biologia.bio.brbiologia.bio.br/curso/biologia geral e evolução/núcleo, divisão... · sua...
TRANSCRIPT






ME = microscópio eletrônico.

RER = retículo endoplasmático rugoso ou granuloso




Análises detalhadas, que incluem reconstituição por computador, permitiram areconstrução de modelos tridimensionais dos complexos de poro. Eles são constituídospor dois anéis com arranjo octogonal e estabelecem o perímetro do poro. Cada anel éancorado na bicamada lipídica, nos pontos em que as membranas interna e externaestão fundidas. A eles se conectam oito fibrilas radiais que se dirigem ao canal central.Estruturas filamentosas se estendem dos anéis citoplasmático e nuclear, constituindouma estrutura semelhante a uma cesta de basquete apenas no lado nuclear. Acredita-seexistirem cerca de 100 proteínas diferentes constituindo esses componentes docomplexo de poro. Elas são coletivamente chamadas de nucleoporinas. O canal centraltem aproximadamente 40 nm de diâmetro, e uma estrutura em forma de grânulo(grânulo central) costuma ser vista em seu interior. Alguns pesquisadores acreditam queela é um componente intrínseco do complexo de poro que regula o transporte demoléculas através do canal central; outros propõem tratar-se de macromoléculas emtrânsito entre o núcleo e o citoplasma. As proteínas da lâmina nuclear estão associadasà membrana nuclear interna e se interrompem nos poros.A associação do complexo raio-anel leva ao estabelecimento de canais entre os raios,fechando parcialmente a abertura do complexo de poro.

O grânulo central é também denominado de complexo transportador ou núcleo central(central plug).

Dois mecanismos diferentes são utilizados no trânsito de moléculas através dos complexos deporo. Moléculas com até 9 nm de diâmetro atravessam rapidamente o complexo de poro nosdois sentidos. Elas se difundem passivamente através dos canais estabelecidos pelos raios docomplexo de poro. A maioria das proteínas e RNA são, no entanto, grandes demais para sedifundirem por esses canais. Essas macromoléculas atravessam os complexos de poro por umprocesso que consome energia, onde tanto as proteínas quanto os RNAs são reconhecidos etransportados seletivamente. Esse transporte ocorre através de um mecanismo regulado, quepermite a abertura dos canais até um diâmetro aproximado de 25 nm, em resposta a sinaisapropriados. As proteínas próprias do núcleo são sintetizadas no citoplasma com um sinal delocalização nuclear, constituído por um segmento de 4-8 aminoácidos, rico em lisina e arginina.Essas proteínas marcadas para destino nuclear atravessam os complexos de poro por ummecanismo que consome energia fornecida pelo ATP e pelo GTP. O sinal nuclear específico éreconhecido por uma proteína citoplasmática, a importina, que se liga à proteína a sertransportadas e estabelece a sua ligação com o complexo de poro, promovendo a suatranslocação através do poro. Após o transporte, a importina se desliga da proteína e retorna aocitoplasma. O sinal de localização nuclear não é removido depois que a proteína entra nonúcleo, o que permite sua reintrodução quando o envoltório nuclear se refaz após a mitose.A exportação de RNAs do núcleo para o citoplasma ocorre também por um processo ativo,mediado por receptores específicos. Os RNAs transcritos no núcleo que desempenham suafunção no citoplasma (mRNA, tRNA e rRNA) são exportados como complexos RNA-proteínas. Ossinais que dirigem a exportação nuclear podem estar presentes nos próprios RNAs ou nasproteínas. As moléculas de mRNA estão complexadas com cerca de 20 proteínas, formando asribonucleoproteínas nucleares heterogêneas ou hnRNPs. Pelo menos um dessas proteínas podeconter um sinal de exportação nuclear e atuar como transportadora do mRNA durante suaexportação para o citoplasma. Também os rRNAs são exportados do núcleo na forma departículas complexadas com proteínas, ou seja, de subunidades ribossômicas. Sua exportaçãotambém pode ser mediada por sinais presentes nas proteínas ribosômicas. Não se sabe aindacomo ocorre a exportação dos tRNAs.



* As histonas se ligam ao DNA graças à interação de seus radicais amino com os radicaisfosfato do DNA. Nem todos os radicais fosfato estão neutralizados pelas histonas, o queconfere à cromatina um caráter ácido (afinidade por corantes básicos).

Cada cromossomo contém um único duplex de DNA, compactado em uma fibra que se estendecontinuamente ao longo de todo o cromossomo. Assim, no que diz respeito à cromatinainterfásica e ao cromossomo mitótico, podemos explicar a compactação de uma molécula deDNA única e extremamente longa, em uma forma na qual pode ser transcrita e replicada etambém ser ciclicamente mais ou menos comprimida.A disposição da cromatina dentro do núcleo e o seu grau de condensação variam de um tipocelular para outro (são característicos de cada tipo celular). Além disso, a mesma célula podeapresentar a cromatina com vários graus de condensação, de acordo com o estágio funcional ecom o estado de diferenciação em que se encontra.


Os genes localizados na eucromatina podem se expressar ou não, dependendo do tipocelular e de suas necessidades metabólicas. Existem pelo menos duas formas deeucromatina: cerca de 10% na forma de cromatina ativa, que é menos condensada, e orestante na forma de cromatina inativa, que é mais condensada. A heterocromatinaconstitutiva está sempre condensada. Consiste, na maior parte, de DNA repetitivo e éencontrada nos centrômeros e ao redor deles e em algumas outras regiões. Ela passapelo ciclo celular com poucas mudanças no seu grau de condensação. Forma uma sériede massas discretas, mas, frequentemente, as diversas regiões de cromatina agregam-seem um cromocentro densamente corado.A heterocromatina facultativa pode existir tanto na forma geneticamente ativa(descondensada) quanto na forma inativa (condensada), como no caso da inativação docromossomo X em mamíferos (corpúsculo de Barr).A mesma fibra cromatínica pode apresentar regiões eucromáticas contínuas com regiõesheterocromáticas. Assim, o material genético é organizado de modo que diferentesestados de compactação sejam mantidos lado a lado, possibilitando a ocorrência dealterações cíclicas no nível de compactação da cromatina entre a intérfase e a divisão, eentre as diferentes fases da vida da célula.

Os cromossomos normais têm um só centrômero, o qual é visto ao microscópio como uma constriçãoprimária, a região em que as cromátides-irmãs estão unidas. O centrômero é essencial para a segregaçãodurante a divisão celular.Funções:ponto de ligação das fibras de microtúbulos durante a divisão celular;ponto de reunião de cromátides-irmãs;motor mecano-químico responsável pelo movimento dos cromossomos para os pólos da célula econclusão da divisão celular.Durante a prófase tardia, um par de cinetócoros forma-se em cada centrômero, cada um ligado a uma dascromátides-irmãs. Múltiplos microtúbulos ligam-se a cada cinetócoro, ligando o centrômero de umcromossomo aos dois pólos do fuso. Os cinetócoros desempenham um papel fundamental nesseprocesso, controlando a reunião e a separação dos microtúbulos acoplados e, por meio da presença demoléculas motoras, dirigindo o movimento cromossômico.Os telômeros são estruturas especializadas, constituídas por DNA repetitivo (repetições em tandem ouagrupadas) e proteína, que cobrem as extremidades dos cromossomos eucarióticos.Funções prováveis:manter a integridade estrutural do cromossomo: se um telômero for perdido, a extremidadecromossômica resultante fica instável, tendendo a fundir-se com as extremidades de outros cromossomosquebrados ou envolver-se em eventos de recombinação ou ser degradada.garantir a replicação completa das extremidades codificadoras dos cromossomos: durante a replicação doDNA, a síntese da fita atrasada é descontínua e requer a presença de algum DNA à frente para um primerde RNA.auxiliar o estabelecimento da arquitetura tridimencional do núcleo e/ou do pareamento cromossômico:as extremidades dos cromossomos parecem estar ligadas à membrana nuclear, sugerindo que ostelômeros auxiliam no posicionamento dos cromossomos.auxiliar no reconhecimento entre cromossomos homólogos para pareamento na meiose.Origens de replicação: necessárias para que ocorra a replicação do DNA na fase S da intérfase. Sãosequencias de DNA situadas próximas aos pontos onde a síntese de DNA é iniciada, que controlam o inícioda replicação. Nos eucariotos existem várias origens da replicação autônomas ao longo de uma únicamolécula (para cada origem, duas forquilhas de replicação bidirecionais).

As histonas H3 e H4 apresentam sequências idênticas em organismos tão distintosquanto a ervilha e o boi, sugerindo que elas desempenham funções idênticas em todosos eucariontes. Os tipos H2A e H2B possuem também sequências idênticas, comalgumas variações espécie-específicas.Em alguns tecidos, a H1 é substituída por histonas especiais. Por exemplo, em eritrócitosnucleados de aves, a histona H5 é encontrada em substituição à H1.

A porção enovelada da molécula das histonas contém alta percentagem de aminoácidoshidrofóbicos, e sua associação com o DNA deve-se à interações hidrofóbicas. Pode-seseparar, por processos químicos, a molécula de DNA das moléculas de histonas. Masquando o DNA e as histonas são colocados juntos em condições favoráveis, ocorrenovamente a formação espontânea de nucleossomos.

O nucleossomo é uma partícula de forma cilíndrica achatada, com 10 nm de diâmetro e6 nm de altura. Cada nucleossomo é constituído por 200 pb de DNA associados a umoctâmero de histonas e a uma molécula de histona H1. O octâmero é formado por duasmoléculas de cada uma das histonas H2A, H2B, H3 e H4. A molécula de H1 se associaexternamente ao DNA que envolve o octâmero. Cada nucleossomo é formado por umcentro ou cerne, constituído pelo octâmero de histonas H2A, H2B, H3 e H4, em torno doqual se enrola um segmento de DNA de aproximadamente 146 pb. Conectando umcentro de nucleossomo a outro, encontra-se um segmento de DNA não associado aproteínas com 15 até 100 pb, chamado DNA de ligação.


Dois tipos de fibras cromatínicas são encontradas no núcleo interfásico: a fibra de 10 nm dediâmetro ou nucleofilamento e a fibra de 30 nm ou solenóide.A fibra de 10 nm constitui o primeiro nível de compactação da cromatina e é formada pelaassociação de nucleossomos adjacentes. A organização dessa fibra depende da interação entreas histonas H1 de nucleossomos vizinhos. A histona H1 de um nucleossomo liga-se através desua extremidade amino-terminal, à extremidade carboxi-terminal da H1 do nucleossomoadjacente, em um arranjo “cabeça-cauda”. Com essa organização, o DNA de ligação não é maisobservado na fibra de 10 nm.A fibra de 30 nm constitui o segundo nível de organização da cromatina e é formada peloenovelamento da fibra de 10 nm em uma estrutura helicoidal. Cada volta da espiral contém 6nucleossomos organizados radialmente, ficando a histona H1 localizada no interior da fibra. Ahistona H1, juntamente com íons Mg2+ em concentração adequada, tem papel preponderantena formação e estabilização dessa fibra.Durante a intérfase, a cromatina que contém os genes ativamente transcritos é formada, em suamaioria, por fibras de 30 nm, enquanto cerca de 10% estão na forma de fibras de 10 nm,permitindo o acesso às enzimas envolvidas na transcrição.

Ainda no núcleo interfásico, as fibras de 30 nm podem se organizar em grandes alçasque se prendem ao envoltório nuclear através da lâmina nuclear. Cerca de 10% dacromatina interfásica também se encontra em um estado altamente condensado – aheterocromatina.Níveis superiores de compactação da cromatina parecem envolver, principalmente, asproteínas não-histônicas. A histona H1 parece, no máximo, participar também doprocesso de compactação, uma vez que a sua fosforilação, durante a prófase, determinaa condensação dos cromossomos.



MO= microscopia ópticaOs nucléolos são corpúsculos arredondados de aspecto esponjoso, mergulhadosdiretamente no nucleoplasma, uma vez que não possuem membrana envolvente. Aporção fibrilar densa é mais central e é formada por RNAr (RNA ribossômico) e proteínasribossomais. A porção granular é mais periférica e é formada por subunidadesribossômicas em formação.A região organizadora do nucléolo é a cromatina associada ao nucléolo, que na divisãoencontra-se nos satélites dos cromossomos acrocêntricos. Não é uma estruturacompacta, pois nota-se a invasão do nucleoplasma. Forma os ribossomos a partir dasproteínas ribossômicas, que são importadas do citoplasma e se associam com o RNAr.





Cariocinese= divisão do núcleo (fase que também recebe o nome de mitose no livro doJunqueira).Citocinese = divisão do citoplasmaPodemos também definir o ciclo celular como sendo uma sequência de eventoscontrolados que leva ao crescimento e divisão celular.

G0 é um estado quiescente no qual as células adultas maduras podem ficar por tempoindeterminado. Neste estágio, as células, desprovidas de fatores de crescimento,mantêm baixo metabolismo, com baixa velocidade de síntese de macromoléculas. Desseestado, alguns tipos celulares podem disparar para a fase proliferativa mediante umestímulo apropriado (nutrientes, hormônios de crescimento ou estímulo mecânico,como uma lesão, por exemplo). Ou seja, nesse estágio, as células permanecemmetabolicamente ativas, mas não se dividem ou então, se dividem apenas quandoestimuladas por sinais extracelulares, com a finalidade de renovação tecidual apósmorte ou lesão celular (hepatócitos por exemplo, que entram em G0, mas após dano aoórgão podem voltar a G1 e continuar o ciclo celular). G0 depende da história da célula alongo prazo: em cada tipo celular, cada fase do desenvolvimento do animal obedece aleis distintas, o que reflete as diferenças em sua maquinaria de controle interno. Porexemplo, no corpo humano algumas células como os neurônios que não continuam sereplicando e sim se mantendo e criando comunicações intercelulares. O estado G0 estámuito relacionado com a redução progressiva das sequências teloméricas do DNAdurante as sucessivas divisões celulares, sugerindo que G0 pode ser provocado pelaincapacidade de manter o comprimento dos telômeros (ou porque as células sãodeficientes em telomerase).G1 ("Gap 1") – Intervalo após a mitose durante o qual as células se preparam para iniciara síntese de DNA. Este período é caracterizado pela transcrição gênica e tradução,levando à síntese de proteínas necessárias para a síntese de DNA.S – Período no qual ocorre a duplicação do DNA celular.G2 ("Gap 2") – Intervalo após a síntese de DNA, durante o qual as células se preparampara a divisão.M – Período de divisão celular (mitose ou meiose).


Antes que a célula possa se dividir ela tem que crescer até alcançar um tamanhoadequado e constante. Em função disto, cerca de 95% do ciclo são gastos em intérfase,mas o tempo médio total desta fase é variável de tipo celular para tipo celular. Aduração varia também com as condições fisiológicas em que a célula se encontra, comoidade celular, disponibilidade de hormônios e de fatores de crescimento, temperatura,pressão osmótica, pressão hidrostática e pressão de oxigênio externas, e mesmo com oritmo circadiano que ocorre em animais. Existem também notáveis diferenças quanto àduração do ciclo celular entre os organismos. Em geral o ciclo dura 12 horas, em tecidosde mamíferos com crescimento muito rápido, e 24 horas em tecidos com crescimentomais lento. Em eucariotos unicelulares (leveduras, por exemplo) o tempo de geração ébem mais curto (aproximadamente 1 hora e meia para a formação de duas células-filhas). Nas células embrionárias, G1 é ausente ou tem duração negligenciável logo apósa fertilização. Neste caso, não ocorre crescimento celular.De todas as fases do ciclo, G1 é a mais variável na maioria das células de animais eplantas. Em geral ocupa muitas horas, durante as quais as células crescem. Esse períodopode variar individualmente de célula para célula, pois é o que mais sofre influência defatores extracelulares. Também é o período em que vários inibidores e mutações sãocapazes de bloquear a proliferação. Depois que as células entram na fase S, fatoresextracelulares não mais determinam os eventos do ciclo celular, os quais passam adepender de controles disparados intracelularmente. Portanto, as demais etapas dociclo, incluindo a mitose, têm tempos de duração mais constantes. A mitose dura maisou menos 1 hora. G2 em geral tem duração de 2 a 4 horas e o período S dura de 7 a 8horas. Apesar dessas fases serem mais constantes, a duração de cada uma varia entreespécies e também entre diferentes estágios do desenvolvimento de um mesmoindivíduo.Nas células embrionárias, G1 é ausente ou tem duração negligenciável logo após afertilização. Neste caso, não ocorre crescimento celular.

O ciclo pode durar algumas horas em tecidos de renovação rápida e até meses emoutros tipos celulares. Alguns tipos de células (terminalmente diferenciadas) não sedividem e permanecem em G0. Outras entram em G0 e, após um dano ao órgão, voltama G1 e continuam o ciclo celular (ex: células hepáticas)

O equilíbrio entre a proliferação celular e a manutenção do número de células nostecidos é alcançado através da apoptose.



Quando ocorrem mutações, proto-oncogenes tornam-se oncogenes, que sãocarcinogênicos e causam multiplicação celular excessiva. Essas mutações levam o proto-oncogene a expressar em excesso sua proteína estimuladora do crescimento ou aproduzir uma forma mais ativa. Os genes supressores de tumores, em contraste,contribuem para o desenvolvimento de câncer quando são inativados por mutações. Oresultado é a perda da ação de genes supressores funcionais, o que priva a célula decontroles cruciais para a inibição de crescimento inapropriado.

Substâncias de natureza polipeptídica = polipeptídeos e proteínas. Em geral, ospolipeptídeos com 100 ou mais aminoácidos são chamados de proteínas, e aqueles commais de 10 e menos de 100 aminoácidos são denominados polipeptídeos.Antes que a célula possa se dividir ela tem que crescer até alcançar um tamanhoadequado e constante, o que depende, entre outros fatores, da disponibilidade dosfatores de crescimento.Crescimento celular= aumento da massa celular através da síntese de proteínas e outrasmacromoléculas.


Os fatores de crescimento que estimulam a proliferação celular (mitógenos) podem serdivididos em dois grandes grupos: fatores de competência e fatores de progressão. Osfatores de competência, tais como o fator de crescimento de fibroblastos (FGF), fator decrescimento derivado de plaquetas (PDGF), fator de crescimento de hepatócito (HGF) eo fator de crescimento mitógeno de Bischoff para células precursoras de mioblastos, osquais atuam inicialmente e são diretamente responsáveis pela reentrada da célula nociclo de divisão celular, ou seja, pela sua passagem do estágio G0 para G1. Já os fatoresde progressão, como o fator de crescimento semelhante à insulina 1 e 2 (IGF-1 e IGF-2) eo fator de crescimento epidérmico (EGF), atuam posteriormente no ciclo celular eestimulam a fase de replicação do DNA, que representa a passagem da fase G1 para S dociclo. Assim, os fatores de competência estimulam o crescimento de células (aumentode massa celular), promovendo a síntese de proteínas e outras macromoléculas einibindo a sua degradação, enquanto os fatores de progressão controlam a taxa dedivisão celular atuando na fase G1 do ciclo celular. O resultado final é a ativação decomplexos G1/S-Cdk, que superam as barreiras inibidoras que normalmente bloqueiama progressão para a fase G1-S (ver aula sobre Controle e regulação do ciclo celular).

Receptores de superfície:1- Receptores com atividade tirosina quinase intrínseca autofosforilação.2- Receptores sem atividade catalítica intrínseca ativam proteínas citosólicas (tirosinaquinsase) que fosforilam o receptor.3- Receptores associadas a proteína G geram mensageiros secundáriosintracitoplasmáticosSistema de transdução de sinais: processo pelo qual sinais extracelulares são detectadose convertidos em sinais intracelulares o que gera uma resposta celular específica.A célula estável, após a cascata de fosforilação proteica, entra no ciclo celular (sai de G0e entra em G1). O sistema de transdução de sinais transfere a informação para o núcleo,onde ocorre alterações na expressão gênica (transcrição dos genes) controlado forfatores reguladores conhecidos como fatores de transcrição.


Caso a célula não receba os estímulos necessários (fatores de crescimento e hormônios)para romper o ponto de restrição, ela entra em G0.Uma vez que tenha passado pelo ponto de restrição (ponto R), a célula estácomprometida a prosseguir até o final da divisão, mesmo na ausência de estímulosadicionais.

Ponto de checagem G1/S: ponto de restriçãoPonto de checagem de replicação do DNA: transição G2/MPonto de checagem do fuso mitótico: transição metáfase/anáfase



MPF também é conhecido como Fator Promotor da Fase M.




Os centrossomos são constituídos por um par de centríolos (que se denominadiplossomo) e um material pericentriolar amorfo e eletrodenso, a partir do qualemanam fibras de microtúbulos radiais. Os centrossomos mas as fibras radiais compõemo chamado áster.

Com o rompimento do envoltório nuclear, não há mais limites físicos entre o citoplasmae o material nuclear. Esses eventos ocorridos a partir do rompimento do envoltórionuclear são considerados por muitos autores como uma etapa distinta denominadaprometáfase.
















Sinapse cromossômica é o emparelhamento (pareamento) de pares de cromossomoshomólogos possibilitada pela formação do complexo sinaptonêmico, uma elaborada estruturaconstituída por diversas proteínas que formam um longo eixo central e duas barras laterais àsquais se associam cromossomos homólogos. É altamente específico; colocando em contato cadaponto de um cromossomo com o seu correspondente. Essa ligação forma um par decromossomos chamado "bivalente".



Durante a diacinese os quiasmas são mantidos, o que é importante para a distribuiçãocorreta dos cromossomos. A falta de quiasmas pode levar a uma segregação incorretados cromossomos homólogos na anáfase.

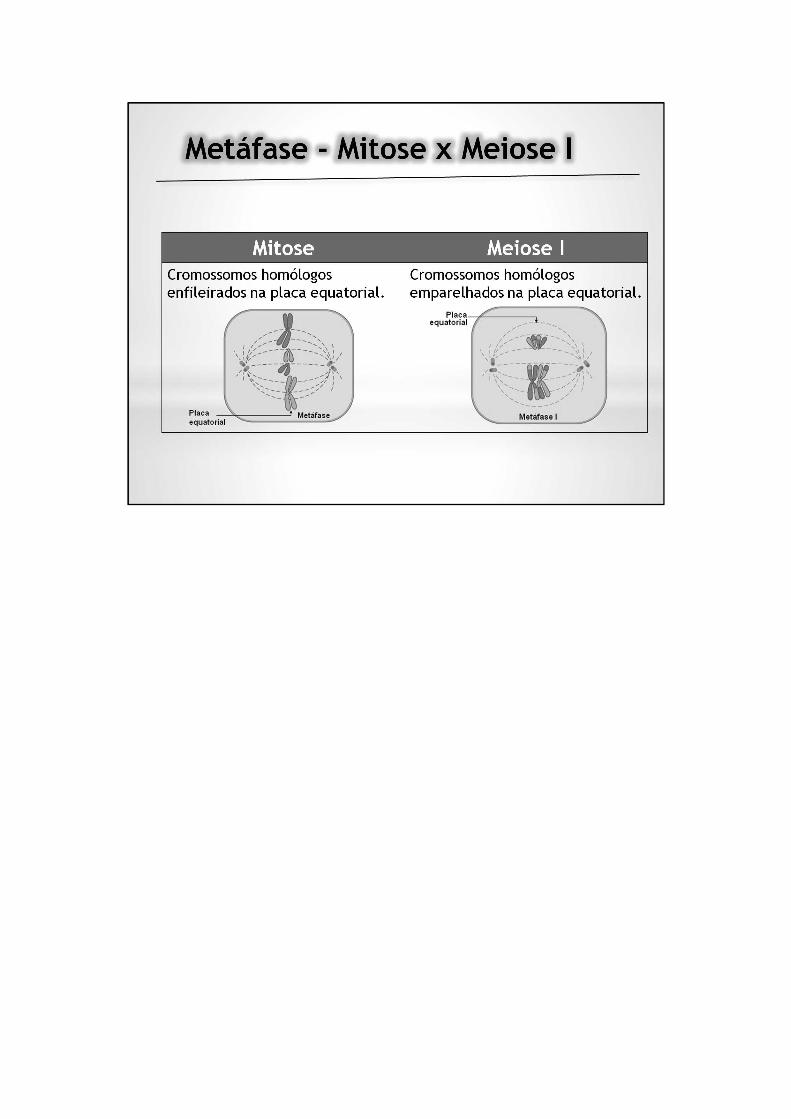









Intercinese: curto intervalo entre a primeira e a segunda divisões da meiose. Nemsempre existe intercinese entre as duas divisões da meiose, ou esta é tão curta quepode ser considerada como inexistente.


Na maioria dos vertebrados, assim como em muitos invertebrados, as divisões meióticaspara a formação do óvulo não são completadas antes que a fertilização ocorra. Comoregra geral, o desenvolvimento é interrompido durante a prófase I da primeira divisãomeiótica. A meiose é retomada e se completa na época da ovulação (maioria dosmamíferos) ou logo após a fertilização (muitos invertebrados, peixes teleósteos, anfíbiose répteis). Em seres humanos, a segunda divisão meiótica somente se completa após apenetração do espermatozoide no ovócito II.




