bioinorgânica do cobre uma diversidade de metaloenzimas de cobre ocorrem em plantas e animais. elas...
TRANSCRIPT

Bioinorgânica do Cobre Prof. Fernando R. Xavier
UDESC 2018

2
Cobre: Características gerais
Ao contrário do ferro, o cobre não estava biodisponível na Terra primitiva. Ao
considerarmos sua atmosfera redutora, os íons cobre permaneceram precipitados na
forma de CuS, um sulfeto bastante insolúvel.
Somente após a produção de O2 em massa pelas cianobactérias, os íons cobre(2+)
começaram a ser utilizados pelos sistemas vivos.

3
Enquanto enzimas que atuam em ambientes redutores trabalham em uma faixa de
potenciais eletroquímicos mais estreita, a presença de oxigênio criou a necessidade
de sistemas que atuassem em uma ampla faixa de potenciais que podem variar de
0 até 0,8 V.
Íons cobre são excelentes candidatos a assumirem este papel. Sendo assim é dito que
se os primeiros estágios da Terra primitiva foram a era bioquímica do ferro,
certamente o estágio seguinte foi a era bioquímica do cobre.
O cobre está presente em um grande número de enzimas, muitas delas envolvidas em
reações de transferência de elétrons, na ativação do dioxigênio e de outras
pequenas moléculas tais como: óxidos de nitrogênio, metano, monóxido de
carbono e ainda na dismutação do H2O2.

4
Os estados de oxidação mais comuns encontrados para íons cobre em sistemas
biológicos são 1+ e 2+. Assim como a forma reduzida do ferro, íons Fe(2+), íons
Cu(1+) podem gerar radicais livres (∙OH) via processo de Fenton.
Considerando outros metais de transição divalentes ad primeira série de transição, o
cobre é o que apresenta complexos mais estáveis sob um ponto de vista
termodinâmico.
Íons Cu(2+) são considerados duros enquanto íons Cu(1+) são macios. Para este
último, fica evidenciada por sua preferência por ligantes contendo enxofre. Ambas as
espécies são cineticamente lábeis.

5
Uma diversidade de metaloenzimas de cobre ocorrem em plantas e animais. Elas são
utilizadas em reações de transferência de elétrons (azurina, plastocianina, lacase),
para reações de oxigenação (tirosinase, ascorbato oxidase) e para transporte de
oxigênio (hemocianina).
Enzimas de cobre
Íons cobre são geralmente coordenados por ligantes enxofre ou nitrogenados.
Metaloenzimas de cobre podem conter íons Cu(1+) d10, Cu(2+) d9, e Cu(3+) d8.
Em termos de números de coordenação, íons Cu(1+) podem variar de 2, 3 ou 4,
enquanto íons Cu(2+) variam entre 4, 5 ou 6. Considerando o NC. 4, íons cobre
divalentes são quadrado-planos enquanto íons monovalentes são tetraédricos.
6
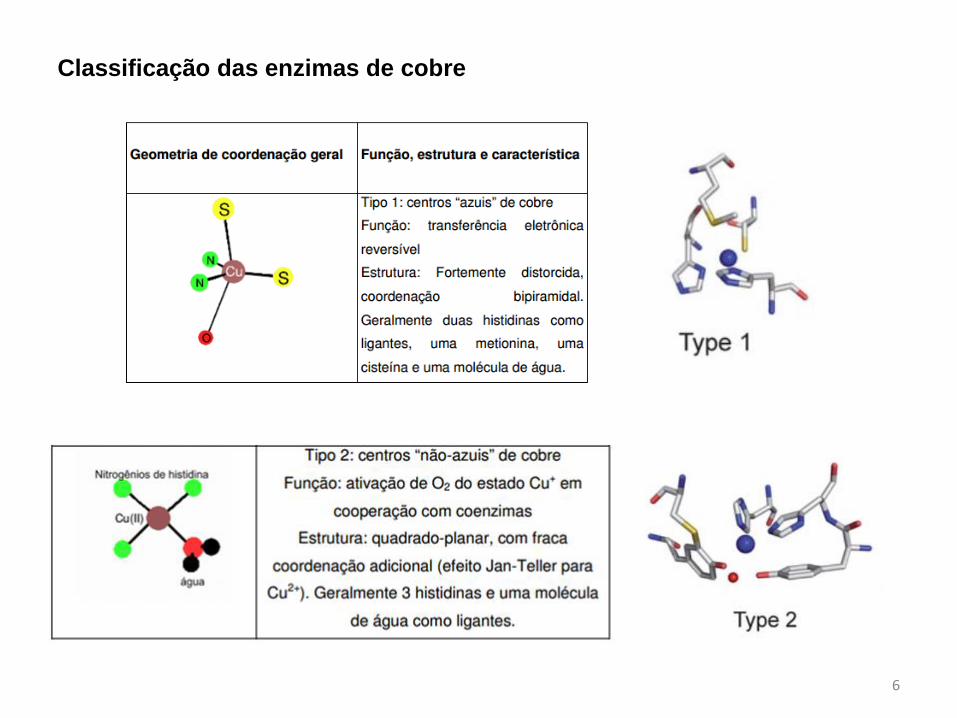
Classificação das enzimas de cobre

7
Mononuclear Binuclear Tetranuclear
Tipo Tipo 1 Tipo 2 Tipo 3 CuA CuZ
UV-Vis Forte ~600 nm e
(em algumas
proteínas)
450 nm
Fraca ~700 nm Fraca ~700 nm Forte ~480 e 530
nm
Forte ~640 nm
Ligantes
comuns
His, Cys His, Asp His His, Cys His, S2-
Geometria sítio
ativo
Tetraédrico
distorcido
Distorcido
tetragonalmente
Tetragonal Trigonal plana cluster
Exemplos Azurina SOD, GAO Catecol Oxidase,
Tirosinase
Citicromo c
oxidase
N2O redutase

8
Enzimas de cobre tipo I (Azuis)

9

10
Tem como função transportar elétrons durante o
processo de fotossíntese. Possui baixa massa molar
(~10,5 kDa) e está presente em plantas superiores.
Plastocianina (Tipo I)
Seu sítio ativo pode ser descrito como uma pirâmide
trigonal distorcida. O plano trigonal é composto por
02 histidinas e uma cisteína enquanto na posição
axial encontra-se uma metionina.
Sua coloração azul (597 nm) é originada de um
processo de TCLM Spπ → Cudx2-y
2.

11
Azurina (Tipo I)
Assim como a plastocianina, tem como função
transportar elétrons durante o processo de
fotossíntese. Possui baixa massa molar (~ 16 kDa)
e está presente em bactérias.
As azurinas estão presentes em processos de
desnitrificação promovidos por bactérias.
Seu sítio ativo pode ser descrito como uma pirâmide
trigonal distorcida. O plano trigonal é composto por
02 histidinas e uma cisteína enquanto na posição
axial encontra-se uma metionina e um grupo cabonil.
Apresenta uma banda de fluorescência intensa em 308 nm.

12
Enzimas de cobre na ativação e redução do O2

13
Enzimas de cobre tipo II (não-azuis)
Enzimas de cobre do tipo II são muito bem caracterizadas do ponto de vista
estrutural, entretanto, para que possam ativar o O2 estas necessitam passar pela
espécie Cu(3+) (improvável) para converter O2 em superóxido.
Para contornar esta possibilidade, atualmente duas soluções são propostas na
literatura:
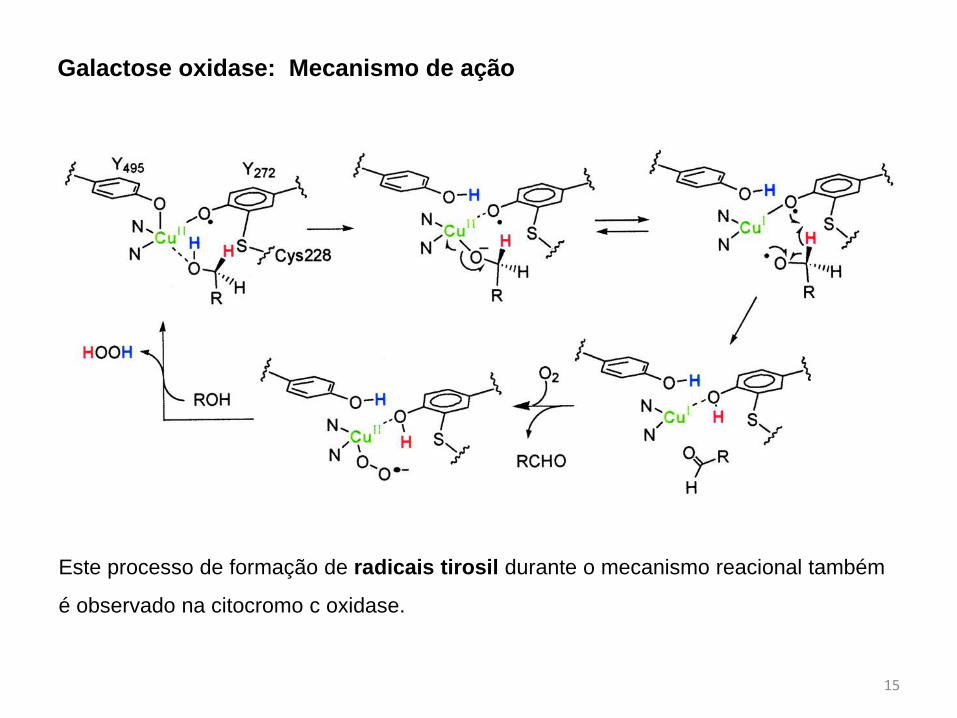
O primeiro caso é da Galactose oxidase (GAO), uma enzima que, na presença de
oxigênio, converte galactose no seu respectivo aldeído com concomitante formação de
H2O2.
Estudos apontam que ao invés de Cu(3+), a GAO é uma metaloenzima radicalar
onde foi detectado a presença da espécie tirosil–Cu(2+) como espécie ativa.

14
Galactose oxidase: Estrutura e sítio ativo
15
Galactose oxidase: Mecanismo de ação
Este processo de formação de radicais tirosil durante o mecanismo reacional também
é observado na citocromo c oxidase.
16
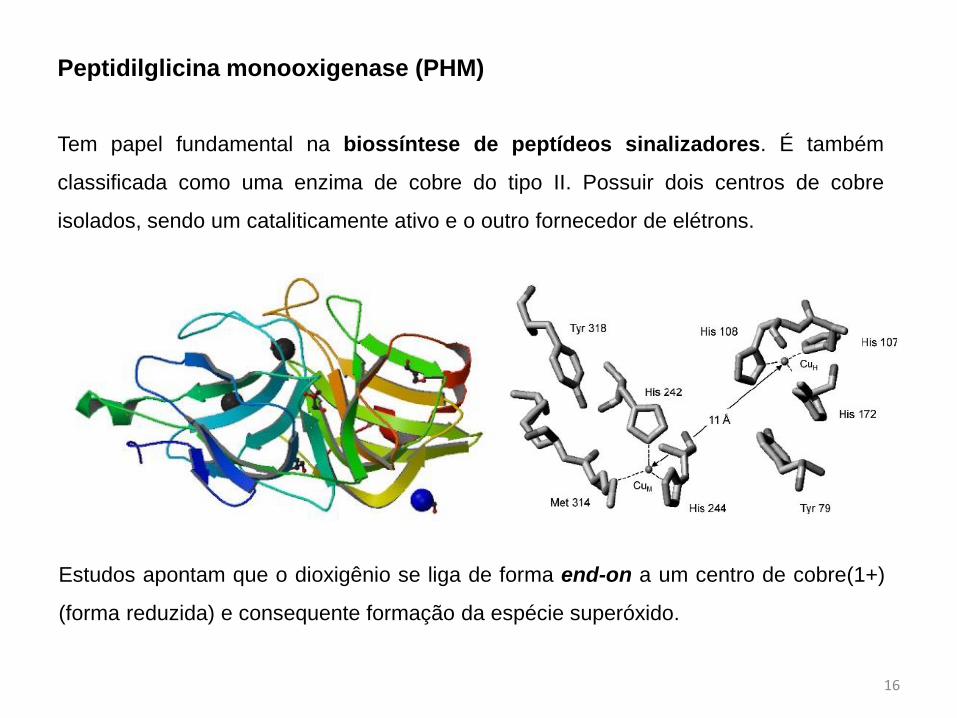
Peptidilglicina monooxigenase (PHM)
Tem papel fundamental na biossíntese de peptídeos sinalizadores. É também
classificada como uma enzima de cobre do tipo II. Possuir dois centros de cobre
isolados, sendo um cataliticamente ativo e o outro fornecedor de elétrons.
Estudos apontam que o dioxigênio se liga de forma end-on a um centro de cobre(1+)
(forma reduzida) e consequente formação da espécie superóxido.

17
Esta forma reduzida é atingida pelo
fornecimento de 1 elétron a longa distância
do outro centro de cobre presente na enzima
através do próprio substrato.

18

19
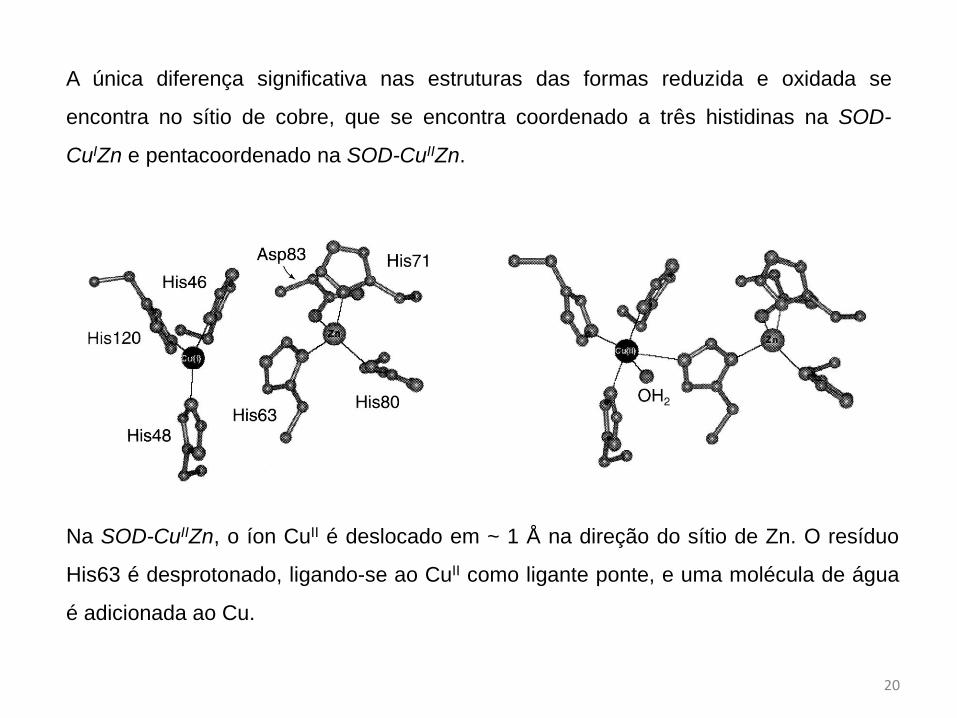
Superóxido dismutase de Cu e Zn
A SOD-CuZn é composta por duas subunidades idênticas, cada uma contendo um cobre
e um zinco no sitio ativo.
As estruturas cristalinas das formas reduzida e oxidada da SOD-CuZn foram
determinadas por cristalografia de raios X.
20
A única diferença significativa nas estruturas das formas reduzida e oxidada se
encontra no sítio de cobre, que se encontra coordenado a três histidinas na SOD-
CuIZn e pentacoordenado na SOD-CuIIZn.
Na SOD-CuIIZn, o íon CuII é deslocado em ~ 1 Å na direção do sítio de Zn. O resíduo
His63 é desprotonado, ligando-se ao CuII como ligante ponte, e uma molécula de água
é adicionada ao Cu.
21
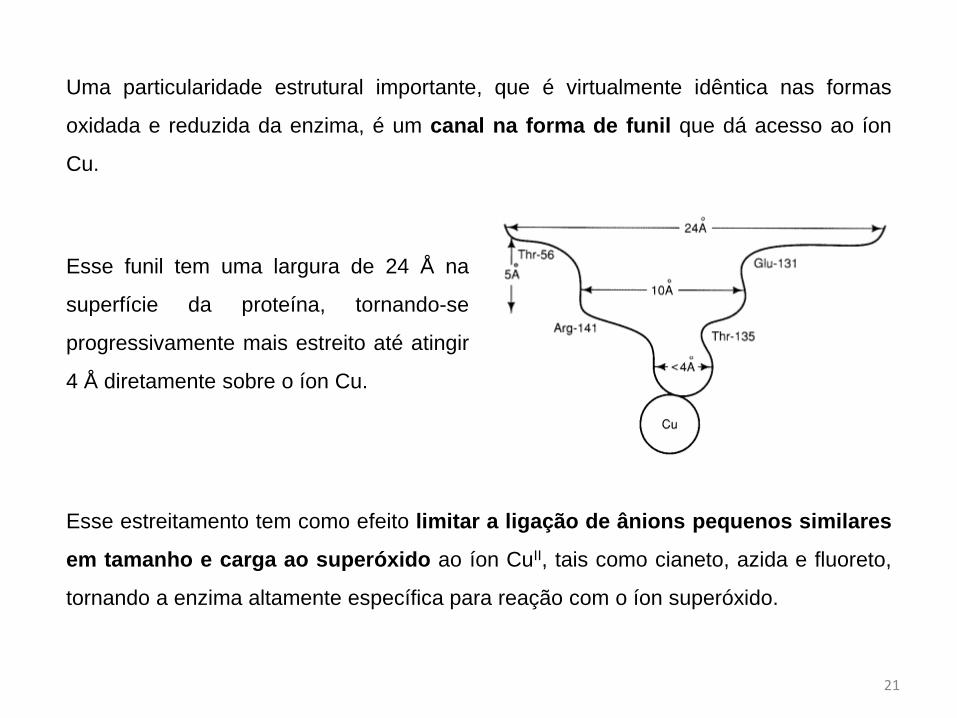
Uma particularidade estrutural importante, que é virtualmente idêntica nas formas
oxidada e reduzida da enzima, é um canal na forma de funil que dá acesso ao íon
Cu.
Esse funil tem uma largura de 24 Å na
superfície da proteína, tornando-se
progressivamente mais estreito até atingir
4 Å diretamente sobre o íon Cu.
Esse estreitamento tem como efeito limitar a ligação de ânions pequenos similares
em tamanho e carga ao superóxido ao íon CuII, tais como cianeto, azida e fluoreto,
tornando a enzima altamente específica para reação com o íon superóxido.

22
O canal que conduz ao sítio ativo também contem uma arginina
conservada ~ 6 Å sobre o íon Cu, cujo papel é gerar o campo
eletrostático que atrai o superóxido até o sítio ativo.
Acredita-se que o grupo guanidina positivamente carregado do resíduo arginina
encontra-se posicionado de forma a guiar o íon superóxido para dentro do canal,
formando ligações de hidrogênio a medida que se aproxima do centro de Cu tanto na
forma reduzida como oxidada.
O canal que dá acesso ao sítio ativo também contém uma cadeia de moléculas de
águas unidas por ligações de hidrogênio que possivelmente participam do
mecanismo da reação fornecendo prótons quando superóxido é reduzido a peróxido.

23
Mecanismo proposto - SOD

24
Redução de O2- para produzir H2O2
A redução de O2- pela SOD-CuIZn parece ocorrer por um mecanismo de esfera externa.
Baseando-se em comparações com compostos modelo de CuI, observou-se que o
arranjo trigonal plano CuI(His)3 possui pouca afinidade para ligar-se a um quarto ligante.
A SOD-CuIZn coordena pequenos ânions, mas aparentemente não ao CuI.

25
Para que o superóxido seja rapidamente reduzido pela SOD-CuIZn, é necessária a
associação de H+ ao O2- ao passo que um elétron é transferido, convertê-lo em
peróxido.
Alternativamente, HOO- aceita um próton da cadeia de moléculas de água conservadas,
formando H2O2 sem que haja formação do complexo Cu-OOH.
Acredita-se que esse próton seja doado pela His63, formando HOO- que possivelmente
se liga ao íon CuII. Uma subsequente protonação ocorre e H2O2 é liberado.

26
Oxidação de O2- para produzir O2
Estudos com ânions como N3-, CN- e F- mostram que esses íons ligam-se diretamente
ao Cu(II) na SOD-CuIIZn.
Com base nesse fato, é provável que O2- também se ligue ao Cu(II) formando o
complexo CuII(O2-) antes que ocorra a transferência de elétron para formar SOD-CuIZn e
O2.
É possível que O2- no canal do sítio ativo da SOD-CuIIZn, interaja com a cadeia de
moléculas de água e Arg conservadas, e finalmente substitua a molécula de água ligada
ao Cu(II).
A transferência de elétrons então ocorre, O2 é liberado e se difunde para forma do canal,
enquanto que a ponte imidazolato é quebrada e a His63 é protonada.

27
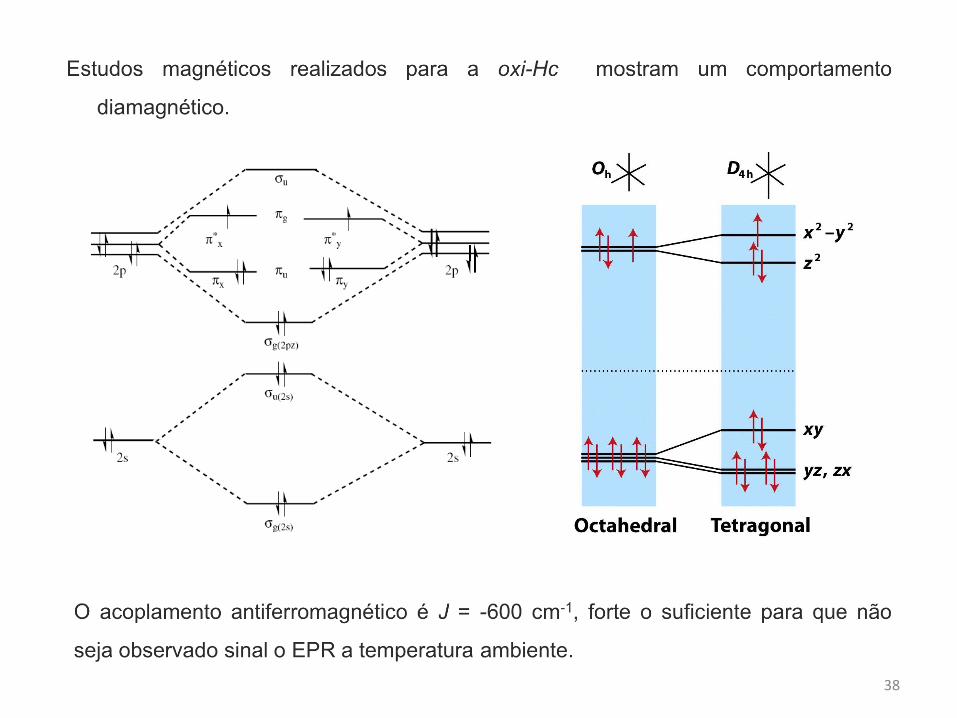
Enzimas de cobre tipo III
Enzimas de cobre do tipo III são caracterizadas por possuírem centros binucleares de
cobre espacialmente próximos e acoplados antiferromagneticamente.
Apesar da diversidade de funções que enzimas desta classe apresentam, a homologia
sequencial e estrutural de seus sítios ativos são consideravelmente preservadas.

28

29
Hemocianina - Hc

30

31

32
33

34
A cada centro estão coordenados três átomos de nitrogênio doadores imidazólicos
correspondentes a resíduos do aminoácido histidina, estando um deles envolvido numa
ligação covalente com um resíduo cisteína.
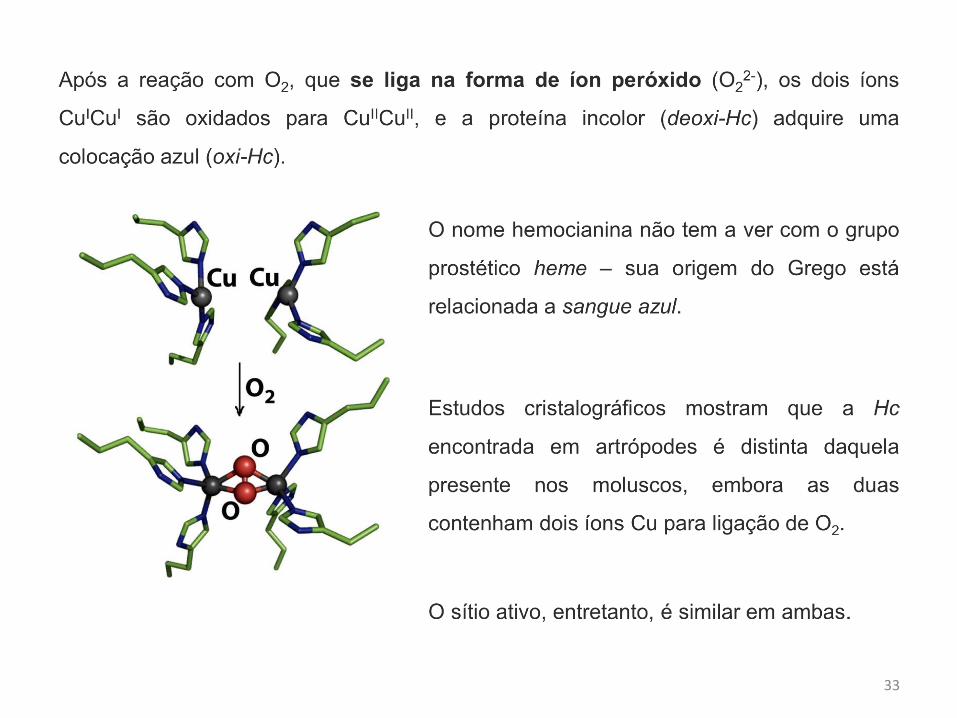
A reação da Hc reduzida com oxigênio resulta numa unidade binuclear de Cu(II)
pentacoordenados separados por uma distância de 3,5 Å e ligados entre si por uma
unidade ponte peroxo, a forma oxi-hemocianina

35

36

37
38

39

40

41

42

43

44

45
46
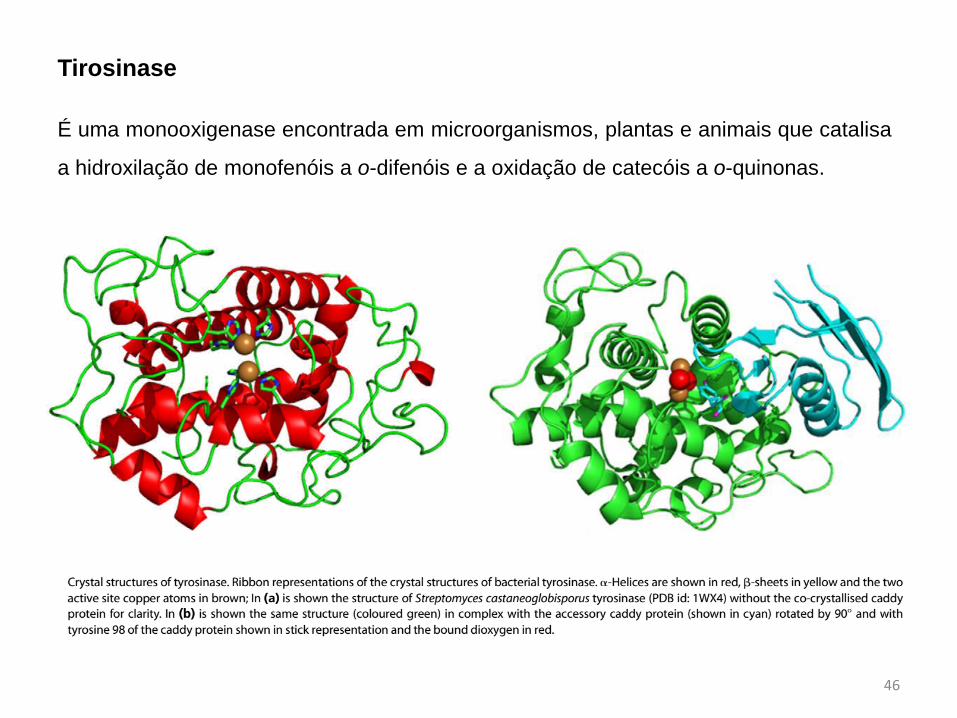
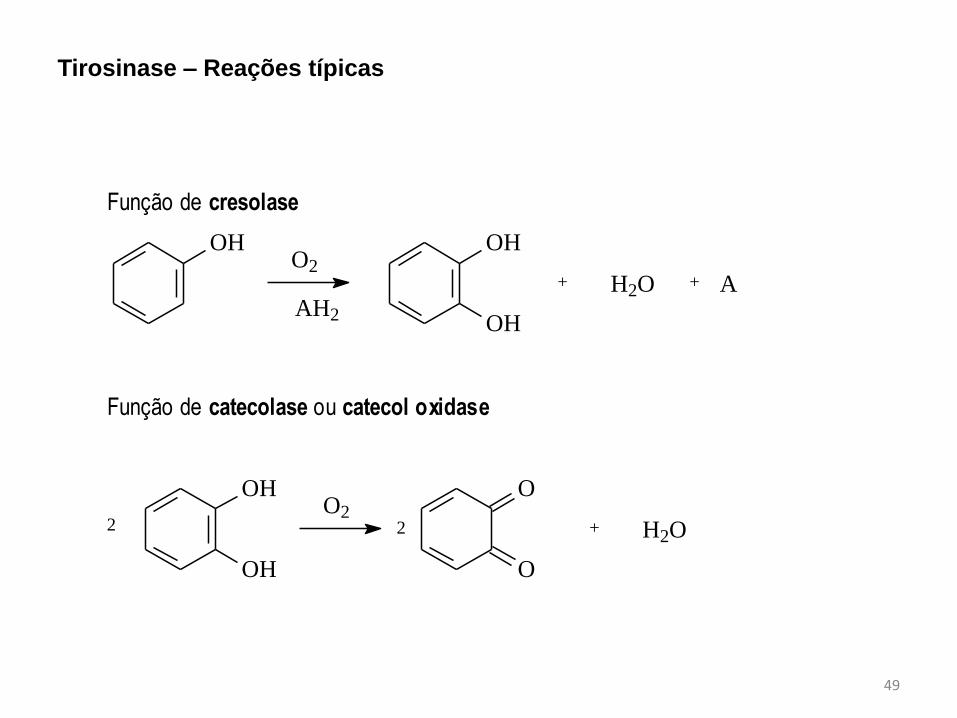
É uma monooxigenase encontrada em microorganismos, plantas e animais que catalisa
a hidroxilação de monofenóis a o-difenóis e a oxidação de catecóis a o-quinonas.
Tirosinase

47
Tirosinase – Tipos e homologia estrutural
• UV - Transições fracas na região de 600-700 nm
• EPR - forte acoplamento antiferromagnético – ausência sinal EPR

48
Tirosinase – Sítio ativo
Comparações das propriedades químicas, EPR, UV-visível, dicroísmo circular e Raman
ressonante revelaram grande semelhança com a hemocianina, permitindo sugerir um
sítio ativo binuclear de cobre que forma um aduto estável com o íon peróxido.
Estudos de EXAFS apontam para
uma distância intermetálica de
3,6 Å para a oxi-tirosinase e 3,4 Å
para a met-tirosinase.
A forma met-tirosinase, também
chamada de estado de repouso,
corresponde a uma espécie
binuclear de cobre(II) que não
contém o íon peróxido
coordenado.
49
i.
ii. Função de cresolase
iii.
O2
OH OH
OHAH2
+ H2O + A
iv.
v. Função de catecolase ou catecol oxidase
vi.
vii.
O2
OH
OH
O
O
+ H2O2 2
Tirosinase – Reações típicas

50

51
Tirosinase – Mecanismo de ação
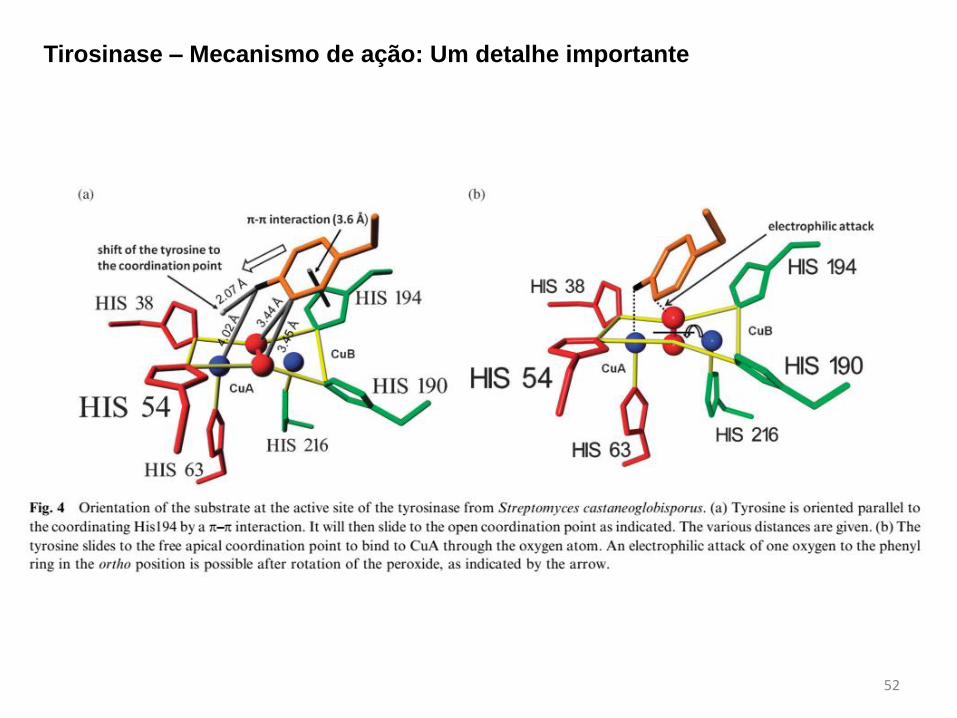
52
Tirosinase – Mecanismo de ação: Um detalhe importante
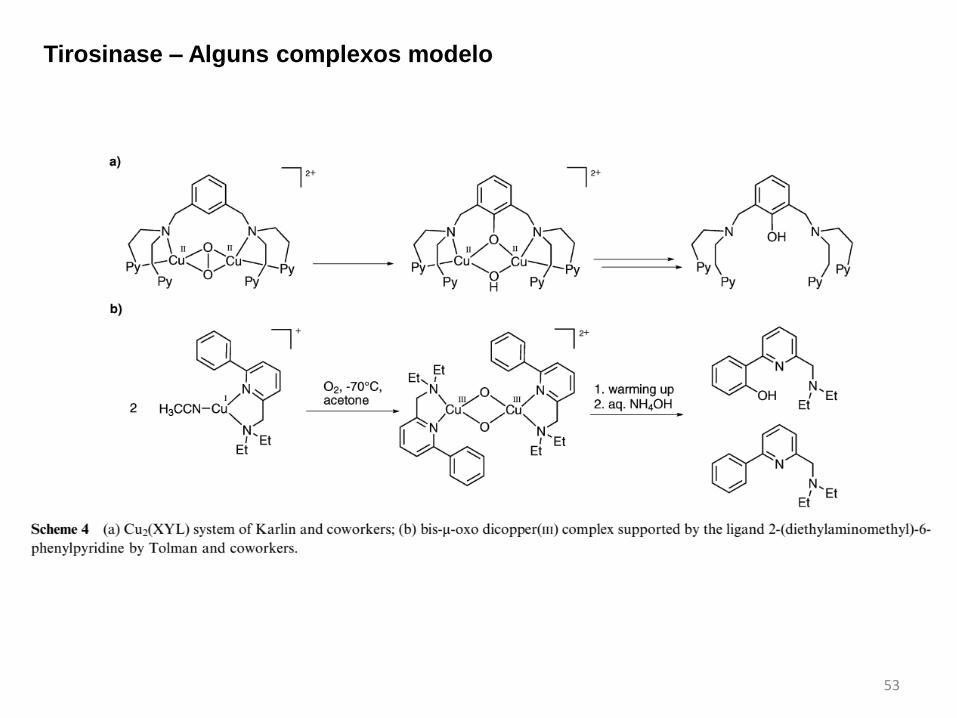
53
Tirosinase – Alguns complexos modelo

54

55

56
Catecol oxidase
A catecol oxidase (CO) é outro membro proeminente do grupo das metaloenzimas de
cobre do tipo III. Catalisa a oxidação de catecóis às correspondentes o-quinonas, sem
apresentar atividade sobre tirosinas.

57
Forma oxidada da CO possui dois centros de Cu(2+) em uma geometria piramidal
trigonal. O ambiente coordenação dos centros de cobre são constituídos por resíduos
histidinas e ponte OH. A distância Cu...Cu é de cerca de 2,9 Å.
Oxi Red
Na forma a reduzida, a distância intermetálica é de 4,4 Å, sem significativas mudanças
conformacionais nos grupos ligantes além da perda da ponte hidróxido.

58
O espectro eletrônico forma oxidada da CO apresenta máximos na faixa de
700-800 nm atribuídos a transições d-d e uma transição na faixa de 310 e 330 nm
atribuída a uma TCLM provavelmente do -hidroxo para os centros de Cu(2+).
Ao coordenar o íon peróxido surgem máximos atribuídos a transições de transferência
de carga do íon peróxido para Cu(II) em 343 nm ( 6500 M-1cm-1) e 580 nm ( 450
M-1 cm-1).
Além disso, os centros binucleares de cobre na catecol oxidase são caracterizados
pelo forte acoplamento antiferromagnético e ausência no espectro de EPR.
Catecol oxidase: Propriedades espectroscópicas

59
Mecanismo de ação proposto por Krebs et. al.

60
Mecanismo de ação proposto por Siegbahn et. al.
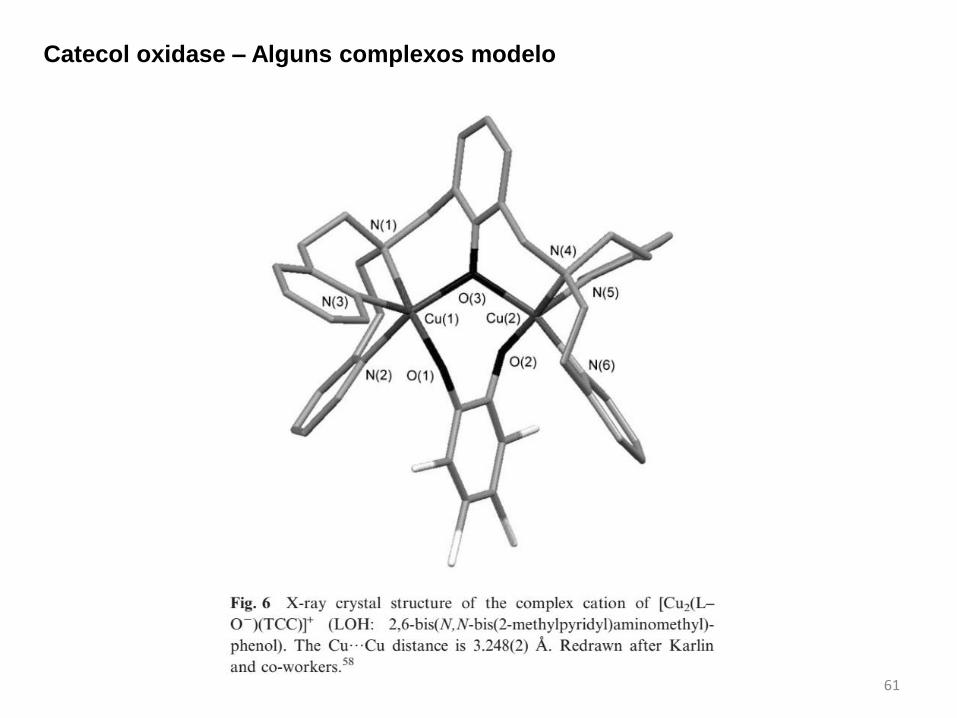
61
Catecol oxidase – Alguns complexos modelo

62

63

64
65