
UNIVERSIDADE BANDEIRANTE DE SÃO PAULO INSTITUTO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE MESTRADO EM FARMÁCIA AVALIAÇÃO DA RUTINA E RUTINA COMPLEXADA COM COBRE(II)
NO METABOLISMO LIPÍDICO DE RATOS INDUZIDOS A HIPERCOLESTEROLEMIA
SÃO PAULO 2012

DANIEL ARTUR FREITAS DE OLIVEIRA
AVALIAÇÃO DA RUTINA E RUTINA COMPLEXADA COM COBRE(II)
NO METABOLISMO LIPÍDICO DE RATOS INDUZIDOS A HIPERCOLESTEROLEMIA
SÃO PAULO 2012
Dissertação apresentada ao programa de Mestrado Profissional em Farmácia da Universidade Bandeirante de São Paulo, como requisito parcial à obtenção do título de mestre. Orientadora: Profa. Dra. Cristina E. Okuyama

Dedico este trabalho a minha orientadora Cristina Eunice Okuyama e todo o pessoal do laboratório no qual passei esses últimos 5 anos.

AGRADECIMENTOS
Agradeço aos meus pais Artur Freitas de Oliveira e Rosângela de Oliveira Lopes que
me educaram e ensinaram os valores morais e éticos.
Agradeço aos meus avôs Ruth, Syllas, Tonica e Mãe que contribuíram de forma
decisiva na minha formação e educação possibilitando, com isso, o meu
desenvolvimento.
Aos meus irmãos Cris e Igo, primos Neto, Andreza, Marina, Bia, Pedro, Fael, Bibo,
Mariana, Dorfo, e tios Zé, Mario, Ana, Júlia, Kito, Rita e Silvano que sempre foram
participativos em minha vida me instruindo e incentivando minhas decisões.
Aos meus amigos espalhados pelo mundo que sempre estiveram por perto nos
momentos de dificuldade. Em especial a turma da graduação e a turma de
Araraquara.
A Dona Rosa e seu Mauro Palomino por terem me “adotado” e terem ajudado em
meu desenvolvimento, bem como agradeço a seus filhos Rafael Renan e André.
A todos do laboratório que sempre me auxiliaram nos meus experimentos e nas
disciplinas do mestrado bem como os conselhos dados nos momentos de
necessidade; Cristina Santos, Ozeraldo Viera, Rafael Temístocles, Dayane Maciel,
Maeli Mosena, Aparecida, Valquiria Losano, Lívia Oyafuso, Renata, Janaina, Carol
Medeiros, Débora, Flávio, Henrique e em especial a Elisa Fortes (bifinho) por ser a
companheira de laboratório e por me auxiliar a fazer as rações dos animais e ao Ivair
Donizete por todo auxílio nestes 5 anos de experimentos.
A todos os professores que sempre foram pacientes com minhas dúvidas e
souberam transmitir um pouco de seus conhecimentos; Sérgio, Niraldo, Susana, Isis,
Claudete.

Agradeço especialmente as professoras Regina e Márcia pelo apoio e os inúmeros
momentos de descontração proporcionados e a minha orientadora Cristina por ter
me “aguentado” por estes anos todos.
Agradeço aos amigos da ETEC Pq. St. Antônio por me auxiliarem nos momentos de
sufoco com aulas e dissertação.
Agradeço a minha baixinha Juliana que tem acompanhado e me motivado nesta
última fase do mestrado.
Ao Dr. André Rennó da UNICAMP pelas análises microscópicas dos tecidos
hepáticos.
Ao apoio financeiro da FAPESP, necessário a realização dos experimentos.

RESUMO
As dislipidemias são alterações fisiopatológicas do metabolismo lipídico, e
suas consequências podem gerar transtornos mais graves, principalmente a
formação da placa de ateroma e consequentemente o enfarto do miocárdio e
acidente vascular encefálico. Alguns estudos demonstram que o flavonóide Rutina,
composto derivado de plantas, tem atividade na regulação do metabolismo lipídico,
porém com baixa atividade. Alguns estudos sugerem que a complexação de
compostos fenólicos com metais de transição pode potencializar o efeito biológico do
composto. No presente trabalho foi investigado o efeito do tratamento de ratos
hipercolesterolêmicos com rutina e rutina complexada a cobre através da avaliação
bioquímica e celular de amostra de sangue e análise anatomo-morfológica dos
órgãos. Os animais foram divididos em sete grupos (n=5): G1 – controle, G2 –
veículo, G3 – sinvastatina 10mg/kg, G4 - rutina 16µmol/Kg, G5 - rutina 160µmol/Kg,
G6 - R-Cu2 16µmol/Kg e G7 - R-Cu2 160µmol/Kg. Os animais receberam dieta
suplementada com colesterol, exceto o grupo controle. O aumento estatístico nos
níveis de colesterol total entre os grupos G1 e G2 confirma a indução da
hipercolesterolemia. Ao se realizar uma análise conjunta dos resultados obtidos após
indução da hipercolesterolemia pode-se perceber que a R-Cu2 teve melhor efeito
que a rutina livre, demonstrando uma tendência na inibição do aumento sérico de
colesterol total, triglicerídeo e LDL, sugerindo assim um possível efeito desse
composto no tratamento das dislipidemias. Na avaliação da peroxidação lipídica,
tanto o tratamento com a rutina livre e quanto a R-Cu2, mesmo com a ingestão de
dieta rica em colesterol, não apresentaram resultados estatisticamente diferentes do
grupo controle (grupo com dieta de ração padrão). Na análise anatomopatológica do
fígado dos animais, os grupos R-Cu2 160µmol/Kg e grupo controle não
demonstraram acumulo de lipídeos, processos inflamatórios e necrose do tecido
hepático enquanto nos demais grupos estas alterações estavam presentes.
Palavra-chave: hipercolesterolemia; flavonóide; peroxidação lipídica.

ABSTRACT
Dyslipidemias are the pathophysiological alterations of lipid metabolism and its
consequences can lead to more serious disorders, especially the formation of
atheromatous plaques and consequently myocardial infarction and stroke. Some
studies show that the flavonoid Rutin, plant-derived compound, has activity in the
regulation of lipid metabolism, but the results are not as impressive. Some studies
suggest that complexation of phenolic compounds with transition metals can enhance
the biological effect of the compound. In the present work was investigated the effect
of treatment of hypercholesterolemic rats with rutin and rutin complexed to copper
thtough biochemical and cellular analysis of blood samples and anatomo-
morphologic evaluation of organs. The animals were divided into seven groups (n =
5): G1 - control, G2 - vehicle, G3 - 10mg/kg simvastatin, G4 - 16μmol/Kg rutin, G5 -
160μmol/Kg rutin, G6 - R-Cu2 16μmol/Kg and G7 - R-Cu2 160μmol/Kg. The animals
were fed a diet supplemented with cholesterol, except the control group. The
statistical difference in total cholesterol levels between groups G1 and G2 confirms
the correct induction of hypercholesterolemia. When conducting an analysis of the
results obtained after induction of hypercholesterolemia can be seen that the R-Cu2
had better effect than the free rutin, showing a strong tendency to inhibit the increase
in serum total cholesterol, triglycerides and LDL, suggesting a possible effect this
compound in dyslipidemias treatment. In the evaluation of lipid peroxidation,
treatment with both free and rutin as the R-Cu2, even with the intake of high-
cholesterol diet, were not statistically different from control group (with standard chow
diet). On pathological examination the highest concentration of the compound R-Cu2
no difference with the control group G1, demonstrating the hepatoprotective activity
of the compound.
Keyword: hypercholesterolemia; flavonoid; lipid peroxidation.

LISTA DE FIGURAS
Figura 1 – Estrutura básica de um ácido graxo.................................................
Figura 2 – Estruturas básicas de um fosfolipídio e um esfingofosfolipídio........
Figura 3 – Estrutura básica de uma molécula de colesterol..............................
Figura 4 - Estrutura de um colesterol................................................................
Figura 5 - O transporte reverso do colesterol....................................................
Figura 6 – Esquema do metabolismo lipídico....................................................
Figura 7 – Estrutura química do flavonoide rutina.............................................
Figura 8 – Possíveis sítios de ligação do metal de transição na molécula de
flavonoide..........................................................................................................
Figura 9 - Níveis séricos de colesterol total em ratos........................................
Figura 10 - Níveis séricos de triglicerídeos em ratos...........................................
Figura 11 - Níveis séricos de HDL em ratos....................................................
Figura 12 - Níveis séricos de LDL em ratos.......................................................
Figura 13 - Avaliação da peroxidação lipídica......................................................
Figura 14 - Avaliação do NO plasmático............................................................
Figura 15 - Contagem de hematócrito em ratos...................................................
Figura 16 - Contagem total de leucócitos em ratos..............................................
Figura 17 - Contagem diferencial de leucócitos em ratos – número de
monócitos.............................................................................................................
Figura 18 - Contagem diferencial de leucócitos em ratos – número de
linfócitos.............................................................................................................
Figura 19 - Contagem diferencial de leucócitos em ratos – número de
neutrófilos.............................................................................................................
Figura 20 - Peso relativo dos rins esquerdo e direito de ratos............................
Figura 21 - Peso relativo de corações provenientes de ratos..............................
Figura 22 - Peso relativo do fígado de ratos.....................................................
Figura 23 - Imagem de fígado obtido após ensaio com dieta rica em colesterol.
Figura 24 - Avaliação microscópica de aorta abdominal obtida após ensaio
com dieta rica em colesterol.................................................................................
16
17
18
19
22
23
29
35
42
43
44
44
47
48
49
50
50
51
51
53
53
54
54
56
57

Figura 25 - Avaliação microscópica da área 1 do tecido hepático de ratos
submetidos a dieta rica em colesterol por 60 dias ............................................
Figura 26 - Avaliação microscópica da área 2 do tecido hepático de ratos
submetidos a dieta rica em colesterol por 60 dias...............................................
Figura 27 - Avaliação microscópica da área 3 do tecido hepático de ratos
submetidos a dieta rica em colesterol por 60 dias...............................................
58
58

LISTA DE TABELAS
Tabela 01 - Grupos de animais divididos de acordo com o tratamento.................
Tabela 02: Análise macroscópica de alguns tecidos dos animais do grupo
controle e dos grupos que receberam dieta suplementada com colesterol...........
38
55

LISTAS DE ABREVIATÚRAS ABCA1 – ATP-binding cassete transportador A1.
ABCG1 - ATP-binding cassete transportador G1.
ADFMChR - 5-allyl-7gen-difluoromethylenechrysin.
Apo – Apoproteína
ATP – Adenosina trifosfato.
CETP – Cholesterol Ester tranfer protein (proteína transportadora de colesterol
esterificado).
CT – Colesterol total.
DNA – Ácido desoxirribonucleico.
IDL – Intermediary Density Lipoprotein (lipoproteína de densidade intermediária)
IMC – Índice de massa corpórea.
IMK – Índice menopausal de Kupperman.
HDL – High Density Lipoprotein (lipoproteína de alta densidade)
HE – Hematoxilina Eosina.
HIV-1 – Vírus da imunodeficiência humana 1.
LCAT – Lecitina colesterol acil transferase.
LDL – Low density lipoprotein (lipoproteína de baixa densidade)
LDLox – Lipoproteínas de baixa densidade oxidadas.
LPL – Lipase Lipoprotéica.
MTT - 3-(4,5- dimethylthiaxolone-2-yl)-2,5-diphenyl tetrazolium bromide.
NK – Natual Killer.
OMS – Organização mundial da saúde.
PLTP – Phospholipids transfer protein (protein transportadora de fosfolipídeos).
Raios U.V. – Raios ultravioleta
R-Cu2 – Rutina complexada a cobre.
RNS – Espécies reativas do nitrogênio.
ROS – Espécies reativas do oxigênio.
R.P.M. – rotações por minuto.
RS+ - Espécie reativa de tióis.
Rut – Rutina.
SOD – Superóxido dismutase.

SR-B1 – Receptor de sequestro classe B tipo 1.
TBA – Ácido tiobarbitúrico.
TBARS – Substãncias reativas ao ácido tiobarbitúrico.
TCA – Ácido Tricloroacético
TG – Triglicérides.
TRC – Transporte Reverso do colesterol.
VLDL – Very Low Density Lipoprotein (lipoproteína de densidade muito baixa)

LISTA DE SÍMBOLOS
H2O2 – Peróxido de hidrogênio.
NO – Óxido Nítrico.
NO+ - Câtion nitrosônio.
•NO – radical óxido nítrico.
NO- - Ânion nitroxila.
O2 – Oxigênio.
O2•- - Ânion superóxido.
•OH – Radical hidroxila.
ONOO- - Peroxinitrito.

SUMÁRIO
1. INTRODUÇÃO........................................................................................................
2. REVISÃO DA LITERATURA..................................................................................
2.1. METABOLISMO LIPÍDICO..................................................................................
2.2. HIPERLIPIDEMIA................................................................................................
2.2.1. Disfunção endotelial e a aterosclerose.............................................................
2.2.2. Radicais livres, estresse oxidativo e peroxidação lipídica................................
2.3. FLAVONÓIDES...................................................................................................
2.3.1. Atividades biológicas dos flavonóides..............................................................
2.3.2. Flavonóides e Metais de Transição..................................................................
3. OBJETIVO GERAL................................................................................................
3.1. OBJETIVOS ESPECÍFICOS...............................................................................
4. MATERIAIS E MÉTODOS....................................................................................
4.1. PREPARAÇÃO DA DROGA COMPLEXADA.....................................................
4.2. ENSAIOS COM DIETA HIPERCOLESTEROLÊMICA........................................
4.2.1. Animais.............................................................................................................
4.2.2. Protocolo Experimental.....................................................................................
4.2.3. Dietas................................................................................................................
4.2.4. Dosagens sanguíneas.......................................................................................
4.2.4.1. Dosagens bioquímicas...................................................................................
4.2.4.2. Avaliação da peroxidação lipídica..................................................................
4.2.4.3. Dosagem plasmática de NO..........................................................................
4.2.4.4. Análise hematológica.....................................................................................
4.2.5. Estudo anatomopatológico................................................................................
4.4. ANÁLISE ESTATÍSTICA......................................................................................
5. RESULTADOS E DISCUSSÃO..............................................................................
5.1. DOSAGENS SANGUÍNEAS................................................................................
5.1.1. Dosagens bioquímicas......................................................................................
5.1.2. Avaliação da peroxidação lipídica....................................................................
5.1.3. Dosagem plasmática de óxido nítrico (NO)......................................................
5.1.4. Análise hematológica........................................................................................
5.2. ESTUDO ANATOMOPATOLÓGICO....................................................................
15
16
16
23
24
26
28
30
34
36
36
37
37
37
37
37
38
39
39
39
40
40
41
41
42
42
42
46
48
49
52

5.2.1. Avaliação macroscópica - Peso relativo dos órgãos.........................................
5.2.2. Avaliação macroscópica – Análise do fígado....................................................
5.2.3. Avaliação microscópica - Análise da aorta........................................................
5.2.4. Avaliação microscópica - Análise do fígado......................................................
6. CONCLUSÃO..........................................................................................................
REFERÊNCIA.........................................................................................................
ANEXOS..................................................................................................................
52
54
55
56
61
62
69

15
1. INTRODUÇÃO
As mudanças nos hábitos de vida decorrentes da transformação da
sociedade na segunda metade do século XX, principalmente nos hábitos alimentares
e num estilo de vida sedentário, acarretaram num aumento expressivo nos índices
de obesidade e consequentemente de doenças associadas ao sobrepeso tal como
as dislipidemias.
Estas são desordens no metabolismo lipídico e tem como característica
laboratorial o aumento dos níveis séricos de colesterol total, principalmente a fração
LDL (lipoproteína de densidade baixa). As dislipidemias podem evoluir para outras
patologias como as doenças cardiovasculares, que segundo estudos da organização
mundial da saúde são as maiores causa de mortes no mundo.
Enquanto esses números tendem a diminuir em países desenvolvidos, no
Brasil a mortalidade decorrente de doenças cardiovasculares tem aumentado.
Estudos do Ministério da Saúde demonstraram que na década de 90,
aproximadamente 32% da população apresentavam sobrepeso (IMC ≥ 25) e que
38% dos homens e 42% das mulheres possuíam os níveis de colesterol total acima
do recomendado (CT> 200mg/dL).
Flavonóides são metabólitos derivados, principalmente, de plantas, estando
presente na dieta diária das pessoas e sendo associados a diversas atividades
biológicas benéficas. Por apresentarem uma característica antioxidante e alguns
estudos sugerirem que compostos flavonóides possam ter uma atividade
hipolipemiante, estes compostos se tornam interessantes no uso terapêutico para a
prevenção ou tratamento de dislipidemias e suas decorrências patológicas. Entre
esses compostos a rutina é um dos mais testados e demonstra ter atividade no
metabolismo lipídico, porém de forma modesta. A complexação de flavonóides a
metais de transição pode potencializar sua atividade biológica, podendo fazer desta
uma molécula interessante para a prevenção das dislipidemias.

16
2. REVISÃO DA LITERATURA
2.1 METABOLISMO LIPÍDICO
Lipídeos é a denominação para um grupo de compostos com estruturas
químicas variadas, relacionados direta ou indiretamente com os ácidos graxos e de
característica hidrofóbica. Os lipídeos mais importantes para o organismo, do ponto
de vista fisiológico, são os triglicérides, fosfolipídeos, o colesterol e o colesterol
esterificado (GENEST, 2003; COBBE; SHEPHERD, 1993).
Os ácidos graxos são cadeias de carbono hidroxiladas e são classificados em
dois grupos, os saturados, sem duplas ligações entre seus átomos de carbono,
mono ou poli-insaturados de acordo com o número de ligações duplas na sua
cadeia. São exemplos de ácidos graxos, o ácido láurico, mirístico, palmítico e
esteárico, todos saturados e os ácidos oléico e linoleico, insaturados.
Quimicamente, os triglicerídeos são compostos por três ácidos graxos ligados
covalentemente a uma molécula de glicerol (figura 1), sendo que estes ácidos
graxos podem variar no seu tipo (número de carbonos e presença ou não de duplas
ligações). O triglicerideo tem características apolares e hidrofóbicas (GENEST, 2003;
COBBE; SHEPHERD, 1993).
Figura 1 – Estrutura básica de um ácido graxo (superior esquerdo), um
glicerol (inferior esquerdo) e de um triglicerídeo (direita).
Fonte: www.ufsc.br (2005)

17
Há dois tipos básicos de fosfolipídeos (figura 2), os mais abundantes no
organismo humano são os constituídos por um esqueleto de glicerol com dois ácidos
graxos e um grupo fosfato ligado a ele, sendo que estes ácidos graxos podem variar
no seu tipo (tipo de ligações e quantidade de carbono em sua cadeia). No grupo
fosfato, podem-se encontrar quatro diferentes moléculas ligadas a ele: colina, dando
origem a lecitina; etanolamina, que origina a fosfatidilinositol; serina que origina a
fosfatilserina; e o inositol, que forma o fosfadilinositol. Os fosfolipídeos estão
presentes em todas as membranas celulares de todas as células e estão envolvidas
nas vias de transdução de sinais após sua hidrólise pela fosfolipase (enzima que
hidrolisa os fosfolipídeos) (COBBE; SHEPHERD, 1993; GENEST, 2003; VON
ECKARDSTEIN et al., 2001).
O outro tipo básico de fosfolipídeo é o esfingofosfolipídeo, quimicamente este
difere dos fosfolipídeos clássicos por não ter a molécula de glicerol, ao invés deste,
a ligação do grupo fosfato e ácidos graxos é feita pela molécula de esfingosina
(figura 2). Este tipo de fosfolipídeo é importante nos processos de manutenção da
estrutura lipoprotéica das membranas, sinalização intracelular, interação entre
células e matriz extracelular, modulação dos receptores de fatores de crescimento
mediando o crescimento celular, diferenciação e morte celular, formação de
microdomínios na membrana plasmática e na formação da bainha de mielina
(GENEST, 2003; MINAMI, 2004; VON ECKARDSTEIN et al., 2001).
Figura 2 – Estruturas básicas de um fosfolipídio e um esfingofosfolipídio.
Fonte: www.fcfar.unesp.br .

18
O colesterol é um álcool monoídrico não saturado da classe dos esteróis,
composto por 27 carbonos (figura 3), e com uma fraca porção polar. É componente
essencial das membranas de células de mamíferos e é substrato para a formação
dos hormônios esteroidais e ácidos biliares. O colesterol é essencial para função das
membranas celulares. A maior parte do colesterol circulante no plasma sanguíneo
está na forma de colesterol esterificado (COBBE; SHEPHERD, 1993; GENEST,
2003).
Figura 3 – Estrutura básica de uma molécula de colesterol
Fonte: www.fcfar.unesp.br.
Devido às inúmeras funções dos lipídeos e o fato dos ácidos graxos
essenciais não poderem ser sintetizados no corpo humano, estes necessitam serem
adquiridos através da dieta. O consumo de ácidos graxos no organismo é alto,
devido estes serem utilizados como fonte de energia, então estes devem ser
ingeridos diariamente para suprir os níveis necessários. E entre os lipídeos, o mais
consumido, na média, são os triglicerídeos, sendo ingerido, aproximadamente, 66g
deste lipídeo. A média de consumo do colesterol é de 250 mg (COBBE;
SHEPHERD, 1993; GENEST, 2003; GINSBERG, 2002).
As características hidrofóbicas dos lipídeos fazem com que estes não possam
ser transportados livremente no plasma (constituído principalmente por água), porém
o metabolismo lipídico evoluiu de forma a conseguir transportar eficientemente estas
moléculas hidrofóbicas de seu sítio de síntese para o sítio de uso (GENEST, 2003;
NOVAK, BYDLOWSKI, 1996).
A maior parte dos lipídeos serão transportados no organismo acrescido de
proteínas específicas, as apoproteínas, formando complexos macromoleculares
chamados de lipoproteínas. As lipoproteínas são constituídas por fosfolipídios e

19
colesterol livre em sua superfície e triglicérides e ésteres de colesterol em seu
núcleo (figura 4) (GENEST, 2003; GINSBERG, 2002; NOVAK, BYDLOWSKI, 1996).
Figura 4: Estrutura de um colesterol
Fonte: modificado de GENEST (2003).
Estas apoproteínas são classificadas como apoA-I, apoA-II, apoA-IV, apoB-
100, apoB-48, apoC-I, apoC-II, apoC-III e apoE e diferem em seu tamanho,
composição química e função na lipoproteína. As quatro funções básicas destas
são: na formação (montagem) e secreção das lipoproteínas; manter a integridade
estrutural das lipoproteínas; sendo co-fator enzimático; e para se ligarem a proteínas
e receptores específicos para a captação celular (NOVAK, BYDLOWSKI, 1996;
GINSBERG, 2002).
Os lipídeos irão apresentar dois trajetos básicos, do local de sua absorção ou
síntese (intestino e fígado respectivamente) para os tecidos, principalmente tecido
muscular e tecido adiposo. E o trajeto das moléculas de colesterol dos tecidos
periféricos para a síntese de membranas, produção de hormônios esteroidais ou
para o fígado para que sejam sintetizados ácidos biliares (GENEST, 2003). Porém,
estes são mais comumente classificados conforme o seu local de origem, podendo
vir da dieta (ciclo exógeno) ou podem ser sintetizados no próprio organismo (ciclo
endógeno) (COBBE; SHEPHERD, 1993).
No ciclo exógeno, ocorre a absorção de lipídeos em nível intestinal. No
intestino, os lipídeos serão emulsificados pelos sais biliares, formando micelas,
depois serão hidrolisados pelas lipases pancreáticas, liberando com isso, ácidos

20
graxos e mono ou diglicerídeos. Estes caem na circulação linfática, e irão formar a
primeira lipoproteína do metabolismo lipídico, o quilomicron. Este irá apresentar a
apoB-48 como apoproteína majoritária, porém será formada também pelas e apoA-I
e apoC-II. Os quilomicrons são constituídos basicamente por triglicerídeos (maior
parte), 9% de fosfolipídeos, 3% de colesterol e 1% de apoB-48. Os quilomicrons
entram rapidamente no compartimento plasmático, onde sofrerá a ação da enzima
lipase lipoprotéica (LPL), que é ativada pela apoC-II, esta enzima está presente na
parede dos vasos sanguíneos e vai hidrolisar 90% dos triglicerídeos. Mediados pela
insulina as moléculas de ácidos graxos serão rapidamente captadas pelas células
musculares para servirem como recurso energético ou então são captados pelo
tecido adiposo para atuarem como reserva energética. Os remanescentes
quilomicrons terão a apoE e devido a isso serão retirados da circulação através de
receptores específicos para essa apo localizados no tecido hepático (GENEST,
2003; NOVAK, BYDLOWSKI, 1996; GINSBERG, 2002). No fígado os lipídeos serão
metabolizados e começará o ciclo endógeno. São 4 os tipos de lipoproteínas
responsáveis por transportar os lipídeos no ciclo endógeno (GUYTON, 1996).
As VLDL (Very low density lipoprotein – Lipoproteínas de densidade muito
baixa): lipoproteínas que apresentam grande concentração de
triglicerídeos.
IDL (intermediary density lipoprotein – lipoproteínas de densidade
intermediária): lipoproteínas que sofreram remoção de boa parte das
moléculas de triglicerídeos e apresentam elevada concentração de
colesterol.
LDL (low density lipoprotein, lipoproteínas de baixa densidade):
lipoproteínas no qual foram removidas todas as moléculas de triglicerídeos
sobrando grande concentração de colesterol.
HDL (high density lipoprotein – lipoproteína de alta densidade): lipoproteína
que apresenta grande concentração de apoproteínas.
Como as necessidades de ácidos graxos são constantes no organismo e
estes não são adquiridos a todo o momento através do ciclo exógeno, o fígado
secreta moléculas de VLDL que irá abastecer os tecidos periféricos com ácidos
graxos. A VLDL é menor que o quilomicron, porém, também apresenta uma grande
concentração de triglicérides e contém a apo-B100 como apoproteína majoritária e

21
as apoproteínas A-I e C-II como secundárias. Os triglicérides serão rapidamente
hidrolisados pela ação da LLP, liberando ácidos graxos para os tecidos. Com a
hidrólise, há uma alteração de apoproteínas e lipídeos presentes na VLDL, ela passa
a conter apo´s C e E e ocorre uma substituição de triglicérides e fosfolipídeos por
ésteres de colesterol, estes são adquiridos da HDL mediada pela ação de duas
enzimas, a cholesterol ester transfer protein (CETP) e pela phospholipids transfer
protein (PLTP), com isso, será formada a IDL, que apresenta a apo B-100 e apo E
como apo´s majoritárias. Esta apresentará dois destinos, ou pode ser recaptada pelo
tecido hepático devido à afinidade de suas apoproteínas com os receptores do
tecido hepáticos ou será processada pela lipase hepática se tornando uma LDL, que
permanecem por longo tempo no plasma (COBBE; SHEPHERD, 1993; GENEST,
2003; GINSBERG, 2002).
A LDL tem um conteúdo apenas residual de triglicerídeos e é composta
principalmente de colesterol e uma única apoproteína, a apo B100, em geral os
triglicerídeos representam somente de 4 a 8% da massa da LDL. Estas lipoproteínas
são responsáveis pela maior parte do transporte de colesterol pelo organismo
abastecendo os tecidos periféricos ou então são removidas pelo fígado, nos dois
casos a mediação ocorre através dos receptores específicos da apoB-100. A
expressão desses receptores é a principal responsável pelo nível de colesterol no
sangue, eles são controlados pela atividade da enzima hidroxi-metil-glutaril CoA
redutase. Quando há uma alta demanda de colesterol pelas células, os receptores
B/E são ativados para aumentar a captação da LDL. Inversamente, quando a célula
está repleta de colesterol, o mecanismo de regulação negativa do receptor protege-a
contra os efeitos deletérios da acumulação deste lipídeo (GENEST, 2003;
GINSBERG, 2002; NOVAK, BYDLOWSKI, 1996).
Este processo é chamado de transporte reverso do colesterol (TRC) e o
mecanismo principal na atividade antiaterogênica do HDL. A HDL é sintetizada no
fígado ou intestino e é primeiramente denominada de HDL nascente, esta contém
pouca apoA-I (apoproteína majoritária da HDL). No plasma, esta HDL nascente
capta colesterol de células periféricas mediado por uma proteína celular específica,
a ATP-binding cassete transportador A1 (ABCA1), com isso ele se trona um HDL
maduro, com um formato esférico. Este colesterol então é esterificado pela enzima
lecitina colesterol acil transferase (LCAT) e se transfere para o núcleo do HDL. Esta
lipoproteína já madura também capta colesterol de células periféricas e de

22
macrófagos, isto pode ocorrer através da ATP-binding cassete transporter G1
(ABCG1) ou receptor de sequestro classe B tipo 1 (SR-BI), aumentando, com isso, o
nível de colesterol esterificado na HDL. Esta pode trocar lipídeos com outras
lipoproteínas, como a IDL e LDL, no caso colesterol esterificado por triglicérides,
esta troca é mediada pela proteína cholesteryl ester transfer protein (CETP) (figura
5). As HDL restantes retornam ao fígado, através da interação com os receptores
SR-BI. No fígado, este colesterol poderá ser utilizado na formação de VLDL´s ou
ácidos biliares (WANG; PENG, 2011; RADER, 2002; DEAN et al., 2001; VON
ECKARDSTEIN et al., 2001).
Figura 5 - O transporte reverso do colesterol. A apo A-I pobre em lipídeos interage com
as células periféricas e captam o colesterol e o fosfolipídeo através do transporte mediado
pelo ABCA1. A HDL madura também capta o colesterol dos macrófagos via SR-B1. O
colesterol no HDL é convertido em éster de colesterol através da enzima LCAT. A HDL
então se liga aos receptores SR-B1 hepáticos e o fígado secreta colesterol diretamente na
bile ou converte-o a ácidos biliares que são secretados pela bile.
Fonte: RADER (2002).
Esta lipoproteína ainda contribui significativamente contra a formação da
placa de ateroma através de mecanismos como a remoção de lipídeos oxidados de
LDL, inibição da fixação de moléculas de adesão e monócitos ao endotélio e
estimulação da liberação de óxido nítrico (REDONDO et al., 2011; VON
ECKARDSTEIN et al., 2001).

23
Figura 6 – Esquema do metabolismo lipídico. Ciclo exógeno mostra formação dos quilomícrons e o
transporte lipídico até o fígado. Ciclo endógeno mostra a formação da VLDL, IDL e LDL e o transporte
dos lipídeos do fígado até os tecidos. A HDL é responsável pelo transporte reverso levando os
lipídeos dos tecidos até o fígado.
Fonte: University of London (2006).
2.2 HIPERLIPIDEMIA
A hiperlipidemia ou hiperlipoproteinemia são dislipidemias e podem ser
definidas como um desequilíbrio nos níveis de lipídeos no organismo, em especial o
triglicerídeo e o colesterol, resultando em um aumento das concentrações
plasmáticas de lipoproteínas não HDL. No organismo sadio há um equilíbrio entre as
concentrações de lipídeos necessários para o metabolismo celular, evitando o
acúmulo excessivo destes. Porém, por razões ainda não definidas totalmente, pode
ocorrer um desequilíbrio que leva à elevação destas moléculas no organismo
(MUSUNURU, 2010).
Hiperlipidemia é um fator de risco para doença cardiovascular. A deposição
de lipídeos no plasma na íntima arterial induz uma resposta inflamatória local e

24
remodelação vascular extensa resultando na formação de placas ateroscleróticas. A
ruptura da placa ou erosão provoca infarto do miocárdio ou acidente vascular
cerebral (POREZ et al., 2012 )
Quanto a sua etiologia elas podem ser classificas em dislipidemias primárias -
decorrentes de fatores genéticos e/ou ambientais como alteração de apoproteínas
ou o sedentarismo, respectivamente - e dislipidemias secundárias - decorrentes de
doenças (diabetes), aos efeitos de medicamentos (anti-hipertensivos, corticóides,
inibidores de proteases) e aos hábitos de vida inadequados (tabagismo, etilismo)
(GONÇALVES et al., 2006).
A hipertrigliceridemia é caracterizada pelo aumento sérico de VLDL, enquanto
que a hipercolesterolemia é consequência do aumento da LDL sérica. Seja com a
hipertrigliceridemia ou a hipercolesterolemia, aumentam-se as chances de
surgimento de doenças, principalmente cardiovasculares. O diagnóstico precoce e
controle dos níveis séricos desses lipídeos, contribui para sensível queda da
incidência e da mortalidade por doença cardiovascular (GENEST, 2003;
MUSUNURU, 2010; GONÇALVES et al., 2006).
As projeções da OMS (Organização Mundial de Saúde) mostram que a
morbidade e a mortalidade causadas por doenças cardiovasculares, nas próximas
décadas, serão a primeira causa de morte em todo o mundo. No Brasil a doença
aterosclerótica coronariana e cerebrovascular já são a maior causa de incapacidade
e morte, correspondendo a 32,1% de todas as causas, concomitantemente, esses
números também vem crescendo em outros países da Europa, Ásia e América do
Sul. Sendo as dislipidemias a principal causa da gênese do mecanismo
fisiopatológico do processo aterosclerótico (MAGALHÃES et al., 2004).
2.2.1 Disfunção endotelial e a Aterosclerose
A aterosclerose é descrita como uma doença inflamatória crônica de origem
multifatorial que ocorre em resposta à agressão endotelial, tendo como
consequência o espessamento da camada sub-íntima macular (ateroma) através do
acúmulo de lipídeos, células inflamatórias e elementos fibrosos, obstruindo total ou
parcialmente o fluxo sanguíneo, acometendo principalmente artérias de médio e

25
grande calibre, sendo a aorta, coronárias, carótidas e cerebrais as de maior
importância clínica (POREZ et al., 2012; BERLINER et al.,1995; VANDERLAAN,
2004).
Consequentemente, a disfunção endotelial aumenta a permeabilidade da
camada íntima, favorecendo a retenção de lipoproteínas. As LDL´s irão sofrer
processo de oxidação e se tornarão imunogênicas proporcionando a quimiotaxia
para macrófagos. Os macrófagos fagocitam as lipoproteínas oxidadas e ficam
retidos na camada íntima do vaso sanguíneo, causando o acúmulo de lipídeos na
parede arterial e consequentemente a formação da placa de ateroma. Estas células
são chamadas de células espumosas, devido a sua grande quantidade de lipídeo
intra-celular (MOURA, 2006; POREZ et al., 2012; BERLINER et al.,1995;
VANDERLAAN, 2004).
Alguns mediadores inflamatórios fazem com que ocorra a migração e
proliferação das células musculares lisas da camada média arterial para a camada
íntima. Nesta camada passam a produzir citocinas e fatores de crescimento, além de
matriz extracelular que formará parte da capa fibrosa da placa aterosclerótica. A
placa de ateroma em estágio desenvolvido é constituída por elementos celulares,
componentes da matriz extracelular e núcleo lipídico. As placas estáveis
caracterizam-se por ter uma capa fibrosa espessa devido ao predomínio de
colágeno, poucas células inflamatórias e uma pequena porção de lipídeos, formando
seu núcleo. As instáveis apresentam uma fina camada fibrótica, atividade
inflamatória intensa, especialmente nas suas bordas laterais, núcleo lipídico bem
proeminente (BERLINER et al.,1995).
A formação da placa de ateroma tem início com lesão endotelial, esta pode
ocorrer por diversos motivos como o aumento sérico das lipoproteínas, em especial
a LDLox (LDL oxidadas), hipertensão arterial e o estresse oxidativo. O principal fator
de risco utilizado na clínica para a prevenção da formação da placa de ateroma tem
sido a avaliação dos níveis séricos de lipoproteínas, principalmente a LDL. Porém, a
LDL nativa, natural, não tem característica aterogênica, esta se torna agressiva
somente quando modificada, principalmente pela ação de radicais livres,
promovendo sua oxidação. Outro importante fator relacionado com a lesão endotelial
é a diminuição da síntese, liberação e atividade do óxido nítrico (NO)
(VANDERLAAN, 2004; BERLINER et al.,1995).

26
O NO é um mediador sintetizado e continuamente liberado pelas células
endoteliais, com importante papel na fisiologia da musculatura lisa vascular
(SPIEKER et al., 2006). Além de ser um vasodilatador potente (IGNARRO et al.,
1981; HUANG et al., 1995), o NO inibe a adesão (RADOMSKI et al., 1987) e
agregação (RADOMSKI et al., 1990) plaquetária e, também, otimiza o fluxo
sanguíneo na microcirculação (MONCADA, HIGGS, 2006).
Uma vez produzido, o NO difunde-se facilmente da célula geradora para a
célula alvo, onde interage rapidamente com moléculas específicas. Sua atuação é
principalmente local, devido ao fato de possuir uma meia-vida curta no espaço
extracelular antes de ser convertido em espécies reativas. Existem relatos que em
condições onde há evidência da deficiência de NO, o processo de aterosclerose está
sendo iniciado ou avançado.
Atualmente, terapias envolvendo o metabolismo das lipoproteínas plasmáticas
ou do colesterol celular, e a intervenção nos processos inflamatórios são importantes
na terapia da aterosclerose (RADER, DAUGHERTY, 2008). O uso de compostos
com atividade antioxidante apresenta grande participação nas terapias, devido ao
sequestro de radicais livres e consequentemente à diminuição de danos celulares.
2.2.2 Radicais livres, estresse oxidativo e peroxidação lipídica
Nos últimos anos, inúmeras evidências têm indicado o papel chave dos
radicais livres e outros oxidantes como grandes responsáveis pelo envelhecimento e
pelas doenças degenerativas associadas ao envelhecimento, como câncer, doenças
cardiovasculares, catarata, declínio do sistema imune e disfunções cerebrais (ATOUI
et al., 2005).
Radicais livres são moléculas instáveis, sendo as mais importantes no
metabolismo humano, as originadas a partir do oxigênio. Este se encontra em
abundância no organismo, sendo gerados, principalmente, no metabolismo
energético (OLSZEWER, 1994).
Um radical livre é definido como qualquer átomo, grupo de átomos ou
molécula contendo um ou mais elétrons desemparelhados ocupando a órbita
externa. O ânion superóxido (O2•-), o radical hidroxila (•OH) e o óxido nítrico (•NO)

27
são exemplos de radicais livres. Existem, entretanto, compostos igualmente reativos
que não possuem elétron desemparelhado na última camada e, portanto, não
podem ser classificados como radicais livres (DRÖGE, 2002). Essas substâncias
são classificadas de maneira mais ampla como espécies reativas, e as mais
estudadas nos sistemas biológicos são as espécies reativas de oxigênio (ROS),
espécies reativas de nitrogênio (RNS), espécies reativas de tióis (RS+), espécies
reativas de cloro, espécies reativas do carbono e complexos de metais de transição
(OLIVEIRA et al., 2009), e incluem o peróxido de hidrogênio (H2O2), o cátion
nitrosônio (NO+), o ânion nitroxila (NO-) e o peroxinitrito (ONOO-) (DRÖGE, 2002).
Em geral, sua instabilidade se dá pelo não pareamento dos elétrons em sua
camada externa. Como tendência natural dos átomos, estes buscam o equilíbrio.
Para isto, necessitam de mais um elétron para conseguir o pareamento de sua
última camada, capturando elétrons de outras moléculas, desestabilizando-as e
provocando um “efeito cascata” (OLSWER, 1994).
Os organismos aeróbios produzem a adenosina trifosfato (ATP) através da
redução completa do O2 por quatro elétrons, através do transporte mitocondrial de
elétrons. Aproximadamente 98% de todo o oxigênio consumido pelas células entram
nas mitocôndrias, onde são reduzidas pela citocromo oxidase. Entretanto, o oxigênio
pode receber menos de quatro elétrons e formar espécies reativas de oxigênio
(RODRIGUES et al., 2003).
Em condições normais, aproximadamente, 5% do oxigênio absorvido pelo
organismo se torna um radical livre, porém, podem ocorrer alterações
fisiopatológicas que alterem este percentual, como por exemplo, o que ocorre nas
inflamações (OLSWER, 1994). Esse aumento do percentual de transformação de O2
em ROS, ou mais amplamente descrevendo, o estresse redox (que pode envolver
vários tipos de espécies reativas), também pode ocorrer devido a hábitos de vida
considerada inadequados como consumo excessivo de álcool, tabagismo, dieta
inadequada, prática exacerbada de exercícios físicos, exposição à radiação
ionizante ultravioleta e outras ondas curtas; condições ambientais impróprias como
aumento da temperatura, e os diversos tipos de poluição; envelhecimento; estados
psicológicos que provoquem estresse emocional; consequência de diversas doenças
como diabetes mellitus, hipertensão arterial, câncer, mal de Alzheimer e mal de
Parkinson (OLIVEIRA et al., 2009).

28
O desequilíbrio entre a produção de ROS/RNS e remoção pelos sistemas de
defesa antioxidantes é denominado estresse oxidativo. O estresse oxidativo é uma
condição celular ou fisiológica de elevada concentração de ROS/RNS que causa
danos moleculares às estruturas celulares, com consequente alteração funcional e
prejuízo das funções vitais, em diversos tecidos e órgãos, tais como músculo,
fígado, tecidos adiposos (BARJA DE QUIROGA, 1992; GOLFARB, 1993), vascular
(DUARTE et al., 1993; FENSTER et al., 2002) e cerebral (HALLIWELL, 1994;
KEYNES, GARTHWAITE, 2004). No entanto, o efeito deletério do estresse oxidativo
varia consideravelmente de acordo com a idade, o estado fisiológico e a dieta
(NIESS et al., 1999).
O aumento do estresse oxidativo pode estar envolvido na patogênese de
diversas doenças cardiovasculares, como a hipertensão (RODRIGO et al., 2007) e
aterosclerose (RADER, DAUGHERTY, 2008; SCHULZ et al., 2008).
Acredita-se que a presença de ROS na parede endotelial pode ocasionar a
formação de LDL-ox, um dos principais fatores para a gênese da placa de ateroma.
Os ácidos graxos da membrana plasmática são alvos dos radicais livres, podendo
desencadear uma reação em cadeia denominada peroxidação lipídica (FONSECA,
2007). Esta é caracterizada por um evento em cadeia tendo malondialdeído como
um de seus subprodutos, este tem a capacidade de se ligar covalentemente a
proteínas provocando alteração em sua molécula. Com isso, o malondialdeído é
utilizado como marcador da peroxidação lipídica em investigações laboratoriais
(FONSECA, 2007). Um dos compostos mais pesquisados devido sua atividade
antioxidante atualmente são os flavonoides.
2.3 FLAVONÓIDES
Os flavonóides são metabólitos secundários dos vegetais, ou seja, apresenta
uma atividade biológica benéfica a espécie, porém não é essencial à vida da planta.
As mais importantes funções dos flavonóides descritas para o metabolismo vegetal
são a proteção contra raios UV; proteção contra insetos, fungos, vírus e bactérias;
atração animal (que propicia a polinização da planta); ação antioxidante; controle da
ação de hormônios; agentes alelopáticos e inibição enzimática (ZUANAZZI;

29
MONTANHA, 2004; DORNAS et al., 2007). Também são responsáveis pela
coloração das flores, frutos e ocasionalmente outras partes da planta (do latim,
Flavus, amarelo) (COSTA, 2002).
Quimicamente eles são classificados como polifenóis, ou seja, possuem um
ou mais núcleos aromáticos contendo substituintes hidroxilados e/ou seus derivados
funcionais. A estrutura padrão da maioria dos flavonóides é composta por 15 átomos
de C (carbono) em seu núcleo fundamental, constituindo de duas fenilas ligadas por
uma cadeia de três C entre elas (figura 7). Nos compostos tricíclicos, as unidades
são chamadas núcleos A, B e C (ZUANAZZI; MONTANHA, 2004).
Figura 7 – Estrutura química do flavonoide rutina.
Do ponto de vista terapêutico, os flavonóides ganharam um impulso devido à
descoberta do “Paradoxo Francês”, no qual foi observado, em populações
francesas, um baixo índice de mortalidade relacionado a doenças cardiovasculares,
mesmo com um hábito alimentar com um alto nível de gordura saturada (NIJVELDT
et al., 2001). Foi observado, que a população francesa apresentava melhores
índices relacionados a doenças cardiovasculares, quando comparados a outra
populações com mesmos hábitos alimentares, o diferencial, porem se apresentava
na ingestão diária de vinho, muito mais consumido pelos franceses. Este fato
sinalizava a possível existência de algum componente cardioprotetor presente no
vinho (PROVIDÊNCIA, 2006). Estudos posteriores indicam que um dos fatores da
proteção cardiovascular provocada pelo vinho se deve ao fato da presença dos
flavonóides (LUZ, 2006).
Os flavonóides apresentam uma variabilidade estrutural muito grande, sendo
classificados em vários grupos, os principais são as flavonas, flavonóis, chalconas,

30
auronas, flavanonas, flavanas, antocianidinas, leucoantocianidinas,
proantocianidinas, isoflavonas e neoflavonóides (DORNAS et al., 2007).
Na dieta, os flavonóides são encontrados em frutas, vegetais e em bebidas
como chá e vinho (NIJVELDT et al., 2001), e seu consumo diário é estimado entre 1
ou 2 g por dia (DORNAS et al., 2007), sendo que um dos flavonóides mais
comumente encontrados é a quercitina, que está presente em frutas, vegetais e
bebidas. Ainda há a luteolina que é encontrada principalmente em pimentões
vermelhos; apegenina no aipo; catequinas em chás, vinhos e chocolate; e
antocininas em frutas vermelhas (sendo o responsável por sua cor), como cerejas,
ameixas, morangos e uvas (SCALBERT; WILLIAMSON, 2000).
Apesar de contida em diversos alimentos (em especial vegetais, frutas e
algumas bebidas), e de sua grande diversidade estrutural, a ingestão de polifenóis é
limitada na dieta da maioria das pessoas. Mesmo assim é a classe de substâncias
antioxidantes mais abundantes em nossa dieta, sendo destes, os flavonóides os
mais ingeridos. Porém a ocorrência das subclasses de flavonóides é restrita a
alguns tipos de alimentos. A maior fonte de flavonóides na alimentação encontra-se
na soja, onde contém, aproximadamente 1 mg de genisteína e dadzeína por grama
de soja (SCALBERT; WILLIAMSON, 2000).
2.3.1 Atividades biológicas dos flavonóides
Os flavonóides são grupos de compostos polifenólicos muito interessantes por
apresentarem uma gama de atividades farmacológicas e serem utilizados no auxilio
ao tratamento de doenças tais como alergia (SHAIK et al., 2006), diabetes mellitus
(HABIBUDDIN et al., 2008), câncer (MILLER et al., 2008), e também nos processos
de infecções virais (DROEBNER et al., 2007), inflamatórios (CHARAMI et al., 2008),
e antioxidantes ou oxidantes (YU et al., 2005; BOOTS et al., 2008).
Os flavonóides kaempferol e kaempferol-3-0-glucoside demonstraram em
ensaios in vivo, terem atividade anti-inflamatória, através dos métodos de edema de
pata e edema de orelha, e também atividade analgésica através do método de
contorções abdominais induzidas por injeção de ácido acético. Nos dois testes que
mensuraram a atividade anti-inflamatória, houve significante inibição inflamatória

31
pelos dois compostos. Acredita-se que seu mecanismo esteja relacionado à inibição
da histamina, serotonina ou da síntese de prostaglandinas. Os dois também
demonstraram efeitos analgésicos no teste de contorções abdominais (PARVEEN et
al., 2007).
Ensaios in vitro, pelo método de viabilidade celular por MTT (3-(4,5-
dimethylthiaxolone-2-yl)-2,5-diphenyl tetrazolium bromide), demonstraram a ação da
5-allyl-7gen-difluoromethylenechrysin (ADFMChR), que é um produto sintético
derivado do flavonóide crisina, no carcinoma hepático humano (linhagem celular
HepG2). Os resultados sugerindo uma indução de apoptose das células
cancerígenas. A análise do efeito do ADFMChR na expressão das proteínas PPARγ,
NF-κB, Bax e Bcl-2 indicou que a apoptose foi induzida pelo aumento da expressão
da proteína PPARγ (TAN et al., 2009).
Alguns trabalhos relatam a atividade antiviral de alguns flavonóides,
principalmente do grupo dos flavonóis, chalconas e seus análogos sintéticos. A
quercetina e seus derivados, demonstraram atividade sobre os vírus herpético
simples do tipo I, vírus respiratório sincial, pseudoraiva, parainfluenza 3, sindbis,
poliomielite do tipo I, coxsackievírus tipo B4 e rinovírus. A diosmina e hesperidina
foram capazes de inibir a replicação do rotavírus, e a baicaleína inibiu a
transcriptase reversa do vírus HIV-1 (ZUANAZZI; MONTANHA, 2004).
A rutina já é utilizada na terapêutica no caso de fragilidade capilar e
hipertensão cardiovascular, sendo, com isso, recomendada na púrpura hemorrágica,
glaucoma, menorragias, hemoptises, hematúrias, varicoses, estados infecciosos,
entre outras. O mecanismo pelo qual isso ocorre é pela inibição da oxidação do
hormônio adrenalina, que é o responsável pela manutenção da resistência capilar
(COSTA, 2002).
Segundo Nijveldt (2001), um importante efeito dos flavonóides é o sequestro
de radicais livres derivados do oxigênio, sendo esta, a atividade mais estudada
destes compostos. São demonstrados também, efeitos na diabetes mellitus, alergia,
câncer, infecções virais, dores de cabeça, úlceras estomacais e duodenais e
inflamações, e grande parte de seus efeitos podem ser explicados devido à sua
ação antioxidante (van ACKER et al., 1998).
Yordanov et al. (2008), demonstraram a atividade de inibição de enzimas
extracelulares secretadas pela Candida albicans, pelos flavonóides apeginina,
kaempferol, berberine e ibogaine. Apesar dos compostos não demonstrarem uma

32
significante atividade candicida, eles foram eficientes em inibir enzimas responsáveis
por aumentar a virulência e facilitar a adesão destes fungos em tecidos no qual não
são considerados de sua flora normal.
No organismo, os antioxidantes podem ser divididos em duas classes: as
substâncias com atividade enzimática e aquelas que não possuem esta atividade.
Na primeira, estão os compostos capazes de bloquear cataliticamente a iniciação
e/ou a propagação da oxidação, ou seja, as enzimas que removem as espécies
reativas de oxigênio. Na segunda classe, estão moléculas que interagem com as
espécies radicalares e são consumidas durante a reação. Nesta classificação,
incluem-se os antioxidantes naturais e sintéticos como os compostos fenólicos.
Dentre as diversas classes de substâncias antioxidantes e anti-inflamatória de
ocorrência natural, os compostos fenólicos têm recebido muita atenção nos últimos
anos, sobretudo por inibirem a peroxidação lipídica e a lipoxigenase in vitro (ATOUI
et al., 2005).
A atividade antioxidante de compostos fenólicos deve-se principalmente às
suas propriedades redutoras. Estas características desempenham um papel
importante no sequestro de radicais livres e quelação de metais de transição, agindo
tanto na etapa inicial quanto no decorrer do processo oxidativo. Os intermediários
formados pela ação de antioxidantes fenólicos são relativamente estáveis, devido à
ressonância do anel aromático presente na estrutura destas substâncias (BOOTS et
al., 2008).
Os flavonóides são potentes agentes antioxidantes, e fazem isso de forma
direta ou indireta, sendo que as atividades que podem interferir na formação de
radicais livres e propagação da ação oxidante são a de quelante de ferro;
sequestrante de radicais livres; inibição das enzimas lipooxigenase, ciclooxigenase,
NADPHoxidase, xantina-oxidase e fosfolipase; estimulação da catalase, superóxido
dismutase e outras enzimas antioxidantes (ZUANAZZI; MONTANHA, 2004).
Alguns desses processos podem ser explicados devido à forma química dos
flavonóides, no qual se acredita que a sua capacidade antioxidante seja devido a
sua estrutura, principalmente as hidroxilas que podem doar elétrons e suportar,
como resultado, a deslocalização em torno do sistema aromático, as hidroxilas
contidas em C3 e C4, atuariam no aumento do potencial antioxidante (DORNAS et
al., 2007), sendo que, quanto maior o número de hidroxilas, maior a atividade como
agente doador de H e de elétrons (BARREIROS et al., 2006).

33
Porém, ainda há muita discussão sobre o mecanismo de ação dos
flavonóides, sendo que seu real desempenho in vivo depende de outros fatores que
não sua composição química e potencial de sequestrar radicais livres in vitro, entre
eles o tipo de radical livre formado. Onde e como são formados esses radicais livres
e doses ideais para obter proteção (BIANCHI; ANTUNES, 1999). Porém, alguns
flavonóides se apresentam mais ativos na atividade de sequestro de radicais livres
do que outros antioxidantes como as vitaminas C e E, tendo as vantagens de quelar
mais metais de transição, estabilidade do flavanoil (radical formado pela reação com
um radical livre) formado e de poderem atuar em ambos os compartimentos
celulares, lipofílico e hidrofílico (BARREIROS et al., 2006).
Os flavonóides miricetina, quercetina e rutina demonstraram melhor atividade
na inibição de danos oxidativos induzidos pela H2O2 no DNA de linfócitos humanos
do que a vitamina C, sendo que a rutina também se demonstrou mais eficiente do
que o manitol na atividade antioxidante sobre o •OH (BIANCHI; ANTUNES; 1999).
Dentre os diversos grupos de flavonóides, se destacam as flavonas e as
catequinas no que se diz respeito ao sequestro de radicais livres. A rutina pertence
ao grupo das flavonas. A rutina é caracterizada como antioxidante, sendo que uma
de suas capacidades é a ação inibitória sobre a enzima xantina oxidase e que esta
sendo relacionada com a oxidação da LDL, processo importante para a gênese
aterosclerótica (NIJVELDT et al., 2001).
Foi demonstrado em estudo que o tratamento com os flavonóides antocianina
e naringina associados reduz os níveis de colesterol total e triglicerídeos em ratos
Wistar (NAGEM et al., 1999).
Estudo com antocianina e naringenina demonstrou que estes flavonóides
proporcionam ganho de peso considerável e redução dos níveis séricos de glicose e
triglicerídeos em coelhos com diabete induzida (RIBEIRO, 2001).
Em pesquisa de Rodrigues et al. (2003), no qual foi avaliada a capacidade da
rutina em induzir variação na glicemia de jejum, alterações séricas na lipoproteína do
colesterol LDL (lipoproína de baixa densidade) e HDL (lipoproteína de alta
densidade) e atividade da enzima superóxido dismutase (SOD), em experimentos
com ratos Wistar, apesar de não terem sidos observadas alterações de peso, da
glicemia e na LDL dos animais, houve uma significante elevação na HDL e na
atividade da enzima SOD, sugerindo um importante efeito na diminuição dos fatores

34
de risco para doenças cardiovasculares. A SOD é responsável por catalisar a
destruição do radical superóxido pela formação de H2O2.
A rutina demonstrou ter a capacidade de reduzir os níveis de colesterol e
triacilglicerídeos, bem como manter elevados os níveis de colesterol HDL em testes
realizados com soro de ratos Wistar hiperlipidêmicos, tratados previamente com
triton (ou tyloxapol, um detergente não aniônico de estrutura polimérica, utilizado
para induzir hiperlipidemia em cobaias) (LIMA et al., 2003).
2.3.2 Flavonóides e Metais de Transição
A relação estrutura-atividade dos flavonóides tem sido extensivamente
estudada na tentativa de se entender quais efeitos estruturais são mais importantes
nas diversas atividades.
Estudos farmacológicos demonstraram que os flavonóides não glicosilados,
como a quercetina e o canferol, possuem maior efeito antioxidante e antiinflamatório
quando na presença de íons de metais de transição como Fe(II) e Cu(II) (van
ACKER et al., 1998). O flavonóide rutina (quercetina glicosilada) também mostrou
maior eficiência na presença de íons de Fe(II) e Cu(II) (AFANAS'EVA et al., 2001).
Entretanto, estes estudos foram realizados in situ, não sendo isolado nenhum
complexo.
A naringina é uma flavona que possui atividade antioxidante, anti-inflamatória
(KORKINA et al., 1997), além de inibir a proliferação in vitro de linhagens de tumores
de mama humanos (AFANAS'EVA et al., 2001). Pereira et al. (2007) demonstraram
que a complexação Naringina-Cu (II), aumenta a atividade antioxidante, anti-
inflamatória e a atividade citotóxica em células tumorais quando comparada ao
flavonóide livre.
Existem estudos que demonstram que flavonóides complexados com metais,
como o ferro, possuem uma efetiva habilidade em promover o sequestro de radicais
livres, e um potente benefício terapêutico para o tratamento de patologias e
disfunções associadas ao estresse oxidativo (KOSTYUK et al., 2007).
Dados preliminares mostram que a rutina quando coordenada ao íon Fe(II),
apresenta uma atividade antioxidante superior à da rutina livre, além de viabilizar o

35
crescimento de células vermelhas em animais anêmicos (PEREIRA et al., 2007;
RETTORI et al., 2008).
Há dois fatores importantes para o planejamento de complexos metálicos
como agentes farmacêuticos: estabilidade e seletividade; e seu sucesso depende do
controle de suas propriedades cinéticas e termodinâmicas, com isso, podendo
encontrar a atividade e melhorar a especificidade biológica minimizando os efeitos
colaterais (BENITE et al., 2007).
A obtenção de compostos com atividade hipolipemiante, antioxidante e anti-
inflamatória complexados com metais de transição, como o ferro, cobre e o zinco,
pode apresentar um interessante papel no combate às patologias relacionadas ao
aumento sérico dos lipídeos e o aumento de radicais livres, como a aterosclerose.
Desta forma, com o presente trabalho pretendeu-se avaliar uma possível
potencialização da atividade da rutina quando complexada ao cobre no metabolismo
lipídico em um modelo de indução de hipercolesterolemia em ratos. Sendo esta a
primeira pesquisa da atividade da rutina complexada a cobre (II) no metabolismo
lipídico, os resultados obtidos são importantes e servem como base para futuras
pesquisas para elucidação dos mecanismos de ação desta molécula.
Figura 8 – Possíveis sítios de ligação do metal de transição na molécula de flavonóide.

36
3. OBJETIVO GERAL
Avaliar a atividade do composto rutina livre e rutina complexada com cobre(II)
no metabolismo lipídico de ratos induzidos a hipercolesterolemia.
3.1. OBJETIVOS ESPECÍFICOS
Avaliar comparativamente entre os compostos rutina e rutina complexada com
cobre(II), os seguintes parâmetros:
• Alterações bioquímicas no sangue de ratos;
• Hemograma;
• Análise anatomopatológica do fígado e aorta dos ratos;
• Análise da peroxidação lipídica do plasma sanguíneo;
• Concentração de NO do plasma sanguíneo.

37
4. MATERIAIS E MÉTODOS
4.1 PREPARAÇÃO DA DROGA COMPLEXADA
O processo de síntese dos compostos rutina complexado a metais de
transição foi realizado no Laboratório de Química de Coordenação da Universidade
Bandeirante de São Paulo (UNIBAN). Estes complexos foram doados pela Profa.
Dra. Regina Mara da Silva Pereira, resultantes do projeto de pesquisa FAPESP
(Processo nº 05/60749-9).
4.2 ENSAIOS COM DIETA HIPERCOLESTEROLÊMICA
4.2.1 Animais
Foram utilizados ratos Wistar machos com idade de 2 meses e peso médio
inicial de 220g, mantidos no biotério da Universidade Bandeirante de São Paulo -
UNIBAN. Os animais foram mantidos à temperatura de 22 ± 3ºC, em ciclo
claro/escuro de 12 horas, com acesso a água e ração ad libitum. Os animais foram
mantidos nos laboratórios por 1 hora antes da realização dos experimentos para
aclimatação sendo que os experimentos foram conduzidos de acordo com o manejo
experimental de animais de laboratório e normas para investigação em animais
conscientes (ZIMMERMMAN, 1983). O procedimento foi analisado e aprovado pela
Comissão de Ética no Uso de Animais da UNIBAN - CEUA/UNIBAN, sob o nº209/11
(ANEXO).
4.2.2 Protocolo Experimental
Os animais foram divididos em sete grupos (n=5), conforme apresentado na
tabela 01.

38
Tabela 01: Grupos de animais divididos de acordo com o tratamento.
GRUPO DOSAGEM
Grupo G1 - controle negativo, ração padrão Água destilada
Grupo G2 - controle negativo, ração suplementada Veículo
Grupo G3 - controle positivo, ração suplementada 10mg/kg de sinvastatina
Grupo G4 - rutina 10, ração suplementada 16µmol/kg de rutina
Grupo G5 - rutina 100, ração suplementada 160µmol/kg de rutina
Grupo G6 - rutina-cobre2 10 (R-Cu2), ração
suplementada 16µmol/kg de R-Cu2
Grupo G7 - rutina-cobre2 100 (R-Cu2), ração
suplementada 160µmol/kg de R-Cu2
Os animais receberam administração das drogas testes pela via oral por
gavagem três vezes por semana, com volume máximo administrado de 1mL/animal.
Devido às características físico-químicas das drogas e para mimetizar uma
possibilidade de futura formulação, os compostos foram preparados em forma de
suspensão utilizando Emulgin a 3%. A sinvastatina foi utilizada como controle
positivo devido a sua atividade no controle do metabolismo através da regulação da
atividade da enzima HMG – CoA redutase.
Durante a indução da hipercolesterolemia e tratamento concomitante, foram
feitas coleta de dados (peso, consumo de ração e água, fezes). Este período teve
duração de 8 semanas, onde os animais receberam tratamento 3 vezes na semana.
Após o período os animais foram submetidos à eutanásia e amostras foram
coletadas para avaliações bioquímicas, hematológicas e morfológicas.
4.2.3 Dietas
Para a indução da hipercolesterolemia foi realizada uma adaptação do
protocolo descrito por Zand et al., (1999). Foi fornecido aos animais, ração
enriquecida com colesterol (2%), ácido cólico (0,5%) por um período de 60 dias. O

39
tiouracil (0,5%) foi adicionado a ração nos primeiros 30 dias. O grupo controle G1
recebeu ração padrão que foi submetida ao mesmo processo de preparo.
4.2.4 Dosagens sanguíneas
Após os 60 dias de tratamento foram realizados as coletas dos materiais para
a análise. Os animais foram mantidos em jejum por 12 horas e então foram
anestesiados com ketamina/xilasina. As amostras de sangue foram coletadas
através da artéria porta-hepática, com auxílio de uma seringa heparinizada.
Parte das amostras de sangue total foi aliquotada em tubos cônicos para a
contagem global de leucócitos, contagem diferencial de leucócitos e hematócrito. A
outra parte foi centrifugada a 2.500 r.p.m. durante 10 minutos, e após, o plasma foi
separado e aliquotado em tubos cônicos. As amostras de sangue total foram
estocadas em geladeira a 8°C, enquanto que o plasma aliquotado foi armazenado
em freezer (-20 a -70ºC) até o momento das dosagens.
4.2.4.1 Dosagens bioquímicas
Foram quantificados os valores de colesterol total, HDL e triglicerídeos. Foram
utilizados Kits de análise enzimático colorimétrico obtido da Labtest Diagnóstica. As
leituras foram realizadas em espectrofotômetro de acordo com as especificações do
laboratório.
O valor de LDL foi calculado matematicamente pela equação de Friedewald:
[LDL = colesterol total – (HDL+TG/5)] de acordo com Nicastro et al (2012).
4.2.4.2 Avaliação da peroxidação lipídica
A avaliação da peroxidação lipídica foi determinada pela verificação de
TBARS (substâncias reativas ao ácido tiobarbitúrico) no plasma através do método
adaptado de Serra et al. (2001). Após o descongelamento das amostras de plasma
coletado dos animais tratados, foram realizados os seguintes procedimentos. Em
tubos cônicos de 1,5mL, contendo 500uL de plasma foram adicionados 500uL de

40
TCA (ácido tricloroacético - 10%). Após homogeneização a mistura foi colocada no
gelo por 10 minutos. Em seguida, os tubos foram centrifugados a 900g por 15
minutos. Após a centrifugação, o sobrenadante foi separado em outro tubo, e foram
acrescentados 500uL de TBA (ácido tiobarbitúrico - 1%). A reação foi incubada a
100oC por 30 minutos. Na sequência, foi realizada a leitura no espectrofotômetro em
535 nm. Paralelamente, foi preparado o “branco” adicionando água no lugar da
amostra.
Após a avaliação das amostras, os resultados (em nmol/uL) foram obtidos
através do cálculo de concentração de TBARS:
C = Absorbância x diluição da amostra/volume de plasma x 0,156= (nmol/ul)
Onde, 0,156 é o fator de concentração para o método.
4.2.4.3 Dosagem plasmática de NO
Para verificação da quantidade de NO (óxido nítrico) pelo efeito da rutina e R-
Cu2 em ratos hipercolesterolêmico foi realizado através da reação de Griess, onde
verificou-se a quantidade de nitrito presente no plasma. Para o preparo da Solução
de Griess foram misturados quantidades iguais (1:1), de sulfanilamida à 1% e N-(1-
naftil) etilenodiamina dihidrocloridrato à 0,1% em ácido fosfórico à 2,5%. Em uma
placa de 96 poços, foram colocados 50uL de plasma e foram adicionados 50uL da
Solução de Griess, deixando reagir por 30min ao abrigo da luz, e a leitura foi
realizada em leitora de placa em 540nm. Nitroprussiato de sódio foi utilizado para a
construção da curva padrão.
4.2.4.4 Análise hematológica
A partir do sangue total foram obtidos os valores de hematócrito, leucócitos
totais e leucócitos diferenciais.
O hematócrito foi obtido através de rotação de capilar contendo o sangue do
animal a 10.000 rpm por 5 minutos em centrífuga de capilar.

41
A contagem total de leucócitos foi quantificada em câmara de Neubauer,
através de diluição de 20µL do sangue total em 380µL de solução de Turk.
A contagem diferencial de leucócitos foi quantificada pela leitura de lâmina
preparada através de coloração panótico.
4.2.5 Estudo anatomopatológico
Foram coletados os rins, fígado, coração e aorta dos animais, todos os órgãos
foram analisados macroscopicamente, limpos, pesados e então guardados em
frascos contendo formol 10% tamponado.
Para avaliação microscópica, as lâminas foram coradas com Hematoxilina e
Eosina (HE) e as fotos obtidas com aumento de 20x. A análise do tecido hepático foi
realizada em três áreas, sendo a área 1 uma região com maior fluxo de sangue
arterial, a área 2 uma região intermediária e a área 3 uma região com um maior fluxo
de sangue venoso, portanto mais propenso a lesões. O estudo anatomopatológico
foi realizado em colaboração com o Doutorando André Rennó do Departamento de
Farmacologia, da Faculdade de Ciências Médicas da UNICAMP.
4.4 ANÁLISE ESTATÍSTICA
Foram realizadas análise de variância One-way ANOVA seguido do Teste de
Bonferroni ou Dunnett, considerando diferenças significativas com P < 0,05. Quando
realizado comparação entre dois grupos foi aplicado Teste t de Student.

42
5. RESULTADOS E DISCUSSÃO
5.1. DOSAGENS SANGUÍNEAS
5.1.1. Dosagens bioquímicas
Segundo a Sociedade Brasileira de Cardiologia (2007) é recomendado que
para definição do perfil lipídico do paciente seja realizada a avaliação de níveis de
colesterol total, HDL, LDL e triglicerídeos. Sendo assim, as amostras sanguíneas
dos animais de todos os grupos, foram quantificadas e obtiveram-se os valores de
colesterol total, HDL, triglicerídeos e LDL.
Os resultados dos níveis séricos de colesterol total (média ± E.P.M) dos
grupos G1, G2, G3, G4, G5, G6 e G7 foram 86,6 ± 4,8, 174,1 ± 23,7, 161,7 ± 31,2,
181,0 ± 10,3, 149,2 ± 9,3, 149,7 ± 21,6, 156,3 ± 23,5mg/dL, respectivamente.
Somente os grupos G2 (veículo) e G4 (rutina 10) apresentaram diferença
significativa em comparação ao grupo G1. Embora exista uma tendência para uma
diminuição do colesterol dos grupos tratados com rutina livre e R-Cu2 (G5, G6 e G7)
em relação ao grupo veículo (G2), no entanto, essas diferenças não foram
significativas (Figura 9).
Figura 9: Níveis séricos de colesterol total em ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Dunnett, considerando diferenças significativas com: ** P<0,01 para a análise estatística do grupo controle (G1) vs demais grupos e,
# P<0,05 para a análise estatística do grupo controle (G2) vs demais grupos.

43
Os níveis de triglicerídeos do grupo controle (G1) apresentavam-se dentro
dos níveis considerados fisiológicos para ratos, segundo a literatura. A
hipertrigliceridemia não foi observada nos ratos submetidos à dieta, como podemos
observar entre os grupos controles. No entanto, o tratamento com R-Cu2 nas
concentrações de 16 e 160µmol/kg (grupos G6 e G7) reduziram significativamente
os níveis de triglicerídeos em aproximadamente 30 e 20%, respectivamente, quando
comparados com os grupos controles G1 e G2 (Figura 10).
0
30
60
90
G1 G2 G3 G4 G5 G6 G7
#
***##
##
Tri
glicerí
deo
s (
mg
/dL
)
Figura 10: Níveis séricos de triglicerídeos em ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 – rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Dunnett, considerando diferenças significativas com: ** e
## P<0,01, análise
estatística de grupos controles (G1 e G2), respectivamente, vs grupos.
Os níveis de HDL do grupo controle (G1) apresentavam-se dentro dos níveis
considerados fisiológicos para ratos, conforme descrito na literatura. Não foram
observadas alterações significativas nos níveis séricos de HDL entre nenhum dos
grupos (Figura 11).

44
0
15
30
45
60
75
G1 G2 G3 G4 G5 G6 G7
#
HD
L (
mg
/dL
)
A Figura 12 mostra que os níveis de LDL estão aumentados em todos os
grupos de animais submetidos à dieta rica em colesterol. O gráfico mostra o
aumento significante de LDL em todos os grupos quando comparado ao grupo
controle (G1). No entanto, os grupos tratados com rutina livre (160µmol/kg – grupo
G5) e R-Cu2 nas concentrações de 16 e 160µmol/kg (grupos G6 e G7) inibiram
significativamente o aumento dos níveis de LDL em 26,1; 28,6 e 23,9% quando
comparados ao grupo veículo (G2).
Figura 11: Níveis séricos de HDL em ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg.
Figura 12: Níveis séricos de LDL em ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Dunnett, considerando diferenças significativas com: ** e ##
P<0,01, análise estatística de grupos controles (G1 e G2), respectivamente, vs grupos.

45
A hiperlipidemia é uma importante causa da aterosclerose e das condições a
ela associadas, tais como coronopatias, doenças cerebrovascular isquêmica e
insuficiência vascular periférica, condições que contribuem para a maior parte da
morbidade e mortalidade entre os adultos idosos e de meia-idade.
Na avaliação dos níveis de colesterol total (CT), pode-se observar um
aumento significativo nos animais dos grupos G1 (ração padrão) e G2 (animais dieta
rica em colesterol) validando o modelo de indução da hipercolesterolemia nos
animais. Além disso, percebe-se que os tratamentos não foram eficazes em inibir o
aumento do colesterol como demonstrado na Figura 9.
Já os níveis de triglicerídeos, embora estejam todos dentro dos níveis normais
(como observado na Figura 10), pode-se verificar uma diminuição dos níveis séricos
nos grupos tratados com R-Cu2 (G6 e G7).
Diversos experimentos têm sido realizados a fim de elucidar a ação dos
flavonóides no metabolismo lipídico. Sudheesh et al. (1997) demonstrou que
flavonóides extraídos de berinjela (Solanum melongena) apresentaram efeito na
redução nos níveis sanguíneos de colesterol total e triacilgliceróis. Isso pode ser
devido à ação da enzima LCAT e LPL, que tiveram um aumento em sua atividade no
experimento. A LCAT é uma enzima responsável por converter o colesterol presente
em quilomícrons, VLDL, LDL e tecidos periféricos em colesterol esterificado,
podendo, assim, ser transportados para o fígado onde serão metabolizados. A LPL é
responsável por hidrolisar os ácidos graxos dos triacilgliceróis presentes em
quilomícrons e VLDL para o tecido adiposo. Também foi observado um aumento nos
níveis de ácidos biliares hepáticos e fecais, bem como esteróis neutros fecais,
indicando uma alta taxa de degradação de colesterol e redução na reabsorção
intestinal de ácidos biliares. Kuppusamy e Das (1992) demonstraram que os
flavonóides têm uma significativa inibição sobre a enzima fosfodiesterase quando
comparado com um potente inibidor desta enzima, sendo que a inibição desta
enzima é relacionada a um efeito lipolítico.
Juzwiak et al. (2005) demonstrou os efeitos hipolipemiantes da quercetina,
associando-os a um efeito de aumento da atividade microssomal, observados pela
elevação do conteúdo de citocromo P450. O aumento da atividade microssomal
também foi demonstrado por MacDonald et al. (1983) com o uso de quercetina e
rutina. Também é relatado o aumento da excreção de sais biliares, aumentando a

46
excreção lipídica. Esses resultados podem elucidar os mecanismos da rutina e rutina
complexada nos seus efeitos no colesterol e triglicerídeos.
Sabe-se que partículas de LDL se acumulam na íntima das artérias
coronárias, isso ocorre principalmente quando a LDL se encontra em altos níveis
sanguíneos ultrapassando as células endoteliais e acumulando na parede dos
vasos. Assim, podem-se desenvolver lesões que levam ao desenvolvimento da
aterosclerose (BERTOLAMI, 2000). Já as partículas de HDL são lipoproteínas
protetoras e reduzem o risco de coronopatias. Para isso, altos níveis de HDL são
desejáveis. Sendo assim, os principais fatores de risco convencionais para doenças
cardiovasculares são LDL elevado, HDL reduzido e alguns fatores de risco
(GOODMAN; GILMAN, 2010).
Os resultados apresentados na Figura 11 demonstram que os níveis séricos
de HDL dos animais tratados com os compostos foram semelhantes ao grupo
veículo. No entanto, o composto R-Cu2 inibiu o aumento significativo dos níveis de
LDL, após indução da hipercolesterolemia (Figura 12). Estes resultados sugerem
que embora o tratamento não tenha aumentado os níveis de HDL, os níveis de LDL
sofreram interferência dos flavonóides.
Ao se realizar uma análise conjunta dos resultados obtidos após indução da
hipercolesterolemia pode-se perceber que a R-Cu2 (nas concentrações de 16 e
160µmol/kg) teve melhor efeito que a rutina livre, demonstrando uma forte tendência
na inibição do aumento sérico de colesterol total, triglicerídeo e LDL, sugerindo
assim um possível efeito contras as dislipidemias, resultado este, também observado
por Kanashiro et al., 2009 com rutina.
5.1.2. Avaliação da peroxidação lipídica
A avaliação da peroxidação lipídica foi determinada pela análise da produção
de TBARS (substâncias reativas ao ácido tiobarbitúrico) no plasma.
Na análise de peroxidação lipídica foi constatada que os animais que
receberam apenas veículo e submetidos à dieta rica em colesterol (grupo G2)
apresentaram aumento de TBARS avaliado. No entanto, os grupos que receberam
tratamentos com sinvastatina, rutina livre e R-Cu2, mesmo com a ingestão de dieta

47
rica em colesterol, não apresentaram resultados estatisticamente diferentes do
grupo controle (grupo com dieta de ração padrão – G1), como podemos observar na
Figura 13.
0
10
20
30
40
50
G1 G2 G3 G4 G5 G6 G7
**
## ##
# #
TB
AR
S (
nm
ol/m
L)
A técnica mais comum usada para medir a lipoperoxidação é o teste do ácido
tiobarbitúrico (TBA), que é um método espectrofotométrico que mede a
concentração dos produtos oriundos da peroxidação dos lipídios. Durante a fase de
iniciação da lipoperoxidação ocorre acúmulo de hidroperóxidos de lipídios na
membrana celular, que se decompõe formando vários aldeídos, sendo o
malonaldeído o principal deles (PATOČKOVÁ et al., 2003).
Embora o modelo de medida de TBARS (substâncias reativas ao ácido
tiobarbitúrico), segundo a literatura, pode ser interferido pela presença de metais de
transição (que podem gerar mais substâncias reativas), não houve interferência na
visualização qualitativa do atual experimento. Pode-se observar que nos grupos
tratados com a droga-teste não houve aumento de TBARS, indicando grandes
chances de estabilidade do complexo flavonóide com o metal. Todos os compostos
testados demonstraram uma atividade no controle da formação de TBARS,
possivelmente devido a atividade antioxidante da rutina e R-Cu2.
Figura 13: Avaliação da peroxidação lipídica. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Dunnett, considerando diferenças significativas com: ** P<0,01 e
# P<0,05, análise estatística de grupos controles (G1 e G2), respectivamente, vs
grupos.
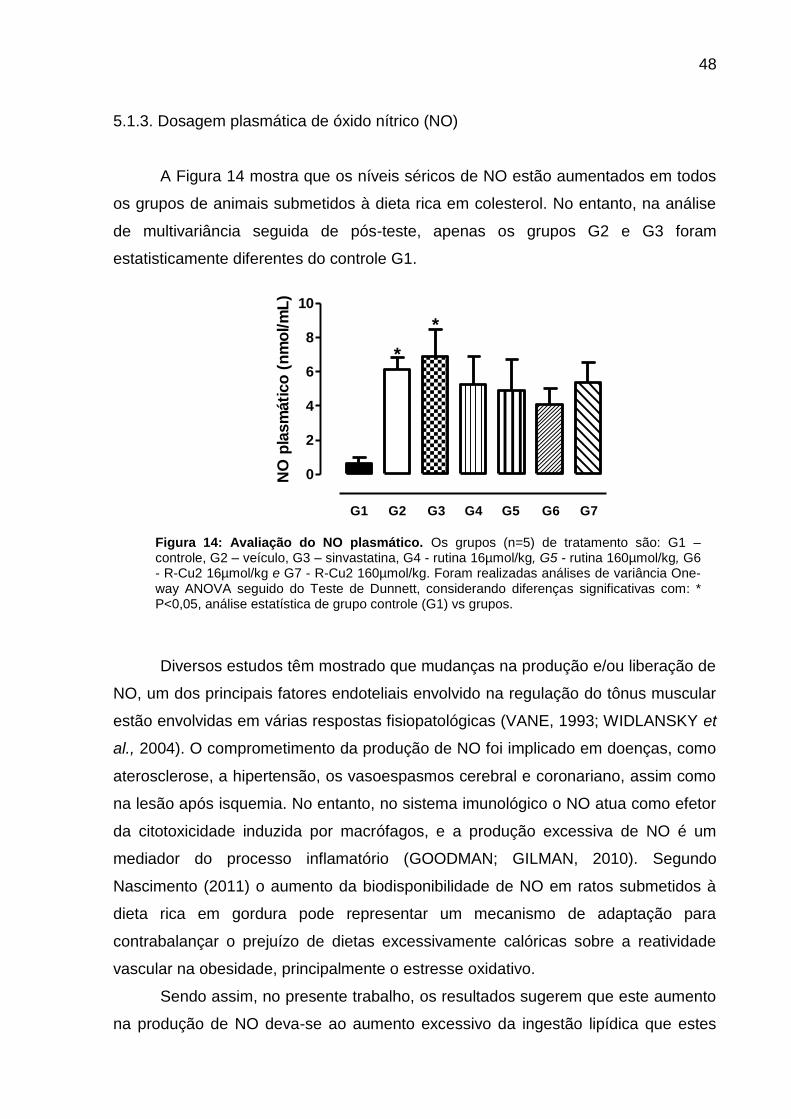
48
5.1.3. Dosagem plasmática de óxido nítrico (NO)
A Figura 14 mostra que os níveis séricos de NO estão aumentados em todos
os grupos de animais submetidos à dieta rica em colesterol. No entanto, na análise
de multivariância seguida de pós-teste, apenas os grupos G2 e G3 foram
estatisticamente diferentes do controle G1.
0
2
4
6
8
10
G1 G2 G3 G4 G5 G6 G7
*
*
NO
pla
sm
áti
co
(n
mo
l/m
L)
Diversos estudos têm mostrado que mudanças na produção e/ou liberação de
NO, um dos principais fatores endoteliais envolvido na regulação do tônus muscular
estão envolvidas em várias respostas fisiopatológicas (VANE, 1993; WIDLANSKY et
al., 2004). O comprometimento da produção de NO foi implicado em doenças, como
aterosclerose, a hipertensão, os vasoespasmos cerebral e coronariano, assim como
na lesão após isquemia. No entanto, no sistema imunológico o NO atua como efetor
da citotoxicidade induzida por macrófagos, e a produção excessiva de NO é um
mediador do processo inflamatório (GOODMAN; GILMAN, 2010). Segundo
Nascimento (2011) o aumento da biodisponibilidade de NO em ratos submetidos à
dieta rica em gordura pode representar um mecanismo de adaptação para
contrabalançar o prejuízo de dietas excessivamente calóricas sobre a reatividade
vascular na obesidade, principalmente o estresse oxidativo.
Sendo assim, no presente trabalho, os resultados sugerem que este aumento
na produção de NO deva-se ao aumento excessivo da ingestão lipídica que estes
Figura 14: Avaliação do NO plasmático. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Dunnett, considerando diferenças significativas com: * P<0,05, análise estatística de grupo controle (G1) vs grupos.

49
ratos foram submetidos. Isso tudo leva a um desequilíbrio, que segundo a literatura
citada anteriormente, força uma compensação adaptativa do metabolismo dos
animais.
5.1.4. Análise hematológica
Nas avaliações dos parâmetros hematológicos foram analisadas o
hematócrito, a contagem global e diferencial de leucócitos.
Na análise hematológica foi observada que os valores dos hematócritos do
grupo controle e dos grupos tratados apresentavam-se dentro dos valores de
referência para ratos. Também foi observado que não houve diferença estatística
entre os grupos estudados, demonstrando que nenhum tratamento interfere neste
parâmetro (Figura 15).
0
20
40
60
G1 G2 G3 G4 G5 G6 G7
Hem
ató
cri
to (
%)
Os valores de leucócitos totais tanto do grupo controle (G1) quanto a dos
grupos tratados também não apresentaram diferença significativa (Figura 16).
Figura 15: Contagem de hematócrito em ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg.

50
0
2
4
6
8
G1 G2 G3 G4 G5 G6 G7
Leu
có
cit
os x
10
6/m
L
Ao analisar os resultados de contagem diferencial de leucócitos foram obtidos
os valores absolutos de monócitos, linfócitos e neutrófilos.
Como demonstrado na Figura 17, o número de monócitos no sangue estava
significativamente reduzido em todos os grupos submetidos à dieta rica em
colesterol.
0.0
0.5
1.0
1.5
2.0
G1 G2 G3 G4 G5 G6 G7
****
**** ** **
Mo
nó
cit
os x
10
6/m
L
Figura 16: Contagem total de leucócitos em ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg.
Figura 17: Contagem diferencial de leucócitos em ratos – número de monócitos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Bonferroni, considerando diferenças significativas com: ** P<0,01, análise estatística de grupo controle (G1) vs grupos.
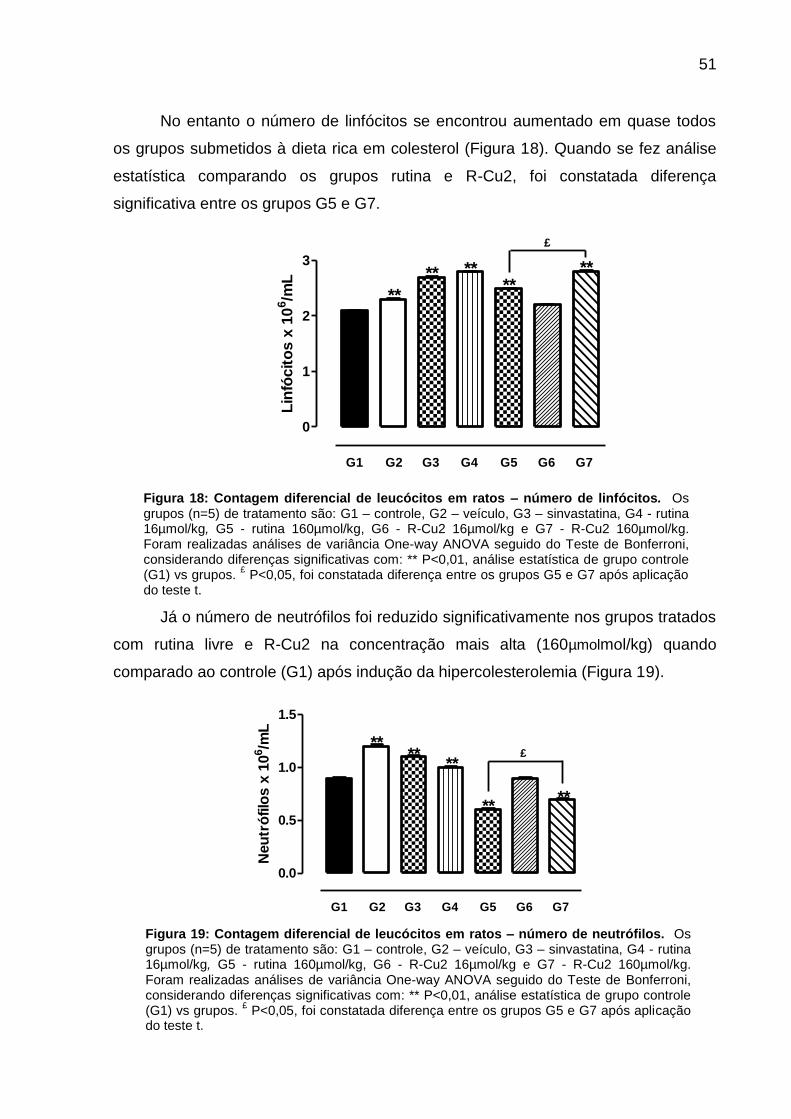
51
No entanto o número de linfócitos se encontrou aumentado em quase todos
os grupos submetidos à dieta rica em colesterol (Figura 18). Quando se fez análise
estatística comparando os grupos rutina e R-Cu2, foi constatada diferença
significativa entre os grupos G5 e G7.
0
1
2
3
G1 G2 G3 G4 G5 G6 G7
****
**
** **
£
Lin
fócit
os x
10
6/m
L
Já o número de neutrófilos foi reduzido significativamente nos grupos tratados
com rutina livre e R-Cu2 na concentração mais alta (160µmolmol/kg) quando
comparado ao controle (G1) após indução da hipercolesterolemia (Figura 19).
0.0
0.5
1.0
1.5
G1 G2 G3 G4 G5 G6 G7
**
****
**
**
£
Neu
tró
filo
s x
10
6/m
L
Figura 18: Contagem diferencial de leucócitos em ratos – número de linfócitos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Bonferroni, considerando diferenças significativas com: ** P<0,01, análise estatística de grupo controle (G1) vs grupos.
£ P<0,05, foi constatada diferença entre os grupos G5 e G7 após aplicação
do teste t.
Figura 19: Contagem diferencial de leucócitos em ratos – número de neutrófilos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Bonferroni, considerando diferenças significativas com: ** P<0,01, análise estatística de grupo controle (G1) vs grupos.
£ P<0,05, foi constatada diferença entre os grupos G5 e G7 após aplicação
do teste t.

52
A aterosclerose associada com hipercolesterolemia é um complexo processo
inflamatório, caracterizado patologicamente pelo recrutamento de monócitos na
parede arterial e deposição de lipoproteínas ricas em colesterol (Murphy et al.,
2011). O recrutamento dos monócitos é um processo mediado por moléculas de
adesão, iniciando com o rolamento, o qual é mediado pela ligação entre E-selectina
em células endoteliais. Todavia, moléculas de adesão participam na interação de
monócitos e células endoteliais apresentando um importante papel na aterogênese.
A ativação das células endoteliais induzida pela hipercolesterolemia aumenta a
expressão dessas moléculas de adesão contribuindo com a aterogênese e a
migração tecidual de monócitos-macrófagos (WU et al., 2009).
Desta forma, a diminuição no número de monócitos circulante (Figura 17)
pode estar associada com as alterações nas células endoteliais provocadas nos
animais que estão hipercolesterolêmicos. Essas alterações fisiológicas induzidas
pelo aumento de lipídio pode justificar também o aumento na quantidade de NO
plasmático (Figura 14).
Além disso, o aumento de linfócitos (Figura 18) pode estar envolvido com a
diminuição de monócitos circulantes e pela indução de um processo inflamatório
crônico.
5.2. ESTUDO ANATOMOPATOLÓGICO
5.2.1. Avaliação macroscópica - Peso relativo dos órgãos
Imediatamente após a coleta dos órgãos, os mesmos foram limpos, pesados
e armazenados. Com o peso dos órgãos, pode-se avaliar o peso relativo dos grupos
estudados. Assim, foi constatado que os pesos relativos dos rins (esquerdo e direito)
não apresentaram diferença significativa entre os grupos, como demonstrada na
Figura 20.

53
0.0
0.1
0.2
0.3
0.4
0.5
G1 G2 G3 G4 G5 G6 G7
G1
G2
G3
G4
G5
G6
G7Peso
rela
tivo
do
rim
esq
uerd
o (
g)
0.0
0.1
0.2
0.3
0.4
0.5
G1 G2 G3 G4 G5 G6 G7
G1
G2
G3
G4
G5
G6
G7Peso
rela
tivo
do
rim
dir
eit
o (
g)
Na Figura 21 está demonstrado o peso relativo de coração proveniente dos
grupos avaliados. Não houve diferença entre estes pesos, sugerindo que não houve
hipertrofia do miocardio em nenhum tratamento.
0.0
0.1
0.2
0.3
0.4
0.5
G1 G2 G3 G4 G5 G6 G7
Peso
rela
tivo
do
co
ração
(g
)
Já na Figura 22, pode-se observar que a ingestão da dieta rica em colesterol
aumentou significativamente o peso do fígado. Um aumento significativo do peso
relativo deste órgão foi verificado em todos os grupos que receberam a dieta.
Nenhum tratamento foi suficiente para evitar a hipertrofia hepática.
Figura 20: Peso relativo dos rins esquerdo e direito de ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg.
Figura 21: Peso relativo de corações provenientes de ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg.

54
0
1
2
3
4
5
G1 G2 G3 G4 G5 G6 G7
* * ** **
G1
G2
G3
G4
G5
G6
G7P
eso
rela
tivo
do
fíg
ad
o (
g)
5.2.2. Avaliação macroscópica – Análise do fígado
O fígado, aparentemente, foi o órgão que mais apresentou alterações
morfológicas. Na Figura 23-A podem ser verificadas as características de um fígado
saudável. Esta imagem foi retirada de um rato do grupo G1 (controle negativo) que
foi alimentado com ração padrão durante todo o período de tratamento. Já a Figura
23-B representa a imagem de um fígado originado de um rato do grupo G2 (veículo)
que recebeu ração suplementada com colesterol durante todo o período de
tratamento. A ilustração demonstra que a dieta rica em colesterol foi responsável
pela alteração de tamanho, coloração e aspecto do órgão. A coloração sugere a
presença de gordura nos hepatócitos. Nos demais grupos G3 a G7, as mesmas
alterações, com diferença de intensidade, foram visualizadas macroscopicamente
como demonstradas na Tabela 2.
Figura 22: Peso relativo do fígado de ratos. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Foram realizadas análises de variância One-way ANOVA seguido do Teste de Bonferroni, considerando diferenças significativas com *P<0,05 quando comparado grupo controle (G1) VS demais grupos.
Figura 23: Imagem de fígado obtido após ensaio com dieta rica em colesterol. A –Fígado obtido de rato do grupo G1 - controle negativo, alimentado com ração padrão. B - Fígado obtido de rato do grupo G2 - veículo, alimentado com ração suplementada com colesterol.
A B

55
Tabela 2: Análise macroscópica de alguns tecidos dos animais do grupo controle e
dos grupos que receberam dieta suplementada com colesterol.
Grupos Aorta Coração Rins Fígado
G1
Sem alterações
Sem alterações
Sem alterações
Sem alterações
G2 Aumento de gorduras
periféricas
Sem alterações
Sem alterações
Alterações de tamanho, coloração muito amarelada e aspecto
esponjoso.
G3 Aumento de gorduras
periféricas
Sem alterações
Sem alterações
Alterações de tamanho, coloração muito amarelada e aspecto esponjoso. *Alguns animais
apresentaram manchas brancas.
G4 Aumento de gorduras
periféricas
Sem alterações
Sem alterações
Alterações de tamanho, coloração muito amarelada e aspecto esponjoso. *Alguns animais
apresentaram manchas brancas
G5 Aumento de gorduras
periféricas
Sem alterações
Sem alterações
Alterações de tamanho, coloração muito amarelada e aspecto
esponjoso.
G6 Aumento de gorduras
periféricas
Sem alterações
Sem alterações
Alterações de tamanho, coloração levemente amarelada e aspecto
esponjoso.
G7 Aumento de gorduras
periféricas
Sem alterações
Sem alterações
Alterações de tamanho, coloração levemente amarelada e aspecto
esponjoso.
5.2.3. Avaliação microscópica - Análise da aorta
Para esta análise, foi retirada a aorta abdominal.
O grupo G1 (controle) não apresentou alteração histológica. Os demais
grupos não apresentaram nenhuma diferença histológica em relação ao grupo G1.
Estes grupos não tiveram alteração de espessamento endotelial e nem presença de
células espumosas (como demonstrado na Figura 24).

56
5.2.4. Avaliação microscópica - Análise do fígado
Para esta análise, foi avaliado o fígado nas zonas I, II e III. Os resultados de
microscopia estão apresentados nas Figuras 25 a 27.
Grupo G1 (controle negativo, ração padrão) – Não foram observadas
alterações histológicas em nenhuma zona avaliada do fígado.
Grupo G2 (controle negativo, ração suplementada) – Foram observadas
alterações micro e macrovacuolares nas 3 zonas avaliadas.
Grupo G3 (controle positivo (10mg/kg de sinvastatina), ração suplementada) -
Foram observadas alterações micro e macrovacuolares nas 3 zonas avaliadas. Além
disso, o tratamento com sinvastatina provocou, em alguns animais, o aparecimento
de área necrótica e foco de inflamação na zona II.
G2 G3 G4
G1
G5 G6 G7
Figura 24: Avaliação microscópica de aorta abdominal obtida após ensaio com dieta rica em colesterol. Os grupos (n=5) de tratamento são: G1 – controle, G2 – veículo, G3 – sinvastatina, G4 - rutina 16µmol/kg, G5 - rutina 160µmol/kg, G6 - R-Cu2 16µmol/kg e G7 - R-Cu2 160µmol/kg. Coloração: hematoxilina e eosina (HE). Fotos: 20 x.

57
Grupo G4 (rutina 16micromol/kg, ração suplementada) - Foram observadas
alterações microvacuolares nas zonas I e II. Além da inflamação na zona II e, em
alguns animais, o aparecimento de área necrótica na zona III.
Grupo G5 (rutina 160micromol/kg, ração suplementada) - Foram observados
poucos focos de alteração macrovacuolares na zona II.
Grupo G6 (rutina-cobre2 (R-Cu2) 16micromol/kg, ração suplementada) -
Foram observadas alterações micro e macrovacuolares nas 3 zonas avaliadas.
Grupo G7 (rutina-cobre2 (R-Cu2) 160micromol/kg, ração suplementada) - Não
foram observadas alterações histológicas em nenhuma zona avaliada do fígado.
Figura 25 – Avaliação microscópica da área 1 do tecido hepático de ratos submetidos a dieta rica em colesterol por 60 dias.

58
Figura 26 – Avaliação microscópica da área 2 do tecido hepático de ratos submetidos a dieta rica em colesterol por 60 dias.
Figura 27 – Avaliação microscópica da área 3 do tecido hepático de ratos submetidos a dieta rica em colesterol por 60 dias.

59
Os principais componentes celulares do parênquima hepático são as células
hepáticas propriamente ditas (hepatócitos) que se dispõem organizadas em lâminas
separadas por sinusóides; células endoteliais, localizadas nos sinusóides; células de
Kupffer localizadas na periferia dos sinusóides. As células de Ito, lipócitos
diferenciados que produzem colágeno e tecido conectivo, responsáveis pela
sustentação do parênquima, juntamente com as células de defesa como as NK
(natural killer) e linfócitos citotóxicos, que também formam o parênquima hepático.
À microscopia de luz, o parênquima hepático é constituído de unidades
funcionais denominadas de ácino ou estrutura acinar hepática. Na unidade acinar, o
sangue é direcionado dos espaços-porta, passando por meio dos sinusóides que
banham as traves de hepatócitos, até a veia centro-lobular.
O parênquima do ácino é dividido em três zonas ou regiões: zona I - mais
próxima dos ramos portais; zona II - zona intermediária; zona III - zona próxima à
veia centro-lobular. Existem algumas diferenças entre as zonas, sobretudo no que
diz respeito à funcionalidade dos hepatócitos. Os hepatócitos da zona I recebem
sangue mais rico em oxigênio, sendo, portanto, mais resistentes à hipoxia e têm
maior capacidade de regeneração. Já os da zona III que recebem nutrição
principalmente de vasos eferentes, pobres em oxigênio, são mais susceptíveis à
lesão isquêmica. São funcionalmente distintos, conferindo ao órgão uma
heterogeneidade funcional. Assim, foi visualizado as diferenças contidas nas três
regiões.
A avaliação histopatológica do fígado possibilita o entendimento das
alterações provocadas com o aumento do peso relativo deste órgão nos grupos de
animais que receberam dieta rica em colesterol. Com isso, vale a pena destacar a
esteatose (acúmulo de triglicerídeos e colesterol dentro dos hepatócitos), uma
alteração extremamente comum em fígados, que pode ser resultado de uma dieta
com excesso de gordura (mas sem ser uma doença propriamente dita).
Pode-se observar nos resultados que o grupo G2 submetido apenas com a
dieta rica em colesterol apresentou alterações morfológicas no fígado em todas as
áreas, comparado ao grupo G1 controle. Observou-se no grupo G2 uma distribuição
difusa dos vacúolos lipídicos nos hepatócitos, caracterizando esteatose. Os
vacúolos são redondos, de limites nítidos e tamanhos variáveis. Os vacúolos
maiores se formam por confluência dos menores. Quando volumosos, deslocam o

60
núcleo e citoplasma do hepatócito para a periferia porque, sendo os lípides (no caso,
triglicerídeos) insolúveis em água, não se misturam com o citoplasma.
No grupo G3 pode-se observar que o tratamento com 10mg/kg de
sinvastatina causou maiores danos hepáticos quando comparado ao grupo G2. Esse
efeito pode estar relacionado, segundo a literatura, com a toxicidade hepática
causada pelos inibidores da HMG-CoA redutase. Entre estes inibidores estão as
estatinas, como exemplo a sinvastatina, que em doses muito elevadas ou
tratamentos prolongados estão associadas ao desenvolvimento de necrose
hepatocelular em animais de laboratório (VAGHASIVA et al., 2009; CHITTURI;
GEORGE, 2002).
O grupo G4 (rutina 16µmol/kg, ração suplementada) também apresentou
alterações hepáticas significativas em alguns animais. Isso pode ser respondido com
o trabalho de Tamura et al (2010) onde relataram que em ratos com consumo
crônico de flavonoide não alterou os parâmetros bioquímicos, embora em 5% do
grupo de ratos machos apresentaram alterações histopatológicas, como a infiltração
celular inflamatória.
No entanto nos grupos submetidos as maiores concentrações de flavonoide
(G5 e G7) apresentaram uma significante diferença na redução dos danos hepáticos
causados pela hipercolesterolemia, quando comparados ao grupo G2. Lima et al
(2003) demonstraram que a rutina possui atividade hipolipidêmica sem apresentar
toxicidade em coelhos submetidos a dieta rica em colesterol. Já outros trabalhos
demonstram que os flavonoides possuem atividade hepatoprotetora em ratos
submetidos a diferentes danos teciduais (SINGAB et al., 2005; YOKOYAMA et al.,
2009). Estes dados da literatura reforçam o interessante resultado obtido,
principalmente com os dados do grupo G7, que não teve diferença com o grupo
controle G1.

61
6. CONCLUSÃO
Ao se realizar uma análise conjunta dos resultados obtidos após indução da
hipercolesterolemia pode-se observar que a R-Cu2 teve melhor efeito que a rutina
livre, demonstrando uma forte tendência na inibição do aumento sérico de colesterol
total, triglicerídeo e LDL, sugerindo assim um possível efeito contras as
dislipidemias.
Na avaliação da peroxidação lipídica, tanto o tratamento com a rutina livre e
quanto a R-Cu2, mesmo com a ingestão de dieta rica em colesterol, não
apresentaram resultados estatisticamente diferentes do grupo controle (grupo com
dieta de ração padrão).
Os resultados também sugerem que o aumento na produção de NO, nos
grupos tratados, deva-se ao aumento excessivo da ingestão lipídica forçando uma
compensação adaptativa do metabolismo dos animais.
No hematológico, ambas as drogas (rutina e rutina complexada)
demonstraram uma diminuição dos monócitos e um aumento dos linfócitos.
Na análise anatomopatológica a maior concentração do composto R-Cu2 não
apresentou diferença com o grupo controle G1, demonstrando a atividade
hepatoprotetora do composto.
Futuramente, pretende-se pesquisar os mecanismos por qual estas drogas
alteram o perfil de colesterol plasmático e elucidar o mecanismo de hepatoproteção
demonstrada pela Rutina complexada ao cobre.

62
7. REFERÊNCIAS AFANAS'EVA, I. B.; OSTRAKHOVITCH, E. A.; MIKHAL'CHIK, E. V.; IBRAGIMOVA, G. A.; KORKINA, L. G. Enhancement of antioxidant and anti-inflammatory activities of bioflavonoid rutin by complexation with transition metals. Biochemical pharmacology. Oxford, v. 15, n. 61, 2001.
ATOUI, A. K.; MANSOURI, A.; BOSKOU, G.; KEFALAS, P. Tea and herbal infusions: Their antioxidant activity and phenolic profile. Food Chem.Barking, v. 89, n. 27, 2005.
BARREIROS, A. L. B. S.; DAVID, J. M.; DAVID, J. P. Estresse oxidativo: relação entre geração de espécies reativas e defesa do organismo. Química Nova, São Paulo, v. 29, n. 1, p. 113-123, out. 2006. BARJA DE QUIROGA G. B. Fat thermogenesis and exercise: two examples of physiological oxidative stress? Free radical biology and medicine, New York, v. 13, n. 4, 1992. BERLINER, J.A.; NAVAB, M.; FOGELMAN, A.M.; FRANK, J.S.; DEMER, L.L.; EDWARDS, P.A.; WATSON, A.D.; LUSIS, A.J. Atherosclerosis: basic mechanisms - oxidation, inflammation and genetics. Circulation, Dallas, v. 91, 1995. BERTOLAMI, M.C. A conexão entre as lipoproteínas e a aterosclerose. Sociedade de Cardiologia do Estado de São Paulo, São Paulo, v.10, 2000. BENITE, A. M. C.; MACHADO, S. P.; BARREIRO, E. J. Uma visão da química bioinorgânica medicinal. Química nova, São Paulo, vol. 30, n. 8, 2007. BIANCHI, M. L. P.; ANTUNES, L. M. G. Radicais livres e os principais antioxidantes da dieta. Revista de Nutrição, Campinas, v. 12, n. 2, maio/ago. 1999.
BOOTS, A.W.; HAENEN, G.R.; BAST, A. Health effects of quercetin: From antioxidant to nutraceutical. European journal of pharmacology, Amsterdam, v. 13, n. 2, 2008. CHARAMI, M.T.; LAZARI, D.; KARIOTI, A.; SKALTSA, H.; HADJIPAVLOU-LITINA, D.; SOULELES, C. Antioxidant and antiinflammatory activities of Sideritis perfoliata subsp. perfoliata (Lamiaceae). Phytotherapy research, London, v. 22, n. 4, 2008. CHITTURI S.; GEORGE J. Hepatotoxicity of Commonly Used Drugs: Nonsteroidal Anti-Inflammatory Drugs, Antihypertensives, Antidiabetic Agents, Anticonvulsants, Lipid-Lowering Agents, Psychotropic Drugs. Seminars in liver disease, v. 22, n. 2, 2002.
COBBE, S.M.; SHEPHERD, J. Cholesterol reduction in the prevention of coronary heart disease: therapeutic rationale and guidelines. British heart journal, v.69, 1993.

63
COSTA, A. F. Farmacognosia, Lisboa, Fundação Calouste Gulbekian, 5 ed., v. 2, 2002, 1032 p. DEAN, M.; HAMON, Y.; CHIMINI, G. The human ATP-binding cassette (ABC) transporter superfamily. Journal of lipid research, Bethesda, v. 42, n. 7, 2001. DORNAS, W. C. et al. Flavonóides: potencial terapêutico no estresse oxidativo. Revista de Ciências Farmacêuticas Básica e Aplicada, Araraquara, v. 28, n. 3, 2008. DROEBNER, K.; EHRHARDT, C.; POETTER, A.; LUDWIG, S.; PLANZ, O. Cystus052, a polyphenol-rich plant extract, exerts anti-influenza virus activity in mice. Antiviral research, Amsterdam, v. 76, n. 1-10, 2007. DRÖGE, W. Free radicals in the physiological control of cell function. Physiological reviews, Bethesda, v. 82, n. 1, 2002. DUARTE, J. A.; APPELL, H. J.; CARVALHO, F.; BASTOS M. L.; SOARES, J. M. Endothelium-derived oxidative stress may contribute to exercise-induced muscle damage. International journal of sports medicine, Stuttgart, v. 14, n. 8, 1993. ECKARDSTEIN, A.V. et al. ATP binding cassette transporter ABCA1 modulates the secretion of apolipoprotein E from human monocyte-derived macrophages. The Federation of American Societies for Experimental Biology Journal, Bethesda, v. 15, 2001. FENSTER, C. P.; WEINSIER, R. L.; DARLEY-USMAR, V. M.; PATEL, R. P. Obesity, aerobic exercise, and vascular disease: the role of oxidant stress. Obesity research, Baton Rouge, v. 10, n. 9, 2002. FONSECA, T. R. Efeito Do exercício físico e do extrato de soja no perfil lipídico, no estresse oxidativo e na aterosclerose em camundongos deficientes do gene para receptor da LDL. 2007. 89 f. Dissertação (Mestre em ciências dos alimentos) – Universidade Federal de Minas Gerais, Belo Horizonte, 2007. GENEST, J. Lipoprotein disorders and cardiovascular risk. Journal of inherited metabolic disease, Lancaster, v. 26, 2003. GINSBERG, H. N. New perspectives on atherogenesis: role of abnormal triglyceride-rich lipoprotein metabolism. Circulation, Dallas, v. 15, 2002. GOLDFARB, G. H. Antioxidants: role of supplementation to prevent exercise-induced oxidative stress. Medicine and science in sports and exercise, Madison, v. 25, n. 2, 1993. GONÇALVES, M. C. R.; DINIZ, M. F. F. M.; BORBA, J. D. C.; NUNES, X. P.; BARBOSA FILHO, J. M. Berinjela (Solanum melongena L.) – mito ou realidade no combate as dislipidemias? Revista Brasileira de Farmacognosia, São Paulo, v. 16, 2006.

64
GOODMAN & GILMAN. Manual de farmacologia e terapêutica. McGrawHill, 2010. GUYTON, A. C.; HALL, J. E. Tratado de Fisiologia Médica. Guanabara, Rio de Janeiro, 9 ed., 1997. HABIBUDDIN, M.; DAGHRIRI, H. A.; HUMAIRA, T.; QAHTANI, M. S.; HEFZI, A. A. Antidiabetic effect of alcoholic extract of Caralluma sinaica L. on streptozotocin-induced diabetic rabbits. Journal of ethnopharmacology, Limerick, v. 117, n. 2, 2008. HALLIWELL, B. Free radicals and antioxidants: a personal view. Nutrition reviews, New York, v. 52, n. 8 (Pt. 1), 1994. HUANG, P.L.; HUANG, Z.; MASHIMO, H.; BLOCH, K.D.; MOSKOWITZ, M.A.; BEVAN, J.A. Hypertension in mice lacking the gene for endothelial nitric oxide synthase. Nature, v. 377, 1995. IGNARRO, L.J.; LIPPTON, H.; EDWARDS, J.C.; BARICOS, W.H.; HYMAN, A.L.; KADOWITZ, P.J. Mechanism of vascular smooth muscle relaxation by organic nitrates, nitrites, nitroprusside and nitric oxide: evidence for the involvement of S-nitrosothiols as active intermediates. Journal of pharmacology and experimental therapeutics, Baltimore, v. 218, 1981. JUZWIAK, S.; WOZCICKI, J.; MOKRZYCKI, K.; MARCHLEWICZ, M.; BIALECKA, M.; WENDA-ROZEWICKA, L.; GAWRONSKA-SZKLARZ, B.; DROZDZIK, M. Effect of quercetin on experimental hyperlipidemia and atherosclerosis in rabbits. Pharmacological reports, Krakow, v. 57, 2005. KANASHIRO, A.; ANDRADE, D. C. O.; KABEYA, L. M.; TURATO, W. M.; FACCIOLI, L. H.; UYEMURA, A. S.; LUCISANO-VALIM, Y. M. Modulatory effects of rutin on biochemical and hematological parameters in hypercholesterolemic Golden Syrian hamsters. Anais da Academia Brasileira de Ciências, Rio de Janeiro, v. 81, n. 1, 2009. KEYNES, R. G.; GARTHWAITE, J. Nitric oxide and its role in ischaemic brain injury. Current molecular medicine, Hilversum, v. 4, n. 2, 2004. KORKINA, L. G.; AFANAS'EV, I. B. Antioxidant and chelating properties of flavonoids. Advances in pharmacology, New York, v. 38, 1997. KOSTYUK, V. A.; POTAPOVICH, A. I.; KOSTYUK, T. V.; CHERIAN, M. G. Metal complexes of dietary flavonoids: evaluation of radical scavenger properties and protective activity against oxidative stress in vivo. Cellular and molecular biology, Elmsford, v. 53, n. 1, 2007. KUPPUSAMY, U. R.; DAS, N. P. Effects of flavonoids on cyclic AMP phosphodiesterase and lipid mobilization in rat adipocytes. Biochemical Pharmacology, Oxford, v. 44, 1992.

65
LIMA, L. P. R.; OLIVEIRA, T. T.; NAGEM, T. J.; PINTO, A. S.; LIMA, E. Q.; SILVA, J. F. Toxidade Aguda de Rutina e Bixina de Bixa orellana. Acta farmaceutica bonaerense: Latin american journal of pharmacy, Buenos Aires, v. 22, n. 1, 2003. LUZ, P. L. Álcool e coração. Arquivos Médicos do ABC, Santo André, v. 31, n. 2, 2006. MAGALHÃES, M. E. C.; BRANDÃO, A. A.; FREITAS, E. V.; POZZAN, R.; BRANDÃO, A. P. Novas perspectivas no tratamento das dislipidemias. Revista da SOCERJ, Rio de Janeiro, v. 17, n. 2, 2004. MacDONALD, I. A.; MADER, J. A.; BUSSARD, R. G. The role of rutin and quercetrin in stimulating flavonol glycosidase activity by cultured cell-free microbial preparations of human feces and saliva. Mutation Research, Amsterdam, v. 122, n. 2, 1983. MILLER, E. G.; PEACOCK, J. J.; BOURLAND, T. C.; TAYLOR, S. E.; WRIGHT, J. M.; PATIL, B. S.; MILLER, E. G. Inhibition of oral carcinogenesis by citrus flavonoids. Nutrition and câncer, Hillsdale, v. 60, n. 1, 2008. MINAMI, L.; MEIRELLES, P. G.; HIROOKA, E. Y.; ONO, E. Y. S. Fumonisinas: efeitos toxicológicos, mecanismos de ação e biomarcadores para avaliação de exposição. Semina: ciências agrárias, Londrina, v. 25, n. 3, 2004. MONCADA, R.; HIGGS, E.A. Nitric oxide and the vascular endothelium. Handbook of Experimental Pharmacology, Berlin, v. 176, 2006. MONCADA, S.; HERMAN, A. G.; VANHOUTTE, P. Endothelium-derived relaxing factor identified as nitric oxide. Trends in pharmacological sciences, Amsterdam, v. 8, 1987. MUSUNURU, K. Atherogenic Dyslipidemia: Cardiovascular Risk and Dietary Intervention. Lipids, v. 45, 2010. NAGEM, T. J.; PEREIRA, W. L.; OLIVEIRA, T. T.; PINTO, A. S.; OLIVEIRA, M. G.; STRINGHETA, P. C. Efeitos de naringina e dos corantes naturais antocianina e carmim no metabolismo lipídico. Revista Brasileira de Farmácia, Rio de Janeiro, v. 80, n. 1, 1999. NAHÁS, E. A. P.; et al. Efeitos da Isoflavona Sobre os Sintomas Climatéricos e o Perfil Lipídico na Mulher em Menopausa. Revista Brasileira de Geriatria e Obstetrícia, Rio de Janeiro, v. 25, n. 5, 2003. NASCIMENTO, T. B.; BAPTISTA, R. F. F.; PEREIRA, P. C.; CAMPOS, D. H. S.; LEOPOLDO, A. S.; LEOPOLDO, A. P. L.; OLIVEIRA JÚNIOR, S. A.; PADOVANI, C. R.; CICOGNA, A. C.; CORDELLINI, A. S. Alterações Vasculares em Ratos Obesos por Dieta Rica em Gordura: Papel da Via L-arginina/NO Endotelial. Arquivos brasileiros de cardiologia, São Paulo, v. 97, n. 1, 2011. NICASTRO, H.; DA LUZ, C. R.; CHAVES, D. F.; DAS NEVES, W.; VALENTE, K. S.; LANCHA JR, A. H. Leucine supplementation combined with resistance exercise

66
improves the plasma lipid profile of dexamethasone-treated rats. Lipids in health and disease, London, v. 11, n. 1, 2012. NIESS, A. M.; DICKHUTH, H. H.; NORTHOFF, H.; FEHRENBACH, E. Free radicals and oxidative stress in exercise--immunological aspects. Exercise immunology review, Champaign, v. 5, 1999. NOVAK, E. M.; BYDLOWSKI, S. P. Biologiamolecular das dislipidemias. Variação genética das apoliproteínas. Arquivos Brasileiros de Cardiologia, São Paulo, v. 67, n. 6, 1996. NIJVELDT, R. J. et al. Flavonóids: A review of probable mechanisms of action and potential applications. The American Journal Of Clinical Nutrition, Bethesda, v. 74, n. 4, 2001. OLIVEIRA, A. C. et al. Fontes vegetais naturais de antioxidantes. Química Nova, São Paulo, v. 32, n. 3, 2009. OLSZEWER, E. O Perigoso Radical Livre. São Paulo: Nova Linha Editorial, 1994. PARVEEN, Z. et al. Antiinflammatory and analgesic activities of Thesium chinense Turcz extracts and its major flavonoids, kaempferol and kaempferol-3-O-glucoside. Yakugaku Zasshi. Journal of the Pharmaceutical Society of Japan, Tokyo, v. 127, n. 8, 2007. PATOCKOVÁ J.; KRSIAK, M.; MARHOL, P.; TŮMOVÁ, E. Cerebrolysin inhibits lipid peroxidation induced by insulin hypoglycemia in the brain and heart of mice. Physiological research, Praha, v. 52, n. 4, 2003. PEREIRA, R. M.; ANDRADES, N. E.; PAULINO, N.; SAWAYA, A. C.; EBERLIN, M. N.; MARCUCCI, M. C.; FAVERO, G. M.; NOVAK, E. M.; BYDLOWSKI, S. P. Synthesis and characterization of a metal complex containing naringin and Cu, and its antioxidant, antimicrobial, antiinflammatory and tumor cell cytotoxicity. Molecules, Basel, v. 12, n. 7, 2007. POREZ G. et al. Bile acid receptors as targets for the treatment of dyslipidemia and cardiovascular disease. Journal of lipid research, Bethesda, v. 53, n. 9, 2012. PROVIDÊNCIA, R. Proteção cardiovascular por bebidas alcoólicas: Bases científicas do Paradoxo Francês. Revista Portuguesa de Cardiologia. Lisboa, v. 25, n. 11, 2006. RADER, D. J.; DAUGHERTY, A. Translating molecular discoveries into new therapies for atherosclerosis. Nature, London, v. 21, 2008. RADER, D. J. High-density lipoproteins and atherosclerosis. American journal of cardiology, New York, v. 90, n. 8A, 2002.

67
RADOMSKI, M. W.; PALMER, R. M.; MONCADA, S. An L-arginine/nitric oxide pathway present in human platelets regulates aggregation. Proceedings of the national academy of sciences of the United States of America, Washington, v. 87, 1990. RADOMSKI, M. W.; PALMER, R. M.; MONCADA, S. Endogenous nitric oxide inhibits human platelet adhesion to vascular endothelium. Lancet, London, v. 2, 1987. REDONDO, S. et al. Emerging therapeutic strategies to enhance HDL function. Lipids in Health and Disease, London, v. 10, 2011. RIBEIRO, J. N. Efeitos de própolis, antocianina e naringenina em coelhos normais e diabéticos. 2001, 130 f. Tese (Magister Scientiae em agroquímica) Universidade Federal de Viçosa, Viçosa, 2001. RODRIGUES, H. G. et al. Suplementação nutricional com antioxidantes naturais: efeito da rutina na concentração de colesterol-HDL. Revista de Nutrição, Campinas, v. 16, n. 3, 2003. RODRIGO, R.; PRAT, H.; PASSALACQUA, W.; ARAYA, J.; GUICHARD, C.; BÄCHLER, J. P. Relationship between oxidative stress and essential hypertension. Hypertension research, Toyonaka, v. 30, n. 12, 2007. SCALBERT, A.; WILLIAMSON, G. Dietary intake and bioavailability of polyphenols. The Journal of Nutrition, Philadelphia, v. 130, n. 1, 2000. SCHULZ, E.; JANSEN, T.; WENZEL, P.; DAIBER, A.; MÜNZEL, T. Nitric oxide, tetrahydrobiopterin, oxidative stress, and endothelial dysfunction in hypertension. Antioxidants and redox signalling, Larchmont, v. 10, n. 6, 2008. SERRA, J. A.; DOMINGUEZ, R. O.; DE LUSTIG, E. S.; et al. Parkinson’s disease is associated with oxidative stress: comparison of peripheral antioxidant profiles in living Parkinson’s, Alzheimer’s and vascular dementia patients. Journal of neural transmission, Wien, v. 108, 2001. SHAIK, Y. B.; CASTELLANI, M. L.; PERRELLA, A.; CONTI, F.; SALINI, V.; TETE, S.; MADHAPPAN, B.; VECCHIET, J.; DE LUTIIS, M. A.; CARAFFA, A.; CERULLI, G. Role of quercetin (a natural herbal compound) in allergy and inflammation. Journal of biological regulators and homeostatic agents, Milano, v. 20, n. 3-4, 2006. SINGAB, A. N.; YOUSSEF, D. T.; NOAMAN, E.; KOTB, S. Hepatoprotective effect of flavonol glycosides rich fraction from Egyptian Vicia calcarata Desf. against CCl4-induced liver damage in rats. Archives of pharmacal research, Seoul, v. 28, n. 78, 2005. SOCIEDADE BRASILEIRA DE CARDIOLOGIA. IV Diretriz Brasileira sobre dislipidemias e prevenção da aterosclerose. Arquivos Brasileiros de Cardiologia. Vol. 88, 2007.

68
SPIEKER, L. E.; FLAMMER, A. J.; LUSCHER, T. F. The vascular endothelium in hypertension. Handbook of Experimental Pharmacology, Berlin, v. 176, part. 2, 2006. SUDHEESH, S.; PRESANNAKUMAR, G.; VIJAYAKUMAR, S., VIJAYALAKSHMI, N.R. Hypolipidemic effect of flavonoids from Solanum melogena. Plant Foods for Human Nutrition, Dordrecht, v.51, 1997. TAMURA, T.; MITSUMORI, K.; MUTO, S.; KASAHARA, H.; KOBAYASHI, S.; OKUHARA, Y.; HAYASHI M; NAGASAWA, T.; ONOZATO, T.; KURODA, J. Fifty-two week chronic toxicity of enzymatically decomposed rutin in Wistar rats. Food and chemical toxicology, Oxford, v. 48, n. 8-9, 2010. TAN, X. W. et al. Induction of apoptosis in human liver carcinoma HepG2 cell line by 5-allyl-7-gen-difluoromethylenechrysin. World Journal of Gastroenterology, Beijing, v. 15, n. 18, 2009. VAGHASIYA, J.; RATHOD, S.; BHALODIA, Y.; MANEK, R.; MALAVIYA, S.; JIVANI N. Protective effect of polyherbal formulation on simvastatin hepatotoxicity in rats. Pharmacology, Basel, v. 1, n. 1, 2009. VAN ACKER, S. A.; VAN BALEN, G. P.; VAN DEN BERG, D. J.; BAST, A.; VAN DER VIJGH, W. J. Influence of iron chelation on the antioxidant activity of flavonoids. Biochemical pharmacology, Oxford, v. 56, n. 8, 1998. VANDERLAAN, A. P.; REARDON, A. C.; GETZ, S. G. Site specificity of atherosclerosis. Arteriosclerosis, Thrombosis and Vascular Biology, Dallas, v.24, 2004. VANE, J. R. Control of the circulation by endothelial mediators. Inaugural G.B. West Memorial Lecture. International archives of allergy and immunology, v. 101, n. 4, 1993. YOKOYAMA, A.; SAKAKIBARA, H.; CROZIER, A.; KAWAI, Y.; MATSUI, A.; TERAO, J.; KUMAZAWA, S.; SHIMOI, K. Quercetin metabolites and protection against peroxynitrite-induced oxidative hepatic injury in rats. Free radical research, Yverdon, v. 43, n. 10, 2009. YORDANOV, M. et al. Inhibition of Candida albicans extracellular enzyme activity by selected natural substances and their application in Candida infection. Canadian Journal of Microbiology, Ottawa, v. 54, n. 6, 2008. YU, J.; WANG, L.; WALZEM, R. L.; MILLER, E. G.; PIKE, L. M.; PATIL, B. S. Antioxidant activity of citrus limonoids, flavonoids, and coumarins. Journal of agricultural and food chemistry, Washington, v. 53, n. 6, 2005. WANG, H.; PENG, D. Q. New insights into the mechanism of low high-density lipoprotein cholesterol in obesity. Lipids in Health and Disease, London, v. 176, 2011.

69
WIDLANSKY, M. E.; SESSO, H. D.; REXRODE, K. M.; MANSON, J. E.; GAZIANO, M. J. Body mass index and total and cardiovascular mortality in men with a history of cardiovascular disease. Archives of internal medicine, Chicago, v. 164, n. 21, 2004. ZAND, T.; HOFFMAN, A. H.; SAVILONIS, B. J.; UNDERWOOD, J. M.; NUNNARI, J. J.; MAJNO, G.; JORIS, I. Lipid deposition in rat aortas with intraluminal hemispherical plug stenosis: A morphological and biophysical study. American journal of pathology, New York, v. 155, n. 1, 1999. ZIMMERMANN, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain, Amsterdam, n. 6, 1983. ZUANAZZI, J. A. S.; MONTANHA, J. A.. Flavonóides. In: SIMÕES, C. M. O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P.; MENTZ, L. A.; PETROVICK, P. R. Farmacognosia da Planta ao Medicamento. Porto Alegre/Florianópolis: UFRGS / UFSC, 5ª ed. Revista e ampliada, primeira reimpressão, p. 577-614. 2004.

70
ANEXO







