desenvolvimento e avaliaÇÃo de mÉtodos …livros01.livrosgratis.com.br/cp055842.pdf · figura...
TRANSCRIPT

INSTITUTO OSWALDO CRUZ
Doutorado em Biologia Celular e Molecular
DESENVOLVIMENTO E AVALIAÇÃO DE MÉTODOS
MOLECULARES PARA O DIAGNÓSTICO DA DENGUE
CELINA DE OLIVEIRA POERSCH
Curitiba 2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
INSTITUTO OSWALDO CRUZ Pós-Graduação em Biologia Celular e Molecular
CELINA DE OLIVEIRA POERSCH
Desenvolvimento e avaliação de métodos moleculares para o diagnóstico da dengue
Tese apresentada ao Instituto Oswaldo Cruz como parte dos requisitos para obtenção do título de Doutor em Biologia Celular e Molecular
Orientador: Prof. Dr. Marco Aurélio Krieger
CURITIBA
2007

Ficha catalográfica elaborada pela
Biblioteca de Ciências Biomédicas/ ICICT / FIOCRUZ - RJ
P745
Poersch, Celina de Oliveira
Desenvolvimento e avaliação de métodos moleculares para o diagnóstico da dengue/ Celina de Oliveira Poersch. – Curitiba, 2007.
xii, 113 f. : il. ; 30 cm. Tese (doutorado) – Instituto Oswaldo Cruz, Biologia Celular e
Molecular, 2007. Bibliografia: f. 99-113
1. Dengue. 2. Dengue – diagnóstico. 3. Aedes. 4. Reação em Cadeia da Polimerase. I. Título.
CDD 614.588 52

iii
INSTITUTO OSWALDO CRUZ Pós-Graduação em Biologia Celular e Molecular
AUTOR: CELINA DE OLIVEIRA POERSCH
DESENVOLVIMENTO E AVALIAÇÃO DE MÉTODOS MOLECULARES PARA O DIAGNÓSTICO DA DENGUE
Orientador: Prof. Dr. Marco Aurélio Krieger
Aprovada em: 21/12/2007
EXAMINADORES:
Prof. Dr. Ricardo Galler - Presidente Prof. Dr. Robrigo Brindeiro Profa. Dra. Cláudia Duarte dos Santos
Curitiba, 21 de dezembro de 2007.

iv
À Clara, que é muito mais importante
do que tudo que eu já fiz ou vou fazer.

v
AGRADECIMENTOS
Aos meus pais que me apóiam em tudo e acreditam em mim mais do que ninguém.
Ao Dr. Marco Aurélio Krieger, que me deu todas as oportunidades e nunca mediu
esforços para que nossos projetos se concretizassem.
Aos diretores do IBMP por me disponibilizarem um ambiente tão estimulante e
acolhedor.
À Anaclete (LACEN-PR) e à Dra. Maria da Glória (IPEC), que gentilmente cederam
as amostras utilizadas nesse trabalho.
A todos da Biometrix pela oportunidade de utilização do Luminex. Um agradecimento
especial ao Maurício Cichon e ao Marcos pela disponibilidade, pela paciência, pelas
dicas e pelo companheirismo.
A todos os colegas do IBMP pela amizade, ajuda, sugestões, ensinamentos,...
Todos vocês, sem exceção, contribuíram direta ou indiretamente para a realização
dessa tese.
À Daniela P. Pavoni pela revisão e, principalmente, pela amizade sincera.
À CAPES, à FIOCRUZ e ao IBMP pelo apoio financeiro.

vi
ÍNDICE
1. INTRODUÇÃO....................................................................................................................................1
1.1. A dengue.......................................................................................................................................1
1.2. Situação atual da doença............................................................................................................3
1.3. O vírus...........................................................................................................................................9
1.4. Aspectos clínicos e patogênese...............................................................................................11
1.5. Viremia e cinética da resposta humoral ao vírus....................................................................13
1.6. Diagnóstico laboratorial.............................................................................................................15
1.6.1. Inibição da hemaglutinação..................................................................................................16
1.6.2. Fixação do complemento......................................................................................................17
1.6.3. Teste de neutralização..........................................................................................................17
1.6.4. ELISA (enzyme-linked immunosorbent assay)..................................................................18
1.6.4.1. MAC-ELISA......................................................................................................................18
1.6.4.2. IgG-ELISA........................................................................................................................19
1.6.4.3. Razão IgM / IgG...............................................................................................................19
1.6.4.4. NS1-ELISA................................................................................................................ ......20
1.6.5. Isolamento viral.....................................................................................................................21
1.6.6. RT-PCR (reverse transcription – polymerase chain reaction)……………………..............22
1.6.6.1. RT-PCR em tempo real....................................................................................................23
1.6.7. Metodologias moleculares inovadoras potencialmente utilizáveis para o diagnóstico da
dengue...........................................................................................................................................25
1.6.7.1. Microarranjo tradicional....................................................................................................25
1.6.7.2. Microarranjos líquidos......................................................................................................27
2. OBJETIVOS.....................................................................................................................................31
3. JUSTIFICATIVAS.............................................................................................................................32
4. MATERIAIS E MÉTODOS................................................................................................................34
4.1. Amostra populacional estudada..............................................................................................34
4.2. Preparo dos padrões para testes de especificidade, sensibilidade e otimização das
reações.............................................................................................................................................35
4.3. Extração do RNA viral...............................................................................................................36
4.4. Desenho dos oligonucleotídeos para PCR em tempo real....................................................36
4.5. Desenho dos oligonucleotídeos para microarranjo líquido..................................................38
4.6. PCR em tempo real....................................................................................................................39
4.6.1. Detecção das seqüências do vírus da dengue por PCR em tempo real.........................43

vii
4.7. Microarranjo líquido..................................................................................................................44
4.7.1. Acoplamento das sondas às micro-esferas de poliestireno............................................46
4.7.2. PCR........................................................................................................................................47
4.7.3. Hibridação do produto de PCR com as sondas acopladas às micro-esferas................48
4.7.4. Detecção das seqüências do vírus da dengue pelo equipamento Luminex®.................49
5. RESULTADOS.................................................................................................................................51
5.1. Avaliação dos diferentes conjuntos de oligonucleotídeos...................................................51
5.2. Otimização da reação de hibridação múltipla (multiplex) para o teste diagnóstico baseado
na metodologia de microarranjo líquido.......................................................................................57
5.3. Otimização da RT-PCR múltipla (multiplex) para o teste diagnóstico baseado na
metodologia de microarranjo líquido............................................................................................59
5.4. Avaliação das diferentes combinações de oligonucleotídeos no ensaio de microarranjo
líquido...............................................................................................................................................60
5.5. Sensibilidade e especificidade dos métodos de diagnóstico molecular desenvolvidos....64
5.6. Aplicação dos métodos desenvolvidos na detecção e sorotipagem do vírus da dengue em
amostras clínicas.............................................................................................................................69
6. DISCUSSÃO.....................................................................................................................................74
7. CONCLUSÕES.................................................................................................................................88
8. PERSPECTIVAS..............................................................................................................................90
ANEXOS...............................................................................................................................................92
REFERÊNCIAS BIBLIOGRÁFICAS.....................................................................................................99

viii
LISTA DE FIGURAS
FIGURA 1.1 – Distribuição do Aedes aegypti durante a década de 30 e nos anos de 1970 e 2006..................................................................................................................3 FIGURA 1.2 – Média anual do número de casos de dengue, reportados à OMS, por década..........................................................................................................................4 FIGURA 1.3 – Número de casos totais de dengue e de febre hemorrágica da dengue reportados à OMS, por ano, na região das Américas..................................................5 FIGURA 1.4 – Número de casos totais de dengue (DF + DHF) e de febre hemorrágica da dengue (DHF) por ano, no Brasil........................................................8 FIGURA 1.5 - Desenho esquemático representando a estrutura do vírus da dengue........................................................................................................................10 FIGURA 1.6 - Resposta imune primária e secundária ao vírus da dengue...............15 FIGURA 4.1 - Representação esquemática das etapas de uma PCR em tempo real..............................................................................................................................41 FIGURA 4.2 – Exemplo de uma curva de dissociação obtida após a amplificação de um segmento de DNA utilizando o corante SYBR Green..........................................43 FIGURA 4.3 – Representação esquemática da detecção de ácidos nucléicos através da técnica de microarranjo líquido..............................................................................46 FIGURA 5.1 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 1 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DEN1 e a sonda TaqMan correspondente...............................................52 FIGURA 5.2 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 1 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DENG e a sonda TaqMan correspondente..............................................53 FIGURA 5.3 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 2 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DEN2 e a sonda TaqMan correspondente...............................................53 FIGURA 5.4 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 2 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DENG e a sonda TaqMan correspondente..............................................54 FIGURA 5.5 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DEN3 e a sonda TaqMan correspondente...............................................54

ix
FIGURA 5.6 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DEN3 e o corante SYBR Green...............................................................55 FIGURA 5.7 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DEN3J e a sonda TaqMan correspondente.............................................55 FIGURA 5.8 – Exemplo de curvas de amplificação obtidas quando o RNA do sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os iniciadores DENG e a sonda TaqMan correspondente..............................................56 FIGURA 5.9 – Valores de fluorescência resultantes da hibridação das diferentes sondas com os produtos de RT-PCRs uniplex complementares a elas e com o branco.........................................................................................................................56 FIGURA 5.10 – Resultados dos ensaios de microarranjo líquido multiplex compostos pelas sondas DEN1, DEN2 e DEN3 hibridadas com os produtos de PCR dos três sorotipos do vírus da dengue, sob diferentes condições...........................................58 FIGURA 5.11 – Hibridação dos produtos da RT-PCR multiplex com a mistura das sondas DEN1, DEN2, DEN3 e DENG........................................................................60 FIGURA 5.12 – Resultados dos ensaios de microarranjo líquido multiplex realizados com diferentes combinações de sondas, para detecção e identificação dos sorotipos 1, 2 e 3 do vírus da dengue........................................................................................61 FIGURA 5.13 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 1 foi testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos sorotipo-específicos DEN1.........................................................................................65 FIGURA 5.14 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 1 foi testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos grupo-específicos DENG............................................................................................65 FIGURA 5.15 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 2 foi testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos sorotipo-específicos DEN2.........................................................................................66 FIGURA 5.16 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 2 foi testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos grupo-específicos DENG............................................................................................66 FIGURA 5.17 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 3 foi testado por PCR em tempo real com o corante SYBR Green utilizando os iniciadores sorotipo-específicos DEN3.........................................................................................67 FIGURA 5.18 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 3 foi testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos sorotipo-específicos DEN3J.......................................................................................67

x
FIGURA 5.19 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 3 foi testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos grupo-específicos DENG............................................................................................68

xi
LISTA DE TABELAS
TABELA 1.1 – Monitoramento da circulação viral, por UF, Brasil, janeiro a julho de 2007..............................................................................................................................7 TABELA 4.1 - Oligonucleotídeos iniciadores e sondas utilizados no ensaio de PCR em tempo real.............................................................................................................38 TABELA 4.2 - Oligonucleotídeos iniciadores e sondas, adaptados de Johnson et al. (2005), utilizados no ensaio de microarranjo líquido..................................................39 TABELA 5.1 – Razão entre os valores de fluorescência (MFI) obtidos nas reações das sondas com os produtos de PCR específicos e não-específicos........................63 TABELA 5.2 – Razão entre o MFI corrigido obtido com a hibridação das sondas DEN1, DEN2J, DEN3 e DENG com os produtos de RT-PCR complementares a elas e o MFI obtido para a mesma sonda na ausência de produto de PCR......................69 TABELA 5.3 - Detecção do RNA do vírus da dengue em soro de paciente, através de dois métodos diagnósticos distintos, utilizando oligonucleotídeos grupo-específicos..................................................................................................................71 TABELA 5.4 - Detecção do RNA do vírus da dengue em soro de paciente, através de dois métodos diagnósticos distintos, utilizando oligonucleotídeos sorotipo-específicos..................................................................................................................71 TABELA 5.5 – Resultados obtidos através da utilização de oligonucleotídeos grupo- e sorotipo-específicos em dois métodos diagnósticos distintos.................................72

xii
INSTITUTO OSWALDO CRUZ
Desenvolvimento e avaliação de métodos moleculares para o diagnóstico da dengue
RESUMO
TESE DE DOUTORADO
Celina de Oliveira Poersch A dengue é hoje a mais importante arbovirose que afeta o homem, constituindo um sério problema de saúde pública no mundo, especialmente em países tropicais onde as condições do meio ambiente favorecem a proliferação dos seus vetores, os mosquitos do gênero Aedes. Atualmente, quase metade da população mundial vive em áreas de risco para a doença e, apesar de sua extensão e gravidade, vacinas ou tratamentos específicos ainda não estão disponíveis. Tendo conhecimento da urgência na melhoria da vigilância epidemiológica da dengue e das deficiências do diagnóstico laboratorial dessa e de outras doenças no país, realizou-se esse trabalho cujo principal objetivo foi o desenvolvimento de métodos diagnósticos baseados em técnicas moleculares de última geração para a detecção e sorotipagem do vírus da dengue. As metodologias escolhidas foram as de PCR em tempo real e de microarranjo líquido, as quais representam os mais recentes avanços na área de diagnóstico molecular. Diferentes conjuntos de oligonucleotídeos sorotipo- e grupo-específicos foram avaliados quanto a capacidade de amplificar e identificar os sorotipos DENV1, DENV2 e DENV3 pelos dois métodos. Dessa forma, desenvolveu-se, baseado na técnica de PCR em tempo real, três ensaios uniplex sorotipo-específicos para dengue e um teste grupo-específico capaz de amplificar todos os sorotipos do vírus. Para o ensaio baseado na técnica de microarranjo líquido os oligonucleotídeos, após testados individualmente, foram combinados em diferentes formatos multiplex, tanto na reação de amplificação (PCR) como na de hibridação. Diferentes protocolos de hibridação foram também avaliados. Quando testada contra curvas padrões obtidas a partir da diluição de quantidades conhecidas dos três sorotipos do vírus, a PCR em tempo real mostrou-se, para todos os ensaios, capaz de amplificar e detectar mesmo a menor quantidade de vírus avaliada, equivalente a 53,57 FFU/ml. Já a sensibilidade do teste de microarranjo líquido multiplex foi de 3.428,48 FFU/ml para DENV1 e 53,57 FFU/ml para DENV2 e DENV3, com os oligonucleotídeos sorotipo-específicos e 214,28 FFU/ml para DENV3 e 53,57 FFU/ml para DENV1 e DENV2, com os oligonucleotídeos grupo-específicos. A especificidade dos dois métodos foi de 100%. A fim de avaliar a capacidade desses testes em diagnosticar e sorotipar o vírus da dengue em amostras clínicas, foram usados 50 soros positivos para IgM anti-dengue, 49 negativos por sorologia e 49 soros de pacientes cuja positividade para dengue foi confirmada por isolamento viral. A concordância entre os dois métodos foi de 92,5% quando comparados os resultados obtidos com os oligonucleotídeos grupo-específicos e de 90,5% para os ensaios sorotipo-específicos. Tomando o grupo positivo por isolamento viral como referência, o método de microarranjo líquido falhou em detectar três amostras que foram consideradas positivas por PCR em tempo real e detectou uma amostra que não foi detectada na PCR. Embora o método baseado em microarranjo líquido tenha apresentado menor sensibilidade do que a PCR em tempo real, as vantagens de permitir a análise simultânea de diversos alvos, a flexibilidade, versatilidade e custo acessível justificam um maior esforço na otimização desse teste. O presente trabalho serviu, acima de tudo, como um estudo de viabilidade técnica das metodologias citadas para o diagnóstico de doenças virais e, acredita-se, não trará apenas benefícios para o diagnóstico da dengue como também para de várias outras doenças prioritárias para o Brasil.

xiii
INSTITUTO OSWALDO CRUZ
Desenvolvimento e avaliação de métodos moleculares para o diagnóstico da dengue
ABSTRACT
TESE DE DOUTORADO
Celina de Oliveira Poersch Nowadays, dengue is the most important arbovirosis affecting humans, being a serious public health problem in the world, especially in tropical regions, where environmental conditions favor the proliferation of its vectors: mosquitoes from the genus Aedes. Currently, almost half of the world’s population lives in areas subject to the disease and, in spite of its breadth and gravity, no vaccines or specific treatments are available. Aware of the need to improve epidemiologic surveillance for dengue and of the deficiencies of laboratorial diagnoses for this and other diseases in the country, this study aimed to develop diagnostic methods based on state-of-the-art molecular techniques for detection and serotype identification of the dengue virus. The selected methods were real-time PCR and liquid microarray, which represent the most recent advances in the field of molecular diagnosis. Different sets of serotype and group-specific oligonucleotides where evaluated for their ability in amplifying and identifying the serotypes DENV1, DENV2 and DENV3 utilizing each of the two methods. In this way, based on the real-time PCR technique, we developed three uniplex serotype-specific assays for dengue and one group-specific test capable of amplifying all virus serotypes. For the assay based on the liquid microarray technique the oligonucleotides, after being individually tested, were combined in different multiplex formats, both in the amplification (PCR) and hybridization reactions. Different hybridization protocols were also evaluated. When tested against standard curves obtained from the dilution of known amounts of the three virus serotypes, real-time PCR proved to be capable, for all assays, of amplifying and detecting even the smallest quantity of virus tested, equivalent to 53,47 FFU/ml. In the other hand, sensibility of detection of the liquid microarray multiplex was 3.428,48 FFU/ml for DENV1 and 53,57 FFU/ml for DENV2 and DENV3, with the serotype-specific oligonucleotides, and 214,28 FFU/ml for DENV1 and 53,57 FFU/ml for DENV2 and DENV3 with the group-specific oligonucleotides. Both methods had 100% of specificity. In order to evaluate these test’s ability in diagnosing and identifying the serotypes of dengue virus in clinical samples, we utilized 50 sera positive for anti-dengue IgM, 49 negative by serology and 49 sera from patients whose infection with dengue virus was confirmed by viral isolation. Agreement between both methods was of 92,5% when comparing results obtained with the group-specific oligonucleotides and of 90,5% for the serotype specific assays. Taking the group confirmed as positive by viral isolation as a reference, the liquid microarray method failed in detecting three samples that were considered positive by the real-time PCR and detected one sample that was not detected by PCR. Although the liquid microarray method showed less sensibility than the real-time PCR, advantages coming from it allowing simultaneous analysis of several targets, its flexibility, versatility and feasible cost justify further work on the optimization of this test. This study served mainly as a test of the technical feasibility of the cited methods for the diagnosis of viral diseases and we believe that it will be beneficial not only for the diagnosis of dengue but also for that of many other priority diseases in Brazil.

1
1. INTRODUÇÃO
1.1. A dengue
Os primeiros relatos de grandes epidemias de uma doença que
possivelmente tratava-se de dengue datam de 1779 e 1780 e foram relatadas em
três continentes (Ásia, África e América do Norte). No entanto, enfermidades com
sintomas muito semelhantes aos da dengue já haviam sido descritas bem antes, em
uma enciclopédia médica chinesa editada no ano de 610 (GUBLER, 2006). Outros
possíveis surtos de dengue ocorreram em 1635 nas Índias Ocidentais e em 1699 no
Panamá (MCSHERRY, 1982). Portanto, é provável que a dengue já apresentasse
uma ampla distribuição geográfica mesmo antes do século XVIII, quando grandes
epidemias ocorreram por todo o mundo. Porém, a dengue foi considerada, por muito
tempo, uma doença benigna, com grandes epidemias acontecendo apenas em
intervalos de 10 a 40 anos (GUBLER e CLARK, 1995).
O sucesso do estabelecimento do ciclo urbano do vírus da dengue (DENV) se
deu, em grande parte, devido à expansão do mosquito africano Aedes aegypti.
Esses mosquitos evoluíram de forma a se tornarem intimamente associados com os
humanos, tornando-se eficientes vetores dos vírus da dengue e da febre amarela
(GUBLER, 1997). A partir do século XVII, devido ao transporte de escravos oriundos
da África, o Aedes aegypti foi levado para os demais continentes e as epidemias de
dengue tornaram-se mais freqüentes e disseminadas. Foi, contudo, após a Segunda
Guerra Mundial que ficou evidente a mudança no comportamento da doença. O
crescimento populacional, a urbanização descontrolada, a falta de programas
efetivos para o controle do vetor e o aumento das viagens comerciais contribuíram

2
para a expansão geográfica do mosquito transmissor e do vírus, permitindo o
estabelecimento de uma pandemia que persiste até hoje (GUBLER, 1997). Além do
aumento da incidência da forma clássica da patogenia (febre da dengue - DF),
relatou-se, em meados do século XX, a emergência de epidemias de uma nova e
mais severa forma da doença, denominada febre hemorrágica da dengue (DHF). A
primeira epidemia de febre hemorrágica ocorreu entre os anos de 1953 e 1954, nas
Filipinas, expandindo rapidamente para vários países asiáticos e ilhas do Pacífico
(WHO, 1997). Durante a década de 70, a DHF tornou-se a principal causa de
hospitalização e morte entre crianças na Ásia (MACKENZIE et al., 2004; WHO,
1997) e, nas décadas subseqüentes, epidemias de febre hemorrágica da dengue
passaram a ocorrer em diversos países.
As Américas Central e do Sul experimentaram as mudanças mais
dramáticas no padrão de ocorrência da doença. Nas décadas de 60 e 70 epidemias
de dengue eram raras devido à erradicação do Aedes aegypti da região. Com a
descontinuidade do programa de erradicação no início dos anos 70, o mosquito re-
invadiu, rapidamente, esses países (figura 1.1) e, desde 1981, grandes epidemias
de febre da dengue e febre hemorrágica da dengue vêm sendo registradas
(GUBLER, 1997; PINHEIRO, 1989).

3
FIGURA 1.1 – Distribuição do Aedes aegypti durante a década de 30 e nos anos de
1970 e 2002.
Fonte: GUBLER, 2006.
1.2. Situação atual da doença
Atualmente, a dengue é considerada a mais importante arbovirose (doença
viral transmitida por artrópodes) do mundo. Nos últimos cinqüenta anos a incidência
da doença aumentou cerca de trinta vezes (MACKENZIE et al., 2004) e,
aproximadamente, 2,5 bilhões de pessoas vivem em áreas de risco para a doença. A
cada década, o número anual de casos de dengue reportados para a Organização
Mundial da Saúde (OMS) cresce exponencialmente. Nos primeiros cinco anos da
década atual, a média anual de casos foi praticamente o dobro daquele reportado na
década anterior (figura 1.2) (NATHAN e DAYAL-DRAGER, 2006). Existem, no
momento, mais de 100 países endêmicos para a dengue e, apesar da fraca
vigilância epidemiológica de muitos países que registram casos da doença, estudos

4
sugerem que mais de 50 milhões de infecções ocorrem anualmente, resultando em
500.000 casos de DHF e 22.000 mortes, principalmente entre crianças das regiões
tropical e subtropical (http://www.who.int/csr/disease/dengue/impact/en/index.html).
FIGURA 1.2 – Média anual do número de casos de dengue, reportados à OMS, por
década.
Fonte: NATHAN e DAYAL-DRAGER, 2006.
A primeira grande epidemia de DHF, que se tem notícia nas Américas,
ocorreu em Cuba, em 1981, com mais de 10.000 casos dessa forma da doença e
158 mortes. De 1981 a 2005, mais de 120.000 casos confirmados de febre
hemorrágica da dengue foram reportados por 28 países da região (GUBLER, 2006;
http://www.paho.org/english/ad/dpc/cd/dengue.htm). Em 2002, foram registrados,
pela primeira vez, mais de 1 milhão de casos totais (DF + DHF) da doença nas
Américas (NATHAN e DAYAL-DRAGER, 2006). Esse número apresentou uma
queda significativa entre 2003 e 2005, voltando, contudo, a crescer em 2006 e 2007.
No ano de 2006 foram registrados 552.141 casos totais de dengue, sendo 14.459 de
908 15497
122174
295554
479848
925896
0100.000200.000300.000400.000500.000
600.000700.000800.000900.000
1.000.000
1955
-195
9
1960
-196
9
1970
-197
9
1980
-198
9
1990
-199
9
2000
-200
5

5
febre hemorrágica. Em 2007, já foram reportados, até o início de outubro, 643.123
casos totais da doença, com mais de 13.000 casos de DHF, resultando em 186
mortes (figura 1.3).
FIGURA 1.3 – Número de casos totais de dengue e de febre hemorrágica da dengue
reportados à OMS, por ano, na região das Américas.
Fonte: http://www.paho.org/english/ad/dpc/cd/dengue.htm
No Brasil, um processo de intensa circulação viral em grandes centros
urbanos iniciou-se somente a partir de 1986, com epidemias explosivas, que foram
se expandindo para todas as regiões brasileiras. No período entre 1986 e 1990,
0
200.000
400.000
600.000
800.000
1.000.000
1.200.000
NÚMERO TOTAL DE CASOS DE DENGUE NAS AMÉRICAS
NÚMERO DE CASOS DE DHF NAS AMÉRICAS
0
2.000
4.000
6.000
8.000
10.000
12.000
14.000
16.000
18.000
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006

6
esses eventos ficaram restritos a alguns estados das regiões Sudeste (Rio de
Janeiro, São Paulo e Minas Gerais) e Nordeste (Pernambuco, Alagoas, Ceará e
Bahia). Inicialmente, foi identificado o sorotipo DENV-1 e, em 1990, o DENV-2 foi
introduzido, passando ambos a circularem intensa e simultaneamente no país. A
partir de 1994, a rápida dispersão do vetor em grande extensão territorial propiciou a
circulação viral em um maior número de estados e municípios, expondo novas
populações, paulatinamente, às infecções. Na década de 90, mesmo em anos não
epidêmicos, a doença registra uma ocorrência de dezenas de milhares de casos por
ano (MS, 1999). O último pico epidêmico ocorreu em 2002, em decorrência da
introdução do sorotipo DENV-3 no país, tendo sido registrados 794.219 mil casos, a
maioria deles no Rio de Janeiro. A dispersão do DENV-3 para os demais estados
tem proporcionado, desde então, o surgimento de surtos e epidemias, que apesar de
graves ainda não atingiram os níveis de 2002. Atualmente, o vetor e o vírus da
dengue encontram-se disseminados pela quase totalidade do território brasileiro e o
país contribui com mais de 80% dos casos registrados nas Américas. O
monitoramento da circulação viral em 2007 (tabela 1.1) demonstrou que o sorotipo
DENV-3 é predominante no país, representando 81% das amostras isoladas.
Entretanto, observa-se, também, um percentual importante (15,2%) de isolamentos
do sorotipo 2 e a persistência do sorotipo 1.

7
TABELA 1.1 – Monitoramento da circulação viral, por UF, Brasil, janeiro a julho de
2007.
Fonte: Secretaria de Vigilância em Saúde (SVS/MS), 2007. *Dados até 31 de Julho de 2007 SI: sem informação
No período de janeiro a julho de 2007, a Secretaria de Vigilância em Saúde
do Ministério da Saúde (SVS/MS) registrou, no Brasil, a ocorrência de 438.949 casos
de dengue clássica (DF), 926 casos de febre hemorrágica da dengue e 98 óbitos
(figura 1.4). Ao compararmos com o ano de 2006, observa-se um aumento de

8
136.488 casos de dengue no país. O Estado de Santa Catarina continua sem
transmissão autóctone de dengue, registrando 420 casos importados. O Estado do
Rio Grande do Sul notificou o primeiro caso confirmado de dengue autóctone, em
abril de 2007, sendo que, no período de janeiro a julho foram notificados 1.016
casos suspeitos de dengue no estado. Para este mesmo período, o Estado do
Paraná registrou 43.691 casos. As transmissões ocorridas no Paraná em 2007
contribuíram para que a Região Sul apresentasse um aumento de 871% no número
de casos quando comparado com o mesmo período de 2006 (MS, 2007).
FIGURA 1.4 – Número total de casos de dengue (DF + DHF) e de febre hemorrágica
da dengue (DHF) por ano, no Brasil.
Fonte: Ministério da Saúde, Secretaria de Vigilância em Saúde, 2007 (MS, 2007).
NÚMERO DE CASOS DE DHF NO BRASIL
0
500
1.000
1.500
2.000
2.500
3.000
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
NÚMERO TOTAL DE CASOS DE DENGUE NO BRASIL
0
100.000
200.000
300.000
400.000
500.000
600.000
700.000
800.000
1980
1983
1986
1989
1992
1995
1998
2001
2004
2007

9
1.3. O vírus
O vírus da dengue (DENV) pertencente à família Flaviviridae e ao gênero
Flavivirus e pode ser classificado em quatro sorotipos distintos, sendo eles DENV-1,
DENV-2, DENV-3 e DENV-4, os quais divergem em, aproximadamente, 30% de
suas seqüências protéicas (HOLMES, 2006). O virion tem formato esférico e
diâmetro entre 40 e 60 nm. O nucleocapsídeo icosaédrico, composto por proteínas C
(capsídeo) e RNA fita simples de polaridade positiva, é envolvido por uma bi-camada
lipídica a qual constitui o envelope. Embebidas na bi-camada do envelope,
encontram-se proteínas E (envelope) e M (membrana), que também pode ser
encontrada em sua forma não processada prM (pré-membrana) (figura 1.5). A
proteína E possui os domínios responsáveis pela interação do vírus com a
membrana da célula hospedeira (CHEN et al., 1996), bem como os epítopos
considerados os principais alvos da resposta humoral, alguns sendo comuns a todos
os flavivírus e outros específicos do grupo ou do sorotipo (GUZMÁN e KOURI,
1996). Após a entrada do vírus na célula por endocitose, a proteína E é, também,
capaz de promover a fusão da membrana do vírus com a membrana do endossomo,
permitindo que a partícula infectante escape dessas vesículas (ROEHRIG, 1997;
STIASNY et al., 2006).

10
FIGURA 1.5 - Desenho esquemático representando a estrutura do vírus da dengue.
Os macrófagos e monócitos são considerados os principais alvos do vírus e
a infecção dessas células ocorre, principalmente, através da interação de receptores
de Fc com partículas virais opsonizadas, sendo, portanto, particularmente importante
em infecções secundárias. Outras células, como, por exemplo, hepatócitos,
linfócitos, células de Kupffer (ROTHMAN, 1997) e células de Langerhans, são, no
entanto, também permissíveis ao vírus, sendo infectadas por um mecanismo
independente de Fc (WU et al., 2000). Nessas células, moléculas de heparan sulfato
presentes em sua superfície interagem com o vírus, facilitando a ligação da proteína
E com receptores específicos, ainda não completamente caracterizados (CHEN et
al., 1997; GERMI et al., 2002; THULLIER et al., 2001).
O genoma do vírus tem um comprimento aproximado de 11 kilobases (kb) e
codifica, além das proteínas estruturais C, prM e E, sete proteínas não estruturais
(NS) denominadas de NS1, NS2A, NS2B, NS3, NS4A, NS4B e NS5. O RNA é
traduzido como uma seqüência única a partir do primeiro ou segundo códon AUG
presente na porção codificadora da molécula, resultando em uma poli-proteína de

11
cerca de 3.400 aminoácidos, que é clivada co- e pós-traducionalmente por proteases
do vírus e do hospedeiro. A região codificadora é flanqueada por regiões não
traduzidas conservadas, fundamentais para a replicação do RNA e para a regulação
da tradução (ALVAREZ et al., 2006). Embora o mecanismo exato de ação de
algumas das proteínas não estruturais ainda não esteja completamente esclarecido,
sabe-se que todas estão envolvidas na replicação ou no processamento da poli-
proteína viral (CHAMBERS et al., 1990; RICE, 2001).
1.4. Aspectos clínicos e patogênese
A infecção por qualquer um dos sorotipos do vírus causa manifestações
clínicas semelhantes que podem variar de febre comum, não diferenciada, a febre
alta associada a fenômenos hemorrágicos, hepatomegalia e aumento da
permeabilidade vascular, caracterizando a febre hemorrágica da dengue (DHF)
(ROTHMAN e ENNIS, 1999). A infecção em humanos por um sorotipo confere
imunidade permanente contra re-infecções subseqüentes pelo mesmo sorotipo, mas
apenas proteção parcial e temporária contra os outros sorotipos. Tem sido
observado que em infecções subseqüentes por diferentes sorotipos do vírus a
chance de desenvolvimento de uma forma mais severa da doença é maior. Esse fato
pode ser explicado pela teoria da “potencialização dependente de anticorpos”
(antibody-dependent enhancement), segundo a qual anticorpos não-neutralizantes,
adquiridos em uma infecção prévia por um vírus heterólogo, ligam-se ao novo
sorotipo infectante e facilitam sua entrada nos macrófagos, por intermédio dos
receptores de Fc (HALSTEAD, 1988; HALSTEAD, 2003; SIMMONS et al., 2006).
Esse mecanismo levaria à rápida ativação de linfócitos T pré-existentes, causando a

12
liberação exacerbada de citocinas e mediadores químicos que causariam os
sintomas característicos da febre hemorrágica (KURANE e ENNIS, 1994). De fato,
os níveis de diversas citocinas, tais como IL-2 (KURANE e INNIS, 1991), IL-6 e
TNFα (KUNO e BAILY, 1994), já foram encontrados elevados em pacientes com
DHF quando comparados com indivíduos com febre da dengue. A hipótese de que a
febre hemorrágica da dengue é causada por fatores predominantemente
imunológicos associados a infecções secundárias é fortemente sustentada pelo fato
de que 90% dos pacientes que desenvolvem a forma mais severa da doença
apresentam evidências sorológicas de uma infecção prévia (SUAYA et al., 2006) e
de que casos de febre hemorrágica ocorrem preferencialmente em áreas onde há a
circulação de múltiplos sorotipos e costuma acometer habitantes dessas regiões,
enquanto que visitantes manifestam a forma clássica da doença (ROTHMAN, 1997).
A observação de que casos de febre hemorrágica acontecem em infecções
primárias e de que nem todas as infecções secundárias resultam em DHF sugere,
no entanto, que outros fatores devem contribuir para a severidade da doença. Entre
eles pode-se citar status imunológico e características genéticas do hospedeiro
(HALSTEAD, 1988; KOURI et al., 1989; SAKUNTABHAI et al., 2005; SIERRA et al,
2006), e alterações na seqüência nucleotídica das cepas, que podem afetar sua
virulência (LEITMEYER et al., 1999; RICO-HESSE et al., 1997; VAUGHN et al.,
2000; WATTS et al., 1999). Esses fatores podem, por exemplo, influenciar na
magnitude da viremia, que apesar de não explicar por si só todos os aspectos da
patogênese parece estar associada com a evolução da doença (LIBRATY et al.,
2002; VAUGHN et al., 2000).
O período de incubação da dengue varia de 3 a 14 dias e o período febril
costuma durar de 2 a 7 dias. Na dengue clássica, a febre é normalmente
acompanhada de mialgia, artralgia, dor retrorbital, fotofobia e linfoadenopatia,

13
podendo ocorrer náusea, vômito, leucopenia, trombocitopenia, erupções cutâneas e
manifestações hemorrágicas. Na primeira fase da febre hemorrágica os sintomas
são muito semelhantes aos da forma clássica. Após a remissão da febre, no entanto,
o aumento da permeabilidade vascular leva à rápida perda de fluidos, que, se não
tratada adequadamente, pode levar a choque e morte (GUBLER, 1998).
A dengue é uma doença de etiologia complexa sendo sua evolução ditada por
interações entre fatores virais e do hospedeiro, e, apesar de alguns avanços, a
elucidação da patogênese depende, ainda, de muita investigação (WHO, 2006).
1.5. Viremia e cinética da resposta humoral ao vírus
A viremia atinge seu pico logo após o aparecimento dos primeiros sintomas,
muitas vezes antes mesmo do paciente apresentar-se doente o suficiente para
recorrer a tratamento médico. Vírus circulantes permanecem, no entanto,
detectáveis geralmente até o quinto dia de doença, coincidindo com o período em
que os níveis de anticorpos começam a elevar-se (VORNDAM e KUNO, 1997).
Anticorpos contra sete das dez proteínas codificadas pelo vírus da dengue já
foram descritos (BRINTON et al., 1998), sendo que aqueles direcionados contra a
proteína E são os únicos neutralizantes e os mais freqüentemente detectados tanto
em respostas primárias quanto em secundárias (VALDÉS et al., 2000).
O isotipo dominante de imunoglobulina em uma infecção primária é o IgM.
Uma pequena porcentagem dos pacientes apresenta níveis detectáveis de IgM já no
primeiro dia da doença e a maioria (cerca de 93%) encontra-se positivo no sexto dia
após o aparecimento dos primeiros sintomas (OPAS, 2004). Os níveis de IgM

14
aumentam rapidamente e atingem seu pico por volta de duas semanas,
permanecendo detectáveis por 2 a 3 meses, o que faz desses anticorpos bons
indicadores de infecções recentes (GUBLER e SATHER, 1988). Anticorpos IgG
começam a aparecer, em uma resposta primária, alguns dias depois dos IgM, sendo
detectáveis a partir do quinto dia de doença. Os títulos de IgG aumentam lentamente
a partir da primeira semana de infecção e permanecem detectáveis por toda a vida.
Indivíduos com imunidade prévia ao vírus da dengue ou mesmo a outro flavivírus
desenvolvem uma resposta secundária caracterizada pelo rápido aumento no título
de IgG quase que imediatamente após o início dos sintomas e pelo alto grau de
reação cruzada, mesmo contra outros flavivírus, apresentado por esses anticorpos
(INNIS et al., 1989). Os níveis de IgM são consideravelmente mais baixos do que na
resposta primária (figura 1.5) (WHO, 1997). A relação entre os títulos de IgM e IgG e
a especificidade dos anticorpos podem ser, portanto, usados na identificação de
respostas primárias e secundárias.
Anticorpos IgM e IgG podem ser específicos para um determinado sorotipo
em algumas infecções primárias, o que geralmente não ocorre em infecções
secundárias. O grau de reação cruzada apresentada por esses anticorpos é variável
e depende do antígeno e do teste diagnóstico utilizado (VORNDAM e KUNO, 1997).
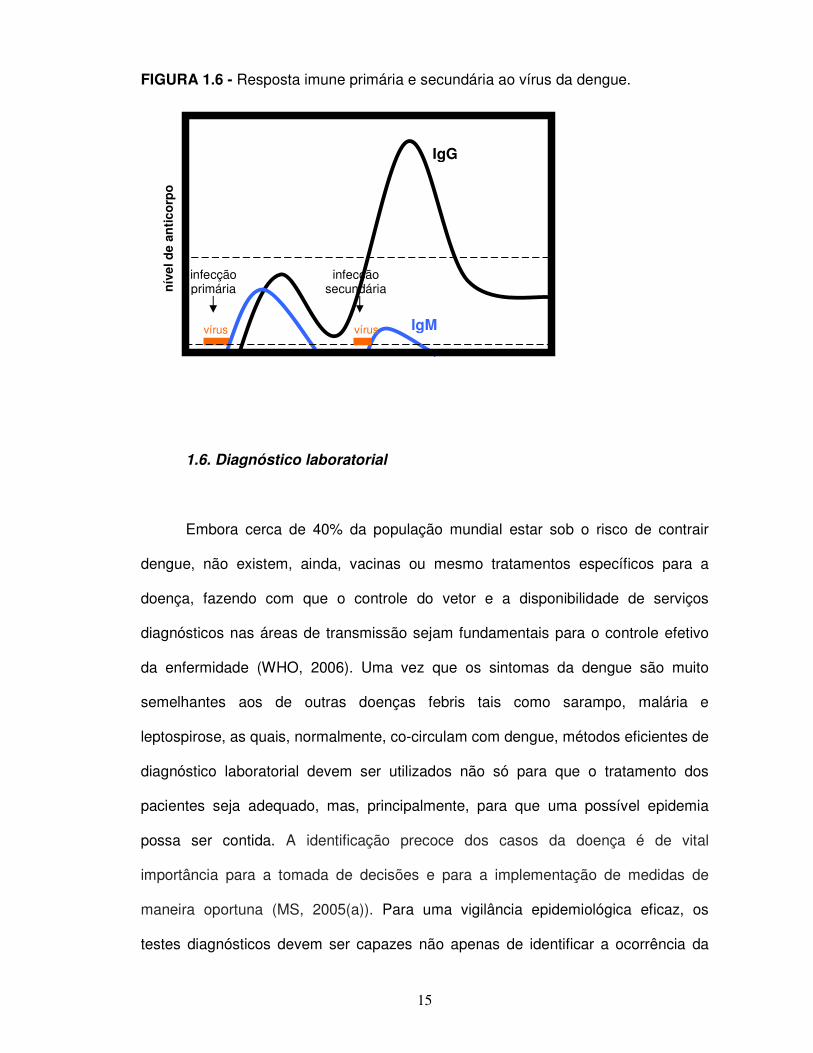
15
FIGURA 1.6 - Resposta imune primária e secundária ao vírus da dengue.
1.6. Diagnóstico laboratorial
Embora cerca de 40% da população mundial estar sob o risco de contrair
dengue, não existem, ainda, vacinas ou mesmo tratamentos específicos para a
doença, fazendo com que o controle do vetor e a disponibilidade de serviços
diagnósticos nas áreas de transmissão sejam fundamentais para o controle efetivo
da enfermidade (WHO, 2006). Uma vez que os sintomas da dengue são muito
semelhantes aos de outras doenças febris tais como sarampo, malária e
leptospirose, as quais, normalmente, co-circulam com dengue, métodos eficientes de
diagnóstico laboratorial devem ser utilizados não só para que o tratamento dos
pacientes seja adequado, mas, principalmente, para que uma possível epidemia
possa ser contida. A identificação precoce dos casos da doença é de vital
importância para a tomada de decisões e para a implementação de medidas de
maneira oportuna (MS, 2005(a)). Para uma vigilância epidemiológica eficaz, os
testes diagnósticos devem ser capazes não apenas de identificar a ocorrência da
infecção primária
infecção secundária n
ível
de
anti
corp
o
IgG
IgM vírus vírus

16
doença, mas também detectar o sorotipo do vírus responsável pela infecção. Além
de contribuir para a melhoria da vigilância epidemiológica e para o manejo clínico
dos pacientes, o uso de testes diagnósticos rápidos e confiáveis são imprescindíveis
para o desenvolvimento e futuras aplicações de anti-virais e vacinas (NIESTERS,
2002).
Tradicionalmente, a dengue tem sido diagnosticada por métodos sorológicos
ou através de isolamento viral. Com os recentes avanços das técnicas moleculares,
no entanto, novos testes têm sido desenvolvidos e devem, em breve, estar
disponíveis comercialmente (BUCHY et al., 2006).
Uma descrição sucinta das técnicas utilizadas para detecção do vírus ou de
anticorpos anti-dengue no soro de pacientes, destacando as principais
características de cada uma dessas abordagens, será feita a seguir.
1.6.1. Inibição da hemaglutinação
Esse ensaio depende da habilidade dos anticorpos em inibir a aglutinação de
hemácias mediada por glicoproteínas virais e, desde o seu desenvolvimento, em
1958, permaneceu praticamente inalterado (VORNDAM e KUNO, 1997). Os soros
devem ser testados em diversas diluições e tratados com acetona ou caolina, a fim
de que inibidores da hemaglutinação sejam removidos. Uma vez que esse teste não
faz a diferenciação entre IgM e IgG, um resultado confiável requer a utilização de
amostras pareadas, coletadas com um intervalo mínimo de 7 dias. Reações
positivas são caracterizadas por um aumento de quatro vezes ou mais entre os
títulos da primeira e segunda amostra, com um título máximo de, pelo menos,
1:1280 em respostas secundárias e menor do que isso em respostas primárias.
Respostas cruzadas com outros flavivírus são comuns, fazendo com que um
diagnóstico preciso não seja possível com base apenas nesse teste (WHO, 1997).

17
1.6.2. Fixação do complemento
Esse teste é pouco utilizado para o diagnóstico da dengue devido a sua baixa
sensibilidade e pouca praticidade (GUBLER, 1998). Ele é baseado no principio de
que moléculas do complemento são consumidas em uma reação antígeno-anticorpo.
Em uma primeira etapa da reação, o soro do paciente, o antígeno e quantidades
conhecidas de complemento são incubados juntos. Se houver a fixação do
complemento, não haverá a lise de hemácias adicionadas em uma segunda etapa
da reação. Uma das vantagens da técnica é que anticorpos fixadores de
complemento aparecem posteriormente aos anticorpos responsáveis pela inibição
da hemaglutinação e persistem por pouco tempo no soro, tornando-a útil para a
detecção de infecções correntes. Além disso, em infecções primárias, os anticorpos
fixadores de complemento são relativamente específicos, permitindo, em alguns
casos, a diferenciação do vírus quanto ao sorotipo (VORNDAM e KUNO, 1997).
1.6.3. Teste de neutralização
Esse teste mede o título de anticorpos neutralizantes em soros de pacientes
convalescentes. A técnica se baseia no princípio de que alguns anticorpos
específicos para o vírus, quando se ligam ao mesmo, impedem sua infecção e
replicação em células de cultura (BUCHY et al., 2006). Diluições do soro,
previamente inativado por calor, são incubadas com quantidades definidas de vírus.
A fração de vírus não neutralizada é adsorvida a uma monocamada de células
suscetíveis e, após um período apropriado de incubação, as placas resultantes são
contadas. O título é determinado como sendo a diluição máxima do soro capaz de
reduzir em 50 a 90% o número de placas. Um aumento de quatro vezes ou mais
entre os títulos de amostras pareadas é indicativo de infecção corrente. Apesar de
ser o método mais sensível de diagnóstico sorológico, é um teste demorado e

18
tecnicamente difícil (CALISHER et al., 1989; GUBLER, 1998). Também, a
interpretação dos resultados é muito influenciada pela linhagem celular e estoque
viral utilizado (BUCHY et al., 2006).
1.6.4. ELISA (enzyme-linked immunosorbent assay)
1.6.4.1. MAC-ELISA
Entre os testes sorológicos para o diagnóstico da dengue, os ensaios imuno-
enzimáticos (ELISAs) são considerados os mais eficientes devido a sua praticidade,
especificidade e sensibilidade. O teste de MAC-ELISA (IgM antibody-capture
enzyme-linked immunosorbent assay), em particular, vem sendo amplamente
utilizado para a confirmação de infecções recentes (WANG et al., 2000). Nesse
teste, anticorpos IgM do soro dos pacientes são capturados não especificamente por
anticorpos anti-IgM fixados em uma placa. Os anticorpos capturados são, então,
testados com antígenos do vírus da dengue e reações positivas são detectadas
utilizando-se um segundo anticorpo anti-dengue conjugado a uma enzima
(VORNDAM e KUNO, 1997). Preparações relativamente grosseiras de antígeno,
como macerado de cérebro de camundongo infectado ou extrato de cultura celular,
podem ser utilizadas, embora testes elaborados com tais reagentes podem
apresentar maior variabilidade intra-lote e são, em geral, mais suscetíveis a
problemas de reação cruzada. O uso de proteínas recombinantes tende a diminuir
ou mesmo a eliminar esses inconvenientes, representando, assim, uma excelente
alternativa para a obtenção de antígenos (CUZZUBBO et al., 2001). A possibilidade
de ocorrência de reações cruzadas deve, no entanto, ser sempre considerada em
testes do tipo ELISA, principalmente quando utilizados em regiões onde existe a co-
circulação de múltiplos flavivírus (SUAYA et al., 2006). Este é o caso do Brasil,
aonde boa parcela da população foi imunizada e, portanto, possui anticorpos

19
circulantes, contra o vírus da febre amarela. Se a reação cruzada entre anticorpos
contra diferentes flavivírus e antígenos da dengue pode acontecer, a reação cruzada
entre anticorpos e antígenos de diferentes sorotipos do vírus da dengue é
praticamente uma regra, impossibilitando a sorotipagem mesmo em casos de
infecção primária (BUCHY et al. 2006).
1.6.4.2. IgG-ELISA
Os testes IgG-ELISA seguem os mesmos princípios e em geral se utilizam
dos mesmos antígenos dos testes de captura de IgM. Porém, por identificarem a
presença de anticorpos do isotipo IgG específicos para o vírus da dengue, são
usados para a detecção de infecções passadas. Em geral, esses testes não são, da
mesma forma, sorotipo-específicos, mas apresentam menos reação cruzada com
outros flavivírus do que os ensaios de MAC-ELISA. Embora os testes de captura de
IgG sejam importantes em estudos de prospecção sorológica, sozinhos não têm
valor para o diagnóstico de infecções agudas ou recentes (BUCHY et al., 2006).
1.6.4.3. Razão IgM / IgG
Uma vez que testes ELISA de captura de IgM e IgG permitem a detecção
quantitativa e qualitativa dos anticorpos, esses dois ensaios podem ser utilizados em
conjunto, a fim de possibilitar a distinção entre respostas primárias e secundárias
(INNIS et al., 1989; MIAGOSTOVICH et al., 1999). A razão IgM / IgG é usada para
este propósito. Razões maiores do que 1,2 são indicativas de respostas primárias ao
vírus e menores do que este valor de respostas secundárias. Esses valores, no
entanto, podem não ser reais em pacientes que apresentam respostas humorais não
clássicas para a doença (FALCONAR et al. 2006). Testes utilizando tiras de
nitrocelulose como suporte sólido, calibrados para detectar IgM em respostas

20
primárias e secundárias e IgG apenas em respostas secundárias, já disponíveis
comercialmente, podem ser usados, da mesma maneira, para diferenciar esses dois
tipos de resposta (CUZZUBBO et al., 2001). A substituição das placas de ELISA por
tiras individuais de nitrocelulose nesses testes permite que eles sejam realizados
sem nenhum equipamento específico. Essa mesma característica, porém, dificulta a
análise de muitas amostras simultaneamente e torna a quantificação muito subjetiva,
já que ela é totalmente dependente de julgamento visual.
1.6.4.4. NS1-ELISA
Atualmente, vários testes de captura de IgM e IgG estão disponíveis
comercialmente, possibilitando a análise rápida e reprodutível de um grande número
de amostras, sem a necessidade de equipamentos sofisticados (LAM et al., 2000;
PALMER et al., 1999; VAUGHN et al., 1999). Apesar das qualidades desses testes e
dos avanços que eles propiciaram na área de diagnóstico, a mais importante
limitação inerente a todos os métodos sorológicos, ou seja, a impossibilidade de
detecção da doença em sua fase inicial, não deve ser menosprezada. A fim de
sobrepujar esse problema e continuar usufruindo os benefícios propiciados pelos
ELISAs, alguns ensaios imuno-enzimáticos para detecção da proteína viral NS1 vêm
sendo desenvolvidos com sucesso (ALCON et al., 2002; YOUNG et al., 2000). O
antígeno NS1 está presente no soro de indivíduos infectados desde o primeiro dia
de sintoma clínico e permanece detectável até o quinto ou sexto dia. Como essa
proteína aparece no início da infecção, antes do surgimento dos primeiros
anticorpos, seu uso como uma ferramenta de detecção precoce da dengue vem
sendo investigado (XU et al., 2006). Alguns estudos, além de avaliarem a detecção
de NS1 como ferramenta para o diagnóstico, identificaram uma correlação entre a

21
severidade da doença e a quantidade desse antígeno no soro (YOUNG et al., 2000;
LIBRATY et al., 2002).
Uma vez que NS1 produz, também, uma forte resposta humoral, o uso dessa
proteína como antígeno em testes de detecção de anticorpos é bastante promissora.
Um ensaio padronizado por SHU et al. (2003) utilizando proteínas NS1
recombinantes mostrou ser sorotipo-específico para soros de fase convalescente de
pacientes com infecção primária e soros de fase aguda de pacientes com infecção
secundária. Esse teste foi, também, capaz de diferenciar respostas primárias de
secundárias, além de apresentar boa correlação com os resultados obtidos através
de testes de neutralização. Outros estudos usando a proteína NS1, no entanto, não
apresentaram os mesmos resultados, sugerindo que mais avaliações são
necessárias a fim de determinar o potencial desses ensaios (BUCHY et al., 2006).
1.6.5. Isolamento viral
Em contraste aos métodos sorológicos, a técnica de isolamento viral é capaz
de diagnosticar a dengue durante a fase aguda da doença, quando os títulos de
anticorpos não atingiram, ainda, níveis detectáveis. O isolamento pode ser feito
inoculando-se soro ou plasma do paciente em cérebro de camundongo, mosquito ou
cultura de células de mamífero ou de inseto. A inoculação em cérebro de
camundongo e em células de mamífero em cultura são técnicas caras, demoradas e
pouco sensíveis e, por isso, não recomendadas (GUBLER, 1998). A inoculação de
mosquito é o método mais sensível, sendo o mais confiável para a confirmação de
casos fatais de febre hemorrágica da dengue. O vírus se replica na maioria dos
tecidos do mosquito e atinge títulos elevados em 4 ou 5 dias. Porém, esse sistema
tem a desvantagem de ser trabalhoso e requerer instalações especiais para a
produção e manutenção dos mosquitos. A utilização de linhagens celulares de

22
insetos é uma boa alternativa à inoculação de mosquitos. Apesar de não ser tão
sensível como esta última técnica e de ser mais suscetível a contaminações, ela é
econômica, requer menos habilidade técnica e permite o processamento de uma
grande quantidade de soros simultaneamente (GUZMÁN e KOURI, 1996; WHO,
2006). O vírus isolado por qualquer um dos métodos acima, ou mesmo presente em
amostras fixadas de tecido, é normalmente detectado através de imunofluorescência
indireta com anticorpos monoclonais sorotipo-específicos (VORNDAM e KUNO,
1997).
No entanto, o vírus pode ser inativado por calor, pH e alguns químicos,
fazendo com que o sucesso do isolamento seja seriamente afetado por problemas
de manipulação e armazenamento das amostras. Outro fator a ser considerado é
que o tempo de duração da viremia é de, apenas, 4 ou 5 dias e algumas cepas
podem manter um título viral abaixo do limite de detecção durante toda a infecção. O
aumento do título de anticorpos faz, também, com que a eficiência do método
diminua rapidamente, o que é particularmente problemático em casos de infecções
secundárias (GUBLER, 1998).
1.6.6. RT-PCR (reverse transcription – polymerase chain reaction)
Uma outra forma de verificar a presença do vírus da dengue é através da
detecção do RNA viral por RT-PCR. A metodologia de PCR, devido sua alta
sensibilidade e rapidez na detecção de, potencialmente, qualquer seqüência
genética, revolucionou a área de diagnóstico de doenças infecciosas (RATCLIFF et
al., 2007). Assim como a técnica de isolamento viral, possui a vantagem, com
relação aos métodos sorológicos, de possibilitar a sorotipagem do vírus infectante e
permitir o diagnóstico da doença em sua fase inicial (DEUBEL, 1992). No entanto,
essa técnica é menos influenciada por problemas de armazenamento e manipulação

23
do soro e pela presença de anticorpos neutralizantes do que o isolamento, além de
ser capaz de fornecer resultados em um período de tempo consideravelmente mais
curto (GUBLER, 1998; SCHILLING et al., 2004).
A RT-PCR vem sendo cada vez mais utilizada como um método diagnóstico
para a dengue (PAULA et al. 2002a). Vários protocolos de RT-PCR para detecção
do RNA do vírus e determinação do seu sorotipo foram desenvolvidos nos últimos
anos, como, por exemplo, o uso de quatro iniciadores sorotipo-específicos em uma
única reação (HARRIS et al., 1998) e reações de amplificação utilizando um único
par de iniciadores universais para dengue seguidas de nested PCR (LANCIOTTI et
al., 1992), análise com enzimas de restrição (PAULA et al., 2002a), ou hibridação
com sondas específicas (HENCHAL et al., 1991).
A RT-PCR permite a detecção de diminutas quantidades de RNA viral mesmo
em amostras onde o vírus está inativo ou associado a anticorpos. No entanto, ela é
altamente sujeita a contaminações, depende de cuidados específicos com a
manipulação do RNA e, assim como ocorre com o isolamento viral, sua aplicação
está restrita ao período de viremia (WHO, 1997). As vantagens da RT-PCR, contudo,
devem contribuir para que essa metodologia substitua, em definitivo, o isolamento
viral como teste para o diagnóstico precoce da infecção.
1.6.6.1. RT-PCR em tempo real
A aceitação da PCR como método diagnóstico e sua implementação na rotina
laboratorial foram grandemente beneficiadas com o advento da PCR em tempo real
(MACKAY et al., 2002; NIESTERS, 2001), o qual ocorreu no início da década de 90
(HIGUCHI et al., 1992; HOLLAND et al., 1991; LEE et al., 1993; LIVAK et al., 1995;).
A PCR em tempo real permite que a detecção dos fragmentos amplificados
(amplicons) seja feita de forma simultânea à reação de amplificação, em oposição

24
aos métodos tradicionais de PCR, aonde o produto de amplificação é detectado
apenas ao final da reação, com a utilização de uma segunda técnica, normalmente
eletroforese em gel. O monitoramento da presença e acúmulo do amplicon em
tempo real é possível graças à marcação de sondas, iniciadores ou do próprio
produto de PCR com moléculas fluorogênicas (MACKAY et al., 2002). A utilização
dessa química, com a conseqüente eliminação de uma etapa de manipulação e
análise após a amplificação, acrescentou ao extremamente sensível método de PCR
maior praticidade, menor tempo para obtenção do resultado e diminuição da
possibilidade de contaminação cruzada (HEID et al., 1996; SHU et al.,2003). Uma
vez que a metodologia de PCR em tempo real permite que os passos de transcrição
reversa (RT), PCR e detecção do produto sejam feitos seqüencialmente, em um
sistema fechado, sem a necessidade de qualquer etapa de manipulação entre eles,
essa técnica é considerada muito mais confiável e reprodutível, além de mais rápida,
do que as técnicas tradicionais de amplificação e, portanto, extremamente
apropriada para o diagnóstico molecular em laboratórios clínicos (BUSTIN, 2002;
VERNET, 2004). Adicionalmente às vantagens já citadas, a PCR em tempo real
possibilita também a quantificação do RNA ou DNA adicionado no início da reação,
e, portanto, a determinação da carga viral, em uma ampla faixa de concentração
(NIESTERS, 2002). Diversos protocolos utilizando essa metodologia para o
diagnóstico ou para a quantificação do RNA do vírus da dengue já foram descritos
(CALLAHAN et al., 2001; CHEN et al., 2001; DROSTEN et al., 2002; HOUNG et al.,
2000; HOUNG et al., 2001; ITO et al., 2004; JOHNSON et al., 2005(b); KONG et al.,
2006; LAI et al., 2007; LAUE et al., 1999; WANG et al., 2002(b); WANG et al., 2003).
A desvantagem da PCR em tempo real em relação à PCR convencional
reside, principalmente, na restrita capacidade multiplex da primeira. A amplificação e
detecção simultânea de alvos distintos é limitada, na PCR em tempo real, pelo

25
número de grupamentos fluorescentes que o equipamento é capaz de detectar (no
máximo cinco) (MACKAY et al., 2002). Esse fato ganha maior importância quando se
pretende identificar agentes que possuem grande variabilidade genética, tornando
os métodos multiplex de detecção tendenciosos e limitados a uma pequena
quantidade de patógenos. GARDNER et al. (2003), por exemplo, estimaram a partir
de dados de alinhamento, que um ensaio de PCR em tempo real apto a detectar
todas as linhagens de HIV com a mesma sensibilidade, deveria conter, no mínimo,
nove pares de iniciadores e sondas. Dessa forma, o caminho natural para aumentar
a capacidade dos testes, permitindo a detecção simultânea de diferentes alvos
moleculares, deve passar, obrigadoriamente, pela introdução de novas plataformas
tecnológicas.
1.6.7. Metodologias moleculares inovadoras potencialmente utilizáveis
para o diagnóstico da dengue
1.6.7.1. Microarranjo tradicional
A técnica de microarranjo consiste na hibridação de diferentes fragmentos de
ácidos nucléicos, marcados com moléculas fluorescentes, com um conjunto de
sondas de oligonucleotídeos imobilizadas em uma superfície sólida (geralmente
vidro, silicone ou plástico). A formação dos híbridos é então detectada através do
mapeamento do sinal fluorescente. As dimensões das áreas cobertas pelas sondas
individuais, denominadas de spot, são microscópicas e organizadas de forma
ordenada, justificando, portanto, o nome de microarranjos. Uma das principais
características dos microarranjos, por terem seus elementos individuais muito
pequenos, é a sua alta densidade, na ordem de milhares de sondas individuais por
centímetro quadrado. A grande quantidade de sondas que pode estar presente em
um microarranjo faz dessa técnica uma poderosa ferramenta para a análise

26
simultânea de diferentes alvos, além de conferir maior sensibilidade e confiabilidade
ao resultado devido à possibilidade de utilização de várias sondas complementares
a um mesmo organismo (BEDNAR, 2000; FREEMAN et al., 2000; HELLER, 2002).
Apesar dos microarranjos terem sua principal aplicação na análise da
expressão gênica e identificação de polimorfismos, o diagnóstico de doenças
infecciosas também se beneficiou dessa tecnologia (BEDNAR, 2000; STRIEBEL et
al., 2003). Microarranjos destinados à identificação e genotipagem dos mais diversos
vírus já foram desenvolvidos e alguns encontram-se disponíveis comercialmente (AN
et al., 2003; BORISKIN et al., 2004; CHIZHIKOV et al., 2002; FOLDES-PAPP et al.,
2004; KAWAGUCHI et al., 2003; KLAASSEN et al., 2004; LAASSRI et al., 2003;
LAASSRI et al., 2005; LI et al., 2001; LOVMAR et al., 2003; OH et al., 2004;
PALACIOS et al., 2007; STRIEBEL et al., 2003; WANG et al., 2002(a)). PALACIOS
et al. (2007), por exemplo, propõem-se, com seu microarranjo de 29.500 sondas, a
detectar não apenas a presença dos mais importantes vírus como das mais variadas
espécies de bactérias, fungos e protozoários.
Além de possibilitar a detecção de diferentes organismos em um mesmo
ensaio, essa metodologia pode ser utilizada para caracterizações moleculares mais
finas, como resistência a drogas e genotipagem, através da identificação de
variantes distintas de determinado agente etiológico (AMEXIS et al., 2002; WADE et
al., 2004). CHERKASOVA et al. (2003) e IVSHINA et al., (2004) propuseram, por
exemplo, um sistema de monitoramento, por microarranjo, da evolução viral. Essa
aplicação é muito importante para estudos evolutivos e de acompanhamento
epidemiológico, em especial para vírus que apresentam evolução rápida, através de
mutação ou recombinação de segmentos genômicos. A genotipagem também é
importante em casos onde o genótipo do patógeno está relacionado com diferenças
na progressão da doença ou na resposta ao tratamento, como é o caso do vírus da

27
hepatite C (HCV) e do papilomavírus humano (HPV), entre outros. Também de
grande importância é a capacidade dessa ferramenta em identificar patógenos em
doenças de etiologia ainda desconhecida (GARDNER et al., 2003; WANG et al.,
2002(a)).
A grande vantagem dessa metodologia reside, então, na sua capacidade de
atender a uma forte demanda do segmento de diagnósticos por testes simultâneos,
onde em apenas um procedimento pode-se avaliar uma grande quantidade de
seqüências alvo. Porém, quando comparado à PCR em tempo real, os ensaios de
microarranjo geralmente perdem em sensibilidade, custo e tempo necessário para
obtenção dos resultados (GARDNER et al., 2003).
1.6.7.2. Microarranjos líquidos
Uma das metodologias que vem ganhando popularidade na área de
diagnóstico faz uso da citometria de fluxo e de micro-partículas esféricas (KELLAR e
IANNONE, 2002). Esse sistema, denominado aqui de microarranjo líquido, consiste
no emprego de uma mistura de diferentes tipos de micro-esferas, uniformes em
tamanho, mas que emitem intensidades de fluorescência distintas, como suporte
sólido para diferentes moléculas de captura, sejam elas anticorpos, antígenos,
peptídeos ou ácidos nucléicos. A ligação das moléculas de captura, já acopladas a
cada uma das micro-esferas, a seus alvos é detectada através da utilização de um
conjugado (molécula repórter) também fluorescente (FULTON et al., 1997;
MCHUGH, 1994; VIGNALI, 2000). Apesar de possível há mais de vinte e cinco anos,
desde o desenvolvimento dos primeiros citômetros de fluxo comerciais, essa técnica
só foi difundida há, aproximadamente, dez anos, graças ao desenvolvimento do
equipamento Luminex® e de um arranjo de cem micro-esferas distintas pela empresa
LuminexCorp. Este equipamento consiste em um citômetro de fluxo compacto,

28
composto por um processador de sinal digital e dois canais de lazer capazes de
excitar os fluorocromos das micro-esferas e do repórter (KELLAR e IANNONE,
2002). Por permitir a detecção de ácidos nucléicos e proteínas, a metodologia de
microarranjo líquido serve como substituta tanto para a técnica de microarranjo
tradicional como para os testes imuno-enzimáticos, com a vantagem, sobre este
último, de possibilitar análise de diversos parâmetros simultaneamente, utilizando
um volume reduzido de amostra.
Apesar de permitir, atualmente, a análise de apenas cem alvos ao mesmo
tempo, contra os milhares que podem ser avaliados por microarranjo tradicional, a
técnica de microarranjo líquido apresenta como benefícios sobre o primeiro o custo
reduzido, maior facilidade na interpretação dos resultados, maiores velocidade de
execução e sensibilidade, devido à melhoria na cinética de hibridação, e maior
flexibilidade (DUNBAR, 2006). E conforme ressaltado por YANG et al. (2001),
enquanto os microarranjos tradicionais são apropriados para a avaliação de
genomas inteiros e poucas amostras e a PCR em tempo real é útil para a análise de
muitas amostras mas de poucos alvos, os microarranjos líquidos são ideais para a
avaliação de dezenas de alvos em muitas amostras.
Nos últimos anos, essa tecnologia vem sendo cada vez mais utilizada e os
inúmeros trabalhos publicados demonstram a aptidão dessa plataforma para a
detecção dos mais variados analitos e a forte tendência em direção à sua adoção
como método diagnóstico. A eficiência da metodologia já pôde ser comprovada em
estudos para a detecção de anticorpos contra antígenos MHC de classe I e II (PEI et
al., 1999; PIAZZA et al., 2006), auto-anticorpos contra cardiolipina, fosfatidilserina e
antígenos nucleares (MARTINS et al., 2004; PIERANGELI et al., 1999), contra toxina
anthrax (BIAGINI et al.; 2004), tétano, difteria e Haemophilus influenzae tipo B
(PICKERING et al., 2002), contra o vírus da hepatite C (MCHUGH et al., 1997), vírus

29
da AIDS (FAUCHER et al., 2006; TOELLNER et al., 2006), papilomavirus humano
(OPALKA et al., 2003), flavivírus (WONG et al., 2004), vírus West Nile e da
encefalite de Saint Louis (JOHNSON et al., 2005(a)) e vírus respiratórios (MARTINS
2002).
Métodos multiplex já foram, também, desenvolvidos por vários autores para a
quantificação de diferentes citocinas secretadas no soro (ALLEN et al., 2007;
CAMILLA et al., 2001; CARSON e VIGNALI, 1999; OLIVER et al., 1998; MORGAN et
al., 2004; SHURIN et al., 2007; YURKOVETSKY et al., 2007). Carson e Vignali
(1999), por exemplo, usaram a tecnologia de microarranjo líquido para o
desenvolvimento de um teste para a detecção simultânea de 15 diferentes citocinas.
Morgan e colaboradores (2004) demonstraram a capacidade de kits comerciais (BD
Biosciences) em avaliar o nível de citocinas associadas à resposta Th1/Th2 em
amostras de lágrima e lavado nasal, de mediadores inflamatórios derivados do
complemento em plasma, de moléculas sinalizadoras em células T, de fatores de
transcrição fosforilados e de marcadores da apoptose. Esse mesmo trabalho
exemplifica como a metodologia pode ser aplicada para a detecção e quantificação
de anticorpos para diferentes antígenos. Outros exemplos da utilização da técnica
são na quantificação simultânea de peptídeos β-amilóide, proteína tau total e formas
hiperfosforiladas desta proteína em fluido cérebro-espinhal, como fator diagnóstico
para Alzheimer (OLSSON et al., 2005) e na determinação dos níveis do antígeno X
do vírus da hepatite B como marcador prognóstico para a doença (PAL et al., 2005).
Em relação à detecção de ácidos nucléicos, a principal aplicação dos testes
de microarranjo líquido tem sido na identificação de polimorfismos dos genes do
MHC e de receptores de células killer (FULTON et al., 1997; NONG et al., 2007) e de
mutações relacionadas ao desenvolvimento de doenças tais como doenças
neurodegenerativas (CHEN et al., 2007; PYATT et al., 2007), fibrose cística

30
(DUNBAR e JACOBSON, 2000; HADD et al., 2004) e leucemia linfoblástica
pediátrica (WALLACE et al., 2003).
Como método de diagnóstico molecular já foi utilizado, entre outras coisas,
para a identificação de bactérias contaminantes ambientais (BAUMS et al., 2007;
SPIRO et al., 2000; SPIRO e LOWE, 2002) e alimentares (DUNBAR et al., 2003),
para a identificação e diferenciação de espécies de bactérias do gênero Vibrio
(TRACZ et al., 2007), para a genotipagem de Mycobacterium tuberculosis (COWAN
et al., 2004), para detecção e classificação de fungos (DIAZ e FELL, 2004), detecção
molecular do vírus da hepatite C (HCV) e vírus herpes simplex (HSV) (SMITH et al.,
1998), genotipagem do HPV (OH et al., 2007) e quantificação do HIV (VAN CLEVE
et al., 1998). MAHONY et al. (2007) utilizaram a técnica para o desenvolvimento de
um teste multiplex para detecção e diferenciação de 20 diferentes vírus respiratórios.
O uso da técnica de microarranjo líquido em estudos de expressão gênica
(WEDEMEYER et al., 2000; YANG et al., 2001) também já foi descrito. Sistemas
multiplex comerciais baseados em microarranjo líquido já são disponibilizados por
empresas como Millipore, Bio-Rad e Qiagen. Espera-se que à medida que essa
tecnologia amadureça muitos outros ensaios sejam ofertados comercialmente e que
a tecnologia passe a ser extensivamente utilizada para detecção de ácidos
nucléicos, ensaios sorológicos e estudos de expressão gênica (DUNBAR, 2006).

31
2. OBJETIVOS
• desenvolver um método de diagnóstico molecular para a dengue
baseado na técnica de PCR em tempo real;
• desenvolver um método de diagnóstico molecular para a dengue
baseado na técnica de microarranjo líquido;
• avaliar os dois métodos desenvolvidos quanto a capacidade em
detectar o RNA do vírus da dengue em amostras clínicas;
• comparar os dois métodos desenvolvidos quanto a sensibilidade e a
especificidade;
• adquirir competência em metodologias que tendem a dominar o setor
de diagnóstico em um futuro próximo.

32
3. JUSTIFICATIVAS
À medida que a incidência da dengue aumenta e considerando a inexistência
de vacinas ou tratamentos específicos para a doença, métodos eficientes de
diagnóstico, juntamente com medidas de controle do inseto vetor, são de
fundamental importância. Testes de diagnóstico laboratoriais, que permitam a
detecção precoce dos casos e forneçam dados acerca do sorotipo do vírus
infectante, são essenciais não apenas para possibilitar o correto manejo clínico,
mas, principalmente, para a melhoria da vigilância epidemiológica, o que levará a um
maior entendimento do comportamento da doença e auxiliará na contenção de
epidemias futuras.
Apesar do Brasil ser endêmico para a dengue, o diagnóstico laboratorial da
doença no país ainda é falho, pois se baseia, quase que exclusivamente, em testes
sorológicos, que não permitem o diagnóstico na fase aguda da infecção e nem a
determinação do sorotipo viral. Apesar dos avanços nas técnicas de biologia
molecular, essas metodologias são ainda pouco aplicadas no setor de diagnóstico
do país. Para dengue, assim como para diversas doenças virais, o isolamento
continua sendo o teste de escolha para a sorotipagem e o diagnóstico na fase aguda
(BUCHY et al., 2006; LAI et al., 2007). O alto custo, necessidade de instalações
especiais, demora para confirmação dos resultados e baixa processividade (HARRIS
et al., 1998; YONG et al. 2007), no entanto, fazem com que essa metodologia não
seja capaz de abranger a extensão necessária para auxiliar, de forma efetiva, no
controle da doença.

33
Melhorias na vigilância epidemiológica da dengue e nos cuidados aos
pacientes só serão possíveis através da introdução definitiva de testes que, além de
permitirem a sorotipagem e diagnóstico precoce da infecção, possam ser
amplamente disponibilizados e forneçam resultados precisos, confiáveis e em um
curto período de tempo. Tendo conhecimento da importância e da necessidade de
popularização dos testes de base molecular e pensando em suprir o atraso
tecnológico do país no que diz respeito à área de diagnóstico, realizou-se esse
trabalho, através do qual propomos a adequação de metodologias de última geração
e capazes de atender as demandas impostas pelo setor de diagnóstico, para a
detecção e sorotipagem do vírus da dengue em amostras clínicas. Com a escolha
das metodologias de PCR em tempo real e microarranjo líquido estamos
empregando os conhecimentos tecnológicos mais avançados disponíveis,
assegurando a possibilidade de aplicação e uso do produto e considerando,
sobretudo, o aspecto custo-benefício, o que, certamente, resultará em grande ganho
para o setor de diagnóstico no país. Além disso, a grande capacidade de
processamento e flexibilidade desses sistemas torna fácil sua adequação para a
análise de outros alvos que futuramente poderão ser inseridos dentro das mesmas
plataformas tecnológicas.
Acreditamos que utilizando a dengue como modelo seremos capazes de
entender e validar técnicas que poderão ser empregada para o diagnóstico
molecular e sorológico de doenças estratégicas para a saúde pública, suprindo,
dessa forma, a carência do setor de diagnóstico e a dependência do país por
insumos produzidos no exterior.

34
4. MATERIAIS E MÉTODOS
4.1. Amostra populacional estudada
Um total de 170 amostras coletadas entre os anos de 2001 e 2007 foi usado
na avaliação dos dois métodos de diagnóstico molecular desenvolvidos. Do total de
amostras, 49 representavam soros de fase aguda, negativos para IgM, a partir dos
quais o vírus da dengue foi isolado em cultura de células C6/36 e sorotipado através
do uso de anticorpos monoclonais sorotipo-específicos. Destas 49, 14 foram
sorotipadas como DENV1, 7 como DENV2 e 28 como DENV3. Essas amostras
foram gentilmente doadas pelo Instituto de Pesquisa Clínica Evandro Chagas (IPEC)
da Fundação Oswaldo Cruz (FIOCRUZ) do Rio de Janeiro, que também realizou o
isolamento viral. Outras 98 amostras foram obtidas junto ao Laboratório Central
(LACEN) do Estado do Paraná e consistiam em soros provenientes de indivíduos
suspeitos de infecção por dengue, que foram coletados durante o último surto da
doença no Estado, ocorrido nos primeiros meses de 2007. Todos esses soros já
haviam sido testados por um ensaio de MAC-ELISA (IgM antibody capture - enzyme-
linked immunosorbent assay), resultando em 48 amostras negativas e 50 amostras
positivas para IgM anti-dengue. As 23 amostras restantes compreendiam controles
negativos para a doença e incluiam dois soros de pacientes positivos para o vírus da
hepatite C (HCV), três positivos para o vírus da hepatite B (HBV), três positivos para
citomegalovírus (CMV), um sobrenadante de cultura do vírus da febre amarela (YFV)
e soro de catorze indivíduos saudáveis, residentes na cidade de Curitiba, que nunca
estiveram em áreas epidêmicas para dengue e nunca apresentaram os sintomas

35
característicos da patogenia. Destes treze, quatro haviam sido vacinados para febre
amarela em torno de cinco dias antes da coleta do sangue.
Após a coleta, os soros foram armazenados a -70 ou a -20oC até seu
processamento para a extração do RNA viral.
4.2. Preparo dos padrões para testes de especificidade, sensibilidade e
otimização das reações
Além das amostras sorológicas, quantidades definidas de vírus da dengue
dos sorotipos DENV1 (cepa BR90), DENV2 (cepa M20558) e DENV3 (isolado local),
previamente purificados e titulados, foram igualmente submetidas à análise pelos
dois métodos diagnósticos. Os RNAs desses padrões de sorotipo e quantidade
conhecidos foram usados nas etapas de padronização dos ensaios, como controle
positivo das reações e para a avaliação da especificidade e sensibilidade de ambos
os métodos desenvolvidos. Para o preparo destes padrões, 122.880 foci forming
units (FFU) de cada um dos três sorotipos do vírus foram adicionados a três
alíquotas distintas de 140 µl de soro negativo para a doença. Após a extração do
RNA viral, curvas padrões foram geradas, a partir de sete diluições seriadas de 1:4
do RNA extraído. Cinco microlitros (5 µl) do ponto mais concentrado da curva foram
usados na fase de otimização dos testes e para a avaliação da especificidade. Para
a verificação da sensibilidade, o mesmo volume (5 µl) de cada um dos oito pontos da
curva (de 2.048 a 0,125 FFU/µl) foram amplificados, em triplicata, por PCR em
tempo real ou submetidos à reação de RT-PCR para posterior análise pela técnica
de microarranjo líquido. A carga viral dos padrões foi calculada em FFU por mililitro
(FFU/ml), com base no volume de soro utilizado para a extração do RNA (140 µl).

36
4.3. Extração do RNA viral
Os RNAs, tanto das amostra de soro de pacientes como dos padrões, foram
isolados diretamente a partir de uma alíquota de 140 µl de cada um dos soros a
serem testados. A extração foi feita com o QIAmp Viral RNA Kit (Qiagen) conforme
instruções do fabricante. Os RNAs eluídos em 60 µl de água livre de RNase foram
armazenados a –70oC até o momento do seu uso.
4.4. Desenho dos oligonucleotídeos para PCR em tempo real
Um total de 208 seqüências que representavam genomas completos do vírus
da dengue dos tipos 1, 2 e 3 presentes no GenBank foram alinhadas através do
software CLUSTAL X (THOMPSON et al., 1997). Regiões do genoma que
apresentavam um alto grau de conservação dentro de um mesmo sorotipo, mas uma
baixa similaridade entre os diferentes sorotipos, foram identificadas visualmente e
selecionadas. A seqüência exata dos oligonucleotídeos iniciadores e sondas
complementares a essas regiões foram determinadas usando o software Primer
Express (Applied Biosystems). O conjunto de oligonucleotídeos que melhor se
adequava aos parâmetros recomendados pelo programa foi escolhido. Dessa forma,
os iniciadores utilizados apresentavam Tm (melting temperature) entre 58 e 60oC e a
sonda uma Tm entre 7 a 10oC superior à dos iniciadores. As sondas não
apresentavam guanina em sua extremidade 5’ e o tamanho do fragmento
amplificado não ultrapassava os 200 pares de base (pb). Buscou-se, também,
minimizar a probabilidade de formação de estruturas secundárias e de dimerização
dos oligonucleotídeos. A aplicação dos mesmos critérios para a seleção dos

37
diferentes oligonucleotídeos permite que todos funcionem sincronicamente sob
condições idênticas.
À extremidade 5’ das sondas foi adicionada a molécula fluorescente FAM (6-
carboxifluoresceína) e à extremidade 3’ ligou-se a molécula não-fluorescente, com
atividade quencher, BHQ1 (black hole quencher 1). Além dos oligonucleotídeos
específicos para cada um dos sorotipos, foi selecionado, a partir de dados obtidos
na literatura, um grupo de iniciadores e sonda complementares a todos os sorotipos
do vírus (DROSTEN et al. 2002). Esses oligonucleotídeos grupo-específicos, que
devem servir como um controle das reações de amplificação e detecção, atendem
os mesmos parâmetros citados acima e são complementares a praticamente todas
as seqüências obtidas no banco de dados. Para o vírus da dengue do tipo 3, um
segundo conjunto de iniciadores e sonda descritos no trabalho de Johnson et al.
(2005(b)) foi, também, testado. A seqüência dos oligonucleotídeos utilizados no
ensaio de PCR em tempo real e suas principais características estão listadas na
tabela 4.1.

38
TABELA 4.1 - Oligonucleotídeos iniciadores e sondas utilizados no ensaio de PCR
em tempo real.
.
Oligonucleotídeo Seqüência (5’ →→→→ 3’) Região genômica Referência
GenBank
Tamanho
do produto
amplificado
DEN1-sense GCTGATGCTGGTGACACCAT M (907 – 926) AF 311958
66 pb DEN1-anti-sense TCGACGAAGTCTCTGTTGCCTAT E (950 – 972) AF311958
DEN1-sonda CCCACGCATCGCATGGCCAT E (929 – 948) AF311958
DEN2- sense ACCATAGGAACGACACATTTCC M (864 – 885) AF 489932
80 pb DEN2- anti-sense CAACGCATTGTCATTGAAGGA M/E (923 – 943) AF 489932
DEN2- sonda AGGGCCTTGATTTTCATCTTACTGACAGC M (888 – 916) AF 489932
DEN3- sense CCCATCCATGACAATGAGATGT M/E (922 – 943) AY 679147
78 pb DEN3- anti-sense TCAACCCACGTAGCTCCTGAT E (979 – 999) AY 679147
DEN3- sonda TGGGAGTAGGAAACAGAGATTTTGTGGAAGG E (945 – 975) AY 679147
DEN3J- sensea GGACTGGACACACGCACTCA M (740 – 759) NC 001475
74 pb DEN3J- anti-sensea CATGTCTCTACCTTCTCGACTTGTCT M (788 – 813) NC 001475
DEN3J- sondaa ACCTGGATGTCGGCTGAAGGAGCTTG M (761 – 786) NC 001475
DENG- senseb GGATAGACCAGAGATCCTGCTGT 3’NCR (10646 – 10668) AF 311958
67 pb DENG- anti-sense1b CATTCCATTTTCTGGCGTTC 3’NCR (10693 – 10712) AF 311958
DENG- anti-sense2b CAATCCATCTTGCGGCGCTC 3’NCR (10607 – 10626) AF 375822
DENG- sondab CAGCATCATTCCAGGCACAG 3’NCR (10674 – 10693) AF 311958
a oligonucleotídeos descritos por Johnson et al. (2005(b)) b oligonucleotídeos descritos por Drosten et al. (2002), complementares a seqüências comuns a todos os sorotipos do vírus da dengue. As bases em azul, nas seqüências dos oligonucleotídeos, diferem em pelo menos uma das cepas analisadas.
4.5. Desenho dos oligonucleotídeos para microarranjo líquido
Os mesmos oligonucleotídeos desenhados para o ensaio de PCR em tempo
real foram, também, utilizados nos testes de microarranjo líquido, com as seguintes
alterações: a molécula fluorescente FAM e o quencher BHQ1 foram removidos das
sondas, os iniciadores anti-sense e as sondas receberam, respectivamente, uma
biotina e uma amina em suas extremidades 5’ e as sondas foram sempre
posicionadas na fita sense, de maneira que suas seqüências eram complementares
à fita extendida a partir do iniciador anti-sense, ou seja, à fita biotinilada. A fim de

39
atender este último requisito, a seqüência descrita na tabela 4.1 para a sonda DEN1,
a qual é complementar à fita anti-sense, foi alterada para a seqüência reversa
complementar da mesma.
Além de todos os oligonucleotídeos já listados na tabela 4.1, os iniciadores e
sondas utilizados por Johnson et al. (2005(b)) para amplificação e detecção dos
vírus da dengue dos tipos 1 e 2 por PCR em tempo real foram, também, adaptados
para a metodologia de microarranjo líquido e avaliados. As seqüências de tais
oligonucleotídeos estão listadas a seguir.
TABELA 4.2 - Oligonucleotídeos iniciadores e sondas, adaptados de Johnson et al.
(2005(b)), utilizados no ensaio de microarranjo líquido.
Oligonucleotídeo Seqüência (5’ →→→→ 3’) Região genômica Referência
GenBank
Tamanho
do produto
amplificado
DEN1J-sense CAAAAGGAAGTCGTGCAATA NS5 (8973 – 8992) AY732480
112 pb DEN1J-anti-sense CTGAGTGAATTCTCTCTACTGAACC NS5 (9060 – 9084) AY732480
DEN1J-sonda CATGTGGTTGGGAGCACGC NS5 (8998 – 9019) AY732480
DEN2J- sense CAGGTTATGGCACTGTCACGAT E (1463 – 1484) DQ181803
77 pb DEN2J- anti-sense CCATCTGCAGCAACACCATCTC E (1519 – 1540) DQ181803
DEN2J- sonda CTCTCCGAGAACAGGCCTCGACTTCAA E (1491 – 1517) DQ181803
As bases em azul, nas seqüências dos oligonucleotídeos, diferem em pelo menos uma das cepas analisadas.
4.6. PCR em tempo real
A fim de permitir um diagnóstico rápido e específico, desenvolveu-se um
ensaio de PCR em tempo real, baseado na hidrólise de sondas fluorescentes, o qual
foi testado na detecção do RNA do vírus da dengue presente no soro de pacientes.
Essa técnica permite a realização da reação de transcrição reversa (RT) e de PCR
em um mesmo tubo, sendo o produto da amplificação detectado imediatamente após
sua síntese, durante o curso de toda a reação de PCR. A detecção em tempo real é

40
possível devido à degradação de uma sonda (sonda TaqMan®), complementar a
uma região interna do segmento amplificado e que apresenta na sua extremidade 5’
uma molécula fluorescente e em sua extremidade 3’ uma molécula quencher, capaz
de, por um fenômeno físico denominado FRET (Fluorescence Ressonance Energy
Transfer), absorver a fluorescência emitida pelo fluoróforo após este ser estimulado
por luz de comprimento de onda específico. O acoplamento de fluoróforos diferentes
a sondas com especificidades distintas permite a realização de ensaios múltiplos
(multiplex), com a detecção de diferentes alvos em uma mesma amostra. Nesse
trabalho foram usados 6-carboxifluoresceína (FAM) e black hole quencher-1 (BHQ1)
como fluoróforo e quencher, respectivamente.
Durante a fase de extensão dos iniciadores, a sonda que estava anelada à
sua seqüência complementar é degradada devido à atividade 5’ – 3’ exonuclease da
enzima Taq DNA polimerase. A degradação ocasiona a separação dos dois
fluoróforos, o que abole o fenômeno FRET, ocasionando o aumento da fluorescência
na reação (figura 1.1) (BUSTIN, 2000). A fluorescência emitida em cada ciclo é
captada pelo sistema óptico do equipamento de PCR em tempo real e representada
em um gráfico. Com base neste gráfico, determina-se, manualmente, um valor
mínimo de fluorescência necessário para que uma amostra seja considerada
positiva. O ciclo em que a fluorescência de certa amostra atinge esse valor mínimo é
denominado de Ct (threshold cycle).
Uma vez que a intensidade da fluorescência emitida é proporcional à
quantidade do produto presente dentro de cada tubo de ensaio, essa técnica
possibilita não só a simples detecção de um fragmento específico de RNA ou DNA
como também a sua quantificação (HEID et al., 1996). A quantificação é feita
amplificando-se amostras padrões, com quantidades conhecidas e variadas de
molde, concomitantemente com amostras cujas quantidades queira-se determinar.

41
Os valores de Ct obtidos para as amostras padrões são utilizados na construção de
uma curva representando o Ct versus o número de cópias iniciais. Os valores de Ct
das amostras desconhecidas são interpolados na curva padrão, sendo possível a
determinação do número de cópias iniciais presentes nessas amostras. Uma vez
que esse método não requer nenhum tipo de processamento após a reação de PCR,
o material amplificado pode ser descartado sem que o tubo seja aberto, evitando a
contaminação de amostras com o produto de uma amplificação anterior.
FIGURA 4.1 - Representação esquemática das etapas de uma PCR em tempo real.
Na etapa de alongamento, a hidrólise da sonda permite a emissão da fluorescência de FAM, que é detectada e registrada a cada ciclo de amplificação.
BHQ1

42
Além da metodologia baseada na degradação das sondas TaqMan®, também
foi avaliado, em alguns casos, a estratégia de PCR em tempo real que se utiliza do
corante SYBR Green para detecção do fragmento amplificado. Esse corante emite
fluorescência quando intercalado a fitas duplas de DNA e, da mesma forma que
ocorre para as sondas TaqMan®, a intensidade da fluorescência emitida é
proporcional a quantidade do material amplificado dentro da reação. Esse método
permite que seja feita, após a etapa de amplificação, uma análise de dissociação do
produto de PCR. Essa análise é feita aumentando-se a temperatura, de 60 até 95oC,
com constante leitura da fluorescência. Quando se atinge uma temperatura igual à
Tm (melting temperature) do amplicon, observa-se uma queda abrupta do sinal.
Esse fenômeno deve ser observado apenas uma vez durante a análise de
dissociação, indicando que ocorreu a amplificação de apenas um alvo e eliminando,
portanto, a possibilidade de ocorrência de amplificações inespecíficas (figura 4.2).
Essa metodologia apresenta as mesmas vantagens do que a sonda no concernente
à rapidez de obtenção dos resultados, redução das chances de ocorrência de
contaminação cruzada e possibilidade de quantificação do material adicionado à
reação. O fato da utilização do SYBR Green eliminar a necessidade de uso da
sonda facilita o desenho do ensaio e reduz seus custos. A desvantagem, no entanto,
com relação à metodologia TaqMan®, reside na impossibilidade da realização de
testes multiplex.

43
FIGURA 4.2 – Exemplo de uma curva de dissociação obtida após a amplificação de
um segmento de DNA utilizando o corante SYBR Green.
A
B
Em A, dado bruto obtido após análise de dissociação de um conjunto de amostras, onde o ponto de inflexão da curva indica a temperatura de desnaturação do amplicon. Em B, o mesmo resultado transformado em derivativa, onde o ponto de inflexão nos dados brutos passa a ser representado como um “pico de dissociação".
4.6.1. Detecção das seqüências do vírus da dengue por PCR em tempo
real
O ensaio de PCR em tempo real foi realizado no sistema de detecção de
seqüência ABI PRISM 7000 (Applied Biosystem), utilizando o kit TaqMan One-Step
RT-PCR Master Mix (Applied Biosystems) para os ensaios com sondas e o kit
SYBR Green Master Mix (Applied Biosystems) para os ensaios com SYBR Green.
Cinco microlitros (5 µl) de RNA extraído de cada amostra foram testados em uma
reação com volume final de 25 µl, contendo 0,2 µM de cada iniciador e os
temperatura
temperatura

44
componentes do kit (master mix e enzima MultiScribe) diluídos conforme
recomendado pelo fabricante. Nas reações baseadas na metodologia TaqMan foram
adicionados, ainda, 0,3 µM de sonda. A ciclagem consistiu de um passo inicial de 30
minutos a 48oC, seguido por 10 minutos a 95oC e 40 ciclos de 30 segundos a 95oC e
1 minuto a 60oC. Para os ensaios utilizando o corante SYBR Green, foi incluído,
após a etapa de amplificação, um passo de análise de dissociação do produto de
PCR. Essa análise consistiu em um passo de desnaturação a 95oC por 1 minuto,
redução da temperatura para 60oC, seguida de um aumento até 95oC, a uma taxa
de, aproximadamente, 1oC/40s com leitura constante da fluorescência. Todas as
amostras foram submetidas, em duplicata, a pelo menos duas reações de
amplificação distintas: uma com os oligonucleotídeos grupo-específicos e outra com
os oligonucleotídeos específicos para o sorotipo mais provável da infecção.
Amostras negativas em qualquer uma das duas reações foram testadas para
os dois outros sorotipos restantes.
4.7. Microarranjo líquido
O príncipio da técnica de microarranjo líquido é a utilização de micro-esferas
de poliestireno como suporte para moléculas de captura que podem incluir qualquer
tipo de ácido nucléico ou proteína. Essas micro-esferas, de 5,6 micrômetros de
diâmetro, são coradas, internamente, com fluoróforos na faixa do vermelho e do
infra-vermelho. O preenchimento de diferentes lotes com proporções distintas dos
dois corantes possibilitou a criação de 100 diferentes grupos de micro-esferas. Cada
grupo carrega, então, uma assinatura única, identificada após a passagem da micro-
esfera por um feixe de laser de determinado comprimento de onda. Diferentes

45
conjuntos de micro-esferas podem ser utilizados em um mesmo ensaio, permitindo,
portanto, a detecção e identificação de até 100 alvos moleculares em um único
volume de reação.
No caso da detecção de ácidos nucléicos, um oligonucleotídeo de captura
(sonda), contendo um grupamento amina em uma de suas extremidade, é ligado
covalentemente à superfície de uma micro-esfera recoberta por grupamentos
carboxila. Alvos moleculares biotinilados, tais como produtos de PCR, ligam-se às
sondas e essa interação é detectada utilizando-se uma molécula repórter também
fluorescente – geralmente streptavidina-R-ficoeritrina – que se ligará à molécula alvo
e será lida por um segundo feixe de laser. A intensidade da fluorescência emitida
pela ficoeritrina é proporcional à concentração do alvo na amostra. Cada micro-
esfera é analisada, individualmente, por um equipamento (tipicamente o Luminex® -
LuminexCorp) capaz de excitar os fluoróforos e detectar o sinal emitido por eles,
classificando as micro-esferas de acordo com sua assinatura espectral (código de
cor) e quantificando a reação ocorrida em sua superfície. Uma representação
esquemática das diversas etapas envolvidas na detecção de ácidos nucléicos
através da técnica de microarranjo líquido encontra-se a seguir.

46
FIGURA 4.3 – Representação esquemática da detecção de ácidos nucléicos através
da técnica de microarranjo líquido.
4.7.1. Acoplamento das sondas às micro-esferas de poliestireno
Cada sonda, contendo uma amina em sua extremidade 5’, foi ligada,
covalentemente, a um conjunto diferente de micro-esferas através do uso de
carbodiimida. Para tal, o estoque de micro-esferas carboxiladas foi vortexado por 2
minutos e 5 x 106 micro-esferas foram transferidas para um tubo de microcentrífuga
de 1,5 ml e centrifugadas por 3 minutos a 8.000g. O sobrenadante foi removido e as
micro-esferas foram ressuspendidas em 50 µl de MES (ácido 2-(N-
morfolino)etanosulfônico) 100 mM (pH 4,5). Adicionou-se 0,2 nmol de sonda e a
suspensão foi vortexada brevemente. Cinco microlitros (5 µl) de uma solução
10 µg/µl de EDC (N-(3-dimetilaminopropil)-N-etilcarbodiimida) foram acrescentados à
mistura, que foi novamente vortexada e incubada a temperatura ambiente, por uma
vírus ou outro patógeno
ácido nucléico extraído
extração do ácido nucléico PCR / RT-PCR
produto amplificado e marcado
hibridação detecção
detecção produto de PCR ligado à sonda

47
hora, ao abrigo da luz. As micro-esferas foram centrifugadas por 3 minutos a 8.000 g
e, após remoção do sobrenadante, lavadas uma vez com 1 ml de Tween-20 0,02%
(v/v). A lavagem compreendeu a suspensão das micro-esferas com o uso de vórtex,
após a adição da solução de Tween-20, centrifugação igual às anteriores e remoção
do sobrenadante. Por fim, as micro-esferas foram ressuspendidas em 100 µl de TE
(Tris-HCl 10 mM, EDTA 1 mM; pH 8,0) e armazenadas a 4oC, ao abrigo da luz.
4.7.2. PCR
A primeira etapa do teste baseado na metodologia de microarranjo líquido
consistiu na amplificação de segmentos específicos dos sorotipos 1, 2 e 3 do vírus
da dengue, através de uma RT-PCR (reverse transcription – polymerase chain
reaction) realizada em dois passos. Primeiramente, foram desenvolvidas reações de
transcrição reversa (RT) e de PCR individuais para cada sorotipo do vírus (uniplex).
Dessa forma, 5 µl de RNA dos controles positivos para cada sorotipo foram,
inicialmente, convertidos em cDNA (DNA complementar) pela ação de uma enzima
transcritase reversa (ImProm II - Promega), em uma reação contendo 2 unidades/µl
desta enzima, 100 µM de cada um dos quatro desoxirribonucleotídeos trifosfatados,
10 mM de ditiotreitol (DTT), 0,3 unidades/µl de inibidor de RNase (RNase OUT -
Invitrogen), 50 mM de Tris-HCl (pH 8,3), 75 mM de KCl, 3 mM de MgCl2 e 0,25 µM
do iniciador anti-sense complementar ao sorotipo em questão em uma reação com
volume final de 20 µl. Após incubação de 1 hora a 42oC, a totalidade do cDNA foi
submetida a uma reação de PCR de 50 µl de volume final, na qual utilizou-se 0,2 µM
do mesmo iniciador anti-sense usado na reação de RT e do iniciador sense
correspondente. Nesta reação foram, também, adicionados 50 mM de KCl, 20 mM
de Tris-HCl (pH 8,4), 1,5 mM de MgCl2, 200 µM de cada um dos quatro
desoxirribonucleotídeos trifosfatados (dNTPs) e 0,025 unidades/µl de enzima Taq

48
DNA polimerase. As amostras foram submetidas a 40 ciclos de desnaturação (94oC
por 30 segundos), anelamento dos iniciadores (60oC por 30 segundos) e extensão
(72oC por 1 minuto), os quais foram antecedidos por 2 minutos a 94oC.
Posteriormente, desenvolveu-se uma reação múltipla (multiplex) de RT-PCR em dois
passos, contendo, portanto, em um mesmo tubo, iniciadores complementares aos
três sorotipos do vírus e os iniciadores grupo-específicos. As concentrações dos
reagentes e condições de amplificação da reação multiplex foram as mesmas
utilizadas para a reação uniplex e já detalhadas acima.
4.7.3. Hibridação do produto de PCR com as sondas acopladas às micro-
esferas
Durante o desenvolvimento inicial, o DNA amplificado por RT-PCR uniplex foi
colocado em contato com as sondas acopladas às micro-esferas em reações de
hibridação também uniplex (apenas uma sonda por tubo de reação). Em uma etapa
posterior, produtos da RT-PCR obtidos da mesma maneira, foram testados em
reações de hibridação multiplex, realizadas sob condições distintas. E por fim, a
condição de hibridação multiplex que forneceu melhores resultados foi testada com
os produtos de PCR obtidos através da RT-PCR multiplex. Utilizando esta última
abordagem, reações contendo combinações diferentes de cada oligonucleotídeo
foram avaliadas, tanto na etapa de amplificação como de hibridação, de maneira que
cada combinação sempre continha um conjunto de oligonucleotídeo específico para
cada sorotipo e os oligonucleotídeos grupo-específicos.
O protocolo de hibridação inicialmente testado consistiu na adição de 5 µl do
produto da RT-PCR e 12 µl de TE (pH 8,0) a 33 µl de uma solução de TMAC 1,5X
(TMAC 4,5 M; sarcosil 0.15% [p/v]; Tris-HCl 75 mM; EDTA 6 mM, pH 8,0) contendo,
aproximadamente, 2.000 micro-esferas de cada código de cor. Essa mistura foi

49
submetida a um passo de desnaturação de 5 minutos a 95oC seguido da etapa de
hibridação de 15 minutos a 55oC. Após a hibridação, as micro-esferas foram
recolhidas por centrifugação a 2.500g por 5 minutos e o sobrenadante removido por
inversão. O produto hibridado foi marcado através da ressuspensão das micro-
esferas em 75 µl de uma solução TMAC 1X (TMAC 3 M; sarcosil 0.1% [p/v]; Tris-HCl
50 mM; EDTA 4 mM, pH 8,0), contendo 3 ng/µl de streptavidina-R-ficoeritrina, e
incubação a temperatura de hibridação por 5 minutos. A reação foi, então, analisada
no equipamento Luminex® (LuminexCorp).
Tomando esse protocolo e os resultados obtidos com ele como referência,
algumas modificações foram propostas e testadas. Avaliou-se, dessa maneira, o
efeito de diferentes temperaturas de hibridação e da introdução de passos de
lavagem após as etapas de hibridação e de marcação. Além da temperatura de
55oC, a hibridação foi também realizada nas temperaturas de 60, 65 e 67oC. As
lavagens, por sua vez, consistiram na adição de 40 µl de Sheath Fluid®
(LuminexCorp) a cada amostra, imediatamente após a hibridação e marcação,
centrifugação a 2.500g por 5 minutos e remoção do sobrenadante por inversão.
Essa seqüência de lavagem foi repetida três vezes após a hibridização e uma vez
após a marcação.
4.7.4. Detecção das seqüências do vírus da dengue pelo equipamento
Luminex®
Após as etapas de hibridização e marcação, a placa contendo até 96
amostras foi posicionada no equipamento Luminex® para contagem das micro-
esferas e quantificação da fluorescência emitida pela ficoeritrina. Cem (100) micro-
esferas de cada especificidade, representando 100 réplicas de cada ensaio, foram
analisadas para cada amostra. O valor mediano da intensidade de fluorescência

50
(MFI) das 100 réplicas foi calculado pelo software do equipamento. Adicionalmente
às 100 medidas feitas para cada amostra, todas as amostras foram analisadas em
duplicata, incluindo o passo de amplificação. Todos os valores medianos de
fluorescência reportados foram corrigidos subtraindo-se dos mesmos o valor de MFI
obtido em uma reação na qual o produto de PCR adicionado na etapa de hibridação
foi substituído por TE (branco). Foi considerada positiva a amostra cuja razão entre o
valor corrigido de MFI da amostra e o valor corrigido de MFI de uma amostra
negativa, presente na mesma placa, foi superior a 4 para DEN1, 70 para DEN2 e 3
para DEN3 e DENG. Esses valores foram determinados com base na análise das
amostras negativas e dos controles positivos. As razões com valores até 15%
inferiores aos citados acima foram classificados como pertencentes à zona cinza do
teste. Amostras que tiveram resultados dentro da zona cinza para as duas réplicas
foram testadas mais duas vezes, sendo que um desses testes foi feito exatamente
como os anteriores e no outro a conversão do RNA em cDNA e a amplificação do
DNA foram feitas em reações uniplex.
A amostra negativa, adicionada a todos os ensaios realizados, consistia no
controle negativo da RT-PCR. Também presente em todos os ensaios estavam o
branco e controles positivos para os sorotipos 1, 2 e 3 do vírus da dengue.

51
5. RESULTADOS
5.1. Avaliação dos diferentes conjuntos de oligonucleotídeos
Na PCR em tempo real, testou-se, inicialmente, os conjuntos de
oligonucleotídeos DEN1, DEN2, DEN3 e DENG (tabela 4.1). Todos forneceram
curvas de amplificação satisfatórias exceto DEN3, que apresentava uma boa
amplificação apenas quando a sonda era excluída da reação e utilizava-se o corante
SYBR Green. Sabendo que somente a metodologia baseada em sonda permitirá o
possível desenvolvimento de um teste multiplex, que inclua, em uma mesma reação,
os oligonucleotídeos específicos para os três sorotipos do vírus, testou-se o grupo
de iniciadores e sonda DEN3J, também específcos para o sorotipo 3 do vírus e que
poderia, então, substituir o conjunto DEN3. Os oligonucleotídeos DEN3J, por sua
vez, mostraram-se eficientes na amplificação do RNA controle para DENV3, através
da metodologia de amplificação e detecção por PCR em tempo real baseada em
sondas TaqMan (figuras 5.1 a 5.8).
A fim de avaliar o desempenho dos sete grupos de oligonucleotídeos no teste
diagnóstico baseado na metodologia de microarranjo líquido, cada conjunto de
iniciadores e sonda foi testado, individualmente, quanto sua capacidade de
amplificar o RNA do vírus da dengue e de detectar o material amplificado,
respectivamente, em reações uniplex de PCR e de hibridação.
Conforme observado na figura 5.9, com exceção do resultado obtido para
DEN1J, as hibridações dos produtos de PCR com as sondas correspondentes,
imobilizadas nas micro-esferas, resultaram em valores medianos de fluorescência

52
(MFI) de, pelo menos, cinco vezes maior do que os valores obtidos quando as
mesmas micro-esferas foram submetidas à hibridação com uma solução que
continha todos os componentes da reação exceto o produto de PCR (branco).
FIGURA 5.1 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 1 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DEN1 e a sonda TaqMan correspondente.
Amplificação dos controles por PCR em tempo real. O eixo x do gráfico de amplificação
representa a intensidade de fluorescência relativa (delta Rn) e o eixo y o ciclo da PCR.

53
FIGURA 5.2 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 1 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DENG e a sonda TaqMan correspondente.
Legenda: ver figura 5.1.
FIGURA 5.3 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 2 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DEN2 e a sonda TaqMan correspondente.
Legenda: ver figura 5.1.

54
FIGURA 5.4 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 2 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DENG e a sonda TaqMan correspondente.
Legenda: ver figura 5.1
FIGURA 5.5 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DEN3 e a sonda TaqMan correspondente.
Legenda: ver figura 5.1.

55
FIGURA 5.6 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DEN3 e o corante SYBR Green.
Legenda: ver figura 5.1.
FIGURA 5.7 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DEN3J e a sonda TaqMan correspondente.
Legenda: ver figura 5.1.

56
FIGURA 5.8 – Exemplo de curvas de amplificação obtidas quando o RNA do
sorotipo 3 do vírus da dengue foi amplificado, por PCR em tempo real, utilizando os
iniciadores DENG e a sonda TaqMan correspondente.
Legenda: ver figura 5.1.
FIGURA 5.9 – Valores de fluorescência resultantes da hibridação das diferentes
sondas com os produtos de RT-PCRs uniplex complementares a elas e com o
branco.
DEN1, DEN2, DEN3, DENG, DEN1J, DEN2J, DEN3J: resultado da hibridação da sonda especificada com o produto de RT-PCR complementar a ela. B: resultado da hibridação da sonda indicada na barra anterior com o branco.
1697
41
2506,5
325
4569
82
1070,5
135,5
707
213,5
5055
194
2527
96,5
0
1000
2000
3000
4000
5000
6000
MFI
DEN1 B DEN2 B DEN3 B DENG B DEN1J B DEN2J B DEN3J B

57
5.2. Otimização da reação de hibridação múltipla (multiplex) para o teste
diagnóstico baseado na metodologia de microarranjo líquido
No processo de desenvolvimento de um teste diagnóstico múltiplo (multiplex)
para dengue, baseado na técnica de microarranjo líquido, quantidades iguais das
micro-esferas acopladas às sondas DEN1, DEN2 e DEN3 (tabela 4.1) foram
combinadas, e essa mistura foi hibridada, separadamente, com os produtos de PCR
obtidos pela amplificação do cDNA dos vírus dos tipos 1, 2 e 3.
Inicialmente, foram testadas duas temperaturas distintas de hibridação: 55 e
60oC. As intensidades de fluorescência medianas (MFI) obtidas com a temperatura
de 60oC foram superiores aquelas conseguidas quando a temperatura de 55oC foi
usada para a hibridação. Porém, mesmo com a temperatura de 60oC, observou-se
reação cruzada entre o produto de PCR de DEN1 e a sonda DEN3, o que motivou a
realização de testes adicionais de hibridação com as temperaturas de 65 e 67oC.
Uma vez que os resultados utilizando estas temperaturas não foram satisfatórios,
desenvolveu-se um protocolo no qual a hibridação foi feita a 60oC, porém, diferente
dos testes anteriores, etapas de lavagem foram incluídas após os passos de
hibridação e de marcação. Este protocolo, por sua vez, forneceu as condições
necessárias para a discriminação entre os três sorotipos, sendo adotado para a
realização de todos os testes cujos resultados serão exibidos a diante. Os sinais de
fluorescência obtidos para cada sonda, em todos os ensaios citados acima podem
ser conferidos nos gráficos que se seguem (figura 5.10).

58
FIGURA 5.10 – Resultados dos ensaios de microarranjo líquido multiplex compostos
pelas sondas DEN1, DEN2 e DEN3 hibridadas com os produtos de PCR dos três
sorotipos do vírus da dengue, sob diferentes condições.
HIBRIDAÇÃO A 55 C
0
1000
2000
3000
4000
5000
MFI
sonda DEN1
sonda DEN2
sonda DEN3
HIBRIDAÇÃO A 60 C
0
1000
2000
3000
4000
5000
MFI
HIBRIDAÇÃO A 65 C
0
1000
2000
3000
4000
5000
PCR DEN1 PCR DEN2 PCR DEN3 branco
MFI

59
Continuação figura 5.10.
5.3. Otimização da RT-PCR múltipla (multiplex) para o teste diagnóstico
baseado na metodologia de microarranjo líquido
Com o objetivo de tornar o teste mais rápido e de mais fácil execução,
desenvolveu-se um protocolo de RT-PCR multiplex, onde os iniciadores específicos
para os diferentes sorotipos do vírus e os iniciadores grupo-específicos foram
adicionados, simultaneamente, a um mesmo tubo de ensaio e avaliados quanto sua
eficiência em realizar, primeiramente, a transcrição reversa dos RNAs controles de
DENV1, DENV2 e DENV3 e, na seqüência, a amplificação dos cDNAs provenientes
dessa retro-transcrição. Após a amplificação, a eficiência do método pôde ser
HIBRIDAÇÃO A 60 C COM LAVAGEM
0
1000
2000
3000
4000
5000
PCR DEN1 PCR DEN2 PCR DEN3 branco
MFI
HIBRIDAÇÃO A 67 C
0
1000
2000
3000
4000
5000
MFI
sonda DEN1
sonda DEN2
sonda DEN3

60
comprovada através da hibridação dos produtos de PCR com as sondas
imobilizadas nas micro-esferas, utilizando as condições definidas no item anterior.
FIGURA 5.11 – Hibridação dos produtos da RT-PCR multiplex com a mistura das
sondas DEN1, DEN2, DEN3 e DENG.
Os valores de MFI apresentados nesse gráfico estão subtraídos do valor de MFI obtido na ausência de produto de PCR (MFI corrigido).
Conforme observado no gráfico acima, a utilização do produto da RT-PCR
multiplex não interferiu na capacidade discriminatória do ensaio e não parece ter
afetado a qualidade do sinal de fluorescência.
5.4. Avaliação das diferentes combinações de oligonucleotídeos no
ensaio de microarranjo líquido
A fim de se obter a melhor intensidade de fluorescência, juntamente com a
melhor discriminação entre os sorotipos, diferentes combinações dos seis conjuntos
de oligonucleotídeos sorotipo-específicos (DEN1, DEN2, DEN3, DEN1J, DEN2J e
0
1000
2000
3000
4000
5000
RNA DEN1 RNA DEN2 RNA DEN3
MFI
sonda DEN1
sonda DEN2
sonda DEN3
sonda DENG

61
DEN3J) foram avaliadas. Cada mistura testada continha apenas um grupo de
oligonucleotídeos específicos para cada sorotipo mais os oligonucleotídeos grupo-
específicos (DENG), em quantidades equimolares. Os RNAs de cada sorotipo do
vírus, após submetidos à transcrição reversa e amplificação multiplex com cada
combinação de iniciadores, foram hibridados, também em uma reação múltipla, com
o conjunto de sondas correspondentes aos iniciadores utilizados na RT-PCR. Os
resultados das hibridações, após leitura no equipamento Luminex®, estão
demonstrados abaixo.
FIGURA 5.12 – Resultados dos ensaios de microarranjo líquido multiplex realizados
com diferentes combinações de sondas, para detecção e identificação dos sorotipos
1, 2 e 3 do vírus da dengue.
COMBINAÇÃO 1
0
1000
2000
3000
4000
5000
MFI
sonda DEN1
sonda DEN2
sonda DEN3
sonda DENG
COMBINAÇÃO 2
0
1000
2000
3000
4000
5000
RNA DEN1 RNA DEN2 RNA DEN3
MFI
sonda DEN1
sonda DEN2J
sonda DEN3
sonda DENG

62
Continuação figura 5.12.
Intensidade mediana de fluorescência obtida através da hibridização dos produtos das RT-PCRs dos RNAs controles de DEN1, DEN2 e DEN3 com as sondas correspondes, em ensaios multiplex. Os valores de MFI apresentados nesse gráfico estão subtraídos do valor de MFI obtido na ausência de produto de PCR (MFI corrigido).
COMBINAÇÃO 3
0
1000
2000
3000
4000
5000M
FI
sonda DEN1
sonda DEN2J
sonda DEN3J
sonda DENG
COMBINAÇÃO 5
0
1000
2000
3000
4000
5000
RNA DEN1 RNA DEN2 RNA DEN3
MFI
sonda DEN1J
sonda DEN2J
sonda DEN3J
sonda DENG
COMBINAÇÃO 4
0
1000
2000
3000
4000
5000
MFI
sonda DEN1J
sonda DEN2J
sonda DEN3
sonda DENG

63
Para facilitar a comparação entre as diferentes combinações de
oligonucleotídeos testadas, foi determinada a razão entre o MFI obtido com a
hibridação de uma sonda com o produto de RT-PCR do sorotipo correspondente e o
MFI obtido quando a mesma sonda foi submetida à hibridação com o produto de RT-
PCR dos outros dois sorotipos não complementares a ela.
TABELA 5.1 – Razão entre os valores de fluorescência (MFI) obtidos nas reações
das sondas com os produtos de PCR específicos e não-específicos.
COMBINAÇÃOa DEN1 DEN2 DEN3 DEN2b DEN3 DEN1 DEN3 DEN1 DEN2
1 (DEN1, DEN2, DEN3, DENG)
68,9 76 6,7 6,5 14,1 26,6 2 (DEN1, DEN2J, DEN3, DENG)
6,7 5,1 5.167,0 5.167,0 3,9 5 3 (DEN1, DEN2J, DEN3J, DENG)
4,7 5,6 40,3 3,4 1,4 1,1 4 (DEN1J, DEN2J, DEN3, DENG)
0 0 42,4 40,9 1,1 0,9 5 (DEN1J, DEN2J, DEN3J, DENG)
0 0 3,3 2,1 1,7 1,1 a combinações de oligonucleotídeos avaliadas. b valores da razão entre o MFI corrigido obtido quando a sonda específica para o sorotipo 1 foi hibridada com o produto de RT-PCR complementar a ela e o MFI corrigido obtido quando a mesma sonda foi hibridada com o produto da RT-PCR do sorotipo 2, para todas as combinações de oligonucleotídeos.
Através da comparação, pode-se concluir que as combinações 1 e 2 foram as
que forneceram os maiores valores de fluorescência e permitiram a melhor
discriminação entre os sorotipos. A combinação 2, composta pelos oligonucleotídeos
DEN1, DEN2J, DEN3 e DENG, foi usada para a execução dos demais ensaios
desse trabalho.

64
5.5. Sensibilidade e especificidade dos métodos de diagnóstico
molecular desenvolvidos
A sensibilidade e a especificidade das duas técnicas de diagnóstico molecular
testadas foram avaliadas utilizando o RNA extraído de alíquotas de soro negativo
acrescidas de quantidades conhecidas dos vírus da dengue dos tipos 1, 2 e 3,
isolados em cultura e purificados em gradiente de sacarose. Após extração, o RNA
foi diluído, resultando em oito pontos distintos, cujas concentrações diferiam quatro
vezes entre si. Foram também usados, para análise da especificidade, RNAs
extraídos de soros negativos para dengue, de soros de indivíduos vacinados contra
febre amarela, do vírus da febre amarela propagado em cultura celular e de soros de
pacientes positivos para o vírus da hepatite C (HCV), hepatite B (HBV) e
citomegalovírus (CMV).
No ensaio de PCR em tempo real, o RNA dos três sorotipos do vírus da
dengue analisados foi detectado em todas as diluições, tanto quando utilizado, para
a amplificação, os oligonucleotídeos sorotipo-específicos (DEN1, DEN2, DEN3 e
DEN3J), como quando se utilizou os oligonucleotídeos complementares a todos os
sorotipos (DENG). Mesmo a concentração mais baixa utilizada (53,57 FFU/ml,
equivalente a 0,625 FFU na reação) permaneceu dentro da faixa de linearidade na
curva-padrão, a qual teve, para todos os casos, um coeficiente de correlação
superior a 0,99 (figuras 5.13 a 5.19).

65
FIGURA 5.13 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 1 foi
testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos
sorotipo-específicos DEN1.
R2 = 0,993
0
5
10
15
20
25
30
35
-1 0 1 2 3 4 5
A quantidade de cópias iniciais acrescentada a cada reação foi de 10.240, 2.560, 640, 160, 40, 10, 2,5 e 0,625, e seu log está representado no eixo x. O valor do Ct (threshold cycle) obtido para cada reação está indicado no eixo y. Os valores representam a média de três reações distintas. R2: coeficiente de correlação
FIGURA 5.14 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 1 foi
testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos
grupo-específicos DENG.
R2 = 0,9982
0
5
10
15
20
25
30
35
40
-1 0 1 2 3 4 5
Legenda: ver figura 5.13.

66
FIGURA 5.15 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 2 foi
testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos
sorotipo-específicos DEN2.
R2 = 0,9986
0
5
10
15
20
25
30
35
40
-1 0 1 2 3 4 5
Legenda: ver figura 5.13.
FIGURA 5.16 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 2 foi
testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos
grupo-específicos DENG.
R2 = 0,9953
0
5
10
15
20
25
30
35
40
-1 0 1 2 3 4 5
Legenda: ver figura 5.13.
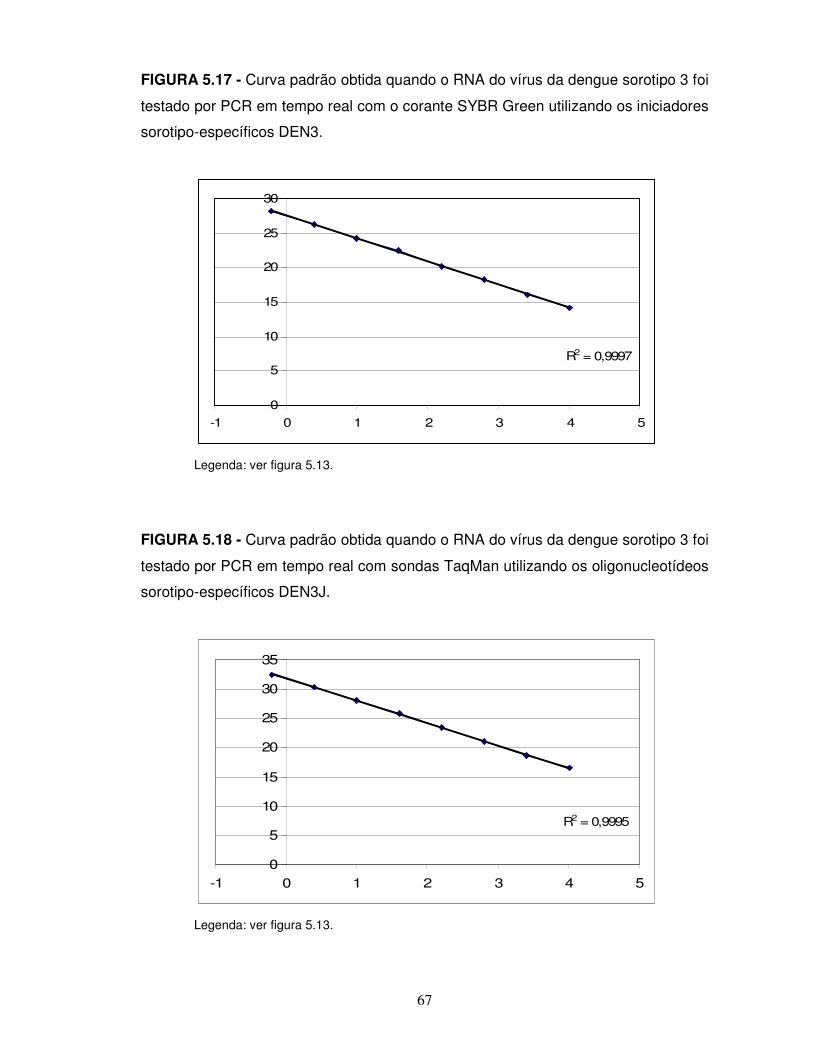
67
FIGURA 5.17 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 3 foi
testado por PCR em tempo real com o corante SYBR Green utilizando os iniciadores
sorotipo-específicos DEN3.
R2 = 0,9997
0
5
10
15
20
25
30
-1 0 1 2 3 4 5
Legenda: ver figura 5.13.
FIGURA 5.18 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 3 foi
testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos
sorotipo-específicos DEN3J.
R2 = 0,9995
0
5
10
15
20
25
30
35
-1 0 1 2 3 4 5
Legenda: ver figura 5.13.

68
FIGURA 5.19 - Curva padrão obtida quando o RNA do vírus da dengue sorotipo 3 foi
testado por PCR em tempo real com sondas TaqMan utilizando os oligonucleotídeos
grupo-específicos DENG.
R2 = 0,9938
0
5
10
15
20
25
30
35
40
-1 0 1 2 3 4 5
Legenda: ver figura 5.13.
Através do ensaio de microarranjo líquido, a menor quantidade de vírus
detectada em todas as réplicas foi de 3.428,48 FFU/ml para DENV1 e 53,57 FFU/ml
para DENV2 e DENV3. Com relação aos oligonucleotídeos grupo-específicos, foram
consideradas como positivas mesmo as amostras contendo a menor quantidade
(53,57 FFU/ml) dos vírus dos tipos DENV1 e DENV2 e, para o sorotipo DENV3,
detectou-se até 214,28 FFU/ml (tabela 5.2).

69
TABELA 5.2 – Razão entre o MFI corrigido obtido com a hibridação das sondas
DEN1, DEN2J, DEN3 e DENG com os produtos de RT-PCR complementares a elas
e o MFI obtido para a mesma sonda na ausência de produto de PCR.
DEN1 DEN2J DEN3 DENG x DEN1a DENG x DEN2b DENG x DEN3c
877.690,88 FFU/ml 13,5 690,6 15,3 11,3 20,0 17,9
219.422,72 FFU/ml 13 576 6,9 16,3 18,3 10,5
54.855,68 FFU/ml 10 497,6 16,4 14 13,4 16,6
13.713,91 FFU/ml 9,6 596,4 10 10,1 9,7 15,4
3.428,48 FFU/ml 5,7 608,5 11,8 6,8 10,8 10,4
857,12 FFU/ml 3,6 862,7 16,9 3,2 8,5 10,1
214,28 FFU/ml 2,3 784,9 7,4 4,7 5,3 3,7
53,57 FFU/ml 1,9 461,9 4,4 5,8 4,3 2,5
a, b, c RNAs controles de DEN1, DEN2 e DEN3, respectivamente, amplificados e detectados pelos oligonucleotídeos grupo-específicos (DENG) Os valores representam a média de três experimentos, que incluem reações de RT-PCR e de detecção distintas. Para serem considerados positivos, os valores devem ser ≥ 4 para DEN1, ≥ 70 para DEN2J e ≥ 3 para DEN3 e DENG.
Vale ressaltar que, tanto nos testes de PCR em tempo real como nos de
microarranjo líquido, nenhuma amostra foi considerada positiva para mais de um
sorotipo e nenhuma amostra negativa para dengue foi detectada, indicando que os
dois métodos apresentam boa especificidade.
5.6. Aplicação dos métodos desenvolvidos na detecção e sorotipagem
do vírus da dengue em amostras clínicas
A fim de avaliar os dois métodos de diagnóstico quanto a sua capacidade em
detectar o RNA do vírus da dengue em amostras clínicas, foram utilizados os RNAs
extraídos de soros de pacientes com suspeita de dengue, submetidos a um teste
sorológico (MAC-ELISA) para a doença, e de soros cuja positividade para a dengue
foi confirmada por isolamento viral. Os resultados obtidos para cada amostra,
através dos diferentes ensaios realizados, encontram-se detalhados no anexo 1.

70
Todos os resultados positivos no grupo de amostras doadas pelo Laboratório
Central do Estado do Paraná (LACEN-PR), provenientes de pacientes supostamente
infectados no início do ano de 2007, foram classificados como sorotipo 3, tanto pela
PCR em tempo real como pelo microarranjo líquido.
Após a realização de todos os ensaios em duplicata, quatro amostras
permaneceram classificadas como “zona cinza” para DEN3 no ensaio de
microarranjo líquido, sendo que duas delas também apresentaram esse resultado
para DENG e duas foram consideradas negativas para dengue grupo-específico.
Estas mesmas quatro amostras foram positivas para DEN3 e DENG na PCR em
tempo real. Além das citadas a cima, outras duas amostras, positivas para DEN3 no
microarranjo e também positivas para DEN3 e DENG na PCR, foram consideradas
zona cinza para DENG, após análise em duplicata pela técnica de microarranjo
líquido. Quando testadas uma terceira vez, as duas amostras, consideradas “zona
cinza” tanto para DEN3 como para DENG, foram classificadas como positivas para
DEN3 e permaneceram dentro da zona cinza para DENG e as outras quatro também
classificadas como “zona cinza” (duas para DEN3 e duas para DENG)
permaneceram dentro dessa faixa. No entanto, todos os valores saíram da zona
cinza e foram considerados positivos quando as amostras foram submetidas a
reações de RT-PCR uniplex.
Para facilitar o entendimento e interpretação dos resultados, criou-se as
tabelas 5.3 e 5.4, onde o número de amostras positivas para dengue grupo-
específico (DENG) e as positivas com os oligonucleotídeos sorotipo-específicos,
foram discriminadas. Conforme observado nestas tabelas, apesar das duas
metodologias de diagnóstico detectarem como positivas aproximadamente o mesmo
número de amostras, houve algumas discordâncias com relação aos resultados
fornecidos pelos dois métodos.

71
TABELA 5.3 - Detecção do RNA do vírus da dengue em soro de paciente, através
de dois métodos diagnósticos distintos, utilizando oligonucleotídeos grupo-
específicos.
no de amostras positivas para dengue grupo-específico no de resultados
discordantes amostras PCR em tempo real microarranjo líquido
IgM negativas (n=48) 18 20 2
IgM positivas (n=50) 8 12 3
isolamento (n=49) 41 41 6
total (n=147) 67 73 11
IgM negativas: grupo de amostras com resultado negativo no teste de MAC-ELISA para dengue; IgM positivas: grupo de amostras com resultado positivo no teste de MAC-ELISA para dengue; isolamento: grupo de amostras positivas para dengue no isolamento viral
TABELA 5.4 - Detecção do RNA do vírus da dengue em soro de paciente, através
de dois métodos diagnósticos distintos, utilizando oligonucleotídeos sorotipo-
específicos.
no de amostras positivas para dengue sorotipo-específico no de resultados
discordantes amostras PCR em tempo real microarranjo líquido
IgM negativas (n=48) 20 21 1
IgM positivas (n=50) 12 12 8
isolamento (n=49) 46 43 5
total (n=147) 78 76 14
Legenda: ver tabela 5.3.
As diferenças entre os resultados obtidos através dos dois métodos ficam
mais claras através da análise da tabela a seguir (tabela 5.5). Nesta tabela, são
apresentados os resultados positivos apenas para os oligonucleotídeos grupo-
específicos, os resultados positivos apenas para os oligonucleotídeos sorotipo-
específicos e os positivos para os dois, simultaneamente, nos dois métodos
diagnósticos.

72
TABELA 5.5 – Resultados obtidos através da utilização de oligonucleotídeos grupo-
e sorotipo-específicos em dois métodos diagnósticos distintos.
no de amostras positivas
PCR em tempo real microarranjo líquido PCR e microarranjo
DEN sorotipo 12 6 4
DEN grupo 1 3 1
DEN sor. + DEN grupo 66 70 62
total 79 79 67
DEN sorotipo: resultados positivos apenas para dengue sorotipo-específico. DEN grupo: resultados positivos apenas para dengue grupo-específico. DEN sor. + DEN grupo: resultados positivos para dengue sorotipo- e grupo-específico, simultaneamente.
Das oito amostras que na PCR em tempo real foram positivas apenas para
dengue sorotipo-específico, três pertencem ao grupo positivo para isolamento viral,
sendo que uma foi sorotipada como DENV1, outra como DENV2 e outra como
DENV3. Quatro amostras pertencem ao grupo positivo para IgM e uma ao grupo
negativo para IgM. Dessas, a amostra de DENV2 (isolamento) é positiva apenas
para DENG no microarranjo, a amostra de DENV1 (isolamento) é positiva para
DENG e DEN1 no microarranjo e a amostra do grupo IgM negativo é positiva para
DEN3 e DENG no microarranjo. As outras cinco amostras (quatro positivas para IgM
e uma para isolamento) são positivas apenas para dengue sorotipo-específico na
PCR em tempo real.
As duas amostras positivas apenas para dengue sorotipo-específico, no
microarranjo líquido, pertencem ao grupo positivo para isolamento viral e são
positivas tanto para dengue grupo- e sorotipo-específico na PCR.
Conforme demonstrado na tabela 5.5, das três amostras positivas somente
para dengue grupo-específico, no microarranjo líquido, uma também apresentou
esse resultado na PCR. Essa amostra pertence ao grupo positivo para isolamento e
foi sorotipada como DENV2. As outras duas também pertencem ao grupo positivo

73
para isolamento viral, sendo que uma (sorotipo DENV2) é positiva apenas para
DEN2 na PCR e a outra (sorotipo DENV1) foi considerada como positiva tanto para
DEN1 como para DENG na PCR.
Das 70 amostras positivas para dengue grupo- e sorotipo-específico no
microarranjo líquido, oito só apresentaram esse resultado para este teste, sendo que
destas, seis são negativas para DENG e dengue sorotipo-específico na PCR (uma é
positiva para isolamento, quatro são positivas para IgM e uma negativa para IgM).
As outras duas (uma IgM negativa e uma IgM positiva) são negativas, na PCR,
apenas para dengue grupo-específico. Das 66 positivas tanto para DENG como para
dengue sorotipo-específico na PCR, quatro não apresentaram o mesmo resultado no
microarranjo. Dessas quatro, todas foram positivas no isolamento, sendo que três
foram positivas, no microarranjo, só para dengue sorotipo-específico e uma,
pertencente ao sorotipo DENV1, só para DENG.

74
6. DISCUSSÃO
As tendências em novas tecnologias para o diagnóstico laboratorial estão
voltadas para testes que permitam a detecção simultânea de diversos patógenos em
um grande número de amostras, que sejam de simples execução, potencialmente
automatizáveis e que forneçam resultados em um curto período de tempo.
Associadas a estas características, ensaios de qualidade devem, certamente, ser
sensíveis e específicos, possibilitando o diagnóstico precoce e diferencial de
doenças diversas. Na área de diagnóstico a necessidade de inovação é constante e
a implantação de metodologias de vanguarda é um requisito obrigatório para o
desenvolvimento da saúde pública. Apesar da enorme importância que o
desenvolvimento tecnológico apresenta nesse segmento, existem, no contexto
nacional, graves limitações de investimentos do setor produtivo privado e grandes
limitações conjeturais no setor produtivo público, que podem aumentar ainda mais a
dependência do país por insumos importados, o que já constitui a realidade para
grande parcela dos testes utilizados no Brasil.
O caso da dengue não é uma exceção. Apesar dos dados alarmantes da
doença no país, pouco tem sido investido para a melhoria do seu diagnóstico e
vigilância epidemiológica. O estabelecimento de um sistema de vigilância embasado
em métodos de diagnóstico laboratorial eficientes, necessita de investimento público
e, portanto, vontade política. Infelizmente, o Brasil sempre se caracterizou por
destinar uma parcela muito pequena do seu orçamento para a saúde pública, a qual,
em geral, é direcionada para programas de saúde mais visíveis. Um dos problemas
que tem acarretado retardo no reconhecimento de uma situação epidêmica,
aumento nos gastos públicos e, indiretamente, diminuição da qualidade da atenção,

75
é, justamente, o uso inadequado das técnicas disponíveis para o diagnóstico da
dengue. O uso dos testes diagnósticos no Brasil não tem sido racional, ocasionando
sobrecarga de trabalho para os laboratórios de saúde pública em períodos
epidêmicos e baixa sensibilidade da vigilância epidemiológica nos períodos não
epidêmicos (MS, 2005(b)), impedindo, dessa forma, que medidas emergenciais
sejam tomadas a tempo de impactar um possível surto. Embora os benefícios de
abordagens públicas a médio e longo prazo para a prevenção e controle da dengue
sejam significativos, o Brasil, assim como outros países endêmicos para a doença,
prefere adotar medidas de controle do vetor apenas durante picos epidêmicos. Essa
estratégia falha e tardia, além de gerar um gasto desnecessário para os cofres
públicos, não resolve de forma alguma o problema da doença. A falta de suporte
laboratorial para a vigilância da dengue é uma realidade em muitos países
endêmicos para a mesma (OOI et al., 2006). No caso do Brasil, apesar da existência
de uma rede laboratorial com potencial para atender a demanda da vigilância,
problemas de infra-estrutura e capacitação técnica devem ser resolvidos. Além
disso, o teste de rotina para o diagnóstico da patologia é um ensaio imuno-
enzimático de captura de IgM (MAC-ELISA), que, apesar de apresentar bons
resultados, só detecta a doença em sua fase tardia, quando os títulos de anticorpos
específicos começam a se elevar, e não permite a identificação do sorotipo
infectante. Ao mesmo tempo, como os níveis de IgM séricos permanecem elevados
por, aproximadamente, três meses após o contágio, esse teste pode classificar como
positivas amostras de pacientes que há muito já resolveram a infecção. Em oposição
à última observação, encontram-se os casos de infecção secundária, onde os níveis
de IgM são, em geral, baixos, podendo não ser detectados nesses testes
(JOHNSON et al., 2005(b)). Como resultado, métodos sorológicos de diagnóstico
exercem menor impacto na vigilância epidemiológica da doença e na orientação da

76
conduta terapêutica (YONG et al., 2007).
A implementação de tecnologias moleculares na virologia clínica resultou não
só na melhoria do diagnóstico de uma série de doenças virais, mas também na
possibilidade de detecção de praticamente qualquer vírus. As técnicas moleculares
mostraram-se especialmente úteis e vantajosas nos casos onde a propagação do
vírus em cultivo celular não é bem sucedida. Devido às inúmeras qualidades, a
implementação de métodos moleculares de diagnóstico na virologia, em substituição
a técnicas tradicionais de isolamento, deve ser maciça e definitiva em um futuro
próximo. Espera-se que nos próximos anos cada vez mais companhias ofereçam
testes diagnósticos baseados em métodos moleculares (VERNET, 2004). Os poucos
laboratórios que fazem a detecção do vírus através de métodos moleculares no
Brasil utilizam ensaios e procedimentos não validados e de desempenho variado,
comprometendo a qualidade dos resultados. Nesse contexto, a implementação e
ampla disponibilização de métodos laboratoriais padronizados para o diagnóstico da
dengue resultarão em benefícios notáveis e devem ser urgentemente consideradas.
A metodologia de PCR em tempo real vem contribuindo para que a PCR seja
cada vez mais aceita como um método diagnóstico (MACKAY et al., 2002;
NIESTERS, 2002). Devido à facilidade de execução dos ensaios, com poucas
possibilidades de manipulação e detecção do alvo pelo próprio equipamento,
durante o curso da reação, essa técnica é considerada muito mais sensível e
reprodutível do que as técnicas tradicionais de amplificação. A ausência de análise
após a etapa de amplificação, além de conferir maior confiabilidade aos resultados,
possibilita, também, a obtenção dos resultados em um curto período de tempo
(LAUE et al., 1999; RATCLIFF et al., 2007; SHU et al., 2003).
Através do ensaio de PCR em tempo real para detecção e sorotipagem do
vírus da dengue desenvolvido nesse trabalho, pôde-se verificar a grande

77
sensibilidade do método. Todos os sorotipos foram detectados mesmo nas menores
concentrações testadas (53,57 FFU/ml ou 0,625 FFU na reação). Esses valores
foram equivalentes (CALLAHAN et al., 2001; HOUNG et al., 2000; HOUNG et al.,
2001; JOHNSON et al., 2005(b)) ou, em alguns casos, superiores (CHEN et al.,
2001; DROSTEN et al., 2002; ITO et al., 2004; KONG et al., 2006; LAUE et al., 1999;
WANG et al., 2002(b); YONG et al., 2007) aos obtidos por outros grupos, em ensaios
baseados na mesma metodologia.
Por possibilitar a realização das etapas de transcrição reversa, amplificação e
detecção em um único tubo, sem a necessidade de intervenções, o ensaio mostrou
ser de fácil otimização. Como em outros testes de base molecular, o principal
desafio encontrado foi na definição das seqüências dos oligonucleotídeos (BAUMS
et al., 2007). Oligonucleotídeos que não só atendam as exigências físico-químicas
impostas pela metodologia, mas que sejam perfeitamente complementares ao maior
número possível de seqüências existentes, são essenciais para o sucesso do
ensaio. Para a determinação das regiões genômicas alvo para os ensaios
desenvolvidos, obteve-se o maior número possível de seqüências do vírus da
dengue junto ao GenBank e levou-se em consideração a conservação genética das
regiões escolhidas e sua ocorrência dentre os isolados brasileiros. Isso garante que
os ensaios sejam mais reprodutíveis, específicos e sensíveis, e principalmente,
capazes de detectar a presença das variantes dos vírus circulantes no país.
A boa qualidade das amplificações e alta sensibilidade da PCR em tempo real
refletiram o cuidado na seleção das seqüências. Mesmo os oligonucleotídeos
obtidos na literatura, que não foram desenhados com base em seqüências
brasileiras, mas são complementares a regiões muito conservadas do vírus,
apresentaram uma grande similaridade com os isolados locais. A seqüência que
mais divergiu das cepas brasileiras foi a da sonda DEN1J, a qual possui duas bases

78
discordantes em relação a um isolado e outras duas em relação a uma segunda
cepa. Apesar do pequeno número (nove) de seqüências completas de isolados
brasileiros presentes no banco de dados não permitir a avaliação da real
significância dessas diferenças, estas podem estar correlacionadas com o fraco
desempenho do conjunto de oligonucleotídeos DEN1J no ensaio de microarranjo
(figura 5.9 e 5.12). O insucesso obtido com a sonda DEN3 no teste de PCR em
tempo real (figura 5.5) só pode ser explicado por problemas na síntese deste
oligonucleotídeo, uma vez que uma sonda de mesma seqüência forneceu bons
resultados no ensaio de microarranjo líquido.
Apesar das reações nos ensaios de microarranjo líquido ocorrerem de forma
mais favorável do que nos microarranjos tradicionais, devido à natureza tri-
dimensional das micro-esferas (DIAZ, 2007), a imobilização das sondas nestas
partículas cria condições menos favoráveis para a interação entre a sonda e seu
alvo do que as condições fornecidas pela PCR em tempo real, que se beneficia de
uma cinética de reação de fase líquida. A existência da etapa de hibridação após a
amplificação dos alvos faz com que a definição da seqüência da sonda para a
técnica de microarranjo líquido requeira especial atenção (DIAZ e FELL, 2004).
O tamanho da sonda está, em geral, diretamente relacionado com a
sensibilidade e inversamente relacionado com a especificidade do ensaio. O
aumento da sensibilidade geralmente observado em ensaios com sondas maiores é
resultado do aumento da eficiência de hibridação dessas sondas, o que pode ser
explicado pelo fato de que a energia livre associada a um mismatch é uma fração da
energia livre total da ligação (ABOU-ELA et al., 1985), fazendo com que o efeito de
um mismatch seja maior quanto menor for o tamanho da sonda (LIVSHITS e
MIRZABEKOV, 1996). No entanto, embora sondas com mais de 50 nucleotídeos
tendam a melhorar a sensibilidade do teste, a possibilidade da formação de

79
estruturas secundárias e da geração de resultados falso-positivos, devido à perda da
especificidade, é maior. Tal efeito é relatado por DIAZ e FELL (2004) e
ARMSTRONG et al. (2000), que observaram uma diminuição substancial da
resolução e, conforme esperado, da especificidade do teste mediante a adição de
bases à seqüência original da sonda.
Por outro lado, sondas pequenas, de 8 a 19 nucleotídeos, como as utilizadas
inicialmente nos ensaios de microarranjo, não permitem o uso de condições muito
estringentes de hibridação, fazendo com que esses fragmentos muito curtos sejam,
em muitos casos, incapazes de fornecer a especificidade necessária ao ensaio. Um
bom equilíbrio entre especificidade e sensibilidade em ensaios de microarranjo tem
sido atingido com sondas de 20 a 30 nucleotídeos. Outro fator que pode afetar a
eficiência de hibridação é o tamanho da seqüência alvo. Melhores resultados são,
em geral, observados quando o tamanho desses fragmentos são inferiores a 300 pb
(revisado por DIAZ, 2007).
As sondas avaliadas nesse trabalho apresentam tamanhos entre 19 e 31
nucleotídeos e os iniciadores utilizados geraram produtos de 66 a 112 pb. As
pequenas diferenças observadas nos tamanhos tanto das sondas como dos
amplicons não puderam ser correlacionadas com as diferenças na qualidade dos
resultados obtidos. Embora sondas com regiões internas de complementaridade
tenham sido evitadas, algumas poucas bases complementares dentro das
seqüências (hairpins) podem ter gerado estruturas secundárias que afetaram a
hibridação dessa molécula de captura com seu alvo. A fim de verificar se essas
interações poderiam estar prejudicando a eficiência do ensaio, a existência de
regiões de complementaridade dentro e entre as seqüências de todos os
oligonucleotídeos foi investigada utilizando o software OligoAnalyzer versão 3.0
(www.idtdna.com/analyzer/Applications/OligoAnalyzer/). O resumo dessas análises

80
encontra-se no anexo 2. Usando o valor de ∆G como medida da estabilidade dessas
interações, verificou-se que todos os possíveis hairpins formados pelos
oligonucleotídeos são desfavoráveis (valores de ∆G positivos) a uma temperatura de
60oC.
A principal vantagem da metodologia de microarranjo líquido, que consiste na
possibilidade de análise simultânea de diferentes alvos, só poderá ser usufruída se
reações de amplificação e hibridação múltiplas (multiplex) forem bem sucedidas.
Para tal, cuidados especiais devem, também, ser tomados para que os
oligonucleotídeos escolhidos interfiram o mínimo possível uns com os outros. Todos
os oligonucleotídeos usados nesse trabalho foram testados, individualmente e dois a
dois, para a possível ocorrência de homo e hetero-dímeros, respectivamente. Dos
resultados obtidos através dessas análises, os quais estão dispostos no anexo 3,
merece especial atenção o fato de que os oligonucleotídeos DENG formam híbridos
estáveis (∆G < -6) com a maioria das seqüências. Na prática, o desempenho desses
oligonucleotídeos foi, de fato, pior do que dos oligonucleotídeos sorotipo-específicos
na PCR em tempo real e, apesar do mesmo não ser observado no teste de
microarranjo líquido, o uso desse conjunto de oligonucleotídeos pode estar
prejudicando o desempenho do ensaio como um todo. Portanto, a não inclusão
dessas seqüências na versão final dos testes multiplex deve ser considerada.
Oligonucleotídeos grupo-específicos seriam relevantes em um ensaio de PCR
em tempo real cujo objetivo não fosse a sorotipagem. Uma reação que permitisse
apenas a detecção do vírus poderia ser incluída em um teste diagnóstico multiplex
para diferentes patógenos ou mesmo em um teste uniplex de triagem para dengue,
o qual direcionaria, por fim, investigações mais detalhadas.
Da mesma maneira que a formação de estruturas secundárias entre os
oligonucleotídeos do grupo DENG e as demais seqüências pode explicar os

81
resultados obtidos na prática, a mesma correlação pode ser feita para os
oligonucleotídeos DEN1 e DEN1J, que formam interações muito estáveis (∆G < -10)
com várias seqüências e cujo desempenho não é tão satisfatório como o de outros
conjuntos de oligonucleotídeos. Outras situações, no entanto, como os altos valores
negativos de ∆G obtidos entre os oligonucleotídeos DEN2J, não estão de acordo
com os bons resultados alcançados com seu uso na prática.
Apesar do pequeno número de alvos desse trabalho permitir a análise
individual de cada oligonucleotídeo e de cada possível par de interação, essa
estratégia, pouco automatizada, não será viável a medida que se aumente a
abrangência do teste. Felizmente, softwares e estratégias que permitem a predição
do comportamento de diversos oligonucleotídeos em reações multiplex e que
auxiliam na determinação das condições mais apropriadas para determinada reação
encontram-se disponíveis gratuitamente e, certamente, serão empregados em
desenvolvimentos futuros (ELNIFRO et al., 2000; HENEGARIU et al., 1997;
SCHOSKE et al., 2003).
Sobre o desenho das sondas para ensaios de microarranjo líquido podemos
concluir, entre outras coisas, que cuidados com relação ao tamanho e seqüência
devem ser tomados, mas que, conforme bem colocado por DIAZ (2007), “apesar dos
esforços em prever o comportamento das sondas baseando-se na posição das
bases complementares, na cinética de hibridação, na formação de estruturas
secundárias e na interação diferencial de regiões com presença ou ausência de
loops na seqüência alvo, até o momento ninguém desenvolveu um conjunto
convincente de regras a seguir”.
Outros fatores que afetam a especificidade e sensibilidade do ensaio são a
temperatura e tempo de hibridação e a quantidade de alvo adicionada à reação
(DUNBAR, 2006). Conforme observado durante o processo de padronização da

82
reação de hibridação, alterações na temperatura ocasionaram grandes mudanças
nos resultados dos ensaios. Como esperado, temperaturas mais baixas diminuíram
o poder de discriminação do teste, gerando resultados falso-positivos e duvidosos.
Já temperaturas muito próximas à Tm das sondas dificultaram a formação de
híbridos entre estas e seus alvos, tornando o ensaio pouco eficiente. A melhor
solução encontrada foi a utilização de uma temperatura intermediária, que apesar de
ainda permitir poucas interações inespecíficas, possibilitou a obtenção de bons
resultados após a introdução de passos de lavagem. Essas etapas de lavagem
reduzem a velocidade de execução do teste, a reprodutibilidade e aumentam a
chance de contaminações e erros. Todos esses inconvenientes, porém, podem ser
suplantados mediante a automação dos procedimentos. Sistemas de pipetagem
automatizados, acoplados a bombas a vácuo ou a separadores magnéticos, já se
encontram amplamente disponíveis comercialmente e forneceriam o nível de
automação e, conseqüentemente, reprodutibilidade e rapidez desejadas ao teste. Do
mesmo modo, diferentes soluções e tempos de lavagem e hibridação devem ser
testados a fim de definir qual o protocolo exato que permitirá a obtenção dos
melhores resultados, em um menor período de tempo e com o mínimo de
intervenções.
A adição de tetrametilamônio clorido (TMAC) ao tampão de hibridação
certamente facilitou a realização desse passo. O uso de TMAC a uma concentração
final de 3 ou 4 M elimina a influência do conteúdo de GC na determinação da Tm da
sonda, fazendo com que as condições de hibridação sejam dependentes do
tamanho da sonda e não da sua composição. A equalização das temperaturas de
dissociação dos pares AT e GC permite que diferentes sondas, com características
distintas, sejam incorporadas em um ensaio multiplex e usadas sob condições de
hibridação idênticas (MASKOS e SOUTHERN, 1992).

83
Já o efeito da quantidade de alvo na hibridação pôde ser observado quando
foi adicionado, à reação, 12 µl de produto de PCR, em vez dos usuais 5 µl usados
em todos os testes. Nesse teste, cujos resultados não foram apresentados,
amostras controles e de pacientes foram sorotipadas erroneamente e algumas ainda
foram consideradas positivas para mais de um sorotipo. Além da perda de
especificidade, sabe-se que a renaturação do produto de PCR é mais evidente
quando o mesmo está presente em concentrações muito elevadas (além do ponto
de saturação) (NOLAN e MANDY, 2001), o que pode ocasionar, também, a perda de
sensibilidade do teste. Quantidades intermediárias, no entanto, podem ser avaliadas,
com o objetivo de se conseguir a máxima sensibilidade sem que a especificidade
seja comprometida.
Mesmo não estando completamente padronizado, as detecções e
sorotipagens feitas com o teste de microarranjo líquido apresentaram boa
concordância com os resultados obtidos por PCR em tempo real. Das 66 amostras
positivas nas PCRs grupo- e sorotipo-específicas, 62 apresentaram o mesmo
resultado no microarranjo líquido, que foi capaz de detectar mais oito amostras. Uma
dessas é sabidamente positiva (resultado confirmado por isolamento viral) e não foi
detectada pela PCR em tempo real. A mesma situação pode, também, estar
ocorrendo para outras duas amostras que foram, igualmente, positivas com os
oligonucleotídeos grupo- e sorotipo-específicos no microarranjo líquido e positivas
somente na PCR sorotipo-específica. Por outro lado, as três amostras positivas para
o isolamento viral que foram, também, positivas para dengue sorotipo-específico na
PCR, porém negativas no microarranjo líquido, sugerem uma menor sensibilidade
desta técnica frente ao teste de PCR em tempo real.
O maior número de resultados positivos obtidos com os oligonucleotídeos
sorotipo-específicos na PCR em tempo real indica que estes oligonucleotídeos

84
apresentam melhor desempenho do que os oligonucleotídeos grupo-específicos, na
detecção do vírus da dengue em amostras clínicas, através desse método. Essa
afirmação ganha confiabilidade quando se toma como referência o grupo de
amostras das quais isolou-se o vírus da dengue. O mesmo não se pode afirmar para
a técnica de microarranjo líquido, pois, também com base nas amostras positivas
para isolamento viral, houve falha na detecção de três amostras através da utilização
dos oligonucleotídeos sorotipo-específicos e de duas amostras com os
oligonucleotídeos grupo-específicos. As cinco amostras positivas exclusivamente na
PCR em tempo real sorotipo-específica podem estar refletindo a maior sensibilidade
deste teste, já que uma dessas amostras pertence ao grupo positivo para isolamento
e as outras quatro pertencem ao grupo positivo para IgM e podem, portanto,
apresentar baixa carga viral. No entanto, a ocorrência de resultados falso-positivos
não podem ser completamente descartados. A maior quantidade de resultados
positivos, fornecidos pelos dois testes moleculares, no grupo de pacientes negativos
para IgM anti-dengue, está de acordo com o fato de que a carga viral é
inversamente proporcional ao título de anticorpo (HALSTEAD, 1988; YONG et al.,
2007). Com relação à sorotipagem, houve concordância de 100% dos resultados,
sugerindo que ambos os testes apresentam boa especificidade.
Um fator importante para a perda da sensibilidade no microarranjo líquido é a
natureza multiplex dessa técnica. A utilização de uma PCR (ou RT-PCR) multiplex
em conjunto com a hibridação também multiplex, nos microarranjos líquidos, é
fundamental para manter a principal característica do teste que é o caráter múltiplo.
Entretanto, a introdução de uma PCR multiplex aumenta a complexidade do
desenvolvimento e pode comprometer a sensibilidade do método. Uma outra
maneira de resolver o problema da perda de sensibilidade, que não implica na
modificação das seqüências dos oligonucleotídeos e nem nas condições de

85
hibridação, pode ser através do uso de abordagens que aumentem a intensidade do
sinal do repórter. Como substitutos dos iniciadores biotinilados, que acrescentam
somente uma molécula de biotina ao alvo, podem ser usados nucleotídeos
biotinilados, ou dendrímeros de DNA (DIAZ, 2007). Segundo LOWE et al. (2004), a
marcação do DNA com dendrímeros resultou em um aumento de dez vezes na
fluorescência em relação ao sinal emitido por uma única molécula de streptavidina-
R-ficoeritrina. Esse nível de sensibilidade permitiu a detecção de uma única
molécula por micro-esfera. Reagentes destinados à amplificação de alvos em
formatos multiplex são comercializados por diferentes empresas e podem, também,
ser empregados.
No presente trabalho, apesar de não terem sido observadas diferenças nos
resultados quando os RNAs controles foram amplificados em RT-PCRs uniplex ou
multiplex, em algumas amostras clínicas, a seqüência do vírus da dengue foi
detectada apenas quando essas foram submetidas a uma RT-PCR uniplex,
sugerindo que a RT-PCR multiplex impõe uma dificuldade a mais ao ensaio, a qual
pode ser relevante em amostras com baixa carga viral.
Apesar de representarem um desafio para o desenvolvimento, metodologias
que permitam a realização de testes multiplex são altamente desejáveis, pois
implicam na redução de custos, trabalho e tempo de execução (DUNBAR, 2006). A
capacidade multiplex da PCR em tempo real é limitada pelos equipamentos
atualmente disponíveis, os quais permitem a detecção simultânea de apenas quatro
fluoróforos distintos. No microarranjo líquido, no entanto, o número de alvos que
podem ser detectados é limitado, apenas, pela capacidade de amplificação de
produtos múltiplos na PCR. Os microarranjos líquidos podem, inclusive, conter
diferentes sondas para o mesmo vírus ou sorotipo. Um certo nível de redundância
pode garantir a acurácia dos resultados além de permitir, através da utilização de

86
sondas complementares a diferentes níveis taxonômicos, a identificação de
patógenos emergentes (DIAZ, 2007; SUMMERBELL et al., 2005).
Conforme se faz perceptível pelo exposto acima, a otimização do ensaio de
microarranjo líquido é mais demorada e trabalhosa do que a da PCR em tempo real.
O fato dessa última metodologia já estar mais consolidada e disseminada, com
reagentes e protocolos bem estabelecidos, contribui para a facilidade de
estabelecimento da mesma. À medida que a técnica de microarranjo líquido torne-
se, da mesma forma, mais corriqueira, protocolos e, principalmente, reagentes
voltados exclusivamente para este fim deverão ser cada vez mais comuns. Mesmo
que de forma mais lenta, o conhecimento atual nessa área permite o
desenvolvimento de testes de excelente qualidade, com a utilização dessa
metodologia. Apesar da quase inexistência de trabalhos comparando o desempenho
de ensaios estabelecidos sobre a plataforma de microarranjo líquido com outras
metodologias, resultados equivalentes entre esta técnica e as técnicas de PCR em
tempo real e microarranjo tradicional foram obtidos por alguns pesquisadores
(ARMSTRONG et al., 2000; BRUNSTEIN e THOMAS, 2006; OH et al, 2007).
Desconsiderando a etapa de extração do RNA, que é comum aos dois
métodos, o tempo necessário para obtenção do resultado através do teste de
microarranjo líquido é, sob as condições atuais, de, aproximadamente, cinco horas.
Com a metade desse tempo, a PCR em tempo real para um sorotipo é concluída.
Considerando que, a fim de investigar uma amostra para a presença dos três
sorotipos do vírus da dengue faz-se necessária a realização de três reações de
amplificação em tempo real, ao passo que com um único ensaio de microarranjo
líquido chega-se a esse resultado, a técnica de microarranjo líquido passa a ser mais
vantajosa com relação ao tempo de execução. Isso se tornará mais evidente à
medida que a capacidade multiplex do microarranjo líquido seja ampliada e um

87
mesmo teste seja utilizado para investigar a presença de diversos patógenos. O
mesmo efeito será observado no custo dos testes. Da maneira como foi executada
nesse trabalho, uma reação de PCR em tempo real custa, aproximadamente, R$
4,50. Ou seja, a investigação dos três sorotipos em uma amostra, por PCR em
tempo real, custaria R$ 13,50 contra R$ 12,00 pelo teste baseado em microarranjo
líquido. Essa diferença aumenta a medida que mais alvos sejam investigados.
Além do menor custo e maior velocidade de execução, decorrentes da sua
forte capacidade em realizar análises múltiplas, uma outra vantagem da técnica de
microarranjo líquido, frente à PCR em tempo real, é a objetividade na análise dos
resultados. Apesar da otimização mais trabalhosa, após padronizado, o microarranjo
é menos subjetivo na interpretação dos resultados do que a PCR, pois a existência
de um valor de corte possibilita a determinação automática das amostras positivas e
negativas através da utilização de algoritmos computacionais simples. Já a PCR em
tempo real depende do ajuste manual do limiar de detecção (threshold) e avaliação
individual de cada curva de amplificação. Além disso, não só o tempo de execução,
como também a chance de erros do ensaio de microarranjo líquido, pode ser
drasticamente reduzido com a introdução de uma reação de RT-PCR em passo
único e a automação do processo, que eliminaria as etapas de centrifugação. Já a
automação da PCR em tempo real, apesar de também dar mais confiabilidade ao
método, não alteraria, de forma significativa, o tempo de execução do teste.
Apesar das particularidades inerentes a cada método, as duas técnicas,
mostraram-se apropriadas para a implementação de testes de diagnóstico molecular
na rotina laboratorial, pois são sensíveis, específicas, de fácil execução, rápidas,
possuem potencial de automação e equipamentos de bancada com flexibilidade
para atender rotinas grandes e pequenas.

88
7. CONCLUSÕES
Através desse trabalho, foram desenvolvidos dois métodos para o diagnóstico
e sorotipagem molecular da dengue. As metodologias escolhidas (PCR em tempo
real e microarranjo líquido) representam os mais recentes avanços na área de
diagnóstico, e ambas provaram ser metodologias de análise em larga escala,
capazes de fornecer resultados confiáveis em um curto período de tempo.
O ensaio de PCR em tempo real mostrou ser de fácil otimização, fácil
execução e altamente sensível. Porém, sua capacidade de efetuar análises múltiplas
é limitada. Ao contrário da técnica de microarranjo líquido, que por envolver várias
etapas, desde a amplificação das amostras até a hibridação e marcação, não é tão
facilmente padronizada, mas possui uma grande capacidade multiplex, que a torna
extremamente atrativa para o setor de diagnóstico. Adicionalmente a sua
capacidade em realizar análises múltiplas, a flexibilidade, possibilidade de
incorporação de vários formatos de ensaios, custo acessível e potencial de
automação são outras das conveniências dessa tecnologia.
O potencial de realizar análises simultâneas de diferentes alvos, da técnica de
microarranjo líquido, foi demonstrado tanto através da hibridação de amplicons
gerados em reações de amplificação uniplex com sondas agrupadas em formato
multiplex, como através da hibridação de amplicons gerados em RT-PCRs multiplex
com sondas agrupadas. Contudo, apesar do grande potencial apresentado pelo
teste de detecção e sorotipagem do vírus da dengue baseado nessa metodologia,
ainda serão necessárias diversas adaptações e validações antes do seu uso
definitivo como um teste diagnóstico.

89
Apesar de apresentarem grande potencial como metodologias diagnósticas, a
automação é essencial para implementação desses ensaios na rotina laboratorial.
Mesmo na PCR em tempo real, que exige menos manipulação, as possibilidades de
erro são elevadas quando um grande número de amostras é manipulado
simultaneamente.
Por fim, esse trabalho não só permitiu o desenvolvimento de novos testes
para o diagnóstico de uma doença de grande importância para a saúde pública do
Brasil, como também possibilitou a aquisição de competência em tecnologias que
devem dominar o setor de diagnóstico. Devido sua grande aplicabilidade, essas
plataformas poderão ser, futuramente, utilizadas para identificação de diferentes
patógenos importantes dentro do contexto epidemiológico brasileiro. O domínio
dessas metodologias poderá resultar em relevantes desdobramentos tecnológicos
aplicáveis a diferentes testes diagnósticos, possibilitando uma maior economia de
divisas e independência tecnológica do país.

90
8. PERSPECTIVAS
Esse trabalho sumariza os avanços alcançados, até o momento, em direção
ao desenvolvimento de testes moleculares para o diagnóstico e sorotipagem da
dengue. Os resultados aqui apresentados nos permitem avaliar as estratégias
empregadas e direcionar pesquisas futuras.
Através dessa análise crítica foi possível inferir que a técnica de microarranjo
líquido representa uma grande promessa para o setor de diagnóstico e, portanto,
mais esforços serão investidos para seu estabelecimento definitivo como método
diagnóstico para dengue e outras patologias. Esses esforços devem se concentrar:
1) na melhoria dos oligonucleotídeos específicos para o sorotipo 1 do
vírus da dengue;
2) no desenvolvimento e otimização de uma reação de RT-PCR em
passo único;
3) na avaliação de diferentes tempos e tampões de hibridação;
4) na avaliação do efeito da adição de diferentes quantidades do
produto de PCR na reação de hibridação;
5) na avaliação de diferentes métodos de marcação dos produtos de
PCR;
6) na automação da metodologia.
A PCR em tempo real comprovou ser uma técnica muito sensível e
específica. Devido suas qualidades como método de detecção de ácidos nucléicos e
por ser uma metodologia já bem estabelecida, a PCR em tempo real, assim como

91
ocorreu nesse trabalho, deverá servir como referência para a padronização de
qualquer outra técnica que se proponha ao mesmo objetivo. A fim de que o ensaio
para o vírus da dengue baseado nessa metodologia fique ainda mais adequado ao
diagnóstico em larga escala, será necessário o estabelecimento de um protocolo que
permita a detecção dos três sorotipos do vírus em um mesmo tubo de ensaio, além
da avaliação de soluções mais automatizadas para o preparo da reação.
A competência adquirida através da realização desse trabalho já está sendo
aplicada para o desenvolvimento de outros testes baseados nas mesmas
metodologias, que deverão ser cada vez mais utilizadas como ferramentas de
diagnóstico.

92
ANEXOS
ANEXO 1 – Resultados obtidos com os testes de PCR em tempo real e de
microarranjo líquido para o diagnóstico da dengue em amostras clínicas.................93
ANEXO 2 – Valores do ∆G mais desfavorável das possíveis estruturas secundárias
internas (hairpins) formadas por cada oligonucleotídeo usado nos ensaios de PCR
em tempo real e microarranjo líquido.........................................................................96
ANEXO 3 – Valores do ∆G mais desfavorável dos possíveis homo e hetero-dímeros
formados pelos oligonucleotídeos usados nos ensaios de PCR em tempo real e
microarranjo líquido................................................................................................... 97

93
ANEXO 1 – Resultados obtidos com os testes de PCR em tempo real e de microarranjo líquido para o diagnóstico da dengue em amostras clínicas.
AMOSTRA COLETA IgM PCR DENG PCR DENESP LUMINEX DENG LUMINEX DENESP ISOLAMENTO
1 - dia 1 NEG NEG NEG NEG NEG NR
2 - dia 1 NEG NEG NEG NEG NEG NR
3 - dia 2 NEG NEG NEG NEG NEG NR
4 - dia 1 NEG NEG NEG NEG NEG NR
5 - dia 3 NEG POS DENV3 POS DENV3 NR
6 - dia 3 NEG POS DENV3 POS DENV3 NR
7 - dia 3 NEG POS DENV3 POS DENV3 NR
8 - dia 4 NEG POS DENV3 POS DENV3 NR
9 - dia 3 NEG NEG NEG NEG NEG NR
10 - dia 1 NEG NEG NEG NEG NEG NR
11 - dia 3 NEG POS DENV3 POS DENV3 NR
13 - NI NEG NEG NEG NEG NEG NR
14 - dia 3 NEG POS DENV3 POS DENV3 NR
15 - dia 0 NEG NEG NEG NEG NEG NR
16 - dia 1 NEG NEG NEG NEG NEG NR
17 - dia 2 NEG NEG NEG NEG NEG NR
18 - dia 2 NEG NEG NEG NEG NEG NR
19 - dia 2 NEG NEG NEG NEG NEG NR
20 - dia 2 NEG NEG NEG NEG NEG NR
21 - dia 2 NEG NEG NEG NEG NEG NR
23 - dia 2 NEG POS DENV3 POS DENV3 NR
24 - dia 2 NEG POS DENV3 POS DENV3 NR
25 - dia 3 NEG NEG DENV3 POS DENV3 NR
26 - dia 6 NEG NEG NEG NEG NEG NR
27 - NI NEG NEG NEG NEG NEG NR
28 - dia 4 NEG NEG NEG NEG NEG NR
29 - dia 3 NEG POS DENV3 POS DENV3 NR
30 - dia 4 NEG POS DENV3 POS DENV3 NR
31 - dia 3 NEG POS DENV3 POS DENV3 NR
32 - dia 3 NEG POS DENV3 POS DENV3 NR
33 - dia 4 NEG POS DENV3 POS DENV3 NR
34 - dia 1 NEG POS DENV3 POS DENV3 NR
35 - dia 5 NEG NEG DENV3 NEG DENV3 NR
36 - dia 1 NEG POS DENV3 POS DENV3 NR
37 - dia 3 NEG NEG NEG NEG NEG NR
38 - dia 2 NEG POS DENV3 POS DENV3 NR
39 - dia 1 NEG POS DENV3 POS DENV3 NR
40 - dia 6 NEG NEG NEG NEG NEG NR
41 - dia 12 NEG NEG NEG NEG NEG NR
42 - dia 7 NEG NEG NEG NEG NEG NR
43 - dia 10 NEG NEG NEG NEG NEG NR
44 - dia 5 NEG NEG NEG NEG NEG NR
45 - dia 4 NEG NEG NEG NEG NEG NR
46 - dia 4 NEG NEG NEG NEG NEG NR
47 - dia 9 NEG NEG NEG POS DENV3 NR
48 - dia 11 NEG NEG NEG NEG NEG NR
49 - dia 8 NEG POS DENV3 POS DENV3 NR
50 - dia 6 NEG NEG NEG NEG NEG NR

94
Continuação anexo 1.
AMOSTRA COLETA IgM PCR DENG PCR DENESP LUMINEX DENG LUMINEX DENESP ISOLAMENTO
1 + dia 9 POS NEG NEG NEG NEG NR
2 + dia 13 POS NEG NEG NEG NEG NR
3 + dia 9 POS NEG NEG NEG NEG NR
4 + dia 11 POS NEG NEG NEG NEG NR
5 + dia 9 POS NEG NEG POS DENV3 NR
6 + dia 10 POS NEG DENV3 NEG NEG NR
7 + dia 7 POS NEG NEG NEG NEG NR
8 + dia 6 POS POS NEG POS NEG NR
9 + dia 7 POS NEG NEG NEG NEG NR
10 + dia 10 POS NEG DENV3 NEG NEG NR
11 + dia 16 POS NEG NEG NEG NEG NR
12 + dia 3 POS POS DENV3 POS DENV3 NR
13 + dia 10 POS NEG NEG NEG NEG NR
14 + dia 6 POS POS DENV3 POS DENV3 NR
15 + dia 9 POS NEG NEG NEG NEG NR
16 + dia 6 POS POS DENV3 POS DENV3 NR
17 + dia 4 POS NEG NEG NEG NEG NR
18 + dia 9 POS NEG NEG NEG NEG NR
19 + dia 7 POS NEG NEG NEG NEG NR
20 + dia 6 POS NEG NEG POS DENV3 NR
21 + dia 4 POS NEG NEG NEG NEG NR
22 + dia 7 POS NEG NEG NEG NEG NR
23 + dia 12 POS NEG NEG NEG NEG NR
24 + dia 5 POS NEG NEG POS DENV3 NR
25 + dia 10 POS NEG NEG NEG NEG NR
26 + dia 7 POS NEG NEG NEG NEG NR
27 + dia 13 POS NEG NEG NEG NEG NR
28 + dia 7 POS NEG DENV3 NEG DENV3 NR
29 + dia 9 POS NEG NEG NEG NEG NR
30 + dia 7 POS POS DENV3 POS DENV3 NR
31 + dia 6 POS NEG NEG NEG NEG NR
32 + dia 7 POS NEG NEG NEG NEG NR
33 + dia 7 POS NEG NEG NEG NEG NR
34 + dia 8 POS NEG NEG NEG NEG NR
35 + dia 10 POS POS DENV3 POS DENV3 NR
36 + dia 7 POS NEG NEG NEG NEG NR
37 + dia 9 POS NEG NEG NEG NEG NR
38 + dia 9 POS NEG DENV3 NEG NEG NR
39 + dia 6 POS NEG NEG NEG NEG NR
40 + dia 8 POS NEG DENV3 NEG NEG NR
41 + dia 5 POS POS DENV3 POS DENV3 NR
42 + dia 5 POS POS DENV3 POS DENV3 NR
43 + dia 5 POS NEG NEG POS DENV3 NR
44 + dia 6 POS NEG NEG NEG NEG NR
45 + dia 9 POS NEG NEG NEG NEG NR
46 + dia 5 POS NEG NEG NEG NEG NR
47 + dia 9 POS NEG NEG NEG NEG NR
48 + dia 7 POS NEG NEG NEG NEG NR
49 + dia 6 POS NEG NEG NEG NEG NR
50 + dia 8 POS NEG NEG NEG NEG NR

95
Continuação anexo 1.
AMOSTRA COLETA IgM PCR DENG PCR DENESP LUMINEX DENG LUMINEX DENESP ISOLAMENTO
5B dia 3 NEG POS DENV1 POS NEG DENV1
6B dia 1 NEG POS DENV1 POS DENV1 DENV1
7B dia 2 NEG POS DENV1 POS DENV1 DENV1
8B dia 7 NEG POS DENV1 POS DENV1 DENV1
9B dia 2 NEG POS DENV1 POS DENV1 DENV1
10B dia 2 NEG POS DENV1 POS DENV1 DENV1
11B dia 3 NEG POS DENV1 POS DENV1 DENV1
12B dia 2 NEG POS DENV1 NEG DENV1 DENV1
13B dia 1 NEG POS DENV1 POS DENV1 DENV1
14B dia 3 NEG POS DENV1 POS DENV1 DENV1
15B dia 3 NEG POS DENV1 POS DENV1 DENV1
16B dia 3 NEG NEG DENV1 POS DENV1 DENV1
17B dia 2 NR POS DENV1 POS DENV1 DENV1
18B dia 2 NR POS DENV1 POS DENV1 DENV1
23B dia 2 NEG NEG NEG NEG NEG DENV2
24B dia 3 NEG POS DENV2 POS DENV2 DENV2
25B dia 4 NEG POS DENV2 NEG DENV2 DENV2
26B dia 1 NEG NEG NEG NEG NEG DENV2
27B dia 1 NEG POS DENV2 POS DENV2 DENV2 28B dia 1 NEG NEG DENV2 POS NEG DENV2 29B dia 2 NEG POS DENV2 POS DENV2 DENV2 33B dia 2 NEG POS DENV3 POS DENV3 DENV3 34B dia 2 NEG POS DENV3 POS DENV3 DENV3 35B dia 5 NEG POS DENV3 POS DENV3 DENV3 36B dia 2 NEG POS DENV3 POS DENV3 DENV3 37B dia 2 NEG POS DENV3 POS DENV3 DENV3 38B dia 4 NEG POS DENV3 POS DENV3 DENV3 39B dia 2 NEG POS DENV3 POS DENV3 DENV3 40B dia 2 NR POS DENV3 POS DENV3 DENV3 42B dia 2 NR POS DENV3 POS DENV3 DENV3 43B dia 99 NEG NEG DENV3 NEG DENV3 DENV3 44B dia 4 NEG POS DENV3 POS DENV3 DENV3 45B dia 0 NR POS DENV3 POS DENV3 DENV3 46B dia 5 NEG POS DENV3 POS DENV3 DENV3 47B dia 0 NR POS DENV3 POS DENV3 DENV3 48B dia 2 NR POS DENV3 POS DENV3 DENV3 49B dia 3 NR POS DENV3 POS DENV3 DENV3 50B dia 3 NR NEG DENV3 NEG NEG DENV3 51B dia 3 NR POS DENV3 POS DENV3 DENV3 52B dia 3 NR NEG DENV3 NEG DENV3 DENV3 53B dia 2 NR NEG NEG POS DENV3 DENV3 54B dia 4 NR POS DENV3 POS DENV3 DENV3 55B dia 3 NR POS DENV3 POS DENV3 DENV3 56B dia 1 NR POS DENV3 POS DENV3 DENV3 57B dia 2 NR POS DENV3 POS DENV3 DENV3 58B dia 4 NEG POS DENV3 POS DENV3 DENV3 59B dia 1 NR POS DENV3 POS DENV3 DENV3 60B dia 1 NR POS DENV3 NEG DENV3 DENV3 61B dia 2 NR POS DENV3 POS DENV3 DENV3
DENG: resultados obtidos com os oligonucleotídeos grupo-específicos DENESP: resultados obtidos com os oligonucleotídeos sorotipo-específicos POS: positivo NEG: negativo NI: não informado NR: não realizado

96
ANEXO 2 – Valores do ∆G mais desfavorável das possíveis estruturas secundárias internas (hairpins) formadas por cada oligonucleotídeo usado nos ensaios de PCR em tempo real e microarranjo líquido.
oligonucleotídeo ∆∆∆∆G oligonucleotídeo ∆∆∆∆G
DEN1F 1,21 DEN1JP 2,2
DEN1R 0,34 DEN2JF 1,84
DEN1P 1,92 DEN2JR 2,53
DEN2F 1,71 DEN2JP 0,68
DEN2R 1,28 DEN3JF 2,31
DEN2P 2,18 DEN3JR 1,41
DEN3F 1,96 DEN3JP 1,35
DEN3R 2,0 DENGF 1,63
DEN3P 2,11 DENGR1 1,66
DEN1JF 2,3 DENGR2 2,34
DEN1JR 2,06 DENGP 2,14

97
ANEXO 3 – Valores do ∆G mais desfavorável dos possíveis homo e hetero-dímeros
formados pelos oligonucleotídeos usados nos ensaios de PCR em tempo real e
microarranjo líquido.
DEN1F
DEN1R DEN1P DEN2F DEN2R DEN2P DEN3F DEN3R DEN3P
DEN1F
-8,32 -3,3 -8,14 -6,37 -6,82 -6,69 -5,02 -5,02 -5,02
DEN1R
-3,3 -6,76 -6,21 -7,1 -5,24 -6,21 -4,75 -5,24 -11,59
DEN1P
-8,14 -6,21 -16,13 -6,5 -3,61 -9,28 -8,45 -5,02 -8,09
DEN2F
-6,37 -7,1 -6,5 -6,59 -4,87 -4,67 -5,37 -6,24 -8,53
DEN2R
-6,82 -5,24 -3,61 -4,87 -3,9 -6,82 -13,67 -6,24 -3,9
DEN2P
-6,69 -6,21 -9,28 -4,67 -6,82 -9,28 -6,6 -6,14 -6,61
DEN3F
-5,02 -4,75 -8,45 -5,37 -13,67 -6,6 -5,38 -3,53 -8,09
DEN3R
-5,02 -5,24 -5,02 -6,24 -6,24 -6,14 -3,53 -6,34 -8,09
DEN3P
-5,02 -11,59 -8,09 -8,53 -3,9 -6,61 -8,09 -8,09 -3,89
DEN1JF
NA NA NA -7,87 -5,37 -6,61 -4,64 -6,91 -7,79
DEN1JR
NA NA NA -3,52 -3,53 -5,47 -4,75 -3,9 -6,35
DEN1JP
NA NA NA -6,72 -5,24 -3,43 -8,09 -9,38 -3,3
DEN2JF
-8,17 -8,16 -9,64 NA NA NA -6,82 -6,62 -4,89
DEN2JR
-8,65 -10,33 -6,5 NA NA NA -8,18 -4,74 -6,22
DEN2JP
-3,55 -8,03 -9,28 NA NA NA -4,75 -6,62 -6,24
DEN3JF
-5,09 -5,09 -5,02 -5,02 -4,87 -3,55 NA NA NA
DEN3JR
-4,87 -6,76 -5,38 -5,19 -8,19 -4,87 NA NA NA
DEN3JP
-5,02 -6,53 -6,21 -8,48 -4,87 -6,69 NA NA NA
DENGF
-7,96 -8,3 -5,09 -6,24 -6,24 -8,03 -6,12 -4,74 -6,24
DENGR1
-5,02 -6,21 -8,16 -8,47 -10,04 -6,21 -5,37 -5,02 -8,54
DENGR2
-5,02 -6,21 -8,7 -4,64 -8,7 -6,21 -6,6 -4,74 -6,6
DENGP
-13,65 -9,76 -6,21 -6,59 -4,64 -7,81 -6,95 -6,62 -8,54
As letras F, R e P, após a sigla dos oligonucleotídeos, são referentes à forward (sense), reverse (anti-sense) e probe (sonda), respectivamente. NA: não aplicável.

98
Continuação anexo 3.
DEN1JF
DEN1JR DEN1JP DEN2JF DEN2JR DEN2JP DEN3JF DEN3JR DEN3JP
DEN1JF
-7,05 -3,52 -10,05 -8,48 -7,05 -11,65 -6,91 -10,07 -4,65
DEN1JR
-3,52 -8,51 -6,36 -6,36 -3,55 -6,69 -8,42 -3,52 -3,52
DEN1JP
-10,05 -6,36 -5,38 -3,61 -6,37 -6,24 -5,25 -5,38 -4,74
DEN2JF
-8,48 -6,36 -3,61 -3,61 -6,5 -6,21 -4,87 -6,01 -7,96
DEN2JR
-7,05 -3,55 -6,37 -6,5 -10,24 -4,75 -5,09 -3,9 -6,69
DEN2JP
-11,65 -6,69 -6,24 -6,21 -4,75 -12,47 -4,64 -10,3 -8,26
DEN3JF
-6,91 -8,42 -5,25 -4,87 -5,09 -4,64 -3,61 -4,87 -4,87
DEN3JR
-10,07 -3,52 -5,38 -6,01 -3,9 -10,3 -4,87 -6,76 -8,19
DEN3JP
-4,65 -3,52 -4,74 -7,96 -6,69 -8,26 -4,87 -8,19 -6,34
DENGF
-6,24 -6,35 -6,69 -6,62 -8,65 -6,62 -6,62 -6,35 -6,62
DENGR1
-6,59 -4,99 -8.09 -6,5 -5,02 -6,21 -8,09 -3,61 -6,6
DENGR2
-7,04 -3,42 -6,75 -6,5 -7,04 -6,68 -8,7 -3,9 -8,07
DENGP
-6,59 -4,99 -5,25 -5,02 -5,09 -7,81 -8,2 -4,67 -11,27
Continuação anexo 3.
DENGF
DENGR1 DENGR2 DENGP
DENGF
-6,12 -8,2 -6,12 -8,65
DENGR1
-8,2 -5,02 -6,75 -6,62
DENGR2
-6,12 -6,75 -9,86 -5,09
DENGP
-8,65 -6,62 -5,09 -3,14

99
REFERÊNCIAS BIBLIOGRÁFICAS
ABOU-ELA F, Koh D, Tinoco I Jr, Martin FJ. Base-base mistmatches. Thermodynamics of double helix formation for dCA3XA3G _ dCT3YT3G (X, Y A,C,G,T). Nucleic Acids Res. 1985 13:3944–3948. ALCON S, Talarmin A, Debruyne M, Falconar A, Deubel V, Flamand M. Enzyme-linked immunosorbent assay specific to dengue virus type 1 nonstructural protein NS1 reveals circulation of the antigen in the blood during the acute phase of disease in patients experiencing primary or secondary infections. J Clin Microb. 2002;40(2):376-81. ALLEN C, Duffy S, Teknos T, Islam M, Chen Z, Albert PS, Wolf G, Van Waes C. Nuclear factor-kappaB-related serum factors as longitudinal biomarkers of response and survival in advanced oropharyngeal carcinoma. Clin Cancer Res. 2007 Jun 1;13(11):3182-90. ALVAREZ DE, Lodeiro MF, Filomatori CV, Fucito S, Mondotte JA, Gamarnik AV. Structural and functional analysis of dengue virus RNA. Novartis Found Symp. 2006;277:120-32. AN HJ, Cho NH, Lee SY, Kim IH, Lee C, Kim SJ, Mun MS, Kim SH, Jeong JK. Correlation of cervical carcinoma and precancerous lesions with human papillomavirus (HPV) genotypes detected with the HPV DNA chip microarray method. Cancer. 2003 Apr 1;97(7):1672-80. ARMSTRONG B, Stewart M, Mazumder A. Suspension arrays for high throughput, multiplexed single nucleotide polymorphism genotyping. Cytometry. 2000 Jun 1;40(2):102-8. BAUMS IB, Goodwin KD, Kiesling T, Wanless D, Diaz MR, Fell JW. Luminex detection of fecal indicators in river samples, marine recreational water, and beach sand. Mar Pollut Bull. 2007 May;54(5):521-36. BEDNÁR M. DNA microarray technology and application. Med Sci Monit. 2000 Jul-Aug;6(4):796-800. BIAGINI RE, Sammons DL, Smith JP, MacKenzie BA, Striley CA, Semenova V, Steward-Clark E, Stamey K, Freeman AE, Quinn CP, Snawder JE. Comparison of a multiplexed fluorescent covalent microsphere immunoassay and an enzyme-linked immunosorbent assay for measurement of human immunoglobulin G antibodies to anthrax toxins. Clin Diagn Lab Immunol. 2004 Jan;11(1):50–55. BORISKIN YS, Rice PS, Stabler RA, Hinds J, Al-Ghusein H, Vass K, Butcher PD. DNA microarrays for virus detection in cases of central nervous system. J infection. Clin Microbiol. 2004 Dec;42(12):5811-8.

100
BRINTON MA, Kurane I, Mathew A, Zeng L, Shi PY, Rothman A, Ennis FA. Immune mediated and inherited defenses against flaviviruses. Clin Diagn Virol. 1998;10:129-39. BRUNSTEIN J, Thomas E. Direct screening of clinical specimens for multiple respiratory pathogens using the Genaco Respiratory Panels 1 and 2. Diagn Mol Pathol. 2006 Sep;15(3):169-73. BUCHY P, Yoksan S, Peeling RW, Hunsperger E. Laboratory tests for the diagnosis of dengue virus infection. Report of the Scientific Working Group meeting on Dengue; 2006 Oct 1–5; Geneva, Switzerland. Geneva: World Health Organization; 2007. p.35-49. BUSTIN SA. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinology. 2000;25:169-93. BUSTIN SA. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): trends and problems. J Mol Endocrinology. 2002;29:23-39. CALISHER CH, Karabatsos N, Dalrymple JM, Shope RE, Porterfield JS, Westaway EG, Brandt WE. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J Gen Virol. 1989 Jan;70 ( Pt 1):37-43. CALLAHAN JD, Wu, S-JL, Dion-Shultz A, Mangold BE, Peruski LF, Watts DM, Porter KR, Murphy GR, Suharyono W, King C-C, Hayes CG, Temenak JJ. Development and evaluation of serotype- and group-specific fluorogenic reverse transcriptase PCR (TaqMan) assays for dengue virus. J Clin Microb. 2001;39(11):4119-24. CAMILLA C, Mély L, Magnan A, Casano B, Prato S, Debono S, Montero F, Defoort JP, Martin M, Fert V. Flow cytometric microsphere-based immunoassay: analysis of secreted cytokines in whole-blood samples from asthmatics. Clin Diagn Lab Immunol. 2001 Jul;8(4):776-84. CARDOSA MJ, Baharudin F, Hamild S, Hool TP, Nimmannitya S. A nitrocellulose membrane based IgM capture enzyme immunoassay for etiological diagnosis of dengue virus infections. Clin Diagn Virol. 1995;3:343-50. CARSON RT, Vignali DA. Simultaneous quantitation of 15 cytokines using a multiplexed flow cytometric assay. J Immunol Methods. 1999 Jul 30;227(1-2):41-52. CHAN YC, Kautner IM, Lam SK. The influence of antibody levels in dengue diagnosis by polymerase chain reaction. J Virol Methods. 1994;49:315-22. CHEN C, Nan B, Lin P, Yao Q. C-reactive protein increases plasminogen activator inhibitor-1 expression in human endothelial cells.Thromb Res. 2007 Oct 17; [Epub ahead of print]. CHEN J, Iannone MA, Li MS, Taylor JD, Rivers P, Nelsen AJ, Slentz-Kesler KA, Roses A, Weiner MP. A microsphere-based assay for multiplexed single nucleotide polymorphism analysis using single base chain extension. Genome Res. 2000 Apr;10(4):549-57.

101
CHEN R-F, Yeh W-T, Yang M-Y, Yang KD. A model of the real-time correlation of viral titers with immune reactions in antibody-dependent enhancement of dengue-2 infections. FEMS Immunol Med Microb. 2001;30:1-7. CHEN Y, Maguire T, Hileman RE, Fromm JR, Esko JD, Linhardt RJ, Marks RM. Dengue virus infectivity depends on envelope protein binding to target cell heparan sulfate. Nat Med. 1997;3:866-71. CHEN Y, Maguire T, Marks RM. Demonstration of binding of dengue virus envelope protein to target cells. J Virol. 1996;70(12):8765-72. CHERKASOVA E, Laassri M, Chizhikov V, Korotkova E, Dragunsky E, Agol VI, Chumakov K. Microarray analysis of evolution of RNA viruses: evidence of circulation of virulent highly divergent vaccine-derived polioviruses. Proc Natl Acad Sci U S A. 2003 Aug 5;100(16):9398-403. CHIZHIKOV V, Wagner M, Ivshina A, Hoshino Y, Kapikian AZ, Chumakov K. Detection and genotyping of human group A rotaviruses by oligonucleotide microarray hybridization. J Clin Microbiol. 2002 Jul;40(7):2398-407. COWAN LS, Diem L, Brake MC, Crawford JT. Transfer of a Mycobacterium tuberculosis genotyping method, Spoligotyping, from a reverse line-blot hybridization, membrane-based assay to the Luminex multianalyte profiling system. J Clin Microbiol. 2004 Jan;42(1):474-7. CRAIG S, Thu HM, Lowry K, Wang XF, Holmes EC, Aaskov J. Diverse dengue type 2 virus populations contain recombinant and both parental viruses in a single mosquito host. J Virol. 2003;77(7):4463-7. CUZZUBBO AJ, Endy TP, Nisalak A, Kalayanarooj S, Vaughn DW, Ogata SA, Clements DE, Devine PL. Use of recombinant envelope proteins for serological diagnosis of dengue virus infection in an immunocromatographic assay. Clin Diag Lab Immunol. 2001;8(6):1150-5. DEUBEL V. Recent advances and prospective researches on molecular epidemiology of dengue viruses. Mem Inst Oswaldo Cruz. 1992;87 Suppl 5:133-6. DIAZ MR. Bead Suspension Arrays for Identifying Fungal Pathogens. Microbe. 2007;2(2):74-81. DIAZ MR, Fell JW. High-throughput detection of pathogenic yeasts of the genus trichosporon. J Clin Microbiol. 2004 Aug;42(8):3696-706. DOLEZALOVA R, Lacinova Z, Dolinkova M, Kleiblova P, Haluzikova D, Housa D, Papezova H, Haluzik M. Changes of endocrine function of adipose tissue in anorexia nervosa: comparison of circulating levels versus subcutaneous mRNA expression. Clin Endocrinol (Oxf). 2007 Nov;67(5):674-8. DROSTEN C, Götting S, Schilling S, Asper M, Panning M, Schmitz H, Günther S. Rapid detection and quantification of RNA of Ebola and Marburg viruses, Lassa virus, Crimean-Congo hemorrhagic fever virus, Rift Valley fever virus, dengue virus, and

102
yellow fever virus by real-time reverse transcription-PCR. J Clin Microb. 2002;40(7):2323-30. Dunbar SA. Applications of Luminex xMAP technology for rapid, high-throughput multiplexed nucleic acid detection. Clin Chim Acta. 2006 Jan;363(1-2):71-82. DUNBAR SA, Jacobson JW. Application of the luminex LabMAP in rapid screening for mutations in the cystic fibrosis transmembrane conductance regulator gene: A pilot study. Clin Chem. 2000 Sep;46(9):1498-500. DUNBAR SA, Vander Zee CA, Oliver KG, Karem KL, Jacobson JW. Quantitative, multiplexed detection of bacterial pathogens: DNA and protein applications of the Luminex LabMAP system. J Microbiol Methods. 2003 May;53(2):245-52. ELNIFRO EM, Ashshi AM, Cooper RJ, Klapper PE. Multiplex PCR: optimization and application in diagnostic virology. Clin Microbiol Rev. 2000 Oct;13(4):559-70. FALCONAR AK, de Plata E, Romero-Vivas CM. Altered enzyme-linked immunosorbent assay immunoglobulin M (IgM)/IgG optical density ratios can correctly classify all primary or secondary dengue virus infections 1 day after the onset of symptoms, when all of the viruses can be isolated. Clin Vaccine Immunol. 2006 Sep;13(9):1044-51. FAUCHER S, Martel A, Sherring A, Bogdanovic D, Malloch L, Kim JE, Bergeron M, Sandstrom P, Mandy FF. A combined HIV-1 protein bead array for serology assay and T-cell subset immunophenotyping with a hybrid flow cytometer: a step in the direction of a comprehensive multitasking instrument platform for infectious disease diagnosis and monitoring. Cytometry B Clin Cytom. 2006 May;70(3):179-88. FREEMAN WM, Robertson DJ, Vrana KE. Fundamentals of DNA hybridization arrays for gene expression analysis. Biotechniques. 2000 Nov;29(5):1042-6, 1048-55. FULTON RJ, McDade RL, Smith PL, Kienker LJ, Kettman JR Jr. Advanced multiplexed analysis with the FlowMetrix system. Clin Chem. 1997 Sep;43(9):1749-56. FUNASA (Fundação Nacional de Saúde). Guia de Vigilância Epidemiológica. 5 ed. Brasília: FUNASA; 2002. GARDNER SN, Kuczmarski TA, Vitalis EA, Slezak TR. Limitations of TaqMan PCR for detecting divergent viral pathogens illustrated by hepatitis A, B, C, and E viruses and human immunodeficiency virus. J Clin Microbiol. 2003 Jun;41(6):2417-27. GERMI R, Crance J-M, Garin D, Guimet J, Lortat-Jacob H, Ruigrok RWH, Zarski J-P, Drouet E. Heparan sulfate-mediated binding of infectious dengue virus type 2 and yellow fever virus. Virology. 2002;292:162-8. GIBBONS R, Vaughn DW. Dengue: an escalating problem. BMJ. 2002;324:1563-6. GOFF LA, Yang M, Bowers J, Getts RC, Padgett RW, Hart RP Rational probe optimization and enhanced detection strategy for microRNAs using microarrays. RNA Biol. 2005 Jul-Sep;2(3):93-100.

103
GUBLER DJ. Dengue and dengue hemorrhagic fever: its history and resurgence as a global public health problem. In: Gubler DJ, Kuno G. Dengue and Dengue Hemorrhagic Fever. New York: CAB International; 1997. GUBLER DJ. Dengue and dengue hemorrhagic fever. Clin Microb Rev. 1998;11(3):480-96. GUBLER DJ, Clark GG. Dengue/dengue hemorrhagic fever: the emergence of a global health problem. Emerging Infect Dis.1995;1(2):55-7. GUBLER DJ. Dengue/dengue haemorrhagic fever: history and current status. Novartis Found Symp. 2006;277:3-16. GUBLER DJ, Kuno G, Sather GE, Waterman SH. A case of natural concurrent human infection with two dengue viruses. Am J Trop Med Hyg. 1985;34(1):170-3. GUBLER DJ, Sather G. Laboratory diagnosis of dengue and dengue hemorrhagic fever. Proceedings of the International Symposium on Yellow Fever and Dengue; 1988; Rio de Janeiro, Brasil. GUZMÁN MG, Kouri G. Advances in dengue diagnosis. Clin Diagn Lab Immunol. 1996;3(6):621-7. HADD AG, Laosinchai-Wolf W, Novak CR, Badgett MR, Isgur LA, Goldrick M, Walkerpeach CR. Microsphere bead arrays and sequence validation of 5/7/9T genotypes for multiplex screening of cystic fibrosis polymorphisms. J Mol Diagn. 2004 Nov;6(4):348-55. HALSTEAD SB. Pathogenesis of dengue: challenges to molecular biology. Science. 1988;239:476-81. HALSTEAD SB. Neutralization and antibody-dependent enhancement of dengue viruses. Adv Virus Res. 2003;60:421-67. HARRIS E, Roberts TG, Smith L, Selle J, Kramer LD, Valle S, Sandoval E, Balmaseda A. Typing of dengue viruses in clinical specimens and mosquitoes by single-tube multiplex reverse transcriptase PCR. J Clin Microb. 1998;36(9):2634-9. HEID CA, Stevens J, Livak KJ, Williams PM. Real time quantitative PCR. Genome Res. 1996 Oct;6(10):986-94. HELLER MJ. DNA microarray technology: devices, systems, and applications. Annu Rev Biomed Eng. 2002;4:129-53. HENCHAL EA, Polo SL, Vorndam V, Yaemsiri C, Innis BL, Hoke CH. Sensitivity and specificity of a universal primer set for the rapid diagnosis of dengue virus infections by polymerase chain reaction and nucleic acid hybridization. Am J Trop Med Hyg. 1991;45(4):418-28. HENEGARIU O, Heerema NA, Dlouhy SR, Vance GH, Vogt PH. Multiplex PCR: critical parameters and step-by-step protocol. Biotechniques. 1997 Sep;23(3):504-11.

104
HIGUCHI R, Dollinger G, Walsh PS, Griffith R. Simultaneous amplification and detection of specific DNA sequences. Biotechnology (N Y). 1992 Apr;10(4):413-7. HOLMES EC. The evolutionary biology of dengue virus. Novartis Found Symp. 2006;277:177-87. HOUNG HH, Hritz D, Kanesa-Thasan N. Quantitative detection of dengue 2 virus using fluorogenic 3’-noncoding sequence. J Virol Methods. 2000;86(1):1-11. HOUNG HS, Chung-Ming Chen R, Vaughn DW, Kanesa-thasan N. Development of a fluorogenic RT-PCR system for quantitative identification of dengue virus serotypes 1-4 using conserved and serotype-specific 3' noncoding sequences. J Virol Methods. 2001 Jun;95(1-2):19-32. INNIS BL, Nisalak A, Nimmannitya S, Kusalerdchariya S, Chongswasdi V, Suntayakorn S, Puttisri P, Hoke H. An enzyme-linked immunosorbent assay to characterize dengue infections where dengue and japanese encephalitis co-circulate. Am J Trop Med Hyg. 1989;40(4):418-27. ITO M, Takasaki T, Yamada K, Nerome R, Tajima S, Kurane I. Development and evaluation of fluorogenic TaqMan reverse transcriptase PCR assays for detection of dengue virus types 1 to 4. J Clin Microbiol. 2004 Dec;42(12):5935-7. IVSHINA AV, Vodeiko GM, Kuznetsov VA, Volokhov D, Taffs R, Chizhikov VI, Levandowski RA, Chumakov KM. Mapping of genomic segments of influenza B virus strains by an oligonucleotide microarray method. J Clin Microbiol. 2004 Dec;42(12):5793-801. JOHNSON AJ, Noga AJ, Kosoy O, Lanciotti RS, Johnson AA, Biggerstaff BJ. Duplex microsphere-based immunoassay for detection of anti-West Nile virus and anti-St. Louis encephalitis virus immunoglobulin m antibodies. Clin Diagn Lab Immunol. 2005(a) May;12(5):566-74. JOHNSON BW, Russell BJ, Lanciotti RS. Serotype-specific detection of dengue viruses in a fourplex real-time reverse transcriptase PCR assay. J Clin Microbiol. 2005(b) Oct;43(10):4977-83. KAWAGUCHI K, Kaneko S, Honda M, Kawai HF, Shirota Y, Kobayashi K. Detection of hepatitis B virus DNA in sera from patients with chronic hepatitis B virus infection by DNA microarray method. J Clin Microbiol. 2003 Apr;41(4):1701-4. KELLAR KL, Iannone MA. Multiplexed microsphere-based flow cytometric assays. Exp Hematol. 2002 Nov;30(11):1227-37. KLAASSEN CH, Prinsen CF, de Valk HA, Horrevorts AM, Jeunink MA, Thunnissen FB. DNA microarray format for detection and subtyping of human papillomavirus. J Clin Microbiol. 2004 May;42(5):2152-60. KONG YY, Thay CH, Tin TC, Devi S. Rapid detection, serotyping and quantitation of dengue viruses by TaqMan real-time one-step RT-PCR. J Virol Methods. 2006 Dec;138(1-2):123-30.

105
KOURI GP, Guzmán MG, Bravo JR, Triana C. Dengue haemorrhagic fever/dengue shock syndrome: lessons from the Cuban epidemic, 1981. Bull World Health Organ. 1989;67(4):375-80. KUNO G, Baily RE. Cytokine responses to dengue infection among Puerto Rican patients. Mem Inst Oswaldo Cruz. 1994;89:179-82. Kurane I, Ennis FA. Cytokines in dengue virus infections: role of cytokines in pathogenesis of dengue hemorrhagic fever. Sem Virol. 1994;5:443-8. KURANE I, Innis BL, Nimmannitya S, Nisalak A, Meager A, Jannus J, Ennis FA. Activation of T lymphocytes in dengue virus infections: high levels of soluble interleukin 2 receptor, soluble CD4, soluble CD8, interleukin 2 and interferon gamma in sera of children with dengue. J Clin Invest. 1991;88:1473-80. LAI YL, Chung YK, Tan HC, Yap HF, Yap G, Ooi EE, Ng LC. Cost-effective real-time reverse transcriptase PCR (RT-PCR) to screen for Dengue virus followed by rapid single-tube multiplex RT-PCR for serotyping of the virus. J Clin Microbiol. 2007 Mar;45(3):935-41. LAM SK, Ew CL, Mitchell JL, Cuzzubbo AJ, Devine PL. Clin Diagn Lab Immunol 2000;7(5):850-2. LANCIOTTI RS, Calisher CH, Gubler DJ, Chang G-J, Vorndam V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J Clin Microb. 1992;30(3):545-51. LAASSRI M, Chizhikov V, Mikheev M, Shchelkunov S, Chumakov K. Detection and discrimination of orthopoxviruses using microarrays of immobilized oligonucleotides. J Virol Methods. 2003 Sep;112(1-2):67-78. LAASSRI M, Dragunsky E, Enterline J, Eremeeva T, Ivanova O, Lottenbach K, Belshe R, Chumakov K. Genomic analysis of vaccine-derived poliovirus strains in stool specimens by combination of full-length PCR and oligonucleotide microarray hybridization. J Clin Microbiol. 2005 Jun;43(6):2886-94. LAUE T, Emmerich P, Schmitz H. Detection of dengue virus RNA in patients after primary or secondary dengue infection by using the TaqMan automated amplification system. J Clin Microb. 1999;37(8):2543-7. LEE LG, Connell CR, Bloch W. Allelic discrimination by nick-translation PCR with fluorogenic probes. Nucleic Acids Res. 1993 Aug 11;21(16):3761-6. LEITMEYER KC, Vaughn DW, Watts DM, Salas R, Chacon IV, Ramos C, Rico-Hesse R. Dengue virus structural differences that correlate with pathogenesis. J Virol. 1999;73(6):4738-47. LIBRATY DH, Young PR, Pickering D, Endy TP, Kalayanarooj S, Green S, Vaughn DW, Nisalak A, Ennis FA, Rothman AL. High circulating levels of the dengue virus nonstructural protein NS1 early in dengue illness correlate with the development of dengue hemorrhagic fever. J Infect Dis. 2002 Oct 15;186(8):1165-8.

106
LIVAK KJ, Flood SJ, Marmaro J, Giusti W, Deetz K. Oligonucleotides with fluorescent dyes at opposite ends provide a quenched probe system useful for detecting PCR product and nucleic acid hybridization. PCR Methods Appl. 1995 Jun;4(6):357-62. LIVSHITS MA, Mirzabekov AD. Theoretical analysis of the kinetics of DNA hybridization with gel-immobilized oligonucleotides. Biophys J. 1996 Nov;71(5):2795-801. LOVMAR L, Fock C, Espinoza F, Bucardo F, Syvänen AC, Bondeson K. Microarrays for genotyping human group a rotavirus by multiplex capture and type-specific primer extension. J Clin Microbiol. 2003 Nov;41(11):5153-8. LOWE M, Spiro A, Zhang YZ, Getts R. Multiplexed, particle-based detection of DNA using flow cytometry with 3DNA dendrimers for signal amplification. Cytometry A. 2004 Aug;60(2):135-44. LUND-JOHANSEN F, Davis K, Bishop J, de Waal Malefyt R. Flow cytometric analysis of immunoprecipitates: high-throughput analysis of protein phosphorylation and protein-protein interactions. Cytometry. 2000 Apr 1;39(4):250-9. MACKAY IM, Arden KE, Nitsche A. Real-time PCR in virology. Nucleic Acids Res. 2002 Mar 15;30(6):1292-305. MACKENZIE JS, Gubler DJ, Petersen LR. Emerging flaviviruses: the spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat Med. 2004 Dec;10(12 Suppl):S98-109. MAHONY J, Chong S, Merante F, Yaghoubian S, Sinha T, Lisle C, Janeczko R. Development of a respiratory virus panel test for detection of twenty human respiratory viruses by use of multiplex PCR and a fluid microbead-based assay. J Clin Microbiol. 2007 Sep;45(9):2965-70. MARTINS TB. Development of internal controls for the Luminex instrument as part of a multiplex seven-analyte viral respiratory antibody profile. Clin Diagn Lab Immunol. 2002 Jan;9(1):41-5. MARTINS TB, Burlingame R, von Mühlen CA, Jaskowski TD, Litwin CM, Hill HR. Evaluation of multiplexed fluorescent microsphere immunoassay for detection of autoantibodies to nuclear antigens. Clin Diagn Lab Immunol. 2004 Nov;11(6):1054-9. MASKOS U, Southern EM. Parallel analysis of oligodeoxyribonucleotide (oligonucleotide) interactions. I. Analysis of factors influencing oligonucleotide duplex formation. Nucleic Acids Res. 1992 Apr 11;20(7):1675-8. MCHUGH TM, Viele MK, Chase ES, Recktenwald DJ. The sensitive detection and quantitation of antibody to HCV by using a microsphere-based immunoassay and flow cytometry. Cytometry. 1997 Oct 1;29(2):106-12. MCHUGH TM. Flow microsphere immunoassay for the quantitative and simultaneous detection of multiple soluble analytes. Methods Cell Biol. 1994;42 (Pt B):575-95.

107
MCSHERRY JA. Some medical aspects of the Darien scheme: was it dengue? Scott Med J. 1982 Apr;27(2):183-4. MIAGOSTOVICH MP, Nogueira RMR, Dos Santos FB, Schatzmayr HG, Araújo ESM, Vorndam V. Evaluation of an IgG enzyme-linked immunosorbent assay for dengue diagnosis. J Clin Virol. 1999;14:183-9. MORGAN E, Varro R, Sepulveda H, Ember JA, Apgar J, Wilson J, Lowe L, Chen R, Shivraj L, Agadir A, Campos R, Ernst D, Gaur A. Cytometric bead array: a multiplexed assay platform with applications in various areas of biology. Clin Immunol. 2004 Mar;110(3):252-66. MS (Ministério da Saúde). Boletim epidemiológico – edição especial. Brasília: Ministério da Saúde, Fundação Nacional de Saúde; 1999. MS (Ministério da Saúde). Balanço dengue janeiro a julho de 2007. Brasília: Ministério da Saúde, Secretaria de Vigilância em Saúde; 2007. MS (Ministério da Saúde, Secretaria de Vigilância em Saúde). Dengue: diagnóstico e manejo clínico. Ministério da Saúde, Secretaria de Vigilância em Saúde, Diretoria Técnica de Gestão. – 2. ed. – Brasília: Ministério da Saúde, 2005(a). MS (Ministério da Saúde, Secretaria de Vigilância em Saúde). Dengue: roteiro para capacitação de profissionais médicos no diagnóstico e tratamento. Ministério da Saúde, Secretaria de Vigilância em Saúde, Diretoria Técnica de Gestão. – 2. ed. – Brasília: Ministério da Saúde, 2005(b). NATHAN MB, Dayal-Drager R. Recent epidemiological trends, the global strategy and public health advances in dengue. Report of the Scientific Working Group meeting on Dengue; 2006 Oct 1–5; Geneva, Switzerland. Geneva: World Health Organization; 2007. p.30-34. NIESTERS HG. Standardization and quality control in molecular diagnostics. Expert Rev Mol Diagn. 2001 Jul;1(2):129-31. NIESTERS HG. Clinical virology in real time. J Clin Virol. 2002 Dec;25 Suppl 3:S3-12. NOLAN JP, Mandy FF. Suspension array technology: new tools for gene and protein analysis. Cell Mol Biol (Noisy-le-grand). 2001 Nov;47(7):1241-56. NONG T, Saito K, Blair L, Tarsitani C, Lee JH. KIR genotyping by reverse sequence-specific oligonucleotide methodology. Tissue Antigens. 2007 Apr;69 Suppl 1:92-5. OH TJ, Kim CJ, Woo SK, Kim TS, Jeong DJ, Kim MS, Lee S, Cho HS, An S. Development and clinical evaluation of a highly sensitive DNA microarray for detection and genotyping of human papillomaviruses. J Clin Microbiol. 2004 Jul;42(7):3272-80. OH Y, Bae SM, Kim YW, Choi HS, Nam GH, Han SJ, Park CH, Cho Y, Han BD, Ahn WS. Polymerase chain reaction-based fluorescent Luminex assay to detect the presence of human papillomavirus types. Cancer Sci. 2007 Apr;98(4):549-54.

108
OLIVER KG, Kettman JR, Fulton RJ. Multiplexed analysis of human cytokines by use of the FlowMetrix system. Clin Chem. 1998 Sep;44(9):2057-60. Olsson A, Vanderstichele H, Andreasen N, De Meyer G, Wallin A, Holmberg B, Rosengren L, Vanmechelen E, Blennow K. Simultaneous measurement of beta-amyloid(1-42), total tau, and phosphorylated tau (Thr181) in cerebrospinal fluid by the xMAP technology. Clin Chem. 2005 Feb;51(2):336-45. OOI EE, Gubler DJ, Nam VS. Dengue research needs related to surveillance and emergency response. Report of the Scientific Working Group meeting on Dengue; 2006 Oct 1–5; Geneva, Switzerland. Geneva: World Health Organization; 2007. p.124-133. OPALKA D, Lachman CE, MacMullen SA, Jansen KU, Smith JF, Chirmule N, Esser MT. Simultaneous quantitation of antibodies to neutralizing epitopes on virus-like particles for human papillomavirus types 6, 11, 16, and 18 by a multiplexed luminex assay. Clin Diagn Lab Immunol. 2003 Jan;10(1):108-15. OPAS (Organização Pan-america de Saúde). Dengue and dengue hemorrhagic fever in the Americas: guidelines for prevention and control (Scientific Publication No. 548), 1994. Washington DC, Pan American Health Organization. PÁL J, Pálinkás L, Nyárády Z, Czömpöly T, Marczinovits I, Lustyik G, Saleh Ali Y, Berencsi G, Chen R, Varró R, Pár A, Németh P. Sandwich type ELISA and a fluorescent cytometric microbead assay for quantitative determination of hepatitis B virus X antigen level in human sera. J Immunol Methods. 2005 Nov 30;306(1-2):183-92. PALACIOS G, Quan PL, Jabado OJ, Conlan S, Hirschberg DL, Liu Y, Zhai J, Renwick N, Hui J, Hegyi H, Grolla A, Strong JE, Towner JS, Geisbert TW, Jahrling PB, Büchen-Osmond C, Ellerbrok H, Sanchez-Seco MP, Lussier Y, Formenty P, Nichol MS, Feldmann H, Briese T, Lipkin WI. Panmicrobial oligonucleotide array for diagnosis of infectious diseases. Emerg Infect Dis. 2007 Jan;13(1):73-81. PALMER CJ, King SD, Cuadrado RR, Perez E, Baum M, Ager AL. Evaluation of the MRL diagnostics dengue fever virus IgM capture ELISA and the PanBio rapid immunochromatographic test for diagnosis of dengue fever in Jamaica. J Clin Microb. 1999;37(5):1600-01. PAULA SO, Lima DM, Fonseca BAL. Detection and identification of dengue-1 virus in clinical samples by a nested-PCR followed by restriction enzyme digestion of amplicons. J Med Virol. 2002(a);66:529-34. PAULA SO, Neto RJP, Corrêa JACT, Assumpção SR, Costa MLS, Lima DM, Fonseca BAL. The use of reverse transcription-polymerase chain reaction (RT-PCR) for the rapid detection and identification of dengue virus in an endemic region: a validation study. Transact Royal Society Trop Med Hyg. 2002(b);96:266-9. PEI R, Lee J, Chen T, Rojo S, Terasaki PI. Flow cytometric detection of HLA antibodies using a spectrum of microbeads. Hum Immunol. 1999 Dec;60(12):1293-302.

109
PIAZZA A, Poggi E, Ozzella G, Borrelli L, Monaco PI, Scornajenghi A, Tisone G, Adorno D. Public epitope specificity of HLA class I antibodies induced by a failed kidney transplant: alloantibody characterization by flow cytometric techniques. Transplantation. 2006 May 15;81(9):1298-305. Pickering JW, Martins TB, Schroder MC, Hill HR. Comparison of a multiplex flow cytometric assay with enzyme-linked immunosorbent assay for auantitation of antibodies to tetanus, diphtheria, and Haemophilus influenzae Type b. Clin Diagn Lab Immunol. 2002 Jul;9(4):872-6. PIERANGELI SS, Silva LK, Harris EN. A flow cytometric assay for the detection of antiphospholipid antibodies. Am Clin Lab. 1999 Jul;18(6):18-9. PINHEIRO FP. Dengue in the Americas. 1980-1987. Epidemiol Bull. 1989;10(1):1-8. PYATT RE, Mihal DC, Prior TW. Assessment of liquid microbead arrays for the screening of newborns for spinal muscular atrophy. Clin Chem. 2007 Nov;53(11):1879-85. RAENGSAKULRACH B, Nisalak A, Maneekarn N, Yenchitsomanus P-T, Limsomwong C, Jairungsri A, Thirawuth V, Green S, Kalayanarooj S, Suntayakorn S, Sittisombut N, Malasit P, Vaughn DW. Comparison of four reverse transcription-polymerase chain reaction procedures for the detection of dengue virus in clinical specimens. J Virol Methods. 2002;105:219-32. RATCLIFF RM, Chang G, Kok T, Sloots TP. Molecular diagnosis of medical viruses. Curr Issues Mol Biol. 2007 Jul;9(2):87-102. RICE CM. Flaviviridae: the viruses and their replication. In: Knipe DM, Howley PM. Fields Virology. 4 ed. Philadelphia: Lippincott Williams and Wilkins; 2001. Rico-Hesse R, Harrison LM, Salas RA, Tovar D, Nisalak A, Ramos C, Boshell J, Mesa MTR, Nogueira RMR, Rosa AT. Origins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology. 1997;230:244-51. ROEHRIG JT. Immunochemistry of dengue viruses. In: Gubler DJ, Kuno G. Dengue and Dengue Hemorrhagic Fever. New York: CAB International; 1997. ROTHMAN AL. Viral pathogenesis of dengue infections. In: Gubler DJ, Kuno G. Dengue and Dengue Hemorrhagic Fever. New York: CAB International; 1997. ROTHMAN AL, Ennis FA. Immunopathogenesis of dengue hemorrhagic fever. Virology. 1999;257:1-6. SAKUNTABHAI A, Turbpaiboon C, Casadémont I, Chuansumrit A, Lowhnoo T, Kajaste-Rudnitski A, Kalayanarooj SM, Tangnararatchakit K, Tangthawornchaikul N, Vasanawathana S, Chaiyaratana W, Yenchitsomanus PT, Suriyaphol P, Avirutnan P, Chokephaibulkit K, Matsuda F, Yoksan S, Jacob Y, Lathrop GM, Malasit P, Desprès P, Julier C. A variant in the CD209 promoter is associated with severity of dengue disease. Nat Genet. 2005 May;37(5):507-13. SCHILLING S, Ludolfs D, Van An L, Schmitz H. Laboratory diagnosis of primary and secondary dengue infection. J Clin Virol. 2004 Nov;31(3):179-84.

110
SCHOSKE R, Vallone PM, Ruitberg CM, Butler JM. Multiplex PCR design strategy used for the simultaneous amplification of 10 Y chromosome short tandem repeat (STR) loci. Anal Bioanal Chem. 2003 Feb;375(3):333-43. SHU PY, Chang SF, Kuo YC, Yueh YY, Chien LJ, Sue CL, Lin TH, Huang JH. Development of group- and serotype-specific one-step SYBR green I-based real-time reverse transcription-PCR assay for dengue virus. J Clin Microbiol. 2003 Jun;41(6):2408-16. SHU PY, Chen LK, Chang SF, Yueh YY, Chow L, Chien LJ, Chin C, Lin TH, Huang JH. Comparison of capture immunoglobulin M (IgM) and IgG enzyme-linked immunosorbent assay (ELISA) and nonstructural protein NS1 serotype-specific IgG ELISA for differentiation of primary and secondary dengue virus infections. Clin Diagn Lab Immunol. 2003 Jul;10(4):622-30. SHURIN GV, Yurkovetsky ZR, Chatta GS, Tourkova IL, Shurin MR, Lokshin AE. Dynamic alteration of soluble serum biomarkers in healthy aging. Cytokine. 2007 Aug;39(2):123-9. SIERRA B, Kouri G, Guzmán MG. Race: A risk factor for dengue hemorrhagic fever. Archives of Virology. 2006;xxx SIMMONS CP, Halstead SB, Rothman A, Harris E, Screaton G, Rico-Hesse R, Vaughn D, Holmes E, Guzman M. Understanding pathogenesis , immune response and viral factors. Report of the Scientific Working Group meeting on Dengue; 2006 Oct 1–5; Geneva, Switzerland. Geneva: World Health Organization; 2007. p.54-60. SMITH PL, WalkerPeach CR, Fulton RJ, DuBois DB. A rapid, sensitive, multiplexed assay for detection of viral nucleic acids using the FlowMetrix system. Clin Chem. 1998 Sep;44(9):2054-6.
SOLOMON T, Mallewa M. Dengue and other emerging flaviviruses. J Infection. 2001;42:104-15. SPIRO A, Lowe M. Quantitation of DNA sequences in environmental PCR products by a multiplexed, bead-based method. Appl Environ Microbiol. 2002 Feb;68(2):1010-3. SPIRO A, Lowe M, Brown D. A bead-based method for multiplexed identification and quantitation of DNA sequences using flow cytometry. Appl Environ Microbiol. 2000 Oct;66(10):4258-65. STIASNY K, Kiermayr S, Heinz FX. Entry functions and antigenic structure of flavivirus envelope proteins. Novartis Found Symp. 2006;277:57-65. STRIEBEL HM, Birch-Hirschfeld E, Egerer R, Földes-Papp Z. Virus diagnostics on microarrays. Curr Pharm Biotechnol. 2003 Dec;4(6):401-15. SUAYA JA, Shepard DS, Beatty ME. Dengue: burden of disease and costs of illness. Report of the Scientific Working Group meeting on Dengue; 2006 Oct 1–5; Geneva, Switzerland. Geneva: World Health Organization; 2007. p.35-49.

111
SUMMERBELL RC, Lévesque CA, Seifert KA, Bovers M, Fell JW, Diaz MR, Boekhout T, de Hoog GS, Stalpers J, Crous PW. Microcoding: the second step in DNA barcoding. Philos Trans R Soc Lond B Biol Sci. 2005 Oct 29;360(1462):1897-903. THOMPSON JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;24:4876-82. THULLIER P, Demangel C, Bedouelle H, Mégret F, Jouan A, Deubel V, Mazié J-C, Lafaye P. Mapping of a dengue virus neutralizing epitope critical for the infectivity of all serotypes: insight into the neutralization mechanism. J Gen Virol. 2001;82:1885-92. TOELLNER L, Fischlechner M, Ferko B, Grabherr RM, Donath E. Virus-coated layer-by-layer colloids as a multiplex suspension array for the detection and quantification of virus-specific antibodies. Clin Chem. 2006 Aug;52(8):1575-83. TOLOU HJG, Couissinier-Paris P, Durand J-P, Mercier V, Pina J-J, Micco P, Billoir F, Charrel RN, Lamballerie X. Evidence for recombination in the natural populations of dengue virus type 1 based on the analysis of complete genome sequences. J Gen Virol. 2001;82:1283-1290. TRACZ DM, Backhouse PG, Olson AB, McCrea JK, Walsh JA, Ng LK, Gilmour MW. Rapid detection of Vibrio species using liquid microsphere arrays and real-time PCR targeting the ftsZ locus. J Med Microbiol. 2007 Jan;56(Pt 1):56-65. TWIDDY SS, Holmes EC. The extent of homologous recombination in members of the genus Flavivirus. J Gen Virol. 2003;84:429-40. VALDÉS K, Alvarez M, Pupo M, Vásquez S, Rodríguez R, Guzmán M. G. Human dengue antibodies against structural and nonstructural proteins. Clin Diagn Lab Immunol. 2000;7(5):856-7. VAN CLEVE M, Ostrerova N, Tietgen K, Cao W, Chang C, Collins ML, Kolberg J, Urdea M, Lohman K. Direct quantitation of HIV by flow cytometry using branched DNA signal amplification. Mol Cell Probes. 1998 Aug;12(4):243-7. Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, Suntayakorn S, Endy TP, Raengsakulrach B, Rothman AL, Ennis F, Nisalak A. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. The J Infec Dis. 2000;181:2-9. VAUGHN DW, Nisalak A, Solomon T, Kalayanarooj S, Dung NM, Kneen R, Cuzzubbo A, Devine P. Rapid serologic diagnosis of dengue virus infection using a commercial capture ELISA that distinguishes primary and secondary infections. Am J Trop Med Hyg. 1999;60(4):693-8. VERNET G. Molecular diagnostics in virology. J Clin Virol. 2004 Dec;31(4):239-47. VIGNALI DA. Multiplexed particle-based flow cytometric assays. J Immunol Methods. 2000 Sep 21;243(1-2):243-55.

112
VORNDAM V, Kuno G. Laboratory diagnosis of dengue virus infections. In: Gubler, DJ, Kuno G. Dengue and dengue hemorrhagic fever. New York: CAB International; 1997. WADE MM, Volokhov D, Peredelchuk M, Chizhikov V, Zhang Y. Accurate mapping of mutations of pyrazinamide-resistant Mycobacterium tuberculosis strains with a scanning-frame oligonucleotide microarray. Diagn Microbiol Infect Dis. 2004 Jun;49(2):89-97. WALLACE J, Zhou Y, Usmani GN, Reardon M, Newburger P, Woda B, Pihan G. BARCODE-ALL: accelerated and cost-effective genetic risk stratification in acute leukemia using spectrally addressable liquid bead microarrays. Leukemia. 2003 Jul;17(7):1404-10. WANG D, Coscoy L, Zylberberg M, Avila PC, Boushey HA, Ganem D, DeRisi JL. Microarray-based detection and genotyping of viral pathogens. Proc Natl Acad Sci U S A. 2002(a) Nov 26;99(24):15687-92. WANG HL, Lin KH, Yueh YY, Chow L, Wu YC, Chen HY, Sheu MM, Chen WJ. Efficient diagnosis of dengue infections using patients’ peripheral blood leukocytes and serum/plasma. Intervirology. 2000;43:107-11. WANG WK, Chao DY, Kao CL, Wu HC, Liu YC, Li CM, Lin SC, Ho ST, Huang JH, King CC. High levels of plasma dengue viral load during defervescence in patients with dengue hemorrhagic fever: implications for pathogenesis. Virology. 2003;305(2):330-8. WANG WK, Sung TL, Tsai YC, Kao CL, Chang SM, King CC. Detection of dengue virus replication in peripheral blood mononuclear cells from dengue virus type 2-infected patients by a reverse transcription-real-time PCR assay. J Clin Microb. 2002(b);40(12):4472-8. WATTS DM, Porter KR, Putvatana P, Vasquez B, Calampa C, Hayes CG, Halstead SB. Failure of secondary infection with American genotype dengue 2 to cause dengue hemorrhagic fever. The Lancet. 1999;354:1431-4. WEDEMEYER N, Göhde W, Pötter T. Flow cytometric analysis of reverse transcription-PCR products: quantification of p21(WAF1/CIP1) and proliferating cell nuclear antigen mRNA. Clin Chem. 2000 Aug;46(8 Pt 1):1057-64. WHO (World Health Organization). Dengue haemorrhagic fever: diagnosis, treatment, prevention and control. 2 ed. Geneva: World Health Organization; 1997. WHO (World Health Organization). Report of the Scientific Working Group meeting on Dengue; 2006 Oct 1–5; Geneva, Switzerland. Geneva: World Health Organization; 2007. WONG SJ, Demarest VL, Boyle RH, Wang T, Ledizet M, Kar K, Kramer LD, Fikrig E, Koski RA. Detection of human anti-flavivirus antibodies with a west nile virus recombinant antigen microsphere immunoassay. J Clin Microbiol. 2004 Jan;42(1):65-72.

113
WOROBEY M, Rambaut A, Holmes EC. Widespread intra-serotype recombination in natural populations of dengue virus. Proc Natl Acad Sci U S A. 1999;96:7352-7. WU S-JL, Grouard-Vogel G, Sun W, Mascola JR, Brachtel E, Putvatana R, Louder MK, Filgueira L, Marovich MA, Wong HK, Blauvelt A, Murphy GS, Robb ML, Innes BL, Birx DL, Hayes CG, Frankel SS. Human skin Langerhans cells are targets of dengue virus infection. Nat Med. 2000;6(7):816-20. XU H, Di B, Pan YX, Qiu LW, Wang YD, Hao W, He LJ, Yuen KY, Che XY. Serotype 1-specific monoclonal antibody-based antigen capture immunoassay for detection of circulating nonstructural protein NS1: Implications for early diagnosis and serotyping of dengue virus infections. J Clin Microbiol. 2006 Aug;44(8):2872-8. YANG L, Tran DK, Wang X. BADGE, Beads Array for the Detection of Gene Expression, a high-throughput diagnostic bioassay. Genome Res. 2001 Nov;11(11):1888-98. YONG YK, Thayan R, Chong HT, Tan CT, Sekaran SD. Rapid detection and serotyping of dengue virus by multiplex RT-PCR and real-time SYBR green RT-PCR. Singapore Med J. 2007 Jul;48(7):662-8. YOUNG PR, Hilditch PA, Bletchly C, Halloran W. An antigen capture enzyme-linked immunosorbent assay reveals high levels of the dengue virus protein NS1 in the sera of infected patients. J Clin Microbiol. 2000;38(3):1053-57. YURKOVETSKY ZR, Kirkwood JM, Edington HD, Marrangoni AM, Velikokhatnaya L, Winans MT, Gorelik E, Lokshin AE. Multiplex analysis of serum cytokines in melanoma patients treated with interferon-alpha2b. Clin Cancer Res. 2007 Apr 15;13(8):2422-8.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo