determinaÇÃo e quantificaÇÃo de protozoÁrios...
TRANSCRIPT

UNIVERSIDADE TECNOLÓGICA FEDERAL DO PARANÁ
DEPARTAMENTO DE ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA
LETÍCIA WLODARSKI
DETERMINAÇÃO E QUANTIFICAÇÃO DE PROTOZOÁRIOS
CILIADOS E BACTÉRIAS DO RÚMEN DE BOVINOS EM PASTAGENS
TEMPERADAS E TROPICAIS
DISSERTAÇÃO
DOIS VIZINHOS
2017

LETÍCIA WLODARSKI
DETERMINAÇÃO E QUANTIFICAÇÃO DE PROTOZOÁRIOS
CILIADOS E BACTÉRIAS DO RÚMEN DE BOVINOS EM PASTAGENS
TEMPERADAS E TROPICAIS
Dissertação apresentada ao Programa de Pós-
Graduação em Zootecnia da Universidade
Tecnológica Federal do Paraná, Campus Dois
Vizinhos, como requisito parcial à obtenção do
título de Mestre em Zootecnia – Área de
Concentração: Produção e Nutrição Animal.
Orientador: Emilyn Midori Maeda
DOIS VIZINHOS
2017

W836d Wlodarski, Letícia.
Determinação e quantificação de protozoários ciliados e
bactérias do rúmen de bovinos alimentados em pastagem
temperadas e tropicais. / Letícia Wlodarski – Dois Vizinhos,
2017.
67f.
Orientadora: Emilyn Midori Maeda.
Dissertação (Mestrado) - Universidade Tecnológica
Federal do Paraná, Programa de Pós-Graduação em
Zootecnia, Dois Vizinhos, 2017.
Bibliografia p.59-61
1. Microbiotal Runinal 2. Ruminantes – Nutrição
I. Maeda, Emilyn Midori, orient. II. Universidade
Tecnológica Federal do Paraná – Dois Vizinhos III. Título
CDD: 636.20852
Ficha catalográfica elaborada por Keli Rodrigues do Amaral Benin CRB: 9/1559
Biblioteca da UTFPR-Dois Vizinhos

Ministério da Educação
Universidade Tecnológica Federal do Paraná
Câmpus Dois Vizinhos
Diretoria de Pesquisa e Pós-Graduação
Programa de Pós-Graduação em Zootecnia
TERMO DE APROVAÇÃO
Título da Dissertação n° 074
Determinação e quantificação de protozoários ciliados e bactérias do rúmen de bovinos
em pastagens temperadas e tropicais
Letícia Wlodarski
Dissertação apresentada às oito horas e trinta minutos do dia quinze de fevereiro de dois mil e
dezessete, como requisito parcial para obtenção do título de MESTRE EM ZOOTECNIA,
Linha de Pesquisa – Produção e Nutrição Animal, Programa de Pós-Graduação em Zootecnia
(Área de Concentração: Produção animal), Universidade Tecnológica Federal do Paraná,
Câmpus Dois Vizinhos. A candidata foi arguida pela Banca Examinadora composta pelos
professores abaixo assinados. Após deliberação, a Banca Examinadora considerou o trabalho
........................................
Banca examinadora:
Emilyn Midori Maeda
UTFPR-DV
Ana Carolina Fluck
UTFPR-DV
Isabel Martinele Corrêa
UFJF-MG
Prof. Dr. Douglas Sampaio Henrique
Coordenador do PPGZO
*A Folha de Aprovação assinada encontra-se na Coordenação do Programa de Pós-Graduação
em Zootecnia.

AGRADECIMENTOS
A Deus, que todos os dias da minha vida me deu forças para nunca desistir.
Aos meus pais Polan e Bernadete e ao meu irmão Leandro, pelo amor, incentivo e apoio
financeiro e emocional.
A minha orientadora Prof. Dra. Emilyn Midori Maeda, pela paciência, apoio e confiança, sem
dúvidas uma pessoa maravilhosa...
À Universidade Tecnológica Federal do Paraná, em especial ao departamento de Zootecnia,
pela oportunidade de realização do mestrado.
Especial agradecimento à Priscila Fregulia (mestranda da UFJF), pela paciência e tempo
dedicado em me ajudar na identificação dos protozoários, e pela amizade que construímos.
Agradeço também a graduanda da UFJF, Elen Furtado, pela hospedagem e confiança, serei
eternamente grata.
A Universidade Federal de Juiz de Fora e a equipe do Laboratório de Protozoologia, pela
amizade e paciência e em especial a professora Marta Tavares d´Agosto pela oportunidade e
disponibilidade em ajudar.
Ao professor Dr. Elias Nunes Martins, a pós-doutoranda Ana Carolina Fluck e ao doutorando
Olmar Antonio Denardin Costa pela ajuda nas análises estatísticas.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela bolsa de
estudos concedida. À Miolar Alimentos Ltda e o Frigorífico Orfimar por abrir as portas para
a realização das coletas e em especial aos colaboradores envolvidos.
Aos meus amigos, por fazerem parte da minha vida.
Muito obrigada!

WLODARSKI, Letícia. Determinação e quantificação de protozoários ciliados e bactérias
do rúmen de bovinos alimentados em pastagens temperadas e tropicais. 2017. 67 folhas.
Dissertação (Mestrado em Zootecnia) – Programa de Pós Graduação em Zootecnia,
Universidade Tecnológica Federal do Paraná. Dois Vizinhos, 2017.
RESUMO
O objetivo do presente trabalho foi identificar e quantificar os protozoários ciliados e
bactérias do rúmen de bovinos cruzados (Europeu x Nelore), com média de 2,5 anos,
alimentados em pastagens de inverno (Azevém e Aveia + Azevém) e verão (Brachiaria spp. e
Brachiaria spp. + Esrela Africana). Amostras de conteúdo ruminal foram obtidas do centro da
massa ruminal, após o abate dos animais. A contagem de bactérias totais (CBT), para as
frações sólida e para líquida, foi realizada em placas com auxílio de contador de colônias
manual. Para a análise morfológica empregou-se a técnica de coloração de Gram e a leitura
das lâminas foi feita em microscópio óptico com objetiva de 100x, sendo as bactérias
classificadas em cocos e bacilos, gram positivos e gram negativos. A avaliação da atividade
fermentativa foi evidenciada pelo crescimento de colônias com coloração rósea atravéz de
uma fonte de carboidrato. A quantificação e identificação dos gêneros de ciliados foram
realizadas em câmara Sedgewick-Rafter em microscopia ótica. Foi determinado o teor de
Matéria Seca, Proteína Bruta, Matéria Mineral, Fibra em Detergente Neutro e Fibra em
Detergente Ácido, Digestibilidade In Vitro na Matéria Seca, Lignina, Celulose, Hemicelulose.
O delineamento experimental foi inteiramente casualizado, com quatro tratamentos e 10
repetições para cada tratamento. Os dados foram submetidos à análise por meio da
metodologia de Modelos Lineares, generalizados, com distribuição Poisson (1%) e foi
utilizado o procedimento GENMOD. Foram observadas bactérias Gram positivas e negativas,
nas formas cocos e bacilos e 11 gêneros de protozoários ciliados, sendo Diplodinium o gênero
predominante. A maior concentração de bactérias foi encontrada nas forrageiras de inverno
(494,8 x1010
mL-1
), com maior intensidade na pastagem de Azevém. Nos protozoários
ciliados, maior concentração foi observada no verão (200,70 x104 mL
-1), principalmente na
pastagem de Brachiaria spp.
Palavras-chave: microbiota ruminal, Ophryoscolecidae, estacionalidade, forragem,
ruminantes

WLODARSKI, Letícia. Determination and quantification of ciliate protozoa and rumen
bacteria of cattle fed in temperate and tropical pastures. 2017, 67 pages. Dissertation
(Master of Animal Science) - Universidade Tecnológica Federal do Paraná. Dois Vizinhos,
2017.
ABSTRACT
The objective of the present work was to identify and quantify the ciliate protozoa and cross-
bovine rumen bacteria (European x Nellore), with an average of 2.5 years, fed on winter
pastures (Azevém and Aveia + Azevém) and summer (Brachiaria spp And Brachiaria spp. +
Esrala Africana). Samples of ruminal contents were obtained from the center of the ruminal
mass, after the slaughter of the animals. The total bacterial count (CBT), for solid and liquid
fractions, was performed in plates with the aid of a manual colony counter. For the
morphological analysis the Gram staining technique was used and the slides were read under
an optical microscope with objective of 100x, being the bacteria classified in cocci and bacilli,
gram positive and gram negative. The evaluation of the fermentative activity was evidenced
by the growth of colonies with pink coloration through a carbohydrate source. The
quantification and identification of ciliate genera were performed in Sedgewick-Rafter
chamber under optical microscopy. The content of Dry Matter, Crude Protein, Mineral
Matter, Neutral Detergent Fiber and Acid Detergent Fiber, In Vitro Digestibility in Dry
Matter, Lignin, Cellulose and Hemicellulose were determined. The experimental design was
completely randomized, with four treatments and 10 replicates for each treatment. The data
were submitted to analysis using the methodology of Linear Models, generalized with Poisson
distribution (1%) and the GENMOD procedure was used. Gram positive and negative bacteria
were observed, in the forms cocci and bacilli and 11 genera of ciliate protozoa, being
Diplodinium the predominant genus. The highest concentration of bacteria was found in
winter forages (494.8 x1010 mL-1
), with higher intensity in the Azevém pasture. In the ciliate
protozoa, higher concentration was observed in the summer (200.70 x 10 4 mL-1
), mainly in
Brachiaria spp.
Keywords: ruminal microbiota, Ophryosclerosis, seasonality, forage, ruminants

LISTA DE TABELAS
Capítulo único: Determinação e quantificação de protozoários ciliados e bactérias do
rúmen de bovinos alimentados em pastagens temperadas e tropicais.
Tabela 1: Composição nutricional das pastagens de inverno e verão ingeridas pelos bovinos
cruzados estudados....................................................................................................................45
Tabela 2: Tabela 2 – Composição do meio de cultura utilizado para a atividade fermentativa
do carboidrato Carboximetilcelulose........................................................................................47
Tabela 3: Médias (x1010
mL-1
) da contagem de bactérias totais (CBT) da fração sólida e
líquida de amostras extraídas de bovinos cruzados (Nelore x Europeu) submetidos ao efeito da
estacionalidade de pastagens temperadas (Azevém e Azevém + Aveia) e tropicais (Brachiaria
spp. e Brachiaria spp + Estrela africana)..................................................................................48
Tabela 4: Flora ruminal e atividade celulolítica da fração sólida e líquida de amostras
extraídas de bovinos cruzados (Nelore x Europeu) submetidos ao efeito da estacionalidade de
pastagens temperadas (Azevém e Azevém + Aveia) e tropicais (Brachiaria spp. e Brachiaria
spp + Estrela africana)..............................................................................................................50
Tabela 5: Ordem, família e gênero de protozoários ciliados em bovinos cruzados (Nelore x
Europeu) submetidos ao efeito da estacionalidade de pastagens temperadas (Azevém e
Azevém + Aveia) e tropicais (Brachiaria spp. e Brachiaria spp + Estrela
africana)....................................................................................................................................51
Tabela 6: Prevalência (%) dos gêneros de protozoários ciliados do rúmen de bovinos
cruzados (Nelore x Europeu) submetidos ao efeito da estacionalidade de pastagens
temperadas (Azevém e Azevém + Aveia) e tropicais (Brachiaria spp. e Brachiaria spp +
Estrela africana.........................................................................................................................52.
Tabela 7: Médias (x104 mL
-1) estimadas para o total dos gêneros de protozoários ciliados do
rúmen encontrados de amostras extraídas de bovinos cruzados submetido submetidos ao
efeito da estacionalidade de pastagens temperadas (Azevém e Azevém + Aveia) e tropicais
(Brachiaria spp. e Brachiaria spp + Estrela africana).............................................................53
Tabela 8: Médias (x104 mL
-1) estimadas para os gêneros de protozoários ciliados em bovinos
cruzados do rúmen encontrados de amostras extraídas de bovinos cruzados submetido
submetidos ao efeito da estacionalidade de pastagens temperadas (Azevém e Azevém +
Aveia) e tropicais (Brachiaria spp. e Brachiaria spp + Estrela africana)..............................55

LISTAS DE FIGURAS
Capítulo único: Determinação e quantificação de protozoários ciliados e bactérias do
rúmen de bovinos alimentados em pastagens temperadas e tropicais.
Figura 1. Desenhos esquemáticos de protozoários ciliados encontrados em ruminantes
domésticos.................................................................................................................................24

LISTA DE ANEXOS
Capítulo único: Determinação e quantificação de protozoários ciliados e bactérias do
rúmen de bovinos alimentados em pastagens temperadas e tropicais.
Anexo A: Comitê de ética..................................................................................................63-64
Anexo B: Bovinos utilizados para a amostragem do conteúdo ruminal..................................65
Anexo C: Abertura do compartimento ruminal para a coleta das amostras............................65
Anexo D: Coleta das amostras para determinação e quantificação de bactérias e protozoários
...................................................................................................................................................65
Anexo E: Inoculação das alíquotas de líquido ruminal em placas de petri estéril para a análise
da contagem bacteriana total (CBT).........................................................................................66
Anexo F: A: Bactéria Gram-negativa; B: Bactéria Gram-positiva e contagem de colônias de
bacterianas ruminais..................................................................................................................66
Anexo G: Gêneros de protozoários ciliados em bovinos cruzados em pastagens de inverno e
verão, Azevém, Azevém e Aveia, Brachiaria spp. e Brachiaria spp e Estrela africana. A:
Diplodinium; B: Eremoplastron; C: Eudiplodinium; D: Dasytricha; E: Eodinium; F:
Ostracodinium; G: Metadinium; H: Polyplastron; I: Epidinium; J: Isotricha; K:
Entodinium................................................................................................................................67

SUMÁRIO
1. INTRODUÇÃO ............................................................................................................... 11
2. REVISÃO BIBLIOGRÁFICA ......................................................................................... 13
2.1 Caracterização ruminal e microbiota .............................................................................. 13
2.2 Bactérias ruminais .......................................................................................................... 15
2.3 Protozoários ruminais ..................................................................................................... 18
2.4 Interação microrganismo – hospedeiro .......................................................................... 24
2.5 Interação entre microrganismos ruminais....................................................................... 25
2.6 Efeito da dieta na microbiologia ruminal ....................................................................... 27
2.7 Técnicas de identificação da microbiota ruminal ........................................................... 31
4. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 33
3. ARTIGO: .............................................................................................................................. 42
INTRODUÇÃO ........................................................................................................................ 43
MATERIAL E MÉTODOS ...................................................................................................... 44
RESULTADOS E DISCUSSÃO ............................................................................................. 48
CONCLUSÃO .......................................................................................................................... 57
CONSIDERÇÕES FINAIS ...................................................................................................... 58
REFERÊNCIAS ....................................................................................................................... 59
5. ANEXOS .............................................................................................................................. 62

11
1. INTRODUÇÃO
As relações mutualísticas entre as populações microbianas apresentam-se como um
dos elementos essenciais no aproveitamento dos nutrientes pelo hospedeiro. A compreensão
dos mecanismos que estão relacionados ao compartimento ruminal e a estrutura da microbiota
possibilitam novas alternativas e tecnologias para a sustentabilidade da produção.
O rúmen representa um microbioma complexo, em que populações de microrganismos
anaeróbios como fungos, bactérias e protozoários realizam diferentes funções físico-químicas,
essenciais para a mantença dos ruminantes. A relação entre microrganismos são específicas e
inter-específicas, provendo energia ao hospedeiro e, sobretudo, no metabolismo de nutrientes
através da degradação da celulose das plantas (CHOUDHURY et al., 2012). Dentre os fatores
que afetam as populações microbianas, dieta, idade e condições de saúde do animal
hospedeiro são os mais relevantes (WELKIE et al., 2010).
Os microrganismos ruminais caracterizam-se pelo enorme número e diversidade de
espécies. Uma espécie é considerada pertencente ao microbioma ruminal quando é anaeróbia,
de crescimento contínuo e é capaz de gerar produtos, ou seja, fornecer condições adequadas
para a mantença do ambiente ruminal. Protozoários ciliados e bactérias representam quase que
em totalidade os microrganismos de maior importância da microbiota ruminal (FONSECA;
DIAS-DA-SILVA, 2001).
As bactérias representam um dos principais microrganismos presentes no conteúdo
ruminal, sendo os seres mais diversos taanto em número de espécies quanto em atividade
metabólica, diversificação na ingestão de alimento e capacidade de adaptação ruminal. São de
suma importância no processo de fermentação e degradação da lignina, celulose,
hemicelulose, proteína, amido e óleo contido nos alimentos. No conteúdo ruminal, o tamanho
das bactérias pode variar de 1 a 5 µm, aproximadamente 1010 células/grama (TEIXEIRA,
2001).
Os protozoários ruminais são unicelulares, anaeróbios, não patogênicos e cujo
tamanho varia de 20 a 200 μm, com densidade aproximada de 104 ciliados por mililitro de
conteúdo ruminal. Estes microrganismos são importantes no processo de fermentação ruminal
e são habilitados na digestão da maioria dos componentes dos alimentos, sobretudo na
digestibilidade da fibra, sendo responsáveis por até 34% da degradação do material fibroso
(KAMRA, 2005).

12
Para que o desenvolvimento de protozoários e bactérias seja de forma desejável, é
necessária a disponibilidade de nitrogênio e energia, onde a quantidade de nitrogênio exigido
está baseada na energia disponibilizada no rúmen (MARTIN et al., 1990). Diante disto, o
efeito da dieta exerce significativa importância nos estudos ruminais, sobretudo em dietas
onde a base é forragem em que os valores nutricionais podem ser alterados devido ao período
estacional. As forragens representam uma das principais fontes de nutrientes para os
herbívoros, desta forma, o conhecimento gerado e as informações disponibilizadas a cerca
deste assunto permitem condições para a otimização da dieta fornecida, principalmente em
relação à digestão e fermentação dos alimentos.
Tendo em vista a importância da comunidade ruminal e sua relação com os bovinos, o
objetivo deste trabalho foi identificar e quantificar os protozoários e bactérias do rúmen de 40
bovinos de corte submetidos à alimentação a pasto: Azevém, Aveia + Azevém, Brachiaria
spp. e Brachiaria spp + Estrela Africana.

13
2. REVISÃO BIBLIOGRÁFICA
2.1 Caracterização ruminal e microbiota
O rúmen compreende um ecossistema complexo, com inúmeras características que
tem por finalidade a fermentação do alimento consumido pelo animal. Esse processo ocorre
pela intervenção dos microrganismos ruminais, os quais transformam o produto ingerido em
ácidos graxos voláteis, que tem por finalidade fornecer energia ao hospedeiro; além da síntese
da proteína, sendo esta realizada pela massa microbiana ruminal (WEIMER, 2009).
O ruminante possui câmeras fermentativas em seu trato digestivo, o que permite a
estes animais maior eficiência no aproveitamento de alimentos fibrosos e produtos da
fermentação, principalmente nos animais mantidos em pastejo. Seu estômago multicavitário
deriva embrionariamente do estômago simples, e é dividido em quatro partes: rúmen, retículo,
omaso e abomaso (OLIVEIRA, 2013).
O compartimento ruminal abriga um ecossistema com características complexas, as
quais podem ser benéficas aos animais, desde que sua capacidade de fermentar alimentos e
transformar os produtos finais dessa fermentação em nutrientes de alto valor nutritivo e
absorvíveis no intestino seja maximizada. O rúmen é composto de partículas, agregados de
alimentos e microrganismos anaeróbios, como protozoários, bactérias, micoplasmas e
bacteriófagos, que convivem de forma interativa entre si. Protozoários ciliados e bactérias
representam quase que em totalidade, os microrganismos de maior importância da microbiota
ruminal (FONSECA; DIAS-DA-SILVA, 2001). A ingestão de alimentos sólidos é a principal
forma de modificar a anatomia e fisiologia do trato digestivo dos ruminantes jovens,
principalmente do rúmen (OLIVEIRA et al., 2007).
A anaerobiose apresenta-se como uma das características peculiares do rúmen, sendo
um dos fatores limitantes para a mantença do ecossistema microbiano, como também auxilia
na mantença da energia utilizada pelo animal. A conservação das condições anaeróbias do
ambiente ruminal é mantida através dos gases oriundos da fermentação, como o gás
carbônico, metano e traços de hidrogênio (KAMRA, 2005).
Entre as características do rúmen tem-se um sistema isotérmico (38º a 42ºC) o qual é
regulado pelo animal hospedeiro, devido ao metabolismo homeotérmico deste; o pH apresenta
variação de acordo com a dieta fornecida, no entanto o ambiente ruminal tende a se manter
neutro em virtude da remoção dos ácidos pela fermentação, que são absorvidos e
neutralizados pela ação tamponante da saliva animal e a baixa quantidade de oxigênio
presente no rúmen, que contribui para o desenvolvimento de microrganismos anaeróbios, e

14
alguns gêneros de bactérias facultativas (TEIXEIRA, 2001; LANA, 2005; KOZLOSKI,
2011).
Durante o processo evolutivo, os ruminantes desenvolveram a capacidade de
aproveitar de forma eficiente carboidratos estruturais como fonte energética e compostos
nitrogenados não-proteicos como fonte de proteína (VALADARES FILHO; PINA, 2006).
Os ruminantes caracterizam-se pela diversidade de microrganismos presentes no
rúmen e intestino, sendo estes responsáveis pela fermentação e digestão dos alimentos,
através da endosimbiose entre as populações existentes, que as permite sobreviver com uma
dieta, em que a principal fonte de carboidrato é a celulose (TEIXEIRA, 2001). Os
microrganismos ruminais são capazes de digerir aproximadamente 70 a 85% da matéria seca
digestível da ração (COELHO DA SILVA; LEÃO, 1979).
Estudos relacionados à microbiologia do rúmen, juntamente com a produção animal,
contribuem para melhorar a eficiência do uso de diferentes alimentos, devido à
disponibilidade de informações a cerca da biodiversidade das populações microbianas e as
relações mutualísticas entre estas. O conhecimento gerado com estas pesquisas permite
condições para a otimização da dieta fornecida, principalmente em relação à digestão e
fermentação dos alimentos (WEIMER, 2011).
A comunidade de microrganismos no rúmen é bastante diversa devido às inúmeras
espécies existentes, sua classificação pode variar de acordo com os diferentes autores. Para
uma espécie ser considerada pertencente à microbiota ruminal, esta deve ser anaeróbia, gerar
subprodutos no rúmen e apresentar crescimento ativo, ou seja, apresentar metabolismo
conciliável com as condições ambientais de um rúmen normal (STEWART et al., 1997).
De acordo com Pedersen e Tannock (1989), a colonização do trato digestório pelos
microrganismos é influenciada por diferentes fatores e fenômenos concomitantes. As
bactérias realizam a colonização de forma mais ágil que os demais microrganismos do rúmen,
sendo os Lactobacilos as primeiras espécies colonizadoras do trato gastrintestinal (CHAVES
et al., 1999).
Para Mcallister et al. (1994), os microrganismos do rúmen que tem capacidade de
interagir com os fragmentos do alimento, podem ser caracterizados em: livres, associados ao
conteúdo ruminal; associado a partículas de forma fraca; e estritamente aderidos as partículas
de alimentos.
A pesquisa voltada a microbiologia ruminal visa constatar como as espécies
microbianas agem na digestão dos alimentos, como os transformam e sintetiza-os. Para que o

15
desenvolvimento de protozoários e bactérias apresente-se de forma desejável, torna-se
necessário a disponibilidade de nitrogênio e energia para estes seres, assim, a quantidade de
nitrogênio exigido está baseada na energia disponibilizada no rúmen. (MARTIN et al., 1990).
A microbiologia ruminal possibilitou a ampliação dos conhecimentos relacionados à
população microbiana e através destes dados e características conhecidas, permitiu melhorias
em relação à alimentação de ruminantes, garantindo melhor eficiência e aproveitamento dos
nutrientes da dieta, o que gera ganhos no desempenho dos animais (MORGAVI et al., 2013).
2.2 Bactérias ruminais
As bactérias representam a principal população de microrganismos presentes no
conteúdo ruminal, sendo os seres mais diversos tanto em número quanto em atividade
metabólica. São de suma importância no processo de fermentação e na degradação da lignina,
celulose, hemicelulose, como também a proteína, o amido e o óleo contido nos alimentos. A
simbiose entre as bactérias e entre outros microrganismos promove a formação de ácidos
graxos voláteis e a proteína microbiana ruminal (TEIXEIRA, 2001).
Os primeiros estudos relacionados à identificação e metabolismo das bactérias
surgiram após a Segunda Guerra Mundial, com a ampliação de pesquisas e técnicas de cultivo
bacteriano em ambiente anaeróbio (HUNGATE, 1966). Anteriormente a este período, os
estudos relacionados à microbiota e a fermentação ruminal eram escassos.
As bactérias constituem de 60 a 90% da biomassa microbiana com cerca de 200
espécies distintas (KOZLOSKI, 2011). No conteúdo ruminal, as bactérias podem variar de 1 a
5 µm, aproximadamente 1011 células/grama (WRIGHT; KLIEVE, 2011). As espécies de
bactérias também podem variar quanto à forma, podendo apresentar-se como cocos, bacilos,
entre outras. As bactérias gram-positivas apresentam uma membrana externa menos
complexa, sendo esta preservada por uma camada glicopeptídica, já as gram-negativas
apresentam dupla membrana externa, as quais são conectadas por uma camada de
glicopeptídeos (KOZLOSKI, 2011). O número exato de espécies de bactérias ruminais ainda
é desconhecido, devido a enorme variedade e a complexidade destes microrganismos. No
entanto, já foram isoladas mais de 400 espécies oriundas do trato digestório de animais
distintos (KRAUSE; RUSSEL, 1996).
As bactérias representam a população mais distinta e numerosa do rúmen, isto se deve
a intensa atividade metabólica, diversificação na ingestão de alimento e capacidade de

16
adaptação ruminal. A classificação das bactérias comumente usada pelos pesquisadores é
fundamentada no tipo de substrato em que os microrganismos agem e nos produtos finais da
fermentação. Desta forma, são denominadas como fermentadoras de carboidratos estruturais
(celulolíticas), fermentadoras de carboidratos não-estruturais, sendo estas as amilolíticas e
pectinolíticas, como também metanogênicas, proteolíticas, lipolíticas e láticas (ARCURI et
al., 2006)
Segundo Kamra (2005), a maioria das bactérias degradadoras de carboidratos
estruturais são gram-positivas. Estas degradam a celulose formando o acetato, disponibilizam
substratos para as demais bactérias, sobretudo as metanogênicas (Gram-positivas), que
utilizam CO2 e H2 para a produção do metano. Já as bactérias amilolíticas (Gram-negativas)
digerem o amido e formam o propionato, o qual é precursor gliconeogênico nos ruminantes.
Também são encontradas outras bactérias responsáveis pela degradação de lipídeos,
principalmente as proteolíticas (Gram-positivas), que geram o nitrogênio em forma de amônia
para as bactérias celulolíticas (ROBSON; STEWART, 1997). Segundo o mesmo autor
também existem as bactérias Gram-variáveis, as quais alteram as características da membrana
de acordo com o ambiente proporcionado. As bactérias fermentadoras de carboidratos
estruturais são hábeis a produzir a enzima extracelular, por meio da hidrólise da celulose. As
celulases, na grande maioria, encontram-se coligadas às células que se aderem à porção
fibrosa do conteúdo ruminal. Esta junção ocorre por meio do glicocálice, que é formado
quando a celulase inicia a degradação da fibra. Posteriormente a formação da matriz de
glicocálise, ocasionado pelo envelopamento celular, as celulases iniciam a digestão da
celulose (CHENG; COSTERTON, 1986). Para o desenvolvimento das bactérias celulolíticas,
os ácidos graxos de cadeia ramificada são fundamentais (DEHORITY, 1987). Os principais
produtos formados pelos organismos celulolíticos são o acetato, succinato, formato, butirato,
H2 e CO2. Também ocorre a liberação de lactato e etanol (HUNGATE, 1966). Entre as
espécies de grande importância tem-se a Butyrivibrio fibrisolvens, Ruminococcus albus,
Ruminococcus flavefaciens e Bacteroides succinogenes (DEHORITY, 1987).
Nas bactérias celulolíticas, a diminuição do pH ruminal (<6,0) minimiza a atividade
destes microrganismos e quando os níveis decrescem mesmo em pequenas quantidades, pode
ocorrer a inibição da digestão da celulose. Estudos comprovam que em baixo pH intracelular,
estes microrganismos cessam a atividade de crescimento, isto ocorre devido a instabilidade no
gradiente de pH, ocasionando uma toxicidade na célula bacteriana pelo acúmulo de ânions
(RUSSEL & WILSON, 1996). A atividade celulolítica também pode ser inibida devido à

17
inclusão de amido na alimentação, o que induz o desenvolvimento de bactérias amilolíticas e
altera a digestão da fibra, em decorrência à competição entre os distintos grupos
(STEENBAKKERS et al., 2008).
Koike et al. (2003) observando a cinética da ligação das bactérias do rúmen à fibra,
constatou que a Fibrobacter succinogenes fixa-se ao feno de forma mais rápida em
comparação as bactérias ruminococos. A aderência da bactéria a partícula do alimento pode
variar aproximadamente 24 horas Brulc et al. (2009) e para Kim et al. (2005) afirmaram que a
partir das 24 horas ocorre a movimentação mecânica das bactérias, isto de forma desuniforme.
Assim pode-se dizer que os distintos microrganismos celulolíticos degradam a fibra em taxas
distintas.
Bactérias que degradam carboidratos de natureza não estrutural são fundamentais na
quebra ruminal do amido, sendo este processo realizado pela enzima amilase. A fermentação
do amido é realizada por espécies do gênero Bacteroides, principalmente a Bacteroides
amylophilus. As bactérias Streptococus bovis e Selenomonas ruminantium produzem através
da fermentação do amido e açúcares solúveis, o acetato. Se as quantidades destes carboidratos
forem elevadas, podem ser sintetizados o acetato, formato, etanol e propionato (RUSSELL,
1988).
Dentre as bactérias proteolíticas ruminais, a maioria tem capacidade de degradar a
proteína. Bacteroides amylophilus e Bacteroides ruminicola utilizam os aminoácidos como
principal fonte de energia. Quando degradada a proteína proveniente da dieta, por parte dos
microrganismos, ocorre a formação de ácidos graxos e amônia. Se houver elevada degradação
de proteína, o hospedeiro reduz a retenção de N (TEIXEIRA, 1991).
A partir da década de 80, foram identificadas três espécies de bactérias fermentadoras
de aminoácidos (Clostridium aminophilum, C. stiklandii e Peptostreptococcus sp.), estas não
empregam os carboidratos como principal fonte de energia e caracterizam-se por desanimar
aminoácidos em proporções superiores as demais bactérias, cerca de 20 vezes a mais (LIMA
et al., 2009). Este grupo torna-se importante em virtude da proteína dietética fornecer amônia
e Ácidos Graxos de Cadeia Curta, quando degradada pelos organismos de rúmen. Dessa
forma, se o nível de desaminação sobressair o nível de aproveitamento da amônia para a
síntese microbiana pode acarretar uma inversão e diminuição na conversão alimentar
(TEDESCHI et al., 2000).
Outro grupo de microrganismos bastante relevante para o ambiente ruminal são as
archeas metanogênicas, as quais auxiliam na manutenção e estabilização das atividades do

18
rúmen, sendo representadas por uma pequena parcela da biomassa ruminal (JANSSEN &
KIRS, 2008). Quando ocorre uma elevada produção de H2 no rúmen, as metanogênicas,
principalmente o gênero Methanobacterium, utilizam este composto para minimizar o CO2 e
sintetizar o CH4, contribuem desta forma, para a reciclagem do NAD oxidado e auxiliam na
continuidade nos processos fermentativos (KOZLOSKI, 2011).
Para que a produtividade e a saúde dos ruminantes sejam mantidas, as bactérias
ruminais são indispensáveis (WELKIE et al., 2010). As alterações no ambiente ruminal e a
simbiose entre as populações microbianas ainda são pouco conhecidas, principalmente para
bovinos alimentados exclusivamente a pasto. Desta forma, o conhecimento das espécies
presentes, bem como a característica de cada, torna-se essencial para garantir resultados
satisfatórios aos rebanhos. A dieta compreende um dos principais fatores que alteram a
microbiota ruminal, assim o estudo a cerca deste assunto, pode facilitar o entendimento da
interação entre os organismos presentes, e a interação destes com a dieta fornecida
(VALADARES FILHO; PINA, 2011).
2.3 Protozoários ruminais
Os protozoários ciliados do rúmen desempenham diversas funções interligadas, sendo
estas, bioquímicas e fisiológicas de grande importância para os ruminantes, sobretudo no
metabolismo de nutrientes (COALHO et al., 2003). São seres unicelulares, anaeróbios, não
patogênicos e variam de 20 a 200 μm, sendo de 10 a 100 vezes maiores que as bactérias
(DEHORITY, 1993). Segundo Teixeira (1991), a população desses microrganismos pode
variar de 100.000 a 1.000.000 células/mL de conteúdo ruminal. Calcula-se que a constituição
dos protozoários representa aproximadamente 2% dos produtos finais da fermentação
(YOKOYAMA; JOHNSON, 1988).
Gruby & Delafond (1843) realizaram os primeiros estudos relacionados aos
protozoários ciliados encontrados no ambiente ruminal, quando estes foram considerados
importantes para os animais ruminantes em relação ao seu metabolismo e nutrição. Colin
(1954) citou a existência dos protozoários através de figuras, as quais representavam as
diferentes espécies existentes. Stein (1858, 1859) caracterizou algumas espécies de ciliados no
compartimento estomacal de carneiros e bovinos, as quais foram designadas como Isotricha
intestinalis, Ophryoscolex purkynjei, Ophryoscolex inermis, Entodinium bursa, Entodinium
dentatum e Entodinium caudatum. Posteriormente, em estudos mais detalhados sobre as

19
espécies de protozoários, o mesmo autor, em estudo sobre Isotricha, relatou e apresentou mais
uma espécie, sendoclassificada como Isotricha prostoma.
No decorrer do tempo, novos estudos sobre a microbiologia do rúmen foram se
tornando evidentes, principalmente em relação aos protozoários. Outros autores, como
Schuberg (1888), passaram a estudar detalhadamente as espécies já identificadas e em seguida
instituiu-se novos gêneros Buetschlia e Dasytricha, e desmembramento do gênero Entodinium
em Entodinium e Diplodinium.
A classificação dos protozoários ciliados do ambiente ruminal (Alveolata, Ciliophora)
é apresentada como filo Ciliophora, e está presente na classe Litostomatea, subclasse
Trichostomatia e divididos em duas ordens: Vestibuliferida Puytorac et al., 1974 e
Entodiniomorphida Reichenow in Doflein and Reichenow, 1929.
Segundo Lynn (2008), a ordem Vestibuliferida inclui cinco famílias de protozoários
ciliados e nos ruminantes é caracterizada pela família Isotrichidae, aqual abrange os gêneros:
Dasytricha, que apresenta características morfológicas como cílios, cobrindo o corpo em
cinécias oblíquas e também fissura do vestíbulo localizado na região posterior do corpo, com
comprimento de 60-100 µm e 38-53 µm de largura. Isotricha, com corpo coberto por cílios
dispostos em cinécias longitudinais e fissura do vestíbulo situado em arranjos distintos,
dependendo da espécie. Oligoisotricha, desprovido de cílios somáticos na parte posterior do
corpo, comprimento de 105-207 µm e largura de 73-128 µm.
Os isotriquídeos apresentam maior propensão por alimentos solúveis, no entanto, não
possuem grande resistência nas condições químicas do ambiente ruminal. Exibem como
característica marcante a capacidade de se fixar à parede do retículo e imigrar ao rúmen
posteriormente a alimentação, isso se deve supostamente devido à ocorrência de açúcares
solúveis. Apresentam películas mais flexíveis e são cobertos de cílios em torno de todo o
corpo (DEHORITY; TIRABASSO, 2000).
Após alguns anos de pesquisa, os ciliados que continham cutícila firme e cílios
localizados na região oral, foram congregados em Entodinomorphida. Os entodinomorfos são
bastante adaptados ao compartimento ruminal e se aderem com facilidade às partículas do
alimento (DEHORITY; TIRABASSO, 2000).
A ordem Entodiniomorphida abrange três subordens, sendo as duas primeiras,
Archistomatina e Blepharocorythina, caracterizadas pelas famílias Buetschliidae e
Blepharocorythidae, que compreendem os gêneros Buetschlia e Charonina. O gênero
Buetschlia, apresenta o corpo em formato oval com duas regiões contendo cílios, na parte

20
anterior e posterior, sendo a anterior composta por uma coroa ciliar. Possui um macronúcleo
elíptico com um vacúolo contrátil (região posterior) e vacúolo consolidado (região posterior),
o comprimento varia de 30-35 µm e largura 20-30 µm. Já o gênero Charonina possui ciliatura
restrita às regiões anterior e posterior do corpo, o vestíbulo é bastante acentuado, o
comprimento pode variar entre 28-46 µm e a largura 9-15 µm (LYNN, 2008).
A caracterização da terceira subordem Entodiniomorphina inclui a família
Ophryoscolecidae. Os componentes desta família são comumente encontrados em ruminantes,
são dotados por regiões com ciliatura retrátil e contém macronúcleo longo. Incluem-se os
gêneros: Entodinium, Eodinium, Diplodinium, Diploplaston, Eremoplastron, Eudiplodinium,
Ostracodinium, Metadinium, Enoploplastron, Elytroplastron, Polyplastron, Epidinium e
Ophryoscolex (LYNN, 2008).
Gênero Entodinium: apresentam uma única região ciliar retrátil, a qual envolve a
fissura do vestíbulo. O comprimento varia de 22-115 µm e a largura 14-90 µm.
Gênero Diplodinium: duas regiões ciliares retráteis dispostas no mesmo plano
corporal. Não possuem placas esqueléticas e o macronúcleo apresenta-se com o terço anterior
inclinado a região ventral. Os vacúolos são contráteis, o comprimento varia de 30-120 µm e a
largura 20-80 µm.
Gênero Eodinium: as regiões ciliares retráteis estão situadas em um mesmo plano
corporal. Não possuem placas esqueléticas e o macronúcleo apresenta-se em formato de
bastão localizado na região dorsal. Os vacúolos são contráteis, o comprimento varia de 40-70
µm e largura 20-40 µm.
Gênero Eremoplastron: as regiões ciliares retráteis estão situadas em um mesmo plano
corporal. Possuem somente uma única placa esquelética estreita e o macronúcleo apresenta-se
em formato de bastão ligeiramente inclinado a região ventral. Os dois vacúolos são contráteis,
os comprimento dos pertencentes a este gênero varia de 35-110 µm e a largura 25-70 µm.
Gênero Eudiplodinium: as regiões ciliares retráteis estão situadas em um mesmo plano
corporal. Possuem uma única placa esquelética alongada, sendo esta alargada na parte anterior
situada próxima ao macronúcleo. O macronúcleo apresenta-se em forma de gancho e os
vacúolos são contráteis. As medidas de comprimento variam de 120-200 µm e a largura 80-50
µm.
Gênero Diploplastron: as duas regiões com presença de cílios retráteis estão situadas
no mesmo plano corporal. As placas esqueléticas são delgadas, sendo o macronúcleo em

21
formato de bastão. Os vacúolos são contráteis, com comprimento de 90-128 µm e largura de
65-87 µm.
Gênero Polyplastron: apresenta duas regiões ciliares retráteis, situadas no mesmo
plano corporal. Possui cinco placas esqueléticas, duas situadas na superfície esquerda e outras
três na superfície direita e cinco vacúolos contráteis. O comprimento varia de 122-210 µm e a
largura 97-130 µm.
Gênero Ostracodinium: exibe duas regiões ciliares situadas no mesmo plano corporal.
Possui somente uma placa esquelética na superfície esquerda. O macronúcleo apresenta-se
lobado, os vacúolos são contráteis que se diferenciam de acordo com a espécie. O
comprimento varia de 50-128 µm e a largura de 36-70 µm.
Gênero Elytroplastron: exibe duas regiões ciliares situadas no mesmo plano corporal.
Possui quatro placas esqueléticas, duas situadas na superfície esquerda e outras duas na
superfície direita e quatro vacúolos contráteis. O macronúcleo se apresenta na forma de
bastão, o comprimento varia de 100-160 µm e a largura 76-100 µm.
Gênero Metadinium: exibe duas regiões ciliares situadas no mesmo plano corporal.
Possui duas placas esqueléticas largas unidas ou não na região posterior. Macronúcleo se
dispõe no formato de ‘E’ ou ‘F’ e os dois vacúolos são contráteis. As medidas de
comprimento variam de 110-210 µm e a largura 80-140 µm.
Gênero Enoploplastron: exibe duas regiões ciliares situadas no mesmo plano corporal.
Apresenta três placas esqueléticas afastadas ou parcialmente acopladas. O macronúcleo se
apresenta na forma de bastão e dois vacúolos contráteis. O comprimento varia de 80-110 µm e
a largura de 60-78 µm.
Gênero Ophryoscolex: apresenta duas regiões ciliares situadas em planos diferentes no
corpo. Possui três placas esqueléticas e uma coroa de espinhos que se inicia na região
mediana até a área posterior do corpo. Exibe inúmeros vacúolos contráteis, o comprimento
varia de 140-190 µm e a largura 80-110 µm.
Gênero Epidinium: duas regiões ciliares retráteis situadas em planos diferentes no
corpo. Apresenta três placas esqueléticas e podem apresentar espinhos na parte posterior. Os
dois vacúolos são contráteis, o comprimento varia de 80-150 µm e a largura 40-70 µm.
Os protozoários do gênero Entodinium spp. são descritos por alguns autores como
predominantes (~90%) em animais alimentados a base de concentrado (HUNGATE, 1966;
NOGUEIRA FILHO et al.,1992). Já os protozoários pertencentes aos gêneros Diplodinium
spp. e Epidinium spp., são comumente encontrados em dietas com altos teores de fibra, pois

22
apresentam concentrações de celulase a qual auxilia na degradação do material fibroso
(JOUANY, 1996).
Segundo Oliveira et al. (1992), quando animais são acondicionados em sistema de
pastejo, adicionado feno ou cana de açúcar, os gêneros Isotricha spp. e Dasytricha spp, são
encontrados em número significativo, visto que, degradam os polissacarídeos não estruturais e
carboidratos solúveis encontrados nas plantas.
Quando observado a diversidade e estrutura da comunidade de protozoários em
bovinos alimentados com diferentes dietas (feno e silagem/grãos), constatou-se maior
predominância da espécie Entodinium spp. quando os animais eram submetidos a alimentação
a base de silagem e grãos e maior foi o diversidade de espécies quando fornecido feno
(TYMENSEN et al., 2012). Segundo o mesmo autor, outros gêneros, Isotricha, Dasytricha,
Ostracodinium, Diplodinium e Diploplastron foram comuns a ambas as dietas. Já para a dieta
a base de feno, os gêneros Dasytricha, Ostracodinium e Diploplastron predominaram. No
entanto, os gêneros Eudiplodinium e Epidinium, foram observados somente nos animais
alimentados com feno enquanto que Ophryoscolex e Polyplastron foram encontrados somente
na dieta contendo silagem/grãos. Dados semelhantes também foram observados em bovinos
em que a predominância dos gêneros foi Entodinium (~ 67%, predominantemente E.
caudatum), Dasytricha (~ 6%), Isotricha (~ 9%) e Diplodinium (~ 18%) (LENG et al., 2011).
Os protozoários ciliados do rúmen apresentam algumas características e peculiaridades
que os tornam importantes para a microbiota ruminal. Segundo Ruiz (1992), estes
microrganismos tem a capacidade de retenção ruminal maior do que quando contrastados com
as bactérias. Após a ingestão das bactérias pelos protozoários, ocorre à transformação da
proteína microbiana em proteína oriunda de protozoários, e com a morte dos protozoários o
nitrogênio originado passa a ser utilizado pelas bactérias.
Segundo Kozloski (2011), os protozoários realizam o engolfamento das bactérias a fim
de utilizar os aminoácidos e ácidos nucleicos presentes. Dietas contendo maior concentração
de grãos torna este processo mais intenso e simplificado do que comparado com dietas
oriundas de pastagens, pois as bactérias aderem-se a sítios que bloqueiam o engolfamento
realizado pelos protozoários.
Em relação ao substrato fermentativo, alguns protozoários apresentam características
celulolíticas, no entanto os substratos elementares utilizados são o amido e os açúcares, pois
são rapidamente fermentáveis e armazenados em amilopectina ou amido protozoário. A
capacidade tamponante também é uma característica relevante dos protozoários, uma vez que

23
as bactérias não tem disponibilidade de grandes quantidades de substratos rapidamente
fermentáveis (WILLIANS, 1986). O quimiotactismo, habilidade de locomoção em gradientes
de concentração contendo açúcares, também é uma particularidade dos protozoários
(ARCURI et al., 2006). Segundo Dehority e Tirabasso (1989), afirmam que os protozoários
pertencentes à Ordem Vestibuliferida, sobretudo os Isotrichidade, aderem-se a parede do
retículo e desta forma migra até o rúmen, posteriormente a ingestão da dieta. Assim ocorre
uma seleção de protozoários, em que os microrganismos maiores ficam reclusos no
compartimento ruminal, o que porporciona a estes seres sobreviver neste ambiente, pois o
tempo de geração é bastante longo (LENG, 1989).
Nos ruminantes, algumas espécies de protozoários são encontradas nas primeiras seis
semanas de vida do animal, principalmente os do gênero Entodinium, sendo que a colonização
destes microrganismos varia conforme a dieta disponibilizada (ARCURI et al., 2006). A
saliva é uma das principais vias de transmissão de protozoários ciliados nos ruminantes, o
contato direto entre os animais faunados contribui para a migração destes microrganismos até
o compartimento ruminal, como também podem ser encontradosem maior quantidade (74%)
retidos no espaço retículo-rúmen (FOULKES; LENG, 1989). A população de protozoários
apresenta variação em relação ao período do dia, sendo a espécie e número de protozoário,
influenciados pela ingestão de alimento e água, característica física e nutricional da dieta e
produção de saliva (ARCURI et al., 2006).
O estudo sobre os protozoários se torna importante a fim de classificar os principais
gêneros e verificar a influência da alimentação sobre as populações destes microrganismos.
Como também o efeito benéfico que podem proporcionar em termos de desempenho e
eficiência alimentar.

24
Figura 1. Desenhos esquemáticos de protozoários ciliados encontrados em ruminantes domésticos. Ordem
Vestibuliferida: Gêneros A: Isotricha (A’- Isotricha prostoma; A’’- Isotricha intestinalis), B: Dasytricha.
Ordem Entodiniomorphida: Gêneros: C: Charonina; D: Entodinium; E: Diplodinium; F: Eodinium; G:
Eremoplastron; H: Ostracodinium; I: Eudiplodinium; J: Diploplastron; K: Metadinium; L: Enoploplastron; M:
Polyplastron (M’: Lado esquerdo; M’’: Lado direito); N: Epidinium e O: Ophryoscolex. Fonte: D’AGOSTO
(1995).
2.4 Interação microrganismo – hospedeiro
O ruminante apresenta grande habilidade de aproveitar uma gama de alimentos para a
obtenção de nutrientes. Essa capacidade se deve a relação simbiótica existente entre as
populações microbianas que habitam o compartimento ruminal e o hospedeiro. Os
microrganismos e o hospedeiro agem sinergicamente convertendo substâncias indigeríveis,
como amônia, celulose e hemicelulose, em produtos finais do processo fermentativo, ácidos
graxos voláteis, vitaminas, aminoácidos e demais componentes responsáveis pela produção de
carne, lã e leite (KAMRA, 2005).
Os herbívoros não contêm enzimas responsáveis pela digestão das fibras presentes na
dieta. Dessa forma, o compartimento ruminal possui características físico-químicas que
possibilitam condições adequadas para o processo fermentativo do alimento, sendo este
realizado pela população microbiana. Entre os microrganismos colonizadores do rúmen, os
protozoários são caracterizados pelo maior tamanho, assim, representam significativa fração
da microbiota ruminal (MONÇÃO et al., 2013).

25
Os ruminantes destinados à produção diferentemente de outros mamíferos realizam
uma simbiose com os microrganismos ruminais, utilizando os nutrientes disponibilizados nos
compostos não-proteicos e carboidratos estruturais. Desta forma, através do metabolismo das
populações microbianas, as enzimas realizam a degradação destes compostos (VALADARES
FILHO; PINA, 2006).
Os bovinos realizam uma interação associativa com as populações microbianas do
rúmen, sendo estes microrganismos peças fundamentais no processo de degradação da fibra.
Disponibilizam produtos do processo fermentativo e nutrientes para o metabolismo animal. A
microbiota ruminal pode ser alterada devido a uma série de fatores, como a dieta, idade e
saúde do animal. Constata-se então, que em bovinos saudáveis, a dieta é a principal
responsável pela composição estrutural da comunidade microbiana ruminal (WELKIE et al.,
2010).
Segundo Brulc et al. (2009), há evidências de que o microbioma bacteriano,
juntamente com os mecanismos metabólicos ruminais são diferentes em relação a dieta e ao
hospedeiro, pois cada animal absorve distintas quantidades de nutrientes, conforme a
necessidade.
Atualmente, as pesquisas voltadas a nutrição de ruminantes têm como um dos focos
principais os microrganismos do rúmen, os quais apresentam diferentes funções no processo
de digestão, além de ter função protetora no controle de microrganismos patogênicos. A
população microbiana é influenciada pelas características ruminais, sendo estas influenciadas
pelo animal hospedeiro, processo de mastigação e ruminação, produtos oriundos da
fermentação, características químicas da saliva, pH e temperatura (MORGAVI et al., 2013).
Para que os microrganismos do rúmen possam desenvolver suas habilidades
específicas, é necessário fornecer condições adequadas para sua mantença no ambiente, sendo
estes responsáveis pela contribuição para o incremento na eficiência alimentar. Assim, a
interdependência entre o hospedeiro e os microrganismos deve ser estudada para que possam
ser observadas as características e funções de cada população existente no compartimento
ruminal.
2.5 Interação entre microrganismos ruminais
O compartimento ruminal possui características de suma importância para a mantença
das populações microbianas (KOZLOSKI, 2011). Assim, apresenta um processo de

26
degradação constante, que, após a ingestão do alimento pelo hospedeiro, pode exibir
transformações ao longo de sua extensão. No entanto, Rankin et al. (1997) citam que as
informações contidas na literatura, referente aos microrganismos ruminais, na maioria das
vezes exibem dados contraditórios. Essas controvérsias podem ser explicadas pelo fato de que
as metodologias empregadas no estudo das interações entre os microrganismos e as espécies
existentes, são inapropriadas.
Pesquisas voltadas ao estudo da microbiologia do rúmen possibilitam um maior
conhecimento a cerca das interações entre as populações microbianas, e a partir destas
informações, novas metodologias poderão ser utilizadas e desta forma possibilitarão em um
futuro próximo, fornecer incrementos que auxiliarão na degradação da porção fibrosa das
plantas forrageiras, como também permitirão o uso de subprodutos oriundos de agroindústrias
(RUSSEL; MANTOVANI, 2002).
Dehority (2004) descreve que as interações entre as populações de microrganismos
ruminais podem ser de parasitismo, onde um microrganismo é favorecido perante algum dano
causado a outro; mutualismo, quando os dois são beneficiados e comensalismo, quando
somente um microrganismo se beneficia e o outro não sofre efeitos.
A predação, peculiaridade dos protozoários, reduz a biomassa bacteriana encontrada
na forma livre no ambiente ruminal (HSU et al., 1991). Esta atividade intensifica a reciclagem
e a eliminação de N pelo hospedeiro, diminui a passagem da proteína microbiana para o
intestino delgado, pelo decréscimo dos microrganismos bacterianos e acúmulo de
protozoários ruminais. Assim, a predação dos protozoários interfere no número e totalidade
das bactérias ruminais (KARNATI et al., 2009).
Os microrganismos de menor tamanho, principalmente o gênero Entodinium, são
predados por espécies de maior tamanho, como Diplodinium, Polyplastrom e Ostracodinium
(FRANZOLIN; DERORITY, 1999). Devido a essa característica predatória dos protozoários,
autores indicam que esta seria a causa das distintas espécies existentes no ambiente ruminal e
que os protozoários também realizam a predação de fungos ruminais (DERORITY; ORPIN,
1997).
Segundo Finlay et al. (1994), os protozoários realizam a predação das bactérias pelo
fato que estas são a principal fonte de compostos nitrogenados. Estudos realizados com
animais faunados e defaunados, comprovam que o número de bactérias em animais faunados
é menor do que quando comparados com animais defaunados. No entanto, esta atividade
predatória pode variar de acordo com a espécie e a quantidade populacional de bactérias. As

27
bactérias ruminais fermentadoras de metano, utilizam H2 oriundo da degradação de polímeros
de plantas, obtendo energia no processo de redução de CO2 em CH4. Essas bactérias
apresentam-se na forma livre no rúmen ou alocados no citoplasma de protozoários ciliados.
As bactérias fermentadoras de carboidratos estruturais agem sinergicamente com os
fungos, e desta forma proporcionam maior intensificação na atividade de degradação
(GORDON; PHILLIPS, 1998). Os fungos estão relacionados com as bactérias devido à
nutrição cruzada, desta forma, disponibilizam açúcares para seu metabolismo (WILLIAMS,
1994).
As bactérias são indispensáveis aos fungos, pois através da interação entre ambos, as
exigências de vitaminas do complexo B, outros substratos e aminoácidos são supridas
(WILLIAMS, 1994). No entanto, estudos mais recentes evidenciaram que algumas espécies
bacterianas, principalmente as espécies Ruminococcus albus e Ruminococcus flavefaciens,
impedem a digestão da celulose pelos fungos anaeróbios ou cessam o crescimento de algumas
estirpes através de uma proteína solúvel (Kamra, 2005).
Na interação entre os microrganismos, alguns são extremamente dependentes de
outros, principalmente em relação ao suprimento de nutrientes, como também agem de forma
antagonista entre si liberando componentes antimicrobianos (WRIGHT et al., 2004; KAMRA,
2005). As interações e interelações entre as populações de microrganismos ruminais podem
apresentar efeito positivo ou negativo, o qual interfere diretamente no hospedeiro,
principalmente sobre a digestibilidade e aproveitamento dos nutrientes ingeridos. Assim,
equilíbrio entre essas populações deve ser estudado a fim de garantir maior eficiência na
fermentação ruminal (DEHORITY, 1998).
2.6 Efeito da dieta na microbiologia ruminal
O gargalo da nutrição de ruminantes está baseado no aumento da capacidade de
ingestão de alimento com o intuito de manter as exigências nutricionais sem alterar as
condições fisiológicas do compartimento ruminal, isto é, garantir a atividade de ruminação e
adequado consumo de volumoso (VELHO et al., 2007).
Dutta et al. (2009) sugerem que uma dieta que consiga balancear teores proteicos e
energéticos resultará no beneficiamento do crescimento microbiano e desempenho
reprodutivo dos animais. Barrière et al. (2003) afirmam que a forragem limita a taxa de

28
digestão do rúmen pela fração fibrosa, pois possuem altos teores de parede celular, podendo
interferir no desempenho de animais de alta produtividade.
Após a ingestão do alimento, este passa primeiramente pelos pré-estômagos e assim os
nutrientes são disponibilizados para o hospedeiro de forma benéfica, pois são gerados
compostos que serão utilizados pelo animal. O rúmen é o compartimento onde ocorrem com
maior intensidade as reações e processos fisiológicos, desta forma a dieta torna-se a principal
responsável pelos efeitos ocasionados na microbiota ruminal, tendo como consequência a
alteração nos processos digestivos, na produção animal e eficiência alimentar. O estudo dos
microrganismos do rúmen e os processos fermentativos ocorridos neste compartimento são
um dos pontos relevantes na nutrição de ruminantes na atualidade (BERCHIELLI et al.,
2006).
Ao longo do tempo, os ruminantes desenvolveram uma elevada capacidade de
ingestão de alimento, juntamente com uma relação mutualística com os diferentes
microrganismos responsáveis pela digestão da fibra, desta forma, as forrageiras compreendem
uma das principais fontes de nutrientes para os herbívoros. A proteína e a energia são os
componentes principais da ração, pois contribuem para a adequada mastigação, ruminação e
demais condições para o funcionamento do rúmen (CASTAGNARA et al., 2012). Dessa
forma, o estudo das diferentes forrageiras e a interação destas com os microrganismos
ruminais, proporciona um maior entendimento dos processos fisiológicos ocorridos no
ambiente ruminal e a influência destes na produção animal (VAN SOEST, 1994).
Salva a aptidão dos microrganimos ruminais de digerirem carboidratos estruturais,
fatores inerentes às plantas como os constituintes da parede celular, sobretudo as ligações
entre a matriz de hemicelulose e lignina e alguns fatores ligados ao animal, como a
mastigação, salivação e pH ruminal, serão responsáveis por limitar a extensão da digestão no
rúmen, por representarem uma barreira física aos processos fibrolíticos (MARTINS et al.,
2006).
Segundo Van Soest (1994), alguns fatores alteram a microbiota ruminal em virtude da
gramínea utilizada. Dietas contendo gramíneas tropicais podem apresentar deficiência em
nitrogênio e alguns nutrientes. Devido a este fator, a ingestão do alimento tende a ser limitada,
pois a digestão torna-se mais lenta e ocorre uma sensação de saciedade no animal. Baixos
teores proteicos na dieta de ruminantes irão afetar o crescimento microbiano e,
consequentemente, a produção de proteínas microbianas, digestão ruminal disponibilidade de
nitrogênio e energia (REYNAL; BRODERICK, 2005).

29
Os microrganismos ruminais exigem uma determinada quantidade de nitrogênio, o
qual é obtido a partir da energia disponibilizada no rúmen, as bactérias e protozoários
necessitam destes dois componentes para que a proliferação das espécies seja adequada
(LUCCI, 1997). Desta forma, alguns microrganismos não tem acesso a determinados
substratos, como também as quantidades disponíveis tendem a diminuir, como consequência
ocorre o desaparecimento ou morte dos microrganismos e a diminuição na produção animal.
Os carboidratos fibrosos representam uma das principais fontes de energia para o
rúmen. As pastagens tropicais fornecem teores médios ou baixos de proteína, assim a
disponibilidade de N-NH3 no compartimento ruminal pode ser uma condição limitante para o
crescimento das populações microbianas (SILVA et al., 2009).
Van Soest (1987) descreve que os maiores teores proteicos da planta são encontrados
nas folhas. O estadio de desenvolvimento da planta irá influenciar diretamente na taxa de
passagem do alimento, características ruminais como pH e microbiota, pois , conforme Grise
et al. (2001), maior conteúdo celular depositado na planta irá resultar em melhor
digestibilidade da pastagem e maior consumo de matéria seca.
Pesquisas relacionadas à influência da estacionalidade e do tipo de pastagem sobre a
microbiota ruminal são bastante escassas. No entanto a literatura apresenta alguns dados e
estudos relacionados ao fornecimento de forrageiras, principalmente em relação aos
protozoários ciliados do rúmen. Nogueira Filho et al. (1992) ao avaliar o fornecimento de
forragem em distintos estágios vegetativos sobre a população de ciliados do rúmen de
bovinos, observaram que os gêneros decrescem em virtude da idade da gramínea. Os
principais gêneros encontrados foram Entodinium spp., Diplodinium spp. e Polyplastron spp.
Segundo os autores, a principal causa para o decréscimo dos ciliados seria a redução dos
açúcares solúveis e o aumento do material fibroso da planta, devido à maturação.
Para Silva et al. (2014) existe uma variação das populações de protozoários em relação
a estação do ano. O mesmo autor, ao estudar os ciliados do rúmen de Bos indicus, observou
maior número de microrganismos nos períodos úmidos. Esta variação pode ser explicada
devido à qualidade nutricional das gramíneas, a qual é alterada pelo efeito da estacionalidade.
Segundo Manella & Lourenço (2004), a variação de protozoários decorrente da
estação do ano pode estar relacionado com as características da pastagem. Estas mudanças
ocorrem devido ao tipo de forragem utilizada, o ciclo fisiológico, a quantidade de matéria
seca e os nutrientes disponibilizados pela planta (ARAÚJO FILHO et al., 2002a).

30
DEHORITY & ODENYO (2003) ao confrontar ruminantes africanos, sendo estes
selvagens e domésticos, verificaram maior número de protozoários em animais oriundos de
um sistema exclusivamente a pasto, do que quando comparado a animais submetidos à
inclusão de níveis de concentrado. Segundo estes autores, o gênero Ophryoscolecidae ocorre
com maior frequência em animais quando a dieta se restringe somente a forrageiras.
O rúmen pode sofrer alterações resultantes da microbiota ruminal, sobretudo em
relação à predação entre os protozoários ciliados e as bactérias. Pois a partir dessa relação
simbiótica, as bactérias tornam-se fonte de ácidos nucleicos e aminoácidos (EADIE, 1967).
Segundo Kozloski (2011), quando os animais recebem como dieta principal as forrageiras, as
bactérias aderem-se ao material fibroso a partir de sítios, o que dificulta o processo de
engolfamento realizado pelos protozoários.
A existência dos protozoários ciliados contribui para o aumento da digestão da
celulose e hemicelulose (FONDEVILA; DEHORITY, 2000). Os ciliados da ordem
Entodiniomorphida, sobretudo das subfamílias Diplodiniinae e Ophyoscolecinae apresentam
maior propensão a dietas contendo elevada quantidade de fibra. São microrganismos menos
resistentes a reduções do pH, quando fornecido dietas contendo níveis elevados de
concentrado, assim são encontrados de forma mais numerosa em alimentos fibrosos
(WILLIAMS; COLEMAN, 1992).
Para Hungate (1966), a família Isotrichidae ocorre com amor frequência em animais
sob pastejo ou recebendo feno, pois apresentam maior capacidade de digestão de
polissacarídeos não estruturais e os carboidratos solúveis da forrageira. Já a família
Ophryoscolecidae é encontrada de forma menos expressiva em dietas contendo somente
grãos. Martinele et al. (2008a) observaram que o gênero Entodinium não apresenta variação
em decorrência da inclusão de concentrado sob dietas a base de capim-elefante, nas
proporções de 20 e 40% em vacas mestiças holandês-zebu e quando fornecido a mesma
quantidade de concentrado para os animais, o número de ciliados passa a sofrer alterações.
Alguns fatores explicam esta alteração, como o período em que o alimento fica retido, valor
de pH e processos fisiológicos de cada hospedeiro (MICHALOWSKI, 1977).
Segundo Dehority (2003) o rúmen apresenta pH entre 5,5 e 7,0 em condições normais,
e o menor valor ocorre duas a seis horas após a alimentação, em virtude da maior produção de
ácidos decorrente da fermentação do alimento, o que tem como consequência uma diminuição
da população de protozoários. Oliveira et al. (1987) ao estudar a concentração de protozoários
ciliados em ovinos submetidos ao pastejo, verificaram que o pH tende a se manter mais

31
estável, 6,9. Quando fornecido dietas com elevada quantidade de celulose e carboidratos
solúveis, o pH não tem uma diminuição exacerbada, isso se deve ao desenvolvimento de
bactérias celulolíticas, pois sua atividade é comprometida em pH 6,0 a 6,1 e inibida abaixo de
5,9 (RUSSEL; DOMBROWSKI, 1980).
A nutrição representa uma importante ferramenta para manipular os microrganismos e
otimizar o ecossistema ruminal, a fim de garantir a produção de carne de qualidade para
atender as demandas de uma população humana crescente. Os conhecimentos gerados pelo
estudo da microbiota ruminal, podem gerar ganhos a médio e longo prazo a partir da
integração dos dados taxonômicos e funcionais das populações microbianas e a relação destes
com a digestão do rúmen, desempenho e metabolismo do hospedeiro (MORGAVI et al.,
2013).
2.7 Técnicas de identificação da microbiota ruminal
O rúmen abriga uma grande variedade de microrganismos, o que torna os estudos de
identificação, caracterização e quantificação bastante complexa. Segundo Shin et al. (2004),
acredita-se que somente 5-10% da microbiota ruminal é conceituada, sendo que existem
aproximadamente 1000 espécies de fungos, protozoários e bactérias.
Ao longo dos anos várias metodologias têm sido empregadas na busca de observar a
microbiota ruminal, desde técnicas tradicionais, como roll-tube technique (HUNGATE, 1969)
ou a utilização da técnica dos números mais prováveis (DEHORITY; TIRABASSO; GRIFO,
1989). Contudo, algumas populações microbianas não podem ser identificadas devido ao
cultivo destas ser in vitro.
Experimentação in vivo apresenta uma série de obstáculos para estimar a síntese
microbiana no rúmen (REYNAL et al., 2003) .Segundo Fernando et al. (2010), os
microrganismos já identificados representam somente uma parte da população microbiana,
sendo este valor muitas vezes subestimado. Em relação à determinação e quantificação de
protozoários e bactérias, várias técnicas são utilizadas, como contagens através de câmaras, no
caso dos protozoários, e metodologias baseadas na utilização de observações feitas em
microscópios. Procedimentos e técnicas baseadas em Dehority (1984) e Ogimoto & Imai
(1981). Para a contagem da massa microbiana, diversas metodologias são empregadas, como
as distintas frações existentes e determinação de bactérias gram-negativas e gram-positivas
(MARTIN et al.,1994).

32
Atualmente, o estudo dos microrganismos do rúmen, sobretudo das bactérias, está
cada vez mais evidente, principalmente em relação à biologia molecular, sendo que através
desta técnica, é possível quantificar e identificar os microrganismos sob qualquer forma de
cultivo. Há pouco tempo, o conhecimento pertinente aos microrganismos ruminais, era
oriundo de pesquisas de cultivo, em que se realizava técnicas de isolamento, enumeração
celular e também a tipificação nutricional. No entanto, o acesso à diversidade de
microrganismos era bastante limitado. Posteriormente surgiram pesquisas complementares
com a utilização de técnicas moleculares, as quais fornecem resultados mais acurados e maior
rapidez nas análises (MCSWEENEY; MACKIE, 2012).
A caracterização da população de microrganismos procariotos baseia-se na utilização
do sequenciamento do gene 16S rRNA, através deste é possível estimar a diversidade e
classificação filogenética, a fim de identificar e quantificar organismos não cultivados
(MAKKAR; CAMEOTRA, 2002; KANG et al., 2009).
A técnica da Reação em Cadeia pela Polimerase (PCR) caracteriza-se por permitir a
multiplicação de determinados trechos do DNA com a utilização do primer de PCR, DNA
polimerase termoestável (enzima Taq polimerase), posteriormente quando se tem sua máxima
concentração em solução específica, torna-se possível sua aplicabilidade em métodos de
isolamento e identificação de substâncias. Essa técnica pode ser obtida por análise tradicional
ou em Tempo Real, sendo que em Tempo Real é possível comparar as amostras e realizar a
mensuração gênica, diferentemente da técnica tradicional em que somente se detecta a
amplificação do trecho de PCR no final da reação (MCSWEENEY et al., 2006)
Outro procedimento aplicável na microbiologia ruminal é a qPCR, um procedimento
simples e que fornece resultados confiáveis. Essa técnica permite o monitoramento contínuo
da síntese dos produtos de PCR, e possibilita a quantificação das bactérias alvo com maior
acurácia, como também tem sido utilizada na quantificação de protozoários (SKILLMAN et
al., 2006), fungos (BOOTS et al., 2012) e bactérias (WANAPAT; CHERDTHONG, 2009) do
rúmen.
Quimiostatos também tem sido utilizado no estudo da eficiência microbiana, os dados
obtidos por essa técnica apresentam alta precisão e confiabilidade, quando aplicado em meios
de cultura, sendo que as taxas de crescimento microbiano estão relacionadas às taxas de
diluição aplicadas (ARCURI et al., 2006).
Conforme Krause et al. (2013), as pesquisas em microbiologia ruminal baseadas em
técnicas moleculares, como a metagenômica, permitem a formulação de probióticos

33
específicos, a inserção de microrganismos no rúmen, otimização da fermentação ruminal e o
desenvolvimento de vacinas para as bactérias produtoras de metano. Estes estudos juntamente
com o avanço das técnicas de pirosequnciamento podem esclarecer as interações entre as
populações microbianas com o hospedeiro, a fim de otimizar a eficiência nutricional e a saúde
dos animais.
As pesquisas voltadas a microbiota ruminal são se suma importância para a nutrição
animal. Visto que, o desenvolvimento da pecuária está intimamente ligado ao conhecimento
sobre os diversos microrganismos que compreendem o compartimento ruminal, bem como os
parâmetros e fatores que afetam o sistema (RUSSELL, 2002).
4. REFERÊNCIAS BIBLIOGRÁFICAS
ARAÚJO-FILHO, J.A.; F.C. CARVALHO; N.L. SILVA. Fenologia y valor nutritivo de
follajes de algunas especies forrajeras de la Caatinga. Agroforestía en las Américas, Costa
Rica, 9: 33-37. 2002.
ARCURI, P. B.; LOPES, F. C. F.; CARNEIRO, J. C. Microbiologia do rúmen. In:
BERCHIELLI, T. T.; PIRES, A. V.; OLIVEIRA, S. G. de. Nutrição de Ruminantes,
Jaboticabal: Funep, p. 111-116, 2006.
BARRIÈRE, Y.; C. GUILLET; GOFFNER, D. e PICHON, M. Genetic variation and
breeding strategies for improved cell wall digestibility in annual forage crops. Anim. Res.
52:193-228, 2003.
BERCHIELLI, T.T. et al. Nutrição de ruminantes. Jaboticabal: Funep, 2006. 583p.
BOOTS, B. et al. Responses of anaerobic rumen fungal diversity (phylum
Neocallimastigomycota) to changes in bovine diet. Journal of Applied Microbiology, v.
114, p. 626-635, 2012.
CASTAGNARA, D. D. et al. Effect of boron and zinc fertilization on white oats grown in soil
with average content of these nutrients. Revista Brasileira de Zootecnia, Viçosa, v. 41, n. 7,
p. 1598 - 1607, 2012.
CHAVES, A. H. et al. Seleção de isolados de Lactobacillus acidophilus usados como
probiótico em bezerros. Revista Brasileira de Zootecnia, 28:1093, 1999.
CHENG, K.H.; COSTERTON. J.W. Microbial adhesion and colonization within the digestive
tract. In: BARNES, E.M.; MEAD, G.C.Anaerobic bacteria in habitats other than man.
LONDON: BlackwELL Sci. PuBL., p. 239-262. 1986.

34
Choudhury, P.K. et al. Harnessing the diversity of rumen microbes using molecular
approaches. In: Sirohi SK, Walli TK, Singh B, Singh N (eds) Livestock Green House Gases:
emis-sion and options for mitigation. Satish Serial Publishing House, Delhi, Section-B,
Chapter-6, pp 65–82. ISBN 978-93-81226-52-065, 2012.
COALHO, M. R.; NOGUEIRA FILHO, J. C. M.; CUNHA, J. A.; LIMA, C. G De. Estudo
dos protozoários ciliados em bovinos consumindo dietas com diferentes níveis de proteína
não degradável no rúmen. Acta Scientiarum. Animal Sciences, Maringá, v. 25, n. 1, p. 193-
199, 2003.
COELHO DA SILVA, J.F.; LEÃO, M.I. Fundamentos de nutrição dos ruminantes.
Piracicaba: Livroceres, 380p. 1979.
COLEMAN, GS; HALL, FJ. Digestion ofEpidinium ecaudatum caudatum by the rumen
ciliate Polyplastron multivesiculatum as shown by studies in the electron microscope. Soc
Gen Microbiol Q 6:29–30, 1978.
COLIN, 1954. In: Cellular domains. NABI, I. R. Wiley-Blackwell, 2011.
DEHORITY, B. A. Microbial interactions in the rumen. Revista de La Facultad de
Agronomía Luz, v. 15, p. 69-86, 1998.
DEHORITY, B. A. Rúmen microbiology. Wooster: ohio state university, 239p. 1987.
DEHORITY, B. A.; ORPIN, G.C. Development of, and natural fluctuations in, rumen
microbial populations. The Rumen Microbial Ecosystem. P. N. Hobson and C. S. Stewart.
London, Blackie Academic, v. 2, p.196-245, 1997.
DEHORITY, B. A.; Rumen microbiology. Nottingham: Nottingham University Press, 2nd
ed. 2004. 372 p.
DEHORITY, B. A.; TIRABASSO, P. A. Antibiosis between ruminal bacteria and ruminal
fungi. Applied and Environmental Microbiology, v. 66, N. 7, p. 2921–2927, July, 2000.
DEHORITY, B. A.; TIRABASSO, P. A.; GRIFO, A. P. Most-probable-number procedures
for enumerating ruminal bacteria, including the simultaneous estimation of total and
cellulolytic numbers in one medium. Applied Environement Microbiology, Washington, v.
55, n. 11, p. 2789–2792, 1989.
DEHORITY, B.A.; ODENYO, A.A. ‘Influence of diet on the rumen protozoal fauna of
indigenous African wild ruminants’, Journal of Eukaryotic Microbiology 50, 220–223,
2003.
DEHORITY, B.A. 1993. Laboratory manual for classification and morphology of rumen
ciliate protozoa. Florida: CRC Press Inc. 96p.
DEHORITY, B.A. Evaluation of subsampling and fixationproceduresused for counting rumen
protozoa. Applied and Environmental Microbiology, v.48, p.182-185, 1984.

35
DEHORITY, B.A. Rumen microbiology. Thrumpton, Nottingham: Nottingham University
Press, 372p., 2003.
DEHORITY, B.A., TIRABASSO, P.A. 1989. Factors affecting the migration and
sequestration of rumen protozoa in family Isotrichiidae. J. Gen. Microbiol., 135(3): 539-548.
DUTTA T K, AGNIHOTRI M K, SAHOO P K, RAJKUMAR V, DAS A K. Effect of
different protein-energy ratio in pulse by-products and residue based pelleted feeds on growth,
rumen fermentation, carcass and sausage quality in Barbari kids. Small Ruminant Research,
v.85, p.34–41, 2009.
EADIE, M. Studies on the ecology of certain rumen ciliate protozoa. Journal General
Microbiology 49: 175-194, 1967.
FERNANDO, S. C.; PURVIS II, H. T.; NAJAR, F. Z.; SUKHARNIKOV, L. O.;
KREHBIEL, C. R.; NAGARAJA, T. G.; ROE, B. A.; DESILVA, U. Rumen Microbial
Population Dynamics during Adaptation to a High-Grain Diet. Applied and Environmental
Microbiology, Oklahoma, v. 76, n. 22, p. 7482-7490, 2010.
FFOULKES, D.; LENG, R.A. Dynamics of protozoa in the rumen of cattle. British Journal
of Nutrition 59, 429–436. 1989.
FINLAY, B.J.; G.ESTEBAN; K.J.CLARKE & A.G.WILLIAMS. Some rumen ciliates have
endosymbiotic methanogens. FEMS Microbiology Letters, 117: 157-162, 1994.
FONDEVILA M., DEHORITY B.A., In vitro culture of Entodinium exiguum and E.
caudatum, with or without rumen bacteria, Journal Animal Science. 78 Suppl. 1 290, 2000.
FONSECA, A.J.M; DIAS-DA-SILVA, A.A. Efeitos da eliminação dos protozoários do
rúmen no desempenho produtivo de ruminantes-Revisão. Revista Portuguesa de Ciências
Veterinárias, Lisboa, 96 (538): 60-64. 2001.
FRANZOLIN, R.; DEHORITY, B. A. Comparision of protozoal populations and digestion
rates between water buffalo and cattle fed an all forage diet. Journal Applied Animal
Research, 16:33, 1999.
GORDON, G. L. R.; PHILLIPS, M. W. The role of anaerobic gut fungi in ruminants.
Nutrition Research Reviews, v. 11, p. 133-168, 1998.
GRISE, M. M.; et al. Avaliação da composição química e da digestibilidade in vitro da
mistura aveia IAPAR 61 (Avena strigosa Schreb) + ervilha forrageira (Pisum arvense L.) em
diferentes alturas sob pastejo. Revista Brasileira de Zootecnia, v.30, n.3, p.659-665, 2001.
GRUBY; DELAFOND, 1843. In: Cellular domains. NABI, I. R. Wiley-Blackwell, 2011.
HSU, J. T. et al. Effects of defaunation and various nitrogen supplementation regimens on
microbial numbers and activity in the rumen of sheep. Journal of Animal Science, 69:1279,
1991.

36
HUNGATE, R. E. A Roll Tube Method for Cultivation of Strict Anaerobes. In: Methods in
Microbiology, Norris, J.R. and D.W. Ribbons (Ed). New York Academic Press, 1969, p. 117 -
132.
HUNGATE, R. E. The rumen and its microbes. New York: Academic Press, 533 p. 1966.
IVAN, M. et al. Effects of Isotricha, Dasytricha, Entodinium, and total fauna on ruminal
fermentation and duodenal flow in wethers fed different diets. J. Dairy Sci.Selangor, 83,
776–787, 2000.
JANSSEN, P. H.; KIRS, M. Structure of the archaeal community of the rumen. Appl.
Environ. Microbiol. 74, 3619–3625. 10.1128/AEM.02812-07, 2008.
JOUANY, J. P. Effect of the rumen protozoa on nitrogem utilization bu ruminats. J. Nutr.,
Bethesda, v. 126 (suppl.), p. 1335-1346, 1996.
KAMRA, D.N. Rumen microbial ecosystem. Current Science, v.89, n.1, p.124-134, 2005.
KANG, S.; DENMAN, S. E.; McSWEENEY, C. S. The use of molecular tools for the study
of rumen ecology. In: Simpósio internacional avanços em técnicas de pesquisa em nutrição de
ruminantes, 2. Pirassununga. Anais...p.179-194, 2009.
KARNATI, S.K.R.; YU, Z.; FIRKINS, J.L. Investigating unsaturated fat, monensin, or
bromoethanesulfonate in continuous cultures retaining ruminal protozoa. II. Interaction of
treatment and presence of protozoa on prokaryotic communities. Journal of Dairy Science,
submitted, 2009.
KOIKE, S. et al. Kinetics of in sacco fiber-attachment of representative ruminal cellulolytic
bacteria monitored by competitive PCR. J. Dairy Sci. 2003;86:1429–1435.
KOZLOSKI, G.V. Bioquímica dos ruminantes. 3ª Ed. UFSM, Santa Maria , 212 p. 2011.
KRAUSE, D. O.; NAGARAJA, T. G.; WRIGHT A. D. G.; Callaway T. R. Board-invited
review: Rumen microbiology: Leading the way in microbial ecology. Journal of Dairy
Science, v. 91, p. 331-341, 2013.
KRAUSE, D. O.; RUSSEL, J. B. How many ruminal bacteria are there? Journal of Dairy
Science, 79: 1467, 1996.
LANA, R.P. Nutrição e alimentação animal. 1 ed. Viçosa, Universidade Federal de Viçosa.
2005. 343p.
LENG, J. et al. Assessment of protozoa in Yunnan Yellow cattle rumen based on the 18S
rRNA sequences. Mol. Biol. Rep. 38: 577–85, 2011.
LENG, R.A. Dynamics of protozoa in the rumen. In: J.V. Nolan, R.A. Leng e D.I. Demeyer
(Editores), The roles of protozoa and fungi in ruminant digestion. Penambul Books,
Armidale, NSW, pp. 51-58, 1989..

37
LIMA, J. R. et al. Bovicin HC5 inhibits wasteful amino acid degradation by mixed ruminal
bacteria in vitro. FEMS Microbiology Letters, 292:78, 2009.
LOWE, S. E.; THEODOROU, M. K.; TRINCI, A. P. Cellulases and xylanase of an anaerobic
rumen fungus grown on wheat straw, wheat straw holocellulose, cellulose, and xylan.
Applied and Environmental Microbiology, v. 53, n. 6, p. 1216-1223, 1987.
LUCCI, C.S. Nutrição e manejo de bovinos leiteiros. São Paulo: Manole, 1997. 169p.
LYNN, D.H. The ciliated protozoa. Characterization, classification and guide to the literature,
3rd ed. Springer, Dordrecth, 2008.
MAKKAR, R. S.; CAMEOTRA S. S. An update on the use of unconventional substrates for
biosurfactant production and their new applications. Applied Microbiology and
Biotechnology, v. 58, p. 428-34, 2002.
MANELLA, M.Q.; LOURENÇO, A.J. População de protozoários ciliados no rúmen de
bovinos nelore em pastos de Brachiaria brizantha marandu recebendo suplemento protéico ou
com livre acesso a banco de proteína de Leucena leucocephala nas diferentes estações do ano.
Boletim da Indústria Animal, n.1, p.1-11, 2004.
MARTIN, C.; WILLIAMS, A.G.; MICHALET-DOREAU, B. Isolation and Characteristics
of the Protozoal and bacterial fractions from bovine ruminal contents. Journal of Animal
Science, v.72, n.11, p.2962-2968, 1994.
MARTIN, S. A. Influence of Aspergillus oryzae fermentation extract (Amaferm) on ruminal
microorganisms. In: Biozyme tech. Symposium, 1990, Rochester. Proceedings… Rochester,
p. 2-19, 1990.
MARTINELE, I. SIQUEIRA-CASTRO, I. C. V. D´AGOSTO, M. Protozoários ciliados no
rúmen de bovinos alimentados com dietas de capim elefante e com dois níveis de
concentrado. Rev. Bras. Saúde Prod. An., v.9, n.1, p. 74-81, 2008 (a).
MARTINS, A.S. et al. Taxa de passagem e parâmetros ruminais em bovinos suplementados
com enzimas fibrolíticas. Revista Brasileira de Zootecnia, v.35, n.3, p.1186-1193, 2006.
MCALLISTE, K. A. et al. Genetic heterogeneity in hereditary haemorrhagic telangiectasia:
possible correlation with clinical phenotype. J. Med. Genet. 31: 927-932, 1994.
McSWEENEY, C. S.; MACKIE, R. Commission on Genetic Resources for Food and
Agriculture. Micro-organisms and ruminant digestion: State of knowledge, trends and future
prospects. Background Study Paper (FAO), n. 61, p. 1-62, 2012.
MCSWEENEY, C.S. et al. Application of recent DNA/RNA-based techniques in rumen
ecology. AAAP Conference, Korea. Asian-Australasian Journal of Animal Science, v.20,
p.283-294, 2006.

38
MICHALOWSKI, T. Diurnal changes in concentration or rumen ciliates and in occurrence of
dividing forms in water buffalo (Bubalus bubalus) fed once daily. Applied Environmental
Microbiology, Washington, 33 (4): 802-804. 1977.
MONÇÃO, F. P. P. et al. Desenvolvimento da microbiota ruminal de bezerros: revisão de
literatura. Revista Unimontes Científica, Montes Claros, v. 15, n. 1 - jan. 2013.
MORGAVI D. P. et al. Rumen microbial (meta) genomics and its application to ruminant
production. Animal 7(s1):184–201, 2013.
NOGUEIRA-FILHO et al. Contagens diferenciais de protozoários ciliados em rúmen de
bovinos arraçoados com capim elefante Napier (Penissetum Purpureum Schum), em vários
estádios de crescimento vegetativo. Braz. J. vet. Res. Anim. Sci., São Paulo, v. 27, n 2, p
215-222, 1992.
OGIMOTO, K.; IMAI, S. Atlas of rumen microbiology. Tokyo: Japan ScientificvSocieties,
231p. 1981.
OLIVEIRA, E. R. et al. Degradação ruminal da fibra em detergente neutro de gramíneas do
gênero Cynodon spp. em quatro idades de corte. Agrarian, v. 6, n. 20, p. 205-214, 2013.
OLIVEIRA, J. S.; ZANINI, A. M.; SANTOS, E. M. Fisiologia, manejo e alimentação de
bezerros de corte. Arquivos de Ciências Veterinária e Zootecnia. Unipar, Umuarama, v.10,
n.1, p. 39-48, 2007.
OLIVEIRA, M. E. M., WATANABE, I.; NOGUEIRA FILHO, J. C. M. Ultra estrutura de
membrana celular de Dasytricha ruminantium encontrada no líquido ruminal de bubalinos
Bubalus bubalis da raça Mediterrânea. In: SIMPOSIO TÉCNICO ESP.
ULTRAESTRUTURA DE MENBRANAS, p. 63-64. 1992.
OLIVEIRA, M.E.M.; J.C.M. NOGUEIRA-FILHO; C.S. LUCCI; W. DUPAS & C.G. LIMA.
Desenvolvimento de populações de protozoários ciliados no rúmen de ovinos (Ovis Aires L.)
criados em Itapetininga, São Paulo. Revista da Faculdade de Medicina Veterinária e
Zootecnia da Universidade de São Paulo, São Paulo, 24 (2): 225-232, 1987.
PEDERSEN, K.; TANNOCK, G. W. Colonization of the porcine gastrointestinal tract by
Lactobacilli. Applied and Environmental Microbiology, 55:179, 1989.
REYNAL, S. M., G. A. BRODERICK, S. AHVENJARVI E HUHTANEN, P. Effect of
feeding protein supplements of differing degradability on omasal flow of microbial and
undegraded protein. J. Dairy Sci. 86:1292–1305, 2003.
REYNAL; S. M.; BRODERICK, G. A. Effect of dietary level of rumen-degraded protein on
production and nitrogen metabolism in lactating dairy cows. J. Dair. Scien., v. 88, p. 4045 –
4064, 2005.
ROBSON, P. N.; STEWART, C. S. The rumen microbial ecosystem. ed. 2, p.523 – 632,
1997.
RUIZ, R. L. Microbiologia zootécnica. São Paulo: Ed. Roca, 1992. 314p.

39
RUSSEL, J. B. Rumen microbiology and its role in ruminant nutrition. Ithaca, NY: Cornell
University Press, p. 121, 2002.
RUSSEL, J. B.; MANTOVANI, H. C. The Bacteriocins of ruminal bactéria and their
potencial as na alternative to antibiotics. Journal of Molecular Microbiology and
Biotechnology, 4:347, 2002.
RUSSEL, J. B.; WILSON, D. B. Why are ruminal cellulolytic bacteria unable to digest
cellulose at low pH? Journal of Dairy Science, 79:1503, 1996.
RUSSEL, J.B. Ecology of rumen microorganisms: energy use. In: DOBSON, A .;DOBSON,
M. Aspect of digestive physiology in ruminants. Ithaca: Comstock Publ. Assoc., p. 74 -98.
1988.
RUSSELL, J.B.; DOMBROWSKI, D.B. Effect of pH on the efficiency of growth by pure
cultures of rumen bacteria in continuous culture. Applied Environmental Microbiology,
v.39, p.604-610, 1980.
SCHUBERG, 1888. In: Cellular domains. NABI, I. R. Wiley-Blackwell, 2011.
SHIN, E. C. et al. Phylogenetic analysis of archaea in three fractions of cow rumen based on
the 16S rDNA sequence. Anaerobe, London, v. 10, n. 6, p. 313 - 319, 2004.
SILVA et al. Protozoários ruminais de novilhos de corte criados em pastagem tropical
durante o período seco. Ciência Animal Brasileira. v. 15, n. 3, 2014.
SILVA, F.F. et al. Suplementação a pasto: disponibilidade e qualidade x níveis de
suplementação x desempenho. Revista Brasileira de Zootecnia, v.38, 371-389, 2009.
SKILLMAN, L. C.; TOOVEY, A. F.; WILLIAMS, A. J.; WRIGHT, A. D. Development and
validation of a real-time PCR method to quantify rumen protozoa and examination of
variability between entodinium populations in sheep offered a hay-based diet. Applied and
Environmental Microbiology, v. 72, p. 200206, 2006.
STEENBAKKERS, P. J. et al. A serpin in the cellulosome of the anaerobic fungus Piromyces
sp. Strain E2. Mycological Research, 112:999, 2008.
STEIN, 1858 e 1859. In: Kıbrıs Evcil Keçilerinde Yaşayan İşkembe Siliyatı Ophryoscolex
Purkynjei Stein, 1858 (Ciliophora: Entodiniomorphida)’de Morfometrik Varyasyonlar.
MERMER, A.; GÖÇMEN, B.; ÇİÇEK, K.; ERGEN, G.; SEZGİN, Y.; YILDIZ, M. Z.
Türkiye Parazitoloji Dergisi, 30 (3): 252-257, 2006.
STEWART, C. S.; FLINT, H. J.; BRYANT, M. P. The rumen bacteria. In: HOBSON, P. N.;
STEWART, C. S. (Eds.). The Rumen Microbial Ecosystem. London: Blackie Academic, v. 2,
p. 10:72, 1997.
TEIXEIRA, J. C. Nutrição de ruminantes, Lavras, MG: ESAL/ FAEPE, 1991.

40
TEIXEIRA, J.C. Nutrição de ruminantes. Lavras: UFLA/ FAEPE, 182 p. Curso de Pós
Graduação Latu Sensu (Especialização) à distância: Produção de Ruminantes, 2001.
TYMENSEN, L.; BARKLEY, C.; MCALLISTER, T.A. Relative diversity and community
structure analysis of rumenprotozoa according to T-RFLP and microscopic methods. J.
Microbiol. Meth. 88: 1–6, 2012.
VALADARES FILHO, S.C.; PINA, D.S. Nutrição de ruminantes. Jaboticabal: FUNEP, v.1,
583p. 2006.
VALDARES FILHO, S.C.; PINA, D.S. Fermentação ruminal. In: BERCHIELLI, T. T.;
PIRES, A. V.; OLIVEIRA, S. G. (Ed). Nutrição de Ruminantes. 2 ed. Jaboticabal: Funep,
2011, p. 161-189.
VAN SOEST, P.J. Interactions of feeding behavior and forage composition. In:
INTERNATIONAL CONFERENCE ON GOATS, 4, 1987, Brasília. Proceedings ... Brasília,
1987, p.971-87.
VAN SOEST, P.J. Nutritional Ecology of the Ruminant. Ithaca: Cornell University Press,
476p. 1994.
VELHO, J.P. et al. Composição bromatológica de silagens de milho produzidas com
diferentes densidades de compactação. Revista Brasileira de Zootecnia, v.36, n.5, p.1532-
1538, 2007.
WANAPAT, M.; CHERDTHONG, A. Use of real-time PCR technique in studying rumen
cellulolytic bacteria population as affected by level of roughage in swamp buffalo. Current
Microbiology, v. 58, p. 294299, 2009.
WEIMER, P. J. End product yields from the extraruminal fermentation of various
polysaccharide, protein and nucleic acid components of biofuels feedstocks. Bioresource
Technology, v.102. n.3, p.3254-9, 2011.
WEIMER, P.J.; RUSSELL, J.B.; MUCK, R.E. Lessons from the cow: what the ruminant
animal can teach us about consolidated bioprocessing of cellulosic biomass. Bioresource
Technology, v.100, p.5323-5331, 2009.
WELKIE, D.G.; STEVENSON, D.M.; WEIMER, P.J. analysis of ruminal bacterial
community dynamics in lactating dairy cows during the feeding cycle. Anaerobe, London,
v.16, n.2, p.94–100, 2010.
WILLIAMS, A . G. Rumen holotrich cilliate protozoa. Microbiological Review,
Washington, v.50, p. 25, 1986.
WILLIAMS, A. P. Amino acid requeriments of the veal calf and beef steers. In:
D’MELLO, J. P. F. (Ed.) Amino acids in farm animal nutrition. Wallingford: CAB. 1994. p.
329-350.

41
WILLIAMS, A.G.; G.S. COLEMAN. The Rumen Protozoa. London: Springer-Verlag,
New York Inc, 423 p. 1992.
WRIGHT A-DG, KLIEVE AV. Does the complexity of the rumen microbial ecology
preclude methane mitigation? Anim Feed Sci Technol 166–167:248–253, 2011.
WRIGHT, A.D.G. et al. Reducing methane emissions in sheep by immunization against
rumen methanogens. Vaccine, v.22, p.3976-3985, 2004.
YOKOYAMA, M. T.; JOHSON, K. A. Microbiologia del rumen e intéstino. In: CHURCH,
C.D. El Rumiante: fisiología digestiva e nutrición. Zaragoza: Editorial Acribia S.A., 1988.
p.137-157.

42
3. ARTIGO:
DETERMINAÇÃO E QUANTIFICAÇÃO DE PROTOZOÁRIOS CILIADOS E
BACTÉRIAS DO RÚMEN DE BOVINOS EM PASTAGENS DE INVERNO E VERÃO

43
INTRODUÇÃO
As forragens representam uma das principais fontes de nutrientes para os herbívoros.
Dietas exclusivamente a pasto podem apresentar grande variação na composição nutricional
para o uso microbiano, em virtude da estacionalidade e da gramínea utilizada. O tipo de
alimento, a qualidade nutricional da dieta e interações entre as populações microbianas
representam os principais fatores relacionados à proporção de espécies no ambiente ruminal
(WILIANMS; COLEMAN, 1991). A composição da comunidade microbiana ruminal é
representada por distintas espécies de fungos, bactérias e protozoários e, através das funções
físico-químicas realizadas por estes microrganismos, sintetizam enzimas degradadoras de
carboidratos estruturais, fornecendo desta forma, energia ao animal hospedeiro (VIDAL et al.,
2007).
A alimentação animal a base de forragem promove condições favoráveis ao
desenvolvimento do rúmen, como também das estruturas internas deste compartimento. A
ingestão de alimentos fibrosos proporciona o surgimento de uma diversidade de espécies
microbianas, sendo estas responsáveis pela formação dos produtos finais da fermentação,
ácidos graxos voláteis, pH do rúmen adequado, condições que promovem intensa atividade
metabólica no ambiente ruminal, sendo estas condições adequadas para o estabelecimento de
protozoários ciliados e bactérias celulolíticas (OLIVEIRA et al., 2007).
Os carboidratos fibrosos representam uma das principais fontes de energia para o
rúmen, contudo, alguns fatores alteram a microbiota ruminal em virtude da gramínea
utilizada. As pastagens tropicais fornecem teores médios ou baixos de proteína, assim a
disponibilidade de N-NH3 no compartimento ruminal é uma condição limitante para o
crescimento das populações microbianas (SILVA et al., 2009). Devido a este fator, a ingestão
do alimento tende a ser limitada, pois a digestão é mais lenta e ocorre sensação de saciedade
ao animal. Baixos teores proteicos e energéticos na dieta de ruminantes podem afetar o
crescimento microbiano e, consequentemente, a produção de proteínas microbianas, digestão
ruminal, disponibilidade de nitrogênio e energia. Já as gramíneas de inverno normalmente
apresentam alta digestibilidade e elevado teor de N degradável, o que contribui para o
aumento da comunidade e diversificação da microbiota ruminal.
Pesquisas relacionadas à microbiologia do rúmen, principalmente voltadas aos
protozoários e bactérias em animais mantidos exclusivamente a pasto, possibilitam um maior
conhecimento das interações entre as populações microbianas e a interação destes com a dieta

44
fornecida. A partir destas informações, novas metodologias poderão ser utilizadas a fim de
auxiliar na compreensão da degradação da porção fibrosa das plantas forrageiras, como
também permitirão o uso de subprodutos oriundos de agroindústrias (RUSSEL;
MANTOVANI, 2002).
Os protozoários ciliados e bactérias ruminais são influenciados por diversos fatores,
como a estacionalidade, características nutricionais e morfológicas da pastagem, concentração
de açúcares e teor de fibra. A complexidade das comunidades microbianas ainda é pouco
conhecida, o que dificulta o avanço de pesquisas que visam otimizar o funcionamento
ruminal. Portanto, a compreensão deste ambiente possibilitará o conhecimento a cerca dos
alimentos fornecidos aos animais, como também a influência destes nos processo
fermentativo (JAMI; MIZRAHI, 2012).
O objetivo foi identificar e quantificar os protozoários e bactérias do rúmen de 40
bovinos de corte submetidos à alimentação à pasto Azevém, Aveia+Azevém, Brachiaria e
Brachiaria+Estrela Africana.
MATERIAL E MÉTODOS
Coleta das amostras ruminais e período experimental
As amostras foram coletadas do conteúdo ruminal de bovinos cruzados (Nelore x
Europeu), com média de 2,5 anos, abatidos em frigoríficos comerciais, oriundos de diferentes
propriedades do sudoeste do Paraná, no período de setembro de 2015 a julho de 2016. Foram
coletadas amostras de 10 diferentes animais de cada pastagem, totalizando 40 amostras. Os
animais foram submetidos a 12 horas de jejum alimentar anteriormente ao abate. Após a
chegada do rúmen na área suja do frigorífico, procedeu-se a lavagem do mesmo para evitar a
contaminação da amostra. Em seguida foi realizada a abertura deste compartimento com o
auxílio de uma faca esterilizada. As amostras foram coletadas da massa central do conteúdo
ruminal, homogeneizadas e acondicionadas em potes, sendo destinadas a determinação de
protozoários e bactérias (Anexos C e D). As coletas foram realizadas nas dependências dos
frigoríficos Miolar Alimentos Ltda, situado no Município de Dois Vizinhos e frigorífico
Orfimar, localizado em Itapejara do O’este, sob o protocolo nº 2015-24 da Comissão de
Ética do Uso de Animais (CEUA).
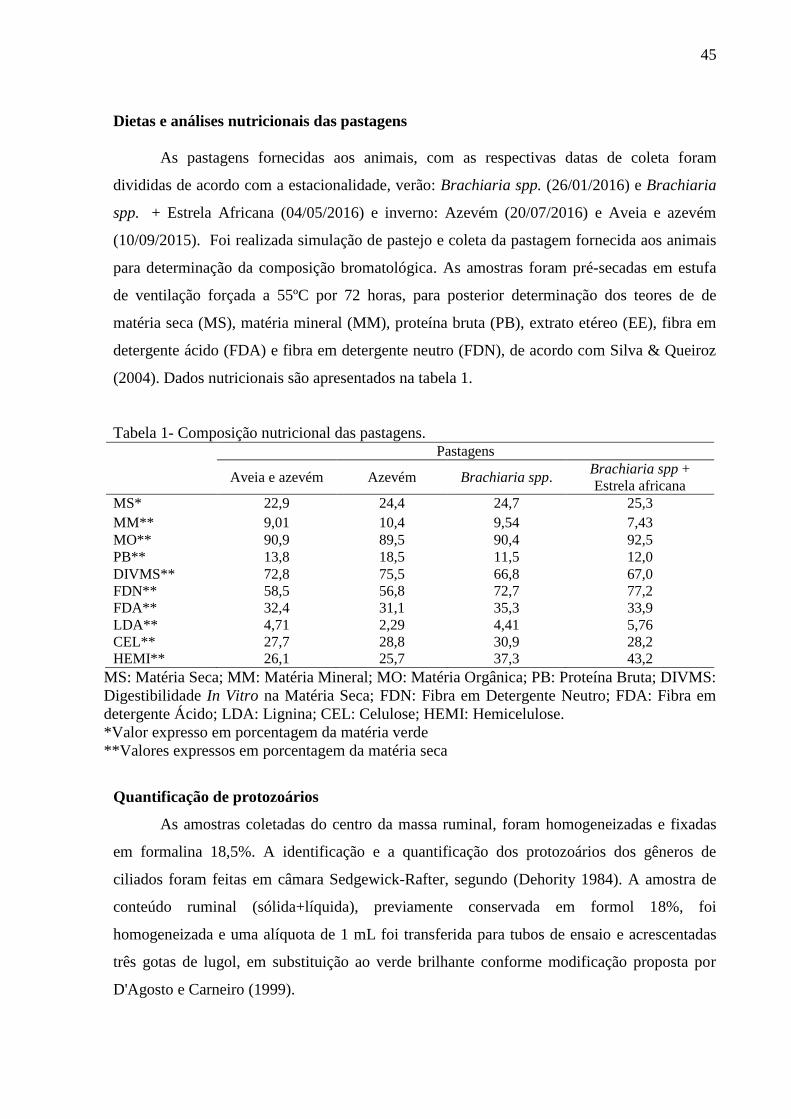
45
Dietas e análises nutricionais das pastagens
As pastagens fornecidas aos animais, com as respectivas datas de coleta foram
divididas de acordo com a estacionalidade, verão: Brachiaria spp. (26/01/2016) e Brachiaria
spp. + Estrela Africana (04/05/2016) e inverno: Azevém (20/07/2016) e Aveia e azevém
(10/09/2015). Foi realizada simulação de pastejo e coleta da pastagem fornecida aos animais
para determinação da composição bromatológica. As amostras foram pré-secadas em estufa
de ventilação forçada a 55ºC por 72 horas, para posterior determinação dos teores de de
matéria seca (MS), matéria mineral (MM), proteína bruta (PB), extrato etéreo (EE), fibra em
detergente ácido (FDA) e fibra em detergente neutro (FDN), de acordo com Silva & Queiroz
(2004). Dados nutricionais são apresentados na tabela 1.
Tabela 1- Composição nutricional das pastagens. Pastagens
Aveia e azevém Azevém Brachiaria spp.
Brachiaria spp +
Estrela africana
MS* 22,9 24,4 24,7 25,3
MM** 9,01 10,4 9,54 7,43
MO** 90,9 89,5 90,4 92,5
PB** 13,8 18,5 11,5 12,0
DIVMS** 72,8 75,5 66,8 67,0
FDN** 58,5 56,8 72,7 77,2
FDA** 32,4 31,1 35,3 33,9
LDA** 4,71 2,29 4,41 5,76
CEL** 27,7 28,8 30,9 28,2
HEMI** 26,1 25,7 37,3 43,2
MS: Matéria Seca; MM: Matéria Mineral; MO: Matéria Orgânica; PB: Proteína Bruta; DIVMS:
Digestibilidade In Vitro na Matéria Seca; FDN: Fibra em Detergente Neutro; FDA: Fibra em
detergente Ácido; LDA: Lignina; CEL: Celulose; HEMI: Hemicelulose.
*Valor expresso em porcentagem da matéria verde
**Valores expressos em porcentagem da matéria seca
Quantificação de protozoários
As amostras coletadas do centro da massa ruminal, foram homogeneizadas e fixadas
em formalina 18,5%. A identificação e a quantificação dos protozoários dos gêneros de
ciliados foram feitas em câmara Sedgewick-Rafter, segundo (Dehority 1984). A amostra de
conteúdo ruminal (sólida+líquida), previamente conservada em formol 18%, foi
homogeneizada e uma alíquota de 1 mL foi transferida para tubos de ensaio e acrescentadas
três gotas de lugol, em substituição ao verde brilhante conforme modificação proposta por
D'Agosto e Carneiro (1999).

46
Após 15 minutos, foi adicionado 9 mL de glicerina a 30%. Para a quantificação dos
protozoários, foi pipetado em cada tubo de ensaio 1 mL de amostra previamente coradas com
lugol e fixadas com glicerina, para preencher a câmara de Sedgewick-Rafter. Foi utilizada
uma grade de contagem em uma das oculares, onde foram quantificados os ciliados presentes
em 50 campos e, posteriormente, após rotação da câmara em 180º, mais 50 campos foram
analisados. O cálculo do número total de ciliados por mililitro de amostra foi realizado
multiplicando-se o número de protozoários ciliados encontrados por 80 e por 20, que
correspondem à superfície total da câmara de contagem e à diluição, respectivamente
(DEHORITY, 1984).
Coleta das amostras – Bactérias ruminais
As amostras do conteúdo ruminal foram coletadas em frasco estéril plástico, com
capacidade para 80 mL. Duas amostras foram coletadas, uma da parte líquida da porção
ruminal e outra da parte sólida. Após a coleta, o material foi armazenado em caixa isotérmica
a 10 oC e encaminhado ao setor de Microbiologia da Faculdade União de Ensino do Sudoeste
do Paraná – UNISEP, para posteriores análises.
Contagem total de bactérias (CBT)
Para a contagem total de bactérias, tanto para a fração sólida quanto líquida, foram
utilizadas 25g de amostra e, em seguida, adicionado 225 mL de solução peptonada tamponada
a 1%, sendo esta a diluição 10-1
. A partir desta, foram realizadas diluições até a proporção 10-
10, em solução peptonada conforme descrito anteriormente. De cada diluição foi retirada uma
alíquota de 1 mL e inoculada em placa de petri estéril, adicionando uma camada de
aproximadamente 30 mL de ágar padrão (plate count agar). Posteriormente, essas placas
foram transferidas para estufa bacteriológica a 36 oC por 24 horas. A leitura das placas foi
realizada com auxílio de contador de colônias manual (adaptado de Koneman, 2008,
metodologia utilizada ma Microbiologia Geral).
Análise morfológica das bactérias
Para a análise morfológica empregou-se a técnica de coloração de Gram, na qual foi
utilizado 100 μL de uma suspenção de bactérias isoladas em ágar padrão, na concentração 0,5
da escala de McFarland. Após, foi adicionado em lâmina de vidro e procedeu-se a
metodologia padrão da coloração, utilizando os corantes violeta giensiana, lugol, solução

47
descorante de álcool cetona e fucsina (adaptado de Koneman, 2008). A leitura das lâminas foi
feita em microscópio óptico em objetiva de 100x. As bactérias foram classificadas em cocos e
bacilos, gram positivos e gram negativos.
Fermentação de carboidratos
Para cada amostra foi usada uma placa de meio de cultura com fonte única de
carboidrato, inoculando 1 mL de suspensação de bactérias na concentração 0,5 da escala de
McFarland sobre a placa, contendo o meio, e, subsequentemente, foi incubada em estufa
bacteriológica a 36 oC por 24 horas.
A composição básica do meio de cultura está representada na tabela abaixo, sendo que
o carboidrato testado foi o carboximetilcelulose. A avaliação da atividade fermentativa é
evidenciada pelo crescimento de colônias com coloração rósea (adaptado segundo Koneman,
2008).
Tabela 2 – Composição do meio de cultura utilizado para a atividade fermentativa do
carboidrato Carboximetilcelulose (Koneman, 2008).
COMPONETES CONCENTRAÇÃO (g.L-1
)
NaNO3 3,0
K2HPO4 1,0
MgSO4 0,5
KCl 0,5
FeSO4 . 7H20 0,01
Ágar padrão 20,0
Carboximetilcelulose 5,0
Análise estatística
O delineamento experimental foi o inteiramente casualizado, com quatro tratamentos,
e 10 repetições.
Os dados foram submetidos à análise por meio da metodologia de Modelos Lineares,
generalizados, assumindo distribuição Poisson com função de ligação logarítmica, com nível
de significância 1%. Foi utilizado o procedimento GENMOD do pacote estatístico SAS.
em que:
: é a observação da contagem do número de espécimes no animal, submetido à dieta i;
: base do logaritmo neperiano
η: função linear dos parâmetros na formula em que:
µ: é o intercepto,

48
: é o efeito da dieta i; i = 1;2;3;4;
: efeito da interação entre dieta e período;
: erro aleatório associado a cada observação .
RESULTADOS E DISCUSSÃO
Na pastagem de Azevém as médias da contagem de bactérias totais encontradas
foram 470,1 x1010
mL-1
(fração líquida) e 24,7 x1010
mL-1
(fração sólida) e Brachiaria spp. +
Estrela Africana 105,2 e 61,2, respectivamente (Tabela 2). Para as gramíneas Azevém +
Aveia e Brachiaria spp. a contagem das bactérias totais não foi possível através da
metodologia empregada devido a alta concentração encontrada nessas amostras, sendo
superior ao limite máximo (>250), o que dificultou a identificação do número de colônias.
Tabela 3- Médias (x1010
mL-1
) da contagem de bactérias totais (CBT) da fração sólida e
líquida de amostras extraídas de bovinos cruzados (Nelore x Europeu) submetidos ao efeito da
estacionalidade de pastagens temperadas (Azevém e Azevém + Aveia) e tropicais (Brachiaria
spp. e Brachiaria spp + Estrela africana).
Tratamentos/Pastagens
CBT
Fração líquida σ Fração sólida σ
Temperadas
Azevém 470,1 742,9 24,7 33,6
Azevém + Aveia >250 0 >250 0
Tropicais
Brachiaria spp. >250 0 >250 0 0
Brachiaria spp. + Estrela
africana 105,2 109,4 61,2 80,5
σ: Desvio padrão.
Foi encontrada maior concentração de bactérias totais na fração líquida das amostras
ruminais de animais em pastejo. Este resultado se explica pelo fato de que na maioria das
pesquisas são encontrados dados oriundos de animais fistulados, em que estes não são
submetidos a longos períodos sem alimentação. Já nos resultados encontrados, foram obtidas
amostras de animais abatidos, os quais foram mantidos em jejum pré abate por
aproximadamente 12 horas, o que justifica a alteração da concentração do número de
bactérias ruminais. Os resultados encontrados são contraditórios aos trabalhos de Wanapat e
Cherdthong (2009), em que foi observada maior prevalência de bactérias celulolíticas,

49
principais degradadoras da fibra, em dietas a base de forragens na fração sólida do conteúdo
ruminal quando comparado com a fase líquida.
A disponibilidade do substrato utilizado pelos microrganismos decresce devido à
escassez de alimento, principalmente no período de jejum pré-abate, impactando no número
de bactérias na porção líquida da amostra ruminal. Segundo Wanapat e Cherdthong (2009),
quando os animais ficam por um longo período sem alimentação, ocorre um maior
desenvolvimento de células e um acréscimo populacional na fase líquida do conteúdo ruminal
ou maior aderência as micropartículas existentes.
Houve maior concentração de bactérias nas gramíneas de inverno quando comparadas
com forrageiras tropicais. Este resultado é justificado pela característica anatômica das
forrageiras, pois as plantas de verão (C4) apresentam células arranjadas de forma mais densa,
externamente aos feixes vasculares, designada arranjo tipo Kranz, sendo este arranjo ausente
nas gramíneas de inverno (C3). Os dados encontrados corroboram com Wilson (1997), que ao
contrastar os tecidos de plantas (C3) e (C4), observou menor prevalência de mesófilo (tecido
caracterizado pela acelerada degradação) em gramíneas de verão, o que contribuiu para a
menor ação e concentração de microrganismos nestas plantas.
De acordo com Kozloski (2011), a aderência e colonização bacteriana podem ser
influenciadas por diversos fatores como capacidade de degradação do microrganismo, o
substrato disponível e o ambiente ruminal em que estes se encontram. Os tecidos das plantas
exercem grande influência na acessibilidade das bactérias ao material fibroso, sendo que as
gramíneas de clima temperado apresentam maior aderência bacteriana, como também são
mais extensamente degradadas, quando comparadas com as forrageiras tropicais.
Em relação à flora ruminal, foi observada a presença de bactérias Gram positivas e
negativas nas pastagens, como também grande diversidade nas formas bacterianas (Tabela 3).
Na pastagem de Azevém se constatou maior abundância nas populações de bactérias
ruminais, Gram positivas e negativas, Cocos e Bacilos, resultados que corroboram com
Dirksen (1993) ao estudar a diversidade da microbiota ruminal de ovinos criados a pasto, o
qual observou maior quantidade de bactérias ruminais nos animais submetidos ao pastejo de
gramíneas de inverno ao comparar com forrageiras de verão. Na pastagem de Brachiaria spp.
não observou-se Bacilos Gram negativos.
Em todas as amostras houve a presença de bactérias Gram positivas, principalmente
nas pastagens de Aveia + Azevém e Brachiaria spp. + Estrela Africana. Este resultado se
explica pela característica do substrato que estas utilizam, pois são degradadoras de

50
carboidratos estruturais e apresentam grande propensão por dietas compostas por alimentos
fibrosos (KAMRA, 2005). As Gram positivas, ao degradar a celulose presente na parede
celular das plantas, formam o acetato, assim, disponibilizam substratos para as demais
bactérias, sobretudo as metanogênicas, as quais também são Gram positivas. As bactérias
proteolíticas também são designadas Gram positivas, estas geram nitrogênio na forma de
amônia paras as celulolíticas (ROBSON; STEWART, 1997). Como houve atividade
celulolítica em todas as amostras estudadas, este seria um fator contribuinte para a maior
densidade de Gram positivas, assim como a maior atividade destes microrganismos em dietas
contendo volumoso.
Tabela 4 - Flora ruminal e atividade celulolítica da fração sólida e líquida de amostras
extraídas de bovinos cruzados (Nelore x Europeu) submetidos ao efeito da estacionalidade de
pastagens temperadas (Azevém e Azevém + Aveia) e tropicais (Brachiaria spp. e Brachiaria
spp + Estrela africana). Flora Bacteriana Atividade celulolítica
Frações do líquido ruminal
Líquida Sólida Líquida Sólida
Temperadas
Azevém CG+;CG-;BG+;BG- CG+; CG-; BG+;BG- + +
Azevém + Aveia CG+;BG+ CG+;BG+ + +
Tropicias
Brachiaria spp. CG+;CG-;BG+ CG+;CG-;BG+ + +
Brachiaria spp. +
Estrela africana CG+;BG+ CG+;BG+ + +
C: Cocos; B: Bacilos; G+: Gram-positivas; G-: Gram-negativas
Em relação aos protozoários ciliados do rúmen, foi observada a ocorrência dos
ciliados nas ordens Vestibuliferida e Entodiniomorphida (Tabela 5). Na ordem
Vestibuliferida, constata-se a presença da família Isotrichidae, composta pelos gêneros
Dasytricha e Isotricha. e na ordem Entodiniomorphida, protozoários da família
Ophryoscolecidae, caracterizada pelos gêneros Entodinium, Diplodinium, Eodinium,
Epidinium, Eremoplastron, Eudiplodinium, Metadinium, Ostracodinium e Polyplastron.
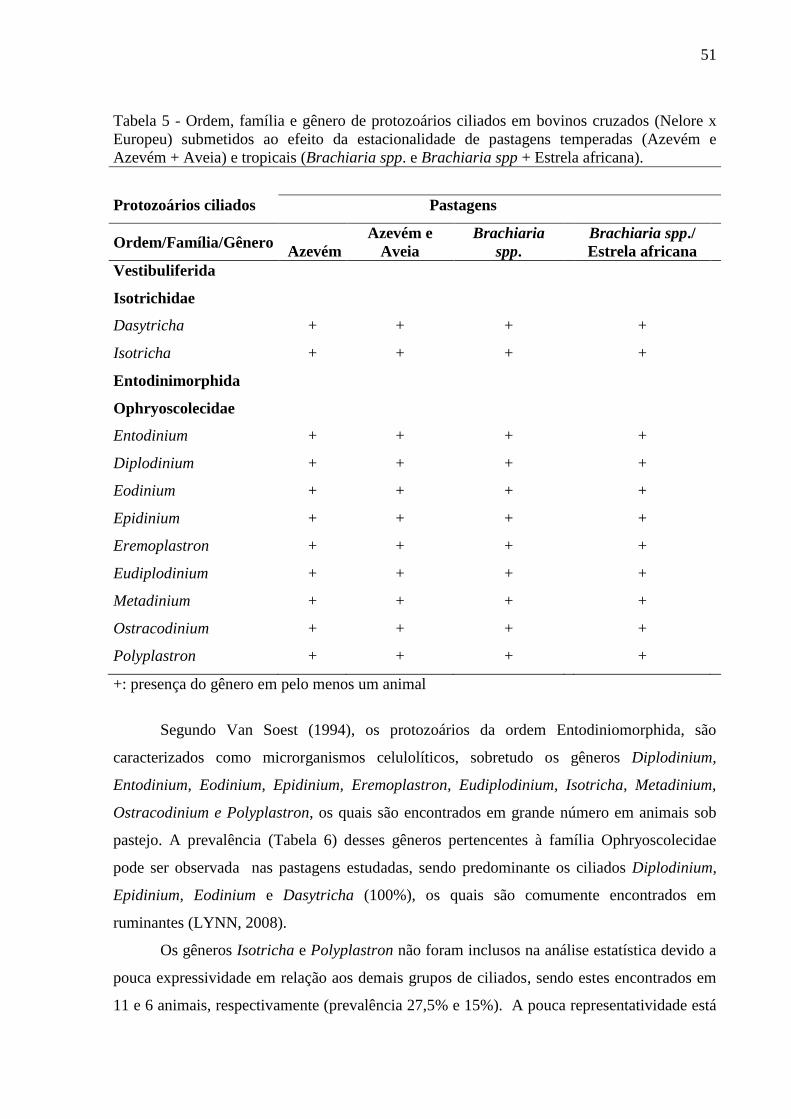
51
Tabela 5 - Ordem, família e gênero de protozoários ciliados em bovinos cruzados (Nelore x
Europeu) submetidos ao efeito da estacionalidade de pastagens temperadas (Azevém e
Azevém + Aveia) e tropicais (Brachiaria spp. e Brachiaria spp + Estrela africana).
Protozoários ciliados Pastagens
Ordem/Família/Gênero Azevém
Azevém e
Aveia
Brachiaria
spp.
Brachiaria spp./
Estrela africana
Vestibuliferida
Isotrichidae
Dasytricha + + + +
Isotricha + + + +
Entodinimorphida
Ophryoscolecidae
Entodinium + + + +
Diplodinium + + + +
Eodinium + + + +
Epidinium + + + +
Eremoplastron + + + +
Eudiplodinium + + + +
Metadinium + + + +
Ostracodinium + + + +
Polyplastron + + + +
+: presença do gênero em pelo menos um animal
Segundo Van Soest (1994), os protozoários da ordem Entodiniomorphida, são
caracterizados como microrganismos celulolíticos, sobretudo os gêneros Diplodinium,
Entodinium, Eodinium, Epidinium, Eremoplastron, Eudiplodinium, Isotricha, Metadinium,
Ostracodinium e Polyplastron, os quais são encontrados em grande número em animais sob
pastejo. A prevalência (Tabela 6) desses gêneros pertencentes à família Ophryoscolecidae
pode ser observada nas pastagens estudadas, sendo predominante os ciliados Diplodinium,
Epidinium, Eodinium e Dasytricha (100%), os quais são comumente encontrados em
ruminantes (LYNN, 2008).
Os gêneros Isotricha e Polyplastron não foram inclusos na análise estatística devido a
pouca expressividade em relação aos demais grupos de ciliados, sendo estes encontrados em
11 e 6 animais, respectivamente (prevalência 27,5% e 15%). A pouca representatividade está

52
relacionada aos fatores intrínsecos do animal hospedeiro. Dessa forma, averigua-se que,
apesar da dieta ser um fator de grande relevância na mantença das condições da microbiota
ruminal, a característica individual e o metabolismo do hospedeiro, exercem grande influência
na população dos protozoários ciliados (GÖÇMEN et al., 2002).
Tabela 6 – Prevalência (%) dos gêneros de protozoários ciliados do rúmen de bovinos
cruzados (Nelore x Europeu) submetidos ao efeito da estacionalidade de pastagens
temperadas (Azevém e Azevém + Aveia) e tropicais (Brachiaria spp. e Brachiaria spp +
Estrela africana).
Gêneros Azevém Azevém +
Aveia Brachiaria spp.
Brachiaria spp. + Estrela
africana
Entodinium 70% 90% 100% 100%
Diplodinium 100% 100% 100% 100%
Eudiplodinium 50% 90% 80% 90%
Eremoplastron 60% 90% 100% 90%
Metadinium 40% 80% 70% 80%
Ostracodinium 30% 40% 100% 80%
Epidinium 100% 100% 100% 100%
Eodinium 100% 100% 100% 100%
Dasytricha 100% 100% 10% 100%
Foi constatado que o gênero predominante foi o Diplodinium, 40,38 x104 mL
-1, sendo
esta densidade também observada em relação às pastagens estudadas, principalmente na
gramínea Azevém (Tabela 8). Os demais gêneros Dasytricha, Entodinium, Eodinium e
Epidinium, também foram observados em concentrações elevadas, 13,62; 11,81; 11,75 e
10,86 (x104 mL
-1), respectivamente.
Segundo Purse e Moir (1966), quando a dieta contém grande quantidade de volumoso
ocorre um aumento nas populações dos ciliados pertencentes ao gênero Diplodinium. A maior
concentração nas populações deste gênero ocorre pela presença da enzima celulase, a qual
favorece a degradação da celulose presente na parede celular das plantas (WILLIANMS;
COLEMAN, 1992). A composição nutricional das dietas também pode afirmar que as
pastagens forneciam condições adequadas para serem consideradas de boa qualidade (Van
Soest, 1994), sendo este um fator contribuinte para o aumento das populações de
Diplodinium.

53
Tabela 7 - Médias (x104 mL
-1) estimadas para o total dos gêneros de protozoários ciliados do
rúmen encontrados de amostras extraídas de bovinos cruzados submetido submetidos ao
efeito da estacionalidade de pastagens temperadas (Azevém e Azevém + Aveia) e tropicais
(Brachiaria spp. e Brachiaria spp + Estrela africana).
Gênero Média Desvio padrão
Entodinium 11,81 23,48
Diplodinium 40,38 22,9
Eudiplodinium 0,43 0,43
Eremoplastron 1,15 1,51
Metadinium 0,42 0,49
Ostracodinium 0,48 0,85
Epidinium 10,86 11,05
Eodinium 11,75 8,36
Dasytricha 13,62 14,36
Os ciliados do gênero Diplodinium apresentam elevada taxa de divisão, a qual é
intensa aproximadamente 8 horas após a ingestão de alimento (MICHALOWSKI, 1977).
Este fator contribuiu para a elevada concentração destes microrganismos, pois os animais
podem ter pastejado por um período maior anteriormente ao abate,assim como o tempo de
transporte pode ter interferido, pois os animais foram adquiridos de diferentes propriedades da
região Sudoeste do Paraná. A variação no número de ciliados deste gênero também foi
observada nas diferentes pastagens avaliadas (Tabela 8).
Na tabela 8 pode ser observada uma significativa concentração dos gêneros Epidinium
e Dasytricha em todas as pastagens, sendo na gramínea Azevém encontrada as maiores
populações, 22,22 e 21,47 x104 mL
-1, respectivamente. De modo geral, o número de ciliados
do gênero Dasytricha apresentou valores expressivos em todas as forrageiras. Estes resultados
são explicados pelo alto teor de celulose nas gramíneas (Tabela 1). Hungate (1966) ao testar
diferentes proporções de volumoso na dieta, observou que quando fornecido maiores níveis de
V:C (70:30), as populações de Dasytricha foram beneficiadas, o que é justificado segundo o
autor, pela capacidade de degradação celulolítica. Marinho (1983) notou um número
significativo de ciliados Dasytricha em amostras ruminais de ovinos abatidos oriundos de
pastagem, corroborando com os resultados, em que foi constatada uma elevada concentração
deste gênero em todas as forrageiras.
O gênero Epidinium é descrito por Manella e Lourenço (2004) e Williamns e
Coleman (1992) como um dos protozoários que participa diretamente e de forma efetiva na

54
degradação dos componentes da parede celular das plantas, sobretudo da celulose.
Bonhomme-Florentin et al. (1978) verificaram o efeito da sazonalidade no número de
protozoários ciliados em bovinos e constataram a diminuição do gênero Epidinium durante
períodos secos. Informações semelhantes aos resultados encontrados, pois houve uma
diminuição na concentração deste gênero quando os animais receberam forrageiras tropicais,
assim como quando em pastejo de Aveia + Azevém, sendo que esta gramínea se encontrava
em final de ciclo (Tabela 8). O teor de fibra das forrageiras (Tabela 1) foi um fator que
contribuiu para o decréscimo na concentração de protozoários, pois em virtude da maturação
da planta, ocorre a diminuição dos açúcares solúveis e aumento da fibra. Segundo Williamns
e Coleman (1992) a espécie Epidinium caudatum possui elevada quantidade de celulase
presente no citoplasma, o que auxilia na degradação do material fibroso.
Os ciliados pertencentes ao gênero Entodinium apresentaram média de 11, 81 x104
mL-1
do total dos gêneros de protozoários das amostras estudadas (Tabela 7) e em relação as
pastagens (Tabela 8), foi encontrado em maior concentração nas forrageiras de verão,
Brachiaria spp.(28,68x104 mL
-1) e Brachiaria spp. + Estrela Africana (11,05x10
4 mL
-1).
Diversos autores demonstram que os Entodinium são caracterizados como ciliados que
apresentam grande propensão por alimentos com elevados teores de energia (D’AGOSTO;
GUEDES, 2000; FRANZOLIN; FRANZOLIN, 2000; NOGUEIRA FILHO et al., 1992).
Tymensen et al. (2012) ao observar a estrutura da comunidade de protozoários em bovinos
alimentados com diferentes dietas (feno e silagem/grãos), constatou maior predominância da
espécie Entodinium spp. quando os animais foram submetidos a alimentação a base de
silagem e grãos e maior foi o diversidade de espécies quando fornecido feno. Apesar das
dietas serem somente a pasto, a predominância deste gênero foi bastante significativa, sendo a
qualidade bromatológica da dieta o fator contribuinte para os resultados encontrados, pois a
pesar de essas pastagens apresentarem elevado teor de FDN e Lignina, a digestibilidade das
mesmas foi alta.

55
Tabela 8 - Médias (x104 mL
-1) estimadas para os gêneros de protozoários ciliados em bovinos cruzados do rúmen encontrados de amostras
extraídas de bovinos cruzados submetido submetidos ao efeito da estacionalidade de pastagens temperadas (Azevém e Azevém + Aveia) e
tropicais (Brachiaria spp. e Brachiaria spp + Estrela africana).
σ: Desvio padrão.
Azevém σ Aveia+Azevém σ Brachiaria spp. σ
Brachiaria spp.+Estrela
Africana σ
Entodinium 5,61 11,71 1,888 1,64 28,688 40,78 11,056 10,85
Diplodinium 48,72 29,95 30,352 15,69 46,096 25,41 36,368 16,05
Eudiplodinium 0,25 0,32 0,496 0,54 0,48 0,43 0,496 0,42
Eremoplastron 0,32 0,32 0,768 0,87 1,312 1,16 2,208 2,35
Metadinium 0,28 0,55 0,272 0,22 0,672 0,63 0,48 0,43
Ostracodinium 0,72 1,57 0,096 0,13 0,576 0,27 0,528 0,58
Epidinium 22,22 16,83 5,568 1,85 9,424 5,61 6,256 3,24
Eodinium 7,07 6,36 13,264 9,43 12,928 8,11 13,744 8,66
Dasytricha 21,47 13,49 3,616 2,26 24,4 17,47 4,992 3,19
Total de ciliados inverno= 162,98 Total de ciliados verão= 200,70

56
O período estacional influenciou na composição dos gêneros de protozoários do rúmen
dos animais oriundos do pastejo de diferentes gramíneas (Tabela 8). Constatou-se maior
concentração de ciliados no verão (200,70 x104 mL
-1). Resultado explicado pela atividade
fibrolítica dos ciliados, os quais aumentam a densidade em pastagens com elevado teor de
fibra e em períodos úmidos (ARAÚJO FILHO, 2002; SILVA et al., 2014).
A característica nutricional das pastagens de verão é um dos fatores que influencia na
taxa de passagem do alimento e consumo animal. Nogueira Filho et al. (1992) afirmam que
dietas a base de volumoso podem apresentar variação na disponibilidade de nutrientes, pois os
diferentes tecidos da planta interferem na degradação microbiana, devido a quantidade de
hemicelulose, celulose e lignina. Esta informação se confirma pelos dados referentes ao
número de protozoários encontrados em relação às diferentes gramíneas, em que a forrageiras
Brachiaria spp. e Brachiaria spp.+ Estrela africana apresentaram maior concentração de
ciliados, bem como os maiores teores de celulose, hemicelulose e lignina, demonstrando que a
atividade microbiana foi mais intensa. Os resultados obtidos também corroboram com os
estudos de Christiansen et al. (1964) e Dehority e Odenyo (2003) em que encontraram grande
diversidade de protozoários ciliados, principalmente da família Ophryoscolecidae, em animais
submetidos ao pastejo de forrageiras de elevado teor fibroso, o que é explicado pelo
decréscimo na taxa de passagem do alimento pelo sistema digestório, o que ocasiona o
aumento dos níveis de pH ruminal, sendo estas condições favoráveis a colonização por parte
destes protozoários.
Apesar de a dieta ter um papel de grande relevância na concentração das populações
de protozoários ciliados do rúmen, outros aspectos também podem ser atribuídos para explicar
a ocorrência dos protozoários, como as características individuais do hospedeiro em relação
aos animais da mesma espécie e a taxa de passagem do alimento (FRANZOLIN;
DEHORITY, 1996). Particularidades metabólicas do hospedeiro, relações mutualísticas entre
os microrganismos e o antagonismo entre as espécies de protozoários ciliados, também
contribuem para a alteração das populações de protozoários (D’AGOSTO; SANTA-ROSA,
1998).
Pesquisas recentes certificam que os protozoários ciliados do rúmen estão ligados
efetivamente nas atividades celulolíticas e hemicelulolíticas do ambiente ruminal. Os ciliados
colonizam as frações da fibra, estimulam a ruminação e secreção salivar, o que mantém o pH
estável para a colonização de microrganismos celulolíticos, como também auxiliam na
degradação dos tecidos da planta e na ação bacteriana (KOZLOSKI, 2011; DEHORITY,

57
2003). Portanto, são de suma importância em bovinos criados em pastagens devido à atuação
destes microrganismos tem-se um maior aproveitamento das forrageiras, proporcionando
maior eficiência alimentar.
CONCLUSÃO
O período estacional influenciou nas populações microbianas do rúmen. A Contagem
Bacteriana Total (CBT) foi maior no inverno, sobretudo na pastagem de Azevém (494,8 x1010
mL-1
), e na fração líquida do conteúdo ruminal (470,1 x1010
mL-1
). Foi constatada a presença
das bactérias gram-positivas e gram-negativas nos bovinos criados a pasto e grande
diversidade nas formas bacterianas, sendo predominantes as formas de cocos e bacilos.
A maior diversidade e concentração dos gêneros de protozoários ciliados foi
encontrada nos animais em pastagens de verão (200,70 x104 mL
-1), sendo predominante o
Diplodinium (40,38 x104 mL
-1). A concentração de ciliados foi mais intensa na pastagem de
Brachiaria spp.

58
CONSIDERÇÕES FINAIS
O maior número de bactérias ruminais foi encontrado nas pastagens de inverno,
principalmente devido à ausência dos feixes vasculares, devido ao metabolismo das plantas
C3, maior prevalência de mesófilo nas plantas, intensa aderência e colonização bacteriana, as
quais são mais extensamente degradas.
Os protozoários ciliados foram encontrados em maior número nas pastagens de verão,
devido a maior atividade destes microrganismos, os quais realizam intensa degradação dos
carboidratos estruturais, celulose, hemicelulose e lignina. A menor taxa de passagem do
alimento favoreceu a degradação da fibra, provavelmente, pelo vível de pH neutro.

59
REFERÊNCIAS
ABRÃO, F. O.; DUARTE, E. R.; NIGRI, A. C. de A.; SILVA, M. L. F.; RIBEIRO, I. C. O.;
SILVA, K. L. da; ROSA, C. A.; RODRIGUEZ, N. M. Caracterização físico-química e
microbiológica e população de fungos no conteúdo ruminal de novilhos de corte hígidos ou
com acidose ruminal. Revista Brasileira de Medicina Veterinária, 37(1):7-14, jan/mar
2015.
BERCHIELLI, T. T.; PIRES, A. V.; OLIVEIRA, S. G. de. Nutrição de Ruminantes,
Jaboticabal: Funep, p. 111-116, 2006.
BONHOMME-FLORENTIN, A, BLANCOU, J, LATTEUR, B, E’tude des variations
saisonnières de la microfaune du rumen de zebus. Protistologica, 14:283-289, 1978.
CHRISTIANSEN, W.C., WOODS, W., BURROUGHS, W. Ration characteristics influencing
rumen protozoal population. Journal of Animal Science, 23(4):984-988. 1964.
D’AGOSTO, M. & SANTA-ROSA, M.R de. Influência do hospedeiro no perfil populacional
e nas populações de ciliados do rúmen de bovinos. Revista Brasileira de Zoologia, Curitiba,
v. 15, p. 389-396, 1998.
D’AGOSTO, M.; GUEDES, P.M.M. Caracterização das populações de ciliados (Protista,
Ciliophora) do rúmen de bovinos de corte no estado de Minas Gerais, Brasil. Revista
Brasileira de Zoociências, v.2, p.81-90, 2000.
D'AGOSTO, M.; CARNEIRO, M.E. Evaluation of lugol solution used for counting rumen
ciliates. Revista Brasileira de Zoologia, v.16, p.725-729, 1999.
DEHORITY, B.A. Evaluation of subsampling and fixationproceduresused for counting rumen
protozoa. Applied and Environmental Microbiology, v.48, p.182-185, 1984.
DEHORITY, B.A. Rumen microbiology. Thrumpton, Nottingham: Nottingham University
Press, 372p., 2003.
DEHORITY, B.A.; ODENYO, A.A. ‘Influence of diet on the rumen protozoal fauna of
indigenous African wild ruminants’, Journal of Eukaryotic Microbiology 50, 220–223,
2003.
DIRKSEN, G. Sistema Digestivo. In: DIRKSEN, G.; GRÜNDER, H.D.; STÖBER, M.
Rosenberger. Exame Clínico dos Bovinos. Guanabara Koogan: Rio de Janeiro, p. 166- 228,
1993.
FRANZOLIN, R., DEHORITY, B.A. Effect of prolonged high-concentrate feeding on
ruminal protozoa concentrations. Journal of Animal Science, 74(11):2803-2809, 1996.
FRANZOLIN, R.; FRANZOLIN, M.H.T. População de protozoários ciliados e
degradabilidade ruminal em búfalos e bovinos zebuínos sob dieta à base de cana de açúcar.
Revista Brasileira de Zootecnia, v.29, p.1853-1861, 2000.

60
GÖÇMEN, B.; DEHORITY, B.A.; RASTGELDI, S. The occurrence of rumen ciliate
Metadinium banksi Dehority, 1985 (Ophryoscolecidae, Entodiniomorphida) from domestic
goats (Capra hircus L.) in Southeastern Turkey. Turkish Journal of Zoology 26, 367-370,
2002.
HSU, J. T.; FAHEY, G. C.; MERCHEN, N.R.; MACKIE, R. Effects of defaunation and
various nitrogen supplementation regimens on microbial numbers and activity in the rumen of
sheep. Journal of Animal Science, 69:1279, 1991.
JAMI, E.; MIZRAHI, I. Composition and smilarity of bovine rumen microbiota across
individual animals. Plos One 7(3): e33306. doi:10.1371/journal.pone.0033306, 2012.
KAMRA, D.N. Rumen microbial ecosystem. Current Science, v.89, n.1, p.124-134, 2005.
KARNATI, S.K.R.; YU, Z.; FIRKINS, J.L. Investigating unsaturated fat, monensin, or
bromoethanesulfonate in continuous cultures retaining ruminal protozoa. II. Interaction of
treatment and presence of protozoa on prokaryotic communities. Journal of Dairy Science,
submitted, 2009.
KONEMAN, E. W. Diagnóstico microbiológico: textos e atlas colorido. Vol. 1. 1ª ed., Rio
de Janeiro: Guanabara? Koogan, 2008.
KOZLOSKI, G.V. Bioquímica dos ruminantes. 3ª Ed. UFSM, Santa Maria , 212 p. 2011.
MANELLA, M.Q.; LOURENÇO, A.J. População de protozoários ciliados no rúmen de
bovinos nelore em pastos de Brachiaria brizantha marandu recebendo suplemento protéico ou
com livre acesso a banco de proteína de Leucena leucocephala nas diferentes estações do ano.
Boletim da Indústria Animal, n.1, p.1-11, 2004.
MARINHO, A. A. M. Ciliate protozoa in the rumen of grazing sheep. Revista Portuguesa de
Ciências Veterinárias, v. 78, p. 157-165, 1983.
MARTINELE, I.; SIQUEIRA-CASTRO, I. C. V.; D´AGOSTO, M. Protozoários ciliados no
rúmen de bovinos alimentados com dietas de capim elefante e com dois níveis de
concentrado. Revista Brasileira de Saúde e Produção Animal, v.9, n.1, p. 74-81, 2008 (a).
MICHALOWSKI, T. Diurnal changes in concentration or rumen ciliates and in occurrence of
dividing forms in water buffalo (Bubalus bubalus) fed once daily. Applied Environmental
Microbiology, Washington, 33 (4): 802-804. 1977.
NOGUEIRA-FILHO, J. C.; LUCCI, C. de S.; MELOTTI, L.; OLIVEIRA, M. E. M. de;
LIMA, C. G. de; CUNHA, J. A. da. Contagens diferenciais de protozoários ciliados em rúmen
de bovinos arraçoados com capim elefante Napier (Penissetum Purpureum Schum), em vários
estádios de crescimento vegetativo. Brazilian Journal of Veterinary Research and Animal
Science, São Paulo, v. 27, n 2, p 215-222, 1992.
REIS, Cândida C. dos: Protozoários ciliados no rúmen de bovinos Nelore e Cruzados
Nelore x Europeu sob diferentes sistemas de alimentação, 2015, 64 f. Dissertação

61
(Mestrado em Zootecnia) – Programa de Pós Graduação em Zootecnia, Universidade
Tecnológica Federal do Paraná. Dois Vizinhos, 2015.
RUSSEL, J. B.; MANTOVANI, H. C. The Bacteriocins of ruminal bactéria and their
potencial as na alternative to antibiotics. Journal of Molecular Microbiology and
Biotechnology, 4:347, 2002.
SILVA, D.J.; QUEIROZ, A.C. Análises de alimentos (métodos químicos e biológicos).
3.ed. Viçosa, MG: Editora UFV, 235p. 2004.
SILVA, F.F.; SÁ, J. F. de.; SCHIO, A. R.; ÍTAVO, L. C. V.; SILVA, R. R.; MATEUS, R. G.
Suplementação a pasto: disponibilidade e qualidade x níveis de suplementação x desempenho.
Revista Brasileira de Zootecnia, v.38, 371-389, 2009.
STATISTICAL ANALYSIS SYSTEM - SAS. System for Microsoft Windows: release 9.0.
Cary: 2004. 1 CD-ROM.
TYMENSEN, L.; BARKLEY, C.; MCALLISTER, T.A. Relative diversity and community
structure analysis of rumenprotozoa according to T-RFLP and microscopic methods. Journal
of Microbiological Methods, 88: 1–6, 2012.
VAN SOEST, P.J. Nutritional Ecology of the Ruminant. Ithaca: Cornell University Press,
476p. 1994.
VIDAL, M. J.; PAIVA, A. C. P.; ARCURI, P. B.; ARCURI, P. B.; LOPES, F.C.F.;
ZAQUINI, L. Efeito de diferentes doses de enxofre no consumo voluntário e nas populações
de protozoários do rúmen de novilhas mestiças alimentadas com capim-elefante de baixa
qualidade. Ciência Agrotécnica, Lavras, v. 31, n. 1, p. 218-222, 2007.
VIEIRA, A.C.S.; AFONSO, J.A.; MENDONÇA, C.L. Características do fluido ruminal de
ovinos Santa Inês criados extensivamente em Pernambuco. Pesquisa Veterinária Brasileira,
27:110-114, 2007.
WANAPAT, M.; CHERDTHONG, A. Use of Real-Time PCR technique in studying rumen
cellulolytic bacteria population asaffected by level of roughage in swamp buffaloes, Current
Microbiology, Vol.58, pp. 294–299, 2009.
WILLIAMS, A.G.; G.S. COLEMAN. The Rumen Protozoa. London: Springer-Verlag, New
York Inc, 423 p. 1992.
WILSON, JR. Structural and anatomical traits of forages influencing their nutritive value for
ruminants. In: Anais do Simpósio Internacional sobre Produção Animal em Pastejo. Viçosa.
Viçosa: Universidade Federal de Viçosa; p. 173-208, 1997.

62
5. ANEXOS

63

64
Anexo A: Comitê de ética

65
Anexo C: Abertura do compartimento ruminal para a coleta das amostras. (Fonte: Autor)
Anexo D: Coleta das amostras para determinação e quantificação de bactérias e protozoários. (Fonte: Autor)
Anexo B: Bovinos utilizados para a amostragem do conteúdo ruminal (Fonte: Autor)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)
(Fonte: Letícia Wlodarski)

66
Anexo E: Inoculação das alíquotas de líquido ruminal em placas de petri estéril para a análise
da contagem bacteriana total (CBT). (Fonte: Autor)
Anexo F: A: Bactéria Gram-negativa; B: Bactéria Gram-positiva e contagem de colônias de
bacterianas ruminais. (Fonte: Autor)
A
B

67
Anexo G: Gêneros de protozoários ciliados em bovinos cruzados em pastagens de inverno e
verão, Azevém, Azevém e Aveia, Brachiaria spp. e Brachiaria spp e Estrela africana. (Fonte:
Autor) A: Diplodinium; B: Eremoplastron; C: Eudiplodinium; D: Dasytricha; E: Eodinium; F:
Ostracodinium; G: Metadinium; H: Polyplastron; I: Epidinium; J: Isotricha; K:
Entodinium.