estrutura da comunidade meiofaunÍstica de ......figura 16: abundancia dos gêneros de nematoda:...
TRANSCRIPT

Universidade Federal de Pernambuco
Centro de Ciências Biológicas
Departamento de Zoologia
Pós-Graduação em Biologia Animal
ESTRUTURA DA COMUNIDADE MEIOFAUNÍSTICA DE
SUBSTRATO INCONSOLIDADO DE DOIS RECIFES
TROPICAIS, COM E SEM PROTEÇÃO AMBIENTAL,
EM TAMANDARÉ (PERNAMBUCO – BRASIL).
ÉRIKA CAVALCANTE LEITE DOS SANTOS
RECIFE
2010

Érika Cavalcante Leite dos Santos
ESTRUTURA DA COMUNIDADE MEIOFAUNÍSTICA DE
SUBSTRATO INCONSOLIDADO DE DOIS RECIFES
TROPICAIS, COM E SEM PROTEÇÃO AMBIENTAL,
EM TAMANDARÉ (PERNAMBUCO – BRASIL).
Dissertação apresentada ao programa de Pós-Graduação em Biologia Animal da Universidade
Federal de Pernambuco como requisito para a obtenção
do título de mestre.
Orientador: Prof. Dr. José Roberto Botelho de Souza.
Co-Orientadora: Profa. Dra. Clélia Márcia Cavalcanti da Rocha.
Recife 2010

Santos, Érica Cavalcante Leite dos Estrutura da comunidade meiofaunística de substrato inconsolidade de dois recifes tropicais, com e sem proteção ambiental em Tamandaré (Pernambuco-Brasil)/ Érica Cavalcante Leite dos Santos. – Recife: O Autor, 2011. 93 folhas : il., fig., tab.
Orientador: José Roberto Botelho de Souza Co-Orientador: Clélia Márcia Cavalcanti da Rocha
Dissertação (mestrado) – Universidade Federal de Pernambuco. Centro de Ciências Biológicas, Biologia Animal, 2011.
Inclui bibliografia
1. Nematoda 2. Polychaeta 3. Pernambuco I. Título.
578.778 9 CDD (22.ed.) UFPE/CCB-2011-161


AGRADECIMENTOS
Ao CNPq pela concessão da bolsa de Mestrado; Ao IBAMA e ao ICM-Bio pela autorização dada para a execução deste trabalho; Ao CEPENE – ICM-Bio por ceder seu alojamento durante as coletas; Ao Projeto Recifes Costeiros, na pessoa do Prof. Dr. Mauro Maida, por todo apoio logístico dado nas coletas, sem vocês esse trabalho não teria saído, muito obrigado; Ao Prof. Dr. José Roberto Botelho por ter me aceitado como sua orientanda, pelo apoio dado durante a execução deste trabalho e pela amizade; À Profa. Dra. Clélia da Rocha, por ter continuado comigo como co-orientadora, por estar sempre presente com seus conselhos, me orientando na vida, e pela amizade; Aos integrantes da banca, por terem aceitado o convite e pelas contribuições dadas a este trabalho; Aos meus ajudantes de coleta Clélia da Rocha, José Roberto Botelho, Eduardo Cavalcante, Kyllderes Lima, Henrique Maranhão, Juliana Rocha, Danielle Lopes, Selma Vasconcelos, Glória Maria, Kamylla Mayara e Adriane Mendes, muito obrigada, a ajuda de vocês foi imprescindível; Aos barqueiros “Inho” e “Seu Ruben” pela disponibilidade, paciência, cuidados, ajudas e conversas durante as coletas; À Profa. Dra. Fernanda Amaral e ao Prof. Dr. Carlos Perez pelo empréstimo de equipamento de mergulho; Ao Prof. Dr. Paulo Santos pelo empréstimo de cubeta para medição do microfitobentos; A Priscila Porchat pelo apoio e conselhos sobre os microfitobentos; À Profa. Dra. Virág Venekey pela identificação dos Nematoda, disponibilidade e conselhos a este trabalho; Ao Laboratório de Porífera, especificamente a Profa. Dra. Josivete Pinheiro e ao Prof Dr. Ulisses Pinheiro por me permitir usar suas dependências sempre que precisei; Ao Laboratório de Fisiologia Vegetal, na pessoa da Profa. Dra. Jarcilene pelo uso de suas dependências e equipamento; Ao Laboratorio de Ecotoxicologia, na pessoa do Prof. Dr. Paulo Carvalho por me permitir usar seus equipamentos e dependências;

A todos que fazem parte do Laboratório de Meiofauna pela disponibilidade sempre que precisei e pelas boas conversas; A todos que fazem ou fizeram parte do Laboratório de Comunidades Marinhas – LACMAR durante esta etapa: Selma Vasconcelos, Adriane Mendes, Glória Maria, Paulo Bonifácio, Danielle Lopes, Cristiana Sette, Kamylla Mayara, Daniele Maciel, Ravi Luna, Mariana e Natália, vocês fizeram este trabalho se tornar mais leve e nossas tardes muito mais alegres, obrigada pela amizade e carinho que tem por mim; Ao enfim Laboratório de Porífera: Josivete Pinheiro, Ulisses Pinheiro, Elielton, Suzane, Paulo Bonifácio, Helcy, Gil, Renato, Juliana e Lucas pela amizade;
À minha turma de mestrado, pela amizade que construímos, pelos momentos felizes que passamos juntos e finalmente por ter a certeza que seremos amigos de longa data; À Daniel de Oliveira, Débora Barbosa e Mônica Marinho, por sermos tão unidos desde o começo, na UFRPE, pelo apoio mútuo nessa etapa de nossas vidas acadêmicas e pela amizade que aprendemos a cultivar; À todos os meus amigos que contribuíram direta e indiretamente para minha sanidade mental, sem dúvida nenhuma fundamental para a execução deste trabalho; À Daniel Lago pela companhia em todos os momentos, pelas palavras e demonstração de apoio e carinho, À minha família, especialmente a minha mãe Adeilda Leite e ao meu pai Atayde dos Santos que aprenderam a aceitar minha escolha em ser bióloga, e hoje se mostram meus fãs mesmo que timidamente, muito obrigada, este apoio é muito importante para mim. Muito Obrigada.

RESUMO
O objetivo deste trabalho foi conhecer a estrutura de comunidade da meiofauna de substrato inconsolidado em ambiente recifal, suas interações com fatores ambientais e relacioná-la com a proteção ambiental e a influência do recife sobre a mesma. Foram feitas coletas nos meses de abril e setembro de 2008, dentro e fora de uma área de proteção ambiental. As duas áreas são compostas por areia grossa, com baixo teor de matéria orgânica. Foram encontrados 15 táxons da meiofauna, 56 gêneros de Nematoda e 18 gêneros de Polychaeta. A meiofauna apresentou diversidade foi maior na área protegida e na área mais afastada do recife, sendo estes resultados significativos. Os Nematoda não apresentaram diferença significativa de diversidade entre as áreas, tanto em relação a proteção quanto em relação ao afastatamento do recife. Os Polychaeta também não apresentou diferença significativa na diversidade entre as áreas comparadas, apesar da riqueza ser significativamente maior na área mais afastada do recife. As diferenças encontradas em relação aos parâmetros sedimentológicos e em relação aos parâmetros de estrutura biológica foram pequenas, indicando que algum fator foi subestimado teria influencia maior que os parâmetros aferidos. Ainda assim a influencia do recife sobre a comunidade foi claramente constatada, existindo ainda a necessidade de mais estudos, tanto em termos de variabilidade espacial, como em relação aos estudos de comunidade e os efeitos de cadeia sobre elas.
Palavras-chave: Nematoda, Polychaeta, Pernambuco

ABSTRACT
The aim of this study was knowing the meiofauna community structure of soft sediment
in a coastal reef and its interactions with environmental factors and the relationship with marine protected area and reef influence. The samples were taken about April and September 2008 inside and outside a marine protected area. The both area are compound by coarse sand with low organic matter taxes. We found 15 taxa of meiofauna what 56 genus of Nematoda and 18 genus of Polychaeta. The meiofauna diversity showed higher and significant values inside the marine protected area and away from the reef . The Nematoda diversity values didn’t show significant differences between the both areas. The Polychaeta diversity values didn’t show significant difference too. But the Polychaeta richness values was higher significant away from the reef. The differences between sediment parameters and biological structure parameters were low and we think that some factor was underestimated and this factor been the biggest influence on the meiofauna community structure. Even so the reef influence over the community was clearly found and we think what the study improvement about spacial variance and the effects of top-down processes over the community are need.
Keyword: Nematoda, Polychaeta, Pernambuco

LISTA DE FIGURAS
Figura 01: Mapa da localização da cidade de Tamandaré, litoral sul de Pernambuco. (Sobral, 2009)
20
Figura 02: Localização das áreas de coleta: pontos 1 e 2 na Ilha da Barra (8o46’02”S; 35o05’39”W) e pontos 1 e 2 no Pirambu (8o45’26”S; 35o05’12”W). A marcação se refere à área protegida da pesca. Fonte: Modificado de Google Earth. (figura meramente ilustrativa)
21
Figura 03: A) ANOVA da quantidade de Clorofila em Tamandaré no mês de setembro; B) ANOVA da quantidade de Feopigmento em Tamandare no mês de setembro. Ilha da Barra ( ) Pirambú ( ).Barra vertical indica 95% do intervalo de confiança, N= 48.
26
Figura 04: Indices de chuva acumulada do ano de 2008 (colunas) e média pluviométrica de período 1961-1990 (linha) na estação meteorológica do Curado, PE. Fonte: INMET
26
Figura 05: A) Tamanho médio do grão dentro e fora da área de restrição à pesca no mês de setembro de 2008 em Tamandaré; B) Tamanho médio do grão próximo e afastado do recife no mês de setembro de 2008 em Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 14.
27
Figura 06: Curva de dominância acumulada dos principais grupos meiofaunais capturados no sedimento inconsolidado do ambiente recifal de Tamandaré, PE no ano de 2008.
28
Figura 07: Abundância nos meses de abril e setembro na área recifal de Tamandaré, representando a meiofauna restante, e destacando a nematofauna e poliquetofauna. Barra vertical indica erro padrão, N= 128.
29
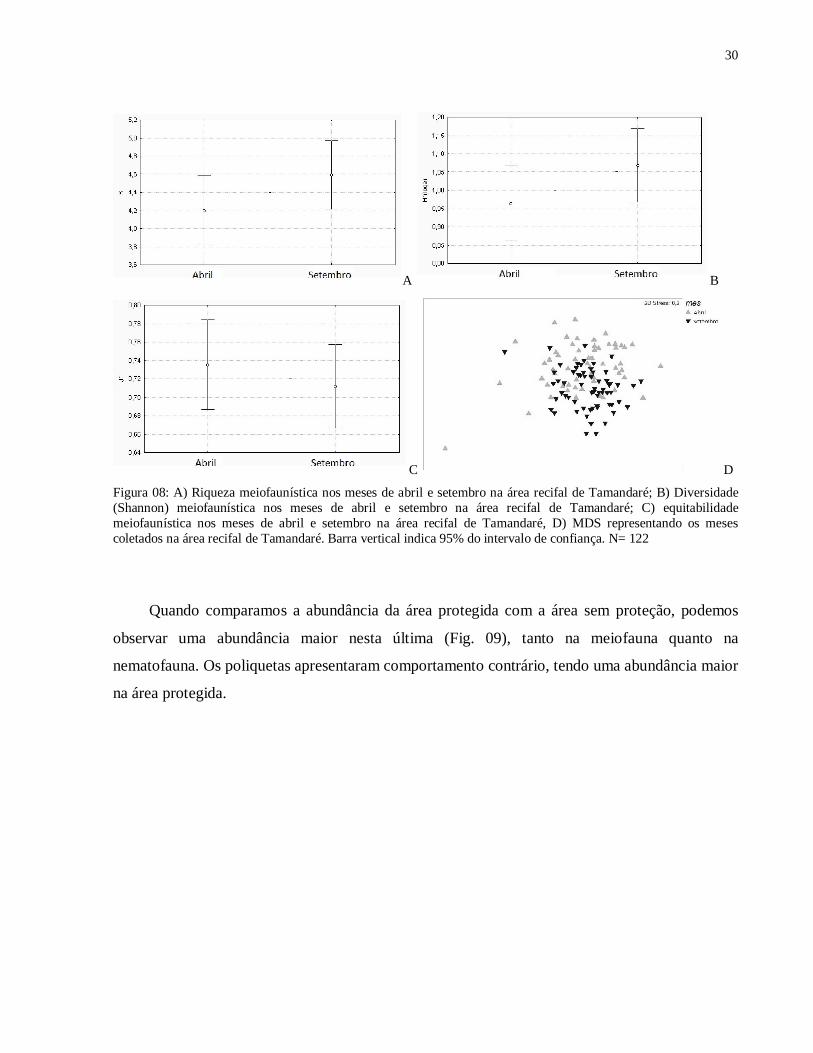
Figura 08: A) Riqueza meiofaunística nos meses de abril e setembro na área recifal de Tamandaré; B) Diversidade (Shannon) meiofaunística nos meses de abril e setembro na área recifal de Tamandaré; C) equitabilidade meiofaunística nos meses de abril e setembro na área recifal de Tamandaré, D) MDS representando os meses coletados na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 122
30
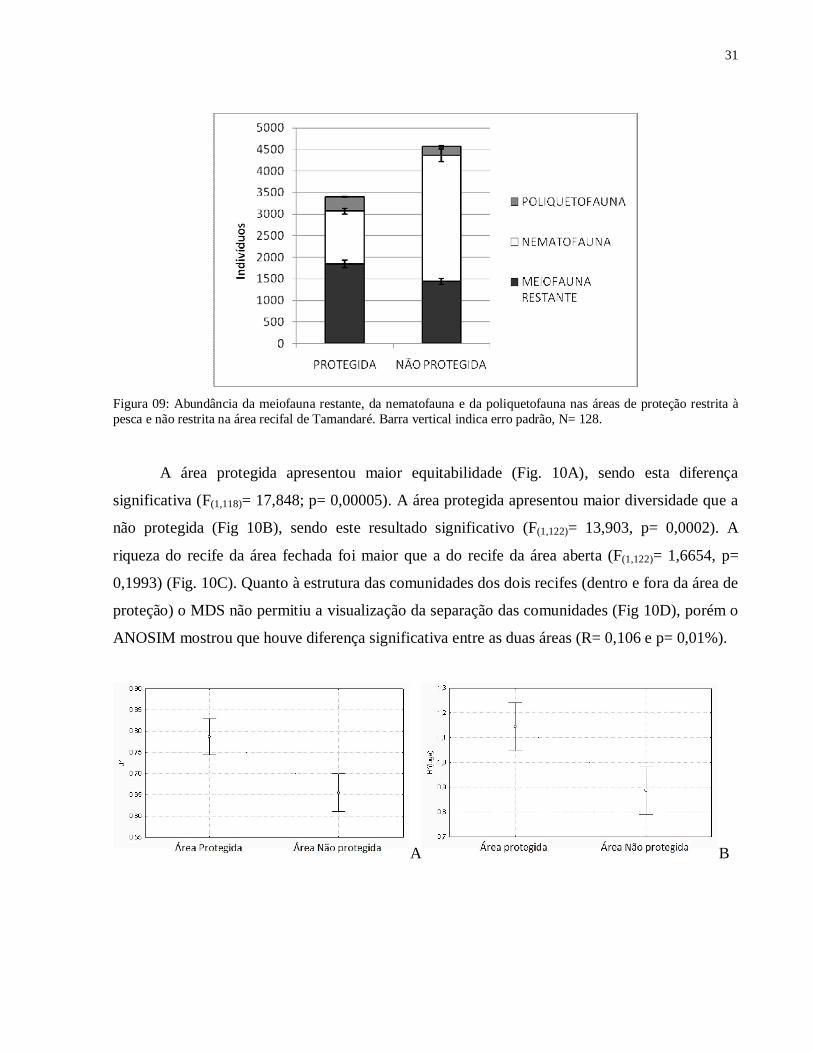
Figura 09: Abundância da meiofauna restante, da nematofauna e da poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica erro padrão, N= 128.
31
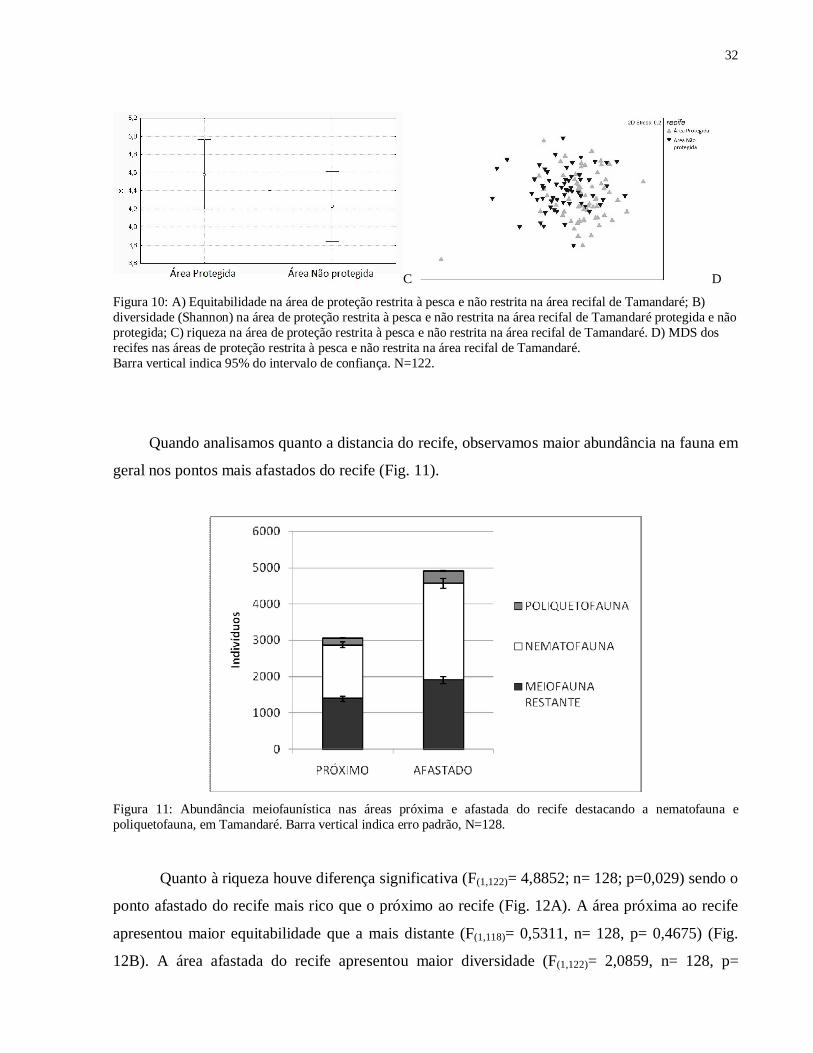
Figura 10: A) Equitabilidade na área de proteção restrita à pesca e não restrita na área recifal de Tamandaré; B) diversidade (Shannon) na área de proteção restrita à pesca e não restrita na área recifal de Tamandaré protegida e não protegida; C) riqueza na área de proteção
32

restrita à pesca e não restrita na área recifal de Tamandaré. D) MDS dos recifes nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N=122. Figura 11: Abundância meiofaunística nas áreas próxima e afastada do recife destacando a nematofauna e poliquetofauna, em Tamandaré. Barra vertical indica erro padrão, N=128.
32
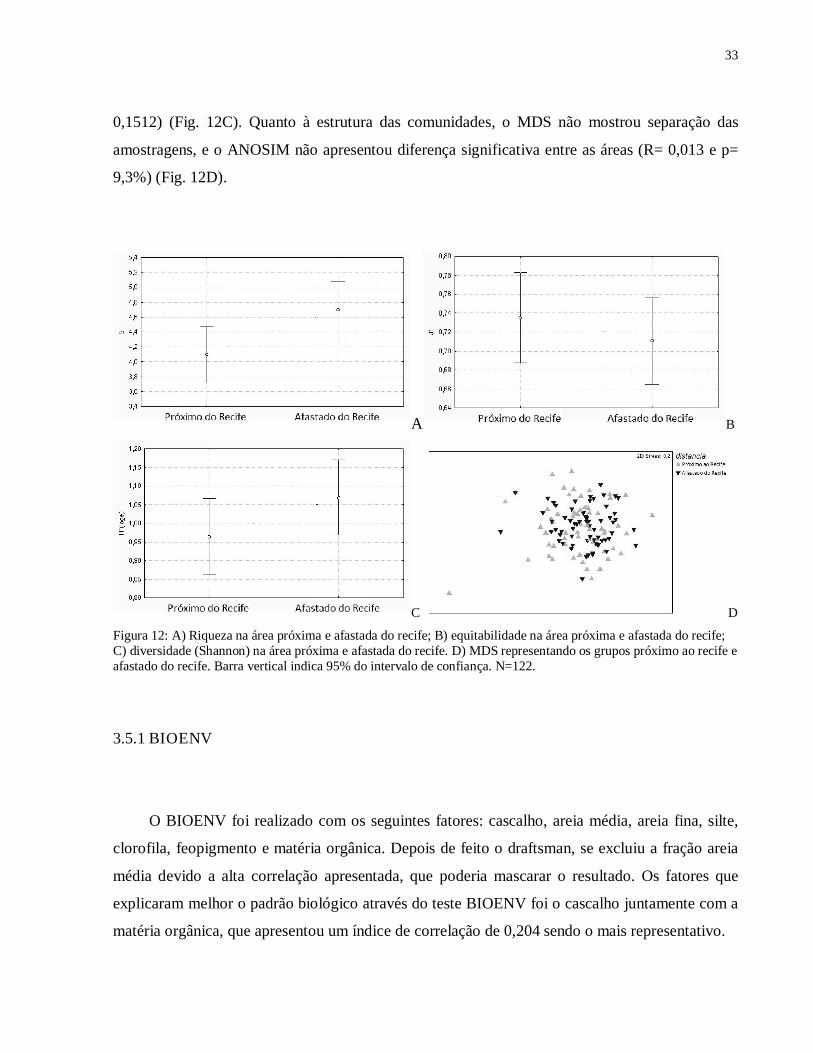
Figura 12: A) Riqueza na área próxima e afastada do recife; B) equitabilidade na área próxima e afastada do recife; C) diversidade (Shannon) na área próxima e afastada do recife. D) MDS representando os grupos próximo ao recife e afastado do recife. Barra vertical indica 95% do intervalo de confiança. N=122.
33
Figura 13: Densidades médias dos gêneros de Nematoda mais abundantes nas amostras coletadas em abril e setembro de 2008, nas duas áreas de coleta, na baía de Tamandaré. Barra vertical indica erro padrão. Fonte: Modificado de WARWICK et al. (1988).
38
Figura 14: Abundancia total dos gêneros de Nematoda: Daptonema, Gomphionema, Theristus, Marylynnia e Setosabatieria, nos meses de abril e setembro de 2008 na área recifal da baía de Tamandaré. Barra vertical indica erro padrão, N= 96.
38
Figura 15: A) Riqueza da nematofauna nos meses de abril e setembro na área recifal de Tamandaré, B) Diversidade da nematofauna nos meses de abril e setembro na área recifal de Tamandaré, C) Equitabilidade na nematofauna nos meses de abril e setembro na área recifal de Tamandaré, D) MDS mostrando a diferença entre os meses amostrados na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 92)
39
Figura 16: Abundancia dos gêneros de Nematoda: Daptonema, Gomphionema, Theristus, Marylynnia e Setosabatieria, áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica erro padrão, N= 96.
40
Figura 17: A) Riqueza da nematofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, B) Diversidade da nematofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, C) Equitabilidade na nematofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, D) MDS comparando recifes nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 92.
41
Figura 18: Abundancia dos gêneros de Nematoda: Daptonema, Gomphionema, Theristus, Marylynnia e Setosabatieria, na área próxima ao recife e afastado deste, na baía de Tamandaré em 2008. Barra vertical indica erro padrão, N= 96.
41
Figura 19: A) Riqueza da nematofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, B) Diversidade da nematofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, C) Equitabilidade na nematofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, D) MDS comparando as comunidades proximo e afastado do recife na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 92.
42

Figura 20: Abundância do gênero Sphaerosyllis nos meses de abril e setembro, na baía de Tamandaré em 2008. Barra vertical indica erro padrão, N= 90.
44
Figura 21: a) Riqueza da poliquetofauna nos meses de abril e setembro na área recifal de Tamandaré, b) Equitabilidade na poliquetofauna nos meses de abril e setembro na área recifal de Tamandaré, c) Diversidade da poliquetofauna nos meses de abril e setembro na área recifal de Tamandaré d) MDS mostrando a diferença entre os meses amostrados na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 71.
45
Figura 22: Abundância de Sphaerosyllis nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré em 2008. Barra vertical indica erro padrão, n= 90.
46
Figura 23: a) Riqueza da poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, b) Equitabilidade na poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, c) Diversidade da poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, d) MDS comparando recifes nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N=71.
47
Figura 24: Abundancia de Sphaerosyllis na área próxima ao recife e afastado deste na área recifal de Tamandaré em 2008. Barra vertical indica erro padrão, N= 90.
47
Figura 25: a) Riqueza da poliquetofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, b) Equitabilidade na poliquetofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, c) Diversidade da poliquetofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, d) MDS comparando as comunidades proximo e afastado do recife na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N=71.
48

LISTA DE TABELAS
Tabela 1: Quantidade média de clorofila e feopigmento encontrada nos pontos de coleta no mês de setembro em Tamandaré. Desvio padrão entre parênteses.
25
Tabela 2: Análise SIMPER para os grupos de acordo com os fatores pré-estabelecidos, apresentando os táxons/gênero que mais constribuíram com as dissimilaridades entre as áreas analisadas em Tamandaré.
34

SUMÁRIO
1 INTRODUÇÃO
1.1 Hipótese
1.2 Objetivos
1.2.1 Objetivo Geral
1.2.2 Objetivos Específicos
2 MATERIAL E MÉTODO
2.1 Área de Estudo
2.2 Métodos
2.2.1 Em campo
2.2.2 Em Laboratório
2.2.3 Análise dos Dados
3 RESULTADOS
3.1 Microfitobentos
3.2 Pluviometria
3.3 Salinidade
3.4 Granulometria
3.5 Meiofauna
3.5.1 BIOENV
3.5.2 SIMPER
3.6 Nematofauna
3.6.1 Correlação
3.7 Poliquetofauna
4 DISCUSSÃO
5 CONCLUSÕES
6 REFERÊNCIAS BIBLIOGRÁFICAS

12
1 INTRODUÇÃO
As áreas de proteção marinha são ferramentas largamente utilizadas para o manejo de
áreas naturais. Os propósitos de seu uso podem incluir conservação (manutenção da diversidade
biológica), recreação (áreas turisticas), histórico (locais históricos e culturais), pesquisas
(monitoramento de impactos) e comercial (conservação de áreas importantes para espécies de
importância econômica) (DAVIS & TISDEL, 1995).
Áreas fechadas em áreas de proteção marinha com grande diversidade e complexidade
podem ter efeito positivo no estoque pesqueiro, quando comparadas com áreas com limitada
proteção ou habitats pobres em qualidade (FRIEDLANDER, 2001). Associações de peixes
recifais também tendem a ser diferentes quando comparadas áreas fechadas com áreas onde
vários níveis de pesca são permitidos (FRIEDLANDER et al. 2003). Estas áreas com restrições
têm altos valores em riqueza, abundância e diversidade, seguidos pela parcialmente protegida e
por fim, pelas áreas de acesso livre (FRIEDLANDER et al. 2003).
Existem 728 unidades de conservação (UC) criadas por decretos federais. Dessas, 429 são
reservas particulares, sendo nenhuma marinha. Das 299 restantes, 45 são unidades de
conservação marinhas ou mistas, onde: quatro são estações ecológicas, oito são Parque
Nacionais, três são reservas biológicas, dez são áreas de proteção ambiental, duas são área de
relevante interesse ecológico, uma é floresta nacional e 17 são reservas extrativistas. Destas 45
unidades de conservação, 17 estão no nordeste e três em Pernambuco (IBAMA, 2009). O
Governo do Estado de Pernambuco criou e mantêm 54 unidades de conservação, onde 14 são
áreas de proteção ambiental (APA) e todas são costeiras ou mistas (SECTMA, 2009). Destas 14
áreas de proteção apenas uma tem plano de manejo, a APA de Guadalupe, porém este plano de
manejo ainda não envolve o ecossistema marinho (trata-se de uma UC mista).
Em levantamento realizado em 2006, foram identificadas 980 áreas de proteção marinhas,
cobrindo 968.650 km2 (18,7%) dos recifes de coral do mundo. No Brasil existem 5 unidades de
conservação marinhas envolvendo recifes, sendo uma avaliada como limitada (APA dos Corais),
duas como parciais, e duas como adequadas (MORA et al. 2006) de acordo com a área ocupada
por corais que é acobertada pela proteção.

13
A Área de Proteção Ambiental Costa dos Corais foi criada por decreto federal em
Outubro de 1997 com o objetivo de ordenar o uso do ecossistema recifal localizado nos 130 km
de costa entre os municípios de Tamandaré - PE e Paripueira – AL. Com a criação da APA dos
corais e do projeto recifes costeiros, sugeriu-se um acompanhamento destes ambientes no estado
de Pernambuco e a criação de uma área fechada a pescadores e recreações náuticas como a caça
subaquática e trânsito constante de embarcações (apenas travessia para mar aberto). Após a
implantação da APA já foi observado um aumento na população de peixes e o aparecimento mais
freqüente de peixes anteriormente raros, como o mero e a caranha (FERREIRA et al., 1995 e
2001). A pesca em Tamandaré, fora da área restrita à pesca, se dá de forma artesanal, sendo as
mais utilizadas as pescas de linha, com arpão, com bicheiro e com rede caceia (FERREIRA et al.,
2001).
Áreas de proteção marinhas com exclusão da pesca são ferramentas importantes para a
conservação do ambiente marinho. Entretanto, o efeito desta medida sobre os invertebrados
existentes no ambiente é pouco conhecido. A APA dos corais em Tamandaré possui um trecho
com exclusão da pesca, que possibilita a realização de trabalhos comparativos com área
semelhantes, mas abertas à pesca, nas proximidades. Padrões encontrados na fauna adjacente,
como variação na abundância e riqueza de espécies, sugerem a interação entre estes dois
ambientes. A heterogeneidade deste ambiente aquático afeta o padrão da distribuição espacial de
populações aquáticas (DOWNING & CYR, 1985), principalmente, os grupos que dão
sustentação a comunidades bentônicas. Vários estudos sugerem que os modelos de estrutura de
comunidades precisam considerar os diferentes aspectos da fauna em suas investigações
(BARROS, 2005; NETTO et al., 2003).
Os ambientes associados a substratos consolidados são considerados de alta produtividade
(FERREIRA et al., 1995; COELHO et al., 2004). A complexidade destes ambientes tem sido
apontada como um dos responsáveis por uma alta biodiversidade (BIRKELAND, 1997). A
própria heterogeneidade do substrato quebra, por exemplo, o gradiente natural de situações de
luminosidade e hidrodinâmica decorrente da profundidade, criando situações de infralitoral
dentro do médio-litoral. Associadas a esses fatores devem ainda ser consideradas as interações
biológicas e os processos de sucessão (BRAWLEY & ADEY, 1981).
Poucos estudos investigam a influencia dos recifes sobre as estruturas das comunidades
adjacentes. A presença do substrato duro deve permitir a regulação de uma amplitude de

14
invertebrados bentônicos, que de outra maneira não poderiam encontrar habitats aptos para sua
sobrevivência pós-recrutamento (DANOVARO & SIMONETTA, 2002). Davis et al. (1982)
observaram que o substrato duro tem um impacto nas comunidades bênticas que vivem no
sedimento adjacente, uma vez que mudam a velocidade e a direção da corrente, a erosão do
sedimento, a taxa de sedimentação, a distribuição do grão e a quantidade de matéria orgânica.
Alguns trabalhos descrevem a meiofauna no ambiente recifal natural (ST. JOHN et al.,
1989, MARANHÃO, 2003, NETTO et al., 2003, GUZMAN, et al., 1987) e também em
ambientes recifais artificiais (DANOVARO et al., 2002; FONSÊCA-GENEVOIS et al., 2006).
Porém, não existe informação sobre a influência dos recifes na comunidade meiofaunística. A
meiofauna compreende organismos de 0,044 mm a 0,5mm de tamanho (MARE, 1942), formando
associações de metazoários bênticos. Estes animais possuem morfologia, fisiologia e ciclos de
vida característicos, habitando o interstício dos sedimentos, também sendo encontrados
associados a outros organismos como algas, cnidários, poliquetas, entre outros (ALONGI, 1985;
VENEKEY et al. 2008). Warwick (1984) a considera uma unidade funcional cujas espécies
desempenham estratégias de sobrevivência diferentes das empreendidas pela macrofauna. A
meiofauna é composta por aproximadamente 30 filos, sendo alguns grupos tipicamente
meiofaunais (Nematoda, Copepoda Harpaticoida, Tardigrada e Gastrotricha), enquanto outros
compõem a meiofauna apenas numa parte do seu ciclo de vida (Gastropoda, Holothuroidea,
Nemertinea e alguns Polychaeta). Esses organismos participam da cadeia alimentar como
alimento (GEE, 1989) e como consumidores (MONTAGNA, 1995) constituindo uma das
maiores fatias no fluxo de energia dos sistemas bentônicos (STEAD et al. 2004). Copepoda e
Ostracoda são grandes consumidores do microfitobentos (MONTAGNA, 1995; CARMAN et al.
1997) e são importantes presas para pequenos instares de peixes,para grandes invertebrados e
para a própria meiofauna (COULL, 1988; MCCALL & FLEEGER, 1995; SCHMID-ARAYA et
al. 2002). Atuam também na remineralização de detritos orgânicos (TENORE et al., 1977) e na
evidente relação na distribuição de biomassa do bentos marinho.
A distribuição da meiofauna é frequentemente relacionada a fatores químicos e físicos
limitantes. A salinidade, a temperatura, o teor de oxigenio dissolvido, caracteristicas do
sedimento, o regime de ondas, as correntes e a amplitude de maré são fatores importantes na
modelagem da estrutura da comunidade meiofaunistica em relação ao substrato, uma vez que as
propriedades do sedimento e a sua distribuição controlam as dimensões do espaço intersticial,

15
interferindo no grau de seleção, tamanho do grão, porosidade e permeabilidade (GIERE, 1993 e
MCLACHLAN & TURNER, 1994). Hullings e Gray (1976) concordando com Jansson (1967)
citam o tamanho do espaco intersticial como o principal fator para a presença da meiofauna.
Porém outros fatores como alimento disponível, estruturas biogênicas de plantas e animais, e
comportamento socio-reprodutivo (ESTEVES, 1995) podem interferir na distribuição da
meiofauna no sedimento.
Alguns estudos mostram que a predação é um fator biológico importante como processo
regulador da estrutura da comunidade bentônica (BELL & COULL, 1978; BARROS, 2005;
MORTON, 2005), pois é sabido que invertebrados da macrofauna (BELL & COULL, 1978;
KENNEDY, 1993; REISE, 1979), peixes (ST. JOHN et al., 1989; MORAIS & BODIOU, 1984) e
até aves (SUTHERLAND et al., 2000) predam de forma intensa os organismos da meiofauna.
Virnstein (1977) observou o aumento da abundância na ausência do predador, e sugeriu que o
tamanho da população é limitado pela predação.
As associações meiofaunais têm sido usadas para descrever variações ambientais, sendo
uma ferramenta importante para descrever os diferentes impactos que atingem os organismos
bênticos. Para este trabalho foram escolhidos os grupos Nematoda e Polychaeta para serem
observados com mais detalhe.
Assim como a meiofauna de forma geral, os Nematoda possuem gerações curtas
permitindo uma rápida resposta, ubiquidade, facilidade de coleta e pequenas amostras são
suficientes para obter dados representativos, características que os tornam bons indicadores
ambientais (PLATT et al. 1984). São considerados o táxon meiofaunístico mais estudado, além
de serem os organismos multicelulares mais abundantes no mundo, com mais de 26.000 espécies
distribuídas em 250 famílias. Apenas entre 4.000 e 5.000 espécies são de vida livre
(LORENZEN, 1994). Os Nematoda de vida livre foram encontrados em todos os ambientes
conhecidos e geralmente são encontrados em quantidade maior que os outros grupos da
meiofauna (PLATT et al., 1984). São considerados fundamentais para a manutenção do fundo
marinho (PLATT & WARWICK, 1980).
Os representantes do filo Nematoda que são marinhos de vida livre geralmente
apresentam maior densidade em sedimentos finos ou lamosos, porém a diversidade
frequentemente é maior em sedimentos grossos (PLATT & WARWICK, 1980). Seu sucesso
adaptativo se deve principalmente ao formato do seu corpo, que é cilindrico, apresentando uma

16
alta pressão dos fluidos internos, lhe permitindo colonizar diversos tipos de habitats
(SCHIEMER, 1987). Além de sua diversidade bucal (WIESER, 1953, 1960; WIESER &
KANWISHER 1961) que proporciona diferentes tipos de consumo.
As primeiras pesquisas em Nematoda no Brasil datam de 1920, com registro de 3 espécies
por Cobb e depois na década de 50 com o Dr. Sebastian Gerlach (VENEKEY, 2007). Atualmente
no Brasil a pesquisa em Nematoda marinho de vida livre concentra-se principalmente na UFPE,
onde existe um núcleo bastante atuante, com inserção internacional.
Os representantes do filo Polychaeta na meiofauna estão entre os grupos mais abundantes,
sendo frequentemente coletados (BOECKNER et al. 2009). Cerca de 10.000 espécies distribuídas
em 82 famílias são conhecidas (ROUSE & PLEIJEL, 2001), sendo destas 250 espécies
conhecidas como meiofaunais (GIERE 1993; WESTHEIDE, 2002). Entre os poliquetas existem
alguns animais que participam da meiofauna apenas na sua fase juvenil e outros que são
considerados permanentes na meiofauna, pois seu ciclo de vida inteiro se passa na meiofauna.
Diferente dos Polychaeta da macrofauna, que lançam sua larva trocófora na coluna d’água, os
Polychaeta da meiofauna não possuem larva trocófora, tendo desenvolvimento direto, onde o
juvenil é geralmente uma versão mais simples do indivíduo adulto (GIERE 1993).
Os poliquetas da meiofauna geralmente são encontrados em sedimentos com grãos
médios a grosseiros (VILLORA-MORENO, 1997), formando associações e seguindo padrões de
zonação no infralitoral (WESTHEIDE, 1972; VILLORA-MORENO et al., 1991). Um ambiente
com alta heterogeneidade formará vários micro-habitats, aumentando a diversidade dos
poliquetas intersticiais (VILLORA-MORENO, 1997). Assim como para a meiofauna de forma
geral, o principal fator para a presença de poliquetas num ambiente seria a disponibilidade de
espaço intersticial (LEE et al., 2006).
Boa parte dos trabalhos que tratam sobre Polychaeta intersticiais no Brasil tem enfoque
taxonômico (SANTOS & LANA, 2003; ASSIS et al. 2007; ZANOL et al. 2000; SANTOS, et al.
2004; SANTOS & MACKIE, 2008), sendo menos frequentes estudos sobre a sua biologia e
ecologia (DI DOMENICO et al. 2009) . Tratando-se de Polychaeta pertencentes à meiofauna, os
estudos taxonômicos não são comuns e os ecológicos são ainda mais raros. Apesar dos
Polychaeta serem abundantes nas amostragens de meiofauna raramente são o objeto de estudo,
sendo geralmente tratados em análises da comunidade meiofaunistica de modo geral.

17
Estudos vêm sendo desenvolvidos com o objetivo de conhecer o papel da meiofauna em
ambientes sujeitos as perturbações naturais (HULINGS & GRAY 1976, PLATT et al. 1984). Este
trabalho vem contribuir com o preenchimento desta lacuna, ao apresentar informações sobre a
estrutura da comunidade meiofaunística em substrato inconsolidado de ecossistema recifal
abordando duas áreas, sendo uma protegida da ação da pesca, analisando ainda a influência do
recife sobre esta comunidade.
1.1 Hipótese
Este trabalho foi desenvolvido sob as hipóteses alternativas de que há diferenças entre as
estruturas das comunidades meiofaunísticas presentes na área de ambiente recifal da APA dos
Corais (PE) e na área de ambiente recifal exposta ao efeito antrópico em Tamandaré, assim como
de que a presença do recife influencia na estrutura da comunidade meiofaunística presente nos
sedimentos adjacentes.
1.2 Objetivos
1.2.1 Objetivo Geral
Analisar a estrutura da comunidade meiofaunística do sedimento inconsolidado próximo a
afastado da parte interna de duas áreas recifais, numa protegida legalmente da pesca e outra não.
1.2.2 Objetivos Específicos

18
- Descrever as associações meiofaunais presentes no substrato inconsolidado adjacente à
parte interna de recifes da praia de Tamandaré em Pernambuco;
-Analisar os padrões de comunidade encontrados com as variáveis ambientais (e.g.,
granulometria, feopigmentos e clorofila a dos sedimentos, salinidade);
- Relacionar os resultados das associações encontrados com a proteção legal à pesca e à
distância do recife.

19
2 MATERIAL E MÉTODOS
2.1 Área de Estudo
O litoral do município de Tamandaré (PE), localizado no extremo norte da APA Costa
dos Corais, tem 14 km de extensão (8º 45' S; 35º 05' W) (Fig. 01), sendo limitado pela
desembocadura do rio Formoso ao norte e o rio Mamucaba ao sul. Localiza-se a 110 km de
Recife, Pernambuco. Possui clima tropical úmido, sendo classificado com tipo As’ de acordo
com Koeppen. A época mais chuvosa do ano é o mês de julho e o período mais seco no mês de
dezembro. A temperatura média é de 24°C podendo atingir uma máxima de 32°C e mínima de
18°C e é dominada por ventos alísios de SE e NE (CPRH, 2003). De acordo com Maida e
Ferreira (1997) a temperatura média da água é de 24°C no inverno e 30,5°C no verão, com
visibilidade de 1m a 20m dependendo da época do ano, quantidade de chuvas, ventos e correntes.
As ondas são consideradas de baixa intensidade, sendo amortecidas pelos recifes, e chegando ao
litoral com 0,66m de altura (FARIAS, 2002).
Assim como em todo litoral de Pernambuco onde se fazem presentes, os recifes em
Tamandaré se distribuem em três linhas de recife em tipo franja, dispostas paralelamente à linha
de praia. Na baía de Tamandaré a laguna apresenta profundidade máxima de 8m (PINHEIRO
2006).

20
Figura 01: Mapa da localização do município de Tamandaré, litoral sul de Pernambuco.
2.2 Métodos
2.2.1 Em campo
As coletas no ecossistema recifal de Tamandaré foram realizadas através de 2
amostragens que ocorreram nos meses de abril e setembro de 2008. No local foram escolhidas
duas áreas com recifes sendo uma dentro da área de proteção restrita, Ilha da Barra, e a outra fora
desta, Pirambú, que está incluído na área de proteção ambiental, porém não esta incluído na área
de restrição a pesca. Em cada área foram traçadas 4 linhas transversais à costa, distantes entre si
mais de 100m e dispostas aleatoriamente, com 2 pontos em cada: um adjacente ao recife e o outro
a 10 metros do primeiro ponto. Em cada ponto foram tomadas 4 amostras (réplicas) biológicas
(totalizando 32 amostras por área), além de 1 amostra para análise de sedimento e 3 para análise
de microfitobentos. A profundidade dos pontos de coleta foi entre 3m e 6m de profundidade.

21
As amostras foram coletadas com auxílio do equipamento autônomo de mergulho,
utilizando-se um amostrador de PVC com 2,5cm de diametro 10 cm2 de área (HULLINGS &
GRAY, 1976) que foi colocado até 10 cm de profundidade no sedimento. O amostrador foi
fechado com um tampo (cap) para exercer a pressão necessária para que o sedimento ficasse
retido no tubo. Assim que retirado do fundo o amostrador foi colocado em saco plástico e lacrado
com liga de borracha, sendo fixado em formol a 4% tamponado com água do mar.
Figura 02: Localização das áreas de coleta: pontos 1 e 2 na Ilha da Barra (8o46’02”S; 35o05’39”W) e pontos 1 e 2 no Pirambu (8o45’26”S; 35o05’12”W). A marcação se refere à área protegida da pesca. Fonte: Modificado de Google Earth. (figura meramente ilustrativa)
A coleta das amostras para análise do microfitobentos foi realizada no mês de setembro
com auxílio de uma seringa de 1,13cm² de área, inserida nos dois primeiros centímetros do
sedimento. Em seguida as amostras foram acondicionadas em frascos âmbar e conservadas em
gelo ao abrigo da luz (COLIJN & DIJKEAMA, 1981) e posteriormente congeladas até o
momento em que foram processadas.
Em cada ponto de coleta foi colhido paralelamente uma amostra sedimentar para análises
granulométricas, apenas no mês de setembro. Amostras de água do mar foram coletadas para
medições dos teores de salinidade locais através de refratômetro portátil.

22
2.2.2 Em laboratório
A extração da fauna foi feita em laboratório, através da lavagem das amostras em peneiras
de 0,5mm e 0,044mm e posterior centrifugação manual (ELMGREN, 1966). O sobrenadante foi
então passado para uma placa de Dolffus, composta de 200 quadrados de 0,25cm2 cada um, para
triagem do material. A meiofauna foi identificada em níveis taxonômicos altos, sendo os
Nematoda e Polychaeta identificados na melhor resolução taxonômica possível. Para isso foram
montadas lâminas para microscopia, através de uma adaptação das técnicas referidas por Cobb
(1917) e De Grisse (1969) para Nematoda. Para a identificação e elaboração da lista foi usada a
classificação de Lorenzen (1994). Para a elaboração de lâminas de Polychaeta foi utilizado o
meio Gel de Glicerina, que é a mistura de 15g de gelatina, 100 ml de água destilada, 100ml de
glicerina pura e 1g de fenol. Foi feita a estimativa total das espécies identificadas.
Após a observação das lâminas, os Nematoda podem ser classificados segundo Wieser
(1953, 1960) e Wieser & Kanwisher (1961) em 1A: apresentando cavidade bucal reduzida ou
ausente, sendo chamado de detritívoro seletivo; 2A: apresenta dentes de tamanhos reduzidos,
sendo conhecidos como raspadores; 1B: apresenta cavidade ampla e sem dentes, tratando-se de
um comedor de depósito não-seletivo; ou 2B: apresenta dentes fortes ou mandíbulas na cavidade
bucal, sendo chamados de predadores onívoros.
Devido ao fato das amostras de microfitobentos terem sido coletadas no meio aquoso e
por conseqüência com bastante água, foram submetidas à decantação durante o descongelamento
antes da extração dos pigmentos. As amostras foram retiradas do freezer cerca de 8h antes da
adição de acetona, para descongelar e decantar no escuro. Após este processo, o sobrenadante foi
retirado com pipeta de Pasteur com cuidado para não tocar o fundo nem suspender a amostra. Em
seguida, foi realizada a extração dos pigmentos.
Para a extração dos pigmentos fotossintéticos foi adicionado às amostras 10 ml de acetona
a 100% (PA). Em seguida foram homogeneizadas e colocadas em refrigeração a 4ºC por 20h no
escuro. As concentrações de clorofila-a e feopigmentos foram determinadas por
espectrofotometria, pela leitura nos comprimentos de onda 665 e 750nm, antes e depois de
acidificadas com 50 microlitros de HCl a 0,05N (COLIJN & DIJKEAMA, 1981), e calculadas
através das equações de Lorenzen (1967).

23
O sedimento coletado foi levado à estufa com temperatura de 75oC num período de 48h.
Foram separados 100 g, lavado o material, colocado em estufa por mais 48 horas e estimada a
massa novamente. O sedimento passou, então, por uma série de peneiras em intervalos de 1 phi,
auxiliado por um agitador mecânico, durante 10 minutos, para classificar o material arenoso
(≥0,062 mm) (SUGUIO, 1973).
Os dados pluviométricos referentes ao ano de 2008 foram fornecidos pelo Ministério da
Agricultura – Instituto Nacional de Meteorologia (INMET).
2.2.3 Análise dos Dados
A homogeneidade dos dados foi comprovada pelo teste de Bartlett. Sendo assim foi
possível utilizar a análise de variância, tratando os dados de maneira independente.
Os parâmetros de sedimento de Folk e Ward (1957) foram obtidos através do programa
SYSGRAN. As diferenças de concentração de clorofila, feopigmentos, tamanho médio do grão,
riqueza de espécies por amostra, diversidade (H’) e equitabilidade, entre as duas áreas e em
relação à distância do recife, foram analisados com testes ANOVA. Estas análises foram feitas
com o programa STATISTICA 6.0;
A proporção de matéria orgânica foi analisada através de ANOVA (um fator) não
paramétrica (Kruskal-Wallis), utilizando o programa BIOESTAT 5.0. A analise de correlação de
Pearson entre os Nematoda e Polychaeta mais abundantes com os parâmetros ambientais foi
realizada com o programa STATISTICA 6.0. Foram utilizados o índice de diversidade de
Shannon e equitabilidade de Pielou (KREBS, 1999) para a analises da estrutura da meiofauna. Os
dados das espécies foram transformados (log x+1) e foi utilizado o coeficiente de Bray Curtis
para a análise de escalonamento multidimensional (MDS), realizada com o programa PRIMER
(CLARKE & WARWICK, 1994), e o resultado gráfico gerado pelo MDS foi confirmado através
do ANOSIM também no mesmo programa.
Foram feitas correlações (BIOENV) objetivando aferir a interferência dos dados
ambientais sobre a comunidade da meiofauna. Antes de realizar o teste BIOENV, foi feito um
draftsman (correlação múltipla utilizando o r de Spearman) para observar quais parâmetros

24
possuíam alta correlação. A fração areia apresentou forte correlação com a fração cascalho, logo
este fator foi retirado antes da análise de acordo com a recomendação de Clarke & Warwick
(2001). Os parâmetros utilizados foram: clorofila, feopigmentos, cascalho, areia e silte.. Para
estes testes foi utilizado o pacote estatístico PRIMER 6.0.
Foi feito também um teste SIMPER para avaliar a influência dos grupos amostrados na
dissimilaridade entre os fatores estabelecidos. Foram utilizados para a realização do SIMPER em
nível de grandes grupos.

25
3 RESULTADOS
3.1 Microfitobentos
A maior concentração média total de clorofila nos sedimentos foi encontrada no Pirambú
(Tab. 1), mas esta diferença não foi significativa (Fig. 03). Os pontos afastados do Recife
apresentaram maior concentração média total, sem diferença significativa também (Fig. 03).
Quanto aos feopigmentos, a maior concentração foi encontrada na área protegida em ponto
próximo ao recife (Tab. 1). A menor concentração ocorreu no Pirambu (afastado do recife) (Tab.
1). A diferença de feopigmentos foi significativa em relação a distância do recife, apenas no
Pirambú (Fig. 03).
Tabela 1: Quantidade média de clorofila e feopigmento encontrada nos pontos de coleta no mês de setembro em Tamandaré. Desvio padrão entre parênteses.
Clorofila µg/ml Protegida Não protegida Média Total
Próximo do Recife 0,9725 (0,44) 0,8825 (0,67) 0,9275 (0,53)
Afastado do Recife 1,595 (1,01) 1,8625 (0,97) 1,7287 (0,97)
Média Total 1,2837 (0,84) 1,3725 (0,93)
Feopigmento µg/ml
Próximo do Recife 16,94 (17,79) 8,1425 (11,46) 12,5412 (14,63)
Afastado do Recife 4,7625 (6,81) 2,17 (1,25) 3,4662 (4,74)
Média Total 10,8512 (14,06) 5,1562 (8,19)
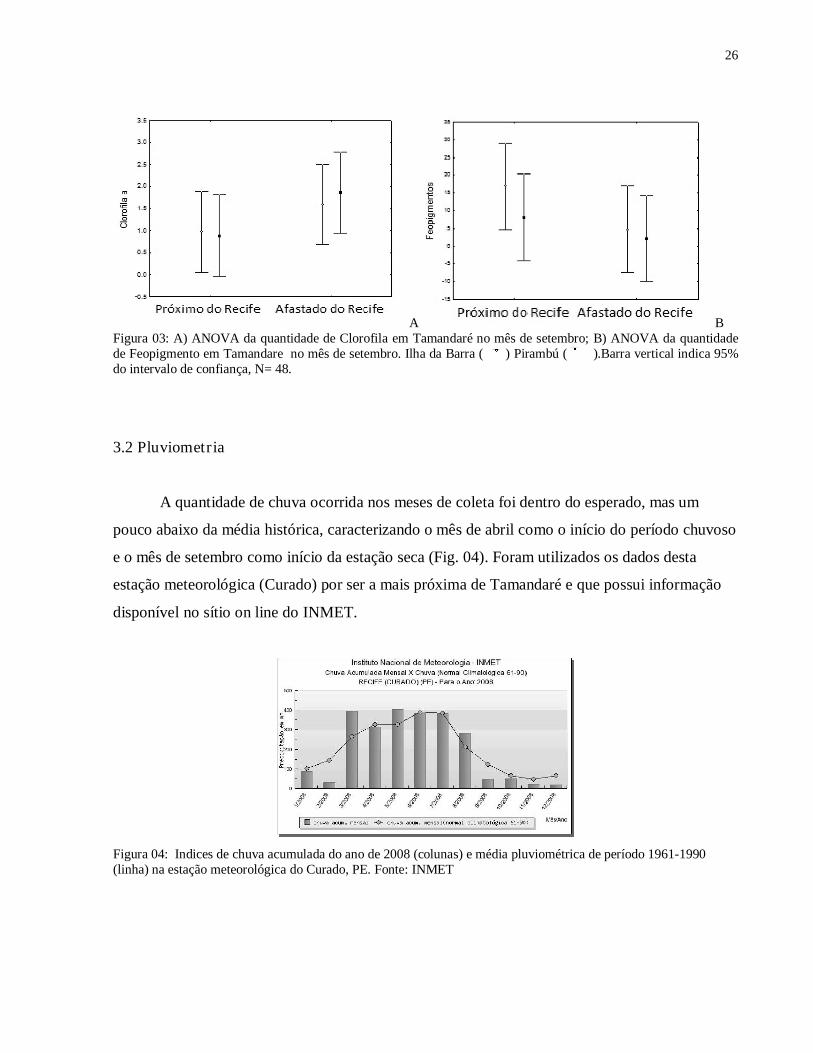
26
A B Figura 03: A) ANOVA da quantidade de Clorofila em Tamandaré no mês de setembro; B) ANOVA da quantidade de Feopigmento em Tamandare no mês de setembro. Ilha da Barra ( ) Pirambú ( ).Barra vertical indica 95% do intervalo de confiança, N= 48.
3.2 Pluviometria
A quantidade de chuva ocorrida nos meses de coleta foi dentro do esperado, mas um
pouco abaixo da média histórica, caracterizando o mês de abril como o início do período chuvoso
e o mês de setembro como início da estação seca (Fig. 04). Foram utilizados os dados desta
estação meteorológica (Curado) por ser a mais próxima de Tamandaré e que possui informação
disponível no sítio on line do INMET.
Figura 04: Indices de chuva acumulada do ano de 2008 (colunas) e média pluviométrica de período 1961-1990 (linha) na estação meteorológica do Curado, PE. Fonte: INMET
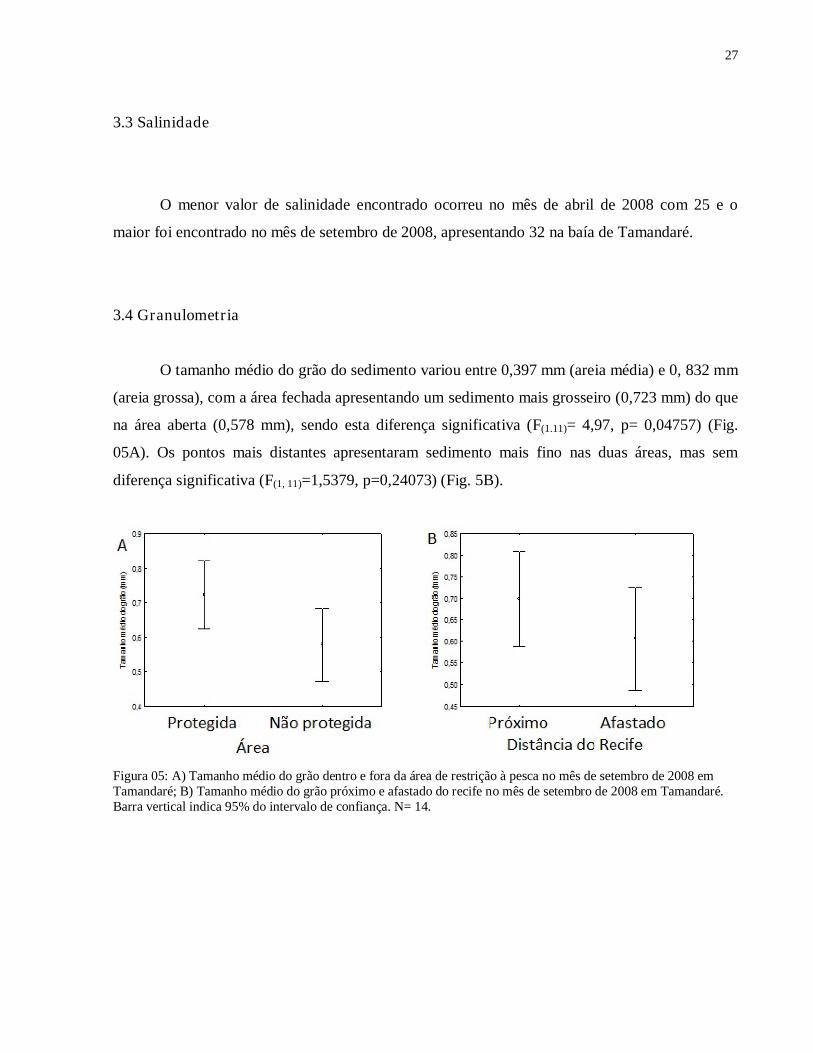
27
3.3 Salinidade
O menor valor de salinidade encontrado ocorreu no mês de abril de 2008 com 25 e o
maior foi encontrado no mês de setembro de 2008, apresentando 32 na baía de Tamandaré.
3.4 Granulometria
O tamanho médio do grão do sedimento variou entre 0,397 mm (areia média) e 0, 832 mm
(areia grossa), com a área fechada apresentando um sedimento mais grosseiro (0,723 mm) do que
na área aberta (0,578 mm), sendo esta diferença significativa (F(1.11)= 4,97, p= 0,04757) (Fig.
05A). Os pontos mais distantes apresentaram sedimento mais fino nas duas áreas, mas sem
diferença significativa (F(1, 11)=1,5379, p=0,24073) (Fig. 5B).
Figura 05: A) Tamanho médio do grão dentro e fora da área de restrição à pesca no mês de setembro de 2008 em Tamandaré; B) Tamanho médio do grão próximo e afastado do recife no mês de setembro de 2008 em Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 14.
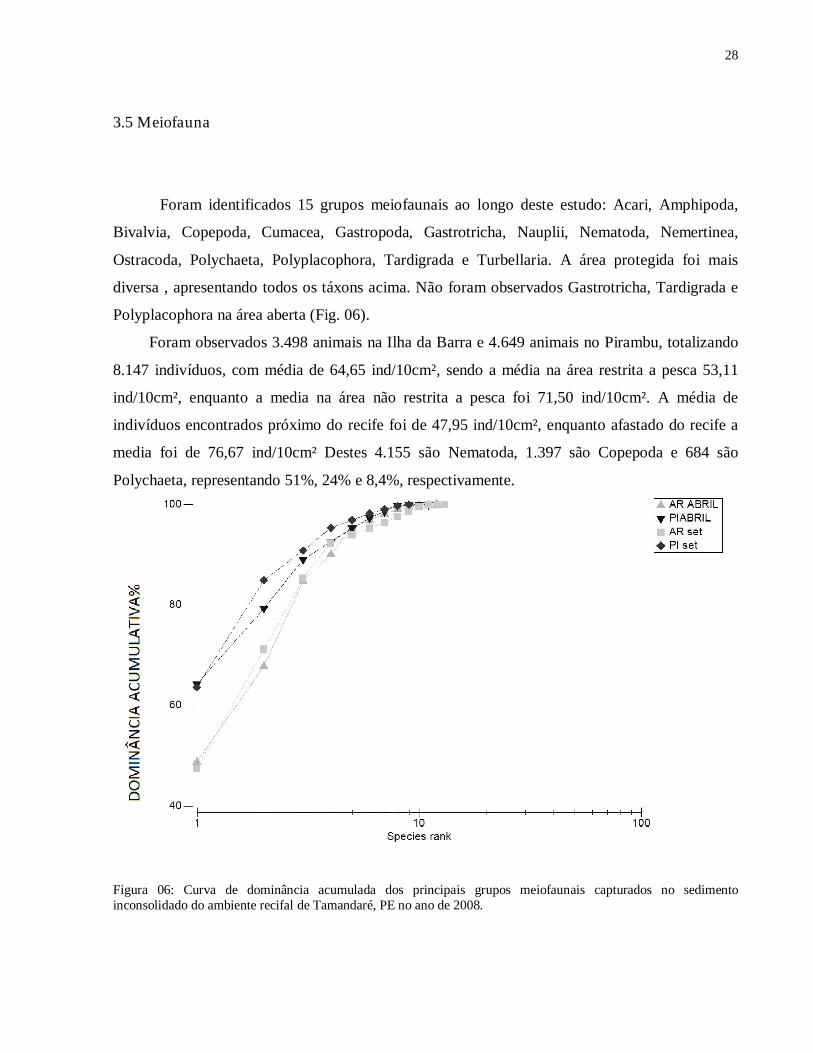
28
3.5 Meiofauna
Foram identificados 15 grupos meiofaunais ao longo deste estudo: Acari, Amphipoda,
Bivalvia, Copepoda, Cumacea, Gastropoda, Gastrotricha, Nauplii, Nematoda, Nemertinea,
Ostracoda, Polychaeta, Polyplacophora, Tardigrada e Turbellaria. A área protegida foi mais
diversa , apresentando todos os táxons acima. Não foram observados Gastrotricha, Tardigrada e
Polyplacophora na área aberta (Fig. 06).
Foram observados 3.498 animais na Ilha da Barra e 4.649 animais no Pirambu, totalizando
8.147 indivíduos, com média de 64,65 ind/10cm², sendo a média na área restrita a pesca 53,11
ind/10cm², enquanto a media na área não restrita a pesca foi 71,50 ind/10cm². A média de
indivíduos encontrados próximo do recife foi de 47,95 ind/10cm², enquanto afastado do recife a
media foi de 76,67 ind/10cm² Destes 4.155 são Nematoda, 1.397 são Copepoda e 684 são
Polychaeta, representando 51%, 24% e 8,4%, respectivamente.
Figura 06: Curva de dominância acumulada dos principais grupos meiofaunais capturados no sedimento inconsolidado do ambiente recifal de Tamandaré, PE no ano de 2008.
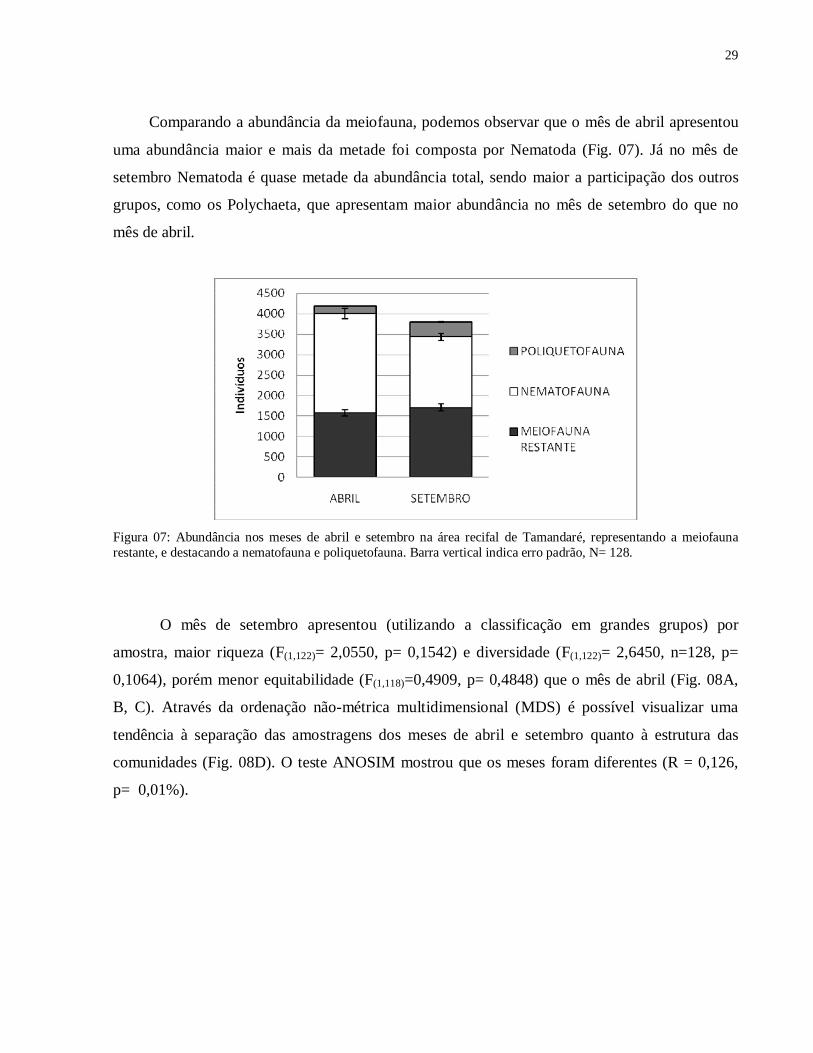
29
Comparando a abundância da meiofauna, podemos observar que o mês de abril apresentou
uma abundância maior e mais da metade foi composta por Nematoda (Fig. 07). Já no mês de
setembro Nematoda é quase metade da abundância total, sendo maior a participação dos outros
grupos, como os Polychaeta, que apresentam maior abundância no mês de setembro do que no
mês de abril.
Figura 07: Abundância nos meses de abril e setembro na área recifal de Tamandaré, representando a meiofauna restante, e destacando a nematofauna e poliquetofauna. Barra vertical indica erro padrão, N= 128.
O mês de setembro apresentou (utilizando a classificação em grandes grupos) por
amostra, maior riqueza (F(1,122)= 2,0550, p= 0,1542) e diversidade (F(1,122)= 2,6450, n=128, p=
0,1064), porém menor equitabilidade (F(1,118)=0,4909, p= 0,4848) que o mês de abril (Fig. 08A,
B, C). Através da ordenação não-métrica multidimensional (MDS) é possível visualizar uma
tendência à separação das amostragens dos meses de abril e setembro quanto à estrutura das
comunidades (Fig. 08D). O teste ANOSIM mostrou que os meses foram diferentes (R = 0,126,
p= 0,01%).
30
A B
C D
Figura 08: A) Riqueza meiofaunística nos meses de abril e setembro na área recifal de Tamandaré; B) Diversidade (Shannon) meiofaunística nos meses de abril e setembro na área recifal de Tamandaré; C) equitabilidade meiofaunística nos meses de abril e setembro na área recifal de Tamandaré, D) MDS representando os meses coletados na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 122
Quando comparamos a abundância da área protegida com a área sem proteção, podemos
observar uma abundância maior nesta última (Fig. 09), tanto na meiofauna quanto na
nematofauna. Os poliquetas apresentaram comportamento contrário, tendo uma abundância maior
na área protegida.
31
Figura 09: Abundância da meiofauna restante, da nematofauna e da poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica erro padrão, N= 128.
A área protegida apresentou maior equitabilidade (Fig. 10A), sendo esta diferença
significativa (F(1,118)= 17,848; p= 0,00005). A área protegida apresentou maior diversidade que a
não protegida (Fig 10B), sendo este resultado significativo (F(1,122)= 13,903, p= 0,0002). A
riqueza do recife da área fechada foi maior que a do recife da área aberta (F(1,122)= 1,6654, p=
0,1993) (Fig. 10C). Quanto à estrutura das comunidades dos dois recifes (dentro e fora da área de
proteção) o MDS não permitiu a visualização da separação das comunidades (Fig 10D), porém o
ANOSIM mostrou que houve diferença significativa entre as duas áreas (R= 0,106 e p= 0,01%).
A B
32
C D
Figura 10: A) Equitabilidade na área de proteção restrita à pesca e não restrita na área recifal de Tamandaré; B) diversidade (Shannon) na área de proteção restrita à pesca e não restrita na área recifal de Tamandaré protegida e não protegida; C) riqueza na área de proteção restrita à pesca e não restrita na área recifal de Tamandaré. D) MDS dos recifes nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N=122.
Quando analisamos quanto a distancia do recife, observamos maior abundância na fauna em
geral nos pontos mais afastados do recife (Fig. 11).
Figura 11: Abundância meiofaunística nas áreas próxima e afastada do recife destacando a nematofauna e poliquetofauna, em Tamandaré. Barra vertical indica erro padrão, N=128.
Quanto à riqueza houve diferença significativa (F(1,122)= 4,8852; n= 128; p=0,029) sendo o
ponto afastado do recife mais rico que o próximo ao recife (Fig. 12A). A área próxima ao recife
apresentou maior equitabilidade que a mais distante (F(1,118)= 0,5311, n= 128, p= 0,4675) (Fig.
12B). A área afastada do recife apresentou maior diversidade (F(1,122)= 2,0859, n= 128, p=
33
0,1512) (Fig. 12C). Quanto à estrutura das comunidades, o MDS não mostrou separação das
amostragens, e o ANOSIM não apresentou diferença significativa entre as áreas (R= 0,013 e p=
9,3%) (Fig. 12D).
A B
C D
Figura 12: A) Riqueza na área próxima e afastada do recife; B) equitabilidade na área próxima e afastada do recife; C) diversidade (Shannon) na área próxima e afastada do recife. D) MDS representando os grupos próximo ao recife e afastado do recife. Barra vertical indica 95% do intervalo de confiança. N=122.
3.5.1 BIOENV
O BIOENV foi realizado com os seguintes fatores: cascalho, areia média, areia fina, silte,
clorofila, feopigmento e matéria orgânica. Depois de feito o draftsman, se excluiu a fração areia
média devido a alta correlação apresentada, que poderia mascarar o resultado. Os fatores que
explicaram melhor o padrão biológico através do teste BIOENV foi o cascalho juntamente com a
matéria orgânica, que apresentou um índice de correlação de 0,204 sendo o mais representativo.
34
3.5.2 SIMPER
De acordo com a análise SIMPER, que verifica a contribuição dos grupos analisados para
as dissimilaridades ou diferenças na estrutura da comunidade meiofaunística presente nos pontos
amostrados, foi observado que 5 grupos foram responsáveis por mais de 50% das
dissimilaridades, sendo Copepoda, Polychaeta, Nematoda, Ostradoda e Turbellaria os que
contribuiram mais para as dissimilaridades entre os grupos amostrados (Tab. 3).
Tabela 2: Análise SIMPER para os grupos de acordo com os fatores pré-estabelecidos, apresentando os táxons/gênero que mais constribuíram com as dissimilaridades entre as áreas analisadas em Tamandaré.
Grupo Dissimilaridade
Média (%)
Táxon/Gênero % de
Contribuição
% Acumulada
Área restrita à pesca e Área não restrita
34,31 Copepoda Polychaeta Nematoda Ostracoda Turbellaria
17,95 13,93 12,53 12,23 9,28
17,95 31,88 44,41 56,64 65,92
Próximo e Afastado do recife
31,03 Copepoda Polychaeta Ostracoda Turbellaria Nematoda
15,68 14,20 12,35 10,80 10,73
15,68 29,87 42,23 53,03 63,75
3.6 Nematofauna
Foram encontrados 56 gêneros de Nematoda nas amostras coletadas no ambiente recifal de
Tamandaré, listados abaixo junto a sua abundância.
Nematoda
Enoplia
Enoplida

35
Enoplina
Thoracostomopsidae
Enoploides Ssaweljev, 1912 (1)
Epacanthion Wieser, 1953 (16)
Anoplostomatidae
Chaetonema Filipjev, 1927 (11)
Phanodermatidae
Crenopharyncx Filipjev, 1934 (1)
Ironidae
Dolicholaimus De Man, 1888 (24)
Leptosomatidae
Platycoma Cobb, 1894 (2)
Oncholaimidae
Oncholaimus Dujardin, 1845 (42)
Viscosia De Man, 1890 (14)
Enchelidiidae
Eurystomina Filipjev, 1921 (5)
Chormadoria
Chromadorida
Chromadorina
Chromadoridae
Actinonema Cobb, 1920 (2)
Chromadorella Filipjev, 1918 (3)
Chromadorita Flipjev, 1922 (2)
Ptycholaimellus Cobb, 1920 (4)
Rhips Cobb, 1920 (4)
Spilophorella Filipjev, 1917 (1)
Ethmolaimidae
Filitonchus Platt, 1982 (1)
Gomphionema Wieser & Hopper, 1966 (247)
Nannolaimus Cobb, 1920 (1)

36
Cyatholaimidae
Cyatholaimus Bastian, 1865 (1)
Longicyatholaimus Micoletky, 1924 (12)
Marylynnia Hopper, 1977 (119)
Nannolaimoides Ott, 1972 (3)
Paracanthonchus Micoletzky, 1924 (9)
Pomponema Cobb, 1917 (51)
Selachinematidae
Cheironchus Cobb, 1917 (2)
Halichoanolaimus De Man, 1886 (5)
Latronema Wieser, 1954 (16)
Synonchiella Cobb, 1933 (2)
Desmodoridae
Acanthopharynx Marion, 1870 (1)
Catanema Cobb, 1920 (1)
Desmodora De Man, 1889 (20)
Eubostrichus Greef, 1869 (1)
Metachromadora Filipjev, 1918 (4)
Spirinia Gerlach, 1963 (27)
Draconematidae
Dracograllus Allen & Noffsinger, 1978 (1)
Microlaimidae
Calomicrolaimus Lorenzen, 1971 (2)
Leptolaimina
Leptolaimidae
Deontolaimus De Man, 1880 (5)
Desmoscolecina
Desmoscolecidae
Tricoma Cobb, 1893 (18)
Monhysterida
Xyalidae

37
Daptonema Cobb, 1920 (371)
Promonhystera Wieser, 1956 (5)
Pseudosteirenia Wieser, 1956 (7)
Steirenia Micoletzky, 1922 (28)
Theristus bastian, 1865 (150)
Trichotheristus Wieser, 1956 (4)
Sphaerolaimidae
Parasphaerolaimus Ditlevsen, 1918 (1)
Linnhomoeidae
Desmolaimus De Man, 1880 (3)
Metalinhomoeus De Man, 1907 (2)
Terschellingia De Man, 1888 (95)
Axonolaimidae
Parodontophora Timm, 1963 (53)
Diplopeltidae
Southerniella Allgén, 1932 (1)
Comesomatidae
Comesoma Bastian, 1865 (20)
Dorylaimopsis Ditlevsen, 1918 (35)
Paracomesoma Hope & Murphy, 1972 (14)
Paramesonchium Hopper, 1967 (12)
Sabatieria Rouville, 1903 (43)
Setosabatieria Platt, 1985 (125)
Foram analisados 1.650 indivíduos, entre estes os mais abundantes foram os gêneros
Daptonema, Gomphionema, Theristus, Setosabatieria e Maylynnia respectivamente. O gênero
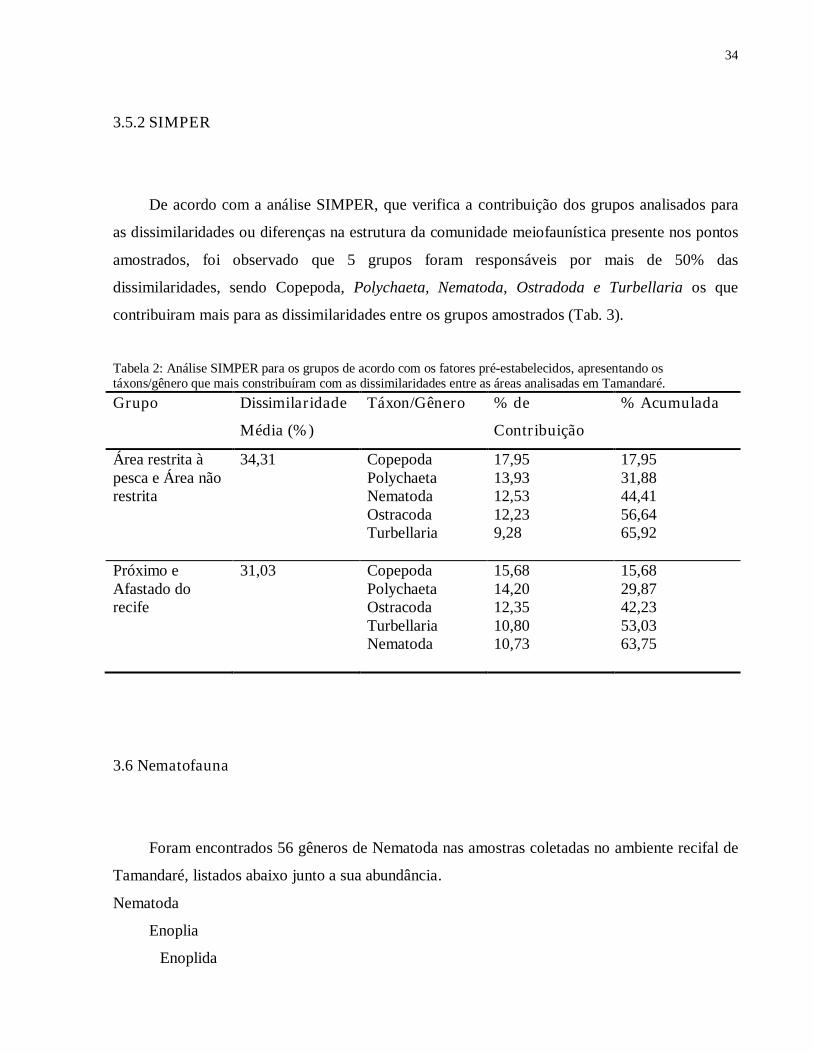
Daptonema (fig.13) e o gênero Theristus (fig. 13) são classificados segundo Wieser (1953, 1960)
e Wieser & Kanwisher (1961) como do tipo 1B. Gomphionema (fig.13) é classificado como do
tipo 2B. O gênero Setosabatieria (fig 13) é classificado como do tipo 1A. E Marylynnia (fig. 13)
é classificado como 2A (fig. 13).
38
Figura 13: Densidades médias dos gêneros de Nematoda mais abundantes nas amostras coletadas em abril e setembro de 2008, nas duas áreas de coleta, na baía de Tamandaré. Barra vertical indica erro padrão. Fonte: Modificado de WARWICK et al. (1988).
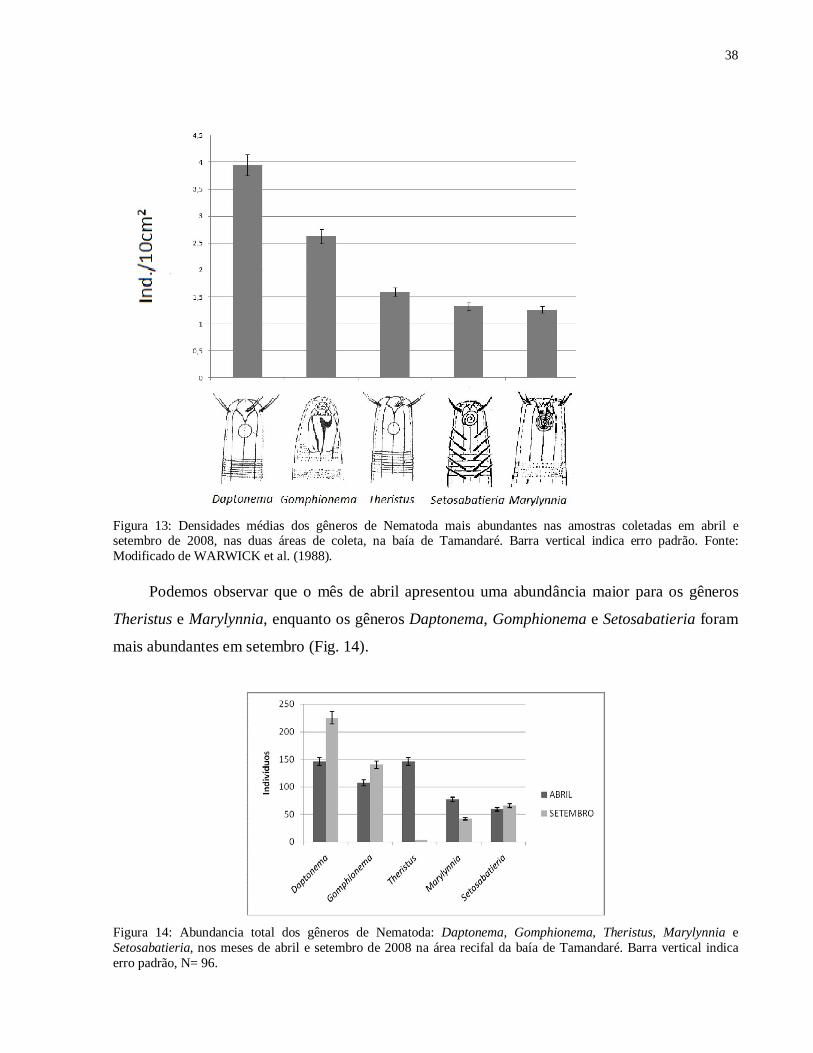
Podemos observar que o mês de abril apresentou uma abundância maior para os gêneros
Theristus e Marylynnia, enquanto os gêneros Daptonema, Gomphionema e Setosabatieria foram
mais abundantes em setembro (Fig. 14).
Figura 14: Abundancia total dos gêneros de Nematoda: Daptonema, Gomphionema, Theristus, Marylynnia e Setosabatieria, nos meses de abril e setembro de 2008 na área recifal da baía de Tamandaré. Barra vertical indica erro padrão, N= 96.
39
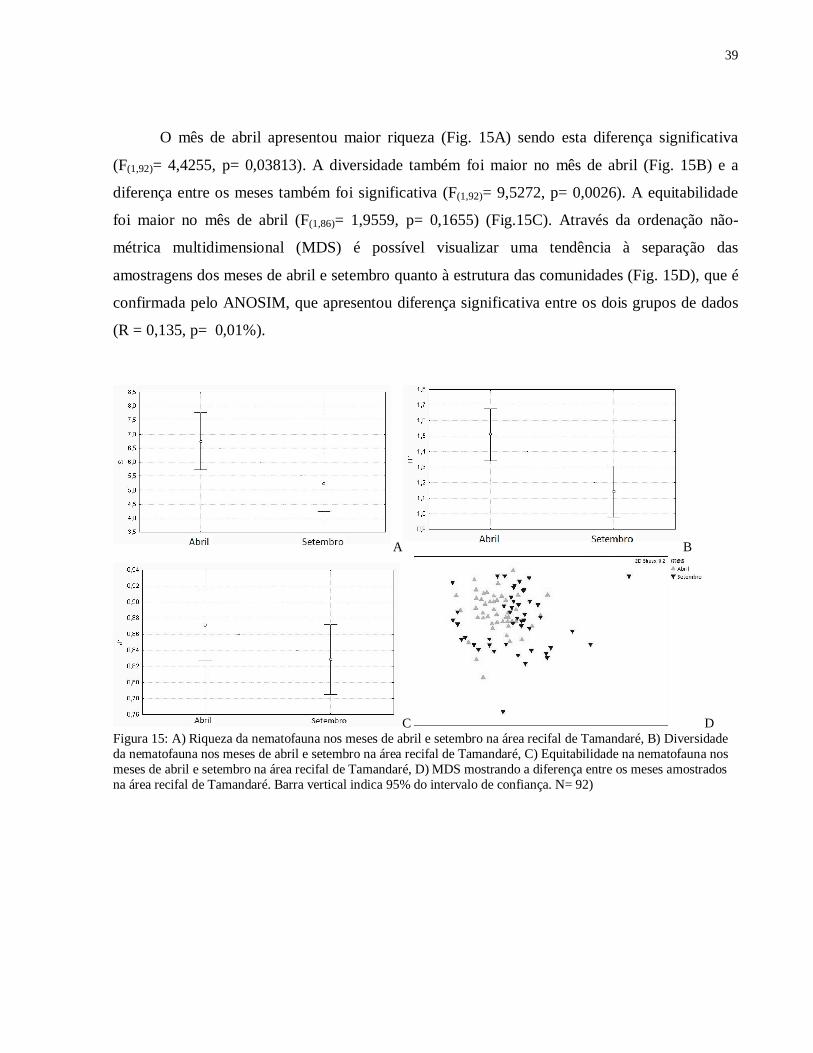
O mês de abril apresentou maior riqueza (Fig. 15A) sendo esta diferença significativa
(F(1,92)= 4,4255, p= 0,03813). A diversidade também foi maior no mês de abril (Fig. 15B) e a
diferença entre os meses também foi significativa (F(1,92)= 9,5272, p= 0,0026). A equitabilidade
foi maior no mês de abril (F(1,86)= 1,9559, p= 0,1655) (Fig.15C). Através da ordenação não-
métrica multidimensional (MDS) é possível visualizar uma tendência à separação das
amostragens dos meses de abril e setembro quanto à estrutura das comunidades (Fig. 15D), que é
confirmada pelo ANOSIM, que apresentou diferença significativa entre os dois grupos de dados
(R = 0,135, p= 0,01%).
A B
C D Figura 15: A) Riqueza da nematofauna nos meses de abril e setembro na área recifal de Tamandaré, B) Diversidade da nematofauna nos meses de abril e setembro na área recifal de Tamandaré, C) Equitabilidade na nematofauna nos meses de abril e setembro na área recifal de Tamandaré, D) MDS mostrando a diferença entre os meses amostrados na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 92)
40
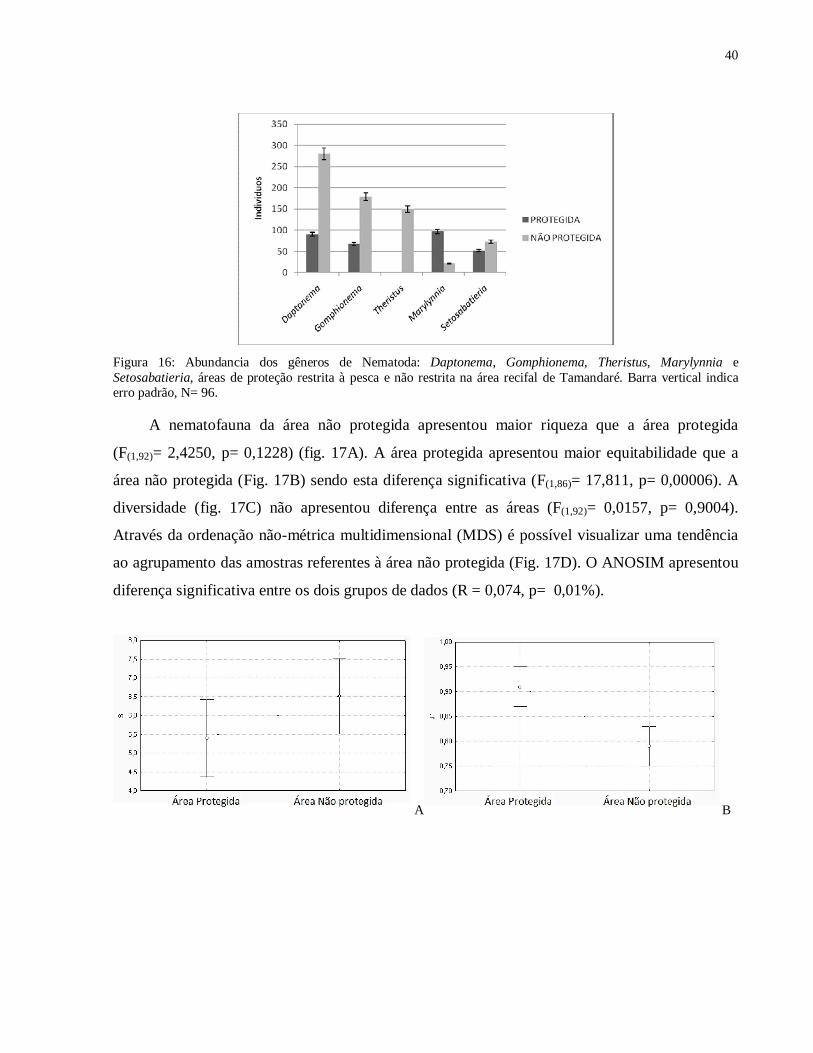
Figura 16: Abundancia dos gêneros de Nematoda: Daptonema, Gomphionema, Theristus, Marylynnia e Setosabatieria, áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica erro padrão, N= 96.
A nematofauna da área não protegida apresentou maior riqueza que a área protegida
(F(1,92)= 2,4250, p= 0,1228) (fig. 17A). A área protegida apresentou maior equitabilidade que a
área não protegida (Fig. 17B) sendo esta diferença significativa (F(1,86)= 17,811, p= 0,00006). A
diversidade (fig. 17C) não apresentou diferença entre as áreas (F(1,92)= 0,0157, p= 0,9004).
Através da ordenação não-métrica multidimensional (MDS) é possível visualizar uma tendência
ao agrupamento das amostras referentes à área não protegida (Fig. 17D). O ANOSIM apresentou
diferença significativa entre os dois grupos de dados (R = 0,074, p= 0,01%).
A B
41
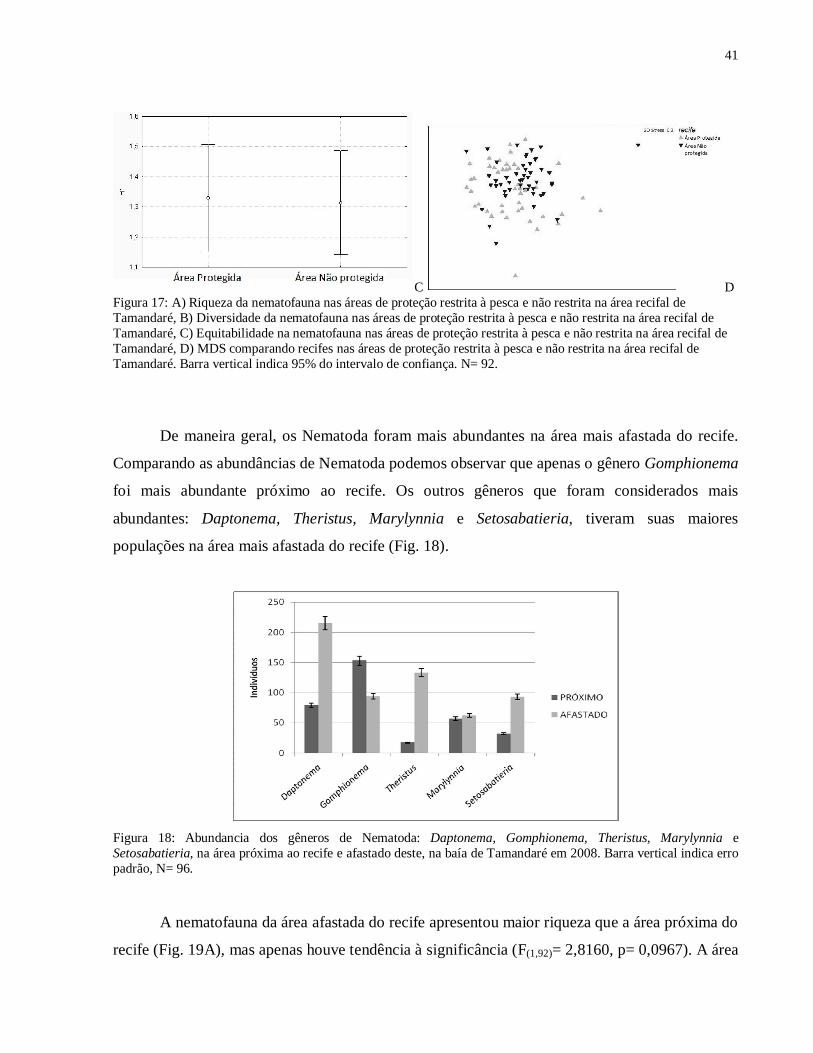
C D Figura 17: A) Riqueza da nematofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, B) Diversidade da nematofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, C) Equitabilidade na nematofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, D) MDS comparando recifes nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 92.
De maneira geral, os Nematoda foram mais abundantes na área mais afastada do recife.
Comparando as abundâncias de Nematoda podemos observar que apenas o gênero Gomphionema
foi mais abundante próximo ao recife. Os outros gêneros que foram considerados mais
abundantes: Daptonema, Theristus, Marylynnia e Setosabatieria, tiveram suas maiores
populações na área mais afastada do recife (Fig. 18).
Figura 18: Abundancia dos gêneros de Nematoda: Daptonema, Gomphionema, Theristus, Marylynnia e Setosabatieria, na área próxima ao recife e afastado deste, na baía de Tamandaré em 2008. Barra vertical indica erro padrão, N= 96.
A nematofauna da área afastada do recife apresentou maior riqueza que a área próxima do
recife (Fig. 19A), mas apenas houve tendência à significância (F(1,92)= 2,8160, p= 0,0967). A área
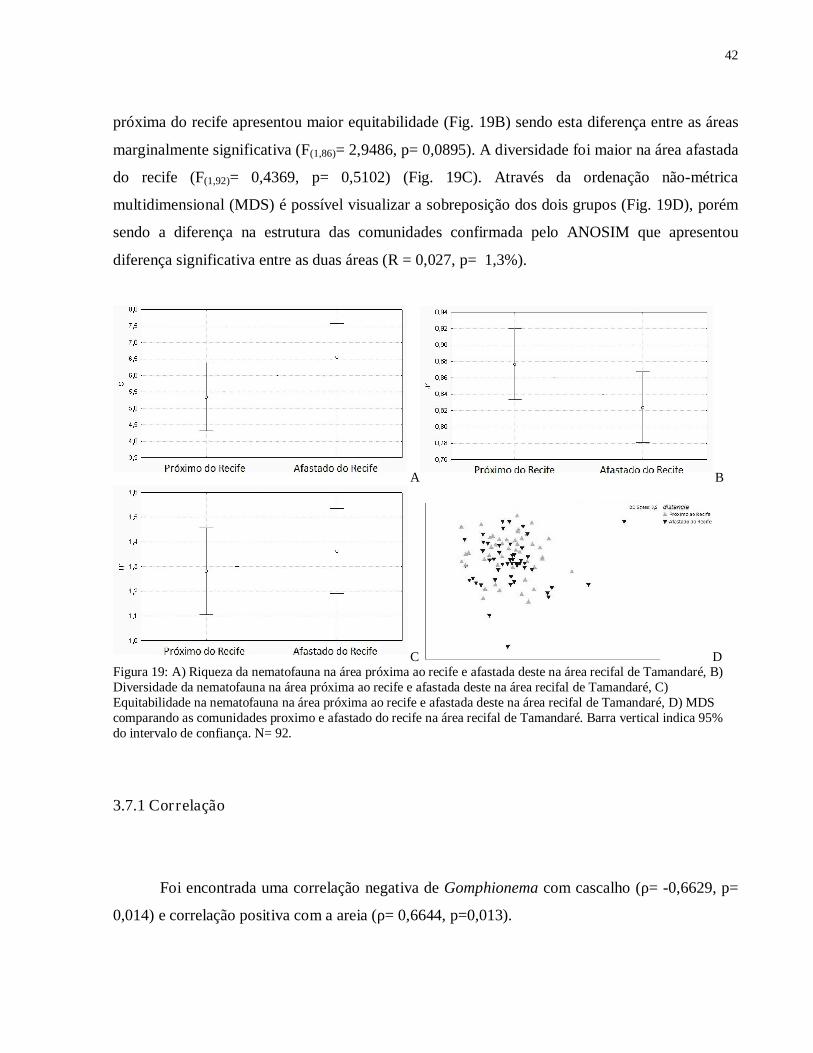
42
próxima do recife apresentou maior equitabilidade (Fig. 19B) sendo esta diferença entre as áreas
marginalmente significativa (F(1,86)= 2,9486, p= 0,0895). A diversidade foi maior na área afastada
do recife (F(1,92)= 0,4369, p= 0,5102) (Fig. 19C). Através da ordenação não-métrica
multidimensional (MDS) é possível visualizar a sobreposição dos dois grupos (Fig. 19D), porém
sendo a diferença na estrutura das comunidades confirmada pelo ANOSIM que apresentou
diferença significativa entre as duas áreas (R = 0,027, p= 1,3%).
A B
C D Figura 19: A) Riqueza da nematofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, B) Diversidade da nematofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, C) Equitabilidade na nematofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, D) MDS comparando as comunidades proximo e afastado do recife na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 92.
3.7.1 Correlação
Foi encontrada uma correlação negativa de Gomphionema com cascalho (ρ= -0,6629, p=
0,014) e correlação positiva com a areia (ρ= 0,6644, p=0,013).

43
3.7 Poliquetofauna
Foram identificadas 9 famílias de Polychaeta e 18 gêneros no ambiente recifal de
Tamandaré, listados abaixo junto à sua abundância.
Orbinida
Parainidae
Aparaonis Hartman, 1965 (7)
Aedicira Hartman, 1957 (8)
Spionida
Spionidae
Prionospio Malmgren, 1867 (7)
Cirratulidae (1)
Phyllodocida
Sigalionidae (7)
Hesionidae
Alikunhia Hartman, 1959 (5)
Gen 1 (1)
Syllidae
Autolytinae
Proceraea Ehlers, 1864 (1)
Gen 2 (1)
Exogoninae
Brania Quatrefages, 1866 (1)
Sphaerosyllis Clarapede, 1863 (180)
Sphaerosyllis annulata Nogueira, San Martin & Fukuda, 2003
Sphaerosyllis centroamericana Hartmann-Schroeder, 1959
Parapionosyllis Fauvel, 1923 (1)
Gen 3 (3)
Eusillynae
44
Syllides Oersted, 1845 (11)
Eudontosyllis Knox, 1960 (2)
Pionosyllis Malmgren, 1867 (3)
Plakosyllis Hartmann-Schroeder, 1956 (1)
Braniella Hartman, 1965 (2)
Gen 4 (1)
Terebellida
Terebellidae (1)
Nerillida
Nerillidae
Meganerilla Boaden, 1961 (1)
Mesonerilla Remane, 1949 (3)
Nerilla Schmidt, 1848 (1)
Psamoriedlia Kersteuer, 1966 (3)
Thalassochaetus Ax, 1954 (1)
Diurodrilidae (1)
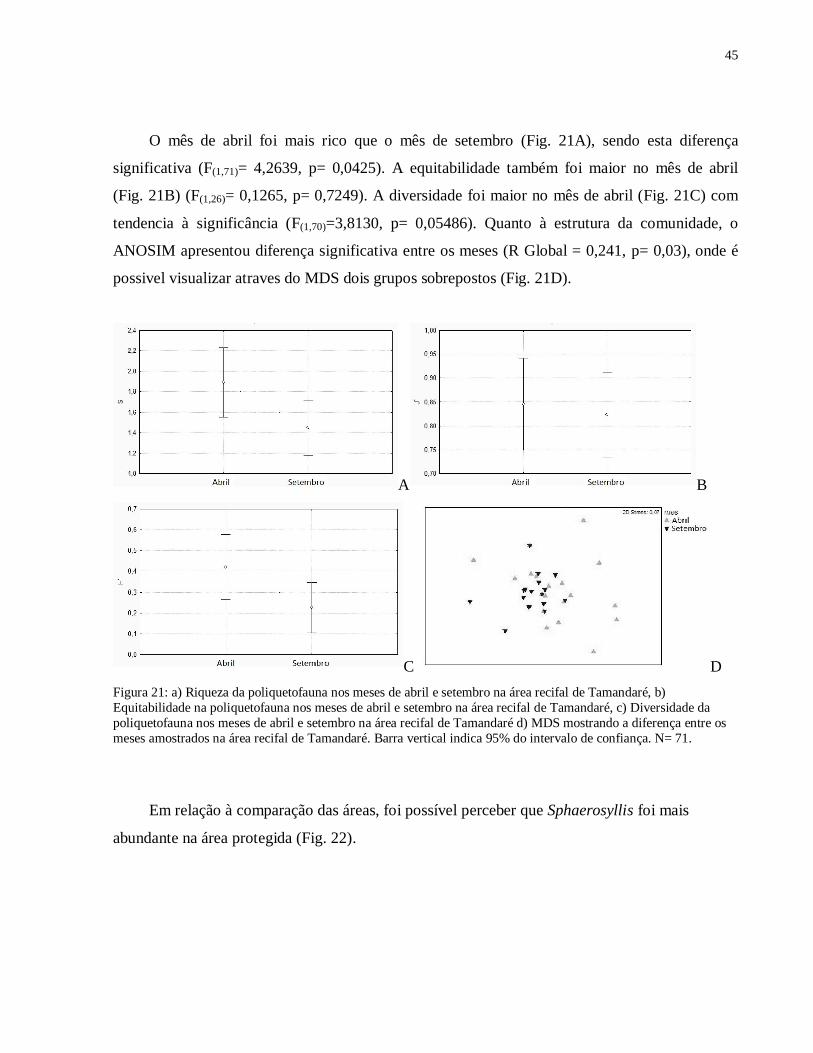
Foram analisados 254 indivíduos, dentre os quais o mais abundante foi Sphaerosyllis, que
pertence à família Syllidae. Em relação à abundância dos Sphaerosyllis, podemos observar que
o mês de setembro apresentou abundância maior que o mês de abril (Fig 20).
Figura 20: Abundância do gênero Sphaerosyllis nos meses de abril e setembro, na baía de Tamandaré em 2008. Barra vertical indica erro padrão, N= 90.
45
O mês de abril foi mais rico que o mês de setembro (Fig. 21A), sendo esta diferença
significativa (F(1,71)= 4,2639, p= 0,0425). A equitabilidade também foi maior no mês de abril
(Fig. 21B) (F(1,26)= 0,1265, p= 0,7249). A diversidade foi maior no mês de abril (Fig. 21C) com
tendencia à significância (F(1,70)=3,8130, p= 0,05486). Quanto à estrutura da comunidade, o
ANOSIM apresentou diferença significativa entre os meses (R Global = 0,241, p= 0,03), onde é
possivel visualizar atraves do MDS dois grupos sobrepostos (Fig. 21D).
A B
C D
Figura 21: a) Riqueza da poliquetofauna nos meses de abril e setembro na área recifal de Tamandaré, b) Equitabilidade na poliquetofauna nos meses de abril e setembro na área recifal de Tamandaré, c) Diversidade da poliquetofauna nos meses de abril e setembro na área recifal de Tamandaré d) MDS mostrando a diferença entre os meses amostrados na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N= 71.
Em relação à comparação das áreas, foi possível perceber que Sphaerosyllis foi mais
abundante na área protegida (Fig. 22).
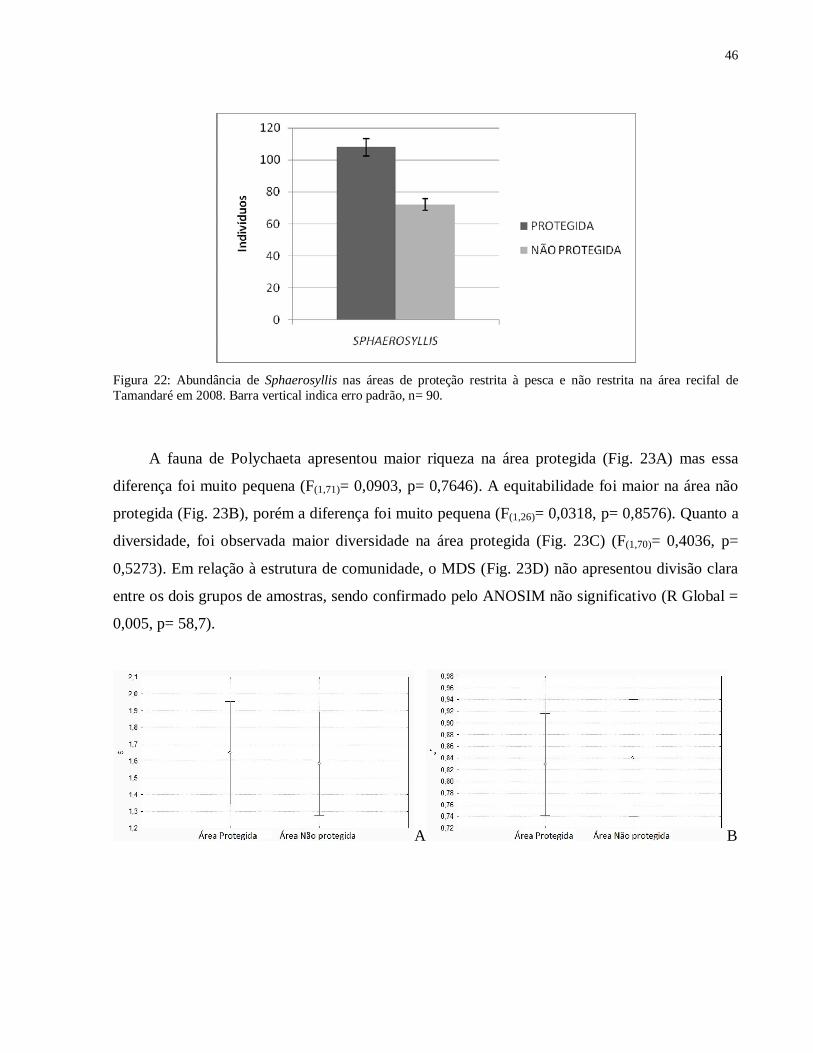
46
Figura 22: Abundância de Sphaerosyllis nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré em 2008. Barra vertical indica erro padrão, n= 90.
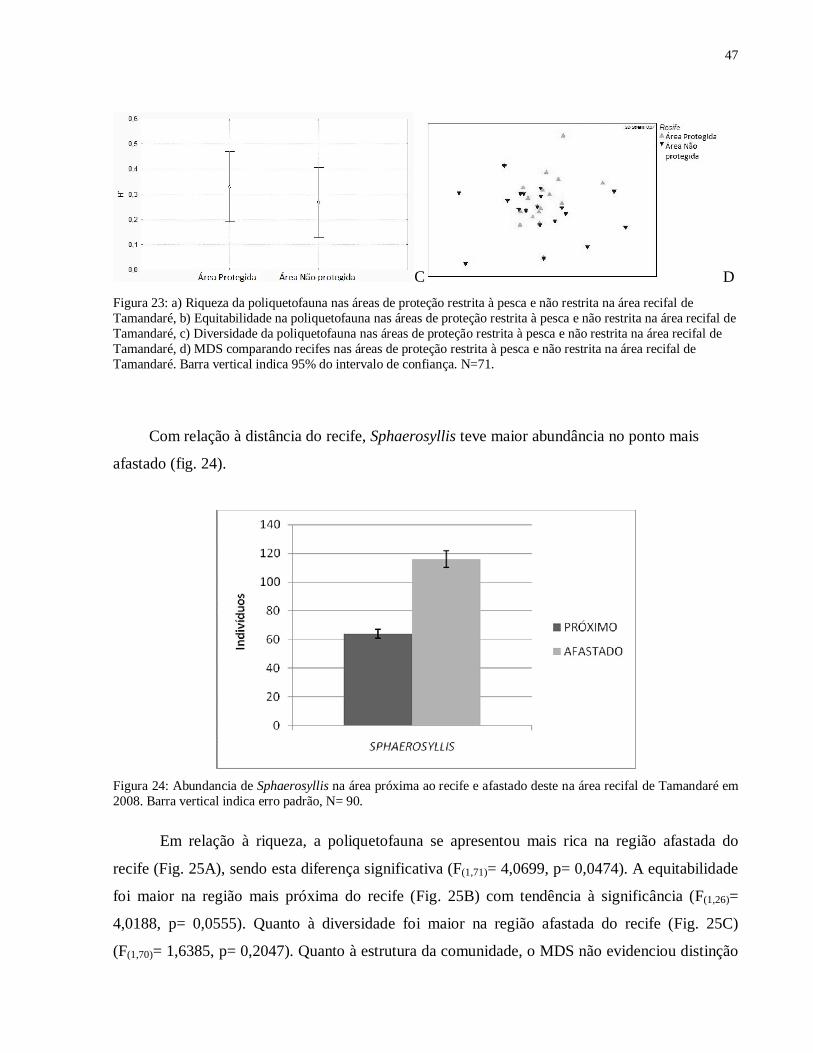
A fauna de Polychaeta apresentou maior riqueza na área protegida (Fig. 23A) mas essa
diferença foi muito pequena (F(1,71)= 0,0903, p= 0,7646). A equitabilidade foi maior na área não
protegida (Fig. 23B), porém a diferença foi muito pequena (F(1,26)= 0,0318, p= 0,8576). Quanto a
diversidade, foi observada maior diversidade na área protegida (Fig. 23C) (F(1,70)= 0,4036, p=
0,5273). Em relação à estrutura de comunidade, o MDS (Fig. 23D) não apresentou divisão clara
entre os dois grupos de amostras, sendo confirmado pelo ANOSIM não significativo (R Global =
0,005, p= 58,7).
A B
47
C D
Figura 23: a) Riqueza da poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, b) Equitabilidade na poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, c) Diversidade da poliquetofauna nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré, d) MDS comparando recifes nas áreas de proteção restrita à pesca e não restrita na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N=71.
Com relação à distância do recife, Sphaerosyllis teve maior abundância no ponto mais
afastado (fig. 24).
Figura 24: Abundancia de Sphaerosyllis na área próxima ao recife e afastado deste na área recifal de Tamandaré em 2008. Barra vertical indica erro padrão, N= 90.
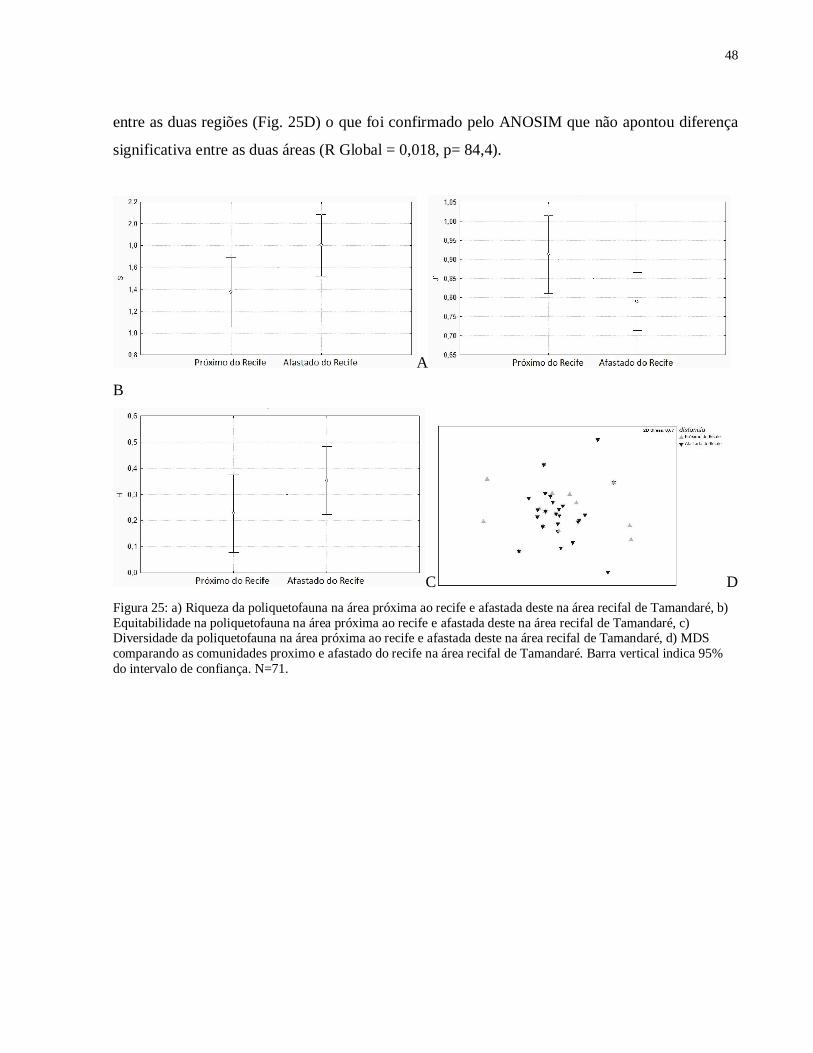
Em relação à riqueza, a poliquetofauna se apresentou mais rica na região afastada do
recife (Fig. 25A), sendo esta diferença significativa (F(1,71)= 4,0699, p= 0,0474). A equitabilidade
foi maior na região mais próxima do recife (Fig. 25B) com tendência à significância (F(1,26)=
4,0188, p= 0,0555). Quanto à diversidade foi maior na região afastada do recife (Fig. 25C)
(F(1,70)= 1,6385, p= 0,2047). Quanto à estrutura da comunidade, o MDS não evidenciou distinção
48
entre as duas regiões (Fig. 25D) o que foi confirmado pelo ANOSIM que não apontou diferença
significativa entre as duas áreas (R Global = 0,018, p= 84,4).
A
B
C D
Figura 25: a) Riqueza da poliquetofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, b) Equitabilidade na poliquetofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, c) Diversidade da poliquetofauna na área próxima ao recife e afastada deste na área recifal de Tamandaré, d) MDS comparando as comunidades proximo e afastado do recife na área recifal de Tamandaré. Barra vertical indica 95% do intervalo de confiança. N=71.

49
4 DISCUSSÃO
Os estudos em áreas de proteção marinhas são de extrema importância para a avaliação
das unidades de conservação, por servirem como coleções vivas devido ao conjunto de
biodiversidade que abrigam e de promover uma situação ímpar que é a pouca influência humana,
fundamental para algumas observações e estudos. Apesar das poucas unidades de conservação
marinha com áreas restritas no Brasil, a APA dos Corais se destaca como a mais bem preservada
do Brasil e através dos trabalhos de pesquisa desenvolvidos no local (Macedo. 2009).
Este trabalho veio contribuir com o conhecimento da ecologia de comunidades do
substrato inconsolidado no ambiente recifal, utilizando a meiofauna para analisar as diferenças
em relação à proteção da pesca e a distancia do recife. Além da contribuição para o conhecimento
de Nematoda e Polychaeta, sendo este grupo raramente abordado de maneira mais detalhada em
estudos de meiofauna.
A área de proteção restrita apresentou os maiores índices de microfitobentos do
sedimento, sendo o feopigmento responsável por este resultado, apresentando o dobro da área não
protegida em média. A média de clorofila diferiu pouco entre as duas áreas amostradas, protegida
e não protegida. Como já citado por MUROLO et al. (2005) o feopigmento geralmente tem maior
concentração que a clorofila a, e ambos tem forte ligação a concentrações de nitrogênio.
microfitobentos .
No geral, as características do sedimento encontradas como a proporção de sedimentos
grosseiros e areia, possuem distribuição unimodal, indicando que os mesmos estão sendo
estruturados por um agente ambiental. As diferenças encontradas nos parâmetros do sedimento
sugerem diferenças pontuais, uma vez que abordam uma pequena área, assim como foi
encontrado por Macedo (2009) para a mesma área.
Apesar de haver diferença significativa entre a granulometria das áreas, o índice de
correlação apresentado pelo BIOENV foi de apenas 0,204 para cascalho e matéria orgânica,
sugerindo que as variações na granulometria e sedimentação explicam muito pouco as diferenças
encontradas para a meiofauna entre as duas áreas.
Assim como neste estudo, Ambrose & Anderson (1990) na Califórnia também
encontraram sedimento mais grosso quando mais próximo do que afastado 10 e 20 metros. Este
padrão também ocorreu na Austrália (BARROS, 2005) e no sul do Brasil (LORENZI, 2004), em

50
recifes artificiais ou naturais, com desenho amostral similar ao deste trabalho. O que mostra a
clara influência do recife no sedimento devido à ação das correntes que se formam pela presença
do recife (LORENZI, 2004). Os recifes de Tamandaré são largos, fazendo com que a onda perca
energia ao passar por eles
Porém na Itália (DANOVARO, 2002) a fração de finos aumentou ou se manteve,
sugerindo uma redução de turbulências e por ser uma área de deposição. Netto et al (1999)
comparando a área externa com a interna do Atol das Rocas, observou sedimento mais grosso na
área externa do que na área interna, neste caso sendo influenciado pelas correntes oceânicas.
Enfim, o sedimento reflete as condições de hidrodinâmicas prevalentes, sendo de grande
importância a intensidade e sentido destas correntes.
Provavelmente dois fatores devem agir nesta área: a hidrodinâmica e a deposição.
Segundo Macedo (2009) a taxa de deposição de finos na baía de Tamandaré é alta, apresentando
média de 662 g/m²/dia. A hidrodinâmica local é conhecida apresentando uma média de 2m de
altura da onda, com pluviometria maior no inverno e com marés de sizígia que cobrem totalmente
o recife levando a remobilização dos sedimentos (MACEDO, 2009). Além da influencia do rio
Una, que deságua em local próximo e com ajuda dos ventos de sul e sudeste, direcionam uma
pluma de sedimentos para a área dos recifes da baía de Tamandaré (MACEDO, 2009).
A salinidade se apresentou dentro do esperado para a região. O menor resultado ocorreu
logo após um mês com chuvas bem acima da média. Enquanto o menos resultado de salinidade
ocorreu num mês com chuvas bem abaixo da média. Talvez por isso os dados de salinidade se
apresentem dessa forma. A pluviosidade também dentro do esperado como já visto por outros
autores sobre a área (ANJOS, 2007; BASTOS, 2006), apenas um pouco abaixo da média
esperada para a época. Porém não há descaracterização do que normalmente ocorre no litoral
pernambucano que é um mês de abril com inicio do aumento das chuvas e setembro como inicio
do período seco.
Maranhão (2003), estudou a meiofauna da parte superior e interna do Recife de Porto de
Galinhas, encontrando 447 ind/cm² no máximo e uma média de 174,6 ind/cm², sendo importante
ressaltar que este trabalho abordou a parte interna do recife e o topo, e na área interna do recife a
média da abundância não ultrapassou 100 ind/cm², semelhante ao encontrado neste trabalho.
Danovaro (2002) encontrou uma média máxima em torno de 380 ind/10cm² e o mínimo próximo
de 100 ind/cm² num recife artificial na Itália. Já Guzman et al (1987) encontrou de 99 a 5.755

51
ind/cm² em um recife de coral do Pacífico. Este trabalho apresentou 64,65 ind/10 cm², com o
máximo de 239 ind/cm², que estaria dentro das médias encontradas por outros autores para
ambiente recifal. Logan e colaboradores (2008), trabalhando no substrato duro de recifes,
encontraram densidades semelhantes às de Tamandaré, 40-290 ind/cm2, embora tenham utilizado
malha de 100 μm, e coletado com ‘air-lift’.
Para Pernambuco são descritos 21 táxons meiofaunísticos, são eles: Nematoda, Copepoda
Turbellaria, Kinorhyncha, Gastrotricha, Polychaeta, Oligochaeta, Nemertinea, Acari, Ostracoda,
Cumacea, Isopoda, Amphipoda, Priapulida, Loricifera, Gastropoda, Bivalvia, Rotifera,
Polyplacophora, Insecta e Tardigrada (DOS SANTOS, 2005, BARBOSA et al 2007, Da Rocha –
Com. Pess., Venekey,et al, 2008). O estudo da meiofauna historicamente tem seu enfoque
principal na taxonomia, sendo a ecologia menos abordada. No litoral de Pernambuco os táxons
melhor estudados são os Nematoda, os Copepoda e os Tardigrada, estes últimos mais
recentemente. Os outros táxons apenas têm sua ocorrência relatada, sendo então carentes de
informações mais detalhadas.
O número de táxons encontrados em trabalhos envolvendo ambiente recifais varia muito.
Em Porto de Galinhas, trabalhando com a meiofauna (MARANHÃO, 2003) obteve 13 táxons,
enquanto Danovaro (2002) relatou apenas 7 táxons em seu trabalho na Itália, também com
meiofauna. Lorenzi (2004) trabalhando com macrofauna intersticial observou 25 táxons em recife
natural no sul do Brasil.
Em Tamandaré foram desenvolvidos estudos meiofaunísticos em vários ambientes como
o ambiente de praia (VENEKEY, 2007), recifal (MARANHÃO, 2000) e estuarino
(VASCONCELOS, 2003). Venekey (2007) relatou 10 táxons, enquanto Vasconcelos (2003) em
área de manguezal do Rio Formoso registrou 14 táxons. Maranhão (2000) registrou apenas 4
táxons na área interna do recife, e 8 táxons no topo recifal, em substrato inconsolidado. Também
em ambiente de praia em Tamandaré, Ribeiro (1999) observou a presença de 14 táxons. Este
trabalho registrou ao todo 15 táxons, números bem maiores que os encontrados por Maranhão
(2000) para o ambiente recifal da área, mas bem próximo dos resultados obtidos nos outros
ambientes do mesmo local.
O mês de setembro apresentou maior riqueza de grandes grupos. Essa diferença temporal
encontrada parece estar relacionada com a mudança marcante que este local sofre no período
chuvoso. Segundo Macedo (2009) a sedimentação juntamente com o maior aporte de águas

52
fluviais ocorrem de forma intensa neste período. Sendo assim, é bem provável que essa grande
modificação ambiental interfira aumentando a complexidade do ambiente, e consequentemente
aumentando a riqueza.
Nematoda foi o grupo dominante, seguido de copepoda. Este padrão foi encontrado para a
maioria dos ambientes estudados em Pernambuco, assim como de outros lugares. De maneira
geral existem 4 grupos que variam na posição de mais abundantes: Nematoda, Copepoda,
Polychaeta e Amphipoda (GIERE, 1993). Em geral é conhecido que em sedimentos finos e
lamosos a dominância seja dos Nematoda e em sedimento grosseiro a dominância seja dos
Copepoda (HEIP et al., 1985; COULL, 1970; BOECKNER et al., 2009). Neste trabalho apesar
do tamanho do grão ser entre médio e grosso, a dominância foi de Nematoda, como geralmente
acontece nos trabalhos da região.
No presente estudo verificou-se que estrutura da comunidade nas duas áreas abordadas foi
similar, embora a diversidade meiofaunística seja maior na área protegida, e a abundância seja
maior fora da área de proteção. É importante ressaltar que grande abundância de Nematoda na
área não protegida refletiu diretamente na abundância da comunidade como um todo. Em relação
à presença do recife, a abundância foi maior nos pontos mais afastados, sendo também nestes
pontos a riqueza significativamente maior.
Lorenzi (2004) em trabalho com macrofauna de ambiente recifal no sul do Brasil, não
encontrou diferenças significativas quanto à riqueza em relação à presença do recife artificial. Em
recife naturais, porém, o autor encontrou riqueza significativamente maior na região afastada do
recife. Posey & Ambrose (1994) também encontraram maior abundância a 75 metros do recife.
Este fator pode ser explicado devido ao “halo trófico” formado adjacente ao recife, sendo mais ou
menos evidente de acordo com tamanho do recife, densidade de predadores e época do ano
(LORENZI, 2004). Apesar disso, alguns autores relatam que a presença do recife no ambiente,
causando modificações nos padrões físicos, seria mais influente na infauna do que fatores como
predação (AMBROSE & ANDERSON, 1994; FABI et al, 2002). Barros (2005) também
encontrou diferença entre a área próxima do recife e a distante dele, sendo a abundancia maior
quando mais afastado do recife, assim como aconteceu neste trabalho.
O distúrbio ambiental exerce um papel critico na heterogeneidade do ambiente e resulta
na diversidade de espécies (SERRANO et al, 2006), já tendo sido observado em comunidades de
substrato duro (SOUSA, 2001). Quando um padrão de baixa riqueza e alta equitabilidade ocorre,

53
parece haver um padrão de baixa complexidade ou presença de ambiente com grande
perturbação. Como foi observado neste trabalho na área próxima do recife houve maior tamanho
do grão, e consequentemente maior hidrodinamismo, e maior quantidade de feopigmentos.
Enquanto que afastado do recife a diversidade foi maior, a quantidade de clorofila foi maior e
maior complexidade (ou menor perturbação).
Observando estudos sobre Nematoda marinhos de vida livre no litoral pernambucano,
verificamos que Castro (2003) encontrou 27 gêneros, na região entre-marés de uma praia
protegida, e Silva (2004) encontrou 56 gêneros em ambientes infralitorais estuarinos. Da Rocha
(2003) encontrou 86 gêneros em ambiente fital. Em trabalho na praia de Tamandaré, Venekey
(2007) encontrou 57 gêneros, em praia aberta, números não muito distante dos aqui apresentados.
Porém dos 56 gêneros deste trabalho, 34 são gêneros diferentes dos encontrados
anteriormente por Venekey 2007. Alguns gêneros como Chaetonema, Deontolaimus,
Dracograllus, Enoploides, Filitonchus, Nannolaimus, que se julgava serem exclusivos de praias
arenosas, e Acantopharyncx, considerado exclusivo de ambiente fital (Venekey, 2007), foram
encontrados no ambiente recifal, porém Dracograllus, Enoploides, Filitonchus, Nannolaimus e
Acantopharyncx apenas um exemplar de cada foi registrado, o que pode indicar um fato
ocasional.
A riqueza da nematofauna foi maior na área sem proteção legal, porém a equitabilidade
entre as espécies foi maior na área protegida, fazendo com que o índice de diversidade (Shannon)
não fosse diferente nas duas áreas. Dos cinco gêneros de Nematoda mais abundantes neste
trabalho, Daptonema, Gomphionema e Theristus são freqüentemente encontrados no conteúdo
estomacal de juvenis de gerreídeos e gobiídeos (SILVA, 2004). Isto deve explicar a menor
abundância destes gêneros na área protegida, onde se supõe haver maior quantidade de peixes
predadores da meiofauna. Marylynnia e Setosabatieria já foram encontrados em conteúdo
estomacal de alguns poucos peixes das famílias acima citadas (SILVA, 2004), porém foram
considerados raros por isto talvez a influência da proteção legal na abundância da nematofauna
não tenha sido tão evidente. Foi possível inclusive observar uma população maior de Marylynnia
na área protegida, isso pode ser explicado pela maior quantidade de microfitobentos na área
protegida do que na área não protegida, uma vez que é um animal herbívoro. Gomphionema
apresentou correlação negativa com cascalho e positiva com areia, o que pode explicar sua menor
abundância na área protegida, uma vez que lá o sedimento se apresentou mais grosso. Esse

54
evento pode ainda ter sido amplificado devido a interações biológicas (parece ser item comum na
dieta de alguns peixes - SILVA, 2004).
Em relação à presença do recife, a riqueza e diversidade foram maiores no ponto mais
afastado, porém a equitabilidade da nematofauna foi menor neste ponto, o que indica uma co-
dominancia de alguns gêneros de Nematoda, confirmado pelos dados de abundância. Apesar de
Nematoda ter sido o táxon mais abundante neste estudo, houve diferença no padrão de
abundância nas duas áreas, com a área protegida apresentando uma equitabilidade maior, este
padrão foi similar nos dois períodos de coleta. Este resultado parece corroborar com o tema da
complexidade do ambiente e hidrodinamismo abordado anteriormente.
O gênero Marylynnia, apesar de não apresentar correlação com microfitobentos, estão
intimamente ligados a eles devido ao hábito alimentar e estes foram encontrados em maior
quantidade na área protegida como citado anteriormente.
Devido ao fato de haver poucos estudos de Polychaeta na região Nordeste, mais
precisamente os indivíduos intersticiais, e nenhum estudo de Polychaeta da meiofauna, muitos
são primeira ocorrência para o nordeste ou para o Brasil. Da família Paraonidae, o gênero
Aparaonis é nova ocorrência para o Brasil e Aedicira é nova ocorrência para o nordeste. O gênero
Alikunhia da família Hesionidae é nova ocorrência para o Brasil. Da família Syllidae os gêneros
Sphaerosyllis, Syllides e Pionosyllis são novas ocorrências para o nordeste; logo as espécies S.
Annulata e S. centro-americana são novas ocorrências para o nordeste. E todos os gêneros
apresentados pertencentes à família Nerillidae e ocorrência da família Diurodrilidae são novas
ocorrências para o Brasil.
Kohn & Lloyd (1973) já citavam a família Syllidae como especialmente abundante em
ambientes recifais, e neste trabalho não foi diferente, onde aproximadamente 70% dos Polychaeta
foram da família Syllidae. Sphaerosyllis foi o gênero mais abundante, são considerados
consumidores de depósito, oportunistas e também se alimentam de diatomáceas (FAUCHAULD
& JUMARS, 1979), vindo dai seu provável sucesso na ocupação deste ambiente. Sphaerosyllis
tem ocorrência nas Ilhas Canárias, Nordeste do Mediterrâneo, Mar Vermelho, Atlântico norte,
Ilhas Galápagos, Mar Negro, Austrália (NUNEZ et al, 1992) , Sudeste do Brasil (AMARAL et al,
2005).
O gênero Sphaerosyllis apresentou maior abundância e a diversidade de Polychaeta foi
maior na área protegida, porém apresentou uma diferença pequena. Devido estes animais serem

55
possivelmente alvo de predação, provavelmente outro fator que não a proteção ambiental deve ter
maior influência na sua abundância, como maior oferta de matéria orgânica na área protegida e
seu hábito oportunista. Devido à pequena diferença na diversidade das duas áreas, pode ser que
em ambas as áreas, estes organismos estejam pressionados pelo mesmo fator ambiental e este
independe da área protegida ou não. Em relação à distância do recife, a riqueza foi maior quando
afastado, e a equitabilidade maior quando perto do recife. A diversidade foi maior afastado do
recife e a abundância também, assim como foi para a meiofauna de forma geral.
Em relação à distância do recife, tanto a abundância quanto a riqueza e diversidade foram
maiores na situação afastada do recife, sendo evidente a influência da presença do recife sobre a
comunidade, como já citado tanto para a meiofauna de maneira geral quanto para a nematofauna.
Os organismos que mais determinaram na diferença entre as áreas amostradas nas duas
situações abordadas neste trabalho foram os Copepoda e em segundo os Polychaeta. Apesar de
Polychaeta não ter sido tão abundante quanto Copepoda e a diferença entre os valores do
SIMPER é pequena. Logo, estes organismos parecem ter bastante importância na manutenção da
comunidade, sendo provavelmente mais afetado em relações bottom-up e top-down, apesar dos
resultados deste trabalho não mostrarem isso com clareza.
Apesar de não ter havido correlação de nenhum fator ambiental com a meiofauna de
maneira geral, esses fatores devem ser considerados, uma vez que estes animais estão
intimamente ligados ao substrato e normalmente o tamanho do grão é o que determina a estrutura
da comunidade. Alguns Nematoda obtiveram correlação com alguns fatores ambientais, já
abordados nesta discussão.
Foi possível observar que tanto a ANOVA usando a meiofauna como descritor conseguiu
diferenciar as áreas quanto à proteção ambiental, porém o mesmo não ocorreu quando utilizada a
nematofauna ou a poliquetofauna como descritor. O ANOSIM e o MDS não indicaram diferença
significativa entre a estrutura desta comunidade das duas áreas abordadas em relação à presença
ou ausência de proteção legal, e nenhum dos grupos abordados funcionou como descritor da
estrutura da comunidade.
Devido ao pouco conhecimento sobre a meiofauna de ambiente recifal, juntamente com
fatores ambientais que podem ser mais determinantes que a proteção ambiental sobre a
comunidade, ainda não é possível determinar um organismo da meiofauna como bio-indicador de

56
qualidade ambiental para este tipo de situação. Porém o fator que pode ser usado como descritor
de área protegida seria o índice de diversidade da meiofauna em geral.
A menor abundância tanto na meiofauna quanto de alguns gêneros de Nematoda,
especificamente os conhecidos como consumidos com freqüência por peixes juvenis devem ser
levados em consideração como descritores. A abundância de Sphaerosyllis poderia ser utilizada
também para ambientes recifais, porém devido ao hábito oportunista deve-se conhecer melhor
sua fisiologia e comportamento diante de áreas realmente degradadas, que não é o caso do local
estudado.
Foi possível observar que a influência do recife sobre a comunidade meiofaunística foi
evidente, atuando tanto na abundância quanto nos índices de riqueza, equitabilidade e
diversidade. Apenas Gomphionema (Nematoda) não seguiu este padrão, apresentando maior
abundância próximo ao recife.
A área de proteção ambiental influenciou pouco a estrutura da comunidade
meiofaunística, sendo necessárias mais pesquisas em áreas de proteção ambiental e em áreas
recifais, para conhecer melhor a taxonomia e interações ecológicas nesses ambientes. A presença
dos recifes influencia a meiofauna, seja pela modificação do substrato, seja pela presença de
animais consumidores da meiofauna, mesmo assim estes efeitos parecem ser pouco conhecidos, e
qual efeito seria mais determinante devem ser objetos de estudos futuros. Através deste estudo,
foi possível conhecer um ambiente pouco explorado por estudos da meiofauna, aumentando
assim o conhecimento sobre a ocorrência e ecologia destes animais de grande importância para a
manutenção da teia trófica marinha.

57
5 CONCLUSÕES
• Em relação a estrutura da comunidade foi possível observar que há diferença tanto entre a
área restrita à pesca quanto na área não protegida da pesca, e também há diferença em relação à
distância do recife.
• Neste trabalho a meiofauna não apresentou correlação com nenhum fator ambiental, mas
alguns de seus componentes sim. Não pode ser descartada a possibilidade de algum fator
ambiental, levantado por este trabalho ou não, ser mais determinante para a meiofauna do que a
proteção ambiental.
• Apesar da diferença de abundância e de diversidade, não foi possível apontar um descritor
da comunidade ou um bio-indicador de qualidade ambiental
• A presença do recife atua significativamente na estrutura das comunidades meiofaunística,
nematofaunística e poliquetofaunística.

58
6 REFERÊNCIAS BIBLIOGRÁFICAS
AGARDY, T. 1997. Marine protected areas an ocean conservartion, Landes Press, Texas, 244p. ALONGI, D. M. 1985. Microbes, meiofauna, and bacterial productivity on tubes construted by the polychaete Capitella capitata. Marine Ecology – Progress series, 23: 207-208. ALONGI, D. M. 1989. Benthic processes across mixed terrigenous-carbonate sedimentary facies on the central Great Bamer Reef continental shelf. Cont. Shelf. Res. 9: 629-663 AMARAL, A. C. Z.; RIZZO, A. E.; ARRUDA, E. P. 2005. Manual de identificação dos invertebrados marinhos da região sudeste-sul do Brasil. EDUSP, Volume 1, 288p. AMARAL, A. C. Z.; NALLIN, S. A. H. & STEINER, T. M. 2006. Catálogo das espécies de Annelida Polychaeta do Brasil. HTTP://www.ib.unicamp.br/destaques\biota\bentos_marinho\prod_cien\texto_poli.pdf (consultado pela última vez em 27/I/2010). AMBROSE, R. F. & ANDERSON, T. W. 1990. Influence of an artificial reef on the surrounding infaunal community. Marine Biology, 107: 41-52. ANJOS, D. L. 2007. Inter-relação da Pluviometria com a Biomassa Fitoplanctônica dos Estuários de Pernambuco (Brasil). Recife, Monografia (Graduação) Universidade Federal de Pernambuco, 103p. ASSIS, J. E.; SAMIGUEL, C. A. & CHRISTOFFERSEN, M. L. 2007. Two new species of Nicomache (Polychaeta: Maldanidae) from the Southwest Atlantic. Zootaxa 1454:27-37. BARBOSA, D. F.; OLIVEIRA, D. A. S.; VERÇOSA, .M .M.; DOS SANTOS, É. C. L.; SOUZA, J. R. B.; DA ROCHA, C. M. C. 2007. Estrutura da comunidade meiofaunística associada a macrófitas na praia do Cupe, litoral sul do estado de Pernambuco. In: ANDRADE, C. C. Oceanografia e Mudanças Globais. São Paulo.. BARROS, F. 2005. Evaluating the importance of predation on subtidal benthic assemblages in sandy habitats around rocky reefs. Acta Oecologica 27 (2005) 211-223. BASTOS, R. B. 2006. Estrutura da comunidade fitoplantônica e variáveis ambientais no estuário do Rio Una-Pernambuco - Brasil. Recife, Dissertação (Mestrado) – Universidade Federal de Pernambuco. BELL, S. S. & COULL, B. C. 1978. Field evidence that shrimp predation regulates meiofauna. Oecologia, v. 35, p. 141-148.

59
BIRKELAND, C. 1997. Introduction. Chapter 1. In: Life and death of coral reefs. Birkeland, C. (ed.) Chapman & Hall, New York, 535p. BOECKNER, M. J. ; SHARMA, J. ; PROCTOR, H. C. 2009. Revisiting the meiofauna paradox : dispersal and colonization of nematodes and other maiofaunal organisms in low- and high-energy environments. Hydrobiologia, 624 : 91-106. BRAWLEY, S. H. & ADEY, W. H. 1981. The effect of micrograzers on algal community structure in a coral reef microcosm. Marine Biology, vol. 61, p 2-3.. CARMAN, K. R.; FLEEGER, J. W., POMARICO, S. M. 1997. Response of benthic food web to hydrocarbon contamination. Limnol. Oceanogr., 42: 561-571. CASTRO, F. J. V. 2003. Variação temporal da meiofauna e da nematofauna em uma área mediolitorânea da bacia do Pina (Pernambuco, Brasil). Recife, Tese (Doutorado) Universidade Federal de Pernambuco, 116p. CLARKE, R. & WARWICK R. M. 1994. Changes in marine communities: an approach to statistical analysis and interpretation. Plymouth. NERC, 187p. COBB, N.A. 1917. Note on nemas. Contr. Sci. Nematol. n.5, p.117-128. COELHO, P. A. ; SANTOS, M. DO C. F. ; FREITAS, A. E. T. S. 2004. Crescimento do aratu-da-pedra, Plagusia depressa (Fabricius, 1775) (Crustacea: Decapoda: Plagusiidae) em Tamandaré - Pernambuco. Boletim técnico científico do CEPENE, Tamandaré, v. 12, n. 1, p. 73-79. COLIJN, F & DIJKEMA, KS. 1981. Species composition of benthic Diatoms and distribution of chlorophyll-a on an intertidal flat in the Dutch Waaden Sea. Mar. Ecol. Prog. Ser., 4: 9-21. COULL, B. C. 1988. Ecology of the marine meiofauna. In: Higgins, R. P. & Thiel, H., eds Introductio to the study of meiofauna. Washngton, D. C. Smithsonian Institution Press. p.18 - 38.. CPRH, 2003. Companhia Pernambucana do Meio Ambiente – Diagnóstico Sócio-Ambiental do Litoral Sul de Pernambuco. 87p. DA ROCHA, C. M. C. 2003. Efeito do substrato fital na comunidade maiofaunística associada, com ênfase aos Nematoda livres. Recife, tese (Doutorado) Universidade Federal de Pernambuco, 120p. DANOVARO, R. & SIMONETTA, F. 2002. Meiofaunal vertical zonation on hard-bottoms: comparison with soft-bottom meiofauna. Marine Ecology – Progress series, 230: 159-169. DANOVARO, R.; GAMBI, C. MAZOLLA, A.; MIRTO, S. 2002. Influence of artificial feefs on the surrounding infauna: analysis of meiofauna. Journal of Marine Science, 59: S356-S362.

60
DAVIS, D & TISDEL, C. 1995. Recreational scuba-diving and carrying capacity in marine protected areas. Ocean & Coastal Management, v. 26, n. 1, p. 19-40. DAVIS,N.,VANBLARICOM,G.R., & DAYTON,P.K.1982. Man-mad estructures on marine sediments: effects on adjacent benthic communities. Marine Biology,70:295–303. DE GRISSE, A.T. 1969. Redescription ou modification de qulques tecniques utilisé dans l’etude des nematodes phytoparasitaries. Mededelingen Rijkfakuteit Landwwetenschappen Gent 34 :351-369. DEAN, W. E. 1974. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignitions : comparison with other methods. Journal of Sedimentary Petrology, v. 44, p. 141-148. DI DOMENICO, M. ; LANA, P. C. ; & GARRAFFONI, A. R. S. 2009. Distribution patterns of interstitial polychaetes in sandy beaches of southern Brazil. Marine Ecology, 30 : 47-62. DOS SANTOS, É. C. L. 2005. Distribuição de Batillipes pennaki Marcus, 1946 (Tardigrada, Heterotardigrada) na margem Sul da Ilha de Itamaracá em período de estiagem. Recife, Monografia, Universidade de Pernambuco, 48p. DOWNING, J.A. & CYR, H. 1985. The quantitative estimation of epiphytic invertebrate populations. Can J Fish Aquat Sci 42:1570-1579 ELMGREN, R. 1966. Baltic benthos communities and the role of meiofauna. Contr. Asko Lab. Univ. of Stockolm, 14: 1-31. ESTEVES, A. M. 1995. Microdistribuição espacial da meiofauna na Coroa do Avião, Pernambuco. Recife. Dissertação (Mestrado). Universidade Federal de Pernambuco. Recife-PE. 74 pp. FABI, G.; LUCCARINI, F.; PANFILI, M.; SOLUSTRI, C.; SPAGNOLO, A. 2002. Effects of na artificial reef on the surrounding soft-bottom community (central Adriatic Sea). Journal of Marine Science, 59:S343-S349. FARIAS, F. S. 2002. Tamoindaré – a dinâmica de suas praias frente às novas tribos repovoadoras do século XX. Recife, Dissertação (Mestrado) Universidade Federal de Pernambuco, 124 p. FAUCHAULD, K & JUMARS, P. A. 1979. The diet of worms: A study of polychaete feeding guilds. Oceanography and Marine Biology Annual Review, 17: 193-284. FERREIRA, B. P.; MAIDA, M.; SOUZA, A. L. T. 1995. Levantamento inicial das comunidades de peixes recifais da região de Tamandaré – PE. Bol. Boletim técnico científico do CEPENE, Tamandaré, v. 3, n. 1, p. 211-230.

61
FERREIRA, B. P.; MAIDA, M. and CAVA, F. C. 2001. Características e perspectivas para o manejo da pesca na APA Marinha Costa dos Corais. In: Congresso Brasileiro de Unidades de Conservação, Campo Grande, MS. Proceedings... Campo Grande. pp. 50-58. FOLK, R. L. & WARD, W. C. 1987. Brazos river bar: a study of significance of grain size parameters. J. Sediment Petrol. 27: 3-26. FONSECA-GENEVOIS, V.; SOMERFIELD, P. J.; NEVES, M. H.B.; COUTINHO, R.; MOENS, T. 2006. Colonization and early succession on artificial substrata by meiofauna. Marine Biology, 148: 1039-1050. FRIEDLANDER, A. M. 2001. Essential fish habitat in the effective design of marine reserves: applications for marine ornamental fishes. Aquarium Sci Conserv 3:135-150. FRIEDLANDER, A. M.; BROWN, E. K.; JOKIEL, P. L.; SMITH, W. R.; RODGERNS, K. S. 2003. Effects of habitat, wave exposure, and marine protected area status on coral reef fish assemblages in the Hawaiian archipelago. Coral Reefs, V. 22, 3:291-305. GEE, J. M. 1989. An ecological economic review of meiofauna as food for fish. Zool. J. Linn. Soc., 96:243-261. GIERE, O. 1993. Meiobenthology: The microscopic fauna in aquatic sediments. Springer-Verlag, Berlin. 328 pp. GUZMAN, H. M.; OBANDO, V. L.; CORTES, J. 1987. Meiofauna associated with a Pacific coral reef in Costa Rica. Coral Reefs, v. 6, p. 107-112. HULINGS, N. C. & GRAY J. S. 1976. Physical factors controlling abundance of meiofauna on tidal and atidal beaches. Marine Biology, Springer-Verlag, 34, 77-83. IBAMA, 2009 http://www.ibama.gov.br/siucweb/listaUc.php. Último acesso 17/I/ 2010. JANSSON, B. 1967. The significance of grain size and pore water content for the intersticial fauna of sandy beaches. Oikos, 18:311-322. KELLEHER, G. & KENCHINGTON, R. 1992. Guidelines for establishing marine protected areas. A marine conservation and development report. IUCN. Switzerland, 79p. KENNEDY, A. D. 1993. Minimal predation upn meiofauna by endobenthic macrofauna in the Exe Estuary, south west England. Mar. Biol. 117: 311-319. KOHN, A. J. & LLOYD, M. C. 1973. Polychaetes of truncated reef limestone substrates on eastern Indian Ocean coral reefs: diversity, abundance and taxonomy. Internationale Revue der gesamten Hydrobiologie und Hydrographie, v. 58, p. 369-400.

62
KREBS, C.J. 1998. Ecological Methodology. Harper & Row, New York, 654p. LEE, M. R.; CORREA, J. A.; SEED, R. 2006. A sediment quality triad assessment of the impact of copper mine tailings disposal on the litoral sedimentary environment on the Atacama region of Northern Chile. Marine Pollution Bulletin, 52:1389-1395. LOGAN, D.; TOWNSEND, K.A.; TOWSEND, K. & TIBBETS, I.R. 2008. Meiofauna sediment relations in leeward slope turf algae of Heron Island reef. Hydrobiologia, 610:269-276. LORENZEN, C. J. 1967. Determination of Chlorophyll and Pheo-Pigments: Spectrophotometric Equations. Limnology and Oceanography, Vol. 12, No. 2, pp. 343-346. LORENZEN, S. 1994. The phylogenetic systematics of freeliving nematodes. The Ray Society Institute, 383p. LORENZI, L. 2004. Estrutura das associações infaunais sublitorais de substrato inconsolidado adjacente a recifes artificiais e naturais (Paraná, Brasil). Curitiba, Tese (Doutorado) Universidade Federal do Paraná, 199p. MACEDO, E. C. 2009. Um ensaio sobre a sedimentação e suas implicações ecológicas nos recifes costeiros da baía de Tamandaré/PE. Recife, Dissertação (Mestrado) Universidade Federal de Pernambuco, 121p. MAIDA, M. & FERREIRA, B. P. 1997. Coral reefs of Brazil: an overview. Proc. 8th Int. Coral Reef Symp. Panamá, 1:263-274. MARANHÃO, G. M. B.; FONSECA-GENEVOIS, V.; PASSAVANTE, J. Z. O. 2000. Meiofauna da Área Recifal da Baía de Tamandaré (Pernambuco, Brasil). Trab. Oceanog. Univ. Fed. PE, Recife, v. 28, n. 1, p. 47-59. MARANHÃO, G. M. B. 2003. Distribuição espaço-temporal da meiofauna e da nematofauna no ecossistema recifal de Porto de Galinhas, Ipojuca, Pernambuco, Brasil. Recife, Dissertação (Mestrado) Universidade Federal de Pernambuco, 116p. MARE, M. F. 1942. A study of marine benthic community with special reference to the micro-organisms. J. Mar. Biol. Ass. UK, 25:517-554. MCCALL, J. N. and FLEEGER, J.W. 1995. Predation by juvenile fish on hyperbenthic meiofauna: a review with dat on post-larval Leiostomus xanturus. Vie Milieu, 45(1): 61-73. MCLACHLAN, A & TURNER, I. 1994. The interstitial environment of sandy beaches. P.S.Z.N.I: Mar. Ecol.,15(3/4): 177-211. MONTAGNA, P. A. 1995. Rates of meiofaunal microbivory: a review. Vie et Millieu, 45: 1-10.

63
MORA, C.; ANDREFOUET, S.; COSTELLO, M.J.; KRANENBURG, C.; ROLLO, A.; VERON, J.; GASTON, K. J.; MYERS, R. A. 2006. Coral reefs and the global network of marine protected areas. Science, v. 312, n. 5781, p. 1750-1751. MORAIS, L. T.; & BODIOU, J. Y. 1984. Predation on meiofauna by juvenile fish in a western Mediterranean flatfish nursery ground. Marine Biology, v. 82, p. 209-215. MORTON, B. 2005. Predator-prey interactions between Lepsiella (Bedeva) paivae (Gastropoda: Muricidae) and Katelysia scalarina (Bivalvia: Veneridae) on Pricess Royal Harbour, western Autralia. Journal of Molluscan Studies 71: 371–378. NETTO, S. A.; ATTRIL, M. J.; WARWICK, R. M. 1999. The effect of a natural water movement related disturbance on the estructure of meiofauna and macrofauna communities in the intertidal sand flat of Rocas Atoll (N.E. Brasil). J. Sea Res. 42: 291-302. NETTO, S. A.; ATTRILL, J. M.; WARWICK, R. M. 2003. The relationship between benthic fauna, carbonate sediments and reef morphology in reef-flat tidal pools of Rocas Atoll (north-east Brazil). Journal of the Marine Biological Association of the UK, 83:2:425-432 NUNEZ, J.; SAN MARTIN, G. & BRITO, M. C. 1992. Exogoninao (Polychaeta, Syllidae) from the Canary Island. Scientia Marina, 56 (1):43-52. PINHEIRO, B. R. 2006. Recrutamento de corais no recife da Ilha da Barra – Tamandaré/PE. Recife, Dissertação (Mestrado) Universidade Federal de Pernambuco, 82p. PLATT, H. M. & WARWICK, R. M. 1980. The significance of free-living nematodes to the littoral ecosystem. Systematic Association Special, v. 7, p. 729-759. PLATT, H. M.; SHAW, K. M. & LAMBSHEAD, P. J. D. 1984. Nematode species abundance patterns and their use in the detection of environmental pertubations. Hydrobiologia 118, 59-66. POSEY, M. H. & AMBROSE JR, W. G. 1994. Effects of proximity to an offshore hard-bottom reef on infaunal abundances. Marine Biology, 118: 745-753. REISE, K. 1979. Moderate predation on meiofauna by the macrobenthos of the Wadden Sea. Helgoland Marine Research, v. 32, p. 453-465. RENAUD-DEBYSER,J. ; SALVAT, B., 1963. Eleménts de prosperité des biotopes des sediments meubles intertidaux et écologie de leurs poputations em microfaune et macrofaune. Vie et Millieu, n. 14, p. 463-550. RIBEIRO, V. S. S. 1999. Relação dos fatores ambientais com a meiofauna na praia de Tamandaré, PE-Brasil. Recife, dissertação (Mestrado) Universidade Federal de Pernambuco, 76p. ROUSE, G. W & PLEIJEL, F. 2001. Polychetes. Oxford University Press, 354 p.

64
SANTOS, C. S. G & LANA, P. C. 2003. Nereididae (Polychata) da costa nordeste do Brasil. III. Gêneros Ceratonereis e Nereis. Iheringia, Sér. Zool., Porto Alegre, 93(1):5-22. SANTOS, C. S. G. & MACKIE, A. S. Y. 2008. New species of Poecilochaetus Claparède, 1875 (Polychaeta, Spionida, Poecilochaetidae) from Paranaguá Bay, southern Brazil. Zootaxa, 1790: 53-68. SANTOS, C. S. G.; NONATO, E. F. & PETERSEN, M. E. 2004. Two new species of Opheliidae ( Annelida: Polychaeta): Euzonus papillatus sp. n. from a northeastern brazilian sandy beach and Euzonus mammillatus sp. n. from the continental shelf of south-eastern Brazil. Zootaxa, 478: 1-12. SCHIEMER, F. 1987. Nematoda. In: PANDIAN, T. J. & VERNBERG, F. J. (Eds), Animal Energetics. Academic Press, v. 1, 186-211. SCHMID-ARAYA, J. M.; HILDREW, A. G.; ROBERTSON, A.; SCHMID, P. E.; WINTERBOTTOM, J. 2002. The importance of meiofauna in food webs: evidence from acid stream. Ecology, 83(5): 1271-1785. SECTMA, 2009. Programa de conservação da biodiversidade de Pernambuco: Criação e implementação de Unidades de Conservação. Recife, 49p. SERRANO, A.; SAN MARTIN, G. & LOPEZ, E. 2006. Ecology of Syllidae (Anelida, Polychaeta) from shallow rocky environments in the Cantabrian Sea (South Bay of Biscay). Scientia Marina, 70S3, p. 225-235. SILVA, M. C. 2004. A meiofauna como estoque alimentar para peixes juvenis (Gobiidae e Gerreidae) do Canal de Santa Cruz, Itamaracá, Pernambuco com ênfase aos Nematoda livres. Recife, dissertação (Mestrado) Universidade Federal de Pernambuco, 101p. SOUSA, W.P. – 2001. Natural disturbance and the dynamics of marine benthic communities. In: M.D. Bertness, S.D. Gaines and M.E. Hay (eds.), Marine Community Ecology. Sinauer Ass. p. 85-130 STEAD, T. K.; SCHIMID-ARAYA, J. M.; HILDREW, A. G. 2004. The contribution os surface to benthic density and biomass in a gravel stream. Arch. Hydrobiol. 160: 171-191. ST. JOHN, J.; JONS, G. P.; SALE, R. F. 1989. Distribution and abundance of soft-sediment meiofauna and a predatory goby in a coral reef lagoon. Coral Reefs, v. 8 2:51-57. SUGUIO, K. 1973, Introdução a Sedimentologia. Ed. Edgard Blucher, EDUSP, São Paulo, 317p. SUTHERLAND, T. F.; SHEPHERD, P. C. F.; ELNER, R. W. 2000. Predation on meiofaunal and macrofaunal invertebrates by western sandpipers (Calidris mauri): evidence for a dual foraging modes. Marine Biology, 137: 983-993.

65
TENORE, K. R.; TIEJEN, J. H. & LEE, J. J. 1977. Effect of meiofauna in incorporation of aged eelegrass, Zoostera marinha, detritus by the polychaete Nephtys incisa. Journal of Fish Research. Bd Canada, 34: p.563 – 567. VASCONCELOS, D. M. 2003. Distribuição espacial da comunidade da meiofauna e diversidade de Copepoda Harpacticoida no estuário do Rio Formoso, Pernambuco. Recife, Dissertação (Mestrado) Universidade Federal de Pernambuco, 74p. VENEKEY, V. 2007. Atualização do conhecimento taxonômico dos Nematoda na costa brasileira e sua ecologia na praia de Tamandaré-PE (Brasil). Recife, Tese (Doutorado) Universidade Federal de Pernambuco, 165p. VENEKEY, V.; FONSECA-GENEVOIS, V. G., DA ROCHA, C. M. C., SANTOS, P. J. P. 2008. Distribuição espaço-temporal da meiofauna em Sargassum polyceratium Montagne (FUCALES, SARGASSACEAE) de um costão rochoso do nordeste do Brasil. Revista Atlântica, Rio Grande, 30 (1) 53-67. VILLORA-MORENO, S. 1997. Environmental heterogeneity and the biodiversity of interstitial Polychaeta. Bulletin of Marine Science, v. 60, p. 494-501. VILLORA-MORENO, S.; CAPACCIONI-AZZATI, R.; GARCIA-CARRASCOSA, A. M. 1991. Meiobenthos of Sandy beaches from de Gulf of Valencia (Western Mediterranean): Ecology of interstitial polychaetes. Bulletin of Marine Science, v. 48, p. 376-385. VIRNSTEIN, R. W. 1977. The importance of predation by crabs and fishies on benthic infauna in Chesapeake Bay. Ecology 58, 1199-1217. WARWICK, R. M. 1984. Species size distribution in marine benthic communities. Oecology, Berlin 61, p.32 - 41. WARWICK, R. M.; PLATT, H. M. & SOMERFIELD, P. J. 1988. Free-living marine nematodes. Part III. The Linnean Society of London, n. 38, 297p. WESTHEIDE, W. 1972. La faune des Polychetes et des Archiannelides dans les plages a Ressac de la cote mediterraneenne de la Tunisie. Bulletin Institut National des Sciences et Technologie de la mer, 2(3), p. 449-468. WESTHEIDE, W. 2002. Polychaetes intersticial families. The Linnean Society of London, 152 p. WETZEL, R. G.; LIKENS, G. E. 1991. Limnological Analyses. Second Edition. Springer-Verlag, New York. 391p. WIESER, W. 1953. Die Beziehung zwischen Mundhoehlengstalt, Ernaehrungsweise und Vorkommem bei frelebenden marinen Nematoden. Eine oekologisch – morphologisch studie. Arkive Zoologische, ser. II, 4, 439-484.

66
WIESER, W. 1960. Benthic studies in Buzzards Bay. II. The meiofauna. Limnology and Oceanography, n 5, 121-137. WIESER, W. & KANWISHER, J. 1961. Ecological and physiological studies on marine nematodes from a small salt marsh near Woods Hole, Massachusetts. Limnology and Oceanography, n 6, 262-270. ZANOL, J.; PAIVA, P. C.; ATTOLINI; F. S. 2000. Eunice and Palola (Eunicidae: Polychaeta) from the eastern brazilian coast (13°00´- 22°30´S). Bulletin of marine science, 67(1): 449-463.