2004), (Figura 4).
A B
Figura 4. Síntomas causados por Xanthomonas campestris pv. vesicatoria en cultivos de
tomate. A) manchas necróticas en tejido foliar. B) heridas necróticas en fruto.
10

Rivera Soto Fatima del Rosario
4.5. Estructura y organización del genoma. Xanthomonas campestris pv. vesicatoria también designado Xanthomonas
axonopodis pv. vesicatoria (Bonas, 1993) o Xanthomonas euvesicatoria (Jones, 2004) tiene un alto contenido genómico de G + C (Leyns, 1984). Xanthomonas
campetris pv vesicatoria contiene un simple cromosoma circular de 5, 178,466 pb
y cuatro elementos extracromosomales. Contiene dos operones rRNA
organizados en el orden 16S-23S-5S y localizados en una región de
aproximadamente 500 kb (entre 4,600,000 pb y 5,100,000 pb) (Thieme et al., 2005).
El alto número de elementos móviles y desviaciones en contenidos de G +
C proporcionan la flexibilidad del genoma en Xanthomonas campestris pv.
vesicatoria. Esto es una ventaja para la evolución de los patógenos por la
necesidad de adaptación contínua para suprimir los mecanismos de defensa del
hospedero (Thieme et al., 2005). Xanthomonas campestris pv. vesicatoria posee DSF (factor de difusión de
señal), constituidos por ácidos grasos alfa, β-insaturados los cuales están
involucrados en la regulación de síntesis de enzimas extracelulares y
exopolisacáridos. Ocho genes (rpfA a H) involucrados en la producción y
percepción de DSF, se están dentro del genoma de X. campestris pv. vesicatoria.
Posee también el factor difusible (DF), el cual es una butirolactona y está
involucrada en la regulación de pigmentos (xanthomonadinas) y en la síntesis de
exopolisacaridos.
El DF es encontrado en el genoma de X. campestris pv. campestris y se
cree que está dentro del genoma de X. campestris pv. vesicatoria por la presencia
de un pigB locus que está implicado en la síntesis de DF en Xanthomonas
campestris pv. campestris (Poplawsky et al., 1997). Tiene una región de 147-kb en el cromosoma (116 genes, XCV1929 a
XCV2044) el cual es únicamente desarrollado para la biosíntesis flagelar (Thieme et al., 2005).
En X. campestris pv. vesicatoria algunos operones codifican componentes
de tipo 4 pili, que le permite a la bacteria la retracción y la adhesión al tejido de la
11

Rivera Soto Fatima del Rosario
planta (Van Doorn et al., 1994). Algúnas bacterias Gram-negativas tienen polisacáridos fijos a su superficie,
las cuales contribuyen a su patogenicidad y a su interacción con las células de
plantas y animales (D,Haeze et al., 2004). El típico exopolisacárido del género
Xanthomonas es el xanthano (Becker et al., 1998). El gen gum controla la
biosíntesis de xhantano en X. campestris pv. campestris y es conservado en X.
campestris pv. vesicatoria. Estudios en X. campestris pv. campestris demuestran
que xhantano gum no es esencial para la patogenicidad pero contribuye en la
agresividad contra el hospedero (Katzen et al., 1998). El sistema de secreción bacterial es importante para la interacción del
patógeno con el hospedero (Buttner et al., 2002). X. campestris pv. vesicatoria
contiene genes para todos los sistemas de transporte de proteinas conocidos en
bacterias gram-negativa. Se conoce el “Sec pathway” el cual es importante para el
transporte de muchas proteínas en el espacio periplasmico. El sistema TAT (tatA-
tatC) manda una ruta alternativa de proteinas al periplasma (Robinson et al., 2001). El sistema de secreción tipo II de patogenicidad de la baceria consiste en
un gran número de proteínas incluyendo posibles determinantes de avirulencia, al
espacio intercelular de la planta (Sandkvist et al., 2001). El principal determinante de la virulencia dentro de X. campestris pv.
vesicatoria es el sistema de secreción tipo III (TTSS), que es codificado por el
grupo de genes de la patogenicidad hrp. Localizado a 35.3-kb del cromosoma
genómico (459,555 pb – 494, 868 pb) (Thieme et al., 2005). 4.6. Importancia del gen hrp.
El gen hrp está altamente conservado entre un gran número de bacterias
fitopatógenas. El gen hrp (reacción de hipersensibilidad y patogenicidad, es
esencial para una evidente interacción con la planta, para la patogenicidad en
plantas susceptibles y la inducción de la Reacción de hipersensibilidad (HR) en
plantas resistentes (Fenselau et al., 1992). El gen hrp determina la patogenicidad, muchos genes hrp son organizados
en grandes grupos que contienen dos o más genes y se pueden aislar de muchas
12

Rivera Soto Fatima del Rosario
bacterias fitopatógenas gram negativas, tales como: patovares de Pseudomona
syringe, Ralstonia solanacearum, Erwinia amylovora y patovares de Xanthomonas
campetris (Bonas, 1994). Los genes hrp de las bacterias fitopatógenas son también muy similares a
los genes que están involucrados en la secreción de factores de patogenicidad
para bacterias patógenas de mamíferos (Fenselau et al., 1992). Bacterias no-
patógenas no pueden producir síntomas en hospederos susceptibles y reacción
de hipersensibilidad en no-hospederos y se cree que no poseen secuencias del
DNA similar al gen hrp (Stall et al., 1990). Muchos genes hrp están involucrados
en una región cromosomal 23 kb y están organizados en seis operones que van
de hrpA a hrpF (Bonas et al., 1991). El gen hrp que determina la hipersensibilidad
y patogenicidad es apropiado para los análisis de detección e identificación de
bacterias fitopatógenas (Willis et al., 1990). Los oligonucleotidos diseñados en el
gen hrp pueden ser usados para determinar la patogenicidad desconocida de
Xanthomonas. La presencia de cepas fitopatógenas en muestras puede ser
determinada por la amplificación de fragmentos hrp sin la necesidad de métodos
de aislamiento del organismo e inoculación en plantas hospederas (Leite et al., 1994). 4.7. Importancia del Cultivo de tomate en Sinaloa.
Sinaloa es considerado en México y en el mundo como una entidad
eminentemente agrícola, donde se concentra gran parte de la producción de
hortalizas del país. A lo largo del último siglo se ha dado un desarrollo de la
horticultura como consecuencia principalmente de la variación que ha
experimentado la demanda, tanto en el mercado nacional como en el mercado
externo (Gonzáles, 1998). Sinaloa obtuvo una producción total de 537,177 ton.
con un rendimiento promedio de 32.735 ton/ha en el ciclo otoño-invierno 2005-
2006. Los municipios con mayor superficie sembrada de tomate en Sinaloa son:
Culiacán (7399 has), Los Mochis (7210), Guasave (7093 has), Mazatlán (1697
has), La Cruz (1567) y Guamúchil (802). Alcanzando una superficie de 26, 530
has en el ciclo 2005-2006, con un valor comercial de 412.6 millones de dólares,
13

Rivera Soto Fatima del Rosario
representando el 37 % del valor total de las exportaciones de legumbres y
hortalizas, siendo una fuente importante de mano de obra ya que se requieren
hasta 254 jornales por hectárea (SAGARPA, 2004). Esta situación favorable de la
horticultura Sinaloense se debe, entre otros factores a la posición geográfica
estratégica, un clima favorable, sus notables avances tecnológicos y gran
capacidad de innovación, además sobre salen la organización eficiente de los
productores, la disposición de tierra cultivable, una enorme infraestructura
hidráulica, disponibilidad de mano de obra, tradición en sus relaciones con las
distribuidoras estadounidenses, facilidades en telecomunicaciones y transporte y
la calidad de sus productos hortícolas (Gonzáles, 1998) . 4.8. Importancia de la mancha bacteriana en tomate.
El género Xanthomonas comprende un grupo de bacterias fitopatógenas de
gran importancia económica, ya que sus especies tienen como hospedero a más
de 124 especies de plantas monocotiledóneas y 268 dicotiledóneas son
hospedantes de Xanthomonas (Leyns et al., 1984). Entre las especies de
Xanthomonas campestris es una de las más importantes pues consta de al menos
125 diferentes patovares que son diferenciados por el hospedante al que afectan
(Bradbury, 1984). La mancha bacteriana es una de las enfermedades más importantes del
tomate y chile en el Estado de Sinaloa, por su amplia distribución y severidad esta
considerada como una de las enfermedades más importantes en la actualidad. El
clima caluroso y alta humedad de esta región costera de México, combinado con
lluvias en invierno y neblinas, constituyen un ambiente ideal para la infección de
hojas pecíolos, tallos y frutos (Cruz et al., 1998). La enfermedad causa la caída de flores, reflejandose en la producción y su
rápida diseminación en favorecida por altas temperaturas (24-30°C). Las pérdidas
causadas por la mancha bacteriana también repercuten en el costo de los
productos químicos utilizados como estrategias de control, como lo es el uso de
fungicidas cúpricos (Lopes & Quezado-Soares, 1997; Goode & Passer, 1980; Jones, 1997).
14

Rivera Soto Fatima del Rosario
Algunos factores que explican el desarrollo de fuertes epidémias por la
mancha bacteriana en el cultivo de tomate son: variabilidad en el control químico y
la ausencia de fuentes de resistencia adecuada (Goode & Passer, 1980; Jones, 1997).
4.9. Ciclo de la enfermedad. La bacteria penetra dentro de las hojas a través de los estómas. En los
frutos, se introduce mediante heridas provocadas por insectos picadores como la
chinche verde (Nezara viridula) por la acción del granizo, o bien por el granizo y
por partículas de arena muy finas arrastradas por vientos muy fuertes (Jones, et
al., 1991). Una vez dentro de la cavidad estomatal la bacteria provoca una
hinchazón y rompimiento de la epidermis. Después se extiende hacia otros
espacios intercelulares adyacentes, y destruye el tejido. Posteriormente la mancha
se seca y el tejido se colapsa (Jaime, 1976). En tejidos jóvenes de hojas y frutos estimulan la división celular; la
hipertrofia es ocasionada por cantidades muy pequeñas de amonio, que es
producido por la bacteria.
Las bacterias se diseminan a través de la semilla infestada, por la lluvia y
durante las labores de cultivos. Sobrevive en residuos de plantas muertas durante
dos años, en semillas secas de tomate durante 16 meses, en plantas de tomate y
en la rizósfera de algúnos cereales de invierno; la bacteria no sobrevive en el
suelo como células individuales (Peterson, 1965; Jones et al., 1991). Las temperaturas de 24 a 30°C, así como la humedad relativa
especialmente gotitas de agua son necesarias para el desarrollo de la enfermedad
(Jones et al., 1991). 4.10. Control de la enfermedad. La desinfección de la semilla es importante para un buen inicio de la
producción de plántulas libres de las bacterias. Se recomienda la desinfección de
la semilla con agua caliente a 50°C durante 30 minutos o tratar la semilla con
productos químicos como el Arasan 75, en una relación 1:2000 durante 5 minutos
15

Rivera Soto Fatima del Rosario
seguido de un lavado de la semilla (León, 1982; Haward y Waterson, 1964). En plántulas que se producen en invernaderos con cubierta de plástico, se
debe de regar solamente cuando sea necesario. Tratamientos al agua con
bactericidas/algicidas resultan de gran utilidad y a muy bajo costo si se
implementan en forma contínua. Tambien conviene aplicar cualquier metodo que
ayude a reducir la temperatura dentro del invernadero, sobre todo durante los
meses más calurosos. Lo anterior no favorece el desarrollo de la enfermedad
(Sánchez, 1986). Las aspersiones con productos químicos a base de cobre mezclados con
antibióticos completan el manejo de la enfermedad.
En el campo, la alternativa de control es el uso de fungicidas a base de
cobre mezclados con ditiocarbamatos de manganeso, así como la aspersión de
antibióticos como el sulfato de estreptomicina.
La rotación de cultivos es recomendable para eliminar las fuentes de
inóculo (residuos de cosecha y plantas muertas) en el campo (Jones et al., 1991). B. Caracterización fisiológica de Xanthomonas campestris pv. vesicatoria.
Por más de 70 años se creía que la mancha bacteriana era causada por
una sola especie, identificada como Xanthomonas campestris pv. vesicatoria
(Jones et al.,1998). Varios estudios se han realizado con el objeto de esclarecer
la etiología de la enfermedad, a través de características fenotípicas y genotípicas.
Las pruebas bioquímicas, son pruebas simples que se han desarrollado
para demostrar la presencia o ausencia de una determinada actividad enzimática,
grupo de enzimas o determinada vía metabólica, crecimiento a una determinada
temperatura, crecimiento en presencia de inhibidores, entre otros. No significan de
ninguna manera un estudio profundo del metabolismo bacteriano (Montealegre, 2002). 1. Caracterización a nivel género.
La capacidad de hidrolizar almidón (actividad amilolítica) fue una de las
primeras características fenotípicas que mostró diferencias entre los aislados de
16

Rivera Soto Fatima del Rosario
Xanthomonas spp. asociados con la mancha bacteriana en tomate y chile. Actualmente se han caracterizado los grupos “A”, “B”, “C” y “D” de Xanthomonas
campetris pv. vesicatoria como agentes causales de la mancha bacteriana. (Jones et al., 1998). Se han reconocido cerca de 100 especies de Xanthomonas como
patógenos de distintas plantas. En el Cuadro 2, se definen algunas de las pruebas
bioquímicas empleadas para la caracterización de 13 generos de bacterias, dentro
de los cuales se ubica Xanthomonas.
Cuadro 2. Características usadas para diferenciar géneros de bacterias
patógenas en plantas capaces de crecer en medio estándar.
Caracter
Erw
inia
Pan
toea
Aci
divo
rax
Pse
udom
onas
Ral
ston
ia
Bur
khol
deria
Xant
hom
onas
Xyl
ophi
lus
Agr
obac
teriu
m
Cla
viba
cter
Clo
strid
ium
Bac
illus
Stre
ptom
yces
Gram - - - - - - - - - + + + +
Crecimiento anaerobio + + - - - - - - - - + + -
Crecimiento aerobio + + + + + + + + + + - + +
Colonias amarillas o
naranjas en YDC o NBY - + - - - - + + -
+ - - -
Colonias mucoides en YDC - - + - + - + - + + ND ND -
Fluorescencia en B de K - - - + - - - - - - - - -
Ureasa - - + - + v - + ND - ND ND ND
Oxidasa - - + - + + - - + - - V +
(Schaad, 2000)
1.1. Fluorescencia. La fluorescencia es la capacidad que tienen ciertos cuerpos de
transformar la luz que reciben en radiaciones de mayor longitud de onda. El medio
B de King, especialmente pobre en hierro, pone de manifiesto la presencia de un
pigmento fluosrescente con luz ultravioleta de 260 nm. La bacteria Xanthomonas
17

Rivera Soto Fatima del Rosario
campestris pv. vesicatoria es una bacteria que no emite luz al crecer en dicho
medio de cultivo y ser expuesta a luz UV (Lehninger, 1995).
1.2. Tinción de Gram La tinción de Gram fue diseñada empíricamente en 1884 por el médico
danes Cristian Gram, quien dividió a las bacterias en organismos Gram-positivos
(Figura 5) y Gram-negativos según su reacción frente al teñido con cristal violeta y
yodo y después de ser decoloradas y tratadas con safranina (Lehninger, 1995). La tinción de Gram es la más importante de las tinciones diferenciales (http://edicion-
micro.usal.es/web/identificacion/AyudaPruebas.html).
Características de las bacterias Gram positivas y Gram negativas:
a) Gram positivas
Ácido Teicoico Peptidoglicano
Pared celular
Membrana Citoplasmática
Ácido Lipoteicoico
Figura 5. Estructura de una bacteria Gram positiva.
En el interior de la célula se encuentra el citoplasma que está constituido
por ribosomas y centralmente se ubica el nucleoide bacteriano que semeja una
madeja de hebras y cuyo principal componente es el DNA.
El perfil de la envoltura externa de la pared celular de la bacteria Gram
positiva, posee como base química fundamental el péptidoglucano y que
corresponde a un polímero de N-acetil glucosamina, unido en posición B-1-4 con
N-acetil murámico, a éste se agregan por el grupo lactilo 4 ó más aminoácidos.
18

Rivera Soto Fatima del Rosario
Esta molécula se polimeriza gran cantidad de veces, de modo que se forma una
malla especial, llamada sáculo de mureína. Dicho compuesto es de vital
importancia para conservar la forma y darle rigidez a la célula bacteriana; si este
compuesto no existiera, la célula reventaría debido a su gran presión interna.
b) Gram negativas
Las células presentan una doble capa denominada membrana externa,
(Figura 6), luego se observa una capa más densa que corresponde al
peptidoglucano, estructura que conforma la pared celular de la bacteria Gram
positiva y que también es responsable de la rigidez y resistancia de la célula Gram
negativa.
Lipopolisacárido Poro
Membrana externa
Lipoproteína Espacio Periplásmico
Peptidoglicano Membrana citoplásmica
Figura 6. Estructura interna de una bacteria gram negativa.
Luego se encuentra el espacio periplasmático y la membrana
citoplasmática que posee similar característica a la de la bacteria Gram positiva.
En el interior se encuentran el citoplasma y nucleoide, que aparecen menos
compactos que las bacterias Gram positivas, pero que son estructuralmente
semejantes.
En las bacterias Gram negativas, como es el caso de Xanthomonas
campestris pv. vesicatoria, existe una membrana externa que no poseen las
bacterias Gram positivas y cuya estructura es semejante a la membrana
citoplasmática. La membrana externa se presenta como un triple perfil en el que
19

Rivera Soto Fatima del Rosario
las líneas densas a los electrones corresponden a proteínas que fijan los
compuestos de metales pesados (para la microscopía electrónica). El perfil
transparente a los electrones tiene una composición química compleja; al exterior
de la bacteria se encuentran polímeros de heterosacáridos o azúcares que están
unidos a un complejo de glúcidos fosfolípidos que se han denominado
lipopolisacáridos (Montealegre, 2002).
1.3. Catalasa. La porción no aminoácido de una proteína conjugada se denomina grupo
prostético. La catalasa es una hemo-enzima que posee un espectro de absorción
característico debido a su grupo prostético hemo (ferroprotoporfirina) y muestra
cambios transitorios en su espectro cuando se muestra con su sustrato, el
peróxido de hidrógeno, lo cual es el reflejo de la descomposición de su complejo
enzima-sustrato. El peróxido de hidrógeno formado por la superóxido dismutasa,
hemoenzima que cataliza la reacción.
H2O2→ H2O + ½ O2
La detección de la catalaza se realiza mediante la identificación de burbujas
de oxígeno formadas, dicha reacción se observa en el caso de Xanthomonas
campestris pv. vesicatoria debido a la acción de dicha enzima cuando se pone en
contacto con su sustrato el peroxido de hidrógeno (Lehninger, 1995).
1.4. Oxidasa. Los citocromos son proteínas transferidoras de electrones que contienen
grupos ferro-porfirina; se encuentran únicamente en las células aeróbicas.
Algunas se hallan localizadas en la membrana mitocondrial interna, donde actúan
secuencialmente para transportar los electrones originados en varios sistemas
deshidrogenasas, hasta el oxigeno molecular. Otros citocromos se encuentran en
el retículo endoplasmático en el que desempeñan un papel en las reacciones de
hidroxilación especializadas. Todos ellos experimentan cambios reversibles de
valencia Fe (II) Fe (III) durante sus ciclos catalíticos. Sus formas reducidas no
20

Rivera Soto Fatima del Rosario
pueden ser oxidadas por el oxígeno molecular, con excepción del citocromo
terminal de la respiración mitocondrial, es decir, el citocromo, a3 o citocromo c
oxidasa, que también contiene cobre íntimamente unido a él.
Los citocromos oxidasas tienen una gran afinidad por el oxígeno, mientras
que las oxidasas negativas no. La reacción se efectúa cuando la proteína actúa
sobre el dihidrocloril tetrametil fenilamina que al combinarse con el naftol se
produce un color azúl y agua, esta reacción no sucede con la bacteria
Xanthomonas campestris pv. vesicatoria, dicha reacción se muestra a
continuación:
N (CH3)2 NH2 + Naftol + O2 → N (CH3)2 Azul + H2O
1.5. Crecimiento en YDC (extracto de levadura dextrosa). Este medio de cultivo es rico en calcio y se utiliza para observar el
crecimiento, elevación, consistencia y color de la bacteria Xanthomonas
campestris pv. vesicatoria. Son colonias de Xanthomonas spp. presenta una
consistencia pegajosa y una coloración amarilla o naranja, esto permite diferenciar
entre otros géneros como de Pseudomonas spp. que no presenta estas
caracteristicas en YDC.
2. Caracterización de especies de Xanthomonas. Pocas especies han sido fácilmente distinguidas del tipo de especies de
Xanthomonas y/o cualquier otra, mediante el uso de pruebas bioquímicas usadas
en el manual de determinación bacteriológica de la octava edición de Bergey´s
(Skerman, 1980). Aun no es posible diferenciar las cuatro especies de Xanthomonas
asociadas con la mancha bacteriana en tomate (X. euvesicatoria, X. vesicatoria, X.
perforans y X. gardneri) empleando pruebas bioquímicas. El último “Índice de
bacterias y cambios en la nomenclatura” publicados en la International Journal of
Systemic Bacteriology entre 1980 y 1992 (Moore, 1992), incluyen X. albilineans,
X. axonopodis, X. campestris, X. citri, X. fragaria, X. malthophilia, X. orizae,
21
Rivera Soto Fatima del Rosario
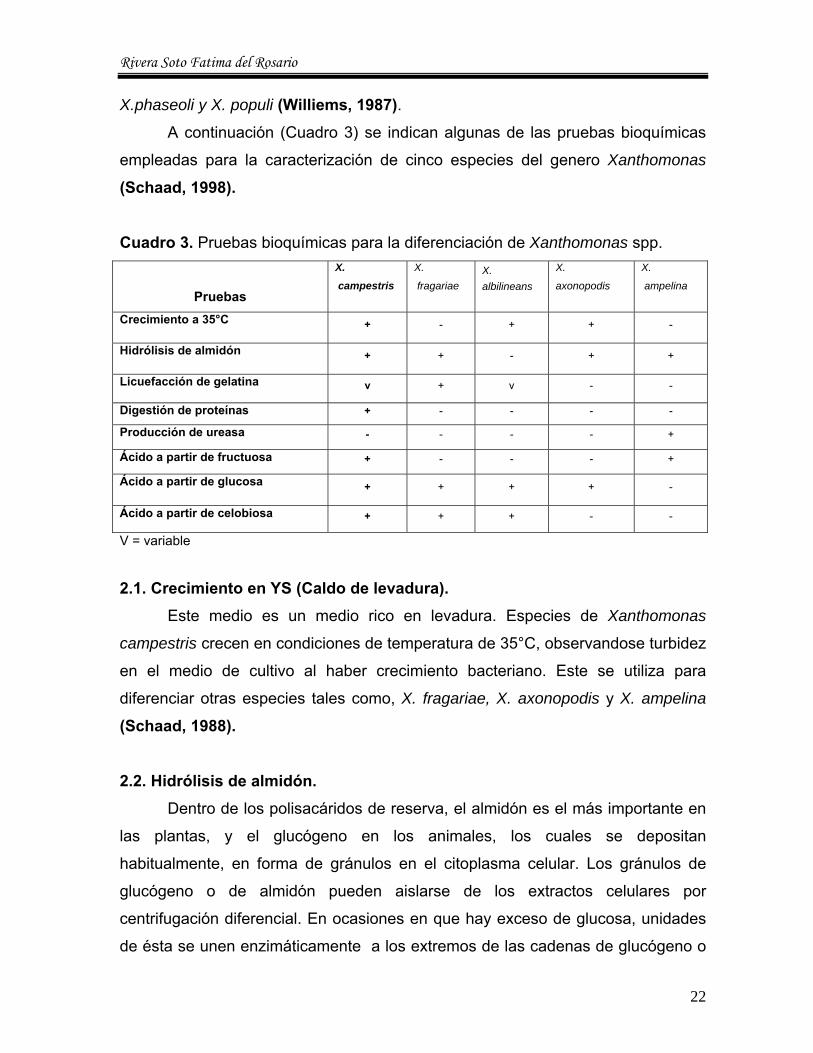
X.phaseoli y X. populi (Williems, 1987). A continuación (Cuadro 3) se indican algunas de las pruebas bioquímicas
empleadas para la caracterización de cinco especies del genero Xanthomonas
(Schaad, 1998).
Cuadro 3. Pruebas bioquímicas para la diferenciación de Xanthomonas spp.
Pruebas
X.
campestris
X.
fragariae X. albilineans
X.
axonopodis
X.
ampelina
Crecimiento a 35°C + - + + -
Hidrólisis de almidón + + - + +
Licuefacción de gelatina v + v - -
Digestión de proteínas + - - - -
Producción de ureasa - - - - +
Ácido a partir de fructuosa + - - - +
Ácido a partir de glucosa + + + + -
Ácido a partir de celobiosa + + + - -
V = variable
2.1. Crecimiento en YS (Caldo de levadura). Este medio es un medio rico en levadura. Especies de Xanthomonas
campestris crecen en condiciones de temperatura de 35°C, observandose turbidez
en el medio de cultivo al haber crecimiento bacteriano. Este se utiliza para
diferenciar otras especies tales como, X. fragariae, X. axonopodis y X. ampelina
(Schaad, 1988). 2.2. Hidrólisis de almidón.
Dentro de los polisacáridos de reserva, el almidón es el más importante en
las plantas, y el glucógeno en los animales, los cuales se depositan
habitualmente, en forma de gránulos en el citoplasma celular. Los gránulos de
glucógeno o de almidón pueden aislarse de los extractos celulares por
centrifugación diferencial. En ocasiones en que hay exceso de glucosa, unidades
de ésta se unen enzimáticamente a los extremos de las cadenas de glucógeno o
22

Rivera Soto Fatima del Rosario
almidón; si existe necesidad metabólica, entonces se liberan de nuevo, también
por acción enzimática y son empleadas como combustible (Lehninger, 1995). El almidón se encuentra en dos formas, la alfa-amilosa y la amilopeptina.
Ciertas bacterias fitopatógenas (Xanthomonas en su gran mayoría), son capaces
de transportar el almidón por medio de enzimas extracelulares conocidas como
alfa y β amilasas, las cuales hidrolizan el almidón a maltosa, de tal forma que esta
ya puede penetrar al interior de la célula y ser atacadas por enzimas
intracelulares para la obtención de energía (Rodríguez, 2001). 2.3. Licuefacción de gelatina.
La gelatina es una proteína simple obtenida de la hidrólisis del colágeno, la
cual puede ser despolimerizada por todas aquellas bacterias que sintetizan la
exoenzima gelatinasa. En el caso de Xanthomonas campestris pv. vesicatoria
produce dicha enzima. Sin embargo, es importante señalar que el medio donde se
va a probar la acción de dicha enzima debe estar exento de carbohidratos, ya que
al fermentarse inhiben la síntesis de la gelatinasa (Rodríguez, 2001).
2.4. Dihidrolasa de arginina. Los aminoácidos básicos en los que los grupos R tienen carga positiva neta
a pH 7, poseen todos seis átomos de carbono. La arginina tiene un grupo
guanídico cargado positivamente.
La detección del sistema enzimático dihidrolasa de arginina, el cual permite a las
bacterias crecer bajo condiciones anaeróbicas, consiste de dos enzimas: la
arginina desmidasa que degrada la arginina a citrulina y NH3 y la citrulina ureidasa
que convierte a la citrulina a ornitina, bióxido de carbono y NH3. (Rodríguez, 2001). 2.5. Utilización de fuentes de carbono (Fructuosa, glucosa y celobiosa). Las fuentes de carbono son tradicionalmente utilizadas para determinar la
actividad de la vía metabólica de muchas bacterias a partir de sustratos (fructousa,
glucosa y celobiosa) que se incorporan en el medio de cultivo y que la bacteria al
23
Rivera Soto Fatima del Rosario
crecer transforma o no. Existen muchas fuentes de carbono para llegar a la
caracterización de Xanthomonas campestris, enlistadas en el manual de Schaad
1988 y 2001. Así, la utilización de fructuosa, glucosa y celobiosa con fuentes de
carbono, permite la diferenciación de X. fragariae, X. albilineans y X. axonopodis,
al utilizar glucosa es diferenciada de X. ampelina y al utilizar celobiosa es
diferenciada de X. axonopodis y X. ampelina de acuerdo al manual de Schaad
1988.
3. Caracterización a nivel patovar. El término patovar no ha sido taxonómicamente estandarizado en el Código
Internacional de Nomenclatura de Bacterias (Sneath, 1992). Desde 1980, se han
propuestos algunos cambios relacionados con la nomenclatura y taxonomía de
Xanthomonas.
El término patovar fue propuesto en 1978 (Young, 1978), como una medida
para preservar el nombre de la planta hospedera, pero los datos fenotípicos no
fueron capaces de diferenciarlas de otras especies.
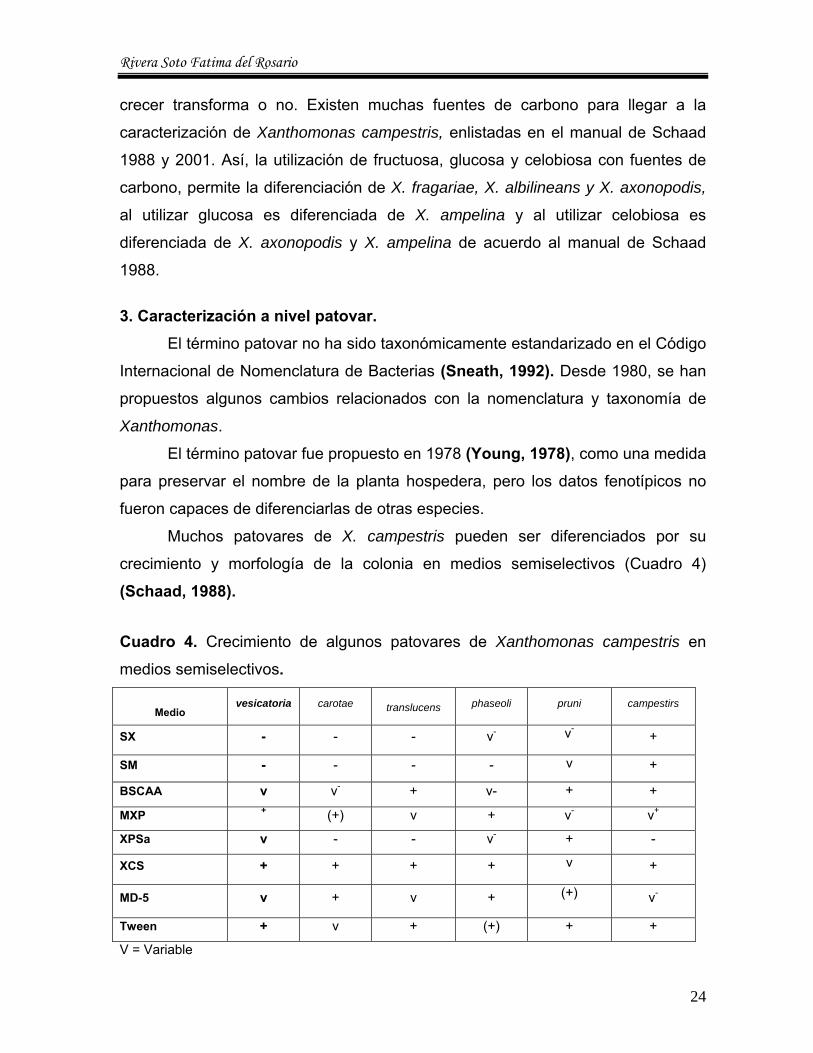
Muchos patovares de X. campestris pueden ser diferenciados por su
crecimiento y morfología de la colonia en medios semiselectivos (Cuadro 4) (Schaad, 1988).
Cuadro 4. Crecimiento de algunos patovares de Xanthomonas campestris en
medios semiselectivos.
Medio vesicatoria carotae translucens phaseoli pruni campestirs
SX - - - v- v-+
SM - - - - v +
BSCAA v v- + v- + +
MXP + (+) v + v- v+
XPSa v - - v- + -
XCS + + + + v +
MD-5 v + v + (+) v-
Tween + v + (+) + +
V = Variable
24

Rivera Soto Fatima del Rosario
3.1. Crecimiento en agar Tween y SX. El medio SX es usado para la caracterización de Xanthomonas spp., en
tejido de plantas y suelo, tambien es usada para semilla. Los medios Tween y SX
son dos de los medios utilizados por Jones et al en 1980 para la caracterización
de pathovares “vesicatoria”, ya que son dos los principales para difrenciar entre el
resto de pathovares de acuerdo al manual de Schaad, 1988.
4. Prueba de patogenicidad. Cuando un organismo se encuentra asociado en forma convincente a una
planta enferma es fundamental determinar un papel como agente causal de la
enfermedad. Para demostrar lo anterior se realizan los postulados de Koch, los
cuales consisten en lo siguiente:
1. El patógeno debe encontrarse asociado con la enfermedad en todas las
plantas enfermas que se examinen.
2. El patógeno debe aislarse y desarrollarse en cultivo (parásitos no
obligados) puro y se deben describir sus características o desarrollarse en
una planta hospedante susceptible (parásito obligado) y registrar su
presencia por los efectos que en ella produzca.
3. El patógeno que se desarrolle en un cultivo puro debe ser inoculado en
plantas sanas de la misma especie y variedad en que apareció la
enfermedad y debe producir la misma enfermedad en la planta inoculada.
4. El patógeno debe registrarse una vez más en cultivo puro y sus
características deben corresponder a las anotadas en el segundo punto
(Agrios, 2004).
Es importante considerar que la infección por bacterias fitopatógenas
ocurre a través de heridas o aberturas naturales, por ello se debe de ayudar a la
bacteria en el proceso de penetración. Con base en lo anterior se han ideado
métodos de inoculación artificial tales como: Aspersión, el cual se usa para inducir
manchas foliares o tizones en los que las bacterias penetran a través de los
25

Rivera Soto Fatima del Rosario
estomas, nectarios o estigmas; inyección ó punción, este método se utiliza en
caso de bacterias que causan marchités o pudrición blanda para ello se realizan
heridas en los cotiledónes y en la axila de la primera hoja verdadera; finalmente, la
inmersión en suspensión bacteriana que es recomendada para R. fascinas, planta
de tabaco, margaritón o semillas de chíncharo y la siembra en suelo esterilizado,
también se recomienda este último método para bacterias que invaden el xilema
(Rodríguez, 2001). Debido al tiempo consumido en la consecución de las pruebas
normales se han ideado pruebas rápidas de patogenicidad (Rodríguez, 2001) (Cuadro 5). C. Caracterización genotípica de Xanthomonas campestris pv. vesicatoria. La detección e identificación de bacterias fitopatógenas dependen del éxito
en el aislamiento de la bacteria en cultivo puro, la realización de pruebas
bioquímicas, serológicas y pruebas de patogencidad (Saettler 1989; Schaad 1989). Algunas veces se requieren medios selectivos para incrementar la
sensibilidad del aislamiento, de las bacterias, lo que inhibe el crecimiento de
bacterias contaminantes asociadas al tejido de la planta (Saettler 1989). Los
antisueros se han usado para la detección e identificación de bacterias, pero los
resultados son variables. Los antisueros policlonales pueden reaccionar con otras
bacterias y pueden ser ineficientes para detectar cepas específicas o patovares de
Xanthomonas campestris. Por lo que el diagnóstico de la mancha bacteriana ha
sido complicado debido a la heterogeneidad del agente causal de la enfermedad
(Bouzar et al., 1994). Actualmente se utilizan técnicas basadas en ácidos
nucleicos para la detección y caracterización de bacterias fitopatógenas
(Bereswill, 1992), por lo tanto, el diagnóstico de enfermedades bacterianas es
más sensible y específico a través de estos métodos (DeBoer, 1996). Para la
caracterización genotípica de Xanthomonas spp. que causan la mancha
bacteriana en plantas de tomate y/o chile se han empleando métodos
dependientes de amplificaciones por PCR (Reacción en cadena de la polimerasa)
(Lows et al., 1999). Se considera que los métodos basados en análisis de ácidos
nucléicos son más confiables y precisos que las características fenotípicas,
26

Rivera Soto Fatima del Rosario
serología, bioquímicas, fisiológicas, composición de ácidos grasos y utilización de
diversas fuentes de carbono, al comparar relaciones genéticas entre
microorganismos.
Cuadro 5. Pruebas rápidas de patogenicidad
Bacteria Parte vegetal
inoculada Método Síntoma esperado
Agrobacterium
rizogenes
Rebanadas de
zanahoria
Punción superficial Raíces adventicias
P. carotovorum Rebanadas de papa
P. fluorescentes Cebolla
X. c. pv. campestris Pepino
Cultivo bacteriano Pudrición suave
E. amylovora Frutos verdes del peral Lesiones necróticas
P. s.pv. syringae Vainas de chíncharo
P. s.pv. pisi Vainas de fríjol Punción con aguja
Lesiones húmedas
P. s.pv. phaseolicola
X. s. pv. phaseolicila
P. grupo syringae
E. amilovora
Hojas de tabaco Infiltración de hojas de
tabaco
Reacción de
hipersensibilidad
R. fascians Plántulas o semillas de
chíncharo dulce
Punción Fasciación
P. s. pv. syringae Frutos de limón Aspersión Necrosis hundida
P.s.pv. lachrymans Frutos de pepino
inmaduro
inyección Lesiones húmedas con
exudado
C. m. Subs.
michiganensis
Lesion levantada
X. vesicatoria Manchas y cancros
P. s. pv. tomato
Plántulas de tomate Inyección y corte de hoja
con navaja Manchas necróticas
pequeñas
C. m. Subs.
michiganensis
Hojas de maravilla Infiltración de
suspensión
Reacción de
hipersensibilidad
C. m. Subs.
sepedonicus
Plántulas de berenjena Inyección axilar Marchitez
A. tumefaciens Plántulas de tomate,
datura, girasol,
caléndula o crisantemo.
Aspersión en heridas en
el tallo
Agallas
27

Rivera Soto Fatima del Rosario
1. Detección de Xanthomonas campestris pv. vesicatoria mediante la Reacción en Cadena de la Polimerasa (PCR).
La técnica de la reacción en cadena de la polimerasa (PCR) desarrollada
por Kary Mullis a mediados de los 80’s revolucionó el campo de la biotecnología
debido a la gran sensibilidad que posee en comparación con otras técnicas tanto
serológicas como convencionales. Básicamente el principio de esta técnica es la
síntesis “in vitro” de una molécula de DNA concreta a miles de moléculas
idénticas en cuestión de minutos. Además la sensibilidad de la técnica depende de
la especificidad de los “primers” u oligonucleótidos ante otros microorganismos
que podrían estar presentes en las muestras (Leite RP et al., 1994). El gen hrp
forma parte de una región altamente conservada del genoma de bacterias
fitopatógenas, por lo que es utilizado para la detección e identificación de un gran
número de especies de Xanthomonas o patovares. Teniendo en cuanta que
bacterias no-patógenas no pueden producir síntomas en hospederos susceptibles
y reacción de hipersensibilidad en no-hospederos y se cree que no poseen
secuencias del DNA similar al gen hrp. El estudio del gen hrp ha permitido la
rápida detección de Xanthomonas spp. mediante el PCR.
Los requerimientos para esta técnica son:
a) Un ADN de doble cadena que actúe como molde o templado.
b) Dos oligonucleótidos sintéticos como iniciadores de la síntesis que deben ser
complementarios a regiones en cadenas opuestas de ADN. La hibridación es
en regiones que flaquean al DNA de interés y con los extremos 3’ orientados
hacia el centro.
c) Una DNA polimerasa termoestable, que resista temperaturas mayores a 95°C,
como la Polimerasa Taq de Thermus aquaticus.
d) Los cuatro dNTPs en un buffer adecuado.
(Balbás, 2002).
28

Rivera Soto Fatima del Rosario
III. HIPÓTESIS. La enfermedad de la mancha bacteriana ocasionada por Xanthomonas
campestris pv. vesicatoria está asociada a cultivos de tomate en Sinaloa.
IV. OBJETIVO. A. Objetivo General.
Detectar y caracterizar aislados de Xanthomonas campestris pv. vesicatoria
asociada con la mancha bacteriana en cultivos de tomate en Sinaloa.
B. Objetivos específicos.
Detectar por PCR la bacteria Xanthomonas campestris pv. vesicatoria en
tomate.
Caracterizar molecular y fisiológicamente aislados de Xanthomonas
campestris pv. vesicatoria asociadas con la mancha bacteriana en tomate.
Determinar la patogenicidad de aislados de Xanthomonas campestris pv.
vesicatoria obtenidos de plantas de tomate.
29

Rivera Soto Fatima del Rosario
V. MATERIALES Y MÉTODOS. 1. Material vegetal.
El muestreo se realizó en plantaciones de tomate del estado de Sinaloa (Ahome, Guasave, Mocorito, Sinaloa de Leyva, Culiacán, Elota, El Fuerte y
Salvador Alvarado), durante los ciclos agrícolas 2004-2006. Las muestras de
plantas sintomáticas se colectaron de lotes comerciales considerando los
síntomas inducidos por el genero de Xanthomonas en tomate. Las muestras se
colectaron en bolsas de polietileno y se trasladaron al laboratorio donde se
conservaron a 4°C paraanalizarse en un periodo no mayor a 48 hr después de su
colecta.
Durante el periodo muestreo se obtuvieron 174 muestras; de ellas 3, 25, 91,
30, 10, 3, 11 y 1 se colectaron en los municipios de Sinaloa de Leyva, Ahome,
Guasave, Culiacán, Mocorito, Salvador Alvarado, Elota y El Fuerte,
respectivamente (Figura 7).
Cabe resaltar que únicamente se colectaron muestras sintomáticas de tejido
foliar de tomate y de los municipios donde existe mayor superficie sembrada. El
número de muestras varió en los municipios debido a la eventualidad de los
síntomas mostrados en campo.
Salvador
Culiacán
Elota
Figura 7. Localización de
muestras de tomate con sínt
Alvarado Ahome
Sinaloa de Leyva
Mocorito
Guasave
los municipios de Sinaloa donde se realizó la colecta de
omas de mancha bacteriana (X. campestris pv. vesicatoria).
30

Rivera Soto Fatima del Rosario
A. METODOLOGÍA. 1. Pruebas simple de identificación para Xanthomonas.
Previo a la caracterización molecular se realizaron pruebas sencillas y
rápidas para eliminar las colonias bacterianas que no presentaban características
morfológicas de Xanthomonas campestris pv. vesicatoria; para ello se
consideraron las características de colonias en AN (agar nutritivo) y la prueba de
fluorescencia en medio B de King, tinción de Gram y forma de las bacterias. Todas
las colonias bacterianas que presentaron características similares al género
Xanthomonas se utilizarón en el desarrollo de este proyecto.
1.1. Aislamiento y condiciones de crecimiento bacterial en AN (agar nutritivo). Para el aislamiento de bacterias a partir de las plantas infectadas se
tomaron pequeños trozos de muestra del borde de la lesión con fin de restringir el
desarrollo posterior de contaminantes. Estos pequeños trozos de muestra se
sumergieron en alcohol al 96%, por un minuto, después se lavaron con agua
destilada estéril 1 minuto y se secaron en papel absorbente, para posteriormente
ser sumergidos en agua destilada estéril (2 ml) en un tubo por un minuto, de la
suspensión resultante se tomó una asada, para sembrar por estriado en cajas
petri con Agar Nutritivo (AN), las cuales se incubaron a 28º C, por 24 a 48 horas.
Las colonias amarillas fueron resembradas en cajas petri con agar nutritivo para
purificación, después de esto fueron preservadas en glicerol al 30% a -70°C.
Todas las cepas se transfirieron a medios de cultivo cada tercer dia ya que era
necesario mantener las colonias jóvenes (no mayores a 48 hr) para la realización
de las pruebas bioquímicas y fisiológicas.
1.2. Tinción Gram. La bacteria se cultivó en placa con AN por 24 horas. Se colocó una gota de
agua destilada estéril en un portaobjeto, y con el asa bacteriológica se tomó una
pequeña porción de una colonia (se recomienda colonias de 24 horas)
mezclandose con el agua para suspender la bacteria y así preparar un frotis, que
31

Rivera Soto Fatima del Rosario
se fijó a la flama de un mechero de alcohol.
Se adicionó una o dos gotas de la solución de cristal violeta, se dejó actuar
un minuto y posteriormente se decantó; se adicionaron una o dos gotas de la
solución de lugol, se dejó actuar por un minuto y luego se decantó. Se decoloró
con etanol hasta quitar el excedente de colorante. Por último se adicionaron dos
gotas de safranina y se dejó actuar por 30 seg y finalmente se enjuagó con agua
corriente para posteriormente observarse al microscopio biológico compuesto
(Carl Zeizz, serie 470801-9097).
1.3. Prueba de Fluorescencia. Después de realizar los aislamientos bacteriológicos en AN (Agar
nutritivo), a las colonias amarillas mucoides se les realizó la prueba de
fluorescencia en medio B de King. Treinta y tres gr de medio B de king se
agregaron en 1000 ml de agua destilada e inmediatamente y se agitaron
vigorosamente, asegurándose de que estuviera completamente disuelto.
Posteriormente se sometió a esterilización en autoclave durante 15 min a 120°C y
15 Lb de presión. El medio de cultivo una vez estéril y a temperatura de 50° C se
dispensó en cajas petri para esperar su solidificación a temperatura ambiente.
Enseguida, en condiciones asépticas se procedió a sembrar la bacteria por estria
en medio AN y posteriormente cuando los cultivos tuvieron 48 horas se
transfirieron al medio B de King. Se utilizó como control positivo una cepa de
Pseudomonas (proporcionada por el Dr. Rubén Félix Gastelum, profesor
investigador de biología de la Universidad de Occidentes). Las cajas sembradas
se incubaron a 27° C de 24 a 48 hr.
B. Caracterización molecular. 1. Extracción de DNA de cepa bacteriana.
La bacteria se inoculó en medio LB y se incubó a 37° C de 16 a 18 horas;
posteriormente se transfirieron 1.5 mL de cultivo bacteriano a un tubo Eppendorf
de 1.5 mL; se centrifugó a 13 000 rpm por 3 min después se decantó el
sobrenadante, se transfirieron nuevamente 1.5 mL de cultivo bacteriano al igual
32

Rivera Soto Fatima del Rosario
que en el primer paso, se centrifugaron y se dacantó. La pastilla se resuspendió
en 467 µL de solución amortiguadora con repetidos pipeteos, enseguida se
añadieron 30 µL de SDS al 10% y 3 µL de proteinasa K (20 mg/mL), se agitó por
inversión y se incubó 1 hora a 37 °C. Se adicionó un volumen igual (500 µL) de cloroformo: alcohol isoamílico (24:1) a -20°C, se mezcló por inversión varias
veces y se centrifugó por 8 min. a 13 000 rpm. Se transfirió la fase superior
acuosa a un tubo nuevo y se adicionaron 15 µL de RNA’sa. Se agitó por inversión
y se incubó a 37° C por 30 min. se adicionaron 500 µL de cloroformo:alcohol
isoamílico (24:1) a -20° C y se centrifugó a 13 000 rpm por 8 min. aquí se transfirió
nuevamente la fase superior acuosa a un tubo nuevo y se añadieron 50 µL de
acetato de sodio 3M pH 5.2 y se agitaron por inversión, se adicionaron 300 µL de
isopropanol a -20° C y se centrifugó por 8 min. a 13 000 rpm. se lavó el DNA con
etanol al 70% a -20°C por 3 min. Y se centrifugó a 13 000 rpm. y se decantó
cuidando de no perder la pastilla. Posteriormente se secó la pastilla y se
resuspendió el DNA en 20 µL de agua bidestilada. El DNA fue almacenado a -
20°C hasta su posterior utilización.
2. Electroforésis para visualizar el DNA genómico. La verificación de la extracción del DNA se llevó a cabo mediante su
visualización en un gel de agarosa por medio de la técnica de electroforesis. Se
utilizó un gel de agarosa al 0.8% con bromuro de etidio a una concentración de 0.2
µL/ml. Primeramente se colocó en un matraz 0.24 gr de agarosa, se adicionaron
30 ml de buffer TAE 1X y se disolvió con temperatura por 1 minuto en horno de
microondas. Posteriormente, se dejó enfriar lo suficiente antes de gelificar, se
agregaron 0.8 µL de bromuro de etidio y por último, se vació en la cámara de
electroforesis hasta enfriarse y gelificar, aproximadamente 10 a 15 min despues.
Una vez gelificado se cargaron 2 µL de la muestra del DNA, junto con 6µL de
agua bidestilada estéril y 2µL de buffer de carga (colorante naranja G). La
electroforesis se corrió a 80 V durante 20-25 min, en una cámara de electroforesis
con el buffer TAE 1X. Finalmente, se visualizó el DNA en un transiluminador de
luz ultravioleta y se colocó en un fotodocumentador de imágenes para registrar los
33

Rivera Soto Fatima del Rosario
resultados (Chemidoc, Biorad).
3. Determinación de la concentración de DNA. Antes de correr la PCR se determinó la concentración del ADN mediante la
lectura de absorbancia en un espectrofotómetro, con una longitud de onda de 260
nm. 5 µL del DNA obtenido fueron diluidos en 450 µL de agua destilada estéril.
Para conocer la concentración del DNA se aplicó la siguiente formula:
Concentración de DNA = (50 ng/µL) (Factor de dilución) (Absorbancia)
De todas las muestras se prepararon alícuotas a una concentración de 20
ng/µL y se utilizaron 2 µL de DNA templado en la reacción de PCR.
4. Detección del patógeno por PCR. La amplificación de la secuencia hrp mediante PCR es una herramienta útil
para la detección e identificación de Xanthomonas. La región hrp no está presente
en bacterias saprofitas y Xanthomonas spp. oportunistas, lo que reduce la
posibilidad de obtener falsos positivos.
El par de primer (Cuadro 6) RST65/RST69 amplifican un fragmento de 420
pb de cepas del patovar vesicatoria; estos primers están diseñados en la
secuencia conservada entre los cuatro grupos de Xanthomonas campestris pv.
vesicatoria. (Obradovic et al., 2003). Cuadro 6. Secuencias de los oligonucleótidos usados en la PCR para detectar aislados de Xanthomonas campestris pv. vesicatoria.
Primer Secuencias 5´→ 3’ Referencia
RST65 GTCGTCGTTACGGCAAGGTGGTCG Obradovic et al., 2003
RST69 TCGCCCAGCGTCATCAGGCCATC Obradovic et al., 2003
La PCR se realizó en tubos Eppendorf de 0.2 ml con un volumen total de
reacción de 25 µL el cual contenía 40 ng de DNA total, 1 X de buffer, 2 mM de
34

Rivera Soto Fatima del Rosario
MgCl2, 0.4 pmol de cada primer, 0.2 mM de nucleótidos trifosfatados (dNTP´s) y
2.0 Unidades de taq polimerasa (Invitrogen Cat. 18038-018).
Para la amplificación de los ácidos nucléicos los tubos Eppendorf con la
mezcla de reacción se colocaron en un termociclador (Cycle-BioRad). El proceso
de la PCR se sujetó a las siguientes condiciones: 94°C por 2 min. (primera etapa
de desnaturalización), 94 °C por 30 seg. (segunda etapa de desnaturalización),
61°C por 45 seg. (etapa de alineamiento) y 72°C por 30 seg. (etapa de extensión).
Los últimos 3 pasos se repitieron por 30 ciclos y se dió una extensión de 72°C por
5 min para la polimerización final. Posteriormente los tubos permanecieron a 4°C
para enfriamiento y congelación de las moléculas sintetizadas.
5. Purificación de bandas (Kit Gen Clean III). Los fragmentos positivos para Xanthomonas campestris pv. vesicatoria se
cortaron del gel y se purificaron siguiendo el protocolo del kit comercial, Gen Clean
Kit III (Gibco-BRL Cat. 1001-200). Enseguida, se colocó la banda en tubos
Eppendorf y se adicionaron 500 µL de ioduro de sodio (NaI) y se incubaron a 55°
C, agitando constantemente hasta que se disolviera la banda con agarosa y liberar
el DNA. Una vez disuelta la agarosa, se agregaron 5 µL de perlas de vidrio (Glass
1000) y se incubó por 5 min a temperatura ambiente agitándose cada minuto por
inversión. Posterior a la incubación se centrifugó por 10 seg a 13 000 rpm y se
eliminó el sobre nadante. El presipitado se lavó tres veces con 500 µL de solución
“New Wash”. En cada lavado la suspension se agitó en un vortex y se centrifugó
por 10 seg a 13,000 rpm descartandose el sobrenadante. Posteriormente la
pastilla se secó a temperatura ambiente y se resuspendió en 10 µL de agua ultra
pura y finalmente se incubó a 55° C por 5 min y se centrifugó a 13,000 rpm
durante 30 seg para liberar el DNA de las perlas de vidrio (Glass 1000) y que este
quedara disuelto en el sobrenadante, el cual se recupero con la pipeta cuidando
de no tocar la pastilla.
Para corroborar la recuperación del DNA purificado, se realizó una corrida
electroforética en un gel de agarosa al 1% con 2 µL del producto, 6 µL de agua
bidestilada estéril y 2 µL de buffer de carga (colorante G).
35

Rivera Soto Fatima del Rosario
6 Ligación de productos de PCR en el vector de clonación. Una vez purificados los fragmentos de DNA de Xanthomonas campestris
pv. vesicatoria se ligaron en el vector de clonación pGEM-T Easy Vector System
(Promega Madison WI USA Cat. 157348). Este vector posee en los extremos una
base de timinas y el fragmento amplificado por PCR extremos con adeninas, que
por complementariedad y con la acción de la enzima ligasa permite el
recirculamiento del vector con el fragmento amplificado. Para la reacción de
ligación se mezclaron 4 µL de DNA purificado, 0.5 µL de vector (25 ng/ml), 5.0 µL
de buffer de clonación (2X), 1 µl de T4 DNA ligasa (3 U/µl) y 2.0 µl de agua
bidestilada estéril. Posteriormente la reacción se incubó a 4 °C durante 12 hr. El
producto de la ligación se utilizó en el proceso de transformación de E. coli.
Cuando no se realizó de inmediato la transformación, la reacción de ligación se
conservó a -20 °C hasta su utilización.
7. Transformación de células competentes. Para el proceso de transformación se utilizaron cajas petri (dos por cada
ligación) con LB sólido, ampicilina (100 mg/ml), IPTG (200ng/ml) (isopropil-β D-
tiogalactopiranosida) y X-Gal (20 ng/ml) (5-bromo-4cloro-3i-ndoli- β-D-
galactopiranosida). Las células competentes de E. coli (JIM 109, Cat. N.-L2001,
Promega) se descongelaron sobre hielo agitándose cuidadosamente por
inversión. En un tubo Eppendorf se colocan entre 2 y 5 µl de la reacción de
ligación (dependiendo de la intensidad de la banda purificada). Posteriormente, se
adicionaron 50 µl de células competentes. La reacción se mezcló golpeando
ligeramente los tubos con los dedos. Rápidamente los tubos fueron colocados en
hielo por 20 min. Posteriormente se procedió al choque térmico de las células por
aproximadamente 45-50 seg en baño maria a 42 °C (sin agitación).
Posteriormente los tubos se regresaron al hielo por dos min. Se agregaron 900 µl
de medio SOC (1% de tristona, 0.5 % de extracto de levadura, 8.5 mM de NaCl,
2.5 mM de KCl, 0.01 mM de MgCl2, 0.02 mM de glucosa 1000 ml de agua
bidestilada) a los tubos que contenían las células transformadas con las
reacciones de ligación y por último se incubó a 37° C con agitación a 160 rpm por
36

Rivera Soto Fatima del Rosario
1.5 hrs.
En la cámara de flujo laminar se adicionaron 100 µl de la reacción de
transformación se distribuyéndose uniformemente a las cajas petri con
LB/ampicilina. El resto del transformado se centrifugó para obtener un
concentrado de las células. Se descartaron aproximadamente 800 µl de LB a otra
caja. Las cajas petri se incubaron por 16 horas a 37 °C. Las colonias
recombinantes con el fragmento de DNA insertado exhiben un color blanco, lo cual
se debe a que la inserción del DNA amplicado interrumpe el marco de lectura de
β-galactosidasa haciéndola no funcional y la hace incapaz de emplear extracto X-
gal para producir la coloración azul que muestra en células no transformadas.
8. Extracción de DNA plasmídico (minipreps). Para realizar la extracción del DNA plasmídico se crecieron las células
recombinantes (de color blanco) por 16 horas en LB líquido con ampicilina a 37°C
y agitación a 160 rpm. Posteriormente se procedió a la extracción del DNA
plasmídico, de acuerdo a las recomendaciones del fabricante del kit Rapid
Plasmid Miniprep Purification Systems (Marlingen Bioscience 50 cat. No 11453-
016 Germany/USA).
Para verificar si se tenía el fragmento insertado en el vector se realizó
digestión con la enzima EcoRI, y en algunos casos el DNA plasmídico de cada
clona se amplificó por PCR con el par de oligonucleótidos RST65/RST69 y es
visualizó en un gel de agarosa al 0.8%.
9. Análisis de restricción del gen hrp amplificado por PCR (RFLP-PCR). Para determinar la presencia de los grupos (“A”, “B”, “C” y “D”)de
Xanthomonas campestris pv. vesicatoria, los productos de PCR fueron digeridos
con la enzima HaeIII que permite observar diferencias entre dichos grupos
(Obradovic et al., 2003). Los análisis de restricción de regiones amplificadas del
gen hrp pueden realizarse para cualquier grupo patógeno de Xanthomonas,
facilitandose la identificación y clasificación de cepas bacterianas (Leite et al., 1994). Todas las amplificaciones por PCR de las muestras fueron clonadas y
37

Rivera Soto Fatima del Rosario
secuenciadas y mediante la técnica de RFLP-PCR se compararon los patrones
obtenidos.
Para el análisis de restricción de los fragmentos amplificados por PCR con
los primers RST65/RST69 que amplifican un fragmento de 420 pb, se utilizó la
enzima de restricción Hae III (Invitrogen). Para la digestión se mezclaron 10 µl del
producto de PCR 2.0 Unidades de enzima, buffer 1X y se sometió a un periodo de
incubación de 2 horas a 37°C. Los fragmentos obtenidos mediante restricción se
separaron en un gel de agarosa al 2%, separados por electroforesis con un voltaje
de 70 V durante 1 hr. El gel de agarosa fue teñido con una solución de bromuro de
etidio, los fragmentos de DNA se visualizaron con luz UV en un transluminador
digitalizado (Chemidoc, Biorad Cat. 170-8070 USA).
10. Secuenciación, análisis y comparación de secuencias.
La secuenciación de clonas y productos directos de PCR se realizó en el
laboratorio de Química de DNA del CINVESTAV-IPN Unidad Irapuato. Se utilizó el
kit Dye Terminator Cycle Sequencing Ready Reaction empleando un secuenciador
ABI PRISM 377 PERKIN-ELMER (CETUR, Norwalk, CT).
Las secuencias obtenidas fueron comparadas con secuencias reportadas
en el banco de genes (Gen Bank) del centro Nacional para la Información de
Biotecnología (NCBI- Nacional Center for Biotechnology Information
htt://www.ncbi.nibh.gov/), empleó el programa BLAST-N de dicho sitio para
conocer el nivel de identidad genética existente con secuencias ya reportadas y
depositadas en el banco.
La comparación de las secuencias del gen hrp y el árbol filogenético fueron
realizados con el programa MegAling y los mapas de restricción se realizaron con
el programa MapDraw mediante el software DNASTAR (versión 2.0 Madison,
Wisconsin, USA). Los mapas fueron comparados con los perfiles de referencia de
Serbia: 93-1 (raza de chile grupo A), 75-3 (raza de tomate 1 grupo A), XV56
(tomate raza 2, grupo B), 91118 (tomate raza 3, grupo C) y (X. gardneri, grupo D)
(Jones et al., 2000) (Figura 8).
38

Rivera Soto Fatima del Rosario
Figura 8. Restricción de fragmentos de 420 pb del gen hrp de Xanthomonas spp.
con la enzima HaeIII con los primers RST65/69. Carriles: A) 93-1 (raza de chile
grupo A); A1) 75-3 (raza de tomate 1 grupo A); B) XV56 (tomate raza 2, grupo B);
C) 91118 (tomate raza 3, grupo C); D) X. gardneri, grupo D.
C. Caracterización fisiológica. Para determinar las características morfológicas de la bacteria asociada a
la mancha bacteriana del tomate se realizaron aislamientos en placas con AN; en
este caso se registraron la forma, color, elevación y superficie de las colonias. A
los diferentes aislamientos se le realizaron pruebas fisiológicas y bioquímicas para
su identificación. Durante esta investigación sólo se realizaron algunas de las
pruebas más comunes para la caracterización Xanthomonas campestris pv.
vesicatoria.
1. Caracterización fisiológica del genero Xanthomonas.
1.1. Catalasa. La catalasa es una enzima que protege a las células frente al peroxido de
hidrógeno y se determina mediante la producción de oxígeno al mezclar la
bacteria con peróxido de hidrógeno. Para realizar esta prueba, se tomó un poco
de colonia bacteriana, el cual se colocó en un portaobjetos; posteriormente se
39

Rivera Soto Fatima del Rosario
añadió a la muestra una gota de peróxido de hidrógeno. La reacción se consideró
positiva a catalaza cuando se desprendieron burbujas de oxígeno.
1.2. Oxidasa. Con una asa de platino se tomó un poco de colonia bacteriana y se colocó
sobre una tira de papel filtro impregnado con la amina (dihidrocloride
phenylenediamine). Cuando el papel viraba del color blanco a purpura en
aproximádamente 10-60 seg, se consideró a la bacteria en estudio oxidasa
positiva.
1.3. Crecimiento en YDC (Extracto de levadura dextrosa). En un matraz se agregaron 500 ml de agua destilada y se añadieron en
orden de aparición 5 gr de extracto de levadura, 10 gr de dextrosa, 10 gr de
carbonato de calcio y 7.5 gr de agar. Se agitó el matraz vigorosamente.
Posteriormente se sometió a esterilización en autoclave durante 15 min a 120°C y
15 Lb de presión. El medio de cultivo una vez estéril y a temperatura de 50° C se
dispensó en cajas petri para esperar su solidificación a temperatura ambiente.
A partir de colonias bacterianas de 24-48 horas desarrolladas AN (agar
nutritivo), se tomaron bacteria con una asa y se colocaron en 5 puntos
equidistantes sobre el medio. Después de la siembra cajas se incubaron a 27° C
de 24 a 48 hr. Colonias amarillas o naranjas y mucoides en este medio de cultivo
son características de Xanthomonas.
2. Caracterización a nivel especie. Las cajas bacterianas que fueron identificadas como Xanthomonas, se
caracterizaron para diferenciar su especie. Se realizaron las siguientes pruebas:
2.1. Crecimiento en YS (Caldo de levadura). Esta prueba se realiza para determinar el crecimiento de la bacteria a 35
°C. Para ello se utilizó el medio líquido YS (Caldo de levadura), Se agregaron, en
1000 ml de agua destilada, 0.5 gr de NH4H2PO4, 0.5 gr de K2HPO4, 0.2 gr de
40

Rivera Soto Fatima del Rosario
MgSO4.7H2O, 5.0 gr de NaCl y 0.5 gr de extracto de levadura. Posteriormente se
agitó vigorosamente y se vaciaron 9 ml en tubos de ensayo los cuales se taparon
perfectamente con un tapón de algodón y se esterilizaron por 15 min a 120 ° C y
a 15 Lb de presión. La inoculación del medio de cultivos se realizó cuando este se
igualó a la temperatura ambiente.
Las cepas en estudio se cultivaron previamente, en medio AN durante 24-
48 horas. A partir de estas colonias jovenes, se tomó una porción de bacterias
mediante el asa y se transfirieron en los tubos con medio líquido estéril. Por último
se colocaron en la incubadora a 35°C por 10-12 días.
2.2. Hidrólisis de almidón. En un matraz Erlenmeyer, se colocaron 500 ml de agua destilada
posteriormente se le adicionaron 10 gr de agar y 5 gr de almidón, se agitaron
vigorosamente asegurándose de que estuvieran disueltos. Posteriormente el
medio de cultivo se esterilizó y se dispensó en cajas de Petri.
Después se procedió a sembrar la bacteria colocando crecimiento
bacteriano en cinco puntos equidistantes equidistantes en cajas de petri
conteniendo el medio de cultivo. Después de la siembre los cultivos se incubaron
por cinco días a 27° C.
Finalmente, las cajas de Petri con la siembre se inundaron con lugol (yodo
1 gr; yoduro de potasio 2 gr; agua destilada 100 ml). La aparición de zonas claras
en los márgenes o debajo de las colonias indicó que el almidón fue hidrolizado
indirectamente por las enzimas alfa y β amilasas.
2.3. Licuefacción de gelatina. En un matraz que contenía 1000 ml de agua destilada se disolvieron 3 gr
de extracto de carne, 5 gr de peptona y 120 gr de gelatina; se agitaron hasta que
todos los componentes se mezclaran; enseguida se colocaron 10 ml de este
medio en tubos de cultivo y se esterilizaron, (como se describe en el apartado
2.1.). El medio de cultivo se conservó a 4°C y se inoculó por punción con colonias
jovenes (24-48 hr). Por ultimo, los tubos inoculados se incubaron a 28°C y se
41

Rivera Soto Fatima del Rosario
determinó la licuefacción del medio por cambio del color a los 5 días.
2.4. Dihidrolasa de arginina. Se prepara el medio Thornley´2A (1000 ml de agua destilada, 1.0 gr de
peptona, 5.0 gr de NaCl, 3.0 gr de K2HPO4, 3.0 gr de agar; 0.01 gr de rojo fenol y
10.0 gr de monoclorhidrato de arginina y el pH se ajustó a 7.2), posteriormente se
vaciaron 9 ml de medio de cultivo a tubos de ensayo y se esterilizaron (como se
describe en el apartado 2.1.).
Una vez que la temperatura del medio de cultivo se igualó a la del
laboratorio la bacteria se inoculó. Para cada aislamiento bacteriano se utilizaron
tres tubos del medio antes señalado. La inoculación del medio se realizó mediante
punción y se utilizó un tubo con el mismo medio como testigo. Una vez inoculada
la bacteria, se adicionó a cada tubo una capa de 1 cm de aceite mineral y se
incubaron durante 72 hr a 28°C.
2.5. Fuentes de carbono (Fructuosa, glucosa y celobiosa). En esta prueba se utilizó medio C de Dye. En un matraz de 1000 ml, se
agregaron 500 ml de agua estéril se agregaron 0.5 gr de NH4H2PO4, 0.5 gr de
K2HPO4, 0.2 gr de MgSO4.7H2O, 5.0 gr de NaCl, 1.0 gr de extracto de levadura,
12.0 gr de agar y 0.7 ml de lila bromocrisol (1.5% en solución de alcohol),
después se agitó vigorosamente, se ajustó el pH a 6.8 por ultimo se esterilizó
(como se describió en el apartado 2.1.). Una vez que el medio se enfrió seagregó
la fuente de carbono filtrada y estéril al 0.5% v/v, se agitó y se vació en cajas
Petri. Cabe mencionar que se preparó un matraz con medio C de Dye para cada
fuente de carbono realizadas (celobiosa, glucosa y fructuosa), los cuales se
dispensaron en cajas de Petri y se inocularon en un periodo no mayor de 48 hr
después de su preparación. La bacteria fue inoculada por estria a partir de
colonias jovenes (24-48 hr) cultivadas en AN (agar nutritivo) y enseguida fueron
incubadas a 28 °C por 48 hr. La fuente de carbono es positiva si existe un cambio
de color en el medio que fue inoculado.
42

Rivera Soto Fatima del Rosario
3. Caracterización a nivel patovar. 3.1. Crecimiento en medio Tween.
El medio Tween es un medio selectivo para X. campestris pv. vesicatoria.
Para esta prueba, en un matraz que contenía 1000 ml de agua destilada se
agregaron 10.0 gr de peptona, 10.0 gr de bromuro de potasio, 0.25 gr de CaCl2 y
15.0 gr de agar. Se agitó vigorosamente hasta que se homogenizó perfectamente,
posteriormente se esterilizó. A una temperatura de aproximadamente de 45°C,
cuidando de que no solidificara, se agregaron los siguientes reactivos: 10.0 ml de
tween, 75.0 mg de cyclohexamida, 25.0 mg de cephalexina, 6.0 mg de 5-
fluorouracil y 0.4 mg de tobramicina. Se agitó suavemente y se vació en cajas petri
(como se mencionó en el apartado 2.1.4.).
La inoclulación se llevó a cabo sembrando la bacteria por estría, partiendo
de colonias de las cepas crecidas en AN por 24-48 horas. Después de la siembra
las cajas se incubaron a 27° C de 24 a 48 hr.
3.2. Crecimiento en medio SX. Este es medio sólido selectivo para X. campestris pv. vesicatoria. Para esta
prueba en un matraz que contenía 500 ml de agua destilada estéril se agregaron
5.0 gr de almidón, 0.5 gr de extracto de carne, 2.5 gr de cloruro de amonio, 1.0 gr
de fosfato de potasio, 0.5 ml de violeta metil 2B (solución al 1% en etanol al 20%),
1.0 ml de verde metil (solución acuosa al 1%) y 6.0 gr de agar, el medio se
esterilizó como se mencionó anteriormente. Una vez que el medio se enfrío se
adicionaron 2.5 ml de una suspensión de cycloexamida. (se adicionaron 5.0 gr a
5.0 de metanol y se aforó con agua estéril), se homogenizó vigorosamente.
Después se vació en cajas petri.
4. Pruebas de patogenicidad. Para las pruebas de patogencidad se usaron tres plantas por aislamiento,
este se llevó a cabo en plantas de tomate con 2 meses de edad. Las plantas se
desarrollaron en vasos de poliestireno de 1.0 L conteniendo una mezcla de muzgo
y vermiculita (1:1) y se fertilizaron con triple 17 cada 10 días. La inculación con el
43

Rivera Soto Fatima del Rosario
aislamiento de Xanthomonas, obtenidas de tomate, se efectuó en plantas en la
etapa de seis hojas verdaderas. El inóculo consistió en una suspensión bacteriana
(1 x 108 células/ml) de 48 hr, la cual se obtuvo a partir de crecimiento bacteriano
en AN. Previo a la inoculación, las plantas se frotaron con un agitador de vidrio
para provocar el rompimiento de tricomas y facilitar la penetración de la bacteria,
la cual se frotó en hojas, peciolos y tallos con algodón impregnado con la misma;
como testigo se utilizó algodón impregnado con agua destilada estéril. Después de
la inoculación, las plantas se colocaron en el invernadero y se cubrieron con una
bolsa de polietileno para mantener la humedad. Las plantas estuvieron en
observación diaria para mantener las condiciones de humedad y temperatura
óptimas para Xanthomonas campestris pv. vesicatoria.
VI. RESULTADOS Y DISCUSIONES.
44

Rivera Soto Fatima del Rosario
A. Sintomatología. La bacteria Xanthomonas campestris. pv. vesicatoria se presenta en toda la
etapa de crecimiento del cultivo de tomate, afectando hojas, tallos y frutos. Se
pueden presentar síntomas más severos cuando el clima es cálido y con alta
humedad; sí este es el caso las lesiones pueden unirse y formar manchas grandes
necróticas irregulares en follaje, causando defoliacion. En algunos casos puede
afectar la floración y causar el aborto de flores. Durante este estudio las muestras
colectadas en las diferentes regiones del estado de Sinaloa (Ahome, Guasave,
Mocorito, Sinaloa de Leyva, Culiacán, Elota, El Fuerte y Salvador Alvarado)
presentaron sintomatología típica de la bacteria. Se observó únicamente manchas
necróticas en tejido foliar, también en las partes apicales de la planta, en la
mayoría de las plantas se observó la necrósis en el envés y por los bordes de las
hojas. También se encontraron síntomas más severos como manchas necróticas
extensivas e irregulares causando defoliación. Cabe mencionar que únicamente
se colectaron plantas sintomáticas (Figuras 9-10).
B
FS
A
igura 9. Síntomas de mancha bacteriana observados en cultivos de tomate en
inaloa. A) Manchas necróticas en toda el área foliar B) y bordes de un foliolo.
A B
45

Rivera Soto Fatima del Rosario
Figura 10. Síntomas observados en cultivo de tomate. A) Defoliación. B) Manchas
necróticas severas e irregulares.
B. Pruebas simples de identificación de Xanthomonas.
Para efectuar la caracterización molecular se realizaron aislamientos de
bacterias asociados a 174 muestras sintomáticas del tomate, colectadas en los
diferentes municipios del estado de Sinaloa. Pruebas fisiologicas sensillas se
llevaron a cabo para determinar las características más importantes de la bacteria
de interes en el presente estudio. Se ignoraron 127 aislados que no presentaron
caracteristicas importantes para este proyecto quedando un total de 47
aislamientos correspondieron al genero de Xanthomonas campestris pv.
vesicatoria, pues presentaron caracteristicas importantes (Figura 11) para estudio.
Una de las pruebas sencillas para la selección de aislados que posteriormente
serian analizados por técnicas moleculares fue la observación de la morfología
colonial en AN (agar nutritivo) y tinción Gram y la prueba de fluorescencia.
46

Rivera Soto Fatima del Rosario
Figura 11. Aislados bacteriológicos obtenidos de tejido foliar sintomático de plantas de
tomate, con mancha bacteriana.
1. Tinción Gram. Anteriormente hablamos de microorganismos Gram-positivos cuando
aparecen teñidos de color azul-violeta y de Gram negativos cuando se visualizan
de color rojo-rosado. En este estudio los 15 aislados bacteriológicos fueron Gram
negativo y de forma bacilar, lo cual es característico del género Xanthomonas (Figura 12).
Figura 12. Fotografía con microscopio de la bacteria Gram negativa y forma bacilar de
aislados bacteriológicos de plantas de tomate.
De acuerdo a la bibliografía, las células Gram positivas retienen el cristal
violeta cuando se tratan con etanol mientras que las Gram negativas no lo tiñen y
se tiñen entonces del color rojo de la safranina
esto se debe a que las células Gram negativas tienen un contenido graso muy
47

Rivera Soto Fatima del Rosario
elevado en su pared celular y al ser decoloradas pierden el complejo formado por
cristal violeta y el iodo tornándose incoloras y posteriormente al ser tratadas con
safranina se tornan rojas y las Gram positivas de lo contrario retienen el complejo
debido a la poca cantidad de lípidos en su pared.
2. Fluorescencia. De los 47 aislados en estudio, sólo en dos se observó pigmento
fluorescente. Lo cual sugirió que estas cepas pertenecen al género Pseudomonas,
que son comúnmente confundidas con Xanthomonas spp. ya que presentan la
misma morfología colonial en AN, pero Pseudomonas spp. es fluorescente en
medio B de King (Figura 13). Se obtuvieron un total de 15 aislados con
características amarillas mucoides no fluorescentes sobre el medio B de King,
para seguir con la caracterización molecular y posteriormente correr la
caracterización fisiológica.
A B
Figura 13. Prueba de fluorescencia en medio B de King, con exposición a luz ultravioleta
(260 nm): A) aislado de tomate, B) Testigo positivo (Pseudomona syringae pv. tomato)
proporcionada por el Dr. Rubén Félix Gastelum.
C. Caracterización molecular 1. Extracción de DNA.
Se estandarizó una técnica de extracción de DNA de cepas bacterianas
(Figura 14) ya que es de gran importancia la obtención de DNA de buena calidad
para la detección de fitopatógenos (Chavez-Medina, 2006).
48

Rivera Soto Fatima del Rosario
1 2 3 4 5 6 7
Figura 14. DNA de aislados bacteriológicos de tejido de tomate (carriles 1 – 7).
2. Detección de bacterias por PCR. A todas las extracciones de DNA de colonias amarillas mucoides, se les
realizó la detección por PCR. Se amplificaron fragmentos de 420 pb de la
secuencia hrp de Xanthomona campestris pv. vesicatoria con los primers
RST65/RST69 (Figura 15). El primer fragmento amplificado por PCR fue clonado
y secuenciado para ser usado como control positivo para todas las detecciones
posteriores.
Se detectó la presencia de Xanthomonas en 15 muestras de las 47
analizadas. La distribución de las muestras positivas fue la siguiente: se
detectaron 2 de 3 en Sinaloa de Leyva, 4 de 25 en Ahome, 2 de 91 en Guasave,
3 de 30 en Culiacán, 1 de 10 en Mocorito, 1 de 3 en salvador alvarado, 1 en El
Fuerte y 1 de 11 en Elota.
Se ha demostrado la presencia de Xanthomonas campestris pv. vesicatoria
en cultivos de tomate en el estado de Sinaloa (Carrillo-Fasio et al., 2001). Se han
hecho investigaciones en otras partes del mundo como El Caribe, América Central
(Bouzar et al., 1999), Norte América, Sur América, Europa, Australia, África y
Estados Unidos (Bouzar et al., 1994).
49

Rivera Soto Fatima del Rosario
1 2 3 4 5 M (-) (+)
1 2 3 4 5 M (-) (+)
uliacan, (M) marcador de peso
olecular 1 kb, (-) control negativo, (+) control positivo.
A
420 pb
B
420 pb
Figura 15. Productos amplificados por PCR del gen hrp de Xanthomonas spp. con el par
de primers RST65/RST69. A) Carriles 1-5 tejido foliar de tomate del municipio de Ahome,
(M) marcador de peso molecular 1 Kb, (-) control negativo, y (+) control positivo. Panel B) carril 1-5 tejido foliar de tomate del municipio de C
m
50

Rivera Soto Fatima del Rosario
3. Identificación de Xanthomonas spp. mediante el análisis por RFLP-PCR. El análisis de restricción de fragmentos amplificados es una herramienta
muy importante para la identificación de grupos de cepas fitopatógenas y
patovares de X. campestris (Leite et al., 1994). Se utilizó la enzima HaeIII para
digerir los fragmentos correspondientes al gen hrp de Xanthomonas spp.
amplificados a partir de muestras de tomate. El análisis de los patrones de
restricción muestra que las Xanthomonas spp. detectadas en esta investigación
pertenecen al grupo A (X. euvesicatoria) y C (X. perforans) ambas nombradas
de bandas,
aso que no sucede con los patrones de bandas de los grupos B y D.
iones de Sinaloa digeridos con la enzima HaeIII. M. Marcador de peso
anteriormente Xanthomonas axonopodis pv. vesicatoria (Jones et al., 2004). Los perfiles de restricción obtenidos con la enzima HaeIII de las 15
muestras de tomate de Sinaloa fueron idénticos a los patrones de bandas del
grupo A y C obtenidos por Obradovic et al (2003) (Figura 16). Con esta enzima es
difícil diferenciar los grupos A y C, ya que presentan el mismo patrón
c
M
123pb
51 pb
135pb
69 pb
1 2 3 4 5 7 8 9 10 11 12
Figura 16. RFLP-PCR. Análisis de restricción de los fragmentos amplificados por PCR
con los primers RST65/RST69 (420 pb). Carriles 1-12 Xanthomonas spp. de tomate de
los diferentes reg
molecular 50 pb.
51

Rivera Soto Fatima del Rosario
4. Análisis de secuencias. Los productos amplificado por PCR de la región hrp de 420 pb con los
primers RST65/RST69 fueron clonados y secuenciados. Cabe señalar que se
clonaron todos los fragmentos amplificados por PCR para comparar las
secuencias en el programa Map Draw y observar diferencias en base a lo
publicado por Obradovic y colaboradores (2003). Durante este trabajo se lograron
encias 147-3 Ahome y 166-4
detectar, clonar y secuenciar 15 fragmentos de Xanthomonas spp (Cuadro 7-8)
El grupo de secuencias de las cepas 161-3 de Culiacán, 154-4 de Ahome,
125-5 Mocorito, 167-4 Ahome, 162-1 Culiacán, G5 Salvador Alvarado, 159-1
Guasave, SE59 Sinaloa de Leyva, 145-1 Elota y 143-C, presentaron una
homología del 100 % entre ellas. El grupo de secu
Ahome tienen una homología del 100 % entre ellas.
Las secuencias de las Xanthomonas spp. en estudio se compararon con
otras secuencias reportadas en el banco de datos (mediante el programa Blast
Search del Nacional Center for Biotechology Information (NCBI), http:// www.
ncbi.nlm.nij.gov/blast), obteniéndose los siguientes resultados: las 15 secuencias
de los diferentes aislados de los municipios de Sinaloa presentan una alta
homología con la secuencia del grupo A (AMO39952) de tomate reportada en el
banco de genes. Las secuencias de Xanthomonas campestrsi pv. vesicatoria de
tomate A154-4, A 167-4, A 147-3 y A166 del municipio de Ahome tienen una
homología del 99.5 al 100 % con la cepa 75-3 (Grupo A), siendo la cepa A166 y la
147-3 las únicas de todas las secuencias obtenidas que presentan una homología
del 100% con la reportada en el banco de genes, las secuencias 161-3, C162-1 y
163-3 del municipio de Culiacán, tienen una homología del 99.3 al 99.5 % con la
cepa 75-3 (Grupo A), del municipio de Mocorito sólo se obtúvo la secuencia M125-
5, la cual presenta una homología del 99.5 % con la secuencia de la cepa 75-3
(Grupo A), las dos secuencias obtenidas del municipio de Sinaloa de Leyva, SE59
y 143-C, ambas presentan una homología del 99.5 % con respecto a la secuencia
de la cepa 75-3 (Grupo A), las secuencias de las cepas del municipio de Guasave,
G158-3 y G159-1, ambas presentan una homología del 99.3 al 99.5% con
respecto a la cepa 75-3 (Grupo A), del municipio de Salvador Alvarado se obtuvo
52

Rivera Soto Fatima del Rosario
53
cepa 75-3 (Grupo A).
una secuencia, G5, con una homología del 99.5 % con respecto a la cepa 75-3
(Grupo A) y por último la secuencia del municipio de Elota, E145-1 presenta una
homología del 99.5 % con la

Rivera Soto Fatima del Rosario
Cuadro 7. Secuencias del gen hrp de Xanthomonas campestris pv. vesicatoria obtenidas de los cultivos de tomate de
diferentes cultivos de Sinaloa.
Número Clona de Xanthomonas spp.
Origen Similitud con otras secuencias
1 A154 AhomeXanthomonas campestris pv. vesicatoria complete genome AMO39952 99%
2 167-4 AhomeXanthomonas campestris pv. vesicatoria complete genome AMO39952 99%
3 147-3 AhomeXanthomonas campestris pv. vesicatoria complete genome AMO39952 99%
4 166-4 Ahome
Xanthomonas campestris pv. vesicatoria complete genome AMO39952 100% 5 G5 Guamúchil
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 6 C161-3 Culiacán
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 7 C162 Culiacán
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 8 C163 Culiacán
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 9 E145-1 Elota
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 10 SL143C Sinaloa de Leyva
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 11 SLSE59-2 Sinaloa de Leyva
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 12 G158-3 Guasave
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 13 G159-1 Guasave
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 14 F169 El Fuerte
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99% 15 M125 Mocorito
Xanthomonas campestris pv. Vesicatoria complete genome AMO39952 99%
54
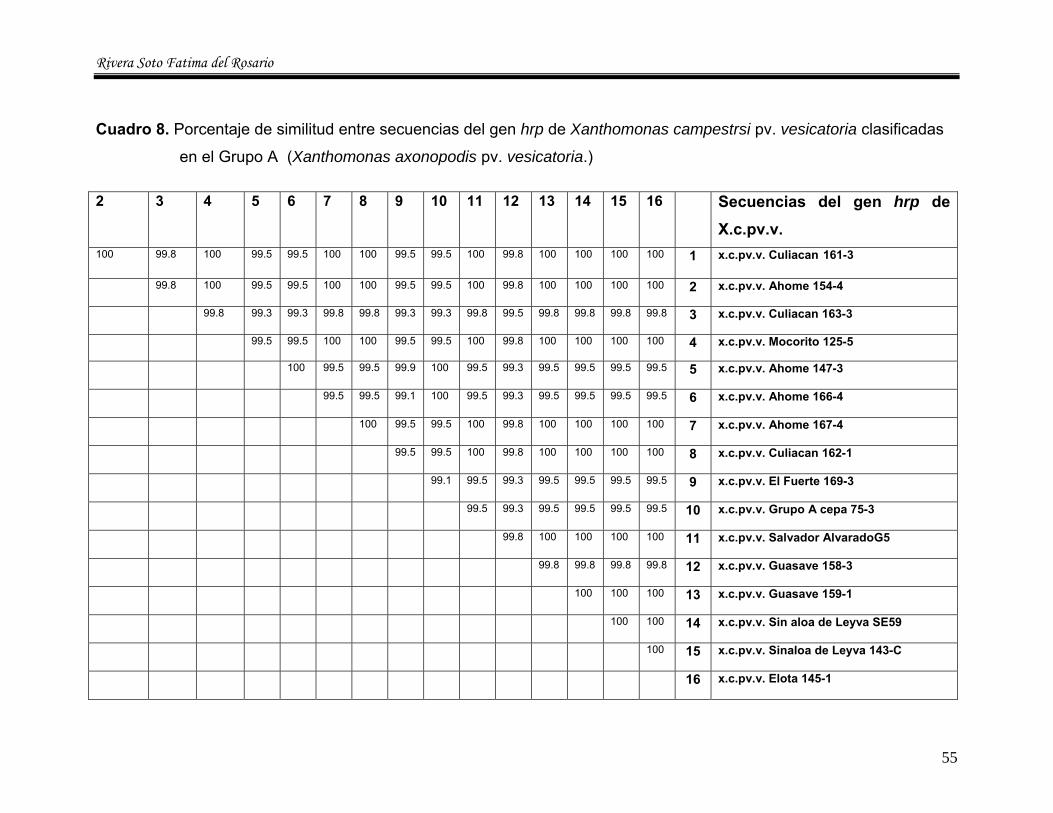
Rivera Soto Fatima del Rosario
55
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Secuencias del gen hrp de X.c.pv.v.
100 99.8 100 99.5 99.5 100 100 99.5 99.5 100 99.8 100 100 100 100 1 x.c.pv.v. Culiacan 161-3
99.8 100 99.5 99.5 100 100 99.5 99.5 100 99.8 100 100 100 100 2 x.c.pv.v. Ahome 154-4 99.8 99.3 99.3 99.8 99.8 99.3 99.3 99.8 99.5 99.8 99.8 99.8 99.8 3 x.c.pv.v. Culiacan 163-3 99.5 99.5 100 100 99.5 99.5 100 99.8 100 100 100 100 4 x.c.pv.v. Mocorito 125-5
100 99.5 99.5 99.9 100 99.5 99.3 99.5 99.5 99.5 99.5 5 x.c.pv.v. Ahome 147-3
99.5 99.5 99.1 100 99.5 99.3 99.5 99.5 99.5 99.5 6 x.c.pv.v. Ahome 166-4
100 99.5 99.5 100 99.8 100 100 100 100 7 x.c.pv.v. Ahome 167-4
99.5 99.5 100 99.8 100 100 100 100 8 x.c.pv.v. Culiacan 162-1
99.1 99.5 99.3 99.5 99.5 99.5 99.5 9 x.c.pv.v. El Fuerte 169-3
99.5 99.3 99.5 99.5 99.5 99.5 10 x.c.pv.v. Grupo A cepa 75-3
99.8 100 100 100 100 11 x.c.pv.v. Salvador AlvaradoG5
99.8 99.8 99.8 99.8 12 x.c.pv.v. Guasave 158-3
100 100 100 13 x.c.pv.v. Guasave 159-1
100 100 14 x.c.pv.v. Sin aloa de Leyva SE59
100 15 x.c.pv.v. Sinaloa de Leyva 143-C
16 x.c.pv.v. Elota 145-1
Cuadro 8. Porcentaje de similitud entre secuencias del gen hrp de Xanthomonas campestrsi pv. vesicatoria clasificadas
en el Grupo A (Xanthomonas axonopodis pv. vesicatoria.)

Rivera Soto Fatima del Rosario
5. Análisis de Restricción. Las 15 secuencias obtenidas del gen hrp de Xanthomonas
campestris pv. vesicatoria de tomate fueron analizadas con el programa Map
Draw del programa DNASTAR. Para la obtención de los mapas de restricción
de las secuencias, se utilizaron las enzimas HaeIII y CfoI. Ambas enzimas
fueron estudiadas por Obradovic et al (2003) además de la enzimas TaqI pero él
solo propone la enzima HaeIII para diferenciar entre los cuatro grupos de
Xanthomonas campestris pv. vesicatoria (A, B, C y D). En este estudio se
analizaron las secuencias en el programa MapDraw usando las enzimas HaeIII y
CfoI. Se tiene secuenciado el genoma completo de la cepa 75-3 de tomate que
corresponde al grupo A (X. euvesicatoria), pero no están aún reportadas las
secuencias del gen hrp de los grupos B (X. vesicatoria), grupo C (X. perforans) y
grupo D (X. gardneri), de acuerdo a su actual reclasificación (Jones et al., 2004). Las secuencias de Xantomonas campestris pv. vesicatoria de tomate del
municipio de Ahome (A 167-4, A 154-4 y A 166-4) presentaron el mismo patron de
restricción con el analisis RFLP-PCR con la enzima HaeIII. Las cepas de tomate
de C163-3, C161-1 (Culiacán ), A154-4, A167-4 (Ahome), M125-5 (Mocorito), G5
(Salvador Alvarado), SLSE59, SL143C (Sinaloa de Leyva) y E145-1 (Elota) tiene
una mismo patron de restricción con la enzima HaeIII y Cfol, pues presentaron
una homología del 100% entre ellas y del 99.5 % con el grupo A de
Xanthomonas campestris pv vesicatoria. Las secuencias C163-3 (Culiacan),
F169-3 (El Fuerte) y G158-3 (Guasave) tienen un mismo patron de restricción
que las secuencias mencionadas con las enzimas HaeIII y Cfol a pesar de las
diferencias en sus secuencias ya que tienen una homología del 99.3 al 99.5%
entre ellas y del 99.1 al 99.3% con el grupo A de Xanthomonas campestris pv.
vesicatoria. El analisis de los sitios de restricción con enzima HaeIII para las
secuencias A 147-3 y A-166-4 (Ahome) presentan un patron de restricción
idéntico al patron del la cepa 75-3 perteneciente al grupo A de Xanthomonas ya
que tienen una homología del 100 % entre ellas y con la cepa del Grupo A. Las
secuencias A 147-3 y A-166-4 (Ahome), tienen un sitio de corte con la enzima
56

Rivera Soto Fatima del Rosario
CfoI en la posición 279 que no presentan el resto de las secuencias, este sitio de
corte tambien lo presenta la secuencia de la cepa del Grupo A reportada en el
“GenBank”. Se puede observar que todas las secuencias presentan los mismos
sitios de corte con la enzima HaeIII estudiada por Obradovic et al (2003) para la
diferenciación entre grupos y comparada con la secuencia del grupo A, las
Xanthomonas spp. aisladas en el estado de Sinaloa pueden pertenecer a este
grupo (Figura 17).
Las diferencias entre secuencias puede ser por reconocimientos de las
enzimas de restricción por diferencias en la secuencia de nucleótidos; aunque la
clasificación de Xanthomonas spp. se puede realizar mediante enzimas de
restricción es importante conocer las secuencias de grupos de Xanthomonas
spp. para realizar una comparación completa.
Se tiene secuenciado el genoma completo de la cepa 75-3 de tomate que
corresponde al grupo A (X. euvesicatoria), pero no se conocen aún las
secuencias del gen hrp de los grupos B (X. vesicatoria), grupo C (X. perforans) y
grupo D (X. gardneri), de acuerdo a su actual reclasificación (Jones et al., 2004).
57

Rivera Soto Fatima del Rosario
Grupo A de Xcv (cepa 75-3)
Xcv C162-1 Xcv C161-3 Xcv C163-3 Xcv A167-4 Xcv A154-4 Xcv G5 Xcv G158-3 Xcv G159-1 Xcv SLSE59 Xcv SL143-C Xcv E145-1 Xcv M125-5 Xcv F169-3
Xcv A147-3 Xcv A166-4
Figura 17. Análisis de sitios de restricción de las secuencias del gen hrp de Xanthomonas campestris pv. vesicatoria Los mapas fueron realizados utilizando la opción de MapDraw del programa DNASTAR. Grupo A de X.c. pv. v. (cepa 75-3 AMO39952 ), C 162-1, C 161-3 y C163-3 (tomate de Culiacán), A 167-4, y A 154-4 (tomate de Ahome), G5 (tomate de Salvador Alvarado), G 158-3 y G 159-1 (tomate de Guasave), SL SE59, SL 143-C (tomate de Sinaloa de Leyva), E145-1 (tomate de Elota), M 125-5 (tomate de Mocorito), F 169-3 (tomate de El Fuerte). La flecha indica la ausencia y los rectangulos la presencia de un sitio de restricción distinto.
100 bp 200 bp 300 bp 400 bp
100 bp 200 bp 300 bp 400 bp
Cfo
I H
ae II
I C
fo I
Hae
III
Hae
III
Hae
III
Cfo
I
Hae
III
Cfo
I
Hae
III
Hae
III
Cfo
I
Cfo
I
Cfo
I
Cfo
I
Hae
III
Hae
III
Hae
III
Hae
III
Hae
III
Cfo
I
Cfo
I
Cfo
I
Cfo
I
Cfo
I
Cfo
I
Hae
III
Hae
III
Hae
III
Hae
III
Hae
III
Hae
III
Cfo
I
Cfo
I
Cfo
I
Cfo
I
Cfo
I
Cfo
I
58

Rivera Soto Fatima del Rosario
6. Análisis Filogenético.
Con base en el análisis de la secuencia del gen hrp, se construyó un árbol
filogen
os aislamientos de Xanthomonas campestris pv. vesicatoria de tomate
C162-1
ético de las 15 secuencias de Xanthomonas campestris pv. vesicatoria
caracterizadas en este estudio. (Figura 18). En este proceso se consideró la
secuencia del Grupo A de Xanthomonas campestris pv. vesicatoria del banco de
genes.
L
, C 161-3, C163-3, A 154-4, A 167-4, M 125-5, G5, G 159-1, G 158-3, F
169-3, SLSE59, SL 143C y E 145-1 se agrupan en una misma rama confirmando
su alta homología tal como se demostró en el análisis de restricción con las
enzimas HaeIII y CfoI donde presentaron patrones de restricción idénticos;
mientras que las secuencias de los aislamientos de las cepas A147-3, A 166-4
se agrupan en una misma rama confirmando su alta homología como se observa
en el analisis de restricción con las enzimas HaeIII y Cfol donde presentan
patrones de restricción idénticos al Grupo A (X. aeuvesicatoria). En base a lo
anterior podemos decir que las cepas aisladas de los municipios de Ahome,
Guasave, El Fuerte, Mocorito, Salvador Alvarado, Sinaloa de Leyva, Culiacán y
Elota estan dentro del grupo A (Xanthomonas euvesicatotia) (a pesar de que
existen pequeñas diferencias entre ellas).
59
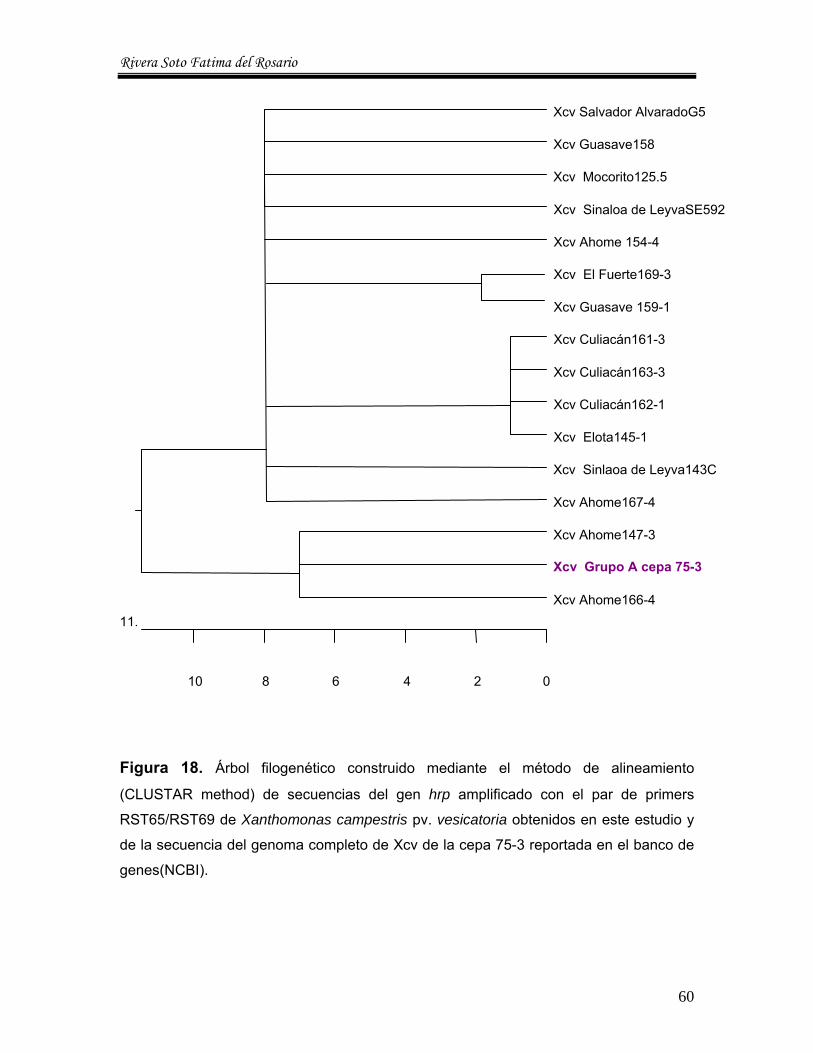
Rivera Soto Fatima del Rosario
Xcv Salvador AlvaradoG5
Xcv Guasave158 Xcv Mocorito125.5 Xcv Sinaloa de LeyvaSE592 Xcv Ahome 154-4
Xcv El Fuerte169-3 Xcv Guasave 159-1 Xcv Culiacán161-3 Xcv Culiacán163-3 Xcv Culiacán162-1 Xcv Elota145-1 Xcv Sinlaoa de Leyva143C
Xcv Ahome167-4 Xcv Ahome147-3 Xcv Grupo A cepa 75-3 Xcv Ahome166-4
11.
10 8 6 4 2 0 Figura 18. Árbol filogenético construido mediante el método de alineamiento
(CLUSTAR method) de secuencias del gen hrp amplificado con el par de primers
RST65/RST69 de Xanthomonas campestris pv. vesicatoria obtenidos en este estudio y
de la secuencia del genoma completo de Xcv de la cepa 75-3 reportada en el banco de
genes(NCBI).
60

Rivera Soto Fatima del Rosario
D.Caracterización fisiológica. Sin duda es importante el poder reafirmar este estudio mediante
metodos convencionales como lo son las pruebas bioquímicas, las cuales fueron
realizadas a los 15 aislados caracterizados molecularmente. Este estudio
tambien se realizó con el fin de conocer y observar el comportamiento del
patogeno, partiendo con su caracterización a nivel género, seguido por especie
y culminando con patovar.
1. Caracterización a nivel genero. 1.1. Catalasa. Esta reacción fue catalasa postivos para los 15 aislados. Por otra parte
la presencia de oxidasa va ligada a la producción de catalasa, ya que esta
degrada el peróxido de hidrógeno al presentar una formación de burbujas al
adicionarlo a la bacteria.
1.2. Oxidasa. Los 15 aislados fueron oxidasa negativos por no presentar coloración
azul al adicionar tetrametil. Según la literatura estos resultados se deben a que
en las células aeróbicas de la bacteria se encuentran los citocromos los cuales
son proteínas transferidoras de electrones, los citocromos oxidasa tienen una
gran afinidad por el oxígeno, mientras que las oxidasas negativas no, es por ello
que al combinarse la proteína (citocromo) con el tetrametil y el naftol se produce
un color azúl, lo cual no sucedió en este caso. Estos resultados demuestran que
la bacteria puede pertenecer al genero Xanthomonas que es una bacteria
aerobia y que no contiene citocromos en su celulas, pues no se presento una
reacción de color azul.
1.3. Crecimiento en YDC (Extracto de levadura dextrosa). A las 48 hrs de incubación en YDC, los 15 aislados presentaron una
coloración mucoide, en cinco una coloración naranja y en 10 una coloración
61

Rivera Soto Fatima del Rosario
amarilla. De acuerdo al manual de Schaad de 2001, la reacción fue positiva
para todas las cepas analizadas en este trabajo (Figura 19).
Figura 19. Colonia de consistencia mucoide, de color amarillo, desarrollado en YDC,
correspondiente a la cepa del Estado de Sinaloa a las 48 horas de incubación.
Los resultados obtenidos de las cinco pruebas fisiológicas realizadas,
coinciden en que las cepas en estudio pertenecen al genero Xanthomonas.
Posteriormente se corrieron pruebas más específicas, para determinación
especies y patovar. 2. Caracterización a nivel especie. 2.1. Crecimiento en YS (Caldo de levadura). Después de 48 hrs de incubación de los aislamientos a 35°C en caldo
YDC se observó crecimiento bacteriano, al ser comparado con el control
negativo se corrobora turbidéz en los 15 aislados. Siendo este el resultado
esperado para especies de Xanthomonas campestris (Figura 20).
62

Rivera Soto Fatima del Rosario
B A
Figura 20. A) Crecimiento de un aislado del Estado de Sinaloa de Xanthomonas
campestris pv. vesicatoria, después de 48 horas de incubación en YS a 35 °C. B) Testigo sin inocular.
2.2. Hidrólisis de almidón. Los 15 aislados de los diferentes municipios de Sinaloa fueron positivos
para la hidrólisis de almidón. Sin embargo, algunos aislados reaccionaron
débilmente por presentar halos poco definidos, sin embargo, se consideraron
positivos para dicha reacción (Figura 21). Esta reacción se debe a que ciertas
bacterias del genero Xanthomonas en su mayoría, son capaces de polimerizar el
almidón por medio de enzimas extracelulares, conocidas como amilasas
(Rodríguez, 2001). La capacidad de hidrolizar almidón (actividad amilolítica) fue
una de las primeras características fenotípicas que muestran diferencias entre
los aislados de la Xanthomonas spp. asociados con la mancha bacteriana en
tomate y chile (Jones et al., 1998). Los aislados asociados a la mancha
bacteriana en tomate son capaces de degradar almidón a diferencia de los
aislados de chile.
63

Rivera Soto Fatima del Rosario
B A
Figura 21. Prueba de hidrólisis de almidón. A) Hidrólisis de almidón positiva el aislado
de Xanthomonas de tomate del municipio de Mocorito. B) Control negativo, aislado de
Xanthomonas campestris pv. vesicatoria de chile proporcionada por el Dr. Rubén Félix
Gastelum (Prof. Investigador de la Universidad de Occidente).
2.3. Licuefacción de gelatina. En esta prueba, ninguno de los 15 aislados en estudio causó
licuefacción de gelatina (Figura 22). La literatura reporta que las especies X.
campestris presenta resultados variables en cuanto a la producción de
gelatinasa. Esta reacción ayuda a la identificación de Serratia, Pseudomonas,
Flavobacterium y Clostridium, tambien ha sido usada por Obradovic et al, en el
2003 para la caracterización de Xanthomonas spp.
A B
Figura 22. Prueba de licuefacción de gelatina. A) Control negativo, sin inocular. B)
Reacción negativa para gelatinasa, de un aislado de Xanthomonas obtenido de tomate
del municipio de Mocorito.
64

Rivera Soto Fatima del Rosario
2.4. Hidrolasa de arginina. En la prueba de dihidrolasa de arginina los 15 aislados dieron reacción
negativa, para la producción de esta enzima, ya que las bacterias no crecieron
bajo condiciones anaerobias, en ninguno de los tubos inoculados se presentó un
cambio de color del medio, lo que indica la carencia del sistema enzimático
dihidrolasa de arginina (Figura 23).
A B C
Figura 23. Prueba de arginina en medio de Thornley 2A. A) y B) Reacción negativa de
la hidrolasa de arginina del aisl do de la planta de tomate con mancha bacteriana
originada de Culiacán. C) Contro
2.5. Fuentes de carbono. En los 15 aislamientos
de incubar la bacteria sobre e
26). Estos resultados nos indic
ácido a partir de carbohidratos
establecido con especies de l
de Schaad 1988. Los 15 aisla
género, especie y se confi
resultados característicos de la
(Cuadro 9).
aA
l negativo, sin inocular.se observó el cambio de color del medio después
l medio base Dye durante 48 hrs. (Figura 24, 25 y an que los 15 aislados fueron capaces de producir
. Los resultados obtenidos están de acuerdo a lo
a bacteria X. campestris establecido en el manual
dos bacteriológicos fueron caracterizados a nivel
rmó su patovar con medios selectivos dando
bacteria Xanthomonas campestris pv. vesicatoria
65

Rivera Soto Fatima del Rosario
Figura 24. Producción de ácido a partir fructuosa como fuente de carbono. A y B
Aislados del municipio de Sinaloa de Leyva mostraron cambios de color en el medio
Dye lo que indica que el azúcar fue utilizado.
B
A
A
B
FiguAisla
reac
A
ra 25. Producción de ácido a partir de la fuente de carbono Glucosa. A) y B).
dos del municipio de Sinaloa de Leyva cambiaron el color del medio lo que indica
ción positiva para la utilización de Glucosa.
66
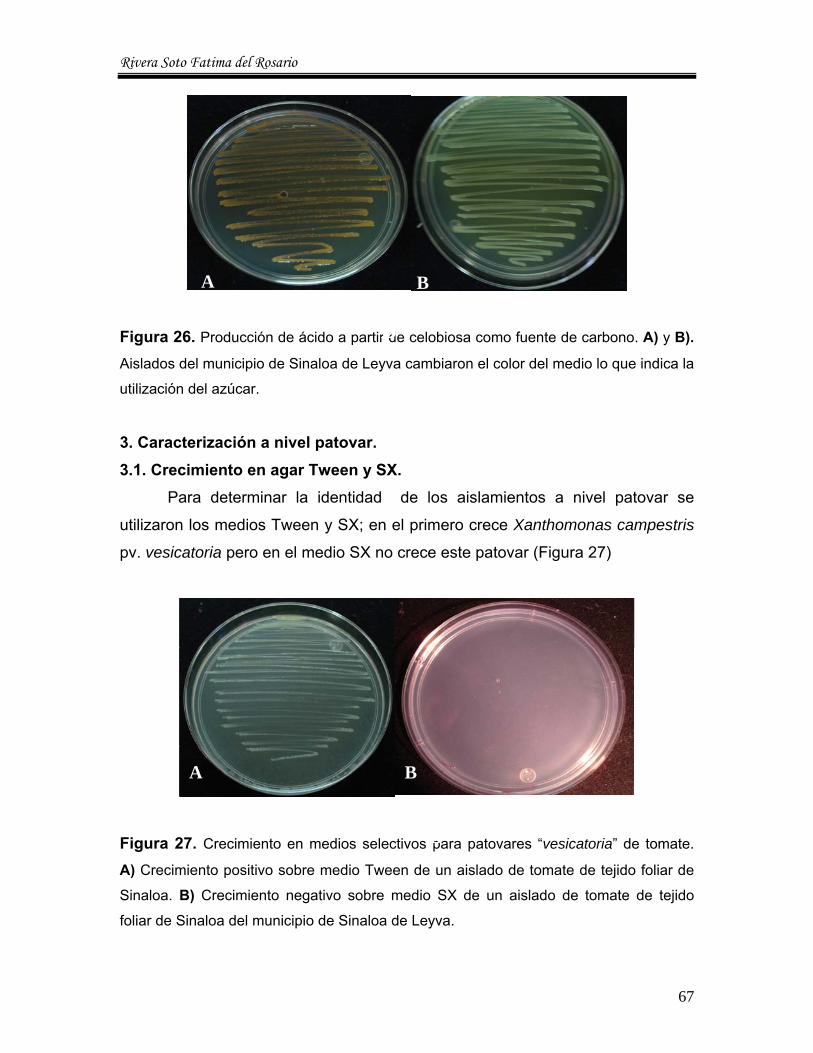
Rivera Soto Fatima del Rosario
B A
Figura 26. Producción de ácido a part de celobiosa como fuente de carbono. A) y B).
Aislados del municipio de Sinaloa de Leyva cambiaron el color del medio lo que indica la
utilización del azúcar.
3. Caracterización a nivel patovar. 3.1. Crecimiento en agar Tween y SX. Para determinar la identidad de los aislamientos a nivel patovar se
utilizaron los medios Tween y SX; en el primero crece Xanthomonas campestris
pv. vesicatoria pero en el medio SX no crece este patovar (Figura 27)
A
FAS
f
A
igura 27. Crecimiento en medios selectivos para patovares “vesicatoria” de tomate.) Crecimiento positivo sobre medio Tween d
inaloa. B) Crecimiento negativo sobre medi
oliar de Sinaloa del municipio de Sinaloa de Le
B
B
irB
e un aislado de tomate de tejido foliar de
o SX de un aislado de tomate de tejido
yva.
67
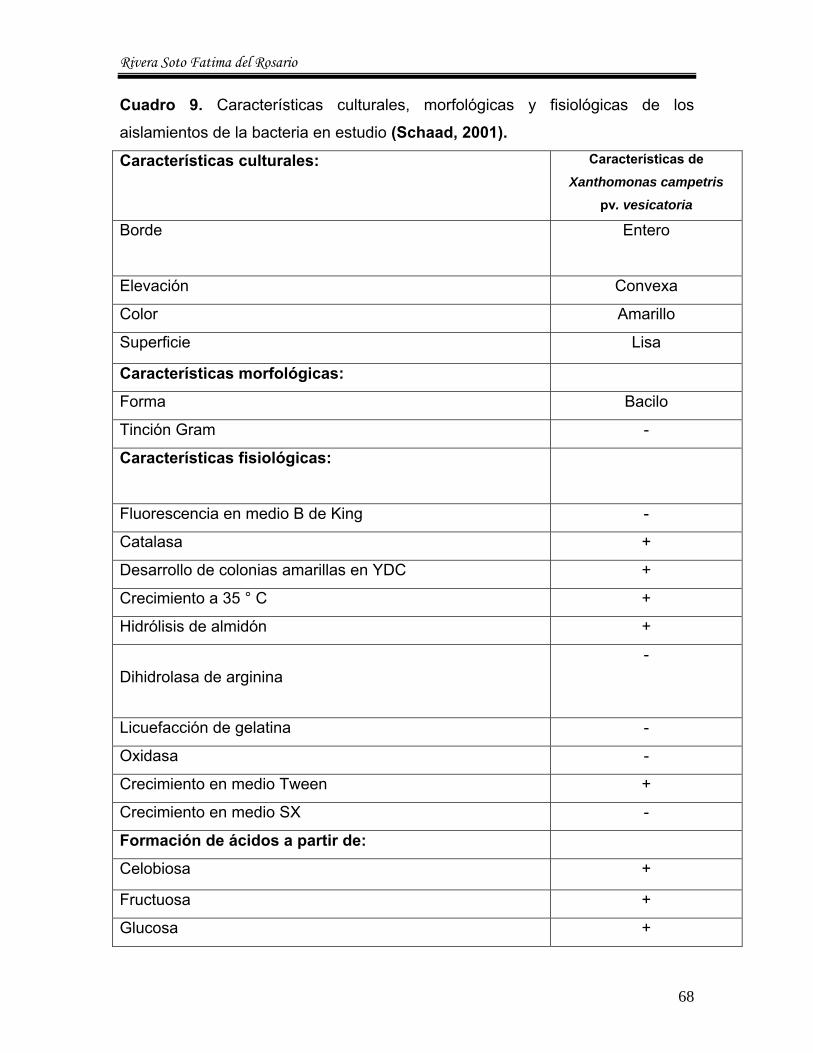
Rivera Soto Fatima del Rosario
Cuadro 9. Características culturales, morfológicas y fisiológicas de los
aislamientos de la bacteria en estudio (Schaad, 2001).
Características culturales: Características de Xanthomonas campetris
pv. vesicatoria
Borde
Entero
Elevación Convexa
Color Amarillo
Superficie Lisa
Características morfológicas:
Forma Bacilo
Tinción Gram -
Características fisiológicas:
Fluorescencia en medio B de King -
Catalasa +
Desarrollo de colonias amarillas en YDC +
Crecimiento a 35 ° C +
Hidrólisis de almidón +
Dihidrolasa de arginina -
Licuefacción de gelatina -
Oxidasa -
Crecimiento en medio Tween +
Crecimiento en medio SX -
Formación de ácidos a partir de:
Celobiosa +
Fructuosa +
Glucosa +
68

Rivera Soto Fatima del Rosario
E. Pruebas de patogenicidad. Una vez que los 15 aislados fueron caracterizados molecularmente y
fisiológicamente, se les realizó la prueba de patogenicidad esto fue para
comprobar su relación patogénica con la mancha bacteriana del tomate en las
diferentes plantaciones en Sinaloa. En este estudio se demostró que los 15
aislados fueron patogénicos. Lo anterior pudo deberse a que todos los islados
en estudio se transfirieron constantemente, lo que pudo inducir a que se
debilitaran y que en consecuencia no ejercieran una fuerte actividad patogénica.
El método de inoculación fue el adecuado y las condiciones de incubación
fueron favorables al desarrollo de mancha bacteriana, lo cual se evidencio en
que las plantas (Figura 28) como controles positivos mostraron síntomas más
severos y característicos de la bacteria Xanthomonas campestris pv.
vesicatoria. Esta cepa es un aislamiento reciente y poco manipulado de ahí que
afectó severamente al tomate. Para comprobar que los síntomas que en las
plantas de tomate eran ocasionados por la bacteria de interes, se pudo aislar el
patógeno y observar crecimiento de colonias amarillas mucoides; además dichas
colonias fueron bacilos Gram negativos. Con esto se cumplieron los postulados
de Koch.
A B
Figura 28. Pruebas de patogenicidad en cultivo de tomate. A) Planta de tomate
inoculada artifialmente con Xanthomonas campestris pv. vesicatoria aislada de
cultivo de tomate del municipio de Culiacán. B) Control positivo, planta inoculada
con un aislamiento de Xanthomonas campestris pv. vesicatoria proporcionada por
el Dr, Rubén Félix.
69

Rivera Soto Fatima del Rosario
CONCLUSIONES
♦ Las pruebas bioquímicas permitieron la caracterización a nivel genero, especie
y pathovar 15 aislados de Xanthomonas campestris pv. vesicatoria de cultivos
de tomate de los municipios de Guasave, Ahome, Mocorito, Salvador Alvarado,
Elota, El Fuerte, Sinaloa de Leyva y Culiacán.
♦ Los aislamientos de Xanthomonas campestris pv. vesicatoria obtenidas de las
15 muestras sintomáticas de tomate en Sinaloa correponden al grupo A
(Xanthomonas euvesicatoria).
♦ Los 15 aislamientos de Xanthomonas campestris pv. vesicatoria de tomate de
los municipios de Sinaloa de Leyva, Culiacán, Elota, Salvador Alvarado,
Guasave, Ahome, Mocorito y El Fuerte resultaron patogénicos en el mismo
cultivo, en condiciones de invernadero.
70

Rivera Soto Fatima del Rosario
BIBLIOGRAFIA Agrios, G. N. 2004. Fitopatología. Segunda edición. Ed. Limusa
México. 838 pag.
Balbás, P. 2002. De la Biología molecular a la biotecnología. Editorial
Trillas. México, D. F. 324 pp.
Becker, A., F. Katzen, A. Pühler, and L. Ielpi. 1998. Xanthan gum
biosynthesis and application: a biochemical/genetic perspective. Appl.
Microbiol. Biotechnol. 50:145-152.
Beer, S. V., D. W. Bauer, X. H. Jiang, R. J. Laby, B. J. Sneath, Z.-M. Wei, D. A. Wilcox, and C. H. Zumoff. 1991. The hrp gene cluster of
Erwinia amylovora. P. 53-60. In H.- Hennecke and D. P. S. Verma
(ed.), Advances in molecular genetics of plantmicrobe interactions, vol.
1. Kluwer Academic Publishers, Dordrecht, The Netherlands. Bereswill, S., A. Pahl, P. Bellemann, W. Zeller, and K. Geider.
1992. Sensitive and species-specific detection of Erwinia amylovora by
polymerase chain reaction analysis. Appl. Environ. Microbiol. 58:3522-
3526.
Blancard, D. 1990. Enfermedades del tomate, observar, identificar,
luchar, Ed. Mundi Prensa, Madrid.
Bonas U., Conrads-Strauch, J., Balbo, I. 1993. Resistance in tomato
to Xanthomonas campestris pv. vesicatoria is determined by alleles of
the pepper-specific avirulence genes avrBs3. Molecular General
Genetics, v.238, p.261-269. Bonas, U., R. Schulte, S. Fenselau, G. V. Minsivage, B. J.
Staskawicz, and R. E. Stall. 1991. Isolation of a gene cluster from
Xanthomonas campestris pv. vesicatoria that determines pathogenicity
and the hypersensitive response on pepper and tomato Mol. Plant
Mocrobe Interact. 4:81-88. Bonas, U. 1994. hrp genes of phytopatogenic bacteria. Curr. Top.
Microbiol. Immunol. 192:79-98. Boucher, C. A., F. Van Gijsegem, P. A. Barberis, M. Arlet, and C.
71

Rivera Soto Fatima del Rosario
Zischek. 1987. Pseudomonas solanacearum genes controlling both
pathogenicity and hypersensivity are tabacco are clustered. J.
Bacteriol. 169:5626-5632.
Bouzar H, Jones JB, Stall RE Hodge NC, Minsavage GV, et al. 1994. Physiological, chemical, serological, and pathogenic analyses of
a worldwide collection of Xanthomonas campestris pv. vesicatoria
strains. Phytopathology 84: 663-71
Bouzar H, Jones JB, Stall RE, Louws FJ, Schneider M, Rademaker JLW, de Bruijn FL and Jackson LE. 1999. Multiphasic analysis of
xanthomonads causing bacterial spot disease an tomato and pepper in
the Caribbean and Central America: Evidence for common lineages
within and between countries. Phytopathology 89: 328-335
Bradbury, J. F. 1984. Genus II. Xanthomonas Dowson 1929, 187, p.
199-210. In N. R. Krieg and J. C. holt (ed), Bergey´s manual of
systematic bacteriology, vol. 1. The Williams & Wilkins Co., Baltimore.
Búttner, D., and U. Bonas. 2002. Getting across-bacterial type III
effector proteins on their way to the plant cell. EMBO J. 21:5313-5322.
Canteros BI, Minsavage GV, Bonas U, Pring D. and Stall RE. 1991. A gene from Xanthomonas campestris pv vesicatoria that determines
avirulence in tomato es related to avrBs3. Mol Plant-Microbe Interact.
4: 628-32.
Carrillo-Fasio, J.A. 2001. Razas de Xanthomonas campestris pv.
vesicatoria (Doidge) Dye presentes en el Estado de Sinaloa, México.
Revista Mexicana de Fitopatología, vol. 19.
Chavez medina, 2006. Detección y caracterización de fitoplasmas
relacionados en enfermedades de solanaceas en México. Tesis de
Maestría en Recursos Naturales y Medio Ambiente. Centro
Interdisciplinario de Investigación para el Desarrollo Integral Regional,
IPN.
Cruz, O.J.E., García, E. R. S. y Carrillo, F.J.A. 1998. Enfermedades
de las Hortalizas. Universidad Autónoma de Sinaloa. Culiacán,
72

Rivera Soto Fatima del Rosario
Sinaloa. 255 p.
DeBoer, S., Cuppels, D. A., and Gitaitis, R. D. 1996. Detecting latent
bacterial infections. Pages 27-57 in: Advances in Botanical research.
S., DeBoer, J. Andrews, and I. C. Tommerup, eds. Academic Press,
London.
D´Haeze, W., and M. Holsters. 2004. Surface polysaccharides enable
bacteria to evade plant immunity. Trends Microbiol. 12:555-561.
Fenselau, S., I. Balbo, and U. Bonas. 1992. Determinants of
pathogenicity in Xanthomonas campestris pv. vesicatoria are related to
proteins involved in secretion in bacterial pathogens of animals. Mol.
Plant Microbe Interact. 5:390-396.
Garrity, G.M., Holt, J. G., 2000. An overview of the road map to the
manual. Systematic Bacteriology. 2.ed. New York: Springer. 20p.
(Bergey´s Manual).
Gitaitis, R., McCarter, S., Jones, J. B. 1992. Disease control in
tomato transplants produced in Giorgia and Florida. Plant Disease,
v.76, n.7, p.651-656.
Gonzáles, Humberto y Margarita Calleja. 1998. La exportación de
frutas y hortalizas a Estados Unidos de Norte América, Guadalajara,
Secretaria de Agricultura, Ganadería y Desarrollo Rural, CIESAS,
CONACYT.
Goode, M. J., Passer, M. 1980. Prevention - key to controlling
bacterial spot and bacterial speck of tomato. Plant Disease, v.64, n. 7,
p.831-834
Goto, M. 1990. Fundamentals of Bacterial Plant Pathology, Ed.
Academic Press. New York.
Jaime S.F., 1976. Mancha bacteriana del tomate y chile, Seminario fit-
623, Colegio de posgraduados, Chapingo, México, inédito.
Jones, J.B., J.P. Jones, R.E. Stall and T.A. Zitter. 1991. Compendium of tomato disease, APS Press, P. 27.
Jones J.B, Bouzar H, Somodi GC, Stall RE and Pernezny K .1998.
73

Rivera Soto Fatima del Rosario
Evidence for the preemptive nature of tomato race 3 of Xanthomonas
campestris pv. vesicatoria in Florida. Phytopathology 88: 33-38
Jones J.B, Bouzar H, Stall RE, Almira EC, Roberts PD, Bowen BW, Sudberry J, Strickler PM and Chun J. 2000. Systematic analysis of
xanthomonads (Xanthomonas spp.). associated with pepper and
tomato lesions. International Journal of Systematic and Evolutionary
Microbiology 50: 1211-1219.
Jones, J. B., G. H. Lacy, H. Bouzar, R. E. Stall, and N. W. Schaad. 2004. Reclassification of the xanthomonads associated with bacterial
spot disease of tomato and pepper. Syst. Appl. Microbiol. 27:755-762.
Jones J. B. 1997. Bacterial spot In: Jones, J. B., Jones, J. P., Stall, R.
E., Zitter, T.A. (Ed.). Compendium of tomato diseases. St. Paul:
America Phytopatologycal Society Press.p.27.
Katzen, F., D. U. Ferreiro, C. G. Oddo, M. V. Ielmini, A. Becker, A. Pühler, and L. Ielpi. 1998. Xanthomonas campestris pv. campestris
gum mutants: effects on xanthan biosynthesis and plant virulence. J.
Bacteriol. 180:1607-1617.
Leite RP, Minsavage GV, Bonas U, Stall RE. 1994. Detection and
identification of phytopatogenic Xanthomonas strain by amplification of
DNA sequence related to the hrp genes of Xanthomonas campestris
pv. vesicatoria. Appl. Environ. Microbiol. 60:1068-77.
Lehninger, A. L. 1995. Principios de Bioquímica; Ed. Omega. México,
D.F.
Leyns, F., M. De Cleene, J.-G. Swings and J. De Ley. 1984. The
host range of the genus Xanthomonas. Bot Rev. 50:308-356.
Lindgren, P. B., R. C. Peet, and N. J. Panopoulos. 1986. Gene
cluster of Pseudomonas syringae pv. Phaseolicola controls
pathogenicity of bean plants and hypersensitivity on nonhost plants. J.
Bacteriol. 168 :512-522.
Lopes, C. A., Quezado-Soares, A. M. 1997. DoenÇãs bacterianas
das hortaliÇas – diagnose e controle. Brasilia: Embrapa-CNPH. p.70.
74

Rivera Soto Fatima del Rosario
Lows, F. J., Rademaker, J. L. W., Bruijn, F. J. 1999. The three D’s of
PCR-based genomic analysis of phytobacteria: diversity, detection,
and disease diagnosis. Annual Review of Phytopathology, v.37, p.81-
125.
McManus, P., Stockwell, V. 2002. Antibiotics for plant disease
control: silver bullets of rusty sabers? www.APSFeatures/APSFea-
antibiotics.htm. Montealegre, 2002. Bacterias. Limusa, Mexico.
Moore, W. E.C. and L. V. H. Moore. 1992. Index of the Bacterial and
Yeast Nomenclatural Changes. Published in the International Journal
of Systematic Bacteriology Approved Lists of Bacterial Names.
(January 1, 1980 to Journal 1, 1992) American Society for
Microbiology. Washington, D. C.
Obradovic, A., A. Mavridis, K. Rudolph, J. D. Janse, M. Arsenejevic, J.B. Jones, G. V. Minsavage, and J. F. Wang. 2003. Characterization and PCR- based typing of Xanthomonas campestris
pv. vesicatoria from pepper and tomatoes in Serbia. Eur. J. Plant
Pathol. 110:285-292.
Pohronezny, K., Waddill, V. H., Schuster, D. J., and Sonoda, R. M. 1986. Integrated pest management for Florida tomatoes. Plant Dis.
70:92-102.
Poplawsky, A., and W. Chun. 1997. pigB determines a diffusible
factor needed for extracellular polysacch slime and xanthomonadin
production in Xanthomonas campestris pv. campestris. J. Bacteriol.
179:439-444.
Quezado-Soares, A. M., y Lopez, A. C. 1999. Control químico de
mancha- bacteriana em tomaterio para processamento industrial.
1999. Summa Phytopatologica. V.25 n.1, p.21.
Robinson, C. and A. Bolhuis. 2001. Protein targeting by the twin-
arginine translocation Nat. Rev. Mol. Cell Biol. 2:350-356.
Rodríguez M., M. L. 2001. Manual para identificación de bacterias
75

Rivera Soto Fatima del Rosario
fitopatógenas. 2da. Edición. Parasitología Agrícola. Chapingo,
México.119. pp.
Schaad, N. W. , A. K. Vidaver, G. H. Lacy, K. Rudolph, and J. B. Jones 2000. Evaluation of proposed amended names of several
pseudomonads and xanthomonads and recommendations.
Phytopathology 90:208-213.
Schaad, N. W. 1988. Laboratory guide for identification of plant
pathogenic bacteria. American Phytopatological Society. St. Paul.
Minnesota, USA. 158 p.
Schaad, N. W. 1989. Detection of Xanthomonas campestris pv.
campestris in Crucifers. Pages 68-75 in: A. W. Schaad, and D. A.
Roth, eds., Detection of Bacteria in Seed and Other Planting Material.
APS Press. St. Paul, MN.
Schaad, N. W. 2001. Laboratory guide for identification of plant
pathogenic bacteria. American Phytopatological Society. St. Paul.
Minnesota, USA.
Saettler, A. W., N. W. Schaad, and D. A. Roth (eds.). 1989. Detection of bacterial in seed and other planting material. American
Phytopatological Society, St. Paul, Minn.
Sahin F, Miller SA. 1997. A source of resistance in Capsicum spp.
accessions to pepper race 6 of Xanthomonas campestris pv.
vesicatoria. Phytopathology 87: S84 (Abstr.).
Sandkvist, M. 2001. Type II secretion and pathogenesis. Infect.
Immun. 69:3523-3535
Skeman, V. B. D., V. McGowan, and P. H. A. Sneath. 1980. Approved List of Bacterial Names. Int. J. Syst. Bacteriol. 30:225-420.
Sneath, P. H. A. 1992. International Code of Nomenclature of Bacteria
and Statutes of the International Committee on Systemic Bacteriology
and Statutes the Bacteriology and Applied Microbiology Section of the
International Union of Microbiological Societies. Bacteriological Code.
1990 Revision. Amer. Soc. for Microbiol., Washington, D. C.
76

Rivera Soto Fatima del Rosario
Stall, R. E., and G. V. Minsavage. 1990. The use of hrp genes to
identify opportunistic xanthomonads, p. 360-374. Klement (ed.), Proc.
7th Int. Conf. Plant Pathog. Bacteria 1989. Akadémiai Kiadó, Budapest.
Stall, R. E. 1993. Xanthomonas campestris pv vesicatoria: cause of
bacterial spot of tomato and pepper. Pages 57-60 in: Xanthomonas. J.
G. Swings and E. L. Civerolo, eds. Chapman and Hall, London, UK.
Sutic D. 1957. Tomato bacteriosis. Special Edition of Institute for Plant
Protection, Belgrade 6: 1-65. English summary: Review of Applied
Mycology 36:734-735.
Thieme F, Koebnik R, Bekel T, Berger C, Boch J, Büttner D. 2005. Insights into genome plasticity and Pathogenicity of the plant
pathogenic bacterium Xanthomonas campestris pv. vesicatoria
revealed by the complete genome sequence. Journal of bacteriology
187:7254-7266.
Van Doorn, J., P. M. Boonekamp, and B. Oudega. 1994. Partial
characterization of fimbriae of Xanthomonas campestris pv. hyacinthi.
Mol. Plant. Microbe Interact. 7:334-344.
Vauterin L, Hoste, Kersters K and Swings J. 1995. Reclassification of Xanthomonas. International Journal of Systematic Bacteriology 3:
472-489.
Willems, A., M. Gillis, K. Kersters, L. Van de Broecke, and J. De Ley. 1987. Transfer of Xanthomonas ampelina Panagopoulos to a new
genus, Xylophilus gen. nov., as Xylophilus ampelinus Panagopoulos
(1969) comb. Nov. Int. J. Syst. Bacteriol. 37:422.430 Wills, K., J.J. Rich, and E. M. Hrabak. 1990. The hrp genes of
phytopathogenic bacteria. Mol. Plant Microbe Interact. 4:132-138.
Young, J. M., Bull, C. T., De Boer, S. H. Firrao, G., Gardan, L., Saddler, G. E., Stead, D. E., Takikawa, Y. 2001. Classification,
Nomenclature, and plant pathogenic bacteria – a clarification.
Phytopathology. 91:617-620.
Young, J. M., D. W. Dye, J. F. Bradbury, C G. Panagopoulos and C.
77

Rivera Soto Fatima del Rosario
F. Robbs. 1978. A proposed nomenclature and classification for plant
pathogenic bacteria. N. Z. J. Agric. Res. 21:153-177.
Internet http://www.monografias.com/trabajos/bacterias/bacterias.html http://edicion-micro.usal.es/web/identificacion/AyudaPruebas.html
http://www.sagarpa.gob.mx. http://www.uphs.upenn.edu/bugdrug/antibiotic_manual/gram.htm
hpp:// www. ncbi.nlm.nij.gov/blast
78