distribuiÇÃo espacial de insetos fitÓfagos e de...
TRANSCRIPT

UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA E
MUCURI – UFVJM
Joyce Gomes Damascena
DISTRIBUIÇÃO ESPACIAL DE INSETOS FITÓFAGOS E DE
INIMIGOS NATURAIS EM LEUCAENA LEUCOCEPHALA UTILIZADA
COMO QUEBRA-VENTO EM REGIÃO DE CERRADO NO BRASIL
DIAMANTINA - MG
2016

JOYCE GOMES DAMASCENA
DISTRIBUIÇÃO ESPACIAL DE INSETOS FITÓFAGOS E DE
INIMIGOS NATURAIS EM LEUCAENA LEUCOCEPHALA UTILIZADA
COMO QUEBRA-VENTO EM REGIÃO DE CERRADO NO BRASIL
Dissertação apresentada ao Curso de Pós-
Graduação Stricto Sensu em Produção Vegetal da
Universidade Federal dos Vales do Jequitinhonha
e Mucuri, como parte das exigências do Programa
de Pós-Graduação em Produção Vegetal, área de
concentração Produção Vegetal, para obtenção do
título de “Mestre”.
Orientador
Prof. Dr. Germano Leão Demolin Leite
Coorientador
Prof. Dr. Marcus Alvarenga Soares
DIAMANTINA - MG
2016

Ficha Catalográfica
Preparada pelo Serviço de Biblioteca/UFVJM Bibliotecário
responsável: Gilson Rodrigues Horta – CRB6 nº 3104
D155d
2016
Damascena, Joyce Gomes.
Distribuição espacial de insetos fitófagos e de inimigos naturais em
leucaena leucocephala utilizada como quebra-vento em região de
cerrado no Brasil. / Joyce Gomes Damascena. Diamantina: UFVJM,
2016.
40 f.
Dissertação (Mestrado) – Universidade Federal dos Vales do
Jequitinhonha e Mucuri. Programa de Pós-Graduação em Produção
Vegetal, 2016.
Orientador: Prof. Dr. Germano Leão Demolin Leite.
Coorientador: Prof. Dr. Marcus Alvarenga Soares.
1. Estratificação. 2. Pontos cardeais. 3. Dossel. 4. Faces foliares.
I. Título.
CDD: 570

JOYCE GOMES DAMASCENA
DISTRIBUIÇÃO ESPACIAL DE INSETOS FITÓFAGOS E DE
INIMIGOS NATURAIS EM LEUCAENA LEUCOCEPHALA UTILIZADA
COMO QUEBRA-VENTO EM REGIÃO DE CERRADO NO BRASIL
DIAMANTINA

OFEREÇO O Senhor me deu força quando achava que não agüentaria; porque o Senhor colocou em
minha vida, familiares e amigos que me apoiaram e incentivaram. Colocou-me junto a
colegas, todos vindos de lugares, criações, sonhos, idéias e estilos de vida tão diferentes.
Deu-me não só mestres, mas também conselheiros; fez com que nesse tempo eu aprendesse
muitas coisas, não só referentes a uma matéria ou outra, mas à vida que me espera. Muito
obrigado DEUS!
DEDICO
Aos meus pais, Helena G. Damascena e Joaquim Damascena, pelo amor, educação, paciência e
todo apoio necessário para ultrapassar barreiras e alcançar as metas que se apresentam em
minha vida, e as minhas madrinhas, Lora, Dinha e Prêta, In Memorian.

AGRADECIMENTOS
A Deus, que está no centro da minha vida; da minha família, dos meus amigos,
das minhas atividades, dos meus pensamentos, ... Tudo está ligado a Ele. Minha segurança!
Agradeço também ao meu noivo João Paulo, que de forma especial e carinhosa
me deu força e coragem me apoiando nos momentos de dificuldades.
Meus irmãos, Ellen e Felipe, pela amizade e por estarem sempre ao meu lado em
momentos importantes da minha vida.
À UFVJM, pelo acolhimento e incentivo e à Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES), pela concessão de bolsa de estudo.
Aos professores do PPGPV pelo compartilhamento de conhecimento e
experiências, contribuindo para minha formação acadêmica e profissional. Em especial ao
professor Evander Ferreira, pela atenção e paciência.
Ao meu orientador Germano Leão Demolin Leite, que me aceitou como orientada,
pelos seus conhecimentos compartilhados e acolhimento, e ao professor Marcus Alvarenga
Soares, que de forma muito carinhosa aceitou-me coorientar.
Agradeço, aos meus colegas que já se tornaram amigos, que vêem me
acompanhando desde a graduação, Adriana, Aline, Márcio, Luan, Igor, pois acreditaram nos
meus sonhos, estiveram sempre ao meu lado, principalmente nos meus estresses,
compreendendo e acompanhando com amor e paciência.
Enfim, a todos que diretamente ou indiretamente me ajudaram e me apoiaram,
principalmente com conselhos que me ajudaram a crescer.

i
RESUMO
DAMASCENA, Joyce Gomes. DISTRIBUIÇÃO ESPACIAL DE INSETOS
FITÓFAGOS E DE INIMIGOS NATURAIS EM LEUCAENA LEUCOCEPHALA
UTILIZADA COMO QUEBRA-VENTO EM REGIÃO DE CERRADO NO
BRASIL. 2016. 40p. (Dissertação - Mestrado em Produção Vegetal) – Universidade
Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, 2016.
Leucaena leucocephala (Lam.) de Wit. (Fabaceae: Mimosoideae), conhecida como
leucena, é bastante cultivada no mundo pelo valor nutricional de suas folhas e ramos finos,
sendo considerado um alimento completo para ruminantes e monogástricos, por isso
utilizada em sistemas agrossilvipastoris. Espécie que também é empregada para
reflorestamento de áreas degradadas podendo controlar erosões hídricas e eólicas, além de
melhorar as condições ambientais. Além disso, leucena pode ser usada em quebra-vento, o
que poderá influenciar na distribuição de pragas e inimigos naturais e dificultar a
localização de plantas hospedeiras. A distribuição desses insetos na planta poderá ocorrer
de acordo com fatores extrínsecos (ex.; vento, insolação), o que varia com a região que a
cultura foi cultivada, e intrínseca (ex.; componentes físico-químicos) relacionados com
genética da planta. O objetivo deste trabalho foi avaliar durante dois anos, a distribuição
espacial de insetos fitófagos e de seus inimigos naturais ao longo da estratificação vertical
do dossel (partes apical, média, basal e no tronco) e horizontal (orientação dos galhos
quanto à exposição do sol nas faces norte, sul, leste e oeste) e da face foliar (adaxial e
abaxial) em L. leucocephala. As avaliações foram realizadas estando L. leucocephala em
quebra-vento com 100 metros de comprimento, disposto com duas fileiras centrais de
Acacia mangium Willd. (Leguminosae) (espaçamento 3 x 3 m) e duas fileiras de L.
leucocephala, uma de cada lado da A. mangium, com espaçamento de 3 m entre fileiras e 2
m entre plantas. Foram avaliados os números de insetos fitófagos, polinizadores e inimigos
naturais, contados quinzenalmente, em 20 árvores de L. leucocephala (16 meses de idade)
durante o período experimental. Essa contagem foi realizada por visualização direta, nas
faces foliares (adaxial e abaxial)/folha, na parte apical, média e basal do dossel e nos
galhos posicionados nas faces norte, sul, leste e oeste, entre 7 e 11 h da manhã. Além disso,
os insetos foram contabilizados no tronco das 20 árvores/avaliação. Avaliaram-se os
índices ecológicos (número de indivíduos, riqueza, diversidade e abundância de espécies).
A fórmula de Colina foi usada para calcular a diversidade. Abundância e riqueza de
espécies (S) foram calculadas pela fórmula de Simpson. Dominância-K foi calculada

ii
traçando a abundância percentual cumulativo contra log classificação de espécies. Os
valores de dominância-K indica a distribuição de dominância e uniformidade dos
indivíduos entre as espécies. As frequências de cada espécie de inseto nas amostras foram
classificadas como constante, comum e rara. No geral, foi observado baixos índices de
insetos em L. leucocephala, sendo encontradas 18 espécies raras, 3 comuns e 1 constante.
Os maiores números de indivíduos tanto de fitófagos, quanto de inimigos naturais e
polinizadores quanto à estratificação horizontal do dossel, foram observados na posição
norte dos ramos. Para os índices de riqueza específica e diversidade de fitófagos foram
observados maiores valores nos galhos voltados a oeste e, para os inimigos naturais, nos
galhos voltados para norte/sul e oeste, respectivamente. Para as variáveis, número de
indivíduos, riqueza específica e índice de diversidade quanto à estratificação vertical do
dossel, os maiores valores para fitófagos foram observados na parte apical de L.
leucocephala; e de inimigos naturais nas partes apicais e medianas, respectivamente. Os
maiores valores de indivíduos, riqueza específica e índice de diversidade tanto para os
fitófagos como os inimigos naturais, foram observados na face abaxial de L. leucocephala.
Quanto à variável abundância, fitófagos mastigadores obtiveram maior índice na posição
norte dos ramos, já sugadores, na posição oeste. Maior abundância de formigas foi
encontrada na posição norte, e maior abundância de abelhas da espécie Tetragonisca
angustula Latreille (Hymenoptera: Apidae) na posição oeste dos ramos do dossel de L.
leucocephala. Quanto à estratificação vertical do dossel, maiores valores de abundância de
fitófagos mastigadores e sugadores foi na região apical do dossel, o que também foi para
formigas, já as abelhas T. angustula em maior abundância na região mediana do dossel de
L. leucocephala. Quanto à face foliar, no geral, para todos os índices avaliados, os maiores
valores foram encontrados na face abaxial das folhas de L. leucocephala. Com este estudo,
podem-se estabelecer estratégias de monitoramento, sabendo-se que fitófagos, inimigos
naturais e polinizadores estão em maior número de indivíduos na posição norte da região
apical do dossel. Já a riqueza específica e diversidade, encontram-se maiores índices de
fitófagos, inimigos naturais e polinizadores na posição oeste da região apical do dossel e
no geral, encontrados na face abaxial das folhas de L. leucocephala.
Palavras-chave: leucena, artrópodes, diversidade.

iii
ABSTRACT
DAMASCENA, Joyce Gomes. SPATIAL DISTRIBUTION OF PHYTOPHAGOUS
INSECTS AND NATURAL ENEMIES IN LEUCAENA LEUCOCEPHALA
UTILIZED AS WINDBREAK IN CERRADO REGION OF BRAZIL. 2016. 40p.
(Dissertation– Master’s degree in Vegetal Production) – Federal University of
Jequitinhonha and Mucuri Valleys, Diamantina, 2016.
Leucaena leucocephala (Lam.) of Wit.(Fabaceae: Mimosoideae), known as leucaena, is
widely cultivated around the world due to the nutritional value of its leaves and thin
branches, being considered a complete ruminant and monogastric feed, hence its use in
agroforestry systems. This species is also used in the reforestation of degraded areas for it
can control hydrous and aeolian erosion and also improve the environmental conditions.
Above all, leucaena can be used in windbreaker, which may influence pests and enemies
distribution and difficult the location of host plants. The distribution of those insects on the
plant may occur according to extrinsic factors (e.g.: wind, insolation), which varies with
the area where the culture was cultivated, and intrinsic (e.g.: physical and chemical
components) related to the plant genetics. The objective of this paper was to evaluate
during two years the spatial distribution of phytophagous insects and their natural enemies
along the vertical canopy stratification (apical, medium and basal parts and also the trunk)
and horizontal (orientation of the branches in relation to the exposition to the sun on north,
south, east and west faces) and of foliar face (adaxial and abaxial) in L. leucocephala. The
evaluations were performed with L. leucocephala being in windbreak with 100 meters in
length, disposed in two central rows of Acacia mangium Willd. (Leguminosae) (spacing of
3 x 3 meters) and two rows of L. leucocephala, one of each side of A. mangium, with
spacing of 3 meters between the rows and 2 meters between the plants. We evaluated the
number of phytophagous insects, pollinator insects and natural enemies, counted every 15
days, in 20 trees of L. leucocephala (16-month old) during the experimental period. This
count was performed by direct visualization on the foliar faces (adaxial and abaxial)/leaf,
on the apical, medium and basal parts of the canopy and on branches placed on the north,
south, east and west faces, between 7hrs and 11hrs in the morning. Above all, the insects
were counted on the trunk of 20 trees/evaluation. The ecological indicators (number of
individuals, affluence, diversity and abundance of species). The formula of Colina was
used to calculate the diversity. Abundance and affluence of species (S) were calculated by
the formula of Simpson. Dominance-K was calculated tracing cumulative percentage

iv
abundance against log classification of species. The values of dominance-K indicate the
dominance and uniformity distribution of individuals between the species. The frequency
of each insect species in the samples was classified as constant, common and rare. In
general, it was observed low indices of insects in L. leucocephala, and 18 rare, 3 common
and 1 constant species were found. The greatest number of phytophagous individuals,
natural enemies and pollinators related to the horizontal canopy stratification were
observed on the north side of the branches. For specific affluence and phytophagous
diversity indices we observed greater values on the branches facing west, and for the
bioregulators, on the branches facing north/south and west, respectively. For the variables,
number of individuals, specific affluence, and diversity index in relation to the vertical
canopy stratification, the greater values for phytophagous were observed on the apical part
of L. leucocephala; and of bioregulators on the apical and medium parts, respectively. The
greatest values of individuals, specific affluence and biodiversity index were observed on
the abaxial face of L. leucocephala for both phytophagous and bioregulators. As for the
abundance variable, chewing phytophagous acquired greater index on the north position of
the branches, while sucking phytophagous, on west position. Greater abundance of ants
was found on the north position, and greater abundance of bees of the Tetragonisca
angustula Latreille (Hymenoptera: Apidae) species on the west position of the canopy
branches of L. leucocephala. Regarding the vertical canopy stratification, greater
abundance values of chewing and sucking phytophagous were found on the apical canopy
region, the same was true for ants, whereas the T. angustula bees were more abundant on
the medium canopy region of L. leucocephala. As for the foliar face, in general for all
evaluated indexes the greatest values were found on the abaxial face of L. leucocephala
leaves. With this paper, we can establish monitoring strategies, knowing that
phytophagous, natural enemies and pollinators are in greater number of individuals on the
north position of the apical canopy region. As for the specific affluence and diversity,
greater phytophagous, natural enemies and pollinators were found on the west position of
the apical canopy region and in general, were also found on the abaxial face of L.
leucocephala leaves.
Key-words: leucena, Arthropods, diversity.

v
LISTA DE TABELAS
ARTIGO CIENTÍFICO Pág.
Tabela 1 36
Tabela 2 37
Tabela 3 38
Tabela 4 39
Tabela 5 40

Sumário RESUMO ........................................................................................................................... i ABSTRACT .................................................................................................................... iii LISTA DE TABELAS ..................................................................................................... v INTRODUÇÃO GERAL ................................................................................................ 1 REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 5
ARTIGO CIENTÍFICO .................................................................................................. 9 Resumo ............................................................................................................................. 9 Abstract .......................................................................................................................... 10
1 Introdução ................................................................................................................... 11 2 Material e Métodos ..................................................................................................... 13 3 Resultados ................................................................................................................... 15 4 Discussão ..................................................................................................................... 17 5 Conclusões ................................................................................................................... 24
Agradecimentos ............................................................................................................. 24
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................ 25

INTRODUÇÃO GERAL
Leucaena leucocephala (Lam.) de Wit. (Fabaceae: Mimosoideae) é a principal
espécie do gênero Leucaena Benth (HUGHES, 1998) compreende 22 espécies, distribuídas
desde o sul do Texas até o Peru (SCHIFINO-WITTMANN, 2004). Durante todo o ano, a
leucena pode apresentar floração, sendo nos meses chuvosos a maior produção de flores.
Quanto ao porte da árvore, a leucena pode ser disposta em três grupos principais: 1) tipo
Havaiano, comumente encontrado na costa do México é arbustivo de até cinco metros de
altura, 2) tipo Salvador, originário do interior da América Central é arbóreo com até vinte
metros de altura e 3) o tipo médio, conhecido como tipo Peru, cujo emprego ainda é
restrito ao Hawaí, México e Austrália, (PEIXOTO et al., 2002) normalmente mais folhoso
e engalhado, sendo o mais utilizado em forrageira e por isso de mais fácil alcance pelos
animais em pastejo (BARRETO et al., 2010). A Leucena possui ráquis pubescente, seus
folíolos são oblongo-lineares (10 a 15 pares) e suas folhas bipinadas, de 4 a 8 pares de
pinas. Apresenta vagens achatadas e finas, contendo de 15 a 25 sementes de cor marrom
brilhante (ALCÂNTARA e BUFARAH, 1979). Suas flores são brancas e numerosas
agrupando-se em capítulo globular de 1,5 cm a 3 cm de diâmetros (PEIXOTO et al., 2002).
Leucaena leucocephala além de ser amplamente cultivada para reflorestamento de áreas
degradadas, também serve como fonte de proteína para alimentação animal em sistemas
agrossilvipastoris, por possuir excelente valor nutritivo e alta produção de matéria seca
(ATAWODI et al., 2008). Devido a sua adaptação aos diversos tipos de solos e diferentes
níveis de precipitação, insolação e resistência à salinidade é bastante cultivada no mundo
(LINS et al., 2007). Pode controlar erosões hídricas e eólicas, melhorando assim as
condições ambientais (HAI, 2007), além de poder ser usada em quebra-ventos.
Árvores usadas como quebra-ventos podem influenciar a distribuição de pragas e
de inimigos naturais e dificultar a localização de plantas hospedeiras (RAO et al., 2000;
BRANDLE et al., 2004). Sendo que, a posição dos ramos de uma árvore quanto ao
caminhamento do sol pode afetar a abundância e riqueza de insetos devido a: 1) direção e
intensidade dos ventos, podendo afetar a dispersão de muitos insetos na copa da planta
hospedeira (LEITE et al., 2011a, b, c) e ocasionar dessecamento das folhas e a queda de flores
e frutos, o que afeta a comunidade de insetos (ex.: menor quantidade de recurso ou de pior
qualidade) (LEITE et al., 2006, 2009, 2011c); 2) exposição ao sol, o que pode influenciar a
qualidade dos tecidos da planta hospedeira (ex.: folhas mais duras) (UNSICKER e MODY,
2005); 3) microclima. Folhas expostas ao sol apresentam um maior dano foliar devido à

2
presença de insetos galhadores, comparado a folhas em camadas intermediárias e sombreadas
(BASSET, 1991; LEITE et al., 2011a), e 4) preferência de herbívoros por partes das plantas
com menor número de inimigos naturais (espaço livre-inimigos) (UNSICKER e MODY, 2005;
LEITE et al., 2012). Além disso, quanto a estratificação vertical, o dossel dessas plantas
oferece recursos para a manutenção da diversidade, onde seus ramos e folhas alteram o
ambiente pelo sombreamento afetando a umidade, evapotranspiração e servindo de refúgio
para insetos herbívoros e inimigos naturais (BRANDLE et al., 2004). O dossel superior está
diretamente exposto a maior radiação solar, onde também se concentra a maior densidade de
folhas, em geral, as mais novas, além de flores e sementes, quando se compara a região inferior
(PARKER, 1995). Além de que, características estruturais e químicas das folhas, também
influenciam a distribuição e o ataque dos fitófagos (PEETERS, 2002). Em geral, as folhas mais
novas de uma planta, possuem maior palatabilidade aos insetos, e por possuírem menor
conteúdo de fibras, serem mais macias e mais nutritivas, são mais predadas. Contudo, essas
folhas novas possuem uma maior concentração de compostos secundários, o que pode explicar
a preferência por insetos herbívoros especialistas (PEETERS, 2002). Já insetos mais
generalistas preferem as folhas maduras. De modo geral, os insetos preferem a face abaxial
da folha por fatores como: (1) essa região se apresentar mais tenra; (2) nervuras mais
salientes; (3) córtex delgado e (4) epiderme mais fina. A face abaxial da folha ainda pode
servir de refúgio para os insetos, seja por predadores e intempéries, além de proteger a
oviposição (ANTÔNIO et al., 2002).
Além de esses fatores externos influenciarem na distribuição e população dos
insetos, é evidente a influência das características físico-químicas na interação planta-
inseto herbívoro. A leucena apresenta alguns atributos que podem favorecer ou não a
presença de insetos: (1) contém taninos flabofênicos (CAVALCANTE et al., 2006), os
quais reduzem significativamente o crescimento e a sobrevivência de insetos, por
inativarem enzimas digestivas e criarem um complexo de taninos-proteínas de difícil
digestão (MELLO e SILVA-FILHO, 2002) e (2) apresenta nectários extraflorais, que
atuam como defesas indiretas e induzidas contra herbívoros. Nectários extraflorais são
estruturas que secretam néctar, estando presentes em órgãos aéreos das plantas (FAHN,
1979). Vários estudos mostram que em troca de néctar, as formigas protegem as plantas
que possuem os nectários extraflorais, atacando os herbívoros (mutualismo defensivo)
(COSTA et al., 1992; DE LA FUENTE e MARQUIS, 1999; CUAUTLE e RICO-GRAY,
2003). Algumas espécies de plantas conseguem aumentar a produção de néctar extrafloral
após sofrerem ataques de herbivoria, por isso, atraindo mais formigas aos ramos cometidos

3
(NESS, 2003; RIOS et al., 2008), já outras espécies de plantas produzem mais néctar nas
folhas novas após danos mecânicos (MONDOR e ADDICOTT, 2003).
Nos lugares onde a leucena é cultivada, a principal praga é o psilídeo
Heteropsylla cubana Crawford (Hemiptera: Psyllidae) (SANTANA, 2008) a qual sua
dispersão é em função do movimento dos ventos (ELDER et al., 1998; RAO et al., 2000).
Este inseto é responsável por graves danos a leucena em várias partes do mundo, pois
podem se alimentar intensamente dessa árvore a ponto de ocasionar desfolhação total, com
consequente morte da planta (SANTANA, 2008). No Brasil, foi registrado primeiramente
em 2000, na região do porto de Tubarão-ES. Em 2005, foi observado em Minas Gerais
(Patos de Minas, Vazante e Uberlândia) e no Paraná (Londrina). Em 2006, foi detectada na
Bahia (Salvador) (SANTANA, 2008). Outros insetos também são comumente encontrados
em associação com L. leucocephala como: Acanthoscelides macrophthalmus (Schaeffer)
(Chrysomelidae: Bruchinae) (RAGHU et al., 2005) e Coccus longulus (Douglas)
(Hemiptera: Coccidae) (ELDER et al., 1998), sendo esses dois insetos, com registro no
Brasil. Em alguns trabalhos foi verificado que Apis mellifera L. e Trigona spinipes
Fabricius (Hymenoptera: Apidae) são os principais visitantes de L. leucocephalla
(CARVALHO e MAÊDA, 1997; MELO-SILVA et al., 2014). Essa visita ocorre em
horário semelhante, a qual o maior pico de visitação acontece de 8-9 horas, o que sugere
ser o melhor horário com maior disponibilidade de recursos florais (MELO-SILVA et al.,
2014). A diminuição das visitas ocorre após as 10h devido à falta de recursos na flor e não
devido à temperatura (ARAUJO et al., 2006). Lembrando que a depender da cultura, T.
spinipes pode influênciar na polinização, diminuindo ou inibindo a frutificação (SERRA,
2010),
No entanto, não há estudos sobre a distribuição de insetos fitófagos e de seus
inimigos naturais em L. leucocephala no Brasil. O conhecimento das distribuições
espaciais de insetos-praga e seus inimigos naturais são importantes não
só por causa de sua importância para melhorar a compreensão da dinâmica populacional de
insetos em geral, mas também porque é fundamental para futuros planejamentos de
estratégias de planos de amostragem para tomadas de decisão sobre o controle de pragas
(TOLEDO et al., 2006). Diante do exposto, objetivou-se identificar os insetos fitófagos,
inimigos naturais e polinizadores. Além disso, as suas distribuições espaciais destes ao
longo da estratificação vertical do dossel (partes apical, média, basal e tronco) e horizontal

4
(orientação dos galhos quanto à exposição do sol nas faces norte, sul, leste e oeste) e da
face foliar (adaxial e abaxial) em plantas de L. leucocephala.

5
REFERÊNCIAS BIBLIOGRÁFICAS
ALCÂNTARA, P.N.; BUFARAH, G., 1979. Plantas forrageiras: gramíneas e
leguminosas, São Paulo, 150p.
ANTÔNIO, A. C. et al. Ovoposição de Diaphania hyalinata L. (Lepidoptera, Pyralidae)
afetada pela face foliar e tricomas. Acta Scientiarum, Maringá, v. 24, n. 2, p. 359-362,
2002.
ARAUJO, V. A.; ANTONINI, Y.; ARAUJO, A. P. A. Diversity of bees and their floral
resources at. Altitudinal areas in the Southern Espinhaço Range, Minas Gerais, Brazil.
Neotropical Entomology, Londrina, v. 35, n. 1, p. 30-40, 2006.
ATAWODI, S. E. et al. Assessment of Leucaena leucocephala leaves as feed supplement
in laying hens. African Journal of Biotechnology, Nairobi, v. 7, n. 3, p. 317-321, 2008.
BARRETO, M. L. DE JESUS. et al. Utilização da leucena (Leucaena leucocephala) na
alimentação ruminantes. Revista Verde, Mossoró, v.5, n.1, p. 07-16, 2010.
BASSET, Y. The spatial distribution of herbivory, mines, and galls within an Australian
rain forest tree. Biotropica, Washington, v. 23, p. 271–281, 1991.
BRANDLE, J. R.; HODGES, L.; ZHOU, X. H. Windbreaks in North American
agricultural systems. Agroforestry Systems, University of Nebraska-Lincoln, v. 61, p. 65–
78, 2004.
CAVALCANTE, G. M.; MOREIRA, A. F. C.; VASCONSELOS, S. D. Potencialidade
inseticida de extratos aquosos de essências florestais sobre mosca-branca. Pesquisa
Agropecuária brasileira, Brasília, v.41, n.1, p.9-14, jan. 2006.
COSTA, F. M. C. B.; OLIVEIRA-FILHO, A. T. and OLIVEIRA, P. S. The role of
extrafloral nectaries in Qualea grandiflora (Vochysiaceae) in limiting herbivory: an
experiment of ant protection in Cerrado vegetation. Ecology Entomology, London, v. 17,
p. 362-365, 1992.
CUAUTLE, M. and RICO-GRAY, V. The effect of wasps and ants on the reproductive
success of the extrafloral nectaried plant Turnera ulmifolia (Turneraceae). Functional
Ecology, Oxford, v. 17, p. 417-423, 2003.

6
CARVALHO, A. G.; MAÊDA, J. M. Biologia floral de L. leucocephala (Leguminosae,
Mimosoidae). Floresta e Ambiente, Seropédica, v. 4, n. 1, p. 25-29, 1997.
DE LA FUENTE, M. A. S. and MARQUIS, R. J. The role of ant-tended extrafloral
nectaries in the protection and benefit of a Neotropical rainforest tree. Oecologia, Berlin,
v. 118, p. 192-202, 1999.
ELDER, R. J.; MIDDLETON, C. H.; BELL, K. L. Heteropsylla cubana Crawford
(Psyllidae) and Coccus longulus (Douglas) (Coccidae) infestations on Leucaena species
and hybrids in coastal central Queensland. Australian Journal of Entomology, Canberra,
v. 37, p. 52-56, 1998.
FAHN, A. Ultra structure of nectaries in relation to nectar secretion. American Journal
of Botany, Columbus – Ohio, v. 66, p. 977-985, 1979.
HAI, R. et al. Changes in biodiversity and ecosystem function during the restoration of a
tropical forest in south China. Science in China Series C: Life Sciences, Beijing, v. 50, n.
2, p. 277-284, 2007.
LEITE, G. L. D. et al. Artrópodes (Arthropoda) associados a mudas de pequizeiro
Caryocar brasiliense Cambess. (Caryocaraceae). Arquivo Instituto Biológico, São Paulo,
v. 73, n.3, p. 365-370, 2006.
LEITE, G. L. D. et al. Within tree distribution of a gall-inducing Eurytoma
(Hymenoptera, Eurytomidae) on Caryocar brasiliense (Caryocaraceae). Revista
Brasileira de Entomologia, Curitiba, v. 53, p. 643-648, 2009.
LEITE, G. L. D. et al. Factors affecting Constrictotermes cyphergaster (Isoptera:
Termitidae) nesting on Caryocar brasiliense trees in the Brazilian savanna.
Sociobiology, Chico, v. 57, p. 165-180, 2011a.
LEITE, G. L. D. et al. Distribution of a leaf vein gall in Caryocar brasiliense
(Caryocaraceae) Tree. Revista Caatinga, Mossoró, v. 24, p. 186-190, 2011b.
LEITE, G. L. D. et al. Within tree distribution of a discoid gall on Caryocar brasiliense.
Revista Colombiana de Entomología, Santa Fé de Bogotá, v. 37, p. 289-293, 2011c.

7
LEITE, G. L. D. et al. Seasonal abundance of hemipterans on Caryocar brasiliense
(Malpighiales: Caryocaraceae) trees in the Cerrado. Florida Entomologist, Gainesville,
p.862-872, 2012.
LINS, C. E. DE LIMA. et al. Efeito de fungos micorrízicos arbusculares no crescimento de
mudas de Leucaena leucocephala (lam.) de Wit. em solos de caatinga sob impacto de
mineração de cobre. Revista árvore, viçosa, v. 31, n. 2, 2007.
MELLO, M. O.; SILVA-FILHO, M. C. Plant-insect interactions: an evolutionary arms
race between two distinct defense mechanisms. Brazilian Journal of Plant Physiology,
Campos dos Goytacazes, v.14, p.71-81, 2002.
MELO-SILVA, C. et al. Reproductive biology of L. leucocephala (Lam.) R. de Wit
(Fabaceae: Mimosoideae): successo fanin vasive species. Neotropical Biology and
Conservation, São Leopoldo, v. 9, n. 2, p. 91-97, 2014.
MONDOR, E. B. and ADDICOTT, J. F. Conspicuous extrafloral nectarines are inducible
in Vicia faba. Ecology Letters, Oxford, v. 6, p. 495-497, 2003.
NESS, J. H. Catalpa bignonioides alters extrafloral nectar production after herbivory and
attracts ant bodyguards. Oecologia, Berlin, v. 134, p. 210-218, 2003.
PARKER, G. G. Structure and microclimate of forest canopies. Forest Canopy (ed. by
M.D. Lowman and N.M. Nadkarni), San Diego, p. 431-455, 1995.
PEETERS, P. J. Correlations between leaf structural traits and the density of herbivorous
insect guilds. Biological Journal of the Linnean Society, United Kingdom , v.77, p. 43-65,
2002.
PEIXOTO, A.M. et al. Enciclopédia Agrícola Brasileira, São Paulo, p. 272.
RAO, M. R.; SINGH, M.P.; DAY, R. Insect pest problems in tropical agroforestry
systems: Contributory factors and strategies for management. Agroforestry Systems, New
York, v. 50, p. 243–277, 2000.
RAGHU, S.; WILTSHIRE, C.; DHILEEPAN, K. Intensity of pre-dispersal seed predation
in the invasive legume Leucaena leucocephala is limited by the duration ofpod retention.
Austral Ecology, Hoboken, v. 30, p. 310–318, 2005.

8
RIOS, R. S.; MARQUIS, R. J. and FLUNKER, J. C. Population and variation in plants
traits associated with ant attraction and herbivory in Chamaecrista fasciculata (Fabaceae).
Oecologia, Berlin, v. 156, p. 577-588, 2008.
SANTANA, D. L. Q. Psilídeos no Brasil: Heteropsylla cubana (Psilídeo da leucena)
Hemiptera: Psyllidae. Colombo-PR: Embrapa Florestas, Comunicado Técnico, n. 212, p.
4, 2008.
SCHIFINO-WITTMANN, M. T. Citogenética do gênero Leucaena Benth. Ciência Rural,
Santa Maria, v. 34, n. 1, p. 309-314, 2004.
SERRA, B. D. V; DE CAMPOS, L. A. Polinização entomófila de abobrinha, Cucurbita
moschata (Cucurbitacea), Neotropical Entomology, Londrina, v. 39, ed. 2, p. 153-159,
2010.
TOLEDO, F. R.; BARBOSA, J. C.; YAMAMOTO, P. T. Distribuição espacial de
Toxoptera citricida (Kirkaldy) (Hemiptera: Aphididae) na cultura de citros. Revista
Brasileira de Fruticultura, Jaboticabal, v. 28, n. 2, p. 194-198, 2006.
UNSICKER, S. B.; MODY, K. Influence of tree species and compass bearing on insect
folivory of nine common tree species in the West African savanna. Journal of Tropical
Ecology, New York, v.21, n.2, p. 227-231, 2005.

9
ARTIGO CIENTÍFICO
DISTRIBUIÇÃO ESPACIAL DE INSETOS FITÓFAGOS E DE INIMIGOS
NATURAIS EM LEUCAENA LEUCOCEPHALA UTILIZADA COMO QUEBRA-
VENTO EM REGIÃO DE CERRADO NO BRASIL
Resumo - Leucaena leucocephala (Lam.) de Wit. (Fabaceae: Mimosoideae) é uma espécie
amplamente cultivada para reflorestamento de áreas degradadas e como fonte de proteína
para alimentação animal em sistemas agrossilvipastoris, sendo às vezes atacada por pragas.
O objetivo foi avaliar durante dois anos, a distribuição espacial de insetos fitófagos e seus
inimigos naturais ao longo da estratificação vertical do dossel (partes apical, média, basal e
no tronco) e horizontal (orientação dos galhos quanto à exposição do sol nas faces norte,
sul, leste e oeste) e da face foliar (adaxial e abaxial) em L. leucocephala de acordo com os
seguintes indicadores de diversidade: número de indivíduos, riqueza (S), diversidade (H’),
frequência, dominância-K e abundância de espécies. Os maiores números de indivíduos
fitófagos e inimigos naturais foram observados em galhos voltados para a face norte.
Contudo, a maior riqueza específica e índice de diversidade de fitófagos foram observados
nos galhos voltados a oeste e para os inimigos naturais, nos galhos voltados para norte/sul
e oeste, respectivamente. Os maiores valores de indivíduos, riqueza específica e índice de
diversidade de fitófagos e inimigos naturais foram observados na parte apical e mediana do
dossel de L. leucocephala. Os maiores valores, tanto para os fitófagos como para os
inimigos naturais, de indivíduos, riqueza específica e índice de diversidade foram
observados na face abaxial de L. leucocephala. Os fitófagos e os inimigos naturais mais
abundantes e com maior dominância-K observados em maior frequência nas amostras
(espécies comuns/constantes) em L. leucocephala foram Trigona spinipes Fabricius
(Hymenoptera: Apidae, Meliponinae), Tropidacris collaris Stoll (Orthoptera: Romaleidae),
Camponotus sp.2 (Hymenoptera: Formicidae) e Brachymyrmex sp. (Hymenoptera:
Formicidae), respectivamente.
Palavras-chave: estratificação, pontos cardeais, dossel, faces foliares.

10
Abstract - Leucaena leucocephala (Lam.) of Wit. (Fabaceae: Mimosoideae) is a widely
cultivated species used for reforestation of degraded areas and as source of protein for
animals in agroforestry systems, sometimes being attacked by pests. The objective was to
evaluate for two years the spatial distribution of phytophagous insects and their natural
enemies along the vertical canopy stratification(apical, medium and basal parts and also
the trunk) and horizontal (orientation of the branches related to the position of the sun on
the north, south, east and west faces) and the foliar face (adaxial and abaxial) in L.
leucocephala according to the following diversity indicators: number of individuals,
affluence (S), diversity (H’), frequency, dominance-K and abundance of the species. The
greater number of phytophagous and bioregulators individuals were observed on branches
facing north. However, the greatest specific affluence and diversity index of phytophagous
was observed on branches facing west and as for the bioregulators, on the branches facing
north/south and west, respectively. The greatest values of individuals, specific affluence
and diversity index of phytophagous and bioregulators were observed in the apical and
medium parts of the canopy of L. leucocephala. The greatest values of individuals, specific
affluence and diversity index for both phytophagous and bioregulators were observed on
the abaxial face of L. leucocephala. The most abundant and with greatest k-dominance
phytophagous and bioregulators observed in the samples (common/constant species) in L.
leucocephala were Trigona spinipes Fabricius (Hymenoptera: Apidae, Meliponinae),
Tropidacris collaris Stoll (Orthoptera: Romaleidae), Camponotus sp.2 (Hymenoptera:
Formicidae) and Brachymyrmex sp. (Hymenoptera: Formicidae), respectively.
Key-words: Stratification, cardinal points, canopy, foliar faces.

11
1 Introdução
Leucaena leucocephala (Lam.) de Wit. (Fabaceae: Mimosoideae) é uma
espécie de crescimento rápido, fixadora de nitrogênio, tem despontado como alternativa
promissora para recuperação de áreas degradadas (DA COSTA e DURIGAN, 2010).
Quando constatado que não está se comportando como invasora de ecossistemas naturais, a
leucena pode enquadrar-se melhor como ruderal (DA COSTA e DURIGAN, 2010), ou
seja, uma espécie vegetal que prolifera apenas em áreas degradadas ou perturbadas
dificultando o estabelecimento de outras espécies, incluindo nativas (WOITKE e DIETZ,
2002). A L. leucocephala pode controlar erosões hídricas e eólicas, melhorando as
condições ambientais (HAI, 2007) e pode ser usada em quebra-ventos.
Árvores usadas como quebra-ventos podem influenciar a distribuição de pragas
e de inimigos naturais e dificultar a localização de plantas hospedeiras (RAO et al., 2000;
BRANDLE et al., 2004). A estratificação horizontal dos ramos quanto aos pontos cardeais
podem afetar a abundância e riqueza de insetos. Isso ocorre devido a direção e intensidade
dos ventos (LEITE et al., 2011a, b, c); insolação (UNSICKER e MODY, 2005);
microclima formado pelo dossel da planta (BASSET, 1991; LEITE et al., 2011a) e
preferência de herbívoros por espaços livres de inimigos naturais (UNSICKER e MODY,
2005; LEITE et al., 2012). Quanto a estratificação vertical, o dossel dessas plantas oferece
grande muitos recursos para a manutenção da biodiversidade de insetos e seus ramos e
folhas alteram o ambiente pelo sombreamento afetando a umidade, evapotranspiração e
servindo de refúgio para insetos herbívoros e inimigos naturais (BRANDLE et al., 2004).
Já as folhas podem influenciar nessa distribuição dos insetos, devido as suas características
físico-químicas (PEETERS, 2002). A variação na intensidade luminosa pode influenciar as
taxas de herbivoria, aumentando a espessura foliar com consequência maior dureza dessa
folha, da epiderme, parênquima paliçádico, cutícula espessada, esclerênquima (ALMEIDA
CORTEZ, 2005). Além de que, a composição química da folha, como conteúdo de lignina,
celulose, fibras e presença de taninos e nectários extraflorais podem influenciar na
distribuição dos insetos e na taxa de herbivoria (PEETERS, 2002). Já quanto à face foliar,
os insetos, em geral, preferem a face abaxial da folha por essa região apresentar-se mais
tenra, onde as nervuras são mais salientes, córtex delgado e epiderme mais fina, e poder
servir de refúgio para os insetos e proteger as suas oviposições contra predadores e
intempéries (ANTÔNIO, 2002). Aliado ao já discutido, uma epiderme com textura rígida,

12
que possua alto teor de sílica e/ou lignina, é uma barreira mecânica que reduz a oviposição
por alguns insetos (LARA, 1991).
Os insetos mais comumente encontrados em associação com L. leucocephala
são: Acanthoscelides macrophthalmus (Schaeffer) (Chrysomelidae: Bruchinae) (RAGHU
et al., 2005) predando sementes, Coccus longulus (Douglas) (Hemiptera: Coccidae)
(ELDER et al., 1998) e o psilídeo Heteropsylla cubana Crawford (Hemiptera: Psyllidae),
sugando seiva. Esta última responsável por graves danos a essa planta em várias partes do
mundo, sendo a sua dispersão em função do movimento dos ventos (ELDER et al., 1998;
RAO et al., 2000). A H. cubana apresenta maior importância para a cultura da leucena,
uma vez que podem se alimentar intensamente dessa árvore a ponto de ocasionar
desfolhação total, com consequente morte da planta (SANTANA, 2008; DRUMOND e
RIBASKI, 2010). No Brasil, foi registrado primeiramente em 2000, na região do porto de
Tubarão-ES. Em 2005, foi observado em Minas Gerais (Patos de Minas, Vazante e
Uberlândia) e no Paraná (Londrina). Em 2006, foi detectada na Bahia (Salvador)
(SANTANA, 2008).
No entanto, não há estudos sobre a distribuição de insetos fitófagos e seus
inimigos naturais em L. leucocephala no Brasil. O conhecimento das distribuições
espaciais de insetos-praga e seus inimigos naturais são importantes para o estabelecimento
de planos de amostragem e na decisão sobre o controle de pragas (TOLEDO et al., 2006).
A predação é um dos fatores que afetam a distribuição e a abundância das espécies
(BEGON et al., 2006).
Diante do exposto, objetivou-se: 1) identificar os insetos fitófagos, inimigos
naturais e polinizadores; 2) as suas distribuições quanto à estratificação vertical do dossel
(partes apical, média, basal e tronco); 3) horizontal (orientação dos galhos quanto à
exposição do sol nas faces norte, sul, leste e oeste) e 4) também nas faces foliares (adaxial
e abaxial) em plantas de L. leucocephala.

13
2 Material e Métodos
Este trabalho foi desenvolvido em área de pastagem no Instituto de Ciências
Agrárias da Universidade Federal de Minas Gerais (ICA/UFMG), município de Montes
Claros, Minas Gerais, de janeiro de 2014 a dezembro de 2015 em região com clima Aw:
tropical de savana de acordo com classificação de Köppen com inverno seco e verão
chuvoso e Latossolo Vermelho Amarelo Distrófico.
O quebra-vento com 100 metros de comprimento foi disposto com duas fileiras
centrais de Acacia mangium Willd. (Leguminosae) (espaçamento 3 x 3 m) e duas fileiras
de L. leucocephala, uma de cada lado da A. mangium, com espaçamento de 3 m entre
fileiras e 2 m entre plantas. As mudas foram preparadas em viveiro e plantadas no local
definitivo em setembro de 2012, em covas de 30 x 30 x 30 cm, misturando-se ao substrato,
fosfato natural reativo na dosagem de 160 g.
Os insetos (fitófagos, inimigos naturais e polinizadores) foram contados, por
visualização direta, nas faces foliares (adaxial e abaxial) na partes apicais, médias e basais
do dossel posicionadas nas faces norte, sul, leste e oeste, totalizando 12 folhas por parte do
dossel/avaliação, entre 7 e 11 h da manhã, em 20 árvores de L. Leucocephala, inicialmente
com 16 meses de idade, por um período de dois anos. Além disso, foram avaliados os
insetos presentes nos troncos (altura do peito) das 20 árvores/avaliação. Os insetos
capturados foram armazenados em frasco com álcool 70%, separados em morfoespécies e
enviados para identificação.
Os índices ecológicos (número de indivíduos, riqueza, diversidade e
abundância de espécies) foram calculados para os insetos identificados e medidos por meio
do cálculo do conjunto de dados de taxa de amostras da diversidade Proversão software 2.
A fórmula de Colina (HILL, 1973) foi usada para calcular a diversidade - diversidade Hill
de primeira ordem: N1 = exp (H') -, que calcula a diversidade em termos do número
efetivo de espécies (HILL, 1973; JOST, 2006). Abundância e riqueza de espécies (S)
foram calculadas pela fórmula de Simpson: D=(ni /N) *100, onde: D=Ni/N; ni= número de
indivíduos por espécie; N= número total de indivíduos; S = riqueza (número de espécies
presentes) (TOWNSEND et al., 2006;. LAZO et al., 2007). Dominância-K foi calculada
traçando a abundância percentual cumulativo contra log classificação de espécies
(LAMBSHEAD et al., 1983). Os valores de dominância-K indica a distribuição de
dominância e uniformidade dos indivíduos entre as espécies (GEE et al., 1985). As

14
frequências de cada espécie de inseto nas amostras foram classificadas como: a) constante
(presença ≥ 50%), b) comum (presença entre 11 e 49%) e c) rara (presença ≤ 10%)
(adaptado por SIQUEIRA et al., 2008).

15
3 Resultados
Observaram-se 11 espécies raras – Euxesta sp. (Diptera: Otitidae), Episyrphus
balteatus De Geer (Diptera: Syrphidae), Aethalion reticulatum L. (Hemiptera:
Aetalionidae), Mahanarva posticata Stal (Hemiptera: Cercopidae), Ferrariana trivitata
Signoret (Hemiptera: Cicadellidae), Dalbulus maidis De long e Wolcott (Hemiptera:
Cicadellidae), Tetragonisca angustula Latreille (Hymenoptera: Apidae), Camponotus sp.1
(Hymenoptera: Formicidae), Camponotus sp.3 (Hymenoptera: Formicidae), Polistes sp.
(Hymenoptera: Vespidae), Mantis religiosa L. (Mantodea: Mantidae) e sete espécies raras
não identificadas. Também encontradas três espécies comuns – Trigona spinipes Fabricius
(Hymenoptera: Apidae, Meliponinae), Brachymyrmex sp. (Hymenoptera: Formicidae) e
Tropidacris collaris Stoll (Orthoptera: Romaleidae) – e uma constante – Camponotus sp.2
(Hymenoptera: Formicidae) em árvores de L. leucocephala. Em relação aos fitófagos,
foram notados que a ordem Hemiptera apresentou maior número famílias (5) e de espécies
(4). Já os inimigos naturais, a ordem Hymenoptera se destacou entre as demais com três
famílias, três morfoespécies, quatro gêneros e uma espécie identificada (Tabela 1).
Os maiores números de indivíduos fitófagos, inimigos naturais e polinizadores
foram observados em galhos voltados para a face norte. Contudo, a maior riqueza
específica e índice de diversidade de fitófagos foram observados nos galhos voltados a
oeste e para os inimigos naturais nos galhos voltados para norte/sul e leste
respectivamente. Os maiores valores de indivíduos, riqueza específica e índice de
diversidade de fitófagos foram observados na parte apical de L. leucocephala; e de
inimigos naturais nas partes apicais e medianas, respectivamente. Os maiores valores, tanto
para os fitófagos como os inimigos naturais, de indivíduos, riqueza específica e índice de
diversidade foram observados na face abaxial de L. leucocephala (Tabela 2).
Quanto ao efeito cardinal, fitófagos mastigadores obtiveram maior abundância
(11) na posição norte dos ramos e com três diferentes espécies. Já os sugadores, maior
abundância (5) na posição oeste, com quatro espécies diferentes. Maior abundância de
formigas (28) na posição norte, com quatro espécies, e maior abundância de abelhas
Tetragonisca angustula Latreille (Hymenoptera: Apidae) (4) na posição oeste dos ramos
do dossel de L. leucocephala (Tabela 3).
Quanto à estratificação vertical do dossel, observou-se que a maior abundância
de fitófagos mastigadores (11) e sugadores (6) foram na região apical, com quatro e seis

16
espécies diferentes, respectivamente. Formigas foram encontradas em maior abundância
(42) também na região apical, com três espécies. Já as abelhas T. angustula em maior
abundância (6) na região mediana do dossel de L. leucocephala (Tabela 4).
As espécies T. spinipes e T. collaris (fitófagos), Camponotus sp.2 e
Brachymyrmex sp. (inimigos naturais) apresentaram-se mais abundantes e com maior
dominância-K na face inferior das folhas de L. leucocephala (Tabela 5).

17
4 Discussão
Foram observadas doze espécies de insetos identificadas, três morfoespécies,
mas, em geral, em baixas densidades em árvores de L. leucocephala. Trigona spinipes e T.
collaris foram os insetos fitófagos com maior frequência e status de espécie comum. Esses
insetos são considerados prejudiciais a diversas culturas, sendo o primeiro, responsável por
danos a brotos para obtenção de fibras utilizadas na construção dos ninhos (SILVA et al.,
1997). Os prejuízos relacionados a essa abelha, foram observados especialmente em
frutíferas, florígenas e leguminosas florestais (ZUCCHI et al, 1993) Além disso, T.
spinipes pode influênciar na polinização, diminuindo ou inibindo a frutificação. Em
Cucurbita moschata D. (Cucurbitaceae), flores que foram visitadas apenas uma vez por T.
spinipes, não produziram frutos, devendo-se, talvez, às suas operárias transportarem pólen
insuficiente (SERRA, 2010). Outro fator relevante é que devido ao seu pequeno tamanho
corporal, essas abelhas tocam os estames e estigmas apenas ocasionalmente e, ao se
permitir a visita de apenas um indivíduo à flor, eliminou-se o comportamento de visita em
"bando", padrão de forrageamento característico dessas espécies (SERRA, 2010). Sendo
assim, esse comportamento de polinizadoras ocasionais, pouco contribui para produção de
frutos, o que poderá variar entre culturas (SERRA, 2010). Já T. collaris pode atacar
diversas espécies silvestres, palmeiras e também plantas úteis ao homem, como frutíferas e
grãos (COSTA et al., 2003). Os inimigos naturais mais frenquentes foram Camponotus sp.2
(constante) e Brachymyrmex sp. (comum).
Coleoptera, Lepidoptera e Hymenoptera são as ordens mais abundantes de
insetos mastigadores no cerrado (PINHEIRO et al., 2002; ZANUNCIO et al., 2002;
ZANETTI et al., 2003). No entanto, em L. leucocephala os insetos mais abundantes
encontrados foram das ordens Hemiptera e Hymenoptera. Para insetos que se alimentam de
seiva, um fator importante é a seleção da planta hospedeira e sua qualidade nutricional
(COOK e DENNO ,1994). Assim, insetos herbívoros sugadores, podem apresentar maior
especificidade ao hospedeiro do que mastigadores (RIBEIRO e PIMENTA, 1991;
STYRSKY e EUBANKS, 2007). Alguns estudos demonstram sincronismo entre
herbívoros sugadores e a fenologia da planta hospedeira (RIBEIRO e PIMENTA, 1991),
sugerindo menor efeito do habitat para os sugadores.
A ligação entre taninos e proteínas ocorre provavelmente através de pontes de
hidrogênio entre os grupos fenólicos dos taninos e determinados sítios das proteínas,

18
emprestando uma duradoura estabilidade a estas substâncias (MONTEIRO et al., 2005),
devido a isso, pode-se sugerir que o maior encontro de sugadores que mastigadores neste
trabalho, pode estar associados aos seguintes fatos. O primeiro fator é que em leucena, o
tanino é um dos fatores antinutricionais. Seu tipo mais comum é a proantocianidina,
também conhecida como tanino condensado (TC) (MONTEIRO et al., 2005). Os TC
localizam-se no interior das células vegetais e/ou no tegumento das sementes, sendo
liberados rapidamente durante a mastigação ou dano mecânico (MONTEIRO et al., 2005).
Nesse primeiro momento, os taninos conferem sensação de adstringência, prejudicando
imediatamente a palatabilidade das folhas (MONTEIRO et al., 2005). Uma vez que o
grupo de insetos herbívoros mastigadores generalistas contribuírem com 75% ou mais de
todo consumo anual de folhas (COLEY e BARONE, 1996), pode ser atribuído ao fato de
ter sido encontrado em menor abundância, quando se compara aos sugadores. Porém Silva
et al. (2009) não encontraram nenhuma relação significativa entre riqueza e abundância de
insetos herbívoros em função da concentração de taninos. Esses mesmos autores
encontraram que a concentração de taninos não variou significativamente entre os habitats
de Cerrado, Mata Ciliar e Floresta Decidual. Compostos secundários não foram medidos
neste estudo e por isso, devemos levar em consideração o fato de que os taninos são
compostos de defesa química constitutiva cujo efeito é dependente da dosagem ( SILVA et
al., 2009). O segundo fator é que os taninos, além de tóxicos, podem ligar-se às proteínas e
outras macromoléculas. Ayres et al. (1997) verificaram que a rápida mortalidade de insetos
tratados com taninos condensados parece ser devido à atividade tóxica destes compostos e
não pela inibição da digestibilidade. Por último, a maior parte dos insetos fitófagos
dependem para viver de substâncias solúveis, tais como açúcares redutores e aminoácidos
livres (OLIVEIRA, 2009). Espécies de tripes, pulgões, cochonilhas, cigarrinhas, cigarras, e
outros insetos raspadores e sugadores, possuem um organismo muito simples e aparelho
digestivo com baixa capacidade de digestão tornando-os incapazes de desdobrar proteínas
em aminoácidos para serem posteriormente recombinados a conveniência de cada um, por
isso, eles dependem de aminoácidos livres existentes na seiva das plantas ou no suco
celular (PARRA, 1991; BURG e MAYER, 2001; SOUZA e RESENDE, 2003), não
ocorrendo a digestão tanino-proteína, que poderiam inibir mais a alimentação de
mastigadores do que de sugadores na leucena.
Há três possibilidades que podem justificar a baixa quantidade de insetos em L.
Leucocephala. A primeira é por se tratar de uma espécie exótica, espera-se que haja menor

19
abundância de artrópodes de espécies nativas como previsto pela hipótese de liberação de
inimigos naturais (KEANE e CRAWLEY, 2002). Essa hipótese propõe que, quando uma
espécie de planta é transferida para fora de sua distribuição geográfica deixa para trás suas
pragas especializadas e, uma vez fora dessa distribuição, essas plantas terão uma vantagem
competitiva sobre espécies nativas (KEANE e CRAWLEY, 2002). Segundo Veloso
(1997), a produção de uma cultura comercial com a utilização de espécies nativas é
limitada pela ocorrência de pragas. A segunda possibilidade é que, apesar de L.
leucocephala apresentar teor relativamente alto de N, baixa lignina e baixo teor de
hemicelulose (BELTRÁN e WUNDERLE JR, 2014), ela contém taninos flabofênicos
(CAVALCANTE et al., 2006). Taninos são classificados como substâncias quantitativas,
por serem redutores digestivos, com efeito proporcional à concentração (STRONG et al.,
1984), reduzem significativamente o crescimento e a sobrevivência de insetos, uma vez
que inativam enzimas digestivas e criam um complexo de taninos-proteínas de difícil
digestão (MELLO e SILVA-FILHO, 2002). E a última possibilidade é a associação entre
plantas e formigas, que em geral, são interações mutualísticas obrigatórias
(HÖLLDOBLER E WILSON, 1990; BRONSTEIN, 1998). A resposta mutualística,
normalmente, é feita pela pressão seletiva e este mutualismo acontece frequentemente,
sendo variado em diferentes espécies de insetos. As formigas visitantes podem predar ou
injuriar insetos herbívoros que encontram sobre as plantas, reduzindo significativamente as
taxas de herbivoria sobre as folhas, botões e flores. Considerando que pelo menos parte dos
indivíduos predados é herbívoro potencial, as formigas acabam exercendo um efeito
protetor sobre a planta (JUNQUEIRA, 2001). O néctar extrafloral é justamente o
mecanismo responsável por manter formigas constantemente forrageando pelas folhas da
planta (DÁTTILO, 2009). Hughes (1998) indica que L. leucocephala apresenta um
nectário extrafloral no pecíolo da folha com forma elíptica e côncava. Maior visitação de
formiga a nectários extraflorais podem favorecer a produção de flores ou de frutos desta
planta devido a diminuição da hervoria reduzindo os danos para árvores de L. leucocephala
por pragas, além de que, brotação das folhas e desenvolvimento de flores antes do período
chuvoso é fato comum em plantas perenes em Cerrado (ALMEIDA et al., 1998; FELFINI
et al., 1999; PEDRONI et al., 2002; ALMEIDA et al., 2006; LEITE et al., 2006). Isso
permite que as plantas aumentem sua área fotossintética quando a eficiência de herbivoria
por insetos é menor (LEITE et al., 2012).

20
Mastigadores herbívoros e formigas foram encontrados em maior abundância
na posição norte dos ramos do dossel de L. leucocephala. Esse fato pode ter ocorrido
devido à qualidade dos recursos alimentares nesse ponto, pois nectários foliares e flores
expostas a correntes de vento (vento predominante na região norte/leste) podem apresentar
maior evaporação e, portanto, aumento de suas concentrações de néctar (DEBRUIJN e
SOMMEIJER, 1997). Já para insetos sugadores foi encontrada maior abundância na
posição oeste dos ramos do dossel, local que recebe menor intensidade de vento e por isso,
pode apresentar umidade mais elevada e temperatura mais baixa, diminuindo a dessecação
desses insetos, melhorando as condições de sobrevivência (RAO et al., 2000). A
abundância da maioria das espécies de sucção nas folhas, flores e frutos em árvores de
Caryocar brasiliense Camb. (Malpighiales: Caryocaraceae) foram semelhantes entre os
pontos cardeais (LEITE et al., 2015). Também T. angustula obteve maior índice de
abundância na face oeste. Duas hipóteses podem explicar essa tendência. A primeira é o
sentido de vento. Este pode influenciar na dispersão dos insetos (FENG et al., 2005;
LEITE et al., 2011a) nas suas migrações (PATHAK et al., 1999; TIXIER et al., 2000;
AUSLANDER et al., 2003; SCHOOLEY e WIENS, 2003; FENG et al., 2005; LEITE et
al., 2009, 2011b) e polinização. Em ecossistemas naturais, as abelhas T. angustula alojam-
se em abrigos devidamente seguros, isto é, protegidos das chuvas, do sol direto e das
correntes de ar (VILLAS-BÔAS, 2012). As abelhas sem ferrão têm porte pequeno, em
geral menor do que 0,8 cm, e por isso se mostram mais sensíveis ao vento (HILÁRIO,
2007). A preferência pelos galhos a oeste pode ser devido à proteção do vento o que
favorece a estabilidade de vôo quando forem à procura de néctar em regiões da planta ou
mesmo quando estas abelhas estão pousadas sobre as flores. Vários estudos (por exemplo,
HEINRICH e RAVEN, 1972; LAROCA et al., 1982; RAMALHO et al., 1991; MORATO
e CAMPOS, 2000; ANTONINI et al., 2005) relatam que as características corporais e
limitações fisiológicas de abelhas, especialmente os menores em tamanho do corpo, podem
gerar padrões de visita que favorecem o vôo em condições que dependem de fatores
ambientais. Neste caso, as abelhas geralmente preferem visitar em períodos de altas
temperaturas e luminosidade e baixa umidade relativa do ar e velocidade do vento
(POLLATO et al., 2012). A segunda hipótese é a fuga de espécies mais agressivas.
Abelhas T. angustula e T. spinipes são abelhas sem ferrão, porém pertecem a espécies e
gêneros diferentes. Apesar de abelhas T. angustula e T. spinipes estarem na mesma região
do dossel (região mediana), elas se encontram em pontos cardeais diferentes (T. angustula

21
ao oeste e T. spinipes ao norte dos ramos em L. leucoceplhala). Isso pode ter ocorrido
porque as T. spinipes são extremamente agressivas, suas operárias apresentam um
comportamento agressivo contra operárias de Apis mellifera L. (Hymenoptera: Apidae)
(BRIZOLA-BONACINA, 2012). Por causa dessa agressividade, a abelha T. angustula,
pode estar se protegendo, evitando regiões onde a T. spinipes se encontra. Apesar das
abelhas T. spinipes possuírem estratégia de forrageamento em horários diferentes das T.
angustula (BRIZOLA-BONACINA, 2012), o gênero Trigona forragea em grupos,
impedindo que outras espécies habite nas flores por eles ocupadas (SERRA, 2010).
Mastigadores, sugadores e formigas, de modo geral, foram encontrados em
maior abundância na região apical do dossel. Para isso, há três possibilidades: a primeira é
que a parte apical do dossel apresenta-se com mais folhas novas, melhor alimento para
herbivoros e portanto, folhas mais tenras e com maior concentração de nutientes essenciais
como sais minerais, aminoácidos, vitaminas e não essenciais como os carboidratos,
lipídeos e esteróis. Muitos aspectos relacionados à biologia dos insetos, como:
comportamento, fisiologia e ecologia, estão de alguma forma incluídos em um contexto
nutricional (PANIZZI e PARRA, 1991). Em folhas jovens e sementes maduras da leucena
armazenam mimosina alcalóides ou leucenol (MASCRÉ, 1937; ADAMS et al., 1945).
Mimosina é um aminoácido não-protéico econtrado em leucena, seu efeito é antimitótico,
inibidor de apoptose neural, além de excluir plantas que crescem nas proximidades via
alelopatia (PAIM, 2005). O acúmulo deste aminoácido é estimulado por dano mecânico,
ácido saicílico que é um sinalizador de ataque de patogénos e pela auxina ácido naftaleno
acético (ANA) (PAIM, 2005). Na folhagem de leucena, mimosina contribui para a sua alta
conteúdo total de N, e juntamente com taninos, podem servir como uma defesa contra
alguns insetos herbívoros (HAMMOND, 1995; BELL, 2003). No entanto, alguns
herbívoros não são afetados por esses compostos, como os sugadores, psilídeo da
Leucaena, que podem quebrar mimosina (KAMADA et al., 1996). Também, as abelhas
Apis mellifera não sentem os efeitos tóxicos da mimosina (PEREIRA, 2005). Elevadas
biomassas de larvas de lepidóptera encontradas em associação com a leucena, sugere que
eles também podem ser capazes de quebrar os compostos secundários (NISHIDA, 2003).
Segundo Coley e Barone (1996), devido à maior qualidade nutricional de
folhas novas, as taxas de herbivoria em espécies tropicais podem ser de 5 a 25 vezes
maiores que em folhas maduras. Esse fato pode ter influenciado a população de inimigos
naturais, uma vez que estes estariam onde há maior quantidade de presas (fitófagos). A

22
segunda possibilidade é que regiões de crescimento da copa possuem uma maior
probabilidade de encontro de folhas jovens (BELL et al.,1999; BARRIOS, 2003) e
consequentemente nectários extraflorais mais novos, isto é, mais cheios de susbstâncias
açucaradas, o que leva a maior presença de formigas nesse local. E a terceira possibilidade
é que provavelmente a preferência dos artrópodes fitófagos para as distribuições em
regiões mais altas do dossel está relacionada a um menor risco de parasitismo
(RAMANAND e OLCKERS, 2013) ou predação por inimigos naturais, tais como formigas
(ELBANNA, 2011).
O que pode explicar a maior presença de fitófagos e formigas em um mesmo
local, é a presença de nectários extraflorais. Isso faz com que seja um valioso recurso
alimentar para muitos artrópodes (RUHREN e HANDEL, 1999), principalmente formigas
das subfamílias Myrmicinae, Formicinae e Dolichoderinae (OLIVEIRA e BRANDÃO,
1991). O modo pelo qual ocorre esse mutualismo entre formigas e plantas, de acordo com
Beattie (1985) é um processo muito sutil e complexo. Segundo esse autor, há quatro
interações básicas entre formigas e plantas: (1) as formigas protegem a planta de
herbívoros e outros inimigos, (2) formigas alimentam-se de nutrientes essenciais para
plantas, (3) formigas ajudam com a dispersão de sementes de plantas e frutos e (4) ajudam
na polinização.
Apesar de mastigadores, de forma geral, serem encontrados em maior
quantidade na região apical do dossel, a maior abundância da abelha T. spinipes foi na
região mediana do dossel, o que também foi observado para a abelha T. angustula. Isto se
deve provavelmente, à maior disponibilidade de recursos alimentares nessa região que a
parte basal e o tronco de árvores (BASSET et al., 2003). As abelhas sem ferrão iniciam sua
atividade de coleta diária de néctar às 5:20 h, e permanece mais ativa até as 10:00h
(NEVES e VIANA, 2002). Essa estratégia de forrageamento possui três possíveis
vantagens: (1) evita que as abelhas sofram coma alta temperatura e a baixa umidade do ar;
(2) possibilita a coleta de recursos tróficos não alterados, o néctar, por exemplo, pode
aumentar sua concentração devido à evaporação ao longo do dia (PROCTOR et al.,1996);
e (3) encontram maior quantidade de alimento, fator limitante para essas abelhas que
possuem colônias muito populosas e ativas durante todo o ano. A aclimatação para altas
temperaturas parece ser imprescindível à sobrevivência dos insetos em condições naturais,
pois as altas temperaturas sazonais e diárias são precedidas por um gradiente de

23
temperaturas mais baixas, e vice-versa (ROMOSER e STOFFOLANO JR., 1994). Esse
resultado concorda com o encontrado por Silva et al. (2015) em A. mangium.
Número de indíviduos, riqueza específica, diversidade, abundância e
dominância-K tanto de fitófagos, quanto de inimigos naturais e polinizadores, obtiveram
índices mais altos na superfície abaxial de folhas. Destacando-se as espécies T. spinipes e
T. collaris (fitófagos) e Camponotus sp.2 e Brachymyrmex sp. (inimigos naturais) nesta
face foliar, isso pode ter ocorrido devido a alguns fatos. O primeiro fato é que na face
abaxial, as folhas apresentam-se mais tenras. Fitófagos, em geral, dão preferência onde às
folhas são mais novas e tenras (o que favorece a alimentação). A segunda possibilidade é
que na região abaxial as nervuras são mais salientes e córtex mais delgado, o que favorece
a alimentação principalmente de insetos sugadores. Além disso, essa região da folha é de
melhor qualidade nutricional do que a da face adaxial e de folhas dos terços mais
inferiores. Insetos succívoros e ácaros preferem, em geral, a face abaxial de folhas apicais
(ou pontos de crescimento) (MIRANDA et al., 1998; SILVA et al., 1998; LEITE et al.,
1999; 2002 e 2003). Essa maior facilidade de sucção da seiva se deve ao floema estar mais
exposto na face abaxial e, também, a resistência do tecido a ser perfurado nesta face
(parênquima lacunoso) ser menor do que na face adaxial (parênquima paliçádico) (CHU et
al., 1995, 2007; GLÓRIA e GUERREIRO, 2003). Em geral, as densidades de insetos
fitófagos são negativamente correlacionadas com o trabalho e a força que exercem ao
rasgar as folhas (PEETERS et al., 2007). Sendo a superfície abaxial das folhas, onde a
epiderme é mais fina, favorece a alimentação de insetos e consequentemente, uma maior
aptidão na atividade de herbivorismo (FIENE et al., 2013). Por último, a face abaxial da
folha confere aos fitófagos maior proteção contra inimigos naturais e intemperies. A face
foliar pode influenciar a oviposição dos insetos devido à proteção que pode proporcionar
contra intempéries climáticas e ação de inimigos naturais. Esta proteção é maior quando os
ovos são colocados na face abaxial, que estando voltada para baixo faz com que estes
fiquem menos expostos a estes agentes do controle natural (ANTÔNIO et al., 2002). Nas
estratégias comportamentais a busca por refúgios realizada pelas presas, pode reduzir o
risco de predação, principalmente quando realizada por predadores visualmente orientados
(KREBS e DAVIES, 1993). Esses refúgios podem ser localizados em folhiços, estruturas
dos troncos e faces foliares abaxiais. Lagartas podem se localizar em partes específicas das
plantas como na face abaxial das folhas. Esse comportamento pode reduzir o risco de
predação já que a probabilidade de detecção de um predador visualmente orientado é

24
menor na face abaxial do que na face adaxial da folha (KREBS e DAVIES, 1993).
Resultados semelhantes foram encontrados por Leite et al. (2006) em C. brasiliense com
sugadores e forrmigas, e por Silva et. al (2014) em A. Mangium, com fitófagos, inimigos
naturais e polinizadores.
5 Conclusões
As espécies T. spinipes e T. collaris têm maior potencial para se tornarem
pragas em plantas de L. leucocephala por serem os mais frenquentes e abundantes e por
terem histórico em outras culturas. Esses resultados podem apoiar programas de controle
de pragas e manutenção de inimigos naturais e polinizadores como os inimigos naturais
mais frenquentes, Camponotus sp.2 e Brachymyrmex sp. em futuras plantações de L.
leucocephala, consorciada (ex.: sistemas agrossilvipastoris) ou não (monocultura).
A maneira pela qual os insetos estão distribuídos na planta, pode-se efetivar
uma amostragem mais correta. Logo, a partir desses resultados, se o objetivo for
estabelecer estratégias de monitoramento, entende-se que fitófagos, inimigos naturais e
polinizadores estão em maior quantidade na posição norte da região apical do dossel. Se o
objetivo for manutenção de inimigos naturais e polinizadores, a riqueza específica e
diversidade, encontram-se em maiores índices na posição oeste da região apical do dossel.
No geral, encontrados na face abaxial das folhas de L. leucocephala.
Agradecimentos
Aos taxonomistas Drs. Ayr de Moura Bello (Coleoptera), Ivan Cardoso
Nascimento (EMBRAPA-ILHÉUS Centro de Pesquisas do Cacau, CEPLAC, Itabuna, BA)
(Formicidae), Paulo Sérgio Fiuza Ferreira (Hemiptera) (UFV) e Luci Boa Nova Coelho
(Universidade Federal do Rio de Janeiro (Cicadellidae) pelas identificações dos espécimes.
A CAPES pela concessão de bolsa de mestrado.

25
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, R.; CRISTOL, S. J.; ANDERSON, A. A.; ALBERT, A. A. The structureof
leucenol, 1. Journal of the American Chemical Society, Washington, v. 67, p. 89-92,
1945.
ALMEIDA, S. P. Frutas nativas do cerrado: Caracterização físico-química e fonte de
potencial de nutrientes, In, SANO, S. M.; ALMEIDA, S. P. Cerrado ambiente e flora.
EMBRAPA/CPAC, Planaltina, p. 247-28, 1998.
ALMEIDA, C. I. M. et al. Fenologia e artrópodes de Copaifera langsdorfii no
cerrado. Revista Brasileira de Plantas Medicinais, Botucatu,v. 8, n. 2, p. 64-70,
2006.
ALMEIDA CORTEZ, J. "Herbivoria e mecanismos de defesa vegetal". In: NOGUEIRA,
R. J. M. C; ARAÚJO, E. L.; WILLADINO, L. G.; CAVALCANTE, U. M. T (Org.).
Estresses ambientais: danos e benefícios em plantas, Recife. p. 389-396. 2005.
ANTÔNIO, A. C. et al. Ovoposição de Diaphania hyalinata L. (Lepidoptera, Pyralidae)
afetada pela face foliar e tricomas. Acta Scientiarum, Maringá, v. 24, n. 2, p. 359-362,
2002
ANTONINI, Y. et al. Diversidade e comportamento dos insetos visitantes florais de
Stachytarpheta glabra Cham. (Verbenaceae), em uma área de campo ferruginoso.
Neotropical Entomology, Ouro Preto, v . 34, p. 555–564, 2005.
AUSLANDER, M.; NEVO, E.; INBAR, M. The effects of slope orientation on plant
growth, developmental instability and susceptibility to herbivores. Journal of
Arid Environments, Filadélfia, V. 55, p. 405-416, 2003.
AYRES, M. P.; CLAUSEN, T. P.; MACLEAN, S. F.; REDMAN, A. M.; REICHARDT,
P. B. Ecology, Washington, v. 78, p. 1696, 1997.
BARRIOS, H. Insect herbivores feeding on conspecific seeslings and trees. Arthropods of
Tropical Forests: Spatio-Temporal Dynamics and Resource Use in the Canopy (ed. By
BASSET, Y. et al.) Cambridge University Press, Cambridge, United Kingdom, p. 282-290,
2003.

26
BASSET, Y. The spatial distribution of herbivory, mines, and galls within an Australian
rain forest tree. Biotropica, Washington, v. 23, p. 271–281, 1991.
BASSET, Y. et al. Arthropods of tropical forests: spatio-temporal dynamics and
resource use in the canopy. Cambridge University Press, Cambridge, p. 474, 2003.
BEATTIE, A. J. The evolutionary ecology ant-plant mutualisms. Cambridge, Cambridge
University Press. p. 182, 1985.
BEGON, M.; TOWNSEND, C. R.; HAPER, J. L. Ecology: from individuals to
ecosystems. Oxford, 2006.
BELL, A. D.; BELL, A. E.; DINES, T. D. Branch construction and bud defence status at
the canopy surface of a West African rainforest. Biological Journal of the Linnean
Society, United Kingdom, v. 66, p. 481-499, 1999.
BELL, E. A. Nonprotein amino acids of plants: significance in medicine, nutrition and
agriculture. Journal of Agricultural Food Chemistry, Washington, v. 51, p. 2854-2865,
2003.
BELTRÁN, W.; WUNDERLE JR, J. M. Temporal dynamics of arthropods on six tree
species in dry woodlands on the Caribbean island of Puerto rico. Journal of Insect
Science, Oxford University, v. 14 p. 199, 2014.
BRANDLE, J. R.; HODGES, L.; ZHOU, X. H. Windbreaks in North American
agricultural systems. Agroforestry Systems, University of Nebraska-Lincoln, v. 61, p. 65–
78, 2004.
BRIZOLA-BONACINA, A. K. et al. Bee visitors of Quaresmeira flowes (Tibouchina
granulosa Cogn.) in the region of Dourados ( MS-Brasil). Sociobiology, Chico, v. 59, ed.
4, p. 1253-1267. 2012.
BRONSTEIN, J. L. The contribution of ant-plant protection studies to our understanding
of mutualism. Biotropica, Washington, v. 30, p. 150-161, 1998.
BURG, I. C. e MAYER, P. H. Alternativas ecológicas para prevenção e controle de pragas
e doenças. Grafit, Francisco Beltrão, p.154, 2001.

27
CAVALCANTE, G. M.; MOREIRA, A. F. C.; VASCONSELOS, S. D. Potencialidade
inseticida de extratos aquosos de essências florestais sobre mosca-branca. Pesquisa
Agropecuária brasileira, Brasília, v.41, n.1, p.9-14, jan. 2006.
CHU, C. C.; HENNEBERRY, T. J.; COHEN, A. C. Bemisia argentifolli ( Homoptera:
Aleyrodidae): host preference and factors affecting oviposition and feeding site preference.
Environmental Entomology, Lanham, v. 24, p. 254-360, 1995.
COLEY, P. D.; BARONE, J. A. Herbivory and plant defensesin tropical forests. Annual
Review of Ecology and Systematics, Palo Alto, v. 27, p.305–335, 1996.
COOK, A. G e DENNO ,R. F. Planthopper/plant interactions: feeding behavior, plant
nutrition, plant defense, and host plant specialization. Pp. 114-139 en: Denno, R. F e TJ
Perfect (eds.). Planthoppers: Their Ecology and Management. Chapman & Hall, p. 799,
1994.
COSTA, M. K. C. M. et al. Descrição histológica do estomedeu de Tropidacris collaris
(Stoll, 1813) (Orthoptera: Romaleidae). Arquivos do Instituto Biológico, São Paulo, v.
70, n. 3, p. 259-263, 2003.
DA COSTA, J. N. M. N.; DURIGAN, G. Leucaena leucocephala (Lam.) de Wit
(Fabacea): Invasora ou ruderal? Revista Árvore, Viçosa, v. 34, n. 5, 2010.
DÁTTILO, W. et al. Interações mutualísticas entre formigas e plantas. Publicação do
Projeto Entomologistas do Brasil, Capinzal, v. 2, p. 32-36, 2009.
DEBRUIJN, L. L. M., AND SOMMEIJER, M. J. Colony foraging in different species of
stingless bees (Apidae, Meliponinae) and the regulation of individual nectar foraging.
Insectes Sociaux, Paris, v. 44, p. 35-47, 1997.
DRUMOND, M. A.; RIBASKI, J. Leucena (Leucaena leucocephala): leguminosa de uso
múltiplo para o semiárido brasileiro. Embrapa Semiárido, Petrolina, p. 9984, 2010.
ELBANNA, S. M. Ant-Acacia interaction: Chemical or physical defense? Entomological
Research, London, v. 41, p. 135-141, 2011.
ELDER, R. J.; MIDDLETON, C. H.; BELL, K. L. Heteropsylla cubana Crawford
(Psyllidae) and Coccus longulus (Douglas) (Coccidae) infestations on Leucaena species

28
and hybrids in coastal central Queensland. Australian Journal of Entomology, Canberra,
v. 37, p. 52-56, 1998.
FENG, H. G. et al. High-altitude windborne transport of Helicoverpa armigera
(Lepidoptera: Noctuidae) in mid-summer in northern China. Journal of Insect Behavior,
Riverside, ed. 18, v. 3, p. 335-349, 2005.
FELFINI, J. M. et al. Estudo fenológico de Stryphnodendron adstringens (Mart.)
Coville no cerrado sensus tricto da Fazenda Água Limpa no Distrito Federal, Brasil.
Revista Brasiliera de Botânica, São Paulo, v. 22, p. 83-90, 1999.
FIENE, J. et al. Foraging on individual leaves by an intracellular feeding insect is not
associated with leaf biomechanical properties or leaf orientation. Plos One, San Francisco
ed. 8, v. 11, 2013.
FREITAS, L. H. C.; SCHIFINO-WITTMANN, M. T.; PAIM, N. R. Variabilidade intra e
intergenotípica em uma população de híbridos entre Leucaena leucocephala
e L. diversifolia. Pesquisa Agropecuária Brasileira, Brasília, v. 36, n. 8, p. 1069-1076,
2001.
GEE, J. M. et al. Effects of organic enrichment on meiofaunal abundance and community
structure in sublittoral soft sediments. Journal of Experimental Marine Biology and
Ecology, Netherlands, v. 91, p. 247-262, 1985.
GLÓRIA, B. A. D.; GUERREIRO, S. M. C. Anatomia Vegetal, Viçosa, p. 438, 2003.
HAI, R. et al. Changes in biodiversity and ecosystem function duringthe restoration of a
tropical forest in south China. Science in China Series C: Life Sciences, Beijing, v. 50, n.
2, p. 277-284, 2007.
HAMMOND, A. C. Leucaena toxicosis and its control in ruminants. Journal of Animal
Science, Champaing, v. 73, p. 1487-1492, 1995.
HEINRICH, B.; RAVEN, P. H. Energetics and pollination ecology. Science, London, v.
176, n. 4035, p. 597–602, 1972.
HILÁRIO, S. D; RIBEIRO, M. F; FONSECA, V. L. I. Efeito do vento sobre a atividade de
vôo de Plebeia remota (Holmberg, 1903) (Apidae, Meliponini). Biota Neotropica,
Campinas, v. 7, n. 3, 2007.

29
HILL, M. O. Diversity and evenness: aunifying notation and its consequences. Ecology,
Washington, v. 54, n. 2, p. 427-432, 1973.
HÖLLDOBLER, B. e E. O. WILSON. “The ants”. Journal of Evolutionary Biology,
Springer, Berlin, v. 5, p. 169-171, 1990.
HUGHES, C. E. Leucaena: manual de recursos genéticos. Oxford : Oxford Forestry
Institute, (Tropical Papers, 37) p. 280, 1998.
JOST, L. Entropy and diversity. Oikos, Hoboken, v. 113, p. 363–375, 2006.
JUNQUEIRA, L. K.; DIEHL, E.; DIEHL-FLEIG, E. Formigas (Hymenoptera:
Formicidae) visitants de Ilex paraguariensis (Aquifoliaceae). Neotropical Entomology,
Londrina, v. 30 n. 1, 2001.
KAMADA, Y. Degradation of toxic mimosine by a hydrolytic enzyme of Leucaena
psyllids (Jumping plant lice). Bioscience Biotechnology Biochemistry,London, v. 60, p.
351-352, 1996.
KEANE, R. M.; CRAWLEY, M. J. Exotic plant invasions and the enemy Release
hypothesis. Trends in Ecology and Evolution, Cambridge, v. 17, p. 164–169, 2002.
KREBS, J. R.; DAVIES, N. B. An introduction to Behavioural Ecology. Oxford:
Blackwell Publishing, 1993.
LAMBSHEAD, P. J. D.; PLATT, H. M.; SHAW, K. M. The detection of differences
among assemblages of marine benthic species based on an assessment of dominance and
diversity. Journal of Natural History, United Kingdom, v. 17, p. 859-874, 1983.
LARA, F. M. Princípios de resistência de plantas a insetos. 2 ed. Ícone, São Paulo, p. 331,
1991.
LAROCA, S.; CURE JR; BORTOLI C. A associação das abelhas silvestres (Hymenoptera,
Apoidea) de uma área restrita no interior da cidade de Curitiba (Brasil): uma abordagem
biocenótica. Dusenia, São Paulo, v. 13, n. 3, p. 93–117, 1982.
LAZO, J. A. et al. Diversidad zoológica asociada a um silvopastoreo leucaena-guineacon
diferentes edades de establecimiento. Pesquisa Agropecuária Brasileira, Brasília, v. 42,
n. 12, p. 1667-1674, 2007.

30
LEITE, G. L. D. et al. Influence of canopy height and fertilization levels to Aculops
lycopersici (Acari: Eriophyidae). Experimental and Applied Acarology, Amsterdam, v.
23, n. 8, p. 633-642, 1999.
LEITE, G. L. D. et al. Effects of leaf compounds, climate and natural enemies on the
incidence of thrips in cassava. Pesquisa Agropecuária Brasileira, Brasília, v. 37, p.
1657-1662, 2002.
LEITE, G. L. D. et al. Natural factors affecting the whitefly infestation on cassava.
Acta Scientiarum Agronomy, Maringá, v. 25, p. 291-297, 2003.
LEITE, G. L. D. et al. Artrópodes (Arthropoda) associados a mudas de pequizeiro
Caryocar brasiliense Cambess. (Caryocaraceae). Arquivo Instituto Biológico, São Paulo,
v. 73, n.3, p. 365-370, 2006.
LEITE, G. L. D. et al. Within tree distribution of a gall-inducing Eurytoma
(Hymenoptera, Eurytomidae) on Caryocar brasiliense (Caryocaraceae). Revista
Brasileira de Entomologia, Curitiba, v. 53, p. 643-648, 2009.
LEITE, G. L. D. et al. Factors affecting Constrictotermes cyphergaster (Isoptera:
Termitidae) nesting on Caryocar brasiliense trees in the Brazilian savanna.
Sociobiology, Chico, v. 57, p. 165-180, 2011a.
LEITE, G. L. D. et al. Distribution of a leaf vein gall in Caryocar brasiliense
(Caryocaraceae) Tree. Revista Caatinga, Mossoró, v. 24, p. 186-190, 2011b.
LEITE, G. L. D. et al. Within tree distribution of a discoid gall on Caryocar brasiliense.
Revista Colombiana de Entomología, Santa Fé de Bogotá, v. 37, p. 289-293, 2011c.
LEITE, G. L. D. et al. Seasonal abundance of hemipterans on Caryocar brasiliense
(Malpighiales: Caryocaraceae) trees in the Cerrado. Florida Entomologist, Gainesville,
p.862-872, 2012.
LEITE, G. L. D. et al. Cardinal distribution of sucking insects in Caryocar brasiliense
(Caryocaraceae) in e Cerrado (Brazil). Revista Colombiana de Entomología, Bogotá, v.
41, p. 105-111, 2015.
MASCRÉ, M. Compt rend, p. 204-890, 1937.

31
MIRANDA, M. M. M. et al. Ecological life table of Tuta absoluta (Meyrick) (Lepidoptera:
Gelechiidae). Biocontrol Science and Technology, Abingdon, v. 8, n. 4, p. 597-606,
1998.
MELLO, M. O.; SILVA-FILHO, M. C. Plant-insect interactions: an evolutionary arms
race between two distinct defense mechanisms. Brazilian Journal of Plant Physiology,
Campos dos Goytacazes, v.14, p.71-81, 2002.
MONTEIRO, J. M. et al. Taninos: uma abordagem da química à ecologia. Química Nova,
São Paulo, v. 28, n. 5, 2005
MORATO, E. F.; CAMPOS, L. A. O. Partição de recursos florais de espécies de Sida
linnaeus e Mauvastrum coromandelianum (Linnaeus) Garck (Malvaceae) entre
Cephalurgus anomalus Moure & Oliveira (Hymenoptera, Andrenidae, Panurginae) e
Melissoptila cnecomala (Moure) (Hymenoptera, Apidae, Eucerini). Revista Brasileira de
Zoologia, Curitiba, v. 17, p. 705–727, 2000.
NEVES, E. L.; VIANA, B. F. As abelhas eussociais (Hymenoptera, Apidae) visitantes
florais em um ecossistema de dunas continentais no médio Rio São Francisco, Bahia,
Brasil. Revista Brasileira de Entomologia, Curitiba, ed. 46, v. 4, p. 571-578, 2002.
NISHIDA, R. Sequestration of defensive substances from plants by Lepdoptera. Annu
Review Entomology, Palo Alto, v. 47, p. 57-92, 2003.
OLIVEIRA, P. S.; BRANDÃO, C. R. F. The ant community associated with extrafloral
nectaries in the Brazilian cerrado. In: Cutler, D.F.; Huxley, C.R. Ant- Plant Interactions.
Oxford Univ. Press, Oxford, p. 601, 1991.
PAIM, D. C. Acúmulo de miosina em Leucaena leucocephala em resposta a etileno,
diferentes auxinas e nitrato de prata. Livro de resumos, Porto Alegre, p. 17-21, 2005.
PANIZZI, A. R.; PARRA, J. R. P. Introdução à ecologia nutricional de insetos. In
PANIZZI, A. R.: PARRA, J. R. P. Ed. Ecologia nutricional de insetos e suas implicações
no manejo integrado de pragas, São Paulo, p. 1-7, 1991.
PATHAK, S. C. et al. Insect drift over the northern Arabian Sea in early summer.
Journal of Biosciences, Bangalore, ed. 24, v. 2, p. 233-240, 1999.

32
PARRA, J. R. P. Consumo e utilização de alimentos por insetos. In: PANIZZI, A. R.,
PARRA, J. R. P. (Eds.). Ecologia nutricional de insetos e suas implicações no manejo de
pragas. Manole, São Paulo, pp. 9-65, 1991.
PEDRONI, F.; SANCHEZ, M.; SANTOS, F. A. M. Fenologia da copaíba (Copaifera
langsdorffii Desf.- Leguminosae, Caesalpinioideae) em uma floresta semidecídua no
sudeste do Brasil. Revista Brasileira Botânica, São Paulo, v. 25, p. 183-194, 2002.
PEETERS, P. J. Correlations between leaf structural traits and the density of herbivorous
insect guilds. Biological Journal of the Linnean Society, London, v. 77, p. 43-65, 2002.
PEETERS, P. J.: SANSON, G.; READ, J. Leaf biomechanical properties and the densities
of her- bivorous insect guilds. Functional Ecology, Oxford, v. 21, p. 246-255.2007.
PEREIRA, F. DE MELLO. Desenvolvimento de ração proteica para abelhas Apis
mellifera utilizando produtos regionais no nordeste brasileiro. Tese (Doutorado em
Zootecnia-Doutorado integrado em zootecnia) Universidade Federal do Ceará, Fortaleza,
2005.
PEREIRA, A. R. Como selecionar plantas para áreas degradadas e controle de erosão.
Belo Horizonte, p. 239, 2005.
PINHEIRO, F. et al. Seasonal pattern of insect abundance in the Brazilian cerrado. Austral
Ecology, Hoboken, ed. 27, v. 2, p. 132-136, 2002.
POLATTO, L. P. et al. Exploitation of floral resources on Sparattosperma leucanthum
(Bignoniaceae): foraging activity of the pollinators and the nectar and pollen thieves. Acta
Ethologica, Berlin, v. 15, p. 119-126, 2012.
PROCTOR, M.; YEO, P.; LACK, A. The Natural History of Pollination. London, Haper
Collins Publishers, p. 479, 1996.
RAGHU, S.; WILTSHIRE, C.; DHILEEPAN, K. Intensity of pre-dispersal seed predation
in the invasive legume Leucaena leucocephala is limited by the duration of pod retention.
Austral Ecology, Hoboken, v. 30, p. 310–318, 2005.
RAMALHO M.; IMPERATRIZ-FONSECA, V. L.; KLEINERT-GIOVANNINI, A.
Ecologia nutricional de abelhas sociais. In: PANIZZI, A. R.; PARRA, J. R. P (eds).

33
Ecologia nutricional de insetos e suas implicações no manejo de pragas. Ed. Manole,
Tamboré, Barueri, pp 225–252, 1991.
RAMANAND, H.; OLCKERS, T. Does height of exposure in the canopy influence egg
mortality in Acanthoscelides macrophthalmus (Coleoptera: Chryso- melidae), a biological
control agent of Leucaena leucocephala in South Africa? Biocontrol Science Technology,
Abingdon, v. 23, p. 545-554, 2013.
RAO, M. R.; SINGH, M.P.; DAY, R. Insect pest problems in tropical agroforestry
systems: Contributory factors and strategies for management. Agroforestry Systems, New
York, v. 50, p. 243–277, 2000.
RIBEIRO, S. P e PIMENTA, H. R. Padrões de abundância e de distribuição temporal de
herbívoros de vida livre em Tabebuia ochracea (Bignoniaceae). Anais da Sociedade
Entomológica do Brasil, v. 20, p. 428-448, 1991.
ROMOSER, W. S.; STOFFOLANO, J. G. J. R. The Science of Entomology, Dubuque,
Brown Communications, ed. 3, p. 532, 1994.
HUGHES, C. E. Leucaena: manual de recursos genéticos. Oxford : Oxford Forestry
Institute, (Tropical Papers, 37) p. 280, 1998.
RUHREN, S.; HANDEL, S. N. Jumping spiders (Salticidae) enhance the seed production
of a plant with extrafloral nectaries. Oecologia, Berlin, v. 119, p. 227-230, 1999.
SANTANA, D. L. Q. Psilídeos no Brasil: Heteropsylla cubana (Psilídeo da leucena)
Hemiptera: Psyllidae. Colombo-PR: Embrapa Florestas, Comunicado Técnico, n. 212, p.
4, 2008.
SCHOOLEY, R. L; WIENS, J. A. Finding habitat patches and directional connectivity.
Oikos, Rio de Janeiro, v. 102, n.3 p. 559-570, 2003.
SERRA, B. D. V; DE CAMPOS, L. A. Neotropical Entomology, Londrina, v. 39, ed. 2,
p. 153-159, 2010.
SILVA, M. M. et al. Influência de Trigona spinipes Fabr. (Hymenoptera: Apidae) na
polinização do maracujazeiro amarelo. Anais da Sociedade Entomológica do Brasil, São
Paulo, v. 26, n. 2, p. 217-221, 1997.

34
SILVA, S. J. R. Recursos tróficos de abelhas Apis mellifera L. (Himenóptera, Apidae) em
uma área de savana do Estado de Roraima: fontes de néctar e pólen. Dissertação
(Mestrado em Ciências Biológicas) - Instituto Nacional de Pesquisas da Amazônia,
Manaus, p. 88, 1998.
SILVA, J. O. et al. Esclerofilia, taninos e insetos herbívoros associados a Copaifera
lagsdorffii Desf. (Fabaceae: Caesalpinioideae) em área de transição Cerrado-Caatinga no
Brasil. Ecologia Austral, Códoba, v. 19, n. 3, 2009.
SILVA, F. W. S. et al. Spatial Distribution of Arthropods on Acacia mangium ( Fabales:
Fabaceae) Trees as Windbreaks in the Cerrado. Florida Entomologist, Gainesville, ed. 97,
v. 2, p. 631-638, 2015.
SIQUEIRA, K. M. M. Estudo comparativo da polinização de Mangifera indica L.
em cultivo convencional e orgânico na região do Vale do Submédio do São
Francisco. Revista Brasileira de Fruticultura, Jaboticabal, v. 30, p. 303-310, 2008.
SOUZA, J. L. E RESENDE, P. Manual de horticultura orgânica. Aprenda Fácil, Viçosa,
p. 564, 2003.
STYRSKY, J. D e EUBANKS, M. D. Ecological consequences of interactions between
ants and honeydew-producing insects. Proceedings of the Royal Society of London.
Series B, Biological Sciences, v. 274, p. 151-164, 2007.
STRONG, D. R.; LAWTON, J. H.; SOUTHWOOD, T. R. E. Insects on plants: community
patterns and mechanisms. Blackwell Scientific, London, p. 313, 1984.
TIXIER, M. S.; KREITER, S.; AUGER, P. Colonization of vine-yards by phytoseiid mites:
their dispersal patterns in the plot and their fate. Experimental and Applied Acarology,
Amsterdam, ed. 24 v. 3, p. 191-211, 2000.
TOLEDO, F. R.; BARBOSA, J. C.; YAMAMOTO, P. T. Distribuição espacial de
Toxoptera citricida (Kirkaldy) (Hemiptera: Aphididae) na cultura de citros. Revista
Brasileira de Fruticultura, Jaboticabal, v. 28, n. 2, p. 194-198, 2006.
TOWNSEND, C. R.; BERGON, M.; HARPER, J. L. Fundamentos em ecologia. Porto
Alegre, 2006.

35
UNSICKER, S. B.; MODY, K. Influence of tree species and compass bearing on insect
folivory of nine common tree species in the West African savanna. Journal of Tropical
Ecology, Cambridge, v. 21, n. 2, p. 227-231, 2005.
VELOSO, V. R. S. Dinâmica populacional de Anastrepha spp. e Ceratitis capitata
(Weid., 1824)(Diptera, Tephritidae) nos cerrados de Goiás. 1997. 115 p. Tese
(Doutorado em Agronomia- Produção Vegetal) Escola de Agronomia da Universidade
Federal de Goiás, Goiânia, 1997.
VILLAS-BÔAS, J. Manual Tecnológico: Mel de abelhas sem Ferrão. 1ª edição. Brasília,
2012.
WOITKE, M. e DIETZ, H. Shifts in dominance of native and invasive plants in
experimental patches of vegetations. Perpectives in Plant Ecology, evolution and
Systematics, v. 5, n. 3, p. 165-184, 2002.
ZANETTI, R. et al. Level of economic damage for leaf-cutting ants in Eucalyptus
plantations in Brazil. Sociobiology, Chico, ed. 42, v. 2, p. 433-442, 2003.
ZANUNCIO, J. C. et al. Spatial distribution of nests of the leaf cutting ant Atta sexdens
rubropilosa (Hymenoptera: Formicidae) in plantations of Eucalyptus urophylla in Brazil.
Sociobiology, Chico, ed. 39, v. 2, p. 231-242, 2002.
ZUCCHI, R. A., S. S. Neto e O. Nakano. Guia de identificação de pragas agrícolas.
Piracicaba, FEAEQ, p.139, 1993.
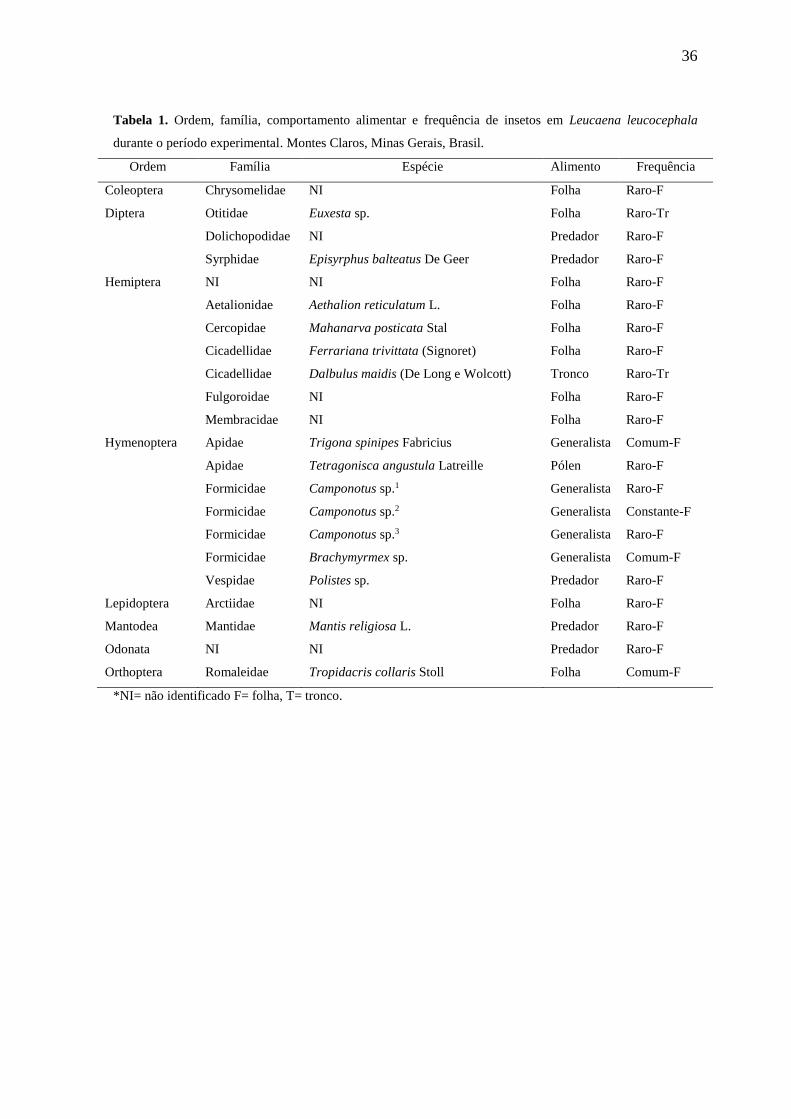
36
Tabela 1. Ordem, família, comportamento alimentar e frequência de insetos em Leucaena leucocephala
durante o período experimental. Montes Claros, Minas Gerais, Brasil.
Ordem Família Espécie Alimento Frequência
Coleoptera Chrysomelidae NI Folha Raro-F
Diptera Otitidae Euxesta sp. Folha Raro-Tr
Dolichopodidae NI Predador Raro-F
Syrphidae Episyrphus balteatus De Geer Predador Raro-F
Hemiptera NI NI Folha Raro-F
Aetalionidae Aethalion reticulatum L. Folha Raro-F
Cercopidae Mahanarva posticata Stal Folha Raro-F
Cicadellidae Ferrariana trivittata (Signoret) Folha Raro-F
Cicadellidae Dalbulus maidis (De Long e Wolcott) Tronco Raro-Tr
Fulgoroidae NI Folha Raro-F
Membracidae NI Folha Raro-F
Hymenoptera Apidae Trigona spinipes Fabricius Generalista Comum-F
Apidae Tetragonisca angustula Latreille Pólen Raro-F
Formicidae Camponotus sp.1 Generalista Raro-F
Formicidae Camponotus sp.2 Generalista Constante-F
Formicidae Camponotus sp.3 Generalista Raro-F
Formicidae Brachymyrmex sp. Generalista Comum-F
Vespidae Polistes sp. Predador Raro-F
Lepidoptera Arctiidae NI Folha Raro-F
Mantodea Mantidae Mantis religiosa L. Predador Raro-F
Odonata NI NI Predador Raro-F
Orthoptera Romaleidae Tropidacris collaris Stoll Folha Comum-F
*NI= não identificado F= folha, T= tronco.

37
Tabela 2. Número de indivíduos (NI), riqueza específica (S) e índice de diversidade (D) de fitófagos,
inimigos naturais e polinizadores em Leucaena leucocephala em função da posição dos ramos aos pontos
cardeais, no dossel e na face foliar em dois anos durante o período experimental por planta. Montes Claros-
MG.
Posição do ramo quanto aos pontos cardeais
Fitófagos Norte Sul Leste Oeste
NI 12 3 3 10
S 4 3 1 6
D 7,33 7,04 1,44 14,71
Inimigos naturais e
polinizadores
NI 33 23 25 24
S 7 7 6 3
D 7,17 7,54 8,10 4,59
Parte do Dossel
Apical Médio Basal Tronco
Fitófagos
NI 17 11 0 2
S 9 4 0 2
D 21,39 8,64 0 3,92
Inimigos naturais e
polinizadores
NI 46 35 24 17
S 7 5 4 3
D 6,60 7,20 4,68 5,75
Face foliar
Abaxial Adaxial
Fitófagos
NI 19 9
S 7 5
D 14,28 11,94
Inimigos naturais e
polinizadores
NI 70 35
S 9 3
D 9,68 3,12

38
Tabela 3. Dominância-K e abundância (entre parênteses) dos insetos fitófagos, inimigos naturais e
polinizadores em Leucaena leucocephala em função da posição dos ramos aos pontos cardeais em dois anos
durante o período experimental por planta. Montes Claros-MG.
Fitófagos
Norte Sul Leste Oeste
Trigona spinipes 50.0(6) Tropidacris
colaris
33.3(1) F. trivittata 100(3) Tropidacris
colaris
40(4)
Tropidacris
Colaris
83.3(4) Trigona spinipes 66.7(1) A. reticulatum 100(0) M. posticata 60(2)
Arctiidae 91.7(1) Membracidae 100(1) Tropidacris
colaris
100(0) A. reticulatum 70(1)
Membracidae 100(1) A. reticulatum 100(0) Fulgoroidea 100(0) Fulgoroidea 80(1)
A. reticulatum 100(0) F. trivittata 100(0) Arctiidae 100(0) Chrysomelidae
90(1)
F. trivittata 100(0) Fulgoroidea 100(0) M. posticata 100(0) Hemiptera 100(1)
Inimigos naturais e polinizadores
Camponotus sp.2 69.7(23) Camponotus sp. 2 69.6(16) Camponotus sp.2 56.0(14) Camponotus sp. 2 70.8(17)
T. angustula 78.8(3) Brachymyrmex sp. 78.3(2) Brachymyrmex sp. 84.0(7) T. angustula 87.5(4)
Brachymyrmex
sp.
87.9(3) Camponotus sp. 3 82.6(1) Camponotus sp. 3 88. 0(1) Brachymyrmex sp. 100(3)
Camponotus sp. 3 90.9(1) E. balteatus 87.0(1) Dolichopodidae 92.0(1) Camponotus sp. 3 100(0)
Mantis religiosa 93.9(1) Odonata 91.3(1) T. angustula 96.0(1) Mantis religiosa 100(0)
Camponotus sp. 1 97.0(1) T. angustula 95.7(1) Polistes sp. 100(1) Camponotus sp. 1 100(0)
Polistes sp. 100(1) Polistes sp. 100(1) Mantis religiosa 100(0) E. balteatus 100(0)

39
Tabela 4. Dominância-K e abundância (entre parênteses) dos insetos fitófagos, inimigos naturais e
polinizadores em Leucaena leucocephala em função do dossel em dois anos durante o período experimental
por planta. Montes Claros-MG.
Fitófagos
Apical Média Basal Tronco
Tropidacris
colaris
41.2(7) Trigona spinipes 45.5(5) Trigona spinipes 100(0) Euxesta sp. 50(1)
M. posticata 52.9(2) F. trivittata 72.7(3) F. trivittata 100(0) Dalbulus maidis 100(1)
Trigona spinipes 64.7(2) Tropidacris
colaris
90.9(2) Tropidacris
colaris
100(0) A. reticulatum 100(0)
A. reticulatum 70.6(1) Membracidae 100(1) Membracidae 100(0) Tropidacris
colaris
100(0)
Fulgoroidea 76.5(1) A. reticulatum 100(0) A. reticulatum 100(0) F. trivittata 100(0)
Arctiidae 82.4(1) Fulgoroidea 100(0) Fulgoroidea 100(0) Fulgoroidea 100(0)
Chrysomelidae 88.2(1) Arctiidae 100(0) Arctiidae 100(0) Arctiidae 100(0)
Hemiptera 94.1(1) M.posticata 100(0) M. posticata 100(0) M. posticata 100(0)
Membracidae 100(1) Chrysomelidae 100(0) Chrysomelidae 100(0) Chrysomelidae 100(0)
Inimigos naturais e polinizadores
Camponotus sp. 2 67.4(31) Camponotus sp. 2 60.0(21) Camponotus sp. 2 75(18) Camponotus sp. 2 58.8(10)
Brachymyrmex
sp.
87.0(9) T. angustula 77.1(6) Polistes sp. 87.5(3) Brachymyrmex sp. 82.4(4)
Camponotus sp. 3 91.3(2) Brachymyrmex sp. 94.3(6) T. angustula 95.8(2) Camponotus sp. 3 100(3)
Mantis religiosa 93.5(1) Camponotus sp. 3 97.1(1) E. balteatus 100(1) Mantis religiosa 100(0)
Dolichopodidae 95.7(1) Camponotus sp. 1 100(1) Camponotus sp. 3 100(0) Camponotus sp. 1 100(0)
Odonata 97.8(1) Mantis religiosa 100(0) Mantis religiosa 100(0) E. balteatus 100(0)
T. angustula 100(1) E. balteatus 100(0) Camponotus sp. 1 100(0) Dolichopodidae 100(0)
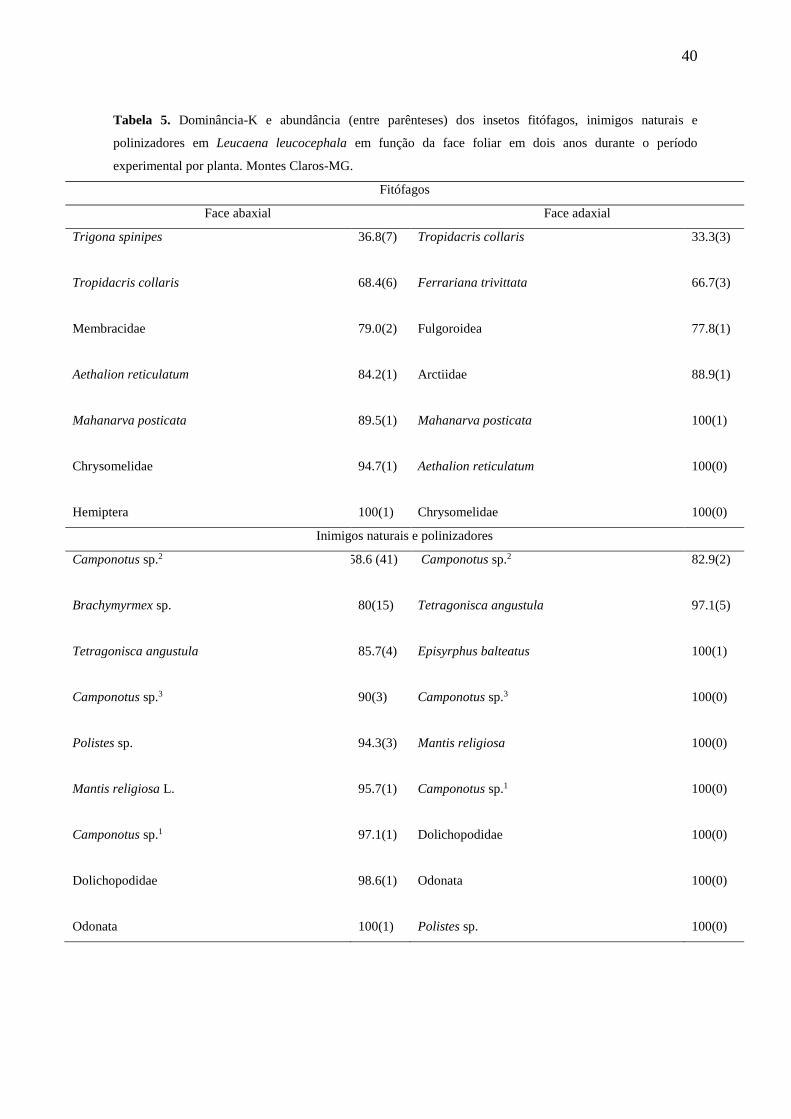
40
Tabela 5. Dominância-K e abundância (entre parênteses) dos insetos fitófagos, inimigos naturais e
polinizadores em Leucaena leucocephala em função da face foliar em dois anos durante o período
experimental por planta. Montes Claros-MG.
Fitófagos
Face abaxial Face adaxial
Trigona spinipes 36.8(7) Tropidacris collaris 33.3(3)
Tropidacris collaris 68.4(6) Ferrariana trivittata 66.7(3)
Membracidae 79.0(2) Fulgoroidea 77.8(1)
Aethalion reticulatum 84.2(1) Arctiidae 88.9(1)
Mahanarva posticata 89.5(1) Mahanarva posticata 100(1)
Chrysomelidae 94.7(1) Aethalion reticulatum 100(0)
Hemiptera 100(1) Chrysomelidae 100(0)
Inimigos naturais e polinizadores
Camponotus sp.2 58.6 (41) Camponotus sp.2 82.9(2)
Brachymyrmex sp. 80(15) Tetragonisca angustula 97.1(5)
Tetragonisca angustula 85.7(4) Episyrphus balteatus 100(1)
Camponotus sp.3 90(3) Camponotus sp.3 100(0)
Polistes sp. 94.3(3) Mantis religiosa 100(0)
Mantis religiosa L. 95.7(1) Camponotus sp.1 100(0)
Camponotus sp.1 97.1(1) Dolichopodidae 100(0)
Dolichopodidae 98.6(1) Odonata 100(0)
Odonata 100(1) Polistes sp. 100(0)
