caracterização das neurotransmissões gabaérgica e...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO FACULDADE DE MEDICINA DE RIEIRÃO PRETO – FMRP-USP
DEPARTAMENTO DE FISIOLOGIA
Caracterização das neurotransmissões GABAérgica e glutamatérgica em um núcleo envolvido no processamento auditivo em pássaros canoros
André Luiz Andreotti Dagostin
Dissertação apresentada ao Departamento de Fisiologia da Faculdade de Medicina de Ribeirão Preto como parte das exigências para a obtenção do título de Mestre em Fisiologia.
Orientador: Prof. Dr. Ricardo Maurício Xavier Leão.
RIBEIRÃO PRETO – SP 2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

FICHA CATALOGRÁFICA
Dagostin, André Luiz Andreotti
Caracterização das neurotransmissões GABAérgica e glutamatérgica em um núcleo envolvido no processamento auditivo em pássaros canoros 87p.:
Dissertação apresentada ao Departamento de Fisiologia da Faculdade de Medicina de Ribeirão Preto como parte das exigências para a obtenção do título de Mestre em Fisiologia. Orientador: Leão, Ricardo Maurício Xavier
1. Neurotransmissão; 2. Eletrofisiologia; 3. GABA; 4. Glutamato; 5.
Mandarim; 6. Canto

FOLHA DE APROVAÇÃO
André Luiz Andreotti Dagostin
Caracterização das neurotransmissões GABAérgica e glutamatérgica em um núcleo envolvido no processamento auditivo em pássaros canoros
Dissertação apresentada ao Departamento de Fisiologia da Faculdade de Medicina de Ribeirão Preto como parte das exigências para a obtenção do título de Mestre em Fisiologia.
Aprovado em: ___/___/___
Banca examinadora
Prof. Dr. Ricardo Maurício Xavier Leão (Orientador)
FMRP-USP
Assinatura___________________________________________
Profa. Dra. Eliane Comoli
FMRP-USP
Assinatura___________________________________________
Prof. Dr. Christopher Kushmerick
ICB-UFMG
Assinatura___________________________________________

ii
DEDICATÓRIA
Aos meus heróis, meus ídolos: meus pais.

iii
AGRADECIMENTOS
A Deus,pela vida e por colocar em meus caminhos pessoas maravilhosas, que me
apoiaram e andaram comigo em algum momento na caminhada da vida, algumas das quais
encontram-se abaixo.
Pai, a você devo tudo o que sou, meus valores, caráter, idéias e ambições. Não existem
palavras para expressar a minha gratidão e admiração por você. Se um dia me tornar metade do
que és, dar-me-ei por feliz.
Mãe, obrigado por todas as idéias, conselhos, cafunés e por que não, pelas broncas. Para
mim, és um exemplo de dedicação, perseverança e aplicação no labor, tanto em casa quanto fora.
João e Eloisa, meus irmãos amados. Fontes de alegria e felicidade. Nenhum de nós é
santo, mas ter vocês em minha vida, me dá um porquê a mais para continuar a batalhar e deixá-los
orgulhosos de mim, da mesma forma que sou de ambos.
Família Andreotti e Dagostin, obrigado por me acolherem e constituírem ninhos
aconchegantes aos quais vôo de volta sempre que possível para um alento a mais.
Minha namorada Ana Flávia, que viu todo o processo desde a entrada no mestrado até a
sua conclusão. Que passou pelos bons e maus momentos de minha vida nesses anos que estamos
juntos, pela compreensão e pelo amor e companheirismo em todos os momentos.
Família Winz e Nery, obrigado por me aceitarem em seu meio e serem comigo pessoas tão
queridas.
Rato, Felipe, Célio, Juju, Becker, Darlan, Fran, Chile. Obrigado por todas as festas, folias,
churrascos e risadas, mas principalmente pela amizade que sempre dispensaram a mim. Foz não
seria tão bonita se vocês não estivessem lá.
Cláudia, Clodoaldo, Alexandre e Henrique. Grandes amigos de colégio que moram em
meu coração. Sei do apoio de vocês, mesmo nos vendo uma vez a cada muitos anos.
Carla Straub. Minha amiga e confidente desde a graduação.
Forno e AZ... Companheiros de bons momentos, maus momentos, presepadas, folias,
vídeo game. Todos seguimos os caminhos da pós-graduação, e ter amigos como vocês é oq eu faz
da vida uma experiência sem igual.
Alessandro, Kiti, Éder e Ângela. Obrigado pelo companheirismo, conselhos, pizzas, jogos,
futebol e, principalmente, amizade incondicional.

iv
Povo das “amarelinhas”: vocês fizeram daquelas quitinetes safadas o melhor lugar para se
viver em toda Cascavel!
Ernane, Augusto, Carlos e Berel. A convivência no lar deve ser harmoniosa para que
nossa casa seja um porto seguro em momentos tempestuosos. Obrigado por serem as pessoas que
fazem isso acontecer.
Ernane e Lígia, obrigado pelas vezes que me ouviram, que riram comigo, me apoiaram e
me ajudaram. Vocês são duas pessoas que vou levar no coração para o resto da vida.
Meninão, João Paulo, João Henrique, Sevi, Renata Maria, Fabi, Fernandinha, Lys,
Augusto, Ernane, Lígia, Cadu, Thiago, Elaine, Paula, Dani, Chefe, Lílian, Dezão. Kusuda,
Marcelo, Roberta, Dawitt, Edu, Ângelo, Adriano, Ricardinho, Renato Rizzo, Renato Soriano e
Roberta Ribeiro. Se rir é um santo remédio, viverei até os 150 anos ao lado desse povo.
Márcio, Mateus, Bruno, Leandro, Zoca, Berel, Thiago Malardo, Zé, “Meu Amigo”, Zé (de
novo), Bebinho, Rodrigo, Rafa e os distantes Valtão e Felipe. Esse grupo faz a quarta feira ser o
dia mais esperado da semana! A pelada no seu Luis é o highlight da semana e estes caras,
jogadores de fibra com os quais tenho o prazer e orgulho de dividir a quadra!
Fernandinho, você eu só tenho a agradecer. Tu és o homem que faz chover! Obrigado
pelas dicas e ajudas na hora das “adaptações técnicas”. Juntamente com Renatão, obrigado pelo
companheirismo nas pedaladas dominicais.
Cláudia B. Vanzela, Elisa M. Aleixo, Fernando C. Rastello e Carlos Belini, funcionários
dedicados que quebram todos os galhos e desfazem todos os nós burocráticos nessa jornada da
pós-graduação.
Bioteristas de Departamento de Fisiologia, Leonardo Sidelis Filho e Eduardo Gomes,
pelos cuidados dispensados aos animais.
Professores do departamento de Fisiologia, pelas aulas ministradas e conhecimento
repassado.
Professor Wamberto Varanda por nos ajudar com a estrutura inicial para nossos
experimentos, pelos ensinamentos repassados nesses anos de convivência e pelas memoráveis
reuniões de laboratório em sua casa.
Marcelo Cairrão, obrigado pela amizade, alto astral no laboratório, pelos ensinamentos
sobre hiopocampo e por confiar a mim os experimentos com a carambola!

v
Cadu, companheiro de laboratório, de risadas, de histórias, de almoços. Você é uma
pessoa especial, meu caro. Se as pessoas fossem prestativas, compreensivas e companheiras uma
centena de vezes menos que você, esse mundo certamente seria um lugar melhor.
Thiago, a organização é uma necessidade dentro do ambiente de trabalho, mas tudo que é
em demasia, faz mal... Mas nem por isso deixo de gostar de você.
Dani, Você me ensinou muita coisa desde o momento que entrei no laboratório e me deu
muitas dicas e conselhos a respeito de tudo dentro da universidade. Obrigado por todo o apoio e
atenção!
Chefe... Deixei você por último pra fechar com chave de ouro. Você me ensinou muita
coisa, me deu uma liberdade de me expressar e trabalhar que poucos dão. Foi alguém que me
incentivou a sempre buscar um pouco mais, que me questionou, elogiou e confiou muito na minha
pessoa. Vou sempre levar você na minha memória exatamente como você se descreveu para mim
a algum tempo atrás “Não sou seu chefe, sou seu amigo que sempre tem razão”. Ricardo, muito
obrigado!

vi
CONTEÚDO
LISTA DE ABREVIATURAS ................................................................................................... VIII
RESUMO ............................................................................................................................... XI
ABSTRACT ...........................................................................................................................XIII
1 – INTRODUÇÃO................................................................................................................ 18
O NIDOPÁLIO CAUDOMEDIAL (NCM)..................................................................................... 25
2 - OBJETIVO....................................................................................................................... 29
3 - METODOLOGIA ............................................................................................................. 31
3.1 - ANIMAIS ....................................................................................................................... 32 3.2 - ESTÍMULO ...................................................................................................................... 32 3.3 - CONFECÇÃO DAS FATIAS ............................................................................................... 33 3.4 - ELETROFISIOLOGIA ......................................................................................................... 35 3.4.1 - VISUALIZAÇÃO DOS NEURÔNIOS ..........................................................................................35 3.4.2 - WHOLE CELL PATCH CLAMP...................................................................................................35 3.4.3 - REGISTRO DAS CORRENTES SINÁPTICAS...................................................................................36 3.5 - ANÁLISE DOS DADOS ..................................................................................................... 37 3.6 - IMUNOCITOQUÍMICA...................................................................................................... 39
4 - RESULTADOS ................................................................................................................. 40
4.1 – EXPRESSÃO DA PROTEÍNA ZENK EM RESPOSTA AO CANTO. .............................................. 41 4.2 – CARACTERIZAÇÃO DAS CORRENTES SINÁPTICA ESPONTÂNEAS (SPSCS) REGISTRADAS S NO NCM.................................................................................................................................... 43 4.2.1 - CORRENTES PÓS-SINÁPTICAS MINIATURA (MPSCS) ..............................................................46 4.2.2 - CARACTERIZAÇÃO DOS SIPSCS E DOS SEPSCS QUANTO AO POTENCIAL DE REVERSÃO. ........50 4.3 – EFEITO DA ESTIMULAÇÃO CANORA NA NEUROTRANSMISSÃO NO NCM ............................. 54 4.3.1 SEGREGAÇÃO SEXUAL DOS PÁSSAROS CONTROLE E ESTIMULADOS............................................56 4.3.2 – ISOLAMENTO DA TRANSMISSÃO GABAÉRGICA PELO DNQX EM PÁSSAROS CONTROLE E ESTIMULADOS. ..................................................................................................................................59 4.3.3 - ISOLAMENTO DA TRANSMISSÃO GLUTAMATÉRGICA...............................................................61 4.3.4- EXCITABILIDADE AUMENTADA DOS NEURÔNIOS GLUTAMATÉRGICOS .....................................63 4.4- NEUROTRANSMISSÃO NO NCM DE FÊMEAS SUBMETIDAS A UM ISOLAMENTO PROLONGADO DO ESTÍMULO CANORO. ............................................................................................................... 65
5 - DISCUSSÃO.................................................................................................................... 68
5.1 – EXPRESSÃO DA PROTEÍNA ZENK EM RESPOSTA AO CANTO. .............................................. 69 5.2 – CARACTERIZAÇÃO DAS CORRENTES SINÁPTICA ESPONTÂNEAS (SPSCS) REGISTRADAS S NO NCM.................................................................................................................................... 69

vii
5.3 – EFEITO DA ESTIMULAÇÃO CANORA NA NEUROTRANSMISSÃO NO NCM ............................. 73 5.4-DIFERENÇAS SEXUAIS NA RESPOSTA AO CANTO NO NCM. ................................................. 75 5.5-CONCLUSÕES. ................................................................................................................. 76
6 – TABELAS ........................................................................................................................ 77
7 – BIBLIOGRAFIA................................................................................................................ 82

viii
LISTA DE ABREVIATURAS
2-AG 2-araquidonoiglicerol
aCSF Fluido Cefalorraquidiano Atificial
AMPA Ácido α-amino-3-3hidroxi-5-metil-4-isoxazolepropiônico
ANOVA Análise de Variância
ATP-Mg Adenosina Trifosfato, sal de Magnésio
BSA Albumina de Soro Bovino
CB1 Receptor Canabinóide Tipo 1
Cm Capacitância da Membrana Celular
CsMSO4 Metanosulfonato de Césio
DAB Diaminobenzidina
DIC Contraste por Interferência Diferencial
DNAÁcido Desoxirribonucléico
DNQX 6,7-Dinitroquinoxalina-2,3-Diona
DSE Supressão da Excitação por Despolarização
DSI Supressão da Inibição por Despolarização
EGTA Ácido Etileno Glicol-bis (2-aminoetileter)-Tetra acético
EPM Erro Padrão da Média
FC Fêmeas Controle
FE Fêmeas Estimuladas
GABA Ácido Gama-Aminobutírico
GTP Guanosina Trifosfato
HEPES Ácido 4-(2-hidroxietil)-1-Piperazino Etano Sulfônico
HW Half Width
ICC Imunocitoquímica
IEG Gene de Expressão Imediata
KGlu Glucontao de Potássio
LTD Depressão a Longo Prazo
LTP Potenciação a Longo Prazo
MC Machos Controle
ME Machos Estimulados
mEPSC Corrente Excitatória Pós-sináptica em miniatura
mGluR1 Receptor Metabotrópico de Glutamato Tipo 1
mGluR5 Receptor Metabotrópico de Glutamato Tipo 5

ix
mIPSC Corrente Inibitória Pós-sináptica em miniatura
PA Potencial de Ação
PBS Tampão Fosfato com Salina
PCA-Na Fosfocreatina, sal de Sódio
PLCβ Fosfolipase Cβ
RNAm Ácido Ribonucléico Mensageiro
RS Resistência em Série
sEPSC Corrente Excitatória Pós-sináptica Espontânea
sIPSC Corrente Inibitória Pós-sinápticas Espontâneas
sPSC Corrente Pós-sináptica Espontânea
TTX Tetrodotoxina
ZENK
Núcleos do cérebro de pássaros canoros:
Av Núcleo Avalanche
B Núcleo Basolateral
CLM Mesopálio Caudolateral
CMM Mesopálio Caudomedial
CN Núcleo Coclear
DLM Núcleo Dorsolateral do Tálamo Medial
DM Núcleo Medial Dorsal
E Núcleo Entopalial
HVC Centro Vocal Superior
L (1,2,3) Campo L (porções 1, 2 e 3)
L Area X Porção Lateral da Área X do Estriado
LLD Núcleo Dorsal – Lemnisco Lateral
LLI Núcleo Intermediário - Lemnisco Lateral
LLV Núcleo Ventral - Lemnisco Lateral
LMAN Núcleo Magnocelular Lateral do Nidopálio Anterior
LMO Núcleo Oval do Mesencéfalo
MLd Núcleo Dorsolateral do Mesencéfalo
NCM Nidopálio Caudomedial
Nif Núcleo Interfacial do Nidopálio
nXIIts Porção Traqueosiringial do Núcleo do Hipoglosso

x
Ov Ovoidalis
PAm Paraambíguo
RA Núcleo Robusto do Arcopálio
RAm Retroambíguo
SO Oliva Superior
TSM Trato Septomesencefálico
Uva Núcleo Uviforme

xi
RESUMO

xii
DAGOSTIN, A.L.A., CARACTERIZAÇÃO DAS NEUROTRANSMISSÕES GABAÉRGICA E
GLUTAMATÉRGICA EM UM NÚCLEO ENVOLVIDO NO PROCESSAMENTO AUDITIVO
EM PÁSSAROS CANOROS. Faculdade de Medicina de Ribeirão Preto, Universidade de São
Paulo, Ribeirão Preto, 2008.
O canto em pássaros canoros é usado para a demarcação territorial e para o acasalamento.
Assim, para que um indivíduo seja bem sucedido em seu contexto social, é necessário que
aprenda a cantar e reconhecer o canto de sua espécie de maneira adequada. A habilidade de
aprender o canto não é característica geral dos pássaros, mas sim, de um grupo específico: os
pássaros canoros (subordem oscine). O circuito telencefálico responsável pelo reconhecimento do
canto é presente tanto em machos quanto em fêmeas, sendo o nidopálio caudomedial (NCM) um
núcleo proeminentemente relacionado a esse reconhecimento e à memória auditiva. O NCM
responde especificamente ao canto da espécie induzindo a expressão do gene de expressão
imediata (IEG) zenk, um fator de transcrição responsável pela expressão de diversos genes alvo.
Porém, os mecanismos neurais e sinápticos no NCM e como eles respondem ao estímulo canoro
são pouco conhecidos. O objetivo deste trabalho é a investigação das características da
neurotransmissão dos neurônios do NCM em fatias frescas. Para isso, mandarins (Taeniopyia
guttatta) adultos machos e fêmeas foram estimulados com canto específico da espécie ou
mantidos em isolamento. O registro da atividade sináptica foi feito utilizando-se a técnica de
patch-clamp em modo voltage clamp para registros de correntes pós-sinápticas espontâneas
(sPSCs). Em nossos registros, observamos que as correntes que compõem a neurotransmissão no
NCM são geradas pelos neurotransmissores GABA (sIPSCs) e glutamato (sEPSCs), sendo os
sIPSCs muito mais abundantes que os sEPSCs. Os registros de correntes em miniatura (na
presença de TTX) mostraram que as sEPSCs correspondem a mEPSCs, diferentemente dos
sIPSCs, que são bastante sensíveis à TTX. A estimulação pelo canto aumentou a excitabilidade da
rede glutamatérgica evidenciada pelo aparecimento de bursts de correntes glutamatérgicas após a
desinibição por bicuculina. Em machos estimulados, essas correntes apareceram mesmo sem a
inibição da transmissão GABAérgica e provavelmente são a causa do aumento da freqüência dos
sISPCs no NCM desses pássaros. Experimentos em fêmeas isoladas de machos por 7 a 30 dias
sugerem que a conectividade da rede glutamatérgica no NCM dependa de um contínuo estímulo
canoro. Concluímos que o estímulo canoro desencadeia um aumento da excitabilidade dos
neurônios glutamatérgicos no NCM que poderia representar uma forma de plasticidade induzida
pelo canto relevante para o reconhecimento vocal do pássaro.

xiii
ABSTRACT

xiv
DAGOSTIN, A.L.A, CHARACTERIZATION OF THE GABAERGIC AND
GLUTAMATERGIC NEUROTRANSMISSIONS WITHIN A NUCLEUS INVOLVED WITH
THE AUDITORY PROCESSING IN SONGBIRDS. School of Medicine of Ribeirão Preto,
University of São Paulo, Ribeirão Preto, 2008
The bird song is mainly used for territory delimitation and mating. Thus, to biologically
succeed, the bird must learn how to sing and recognize the song from its species properly. This
skill is not a general characteristic of the birds; instead it is present within one specific group: the
songbirds (oscine suborder). The telencephalic circuit responsible for recognition of the song is
present in both males and females, being the caudal medial nidopallium (NCM) a prominent area
related to this recognition and to auditory memory. The NCM responds to the song specifically,
evidenced by the expression of the immediate expression gene (IEG) zenk, a transcriptional factor
responsible for the expression of a variety of genes. Notwithstanding, the neural and synaptic
mechanisms of the NCM and how they behave under song stimulation are poorly understood. The
aim of this work is to investigate the characteristics of the neurotransmission of the neurons
within the NCM in fresh brain slices. For this, adult zebra finches (Taeniopyia guttatta) males and
females were stimulated with a species-specific song or kept in isolation. The synaptic activity
record was made using the patch-clamp technique in voltage-clamp mode to acquire data about
the spontaneous post synaptic currents (sPSCs). We observed that the neurotransmission within
the NCM is generated by the neurotransmitters GABA (sIPSCs) and glutamate (sEPSCs), being
the sIPSCs much more abundant than the sEPSCs. The miniature currents records (TTX present)
showed that the sEPSCs actually correspond to the mEPSCs, on the other hand, it showed that the
sIPSCs are quite sensitive to TTX. The song stimulation increased the glutamatergic network
excitability, evidenced by the appearance of glutamatergic current bursts after disinhibition by
bicuculine. In stimulated males, sometimes, these currents appeared even with the GABAergic
inhibition of the neurotransmission present. This is probably the cause of the observed increase in
the sISPCs frequency in the NCM from these birds. Experiments with prolonged male-isolated
females (7-30 day isolation) suggest that the connectivity of the glutamatergic network within
NCM relies on a continuous song stimulus. We conclude that the song stimulus triggers an
increase of the excitability of the glutamatergic neurons within the NCM that could represent
some kind of song-induced plasticity relevant for the bird’s vocal recognition.

18
1 – INTRODUÇÃO

19
A vocalização é uma forma de comunicação amplamente usada por diversos animais,
atingindo, na espécie humana, níveis extremos de complexidade tanto na produção de sons quanto
nos significados embutidos neles. É evidente a importância do correto reconhecimento do padrão
de sons da voz para a comunicação precisa entre dois indivíduos, bem como as características
tonais e de timbre da voz, as quais representam uma assinatura específica importante para o
reconhecimento individual num contexto social. A habilidade de comunicação através da
vocalização, apesar de ser amplamente utilizada em mamíferos, contrasta com a habilidade restrita
de seu aprendizado entre os indivíduos dessa classe, já que apenas uma espécie de primatas
(humanos), os cetáceos (baleias e golfinhos - Guinee & Payne, 1988) e duas espécies de
morcegos (Boughman, 1998) possuem mecanismos que os permitem aprender padrões complexos
de vocalização. Por outro lado, na classe das aves, três grandes grupos possuem essa capacidade:
papagaios, beija-flores e passeriformes pertencentes à sobordem oscine (também conhecidos
como pássaros canoros).
A subordem dos oscinos (ordem passeriformes) é o grupo mais amplamente utilizado para
pesquisas em neurobiologia do canto, sendo os canários (Serinus canaria), mandarins
(Taeniopygia guttata) e manons (Lonchura domestica) os mais largamente utilizados (Brenowitz
& Beecher, 1995). Esses animais possuem uma forma similar de aprendizado do canto, onde um
modelo, como por exemplo um tutor ou até mesmo um canto gravado, servem para ensinar o
animal a vocalizar, porém, cada espécie tem suas preferências inatas em relação à estrutura do
canto que preferem aprender. Testes onde animais são treinados para pressionar diferentes botões,
sendo que cada um dos botões é responsável pela produção de um canto, entre os quais, o do
tutor, mostram que o botão relacionado ao canto do tutor é pressionado muitas vezes mais (cerca
de 60%) do que os demais, evidenciando a preferência inata do animal (Marler, 1989).
O canto aprendido tem em geral uma duração longa que pode variar de alguns segundos
até algumas dezenas de segundos, chegando até mesmo a minutos, e é diferenciado dos chamados
inatos, que são vocalizações curtas e normalmente não aprendidas (Doupe & Kuhl, 1999). O canto
possui uma estrutura temporal em que componentes silábicos se agrupam em motivos ou frases,
que são séries de sílabas iguais ou diferentes. Diferentes espécies de pássaros podem agrupar
diferentes frases em diferentes ordens ou vocalizar sílabas em várias ordens. Essa estrutura se
assemelha à sintaxe (entenda-se sintaxe simplesmente como a ordem das sílabas dentro das frases)
da linguagem humana, evidentemente sem as regras complexas de gramática que a regem. Porém,
diferentemente da fala humana, o canto dos pássaros não possui semântica, ou seja, significado
abstrato, servindo basicamente para demarcação de território e acasalamento, porém, este assunto
é ainda controverso, pois existe evidência de que o canto ou outras vocalizações também podem

20
conferir significado num sentido mais abstrato. Por exemplo, o número de chamados no chapim-
de-cabeça-negra (Poecile atricapillus), correlacionado com aparecimento de predadores, parece
anunciar a presença e ameaça que diferentes predadores representam (Williams, 2004), podendo
usar diferentes cantos para essas funções (Catchpole, 1983). Ao fazermos um paralelo com
humanos, vemos que existem diferenças marcantes, como a produção do canto ser, na maioria das
vezes, restrita aos machos, pois apenas eles possuem um aparato neural específico para essa
função (ver mais abaixo). Por outro lado, semelhanças expressivas existem, como a habilidade de
discriminar sons diversos e memorizar os sons específicos da espécie, aspectos essenciais para a
comunicação vocal e, da mesma forma que em humanos, diferentes estruturas cerebrais de
pássaros canoros têm sido descritas como participantes na discriminação de sons.
Uma das peculiaridades no uso de pássaros canoros para o estudo do reconhecimento
vocal é a distância filogenética existente entre pássaros e mamíferos, o que culmina em diferenças
pronunciadas entre estrutura do cérebro de um e de outro. Devido a isso, interessa-nos discutir
alguns estudos a respeito da homologia entre o sistema nervoso central de humanos e outros
animais. Esses estudos foram iniciados por Edinger entre 1885 e 1908, usando como base de
estudos “A Origem das Espécies” de Darwin (Jarvis et al., 2005). Posteriormente, uma teoria
sobre a evolução do cérebro de vertebrados formulada por estudiosos como J.B. Johnston, G.C.
Huber, E.C. Crosby, C.U. Ariëns-Kappers, e C.J. Herrick dizia que as funções cerebrais se
transferiam gradualmente entre diferentes áreas no decorrer da evolução (Reiner et al., 2004).
Tanto é que, usando os humanos e seu córtex evoluído como base para comparação com outros
indivíduos, postulou-se, no começo do século 20, que pássaros seriam seres inferiores no que diz
respeito à inteligência, pois teriam suas ações embasadas em impulsos instintivos provenientes de
seus gânglios da base bastante desenvolvidos, faltando-lhes base neural para cognição e
aprendizado (Reiner, 2005). Nessa linha de raciocínio, acreditava-se que em répteis, apenas uma
pequena porção cortical era presente, enquanto os gânglios da base ocupavam um volume
desproporcionalmente maior que nos mamíferos, assim, avançando na filogenia animal, essa
porção cortical haveria hipertrofiado para a formação do córtex dos mamíferos e essa proporção
manter-se-ia dependendo da posição da classe dentro da linha evolutiva (Reiner et al., 2004).
Devido a essas idéias inicialmente equivocadas, por muito tempo se pensou que o cérebro do
pássaro seria composto basicamente de gânglios da base e seu comportamento seguiria apenas
padrões pré-estabelecidos e estáticos. Entretanto, no decorrer do século XX, um grande número de
evidências mostrou que o cérebro dos pássaros possui um grande território palial com funções
similares ao do córtex dos mamíferos (Figura 1.1).

21
Figura 1.1: Em A, interpretação e nomenclatura defasadas do cérebro de pássaro (pombo); em B, interpretação e
nomenclatura aceitas atualmente para o cérebro de pássaro (pombo) e em C, as áreas correspondentes em mamífero
(rato). Adaptado de Jarvis et al, 2004.
Porém, anatomicamente o pálio das pássaros constitui-se de uma maneira um pouco
diferente (Figura 1.2), organizando-se de forma globular (em núcleos), e seu homólogo mamífero,
o córtex, organizado em estrutura laminar (Jarvis et al., 2005).
Fig 1.2: O cérebro de Mandarim (Taeniopygia guttata) em corte parassagital, sendo a região do pallium destacada em
cinza e a correspondente ao NCM, listrada (Adaptado de AvianBrain.org).
Com essa visão de um pallium grande e bem desenvolvido que processa informações da
mesma forma que o córtex mamífero, uma reavaliação das habilidades cognitivas das aves desde
os anos 50 mostrou de maneira progressiva que estas são bem mais complexas do que
inicialmente imaginado (Marler, 1955); (Thorpe, 1958). Exemplos dessas habilidades são os
Ven
tral
Dor
sal
Medial Lateral
Caudal Rostral
Ven
tral
Dor
sal

22
pombos, que conseguem memorizar até 725 padrões visuais (Von Fersen & Elius, 1989) ou
corvos que fazem uso de galhos e folhas como ferramentas para obtenção de alimento e passam
esse conhecimento para outros de sua espécie (Pollok et al., 2000; Hunt & Gray, 2003). Outros
pássaros, como Aphelocoma californica (membro da família dos corvos) mostram memória
episódica, habilidade antes atribuída apenas a humanos (Clayton & Dickinson, 1998), corujas
possuem uma capacidade de localização sonora altamente sofisticada desenvolvida através da
aprendizagem (Knudsen, 2002) e, finalmente, pássaros como papagaios, beija-flores e da
subordem oscine que possuem a habilidade de aprendizagem vocal (Jarvis et al., 2000). O estudo
do canto desses pássaros que aprendem a vocalizar (pássaros canoros) começou em 1958 com
William Thorpe, que mostrou a necessidade do tentilhão (Fringilla coelebs) de ouvir a canção de
um macho tutor adulto para a produção posterior de um canto individual próprio. Outros
investigadores, como Peter Marler, Masakazu Konishi e Fernando Nottebohm seguiram a linha de
Thorpe e abriram caminhos para uma geração de estudiosos contribuindo no estudo do
comportamento e controle neural do canto (revisto em:(Brenowitz et al., 1997).
Pássaros canoros tem sido um sistema atrativo para o estudo das bases neurais do
aprendizado devido à maneira com a qual aprendem a cantar imitando o pai ou um tutor, criando
um padrão de canto próprio, que dura para o resto da vida do animal. O canto possui algumas
características que o tornam um modelo versátil para o estudo da neurobiologia comportamental
(Doupe & Kuhl, 1999; Williams, 2004), como (1) sua natureza estereotipada e mensurável; (2) a
similaridade entre seu aprendizado e o aprendizado da fala humana, onde ambos aprendem essa
vocalização complexa no início da vida, sendo ela dependente da escuta de um modelo (adulto)
para que possa ser copiada e da escuta do seu próprio canto para que haja refino e manutenção
deste na vida adulta e, para discernir qual o canto a imitar, existe uma predisposição inata para o
aprendizado dos sons produzidos por indivíduos da própria espécie em detrimento daqueles
produzidos por outros animais; (3) a caracterização precisa de um circuito dedicado
exclusivamente à aquisição e outro à produção do canto onde centros motores e auditivos no
telencéfalo interagem para controle da vocalização e (4) um período crítico para a aprendizagem
desta. Em mandarins, essa aprendizagem do canto ocorre entre 30 e 95 dias, período conhecido
como sensível para a aprendizagem vocal, porém, o desenvolvimento do canto acontece apenas
nos machos, pois nesta espécie (como em vários outros oscinos), o tamanho dos núcleos do
sistema do canto dos machos é aumentado quando comparados aos das fêmeas, que são bastante
atrofiados (Nottebohm & Arnold, 1976); (MacDougall-Shackleton & Ball, 1999). Esse
desenvolvimento depende de um tutor (sempre um macho) e é dividido em fases. Inicialmente,
pássaros jovens apenas escutam o tutor para aquisição de um modelo de canto (aquisição de
memória ou sensória). Em seguida, vem a fase sensório-motora, onde o animal começa a

23
vocalizar e escutar seu próprio canto. Essa retroalimentação (escutar o próprio canto) é necessária
para a comparação do canto produzido com o canto aprendido (daí o nome “sensório-motora”).
Esse canto inicial se desenvolve até um ponto bem estruturado, porém ainda variável. O processo
estará apenas totalmente concluído quando o canto for cristalizado e se apresentar de forma
estereotipada semelhante ao modelo apresentado pelo tutor. Esse processo de cristalização
depende de testosterona para que seja completado (Brenowitz et al., 1997) e também da síntese
local de ácido retinóico no núcleo HVC (Denisenko-Nehrbass et al., 2000).
Nos pássaros canoros, uma extensa rede de núcleos interconectados foi identificada como
a responsável pelo reconhecimento, aprendizado e produção do canto, adequadamente chamada
de “sistema do canto” (Nottebohm, 2000) que inexiste nos demais pássaros. O sistema do canto
dos pássaros canoros consiste de três circuitos: um auditivo e dois ligados ao canto (um posterior
e outro anterior). Os circuitos auditivo e vocais se comunicam sinapticamente via conexões da
área auditiva do mesopálio caudolateral (CLM) para as áreas vocais do centro vocal superior
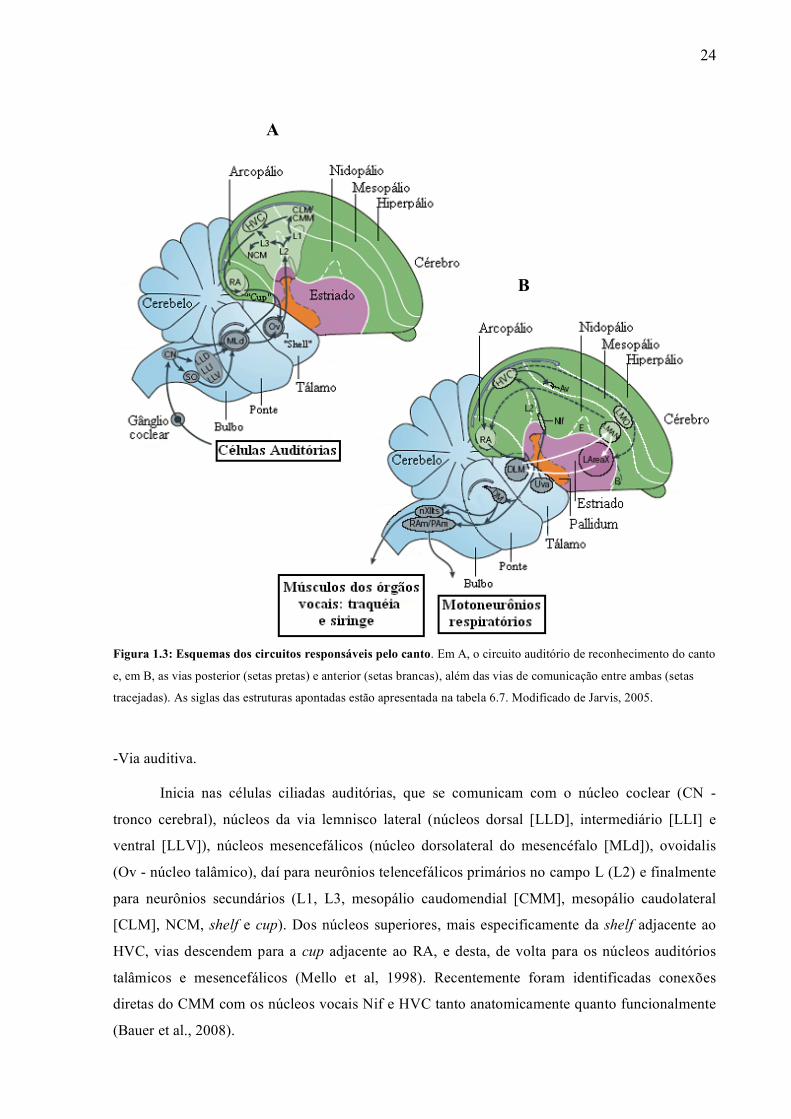
(HVC) e núcleo interface (Nif) (Bauer et al., 2008) (ver tabela 6.7 para a definição das siglas).
-Vias ligadas ao canto:
As vias do canto destinadas à sua produção são exclusivas de pássaros que aprendem a
vocalizar (oscinos, piscitaciformes e troquiliformes) (Jarvis et al., 2000). Nos pássaros canoros, a
via posterior inicia-se no HVC, com neurônios projetando para o núcleo robusto do arcopálio
(RA). Deste, projeções atingem o mesencéfalo no núcleo medial dorsal (DM) e tronco cerebral na
parte traqueosiringeal do núcleo do hipoglosso (nXIIts), núcleos que controlam o órgão vocal
(siringe) e os motoneurônios respiratórios. Nos pássaros canoros, esta via é responsável pela
produção de vocalizações aprendidas (Nottebohm et al., 1976). A via anterior forma uma volta,
iniciando na região palial, mais especificamente no núcleo magnocelular lateral do nidopálio
anterior (LMAN), gânglios da base (área X), região talâmica (núcleo dorsolateral do tálamo
medial - DLM) e de volta ao LMAN. Esta via é responsável pelo aprendizado vocal (Bottjer et al.,
1984; Scharff & Nottebohm, 1991) e parece ter papel no contexto social, sintaxe e manutenção do
canto adulto (Jarvis et al., 1998; Hessler & Doupe, 1999; Williams & Mehta, 1999; Brainard &
Doupe, 2000; Kobayashi et al., 2001) (Figura 1.3A).
24
Figura 1.3: Esquemas dos circuitos responsáveis pelo canto. Em A, o circuito auditório de reconhecimento do canto
e, em B, as vias posterior (setas pretas) e anterior (setas brancas), além das vias de comunicação entre ambas (setas
tracejadas). As siglas das estruturas apontadas estão apresentada na tabela 6.7. Modificado de Jarvis, 2005.
-Via auditiva.
Inicia nas células ciliadas auditórias, que se comunicam com o núcleo coclear (CN -
tronco cerebral), núcleos da via lemnisco lateral (núcleos dorsal [LLD], intermediário [LLI] e
ventral [LLV]), núcleos mesencefálicos (núcleo dorsolateral do mesencéfalo [MLd]), ovoidalis
(Ov - núcleo talâmico), daí para neurônios telencefálicos primários no campo L (L2) e finalmente
para neurônios secundários (L1, L3, mesopálio caudomendial [CMM], mesopálio caudolateral
[CLM], NCM, shelf e cup). Dos núcleos superiores, mais especificamente da shelf adjacente ao
HVC, vias descendem para a cup adjacente ao RA, e desta, de volta para os núcleos auditórios
talâmicos e mesencefálicos (Mello et al, 1998). Recentemente foram identificadas conexões
diretas do CMM com os núcleos vocais Nif e HVC tanto anatomicamente quanto funcionalmente
(Bauer et al., 2008).
A
B

25
O Nidopálio Caudomedial (NCM)
O NCM (estrutura listrada na figura 1.2), juntamente com as demais áreas auditórias
aferentes ao HVC e o resto do sistema do canto (representado na figura 1.3) é, em pássaros,
considerado análogo às porções supragranulares dos córtices auditivo primário e de associação em
mamíferos (Doupe & Kuhl, 1999). O NCM é facilmente identificado por ser uma região grande,
com bordas ventral, dorsal e caudal definidas pela zona periventricular. Na parte rostral, seu limite
é o Campo L2, porém lateralmente, seus limites não são distintos (esquema de cortes
evidenciando o NCM na figura 3.2 – metodologia) (Mello & Clayton, 1994).
Quando um pássaro canoro recebe um estímulo acústico de um canto espécie-específico,
seu telencéfalo é estimulado, e é o NCM a região que exibe a expressão mais robusta do gene de
expressão imediata (IEG) zenk (também conhecido como zif-268, erg-1, NGFI-A e Krox-24
(Milbrandt, 1987; Christy et al., 1988; Lemaire et al., 1988; Sukhatme et al., 1988; Mello et al.,
1992)). O IEG zenk é considerado um gene dependente de atividade neural e é expresso em
resposta à despolarização neuronal (Milbrandt, 1987; Sukhatme et al., 1988). Ele é um fator de
transcrição do tipo zinc-finger, que se liga ao seu sítio específico no DNA presente em genes
distintos (Christy & Nathans, 1989; Pavletich & Pabo, 1991; Swirnoff & Milbrandt, 1995). Em
mamíferos, a indução do zenk é relacionado com o fenônemo de potenciação a longo prazo (LTP)
e formação de memória (Cole et al., 1989; Wisden et al., 1990; Abraham et al., 1993; Roberts et
al., 1996; Jones et al., 2001). Além do zenk, o NCM expressa outros IEGs em resposta ao canto,
como Arc, c-fos e c-jun (Mello & Clayton, 1994; Nastiuk et al., 1994; Bolhuis et al., 2000; Bailey
& Wade, 2003; Velho et al., 2005). Para espressão de zenk e Arc no NCM, a via responsável é a
iniciada com a ativação da MAP quinase (Velho et al., 2005). A expressão do zenk ocorre rápida e
transientemente após o estímulo do canto (Mello et al., 1992; Mello & Clayton, 1994) tanto em
machos quanto em fêmeas (Mello et al., 1992; Bailey & Wade, 2005) e é abolida quando o animal
é ensurdecido (Jarvis & Nottebohm, 1997) e a expressão do mRNA zenk atinge seu pico em 30
minutos após o início do estímulo, ao passo que o da proteína ZENK ocorre entre 1 e 2 horas após
o início do estímulo (Mello & Ribeiro, 1998).
Além da marcação do RNAm do zenk por hibridização in situ e da proteína por
imunocitoquímica, dados eletrofisiológcos de registro multiunitário confirmam o fato de que o
NCM é bastante responsivo à estímulos auditivos, em especial, àqueles específicos da espécie, o
que confirma sua participação no processamento da informação sonora (Chew et al., 1995; Chew
et al., 1996a; Stripling et al., 1997; Stripling et al., 2001). Evidências recentes sugerem que o
NCM contenha a representação neural do canto aprendido (Bolhuis et al., 2000; Terpstra et al.,
26
2004; Phan et al., 2006). Recentemente, Gobes & Bolhuis (2007) mostraram que a lesão bilateral
do NCM leva à uma deficiência no reconhecimento desse canto do tutor, sem prejudicar a
produção do canto do animal
Quando um canto diferente é apresentado ao pássaro (seja um som reproduzido em
laboratório ou o canto de um indivíduo na natureza), os neurônios do NCM são ativados e dois
fenômenos ocorrem: uma resposta vigorosa de aumento da atividade neuronal no NCM, como
visto por registros multiunitários (Chew et al., 1995) e um aumento na expressão de zenk (Mello
et al., 1992; Mello et al., 1994). Isso se deve ao fato de que neurônios do NCM de mandarins são
mais responsivos a cantos de pássaros da mesma espécie do que outras classes de sons, sejam eles
sons artificiais ou mesmo cantos de outras pássaros (Chew et al., 1996a), ao contrário de outros
núcleos do sistema do canto, como o HVC (Margoliash, 1983), que mostram amplitudes de
registro de campo in vivo similares para todos os sons testados, corroborando a hipótese de que o
NCM tem função no reconhecimento do canto específico da espécie.
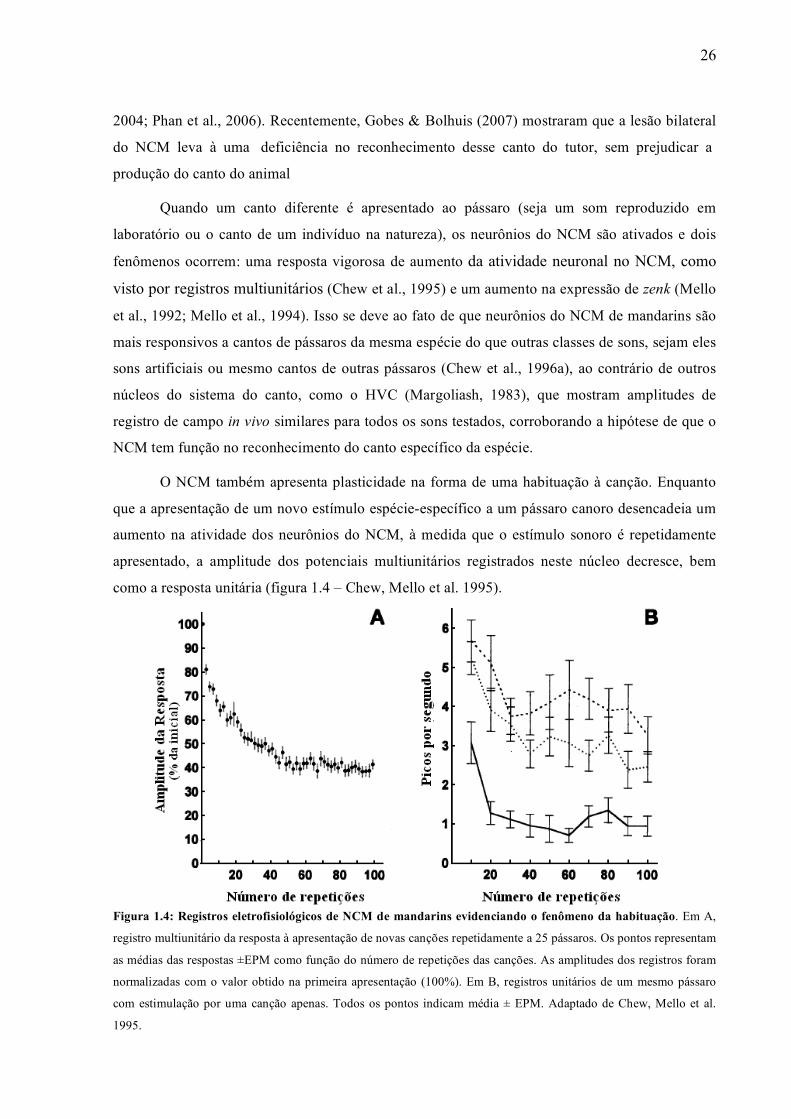
O NCM também apresenta plasticidade na forma de uma habituação à canção. Enquanto
que a apresentação de um novo estímulo espécie-específico a um pássaro canoro desencadeia um
aumento na atividade dos neurônios do NCM, à medida que o estímulo sonoro é repetidamente
apresentado, a amplitude dos potenciais multiunitários registrados neste núcleo decresce, bem
como a resposta unitária (figura 1.4 – Chew, Mello et al. 1995).
Figura 1.4: Registros eletrofisiológicos de NCM de mandarins evidenciando o fenômeno da habituação. Em A,
registro multiunitário da resposta à apresentação de novas canções repetidamente a 25 pássaros. Os pontos representam
as médias das respostas ±EPM como função do número de repetições das canções. As amplitudes dos registros foram
normalizadas com o valor obtido na primeira apresentação (100%). Em B, registros unitários de um mesmo pássaro
com estimulação por uma canção apenas. Todos os pontos indicam média ± EPM. Adaptado de Chew, Mello et al.
1995.
27
Isso constitui o processo da habituação, que ocorre especificamente para cada um dos
novos estímulos aplicados ao animal, ou seja, para cada canto novo, uma atividade aumentada é
observada, seguida de decréscimo da magnitude da resposta ao estímulo conforme o número de
repetições. Essas alterações podem durar até 20 horas para as canções espécie-específicas e até 4
horas para os demais sons (Chew et al., 1995) e são dependentes da síntese de proteína para a sua
manutenção (Chew et al., 1996a).
Além das alterações eletrofisiológicas que ocorrem no NCM no processo de habituação,
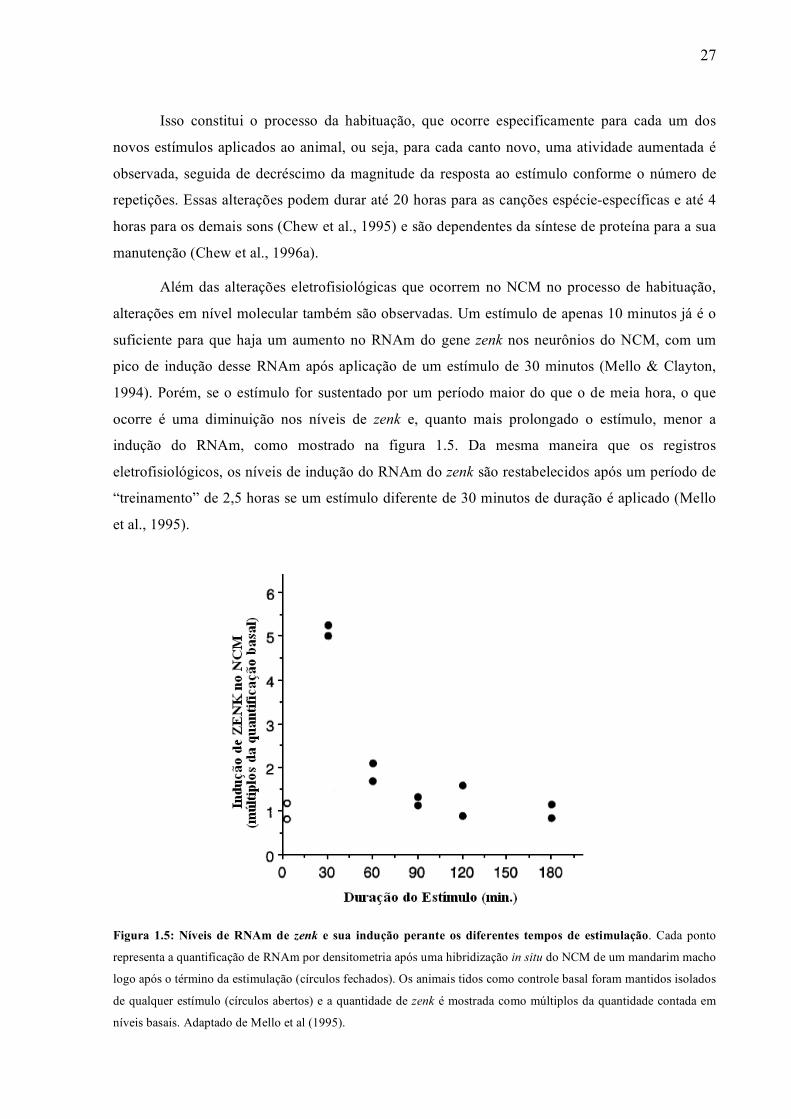
alterações em nível molecular também são observadas. Um estímulo de apenas 10 minutos já é o
suficiente para que haja um aumento no RNAm do gene zenk nos neurônios do NCM, com um
pico de indução desse RNAm após aplicação de um estímulo de 30 minutos (Mello & Clayton,
1994). Porém, se o estímulo for sustentado por um período maior do que o de meia hora, o que
ocorre é uma diminuição nos níveis de zenk e, quanto mais prolongado o estímulo, menor a
indução do RNAm, como mostrado na figura 1.5. Da mesma maneira que os registros
eletrofisiológicos, os níveis de indução do RNAm do zenk são restabelecidos após um período de
“treinamento” de 2,5 horas se um estímulo diferente de 30 minutos de duração é aplicado (Mello
et al., 1995).
Figura 1.5: Níveis de RNAm de zenk e sua indução perante os diferentes tempos de estimulação. Cada ponto
representa a quantificação de RNAm por densitometria após uma hibridização in situ do NCM de um mandarim macho
logo após o término da estimulação (círculos fechados). Os animais tidos como controle basal foram mantidos isolados
de qualquer estímulo (círculos abertos) e a quantidade de zenk é mostrada como múltiplos da quantidade contada em
níveis basais. Adaptado de Mello et al (1995).

28
Pouco se sabe quanto à fisiologia celular e sináptica dos neurônios do NCM. Pinaud et al.
(2004) mostraram que aproximadamente metade dos neurônios do NCM são GABAérgicos, e que
podem expressar zenk em resposta à canção. Esses neurônios parecem estar inibindo uma
atividade glutamatérgica forte, pois a inibição das correntes sinápticas por bicuculina levou ao
aparecimento de correntes glutamatérgicas espontâneas não vistas previamente à aplicação da
droga (Pinaud et al., 2004). Alem disso, o NCM recebe intensa inervação noradrenérgica (Mello
et al., 1998) e expressa a enzima aromatase em sua parte caudal (Pinaud et al., 2006). Entretanto,
os substratos sinápticos da fisiologia do NCM ainda são desconhecidos.

29
2 - OBJETIVO

30
Apesar da quantidade de informações obtidas por análise de expressão gênica e
eletrofisiologia in vivo no NCM, nada se sabe sobre a fisiologia da neurotransmissão nesse núcleo.
Tendo em vista a relevância desse núcleo para o reconhecimento da memória auditiva em
pássaros e das potenciais correlações que podemos fazer com esses processos em mamíferos,
resolvemos investigar usando técnicas eletrofisiológicas in vitro a fisiologia da neurotransmissão
em fatias a fresco do NCM de mandarins (Taenopyigia guttatta) (o modelo mais utilizado no
estudo da neurobiologia do canto) e sua plasticidade em resposta ao estímulo biológico, no caso, o
canto específico do pássaro. Para isso, usamos pássaros em duas condições: isolados
acusticamente por 12 horas (controles) e estimulados pelo canto (estimulados). Nosso paradigma
é que a estimulação canora ativaria neurônios específicos do NCM (evidenciados pela expressão
do IEG zenk) e alteraria sua fisiologia gerando um substrato para o fenômeno de plasticidade da
habituação ao canto.

31
3 - METODOLOGIA

32
3.1 - Animais
Os animais utilizados nos experimentos foram mandarins machos e fêmeas (Taeniopygia
guttata) (Figura 3.1), pássaros pertencentes à ordem passeriforme, subordem passeri (oscine),
família passeridae e subfamília estrildinae, nativos da região da Austrália, Timor Leste e
Indonésia, porém criados como pássaros ornamentais em todo o mundo.
Figura 3.1: Espécimes de mandarins. À esquerda, uma fêmea e à direita, um macho. Notar o dimorfismo no fenótipo
da plumagem dos animais.
Todos os espécimes foram adquiridos de criadores locais, mantidos no biotério do
Departamento de Fisiologia da Faculdade de Medicina de Ribeirão Preto em gaiolas com água e
comida à vontade e mantidas junto à janela para um ciclo claro/escuro com luz naturalPara cada
experimento, um animal foi usado por vez. Um dia antes do experimento, o animal escolhido foi
retirado do biotério e transferido para uma câmara acústica (Insight Instrumentos) dentro do
Laboratório de Fisiologia e Plasticidade Sináptica e mantidos dentro de uma gaiola com água e
comida à vontade pelo período de uma noite. Um temporizador automático acoplado a uma
luminária foram usados para ditar o ciclo claro/escuro da câmara.
3.2 - Estímulo
Na manhã do dia seguinte ao isolamento, os animais tiveram um dos dois destinos:
_Animais controle: os mandarins foram sacrificados por decapitação cerca de 15 minutos
após o acender das luzes da caixa de isolamento acústico.
_ Animais estimulados: os mandarins foram estimulados acusticamente com uma
reprodução de 7 canções espécie-específicas jamais ouvidas por estes animais durante 15 s em
intervalos de 45 s por aproximadamente meia hora. Após uma hora do término das canções (esse

33
intervalo após o estímulo corresponde ao tempo necessário para que ocorra o pico de expressão do
ZENK (Mello & Ribeiro, 1998)) os animais foram então sacrificados por decapitação.
O estímulo acústico foi aplicado aos animais pela reprodução de um CD contendo as
canções dos mandarins por um tocador de CDs portátil conectado a duas caixas acústicas
localizadas dentro da caixa de isolamento. O nível de intensidade acústica (72 - 76 dB) foi medido
por um medidor de decibéis portátil (Radio Shack). As canções foram obtidas no zebrafinch song
archive (http://wso.williams.edu/~hwilliam/ZFsongs/) na página da Internet da Dra. Heather
Williams (Departamento de Biologia, Williams College, Williamstowm, MA. EUA). O protocolo
de estimulação usado seguiu os parâmetros dos protocolos utilizados para a estimulação da
expressão da proteína ZENK (Mello & Clayton, 1994).
Todo o procedimento foi aprovado de acordo com o protocolo do comitê de ética no
024/2003 aprovado pelo CETEA-FMRP.
3.3 - Confecção das fatias
Os cérebros dos mandarins foram cuidadosamente removidos após a decapitação e
imediatamente colocados em solução de corte, o aCSF (artificial cerebrospinal fluid) modificado
para corte (com alta sacarose – Tabela 6.2), gelado e pré-borbulhado com mistura carbogência
(95%O2 e 5% CO2) para ajuste do pH (7,3 – 7,4). Depois de mergulhados no aCSF, os
hemisférios foram separados com a ajuda de uma lâmina de bisturi e colados independentemente
em uma plataforma, apoiados por um bloco de agar 4% para corte em vibrátomo (Vibratome,
série 1000 - manual), onde foram feitos cortes parassagitais de 200 µm de espessura orientados da
porção medial para a porção lateral.
O NCM não possui limites claramente identificáveis em sua porção lateral. Devido a essa
característica, em nosso protocolo de corte, retiramos de cada um dos hemisférios no máximo 4 –
5 fatias de 200 µm, como ilustrado pelos desenhos em câmara clara da figura 3.2, onde o esquema
do NCM usado em nossos registros eletrofisiológicos está pintado em cinza.
Depois de cortadas as fatias, transferimo-las para um becker com a mesma solução de
corte borbulhada com mistura carbogência, porém à temperatura ambiente, onde ficaram
descansando por pelo menos uma hora antes dos registros.

34
Figura 3.2: Desenho em câmara clara de diferentes níveis do cérebro de mandarim. Os cortes são mostrados
iniciando na porção medial (mais acima) e terminando na porção lateral (abaixo). A estrutura pintada em cinza e
apontada como “1” é a porção do NCM utilizável em experimentos. 1 – NCM; 2 – Terceiro ventrículo; 3 –
Hipocampo; 4 – Cerebelo; 5 – Campo L. Desenho: Dr. Eric Jarvis, Duke University, NC-USA, disponibilizado em
http://avianbrain.org/.

35
3.4 - Eletrofisiologia
3.4.1 - Visualização dos neurônios
Os neurônios do NCM foram visualizados através de um microscópio Olympus BX51WI
com sistema de Contraste por Interferência Diferencial (DIC) acoplado a uma câmera Hamamatsu
(modelo C7500-50) e um monitor monocromático SONY. Ao visualizar o NCM no monitor, este
não apresenta uma organização celular distinta, sendo composto de neurônios homogeneamente
distribuídos, como pode ser visto na figura 3.3. Todo o sistema é permanentemente mantido sobre
uma mesa pneumática (TMC) para o isolamento mecânico do ambiente.
Figura 3.3: Imagem capturada com sistema DIC com uma objetiva de 60x de neurônios do NCM.
3.4.2 - Whole cell patch clamp
O sistema de registro é composto por um amplificador HEKA, modelo EPC-10, com
ligação direta via fibra óptica a uma placa de aquisição integrada (LIH 1600) acoplada a um
computador tipo PC para processamento e armazenamento dos dados no disco rígido para
posterior análise. A configuração usada para todos os registros foi a de whole cell patch clamp,
com fixação da voltagem para visualização das correntes sinápticas – voltage clamp.
Pipetas de borossilicato (DE: 1,5 mm; DI: 0,86 mm) com filamento interno (Sutter
Instruments) foram confeccionadas com um puxador de pipetas Sutter P-97 e preenchidas com
solução interna de pipeta contendo cloreto de césio como soluto principal (Tabela 6.3) ou

36
metanosulfonato de césio (Tabela 6.4), filtradas com filtros de 20µm (Millipore), resultando em
pipetas com resistências entre de 3-6MΩ aproximadamente quando mergulhadas na solução de
registro. A solução de perfusão da fatia usada foi o aCSF (Tabela 6.1) borbulhado com mistura
carbogênica (95%O2 e 5% CO2). As soluções e drogas foram aplicadas às fatias por um sistema
de múltiplas vias impelido por força gravitacional, confeccionado no próprio laboratório. A
remoção das soluções foi toda feita através de pressão negativa por bomba de sucção.
Depois da pipeta preenchida e acoplada ao probe com um eletrodo de prata cloretado, esta
foi então mergulhada no aCSF do banho e uma pressão positiva aplicada através de uma seringa
de 10 mL. A aproximação da pipeta ao neurônio foi feita com um micromanipulador hidráulico
(Syskiu Instruments). Quando a ponta da pipeta tocava o neurônio escolhido, a pressão era
inicialmente igualada à atmosférica e em seguida, aplicava-se uma leve sucção. Com isso,
resistência da pipeta aumentava para algumas centenas de MΩ e a configuração cell attached era
formada e a capacitância da pipeta eletronicamente cancelada. Com um pouco mais de sucção, o
retalho da membrana na boca da pipeta era rompido e o modo whole cell, configurado. Os
neurônios foram mantidos a -70 mV e os valores de capacitância de membrana (Cm) e resistência
em séria (RS) obtidos. Os valores de RS foram constantemente monitorados e, registros com
valores de RS superiores à 30 MΩ foram descartados. A maioria dos registros teve sua RS
compensada digitalmente (lag de 10 µs) em até 80%.
3.4.3 - Registro das correntes sinápticas
As correntes foram filtradas low-pass a 3 KHz e adquiridas a 10 kHz. Dois registros de 60
segundos cada um foram adquiridos por condição (com ou sem droga). Os dados de corrente
foram adquiridos com o software PULSE (HEKA). Todos os registros foram iniciados após 5
minutos da configuração de whole cell ter sido estabelecida, para estabilização do sistema. Para os
protocolos de aplicação de droga, o registro foi feito apenas depois de 5 mL de solução com droga
ter passado pelo banho (aproximadamente 4 minutos).
As correntes inibitórias pós-sinápticas espontâneas (sIPSCs) foram isoladas através da
aplicação de 6,7-dinitroquinoxalina-2,3-diona (DNQX) numa concentração de 20 µM. Os
registros foram feitos em um holding de -70 mV (ao menos quando indicado um valor diferente) e
solução interna de pipeta com CsCl como soluto principal. As correntes excitatórias pós-
sinápticas espontâneas (sEPSCs), foram isoladas pela aplicação de bicuculina 10 µM e todos os
registros foram feitos a -70 mV quando o CsCl foi usado como soluto principal da solução interna
de pipeta. Alguns experimentos foram conduzidos com solução interna contendo metanosulfonato

37
de césio como principal sal a fim de medirmos as correntes sinápticas em uma situação de baixo
cloreto intracelular, assim como alguns neurônios foram registrados em current-clamp usando
solução interna de pipeta com gluconato de potássio como principal soluto (tabela 6.5).
As correntes pós-sinápticas miniatura (mPSCs) foram registradas na presença de
tetrodotoxina (TTX) 1 µM ao banho associada a um dos dois antagonistas (DNQX ou bicuculina)
nas concentrações descritas.
3.5 - Análise dos dados
Todos os gráficos e análises estatísticas foram realizadas no programa Prism (GraphPad;
versão 4.02), com exceção da análise das curvas de fração cumulativa, as quais foram feitas no
Microsoft Excel. As análises consistiram de Análise de Variância (ANOVA) de duas vias quando
duas variáveis foram confrontadas, teste t pareado e não pareado quando apenas uma variável foi
levada em consideração e parâmetros “D” (maior diferença entre as duas distribuições) e mediana
para comparação de curvas de fração cumulativa.
As análises das correntes espontâneas foram feitas no programa Mini Analysis
(Synaptosoft). Os registros foram primeiramente escolhidos e transformados em um formato
legível pelo sistema (ABF), em seguida, os parâmetros de análise do programa foram ajustados
para cada arquivo individual e as correntes foram visualizadas uma a uma e marcadas
manualmente. O programa nos retornou os dados de interesse (freqüência, amplitude e half-width
[medida correspondente à largura do sPSC no ponto médio de sua amplitude]) em forma de uma
tabela com as médias de cada um dos registros, de onde foram anotados e organizados em
planilhas no Microsoft Excel.
A análise da cinética das correntes sinápticas em miniatura foi feita no programa Mini
Analysis usando-se a média de todos os eventos registrados em um arquivo de um minuto após o
isolamento farmacológico dos sIPSCs ou sEPSCs e aplicação de TTX, como mostrado na figura
2.3. A partir do traçado médio, os valores de constante de tempo (τ), decaimento e half width
foram calculados e os gráficos montados no GraphPad-Prism.

38
Figura 2.3: Traçados individuais agrupados (A) e a média dos mesmos (B) calculados na interface de análise de grupo
do programa Mini Analysis.
Os gráficos de fração cumulativa foram construídos no GraphPad-Prism como média de
vários traçados distintos gerados pelo programa Mini Analysis (Figura 2.4).
Figura 2.4: Exemplo de construção do gráfico das médias das curvas de fração cumulativa.
O nível de significância dos testes realizados foi de 0,05 e os resultados em barra são
apresentados como média ± erro padrão da média (EPM).
A B

39
3.6 - Imunocitoquímica
Usamos a imunocitoquímica (ICC) para evidenciar a expressão do gene zenk através da
marcação com diaminobenzidina (DAB) de seu transcrito ZENK. O material que utilizamos para
tal foram fatias obtidas através do mesmo protocolo de corte daquelas utilizadas nos experimentos
de eletrofisiologia, as quais retiramos diretamente do vibrátomo para o paraformaldeído (4%),
onde permaneceram por meia hora e depois as lavamos em tampão fosfato com salina (PBS –
Tabela 6.6 da lista de soluções) por 3 vezes com duração de 20 minutos cada lavagem. Depois de
fixadas as fatias, as estocamos em geladeira por até quatro dias para que as reações de ICC das
demais fatias coletadas durante a semana fossem desenvolvidas em conjunto.
Iniciamos as ICCs incubando as diferentes fatias em poços numa placa de cultura de
acrílico e mergulhando-as em solução de H2O2 a 0,3% por 10 minutos, seguida de lavagem em
PBS, 3x por 20 minutos cada. Um bloqueador, a albumina de soro bovina (bovine serum albumine
- BSA) foi adicionado e mantido por uma hora. Após essa etapa, incubamos as fatias com o
anticorpo primário para ZENK (Anti-erg1, Santa Cruz Biotechnology – SC189) a uma diluição de
1:500 (estoque: 200µg/mL) em geladeira por dois dias. O passo seguinte foi a lavagem das fatias
em PBS 3x por 15 minutos seguido da aplicação do anticorpo secundário biotinilado goat-anti-
rabbit (Chemicon International ap132b – 2mL) diluído 1:50 durante duas horas. O preparo do
complexo amplificador avidina-biotina (Vectastain Elite ABC Kit) feito pelo menos meia hora
antes de sua adição, consistiu na reação de 40 µL da solução com avidina combinada com 40µL
da solução com biotina com volume completado pra 1mL. Após a ligação do antígeno com o
anticorpo secundário, as fatias foram lavadas em PBS 3x por 30 minutos e colocadas para reagir
com o complexo AB por duas horas, com posterior lavagem com PBS 3x de 15 minutos. A reação
de coloração com DAB foi feita pingando-se a solução previamente preparada (1 mg/mL de DAB,
H2O2 (30%) 1:1000 em PBS) sobre as fatia montadas em uma lâmina super frost por pelo menos
5 minutos.
A montagem das lâminas com Etellan foi feita no dia seguinte, após desidratação com
lavagem dos espécimes em uma série de soluções alcoólicas (70%, 90% e 100%) e deslipidização
em Xilol.

40
4 - RESULTADOS

41
4.1 – Expressão da proteína ZENK em resposta ao canto.
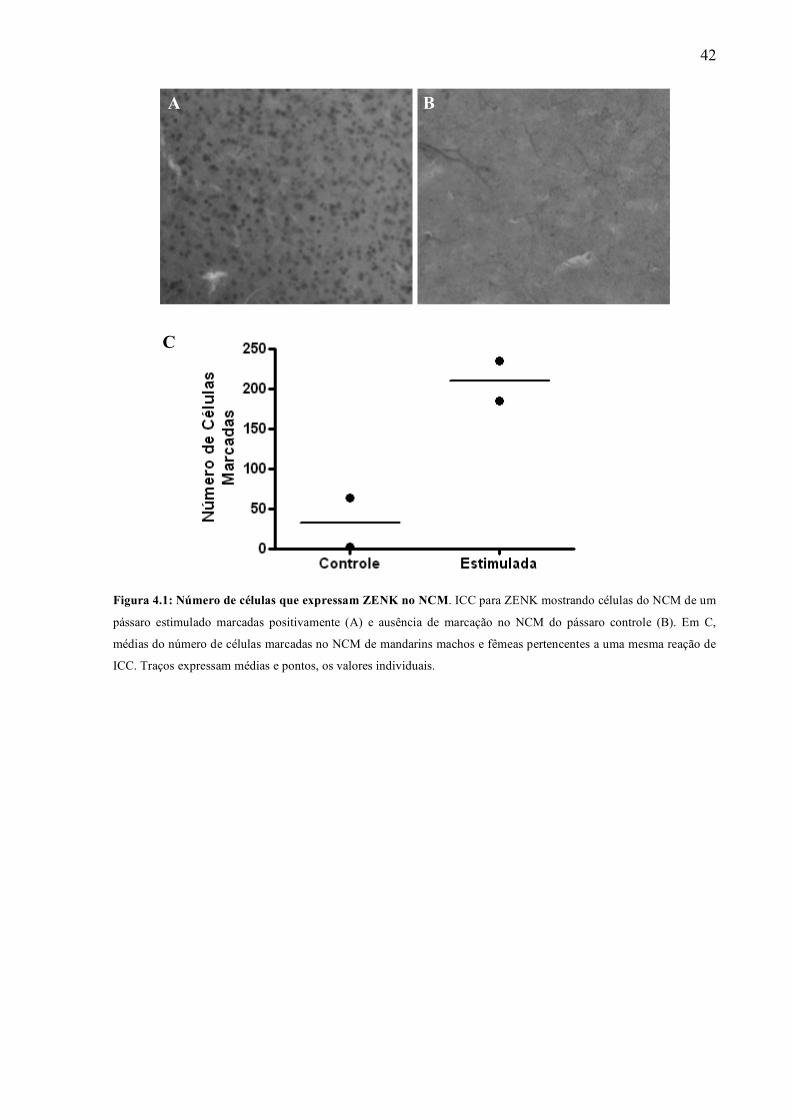
A expressão do gene zenk pode ser evidenciada pela marcação por imunocitoquímica de
seu transcrito ZENK. Como essa proteína é expressa quando o pássaro é estimulado por um canto
co-específico, sua expressão nas fatias serve como um controle da eficácia do protocolo de
estimulação e do isolamento acústico do pássaro. Após um protocolo de estimulação, espera-se
que a proteína ZENK seja expressa em grande quantidade; por outro lado, se o animal é mantido
sem estímulo, sua expressão é esperada ser baixa.
Na figura 4.1B, observamos que no NCM de pássaros controle a expressão de ZENK é
baixa, enquanto que no NCM dos pássaros estimulados é claramente maior (figura 4.1A)
(pássaros controle: 85 ± 43 células marcadas, n = 5; pássaros estimulados: 411 ± 196 células
marcadas, n = 4). Dentro de nossas estimativas observamos uma grande variação da resposta ao
zenk intra-grupo e, um possível motivo de variação pode ter sido diferentes qualidades da
marcação. Para isso comparamos 4 fatias (2 controle e 2 estimuladas) que foram submetidas ao
mesmo protocolo de marcação por DAB na mesma lâmina (Figura 4.1C). Nessas fatias, fica claro
que mesmo com essas variações, os pássaros estimulados apresentaram uma expressão de ZENK
maior do que os controles. Esses dados mostram que tanto o isolamento acústico quanto a
estimulação canoro à qual submetemos os pássaros são eficientes e que o procedimento de
sacrifício do animal e obtenção das fatias não estimula a expressão de ZENK.
42
Figura 4.1: Número de células que expressam ZENK no NCM. ICC para ZENK mostrando células do NCM de um
pássaro estimulado marcadas positivamente (A) e ausência de marcação no NCM do pássaro controle (B). Em C,
médias do número de células marcadas no NCM de mandarins machos e fêmeas pertencentes a uma mesma reação de
ICC. Traços expressam médias e pontos, os valores individuais.
C
A B

43
4.2 – Caracterização das correntes sináptica espontâneas (sPSCs) registradas s no NCM.
Quando as células do NCM dos mandarins foram mantidas em voltage clamp a -70mV,
observamos quase sempre a presença de correntes sinápticas espontâneas, representadas por
deflexões rápidas da corrente basal com um retorno à linha de base mais lento (Figura 4.2). Essas
correntes refletem a liberação de neurotransmissores pelos neurônios que fazem sinapses com o
neurônio registrado.
Figura 4.2: Exemplo de registro de sPSCs em voltage clamp de neurônio do NCM de mandarim. Registro feito a
-70 mV com solução interna de pipeta de CsCl.
Para que pudéssemos observar quais os transmissores responsáveis pelos sPSCs no NCM,
aplicamos antagonistas farmacológicos dos receptores ionotrópicos glutamatérgicos
AMPA/kainato (DNQX) e GABAA (bicuculina) no NCM de pássaros controle. A aplicação de
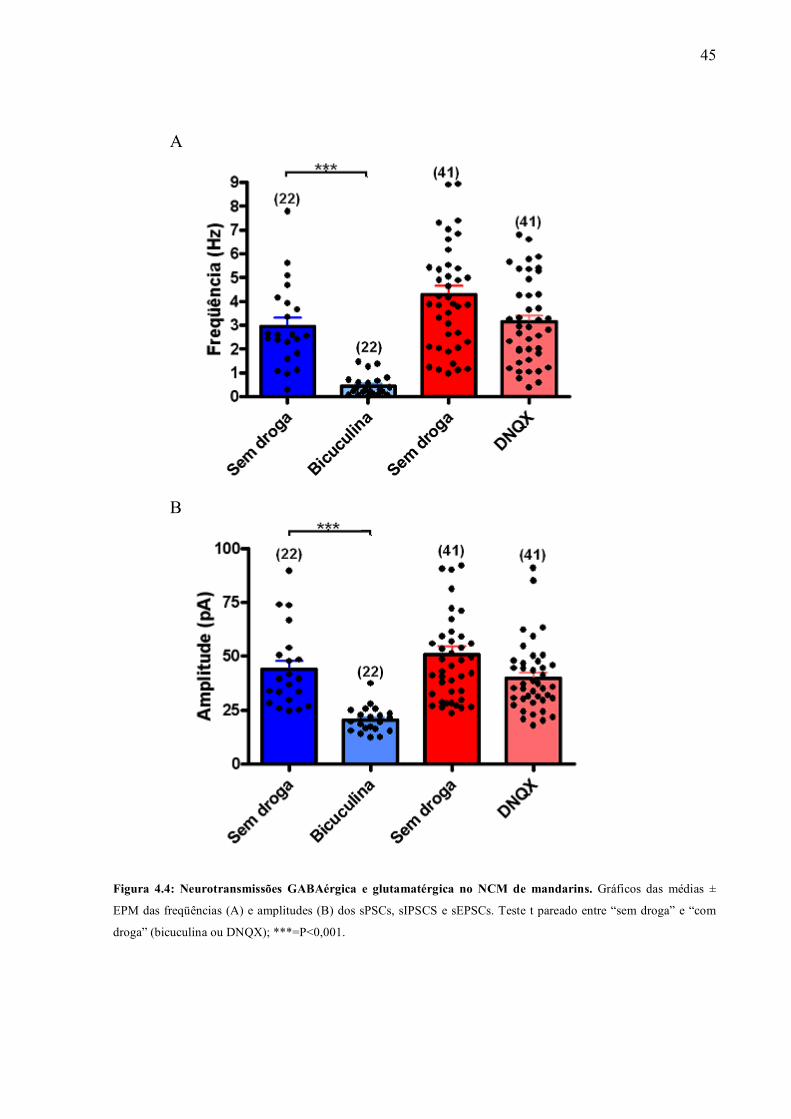
bicuculina ao banho produziu uma redução significativa tanto na freqüência (de 3,0 ± 0,31 Hz
para 0,5 ± 0,1 Hz [85 ± 3 %]; n = 22; p<0,001) quanto na amplitude (de 44 ± 4 pA para 25,5 ± 1,3
pA [20,5 ± 1,3 %]; n = 22; p<0,001) dos sPSCs (Figuras 4.3A [traçado] e 4.4 [gráfico]). Já a
aplicação de DNQX teve um pequeno efeito inibitório, porém não significativo, sobre a
freqüência e a amplitude dos sPSCs (de 4,3 ± 0,4 Hz para 3,14 ± 0,3 Hz [21,6 ± 5,5 %] e de 50,5
± 4 pA para 40 ± 2,5 pA [16,2 ± 3,2 %] respectivamente; n = 41; p<0,05 – Figuras 4.3B [traçado]
e 4.43 [gráfico]). Quando aplicamos ambos DNQX e bicuculina ao banho, os sPSCs foram
totalmente bloqueados (Figura 4.3C). Assim, concluímos que os sPSCs registrados nos neurônios
do NCM são gerados pela ativação de receptores glutamatérgicos e GABAérgicos, e que a
transmissão GABAérgica é mais abundante do que a glutamatérgica.

44
Figura 4.3: Efeito dos antagonistas glutamatérgico (DNQX) e GABAérgico (bicuculina) sobre os sPSCs no NCM
de mandarins. Em A, exemplo de registro em voltage clamp (-70 mV) de sPSCs (controle) seguido da aplicação de
bicuculina. Em B, exemplo de registro em voltage clamp (-70 mV) de sPSCs seguido da aplicação de DNQX. Em C, o
efeito da bicuculina e DNQX aplicados juntos ao banho. A figura D mostra a fração cumulativa referente às
freqüências em situação controle (sem droga), com aplicação de DNQX (GABA) e aplicação de bicuculina
(Glutamato).
D
45
Figura 4.4: Neurotransmissões GABAérgica e glutamatérgica no NCM de mandarins. Gráficos das médias ±
EPM das freqüências (A) e amplitudes (B) dos sPSCs, sIPSCS e sEPSCs. Teste t pareado entre “sem droga” e “com
droga” (bicuculina ou DNQX); ***=P<0,001.
A
B
46
4.2.1 - Correntes Pós-sinápticas Miniatura (mPSCs)
Visto que GABA e glutamato são os responsáveis pela transmissão espontânea que
observamos no NCM, decidimos estudar as características cinéticas das correntes sinápticas
inibitórias e excitatórias no NCM. Para isso, utilizamos as correntes sinápticas miniaturas
registradas na presença do bloqueador de canais de sódio tetrodotoxina (TTX). As correntes
miniaturas refletem a liberação de uma ou poucas vesículas por vez (quanta) e por isso são ideais
para o estudo da cinética dos receptores presentes na fenda sináptica.
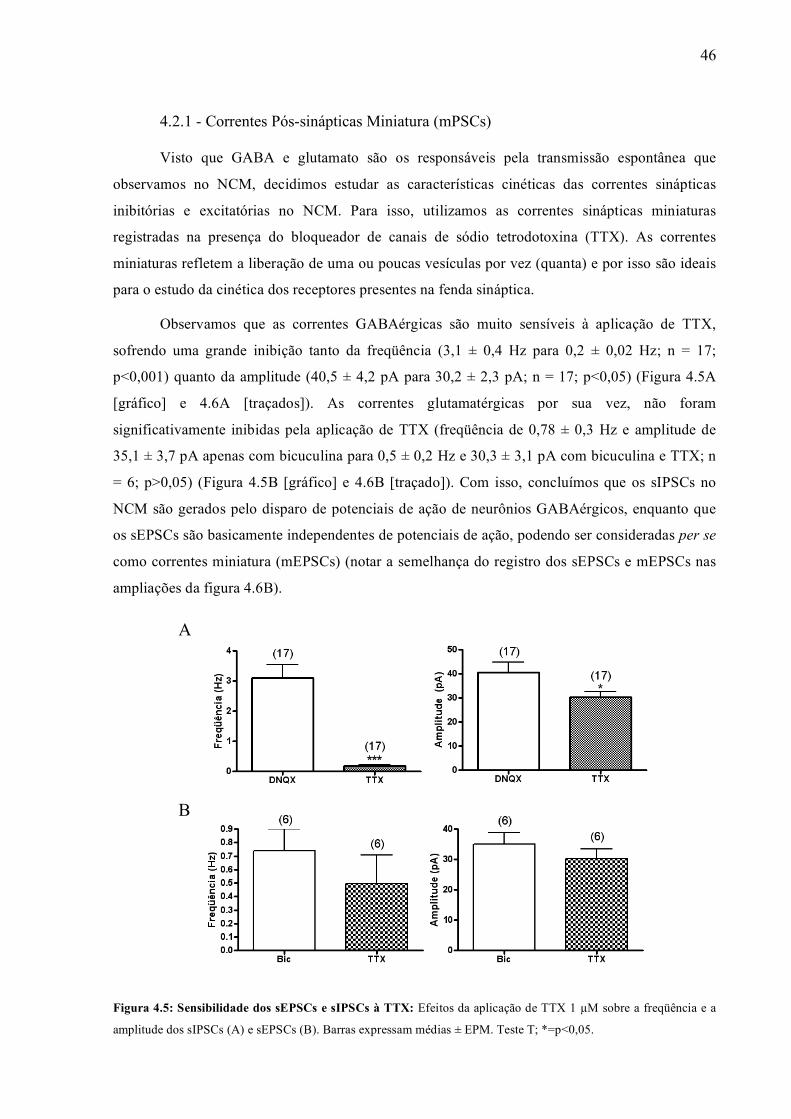
Observamos que as correntes GABAérgicas são muito sensíveis à aplicação de TTX,
sofrendo uma grande inibição tanto da freqüência (3,1 ± 0,4 Hz para 0,2 ± 0,02 Hz; n = 17;
p<0,001) quanto da amplitude (40,5 ± 4,2 pA para 30,2 ± 2,3 pA; n = 17; p<0,05) (Figura 4.5A
[gráfico] e 4.6A [traçados]). As correntes glutamatérgicas por sua vez, não foram
significativamente inibidas pela aplicação de TTX (freqüência de 0,78 ± 0,3 Hz e amplitude de
35,1 ± 3,7 pA apenas com bicuculina para 0,5 ± 0,2 Hz e 30,3 ± 3,1 pA com bicuculina e TTX; n
= 6; p>0,05) (Figura 4.5B [gráfico] e 4.6B [traçado]). Com isso, concluímos que os sIPSCs no
NCM são gerados pelo disparo de potenciais de ação de neurônios GABAérgicos, enquanto que
os sEPSCs são basicamente independentes de potenciais de ação, podendo ser consideradas per se
como correntes miniatura (mEPSCs) (notar a semelhança do registro dos sEPSCs e mEPSCs nas
ampliações da figura 4.6B).
Figura 4.5: Sensibilidade dos sEPSCs e sIPSCs à TTX: Efeitos da aplicação de TTX 1 µM sobre a freqüência e a
amplitude dos sIPSCs (A) e sEPSCs (B). Barras expressam médias ± EPM. Teste T; *=p<0,05.
B
A

47
Figura 4.6: Ação da TTX sobre os sPSCs. Em B, exemplos de traçados em Voltage Clamp onde a TTX
1µM é aplicada junto com DNQX (A) ou bicuculina (B). Os quadros em B mostram em maior detalhe os formatos dos
sIPSCs e mIPSCs.
A
B
48
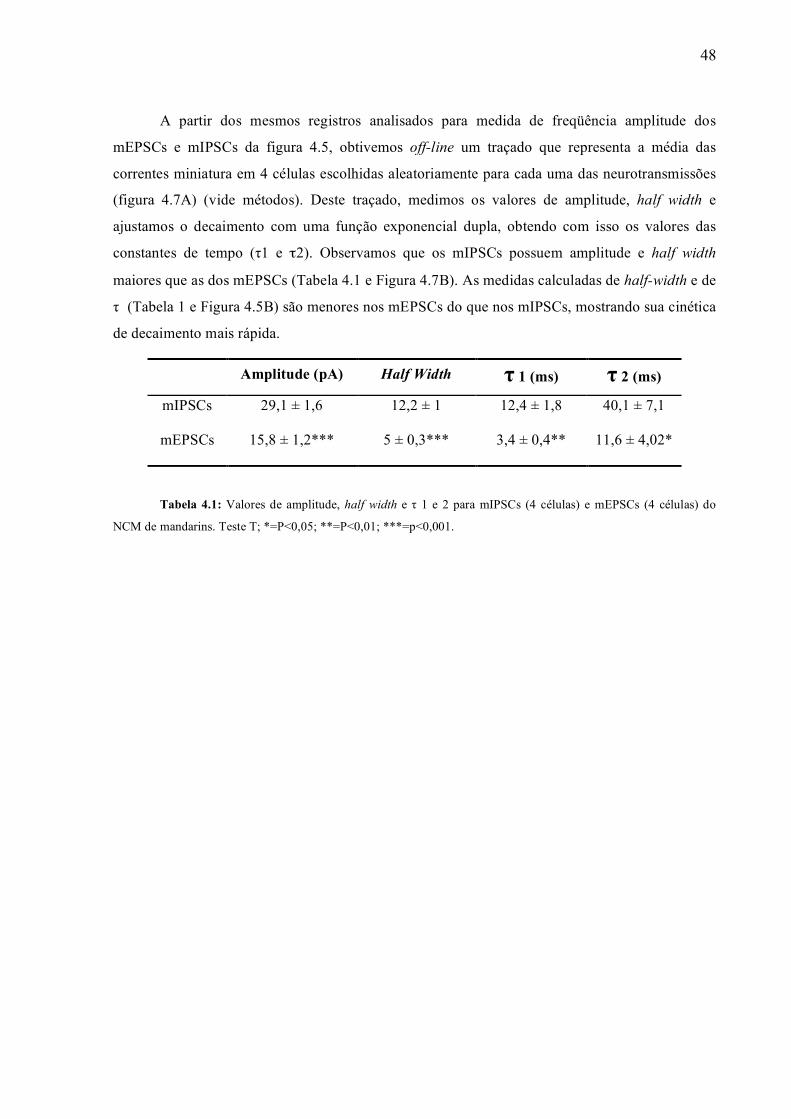
A partir dos mesmos registros analisados para medida de freqüência amplitude dos
mEPSCs e mIPSCs da figura 4.5, obtivemos off-line um traçado que representa a média das
correntes miniatura em 4 células escolhidas aleatoriamente para cada uma das neurotransmissões
(figura 4.7A) (vide métodos). Deste traçado, medimos os valores de amplitude, half width e
ajustamos o decaimento com uma função exponencial dupla, obtendo com isso os valores das
constantes de tempo (τ1 e τ2). Observamos que os mIPSCs possuem amplitude e half width
maiores que as dos mEPSCs (Tabela 4.1 e Figura 4.7B). As medidas calculadas de half-width e de
τ (Tabela 1 e Figura 4.5B) são menores nos mEPSCs do que nos mIPSCs, mostrando sua cinética
de decaimento mais rápida.
Amplitude (pA) Half Width τ 1 (ms) τ 2 (ms)
mIPSCs 29,1 ± 1,6 12,2 ± 1 12,4 ± 1,8 40,1 ± 7,1
mEPSCs 15,8 ± 1,2*** 5 ± 0,3*** 3,4 ± 0,4** 11,6 ± 4,02*
Tabela 4.1: Valores de amplitude, half width e τ 1 e 2 para mIPSCs (4 células) e mEPSCs (4 células) do
NCM de mandarins. Teste T; *=P<0,05; **=P<0,01; ***=p<0,001.

49
B i
iii
ii
A i ii
Figura 4.7: Cinética dos mPSCs do NCM de
machos. Em Ai, o registro médio de 27 mIPSCs;
em Aii, registro médio de 20 mEPSCs, ambos de
uma única célula. Rise time (10-90%): pontos
amarelos; decay (90-37%): pontos lilás; pico: “X”
vermelho: linha de base: “X” e linha verdes;
curva de ajuste da dupla exponencial: linha
vermelha [equação de ajuste: y = A1 * exp ( -x /
tau1) + A2 * exp (-x / tau2) + Baseline]. Em Bi, as
medidas de amplitude dos mIPSCs e mEPSCs, em
Bii, suas half width e em Biii seus respectivos τ1 e
τ2. Barras expressam médias ± EPM. Teste T.
*=p<0,05; **=p<0,01; ***=p<0,001.

50
4.2.2 - Caracterização dos sIPSCs e dos sEPSCs quanto ao potencial de reversão.
Com uma solução interna de pipeta contendo CsCl como íon principal, onde a
concentração de cloreto é alta, as correntes inibitórias e excitatórias exibem padrões de sentido e
reversão similares (correntes para dentro e reversão em 0mV) devido à força eletromotriz que age
sobre os íons, puxando os cátions para dentro da célula e empurrando os ânions para fora dela
com a mesma intensidade. Para investigar o comportamento das diferentes correntes
isoladamente, sem a intervenção farmacológica, utilizamos uma solução interna de pipeta
contendo, como íon principal, o metanosulfonato de césio (CsMSO4). Com ela, separamos os
sIPSCs e sEPSCs de acordo com seus respectivos potenciais de reversão. Nessa solução
obtivemos um potencial de reversão calculado para o cloreto de –52 mV, descontando um
potencial de junção estimado em 14 mV (estimado usando-se a mobilidade do gluconato), um
valor próximo ao observado experimentalmente, de -47 mV.
Para registrarmos os sEPSCs, mantivemos o neurônio no potencial de reversão observado
para o cloreto (-47 mV) (Figura 4.8Ai). Para registrarmos os sIPSCs, mantivemos o neurônios a -
20 mV onde os sIPSCs apresentavam polaridade positiva e sEPSCs polaridade negativa (Figura
4.8Aii). Inicialmente, observamos que as freqüências dos sIPSCs e dos sEPSCs foram pouco
alteradas após a aplicação de droga (Figura 4.6B). Porém, observamos uma redução não
significativa da freqüência dos sIPSCs em -20mV comparados com os valores a -70mV (Tabela
4.2). O mesmo não foi observado com os sEPSCs nos diferentes potenciais. Com isso, concluímos
que a interação entre as neurotransmissões (estimulação da neurotransmissão GABAérgica pela à
glutamatérgica), se existir, deve ser pequena.
Tabela 2: Freqüências e amplitudes dos sIPSCs e sEPSCs registrados em diferentes potenciais antes e após a aplicação
de DNQX (sIPSCs) ou bicuculina (sEPSCs). Resultados mostrados como média ± EPM. Testes t; p>0,05.
Grupo (n) Freqüência (Hz)
sIPSCs -70 mV (n = 43) 3,2 ± 1,9
sIPSCs -20 mV (n = 21) 2,0 ± 0,3
sIPSCs -20 mV [DNQX] (n = 21) 2,1 ± 0,4
sEPSCs -70 mV (n = 27) 0,5 ± 0,4
sEPSCs -47 mV (n = 3) 0,5 ± 0,1
sEPSCs -47 mV [Bicuculina] (n = 3) 0,6 ± 0,2

51
A
-47 mV
ii
-20 mV
i
Figura 4.8: Correntes registradas com uma solução interna de baixo cloreto (metanosulfonato de césio). Em
A, (i) registro a -47 mV, onde a corrente glutamatérgica é observada e a GABAérgica encontra-se no seu ponto de
reversão e (ii) a -20 mV, onde as correntes GABAérgicas (correntes de saída) são observadas junto às glutamatérgicas
(correntes de entrada) num mesmo registro. Em B, (i) freqüência dos sIPSCs num potencial de -20mV antes e após a
aplicação de DNQX 20 µM e (ii) freqüência dos sEPSCs num potencial de -47mV antes e após a aplicação de
bicuculina. Barras expressam médias ± EPM. ANOVA de duas vias. P>0,05.
B

52
Como é sabido que sinalização retrógrada por neurotransmissores liberados por
despolarização neuronal inibem a neurotransmissão GABAérgica quando o neurônio pós-
sináptico é despolarizado (Kreitzer & Regehr, 2001; Ohno-Shosaku et al., 2001; Wilson & Nicoll,
2001), decidimos, estudar porque a -20mV a freqüência observada dos sIPSCs era menor do que a
observada a -70mV.
Assim, testamos se o potencial do neurônio registrado teria influência na freqüência dos
sIPSCs. Para tal, registramos os sIPSCs (DNQX presente) em dois potencias simétricos (-50mV e
+50mV – Figura 4.9A). Observamos que a freqüência dos sIPSCs foi significativamente menor no
potencial positivo do que no potencial negativo (-50 mV = 2,1 ± 0,4 Hz; +50mV = 1,1 ± 0,3 Hz; n
= 6; p<0,05) (Figura 4.9Bi), enquanto que a amplitude não foi alterada (-50 mV = 36 ± 5,7 e +50
mV = 33,2 ± 7,8; p>0,05). Com isso, concluímos que o potencial da membrana do neurônio pós-
sináptico inibe freqüência dos sIPSCs no NCM possivelmente por um mecanismo de sinalização
retrógrada semelhante ao observado no hipocampo e cerebelo de roedores.

53
Figura 4.9: Diferença na freqüência
dos sIPSCs em potenciais de
membrana com mesma força
eletromotriz e sentidos opostos. Em A,
exemplos dos registros feitos a -50 mV e
a +50 mV em um dos neurônios do
NCM. As setas indicam em A os sIPSCs
observados a +50 mV. B, valores de
freqüência (i) e amplitude (ii) nos
potenciais de membrana de -50 mV e
+50 mV. Barras expressam médias ±
EPM. n = 6 (4 machos e 2 fêmeas).
Teste t; *=p<0,05.
B
ii
i
(6)
(6)
(6) (6)
500 ms 50 p
A
-50 mV
+50 mV
A ↓ ↓↓ ↓

54
4.3 – Efeito da estimulação canora na neurotransmissão no NCM
O principal interesse em nossa investigação foi saber se a apresentação do canto pode
resultar em alterações nas propriedades da neurotransmissão GABAérgica e glutamatérgica no
NCM. Dessa forma, registramos os sPSCs do NCM dos mandarins em duas situações de estímulo:
pássaro em situação controle (isolado e sem qualquer estímulo auditivo apresentado) e pássaro
estimulado (quando lhe foi apresentado um canto espécie-específico), sacrificados no tempo
correspondente ao pico da expressão do ZENK. (Mello & Ribeiro, 1998).
Observamos que os valores de freqüência dos sPSCs no grupo controle foi menor, mas
não significativo (p>0,05), do que o observado no estimulado (3,8 ± 0,4 Hz (n = 38) e 4,5 ± 0,6
Hz (n = 29) respectivamente; p>0,05) (Figura 4.10A). As amplitudes dos sPSCs do grupo controle
também não diferiram significativamente das observadas no grupo estimulado (54,3 ± 5,7 pA e
48,1 ± 3,5 pA respectivamente; p>0,05) (Figura 4.10B).

55
(38)
Figura 4.10: Freqüências e amplitudes de sPSCs do NCM de pássaros controle e estimulados registradas a
-70mV. Em A, freqüências de sPSCs mostradas individualmente (pontos negros) e como médias (barras). Em B,
amplitudes também mostradas individualmente (pontos negros) sobrepostos às barras. Barras expressam médias ±
EPM. Teste T, p>0,05.
Controle Estimuladas 0
5
10
15
Freq
üênc
ia (H
z)
A
(38)
(29)
Controle Estimuladas 0
50
100
150
200
Am
plitu
de (p
A)
B
(29)
(38)

56
4.3.1 Segregação sexual dos pássaros controle e estimulados
O estímulo sonoro espécie-específico não produz efeito significativo na freqüência ou
amplitude dos sPSCs no NCM, porém, o canto não é o único fator que nós levamos em
consideração. Estudos recentes evidenciaram que o NCM pode sofrer influência do sexo do
animal (Pinaud et al., 2006; Tersptra et al., 2004, 2006) e, como o NCM é bem desenvolvido e
presente em pássaros de ambos os sexos, (diferentemente dos núcleos envolvidos com a produção
do canto, desenvolvidos apenas nos machos (Nottebohm & Arnold, 1976; MacDougall-
Shackleton & Ball, 1999)), segregamos os resultados de acordo com estímulo (controle e
estimulados) e sexo do animal para observar se o NCM de machos e fêmeas responde de forma
diferente ao canto.
Neste novo arranjo dos dados, notamos uma maior freqüência dos sPSCs nos machos
estimulados (MEs) (5,9 ± 1,0 Hz, n = 14; p<0,05), em relação aos demais grupos (fêmeas
estimuladas (FEs): 3,2 ± 0,4 Hz, n = 15; machos controle (MCs): 3,9 ± 0,5 Hz, n = 15; fêmeas
controle (FCs): 3,7 ± 0,5 Hz, n = 23) (Figura 4.11). A ANOVA de duas vias mostrou que o
“gênero” tem efeito significativo sobre a freqüência dos sPSCs (p=0,02) ao contrário da
“condição” (controle versus estimulado; p=0,2) e que existe uma interação entre ambas variáveis
(p=0,04). Um pós-teste de Bonferroni considerou significativa a diferença entre as freqüências de
machos e fêmeas do grupo estimulado.
Os valores de amplitude não apresentaram diferenças significativas entre os grupos (MCs
= 47,4 ± 5,3 pA, n = 15; FCs = 48,8 ± 7,2 pA, n = 3; MEs = 51,7 ± 5,4 pA, n = 14; FEs = 46,2 ±
4,0 pA, n = 15; p>0,05) (Figura 4.9). Esses dados mostram que o sexo do pássaro é um fator que
influencia a resposta do NCM ao canto.

57
Figura 4.11: Segregação por estímulo e sexo dos valores de freqüência e amplitude dos sPSCs do NCM de
mandarins. Em A, freqüência do sPSCs e em B suas respectivas amplitudes. Barras expressam médias ± EPM.
ANOVA de duas vias. ** indica o fator (sexo - machos) que mais tem efeito na comparação, com P= 0,02; *=p<0,05.
Pós-teste de Bonferroni.
(14)
(15)
(15) (23)
(14) (15)
(15) (23)
58
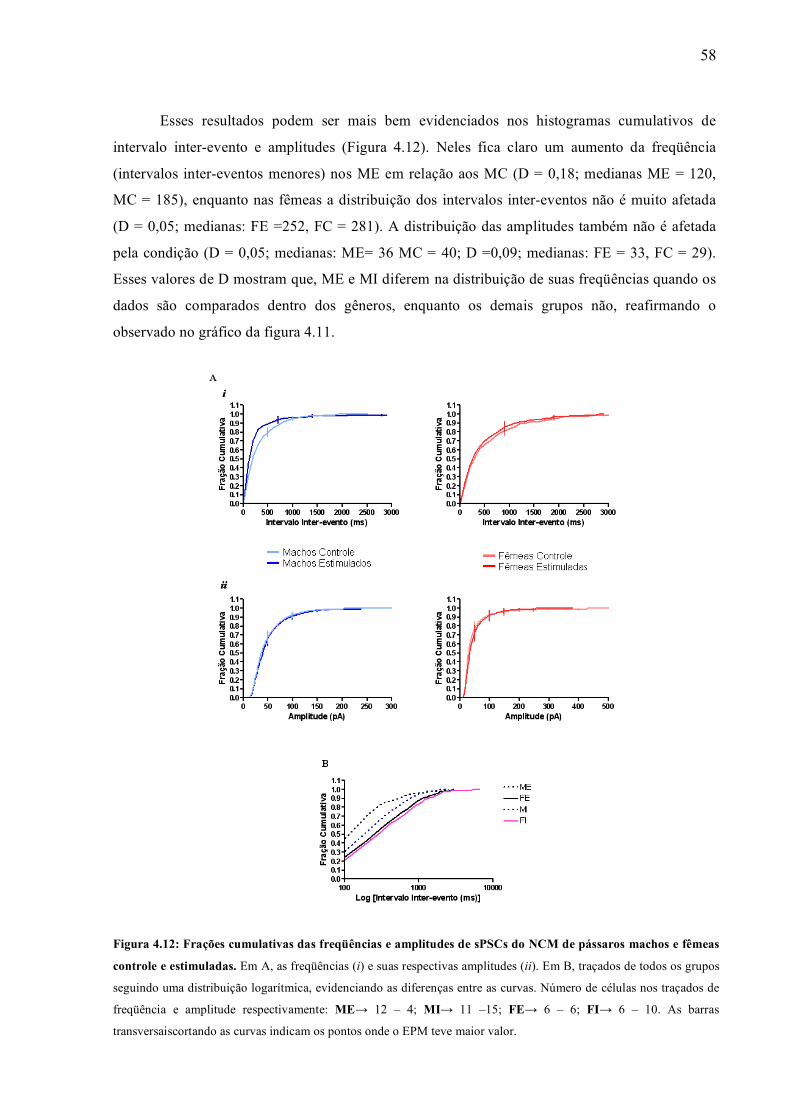
Esses resultados podem ser mais bem evidenciados nos histogramas cumulativos de
intervalo inter-evento e amplitudes (Figura 4.12). Neles fica claro um aumento da freqüência
(intervalos inter-eventos menores) nos ME em relação aos MC (D = 0,18; medianas ME = 120,
MC = 185), enquanto nas fêmeas a distribuição dos intervalos inter-eventos não é muito afetada
(D = 0,05; medianas: FE =252, FC = 281). A distribuição das amplitudes também não é afetada
pela condição (D = 0,05; medianas: ME= 36 MC = 40; D =0,09; medianas: FE = 33, FC = 29).
Esses valores de D mostram que, ME e MI diferem na distribuição de suas freqüências quando os
dados são comparados dentro dos gêneros, enquanto os demais grupos não, reafirmando o
observado no gráfico da figura 4.11.
Figura 4.12: Frações cumulativas das freqüências e amplitudes de sPSCs do NCM de pássaros machos e fêmeas
controle e estimuladas. Em A, as freqüências (i) e suas respectivas amplitudes (ii). Em B, traçados de todos os grupos
seguindo uma distribuição logarítmica, evidenciando as diferenças entre as curvas. Número de células nos traçados de
freqüência e amplitude respectivamente: ME→ 12 – 4; MI→ 11 –15; FE→ 6 – 6; FI→ 6 – 10. As barras
transversaiscortando as curvas indicam os pontos onde o EPM teve maior valor.

59
4.3.2 – Isolamento da transmissão GABAérgica pelo DNQX em pássaros controle e estimulados.
Como a neurotransmissão no NCM é composta por correntes geradas por GABA e
glutamato, fizemos um estudo mais detalhado do que o apresentado na figura 4.3 para observar
seus comportamentos e suas participações individuais nas diferentes condições em ambos os
sexos.
Ao isolarmos as correntes sinápticas GABAérgicas pela aplicação de DNQX, observamos
uma inibição significativa da freqüência dos sPSCs no NCM dos ME (p<0,05), enquanto que nos
outros grupos, a freqüência dos sPSCs dos pássaros não foi significativamente afetada (p>0,05)
(Tabela 4.3). Resultado similar foi observado com as amplitudes, que foram significativamente
inibidas nos ME (p<0,05), mas pouco alteradas nos demais grupos (p>0,05) (Tabela 4.3).
Analisando as freqüências e amplitudes das correntes GABAérgicas isoladas, não
observamos diferença significativa desses parâmetros entre os diversos grupos (ANOVA de duas
vias, p>0,05 – Figura 4.13). Concluímos, com esses experimentos, que o aumento na freqüência
dos sPSCs nos machos estimulados não se deve a um aumento da freqüência dos sIPSCs
GABAérgicos, mas provavelmente a um aumento da excitação glutamatérgica.
Grupo (n) Freqüência (Hz) Amplitude (pA)
Sem Droga DNQX 20µM Sem Droga DNQX 20µM
ME (11) 6,4 ± 1,2 3,9 ± 0,6 * 55,8 ± 6,5 38,1 ± 4 **
MC (11) 4,1 ± 0,6 3,9 ± 0,6 46,3 ± 5,7 43,7 ± 6,2
FE (8) 3,7 ± 1,2 2,8 ± 0,7 53,1 ± 7,9 37,9 ± 4,1
FC (12) 3,8 ± 0,8 3,2 ± 0,2 51,2 ± 10,8 39,1 ± 4,9
Tabela 4.3: Valores de freqüência e amplitude de neurônios de NCM de machos e fêmeas antes e depois da
aplicação de DNQX 20µM. ** = P<0,01; * = P<0,05; teste t pareado.

60
Figura 4.13: Fração GABAérgica dos sPSCs no NCM. Freqüências e amplitudes dos sIPSCs em neurônios do
NCM de machos e fêmeas controle ou estimulados, isolados pela aplicação de DNQX 20 µM ao banho. Barras
expressam médias ± EPM. ANOVA de duas vias; p>0,05.
Controle

61
4.3.3 - Isolamento da transmissão Glutamatérgica.
Em vista da maior fração das correntes sinápticas inibida por DNQX no NCM dos ME,
seria de se esperar que após a aplicação de bicuculina no NCM desses pássaros, observássemos
um maior número de correntes glutamatérgicas (sEPSCs) em relação aos outros grupos.
A bicuculina inibiu fortemente a freqüência e a amplitude das correntes sinápticas no
NCM de todos os grupos de pássaros (Tabela 4.4). Entretanto, após a aplicação de bicuculina, as
freqüências dos sEPSCs foram todas similares. Em experimentos onde medimos as correntes
glutamatérgicas usando uma solução de baixo cloreto e mantivemos o neurônio em um potencial
próximo ao de reversão do cloreto (-47 mV), a freqüência dos sEPSCs nos machos estimulados
(0,63 ± 0,16 Hz; n = 3) foi similar ao observado com CsCl após aplicação de bicuculina (Tabela
4.4). Isso mostra que não é a freqüência dos ePSCs glutamatérgicos a responsável direta pela
diferença observada.
A amplitude foi, entretanto, significativamente menor nos pássaros estimulados em
relação aos pássaros isolados (efeito observado em ambos os sexos) (p<0,05, ANOVA de duas
vias – Figura 4.12), enquanto a freqüência não mostrou essa redução significativa (P>0,05,
ANOVA de duas vias – Figura 4.14).
Grupo (n) Freqüência (Hz) Amplitude (pA)
Sem Droga Bicuculina Sem Droga Bicuculina
ME (4) 3,4 ± 1,5 0,3 ± 0,1 44,2 ± 11,4 21,5 ± 2,7
MC (6) 4 ± 0,8 0,5 ± 0,1** 64,3 ± 14 30,3 ± 3,8*
FE (6) 3,1 ± 0,5 0,6 ± 0,2** 38,5 ± 3,8 18 ± 1,6**
FC (11) 3,3 ± 0,7 0,6 ± 0,1* 60,5 ± 15,4 24 ± 3*
Tabela 4: Freqüências e amplitudes das correntes sinápticas no NCM de mandarins machos e fêmeas antes da
aplicação de bicuculina. (sPSCs - “Sem Droga”) e após a aplicação (“Bicuculina”), isolados ou estimulados. Teste t
pareado. *=p<0.05; **p<0,01.

62
Figura 4.14: Valores de freqüência e amplitude dos sEPSCs. Os valores apresentados dizem respeito apenas aos
registros obtidos em presença da bicuculina de machos e fêmeas controle e estimulados. Em A, as freqüências e em B
as amplitudes. Barras expressam médias ± EPM. ANOVA de duas vias; **=p<0,05.
A
B

63
4.3.4- Excitabilidade aumentada dos neurônios glutamatérgicos
No decorrer dos experimentos, a aplicação de bicuculina eventualmente gerava fenômenos
distintos dos sEPSCs. Esses fenômenos eram bursts de eventos glutamatérgicos de grande
amplitude e duração (figura 4.15A) como descrito anteriormente por Pinaud et al (2004). Estes
bursts apareceram com maior freqüência dependendo do sexo e do protocolo de estimulação
aplicado ao animal, sendo os ME mais suscetíveis ao aparecimento destes (9 em 11 células) e as
FC, menos (nenhum registro em 7 células) (figura 4.15D).
Esses fenômenos, ao contrario dos sEPSCs, são grandes o suficiente para deflagrar o
disparo de potenciais de ação no neurônio pós-sináptico quando registrado com uma solução
interna de pipeta contendo potássio fisiológico (no caso, gluconato de potássio [KGlu]) (Figura
4.15B).
Em 8 neurônios provenientes de machos estimulados, estes bursts glutamatérgicos foram
observados mesmo na ausência de bicuculina, isso pode ser visto em um registro feito com
solução interna de pipeta contendo CsMSO4, onde o burst ocorre ao mesmo tempo em que sIPSCs
são vistos como deflexões da corrente para cima (Figura 4.15C). Em ambos os casos (com ou sem
bicuculina), a aplicação de DNQX eliminou o aparecimento dos bursts, mostrando sua origem
glutamatérgica (não mostrado).

64
C
Figura 4.15: Bursts glutamatérgicos em
pássaros estimulados e controles. Em A,
exemplo de bursts registrados na presença de
bicuculina a -70 mV. Em B, potenciais de ação
(4) sendo disparados no pico de um dos bursts
registrados com KGlu em Current Clamp. Em
C, exemplo de burst ocorrido a -20 mV sem a
aplicação de bicuculina, onde podemos ver o
burst propriamente dito juntamente com
sIPSCs (deflexões para cima). Em D, a
freqüência de aparecimento dos bursts nos
diferentes protocolos. Notar a escala das barras
de escala para noção de amplitude e duração.
A i ii
B
D
C
0
2 5
5 0
7 5
1 0 0 9 / 1 1
3 / 1 0
7 / 1 4
0 / 7
% d
e cé
lula
s
ME
FC
MC
FE

65
4.4- Neurotransmissão no NCM de fêmeas submetidas a um isolamento prolongado do estímulo canoro. Machos estimulados
Todos os pássaros foram mantidos em um mesmo recinto dentro do biotério setorial antes
de serem isolados no dia anterior ao experimento. Com isso, eles ouviram constantemente cantos
dos machos presentes nas gaiolas. Para descobrir se essa exposição crônica ao canto estava ou não
mascarando quaisquer resultados, mantivemos um grupo de fêmeas sem a presença dos machos
por pelo menos uma semana no biotério setorial antes do registro dos sPSCs.
Estas fêmeas prolongadamente isoladas mostraram um aumento significativo da
freqüência dos sPSCs tanto dos pássaros controle (5,4 ± 1,4 Hz; n = 10) quanto dos estimulados
(5,9 ± 0,9 Hz; n = 7; p<0,05) em relação às fêmeas normais (controle = 3,6 ± 0,6 Hz; n = 23 e
estimuladas = 3,2 ± 0,4 Hz; n = 15), sendo que a variável “manutenção” (basais versus normais) é
apontada pela ANOVA de duas vias como o fator que mais influencia na diferença observada
(p=0,0075). As amplitudes não apresentaram diferenças significativas entre os dois protocolos de
manutenção dos pássaros (basais controle = 48,9 ± 0,9 pA; basais estimuladas = 58,3 ± 3,2 pA;
FCs = 48,9 ± 7,2 pA; FEs = 46,2 ± 4 pA; p>0,05 - Figura 4.16A).
Um fenômeno em particular que nos chamou atenção neste protocolo foi o fato de três
células de fêmeas isoladas apresentarem freqüências elevadas depois da aplicação de bicuculina,
como mostrado pelo registro da figura 4.17A e pelo gráfico da figura 4.17B, que mostra as
freqüências e amplitudes antes e após a aplicação da droga. As setas indicam os casos citados. A
half-width dessas correntes foi menor e similar à dos sEPSCs, sugerindo que essas correntes são
de fato glutamatérgicas (Figura 4.17C).
Mesmo nesses pássaros não observamos uma resposta mais pronunciada à canção,
descartando assim, algum efeito de longo prazo de habituação no NCM. Por outro lado, esse
isolamento alterou de alguma forma a fisiologia do núcleo, parecendo que a privação do canto
parece eliminar a conectividade da rede glutamatérgica do NCM, gerando uma atividade
glutamatérgica exacerbada ao invés dos bursts observados anteriormente.

66
Figura 4.16: Análise das correntes sinápticas das pássaros prolongadamente isoladas. Os gráficos representam
freqüência e amplitude dos sPSCs de fêmeas normais e prolongadamente isoladas quando estimuladas ou controle.
Barras expressam médias ± EPM. ANOVA de duas vias. * = P<0,05; **=p<0,01.

67
C
Figura 4.17: Fêmeas com isolamento prolongado: Em A, registros de sPSCs antes e sEPSCs após a aplicação de
bicuculina. Traçados em correspondem à célula do dia 11/12/2006. Em B, os valores individuais de freqüência e
amplitude antes e após a aplicação de bicuculina com as setas indicando os casos de alta freqüência após essa
aplicação. Em C, médias das half widths dos três casos isolados antes e após a aplicação de bicuculina.
A
B

68
5 - DISCUSSÃO

69
5.1 – Expressão da proteína ZENK em resposta ao canto.
Mello et al (1998) mostraram que a expressão de zenk ou de sua proteína no NCM ocorre
de maneira acentuada em resposta a canções espécie-específicas tanto em machos quanto em
fêmeas. Os resultados obtidos em nosso laboratório com a ICC do ZENK estão de acordo com os
descritos na literatura.
Os protocolos para a ICC foram desenvolvidos à medida que os experimentos de
eletrofisiologia foram realizados, pois as fatias que marcamos vinham dos mesmos animais
utilizados nos registros. Para normalizar as variações do processo de marcação com DAB,
mostramos na figura 4.1 apenas quatro espécimes de uma mesma reação (2 controle e 2
estimulado) para comparação. Os resultados dessa ICC mostram que os protocolos de isolamento
e estimulação são eficazes, exibindo uma marcação da proteína muito mais alta em animais
estimulado comparada àquela dos animais isolados. Tal é a diferença que, por área estimada,
alguns animais isolados apresentam 2 ou 4 células marcadas, enquanto animais estimulados
apresentaram contagens tão grandes quanto 997 células.
Uma preocupação nossa foi a indução da expressão de ZENK em resposta aos demais
protocolos aplicados ao tecido nervoso do animal, como corte, manipulação e tempo de espera na
solução de corte antes do registro. Porém, nenhum desses itens foi estímulo para mudança da
expressão da proteína.
5.2 – Caracterização das correntes sináptica espontâneas (sPSCs) registradas s no NCM.
Quando fizemos registros em voltage clamp dos neurônios do NCM, observamos sPSCs
com facilidade. Isolando farmacologicamente os neurotransmissores responsáveis pelos sPSCs,
vimos que dentro do NCM, apenas GABA e glutamato dominam a neurotransmissão espontânea
presente em nossos registros e que, em termos de porcentagem, a contribuição das correntes
GABAérgicas é muito maior do que a das glutamatérgicas. Os potenciais de reversão por sua vez,
quando da utilização da solução interna com baixo cloreto, reverteram de acordo com o esperado,
com a corrente GABAérgica em aproximadamente -47 mV, e a glutamatérgica em 0 mV .
Curiosamente, o RNAm dos receptores glicinérgicos GlyRα2 e GlyRβ foram
identificados no NCM de mandarins (Mello et al., 2007). Sua ação foi observada pela injeção
unilateral de estriquinina no NCM de mandarins (antagonista dos receptores glicinérgicos), o que
aumentou a atividade espontânea de bursting e o tamanho da resposta fásica ao canto registrada

70
no lado injetado. Já a microinjeção de glicina, como seria de se esperar, teve efeito oposto no
NCM. Entretanto em nossos experimentos não observamos nenhuma corrente que não fosse
completamente antagonizada pela aplicação concomitante de bicuculina e DNQX. É possível que
esses receptores glicinérgicos sejam extra-sinápticos, porém a origem da glicina que ativaria
fisiologicamente esses receptores ainda é obscura.
As correntes GABAérgica e glutamatérgica, quando analisadas através das correntes
miniatura, foram nitidamente diferentes uma da outra. Quanto à cinética das correntes, os
parâmetros utilizados para comparação se mostraram muito distintos. Os valores de amplitude,
half width e τ dos mIPSCs evidenciaram características de uma corrente mais lenta (em termos de
decaimento) do que a corrente deflagrada pelos mEPSCs, da mesma maneira que normalmente se
observa em preparações de mamíferos.
Ao contrário da relação entre ePSCs e iPSCs, os mEPSCs foram mais abundantes no
NCM do que os mIPSCs, sendo os valores de freqüência e amplitude dos mEPSCs muito
similares aos observados nos sEPSCs. Esta observação nos permite afirmar que os sEPSCs são, na
verdade, eventos quantais, que ocorrem sem a necessidade de um potencial de ação (PA). Já os
sIPSCs por sua vez, foram significativamente afetados pela TTX, evidência que prova que os
sIPSC são dependentes do disparo de potenciais de ação dos neurônios GABAérgicos para
ocorrerem. Outra característica relacionada aos neurônios GABAérgicos é a falta de efeito do
DNQX em inibir os sPSCs. Isso sugere que a excitação glutamatérgica não é necessária para a
atividade dos neurônios GABAérgicos (pelo menos nas condições basais). Experimentos
anteriores de nosso grupo em pássaros retirados diretamente do aviário obtiveram o mesmo
resultado.
Esses dados sugerem que a transmissão glutamatérgica no NCM está sob controle da
ativação dependente de estímulo das aferências (como do campo L), enquanto que a transmissão
GABAérgica possui uma atividade intrínseca independente (em condições basais) da estimulação
glutamatérgica. Observamos, entretanto, que em machos, principalmente após a estimulação
canora, a desinibição pela bicuculina produziu uma atividade de bursting glutamatérgico
espontâneo, o que sugere que os neurônios glutamatérgicos podem ter a capacidade de gerar
atividade espontânea e que devem estar fortemente conectados entre si (ver discussão posterior).
Para estudarmos isoladamente as transmissões glutamatérgica e GABAérgica sem a
necessidade de aplicação de antagonistas, fizemos registros das correntes sinápticas em solução de
baixo cloreto em diferentes potenciais. No caso, -47 mV para observar as correntes
glutamatérgicas (reversão do cloreto) e a -20mV para observar as correntes GABAérgicas (acima
da reversão do cloreto, produzindo correntes positivas). Os antagonistas bicuculina e DNQX

71
foram posteriormente aplicados para analisarmos seus efeitos sobre a neurotransmissão. Nessas
condições, bicuculina e DNQX tiveram o efeito esperado sobre as correntes GABAérgicas e
glutamatérgicas respectivamente, mas não afetaram as correntes remanescentes, confirmando que
o bloqueio de um tipo de corrente sináptica não afeta o comportamento do outro (em condições
basais).
As freqüências dos sEPSCs mantiveram-se inalteradas quando comparamos registros a -
70mV e -47mV na presença de bicuculina, porém, os valores de freqüência dos sIPSCs obtidos a -
20mV foram menores do que aqueles observados a -70 mV na presença de DNQX. Para
confirmar essa observação e saber se o potencial da membrana pós-sináptica dos neurônios do
NCM pode interferir com a liberação de GABA, realizamos experimentos usando solução interna
de CsCl em potenciais de membrana simétricos (-50 mV e 50 mV) e registramos os sIPSCs na
presença de DNQX. Nessas condições, o módulo da força eletromotriz para o cloreto é igual, mas
de polaridade oposta, para os dois potenciais de membrana. A partir desse protocolo, observamos
que os sIPSCs foram significativamente menos freqüentes a 50 mV do que a -50mV, sugerindo
uma retroinibição da liberação de GABA pelo neurônio pós-sináptico. Esse tipo de inibição
retrograda é associado à liberação de endocanabinóides pelos neurônios pós-sinápticos.
No sistema nervoso central são dois os tipos de endocanabinóides encontrados: a
anandamida (N-araquidonoiletanolamida) (Devane et al., 1992), que é sintetizada a partir de
lipídio da membrana pela N-acetiltransferase e fosfolipase D (Di Marzo et al., 1994; Sugiura et
al., 1996a; Sugiura et al., 1996b) e o 2AG (2-araquidonoiglicerol) (Mechoulam et al., 1995;
Sugiura et al., 1995), o qual é presente numa quantidade maior que a anandamida no cérebro e é
sintetizado também a partir de lipídios da membrana através da fosfolipase C juntamente com a
diacilglicerol lipase (Freund et al., 2003).
Quando sua via de produção é ativada, os endocanabinóides são sintetizados pelos
neurônios pós-sinápticos e se difundem retrogradamente até o neurônio pré-sináptico, onde se
ligam aos receptores CB1 (Matsuda et al., 1990) que são acoplados à proteína Gi/o
(Hashimotodani et al., 2007), inibindo correntes de cálcio pré-sinápticas (Kushmerick et al., 2004)
e conseqüentemente a liberação de neurotransmissores pelo terminal pré-sináptico (Kreitzer &
Regehr, 2001; Ohno-Shosaku et al., 2001; Wilson & Nicoll, 2001). Essa supressão da liberação de
neurotransmissores devido à influência do neurônio pós-sináptico foi primeiramente evidenciada
pelo fenômeno conhecido como “supressão da inibição por despolarização” (depolarization
supression of inhibition – DSI), descrito por (Llano et al., 1991) no cerebelo e em seguida por
(Pitler & Alger, 1992) no hipocampo. O fenômeno consiste na redução da freqüência de sIPSCs
registrada logo após a aplicação de um pulso despolarizante, desencadeado pelo influxo de cálcio

72
nos terminais pós-sinápticos (Pitler & Alger, 1992). O aumento na concentração de cálcio
citoplasmático inicia, por uma via ainda desconhecida (Hashimotodani et al., 2007), a produção
de endocanabinóides, que se difundem retrogradamente pela fenda sináptica e agem sobre seus
receptores, reduzindo transitoriamente a liberação de neurotransmissores. Prova disso é a
supressão do fenômeno pela aplicação de antagonistas CB1 (AM281 e SR141716A) em neurônios
de hipocampo em cultura (Ohno-Shosaku et al., 2001) ou fatia (Wilson & Nicoll, 2001).
Da mesma forma que o DSI ocorre em sinapses inibitórias, um fenômeno similar ocorre
em sinapses excitatórias, a supressão da excitação por despolarização (depolarization supression
of excitation – DSE), descrito por Kreitzer e Regehr (2001) em células de Purkinje no cerebelo.
Sua indução, assim como a do DSI, é mediada por endocanabinóides e é suprimida na presença de
antagonista CB1.
Além da via ativada pela entrada de cálcio no termina pós-sináptico, a ativação dos
receptores metabotrópicos de glutamato 1 e 5 (mGluR1 e mGluR5) também pode estimular a
síntese de endocanabinóides. Estes receptores são acoplados à proteína Gq e, quando ativados,
estimulam as fosfolipases Cβ (PLCβ), que produzem diacilglicerol, o qual é convertido pela
diacilglicerol lipase em 2-AG que finalmente se difunde para a fenda sináptica e age sobre seu
receptor (revisto por Hashimotodani, 2007). Essa inibição retrógrada da transmissão sináptica tem
sido descrita em sinapses por todo o cérebro, sugerindo uma abrangência difusa desse mecanismo
de regulação sináptica (Kreitzer & Regehr, 2002; Wilson & Nicoll, 2002).
Baseado nisso, seria então plausível supor que o fenômeno da DSI poderia ocorrer
também no NCM. Porém, experimentos preliminares por nós iniciados tentando reproduzir o DSI
no NCM usando o mesmo protocolo aplicado no hipocampo por Pitler e Alger (1992) para a
indução do fenômeno, não produziram a inibição transitória típica do DSI. Possivelmente, no
NCM, uma despolarização mais prolongada seja necessária para promover esse fenômeno,
diferente das despolarizações de 2 segundos usadas para promover o DSI. Nosso grupo pretende
continuar a investigação deste fenômeno, especialmente porque receptores CB1 são encontrados
no NCM dos mandarins (Soderstrom & Johnson, 2000) e sua ativação inibe a expressão do zenk
(Whitney et al., 2003). Finalmente, deve-se salientar que esse efeito do potencial da membrana
pós-sináptica sobre a neurotransmissão GABAérgica gera problemas na interpretação dos
resultados registrados em potenciais despolarizados, fato que deve ser levado em conta.

73
5.3 – Efeito da estimulação canora na neurotransmissão no NCM
Em nossos experimentos, observamos que a apresentação do canto gerou 3 efeitos sobre a
neurotransmissão no NCM: (i) um aumento, dependente da ativação de receptores
glutamatérgicos, da freqüência dos sIPSCs GABAérgicos nos machos; (ii) um aumento na
probabilidade do aparecimento de bursts glutamatérgicos em resposta à desinibição por bicuculina
ou espontaneamente, em especial nos machos e (iii) uma diminuição na amplitude dos sEPSCs no
NCM de pássaros de ambos os sexos.
Como não foi observado um aumento na freqüência ou na amplitude dos sEPSCs no NCM
dos machos estimulados, acreditamos que o aumento da atividade de bursts glutamatérgicos
espontâneos pelo canto deva ser responsável por esse aumento na transmissão GABAérgica. Esses
bursts poderiam ativar tanto receptores sinápticos como extra-sinápticos por spill-over (Kullmann
& Asztely, 1998) nos neurônios GABAérgicos, potenciando o disparo desses. É interessante notar
que, olhando individualmente alguns registros de machos estimulados, observamos que o aumento
da freqüência dos sPSCs não foi homogêneo, mas localizados temporalmente (intervalos de pouco
segundos com alta freqüência de sPSCs durante o registro), o que foi atenuado pela aplicação de
DNQX. Portanto, o aumento da freqüência dos sIPSCs nos machos estimulados é provavelmente
uma conseqüência do aumento da neurotransmissão glutamatérgica em bursts.
Por outro lado, não podemos descartar a hipótese de que essas sinapses GABAérgicas
potenciadas sejam provenientes de neurônios de projeção de outros núcleos (como o CMM, o qual
também é responsável pelo processamento de informações complexas relacionadas ao canto e
possui comunicação recíproca com o NCM (Vates et al., 1996) ou o campo L2 (origem das
aferências primárias do NCM (Vates, Broome et al., 1996))), que estariam sendo ativadas por
esses neurônios glutamatérgicos os quais estariam, por sua vez, sendo ativados pelo canto.
Entretanto, como não se sabe se esses neurônios de projeção GABAérgicos realmente existem e
devido ao número grande de neurônios GABAérgicos no NCM (Pinaud et al., 2004),
consideramos essa hipótese menos plausível.
O aumento da neurotransmissão glutamatérgica em bursts foi, em nossa opinião, o efeito
mais marcante do canto sobre o NCM. Isso sugere veementemente que os neurônios
glutamatérgicos se encontram conectados formando redes locais ou globais de excitação neural,
sendo esse efeito curiosamente mais intenso nos machos do que nas fêmeas. Correntes de cálcio
de baixo limiar (tipo T) e correntes catiônicas de entrada do tipo Ih são associadas com atividade
auto-regenerativa e disparo em bursts em neurônios (McCormick, 2004) e, um aumento de sua
expressão ou uma regulação positiva dessas correntes pelo canto poderia ser o mecanismo de

74
geração da atividade em bursts. Exemplos de alterações da excitabilidade da membrana em
resposta à experiência já foram reportadas o sistema nervoso de vertebrados e invertebrados
(Zhang & Linden, 2003). Recentemente alterações na expressão da corrente catiônica retificadora
de entrada Ih foram observadas em conjunto com a indução de LTP e LTD em neurônios
hipocampais (Fan et al., 2005; Brager & Johnston, 2007), sendo que LTP induziu um decremento
da Ih e LTD um aumento. Esses resultados suportam nossa hipótese que a ativação de neurônios
do NCM pelo canto pode alterar a excitabilidade neuronal.
O fato de o canto desencadear o disparo de neurônios do NCM é um forte indicativo de
um fenômeno de plasticidade neural que deve ter implicações fisiológicas nesse núcleo. Esses
neurônios após serem ativados pelo canto, teriam então uma excitabilidade aumentada, facilitando
o disparo de PAs apenas em resposta ao canto que estimula a via potenciada específica em
detrimento a qualquer outra.
Chamou nossa atenção que em 3 neurônios do NCM de fêmeas isoladas prolongadamente
dos machos, a aplicação de bicuculina ao invés de produzir o aparecimento de bursts
glutamatérgicos induziu um aumento da freqüência dos sEPSCs, fenômeno que nunca observamos
em nenhum outro protocolo, seja no NCM das fêmeas mantidas com machos ou no NCM dos
próprios machos. Esse dado sugere que nessas fêmeas a falta do estímulo canoro desorganizou a
rede glutamatérgica no NCM. Ou que possíveis alterações nos níveis de estrógeno devido ao
isolamento dos machos tenha alterado a rede glutamatérgica do NCM. Suportando essa hipótese
foi observado que em fêmeas pardais de garganta branca (Zonotrichia albicollis) a resposta à
canção no NCM (medida pela expressão de ZENK) só foi específica em fêmeas com níveis de
estrógeno suficientes para gerar resposta copulatória (Maney et al., 2006).
Uma alternativa à hipótese de um aumento da excitabilidade neuronal glutamatérgica seria
que os neurônios glutamatérgicos já tivessem propriedades de membrana que levassem a disparos
de potenciais de ação espontâneos e que em resposta ao canto e estes se tornassem mais
conectados, talvez por um fenômeno semelhante a potenciação em longo prazo (LTP), gerando
assim os bursts. Consideramos essa hipótese menos provável pelas seguintes razões: (i) os
sEPSCs registrados após a aplicação de bicuculina foram equivalentes às correntes miniatura; (ii)
não observamos um aumento da freqüência dos sEPSCs, o que poderia ser sugestivo do
aparecimento de sinapses antes “silenciosas” (Liao et al., 1995; Liao et al., 2001; Poncer &
Malinow, 2001) ou de LTP pré-sináptico (Dolphin et al., 1982; Malinow & Tsien, 1990) e (iii) a
amplitude dos sEPSCs foi de fato diminuída pelo canto, o contrário do que seria esperado no LTP
(Barrionuevo et al., 1986), sendo mais parecido a um fenômeno que pode ser relacionado à
depressão a longo prazo (LTD) do hipocampo (Lee et al., 1998).

75
Curiosamente, o NCM apresenta habituação à repetição do estímulo do canto, o que
contrasta com o que observamos. Isso pode significar que o efeito final do aumento da
excitabilidade glutamatérgica seja uma diminuição da atividade neuronal, talvez devido a
neurônios pós sinápticos com excitabilidade reduzida ou talvez por aumento da inibição
GABAérgica, como vimos nos machos estimulados. Como neurônios GABAérgicos são ativados
pelo canto (identificados pela expressão de ZENK; Pinaud et al., 2004) essa hipótese não é
improvável. É possível, dessa maneira, pensar que a habituação ocorra em neurônios inervados
pelos neurônios glutamatérgicos que foram potenciados pelo canto. Adicionalmente a inibição da
amplitude dos sEPSCs observada nas aves estimuladas poderia também contribuir com a
expressão da habituação.
5.4-Diferenças sexuais na resposta ao canto no NCM.
Ao contrário das vias envolvidas com a produção e o aprendizado do canto, as vias
auditivas telencefálicas são presentes em pássaros de ambos os sexos. Normalmente, assume-se
que não existam diferenças sexuais na fisiologia dessas áreas, o que torna nossos resultados, a
princípio, inusitados. No seu estudo pioneiro sobre a expressão do zenk no NCM em resposta à
canção, Mello et al, (1992) não encontraram nenhuma diferença quantitativa na expressão do zenk
no NCM de machos e fêmeas. Posteriormente Bailey e Wade (2005) não observaram diferença na
expressão de ZENK e FOS no NCM e CMM de mandarins machos e fêmeas juvenis (p45 -
correspondente à fase sensorimotora dos machos). A resposta eletrofisiológica à canção também
foi semelhante em mandarins de ambos os sexos (Chew et al., 1996b). Por outro lado, Pinaud et
al, (2004) encontraram um maior número de neurônios GABAérgicos que expressam calbidina no
NCM de mandarins machos do que no NCM das fêmeas. Além disso, Terpstra et al. (2006)
observaram que o NCM de mandarins fêmeas apresentava uma reposta menor à apresentação da
canção do macho tutor (a fêmea, apesar de não cantar, também precisa aprender a reconhecer o
canto da espécie por um tutor durante seu desenvolvimento) (Miller, 1979; Clayton & Dickinson,
1998) do que o CMM, enquanto nos machos essa preferência não foi observada (Terpstra et al.,
2004). Esses dados sugerem que o NCM não obstante não apresentar um claro dimorfismo sexual,
deve possuir diferenças sexuais mais sutis que influem em sua fisiologia.
Nossos dados mostram um aumento na excitabilidade glutamatérgica mais acentuada ao
canto no NCM de machos do que de fêmeas, sugerindo que o NCM dos machos seja mais
responsivo ao canto do que o das fêmeas. É interessante notar que o NCM de mandarins machos e
fêmeas expressa em sua porção caudal a enzima aromatase, que converte a testosterona em

76
estrógeno (Pinaud et al., 2004), sugerindo que o NCM receba influência dos níveis de hormônios
sexuais. Entretanto, é prematuro afirmar que essa diferença seja relacionada às diferenças na
resposta fisiológica ao canto no NCM antes que controles mais rigorosos sejam feitos para
sabermos se houve alguma auto-estimulação dos machos pela canção antes do sacrifício, já que
este ocorreu num intervalo de cerca de 10 minutos após o acender das luzes e também pelo fato de
que nos machos estimulados, a própria produção do canto durante a estimulação pode ter
aumentado o número de neurônios ativados no NCM em relação às fêmeas estimuladas.
5.5-Conclusões. Nossos experimentos são pioneiros na investigação da fisiologia sináptica do NCM de sua
plasticidade ao canto. Concluímos que o NCM aumenta a excitabilidade de sua rede
glutamatérgica em resposta a audição do canto específico do pássaro e que a amplitude dos
sEPSCs é diminuída pela estimulação canora. Esses fenômenos podem estar implicados na
memória auditiva do canto e podem ser um substrato fisiológico do fenômeno da habituação ao
canto observado no NCM.

77
6 – TABELAS

78
Tabela 6.1
aCSF
Soluto Concentração (mM)
NaCl 125
KCl 2,5
NaHCO3 25
NaHPO4 1,25
Glicose 25
Mio-Inositol 3
CaCl2 2
MgCl2 1
Piruvato de sódio 2
Ácido ascórbico 0,4
Tabela 6.2
aCSF modificado para corte
Soluto Molaridade
NaCl 87
KCl 2,5
NaHCO3 25
NaHPO4 1,25
Sacarose 75
Glicose 25
Mio-Inositol 3
CaCl2 0,2
MgCl2 7
Piruvato de sódio 2
Ácido ascórbico 0,4

79
Tabela 6.3
Solução interna de Cloreto de Césio
Soluto Concentração (mM)
CsCl 140
EGTA 5
HEPES 10
ATP-Mg 4
PCA-Na* 20
GTP 0,3
pH 7,2-7,2 com CsOH
310 mOsm
*Fosfocreatina - sal dissódico
Tabela 6.4
Solução interna de Metanosulfonato de Césio
Soluto Concentração (mM)
CsCMeSO4 130
KCl 10
EGTA 5
HEPES 10
ATP-Mg 4
PCA-Na* 20
GTP 0,3
pH 7,2-7,2 com CsOH
310 mOsm
*Fosfocreatina - sal dissódico

80
Tabela 6.5
Solução Interna de Gluconato de Potássio
Soluto Concentração (mM)
Gluconato de Potássio 130
EGTA 5
HEPES 10
KCl 20
ATP-Mg 2
GTP-Na 0,2
PCA-Na* 5
pH = 7,3 com KOH
310 mOsm
*Fosfocreatina - sal dissódico
Tabela 6.6
PBS
Soluto Concentração (mM)
NaCl 142
KCl 2,7
KH2PO4 1,5
NaHPO4 8,1
pH 7,4

81
Tabela 6.7
Núcleos do sistema do canto dos pássaros canoros. Sigla Nome
Av Núcleo Avalanche
B Núcleo Basorostral
CLM Mesopálio Caudolateral
CMM Mesopálio Caudomedial
CN Núcleo Coclear
DLM Núcleo Dorsolateral do Tálamo Medial
DM Núcleo Medial Dorsal
E Núcleo Entopalial
HVC Centro Vocal Superior
L (1,2,3) Campo “L” (porções 1, 2 e 3)
LArea X Porção Lateral da Área X do Estriado
LLD Lemnisco Lateral – núcleo dorsal
LLI Lemnisco Lateral – núcleo intermediário
LLV Lemnisco Lateral – núcleo ventral
LMAN Núcleo Magnocelular Lateral do Nidopálio Anterior
LMO Núcleo Oval do Mesopálio
MLd Núcleo Dorsolateral do Mesencéfalo
NCM Nidopálio caudomedial
Nif Núcleo Interfacial do Nidopálio
nXIIts Parte Traqueosiringeal do Núcleo do Hipoglosso
Ov Ovoidalis
PAm Paraambíguo
RA Núcleo Robusto do Arcopálio
RAm Retroambíguo
SO Oliva Superior
TSM Trato septomesencefálico
Uva Núcleo Uviforme

82
7 – BIBLIOGRAFIA

83
ABRAHAM, W. C., MASON, S. E., DEMMER, J., WILLIAMS, J. M., RICHARDSON, C.
L., TATE, W. P., LAWLOR, P. A. e DRAGUNOW, M. (1993). Correlations between immediate early gene induction and the persistence of long-term potentiation. Neuroscience 56(3): 717-27.
BAILEY, D. J. & WADE, J. (2003). Differential expression of the immediate early genes FOS and ZENK following auditory stimulation in the juvenile male and female zebra finch. Brain Res Mol Brain Res 116(1-2): 147-54.
BAILEY, D. J. & WADE, J. (2005). FOS and ZENK responses in 45-day-old zebra finches vary with auditory stimulus and brain region, but not sex. Behav Brain Res 162(1): 108-15.
BARRIONUEVO, G., KELSO, S. R., JOHNSTON, D. e BROWN, T. H. (1986). Conductance mechanism responsible for long-term potentiation in monosynaptic and isolated excitatory synaptic inputs to hippocampus. J Neurophysiol 55(3): 540-50.
BAUER, E. E., COLEMAN, M. J., ROBERTS, T. F., ROY, A., PRATHER, J. F. e MOONEY, R. (2008). A synaptic basis for auditory-vocal integration in the songbird. J Neurosci 28(6): 1509-22.
BOLHUIS, J. J., ZIJLSTRA, G. G., DEN BOER-VISSER, A. M. e VAN DER ZEE, E. A. (2000). Localized neuronal activation in the zebra finch brain is related to the strength of song learning. Proc Natl Acad Sci U S A 97(5): 2282-5.
BOTTJER, S. W., MIESNER, E. A. e ARNOLD, A. P. (1984). Forebrain lesions disrupt development but not maintenance of song in passerine birds. Science 224(4651): 901-3.
BOUGHMAN, J. W. (1998). Vocal learning by greater spear-nosed bats. Proc Biol Sci 265(1392): 227-33.
BRAGER, D. H. & JOHNSTON, D. (2007). Plasticity of intrinsic excitability during long-term depression is mediated through mGluR-dependent changes in I(h) in hippocampal CA1 pyramidal neurons. J Neurosci 27(51): 13926-37.
BRAINARD, M. S. & DOUPE, A. J. (2000). Interruption of a basal ganglia-forebrain circuit prevents plasticity of learned vocalizations. Nature 404(6779): 762-6.
BRENOWITZ, E. A. & BEECHER, M. D. (1995). Song learning in birds: diversity and plasticity, opportunities and challenges. TRENDS in Neurosciences 28(3): 127-132.
BRENOWITZ, E. A., MARGOLIASH, D. e NORDEEN, K. W. (1997). An introduction to birdsong and the avian song system. J Neurobiol 33(5): 495-500.
CATCHPOLE, C. K. (1983). Variation in the song of the great reed warbler Acrocephalus arundinaceus in relation to mate attraction and territorial defense. Animal Behaviour 31: 1217-1225.
CHEW, S. J., MELLO, C., NOTTEBOHM, F., JARVIS, E. e VICARIO, D. S. (1995). Decrements in auditory responses to a repeated conspecific song are long-lasting and require two periods of protein synthesis in the songbird forebrain. Proc Natl Acad Sci U S A 92(8): 3406-10.
CHEW, S. J., VICARIO, D. S. e NOTTEBOHM, F. (1996a). A large-capacity memory system that recognizes the calls and songs of individual birds. Proc Natl Acad Sci U S A 93(5): 1950-5.
CHEW, S. J., VICARIO, D. S. e NOTTEBOHM, F. (1996b). Quantal duration of auditory memories. Science 274(5294): 1909-14.
CHRISTY, B. & NATHANS, D. (1989). DNA binding site of the growth factor-inducible protein Zif268. Proc Natl Acad Sci U S A 86(22): 8737-41.

84
CHRISTY, B. A., LAU, L. F. e NATHANS, D. (1988). A gene activated in mouse 3T3 cells by serum growth factors encodes a protein with "zinc finger" sequences. Proc Natl Acad Sci U S A 85(21): 7857-61.
CLAYTON, N. S. & DICKINSON, A. (1998). Episodic-like memory during cache recovery by scrub jays. Nature 395(6699): 272-4.
COLE, A. J., SAFFEN, D. W., BARABAN, J. M. e WORLEY, P. F. (1989). Rapid increase of an immediate early gene messenger RNA in hippocampal neurons by synaptic NMDA receptor activation. Nature 340(6233): 474-6.
DENISENKO-NEHRBASS, N. I., JARVIS, E., SCHARFF, C., NOTTEBOHM, F. e MELLO, C. V. (2000). Site-specific retinoic acid production in the brain of adult songbirds. Neuron 27(2): 359-70.
DEVANE, W. A., HANUS, L., BREUER, A., PERTWEE, R. G., STEVENSON, L. A., GRIFFIN, G., GIBSON, D., MANDELBAUM, A., ETINGER, A. e MECHOULAM, R. (1992). Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 258(5090): 1946-9.
DI MARZO, V., FONTANA, A., CADAS, H., SCHINELLI, S., CIMINO, G., SCHWARTZ, J. C. e PIOMELLI, D. (1994). Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 372: 686-691.
DOLPHIN, A. C., ERRINGTON, M. L. e BLISS, T. V. (1982). Long-term potentiation of the perforant path in vivo is associated with increased glutamate release. Nature 297(5866): 496-8.
DOUPE, A. J. & KUHL, P. K. (1999). Birdsong and human speech: common themes and mechanisms. Annu Rev Neurosci 22: 567-631.
FAN, Y., FRICKER, D., BRAGER, D. H., CHEN, X., LU, H. C., CHITWOOD, R. A. e JOHNSTON, D. (2005). Activity-dependent decrease of excitability in rat hippocampal neurons through increases in I(h). Nat Neurosci 8(11): 1542-51.
FREUND, T. F., KATONA, I. e PIOMELLI, D. (2003). Role of endogenous cannabinoids in synaptic signaling. Physiol Rev 83(3): 1017-66.
GUINEE, L.N. & PAYNE, K.B. (1988). Rhyme-like repetitions in songs of Humpback Whales. Ethology 79: 295-306
HASHIMOTODANI, Y., OHNO-SHOSAKU, T. e KANO, M. (2007). Endocannabinoids and synaptic function in the CNS. Neuroscientist 13(2): 127-37.
HESSLER, N. A. & DOUPE, A. J. (1999). Social context modulates singing-related neural activity in the songbird forebrain. Nat Neurosci 2(3): 209-11.
HUNT, G. R. & GRAY, R. D. (2003). Diversification and cumulative evolution in New Caledonian crow tool manufacture. Proc Biol Sci 270(1517): 867-74.
JARVIS, E. D., GUNTURKUN, O., BRUCE, L., CSILLAG, A., KARTEN, H., KUENZEL, W., MEDINA, L., PAXINOS, G., PERKEL, D. J., SHIMIZU, T., STRIEDTER, G., WILD, J. M., BALL, G. F., DUGAS-FORD, J., DURAND, S. E., HOUGH, G. E., HUSBAND, S., KUBIKOVA, L., LEE, D. W., MELLO, C. V., POWERS, A., SIANG, C., SMULDERS, T. V., WADA, K., WHITE, S. A., YAMAMOTO, K., YU, J., REINER, A. e BUTLER, A. B. (2005). Avian brains and a new understanding of vertebrate brain evolution. Nat Rev Neurosci 6(2): 151-9.
JARVIS, E. D. & NOTTEBOHM, F. (1997). Motor-driven gene expression. Proc Natl Acad Sci U S A 94(8): 4097-102.
JARVIS, E. D., RIBEIRO, S., DA SILVA, M. L., VENTURA, D., VIELLIARD, J. e MELLO, C. V. (2000). Behaviourally driven gene expression reveals song nuclei in hummingbird brain. Nature 406(6796): 628-32.

85
JARVIS, E. D., SCHARFF, C., GROSSMAN, M. R., RAMOS, J. A. e NOTTEBOHM, F. (1998). For whom the bird sings: context-dependent gene expression. Neuron 21(4): 775-88.
JONES, M. W., ERRINGTON, M. L., FRENCH, P. J., FINE, A., BLISS, T. V., GAREL, S., CHARNAY, P., BOZON, B., LAROCHE, S. e DAVIS, S. (2001). A requirement for the immediate early gene Zif268 in the expression of late LTP and long-term memories. Nat Neurosci 4(3): 289-96.
KNUDSEN, E. I. (2002). Instructed learning in the auditory localization pathway of the barn owl. Nature 417(6886): 322-8.
KOBAYASHI, K., UNO, H. e OKANOYA, K. (2001). Partial lesions in the anterior forebrain pathway affect song production in adult Bengalese finches. Neuroreport 12(2): 353-8.
KREITZER, A. C. & REGEHR, W. G. (2001). Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. Neuron 29(3): 717-27.
KREITZER, A. C. & REGEHR, W. G. (2002). Retrograde signaling by endocannabinoids. Curr Opin Neurobiol 12(3): 324-30.
KULLMANN, D. M. & ASZTELY, F. (1998). Extrasynaptic glutamate spillover in the hippocampus: evidence and implications. Trends Neurosci 21(1): 8-14.
KUSHMERICK, C., PRICE, G. D., TASCHENBERGER, H., PUENTE, N., RENDEN, R., WADICHE, J. I., DUVOISIN, R. M., GRANDES, P. e VON GERSDORFF, H. (2004). Retroinhibition of presynaptic Ca2+ currents by endocannabinoids released via postsynaptic mGluR activation at a calyx synapse. J Neurosci 24(26): 5955-65.
LEE, H. K., KAMEYAMA, K., HUGANIR, R. L. e BEAR, M. F. (1998). NMDA induces long-term synaptic depression and dephosphorylation of the GluR1 subunit of AMPA receptors in hippocampus. Neuron 21(5): 1151-62.
LEMAIRE, P., REVELANT, O., BRAVO, R. e CHARNAY, P. (1988). Two mouse genes encoding potential transcription factors with identical DNA-binding domains are activated by growth factors in cultured cells. Proc Natl Acad Sci U S A 85(13): 4691-5.
LIAO, D., HESSLER, N. A. e MALINOW, R. (1995). Activation of postsynaptically silent synapses during pairing-induced LTP in CA1 region of hippocampal slice. Nature 375(6530): 400-4.
LIAO, D., SCANNEVIN, R. H. e HUGANIR, R. (2001). Activation of silent synapses by rapid activity-dependent synaptic recruitment of AMPA receptors. J Neurosci 21(16): 6008-17.
LLANO, I., LERESCHE, N. e MARTY, A. (1991). Calcium entry increases the sensitivity of cerebellar Purkinje cells to applied GABA and decreases inhibitory synaptic currents. Neuron 6(4): 565-74.
MACDOUGALL-SHACKLETON, S. A. & BALL, G. F. (1999). Comparative studies of sex differences in the song-control system of songbirds. Trends in neuroscience 22: 432-436.
MALINOW, R. & TSIEN, R. W. (1990). Presynaptic enhancement shown by whole-cell recordings of long-term potentiation in hippocampal slices. Nature 346(6280): 177-80.
MANEY, D. L., CHO, E. e GOODE, C. T. (2006). Estrogen-dependent selectivity of genomic responses to birdsong. Eur J Neurosci 23(6): 1523-9.
MARGOLIASH, D. (1983). Acoustic parameters underlying the responses of song-specific neurons in the white-crowned sparrow. J Neurosci 3(5): 1039-57.
MARLER, P. (1955). Characteristics of Some Animal Calls. Nature 176: 6-8. MARLER, P. (1989). Learning by Instinct: birdsong. American Speech-Language-Speech
Association 31: 75-79.

86
MATSUDA, L. A., LOLAIT, S. J., BROWNSTEIN, M. J., YOUNG, A. C. e BONNER, T. I. (1990). Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 346(6284): 561-4.
MECHOULAM, R., BEN-SHABAT, S., HANUS, L., LIGUMSKY, M., KAMINSKI, N. E., SCHATZ, A. R., GOPHER, A., ALMOG, S., MARTIN, B. R., COMPTON, D. R. e ET AL. (1995). Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol 50(1): 83-90.
MELLO, C., NOTTEBOHM, F. e CLAYTON, D. (1995). Repeated exposure to one song leads to a rapid and persistent decline in an immediate early gene's response to that song in zebra finch telencephalon. J Neurosci 15(10): 6919-25.
MELLO, C. V. & CLAYTON, D. F. (1994). Song-induced ZENK gene expression in auditory pathways of songbird brain and its relation to the song control system. J Neurosci 14(11 Pt 1): 6652-66.
MELLO, C. V., LOVELL, P. V., LU, K. e VICARIO, D. S. (2007). Glycinergic transmission in the avian auditory telencephalon. Society for Neuroscience Abstract. San Diego.
MELLO, C. V., PINAUD, R. e RIBEIRO, S. (1998). Noradrenergic system of the zebra finch brain: immunocytochemical study of dopamine-beta-hydroxylase. J Comp Neurol 400(2): 207-28.
MELLO, C. V. & RIBEIRO, S. (1998). ZENK protein regulation by song in the brain of songbirds. J Comp Neurol 393(4): 426-38.
MELLO, C. V., VICARIO, D. S. e CLAYTON, D. F. (1992). Song presentation induces gene expression in the songbird forebrain. Proc Natl Acad Sci U S A 89(15): 6818-22.
MILBRANDT, J. (1987). A nerve growth factor-induced gene encodes a possible transcriptional regulatory factor. Science 238(4828): 797-9.
MILLER, D. B. (1979). Long-term recognition of father's song by female zebra finches. Nature 280: 389-391.
NASTIUK, K. L., MELLO, C. V., GEORGE, J. M. e CLAYTON, D. F. (1994). Immediate-early gene responses in the avian song control system: cloning and expression analysis of the canary c-jun cDNA. Brain Res Mol Brain Res 27(2): 299-309.
NOTTEBOHM, F. (2000). The Design of Animal Communication. Cambridge, MIT Press. NOTTEBOHM, F. & ARNOLD, A. P. (1976). Sexual dimorphism in vocal control areas of
the songbird brain. Science 194(4261): 211-3. NOTTEBOHM, F., STOKES, T. M. e LEONARD, C. M. (1976). Central control of song in
the canary, Serinus canarius. J Comp Neurol 165(4): 457-86. OHNO-SHOSAKU, T., MAEJIMA, T. e KANO, M. (2001). Endogenous cannabinoids
mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron 29(3): 729-38.
PAVLETICH, N. P. & PABO, C. O. (1991). Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 A. Science 252(5007): 809-17.
PHAN, M. L., PYTTE, C. L. e VICARIO, D. S. (2006). Early auditory experience generates long-lasting memories that may subserve vocal learning in songbirds. Proc Natl Acad Sci U S A 103(4): 1088-93.
PINAUD, R., FORTES, A. F., LOVELL, P. e MELLO, C. V. (2006). Calbindin-positive neurons reveal a sexual dimorphism within the songbird analogue of the mammalian auditory cortex. J Neurobiol 66(2): 182-95.
PINAUD, R., VELHO, T. A., JEONG, J. K., TREMERE, L. A., LEAO, R. M., VON GERSDORFF, H. e MELLO, C. V. (2004). GABAergic neurons participate in the brain's response to birdsong auditory stimulation. Eur J Neurosci 20(5): 1318-30.
PITLER, T. A. & ALGER, B. E. (1992). Postsynaptic spike firing reduces synaptic GABAA responses in hippocampal pyramidal cells. J Neurosci 12(10): 4122-32.

87
POLLOK, B., PRIOR, H. e GUNTURKUN, O. (2000). Development of object permanence in food-storing magpies (Pica pica). J Comp Psychol 114(2): 148-57.
PONCER, J. C. & MALINOW, R. (2001). Postsynaptic conversion of silent synapses during LTP affects synaptic gain and transmission dynamics. Nat Neurosci 4(10): 989-96.
REINER, A. (2005). A new avian brain nomenclature: why, how and what. Brain Res Bull 66(4-6): 317-31.
REINER, A., PERKEL, D. J., MELLO, C. V. e JARVIS, E. D. (2004). Songbirds and the revised avian brain nomenclature. Ann N Y Acad Sci 1016: 77-108.
ROBERTS, L. A., HIGGINS, M. J., O'SHAUGHNESSY, C. T., STONE, T. W. e MORRIS, B. J. (1996). Changes in hippocampal gene expression associated with the induction of long-term potentiation. Brain Res Mol Brain Res 42(1): 123-7.
SCHARFF, C. & NOTTEBOHM, F. (1991). A comparative study of the behavioral deficits following lesions of various parts of the zebra finch song system: implications for vocal learning. J Neurosci 11(9): 2896-913.
SODERSTROM, K. & JOHNSON, F. (2000). CB1 cannabinoid receptor expression in brain regions associated with zebra finch song control. Brain Res 857(1-2): 151-7.
STRIPLING, R., KRUSE, A. A. e CLAYTON, D. F. (2001). Development of song responses in the zebra finch caudomedial neostriatum: role of genomic and electrophysiological activities. J Neurobiol 48(3): 163-80.
STRIPLING, R., VOLMAN, S. F. e CLAYTON, D. F. (1997). Response modulation in the zebra finch neostriatum: relationship to nuclear gene regulation. J Neurosci 17(10): 3883-93.
SUGIURA, T., KONDO, S., SUKAGAWA, A., NAKANE, S., SHINODA, A., ITOH, K., YAMASHITA, A. e WAKU, K. (1995). 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun 215(1): 89-97.
SUGIURA, T., KONDO, S., SUKAGAWA, A., TONEGAWA, T., NAKANE, S., YAMASHITA, A., ISHIMA, Y. e WAKU, K. (1996b). Transacylase-mediated and phosphodiesterase-mediated synthesis of N-arachidonoylethanolamine, an endogenous cannabinoid-receptor ligand, in rat brain microsomes. Comparison with synthesis from free arachidonic acid and ethanolamine. Eur J Biochem 240: 53-62.
SUGIURA, T., KONDO, S., SUKAGAWA, A., TONEGAWA, T., NAKANE, S., YAMASHITA, A. e WAKU, K. (1996a). Enzymatic synthesis of anandamide, an endogenous cannabinoid receptor ligand, through N-acylphosphatidylethanolamine pathway in testis: involvement of Ca(2� )-dependent transacylase and phosphodiesterase activities. Biochem Biophys Res Commun 218: 113-117.
SUKHATME, V. P., CAO, X. M., CHANG, L. C., TSAI-MORRIS, C. H., STAMENKOVICH, D., FERREIRA, P. C., COHEN, D. R., EDWARDS, S. A., SHOWS, T. B., CURRAN, T. e ET AL. (1988). A zinc finger-encoding gene coregulated with c-fos during growth and differentiation, and after cellular depolarization. Cell 53(1): 37-43.
SWIRNOFF, A. H. & MILBRANDT, J. (1995). DNA-binding specificity of NGFI-A and related zinc finger transcription factors. Mol Cell Biol 15(4): 2275-87.
TERPSTRA, N. J., BOLHUIS, J. J. e DEN BOER-VISSER, A. M. (2004). An analysis of the neural representation of birdsong memory. J Neurosci 24(21): 4971-7.
THORPE, W. H. (1958). The learning of song patterns by birds, with special reference to the song of the chaffinch, Fringilla coelebs. Ibis 100: 535-570.
VATES, G. E., BROOME, B. M., MELLO, C. V. e NOTTEBOHM, F. (1996). Auditory pathways of caudal telencephalon and their relation to the song system of adult male zebra finches. J Comp Neurol 366(4): 613-42.

88
VELHO, T. A., PINAUD, R., RODRIGUES, P. V. e MELLO, C. V. (2005). Co-induction of activity-dependent genes in songbirds. Eur J Neurosci 22(7): 1667-78.
VON FERSEN, L. & ELIUS, J. D. (1989). Long-term retention of many visual patterns by pigeons. Ethology 82: 141-155.
WHITNEY, O., SODERSTROM, K. e JOHNSON, F. (2003). CB1 cannabinoid receptor activation inhibits a neural correlate of song recognition in an auditory/perceptual region of the zebra finch telencephalon. J Neurobiol 56(3): 266-74.
WILLIAMS, H. (2004). Birdsong and singing behavior. Annals of the New York Academy Science 1016: 1-30.
WILLIAMS, H. & MEHTA, N. (1999). Changes in adult zebra finch song require a forebrain nucleus that is not necessary for song production. J Neurobiol 39(1): 14-28.
WILSON, R. I. & NICOLL, R. A. (2001). Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 410(6828): 588-92.
WILSON, R. I. & NICOLL, R. A. (2002). Endocannabinoid signaling in the brain. Science 296(5568): 678-82.
WISDEN, W., ERRINGTON, M. L., WILLIAMS, S., DUNNETT, S. B., WATERS, C., HITCHCOCK, D., EVAN, G., BLISS, T. V. e HUNT, S. P. (1990). Differential expression of immediate early genes in the hippocampus and spinal cord. Neuron 4(4): 603-14.
ZHANG, W. & LINDEN, D. J. (2003). The other side of the engram: experience-driven changes in neuronal intrinsic excitability. Nat Rev Neurosci 4(11): 885-900.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo