flutuações populacionais de espécies de amebas testáceas...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE MARINGÁ CURSO DE PÓS-GRADUAÇÃO EM
ECOLOGIA DE AMBIENTES AQUÁTICOS CONTINENTAIS
Flutuações populacionais de espécies de amebas testáceas (Rhizopoda) no plâncton de distintos ambientes
da planície de inundação do alto rio Paraná.
GEZIELE MUCIO ALVES
Maringá – PR
03/2006
1

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

GEZIELE MUCIO ALVES
Flutuações populacionais de espécies de amebas testáceas (Rhizopoda) no plâncton de distintos ambientes
da planície de inundação do alto rio Paraná.
Dissertação de mestrado apresentada ao curso de Pós-Graduação em Ecologia de Ambientes Aquáticos Continentais, Departamento de Biologia da Universidade Estadual de Maringá, como parte dos requisitos para obtenção do título de mestre em Ciências Ambientais.
Orientador: Prof. Dr. Fábio Amodêo Lansac-Tôha
Maringá – PR
03/2006
2

Agradecimentos
À Deus, por estar comigo em todas as etapas da minha vida. Ao meu orientador, Fábio Amodêo Lansac-Tôha, pela amizade, paciência, orientação
e confiança em meu trabalho. À Luiz Felipe Machado Velho, pela carinho, pelas sugestões e auxílio na elaboração
desse trabalho. À Claudia Costa Bonecker, pela amizade e por me atender com carinho nos
momentos de dificuldades. Aos professores, Alberto Prioli, Luis Carlos Gomes e Luis Maurício Bini pelas
sugestões e auxílio nas análises estatísticas. Com muito carinho, à Érica, pela amizade, companheirismo, por ser responsável por
minhas primeiras noções de sincronia e pela imensa contribuição em me ensinar a maior parte das análises deste estudo.
À amigas Marisa e Fabiana, por me apoiarem em todas as etapas deste trabalho e
fazerem do laboratório minha segunda família. Aos amigos de laboratório Gustavo, Leandro, Fábio, Jú, Flávia, Anderson, Deise,
Renata, Ana Paula, Ciro, Dani, Ariane, por tornarem os momentos de trabalho mais descontraídos.
Ao pessoal da biblioteca do curso, ao programa de Pós-Graduação em Ecologia de
Ambientes Aquáticos Continentais e ao CNPq pela concessão da bolsa. À minha família, meu pai, minha irmã e em especial minha mãe que, sem o auxílio
dela, a realização desse estudo seria muito mais difícil. À minha paixão, minha filha Natália e ao meu esposo Clodoaldo pelo amor, apoio e
incentivo em meus estudos. À todos que de alguma forma fizeram parte desse período de trabalho.
3

Este trabalho foi elaborado de acordo com as regras da revista
Hydrobiologia
4

Resumo Flutuações temporais e espaciais da abundância são características marcantes de todas as populações animais. A sincronia regional na dinâmica de populações locais tem sido verificada em vários estudos. Dessa forma, este trabalho teve como objetivo investigar a existência de padrões coerentes de flutuação temporal da abundância das principais populações de tecamebas no plâncton da planície de inundação do alto rio Paraná. As amostragens de tecamebas foram realizadas trimestralmente, entre fevereiro e novembro, no período de 2000 a 2002, e semestralmente, em 2003, em 12 ambientes associados a diferentes rios. As coletas foram obtidas à subsuperfície da região pelágica com auxílio de uma moto-bomba, sendo filtrados 600 litros de água por amostra em uma rede de plâncton com 68 µm de abertura de malha. O material coletado foi fixado com solução de formaldeído (4%), tamponada com carbonato de cálcio. Foram realizadas sub-amostragens com auxílio de uma pipeta tipo Hensen-Stempell (2,5mL), para a determinação da abundância, e as contagens realizadas em câmaras de Sedgwick-Rafter. Para a identificação, os exemplares foram triados de cada amostra e montados em lâminas em glicerina. Foram registrados 93 táxons de tecamebas, pertencentes a 8 famílias, sendo Difflugiidae, Arcellidae e Centropyxidae as mais representativas. Arcella conica, A. discoides, A. vulgaris, Centropyxis aculeata, C. ecornis, Difflugia gramen, D. pseudogramen e Cucurbitella dentata foram as espécies mais abundantes, sendo estas selecionadas para as análises de sincronia espacial. Não foi registrado padrão coerente de flutuação da abundância para as principais espécies considerando todos os ambientes estudados da planície. Entretanto, analisando-se somente os canais, foi registrada sincronia espacial para Centropyxis ecornis. A estatística de Mantel demonstrou que o nível de sincronia, identificado pelos coeficientes de correlação de Spearman e Pearson, não esteve correlacionado com a distância geográfica entre os ambientes. Nesse sentido, pode-se concluir que, na planície de inundação do alto rio Paraná, os fatores regionais não são preponderantes sobre a dinâmica populacional de tecamebas Palavras chaves: zooplâncton, Protozoa, heterogeneidade espacial e temporal, sincronia espacial.
5

Abstract Temporal and spatial abundance fluctuations are remarkable features of all animal populations. Regional synchrony on local populations dynamics have been verified in several researches. In this way, this study aimed to investigate the existence of coherent patterns in abundance temporal fluctuations of principal testate amoebae populations in plankton from upper Paraná river floodplain. Testate amoebae sampling were carried out every three months, between February and November, during 2000 and 2002, and semestrially in 2003, in 12 environments associated to different rivers. Samples were obtained at subsurface of pelagic region, using a motorized pump and a 68 µm mesh plankton net. Collected material was preserved with formaldehyde (4%) buffered with calcium carbonate. Subsampling was made using a Hensen-Stempell pipette for abundance determination. For species identification, individuals were removed from each sample and put on a glycerin slide. Ninety three testate amoebae taxa were identified, which belongs to 8 families. Among them Difflugiidae, Arcellidae and Centropyxidae were the most specious ones. Arcella conica, A. discoides, A. vulgaris, Centropyxis aculeata, C. ecornis, Difflugia gramen, D. pseudogramen and Cucurbitella dentata were the most abundant species, and they were chosen to spatial synchrony analysis. It was not registered any coherent pattern of abundance fluctuation, considering all floodplain studied environments. Nevertheless, when analyzing only the channels, it was observed spatial synchrony to Centropyxis ecornis. Mantel statistic results showed that synchrony level, identified by Spearman and Pearson correlation's, was not correlated to geographic distance among environments. So, we can conclude that on upper Paraná river floodplain, regional factors were not preponderant on testate amoebae population dynamic. Key words: zooplankton, Protozoa, spatial and temporal heterogeneity, spatial synchrony.
6

Introdução
Planícies de inundação são ecossistemas formados por rios, lagoas e canais ligados
ao rio principal, apresentando comunicação de pequena ou grande extensão, em função da
presença de canais permanentes e das alterações do nível fluviométrico (Agostinho &
Zalewski, 1996). As modificações no nível fluviométrico provocam alterações nas
características físicas e químicas ambientais e, conseqüentemente, a biota responde com
adaptações morfológicas, fisiológicas ou etológicas, ou ainda, alterando a estrutura das
comunidades (Junk et al., 1989; Neiff, 1990).
Heino et al. (1997) afirmaram que flutuações temporais e espaciais da abundância
são características marcantes de todas as populações animais. Moran (1953) propôs que
fatores independentes da densidade (geralmente fatores climáticos, de acordo com Royama,
1992), se estiverem correlacionados entre as regiões, podem induzir a sincronia espacial em
duas ou mais populações que apresentam a mesma estrutura intrínseca (dependente da
densidade, como predação ou competição). Esse fenômeno foi denominado por Royama
(1992) de “teorema de Moran” ou “efeito Moran”.
De acordo com Ranta et al. (1999), as características de dinâmica sincronizada das
populações englobam o nível global de sincronia, a tendência central da sincronia, a
dependência da distância e as inter-relações entre essas variáveis. Esses mesmos autores
consideram que a distância que separa as populações locais está inversamente relacionada à
sincronia. Entretanto, em estudos com a comunidade zooplanctônica, Cottenie et al. (2001)
afirmaram que ambientes próximos e mesmo conectados podem ser distintos em relação à
estrutura dessa comunidade e essas diferenças estão correlacionadas com as diferenças na
estrutura trófica e interações bióticas presentes nesses ambientes.
Segundo Kendall et al. (2000), a sincronia regional na dinâmica de populações
locais tem sido verificada em vários estudos, como de parasitas, insetos, peixes, pássaros e
mamíferos. O entendimento das causas de sincronia em ampla escala representa,
recentemente, uma das preocupações em estudos de ecologia de populações devido à
tendência de redução de metapopulações com o aumento da correlação regional. Muitos
aspectos de reservas naturais e a efetiva conservação de espécies ameaçadas dependem do
nível de sincronia regional na dinâmica dessas populações (Kendall et al., 2000).
7

Padrões regionais similares de variabilidade populacional têm como evidência que
as abundâncias das populações são determinadas, de alguma forma, por fatores operando
em escala regional. Por outro lado, a ausência de sincronia implica na ação predominante
de reguladores em escala local (Kratz et al., 1987). O grau de coerência temporal pode,
portanto, indicar se as populações, em determinada região, são mais influenciadas por
fatores locais (intrínsecos) ou por fatores regionais (extrínsecos) (Nicholson, 1933;
Andrewartha & Birch, 1954).
Em ecossistemas aquáticos, pertencentes à planície de inundação do alto rio
Paraná, Lansac-Tôha et al. (1997, 2004) mostraram que a comunidade zooplanctônica
apresenta alterações na abundância em decorrência de mudanças morfométricas do
ambiente, do tempo de inundação e das características peculiares do rio que ocasiona esta
inundação. Dentre os grupos zooplanctônicos estudados nesses ambientes aquáticos,
encontram-se as amebas testáceas ou tecamebas (Lansac-Tôha et al., 1997, 2004; Velho et
al., 1999, 2003, 2004; Bini et al., 2003). Segundo Hardoim (1997), esses organismos
possuem uma série de características que os tornam interessantes nas investigações
ecológicas, como, sua alta abundância em muitos ecossistemas aquáticos continentais,
grande produção de biomassa e seu tamanho e tempo de geração, que permitem
investigações sobre processos demográficos tanto em escala espacial como temporal.
Lena & Zaidenwerg (1975) acreditam que a presença de tecamebas no plâncton
deva ser atribuída apenas a processos hidrodinâmicos, como o arraste de organismos do
sedimento e vegetação marginal. Entretanto, estudos têm mostrado que pelo menos parte de
seu ciclo de vida deve ocorrer no ambiente planctônico de forma a garantir o sucesso dessas
populações (Velho et al., 1999, 2003). Esses mesmos autores, afirmaram que a
hidrodinâmica possui maior importância na determinação da estrutura das assembléias de
tecamebas que a variabilidade temporal.
Considerando que alterações na composição e abundância de tecamebas podem ser
preditas pela heterogeneidade temporal e espacial do compartimento planctônico,
pressupõe-se que as populações mais abundantes de tecamebas apresentam o mesmo
padrão de flutuação em escala regional nos diferentes ambientes da planície de inundação e
que essa sincronia seja maior quando considerados ambientes próximos, associados ao
8
mesmo rio (Paraná, Ivinheima ou Baía), ou pertencentes a mesma categoria (rio, canal,
lagoa aberta ou lagoa fechada).
Material e métodos
Área de estudo
A trabalho foi desenvolvido na “Área de Proteção Ambiental das Ilhas e Várzeas
do Rio Paraná”, criada pelo Governo Federal em 1997, na planície de inundação do alto rio
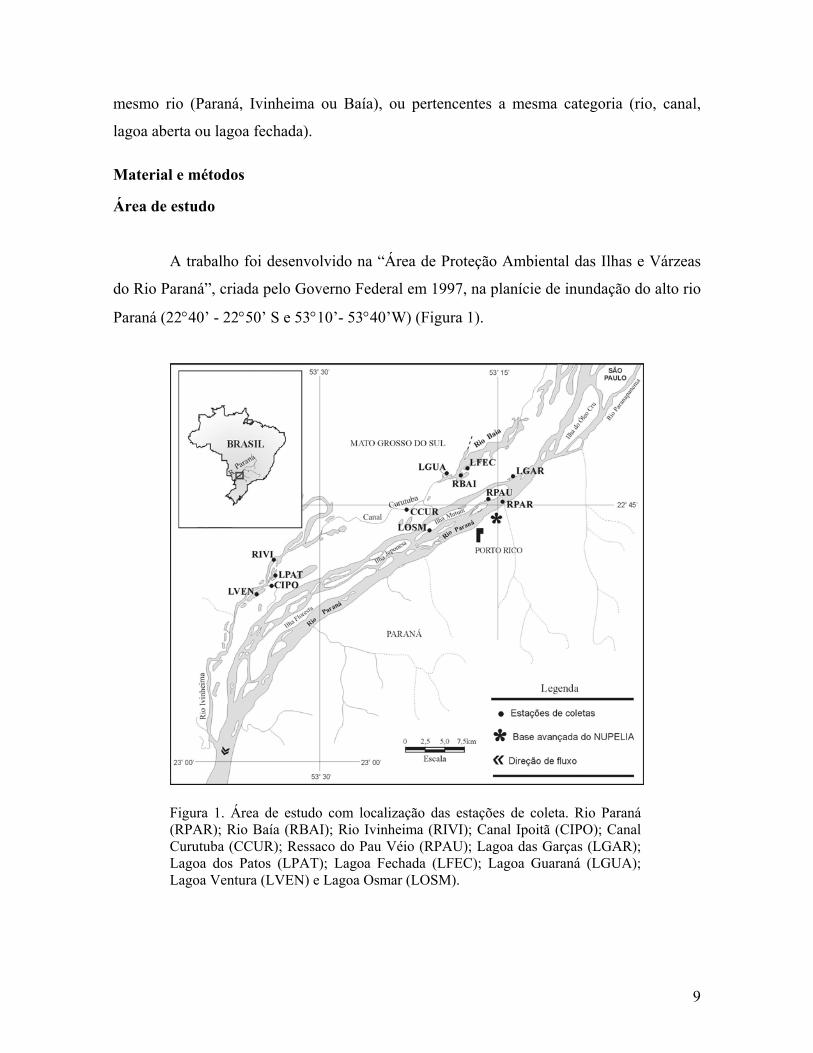
Paraná (22°40’ - 22°50’ S e 53°10’- 53°40’W) (Figura 1).
Figura 1. Área de estudo com localização das estações de coleta. Rio Paraná (RPAR); Rio Baía (RBAI); Rio Ivinheima (RIVI); Canal Ipoitã (CIPO); Canal Curutuba (CCUR); Ressaco do Pau Véio (RPAU); Lagoa das Garças (LGAR); Lagoa dos Patos (LPAT); Lagoa Fechada (LFEC); Lagoa Guaraná (LGUA); Lagoa Ventura (LVEN) e Lagoa Osmar (LOSM).
9

O sistema Paraná é formado pelo rio Paraná e lagoas de várzea associadas
presentes nas ilhas e várzeas. O trecho estudado do rio Paraná (22º45’S – 53º15’W) tem
padrão multicanal, apresentando velocidade média de corrente relativamente alta, com
largura variada e presença de extensas ilhas e barras. Neste trecho, o rio apresenta
profundidade média de 4,0 m, podendo atingir 15,0 m de profundidade máxima (Thomaz et
al., 1992).
O sistema Baía é formado pelo rio Baía ( trecho 22º43’S – 53º17’W) e um grande
número de lagoas associadas ao longo de seu curso. Esse rio comunica-se com o rio Paraná
através de um canal de ligação em seu trecho inferior. Apresenta largura variada e
profundidade média de 3,2 m. Trata-se de um rio sinuoso, considerado como um ambiente
semilótico, e diretamente influenciado pelo regime hidrológico do rio Paraná. A sua baixa
velocidade de corrente e pequena declividade do leito permite que seja caracterizado como
um rio de planície. A reduzida velocidade de corrente possibilita a ocorrência de
estratificação térmica e química (Thomaz et al., 1991).
O sistema Ivinheima é formado por lagoas de planície de inundação associadas ao
rio Ivinheima (trecho 22º47’S – 53º32’W) e pelo próprio rio. Este é um dos principais
tributários da margem direita do rio Paraná, com profundidade média de 3,9 m. Caracteriza-
se por apresentar águas turbulentas, escoando sobre a planície. Corre paralelo ao rio Paraná
em seu curso inferior, conectando-se ao rio Baía pelo canal Curutuba e ao rio Paraná pelo
canal Ipoitã e dois outros canais (Thomaz et al., 1992).
Para a realização desse estudo, foram consideradas 12 estações de amostragem,
abrangendo três rios (Paraná, Baía e Ivinheima), três lagoas com comunicação permanente
com o rio principal (lagoas abertas), três lagoas sem comunicação com o rio principal,
associadas a ele somente pelo lençol freático (lagoas fechadas), dois canais e um ressaco
(ambiente lêntico formado por depósito de sedimento em uma das margens do rio) (Figura
1; Tabela 1).
10
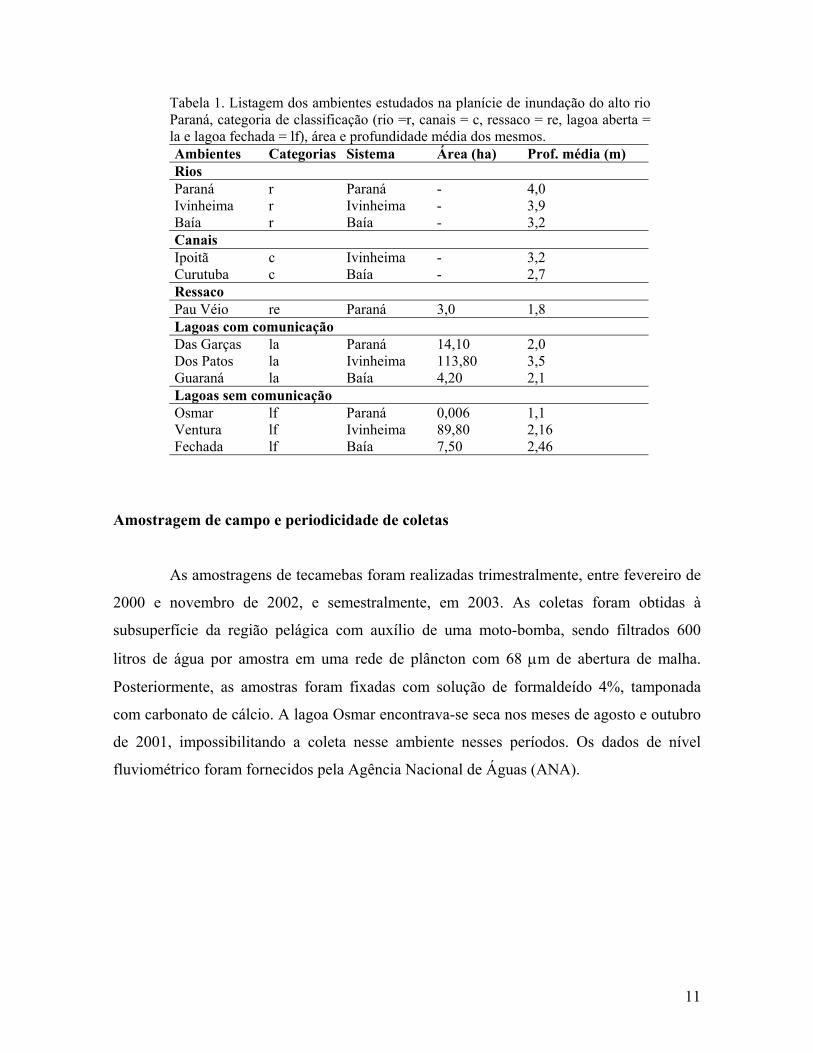
Tabela 1. Listagem dos ambientes estudados na planície de inundação do alto rio Paraná, categoria de classificação (rio =r, canais = c, ressaco = re, lagoa aberta = la e lagoa fechada = lf), área e profundidade média dos mesmos. Ambientes Categorias Sistema Área (ha) Prof. média (m) Rios Paraná r Paraná - 4,0 Ivinheima r Ivinheima - 3,9 Baía r Baía - 3,2 Canais Ipoitã c Ivinheima - 3,2 Curutuba c Baía - 2,7 Ressaco Pau Véio re Paraná 3,0 1,8 Lagoas com comunicação Das Garças la Paraná 14,10 2,0 Dos Patos la Ivinheima 113,80 3,5 Guaraná la Baía 4,20 2,1 Lagoas sem comunicação Osmar lf Paraná 0,006 1,1 Ventura lf Ivinheima 89,80 2,16 Fechada lf Baía 7,50 2,46
Amostragem de campo e periodicidade de coletas
As amostragens de tecamebas foram realizadas trimestralmente, entre fevereiro de
2000 e novembro de 2002, e semestralmente, em 2003. As coletas foram obtidas à
subsuperfície da região pelágica com auxílio de uma moto-bomba, sendo filtrados 600
litros de água por amostra em uma rede de plâncton com 68 µm de abertura de malha.
Posteriormente, as amostras foram fixadas com solução de formaldeído 4%, tamponada
com carbonato de cálcio. A lagoa Osmar encontrava-se seca nos meses de agosto e outubro
de 2001, impossibilitando a coleta nesse ambiente nesses períodos. Os dados de nível
fluviométrico foram fornecidos pela Agência Nacional de Águas (ANA).
11

Análise laboratorial
Para a análise qualitativa, os exemplares foram triados de cada amostra e
montados em lâminas com glicerina para posterior identificação. A identificação foi
baseada na seguinte bibliografia: Deflandre (1928, 1929); Gauthier Liévre & Thomaz
(1958, 1960), Vucetich (1973), Ogden & Hedley (1980), Velho et al. (1996), Velho &
Lansac-Tôha (1996) e Hardoim (1997).
A análise quantitativa foi realizada em câmara de Sedgwick-Rafter sob
microscópio óptico. Para a determinação da abundância foram realizadas subamostragens
com pipeta tipo Hensen-Stempel (2,5mL). As amostras foram coradas com rosa de bengala
e apenas os organismos que apresentavam protoplasma corado foram contados e
identificados, assumindo-se que estavam vivos no momento da coleta. A contagem dos
organismos foi baseada na metodologia de Bottrell et al. (1976), tendo sido estabelecidas
três subamostras subseqüentes para cada amostra coletada. A abundância foi expressa em
ind. m-3.
Análise dos dados
Dentre os táxons registrados, no conjunto de n amostras analisadas (diversidade γ),
foram selecionadas, para a análise de sincronia espacial, as espécies mais abundantes, que
perfizeram 82,3% da abundância total.
Foi empregado o coeficiente de correlação intraclasse (ri), o qual quantifica o grau
de sincronia da flutuação de abundância entre duas ou mais populações ao longo do tempo,
visando avaliar a existência de coerência temporal entre as populações de cada espécie
entre os ambientes. Comparando duas populações, esse coeficiente varia de –1 a 1, e o
ponto onde os fatores extrínsecos (regionais) são mais importantes corresponde ao valor
crítico de ri (onde p=0,05 para uma diferença significativa de 0). Com mais de duas
populações, ri é semelhante à “correlação média” entre todas as populações. Assim, o valor
crítico e o limite inferior para ri mudam quanto mais populações forem analisadas. Como
resultado, a coerência temporal pode ser visualizada como tendo duas dimensões (Figura
2), onde ri e o número de populações n são eixos ortogonais que definem o plano de
12

coerência. Influências extrínsecas e intrínsecas são balanceadas ao longo de uma curva que
une os valores críticos de ri para diferentes números de populações. A espécie mais
coerente regionalmente será a mais distante do valor crítico (e distante do 0) e inclui o
maior número de populações.
Figura 2. Diagrama representando as dimensões da coerência temporal. A curva pontilhada une os valores críticos de coerência (p=0,05) para comparações envolvendo diferentes números de populações e divide o plano em áreas de predominância de fatores intrínsecos e extrínsecos. A curva sólida une pontos que delineiam os limites inferiores de coerência negativa para diferentes números de populações. Em uma comparação entre n ambientes, o valor mínimo possível de coerência é -1/(n-1). Valores à esquerda desta curva não são possíveis. (Modificado de Rusak et al., 1999).
Em uma comparação entre n ambientes, o coeficiente de correlação intraclasse (ri)
foi estimado como:
ri= MSM - MSE / MSM + (n-1)MSE,,
onde MSM e MSE são os quadrados médios para os meses e para o erro de uma ANOVA
bifatorial (ambiente e meses) sem replicação, respectivamente. Utilizando a ANOVA
bifatorial, decompõe-se em fatores um componente da variação devido à diferenças entre os
ambientes. Quando a sincronia aumenta, o MSE aproxima-se de zero e ri aproxima-se de 1.
Quando as flutuações populacionais são assíncronas, a variância atribuída ao erro
aumentará até ser maior que MSM (há mais variabilidade dentro do mês do que entre os
13

meses) e a correlação se tornará negativa. Devido a essa relação, a correlação intraclasse
pode diminuir até -1, quando n=2. Esse coeficiente de correlação está descrito em Rusak et
al. (1999). Para essa análise, os dados de abundância foram previamente log transformados
(log x+1).
Para investigar a existência de padrões coerentes de flutuação temporal, em escala
regional, na planície de inundação, foram analisados os cinco diferentes tipos de ambientes
amostrados (rios, canais, ressaco, lagoas abertas e fechadas). Para verificar a existência de
sincronia espacial em ambientes próximos associados ao mesmo rio, foram avaliados três
tipos de ambientes para cada sistema, sendo eles, o rio principal, uma lagoa aberta e uma
lagoa fechada. Para verificar a existência dessa autocorrelação espacial em ambientes
pertencentes a mesma categoria, foram analisados separadamente os canais, rios, lagoas
abertas e lagoas fechadas.
Visando verificar a existência de correlação significativa entre a flutuação da
abundância das principais espécies, comparando, par a par, os ambientes amostrados, foram
estimados os coeficientes de correlação de Pearson (r) e de Spearman (R). O primeiro
detecta somente uma relação linear entre a variação da abundância das populações e o
segundo identifica uma relação crescente ou decrescente em formato de curva. Essas
análises foram realizadas utilizando o pacote estatístico Statistica 5.5 (Statsoft Inc., 1996).
Para correlacionar o grau de sincronia estimado (Pearson e Spearman) e a distância
geográfica (Km) entre os ambientes analisados foi utilizada a estatística de Mantel (z). O
teste faz comparação entre duas matrizes, uma com as distâncias geográficas (dij) entre os
ambientes e outra com o coeficiente de correlação (cij), calculado através do coeficiente de
Pearson ou Spearman, utilizando a fórmula:
Z = ΣiΣj cij dij
A estatística de Mantel testa a significância da correlação entre os valores de cij e
dij (Shanker e Sukumar, 1999). Para essa análise foi utilizado o pacote NTSYS-PC versão
2.1 (Rohlf, 2000).
Para realização dos coeficientes de correlação de Pearson e Spearman, os dados
não foram transformados. Todos os resultados foram testados para o nível de significância
de p< 0,05.
14

Resultados
No ano de 2000, o rio Paraná não apresentou períodos nitidamente diferenciados
de cheia e seca, sendo registrado apenas um pulso de curta duração (aproximadamente 15
dias) nos meses de março e abril (Figura 3). O ano de 2001 não foi caracterizado por uma
cheia expressiva, mas foram observados períodos distintos de variação do nível
fluviométrico, sendo o primeiro de janeiro a junho, com um período seco em março, um
período mais seco (valores inferiores a 1,7 m), de julho a novembro e elevação novamente
do nível no mês de dezembro. Por outro lado, em 2002 e 2003, entre janeiro e março,
prevaleceram altos valores de nível fluviométrico, caracterizando um período de cheia e,
posteriormente, foram observadas oscilações de curto período e de valores inferiores aos
registrados no início do ano (Figura 3).
Nív
el F
luvi
omét
rico
(m)
0.5
1.5
2.5
3.5
4.5
5.5
6.5
jan0
0m
arm
aio jul
set
nov
jan0
1m
arm
aio jul
set
nov
jan0
2m
arm
aio jul
set
nov
jan0
3m
arm
ai jul
set
nov
Figura 3. Nível fluviométrico do rio Paraná medido diariamente em Porto São José. As setas indicam os períodos de coleta. A linha horizontal indica o nível de transbordamento.
Durante o período estudado, nos distintos ambientes amostrados, foram registrados
93 táxons de tecamebas, pertencentes a 8 famílias, sendo Difflugiidae (46 táxons),
Arcellidae (21 táxons) e Centropyxidae (12 táxons), as mais representativas (Tabela 2).
15

Tabela 2. Inventário faunístico de tecamebas registradas em diferentes ambientes da planície de inundação do alto rio Paraná no período de 2000 a 2003. Arcellidae Arcella arenaria Greeff, 1866 A. hemisphaerica f. undulata Deflandre, 1928 A. brasiliensis Cunha, 1913 A. megastoma Pénard, 1902 A. catinus Pénard, 1890 A. mitrata Leidy, 1879 A. conica (Playfair, 1917) A. mitrata var. spectabilis Deflandre, 1928 A. costata Ehrenberg, 1847 A. nordestina Vucetich, 1973 A. crenulata Deflandre, 1928 A. rota Daday, 1905 A. dentata Ehrenberg, 1838 A. vulgaris Ehrenberg, 1830 A. discoides Ehrenberg, 1843 A. vulgaris f. elegans Deflandre, 1928 A. gibbosac Pénard, 1890 A. vulgaris var. penardi Deflandre, 1928 A. gibbosa var. mitriformis Deflandre, 1928 A. vulgaris f. undulata Deflandre, 1928 A. hemisphaerica Perty, 1852 Centropyxidae Centropyxis aculeata (Ehrenberg, 1838) C. ecornis (Ehrenberg, 1841) C. aerophila Deflandre, 1929 C. gibba Deflandre, 1929 C. aerophila var. sphagnicola Deflandre, 1929 C. hirsuta Deflandre, 1929 C. cassis (Wallich, 1864) C. marsupiformis (Wallich, 1864) C. constricta (Ehrenberg, 1841) C. platystoma (Pénard, 1902) C. discoides (Pénard, 1890) C. spinosa (Cash, 1905) Difflugiidae Cucurbitella crateriformis Gauthier-Lièvre & Thomas, 1960 D. lobostoma Leidy, 1879 C. dentata f. crucilobata Gauthier-Lièvre & Thomas, 1958 D. lobostoma var. cornuta Gauthier-Lièvre & Thomas, 1958 C. dentata f. quinquilobata Gauthier-Lièvre & Thomas, 1960 D. lobostoma var. multilobata Gauthier-Lièvre & Thomas, 1958 C. dentata f. trilobata Gauthier-Lièvre & Thomas, 1960 D. lobostoma var. tuberosa Gauthier-Lièvre & Thomas, 1958 C. madagascariensis Gauthier-Lièvre & Thomas, 1960 D. microclaviformis (Kourov, 1925) C. mespiliformis Pénard, 1902 D. muriformis Gauthier-Lièvre & Thomas, 1958 C. mespiliformis var. africana Gauthier-Lièvre & Thomas, 1958 D. muriformis f. crucilobata Gauthier-Lièvre & Thomas, 1958 Difflugia acuminata Ehrenberg, 1838 D. nebeloides Gauthier-Lièvre & Thomas, 1958 D. acutissima Deflandre, 1931 D. oblonga Ehrenberg, 1838 D. amphoralis var. globosa Gauthier-Lièvre & Thomas, 1958 D. parva (Thomas, 1954) D. angulostoma Gauthier-Lièvre & Thomas, 1958 D. pleustonica Dioni, 1970 D. bicruris Gauthier-Lièvre & Thomas, 1958 D. pseudogramen Gauthier-Lièvre & Thomas, 1960 D. corona Wallich, 1864 D. stellastoma Vucetich, 1989 D. corona f. tuberculata Vucetich, 1973 D. urceolata Carter, 1864 D. curvicaulis Pénard, 1899 D. ventricosa Deflandre, 1926 D. difficilis Thomas, 1955 D. ventricosa f. recticaulis Dioni, 1970 D. elegans Pénard, 1890 Difflugia sp1 D. globulosa Dujardim, 1837 Difflugia sp2 D. gramen Pénard, 1902 Difflugia sp3 D. helvetica var. multilobata Gauthier-Lièvre & Thomas, 1958 Difflugia sp4 D. kempnyi Stepánek, 1953 Difflugia sp5 D. limnetica (Levander, 1900) Pontigulasia compressa (Carter, 1864) D. lithophila Pénard, 1902 Suiadifflugia multipora Green, 1975 Euglyphidae Euglypha acanthophora (Ehrenberg, 1841) Lesquereusiidae Lesquereusia mimetica Pénard, 1902 Netzelia oviformis (Cash, 1909) L. modesta Rhumbler, 1896 N. wailesi (Ogden, 1980) L. spiralis (Ehrenberg, 1840) Hyalosphenidae Heleopera sp. Nebela sp. .Nebela penardiana Deflandre, 1936 Plagiopyxidae Plagiopyxis callida Pénard, 1910 Hoogenraadia cryptostoma Gauthier-Lièvre & Thomas, 1958 Trigonopyxidae Cyclopyxis eurystoma (Deflandre, 1929) C. khali (Deflandre, 1929) C. impressa (Daday, 1905)
16

As espécies mais abundantes foram Difflugia gramen (30,1%), D. pseudogramen
(17%), Centropyxis aculeata (14,8%), Cucurbitella dentata (9,9%), Arcella discoides
(3,5%), Centropyxis ecornis (2,9%), A. conica (2,9%) e A. vulgaris (1,6%). As demais
espécies (85) somaram 17,7% da abundância total (Figura 4).
����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
D. gramen (30,1%)
D. pseudogramen (17%)
���������������Outras (17,7%)
������������������������������������������������������������������������������������������
A. vulgaris (1,6%)
������������A. conica (2,3%) ��������������������������������
��������������������������������C. ecornis (2,9%) ����������������������������������������������������������������A. discoides (3,5%)�����������������������������������������������������������������������������������������������������������������������������������������������������������
C. dentata (9,9%)
����������������
C. aculeata (14,8%)
Figura 4. Abundância relativa de tecamebas durante o período de estudo em diferentes ambientes da planície de inundação do alto rio Paraná.
Analisando-se a flutuação temporal da abundância, das principais espécies, em
todos os ambientes amostrados da planície de inundação, foi observada a existência de
várias linhas assíncronas, indicando a ausência de um padrão temporal (Figura 5). Difflugia
pseudogramen apresentou uma dinâmica temporal mais ordenada, contudo, não foram os
mesmos ambientes, com flutuações semelhantes, durante todo o período.
17

meses
log(
ind.
m-3+1
)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
Arcella conicari= -0,084p= 0,261
meses
log(
ind.
m-3+1
)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 set
A. discoidesri= -0,090p= NS
meses
log(
ind.
m-3
+1)
������������������������������������������
������������������������
����������������������������
����������������������������
��������������������������
��������������
����������������������������
��������������������������������������
���������������������
������
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
A. vulgarisri= -0,091p= NS
meses
log(
ind.
m-3+1
)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
Centropyxis aculeatari= -0,091p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. ecornisri= -0,091p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
Difflugia gramenri= -0,086p= 0,350
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag no fe/02 ma ag no mç/03 se
D. pseudogramenri= -0,090p= 0,735
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
Cucurbitella dentatari= -0,090p= NS
Figura 5. Flutuação temporal da abundância das principais espécies de tecamebas nos distintos ambientes da planície de inundação do alto rio Paraná (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
Considerando-se apenas os ambientes associados ao rio Paraná, não foi observada
tendência à sincronia para nenhuma espécie analisada. Essa assincronia temporal pode ser
verificada pela ausência de linhas paralelas dos padrões de flutuação da abundância entre os
18
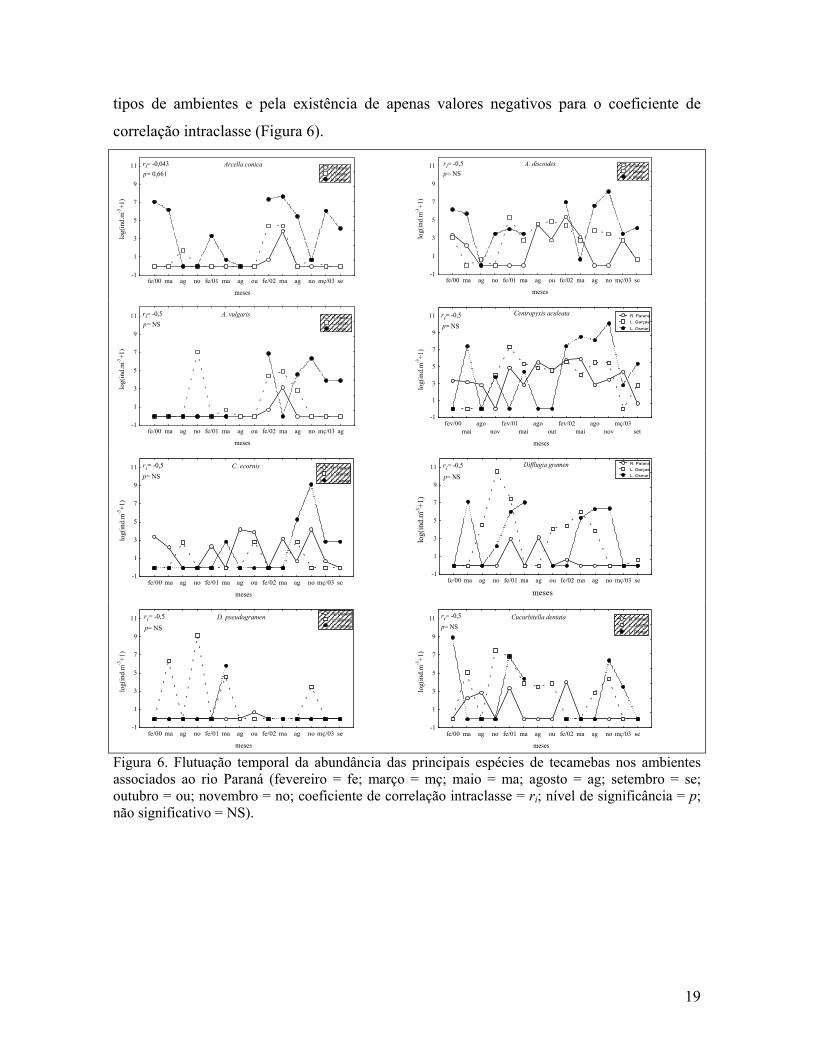
tipos de ambientes e pela existência de apenas valores negativos para o coeficiente de
correlação intraclasse (Figura 6).
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
��������������������������
R. ParanáL. GarçasL. Osmar
Arcella conicari= -0,043p= 0,661
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
���������������������������������������
R. ParanáL. GarçasL. Osmar
A. discoidesri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 ag
��������������������������R. Paraná
L. GarçasL.Osmar
A. vulgarisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fev/00mai
agonov
fev/01mai
agoout
fev/02mai
agonov
mç/03set
R. ParanáL. GarçasL. Osmar
Centropyxis aculeatari= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
����������������������������R. Paraná
L. GarçasL. Osmar
C. ecornisri= -0,5p= NS
meses
log(
ind.
m-3+1
)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáL. GarçasL. Osmar
Difflugia gramenri= -0,5
p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
����������������������������
R. ParanáL. GarçasL. Osmar
D. pseudogramenri= -0,5
p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
����������������������������R. Paraná
L. GarçasL. Osmar
Cucurbitella dentatari= -0,5p= NS
Figura 6. Flutuação temporal da abundância das principais espécies de tecamebas nos ambientes associados ao rio Paraná (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
19
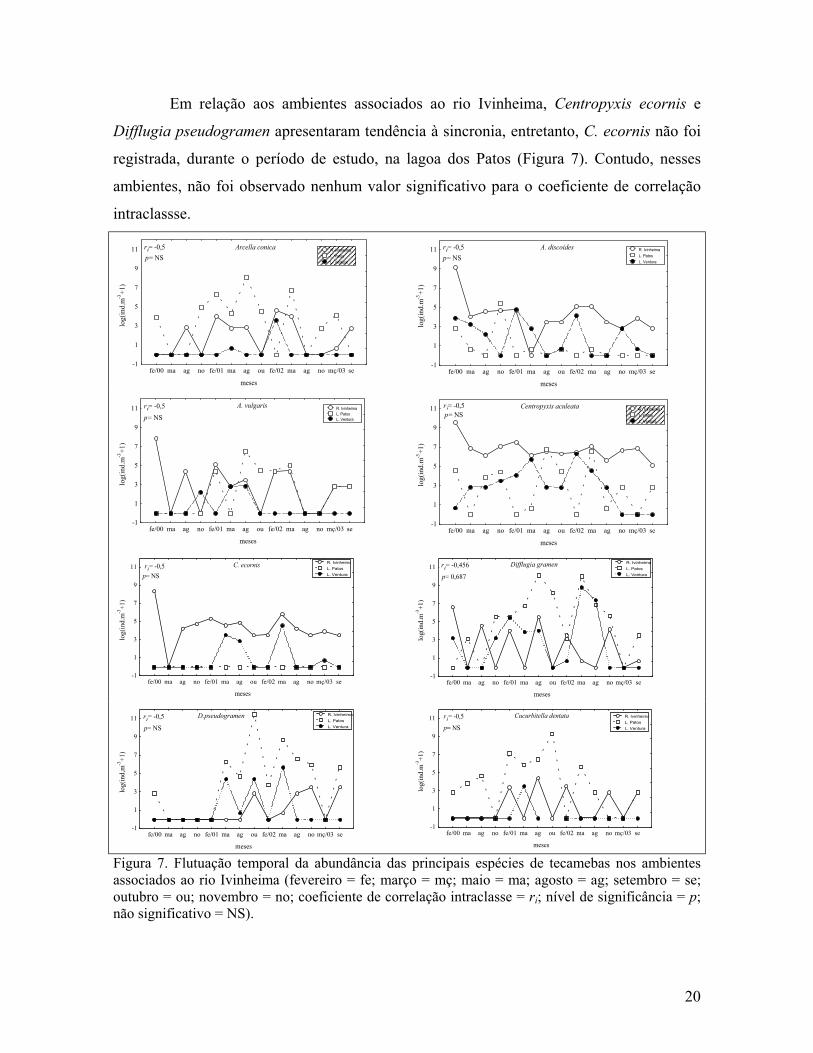
Em relação aos ambientes associados ao rio Ivinheima, Centropyxis ecornis e
Difflugia pseudogramen apresentaram tendência à sincronia, entretanto, C. ecornis não foi
registrada, durante o período de estudo, na lagoa dos Patos (Figura 7). Contudo, nesses
ambientes, não foi observado nenhum valor significativo para o coeficiente de correlação
intraclassse.
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
��������������������������������R. Ivinheima
L. PatosL. Ventura
Arcella conicari= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. IvinheimaL. PatosL. Ventura
A. discoidesri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. IvinheimaL. PatosL. Ventura
A. vulgarisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
��������������������������������R. Ivinheima
L. PatosL. Ventura
Centropyxis aculeatari= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. IvinheimaL. PatosL. Ventura
C. ecornisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. IvinheimaL. PatosL. Ventura
Difflugia gramenri= -0,456
p= 0,687
meses
log(
ind,
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. IvinheimaL. PatosL. Ventura
D.pseudogramenri= -0,5
p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. IvinheimaL. PatosL. Ventura
Cucurbitella dentatari= -0,5
p= NS
Figura 7. Flutuação temporal da abundância das principais espécies de tecamebas nos ambientes associados ao rio Ivinheima (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
20
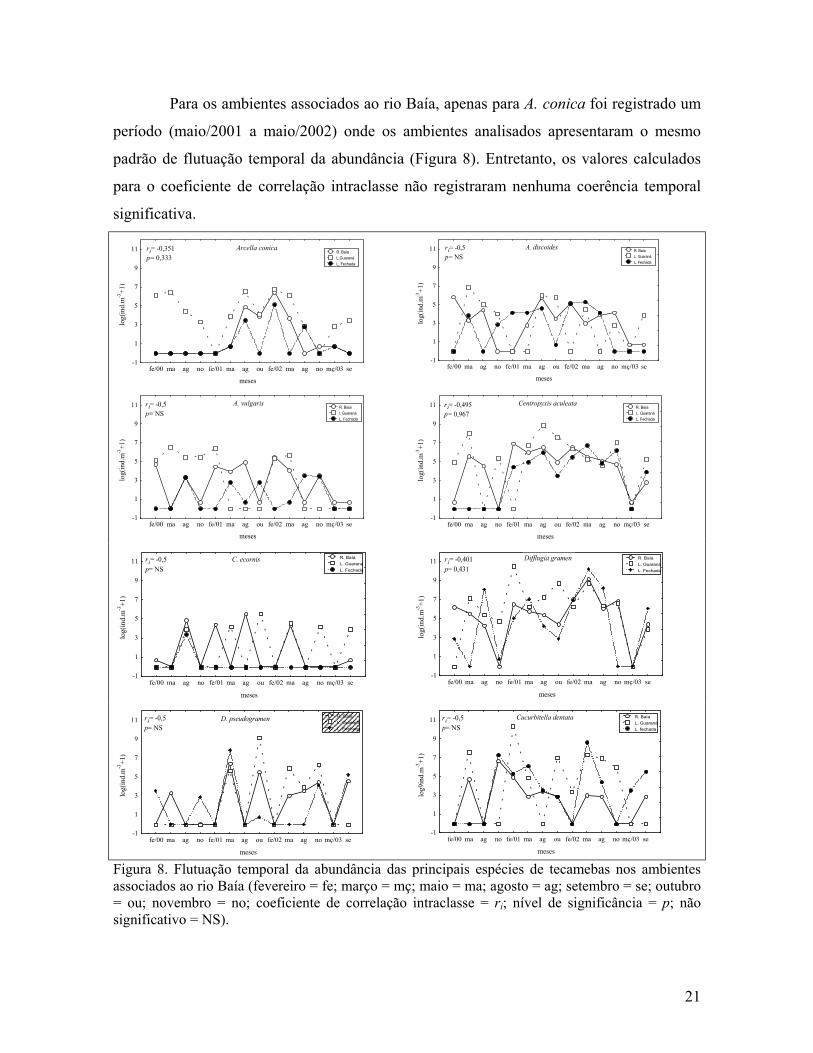
Para os ambientes associados ao rio Baía, apenas para A. conica foi registrado um
período (maio/2001 a maio/2002) onde os ambientes analisados apresentaram o mesmo
padrão de flutuação temporal da abundância (Figura 8). Entretanto, os valores calculados
para o coeficiente de correlação intraclasse não registraram nenhuma coerência temporal
significativa.
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. BaíaL.GuaranáL. Fechada
Arcella conicari= -0,351p= 0,333
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. BaíaL. GuaranáL. Fechada
A. discoidesri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. Baíal. GuaranáL. Fechada
A. vulgarisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. BaíaL. GuaranáL. Fechada
ri= -0,495p= 0,967
Centropyxis aculeata
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. BaíaL. GuaranáL. Fechada
C. ecornisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. BaíaL. GuaranáL. Fechada
Difflugia gramenri= -0,401p= 0,431
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
���������������������������������������������
R. BaíaL. GuaranáL. Fechada
D. pseudogramenri= -0,5p= NS
meses
log9
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. BaíaL. GuaranáL. fechada
Cucurbitella dentatari= -0,5p= NS
Figura 8. Flutuação temporal da abundância das principais espécies de tecamebas nos ambientes associados ao rio Baía (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
21
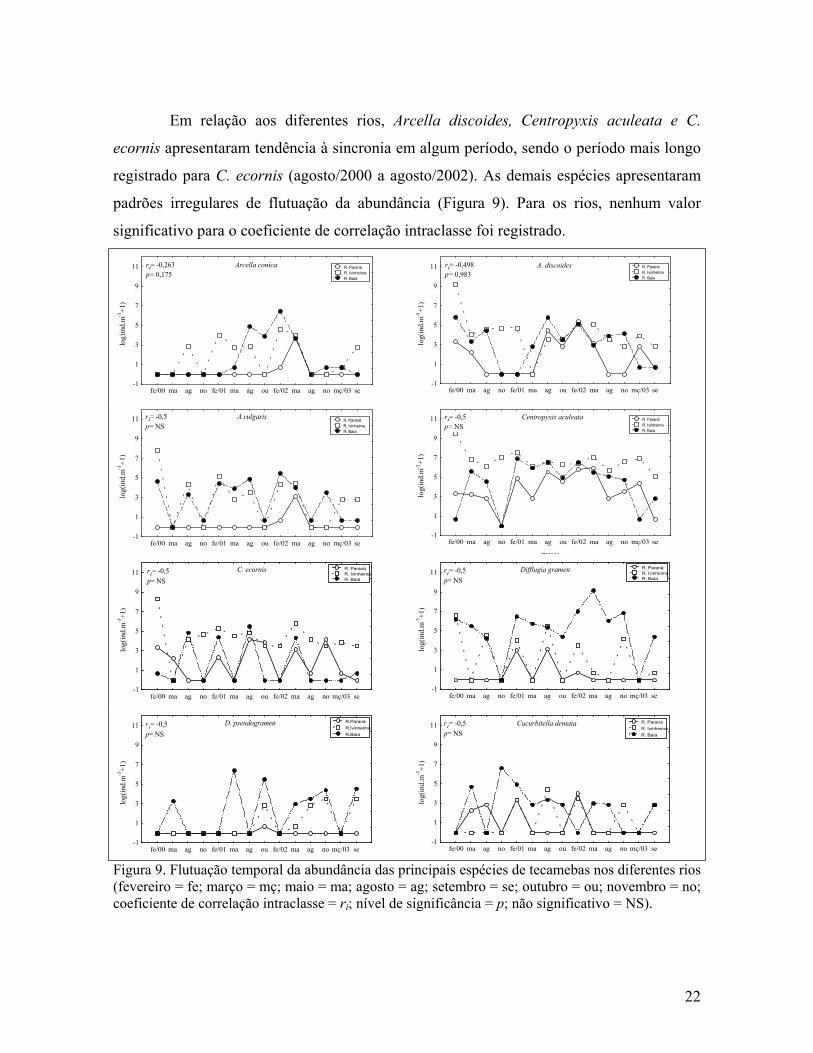
Em relação aos diferentes rios, Arcella discoides, Centropyxis aculeata e C.
ecornis apresentaram tendência à sincronia em algum período, sendo o período mais longo
registrado para C. ecornis (agosto/2000 a agosto/2002). As demais espécies apresentaram
padrões irregulares de flutuação da abundância (Figura 9). Para os rios, nenhum valor
significativo para o coeficiente de correlação intraclasse foi registrado.
log)
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
Arcella conicari= -0,263p= 0,175
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
ri= -0,498p= 0,983
A. discoides
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
A.vulgarisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
Centropyxis aculeatari= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
C. ecornisri= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
Difflugia gramenri= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R.ParanáR.IvinheimaR.Baía
D. pseudogramenri= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
R. ParanáR. IvinheimaR. Baía
Cucurbitella dentatari= -0,5p= NS
mesesFigura 9. Flutuação temporal da abundância das principais espécies de tecamebas nos diferentes rios (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
22

Analisando-se apenas os canais, Centropyxis aculeata apresentou tendência `a
sincronia, com valor positivo do coeficiente de correlação, porém não significativo.
Somente C. ecornis apresentou padrões congruentes de flutuação da abundância com valor
significativo de ri (Figura 10).
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. IpoitãC. Curutuba
Arcella conicari= -1p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. IpoitãC. Curutuba
A. discoidesri= -1p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. IpoitãC.Curutuba
A. vulgarisri= -0,076p= 0,117
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. IpoitãC. Curutuba
Centropyxis aculeatari= 0,105p= 0,070
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. IpoitãC. Curutuba
C. ecornisri= 0,600p= 0,019
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fev/00 mai ago nov fev/01 mai ago out fev/02 mai ago nov mar/03 set
C. IpoitãC. Curutuba
Difflugia gramenri= -1p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
IpoitãCurutuba
D. pseudogramenri= -1p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
C. IpoitãC. Curutuba
Cucurbitella dentatari= -0,342p= 0,242
Figura 10. Flutuação temporal da abundância das principais espécies de tecamebas nos canais estudados (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
23

Considerando-se as lagoas abertas, nenhuma das espécies analisadas apresentou
qualquer tendência à sincronia, apesar de Diffugia gramen apresentar a mesma flutuação da
abundância, para as diferentes lagoas, entre fevereiro e agosto de 2002 (Figura 11). Assim,
todos os valores de ri calculados foram negativos.
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
Arcella conicari= -0,5p= NS
meses
log9
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
A. discoidesri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
A. vulgarisri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
Centropyxis aculeatari= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
C. ecornisri= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
Difflugia gramenri= -0,5p= NS
meses
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
D. pseudogramenri= -0,450p= 0,652
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
GarçasPatosGuaraná
Cucurbitella dentatari= -0,490p= 0,919
mesesFigura 11. Flutuação temporal da abundância das principais espécies de tecamebas nas diferentes lagoas abertas (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
24

Para as lagoas fechadas nenhum padrão de flutuação de abundância foi encontrado
(Figura 12). Da mesma forma, não foi registrado valor significativo para ri. lo
g(in
d.m
-3+1
)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
Arcella conicari= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
A.discoidesri= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
A. vulgarisri= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
Centropyxis aculeatari= -0,5p= NS
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
C. ecornisri= -0,5p= NS
log9
ind.
m-3+1
)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
Difflugia gramenri= -0,350p= 0,297
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
D. pseudogramenri= -0,413p= 0,527
log(
ind.
m-3
+1)
-1
1
3
5
7
9
11
fe/00 ma ag no fe/01 ma ag ou fe/02 ma ag no mç/03 se
OsmarVenturaFechada
Cucurbitella dentatari= -0.5p= NS
mesesFigura 12. Flutuação temporal da abundância das principais espécies de amebas testáceas nas diferentes lagoas fechadas (fevereiro = fe; março = mç; maio = ma; agosto = ag; setembro = se; outubro = ou; novembro = no; coeficiente de correlação intraclasse = ri; nível de significância = p; não significativo = NS).
As espécies apresentaram grau de coerência semelhante, comparando-se as
distintas unidades, e com valores na região de predomínio de fatores intrínsecos sobre os
padrões de abundância. Somente Centropyxis ecornis, analisando-se apenas os canais,
25

apresentou correlação positiva e significativa, ou seja, com valor na região de predomínio
de fatores extrínsecos (Figura 13). Arcella conica
coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,0 -0,8 -0,6 -0,4 -0,2 0,0 0,2 0,4 0,6 0,8 1,0
TOD
CA
SILALF PR
SB
RI
A. discoides
Coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9 1,1
TOD
CA
SIPRSB
LALFRI
A. vulgaris
Coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9 1,1
TOD
CA
SIPRSB
LALFRI
Centropyxis aculeata
CoerênciaN
úmer
o de
am
bien
tes
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9 1,1
CA
TOD
SIPRSB
LALFRI
C. ecornis
Coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9
TOD
CA
SIPRSB
LALFRI
Difflugia gramen
Coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9 1,1
TOD
CA
LARIPR SI
SB
LF
D. pseudogramen
Coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9
TOD
CA
PRSISBRI
LA
LF
Cucurbitella dentata
Coerência
Núm
ero
de a
mbi
ente
s
0
2
4
6
8
10
12
14
-1,1 -0,9 -0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9
TOD
CA
SIPRBA
RILALF
Figura 13. Coerência temporal das principais espécies de tecamebas em todos os ambientes estudados da planície de inundação e em suas subunidades.
Os resultados das correlações de Spearman evidenciaram que Difflugia
pseudogramen apresentou os valores mais altos do coeficiente e a maior quantidade (19) de
correlações significativas (Tabela 3). Os ambientes correlacionados com maiores valores de
R foram o canal Ipoitã com a lagoa Ventura (0,846) e a lagoa dos Patos com a lagoa
Guaraná (0,840). Na primeira correlação, os ambientes são pertencentes ao sistema
26

Ivinheima, e na segunda, pertencentes a sistemas diferentes, porém com características
hidrodinâmicas semelhantes (lagoas abertas).
Tabela 3: Correlações significativas (p<0,05) de Spearman (R) para as principais espécies de tecamebas nos diferentes ambientes da planície de inundação do alto rio Paraná (Par=Rio Paraná; Ivi= R. Ivinheima; Ba=R. Baía; Ipo= Canal Ipoitã; Cur= C.Curutuba; Pau= Ressaco do Pau Véio; Gar= Lagoa Garças; Pat=L.Patos; Gua=L.Guaraná; Osm=L.Osmar; Vem=L.Ventura; Fec=L.Fechada).
Arcella conica
A discoides
A. vulgaris
Centropyxis
C. ecornis
Difflugia
D. pseudogram
C. dentata
Par x Ivi 0,571 - - - - - - - Par x Ba - - - 0,566 - - - - Par x Ipo - - - - - - 0,735 0,538 Par x Pau - - - - - - 0,638 - Par x Gar 0,763 - 0,592 - - - - - Par x Osm 0,633 - - - - - - - Par x Fec - - - 0,535 - - - - Par x Gua - - - - - - 0,565 - Par x Ven - - - - - - 0,575 - Ivi x Cur - 0,534 - - 0,541 - 0,629 0,536 Ivi x Ba - - 0,769 - 0,572 - 0,672 - Ivi x Pat - - - - - - 0,698 - Ivi x Gua - - - - - - 0,637 - Ba x Cur - - - - 0,568 0,568 - - Ba x Ipo - - - - - - 0,674 - Ba x Gar - - - 0,670 - - - 0,667 Ba x Vent - - - 0,648 - - - - Ba x Fec - - - 0,590 - - 0,566 0,607 Ba x Pat - - - - - - 0,717 - Ba x Gua - - - - - - 0,724 - Ipo x Cur - - - 0,643 - - - - Ipo x Fec - - - - 0,534 0,534 - - Ipo x Gua 0,539 - - - - - 0,704 - Ipo x Osm 0,567 - - - - - - - Ipo x Pat - 0,588 - - - - 0,593 - Ipo x Ven - - - - - - 0,846 - Pau x Gua - - - - 0,599 0,599 - 0,593 Cur x Osm 0,563 - - - - - - - Cur x Fec - 0,587 - 0,595 - - - - Cur x Gua - - 0,707 - - - - Cur x Pat - - - - 0,692 0,692 - - Pat x Gua - - - - 0,731 0,731 0,840 - Pat x Ven - - - - 0,553 0,553 0,674 - Vent x Osm - - - - - - 0,604 - Ven x Fec 0,574 - - 0,571 0,571 - - Gar x Fec - - - 0,706 - - - - Gua x Vent - - - - - - 0,670 -
27

Difflugia pseudogramen também apresentou os maiores valores para a correlação
de Pearson (Tabela 4). Maiores valores foram registrados para ambientes pertencentes a
sistemas diferentes, ou seja, ambientes distantes e, em alguns casos, com hidrodinâmicas
diferentes. Essa análise evidenciou a correlação significativa, para Centropyxis ecornis
(0,839), entre os canais Ipoitã e Curutuba, identificada anteriormente pelo coeficiente de
correlação intraclasse, demonstrando a natureza linear dessa correlação.
Tabela 4: Correlações significativas (p<0,05) de Pearson (r) para as principais espécies de tecamebas nos diferentes ambientes da planície de inundação do alto rio Paraná (Par=Rio Paraná; Ivi= R. Ivinheima; Ba=R. Baía; Ipo= Canal Ipoitã; Cur= C.Curutuba; Pau= Ressaco do Pau Véio; Gar= Lagoa Garças; Pat=L.Patos; Gua=L.Guaraná; Osm=L.Osmar; Vem=L.Ventura; Fec=L.Fechada).
Arcella conica
A discoides
A. vulgaris
Centropyxis aculeata
C. ecornis
Difflugia gramen
D. pseudogramen
C. dentata
Par x Ivi - - - - - - - 0,765 Par x Ba - - - 0,545 - - - - Par x Ipo - - - 0,794 - - - 0,955 Par x Pau - 0,861 - - - - - - Par x Gar 0,700 - - - - - - - Par x Osm 0,710 - - - 0,538 - - - Par x Fec - - - 0,722 - - - - Par x Gua - - - - - 0,973 - - Par x Pat - - 0,787 0,679 - - - - Par x Cur - - - 0,845 - - - - Ivi x ipo - - - - - - - 0,827 Ivi x Cur - - - - - - 0,836 - Ivi x Baia 0,770 0,885 - - - - - - Ivi x Gar 0,811 - - - - - - - Ivi x Osm 0,564 - - - - - - - Ivi x Fec 0,732 - - - - - - - Ivi x Ven 0,761 - - - - - - - Ba x Cur - - - - - 0,993 - 0,732 Ba x Ipo - - - - - - 0,983 - Ba x Pau - - - - - - - 0,755 Ba x Gar 0,679 - - 0,739 - - 0,961 0,842 Ba x Ven 0,976 - - - - 0,965 0,983 - Ba x Fec 0,993 - - - - 0,975 0,992 - Ba x Pat - - - - - 0,970 - 0,723 Ba x Gua 0,640 - - - - - - 0,794 Ba x Osm - - 0,712 - - - 0,983 - Ipo x Cur - - - 0,942 0,839 - - - Ipo x Fec - - - 0,666 - - 0,997 - Ipo x Pat - 0,796 - - - - - - Ipo x Vem - - - - 0,793 - - - Ipo x Gar - - 0,900 - - - 0,945 - Cur x Vent - - - 0,604 0,882 0,972 - - Cur x Pau - - 0,655 - - - - 0,984 Cur x Gar - - - - - - - 0,974 Cur x Fec - - - 0,723 - 0,984 - - Cur x Gua - - 0,634 - - - 0,688 0,982 Cur x Pat - - - - - 0,984 - 0,914 Pau x Gar - - - - - - - 0,978 Pau x Pat - - - - - - - 0,952 Pau x Gua - - 0,590 - - - - 0,997 Pat x Gua - - - 0,599 - - - - Pat x Ven - - - - - 0,961 - - Pat x Fec - - - 0,704 - 0,969 - - Gar x Pat - - - - - - - 0,925 Gar x Gua - - - - - - 0,682 0,989 Gar x Fec 0,630 - - - - - 0,946 - Gar x Osm 0,860 - - - - - 0,945 -
28
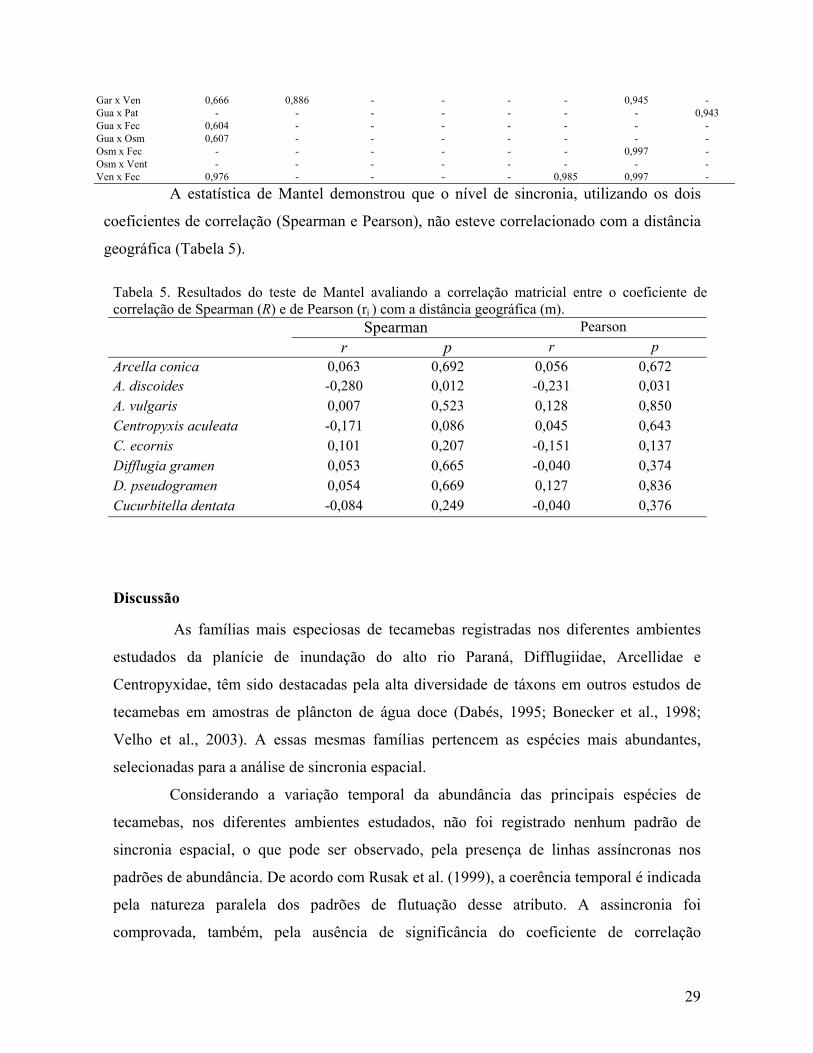
Gar x Ven 0,666 0,886 - - - - 0,945 - Gua x Pat - - - - - - - 0,943 Gua x Fec 0,604 - - - - - - - Gua x Osm 0,607 - - - - - - - Osm x Fec - - - - - - 0,997 - Osm x Vent - - - - - - - - Ven x Fec 0,976 - - - - 0,985 0,997 -
A estatística de Mantel demonstrou que o nível de sincronia, utilizando os dois
coeficientes de correlação (Spearman e Pearson), não esteve correlacionado com a distância
geográfica (Tabela 5).
Tabela 5. Resultados do teste de Mantel avaliando a correlação matricial entre o coeficiente de correlação de Spearman (R) e de Pearson (ri ) com a distância geográfica (m). Spearman Pearson r p r p Arcella conica 0,063 0,692 0,056 0,672 A. discoides -0,280 0,012 -0,231 0,031 A. vulgaris 0,007 0,523 0,128 0,850 Centropyxis aculeata -0,171 0,086 0,045 0,643 C. ecornis 0,101 0,207 -0,151 0,137 Difflugia gramen 0,053 0,665 -0,040 0,374 D. pseudogramen 0,054 0,669 0,127 0,836 Cucurbitella dentata -0,084 0,249 -0,040 0,376
Discussão
As famílias mais especiosas de tecamebas registradas nos diferentes ambientes
estudados da planície de inundação do alto rio Paraná, Difflugiidae, Arcellidae e
Centropyxidae, têm sido destacadas pela alta diversidade de táxons em outros estudos de
tecamebas em amostras de plâncton de água doce (Dabés, 1995; Bonecker et al., 1998;
Velho et al., 2003). A essas mesmas famílias pertencem as espécies mais abundantes,
selecionadas para a análise de sincronia espacial.
Considerando a variação temporal da abundância das principais espécies de
tecamebas, nos diferentes ambientes estudados, não foi registrado nenhum padrão de
sincronia espacial, o que pode ser observado, pela presença de linhas assíncronas nos
padrões de abundância. De acordo com Rusak et al. (1999), a coerência temporal é indicada
pela natureza paralela dos padrões de flutuação desse atributo. A assincronia foi
comprovada, também, pela ausência de significância do coeficiente de correlação
29

intraclasse, estimado para os ambientes analisados. Assim, pode-se afirmar que não houve a
ação de um evento regional capaz de atuar nos diferentes ambientes da planície, de forma
que os mesmos apresentassem características homogêneas e induzissem as populações de
tecamebas flutuarem sincronicamente. Takahashi (2004) também não encontrou padrões
congruentes para a flutuação populacional de cladóceros nessa mesma planície.
Um fator atuante em escala espacial implica na similaridade da dinâmica, dentro
da área de influência (Bjornstad et al., 1999). Nesse sentido, esperava-se que, em ambientes
próximos, as tecamebas possuiriam padrões congruentes de flutuação temporal. Contudo,
analisando o coeficiente de correlação intraclasse para ambientes próximos associados ao
mesmo rio, bem como os resultados da estatística de Mantel, pode-se inferir que a
proximidade não influenciou os padrões de flutuação da abundância desses organismos.
Dessa forma, os padrões coerentes de flutuação da abundância, identificados através dos
coeficientes de correlação de Spearman e Pearson, para ambientes pertencentes a sistemas
diferentes, e em alguns casos, à categorias diferentes, foram, provavelmente, determinados
pela associação de fatores regionais influenciando os atributos químicos e físicos e
interações biológicas (fatores locais). Entretanto, alguns trabalhos têm demonstrado que a
sincronia espacial diminuiu significativamente com o aumento da distância entre os locais,
como para algumas espécies de insetos (Hanski & Woiwod, 1993), aves (Ranta et al., 1995)
e peixes (Domingues et al., 2005).
Os resultados da análise do coeficiente de correlação intraclasse, considerando
ambientes pertencentes a mesma categoria (rio, canal, lagoa aberta ou lagoa fechada),
evidenciaram valor significativo apenas para Centropyxis ecornis, nos diferentes canais.
Essa sincronia sugere que uma parte da variação temporal da abundância foi produzida por
fatores operando em uma escala maior que a local (Rusak et al., 1999), ou seja, as
características próprias dos canais, como a morfometria (profundidade, área e “fetch”),
associadas a fatores regionais, como a alteração do nível fluviométrico, determinaram o
padrão sincrônico registrado para as populações de C. ecornis. Dessa forma, pode-se
sugerir que apenas o tipo de ambiente, com características hidrodinâmicas semelhantes, não
funciona como regulador da flutuação da abundância de tecamebas.
Para existir sincronia espacial, conforme proposto pelo “efeito Moran”, os fatores
extrínsecos (regionais) deveriam estar correlacionados e as populações precisariam
30

apresentar a mesma estrutura intrínseca (predação e competição) (Moran, 1953). Em
regiões tropicais, a alteração do nível fluviométrico atua como principal fator regional
responsável pela variação temporal dos fatores físicos, químicos e bióticos em sistemas
rios-planície de inundação. Entretanto, a temperatura, a ação do vento, a radiação
subaquática e a precipitação pluviométrica também afetam localmente os hábitats aquáticos
e transicionais desses ambientes (Tundisi, 1983; Thomaz et al., 1992).
Embora os resultados desse estudo não corroborem o pressuposto de sincronia
populacional de tecamebas em todos ambientes da planície de inundação, bem como em
suas subunidades (em sistemas e categorias semelhantes), esses mesmos resultados
corroboram alguns padrões encontrados por outros autores, nessa mesma planície, os quais
evidenciaram que os fatores locais, como mudanças morfométricas e hidrodinâmicas
diferenciadas, são mais importantes que a variabilidade temporal na determinação dos
padrões de abundância desse grupo de protozoários, refutando a importância do pulso de
inundação, considerado como fator preponderante na determinação de vários processos e
padrões ecológicos em sistemas de planície de inundação (Lansac-Tôha et al. 1997, 2004;
Velho et al., 2003), visto que os dados registraram a predominância de fatores locais
atuando sobre a dinâmica de tecamebas.
Deve-se considerar, no entanto, que a duração e o período de início de águas altas
variaram consideravelmente entre os anos de 2000 e 2003. Essa variação está
provavelmente relacionada à construção de barragens, como a de Porto Primavera,
localizada a montante da área de estudo. A operação das barragens tem atuado na redução
da amplitude de variação dos níveis hidrométricos do alto rio Paraná, o que pode estar
reduzindo a influência desses eventos sobre a dinâmica das comunidades aquáticas na
planície de inundação do alto rio Paraná (Agostinho & Zalewsky, 1996; Souza Filho &
Stevaux, 2004). Além disso, os eventos regionais podem agir de forma distinta dependendo
das características locais, considerando a heterogeneidade espacial da planície, em que a
morfometria, grau de comunicação e hidrodinâmica conferem, a cada um dos hábitats,
características limnológicas próprias, que interferem na presença e distribuição dos
organismos (Thomaz et al., 1997). Webster et al. (2000), analisando a dinâmica temporal de
variáveis limnológicas, concluíram que a sincronia tende a ser maior em lagos com
hidrologia relativamente simples.
31

Populações que não apresentam dinâmicas correlacionadas necessitam ser
monitoradas em um número maior de ambientes, visto que diferenças locais são
preponderantes. Caso tivesse sido registrada a sincronia espacial para a flutuação da
abundância de tecamebas, os resultados obtidos de um único ambiente poderiam ser
extrapolados para outros ambientes pertencentes a mesma planície, de modo a usá-lo como
site sentinela, acarretando em uma redução do esforço de amostragem.
Dessa forma, os resultados não permitiram aceitar a hipótese predita, considerando
a ausência de sincronia registrada para as flutuações de abundância das principais espécies
de tecamebas, quando analisados todos os ambientes estudados da planície, bem como,
quando comparados os ambientes próximos e, ainda, ao se analisar as diferentes categorias,
com exceção do padrão coerente registrado para Centropyxis ecornis, para os diferentes
canais. Desse modo, pode-se concluir que, na planície de inundação do alto rio Paraná, os
fatores regionais não são preponderantes na determinação da dinâmica populacional de
tecamebas.
Referências
Agostinho, A. A. & M. Zalewski, 1996. A planície alagável do alto rio Paraná: importância e preservação. Eduem, Maringá, 100pp.
Andrewartha, H. G. & L. C. Birch, 1954. The distribution and abundance of animals. University of Chicago Press, Chicago.
Bini, L. M., L. F. M. Velho & F. A. Lansac-Tôha, 2003. The effect of connectivity on the relationship between local and regional species richness of testate amoebae (Protozoa, Rhizopoda) in floodplain lagoons of the upper Paraná river, Brazil. Acta Oecologica 24:145-151.
Bjornstad, O. N., C. N. Stenseth & T. Saitoh, 1999. Synchrony and scaling in dynamics of voles and mice in Northern Japan. Ecological Society of America. Ecology 80(2): 622-637.
Bonecker, C. C., F. A. Lansac-Tôha & L. M. Bini, 1998. Composition of zooplanckton in different environments of Mato Grosso Pantanal, Mato Grosso, Brazil. In: Anais do VIII Seminário Regional de Ecologia. Universidade Federal de São Carlos, São carlos, 1123-1135.
Bottrel, H. H., A. Duncan, Z. Gliwicz, E. Grygierek, A. Herzig, A. Hillbricht-Illkowska, H. Kurasawa, P. Larsson & T. Weglenska, 1976. A review of some problems in zooplankton production studies. Norwegian Journal of Zoology 24: 419-456.
Cottenie, K., N. Nuytten, E. Michels & L. De Meester, 2001. Zooplankton community structure and environmental conditions in a set of interconnected ponds. Hydrobiologia 442: 339-350.
32

Dabés, M. B. G. S., 1995. Composição e descrição do zooplâncton de 5 (cinco) lagoas marginais do rio São Francisco, Pirapora, Três Marias, Minas Gerais-Brasil. Revista Brasileira de Biologia 55: 831-845.
Deflandre, G., 1928. Le genre Arcella Ehrenberg. Archiv für Protistenkuned 64: 152-287. Deflandre, G., 1929. Le genre Centropyxis Stein. Archiv für Protistenkuned 67: 322-375. Domingues, W.M., L. M. Bini & A. A. Agostinho, 2005. Spatial synchony of a highly
endemic fish assemblage (Segredo reservoir, Iguacu river, Parana state, Brazil. Brazilian Journal of Biology 65(3): 439-449.
Gauthier-Lievre, L. & R. Thomas, 1958. Le genre Difflugia, Pentagonia, Maghrebia et Hoogenraadia (Rhizopodes Testacès) en Afrique. Archiv für Protistenkuned 103: 1-370.
Gauthier-Lievre, L. & R. Thomas, 1960. Le genres Cucurbitella Pènard. Archiv für Protistenkuned 104(4): 569-60.
Hanski, I. & I. P. Woiwod, 1993. Spatial synchrony in the dynamics of moth and aphid populations. Journal of Animal Ecology 62: 656-668.
Hardoim, E. L., 1997. Taxonomia e ecologia de Testacea (Protista, Rhizopoda) do Pantanal do Poconé – rio Bento Gomes e vazante Birici, Mato Grosso, Brasil. Tese de Doutorado. São Carlos. Universidade Federal de São Carlos, São Carlos.
Heino, M., V. Kaitala, E. Ranta & J. Lindströn, 1997. Synchronous dynamics and rates of extinction in spatially structured populations. Proceedings of the Royal Society of London Series B-Biological Sciences 264: 481-486.
Junk, W. J., P. B. Bayley & R. E. Sparks, 1989. The flood pulse concept in river-floodplain systems. Canadian Special Publication of Fisheries and Aquatic Sciences 106: 110-127.
Kendall, E. B., O. N. Bjornstad, J. Bascompte, T. H. Keitt & W. F. Fagan, 2000. Dispersal, environmental correlation and spatial synchrony in population dynamics. American Naturalist 155(5): 628-636.
Kratz, T. K., P. A. Soranno, & J. J. Magnuson, 1987. Inferences from spatial and temporal variability in ecosystems: long term data from lakes. American Naturalist 129: 830-846.
Lansac-Tôha, F. A., C. C. Bonecker,. L. F. M. Velho &. A. F Lima, 1997. Composição, distribuição e abundância da comunidade zooplanctônica. In: Vazzoler, A. E. A.M., A. A., Agostinho & N. S Hahn (ed.). A planície de inundação do alto rio Paraná: Aspectos físicos, biológicos e socioeconômicos, Eduem, Maringá, 117-155.
Lansac-Tôha, F. A., C. C. Bonecker & L. F. M. Velho, 2004. Composition, species richness and abundance of the zooplankton community. In Thomaz, S. M., A. A. Agostinho & N. S. Hahn (eds), The Upper Paraná River and its floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden: 145-190.
Lena, H.; & S. J. Zaindenwerg, 1975. Tecamebas del delta del Paraná (Argentina). Revista Española de Micropaleontologia 7(3): 519-537.
Moran, P. A. P., 1953. The statistical analysis of the Canadian lynx cycle. II. Synchronization and meteorology. Australian Journal of Zoology 1: 291-298.
Neiff, J. J., 1990. Ideas para la interpretación ecologica del Paraná. Interciencia 15(6): 424-441.
Nicholson, A. J., 1933. The balance of animal populations. Journal of Animal Ecology 2: 132-178.
33

Ogden, C. G.; & R. H. Hedley, 1980. An atlas of freshwater testate amoebae. University Press, Oxford: 222pp.
Ranta E., V Kaitala, J Lindström & H. Lindén, 1995. Synchrony in tetraonid population dynamics. Journal of Animal Ecology 64: 767-776.
Ranta, E., V. Kaitala, J. Lindströn & H. Lindén, 1999. Spatially autocorrelated disturbances and patterns in population synchrony. Proceedings of the Royal Society of London Series B-Biological Sciences 266: 1851-1856.
Rohlf, F. J. 1989. NTSYS: numerical taxonomy and multivariate analysis system. Exeter Softwares, New York.
Royama, T., 1992. Analytical population dynamics, Chapman & Hall, London. Souza Filho, E. E., P. C. Rocha, E. Comunello & J. C. Stevaux, 2004. Effects of the Porto
Primavera Dam on physical environment of the downstream floodplain. In Thomaz, S. M., A. A. Agostinho & N. S. Hahn (eds), The Upper Paraná River and its floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden: 55-74.
Rusak, J. A., N. D. Yan, K. M. Somers & D. J. McQueen, 1999. The temporal coherence of zooplankton population abundances in neighboring north-temperate lakes. American Naturalist 153: 46-58.
Shanker, K. & R. Sukumar, 1999. Synchrony in small mammmal populations of montante forest patches in southern India. Journal of Animal Ecology 68: 50-59.
Statsoft Inc., 1996. Tulsa: Statistica. 3v. Takahashi, E. M., 2004.Flutuações populacionais de cladóceros planctônicos na planície de
inundação do alto rio Paraná (PR/MS).. Dissertação de Mestrado, Universidade Estadual de Maringá, UEM: Maringá.
Thomaz, S. M., M. C. Roberto, F .A. Lansac-Tôha, F. A. Esteves & A. F. Lima, 1991. Dinâmica temporal dos principais fatores limnológicos do rio Baía - planície de inundação do alto rio Paraná, MS, Brasil. Revista Unimar 13: 299-312.
Thomaz, S. M., M. C. Roberto, F .A. Lansac-Tôha, A. F. Lima &, F. A. Esteves, 1992. Característica limnológicas de uma estação de amostragem do alto rio Paraná e outra do baixo rio Ivinheima- (PR, MS-Brasil). Acta Limnologica Brasiliensia 4: 32-51.
Thomaz, S. M, M. C. Roberto & L. M. Bini, 1997. Caracterização Limnológica dos ambientes aquáticos e influência dos níveis fluviométricos. In: Vazzoler, A. E. A.M., A. A., Agostinho & N. S Hahn (ed.). A planície de inundação do alto rio Paraná: Aspectos físicos, biológicos e socioeconômicos, Eduem, Maringá, 73-102.
Tundisi, J. G. 1983. A review of basic ecological process interacting with production and stading-stock of phytoplankton in lakes and reservoir in Brazil. Hydrobiologia 100: 223-243.
Velho, L. F. M. & F.A Lansac-Tôha, 1996. Testate amoebae (Rhizopodea-Sarcodina) from zooplankton of the high Paraná river floodplain, State of Mato Grosso do Sul, Brazil: II. Family Difflugidae. Studies Neotropical Fauna and Environment 31: 179-192.
Velho, L. F. M., F.A Lansac-Tôha & M. Serafim-Junior, 1996. Testate amoebae (Rhizopodea- Sarcodina) from zooplankton of the high Paraná river floodplain, State of Mato Grosso do Sul, Brazil. I. Families Arcellidae and Centropyxidae. Studies Neotropical Fauna and Environment 31: 135-150.
Velho, L. F. M., F.A Lansac-Tôha & L. M. Bini, 1999. Spatial and temporal variation in densities of testate amoebae in the plankton of the upper Paraná river. Hydrobiologia 411: 103-113.
34

Velho, L. F. M., F. A Lansac-Tôha & L. M. Bini, 2003. Influence of environmental heterogeneity on the structure of testate amoebae (Protozoa, Rhizopoda) assemblages in the plankton of the upper Paraná river floodplain, Brazil. International Review of Hydrobiology 88: 154-166.
Velho, L. F. M., L. M. Bini & F.A Lansac-Tôha, 2004. Testate amoebae (Rhizopodea-Sarcodina) diversity in plankton of the upper Paraná River floodplain, Brazil. Hydrobiologia 523: 103-111.
Vucetich, M. C., 1973. Estudio de tecamebianos argentinos, en especial los del dominio pampasico. Revista del Museu de La Plata, serie Zoologia 11(108): 287-332.
Webster, K. E., P. A. Soranno, S. B. Baines, T. K. Kratz, C. J. Dillon, P. Campbell, E. J. Fee & R. E. Hecky, 2000. Structuring features of lake districts: landscape controls on lake chemical responses to drought. Freshwater Biology 42: 499-515.
35

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo