desempenho e digstibilidade em ovinos … o ensaio de digestibilidade e balanço nitrogenado foram...
TRANSCRIPT

DESEMPENHO E DIGSTIBILIDADE EM OVINOS DA RAÇA SANTA INÊS
ALIMENTADOS EM DIFERENTES CODIÇÕES DE BALANÇOS DE PROTEÍNA DEGRADÁVEL NO RUMEN E PROTEÍNA
METABOLIZÁVEL
FLÁVIO MORENO SALVADOR
2007

FLÁVIO MORENO SALVADOR
DESEMPENHO E DIGESTIBILIDADE EM OVINOS DA RAÇA SANTA INÊS ALIMENTADOS COM DIFERENR]TES CONDIÇÕES DE
BALANÇOS DE PROTEÍNA DEGRADÁVEL NO RUMEN E PROTEÍNA METABOLIZÁVEL
Tese apresentada à Universidade Federal de Lavras, como parte das exigências do Programa de Pós-graduação em Zootecnia, área de concentração em Nutrição de Ruminantes, para obtenção do título de “Doutor”. Orientador
Prof. Dr. Juan Ramón Olalquiaga Pérez
LAVRAS MINAS GERAIS - BRASIL
2007

SUMÁRIO
Página
RESUMO i
1 INTRODUÇÃO 1
2 REFERÊNCIAL TEÓRICO 3
2.1. A fermentação dos alimentos no rúmen 3
2.2. Os sistemas de alimentação de ruminantes 4
2.2.1. Os sistemas britânicos ARC e AFRC 9
2.2.2. A proteína dietética e seu fracionamento de acordo com os sistemas de alimentação de ruminantes 13
2.3. A proteína de origem microbiana na nutrição protéica dos ruminantes 16
2.3.1. Aspectos interferentes na síntese de proteína microbiana 19
2.3.2. A sincronia no rúmen 20
2.3.3. O tamanho corporal e sua relação com a síntese de proteína microbiana 24
2.4. Considerações acerca do não atendimento das demandas para otimização da síntese de proteína microbiana 30
3 MATERIAL E MÉTODOS 34
3.1. ENSAIO DE DESEMPENHO 34
3.1.1. Local, instalações e período de realização 34
3.1.2. Animais e alimentos 35
3.1.3. Elaboração das dietas e manejo alimentar 35
3.1.4. Tratamentos 38
3.1.5. Coleta de alimentos e sobras 43
3.1.6. Análises bromatológicas 43
3.1.7. Delineamento experimental 44
3.2. ENSAIO DE DIGESTIBILIDADE E BANAÇO NITROGENADO 45
3.2.1. Local, instalações e período de realização 45

3.2.2. Animais e alimentos 46
3.2.3. Elaboração das dietas e manejo alimentar 46
3.2.4. Tratamentos 49
3.2.5. Coletas de alimentos, sobras, fezes e urina 53
3.2.6. Análises químico-bromatológicas 53
3.2.7. Cálculos da digestibilidade e do balanço nitrogenado 54
3.2.8. Delineamento experimental 55
4 RESULTADOS E DISCUSSÃO 57
4.1. ENSAIO DE DESEMPENHO 57
4.1.1 Consumo de matéria seca e de nutrientes 58
4.1.2. Ganho de peso e conversão alimentar 80
4.2. ENSAIO DE DIGESTIBILIDADE E BALANÇO DE NITROGÊNIO 89
4.2.1. Consumo e digestibilidade aparente dos nutrientes 89
4.2.2 Balanço de nitrogênio 100
5 CONCLUSÕES 108
REFERÊNCIAS BIBLIOGRÁFICAS 110

i
RESUMO
SALVADOR, Flávio Moreno. Desempenho e digestibilidade em ovinos da raça Santa Inês alimentados em diferentes condições de balanços de proteína degradável no rúmen e proteína metabolizável. Lavras: UFLA, 2007. 130p. (Teses - Doutorado em Zootecnia).1 Foram conduzidos dois experimentos para avaliar se o atendimento ou não da demanda do rúmen em proteína degradável (PDR), em associação com condições de atendimento ou superávit de proteína metabolizável (PM), afetavam o desempenho, a ingestão de matéria seca e a digestibilidade dos nutrientes em ovinos alimentados com volumoso tropical, de acordo com as recomendações do sistema AFRC. No experimento de desempenho foram utilizados 16 cordeiras da raça Santa Inês, com peso médio inicial de 21,5 kg, em delineamento inteiramente casualizado. Para o ensaio de digestibilidade e balanço nitrogenado foram utilizadas oito borregas Santa Inês, distribuídas em dois quadrados latinos 4x4. Os tratamentos foram: A) atendimento da exigência de PDR e condição superavitária de PM, com uso de fonte de proteína verdadeira; B) não atendimento da demanda em PDR, porém, atendimento das exigências em PM; C) atendimento da exigência de PDR e condição superavitária de PM, com uso nitrogênio não-protéico; D) não atendimento da demanda em PDR, porém com condição superavitária de PM, por meio do uso de fonte de proteína não degradável no rúmen. Os tratamentos não promoveram diferenças (P<0,05) no tocante às ingestões de matéria seca, conversão alimentar e ganhos de peso. Este se situou dentro de um nível relativamente modesto (116 g/dia), levando em consideração a espécie, sexo e categoria animal. As digestibilidades da matéria seca e das frações fibrosas (FDN e FDA) não foram afetadas pelas distintas condições de equilíbrios entre a PDR e PM. O sistema AFRC não permitiu boa precisão na consecução dos níveis de desempenho pré-determinados, mostrando-se não ser ajustado às condições de volumoso e raça tropicais.

1
1 INTRODUÇÃO
A proteína tem sido um dos nutrientes mais pesquisados na nutrição de
ruminantes, sendo que, em razão de sua natureza diversificada, têm sido
atribuída a ela ganhos diferenciados no desempenho animal, bem como a
possibilidade da melhor extração de energia das porções fibrosas dos alimentos
volumosos devido ao atendimento das demandas microbianas por nitrogênio.
Por estas razões, a adequação nos teores de proteína nas dietas animais pode
contribuir com a otimização da utilização do nitrogênio permitindo maior
economicidade nos sistemas de produção, além de poder reduzir a contaminação
ambiental devido a menor excreção deste elemento.
Como resultado do grande volume de informações resultante das
pesquisas, não somente acerca da proteína, mas também relacionadas à nutrição
energética de animais ruminantes, propostas de sistemas de alimentação têm
sido formuladas e a característica que estes sistemas tem em comum entre si, e
que os difere das propostas voltadas para outras espécies animais reside na
intima relação entre a energia e a proteína na predição do rendimento
microbiano a partir da ingestão de nitrogênio degradável e matéria orgânica
fermentável no rúmen.
O princípio básico destes sistemas é a busca em atender a exigência de
nitrogênio da microbiota ruminal, maximizando seu crescimento e,
posteriormente, quantificar o aporte de nutrientes disponíveis para ser digerido,
absorvido e utilizado pelo animal. Estes modelos idealizam o sincronismo entre
a digestão ruminal de proteínas e carboidratos e esta premissa de otimização do
crescimento microbiano ruminal condicionará, segundo os modelos de equilíbrio
entre energia e proteína no rúmen, sobra de proteína metabolizável em relação à
respectiva exigência animal. Ainda segundo os mesmos modelos, esta sobra de

2
proteína metabolizável tem como conseqüência gasto energético para sua
metabolização.
Este aspecto gera alguns questionamentos dentro do enfoque da nutrição
protéica dos ruminantes, principalmente relacionados à natureza da proteína,
com possíveis limitações ao crescimento microbiano. Até que ponto o não
atendimento destes fatores pode prejudicar o desempenho e a digestibilidade das
dietas são questões pouco abordadas na literatura científica.
O objetivo deste trabalho foi verificar se, e em que extensão, o déficit de
proteína degradável no rúmen (PDR) associado ou não a um superávit de
proteína metabolizável (PM) afetam o desempenho, a ingestão de matéria seca e
de nutrientes e a digestibilidade dos componentes dietéticos em dietas para
ovinos da raça Santa Inês, alimentados com volumoso tropical, segundo as
condições propostas pelo sistema britânico AFRC (1993).

3
2 REFERENCIAL TEÓRICO
2.1. A fermentação dos alimentos no rúmen
Os processos microbianos do rúmen permitem converter alimentos
fibrosos e proteínas de baixa qualidade, e mesmo compostos nitrogenados não-
protéicos, em nutrientes valiosos para o animal ruminante. Beever & Moud
(2000) salientam que “a principal razão pela qual monogástricos e pré-
ruminantes são incapazes de utilizar quantidades significativas de forragens, é
que, assim como outros mamíferos, eles não possuem enzimas capazes de
quebrar os polímeros complexos que formam as paredes celulares dos vegetais.
Nos ruminantes, no entanto, o principal sítio de digestão, com relação às
forragens, é o rúmen, local onde o alimento é retido por períodos substanciais e
sujeito á fermentação microbiana extensiva, sob condições anaeróbicas”.
Dentre os compostos resultantes dos processos fermentativos no rúmen,
Owens & Goetsch (1988) destacam os ácidos graxos voláteis, a proteína
microbiana e vitaminas K e do grupo B como sendo de grande importância para
a nutrição dos animais ruminantes. Bergman (1990) salienta que os produtos
finais da fermentação são parcialmente determinados pela natureza da dieta,
posto que esta pode mudar a atividade metabólica dos microrganismos,
provendo novos ou diferentes substratos que influenciam a qualidade e a
natureza desses produtos. Além disso, as quantidades dos compostos resultantes
dos processos fermentativos são elaboradas não somente em função das
composições químicas dos alimentos ingeridos, mas são também dependentes da
quantidade e do tempo de permanência dos alimentos nos compartimentos pré-
gástricos (Hungate, 1966), uma vez que a quantidade e a composição da dieta

4
são variáveis externas que afetam a ingestão, a taxa de digestão, a taxa de
passagem e, desta maneira, o turnover do conteúdo ruminal (Van Soest, 1994).
Assim, têm-se ressaltado o aspecto de que a dieta é, provavelmente, o
fator mais importante que influencia o número e a proporção relativa das
diferentes espécies de microrganismos ruminais e, por conseguinte, os produtos
finais da fermentação. A mudança da dieta de um animal resulta num período de
transição na população ruminal, em que a proporção das diferentes espécies
ruminais varia para um novo balanço, que melhor se ajuste às mudanças
dietéticas (Valadares Filho & Pina, 2006).
Portanto, a fermentação pré-gástrica a que os alimentos são submetidos
ao serem ingeridos por animais com rúmen funcional pode contribuir para a
ocorrência de erros na predição do desempenho animal ao se levar em conta
unicamente os componentes da dieta (Russell et al., 1992).
2.2. Os sistemas de alimentação de ruminantes
O desenvolvimento dos diversos sistemas de alimentação de ruminantes
sempre caminhou paralelamente ao aprimoramento das técnicas de avaliação e
caracterização dos alimentos.
O sistema de Weende para análise dos alimentos (também chamado
método proximal), estabelecido há mais de 100 anos, baseia-se na determinação
das proporções de umidade, compostos nitrogenados, cinzas, extratos solúveis
em éter e porções fibrosas insolúveis em ácido e álcali e foi a primeira base para
a estimação de valores de energia dos alimentos, mais precisamente, do total de
nutrientes digestíveis (NDT). Entretanto, tem sua utilização muitíssimo limitada
quando se busca conhecer mais precisamente o valor nutritivo dos alimentos, as
exigências nutricionais e assim predizer o desempenho animal.

5
Neste contexto, Sniffen et al. (1992) enfatizaram que para se alcançar
um melhor ajuste na formulação de dietas para ruminantes, obtendo-se
concomitantemente redução em custos, faz-se necessário que os alimentos
utilizados pelos animais tenham suas características químicas bem definidas,
com suas porções componentes fracionadas no sentido de melhor caracterizá-las
sob a óptica da alimentação animal.
Na busca por atender esta necessidade, o sistema proposto por Goering
& Van Soest (1970), baseado na solubilidade dos alimentos em soluções
detergentes, permitiu a identificação e classificação de diferentes porções dos
alimentos, fazendo inferências às suas disponibilidades biológicas. Graças a este
sistema, tornou-se possível a melhor caracterização, principalmente dos
carboidratos, representados até então pela fibra bruta e extrativos não-
nitrogenados, e também do componente nitrogenado (proteína bruta) dos
alimentos.
Paralelamente, conceitos sobre a nutrição de ruminantes têm evoluído de
forma considerável nas últimas duas décadas. Até o final da década de 70, as
estimativas das exigências energéticas e protéicas eram obtidas a partir de
ensaios de desempenho e digestibilidade. O desenvolvimento e aprimoramento
dos ensaios de metabolismo, a partir da década de 80, possibilitaram o
desenvolvimento do método fatorial de exigências utilizado até o momento pelos
principais sistemas disponíveis.
Os requerimentos de proteína determinados pela abordagem fatorial
consideram separadamente as demandas de mantença (sendo nestas incluídas as
perdas endógenas de nitrogênio através das fezes e urina e por intermédio da
descamação da pele) e as exigidas para a consecução da produção animal (NRC,
2006). O atendimento das exigências totais de proteína pelos tecidos dos animais
ruminantes é obtido pela absorção, ao nível do intestino delgado, dos produtos
finais resultantes dos processos digestivos sofridos por compostos nitrogenados,

6
em especial pelo montante de aminoácidos disponibilizados para a absorção. As
fontes de proteína que chegam ao intestino dos ruminantes são a proteína
microbiana, a proteína dietética que não sofreu a ação da microbiota ruminal
(durante sua permanência nas câmaras pré-gástricas) e a proteína endógena. Ao
“pool” dos aminoácidos provenientes da digestão dessas fontes e absorvidos no
intestino dá-se o nome de proteína metabolizável. (Burroughs et al., 1975a;
ARC, 1980).
Os sistemas de alimentação evoluíram das determinações de exigências
em proteína bruta para os atuais modelos de proteína metabolizável, que
permitem adequar as exigências da população microbiana ruminal em
compostos nitrogenados, assim como as exigências do ruminante em proteína
metabolizável. Os sistemas de proteína metabolizável têm estimulado e
permitido avanços no conhecimento das exigências em aminoácidos dos
ruminantes e no balanceamento do perfil de aminoácidos essenciais da proteína
metabolizável (Santos, 2006).
Estes avanços no conhecimento da caracterização e composição química
dos alimentos bem como na determinação das demandas nutricionais dos
animais, teve sua integração facultada e ampliada graças ao advento da
tecnologia dos computadores, cujo impulso de desenvolvimento iniciou-se por
volta dos anos 60. Graças a esta tecnologia, a habilidade dos cientistas em
descrever matematicamente as relações biológicas foi grandemente
incrementada e assim, modelos matemáticos passaram a ser construídos
objetivando descrever vários aspectos relacionados à nutrição e alimentação
animal (Russell, 2002).
Define-se um modelo como uma representação simplificada, abstrata e
idealizada de determinada realidade. Qualquer tipo de modelo, por definição,
deve basear-se em argumentações ordenadas, lógicas e justificáveis,
pressupostas a partir do conhecimento científico existente sobre o assunto em

7
questão. Um modelo matemático nada mais é do que uma equação ou um
conjunto de equações que representam o comportamento de um sistema, cuja
resolução implica a predição de mudanças que podem ocorrer na realidade; é a
conseqüência ou o resultado direto de empreendimentos analíticos para a
abstração e definição do mundo real, em termos matemáticos precisos (Mertens,
1976).
Os modelos matemáticos são essencialmente classificados como
‘determinísticos’ ou ‘estocásticos’, ‘dinâmicos’ ou ‘estáticos’ e ‘empíricos’ ou
‘mecanísticos’. Modelos determinísticos são baseados na assunção de que as
soluções obtidas derivam de equações ou funções exatas, entretanto é sabido que
dados biológicos freqüentemente tem elevado grau intrínseco de variabilidade.
Os modelos estocásticos (também ditos probabilísticos) se valem de relações
estatísticas no cômputo das variações, tendo estas a amplitude da variância de
cada um de seus componentes. Modelos estáticos ignoram o efeito do tempo,
mas modelos dinâmicos descrevem relações tempo-dependente dos fatores que
tenham esta característica (Baldwin & Donovan, 2000; Russell, 2002).
Modelos empíricos são construídos a partir da descrição da observação
de dados, os quais são ajustados a uma equação (ou conjunto de equações
matemáticas). Já os modelos mecanísticos (também denominados ‘teóricos’)
buscam promover a descrição de um sistema com a compreensão dos fatores
causais concernentes com os mecanismos envolvidos no sistema em estudo.
Estes modelos são construídos através do exame da estrutura do sistema,
compartimentalizando-o e analisando o comportamento de todo o sistema em
termos dos componentes individuais (compartimentos) e das interações entre
eles (Dijkstra & France, 1995).
Os sistemas de alimentação de ruminantes atualmente em uso se valem
principalmente de equações empíricas (AFRC, CSIRO, INRA, NRC,
DVB/OEB-System - Dijkstra et al., 1998). Embora difiram entre si, quanto à

8
terminologia e detalhamento, estes sistemas são conceitualmente similares em
seus objetivos de prever o fluxo de energia disponibilizado aos animais e
microrganismos ruminais, a quantidade de N passível de ser utilizada por
aqueles seres, pela estimativa de síntese de proteína microbiana e pelo
conseqüente N-aminoácido microbiano que seja disponibilizado ao ruminante
hospedeiro, pelos aspectos cinéticos dos nutrientes no rúmen, pelo ‘rendimento’
de nutrientes que alcança o intestino delgado e conseqüentemente, pelo próprio
desempenho animal.
Algumas propostas de modelos mecanísticos existentes, bem menos
notórias (Dijkstra et al., 1992; Lescoat & Sauvant, 1995, entre outras),
descrevem, por exemplo, a síntese de proteína microbiana baseada não somente
na quantidade, mas no tipo de matéria orgânica disponível, na utilização desta
nos processos de crescimento e de não crescimento microbiano, nas interações
entre as classes de microrganismos, etc.
Comparativamente aos modelos empíricos, os conceitos adotados nos
modelos mecanísticos são mais variáveis posto que os objetivos da modelagem
mecanística e as hipóteses subjacentes são completamente diferentes. O
principal objetivo dos modelos mecanísticos de rúmen é o de prover um
conhecimento integrado dos aspectos envolvidos e permitir predizer o perfil de
nutrientes (incluindo a proteína microbiana), disponível para absorção. Por
exemplo, nenhum dos sistemas empíricos utilizados na avaliação de proteína
para ruminantes considera explicitamente as interações existentes entre as
diferentes populações de microrganismos existentes no rúmen e os variáveis
efeitos de suas atividades na degradação da matéria orgânica presente. Outro
aspecto também desconsiderado nos atuais modelos empíricos refere-se à
consideração dada às variações ocorrentes quanto ao tempo, relevando assim as
oscilações entre dias e dentro de cada dia, fazendo com que os modelos
empíricos observem o animal segundo uma condição ‘steady-state’, o que

9
absolutamente não condiz com a realidade, classificando assim estes sistemas
como empíricos e estáticos (Sniffen & Robinson, 1987).
Tem-se ainda que os modelos podem conter tanto elementos empíricos
como mecanísticos em sua construção, sendo assim chamados de modelos
mistos, como é o caso do sistema de Cornell (CNCPS). Para exemplificar esta
característica, pode-se citar que este sistema relaciona a disponibilidade de
substratos à utilização da energia em processos de crescimento e não-
crescimento microbiano (enfoque mecanístico), mas a degradação dos substratos
é representada sem considerar a interação dos efeitos entre carboidratos e
nitrogênio ou os efeitos das atividades microbianas sobre a degradação da
matéria orgânica no rúmen (enfoque empírico).
Seja como for, cada abordagem assumida de modelagem possui
vantagens e desvantagens, dependendo do objetivo específico. Um modelo
empírico baseado diretamente em um conjunto definido de dados pode, dentro
deste limite, prover respostas bastante acuradas em sua predição. Em
contrapartida, modelos mecanísticos, que tiveram seus componentes e
parâmetros advindos de numerosos e variados bancos de dados, podem não
fornecer predições tão exatas, entretanto permitem um entendimento melhor do
comportamento do sistema como um todo. Outro aspecto importante reside no
fato de que os modelos empíricos são normalmente mais simples e, portanto,
mais facilmente utilizáveis, constituindo por esta razão práticas ferramentas
utilizadas inclusive na elaboração de dietas.
2.2.1. Os sistemas britânicos ARC e AFRC
Os sistemas de nutrição e alimentação de ruminantes de origem britânica
(ARC, 1980 e AFRC, 1993) trouxeram relevantes contribuições sobre a
utilização dos nutrientes por estes animais. O conceito de energia metabolizável

10
assumido pelo ARC (1980) tem como base a relação entre os consumos de
energia metabolizável (nos alimentos ou dieta) com a retenção da energia líquida
nos produtos e no metabolismo animal. A ingestão de energia metabolizável
refere-se à energia bruta ingerida menos a energia bruta contida nas fezes, urina
e gases de combustão (majoritariamente metano).
Especificamente no que diz respeito ao aproveitamento da energia, o
ARC (1980) estabeleceu o conceito da metabolizabilidade (q), definida como a
energia metabolizável do alimento dividida por sua energia bruta. A
metabolizabilidade da energia à mantença é simbolizada por qm e, em qualquer
outro nível de alimentação, qL. A eficiência de utilização da energia
metabolizável (simbolizada por k) é definida como o aumento na retenção de
energia que ocorre por unidade de incremento de energia metabolizável
oferecida, tendo por isso estreita relação com a metabolizabilidade da energia
em cada alimento. A eficiência de utilização da energia metabolizável foi então
apresentada como função linear da metabolizabilidade da energia, sendo
específica quanto à função fisiológica de interesse (mantença, ganho de peso
corporal, lactação, etc.).
Por convenção, os sistemas de alimentação consideram que quando a
retenção de energia corporal é zero, diz-se que o animal está em manutenção, ou
seja, consome e dissipa energia para a manutenção dos processos vitais e
metabólicos básicos. Já a retenção de energia refere-se à taxa de deposição
energética corporal, que pode ser obviamente negativa quando o nível de
ingestão energética está abaixo da mantença. Assim posto, considerando que a
energia metabolizável de cada alimento possui uma metabolizabilidade que lhe é
peculiar e a eficiência com que é utilizada é variável em função do objetivo de
seu uso, faz-se imperativo que, para se estimarem as exigências de energia
metabolizável de mantença e ganho de peso seja necessário, primeiramente,

11
estimar a exigência líquida de energia para mantença e para ganho,
respectivamente.
O ARC (1980) relacionou ainda a quantidade de energia provinda do
alimento consumido e a exigência energética dos animais, auferindo quantas
vezes a energia exigida para mantença é ingerida, e a este valor foi dado o nome
de nível de produção, representado por L. O sistema AFRC (1993) também
assume este postulado. Este valor calculado, enquanto não considerado para o
computo das demandas energéticas, é levado em conta para o cálculo do
potencial de crescimento microbiano ruminal.
A exigência de energia metabolizável para ganho de peso, como o
sistema AFRC (1993) o apresenta, é dependente do tamanho do ganho de peso
buscado, da composição energética corporal dos animais (Eg) e da eficiência de
utilização da energia metabolizável para ganho de peso (kg).
A ingestão de energia metabolizável (IEM), menos a exigência
energética de mantença (EMm), resulta na sobra de energia metabolizável
disponível para ganho de peso (EMg) que será utilizada com a eficiência kg. Para
o balanceamento de dietas, o sistema propõe a seguinte função para o balanço
energético:
IMS x [EM]dieta = EMm + EMg
em que IMS é a ingestão de matéria seca (kg/dia); [EM]dieta é a concentração de
energia metabolizável da dieta (Mcal/kg MS); EMm é a exigência de energia
metabolizável para mantença (Mcal/dia) e EMg é a energia metabolizável de
ganho de peso (Mcal/dia).
Nos sistemas britânicos, a exigência de proteína metabolizável,
igualmente ao que se dá no tocante à energia, é resultante do somatório das
demandas de mantença e produção, sendo considerada a parte da demanda para

12
mantença as perdas derivadas da descamação da pele e do crescimento de lã e
pelos.
Para estimar a síntese de proteína microbiana, o sistema considera
primeiramente o potencial de crescimento microbiano (YPBmic) possível de ser
obtido, sendo este definido como função do nível de ingestão de energia
metabolizável, ou melhor, pelo nível de produção (L), conforme a equação
YPBmic = 7 + 6 x (1 - e (-0,35 x L))
Assim, tem-se que o crescimento microbiano (Y) é obtido, segundo o
AFRC (1993), a partir de:
Y = IEMfe x YPBmic
em que (YPBmic) é o potencial de crescimento microbiano expresso em g PB
microbiana/Mcal IEMfe; (L) é o nível de produção (ingestão de EM em relação à
exigência de mantença) e IEMfe representa a ingestão de energia metabolizável
fermentável (em Mcal/dia).
A inclusão de proteína dietética se desenvolve em uma segunda etapa do
balanceamento conforme apresentada para o ARC (1980) e AFRC (1993). A
partir da definição da concentração energética da dieta, que é função de
consumo, peso vivo, composição e intensidade de ganho de peso, busca-se
incluir proteína degradável na dieta a fim de atender prioritariamente à demanda
de proteína para crescimento microbiano ruminal. Desta forma, a inclusão de
uma dada fonte de proteína verdadeira ou de NNP dependerá da escala de
crescimento microbiano que o balanço energético da dieta permite.

13
2.2.2. A proteína dietética e seu fracionamento de acordo com os sistemas de alimentação de ruminantes
O sistema britânico ARC (1980) basicamente subdivide os compostos
nitrogenados presentes nos alimentos em duas porções, uma passível de ser
degradada no rúmen e outra indegradável (RDN e UDN). Por outro lado, Van
Soest (1994) chamou a atenção para o aspecto de que simplesmente classificar o
nitrogênio dietético como N solúvel e N insolúvel no rúmen não é adequado
para o entendimento de seu valor nutricional e de sua dinâmica no trato
digestivo dos ruminantes. A razão para tal reside no fato de que esta divisão
simplista não distingue nitrogênio não-protéico (NNP) de proteína verdadeira,
ou ainda, não leva em conta a indisponibilidade, mesmo que parcial, da fração
insolúvel, além de assumir que a insolubilidade confere características de baixa
velocidade de degradação. A proteína presente nas folhas da alfafa (Medicago
sativa) é desnaturada (tornando menos solúvel) durante o processo de fenação,
porém, continua rapidamente degradável, enquanto que a albumina sérica
solúvel fermenta muito lentamente.
O sistema americano (NRC, 1985) aperfeiçoou mais o fracionamento,
dividindo a proteína dos alimentos em três frações, sendo uma rapidamente
degradada no rúmen, uma potencialmente degradável e outra indegradável.
Russell et al. (1992) e Sniffen et al. (1992), entretanto, consideraram este
modelo ainda limitado e sugeriram novas subdivisões.
Para que o processo de degradação das proteínas no rúmen fosse mais
bem explorado, e avaliadas as contribuições da proteína dietética e microbiana
para a quantidade e qualidade dos aminoácidos absorvidos ao nível do intestino
delgado, fizeram-se necessárias investigações mais acuradas da disponibilidade
potencial das diferentes frações que compõem a proteína. Progressos
significativos foram obtidos no desenvolvimento de metodologias visando
relacionar a solubilidade de diferentes frações da proteína com sua

14
suscetibilidade à degradação enzimática (Krishnamoorthy et al., 1982; 1983).
Assim, o sistema Cornell Net Carbohydrate and Protein System - CNCPS
(detalhadamente descrito por Fox et al., 2003; Fox et al., 2004), foi desenvolvido
desde o início dos anos 90, caracterizando o componente protéico dos alimentos
como dividido em três frações básicas: nitrogênio não-protéico (fração A),
proteína verdadeira (fração B) e nitrogênio indisponível (fração C). A fração B,
por sua vez, foi subdividida em três outras sub-frações, conforme a velocidade
de degradação (taxa de degradação) dentro do rúmen: B1 (proteína solúvel,
rapidamente degradada no rúmen), B2 (proteína insolúvel, moderadamente
degradada no rúmen) e B3 (proteína insolúvel, lentamente degradada no rúmen).
Na Tabela 1A, apresentada nos anexos, pode ser visualizado um esquema das
frações protéicas dos alimentos e a forma de se obter tais frações. Esta maneira
de fracionar os constituintes nitrogenados dos alimentos, em associação com o
devido fracionamento dos carboidratos, permite maximizar a sincronização das
atividades microbianas sobre a proteína e carboidratos no rúmen, maximizando a
produção microbiana, a extração de energia dos alimentos e também minimizar
perdas nitrogenadas (Pereira, 1999).
Dentre as frações (A, B e C), a fração B é a mais variável, quanto ao
intervalo de degradação dentro do rúmen, uma vez que se assume que a fração A
é completamente degradada e que a fração C é completamente indisponível.
Assim ressalta-se que o fracionamento do nitrogênio pura e simplesmente não
implica em condições de se poder predizer todo o montante de N que estará
disponibilizado para a população microbiana. É fundamental que associado à
caracterização da fração protéica do alimento sejam auferidas também as
velocidades de degradação de cada uma das frações a fim de permitir estimar o
montante efetivo de proteína degradável (PDR) e não degradável no rúmen
(PNDR). Várias propostas têm sido feitas no sentido de se tentar combinar
estimativas da taxa de desaparecimento (Kd) da fração B no rúmen, com a taxa

15
de passagem (Kp) com o intuito de se estimar a PDR e PNDR. O NRC (2001),
baseando-se em dados de 190 experimentos, propôs das equações conforme:
PDR = A + {Bi x [ KdBi / (KdBi + KpBi)]}
PNDR = Bi x [ KpBi / (KpBi + KdBi)] + C
Onde:
PDR: proteína degradável no rúmen
PNDR: proteína não degradável no rúmen
A: fração A da proteína (% da PB)
Bi: fração Bi da proteína, sendo i = 1, 2 e 3 (% da PB)
C: fração C da proteína (% da PB)
KdBi; taxa de degradação da fração Bi (% por hora)
KpBi; taxa de passagem da fração Bi (% por hora)
O modelo proposto pelo ARC (1980) e adaptado pelo AFRC (1993),
descrito anteriormente, pode ser utilizado para predizer dietas para os ovinos. No
entanto, os componentes do modelo do AFRC (1993) que atualmente podem ser
considerados inadequados para os ovinos, frente aos novos trabalhos científicos,
foram mais adequadamente ajustados com base em uma extensiva revisão
bibliográfica para melhorar a precisão dos modelos, conforme pode ser proposto
por Cannas et al. (2004). Este sistema tem como base a estrutura do CNCPS e
prevê o efeito do nível de alimentação sobre a utilização da dieta, tendo sido
apresentado à comunidade científica como a versão do sistema de Cornell para a
espécie ovina (denominado por isso, CNCPS-S). O modelo teve ênfase especial
em ovinos leiteiros.
Assim é que modelos mais modernos de alimentação de ovinos vêm se
desenvolvendo com o objetivo de melhorar a acurácia de suas estimativas.

16
Exemplos nesta direção são o próprio CNCPS-S, de origem norte-americana e o
MIPAF (italiano), citado por Cannas & Atzori (2005).
2.3. A proteína de origem microbiana na nutrição protéica dos ruminantes
Na nutrição de animais ruminantes é fato conhecido que uma substancial
proporção dos aminoácidos absorvidos pelos ruminantes (mais de 50% e não
raras vezes este montante pode chegar de dois terços a três quartos) é
proveniente da proteína microbiana sintetizada no rúmen (AFRC, 1993). Esta é
sintetizada no processo fermentativo de degradação ruminal a partir de proteína
dietética, proteína microbiana reciclada, nitrogênio reciclado via saliva e sangue
ou mesmo fontes de nitrogênio não protéico (Teixeira & Salvador, 2004).
A proteína microbiana é considerada fonte de boa qualidade, em relação
à sua digestibilidade intestinal (entre 80 a 85%) e ao seu perfil de aminoácidos
(NRC, 2000), sendo sua composição aminoacídica similar à da proteína dos
tecidos do próprio corpo do animal, bem como da proteína encontrada no leite
(Schwab, 1996). Em comparação à composição da proteína de concentrados
protéicos de origem vegetal, a proteína microbiana contém maior proporção de
metionina e lisina e (após a proibição da utilização de alimentos de origem
animal em dietas destinadas a ruminantes no Brasil) não existem fontes que
atendam melhor aos requerimentos aminoacídicos do animal que a proteína
microbiana (Verbic, 2002).
Foi graças à qualidade desta fonte protéica que Virtanen (1966)
conseguiu demonstrar que vacas com moderadas produções leiteiras
conseguiram a satisfação de suas demandas nutricionais em aminoácidos tendo
como fonte dietética de nitrogênio (N) apenas a uréia.

17
Assim, a proteína microbiana ruminal deve ser considerada como uma
importante fonte protéica e de fato, as exigências de proteína metabolizável dos
ruminantes são atendidas mediante a absorção no intestino delgado da proteína
verdadeira microbiana e proteína dietética não degradada no rúmen. O
suprimento de proteína metabolizável derivada da proteína microbiana é similar
àquela advinda das silagens de gramíneas (aproximadamente 64% - AFRC,
1993) e apresenta bom perfil de aminoácidos, apesar de variável (Storm &
Ørskov, 1983).
Embora o rúmen apresente vantagens, particularmente quando aos
animais são oferecidos alimentos de baixa qualidade, ele pode, por outro lado,
vir a promover a ineficiência de utilização de nitrogênio pelos ruminantes. Por
exemplo, quando vacas leiteiras receberam concentrado com níveis e
características clássicas em associação a 16 diferentes tipos de silagens de
gramíneas, a eficiência de conversão de nitrogênio alimentar em nitrogênio do
leite variou de 23 a 32% (Dewhurst et al. 1996).
Condições em que ocorra a ingestão excessiva de compostos
nitrogenados, sem o devido aporte de energia disponível podem favorecer o
comprometimento do desempenho produtivo e reprodutivo dos animais,
aumentar as exigências energéticas, elevar significativamente os custos de
produção, além de poder ser fator de agravamento de poluição ambiental devido
ao incremento na excreção do N excedente (NRC, 2000). Portanto, o
desconhecimento de como o rúmen desempenhará em uma dada situação
constitui-se na principal causa do uso ineficiente da proteína dietética. Torna-se
então essencial predizer ou estimar o montante de proteína microbiana produzida
a fim de corrigir problemas bem como otimizar o uso do nitrogênio da forragem
e das fontes protéicas, geralmente de custos elevados. Assim, conseguir-se
determinar o tamanho do crescimento microbiano ocorrente no rúmen e a
respectiva absorção do pool de aminoácidos ali elaborado, ao nível do intestino

18
delgado, é fundamental para permitir a adequação na formulação de dietas de
ruminantes.
O NRC (1985) discute o crescimento microbiano sob três contextos: a
eficiência microbiana, a massa microbiana e o fluxo microbiano. A síntese de
proteína microbiana, sob o aspecto da eficiência microbiana, tem sido expressa
de maneiras diferentes, segundo diferentes sistemas de alimentação: em função
da quantidade de nutrientes digestíveis totais - NDT (NRC, 1985), em relação à
matéria orgânica degradada no rúmen - MODR (ARC, 1980) ou em função da
quantidade de carboidratos degradados no rúmen - CHODR (sistema Cornell -
CNCPS - Sniffen et al., 1992). Este último enfoque motivou a proposição de
uma classificação simplista pelo CNCPS dos microrganismos ruminais de
acordo com o tipo de carboidrato utilizado como fonte de energia (Russell,
1984; Russell et al., 1992). Valadares Filho (1995) salienta considerar que a
melhor forma de expressar a eficiência microbiana seria em relação aos
CHODR, comparativamente à MODR e ao NDT, posto que são os carboidratos
que definitivamente constituem a principal fonte de energia para os
microrganismos ruminais, quando comparados aos lipídios e à proteína bruta.
O sistema AFRC (1993), porém, definiu o conceito de energia
metabolizável fermentável (EMfe), por meio do qual se busca refletir a
disponibilidade de energia metabolizável dos alimentos para os microrganismos
ruminais. Assim, a participação energética da gordura é desprezada pelo sistema,
que não considera o uso da energia proveniente da gordura pela flora microbiana
ruminal.
Se por um lado torna-se evidente as vantagens obtidas ao poder se
estimar a síntese de proteína microbiana, a literatura a este respeito tem sido
revisada em diversas ocasiões e resulta em poucos prognosticadores totalmente
confiáveis acerca da síntese de proteína microbiana ou acerca da eficiência da
mesma (Broderick & Merchen, 1992; Firkins, 1996). Isto se dá tanto em função

19
da complexidade do sistema do rúmen como também em função das
dificuldades técnicas de estimar a síntese de proteína microbiana principalmente
in vivo. Existem dois problemas distintos, que são determinar qual a quantidade
de material que deixa o rúmen e então estimar qual é a proporção de material
protéico de origem microbiana dentre este material. Estas estimativas têm
requerido o uso de animais canulados os quais tendem a somar a estes
problemas, baixa repetibilidade. Titgemeyer (1997) revisando a literatura
demonstrou que, em média, grupos de 12 animais por tratamento são necessários
para identificar diferenças significativas entre tratamentos da ordem de 10%
quanto à eficiência de síntese de proteína microbiana.
2.3.1. Aspectos interferentes na síntese de proteína microbiana
Em condições normais os fatores que afetam de modo marcante a síntese
de proteína microbiana são de foro alimentar. Vários trabalhos foram citados por
Ribeiro et al. (2001) os quais mencionam que a síntese de proteína microbiana
no rúmen é variável e dependente das concentrações e da qualidade das fontes
de energia e de nitrogênio dietéticos no rúmen, da taxa de diluição ruminal, da
freqüência de alimentação, do consumo de alimentos, da proporção entre
volumosos e concentrados (que obviamente está relacionada à qualidade e
quantidades das fontes de N e energia), do processamento pelo qual o volumoso
pode vir a passar, da presença de determinados aditivos e/ou ionóforos e da
presença de minerais (especialmente fósforo, magnésio e enxofre). O fluxo
microbiano, discutido pelo NRC (1985), é um dos importantes aspectos
relacionados ao crescimento microbiano e este é, por sua vez, dependente das
relações entre o tamanho das partículas, do volume e da taxa de passagem no
rúmen. Este último aspecto tem importância relevante posto que a taxa de
passagem acaba interferindo na redução da idade média da população

20
microbiana (em função da remoção de organismos maduros) e assim reduzem a
demanda energética de mantença desta microbiota, elevando assim a eficiência
de uso da energia do sistema para crescimento microbiano (Polan, 1988).
Entretanto, a literatura concernente à síntese de proteína microbiana é até
certo ponto confusa e não raramente contraditória e isto decorre em razão tanto
do aspecto dos complexos fatores envolvidos bem como decorrente das
dificuldades de mensuração desta síntese. Existem numerosos tratamentos que
têm apresentados claros efeitos que podem estar relacionados aos componentes
dos modelos clássicos de crescimento microbiano, modelos estes que relacionam
o crescimento à presença e/ou abundância de substrato (em especial N) e à
disponibilidade de energia dos microrganismos (Clark et al., 1992; Jetana et al.,
2000).
2.3.2. A sincronia no rúmen
O conceito de sincronia no rúmen tem sido proposto como sendo
fundamental para simplificação da descrição do suprimento de energia e proteína
aos microrganismos ruminais. É considerado que a síntese de proteína
microbiana será maximizada por intermédio da sincronia entre a
disponibilização de energia fermentável e de nitrogênio degradável no rúmen.
Rooke et al. (1987) demonstraram o principio de incrementar a síntese de
proteína microbiana quando glicose, acompanhada ou não por caseína, foi
infundida dentro do rúmen de vacas alimentadas com dietas à base de silagem de
gramíneas. Nas condições particulares em que a caseína foi adicionada,
verificou-se incremento da síntese de proteína microbiana. Desde então diversas
outras tentativas testaram a hipótese da ‘sincronia’ e um resumo de alguns
trabalhos, relativos ao modo em como a sincronia e o assincronismo foram
obtidos experimentalmente são citados na revisão de Dewhurst et al. (2000).

21
É possível alterar a sincronização de dietas, tanto por intermédio da
alteração dos componentes da dieta (mudança de ingredientes) como por meio
da modificação das proporções entre os ingredientes ou ainda dosando formas
específicas de energia e N dentro do rúmen, ou a combinação entre as diversas
maneiras descritas. Porém, não raro ocorre impossibilidade de identificação se
um incremento na eficiência de síntese de proteína microbiana foi obtido em
função do processo de sincronização de energia e nitrogênio disponíveis no
rúmen ou se é efeito associado à manipulação de ingredientes (nível e tipo)
(Herrera-Saldana et al., 1990; Sinclair et al., 1993). Uma outra falha potencial de
experimentos onde as taxas de degradação/ fermentação das frações de proteínas
e carboidratos são pré-determinadas em estudos in sacco com o objetivo de
calcular um ‘índice de sincronização’ reside no fato de que a secagem e a
moagem de substratos altera as características e assim, a disponibilidade dos
componentes dietéticos que são utilizados para determinar estes ‘índices’.
Fornecendo os mesmos ingredientes de acordo com diferentes padrões
alimentares ou de infusão de nutrientes, diretamente dentro do rúmen, constitui-
se em uma forma mais robusta de teste. Entretanto, mesmo neste tipo de ensaio
pequenas evidências conclusivas têm sido apresentadas as quais mostram efeitos
positivos na síntese de proteína microbiana (Henning et al., 1993) ou na
produção de leite (Kolver et al., 1998). Henning et al. (1993) demonstrou que a
contínua infusão exclusiva de açúcar aumentou a eficiência de crescimento
microbiano e eles concluíram que a manipulação dietética deveria objetivar em
prover tanto um suprimento de energia enquanto houvesse um suprimento
adequado de quantidades de N degradável no rúmen. Por outro lado, Henning et
al (1991), citados por Henning et al. (1993), demonstraram que, sob condições in
vitro, uma dose única de glicose foi superior a outros padrões de fontes
energéticas em elevar a síntese microbiana e a eficiência. Dewhurst et al. (2000)
relata que pesquisadores do Instituto de Pesquisa em Meio-ambiente e

22
Pastagens, de Aberystwyth (Reino Unido) forneceram dietas formuladas para
serem tanto altamente sincrônicas ou altamente assíncronas, em um sistema de
cultura contínuo simulando o rúmen. As dietas sincrônicas permitiram maiores
rendimentos microbianos, apesar do efeito assíncrono ter sido praticamente
eliminado em razão da alimentação continua nestes sistemas.
Efeitos mais evidentes os quais são atribuídos ao sincronismo podem ser
resultados de efeitos específicos de nutrientes individuais, particularmente de
frações da energia e/ou proteína. Neste contexto, é útil considerar a hipótese da
‘sincronia’ e sua interpretação a partir do ponto de vista de dois sistemas de
alimentação bastante distintos, por exemplo, baseados em pastejo de forragens
frescas e alimentação a partir de forragens conservadas (tais como silagens e
fenos). Chamberlain & Choung (1995) apresentaram o caso de ‘assincronismo’
com dietas baseadas em forragens conservadas, onde um grande desequilíbrio
pode surgir, com pouca energia prontamente disponível, na presença de
abundantes compostos derivados da degradação protéica (peptídeos,
aminoácidos e amônia). De fato, contrariamente à maioria dos achados
discutidos anteriormente, Kim et al. (1999a) observaram aumento no fluxo de N
microbiano em vacas leiteiras quando foram criadas condições sincrônicas no
rúmen por meio da infusão de açúcares em diferentes tempos tendo como dieta
silagem e concentrados. Porém, em outro estudo semelhante, onde a silagem foi
fornecida sem o concentrado (Kim et al., 1999b), não houve efeito da infusão.
Os autores sugeriram que o grau de sincronia apenas influenciará a síntese de
proteína microbiana em dietas que contenham elevadas concentrações de
carboidratos prontamente fermentáveis, embora isto apenas seja provável se a
capacidade dos microrganismos em armazenar amido for excedida. Torna-se
difícil explicar estes achados contraditórios visto que respostas positivas seriam
mais prováveis constatar em condições onde as vacas recebessem
exclusivamente silagens.

23
Por outro lado, durante pastejos de gramíneas frescas, níveis
relativamente altos de açúcares solúveis e compostos resultantes de degradação
protéica se tornarão disponíveis por períodos extensos, em razão das ações de
enzimas vegetais e microbianas. Além disso, se excessos de açúcar estiverem
disponíveis durante o pastejo de forragens frescas e existir uma deficiência
transitória de N, as bactérias ruminais podem sintetizar e armazenar amido (até
75% da matéria seca celular), o qual pode ser armazenado para utilização
posterior, quando o suprimento de N for recomposto. Assim, assincronismo
entre o suprimento de energia e N pode não representar grande problema durante
períodos de pastejo de forragens frescas bem como quando as dietas forem
baseadas em silagens de gramíneas. Fatores tais como o nível, o tipo e o
equilíbrio de diferentes fontes de carboidratos podem assumir maior
importância, particularmente quando variações no conteúdo de açúcares solúveis
(e outros nutrientes) são consideradas para forragens pastejadas na primavera ou
verão. Seria então digno de proposição que, para o caso de silagens o termo
‘equilíbrio’ o qual abarca todos estes requerimentos, deveria substituir o termo
‘sincronia’ quando descrever o suprimento de energia e de N. Na realidade, tem
sido demonstrado que açúcares solúveis como a sacarose, lactose e frutose são
superiores ao amido (cereais são usualmente utilizados para suplementações
práticas, ao invés da sacarose) como fontes de energia para a fixação do N
microbiano no rúmen. Este dado pode ter significância para a síntese microbiana
em animais em pastejo, uma vez que os níveis de amido são muitíssimos baixos
em gramíneas frescas a frutana, polissacarídeo de armazenagem, pode
representar ao redor de 70% do total de carboidratos hidrossolúveis. E sobre ela
muito pouco tem sido reportado, no que diz respeito às características de
fermentação e o uso desta pelos microrganismos. Porém, a taxa e a extensão de
produção de gás a partir das frutanas sob condições simuladas de rúmen in vitro
tem demonstrado que este açúcar se comporta de forma semelhante ao amido e é

24
fermentada consideravelmente mais lentamente que a sacarose ou a glicose
(Dewhurst et al., 2000).
O uso de novas variedades de gramíneas, desenvolvidas para conter
elevadas concentrações de carboidratos hidrossolúveis, objetivando equilibrar o
suprimento energético ao rúmen e melhorar s produção leiteira em vacas, foi
avaliado por Miller et al. (1999), citados por Dewhurst et al., 2000. Esta
abordagem pode solucionar o problema de suprimento energético a partir do
ponto de vista do tipo e do nível de açúcar, posto que a maioria dos açúcares
presentes nestas linhagens encontra-se na forma de polímeros de frutanas.
Aqueles autores obtiveram aumentos na produção leiteira quando vacas se
alimentaram destas forragens ricas nestes tipos de carboidratos hidrossolúveis,
comparativamente às forragens controle (baixos níveis de frutanas) e assim
sugeriram que a principal causa foi decorrente provavelmente de um aumento da
ingestão de matéria seca digestível. Entretanto, os estudos da digestão no rúmen
não foram levados a efeito e um efeito na síntese microbiana também é possível.
A manipulação de linhagens forrageiras para características que resultem em
maiores concentrações de carboidratos hidrossolúveis e redução da
degradabilidade da proteína são caminhos a avançar e tem o potencial de
coadunar qualidade da forragem aos requerimentos ruminais em açucares, N e
muitos outros nutrientes.
2.3.3. O tamanho corporal e sua relação com a síntese de proteína microbiana
A predição da síntese de proteína microbiana em determinadas
condições de alimentação, sob a óptica de modelos empíricos, permite a
constatação de comportamentos característicos. Por exemplo, ao se lançar mão
das equações propostas segundo o sistema AFRC (1993), e realizando simples
exercícios algébricos, poderá ser verificado que o rendimento microbiano se dá
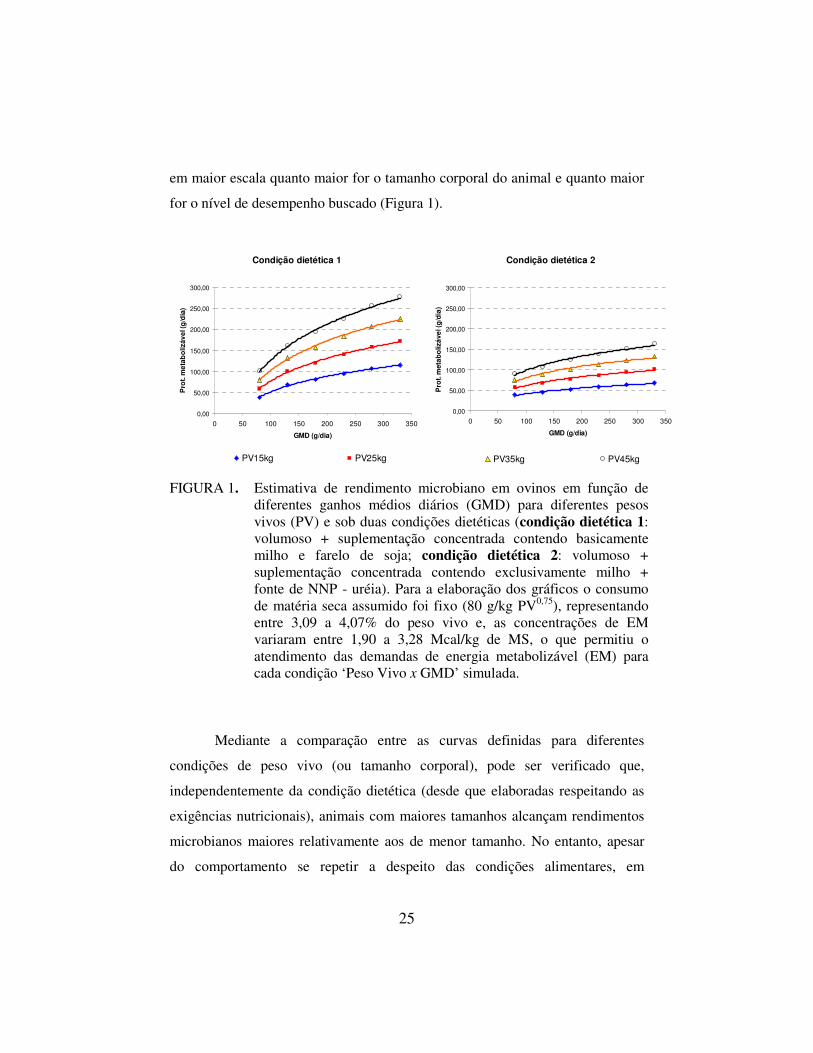
25
em maior escala quanto maior for o tamanho corporal do animal e quanto maior
for o nível de desempenho buscado (Figura 1).
Condição dietética 1
0,00
50,00
100,00
150,00
200,00
250,00
300,00
0 50 100 150 200 250 300 350
GMD (g/dia)
Pro
t. m
eta
bo
lizáv
el (
g/d
ia)
PV15kg PV25kg
Condição dietética 2
0,00
50,00
100,00
150,00
200,00
250,00
300,00
0 50 100 150 200 250 300 350
GMD (g/dia)P
rot.
meta
bo
lizá
ve
l (g
/dia
)
PV35kg PV45kg
FIGURA 1. Estimativa de rendimento microbiano em ovinos em função de diferentes ganhos médios diários (GMD) para diferentes pesos vivos (PV) e sob duas condições dietéticas (condição dietética 1: volumoso + suplementação concentrada contendo basicamente milho e farelo de soja; condição dietética 2: volumoso + suplementação concentrada contendo exclusivamente milho + fonte de NNP - uréia). Para a elaboração dos gráficos o consumo de matéria seca assumido foi fixo (80 g/kg PV0,75), representando entre 3,09 a 4,07% do peso vivo e, as concentrações de EM variaram entre 1,90 a 3,28 Mcal/kg de MS, o que permitiu o atendimento das demandas de energia metabolizável (EM) para cada condição ‘Peso Vivo x GMD’ simulada.
Mediante a comparação entre as curvas definidas para diferentes
condições de peso vivo (ou tamanho corporal), pode ser verificado que,
independentemente da condição dietética (desde que elaboradas respeitando as
exigências nutricionais), animais com maiores tamanhos alcançam rendimentos
microbianos maiores relativamente aos de menor tamanho. No entanto, apesar
do comportamento se repetir a despeito das condições alimentares, em

26
concordância com a afirmação de Ribeiro et al. (2001), condições alimentares
diferentes podem promover rendimentos microbianos distintos em termos de
grandezas numéricas e as premissas do modelo do AFRC (1993) parecem
acomodar este aspecto.
Este comportamento de simulação se verifica também em outros
sistemas de alimentação, posto que as equações consideradas levam em conta os
mesmos aspectos, ou seja, o potencial de crescimento microbiano frente à
disponibilidade energética (seja expressa como NDT, MODR ou CHODR) e de
compostos nitrogenados.
Desta forma, certas condições dietéticas demonstrarão a pressuposição
da existência de tamanhos corporais ‘limites’, abaixo dos quais a síntese de
proteína microbiana per si não é capaz de prover volume suficiente para o
atendimento das demandas em proteína metabolizável, não permitindo, por
conseguinte, o alcance de níveis de desempenhos acima de determinados
patamares. Ao se efetuar simulações de síntese de proteína microbiana em
ovinos com pesos vivos e ganhos distintos, determinando curvas que
representem tanto a exigência de proteína metabolizável bem como o montante
de proteína metabolizável passível de ser alcançado por meio da dieta e da
síntese microbiana ruminal, verifica-se que em circunstâncias em que dieta
provê energia e apenas uréia como fonte de nitrogênio, esta pressuposição
definitivamente se configura enquanto que sob condições dietéticas diferentes
(onde, por exemplo, a fonte de N seja o farelo de soja), esta ocorrência não se dá
(Figura 2).

27
GMD = 180g/dia (cond. diet. 1)GMD = 180g/dia (cond. diet. 1)GMD = 180g/dia (cond. diet. 1)GMD = 180g/dia (cond. diet. 1)
0,00
50,00
100,00
150,00
200,00
250,00
Pro
t. m
eta
bo
lizá
ve
l (g
/dia
)
GMD = 180g/dia (cond. diet. 2)GMD = 180g/dia (cond. diet. 2)GMD = 180g/dia (cond. diet. 2)GMD = 180g/dia (cond. diet. 2)
0,00
50,00
100,00
150,00
200,00
250,00
Pro
t. m
eta
bo
lizá
ve
l (g
/dia
)
GMD = 280g/dia (cond. diet. 1)GMD = 280g/dia (cond. diet. 1)GMD = 280g/dia (cond. diet. 1)GMD = 280g/dia (cond. diet. 1)
0,00
50,00
100,00
150,00
200,00
250,00
300,00
Pro
t. m
eta
bo
lizá
ve
l (g
/dia
)
GMD = 280g/dia (cond. diet. 2)GMD = 280g/dia (cond. diet. 2)GMD = 280g/dia (cond. diet. 2)GMD = 280g/dia (cond. diet. 2)
0,00
50,00
100,00
150,00
200,00
250,00
300,00
Pro
t. m
eta
bo
lizá
ve
l (g
/dia
)
GMD = 330g/dia (cond. diet. 1)GMD = 330g/dia (cond. diet. 1)GMD = 330g/dia (cond. diet. 1)GMD = 330g/dia (cond. diet. 1)
0,00
100,00
200,00
300,00
400,00
0 10 20 30 40 50 60
Peso Vivo (kg)
Pro
t. m
eta
bo
lizá
ve
l (g
/dia
)
GMD = 330g/dia (cond. diet. 2)GMD = 330g/dia (cond. diet. 2)GMD = 330g/dia (cond. diet. 2)GMD = 330g/dia (cond. diet. 2)
0,00
100,00
200,00
300,00
400,00
0 10 20 30 40 50 60
Peso Vivo (kg)
Pro
t. m
eta
bo
lizá
ve
l (g
/dia
)
PMet dietaExig PMet
FIGURA 2. Curvas de exigência e saldo de proteína metabolizável para diferentes condições de pesos vivos e ganhos de peso (GMD). Condições dietéticas 1 e 2 conforme descritas na Figura 1.
Para todas as circunstâncias simuladas, as curvas de exigência e saldo
dietético são divergentes, cruzando-se em pontos distintos, dependendo do
patamar de desempenho proposto (ganho de peso).
28
Conforme já mencionado, nas condições dietéticas em que fontes
verdadeiras de proteína foram consideradas, não se observa incapacidade no
atendimento das exigências e, portanto, não se sustenta a hipótese de tamanho
limite. Em contrapartida, nas condições dietéticas onde a proteína metabolizável
deriva eminentemente da síntese ruminal de proteína microbiana verifica-se que
para qualquer patamar de desempenho apenas em ovinos acima de 21 kg de peso
vivo, ingerindo volumosos tropicais de boa qualidade (considerado nas
simulações), a maximização da síntese de proteína microbiana contribuirá
contundentemente para o atendimento das demandas em proteína metabolizável
sendo, na realidade, obtido volumes que excedem em muito às exigências.
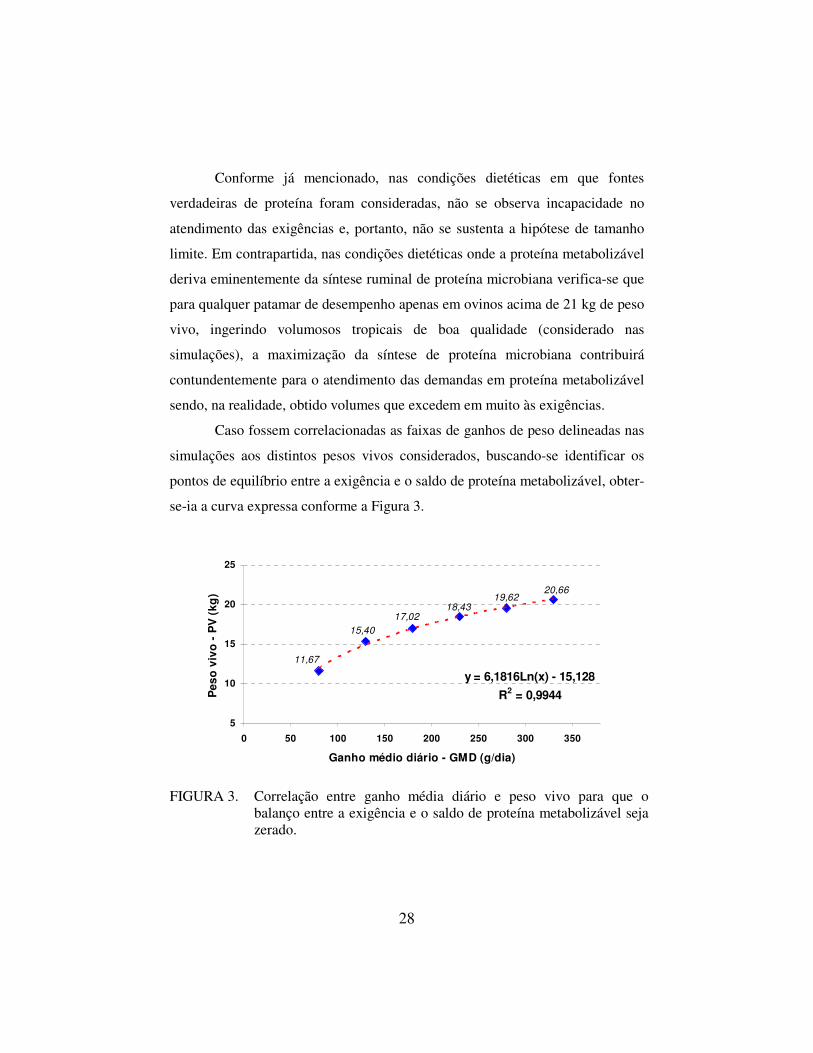
Caso fossem correlacionadas as faixas de ganhos de peso delineadas nas
simulações aos distintos pesos vivos considerados, buscando-se identificar os
pontos de equilíbrio entre a exigência e o saldo de proteína metabolizável, obter-
se-ia a curva expressa conforme a Figura 3.
20,6619,62
18,4317,02
15,40
11,67
y = 6,1816Ln(x) - 15,128
R2 = 0,9944
5
10
15
20
25
0 50 100 150 200 250 300 350
Ganho médio diário - GMD (g/dia)
Pe
so
viv
o -
PV
(k
g)
FIGURA 3. Correlação entre ganho média diário e peso vivo para que o balanço entre a exigência e o saldo de proteína metabolizável seja zerado.

29
Um aspecto a ressaltar é que a conseqüente elevação nas demandas
nutritivas de energia metabolizável em função de maiores tamanhos corporais e
níveis de desempenho, implica em incrementos na demanda dietética por
proteína degradável no rúmen, visando otimizar o crescimento microbiano.
Assim, o total de proteína microbiana obtida quando se permite sua otimização,
somado à fração não degradada da proteína dietética digestível, resultam em um
montante de proteína metabolizável dietética superior à respectiva demanda para
um dado nível de desempenho, gerando assim, absorção de volumes maiores de
aminoácidos além do atendimento das exigências.
Harmeyer & Martens (1980) comentam que tanto a amônia absorvida
através do epitélio ruminal, devido a ingestão de proporções elevadas de
compostos nitrogenados rapidamente degradados no rúmen, quanto o nitrogênio
resultante do ‘desmonte’ de aminoácidos ao nível hepático, provenientes de
elevadas ingestões a partir do intestino delgado, exercem o mesmo efeito
estimulatório sobre a síntese de uréia no fígado, sendo apenas diferente a
velocidade com que este processo se inicia. Isto implica que incrementos no
montante de aminoácidos absorvidos podem levar a sobrecarga hepática e isto
gera um custo metabólico, visto que o processo de reconversão de amônia em
uréia no fígado custa cerca de 12 kcal/g de nitrogênio (Van Soest, 1994). Nas
condições em que o metabolismo de aminoácidos gerar amônia, boa parte deste
custo energético pode vir a ser suprido pelo próprio esqueleto carbônico
derivado do ‘desmonte’ dos aminoácidos (Harris & Lobley, 1991).
Deste modo, tem-se que a busca por equilíbrio entre o aporte de energia
disponível e de compostos nitrogenados ao nível de rúmen, objetivando otimizar
a síntese de proteína microbiana, na tentativa de não comprometer o
desempenho produtivo e reprodutivo e aumentar as exigências energéticas
(Pereira, 1999; NRC, 200), implica em favorecer excedentes de aminoácidos

30
para absorção no intestino delgado, podendo incorrer assim nas mesmas
conseqüências que se intenta evitar ao equilibrar o rúmen.
2.4. Considerações acerca do não atendimento das demandas para otimização da síntese de proteína microbiana
A capacidade dos animais de consumir alimentos em quantidades
suficientes para alcançar suas exigências de mantença e produção é um dos
fatores mais importantes em sistemas de produção, principalmente se estes
forem em grande parte dependentes de volumosos (Sniffen et al., 1993). Illius &
Jessop (1996) afirmaram que a predição do consumo é o ponto critico de todos
os métodos e modelos atuais de formulação de dietas.
Em condições de consumo de forragem de baixo valor nutritivo, uma
séria consideração a ser feita e preconizada por todos os sistemas de alimentação
de ruminantes vigentes, é a busca pelo atendimento das necessidades ruminais
de nitrogênio para assegurar a digestão da porção fibrosa e, por conseguinte, do
consumo. A indigestibilidade da matéria seca é o principal fator que diminui o
consumo de alimentos em ruminantes (Mertens, 1994; Van soest, 1994; Allen,
1996) e esta tem, por sua vez, elevada relação com a proporção de parede
celular, ou mais diretamente, com o conteúdo em fibra insolúvel em detergente
neutro (FDN), sendo este constituinte um importante definidor da qualidade da
dieta. Esta característica é particularmente mais contundente em forrageiras
típicas de regiões tropicais uma vez que as plantas que se desenvolvem sob tais
condições apresentam composição nutricional bastante diferente daquela obtida
em regiões de clima temperado (Van Soest, 1994).
Assim, a necessidade de proteína degradável no rúmen para efetiva
atuação da flora microbiana, conforme recomendação do NRC (1985) está em
torno de 13% dos nutrientes digestíveis totais (NDT). Esse valor também é

31
ratificado pelo sistema AFRC (1993), sendo pouco menor que os valores
descritos para animais em crescimento e lactação. Ainda, segundo o AFRC
(1993), o requerimento de proteína degradável no rúmen é estimado em função
da quantidade de matéria orgânica que é aparentemente degradada no rúmen
(MODR).
Diante da qualidade da natureza da proteína microbiana já salientada, e
considerando o fato de poder se incrementar sua síntese a custos reduzidos, por
intermédio do uso fontes de nitrogênio não-protéico, como, por exemplo a uréia
(Salvador, 2001), tem sido objetivo da alimentação de ruminantes maximizar o
fluxo de proteína microbiana para o intestino delgado, aumentando assim a
eficiência produtiva. Klopfenstein (1996) afirma que os requerimentos de
proteína degradável para crescimento microbiano devem ser atendidos, antes que
uma resposta à proteína de escape possa ser manifestada.
Nesta direção, faz-se necessário quantificar a contribuição da síntese
ruminal de proteína microbiana para um melhor entendimento do processo de
conversão dos nutrientes dietéticos em proteína microbiana e dos fatores que o
afetam. Sniffen & Robinson (1987) enfatizam que a acurácia na predição do
crescimento da população de microrganismos no rúmen e o fluxo da proteína
microbiana para o intestino delgado têm importante papel na predição da
utilização de carboidratos e proteína pelos ruminantes, bem como no
desenvolvimento e aprimoramento de sistemas alimentares destes animais.
Noller et al. (1996) comentam que a parcela da proteína dietética que
escapa à degradação ruminal torna-se mais importante à medida que a produção
animal aumenta, embora um adequado suprimento de proteína degradável seja
necessário para manter a função ruminal, que é essencial para maximizar o
consumo de forragem e a digestibilidade no rúmen. Reis et al. (1996) chamam a
atenção de que a utilização de fontes de proteína de baixa degradabilidade é

32
adequada quando a disponibilidade de forragem é alta, mas com baixo conteúdo
de PB (menor que 7% na base seca).
Interessantemente, uma das primeiras publicações em língua portuguesa
a respeito de zootecnia, datada de 1878, descrevia que em Portugal, os bovinos
até então, utilizados para tração animal e que tinham acesso a farinha de peixe
(fonte protéica de baixa degradabilidade ruminal e alta qualidade), tinham o
apetite estimulado, e segundo ele apresentavam “o estômago fortalecido” para a
digestão das palhas e outras forragens que consumiam, fato que lhes permitia
melhores condições de crescimento e desenvolvimento (Baganha, 1878).
Relatos mais recentes apontam também nesta direção, como é o caso do
trabalho de Gutierrez-Ornelas & Klopfenstein (1994), que trabalhando com
novilhos consumindo palha, observaram ganhos de 400 g/dia quando
suplementados com concentrado contendo 33,4% de proteína de escape (farinha
de sangue e glúten de milho), comparado a um ganho de 260 g/dia do grupo sem
suplementação. Richards et al. (1995) suplementaram bezerros de corte com 8
meses de idade, em regime de pasto, com glúten de milho e comparam os
resultados com outros dois lotes que receberam apenas milho e milho mais uréia.
Descreveram resultados bastante favoráveis para o primeiro lote, com diferença
de 11% no que se refere ao ganho de peso. Porém, não foi observada diferença
significativa na ingestão de forragem. Os autores atribuíram esta resposta ao fato
de não ter havido limitação quanto à ingestão de proteína metabolizável entre os
tratamentos.
Badra (1996), fornecendo 770 g/animal/dia de suplemento com proteína
de escape ruminal para novilhos nelore, em pastagem de Brachiaria brizantha
cv. Marandu vedada para seca, observou resultados animadores, ou seja, 0,825
kg/animal/dia e 0,260 kg/animal/dia para os grupos suplementados e não
suplementados, respectivamente. Da mesma forma, avaliando a estratégia de
suplementação protéica, quanto a degradabilidade ruminal da proteína, Bispo

33
(2000), em condições semelhantes às apresentadas por Badra (1996), observou
ganhos de 0,532; 0,405 e 0,254 kg/animal/dia para animais que consumiam os
suplementos com proteína não degradável no rúmen (PNDR), proteína
degradável no rúmen (PDR) e sem suplementação, respectivamente.
Siqueira (2001) relatou que o não atendimento dos níveis de proteína
degradável no rúmen não comprometeu o desempenho de bovinos para o nível
de ganho estudado (0,670 kg/dia).

34
3 MATERIAL E MÉTODOS
O experimento constou de dois ensaios, sendo um de desempenho e
outro de digestibilidade e balanço nitrogenado. Ambos foram realizados nas
instalações do Setor de Ovinocultura da Universidade Federal de Lavras
(UFLA), situada no município de Lavras - MG. A cidade está situada a 21º14’
de latitude sul, 45º00’ de longitude W.Gr. e altitude de 918 m.
3.1. ENSAIO DE DESEMPENHO
3.1.1. Local, instalações e período de realização
O ensaio de desempenho foi realizado nas dependências do Setor de
Ovinocultura da UFLA, no período compreendido entre 09/05/2005 a
13/09/2005, sendo os primeiros 14 dias (09/05/2005 a 23/05/2005) reservados à
adaptação dos animais às instalações, manejo e ajuste das dietas e os 112 dias
seguintes (24/05/2005 a 13/09/2005) compuseram o período experimental
propriamente.
Para a realização deste ensaio, os animais experimentais foram
instalados em baias individuas com área de 1,3 m2, com o piso recoberto por
maravalha de madeira (“cama”), contendo cada baia um cocho de alimentação e
um bebedouro individuais.

35
3.1.2. Animais e alimentos
Foram utilizadas dezesseis borregas da raça Santa Inês, com peso médio
e erro padrão iniciais de 21,55 ± 1,03 kg (início do período experimental, em
24/05/2005). Dois dias após terem sido instaladas nas baias individuais
(11/05/2005), as borregas foram vermifugadas.
A alimentação dos animais consistiu de feno de capim coastcross
(Cynodon dactylon L. Pers.) moído e ração concentrada. Foi utilizado um
moinho de martelos para proceder à moagem do feno, reduzindo-o a partículas
com tamanho de aproximadamente um (01) cm. Para o preparo dos concentrados
experimentais foram utilizados milho moído, farelo de soja, glutenose, uréia
(como fonte de nitrogênio não-protéico) e suplemento mineral.
A caracterização nutritiva dos ingredientes utilizados na elaboração das
dietas experimentais do ensaio de desempenho encontra-se na Tabela 1.
3.1.3. Elaboração das dietas e manejo alimentar
Após os animais terem sido pesados no inicio do período de adaptação,
estes foram sorteados aos tratamentos e de posse de seus pesos vivos, foram
calculadas as dietas que foram elaboradas segundo os princípios e
recomendações estabelecidos pelo sistema britânico AFRC (1993).
Para fins de estimação das demandas diárias de energia metabolizável,
foi proposto um ganho de peso médio diário da ordem de 160 g/animal, o que
correspondeu, em média, a aproximadamente 0,74% do peso vivo inicial dos
animais.
Para se proceder à elaboração das dietas, os animais tiveram seus
consumos voluntários avaliados durante o período de adaptação (09/05/2005 a
23/05/2005). O objetivo deste procedimento foi auferir a capacidade dos animais

36
TABELA 1. Caracterização dos alimentos utilizados na elaboração das dietas experimentais do ensaio de desempenho.
EM1 MS2 PB2 PDR3 PM4 FDNN2 FDAN
2 Frações (% da PB)2 ; 5 Alimentos
(Mcal/kgMS) (%) (% da MS) A B C
Feno Cynodon 1,53 82,66 7,75 3,28 4,77 81,43 40,59 19,51 67,81 12,68
Milho 3,30 83,05 9,63 4,71 6,98 14,871 2,08 3,91 87,50 8,59
Farelo soja 3,16 83,14 56,38 38,30 41,04 16,02 7,06 9,60 89,87 0,53
Glutenose 3,22 88,05 77,30 34,21 63,34 1,96 0,17 5,05 94,12 0,83
Uréia - 100,00 281,00 281,00 143,31 - - 100 - - 1 Energia metabolizável segundo AFRC (1993) 2 Resultados obtidos através de análises realizadas no laboratório do Depto de Zootecnia - UFLA 3 PDR: proteína degradável no rúmen - concentrações médias obtidas a partir dos valores das frações da PB de cada alimento,
segundo o sistema CNCPS (2003) 4 PM: proteína metabolizável - concentrações médias obtidas a partir dos valores das frações da PB de cada alimento, segundo o
sistema CNCPS (2003) 5 Valores para a fração B resultantes da soma das sub-frações B1, B2 e B3.

37
em ingerir os alimentos que seriam fornecidos, em especial a capacidade de
consumo possível de ser alcançada quanto ao alimento volumoso (feno moído).
As dietas fornecidas durante este período pré-experimental (adaptação) foram
elaboradas seguindo os princípios que regeram a definição dos tratamentos.
Inicialmente foram realizadas ofertas de alimentos considerando ingestões de
matéria seca da ordem de 70 g/kg PV0,75, as quais foram elevadas até que houve
a estabilização da ingestão, que se situou em torno de 90 g MS /kg PV0,75, sendo
então este valor assumido para a elaboração das mesmas dietas (tratamentos) no
período experimental.
As dietas experimentais foram calculadas de forma individual, isto é,
para cada animal foi elaborada a dieta específica ao seu peso vivo, de acordo
com os princípios do tratamento ao qual pertencia. Assim, não foi levada a efeito
a confecção de dietas considerando o peso médio de cada grupo de animais que
compunha as parcelas de cada tratamento. Este procedimento foi assumido para
que se impusesse a cada animal, dentro do tratamento do qual era integrante, a
condição alimentar estritamente exata aos princípios do tratamento aplicado.
Desta forma, buscou-se permitir que a avaliação do desempenho animal
ocorresse em cada animal nas condições dietéticas exatas do tratamento.
A alimentação dos animais se deu em duas refeições diárias, às 08:00 h e
16:00 h. Todo o volumoso (feno moído) foi fornecido de uma única vez, na
refeição da manhã, sendo o concentrado fornecido 50% pela manhã e 50% à
tarde.
Os animais também receberam, adicionado ao concentrado, uma mistura
mineral completa1 (macro e microminerais) em quantidade suficiente para
garantir o consumo de 15 g/animal/dia.
1 Cada 1000g de suplemento continha: P 65g; Ca 120g; Na 152g; Mg 5g; S 25g; Zn 2.000mg; Cu 1.500mg; Fe 1.200mg; I 120mg; Co 80mg; Se 12mg; F (máx) 650mg.

38
Cada animal teve à sua disposição água limpa e fresca em tempo
integral, em baldes plásticos adequados para este propósito.
3.1.4. Tratamentos
Os tratamentos consistiram de quatro dietas cujas elaborações levaram
em consideração condições que tanto priorizassem o atendimento da demanda de
nitrogênio no rúmen, otimizando a síntese de proteína microbiana, bem como,
estabelecendo condições em que o atendimento das exigências nitrogenadas
focasse apenas as demandas dos animais, não necessariamente satisfazendo o
potencial para incrementar a síntese de proteína microbiana ruminal.
Os tratamentos podem ser assim definidos:
Tratamento “A”: Ajuste na oferta de proteína degradável no rúmen em
função do potencial de crescimento microbiano face à
quantidade de energia fermentável advinda da dieta,
segundo o sistema AFRC (1993). Nesta circunstancia,
segundo o próprio sistema, haverá sobra de proteína
metabolizável em relação à respectiva exigência
animal. Utilizaram-se no preparo dos concentrados
apenas fontes protéicas verdadeiras (milho e farelo de
soja);
Tratamento “B”: Atendimento apenas das demandas protéicas dos
animais (atendimento da exigência em proteína
metabolizável), buscando minimizar a disponibilização
de proteína degradável no rúmen. Foi utilizada fonte de
proteína não degradável no rúmen (glutenose);

39
Tratamento “C”: Ajuste da dieta do Tratamento B na direção do
atendimento da premissa assumida para o Tratamento
A (satisfação da demanda de N no rúmen para
otimização da síntese de proteína microbiana),
entretanto, fazendo uso de fonte de nitrogênio não-
protéico (uréia);
Tratamento “D”: Fornecimento de proteína metabolizável nas mesmas
proporções da dieta A, porém, sem lançar mão da
otimização do crescimento microbiano ruminal (sem
ajuste do potencial de crescimento à energia
disponível), se valendo para isto de fornecimento de
maiores proporções na dieta de proteína não degradável
no rúmen (glutenose).
Para a construção das dietas experimentais do ensaio de desempenho, a
partir das equações estabelecidas pelo sistema AFRC (1993), foi efetuado
primeiramente o cálculo das demandas energéticas de cada um dos animais em
função das necessidades de mantença (EMm) e de ganho de peso vivo
estabelecido (160g/dia). Uma vez tendo sido definida também a ingestão de
matéria seca por kg de PV0,75 (90g), obteve-se uma densidade energética
dietética comum a todas as dietas (isoenergéticas) da ordem de 2,51 Mcal/kg MS
(equivalendo a 10,51 MJ/kg MS ou ainda 69,50% NDT). A partir da estimativa
de ingestão de energia metabolizável, foi realizada a estimativa de aporte de
energia metabolizável fermentável (EMfe) e, assim, pôde ser determinado o
potencial de síntese de proteína microbiana a partir da equação proposta pelo
sistema AFRC (1993), conforme (valores expressos em g PB/Mcal de EMfe
inferida):

40
YPBmic = (7 + 6 x (1 - e (-0,35 x L)),
em que L refere-se, segundo o sistema ARC (1980), ao nível de produção e é
resultante da razão entre a ingestão total de energia metabolizável fermentável e
a exigência de energia metabolizável para mantença, conforme:
(L = IEMfe / EMm).
A obtenção dos valores de EMfe está relacionada com as concentrações
de energia metabolizável e de extrato etéreo (EE) dos alimentos e é possibilitada
por meio da equação:
EMfe = EM (em Mcal/kgMS) - (8,37 x EE (em kg/kgMS)).
(valores expressos em Mcal)
O montante de proteína microbiana possível de síntese (Y - expresso em
g/dia), em função do aporte de energia, é obtido pela expressão:
Y = IEMfe x YPBmic.
Posto que o sistema AFRC (1993) assume que a exigência de PDR
efetiva seja igual a 1 (100%) do crescimento microbiano estimado, tem-se que:
Y (em g/dia) = Exigência de PDR (em g/dia).
Na Tabela 2 estão apresentadas as proporções dos ingredientes na
elaboração dos concentrados experimentais e suas composições nutricionais e na
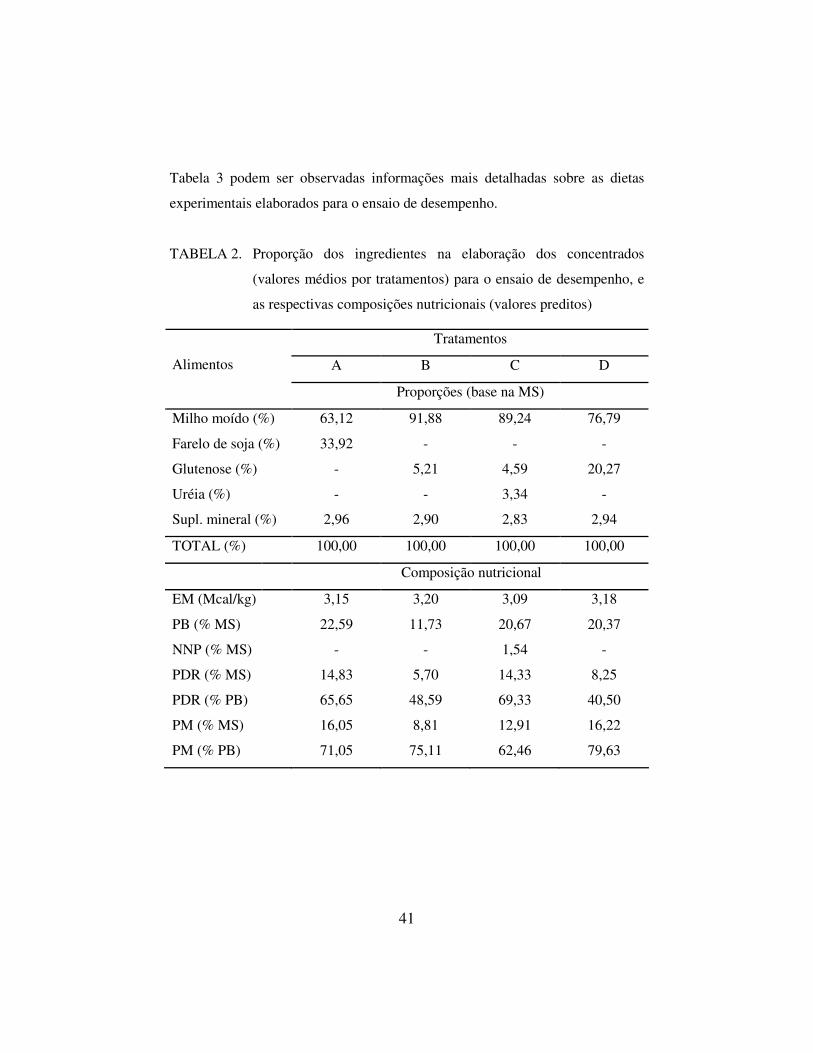
41
Tabela 3 podem ser observadas informações mais detalhadas sobre as dietas
experimentais elaborados para o ensaio de desempenho.
TABELA 2. Proporção dos ingredientes na elaboração dos concentrados
(valores médios por tratamentos) para o ensaio de desempenho, e
as respectivas composições nutricionais (valores preditos)
Tratamentos
A B C D Alimentos
Proporções (base na MS)
Milho moído (%) 63,12 91,88 89,24 76,79
Farelo de soja (%) 33,92 - - -
Glutenose (%) - 5,21 4,59 20,27
Uréia (%) - - 3,34 -
Supl. mineral (%) 2,96 2,90 2,83 2,94
TOTAL (%) 100,00 100,00 100,00 100,00
Composição nutricional
EM (Mcal/kg) 3,15 3,20 3,09 3,18
PB (% MS) 22,59 11,73 20,67 20,37
NNP (% MS) - - 1,54 -
PDR (% MS) 14,83 5,70 14,33 8,25
PDR (% PB) 65,65 48,59 69,33 40,50
PM (% MS) 16,05 8,81 12,91 16,22
PM (% PB) 71,05 75,11 62,46 79,63
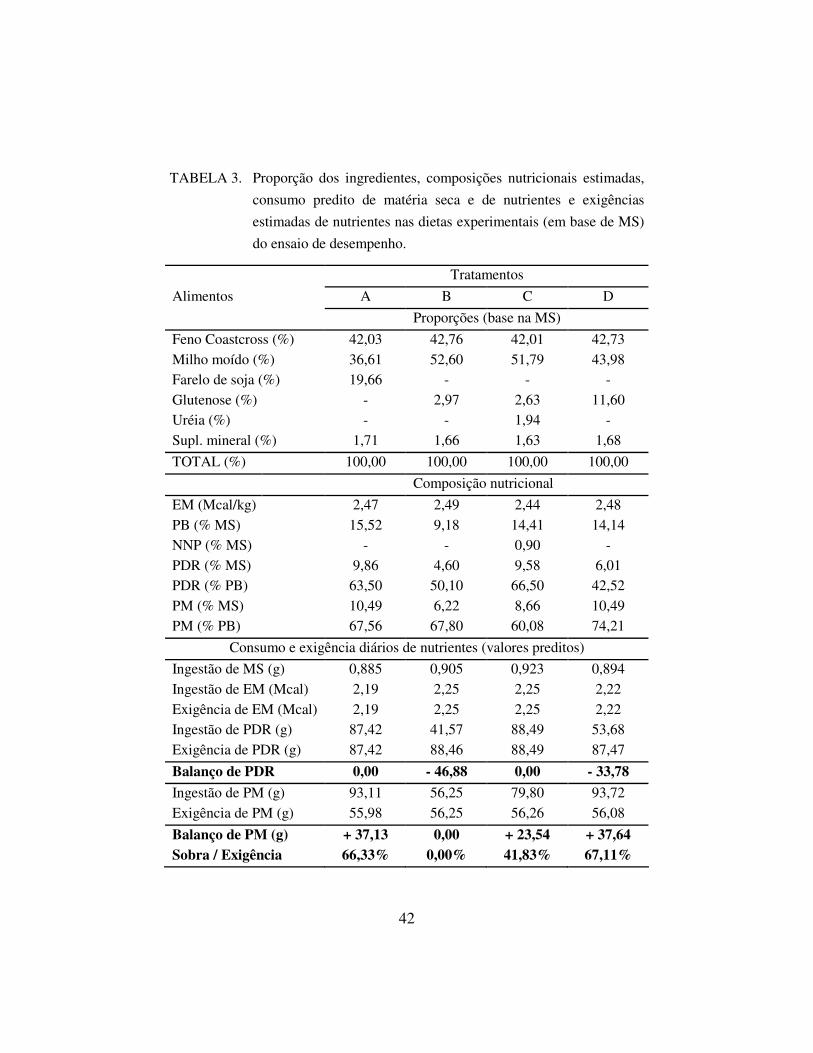
42
TABELA 3. Proporção dos ingredientes, composições nutricionais estimadas,
consumo predito de matéria seca e de nutrientes e exigências
estimadas de nutrientes nas dietas experimentais (em base de MS)
do ensaio de desempenho.
Tratamentos
Alimentos A B C D
Proporções (base na MS)
Feno Coastcross (%) 42,03 42,76 42,01 42,73 Milho moído (%) 36,61 52,60 51,79 43,98 Farelo de soja (%) 19,66 - - - Glutenose (%) - 2,97 2,63 11,60 Uréia (%) - - 1,94 - Supl. mineral (%) 1,71 1,66 1,63 1,68
TOTAL (%) 100,00 100,00 100,00 100,00
Composição nutricional
EM (Mcal/kg) 2,47 2,49 2,44 2,48 PB (% MS) 15,52 9,18 14,41 14,14 NNP (% MS) - - 0,90 - PDR (% MS) 9,86 4,60 9,58 6,01 PDR (% PB) 63,50 50,10 66,50 42,52 PM (% MS) 10,49 6,22 8,66 10,49 PM (% PB) 67,56 67,80 60,08 74,21
Consumo e exigência diários de nutrientes (valores preditos)
Ingestão de MS (g) 0,885 0,905 0,923 0,894 Ingestão de EM (Mcal) 2,19 2,25 2,25 2,22 Exigência de EM (Mcal) 2,19 2,25 2,25 2,22 Ingestão de PDR (g) 87,42 41,57 88,49 53,68 Exigência de PDR (g) 87,42 88,46 88,49 87,47
Balanço de PDR 0,00 - 46,88 0,00 - 33,78
Ingestão de PM (g) 93,11 56,25 79,80 93,72 Exigência de PM (g) 55,98 56,25 56,26 56,08
Balanço de PM (g) + 37,13 0,00 + 23,54 + 37,64 Sobra / Exigência 66,33% 0,00% 41,83% 67,11%

43
3.1.5. Coleta de alimentos e sobras
Os alimentos fornecidos foram amostrados semanalmente e as amostras
foram posteriormente homogeneizadas, formando uma única amostra composta
por alimento. Diariamente, o alimento recusado (sobras) era recolhido antes do
fornecimento da refeição matutina, pesado e amostrado (em torno de 35% da
sobra total).
Todas as amostragens feitas dos alimentos ofertados e das sobras foram
congeladas a -20 ºC para posteriores análises químico-bromatológicas.
3.1.6. Análises bromatológicas
Para a determinação da matéria pré-seca dos alimentos utilizou-se estufa
com circulação forçada de ar com temperatura regulada para 60 ºC por 72 horas.
Após a pré-secagem, as amostras foram moídas em moinho tipo Willey com
peneira de um (01) mm.
Todas as amostras de alimentos e de sobras foram analisadas para
determinação da matéria seca total (MS) e da proteína bruta (PB) segundo as
metodologias descritas por Silva & Queiroz (2002). Também foram analisadas
para determinação das concentrações de fibras insolúveis em detergente neutro
(FDN) e ácido (FDA), segundo os procedimentos recomendados por Goering &
Van Soest (1970) e foi efetuado o fracionamento da PB de todas as amostras
(alimentos e sobras) segundo metodologias descritas por Malafaia & Vieira
(1994) e Reis (2005).

44
3.1.7. Delineamento experimental
Considerando o fato de que o crescimento animal não ocorre de forma
linear, e sim sigmoidal, objetiva-se investigar o efeito dos tratamentos em
momentos distintos do desenvolvimento e crescimento dos animais. Assim, as
variáveis investigadas no ensaio de desempenho o foram considerando duas
fases do desenvolvimento corporal dos animais, a saber, dos 21 aos 28 kg e de
28 a 36 kg de peso vivo.
Deste modo, o ensaio de desempenho ocorreu em um delineamento
experimental inteiramente casualizado, em um esquema fatorial 2x4, sendo duas
fases do crescimento e quatro tratamentos, com quatro repetições por tratamento.
As variáveis analisadas neste ensaio foram o ganho de peso médio
diário, a conversão alimentar e a ingestão de alimentos.
Os dados obtidos para tais variáveis foram submetidos à análise de
variância e as médias foram comparadas pelo teste de Tukey a 5% de
probabilidade, utilizando o software estatístico “SISVAR” (Ferreira, 2000).
O modelo estatístico para este estudo é:
Yijk = µµµµ + Fi + Dj + FDij + eijk ,
em que:
Yijk representa o k-ésimo valor observado correspondente à fase i da
dieta j;
µµµµ é uma constante associada a todas as observações;
Fi é o efeito da fase i de crescimento animal, com i = 1 e 2;
Dj é o efeito da dieta j estabelecido, com j = 1,2, 3 e 4;
FDij é efeito da interação entre a fase i e a dieta j; e

45
eijk é o erro experimental associado a Yijk, com k = 1,2,3 e 4; e que por
hipótese tem distribuição normal com média zero e variância δ2.
Todos os dados auferidos no ensaio passaram inicialmente por teste de
normalidade (teste de Shapiro-Wilk), tendo sido com auxílio do software
Statistical Analysis System (SAS, 1991).
3.2. ENSAIO DE DIGESTIBILIDADE E BALANÇO NITROGENADO
3.2.1. Local, instalações e período de realização
O ensaio de digestibilidade e balanço nitrogenado foi realizado nas
instalações do Setor de Ovinocultura da UFLA, ocorrendo no período
compreendido entre 10/06/2005 a 25/09/2005 (107 dias), sendo que os primeiros
39 dias (10/06/2005 a 19/07/2005) foram destinados a permitir a adaptação dos
animais principalmente às instalações (gaiolas) e manejo. Os 66 dias
subseqüentes foram distribuídos em quatro rodadas (períodos) dentro de um
delineamento em quadrados latinos, com média de 10 dias para adaptação às
dietas (fase pré-experimental) e cinco (5) dias de coleta (fase experimental).
Os animais experimentais foram instalados em gaiolas metálicas
individuais adequadas para ensaios de digestibilidade in vivo, providas de
comedouro e bebedouro individuais.
Cada gaiola metabólica possuía, acoplado ao assoalho, um sistema de
captação de fezes e urina. As fezes foram recolhidas em bandejas plásticas e a
urina foi recolhida em baldes plásticos, adaptados com uma tela separadora,
evitando que as fezes e a urina se misturassem. Em cada balde foram colocados

46
100 mL de solução de H2SO4 a 10% para acidificar a urina, evitando perdas por
volatilização.
3.2.2. Animais e alimentos
Foram utilizadas dois grupos com quatro borregas cada, (fêmeas em
crescimento) da raça Santa Inês, sendo o primeiro grupo composto de animais
com mais idade em relação ao segundo, com peso médio e desvio padrão iniciais
de 29,4 ± 1,13 kg; e o segundo grupo, constituído por animais um pouco mais
jovens e com peso médio e desvio padrão iniciais de 23,8 ± 0,95 kg.
Os animais foram vermifugados com endoparasiticida oral dois dias após
terem sido alojados nas gaiolas metabólicas e a vermifugação foi efetuada
apenas uma única vez em todo o ensaio.
A alimentação dos animais consistiu de feno de capim coastcross
(Cynodon dactylon L. Pers.) moído e ração concentrada. Foi utilizado um
moinho de martelo para proceder à moagem do feno, reduzindo-o a partículas
com tamanho de aproximadamente um (01) cm. Para o preparo dos concentrados
experimentais foram utilizados milho moído, farelo de soja, glutenose, uréia
(como fonte de nitrogênio não-protéico) e suplemento mineral.
A caracterização nutritiva dos alimentos utilizados na elaboração das
dietas experimentais do ensaio de digestibilidade encontra-se apresentada na
Tabela 4.
3.2.3. Elaboração das dietas e manejo alimentar
A primeira pesagem dos animais se deu em 10/06/2005, quando então os
animais foram sorteados aos tratamentos e instalados nas gaiolas metabólicas.
De posse dos valores de seus pesos vivos, foram calculadas as dietas, elaboradas

47
TABELA 4. Caracterização dos alimentos utilizados na elaboração das dietas experimentais do ensaio de digestibilidade
e balanço nitrogenado.
EM1 MS2 PB2 PDR3 PM4 FDNN2 FDAN
2 Alimentos
(Mcal/kgMS) (%) (% da MS)
Feno Cynodon 1,53 82,59 8,02 3,39 4,94 76,91 47,22
Milho 3,30 79,89 9,33 4,56 6,76 14,24 3,90
Farelo soja 3,16 84,71 56,01 38,05 40,77 15,98 8,73
Glutenose 3,22 85,49 77,23 34,17 63,28 1,46 0,39
Uréia - 98,00 281,00 281,00 143,31 - - 1 Energia metabolizável segundo AFRC (1993) 2 Resultados obtidos através de análises realizadas no laboratório do Depto de Zootecnia - UFLA 3 PDR: proteína degradável no rúmen - concentrações médias obtidas a partir dos valores das frações da PB de cada
alimento, segundo o sistema CNCPS (2003) 4 PM: proteína metabolizável - concentrações médias obtidas a partir dos valores das frações da PB de cada alimento,
segundo o sistema CNCPS (2003)

48
segundo os princípios e recomendações estabelecidos pelo sistema AFRC
(1993).
O procedimento na elaboração das dietas foi semelhante ao efetivado no
ensaio de desempenho, ou seja, foram estimadas inicialmente as demandas
diárias de energia metabolizável, sendo considerado um ganho de peso vivo de
160 g/dia, correspondendo, em média, a aproximadamente 0,54% do peso vivo
para o grupo de borregas mais pesadas, e 0,67% do peso vivo dos animais mais
leves, no início do ensaio.
Para se proceder à elaboração das dietas, os animais tiveram seus
consumos voluntários avaliados durante o período de adaptação (de 10 a 21/06
de 2005 - 11 dias). O objetivo deste procedimento foi auferir a capacidade dos
animais em ingerir os alimentos que seriam fornecidos, em especial a capacidade
de consumo possível de ser obtida quanto ao alimento volumoso (feno moído).
As dietas fornecidas durante este período pré-experimental foram elaboradas
segundo os princípios que regiram a definição dos tratamentos. Inicialmente
foram realizadas ofertas de alimentos considerando ingestões de matéria seca da
ordem de 75g/kg PV0,75, as quais foram elevadas até que houvesse a
estabilização da ingestão, que se situou em 90 g MS /kg PV0,75, sendo então este
valor o assumido para a elaboração das mesmas dietas (tratamentos) no período
experimental. Em 21 de junho de 2005 os animais foram novamente pesados e
então foi levado a efeito o mesmo procedimento, ou seja, auferiram-se os pesos
vivos médios de cada animal e elaboraram-se as dietas experimentais com base
no consumo de 90 g/kg PV0,75, sendo daquele momento em diante considerado
período pré-experimental.
Da mesma forma como ocorreu no ensaio de desempenho, as dietas
experimentais foram calculadas de forma individual, isto é, para cada animal foi
elaborada a dieta específica ao seu peso vivo, de acordo com os princípios do
tratamento ao qual pertencia.

49
Os animais receberam alimentação em duas refeições diárias, às 08:00 h
e 16:00 h. Cada animal recebeu todo o volumoso (feno moído) pela manhã, de
uma única vez, sendo o concentrado fornecido 50% em cada refeição.
Cada animal teve à sua disposição água limpa e fresca ad libitum,
disponibilizada em baldes plásticos adequados para este fim. O suplemento
mineral utilizado no preparo dos concentrados apresentava tanto macro como
microminerais2.
3.2.4. Tratamentos
Os tratamentos consistiram de quatro dietas elaboradas segundo o
efetuado no ensaio de desempenho. Conforme já descrito, as dietas levaram em
consideração condições que tanto priorizassem o atendimento da demanda de
nitrogênio no rúmen, otimizando a síntese de proteína microbiana, bem como,
condições em que o atendimento das exigências nitrogenadas focasse apenas as
demandas dos animais, não necessariamente satisfazendo o potencial para
incrementar a síntese de proteína microbiana ruminal. A definição mais
detalhada de cada tratamento pode ser verificada no tópico 3.1.4.
Da mesma forma como já descrito no ensaio de desempenho, utilizando
as equações e recomendações do sistema AFRC (1993) foi efetuada a construção
das dietas experimentais do ensaio de digestibilidade, sendo primeiramente
realizado o cálculo das demandas energéticas de cada um dos animais em função
das necessidades de mantença (EMm) e de ganho de peso vivo estabelecido
(160g/dia). Uma vez tendo sido definida também a ingestão de matéria seca por
kg de PV0,75 (90g), obteve-se uma densidade energética dietética comum a todas
as dietas (isoenergéticas) da ordem de 2,55 Mcal/kg MS (equivalendo a 10,67
2 Cada 1000g de suplemento continha: P 65g; Ca 120g; Na 152g; Mg 5g; S 25g; Zn 2.000mg; Cu 1.500mg; Fe 1.200mg; I 120mg; Co 80mg; Se 12mg; F (máx) 650mg.

50
MJ/kg MS ou ainda 70,5% NDT). A partir da estimativa de ingestão de energia
metabolizável, foi realizada a estimativa de aporte de energia metabolizável
fermentável (EMfe) e, assim, pôde ser determinado o potencial de síntese de
proteína microbiana a partir da equação proposta pelo sistema AFRC (1993),
conforme (valores expressos em g PB/Mcal de EMfe inferida):
YPBmic = (7 + 6 x (1 - e (-0,35 x L)),
em que L refere-se, segundo o sistema ARC (1980), ao nível de produção e é
resultante da razão entre a ingestão total de energia metabolizável fermentável e
a exigência de energia metabolizável para mantença, conforme:
(L = IEMfe / EMm).
A obtenção dos valores de EMfe está relacionada com as concentrações
de energia metabolizável e de extrato etéreo (EE) dos alimentos e é possibilitada
por meio da equação:
EMfe = EM (em Mcal/kgMS) - (8,37 x EE (em kg/kgMS)).
(valores expressos em Mcal)
O montante de proteína microbiana possível de síntese (Y - expresso em
g/dia), em função do aporte de energia, é obtido pela expressão:
Y = IEMfe x YPBmic.
Posto que o sistema AFRC (1993) assume que a exigência de PDR
efetiva seja igual a 1 (100%) do crescimento microbiano estimado, tem-se que:
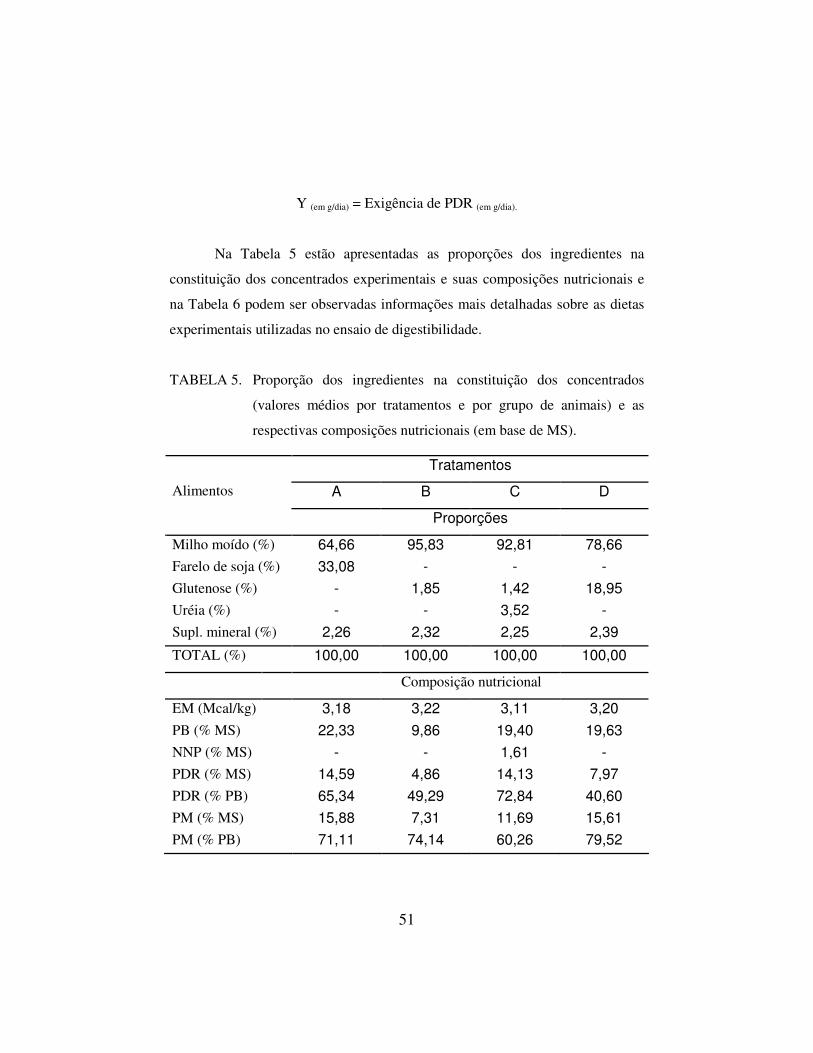
51
Y (em g/dia) = Exigência de PDR (em g/dia).
Na Tabela 5 estão apresentadas as proporções dos ingredientes na
constituição dos concentrados experimentais e suas composições nutricionais e
na Tabela 6 podem ser observadas informações mais detalhadas sobre as dietas
experimentais utilizadas no ensaio de digestibilidade.
TABELA 5. Proporção dos ingredientes na constituição dos concentrados
(valores médios por tratamentos e por grupo de animais) e as
respectivas composições nutricionais (em base de MS).
Tratamentos
A B C D Alimentos
Proporções
Milho moído (%) 64,66 95,83 92,81 78,66
Farelo de soja (%) 33,08 - - -
Glutenose (%) - 1,85 1,42 18,95
Uréia (%) - - 3,52 -
Supl. mineral (%) 2,26 2,32 2,25 2,39
TOTAL (%) 100,00 100,00 100,00 100,00
Composição nutricional
EM (Mcal/kg) 3,18 3,22 3,11 3,20
PB (% MS) 22,33 9,86 19,40 19,63
NNP (% MS) - - 1,61 -
PDR (% MS) 14,59 4,86 14,13 7,97
PDR (% PB) 65,34 49,29 72,84 40,60
PM (% MS) 15,88 7,31 11,69 15,61
PM (% PB) 71,11 74,14 60,26 79,52

52
TABELA 6. Proporção dos ingredientes, composições nutricionais estimadas, consumo predito de matéria seca e de nutrientes e exigências estimadas de nutrientes nas dietas experimentais (valores médios por tratamentos e por grupo de animais) do ensaio de digestibilidade (base na MS).
Tratamentos
Alimentos A B C D
Proporções
Feno Coastcross (%) 37,05 38,63 37,92 38,88 Milho moído (%) 40,70 58,81 57,61 48,08 Farelo de soja (%) 20,83 - - - Glutenose (%) - 1,14 0,88 11,58 Uréia (%) - - 2,19 - Supl. mineral (%) 1,42 1,42 1,40 1,46
TOTAL (%) 100,00 100,00 100,00 100,00
Composição nutricional
EM (Mcal/kg) 2,57 2,57 2,51 2,55 PB (% MS) 16,20 8,28 14,23 14,25 NNP (% MS) - - 1,00 - PDR (% MS) 10,30 4,19 9,91 6,04 PDR (% PB) 63,55 50,54 69,67 42,41 PM (% MS) 11,04 5,56 8,31 10,63 PM (% PB) 68,12 67,09 58,42 74,61
Consumo e exigência diários de nutrientes (valores preditos)
Ingestão de MS (g) 1,054 1,056 1,074 1,027 Ingestão de EM (Mcal) 2,71 2,71 2,70 2,62 Exigência de EM (Mcal) 2,71 2,71 2,70 2,62 Ingestão de PDR (g) 108,51 44,19 106,48 62,05 Exigência de PDR (g) 108,51 107,01 106,48 103,99
Balanço de PDR 0,00 - 62,82 0,00 - 41,95
Ingestão de PM (g) 116,35 58,62 89,21 109,21 Exigência de PM (g) 58,60 58,62 58,56 58,17
Balanço de PM (g) + 57,75 0,00 + 30,65 + 51,04 Sobra / Exigência 98,56% 0,00% 52,35% 87,75%

53
3.2.5. Coleta de alimentos, sobras, fezes e urina
Os alimentos fornecidos foram amostrados todas as semanas e as
amostras foram posteriormente homogeneizadas, formando uma única amostra
composta por alimento.
O alimento recusado (sobras) foi recolhido diariamente, antes do
fornecimento da refeição matutina, pesado e amostrado (em torno de 35% da
sobra total).
As fezes e a urina foram recolhidas diariamente pela manhã. A coleta de
fezes foi total, seus pesos foram anotados, estas foram amostradas (20% do total
diário) e, então, acondicionadas em sacos plásticos devidamente identificados.
A urina produzida por cada animal teve seu volume (mL) também
registrado e foram efetuados amostragem (10% do volume diário) e
acondicionamento das amostras em vidro âmbar devidamente identificado para
cada animal.
Todas as amostragens feitas do alimento ofertado, das sobras, das fezes e
da urina e, após o seu devido acondicionamento para armazenagem (sacos
plásticos ou vidros), foram congeladas a -20 ºC para posteriores análises
químico-bromatológicas.
3.2.6. Análises químico-bromatológicas
Para a determinação da matéria pré-seca dos alimentos (feno e
concentrados), sobras e fezes, utilizou-se estufa com circulação forçada de ar
com temperatura regulada para 60 ºC por 72 horas. Após a pré-secagem, as
amostras foram moídas em moinho tipo Willey com peneira de 1 mm.
Para a análise das urinas colhidas durante as fases de coleta, que se
encontravam armazenadas em frascos de vidro âmbar, foi realizado o

54
descongelamento destas, as quais foram, então, homogeneizadas por agitação e
filtradas com uso de algodão hidrófilo e funil. As amostras foram analisadas
para N total.
As amostras dos ingredientes utilizados no preparo dos concentrados
experimentais, do feno, das sobras e das fezes foram analisadas para MS e PB
segundo as metodologias descritas por Souza & Queiroz (2002). Também foram
analisadas para a determinação das concentrações de FDN e FDA, segundo os
procedimentos recomendados por Goering & Van Soest (1970).
3.2.7. Cálculos da digestibilidade e do balanço de N
Os valores de digestibilidade aparente (DIG) dos nutrientes foram
obtidos pela fórmula proposta por Coelho da Silva & Leão (1979), apresentada a
seguir:
DIG = [(ING x %ING) - (SOB x %SOB)] - (FEZ x %FEZ) x 100
(ING x %ING) - (SOB x %SOB)
em que:
ING = quantidade de alimento consumido;
%ING = teor do nutriente no alimento fornecido;
SOB = quantidade de sobras retiradas;
%SOB = teor do nutriente nas sobras;
FEZ = quantidade de fezes coletadas;
%FEZ = teor do nutriente nas fezes.

55
O balanço de N é obtido subtraindo-se o total de N excretado nas fezes e
na urina do total de N ingerido, representando o total de N que efetivamente
ficou retido no organismo animal, conforme:
N RETIDO = (N Fornecido - N Sobras) - (N Fezes + N Urina).
Os valores obtidos a partir da subtração do total de N ingerido, menos o
N contido nas fezes, referem-se ao N absorvido, conforme:
N ABSORVIDO = (N Fornecido - N Sobras) - N Fezes.
Os valores de N (ingerido e excretado nas fezes e urina) foram obtidos a
partir das análises químicas realizadas, conforme já mencionado.
3.2.8. Delineamento experimental
O delineamento experimental utilizado foi o de quadrados latinos, em
um esquema change-over, tendo sido utilizado dois (2) quadrados 4x4, com
quatro animais por quadrado e quatro tratamentos (dietas). Cada quadrado foi
composto por um dentre os dois grupamentos de animais (diferentes em relação
aos pesos vivos).
O ensaio objetivou o estudo do consumo e digestibilidade aparente da
MS, MO, PB, FDNN e FDAN e também do balanço de nitrogênio, o que resultou
em um total de trinta e duas parcelas experimentais ao serem considerados os
dois quadrados.
Os dados obtidos para tais variáveis foram submetidos à análise de
variância e as médias foram comparadas pelo teste de Tukey a 5% de
probabilidade, utilizando o software estatístico “SISVAR” (Ferreira, 2000).

56
O modelo estatístico para o estudo de consumos, digestibilidade dos
nutrientes mencionados e balanço nitrogenado foi:
Yijklr = µ + Qi + P(Qi)j + A(Qi)k + Tl + TQil + eijklr ,
em que:
Yijklr representa o r-ésimo valor observado correspondente ao quadradro
i no período j pelo animal k no tratamento (dieta) l;
µ é uma constante associada a todas as observações;
Qi é o efeito do quadrado latino i, com i = 1 e 2;
P(Qi)j é o efeito do período j dentro do quadrado latino i, com j = 1, 2, 3
e 4;
A(Qi)k é efeito do animal k dentro do quadrado latino i, com k variando
de 1 a 8;
eijklr é o erro experimental associado a Yijklr, que por hipótese tem
distribuição normal com média zero e variância δ2.

57
4 RESULTADOS E DISCUSSÃO
Os resultados obtidos para todas as variáveis analisadas, tanto no ensaio
de desempenho como no de digestibilidade e balanço nitrogenado, foram
submetidos ao teste de normalidade (Shapiro-Wilk) e foi confirmada a
distribuição normal para os valores obtidos.
4.1. ENSAIO DE DESEMPENHO
No ensaio de desempenho foram avaliadas as ingestões de matéria seca
(MS), de fibra insolúvel em detergente neutro corrigida para nitrogênio (FDNN),
de fibra insolúvel em detergente ácido corrigida para nitrogênio (FDAN), de
proteína bruta (PB), das frações da PB segundo o sistema de Cornell, da proteína
degradável no rúmen e da proteína metabolizável. Foram também analisados o
ganho médio de peso diário e a conversão alimentar.
Na Tabela 7 observam-se diferenças na participação dos alimentos nas
dietas em relação ao proposto (Tabela 3), que se devem às variações ocorridas
nos valores das concentrações de matéria seca e nutrientes efetivamente
alcançados nas análises dos alimentos utilizados. Pode ser constatado que, em
função destas variações, as concentrações nutritivas também sofreram pequenas
variações.
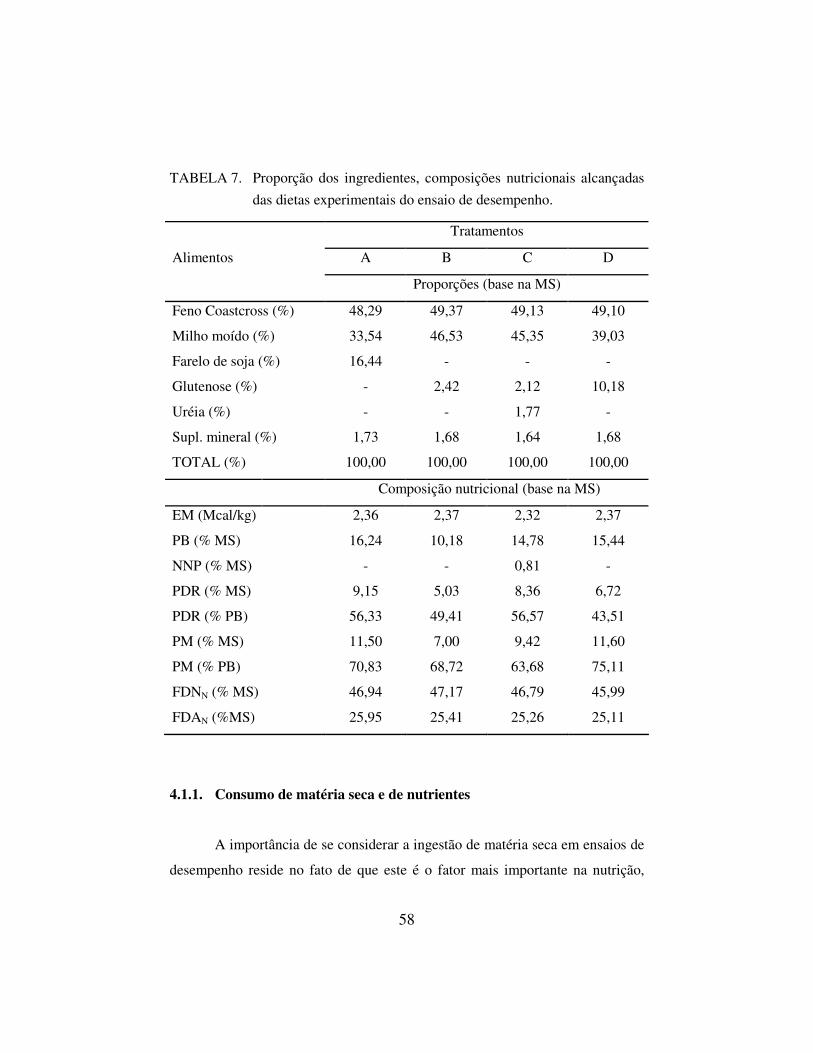
58
TABELA 7. Proporção dos ingredientes, composições nutricionais alcançadas
das dietas experimentais do ensaio de desempenho.
Tratamentos
Alimentos A B C D
Proporções (base na MS)
Feno Coastcross (%) 48,29 49,37 49,13 49,10
Milho moído (%) 33,54 46,53 45,35 39,03
Farelo de soja (%) 16,44 - - -
Glutenose (%) - 2,42 2,12 10,18
Uréia (%) - - 1,77 -
Supl. mineral (%) 1,73 1,68 1,64 1,68
TOTAL (%) 100,00 100,00 100,00 100,00
Composição nutricional (base na MS)
EM (Mcal/kg) 2,36 2,37 2,32 2,37
PB (% MS) 16,24 10,18 14,78 15,44
NNP (% MS) - - 0,81 -
PDR (% MS) 9,15 5,03 8,36 6,72
PDR (% PB) 56,33 49,41 56,57 43,51
PM (% MS) 11,50 7,00 9,42 11,60
PM (% PB) 70,83 68,72 63,68 75,11
FDNN (% MS) 46,94 47,17 46,79 45,99
FDAN (%MS) 25,95 25,41 25,26 25,11
4.1.1. Consumo de matéria seca e de nutrientes
A importância de se considerar a ingestão de matéria seca em ensaios de
desempenho reside no fato de que este é o fator mais importante na nutrição,

59
pois estabelece as quantidades de nutrientes disponíveis para a saúde e produção
animal (NRC, 2001).
Na Tabela 8 podem ser visualizados os resultados concernentes aos
consumos de matéria seca. Os valores estão apresentados em termos de
ingestões totais (g/dia), em relação ao peso vivo (%PV) ou ainda em relação ao
peso metabólico dos animais (g/kg PV0,75). Não houve efeito de entre interação
tratamento e fase (P>0,05).
TABELA 8. Valores médios1 de ingestão de matéria seca, apresentados em
termos do total (IMST), em relação ao peso vivo (IMS%PV) e em
relação ao peso metabólico (IMSPV0,75), em função dos
tratamentos e das fases consideradas para ensaio de desempenho.
IMST IMS%PV IMSPV0,75 Tratamentos
(g/dia) (%) (g/kg PV0,75)
A 951,65 3,50 a 79,76
B 896,72 3,22 ab 73,80
C 885,51 3,15 ab 72,47
D 891,60 3,12 b 71,94
Estudos entre as fases do ensaio 2
1ª Fase 817,81 b 3,31 73,68
2ª Fase 994,94 a 3,19 75,31
CV (%) 13,15 7,97 8,65
1 Médias de tratamentos seguidas de mesma letra não diferem entre si pelo teste Tukey com nível de significância de 5%. 2 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.

60
Embora a ingestão verificada para o tratamento A, em termos de valores
totais (g/dia), tenha sido superior numericamente aos resultados dos demais
tratamentos, não diferiu estatisticamente destes outros. Valadares et al. (1997a)
comentam que avaliar ingestão de matéria seca ou outro nutriente quando
expressa em termos totais não parece adequado por dificultar comparações entre
experimentos e mesmo dentro de experimentos em face da variação do peso vivo
entre os animais. Geralmente, ocorre aumento de consumo com o aumento de
peso corporal, o que indica ser mais conveniente expressar consumos em função
do peso corporal.
Apesar de terem sido identificadas diferenças significativas para a
ingestão de matéria seca quando considerada em função da porcentagem de peso
vivo, a outra forma de expressar o consumo em relação ao peso vivo (g/kg
PV0,75) não apontou diferenças entre os tratamentos (P>0,05). A comparação de
médias da IMS%PV efetuada por testes estatísticos que não sobrepõem médias,
como o teste de Scott-Knott, apresenta a média do tratamento A (3,50%) como
superior (P<0,05) às demais, sendo todas estas iguais entre si (P>0,05).
A diferença verificada entre as fases, considerada do ponto de vista da
ingestão total (g/cab/dia) é perfeitamente compreensível posto que as
capacidades volumétricas dos pré-estômagos sabidamente aumentam com a
majoração do tamanho animal - conseqüência do próprio crescimento (Kolb,
1984). Assim, uma vez que na segunda fase os animais encontravam-se maiores,
portanto aptos a conseguir ingestões maiores, isto de fato se deu. Esta
ocorrência, no entanto, não é verifica quando se ‘corrige’ o efeito do tamanho
animal, ou seja, quando se avalia a ingestão em termos relativos ao peso vivo
(Valadares et al., 1997a).
Ao se avaliar a ingestão da matéria seca em termos do peso metabólico,
verifica-se que os valores alcançados (média geral em torno de 74,50 g/kg
PV0,75) não estão tão próximos do considerado por ocasião da elaboração das

61
dietas (90 g/kg PV0,75). Este aspecto deve ser considerado pois é possível que,
dentro das recomendações do sistema AFRC (1993) adotado, as dietas
elaboradas talvez não tenham sido efetivamente aplicadas aos animais,
particularmente no que concerne ao atendimento energético. Este aspecto será
discutido mais adequadamente em oportunidade adiante.
O efeito positivo do uso suplementar dos concentrados sobre as dietas
pode ser evidenciado ao se comparar os níveis de ingestão obtidos, em termos de
percentual do peso vivo ou em relação ao peso metabólico, com os dados de
Rodrigues et al. (1998) e Moreira et al. (2001), que alimentaram ovinos
exclusivamente com feno de capim coastcross e auferiram ingestões de matéria
seca da ordem de 2,1 e 2% do peso vivo ou 50,65 e 51,64 g/kg PV0,75,
respectivamente. Mesmo em circunstâncias em que dietas à base de feno de
coastcross foram suplementadas exclusivamente com fonte de nitrogênio-não
protéico (amiréia), permitindo que as concentrações dietéticas finais de PB
estivessem por volta de 13%, não permitiram que ovinos alcançassem patamares
de ingestões verificadas no presente estudo (Salvador et al., 2004).
Na estimativa do consumo, Mertens (1992) sugere que limitações
relativas ao animal, ao alimento e às condições do manejo alimentar sejam
consideradas. Sob o aspecto do animal, fatores de ordem física, relacionados à
capacidade de distensão do próprio rúmen, têm papel importante na regulação do
consumo e neste sentido, o pesquisador salienta que a fibra, melhor representada
pela FDN, está intimamente ligada a este aspecto e à fermentação ruminal. Por
conseguinte, a digestibilidade e o valor energético da dieta também se
relacionam à sua porção fibrosa, uma vez que este componente é o que melhor
representa os constituintes de baixa degradação. Tem-se então que, dietas que
possam proporcionar uma mais ampla fermentação da fração fibrosa podem ter
como resultado o incremento da ingestão (Conrad, 1966; Mertens, 1994; Van
Soest, 1994; Allen, 1996). Roseler et al. (1993) comentam que a concentração e

62
a qualidade da proteína da dieta podem modificar o consumo pelos animais
ruminantes, alterando tanto o mecanismo físico, como o fisiológico. Reduções
no teor de PB da dieta para níveis abaixo de 12%, ou na disponibilidade de
nitrogênio, podem reduzir a digestão da fibra e conseqüentemente, restringir o
consumo.
Considerando que as dietas deste trabalho apresentavam concentrações
energéticas e de FDNN semelhantes entre si (em torno de 2,35 Mcal/kg e
46,72%, respectivamente), seria esperado que os tratamentos B e D pudessem
resultar em ingestões de matéria seca menores, uma vez que estes não priorizam
o atendimento das demandas nitrogenadas da população microbiana, por não
oferecerem proporções de N disponível que promovessem seu crescimento e, no
caso específico do tratamento B, os valores de PB dietéticos foram inferiores a
12%, como preconizado por Roseler et al. (1993). Wilson & Kennedy (1996)
sustentam que o não atendimento dos requerimentos microbianos em nitrogênio
repercute na limitação de seu crescimento e depressão da digestão da parede
celular, resultando finalmente em diminuição do consumo. Deste modo, tem-se
que o fornecimento de nitrogênio disponível ao pleno desenvolvimento da
população microbiana melhora a digestão da matéria orgânica dietética,
resultando em incremento na taxa de passagem e permitindo aos animais
consumirem mais alimento (Romney & Gill, 2000). No entanto, no presente
trabalho estas afirmações não se consolidaram verdadeiras, posto que não se
verificaram diferenças entre as ingestões de matéria seca a despeito de se buscar
ou não o atendimento da demanda do rúmen por nitrogênio disponível.
Por outro lado, Ørskov (1992) afirmou que o nitrogênio suplementar
pode não influi no consumo de matéria seca quando a dieta apresentar teores de
proteína superiores a 12%. Este fato foi verificado no trabalho de Zundt et al.
(2002), que alimentaram cordeiros e cordeiras ‘three-cross’ (½ Texel + ¼
Bergamácia + ¼ Corriedale), pesando em média 30 kg, com níveis crescentes de

63
proteína bruta na dieta (12 a 24%) e não observaram incrementos na ingestão
alimentar. No presente estudo, os tratamentos A e D apresentaram concentrações
protéicas similares (16,24 e 15,44%, respectivamente) e, embora entre os dois,
somente o tratamento A tenha proporcionado a satisfação das demandas em
proteína degradável (PDR), o fato dos níveis de PB serem semelhantes
poderiam, de acordo com a declaração de Ørskov, explicar a igualdade entre os
consumos de MS.
Existem, entretanto, dados na literatura que não assentem com a
afirmação de Ørskov. Ortiz et al. (2005), alimentando cordeiros machos
lactentes da raça Suffolk com dietas contendo 15, 20 ou 25% de PB em sistema
de creep feeding (elaboradas apenas com milho, farelo de trigo e farelo de soja)
observaram elevações no consumo de MS de quase o dobro (0,197 kg/cab/dia
para 15%PB a 0,386 kg/cab/dia, para 25%PB). Outras pesquisas também
constataram o efeito de incrementos na ingestão de matéria seca com ovinos
(Meherez & Ørskov, 1978; Viera et al., 1980; Huston et al. 1988; Fluharty &
McClure, 1997) e também com bovinos (Valadares et., 1997a; Ítavo et al., 2002;
Rennó, 2003; Cavalcante et al. 2005; Obeid et al., 2006).
Ainda no aspecto relativo à concentração de PB e a ingestão de matéria
seca, salienta-se a afirmação de que dietas com concentrações de proteína bruta
inferiores a 7% têm suas ingestões limitadas por aportes insuficientes de
nitrogênio no rúmen (Martin et al., 1981). Esta circunstância não se dá neste
trabalho uma vez que todos os tratamentos apresentaram PB em concentrações
superiores a 10%. Porém, deve-se atentar para o fato de que a oferta efetiva de N
no rúmen refere-se eminentemente à concentração de proteína degradável (PDR)
e sob este aspecto, o tratamento B contou com níveis de PDR inferiores a 6%, e
ainda assim este tratamento não promoveu valores de ingestões de matéria seca
menores que os demais.

64
Morrison & Mackie (1996), discutindo as rotas bioquímico-enzimáticas
da assimilação microbiana da amônia no rúmen, sugerem a possibilidade de que
as concentrações ótimas de amônia para permitir a digestão ruminal da fibra e as
necessárias para otimizar a síntese de proteína microbiana possam não se situar
nos mesmos patamares. Este fato pode explicar os resultados de Araújo et al.
(1998) e de Signoretti et al. (1999), que alimentaram bezerros com proporções
crescentes de volumoso nas dietas (de 10 a 90% e 10 a 55%, respectivamente),
reduzindo os teores dietéticos de NDT em termos totais entre 12 a 20 pontos
percentuais, respectivamente, e mesmo assim não verificaram redução na
digestibilidade da fração fibrosa (FDN e FDA), apesar de se observar
diminuição da ingestão de matéria seca (em termos de percentagem do peso
vivo) para o primeiro autor e manutenção do nível de consumo de MS, para o
segundo autor. Ou seja, condições de dietas menos energéticas embora
isonitrogenadas vieram a promover a pequena redução ou mesmo manutenção
da ingestão de matéria seca, mas não interferiram na digestão microbiana da
fibra.
Um aspecto a se considerar na avaliação das ingestões de matéria seca
das dietas, diz respeito à inclusão de uréia nas dietas, que no presente estudo se
dá somente no tratamento C. Um dos trabalhos pioneiros na investigação da
influência da presença da uréia nas dietas de ruminantes foi o de Huber & Cook
(1972), no qual se dá a inclusão desta fonte nitrogenada na ordem de 1 a 3% da
MS do concentrado e sob três diferentes maneiras de administração (oral,
ruminal e abomasal). Os autores verificaram efeito depressivo no consumo de
alimento quando a inclusão na dieta se deu em níveis mais elevados e quando se
processou por via oral, considerando que o sabor indesejável da uréia foi o
aspecto preponderante para promover a redução de consumo. Este aspecto foi
também salientado por Borges (1999) e Silva et al. (2001).

65
Huber & Kung (1981) apontaram a redução nas ingestões de alimentos
quando a concentração de uréia na MS da dieta se situar acima de 1,5%, mesmo
em condições de animais adaptados fisiologicamente à presença deste composto
nas dietas.
No presente estudo, a concentração uréia na dieta do tratamento C
situou-se acima do nível preconizado como limite (1,5%), e, embora os valores
de ingestão observados para este tratamento tenham sido ligeiramente inferiores
em termos numéricos, não se diferenciaram (P>0,05) dos demais tratamentos.
Na realidade, encontram-se na literatura estudos que levaram a efeito
substituições parciais de farelos proteinosos por uréia na dieta total, obtendo
assim crescimento da participação da uréia na composição das dietas, sem que
fossem constatadas reduções significativas na ingestão de matéria seca e de
outros nutrientes em ovinos (Siqueira et al., 1981; Lavezzo et al., 1996;
Fagundes Neto et al., 2001), em vacas lactantes (Guidi, 1999; Imaizumi, 2000;
Carmo et al. 2001) ou em bovinos de corte (Ferreira et al., 1996; Magalhães et
al., 2002; Souza et al., 2002; Rennó et al., 2005; Paixão et al., 2006). Ressalta-se
ainda o fato de que ao se comparar dietas isonitrogenadas cujos concentrados
sejam elaborados com milho (cuja PB é considerada eminentemente de escape
[entre 55 a 60% - Sindt et al., 1993]) e acrescidas de farelo de soja ou uréia,
similares aos tratamentos A e C da presente pesquisa, não têm apresentado
diferenças quanto à ingestão de MS (Clark et al., 1970; Greathouse et al., 1974 e
Plegge et al., 1983, citados por Shain et al., 1998).
Apesar de não ser objetivo de investigação deste trabalho verificar as
influências e resultados advindos do uso de inclusões crescentes de uréia ou
averiguar os efeitos decorrentes do uso de concentrações maiores de PB em
dietas de ruminantes, a abordagem de pesquisas envolvidas nestes assuntos
decorre do fato de que inclusões crescentes de uréia (ou de qualquer outra fonte
de NNP) ou ainda de que o incremento no teor de PB das dietas, resulta em

66
modificações de aportes de N degradável no rúmen e de montantes diferentes de
proteína de origem microbiana e dietética não degradada no contexto ruminal,
que atingem o intestino delgado (proteína metabolizável de ‘naturezas’ distintas,
quanto ao perfil de aminoácidos). Ou sejam, fornecimento de quantidades
variáveis de proteína ou de NNP nas dietas podem vir a estimular mais ou
menos a síntese de proteína microbiana, conseqüentemente, gerar saldos de
proteína metabolizável (em função da proteína microbiana). Mas, em todas estas
circunstâncias, o que normalmente se verifica são condições de favorecimento
da síntese de proteína microbiana.
Nos trabalhos de Shain et al. (1998) e de Franco (2001), condições
relativas ao balanço entre o consumo e a satisfação das demandas em PDR e em
proteína metabolizável (PM) foram consideradas, como se dá no presente
estudo.
No trabalho de Shain et al. (1998), os autores trabalharam com novilhos
cruzados em terminação (em torno de 350 kg de peso vivo) recebendo dietas
ricas em concentrados, sendo que 79,5% da dieta era constituída de milho
laminado (‘dry-rolled corn’) e a relação volumoso:concentrado foi de 10:90,
portanto, dietas altamente energéticas. Os autores, por intermédio de inclusões
crescentes de uréia (0; 0,88; 1,34 e 1,96% da MS dietética), verificaram a
satisfação das exigências em PDR (com exceção da dieta sem adição de uréia) e
condições superavitárias no tocante às demandas de proteína metabolizável,
conforme se investiga também neste estudo. No ensaio de desempenho, os
autores não observaram diferenças entre os tratamentos quanto ao consumo de
MS, inclusive para o tratamento onde se promoveu deficiência de PDR (como é
o caso do tratamento B, presente).
Franco (2001) também trabalhou com novilhos cruzados com peso
médio de 315 kg, em regime de confinamento e promoveu combinações de
diferentes alimentos concentrados (milho, polpa cítrica, farelo de soja, uréia e

67
farelo de glúten de milho), de modo a elaborar dietas para crescimento que
resultassem em balanços de PDR e de proteína metabolizável em diferentes
situações de superávit ou déficit. O autor verificou que nas distintas condições
nutricionais propostas a ingestão de matéria seca não foi diferente entre os
tratamentos, independentemente da maneira de expressá-la (em termos totais,
em relação ao peso vivo [%] ou peso metabólico [PV0,75]). Deve ser salientado
que nestes dois trabalhos, a performance buscada em termos de ganhos de peso
foram elevadas (1,54 kg/dia e 1,12 kg/dia, para Shain et al. [1998] e Franco
[2001], respectivamente), considerando o tipo animal experimental, enquanto
que no presente estudo, os ganhos propostos para cordeiras da raça Santa Inês
(160 g/dia) são relativamente modestos frente aos apresentados por Furusho-
Garcia (2001), por exemplo.
A Tabela 9 apresenta os consumos de fibra em detergente neutro
corrigida para o nitrogênio (FDNN). Assim como efetuado com relação à MS, os
valores estão apresentados em termos de ingestões totais (g/dia), em relação ao
peso vivo (%PV) ou ainda em relação ao peso metabólico dos animais (g/kg
PV0,75). Não houve efeito de interação entre tratamentos e fases.
Não se verificaram diferenças entre os tratamentos (P>0,05) no tocante
às ingestões de FDNN. Considerando o fato de que todos as dietas foram
elaborados mantendo a relação volumoso:concentrado em aproximadamente
49:51, e sendo unicamente o feno de coastcross o alimento volumoso utilizado,
em razão de não terem sido identificadas diferenças quanto aos consumos de
MS, já seria esperado que as ingestões das porções fibrosas não viessem a ser
distintas.
Pode ser constatado que as ingestões observadas para a segunda fase
foram superiores em relação à primeira, ocorrência esperada e relacionada à
maior ingestão de MS ocorrida na segunda fase comparativamente à primeira.
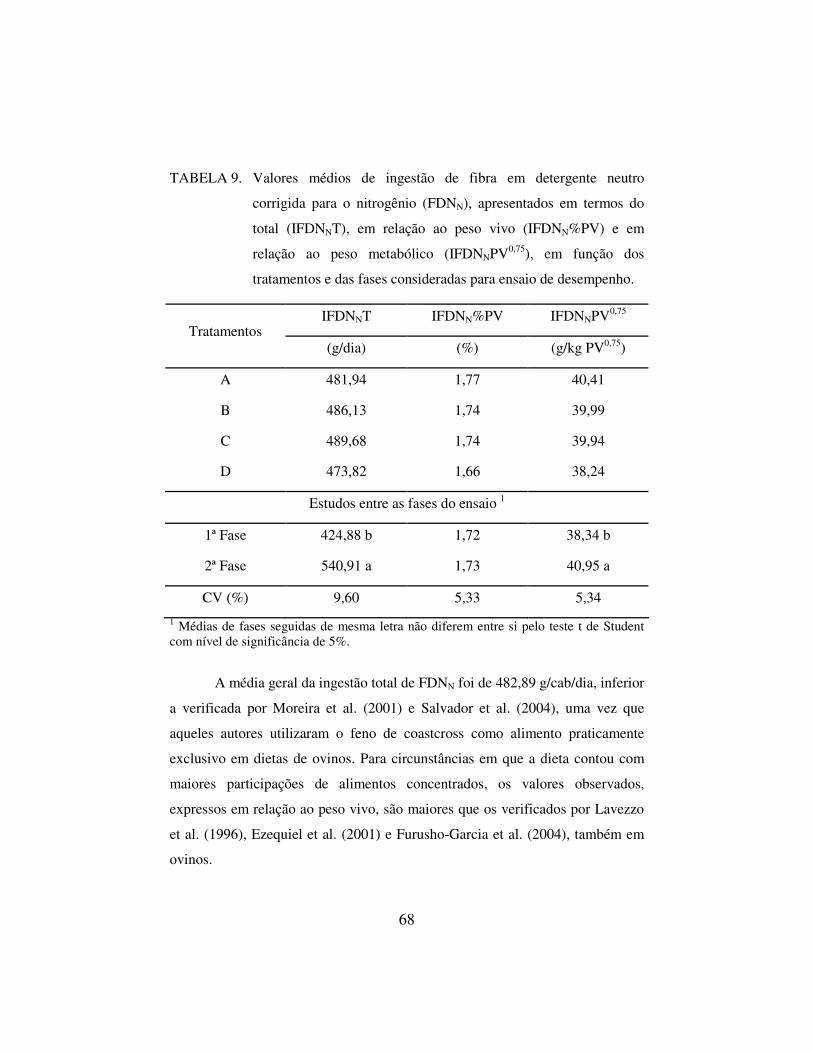
68
TABELA 9. Valores médios de ingestão de fibra em detergente neutro
corrigida para o nitrogênio (FDNN), apresentados em termos do
total (IFDNNT), em relação ao peso vivo (IFDNN%PV) e em
relação ao peso metabólico (IFDNNPV0,75), em função dos
tratamentos e das fases consideradas para ensaio de desempenho.
IFDNNT IFDNN%PV IFDNNPV0,75 Tratamentos
(g/dia) (%) (g/kg PV0,75)
A 481,94 1,77 40,41
B 486,13 1,74 39,99
C 489,68 1,74 39,94
D 473,82 1,66 38,24
Estudos entre as fases do ensaio 1
1ª Fase 424,88 b 1,72 38,34 b
2ª Fase 540,91 a 1,73 40,95 a
CV (%) 9,60 5,33 5,34
1 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.
A média geral da ingestão total de FDNN foi de 482,89 g/cab/dia, inferior
a verificada por Moreira et al. (2001) e Salvador et al. (2004), uma vez que
aqueles autores utilizaram o feno de coastcross como alimento praticamente
exclusivo em dietas de ovinos. Para circunstâncias em que a dieta contou com
maiores participações de alimentos concentrados, os valores observados,
expressos em relação ao peso vivo, são maiores que os verificados por Lavezzo
et al. (1996), Ezequiel et al. (2001) e Furusho-Garcia et al. (2004), também em
ovinos.

69
Apesar do presente trabalho utilizar dietas com teores médios de FDNN
ao redor de 46,72%,, em razão da presença de alimentos concentrados
compondo as referidas dietas (aproximadamente 51%), as ingestões de fibra
expressas em relação ao peso vivo animal têm como média geral o valor de
1,73%, valor superior aos observados pelos autores supra citados (1,40 e 1,53%
do peso vivo, respectivamente para Moreira et al., 2001 e Salvador et al., 2004),
em cujos trabalhos o feno era alimento exclusivo.
Este aspecto pode ser um indicativo de que a ingestão diária de matéria
seca não teve a fibra como fator limitador, e sim a ingestão de energia. Deste
modo, a suposição anteriormente mencionada de que haveria a possibilidade de
não ter ocorrido o atendimento energético dos animais pode não ter fundamento.
Segundo o NRC (2006), o potencial de ingestão de alimentos pelos
animais é resultado da combinação de sua demanda por energia e por sua
capacidade física de ingerir a dieta, ambas relacionadas com o tamanho corporal
e com o peso à maturidade esperado. Mertens (1994) salienta ainda que fatores
psicogênicos também têm um papel importante no controle da ingestão,
referindo-se à resposta comportamental dos animais frente a aspectos
estimuladores ou inibidores presentes no alimento ou advindos do manejo ou
ambiente. Para Conrad (1966), dietas com digestibilidade inferior a 66% têm o
controle exercido prioritariamente por fatores físicos que são os resultantes da
constituição fibrosa da dieta, capazes de promoverem a distensão física do
complexo rúmen-retículo (“rumen-fill”). Ao se avaliar a digestibilidade da MS
das dietas da presente pesquisa poderá se constatar que os valores observados
estão muito próximos dos mencionados por Conrad.
A Tabela 10 apresenta os consumos de fibra em detergente ácido
corrigida para o nitrogênio (FDAN). Os valores estão apresentados em termos de
ingestões totais (g/dia), em relação ao peso vivo (%PV) ou ainda em relação ao

70
peso metabólico dos animais (g/kg PV0,75) e também não se constatou efeito de
interação entre tratamentos e fases.
TABELA 10. Valores médios1 de ingestão de fibra em detergente ácido
corrigida para o nitrogênio (FDAN), apresentados em termos do
total (IFDANT), em relação ao peso vivo (IFDAN%PV) e em
relação ao peso metabólico (IFDANPV0,75), em função dos
tratamentos e das fases consideradas para ensaio de desempenho.
IFDANT IFDAN%PV IFDANPV0,75 Tratamentos
(g/dia) (%) (g/kg PV0,75)
A 266,33 0,98 a 22,33
B 261,97 0,94 ab 21,55
C 263,30 0,93 ab 21,48
D 257,30 0,90 b 20,77
Estudos entre as fases do ensaio 2
1ª Fase 231,14 b 0,94 20,86 b
2ª Fase 293,32 a 0,94 22,21 a
CV (%) 9,96 5,61 5,69
1 Médias de tratamentos seguidas de mesma letra não diferem entre si pelo teste Tukey com nível de significância de 5%. 2 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.
Os consumos expressos em base g/kg PV0,75 verificados foram
superiores aos alcançados por Coutinho Filho et al. (1995) (média gera de 21,53
g/kg PV0,75 no presente estudo versus 14,65 g/kg PV0,75, de Coutinho Filho et al.,
1975). Aqueles autores alimentaram cordeiros da raça Polwarth pesando
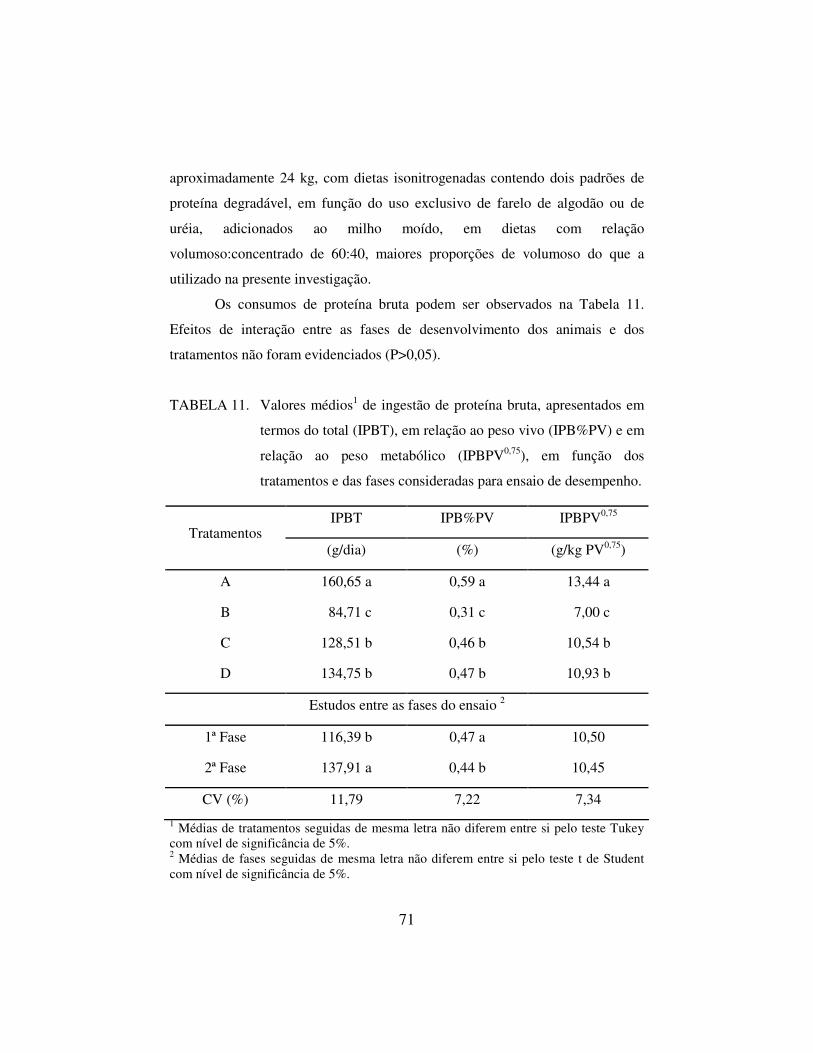
71
aproximadamente 24 kg, com dietas isonitrogenadas contendo dois padrões de
proteína degradável, em função do uso exclusivo de farelo de algodão ou de
uréia, adicionados ao milho moído, em dietas com relação
volumoso:concentrado de 60:40, maiores proporções de volumoso do que a
utilizado na presente investigação.
Os consumos de proteína bruta podem ser observados na Tabela 11.
Efeitos de interação entre as fases de desenvolvimento dos animais e dos
tratamentos não foram evidenciados (P>0,05).
TABELA 11. Valores médios1 de ingestão de proteína bruta, apresentados em
termos do total (IPBT), em relação ao peso vivo (IPB%PV) e em
relação ao peso metabólico (IPBPV0,75), em função dos
tratamentos e das fases consideradas para ensaio de desempenho.
IPBT IPB%PV IPBPV0,75 Tratamentos
(g/dia) (%) (g/kg PV0,75)
A 160,65 a 0,59 a 13,44 a
B 84,71 c 0,31 c 7,00 c
C 128,51 b 0,46 b 10,54 b
D 134,75 b 0,47 b 10,93 b
Estudos entre as fases do ensaio 2
1ª Fase 116,39 b 0,47 a 10,50
2ª Fase 137,91 a 0,44 b 10,45
CV (%) 11,79 7,22 7,34
1 Médias de tratamentos seguidas de mesma letra não diferem entre si pelo teste Tukey com nível de significância de 5%. 2 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.

72
Na elaboração das dietas não se objetivou delinear condições que
buscassem concentrações protéicas dietéticas idênticas (“isonitrogenadas”),
posto que a atenção esteve voltada principalmente para a relação entre
exigências de proteína metabolizável, exigências do rúmen por proteína
degradável e balanços no atendimento ou não destas demandas. Assim, seria
esperado que pudesse haver diferenças nas concentrações de proteína bruta dos
tratamentos, embora os teores de PB das dietas dos tratamentos A, C e D,
tenham sido próximos (16,24; 14,78 e 15,44%, respectivamente [Tabela 3]).
Trabalhos que enfocaram o balanço entre as exigências e aportes de PDR e/ou
proteína metabolizável, tais como o de Shain et al. (1988), de Franco (2001) e o
de Costa (2001) também apresentam teores de PB distintos entre si para os
tratamentos testados.
As ingestões de PB também diferiram entre si (P<0,05), sendo que as
médias obtidas para qualquer das formas de expressar os consumos,
apresentaram a mesma ordem de posição das concentrações dietéticas de PB.
Isto é, as ingestões acompanharam a mesma ordenação de seqüência (maior para
menor) verificada para os teores de PB: tratamentos A, D, C e B, embora as
médias de consumo dos tratamentos D e C tenham sido semelhantes (P>0,05).
Em todos os alimentos e sobras foi realizada a análise para
fracionamento da PB (segundo o modelo do sistema CNCPS). Assim,
considerando os valores verificados das frações, das taxas de degradação
estimadas pelo modelo CNCPS e das taxas de passagem auferidas pelas
equações do sistema AFRC (1993) para cada alimento, foram feitas estimativas
das ingestões de proteína degradável no rúmen e de proteína metabolizável.
As Tabelas 12 e 13 apresentam os valores de ingestão da fração A
(correspondendo ao NNP) e fração B (proteína verdadeira degradável no rúmen)
da PB.

73
Pela análise estatística dos consumos da fração A foi verificado existir
interação entre os tratamentos e as fases de desenvolvimento dos animais.
TABELA 12-1. Valores médios1 de ingestão da fração A da proteína bruta,
apresentados em termos do total (g/dia), em função dos
tratamentos e das fases consideradas para ensaio de
desempenho (CV = 9,88%).
Fases2 Tratamentos1
1ª 2ª Médias
A 15,15 b B 19,21 a B 17,16 B
B 7,68 a C 8,87 a C 8,27 D
C 49,54 b A 66,43 a A 57,98 A
D 10,85 a BC 13,00 a C 11,92 C
Médias 20,80 b 26,87 a
1Médias seguidas de mesma letra maiúscula, na coluna, não diferem entre si pelo teste Tukey com um nível nominal de significância de 5%. 2Médias seguidas de mesma letra minúscula, na linha, não diferem entre si pelo teste t de Student com um nível nominal de significância de 5%.
Como pode ser observado em relação às médias totais por tratamento, o
maior valor é o verificado para o tratamento C (57,98 g/dia), justamente pelo
fato de ser o único que contempla uma fonte de NNP (uréia) em sua formulação,
daí ter propiciado os maiores consumos nesta fração. Quando se analisam os
valores observados para cada tratamento considerando o comportamento dentro
de cada fase, percebe-se que para o tratamento C, os consumos foram diferentes
entre a fases, sendo maiores na segunda fase, muito certamente em razão dos
aportes terem sido maiores simplesmente pelo fato dos animais estarem em

74
maior porte e também mais adaptados a inclusão deste insumo em suas dietas,
ingerindo-as mais facilmente a despeito de aspectos relativos à pouca
palatabilidade que a uréia possa eventualmente promover.
Observa-se também que as menores ingestões, para ambas as fases, se
verificaram para os tratamentos B e D, justamente os que contaram com a
inclusão de glutenose em suas composições, ingrediente cuja proteína é
reconhecidamente pouco degradável no rúmen.
A Tabela 12-2 apresenta esta mesma variável porém, relativa ao peso
metabólico dos animais.
TABELA 12-2. Valores médios1 de ingestão da fração A da proteína bruta,
apresentados em relação ao peso vivo metabólico (g/kg PV0,75),
em função dos tratamentos e das fases consideradas para ensaio
de desempenho (CV = 6,04%).
Fases2 Tratamentos1
1ª 2ª Médias
A 1,39 a B 1,49 a B 1,44 B
B 0,69 a D 0,67 a D 0,68 D
C 4,44 b A 5,00 a A 4,72 A
D 0,97 a C 0,93 a C 0,96 C
Médias 1,87 b 2,03 a
1Médias seguidas de mesma letra maiúscula, na coluna, não diferem entre si pelo teste Tukey com um nível nominal de significância de 5%. 2Médias seguidas de mesma letra minúscula, na linha, não diferem entre si pelo teste t de Student com um nível nominal de significância de 5%.

75
TABELA 13. Valores médios1 de ingestão da fração B da proteína bruta,
apresentados em termos do total (IFrBT) e em relação ao peso
metabólico (IFrBPV0,75), em função dos tratamentos e das fases
consideradas para ensaio de desempenho.
IFrBT IFrBPV0,75 Tratamentos
(g/dia) (g/kg PV0,75)
A 135,83 a 11,36 a
B 68,01 c 5,63 c
C 62,62 c 5,17 c
D 114,83 b 9,33 b
Estudo entre as fases do ensaio2
1ª Fase 88,49 b 7,99
2ª Fase 102,22 a 7,76
CV (%) 13,01 7,95
1 Médias de tratamentos seguidas de mesma letra não diferem entre si pelo teste Tukey com nível de significância de 5%. 2 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.
Os consumos de proteína degradável no rúmen e de proteína
metabolizável estão apresentados nas Tabelas 14 e 15, respectivamente. Os
tratamentos proporcionaram ingestões diferentes (P<0,05) entre si, tanto para o
consumo de proteína degradável no rúmen (PDR) como para a proteína
metabolizável (PM), e deve ser salientado que as dietas ao serem elaboradas não
buscavam permitir ingestões semelhantes entre alguns tratamentos. O objetivo
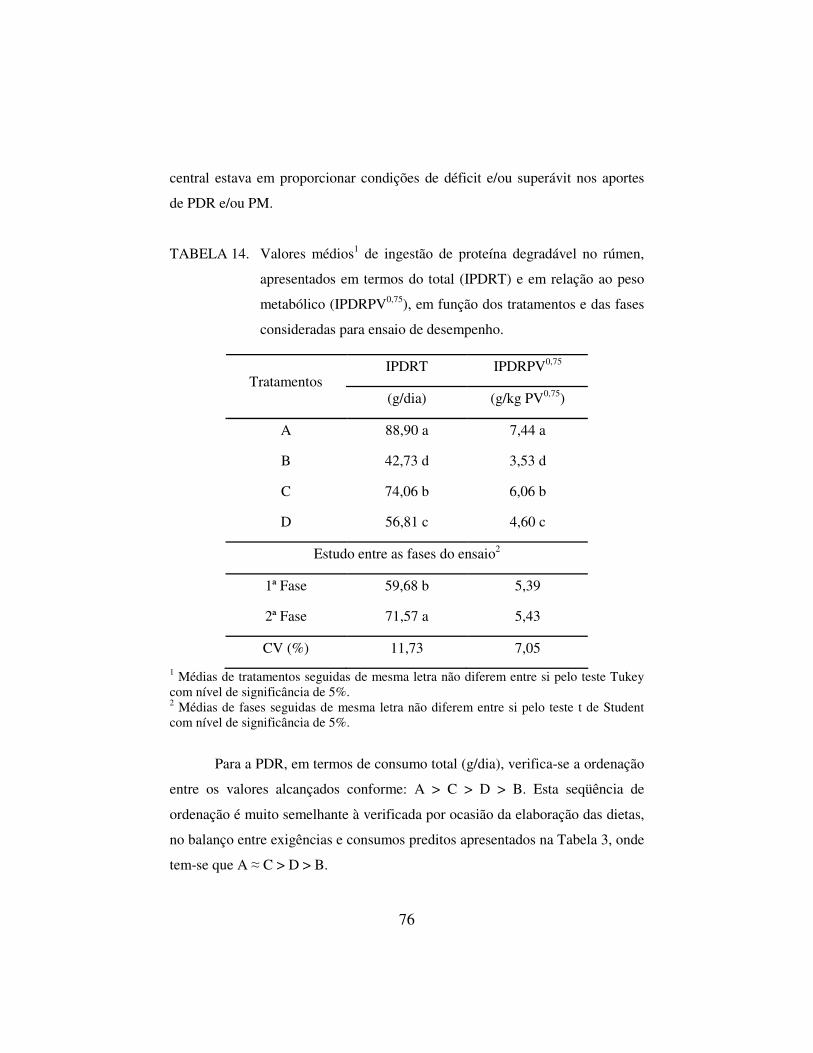
76
central estava em proporcionar condições de déficit e/ou superávit nos aportes
de PDR e/ou PM.
TABELA 14. Valores médios1 de ingestão de proteína degradável no rúmen,
apresentados em termos do total (IPDRT) e em relação ao peso
metabólico (IPDRPV0,75), em função dos tratamentos e das fases
consideradas para ensaio de desempenho.
IPDRT IPDRPV0,75 Tratamentos
(g/dia) (g/kg PV0,75)
A 88,90 a 7,44 a
B 42,73 d 3,53 d
C 74,06 b 6,06 b
D 56,81 c 4,60 c
Estudo entre as fases do ensaio2
1ª Fase 59,68 b 5,39
2ª Fase 71,57 a 5,43
CV (%) 11,73 7,05
1 Médias de tratamentos seguidas de mesma letra não diferem entre si pelo teste Tukey com nível de significância de 5%. 2 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.
Para a PDR, em termos de consumo total (g/dia), verifica-se a ordenação
entre os valores alcançados conforme: A > C > D > B. Esta seqüência de
ordenação é muito semelhante à verificada por ocasião da elaboração das dietas,
no balanço entre exigências e consumos preditos apresentados na Tabela 3, onde
tem-se que A ≈ C > D > B.

77
TABELA 15. Valores médios1 de ingestão de proteína metabolizável,
apresentados em termos do total (IPMT) e em relação ao peso
metabólico (IPMPV0,75), em função dos tratamentos e das fases
consideradas para ensaio de desempenho.
IPMT IPMPV0,75 Tratamentos
(g/dia) (g/kg PV0,75)
A 111,48 a 9,33 a
B 58,82 c 4,87 d
C 80,54 b 6,61 c
D 97,16 a 7,87 b
Estudo entre as fases do ensaio2
1ª Fase 80,28 b 5,39
2ª Fase 93,72 a 5,43
CV (%) 12,66 7,90
1 Médias de tratamentos seguidas de mesma letra não diferem entre si pelo teste Tukey com nível de significância de 5%. 2 Médias de fases seguidas de mesma letra não diferem entre si pelo teste t de Student com nível de significância de 5%.
Para a ingestão de PM, por sua vez, tem-se que a ordenação observada é
A > D > C > B, também semelhante à proposta por ocasião da elaboração das
ditas, conforme exposto na Tabela 3, a saber: A ≈ D > C > B.
Na Tabela 16, apresentada a seguir, podem ser visualizados os balanços
de PDR e PM estimados a partir das dietas efetivamente aplicadas aos animais,
conforme apresentadas na Tabela 7.

78
Conforme já mencionado anteriormente, em razão dos resultados das
análises dos alimentos utilizados na composição das dietas terem sido distintos
dos considerados por ocasião da elaboração destas, alterações nas concentrações
nutritivas das dietas também se efetivaram. Entretanto, estas variações não
comprometeram o objetivo inicial da pesquisa, que era de combinar situações de
déficit ou superávit de PDR e PM. Como pode ser verificado, os balanços de
PDR e PM verificados estão bastante próximos aos inicialmente propostos e
apresentados na Tabela 3.
TABELA 16. Balanço entre exigências nutricionais estimadas e ingestões
efetivados de nutrientes nas dietas experimentais (em base de
MS) do ensaio de desempenho.
Tratamentos
Nutrientes A B C D
Consumos realizados e exigências de nutrientes
Exigência de EM (Mcal) 2,19 2,25 2,25 2,22
Ingestão estimada EM1 2,25 2,12 2,05 2,11
Balanço de EM (Mcal) + 0,06 -0,13 - 0,20 -0,11
Exigência de PDR (g) 87,42 88,46 88,49 87,47
Ingestão de PDR (g) 88,90 42,73 74,06 56,81
Balanço de PDR (g) + 1,48 -45,73 - 14,43 -30,66
Exigência de PM (g) 55,98 56,25 56,26 56,08
Ingestão de PM (g) 111,48 58,82 80,54 97,16
Balanço de PM (g) + 55,51 + 2,57 + 24,27 + 41,07
Sobra / Exigência 99,16% 4,57% 43,14% 73,23%

79
Embora as dietas tenham sido elaboradas segundo os princípios e
recomendações do sistema britânico AFRC (1993), considerando a categoria
animal utilizado no presente ensaio (cordeiras com aproximadamente 7 meses de
idade, pesando em média ao redor de 21,5 kg e com ganho de peso proposto de
160 g/dia), quando se verificam os valores da caracterização nutricional das
dietas efetivamente administradas aos animais (Tabela 7) e as ingestões
estimadas de nutrientes (Tabela 16), e se efetua a comparação com as demandas
apresentadas em edição recente do National Research Council para ovinos
(NRC,2006), têm-se que a publicação norte-americana propõe valores maiores
de exigências em proteína metabolizável do que o AFRC, embora as
recomendações para energia metabolizável sejam muito similares entre as duas
publicações.
Uma vez que o balanço da ingestão estimada de energia metabolizável
apresentado na Tabela 16 (média geral de 2,13 Mcal/dia) resulta praticamente no
equilíbrio entre as necessidades animais e os consumos, associado ao fato de que
as estimativas de exigências deste nutriente para a presente pesquisa estão em
conformidade com publicação mais recente (NRC, 2006), indicam que houve a
satisfação dos requerimentos energéticos dos animais e que muito certamente o
consumo de alimento teve a energia como balizador. Alves et al. (2003),
estudando níveis crescentes de energia metabolizável (2,42; 2,66 e 2,83 Mcal/kg
MS) na dieta de animais da raça Santa Inês (peso e idade similares aos do
presente trabalho, porém tratou-se serem machos inteiros) não verificaram
diferenças nos ganhos de peso e conversão alimentar entre animais e concluíram
que os consumos das dietas tiveram o aporte energético como fator regulador.
Naquele trabalho, os ganhos de peso médios observados para os animais
alimentados com as dietas de menores densidades energéticas (2,42 Mcal/kg S)
foram de 0,123 kg/cab/dia, ligeiramente superiores aos do presente trabalho
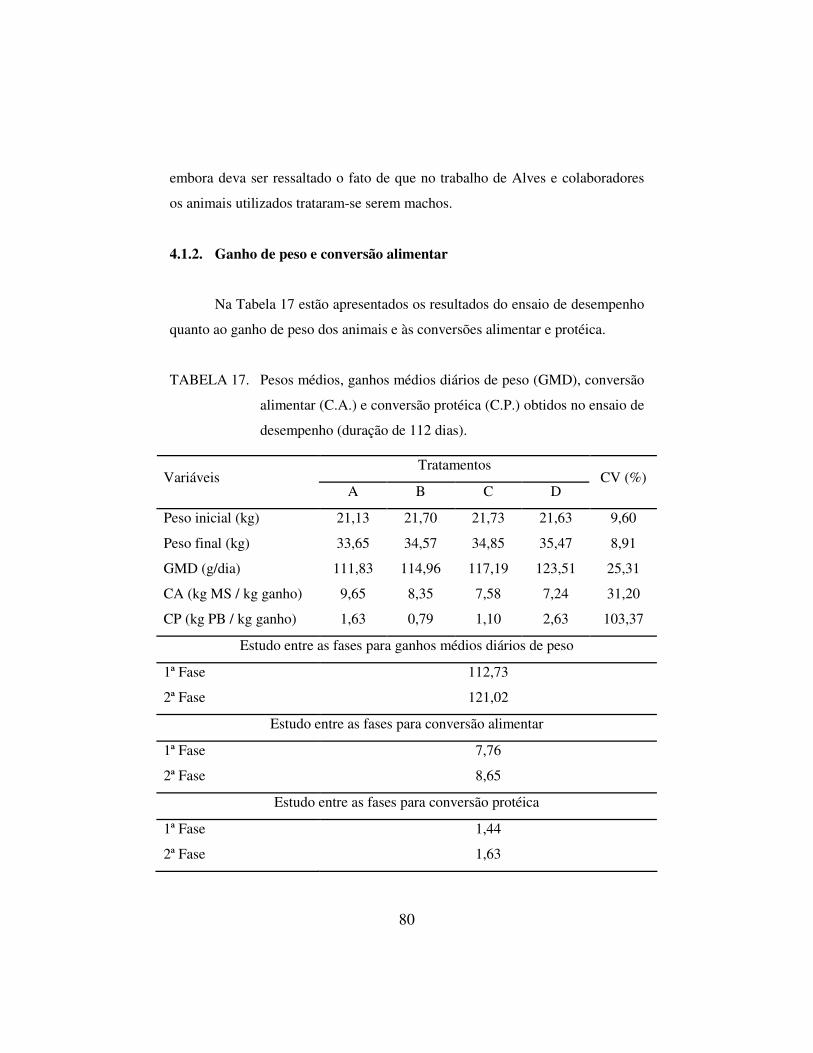
80
embora deva ser ressaltado o fato de que no trabalho de Alves e colaboradores
os animais utilizados trataram-se serem machos.
4.1.2. Ganho de peso e conversão alimentar
Na Tabela 17 estão apresentados os resultados do ensaio de desempenho
quanto ao ganho de peso dos animais e às conversões alimentar e protéica.
TABELA 17. Pesos médios, ganhos médios diários de peso (GMD), conversão
alimentar (C.A.) e conversão protéica (C.P.) obtidos no ensaio de
desempenho (duração de 112 dias).
Tratamentos Variáveis
A B C D CV (%)
Peso inicial (kg) 21,13 21,70 21,73 21,63 9,60
Peso final (kg) 33,65 34,57 34,85 35,47 8,91
GMD (g/dia) 111,83 114,96 117,19 123,51 25,31
CA (kg MS / kg ganho) 9,65 8,35 7,58 7,24 31,20
CP (kg PB / kg ganho) 1,63 0,79 1,10 2,63 103,37
Estudo entre as fases para ganhos médios diários de peso
1ª Fase 112,73
2ª Fase 121,02
Estudo entre as fases para conversão alimentar
1ª Fase 7,76
2ª Fase 8,65
Estudo entre as fases para conversão protéica
1ª Fase 1,44
2ª Fase 1,63

81
Em nenhuma destas variáveis foram identificadas diferenças entre os
tratamentos e entre fases (P>0,05).
Seria esperado que os ganhos de peso verificados na primeira fase
viessem a ser superiores aos da segunda fase, devido ao ‘impulso’ fisiológico
maior para crescimento admissível para o primeiro momento de crescimento,
entretanto isto não se deu embora os índices relativos à eficiência biológica
(conversão alimentar e conversão protéica) apontem superioridade para a
primeira fase em relação à segunda, ainda que apenas numericamente.
Os ganhos de peso observados no presente trabalho (média geral de
116,87 g/cab/dia) ficaram aquém do objetivado por ocasião das formulações das
dietas (160 g/cab/dia). Quintão (2006) também se valeu do sistema AFRC para
elaborar dietas para fêmeas Santa Inês, mais pesadas que as utilizadas neste
ensaio, propondo ganhos de peso da ordem de 180 g/dia e verificou valores
médios de 132 g/dia, também menores do que os propostos. Considerando o fato
de que a base de informações que foram utilizados para a construção e
desenvolvimento daquele sistema tenha origem em pesquisas desenvolvidas com
animais, alimentos e condições ambientais e experimentais principalmente
vigentes no Reino Unido, devem constituir provavelmente a razão para este
desvio entre os ganhos propostos e os efetivamente verificados.
Furusho-Garcia (2001), trabalhando com fêmeas em crescimento da raça
Santa Inês (e com pesos variando entre 25 a 35 kg), encontraram valores de
ganho de peso (0,143 kg/dia) superiores aos obtidos no presente trabalho (0,117
kg/dia), entretanto, deve ser salientado que uma importante diferença entre o
trabalho daquele pesquisador e esta investigação é de que a densidade energética
(EM) assumida para o primeiro foi de 2,64 Mcal/kg de MS, contra 2,35 Mcal/kg
no segundo e isto se deve basicamente ao fato da diferença na relação

82
volumoso:concentrado assumido para cada uma das duas pesquisas: 20:80 para
Furusho-Garcia (2001) versus 49:51 para a presente.
O ponto a ser ressaltado nesta comparação é de que os ganhos de peso
obtidos neste presente trabalho não tiveram déficits energéticos como fator
limitador para a manifestação de desempenho animal, ou ainda para que se
permitisse que as diferenças entre os tratamentos (condições de balanços
diferentes de PDR e PM ) pudessem ser evidenciadas.
Owens & Zinn (1988) afirmaram que as exigências de aminoácidos pelo
animal dependem do potencial de produção. Assim, para mantença e
crescimento lento, o suprimento de aminoácidos provindos da proteína dos
microrganismos ruminais é adequado, mas para altas taxas de produção de leite
ou de crescimento, a proteína microbiana como única fonte de aminoácidos
torna-se insuficiente. Enquanto esta afirmação pode explicar e parte a ausência
de diferença entre os resultados de ganhos de peso da presente pesquisa, esta
asserção entretanto, não encontra respaldo nos resultados de Andriguetto &
Cavassin (2002), que alimentaram cordeiros em confinamento utilizando
proteína de soja submetida a três formas de tratamentos de forma a proporcionar
níveis decrescentes de degradabilidade em suas frações protéicas. As dietas
foram estabelecidas para atingir concentrações protéicas e energéticas
semelhantes entre si (18% de PB e 2,37 Mcal de EM/kg MS) e os autores não
verificaram diferença entre as diferentes quantidades de PDR ofertada para as
variáveis consumo de MS, conversão alimentar e ganho de peso, sendo este
último considerado bastante elevado para cordeiros (ao redor de 400 g/cab/dia).
Cervieri et al. (2001) avaliaram as respostas ao fornecimento de dietas
com diferentes níveis de PDR durante as fases de crescimento e terminação de
bovinos em confinamento. Os autores estudaram aspectos de desempenho e de
qualidade de carcaça. Os tratamentos foram assim dispostos:

83
Crescimento Terminação
Trat 1: 69% da PB como PDR 69% da PB como PDR
Trat 2: 77% da PB como PDR 69% da PB como PDR
Trat 3: 61% da PB como PDR 69% da PB como PDR
As dietas eram isonitrogenadas e isoenergéticas e no preparo destas
lançou-se mão também de fontes protéicas de baixa degradabilidade (farinha de
sangue). Os autores identificaram diferenças significativas para características
de desempenho (ganho diário e CA) apenas na fase de crescimento, apontando
superioridade para os tratamentos 1 e 3. Quando foi considerada toda a atividade
de confinamento (as duas fases juntas), não foi identificada superioridade para
nenhum dos tratamentos. Segundo os autores, os melhores resultados obtidos
quanto ao ganho médio de peso para os animais do tratamento T1 e conversão
alimentar para os dos tratamentos T1 e T3 durante a Fase 1 podem ser atribuídos
ao atendimento adequado da maior exigência de proteína do ganho dos animais,
já que bezerros não castrados e recém-desmamados apresentam maior
crescimento muscular em comparação com o comportamento de deposição do
tecido adiposo durante o período inicial de confinamento e, assim, a necessidade
de proteína metabolizável que é mais elevada nesta fase teria sua demanda
atendida quando a proteína dietária (oriunda dos ingredientes básicos ou de
suplementos protéicos) escapasse da degradação ruminal e se tornasse
disponível para absorção no intestino delgado.
Outros trabalhos onde desequilíbrios nos balanços de PDR e proteína
metabolizável foram considerados também não identificaram diferenças nos
ganhos de peso ou de eficiência alimentar para animais em ensaios de
desempenho.
Shain et al. (1998) alimentaram novilhos cruzados em sistemas de
confinamento onde, através de inclusões crescentes de uréia (0; 0,88; 1,34 e

84
1,96% na MS), promoveram condições em que em pelo menos dois tratamentos
houve déficit no atendimento das demandas de PDR do rúmen, apesar de todos
os tratamentos propiciarem o atendimento dos requerimentos em proteína
metabolizável (superávit médio de 31,5%).
Os autores verificaram igualdade nos consumos, entretanto, nas
condições onde o déficit de proteína degradável no rúmen (PDR) foi mais crítico
(-39,2% ─ tratamento com 0% de uréia), os ganhos de peso e a eficiência
alimentar foram inferiores em relação aos outros três tratamentos (6,6% e 5,4%
inferiores, respectivamente para o GMD e CA). Para o tratamento onde o déficit
de PDR foi menor (apenas -7,3% das demandas ─ tratamento com 0,88% de
uréia), os autores argumentam que provavelmente a reciclagem do nitrogênio
facultou o atendimento da demanda em PDR ou que a própria estimativa de
exigência possa estar superestimada. As equações do NRC (1985) para a
reciclagem de N indicam que, conforme se dá a redução das concentrações
protéicas dietéticas ocorre um incremento na porcentagem de reciclagem do N
(em termos de N ingerido), no entanto, em circunstâncias de animais altamente
produtivos, quando as condições de ritmo de crescimento são elevadas, a síntese
de tecidos altamente ativada atua como um verdadeiro ‘escoadouro’ de
nitrogênio, reduzindo deste modo sobremaneira a síntese de uréia e
concomitantemente diminuindo a possibilidade de se dar reciclagem. Para as
condições do trabalho de Shain e colaboradores é provável que devido ao déficit
de PDR para o tratamento com 0,88% de uréia na MS ser bastante diminuto, a
ainda que pequena reciclagem possa ter dado cabo de suprir a demanda.
No presente estudo, no tratamento onde se estimou um déficit de PDR
em relação às exigências da ordem de -45,73% sem entretanto que se fizesse
manifesto esta deficiência em face ao desempenho animal, parece pouco
provável que a reciclagem do nitrogênio tenha podido cumprir a

85
complementaridade dos requerimentos, dado a magnitude do desequilíbrio
(quase 50% da demanda).
A questão toma conotação mais intrigante quando são trazidos a tona os
diversos autores que enfatizam a redução da capacidade ingestiva e digestiva da
matéria seca (mormente da fibra) e, conseqüentemente, piora do desempenho
quando não são satisfeitas as demandas microbianas por nitrogênio (Burroughs
et al, 1975b; Mertens, 1994; Van soest, 1994; Allen, 1996). Shain et al (1998)
afirmaram, com base em seus resultados de investigação, que:dietas com PDR
menor de 6,4% na MS reduzem o ganho de peso e a conversão alimentar.
Vale ressaltar no entanto, que Stern & Hoover (1979) indicaram que a
concentração ótima de amônia no rúmen requerida para máximas taxas de
fermentação pode não necessariamente ser a mesma indicada para proporcionar
máximas sínteses de proteína microbiana. Milton & Brandt (1994) verificaram
que a digestibilidade do amido foi elevada quando maiores aportes de N
degradável (uréia) foram administrados, porém os autores não observaram
aumentos na síntese de proteína microbiana e Shain et al. (1998) não
encontraram aumento nas concentrações de ácidos graxos voláteis ao elevarem o
aporte de N degradável no rúmen.
Outro trabalho interessante é o de Franco (2001), onde o autor,
diferentemente do objetivado por Shain et al. (1998), deliberadamente promove
combinações de condições de déficit e/ou superávit de PDR e/ou PM,
semelhantemente ao pretendido no presente estudo. O pesquisador estudou o
desempenho de novilhos cruzados em 112 de confinamento (ganho diário
pretendido de 1,20 kg /animal) utilizando dietas contendo a silagem de milho
como volumoso e milho e polpa cítrica como farelos energéticos. A combinação
do uso de uréia; farelo de soja e farelo de glúten de milho (glutenose), objetivou
buscar condições de balanço de proteína degradável no rúmen e de proteína
86
metabolizável que podem ser esquematicamente assim apresentados (e
permitindo a comparação com o presente ensaio):
Franco (2001) Presente Ensaio
Trat PDR PM Trat PDR PM
1: “0” Defic A: “0” Super
2: Super “0” B: Defic “0”
3: Defic “0” C: “0” Super
4: “0” “0” D: Defic Super
onde “0” indica balaço zerado (ingestão igual demanda), “Super” indica
condição superavitária (ingestão maior que demanda) e “Defic” indica condição
deficitária (ingestão menor que requerimento). Somente há similaridade entre os
ensaios quanto ao tratamento 3 de Franco (2001) e o tratamento B no presente
estudo, ambos considerando apenas o uso de farelo de glúten de milho
(glutenose) como fonte protéica.
O aspecto a salientar está relacionado ao fato de que no ensaio da
presente investigação, os ganhos propostos para a espécie utilizada (ovinos) são
relativamente modestos (proposição de 160 g/cab/dia, tendo se atingido a média
geral de aproximadamente 117g/cab/dia) enquanto que o nível de performance
almejado por Franco foi de 1.200 g/cab/dia, patamar já considerado elevado em
se tratando de terem sido utilizados bovinos machos cruzados como animais
experimentais. Assim, averiguar se a intensidade de desempenho proposto possa
ter efeito na manifestação de respostas significativas, é um aspecto importante a
ser levado em conta.
Franco obteve bom nível de performance em ganho de peso, embora
ligeiramente menor do que o pretendido (média geral alcançada de 1.120
g/cab/dia), porém não se verificou efeito das diferentes condições de

87
atendimento ou não dos requerimentos em PDR e/ou PM, tanto para o consumo
de matéria seca bem como para os ganhos de peso. Esta ocorrência de certa
forma vem confrontar à afirmação de Parsons & Allison (1991), segundo os
quais o fornecimento de proteína não degradável só deve ser realizado depois da
satisfação das demandas do rúmen e quando o suprimento de proteína
microbiana for insuficiente para proporcionar o atendimento da demanda por
proteína metabolizável.
Outro trabalho também considerando condições diferenciadas de
atendimento das exigências de proteína degradável no rúmen e proteína
metabolizável é o de Costa (2001), que foi levado a efeito com novilhos
cruzados jovens (peso médio inicial de 150 kg), recriados a pasto (Brachiaria
decumbens) e recebendo suplementação em quantidades fixas (0,5% do PV),
sendo os suplementos formulados com o objetivo de permitir ganhos de peso de
500g/cab/dia. Os tratamentos adotados pelo pesquisador podem ser assim
esquematizados, com relação aos balanços de PDR e PM:
Trat PDR PM
1: Sem nenhuma suplementação
2: Déficit Atendimento OK
3: Atendimento OK Superávit
4: Déficit Superávit
5: Superávit Atendimento OK
6: Suplementação ad libitum (ganho máximo)
Diferentemente do observado por Franco (2001) e na presente pesquisa,
Costa (2001) verificou os maiores ganhos de peso para o tratamento 6
(suplementação ad libitum), auferindo média de 0,734 kg/cab/dia, o que
realmente foi esperado pelo pesquisador. Entretanto, inusitadamente, não foram
verificadas diferenças entre o tratamento 1 (testemunha sem suplementação

88
alguma) e os tratamentos 2, 3 e 5, enfatizando que nos tratamentos 3 e 5 tanto os
requerimentos em PDR como em PM estavam atendidos (ou em superávit).
Ocupando a posição de segundo melhor resultado, está o proporcionado pelo
tratamento 4, que não atendeu as demandas de N do rúmen, em razão do uso em
grande proporção de farelo de glúten de milho, reconhecida fonte de proteína de
baixa degradabilidade. O pesquisador concluiu que a busca em prover condições
de ajuste de N degradável no rúmen, conforme preconizado pelos principais
sistemas de alimentação de ruminantes, não é limitante para o desenvolvimento
do ruminante e o cumprimento de suas funções produtivas, não sendo a única via
de promover efetivo crescimento nos ruminante, uma vez atendidas as
necessidades de proteína metabolizável, por meio do uso de fontes de baixa
degradabilidade.
Nesta direção, Andrade & Alcalde (1995) sugeriram que em gramíneas
tropicais pobres em proteína e com alta degradabilidade pode haver resposta à
suplementação com proteína de alta qualidade e baixa degradabilidade.
Entretanto, esta perspectiva não é avalizada por Cochran et al. (1998), que
defenderam que alimentos normalmente utilizados em programas de
suplementação de ruminantes, particularmente sob condições tropicais, não
devem contemplar o uso de proteína de baixa degradabilidade, pois esta fração
do alimento pode não contribuir diretamente para a produção microbiana e a
digestão primária da forragem, ocorrente no rúmen.
Por fim, um aspecto a salientar refere-se à grande amplitude verificada
nos coeficientes de variação para os índices de conversão. Neste aspecto é
importante ressaltar que segundo Guidoni (1994), citado por Zadra (2007), tanto
a conversão alimentar, como a conversão protéica e a eficiência alimentar,
resultam em índices viesados para a estimação do desempenho nutricional, e
portanto sujeitos a intervalos de amplitudes maiores do que os observados para o
consumo e o ganho de peso, separadamente.

89
4.2. ENSAIO DE DIGESTIBILIDADE E BALANÇO DE NITROGÊNIO
No ensaio de digestibilidade foram avaliadas as ingestões e a
digestibilidade aparente de matéria seca (MS) e matéria orgânica (MO), da fibra
insolúvel em detergente neutro corrigida para nitrogênio (FDNN), da fibra
insolúvel em detergente ácido corrigida para nitrogênio (FDAN) e da proteína
bruta (PB). Foi também efetuado o balanço nitrogenado dos animais em relação
aos tratamentos, avaliando-se as ingestões, excreções e retenções do nitrogênio.
Na Tabela 18 observam-se diferenças na participação dos alimentos nas
dietas em relação ao proposto (Tabela 6), que se devem às variações ocorridas
nos valores das concentrações de matéria seca e nutrientes efetivamente
alcançados nas análises dos alimentos utilizados. Pode ser constatado que, em
função destas variações, as concentrações nutritivas também sofreram pequenas
variações.
4.2.1. Consumo e digestibilidade aparente dos nutrientes
As médias referentes ao consumo de MS, MO, FDNN, FDAN e PB,
expressas em gramas por animal por dia, porcentagem do peso vivo (%PV) e
gramas por kg de peso metabólico (g/kg PV0,75), bem como os coeficientes de
digestibilidade observados, estão apresentados na Tabela 19.
De forma parecida ao ocorrido no ensaio de desempenho, as ingestões
dos nutrientes verificadas em termos de valores totais (g/dia) para o tratamento
A, foram superiores numericamente aos resultados dos demais tratamentos, sem
que houvesse, entretanto, diferença estatística entre eles, com exceção do
consumo de PB. Excetuando-se as ingestões de PB, quando a avaliação é
expressa em função do peso corporal (em %PV ou em g/kg PV0,75), conforme
sugerido por Valadares et al. (1997a), tem-se que apenas para a MS, expressa em
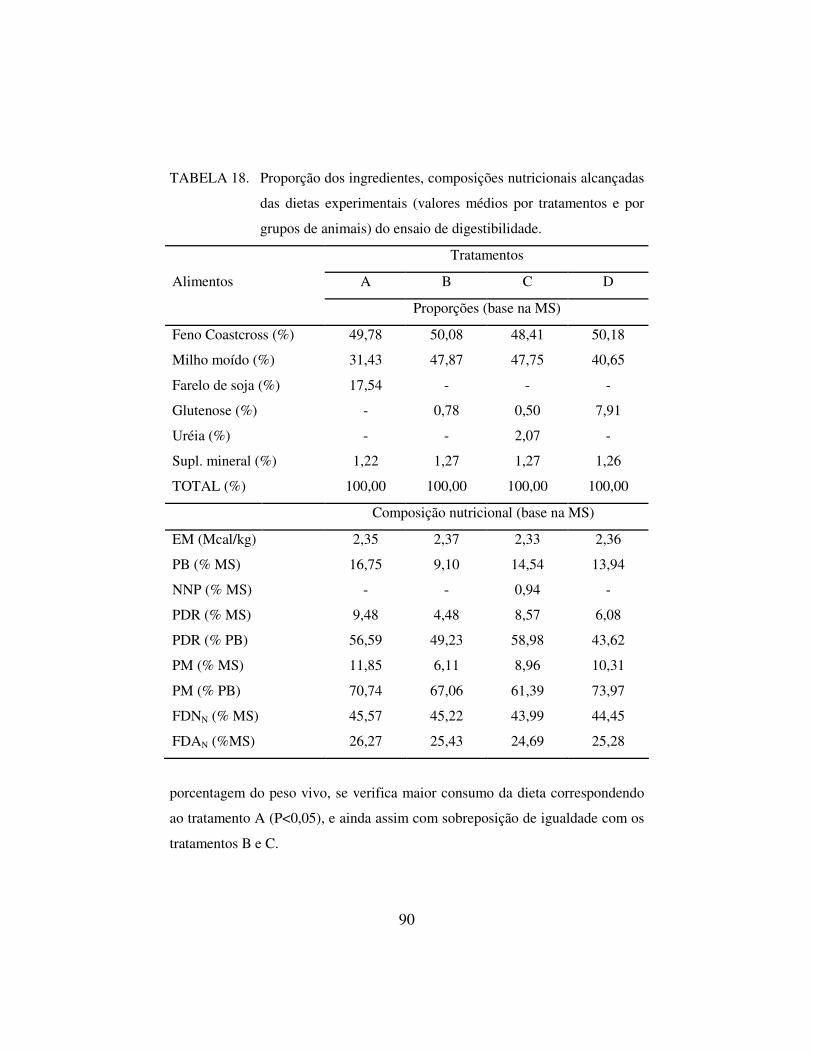
90
TABELA 18. Proporção dos ingredientes, composições nutricionais alcançadas
das dietas experimentais (valores médios por tratamentos e por
grupos de animais) do ensaio de digestibilidade.
Tratamentos
Alimentos A B C D
Proporções (base na MS)
Feno Coastcross (%) 49,78 50,08 48,41 50,18
Milho moído (%) 31,43 47,87 47,75 40,65
Farelo de soja (%) 17,54 - - -
Glutenose (%) - 0,78 0,50 7,91
Uréia (%) - - 2,07 -
Supl. mineral (%) 1,22 1,27 1,27 1,26
TOTAL (%) 100,00 100,00 100,00 100,00
Composição nutricional (base na MS)
EM (Mcal/kg) 2,35 2,37 2,33 2,36
PB (% MS) 16,75 9,10 14,54 13,94
NNP (% MS) - - 0,94 -
PDR (% MS) 9,48 4,48 8,57 6,08
PDR (% PB) 56,59 49,23 58,98 43,62
PM (% MS) 11,85 6,11 8,96 10,31
PM (% PB) 70,74 67,06 61,39 73,97
FDNN (% MS) 45,57 45,22 43,99 44,45
FDAN (%MS) 26,27 25,43 24,69 25,28
porcentagem do peso vivo, se verifica maior consumo da dieta correspondendo
ao tratamento A (P<0,05), e ainda assim com sobreposição de igualdade com os
tratamentos B e C.
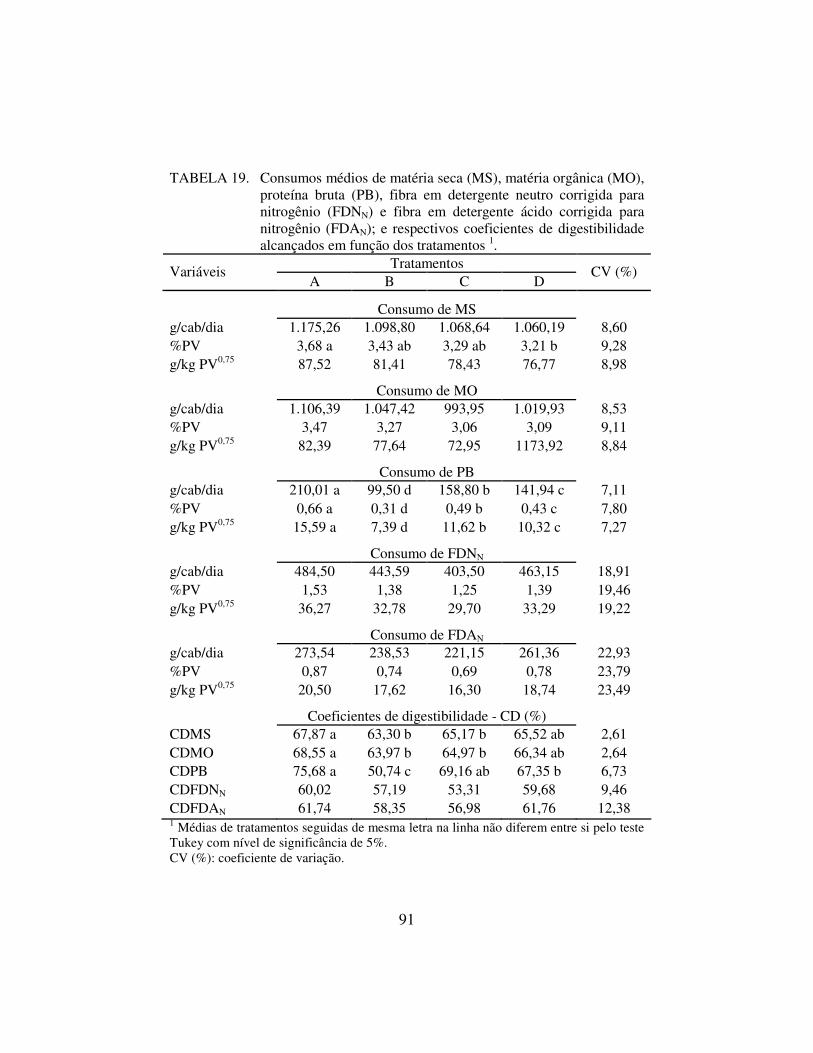
91
TABELA 19. Consumos médios de matéria seca (MS), matéria orgânica (MO), proteína bruta (PB), fibra em detergente neutro corrigida para nitrogênio (FDNN) e fibra em detergente ácido corrigida para nitrogênio (FDAN); e respectivos coeficientes de digestibilidade alcançados em função dos tratamentos 1.
Tratamentos Variáveis
A B C D CV (%)
Consumo de MS
g/cab/dia 1.175,26 1.098,80 1.068,64 1.060,19 8,60 %PV 3,68 a 3,43 ab 3,29 ab 3,21 b 9,28 g/kg PV0,75 87,52 81,41 78,43 76,77 8,98
Consumo de MO
g/cab/dia 1.106,39 1.047,42 993,95 1.019,93 8,53 %PV 3,47 3,27 3,06 3,09 9,11 g/kg PV0,75 82,39 77,64 72,95 1173,92 8,84
Consumo de PB
g/cab/dia 210,01 a 99,50 d 158,80 b 141,94 c 7,11 %PV 0,66 a 0,31 d 0,49 b 0,43 c 7,80 g/kg PV0,75 15,59 a 7,39 d 11,62 b 10,32 c 7,27
Consumo de FDNN
g/cab/dia 484,50 443,59 403,50 463,15 18,91 %PV 1,53 1,38 1,25 1,39 19,46 g/kg PV0,75 36,27 32,78 29,70 33,29 19,22
Consumo de FDAN
g/cab/dia 273,54 238,53 221,15 261,36 22,93 %PV 0,87 0,74 0,69 0,78 23,79 g/kg PV0,75 20,50 17,62 16,30 18,74 23,49
Coeficientes de digestibilidade - CD (%)
CDMS 67,87 a 63,30 b 65,17 b 65,52 ab 2,61 CDMO 68,55 a 63,97 b 64,97 b 66,34 ab 2,64 CDPB 75,68 a 50,74 c 69,16 ab 67,35 b 6,73 CDFDNN 60,02 57,19 53,31 59,68 9,46 CDFDAN 61,74 58,35 56,98 61,76 12,38 1 Médias de tratamentos seguidas de mesma letra na linha não diferem entre si pelo teste Tukey com nível de significância de 5%. CV (%): coeficiente de variação.

92
A ausência de diferença verificada corrobora os resultados obtidos por
Cecava et al. (1990; 1991), Christensen et al. (1993) e Branco et al. (2004),
sendo coerente com a afirmação de Mertens (1994), que salientou a dificuldade
de se estabelecer um mecanismo pelo qual o aumento da concentração de
proteína na dieta, ou mesmo, o aumento da disponibilização de N no rúmen,
resultasse em aumento na ingestão, sem incluir a teoria da regulação de energia.
Um aspecto que deve ser lembrado reside no fato de que as ofertas de
alimentos não se procederam de forma restrita, conforme adotado por Salvador
et al. (2004) e Zeoula et al. (2006), que optaram por este procedimento para se
evitar efeitos sobre os coeficientes de digestibilidade e manter a imposição das
condições dietéticas dos tratamentos, principalmente levando em conta a
presença nos concentrados que ingredientes que poderiam interferir nos
consumos. Assim, sugere-se que a igualdade entre as ingestões é resposta aos
próprios tratamentos.
Ressalta-se o aspecto de que algumas dietas contatam em suas
composições com ingredientes que sabidamente podem interferir naturalmente
na ingestão, como a uréia por exemplo (tratamento C). Entretanto, curiosamente
o tratamento em que a ingestão de matéria seca foi numericamente menor foi o
tratamento D, e este é caracterizado pela maior inclusão da glutenose (farelo de
glúten de milho) na composição dos concentrados, relativamente aos demais
tratamentos. Costa (2001), pesquisando diferentes condições de balanço entre
ingestões e requerimentos de proteína degradável no rúmen (PDR) e proteína
metabolizável (PM) em novilhos em confinamento, observaram certa rejeição
por parte dos animais em ingerir o tratamento que também contava com maior
participação da glutenose no preparo dos concentrados, sendo inclusive
necessário promover a substituição de alguns animais em função da recusa ao
suplemento.

93
Embora já tenha sido bastante reportado que a quantidade e qualidade da
proteína dietética pode modificar o consumo pelos animais ruminantes,
alterando tanto o mecanismo físico como o fisiológico do controle da ingestão
(Roseler et al., 1993; Mertens, 1994; Allen, 1996), não foram constatados estes
efeitos no presente estudo para os consumos de MS, MO e porções fibrosas,
verificando-se apenas diferenças quanto às ingestões de PB. Pode ser verificado
por meio da Tabela 18, que as concentrações de PB e de PDR (como % da PB)
chegaram a variar em até 35 e 84%, respectivamente, sem que esta variação
promovesse diferenças significativas nas ingestões dos nutrientes supra
mencionados.
Praticamente todas as considerações feitas quanto às ingestões de
nutrientes apresentadas ao ser discutido o ensaio de desempenho são
perfeitamente pertinentes e cabíveis no presente momento, no que concerne ao
ensaio de digestibilidade. Entretanto, enquanto já foi sugerido que o consumo de
matéria seca (e, por conseguinte, das frações fibrosas) não tenha tido o
enchimento físico do rúmen como fator limitador e sim o consumo energético,
outra consideração concernente ao aspecto energético deve ser levada em conta,
que se relaciona ao fato de ter ou não sido efetivada a respectiva otimização da
síntese de proteína microbiana, objetivada nos tratamentos A e C, e que está
relacionada à disponibilidade ruminal não somente de N (modulado pelos
tratamentos) mas também da disponibilidade de energia no rúmen para este fim.
Preston & Leng (1987) afirmaram que um dos fatores que controla o
consumo de ruminantes é a absorção de aminoácidos no intestino. Este aspecto
acerca do controle de ingestão já foi salientado por outros pesquisadores como
Minson (1990) e Reis et al. (1996), segundo os quais as melhores respostas à
suplementação protéica têm sido observadas com forragens de baixa qualidade,
mas com alta disponibilidade, através do uso de proteína de escape. Não se deve
considerar esta abordagem como excludente dos outros mecanismos mais

94
freqüentemente considerados ao se ponderar a respeito do controle da ingestão,
mas se for considerada a possibilidade de que a ausência de diferenças entre os
consumos de MS ocorrente entre os tratamentos possa ter tido como uma das
causas a mesma quantidade de aminoácidos disponibilizados para absorção no
intestino delgado, ter-se-á que não se deu efetivamente a aplicação dos
tratamentos aos animais, posto que seria possível supor que todos os saldos de
proteína metabolizável no intestino delgado foram semelhantes, o que não
igualaria os tratamentos entre si.
Este foco da discussão tem como base de argumentação o papel do
componente majoritário dos concentrados (e o segundo ingrediente em maior
proporção nas dietas) como fonte de carboidrato não-fibroso fermentável no
rúmen: o milho.
Martins et al. (1999) classificaram o milho como alimento de lenta
degradação ruminal, comparativamente a outras fontes de amido como, por
exemplo, a raspa de mandioca. Deve ser lembrado que a velocidade de
degradação ruminal, produzida pela ação microbiana sobre as diferentes frações
dos alimentos, tem ação sobre a dinâmica e o equilíbrio dos fluxos de substratos
disponíveis para os microrganismos do rúmen (McCarthy et al., 1989) e que, por
isso, variando-se a fonte e a degradabilidade dos carboidratos não-fibrosos nas
dietas, pode-se otimizar a síntese de proteína microbiana no rúmen e a eficiência
de utilização de proteína não degradável no rúmen (Casper & Schingoethe,
1989). Galyean & Owens (1991) afirmaram ainda que efetuar alteração da fonte
protéica quanto à degradabilidade de sua proteína tem pequeno efeito sobre a
digestão ruminal da MO ou da fibra, quando os carboidratos fermentáveis
presentes na dieta são de lenta degradação, como nas condições de dietas com
elevadas proporções de volumosos de baixa qualidade.
Alguns pesquisadores avaliaram a disponibilidade do N liberado no
rúmen com fontes de amido de baixa (milho moído) (Caldas Neto et al., 2007) e

95
alta (farinha de varredura de mandioca) degradabilidade ruminal (Prado et al.,
2004; Caldas Neto et al., 2007). Em estudos de digestibilidade da MS in vitro, os
resultados indicaram a existência de um grau de sincronização entre os teores de
PDR e a fonte de amido de alta degradabilidade ruminal (Prado et al., 2004), não
ocorrendo o mesmo quando a fonte de amido foi o milho (Prado et al., 2004;
Caldas Neto et al., 2007). Zeoula et al. (2006) sugeriram que para teores de PDR
variando de 47 a 70% da PB, quando em associação ao milho moído, a energia
disponibilizada para a síntese de proteína microbiana parece ser limitante. As
dietas deste presente estudo encontram-se dentro desta faixa (Tabela 18).
Prado et al. (2004) estudaram a digestibilidade aparente da MS em
ovinos alimentados com dietas de diferentes proporções de PDR, tendo a farinha
de varredura de mandioca (alta degradabilidade) como fonte de carboidratos
não-fibrosos, enquanto que Zeoula et al. (2006) desenvolveram trabalho muito
similar, porém utilizando o milho moído como fonte de amido. Confrontando os
dois trabalhos (amido de alta degradabilidade vs amido de baixa degradabilidade
- Prado e al., 2004 vs Zeoula e al., 2006, respectivamente) constata-se que, da
mesma forma como se deu no presente estudo, não houve diferença entre as
ingestões de nutrientes para nenhuma das pesquisas mencionadas, porém, a
digestibilidade aparente da MS diferiu no trabalho de Prado e colaboradores,
tendo comportamento linear crescente ao se elevar o nível de PDR, e não diferiu
no de Zeoula e colaboradores. O presente estudo, embora não voltado
essencialmente para avaliação de níveis de PDR nas dietas, verificou diferenças
na digestibilidade da MS e MO, tendo utilizado o milho como fonte de amido
fermentável no rúmen, contrapondo-se assim, de certa forma, aos resultados
colhidos por Zeoula et al. (2006), Fu et al. (2001) e Araújo et al. (1995),
concordando porém, com o verificado por King et al. (1990) que também
trabalharam com dietas elaboradas com fontes de proteína de diferentes
degradabilidades tendo o milho como fonte de amido, e verificaram diferenças

96
na digestibilidade da matéria seca, nas circunstancias em que os valores de PDR
dietéticos foram maiores.
MacCarthy et al. (1989) observaram que rações compostas de milho,
independentemente da fonte de proteína (farinha de peixe ou farelo de soja -
lenta e rápida degradabilidade, respectivamente), propiciaram maiores consumos
de MS, MO e amido e maior produção de leite em vacas Holandesas multíparas,
em início de lactação, quando comparadas com rações que continham a cevada
(carboidrato de rápida degradação ruminal) como fonte de amido. Ou seja, estes
autores evidenciaram respostas quanto aos consumos de MS e MO ao variarem a
combinação entre fontes de carboidratos e de proteínas de alta e baixa
degradabilidade, constatação não colhida nos outros estudos já mencionados,
inclusive na presente pesquisa. Além disso, observaram efeito sobre o consumo
quando a fonte de amido foi de baixa degradabilidade (milho), não verificando
efeitos quando a fonte de amido foi a cevada (alta degradabilidade). Deste modo
não parece ainda completamente esclarecido estes aspectos referentes à
sincronização ou otimização da síntese de proteína microbiana em relação ao
arranjo combinatório entre a qualidade da fonte protéica e a dos carboidratos
não-fibrosos.
A pouca concordância entre os resultados verificados na literatura
sugerem que as respostas referentes às ingestões e digestibilidade de nutrientes
auferidas na presente pesquisa podem ser assumidas como complementares aos
dados já existentes e leva a crer definitivamente que os tratamentos deste
presente estudo foram aplicados aos animais de forma efetiva.
Quanto à PB, uma vez que os consumos de PB foram todos diferentes
entre si (P<0,05) e, ao se considerar o fato de que as ingestões de MS não
diferiram, tem-se que as concentrações de PB dietéticas entre os tratamentos
foram também significativamente diferentes. Pode ser observado que a

97
ordenação (‘ranqueamento’) dos consumos de PB segue a mesma seqüência dos
níveis dietéticos de proteína bruta.
Chama a atenção o fato de que as menores ingestões de PB
proporcionadas pelo tratamento B (7,39 g/kg PV0,75), correspondendo a 47% da
maior ingestão colhida (tratamento A), e ainda apresentando menor proporção
de sua proteína como PDR, em relação ao tratamento A, não chegaram a
interferir na digestibilidade das frações fibrosas (FDNN e FDAN). Sabidamente, a
quebra das estruturas que compõem as paredes celulares vegetais é uma tarefa só
facultada eminentemente graças à ação de enzimas microbianas especificas e,
portanto, em condições dietéticas em que seja possível incrementar a atividade e
o crescimento das populações microbianas é esperado que melhoras nos
coeficientes de digestibilidade da MS e/ou das porções fibrosas sejam colhidas.
Entretanto, neste sentido é oportuno apontar a completa falta de relação
entre concentrações de N-amônia no rúmen e eficiência da síntese microbiana,
verificada por Bach et al (2005), demonstrando que a eficiência da síntese
microbiana é incapaz de estimar a eficiência com que os microrganismos captam
o N disponível no rúmen e o utilizam para incremento de seus crescimentos e
metabolismos. Além disso, vale ressaltar também os estudos de Stern & Hoover
(1979), que indicaram que a concentração ótima de amônia no rúmen requerida
para máximas taxas de fermentação pode não necessariamente ser a mesma
indicada para proporcionar máximas sínteses de proteína microbiana; e ainda, as
investigações de Morrison & Mackie (1996), que sugeriram a possibilidade de
que as concentrações ótimas de amônia para permitir a digestão ruminal da fibra
e as necessárias para otimizar a síntese de proteína microbiana possam não se
situar nos mesmos patamares.
Quanto a digestibilidade auferida para PB, os valores encontrados
apresentam amplitudes bastante grandes, variando de 75,68% (tratamento A) até
57,19% (tratamento B). Estes dois tratamentos apresentam também níveis de PB

98
dietéticos que ocupam os justamente os extremos do intervalo de variação entre
todos os tratamentos (16,75 e 9,10%, respectivamente para os tratamentos A e
B). Segundo Owens & Zinn (1988), um dos fatores que afetam a digestibilidade
aparente da proteína é a quantidade consumida deste nutriente. Este fato pode
ser ilustrado pelos trabalhos de Ezequiel (1987), Klusmeyer et al. (1990) e
Valadares et al. (1997b), segundo os quais a digestibilidade aparente da PB
observada aumentou com a elevação do teor de N nas dietas. A razão apontada
para tal fenômeno, segundo Stallcup et al. (1975), deve-se ao fato de que, à
medida que o conteúdo de N da dieta se eleva, há uma diminuição proporcional
do N endógeno nos compostos nitrogenados fecais.
No presente estudo, as ingestões de PB foram, conforme já mencionado,
diferentes (P<0,05), entretanto, as excreções fecais não somente foram iguais
estatisticamente (P>0,05), como foram numericamente muito próximas (Tabela
20), e assim, como conseqüência do próprio método para se efetuar o cálculo do
coeficiente de digestibilidade, tem-se que nas condições de consumos de PB
maiores os coeficientes de digestibilidade também serão maiores e vice-versa,
uma vez que as excreções são praticamente as mesmas.
TABELA 20. Excreção fecal de proteína bruta (PB) observada em função dos
tratamentos.
Tratamentos Variáveis
A B C D CV (%)
Consumo de MS
g/cab/dia 51,04 48,90 48,85 46,03 11,06
%PV 0,16 0,15 0,15 0,14 12,56
g/kg PV0,75 3,80 3,63 3,59 3,36 12,18

99
Em outro trabalho (Silva et al., 2007a), novilhos mestiços foram
alimentados com dietas isonitrogenadas apresentando níveis de PB dietéticos
relativamente baixos (apenas 7% na MS), em razão de ter sido adotada relação
volumoso:concentrado de 94:6, com dietas à base de feno de Tifton 85 e
suplementadas com diferentes fontes protéicas: uréia, farelo de soja e farelo de
glúten de milho. Naquele trabalho foram colhidos coeficientes de digestibilidade
da PB elevados e estatisticamente iguais (média de 77,34%) o que se contrapõe à
argumentação relacionada ao fato de que consumos baixos de PB resultam em
coeficientes de digestibilidade também baixos.
Em pesquisas onde os níveis de PDR dietéticos foram elevados (maiores
que 55% da PB), normalmente os coeficientes de digestibilidade da PB também
foram elevados como os observados na presente pesquisa (tratamentos A e C, ao
redor de 70%), como é o caso de Lizieire et al. (1990); Araújo et al. (1994);
Rennó. (2003) e Zeoula et al. (2006). Entretanto, a literatura também apresenta
resultados de digestibilidade reduzidos, em torno de 50%, mesmo quando as
condições dietéticas envolvam fontes protéicas de alta e de baixa
degradabilidade, como pode ser constatado nos trabalhos de Dutra et al. (1997 -
farelo de soja vs glutenose+farinha de sangue, e CDPB menores que 53%);
Sampaio et al. (2000 - farelo de algodão vs levedura de cana-de-açúcar vs uréia,
CDPB médio de 53%) e Fregadolli et al. (2001b - levedura de cana-de-açúcar vs
farelo de algodão + farinha de carne e ossos, CDPB médio de 54%)
O trabalho de Fregadolli et al. (2001b) avaliou combinações entre fontes
de amido de baixa (milho) e alta (casca de mandioca desidratada)
degradabilidade, associadas a fontes protéicas de alta (levedura de cana-de-
açúcar) e baixa (farelo de algodão + farinha de carne e ossos) degradabilidade
em novilhos Holandês. Os autores não constataram interferência dos tratamentos
quanto aos consumos de MS e MO, porém constatou-se interação para as
ingestões de PB e é especialmente interessante que, no tocante à digestibilidade

100
da PB, não se verificaram diferenças entre as fontes de amido ou de proteína,
mas foi significativa a interação entre as combinações de degradabilidade.
Quando a fonte protéica foi de alta degradabilidade (levedura), a melhor
digestibilidade da PB foi alcançada com fonte de amido de baixa
degradabilidade (milho) e vice-versa. Esta resposta vem de encontro a diversos
trabalhos que apontam que para se alcançar aumentos na eficiência microbiana
e, conseqüente maior fluxo de proteína ao intestino delgado, deve se priorizar a
utilização de fontes de proteína de alta degradabilidade em associação a fontes
de amido também de alta degradabilidade (Rooke et al., 1987; Herrera-Saldana
et al., 1990; Poore et al., 1993; Kim et al., 1999a).
Embora não tenha sido objeto de investigação deste presente estudo,
avaliar e quantificar a síntese de proteína microbiana, trabalhos que mensuraram
este parâmetro em condições dietéticas em que se combinaram fontes de energia
e de nitrogênio de diferentes degradabilidades (Fregadolli et al., 2001a;
Valkeners et al., 2006), bem como diferentes condições de níveis de PDR
dietéticos, por intermédio do uso de fontes protéicas diferentes (Magalhães et al.,
2005; Pina et al., 2006; Silva et al., 2007b), não verificaram efeito dos
tratamentos aplicados sobre a síntese de proteína microbiana ao nível de rúmen.
Deste modo, não seria inadmissível crer, diante dos resultados verificados neste
ensaio, que é muito provável não ter havido intensidades distintas na síntese de
proteína microbiana ruminal.
4.2.2. Balanço de nitrogênio
Na Tabela 21 constam os resultados obtidos no estudo do balanço de
nitrogênio auferido em função dos tratamentos. O balanço refere-se ao saldo
líquido de nitrogênio retido, após terem sido deduzidas do montante ingerido, as
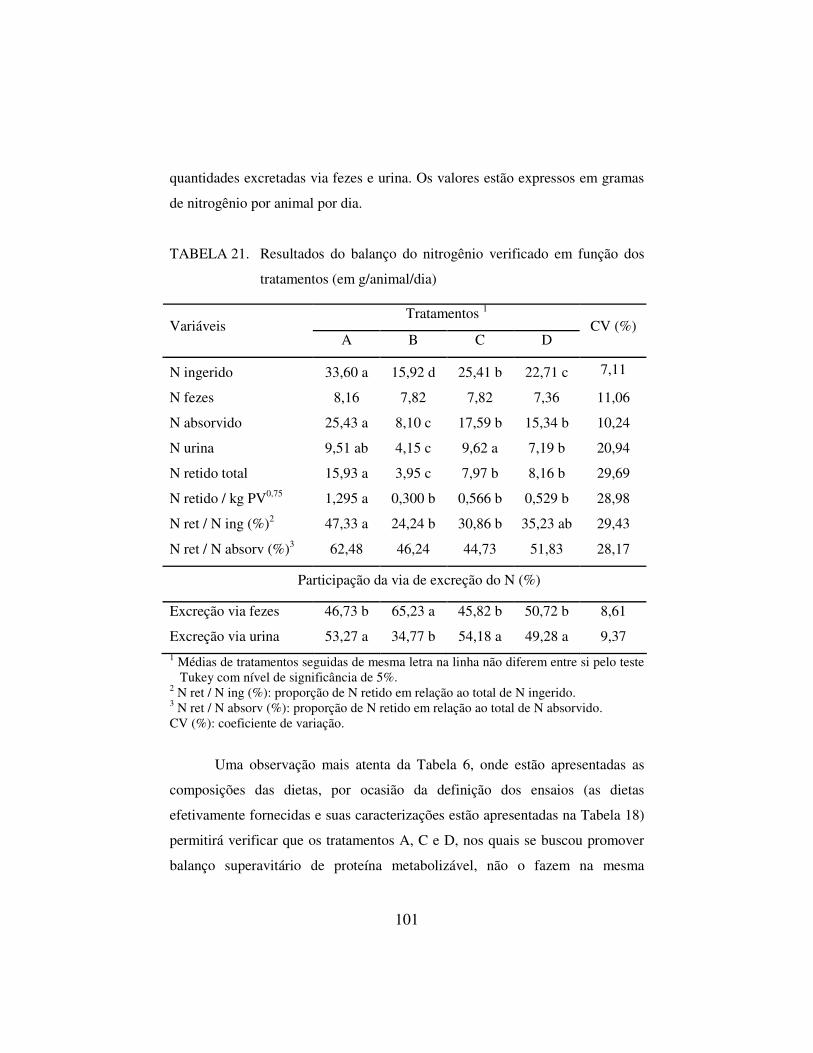
101
quantidades excretadas via fezes e urina. Os valores estão expressos em gramas
de nitrogênio por animal por dia.
TABELA 21. Resultados do balanço do nitrogênio verificado em função dos
tratamentos (em g/animal/dia)
Tratamentos 1 Variáveis
A B C D CV (%)
N ingerido 33,60 a 15,92 d 25,41 b 22,71 c 7,11
N fezes 8,16 7,82 7,82 7,36 11,06
N absorvido 25,43 a 8,10 c 17,59 b 15,34 b 10,24
N urina 9,51 ab 4,15 c 9,62 a 7,19 b 20,94
N retido total 15,93 a 3,95 c 7,97 b 8,16 b 29,69
N retido / kg PV0,75 1,295 a 0,300 b 0,566 b 0,529 b 28,98
N ret / N ing (%)2 47,33 a 24,24 b 30,86 b 35,23 ab 29,43
N ret / N absorv (%)3 62,48 46,24 44,73 51,83 28,17
Participação da via de excreção do N (%)
Excreção via fezes 46,73 b 65,23 a 45,82 b 50,72 b 8,61
Excreção via urina 53,27 a 34,77 b 54,18 a 49,28 a 9,37 1 Médias de tratamentos seguidas de mesma letra na linha não diferem entre si pelo teste
Tukey com nível de significância de 5%. 2 N ret / N ing (%): proporção de N retido em relação ao total de N ingerido. 3 N ret / N absorv (%): proporção de N retido em relação ao total de N absorvido. CV (%): coeficiente de variação.
Uma observação mais atenta da Tabela 6, onde estão apresentadas as
composições das dietas, por ocasião da definição dos ensaios (as dietas
efetivamente fornecidas e suas caracterizações estão apresentadas na Tabela 18)
permitirá verificar que os tratamentos A, C e D, nos quais se buscou promover
balanço superavitário de proteína metabolizável, não o fazem na mesma

102
proporção. Enquanto o tratamento A estabelece um superávit de PM de quase
99% em relação à demanda, os tratamentos C e D o fazem, respectivamente, em
proporção de 52 e 88% relativamente às exigências. Esta particularidade, aliada
às próprias características dos animais por ocasião da formulação das dietas
(pesos vivos), aos quais foram sorteados os tratamentos, resultou em
concentrações de proteína bruta diferentes para estes três tratamentos. Deste
modo, posto que os consumos de matéria seca em termos totais não diferiram
entre os tratamentos, tem-se que as diferenças verificadas nas ingestões de
nitrogênio estão relacionadas na verdade à concentração de PB presente nas
dietas. Conforme já mencionado, os tratamentos não foram delineados com
vistas a promover gradientes de concentração de PB ou de PDR. Esta ocorrência
é, na realidade, conseqüência do princípio que norteou a definição dos
tratamentos (balanços diferenciados de PDR e/ou PM).
A igualdade verificada entre as excreções fecais observadas (P>0,05) é
fato bastante comum em avaliações de balanços de nitrogênio. Van Soest (1994)
afirmou que as perdas fecais de nitrogênio são menos flexíveis que as urinárias e
que correspondem em média a 0,6% do total de matéria seca ingerida e entre 3 a
4% do total de PB ingerida. Neste presente estudo, a excreção fecal de N média
situou-se ligeiramente acima destes patamares (0,71% e 5,11% da MS e PB
ingeridas, respectivamente).
Trabalhos que levaram em consideração a investigação de concentrações
crescentes de PB na dieta (Lizieri et al., 1990; Rennó, 2003; Cavalcante et al.,
2006), ou que estabeleceram condições de dietas isonitrogenadas porém
elaboradas com fontes protéicas diferentes (o que pode resultar em diferentes
proporções de PDR na dieta), como o de Salman et al. (1997), ou ainda,
pesquisas que avaliaram o balanço nitrogenado em condições de gradientes de
concentrações de PDR na dieta (Lizieri et al., 1990; Oliveira Jr et al. 2004;
Zeoula et al., 2006) não identificaram excreções fecais de nitrogênio diferentes.

103
Por outro lado, existem também pesquisas que identificaram excreções fecais de
N diferentes entre os tratamentos quando estes eram caracterizados por níveis
diferentes de PB (Valadares et al., 1997b) ou pelo uso de diferentes fontes
protéicas (Ezequiel et al. 2000; Sampaio et al., 2000) ou ainda pela elevação da
proporção de PDR dietético (Araújo et al., 1994).
As excreções do nitrogênio pela via urinária foram diferentes (P<0,05),
sendo que os tratamentos C e A apresentaram os maiores valores, justamente
aqueles onde a participação da PDR foi maior (Tabela 18 ─ 58,98 e 56,59% da
PB, respectivamente). Chalupa et al. (1970) comentam que maiores excreções
nitrogenadas via urina são decorrentes de excesso de N solúvel na dieta ou da
ineficiência no aproveitamento deste pelos microrganismos ruminais,
principalmente quando em condições de excessos de PB ou do uso excessivo de
fontes de NNP na dieta. De fato, os dados de trabalhos onde se promoveu
elevação da concentração de PB ou da participação da PDR na dieta mostram
aumentos na excreção do nitrogênio por meio da urina (Lizieri et al., 1990;
Valadares et al., 1997b; Cavalcante et al., 2006).
Porém, na literatura podem ser encontrados resultados de igualdade na
excreção urinária de N em condições experimentais de gradientes de PDR
dietéticos (Lavezzo et al., 1996; Zeoula et al., 2006) ou em situações de dietas
isonitrogenadas elaboradas com fontes protéicas diferentes (Ezequiel et al.,
2000; Sampaio et al., 2000).
Cecava & Hancock (1994) verificaram que a excreção de N urinário
foram maiores em novilhos alimentados com dieta contendo uréia (1,35% na
MS) que naqueles que receberam combinações de farelo de soja e farinha de
penas na dieta. Rennó (2003) também observou aumento linear na excreção
média de uréia via urina, em função dos níveis dietéticos de uréia (0; 0,65; 1,30
e 1,95% na MS).

104
No presente estudo, onde foi feito uso de uréia (NNP) na composição da
dieta (tratamento C), a participação deste componente encontra-se dentro de uma
faixa um pouco acima dos 1,5% sugerido por Huber & Kung (1981), porém já
foi devidamente comentado que a oferta de N disponível ao rúmen se deu em
função da energia disponibilizada e do potencial de crescimento microbiano, o
que leva a crer não ter havido ingestão excessiva de nitrogênio não-protéico que
justificassem os valores majorados. Entretanto, ao se calcular as proporções de
N urinário em relação ao consumo total de N obtém-se os resultados de 28,30;
26,07; 37,86 e 31,66% para os tratamentos A, B, C e D, respectivamente. Ou
seja, o tratamento que considerou a inclusão de NNP promoveu uma maior
excreção de N em quase 10 unidades percentuais relativamente ao tratamento
que otimizou a síntese de proteína microbiana por intermédio de fonte protéica
verdadeira (tratamento A - farelo de soja). É provável que, em virtude da maior
solubilidade da uréia no rúmen, esta venha a permitir uma maior absorção
através das paredes ruminais, contribuindo para que haja a redução das
concentrações de n nas fezes e que a via de excreção predominante para todo e
qualquer excesso seja a urinária (Harmeyer & Martens, 1980).
Contrariando estas proporções, pode-se citar o trabalho de Oliveira Jr
(2002), que verificou em novilhos que dieta com participação de uréia
promoveram menor excreção urinária do que aquelas elaboradas exclusivamente
com farelo de soja como fonte de PB.
Interessante verificar que, sendo a perda de N por meio das fezes menos
flexível comparativamente à excreção urinária, compete a esta última exercer
um mais importante papel na retenção do nitrogênio no organismo animal. Isto
explica a participação mais efetiva da via urinária para os tratamentos A, C e D,
nos quais as concentrações de PB eram todas iguais ou superiores a 14%, e, por
outro lado, a excreção do N por via fecal ter majoritária para o tratamento B.

105
Em função da igualdade verifica entre as excreções fecais (P>0,05), da
excreção urinária ter ocorrido em intensidades diferentes (P<0,05) em um
contexto de ingestões também diferentes, tem-se que o balanço nitrogenado,
expresso como N retido, também foi diferentes para os tratamentos. A menor
retenção de N verificada foi a obtida pelo tratamento B, onde a concentração
protéica na dieta, além de ser a menor, também foi caracterizada por ser baixa
degradabilidade. O tratamento D apresenta a menor concentração de PDR (como
% da PB), no entanto este tratamento apresenta concentração protéica cerca de
53% maior que a do tratamento B.
Entretanto, quando se observa a proporção do N retido em relação ao N
absorvido (alguns autores [Salman et al., 1997; Oliveira Jr, 2002] denominam
este parâmetro de “valor biológico”, o que não parece ser uma terminologia
adequada), não se constata diferença (P>0,05) entre os tratamentos, embora o
tratamento A tenha apresentado maior valor numérico.
Interessantemente, apesar da menor retenção proporcionada pelo
tratamento B, correspondendo em torno de apenas 25% da verificada para o
tratamento A e em cerca de 49% da alcançada pelos tratamentos C e D, esta
ocorrência não promoveu pior performance em termos de crescimento e
capacidade de aproveitamento dos alimentos. Ao menos para a espécie animal
em questão (ovinos), nas características raciais, de sexo e nos patamares de
desempenho alcançados (fêmeas Santa Inês, com ganhos de peso modestos - ao
redor de 120 g/cab/dia).
Oldham (1984) asseverou que inter-relações entre os nutrientes que
geram aporte energético e os que resultam em compostos nitrogenados, tanto
dentro do rúmen bem como no corpo do animal, podem ter grandes efeitos no
padrão geral de utilização dos nutrientes. Embora grande ênfase têm sido dada
durante anos às inter-relações entre estes dois grupos de nutrientes dentro do
contexto do rúmen, não menos importante, porém menos enfatizada, é o

106
conjunto de inter-relações entre estes sob o âmbito do corpo animal. A
característica comum entre todos os sistemas de alimentação de ruminantes no
tocante à nutrição energético-protéica, e que os diferencia dos sistemas voltados
para outras espécies e mesmo para outros grupos de nutrientes, é que as
necessidades de energia e proteína estão intimamente ligadas de uma maneira
em uma relação formal de predições e ‘feedback’. Este é o principal passo para
estabelecer uma base quantitativa nestas relações entre a utilização de compostos
nitrogenados e compostos que possam fornecer energia. Porém, enquanto estas
relações tem sido efusivamente investigadas no ambiente ruminal, em menor
profusão tem se explorado estas relações no que diz respeito ao metabolismo
intermediário.
O suprimento de nutrientes aos tecidos interage tanto com os sistemas de
controle de mantém a homeostase metabólica bem como com o sistema que
mantém uma dada condição particular fisiológica (homeorrese). As quantidades
relativas de compostos energéticos e nitrogenados supridos aos animais são,
entretanto, aptas para determinar a eficiência líquida de uso dos nutrientes
absorvidos e esta abordagem conceitual não é recente, posto que em 1946,
Phillipson (citado por Oldham & Smith, 1982) afirmou que “os ruminantes
vivem não somente a partir de ácidos graxos e microrganismos, mas também de
todo alimento, amido ou proteína que deixa o rúmen sem ter sido afetado pela
ação microbiana e a resposta final depende das quantidades destes materiais que
ficam disponibilizados ao animal”.
È importante lembrar os conceitos de controle homeorrético da
utilização de nutrientes descritos por Bauman & Currie (1980), os quais
ressaltaram que, para fêmeas em lactação, nutrientes são requeridos para manter
diferentes estados fisiológicos no decorrer de um ciclo lactacional e que
portanto, implica que o uso dos nutrientes absorvidos também se dê de forma
modificada no transcorrer da lactação, assim como ocorre com a capacidade

107
ingestiva, a capacidade de mobilização de reservas orgânicas, etc. Assim, não é
digno de desconsideração supor que para a atividade de crescimento, esta
modulação do uso dos nutrientes também venha a ocorrer. Oldham (1984)
enfatiza que mais alta eficiência de utilização da proteína tem sido conseguida
em dietas onde a proteína é mais severamente limitada sem que isto implique em
melhoria da performance ou da lucratividade da exploração.
Anteriormente, muitas respostas animais “inexplicáveis” diante do
fornecimento de nutrientes (tipo e nível) eram descritas como resultantes de
interferências advindas do potencial genético dos animais, da ingestão alimentar,
da qualidade organoléptica dos alimentos, etc. Pouca consideração era dada à
natureza da proteína suprida pela dieta, por exemplo, em termos de sua
degradabilidade no rúmen. Entretanto, com a ampliação do conhecimento acerca
de muitos aspectos fisiológicos, e principalmente, acerca da caracterização mais
precisa das frações dos alimentos e do papel destas no metabolismo microbiano
e animal, permitiu um melhor entendimento da manifestação de algumas
respostas animais. Assim, um maior investimento de recursos e esforços na
investigação de aspectos do metabolismo intermediário se mostra necessário
para permitir um maior entendimento das respostas fisiológicas animais e o
surgimento ou desenvolvimento de modelos e sistemas de maior completude.

108
5 CONCLUSÃO
O não atendimento da demanda do rúmen em termos de proteína
degradável, visando otimizar o conseqüente crescimento microbiano, bem como
a promoção de condições superavitárias de proteína metabolizável, por
intermédio tanto da otimização da síntese de proteína microbiana como da
utilização em maior escala de proteína não degradável no rúmen, não
promoveram alterações nas ingestões de matéria seca nem mesmo no ganho de
peso e conversão alimentar em cordeiras Santa Inês, sob condições de
desempenhos moderados.
A presença de proteína degradável no rúmen em concentrações
suficientes para que se obtenha o atendimento da exigência em proteína
metabolizável de cordeiras em ritmos de crescimento moderado é suficiente para
permitir que as digestibilidades da matéria seca e das porções fibrosas não sejam
comprometidas, embora possa proporcionar menores retenções de nitrogênio.
A utilização do sistema britânico AFRC (1993) para ajuste da
alimentação de ovinos de raças origem tropical, alimentadas com forrageiras
também tropicais, não permite boa precisão na consecução dos níveis de
desempenho almejados quanto ao desenvolvimento ponderal.
A informação do teor de proteína bruta bem como da proporção de
proteína degradável no rúmen não permite uma predição acurada na forma como
os animais vão utilizar este nutriente ou todo o alimento.
Pesquisas semelhantes, que venham contemplar o uso de fontes de
carboidratos não-fibrosos de mais alta degradabilidade ruminal devem ser
levadas a efeito, inclusive considerando patamares mais altos de ganhos de peso.
Investigações futuras devem levar em considerem a mensuração da
degradabilidade da matéria seca e de nutrientes, a estimativa de produção

109
microbiana e a quantificação de proteína metabolizável disponibilizada ao nível
do intestino, objetivando proporcionar uma maior completude no conhecimento
dos efeitos advindo do atendimento ou não da demanda em proteína degradável
no rúmen sob diferentes condições de balanço de proteína metabolizável.

110
REFERÊNCIAS BIBLIOGRÁFICAS
AGRICULTURAL RESEARCH COUNCIL. Nutrients requirements of ruminant livestock. Wallingford: CAB International, 1980. 351p.
AGRICULTURAL AND FOOD RESEARCH COUNCIL. Energy and protein requirements of ruminants. Wallingford: CAB International, 1993. 159p.
ALLEN, M. S. Physical constraints on voluntary intake of forage by ruminants. Journal of Animal Science, Champaign, v. 74, n. 12, p. 3063-3075, Dec. 1996.
ALVES, K. S.; CARVALHO, F. F. R. de; VÉRAS, A. S. C.; ANDRADE, M. F. de; COSTA, R. G.; BATISTA, A. M. V.; MEDEIROS, A. N. de; SOUTO MAIOR Jr, R. J. de; ANDRADE, D. K. B. de. Níveis de energia em dietas para ovinos Santa Inês: desempenho. Revista Brasileira de Zootecnia, Viçosa, v. 32, n. 6 (Supl. 2), p. 1937-1944, 2003.
ANDRADE, P.; ALCALDE, C. Nutrição e alimentação do novilho precoce. In; ENCONTRO NACIONAL SOBRE NOVILHO PRECOCE. 1995, Campinas. Anais... Campinas: CATI, 1995, p. 93-109.
ANDRIGUETTO, J. L.; CAVASSIN, E. Proteína protegida de soja e desempenho de cordeiros em confinamento. Archives of Veterinary Science, Curitiba, v. 7, n. 1, p. 49-55, 2002.
ARAÚJO, G. G. L. de; COELHO DA SILVA, J. F.; VALADARES FILHO, S. de C.; LEÃO, M. I.; CASTRO, A. C. G.; QUEIROZ, A. C. de. Efeito da degradabilidade da proteína sobre o consumo e digestão de matéria seca, matéria orgânica e carboidratos estruturais em vacas lactantes. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v. 24, n. 3, p. 371-381, 1995.
ARAÚJO, G. G. L. de; COELHO DA SILVA, J. F.; VALADARES FILHO, S. de C.; LEÃO, M. I.; VALADARES, R. F. D.; ALMEIDA, G. A. P. de. Efeito da degradabilidade da proteína sobre o consumo e digestão da proteína bruta, do extrato etéreo e balanço do nitrogênio de vacas lactantes. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v. 24, n. 2, p. 258-267, 1994.
ARAÚJO, G. G. L. de; COELHO DA SILVA, J. F.; VALADARES FILHO, S. de C.; CAMPOS, O. F. de; CASTRO, A. C. G. de; SIGNORETTI, R. D.; TURCO, S. H. N.; HENRIQUES, L. T. Consumo e digestibilidade total dos

111
nutrientes de dietas contendo diferentes níveis de volumosos, em bezerros. Revista Brasileira de Zootecnia, Viçosa, v. 27, n. 2, p. 345-354, 1998.
BADRA, A. Suplementação protéica de bovinos em pastagens de Brachiaria
brizantha (Hochst) Stapf cv. Marandu. 1996. 36p. Monografia (Trabalho de Graduação) - Faculdade de Ciências Agrárias e Veterinárias - UNESP, Jaboticabal.
BAGANHA, A. Tratado elementar de hygiene pecuária e zootecnia moderna. Porto, 1878, 262p.
BALDWIN, R. L.; DONOVAN, K. .C. Modelling the lactating dairy cow. In: THEODOROU, M. K.; FRANCE, J. (Eds). Feeding systems and feed evaluation models. Wallingford: CAB International, 2000. p. 323-342.
BAUMAN, D. E.; CURRIE, W. B. Partitioning of nutrients during pregnancy and lactation: a review of mechanisms involving homeostasis and homeorhesis. Journal of Dairy Science, Champaign, v. 63, n. 9 p. 1514-1529, 1980.
BEEVER, D. E.; MOUD, L. F. Forage evaluation for efficient ruminant livestock production. In: GIVENS, D. I.; OWENS, E.; AXFORD, R. F. E.; OMED, H. E. (Ed.) Forage evaluation in ruminant nutrition. Wallingford: CAB Publishing, 2000, cap.2, p. 15-42.
BERGMAN, E. N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiological Reviews, v. 70, n. 2, p. 567-590, 1990.
BISPO, A. R. Efeito da uréia e da glutenose na suplementação de tourinhos nelore em pastagens de Brachiaria brizantha (Hoscht) Stap cv. Marandu. 2000. 73p. Dissertação (Mestrado em Zootecnia) - Universidade Estadual Paulista / FCAV, Jaboticabal.
BORGES. A. L. C. C. Controle da ingestão de alimentos. Caderno Técnico da Escola de Veterinária da UFMG, Belo Horizonte, v. 27, p. 67-79, 1999.
BRANCO, A. F.; MOURO, G. F.; HARMON, D. L.; RIGOLON, L. P.; ZEOULA, L. M.; MAIA, F. J.; CONEGLIAN, S. M. Fontes de proteína,ingestão de alimentos e fluxo esplâncnico de nutrientes em ovinos. Revista Brasileira de Zootecnia, Viçosa, v. 33, n. 2, p. 444-452, 2004.

112
BRODERICK, G. A.; MERCHEN, N. R. Markers for quantifying microbial protein synthesis in the rumen. Journal of Dairy Science, Champaign, v. 75, n. 9, p. 2618-2632, 1992.
BURROUGHS, W.; NELSON, D. K.; MERTENS, D. R. Evaluation of protein nutrition by metabolizable protein and urea fermentation potential. Journal of Dairy Science, Champaign, v. 58, p. 611-219, 1975a.
BURROUGHS, W.; NELSON, D. K.; MERTENS, D. R. Protein physiology and its implications in the lactating cow: The metabolizable protein feeding standard. Journal of Animal Science, Champaign, v. 41, p. 933-944, 1975b.
CALDAS NETO, S. F.; ZEOULA, L. M.; KAZAMA, R.; PRADO, I. N. do; GERON, L. J. V.; OLIVEIRA, F. C. L. de; PRADO, O. P. P. do. Proteína degradável no rúmen associada a fontes de amido de alta ou baixa degradabilidade: digestibilidade in vitro e desempenho de novilhos em crescimento. Revista Brasileira de Zootecnia, Viçosa, v. 36, n. 2, p. 452-460, 2007.
CALDAS NETO, S. F.; ZEOULA, L. M.; PRADO, I. N. do; KAZAMA, R.; PRADO, O. P. P. do; OLIVEIRA, F. C. L. de; GERON, L. J. V. Avaliação do efeito do nível de proteína degradável no rúmen em rações contendo fontes de amido de alta degradabilidade ruminal. 1. Digestibilidade in vitro. In: RUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002. Anais... Recife: SBZ, 2002. (1 CD-ROOM)
CANNAS, A.; ATZORI, A. S. Development and evaluation of a model to predict sheep nutrient requirements and feed utilization. Italian Journal of Animal Science, Bologna, v. 4, suppl. 1, p. 15-33, 2005.
CANNAS, A.; TEDESCHI, L. O.; FOX, D. G.; PELL, A. N.; VAN SOEST, P. J. A mechanistic model for predicting the nutrient requirements and feed biological values for sheep. Journal of Animal Science, Champaign, v. 82, n. 1, p. 149-169, 2004.
CARMO C. A.; SANTOS, F. A. P.; SCOTON, R. A.; FERNANDES, R. H. R.; NUSSIO, L. G.; PIRES, A. V. Substituição do farelo de soja por uréia ou amiréia em dietas para vacas em final de lactação. 2. Metabolismo. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: SBZ, 2002. 1 CD-ROM.

113
CASPER, D. P.; SCHINGOETHE, D. J. Lactational response of dairy cows to diets varying in ruminal solubilities of carbohydrate and crude protein. Journal of Dairy Science, Champaign, v. 72, n. 2, p. 928-936, 1989.
CAVALCANTE, M. A. B.; PEREIRA, O. G.; VALADARES FILHO, S. de C.; RIBEIRO, K. G. Níveis de proteína bruta em dietas para bovinos de corte: Consumo, digestibilidade total e desempenho produtivo. Revista Brasileira de Zootecnia, Viçosa, v. 34, n. 3, p. 711-719, 2005.
CAVALCANTE, M. A. B.; PEREIRA, O. G.; VALADARES FILHO, S. de C.; RIBEIRO, K. G.; PACHECO, L. B. B.; ARAÚJO, D.; LEMOS, V. M. C. Níveis de proteína bruta em dietas para bovinos de corte: Parâmetros ruminais, balanço de compostos nitrogenados e produção de proteína microbiana. Revista Brasileira de Zootecnia, Viçosa, v. 36, n. 1, p. 203-210, 2006.
CERVIERI, R. da C.; ARRIGONE, M. B.; OLIVEIRA, H. N. de; SILVEIRA, A. C.; CHARDULO, L. A. L.; MARTINS, C. L. Desempenho e características de carcaça de bezerros confinados recebendo dietas com diferentes degradabilidades da fração protéica. Revista Brasileira de Zootecnia, Viçosa, v. 30, n. 5, p. 1590-1599, 2001
CECAVA, M. J.; HANCOCK, D. L. Effects of anabolic steroids on nitrogen metabolism and growth of steers fed corn silage and corn-based diets supplemented with urea or combination of soybean meal and feather meal. Journal of Animal Science, Champaign, v. 72, n. 9, p. 515-522, 1994.
CECAVA, M. J.; MERCHEN, N. R.; BERGER, L. L.; FAHEY JR., G. C. Intestinal supply of amino acids in sheep fee alkaline hydrogen peroxide-treated wheat straw-based diets supplemented with soybean meal or combination of corn gluten meal and blood meal. Journal of Animal Science, Champaign, v. 68, n. 2, p. 467-477, 1990.
CECAVA, M. J.; MERCHEN, N. R.; BERGER, L. L.; MACKIE R. I.; FAHEY JR., G. C. Effects of dietary energy level and protein source on nutrient digestion and ruminal nitrogen metabolism in steers. Journal of Animal Science, Champaign, v. 69, n. 5, p. 2230-2243, 1991.
CHALUPA, W.; CLARK, J.; OPLIGER, P.; LAVKER, R. Detoxication of ammonia in sheep fed soy protein or urea. Journal of Nutrition, Bethesda, v. 100, n. 2, p. 170-176, 1970.

114
CHALUPA, W.; SNIFFEN, C. J.; FOX, D. G.; VAN SOEST, P. J. Model generated protein degradation nutritional information. In: CORNELL NUTRITION CONFERENCE FOR FEED MANUFACTORERS. 1991, Ithaca. Proceedings... Ithaca: Cornell University, 1991. p. 44-51.
CHAMBERLAIN, D. G.; CHOUNG, J. J. The importance of rate of ruminal fermentation of energy sources diets of dairy cows. In: GARNSWORTHY, P. C.; COLE, D.J. A (Eds), Recent Advances in Animal Nutrition. Nottingham: Nottingham University Press, 1995, p. 3-27.
CHRISTENSEN, R. A.; CAMERON, M. R.; KLUSMEYER, T. H.; ELLIOTT, P.; CLARK, J. H.; NELSON, D. R.; YU, U. Influence of amount and degradability os dietary protein on production of milk components by lactating Holstein cows. Journal of Dairy Science, Champaign, v. 76, n. 11, p. 3497-3513, 1993.
CLARK, J. H.; KLUSMEYER, T. H.; CAMERON, M. R. Microbial protein synthesis and flow of nitrogen fractions to the duodenum of dairy cows. Journal of Dairy Science, Champaign, v. 75, n. 8, p. 2304-2323, 1992.
COCHRAN, R. C.; KOSTER, H. H.; OLSON, K. C. Supplemental protein for grazing cattle. Feedstuffs, v. 70, n. 7, p. 12-19, 1998.
COELHO DA SILVA, J. F.; LEÂO, M. I. Fundamentos de nutrição dos ruminantes. Piracicaba: Livroceres, 1979. 380p.
CONRAD, H. R. Symposium on factors influencing the voluntary intake of herbage for ruminants: Physiological and physical factors limiting feed intake. Journal of Animal Science, Champaign, v. 25, n. 1, p. 227-235, 1966.
COSTA, R. M. Avaliação de suplementos com proteína degradável e de escape ruminal para recria de bovinos. 2001. 47p. Dissertação (Mestrado em Zootecnia) - Universidade Estadual Paulista / FCAV, Jaboticabal.
COUTINHO FILHO, J. L. V.; SAMPIO, A. A. M.; EZEQUIEL, J. M. B.; OLIVEIRA, M. D. S.; VIEIRA, P. de F. Efeito de fontes de nitrogênio sobre a ingestão e digestibilidade aparente de diferentes rações. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v. 24, n. 6, p. 1038-1044, 1995.
DEWHURST, R. J.; DAVIS, D. R.; MERRY, R. J. Microbial protein supply from the rumen. Animal Feed Science and Technology, Amsterdam, v. 85, n. 1, p. 1-21, 2000.

115
DEWHURST, R. J.; MITTON, A. M.; OFFER, N. W. THOMAS, C. Effects of the composition of grass silages on milk production and nitrogen utilization by dairy cows, Animal Science, v. 62, p. 25-34, 1996.
DIJKSTRA, J.; FRANCE, J. Modeling and methodology in animal science. In: DANFAER, A.; LESCOAT, P. (Eds.) FOURTH INTERNATIONAL WORKSHOP ON MODELING NUTRIENT UTILIZATION IN FARM ANIMALS, 1995, Foulum. Proceedings…, Foulum, Denmark: National Institute of Animal Science, 1995, p. 9-18.
DUTRA, A. R.; QUEIROZ, A. C. de; PEREIRA, J. C.; VALADARES FILHO, S. de C.; THIÉBAUT, J. T. L.; MATOS, F. N.; RIBEIRO, C. V. D. M. Efeitos dos níveis de fibra e das fontes de proteína sobre o consumo e digestão dos nutrientes em novilhos. Revista Brasileira de Zootecnia, Viçosa, v. 26, n. 4, p. 787-796, 1997.
EZEQUIEL, J. M. B. Exigências de proteína e minerais em bovinos: frações endógenas. 1987. 131p. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa, Viçosa.
EZEQUIEL, J. M. B.; SAMPAIO, A. A. M.; SEIXAS, J. R. C.; OLIVEIRA, M. M. de. Balanço de nitrogênio e digestão total da proteína e da energia de rações contendo farelo de algodão, levedura de cana-de-açúcar ou uréia, em ovinos. Revista Brasileira de Zootecnia, Viçosa, v, 29, n. 6 (supl 2), p. 2332-2337, 2000.
EZEQUIEL, J. M. B.; SAMPAIO, A. A. M.; SEIXAS, J. R. C.; OLIVEIRA, M. M. de. Digestibilidade aparente da energia e da fibra de dietas para ovinos contendo uréia, amiréia ou farelo de algodão. Revista Brasileira de Zootecnia, Viçosa, v, 30, n. 1, p. 231-235, 2001.
FAGUNDES NETO, J. C.; BRAGA, A. P.; RIBEIRO, H. U.; BARRA, P. B.; VASCONCELOS, S. H. L.; SILVA, A. C. da; LACERDA, P. M. de O.; LEITE, W. de P.; BRAGA, Z. C. A. da C. Substituição do farelo de soja pela mistura milho/uréia sobre o desempenho de ovinos mestiços de Santa Inês. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38., 2001, Piracicaba. Anais... Piracicaba: SBZ, 2001. 1 CD-ROM.
FERREIRA, D. F. Análises estatísticas por meio do Sisvar para Windows versão 4.0. In: REUNIÃO ANULA DA REGIÃO BRASILEIRA DA SOCIEDADE INTERNACIONAL DE BIOMETRIA, 2000, São Carlos. Resumos... São Carlos: RBRAS/UFSCar, 2000. p. 255-258.

116
FERREIRA, J. J.; SALGADO, J. G.; CARNEIRO, J. C. Efeitos de diferentes fontes e níveis de substituição de proteína por uréia na dieta de bovinos confinados. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 33., 1996, Fortaleza. Anais... Fortaleza: SBZ, 1996, p. 31.
FIRKINS, J.L. Maximizing microbial protein synthesis in the rumen. Journal of Nutrition, Pennsylvania, v. 126, p. 1347S-1354S, 1996.
FLUHARTY, F. L.; McCLURE, K. E. Effects of dietary energy intake and protein concentration on performance and visceral organ mass in lambs. Journal of Animal Science, Champaign, v. 75, n. 3, p. 604-610, 1997.
FOX, D. G.; TEDESCHI, L. O.; TYLUTKI, T. P.; RUSSEL, J. B.; VAN AMBURGH, M. E.; CHASE, L. E.; PELL, A. N.; OVERTON, T. R. The Cornell net carbohydrate and protein system model for evaluating herd nutrition and nutrient excretion. Animal Feed Science and Technology, Amsterdam, v. 112, n. 1/4, p. 29-78, 2004.
FOX, D. G.; TYLUTKI, T. P.; TEDESCHI, L. O.; VAN AMBURGH, M. E.; CHASE, L. E.; PELL, A. N.; OVERTON, T. R.; RUSSELL, J. B. Sistema de carboidratos e proteínas “líquidos” para avaliação da nutrição de rebanhos e excreção de nutrientes (CNCPS, versão 5.0): documentação do Modelo CNCPS./ Danny Gene Fox... [et al.]; tradução de Fernando César Ferraz Lopes...[et al] - Juiz de fora: Embrapa Gado de Leite, 2003. 202p.
FRANCO, G. F. Avaliação da proteína degradável no rúmen e da proteína metabolizável segundo o sistema de Cornell em rações de novilhos confinados. 2001. 74p. Tese (Doutorado em Zootecnia) - Universidade Estadual Paulista / FCAV, Jaboticabal.
FREGADOLLI, F. L.; ZEOULA, L. M.; BRANCO, A. F.; PRADO, I. N. do; CALDAS NETO, S. F.; GUIMARÃES, K. C.; KASSIES, M. P.; DALPONTE, A. O. Efeito das fontes de amido e nitrogênio de diferentes degradabilidade ruminais. 2. pH, concentração de amônia no liquido ruminal e eficiência de síntese microbiana. Revista Brasileira de Zootecnia, Viçosa, v. 30, n. 3, p. 870-879, 2001a.
FREGADOLLI, F. L.; ZEOULA, L. M.; PRADO, I. N. do; BRANCO, A. F.; CALDAS NETO, S. F.; KASSIES, M. P.; DALPONTE, A. O. Efeito das fontes de amido e nitrogênio de diferentes degradabilidade ruminais. 1. Digestibilidades parcial e total. Revista Brasileira de Zootecnia, Viçosa, v. 30, n. 3, p. 858-869, 2001b.

117
FU, C. J.; FELTON, E. E.; LEHMKUHLER, J. W.; KERLEY, M. .S. Ruminal peptide concentration required to optimize microbial growth and efficiency. Journal of Animal Science, Champaign, v. 79, n. 5, p. 1305-1312, 2001.
FURUSHO-GARCIA, I. F. Desempenho, características de carcaça, alometria dos cortes e tecidos e eficiência da energia, em cordeiros Santa Inês e cruzas com Texel, Ile de France e Bergamácia. 2001. 316 p. Tese (Doutorado em Zootecnia) - Universidade Federal de Lavras, Lavras.
FURUSHO-GARCIA, I. F.; PÉREZ, J. R. O.; BONAGURIO, S.; ASSIS, R. M.; PEDREIRA, B. C.; SOUZA, X. R. de. Desempenho de cordeiros Santa Inês puros e cruzas Santa Inês com Texel, Ile de France e Bergamácia. Revista Brasileira de Zootecnia, Viçosa, v. 33, n. 6, p. 1591-1603, 2004.
GALYEAN, M. L.; OWENS, F. N. Effects of diet composition and level of feed intake on site and extent of digestion in ruminants. In: TSUDA, T.; SASAKI, Y.; KAWASHIMA, R. (Eds) Physiological aspects of digestion and metabolism in ruminants: proceedings of the seventh International Symposium on Ruminant Physiology. San Diego: Academic Press Inc., 1991. p. 483-514.
GOERING, H. K.; VAN SOEST, P. J. Forage fiber analysis (apparatus, reagents, procedures and some applications). Washington: USDA/ARS. 1970. 379p.
GUIDI, M. T. Efeito de teores e fontes de proteína sobre o desempenho de vacas de leite e digestibilidade dos nutrientes. 1999. 92p. Dissertação (Mestrado em Agronomia) - Escola Superior de Agricultura “Luiz de Queiroz”, Piracicaba.
GUTIERREZ-ORNELAS, E.; KLOPFENSTEIN, T. Alfalfa and escape protein supplements for grazed corn residues. Journal of Animal Science, Champaign, v. 72, n. 12, p. 3042-3048, 1994.
HARMEYER, J.; MARTENS, H. Aspects of urea metabolism in ruminants with reference to the goat. Journal of Dairy Science, Champaign, v. 63, n. 10, p. 1707-1728, 1980.
HARRIS, P. M.; LOBLEY, G. E. Amino acid and energy metabolism in peripheral tissues of ruminants. In: TSUDA, T.; SASAKI, Y.; KAWASHIMA, R. (Eds) Physiological aspects of digestion and metabolism in ruminants:

118
proceedings of the seventh International Symposium on Ruminant Physiology. San Diego: Academic Press Inc., 1991. p. 201-230.
HENNING, P. H.; STEYN, D. G.; MEISSNER, H. H. Effect of synchronization of energy and nitrogen supply on ruminal characteristics and microbial growth. Journal of Animal Science, Champaign, v. 71, n. 9, p. 2516-2528, 1993.
HERRERA-SALDANA, R.; GOMEZ-ALARCON, R.; TORABI, M.; HUBER, J. T. Influence of synchronizing protein and starch degradation in the rumen on nutrient utilization and microbial protein synthesis. Journal of Dairy Science, Champaign, v. 73, n. 1, p. 142-148, 1990.
HUBER, J. T.; COOK, R. M. Influence of site of administration of urea on voluntary intake of concentrate by lactating cows. Journal of Dairy Science, Champaign, v. 55, n. 10, p.1470-1473, 1972.
HUBER, J. T.; KUNG, L. Protein and non-protein nitrogen utilization in cattle. Journal of Animal Science, Champaign, v. 64, n. 6, p.1170-1195, 1981.
HUNGATE, R. E. The rumen and its microbes. Academic Press, New York, 1966. 533p.
HUSTON, J. E.; ENGDAHL, R. S.; BALES, K. M. Intake and digestibility in sheep and goats fed three forages with different levels of supplemental protein. Small Ruminant Research, Amsterdam, v. 1, n, 1, p.81-88, 1988.
ILLIUS, A. W.; JESSOP, N. S. Metabolic constraints on voluntary intake in ruminants. Journal of Animal Science, Champaign, v. 74, n. 12, p. 3052-3062, 1996.
IMAIZUMI, H. Avaliação de diferentes fontes e teores de proteína degradável no rúmen sobre o desempenho e parâmetros ruminais e sangüíneos de vacas holandesas em final de lactação. 2000. 69p. Dissertação (Mestrado em Agronomia) - Escola Superior de Agricultura “Luiz de Queiroz” - Piracicaba.
ITAVO, L. C. V.; VALADARES FILHO, S. de C.; SILVA, F. F. da; VALADARES, R. F. D.; CECON, P. R. Níveis de concentrado e proteína bruta na dieta de bovinos Nelore nas fases de recria e terminação: Consumo e digestibilidade. Revista Brasileira de Zootecnia,Viçosa, v. 31, n. 2 (Supl.), p. 1033-1041, 2002.

119
JETANA, T., ABDULLAH, N.; HALIM, JALALUDIN, R. A.; HO, Y. W. Effects of energy and protein supplementation on microbial-N synthesis and allantoin excretion in sheep fed guinea grass. Animal Feed Science and Technology, Amsterdam, v. 84, p. 167-181, 2000.
KIM, K. H.; CHOUNG, J. J.; CHAMBERLAIN, D. G. Effects of varying degrees of synchrony of energy and nitrogen release in the rumen on the synthesis of microbial protein in cattle consuming a diet of grass silage and cereal-based concentrate. Journal of the Science of Food and Agriculture, v. 79, p. 1441-1447, 1999a.
KIM, K. H.; OH, Y. G.; CHOUNG, J. J.; CHAMBERLAIN, D. G. Effects of varying degrees of synchrony of energy and nitrogen release in the rumen on the synthesis of microbial protein in cattle consuming a diet of grass silage. Journal of the Science of Food and Agriculture, v. 79, p. 833-838, 1999b.
KING, K. J.; HYBER, J. T.; SADIK, M.; BERGEN, W. G.; GRANT, A. L.; KING, V. L. Influence of dietary protein sources on the amino acids profiles available for digestion and metabolism in lactating cows. Journal of Dairy Science, Champaign, v. 73, n. 11, p. 3208-3216, 1990.
KLOPFENSTEIN, T. Need for escape protein by grazing cattle. Animal Feed Science and Technology, v. 60, n. 3-4, p. 191-199, 1996.
KOLB, E. Fisiologia veterinária. Guanabara Koogan, Rio de Janeiro, 1984. 612p.
KOLVER, E.; MULLER, L. D.; VARGA, G. A.; CASSIDY, T. J. Synchronization of ruminal degradation of supplemental carbohydrate with pasture nitrogen in lactating dairy cows. Journal of Dairy Science, Champaign, v. 81, n. 8, p. 2017-2028, 1998.
KRISHNAMOORTHY, U.; MUSCATO, T. V.; SNIFFEN, C. J.; VAN SOEST, P. J. Nitrogen fractions in selected feedstuffs. Journal of Dairy Science, Champaign, v. 65, n. 1, p. 217-225, 1982.
KRISHNAMOORTHY, U.; SNIFFEN, C. J.; STERN, M. D.; VAN SOEST, P. J. Evaluation of a mathematical model of rumen digestion and in vitro simulation of rumen proteolysis to estimate the rumen-undegraded content of feedstuffs. British Journal of Nutrition, Cambridge, v. 50, p. 555-568, 1983.

120
KLUSMEYER, T. H. ; McCARTHY JR, R. D.; CLARK, J. H.; NELSON, D. R. Effects of source and amount of protein on ruminal fermentation and passage of nutrients to the small intestine of lactating cows. Journal of Dairy Science, Champaign, v. 73, n. 12m p. 3526-3537, 1990.
LAVEZZO, O. E. N. M.; LAVEZZO, W.; BURINI, R. C. Efeitos nutricionais da substituição parcial do farelo de soja por uréia, em dietas de ovinos. Comparação da digestibilidade aparente e balanço de nitrogênio com a cinética do metabolismo da 15N-glicina. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v. 25, n. 2, p. 282-297, 1996.
LIZIEIRE, R. S.; COELHO DA SILVA, J. .F; LEÃO, M. I.; VALDARES FILHO, S. de C.; CAMPOS, O. F. de. Níveis crescentes de proteína degradável no rúmen de cabras. 1. Efeitos sobre o consumo, digestibilidade aparente e balanço de nitrogênio. Revista da Sociedade Brasileira de Zootecnia, Viçosa, v. 19, n. 6, p. 552-561, 1990.
MAGALHÃES, K. A.; VALADARES FILHO, S. de C.; VALADARES, R, F. D.; PAULINO, M. F.; PAULINO, P. V. R.; PORTO, M. O.; CHIZZOTTI, M. L.; ANDREATTA, K.; ADÃO, L. A. Níveis de uréia em substituição ao farelo de soja na dieta de novilhos de origem leiteira em confinamento. 1. Desempenho. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: SBZ, 2002. 1 CD-ROM.
MAGALHÃES, K. A.; VALADARES FILHO, S. de C.; VALADARES, R, F. D.; PAIXÃO, M. L.; PINA, D. dos S.; PAULINO, P. V. R.; CHIZZOTTI, M. L.; MARCONDES, M. I.; ARAÚJO, A. M.; PORTO, M. O. Produção de proteína microbiana, concentração plasmática de uréia e excreções de uréia em novilhos alentados com diferentes níveis de uréia ou casca de algodão. Revista Brasileira de Zootecnia, Viçosa, v. 34, n. 4, p. 1400-1407, 2005.
MALAFAIA, P. A. M.; VIEIRA, R. A. M. Técnicas de determinação e avaliação dos compostos nitrogenados em alimentos para ruminantes. In: SIMPÓSIO INTERNACIONAL DE DIGESTIBILIDADE EM RUMINANTES,1997, Lavras. Anais... Lavras: UFLA, 1997. p. 29-54.
MARTIN, L. C.; AMMERMAN, C. B.; HENRY, P. R.; LOGGINS. P. E. Effect of level and form of supplemental energy and nitrogen on utilization of low quality roughages by sheep. Journal of Animal Science, Champaign, v. 53, n. 2, p. 479-488, 1981.

121
MARTINS, A. S.; ZEOULA, L. M.; PRADO, I.N. do; MARTINS, E. N.; LOYOLA, V. R. Degradabilidade ruminal in situ d matéria seca e proteína bruta das silagens de milho e sorgo e de alguns concentrados. Revista Brasileira de Zootecnia, Viçosa, v. 28, n. 5, p. 1109-1117, 1999.
McCARTHY, JR., R. D., KLUSMEYER, T. H., VICINI, J. L., CLARK, J. H., NELSON, D. R.. Effects of source of protein and carbohydrate on ruminal fermentation and passage of nutrients to the small intestine of lactating cows. Journal of Animal Science, Champaign, v. 70, n. 8, p. 2002-2016, 1989.
MEHEREZ, H. Z.; ØRSKOV, E. R. Protein degradation and optimum urea concentration in cereal based diets for sheep. British Journal of Nutrition, Cambridge, v. 40, n. 2, p. 437-445, 1978.
MERTENS, D. R. Analysis of fiber in feeds and its uses in feed evaluation and ration formulation. In: SIMPÓSIO INTERNACIONAL DE RUMINANTES. REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 29., 1992, Lavras. Anais… Lavras: SBZ, 1992, p. 188-219.
MERTENS, D. R. Principles of modeling and simulation in teaching and research. Journal of Dairy Science, Champaign, v. 60, n. 7, p. 1176-1186, 1976.
MERTENS, D. R. Regulation of forage intake. In: FAHEY JR., G. C. (Ed.) Forage quality, evaluation and utilization. Madison: American Society of Agronomy, 1994. p. 450-493.
MILTON. C. T.; BRANDT, R. T. Level of urea in high grain diets: nutrient digestibility, microbial production and rumen metabolism. In: CATTLE FEEDERS DAY PROGRAM, 704., 1994, Kansas. Report... Kansas: Kansas State University, 1994, p. 4-6.
MINSON, D. J. Forage in ruminant nutrition. New York: Academic Press, 1990. 483p.
MOREIRA, A. L.; PEREIRA, O. G.; GARCIA, R.; VALADARES FILHO, S. de C.; CAMPOS, J. M. de S.; MORAES, S. A.; ZERVOUDAKIS, J. T. Consumo e digestibilidade aparente dos nutrientes da silagem de milho e dos fenos de alfafa e de capim coastcross, em ovinos. Revista Brasileira de Zootecnia, Viçosa, v. 30, n. 3, p. 1099-1105, 2001 (Supl.1).

122
MORRISON, M.; MACKIE, R. I. Nitrogen metabolism by ruminal microorganisms: current understanding and future perspectives. Australian Journal of Agricultural Research, Victoria, v. 47, n. 2, p. 227-246, 1996
NATIONAL RESEARCH COUNCIL. Nutrient requirements of beef cattle. 7th ed. Washington, D. C.: National Academy Press, 2000. 242p.
NATIONAL RESEARCH COUNCIL. Nutrient requirements of dairy cattle. 7th ed. Washington, D. C.: National Academy Press, 2001. 381p.
NATIONAL RESEARCH COUNCIL. Nutrient requirements of small ruminants. 1st ed. Washington, D. C.: National Academy Press, 2006. 362p.
NATIONAL RESEARCH COUNCIL. Ruminant nitrogen usage. Washington, D. C.: National Academy Press, 1985. 138p.
NOLLER, C. H.; NASCIMENTO JR., D. do; QUEIROZ, D. S. Exigências nutricionais de animais em pastejo. In: PEIXOTO, A. M.; MOURA, J. C. de; FARIA, V. P. de (Eds.) SIMPÓSIO SOBRE MANEJO DA PASTAGEM: Produção de bovinos a pasto, 13. 1996, Piracicaba. Anais... Piracicaba: FEALQ, 1996, p. 319-352.
OBEID, J. A.; PEREIRA, O. G.; PEREIRA, D. H.; VALADARES FILHO, S. de C.; CARVALHO, I. P. C. de; MARTINS, J. M. Níveis de proteína bruta em dietas para bovinos de corte: consumo , digestibilidade e desempenho produtivo. Revista Brasileira de Zootecnia, Viçosa, v. 35, n. 6, p. 2434-2442, 2006.
OLIVEIRA JR, R. C. de; PIRES, A. V.; FERNANDES, J. J. de R.; SUSIN, I.; SANTOS, F. A. P.; ARAÚJO, R. C. de. Substituição total do farelo de soja por uréia ou amiréia em dietas com alto teor de concentrado, sobre a amônia ruminal, os parâmetros sangüíneos e o metabolismo do nitrogênio em bovinos de corte. Revista Brasileira de Zootecnia, Viçosa, v. 33, n. 3, p. 738-748, 2004.
OLIVEIRA JR, R. C. de. Substituição do farelo de soja por uréia ou amiréia em dietas de bovinos de corte. I. Digestibilidade de nutrientes, balanço de nitrogênio, parâmetros ruminais e sangüíneos; II. Desempenho e; III. Avaliação de indicadores de digestibilidade. 2002. 198p. Tese (Doutorado em Agronomia) - Escola Superior de Agricultura “Luis de Queiroz”, Piracicaba.
OLDHAM, J. D.; SMITH, T. Protein-energy interrelationships for growing and for lactating cattle. In: MILLER, E. L.; PIKE, I. H.; VAN ES (Ed.) Protein contribution of feedstuffs for ruminants. Butterworths: London, 1982, p.103.

123
OLDHAM, J. D. Protein-energy interrelationship in dairy cows. Journal of Dairy Science, Champaign, v. 67, n.
ØRSKOV, E. R. Protein nutrition in ruminants. 2nd Ed. London: Academic Press, 1992. 175p.
ORTIZ, J. S.; COSTA, C.; GARCIA, C. A.; SILVEIRA, L. V. de A. Efeito de diferentes níveis de proteína bruta na ração sobre o desempenho e as características de carcaça de cordeiros terminados em creep feeding. Revista Brasileira de Zootecnia, Viçosa, v. 34, n. 6 (Supl.), p. 2390-2398, 2005.
OWENS, F. N.; GOETSCH, A. L. Ruminal fermentation. In: CHURCH, D. C. The ruminant animal - digestive physiology and nutrition. Simon & Schuster Englewood Cliffs, Waveland Press: Illinois, 1988, p. 145-171.
OWENS, F. N.; ZINN, R. Protein metabolism in ruminants. In: CHURCH, D. C. The ruminant animal - digestive physiology and nutrition. Simon & Schuster Englewood Cliffs, Waveland Press: Illinois, 1988, p. 255-281.
PAIXÃO, M. L.; VALADARES FILHO, S. de C.; LEÃO, M. I.; VALADARES. R. F. D.; PAULINO, M. F.; MARCONDES, M. I.; FONSECA. M. A.; SILVA, P. A.; PINA, D. dos S. Uréia em dietas para bovinos: Consumo, digestibilidade dos nutrientes, ganho de peso, características de carcaça e produção microbiana. Revista Brasileira de Zootecnia, Viçosa, v. 35, n. 6, p. 2451-2460, 2006.
PARSONS, S. D.; ALLISON, C. D. Grazing management as it affects nutrition, animal production and economics of beef production. In: MASS. J. (Ed). Veterinary Clinics of North America. Saunders Company: Philadelphia. 1991, p. 77-97.
PEREIRA, E. S. Dinâmica ruminal e pós-ruminal da proteína e de carboidratos: aplicação de um modelo matemático para avaliação de dietas à base de cana-de-açúcar. 1999. 95p. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa, Viçosa.
PINA, D. dos S.; VALADARES FILHO, S. de C.; VALADARES, R. F D.; DETMANN, E. CAMPOS, J. M. de S.; FONSECA, M. A.; TEIXEIRA, R. M. A.; OLIVEIRA, A. S. de. Síntese de proteína microbiana e concentração de uréia em vacas alimentadas com diferentes fontes de proteína. Revista Brasileira de Zootecnia, Viçosa, v. 35, n. 4, p. 1552-1559, 2006.

124
POLAN, C. E. Dietary protein and microbial protein contribution. Journal of Nutrition, Pennsylvania, v. 18, p. 242-248, 1988.
POORE, M. H.; MOORE, J. A.; ECK. T. P.; Effect of fiber source and ruminal starch degradability on site and extension of digestion in dairy cows. Journal of Dairy Science, Champaign, v. 76, n. 10, p. 2244-2259, 1993.
PRESTON, T. R.; LENG, R. A. Matching ruminant production systems with available resources in the tropics and subtropics. Penambul Books: Armidale, Australia, 1987. 245p.
PRADO, O. P. P. do; ZEOULA, L. M.; GERON, L. J. V.; CALDAS NETO, S. F.; MIDORI, E. M.; FERELI, F.; KAZAMA, R.; OLIVEIRA, F. C. L. de. Balanço do nitrogênio e digestibilidade da energia bruta de rações com diferentes teores de proteína degradável no rúmen e fonte de amido de alta degradabilidade ruminal em ovinos. In; REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41., 2004. Campo Grande. Anais... Campo Grande: SBZ, 2004 (1 CD-ROOM)
QUINTÃO, F. A. Valor nutritivo de dietas à base de feno de “coastcross” suplementadas com uréia ou amiréia no desempenho de ovelhas da raça Santa Inês. 2006. 99p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Lavras - Lavras.
REIS, R. A.; RODRIGUES, L. R. A. Suplementação como estratégia de manejo da pastagem. In: PEIXOTO, A. M.; MOURA, J. C. de; FARIA, V. P. de (Eds.) SIMPÓSIO SOBRE MANEJO DA PASTAGEM: Produção de bovinos a pasto, 13. 1996, Piracicaba. Anais... Piracicaba: FEALQ, 1996, p. 123-150.
REIS, S. T. dos. Fracionamento e degradabilidade ruminal de proteínas e carboidratos de forrageiras do gênero Cynodon. 2005. 70p. Tese (Doutorado em Zootecnia) - Universidade Federal de Lavras, Lavras.
RENNÓ. L. N. Consumo e digestibilidade total e parcial, produção microbiana, parâmetros ruminais e excreções de uréia e creatinina em novilhos alimentados com dietas contendo quatro níveis de uréia ou dois níveis de proteína. 2003. 252p. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa, Viçosa.
RENNÓ, L. N.; VALADARES FILHO, S. de C.; VALADARES, R. F. D.; CECON, P. R.; BACKES, A. A.; RENNÓ, F. P.; ALVES, D. D.; SILVA. P. A. Níveis de uréia na ração de novilhos de quatro grupos genéticos: consumo e

125
digestibilidade totais. Revista Brasielira de Zootecnia, Viçosa, v. 34, n. 5, p. 1775-1785, 2005.
RIBEIRO, K. G.; GARCIA, R.; PEREIRA, O. G.; VALADARES FILHO, S. de C.; CECON, P. R. Eficiência microbiana, fluxo de compostos nitrogenados no abomaso, amônia e pH ruminais em bovinos recebendo dietas contendo feno de capim-Tifton 85 de diferentes idades de rebrota. Revista Brasileira de Zootecnia, Brasília, v. 30, n. 2, p. 581-588, 2001.
RICHARDS, C.; SHAIN, D.; STOCK, R.; KLOPFENSTEIN, T. Effect of wet corn gluten feed and supplemental protein on calf finishing performance. In: Nebraska Beef Reports, MP 62-A, p. 26-27, 1995.
RODRIGUES, P. H. M.; RODRIGUES, R. R.; FERNADES, J. I. M. ; PASSINI, R.; MELOTTI, L.; FERRAZ, E.; CASTRO, A. L. de. Digestibilidade aparente com ovinos de duas gramíneas do gênero Cynodon [Cynodon dactylon (L.) Pers]. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998,Botucatu. Anais… Botucatu: SBZ, 1998. p.503.
ROMNEY, D. L.; GILL, M. Intake of forages. In: GIVENS, D. I.; OWEN, E.; AXFORD, R. F. E.;OMED, H. M. (Edit). Forage evaluation in ruminant nutrition. London: CABI Publishing, 2000. p. 43-62.
ROOKE, J. A.; LEE, N. H.; ARMSTRONG, D. G. The effect of intraruminal infusions of urea, casein, glucose syrup and a mixture of casein and glucose syrup on nitrogen digestion in the rumen of cattle receiving grass-silage diets. British Journal of Nutrition, Cambridge, v. 57, n. 1, p. 89-98, 1987.
ROSELER, D. K.; FERGUSON, J. D.; SNIFFEN. C. J.; HERREMA, J. Dietary protein degradability effects on plasma and milk urea nitrogen and milk nonprotein nitrogen in Holstein cows. Journal of Dairy Science, Champaign, v. 76, n. 2, p. 525-534, 1993.
RUSSELL, J. B. Factors influencing competitions and compositions of rumen bacterial flora. In: HERBIVORE NUTRITION IN SUB-TROPICS AND TROPICS SYMPHOSIUM. 1984, Graighall. Proceedings… Graighal: The Science Press, South Africa, 1984. p. 313.
RUSSELL, J. B.; O’CONNOR, J. D.; FOX, D. G.; VAN SOEST, P. J.; SNIFFEN, C. J. A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. Journal Animal Science, Champaign, v. 70, n. 11, p. 3551-3561, 1992.

126
RUSSELL, J. B. Rumen microbiology and its role in ruminant nutrition. Ithaca, NY, 2002, 119p.
SALMAN, A. K. D.; MATARAZZO, S. V.; EZEQUIEL, J. M. B.; KRONKA, S. N.; SEIXAS, J. R. C.; SOARES, W. V. B.; MARTINS JR., A. P. Estudo do balanço nitrogenado e da digestibilidade da matéria seca e proteína de rações para ovinos, suplementadas com amiréia, uréia ou farelo de algodão. Revista Brasileira de Zootecnia, Viçosa, v. 26, n. 1, p. 179-185, 1997.
SALVADOR, F. M.; TEIXEIRA, J. C.; PÉREZ, J. R. O.; EVANGELISTA, A. R.; MUNIZ, J. A. Utilização de amiréias (produto da extrusão amido + uréia) com diferentes proporções de uréia: 1. Consumo e digestibilidade aparente da matéria seca, fibra em detergente neutro e fibra em detergente ácido. Ciência e Agrotecnologia Lavras, v. 28, n. 1, p. 199-205, 2004
SALVADOR, F. M. Utilização de amiréias (produto da extrusão amido + uréia) em ovinos alimentados com feno de Coastcross. 2001. 99p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Lavras, Lavras.
SAMPAIO. A. A. M.; VIEIRA, P. de F.; BRITO, R. M. de. Digestão total e parcial de nutrientes em bovinos alimentados com rações contendo levedura, uréia ou farelo de algodão. Revista Brasileira de Zootecnia, Viçosa, v. 29, n. 2, p. 589-597, 2000.
SANTOS. F. A. P. Metabolismo de proteínas. In; BERCHIELLI, T. T.; PIRES, A. V.; OLIVEIRA, S. G. de.(Ed.) Nutrição de ruminantes. Jaboticabal: FUNEP, 2006, p. 255-286.
SAS INSTITUTE INC. SAS User’s guide: statistics. 5th ed. Cary: NC, 1991. 1290p.
SCHWAB, C. G. Amino acid nutrition of the dairy cow: current status. In: CORNELL NUTRITION CONFERENCE FOR FEED MANUFACTORERS 1996, Ithaca. Proceedings... Ithaca: Cornell University, 1996. p. 184-198.
SHAIN, D. H.; STOCK, R. A.; KLOPFENSTEIN, T. J.; HEROLD, D. W. Effect of degradable intake protein level on finishing cattle performance and ruminal metabolism. Journal of Animal Science, Champaign, v. 76, n. 1, p. 242-248, 1998.
SIGNORETTI, R. D.; COELHO DA SILVA, J. F.; VALADARES FILHO, S. de C.; PEREIRA, J. C.; CECON, P. R.; QUEIROZ, A. C. de; ARAÚJO, G. G.

127
L. de; ASSIS, G. M. L. de Consumo e digestibilidade aparente em bezerros da raça holandesa alimentados com dietas contendo diferentes níveis de volumoso. Revista Brasileira de Zootecnia, Viçosa, v. 28, n. 1, p. 169-177, 1999.
SILVA, D. J.; QUEIROZ, A. C. de; Análise de alimentos (Métodos químicos e biológicos). Viçosa: UFV, 2002. 235p.
SILVA, R. M. N. da; VALADARES, R, D, F.; VALADARES FILHO, S. de C.; CECON, P. R.; CAMPOS, J. M. de S.; OLIVEIRA, G. A. de; OLIVEIRA, A. S. Uréia para vacas em lactação. 1. Consumo, digestibilidade, produção e composição do leite. Revista Brasileira de Zootecnia, Viçosa, v. 30, n. 5, p. 1639-1649, 2001.
SILVA, E. A. da; BERCHIELLI, T. T.; REIS, R. A.; FERNANDES, J. J. de R.; SATO, K. J.; PAES, J. M. V. Teores de proteína bruta para bovinos alimentados com feno de tifton 85: consumo e digestibilidades total e parcial. Revista Brasileira de Zootecnia, Viçosa, v. 36, n. 1, p. 237-245, 2007a.
SILVA, E. A. da; BERCHIELLI, T. T.; REIS, R. A.; PIRES, A. V.; SATO, K. J.; PAES, J. M. V.; LOPES, A. D. Teores de proteína bruta para bovinos alimentados com feno de tifton 85: parâmetros ruminais, eficiência de síntese microbiana e degradabilidade in situ. Revista Brasileira de Zootecnia, Viçosa, v. 36, n. 1, p. 225-236, 2007b.
SINCLAIR, L. A.; GARNSWORTHY, P. C.; NEWBOLD, J. R.; BUTTERY, P. J. Effect of synchronizing the rate of dietary energy and nitrogen release on rumen fermentation and microbial protein synthesis in sheep. Journal of Agricultural Science, v. 120, p. 251-263, 1993.
SINDT, M. H.; STOCK, R.; KLOPFENSTEIN, T. J.; SHAIN, D. H. Protein sources for finishing calves as affected by management system. Journal of Animal Science, Champaign, v. 71, n. 5, p. 1047-1056, 1993.
SIQUEIRA, E, R. Efeito da uréia como fonte de NNP sobre o consumo e digestibilidade em rações de ovinos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 18., 1981, Goiânia. Anais... Goiânia: SBZ, 1981, p. 363.
SIQUEIRA, G. B. de Efeito da suplementação protéica sobre o desempenho, ingestão voluntária e eficiência alimentar de bovinos de corte consumindo volumosos de baixa qualidade. 2001. 50p. Dissertação (Mestrado em Zootecnia) - Universidade Estadual Paulista, Ilha Solteira.

128
SNIFFEN, C. J.; BEVERLY, R. W.; MOONEY, C. S.; ROE, M. B.; SKIDMORE, A. L.; BLACK. J. R. Nutrient requirements versus supply in the dairy cow: strategies to account for variability. Journal of Dairy Science, Champaign, v. 76, n. 10, p. 3160-3178, 1993.
SNIFFEN, C. J.; .O’CONNOR, J. D.; VAN SOEST, P. J.; FOX, D. G.; RUSSELL, J. B. A net carbohydrate and protein system for evaluating cattle diets. II. Carbohydrate and protein availability. Journal of Animal Science, Champaign, v. 70, n. 11, p. 3562-3577, 1992.
SNIFFEN, C. J.; ROBINSON, P. H. Protein and fiber digestion, passage and utilization in lactating cows: microbial growth and flow as influenced by dietary manipulations. Journal of Dairy Science, Champaign, v. 70, n. 2, p. 425-441, 1987.
SOUZA, V. G. PEREIRA, O. G.; VALDARES FILHO, S. de C.; RIBEIRO, K. G.; SILVA, B. C. da; ZGO, R. F. Consumo e digestibilidade de bovinos de corte recebendo dietas com diferentes níveis de uréia. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: SBZ, 2002. 1 CD-ROM.
STALLCUP, O. T.; DAVIS, G. V.; SHIELDS, L. Influence of dry matter and nitrogen intakes on fecal nitrogen losses in cattle. Journal of Dairy Science, Champaign, v. 58, n. 9, p. 1301-1307, 1975.
STORM, E.; ØRSKOV, E. R. The nutritive value of rumen micro-organisms in ruminants. 1. Large-scale isolation and chemical composition of rumen micro-organisms. British Journal of Nutrition, Cambridge, v. 50, p. 463-470, 1983.
TEIXEIRA, J. C.; SALVADOR, F. M. Amiréia: uma revolução na nutrição de ruminantes. Lavras: UFLA, 2004. 174 p.
TITGEMEYER, E. C. Design and interpretation of nutrient digestion studies. Journal of Animal Science, Champaign, v.75, n. 8, p. 2235-2247, 1997.
VALADARES FILHO, S. de C. Eficiência de síntese de proteína microbiana, degradação ruminal e digestibilidade intestinal da proteína bruta, em bovinos. In: SIMPÓSIO INTERNACIONAL SOBRE EXIGÊNCIAS NUTRICIONAIS DE RUMINANTES, 1995, Viçosa. Anais... Viçosa: UFV/ DZO, 1995, p. 355-388.

129
VALADARES FILHO, S. de C.; PINA, D. dos S. Fermentação ruminal. In: BERCHIELLI, T. T.; PIRES, A. V.; OLIVEIRA, S. G. de.(Ed.) Nutrição de ruminantes. Jaboticabal: FUNEP, 2006, p. 151-182.
VALADARES, R. F. D.; GONÇALVES, L. C.; RODRIGUEZ, N. M.; SAMAPAIO, I. B.; VALADARES FILHO, S. de C.; QUEIROZ, A. C. Níveis de proteína em dietas de bovinos. 1. Consumo e digestibilidades aparentes totais e parciais. Revista Brasileira de Zootecnia, Viçosa, v. 26, n. 6, p. 1252-1258, 1997a.
VALADARES, R. F. D.; GONÇALVES, L. C.; SAMAPAIO, I. B.; RODRIGUEZ, N. M.; COELHO DA SILVA, J. F. Níveis de proteína em dietas de bovinos. 2. Consumo, digestibilidades e balanço de compostos nitrogenados. Revista Brasileira de Zootecnia, Viçosa, v. 26, n. 6, p. 1259-1263, 1997b.
VALKENERS, D.; THÉWIS, A.; AMANT, S.; BECKERS, Y. Effect of various levels of imbalance between energy and nitrogen release in rumen on microbial protein synthesis and nitrogen metabolism in growing double-muscled Belgian Blue bulls fed a corn silage-based diet. Journal of Animal Science, Champaign, v. 84, n. 4, p. 877-885, 2006.
VAN SOEST, P. J. Nutritional ecology of the ruminant. 2. ed. Ithaca: Cornell University, 1994. 476p.
VERBIC, J. Factors affecting microbial protein synthesis in the rumen with emphasis on diets containing forages. Viehwirtschaftliche Fachtagung, 24-25. Apr, 2002.
VIEIRA, D. M.; McLEOD, G. K.; BURTON, J. H. et al. Nutrition of the weaned Holstein calf. 1. Effect of dietary protein levels on rumen metabolism. Journal of Animal Science, Champaign, v. 50, n. 5, p.937-944, May. 1980.
VIRTANEN, A. I. Milk production of cows on protein-free feed. Science, v. 153, p. 1603, 1966
WILSON, J. R.; KENNEDY, P. M. Plant and animal constraints to voluntary feed intake associated with fiber characteristics and particle breakdown and passage in ruminants. Australian Journal of Agricultural Research, Victoria, v. 47, n. 1, p. 199-225, 1996.

130
ZADRA, A. Eficiência bioeconômica de grupos genéticos no confinamento: zebuínos x taurinos. In: ENCONTRO SOBRE CONFINAMENTO, 2., 2007, Jaboticabal. Anais... Jaboticabal: FCAV-UNESP, 2007, p.133-154.
ZEOULA, L. M.; FERELI, F.; PRADO, I. N. do; GERON, L. J. V.; CALDAS NETO, S. F.; PRADO, O. P. P. do; MAEDA, E. M. Digestibilidade e balanço de nitrogênio de rações com diferentes teores de proteína degradável no rúmen e milho moído como fonte de amido em ovinos. Revista Brasileira de Zootecnia, Viçosa, v. 35, n. 5, p. 2179-2186, 2006.
ZUNDT, M.; MACEDO, F. de A. F.; MARTINS, E. N.; MEXIA, A. A.; YAMAMOTO, S. M. Desempenho de cordeiros alimentados com diferentes níveis protéicos. Revista Brasileira de Zootecnia, Viçosa, v. 31, n. 3, p. 1307-1314, 2002.