caracterização ecológica dos peixes recifais do atol das rocas
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS, DEPARTAMENTO DE
OCEANOGRAFIA E LIMNOLOGIA. PROGRAMA DE PÓS-GRADUAÇÃO EM BIOECOLOGIA
AQUÁTICA
Caracterização ecológica dos peixes recifais do Atol das Rocas
Igor Emiliano Gomes Pinheiro
Natal – RN 2006

ii
Caracterização ecológica dos peixes
recifais do Atol das Rocas
Igor Emiliano Gomes Pinheiro
Orientador: Prof. Dr. Jorge Eduardo Lins de Oliveira
Dissertação apresentada ao Departamento de Oceanografia e Limnologia do Centro de Biociências da Universidade Federal do Rio Grande do Norte, como parte dos requisitos exigidos para obtenção do título de mestre em Bioecologia Aquática.
Natal - RN
Dezembro, 2006

iii
Caracterização ecológica dos peixes recifais do Atol das Rocas
Igor Emiliano Gomes Pinheiro
Banca Examinadora:
Prof. Dr. Fúlvio Aurélio de Morais Freire (UFERSA)
Prof. Dr. Jorge Eduardo Lins de Oliveira (UFRN)
Prof. Dr. Jorge Pablo Castello (FURG)
Prof. Dr. Ricardo Clapis Garla (UFRN)
Natal – RN
2006

iv
Divisão de Serviços Técnicos
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede Pinheiro, Igor Emiliano Gomes. Caracterização ecológica dos peixes recifais do Atol das Rocas / Igor Emiliano Gomes Pinheiro. – Natal, RN, 2006. 84 f. : il. Orientador : Jorge Eduardo Lins. Dissertação (Mestrado) - Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-graduação em Bioecologia Aquática.
1. Peixes recifais - Dissertação. 2. Ecologia - Dissertação. 3.Unidade de conservação – Dissertação. 4.Complexidade estrutural – Dissertação. I. Lins, Jorge Eduardo. II. Título.
RN/UF/BCZM CDU 567

v
Agradecimentos
A meus pais por todo incentivo, apoio e por tudo que eles sempre
fizeram por mim para que tudo caminhasse da melhor maneira possível.
Ao Prof. Dr. Jorge Lins pela oportunidade de trabalhar no LABIPE e pela
confiança e apoio sempre demonstrados em relação ao trabalho por mim
desenvolvido.
A minha amiga e namorada Lorena Candice, primeiramente pela sua
companhia desde minha chegada a Natal, e também pelas inúmeras edições
de imagens, sugestões e correções.
A todos os integrantes do Laboratório de Biologia Pesqueira (LABIPE),
em especial a José Garcia pelas sugestões desde o plano de amostragem até
a conclusão desta dissertação.
Ao Prof. Fúlvio pelas contribuições e indispensável ajuda com relação às
análises estatística. Agradeço também Dr. Ricardo Garla e ao Prof. Dr. Jorge
Pablo Castello pelas importantes contribuições.
Aos companheiros “atoleiro” Risonaldo (Magão), Jariam, Neílton e
Janaina. Um agradecimento especial a minha amiga Janaína pelas centenas
de quadrantes feitos durante nossa estadia no Atol das Rocas.
A meus amigos de longa data Arthur (Tony), Augusto (Preto), Aline e aos
amigos feitos durante o mestrado Vinicius (Venécius), Rodrigo, Eduardo,
Abraão, Meireles e Guilherme.
A bibliotecária Izamar da Silva Freitas, Bibliotecária-Chefe da Faculdade
de Odontologia do Câmpus de Araçatuba/UNESP - Universidade Estadual
Paulista Júlio de Mesquita Filho pelos inúmeros artigos enviados. Sua a ajuda
foi essencial, muito obrigado.
Meus sinceros agradecimentos a Zélia Brito chefe na Reserva Biológica
do Atol das Rocas pela oportunidade e logística cedida para que assim se
tornasse possível à realização deste trabalho.

1
Sumário Lista de Figuras ............................................................................................. 2 Lista de Tabela .............................................................................................. 4 Resumo ............................................................................................................ 5 Abstract ............................................................................................................ 6 1)Introdução.................................................................................................... 7
1.2) Objetivos específ icos...................................................................... 11 2) Materiais e métodos .............................................................................. 12
2.1) Área de estudo ................................................................................. 12 2.2) Expedições ........................................................................................ 15
2.2.1) Estudo piloto............................................................................... 15 2.2.2) Expedição para amostragens ................................................. 17
2.3) Piscinas amostradas ....................................................................... 18 2.3.1) Barretinha.................................................................................... 19 2.3.2) Barreta Falsa .............................................................................. 20 2.3.3) Cemiteriozinho ........................................................................... 21 2.3.4) Âncoras ........................................................................................ 22
2.4) Metodologia de Amostragem ........................................................ 24 2.5) Análise dos dados ........................................................................... 29
3) Resultados ............................................................................................... 33 3.2.1) Barretinha.................................................................................... 36 3.2.2) Barreta falsa ............................................................................... 38 3.2.3) Cemiteriozinho ........................................................................... 40 3.2.4) Âncoras ........................................................................................ 42
3.3) Piscinas Abertas x Piscinas Fechadas....................................... 44 3.3.1) Abundância Relativa................................................................. 44 3.3.2) Abundância relativa nos diferentes t ipos de fundo .......... 46 3.3.3) Índices de diversidade ............................................................. 49
3.4) Análises de agrupamento .............................................................. 52 3.5) Relações entre peixes e as variáveis de substrato ................ 55
4) Discussão ................................................................................................. 58 4.1) Metodologia ....................................................................................... 58 4.2) Piscinas Abertas x Piscinas Fechadas....................................... 60
4.2.1) Abundância relativa .................................................................. 60 4.2.2) Abundância relativa nos diferentes t ipos de fundo .......... 61 4.2.3) Índices de diversidade ............................................................. 62
4.3)Análises de agrupamento................................................................ 65 4.5) Complexidade estrutural ................................................................ 69
5)Conclusão .................................................................................................. 73 6) Referências Bibliográficas ................................................................... 74

2
Lista de Figuras Figura 1: Localização do Atol das Rocas .................................................... 12 Figura 2: Desenho esquemático das piscinas do Atol das Rocas. B. Falsa = Barreta Falsa e Cemit.= Cemiteriozinho .................................... 18 Figura 3: Piscina denominada Barretinha. A seta aponta a barreta por onde ocorre a l igação com mar aberto. ............................................... 19 Figuras 4: Piscina denominada de Barreta Falsa. A seta aponta a barreta por onde ocorre a l igação com mar aberto ................................ 20 Figura 5: Piscina denominada de Cemiteriozinho. ................................. 21 Figura 6: Piscina denominada das Ancoras ............................................... 23 Figura 7: Desenho amostral. Q1,Q2, Q3 e Q4 são os quadrados amostrados e o quadrante central marca a estação. ............................ 25 Figura 8: Porcentagens dos tipos de substratos amostrados, da abundância das espécies em cada tipo de substrato e da abundância por unidade de área na piscina denominada Barretinha. No período de 22/05 à 21/06/2006. ...................................... 36 Figura 9: Abundância relativa das famílias mais representativas na piscina Barretinha. No período de 22/05 à 21/06/2006. ...................... 37 Figura 10: Porcentagens dos t ipos de substratos amostrados, da abundância das espécies em cada tipo de substrato e da abundância por unidade de área na piscina denominada Barreta Falsa. No período de 22/05 à 21/06/2006. ................................................. 38 Figura 11: Abundância relativa das famílias mais representativas na piscina Barreta Falsa. No período de 22/05 à 21/06/2006. ......... 39 Figura 12: Porcentagens dos t ipos de substratos amostrados, da abundância das espécies em cada tipo de substrato, e a abundância por unidade de área na piscina denominada Cemiteriozinho. No período de 22/05 à 21/06/2006. ............................. 40 Figura 13: Famílias mais representativas observadas na piscina Cemiteriozinho. No período de 22/05 à 21/06/2006. ............................. 41 Figura 14 : Porcentagens dos tipos de substratos amostrados, da abundância das espécies em cada tipo de substrato e a abundância por unidade de área na piscina denominada das Ancoras. No período de 22/05 à 21/06/2006. ........................................... 42

3
Figura 15 : Famílias mais representativas observadas na piscina das Ancoras. No período de 22/05 à 21/06/2006. .................................. 43 Figura 16: Abundância relativa das famílias mais representativas do Atol das Rocas. No período de 22/05 à 21/06/2006. ...................... 44 Figura 17: Abundância relativa das espécies mais representativas do Atol das Rocas. No período de 22/05 à 21/06/2006. ...................... 45 Figura 18: Abundância relativa das espécies dominantes em cada uma das piscinas amostradas. No período de 22/05 à 21/06/2006......................................................................................................................................... 46 Figura 19: Porcentagens de cada tipo de substrato amostrado, da abundância das espécies em cada tipo de substrato e abundância por unidade de área (Abund/área), para Fundo Rochoso (FR), fundo Arenoso (FA) e fundo de Cascalho (FC); nas piscinas amostradas. No período de 22/05 à 21/06/2006. .................................... 47 Figura 20: Valores dos índices de diversidade e equitatividade calculados para as 4 piscinas amostradas. No período de 22/05 à 21/06/2006. ............................................................................................................... 49 Figura 21: Valores dos índices de diversidade e equitatividade nos diferentes t ipos de fundo das piscinas amostradas. (FR) Fundo Rochoso, (FA) Fundo Arenoso, Fundo de Cascalho (FC). No período de 22/05 à 21/06/2006. ...................................................................... 50 Figura 22: Fundo Rochoso (FR), Fundo de Arenoso (FA), Fundo de Cascalho (FC) .......................................................................................................... 52 Figura 23: Fundo de Rochoso (FR), Fundo de Areia (FA), Fundo de Cascalho (FC). ......................................................................................................... 53 Figura 24: Riqueza (S) de espécies nos diferentes t ipos de substrato das piscinas amostradas. A1, A2, B1 e B2 são as legendas dos grupos formados pelo Cluster. ............................................ 54 Figura 25: Resultados obtidos através de ACC com as variáveis de substrato e abundância das espécies: A.sax, Abudefduf saxati l is; A.chi, Acanthurus chirurgus; A.coe, Acanthurus coeruleus; C.ful, Cephalopholis fulva; C.ocel, Chaetodon ocellatus; C. gla, Coryphopterus glaucofraenum; G.tho, Gnatholepis thompsoni; H.rad, Halichoeres radiatus; H.ads, Holocentrus adscensionis; M.tri, Malacoctenus tr iangulatus; S.amp, Sparisoma amplum; S.axi, Sparisoma axil lare; S.fro, Sparisoma frondosum; S.roc, Stegastes rocasensis; T. nor, Thalassoma noronhanum. Eigenvalues: eixo 1= 0,06; eixo 2= 0,07. Os dois primeiros eixos respondem a 14% das variâncias. O teste de Monte-Carlo para todos os eixos canônicos foram signif icativos (p < 0,05). ........................................................................ 56

4
Lista de Tabela Tabela 1: Famílias e espécies identif icadas durante o estudo piloto. .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16 Tabela 2: Escore de avaliação de habitat (EAH) (GRATWICKE & SPEIGHT, 2005), impresso na parte posterior da prancheta de anotações subaquáticas.... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 Tabela 3: Famílias e espécies observadas durante as amostragens e suas respectivas Categorias Tróficas (C.T) .São marcadas com ( X ) as piscinas em que cada espécies foi observada. Espécies que só ocorreram em piscinas abertas (PA) ou piscinas fechadas (PF), também são marcadas com ( X ). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34 Tabela 4: Riqueza de espécies (S), total de observações (N), medida de equitatividade (J) e diversidade de Shannon-Wiener (H’), na piscina denominada Barretinha. ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 Tabela 5: Riqueza de espécies (S), total de observações (N), medida de equitatividade (J) e diversidade de Shannon-Wiener (H), na piscina denominada Barreta Falsa. ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 Tabela 6: Riqueza de espécies (S), total de observações (N), medida de equitatividade (J) e diversidade de Shannon-Wiener (H), na piscina denominada Cemiteriozinho. ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 Tabela 7: Riqueza de espécies (S), total de observações (N), medida de equitatividade (J) e diversidade de Shannon-Wiener (H), na piscina denominada Ancoras. ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43 Tabela 8: Resultados do teste de Tukey. São marcados com (*) os t ipos de fundo com diferenças signif icativas a nível de (p < 0,05)... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 Tabela 9: Valores de porcentagem (%) das espécies que mais contribuíram para a dissimilaridade entre os diferentes t ipos de fundo. ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 Tabela 10: Valores de Fisher (F) e da probabil idade (p) calculados nas comparações entre os diferentes t ipos de fundo. São marcados com (*) os t ipos de fundo com diferenças signif icativas ao nível de p < 0,05. O grau de l iberdade para essas análises foi 8. ... . . . . . . . . . 51 Tabela 11: Contribuição de todas as variáveis do EAH para as variações na abundância e diversidade de espécies. São apresentadas as correlações e os níveis de signif icância para cada uma das cinco variáveis. São marcados com (*) as variáveis de substrato com diferenças signif icativas a nível de (p < 0,05). ... . . 57

5
Resumo Este trabalho realizado no Atol das Rocas (3º 51’ S; 33º 49’
W), o único atol do Atlântico Sul e a primeira área marinha
protegida brasileira, foi norteado pela hipótese de que: a
composição das comunidades de peixes varia de acordo com a
posição das piscinas; ou seja, com ou sem conexão permanente
com o oceano. Para testar a validade desta hipótese foram
realizadas estimativas da abundância de peixes em piscinas
conectadas (Barretinha/Barreta Falsa) e não conectadas com
oceano (Cemiteriozinho/Âncoras), realizando uma caracterização
ecológica dos peixes que habitam estas piscinas. Adicionalmente a
complexidade estrutural dos locais amostrados também foi
avaliada, com intuito de verif icar variações da abundância e
diversidade de peixes em função deste fator. Pelo fato desta
pesquisa ter sido realizada dentro dos l imites de uma unidade de
conservação, as amostragens foram realizadas através do censo
visual. Um total de 56 espécies foram identif icadas no estudo
piloto e durante as amostragens 48 destas foram observadas. Os
resultados gerados através de técnicas analít icas uni e
mult ivariadas permitiram constatar que o fator determinante nas
variações de densidade, riqueza e diversidade dos peixes é o t ipo
de substrato (consolidados/não consolidados) e não fato das
piscinas estarem ou não conectada permanentemente com oceano.
Com relação à complexidade estrutural; 58% das variações na
diversidade de peixes são atribuídas a mudanças na complexidade
estrutural enquanto que 12% das variações na abundância são
atribuídos a este fator.

6
Abstract
The present work was carried out at the Atol das Rocas (3º
51’ S; 33º 49’ W), the only atoll of the South Atlant ic and the first
Brazil ian marine protected area. It was guided by the following
hypothesis: the composit ion of the f ish communities varies in
agreement with the posit ion of the pools; in other words, with or
without permanent ocean connection. To test the validity of this
hypothesis, the f ish abundance was estimated in the connected
pools (Barretinha/Barreta Falsa) and unconnected ones
(Cemiteriozinho/Âncoras), carrying an ecological characterization
of the f ishes that inhabit these pools. Addit ionally, the structural
complexity of the sampled places was also evaluated intending to
verify the variations of the abundance and diversity of f ishes in
function to this factor. By the fact of this research was being
carried out through the l imits of a conservation unit, the samples
was realized using visual census techniques. The results generated
through uni and mult ivariate analytic techniques allowed the
evidence that decisive factor in the density, r ichness and diversity
variations of f ishes are l inked to the substratum type (Hard / Soft
bottom) and not by the fact that the pool are or are not connected
permanently to the ocean. In relation to the structural complexity;
58% of the variations in the diversity of f ishes were attr ibuted to
changes in the structural complexity, while 12% of the variations in
the abundance were attr ibuted to the structural complexity.

7
1)Introdução
A característica mais marcante dos peixes recifais é sua
diversidade em termos de espécies e morfologia (SALE, 1991).
Dentre os peixes marinhos, os peixes recifais representam a maior
diversidade, correspondendo de 30 a 40% do total das espécies.
Em certas regiões, como no Indo-Pacífico, podem ser encontradas
cerca de 4.000 espécies (CHOAT & BELLWOOD, 1991), quase
2.000 somente na grande barreira de corais da Austrália.
Associada a esta grande diversidade, a ict iofauna recifal é uma
importante fonte de alimento para milhões de pessoas que vivem
em regiões costeiras tropicais de todo o mundo (RUSS, 1991;
MUNRO, 1996).
Atualmente, muitos ecologistas expressam consenso de que
as capturas globais de peixes estão declinando (PAULY et al.,
2002; MCCLANAHAN, 2002); as numerosas populações de animais
marinhos estão desmoronando (HUTCHINGS, 2000; MYERS &
WORN, 2003) e os recifes de corais estão sendo extensivamente
danif icados ou destruídos (PANDOLFI et al. , 2003). Como forma de
preservar os hábitats e minimizar a perda de biodiversidade,
assegurando a continuidade dos recursos vivos para as futuras
gerações, tem-se recorrido ao estabelecimento de Unidades de
Conservação (UCs), ou seja, “superfícies de terra ou mar
consagradas à proteção e manutenção da diversidade biológica”
(IUCN, 1994).
Recentemente, as Áreas Marinhas Protegidas (AMP) têm se
tornado o foco principal da conservação dos ambientes marinhos
(DENNY & BABCOCK, 2003), e um grande número de áreas como
estas foram estabelecidas em todo mundo durante o últ imo século
(JONES et al., 1993 apud LIZASO et al., 2000). Atualmente,
existem mais de 1.300 AMPs no mundo (KELLEHER et al., 1995), e
outras estão sendo criadas aceleradamente (RODWELL &
ROBERTS, 2000).

8
O primeiro esforço dirigido à criação de uma AMP, foi no
inicio do século XX, na Grande Barreira de corais, Austrália
(MORNING POST, 1906), porém, a proposta foi rejeitada pelos
pescadores. A primeira AMP declarada usando legislação
específica, incluindo provisões para a gerência de águas
circunvizinhas e de sua biota, foi declarada em 1935 no
monumento nacional de Jefferson Fort, Florida (RANDALL, 1968).
No Brasil, a primeira unidade de conservação marinha
federal, a ser estabelecida, pelo Decreto Lei n° 83.549 de cinco de
julho de 1978 foi a Reserva Biológica do Atol das Rocas. Reservas
Biológicas (REBIO) são Unidades de Conservação destinadas à
preservação integral da biota e demais atributos naturais
existentes em seus l imites, sem interferência humana direta ou
modificações ambientais, excetuando-se as medidas de
recuperação de seus ecossistemas alterados e as ações de manejo
necessárias para recuperar e preservar o equilíbrio natural, a
diversidade biológica e os processos ecológicos naturais. A
legislação permite apenas pesquisas científ icas devidamente
autorizadas de acordo com seu plano de manejo, sendo, qualquer
outra atividade confl it iva com a legislação existente. (Sistema
Nacional de Unidades de Conservação, 2000).
Até o início da década de 90, apenas aspectos biológicos do
Atol das Rocas tinham sido mais intensivamente investigados. Os
primeiros estudos, que se têm registros foram realizados por
FARIA & SILVA (1937a - b), sobre a ictiofauna e sobre
Palinurídeos (Crustacea-Macrura); posteriormente, alguns
trabalhos foram publicados sobre macroalgas (OLIVEIRA FILHO,
1972; OLIVEIRA FILHO & MENEZES, 1972; OLIVEIRA FILHO &
UGADIM, 1974; 1976).
O estabelecimento de uma estação permanente na reserva
em 1994 permitiu o desenvolvimento de diversos projetos
científ icos. São exemplos de alguns destes estudos os
desenvolvidos por KIKUCHI (1994) com geomorfologia,
estratigrafia e sedimentologia do Atol das Rocas e ECHEVERRÍA

9
et al. (1996) com cnidários. ROSA & MOURA (1997) realizaram um
estudo sobre a estrutura da comunidade de peixes recifais em
diferentes áreas do Atol. Contudo até o presente, nenhum estudo
buscou caracterizar e comparar a ecologia (abundância, r iqueza,
diversidade e estrutura trófica) da ictiofauna em áreas com
características contrastantes no Atol das Rocas.
A grande maioria dos estudos sobre as comunidades de
peixes recifais se concentram na Austrália e no Caribe (SALE,
1991). O Atlântico Sul ocidental possui umas das comunidades
recifais menos conhecidas do mundo (SPALDING et al., 2001;
MMA, 2002). No Brasil no que se refere à ict iofauna recifal, a partir
da metade da década de 90, um número crescente de trabalhos
foram realizados nas regiões costeiras, abrangendo tanto aspectos
zoogeográficos (FLOETER & GASPARINI, 2000; FLOETER et al.,
2001; ROCHA, 2003; FERREIRA et al. , 2004; FLOETER et al. ,
2004; FEITOZA et al. , 2005) como descrit ivos (MOURA, 1995;
ROCHA et al., 1998; GASPARINI et al., 1999; FERREIRA et al.,
2001; ROCHA & ROSA, 2001; MOURA et al. 2003, GUIMARÃES et
al. 2004). Apesar dos trabalhos de âmbito zoogeográficos
incluírem às i lhas oceânicas, estes não buscam caracterizar a
ictiofauna destas regiões e sim compará-las com outras províncias
zoogeográficas. Trabalhos publicados voltados à estrutura das
comunidades de peixes das i lhas oceânicas ainda permanecem
escassos (LUBBOCK & EDWARDS 1981; ROSA & MOURA, 1997;
GASPARINI & FLOETER, 2001; FEITOZA et al., 2003).
Um importante aspecto que é abordado neste trabalho e que
recentemente vem ganhando atenção no Brasil, refere-se às
relações entre a ictiofauna e a complexidade estrutural do
ambiente. Na costa brasileira, o único trabalho publicado que
buscou verif icar especif icamente essas relações foi realizado por
FERREIRA et al. (2001) nos costões rochosos de Arraial do Cabo,
no Estado do Rio de Janeiro. Neste estudo os peixes foram
contabil izados através de transectos e as características do
substrato foram analisadas através de diferentes técnicas. Os

10
autores observaram que a diversidade assim como o número de
espécies relacionam-se posit ivamente com a complexidade
estrutural.
Em diversas partes do mundo, estes aspectos já foram
intensamente estudados e, frequentemente, a complexidade
estrutural relaciona-se de maneira signif icativa com diversidade de
peixes (MCCLANAHAN 1994, MCCORMICK, 1994; REÑONES et al.
1997, GARCIA CHARTON & PEREZ RUZAFA, 1998; GRATWICKE
& SPEIGHT, 2005). Em contraste, a abundância total de peixes é
menos influenciada por variações na complexidade estrutural com
estudos mostrando relações fracas ou até mesmo ausência de
relações entre estes fatores (MCCLANAHAN, 1994; MCCORMICK,
1994; FRIEDLANDER & PARRISH, 1998; GRATWICKE & SPEIGHT,
2005).
Na Reserva Biológica do Atol das Rocas, até o presente,
somente três estudos foram voltados a ictiofauna recifal (ROSA &
MOURA, 1997; MOURA 1998; MARQUES, 2003) e nenhum deles
visou caracterizar e comparar a ecologia da ictiofauna em locais
com características contrastantes, verif icando as relações dos
peixes com os diferentes t ipos de substratos. Desta forma, visando
contribuir para um melhor entendimento sobre a estrutura das
comunidades de peixes recifais desta região, este estudo foi
executado com o objetivo de se realizar uma análise ecológica das
comunidades de peixes que habitam as piscinas1 existentes no
anel recifal do Atol das Rocas durante a maré baixa.
1 Piscinas referem-se a depressões de dimensões variadas que ocorrem no anel recifal do Atol das Rocas

11
1.2) Objetivos específicos
• Estimar a abundância, a riqueza, a diversidade e a
equitabil idade ictiológica, comparando estes índices entre piscinas
abertas2 e fechadas3.
• Analisar a relação entre o t ipo de substrato, a abundância e a
diversidade de peixes que ocorrem nas piscinas.
2 - Piscina abertas localizam-se nas bordas do anel recifal do Atol das Rocas, sendo que estas se mantêm permanentemente ligadas com oceano. 3 - Piscinas fechadas localizam-se no interior do anel recifal e durante os períodos de maré baixa estas permanecem isoladas da frente recifal e da laguna.

12
2) Materiais e métodos
2.1) Área de estudo O Atol das Rocas (Figura 1) está localizado no topo de uma
cadeia de montanhas submarinas no oceano Atlântico Sul, sendo
que sua base encontra-se a 4.000 metros de profundidade no leito
oceânico (GHERARDI & BOSENCE, 1999). Sua área é definida
pelas coordenadas 3º 51’ S Latitude e 33º 49’ W Longitude,
distante 266 km da cidade de Natal, no nordeste brasileiro
(GHERARDI & BOSENCE, 2005). A Reserva Biológica do Atol das
Rocas, inclui o Atol e as águas que o circundam até a isóbata de
1.000 metros, abragendo uma área de 360 Km2.
Figura 1: Local ização do Atol das Rocas
O Atol das Rocas é o único do Oceano Atlântico Sul e um
dos menores do mundo (LEÃO & DOMINGUEZ, 2000). Seu formato
elíptico tem o eixo maior (E W) com 3,5 km de comprimento e o
eixo menor (N-S) com 2,5 km de comprimento (SOARES-GOMES et

13
al., 2001, GHERARDI & BOSENCE, 2001). Os recifes deste atol
são construídos por uma assembléia de algas coralíneas
incrustantes, gastrópodes vermetídeos e foraminíferos incrustantes
(GHERARDI & BOSENCE, 2005). O principal construtor da
estrutura recifal são algas coralíneas da espécie Porolithon cf.
pachydermum, que ocorrem associados com outros quatro gêneros
de algas coralíneas: Porolithon sp., Lithophyllum sp., Lithoporella
sp., e Sporolithon sp . (GHERARDI & BOSENCE, 2001).
Oceanograficamente, a região da REBIO do Atol das Rocas
recebe influência da Corrente Sul Equatorial de direção leste-
oeste, que se desloca da costa africana para costa brasileira
paralelamente ao equador geográfico (JOP’S II, 1996; GOES,
2005) . A Cadeia de Fernando de Noronha, na qual o Atol está
inserido, tem um importante papel na região; pois devido ao seu
relevo submarino, as correntes oceânicas locais podem interagir
com as feições f isiográficas, como as i lhas e os bancos
submersos, originando um complexo sistema de circulação
(RODEN, 1987). Tal sistema pode promover a elevação das
isotermas (ressurgência) e a formação de colunas de Taylor,
transportando verticalmente os nutrientes (RODEN, 1987). Este
aporte de nutrientes para a zona eufótica é necessário para o
aumento da produtividade primária nestas águas, que são
predominantemente oligotróficas (TRAVASSOS et al., 1999).
A temperatura da água tem média de 27°C, contudo nas
piscinas fechadas dentro do anel recifal durante a maré baixa,
pode chegar a 39°C (LEÃO & DOMINGUEZ, 2000). A precipitação
é distribuída irregularmente ao longo do ano, com uma média
mensal variando de 860 mm (abri l/1992) a 2.663 mm (agosto/1992)
(KIKUCHI, 1994). Os ventos do quadrante ESE são dominantes
durante o ano todo, com velocidades variando de 6 a 10 m/s
(LEÃO & DOMINGUEZ, 2000). Porém tais parâmetros possuem
relativa sazonalidade; nos meses de inverno (de junho a agosto)
observa-se uma freqüência 35 % de ventos SE e 15 % de ventos E;
com velocidades variando entre 11,0 e 15,0 m/s. Já no verão (de

14
dezembro a abri l), a freqüência f ica em torno de 20 % para os
ventos SE e E, com velocidade freqüentemente acima de 20,0 m/s
(KIKUCHI, 1994). A visibi l idade da água, em condições de tempo
bom é geralmente superior a 20 metros (LEÃO & DOMINGUEZ,
2000), fator importante que foi levado em consideração visto que
as amostragens foram realizadas através do mergulho.

15
2.2) Expedições
2.2.1) Estudo piloto Visando um reconhecimento prévio do local foi realizado um
estudo piloto no período de 4 de novembro a 5 de dezembro de
2005. Este estudo teve como objetivo definir as estações
amostrais, o método de amostragem e captar imagens para
identif icação das diferentes espécies de peixes que ocorrem na
região. Para isso foram realizados 33 horas de observações
subaquáticas através do mergulho autônomo e l ivre, em diferentes
áreas do anel recifal.
As imagens e fotografias captadas auxil iaram na
identif icação de 36 diferentes famílias de peixes, totalizando 56
espécies (tabela 1). A identif icação das famílias e espécies foi
realizada segundo HUMMAN & DELOACH (2003), ARAÚJO et al.
(2004) e FROESE & PAULY (2005). Os descritores das espécies
estão de acordo com FROESE & PAULY (2005).

16
Tabela 1: Famílias e espécies identificadas durante o estudo piloto.
Famílias Espécies Acanthurus chirurgus (Bloch, 1787) ACANTHURIDAE Acanthurus coeruleus (Bloch & Schneider, 1801)
ALBULIDAE Albula vulpes (Linnaeus, 1758) Apogon ssp APOGONIDAE Phaeoptyx pigmentaria (Poey, 1860) Balistes vetula ( Linnaeus, 1758) BALISTIDAE Melichthys niger (Bloch, 1786)
BELONIDAE Platybelone argalus (Lesueur, 1821) Ophioblennius trinitatis (Valenciennes, 1836) BLENNIIDAE Entomacrodus vomerinus (Valenciennes, 1836)
BOTHIDAE Bothus lunatus (Linnaeus, 1758) Caranx latus (Agassiz, 1831) CARANGIDAE Caranx bartholomaei (Cuvier, 1833)
CARCHARHINIDAE Negaprion brevirostris (Poey, 1868) CHAETODONTIDAE Chaetodon ocellatus (Bloch, 1787)
CLUPEIDAE Harengula clupeola (Cuvier, 1829) DACTYLOPTERIDAE Dactylopterus volitans (Linnaeus, 1758)
DASYATIDAE Dasyatis americana (Hildebrand & Schroeder, 1928)
DIODONTIDAE Diodon hystrix (Linnaeus, 1758) GINGLYMOSTOMIDAE Ginglymostoma cirratum (Bonnaterre, 1788)
Coryphopterus glaucofraenum (Gill, 1863) Gnatholepis thompsoni (Jordan, 1904) GOBIIDAE Bathygobius soporator (Valenciennes, 1837) Haemulon chrysargyreum (Günther, 1859) HAEMULIDAE Haemulon parra (Desmarest, 1823) Holocentrus adscensionis (Osbeck, 1765) HOLOCENTRIDAE Myripristis jacobus (Cuvier, 1829)
KYPHOSIDAE Kyphosus sectatrix (Linnaeus, 1758) Halichoeres radiatus (Linnaeus, 1758) LABRIDAE Thalassoma noronhanum (Boulenger, 1890)
LABRISOMIDAE Malacoctenus triangulatus (Springer, 1959) LUTJANIDAE Lutjanus jocu (Bloch & Schneider, 1801)
MALACANTHIDAE Malacanthus plumieri (Bloch, 1786) MONACANTHIDAE Aluterus scriptus (Osbeck, 1765)
MULLIDAE Mulloidichthys martinicus (Cuvier, 1829) Echidna catenata (Bloch, 1795) Enchelycore nigricans (Bonnaterre, 1788) Gymnothorax funebris (Ranzani, 1840) Gymnothorax miliaris (Kaup, 1856) Gymnothorax vicinus (Castelnau, 1855)
MURAENIDAE
Muraena pavonina (Richardson, 1845) OPHICHTIDAE Myrichthys ocellatus (Lesueur, 1825) OSTRACIIDAE Lactophrys trigonus (Linnaeus, 1758)
POMACANTHIDAE Pomacanthus paru (Bloch, 1787) Abudefduf saxatilis (Linnaeus, 1758) Chromis multilineata (Guichenot, 1853) Stegastes pictus (Castelnau, 1855)
POMACENTRIDAE
Stegastes rocasensis (Emery, 1972) Sparisoma amplum (Ranzani, 1842) Sparisoma axillare (Steindachner, 1878) SCARIDAE Sparisoma frondosum (Agassiz, 1831) Cephalopholis fulva (Linnaeus, 1758) Paranthias furcifer (Valenciennes, 1828) SERRANIDAE Rypticus saponaceus (Bloch & Schneider, 1801)
SPHYRAENIDAE Sphyraena barracuda (Edwards, 1771) SYNODNTIDAE Synodus intermedius (Spix & Agassiz, 1829)

17
2.2.2) Expedição para amostragens A segunda expedição foi realizada no período de 22 de maio
a 21 de junho de 2006. Durante esta expedição foram amostradas
4 diferentes piscinas sendo duas conectadas com o oceano
(abertas) e duas no interior do anel recifal, sem conecção
permanente com o oceano (fechadas). Ao total, foram amostradas
25 estações nas piscinas abertas e 25 em piscinas fechadas,
totalizando 50 censos para análise. Todas as amostragens foram
realizadas através do mergulho l ivre, visto que as piscinas
definidas para amostragens têm profundidade máxima de 4,5
metros e também pelo fato de que a logística necessária para que
todas as amostragens fossem feitas por meio do mergulho
autônomo seria inviável.
Devido a dif iculdade de amostragens noturnas, todos os
censos foram realizados entre 8 e 16 horas, período em que a
maior parte dos peixes permanecem ativas no Atol (cf. MOURA
1998).

18
2.3) Piscinas amostradas
Foram definidas 4 piscinas para amostragem (Figura 2),
sendo duas conectadas permanentemente com oceano (Barretinha
e Barreta Falsa) e duas sem conexão permanente com oceano
(Cemiteriozinho e Âncoras). As piscinas determinadas para
amostragem tiveram suas áreas estimadas com uso de um GPS
(Garmin Etrex).
Figura 2: Desenho esquemát ico das piscinas do Atol das Rocas. B. Falsa = Barreta Falsa e Cemit.= Cemiter ioz inho
O critério usado para definição das piscinas foi à
diversidade de substratos encontrados, a área das piscinas e a
viabil idade de realização das amostragens, visto que as
amostragens foram feitas sempre na maré baixa e o deslocamento
entre as estações era feito caminhando. Esta l imitação não
permitiu constatações a cerca das variações na estrutura da
comunidade de peixes durante as variações de maré (maré alta).
As descrições das piscinas amostradas são apresentadas nos
itens abaixo.

19
2.3.1) Barretinha Piscina permanentemente conectada com oceano (Figura 3),
localizada no setor oeste do anel recifal do Atol das Rocas. No
período de preamar uma forte correnteza se estabelece em direção
ao mar aberto diminuindo de intensidade durante a maré baixa.
Neste período a profundidade máxima nesta piscina é 4,5 metros
sendo que a profundidade média dos quadrados amostradas foi 2,6
metros.
Figura 3: Piscina denominada Barret inha. A seta aponta a barreta por onde ocorre a l igação com mar aberto.
Recifes em manchas (patch reefs) de dimensões variadas
ocorrem em toda a extensão da piscina dominando grande parte do
substrato. Os fundos não consolidados ocupam uma parcela
signif icativa da piscina recifal: cerca de 20% é ocupado por fundos
arenosos e 10% por fundos de cascalho. A parede recifal que
delimita a piscina tem inclinação pouco acentuada, sendo
constituída por algas calcárias, foraminíferos, moluscos,
vermitídeos e corais hematípicos; composição semelhante à de
todo atol. A cobertura, tanto da parede recifal como dos fundos
consolidados é constituída por algas pardas, vermelhas e verdes,
além de moluscos, corais, zoantídeos e outros cnidários.

20
A área total determinada para amostragem foi de
aproximadamente 1.120 metros quadrados, localizados na porção
mais próxima a barreta (canal de comunicação com mar). Esta
área foi dividida em 14 estações de aproximadamente 80 metros
quadrados, tendo 10 metros de comprimento por 8 de largura (10m
x 8m) cada estação. Nesta piscina foram sorteadas 11 estações
para amostragem onde a visibi l idade variou entre 7 e 15 metros.
2.3.2) Barreta Falsa
Piscina com conexão permanente com mar aberto (Figura
4), localizada no setor oeste do anel recifal do Atol das Rocas. No
período de maré alta observa-se o aumento no batimento de ondas
e nota-se o aparecimento de uma forte correnteza que corre em
direção ao mar aberto. Na maré baixa a Barreta Falsa
freqüentemente proporciona condições favoráveis ao mergulho
com águas calmas e com visibi l idade variando entre 5 e 15 metros;
a profundidade máxima na piscina neste período é 4,5 metros. A
profundidade média nos quadrados amostrados foi 2,7 metros.
Figuras 4: Piscina denominada de Barreta Falsa. A seta aponta a barreta por onde ocorre a l igação com mar aberto
As paredes que delimitam a piscina têm declividade
acentuada e são constituídas por substrato consolidado de
composição semelhante à de todo atol. Fundos consolidados

21
predominam nesta piscina, dominando aproximadamente 90% da
piscina recifal. Fundos não consolidados ocorrem na porção mais
próxima ao mar aberto, porém ocupam uma pequena parcela da
área amostrada. A cobertura da parede recifal e dos fundos
consolidados é semelhante a da Barretinha, porém nesta piscina
ocorre uma maior abundância de algas.
A área total determinada para amostragem foi de
aproximadamente 1.280 metros quadrados, abragendo
praticamente toda a área da piscina. Esta área foi dividida em 16
estações, tendo 10 metros de comprimento por 8 de largura (10m x
8m) cada estação. Destas 16 estações foram sorteadas 14 para
amostragem.
2.3.3) Cemiteriozinho
Piscina recifal sem conexão permanente com mar aberto
(Figura 5), localizada no setor oeste do anel recifal do Atol das
Rocas. Esta piscina é formada por uma depressão no anel recifal,
sendo que este local permanece isolado da frente recifal durante a
maré baixa. No período de maré alta é estabelecida uma
comunicação com mar aberto, sendo observado o aumento do
batimento das ondas e o aparecimento de uma correnteza que
corre em direção ao mar aberto.
Figura 5: Piscina denominada de Cemiter ioz inho.

22
As paredes que delimitam a piscina apresentam inclinação
negativa, formando cavernas próximas à interface com a areia.
Recifes em manchas (“patch reefs”) ocorrem em algumas partes da
piscina recifal, porém o substrato dominante é o não consolidado.
A planície arenosa ocupa cerca de 30% da piscina. A areia calcária
de coloração branca é recoberta por uma fina e inconspícua
camada de algas f i lamentosas verdes (MOURA, 1998). Apesar de
parecer pobre em recursos alimentares a planície arenosa abriga
uma rica fauna de invertebrados enterrada no substrato (HOBSON
& CHESS, 1986). Os fundos de cascalho ocupam cerca 40% da
piscina recifal e os fundos consolidados 30%.
A área total determinada para amostragem foi de
aproximadamente 640 metros quadrados, que abragem toda a área
da piscina. Esta área foi dividida em 8 estações de 80 metros
quadrados, tendo 10 metros de comprimento por 8 de largura (10m
x 8m) cada estação. Nesta piscina foram sorteadas 6 estações
para amostragem, com profundidade máxima de 5 metros onde a
visibi l idade variou entre 5 e 15 metros. A profundidade média nos
quadrados amostrados foi 2,7 metros.
2.3.4) Âncoras
Piscina recifal sem conexão permanente com mar aberto
(Figura 6), localizada no setor leste do anel recifal do Atol das
Rocas. No período de maré alta é estabelecida uma conexão entre
a laguna permanente e a frente recifal, sendo observado o
aumento no batimento das ondas e o aparecimento de uma forte
correnteza que corre para oeste. Na maré baixa a piscina das
Âncoras freqüentemente proporciona condições favoráveis ao
mergulho com águas calmas e ótima visibi l idade.

23
Figura 6: Piscina denominada das Ancoras
As paredes que delimitam esta piscina apresentam
inclinação negativa em todo lado norte, formando cavernas
próximas à interface com a areia; já no lado sul esta inclinação é
pouco acentuada. Recifes em manchas (“patch reefs”) de
dimensões variadas ocorrem em toda a extensão da piscina recifal,
sendo mais abundantes no lado sul; neste mesmo lado da piscina
foram observados alguns restos de naufrágio. A planície arenosa
ocupa cerca de 30% da área total amostrada, sendo mais
abundante na porção norte. Já os fundos de cascalho ocupam
cerca de 10% da porção oeste da piscina. A cobertura dos
substratos consolidados e não consolidados são semelhantes à
das outras piscinas escolhidas para amostragem.
A área total determinada para amostragem foi de 1.760
metros quadrados, que abrangem quase a toda a área da piscina.
Esta área foi dividida em 22 estações de 80 metros quadrados,
tendo 10 metros de comprimento por 8 de largura. Nesta piscina
foram sorteadas para amostragem 19 estações com profundidade
máxima de 4,5 metros, onde a visibi l idade variou entre 10 e 20
metros. A profundidade média nos quadrados amostrados foi 2,8
metros.

24
2.4) Metodologia de Amostragem
O fato desta pesquisa ter sido realizada dentro dos l imites de
uma Unidade de Conservação (UN), estimula o uso de métodos
amostrais que permitam estimar a abundância das populações de
peixes sem afetá-las (MESA & VACCHI 1999). Sendo assim o
presente trabalho contou com amostragens visuais “ in situ”, da
fauna ictiológica, ou seja, as amostragens foram realizadas através
do censo visual buscando, assim, causar o mínimo de
perturbações signif icativas à comunidade.
As técnicas de censo visual subaquático são amplamente
usadas em estudos populacionais de ictiofauna (SAMOILYS &
CARLOS, 2000). Este t ipo de metodologia foi primeiramente
descrita por BROCK (1954), e é aplicada para estimativas quali-
quantitativas, pois não somente provê informações a respeito da
presença da espécie, como também permite que se estime a atual
abundância da população por unidade de área pesquisada.
A metodologia empregada neste estudo foi descrita por
GRATWICKE & SPEIGHT, 2005. Para cada estação visitada foram
amostrados 4 quadrantes, com tamanho de 2,5 x 2,5 m. Os
quadrantes foram definidos a partir de um quadrante central não
amostrado (Figura 7). Assim para cada estação, 25 m2 foram
amostrados.
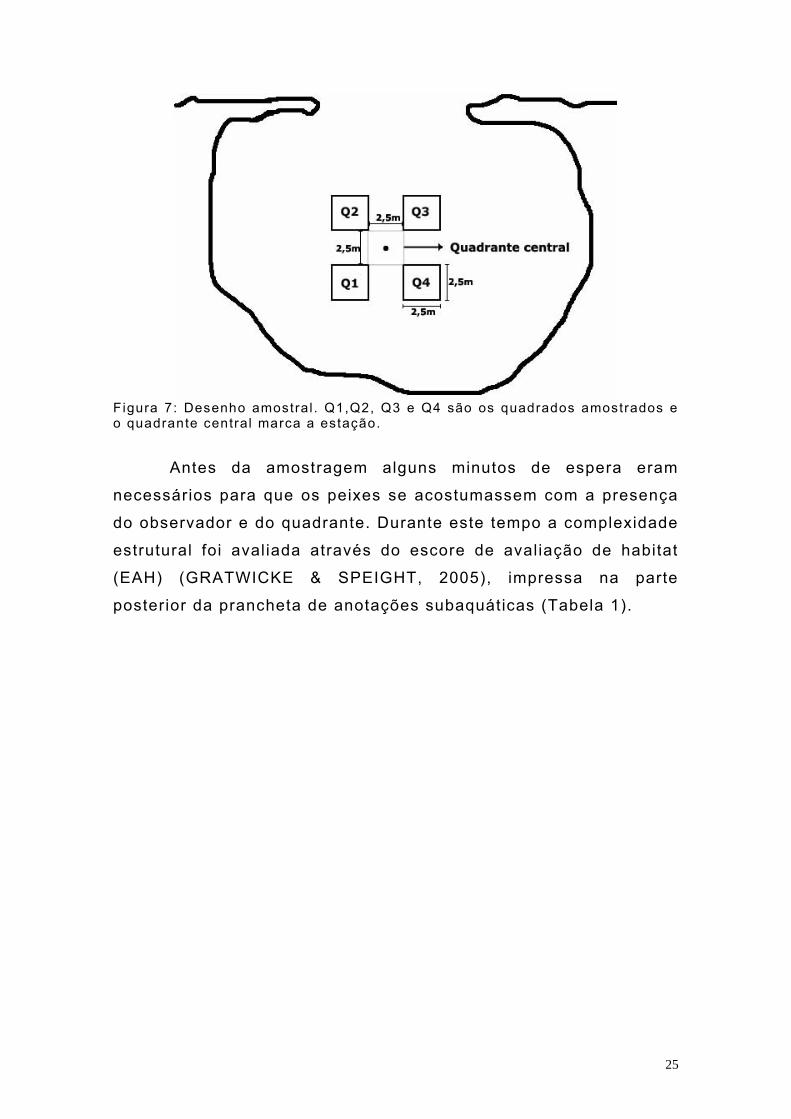
25
F igura 7: Desenho amostral . Q1,Q2, Q3 e Q4 são os quadrados amostrados e o quadrante central marca a estação.
Antes da amostragem alguns minutos de espera eram
necessários para que os peixes se acostumassem com a presença
do observador e do quadrante. Durante este tempo a complexidade
estrutural foi avaliada através do escore de avaliação de habitat
(EAH) (GRATWICKE & SPEIGHT, 2005), impressa na parte
posterior da prancheta de anotações subaquáticas (Tabela 1).

26
Tabela 2: Escore de aval iação de habi tat (EAH) (GRATWICKE & SPEIGHT, 2005), impresso na parte poster ior da prancheta de anotações subaquát icas.
ESCALA DE QUANTIFICAÇÃO DA COMPLEXIDADE ESTRUTURAL DO HÁBITAT 1 2 3 4 5
Rugosidade (topografia visual
estimada do substrato de cada
quadrante)
Variedade de formas de
crescimento (lobosa,
filamentosa, em forma de fita,
maciça, ramificada, cilíndrica, tubular,
em forma de placa, incrustante)
<2 3 a 4 5 a 6 7 a 8 9 a 10
Altura (estimativa visual da altura média do ponto mais baixo da arquitetura do hábitat) (cm)
0 a 9 10 a 19 20 a 39 40 a 79 >80
Categorias de tamanho dos
refúgios (buracos ou aberturas na arquitetura do
hábitat das seguintes
categorias: 1-5, 6-15, 16-30, 31-50 e
>50cm)
0 e 1 2 3 4 5
Cobertura viva (porcentagem total
da cobertura de corais, raiz de
mangue, fanerógamas
marinhas, banco de algas e esponjas)
0 a 9 10 a 19 20 a 39 40 a 79 80 a 100
Substrato duro (%) 0 a 19 20 a 39 40 a 59 60 a 79 80 a 100
Cada quadrante que era demarcado com auxílio de uma trena
foi observado por 5 minutos, a uma distância de 1 metro, onde
todos os peixes na coluna d’água ou que eram facilmente
observados sobre o substrato foram identif icados e contados.
Posteriormente eram gastos de 1 a 5 minutos a procura de peixes
que se encontram entocados ou camuflados no substrato. A
porcentagem de cobertura também é anotada na prancheta. As

27
categorias usadas são; areia4, cascalho5 e rocha6. Para cada
quadrante amostrado foi f ixada uma categoria dependendo do
substrato dominante. Por exemplo, um quadrante com 30% de
cascalho, 20% de algas e 40% de rochas, foi classif icado como
rocha.
O perfi l do substrato feito através do quadrante de 2,5 x 2,5m
foi examinado para avaliar o escore rugosidade (neste estudo os
substratos referenciam a planície arenosa, cascalho, recifes em
manchas e parede recifal). Isto signif ica que a planície de areia
seria classif icada em escore 1. Substratos planos com poucos
recifes em manchas foram classif icados em escore 2, enquanto
que o substrato com perfi l mais complexo foi o preenchido por
rochas e corais sendo classif icado com escore 5.
O termo ”formas de crescimento” foi aplicado para
organismos vivos como algas coralíneas, corais e algas. O objetivo
deste escore é avaliar a diversidade de atributos estruturais do
habitat que possam prover recursos alimentares para diferentes
espécies de peixe.
A média de altura da arquitetura do habitat foi estimada
visualmente buscando distinguir hábitats altos e baixos. A media
de altura do recife é usualmente baseada no menor ponto do
quadrado.
A categoria tamanho dos refúgios de ROBERTS & ORMOND
(1987) foi usada neste estudo. A cobertura viva foi estimada
através de observações do substrato, avaliando a porcentagem da
área coberta por corais, algas ou algum outro substrato vivo. O
substrato duro refere-se à porcentagem de substrato que não era
areia ou cascalho.
4 Os fundos arenosos são compostos de areia calcária de coloração branca, geralmente recoberta por uma fina e inconspícua camada de algas filamentosas verdes (MOURA, 1998). 5 Os fundos de cascalho formados por restos de corais e fragmentos do recife, que se desprende do seu lugar de origem em eventos como tempestades ou mesmo pela ação antrópica (RASSER & RIEGL, 2002). 6 Fundos rochosos ou consolidados são formados por algas calcárias e foraminíferos incrustrantes (ROSA & MOURA 1997).

28
Nota-se que estas categorias são aproximadas e foram
usadas com intuito de dar somente uma impressão a cerca de que
o t ipo dominante do substrato que corresponde ao EAH. Assim é
possível ter uma rápida avaliação da complexidade estrutural do
habitat.

29
2.5) Análise dos dados
Os dados de abundância relativa, obtidos por intermédio do
censo visual foram analisados através de técnicas univariadas e
multivariadas.
Foram empregados os índices de diversidade de Shannon-
Wiener (H`) (SHANNON & WEAVER, 1949) (Equação 1) e de
eqüitatividade de Pielou (J) (PIELOU, 1974) (Equação2).
H’ = - ∑ p .( log2 p) (Equação 1) Onde (p) é a proporção total da amostragem pertencente a
cada espécie.
J = H’/ Hmáx (Equação 2)
Onde H max.= é a máxima diversidade de espécies, calculado
pelo log2 (S).
S = é o total de espécies avistadas em cada estação.
Os índices de diversidade agregam riqueza e equitatividade
em um único descritor numérico, sendo frequentemente uti l izados
na l i teratura sobre diversidade biológica (e.g., RODDA, 1993 apud
MOURA, 2003). Diversos índices já foram propostos (MAGURRAM,
1988), porém os mais amplamente empregados são os índices
Shannon-Wiener (H’) e de Pielou (J) (KREBS, 1989). Estes
índices foram calculados para cada piscina amostrada, para todos
os t ipos de substrato amostrados em cada piscina e para todos os
quadrados amostrados.
Buscando ponderar as abundâncias nos diferentes t ipos de
fundo das piscinas amostradas, foi calculado a abundância por
unidade de área (equação 3) para todos os t ipos de fundo das
quatro piscinas.
Abund./área = (T. O. / n) x 100 (Equação 3)

30
onde: T.O = total de observações no determinado tipo de
fundo
n = número de quadrados amostrado no tipo de fundo
determinado.
As comparações da abundância de peixes entre as piscinas
abertas e fechadas, e entre os diferentes t ipos de fundo foram
realizadas por meio de análises de variância de um fator (One-way
ANOVA) com intervalo de confiança de 95% (α< 0,05). Nas
análises de variância quando observada diferença signif icativas
aplicou-se o teste a posteriori de Tukey (p < 0,05). Adicionalmente,
foi também aplicada à rotina analít ica SIMPER (Similarity
percentages - species contributions) buscando identif icar as
espécies que mais contribuíram para dissimilaridade entre os
grupos definidos para análise (espécies mais discriminantes)
(CLARKE & WARWICK, 1994).
A normalidade foi testada através do teste de Kolmogorov. A
homogeneidade foi testada através do teste de Levene. Nos casos
onde os dados não se mostraram normais ou homogêneos estes
foram transformados pela raiz quadrada para assumirem os pré-
requisitos para análise.
A partir dos dados qualitativos (presença/ausência), foram
geradas planilhas de similaridade através do índice de Bray-Curtis
e posteriormente foram feitas análises de agrupamento
(Hierarchical Cluster). Como procedimento de aglomeração foi
uti l izado o UPGMA (“Unweighted Pair-Group Mean Average”) para
verif icar a determinação dos grupos hierárquicos.
As análises de agrupamento são técnicas analít icas criadas
para reconhecer subgrupos semelhantes de indivíduos ou objetos.
Especif icamente, as análises de agrupamento são técnicas
mult ivariadas que têm por objetivo classif icar uma amostragem de
indivíduos ou objetos, em grupos menores mutuamente exclusivos,
baseados na similaridade entre estes grupos (HAIR et al., 1998).
Esta análise foi empregada a f im de avaliar a similaridade com

31
relação à presença e ausência das espécies entre os diferentes
t ipos de fundo das piscinas amostradas.
Relações estatísticas entre a abundância de peixes com
EAH, foram quantif icadas via análise de correspondentes canônica
(ACC) e de regressão múlt ipla (α< 0,05). Para a realização destas
análises foram consideradas apenas as espécies que ocorreram
em ¼ dos quadrados amostrados, ou seja, espécies que ocorreram
em pelo menos 1 quadrado de cada estação amostradas.
Antes da realização das análises de regressão múlt ipla foi
calculado para as variáveis abióticas, o fator inf lação da variância
(FIV) através do programa estatíst ico SPSS versão 13.0. Este fator
indica o efeito que as variáveis independentes têm sobre o erro
padrão do coeficiente de regressão. Valores altos de FIV indicam
alto grau de colinearidade ou multicolinearidade entre variáveis
independentes (HAIR et al. , 1998), o que pode afetar os
resultados. Variáveis independentes com valores altos de FIV são
excluídas dessas análises.
Para a (ACC), todos os dados foram padronizados através do
método de Hil l (HILL,1979). Para testar a signif icância das
correlações entre os fatores bióticos e abióticos foi uti l izado o
teste de simulação de Monte Carlo (100 permutações), que se
baseia na geração de números aleatórios provenientes de uma
distribuição de probabil idade, a partir das 2 matrizes selecionadas
das espécies e das variáveis ambientais. A hipótese de nulidade
(Ho) levanta a ausência de relação entre as matrizes. Para essa
hipótese, os valores das l inhas da segunda matriz são misturados
aleatoriamente. Isso destrói a relação entre a primeira matriz e a
segunda, mas mantém a correlação dentro da segunda matriz
(MCCUNE & GRACE, 2002)
A ACC foi usada para descrever o relacionamento de cada
uma das espécies de peixes com as variáveis de substrato.
Contudo esta análise não permite que predições sejam feitas sobre
quais variáveis de substrato influenciam a abundância de todas as
espécies de peixes.

32
Buscando avaliar este fator foram geradas as análises de
regressão múlt ipla, buscando verif icar relações entre a abundância
e diversidade de peixes com as variáveis de substrato. Para esta
análise os dados uti l izados foram transformados para serem
aceitos de acordo com as premissas exigidas de normalidade e
homocedasticidade.
Todos os procedimentos analít icos, com exceção do calculo
do FIV foram realizados através dos programas estatísticos
Statist ica, versão 6.0, Primer (Plymouth Routines In Multi
Ecological Research), versão 5.1.2, e PC-ORD versão 4.0.
Buscando diminuir possíveis fontes de interferência durante
as análises foi excluída dos cálculos percentuais a espécie
Harengula clupeola (Clupeidae), pois um grande cardume com
milhares de indivíduos foi observado em um único evento, portanto
esta foi excluída das análises.
Famílias e espécies que tiveram suas abundâncias relativas
superiores a 1%, foram consideradas como mais representativas.

33
3) Resultados
Os dados obtidos neste trabalho são resultados de 62 dias de
permanência no Atol das Rocas onde foram realizados 75 horas de
observações subaquáticas. Realizaram-se 50 censos, sendo 25 em piscinas conectadas
com mar e 25 em piscinas sem conexão permanente com mar.
Durante as amostragens, foram observadas 48 espécies das 56
identif icadas no estudo piloto (Tabela 3).

34
Tabela 3: Famíl ias e espécies observadas durante as amostragens. São marcadas com ( X ) as p isc inas em que cada espécies fo i observada. Espécies que só ocorreram em pisc inas abertas (PA) ou pisc inas fechadas (PF), também são marcadas com ( X ) .
Famílias Espécies Barretinha Barreta Falsa PA Ancoras Cemiteriozinho PF
Acanthurus chirurgus x x x x ACANTHURIDAE Acanthurus coeruleus x x x x
ALBULIDAE Albula vulpes x x APOGONIDAE Phaeoptyx pigmentaria x x BALISTIDAE Balistes veluta x x BELONIDAE Platybelone argalus x x BLENNIIDAE Ophioblennius trinitatis x x x BOTHIDAE Bothus ocellatus x x x x
Caranx latus x x x CARANGIDAE Caranx bartholomaei x x x
CARCHARHINIDAE Negaprion brevirostris x x CHAETODONTIDAE Chaetodon ocellatus x x x x DACTYLOPTERIDAE Dactylopterus volitans x x
DASYATIDAE Dasyatis americana x x GINGLYMOSTOMIDAE Ginglymostoma cirratum x x
Coryphopterus glaucofraenum x x x x GOBIIDAE Gnatholepis thompsoni x x x x Haemulon chrysargyreum x x x HAEMULIDAE Haemulon parra x x x Holocentrus adscensionis x x x x HOLOCENTRIDAE Myripristis jacobus x x x x
KYPHOSIDAE Kyphosus sectatrix x x Halichoeres radiatus x x x x LABRIDAE Thalassoma noronhanum x x x x
LABRISOMIDAE Malacoctenus triangulatus x x x x LUTJANIDAE Lutjanus jocu x x x x

35
Tabela 3: Continuação
MALACANTHIDAE Malacanthus plumieri x x MONACANTHIDAE Aluterus scriptus x x
MULLIDAE Mulloidichthys martinicus x x x Gymnothorax funebris x x Gymnothorax miliaris x x x x Gymnothorax vicinus x x x
MURAENIDAE
Muraena pavonina x x OPHICHTIDAE Myrichthys ocellatus x x OSTRACIIDAE Lactophrys trigonus x x
POMACANTHIDAE Pomacanthus paru x x x x Abudefduf saxatilis x x x x Chromis multilineata x x Stegastes pictus x x x x
POMACENTRIDADE
Stegastes rocasensis x x x x Sparisoma amplum x x x x Sparisoma axillare x x x x SCARIDAE Sparisoma frondosum x x x x Cephalopholis fulva x x x x
SERRANIDAE Paranthias furcifer x x SPHYRAENIDAE Sphyraena barracuda x x SYNODNTIDAE Synodus intermedius x x x

36
3.2.1) Barretinha Na Barretinha realizaram-se 11 censos, que foram sorteados
aleatoriamente entre as 14 estações. Dos 44 quadrados
amostrados nesta piscina, 68% estavam sobre fundos rochosos,
27% sobre fundos arenosos e 5% sobre fundos de cascalho; tal
fato demonstra o predomínio de substrato consolidado nesta
piscina. A f igura 8 mostra a porcentagem dos tipos de substratos
amostrados, a porcentagem da abundância das espécies em cada
substrato e a abundância por unidade de área.
Figura 8: Porcentagens dos t ipos de substratos amostrados, da abundância das espécies em cada t ipo de substrato e da abundância por unidade de área na piscina denominada Barret inha. No período de 22/05 à 21/06/2006.
Foram observadas 26 famílias e 37 diferentes espécies de
peixes (Tabela 3), totalizando 2.679 observações. A tabela 4
mostra os valores dos índices ecológicos e o total de observações
nos diferentes t ipos de substratos na Barretinha.

37
Tabela 4: Riqueza de espécies (S), tota l de observações (N), medida de equi tat iv idade (J) e divers idade de Shannon-Wiener (H’) , na piscina denominada Barret inha.
S N J H' Substrato rochoso 35 2180 0,6398 2,28 Substrato arenoso 20 410 0,731 2,01
Substrato de cascalho
11 89 0,7706 1,87
O índice de diversidade (H’) calculado para esta piscina foi
2,32 e a medida de equitatividade (J) 0,643. Os valores das
abundâncias relativas das famílias mais representativas são
apresentados na figura 9.
41,7
16,910,8
7,54,2 4,0 3,8 2,8 2,1 1,9 1,4 1,2
05
1015202530354045
Labri
dae
Pomac
entrid
ae
Gobiid
ae
Acanth
uridae
Blenniid
ae
Scarid
ae
Serranid
ae
Labri
somida
e
Albulida
e
Carangid
ae
Holoce
ntridae
Chaetod
ontid
ae
Famílias
%
Figura 9: Abundância relat iva das famíl ias mais representat ivas na piscina Barret inha. No período de 22/05 à 21/06/2006.
As cinco espécies mais abundantes foram: Thalassoma
noronhanum (39,75%), Stegastes rocasensis (9,26%),
Coryphopterus glaucofraenum (8,70%), Abudefduf saxati l is (7,39%)
e Acanthurus chirurgus (6,57%). Espécies como o Albula vulpes ,
Aluterus scriptus, Balistela veluta, Dactylopterus volitans ,
Ginglymostoma cirratum e Negaprion brevirostris só foram
observadas nesta piscina.
38
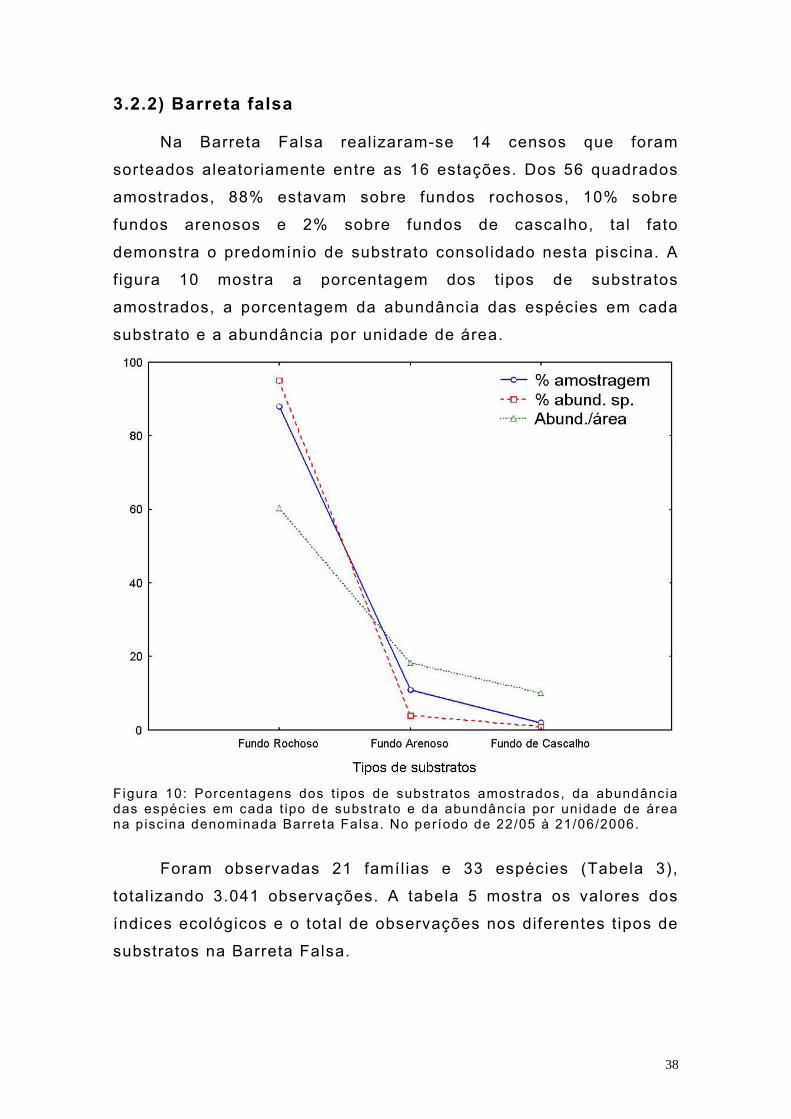
3.2.2) Barreta falsa Na Barreta Falsa realizaram-se 14 censos que foram
sorteados aleatoriamente entre as 16 estações. Dos 56 quadrados
amostrados, 88% estavam sobre fundos rochosos, 10% sobre
fundos arenosos e 2% sobre fundos de cascalho, tal fato
demonstra o predomínio de substrato consolidado nesta piscina. A
f igura 10 mostra a porcentagem dos t ipos de substratos
amostrados, a porcentagem da abundância das espécies em cada
substrato e a abundância por unidade de área.
Figura 10: Porcentagens dos t ipos de substratos amostrados, da abundância das espécies em cada t ipo de substrato e da abundância por unidade de área na piscina denominada Barreta Falsa. No período de 22/05 à 21/06/2006.
Foram observadas 21 famíl ias e 33 espécies (Tabela 3),
totalizando 3.041 observações. A tabela 5 mostra os valores dos
índices ecológicos e o total de observações nos diferentes t ipos de
substratos na Barreta Falsa.

39
Tabela 5: Riqueza de espécies (S), tota l de observações (N), medida de equi tat iv idade (J) e d iversidade de Shannon-Wiener (H), na piscina denominada Barreta Falsa.
S N J H' Substrato rochoso 31 2902 0,6757 2,32 Substrato arenoso 15 109 0,6902 1,815
Substrato de cascalho
6 30 0,7602 1,362
O índice de diversidade (H’) calculado para esta piscina foi
2,31 e a medida de equitatividade (J) 0,662. Os valores das
abundâncias relativas das famílias mais representativas são
apresentados na figura 11.
38,90
20,55
10,926,22 4,54 3,85 3,78 3,26 2,20 1,74 1,48 1,35
05
1015202530354045
Labri
dae
Pomac
entrid
ae
Gobiid
ae
Acanthuri
dae
Serran
idae
Holoce
ntrida
e
Labri
somida
e
Scarid
ae
Haemuli
dae
Caran
gidae
Blennii
dae
Chaeto
donti
dae
Famílias
%
Figura 11: Abundância re lat iva das famíl ias mais representat ivas na pisc ina Barreta Falsa. No período de 22/05 à 21/06/2006.
As cinco espécies mais abundantes foram: Thalassoma
noronhanum (36,96%), Stegastes rocasensis (12,40%),
Coryphopterus glaucofraenum (9,40%), Abudefduf saxati l is (7,43%)
e Acanthurus chirurgus (5,72%). Espécies como o Kyphosus
sectatrix, Muraena Pavonina, Myrichthys ocellatus e Phaeoptyx
pigmentariam só foram observadas nesta piscina.

40
3.2.3) Cemiteriozinho Na piscina do Cemiteriozinho realizaram-se 6 censos que
foram sorteados aleatoriamente entre as 8 estações. Dos 24
quadrados amostrados, 29% estavam sobre fundos rochosos, 29 %
sobre fundos arenosos e 42% sobre fundos de cascalho. Tal fato
demonstra o predomínio de substratos não consolidados nesta
piscina. A f igura 12 mostra a porcentagem dos tipos de substratos
amostrados, a porcentagem da abundância das espécies em cada
substrato e a abundância por unidade de área.
Figura 12: Porcentagens dos t ipos de substratos amostrados, da abundância das espécies em cada t ipo de substrato, e a abundância por unidade de área na piscina denominada Cemiter ioz inho. No período de 22/05 à 21/06/2006.
Foram observadas 14 famílias e 23 espécies (Tabela 3),
totalizando 895 observações. A tabela 6 mostra os valores dos
índices ecológicos e o total de observações nos diferentes t ipos de
substratos na piscina do Cemiteriozinho.
41
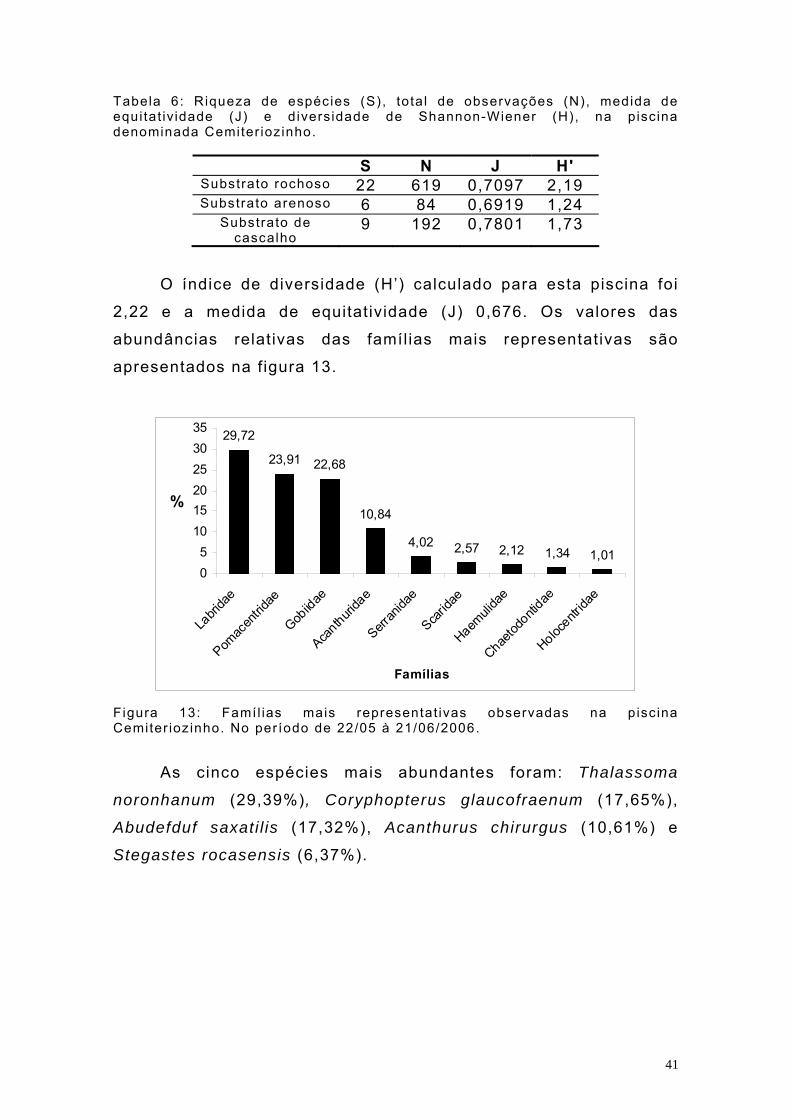
Tabela 6: Riqueza de espécies (S), tota l de observações (N), medida de equi tat iv idade (J) e d iversidade de Shannon-Wiener (H), na piscina denominada Cemiter ioz inho.
S N J H' Substrato rochoso 22 619 0,7097 2,19 Substrato arenoso 6 84 0,6919 1,24
Substrato de cascalho
9 192 0,7801 1,73
O índice de diversidade (H’) calculado para esta piscina foi
2,22 e a medida de equitatividade (J) 0,676. Os valores das
abundâncias relativas das famílias mais representativas são
apresentados na figura 13.
29,72
23,91 22,68
10,84
4,02 2,57 2,12 1,34 1,0105
101520253035
Labri
dae
Pomac
entrid
ae
Gobiid
ae
Acanthuri
dae
Serran
idae
Scarid
ae
Haemuli
dae
Chaetod
ontidae
Holocentr
idae
Famílias
%
Figura 13: Famíl ias mais representat ivas observadas na pisc ina Cemiter ioz inho. No período de 22/05 à 21/06/2006.
As cinco espécies mais abundantes foram: Thalassoma
noronhanum (29,39%), Coryphopterus glaucofraenum (17,65%),
Abudefduf saxati l is (17,32%), Acanthurus chirurgus (10,61%) e
Stegastes rocasensis (6,37%).

42
3.2.4) Âncoras Na piscina das Âncoras realizaram-se 19 censos que foram
sorteados aleatoriamente entre as 22 estações. Dos 76 quadrados
amostrados nesta piscina, 53% estavam sobre fundos rochosos,
39% estavam sobre fundos arenosos e 8% sobre fundos de
cascalho, tal fato demonstra uma equivalência entre os substratos
consolidados e não consolidados nesta piscina. A f igura 14 mostra
a porcentagem dos tipos de substratos amostrados, a porcentagem
da abundância das espécies em cada substrato e a abundância por
unidade de área.
Figura 14 : Porcentagens dos t ipos de substratos amostrados, da abundância das espécies em cada t ipo de substrato e a abundância por unidade de área na piscina denominada das Ancoras. No período de 22/05 à 21/06/2006.
Foram observadas 22 famíl ias e 35 espécies (Tabela 3),
totalizando 2.991 observações. A tabela 7 mostra os valores dos
índices ecológicos e o total de observações nos diferentes t ipos de
substratos na piscina das Ancoras.

43
Tabela 7: Riqueza de espécies (S), tota l de observações (N), medida de equi tat iv idade (J) e d iversidade de Shannon-Wiener (H), na piscina denominada Ancoras.
S N J' H' Substrato rochoso 32 2184 0,7175 2,487 Substrato arenoso 23 695 0,7474 2,043
Substrato de cascalho
15 112 0,7629 1,907
O índice de diversidade (H’) calculado para esta piscina foi
2,52 e a medida de equitatividade (J) foi 0,7606. Os valores das
abundâncias relativas das famílias mais representativas são
apresentados na figura 15.
31,47
21,2015,28
11,576,32 4,11 2,64 1,81 1,57
05
101520253035
Pomac
entrid
ae
Labri
dae
Gobiid
ae
Acanth
urida
e
Serran
idae
Scarid
ae
Haemuli
dae
Holoce
ntrida
e
Chaeto
donti
dae
Famílias
%
Figura 15 : Famíl ias mais representat ivas observadas na pisc ina das Ancoras. No período de 22/05 à 21/06/2006.
As cinco espécies mais abundantes foram: Thalassoma
noronhanum (19,53%), Stegastes rocasensis (15,22%),
Coryphopterus glaucofraenum (14,65%), Abudefduf saxati l is
(12,61%) e Acanthurus chirurgus (10,10%). Espécies como
Chromis multi l ineata, Dasyatis americana, Lactophrys tr igonus e
Paranthias furcifer só foram observadas nesta piscina.

44
3.3) Piscinas Abertas x Piscinas Fechadas
3.3.1) Abundância Relativa Uma primeira análise destes dados revela a maior
abundância relativa das famílias Labridae, Pomacanthidae,
Gobiidae e Acanthuridae em todas as estações, com exceção da
piscina das Âncoras onde a família Pomacanthidae é a mais
abundante. A abundância relativa total das famílias e das espécies
mais representativas das piscinas amostradas são apresentados
nas figuras 16 e 17, respectivamente.
33,33
23,65
13,368,67
4,84 3,67 2,23 2,11 1,79 1,65 1,35 1,3305
101520253035
Labri
dae
Pomac
entrid
ae
Gobiid
ae
Acanthuri
dae
Serran
idae
Scarid
ae
Labri
somidae
Holocentr
idae
Haemuli
dae
Blennii
dae
Chaetod
ontidae
Carangida
e
Famílias
%
Figura 16: Abundância relat iva das famíl ias mais representat ivas do Atol das Rocas. No período de 22/05 à 21/06/2006.

45
1,11
1,34
1,35
1,56
1,65
1,71
1,73
1,88
2,23
4,74
7,80
9,95
11,63
11,85
31,63
0 5 10 15 20 25 30 35
Haemulon chrysargyreum
Sparisoma axillare
Chaetodon ocellatus
Sparisoma amplum
Ophiobleninus trinitatis
Halichoeres radiatus
Gnatholepis thompsoni
Holocentrus ascensionis
Malacoctenus triangulatus
Cephalopholis fulva
Acanthurus chirurgus
Abudefduf saxatilis
Coryphopterus glaucofraenum
Stegastes rocasensis
Thalassoma noronhanum
Esp
écie
s
%
Figura 17: Abundância relat iva das espécies mais representat ivas do Atol das Rocas. No período de 22/05 à 21/06/2006.
A aplicação da análise de variância não identif icou efeitos
signif icativos (Anova; G.L=28; F= 3,189 p= 0,079) na comparação
da abundância relativa das espécies mais representativas entre as
piscinas abertas e fechadas.
A abundância relativa das 5 espécies (Thalassoma
noronhanum, Stegastes rocasensis, Coryphopterus glaucofraenum,
Abudefduf saxati l is e Acanthurus chirurgus) que abrageram a maior
parte da abundância relativa total nas piscinas amostradas são
apresentadas na figura 18.

46
Figura 18: Abundância relat iva das espécies dominantes em cada uma das pisc inas amostradas. No período de 22/05 à 21/06/2006. Nota-se a maior abundância do T.noronhanum e do S.
rocasensis em todas as piscinas amostradas com exceção da
piscina do Cemiteriozinho onde o C. glaucofraenum é a segunda
espécie mais abundante.
3.3.2) Abundância relativa nos diferentes tipos de fundo
Dos 9.606 peixes observados, 82% estavam sobre fundos
rochosos, 13% sobre fundos arenosos e 5% sobre fundos de
cascalho. A f igura 19 mostra a abundância por unidade de área, a
porcentagem da abundância das espécies e a porcentagem dos
tipos de fundo amostrados nas quatro piscinas (os valores de
porcentagem foram calculados separadamente para cada piscina).

47
Figura 19: Porcentagens de cada t ipo de substrato amostrado, da abundância das espécies em cada t ipo de substrato e abundância por unidade de área (Abund/área), para Fundo Rochoso (FR), fundo Arenoso (FA) e fundo de Cascalho (FC); nas pisc inas amostradas. No período de 22/05 à 21/06/2006.
Nota-se que as maiores abundâncias foram verif icadas sobre
substratos rochosos, em todas as piscinas amostradas,
independente da piscina ser ou não conectada permanentemente
com oceano. Os fundos de cascalho se mostraram menos
abundantes na maioria das piscinas, com exceção da piscina do
Cemiteriozinho.
A abundância por unidade de esforço manteve o mesmo
padrão em todas as piscinas com os maiores valores sendo
verif icados nos fundos rochosos e os menores nos fundos de
cascalho (Figura 19).
A aplicação da análise de variância não identif icou diferenças
signif icativas na comparação da abundância por unidade de área
entre os fundos rochosos das piscinas abertas e fechadas
(ANOVA; G.L= 2;F=1,378; p = 0,362), o mesmo ocorreu com os

48
fundos de areia (ANOVA; G.L=2;F=0,644; p =0,506) e cascalho
(ANOVA;G.L=2;F=0,078; p =0,806).
Contudo quando comparado à abundância por unidade de
esforço nos diferentes t ipos de fundo, diferenças signif icativas
foram observadas (ANOVA;G.L=9;F=43,26; p = 0,000). A aplicação
do teste de Tukey mostrou que as diferenças signif icativas
ocorreram entre os fundos consolidados e não consolidados (Tabela 8).
Tabela 8: Resultados do teste de Tukey. São marcados com (*) os t ipos de fundo com di ferenças s igni f icat ivas ao nível de p < 0,05.
Fundo Rochoso
Fundo Arenoso
Fundo de cascalho
Fundo Rochoso --- 0,000354* 0,000196* Fundo Arenoso 0,000354* --- 0,185860 Fundo de cascalho 0,000196* 0,185860 ---
O método analít ico SIMPER mostrou que as espécies que
mais contribuíram para a constatação destas diferenças foram o
Thalassoma noronhanum, Stegastes rocasensis, Coryphopterus
glaucofraenum e Abudefduf saxati l is como apresentado na tabela
9.
Tabela 9: Valores de porcentagem (%) das espécies que mais contr ibuíram para a dissimi lar idade entre os di ferentes t ipos de fundo.
Rocha x Areia Rocha x Cascalho Espécie Contrib. % Contrib. % Thalassoma noronhanum 32,7 33,06 Stegastes rocasensis 12,46 12,3 Coryphopterus glaucofraenum 8,39 9,7 Abudefduf saxati l is 8,35 9,29 Total 61,9 64,35

49
3.3.3) Índices de diversidade
Foram calculados os índices de diversidade das quatro (4)
piscinas amostradas separadamente, dos diferentes t ipos de fundo
e de todas as estações amostradas em cada piscina.
Os índices de diversidade calculados para cada piscina
t iveram o mínimo de 2,22 e máximo de 2,52. O maior índice de
diversidade foi verif icado na piscina das Âncoras seguido da
Barretinha, Barreta Falsa e Cemiteriozinho. Com relação à medida
de equitatividade, esta se mostrou maior nas piscinas sem conexão
permanente com oceano, como mostra a Figura 20.
Figura 20: Valores dos índices de diversidade e equi tat iv idade calculados para as 4 piscinas amostradas. No período de 22/05 à 21/06/2006.
Com relação aos índices de diversidade calculados para cada
tipo de fundo das piscinas amostradas, verif icou-se que os maiores
valores foram observados sobre substratos rochosos, seguido dos
substratos arenosos e de cascalho, tanto nas piscinas conectadas
como não conectadas com o oceano. A única exceção é a piscina

50
do Cemiteriozinho onde a diversidade no fundo de cascalho foi
maior. Com a medida de Equitatividade, ocorreu o inverso do que
foi observado com os índices de diversidade, ou seja, os maiores
valores de equitatividade foram observados nos fundos de
cascalho, areia e rocha respectivamente como mostra a Figura 21.
Figura 21: Valores dos índices de divers idade e equi tat iv idade nos di ferentes t ipos de fundo das pisc inas amostradas. (FR) Fundo Rochoso, (FA) Fundo Arenoso, Fundo de Cascalho (FC). No período de 22/05 à 21/06/2006.
A aplicação da análise de variância não identif icou diferenças
signif icativas na comparação da diversidade de espécies
conferidas nas diferentes estações amostradas das piscinas
abertas e fechadas ANOVA;G.L=48;F=0,05; p = 0,829). O mesmo
ocorreu na comparação da diversidade de espécies entre os t ipos
de fundo entre as piscinas amostradas (ANOVA;G.L=8;F=1,678; p
= 0,248).

51
Porém, quando comparados os índices de diversidade entre
os fundos de areia, cascalho e rocha diferenças signif icativas
foram observadas (Tabela10).
Tabela 10: Valores de Fisher (F) e da probabi l idade (p) calculados nas comparações entre os di ferentes t ipos de fundo. São marcados com (*) os t ipos de fundo com diferenças s igni f icat ivas ao nível de p < 0,05. O grau de l iberdade para essas anál ises foi 8.
Fundo de pedra Fundo de areia Fundo de cascalho
F p F p F p Fundo de pedra 2,7792 0,146542 9,5206* 0,021510*Fundo de areia 2,7792 0,146542 0,1345 0,726359
Fundo de cascalho
9,5206* 0,021510* 0,1345 0,726359

52
3.4) Análises de agrupamento
A análise de agrupamento gerada com os dados de presença
ou ausência das espécies observadas nos diferentes tipos de
fundo das piscinas abertas e fechadas separou 2 grupos: o
primeiro formado por fundos consolidados e o segundo formados
por fundos não consolidados (Figura22).
Figura 22: Fundo Rochoso (FR), Fundo de Arenoso (FA), Fundo de Cascalho (FC)
Nota-se o agrupamento dos fundos consolidados e não
consolidados, independente de serem áreas com características
contrastantes, ou seja, piscina com conexão permanente com o
oceano e sem conexão.
Outra analise de agrupamento gerada com os dados de
presença e ausência das espécies nos diferentes t ipos de fundo
das piscinas amostradas, separou 2 grandes grupos e 4 subgrupos
(Figura 23).
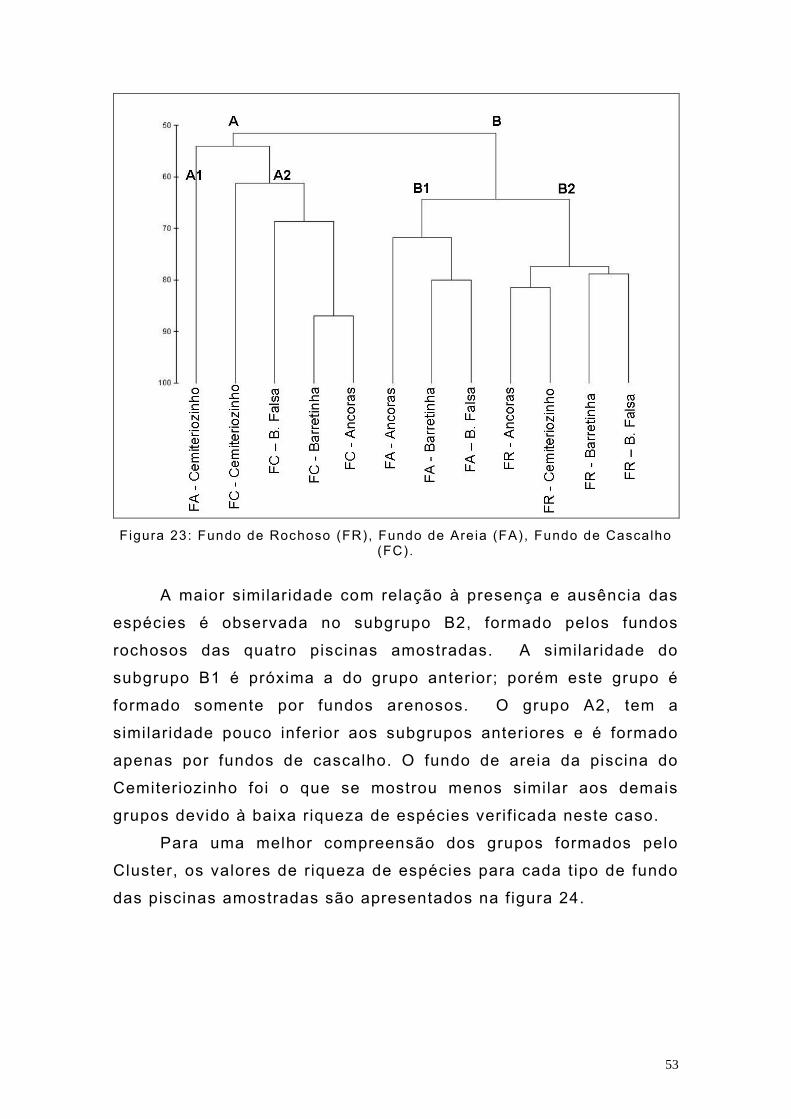
53
Figura 23: Fundo de Rochoso (FR), Fundo de Areia (FA), Fundo de Cascalho
(FC).
A maior similaridade com relação à presença e ausência das
espécies é observada no subgrupo B2, formado pelos fundos
rochosos das quatro piscinas amostradas. A similaridade do
subgrupo B1 é próxima a do grupo anterior; porém este grupo é
formado somente por fundos arenosos. O grupo A2, tem a
similaridade pouco inferior aos subgrupos anteriores e é formado
apenas por fundos de cascalho. O fundo de areia da piscina do
Cemiteriozinho foi o que se mostrou menos similar aos demais
grupos devido à baixa riqueza de espécies verif icada neste caso.
Para uma melhor compreensão dos grupos formados pelo
Cluster, os valores de riqueza de espécies para cada tipo de fundo
das piscinas amostradas são apresentados na figura 24.

54
Figura 24: Riqueza (S) de espécies nos di ferentes t ipos de substrato das pisc inas amostradas. A1, A2, B1 e B2 são as legendas dos grupos formados pelo Cluster . Nota-se os maiores valores de riqueza de espécies nos
fundos rochosos (grupo B2), seguido dos fundos arenosos (B1) e
de cascalho (A2). O fundo de areia da piscina do Cemiteriozinho
se inclui num grupo a parte dos demais (A1).

55
3.5) Relações entre peixes e as variáveis de substrato
O substrato duro foi a variável de substrato com maior grau
de mult icolinearidade o que indica que variações das demais
variáveis estão associadas a variações na porcentagem de
substrato duro. Isto pode causar problemas de mult icolinearidade,
portanto o substrato duro foi excluído das análises.
A análise de correspondentes canônica formou três
diferentes grupos de espécies. O grupo 1 formado por espécies
associadas à maior complexidade estrutural: Cephalopholis fulva,
Chaetodon ocellatus, Halichoeres radiatus, Holocentrus
adscensionis, Malacoctenus tr iangulatus, Sparisoma amplum e
Stegastes rocasensis. O grupo 2 é formado por espécies que
ocorrem tanto em ambientes de maior complexidade estrutural
como em ambientes menos complexos. Este grupo é formado pelas
seguintes espécies: Acanthurus chirurgus, Acanthurus coeruleus,
Thalassoma noronhanum, Sparisoma axil lare, Sparisoma
frondosum e Abudefduf saxati l is. Por últ imo o grupo 3 formado por
espécies associados a ambientes de baixa complexidade
estrutural: Coryphopterus glaucofraenum e Gnatholepis thompsoni
(Figura 25).

56
Figura 25: Resultados obt idos através de ACC com as var iáveis de substrato e abundância das espécies: A.sax, Abudefduf saxat i l is ; A.chi , Acanthurus chirurgus; A.coe, Acanthurus coeruleus; C.ful , Cephalophol is fu lva; C.ocel , Chaetodon ocel latus; C. gla, Coryphopterus glaucofraenum; G.tho, Gnatholepis thompsoni ; H.rad, Hal ichoeres radiatus; H.ads, Holocentrus adscensionis; M.tr i , Malacoctenus tr iangulatus; S.amp, Sparisoma amplum; S.axi , Spar isoma axi l lare; S.fro, Spar isoma frondosum; S.roc, Stegastes rocasensis; T. nor, Thalassoma noronhanum. Eigenvalues: eixo 1= 0,06; eixo 2= 0,07. Os dois pr imeiros eixos respondem a 14% das var iâncias. O teste de Monte-Car lo para todos os eixos canônicos foram signi f icat ivos (p < 0,05).
A ACC mostra as diferentes relações de cada espécie com as
variáveis de substrato. Por exemplo, o M. triangulatus (Grupo1),
prefere áreas com maior porcentagem de cobertura viva (fundos
rochosos com grande cobertura viva). Enquanto que C.
glaucofraenum tem preferências por ambientes com menor
cobertura viva e menor complexidade estrutural (fundos arenosos).
A análise de regressão múltipla mostrou que as combinações
das variáveis de substrato contribuem com cerca de 58% da
variação na diversidade de peixes e com 12% das variações na
abundância das espécies.

57
Duas variáveis relacionaram-se de maneira signif icativa com
a diversidade, são essas: Rugosidade e formas de crescimento.
Enquanto que para abundância a única variável que se relacionou
de maneira signif icativa foi a rugosidade (Tabela11).
Tabela 11: Contr ibuição de todas as var iáveis do EAH para as var iações na abundância e divers idade de espécies. São apresentadas as correlações e os níveis de s igni f icância para cada uma das c inco var iáveis. São marcados com (*) as var iáveis de substrato com di ferenças signi f icat ivas a nível de (p < 0,05).
Abundância Diversidade Correlação parcial p Correlação parcial p Rugosidade 0,22 0,000* 0,43 0,000*Formas de Crescimento 0,05 0,094 0,37 0,000*Altura 0,01 0,840 0,07 0,475Tamanho dos refúgios 0,04 0,272 0,12 0,223Cobertura viva 0,05 0,158 -0,16 0,091Regressão multipla r = 0,12 r = 0,58

58
4) Discussão
4.1) Metodologia Dentre as diferentes técnicas empregadas atualmente em
estudos de peixes em ambientes recifais, o censo visual é o mais
amplamente empregado em ambientes rasos, com substratos
heterogêneos tal como recifes de corais e costões rochosos (DE
GIROLAMO & MAZZOLDI 2001, SAMOILYS & CARLOS, 2000). Isto
deve-se ao seu baixo custo relativo, caráter não destrutivo e não
invasivo (MOURA 2003); grande parte do conhecimento acumulado
sobre ecologia de peixes em ambientes recifais é derivada de
amostragens diretas feitas através de censos visuais (SAMOILYS
& CARLOS, 2000; ACKERMAN & BELLWOOD, 2000).
Entretanto, assim como outros métodos de amostragens o
censo visual apresenta possíveis fontes de erro (SALE, 1997,
HARMELIN-VIVIEN et al., 1985 apud DE GIROLAMO & MAZZOLDI
2001).
Diversos autores (BROCK, 1982, BELLWOOD & ALCALA,
1988, LINCOLN-SMITH, 1989, ACKERMAN E BELLWOOD, 2000,
WILLIS, 2001) relatam que as técnicas de censo visual,
normalmente empregadas nos estudos da ictiofauna em ambientes
recifais geralmente subestimam peixes com hábitos crípticos. No
presente trabalho, visando diminuir esta possível fonte de erro,
espécies que são facilmente avistadas foram contabil izadas
separadas das espécies com hábitos de entocamento ou de
camuflagem, como citado no item 2.4. A escolha deste desenho
amostral apóia-se no fato de que os censos onde espécies com
diferentes hábitos (pelágicos ou crípticos) são amostradas
separadamente produzem estimativas mais acuradas (BOHNSACK
& BANNEROT 1986, DE GIROLAMO & MAZZOLDI 2001, MOURA
2003).
Alguns autores (STANLEY & WILSON, 1995, DE GIROLAMO
& MAZZOLDI 2001) relatam que em amostragens feitas através do

59
censo visual, alguns peixes podem ser afetados posit ivamente ou
negativamente pela presença do observador. Buscando minimizar
esta possível fonte de influência antes de cada censo, foram
gastos cinco minutos de espera para que os peixes se
acostumassem com a presença do observador e do quadrado. DE
GIROLAMO & MAZZOLDI (2001), comparando desenhos amostrais
baseados na metodologia de transectos, observaram que não há
diferenças signif icativas em censos realizados com intervalos de
espera de 5, 10 e 15 minutos (DE GIROLAMO & MAZZOLDI, 2001),
baseados nesta constatação, foi estabelecido o intervalo de 5
minutos.
Amostragens diretas feitas através do mergulho envolvem
fontes de influência adicionais como o comportamento e
experiência do observador, além da tomada de decisões
subjetivas. Ressalta-se que dif ici lmente todas as fontes de
influência possam ser eliminadas, o grande desafio é reconhecê-
las tentando assim minimizá-las (EDGARD et al. 2004). Sendo
assim a melhor metodologia a ser aplicada vai depender dos
objetivos do estudo, da logística disponível, das características do
ambiente, da profundidade, da velocidade da corrente, da
visibi l idade da água, das ondas, dentre outros fatores (EDGARD et
al. 2004). Da mesma forma, o melhor delineamento da amostragem
(número de amostragens, horários das amostragens, período de
maré, etc) também varia conforme os fatores supracitados.
No presente estudo a metodologia empregada mostrou-se
eficiente, pois foram observadas durante os censos 48 diferentes
espécies de peixes, número semelhante ao observado nos
trabalhos realizados por ROSA & MOURA (1997) e MOURA (1998);
sobre a estrutura da comunidade de peixes do Atol das Rocas,
onde outra metodologia de amostragem visual foi uti l izada.

60
4.2) Piscinas Abertas x Piscinas Fechadas
4.2.1) Abundância relativa No geral, características da comunidade de peixes do Atol
das Rocas, como a composição taxonômica e a dominância de
determinadas espécies foram verif icadas em todas as piscinas
tanto conectadas como não conectadas com mar aberto durante a
maré baixa.
Espécies como o Thalassoma noronhanum, Stegastes
rocasensis, Coryphopterus glaucofraenum, Acanthurus chirurgus e
Abudefduf saxati l is foram as mais abundantes em todas as
piscinas.
Como já comentado o Thalassoma noronhanum foi a espécie
mais abundante em todas as piscinas. A segunda espécie mais
abundante nas piscinas amostradas foi o Stegastes rocasensis,
com exceção da piscina do Cemiteriozinho, onde o Coryphopterus
glaucofraenum foi à segunda espécie mais abundante; fato
atribuído a maior extensão da planície arenosa nesta piscina. O C.
glaucofraenum tem preferência por fundos arenosos próximo a
recifes (HUMANN & DELOACH, 2003), ao contrário do S.
rocasensis que tem preferência por águas rasas e fundos
consolidados (SMITH, 2004). Na piscina do Cemiteriozinho o S.
rocasensis foi a terceira espécie mais abundante.
As cinco espécies abrangeram, 72,86 % do total das
observações, o que indica que estas são as mais abundantes no
interior do anel recifal do Atol das Rocas. É importante ressaltar
que as duas espécies que abrageram a maior parcela da
abundância relativa (Thalassoma noronhanum e Stegastes
rocasensis) e que, conseqüentemente, são as espécies
dominantes; são espécies de distr ibuição restrita na costa
brasileira. A mais abundante, o Thalassoma noronhanum distribui-
se entre a costa do Estado do Maranhão ate o Estado de São
Paulo. Já o Stegastes rocasensis é endêmico do conjunto insular

61
oceânico Rocas/Fernando de Noronha (MENEZES et al. , 2003).
Juntas estas duas espécies abrageram 43,48% do total de
observações, porcentagem muito semelhante à observada por
ROSA & MOURA (1997) onde o total de observações feitas para
essas espécies foi 42,84% .
O fato da comparação das abundâncias relativas das
espécies entre piscinas com e sem conexão com oceano não terem
resultado em diferenças signif icativas, sugere que o fator
conectada /não conectada não influência a abundância de peixes
nas piscinas amostradas durante a maré baixa.
4.2.2) Abundância relativa nos diferentes tipos de fundo
A maior parte dos censos realizados nas piscinas
conectadas permanentemente com o oceano foram realizados
sobre substratos rochosos. Nas piscinas sem conexão permanente
com o oceano uma parcela maior de substratos não consolidados
foi amostrada se comparado com as piscinas conectadas com o
oceano. Ambos os fatos são corroborados por ROSA & MOURA
(1997).
Apesar de haver um maior esforço amostral sobre
substratos rochosos nas piscinas das Âncoras, Barretinha e
Barreta Falsa, observa-se que em todos esses casos a
porcentagem da abundância foi maior que a porcentagem de
amostragens. O contrario ocorre nos fundos não consolidados. Na
piscina do Cemiteriozinho, mesmo tendo um maior esforço
amostral sobre fundos não consolidados, as maiores abundâncias
de peixes foram observadas sobre substratos rochosos. Tais fatos
sugerem a maior abundância de peixes sobre fundos consolidados,
independente da piscina ser ou não conectada permanentemente
com oceano. De fato, quando aplicada análise de variância,
buscando comparar as abundâncias por unidade de área nos
diferentes t ipos de fundo das piscinas com e sem conexão
permanente com oceano, não foram observadas diferenças

62
signif icativas. Porém, quando comparada à abundância por
unidade de área nos distintos t ipos de fundo, diferenças
signif icativas foram observadas entre os fundos consolidados e
não consolidados.
A estrutura dos substratos consolidados fornece abrigo e
locais para alimentação da maior parte das espécies de pequeno
porte, que compõem a maior parcela, em número de espécies e
indivíduos, das comunidades de peixes recifais (SALE, 1991).
Este t ipo de substrato, por possuir arquitetura mais complexa que
os campos de algas, substratos arenosos e de cascalho,
frequentemente estão relacionados à maior abundância de peixes
(NAGELKERKEN et al., 2000; FERREIRA et al. , 2001; MATEO &
TOBIAS, 2001; GRATWICKE & SPEIGHT, 2005), fato também
constatado no presente estudo.
Verif icou-se, por meio de analises de SIMPER, que as
espécies que mais contribuíram para constatação dessas
diferenças foram: Thalassoma noronhanum, Stegastes rocasensis,
Coryphopterus glaucofraenum e Abudefduf saxati l is. Todas as
espécies acima citadas têm grande abundância no Atol, e habitam
preferencialmente fundos consolidados, com exceção do
Coryphopterus glaucofraenum que vive na interface areia / rocha
(HUMMAN & DELOACH, 2003; SMITH, 2004).
Essas verif icações vêm sugerir que o fator que mais
influência na abundância dos peixes é o t ipo de substrato e não o
fato da piscina ter ou não conexão permanente com oceano.
4.2.3) Índices de diversidade O maior valor no índice H’ verif icado na piscina das Âncoras
(2,52) pode estar relacionado à variedade de atributos estruturais
existentes nesta piscina. Como descrito no item 2.3.1, esta piscina
possui cavernas na interface rocha areia, “patch reef” e fundos não
consolidados; cada um destes fatores propicia habitat favorável a
uma determinada gama de espécies o que acarretou num maior

63
índice de diversidade nesta piscina. No estudo realizado por
CHEROSKE et al. (2000) a diversidade de peixes, também
apresentou valores mais altos devido às variações nas estruturas
do recife. Apesar dos atributos estruturais citados estarem
presentes nas outras piscinas, estes não são bem representados
como na piscina das Âncoras, como descrito no item 2.3.
Outro fator que pode ter contribuído para a maior
diversidade da piscina das Âncoras é a sua localização, situada a
barlavento do Atol das Rocas. O anel recifal do Atol é subdividido
num arco de barlavento e num arco de sotavento (KIKUCHI, 1999).
A região onde a piscina das Âncoras está localizada é a de maior
energia no Atol (KIKUCHI, 1994) e é por onde grande parte da
água entra durante a preamar. Este fator pode acarretar numa
maior concentração de organismos planctônicos nas piscinas
situadas a barlavento do Atol. Uma constatação que pode vir a
comprovar essa maior concentração de plâncton nesta região, são
as espécies Chromis multi l ineata e Paranthias furcifer, ambas
planctófagas (HUMMAN & DELOACH, 2003, SMITH, 2004), que
foram observadas com freqüência somente na piscina das Âncoras.
Entre as demais piscinas os índices H’ mostraram-se
semelhantes, com o maior valor sendo observado na Barretinha,
seguido da Barreta Falsa e Cemiteriozinho. Os maiores valores
observados nas piscinas abertas estão relacionados à maior
extensão do substrato rochoso e a presença signif icativa de “patch
reef”. Tais fatores proporcionam uma maior heterogeneidade de
nichos e consequentemente uma maior diversidade (SALE, 1991).
A grande extensão da planície arenosa no Cemiteriozinho
contribuiu para a menor diversidade nesta piscina. De fato, a
heterogeneidade dos substratos é um importante fator para
explicar a diversidade de espécies (GARCIA-CHARTON & PEREZ-
RUZAFA, 1998; ABURTO-OROPEZA & BALART, 2001).
Com relação à medida de equitatividade, os maiores valores
foram encontrados nas piscinas sem conexão permanente com
oceano, o que indica uma maior homogeneidade entre as espécies

64
nestas piscinas. Esta verif icação demonstra que as espécies estão
mais próximas de estarem com densidades igualmente distribuídas
em piscinas fechadas. Os maiores valores de equitatividade estão
relacionados com a menor freqüência de perturbações físicas
(hidrodinâmica) e biológicas (predação) (HUSTON, 1979) nas
piscinas sem conexão permanente com oceano.
Os índices de diversidade calculados para os diferentes
t ipos de fundo tiveram seus maiores valores sobre fundos
consolidados, fato corroborado por ROSA & MOURA (1997).
Os fundos consolidados, além de concederem abrigo
também provêm locais para alimentação de um grande número de
espécies (GUIDETTI, 2000). A maior complexidade estrutural dos
substratos consolidados suporta uma maior diversidade de peixes
devido a características destes ambientes (rugosidade, cobertura
viva, fendas, etc). Já os fundos não consolidados, devido à
escassez de refúgios e de alimentos, se comparado aos fundos
consolidados freqüentemente são associados a menores valores
de diversidade (SALE, 1991; GUIDETTI, 2000; ABURTO-OROPEZA
& BALART, 2001).
Os maiores valores de diversidade, observados nos fundos
arenosos em comparação com os fundos de cascalho, podem estar
relacionado ao freqüente uso do substrato arenoso por diferentes
espécies, que uti l izam este ambiente para alimentação (ROSA &
MOURA, 1997). Os fundos de cascalho são formados por restos
de corais e fragmentos do recife, que se desprendem de seu lugar
de origem em eventos como tempestades ou mesmo pela ação
antrópica. O resultado da ação deste agente é a formação de um
substrato inadequado à regeneração e ao desenvolvimento de
organismos bentônicos (RASSER & RIEGL, 2002) o que faz como
que este substrato seja pouco acessado por peixes em busca de
alimento.
Com relação à medida de equitatividade, de maneira geral,
mostrou certo padrão entre as piscinas, tendo sempre os menores
valores sobre fundo rochosos e maiores valores sobre fundos de

65
cascalho, o inverso do observado com índice H’. Os menores
valores de abundância e diversidade sobre os fundos de cascalhos
contribuíram para a constatação dos maiores valores de
equitatividade neste habitats.
A não constatação de diferenças signif icativas na
comparação dos índices de diversidade entre as piscinas sugere
que o fato das piscinas terem ou não conexão permanente com o
oceano, não influência de maneira signif icativa a diversidade de
espécies das piscinas amostradas. Porém, na comparação entre os
t ipos de substrato, diferenças signif icativas foram verif icadas com
relação aos fundos rochosos e de cascalho. De maneira geral os
fundos de cascalho foram os que apresentaram os menores valores
de diversidade nas piscinas amostradas, fato atribuído à baixa
complexidade estrutural e a escassez de alimentos sobre estes
fundos, ao contrario dos fundos rochosos (RASSER & RIEGL,
2002; GRATWICKE & SPEIGHT, 2005).

66
4.3)Análises de agrupamento Através dessa análise, foi possível observar segregação de
dois grupos no Cluster: o primeiro formado pelos fundos
consolidados e o segundo formados por fundos não consolidados. A separação destes grupos ocorreu devido ao fato de que
algumas espécies somente foram observadas sobre fundos
consolidados (grupo 1). São exemplos dessas espécies:
Gymnothorax funebris, Gymnothorax mil iaris, Gymnothorax vicinus,
Muraena pavonina, Myripristis jacobus, Phaeoptyx pigmentaria e
Pomacanthus paru. Todas as espécies citadas têm hábitos
preferencialmente de viverem sobre fundos rochosos (HUMMAN &
DELOACH, 2003).
O grupo 2, formado por substratos não consolidados, a
exemplo do grupo 1, têm espécies que somente foram avistadas
nesse ambiente: Bothus ocelatus, Dactylopterus volitans, Dasyatis
americana e Lactophrys tr igonus. Todas essas espécies segundo
HUMMAN & DELOACH (2003) têm como hábito viverem
preferencialmente sobre fundos não consolidados.
Visto que a geração do cluster é baseada nos dados de
presença e ausência das espécies, e que os grupos delimitados
pelo cluster são mutuamente exclusivos, pode-se constatar que a
formação dos grupos ocorreu devido às diferentes assembléias de
peixes que são observadas sobre fundos consolidados e não
consolidados. Os grupos formaram-se de acordo com o tipo de
substrato independente da piscina ser ou não conectada
permanentemente com mar aberto.
Pela análise do Cluster foi possível observar grupos
diferentes de substratos consolidados e não consolidados. O
grupo B é formado pelos t ipos de fundo onde foram conferidos os
maiores valores de riqueza de espécies, ao contrário do grupo A.
O subgrupo B2, o de maior similaridade e também o de
maior r iqueza, é formado apenas por fundos rochosos, fato
explicado pela ocorrência de algumas espécies somente em fundos

67
consolidados. As maiores riquezas observadas, dentro do grupo
B2, são atribuídas a maior complexidade estrutural nos fundos
consolidados, resultados corroborados por ROSA & MOURA
(1997), GRATWICKE & SPEIGHT (2005).
Outros autores, trabalhando em ambientes marinhos,
estuarinos e de água doce (FERRELL & BELL, 1991, WEST &
KING, 1996, JENKINS & WHEATLEY, 1998, GUIDETTI, 2000),
observaram que a similaridade entre hábitats com maior
complexidade tende a ter similaridade superior em relação a
hábitats de menor complexidade estrutural, fato também observado
neste estudo.
Com relação subgrupo B1, que possui similaridade pouco
inferior ao subgrupo citado anteriormente, é formado por fundos
arenosos. Neste subgrupo observam-se riquezas relativamente
altas, devido à presença de substratos consolidados sobre os
fundos arenosos os chamados “patch reefs”. A presença destas
manchas fez com que aumentasse a riqueza nos fundos de areia,
nas piscinas amostradas com exceção da piscina do
Cemiteriozinho onde os “patch reefs” são mais esparsos, como já
descrito no item 2.3.3. O grupo A é formado, em sua maioria, por
fundos de cascalho com exceção do fundo de areia na piscina do
Cemiteriozinho. Neste grupo são verif icados os menores valores de
riqueza de espécies. A região de substrato não consolidado, no
Atol das Rocas, é a que apresenta a menor riqueza de espécies de
peixes (ROSA & MOURA, 1997), relacionada à menor
complexidade estrutural deste ambiente.
As maiores riquezas observadas nos fundos arenosos, se
comparadas aos fundos de cascalho, pode estar relacionado ao
fato de que a planície arenosa é frequentemente usada para
alimentação por uma parcela signif icativa dos peixes estudados
neste trabalho. Espécies como D. americana, M. ocellatus, C.
fulva, M. plumiere, C. ruber, L. jocu, H.chrysargyreum, H. parra, M.
martinicus, A. saxati l is, H.radiatus, C. glaucofraenum, G.
thompsoni, A. chirurgus, A.coeruleus e L. tr igonu citadas; no

68
estudo realizado MOURA (1998) foram observadas diariamente
sobre a planície arenosa em atividade de alimentação. Destas, o
C. ruber, D. americana, H. chrysargyreum, H. parra e L. tr igonus,
somente foram observadas no presente estudo sobre substratos
arenosos e não sobre substratos de cascalho. Apesar da aparente
escassez de recursos alimentares no fundo arenoso, uma rica
fauna de invertebrados vive enterrada neste substrato (HOBSON &
CHESS, 1986) e servindo de alimento para uma gama de espécies.
Tais fatos vêm confirmar que o fator que mais influencia a
composição e consequentemente a riqueza de espécies das
assembléias de peixes são os t ipos de fundo. Os fundos rochosos,
por possuírem uma arquitetura mais complexa que os fundos
arenosos e de cascalho, frequentemente estão relacionados à
maior riqueza de espécies (GRATWICKE & SPEIGHT, 2005), fato
também observado neste estudo.

69
4.5) Complexidade estrutural
Os ambientes marinhos mais complexos suportam um maior
número de espécies de peixes do que ambientes menos
complexos. (ROBERTS & ORMONT, 1987; MCCLANAHAN 1994;
MCCORMICK, 1994; GARCIA CHARTON & PEREZ RUZAFA, 1998;
FERREIRA et al. 2001, GRATWICKE & SPEIGHT, 2005).
O EAH empregado neste estudo, respondeu a uma parcela
signif icativa das variações na diversidade de peixes (58%) e a uma
parcela menor, porém signif icativa nas variações na abundância
(12%). De fato, variações na abundância relacionadas à
complexidade estrutural são menos consistentes se comparada
com as variações na riqueza e diversidade de peixes
(FRIEDLANDER & PARRISH, 1998; GRATWICKE & SPEIGHT,
2005).
Embora a maioria das espécies seja observada em diferentes
ambientes, a densidade delas é determinada pelas características
do hábitat (ABURTO-OROPEZA & BALART, 2001).
Das espécies envolvidas nestas análises (ACC e regressão
múlt ipla), algumas são generalistas, com ampla distribuição nos
diferentes ambientes (grupo 2), outras têm hábitos mais
específicos (grupo 1 e 3). Por exemplo: H. adscensionis que
durante o dia permanece dentro de locas e geralmente em locais
sombreados (HUMMAN & DELOACH 2003), é a espécie com maior
relação posit iva com a variável tamanho dos refúgios. Enquanto
que espécies como T. noronhanum, A saxati l is e A. chirurgus que
tem ampla distribuição no Atol das Rocas, estão menos
relacionadas com a variável supracitada e também com as demais
variáveis de substrato (Figura 27).
Já o C. glaucofraenum e o G. thompsoni , espécies que tem
preferência por ambientes de baixa complexidade estrutural como
os fundos arenosos próximo a recifes, (HUMMAN & DELOACH,
2003) são as espécies com maiores relações negativas com as
variáveis de substrato (Figura 27). Em outras palavras, existe uma

70
relação inversamente proporcional entre a abundância destas
espécies e a complexidade estrutural.
A análise de regressão múlt ipla mostrou que das cinco
variáveis de substrato analisadas neste estudo, duas relacionaram-
se de maneira signif icativa com a diversidade (Rugosidade e
formas de crescimento) e uma com abundância (Rugosidade).
A rugosidade foi a variável mais signif icante respondendo por
22% das variâncias na abundância e com 43% das variâncias na
diversidade de peixes. Esta constatação decorre do fato de que os
substratos com maior rugosidade proporcionam área de
assentamento para diferentes organismos que por sua vez
proporcionam nichos específicos para uma variada gama de
espécies. Tal fato tem uma grande importância biológica, pois
muitas espécies de peixes alimentam-se destes organismos
(FERREIRA et al. 2001). Adicionalmente o substrato rugoso pode
oferecer um maior número de abrigos contra predadores (ALMANY,
2004). Tais fatores acarretam numa relação posit iva e signif icativa
da rugosidade com a diversidade e abundância de peixes.
A variedade de formas de crescimento foi a segunda variável
mais importante do EAH. Esta respondeu por 36% das variâncias
na diversidade; contudo para as variâncias na abundância esta
variável não foi signif icativa respondendo a apenas 5% das
variâncias. A variedade de formas de crescimento é uma indicação
dos tipos diferentes de microhabitats. Uma maior variedade destes
microhabitats oferece uma fonte potencial de recursos alimentares,
de camuflagem e de refúgio; que por sua vez propicia ambientes
favoráveis a diferentes espécies peixes. O conceito de nicho
proposto por HUTCHINSON (1957), prediz que mais espécies
podem coexistir em áreas com maior variedade de recursos,
porque haveria nichos potenciais que poderiam ser ocupados por
diferentes espécies (HUTCHINSON, 1957).
Outros autores usando métodos diferentes para acessar a
variedade de formas de crescimento, verif icaram relações posit ivas
entre a riqueza e diversidade de peixes com a riqueza de espécies

71
de corais ou a diversidade de substratos (ROBERTS & ORMONT,
1987; CHABANET et al. 1997; OHMAN & RAJASURIYA 1998). No
entanto, cabe aqui ressaltar que alguns autores já verif icaram a
inexistência desta correlação (RISK, 1972; LUCKHURST &
LUCKHURST, 1978 apud GRATWICKE & SPEIGHT, 2005).
A altura do substrato, não foi um fator signif icante das
variâncias na abundância e diversidade de peixes. Porém, foram
observadas relações posit ivas para ambos os casos. De fato,
existe uma tendência de que peixes aglomerem-se no entorno de
hábitats com características proeminentes (BEETS, 1989). Contudo
neste estudo esta variável não contribuiu de forma signif icativa
para as variâncias dos fatores biológicos analisados.
Com relação ao tamanho dos refúgios, teoricamente o
aumento no número de refúgios acarretaria no aumento tanto da
abundância como da diversidade de espécies. A metodologia
empregada foi descrita por ROBERTS & ORMOND (1987), onde os
autores observaram que o tamanho dos refúgios relaciona-se de
maneira signif icativa com abundância de peixes. No entanto, no
presente estudo outras variáveis de substrato (rugosidade e
formas de crescimento) foram mais importantes nas variações na
abundância e na diversidade.
A porcentagem de cobertura não foi uma variável signif icativa
para explicar as variâncias na abundância e diversidade de peixes.
Esta variável se relaciona posit ivamente com abundância e
negativamente com a diversidade. Tal fato indica que áreas com
maior porcentagem de cobertura viva abrigam uma maior
abundância de peixes, mas não uma maior diversidade. Diversos
autores já observaram relações posit ivas da cobertura viva com a
abundância, r iqueza e diversidade de peixes (FERREIRA et al.
2001, GRATWICKE & SPEIGHT, 2005); embora outros autores já
tenham observado a ausência destas relações (ROBERTS &
ORMOND, 1987; OHMAN & RAJASURIYA 1998). As relações
negativas observadas no presente estudo com relação à cobertura
viva, podem parecer contraditórios com os demais trabalhos que

72
abordam esta questão. Contudo tais diferenças possam ser
recorrentes das diferentes metodologias empregadas além das
diferentes características ecológicas do ambientes estudados.
A metodologia descrita por GRATWICKE & SPEIGHT (2005)
para avaliar a complexidade estrutural, é um método direto de
avaliação das variáveis de substrato. Considerando que este
método é robusto em comparação com os demais métodos de
avaliação da complexidade estrutural, este se mostrou eficiente
explicando uma parcela signif icativa das variações na diversidade
(58%) e na abundância (12%) de peixes. Sendo assim, esta
metodologia é de grande uti l idade para estudos relacionados a
esta área de conhecimento, pois além do baixo custo relativo, este
não tem como pré-requisito a experiência com relação à taxonomia
de algas e corais, sendo um método simples e rápido se aplicado
para avaliar a complexidade estrutural de ambientes recifais.

73
5)Conclusão
Constatações geradas neste trabalho permitem concluir que:
As observações realizadas indicam que o fato das piscinas
estarem ou não conectadas permanentemente com o oceano não é
o fator determinante na composição das comunidades de peixes
recifais no Atol das Rocas. O fator de maior influência sobre a
abundância, r iqueza e diversidade de peixes são os diferentes
t ipos de substrato.
O único fator que mostrou certa tendência com relação ao
fator conectada/não conectada foi à medida de equitatividade. Os
maiores valores de equitatividade, observados em piscinas sem
conexão permanente com oceano, indicam que esta característica
(sem conexão) favorece uma melhor distribuição das assembléias
de peixes.
Com relação à complexidade estrutural, a variável de
substrato com maior influência sobre as variações da abundância e
diversidade de peixes no Atol das Rocas é a rugosidade seguida
da variável formas de crescimento. Ambas as variáveis
responderam de maneira signif icativas as variações na diversidade
de espécies enquanto que somente a rugosidade se relaciona da
maneira signif icativa com abundância.

74
6) Referências Bibliográficas ABURTO-OROPEZA, A. & BALART, E. F. 2001. Community
Structure of reef f ish in several habitats of rocky reef in the Gulf of California. Mar. Ecol, 22 (4):283-305.
ACKERMAN, J.L. & BELLWOOD, D.R. 2000. Reef f ish
assemblages: A re-evaluating using enclosed rotenone stations. Mar. Ecol. Prog. Ser. 206: 227-237.
ALMANY, G. R. 2004. Does increased habitat complexity reduce
predation and competit ion in coral reef f ish assemblages?. Oikos 106: 275-284.
ARAÚJO, M.E.; TEIXEIRA, J.M.C. & OLIVEIRA, A.M.E. 2004.
Peixes estuarinos do nordeste brasileiro. Guia Ilustrado. Edições UFC, Fortaleza. 260p.
BEETS, J. P. 1989. Experimental evaluation of fish recruitment
to combinations of fish aggregating devices and benthic artif icial reefs. Bulletin of Marine Science 44, 973–983.
BELLWOOD, D. R. & ALCALA, A. C. 1988. The effect of a
minimum length specification on visual estimates of density and biomass of coral reef fishes. Coral Reefs 1998 (7):23-27.
BOHNSACK, J. A. & BANNEROT, S. P. 1986. A stationary
visual census technique for quantitatively assessing community structure of coral reef fishes. NOAA Tech. Rep. NMFS 41, 15p.
BROCK, V. E. 1954. A preliminary report on a method of
estimating reef fish populations. J. Wildl. Man., 18 (3):297-308.
CHABANET, P.; RALAMBONDRAINY, H.; AMANIEU, M.;
FAURE, G. & GALZIN, R. 1997. Relationship between coral reef substrata and fish. Coral Reefs 16, 93–102.
CHEROSKE, A. G.; WILLIAMS, S. L. & CARPENTER, R. C.
2000, Effects of physical and biological disturbances on algal turfs in Kaneohe Bay, Hawaii. Journal of Experimental Marine Biology and Ecology, 248(1): 1-34.

75
CHOAT, J. H. & BELLWOOD, D. R. 1991. Reef f ishes: their history
and evolution. In: Sale, P.E. The ecology of f ishes on Coral Reefs. New York : Academic Press, San Diego. pp. 39-68.
CHOAT, J.H., CLEMENTS, K.D. & ROBBINS, W.D. 2002. The
trophic status of herbivorous f ishes on coral reefs. 1: dietary analyses. Marine Biology, 140, 613–623.
CLARKE, K. R. &. WARWICK, R. W. 1994. Change in marine
communities: an aproach to statist ical analysis and interpretation. Bournemouth, Bourne. 859 p.
DE GIROLAMO M. & MAZZOLDI C. 2001. The application of
visual census on Mediterranean rocky habitats. Mar. Envir. Res. 51 (2001): 1-16.
DENNY, C. M. & BABCOCK, R. C. 2003. Do partial marine
reserves protect reef fish assemblages? Biological Conservation, April (2003)
ECHEVERRÍA, C. A.; PIRES, D. O.; MEDEIROS, M. S. &
CASTRO, C. B. 1996. Cnidarians of the Atol das Rocas. Proc. 8th Int. Coral Reef Sym, Panama, Allen Press.
EDGAR, G. J.; BARRETT, N. S. & MORTON, A. J. 2004. Biases
associated with the use of underwater visual census techniques to quantity the density and size-structure of f ish populations. J.Exper. Mar. Biol. Ecol. 208: 269-290.
FARIA, A. & SILVA, D. 1937a. A fauna ictiológica de Roccas.
Observações sobre a maturidade dos órgãos genitais e freqüência das principais espécies. Revista D.N.P.A. Ministério da Agricultura IV(1-2-3): 1-38
FARIA, A. & SILVA, D. 1937b. Os Palinurídeos do Brasil
(Crustacea-Macrura). Revista D.N.P.A. Ministério da Agricultura IV(4-5-6): 1-47.
FEITOZA, B. M.; ROCHA, L. A.; LUIZ JUNIOR, O. J.;
FLOETER, S. R. & GASPARINI, J. L. 2003. Reef fish of St. Paul’s Rocks: news record and notes on biology and zoogeography. Aqua. 7(2): 61-82.
FEITOZA, B. M.; ROSA, R. S. & ROCHA, L. A. 2005. Ecology
and zoogeography of deep-reef fishes in northeastern Brazil. Bulletin of Marine Science, 76(3): 725-742.

76
FERREIRA, C. E. L.; GONÇALVES, J. E. A. & COUTINHO, R. 2001. Community structure of fishes and habitat complexity on a tropical rocky shore. Environmental Biology of Fishes 61: 353–369.
FERREIRA, C. E. L.; FLOETER, S. R; GASPARINI, J. L.;
FERREIRA, B. P. & JOYEUX, J. C. 2004. Trophic structure patterns of Brazilian reef fishes: a latitudinal comparison. Journal of Biogeography (31), 1093–1106.
FERRELL, D. J. & BELL, J. D. 1991 Differences among
assemblages of fish associated with Zostera capricorni and bare sand over a large spatial scale. Marine Ecology Progress Series 72, 15–24.
FLOETER, S. R & GASPARINI, J. L. 2000. The southeastern
Atlantic reef-fish fauna: composition and zoogeographic patterns. J. Fish Biol. 56:1099 -1114.
FLOETER, S. R.; GUIMARÃES, R. Z. P.; ROCHA, L. A.;
FERREIRA, C. E. L.; RANGEL, C. A. & GASPARINI, J. L., 2001, Geographic variation in reef-fish assemblages along the Brazilian coast. Global Ecology and Biogeography, 10(4): 423-431.
FLOETER, S. R.; FERREIRA, C. E. L.; DOMINICI-
AROSEMENA, A. & ZALMON, I. R. 2004. Latitudinal gradients in Atlantic reef fish communities: trophic structure and spatial use patterns. Journal of Fish Biology 64, 1680–1699.
FRIEDLANDER, A. & PARRISH, J.D. (1998) Habitat
characteristics affecting fish assemblages on a Hawaiian coral reef. Journal of Experimental Marine Biology and Ecology 224: 1–30.
FROESE, R. & PAULY, D. 2005. Eds. FishBase. World Wide
Web electronic publication.www.fishbase.org, version (07/2006).
GARCIA CHARTON, J. A. & PEREZ RUZAFA, A. 1998.
Correlation between habitat structure and a rocky reef fish assemblage in the Southwest Mediterranean. Mar. Ecol. 19:111–128.

77
GASPARINI, J. L.; MOURA, R. L. & SAZIMA, I. 1999. Stegastes trindadensis n.sp. (Pisces: Pomacentridae), a new damselfish from Trindade Island, off Brazil. Boletim do Museu de BIologia Mello Leitão, (N. Ser.), v.10, p.3-11, 1999.
GASPARINI, J. L. & FLOETER, S. R. 2001. The shore fishes of
Trindade Island, western South Atlantic. Journal of Natural History, 2001, 35, 1639–1656
GHERARDI, D. F. M. E BOSENCE, D. W. J. 1999. Modeling of
the ecological succession of encrusting organisms in recent coralline-algal frameworks from Atol das Rocas, Brazil. Palaios 14(2): 145-158.
--- --- --- (2001). Composition and community structure the
coralline alga reefs from Atol das Rocas, South Atlantic, Brazil. Coral Reefs (2001) 19:205-219.
--- --- --- (2005). Late Holocene reef growth and relative sea-
level changes in Atol das Rocas, equatorial South Atlantic. Coral Reefs (2005) 24: 264–272
GOES, C. A., 2005. Correntes superficiais no Atlântico Tropical,
obtidas por dados orbitais, e sua influencia na dispersão de larvas de lagosta. Dissertação de Mestrado em Sensoriamento Remoto. São José dos Campos: INPE, 2005. 35p. - (INPE-1111-TDI/111)
GRATWICKE, B. & SPEIGHT, M. R. 2005. The relationship between
fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. Journal of Fish Biology (2005) 66, 650–667.
GUIDETTI, P. 2000. Differences among fish assemblages
associated with nearshore Posidonia oceanica seagrass beds, rocky-algal reefs and unvegetated sand habitats in the Adriatic Sea. Estuarine Coastal and Shelf Science 50, 515–529.
GUIMARÃES, R. Z. P.; GASPARINI, J. L. & ROCHA, L. A. 2004. A
new cleaner goby of the genus Elacatinus (Teleostei: Gobiidae), from Trindade Island, off Brazil. Zootaxa, v.770, p1-8.

78
HAIR JR, J. F., ANDERSON, R. E., TATHAN, R. L., BLCACK, W.C., 1998. Multivariate Data Analysis. Fifth Edit ion, Prentice Hall. 730p.
HILL, M.O. DECORANA 1979. A FORTRAN program for detrented
correspondence analysis and reciprocal averaging. Ecology and Systematics, Cornell University, Ithaca, New York. 52pp.
HOBSON, E. S. & CHESS, J. R. 1986. Relationship among fishes
and their prey in a nearshore sand community off southern California. Env. Biol. Fish., 17 (3): 201-206.
HUMANN, P., & DELOACH, N., 2003. Reef f ish identif ication.
Florida, Caribbean, Bahamas. New world publications, Inc. Florida, USA. 481p.
HUSTON, M. 1979. A general hypothesis of species diversity.
American Naturalist , 113, 81-101. HUTCHINSON, G. E. 1957. Concluding remarks, Cold Spring
Harbor Symposium. Quantitative Biology 22, 415–427. HUTCHINGS, J. A. 2000. Collapse and recovery of marine f ishes .
Nature 406, 882–884 IUCN. 1994. CPNAP. CMMC. Directr ices para las categorías de
manejo de las areas protegidas. Gland, 1994. 261p. JENKINS, G. P. & WHEATLEY, M. J. 1998. The influence of habitat
structure on nearshore f ish assemblages in a southern Australian embayment: comparison of shallow seagrass, reef-algal and unvegetated sand habitats, with emphasis on their importance to recruitment. Journal of Experimental Marine Biology and Ecology 221, 147–172.
JOP’S II, 1996. Joint Oceanographic Projects II (Cruise report and
first results). Sedimentation processes and productivity in the continental shelf waters off East and Northeast Brazil. Werner Ekau e Bastiaan Knoppers (eds.). Bremen: Center for Tropical Marine Ecology. 151 pp.
KELLEHER, G.; BLEAKLEY, C. & WELLS, S. 1995. (Eds.) A global
representative system of marine protected areas. Washington: Great Barrier Reef Marine Park Authority, World Bank and World Conservation Union (IUCN). 4 v.
KIKUCHI, R. K. P. Geomorfologia, Estratigrafia e Sedimentologia
do Atol das Rocas (Rebio-IBAMA/RN). 1994. 144f. Dissertação (Mestrado em Geologia), Universidade Federal da Bahia, Salvador, 1994

79
KIKUCHI, R. K. P. 1999. Atol das Rocas, Atlântico sul equatorial
ocidental, Brasil. In: Schobbenhaus,C.; Campos,D.A.; Queiroz,E.T.; Winge,M.; Berbert-Born,M. (Edit.) Sít ios Geológicos e Paleontológicos do Brasil. Publicado na Internet no endereço http://www.unb.br/ig/sigep/sit io033/sit io033.htm.
KREBS, C. J. 1999. Ecological methodology. Second Edit ion.
Benjamin/Cummings. Menlo Park, CA. 581p LEÃO, Z. M. A. N. & DOMINGUEZ, J. M. L., 2000, Tropical coast of
Brazil. Marine Pollution Bulletin, 41(1-6): 112-122. LINCOLN-SMITH, M. P. 1989. Improving multiespecies rocky reefs
f ish censuses by counting different groups of species using different procedures. Env. Biol. Fish. 26: 29-37.
LIZASO, J. L.; GOÑI, R.; REÑONES, O.; CHARTON, J. A. G.;
GALZIN, R.; BAYLE, J. T.; JEREZ, P. S; RUZAFA, A. P.; & RAMOS, A. A. 2000. Density dependence in marine protected populations: a review. Environmental Conservation 27 (2): 144–158.
LUBBOCK, R. & EDWARDS, A. 1981. The fishes of Saint Paul’s
Rocks. Journal of Fish Biology, v.17, p.283-303. MAGURRAN, A. E. 1988. Ecological diversity and its measurement.
University Press, Princeton, New Jersey. MARQUES, L. V. 2003. Influência de peixes herbívoros na
palatabil idade de macrolagas do Atol das Rocas. Dissertação de mestrado (Mestrado em Biologia Marinha). Universidade Federal Fluminense, Rio de Janeiro. 49p.
MATEO, I. & TOBIAS, W. J. 2001. Distribution of shallow water
coral reef f ishes on the northeast coast of St. Croix, USVI. Caribbean Journal of Science 37, 210–226.
MCCLANAHAN, T. R. 1994. Kenyan coral reef lagoon fish: effects
of f ishing, substrate complexity, and sea urchin. Coral reefs 13:231-241
MCCLANAHAN, T. R. 2002. The near future of coral reefs.
Environ.Conserv. 29: 460–483. MCCORMICK, M. I. 1994. Comparison of f ield methods for
measuring surface topography and their association with a tropical reef f ish community. Mar. Ecol. Prog. Ser. 112:87-96.

80
MCCUNE, B. & GRACE, J. R. 2002. Analysis of ecological
communities. Gleneden Beach, Oregon.300 p. MENEZES, N.A.; BUCKUP, P.A.; FIGUEIREDO, J.L. & MOURA,
R.L. 2003. Catálogo das espécies de peixes marinhos do Brasil. Museu de Zoologia da USP, São Paulo. 159p.
MESA G. & VACCHI M. 1999. An Analysis of the Coastal Fish
Assemblage of the Ustica Island Marine Reserve (Mediterranean Sea). Marine Ecology 20 (2): 147-165.
MMA 2002. Avaliação e ações prioritárias para a conservação da
biodiversidade das zonas costeiras e marinhas. Ministério do Meio Ambiente.
MORNING POST, 1906. Green Island recreation reserve. Cairns
Australia. 1p. MOURA, R. L. 1995. A new species of Chromis (Perciformes:
Pomacentridae) from the southeastern coast of Brazil, with comments on other species of the genus. Revue Fr. Aquariol. 21: 91–96.
MOURA, R. L. 1998. Atividade, distribuição e táticas alimentares
de uma comunidade de peixes do Atol das Rocas. Dissertação (Mestrado em Ciências na área de Zoologia), Instituto de Biociências da Universidade de São Paulo, São Paulo. 108p.
_______ 2003. Riqueza de espécies, diversidade e organização de
assembléias de peixes em ambientes recifais: um estudo ao longo do gradiente latitudinal da costa brasileira. Tese (Doutorado em Ciências, na área de Zoologia), Instituto de Biociências da Universidade de São Paulo. 620p.
MOURA, R. L.; FIGUEIREDO, J. L.; MENEZES, N. A. 2003. Família
Labridae. In: MENEZES, N.A.; BUCKUP, P.A.; FIGUEIREDO, J.L.; MOURA, R. L. (ed). Catálogo das espécies de peixes marinhos do Brasil. São Paulo: Museu de Zoologia da Universidade de São Paulo, n.160, p. 92-93.
MUNRO J. L. 1996. The scope of coral reef Fisheries and their
management. In: Polunin N., Roberts C. (ed) Coral reef Fisheries. Chapman and Hall, New York 129 p.
MYERS, R. A. & WORM, B. (2003) Rapid worldwide depletion of
predatory f ish communit ies. Nature 423, 280–283.

81
NAGELKERKEN, I.; DORENBOSCH, M.; VERBERK, W. C. E. P.; COCHERET DE LA MORINIERE, E. & VAN DER VELDE, G. 2000. Importance of shallow-water biotopes of a Caribbean bay for juvenile coral reef f ishes: patterns in biotope association and spatial distribution. Marine Ecology Progress Series 202, 175–192.
OHMAN, M. C. & RAJASURIYA, A. 1998. Relationships between
habitat structure and fish communit ies on coral and sandstone reefs. Environmental Biology of Fishes 53, 19–31.
OLIVEIRA FILHO, E. C. 1972. As algas marinhas bentônicas do
Atol das Rocas. Ciência e Cultura 24(6): 328 (supl). OLIVEIRA FILHO, E. C. & MENEZES, E N. A. 1972. Notas
biológicas sobre o Atol das Rocas. Ciência e Cultura 24(6): 361 (supl).
OLIVEIRA FILHO, E. C. & UGADIM, Y. 1974. New references of
benthic marine algae to Brazil ian f lora. Boletim de Botânica 2: 71-91.
OLIVEIRA FILHO, E. C. & UGADIM, Y. 1976. A survey of the
marine algae of Atol das Rocas (Brazil). Phycologia 15(1): 41-44.
PANDOLFI, J. M.; BRADBURY, R. H.; SALA, E.; HUGHES, T. P.;
BJORNDAL, K. A.; COOKE, R. G.; MCARDLE, D.; MCCLENACHAN, L.; NEWMAN, M. J. H.; PAREDES, G.; WARNER, R. R. & JACKSON, J. B. C. 2003. Global Trajectories of the Long-Term Decline of Coral Reef Ecosystems. Science 301, 955–958
PAULY, D.; CHRISTENSEN, V.; GUENETTE, S.; PITCHER, T. R.;
SUMAILA, U. R.; WALTERS, C. J.; WATSON, R. & ZELLER, D. 2002. Towards sustainabil i ty in world f isheries.Nature 418, 689–695.
PIELOU, E. C., 1974. Population and communication ecology:
Principles and Methods. Gordon and Breach Science, London. 424p.
RANDALL, J., 1968. Conservation in the sea: a survey of marine parks. Oryx, 10(1): 31–37.
RASSER, M.W. & RIEGL, B. 2002. Holocene coral reef rubble and
its binding agents. Coral Reefs (2002) 21:57–72.

82
ROBERTS, C. M. & ORMOND, R. F. G. (1987). Habitat complexity and coral reef f ish diversity and abundance on Red Sea fringing reefs. Marine Ecology Progress Series 41, 1–8.
ROCHA, L.A.; ROSA, I.L. & ROSA, R.S. 1998. Peixes recifais da
costa da Paraíba, Brasil. Revista Brasileira de Zoologia 15(2), 553-566.
ROCHA, L.A. & ROSA, I. L 2001. Baseline assessment of reef f ish
assemblages of Parcel Manuel Luiz Marine State Park, Maranhão, north-east Brazil. Journal of Fish Biology (2001) 58 , 985–998
ROCHA, L.A. 2003. Patterns of distribution and processes of
speciation in Brazil ian reef f ishes. Journal of Biogeography, 30, 1161–1171
RODEN, G. L., 1987. Effects of seamounts and seamounts chains
on ocean circulation and thermocline structure. Geophys. Monogr. 43: 335-354.
RODWELL, L. D. & ROBERTS, C. M., 2000. Economic implications
of marine protected areas for coral reef f isheries. Sweden: Cordio. 18 p.
ROSA, R. S. & MOURA, R. L., 1997, Visual assessment of reef f ish
community structure in the Atol das Rocas Biological Reserve, off northeastern Brasil, pp. 983-986, 8th International Coral Reef Symposium. Smithsonian Tropical Research Institute. Panamá.
RUSS, G. R. 1991. Coral reef f isheries: effects and yields. In: Sale
P. F. (ed) The ecology of f ishes on coral reefs. Academic Press, San Diego, pp 601–635
SALE, P. F. 1991. The ecology of f ishes on coral reefs. Academic
Press, San Diego, CA. 754p SALE, P. F. 1997. Visual census of f ishes: hoe well do we see what
is there? Proc Int. Coral Reef Symp. 8 (2): 1435 – 1440. SAMOILYS, M. A. & CARLOS, G. 2000. Determining methods of
underwater visual census for estimating the abundance of coral reef f ishes. Env.Biol.Fush. 57:289-304.
SHANNON, C.E & WEAVER, W., 1949. The Mathematical Theory of
Communication. University of I l l inois Press, Urbana, I l l inois. 163p.

83
SMITH, C.L. 2004. National Audubon Society. Field guide to tropical marine f ishes of the Caribbean, the Gulf of México, Florida, the Bahamas, and Bermuda. National Audubon Society. Chanticleer Press Edit ion, New York. 718p.
SPALDING, M. D.; RAVILIOUS, C. & GREEN, E. P. 2001. World
atlas of coral reefs. United Nations Environmental program, WCMC.
SISTEMA NACIONAL DE UNIDADES DE CONSERVAÇÃO - SNUC,
LEI No 9.985. Julho de 2000. Capítulo III, art igo 10. SOARES-GOMES, A., VILLAÇA, R. C. & PEZZELLA, C. A. C.,
2001, Atol das Rocas ecossistema único no Atlântico Sul. Ciência Hoje, 29 (172): 32-39.
STANLEY, D. R., & WILSON, C. A. 1995. Effect of scuba divers on
f ish density and target strength estimates from stationary dual-beam hydroacoustics. Transaction of the American Fisheries Society, 124, 946±949.
TRAVASSOS, P.; HAZIN, F. H. V.; ZAGAGLIA, J. R.; ADVÍNCULA,
R. & SCHOBER, J. 1999. Thermohaline structure around seamounts and islands off North-Eastern Brazil. Archieve of Fishery and Marine Research, v. 47, n. 2/3, p. 211-222.
WEST, R. J. & KING, R. J. 1996. Marine, brackish, and freshwater
f ish communities in the vegetated and bare shallows of an Australian River. Estuaries 19, 31–41.

84