efeito de diferentes condições ambientais em áreas de
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SANTA CATARINA CENTRO DE CIÊNCIAS AGRÁRIAS
DEPARTAMENTO DE AQÜICULTURA
Efeito de diferentes condições ambientais em áreas de cultivo
sobre alimentação e biodeposição do mexilhão Perna perna.
JEAN FRANCO SCHMITT
Dissertação apresentada ao curso de Pós-Graduação em Aqüicultura do Centro de Ciências Agrárias da Universidade Federal de Santa Catarina, como requisito parcial à obtenção do título de Mestre em Aqüicultura. Orientador : Prof. Dr. Jaime Fernando Ferreira
Florianópolis (SC) 2002

ii
FICHA CATALOGRÁFICA
biod
Ciên
6.Se
Schmitt, Jean Franco, Efeito de diferentes condições ambientais em áreas de cultivo sobre alimentação e
eposição do mexilhão Perna perna. / Jean Franco Schmitt – 2002. 89 p. Orientador: Dr Jaime Fernando Ferreira Dissertação (Mestrado) – Universidade Federal de Santa Catarina, Centro de
cias Agrárias, Programa de Pós-Graduação em Aqüicultura. 1.Alimentação, 2.Biodeposição, 3.Maricultura, 4.Mexilhão, 5.Perna perna.
ston.

iii
Efeito de diferentes condições ambientais em áreas de cultivo sobre alimentação e biodeposição do mexilhão Perna perna.
Por
JEAN FRANCO SCHMITT
Esta dissertação foi julgada adequada para a obtenção do título de
MESTRE EM AQÜICULTURA
e aprovada em sua forma final pelo Programa de Pós-Graduação em Aqüicultura.
___________________________________ Prof. Jaime Fernando Ferreira, Dr.
Coordenador do Curso
Banca Examinadora:
____________________________________________ Dr. Jaime Fernando Ferreira - Orientador
____________________________________________ Dra. Aimê Rachel Magenta Magalhães
___________________________________________ Dr. Carlos Augusto França Schettini

iv
Dedico este trabalho aos meus familiares pelo
apoio incondicional em todos os momentos,
especialmente à minha mãe pelo exemplo de
garra, perseverança e ao meu pai, in
memorian, pelo exemplo de integridade e de
humildade, de quem jamais esquecerei...

v
AGRADECIMENTOS
Agradeço ao Prof. Dr. Jaime Fernando Ferreira pela orientação durante o mestrado e
em todas as etapas da realização deste tabalho, e principalmente pela paciência...
Ao CNPq pela concessão da bolsa de mestrado durante o curso.
Ao Carlito Klunk e à toda equipe do Departamento de Aqüicultura, sempre muito
eficientes.
Ao parceiro de pesquisa (e praticamente co-orientador) M.Sc. Felipe M. Suplicy
(“Supla”) pelo auxílio fundamental durante os experimentos de campo e análises de
laboratório, pelas discussões enriquecedoras e pelas constantes demonstrações de empenho.
À Profa. Aimê R. M. Magalhães pela disponibilização de bibliografia e de
equipamento fotográfico, sempre muito atenciosa...
Ao pesquisador M.Sc. Guilherme S. Rupp por fornecer os dados semanais de
parâmetros físico-químicos e seston de Porto Belo.
Aos produtores de Enseada do Brito (Sr. Luís) e de Porto Belo (Marcelo) por
disponibilizar desde os mexilhões utilizados nos experimentos até as embarcações e balsas
que possibilitaram sua execução.
Ao Itamar (“Mala”, “Zéguedé”) pelo transporte do equipamento e do pessoal durante
as coletas de campo.
Aos estagiários do LCMM, Kelly, Ricardo, André, Michela e Taís pelo auxílio nas
coletas de campo e nas análises de laboratório.
Aos funcionários do LCMM, Cláudio, Marisa, Chico, Adriana, João, Micheline,
Eduardo, Sino, Juça, Zezé e aos estagiários Ruth, Rodrigo, Luís e Conrado, pela boa vontade
e paciência para comigo.
À colega e amiga Dominique Pires Ibbotson pelo auxílio nas coletas de campo, mas
principalmente pela amizade...
Ao amigo Rodrigo Schveitzer (“Perú”) pela ajuda nas análises estatísticas, por sua
amizade e pela rica convivência nos últimos dois anos.
Aos amigos e grandes vizinhos Marcelo e Vívian, também ao Seu Washington e à
Dona Vilma, pelos churrascos e peixadas, pela amizade, carinho e simpatia.
Aos amigos Franklin, Torigoi e Tânia pelas conversas, sessões de vídeo, tardes na
praia, cafezinhos vespertinos...

vi
Aos amigos colegas de mestrado: Adrianas pernambucana e gaúcha, Alexandre,
Arthur, Carina, Carla, Charles, Daniela, Fabrini, Gabriela, Gilvan, Hugo, Maria Luiza,
Marcão, Marquinhos, Pato, Sandra, Simone por compartilharem dos momentos festivos e de
labuta durante estes dois anos.
Ao meu pai, Romeu Ernesto Willecke (in memorian), que tornou possível seguir esta
carreira graças ao seu apoio moral e financeiro.
E à Deus, por me possibilitar a convivência com todas estas pessoas, e por permitir
que eu vencesse a mais este desafio.

vii
SUMÁRIO
Lista de figuras do Artigo Científico.................................................................................... viii
Lista de tabelas do Artigo Científico.................................................................................... xi
Lista de abreviaturas............................................................................................................. xiii
Resumo................................................................................................................................. xiv
Abstract................................................................................................................................ xv
Introdução............................................................................................................................ 16
Aspectos do cultivo de moluscos bivalves e sua sustentabilidade.................................. 16
Alimentação e fisiologia energética de moluscos bivalves............................................. 18
A biodeposição e os impactos ambientais resultantes do cultivo de bivalves................. 21
ARTIGO CIENTÍFICO - Efeito de diferentes condições ambientais em áreas de cultivo sobre alimentação e biodeposição do mexilhão Perna perna....................................................... 26
Resumo............................................................................................................................ 27
1.Introdução..................................................................................................................... 28
2.Materiais e Métodos...................................................................................................... 30
2.1. Áreas de estudo................................................................................................ 30
2.2. Monitoração de variáveis ambientais............................................................... 32
2.3. Sistema Experimental....................................................................................... 33
2.4. Material Biológico............................................................................................ 34
2.5. Quantificação de matéria orgânica e inorgânica no seston e nos biodepósitos 35
2.6. Determinação das taxas fisiológicas................................................................. 35
2.7. Padronização das taxas fisiológicas.................................................................. 35
2.8. Análises estatísticas........................................................................................... 36
3.Resultados........................................................................................................................ 37
3.1. Variáveis ambientais.......................................................................................... 37
3.2. Experimentos de alimentação e de biodeposição............................................... 42
4.Discussão......................................................................................................................... 58
5.Agradecimentos.................................................................................................................71
6.Referências........................................................................................................................71
CONSIDERAÇÕES FINAIS................................................................................................... 81
REFERÊNCIAS BIBLIOGRÁFICAS DA INTRODUÇÃO................................................... 83

viii
LISTA DE FIGURAS Figura 1. Localização das áreas de estudo, com mapas do Brasil e do Estado de Santa
Catarina, destacando as localidades de Porto Belo (PB) e de Enseada do Brito (EB), no litoral catarinense.................................................................... 30
Figura 2. Localização das áreas de estudo: área experimental em Porto Belo (PB –
27°08’12.67”S; 48°32’23.40”W), nas proximidades da Ilha João da Cunha (Ilha de Porto Belo), no litoral catarinense..................................................... 31
Figura 3. Localização das áreas de estudo: área experimental em Enseada do Brito
(EB – 27°46’30.29”S; 48°37’47.98”W), no litoral catarinense...................... 32
Figura 4.
Variáveis hidrológicas (médias mensais ± desvio padrão) obtidas em monitoração semanal realizada em Enseada do Brito (EB) e em Porto Belo (PB). Parâmetros físico-químicos e seston (fração entre 1,2 e 230 µm). a : temperatura (° C); b : salinidade (PSU); c : oxigênio dissolvido (OD, mg L-
1); d : nível de saturação de oxigênio dissolvido (OD, %); f : clorofila a (µg L-1); e : feopigmentos (µg L-1)........................................................................
39
Figura 5.
Variáveis hidrológicas (médias mensais ± desvio padrão) obtidas em monitoração semanal realizada em Enseada do Brito (EB) e em Porto Belo (PB). Parâmetros físico-químicos e seston (fração entre 1,2 e 230 µm). a : turbidez (NTU); b : matéria particulada total (TPM, mg L-1); c : matéria inorgânica particulada (PIM, mg L-1); d : matéria orgânica particulada (POM, mg L-1); e : fração orgânica do material em suspensão (f, POM:TPM); f : razão clorofila a / feopigmentos (Cla / Feop).......................
40
Figura 6.
Material em suspensão (fração entre 1,2 e 230 µm; mg L-1) e taxas fisiológicas (alimentação e biodeposição; mg h-1) do mexilhão Perna perna em dois locais de cultivo e em três experimentos por local (médias ± desvio padrão) Porto Belo: PB1 = Experimento 1; PB2 = Experimento 2; PB3 = Experimento 3; Enseada do Brito: EB1 = Experimento 1; EB2 = Experimento 2; EB3 = Experimento 3. a : matéria particulada em suspensão (TPM), matéria inorgânica particulada (PIM) e matéria orgânica particulada (POM); b : taxa de clareamento (CR, L h-1); c : taxas de biodeposição (BR), rejeição (RR) e egestão (ER); d : fração orgânica dos biodepósitos (b), das pseudofezes (p) e das fezes (e)...................................................................................................................
48
Figura 7.
Alimentação e produção de biodepósitos do mexilhão Perna perna em dois locais de cultivo e em três experimentos por local (médias ± desvio padrão). Porto Belo: PB1 = Experimento 1; PB2 = Experimento 2; PB3 = Experimento 3; Enseada do Brito: EB1 = Experimento 1; EB2 = Experimento 2; EB3 = Experimento 3. a : taxas de filtração (FR) e de ingestão (IR); b : fração orgânica do seston (f) e eficiência de seleção (SE); c : taxas de ingestão orgânica (OIR) e de absorção (AR); d : fração orgânica do material ingerido (i) e eficiência de absorção (AE)............................................................................................................... 49

ix
Figura 8.
Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo e Enseada do Brito) durante três experimentos por local. a : taxa de clareamento (CR) e matéria particulada em suspensão (TPM); b : taxa de clareamento (CR) e fração orgânica do seston (f); c : taxa de clareamento (CR) e matéria inorgânica particulada (PIM); d : taxa de clareamento (CR) e matéria orgânica particulada (POM); e : taxa de clareamento (CR) e temperatura; f : taxa de clareamento (CR) e salinidade........................................................................ 52
Figura 9.
Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo e Enseada do Brito) durante três experimentos por local. a : taxa de biodeposição (BR) e matéria particulada em suspensão (TPM); b : taxa de biodeposição (BR) e fração orgânica do seston (f); c : taxa de biodeposição (BR) e matéria inorgânica particulada (PIM); d : taxa de biodeposição (BR) e matéria orgânica particulada (POM); e : taxa de biodeposição (BR) e temperatura; f : taxa de biodeposição (BR) e salinidade..................................................... 53
Figura 10.
Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo e Enseada do Brito) durante três experimentos por local. a : taxa de filtração (FR) e matéria particulada em suspensão (TPM); b : taxa de ingestão (IR) e taxa de filtração (FR); c : taxa de rejeição (RR) e matéria particulada em suspensão (TPM); d : taxa de ingestão (IR) e taxa de rejeição (RR); e : taxa de egestão (ER) e matéria particulada em suspensão (TPM); f : taxa de ingestão (IR) e taxa de egestão (ER)......................................................................................
54
Figura 11.
Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo - PB e Enseada do Brito - EB) durante três experimentos por local (EB1, EB2, EB3, PB1, PB2, PB3). a : eficiência de seleção (SE) e matéria particulada em suspensão (TPM); b : eficiência de seleção (SE) e matéria orgânica particulada (POM); c : eficiência de seleção (SE) e fração orgânica do seston (f); d : eficiência de seleção (SE) e taxa de filtração (FR); e : fração orgânica ingerida (i) e eficiência de seleção (SE); f : eficiência de absorção (AE) e eficiência de seleção (SE)...............................................................................
55
Figura 12.
Correlações entre taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo - PB e Enseada do Brito - EB) durante três experimentos por local (EB1, EB2, EB3, PB1, PB2, PB3). a : taxa de ingestão orgânica (OIR) e matéria orgânica particulada (POM); b : fração orgânica ingerida (i) e fração orgânica do seston (f); c : taxa de ingestão orgânica (OIR) e taxa de filtração (FR); d : eficiência de absorção (AE) e fração orgânica do seston (f); e : taxa de ingestão orgânica (OIR) e taxa de ingestão (IR); f : eficiência de absorção (AE) e fração orgânica ingerida (i)
56

x
Figura 13. Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão
Perna perna em dois locais de cultivo (Porto Belo - PB e Enseada do Brito - EB) durante três experimentos por local (EB1, EB2, EB3, PB1, PB2, PB3). a : taxa de absorção (AR) e matéria particulada total (TPM); b : taxa de absorção (AR) e taxa de filtração (FR); c : taxa de absorção (AR) e matéria orgânica particulada (POM); d : taxa de absorção (AR) e taxa de ingestão (IR); e : taxa de absorção (AR) e fração orgânica do seston (f); f : taxa de absorção (AR) e fração orgânica ingerida (i)...................................... 57

xi
LISTA DE TABELAS Tabela 1: Datas, horários e condições ambientais durante os experimentos de
alimentação e de biodeposição com mexilhão Perna perna em Enseada do Brito e em Porto Belo...................................................................................
33
Tabela 2: Material biológico utilizado nos experimentos de alimentação e de biodeposição com mexilhão Perna perna em Enseada do Brito e em Porto Belo..............................................................................................................
34
Tabela 3: Resultados das análises de variância bi-fatorial (ANOVA) avaliando o efeito dos fatores área de cultivo (local) e meses de ano sobre parâmetros hidrológicos. Variáveis físico-químicas da água: temperatura, salinidade, oxigênio dissolvido (OD e OD%) e turbidez; características quali-quantitativas do seston (fração entre 1,2 e 230 µm): matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla), feopigmentos (Feop) e razão clorofila a / feopigmentos (Cla/Feop)....................................................................................................
37
Tabela 4: Parâmetros de qualidade da água do mar das áreas de cultivo estudadas: comparação dos valores médios (médias ± desvio padrão) de monitoramento semanal realizado em Enseada do Brito (EB) e em Porto Belo (PB). Variáveis físico-químicas da água: temperatura, salinidade, oxigênio dissolvido (OD e OD%) e turbidez; características quali-quantitativas do seston (fração entre 1,2 e 230 µm): matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla), feopigmentos (Feop) e razão clorofila a / feopigmentos (Cla/Feop).....................................................................................................
38
Tabela 5: Coeficientes de correlação entre matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla) e feopigmentos (Feop) em Enseada do Brito.........................................................................
41
Tabela 6: Coeficientes de correlação entre matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla) e feopigmentos (Feop) em Porto Belo....................................................................................
42
Tabela 7: Variáveis físico-químicas da água (médias ± desvio padrão) em Enseada do Brito e em Porto Belo durante os experimentos de alimentação e de biodeposição: temperatura (° C), salinidade (PSU), oxigênio dissolvido (OD, mg L-1) e turbidez (NTU).....................................................................
42

xii
Tabela 8: Parâmetros de qualidade da água do mar das áreas de cultivo estudadas:
comparação dos valores médios (médias ± desvio padrão) dos experimentos em Enseada do Brito (EB) e em Porto Belo (PB). Variáveis físico-químicas: temperatura, salinidade, oxigênio dissolvido (OD e OD%) e turbidez...................................................................................................... 43
Tabela 9: Material em suspensão (fração entre 1,2 e 230 µm) em Enseada do Brito e em Porto Belo durante os experimentos de alimentação e biodeposição: médias ± desvio padrão de matéria particulada total (TPM, mg L-1), matéria inorgânica particulada (PIM, mg L-1), matéria orgânica particulada (POM, mg L-1) e fração orgânica do material em suspensão (f)...........................
44
Tabela 10: Parâmetros de qualidade da água do mar das áreas de cultivo estudadas: comparação dos valores médios (médias ± desvio padrão) dos experimentos em Enseada do Brito (EB) e em Porto Belo (PB). Características quali-quantitativas do seston (fração entre 1,2 e 230 µm): matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM) e fração orgânica do material em suspensão (f)..............................................................................................................
45
Tabela 11: Taxas de alimentação (médias ± desvio padrão) do mexilhão Perna perna em Enseada do Brito (EB) e em Porto Belo (PB) durante cada experimento: CR = taxa de clareamento (L h-1); FR = taxa de filtração (mg h-1); IR = taxa de ingestão (mg h-1); OIR = taxa de ingestão orgânica (mg h-1); AR = taxa de absorção (mg h-1)...............................................................................
46
Tabela 12: Taxas de alimentação (médias ± desvio padrão) do mexilhão Perna perna em Enseada do Brito (EB) e em Porto Belo (PB) durante cada experimento: SE = eficiência de seleção; i = fração orgânica do material ingerido; AE = eficiência de absorção............................................................................
46
Tabela 13:
Taxas de produção de biodepósitos (médias ± desvio padrão) do mexilhão Perna perna em Enseada do Brito (EB) e em Porto Belo (PB) durante cada experimento: RR = taxa de rejeição (mg h-1); ER = taxa de egestão (mg h-
1); BR = taxa de biodeposição (mg h-1); razão RR/BR e razão ER/BR....................................................................................................
46
Tabela 14: Taxas de alimentação e de produção de biodepósitos do mexilhão Perna perna. Comparação dos valores médios (médias ± desvio padrão) dos experimentos em Enseada do Brito (EB) e em Porto Belo (PB): CR = taxa de clareamento (L h-1); FR = taxa de filtração (mg h-1); IR = taxa de ingestão (mg h-1); OIR = taxa de ingestão orgânica (mg h-1); AR = taxa de absorção (mg h-1); SE = eficiência de seleção; i = fração orgânica do material ingerido; AE = eficiência de absorção; RR = taxa de rejeição (mg h-1); ER = taxa de egestão (mg h-1); BR = taxa de biodeposição (mg h-1); razão RR/BR e razão ER/BR; p = fração orgânica das pseudofezes; e = fração orgânica das fezes; b = fração orgânica dos biodepósitos.......... 50

xiii
LISTA DE ABREVIATURAS
AE: eficiência de absorção (fração). AR: taxa de absorção (mg h-1). b: fração orgânica dos biodepósitos (fração). BR: taxa de biodeposição (mg h-1). C: Carbono. Cla: clorofila a (µg L-1). Cla / Feop: razão entre concentrações de clorofila a e de feopigmentos (fração). COP: Carbono orgânico particulado. CR: taxa de clareamento (L h-1). f : fração orgânica do seston (fração). Feop: feopigmentos (µg L-1). FR: taxa de filtração (mg h-1). e: fração orgânica das fezes (fração). ER: taxa de egestão (mg h-1). i: fração orgânica ingerida (fração). IBR: taxa de biodeposição inorgânica (mg h-1). IER: taxa de egestão inorgânica (mg h-1). IFR: taxa de filtração inorgânica (mg h-1). IIR: taxa de ingestão inorgânica (mg h-1). IR: taxa de ingestão (mg h-1). IRR: taxa de rejeição inorgânica (mg h-1). MFL: perdas metabólicas fecais. N: Nitrogênio. NOP: Nitrogênio orgânico particulado. OBR: taxa de biodeposição orgânica (mg h-1). OD: oxigênio dissolvido (mg L-1). OD%: saturação de oxigênio dissolvido (%). OER: taxa de egestão orgânica (mg h-1). OFR: taxa de filtração orgânica (mg h-1). OIR: taxa de ingestão orgânica (mg h-1). ORR: taxa de rejeição orgânica (mg h-1). P: Fósforo. p: fração orgânica das pseudofezes (fração). PIM: matéria inorgânica particulada (mg L-1). POM: matéria orgânica particulada (mg L-1). RR: taxa de rejeição (mg h-1). SE: eficiência de seleção (fração). SFG: potencial de crescimento. TPM: matéria particulada total (mg L-1).

xiv
RESUMO Este estudo avaliou a alimentação e a produção de biodepósitos do mexilhão Perna perna em duas áreas de cultivo no sul do Brasil diferenciadas pelos parâmetros do seston, em três experimentos por local. A Enseada do Brito (EB) apresentou altas concentrações de material particulado total (TPM), de matéria inorgânica particulada (PIM) e de matéria orgânica particulada (POM), mas menor fração orgânica no seston (f) em relação a Porto Belo (PB). Em geral, a alimentação e a biodeposição do mexilhão P. perna variaram de acordo com as características do seston, apresentando comportamentos similares aos observados para outras espécies de bivalves filtradores. As taxas de clareamento (CR) foram menores em EB, onde as concentrações de material em suspensão (TPM, PIM e POM) foram maiores. Já as taxas de filtração (FR), de ingestão (IR) e de ingestão orgânica (OIR) foram maiores em EB, aumentando de acordo com o incremento do seston. As taxas de absorção (AR) seguiram as mesmas tendências de FR, IR e OIR, entretanto as ARs médias foram similares entre EB e PB. A eficiência de seleção (SE) e a fração orgânica ingerida (i) aumentaram conforme incrementou o teor orgânico do seston (f), sendo maiores em PB. As eficiências de absorção (AE), por sua vez, incrementaram com SE e i, mas seus valores médios também foram similares entre os locais de estudo. O mexilhão P. perna apresentou uma alta eficiência de seleção mesmo sob condições de alta turbidez em EB, maximizando sua aquisição energética em termos de absorção de alimento (AR e AE). A produção de biodepósitos foi maior em EB do que em PB, com teor orgânico menor nos biodepósitos produzidos, em função das próprias características do seston (maior TPM e menor f), bem como da maior contribuição de pseudofezes, organicamente mais pobres que as fezes.

xv
ABSTRACT This study analysed the feeding and the biodeposits production by the mussel Perna perna at two culture sites in southern Brazil, differentiated by the seston parameters, in three experiments conducted at both sites. Enseada do Brito (EB) was characterised by higher concentrations of total particulate matter (TPM), higher particulate inorganic matter (PIM) and higher particulate organic matter (POM) values, but lower organic content of seston (f) than at Porto Belo (PB). In general, the feeding and the biodeposits production by mussel P. perna changed according to seston variation, and showed similar behaviour than other filter-feeding bivalve molluscs. Clearance rates (CR) were lower at EB, where the concentrations of suspended matter (TPM, PIM and POM) were higher. Filtration rates (FR), ingestion rates (IR) and organic ingestion rates (OIR) were higher at EB, increasing in accordance with the increase of seston concentrations. The absorption rates (AR) followed the same trends of FR, IR and OIR, however the average AR were similar at both sites. Selection efficiency (SE) and organic content of ingested matter (i) increased in accordance with the increase of the organic content of the seston (f), and were higher at PB. The absorption efficiency (AE), in turn, increased with SE and i, but the average values were similar at both sites. The mussel P. perna showed high selection efficiency at EB, even with high turbidity, maximizing his energetic intake in relation to food absorption (AR and AE). The biodeposits production was higher at EB than at PB, with lower organic content of biodeposits at this site, as a function of seston characteristics (higher TPM and lower f values), and higher pseudofaeces production with lower organic content than faeces.

16
INTRODUÇÃO
Aspectos do cultivo de moluscos bivalves e sua sustentabilidade
Os moluscos bivalves representam um dos grupos mais importantes para a aqüicultura,
tanto do ponto de vista produtivo quanto econômico, constituindo-se em um recurso alimentar
altamente nutritivo e com baixo custo de produção, especialmente em relação a peixes e
crustáceos (PAREJO, 1989; NEWKIRK, 1990). Os bivalves alimentam-se filtrando
microalgas e matéria orgânica em suspensão da coluna d’água (JØRGENSEN, 1990) sendo,
portanto, dependentes exclusivamente da produtividade natural para sua alimentação.
O cultivo de bivalves é uma atividade de grande alcance social, gerando empregos,
ajudando a fixar populações nativas litorâneas em seu ambiente tradicional e proporcionando
um complemento de renda para pescadores artesanais (ROSA, 1997; VINATEA ARANA,
1999). Além disso, colabora com a preservação das regiões onde se estabelecem, com a
conservação da qualidade da água costeira, manutenção dos estoques naturais, e incremento
de estoques pesqueiros pelo aumento da biodiversidade no entorno das estruturas de cultivo
(BARG, 1992; VINATEA ARANA, 2000).
No Brasil, a malacocultura marinha é atualmente desenvolvida por 1.600 produtores,
com uma produção no ano de 2000 de 12.500 toneladas de mexilhões e 1,3 milhões de dúzias
de ostras (PROENÇA apud CARVALHO FILHO, 2001). Em Santa Catarina, principal
Estado produtor, houve um grande desenvolvimento na última década, sendo que a produção
do mexilhão Perna perna, a principal espécie cultivada, evoluiu de 190 toneladas em 1990
para 11.359 toneladas em 2000, o que representa mais de 90% da produção nacional
(OLIVEIRA NETO apud CARVALHO FILHO, 2001). Atualmente esta atividade é realizada
em 13 municípios litorâneos, envolvendo 1050 produtores (OLIVEIRA NETO, op. cit.). Tal
êxito é devido a uma eficiente parceria envolvendo os setores de pesquisa, extensão e
produção (FERREIRA & MAGALHÃES, 1992; POLI & LITLLEPAGE, 1998; OLIVEIRA
NETO apud VINATEA ARANA, 1999).
Para que a mitilicultura se estabeleça em bases racionais e sustentáveis, são
necessárias pesquisas para avaliar as características de cultivo e os critérios utilizados para
organizar e desenvolver a atividade (ROSA, 1997), bem como avaliar seus efeitos sobre o
ambiente. Para tanto, informações relacionadas à bioecologia do mexilhão Perna perna são

17
fundamentais, especialmente com relação à sua fisiologia alimentar, que só recentemente
vem sendo estudada com maior intensidade (SUPLICY, 2000).
O processo de alimentação dos mexilhões (MOLLUSCA: BIVALVIA, Família
Mytilidae) pode ser resumido em cinco etapas (baseado em BAYNE & HAWKINS, 1990):
Através do batimento dos cílios branquiais, é criada uma corrente inalante onde
partículas em suspensão na água (seston) são retidas pelo sistema de cílios das
brânquias;
As partículas removidas são envoltas com muco e transportadas até os palpos labiais,
seguindo até a boca;
A partir de certos limites de concentração do seston, ocorre uma seleção através dos
cílios e palpos labiais, por rejeição de partículas agregadas com muco, sem ingeri-las.
Estas partículas agregadas, denominadas pseudofezes, são eliminadas ventralmente
através da região inalante (região ventral anterior);
O material ingerido sofre digestão intra ou extra-celular, sendo em seguida absorvido;
Finalmente, o material ingerido e não absorvido é eliminado na forma de fezes, através
da região exalante, juntamente com uma pequena parcela de material originado das
células e enzimas envolvidas no processo digestivo.
A matéria particulada em suspensão, fonte alimentar dos mexilhões, pode sofrer
amplas variações em quantidade e qualidade na água do mar. Portanto, experimentos que
acompanhem as variações ambientais e as conseqüências destas variações são relevantes para
a compreensão dos impactos gerados pelo cultivo de mexilhões, como organismos filtradores
ou suspensívoros, no fluxo de material particulado em ambientes costeiros (DAME, 1996).
Muitas informações podem ser obtidas a partir de alterações na quantidade e
proporção entre matéria particulada total (seston) e matéria orgânica particulada disponível
para os bivalves, e, nesse contexto, a qualidade de matéria particulada em suspensão na água
é definida como sendo a proporção de matéria orgânica particulada no seston (NAVARRO et
al., 1991; IGLESIAS et al., 1998; MELLO, 1999).
Caracterizando um estoque natural ou uma área de cultivo de mexilhões como um
sistema, pode-se avaliar a taxa de crescimento da população de mexilhões através dos
princípios da fisiologia energética, baseada em equações de balanço energético, possibilitando
analisar como fatores endógenos e exógenos afetam o crescimento de uma espécie (MELLO,
op. cit.). A partir da produção de modelos dos processos de assimilação (absorção alimentar)

18
e gastos energéticos (respiração e excreção) calcula-se o potencial de crescimento do
organismo, também denominado scope for growth (DAME, op. cit.).
O ganho energético em moluscos bivalves é medido através da determinação da taxa
de ingestão alimentar e da eficiência de absorção do alimento ingerido, e, recentemente, vêm
aumentando as pesquisas com o objetivo de analisar a complexa dependência existente entre a
absorção alimentar as características do ambiente onde o alimento é disponível (IGLESIAS et
al., 1998), gerando subsídios para avaliação do potencial de cultivo de uma área (capacidade
de carga). Com essas informações, podem-se definir as densidades máximas de cultivo que
podem ser mantidas em cada área ou ambiente, nas quais os níveis de produção são
maximizados, sem afetar negativamente as taxas de crescimento dos moluscos (MELLO,
1999; SUPLICY, 2000).
Outro aspecto importante a se considerar em termos de capacidade de carga é o
impacto ambiental gerado pelo cultivo de moluscos. Com a instalação dos cultivos, uma das
principais alterações em ambientes costeiros é o aumento da taxa de sedimentação, resultante
das alterações hidrodinâmicas promovidas pelas estruturas de cultivo bem como da
biodeposição decorrente da produção de fezes e pseudofezes pelos bivalves (BARG, 1992).
Em decorrência podem ocorrer alterações biogeoquímicas no sedimento, afetando a
comunidade bentônica (DAHLBÄCK & GUNNARSSON, 1981; OTTMANN & SORNIN,
1982; TENORE et al., 1982; KASPAR et al., 1985; KAUTSKY & EVANS, 1987; GRANT
et al., 1995; STENTON-DOZEY et al., 1999; HAYAKAWA et al., 2001; CHAMBERLAIN
et al., 2001), e, dependendo da intensidade, estes impactos podem causar o comprometimento
de ambientes costeiros inclusive para a mariculturam como assoreamento, eutrofização,
hipoxia, produção de gás sulfídrico, etc. (BARG, 1992).
Alimentação e fisiologia energética de moluscos bivalves
Os primeiros estudos de modelos numéricos para bivalves em termos de fluxo de
energia foram realizados por Thompson & Bayne (1974) em experimentos com Mytilus
edulis.
Experimentos de fisiologia energética têm sido realizados através de medições em
sistemas fechados ou estáticos (método de Coughlan), ou ainda por medições em sistemas
abertos de fluxo contínuo, baseando-se no cálculo da taxa de clareamento, através da
determinação da redução da concentração de alimento que passa por uma câmara com um ou

19
mais indivíduos com controle da velocidade de fluxo (BAYNE et al., 1987; IGLESIAS et al.,
1998).
A determinação experimental da taxa de ingestão alimentar é usualmente feita
medindo-se a taxa de clareamento e multiplicando-a pela concentração de material
particulado na água. A taxa de clareamento é o volume de água do qual o animal filtrou
totalmente as partículas em suspensão, em um determinado intervalo de tempo (NAVARRO,
et al., 1991; IGLESIAS et al., 1998; MELLO, 1999).
A taxa de filtração é obtida com a multiplicação da taxa de clareamento pela
concentração de matéria particulada em suspensão na água disponível para o animal em
estudo. Se há produção de pseudofezes (que se refere à taxa de rejeição), o valor das mesmas
é subtraído da taxa de filtração, obtendo-se então a taxa de ingestão. Não havendo produção
de pseudofezes, a taxa de filtração é considerada igual à taxa de ingestão (BAYNE et al.,
1987; IGLESIAS et al., 1998).
A partir da avaliação de diversas metodologias, Navarro & Thompson (1994)
concluíram que o método de Conover (1966) é o mais adequado para se estimar a eficiência
de absorção alimentar em bivalves filtradores sob condições de alimentação natural (seston).
Este método baseia-se no fato de que a matéria inorgânica particulada ingerida não é
significativamente absorvida, portanto servindo como um marcador inerte para os processos
digestivos. Assim, as taxas de ingestão e de egestão de matéria inorgânica são iguais. Porém,
quando há produção de pseudofezes (rejeição de partículas) pelos bivalves, além de amostras
representativas das fezes (egestão) são necessárias coletas amostrais quantitativas de
pseudofezes (IGLESIAS et al., 1998). A absorção líquida é igual à absorção bruta subtraída
pelas perdas fecais endógenas (produção de enzimas digestivas, muco, etc.) que são
desprezíveis, considerando dietas suficientes para resultar em uma absorção líquida positiva,
mas em condições de alimento de baixa qualidade nutricional, as perdas fecais são
importantes (BAYNE et al., 1988).
Vários estudos de fisiologia energética foram realizados com bivalves a partir de
experimentos de laboratório com dietas artificiais (WINTER, 1973; WINTER & LANGTON,
1976; NAVARRO & WINTER, 1982; BERRY & SCHLEYER, 1983) ou ainda simulando a
composição do seston encontrado no ambiente (WINTER, 1976; BAYNE et al., 1987; 1989;
1993; IGLESIAS et al., 1992; LABARTA et al., 1997; NAVARRO & THOMPSON, 1994;
NAVARRO & ULLOA, 1995; NAVARRO et al., 1996; NAVARRO & WIDDOWS, 1997;
WONG & CHEUNG, 1999). Também no Brasil, Resgalla Jr. et al. (1999) realizaram estudos

20
sobre a variabilidade das taxas fisiológicas (taxa de crescimento, taxa de filtração, taxa de
respiração e taxa de assimilação) do mexilhão Perna perna em dois sítios de cultivo do litoral
norte de Santa Catarina, com dietas artificiais sob condições de laboratório.
No entanto, para uma melhor estimativa das taxas de biodeposição, eficiência de
absorção e tempo de passagem do alimento pelo trato digestivo, considerando diferenças
regionais, variações diárias (ciclo de marés, por exemplo) ou sazonais em termos de
comportamento alimentar e balanço energético, foram também desenvolvidas técnicas de
estudos em campo (BAYNE et al., 1976; BAYNE & WIDDOWS, 1978; NAVARRO et al.,
1991; NAVARRO, 1983; OKUMUS & STIRLING, 1994; CRANFORD & HARGRAVE,
1994; CRANFORD et al., 1998; IGLESIAS et al., 1996; URRUTIA et al., 1996; NAVARRO
& THOMPSON, 1996, 1997; BARILLÉ et al., 1997; POUVREAU et al., 2000; HAWKINS
et al., 1996, 1998a, 1998b, 1999, 2001; WONG & CHEUNG, 2001a, 2001b; GARDNER,
2002).
Nos experimentos de fisiologia energética, um dos métodos para o cálculo da
concentração de partículas nas amostras de água do ambiente e nas amostras de biodepósitos
consiste em submetê-las à filtração em 0,45 ou 1,2 µm (dependendo da eficiência de retenção
de cada espécie), com posterior análise gravimétrica do material retido no filtro, sendo o peso
de material seco relacionado à matéria particulada total, e o peso de material calcinado
relacionado à determinação da proporção entre matéria orgânica e inorgânica (NAVARRO &
WINTER, 1982; PÉREZ CAMACHO & GONZÁLEZ, 1984; BAYNE et al., 1987; 1989;
1993; IGLESIAS et al., 1992; NAVARRO & JARAMILLO, 1994; NAVARRO & ULLOA,
1995; NAVARRO & WIDDOWS, 1997; NAVARRO & THOMPSON, 1996; 1997;
LABARTA et al., 1997).
Navarro et al. (1991) realizaram experimentos em campo, sobre a fisiologia energética
de mexilhões, onde as taxas de alimentação foram estimadas indiretamente a partir das taxas
de biodeposição, utilizando a matéria inorgânica das fezes e pseudofezes como marcador
quantitativo da matéria particulada total filtrada, sem a necessidade de controlar a velocidade
de fluxo de água no sistema. Foram coletadas amostras de água da entrada do sistema e de
fezes e pseudofezes das unidades experimentais a intervalos de tempo determinados.
Calculou-se a taxa de clareamento através da razão entre a matéria inorgânica dos
biodepósitos e a matéria inorgânica da água na entrada do sistema, e a partir destes dados
foram estimadas as taxas de ingestão e absorção, bem como a eficiência de absorção (método
de Conover).

21
Urrutia et al. (1996) testaram a metodologia proposta por Navarro et al. (1991),
denominada como método dos biodepósitos, comparando-a com o método de fluxo contínuo,
obtendo um coeficiente de correlação de 0,86 entre as taxas de clareamento dos dois métodos.
Em relação à metodologia de fluxo contínuo, o método de biodepósitos proposto por
Navarro et al. (1991) apresenta algumas vantagens práticas, como eliminação do controle da
velocidade de fluxo de água, diminuição da quantidade de amostras a serem coletadas no
experimento, além de fornecer um resultado integrado no período de tempo considerado para
o acúmulo de fezes e pseudofezes amostradas, diminuindo assim os erros resultantes de
variações que ocorrem em curta escala de tempo (MELLO, 1999).
Considerando estas vantagens, Mello (op. cit.) adaptou a metodologia proposta por
Navarro et al. (op. cit.) às nossas condições, realizando o primeiro experimento em campo de
fisiologia alimentar do mexilhão Perna perna em condições de cultivo no Brasil, sendo a
elaboração e execução desta pesquisa baseada neste trabalho.
A biodeposição e os impactos ambientais resultantes do cultivo de bivalves
A sedimentação ou deposição é um processo de remoção de material em suspensão da
coluna d’água. Similarmente, invertebrados suspensívoros, tais como mexilhões, ostras,
cracas, copépodos e tunicados, removem partículas, as processam e liberam para a coluna
d’água como pelotas fecais (JARAMILLO et al., 1992). Organismos filtradores são capazes
de remover uma grande quantidade de material particulado do seston, processando partículas
menores que 100 µm e aglomerando-as com muco, para então liberá-las em “pacotes” de
alguns milímetros, favorecendo assim sua sedimentação (DESLOUS-PAOLI et al., 1992;
SCHETTINI et al., 1997; D’AQUINO, 2000). Das partículas filtradas, uma parte passa
através do trato digestivo e sofre eliminação parcial na forma compactada de fezes. Sob certas
circunstâncias, muitos moluscos bivalves rejeitam parte do material filtrado sem mesmo
ingeri-lo na forma de uma massa menos compacta denominada de pseudofezes (HAVEN &
MORALES-ALAMO, 1972). As fezes e pseudofezes compõem os biodepósitos, e o processo
em que este material sedimenta no fundo denomina-se biodeposição (HAVEN & MORALES-
ALAMO, 1966).
Como sedimentam rapidamente, os biodepósitos podem influenciar as condições
físicas e químicas do ambiente no fundo, portanto a produtividade e biodiversidade dos
ecossistemas bentônicos (NAVARRO & THOMPSON, 1997). Após a deposição, as pelotas

22
fecais podem contribuir com alterações físico-químicas e biológicas no sedimento do fundo,
como descrito por Dahlbäck & Gunnarsson (1981). Os biodepósitos produzidos por
organismos marinhos podem ter uma importante função trófica, dependendo de sua
composição nutricional (NAVARRO & THOMPSON, op. cit.). Outros autores estudadaram a
biodeposição por ostras (ITO & IMAI, 1955; TENORE & DUNSTAN, 1973) e discutiram
sua importância na cadeia trófica em sistemas de aqüicultura. Newell (1965) observou que
pelotas fecais de alguns moluscos marinhos continham matéria orgânica assimilável, passíveis
de reingestão pelos próprios organismos que as expeliam ou de ingestão por outras espécies.
Jaramillo et al. (1992) determinaram que as taxas de biodeposição média de
populações de mexilhões Choromytilus chorus e Mytilus chilensis no estuário do rio Queule,
sul do Chile, foram, respectivamente, de 271 e de 234 g de peso seco m-2 . d-1, e que os
biodepósitos continham 22 e 21% de matéria orgânica, 6,4 e 6,0% de carbono, e 0,3 e 0,4 %
de nitrogênio. Navarro (1983) encontrou concentrações similares de matéria orgânica (17-
27%) nos biodepósitos de Mytilus chilensis na baía de Yaldad, calculando que uma balsa de
cultivo (com 300 cordas de 8 m cada, 2.400 mexilhões por corda) produziria 15,8 Kg de peso
seco de fezes por dia. Haven & Morales-Alamo (1966) descreveram uma ampla variação na
produção de biodepósitos por ostras Crassostrea virginica, com uma média semanal de 1,62 g
de peso seco por indivíduo, chegando ao máximo de 3,92 g de peso seco semanal por ostra.
Estes biodepósitos continham de 9 a 23% de matéria orgânica e de 4 a 12% de carbono
orgânico.
Kautsky & Evans (1987) enfatizam que a maior contribuição dos mexilhões para a
rede trófica ocorre nos processos de sedimentação (biodeposição) e ciclagem de nutrientes,
uma vez que grande parte do material ingerido (alimento) é expelido como fezes, pseudofezes
e nutrientes dissolvidos. Além disso, a atividade bacteriana nos biodepósitos é potencializada
pelo aumento na relação susperfície/volume das partículas liberadas, e a matéria orgânica
depositada pode representar uma fração importante de alimento disponível para outros
invertebrados.
Na zona costeira do Mar Báltico, Mytilus edulis tem uma importante função no fluxo
vertical entre os ecossistemas pelágico (coluna d’água) e bentônico (fundo), incrementando a
deposição anual total de carbono, nitrogênio e fósforo em 10%, e pela circulação e
regeneração da demanda anual de 12% de nitrogênio e de 22% de fósforo da produção
primária pelágica naquela área (KAUTSKY & EVANS, op. cit.).

23
Alterações hidrodinâmicas são promovidas com a instalação dos cultivos, com
diminuição da capacidade de transporte de material particulado pelas correntes (advecção),
gerando a formação de depósitos localizados por sedimentação de partículas em suspensão
(OTTMANN & SORNIN, 1982; BARG, 1992; SCHETTINI et al., 1997). Mattsson & Lindén
(1983) verificaram uma sedimentação pelo menos três vezes maior sob instalações de
mitilicultura em sistema de long line em relação a áreas de controle (sem cultivos) na costa
oeste da Suécia. Tal aporte pode provocar mudanças nas características físico-químicas dos
sedimentos e na macroinfauna associada, com conseqüências para a comunidade da área e
para os próprios bivalves cultivados (MATTSSON & LINDÈN, op cit., BERTRÁN et al.,
1988).
Por apresentarem alto conteúdo de água, os biodepósitos são altamente suscetíveis à
ressuspensão logo depois de formados, dando origem a um “biosseston”, que consiste de
agregados bacterianos, células do fitoplâncton fragmentadas, detritos e partículas resultantes
dos processos digestivos (KAUTSKY & EVANS, 1987). Entretanto, o processo natural de
sedimentação e compactação da matéria orgânica depositada no fundo aumenta a estabilidade
do sedimento (coesão, em até 200%), dificultando posterior erosão e transporte (OTTMANN
& SORNIN, 1982).
Supõe-se que a criação de um ambiente redutor no fundo (camada limite, interface
água - sedimento) por eutrofização afete a produtividade dos organismos cultivados,
principalmente em áreas rasas e com baixa hidrodinâmica, ou seja, pouca circulação e
renovação de água e de seus constituintes físico-químicos, particulados e dissolvidos
(OTTMANN & SORNIN, op. cit.; BARG, 1992).
Schettini et al. (1997), em um estudo preliminar, avaliaram a sedimentação sob
cultivos de bivalves na Armação do Itapocoroy, no litoral norte de Santa Catarina, e
obtiveram taxas relativamente altas (118 ± 65 g m-2 dia-1), mas com baixas porcentagens de
sedimentos finos (predomínio de areia e alto teor de carbonatos) e 17,5% de matéria orgânica
(3,1± 0,6 % de carbono orgânico particulado). Em outro levantamento realizado na mesma
área, D’Aquino (2000) verificou uma taxa de sedimentação média de 262 g m-2 dia-1, com
valores mínimo e máximo, de 63 g m-2 dia-1 e de 1.050 g m-2 dia-1, respectivamente. O teor de
matéria orgânica médio foi de 2,3% (mínimo de 1% e máximo de 4%).
A partir de informações sobre a hidrodinâmica da enseada de Armação do Itapocoroy
(SCHETTINI et al., 1999) e seu papel na remobilização de sedimentos finos (D’AQUINO,
2000), concluiu-se que este material sofre constante ressuspensão do fundo por atuação de

24
ondas, resultando na dispersão do material em suspensão por advecção (transporte por
correntes). Ainda neste local, Proença & Schettini (1998) avaliaram o fluxo vertical de
fitodetritos e observaram taxas médias 5 vezes superiores em relação a uma área de referência
e, dado esse expressivo fluxo vertical comparado à pequena biomassa na coluna d’água,
deduziram que o fitoplâncton alóctono trazido por advecção é uma importante fonte alimentar
para a manutenção dos moluscos cultivados na enseada.
Portanto, as características hidrodinâmicas das áreas de cultivo, envolvendo a atuação
das marés, ventos e ondas, como agentes promotores da circulação de massas d’água em
ambientes costeiros, têm importante papel na sustentabilidade destes locais, atuando no
enriquecimento destas áreas com fitoplâncton e nutrientes, no fornecimento de oxigênio
dissolvido, no transporte e dispersão de biodepósitos (PROENÇA & SCHETTINI, op. cit.;
PROENÇA et al., 1999; CHEVARRIA, 1999; D’AQUINO, 2000).
Um exemplo dos impactos ambientais negativos gerados pelo cultivo de moluscos
marinhos é relatado por Ottmann & Sornin (1982). Nas extensas planícies de maré da costa
atlântica francesa, mais especificamente nas baías de Mont-Saint-Michel e de Aiguillon, e na
enseada de Marennes-Oleron, onde tradicionalmente são cultivados mexilhões e ostras em
sistemas fixos (estacas e mesas, respectivamente), observou-se um aumento nas taxas de
sedimentação em até 1,5 Kg m-2.dia-1, gerando uma taxa de assoreamento de
aproximadamente 0,7 m por ano. Além da diminuição da profundidade destas áreas, outra
conseqüência é a eutrofização promovida pela biodeposição (produção de fezes e
pseudofezes). Nestas condições, a hipoxia e a produção de gás sulfídrico aumentam a
mortalidade dos bivalves, interferindo negativamente na produtividade.
No Japão houve declínio na produtividade de cultivos de ostra, o que foi atribuído a
deterioração da qualidade das áreas de cultivo induzida pela atividade filtradora dos bivalves
(ITO & IMAI, 1955; KUSUKI, 1978). Segundo Mariojouls & Kusuki (1987), na Baía de
Hiroshima cada balsa de cultivo produzia 78,7 Kg dia-1 de biodepósitos (fezes e pseudofezes)
e a proporção de deposição total devida a sedimentação natural foi de 31 a 48% sob as balsas
de cultivo comparada a 89% nas áreas fora dos cultivos.
Por outro lado, nas rias espanholas, a produção de biodepósitos resultante da
mitilicultura tem resultado em benefícios ambientais e econômicos. De acordo com Tenore et
al. (1982), as características hidrodinâmicas destas áreas favorecem o transporte destes
biodepósitos para a plataforma continental adjacente, proporcionando uma fonte adicional de
alimento para a comunidade bentônica epifaunal. Alimentando-se dos detritos oriundos dos

25
cultivos, esta macrofauna, composta por crustáceos e poliquetos, evita o processo de
eutrofização no fundo. Além disso, o incremento desta biomassa bentônica potencializou a
captura de peixes demersais, seus predadores, que são importantes recursos pesqueiros
naquela região. Outro recurso pesqueiro local beneficiado indiretamente por este aporte de
matéria orgânica oriundo da mitilicultura galega é a lagosta Nephrops norvegicus que, como a
ictiofauna, preda sobre a infauna bentônica que por sua vez alimenta-se dos detritos oriundos
dos cultivos (LÓPEZ-JAMAR et al., 1992). Mattsson & Lindén (op. cit.) também relatam que
houve aumento da pesca nas áreas próximas aos cultivos de mexilhões na costa ocidental
sueca, especialmente no primeiro ano de instalação e operação dos cultivos.
Adaptações a condições ambientais adversas tendem a provocar estresse metabólico
nos animais, cujo resultado em um cultivo refletiria na queda de produtividade. Portanto,
identificar e quantificar parâmetros bióticos e abióticos, endógenos e exógenos, que
influenciem negativamente na produtividade de organismos cultivados se faz necessário, a
fim de prevenir e mitigar estes efeitos (MELLO, 1999).
O presente estudo pretende gerar subsídios para avaliação dos potenciais de cultivo de
duas áreas distintas do litoral de Santa Catarina, tanto em termos da produtividade de cada
local como do impacto resultante da atividade. Neste sentido, serão analisadas as taxas
fisiológicas de alimentação (taxas de clareamento, eficiências de seleção, eficiências de
absorção) e de biodeposição (taxas de produção de biodepósitos e sua composição em termos
de matéria orgânica e inorgânica) do mexilhão Perna perna. Assim, teremos susbsídios para o
desenvolvimento de modelos de capacidade de carga para as nossas condições de cultivo,
possibilitando assim o estabelecimento da capacidade de suporte para implantação de novos
parques aqüícolas marinhos (SUPLICY, 2000).

26
ARTIGO CIENTÍFICO
Efeito de diferentes condições ambientais em áreas de cultivo
sobre alimentação e biodeposição do mexilhão Perna perna.
J. F. Schmitt1*, J. F. Ferreira1* & F. M. Suplicy2
1Laboratório de Cultivo de Moluscos Marinhos, Departamento de Aqüicultura, Universidade
Federal de Santa Catarina, Caixa Postal 10.138, Florianópolis, Santa Catarina, CEP 88062-
970, Brasil.
2School of Aquaculture, University of Tasmania, Locked-bag 1-370, Launceston, Tasmania,
7250, Australia.
*Correspondências para [email protected] ou para [email protected]

27
Resumo
Este estudo avaliou o efeito de diferentes condições ambientais sobre a alimentação e a produção de biodepósitos do mexilhão Perna perna em duas áreas de cultivo no sul do Brasil, a partir de experimentos realizados durante verão e outono. As principais características ambientais que diferenciam estes locais são os parâmetros do seston: Enseada do Brito (EB) apresenta alta turbidez (maiores concentrações de material particulado total, TPM, de matéria inorgânica particulada, PIM, e de matéria orgânica particulada, POM) em relação a Porto Belo (PB), que por sua vez apresenta maiores níveis de fração orgânica do seston (f). Em geral, a alimentação e a produção de biodepósitos pelo mexilhão Perna perna variou de acordo com as características do seston, apresentando tendências similares àquelas observadas para outras espécies de bivalves filtradores. As taxas de clareamento (CR) foram menores em Enseada do Brito, sob condições de maior oferta alimentar (POM). As taxas de filtração (FR), de ingestão (IR) e de ingestão orgânica (OIR) foram maiores em EB, acompanhando o aumento do seston. As taxas de absorção (AR) seguiram a mesma tendência de FR, IR e OIR, entretanto as ARs médias foram similares entre os locais estudados. A eficiência de seleção (SE) e a fração orgânica ingerida (i) aumentaram conforme incrementou o teor orgânico do seston (f), sendo maiores em PB. As eficiências de absorção (AE), por sua vez, incrementaram com SE e i, mas seus valores médios também foram similares entre os locais de estudo. O mexilhão Perna perna apresentou uma alta eficiência de seleção mesmo sob condições de alta turbidez resultante da ressuspensão em EB no verão, maximizando sua aquisição energética em termos de absorção de alimento (AR e AE). A biodeposição foi maior em EB do que em PB, em decorrência dos níveis mais elevados de material em suspensão (TPM), com teor orgânico menor de biodepósitos produzidos em EB em função das próprias características do seston (menor f), bem como da maior contribuição de pseudofezes, organicamente mais pobres que as fezes.
Palavras chaves: alimentação, biodeposição, maricultura, mexilhão, Perna perna, seston.

28
1. Introdução
No Brasil, a malacocultura marinha gerou em 2000 uma produção de 12.500 toneladas
de mexilhões e 1,3 milhões de dúzias de ostras (Proença apud Carvalho Filho, 2001). Em
Santa Catarina, principal Estado produtor, houve um grande desenvolvimento na última
década, sendo que a produção de mexilhões, evoluiu de 190 toneladas em 1990 para 11.359
toneladas em 2000 (Oliveira Neto apud Carvalho Filho, 2001), o que representa mais de 90%
da produção nacional. Perna perna, a principal espécie cultivada, é o maior mitilídeo
brasileiro, atingindo o tamanho comercial (7 cm) com 6 a 9 meses de cultivo (Ferreira &
Magalhães, 1997). Para que a mitilicultura se estabeleça em bases racionais e sustentáveis,
são necessárias pesquisas para avaliar as características de cultivo e os critérios utilizados
para organizar e desenvolver a atividade (Rosa,1997), bem como avaliar seus efeitos sobre o
ambiente. Para tanto, informações relacionadas à bioecologia do mexilhão Perna perna são
fundamentais, especialmente com relação às condições ambientais em áreas de cultivo do
litoral brasileiro.
A matéria particulada em suspensão, fonte alimentar dos mexilhões, pode sofrer
amplas variações em quantidade e qualidade na água do mar. O estudo destas variações
ambientais e suas conseqüências são indispensáveis para a compreensão dos impactos gerados
pelo cultivo de mexilhões, como organismos filtradores ou suspensívoros, no fluxo de
material particulado em ambientes costeiros (Dame, 1996).
Caracterizando um estoque natural ou uma área de cultivo de mexilhões como um
sistema, pode-se avaliar a taxa de crescimento da população de mexilhões através dos
princípios da fisiologia energética. A partir da produção de modelos dos processos de
assimilação (absorção alimentar) e gastos energéticos (respiração e excreção) calcula-se o
potencial de crescimento do organismo, também denominado “scope for growth” (Bayne &
Newell, 1983; Dame, 1996).
O ganho energético em moluscos bivalves é medido através da determinação da taxa
de ingestão alimentar e da eficiência de absorção do alimento ingerido e recentemente vêm
aumentando as pesquisas com o objetivo de analisar a complexa dependência existente entre a
absorção alimentar as características do ambiente onde o alimento é disponível (Iglesias et al.,
1998), gerando subsídios para avaliação do potencial de cultivo de uma área (capacidade de
carga).

29
Outro aspecto importante a se considerar em termos de capacidade de carga é o
impacto ambiental gerado pelo cultivo de moluscos. Com a instalação dos cultivos ocorre
aumento da taxa de sedimentação, resultante das alterações hidrodinâmicas promovidas pelas
estruturas de cultivo bem como da biodeposição oriunda da produção de fezes e pseudofezes
pelos bivalves (Barg, 1992). Em decorrência podem haver alterações biogeoquímicas no
sedimento que afetam a comunidade bentônica (Dahlbäck & Gunnarsson, 1981; Ottmann &
Sornin, 1982; Tenore et al., 1982; Kaspar et al., 1985; Kautsky & Evans, 1987; Grant et al.,
1995; Stenton-Dozey et al., 1999; Hayakawa et al., 2001; Chamberlain et al., 2001) e,
dependendo da intensidade, estes impactos podem causar o comprometimento dos ambientes
costeiros inclusive para a maricultura (assoreamento, eutrofização, hipoxia, produção de gás
sulfídrico, etc.) (Barg, 1992).
No Brasil, são recentes os estudos avaliando impactos ambientais da maricultura,
como taxas de sedimentação e fluxos verticais de fitodetritos em áreas de cultivo (Schettini et
al., 1997; Proença & Schettini, 1998). Em termos de fisiologia energética alimentar, Resgalla
Jr. et al. (1999) realizaram estudos sobre a variabilidade das taxas fisiológicas (taxa de
crescimento, taxa de filtração, taxa de respiração e taxa de assimilação) de mexilhões Perna
perna provenientes de dois sítios de cultivo do litoral norte de Santa Catarina, sob condições
de laboratório e com dietas artificiais. No entanto, o primeiro estudo de campo possibilitando
analisar a fisiologia alimentar e biodeposição de Perna perna sob condições naturais foi
realizado por Mello (1999), a partir do Método dos Biodepósitos desenvolvido por Navarro
et al. (1991).
Este experimento pretende avaliar o efeito de duas áreas de cultivo do litoral
catarinense que apresentam condições ambientais e de cultivo distintas, Ilha João da Cunha,
na enseada de Porto Belo (município de Porto Belo), e Enseada do Brito (município de
Palhoça), sobre a alimentação (taxas de filtração, rejeição, egestão e absorção, bem como
eficiências de seleção e de absorção) e a biodeposição (taxas de produção de biodepósitos e
sua composição em termos de matéria orgânica e inorgânica) do mexilhão Perna perna,
gerando subsídios para determinação da capacidade carga destes parques aqüícolas.

30
2. Materiais e métodos
2.1. Áreas de estudo
O trabalho foi realizado em duas áreas de cultivo de moluscos, no litoral de Santa
Catarina, Brasil (Fig. 1):
Brasil, com o Estado de Santa Catarina (SC) em
destaque:Porto Belo
Enseada do Brito
Figura 1: Localização das áreas de estudo, com mapas do Brasil e do Estado de Santa Catarina, destacando as localidades de Porto Belo (PB) e de Enseada do Brito (EB), no litoral catarinense (adaptado de http://www.brasil-turismo.com/santa-catarina/mapas-sc.htm).
Enseada de Porto Belo, proximidades da Ilha João da Cunha (ou de Porto Belo),
(27°08’S, 48°32’W), município de Porto Belo (Fig. 2): área de cultivo localizada em
enseada semi-abrigada, de águas com baixa turbidez, substrato de fundo
predominantemente arenoso e profundidade de 3 a 5 metros, com poucos rios
pequenos desaguando na região como os rios Perequê e Porto Belo, e menor
concentração de cultivos: 2,9 ha. de área cultivada (Santos, comunicação pessoal);
Enseada do Brito (27°46’S, 48°37’W), Baía Sul da Ilha de Santa Catarina (face
continental), município de Palhoça (Fig. 3): área de cultivo situada em enseada
abrigada, de águas normalmente mais turvas, substrato de fundo lodoso, com
profundidades entre 1 e 3 metros, com vários rios desaguando na região da enseada
(rios Brito e Cambirela) e da baía sul (rios Tavares, Defuntos e Massiambu), e maior
concentração de cultivos: 36,4 ha. de área cultivada (Quadros, comunicação pessoal).

31
Na região das áreas de estudo, as marés são do tipo micromaré com altura média de 0,8 m
(0,4 m em quadratura e 1,2 m em sizígia), de regime misto com predominância de marés
de ciclo semi-diurno (Schettini et al., 1996, 2002). Os ventos predominates sopram do
quadrante NE, levando a formação de ondulações do tipo sea, mais comuns em baías e
enseadas protegidas. Tanto a Enseada de Porto Belo quanto da Enseada do Brito são
abrigadas do ataque de ondas de maior energia (do tipo swell) oriundas dos quadrantes
Leste e Sul.
Figura 2: Localização das áreas de estudo: área experimental em Porto Belo (PB – 27°08’12.67”S; 48°32’23.40”W), nas proximidades da Ilha João da Cunha (Ilha de Porto Belo), no litoral catarinense.

32
Figura 3: Localização das áreas de estudo: área experimental em Enseada do Brito (EB – 27°46’30.29”S; 48°37’47.98”W), no litoral catarinense.
2.2. Monitoração de variáveis ambientais
Nas áreas de estudo foram monitorados semanalmente, de janeiro a junho de 2001,
parâmetros físico-químicos da água (“in situ”): salinidade (PSU), temperatura (° C), e
oxigênio dissolvido (mg L-1 e % de saturação) com aparelho multianalisador Multi Line P4. A
partir de amostras de água foram determinadas em laboratório as concentrações de matéria
particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada
(POM), clorofila a (Cla), feopigmentos (Feop) e fração orgânica do seston (f), além da
turbidez (NTU) através de turbidímetro La Motte 2020.
Para as análises de seston, as amostras de água (pré-filtradas em malha de 230 µm)
foram filtradas em filtros Fisher G4 (previamente lavados com água destilada, calcinados e
pesados). Ao final de cada filtragem os filtros com amostra foram enxaguados com água
destilada (cerca de 50 mL) para retirada dos sais. Os pesos (mg L-1) de matéria particulada
total foram obtidos após secagem de filtros contendo as amostras em em estufa a 60 °C por 48

33
horas. Os pesos de matéria inorgânica (mg L-1) foram obtidos calcinando-se as amostras de
matéria particulada total a 450°C por 4 horas, e a matéria orgânica (POM) foi determinada
pela diferença entre TPM e PIM (mg L-1). O índice f foi obtido pela razão POM/TPM. Para
cada amostra de água foram filtradas três alíquotas e, a cada vinte filtros com amostras, foram
usados três filtros como “branco” (filtrados com água destilada). A média dos pesos dos
filtros utilizados como “branco” foi subtraída dos valores obtidos com as amostras (adaptado
de Strickland & Parsons, 1972).
Para as análises de clorofila a (Cla) e feopigmentos (Feop), as amostras de água foram
filtradas em filtros Wathman 934-AH, sendo os filtros com material retido submetidos a
extração em solução de acetona 90% a 4°C, no escuro, por até 20 horas. O extrato foi
centrifugado (3000 rpm, 10 min.) e submetido à determinação por fluorimetria (fluorímetro
TURNER AU10), com duas leituras: uma anterior e outra posterior à acidificação com HCl
(0,003 N), segundo método proposto por Barroso & Littlepage (1998).
2.3. Sistema experimental
Os experimentos foram realizados na enseada de Porto Belo (Ilha João da Cunha/Ilha
de Porto Belo) em 07/02/2001 (PB1), em 31/03/2001 (PB2) e em 06/07/2001 (PB3); e na
Enseada do Brito em 14/03/2001 (EB1), em 14/04/2001 (EB2) e em 05/06/2001 (EB3),
caracterizando três experimentos em cada local (detalhes na Tab. 1).
Tabela 1: Datas, horários e condições ambientais durante os experimentos de alimentação e de biodeposição com mexilhão Perna perna em Enseada do Brito e em Porto Belo. Experimento Data Horário Maré Vento Ondulação Turbidez
(média ± d.p.) EB1 14/03/2001 9:55 – 13:55 Enchente de
quadratura NE forte a
médio Sea forte a
média 17,2 ± 7,3
EB2 14/04/2001 10:15 – 15:15 Enchente de quadratura
NE fraco a médio
Sea fraca a média
7,7 ± 1,8
EB3 05/06/2001 11:00 – 16:00 Enchente e vazante de
sizígia
NE médio a fraco
Sea média a fraca
4,6 ± 1,7
PB1 07/02/2001 10:20 – 14:40 Enchente de sizígia
Sem ventos a NE fraco
Sem ondas a Sea fraca
0,6 ± 0,2
PB2 31/03/2001 10:10 – 15:10 Enchente e vazante de quadratura
NE fraco a médio
Sea fraca a média
1,0 ± 0,2
PB3 06/07/2001 10:45 – 15:45 Enchente de sizígia
Sem ventos
Sem ondas 0,3 ± 0,1

34
O sistema experimental foi montado sobre balsas de trabalho utilizadas pelos
produtores nas atividades de manejo dos moluscos, situadas nas áreas de estudo, e constituiu-
se de uma bandeja compartimentada em câmaras individuais para cada animal, com água
circulante bombeada do próprio ambiente de cultivo em estudo (Mello, 1999). A água de
entrada era distribuída homogeneamente pelos compartimentos e escoada por uma calha
central, havendo ainda um mecanismo de controle do fluxo de água no sistema para evitar a
ressuspensão dos biodepósitos. O fluxo variou entre 0,14 e 0,39 L s-1.
Foram coletadas periodicamente amostras da água de entrada do sistema (1 L; no
início, meio e fim de cada bateria), e dos biodepósitos de cada unidade experimental (em
intervalos de 1 hora, ao longo de 4 horas) com uso de micropipeta.
Durante os experimentos de campo também foram analisados “in situ”, salinidade
(PSU), temperatura (° C) e oxigênio dissolvido (mg L-1) na água, com aparelho
multianalisador Multi Line P4.
2.4. Material biológico
Os mexilhões da espécie Perna perna utilizados nos experimentos de alimentação e
biodeposição (detalhes na Tab. 2) foram coletados nas próprias áreas de estudo sendo
previamente limpos, através de escovação e retirada de organismos epibiontes, e aclimatados
no sistema experimental por 1 hora. Em cada experimento foram realizadas medições de
comprimento de concha (cm) e de peso seco (60 °C por 48 hs.) das partes moles (g) de cada
animal experimental.
Tabela 2: Material biológico utilizado nos experimentos de alimentação e de biodeposição com mexilhão Perna perna em Enseada do Brito e em Porto Belo.
Experimento n comprimento (cm) peso (g) EB1 15 6,58 ± 1,42 1,37 ± 0,76
EB2 12 7,05 ± 1,01 1,26 ± 0,57
EB3 8 6,92 ± 0,95 1,23 ± 0,67
PB1 8 6,60 ± 1,94 1,85 ± 1,34
PB2 9 6,42 ± 1,30 1,69 ± 0,81
PB3 9 7,26 ± 0,94 1,82 ± 0,83

35
2.5. Quantificação de matéria orgânica e inorgânica no seston e nos biodepósitos
As amostras de água foram filtradas em filtros Fisher G4 e os biodepósitos (fezes e
pseudofezes) em filtros Wathman GF/C e processadas de acordo com o método descrito
anteriormente para as análises de seston (item 2.2.).
2.6. Determinação das taxas fisiológicas
A partir dos dados de matéria particulada total, inorgânica e orgânica das amostras de
água (TPM, PIM, POM) e dos biodepósitos (fezes e pseudofezes), foram obtidas as seguintes
taxas fisiológicas: das pseudofezes obtivemos (em mg h-1) as taxas de rejeição (RR), rejeição
inorgânica (IRR), rejeição orgânica (ORR); das fezes obtivemos (em mg h-1) as taxas de
egestão (ER), egestão inorgânica (IER), egestão orgânica (OER); taxas de biodeposição (BR
= RR + ER; em mg h-1), biodeposição inorgânica (IBR = IRR + IER; em mg h-1) e
biodeposição orgânica (OBR = ORR + OER; em mg h-1); taxa de clareamento (CR =
IBR/PIM, em L h-1); taxas de filtração (FR = TPM * CR; em mg h-1), filtração inorgânica
(IFR = PIM * CR; em mg-1) e filtração orgânica (OFR = POM * CR; em mg h-1); as taxas de
ingestão (IR = FR - RR; em mg h-1), ingestão inorgânica (IIR = IFR - IRR; em mg h-1) e
ingestão orgânica (OIR = OFR - ORR; em mg h-1); fração orgânica ingerida (i = OIR/IR);
taxa de absorção (AR = OIR - OER; em mg h-1); a eficiência de absorção (AE = AR/OIR);
frações orgânicas de fezes (e = OER/ER), pseudofezes (p = ORR/RR) e biodepósitos (b =
OBR/BR); eficiência de seleção (SE = 1 – (p/f)).
2.7. Padronização das taxas fisiológicas
Como os mexilhões utilizados nos experimentos apresentavam tamanhos variados
(comprimento de concha entre 4 e 9 cm, vide Tab. 2), para eliminar a variabilidade em função
do tamanho as taxas fisiológicas (alimentação e biodeposição) foram padronizadas para um
comprimento específico de 7 cm, já que o comprimento médio de concha (± DP) foi de 6,68 ±
1,38 cm (CV = 20,6%). A opção pela padronização das taxas fisiológicas em função do
comprimento de concha se deve a alta variabilidade dos dados de peso seco dos mexilhões:
1,49 ± 0,86 g (CV = 57,6%). Sendo assim, padronizações em função de peso seco poderiam
levar a resultados incorretos (Iglesias et al., 1996). As padronizações foram efetuadas a partir

36
da fórmula Ys = Y * (7/X)b, onde Ys é a taxa fisiológica padronizada, Y é a taxa fisiológica
observada, 7 é o comprimento padrão (= 7 cm), X é o comprimento do mexilhão experimental
e b é a potência de incremento da taxa fisiológica em relação ao tamanho corpóreo do bivalve.
A potência b, oriunda da equação alométrica Y = aXb (fórmula logarítmica: logY = loga + b*
logX) foi obtida a partir de regressão linear entre logX (comprimento) e logY (taxa
fisiológica) para cada experimento (Microsoft Excel 7.0).
2.8. Análises estatísticas
A normalidade dos dados foi avaliada através do gráfico de probabilidade normal
(“normal probability plot”) e a homogeneidade de variâncias (homocedasticidade) pelo teste
de Levene.
Para aumentar a normalidade dos dados e reduzir sua heterocedasticidade, as variáveis
físico-químicas, concentrações do seston (material em suspensão e pigmentos), bem como as
taxas fisiológicas, foram transformadas para a escala log(x). As variáveis de proporção (f,
Cla/Feop, p, e, b, i, SE, AE) foram transformadas em arcoseno (x1/2).
Para avaliar o efeito dos ambientes de cultivo (locais) e dos meses do ano (tempo)
sobre as variáveis ambientais monitorizadas semanalmente foram realizadas análises de
variância bi-fatorial (ANOVA two-way). Sendo observada interação significativa (p<0,05)
entre fatores (local x tempo), foram avaliadas as diferenças tanto entre locais em cada mês do
ano quanto entre meses do ano para cada local através de teste Tukey HSD para n’s desiguais.
Ainda foram realizados testes de correlação (α = 0,05) entre as diferentes variáveis
ambientais para cada local.
Os experimentos foram comparados entre si em termos de variáveis ambientais, taxas
de alimentação e de biodeposição, através de análise de variância uni-fatorial (ANOVA one-
way), para avaliação do efeito dos locais sobre os resultados. Diferenças significativas
(p<0,05) foram avaliadas pelo teste Tukey HSD para n’s desiguais. Também foram
comparadas as médias dos parâmetros experimentais (variáveis ambientais e taxas
fisiológicas) entre os dois locais utilizando-se o Teste t de Student (α = 0,05) para amostras
independentes (grupos de amostras).
Para as análises estatísticas supracitadas utilizou-se o programa STATISTICA 5.0.
Ainda foram realizados testes de correlação (α = 0,05) entre as variáveis ambientais e
taxas fisiológicas, bem como entre taxas fisiológicas, utilizando as médias experimentais dos

37
dois locais estudados, sem transformação dos dados, com plotação gráfica em planilha
eletrônica Microsoft Excel 7.0.
3. Resultados
3.1. Variáveis ambientais
Os parâmetros hidrológicos neste trabalho podem ser avaliados quanto a sua variação
temporal (entre meses; Figs. 4 e 5, Tab. 3) e geográfica (entre locais; Tabs. 3 e 4; Figs. 4 e 5).
Tabela 3: Resultados das análises de variância bi-fatorial (ANOVA) avaliando o efeito dos fatores área de cultivo (local) e meses de ano sobre parâmetros hidrológicos. Variáveis físico-químicas da água: temperatura, salinidade, oxigênio dissolvido (OD e OD%) e turbidez; características quali-quantitativas do seston (fração entre 1,2 e 230 µm): matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla), feopigmentos (Feop) e razão clorofila a / feopigmentos (Cla/Feop).
Variável Entre Locais Entre Meses Interação Temperatura * *** Ns
Salinidade ns *** Ns
OD ns ** Ns
OD % ns ns Ns
Turbidez *** ns Ns
TPM *** ns Ns
PIM *** ns Ns
POM ** ns Ns
f ** * Ns
Cla *** ns **
Feop *** ns *
Cla/Feop ns ns Ns
ns : diferença não significativa; * : p<0,05; ** : p<0,01; *** : p<0,001.
Na escala de tempo mensal (Tab. 3), foram observadas variações significativas de
temperatura (p<0,001), salinidade (p<0,001) e de oxigênio dissolvido (p<0,01), com

38
tendências similares entre os dois locais. As temperaturas foram mais elevadas nos meses de
verão (janeiro, fevereiro e março) e abril em relação a maio e junho. A salinidade não
apresentou tendência sazonal tão marcada, com picos em janeiro e em abril. O oxigênio
dissolvido apresentou um pico pronunciado em fevereiro.
Em termos de seston, as variações mensais foram claramente observadas somente para
a fração orgânica do material em suspensão (f), com p<0,05, que apresentou picos em
fevereiro e maio em ambos os locais. Já a clorofila a (Cla) apresentou pico em março na
Enseada do Brito (EB), enquanto em Porto Belo (PB) os níveis mais altos ocorreram em
janeiro e fevereiro. Os feopigmentos (Feop) acompanharam as variações de Cla.
Tabela 4: Parâmetros de qualidade da água do mar das áreas de cultivo estudadas: comparação dos valores médios (médias ± desvio padrão) de monitoramento semanal realizado em Enseada do Brito (EB) e em Porto Belo (PB). Variáveis físico-químicas da água: temperatura, salinidade, oxigênio dissolvido (OD e OD%) e turbidez; características quali-quantitativas do seston (fração entre 1,2 e 230 µm): matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla), feopigmentos (Feop) e razão clorofila a / feopigmentos (Cla/Feop).
Variável Enseada do Brito Porto Belo Diferença entre Locais
Temperatura 23,0 ± 6,3 23,9 ± 6,6 Ns
Salinidade 29,2 ± 7,6 29,5 ± 7,9 Ns
OD 6,2 ± 1,7 6,2 ± 1,8 Ns
OD % 83,9 ± 23,1 86,7 ± 24,9 Ns
Turbidez 4,5 ± 3,7 1,0 ± 0,8 ***
TPM 9,2 ± 6,8 2,8 ± 2,0 ***
PIM 7,8 ± 6,0 2,1 ± 1,7 ***
POM 1,4 ± 1,0 0,8 ± 0,6 **
f 0,2 ± 0,1 0,3 ± 0,2 *
Cla 2,6 ± 1,5 1,2 ± 0,8 ***
Feop 1,3 ± 0,7 0,5 ± 0,4 ***
Cla/Feop 2,1 ± 0,8 2,9 ± 2,8 Ns
ns : diferença não significativa; * : p<0,05; ** : p<0,01; *** : p<0,001.

39
15
20
25
30
35
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
a
Tem
pera
tura
(°C
)
EB PB
20
25
30
35
40
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
b
Salin
idad
e (P
SU)
EB PB
4
5
6
7
8
9
10
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
c
OD
(mg
L-1)
EB PB
50
60
70
80
90
100
110
120
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
d
OD
(%)
EB PB
0
1
2
3
4
5
6
7
8
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
e
Clo
rofil
a a
(ug
L-1)
EB PB
0
1
2
3
4
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
f
Feop
igm
ento
s (u
g L-1
)
EB PB
Figura 4: Variáveis hidrológicas (médias mensais ± desvio padrão) obtidas em monitoração semanal realizada em Enseada do Brito (EB) e em Porto Belo (PB). Parâmetros físico-químicos e seston (fração entre 1,2 e 230 µm). a : temperatura (°C); b : salinidade (PSU); c : oxigênio dissolvido (OD, mg L-1); d : nível de saturação de oxigênio dissolvido (OD, %); f : clorofila a (µg L-1); e : feopigmentos (µg L-1).

40
0
5
10
15
20
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
a
Turb
idez
(NTU
)
EB PB
0
5
10
15
20
25
30
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
b
TPM
(mg
L-1)
EB PB
0
5
10
15
20
25
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
c
PIM
(mg
L-1)
EB PB
0
1
2
3
4
5
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
d
POM
(mg
L-1)
EB PB
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
e
f
EB PB
0
2
4
6
8
10
12
Jan/
01
Fev/
01
Mar
/01
Abr
/01
Mai
/01
Jun/
01
f
Cla
:Feo
p
EB PB
Figura 5: Variáveis hidrológicas (médias mensais ± desvio padrão) obtidas em monitoração semanal realizada em Enseada do Brito (EB) e em Porto Belo (PB). Parâmetros físico-químicos e seston (fração entre 1,2 e 230 µm). a : turbidez (NTU); b : matéria particulada total (TPM, mg L-1); c : matéria inorgânica particulada (PIM, mg L-1); d : matéria orgânica particulada (POM, mg L-1); e : fração orgânica do material em suspensão (f, POM:TPM); f : razão clorofila a / feopigmentos (Cla / Feop).

41
Comparando os locais (Tab. 4), constatamos que as variáveis mais importantes em
termos de distinção dos ambientes de cultivo são os parâmetros do seston, ou seja, turbidez,
TPM, PIM, POM, f, Cla e Feop. Diferenças significativas foram observadas quanto a
turbidez, que foi mais alta em Enseada do Brito em relação a Porto Belo (p<0,001). As
concentrações de TPM e PIM (p<0,001), POM (p<0,01) foram sempre mais elevadas em EB,
o mesmo ocorrendo em termos de pigmentos (p<0,001). Já a fração orgânica do seston,
representada pelo índice f, foi superior em PB (p<0,05).
Comparando as matrizes de correlação entre parâmetros do seston (Tabs. 5 e 6),
observamos tendências distintas entre os locais. Em EB houve alta correlação de POM com
TPM (r = 0,84; p<0,001) e PIM (r = 0,73; p<0,001), f apresentou correlação negativa com
PIM (r = - 0,54; p<0,05). Quanto aos pigmentos, Cla e Feop apresentaram correlação com
TPM (r = 0,51; p<0,05 e r = 0,67; p<0,01; respectivamente), PIM (r = 0,51; p<0,05 e r = 0,67;
p<0,01; respectivamente) e POM (r = 0,45 e 0,56; p<0,05). Em PB, POM apresentou
correlação com TPM (r = 0,62; p<0,01), f apresentou correlação negativa com PIM (r = -
0,57; p<0,01) e positiva com POM (r = 0,66; p<0,01), mas Cla e Feop não apresentaram
qualquer correlação com os demais parâmetros, a não ser entre si (Cla x Feop). Tabela 5: Coeficientes de correlação entre matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla) e feopigmentos (Feop) em Enseada do Brito.
Variáveis TPM PIM POM f Cla Feop
TPM 1
PIM 0,98*** 1
POM 0,84*** 0,73*** 1
f - 0,38 - 0,54* 0,18 1
Cla 0,51* 0,51* 0,45* - 0,19 1
Feop 0,67** 0,67** 0,56* - 0,30 0,79*** 1 * : p<0,05; ** : p<0,01; *** : p<0,001.

42
Tabela 6: Coeficientes de correlação entre matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM), fração orgânica do material em suspensão (f), clorofila a (Cla) e feopigmentos (Feop) em Porto Belo.
Variáveis TPM PIM POM f Cla Feop
TPM 1
PIM 0,90*** 1
POM 0,62** 0,25 1
f - 0,17 - 0,57** 0,66** 1
Cla 0,41 0,38 0,31 - 0,04 1
Feop 0,21 0,19 0,18 0,02 0,49* 1 * : p<0,05; ** : p<0,01; *** : p<0,001.
3.2. Experimentos de alimentação e de biodeposição
Variáveis físico-químicas
Os resultados das análises de variância e da comparação de médias dos parâmetros
físico-químicos durante os experimentos de alimentação e de biodeposição estão descritos na
Tabela 7. A comparação das médias destes parâmetros entre locais, com a utilização de Teste
t de Student, é apresentada na Tabela 8.
Tabela 7: Variáveis físico-químicas da água (médias ± desvio padrão) em Enseada do Brito e em Porto Belo durante os experimentos de alimentação e de biodeposição: temperatura (°C), salinidade (PSU), oxigênio dissolvido (OD, mg L-1) e turbidez (NTU). Experimento Temperatura Salinidade OD OD % Turbidez
EB1
EB2
EB3
PB1
PB2
PB3
25,8 ± 0,6bc
25,5 ± 0,5c
22,2 ± 0,3d
29,2 ± 0,3a
26,5 ± 0,4b
18,4 ± 0,1e
33,0 ± 0,1a
32,2 ± 0,2 b
29,0 ± 0,1e
30,8 ± 0,1c
30,3 ± 0,5d
31,2 ± 0,1c
-
7,0 ± 0,2a
6,8 ± 0,3a
-
6,4 ± 0,6ab
5,9 ± 0,3b
-
0,99 ± 0,01a
0,94 ± 0,04a
-
0,92 ± 0,08a
0,76 ± 0,04b
17,2 ± 7,3a
7,7 ± 1,8b
4,6 ± 1,7c
0,6 ± 0,2e
1,0 ± 0,2d
0,3 ± 0,1f
Letras (sobrescrito) diferentes indicam diferenças significativas entre experimentos (p<0,05).
Como observado nos dados obtidos na monitoração semanal, também não foi
detectada diferença estatística para a temperatura média da água durante os experimentos de
alimentação e biodeposição entre Porto Belo (PB) e em Enseada do Brito (EB). Já quando
comparados os experimentos entre si, a temperatura da água foi mais elevada durante o

43
experimento PB1 e menor em PB3 (p<0,05). Em EB1 a temperatura foi estatisticamente
similar a EB2 e a PB2 (p>0,05), porém mais elevada que em EB3 (p<0,05). Entretanto, deve-
se levar em conta que em PB1 o experimento foi realizado no início de fevereiro (alta estação
de verão), quando as temperaturas alcançam seu auge, ao passo que em EB1 o experimento
ocorreu no final da estação. Já os experimentos EB3 e PB3 foram realizados no final do
outono e início do inverno, respectivamente, quando as temperaturas da água já estão mais
frias. No caso dos experimentos EB1, PB2 e EB2, a realização dos experimentos ocorreu no
período de transição entre o final do verão e início do outono, em um intervalo de no máximo
um mês, explicando as temperaturas similares entre os três experimentos.
Tabela 8: Parâmetros de qualidade da água do mar das áreas de cultivo estudadas: comparação dos valores médios (médias ± desvio padrão) dos experimentos em Enseada do Brito (EB) e em Porto Belo (PB). Variáveis físico-químicas : temperatura, salinidade, oxigênio dissolvido (OD e OD%) e turbidez.
Variável EB (média ± desvpad) PB (média ± desvpad) Diferença entre Locais (médias experimentais)
Temperatura 24,7 ± 1,7 25,2 ± 4,5 ns
Salinidade 31,6 ± 1,7 30,5 ± 0,4 ns
OD 6,9 ± 0,2 6,2 ± 0,6 **
OD% 0,97 ± 0,04 0,85 ± 0,10 **
Turbidez 10,3 ± 7,2 0,6 ± 0,3 ***
TPM 18,0 ± 11,8 1,5 ± 0,4 ***
PIM 15,5 ± 9,4 1,0 ± 0,4 ***
POM 2,5 ± 2,9 0,5 ± 0,3 ***
f 0,12 ± 0,05 0,33 ± 0,15 ***
ns : diferença não significativa; * : p<0,05; ** : p<0,01; *** : p<0,001.
Também a salinidade média foi estatisticamante similar entre EB e PB, sendo que
entre os experimentos foi maior em EB1, seguido de EB2 e menor em EB3, com os
experimentos PB3, PB1 e PB2 apresentando valores intermediários (p<0,05), ao passo que
não houve diferença entre experimentos PB1 e PB3. As diferenças encontradas entre os
experimentos podem ser explicadas pela eventual interferência de fatores hidrológicos e
meteorológicos locais, como ocorrências de chuvas e alterações de vazão de rios e córregos
que deságuam nas proximidades das áreas de estudo (por exemplo: rios Tavares, Defuntos,
Massiambu, Brito e Cambirela em Enseada do Brito e rios Perequê e Porto Belo em Porto
Belo).

44
Quanto aos níveis de oxigênio dissolvido na água (OD% e OD em mg L-1), devido a
problemas com o equipamento, só foram realizadas análises durante os experimentos EB2,
EB3, PB2 e PB3. Para o oxigênio dissolvido (mg L-1), em EB2 e em EB3 as concentrações
foram mais elevadas em relação a PB3 (p<0,05), ficando PB2 num patamar intermediário
(p>0,05). Em termos de saturação de oxigênio dissolvido (%), PB3 foi o único experimento
distoante dos demais (p<0,05). Entretanto, todas as concentrações observadas são
consideradas adequadas para atender a demanda metabólica dos bivalvos cultivados.
Para turbidez (NTU), um indicador indireto de concentração do seston, verificou-se
diferença significativa entre todos os experimentos (p<0,05), onde os experimentos de
Enseada do Brito (EB1, EB2 e EB3) apresentaram níveis claramente superiores aos de Porto
Belo (PB1, PB2 e PB3).
Seston
Os resultados das análises de variância e da comparação de médias dos parâmetros do
seston durante os experimentos de alimentação e biodeposição estão descritos na Tabela 9,
com os dados de TPM, PIM, POM e f sendo apresentados graficamente nas Figuras 3.a e 4.b.
A comparação das médias destes parâmetros entre locais, com a utilização de Teste t de
Student, é apresentada na Tabela 10.
Tabela 9: Material em suspensão (fração entre 1,2 e 230 µm) em Enseada do Brito e em Porto Belo durante os experimentos de alimentação e biodeposição: médias ± desvio padrão de matéria particulada total (TPM, mg L-
1), matéria inorgânica particulada (PIM, mg L-1), matéria orgânica particulada (POM, mg L-1) e fração orgânica do material em suspensão (f). Experimento TPM PIM POM f EB1
EB2
EB3
PB1
PB2
PB3
29,6 ± 11,9a
12,5 ± 3,1b
9,9 ± 3,1b
1,7 ± 0,4c
1,6 ± 0,4cd
1,2 ± 0,3d
24,9 ± 9,2a
11,3 ± 2,9b
8,8 ± 3,0b
1,0 ± 0,4cd
1,3 ± 0,3c
0,8 ± 0,2d
4,8± 3,8a
1,2 ± 0,4b
1,0 ± 0,2bc
0,7 ± 0,3c
0,3 ± 0,2d
0,4 ± 0,1cd
0,15 ± 0,06bc
0,10 ± 0,02c
0,11 ± 0,03bc
0,41 ± 0,17a
0,20 ± 0,08b
0,36 ± 0,09a
Letras (sobrescrito) diferentes indicam diferenças significativas entre experimentos (p<0,05).
Comparando as concentrações médias de material em suspensão, em termos de
matéria particulada total (TPM), houve diferença significativa entre locais em ambos os
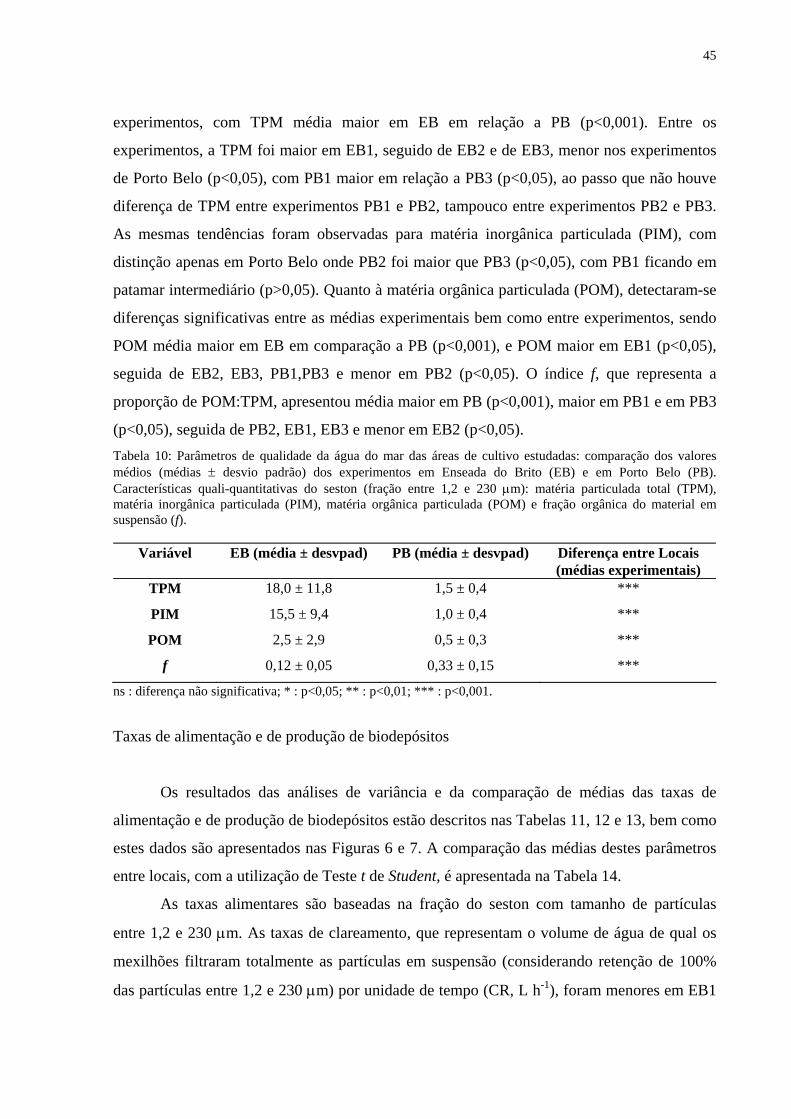
45
experimentos, com TPM média maior em EB em relação a PB (p<0,001). Entre os
experimentos, a TPM foi maior em EB1, seguido de EB2 e de EB3, menor nos experimentos
de Porto Belo (p<0,05), com PB1 maior em relação a PB3 (p<0,05), ao passo que não houve
diferença de TPM entre experimentos PB1 e PB2, tampouco entre experimentos PB2 e PB3.
As mesmas tendências foram observadas para matéria inorgânica particulada (PIM), com
distinção apenas em Porto Belo onde PB2 foi maior que PB3 (p<0,05), com PB1 ficando em
patamar intermediário (p>0,05). Quanto à matéria orgânica particulada (POM), detectaram-se
diferenças significativas entre as médias experimentais bem como entre experimentos, sendo
POM média maior em EB em comparação a PB (p<0,001), e POM maior em EB1 (p<0,05),
seguida de EB2, EB3, PB1,PB3 e menor em PB2 (p<0,05). O índice f, que representa a
proporção de POM:TPM, apresentou média maior em PB (p<0,001), maior em PB1 e em PB3
(p<0,05), seguida de PB2, EB1, EB3 e menor em EB2 (p<0,05). Tabela 10: Parâmetros de qualidade da água do mar das áreas de cultivo estudadas: comparação dos valores médios (médias ± desvio padrão) dos experimentos em Enseada do Brito (EB) e em Porto Belo (PB). Características quali-quantitativas do seston (fração entre 1,2 e 230 µm): matéria particulada total (TPM), matéria inorgânica particulada (PIM), matéria orgânica particulada (POM) e fração orgânica do material em suspensão (f).
Variável EB (média ± desvpad) PB (média ± desvpad) Diferença entre Locais (médias experimentais)
TPM 18,0 ± 11,8 1,5 ± 0,4 ***
PIM 15,5 ± 9,4 1,0 ± 0,4 ***
POM 2,5 ± 2,9 0,5 ± 0,3 ***
f 0,12 ± 0,05 0,33 ± 0,15 ***
ns : diferença não significativa; * : p<0,05; ** : p<0,01; *** : p<0,001.
Taxas de alimentação e de produção de biodepósitos
Os resultados das análises de variância e da comparação de médias das taxas de
alimentação e de produção de biodepósitos estão descritos nas Tabelas 11, 12 e 13, bem como
estes dados são apresentados nas Figuras 6 e 7. A comparação das médias destes parâmetros
entre locais, com a utilização de Teste t de Student, é apresentada na Tabela 14.
As taxas alimentares são baseadas na fração do seston com tamanho de partículas
entre 1,2 e 230 µm. As taxas de clareamento, que representam o volume de água de qual os
mexilhões filtraram totalmente as partículas em suspensão (considerando retenção de 100%
das partículas entre 1,2 e 230 µm) por unidade de tempo (CR, L h-1), foram menores em EB1

46
e em EB3 (p<0,05), intermediárias em EB2, PB1, PB2 e maiores em PB3 (p<0,05), esta
última estatisticamente similar a PB2, sendo que a CR média foi menor em EB (p<0,001).
Tabela 11: Taxas de alimentação (médias ± desvio padrão) do mexilhão Perna perna em Enseada do Brito (EB) e em Porto Belo (PB) durante cada experimento: CR = taxa de clareamento (L h-1); FR = taxa de filtração (mg h-
1); IR = taxa de ingestão (mg h-1); OIR = taxa de ingestão orgânica (mg h-1); AR = taxa de absorção (mg h-1).
Experimento CR FR IR OIR AR EB1
EB2
EB3
PB1
PB2
PB3
3,24 ± 0,83c
5,26 ± 1,71b
3,19 ± 0,51c
5,46 ± 1,50b
5,97 ± 1,66ab
8,35 ± 1,83a
87,86 ± 29,99a
65,52 ± 21,19a
26,29 ± 4,04b
9,64 ± 2,75c
9,63 ± 2,63c
9,16 ± 2,12c
22,16 ± 5,26a
13,55 ± 5,62b
9,05 ± 1,36b
8,76 ± 2,36bc
5,85 ± 1,69c
8,20 ± 1,66bc
11,28 ± 2,65a
2,65 ± 0,99c
1,86 ± 0,34c
4,42 ± 1,29b
1,76 ± 0,57c
3,64 ± 0,76bc
10,31 ± 2,53a
1,46 ± 0,62c
0,89 ± 0,22c
3,59 ± 1,11b
1,05 ± 0,52c
2,98 ± 0,81b
Letras (sobrescrito) diferentes indicam diferenças significativas entre experimentos (p<0,05).
Tabela 12: Taxas de alimentação (médias ± desvio padrão) do mexilhão Perna perna em Enseada do Brito (EB) e em Porto Belo (PB) durante cada experimento: SE = eficiência de seleção; i = fração orgânica do material ingerido; AE = eficiência de absorção.
Experimento SE i AE EB1
EB2
EB3
PB1
PB2
PB3
0,52 ± 0,03ab
0,18 ± 0,03c
0,26 ± 0,05bc
0,68 ± 0,06a
0,42 ± 0,18b
0,61 ± 0,31ab
0,47 ± 0,06a
0,19 ± 0,03c
0,20 ± 0,03c
0,50 ± 0,02a
0,30 ± 0,06b
0,44 ± 0,02a
0,86 ± 0,04a
0,41 ± 0,07c
0,47 ± 0,05bc
0,79 ± 0,02a
0,55 ± 0,10b
0,80 ± 0,07a
Letras (sobrescrito) diferentes indicam diferenças significativas entre experimentos (p<0,05).
Tabela 13: Taxas de produção de biodepósitos (médias ± desvio padrão) do mexilhão Perna perna em Enseada do Brito (EB) e em Porto Belo (PB) durante cada experimento: RR = taxa de rejeição (mg h-1); ER = taxa de egestão (mg h-1); BR = taxa de biodeposição (mg h-1); razão RR/BR e razão ER/BR.
Experimento RR ER BR RR/BR ER/BR EB1
EB2
EB3
PB1
PB2
PB3
69,71 ± 20,10a
51,97 ± 16,43a
17,24 ± 3,98b
0,88 ± 0,49d
3,78 ± 1,80c
0,96 ± 0,57d
11,84 ± 3,74a
12,09 ± 5,07a
8,16 ± 1,26ab
5,17 ± 1,30b
4,80 ± 1,46b
5,36 ± 0,75b
81,55 ± 21,26a
64,06 ± 20,58a
25,40 ± 3,88b
6,05 ± 1,67c
8,58 ± 2,14c
6,22 ± 1,28c
0,85 ± 0,06a
0,81 ± 0,04a
0,67 ± 0,07b
0,14 ± 0,06d
0,43 ± 0,14c
0,15 ± 0,07d
0,15 ± 0,06c
0,19 ± 0,04c
0,33 ± 0,07c
0,86 ± 0,06a
0,57 ± 0,14b
0,87 ± 0,06a
Letras (sobrescrito) diferentes indicam diferenças significativas entre experimentos (p<0,05). As taxas de filtração (FR, mg h-1) apresentaram média maior em EB (p<0,001), sendo
maiores em EB1 e EB2, seguidas de EB3, e menores em PB1, PB2 e PB3 (p<0,05). A taxa de

47
ingestão (IR, mg h-1) foi maior em EB1 e menor em PB2 (p<0,05), sendo IR média maior em
EB que em PB (p<0,001). Já a taxa de ingestão orgânica (OIR, mg h-1) foi maior em EB1
seguida de PB1 e de PB3 (p<0,05), enquanto que entre EB2, EB3 e PB2, com as menores
médias, não houve diferença significativa (p>0,05), mas a OIR média foi maior em EB que
em PB (p<0,05).
As taxas de absorção (AR, mg h-1), que correspondem ao alimento que é efetivamente
convertido para produção de tecido somático e reprodutivo, apresentaram tendências similares
àquelas observadas para OIR, ou seja, AR foi maior em EB1 seguida de PB1 seguida de PB1
e de PB3 (p<0,05), enquanto que entre EB2, EB3 e PB2, com as menores médias, não houve
diferença significativa (p>0,05), mas não foi detectada diferença estatística para a AR média
entre EB e PB (p>0,05).
A eficiência de seleção (SE), um índice representativo da competência dos bivalves
para selecionar e ingerir as partículas orgânicas que compõem o seston, mostrou variação
tanto entre as médias locais como entre os experimentos. A SE foi maior em PB1 e menor em
EB2 (p<0,05), e SE média foi maior em PB do que em EB (p<0,001), seguindo tendência
similar àquela observada para f.
A fração orgânica do material ingerido (i) variou entre os experimentos, sendo maior
para PB1, EB1 e PB3, seguida de PB2, e menor em EB3 e EB2 (p<0,05). A i média foi maior
em PB do que em EB (p<0,001). Em termos proporcionais, a fração de matéria orgânica
particulada absorvida em relação à ingerida (AR:OIR, eficiência de absorção, AE) variou
entre experimentos mas não entre as médias locais. AE foi maior em EB1, em PB3 e em PB1,
seguida de PB2, de EB3 e menor em EB2 (p<0,05).

48
a
0
15
30
45
PB1 PB2 PB3 EB1 EB2 EB3
mg
L-1
TPM PIM POM
b
0
2
4
6
8
10
12
PB1 PB2 PB3 EB1 EB2 EB3
CR
(L h
-1)
c
0
20
40
60
80
100
120
PB1 PB2 PB3 EB1 EB2 EB3
mg
h-1
BR RR ER
d
0
0,05
0,1
0,15
0,2
0,25
PB1 PB2 PB3 EB1 EB2 EB3b p e
Figura 6: Material em suspensão (fração entre 1,2 e 230 µm; mg L-1) e taxas fisiológicas (alimentação e biodeposição; mg h-1) do mexilhão Perna perna em dois locais de cultivo e em três experimentos por local (médias ± desvio padrão). Porto Belo: PB1 = Experimento 1; PB2 = Experimento 2; PB3 = Experimento 3; Enseada do Brito: EB1 = Experimento 1; EB2 = Experimento 2; EB3 = Experimento 3. a : matéria particulada em suspensão (TPM), matéria inorgânica particulada (PIM) e matéria orgânica particulada (POM); b : taxa de clareamento (CR, L h-1); c : taxas de biodeposição (BR), rejeição (RR) e egestão (ER); d : fração orgânica dos biodepósitos (b), das pseudofezes (p) e das fezes (e).

49
a
0
20
40
60
80
100
120
140
PB1 PB2 PB3 EB1 EB2 EB3
mg
h-1
FR IR
b
0,0
0,2
0,4
0,6
0,8
1,0
PB1 PB2 PB3 EB1 EB2 EB3f SE
c
0
5
10
15
PB1 PB2 PB3 EB1 EB2 EB3
mg
h-1
OIR AR
d
0,0
0,2
0,4
0,6
0,8
1,0
PB1 PB2 PB3 EB1 EB2 EB3i AE
Figura 7: Alimentação e produção de biodepósitos do mexilhão Perna perna em dois locais de cultivo e em três experimentos por local (médias ± desvio padrão). Porto Belo: PB1 = Experimento 1; PB2 = Experimento 2; PB3 = Experimento 3; Enseada do Brito: EB1 = Experimento 1; EB2 = Experimento 2; EB3 = Experimento 3. a : taxas de filtração (FR) e de ingestão (IR); b : fração orgânica do seston (f) e eficiência de seleção (SE); c : taxas de ingestão orgânica (OIR) e de absorção (AR); d : fração orgânica do material ingerido (i) e eficiência de absorção (AE).
As taxas de rejeição (RR, mg h-1), que representam a quantidade de partículas
rejeitadas na forma de pseudofezes, foram maiores em EB1 e EB2, seguidas de EB3, de PB2 e
menores em PB3 e em PB1 (p<0,05), com RR média maior em EB (p<0,001). As taxas de
egestão (ER, mg h-1), que correspondem as partículas ingeridas eliminadas nas fezes, foram
sempre maiores em Enseada do Brito e menores em Porto Belo, não havendo diferença entre

50
experimentos para ambos os locais (EB1 = EB2 = EB3 e PB1 = PB2 = PB3),
conseqüentemente o valor da ER média foi maior em EB (p<0,001). Tabela 14: Taxas de alimentação e de produção de biodepósitos do mexilhão Perna perna. Comparação dos valores médios (médias ± desvio padrão) dos experimentos em Enseada do Brito (EB) e em Porto Belo (PB): CR = taxa de clareamento (L h-1); FR = taxa de filtração (mg h-1); IR = taxa de ingestão (mg h-1); OIR = taxa de ingestão orgânica (mg h-1); AR = taxa de absorção (mg h-1); SE = eficiência de seleção; i = fração orgânica do material ingerido; AE = eficiência de absorção; RR = taxa de rejeição (mg h-1); ER = taxa de egestão (mg h-1); BR = taxa de biodeposição (mg h-1); razão RR/BR e razão ER/BR; p = fração orgânica das pseudofezes; e = fração orgânica das fezes; b = fração orgânica dos biodepósitos.
Variável EB (média ± desvpad) PB (média ± desvpad) Diferença entre Locais (médias experimentais)
CR 3,92 ± 1,50 6,64 ± 2,06 ***
FR 66,13 ± 33,18 9,47 ± 2,41 ***
IR 16,21 ± 7,22 7,56 ± 2,24 ***
OIR 6,17 ± 4,85 3,23 ± 1,43 *
AR 5,12 ± 4,86 2,50 ± 1,36 ns
SE 0,34 ± 0,16 0,57 ± 0,23 ***
i 0,32 ± 0,15 0,41 ± 0,09 **
AE 0,62 ± 0,22 0,71 ± 0,14 ns
RR 51,63 ± 26,07 1,91 ± 1,77 ***
ER 11,09 ± 4,13 5,11 ± 1,18 ***
BR 62,72 ± 28,48 6,99 ± 2,05 ***
RR/BR 0,80 ± 0,09 0,24 ± 0,17 ***
ER/BR 0,20 ± 0,09 0,76 ± 0,17 ***
p 0,08 ± 0,01 0,13 ± 0,06 ***
e 0,10 ± 0,02 0,15 ± 0,03 ***
b 0,08 ± 0,01 0,14 ± 0,03 ***
ns : diferença não significativa; * : p<0,05; ** : p<0,01; *** : p<0,001.
As taxas de biodeposição (BR, mg h-1), que correspondem à soma de RR e ER,
apresentaram tendências similares às observadas para ER, ou seja, foram maiores em Enseada
do Brito em ambos os experimentos em relação a Porto Belo (p<0,05), com EB1 igual a EB2
e PB1 igual a PB2 e a PB3, com EB3 apresentando média intermediária, conseqüentemente a
BR média foi maior em EB (p<0,001). Entretanto, as contribuições relativas de fezes e
pseudofezes em termos de biodepósitos variaram marcadamente, especialmente entre os
locais (Fig. 3c). Na Enseada do Brito, as RRs corresponderam com mais de 60% das BRs

51
(85%, 81% e 67% em EB1, EB2 e EB3, respectivamente). Em Porto Belo, esta proporção foi
inferior às ERs (14%, 43% e 15% em PB1, PB2 e PB3, respectivamente). As frações
orgânicas médias das pseudofezes, fezes e biodepósitos (p, e e b, respectivamente) foram
maiores em Porto Belo (p<0,001). A mesma tendência foi observada para a fração p, onde
PB1 e PB2 foram estatisticamente maiores que EB1, EB2 e EB3 (p<0,05), com PB3 sendo a
média intermediária (p>0,05). A fração e foi maior em PB1 e PB2, seguida de PB3, EB3 e
EB2, e menor em EB1 (p<0,05), e b foi maior em PB1, seguido de PB2 e PB3, menor em
EB1, EB2 e EB3 (p<0,05).
As Figuras 8 a 13 apresentam os gráficos de correlação entre as variáveis ambientais e
as taxas e índices fisiológicos, bem como os níveis de correlação entre as diferentes taxas e
índices fisiológicos observados conjuntamento nos exeperimentos realizados em Porto Belo e
em Enseada do Brito.
As taxas de clareamento (CR) apresentaram correlação negativa em relação às
concentrações de matéria particulada em suspensão (TPM, r = - 0,70), matéria inorgânica
particulada (PIM, r = - 0,71) e matéria orgânica particulada (POM, r = - 0,61), correlação
positiva em relação ao teor de matéria orgânica do seston (f, r = 0,65), correlação negativa em
relação à temperatura da água (r = - 0,39) e correlação nula com a salinidade da água (r =
0,00), de acordo com os gráficos apresentados na Figura 8. Entretanto, cabe frisar que
estatisticamente nenhuma destas correlações foi significativa (p>0,05).
De acordo com os gráficos apresentados na Figura 9, as taxas de biodeposição (BR)
apresentaram forte correlação positiva em relação às concentrações de matéria particulada em
suspensão (TPM, r = 0,93; p<0,01), matéria inorgânica particulada (PIM, r = 0,94; p<0,01) e
matéria orgânica particulada (POM, r = 0,84; p<0,05), correlação negativa em relação ao teor
de matéria orgânica do seston (f, r = - 0,67), fraca correlação positiva em relação à
temperatura da água (r = 0,16) e correlação positiva com a salinidade da água (r = 0,72),
frisando que as três últimas correlações não apresentam significância estatística (p>0,05).

52
Figura 8: Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo e Enseada do Brito) durante três experimentos por local. a : taxa de clareamento (CR) e matéria particulada em suspensão (TPM); b : taxa de clareamento (CR) e fração orgânica do seston (f); c : taxa de clareamento (CR) e matéria inorgânica particulada (PIM); d : taxa de clareamento (CR) e matéria orgânica particulada (POM); e : taxa de clareamento (CR) e temperatura; f : taxa de clareamento (CR) e salinidade.

53
Figura 9: Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo e Enseada do Brito) durante três experimentos por local. a : taxa de biodeposição (BR) e matéria particulada em suspensão (TPM); b : taxa de biodeposição (BR) e fração orgânica do seston (f); c : taxa de biodeposição (BR) e matéria inorgânica particulada (PIM); d : taxa de biodeposição (BR) e matéria orgânica particulada (POM); e : taxa de biodeposição (BR) e temperatura; f : taxa de biodeposição (BR) e salinidade.

54
Figura 10: Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo e Enseada do Brito) durante três experimentos por local. a : taxa de filtração (FR) e matéria particulada em suspensão (TPM); b : taxa de ingestão (IR) e taxa de filtração (FR); c : taxa de rejeição (RR) e matéria particulada em suspensão (TPM); d : taxa de ingestão (IR) e taxa de rejeição (RR); e : taxa de egestão (ER) e matéria particulada em suspensão (TPM); f : taxa de ingestão (IR) e taxa de egestão (ER).

55
Figura 11: Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo - PB e Enseada do Brito - EB) durante três experimentos por local (EB1, EB2, EB3, PB1, PB2, PB3). a : eficiência de seleção (SE) e matéria particulada em suspensão (TPM); b : eficiência de seleção (SE) e matéria orgânica particulada (POM); c : eficiência de seleção (SE) e fração orgânica do seston (f); d : eficiência de seleção (SE) e taxa de filtração (FR); e : fração orgânica ingerida (i) e eficiência de seleção (SE); f : eficiência de absorção (AE) e eficiência de seleção (SE).

56
Figura 12: Correlações entre taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo - PB e Enseada do Brito - EB) durante três experimentos por local (EB1, EB2, EB3, PB1, PB2, PB3). a : taxa de ingestão orgânica (OIR) e matéria orgânica particulada (POM); b : fração orgânica ingerida (i) e fração orgânica do seston (f); c : taxa de ingestão orgânica (OIR) e taxa de filtração (FR); d : eficiência de absorção (AE) e fração orgânica do seston (f); e : taxa de ingestão orgânica (OIR) e taxa de ingestão (IR); f : eficiência de absorção (AE) e fração orgânica ingerida (i).

57
Figura 13: Correlações entre variáveis ambientais e taxas fisiológicas do mexilhão Perna perna em dois locais de cultivo (Porto Belo - PB e Enseada do Brito - EB) durante três experimentos por local (EB1, EB2, EB3, PB1, PB2, PB3). a : taxa de absorção (AR) e matéria particulada total (TPM); b : taxa de absorção (AR) e taxa de filtração (FR); c : taxa de absorção (AR) e matéria orgânica particulada (POM); d : taxa de absorção (AR) e taxa de ingestão (IR); e : taxa de absorção (AR) e fração orgânica do seston (f); f : taxa de absorção (AR) e fração orgânica ingerida (i).

58
As taxas de filtração (FR), rejeição (RR) e egestão (ER) apresentaram forte correlação
positiva com as concentrações de matéria particulada da água (TPM), respectivamente r =
0,94 (p<0,01), r = 0,94 (p<0,01) e r = 0,86 (p<0,05). As taxas de ingestão (IR) apresentaram
forte correlação positiva em relação às taxas de filtração (FR, r = 0,94, p<0,01), rejeição (RR,
r = 0,93, p<0,01) e egestão (ER, r = 0,86, p<0,05), conforme verificamos nos gráficos da
Figura 10.
A eficiência de seleção (SE) apresentou correlação positiva significativa apenas com o
teor orgânico do seston (f, r = 0,89, p<0,01), o que não ficou evidenciado em relação aos
demais parâmetros (FR, TPM e POM). Já a fração orgânica ingerida (i) e a eficiência de
absorção (AE) apresentaram correlação positiva significativa com SE (r = 0,96, p<0,01 e r =
0,91, p<0,01, respectivamente), conforme observado na Figura 11.
Como observado na Figura 12, as taxas de ingestão orgânica (OIR) apresentaram forte
correlação positiva com a concentração de matéria orgânica particulada (POM, r = (0,93,
p>0,01), com a taxa de ingestão (IR, r = 0,88, p<0,01) e correlação sem significância
estatística com a taxa de filtração (FR, r = 0,69, p>0,05). Já para a fração orgânica ingerida
(i), houve correlação significativa com f (r = 0,76, p<0,05). A eficiência de absorção (AE)
apresentou correlação positiva significativa com a fração orgânica de ingestão (i, r = 0,98,
p<0,01) e não significativa positiva com o a fração orgânica do seston (f, r = 0,65, p>0,05).
Já as taxas de absorção (AR), de acordo com a Figura 13, apresentaram correlações
positivas significativas com as concentrações de matéria orgânica particulada da água (POM,
r = 0,91, p<0,01) e com a taxa de ingestão (IR, r = 0,86, p<0,01), e correlações positivas não
significativas com TPM (r = 0,78, p>0,05), FR (r = 0,65, p>0,05) e i (r = 0,66, p>0,05).
4. Discussão
A partir dos parâmetros hidrológicos obtidos no monitoramento semanal, podemos
caracterizar as áreas de estudo como bastante distintas com relação às características do
seston: a Enseada do Brito, área com mais elevada concentração de material em suspensão
(concentrações médias mensais de TPM em torno 10 mg L-1, POM em torno de 1,5 mg L-1, f
de aproximadamente 0,2, Cla em torno de 2,7 µg L-1), mais rasa (profundidade média em
torno de 2 metros) e mais suscetível à ressuspensão de partículas do fundo (constituído de
sedimentos finos), principalmente por ação de ondas e ventos. Já Porto Belo apresenta
menores concentrações de seston (concentrações médias mensais de TPM em torno de 3 mg

59
L-1, POM em torno de 0,8 mg L-1, f de aproximadamente 0,3, Cla em torno de 1,2 µg L-1),
mais profunda (profundidade média em torno de 4 metros) e fundo com sedimento de maior
granulação (arenoso), onde a remobilização de material do fundo pode ocorrer
esporadicamente por atuação de ondas oceânicas (“swell”) mas em menor intensidade.
Na Enseada do Brito (EB), as correlações positivas entre POM, TPM e PIM, e a
correlação negativa de f com PIM sugerem que, tanto as partículas inorgânicas quanto as
orgânicas podem ser de originadas da ressuspensão do material de fundo e que o teor orgânico
no seston (f) é altamente dependente da diluição gerada pelo aumento de PIM, entretanto esta
hipótese necessita ser confirmada, pois também pode haver importante contribuição dos rios
que deságuam na região.
A ressuspensão pode causar dramáticas alterações no seston de áreas costeiras,
promovendo o aporte de uma ampla gama de partículas, como sedimentos inorgânicos,
microalgas bentônicas, pelotas fecais, bactérias e detritos orgânicos de várias origens
(fragmentos de macroalgas e macrófitas, etc.), interferindo nas características nutricionais do
material em suspensão e, portanto, na alimentação de organismos filtradores (Fréchette &
Grant, 1991). Considerando as características ambientais de EB, é possível que sua maior
biomassa fitoplanctônica, indicada pelas mais elevadas concentrações de Cla, resulte da
ressuspensão do microfitobentos do fundo, tendo em vista a correlação positiva encontrada
entre Cla e o material em suspensão (TPM, PIM e POM). As microalgas bentônicas, além de
constituírem-se num importante recurso em termos de biomassa e de produção primária,
também contribuem com a produção de oxigênio, especialmente na interface água-sedimento
(Barranguet, 1997). Quanto aos feopigmentos (Feop), seus níveis podem estar refletindo tanto
o efeito da pastagem dos bivalvos e/ou zooplâncton sobre as microalgas como a ressuspensão
do material do fundo (Hawkins et al., 1986; Navarro et al. 1993; Navarro & Jaramillo, 1994;
Barranguet, 1997; Proença & Schettini, 1998).
Em Porto Belo (PB), f também apresentou correlação negativa com PIM, e positiva
com POM, indicando que as variações simultâneas nas concentrações das partículas
inorgânicas e orgânicas regulam a qualidade do material em suspensão. A inexistência de
correlação de Cla e Feop com os demais parâmetros do seston (principalmente POM) indica
que as contribuições em termos de matéria orgânica em suspensão neste local podem estar
sendo reguladas por outros compostos orgânicos em PB, como detritos, microrganismos
heterotróficos, etc.

60
Em termos de alimentação de bivalves, ainda pouco se sabe sobre suas exigências
nutricionais mínimas, especialmente em condições naturais, sendo que só recentemente estão
sendo publicados alguns trabalhos relacionando dados de crescimento e fisiologia energética
(“scope for growth”) com concentrações naturais de POM, Cla, carbono e nitrogênio
particulados (COP e NOP), como nos trabalhos de Hickman et al. (1991); Navarro et al.
(1991); Navarro & Thompson (1996); Hawkins et al. (1999); Gardner (2000); Wong &
Cheung (2001a).
Na Nova Zelândia, Hawkins et al., 1999 avaliaram o comportamento alimentar de
Perna canaliculus, e observaram que o principal fator condicionando o crescimento deste
bivalve é a biomassa fitoplanctônica, medida através dos níveis de Cla. Segundo estes
autores, o período entre 1996 e 1998 quando houve quedas de produção desta espécie nas
áreas de cultivo neozelandesas, por baixa taxa de crescimento e redução nos índices de
condição, estariam relacionadas às baixas concentrações de Cla na água, inferiores a 1 µg L-1.
Hickman et al. (1991) ainda sugeriram que em concentrações de COP inferiores a 200 µg L-1,
Perna canaliculus apresentaria quedas de índice de condição, refletindo em perdas de
produtividade dos mexihões cultivados em Pelorus - Kenepuru Sound. Smaal & van Stralen
(1990) observaram uma forte correlação entre as médias anuais de índice de condição de
Mytilus edulis e a produção primária média anual (g C m -2 ano -1) no estuário de Eastern
Scheldt (Holanda), mas não houve relação do índice de condição com a biomassa
fitoplanctônica (Cla, mg m -3). A partir destes dados os autores sugeriram haver uma relação
direta entre a produção primária local e o crescimento dos mexilhões.
Em outro estudo, Gardner (2000) comparou a fisiologia energética de três mitilídios,
Aulacomya maoriana, Mytilus galloprovincialis e Perna canaliculus, em dois locais no litoral
da Nova Zelândia (Mahanga Bay e Island Bay), e obteve valores negativos de “scope for
growth” (SFG) em condições onde as concentrações de POM foram inferiores a 3 mg L-1 e o
teor orgânico foi inferior ou igual a 30% da TPM (f ≤ 0,30). No Canadá (Logy Bay),
Modiolus modiolus apresentou SFGs negativos quando as concentrações de POM foram
inferiores a 0,70 mg L-1, mesmo correspondendo a um percentual orgânico em torno de 70 %
(f ≈ 0,70) (Navarro & Thompson, 1996). Para Perna viridis em Kat O, Hong Kong, o SFG foi
próximo de zero quando POM ficou em torno de 2 mg L-1 e f em aproximadamente 0,4
(Wong & Cheung, 2001a). Já Navarro et al. (1991), que avaliaram a fisiologia energética de
mexilhões Mytilus galloprovincialis cultivados nas rías galegas (Ria de Arosa, Espanha)
obtiveram valores positivos de SFG (embora relativamente baixos) com concentrações de

61
POM inferiores a 1 mg L-1, embora em uma fração orgânica maior, variando em torno de 50%
(f = 0,50).
Estas diferenças podem estar refletindo a adaptação das diferentes espécies sob
distintas condições ambientais (Hawkins & Bayne, 1992), mas devemos considerar que em
termos nutricionais, a composição do seston deve ser analisada não somente quanto à fração
orgânica e seu peso seco, mas também suas características em termos de número e volume de
partículas, concentração de pigmentos (clorofila, feopigmentos), composição química (níveis
de carbono e nitrogênio particulados) e bioquímica (carboidratos, lipídios e proteínas), uma
vez que estes constituintes podem variar dependendo da origem desta matéria orgânica, que
pode ser detrítica (material em decomposição, bactérias) e/ou constituída de células vivas do
fitoplâncton ou do microfitobentos, variando seu valor nutricional de acordo com sua
digestibilidade e teor energético, entre outros fatores (Navarro et al. 1993; Navarro &
Jaramillo, 1994; Danovaro & Fabiano, 1997).
Outra questão é a composição do plâncton, uma vez que os bivalves filtradores
apresentam capacidade de seleção entre diferentes espécies (em função do tamanho, forma,
etc.), ingerindo preferencialmente aquelas que são passíveis de absorção em quantidades
significativas, adaptando-se à oferta alimentar disponível em cada ambiente (fitoplâncton,
bacterioplâncton, detritos) (Shumway et al., 1985; Navarro & Thompson, 1997; Newell et al.,
1989; Berry & Schleyer, 1983). Sendo assim, análises quali-quantitativas do plâncton também
são importantes para se determinar qual a disponibilidade alimentar das áreas onde os
moluscos são cultivados.
Embora se considere que partículas com tamanho inferior a 1 µm dificilmente são
retidas por bivalves filtradores adultos (Jørgensen, 1990), Berry & Schleyer (op. cit.)
verificaram que exemplares de Perna perna (45 a 95 mm de comprimento) foram capazes de
reter partículas de até 0,46 µm em seu sistema filtrador, com uma eficiência de retenção
variando entre 11 e 64%. Portanto, deve-se ressaltar que em nossos experimentos as partículas
componentes do bacterioplâncton (de tamanho inferior a 1 µm) foram subestimadas em
nossas análises de seston e de biodepósitos devido às características dos filtros utilizados
(porosidade de 1,2 µm).
Nos bivalves, as taxas alimentação e biodeposição podem ser adversamente afetadas
por fatores abióticos tais como temperatura da água (Tsuchyia, 1980) e baixas salinidades,
que resultam em fechamento das valvas (Shumway, 1977), reduzindo a atividade alimentar
(Bohle, 1972; Navarro, 1988) e, conseqüentemente, diminuindo as taxas de biodeposição

62
(Tsuchyia, op. cit.). Por outro lado, altas concentrações de alimento podem incrementar a
suspensivoria (filtração) e portanto as taxas de bideposição (Tenore & Dunstan, 1973a,
1973b).
O mexilhão Perna perna é uma espécie eurihalina, sobrevivendo a salinidades
variando entre 19 e 49 psu (ou partes por mil) (Salomão et al., 1980), com salinidades letais
inferior e superior de 8 e 54 psu respectivamente (Segnini de Bravo et al., 1998), sendo sua
faixa ótima situada entre 34 e 36 psu (Rafael, 1978; Salomão & Magalhães, 1978). Esta
espécie também é euritérmica, tolerando uma variação entre 5 e 30 °C (Zuim & Mendes,
1977), com temperaturas letais inferior e superior de 3 e 34,5 °C respectivamente (Segnini de
Bravo et al., op. cit.) e faixa ótima entre 21 e 28 °C (Vélez & Epifanio, 1981).
No caso do presente estudo, embora a salinidade da água tenha variado
significativamente entre os experimentos, esta variação não parece apresentar uma
significância fisiológica, uma vez que os valores são normalmente superiores a 30 psu, ou
seja, bem próximos da faixa ótima para P. perna, e com diferenças predominantemente
inferiores a 4 psu. Também neste estudo foi constatado que não houve correlação entre
salinidade e taxa de clareamento (CR). Para a taxa de biodeposição (BR) houve correlação
positiva, mas não significativa. Quanto à temperatura da água, esta oscilou
predominantemente em acima dos 20 °C e abaixo dos 30 °C (a maior diferença entre a maior
e a menor temperatura dos experimentos foi de aproximadamente 10 °C). Também no
presente estudo não foi evidenciada correlação entre alimentação (CR) e biodeposição (BR) e
a temperatura. Outros trabalhos avaliando as variações sazonais das taxas de alimentação não
evidenciaram relação com a variação na temperatura (ou esta relação foi fraca), em função
dos bivalves apresentarem independência térmica na alimentação, devido à sua capacidade de
aclimatação (Widdows, 1978; Bayne & Newell, 1983; Jørgensen et al., 1990; Riisgård, 1991;
Navarro & Thompson, 1996; Smaal et al., 1997; Cranford & Hill, 1999).
Portanto, os parâmetros que parecem caracterizar maior influência sobre as taxas de
alimentação e biodeposição são os quali-quantitativos do seston (TPM, PIM, POM e f), a
exemplo do que foi observado por outros autores (Bayne & Widdows, 1978; Widdows, 1978;
Widdows et al., 1979; Prins et al., 1994; Navarro & Thompson, 1996, 1997; Gardner, 2002).
Isto fica evidente quando comparamos a variação das CRs e BRs com relação às
alterações de concentração de TPM, PIM, POM e de f, onde as CRs apresentam tendência de
aumento em função da menor disponibilidade de matéria em suspensão, a despeito da queda
nas temperaturas da água, especialmente no caso de Porto Belo, e as BRs aumentam em forte

63
relação direta com as concentrações do seston (TPM, PIM e POM) e sem relação aparente
com a temperatura da água.
Comparando os dois locais, podemos observar algumas tendências: o POM aumenta
com a TPM, ao mesmo tempo em que a fração orgânica (f) diminui. Assim, EB que apresenta
níveis mais altos de material em suspensão, proporciona maior quantidade de alimento (POM)
que PB, mas a matéria orgânica em PB sofre menor diluição (maior f).
Conforme aumenta a concentração de alimento, os bivalves tendem a diminuir o fluxo
de água com partículas em suspensão através de seu sistema filtrador, ou seja, diminuem a CR
(Bayne & Widdows, 1978; Widdows et al., 1979), daí as menores CRs em EB.
Em experimentos realizados no final da primavera e início do verão na Enseada de
Sambaqui, na Baía Norte da Ilha de Santa Catarina, Mello (1999) estudou a alimentação e
biodeposição de P. perna sob condições ambientais, e verificou que a CR apresentou
correlação positiva com POM (r = 0,88), mas principalmente com f (r = 0,99). No presente
estudo, a tendência de aumento de CR com f também foi observada, mas não foi tão
claramente pronunciada (r = 0,65). Já em relação às concentrações do seston, a CR apresentou
tendência de relação inversa com TPM (r = - 0,70), PIM (r = - 0,71) e POM (r = - 61). Estas
tendências ficaram evidentes graficamente comparando os dois locais, mas não ficaram tão
claras quando comparamos os experimentos em cada local.
De acordo com a literatura, os mitilídeos apresentam uma variedade de respostas
fisiológicas distintas em termos de CR em relação à variações no seston sob condições de
alimentação natural. Na Nova Zelândia, Gardner (2002) estudou os efeitos da variabilidade do
seston sobre as taxas de clareamento e eficiências de absorção em Aulacomya maoriana,
Mytilus galloprovincialis e Perna canaliculus, e observou que as CRs eram principalmente
explicadas por variações em POM e f. Comparando a fisiologia energética destas mesmas
espécies em relação às características do seston de duas baías neozelandesas, Gardner (2000)
já havia observado esta tendência, onde as CRs mais altas foram obtidas nas condições onde
as concentrações de POM eram mais elevadas.
Em populações de mexilhões Mytilus edulis cultivados em duas lagunas costeiras
escocesas (Loch Etive e Loch Leven), Okumus e Stirling (1994) verificaram uma correlação
positiva significativa entre CR e POM, sendo as CRs mais rápidas obtidas onde o índice f foi
mais alto e o TPM mais baixo. Já Hawkins et al. (1996) observaram que a CR de M. edulis
apresenta relação positiva com TPM, independentemente de f, enquanto que Hawkins et al.
(1997) verificaram uma relação inversa entre CR e POM para esta mesma espécie. Para Perna

64
viridis a CR apresenta relação inversa com POM (Hawkins et al., 1998), também com TPM e
relação positiva com f (Wong & Cheung, 2001b). Hawkins et al. (1999) também verificaram
relação inversa entre CR e POM para P. canaliculus, entretanto a combinação das variações
de Cla e de TPM explicaram melhor as oscilações de CR em relação a às alterações de POM
isoladamente.
Ao mesmo tempo em que aumenta a carga de partículas filtradas conforme incrementa
a concentração no seston, ou seja, FR aumenta com TPM, o sistema de seleção pré-ingestiva
atua no sentido de não colapsar o sistema digestivo por repleção da sua capacidade de
processamento, que é limitada quanto ao volume de partículas que podem ser processadas,
através de rejeição de parte do material filtrado na forma de pseudofezes. Por isso, RR
aumenta com FR (tendência também obsevada por Mello, 1999). É o que pudemos verificar
em nossos experimentos com P. perna, onde as maiores FRs e RRs foram observadas nas
condições de maior turbidez, e em Enseada do Brito em relação a Porto Belo, assim como
ocorre em outras espécies de bivalves, tanto em ambientes de clima temperado como de
regiões tropicais (Hawkins et al, 1996, 1998a, 1998b).
As taxas de ingestão (IRs) mantiveram tendência de aumento conforme aumentaram
as FRs, mesmo com o incremento das RRs, a exemplo do que observaram Mello (1999) para
esta espécie (Perna perna), Hawkins et al. (1996, 1998a) em Mytilus edulis e Hawkins et al.
(1999) para Perna canaliculus, e não estabilizaram a partir do início de produção de
pseudofezes como preconizavam Winter (1978), Widdows et al. (1979), Bayne & Hawkins
(1990), quando supostamente ocorreria a repleção do trato digestivo.
Segundo Hawkins et al, (1998a), a eficiência de seleção (SE) tende a aumentar com
FR, isto é, aumenta a proporção de partículas orgânicas seletivamente ingeridas em relação às
inorgânicas que são preferencialmente rejeitadas (Hawkins & Bayne, 1990). Mello (op. cit.)
também obteve relação positiva entre SE e FR sob condições de alta turbidez, com SE
estabilizando em aproximadamente 0,4 quando FR foi superior a 100 mg h-1. Esta tendência
não ficou evidente no presente estudo. Já para SE encontramos relação positiva com o teor
orgânico do seston (f), conforme preconizado por Hawkins et al, (1998a), como ficou
evidente comparando PB com EB e os diferentes experimentos em cada local. Estas
tendências corroboram com observações feitas para Mytilus edulis (Hawkins et al., 1996) e
Perna viridis (Hawkins et al., 1998b).
Em decorrência do incremento simultâneo de FR e SE, o teor orgânico das partículas
ingeridas (i) aumenta em relação direta com as taxas de alimentação (FR, IR), resultando no

65
incremento da eficiência de absorção (AE) (Hawkins et al., 1996, 1998a, 1998b; Mello, op.
cit.). Este conjunto de fatores resulta então no aumento do aproveitamento da matéria
orgânica ingerida (OIR) que foi absorvida (AR). É o que constatamos, em parte, também em
relação a Perna perna no presente estudo, tanto em condições de águas turvas (EB) como em
águas claras (PB), onde i e AE incrementaram com SE, OIR aumentou com IR e AR com IR.
Em termos de AEs, em geral estas foram maiores quando os valores de f foram
maiores, tanto em EB quanto em PB, embora não tenha sido observada correlação
significativa entre AE e f. Quando comparamos os locais, não houve diferença significativa de
AEs (EB = 0,62; PB = 0,71), embora f e i tenham sido maiores em Porto Belo (EB: f = 0,12, i
= 0,32; PB: f = 0,33, i = 0,41). Entretanto, comparando os experimentos, observamos
comportamentos distintos nas relações entre as variáveis do seston e a eficiência de absorção.
É o caso de AE em EB1 (0,86), que foi estatisticamente tão elevada quanto PB3 (0,80) e PB1
(0,79) em relação às demais, enquanto que f foi significativamente mais alta somente em PB1
(0,41) e PB3 (0,36), ao passo que em EB1 (0,15) observou-se apenas a quarta maior média de
f. Já em termos de POM, EB1 (4,8 mg L-1) apresentou a média estatisticamente mais elevada
em relação a PB1 (0,7 mg L-1) e PB3 (0,4 mg L-1). Podemos inferir, portanto, que quando as
diferenças em termos de concentração no seston (principalmente de POM) foram mais
discrepantes, houve maior eficiência de absorção (AE) em condições de águas turvas, mas
quando esta diferença diminuiu em relação às condições de menor turbidez, o fator que talvez
tenha determinado o aumento em AE seja o teor orgânico do seston (f).
Berry & Schleyer (1983) relatam uma relação positiva entre AE e POM para P. perna
na África do Sul, em P. viridis verificou-se uma relação negativa com TPM e positiva com f
(Wong & Cheung, 2001b), M. galloprovincialis e P. canaliculus apresentaram relação
positiva com TPM, enquanto A. maoriana não evidenciou relação alguma de AE com as
variações do seston (Gardner, 2002). Comparando locais distintos, Gardner (2000) observou
que para estas três espécies as AEs foram mais elevadas onde POM e f foram mais altos,
Okumus e Stirling (1994) verificaram a mesma tendência para M. edulis.
A eficiência de absorção, AE (AR:OIR), aumenta em função da fração orgânica
ingerida, i (OIR:IR), que por sua vez aumenta com com f (Hawkins & Bayne, 1990). Em
parte, estas foram tendências observadas neste estudo para Perna perna, no caso de AE em
relação a f, e principalmente i em relação a f e de AE em relação a i. A fração residual, que
corresponde a OER:OIR, representa as perdas metabólicas fecais (MFL, de metabolic faecal
loss) resultantes da digestão (Hawkins & Bayne, op. cit.). Estas perdas fecais são formadas a

66
partir de fragmentos celulares, enzimas e muco, eliminados durante os processos digestivos.
Com o aumento da qualidade do alimento (f), aumenta a proporção absorvida em relação à
eliminada nas fezes, como conseqüência há incremento da AE. Em dietas pobres (baixo teor
orgânico), estas perdas fecais tornam-se quantitativamente importantes (Hawkins & Bayne,
op. cit.), reduzindo AE.
Comparando as taxas de absorção (AR), verificamos que no experimento de verão em
Enseada do Brito (EB1) houve uma alta absorção (10,31 mg L-1), apesar do baixo f (0,15) e
alto TPM (29,6 mg L-1). Durante este experimento observamos uma forte ação turbulenta do
vento que, por ressuspensão, pode ter resultado na alta concentração de material em
suspensão. Isto pode indicar que Perna perna é bastante eficiente em termos de seleção sob
condições de alta turbidez. Também demonstra que as partículas ressuspendidas podem
compor uma fonte nutricional importante para os bivalves cultivados naquela área.
Em termos de fisiologia energética, o “scope for growth” (SFG) representa o balanço
energético líquido de um organismo resultante do cálculo entre ganho e perda de energia,
obtidos da assimilação (A) com subtração dos gastos de respiração (R) e excreção (U), ou
seja, SFG = A - (R + U) (Bayne & Newell, 1983; Dame, 1996).
Valores positivos de SFG representam ganho energético líquido, que poderão ser
alocados para crescimento somático e reprodutivo, portanto este parâmetro permite estimar o
potencial de crescimento do organismo. Valores negativos de SFG normalmente decorrem de
custos respiratórios maiores que a assimilação energética, mas também podem ocorrer em
função de AEs negativas resultantes de perdas fecais (MFL) significativas. Os gastos
decorrentes de excreção são normalmente baixos em relação ao custo energético total (menos
de 10%) (Hawkins & Bayne, 1984, 1992; Bayne & Hawkins, 1990). As AEs (eficiências de
absorção líquida, isto é, com dedução das MFLs) podem variar normalmente entre - 60% e
60% (- 0,6 à 0,6), segundo Bayne & Hawkins (1990).
As AEs obtidas para Perna perna nas condições aqui avaliadas foram, em média, bem
elevadas: 41 a 86% em Enseada do Brito e 55 a 80% em Porto Belo. Mello (1999) obteve
para esta espécie AEs de até 90% em alguns indivíduos em Sambaqui (Baía Norte).
Utilizando dietas artificiais (culturas monoalgais), Resgalla Jr. et al. (1999) obtiveram taxas
de assimilação variando aproximadamente entre 20 e 80% e entre 40 e 80% em mexilhões
Perna perna provenientes de áreas de cultivo em Armação do Itapocoroy e em Zimbros,
respectivamente. Na África do Sul, Berry & Schleyer (1983) obtiveram uma AE média de

67
61% para esta espécie, considerando as partículas do seston entre 2 e 100 µm, com POM
médio de 3,16 mg L -1.
As AEs normalmente apresentam valores negativos quando o teor orgânico do seston
(f) é inferior a 20 ou 10% (Hawkins & Bayne, 1992; Bayne & Hawkins, 1990). Entretanto
estes valores foram obtidos em experimentos com Mytilus edulis sob dietas artificiais,
normalmente utilizando culturas monoalgais, com alto teor orgânico e sem interferência de
componentes orgânicos refratários (não digeríveis) como detritos vegetais, etc., que
normalmente podem ocorrer no seston natural. Cranford & Hill (1999) avaliaram a variação
sazonal na alimentação in situ de Mytilus edulis e Placopecten magellanicus no litoral
canadense (Bedford Basin e Mahone Bay, Nova Scotia), e verificaram AEs inferiores a zero
quando o POM correspondeu a 30% do seston (f = 0,30), com TPM normalmente inferior a 8
mg L -1.
Segundo Gardner (2000), Aulacomya maoriana, Mytilus galloprovincialis e Perna
canaliculus apresentaram AEs médias variando entre 0 e 7% quando o teor orgânico médio do
seston foi de 30% em Island Bay (Nova Zelândia) durante experimentos de verão. Já no
inverno, neste mesmo local, a porcentagem de POM caiu para aproximadamente 21%,
enquanto que as AEs médias para os três mitilídios subiram para valores entre 3 e 34%. Esta
aparente contradição pode ser explicada pelas diferenças em termos de concentração média de
POM (2,8 mg L -1 no verão e 3,0 mg L -1 no inverno), mas principalmente em termos de
número de partículas (em torno de 8.000 por ml no verão e mais de 20.000 por ml no
inverno), o que denota a fração orgânica do seston como apenas um dos vários critérios em
termos de valor nutricional do seston, sendo necessários outros parâmetros para uma
interpretação mais completa da variação do comportamento alimentar dos bivalves sob
condições naturais.
Em termos de alimentação e assimilação, nossos dados para Perna perna nas duas
áreas analisadas são bastante satisfatórios, entretanto são dados preliminares, que necessitam
ainda maiores estudos acompanhando todas as variações sazonais durante o ano bem como as
variações nictemerais (ciclos de marés), além de dados simutâneos de respiração e excreção
para esta espécie sob condições naturais, para que possa ser determinado o balanço energético
em termos de “scope for growth” (SFG), analisando então o potencial de crescimento da
espécie sob condições de cultivo nas diferentes áreas a serem estudadas.
Embora vários trabalhos têm utilizado biodepósitos para avaliar a alimentação em
bivalves, raramente faz-se abordagem quanto a produção de biodepósitos per si, avaliando as

68
taxas de biodeposição em relação às variações do seston, como em Tsuchyia (1980), Navarro
(1983), Navarro & Thompson (1997), Deslous-Paoli (1992), Jaramillo et al. (1997), Mello
(1999), Pouvreau et al. (2000). Na Enseada do Brito, onde as concentrações do seston sempre
foram mais elevadas, a produção de biodepósitos (BR) foi bem mais alta em relação a Porto
Belo, demonstrando claramente a relação entre BR e TPM. Em condições de baixo TPM
(médias de 1,2 - 1,7 mg L-1), a produção de fezes (ER) é sempre maior ou equivalente à
liberação de pseudofezes (RR), como foi observado em Porto Belo. Já em águas mais turvas
(TPMs médias entre 9,9 e 29,6 mg L-1), como na Enseada do Brito, a proporção de
pseudofezes sempre é mais alta, com RR correspondendo em peso seco em até quase 6 vezes
a ER.
Quanto ao teor orgânico dos biodepósitos (b), este foi sempre mais alto conforme
diminuiu a proporção de pseudofezes (RR) em relação à biodeposição total (BR), uma vez
que a fração orgânica das pseudofezes (p) foi predominantemente menor que a fração
orgânica das fezes (e). Isto ocorre pelo fato dos bivalves rejeitarem seletivamente partículas
inorgânicas nas pseudofezes, resultando numa ingestão organicamente mais rica, cuja parcela
não absorvida é eliminada nas fezes juntamente com outros resíduos da digestão (células,
enzimas, etc.) (Hawkins & Bayne, 1990). Além disso, em PB, onde a oferta alimentar foi de
melhor qualidade (f médio variou entre 0,20 e 0,41) em relação a EB (f médio entre 0,10 e
0,15), tanto p quanto e foram mais elevados, incrementando b. Mello (1999) relatou que em
Sambaqui (TPM entre 10,00 e 31,33 mg L-1; f entre 0,10 e 0,17) e a fração orgânica dos
biodepósitos de Perna perna variou entre 8 e 10% (pseudofezes e fezes, respectivamente),
valores bastante similares aos observados neste trabalho em Enseada do Brito (b variou entre
0,8 e 0,9). Já em Porto Belo, b variou entre 0,14 e 0,17.
Em termos de taxas de biodeposição específicas por grama de peso seco (partes
moles), considerando que os mexilhões neste estudo apresentaram em média 1,5 g de peso
seco, para Porto Belo a BR média de Perna perna foi de 4,66 mg h-1 g-1 de peso seco de carne
(111,84 mg por dia), e para Enseada do Brito de 41,81 mg h-1 g-1 de peso seco de carne
(1.003,52 mg por dia). Mello (1999) em Sambaqui obteve para esta espécie uma taxa de
biodeposição média de aproximadamente 80 mg h-1 g-1 de peso seco de carne (1.920 mg por
dia).
Experimentos de biodeposição com outros bivalves (Mytilus edulis, Crassostrea
virginica, Mercenaria mercenaria, Crassostrea gigas) também demonstraram correlação
positiva entre a concentração de material em suspensão e a produção de biodepósitos (Tenore

69
& Dunstan, 1973a; Tsuchyia, 1980; Hildreth, 1980; Deslous-Paoli, 1992), e que a partir de
uma determinada concentração (TPM > 10 mg L-1) a produção de pseudofezes foi igual ou
superior à de fezes, uma vez que a produção de fezes apresentou relativa estabilização
(Tsuchiya, op. cit., Deslous-Paoli, op. cit.).
Sendo assim, podemos concluir que ambientes com alta turbidez apresentam maior
potencial de biodeposição, enquanto que em áreas com águas de baixa turbidez, a proporção
orgânica dos biodepósitos tende a ser mais elevada.
No mar Báltico, a biodeposição por Mytilus edulis pode corresponder a até 141% da
sedimentação natural em termos de peso seco, e até 173% do peso seco livre de cinzas,
incrementando a deposição anual total de carbono (C), nitrogênio (N) e fósforo (P) em 10%
(Kautsky & Evans, 1987).
Jaramillo et al. (1992) determinaram as taxas médias de biodeposição de populações
de mexilhões em um estuário no sul do Chile (rio Queule), onde TPM variou
aproximadamente entre 4 e16 mg L-1 e f entre 0,2 e 0,6. Choromytilus chorus apresentou uma
média anual de biodeposição de 271 g de peso seco m-2 dia-1 (22% de matéria orgânica, 6,4%
de C e 0,3% de N), e Mytilus chilensis de 234 g de peso seco m-2 dia-1 (21% de matéria
orgânica, 6,0% de C e 0,4% de N). Navarro (1983) encontrou concentrações similares de
matéria orgânica (17-27%) nos biodepósitos de uma população de Mytilus chilensis na baía de
Yaldad (TPM entre 2,17 e 5,26 mg L-1; f entre 0,32 e 0,55), com uma produção de 21,94 mg
de peso seco g-1 dia-1 de fezes. Haven & Morales-Alamo (1966) descreveram uma ampla
variação na produção de biodepósitos por ostras Crassostrea virginica, com uma média
semanal de 1,62 g de peso seco por indivíduo, chegando ao máximo de 3,92 g de peso seco
semanal por ostra, e estes biodepósitos continham de 9 a 23% de matéria orgânica e de 4 a
12% de carbono orgânico.
No Japão houve declínio na produtividade de cultivos de ostra, o que foi atribuído à
deterioração da qualidade das áreas de cultivo induzida pela atividade filtradora dos bivalves
(Ito & Imai, 1955; Kusuki, 1978). Segundo Mariojouls & Kusuki (1987), na baía de
Hiroshima cada balsa de cultivo produzia 78,7 Kg dia-1 de biodepósitos (fezes e pseudofezes),
e a proporção de deposição total devida a sedimentação natural foi de 31 a 48% sob as balsas
de cultivo comparada a 89% nas áreas fora dos cultivos.
Para Mytilus chilensis, Navarro (1983) estipulou que uma balsa de cultivo com 300
cordas de 8 metros cada, com 2.400 mexilhões, produz 15,8 Kg de fezes por dia na baía de

70
Yaldad. Mello (1999) calculou que um espinhél com 50 cordas de 1,5 metros cada, com 700
mexilhões, pode produzir até 161,54 Kg de biodepósitos por dia em Sambaqui.
Para estimar a produção diária de biodepósitos em cada local a partir dos dados deste
experimento, vamos considerar as densidades de cultivo recomendadas, com espaçamento
mínimo entre cordas de 0,5 metros, e de 5 metros entre espinhéis (EPAGRI & UFSC, 1994),
obtendo então 20 espinhéis por ha de área cultivada, sendo cada espinhel com comprimento
de 70 metros e cada metro de corda contendo 465 mexilhões. A partir das características de
cultivo de cada local, com Porto Belo apresentando 2,9 ha de área cultivada e cordas com
comprimento médio de 1,5 metros, e Enseada do Brito com 36,4 ha cultivados com cordas de
aproximadamente 1 metro, podemos estipular uma produção diária de 950,14 Kg de
biodepósitos em Porto Belo e de 71.339,43 Kg de biodepósitos em Enseada do Brito.
Entretanto temos que considerar que estas estimativas não consideram os processos de
dispersão e de ciclagem destes biodepósitos, que precisam ser mais bem estudados.
O impacto gerado por esta carga de biodepósitos nas áreas de cultivo depende
basicamente de dois processos: do grau de dispersão destas partículas no ambiente e da
velocidade de degradação deste material pela biota. Hayakawa et al. (2001) observaram taxas
de sedimentação oscilando entre 2,9 e 11 g de peso seco m-2 dia-1 sob cultivos de Crassostrea
gigas no estuário de Ofunato (Japão), quando os valores esperados a partir das taxas de
biodeposição dos bivalves seriam de mais de 260 g de peso seco m-2 dia-1. Estes
pesquisadores atribuíram esta diferença à dispersão dos biodepósitos por correntes marinhas
atuando no transporte deste material. Verificaram ainda que apesar de ocorrer variação
sazonal na sedimentação local, não houve alteração na composição química do sedimento
quanto às concentrações de C, N e P, sugerindo uma degradação rápida dos biodepósitos na
interface água-sedimento.
No presente estudo, os locais avaliados apresentam características fisiográficas
bastante distintas. A enseada de Porto Belo caracteriza-se por apresentar maior interação com
o oceano (área mais exposta), portanto está sujeita a sofrer menor impacto por biodeposição
em função dos processos advectivos que podem atuar na dispersão de partículas. A Enseada
do Brito situa-se dentro de uma baía rasa (Baía Sul de Florianópolis), que se comunica com o
oceano por um canal estreito ao sul, restringindo sua interação com o mar aberto.
Estas características somadas à grande carga de biodepósitos produzidos pelos
mexilhões sugerem que a Enseada do Brito está mais sujeita a sofrer impactos decorrentes da
biodeposição. Entretanto, informações a respeito dos padrões de circulação nestas áreas, taxas

71
de sedimentação e análises de sedimento do fundo sob os cultivos são necessárias para avaliar
os efeitos de dispersão e a velocidade de degradação e ciclagem dos biodepósitos.
5. Agradecimentos
Somos gratos aos produtores de Porto Belo (Marcelo) e de Enseada do Brito (Sr. Luís)
pelo fornecimento dos mexilhões bem como pela disponibilização das balsas para a execução
dos experimentos de alimentação e biodeposição. Também somos gratos aos técnicos
extensionistas da Epagri Milton Francisco de Quadros (Palhoça) e Evaldo Manoel dos Santos
(Porto Belo) pela disponibilização dos dados referentes às áreas de cultivo daquelas
localidades. Agradecemos ainda aos funcionários, estagiários e pesquisadores do LCMM-
UFSC pelo auxílio nas diversas etapas deste trabalho e ao CNPq pela concessão de bolsa de
Mestrado a Jean Franco Schmitt.
6. Referências
Barg, U. C., 1992. Guidelines for the promotion of environmental management of coastal aquaculture development. FAO Fisheries Technical Paper, n. 328.
Barillé, L., Prou, J., Héral, M., Razet, D., 1997. Effects of high natural seston concentrations
on the feeding, selection, and absorption of the oyster Crassostrea gigas (Thunberg). J. Exp. Mar. Biol. Ecol., 212: 149-172.
Barranguet, C., 1997. The role of microphytobenthic primary production in a mediterranean
mussel culture area. Estuarine, Coastal and Shelf Science, 44: 753-765. Barroso, G. F., Littlepage, J., 1998. Protocolo para análise de clorofila a e feopigmentos
pelo método fluorimétrico (Fluorímetro TD-700). Vitória: Programa Brasileiro para Integração da Maricultura/Programa de Monitoramento Ambiental. 19 p.
Bayne, B. L.; Hawkins, A. J. S., 1990. Filter-feeding in bivalve molluscs: controls of energy
balance. In: Mellinger, J. (ed.) Animal nutrition and transport processes. I. Nutrition in wild and domestic animals. Comp. Physiol. Basel. Karger, 5: 70-83.
Bayne, B. L.; Hawkins, A. J. S., Navarro, E., 1987. Feeding and digestion by the mussel
Mytilus edulis L. (Bivalvia: Mollusca) in mixtures of silt and algal cells at low concentrations. J. Exp. Mar. Biol. Ecol., 111: 1-22.
Bayne, B. L.; Hawkins, A. J. S., Navarro, E., 1988. Feeding and digestion in suspension-
feeding bivalve molluscs: the relevance of physiological compensations. Amer. Zool., 28: 147-759.

72
Bayne, B. L.; Hawkins, A. J. S., Navarro, E., Iglesias, I. P., 1989. Effects of seston
concentration on feeding, digestion and growth in the mussel Mytilus edulis. Mar. Ecol. Prog. Ser., 55: 47-54.
Bayne, B. L.; Iglesias, I. P.; Hawkins, A. J. S.; Navarro, E.; Heral, M., Deslous-Paoli, J. M.,
1993. Feeding behaviour of the mussel, Mytilus edulis: responses to variations in quality and organic content of the seston. J. Mar. Biol. Ass. U. K., 73: 813-829.
Bayne, B. L., Newell, R. C., 1983. Physiological energetics of marine molluscs. In: Saleudin,
A. S. M., Wilber, K. M. (eds). The Mollusca, Vol. 4, Physiology. Academic Press, New York, p. 407-515.
Bayne, B. L.; Widdows, J., 1978. The physiological ecology of two populations of Mytilus
edulis L. Oecologia, 37 (2): 137-162. Bayne, B. L.; Widdows, J., Newell, R. I. E., 1976. Physiological measurements on estuarine
bivalve molluscs in the field. In: Keegan, B. F.; Ceidigh, P. O., Boaden, P. J. S. (eds.) Biology of benthic organisms – 11th European Symposium on Marine Biology, Galway, October 1976. Oxford: Pergamon Press. p. 57-68.
Berry, P. F., Schleyer, M. H., 1983. The brown mussel Perna perna on the Natal coast, South
Africa: utilization of available food and energy budget. Mar. Ecol. Prog. Ser., 13: 201-210.
Bertrán, C.; Jaramillo, E.; Bravo, A.; Velásquez, C., 1988. Acuicultura de Mitilidos en un
estuario del centro-sur de Chile: efectos de los biodepositos. 15 p. Separata - Instituto de Zoología, Facultad de Ciencias, Universidad Austral de Chile.
Bohle, B., 1972. Effects of adaptation to reduced salinity on filtration activity and growth of
mussels (Mytilus edulis). J. Exp. Mar. Biol. Ecol., 10: 41-49. Carvalho Filho, J., 2001. Panorama da malacocultura brasileira. Panorama da Aqüicultura.
Vol. 11, n° 64, p. 25-31. Chamberlain, J.; Fernandes, T. F.; Read, P.; Nickell, T. D., Davies, I. M. 2001. Impacts of
biodeposits from suspended mussel (Mytilus edulis L.) culture on the surrouding surficial sediments. ICES Journal of Marine Science, 58: 411-416.
Chevarria, G. G., 1999. Caracterização biogeoquímica de uma área de cultivo de
moluscos - Enseada de Armação do Itapocoroy - Penha/SC. 60 p. Monografia (Graduação em Oceanografia) - Centro de Ciências Tecnológicas da Terra e do Mar, Universidade do Vale do Itajaí.
Cranford, P. J., Hargrave, B. T., 1994. In situ time-series measurement of ingestion and
absorption rates of suspension-feeding bivalves: Placopecten magellanicus. Limnol. Oceanogr., 39(3): 730-738.

73
Cranford, P. J., Hill, P. S., 1999. Seasonal variation in food utilization by the suspension-feeding bivalve molluscs Mytilus edulis and Placopecten magellanicus. Mar. Ecol. Prog. Ser., 190: 223-239.
Cranford, P. J., Emerson, C.W., Hargrave, B. T., Milligan, T. G., 1998. In situ feeding and
absorption responses of sea scallops Placopecten magellanicus (Gmelin) to storm-induced changes in the quantity and composition of the seston. J. Exp. Mar. Biol. Ecol., 219: 45-70.
Dahlbäck, B., Gunnarsson, L. Å. H., 1981. Sedimentation and sulfate reduction under a
mussel culture. Mar. Biol., 63: 269-275. Dame, R. F., 1996. Ecology of marine bivalves: an ecossystem approach. Boca Raton:
CRC Marine Science Series, 254 p. Danovaro, R., Fabiano, M., 1997. Seasonal changes in quality and quantity of food available
for benthic suspension-feeders in the Golfo Marconi (North-western Mediterranean). Estuarine, Coastal and Shelf Science, 44: 723-736.
D’Aquino, C. de A., 2000. Papel das ondas e correntes na remobilização de sedimentos
finos em zonas de cultivo de moluscos marinhos. 46 p. Monografia (Graduação em Oceanografia) - Centro de Ciências Tecnológicas da Terra e do Mar, Universidade do Vale do Itajaí.
EPAGRI & UFSC, 1994. Manual de cultivo do mexilhão Perna perna. Editora da Epagri.
134 p. Fernandes, F. C., 1985. Mitilicultura. Parte A - Enfoque bioecológico. In: Manual de
Maricultura. Rio de Janeiro: Instituto de Pesquisas da Marinha, Cap. V-A, 24 p. Ferreira, J. F., Magalhães, A. R. M., 1992. Cultivo de mexilhões em Santa Catarina.
Panorama da Aqüicultura. Vol. 2, n° 14, p. 10-11. Ferreira, J. F., Grümann, A. 1999. Cultivo de moluscos marinhos. In: CNI/IEL. Tecnologia e
Inovação para a indústria: biotecnologia, novos materiais, tecnologia da informação. Brasília: SEBRAE, p. 26-29.
Ferreira, J. F., Magalhães, A. R. M., 1997. Mexilhões - biologia e cultivo. Florianópolis:
UFSC. 58 p. Fréchette, M., Grant, J., 1991. An in situ estimation of the effect of wind-driven resuspension
on the growth of the mussel Mytilus edulis L. J. Exp. Mar. Biol. Ecol., 148: 210-213. Gardner, J. P. A., 2000. Where are the mussels on Cook Strait (New Zealand) shores? Low
seston quality as a possible factor limiting multi-species distributions. Mar. Ecol. Prog. Ser., 194: 123-132.

74
Gardner, J. P. A., 2002. Effects of seston variability on the clearance rate and absorption efficiency of the mussel Aulacomya maoriana, Mytilus galloprovincialis and Perna canaliculus from New Zealand. J. Exp. Mar. Biol. Ecol., 268: 83-101.
Grant, J., Hatcher, A., Scott, D. B., Pocklington, P., Schafer, C. T., Winters, G. V., 1995. A
multidisciplinary approach to evaluating impacts of shellfish aquaculture on benthic communities. Estuaries, 18(1A): 124-144.
Haven, D. S., Morales-Alamo, R., 1966. Aspects of biodeposition by oyster and other
invertebrate filter feeders. Limnol. Oceanogr., 20: 924-934. Haven, D. S., Morales-Alamo, R., 1972. Biodeposition as a factor in sedimentation of five
suspended solids in estuaries. Mem. Geol. Soc. Am., 133: 121-130. Hawkins, A. J. S., Bayne, B. L., 1992. Physiological interrelations, and the regulation of
production. In: Gosling, E. (ed.), 1992. The Mussel Mytilus : ecology, physiology, genetics and culture. Elsevier, Amsterdan. p. 171-222.
Hawkins, A. J. S., Bayne, B. L., Mantoura, R. F. C., Llewellyn C. A., Navarro, E., 1986.
Chlorophyll degradation and absorption throughout the digestive system of the blue mussel Mytilus edulis L. J. Exp. Mar. Biol. Ecol., 96: 213-223.
Hawkins, A. J. S., Bayne, B. L., Bougrier, S., Héral, M., Iglesias, J. I. P., Navarro, E., Smith,
R .F. M., Urrutia, M. B., 1998a. Some general relationships in comparing the feeding physiology of suspension-feeding bivalve molluscs. J. Exp. Mar. Biol. Ecol., 219: 87-103.
Hawkins, A. J. S., Fang, J. G., Pascoe, P. L., Zhang, J. H., Zhang, X. L., Zhu, M. Y., 2001.
Modelling short-term responsive adjustments in particle clearance rate among bivalve suspension-feeders: separate unimodal effects of seston volume and composition in the scallop Chlamys farreri. J. Exp. Mar. Biol. Ecol., 262: 61-73.
Hawkins, A. J. S., James, M. R., Hickman, R. W., Hatton, S., Wheatherhead, M., 1999.
Modelling of suspension-feeding and growth in the green-lipped mussel Perna canaliculus exposed to natural and experimental variations of seston availability in the Marlborough Sounds, New Zealand. Mar. Ecol. Prog. Ser., 191: 217-232.
Hawkins, A. J. S., Smith, R. F. M., Bayne, B. L., Héral, M., 1996. Novel observations
underlying the fast growth of suspension-feeding shellfish in the turbid environments: Mytilus edulis. Mar. Ecol. Prog. Ser., 131: 179-190.
Hawkins, A. J. S., Smith, R. F. M., Bougrier, S., Bayne, B. L., Héral, M., 1997. Manipulation
of dietary conditions for maximal growth in mussels, Mytilus edulis L., from the Marennes-Oléron Bay, France. Aquat. Living Resour., 10: 13-22.
Hawkins, A. J. S., Smith, R. F. M., Tan, S. H., Yasin, Z. B., 1998b. Suspension-feeding
behaviour in tropical bivalve molluscs: Perna viridis, Crassostrea belcheri, Crassostrea iradelei, Saccostrea cucculata and Pinctada margarifera. Mar. Ecol. Prog. Ser., 166: 173-185.

75
Hayakawa, Y., Kobayashi, M., Izawa. M., 2001. Sedimentation flux from mariculture of
oyster (Crassostrea gigas) in Ofunato estuary, Japan. ICES Journal of Marine Science, 58: 435-444.
Hickman, R. W., Waite, R. P., Illingworth, J., Meredyth-Young, J. L., Payne, G., 1991. The
relationship beween farmed mussels, Perna canaliculus, and available food in Pelorus-Kenepuru Sound, New Zealand, 1983-1985. Aquaculture, 99: 49-68.
Hildreth, D. I., 1980. Bioseston production by Mytilus edulis and its effects in experimental
systems. Mar. Biol. 55: 309-315. Iglesias, J. I. P., Navarro, E., Alvarez Jorna, P., Armentia, I., 1992. Feeding, particle selection
and absortion in cockles Cerastoderma edule (L.) exposed to variable conditions of food concentration and quality. J. Exp. Mar. Biol. Ecol., 162: 177-198.
Iglesias, J. I. P., Pérez Camacho, A., Navarro, E., Labarta, U., Beiras, R.; Hawkins, A. J. S.,
Widdows, J., 1996. Microgeographic variability in feeding, absortion and condition in mussels (Mytilus galloprovincialis Lmk.): an transplant experiment. J. Shellfish Res., 15(3): 673-680.
Iglesias, J. I. P., Urrutia, M. B., Navarro, E., Ibarrola, I., 1998. Measuring feeding and
absorption in suspension-feeding bivalves: an appraisal of the biodeposistion method. J. Exp. Mar. Biol. Ecol., 219: 71-86.
Ito, S., Imai, T., 1955. Ecology of oyster bed. I.: On the decline of productivity due to
repeated culture. Tokoku J. Agric. Res., 5: 251-268. Jaramillo, E., Beltran, C., Bravo, A., 1992. Mussel biodeposition in an estuary in southern
Chile. Mar. Ecol. Prog. Ser., 82: 85-94. Jørgensen, C. B., 1990. Bivalve filter feeding: hydrodinamics, bioenergetics, physiology
and ecology. Fredensborg: Olsen & Olsen. 140 p. Jørgensen, C. B., Larsen, P. S., Riisgård, H. U., 1990. Effects of temperature on the mussel
pump. Mar. Ecol. Prog. Ser., 64: 89-97. Kaspar, H. F., Gillespie, P. A., Boyer, I. C., Mackenzie, A. L., 1985. Effects of mussel
aquaculture on the nitrogen cycle and benthic communities in Kenepuru Sound, Marlborough Sounds, New Zealand. Mar. Biol., 85: 127-136.
Kautsky, N., Evans, S., 1987. Role of biodeposition by Mytilus edulis in the circulation of
matter and nutrients in a Baltic coastal ecosystem. Mar. Ecol. Prog. Ser., 38: 201-212. Kusuki, Y., 1978. Relationships between quantities of faecal material produced and quantities
of suspended matter removed by the Japanese oyster. Bull. Jap. Soc. Sci. Fish., 44: 1183- 1185.

76
Labarta, U.; Fernández-Reiríz, M. J., Babarro, J. M. F., 1997. Differences in physiological energetics between intertidal and raft cultivated mussels Mytilus galloprovincialis. Mar. Ecol. Progr. Ser., 152: 167-173.
Mariojouls, C., Kusuki, Y., 1987. Appréciation des quantités de biodépôts émis par les huîtres
en élevage suspendu dans la baie d’Hiroshima. Haliotis, 16: 221-231. Mattsson, J., Lindén, O., 1983. Benthic macrofauna succession under mussels, Mytilus edulis
L. (Bivalvia), cultured on hanging long lines. Sarsia, 68: 97-102. Mello, L. G. M., 1999. Taxas de biodeposição e clareamento do mexilhão Perna perna em
ambiente de cultivo. 33 p. Dissertação (Mestrado em Aqüicultura) - Departamento de Aqüicultura, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina.
Melo, E., Martins, R. P., Franco, D. 1999. Standing wave tide at Florianópolis Bay (Brazil)
and its influence on bay pollution. Laboratório de Hidrologia Aplicada e Hidráulica Marítima, Departamento de Engenharia Ambiental, Universidade Federal de Santa Catarina. 10 p. Não publicado.
Navarro, E., Iglesias, J. I. P.; Pérez Camacho, A., Labarta, U., Beiras, R., 1991. The
physiological energetics of mussel (Mytilius galloprovincialis Lmk) from different cultivation rafts in the Ria de Arosa (Galicia, NW Spain). Aquaculture, 94: 197-212.
Navarro, E., Iglesias, J. I. P.; Pérez Camacho, A., Labarta, 1996. The effect of diets of
phytoplancton and suspended bottom material on feeding and absortion of rafts mussels (Mytilus galloprovincialis Lmk). J. Exp. Mar. Biol. Ecol., 198: 175-189.
Navarro, J. M., 1983. Oferta alimenticia natural y su efecto sobre los procesos fisiológicos del
bivalvo Mytilus chilensis en la bahía de Yaldad, Chiloé (Sur de Chile). Mems. Asoc. Latioam. Acuicult., 5: 175-187.
Navarro, J. M., 1988. The effects of salinity on the physiological ecology of Choromytilus
chorus (Molina, 1782) (Bivalvia: Mytilidae). J. Exp. Mar. Biol. Ecol., 122: 19-33. Navarro, J. M., Clasing, E., Urrutia, G., Asencio, G., Stead, R., Herrera, C., 1993.
Biochemical composition and nutritive value of suspended particulate matter over a tidal flat of southern Chile. Estuarine, Coastal and Shelf Science, 37: 59-73.
Navarro, J. M., Jaramillo, R., 1994. Evaluacion de la oferta alimentaria natural disponible a
organismos filtradores de la Bahia de Yaldad (43°08’; 73°44’), sur de Chile. Rev. Biol. Mar., Valparaíso, 29(1): 57-75.
Navarro, J. M., Thompson, R. J., 1994. Comparision and evaluation of different techniques
for measuring absortion efficiency in suspension feeders. Limnol. Oceanog., 39(1): 159-164.
Navarro, J. M., Thompson, R. J., 1996. Physiological energetics of the horse mussel Modiolus
modiolus in a cold ocean environment. Mar. Ecol. Prog. Ser., 138: 135-148.

77
Navarro, J. M., Thompson, R. J., 1997. Biodeposition by the horse mussel Modiolus
modiolus (Dillwyn) during the spring diatom bloom. J. Exp. Mar. Biol. Ecol., 209: 1-13. Navarro, J. M., Ulloa, L., 1995. Effects of environmental factors on physiological responses
and fluctuations in the soft tissue weigth of bivalve Aulacomya ater in a bay of southern Chile. In: Proceedings of the 30th European Marine Biological Symposium, Southampton, UK, september, 1995. p. 21-33.
Navarro, J. M., Widdows, J., 1997. Feeding physiology of Cerastodesma edule in response to
a wide range seston concentration. Mar. Ecol. Prog. Ser., 152: 175-186, 1997. Navarro, J. M., Winter, J. E., 1982. Ingestion rate, assimilation efficiency and energy balance
in Mytilus chilensis in relation to body size and different algal concentratons. Mar. Biol., 67: 255-266.
Newell, R. C., 1965. The role of detritus in the nutrition of two marine deposit feeders, the
prosobranch Hydrobia ulvae and the bivalve Macoma balthica. Proc. Zool. Soc. Lond., 144: 25-45.
Newell, R. C., Shumway, S. E., Cucci, T. L., Selvin, R., 1989. The effects of natural seston
particle size and type on feeding rates, feeding selectivity and food resource availability for the mussel Mytilus edulis Linnaeus, 1758 at bottom culture sites in Maine. J. Shellfish Res., 8 (1): 187-196.
Newkirk, G. F., 1990. A look at mollusc culture in developing countries: problems and
potentials. In: Hernandez R., A. (ed.) Cultivo de moluscos en America Latina. Bogotá: Guadalupe/CIID, p. 99-114.
Okumus, I., Stirling, H. P., 1994. Physiological energetics of cultivated mussel (Mytilus
edulis) populations in two Scottish west coast sea lochs. Mar. Biol., 119: 125-131. Ottmann, F., Sornin, J., 1982. Observations on sediment accumulation as a result of mollusk
culture systems in France. In: Proceedings of SIUEC. Vol. 1. Rio Grande (RS) - Brasil, 21 to 27, November, 1982. p. 329 - 337.
Pérez Camacho, A., González, R., 1984. La filtración del mejillón (Mytilus edulis L.) en
laboratorio. In: Actas do Primeiro Seminario de Ciencias do Mar, As Rías Galegas. Cuadernos da Área de Ciencias Mariñas, Seminario de Estudos Galegos, 1: 427-437.
Pouvreau, S., Bodoy, A., Buestel, D., 2000. In situ suspension feeding behaviour of the pearl
oyster, Pinctada margaritifera: combined effects of body size and wheater-related seston composition. Aquaculture, 181: 91-113.
Prins, T. C., Dankers, N., Smaal, A. C., 1994. Seasonal variation in the filtration rates of a
semi-natural mussel bed in relation to seston composition. J. Exp. Mar. Biol. Ecol., 176: 69-86.

78
Proença, L. A. O., Schettini, C. A. F., 1998. Effect of shellfish culture on phytodetritus vertical fluxes in tropical waters - southern Brazil. Rev. Bras. Oceanogr., 46(2): 125-133.
Rafael, P. R. B., 1978. Mitilicultura - resposta do mexilhão Perna perna (Pelecypoda,
Mytilidae) à diferentes salinidades. Resumos da 30a reunião da SBPC. Resgalla Jr., C., Manzoni, G. C. Kuroshima, K. N., Reis Fo., R. W., Laitano, K. S., 1999.
Variabilidade nas taxas fisiológicas do mexilhão Perna perna em dois sítios de cultivo do litoral norte de Santa Catarina. Notas Técnicas da Facimar, 3: 33-40.
Riisgård, H. U., 1991. Fitration rate and growth in the blue mussel, Mytilus edulis Linnaeus,
1758: dependence on algal concentration. J. Shellfish Res., 10: 29-35. Rosa, R. C. C., 1997. Impacto do cultivo de mexilhões nas comunidades pesqueiras de
Santa Catarina. 188 p. Dissertação (Mestrado em Aqüicultura) - Departamento de Aqüicultura, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina.
Salomão, L. C., Magalhães, A. R. M., 1978. Sobrevivência do molusco Perna perna
submetido à diferentes salinidades. Resumos do V Simpósio Latinoamericano de Oceanografia Biológica, 247-248.
Salomão, L. C., Magalhães, A. R. M., Lunetta, J. E., 1980. Influência da salinidade na
sobrevivência de Perna perna. Boletim de Fisiologia Animal, 4: 143-152. Segnini de Bravo, M. I., Chung, K. S., Pérez, J. E., 1998. Salinity and temperature tolerances
of the green and brown mussels, Perna viridis e Perna perna (Bivalvia: Mytilidae). Rev. Biol. Trop., 46(5): 121-125.
Schettini, C.A.F.; Carvalho, J.B.; Jabor, P., 1996. Hidrologia e distribuição de material em
suspensão de quatro estuários de Santa Catarina. In: III Simpósio sobre Oceanografia, 1996, São Paulo. Resumos do III Simpósio sobre Oceanografia. São Paulo: IOUSP.
Schettini, C. A. F., Resgalla Jr., C., Kuroshima, K. N., 1997. Avaliação preliminar da taxa de
sedimentação na região de cultivo de moluscos (Perna perna) na enseada da Armação – SC. Notas Técnicas da Facimar, 1: 1-8.
Schettini, C. A. F., De Carvalho, J. L. B., Truccolo, E. C., 1999. Aspectos hidrodinâmicos da
Enseada da Armação de Itapocoroy, SC. Notas Técnicas da Facimar, 3: 99-109. Schettini, C.A.F.; Santos, M.I.F. & Abreu, J.G.N., 2002. Observações dos sedimentos de
fundo de uma plataforma abrigada sob influência de atividade de dragagem: Saco dos Limões, Florianópolis, SC. Notas Técnicas da Facimar, 6: 165-175.
Shumway, S. E., 1977. The effect of fluctuating salinity on the tissue water content of eight
species of bivalve molluscs. J. Comp. Physiol., 116: 269-285. Shumway, S. E., Cucci, T. L., Newell, R. C., Yentsch, C. M., 1985. Particle selection,
ingestion, and absorption in filter-feeding bivalves. J. Exp. Mar. Biol. Ecol., 91: 77-92.

79
Smaal, A. C., van Stralen, M. R., 1990. Average annual growth and condition of mussels as a function of food source. Hydrobiologia, 195: 179-188.
Smaal, A. C., Vonck, A. P. M. A., Bakker, M., 1997. Seasonal variation in physiological
energetics of Mytilus edulis and Cerastoderma edule of different size classes. J. Mar. Biol. Assoc. UK, 77: 817-838.
Stenton-Dozey, J. M. E., Jackson, L. F., Busby, A. J., 1999. Impact of mussel culture on
macrobenthic community structure in Saldanha Bay, South Africa. Marine Pollution Bulletin, 39 (1-12): 357-366.
Strickland, J. D. H., Parsons, T. R., 1965. A manual of sea water analysis. Bulletin n. 125,
Otawa: Fisheries Research Board of Canada. 202 p. Suplicy, F. M., 2000. A capacidade de suporte nos cultivos de moluscos: está no decreto mas
poucos sabem do que se trata. Panorama da Aqüicultura, v. 10, n. 57, p. 21-24. Tenore, K. R., Corral, J., Gonzalez, N., 1982. Effects of intensive mussel culture on food
chain patterns and production in coastal Galicia. NW Spain. Proceedings of SIUEC. Vol. 1. Rio Grande (RS) - Brasil, 21 to 27, November, 1982. p. 321 - 328.
Tenore, K. R., Dunstan, W. M., 1973a. Comparison of feeding and biodeposition of three
bivalves at different food levels. Mar. Biol., 21: 190-195. Tenore, K. R., Dunstan, W. M., 1973b. Comparison of rates of feeding and biodeposition of
the american oyster, Crassostrea virginica Gmelin, fed different species of phytoplankton. J. Exp. Mar. Biol. Ecol., 12: 19-26.
Thompson, R. J., Bayne, B. L., 1974. Some relationships between growth, metabolism and
food in the mussel Mytilus edulis. Mar. Biol., 27: 317-326. Tsuchiya, M., 1980. Biodeposit production by the mussel Mytilus edulis L. on rocky shores.
J. Exp. Mar. Biol. Ecol., 47: 203-222. Urrutia, M. B., Iglesias, J. I. P., Navarro, E., Prou, J., 1996. Feeding and absortion in
Cerastoderma edule under environmental conditions in the Bay of Marennes – Olerón (Western France). J. Mar. Biol. Assoc. U. K., 76: 431-450.
Widdows, J., 1978. Combined effects of body size, food concentration and season on the
physiology of Mytilus edulis. J. Mar. Biol. Assoc. U. K., 58: 109-124. Widdows, J., Fieth, P., Worral, C. M., 1979. Relationships between seston, available food and
feeding activity in the common mussel Mytilus edulis. Mar. Biol., 50: 195-207. Winter, J. E., 1973. The filtration rate of Mytilus edulis and its dependence on algal
concentration, measured by a continuous automatic recording apparatus. Mar. Biol., 22: 317-328.

80
Winter, J. E., 1976. Feeding experiments with Mytilus edulis L. at small laboratory scale. II. The influence of suspended silt in addition to algal suspensions on growth. In: Persoone, G., Jaspers, E. (eds.) 10th European Symposium of marine Biology, Ostend, Belgium, Sept. 17-23, (1), Wetteren: Universa Press.
Winter, J. E., Langton, R. W., 1976. Feeding experiments with Mytilus edulis L. at small
laboratory scale. I. The influence of the total amount of food ingested and food concentration on growth. In: Persoone, G., Jaspers, E. (eds.) 10th European Symposium of marine Biology, Ostend, Belgium, Sept. 17-23, (1), Wetteren: Universa Press.
Wong, W. H., Cheung, S. G., 1999. Feeding behaviour of the green mussel, Perna viridis (L.):
responses to variation in seston quantity and quality. J. Exp. Mar. Biol. Ecol., 236: 191-207.
Wong, W. H., Cheung, S. G., 2001a. Feeding rates and scope for growth of green mussels,
Perna viridis (L.) and their relationship with food availability in Kat O, Hong Kong. Aquaculture, 193: 123-137.
Wong, W. H., Cheung, S. G., 2001b. Feeding rhythms of the green-lipped mussel, Perna
viridis (Linnaeus, 1758) (Bivalvia: Mytilidae) during spring and neap tidal cycles. J. Exp. Mar. Biol. Ecol., 257: 13-36.
Valenti, W. C., (ed.), 2000. Aqüicultura no Brasil : bases para um desenvolvimento
sustentável. Brasília: CNPq/Ministério da Ciência e Tecnologia. 399 p. Vélez, R. A., Epifanio, C. E., 1981. Effects of temperature and ration on gametogenesis and
growth in the tropical mussel Perna perna (L.). Aquaculture, 22: 21-26. Vinatea Arana, L. A., 1999. Aqüicultura e desenvolvimento sustentável: subsídios para a
formulação de políticas de desenvolvimento da aqüicultura brasileira. Florianópolis: Editora da UFSC. 310 p.
Vinatea Arana, L. A. 2000. Modos de apropriação e gestão patrimonial de recursos
costeiros. Estudo de caso sobre o potencial e os riscos do cultivo de moluscos marinhos na Baía de Florianópolis, Santa Catarina. 245 p. Tese (Doutorado em Ciências Humanas) - Departamento de Ciências Humanas, Centro de Filosofia e Ciências Humanas, Universidade Federal de Santa Catarina, Florianópolis.
Zuim, S. M. F., Mendes, E. G., 1977. Sobrevivência do bivalve Perna perna em diferentes
temperaturas. In: Reunião Anual da SBPC. Resumos. p. 672.

81
CONSIDERAÇÕES FINAIS
Conclusões:
• em relação às distintas condições ambientais encontradas em Enseada do Brito e Porto
Belo, a alimentação e a produção de biodepósitos pelo mexilhão Perna perna variou de
acordo com as características do seston e apresentou tendências similares às observadas
para outras espécies de bivalves filtradores;
• as taxas de clareamento (CR) foram menores sob condições de maior oferta alimentar
(maior concentração de matéria particulada total, TPM, de matéria inorgânica particulada,
PIM, e de matéria orgânica particulada, POM), em Enseada do Brito;
• já as taxas de filtração (FR), de ingestão (IR) e de ingestão orgânica (OIR) foram maiores
em Enseada do Brito, aumentando de acordo com o incremento do seston (TPM, PIM e
POM);
• as taxas de absorção (AR) seguiram as mesmas tendências de FR, IR e OIR, entretanto as
ARs médias foram similares entre Enseada do Brito e Porto Belo;
• a eficiência de seleção (SE) e a fração orgânica ingerida (i) aumentaram conforme
incrementou o teor orgânico do seston (f), sendo maiores em Porto Belo;
• as eficiências de absorção (AE), por sua vez, incrementaram com SE e i, mas seus valores
médios também foram similares entre os locais de estudo;
• o mexilhão Perna perna apresentou uma alta eficiência de seleção, mesmo sob condições
de alta turbidez resultante de ressuspensão em Enseada do Brito no verão, maximizando
sua aquisição energética em termos de absorção de alimento (AR e AE);
• a biodeposição foi maior em Enseada do Brito do que em Porto Belo em decorrência dos
níveis de material em suspensão mais elevados, com teor orgânico menor de biodepósitos
produzidos em Enseada do Brito, em função das próprias características do seston (menor
f), bem como da maior contribuição de pseudofezes, organicamente mais pobres que as
fezes.

82
Recomendações:
• avaliar a variação do seston e de sua composição em termos número, volume e tamanho
unitário de partículas, composição do plâncton, composição química e bioquímica do material
particulado, bem como suas variações em termos sazonais e nictemerais, e em vários pontos e
profundidades dentro de cada área de estudo;
• avaliar a alimentação e a biodeposição dos bivalves em relação à estas variáveis e às
respectivas escalas de variação (temporal e espacial);
• realizar estudos referentes a fatores endógenos (grau de maturação sexual, presença e grau
de infestação por parasitas e/ou outros agentes patógenos, origem da semente, etc.) e sua
influência em relação às taxas fisiológicas dos bivalves;
• realizar estudos quanto à características hidrodinâmicas nas áreas de cultivo de bivalves;
• realizar estudos quanto as taxas de sedimentação nas áreas de cultivo de bivalves;
• avaliar o teor orgânico e a composição química dos sedimentos sob os cultivos e sua
variação ao longo do tempo;
• avaliar a composição biológica em termos de organismos planctônicos e bentônicos a fim
de detectarem-se possíveis alterações decorrentes do cultivo de bivalves.

83
REFERÊNCIAS BIBLIOGRÁFICAS DA INTRODUÇÃO
BARG, U. C. Guidelines for the promotion of environmental management of coastal aquaculture development. FAO Fisheries Technical Paper, n. 328, 1992. BARILLÉ, L; PROU, J.; HÉRAL, M. & RAZET, D. Effects of high natural seston concentrations on the feeding, selection, and absorption of the oyster Crassostrea gigas (Thunberg). J. Exp. Mar. Biol. Ecol., 212: 149-172, 1997. BAYNE, B. L. & HAWKINS, A. J. S. Filter-feeding in bivalve molluscs: controls of energy balance. In: MELLINGER, J. (ed.) Animal nutrition and transport processes. I. Nutrition in wild and domestic animals. Comp. Physiol. Basel. Karger, 5: 70-83, 1990. BAYNE, B. L.; HAWKINS, A. J. S. & NAVARRO, E. Feeding and digestion by the mussel Mytilus edulis L. (Bivalvia: Mollusca) in mixtures of silt and algal cells at low concentrations. J. Exp. Mar. Biol. Ecol., 111: 1-22, 1987. BAYNE, B. L.; HAWKINS, A. J. S. & NAVARRO, E. Feeding and digestion in suspension-feeding bivalve molluscs: the relevance of physiological compensations. Amer. Zool., 28: 147-759, 1988. BAYNE, B. L.; HAWKINS, A. J. S.; NAVARRO, E. & IGLESIAS, I. P. Effects of seston concentration on feeding, digestion and growth in the mussel Mytilus edulis. Mar. Ecol. Prog. Ser., 55: 47-54, 1989. BAYNE, B. L.; IGLESIAS, I. P.; HAWKINS, A. J. S.; NAVARRO, E.; HERAL, M. & DESLOUS-PAOLI, J. M. Feeding behaviour of the mussel, Mytilus edulis: responses to variations in quality and organic content of the seston. J. Mar. Biol. Ass. U. K., 73: 813-829, 1993. BAYNE, B. L. & WIDDOWS, J. The physiological ecology of two populations of Mytilus edulis L. Oecologia, 37 (2): 137-162, 1978. BAYNE, B. L.; WIDDOWS, J. & NEWELL, R. I. E. Physiological measurements on estuarine bivalve molluscs in the field. In: KEEGAN, B. F.; CEIDIGH, P. O. & BOADEN, P. J. S. (eds.) Biology of benthic organisms – 11th European Symposium on Marine Biology, Galway, October 1976. Oxford: Pergamon Press, 1976. p. 57-68. BERRY, P. F. & SCHLEYER, M. H. The brown mussel Perna perna on the Natal coast, South Africa: utilization of available food and energy budget. Mar. Ecol. Prog. Ser., 13: 201-210, 1983. BERTRÁN, C.; JARAMILLO, E.; BRAVO, A. & VELÁSQUEZ, C. Acuicultura de Mitilidos en un estuario del centro-sur de Chile: efectos de los biodepositos. 1988. 15 p. Separata - Instituto de Zoología, Facultad de Ciencias, Universidad Austral de Chile. CARVALHO FILHO, J. Panorama da malacocultura brasileira. Panorama da Aqüicultura. Vol. 11, n° 64, p. 25-31, 2001.

84
CHAMBERLAIN, J.; FERNANDES, T. F.; READ, P.; NICKELL, T. D. & DAVIES, I. M. Impacts of biodeposits from suspended mussel (Mytilus edulis L.) culture on the surrouding surficial sediments. ICES Journal of Marine Science, 58: 411-416, 2001. CHEVARRIA, G. G. Caracterização biogeoquímica de uma área de cultivo de moluscos - Enseada de Armação do Itapocoroy - Penha/SC. 1999. 60 p. Monografia (Graduação em Oceanografia) - Centro de Ciências Tecnológicas da Terra e do Mar, Universidade do Vale do Itajaí. CONOVER, R. J. Assimilation of organic matter by zooplankton. Limnol. Oceanogr., 11: 338-345, 1966. CRANFORD, P. J.; EMERSON, C.W.; HARGRAVE, B. T. & MILLIGAN, T. G. In situ feeding and absorption responses of sea scallops Placopecten magellanicus (Gmelin) to storm-induced changes in the quantity and composition of the seston. J. Exp. Mar. Biol. Ecol., 219: 45-70, 1998. CRANFORD, P. J. & HARGRAVE, B. T. In situ time-series measurement of ingestion and absorption rates of suspension-feeding bivalves: Placopecten magellanicus. Limnol. Oceanogr., 39(3): 730-738, 1994. DAHLBÄCK, B. & GUNNARSSON, L. Å. H. Sedimentation and sulfate reduction under a mussel culture. Mar. Biol., 63: 269-275, 1981. DAME, R. F. Ecology of marine bivalves: an ecossystem approach. Boca Raton: CRC Marine Science Series, 1996. 254 p. D’AQUINO, C. de A. Papel das ondas e correntes na remobilização de sedimentos finos em zonas de cultivo de moluscos marinhos. 2000. 46 p. Monografia (Graduação em Oceanografia) - Centro de Ciências Tecnológicas da Terra e do Mar, Universidade do Vale do Itajaí. FERREIRA, J. F. & GRÜMANN, A. Cultivo de moluscos marinhos. In: CNI/IEL. Tecnologia e Inovação para a indústria: biotecnologia, novos materiais, tecnologia da informação. Brasília: SEBRAE, 1999. p. 26-29. FERREIRA, J. F. & MAGALHÃES, A. R. M. Cultivo de mexilhões em Santa Catarina. Panorama da Aqüicultura. Vol. 2, n° 14, p. 10-11, 1992. GARDNER, J. P. A. Effects of seston variability on the clearance rate and absorption efficiency of the mussel Aulacomya maoriana, Mytilus galloprovincialis and Perna canaliculus from New Zealand. J. Exp. Mar. Biol. Ecol., 268: 83-101, 2002. GRANT, J.; HATCHER, A.; SCOTT, D. B.; POCKLINGTON, P.; SCHAFER, C. T. & WINTERS, G. V. A multidisciplinary approach to evaluating impacts of shellfish aquaculture on benthic communities. Estuaries, 18(1A): 124-144, 1995. HAVEN, D. S. & MORALES-ALAMO, R. Aspects of biodeposition by oyster and other invertebrate filter feeders. Limnol. Oceanogr., 20: 924-934, 1966. HAVEN, D. S. & MORALES-ALAMO, R. Biodeposition as a factor in sedimentation of five suspended solids in estuaries. Mem. Geol. Soc. Am., 133: 121-130, 1972.

85
HAWKINS, A. J. S.; BAYNE, B. L.; BOUGRIER, S.; HÉRAL, M.; IGLESIAS, J. I. P.; NAVARRO, E.; SMITH, R .F. M. & URRUTIA, M. B. Some general relationships in comparing the feeding physiology of suspension-feeding bivalve molluscs. J. Exp. Mar. Biol. Ecol., 219: 87-103, 1998a. HAWKINS, A. J. S.; FANG, J. G.; PASCOE, P. L.; ZHANG, J. H.; ZHANG, X. L. & ZHU, M. Y. Modelling short-term responsive adjustments in particle clearance rate among bivalve suspension-feeders: separate unimodal effects of seston volume and composition in the scallop Chlamys farreri. J. Exp. Mar. Biol. Ecol., 262: 61-73, 2001. HAWKINS, A. J. S.; JAMES, M. R.; HICKMAN, R.W.; HATTON, S. & WHEATERHEAD, M. Modelling of suspension-feeding and growth in the green-lipped mussel Perna canaliculus exposed to natural and experimental variations of seston availability in the Marlborough Sounds, New Zealand. Mar. Ecol. Prog. Ser., 191: 217-232, 1999. HAWKINS, A. J. S.; SMITH, R. F. M.; BAYNE, B. L. & HÉRAL, M. Novel observations underlying the fast growth of suspension-feeding shellfish in the turbid environments: Mytilus edulis. Mar. Ecol. Prog. Ser., 131: 179-190, 1996. HAWKINS, A. J. S.; SMITH, R. F. M.; BOUGRIER, S.; BAYNE, B.L. & HÉRAL, M. Manipulation of dietary conditions for maximal growth in mussels, Mytilus edulis L., from the Marennes-Oléron Bay, France. Aquat. Living Resour., 10: 13-22, 1997. HAWKINS, A. J. S.; SMITH, R. F. M.; TAN, S. H.; YASIN, Z. B. Suspension-feeding behaviour in tropical bivalve molluscs: Perna viridis, Crassostrea belcheri, Crassostrea iradelei, Saccostrea cucculata and Pinctada margarifera. Mar. Ecol. Prog. Ser., 166: 173-185, 1998b. HAYAKAWA, Y.; KOBAYASHI, M. & IZAWA. M. Sedimentation flux from mariculture of oyster (Crassostrea gigas) in Ofunato estuary, Japan. ICES Journal of Marine Science, 58: 435-444, 2001. IGLESIAS, J. I. P.; NAVARRO, E.; ALVAREZ JORNA, P. & ARMENTIA, I. Feeding, particle selection and absortion in cockles Cerastoderma edule (L.) exposed to variable conditions of food concentration and quality. J. Exp. Mar. Biol. Ecol., 162: 177-198, 1992. IGLESIAS, J. I. P.; PÉREZ CAMACHO, A.; NAVARRO, E.; LABARTA, U.; BEIRAS, R.; HAWKINS, A. J. S. & WIDDOWS, J. Microgeographic variability in feeding, absortion and condition in mussels (Mytilus galloprovincialis Lmk.): an transplant experiment. J. Shellfish Res., 15(3): 673-680, 1996. IGLESIAS, J. I. P.; URRUTIA, M. B.; NAVARRO, E. & IBARROLA, I. Measuring feeding and absorption in suspension-feeding bivalves: an appraisal of the biodeposistion method. J. Exp. Mar. Biol. Ecol., 219: 71-86, 1998. ITO, S. & IMAI, T. Ecology of oyster bed. I.: On the decline of productivity due to repeated culture. Tokoku J. Agric. Res., 5: 251-268, 1955.

86
JARAMILLO, E.; BELTRAN, C. & BRAVO, A. Mussel biodeposition in an estuary in southern Chile. Mar. Ecol. Prog. Ser., 82: 85-94, 1992. JØRGENSEN, C. B. Bivalve filter feeding: hydrodinamics, bioenergetics, physiology and ecology. Fredensborg: Olsen & Olsen, 1990. 140 p. KASPAR, H. F.; GILLESPIE, P. A.; BOYER, I. C. and MACKENZIE, A. L. Effects of mussel aquaculture on the nitrogen cycle and benthic communities in Kenepuru Sound, Marlborough Sounds, New Zealand. Mar. Biol., 85: 127-136. 1985. KAUTSKY, N. & EVANS, S. Role of biodeposition by Mytilus edulis in the circulation of matter and nutrients in a Baltic coastal ecosystem. Mar. Ecol. Prog. Ser., 38: 201-212, 1987. KUSUKI, Y. Relationships between quantities of faecal material produced and quantities of suspended matter removed by the Japanese oyster. Bull. Jap. Soc. Sci. Fish., 44: 1183- 1185, 1978. LABARTA, U.; FERNÁNDEZ-REIRÍZ, M. J. & BABARRO, J. M. F. Differences in physiological energetics between intertidal and raft cultivated mussels Mytilus galloprovincialis. Mar. Ecol. Progr. Ser., 152: 167-173, 1997. LÓPEZ-JAMAR, E.; CAL, R. M.; GONZÀLEZ, G.; HANSON, R. B.; REY, J.; SANTIAGO, G. & TENORE, K. R. Upwelling and douwelling effects on the benthic regime of the continetal shelf of Galicia, NW Spain. Mar. Res., 50: 465-488, 1992. MARIOJOULS, C. & KUSUKI, Y. Appréciation des quantités de biodépôts émis par les huîtres en élevage suspendu dans la baie d’Hiroshima. Haliotis, 16: 221-231, 1987. MATTSSON, J. & LINDÉN, O. Benthic macrofauna succession under mussels, Mytilus edulis L. (Bivalvia), cultured on hanging long lines. Sarsia, 68: 97-102, 1983. MELLO, L. G. M. Taxas de biodeposição e clareamento do mexilhão Perna perna em ambiente de cultivo. 1999. 33 p. Dissertação (Mestrado em Aqüicultura) - Departamento de Aqüicultura, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina. NAVARRO, E.; IGLESIAS, J. I. P.; PÉREZ CAMACHO, A.; LABARTA, U. & BEIRAS, R. The physiological energetics of mussel (Mytilius galloprovincialis Lmk) from different cultivation rafts in the Ria de Arosa (Galicia, NW Spain). Aquaculture, 94: 197-212, 1991. NAVARRO, E.; IGLESIAS, J. I. P.; PÉREZ CAMACHO, A. & LABARTA, U. The effect of diets of phytoplancton and suspended bottom material on feeding and absortion of rafts mussels (Mytilus galloprovincialis Lmk). J. Exp. Mar. Biol. Ecol., 198: 175-189, 1996. NAVARRO, J. M. Oferta alimenticia natural y su efecto sobre los procesos fisiológicos del bivalvo Mytilus chilensis en la bahía de Yaldad, Chiloé (Sur de Chile). Mems. Asoc. Latioam. Acuicult., 5: 175-187, 1983. NAVARRO, J. M. & JARAMILLO, R. Evaluacion de la oferta alimentaria natural disponible a organismos filtradores de la Bahia de Yaldad (43°08’; 73°44’), sur de Chile. Rev. Biol. Mar., Valparaíso, 29(1): 57-75, 1994.

87
NAVARRO, J. M. & THOMPSON, R. J. Comparision and evaluation of different techniques for measuring absortion efficiency in suspension feeders. Limnol. Oceanog., 39(1): 159-164, 1994. NAVARRO, J. M. & THOMPSON, R. J. Physiological energetics of the horse mussel Modiolus modiolus in a cold ocean environment. Mar. Ecol. Prog. Ser., 138: 135-148, 1996. NAVARRO, J. M. & THOMPSON, R. J. Biodeposition by the horse mussel Modiolus modiolus (Dillwyn) during the spring diatom bloom. J. Exp. Mar. Biol. Ecol., 209: 1-13, 1997. NAVARRO, J. M. & ULLOA, L. Effects of environmental factors on physiological responses and fluctuations in the soft tissue weigth of bivalve Aulacomya ater in a bay of southern Chile. In: Proceedings of the 30th European Marine Biological Symposium, Southampton, UK, september, 1995. p. 21-33. NAVARRO, J. M. & WIDDOWS, J. Feeding physiology of Cerastodesma edule in response to a wide range seston concentration. Mar. Ecol. Prog. Ser., 152: 175-186, 1997. NAVARRO, J. M. & WINTER, J. E. Ingestion rate, assimilation efficiency and energy balance in Mytilus chilensis in relation to body size and different algal concentratons. Mar. Biol., 67: 255-266, 1982. NEWELL, R. C. The role of detritus in the nutrition of two marine deposit feeders, the prosobranch Hydrobia ulvae and the bivalve Macoma balthica. Proc. Zool. Soc. Lond., 144: 25-45, 1965. NEWKIRK, G. F. A look at mollusc culture in developing countries: problems and potentials. In: HERNANDEZ R., A. (ed.) Cultivo de moluscos en America Latina. Bogotá: Guadalupe/CIID, 1990. p. 99-114. OKUMUS, I. & STIRLING, H. P. Physiological energetics of cultivated mussel (Mytilus edulis) populations in two Scottish west coast sea lochs. Mar. Biol., 119: 125-131. 1994. OTTMANN, F. & SORNIN, J. Observations on sediment accumulation as a result of mollusk culture systems in France. In: Proceedings of SIUEC. Vol. 1. Rio Grande (RS) - Brasil, 21 to 27, November, 1982. p. 329 - 337. PAREJO, C. B. Moluscos : tecnologia de cultivo. Madrid: Mundi - Prensa, 1989. PÉREZ CAMACHO, A. & GONZÁLEZ, R. La filtración del mejillón (Mytilus edulis L.) en laboratorio. In: Actas do Primeiro Seminario de Ciencias do Mar, As Rías Galegas. Cuadernos da Área de Ciencias Mariñas, Seminario de Estudos Galegos, 1: 427-437, 1984. POUVREAU, S.; BODOY, A. & BUESTEL, D. In situ suspension feeding behaviour of the pearl oyster, Pinctada margaritifera: combined effects of body size and wheater-related seston composition. Aquaculture, 181: 91-113, 2000.

88
POLI, C. R. & LITLLEPAGE, J. Desenvolvimento do cultivo de moluscos no Estado de Santa Catarina. In: Anais do Aquicultura Brasil’98. Volume 1, Recife, 2 a 6 de novembro de 1998. p. 163-181. PROENÇA, L. A. O. & SCHETTINI, C. A. F. Effect of shellfish culture on phytodetritus vertical fluxes in tropical waters - southern Brazil. Rev. Bras. Oceanogr., 46(2): 125-133. 1998. RESGALLA Jr., C.; MANZONI, G. C.; KUROSHIMA, K. N.; REIS Fo., R. W. & LAITANO, K. S. Variabilidade nas taxas fisiológicas do mexilhão Perna perna em dois sítios de cultivo do litoral norte de Santa Catarina. Notas Técnicas da Facimar, 3: 33-40, 1999. ROSA, R. C. C. Impacto do cultivo de mexilhões nas comunidades pesqueiras de Santa Catarina. 1997. 188 p. Dissertação (Mestrado em Aqüicultura) - Departamento de Aqüicultura, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina. SCHETTINI, C.A.F.; CARVALHO, J.B.; JABOR, P. Hidrologia e distribuição de material em suspensão de quatro estuários de Santa Catarina. In: III Simpósio sobre Oceanografia, 1996, São Paulo. Resumos do III Simpósio sobre Oceanografia. São Paulo: IOUSP, 1996. SCHETTINI, C. A. F.; RESGALLA Jr., C. & KUROSHIMA, K. N. Avaliação preliminar da taxa de sedimentação na região de cultivo de moluscos (Perna perna) na enseada da Armação – SC. Notas Técnicas da Facimar, 1: 1-8, 1997. SCHETTINI, C. A. F.; De CARVALHO, J. L. B. & TRUCCOLO, E. C. Aspectos hidrodinâmicos da Enseada da Armação de Itapocoroy, SC. Notas Técnicas da Facimar, 3: 99-109, 1999. SCHETTINI, C.A.F.; SANTOS, M.I.F. & ABREU, J.G.N. Observações dos sedimentos de fundo de uma plataforma abrigada sob influência de atividade de dragagem: Saco dos Limões, Florianópolis, SC. Notas Técnicas da Facimar, 6: 165-175, 2002. STENTON-DOZEY, J. M. E.; JACKSON, L. F. and BUSBY, A. J. Impact of mussel culture on macrobenthic community structure in Saldanha Bay, South Africa. Marine Pollution Bulletin, 39 (1-12): 357-366. 1999. STRICKLAND, J. D. H. & PARSONS, T. R. A manual of sea water analysis. Bulletin n. 125, Otawa: Fisheries Research Board of Canada. 1965. 202 p. SUPLICY, F. M. A capacidade de suporte nos cultivos de moluscos: está no decreto mas poucos sabem do que se trata. Panorama da Aqüicultura, v. 10, n. 57, p. 21-24, 2000. TENORE, K. R.; CORRAL, J. & GONZALEZ, N. Effects of intensive mussel culture on food chain patterns and production in coastal Galicia. NW Spain. Proceedings of SIUEC. Vol. 1. Rio Grande (RS) - Brasil, 21 to 27, November, 1982. p. 321 - 328. TENORE, K. R. & DUNSTAN, W. M. Comparison of rates of feeding and biodeposition of the american oyster, Crassostrea virginica Gmelin, fed different species of phytoplankton. J. Exp. Mar. Biol. Ecol., 12: 19-26, 1973.

89
THOMPSON, R. J. & BAYNE, B. L. Some relationships between growth, metabolism and food in the mussel Mytilus edulis. Mar. Biol., 27: 317-326, 1974. URRUTIA, M. B.; IGLESIAS, J. I. P., NAVARRO, E. & PROU, J. Feeding and absortion in Cerastoderma edule under environmental conditions in the Bay of Marennes – Olerón (Western France). J. Mar. Biol. Assoc. U. K., 76: 431-450, 1996. WINTER, J. E. The filtration rate of Mytilus edulis and its dependence on algal concentration, measured by a continuous automatic recording apparatus Mar. Biol., 22: 317-328, 1973. WINTER, J. E. Feeding experiments with Mytilus edulis L. at small laboratory scale. II. The influence of suspended silt in addition to algal suspensions on growth. In: PERSOONE, G. & JASPERS, E. (eds.) 10th European Symposium of marine Biology, Ostend, Belgium, Sept. 17-23, (1), Wetteren: Universa Press, 1976. WINTER, J. E. & LANGTON, R. W. Feeding experiments with Mytilus edulis L. at small laboratory scale. I. The influence of the total amount of food ingested and food concentration on growth. In: PERSOONE, G. & JASPERS, E. (eds.) 10th European Symposium of marine Biology, Ostend, Belgium, Sept. 17-23, (1), Wetteren: Universa Press, 1976. WONG, W. H. & CHEUNG, S. G. Feeding behaviour of the green mussel, Perna viridis (L.): responses to variation in seston quantity and quality. J. Exp. Mar. Biol. Ecol., 236: 191-207, 1999. WONG, W. H. & CHEUNG, S. G. Feeding rates and scope for growth of green mussels, Perna viridis (L.) and their relationship with food availability in Kat O, Hong Kong. Aquaculture, 193: 123-137, 2001a. WONG, W. H. & CHEUNG, S. G. Feeding rhythms of the green-lipped mussel, Perna viridis (Linnaeus, 1758) (Bivalvia: Mytilidae) during spring and neap tidal cycles. J. Exp. Mar. Biol. Ecol., 257: 13-36, 2001b. VALENTI, W. C. (ed.) Aqüicultura no Brasil : bases para um desenvolvimento sustentável. Brasília: CNPq/Ministério da Ciência e Tecnologia. 2000. 399 p. VINATEA ARANA, L. A. Aqüicultura e desenvolvimento sustentável: subsídios para a formulação de políticas de desenvolvimento da aqüicultura brasileira. Florianópolis: Editora da UFSC, 1999. 310 p. VINATEA ARANA, L. A. Modos de apropriação e gestão patrimonial de recursos costeiros. Estudo de caso sobre o potencial e os riscos do cultivo de moluscos marinhos na Baía de Florianópolis, Santa Catarina. 2000. 245 p. Tese (Doutorado em Ciências Humanas) - Departamento de Ciências Humanas, Centro de Filosofia e Ciências Humanas, Universidade Federal de Santa Catarina, Florianópolis.