estudo do efeito de um dispositivo intravaginal de liberao ... · o propósito deste estudo foi...
TRANSCRIPT

1
1-INTRODUÇÃO
Com o uso cada vez mais freqüente da inseminação artificial e da
transferência de embriões na espécie eqüina se faz necessário recorrer ao controle
do ciclo estral. Assim, quando se usa sêmen refrigerado, cuja capacidade
fecundante se limita a dois ou três dias, é altamente desejável que se disponha de
um lote de fêmeas com estro programado e coordenado com a chegada do sêmen
com o objetivo de aumentar a eficiência. Em se tratando do uso do sêmen
criopreservado, o controle do estro e da ovulação é crítico, visto que a inseminação
deve proceder poucas horas antes da ovulação, devido ao fato do sêmen submetido
a esse processo ter uma sobrevida de 6 a 12 horas no trato reprodutivo da fêmea.
Fica evidente a existência de uma grande lacuna na disponibilidade de uma
terapia progestacional eficaz, mais segura e de menor custo, tornando o
desenvolvimento de uma nova intervenção terapêutica na manipulação dos eventos
reprodutivos da égua uma necessidade na criação de eqüinos.
O propósito deste estudo foi justamente o de preencher esta lacuna,
buscando oferecer aos técnicos e criadores um novo protocolo de sincronização de
estros e ovulações em éguas que reúna eficiência, facilidade de execução e custo
acessível.
Objetivou-se, com este estudo, melhorar a eficiência reprodutiva nos
programas de inseminação artificial ou por monta natural otimizando o uso de

2
garanhões com menor número de coberturas e também reduzindo o risco de
instalação de processos infecciosos uterinos delas decorrentes.
Outro fator que serviu de grande motivação à condução deste estudo foi
justamente o fato de não haver sido encontrada na literatura consultada nenhuma
publicação científica brasileira que tenha tido como tema o uso de dispositivos
intravaginais com progesterona em éguas, somente tendo sido realizados trabalhos
com progesterona sob a forma oral e de implantes subcutâneos. Através deste
experimento procurou-se determinar como éguas de uma raça genuinamente
nacional, como a Mangalarga Marchador, iriam se comportar, sob condições
tropicais, ao uso deste dispositivo e qual o grau de eficiência que seria obtido com
um protocolo de sincronização de estros e ovulações que utilize este recurso
terapêutico.
Este estudo teve como objetivo principal testar dois protocolos de
sincronização de estros e ovulações em éguas com a utilização de um dispositivo
intravaginal de liberação controlada de progesterona (CIDR®, Pharmacia Brasil;
Pfizer) associado a outras drogas: benzoato de estradiol, prostaglandina e hCG
(gonadotrofina coriônica humana).
Este método possui várias vantagens em relação ao consagrado
universalmente altrenogest (Regumate®, Intervet, USA) utilizado com esta finalidade
em éguas. Facilidade de aplicação com economia de mão-de-obra e diminuição do
estresse dos animais, além da grande diferença que separa os custos de ambos os
produtos são algumas, entre outras vantagens, que o uso do dispositivo oferece.
Os custos das fontes de progesterona nos diferentes protocolos representam
os seguintes valores em dólares americanos (cotação em 02/2005): 1- Progesterona
repositol oleosa injetável (200mg/dia) por 10 dias: U$ 180,00 (fonte: agri-med, 2005);
2- Altrenogest (Regumate®) 8 ml/dia por 14 dias: U$ 40,00 (fonte: agri-med, 2005) e
3- CIDR® : U$ 12,00 (fonte: Pfizer, 2005) sendo que o dispositivo em bovinos pode
ser reutilizado por 2 vezes, o que diminuiria o custo para U$ 4,00. Não existem
estudos em éguas que permitam determinar a eficiência do dispositivo reutilizado
nos protocolos de sincronização, motivo pelo qual utilizamos o dispositivo uma única
vez.
O objetivo da determinação do estágio do ciclo estral (estro ou diestro) em
que os animais se encontravam no início do estudo foi o de se avaliar se haveria

3
alguma influência nos parâmetros de tempo de aparecimento de estro, tempo de
detecção de um folículo de 35mm e taxa de fertilidade nas diferentes fases do ciclo
estral.
O objetivo do acompanhamento reprodutivo por meio da ultra-sonografia foi
observar as alterações morfológicas do útero e ovários que viessem a ser
desencadeadas por influência da progesterona liberada pelo CIDR® e para realizar o
diagnóstico de gestação com o intuito de avaliar a interferência do CIDR® sobre a
fertilidade quando comparado com um grupo controle, que recebeu o dispositivo
isento de progesterona.
O propósito de se comparar o Lutalyse® (dinoprost) com o Vetecor® (hCG) foi
o de procurar diminuir custos, tornando o protocolo proposto o mais viável
economicamente, visto que o valor de uma dose de hCG pode chegar a custar de 10
a 15 vezes mais do que a de prostaglandina, dependendo do produto comercial a
ser utilizado.
O estudo das possíveis alterações nos parâmetros do hemograma: série
branca, série vermelha e plaquetas objetivou identificar a influência do CIDR® sobre
o metabolismo dos animais submetidos a esta terapia.

4
2 - REVISÃO BIBLIOGRÁFICA
2.1 - Progesterona e suas funções biológicas na égua
A progesterona é o progestágeno natural de maior prevalência, sendo
secretada pelas células luteínicas do corpo lúteo, pela placenta e pelas glândulas
adrenais. A progesterona, assim como os andrógenos e os estrógenos, é
transportada na corrente circulatória por uma proteína de ligação. Sua secreção é
estimulada primariamente pelo LH (Hafez, 2004).
Os efeitos normais da progesterona são vistos apenas se o tecido-alvo tiver
sido anteriormente submetido a um período de estimulação pelo estrógeno. Esta
sensibilização pelo estrógeno leva a um efeito sinérgico. A progesterona age no
útero provocando quietude no miométrio e secreção do leite uterino pelas glândulas
endometriais. As glândulas uterinas aumentam em profundidade, mais
especificamente em ramificações e tortuosidades. A espiralização das glândulas
aumenta sua capacidade funcional, depois que o desenvolvimento do ducto reto
ocorreu sob a influência do estrógeno (Mckinnon et al.., 1997).
Doses elevadas de progesterona inibem a produção de gonadotrofinas pela
glândula hipófise. Em alguns animais em estro isto pode funcionar como regulador
do período de diestro, porque logo que o corpo lúteo pára de secretar progesterona,

5
segue-se súbita secreção de FSH, que causa desenvolvimento de folículos pró-estro
(Mckinnon et al.., 1997).
A meia-vida fisiológica da progesterona é de apenas 22 a 36 minutos na
égua. Isto significa que uma secreção constante é essencial para manter os níveis
séricos adequados. O transporte dos estrógenos, particularmente o estradiol, e
progestágenos, particularmente a progesterona, no plasma é semelhante. Ambos
são fracamente, mas completamente ligados à albumina. O excesso de
progesterona é fortemente ligado à transcortina. Quando os esteróides sexuais
atingem as células alvo são liberados das proteínas transportadoras e penetram no
interior destas, onde atuam. Quando a síntese protéica é induzida, as células alvo
funcionam. Por exemplo, uma célula glandular pode secretar; a mitose pode ser
estimulada em célula epitelial; o estro psíquico pode ser induzido se a célula alvo for
um neurônio particular; ou a hipertrofia pode ser induzida em célula do miométrio
(Mckinnon et al.., 1997).
A progesterona desempenha as seguintes funções: prepara o endométrio
para a implantação e manutenção da prenhez, aumentando a atividade secretora
das glândulas do endométrio e inibindo a motilidade do miométrio; atua
sinergisticamente com os estrógenos na indução do comportamento de estro; auxilia
no desenvolvimento do tecido secretor (alvéolos) da glândula mamária; provoca
inibição do estro e do pico pré-ovulatório do LH quando em níveis elevados,
desempenhando, portanto, papel fundamental na regulação do ciclo estral (Hafez,
2004).
2.2 - Utilização terapêutica da progesterona e progestágenos na égua
Atualmente, a maior parte das terapias com progestágenos requer uma
administração diária, dificultando o seu emprego no manejo dos haras e onerando o
custo do tratamento, além de poder ocasionar efeitos nocivos devido às altas
concentrações de progesterona desnecessárias na delicada fisiologia reprodutiva
das éguas. Isto pode ocorrer, uma vez que a dose diária ultrapassa, nas primeiras
horas, os níveis terapêuticos requeridos para manter o efeito clínico desejado até a
próxima dose (Dumitriu e Dimitriu, 1994). A terapia com progesterona e progestágenos tem uma enorme
aplicabilidade no manejo reprodutivo da égua, atuando na: 1) regulação do estro em

6
éguas no período de transição estacional, isto é, éguas que se encontram no terço
final do período de anestro estacional. Esta intervenção é de extrema importância
na criação da raça PSI (puro sangue inglês), já que o início da estação de monta
oficial (dia 15 de julho) começa muito antes da estação de monta fisiológica da
espécie, impondo, portanto, a antecipação dos eventos reprodutivos destas éguas
devido à vantagem futura do nascimento antecipado de seus produtos no calendário
esportivo; 2) regulação do estro em éguas cíclicas não-lactentes visando a um
melhor manejo na cobertura, segundo a disponibilidade do garanhão, e na
sincronização de estros para programas de inseminação artificial e transferência de
embriões; 3) regulação de estro em éguas cíclicas lactentes visando à postergação
do estro do potro para favorecer a limpeza fisiológica e a involução uterina e,
conseqüentemente, aumentar a taxa de concepção da primeira ovulação pós-parto;
4) supressão do comportamento de estro por tempo prolongado em éguas
participantes de eventos hípicos; 5) na manutenção de prenhez de éguas com corpo
lúteo pouco funcional ou com luteólise prematura e 6) como coadjuvante no
tratamento de retenção de placenta e involução uterina (McKinnon,1996). Outras indicações terapêuticas da progesterona na égua incluem anestro
causado por estresse por mudanças de manejo, principalmente em éguas virgens,
lactação, etc. Convém ressaltar que, antes de se pensar em utilizar a progesterona
como terapêutica, deve-se realizar um diagnóstico diferencial com condições que
também levam à aciclia, porém sem ter como indicação a progesterona, tais como
deficiências nutricionais, doenças debilitantes, corpo lúteo persistente, entre outros
(Hafez, 2004).
A progesterona pode ser utilizada para tratamento de animais na condição
de anestro lactacional, devendo, porém, esta condição ser distinguida daquela
observada em situações onde ocorre um corpo lúteo persistente, geralmente mais
comum do que a primeira condição (Ginther,1992).
Existem várias formas de apresentação da progesterona no mercado. A
preparação progesterona Repositol age através da liberação de hormônio de uma
base de propilenoglicol na circulação sangüínea. Esta preparação deve ser utilizada
numa dose de pelo menos 1000 mg a cada 4 a 7 dias ou 2000 mg a cada 7 dias para
manter a concentração de progesterona acima de 1 ng/ml. Devido às reações
teciduais indesejáveis, esta é a via menos recomendada para uso em éguas, não
estando mais disponível no mercado (Neely, 1988).

7
Um progestágeno sintético, o Altrenogest (ReguMate®), foi desenvolvido
para controlar o estro na égua, sendo o único progestágeno ativo via oral eficaz
nesta espécie, embora tenha sido citado o uso de outros, como o acetato de
melengestrol (MGA – Ovaban®). Quando o MGA foi administrado na dieta, numa
dosagem de 10 ou 20 mg/dia por 9 a 15 dias, falhou na inibição do estro e ovulação
(Loy e Swan, 1966). O acetato de melengestrol (MGA), administrado na dose de 20
mg, foi ineficaz na prevenção do estro (Loy e Swan, 1966). Neely (1979) selecionou
quatro éguas velhas com histórico de morte embrionária e fraco tônus uterino aos 18
a 19 dias pós-cobertura. Destas, duas foram submetidas a uma dose de 0,4 mg de
MGA duas vezes ao dia, considerada eficaz na inibição do estro e ovulação em
vacas (Zimbelman e Smith, 1966), enquanto as restantes serviram de controle. O
tratamento durou quatro meses a partir da cobertura. As éguas tratadas
permaneceram prenhes enquanto uma égua controle reabsorveu o embrião e a
outra permaneceu em gestação. Neely (1988) verificou que altas doses (>100 mg)
de MGA podem ser mais eficazes em suprimir o estro.
O acetato de clormadinona (CAP), embora ainda não disponível no Brasil,
tem sido utilizado na alimentação de éguas na Europa. Infelizmente, o CAP na
ração, numa dosagem de 10 mg/dia, falhou em inibir o estro e a ovulação (Arthur e
Allen, 1972).
Um outro composto progesterônico utilizado na Europa é a proligestona
(Delvosterona), um progestágeno injetável de longa duração que não apresentou
resultados satisfatórios em estudos realizados por Neely (1979).
O acetato de medroxiprogesterona (MAP) (Depo-provera®) tem sido
sugerido como eficaz no tratamento da manutenção de prenhez. Este é,
supostamente, um progestágeno injetável de longa ação administrado a cada 8 a 14
dias numa dose de 200 a 250 mg. No entanto, não foi conduzido um estudo
controlado de avaliação da eficiência deste progestágeno na espécie eqüina. Neely
(1974) administrou 250 mg de MAP em uma égua ovariectomizada de 410 kg que,
previamente, recebeu 2 mg de cipionato de estradiol (ECP®) durante 45 dias para
mantê-la em estado de estro. Foi determinada a resposta à indução diária de estro e
as concentrações de progesterona aos 14 dias posteriores à aplicação do MAP.
Obtiveram-se amostras para biópsia endometrial antes da administração do MAP e
aos 4, 7 e 14 dias após a aplicação. Os resultados mostraram uma resposta
progesterônica no dia 4, uma regressão desta resposta no dia 7 com aparência

8
hipoplásica anéstrica e uma moderada estimulação das glândulas endometriais no
dia 14, tendo a égua exibido o estro todos os dias, exceto no dia 4 após a aplicação
do MAP. A concentração de progesterona se manteve em 0,0 ng/ml. Portanto, 250
mg de MAP causou uma estimulação endometrial passageira no dia 4, seguida por
uma diminuição desta resposta. Esta estimulação endometrial não se compara em
magnitude com a densidade glandular ou duração da resposta do endométrio,
previamente verificada com a aplicação de 1000 ou 2000 mg de progesterona de
liberação lenta administrada a intervalos semanais em éguas ovariectomizadas.
Um produto similar ao MAP, caproato de hidroxiprogesterona (Hyproval),
surgiu como progestágeno de longa ação. No entanto, esta droga mostrou-se
ineficaz na prevenção da perda de gestação e na supressão da conduta sexual em
éguas (Neely, 1988).
Uma outra maneira de provir progesterona por longo período de tempo é
através de implantes (Bristol, 1981; Scheffrahn et al.,1982) ou esponjas (Palmer,
1985). Bristol (1981) verificou que um implante de 6 mg de norgestomet (Synchro-
Mate B®) foi ineficaz no controle do estro. Um outro estudo demonstrou que o
Norgestamet, numa dosagem de 3,0 mg/dia, não teve efeito na atividade folicular,
comportamento de estro ou sobre a concentração sangüínea do hormônio
luteinizante (LH) em éguas em fase transicional tardia (Wiepz et., al 1988). O
Norgestomet mostrou-se ineficaz na supressão do estro (Scheffrahn 1982). Esponjas
impregnadas com 0,5 e 1,0 g de Altrenogest (Regu-Mate®) colocadas na vagina por
sete dias resultou num alto grau de sincronização do estro e ovulação (Palmer,
1985), tendo, contudo, induzido um processo inflamatório com aderência da mucosa
vaginal. Nenhuma destas formas está disponível atualmente no comércio para
eqüinos.
2.3 - Manipulação farmacológica do ciclo estral da égua
Anestro fisiológico ou aciclia é um dos assuntos de maior interesse dos
criadores de cavalo em todas as partes do mundo. Em zonas temperadas, o
problema é sazonal, associado com a curta duração dos dias, sendo visto na maioria
das éguas no inverno e na primavera (Sharp, 1980). Sem qualquer tratamento,
quase todas as éguas reassumirão sua ciclicidade no final da primavera e
continuarão a ciclar normalmente até o outono ou inverno (Ginther, 1992).

9
A incorporação de tratamentos que sincronizem a emergência de ondas
foliculares em programas de sincronização proporciona a presença de um folículo
dominante em crescimento no momento da suspensão da administração de
progesterona ou aplicação de prostaglandina, resultando na sincronização da
ovulação e permitindo a utilização da inseminação artificial ou monta natural com
altas taxas de prenhez (Almeida et al.., 2001).
Os agentes farmacológicos que atualmente são utilizados para controlar o
ciclo estral na égua são os mesmos utilizados nos bovinos, cujos protocolos
algumas vezes são aplicados da mesma forma, sem se considerar a diferença entre
as espécies. Desde a década de 1970 se generalizou o uso de um potente agente
luteolítico, a prostaglandina (PGF2α), pela sua facilidade de aplicação (Loy et al..,
1981).
Outra forma de controlar estros é mediante a administração de agente
progestacionais (progesterona e progestágenos sintéticos), os quais são utilizados
há bastante tempo, mediante várias vias de aplicação: a oral e a intramuscular (Loy
et al.., 1981), subcutânea, bem como esponjas ou dispositivos intravaginais (Loy e
Swann, 1966; Arberter et al.., 1994). A estratégia do uso de progestágenos consiste
em prolongar artificialmente a fase lútea por um tempo superior à duração de um
corpo lúteo normal. O bloqueio do eixo hipotalâmico-hipofisário exercido pelo
progestágeno exógeno inibe a liberação do hormônio luteinizante (LH) durante o
tratamento, impedindo a manifestação do comportamento estral. Com a terminação
brusca do tratamento, produz-se um efeito similar à luteólise, ocorrendo a
manifestação do estro em torno do terceiro dia pós-tratamento. No entanto, o
tratamento com progestágenos não inibe a liberação do hormônio folículo
estimulante (FSH) a partir da adenohipófise, razão pela qual o desenvolvimento
folicular continua. (Van Nieker et. al, 1982; Squires et al.., 1979; Webel e Squires et
al.., 1982). As altas concentrações iniciais de progesterona (4-5 ng/ml) e estrógeno
(2,5-3,0 pg/ml) absorvidos através da mucosa vaginal quando se utilizam os
dispositivos impregnados com progesterona associados com aplicação parenteral
de estrógenos têm efeitos inibitórios sobre a liberação de LH e FSH,
respectivamente, cujo efeito prático é o de inibir a emergência de um folículo
dominante em uma onda folicular presente. Como a alta concentração de estradiol
de origem exógena cai rapidamente, um novo folículo dominante pode desenvolver-

10
se durante o tratamento, porém, este não chegará a ovular, pois os níveis de LH
são insuficientes devido ao bloqueio da progesterona (Wild et al.., 2002).
A diferença da fêmea bovina em relação à égua é que esta última apresenta
uma fase luteal curta e geralmente com uma só onda folicular, motivo pelo qual os
programas baseados no encurtamento do ciclo estral com prostaglandinas tendem a
apresentar altos índices de fracassos. As razões principais destas falhas estão
vinculadas ao tamanho do folículo ovulatório e ao momento da aplicação da
prostaglandina (Bergfelt, 2000). Se o folículo é de tamanho pré-ovulatório, pode
sofrer atresia ou ovular sem manifestação de estro. Por outro lado, se existe um
folículo em crescimento, o estro pode ocorrer em um curto período de tempo, como
em um prazo de 3 dias. Estes fatores representam uma dispersão significativa com
o inconveniente de que a ovulação pode ocorrer imediatamente depois do
tratamento ou mesmo demorar quase duas semanas (Wild et al.., 2002).
2.4 - Dispositivos intravaginais contendo progesterona disponíveis no mercado
Não existe indicação por parte dos laboratórios fabricantes do uso de
dispositivos intravaginais na espécie eqüina, somente para as espécies bovina,
bubalina, caprina e ovina.
Os tipos de dispositivos disponíveis no mercado encontram-se na Tabela 1.
Tabela 1: Dispositivos intravaginais (DIV) disponíveis para controle reprodutivo em
bovinos, bubalinos, caprinos e ovinos.
Tipo Nome comercial
Quantidade de Progesterona
Laboratório Fabricante
DIV CIDR® 1,50 g Pfizer DIV DIB® 1,90 g Sintex S.A-Argentina DIV PRIDTM 1,55 g Sanofi-França DIV Triu-B® 1,30 g Biogenesis-Argentina DIV Cronipres® 1.00 g Biogenesis-Brasil
2.5 - Uso de dispositivos intravaginais em éguas para indução e regulação de estros

11
Newcombe e Wilson (1997) utilizaram um dispositivo intravaginal
impregnado com progesterona de liberação lenta CIDR® (Easy-Breed CIDR®:
InterAg, Hamilton, New Zeland) para indução de estro e ovulação em éguas em
anestro.
Newcombe (1998) utilizou o CIDR® em 87 éguas que haviam falhado em
produzir folículos ovulatórios na ausência de um corpo lúteo funcional (anestro
prolongado) por vários períodos até que um folículo atingisse um diâmetro mínimo
de 30 mm. Noventa e oito por cento (57/58) das éguas que apresentaram um folículo
maior ou igual a 30 mm ao final do tratamento ovularam após uma injeção de 2500
U.I de hCG (gonadotrofina coriônica humana) entre zero e seis dias após a retirada
do dispositivo; sessenta e cinco por cento das éguas (37/58) tiveram sua prenhez
confirmada aos 14-17 dias. Do total de éguas, 26 falharam em desenvolver um
folículo com diâmetro de no mínimo 30 mm mesmo com o prolongamento do
tratamento. Destas 26 éguas, 7 ovularam e 7 falharam em ovular mesmo após a
remoção do dispositivo e aplicação do hCG. As outras 12 éguas receberam um
implante de GnRH (deslorelin) por um curto intervalo de tempo: 9 ovularam, 7 foram
inseminadas e 4 (57%) tornaram-se prenhes. O autor concluiu com estes estudos
que ambos os métodos foram satisfatórios para tratar éguas em anestro persistente.
Morete et al.. (2004) utilizaram o dispositivo intravaginal Triu-B (Biogenesis,
Argentina) com objetivo de determinar os níveis de progesterona sérica (P4), o
conforto do animal e a eficácia do dispositivo em diminuir o período transicional em
éguas. O estudo foi realizado durante o período transicional inverno-primavera.
Foram utilizados 3 grupos de éguas, tendo o primeiro recebido o dispositivo
contendo zero g de P4 (placebo), o segundo 1,0 g e o terceiro 1,3 g (Triu-B®) os
quais permaneceram por um período de 10 dias. Após a retirada dos dispositivos, os
animais tiveram seus ovários monitorados por ultra-sonografia até que o folículo
dominante atingisse um diâmetro mínimo de 35 mm, momento em que era feita uma
aplicação de hCG (gonadotrofina coriônica humana) a fim de induzir a ovulação.
Embora todas as éguas se encontrassem em estro no momento da colocação do
dispositivo, oitenta e nove por cento (16/18) tiveram seu estro suprimido com os
dispositivos contendo progesterona, enquanto somente vinte por cento (1/5)
daquelas que receberam o placebo demonstraram este efeito. Uma maior
porcentagem de ovulações também foi observada no primeiro grupo em relação ao

12
placebo (78 vs 40%, p<0.11). Observou-se, também, um adiantamento na ovulação
de trinta e oito por cento nas fêmeas tratadas comparadas com aquelas que
receberam placebo.
Handler et al.. (2004) realizaram um estudo utilizando um dispositivo
intravaginal PRIDTM (Sanofi,França) em éguas a fim de estudar os efeitos deste
dispositivo nos diferentes estágios do ciclo reprodutivo e a secreção de progesterona
e LH (hormônio luteinizante) nestes animais. As éguas foram divididas em 3 grupos
de acordo com seu estado reprodutivo: estro, diestro e anestro.Todas receberam o
dispositivo e os mesmos foram mantidos por 12 dias. No momento da retirada, foi
feita uma aplicação de 250 microgramas do agente luteolítico cloprostenol
(EstrumateTM, Essex, Alemanha). Foram feitas mensurações diárias dos ovários por
ultra-sonografia e mensurações dos níveis de progesterona e LH. Como resultados
do trabalho, os autores observaram que a maior resposta de estros e ovulações
ocorreu nas éguas que se encontravam em anestro quando comparadas àquelas em
diestro e estro. Os intervalos entre a remoção do dispositivo até o estro/ovulação
variaram de 4,0/21,0+1,0 dia em anestro, 3,3+0.7/7,1+.0.9 dias em diestro e
3,6+0.3/8,8+0,9 dias em éguas em estro (p<0.05). Concentrações médias de
progestágeno depois da colocação do dispositivo variaram de 0,3+0,1 ng/ml em
éguas em anestro e estro e 7,1ng/ml em éguas no diestro (p<0,01). Concentrações
de LH e progesterona mensuradas por 4 horas um dia após a colocação do
dispositivo foram maiores nas éguas em diestro em relação àquelas em anestro e
estro. Os autores concluíram que a presença de um corpo lúteo no momento da
colocação do dispositivo tem um importante impacto na eficácia do tratamento.
Videla et al.. (2002) realizaram um estudo utilizando dispositivos
intravaginais DIB (Syntex, S.A, Argentina) contendo duas quantidades diferentes de
progestágeno; 1,38g e 1,90g. O objetivo do trabalho foi avaliar as concentrações
séricas de progestágeno dos 2 tipos de dispositivos e caracterizar a dinâmica
ovariana durante o tratamento em animais cíclicos. Foram utilizados 2 grupos de
éguas tendo cada um recebido dispositivos com concentrações diferentes de
progestágeno. Os dispositivos permaneceram por 12 dias e durante este período os
animais tiveram a atividade ovariana monitorada por ultra-sonografia e também
foram coletadas amostras de sangue para dosagens de progesterona. Ao final do
estudo os autores concluíram que, para se obter concentrações de progesterona
sérica suficiente para causar inibição do crescimento folicular e ovulação os

13
dispositivos intravaginais utilizados, devem conter, no mínimo, 1,9 gramas de
progestágeno.
Wild et al.. (2002) utilizaram o PRID® em 11 éguas durante 12 dias não
tendo sido evidenciada a manifestação de estro em nenhum dos animais durante
este período. Após a retirada do dispositivo, somente uma égua não apresentou
estro, sendo que esta tinha um histórico de 3 anos de anestro. Das 10 éguas que
exibiram estro, uma (10%) o manifestou 24 horas após a retirada, duas (20%) após
18 horas e cinco após 72 horas. As outras duas restantes apresentaram estro 5 a 6
dias após a retirada do dispositivo. O autor atribuiu esta dispersão ao fato da
progesterona, apesar de apresentar controle eficiente sobre a manifestação de
estro, não apresenta efeito inibitório sobre a secreção de FSH (hormônio folículo
estimulante), razão pela qual pode haver no momento da retirada do dispositivo um
folículo em estágio avançado de desenvolvimento a ponto de ovular, o qual pode
entrar em atresia ou ovular imediatamente após a retirada do dispositivo, sem,
contudo, haver manifestação explícita dos sinais de estro pela égua. Em outros
casos, o folículo presente ao final do tratamento pode ter um escasso
desenvolvimento, fazendo com que a manifestação do estro seja mais tardia
(Squires et al.., 1979; Webel, 1975) atendendo ao fato de que um folículo cresce de
2 a 3 mm por dia durante o estro. Isso levou os autores a considerar que o PRID®
,embora seja eficiente no bloqueio da manifestação do estro, apresenta variações
consideráveis na sincronização da ovulação. Todos os estros apresentados pelas 10
éguas tiveram manifestação normal, a exceção de uma que apresentou um estro
com 15 dias de duração. As éguas receberam uma dose de 2500 UI de hCG
(gonadotrofina coriônica humana) por via endovenosa no momento em que
apresentaram um folículo maior ou igual a 35mm. Das 11 éguas tratadas, 9
tornaram-se prenhes, 8 através da inseminação artificial com sêmen refrigerado e
uma com estro pouco pronunciado através de monta natural, todas realizadas 24
horas após a administração do hCG. A égua com estro prolongado não se tornou
prenhe. Apesar do número reduzido de animais utilizados neste experimento, os
autores consideraram que a taxa de prenhez obtida foi amplamente satisfatória e
coincide com os resultados obtidos por outros autores (Van Niekerk et al.., 1982) e
sugerem que o uso do PRID® não interfere na fertilidade das éguas.

14
2.6 - Contra-indicações do uso da progesterona e progestágenos em éguas
Não se deve administrar nenhum tipo de progestágeno em éguas com
história de infecção uterina. Washburn et al.. (1982) observaram que éguas
ovariectomizadas tratadas com progesterona apresentavam resposta fagocítica
diminuída. Colbern et al.. (1987) demonstraram que éguas ovariectomizadas ou
intactas expostas à contaminação bacteriana mantiveram a infecção uterina pelo
período de tratamento com progesterona. Alexander et al.. (1991) determinaram o
efeito do tratamento com progesterona na suscetibilidade de infecção uterina. Eles
selecionaram 32 éguas na fase transicional, coletaram swabs uterinos de todas as
éguas antes de as dividirem em 2 grupos; 16 controles e 16 submetidas à terapia
com progesterona (150 mg /dia por 12 dias). Quando as éguas exibiram estro foi
coletado um segundo swab. Na coleta inicial, nenhuma égua apresentou o
crescimento bacteriano. No entanto, cinqüenta e seis por cento das éguas tratadas
com progesterona apresentaram swab com contaminação bacteriana versus vinte e
cinco por cento das éguas controle. A partir desse experimento, os autores
concluíram que a progesterona pode ser nociva à saúde do útero, particularmente
em éguas velhas. No entanto, em ambos os estudos (Webel, 1975; Allen et al..,
1980; Squires et al.., 1979 e 1983), baseados em achados clínicos, não foi
observado que o tratamento progesterônico por 12 a 15 dias tenha alterado a
fertilidade reprodutiva em éguas ginecologicamente sadias.
Segundo os fabricantes (Pfizer; Pharmacia), o uso do CIDR® é contra-
indicado em fêmeas impúberes, debilitadas ou portadoras de patologias genéticas
ou adquiridas do sistema genital, como processos infecciosos e/ ou inflamatórios,
visto que o mesmo tende a induzir um processo infeccioso benigno na mucosa
vaginal pela ação mecânica que exerce sobre as paredes deste órgão, bem como é
um fator de consenso entre vários autores que o aparelho genital, quando sob efeito
progesterônico, é mais susceptível à invasão de microorganismos. Cabe ressaltar
que estas contra-indicações são recomendadas para as espécies para as quais o
CIDR® é indicado (bovinos, bubalinos,ovinos e caprinos).

15
3 - MATERIAL E MÉTODOS
3.1 - Ensaio Experimental 1
Na primeira fase do experimento realizado nas instalações de Universidade
Estadual Norte Fluminense Darcy Ribeiro (UENF), localizada no município de
Campos dos Goitacazes – RJ, foram pesquisados os efeitos do CIDR® sobre a
integridade endometrial em éguas. O CIDR® (Pharmacia, Brasil) é um dispositivo de
material plástico em forma de Y impregnado com 1,5 g de progesterona em veículo
de liberação controlada. Para tanto, foram utilizadas cinco éguas mestiças com
média de idade de 5,5 + 1,8 anos (S=0,8 ano), variando de 6 a 10 anos de idade
(média de 7,8 anos) com baixo escore corporal. Estes animais eram mantidos em
pastejo de capim colonião sem suplementação.
A integridade endometrial foi avaliada através de exames de citologia e
biópsia realizados no Laboratório de Anatomia Patológica da UENF. Antes da coleta
das amostras foram realizadas a assepsia das regiões vulvar e perineal, seguida da
imobilização da cauda utilizando-se uma bandagem de gaze. Os animais foram
contidos em um tronco apropriado para a realização dos procedimentos. O exame
citológico foi feito por meio da análise de esfregaços de amostras endometriais
fixadas com propanol 95% coletadas com swab inseridos no interior da cavidade

16
uterina fixados com uma sonda apropriada. A coloração do esfregaço fixado foi
feita, primariamente com solução de May-Grunwald (Merck 1412) durante 45
segundos e depois, com solução de Giemsa (Merck 9204) durante 15 segundos. As
lâminas foram examinadas em microscópico óptico comum. As amostras para
exames de biópsia foram obtidas por meio do uso de uma pinça de YOMAN de 50
cm de comprimento com a embocadura de, aproximadamente, 0,5 X 2,0 cm que,
após prévia flambagem, foi introduzida na cavidade uterina para se obter uma
amostra. As amostras retiradas foram imediatamente colocadas em solução
fixadora de Bouin por 24 horas, tendo sido transferidas, após este período, para
uma solução de formol a 10%. O tecido fixado foi incluído em parafina e corado pela
hematoxilina-eosina.
Todas as coletas de materiais para exame, assim como o acompanhamento
por ultra-sonografia utilizando-se um equipamento Nokia modelo 500 com
transdutor transretal de 5 Mhz, foram realizados em dois momentos: antes da
colocação do CIDR® e no dia da sua retirada, que foi feita 12 dias após o início do
estudo. Após a retirada dos dispositivos, as éguas foram submetidas a uma
inspeção diária com o intuito de verificar o tempo necessário para a completa
resolução clínica do processo de vaginite. Os animais foram considerados
totalmente recuperados no momento em que não foi observada a presença de
qualquer tipo de secreção com aspecto, cor ou odor considerados anormais.
3.2 - Ensaio Experimental 2
A segunda fase do experimento foi realizada no mês de novembro de 2004
no Haras São Joaquim, localizado no município de Cachoeiro de Itapemirim, sul do
estado do Espírito Santo. Foram utilizadas 66 éguas entre 2,5 anos e 10 anos de
idade (média de 6,5 anos), sadias, não-gestantes, não-lactantes da raça
Mangalarga Marchador, mantidas a campo em pastagem de capim-colonião sem
suplementação alimentar, tendo à disposição mistura mineral e água de boa
qualidade. Todos os animais receberam regularmente as vacinas e os vermífugos
preconizados no programa sanitário adotado no haras. As éguas foram submetidas
a um exame de admissão, que incluiu anamnese, exame clínico geral, ginecológico
manual (retal e vaginal) e ultra-sonográfico dos ovários e do útero. Foi utilizado um

17
tronco de contenção coberto apropriado para a realização de todos os
procedimentos constantes no estudo.
Nesta fase do estudo, as 66 éguas foram divididas em 3 grupos: 22 destas
foram submetidas ao protocolo 1 (grupo 1) com a manutenção do dispositivo por 8
dias, 22 éguas foram submetidas ao protocolo 2 (grupo 2) com a manutenção do
dispositivo por 10 dias e o grupo controle composto pelos 22 animais restantes que
receberam o dispositivo isento de progesterona (obtidos após a manutenção dos
mesmos por 7 dias em solução de éter sulfúrico a 5%) e o mantiveram por 10 dias
na cavidade vaginal. Esta divisão dos animais dentro dos respectivos grupos foi
feita de forma totalmente aleatória quanto ao fator indivíduo e fase do ciclo estral.
Todos os animais foram submetidos, além da inserção dos dispositivos na cavidade
vaginal, à aplicação de outras drogas constantes nos protocolos: prostaglandina
(Lutalyse®), benzoato de estradiol (Estrogin®) e hCG (Vetecor®), O estudo foi conduzido durante todo o mês de novembro de 2004.
3.2.1 - Procedimentos que antecederam a instalação dos protocolos
Visando a minimizar fatores que pudessem influenciar os resultados obtidos
nos protocolos utilizados, antes da instalação dos mesmos, foi estabelecida uma
seqüência de procedimentos descritos a seguir:
• Cadastramento dos animais: foi feita uma ficha individual de todas as
éguas na qual constavam informações referentes aos respectivos históricos e
achados no exame clínico geral e reprodutivo.
• Exame da cavidade vaginal: foi feita uma avaliação visual da cavidade
vaginal através do uso de um espéculo vaginal de Polansky, esterilizado por
flambagem e lubrificado com gel estéril à base de água, objetivando observar a
existência de áreas com quaisquer sinais de processos inflamatórios, infecciosos
e/ou neoplásicos, visto que, uma vez apresentando quaisquer destas
anormalidades, o animal era descartado do estudo.
• Avaliação do trato reprodutivo por ultra-sonografia: foram feitas
avaliações por ultra-sonografia do trato reprodutivo de todas as éguas utilizando-se
um equipamento Nokia modelo 500 com transdutor transretal de 5 Mhz, sendo a

18
primeira avaliação no dia da colocação do dispositivo intravaginal e a última no dia
da retirada do mesmo. As imagens obtidas através destes exames foram impressas
(impressora SONY, UP 890-MD) para que fosse feita uma avaliação mais precisa da
dinâmica útero-ovárica no decorrer do experimento.
Por meio destas avaliações as éguas foram agrupadas em dois grupos
distintos: a) éguas em estro: com presença de folículos maiores de 20 mm em um ou
ambos os ovários e início de edematização do endométrio uterino, traduzido pela
presença de dobras do mesmo, tônus uterino aumentado e abertura cervical; b)
éguas em diestro: ausência de folículos maiores de 20 mm, presença de corpo lúteo,
ausência de edematização, traduzida por aparência homogênea do endométrio
uterino, diminuição do tônus uterino e cérvix fechada. Éguas em anestro (sem
presença de folículos maiores de 10 mm e corpo lúteo) e éguas que apresentassem
alguma patologia uterina (endometrites, cistos, tumores) não foram utilizadas no
estudo.
As características da cavidade vaginal e da cérvix ao exame utilizando-se o
espéculo também foram avaliadas e levadas em consideração em conjunto com os
outros parâmetros para a determinação do estágio do ciclo estral em que o animal
se encontrava. Esta avaliação vaginal com espéculo e o exame por ultra-sonografia
não foram realizados com freqüência diária em função dos riscos de contaminação
que exame especular proporciona e as reações de tenesmo vaginal desencadeadas
pelo exame por ultra-sonografia.
• Coleta de amostras de sangue para realização de hemograma: foram
feitas coletas de duas amostras de sangue da veia jugular de todas as éguas
utilizando-se frascos vaccutainer de 10 ml com heparina sódica para realização de
hemograma completo. A primeira coleta foi realizada no dia da colocação do CIDR®
e a segunda no dia da retirada do mesmo.

19
4 - PROTOCOLOS DE SINCRONIZAÇÃO DE ESTRO E OVULAÇÃO
Os procedimentos realizados na instalação dos protocolos seguiram a
mesma seqüência, somente diferindo quanto ao tempo de permanência do
dispositivo e/ou ao tipo de droga utilizada nos diferentes protocolos conforme
descrito a seguir:
• Introdução do dispositivo intravaginal com progesterona (CIDR®): as
éguas foram higienizadas na sua região ano-genital com água e sabão, desinfetadas
com solução contendo 1% dicloroisocianurato de sódio anidro (Agrisept®, Schering
Plought) e secas com papel toalha. Logo após foi feita a introdução do CIDR ®,
previamente lubrificado com solução antibiótica à base de Gentamicina (Gentrin®,
Laboratório Ouro Fino, Brasil). Com o auxílio de um aplicador específico, também
submetido ao tratamento com a mesma solução antibiótica, os dispositivos foram
introduzidos no interior da cavidade vaginal das 66 éguas participantes do estudo
onde os mesmos permaneceram por 8 dias nos animais do grupo 1 e por 10 dias
nos animais do grupo 2 e do grupo controle. Este dispositivo possui em sua
extremidade posterior um cordel que permanecia suspenso sobre a vulva, e que foi
utilizado para a retirada do mesmo nos dias previstos do dois protocolos.

20
• Aplicação do benzoato de estradiol (Estrogin®) e prostaglandina (Lutalyse®): foi feita uma dose de 10 mg de benzoato de estradiol (Estrogin®,
Farmavet, Brasil) via I.M. logo após a colocação do CIDR®. Simultaneamente foi
aplicado 1 ml de Lutalyse® via I.M. (Laboratório Pharmacia, Brasil), equivalente a
5mg de Dinoprost. Este protocolo foi iniciado nos grupos tratados
independentemente do estágio do ciclo estral em que os animais se encontrassem,
estro ou diestro.
• Retirada do CIDR® e aplicação de prostaglandina (Lutalyse®): decorridos
8 dias do início do protocolo nos animais do grupo 1 e 10 dias nos animais do grupo
2 e do grupo controle foram retirados os dispositivos e imediatamente foi aplicado
uma dose de 1 ml de Lutalyse®, via I.M. em todos os animais .
• Exame da cavidade vaginal: após a retirada do CIDR® foi feito um exame
da cavidade vaginal nos animais utilizando-se o mesmo equipamento e os mesmos
procedimentos utilizados no exame que antecedeu àcolocação do dispositivo.
Naqueles animais que apresentaram indícios de processo inflamatório,
traduzidos por presença de secreção mucopurulenta com cor, aspecto e odor
anormais, foi feito um novo exame a cada 24 horas com o intuito de observar o
período de cura clínica espontânea, tal com foi feito na primeira fase do experimento.
• Monitoramento diário por ultra-sonografia para acompanhamento do desenvolvimento folicular e aplicação de indutor de ovulação: foi feito um
acompanhamento diário por ultra-sonografia a partir da retirada do CIDR® em todas
as éguas até que fosse diagnosticada a presença de um folículo com um diâmetro
mínimo de 35 mm, considerado neste estágio como folículo pré-ovulatório. No
momento em que esta condição foi observada, foi feita a indução da ovulação
através da aplicação de uma dose única de 2.500 U.I de hCG (Vetecor®), por via I.V
na metade dos 30 animais que apresentaram esta condição e na outra metade foi
aplicado 1 ml de Dinoprost (Lutalyse®), por via I.M Esta divisão dos animais que
receberam as drogas foi totalmente aleatória, sendo a mesma utilizada também no
grupo controle.

21
• Realização da cópula: as cópulas foram efetuadas 24 horas após ter sido
diagnosticado o folículo pré-ovulatório de 35 mm em um dos ovários e ter sido feita a
aplicação do indutor de ovulação. Foi feito um monitoramento diário ultra-sonográfico
para que se verificasse a ocorrência da ovulação. Nas éguas nas quais não se
observou a ocorrência da ovulação 48 horas após esta cobertura, uma outra foi
efetuada a cada 48 horas até que a ovulação tivesse se processado.
Estas cópulas foram efetuadas por três garanhões da raça Mangalarga
Marchador com idades de cinco, sete e 13 anos, com fertilidade comprovada através
de exames andrológicos e históricos de coberturas anteriores com índices de
prenhez entre 70 e 80% durante a última estação de monta na qual eles foram
utilizados.
Os exames andrológicos foram realizados 30 dias antes do início da estação
de monta. Os garanhões foram coletados com o auxílio de uma vagina artificial
modelo Hannover. Todos apresentaram boa libido quando em contato com a égua
manequim tendo efetuado a monta num tempo máximo de 5 minutos. As amostras
de sêmen coletadas foram examinadas com o auxílio de um microscópio óptico
comum no aumento de 100 vezes para verificação da motilidade. Os resultados
demonstraram que as três amostras apresentaram motilidade progressiva entre 70 e
80%. A concentração espermática foi avaliada utilizando-se para esse propósito uma
câmara hematimétrica (Newbawer) e os resultados mostraram taxas de
concentração variando entre 120 a 140 milhões de células espermáticas por mililitro.
• Diagnóstico de gestação por ultra-sonografia
Foi realizado o exame por meio da ultra-sonografia nas éguas 15 dias após a
última cobertura para se verificar o índice de prenhez. Outras avaliações ultra-
sonográficas adicionais foram realizadas aos 30 e 45 e 60 dias a fim de se observar
a ocorrência de perdas embrionárias precoces.
Um resumo dos procedimentos realizados nos protocolos utilizados pode ser
observado na figura 1 e na figura 2.

22
Colocação do CIDR Retirada do CIDR® e Ind. ovulaçâo
Aplicação de Estrogin® e Aplicação de fol.≥35mm) ⇓
Lutalyse® (Dia 0) ↓ Lutalyse® (Dia 8) (?)
↓ ⇓ ⇓ ⇓ ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐___⏐
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
↑
Início do protocolo DIAS
Figura 1: Organização esquemática dos procedimentos realizados no estudo: manipulação farmacológica do ciclo reprodutivo da égua através da utilização de um dispositivo impregnado com progesterona de liberação controlada (CIDR ®) por 8 dias.
Colocação do CIDR Retirada do CIDR® e Ind. ovulaçâo *
Aplicação de Estrogin® e Aplicação de fol.≥35mm) ⇓
Lutalyse® (Dia 0) ↓ Lutalyse® (Dia 10) (?)
↓ ⇓ ⇓ ⇓ ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐ | ⏐___⏐
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Início do protocolo DIAS
Figura 2: Organização esquemática dos procedimentos realizados no estudo: manipulação farmacológica do ciclo reprodutivo da égua através da utilização de um dispositivo impregnado com progesterona de liberação controlada (CIDR ®) por 10 dias.

23
5 - ANÁLISE ESTATÍSTICA
Foram realizadas análises descritivas dos dados obtidos para as diversas
características avaliadas sendo que, para supressão do estro, reação ao CIDR®,
tempo decorrido entre a retirada do CIDR® até a manifestação do estro, tempo de
surgimento de um folículo ≥ 35 mm, freqüência das éguas ovuladas com Lutalyse® e
Vetecor® e éguas prenhas com Lutalyse e Vetecor – foi utilizada a estatística do Qui-
quadrado (2 א), considerando-se um nível de 5% de probabilidade (p<0,05), no
intuito de verificar a independência entre os dados observados e os diversos
tratamentos aos quais as éguas foram submetidas (Gomes,1998).

24
6 - RESULTADOS E DISCUSSÃO
6.1 - Ensaio experimental 1
A única alteração que pôde ser atribuída ao dispositivo foi a instalação de
um quadro de vaginite com secreção mucopurulenta em 3 dos 5 animais tratados
(60%), mas que apresentou cura espontânea em todos os animais 72 horas após a
sua retirada, não tendo sido necessária a utilização de nenhuma terapia, tal como
relatada por Wild et al.. (2002) e Morete et al.. (2004).
Para a interpretação dos resultados obtidos a partir do exame de citologia
vaginal, foi levado em conta, para se fazer um diagnóstico dos resultados
quantitativos de neutrófilos de um swab, a relação da presença de outras células
(células do epitélio endometrial) dentro de uma contagem total de duzentas células.
O critério utilizado para se considerar um resultado positivo foi aquele proposto por
Ball et al. (1988) que preconiza que a presença de mais de 2% de
polimorfonucleares seja o valor a partir do qual possa se considerar a égua como
portadora de um processo inflamatório.
O resultado dos exames de biópsia endometrial foi baseado na
diferenciação da infiltração celular que ocorre normalmente nas diferentes fases do
ciclo estral em comparação àquela observada nos processos patológicos e o grau
de fibrose das glândulas uterinas (Rickets, 1978).

25
Em todos os exames realizados 72 horas após a retirada dos dispositivos
(ultra-sonografia, citologia e biópsia uterinas), não foi constatada nenhuma alteração
em relação aos resultados obtidos nestes mesmos exames realizados antes da
colocação dos mesmos.
6.2 - Ensaio experimental 2
Os resultados dos valores do teste qui-quadrado ( Pא 2) dos parâmetros
pesquisados nos protocolos de sincronização de estros e ovulação utilizando-se o
CIDR® encontram-se descritos na tabela 2, tabela 3, tabela 4, tabela 5 e tabela 6
(CV=14,5%). Tabela 2: Resultados percentuais dos parâmetros pesquisados em função dos
diferentes protocolos de sincronização de estros e ovulação utilizando-se o CIDR®.
Grupos Parâmetros
1 (CIDR® por 8
dias)
2 (CIDR® por 10
dias)
Controle (CIDR® por 10 dias sem P4)
Valores de 2χ
Estágio do ciclo estral no início do experimento
Estro:100 % Diestro: 0 %
Estro: 72,73 % Diestro: 27,27%
Estro: 81,82 % Diestro: 18,18 %
-
Perdas do CIDR® 9,09% 9,09% 0% -
Supressão do estro
100 % 100 % 11,11 % 47,29*
Reação ao CIDR® (vaginite)
27,38 %
9,09 % 27,38 % 2,90ns
Recuperação da vaginite até 48 hs
100 % 100 % 100 % -
*Teste Qui-quadrado entre vários parâmetros pesquisados, significativo (*) (GL=2,P < 0,05) e não-significativo (ns).
26
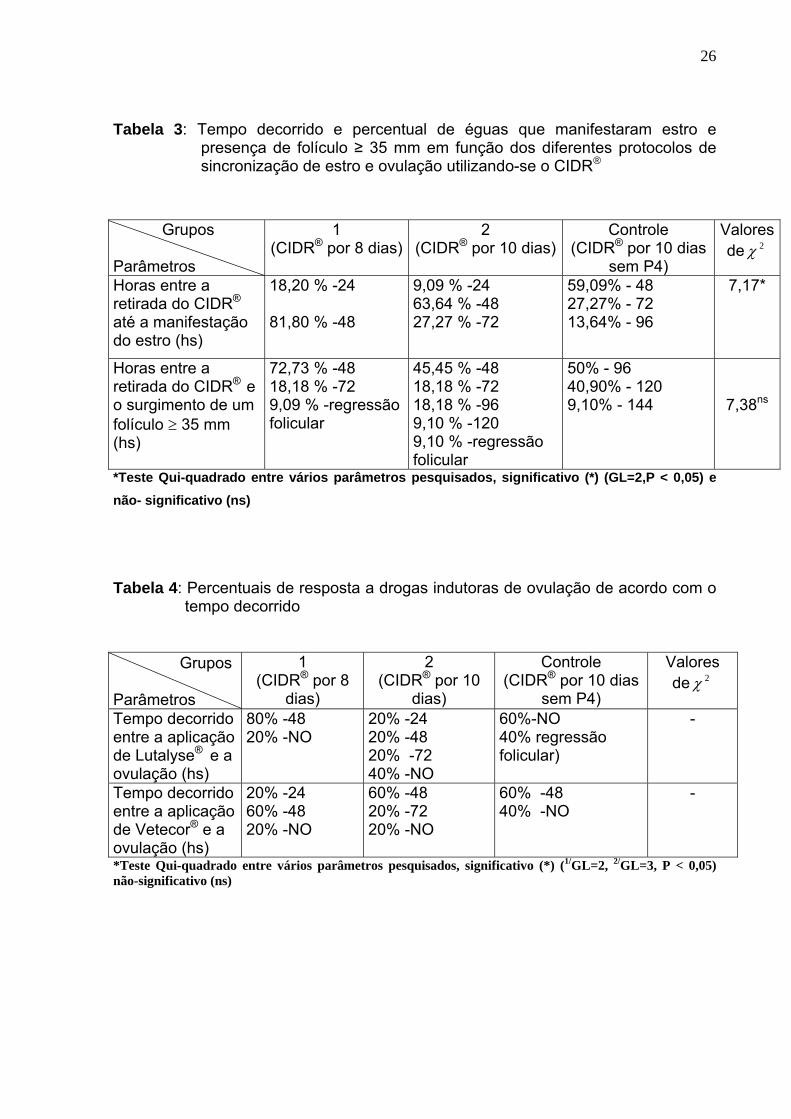
Tabela 3: Tempo decorrido e percentual de éguas que manifestaram estro e
presença de folículo ≥ 35 mm em função dos diferentes protocolos de sincronização de estro e ovulação utilizando-se o CIDR®
Grupos Parâmetros
1 (CIDR® por 8 dias)
2 (CIDR® por 10 dias)
Controle (CIDR® por 10 dias
sem P4)
Valores de 2χ
Horas entre a retirada do CIDR®
até a manifestação do estro (hs)
18,20 % -24 81,80 % -48
9,09 % -24 63,64 % -48 27,27 % -72
59,09% - 48 27,27% - 72 13,64% - 96
7,17*
Horas entre a retirada do CIDR® e o surgimento de um folículo ≥ 35 mm (hs)
72,73 % -48 18,18 % -72 9,09 % -regressão folicular
45,45 % -48 18,18 % -72 18,18 % -96 9,10 % -120 9,10 % -regressão folicular
50% - 96 40,90% - 120 9,10% - 144
7,38ns
*Teste Qui-quadrado entre vários parâmetros pesquisados, significativo (*) (GL=2,P < 0,05) e não- significativo (ns)
Tabela 4: Percentuais de resposta a drogas indutoras de ovulação de acordo com o
tempo decorrido Grupos Parâmetros
1 (CIDR® por 8
dias)
2 (CIDR® por 10
dias)
Controle (CIDR® por 10 dias
sem P4)
Valores de 2χ
Tempo decorrido entre a aplicação de Lutalyse® e a ovulação (hs)
80% -48 20% -NO
20% -24 20% -48 20% -72 40% -NO
60%-NO 40% regressão folicular)
-
Tempo decorrido entre a aplicação de Vetecor® e a ovulação (hs)
20% -24 60% -48 20% -NO
60% -48 20% -72 20% -NO
60% -48 40% -NO
-
*Teste Qui-quadrado entre vários parâmetros pesquisados, significativo (*) (1/GL=2, 2/GL=3, P < 0,05) não-significativo (ns)
27
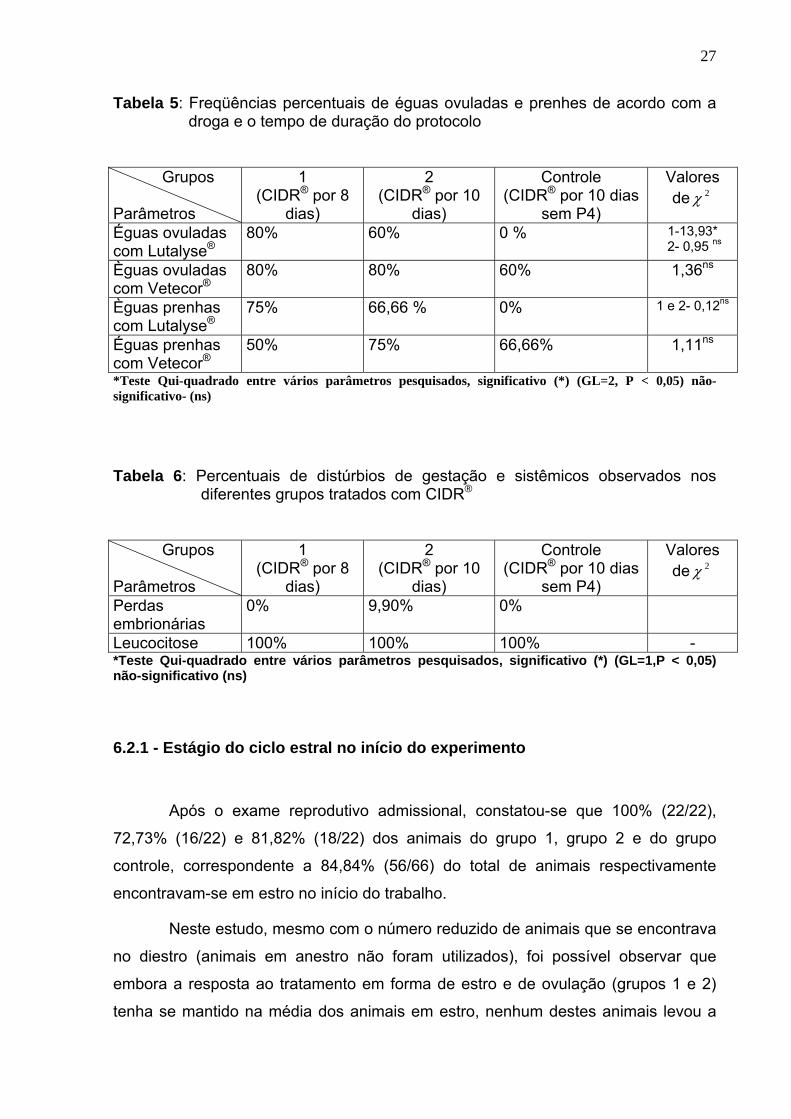
Tabela 5: Freqüências percentuais de éguas ovuladas e prenhes de acordo com a droga e o tempo de duração do protocolo
Grupos Parâmetros
1 (CIDR® por 8
dias)
2 (CIDR® por 10
dias)
Controle (CIDR® por 10 dias
sem P4)
Valores de 2χ
Éguas ovuladas com Lutalyse®
80% 60% 0 % 1-13,93* 2- 0,95 ns
Èguas ovuladas com Vetecor®
80%
80% 60% 1,36ns
Èguas prenhas com Lutalyse®
75% 66,66 %
0% 1 e 2- 0,12ns
Éguas prenhas com Vetecor®
50%
75% 66,66% 1,11ns
*Teste Qui-quadrado entre vários parâmetros pesquisados, significativo (*) (GL=2, P < 0,05) não-significativo- (ns)
Tabela 6: Percentuais de distúrbios de gestação e sistêmicos observados nos
diferentes grupos tratados com CIDR®
Grupos Parâmetros
1 (CIDR® por 8
dias)
2 (CIDR® por 10
dias)
Controle (CIDR® por 10 dias
sem P4)
Valores de 2χ
Perdas embrionárias
0% 9,90% 0%
Leucocitose 100% 100% 100% - *Teste Qui-quadrado entre vários parâmetros pesquisados, significativo (*) (GL=1,P < 0,05) não-significativo (ns) 6.2.1 - Estágio do ciclo estral no início do experimento
Após o exame reprodutivo admissional, constatou-se que 100% (22/22),
72,73% (16/22) e 81,82% (18/22) dos animais do grupo 1, grupo 2 e do grupo
controle, correspondente a 84,84% (56/66) do total de animais respectivamente
encontravam-se em estro no início do trabalho.
Neste estudo, mesmo com o número reduzido de animais que se encontrava
no diestro (animais em anestro não foram utilizados), foi possível observar que
embora a resposta ao tratamento em forma de estro e de ovulação (grupos 1 e 2)
tenha se mantido na média dos animais em estro, nenhum destes animais levou a

28
gestação a termo, ou seja, dos 10 animais em que se iniciou o protocolo que se
encontravam em diestro, apenas 2 tornaram-se gestantes, sendo que ambos,
sofreram processo de reabsorção embrionária detectada por ultra-sonografia aos 30
dias pós-cobertura.
Handler et al. (2004) utilizaram um dispositivo intravaginal PRIDTM (Sanofi,
França) em éguas, objetivando estudar os efeitos deste nos diferentes estágios do
ciclo reprodutivo e a secreção de progesterona e LH (hormônio luteinizante),
concluíram que a presença de um corpo lúteo no momento da colocação do
dispositivo tem um importante impacto na eficácia do tratamento. No presente
estudo provavelmente pode-se ter evitado esta interferência com a incorporação da
aplicação da prostaglandina no início dos protocolos testados.
Os melhores resultados obtidos nos animais em estro talvez tenha sido
devido à aplicação do benzoato de estradiol e da prostaglandina no início do
protocolo, levando à inibição hipotalâmica e a destruição do tecido luteal levando à
uma redução dos níveis sanguíneos do FSH e da progesterona, respectivamente, tal
como afirmaram Squires et al. (1979) e (Webel, 1975).
6.2.2 - Perdas do dispositivo
O percentual de perdas dos dispositivos foi de 9,09% (2/22) nos grupos 1,
9,09% (2/22) no grupo 2 e 0% no grupo controle, importando num total de perdas de
6,06% (04/66). Em razão das perdas, torna-se necessário que novos estudos sejam
realizados visando elaborar um dispositivo mais adaptado à anatomia da égua
desde que, mantenha a sua eficiência em permanecer no local adequado durante o
tempo do protocolo, tal como o observado neste estudo.
Não se dispõem de dados na literatura que estabeleçam valores médios de
perdas de dispositivos intravaginais em éguas, talvez pelo pequeno número de
estudos desenvolvidos com estes dispositivos nesta espécie.
Cabe salientar, entretanto, que o período máximo no qual os animais
permaneceram sem o dispositivo foi de 12 horas, uma vez que, no momento em que
era feita uma das duas inspeções diárias e era constatada a perda do dispositivo,
outro era imediatamente reposto.

29
Acredita-se que estas perdas não tenham causado interferência nos
resultados obtidos, pois os 4 animais que perderam os dispositivos encontravam-se
em estro no início do experimento, tendo interrompido sua manifestação 24 horas
após a sua colocação, só tendo retornado a essa condição após a retirada do
dispositivo. Isto demonstra que o tempo em que o animal ficou sem a influência
hormonal não foi suficiente para causar o desbloqueio hipotalâmico-hipofisário a
ponto de levar o animal a manifestar sinais de estro.
6.2.3 - Supressão do estro
No presente estudo não foi observada manifestação de estro em nenhum
dos animais do grupo 1 (22/22) e do grupo 2 (16/16), conforme demonstrado na
figura 3, o que demonstra que a progesterona contida no dispositivo foi liberada em
quantidades suficientes para promover a inibição dos sinais do estro.
A aplicação do Lutalyse® no primeiro dia do protocolo foi realizada com o
propósito de eliminar qualquer fragmento de tecido luteal que porventura existisse
nos ovários. Com este procedimento, nos certificamos de que não haveria secreção
de progesterona pelo tecido ovariano.
A supressão do estro em 100% dos grupos tratados com o CIDR® com
progesterona coincide com o valor obtido por Wild et al. (2002) utilizando o PRID®
em 11 éguas durante 12 dias não tendo sido evidenciada a manifestação de estro
em nenhum dos animais durante este período.
30
% de Supressão do Estro
100,0 100,0
11,1
0,0
20,0
40,0
60,0
80,0
100,0
120,0
1 2 3
Grupo
Freq
uenc
ias
(%)
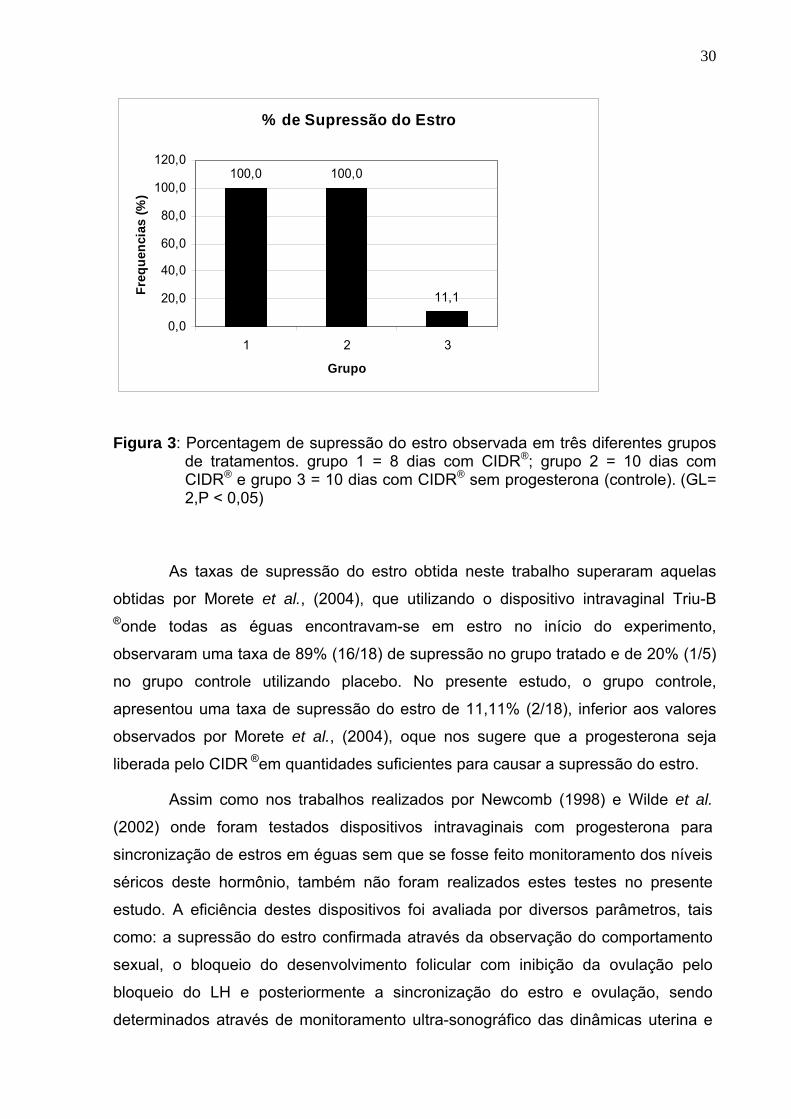
Figura 3: Porcentagem de supressão do estro observada em três diferentes grupos
de tratamentos. grupo 1 = 8 dias com CIDR®; grupo 2 = 10 dias com CIDR® e grupo 3 = 10 dias com CIDR® sem progesterona (controle). (GL= 2,P < 0,05)
As taxas de supressão do estro obtida neste trabalho superaram aquelas
obtidas por Morete et al., (2004), que utilizando o dispositivo intravaginal Triu-B
®onde todas as éguas encontravam-se em estro no início do experimento,
observaram uma taxa de 89% (16/18) de supressão no grupo tratado e de 20% (1/5)
no grupo controle utilizando placebo. No presente estudo, o grupo controle,
apresentou uma taxa de supressão do estro de 11,11% (2/18), inferior aos valores
observados por Morete et al., (2004), oque nos sugere que a progesterona seja
liberada pelo CIDR ®em quantidades suficientes para causar a supressão do estro.
Assim como nos trabalhos realizados por Newcomb (1998) e Wilde et al.
(2002) onde foram testados dispositivos intravaginais com progesterona para
sincronização de estros em éguas sem que se fosse feito monitoramento dos níveis
séricos deste hormônio, também não foram realizados estes testes no presente
estudo. A eficiência destes dispositivos foi avaliada por diversos parâmetros, tais
como: a supressão do estro confirmada através da observação do comportamento
sexual, o bloqueio do desenvolvimento folicular com inibição da ovulação pelo
bloqueio do LH e posteriormente a sincronização do estro e ovulação, sendo
determinados através de monitoramento ultra-sonográfico das dinâmicas uterina e

31
ovariana. O efeito sobre a mucosa vaginal por meio do acompanhamento de
visualização direta da cavidade vaginal foi feito através do uso de espéculo; a
reação inflamatória por meio de exames hematológicos e o efeito sobre as taxas de
fertilidade por meio do acompanhamento ultra-sonográfico para diagnóstico e
manutenção da gestação. Mottershead (2004) afirma ser impossível se obter uma
acurada mensuração do status dos níveis de progesterona séricos em éguas, uma
vez que ocorrem oscilações diárias e dentro de um mesmo dia. O mesmo autor
recomenda que, caso se queira ter uma idéia aproximada sobre os níveis de
progesterona em uma égua deve-se efetuar de 3 a 4 coletas diárias, por no mínimo
4 dias seguidos.
6.2.4 - Reação ao dispositivo e tempo de recuperação da vaginite
Com relação à reação ao dispositivo, análise estatística mostrou não ter
havido significância (p>0,05) entre os grupos tratados e o grupo controle (figura 4) o
que demonstra que a reação inflamatória desencadeada deveu-se à presença
mecânica do dispositivo (trauma físico) e não ao hormônio (progesterona) nele
contido.

32
Reação ao CIDR
0
5
10
15
20
25
30O
corr
ênci
a de
Vag
inite
(%)
Figura 4: P
gc
Dife
apresentara
no presente
torno de 21
7). Esta dife
solução an
dispositivos
éguas.
A n
se, certame
presença d
bovinos. A
consumo, e
agradaria v
entretanto,
embriões o
sentimentos
de produçã
27,38
1 2
Tratamento
9
orcentagem da ocorrênrupos de tratamentos. gom CIDR®; e grupo 3 = 1
rente do observado por
m uma reação inflamató
estudo, observamos u
,22% (14/66) com 100%
rença pode ser explicad
tibiótica a base de gent
antes dos mesmos ter
ão indicação deste dispo
nte, ao fato do mesmo
e secreção mucopurul
diferença reside no fa
nquanto que os eqüinos,
er o animal manifesta
não seria o bastante, v
u simplesmente como m
tão conservadores nos
o ou mesmo instrumento
27,38
Seqüência1
9,0
3
cia de vaginite observadas em três diferentes rupo 1 = 8 dias com CIDR®; grupo 2 = 10 dias 0 dias com CIDR ® sem progesterona (controle.)
Morete et al., (2004), onde 74% dos animais
ria e obtiveram cura espontânea em 72 horas,
ma incidência deste processo inflamatório em
dos animais recuperados em 48 horas (Figura
a pelo fato de ter sido feita uma aspersão com
amicina (Gentrin®, Laboratório Ouro Fino) nos
em sido introduzidos na cavidade vaginal das
sitivo intravaginal para a espécie eqüina deve-
desencadear uma reação inflamatória local com
enta pouco abundante tal como ocorre nos
to de bovinos serem vistos como animais de
na sua maioria, não o são e ao proprietário não
ndo sinais de desconforto. Este argumento,
isto que animais utilizados como receptoras de
atrizes na maioria das vezes, não despertam
seus proprietários, sendo vistos como animais
s para uma atividade profissional ou esportiva.

33
Outro fator relevante reside no fato de que a reação inflamatória
desencadeada foi localizada e restrita à área ocupada pelo dispositivo tendo cura
espontânea dois a três dias após a sua retirada sem comprometimento de outras
partes do aparelho genital, além de não haver comprometimento da fertilidade
conforme verificou Newcombe, (1998). Estes efeitos restritos à cavidade vaginal
foram comprovados na primeira fase do estudo realizado com 05 éguas onde foram
feitas avaliações por meio de ultra-sonografia, exames de citologia e biópsia
uterinas, não tendo sido observada nenhuma alteração na integridade endometrial
que pudesse ser atribuída ao dispositivo.
Em 100% dos animais participantes do estudo, observou-se uma completa
recuperação em 72 horas sem que houvesse necessidade de qualquer tipo de
tratamento. Isso indica que esta vaginite não se constituiu, neste estudo, em um
empecilho ao uso do dispositivo em protocolos de sincronização na espécie eqüina
ao contrário do que muitos alegam, inclusive até os próprios fabricantes do
dispositivo, utilizado no experimento, e de seus similares.
6.2.5-Tempo decorrido entre a retirada do dispositivo até a manifestação do estro
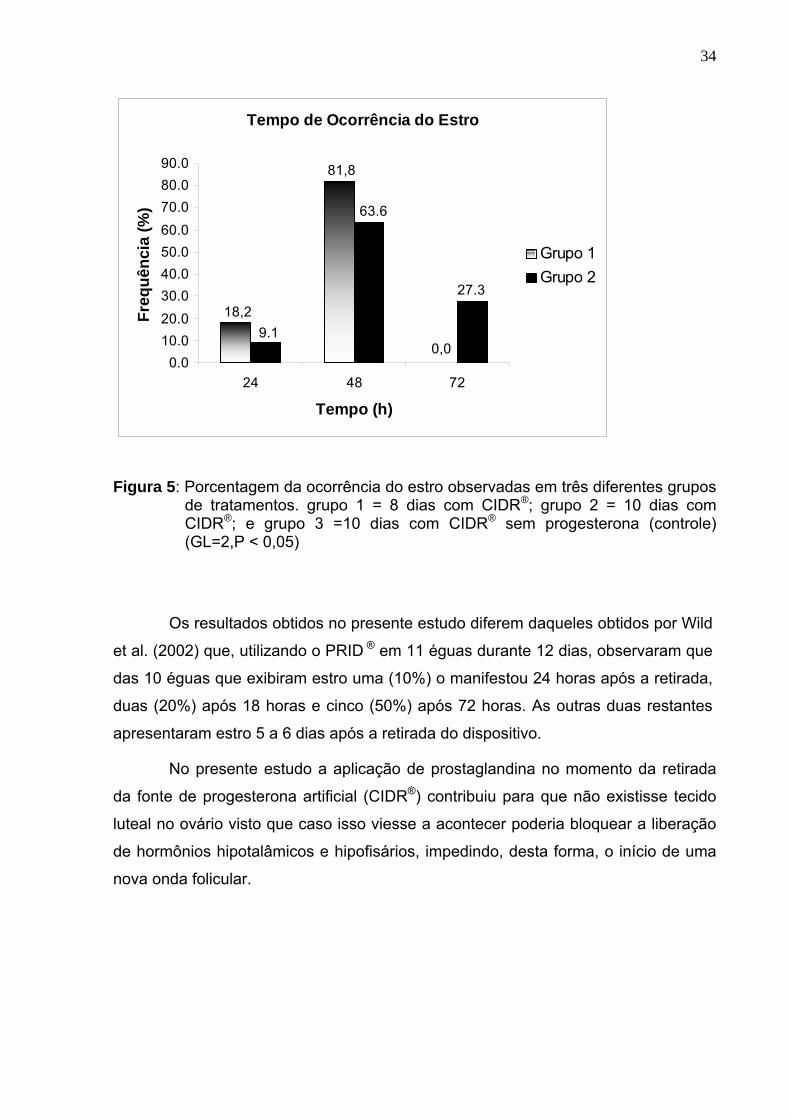
Observou-se uma diferença estatística significativa (p<0,05) entre os grupos
1 e 2 em relação ao tempo decorrido entre a retirada do dispositivo e a manifestação
do estro. Conforme pode ser observado na figura 5, as éguas do grupo 1
apresentaram a maior concentração de estro nas primeiras 48 horas com 81,8%
(18/22), enquanto que as éguas do grupo 2 apresentaram 63,64% (14/22) dos
animais com manifestações de estro até 48 e o restante até 72 horas, ou seja,
27,27% (6/22). Esta diferença pode ser explicada pelo menor tempo de exposição à
progesterona dos animais do grupo 1 que possa ter levado à uma menor
sensibilização do eixo hipotalâmico-hipofisário e que estes animais tenham tido um
menor período refratário à resposta ocasionada no momento da supressão do
fornecimento da progesterona com a retirada do dispositivo ou que a retirada da
fonte de progesterona após um período mais curto tenha ocorrido num momento
mais favorável da onda folicular.
34
Tempo de Ocorrência do Estro
18,2
81,8
0,0
63.6
27.3
9.1
0.010.020.030.040.050.060.070.080.090.0
24 48 72
Tempo (h)
Freq
uênc
ia (%
)
Grupo 1Grupo 2
Figura 5: Porcentagem da ocorrência do estro observadas em três diferentes grupos
de tratamentos. grupo 1 = 8 dias com CIDR®; grupo 2 = 10 dias com CIDR®; e grupo 3 =10 dias com CIDR® sem progesterona (controle) (GL=2,P < 0,05)
Os resultados obtidos no presente estudo diferem daqueles obtidos por Wild
et al. (2002) que, utilizando o PRID ® em 11 éguas durante 12 dias, observaram que
das 10 éguas que exibiram estro uma (10%) o manifestou 24 horas após a retirada,
duas (20%) após 18 horas e cinco (50%) após 72 horas. As outras duas restantes
apresentaram estro 5 a 6 dias após a retirada do dispositivo.
No presente estudo a aplicação de prostaglandina no momento da retirada
da fonte de progesterona artificial (CIDR®) contribuiu para que não existisse tecido
luteal no ovário visto que caso isso viesse a acontecer poderia bloquear a liberação
de hormônios hipotalâmicos e hipofisários, impedindo, desta forma, o início de uma
nova onda folicular.

35
6.2.6-Tempo decorrido entre a retirada do CIDR® até a detecção de um folículo
≥ 35mm
Não foi constatada diferença estatística significativa (p>0,05) entre os
grupos, sendo que a maioria dos animais no grupo 1 (72,73% ; 8/11), e 45,45%
(5/11) do grupo 2 apresentou o folículo ≥ 35 mm até 48 horas após a retirada do
CIDR®, enquanto que o grupo controle apresentou esta condição em até 96 horas
(11/22).
Pode-se concluir a partir destes dados, que o tempo de permanência do
dispositivo não interferiu na taxa de crescimento folicular. Isto, na verdade, já era
esperado uma vez que segundo Wild et al. (2002) a progesterona tem efeito
inibitório sobre a ação do hormônio responsável pela ovulação (LH), mas não
interfere na ação do hormônio responsável pelo crescimento folicular (FSH).
Morete et al. (2004) observou que a média de tempo decorrida entre a
retirada dos dispositivos e a aplicação de hCG foi de 64 + 12 hr (p<0,68) e a
ovulação foi 110 + 14 h para todos os grupos (p<0,82). Apesar de no momento da
retirada do dispositivo 74% das éguas apresentassem vaginite purulenta, este
quadro desapareceu espontaneamente em 72 horas. Os autores concluíram que o
uso dos dispositivos diminuiu a o período de transição e induziu a ovulação em 38%
das éguas tratadas.
A aplicação de uma segunda dose de prostaglandina no momento da
retirada do CIDR® deveu-se ao fato de existir a possibilidade de, no momento da
aplicação da primeira dose de prostaglandina o corpo lúteo presente não
apresentasse maturidade para ser susceptível à ação da prostaglandina, o que só
ocorre naquelas estruturas formadas após 6 dias da ovulação conforme descrito por
Loy et al. (1981).. Outra possibilidade é a de ocorrer ovulação de um folículo mesmo
durante o tratamento, onde se pressupõem ter ocorrido o bloqueio da secreção de
LH através da progesterona, com a formação de um corpo lúteo o que iria interferir
no surgimento de uma nova onda folicular ao final do tratamento caso a
prostaglandina não fosse aplicada.

36
6.2.7-Tempo decorrido entre a aplicação do indutor de ovulação até a ovulação
Com relação ao tempo decorrido entre a aplicação do indutor de ovulação e
a ovulação e considerando somente os animais que responderam à terapia
ovulatória (4 animais do grupo 1; 6 do grupo 2 e 10 do grupo controle não ovularam,
sofrendo luteinização prematura ou apresentaram regressão folicular) todos os 3
grupos apresentaram uma maior concentração de ovulações 48 horas após a
aplicação dos indutores de ovulação com 100% (8/8) para Lutalyse® e 100% (8/8)
para Vetecor® no grupo 1; 66,66% (4/6) para Lutalyse®, 75% (6/8) para Vetecor® no
grupo 2 e 0 % (0/10) para Lutalyse® e 100% (6/6) para Vetecor® no grupo controle.
Estes resultados diferem daqueles obtidos por Newcombe (1997), onde 98% (57/58)
das éguas apresentaram ovulações entre os dias zero e 6 pós-tratamento após a
aplicação de 2.500 U.I de hCG.
O fato de nenhuma das éguas do grupo 3 (controle) ter ovulado após a
aplicação do Lutalyse® pode ser explicado pelo fato de que nenhuma delas
apresentava um folículo com diâmetro mínimo de 40 mm no momento da aplicação
do medicamento. Autores como Mckinnon (1997) e Hafez (2004) descreveram que
somente folículos com diâmetro igual ou superior à 40 mm são responsivos à ação
ovulatória das prostaglandinas. Isto ocorreu devido ao fato de que, quando o
monitoramento foi iniciado todas apresentavam folículos menores que 40 mm (a
aplicação do Lutalyse® era realizada no momento em que o folículo atingia o
diâmetro de 35 mm) enquanto que algumas éguas dos grupos 1 e 2 apresentaram
folículos com diâmetro igual ou superior à 40mm no momento da retirada do CIDR®,
o que poderia explicar a resposta favorável dos animais destes grupos.
Não deve ser deixado de levar em conta que, não só o diâmetro folicular no
momento da aplicação do Lutalyse® é importante para explicar a diferença
observada entre a resposta ao tratamento entre os grupos tratados e o grupo
controle mas também o fato dos grupos responsivos terem sido previamente
sensibilizados por meio do uso da progesterona contida no CIDR®.
Hafez, (2004) relatou a propriedade das prostaglandinas em induzir a
ovulação em folículos com diâmetro igual ou maior de 40 mm. Com o objetivo de
verificar a eficiência desta droga com este intuito, a mesma foi empregada em
metade dos animais que foram utilizados no experimento, sendo que a outra metade
recebeu o hCG, droga universalmente consagrada como sendo aquela que

37
apresenta os melhores resultados quando o objetivo é o de induzir a ovulação em
éguas.
Este procedimento foi adotado visto que o aumento dos índices de fertilidade
proporcionados pela utilização do hCG já foi confirmado por trabalhos científicos de
vários autores como MacDonald (1983) e Palmer (1985) e a não utilização destas no
grupo controle poderia nos levar a conclusões distorcidas.
Diferença significativa quanto ao número de éguas ovuladas com Lutalyse®,
foi observada pelo teste do (p<0,05) quando comparados os três grupos
experimentais. Para validar o motivo de tal diferença, se devido ao tempo de
presença do dispositivo na cavidade vaginal, ou a presença do hormônio, aplicou-se
o referido teste somente nos grupos 1 e 2, o qual demonstrou resultado não-
significativo ( =0,95
2χ
2χ ns). Dessa forma, fica evidenciado que a diferença identificada
se deve a presença do hormônio e não ao tempo de exposição do mesmo.
Assim, com relação à percentagem de éguas ovuladas com Lutalyse®, observou-se uma diferença estatística significativa (p<0,05) entre os grupos tratados
e o controle o que poderia reforçar a nossa recomendação de se utilizar o protocolo
do grupo 1 pelas vantagens já descritas. (figura 9)
Com relação à percentagem de éguas ovuladas com Lutalyse®, observou-se
uma diferença estatística significativa (p<0,05) entre os grupos tratados o que
poderia reforçar a nossa recomendação de se utilizar o protocolo do grupo 1 pelas
vantagens já descritas e pelo mesmo ter apresentado uma maior taxa de ovulação
quando comparado aos outros dois grupos (figura 6).
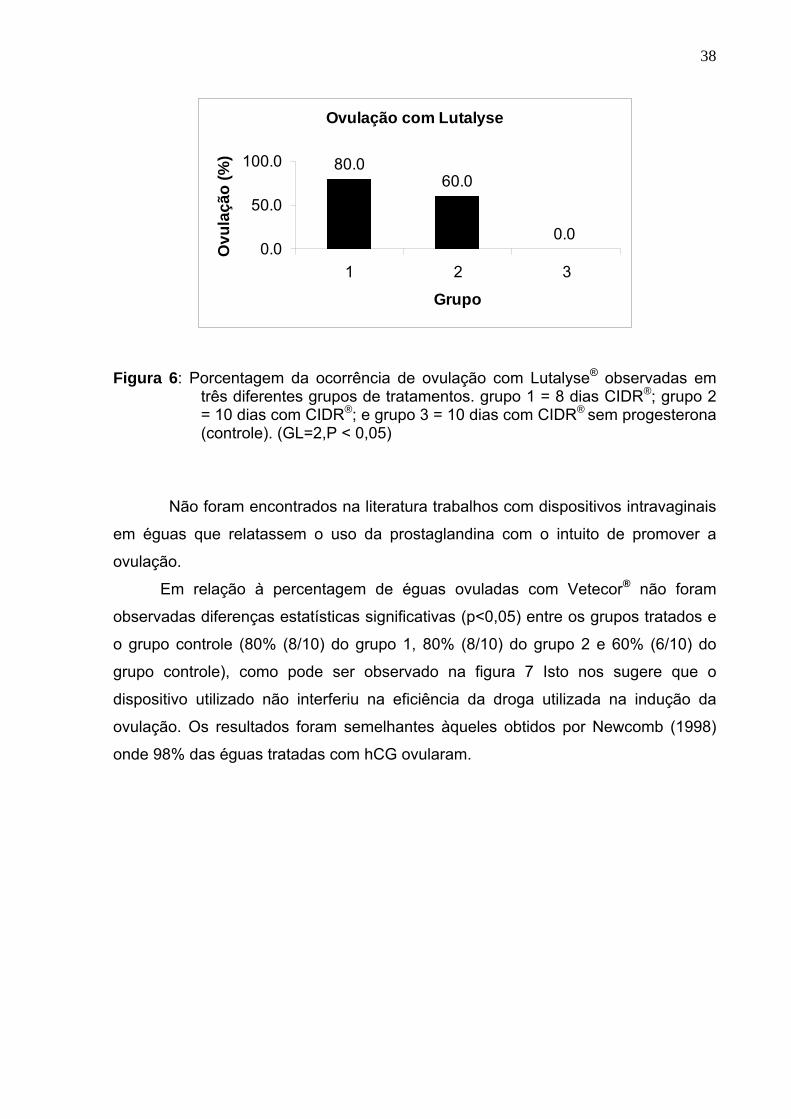
38
Ovulação com Lutalyse
80.060.0
0.00.0
50.0
100.0
1 2 3
Grupo
Ovu
laçã
o (%
)
Figura 6: Porcentagem da ocorrência de ovulação com Lutalyse® observadas em
três diferentes grupos de tratamentos. grupo 1 = 8 dias CIDR®; grupo 2 = 10 dias com CIDR®; e grupo 3 = 10 dias com CIDR® sem progesterona (controle). (GL=2,P < 0,05)
Não foram encontrados na literatura trabalhos com dispositivos intravaginais
em éguas que relatassem o uso da prostaglandina com o intuito de promover a
ovulação. Em relação à percentagem de éguas ovuladas com Vetecor® não foram
observadas diferenças estatísticas significativas (p<0,05) entre os grupos tratados e
o grupo controle (80% (8/10) do grupo 1, 80% (8/10) do grupo 2 e 60% (6/10) do
grupo controle), como pode ser observado na figura 7 Isto nos sugere que o
dispositivo utilizado não interferiu na eficiência da droga utilizada na indução da
ovulação. Os resultados foram semelhantes àqueles obtidos por Newcomb (1998)
onde 98% das éguas tratadas com hCG ovularam.
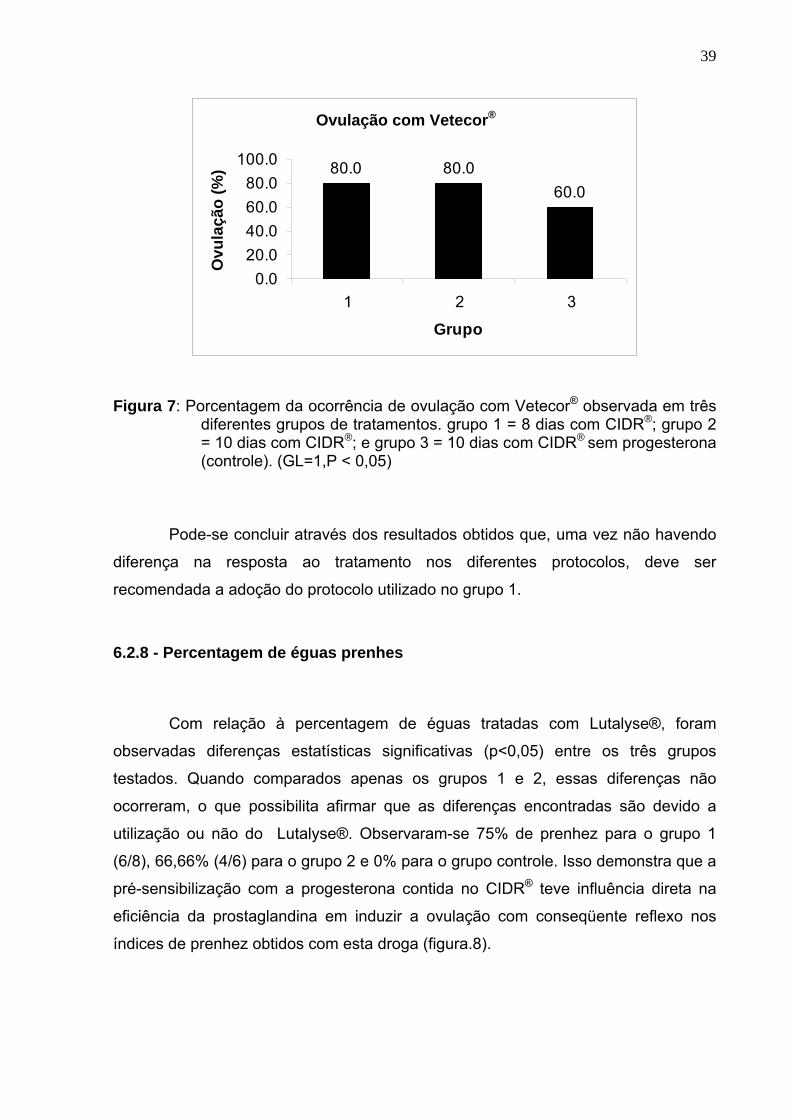
39
Ovulação com Veronal
80.0 80.060.0
0.020.040.060.080.0
100.0
1 2 3
Grupo
Ovu
laçã
o (%
)
Ovulação com Vetecor®
Figura 7: Porcentagem da ocorrência de ovulação com Vetecor® observada em três diferentes grupos de tratamentos. grupo 1 = 8 dias com CIDR®; grupo 2 = 10 dias com CIDR®; e grupo 3 = 10 dias com CIDR® sem progesterona (controle). (GL=1,P < 0,05)
Pode-se concluir através dos resultados obtidos que, uma vez não havendo
diferença na resposta ao tratamento nos diferentes protocolos, deve ser
recomendada a adoção do protocolo utilizado no grupo 1.
6.2.8 - Percentagem de éguas prenhes
Com relação à percentagem de éguas tratadas com Lutalyse®, foram
observadas diferenças estatísticas significativas (p<0,05) entre os três grupos
testados. Quando comparados apenas os grupos 1 e 2, essas diferenças não
ocorreram, o que possibilita afirmar que as diferenças encontradas são devido a
utilização ou não do Lutalyse®. Observaram-se 75% de prenhez para o grupo 1
(6/8), 66,66% (4/6) para o grupo 2 e 0% para o grupo controle. Isso demonstra que a
pré-sensibilização com a progesterona contida no CIDR® teve influência direta na
eficiência da prostaglandina em induzir a ovulação com conseqüente reflexo nos
índices de prenhez obtidos com esta droga (figura.8).

40
Taxa de Prenhêz com Lutalyse
75,066,7
0102030405060708090
1 2
Grupo
Freq
uênc
ia (%
)
Figura 8 Porcentagem de prenhez com Lutalyse® observadas em dois diferentes grupos de tratamentos. Grupo 1 = 8 dias com CIDR®; Grupo 2 = 10 dias com CIDR® (GL=1, P < 0,05)
Não foi observada diferença estatística significativa (p>0,05) entre os grupos
tratados com Vetecor®, sendo 50% para o grupo 1 (04/08), 75% (6/8) para o grupo 2
e 66,66% (04/06) para os tratados com Vetecor® (Figura 9); não havendo
interferência nas taxas de prenhez nem pelo uso do dispositivo, nem tampouco pelo
uso do hCG como droga indutora de ovulação .
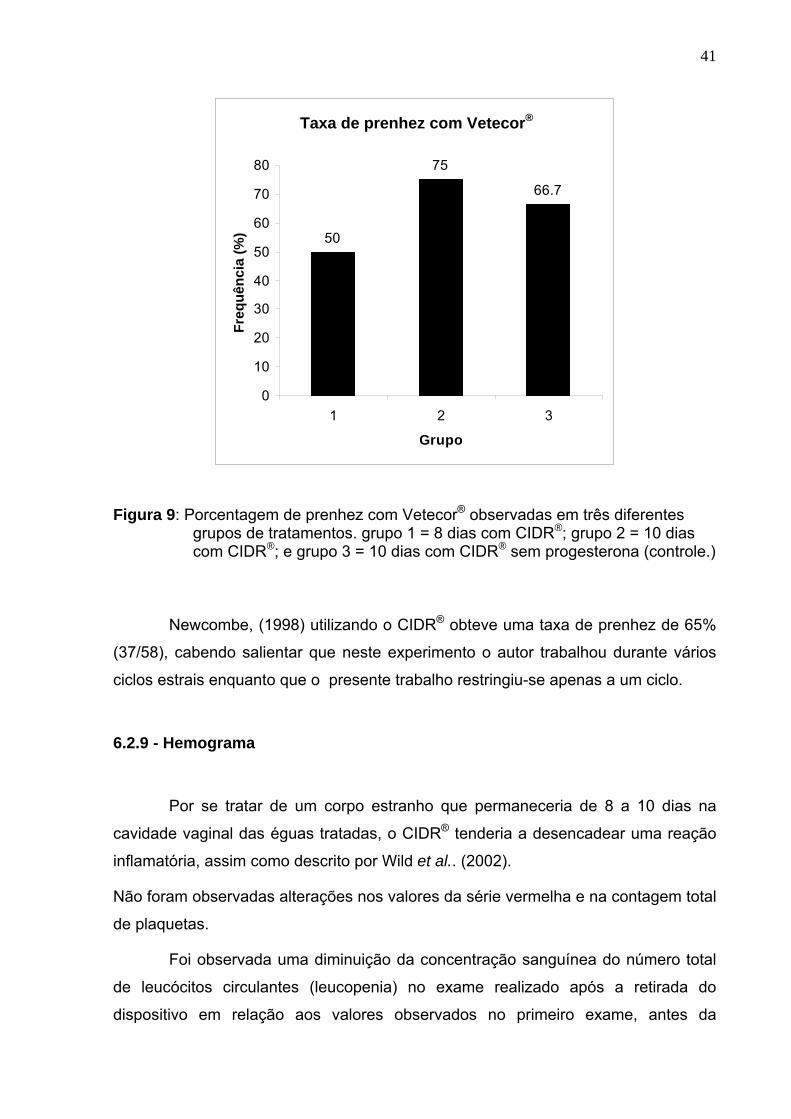
41
Taxa de Prenhêz com Veronal
50
75
66.7
0
10
20
30
40
50
60
70
80
1 2 3
Grupo
Freq
uênc
ia (%
)
Taxa de prenhez com Vetecor®
Figura 9: Porcentagem de prenhez com Vetecor® observadas em três diferentes grupos de tratamentos. grupo 1 = 8 dias com CIDR®; grupo 2 = 10 dias com CIDR®; e grupo 3 = 10 dias com CIDR® sem progesterona (controle.)
Newcombe, (1998) utilizando o CIDR® obteve uma taxa de prenhez de 65%
(37/58), cabendo salientar que neste experimento o autor trabalhou durante vários
ciclos estrais enquanto que o presente trabalho restringiu-se apenas a um ciclo.
6.2.9 - Hemograma
Por se tratar de um corpo estranho que permaneceria de 8 a 10 dias na
cavidade vaginal das éguas tratadas, o CIDR® tenderia a desencadear uma reação
inflamatória, assim como descrito por Wild et al.. (2002).
Não foram observadas alterações nos valores da série vermelha e na contagem total
de plaquetas.
Foi observada uma diminuição da concentração sanguínea do número total
de leucócitos circulantes (leucopenia) no exame realizado após a retirada do
dispositivo em relação aos valores observados no primeiro exame, antes da

42
colocação dos dispositivos intravaginais em 100% dos animais de ambos os grupos
que utilizaram o CIDR® com progesterona (grupo 1 e grupo 2) e do grupo que
utilizou o CIDR® sem progesterona (grupo controle).
Isso demonstra que o processo inflamatório foi desencadeado pelo trauma
mecânico proporcionado pela presença do dispositivo e não pela ação da
progesterona, ao contrário observado por Palmer (1985) que atribuiu à presença do
Altrenogest (Regu-Mate®) impregnado em esponjas colocadas na cavidade vaginal
como agente causal da inflamação e aderência observadas na mucosa vaginal.
A leucopenia desencadeada pelo uso do CIDR® e a observação da cura de
processos inflamatórios endometriais através do uso deste em alguns animais no
decorrer do experimento, apesar das contra-indicações, sugere que a migração
leucocitária ocorra para todo o aparelho reprodutivo e não somente para a cavidade
vaginal.
Não foram encontrados na literatura trabalhos com dispositivos intravaginais
em éguas que tenham investigado a relação do seu uso com possíveis alterações
hematológicas.
6.2.10 - Perdas embrionárias
Foi registrada uma percentagem de 9,9% de perdas embrionárias no grupo 2
(2/22), não tendo sido observada nenhuma perda nos grupos 1 e controle.
Este baixo índice nos leva a concluir não ter havido influência do CIDR® na
fertilidade dos animais tratados.
Não foram encontrados na literatura trabalhos com dispositivos intravaginais
em éguas que relatassem o percentual de perdas embrionárias ocorrido após o uso
destes.

43
7-CONCLUSÕES
O CIDR® mostrou-se eficiente para a sincronização de estros e ovulações,
sendo sugerido a utilização do protocolo de 8 dias pela redução de custos que este
proporciona
O uso do CIDR® não comprometeu os índices de fertilidade, tendo os mesmos
se mantido dentro da média esperada para a espécie eqüina em um ciclo estral.
O uso da prostaglandina mostrou-se eficiente para a indução da ovulação em
previamente tratadas com o CIDR®, constituindo-se na droga de escolha para este
propósito uma vez que o seu preço é expressivamente inferior ao hCG, e pelo fato
adicional de não sensibilizar o organismo animal a produzir anticorpos.
O único efeito sistêmico causado pelo uso do CIDR® foi uma leucocitose, que,
no entanto, não comprometeu o estado de saúde geral dos animais
O tratamento prévio do CIDR® com solução antibiótica a base de gentamicina
parece ter contribuído para a redução da resposta inflamatória, devendo, pois, ser
adotado como rotina.
Esta alta taxa de sincronização observada nos mostra que o CIDR® pode
constituir-se numa ferramenta reprodutiva extremamente útil nos protocolos de
sincronização tanto para a inseminação de matrizes como para éguas receptoras de

44
embriões, reduzindo substancialmente o número destas em um programa de
transferência de embriões.

45
8-REFERÊNCIAS BIBLIOGRÁFICAS
AGRIMED: costs of hormones in oestrus sinchronyzation; http//www.agrimed.com em 02/01/2 0,05
ALEXANDER, S.L. E IRVINE, C.H.G. (1991): Control of onset of breeding season in the mare and its artificial regulation by progesterone treatment. J. Reprod. Fertil. Suppl., 44:307-319.
ALLEN, W.R., URWIN, V., SIMPSON, D.J., GREENWOOD, R.S., CROWHURST, R.C. E ELLIS, D.R (1980).: Preliminary studies on the use of an oral progestagen to induce estrous and ovulation in seasonally anoestrous Thoroughbred mares. Equine Vet. J.,12:141-145.
ALLEN, W. R. (1996).Annual Report of th Thoroughbred Breeders Association, pp 26-30.
ALMEIDA ET AL.; estrus synchronization and follicular dynamics of crioulo mares by norgestomet, melengestrol acetate and altrenogest treatments (2001).. Braz.J.Vet Res. Anim. Sci. Vol 38 No. 6. São Paulo.
ARTHUR, G.H. E ALLEN, W.E. (1972): Clinical observation on reproduction in a pony study. Equine Vet. J., 4:109-117.
BANCUS, K.L. ET AL..: Effect of transportation on early embryonic death in mares. J. Anim. Sci (1990a).., 68:345-351.
BANCUS, K.L. ET AL.. (1990b).Effect of transportation on the estrous cycle and concentration of hormones in mares 1990b. J. Anim. Sci., 68:419-426.

46
BANCUS, K.L. ET AL.. (1990b).: Effect of transportation on early embryonic death in mares. J. Anim. Sci., 68:345-3511990b.
BANCUS, K.L. ET AL.. (1990b): Effect of transportation on the estrous cycle and concentration of hormones in mares. J. Anim. Sci., 68:419-426.
BRISTOL, F. (1981): Study on estrous synchronization in mares. Proc. Soc. Theriogenology, 258-264.
COLBERN, G.T. ET AL.. (1987): Development of a model to study endometritis in mares. J. Equine Vet. Sci. 7:73-7.
DAELS, P.F. ET AL.. (1987): Effect of Salmonella typhimurium endotoxin on PFG2α release and fetal death in the mare. J. Reprod. Fertil. Suppl., 35:485-492,.
DOUGLAS, R.H., BURNS, P.J. E HERSHMAN, L. (1985).: Physiological and commercial parameters for producing progeny for subfertile mares by embrio transfer. Equine Vet. J. Suppl., 3: 111-114,
DUMITRIU, S. E DIMITRIU, M.(1994): Polymeric Drug Carriers. In: Dumitriu, S. (ed) Polymeric Biomaterials. 2.ed. New York: Marcel Dekker. Cap. 14:435-724.
GINTHER, O.J. (1992). Reproductive Biology of the mare. Cross Plains, Wisconsin: Equiservices.
GINTHER, O. (1985) : Embryonic loss in mares: Incidence, time of occurrence and hormonal involvement. Theriogenology, 23:77-83.
GINTHER, O.J. et al. (1977). Estrus, ovulatin and conception following synchronization with progesterone, PGF 2 alfa and HCG in pony mares. J. Anim.Sci 44:431-437.
GOMES, PIMENTEL : Introdução à estatística experimental. Quinta Edição. Editora Globo.
HAFEZ E HAFEZ (2004). Reprodução Animal. Sétima Edição. Editora Manole
HANDLER, S.SCHONLIEB, H. O. HOPPEN, C. AURICH (2004). Effect of endogenous progesterone concentration on the efficacy of PRIDTM treatment in mares. 15th International Congress on Animal Reproduction, Porto Seguro, Brazil.
HAWKINS, D. L., NEELY, D.P., E STABENFELDT, G.H (1979.): Plasma progesterone concentrations derived from the administration of exogenous progesterone to ovariectomized mares. J. Reprod. Fertil. Suppl., 27:211-216.
HINRICHS, K., SERTICH, P.L., CUMMINGS, M.R. E KENNEY, R.M (1985.): Pregnancy in ovariectomized mares achieved by embryo transfer. Equine Vet. J. Suppl., 3:74-75.
HOLTAN, D.W., SQUIRES, E.L., LAPIN, D.R. E GINTHER, O.J (1979).: Effect of ovariectomy on pregnancy in mares. J. Reprod. Fertil. Suppl., 27:457-463.

47
HUGHES, J.P., STABENFELT, G.H. E KENNEDY, P.C. (1980): The estrous cycle and selected functional and pathologic ovarian abnormalities in the mare. Vet. Clin. North Am. (Large Anim. Pract.), 2(2): 225-239..
LOY, R.G. E SWAN, S.M. (1966).: Effects of exogenous progestogens on reproductive phenomena in mares. J. Anim. Sci., 25:811-879.
LOY, R.G. (1980). Characteristics of pastpartum reproduction in mares. VeterinaryClinics of North America: Large Animal Practice 2: 345-359.
LOY, R.G., PEMSTEIN, R., O’CANNA, D. E DOUGLAS,R.H (1981).: Control of ovulation in cycling mares with ovarian steroids and prostaglandin. Theriogenology, 15:191-200,
LOY, R.G., EVANS, M.J., PEMSTEIN, R., E TAYLOR, T.B. (1982).: Effect of injected ovarian steroids on reproductive patterns and performance in postpartum mares. J. Reprod. Fertil. Suppl., 32:199-204.
MCKINNON, O. A., Nobelius, A. M, Tarrida del Marmol Figuera, S.,Sskidmore, J., Vasey, J.R. and Trigg, T.E, (1997) Predictable ovulation in mares treated ith an implant of the GnRh anlogue deslorelin. Equine Veterinary Journal 25: 321-3232.
MCKINNON. O.A, VASEY, J. R., LESCUN, T. B e TRIGG, T. E, (1996). Repeated use of a GnRh anlogue deslorelin (Ovuplant) for hastening ovulation in transitional mares. Equine Veterinary Journal 25: 321-323.
MCKINNON, A.O. et al.. (1987): Diagnostic ultrasonography of uterine pathology in the mare. Proc. Am. Assoc. Equine Pract., pp. 605-622..
MCKINNON, A.O., SQUIRES, E.L., CARNEVALE, E.M. E HERMENET, M.S (1988a).: Ovariectomized steroid-treated mares as embryo transfer recipients and as model to study the role of progestin in pregnancy maintenance. Theriogenology. 29:1011-1063.
MCKINNON, A.O., et al. (1988b): Ultrasonographic studies on the reproductive tract of postpartum mares: Effect of involution and uterine fluid on pregnacy rates in mares with normal and delayed first postpartum ovulatory cycles. J. Am. Vet. Med. Assoc., 192:350-353,.
MCDONALD, L.E (1983).: Hormônios que atuam na reprodução, in: Farmacologia e Terapêutica em Veterinária, Jones, L.M., Boott, N.H. e McDonald, L.E., Ed Guanabara Koogan- Rio de Janeiro, 4°ed.,.
MEINERT, S. SILVA, J.F. S.KROETZ, i. KLUG, E. TRIGG, T.E e HOPPEN, H.O .(1993). Advancing the time of ovulation in the mare with short acting implant releasing the GnRh anlogue deslorelin. Equine Veterinary Journal 24: 65-68.
MEYERS, P, BOWMAN, T.BLODGETT, G.CONBOY, J.S. GIMENEZ, T.REID, MP. TAYLOR, B.C.THAYER, J. JOCHLE, W. e TRIGG T. E (1997). Use of a GnrH analogue, deslorelin acetate, in a slow release implant to accelerate ovulation in oestrus mares. Veterinary Record 140: 249-252

48
MORETE, M.AZURRA, J.M. VEJA, A. L. MIGLIORISI e R.L. DE SOTA (2004). Control of the estrus cycle in mares with an intra-vaginal progestrone-releasing device. 15th International Congress on Animal Reproduction,, Porto Seguro, Brazil.
MOTTERSHEAD. JOS; Inducing early estrus in mare, Equine reproduction articles; www. equine-reproduction.com, em 02/03/2005.
MOTTERSHEAD. JOS; Equine Yahhoo Groups; assunto: regumate; 01/04/2004.
NEELY, D.P. (1988): Progesterone/progestin therapy in the brood-mare. Proc. Am. Assoc. Equine Pract., pp 203-218.
NEELY, D.P (1979): Studies on the control of luteal fuction and prostaglandin release in the mare. PhD Thesis, University of California, Davis.
NEWCOMB JONH R. (1998) Anoestrus in mares: experiences using Ovuplant, a short-term implant of the GnRh analogue deslorelin, following implantation with CIDR-B, a progesterone releasing intravaginal device. Irish Veterinary Journal 51: 582-585.
NEWCOMB JONH R e WILSON, M.C (1997).The use of progesterone releasing intravaginal devices to induce oestrus and ovulation in anoestrous Standardbred mares in Australia. Equine Practice 19: 13-21
PALMER, E. (1985): Recent attempts to improve synchronization of ovulation and to induce superovulation in the mare. Equine Vet, J. Suppl., 3:11-18,.
PARRY-WEEKS, L.C. E HOLTON, D.W. (1987).: Effect of Altrenogest on pregnancy maintenance in unsychronized equine embrio recipients. J. Reprod. Fertil. Suppl., 35:433-438.
PFIZER DO BRASIL-Produtos; http//www.pfizer.com.br. em 20/03/2005
SCHEFFRAHN, NS, ET AL.. (1982): Reproductive hormones secretions in pony mares subsequent to ovulation control durig late winter. Theriogenology, 17:571-585,.
SHARP D.C. (1980). Environmental influences of reproduction in horses. In: Veterinary Clinics of North America: Large Animal Practice. Edited by J.Hughes.Philadelphia: Saunders, pp 207-273.
SHIDELER, R.K. ET AL.. (1982): Progesterone therapy of ovariectomized pregnant mares. J. Reprod. Fertil. Suppl., 32:459-464,.
SHIDELER, R.K. ET AL.: (1983) The effect of Altrenogest, anoral progestin, on hematologic and biochemical parameters in mares. Vet.Hum.Toxicol, 25:250-252.
SHOEMAKER, C.F., SQUIRES, E.L. E. SHIDELER, R.K. (1989) Safety of Altrenogest in pregnant mares and on health and development of offspring. J. Equine Vet. Sci. 9: 69-72.

49
SILVIA, P.J., SQUIRES, E.L. E NETT, T.M. (1986): Changes in the hypothalamic-hypophyseal axis of mares associated with seasonal reproductive recrudescence. Biol. Repod., 35:897-905.
SQUIRES, E.L. ET AL. (1983): Relationship of Altrenogest to ovarian activity, hormone concentrations and fertility of mares. J. Anim. Sci., 56:901-910.
SQUIRES, E.L., STEVENS, W.B., MCGLOTHLIN, D.E. E PICKETT, B.W (1979).: Effect of an oral progestin on the estrous cycle and fertility of mares. J. Anim. Sci., 49:729-735.
SQUIRES, E.L., NETT, T.M., WIEPZ, G.J. E MOCK, E.J. (1885): Use of an enzyme assay for determination of progesterone in broodmares. Proc. Am. Assoc. Equine Pract., pp. 565-570.
SQUIRES, E.L. E GHINTER, O.J. (1975.): Collection technique and progesterone concentration of ovarian and uterine venous blood in mares. 1975. J. Anim. Sci., 40:275-281.
SQUIRES, E.L., SHIDELER, R.K. E MCKINNON, A.O. (1989): Reproductive performance of offspring from mares administered Altrenogest during gestation. J. Equine Vet. Sci. 9: 73-76.
TAYLOR, T.B., PEMSTEIN, R., E LOY, R.G (1982).: Control of ovulation in mares in the early breeding season with ovarian steroids and prostaglandin., J. Reprod. Fertil. Suppl., 32: 219-224.
VAN NIEKERK, C.H. E MORGENTHAL, J.C (1982).: Fetal loss and stress on plasma progesteron levels in pregnant Throgoughbred mares. J. Reprod. Fertil. Suppl., 32:453-457.
VIDELA DORNA, F. GONZALEZ, C. BIANCHI e M.A. ABA (2004).Changes after insertion of intravaginal devices containing two different doses of progesterone in mares. 15th International Congress on Animal Reproduction, Porto Seguro, Brazil.
WASHBURN, S.M., KLESIUS, P.H., GANJAMV.K. E BROWN, B.G. (1982): Effect of estrogen and progesterone on the phagocytic response of ovariectomized mares infested in utero with β-hemolytic Streptococci. Am. J. Vet. Res., 43:1367-1375.
WEBEL, S.K (1975).: Estrous control in horses with a progestin. J. Anim. Sci., 41:385.
WEBEL, S.K. E SQUIRES, E.L (1982): Control of the oestrous cycle in mares with Altrenogest. J. Reprod. Fertil. Suppl., 32: 193-198.
WIEPZ, G.J. SQUIRES, E.L. E CHAMPMAN, P.L. (1988): Effects of norgestomet, Altrenogest and/or estradiol on folicular and hormonal characteristics of late transitional mares. Theriogenology, 30:181-193.
WILD, R. OSCAR ET AL. (2002). Uso de un dispositivo intravaginal para el control del estro en yeguas. Revista Zootecnia Tropical, 20 (4); 483-492.

50
ZIMBELMAN, R.G. E SMITH, L.W. (1966) Control of ovulation in cattle with melengestrol acetate. J. Reprod. Fertil. 11:185-19.