fundaÇÃo universidade federal de rondÔnia … · a região do interflúvio madeira-purus possui...
TRANSCRIPT

FUNDAÇÃO UNIVERSIDADE FEDERAL DE RONDÔNIA
NÚCLEO DE CIÊNCIAS EXATAS E DA TERRA
PROGRAMA DE PÓS-GRADUAÇÃO EM DESENVOLVIMENTO REGIONAL
E MEIO AMBIENTE
BORBOLETAS FRUGÍVORAS DA ESTAÇÃO ECOLÓGICA DE CUNIÃ:
PADRÕES DE DISTRIBUIÇÃO E EFEITOS DE VARIÁVEIS AMBIENTAIS
CAMILA MOURA LEMKE
Porto Velho (RO)
2017

FUNDAÇÃO UNIVERSIDADE FEDERAL DE RONDÔNIA
NÚCLEO DE CIÊNCIAS EXATAS E DA TERRA
PROGRAMA DE PÓS-GRADUAÇÃO EM DESENVOLVIMENTO REGIONAL
E MEIO AMBIENTE
BORBOLETAS FRUGÍVORAS DA ESTAÇÃO ECOLÓGICA DE CUNIÃ:
PADRÕES DE DISTRIBUIÇÃO E EFEITOS DE VARIÁVEIS AMBIENTAIS
CAMILA MOURA LEMKE
Orientador: Prof. Dr. Angelo Gilberto
Manzatto
Dissertação de Mestrado apresentada junto ao
Programa de Pós-Graduação em
Desenvolvimento Regional e Meio Ambiente,
Área de Concentração em Ambiente, Saúde e
Sustentabilidade para obtenção do título de
mestre em Desenvolvimento Regional e Meio
Ambiente.
Porto Velho (RO)
2017

Dados Internacionais de Catalogação na Publicação
Fundação Universidade Federal de Rondônia
Gerada automaticamente mediante informações fornecidas pelo(a) autor(a)
L544b Lemke, Camila Moura.
Borboletas frugívoras da Estação Ecológica de Cuniã : padrões de
distribuição e efeitos de variáveis ambientais / Camila Moura Lemke. -- Porto Velho, RO, 2017.
60 f. : il.
Orientador(a): Prof. Dr. Angelo Gilberto Manzatto
Dissertação (Mestrado em Desenvolvimento Regional e Meio Ambiente) - Fundação Universidade Federal de Rondônia
1.Conservação. 2.RAPELD. 3.Nymphalidae. I. Manzatto, Angelo Gilberto. II. Título.
CDU 502.3(811.1)
Bibliotecário(a) Ozelina do Carmo de Carvalho CRB 11/486

CAMILA MOURA LEMKE
BORBOLETAS FRUGÍVORAS DA ESTAÇÃO ECOLÓGICA DE CUNIÃ: PADRÕES DE DISTRIBUIÇÃO E EFEITOS DE VARIÁVEIS AMBIENTAIS
Comissão Examinadora
Dr. Rodrigo Barros Rocha Membro Titular
Fundação Universidade Federal de Rondônia/Embrapa/RO
União das Escolas Superiores de Rondônia
Porto Velho, 23 de novembro de 2017

D E D I C A T Ó R I A
À minha família.

AGRADECIMENTOS
A Deus e ao Universo por sempre conspirarem ao meu favor.
Ao meu Orientador, Angelo Gilberto Manzatto, muito obrigada pela oportunidade de
orientação, pela paciência e incentivos na construção dessa trajetória.
A meu pai Marcos Lemke pela participação na primeira expedição de campo do mestrado, à
minha mãe Naderge Moura e irmãs Carina Moura Lemke e Catrin Moura Lemke pelo trabalho
de organização da coleção de borboletas e por tudo que representam na minha vida.
Ao meu companheiro, por estar sempre presente, com amor, incentivar os trabalhos, comemorar
as vitórias e aguentar os estresses. E à família Costa pelo carinho.
À Adeilza Sampaio e ao Alex (o leão) Oliveira, pela amizade e apoio em campo e laboratório
nesses dois anos.
À Susamar Pansini e Ricardo Andrade pela ajuda com os dados e análises estatísticas.
À Carolina Brum por dividir o projeto e trabalho.
Ao amigo/irmão Samuel Nienow pelo empurrãozinho à minha vontade de ser borboletóloga e
fazer o mestrado.
À amiga conselheira Carmô, que incentivou o mestrado e trabalho no Cuniã.
Ao Pedro Nascimento, Kamila Miranda, Flaviana Bezerra, Reno e os Carlos que ajudaram nas
coletas de dados.
Ao Marcio Uehara-Prado pelo ensinamento de borboletas.
Ao amigo de trabalho, Paulinho Bonavigo.
A Carolina Augusto, amiga que o mestrado me deu. E a todos os colegas da turma 2015/2.
Universidade Federal de Rondônia (UNIR) e ao Programa de Pós-Graduação Desenvolvimento
Regional e Meio Ambiente (PGDRA) pela oportunidade de capacitação profissional. Em
especial aos Professores Vanderlei Maniesi, Arneide Bandeira, Rodrigo Barros, Maurício
Reginaldo, Artur Moret, Mariangela de Azevedo, Wanderlei Bastos e Ari Ott,
e a secretária do PGDRA, Dona Izabel, por ser extremamente competente, sempre tirar minhas
dúvidas e quebrar uns galhos!
Capes pelo apoio financeiro através da bolsa de estudo concedida.
Ao Laboratório de Biogeoquímica Ambiental Wolfgang C. Pfeiffer pela acolhida e espaço.
A todos que de alguma forma me ajudaram para que esse trabalho fosse possível...

“Bater de asas de uma Borboleta no Brasil, provoca um tornado no Texas?”
Edward Norton Lorenz, 1972.
No Congresso Anual da Associação Americana para o Avanço da Ciência (AAAS).

RESUMO
A ação antrópica vem alterando a paisagem natural e degradando os ecossistemas amazônicos,
assim, inventariar as espécies presentes localmente é a melhor maneira de salvaguardar o
conhecimento. É imprescindível que especialistas compreendam os padrões e processo destas
mudanças, bem como, compreender como as populações biológicas respondem a estas
modificações. E assim poder inferir em tomadas de decisões quanto a biodiversidade,
conservação e uso dos recursos. A Região do interflúvio Madeira-Purus possui características
ecológicas e ambientais relevantes para o entendimento de padrões e processos ecológicos. As
borboletas frugívoras apresentam algumas vantagens que facilitam o estudo de suas populações,
são facilmente capturadas com armadilhas de iscas, são ricas em espécies, são bem conhecidas
taxonomicamente na literatura, há possibilidade mínima de capturas ao acaso, o que não ocorre
com outras metodologias; e proporcionam padronização e simultaneidade do esforço em
diferentes áreas. A família Nymphalidae possui estreita correlação com a riqueza total do
ambiente e podem ser usadas como indicadores rápidos dos diferentes tipos de variações na
paisagem, pois possuem ciclo de vida rápido em relação a outros grupos de animais que
levariam mais tempo para se estabelecer. Além disso, suas espécies representariam outros
grupos na resposta ao distúrbio. Face ao exposto o presente trabalho objetivou caracterizar a
comunidade de borboletas frugívoras da família Nymphalidae da ESEC-Cuniã, reportar a
densidade, determinar os padrões espaciais e temporais e testar os efeitos de variáveis
ambientais nas assembleias de borboletas frugívoras. As densidades das espécies de borboletas
foram determinadas pelo método de amostragem RAPELD. Foram realizadas em 2016 duas
expedições ao campo para coleta de dados em amostragem padronizada seguindo a
sazonalidade da região Amazônica. Utilizando-se armadilhas VSR, iscadas com uma mistura
de caldo de cana e banana fermentada por 48h. Foram amostradas em 30(trinta) parcelas um
total de 667(seiscentos e sessenta e sete) indivíduos representando 4(quatro) subfamílias,
distribuídos em 10(dez) tribos e 73(setenta e três) espécies. A subfamília mais abundante foi
Biblidinae com 479 indivíduos (72% da abundância) com representantes de quatro tribos:
Epicaliini(365), Epiphilini (101), Ageroniini (9) e Callicorini (4). Catonephele acontius foi a
espécie mais abundante e frequente, com 220 espécies. Satyrinae foi a subfamília mais rica com
35 espécies, seguida por Biblidinae, Charaxinae e Nymphalinae com 19, 16, 3 espécies,
respectivamente. A ordenação (MDS) das assembleias de borboletas frugívoras e a formação
de subgrupos sugerem que, a borboletas frugívoras apresentam diversos níveis de similaridade
entre e dentre os subgrupos reportados. Observou-se que os melhores preditores para mudanças
das assembleias de borboletas frugívoras foram gradientes argilosos, alto teor de fósforo e a
estrutura da vegetação estruturam fortemente as assembleias de borboletas frugívoras na região
do interflúvio entre os rios Madeira e Purus. Comprovou-se que a Estação Ecológica de Cuniã
é uma região de refúgio da lepidopterofauna de Rondônia e sul do Amazonas, fundamental para
a manutenção da biota e biodiversidade local e regional.
Palavras-chave: Conservação. RAPELD. Nymphalidae.

ABSTRACT
The anthropic action has altered the natural landscape and degraded the Amazonian ecosystems,
like this, to inventory the species present locally is the best way to safeguard the knowledge. It
is imperative that specialists understand the patterns and process of these changes, as well as
understand how biological populations respond to these changes. And be able to infer in
decisions about biodiversity, conservation and use of resources. The Madeira-Purus interfluvial
region has ecological and environmental characteristics relevant to the understanding of
ecological patterns and processes. Frugivorous butterflies have some advantages that facilitate
the study of their populations, are easily captured with bait traps, are rich in species, are well
known taxonomically in the literature, there is a minimum possibility of random captures,
which is not the case with other methodologies; And provide standardization and concurrency
of effort in different areas. The Nymphalidae family has a close correlation with the total
richness of the environment and can be used as quick indicators of the different types of
variations in the landscape, since they have a fast life cycle in relation to other groups of animals
that would take more time to establish themselves. In addition, their species would represent
other groups in the response to the disorder. In view of the above, the present work aimed to
characterize the community of frugivorous butterflies of the Nymphalidae family of ESEC-
Cuniã, to report the density, to determine the spatial and temporal patterns and to test the effects
of environmental variables on assemblages of frugivorous butterflies. The densities of the
butterfly species were determined by the RAPELD sampling method. Two field trips were
carried out in 2016 to collect data on standardized sampling, following the seasonality of the
Amazon region. Using VSR traps, baited with a mixture of cane juice and fermented banana
for 48h. A total of 667 (sixty - seven) individuals representing 4 (four) subfamilies, distributed
in 10 (ten) tribes and 73 (seventy three) species were sampled in 30 (thirty) plots. The most
abundant subfamily was Biblidinae with 479 individuals (72% of abundance) with
representatives of four tribes: Epicaliini (365), Epiphilini (101), Ageroniini (9) and Callicorini
(4). Catonephele acontius was the most abundant and frequent species, with 220 species.
Satyrinae was the richest subfamily with 35 species, followed by Biblidinae, Charaxinae and
Nymphalinae with 19, 16, 3 species, respectively. The ordering (MDS) of assemblages of
frugivorous butterflies and the formation of subgroups suggest that, to frugivorous butterflies,
there are several levels of similarity between and among the subgroups reported. It was
observed that the best predictors for frugivorous butterfly assemblage changes were clayey
gradients, high phosphorus content and vegetation structure strongly structure the assemblages
of frugivorous butterflies in the region of the interflow between the Madeira and Purus rivers.
It was verified that the Cuniã Ecological Station is a refuge region of the lepidopterofauna of
Rondônia and south of the Amazon, fundamental for the maintenance of biota and local and
regional biodiversity.
Keywords: Conservation. RAPELD. Nymphalidae.

LISTA DE FIGURAS
Figura 1 Estação Ecológica do Cuniã – ESEC Cuniã área em verde- localizada na BR
319 no sentido Porto Velho – Humaitá.
21
Figura 2 Grade padrão para inventários bióticos e abióticos proposto pelo Programa
de Pesquisa em Biodiversidade - PPBio instalada na ESEC-Cuniã localizada na BR
319 no sentido Porto Velho – Humaitá, Rondônia. Os círculos verdes representam as
parcelas terrestres que foram utilizadas neste estudo.
23
Figura 3 Modelo ilustrativo de armadilha: (a) corpo da armadilha confeccionado em
tule; (b) funil interno; (c) plataforma onde a isca é colocada.
24
Figura 4 Representação das parcelas instaladas na grade da ESEC Cuniã (A).
Disposição das 3 armadilhas Van Someren-Rydon ao longo dos 250 m de extensão de
cada parcela. As curvas representam a não linearidade das parcelas, e não desnível do
terreno (B).
25
Figura 5 Coleta em uma das armadilhas instaladas em parcela da ESEC-Cuniã (A).
Amostra coletada em processo de identificação (B).
26
Figura 6 Proporção da abundância das Subfamílias coletadas na ESEC-Cuniã. 31
Figura 7 Abundância das 10 tribos coletadas na ESEC-Cuniã. 31
Figura 8 Dendograma de dissimilaridade das assembleias de borboletas frugívoras das
30 na ESEC Cuniã, Porto Velho, Rondônia.
36
Figura 9 Análise de Ordenação MDS (Configuração Stress de Kruskal (1) = 0,276)
com dados quantitativos (Bray e Curtis) das assembleias de borboletas frugívoras na
ESEC Cuniã, Porto Velho, Rondônia.
36
Figura 10 Densidade relativa de espécies de borboletas frugívoras ao longo do
gradiente de solo: Teores de Argila e Fósforo (P), de peso da massa seca de
Serrapilheira e hidrológico considerando a distância mínima horizontal à rede de
drenagem (DHR).
38
Figura 11 Resultados gráficos do melhor modelo fornecido pelas regressões lineares
múltiplas no qual Teor de Argila, Teor de Fósforo (P), Densidade de Palmeiras,
Florística, MDSBiomassa arbórea são os preditores para mudanças na composição de
borboletas frugívoras.
41

LISTA DE TABELAS
Tabela 1 Lista de espécies de borboletas frugívoras coletadas na ESEC-Cuniã com
sua respectiva abundância.
32
Tabela 2 Seleção do melhor modelo de Regressão Linear, selecionados pela menor
distância de AIC e R² Ajustado
39
Tabela 3 Parâmetros de cada modelo da Regressão Linear. 40

LISTA DE SIGLAS
PPBIO- Programa de Pesquisa em Biodiversidade
MDS- Análise de escalonamento Multidimensional métrico
ESEC- Estação Ecológica
GICJ- Gestão Integrada Cuniã-Jacundá
RAPELD- Pesquisas ecológicas de longa duração com uso de inventários rápidos
para avaliação da complementariedade biótica e uso da terra na Amazônia
VSR- Van Someren-Rydon
REDELEP- Rede Nacional de Pesquisa e Conservação dos Lepidópteros
AIC- Critério de Informação de Akaike
DVL- Distância vertical do lençol freático (verificado no piezômetro)
DHR- Distância mínima horizontal à rede de drenagem (distância do igarapé)
BC- Bray-Curtis
SD- Sorensen-Dice
TA- Teor de Argila;
ES- Espessura de Serrapilheira;.
TP- Teor de Fósforo;
DP- Densidade de Palmeiras;
F- Florística;
MDSBio- MDS Biomassa arbórea;
MS- Peso da Massa seca de Serrapilheira;
DA- Densidade de Árvores;
MDSFlo- MDS Florística;
CA- Cotas Altitudinais do Terreno

SUMÁRIO
INTRODUÇÃO .......................................................................................................
13
1 REFERENCIAL TEÓRICO................................................................................... 16
1.1 RONDÔNIA E O DESFLORESTAMENTO............................................................ 11
1.2 BORBOLETAS FRUGÍVORAS.............................................................................. 12
2 OBJETIVOS.............................................................................................................
20
2.1 OBJETIVO GERAL................................................................................................. 20
2.2 OBJETIVOS ESPECÍFICOS.................................................................................... 20
3 METODOLOGIA...................................................................................................
21
3.1 ÁREA DE ESTUDO................................................................................................. 21
3.2 DESENHO AMOSTRAL......................................................................................... 22
3.3 ESTIMATIVAS DE DENSIDADE DE BORBOLETAS........................................ 23
3.4 VARIÁVEIS AMBIENTAIS.................................................................................... 27
3.5 ANÁLISE DOS DADOS.......................................................................................... 28
4 RESULTADOS.........................................................................................................
31
4.1 BORBOLETAS FRUGÍVORAS DA ESEC CUNIA................................................ 31
4.2 PADRÕES ESPACIAIS E TEMPORAIS DAS ASSEMBLEIAS DE BOBOLETAS
FRUGÍVORAS..................................................................................................................
34
4.3 EFEITO DAS VARIAVEIS AMBIENTAIS............................................................... 37
5 DISCUSSÃO............................................................................................................
42
CONCLUSÃO..........................................................................................................
46
REFERÊNCIAS......................................................................................................
47
APÊNDICE.............................................................................................................. 55

13
INTRODUÇÃO
O bioma amazônico ocupa uma posição de destaque em relação à biodiversidade
mundial (SILVA; RYLANDS; FONSECA, 2005), e ainda exerce um papel importante nos
ciclos globais, principalmente o carbono e também o ciclo da água, os quais afetam outras
regiões, incluindo as áreas mais expressivas para a agricultura brasileira (ANDRADE et al.,
2017; MARENGO, 2007).
Nos últimos anos, tem sido crescente a preocupação com o uso e a conservação dos
recursos florestais e ambientais com base em protocolos de monitoramento padronizados
espacialmente visando ampliar avanços no entendimento de padrões e processos ecológicos
(MAGNUSSON et al., 2005).
Neste contexto, a Amazônia apresenta uma posição de destaque, ocupando um território
de 4,196.943 milhões de km². Apesar disso, a pesquisa na Amazônia ainda é relativamente
incipiente e fragmentada, não exercendo o impacto necessário sobre políticas públicas para uma
região megadiversa tão importante. Estima-se que uma pequena parte foi descrita e catalogada
cientificamente (BRASIL, 2004; PEIXOTO, et al., 2016).
A ação antrópica vem alterando a paisagem natural e degradando os ecossistemas
amazônicos, desta forma, inventariar a biota e relacioná-las a diferentes preditores ambientais
é um bom caminho para propor ações para o uso e o manejo mais adequado de componentes
específicos da biodiversidade.
A maioria desses usos requer a identificação correta de espécies ou outras unidades
taxonômicas, informação sobre a distribuição e abundância dos organismos, e a relação entre
distribuições de organismos e o uso potencial e efetivo da terra. Essas informações são
necessárias para o zoneamento de unidades de conservação, planejamento regional do uso da
terra, avaliação de impactos ambientais, determinação de cotas de extração para indústrias
extrativistas e a identificação do potencial econômico de novos produtos.
Collinge (2001), destaca que esforços de conservação biológica bem-sucedidos exigem
que os biólogos, ecólogos e evolucionistas compreendam simultaneamente padrões e processo
de mudança da paisagem, como perda de habitat e fragmentação, bem como, compreender
como as populações biológicas respondem a estas modificações em larga escala.
As planícies amazônicas são regiões pouco conhecidas e relativamente bem preservadas
(SOUSA, 2007). A Rodovia Manaus-Porto Velho (BR 319) é o único acesso às florestas de
terra-firme na região. Fearnside e Graça (2006) destacam que a região é composta por um
mosaico de unidades de conservação estaduais e federais, concentradas em uma única porção

14
do interflúvio Purus-Madeira e em sua maioria, unidades de uso direto, como reservas
extrativistas ou de uso sustentável.
A Região do interflúvio entre os rios Madeira e Purus é relativamente bem estudada,
foram conduzidos estudos acerca da estrutura da vegetação por Pansini et al., 2016 (palmeiras),
Lima, 2012 (palmeiras), Andrade et al., 2017 (fitossociologia), Sampaio et al., 2016
(Balanophloraceae), Cunha et al., 2014 (Hymenophyllaceae), Galvão, 2016 (Serrapilheira),
Cabral, 2015 (epífitas e hemiepífitas), Ribeiro, 2014 (herbáceas), Magalhães, 2007
(Melastomataceae), Oliveira, 2016 (ervas), bem como outros projetos de estudos da
biodiversidade. E possui informações estruturadas a acerca dos mecanismos mantenedores e
reguladores da biodiversidade local e regional, além de possuir características ecológicas e
ambientais relevantes para o entendimento de padrões e processos ecológicos.
O Interflúvio reúne variações e gradientes ambientais que a distinguem das demais áreas
da Amazônia central. Em particular, o regime pluviométrico e o relevo da região, pouco
variável, com mudanças de altitude muito sutis ao longo do interflúvio (PANSINI, 2011).
Dentre algumas características que devem ser salientadas destacam-se um interessante
gradiente climático entre Manaus e Porto Velho. A duração da estação de menor pluviosidade
varia de 1 a 5 meses ao longo do ano, apresentando precipitações em torno de 100mm. O regime
hidrológico é dependente da precipitação local, com corpos d’água sazonais e lençol freático
superficial.
Tendo em vista a expectativa de ocupação acelerada da região nos próximos anos é
urgente o desenvolvimento de pesquisas que permitam conhecer melhor seus recursos naturais,
procurando dar subsídios para o uso e manejo de recursos florestais e, em paralelo, propor ações
que minimizem o impacto da reabertura da estrada. A repavimentação e recuperação da BR 319
tem gerado um aumento no processo de ocupação de áreas no seu entorno (FEARNSIDE;
GRAÇA, 2006).
Para esses tipos de pesquisas e ou monitoramentos são elencados representantes da fauna
e flora para serem indicadores de qualidade ambiental, estes devem apresentar quatro
características básicas: viabilidade; custo efetivo para amostragem; fácil e confiável
identificação; funcionalidade e responder aos distúrbios de maneira consistente (PEARCE;
VENIER, 2006).
As borboletas têm sido cada vez mais sugeridas como organismos úteis no monitoramento
da qualidade ambiental, pois são sensíveis às mudanças ambientais provenientes de diferentes
tipos de perturbação (DEVRIES et al., 1997; BROWN; FREITAS, 2000; UEHARA-PRADO;
RIBEIRO, 2012).

15
Além de possuírem características favoráveis, como facilidade de amostragem e
identificação, estudos recentes demonstraram que borboletas podem atuar como representantes
de outros grupos de invertebrados e vertebrados em relação às suas respostas às perturbações
antrópicas. (BARLOW et al., 2007; UEHARA-PRADO et al., 2009).
As borboletas frugívoras podem ser indicadores da riqueza de grupos como árvores e
pássaros, podendo ser usadas como táxon substituto (surrogate) em inventários biológicos com
finalidades conservacionistas e biomonitoramento (SCHULZE et al., 2004).
O presente trabalho se propôs, portanto, a responder algumas questões referentes aos
padrões de distribuição de espécies de borboletas frugívoras em um trecho do interflúvio
Madeira-Purus como: (1) Quais espécies ocorrem na ESEC-Cuniã? (2) As espécies possuem
distribuição restrita ao longo dos gradientes ambientais? (3) Quais são os efeitos das variáveis
ambientais em meso-escala em uma floresta de terra-firme na planície do interflúvio Madeira-
Purus?
A resposta a estas questões permitiu ampliar o conhecimento da distribuição de borboletas
frugívoras na região do Interflúvio Madeira-Purus. Auxiliar na condução de futuros planos de
preservação em áreas de conservação e subsidiar políticas públicas do Estado de Rondônia, no
que se refere a tomada de decisões visando à preservação dessas importantes áreas.

16
1 REFERÊNCIAL TEÓRICO
1.1 RONDÔNIA E O DESFLORESTAMENTO
Criado em 22 de dezembro de 1981 - pela Lei Complementar nº 041, correspondente a
cerca 2,86% do território brasileiro e a 6,79% da região norte, o estado de Rondônia possui um
território de 237.590,547km² e se insere na área de abrangência da Amazônia Legal porção
ocidental entre os paralelos 7º 58' e 13º 43' de longitude oeste de Greenwich. (BRASIL, 2010).
O estado de Rondônia historicamente tem seu desenvolvimento a partir de exploração
do meio ambiente. Cemin (2006) afirma que o processo de colonização de Rondônia constitui-
se em prática predatória quanto aos recursos humanos e naturais.
Oliveira (2001) também aborda os processos de apropriação da natureza através dos
programas para o desenvolvimento do estado de Rondônia, este destaca a descoberta de ouro
no rio Corumbiara, no século XVIII; a conquista e o povoamento dos vales do Guaporé,
Mamoré e Madeira; a construção do Real Forte do Príncipe da Beira, no período colonial; o
Primeiro e o Segundo Ciclos da Extração de Látex; a construção da Estrada de Ferro Madeira-
Mamoré em 1912 e a descoberta de minério de estanho (cassiterita) em 1952. O período mais
expressivo do desenvolvimento regional ocorreu a partir da abertura da BR 364, e com
implantação de projetos de colonização, nas décadas de 60, 70 e 80 pelo Governo Federal,
através do INCRA que realizou um programa de povoamento, visando à distribuição de terras
para atrair pessoas dispostas a se instalar e a contribuir para o controle administrativo e
desenvolvimento socioeconômico do território, fato que provocou a entrada de vários
imigrantes vindos da Região Sul do Brasil. Porém, os órgãos governamentais, pouco se
preocupavam com a preservação do meio ambiente, gerando como consequência um
desmatamento desordenado.
A taxa de desmatamento na Amazônia variou ao longo do tempo e tem aumentado nos
últimos anos (FEARNSIDE, 2003), Rondônia está entre os cinco Estados brasileiros com
maiores índices de desmatamento da Amazônia (AZEVEDO et al., 2016).
De acordo com Penin et al. (2011), no Estado ocorrem obras de infraestrutura, em um
ritmo já acelerado de colonização, além do significativo crescimento das últimas décadas que
promete intensificar-se nos próximos anos, com a abertura de estradas, a implementação e
funcionamento das hidrelétricas de Jirau e Santo Antônio, e mais três de grande porte,
projetadas para os rios Mamoré e Machado, e outras 10 Pequenas Centrais Hidrelétricas (PCHs)
em execução, gasodutos, a expansão da pecuária e da monocultura agrocapitalista que ocupam

17
áreas cada vez maiores de nossas florestas. Nas cidades, em crescente ritmo de expansão, a
periferia, antes constituída por terras pouco ocupadas, passa a oferecer o cenário dos novos
bairros.
Durante as últimas décadas, a ação humana transformou as condições naturais da região
e modernizou a economia regional, sem observar as questões ambientais e as disparidades
regionais.
Com o propósito de salvaguardar parte da área existente entre o Interflúvio Madeira-
Purus a ESEC Cuniã, foi criada em 2001, através do decreto de 27 de setembro de 2001 por
meio da Lei 9.985, de 18 de julho de 2.000 (BRASIL, 2001).
Localizada ao norte do Estado de Rondônia e ao sul do Estado do Amazonas, nos
municípios de Porto Velho-RO e Canutama-AM. Atualmente a sua área total é de 186.743,26
hectares, divididos em duas áreas distintas, adjacentes à RESEX Cuniã, denominadas área I e
área II. De relevante importância ecológica, pois é um ambiente rico em lagos e várzeas
formadas pelo rio Madeira e seus afluentes, e promove a proteção de uma parcela significativa
do bioma amazônico e de manchas de transição com o cerrado e campos naturais, somando a
área total de 408 mil hectares. É uma área de terra firme de floresta ombrófila aberta (BRASIL,
2012).
É importante salientar que existe atualmente alta expectativa de ocupação acelerada na
região para os próximos anos devido ao processo de repavimentação e recuperação da BR 319
e construção da ponte sobre o Rio Madeira.
1.2 BORBOLETAS FRUGÍVORAS
Como as ameaças de degradação ambiental encontram-se em ritmo acelerado e partindo
da prerrogativa de que o grande desafio atual é buscar o máximo de conhecimento sobre os
ecossistemas característicos de nossa região e apresentar sugestões de como esse conhecimento
pode ser utilizado para o desenvolvimento sustentável, é urgente a realização de pesquisas que
permitam conhecer melhor seus recursos naturais, dentre eles, as borboletas frugívoras da
família Nymphalidae que estão em crescente utilização como bioindicadores em
monitoramentos em Unidades de Conservação no Brasil (NOBRE et al., 2014).
Estas possuem um valor particular como indicador ecológico, pois além de serem
carismáticas, constituem um grupo de fácil visualização e identificação (MIELKE e
CASAGRANDE, 1997). Borboletas são insetos com amplo apelo em campanhas de

18
conservação (SCHULZE et al., 2004), e possuem a capacidade de indicar alterações ambientais
devido a sua estreita associação aos hábitats (GUTIÉRREZ, 1997; FREITAS et al., 2003).
Quando adultos, esses insetos podem alimentar-se de néctar ou pólen (borboletas
nectarívoras) ou podem adquirir seus nutrientes de matéria orgânica em decomposição (como
frutos, fezes e carcaças) e exsudatos vegetais. Nesse caso, as borboletas são denominadas
frugívoras (DEVRIES, 1987).
A guilda de borboletas frugívoras está representada por espécies das subfamílias
Biblidinae, Charaxinae, Satyrinae e Nymphalinae (tribo Coeini) da família Nymphalidae
(DEVRIES, 1987, para classificação e sistemática da família, consultar WAHLBERG et al.,
2009).
Estudos ecológicos com borboletas frugívoras indicam que padrões de diversidade
desses insetos estão associados a variáveis como precipitação (DEVRIES e WALLA, 2001),
estrutura da vegetação (KOH e SODHI, 2004, PARDONNET et al., 2013), temperatura
(RIBEIRO e FREITAS, 2010), fenologia de folhas e abertura do dossel florestal (BARLOW et
al., 2007).
Além disso, as borboletas frugívoras podem ser indicadores da riqueza de grupos como
árvores e pássaros, podendo ser usadas como táxon substituto (surrogate) em inventários
biológicos com finalidades conservacionistas e biomonitoramento (SCHULZE et al., 2004).
As borboletas frugívoras apresentam algumas vantagens que facilitam o estudo de suas
populações (UEHARA-PRADO et al., 2004; FURLANETTI, SETTE e LOUZADA, 2007) são
facilmente capturadas com armadilhas de iscas, são ricas em espécies, são bem conhecidas
taxonomicamente na literatura, há possibilidade mínima de capturas ao acaso, o que não ocorre
com outras metodologias; e proporcionam padronização e simultaneidade do esforço em
diferentes áreas (UEHARA-PRADO et al., 2004).
A família Nymphalidae possui estreita correlação com a riqueza total do ambiente e
podem ser usadas como indicadores rápidos dos diferentes tipos de variações na paisagem
(BROWN; FREITAS, 2000), pois possuem ciclo de vida rápido em relação a outros grupos de
animais que levariam mais tempo para se estabelecer. Além disso, suas espécies representariam
outros grupos na resposta ao distúrbio (UEHARA-PRADO et al., 2009).
Pyle (1980) sugere o uso da diversidade de borboletas frugívoras como indicador da
diversidade de plantas, pois a co-evolução entre borboletas e suas plantas hospedeiras no estágio
larval tem, em alguns casos levado a uma alta especificidade borboleta-planta. Uehara-Prado et
al. (2009) verificaram que a riqueza de borboletas desta guilda correlacionou-se positivamente

19
à riqueza de espécies arbóreas, sugerindo que independente do ambiente, a riqueza de
borboletas frugívoras pode atuar como indicadora da biodiversidade.
Portanto o conhecimento das espécies ao longo de gradientes hidrológicos e ambientais
podem fornecer informações relevantes sobre o ecossistema.
Para isto se faz necessário inventário de espécies bem como estudos ecológicos
envolvendo este grupo, sendo estas muito sensíveis a mudanças negativas em qualquer dos
fatores ambientais dos quais dependem, são fáceis de serem monitoradas por não-especialistas
até mesmo pessoas com baixo grau de instrução após passarem por treinamento são capazes de
se tornarem monitores de borboletas frugívoras e saberem como está a saúde do ambiente.
Tornando ferramenta útil para conservação desse e de outros locais a partir da
apropriação do processo de conservação pelos atores locais. E gerando informações locais,
subsidiando assim a nível regional para conservação do bioma amazônico.

20
2 OBJETIVOS
2.1 GERAL
Caracterizar a comunidade de borboletas frugívoras da Estação Ecológica de Cuniã em
relação a variáveis ambientais e padrões de distribuição.
2.2 ESPECÍFICOS
• Estabelecer o marco zero de amostragem para o monitoramento das borboletas
frugívoras.
• Reportar a densidade de borboletas frugívoras.
• Determinar os padrões espaciais e temporais das assembleias de borboletas.
• Testar os efeitos das variáveis ambientais ao longo do gradiente distributivo das
borboletas frugívoras.

21
3 METODOLOGIA
3.1 ÁREA DE ESTUDO
O estudo foi conduzido na Estação Ecológica do Cuniã – ESEC Cuniã que é uma unidade
de conservação de proteção integral, segundo SNUC é destinada a realização de pesquisas,
criada por Decreto Federal de 27 de setembro de 2001, atualmente com uma área total de
186.743,26 hectares, localizada ao norte do Estado de Rondônia, no município de Porto Velho-
RO e Canutama/AM. Pela diplomação legal: Dec s/nº de 27 de setembro de 2001/ Dec s/nº de
21 de dezembro de 2007/ Lei n.º 12.249, de 11 de junho de 2010 (BRASIL, 2001; BRASIL,
2012) (ver Figura 1).
Figura 1 Estação Ecológica do Cuniã – ESEC Cuniã área em verde- localizada na BR 319 no sentido Porto
Velho – Humaitá.
Fonte: Fabíola Gomes Vieira, 2013.
O acesso ao lado leste é realizado por via fluvial através do Rio Madeira, sentido Porto
Velho - Manaus, até o Distrito de Nazaré. Já, o acesso ao lado oeste da unidade ocorre pela BR-
319, sentido Porto Velho - Humaitá-AM, possibilitando o deslocamento à área de terra firme
(BRASIL, 2012).
O acesso pela via fluvial permite integração às comunidades ribeirinhas Pau D´Arco e
Belém. A utilização do rio Madeira para deslocamento favorece acesso aos lagos e igarapés da

22
unidade, em especial ao igarapé Aponiã, igarapé Capitari e o Lagos Peixe boi, Barraquinha e
Pau D´Arco (BRASIL, 2012).
A ESEC Cuniã, juntamente com a Reserva Extrativista do Lago do Cuniã e a Floresta
Nacional de Jacundá fazem parte da Gestão Integrada Cuniã-Jacundá (GICJ). A geomorfologia
da região é caracterizada pela presença de grandes interflúvios tabulares com topografia plana
e altitudes variando entre 60 e 90 metros, possui uma extensa rede de igarapés que cortam a
reserva (BRASIL, 2012). Esta condição produz uma rede de água densa bacia formada por
inúmeros córregos e lagos escuros.
A região central do interflúvio é formada por um mosaico de vegetação, desde florestas
de terra firme, florestas aluviais, alguns trechos de "fechado", "Campinas" e vegetação herbácea
em solos arenosos (MALDONADO et. al, 2012).
A vegetação caracteriza-se por área de transição entre as florestas densas, abertas e áreas
de campinaranas. O termo Campinarana geralmente é usado para vegetação que se desenvolve
sobre solos arenosos extremamente pobres (oligotróficos), na maioria dos casos hidromórficos,
e ricos em ácido húmico. No entanto o termo engloba um complexo mosaico de formações não
florestais, porém não savânicas, com ocorrência esporádica, mas frequente em toda a região
Amazônica (PIRES, 1974; PIRES, PRANCE, 1985; RICHARDS, 1996).
As áreas de Campinarana geralmente apresentam subdossel com árvores pequenas e finas
e com escassez de espécies arbóreas o que contrasta com a maioria das florestas Amazônicas.
Este tipo de vegetação exibe uma tendência pronunciada de dominância por uma ou poucas
espécies (ANDERSON, 1981).
3.2 DESENHO AMOSTRAL
O projeto utilizou os módulos de amostragem seguindo as diretrizes do Programa de
Pesquisa em Biodiversidade - PPBio criado pelo Ministério de Ciência e Tecnologia - MCT
para aumentar a eficiência de estudos de monitoramento de biodiversidade na Amazônia. O
PPBio está baseado no uso de protocolos de amostragem padronizados e integrados
espacialmente, usando infraestrutura modular de trilhas e parcelas de amostragem
(MAGNUSSON et.al., 2013). O delineamento amostral segue recomendações da metodologia
RAPELD (MAGNUSSON et.al., 2005).
Os principais aspectos do método podem ser resumidos da seguinte forma: (i) a grade é
uma área de 25 km2 onde foram instaladas 12 trilhas de 5 km de comprimento, 6 no sentido

23
norte-sul e 6 no sentido leste-oeste; (ii) a grade possui 30 parcelas de terra firme permanentes
de 250 m com largura variável adjacentes às trilhas, orientadas norte-sul. O eixo central da
parcela segue a curva de nível; (iii) a amostragem de organismos nas parcelas deve cobrir a
extensão total das parcelas amostradas na grade. Amostragens baseadas nas trilhas devem cobrir
todas as trilhas na grade (ver Figura 2).
Figura 2 Grade padrão para inventários bióticos e abióticos proposto pelo Programa de Pesquisa em
Biodiversidade - PPBio instalada na ESEC-Cuniã localizada na BR 319 no sentido Porto Velho – Humaitá,
Rondônia. Os círculos verdes representam as parcelas de terra firme que foram utilizadas neste estudo.
Fonte: Andrade et al., 2017.
3.3 ESTIMATIVAS DE DENSIDADE DE BORBOLETAS
A amostragem foi realizada nas 30 parcelas instaladas em terra-firme (ver figura 2). As
parcelas possuem 250m de comprimento e são marcadas a cada 10m por piquete (em cano pvc
com placa em alumínio contendo numeração), em cada parcela foram dispostas três armadilhas.
A primeira instalada na marcação entre 0-50m, a segunda entre as marcações de 100m-150m e
a terceira entre as marcações 200m-250m, conforme disponibilidade de uma ou duas árvores
que sustentassem a armadilha, com a distância de 1m do chão (conforme figura 4 e 5). As
armadilhas permaneceram ativas durante seis (6) dias, sendo revisadas a cada 48 horas,
totalizando três (3) visitas para coleta de dados e espécimes. Este procedimento segue as

24
recomendações de Hughes et al. (1998) e Graça (2014) em estudos ecológicos e taxonômicos
de borboletas frugívoras.
Figura 3 Modelo ilustrativo de armadilha(A): (a) corpo da armadilha confeccionado em tule; (b) funil
interno; (c) plataforma onde a isca é colocada. E Armadilha VSR instalada em uma parcela da ESEC-
Cuniã(B).
Fonte: UEHARA-PRADO e RIBEIRO, 2012, Foto: LEMKE, 2016.
a
b
b
B A
c

25
Figura 4 Representação das parcelas instaladas na grade da ESEC Cuniã (A). Disposição das 3 armadilhas Van
Someren-Rydon ao longo dos 250 m de extensão de cada parcela. As curvas representam a não linearidade das
parcelas, e não desnível do terreno(B).
B
A

26
Figura 5 Coleta em uma das armadilhas instaladas em parcela da ESEC-Cuniã(A). Amostra coletada de
Hamadryas alicia (H. Bates, 1865) em processo de identificação (B).
Foto: NASCIMENTO, 2016; LEMKE, 2017
As coletas foram realizadas no ano de 2016, nos períodos de 25 de fevereiro à 04 de
março e 26 de julho a 04 de agosto caracterizando o ciclo de águas altas (AA) e de águas baixas
(AB) respectivamente.
As borboletas foram capturadas utilizando-se armadilhas modelo Van Someren-Rydon
(RYDON, 1964) ou apenas VSR, criada por estes autores e modificadas (detalhes em SHUEY
1997) conforme modelo apresentado em UEHARA-PRADO e RIBEIRO, 2012 (ver detalhes
na Figura 3). Nestas armadilhas foram inseridas na plataforma basal copo plástico de 50 ml
com uma mistura fermentada por 48 horas de caldo de cana e banana na proporção de 1 litro de
caldo para 3 kg de banana amassada, esta composição foi estabelecida no protocolo de
monitoramento de borboletas frugívoras em Unidades de Conservação desenvolvido pela Rede
Nacional de Pesquisa e Conservação de Lepidópteros (RedeLep) (NOBRE et al., 2014).
As borboletas capturadas foram sacrificadas por pressão no tórax e acondicionadas em
envelopes entomológicos confeccionados em papel manteiga e armazenados em sacos plásticos
com fechamento hermético contendo cravos da índia e algodão absorvente visando a
conservação das amostras. A identificação seguiu a classificação taxonômica proposta por
A B

27
Wahlberg et al. (2009), por meio de guias ilustrados Garwood et al., 2009; Vieira et al., 2014,
guia de identificação online (http://butterfliesofamerica.com) e por comparações feitas com
material da Coleção Entomológica da Universidade Federal de Rondônia (UNIR) onde o
material testemunho foi depositado sob o tombamento UFRO.ELE000572 a
UFRO.ELE001238 (Figura 5).
3.4 VARIÁVEIS AMBIENTAIS
As variáveis ambientais utilizadas neste estudo foram realizadas em estudos conduzidos
por Pansini et. al. (2016) e Andrade et. al. (2017). Estes trabalhos incluíram aspectos edáficos,
hidrológicos e da estrutura da vegetação, os quais foram utilizados como variáveis preditoras
para testes inferenciais, os quais seguem uma breve descrição:
(i) A coleta de solo por parcela foi realizada por meio da homogeneização de seis
amostras equidistantes (a cada 50m), na profundidade de 0-10 cm. Em cada ponto, foi retirada
uma amostra de solo superficial de 30 cm x 30 cm x 5 cm com auxílio de trado holandês. Em
seguida foram removidos folhiços e raízes e armazenados em sacos plásticos e conduzidos ao
Laboratório Temático de Solos e Plantas do INPA, onde foram analisados os percentuais de
argila, silte e areia; pH do solo em água; macro e micronutrientes essenciais (mg/kg) Fe2+, Mn2+,
Zn2+ e fósforo disponível (P); bases trocáveis (cmocl/kg) K+, Ca2+, Mg2+ e Al3+. As análises
físico-químicas foram conduzidas de acordo com protocolos estabelecidos pela EMBRAPA
(2009);
(ii) As variáveis hidrológicas utilizadas foram obtidas considerando a distância mínima
horizontal à rede de drenagem (DHR) e vertical do lençol freático (DVL) por parcela. A DHR
refere-se à distância em linha reta ao igarapé mais próximo. Esta medida foi obtida com auxílio
do Software QGis versão 2.18 (2017). A DVL resultou do valor mínimo obtido por medição da
profundidade do lençol freático obtidos dos piezômetros instalados por parcela, ao longo de 12
meses, utilizando-se um medidor digital Solinst Model102. As etapas de perfuração dos poços
e instalação dos piezômetros podem ser conferidas em Pansini (2016);
(iii) As cotas atitudinais por parcela foram obtidas do Repositório de Dados do PPBio
disponibilizados no site https://ppbiodata.inpa.gov.br/metacatui/#view/menger.124.2. As
informações das cotas altitudinais foram obtidas por meio da contratação de um topógrafo
profissional, que mediu a altitude em relação ao nível do mar ao longo das trilhas. O valor de
altitude de cada parcela permanente corresponde à cota de altitude da trilha no ponto inicial da

28
parcela. Como as parcelas seguem uma curva de nível, o valor de altitude é constante em toda
extensão da parcela;
(iv) A espessura da serrapilheira foi medida ao longo do eixo principal da parcela, a
cada dois metros. Um bastão de metal graduado de dois milímetros de diâmetro foi forçado
através da camada de folhas, tomando o devido cuidado para não compactá-las, até atingir a
superfície do solo, os valores foram obtidos em centímetro (cm) (PANSINI, 2011).
(v) Termohigrômetros digitais (modelo 190 Supermedy) foram utilizados para registrar
dados de umidade e temperatura por parcela.
(vi) Os dados referentes a estrutura da vegetação (densidade de palmeiras, árvores,
arvoretas, equações alométricas e fitossociológicas/florísticas da vegetação e peso da massa
seca de serrapilheira foram detalhados em Andrade et al. (2017).
3.5 PREPARAÇÃO E ANÁLISE DOS DADOS
A partir dos dados coletados em campo, aqueles obtidos dos dados/metadados do PPBio
e trabalhos realizados na ESEC-Cuniã por Pansini (2016) e Andrade et al. (2017), foi
constituído um banco de dados em planilha do programa EXCEL® com as informação de
Número de Coleta, Data de Início(data que foi instalada a armadilha) e Data de cada Coleta,
Localização da Parcela, Horário da Revisão, Temperatura, Umidade, Coordenada Geográfica
de cada armadilha, Número da Armadilha, além de dados referentes a identificação taxonômica
como Família, Subfamília, Tribo, Gênero e Espécie, e outros dados referente ao Sexo (quando
a espécie apresentava dimorfismo sexual aparente) para que a partir deste fosse prospectado
todas as demais análises.
Foram elaboradas tabelas dinâmicas, com dados quantitativos (densidade de borboletas
frugívoras por unidade amostral) e com dados qualitativos (presença ou ausência de borboletas
frugívoras). Posteriormente, análises multivariadas foram aplicadas com o propósito de detectar
padrões estruturais, espaciais e temporais.
Para avaliar padrões temporais das assembleias de borboletas frugívoras e verificar se
haveria diferença nas assembleias no período de águas altas e baixas foi realizado teste de
Mantel com a matriz das distâncias geográficas das parcelas e matriz de cada período avaliado.
Ele é usado para testar a hipótese nula de ausência de relação entre duas matrizes simétricas,
calculadas a partir de dois conjuntos de variáveis diferentes medidas nos mesmos objetos. A

29
significância é testada pelo método de randomização (teste de Monte Carlo) (VALENTIN,
2012).
Para descrever e analisar espacialmente as assembleias de borboletas frugívoras foram
aplicadas análises de classificação e escalonamento multidimensional. Na análise de
classificação foi elaborada uma matriz qualitativa onde utilizou-se o índice de similaridade de
Sorensen-Dice (DICE, 1945; SORENSEN, 1957) e o índice de Bray-Curtis como medida de
dissimilaridade (BRAY; CURTIS, 1957). A análise de agrupamento é uma técnica indicada
visando agregar informações mais semelhantes e, através disso, possibilitar comparações entre
os dados (HAIR et al., 2005).
A dimensionalidade das assembleias de borboletas frugívoras foi reduzida por
escalonamento multidimensional métrico (MDS). O MDS reduz as informações de um elevado
número de atributos (espécies), até um pequeno número de variáveis ou eixos, que ordenam os
pontos amostrais baseados numa distância estatística a partir das distâncias originais
(MCCUNE e GRACE, 2002).
As ordenações da composição da comunidade de borboletas frugívoras foram feitas com
base nos dados de presença e ausência (ordenações qualitativas) e com base nas abundâncias
das espécies (ordenações quantitativas). As ordenações quantitativas foram usadas para
capturar os padrões apresentados pelas espécies mais abundantes e, desta forma, evidenciar a
contribuição da abundância relativa das espécies para a diferença entre locais. As duas formas
(qualitativos e quantitativos) foram usadas para se verificar a variabilidade no padrão de
resposta levando em consideração apenas a ocorrência das espécies ou suas abundâncias.
Os resultados obtidos com os dois primeiros eixos (escores) de MDS foram usados
como variáveis dependentes nos testes inferenciais do efeito das variáveis ambientais. Para
determinar a melhor combinação de variáveis ambientais afetando a composição de borboletas,
foram realizadas regressões múltiplas com os preditores granulométricos (areia, silte, argila) e
teor de fósforo (P), hidrológicos considerando a distância mínima horizontal à rede de drenagem
(DHR) e vertical do lençol freático (DVL), a densidade de palmeiras, árvores, arvoretas,
equações alométricas e fitossociológicas/florísticas da vegetação, a espessura de serrapilheira e
o peso da massa seca. E aplicada à transformação quando necessárias, seguindo a equação
proposta por Zar (1999).
O Critério de Informação de Akaike (AIC) foi aplicado, como método de seleção de
modelo para regressão múltipla, visando selecionar um subconjunto de variáveis preditoras para
explicar com mais parcimônia a variação nas variáveis resposta utilizadas (GOTELLI e
ELLISON, 2011).

30
O programa R® (version 1.0.136 – © 2009-2016 RStudio, Inc.) foi usado para gerar a
ordenação simples direta das espécies em função das variáveis ambientais testadas. Estes
mesmos preditores foram utilizados na regressão múltipla com o auxílio do software Action
Stat® e XLSTAT® (ACTION STAT, 2016; ADDINSOFT, 2017).

31
4 RESULTADOS
4.1 BORBOLETAS FRUGÍVORAS DA ESEC CUNIA
Foram amostradas 667 (seiscentos e sessenta e sete) indivíduos representando 4 (quatro)
subfamílias, distribuídos em 10 (dez) tribos e 73 (setenta e três) espécies. A subfamília mais
abundante foi Biblidinae com 479 indivíduos (72% da abundância) (Figura 6) com
representantes de quatro tribos: Epicaliini (365), Epiphilini (101), Ageroniini (9) e Callicorini
(4) contidos na figura7 juntamente com as outras tribos.
Figura 6 Proporção da abundância das Subfamílias coletadas na ESEC-Cuniã.
Figura 7 Abundância das 10 tribos coletadas na ESEC-Cuniã.
Tendo como espécie mais abundante Catonephele acontius com 220 representantes (64
fêmeas e 156 machos) sendo também a mais frequente capturada nas 28 de 30 parcelas e
apresentando a maior abundância em 24 delas. Outras espécies mais abundantes foram,
72%
16%
10%
2%
Biblidinae
Satyrinae
Charaxinae
Nymphalinae
050
100150200250300
350400
Ab
un
dân
cia

32
respectivamente, Catonephele numilia (87), Temenis laothoe (78), Eunica viola (32) e Nica
flavilla (21).
A comunidade total de borboletas foi representada por poucas espécies muito
abundantes e muitas espécies raras (que foram registradas poucas vezes durante o período de
estudo). Quatorze espécies tiveram 6 ou mais indivíduos (n >5). Cerca de 40% das espécies
capturadas (S=30) foram representadas por apenas um indivíduo (singletons). A lista das
espécies é apresentada na Tabela1.
Satyrinae foi a subfamília mais rica com trinta e cinco (35) espécies, seguida por
Biblidinae, Charaxinae e Nymphalinae com dezenove, dezesseis e três espécies,
respectivamente.
Tabela 1
Lista de espécies de borboletas frugívoras coletadas na ESEC-Cuniã com sua respectiva abundância.
SUBFAMÍLIA
TRIBO Espécie
Indivíduos
Águas
altas
Águas
baixas
Total
Geral
BIBLIDINAE
AGERONIINI
Hamadryas alicia (H. Bates, 1865) 1 0 1
Hamadryas arinome (Lucas, 1853) 0 8 8
AGERONIINI
Total 1 8 9
CALLICORINI
Callicore excelsior (Hewitson, 1858) 0 4 4
CALLICORINI
Total 0 4 4
EPICALIINI
Catonephele acontius (Linnaeus, 1771) 32 188 220
Catonephele numilia (Cramer, 1775) 4 83 87
Dynamine pebana (Staudinger, 1885) 0 1 1
Eunica anna (Cramer, 1780) 0 4 4
Eunica malvina (H. Bates, 1864 ) 0 2 2
Eunica veronica (H. Bates, 1864) 0 5 5
Eunica viola (H. Bates, 1864) 1 31 32
Nessaea obrinus (Linnaeus, 1758) 2 3 5
Pyrrhogyra amphiro (H. Bates, 1865 ) 0 1 1
Pyrrhogyra crameri (Aurivillius, 1882) 1 2 3
Pyrrhogyra edocla (E. Doubleday, 1848) 0 2 2
Pyrrhogyra otolais (H. Bates, 1864) 0 3 3
EPICALIINI Total 40 325 365
EPIPHILINI
Asterope leprieuri (Feisthamel, 1835) 0 1 1
Nica flavilla (Godart, 1824) 2 19 21
Temenis laothoe (Cramer, 1777) 2 76 78

33
Continuação Tabela 1
Lista de espécies de borboletas frugívoras coletadas na ESEC-Cuniã com sua respectiva abundância
Temenis pulchra (Hewitson, 1861) 0 1 1
EPIPHILINI Total 4 97 101
BIBLIDINAE Total 45 434 479
CHARAXINAE
ANAEINI
Fountainea ryphea (Cramer, 1775) 0 3 3
Hypna clytemnestra (Cramer, 1777) 0 2 2
Memphis acidalia (C. Felder & R. Felder, 1867) 1 3 4
Memphis glauce (C. Felder & R. Felder, 1862) 0 1 1
Memphis leonida (Stoll, 1782) 0 16 16
Memphis moruus (Fabricius, 1775) 0 9 9
Memphis oenomais (Boisduval, 1870) 1 6 7
Memphis phantes (Hopffer, 1874) 0 1 1
Memphis polycarmes (Fabricius, 1775) 0 1 1
Siderone galanthis (Cramer, 1775) 0 1 1
Zaretis isidora (Cramer, 1779) 1 3 4
Zaretis itys (Cramer, 1777) 0 1 1
ANAEINI Total 3 47 50
PREPONINI
Archaeoprepona amphimachus (Fabricius, 1775) 0 2 2
Archaeoprepona demophon (Stoffel & Descim, 1974) 0 7 7
Prepona dexamenus (Hopffer, 1874) 0 2 2
Prepona pheridamas (Cramer, 1777) 4 2 6
PREPONINI Total 4 13 17
CHARAXINAE
Total 7 60 67
NYMPHALINAE
COEINI
Colobura annulata (Willmott, Constantino & J. Hall,
2011) 1 0 1
Colobura dirce (Linnaeus, 1758) 6 2 8
Tigridia acesta (Linnaeus, 1758) 0 3 3
COEINI Total 7 5 12
NYMPHALINAE
Total 7 5 12
SATYRINAE
BRASSOLINI
Bia actorion (Linnaeus, 1763) 7 9 16
Caligo eurilochus (Cramer, 1775) 1 0 1
Catoblepia berecynthia (Cramer, 1777) 2 10 12
Catoblepia soranus (Westwood, 1851) 3 7 10
Opsiphanes invirae (Hübner, 1808) 0 5 5
Opsiphanes quiteria (Stoll, 1780) 1 0 1
BRASSOLINI
Total 14 31 45
MORPHINI
Morpho achilles (Linnaeus,1758) 0 1 1

34
Continuação Tabela 1
Lista de espécies de borboletas frugívoras coletadas na ESEC-Cuniã com sua respectiva abundância
Morpho helenor (Cramer, 1776) 0 2 2
MORPHINI Total 0 3 3
SATYRINI
Amphidecta calliomma (C. Felder & R. Felder, 1862) 3 0 3
Caeruleuptychia caerulea (A. Butler, 1869) 0 1 1
Chloreuptychia callichloris (A. Butler, 1867) 0 4 4
Chloreuptychia herseis (Godart, 1824) 1 0 1
Chloreuptychia hewitsonii (A. Butler, 1867) 1 0 1
Cissia myncea (Cramer, 1780) 2 0 2
Erichthodes antonina (C. Felder & R. Felder, 1867) 0 3 3
Erichthodes arius (Weymer, 1911) 0 1 1
Euptychia insolata (Butler & Druce, 1872) 1 0 1
Hermeuptychia fallax (C. Felder & R. Felder, 1862) 0 1 1
Magneuptychia antonoe (Cramer, 1775) 0 1 1
Magneuptychia ocnus (A. Butler, 1867) 1 0 1
Pareuptychia binocula (A. Butler, 1869) 0 1 1
Pareuptychia hesionides (Forster, 1964) 1 3 4
Pareuptychia ocirrhoe (Fabricius, 1776) 1 0 1
Pareuptychia summandosa (Gosse,1880) 0 1 1
Paryphthimoides poltys (Prittwitz, 1865) 0 2 2
Pseudodebis euptychidia (A. Butler, 1868) 0 3 3
Pseudodebis marpessa (Hewitson, 1862) 0 4 4
Pseudodebis sp. 0 2 2
Pseudodebis valentina (Cramer, 1782) 1 3 4
Taygetis cleopatra (C. Felder & R. Felder, 1867) 3 5 8
Taygetis leuctra (A. Butler, 1870) 1 2 3
Taygetis sosis (Hopffer, 1874) 0 1 1
Taygetis sp. 1 0 1
Taygetis sylvia (H. Bates, 1866) 0 1 1
Taygetis thamyra (Cramer, 1779) 1 2 3
Taygetis tripunctata (Weymer, 1907) 0 1 1
Taygetis virgilia (Cramer, 1776) 0 1 1
SATYRINI Total 18 43 61
SATYRINAE Total 32 77 109
Total Abundância 91 576 667
Total Riqueza 32 63 73
4.2 PADRÕES ESPACIAIS E TEMPORAIS DAS ASSEMBLEIAS DE BORBOLETAS
FRUGÍVORAS
O agrupamento e ordenação obtidos a partir de dados quantitativos e qualitativos
apresentaram variações quanto à formação de grupos; contudo os padrões evidenciados pelos
dados quantitativos foram mais congruentes, quando comparados com os qualitativos obtidos

35
pela ordenação simples direta (Figura 10). Evidenciando que as borboletas frugívoras
estabelecidas ao longo de gradientes argilosos, hidrológicos, edáficos e estrutura da vegetação
estruturam suas assembleias e determinam os padrões distributivos de suas populações. Os
padrões espaciais e temporais emergentes das assembleias de borboletas frugívoras podem ser
visualizados pelas figuras 08 (dendrograma) e 09 (ordenação).
A análise de classificação apresenta uma decomposição de variação para a classificação
ótima, com 68,51% da variação para os grupos Intra-classe e para os grupos Inter-classes
31,49%, mostrando os diferentes níveis de (dis) similaridade entres as unidades amostrais.
A ordenação por MDS usando dados quantitativos apresentou menor stress = 0,276
(Figura 9) indicando bom ajuste do modelo no tratamento da variabilidade dos dados e indicam
robustez na formação dos grupos encontrados, conforme metodologia detalhada em Minchin
(1987).
A ordenação (MDS) das assembleias de borboletas frugívoras e a formação de
subgrupos com diferentes graus de dissimilaridade (classificação por análise de Cluster)
sugerem que, a borboletas frugívoras apresentam diversos níveis de similaridade entre e dentre
os subgrupos reportados.
Portanto, pode-se afirmar, que a variabilidade nos padrões distributivos das diferentes
populações de borboletas frugívoras constitui-se em níveis funcionais em mesoescala, de
acordo com performance das espécies, as quais contribuem com ampla distribuição e suas
restrições ambientais, as quais determinam a variabilidade das espécies ao longo da estrutura
interna da floresta de terra firme na ESEC Cuniã. As espécies Catonephele acontius,
Catonephele numilia, Eunica viola, Memphis leonida e Temenis laothoe, por exemplo, tiveram
distribuição bem ampla na área de estudo estando presente em todo o gradiente.

36
Figura 8 Dendrograma de dissimilaridade com dados quantitativos (Bray e Curtis) das assembleias de
borboletas frugívoras na ESEC Cuniã, Porto Velho, Rondônia.
Figura 9 Análise de Ordenação MDS (Configuração Stress de Kruskal (1) = 0,276) com dados
quantitativos (Bray e Curtis) das assembleias de borboletas frugívoras na ESEC Cuniã, Porto Velho,
Rondônia.
O teste de Mantel não foi significativo (r(AB.C)=0,027; p=0,422) evidenciando que as
assembleias no período de águas altas (chuva) e águas baixas (seca) na Estação Ecológica do
Cuniã diferem-se quanto a sua composição de espécies. Contudo, quando aplicado para os
T24
T14
T30
T16
T23
T6 T13
T2 T26
T1 T9 T4 T7 T8 T12
T19
T17
T27
T15
T20
T22
T3 T18
T21
T5 T11
T25
T29
T10
T28
0
0,5
1
1,5
2
2,5
3
Dissimilaridad
e
T1
T2
T3
T4
T5
T6
T7T8
T9
T10
T11
T12
T13 T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
-0,6
-0,4
-0,2
0
0,2
0,4
0,6
0,8
-1 -0,8 -0,6 -0,4 -0,2 0 0,2 0,4 0,6 0,8
Dim
2
Dim1

37
dados quantitativos (r(AB.C)=0,127; p=0,001) não indicam diferenças distributivas e /ou
estruturais.
Nas coletas realizadas em duas estações sazonais na Estação Ecológica de Cuniã foi
registrado 91 indivíduos de 32 espécies em fevereiro/março (água altas) e 576 indivíduos de 63
espécies em julho/agosto (águas baixas), as duas estações compartilharam 20 espécies. (Tabela
1).
4.3 EFEITO DAS VARIAVEIS AMBIENTAIS
O padrão de distribuição das assembleias de borboletas frugívoras ao longo do gradiente
argiloso, teor de fósforo (P), peso da massa seca de serrapilheira e hidrológico (considerando a
distância mínima horizontal à rede de drenagem (DHR)) podem ser visualizados na figura 10.
Foram geradas ordenações simples direta de variáveis ambientais com dados quantitativos e
qualitativos; no entanto, devido à convergência de resposta no padrão foram usados os dados
quantitativos.
As assembleias de borboletas frugívoras apresentaram um padrão hierárquico com
acúmulo de espécies na porção do gradiente com as maiores proporções de argila e
concentrações de fósforo (P). Já, para a gradiente de peso da massa seca de serrapilheira e
distância dos corpos d’água, estes foram determinados pelo maior acúmulo de espécies para as
parcelas com menor peso da camada de serapilheira e maior acúmulo de espécies nas áreas
drenadas ou parcialmente drenadas respectivamente.
O gradiente argiloso revelou-se determinante na estruturação das assembleias. A maior
parte das espécies (cerca de 55%) mostraram ter distribuição ampla pelos terrenos argilosos.
Por outro lado, verifica-se que poucas espécies respondem aos extremos do gradiente. Já para
o gradiente hidrológico, a maior parte das espécies mostrou preferência pelos terrenos com
maior proximidade da rede de drenagem, e outro grupo mostrou-se indiferente, revelando
subconjuntos de assembleias de borboletas.

38
Figura 10 Densidade relativa de espécies de borboletas frugívoras ao longo do gradiente de solo: Teores de Argila
e Fósforo (P), peso da massa seca de Serrapilheira (MS) e hidrológico considerando a distância mínima horizontal
à rede de drenagem (DHR).
A
B
C D

39
A partir da contribuição quantitativa o melhor modelo fornecido pelas regressões
lineares múltiplas selecionou o teor de argila (TA) (t = 3,759; p = 0,001), teor de fósforo (TP)
(t = -3,098; p = 0,005), densidade de palmeiras (DP) (t = 2,956; p = 0,007), florística (F) (t =
3,070; p = 0,005) e MDS Biomassa arbórea (MDSBio) (t = -2,335; p = 0,029) como sendo os
preditores a influenciar a composição de borboletas (R² = 73%; AIC = -93,066). Já a influência
da dimensão qualitativa selecionou como preditores para mudança na composição de borboletas
(R² = 62%; AIC = -82,637) os preditores peso da massa seca de serrapilheira (MS) (-t = 2,342;
p = 0,029), cotas altitudinais do terreno (CA) (t = 3,287; p = 0,003) e MDS Biomassa (MDSBio)
(t = -2,337; p = 0,029) (Tabela 2; Tabela 3 e Figura 11).
TA: Teor de Argila; DHR: Distância reta horizontal do dreno (igarapé); ES: Espessura de Serrapilheira; TP: Teor
de Fósforo; DP: Densidade de Palmeiras; F: Florística; MDSBio: MDS Biomassa arbórea; MS: Peso da Massa
seca de Serrapilheira; DA: Densidade de Árvores; MDSFlo: MDS Florística; CA: Cotas Altitudinais do Terreno.
BC: Bray-Curtis usados dados quantitativos; SD: Sorensen-Dice usados dados qualitativos.
Tabela 2
Seleção do melhor modelo de Regressão Linear, selecionados pela menor distância de AIC e R² Ajustado
Índice Eixo
MDS Seleção de modelos AIC R²
R²
Ajustado
BC
EIXO
Dim 1 TA / DHR / ES / TP / DP / F / MDSBio -93,066 0,735 0,654
EIXO
Dim 2 TA / DHR / MS / TP / DP / DA / F / MDSflo / MDSBio -83,370 0,694 0,563
SD
EIXO
Dim 1 DHR /MS / ES / TP/ CA / DA / F / MDSBio -82,637 0,624 0,488
EIXO
Dim 2 TA / MS / ES/ DA / MDSBio -75,136 0,484 0,377

40
TA: Teor de Argila; DHR: Distância reta horizontal do dreno (igarapé); ES: Espessura de Serrapilheira; TP: Teor
de Fósforo; DP: Densidade de Palmeiras; F: Florística; MDSBio: MDS Biomassa arbórea; MS: Peso da Massa
seca de Serrapilheira; DA: Densidade de Árvores; MDSFlo: MDS Florística; BC: Bray-Curtis usados dados
quantitativos; SD: Sorensen-Dice usados dados qualitativos.***P < 0.001 **P < 0.01 *P < 0.05 P < 0.1. †
Tabela 3
Parâmetros de cada modelo da Regressão Linear.
Índice Eixo
MDS Fonte Erro padrão t Pr > |t|
BC
EIXO Dim 1
TA 0,041 3,759 0,001***
DHR 0,039 -1,360 0,187†
ES 0,043 -1,400 0,175†
TP 0,039 -3,098 0,005***
DP 0,040 2,956 0,007***
F 0,060 3,070 0,005***
MDSBio 0,002 -2,335 0,029**
EIXO Dim 2
TA 0,052 1,142 0,266†
DHR 0,062 -2,160 0,043**
MS 0,055 -2,588 0,017**
TP 0,046 1,127 0,273†
DP 0,081 -1,960 0,063*
DA 0,091 -2,637 0,015**
F 0,088 2,262 0,034**
MDSflo 0,456 -3,685 0,001***
MDSBio 0,003 3,305 0,003***

41
Figura 11 Resultados gráficos do melhor modelo fornecido pelas regressões lineares múltiplas no qual Teor
de Argila, Teor de Fósforo (P), Densidade de Palmeiras, Florística, MDSBiomassa arbórea são os
preditores para mudanças na composição de borboletas frugívoras.

42
5 DISCUSSÃO
Na estimativa da densidade de borboletas frugívoras, as espécies Catonephele acontius,
Catonephele numilia, Eunica viola, Memphis leonida e Temenis laothoe, tiveram distribuição
ampla. Catonephele acontius apresentou maior abundância e frequência, possui dimorfismo
sexual bem acentuado, sendo o macho preto com padrões laranja forte e as fêmeas pretas com
bandas transversais amarelas (CANALS, 2003).
Os machos foram mais abundantes e frequentes do que as fêmeas, ambos se alimentam
de frutas no chão e dossel da floresta, os machos também absorvem a umidade rica em minerais
ao longo do gradiente edáfico e hidrológico. C. acontius sendo mais abundante e frequente
apresentou estrita relação com o teor de argila, como variável influenciadora da distribuição
das borboletas.
Contudo, deve-se salientar, o primeiro registro para Rondônia de Hamadryas alicia
Bates 1865 coletada na ESEC-Cuniã em período considerado com maior precipitação. Esta
espécie de predominância florestal é considerada rara por especialistas (D’ABRERA, 1981).
Por consequência da fragmentação florestal é pouco registrada em estudos e monitoramentos.
No portal Global Biodiversity Information Facility (Gbif) foram verificados 9 registros em todo
o mundo, com ocorrência no Peru e Brasil. A ocorrência desta espécie e da diversidade
registrada neste estudo comprova que a ESEC-Cuniã é uma região de refúgio da
lepidopterofauna de Rondônia e sul do Amazonas, fundamental para a manutenção da biota e
biodiversidade local e regional.
Os padrões espaciais e temporais registrados neste estudo evidenciam, quais e como as
variações estruturais e no ambiente podem promover mudanças significativas na distribuição
espacial de borboletas frugívoras.
O teste de Mantel mostrou que a sazonalidade é suficiente para capturar diferenças na
composição de espécies e ainda refletir variações nos padrões estruturais. Corroborando que as
espécies singletons não prejudicam a resposta das assembleias de borboletas frente às variáveis
ambientais, significando que o padrão é dirigido por espécies comuns.
As borboletas frugívoras da Estação Ecológica de Cuniã apresentam uma forte mudança
temporal em sua distribuição. O período de águas baixas (meses julho e agosto), apresentou
maior abundância e riqueza. As borboletas frugívoras demonstram alguma sazonalidade, pois
são influenciadas pela ação direta dos fatores climáticos e estruturais da vegetação, entre outros
(BROWN; FREITAS, 2000).
Segundo Brown Jr. (1992), as borboletas apresentam maior abundância de indivíduos

43
nas épocas mais quentes, com temperaturas mais altas, onde há maior abundância de recursos
alimentares e a distribuição de recursos nutricionais provavelmente o maior fator controlador
da estrutura de borboletas não migratórias em florestas tropicais (GILBERT, 1984).
Corroborando com o resultado evidenciado na comunidade de borboletas frugívoras na ESEC
Cuniã.
Quanto aos efeitos das variáveis ambientais pelas análises das ordenações diretas a
maior parte das espécies mostrou preferência pelos terrenos com maior proximidade da rede de
drenagem, acredita-se que este fator estabeleça diversidade de plantas e proporcione recurso
alimentar às borboletas frugívoras.
Ainda é importante salientar que gradientes argilosos, com concentração de fósforo e
de massa seca da serapilheira atuaram como selecionadores nos padrões distributivos das
borboletas frugívoras na Estação Ecológica do Cuniã.
Gradientes de solos argilosos, com alto teor de Fósforo, massa seca de serrapilheira, a
estrutura da vegetação (biomassa arbórea, diversidade florística e densidade de palmeiras)
parecem atuar como variáveis que estruturam fortemente o gradiente de resposta das espécies
de borboletas frugívoras na Região do Interflúvio. Muitos trabalhos mostram que diversos
grupos bióticos respondem aos fatores edáficos (COSTA et. al., 2005). Este estudo também
demonstrou que as assembleias de borboletas respondem ao gradiente hidrológico, as mudanças
na composição de espécies vegetais, mudanças na cota altitudinal do terreno, de peso da massa
seca de serrapilheira e espessura de serrapilheira. Generalizações a respeito das respostas das
espécies aos gradientes edáficos e hidrológicos devem ser vistos com cautela para regiões
específicas de diferentes interflúvios.
Graça (2014) ressalta que alguns fatores bióticos e abióticos têm sido registrados como
influenciadores dos padrões de riqueza, diversidade e distributivos das assembleias de
borboletas frugívoras na Amazônia Central. Principalmente, a estrutura da vegetação e do
hábitat, os quais determinam variações na estrutura interna da floresta, acarretando mudanças
na composição das assembleias de espécies de borboletas (RAMOS, 2000; KOH e SODHI,
2004; PARDONNET et al., 2013).
A espessura da serapilheira e a disponibilidade de fósforo atuam como fatores limitantes
para as assembleias de borboletas nas florestas tropicais (KASPARI e YANOVIAK, 2008
2009). Fatores e efeitos indiretos associados a espessura da serapilheira merecem destaque, uma
vez que outros grupos de invertebrados que forrageiam no solo, poderiam gerar interações
presa-predador, com as borboletas frugívoras quando estas buscam frutos caídos no chão
florestal (HILL et al., 2001; SOURAKOV et al., 2012), expondo as borboletas em gradiente

44
natural de predação (KASPARI e YANOVIAK 2009).
Os principais preditores atuam como filtros interagindo em estágios diferentes no ciclo
de vida, influenciando os padrões de distribuição sob dois aspectos principais: (i) A composição
de plantas parece atuar nas formas imaturas (larval) devido à especificidade na procura de
plantas hospedeiras, e (ii) as variáveis edáficas definem o tipo de ambiente onde os adultos
podem forragear (machos e fêmeas) e atuam como fatores limitantes para as diferentes
populações.
Graça (2014) destaca que a associação entre borboletas e plantas está intimamente
ligado a dependência delas durante a fase larval e posteriormente na fase adulta na utilização
de fontes diferentes de recursos. Graça (2014), Pardonnet et al (2013) e DeVries, (1987)
destacam, ainda, outras aspectos ligados a história natural desse grupo que merecem destaque:
(i) Diferentes populações de borboletas frugívoras preferem espécies particulares de plantas, o
que podem estar associado a sua palatabilidade e acessibilidade a estes recursos (SPENCER,
1988) (ii) machos e fêmeas podem diferir quanto a acessibilidade e palatabilidade, pois o local
que os machos forrageiam e as fêmeas forrageiam e ovipositam pode diferir daqueles locais
onde não ovipositam e são coletados. Desta forma, diferentes populações de borboletas
preferem espécies particulares de plantas para hospedarem, tanto na sua fase larval, como adulta
e de acordo com o sexo. Assim, à medida que a composição de espécies de plantas muda ao
longo do gradiente ambiental, a disponibilidade de plantas hospedeiras acompanha, o que
provoca a substituição de espécies de borboletas ao longo desse mesmo gradiente.
No entanto, é importante considerar que há ambientes acessíveis e não acessíveis de
acordo com o grupo de interesse (ESTRADA; COATES-ESTRADA, 2002; GRELLE, 2003).
A qualidade de recursos disponíveis é um importante parâmetro e, o qual pode estar associado
a acessibilidade ou não. A estrutura da vegetação inadequada à manutenção do ciclo de vida e
distúrbios causados por atividades antrópicas (ESTRADA; COATES-ESTRADA, 2002;
PARDINI et al., 2009) tem sido sugerido como variáveis importantes a serem testadas.
Com relação ao gradiente de serapilheira e sua associação com as borboletas frugívoras,
Graça (2014) destaca que existem conflitos quanto a interpretação de alguns estudos (KOH e
SODHI 2004; KASPARI e YANOVIAK, 2009). Os principais aspectos são: (i) a relação entre
profundidade de serapilheira vs borboletas frugívoras podem estar associados com diferentes
níveis e graus de perturbação ambiental e da paisagem; (ii) a profundidade de serapilheira
permite que predadores de solo modulem as interações. Essa pressão seletiva ambiental pode
estar selecionando espécies de borboletas com mecanismos de fuga mais eficientes, percepção
do ambiente mais refinada ou respostas mais rápidas a ataques de predadores (Graça, 2014).

45
O Padrão registrado neste estudo assemelha-se ao obtido por Graça (2014) para as
florestas da Amazônia Central, onde predominam regiões onde platôs, vertentes e baixos
predominam, os quais diferem daqueles registrados para a Estação Ecológica do Cuniã.
Os estudos que investigam a associação de variáveis ambientais com a biodiversidade
são essenciais para prever a distribuição de espécies e modelar as mudanças decorrentes de
diversos fatores ambientais e de uso e modificação da paisagem (MAGNUSSON et al, 2005),
estas informações são críticas e, podem indicar, sobretudo, regiões de alta diversidade e detectar
centros de endemismo, tornando o processo de criação de unidades de conservação mais
adequado, fortalecendo ações em âmbito estadual e regional (MESQUITA et al., 2007;
CANDIDO et al., 2007).
Por outro lado, os invertebrados respondem rapidamente a modificações em seu
ambiente, principalmente em função de seu curto ciclo de vida (KREMEN et al., 1993),
portanto, podem funcionar como modelos mais adequados, que outros grupos de representantes
da fauna e da flora, visando testar respostas ecológicas a variabilidade ambiental (MORITZ et
al., 2001; BASSET et al., 2009), sendo que a maioria dos grupos de artrópodes terrestres são
sensíveis às mudanças ambientais (TOURINHO, 2007; SATURNINO, 2007; BRAGAGNOLO
et al., 2007; ASSMANN; JANSSEN, 1999; BROMHAM et al., 1999; PAOLETTI, 1999;
SANTOS et al., 2008; LEWINSOHN et al. 2005).
Neste contexto, o estudo das borboletas frugívoras realizado na Estação Ecológica do
Cuniã localizada na região do interflúvio entre os rios Madeira e o Purus é fundamental para
preencher lacunas no conhecimento da biodiversidade de invertebrados, e ainda se inserir em
estudos que incorporam inventários padronizados, para levantar de forma sistemática, a
ocorrência de borboletas frugívoras na Amazônia Sul Ocidental.
Estas informações pretendem melhorar o conhecimento taxonômico local e regional,
fortalecer coleções de referência, apontar os melhores indicadores de qualidade ambiental, e,
principalmente, indicar estes táxons para inventários rápidos (UEHARA-PRADO et al. 2005,
BARLOW et al. 2007, UEHARA-PRADO et al. 2009).

46
CONCLUSÃO
Com este estudo aumentou-se o conhecimento de borboletas frugívoras que habitam o
Interflúvio. E fornece um aporte para o conhecimento e a conservação da diversidade.
Comprovou-se que a Estação Ecológica de Cuniã é uma região de refúgio da lepidopterofauna
de Rondônia e sul do Amazonas, fundamental para a manutenção da biota e biodiversidade
local e regional.
A análise da densidade, dos padrões espaciais e temporais das assembleias de borboletas
frugívoras da ESEC-Cuniã, permitiu concluir que a variabilidade nos padrões distributivos das
populações de borboletas frugívoras constitui-se em níveis funcionais, de acordo com
performance das espécies, as quais contribuem com ampla distribuição e suas restrições
ambientais determinam a variabilidade das espécies ao longo da estrutura interna da floresta de
terra firme.
Observou-se que os melhores preditores para mudanças das assembleias de borboletas
frugívoras foram gradientes argilosos, alto teor de fósforo, a estrutura da vegetação (biomassa
arbórea, diversidade florística e densidade de palmeiras). Essas variáveis, e mais o gradiente
hidrológico, as mudanças na composição de espécies vegetais, de peso de massa seca de
serrapilheira e espessura da serrapilheira, estruturam fortemente as assembleias de borboletas
frugívoras na região do interflúvio entre os rios Madeira e Purus.

47
REFERÊNCIAS
ACTION STAT, Action stat para excel, versão 3.01. 2016. Disponível em
<http://www.portalaction.com.br> acesso em 12.02.2017.
ADDINSOFT, Xlstat for excel, version 19.02. Addinsoft, New York. 2017.
ANDRADE, R. T. G.; PANSINI, S.; SAMPAIO, A. F.; RIBEIRO, M. S.; CABRAL, G. S.;
MANZATTO, A.G. Fitossociologia de uma floresta de terra firme na Amazônia Sul
Ocidental, Rondônia, Brasil. BIOTA AMAZÔNIA, v. 7, p. 36-43, 2017.
ANDERSON, A. B. White sand vegetation of Brazilian Amazonia. Biotropica 13:199-210.
1981.
ASSMANN, T.; JANSSEN. The effects of habitats changes on the endangered ground beetle
Carabus nitens (Coleoptera, Carabiidae). Journal of Insect Conservation, 3:107-116. 1999.
AZEVEDO, A., ALENCAR, A., MOUTINHO, P., RIBEIRO, V., REIS, T., STABILE, M.,
GUIMARÃES, A. Panorama sobre o desmatamento na Amazônia em 2016. IPAM, Brasília,
DF. 2016. Disponível em
<http://www.observatorioflorestal.org.br/sites/default/files/panorama-desmatamento-
amazonia-2016.pdf > Acesso em 20/04/2017.
BARLOW J, OVERAL WL, ARAUJO IS, GARDNER TA, PERES CA. The value of
primary, secondary and plantation forests for fruitfeeding butterflies in the Brazilian Amazon.
Journal of Applied Ecology, 44:1001–1012. 2007.
BASSET, Y.; HAWKINS, B. & LEATHER, S. Visions for insect conservation and diversity:
spanning the gap between practice and theory. Insect Conservation and Diversity: 2: 1-4.
2009.
BRAGAGNOLO, C.; NOGUEIRA, A.A.; PINTO-DA-ROCHA, R. & PARDINI, R.
Harvestmen in an Atlantic Forest fragmented landscape: evaluating assemblage response to
habitat quality and quantity. Biological Conservation. 139: 389-400. 2007.
BRASIL. Instituto Brasileiro de Geografia e Estatística. Mapa de Biomas do Brasil, primeira
aproximação. Rio de Janeiro: IBGE. 2004. Disponível em <www.ibge.gov.br.> Acesso em
24/02/2017.
BRASIL. Instituto Brasileiro de Geografia e Estatística. 2010. Disponível em
<http://www.ibge.gov.br> acesso em 20.07.2015
BRASIL. MCT. Programa de Pesquisa em Biodiversidade. 2012. Disponível em
<http://www.ppbio.inpa.gov.br> acesso em 22.08.2016
BRASIL. MMA. Instituto Chico Mendes. 2001. Disponível em <http://www.icmbio.gov.br>
acesso em 22.08.2016
BRAY J.R., CURTIS J.T. (1957): An ordination of the upland forest communities of
Southern Wisconsin. Ecological Monographies 27:325-349p. 1957.

48
BROMHAM, L., CARDILLO, M., BENNET, F.A. & ELGAR, M. A. Effects of Stock
Grazing on the Ground Invertebrate. 1999.
BROWN JR., K. S. Borboletas da serra do Japi: diversidade, habitat, recursos alimentares e
variação temporal; p. 142-187 In L. P. C. Morellato (org.)., História Natural da Serra do Japi:
ecologia e preservação de uma área florestal no sudeste do Brasil. Campinas:
Unicamp/Fapesp. 1992.
BROWN JR, K. S; FREITAS A. V. L. Atlantic Forest Butterflies: Indicators for Landscape
Conservation BlOTROPlCA 32(4b): 934-956p. 2000.
CABRAL G. S. Efeitos de variáveis ambientais nos padrões distributivos das assembleias de
epífitas e hemiepífitas no interflúvio entre os rios Purus e Madeiras, Estação Ecológica do
Cuniã, Porto Velho (RO). Monografia (Graduação em Ciências Biológicas) - Universidade
Federal de Rondônia. 2015.
CANALS, G.R. Mariposas de Misiones. L.O.L.A. Buenos Aires, 492p. 2003.
CANDIDO, L.A.; MANZI, A.O.; TOTA, J.; SILVA, P.R.T.; SILVA, M.F.S.; SANTOS,
R.M.N. & CORREIA, F.W.S. O Clima atual e o futuro da Amazônia nos cenários do IPCC: a
questão da savanização. Ciência & Cultura, 59: 44-47. 2007.
CEMIN, Arneide Bandeira. A colonização em Rondônia: imaginário amazônico e projetos de
desenvolvimento - tecnologias do imaginário, dádivas-veneno e violência. Mesa Redonda I:
Rondônia – Tecnologias do Imaginário, fronteiras da colonização e violência. Coordenada por
Juremir Machado da Silva (PUC/RS) no Congresso Internacional - XIV Ciclo de Estudos do
Imaginário – As dimensões Imaginárias da Natureza, promovido pelo Núcleo Interdisciplinar
de Estudos sobre o Imaginário da UFPe, Recife, 29 de outubro a 01 de novembro de 2006.
COLLINGE, S. K. Spatial ecology and biological conservation - introduction. Biological
Conservation, 100: 1–2. 2001.
COSTA, F. R. C.; MAGNUSSON, W. E; LUIZÃO, R. C. Mesoscale distribution patterns of
Amazonian understory herbs in relation to topography, soil and watersheds. Journal of
Ecology, 93: 863-878. 2005.
CUNHA, M. F. M.; SAMPAIO, A. F.; Frigeri, R.B.C.; MANZATTO, A. G.
Hymenophyllaceae (Monilophyta) da Estação Ecológica Cuniã, município de Porto Velho,
Rondônia. Acta Biológica Catarinense, v. 1, p. 46-59-59, 2014.
D'ABRERA, B. Butterflies of the Neotropical Region.Lansdowne Editions. Melbourne, in
association with E.W. Classey. 1981.
DEVRIES, P. J. The butterfl ies of Costa Rica and their natural history: Papilionidae,
Pieridae, and Nymphalidae. Princeton: Princeton University Press, 1987.
DEVRIES, P. J.; MURRAY, D.; LANDE, R. Species diversity in vertical, horizontal, and
temporal dimensions of a fruit-feeding butterfl y community in an Ecuadorian rainforest.
Biological Journal of the Linnean Society. v. 62, p.343-364. 1997.

49
DEVRIES P. J.; WALLA T. R. Species diversity and community structure in neotropical
fruit-feeding butterflies. Biological Journal of the Linnean Society. 74: 1–15. 2001.
DICE, Lee R. Measures of the Amount of Ecologic Association Between Species. Ecology 26
(3): 297-302. doi:10.2307/1932409. JSTOR 1932409. 1945.
ESTRADA, A.; COATES-ESTRADA, R. Bats in continuous forest, forest fragments and in
an agricultural mosaic habitat-island at Los Tuxtlas, Mexico. Biol. Conserv. 103, 237-245.
2002.
FEARNSIDE, P. M. A floresta amazônica nas mudanças globais. Manaus. INPA. 134 p.
2003.
FEARNSIDE, P.M. e GRAÇA, P.M.L.A. BR-319: Brazil’s Manaus-Porto Velho Highway
and the potential impact of linking the arc of deforestation to central Amazonia.
Environmental Management. 38(5): 705-716. 2006.
FREITAS, A.V.L., FRANCINI, R.B. & BROWN JR., K.S. Insetos como indicadores
ambientais. Capítulo 5 In Métodos de estudos em biologia da conservação e manejo da vida
silvestre (L Cullen Jr., C. Valladares-Pádua & R. Rudran, orgs.). Editora da UFPR, p. 125-
151. 2003.
FURLANETTI, P.R.R.; SETTE, I.M.S.; LOUZADA, J.N.C. A comunidade de borboletas
frugívoras em fragmentos florestais conectados e desconectados por corredores: influência na
diversidade, composição e tamanho corpóreo. In: CONGRESSO DE ECOLOGIA DO
BRASIL, 8, 2007, Caxambu. Anais... Minas Gerais: SEB. 2007. Disponível em: <
http://sebecologia.org.br/viiiceb/pdf/1475.pdf> Acesso em 30 ago. 2016.
GALVÃO E. E. Produção de Serapilhera em uma Floresta de Terra Firme na Região do
Interfluvio entre os Rios Purus e Madeira, Porto Velho (RO), Brasil. Monografia (Graduação
em Ciências Biológicas) - Universidade Federal de Rondônia. 2016.
GLOBAL BIODIVERSITY INFORMATION FACILITY (Gbif). 2017. Disponível em
<http://www.gbif.org/occurrence/search?taxon_key=5129513&basis_of_record=PRESERVE
D_SPECIMEN.> Acesso em 20/05/2017
GRAÇA, M. B. C. S. Diversidade, Padrões De Distribuição E Esforço De Coleta De
Borboletas Frugívoras Em Floresta Ombrófila Densa Da Amazônia, Manaus, Brasil.
Dissertação de Mestrado. Programa De Pós-Graduação Em Entomologia. Instituto Nacional
De Pesquisas Da Amazônia – INPA. Editora INPA, Manaus, Amazonas, Brasil. 2014.
GOTELLI, NJ. e ELLISON, AM. Princípios de estatística em ecologia. Porto Alegre:
Artmed. 528 p. 2011.
GRELLE, C.E.V Forest structure and vertical stratification of small mammals in a secondary
Atlantic forest, southeastern Brazil. Studies on Neotropical Fauna and Environment, 38: 81-
85. 2003.
GUTIÉRREZ, D. Importance of historical factors on species richness and composition of

50
butterfly assemblages (Lepidoptera: Rhopalocera) in a northern Iberian mountain range.
Journal of Biogeography, 24: 77-88. 1997.
GARWOOD K., LEHMAN R., CARTER W., CARTER G. Butterflies of Southern
Amazonia. McAllen, Texas. 2009.
GILBERT, L.E. The Biology of butterfly communities. In The Biology of Butterflies (R.I.
Vane-Wright & P.E. Ackery, eds.). Florida: Academic Press Inc., p.41–54. 1984.
HAIR J. F., BLACK, ROLPH E. ANDERSON, RONALD L. TATHAM. Analise
Multivariada de Dados. Editora Bookman, 593 p. 2005.
HILL, J. K., K. C. HAMER, J. TANGAH, AND M. DAWOOD. Ecology of tropical
butterflies in rainforests gaps. Oecologia 128: 294–302. 2001.
HUGHES, J. B.; DAILY, G. C.; EHRLICH, P. R. Use of fruit bait traps for monitoring of
butterflies (Lepidoptera: Nymphalidae). Revista de Biologia Tropical. v. 46, n. 3, p.697-704.
1998.
KASPARI, M.; YANOVIAK, S.P. Biogeography of litter depth in tropical forests: evaluating
the phosphorus growth rate hypothesis. Functional Ecology, 22: 919–923. 2008.
KASPARI M.; SP YANOVIAK, R DUDLEY, M YUAN, AND NA CLAY. Sodium shortage
as a constraint on the carbon cycle in an inland tropical rainforest PNAS 106:19405-19409.
2009.
KOH, L.P.; SODHI, N.S. Importance of reserves, fragments, and parks for butterfly
conservation in a tropical urban landscape. Ecological Applications, 14: 1695–1708. 2004.
KREMEN, C., COLWELL, R.K., ERWIN, T.L., MURPHY, D.D., NOSS, R.F. &
SANJAYAN, M.A. Terrestrial arthropod assemblages: their use in conservation planning.
Conservation Biology, 7: 796-808p. 1993.
LEWINSOHN, T. M.; FREITAS, A.V.; PRADO, P. I. Conservação de invertebrados
terrestres e seus habitats no Brasil. Megadiversidade, Campinas, São Paulo. v. 1. n. 1, p. 1-2.
2005.
LIMA C. B. A. Padrões distributivos das assembleias de palmeiras ao longo de gradientes
ripários na estação ecológica do Cuniã, Interfluvio Purus-Madeira, Rondônia. Dissertação
(Mestrado em Desenvolvimento Regional e Meio Ambiente) - Universidade Federal de
Rondônia. 2012.
MALDONADO, F.D.; KEIZER, E.W.H.; GRAÇA, P.M.L.A.; FEARNSIDE, P.M.; VITEL
C.S. Previsão temporal da distribuição espacial do desmatamento no interflúvio PurusMadeira
até o ano 2050. Pp. 183-196 In: W.C. de Sousa Junior, A.V. Waichman, P.A.A. Sinisgalli,
C.F. de Angelis & A.R. Romeiro (eds.) Rio Purus: Água, Território e Sociedade na Amazônia
Sul-Ocidental. LibriMundi, Goiânia, Goiás. 282 pp. 2012.
MAGALHÃES L. P. Caracterização taxonomica da familia melastomataceae na estação
Ecológica de Cuniã - Esec Cuniã, Porto Velho-RO. Monografia (Graduação em Ciências

51
Biológicas) - Universidade Federal de Rondônia. 2007.
MAGNUSSON W.E., LIMA A.P., LUIZÃO R., LUIZÃO F., COSTA F.R.C., CASTILHO
C.V., KINUPP V.P. 2005. RAPELD: a modification of the gentry method for biodiversity
surveys in long-term ecological research sites. Biota Neotropica. 5(2):1-6. 2005.
MAGNUSSON W., BRAGA-NETO R., PEZZINI F., BACCARO F., O monitoramento da
biodiversidade. Manaus: Áttema Editorial: Assessoria e Design, 352p. 2013.
MARENGO J. A., Mudanças climáticas e seus efeitos sobre a Biodiversidade: Caracterização
do clima atual e definição das alterações climáticas para o Território Brasileiro ao longo do
Século XXI. Series Biodiversidade No. 26, MMA, 2007.
MCCUNE, B. & GRACE J.B. Nonmetric Multidimensional Scaling. Em: Analysis of
Ecological Communities. MJM, Software, Oregon. pp: 125.2002.
MESQUITA, R., MARINELLI, C.E. & PINHEIRO, P.S. Ciência e formulação de políticas de
conservação na Amazônia, p. 239-244. In: Rapp Py-Daniel, L., Deus, C.P., Henriques, A.L.,
Pimpão, D.M. & Ribeiro, O.M. (Eds.). Biodiversidade do médio Madeira: bases científicas
para propostas de conservação. Manaus, INPA, 244p. 2007.
MINCHIN P. R. An evaluation of the relative robustness of techniques for ecological
ordination. Vegetatio 69: 89-107. Dr W. Junk Publishers, Dordrecht - Printed in the
Netherlands 89 1987.
MIELKE O.H.H. e CASAGRANDE M. M., Papillonoidea E Hesperioidea (Lepidoptera) Do
Parque Estadual Do Morro Do Diabo, Teodoro Sampaio, São Paulo, Brasil Revista brasileira
de. Zoologia. 14 (4): 967 -1001p. 1997.
MORITZ, C., RICHARDSON, K.S., FERRIER, S., MONTEITH, G.B., STANISIC, J.,
WILLIAMS, S.E., WHIFfiN, T. Biogeographical concordance and efficiency of taxon
indicators for establishing conservation priority in a tropical rainforest biota. Proceedings of
the Royal Society of London Series B-Biological Sciences 268, 1875–1881p. 2001.
NOBRE, R. de A. et al. Monitoramento da biodiversidade: roteiro metodológico de aplicação.
Brasília: Icmbio, 40 p. 2014.
OLIVEIRA A. E. Efeitos das variáveis ambientais nas assembleias de ervas na Estação
Ecológica do Cuniã (Interflúvio Madeira-Purus - trecho Humaitá (AM) - Porto Velho (RO.
Iniciação científica PIBIC. (Graduação em Ciências Biológicas) - Universidade Federal de
Rondônia. 2016.
OLIVEIRA, O. A. História Desenvolvimento e Colonização do Estado de Rondônia. 4 ed.
Dinâmica. Porto Velho. 2001.
PAOLETTI, M.G. Using bioindicators based on biodiversity to assess landscape
sustainability. Agriculture, Ecosystems and Environment, 74: 1-18. 1999.
PANSINI, S. Efeito Das Variáveis Ambientais Sobre As Assembléias De Palmeiras na
ESEC.Cuniã – Interflúvio Madeira-Purus, Rondônia, Brasil. Dissertação (Mestrado Programa

52
de Pós-Graduação em Desenvolvimento Regional e Meio Ambiente). UNIR. Porto Velho,
Rondônia. 2011.
PANSINI, S. Relação entre hidrologia e assembleia de palmeiras ao longo de gradientes
ambientais na ESEC Cuniã (interflúvio PurusMadeira). Tese (Doutorado em
BIODIVERSIDADE E BIOTECNOLOGIA - REDE BIONORTE) - Universidade Federal do
Amazonas. 2016.
PARDINI, R. FARIA, D. ACCACIO, G.M., LAPS, R.R., MARIANO-NETO, E.,
PACIENCIA, M.L.B., DIXO, M. & BAUMGARTEN, J. The challenge of maintaining
Atlantic forest biodiversity: a multi-taxa conservation assessment of specialist and generalist
species in an agro-forestry mosaic in Southern Bahia. Biol. Conserv., 142: 1178-1190. 2009.
PARDONNET, S.; BECK, H.; MILBERG, P.; BERGMAN, K. Effect of tree-fall gaps on
fruitfeeding nymphalid butterfly assemblage in a Peruvian rain forest. Biotropica, 54: 612–
619. 2013.
PEARCE, J.L.; VENIER, L.A. The use of ground beetles (Coleoptera: Carabidae) and spiders
(Araneae) as bioindicators of sustainable forest managmente: a review. Ecological Indicators.
6: 780–793. 2006.
PEIXOTO, A. L.; LUZ J. R. P.; BRITO M.A.D. Conhecendo a biodiversidade. Brasília:
MCTIC, CNPq, PPBio, 2016.
PENIN, A. et al. O Curso de Bacharelado em Arqueologia na Fundação Universidade Federal
de Rondônia (UNIR). Jornal da Sociedade de Arqueologia Brasileira. v3. Brasil. 2011.
PIRES, J. M. Tipos de vegetação da Amazônia. Brasil Florestal, 5(17): 48-58. 1974.
PIRES, J. M. & G. T. PRANCE. The vegetation types of the Brazilian Amazon. In Key
Environments: Amazonia (G.T. Prance & T. E. Lovejoy, eds.), pp. 109-145. Pergamon Press,
Oxford. 1985.
PYLE, R.M. Butterfly eco-geography and biological conservation in Washington. Atala
8:126. 1980.
R Development Core Team. Version 1.0.136 – © 2009-2016 RStudio, Inc. R: A language and
environment for statistical computing. R Foundation for Statistical Computing, Vienna,
Austria. ISBN 3-90005107-0, URL http://www.R-project.org.
RAMOS, F. A. Nymphalid butterfly communities in an Amazonian forest fragment. Journal
of Research on the Lepidoptera. 35: 29-41. 2000.
RIBEIRO, M. S. Efeito das variáveis ambientais na distribuição de herbáceas na Estação
Ecológica do Cuniã, Interfluvio Madeira-Purus, Rondônia, Brasil. Monografia (Graduação em
Ciências Biológicas) - Universidade Federal de Rondônia. 2014.
RIBEIRO, D. B.; FREITAS A. V. L. Differences in termal responses in a fragmented
landscape: temperature affects the sampling of diurnal, but not nocturnal fruit-feeding
Lepidoptera. The Journal of Reserch on the Lepidoptera, v. 42, p.1-4. 2010.

53
RICHARDS, P.W. The Tropical Rain Forest. 2 ed. Cambridge, Cambridge University Press,
575 p. 1996.
RYDON, A. Notes on the use of butterfly traps in East Africa. Journal of the Lepidopterists’
Society. 18: 51–58. 1964.
SAMPAIO, A. F.; RIBEIRO, M. S.; PANSINI, S.; CUNHA, M. F. M.; MANZATTO, A. G.
Balanophoraceae da Estação Ecológica do Cuniã, Porto Velho, RO, Brasil. Acta Biológica
Catarinense, v. 3, p. 73-79, 2016.
SANTOS, E. M. R.; FRANKLIN, E. & MAGNUSSON, W. Cost-efficiency of Sub-sampling
Protocols to Evaluate Oribatid Mites Communities in an Amazonian Savanna. Biotropica, 40:
728-735. 2008.
SATURNINO, R.F. Efeitos do isolamento e da perda de área de floresta sobre comunidades
insulares de aranhas, Amazônia Central, Brasil. Dissertação de mestrado. Instituto Nacional
de Pesquisas da Amazônia/Universidade Federal do Amazonas, 63pp. 2007.
SCHULZE, C. H.; WALTERT, M.; KESSLER, P. J. A.; PITOPANG, R.; SHAHABUDDIN;
VEDDELER, D.; MÜHLENBERG, M.; GRADSTEIN, S. R.; LEUSCHNER, C.; STEFFAN-
DEWENTER, I.; TSCHARNTKE, T. Biodiversity indicator groups of tropical land-use
systems: comparing plants, birds and insects. Ecological Applications, 14: 1321–1333. 2004.
SHUEY, J. A. An optimizing portable bait trap for quantitative sampling of butterflies.
Tropical Lepidoptera. n. 8, p. 1-4. 1997.
SILVA, J.M.C.; RYLANDS, A.B.; FONSECA, G.A.B. O destino das áreas de endemismo da
Amazônia. Megadiversidade. 1(1):124-131. 2005. Disponível em
<www.scielo.br> acesso em 24/02/2017
SORENSEN, T. A method of establishing groups of equal amplitude in plant sociology based
on similarity of species and its application to analyses of the vegetation on Danish commons.
Kongelige Danske Videnskabernes Selskab 5 (4): 1-34. 1957.
SOURAKOV, A., DUEHL, A., & SOURAKOV, A., Foraging Behavior of the Blue Morpho
and Other Tropical Butterflies: The Chemical and Electrophysiological Basis of Olfactory
Preferences and the Role of Color.– Psyche. 10 pp. 2012.
SOUSA T.E.L. Distribuição de Palmeiras (Arecaceae) ao Longo de Gradientes Ambientais no
Baixo Interflúvio Purus-Madeira, Brasil. Dissertação de Mestrado. Instituto Nacional de
Pesquisas da Amazônia, Manaus. 2007.
TOURINHO, A. L. M. Padrões de distribuição e fatores condicionantes da riqueza e
composição de opiliões na várzea do Rio Amazonas – Brasil (Arachnida: Opiliones). Tese de
Doutorado. Instituto Nacional de Pesquisas da Amazônia/Universidade Federal do Amazonas,
Manaus, 24 pp. 2007.
UEHARA-PRADO, M. Efeitos de Fragmentação Florestal na Guilda de Borboletas
Frugívoras do Planalto Atlântico Paulista. Dissertação de Mestrado. Universidade Estadual de

54
Campinas. Instituto de Biologia. Campinas-São Paulo, Brasil. 2003.
UEHARA-PRADO, M.; FREITAS, A. V. L.; FRANCINI, R. B.; BROWN JR., K. S. Guia
das borboletas frugívoras da Reserva Estadual do Morro Grande e região de Caucaia do Alto,
Cotia (São Paulo). Biota Neotropica. v. 4, p. 1-25. 2004.
UEHARA-PRADO, M.; BROWN, K. S. Jr.; FREITAS, A. V. L. Biological traits of
frugivorous butterflies in a fragmented and a continuous landscape in the South Brazilian
Atlantic Forest. Journal of the Lepidopterists’ Society, v. 59, p. 96-106. 2005.
UEHARA-PRADO Selecting terrestrial arthropods as indicators of small-scale disturbance: A
first approach in the Brazilian Atlantic Forest. Biological Conservation. p.1220–1228. 2009.
UEHARA-PRADO, M. & RIBEIRO D. B. Borboletas em Floresta Atlântica: métodos de
amostragem e inventário de espécies na Serra do Itapeti. Pp. 167-186 In: MORINI, M. S. C.
& V. F. O. MIRANDA(Orgs.). Serra do Itapeti: aspectos históricos, sociais e naturalísticos.
Bauru: Canal6. v. 1. 400 pp. 2012.
VALENTIN, J. Ecologia Numérica: Uma Introdução à Análise Multivariada de Dados
Ecológicos. Interciência, Rio de Janeiro. 2012.
VIEIRA, F. G., Estrutura e Composição da Assembleia de Peixes de Pequeno Porte em
Igarapés de Terra Firme da Estação Ecológica de Cuniã, Porto Velho-RO. Dissertação
(Mestrado Programa de Pós-Graduação em Desenvolvimento Regional e Meio Ambiente).
UNIR. Porto Velho, Rondônia. 2013.
VIEIRA, R.S.; MOTA, C.; AGRA, D.B.; LOPES, L.M.; CEZAR, K.F.S. Guia Ilustrado de
Borboletas da Reserva Florestal Adolpho Ducke. Editora INPA, Manaus, 2014
WAHLBERG, N.; J. LENEVEU; U. KODANDARAMAIAH; C. PEÑA; S. NYLIN; A. V. L.
FREITAS & A. V. Z. BROWER. Nymphalid butterflies diversify following near demise at
the Cretaceous/Tertiary boundary. Proceedingsofthe Royal Society B 276: 4295-4302.p 2009.
ZAR, JH, Biostatistical Analysis. 1999.

55
APÊNDICE
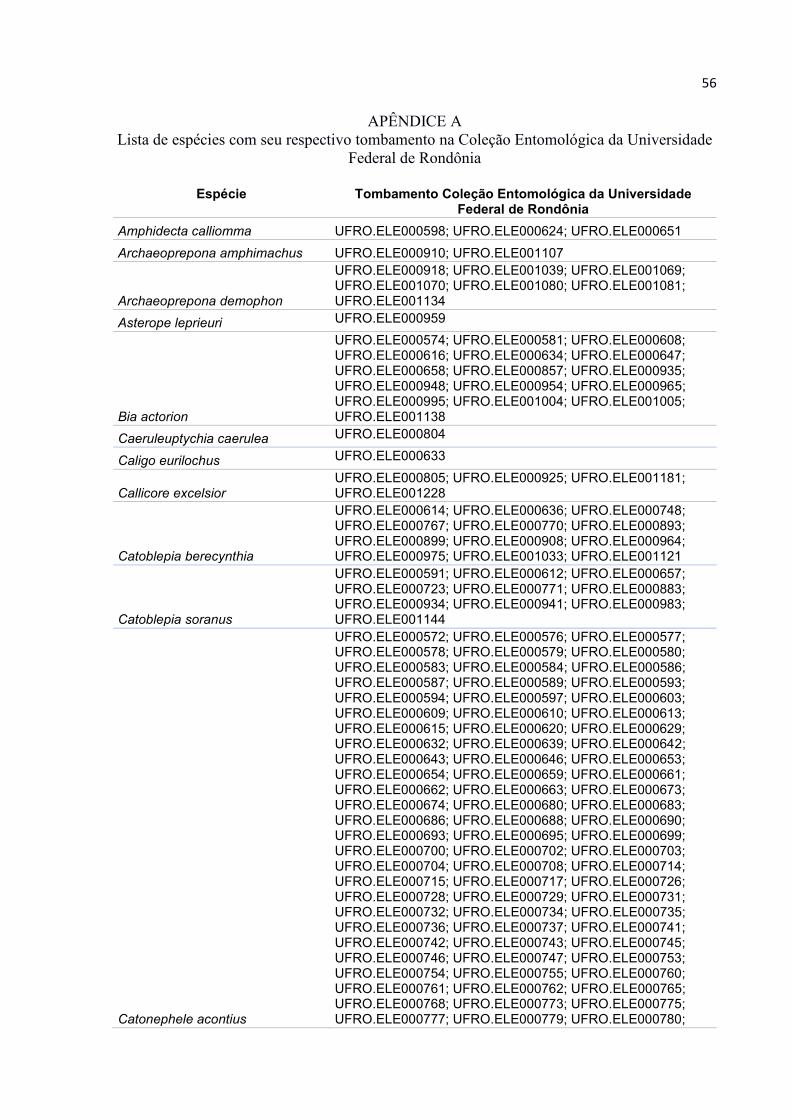
56
APÊNDICE A
Lista de espécies com seu respectivo tombamento na Coleção Entomológica da Universidade
Federal de Rondônia
Espécie Tombamento Coleção Entomológica da Universidade
Federal de Rondônia
Amphidecta calliomma UFRO.ELE000598; UFRO.ELE000624; UFRO.ELE000651
Archaeoprepona amphimachus UFRO.ELE000910; UFRO.ELE001107
Archaeoprepona demophon
UFRO.ELE000918; UFRO.ELE001039; UFRO.ELE001069; UFRO.ELE001070; UFRO.ELE001080; UFRO.ELE001081; UFRO.ELE001134
Asterope leprieuri UFRO.ELE000959
Bia actorion
UFRO.ELE000574; UFRO.ELE000581; UFRO.ELE000608; UFRO.ELE000616; UFRO.ELE000634; UFRO.ELE000647; UFRO.ELE000658; UFRO.ELE000857; UFRO.ELE000935; UFRO.ELE000948; UFRO.ELE000954; UFRO.ELE000965; UFRO.ELE000995; UFRO.ELE001004; UFRO.ELE001005; UFRO.ELE001138
Caeruleuptychia caerulea UFRO.ELE000804
Caligo eurilochus UFRO.ELE000633
Callicore excelsior UFRO.ELE000805; UFRO.ELE000925; UFRO.ELE001181; UFRO.ELE001228
Catoblepia berecynthia
UFRO.ELE000614; UFRO.ELE000636; UFRO.ELE000748; UFRO.ELE000767; UFRO.ELE000770; UFRO.ELE000893; UFRO.ELE000899; UFRO.ELE000908; UFRO.ELE000964; UFRO.ELE000975; UFRO.ELE001033; UFRO.ELE001121
Catoblepia soranus
UFRO.ELE000591; UFRO.ELE000612; UFRO.ELE000657; UFRO.ELE000723; UFRO.ELE000771; UFRO.ELE000883; UFRO.ELE000934; UFRO.ELE000941; UFRO.ELE000983; UFRO.ELE001144
Catonephele acontius
UFRO.ELE000572; UFRO.ELE000576; UFRO.ELE000577; UFRO.ELE000578; UFRO.ELE000579; UFRO.ELE000580; UFRO.ELE000583; UFRO.ELE000584; UFRO.ELE000586; UFRO.ELE000587; UFRO.ELE000589; UFRO.ELE000593; UFRO.ELE000594; UFRO.ELE000597; UFRO.ELE000603; UFRO.ELE000609; UFRO.ELE000610; UFRO.ELE000613; UFRO.ELE000615; UFRO.ELE000620; UFRO.ELE000629; UFRO.ELE000632; UFRO.ELE000639; UFRO.ELE000642; UFRO.ELE000643; UFRO.ELE000646; UFRO.ELE000653; UFRO.ELE000654; UFRO.ELE000659; UFRO.ELE000661; UFRO.ELE000662; UFRO.ELE000663; UFRO.ELE000673; UFRO.ELE000674; UFRO.ELE000680; UFRO.ELE000683; UFRO.ELE000686; UFRO.ELE000688; UFRO.ELE000690; UFRO.ELE000693; UFRO.ELE000695; UFRO.ELE000699; UFRO.ELE000700; UFRO.ELE000702; UFRO.ELE000703; UFRO.ELE000704; UFRO.ELE000708; UFRO.ELE000714; UFRO.ELE000715; UFRO.ELE000717; UFRO.ELE000726; UFRO.ELE000728; UFRO.ELE000729; UFRO.ELE000731; UFRO.ELE000732; UFRO.ELE000734; UFRO.ELE000735; UFRO.ELE000736; UFRO.ELE000737; UFRO.ELE000741; UFRO.ELE000742; UFRO.ELE000743; UFRO.ELE000745; UFRO.ELE000746; UFRO.ELE000747; UFRO.ELE000753; UFRO.ELE000754; UFRO.ELE000755; UFRO.ELE000760; UFRO.ELE000761; UFRO.ELE000762; UFRO.ELE000765; UFRO.ELE000768; UFRO.ELE000773; UFRO.ELE000775; UFRO.ELE000777; UFRO.ELE000779; UFRO.ELE000780;

57
UFRO.ELE000783; UFRO.ELE000785; UFRO.ELE000786; UFRO.ELE000787; UFRO.ELE000788; UFRO.ELE000789; UFRO.ELE000790; UFRO.ELE000791; UFRO.ELE000792; UFRO.ELE000797; UFRO.ELE000798; UFRO.ELE000799; UFRO.ELE000800; UFRO.ELE000801; UFRO.ELE000806; UFRO.ELE000807; UFRO.ELE000808; UFRO.ELE000810; UFRO.ELE000812; UFRO.ELE000813; UFRO.ELE000814; UFRO.ELE000816; UFRO.ELE000819; UFRO.ELE000821; UFRO.ELE000823; UFRO.ELE000825; UFRO.ELE000826; UFRO.ELE000830; UFRO.ELE000833; UFRO.ELE000834; UFRO.ELE000835; UFRO.ELE000836; UFRO.ELE000840; UFRO.ELE000847; UFRO.ELE000850; UFRO.ELE000852; UFRO.ELE000861; UFRO.ELE000866; UFRO.ELE000884; UFRO.ELE000888; UFRO.ELE000892; UFRO.ELE000896; UFRO.ELE000900; UFRO.ELE000901; UFRO.ELE000903; UFRO.ELE000904; UFRO.ELE000907; UFRO.ELE000909; UFRO.ELE000912; UFRO.ELE000913; UFRO.ELE000914; UFRO.ELE000915; UFRO.ELE000924; UFRO.ELE000928; UFRO.ELE000929; UFRO.ELE000930; UFRO.ELE000931; UFRO.ELE000932; UFRO.ELE000937; UFRO.ELE000938; UFRO.ELE000939; UFRO.ELE000952; UFRO.ELE000953; UFRO.ELE000955; UFRO.ELE000958; UFRO.ELE000962; UFRO.ELE000968; UFRO.ELE000969; UFRO.ELE000976; UFRO.ELE000978; UFRO.ELE000979; UFRO.ELE000980; UFRO.ELE000991; UFRO.ELE000992; UFRO.ELE000993; UFRO.ELE000994; UFRO.ELE000997; UFRO.ELE000998; UFRO.ELE000999; UFRO.ELE001011; UFRO.ELE001012; UFRO.ELE001014; UFRO.ELE001018; UFRO.ELE001022; UFRO.ELE001023; UFRO.ELE001027; UFRO.ELE001028; UFRO.ELE001035; UFRO.ELE001036; UFRO.ELE001040; UFRO.ELE001043; UFRO.ELE001044; UFRO.ELE001049; UFRO.ELE001050; UFRO.ELE001053; UFRO.ELE001055; UFRO.ELE001061; UFRO.ELE001062; UFRO.ELE001063; UFRO.ELE001072; UFRO.ELE001077; UFRO.ELE001078; UFRO.ELE001092; UFRO.ELE001095; UFRO.ELE001105; UFRO.ELE001120; UFRO.ELE001126; UFRO.ELE001129; UFRO.ELE001130; UFRO.ELE001131; UFRO.ELE001132; UFRO.ELE001136; UFRO.ELE001141; UFRO.ELE001142; UFRO.ELE001145; UFRO.ELE001146; UFRO.ELE001154; UFRO.ELE001156; UFRO.ELE001157; UFRO.ELE001161; UFRO.ELE001162; UFRO.ELE001163; UFRO.ELE001175; UFRO.ELE001176; UFRO.ELE001185; UFRO.ELE001186; UFRO.ELE001209; UFRO.ELE001210; UFRO.ELE001215; UFRO.ELE001216; UFRO.ELE001220; UFRO.ELE001222; UFRO.ELE001223; UFRO.ELE001224; UFRO.ELE001225; UFRO.ELE001227; UFRO.ELE001230; UFRO.ELE001231; UFRO.ELE001235; UFRO.ELE001236; UFRO.ELE001237; UFRO.ELE001238;
Catonephele numilia
UFRO.ELE000582; UFRO.ELE000601 UFRO.ELE000623; UFRO.ELE000627; UFRO.ELE000665; UFRO.ELE000671; UFRO.ELE000677; UFRO.ELE000679; UFRO.ELE000684; UFRO.ELE000685; UFRO.ELE000694; UFRO.ELE000696; UFRO.ELE000705; UFRO.ELE000706; UFRO.ELE000707; UFRO.ELE000710; UFRO.ELE000711; UFRO.ELE000716; UFRO.ELE000733; UFRO.ELE000752; UFRO.ELE000759; UFRO.ELE000763; UFRO.ELE000776; UFRO.ELE000778; UFRO.ELE000782; UFRO.ELE000793; UFRO.ELE000794; UFRO.ELE000795; UFRO.ELE000796; UFRO.ELE000802; UFRO.ELE000811; UFRO.ELE000815; UFRO.ELE000817; UFRO.ELE000818; UFRO.ELE000820; UFRO.ELE000824;

58
UFRO.ELE000827; UFRO.ELE000828; UFRO.ELE000829; UFRO.ELE000837; UFRO.ELE000838; UFRO.ELE000841; UFRO.ELE000842; UFRO.ELE000843; UFRO.ELE000848; UFRO.ELE000849; UFRO.ELE000853 UFRO.ELE000858; UFRO.ELE000859; UFRO.ELE000860; UFRO.ELE000865; UFRO.ELE000875; UFRO.ELE000887; UFRO.ELE000894; UFRO.ELE000897; UFRO.ELE000906; UFRO.ELE000916; UFRO.ELE000917; UFRO.ELE000922; UFRO.ELE000946; UFRO.ELE000951; UFRO.ELE000966; UFRO.ELE000967; UFRO.ELE000970; UFRO.ELE000971; UFRO.ELE000981; UFRO.ELE000990; UFRO.ELE001016; UFRO.ELE001029;; UFRO.ELE001030; UFRO.ELE001034; UFRO.ELE001045; UFRO.ELE001046; UFRO.ELE001051; UFRO.ELE001064; UFRO.ELE001071; UFRO.ELE001090; UFRO.ELE001147; UFRO.ELE001148; UFRO.ELE001149; UFRO.ELE001155; UFRO.ELE001158; UFRO.ELE001164; UFRO.ELE001165; UFRO.ELE001177; UFRO.ELE001184; UFRO.ELE001199;
Chloreuptychia callichloris UFRO.ELE000895; UFRO.ELE000986; UFRO.ELE000996; UFRO.ELE001114;
Chloreuptychia herseis UFRO.ELE000650;
Chloreuptychia hewitsonii UFRO.ELE000617;
Cissia myncea UFRO.ELE000625; UFRO.ELE000640;
Colobura annulata UFRO.ELE000644;
Colobura dirce
UFRO.ELE000600; UFRO.ELE000626; UFRO.ELE000630; UFRO.ELE000645; UFRO.ELE000649; UFRO.ELE000656; UFRO.ELE001008; UFRO.ELE001102;
Dynamine pebana UFRO.ELE001109;
Erichthodes antonina UFRO.ELE000697; UFRO.ELE000724; UFRO.ELE000781
Erichthodes arius UFRO.ELE001074;
Eunica anna UFRO.ELE001119; UFRO.ELE001123; UFRO.ELE001171; UFRO.ELE001173;
Eunica malvina UFRO.ELE000669; UFRO.ELE001214
Eunica veronica UFRO.ELE001094; UFRO.ELE001108; UFRO.ELE001172; UFRO.ELE001178; UFRO.ELE001189;
Eunica viola
UFRO.ELE000604; UFRO.ELE000670; UFRO.ELE000718; UFRO.ELE000854; UFRO.ELE000867; UFRO.ELE000926; UFRO.ELE000949; UFRO.ELE000950; UFRO.ELE000972; UFRO.ELE000977; UFRO.ELE000984; UFRO.ELE001007; UFRO.ELE001015; UFRO.ELE001017; UFRO.ELE001021; UFRO.ELE001042; UFRO.ELE001060; UFRO.ELE001088; UFRO.ELE001118; UFRO.ELE001135; UFRO.ELE001143; UFRO.ELE001179; UFRO.ELE001187; UFRO.ELE001188; UFRO.ELE001200; UFRO.ELE001201; UFRO.ELE001202; UFRO.ELE001211; UFRO.ELE001212; UFRO.ELE001213; UFRO.ELE001217; UFRO.ELE001229;
Euptychia insolata UFRO.ELE000622;
Fountainea ryphea UFRO.ELE001073; UFRO.ELE001101; UFRO.ELE001133;
Hamadrias arinome UFRO.ELE001197;
Hamadryas alicia UFRO.ELE000655;
Hamadryas arinome
UFRO.ELE000750; UFRO.ELE000881; UFRO.ELE000885; UFRO.ELE000905; UFRO.ELE001037; UFRO.ELE001052; UFRO.ELE001115;
Hermeuptychia fallax UFRO.ELE000689;

59
Hypna clytemnestra UFRO.ELE001079; UFRO.ELE001106;
Magneuptychia antonoe UFRO.ELE001198;
Magneuptychia ocnus UFRO.ELE000619;
Memphis acidalia UFRO.ELE000588; UFRO.ELE001032; UFRO.ELE001048; UFRO.ELE001153;
Memphis glauce UFRO.ELE000982;
Memphis leonida
UFRO.ELE000666; UFRO.ELE000740; UFRO.ELE000851; UFRO.ELE000868; UFRO.ELE000886; UFRO.ELE000902; UFRO.ELE000919; UFRO.ELE000920; UFRO.ELE000945; UFRO.ELE000988; UFRO.ELE001009; UFRO.ELE001024; UFRO.ELE001025; UFRO.ELE001056; UFRO.ELE001091; UFRO.ELE001206;
Memphis moruus
UFRO.ELE000772; UFRO.ELE000870; UFRO.ELE001010; UFRO.ELE001020; UFRO.ELE001057; UFRO.ELE001058; UFRO.ELE001182; UFRO.ELE001221; UFRO.ELE001232;
Memphis oenomais
UFRO.ELE000611; UFRO.ELE000940; UFRO.ELE000942; UFRO.ELE000944; UFRO.ELE000947; UFRO.ELE001001; UFRO.ELE001152;
Memphis phantes UFRO.ELE000987; UFRO.ELE000878;
Morpho achiles UFRO.ELE001003
Morpho helenor UFRO.ELE000880;
Nessaea obrinus UFRO.ELE000607; UFRO.ELE000631; UFRO.ELE000882; UFRO.ELE001006;
Nica flavilla
UFRO.ELE000628; UFRO.ELE000635; UFRO.ELE000667; UFRO.ELE000672; UFRO.ELE000675; UFRO.ELE000682; UFRO.ELE000691; UFRO.ELE000692; UFRO.ELE000709; UFRO.ELE000720; UFRO.ELE000730; UFRO.ELE000739; UFRO.ELE000756; UFRO.ELE000803; UFRO.ELE000831; UFRO.ELE000845; UFRO.ELE000863; UFRO.ELE000898; UFRO.ELE000921; UFRO.ELE001127; UFRO.ELE000698; UFRO.ELE000769; UFRO.ELE000869; UFRO.ELE001054; UFRO.ELE001111;
Pareuptychia binocula UFRO.ELE001093; UFRO.ELE000599; UFRO.ELE001110; UFRO.ELE001125;
Pareuptychia ocirrhoe UFRO.ELE000596
Pareuptychia summandosa
UFRO.ELE001174; UFRO.ELE001183; UFRO.ELE000936; UFRO.ELE000592; UFRO.ELE000602; UFRO.ELE000605; UFRO.ELE000638; UFRO.ELE000725; UFRO.ELE001086; UFRO.ELE001087; UFRO.ELE001067; UFRO.ELE001082; UFRO.ELE001218; UFRO.ELE001068; UFRO.ELE001104; UFRO.ELE000652; UFRO.ELE001038; UFRO.ELE001084; UFRO.ELE001026;
Pyrrhogyra crameri UFRO.ELE001169; UFRO.ELE001170;
Pyrrhogyra edocla UFRO.ELE001059;
Pyrrhogyra otolais UFRO.ELE000923; UFRO.ELE000943; UFRO.ELE001100;
Taygetis cleopatra
UFRO.ELE000637; UFRO.ELE000933; UFRO.ELE000985; UFRO.ELE001066; UFRO.ELE001233; UFRO.ELE000575; UFRO.ELE000722; UFRO.ELE000862; UFRO.ELE000721;
Taygetis sp. UFRO.ELE000641;
Taygetis sylvia UFRO.ELE001112; UFRO.ELE000606; UFRO.ELE001019;
Taygetis tripunctata UFRO.ELE000764;
Taygetis virgilia UFRO.ELE000719; UFRO.ELE000573; UFRO.ELE000585; UFRO.ELE000664; UFRO.ELE000676; UFRO.ELE000678;

60
UFRO.ELE000681; UFRO.ELE000687; UFRO.ELE000712; UFRO.ELE000713; UFRO.ELE000738; UFRO.ELE000744; UFRO.ELE000749; UFRO.ELE000751; UFRO.ELE000757; UFRO.ELE000758; UFRO.ELE000766; UFRO.ELE000774; UFRO.ELE000784; UFRO.ELE000809; UFRO.ELE000844; UFRO.ELE000855; UFRO.ELE000856; UFRO.ELE000864; UFRO.ELE000871; UFRO.ELE000872; UFRO.ELE000874; UFRO.ELE000876; UFRO.ELE000877; UFRO.ELE000879; UFRO.ELE000889; UFRO.ELE000911; UFRO.ELE000956; UFRO.ELE000957; UFRO.ELE000960; UFRO.ELE000961; UFRO.ELE000963; UFRO.ELE000973; UFRO.ELE000974; UFRO.ELE001013; UFRO.ELE001031; UFRO.ELE001041; UFRO.ELE001047; UFRO.ELE001065; UFRO.ELE001075; UFRO.ELE001076; UFRO.ELE001085; UFRO.ELE001089; UFRO.ELE001096; UFRO.ELE001097; UFRO.ELE001098; UFRO.ELE001099; UFRO.ELE001116; UFRO.ELE001117; UFRO.ELE001122; UFRO.ELE001128; UFRO.ELE001137; UFRO.ELE001140; UFRO.ELE001150; UFRO.ELE001151; UFRO.ELE001160; UFRO.ELE001166; UFRO.ELE001167; UFRO.ELE001168; UFRO.ELE001180; UFRO.ELE001190; UFRO.ELE001191; UFRO.ELE001192; UFRO.ELE001193; UFRO.ELE001194; UFRO.ELE001195; UFRO.ELE001196; UFRO.ELE001203 UFRO.ELE001204; UFRO.ELE001205; UFRO.ELE001207; UFRO.ELE001208; UFRO.ELE001219; UFRO.ELE001226;
Temenis pulchra UFRO.ELE000890;
Tigridia acesta UFRO.ELE000832; UFRO.ELE000873; UFRO.ELE000989;
Zaretis isidora UFRO.ELE000590; UFRO.ELE000727; UFRO.ELE000822; UFRO.ELE000839;
Zaretis itys UFRO.ELE001000;