biogeografia américa do sul - padrões e processos 2011
TRANSCRIPT


j j j j j j j j j j j j j J
j j j j j j j
j j j j
j j
j j j j

Q r) l� j\�...r-. J�\
J ,UL
:1 :1 [) r� J 0

NOTA A Editora, os Organizadores e os Colaboradores não se responsabilizam por quaisquer consequências advindas do uso das informações contidas neste livro.
A Editora

60;l��JJ J
r) ( . ) �...:.., P 1
Doutor em Ciências (Entomologia) pela Universidade Federal do Paraná. Professor Titular do Departamento de Zoologia da Universidade Federal do Paraná. Bolsista de Produtividade em Pesquisa do Conselho Nacional de
Desenvolvimento Científico e Tecnológico (CNPq), nível I A.
Doutor em Entomologia pela Comell University. Professor Adjunto do Centro de Ciências Naturais e Humanas da Universidade Federal do ABC.
ROCA

Copyright © 201 1 da 1 ª Edição pela Editora Roca Ltda.
ISBN: 978-85-7241-896-6
Nenhuma parte desta publicação poderá ser reproduzida, guardada pelo sistema "retrieval" ou transmitida de qualquer
modo ou por qualquer outro meio, seja este eletrônico, mecânico, de fotocópia, de gravação, ou outros, sem prévia
autorização escrita da Editora.
Capa GABRIEL ANTONIO REZENDE DE PAULA
CIP-BRASIL. CATALOGAÇÃO-NA-FONTE SINDICATO NACIONAL DOS EDITORES DE LIVROS, RJ.
C322b
Carvalho, Claudio J. B. de Biogeografia da América do Sul : padrões e processos
/ Claudio J. B. de Carvalho, Eduardo A. B. Almeida. - São Paulo : Roca, 2010.
Contém glossário Inclui bibliografia e índice ISBN 978-85-7241 -896-6
1 . B iogeografia - América do Sul. 2. B iodiversidade - América do Sul. I . Almeida, Eduardo A. B. n. Título.
1 0-4 1 68. CDD: 578.098 CDU: 574(8)
2011
Todos os direitos para a língua portuguesa são reservados pela
EDITORA ROCA LTDA. Rua Dr. Cesário Mata Jr., 73
CEP 01221-020 - São Paulo - SP
Tel .: (11) 3331-4478 - Fax: (11) 3331-8653 E-mail: ven [email protected] - www.editoraroca.com.br
Impr esso no Brasil Printed in Brazil

-Clr', <:� ',,�'
Há pouco mais de 30 anos, Léon Croizat iniciou a sua
Biogeografía Analítica y Sintetica ("Panbiogeografía") de las Américas com a seguinte declaração: "Não é de nenhuma forma ousada a afirmação de que não se conhece hoje na Ibero-América uma biogeografia que mereça o título de científica". Continuou, afirmando que não possuímos uma filosofia, uma ideia geral ou um conjunto de noções básicas que nos permita enfrentar os problemas da distribuição geográfica e composição taxonômica de
plantas e animais que ocorrem nas Américas, desde o Rio Grande do Norte até a Terra do Fogo, desde o Estreito de Bering até a pequena ilha de Fernando de Noronha. O que encontramos, com frequência, é uma compilação de modelos, normas, métodos, trabalhos etc. , de zoogeógrafos e fitogeógrafos estrangeiros, que consideramos como mestres irrebatíveis que nunca alcançaremos. Léon Croizat já indicava, naquela época, a necessidade da construção de uma biogeografia desenvolvida por nós mesmos, embasada grandemente pelo reconhecimento e pelas comparações dos padrões de distribuições de animais e plantas.
Ainda hoje não é difícil constatar na literatura a quase
total ausência de livros de Biogeografia genuinamente escritos em português. No Brasil, com sua intensa e profícua produção de conhecimento sobre a biodiversidade, nas suas diferentes formas, faltava um livro com conteúdo amplo e abrangente que divulgasse as atuais ideias biogeográficas para a América do Sul, continente biologicamente híbrido e biogeograficamente muito rico.
O embrião deste livro foi formado há cerca de dez anos, quando se sentiu a necessidade de ter um livro para biogeografia em português. Um livro que reunisse as ideias novas e não apenas uma repetição de antigos chavões biogeográficos, encarquilhados em conceitos antigos e apresentados em traduções pouco recentes . Este embrião foi se desenvolvendo a cada participação em congressos brasileiros de Zoologia, Entomologia, Ecologia ou Conservação. Nestes, percebia-se a falta de um acompanhamento mais crítico dos estudantes nos temas de biogeografia, talvez pela falta de uma obra em português que desmistificasse as teorias e simplificasse
o entendimento dos métodos exibidos e discutidos. Este foi o principal objetivo da concepção deste livro: ter disponível, em língua portuguesa, um livro com condições plenas de iniciar os estudantes nesta ciência dinâmica e de contínua evolução.
A biogeografia encontra-se no meio de um furacão de proposições de mudanças metodológicas e consoli
dação de conceitos. Cada vez mais métodos estão sendo desenvolvidos para o reconhecimento e a explicação dos padrões de distribuição dos organismos. Alguns métodos também estão começando a ser utilizados como ferramentas para resolução de problemas imediatos em áreas de investigação próximas, como de conservação biológica ou de natureza médico-sanitária.
Como qualquer livro com múltiplos capítulos, as ideias expostas estão sob a óptica dos autores. A diversidade de métodos explorados em cada um dos capítulos reflete a variedade de abordagens atualmente disponíveis para a pesquisa biogeográfica. Os autores tiveram a independência para buscarem as ferramentas que consideravam mais apropriadas, de acordo com as suas convicções biogeográficas.
O livro é iniciado por prefácio conciso e seguido de quatro seções - Histórico e Conceitos (quatro capítulos), Métodos e Aplicações (seis capítulos), Evolução Espacial da Região (quatro capítulos) e Padrões e Processos - Estudos de Casos (três capítulos). São 1 7 capítulos confeccionados por 26 autores, jovens em sua maioria, mas com produção biogeográfica já consolidada. O encadeamento de seções e capítulos seguiu de uma visão histórica e conceitual da biogeografia para uma visão das aplicações dos métodos e evolução espacial da região. Na última seção está incluído o estudo de casos de diferentes organismos no continente sul-americano. O livro é finalizado pelo glossário dos principais termos utilizados .
Os organizadores agradecem a Editora Roca pela sensibilidade de produzir uma obra deste tipo; aos revisores dos capítulos, pela leitura criteriosa e atenta (em ordem alfabética do primeiro nome): Camila C. Ribas, Carlos Roberto Brandão, Charles Morphy Dias dos Santos,

VI - Apresentação
Daniela Maeda Takiya, Elaine Della Giustina Soares, Fernanda Werneck, Gabriel Augusto Rodrigues de Melo, Guilherme Schnell e Schuehli, José Alexandre Felizola Diniz Filho, José Francisco de Oliveira Neto, José Maria Cardoso da Silva, Juan J. Morrone, Kirstern Lica F. Haseyama, Lúcia Massutti de Almeida, Luiz Malabarba, Mareio Roberto Pie, Maurício Osvaldo Moura, Paula Posadas, Renata G. Netto, Roberto E. Reis, Rodrigo A.
Torres, Rodrigo C. Marques, Sílvio S . Nihei, Tania Escalante, Valéria C. Muschner; e a Diana Grisales, pela tradução para o português dos manuscritos originalmente escritos em espanhol (Capítulos 2 e 1 1 ) .
Claudio 1. B. de Carvalho Eduardo A. B. Almeida
Novembro de 201 0

T
A América do Sul sempre representou um problema para os biogeógrafos, principalmente em relação a sua fauna. Don Félix de Azara, por exemplo, em 1809, em sua obra Voyages dans l'Amérique Méridionale, após descrever as espécies de mamíferos do Paraguai, comentaval : "Considera-se em geral como uma verdade incontestável que todos os quadrúpedes têm sua origem no velho continente, de onde passaram para a América. Procura-se, em consequência, o lugar por onde essa passagem pôde efetuar-se; e como os continentes se aproximam ao norte mais do que em qualquer outro lugar, crê-se que foi por lá que eles passaram. Não parece difícil aplicar essa ideia àqueles quadrúpedes que povoaram toda a América ou a maior parte desse continente, tais como a anta, os veados, os porcos-da-mato, a onça, a suçuarana, as jaguatiricas e muitos outros, que perfazem uma série ininterrupta desde o norte da América até o sul, e que parecem indicar o caminho que seguiram; embora sejamos levados a crer que jamais tenham existido no antigo continente, pois ali hoje não mais se acham, pode-se presumir que o homem os tenha exterminado".
O naturalista espanhol referia-se à primeira e mais
longeva teoria biogeográfica formulada - a do centro de origem e dispersão da biota, que aparece duas vezes no livro do Gênesis da Bíblia. Como foi admitido durante mais de um milênio, Deus teria criado originalmente todas as espécies (entidades imutáveis, por saírem perfeitas das mãos do Criador) de animais num único ponto da face da Terra - o Jardim do Éden, de onde se dispersaram, após o pecado original do primeiro casal de humanos, para ocupar o resto do planeta. Após o Dilúvio, o Monte Ararat, onde parou a arca de N oé, funcionou como um segundo centro de origem e dispersão dos animais salvos pelo patriarca, que tornaram a povoar toda a superfície de nosso planeta.
Como toda teoria, esta também logo encontrou obstáculos epistemológicos. Como os animais oriundos do Ararat, sem utilidade para o homem, nem capacidade de nadar ou voar, chegaram às ilhas oceânicas, depois de terem repovoado o Velho Mundo? Santo Agostinho (em seu livro De civitate Dei) imunizou a teoria contra esse
teste, admitindo haverem eles sido transportados por anjos para as ilhas do Atlântico Norte2 .
O problema agravou-se com o descobrimento da América e sua estranha biota - existiam animais muito semelhantes aos do Velho Mundo (como os veados), outros bastante diferentes (porcos, macacos) e um terceiro grupo (como os marsupiais, por exemplo) que eram então apenas conhecidos do Novo Mundo. Isto foi claramente percebido e descrito pelo Pe. Joseph d' Acosta em seu livro de 1590. Por que existiam esses diferentes grupos? Como chegaram a este lado do Atlântico? Propôs-se uma ponte intercontinental (a At1ântida) ligando a Europa à América, hipótese derrubada por d' Acosta, ao postular uma passagem pelo estreito de Bering, através do qual os animais provindos do Ararat puderam chegar à América3.
Mas por que alguns permaneceram muito parecidos com seus ancestrais, outros se diferenciaram um tanto e outros ainda ficaram tão diferentes que não podiam ser relacionados a qualquer espécie da Eurásia? Por que certas espécies existiam apenas na América? Buffon, em 1778, após várias tentativas prévias de elucidar o pro
blema, terminou postulando, em seu clássico "Épocas
da Natureza", uma criação separada para os mamíferos da América do Su14.
AzaraI também se ocupou desta questão: "Tão natural quanto pareça esse modo de pensar, podem-se-Ihe fazer várias objeções, e eis aqui a primeira:
1 Q - Parece impossível que o tamanduá-bandeira, o
tamanduá-colete, o ouriço-cacheiro, assim como as várias espécies de marsupiais e de tatus que se acham nas duas Américas possam ter feito uma tão longa via
gem, dadas sua preguiça e poltronice excessivas; e não
se concebe que causa tê-las-ia determinado a viajar.
[ . . .]. 2 Q - A transmigração de algumas espécies parece impossível. Por exemplo, minha capivara e minha lontra
nunca entram na água do mar; e nunca vi nem ouvi
dizer que esses animais se afastem mais de trinta passos
do rio ou do lago onde vivem [ . . .]. 3 Q - O tuco-tuco nunca sai de sua habitação subterrânea [ . . . J; 4Q - Três
espécies de gatos ( . . . ), o cangambá, o ouriço-cacheiro,

VIII- Prefácio
a viscacha, a lebre da Patagônia, os tatus ( .. .) acham-se ao sul dos 26° 30' de latitude ( . . . ) e nenhum ao norte desse paralelo. Como concordar este fato com a passagem desses animais de um continente a outro? [ ... ]. Se, para responder a esta dificuldade, supõe-se que os continentes estavam unidos do lado do sul, e que foi por lá que se efetuou a passagem [uma das hipóteses levantada previamente publicada e depois abandonada por BuffonJ, caímos nos mesmos inconvenientes, pois nenhum desses quadrúpedes existe na África".
O autor só viu uma solução - "cada espécie de insetos e de quadrúpedes não provém de um único casal primordial, mas de vários casais idênticos criados nos diferentes lugares onde os vemos hoje". Pois, acrescentou, "se a criação que concerne à zoologia tivesse sido instantânea e de um só casal de cada espécie, quem teria podido fornecer a alimentação daquelas que vivem às expensas das outras? Elas morreriam de fome ou teriam exterminado a raça daquelas que lhes serviram de alimento . . . " [Objeção já publicada por Eberhardt Zimmermann em seu Specimen Zoologiae geographicae Quadrupedum]5 .
Em 1 820, Augustin Pyramus de Candolle6, num genial artigo intitulado Géographie botanique, propôs pela primeira vez a divisão dos continentes em regiões fitogeográficas ou áreas de endemismo; em 1 838 incluiu várias ilhas oceânicas em seu sistema e propôs 40 regiões fitogeográficas; o que hoje chamamos Região Neotropical incluía as subregiões "México ou América Central", "Antilhas", "Colômbia", "Guianas", "Peru", "Chile", "Brasil tropical", "Brasil austral e Buenos Aires" e "Terras magelânicas"7. Ludwig Karl Schmarda8, em 1 853, confirmou essas divisões para os animais terrestres. James Dwight Dana9, no mesmo ano, ocupou-se da distribuição dos animais marinhos, separando os oceanos em cinco "reinos", cada qual com várias "zonas" biogeográficas. Philip Lutley SclaterlO, em 1 858, propôs uma classificação dos continentes em cinco grandes regiões (que incluíam as áreas de endemismo descobertas por Candolle e Schmarda), sistema seguido até os dias de hoje; considerou tais regiões como "centros de criação", em que Deus teria criado separadamente as espécies.
O advento do paradigma evolutivo de Wallace-Darwin, no século XIX, representou a derrocada do Criacionismo, por admitir a comunidade de descendência das espécies. No entanto, a biogeografia continuou se baseando na antiga teoria do centro de origem e dispersão, admitindo-se agora que à medida que se dispersam, as espécies originam novas espécies descendentes. Continuaram presentes, entretanto, os problemas das barreiras à livre dispersão (o problema pela primei-
ra vez levantado por Santo Agostinho) e da causa das diferenças entre as faunas das diversas regiões . Por estranho que pareça, a Biogeografia, dentro da perspectiva
histórica advinda da teoria da evolução, continuou meramente descritiva (preocupada apenas com os padrões de distribuição), tendo seu apogeu no clássico livro de Wallace1 1 The Geographical Distribution of Animais em 1 876, em que mostrou como os cinco grupos tradicionais de vertebrados estão repartidos nas subregiões das cinco grandes regiões de Sclater. Pouco ou nada contribuiu quanto aos processos de formação desses padrões.
Os notáveis progressos das ciências geológicas no século XX contribuíram para a total reformulação das ideias biogeográficas. A teoria da deriva continental, publicada por Alfred Wegener12 em 1 9 1 5 (Die Enststehung der Kontinente und Ozeane), foi acatada por muitos biólogos - foram os continentes que se moveram, e não os animais. Uma verdadeira "revolução copernicana" no campo da biologia comparada, a magnífica teoria da Tectônica Global veio aperfeiçoar ainda mais essa explicação.
Três grandes revoluções científicas ocorreram no século passado - a Sistemática Filogenética de Willi Hennig, a Pan-biogeografia de Léon Croizat e a Biogeografia por Vicariância, que afetaram profundamente a Biologia Comparada, com extraordinária riqueza de resultados.
Ainda resta muito a fazer em relação à biota da América do Sul - um campo promissor para novas gerações de pesquisadores. É a eles que se dirige esta obra, bastante abrangente e que certamente será de grande importância para incentivar novos estudos sobre esta fascinante ciência - hoje uma feliz e fecunda síntese de todos os campos da História Natural.
Nelson Papavero Museu de Zoologia
Universidade de São Paulo
REFERÊNCIAS BIBLIOGRÁFICAS 1 . AZARA, F. Voyages dans l'Amérique méridionale par Don
Félix de Azara, depuis 1 781 jusqu'en 1801; contenant la description géographique, politique et civile du Paraguay et de la riviere de La Plata; l'histoire de la découverte et de la conquête de ces contrées; des détails nombreux sur leur histoire naturelle, et sur les peuples sauvages qui les habitent; le récit des moyens employés par les Jésuites pour assujetir et civiliser les indigenes, etc. Publiés d' apres les manuscrits de I' auteur avec une notice sur sa vie et ses écrits, par C. A. Walckenaer; enrichis de notes par G. Cuvier. .. Suivis de l'histoire naturelle des Oiseaux du Paraguay et de la Plata, par le même auteur, traduite, d'apres l'original espagnol, et augmentée de notes par M. Sonnini. Accompagné d'un atlas de vingt-cinq planches. Tome Premier. Paris: Dentu, Imprimeur-Libraire, 1 809.

2. PAPAVERO, N . ; TEIXEIRA, D. M . ; LLORENTE-BOUSQUETS, J. História da Biogeografia no Período Pré-evolutivo. São Paulo: Plêiade & Fundação de Amparo à Pesquisa do Estado de São Paulo, 1 997.
3 . D' ACOSTA, J. Historia natural y moral de las Indias, en que se tratan de cosas notables deZ cielo, y elementos, metales, plantas y animales del/as y los ritos, y ceremonias, leyes e gobiernos, y guerras de los Indios. Sevilla: Juan de Léon, 1590.
4. BUFFON, G. L. Histoire naturelle. Supplément V. Epoques de la nature. Paris: Imprirnérie Roya1e, 1778.
5 . ZIMMERMANN, E. Specimen zoologiae geographicae quadrupedum domicilia et migrationes sistens. Leipzig: Lugduni Batavorum, 1777.
6. DE CANDOLLE, A. P. Géographie botanique. In: LEVRAULT, F. C. Dictionnaire des sciences naturel/es 1 9 . Paris: Levrault, 1 820. p. 359-436.
Prefácio - IX
7. DE CANDOLLE, A. P. Statistique de lafamille des Composées. Paris & Strasbourg: Treutel & Würz, 1 838.
8. SCHMARDA, L. K. Die geographische Verbreitung der Thiere, 3 vols. Wien: Carl Gerold & Sohn, 1 853.
9. DANA, J. D. On the isothermal oceanic chart, illustrating the geographical distribution of marine animaIs. Am. J. Sci. Arts, n. 2, v. 1 6 (47), p. 153-1 67 ; 3 1 4-327, 1 853.
10. SCLATER, P. L. On the geographical distribution of the members of the class Aves. J. Prac. Linnean Soe. London (Zoo/.), v. 2, p. 1 30-145, 1 858.
1 1 . WALLACE, A. R. The geographica/ distribution of animaIs; with a study of the relations of living and extinct faunas as elucidating the past changes ofthe Earth 's surface, 2 vols. New York: Harper & Brothers, Publishers, 1 876.
1 2. WEGENER, A. Die Entstehung der Kontinente und Ozeane. Braunschweig: Vieweg, 1 9 1 5 .


(?\� '\ O:-/"J " !v', � ,
: ,,_),
Adrian Antonio Garda. PhD em Zoologia pela University of Oklahoma. Professor Adjunto do Departamento de Botânica, Ecologia e Zoologia da Universidade Federal do Rio Grande do Norte.
Alexandre Cunha Ribeiro. Professor Adjunto do Departamento de Biologia e Zoologia do Instituto de Biociências da Universidade Federal de Mato Grosso.
Charles Morphy D. Santos. Doutor em Entomologia pela Universidade de São Paulo. Professor Adjunto do Centro de Ciências Naturais e Humanas da Universidade Federal do ABC.
Domingos Benício Oliveira Silva Cardoso. Doutorando do Programa de Pós-graduação em Botânica da
Universidade Estadual de Feira de Santana.
Edgardo Ortiz-Jaureguizar. Doutor em Ciências �aturais. Professor Titular da Cátedra de Mastozoologia, Facultad de Ciencias Naturales y Museo, Universidad �acional de La Plata. Investigador Independente do Consejo Nacional de Investigaciones Científicas y Técnicas.
Ernesto Luiz Lavina. Doutor em Ciências pela Cniversidade Federal do Rio Grande do Sul. Professor A.djunto do Curso de Pós-graduação em Geologia da Cniversidade do Vale do Rio dos Sinos. Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível 2.
Felipe de Mello Martins. Doutor em Biologia Genética pela Universidade de São Paulo. Pós-doutorando do Departamento de Zoologia da Universidade de São Paulo.
Fernando César Vieira Zanella. Doutor em Entomologia pela Universidade de São Paulo. Professor .-'1.ssociado da Universidade Federal de Campina Grande. campus de Patos . Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível 2.
Flávio César Thadeo de Lima. Bolsista de Pós-doutorado do Museu de Zoologia da Universidade de São Paulo.
Gerson Fauth. Doutor em Geologia pela Universidade de Heidelberg, Alemanha. Professor Adjunto do
Curso de Pós-graduação em Geologia da Universidade do Vale do Rio dos Sinos.
John R. Grehan. Doutor em Zoologia/Entomologia pela Victoria University ofWellington. Diretor Científico e de Pesquisa do B uffalo Museum of Science, Buffalo, Nova York.
José Alexandre Felizola Diniz Filho. Doutor em Ciências Biológicas (Zoologia) pela Universidade Estadual Paulista Júlio de Mesquita Filho. Professor Titular do Departamento de Ecologia da Universidade Federal de Goiás. Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível IA.
José Maria Cardoso da Silva. PhD em Zoologia pela U niversity of Copenhagen. Vice-presidente para a América do Sul, Conservação Internacional. B olsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível 2 .
Juan J. Morrone. Professor Titular C de tempo completo da Facultad de Ciencias, Universidad Nacional Autónoma de México.
Luciano Paganucci de Queiroz. Professor Titular do Departamento de Ciências Biológicas da Universidade Estadual de Feira de Santana. Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível lD.
Máreia Souto Couri. Doutor em Ciências (Parasitologia) pela Universidade Rural do Rio de Janeiro. Professor Associado do Departamento de Entomologia do Museu Nacional da Universidade Federal do Rio de Janeiro. Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível I A.
Mareio Bernardino DaSilva. Doutor em Zoologia pelo Instituto de Biociências da Universidade de São Paulo. Professor do Departamento de Sistemática e Ecologia da Universidade Federal da Paraíba.
Marcus Vinicius Domingues. Doutor em Zoologia pela Universidade Federal do Paraná. Professor Adjunto da Universidade Federal do Pará - Campus Universitário de Bragança, Instituto de Estudos Costeiros.

XII - Colaboradores
Miguel Bastos Araújo. PhD Geography (Biogeography), University of London, UK. Professor do Departamento de Biodiversidad y Biología Evolutiva, Museo Nacional de Ciencias Naturales, Madri, Espanha; Cátedra Rui Nabeiro - Biodiversidade, CIBIO, Universidade de Évora, Évora, Portugal.
N aércio Aquino Menezes. Professor Titular Aposentado do Museu de Zoologia da Universidade de São Paulo. Bolsista de Produtividade do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível IA.
Paula Posadas. Doutor em Ciências Naturais. Docente da Cátedra de Biogeografía, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Investigadora Adjunta do Consejo Nacional de Investigaciones Científicas y Técnicas.
Peter Lõwenberg Neto. Doutor em Ciências (Entomologia) pela Universidade Federal do Paraná. Professor da Universidade Federal da Integração Latino-americana.
Ricardo Pinto-da-Rocha. Professor do Departamento de Zoologia do Instituto de Biociências da Universidade de São Paulo. Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível ID.
Silvio Shigueo Nihei. Doutor em Ciências Biológicas (Entomologia) pela Universidade Federal do Paraná. Professor Doutor do Departamento de Zoologia do Instituto de Biociências da Universidade de São Paulo. Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico, nível 2.

] Capítulo 1 - Evolução Geológica da América do Sul nos Últimos 250 Milhões de Anos . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
Ernesto Luiz Lavina • Gerson Fauth
Capítulo 2 - América do Sul e Geografia da Vida: Comparação de Algumas Propostas de Regionalização .. .. . . . 14 Juan J. Morrone
Capítulo 3 -Áreas de Endemismo................................................................................................................ 4 1 Claudio J. B. de Carvalho
Capítulo 4 - Lógica da Biogeografia de Vicariância ........ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52 Eduardo A. B . Almeida • Charles Morphy D . Santos
Capítulo 5 - Introdução à Pan-biogeografia: Método e Síntese.... .............. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65 John R. Grehan
Capítulo 6 - Biogeografia Cladística............................................................................................................ 99 Silvio Shigueo Nihei
Capítulo 7 -Modelos de Eventos para Reconstrução Biogeográfica........................................................... 1 23 Eduardo A. B. Almeida
Capítulo 8 - Filogeografia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137 Felipe de Mello Martins • Marcus Vinicius Domingues
Capítulo 9 -Macroecologia e Mudanças Climáticas ................................................................................... 1 5 1 José Alexandre Felizola Diniz Filho • Miguel Bastos Araújo
Capítulo 1 0 - Conservação da Biodiversidade e Biogeografia Histórica...... ........ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162 Peter Lowenberg Neto
Capítulo 1 1 -Evolução da Região Andina da América do Sul ................................................................ .... 175 Paula Posadas • Edgardo Ortiz-Jaureguizar
Capítulo 1 2 - Padrões e Processos Biogeográficos na Amazônia ....... ................................... ...................... 1 89 José Maria Cardoso da Silva • Adrian Antonio Garda
Capítulo 1 3 - Evolução da Biota da Diagonal de Formações Abertas Secas da América do Sul ................ 198 Fernando César Vieira Zanella
Capítulo 1 4 - História Biogeográfica da Mata Atlântica: Opiliões (Arachnida) como Modelo para sua Inferência................................................................................................................. 221 Marcio Bernardino DaSilva • Ricardo Pinto-da-Rocha
Capítulo 15 - Caatinga no Contexto de uma Metacomunidade: Evidências da Biogeografia, Padrões Filogenéticos e Abundância de Espécies em Leguminosas...................................... 241 Domingos Benício Oliveira Silva Cardoso • Luciano Paganucci de Queiroz
Capítulo 1 6 -Biogeografia dos Peixes de Água Doce da América do SuL................. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261 Alexandre Cunha Ribeiro • Flávio César Thadeo de Lima • Naércio Aquino Menezes
Capítulo 1 7 - Biogeografia de Muscidae (Insecta, Diptera) da América do SuL......... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 277 Claudio J. B. de Carvalho • Márcia Souto Couri
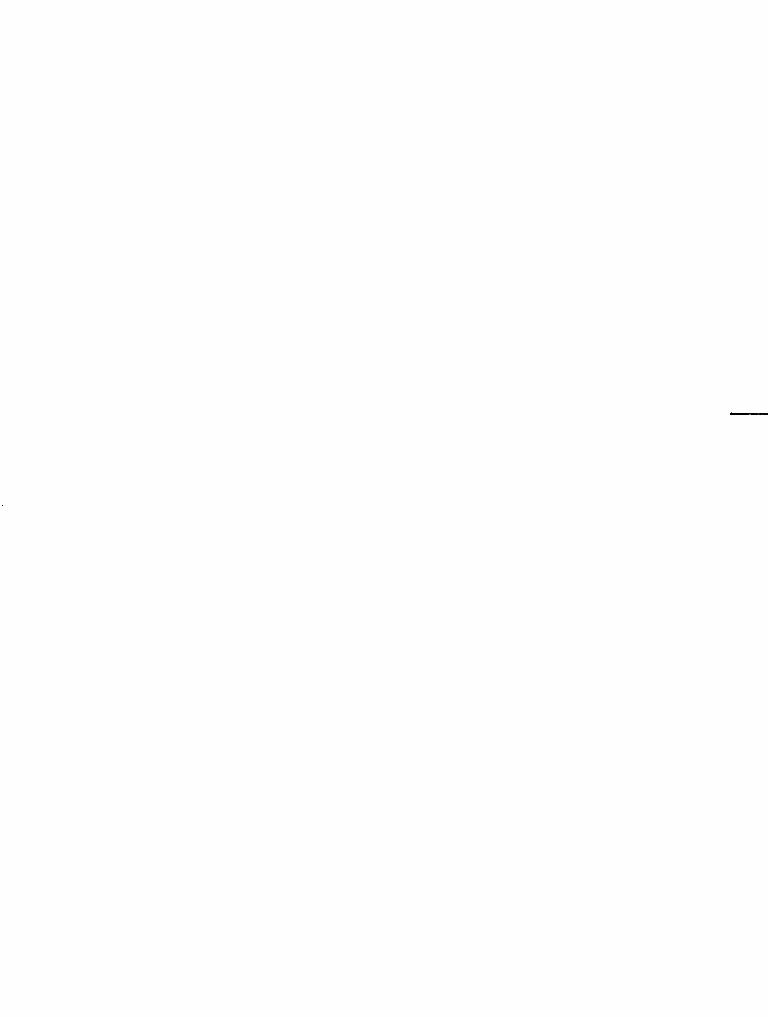

, --------------------------------------------------------------�------�

1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1

,)
Ernesto Luiz Lavina
Gerson Fauth
A história evolutiva da América do Sul nos últimos 250 milhões de anos está relacionada a uma sucessão de grandes eventos geológicos que modificaram tanto os continentes e as bacias oceânicas quanto o clima e a própria evolução das espécies.
A superfície terrestre está em transformação contínua ao longo do tempo. Não existe, em termos de história da Terra, qualquer fator natural que seja constante. A geografia planetária, por exemplo, quando visualizada em termos de tempo geológico, é extraordinariamente mutá\Oel. Cabe destacar que apenas no último bilhão de anos I cerca de 20% do tempo geológico), dois supercontinentes se formaram a partir da reunião de diversos continentes e ilhas, e se fragmentaram dando origem a novos continentes e ilhas CRodínia: 650 a 600 Ma *, e Pangeia: 270 a 220 Ma). Como outros exemplos notáveis de mudanças paleogeográficas, podemos referir a Antárrica, que hoje ocupa a região do polo sul, mas já esteve, juntamente com Austrália c Índia, na região polar ártica I há aproximadamente 700 Ma), ou o atual continente africano, uma série de ilhas há um bilhão de anos, agregadas durante a formação de Rodínia. Intimamente associados às modificações paleogeográficas, o vulcanismo e a fotos síntese adquirem papel fundamental na e\olução do clima. Em verdade, papéis antagônicos, pois o yulcanismo adiciona CO2 à atmosfera, enquanto a
, A sigla Ma sign ifica "mi lhões de anos".
1
fotossíntese o retira. Períodos de formação ou de desagregação de supercontinentes apresentam aumento significativo do vulcanismo, implicando em maior produção de CO2 e, em consequência, amplificando o efeito estufa. Ao contrário, em fases de menor intensidade vulcânica, o fitoplâncton Ce o solo) absorve CO2, diminuindo sua concentração na atmosfera, fazendo com que o planeta esfrie.
Cadeias de realimentação ocorrem naturalmente, se relacionando a diversos fatores, desde a posição geográfica dos continentes (paleolatitude) até as variações na excentricidade da órbita terrestre e na obliquidade do eixo de rotação. Fator fundamental de realimentação climática para períodos frios é a presença de um continente em latitude polar. Uma calota polar posicionada diretamente sobre a água não afeta o nível do mar. Entretanto, se estiver situada sobre um continente, sua expansão acarreta, necessariamente, o rebaixamento do nível do mar. Em períodos mais frios, o gelo polar se expande em área e passa a refletir maior quantidade de luz solar de volta ao espaço, diminuindo a temperatura média do planeta. ° gelo polar cresce às expensas da água do oceano, que pode rebaixar dezenas de metros em poucos milhares de anos. Em períodos de mar baixo, os continentes têm maior dificuldade de reter umidade e tendem a secar na faixa tropical (latitudes inferiores a 30°, onde ocorrem as maiores taxas de insolação e de evaporação). Os desertos se expandem rapidamente nesta faixa e os grãos de quartzo da areia das dunas têm capacidade quase igual à do gelo em refletir a luz solar para o espaço, tomando a Terra ainda mais fria. Uma cadeia de realimentação, como a descrita, é usada para

4 - Histór ico e Conceitos
contextualizar os grandes eventos glaciais, como o ocorrido no final do Pré-cambriano, no qual as evidências sugerem que toda a superfície da Terra tenha congelado, mesmo na faixa equatorial (o continente de Rodínia situava-se na região polar sul), Carbonífero (359 a 299
Ma), quando 25% da superfície da Terra ficaram cobertos por gelo (Gondwana ocupava o polo sul), ou mesmo o período glacial pelo qual passamos. Uma era glacial pode terminar, lentamente, pela movimentação de continentes para fora da região polar, ou abruptamente, em razão de uma fase de intensificação do vulcanismo. Em períodos de maior atividade vulcânica, a situação se inverte, o gelo se retrai e o mar se eleva, invadindo os continentes. Períodos de "mar alto" apresentam tendência a maior uniformidade climática e, com frequência, extensas áreas úmidas e grande desenvolvimento de florestas.
Cabe, portanto, destacar que na história da Terra, o único fator imutável é a continuidade das transformações. Neste sentido, os últimos 250 Ma da história de nosso planeta apresentam muitas particularidades. Durante este tempo, incomensurável para a percepção humana, mas que corresponde a apenas 5 % da história da Terra, surgiram dois novos oceanos (Atlântico e Índico), e o supercontinente Pangeia se dividiu em várias partes, dando origem aos continentes atuais. Associado às imensas variações na geografia planetária, também o clima passou por grandes transformações. O intenso vulcanismo associado à existência da Pangeia, desde a sua formação (Permiano e Triássico) até a fragmentação (Jurássico e Cretáceo), produziu um efeito estufa tão intenso que ocasionou o desaparecimento total do gelo da superfície da Terra, mesmo nas regiões polares. Como consequência, a formação da Pangeia provocou também drásticas alterações na dinâmica dos seres vivos. As biotas empobreceram em gêneros e espécies na passagem do Permiano ao Triássico (250 Ma). Em verdade, o evento de extinção de biomassa, ocorrido neste momento, é considerado a maior crise da história da vida na Terra. As evidências sugerem que mais de 90% das espécies tenham-se extinguido em tempo relativamente curto. A grande extensão da Pangeia, que dificultava ou mesmo impedia a distribuição da umidade para o interior, a progressiva retração areal dos mares rasos epicontinentais, o vulcanismo e o efeito estufa decorrentes, são alguns fatores considerados cruciais para a redução drástica do bioma. Neste tempo, as latitudes altas eram mais favoráveis à vida sobre a Terra1,2. No início do Triássico, florestas e faunas de répteis e répteis mamaliformes se desenvolviam em paleolatitudes tão altas quanto 80° sul (região das atuais montanhas Transantár-
ticas). Durante o Mesozoico, crocodilos e dinossauros habitaram diversas regiões circumpolares do hemisfério norte (Groenlândia, Alasca e Sibéria) e dinossauros viveram na Península Antártica durante o Cretáceo. O efeito estufa sofreu leve arrefecimento ao final do Cretáceo, mas fortaleceu-se novamente no início da era Cenozoica, e somente perdeu intensidade a partir do período entre 45 e 40 Ma, quando a Antártica, que havia saído da região polar durante o Mesozoico, retornou ao � círculo polar. Desde então, o gelo passou outra vez a t;
.:., fazer parte da história da Terra. Os principais eventos t geológicos e climáticos ocorridos na América do Sul são �
� apresentados na Figura 1 . 1 . b, A história acima resumida, envolvendo extraordinárias
mudanças geográficas e climáticas, é uma aquisição relativamente recente, pois apenas a partir da década de 1 970 passou a ser bem entendida. Resulta, porém, de esforços da comunidade científica durante cerca de 300 anos. Pode-se dizer que tenha se iniciado durante os séculos XVIII e XIX, quando houve grandes avanços na cartografia, no detalhamento de mapas e pesquisas em ciências naturais em regiões inóspitas, distantes da Europa. Com certa regularidade, alguns países europeus lançavam grandes expedições científicas aos "novos mundos" no intuito de conhecer e descrever fauna, flora e geografia locais (por exemplo, trabalhos de Humboldt, Darwin e Saint Hilaire). Desde o surgimento dos primeiros mapas mundiais, a coincidência dos contornos da América do Sul e da África intrigou as mentes mais imaginativas sobre o cataclisma que os teria separado (Francis Bacon, 1 620; Antônio Snider-Pellegrini, 1 858; e Marcel Bertrand, Frederick B. Taylor e Edward Suess; na passagem do século XIX ao século XX). Em 1 9 1 2, um meteorologista alemão, Alfred Lothar Wegener, surpreendeu o mundo científico ao apresentar a tese de que os continentes atuais estiveram todos reunidos, no final do Paleozoico e início do Mesozoico, em um único continente, ao qual chamou de Pangeia. Segundo Wegener, a Pangeia teve curta duração, tendo se fragmentado em dois continentes maiores, Gondwana, ao sul, e Laurásia, ao norte. Posteriormente, a Laurásia fragmentou-se em América do Norte e Eurásia (a Groenlândia, em termos geológicos, faz parte da América do Norte) e Gondwana em Índia, Austrália, Antártica, África e América do Sul.
A chave para Wegener montar o quebra-cabeças e sustentar a "teoria da deriva dos continentes" veio da similaridade das linhas de costa dos continentes, do registro fóssil, pois organismos semelhantes eram encontrados em continentes separados por vários milhares de quilômetros, e também da evolução climática. Répteis terrestres do início do Triássico, como os gêneros
150
-
Evolução Geológica da América do Sul nos Últimos 250 M i lhões de Anos - 5
o Início da fragmentação Pangeia
10': - Evol ução paleogeográfica e eventos geológ icos, c l imáticos e a lterações na flora e na fauna da América do Su l �os últimos 250 Mal-26. Ma = mi lhões de anos. As setas ind icam os eventos de grande extinção da vida na Terra.

6 - Histórico e Conce itos
Procolophon, Lystrosaurus e Thrinaxodon, encontrados em quase todos os continentes (se distribuem desde a plataforma siberiana até a região das montanhas Transantárticas), deixam claro a continuidade da superfície emersa, a não existência de barreiras geográficas e a notável uniformidade climática. Todavia, talvez a corroboração mais impressionante da teoria seja a explicação da distribuição geográfica dos carvões e dos depósitos glaciais do período Carbonífero (300 Ma). O carvão foi gerado por florestas de latitudes equatoriais, mas está hoje localizado em regiões subtropicais (Estados Unidos e Europa). Depósitos associados às geleiras do Carbonífero são encontrados em todos os continentes do hemisfério sul e na Índia. Observados no mapa atual, a distribuição do carvão e dos depósitos glaciais é absolutamente inexplicável, pois teríamos de admitir, além de florestas "equatoriais" em latitudes hoje improváveis, geleiras em latitudes baixas, como no Brasil, onde depósitos glaciais ocorrem desde o Rio Grande do Sul até Mato Grosso do Sul e Goiás (Grupo Itararé, da Bacia do Paraná), ou como no caso da Índia, na região equatorial. Quando Wegener reuniu todos estes elementos sobre o mapa da Pangeia, a área sob influência glacial demarcou a região polar (sul) e as florestas ocuparam a região equatorial do planeta. Entretanto, apesar de não haver qualquer elemento falseador da nova teoria, ela foi absolutamente rejeitada pela comunidade científica do início do século xx. E não se pense que os cientistas da época eram refratários às novas ideias; o início do século xx foi o tempo de Albert Einstein, Max Planck, Rutherford, Niels Bohr e Werner Heisenberg; foi o tempo da teoria do átomo, da teoria da relatividade e das mecânicas quântica e ondulatória. Os pesquisadores não conseguiam entender, e Wegener não conseguiu explicar, como os continentes se moviam. Ou, dito de outro modo, como poderiam imensos continentes andar pelo mundo "à deriva", como jangadas ao sabor dos ventos e correntes marítimas? As ideias de Wegener somente começaram a ser aceitas durante a década de 1 960, quando as anomalias magnéticas dos basaltos oceânicos demonstraram, de modo inequívoco, as taxas de geração e espalhamento do assoalho oceânico.
A visão que temos hoje do continente sul-americano, cercado por oceanos e quase isolado, teve início apenas a partir da desagregação da Pangeia (durante o Triássico, há cerca de 230 Ma). De fato, separada relativamente cedo do continente do norte (Laurásia), a América do Sul apresentava ligação apenas com Antártica e Austrália (pela Península Antártica), desenvolvendo biotas autóctones, algumas extraordinárias, como as faunas de marsupiais cenozoicos. A.penas o progressivo soergui-
mento da América Central durante o Cenozoico modificou esta situação, propiciando vários pulsos de imigração e substituição progressiva da fauna ancestral pela originária da América do Norte, em fuga da glaciação que se desenvolvia no hemisfério norte.
N o começo da era Mesozoica, a geografia planetária era dominada pela Pangeia, cercada por um único e imenso oceano (Panthalassa, cuja representação atual é o Oceano Pacífico). A América do Sul, tal como a conhecemos hoje, não possuía qualquer individualização com relação aos demais terrenos gondwânicos. Poder-se-ia seguir por terra desde a região do atual Brasil até a Índia ou Austrália, sem qualquer obstáculo importante. O território sul-americano localizava-se no setor sudoeste da Pangeia, conectando-se diretamente com África e Antártica. Suas rochas mais antigas pertencem aos crátons do Amazonas (escudos de Guaporé e Guiana), São Francisco, Rio da Prata, São Luiz e Luis Alves, e registram uma história geológica anterior a um bilhão de anos, fornecendo elementos importantes para a construção da história pré-cambriana da Terra3. Sobre os crátons, bacias sedimentares, por vezes com grandes dimensões, se desenvolveram durante o Paleozoico. As rochas mais jovens, e especialmente as bacias sedimentares que evoluíram durante o Mesozoico e o Cenozoico (por exemplo, Bacias do Paraná, Parnaíba, Amazonas, Neuquén, e bacias da margem andina), estão relacionadas aos eventos geológicos ocorridos nos últimos 250 Ma.
No Triássico, o clima árido predominava em toda a parte norte, desde o sul da província de Buenos Aires (Argentina) até o norte do Brasil, Guianas, Venezuela e margem oeste do continente. Era decorrente, em boa parte, da grande extensão de terras em latitude tropica12
e do desenvolvimento de um sistema mundial de megamonções4. As condições de maior umidade estavam restritas à parte sul (hoje Terra do Fogo e Patagônia), onde, em muitos momentos, florestas se desenvolveram. N a margem oeste, em vez da Cordilheira dos Andes, um imenso sistema vulcânico, que se tornou ativo na parte superior do Permiano (Formação Choyoi; cerca de 270 Ma), ainda se encontrava em franco desenvolvimento. Produzia grandes volumes de CO2 e se somava ao vulcanismo siberiano na manutenção do efeito estufa planetário. A metade norte da futura América do Sul, durante fases menos secas, era ocupada por uma flora em que dominavam as gimnospermas e faunas de répteis e répteis mamaliformes. Fragmentos deste tempo ficaram

preservados em poucos locais da América do Sul, com destaque para a região central do estado do Rio Grande do Sul (Formações Santa Maria e Caturrita, da Bacia do Paraná) e a Bacia de Ischigualasto, no noroeste da Argentina (Província de San Juan). Nestes locais, as evidências mostram que os cinodontes, ancestrais dos mamíferos, já construíam mecanismos internos cada vez mais eficientes para a regulação da temperatura corporal. Também demonstram, objetivamente, que, emergindo de um grupo inexpressivo de répteis, os primeiros dinossauros caminharam sobre o território sul-americano, espalhando-se, logo depois, por toda a Pangeia.
c
A separação entre continentes envolve longos períodos, estando sempre ligada a episódios vulcânicos importantes, extensas regiões afetadas por terremotos, soerguimentos e rebaixamentos de grandes de áreas, eventos que foram marcantes para a história geológica da América do Sul e que ocorrem até os dias de hoje.
A abertura e o surgimento do Oceano Atlântico teve início após a separação da Pangeia, durante a fase de fragmentação de Gondwana, ocorrida ao final do Triássico e início do Jurássico. A divisão África-América do Sul deu-se a partir dos extremos norte e sul, como duas separações continentais praticamente simultâneas. A primeira iniciou-se há cerca de 220 milhões de anos, na região da Venezuela, e está relacionada à separação entre o norte da África e a América do Norte. O segundo evento de separação começou há 200 milhões de anos ao sul da Argentina (região ao norte das Ilhas Malvinas) e África do Sul. Naquele momento, Gondwana já estava dividido em distintos continentes (América do Sul-África, Índia e Antártica-Austrália, embora ainda existisse comunicação entre América do Sul e Antártica-Austrália). Estas aberturas oceânicas ao norte e ao sul funcionaram como um "zíper", em que a crosta oceânica foi sendo rasgada de leste-oeste na região da Venezuela e sul-norte a partir da Argentina.
A abertura do Oceano Atlântico ocorreu em função y � da ascensão de magma do manto (hot spots), com =' '1 geração de crosta oceânica. A partir da criação de crosta "7 r:;' oceânica na dorsal do Atlântico, as placas tectônicas � sul-americana e africana começaram a se movimentar x S em sentidos opostos. A abertura do Atlântico pode ser
subdividida em cinco fases distintas:
• Processos extensionais: em que a crosta terrestre da região começou a ser estendida e afinada, iniciando o processo de separação entre os dois continentes.
Evolução Geológica da América do Sul nos Últ imos 250 Milhões de Anos - 7
• Falhamentos: na região da separação onde a crosta se tornou mais fina ocorreram grandes falhamentos, pelos quais extravasaram as primeiras lavas. Os sistemas de falhas também movimentaram verticalmente os blocos e o rebaixamento de extensas áreas possibilitou a criação de grandes lagos.
• Afinamento Crustal: rebaixada (estirada e afinada), a crosta recebeu sedimentos continentais e derrames de lava. Ocorreram também as primeiras incursões marinhas.
• Vulcanismo: as rochas vulcânicas são responsáveis pelo acréscimo de crosta. Formaram-se os primeiros depósitos de evaporitos (sais) no proto-oceanoAtlântico, concentrados principalmente nas bacias da região sudeste do Brasil e nas bacias africanas em Angola. Surge a cadeia mesoceânica.
• Oceano aberto: o Oceano Atlântico aumenta de largura e inicia a circulação oceânica e a sedimentação carbonática marinhas.
Antelior ou ao longo do estiramento da crosta terrestre e da formação do Atlântico, algumas feições geológicas diferentes foram produzidas. O atual Rio da Prata encaixou-se em fraturas e falhas geológicas ortogonais à direção da abertura atlântica durante o Cretáceo Inferior6. A Serra da Mantiqueira, já existente desde 500 Ma, foi reativada e soerguida durante o Cretáceo Superior (aproximadamente 80 Ma), fazendo com que a direção do sistema fluvial e, em consequência, do aporte sedimentar das bacias do Paraná e Santos, se alterasse. Outras feições produzidas foram as bacias sedimentares do Recôncavo e Tucano, próximo a Salvador, e São Jorge, no sul da Argentina, que seriam direções onde a ruptura começou, no início do Cretáceo, mas não se desenvolveu (aulacógenos: não houve geração de crosta oceânica). O último elo da ligação física entre América do Sul e África se rompeu no início do Cretáceo ( 1 1 0 Ma) e estava relacionado às rochas antigas da província Borborema7, localizada na região entre os estados de Sergipe e Rio Grande do Norte, no Brasil, e a Nigéria, na África. Exemplo atual para ilustrar como teria ocorrido a separação entre América do Sul e África é encontrado na região leste do continente africano. A visualização de um mapa geográfico revela a existência de uma série de lagos alinhados a grandes sistemas de falhamento. Com efeito, nesta região o magma do manto forçou a crosta, rompendo-a, com abatimento de grandes blocos (rift valleys africanos), que se transformaram em lagos relativamente profundos (por exemplo, Niassa, Tanganica, Kivu e Albert). Mais a norte, o golfo de Áden e o Mar Vermelho exibem o estágio posterior.

8 - H istórico e Conceitos
quando ocorreu formação de crosta oceânica e a crosta continental já está dividida em duas placas (placa Africana, a sul e placa da Arábia, a norte). O rebaixamento crustal decorrente fez com que a água oceânica ocupasse toda a região. O Oceano Atlântico, em seu início, foi semelhante ao Mar Vermelho, apenas com extensão maior. E o Mar Vermelho, no futuro geológico, caso haja o rompimento de uma estreita faixa de terra, poderá se unir ao Mar Mediterrâneo, dando origem a um novo oceano, tal como aconteceu no passado com o Atlântico.
A fragmentação da Pangeia e de Gondwana ao longo do Cretáceo produziu estiramentos crustais, rebaixando a crosta continental, e provocou elevação progressiva do nível do mar. A invasão, pelo mar, de grandes extensões continentais permitiu o estabelecimento de condições úmidas e desenvolvimento de florestas em vários continentes. De um modo geral, porém, após a separação da África, a metade norte da América do Sul continuou sob clima quente e seco até o final do Mesozoico. Em toda esta extensa região, as fases úmidas foram relativamente raras e, ao contrário, em vários locais, são encontradas evidências de condições áridas (por exemplo, Formações Guará [final do Jurássico/início do Cretáceo], Botucatu e Grupo Bauru [Cretáceo] na Bacia do Paraná; Formações Corda e Grajaú [Cretáceo] na Bacia do Parnaíba). Cabe destaque ao "deserto de Botucatu", com idade de 130 Ma (Cretáceo), cujas dunas encobriram desde o norte do Uruguai e Argentina, parte do Paraguai até o sul do Estado do Mato Grosso, sendo comparável ao deselio do Saara.
A Patagônia e o norte da Península Antártica, entretanto, na maior parte do tempo mantiveram-se em condições de clima mais úmido, com extensas florestas onde evoluíram notáveis paleofaunas de dinossauros. As fases marinhas, no grande golfo formado durante o Jurássico (Bacia de Neuquén, Argentina), aberto para o Pacífico, são marcadas por exuberante paleofauna de invertebrados e também por abundantes peixes e répteis aquáticos (ictiossauros, plesiossauros, pliossauros e tartarugas).
.. A faixa andina compreende uma grande cadeia de mon-tanhas que se estende por quase 9.000km de extensão, desde o extremo sul do Chile até o norte da Colômbia, possuindo entre 250 e 900km de largura. A geomorfologia atual dos Andes apresenta basicamente três distintos relevos:
• Região norte: onde os Andes estão segmentados em diferentes cadeias de montanhas.
• Região central: com os maiores cumes e maior largura, com destaque ao cerro Aconcágua com 7 mil metros acima do nível do mar (fronteira entre Argentina e Chile, próximo a Mendoza).
• Região sul: mais estreita e com montanhas de menor altitudé8.
Como mais um notável exemplo da mutabilidade da superfície terrestre, hoje a faixa andina apresenta montanhas com grande altitude, mas no passado, anterior às montanhas, exibia terrenos mais planos, semelhante aos � que hoje existem na margem do Atlântico. t:
.:.., A cordilheira dos Andes evoluiu de distintos modos � ao longo de sua grande extensão, em razão das variações 00
"" nos esforços tectônicos. Nos últimos 200 milhões de 6: anos, dois grandes eventos contribuíram para formar a geografia andina atual. O primeiro foi relacionado à separação da África e à migração do continente sul-americano para oeste. Esta migração fez com que fosse gerada uma tensão no limite com a placa de Nazca (uma das placas tectônicas que compõem o assoalho do Oceano Pacífico), gerando espessamento da crosta e produzindo as primeiras elevações do terreno, associada à cadeia de picos vulcânicos. O segundo e mais forte evento iniciou-se no fim do Mesozoico e esteve relacionado à intensificação dos esforços convergentes no limite com a placa de Nazca, gerando um cordão montanhoso (Patagonides). A principal consequência foi a subducção da placa de Nazca por baixo da placa sul-americana provocando espessamento da crosta sul-americana, em consequência da compressão dos sedimentos da margem continental. Subducção ocorre quando a resistência das rochas é vencida pelo acúmulo de pressão em decorrência de movimentos contrários e, neste caso, a placa com crosta oceânica, mais densa, se desloca para baixo, em sentido ao manto. Esta é a explicação para a existência da fossa do Peru-Chile no bordo oeste do continente. O desenvolvimento de uma zona de subducção no bordo oeste da América do Sul teve grandes repercussões, dando forma ao relevo atual daquela região. Na Patagônia, a pendente regional se inverteu de oeste para leste e, na parte norte, o mar passou a entrar no continente a partir do Atlântico .
No início do Oligoceno (34 Ma) a intensidade dos esforços na borda oeste da América do Sul aumentou e o soerguimento dos Andes foi acelerado. Como consequência, maior volume de rochas da placa de Nazca penetrou sob a placa sul-americana. O ângulo de penetração da placa de Nazca tem variado ao longo do

tempo nas distintas regiões e isto tem reflexo direto sobre o vulcanismo. Na região central do Andes, por exemplo, o ângulo de subducção é baixo (5°) e o vulcanismo é quase inexistente, ao passo que em outras regiões, onde o ângulo é maior, o vulcanismo é mais intens06.
A intensificação do soerguimento dos Andes provocou substanciais mudanças ambientais e climáticas no bordo oeste da placa sul-americana. Na região da Amazônia, a pendente regional foi invertida e a drenagem, que fluía em sentido ao Oceano Pacífico, se inverteu, dirigindo-se, a partir de agora, ao Oceano Atlântic09. A região do pantanal mato-grossense também foi afetada, tendo o escoamento sido dificultando em decorrência da fase mais moderna do soerguimento dos Andes10. No norte da Patagônia Argentina, o mar interior que existia a leste dos Andes desde o início da era Cenozoica desapareceu e o clima tornou-se extremamente árido. A mudança do clima úmido para desértico na Patagônia é explicada pelo "efeito de sombra de barreira", ou seja, antes dos Andes, nas latitudes superiores a 35° sul, a circulação atmosférica dominante dirigia o vento úmido do Oceano Pacífico diretamente para o interior do continente (ventos contra-alísios ou westerlies, relacionados à rotação da Terra). Com o aparecimento das montanhas, o ar úmido do oceano necessita se elevar alguns milhares de metros, esfriando e perdendo capacidade de reter umidade. Deste modo, a umidade fica retida no lado J.ndino do Pacífico (Chile). Ao cruzar a cordilheira, o yento se torna extremamente seco e, quando baixa à Patagônia Argentina, se aquece novamente e passa a retirar umidade do solo, ressecando-o. Ao contrário da Patagônia, o deserto de Atacama, por se situar no lado �este da América do Sul (norte do Chile), em latitude :nferior a 30°, apresenta circulação atmosférica dominaja pelos ventos alísios, que se movem de leste-sudeste ;Cara oeste-noroeste. Têm, portanto, que atravessar todo .) continente sul-americano, deixando boa parte da Jmidade pelo caminho. Depois de subir a cadeia andina, ='00 e completamente seco, o vento induz grande resse: amento em toda a região . Cabe destacar que a : irculação atmosférica terrestre tende, nas latitudes :�opicais, a tornar mais úmido o bordo leste dos conti:-.entes e mais árido o bordo oeste. A situação se inverte
.. :-'Ci faixa subtropical.
:'-Ja parte média do Mioceno ( 14 Ma), em uma fase �elativamente quente e úmida da Terra, o mar encobriu ;rande parte da América do Sul, Europa e Ásia. No :erritório sul-americano, em parte também em conse.:.uência da flexura de uma faixa interna paralela aos -\ndes, um grande braço de mar interior se estendeu
Evolução Geológ ica da América do Sul nos Ú lt imos 250 M i l hões de Anos - 9
desde a Amazônia até a Patagônia, limitado, a oeste, pela cadeia andina, e a leste, pelo escudo brasileirol l . O escudo das Guianas, a norte, e o escudo Brasileiro (Uruguai e regiões sul, sudeste, centro-oeste e nordeste do Brasil) tornaram-se, neste tempo, grandes ilhas.
A América do Sul possui muitos registros de eventos vulcânicos e hoje, na cadeia andina, muitos vulcões estão em atividade. Contudo, durante o Cretáceo, em um momento anterior, portanto, ao soerguimento dos Andes, o continente foi palco de um dos maiores eventos vulcânicos da história geológica da Terra. Deste episódio, de curta duração temporal, resultou a estruturação do que hoje conhecemos como a Serra Geral, região com predominância de relevo do tipo planalto, com altitude moderada a alta, e dominada por rochas de origem vulcânica. Vestígios dos derrames de lava são encontrados na região sudeste da América do Sul, dentro dos limites da bacia sedimentar do Paraná, estendendo-se por ampla região que inclui o sul do Mato Grosso, Goiás e Minas Gerais, oeste de São Paulo, Paraná e Santa Catarina, norte e oeste do Rio Grande do Sul, nordeste da Argentina, oeste do Uruguai e sudeste do Paraguai. As rochas vulcânicas estão espalhadas em área aproximada de 1 ,2 milhão de km2 (equivalente à extensão areal do Peru), podendo atingir até 1 .700m de espessura no oeste do estado de São Paulo. Nos estados de Santa Catarina (Serra do Rio do Rasto) e Rio Grande do Sul (Aparados da Serra) existem taludes verticalizados onde podem ser observados vários derrames.
Eventos vulcânicos são quase sempre associados aos vulcões isolados, cujas erupções geram edifícios vulcânicos em forma de grandes cones. Entretanto, o vulcanismo que ocorreu na margem leste da América do Sul foi diferente. Sua origem está relacionada à grande e praticamente interminável fonte de calor que se instalou abaixo da crosta, na região próxima à divisa entre os estados de Goiás e Mato Grosso. Esta fonte de calor, que pode ser designada como pluma mantélica (hot spot), pode ser explicada como uma bolha de magma com grandes proporções e alta temperatura, originária de região profunda do manto terrestre. Teria subido atravessando o manto e se posicionado na parte inferior da crosta continental sul-americana. Este material forçou e enfraqueceu a crosta até que, por grandes fraturas, o magma ascendeu e extravasou lavas de composição

1 0 - Histórico e Conceitos
básica e ácida. As erupções vulcânicas que formaram a Serra Geral ocorreram no início do Cretáceo, entre 137 e 1 27 Ma. Os derrames se iniciaram na região norte da Bacia do Paraná (divisa de Goiás com Minas Gerais) e foram migrando para sul através do tempo, de modo que os últimos eventos se deram no Uruguai. Em verdade, a pluma mantélica se manteve estacionária e a variação regional do vulcanismo registra o deslocamento do continente para norte durante este períod012.
Ao longo dos últimos 250 milhões de anos, vários eventos de extinção eliminaram diferentes níveis de vida (flora e fauna) em toda a superfície da Terra. Estes eventos, catástrofes com relação à vida, são importantes marcadores de fim e início de eras e períodos da escala do tempo geológico. O conteúdo fossilífero e as características dos estratos sedimentares são estudados visando compreender os fenômenos físicos envolvidos, buscando revelar quais teriam sido as causas potenciais para o desequilíbrio ambiental dos grupos orgânicos afetados (fatores climáticos e/ou geográficos, ou mesmo externos à Terra). A expressão "evento de extinção em massa" é usada quando a extinção acontece em tempo geológico extremamente curto, afetando grande diversidade de organismos que ocupam diferentes ambientes ou nichos ecológicos, e elimina um percentual significativo de famílias, classes ou mesmo filos. Como parâmetro, é importante salientar que se analisa na escala do tempo geológico e, por conseguinte, alta taxa de mortalidade, perceptível na ordem de grandeza de 1 milhão de anos, pode ser considerada como muito rápida.
O estudo sistemático mostrou que várias causas naturais podem ser atribuídas para explicar o desaparecimento de grupos de organismos em curto intervalo de tempo. Entre estas se destacam mudanças climáticas, impactos de meteoros, rebaixamentos do nível do mar em decorrência de eras glaciais, vulcanismo e, em ambientes marinhos, mudança no padrão de circulação das correntes oceânicas e disponibilidade de nutrientes.
A Terra registra cinco grandes eventos de extinção em massa que provocaram sérias alterações na vida da Terra nos últimos 700 Ma. Três destas extinções ocorreram nos últimos 250 Ma:
• Na passagem do Permiano ao Triássico, há 245 Ma (eliminou 95% das espécies).
• Limite Triássico-Jurássico, há 208 Ma (eliminou 76% das espécies).
• Limite Cretáceo-Paleógeno, há 65 Ma (eliminou 75% das espécies) 13 .
A extinção do limite Permiano-Triássico se deu em intervalo de aproximadamente 1 milhão de anos e estaria relacionada à combinação de mudanças na paleogeografia e queda do nível do mar (encurtamentos crustais decorrentes da formação da Pangeia) , ao intenso vulcanismo e às alterações climáticas consequentes (desertificação). Foi a maior crise da história da Terra, afetando linhagens muito antigas de anfíbios e répteis, que desapareceram, restando, no início do Triássico, diversidade faunística muito baixa e nichos ecológicos quase vazios e prontos para serem ocupados por espécies oportunistas 14. Esta extinção constitui-se em um dos principais campos de estudo da ciência geológica, para onde converge a atenção de pesquisadores de muitas especialidades diferentes15 . Por sua vez, a extinção do limite Triássico-Jurássico foi mais seletiva, afetando principalmente as floras Thinnfeldia e Dicroidium, grandes anfíbios e répteis sinápsidos e arcos sáurios (com exceção dos dinossauros) . No final do Triássico, decorridos mais de 30 Ma da grande crise do limite Permo-Triássico, a fauna reptiliana estava novamente muito diversificada. Com a crise do limite Triássico-Jurássico, porém, a diversidade faunística reduziu outra vez. Não tendo sido afetados de modo significativo, os dinossauros, no início do Jurássico, estavam habilitados ao domínio absoluto dos continentes . A "era dos dinossauros", de fato, inicia-se a partir deste evento. Hipóteses para explicar a crise do limite Triássico-Jurássico incluem desde erupções vulcânicas maciças, queda no nível do mar, mudanças no clima até impacto de meteoro, mas até o momento não há consenso sobre o que de fato teria ocorrido. Existe a suspeita de que as alterações na vegetação, com supressão de grupos importantes e expansão das coníferas 1 6, 17, tenha afetado diretamente aos herbívoros e, em consequência, também aos carnívoros.
Durante a passagem Cretáceo-Paleógeno, as extinções aconteceram em curto intervalo de tempo geológico, inferior a 500 mil anos, em razão principalmente da queda de um grande meteoro no golfo do México. No início da década de 1 990, foi encontrada uma grande cratera de impacto na região de Chicxulub, na atual península de Yucatán, com diâmetro de l 70km, fonnada pelo choque de um meteoro com cerca de l Okm de diâmetro. A energia liberada pelo choque deste bólido com a Terra seria equivalente a várias vezes a detonação simultânea de todo atual arsenal atômico da Terra1S . Este impacto provocou drásticas mudanças ambientais rela-

cionadas ao intenso calor gerado no momento do choque, grandes incêndios, noites intermináveis pelo acúmulo de pó na atmosfera superior, ocasionando o bloqueio da radiação solar e, em consequência, frio glacial, mudanças na química das águas do mar e oscilação do nível nos oceanos. Estas mudanças afetaram rapidamente toda a cadeia alimentar. A fotos síntese pode ter deixado de acontecer durante alguns anos, em decorrência do bloqueio da entrada de luz solar1 9•20.
As suspeitas de que a extinção dos dinossauros poderia ter sido ocasionada pela queda de um meteoro se iniciaram a partir do estudo de algumas regiões onde são encontradas rochas do limite Cretáceo-Paleógeno. Nestes locais, no ponto exato do limite existem camadas centimétricas de rochas sedimentares com altos teores do elemento químico irídio, comumente encontrado em meteoros. Ao contrário do que ocorreu com a extinção do Permo-Triássico, no qual as rochas não ficaram preservadas, o limite Cretáceo-Paleógeno pode ser encontrado em uma dezena de locais onde as evidências do evento são abundantes.
N a América do Sul três seções são conhecidas por possuírem rochas bem preservadas da passagem Cretáceo-Paleógeno: Bacia de Neuquén, na Argentina e Bacias de Campos e Pernambuco, no Brasil. O limite Cretáceo-Paleógeno em Pernambuco aflora em uma pedreira localizada a 30km ao norte da cidade de Recife e registra drástica mudança na microfauna marinha (por exemplo, ostracodes, foraminíferos, radiolários).
Além destes grandes eventos de extinção, ocorreram muitos outros episódios de redução na diversidade da nora e da fauna. O início do Eoceno (55 Ma) foi um período de grandes desequilíbrios, com extinção de muitos foraminíferos bentônicos nos oceanos. É bastante provável que a temperatura média global tenha se e levado cerca de 6°C em 20 mil anos21 . O efeito estufa decorrente, conhecido como "máximo termal do Eoce:10", deve ter liberado o metano aprisionado no fundo dos oceanos, injetando ainda mais CO
2 na atmosfera.
Como resultado, houve acidificação das águas e exp3Jl, ão da zona anóxica nos oceanos. Mas, a partir do ='.áximo termal, o plâncton marinho passou por uma fase
= de grande expansão e, nos continentes, as florestas tive-- �::m sua área muito aumentada e os mamíferos modernos
:�iciaram o seu grande desenvolvimento. Ao final do :\Iioceno (5 Ma), 30% das espécies de mamíferos desa:,areceram e existe a possibilidade de que isto tenha �' corrido em tempo muito curto e possa representar e?isódio de extinção de massa. Nas Américas, no ?leistoceno ( l O mil anos), grandes mamíferos desapa-e-:eram22, em percentuais que podem ultrapassar 50%.
Evolução Geológica da América do Su l nos Últ imos 250 M i lhões d e Anos - 1 1
Muitos, como mamutes, mastodontes, tigre dentes de sabre e preguiças gigantes se extinguiram de fato, enquanto outros, como elefantes, cavalos e camelos, desapareceram apenas nos continentes americanos, pois, de modo geral, Europa, Ásia e África foram menos afetadas.
A uniformidade climática do Mesozoico continuou por mais alguns milhões de anos durante a era Cenozoica (pelo menos até o final do Eoceno). A partir do Oligoceno, entretanto, a temperatura média começou a diminuir, primeiro mais lentamente, e com amplas flutuações, depois de modo mais acelerado a partir do início do Pleistoceno (2 Ma). No começo do Oligoceno (32 Ma), a Terra esfriou por curtíssimo espaço de tempo, causando grande queda do nível do mar, e pela primeira vez, geleiras permanentes se formaram na Antártica. Nos mares, a microfauna foi bastante afetada, e na Patagônia, onde este evento climático coincidiu com o soerguimento andino, houve o desaparecimento das floras tropicais e a diminuição das espécies subtropicais.
Há 1 ,5 Ma a calota polar ártica era imensa, encobrindo cerca de dois terços da América do Norte, toda a Groenlândia e o norte da Europa e Sibéria. Os continentes do hemisfério sul, à exceção da Antártica, foram menos atingidos que os do norte, tendo a calota polar sul se expandido sobre o Atlântico Sul. Na América do Sul, Terra do Fogo, Patagônia e metade sul da cadeia andina foram duramente atingidas. Vestígios de antigas geleiras são encontrados em todas estas regiões. Na Terra do Fogo, o canal de Beagle e o estreito de Magalhães são vales escavados por gigantescas geleiras que se dirigiam ao oceano durante o máximo avanço do gelo (entre 35 e 1 8 mil anos). Neste tempo, o volume de gelo sobre os continentes foi tão grande que o nível do mar encontrava-se entre 1 20 e 1 30m mais baixo do que hoje, expondo as plataformas continentais23. O rebaixamento do nível do mar terminou com o isolamento da porção norte da América do Sul e a partir de uma ponte de terras emersas (América Central) passou a ocorrerem trocas faunísticas entre as Américas do Norte e do Sul. Para a América do Sul, a invasão denotou significativa substituição da fauna de mamíferos, hoje composta em grande parte por elementos imigrados. A era glacial ocasionou a redução da área de florestas tropicais do mundo inteiro. Na Amazônia, acredita-se que grande parte da floresta foi alterada para savana por longos períodos, restando apenas núcleos de floresta (ilhas) em

12 - H istórico e Conceitos
setores mais úmidos. A floresta foi gradativamente reconstituída a partir do término da era glacial (entre 1 8 e 15 mil anos)24.
Vivemos em um mundo sob constante transformação, onde a mudança contínua elimina, em termos de tempo geológico, qualquer possibilidade de estabilidade. A mudança pode ocorrer de modo lento, na escala de milhões de anos; pode ser abrupta, envolvendo milhares de anos, ou mesmo instantânea, quando se relaciona aos eventos episódicos, como a queda do grande meteoro que levou os dinossauros à extinção. A contínua movimentação dos continentes por diferentes latitudes gera alterações climáticas de longo prazo, lentas e graduais, porém muitas vezes produzindo condições críticas ou mesmo inóspitas (por exemplo, formação da Pangeia; posição atual da Antártica no polo sul). As eras glaciais são disparadas em curto intervalo de tempo e em poucos milhares de anos o nível do mar pode baixar mais de uma centena de metros. Uma glaciação afeta não só as terras localizadas nas latitudes altas, mas também as das faixas equatoriais, pois diminui a capacidade dos continentes em reter umidade, por vezes ressecando extensas áreas. Ao contrário, épocas de grande atividade vulcânica geram efeito estufa ampliado, que derrete o gelo polar e inunda as terras baixas sobre os continentes, propiciando, muitas vezes, o grande desenvolvimento de florestas (em razão de maior umidade e CO
2 aumen
tado). Erupções vulcânicas catastróficas podem jogar grandes volumes de cinza na atmosfera superior, bloqueando a radiação solar e gerando, instantaneamente, frio glacial por vários anos, até que a atmosfera fique outra vez transparente à luz solar.
A América do Sul, nos últimos 250 Ma, passou por muitas transformações. Experimentou grandes alterações desde sua existência não individualizada no interior, primeiro da Pangeia e depois em Gondwana, em seguida constituindo um continente junto com a África, até finalmente se tornar um continente.
° vulcanismo foi um elemento importante da história geológica do continente, localizado primeiro no bordo oeste, no início da era mesozoica, depois no bordo leste ao final desta era e novamente no bordo oeste com a elevação da cadeia andina durante a era cenozoica. Neste jogo da tectônica de ruptura, que provocou a separação da África, e o consequente tensionamento no bordo oeste, o continente teve sua declividade alterada, produzindo diversos fenômenos, entre eles a inversão
no curso da drenagem (primeiro para oeste e depois para leste) e o sentido das invasões marinhas (do Pacífico no início, depois do Atlântico) .
Embora tenha migrado muito pouco em termos latitudinais (seu principal movimento é para oeste), o continente passou por grandes alterações climáticas. Desertificação nas baixas latitudes e condições mais úmidas e mesmo florestas nas latitudes médias e altas 'D
foram situações comuns na América do Sul ao longo da era Mesozoica. A separação da África e a elevação dos Andes modificaram esta tendência, e hoje temos, na região norte, umidade e floresta na parte central e aridez no lado do Oceano Pacífico, enquanto na região sul, acima dos 30° S, ocorre o inverso, com umidade no bordo oeste e aridez na Patagônia argentina. Durante os últimos 5 milhões de anos, a Terra do Fogo e a Patagônia foram ocupadas pelo gelo, mas a Amazônia também foi muito afetada, com expansão da savana e grande retração da área fiorestada.
A evolução da vida na Terra está relacionada aos eventos geológicos. Durante as grandes crises, a vida sofre redução de diversidade, mas depois, sob novas condições, segue adiante e se expande. Grupos orgânicos surgem, podem inclusive ser favorecidos por uma crise que afeta aos demais, porém desaparecem em algum momento. Seguindo o processo natural, nos próximos milhões de anos, os continentes continuarão em movimento, ocorrerão novos eventos vulcânicos e, como consequência, mudanças climáticas serão decisivas para o surgimento e a extinção de segmentos da vida na Terra26.
Aos professores Wilson Wildner e Cristianini Trescasto Bergue, pelas discussões sobre os diversos eventos ocorridos na América do Sul. À professora Tânia Dutra, pela revisão crítica do original e pelo auxílio na compreensão da evolução da fiora sul-americana.
REFERÊNCIAS BIBLIOGRÁFICAS 1 . DOTT, R. H.; BATTEN, R. L. Evolution of the Earth. 4. ed.
New York: McGraw-Hill Book, 1 988. 555p. 2. HALLAM, A. A. Review of Mesozoic climates. JOllrnal of the
Geological Society (London), v. 142, p. 433-445, 1985. 3 . DE WIT, M. J . ; BRITO NEVES, B . B.; TROUW R. A. J. et a!.
Pre-Cenozoic correlations across the South At1antic region: "the ties that bind". In: PANKHURST, R. J . ; TROUW, R. A. J . ; BRITO NEVES, B . B . et a! . West Gondwana: Pre-cenozoic Correlations Across the South Atlantic Region. Geological Society, London: Special Publications, 2008. capo 1, p. 1 -8 .
4. PARRISH, J. T. Climate of the supercontinent Pangea. Journal of Geology, v. 1 0 1 , p. 215-233, 1 993.
-J 00 00 U> .:.., N -1'0 , cc
'D O' O-

5. MOHRIAK, W; NEMCOK, M.; ENCISO, G. South Atlantic divergent margin evolution: rift-border uplift and salt tectonics in the basins of SE Brazil. In: PANKHURST, R. 1. ; TROUW, R. A 1. ; BRITO NEVES, B. B. et alo West Gondwana: Pre-cenozoic Correlations Across the South Atlantic Region. Geological Society, London: Special Publications, 2008. capo 19, p. 365-398.
6. ORNE, A. R. The tectonic framework of South America. In: VEBLEN, T. T.; YOUNG, K. R.; ORNE, A R. The Physical Geography of SouthAmerica. Oxford: Oxford Press, 2007. capo 2, p. 3-22.
7 . ARTHAUD, M. H.; CABY, R.; FUCK, R. A et a!. Geology of the northern Borborema Province, NE Brazil and its correlation with Nigeria, NW Africa. In: PANKHURST, R. J.; TROUW, R. A 1.; BRITO NEVES, B . B . et a!. West Gondwana: Pre-cenozoic Correlations Across the South Atlantic Region. Geological Society, London: Special Publications, 2008. p. 49-67.
8. NEVES, S. P. Dinâmica do Manto e Deformação Continental. 2. ed. Recife: Universitária, UFPE, 2008. 1 68p.
9. MAPES, R. W.; NOGUEIRA, A C. R.; COLEMAN, D. S . et a!. Evidence for a continent scale drainage inversion in the Amazon Basin since the Lat� Cretaceous. PhiladelphiaAnnual Meeting (Pennsylvania), GSA, paper n. 2 1 4-3, 2006.
10. ASSINE, M. L. A bacia sedimentar do Pantanal Mato-Grossense. In: MATESSO-NETO, Y.; BARTORELLI, A; CARNEIRO, C. R. et a!. Geologia do Continente Sul-americano: evolução da obra de Fernando Flávio Marques de A lmeida. 1 . ed., São Paulo: Beca, 2004. capo 4, p. 6 1 -74.
1 1 . WEBB, S . D. Biological implications of the Middle Miocene Amazon seaway. Science, V. 269, p. 3 6 1 -362, 1995.
1 2. WILDNER, W; SANTOS J. O. S.; HARTMANN, L. A et a!. Clímax final do vulcanismo Serra Geral em 135 Ma: primeiras idades U-Pb em zircão. In: XLIII CONGRESSO BRASILEIRO DE GEOLOGIA, 2006. Aracaju. Anais do XLIII Congresso Brasileiro de Geologia. Aracaju: SBG, 2008. CD ROMAO-480, p. 1 26.
1 .3 . BRENCHLEY, P. J . ; HAERPER, D. A T. Palaecology: ecosystems, environments and evolution. 1 . ed. Londres: Chapman & Hal!, 1 998. 402p.
Evolução Geológica da América do Sul nos Últimos 250 M i lhões de Anos - 1 3
14. BENTON, M. 1. Mass extinction, biotic and abiotic. In: NIERENBERG, W. A Encyclopedia ofEnvironmenta l Biology. San Diego: Academic Press, 1995. V. 2. p. 523-534.
1 5 . LAVINA, E. L. A geologia e o processo histórico (ou, sobre como se constrói um passado a marteladas). Gaea. V. 2. n. 1 , p. 29-39, 2006.
16. WILLIS, K. J.; MCELWAIN, J. C. The Evolution ofPlants. 1 . ed. New York: Oxford University Press, 2002. 378p.
17. DUTRA, T. L.; STRANS, A História das Araucariaceae: a contribuição dos fósseis para o entendimento das adaptações modernas da família no hemisfério sul, com vistas ao seu manejo e conservação. In: RONCH!, L. H. ; COELHO. O. G. (eds.). Tecnologia, Diagnóstico e Planejamento Ambiental. São Leopoldo: Unisinos, 2003. p. 293-3 5 1 .
1 8 . ALVAREZ, W. T. rex and the Crater of Doom. 1 . ed. New Jersey: Princeton University Press, 1997. 1 85p.
19. STANLEY, S . M. Earth System History. 1. ed. New York: Freeman and Company, 1 998. 6 1 5p.
20. LEVIN, H. The Earth Through Time. 8. ed. Hoboken: John Wiley & Sons, 2006. 547p.
2 1 . KENNETT, J. P.; STOTT, L. D. Abrupt deep-sea warming, palaeoceanographic changes and benthic extinctions at the end of the Palaeocene. Nature, V. 353, p. 225-229, 1 99 1 .
22. CARTELLE, C . Pleistocene mammals o f the cerrado and caatinga of Brazil. In: EISENBERG, J. F; REDFORD, K. H. Mammals of the Neotropics: the central neotropics. Chicago: University of Chicago Press, 1 999. p. 27-46.
23. SUGUIO, K.; ANGULO, R. 1 . ; CARVALHO, A. M. et aI. Paleoníveis do mar e paleolinhas da costa. In: SOUZA, C. R. G.; SUGUIO, K. ; OLIVEIRA, A. M. S. et a!. Quaternário do Brasil. I . ed. Ribeirão Preto: Holos, 2005. capo 6, p. 1 14-129.
24. HAFFER, 1. Ciclos de tempo e indicadores de tempos na história da Amazônia. Estudos Avançados, V. 6, p. 7-39, 1992.
25. HAQ, B. U. The Geological Time Table. 6. ed. Amsterdam: Elsevier, 2007.
26. SCOTESE, C. R. PALEOMAP Project. Disponível em: http :// www.scotese.comlearth.htm. Acesso em: 07/011201 0.

_____________________ 2
Juan J. Morrone
A biota da América do Sul tem sido e continua sendo fundamental para a investigação em biogeografia. B asta recordar que Darwin 1 iniciou a Origem das Espécies da seguinte maneira:
"Quando estava a bordo do H.M.S. Beagle como naturalista, fiquei muito impressionado com certos fatos da distribuição dos habitantes da América de Sul e das relações geológicas entre os habitantes atuais e passados desse continente. Parecia-me que estes fatos lançavam alguma luz sobre a origem das espécies - esse mistério dos mistérios, como tem sido chamado por um dos nossos principais filósofos ".
Durante o último século e meio foram levadas a cabo numerosas análises biogeográficas para América de Sul, tanto referentes a propostas de regionalização como a análise de padrões ecológicos e evolutivos. Da análise exaustiva dessas contribuições, com certeza, surgirão chaves para compreender a evolução da pesquisa biogeográfica sul-americana.
As regionalizações biogeográficas constituem elementos fundamentais que resultam da investigação biogeográfica. Elas são utilizadas por investigadores que precisam descrever a distribuição de um determinado táxon, estabelecer comparações bióticas entre áreas diferentes ou referir-se a essas áreas para conservação. Por isso, constituem sistemas de referência que são utilizadas pelas mais diversas disciplinas2.
Neste capítulo, pretendo fazer uma contribuição à historiografia da biogeografia da América do Sul. Do vasto material disponível, selecionei 1 2 propostas de regionalizaçã03-14. Meu objetivo é analisar criticamente e discutir essas regionalizações.
Cabrera e Yepes3, baseando-se na distribuição dos mamíferos sul-americanos, reconheceram as sub-regiões propostas por Sclater15 para a região Neotropical: sub-região guiano-brasileira e sub-região patagônica.
Corresponde à parte tropical da América do Sul. Nela predominam planícies com selvas ou bosque chaquenho e savanas com pequenos planaltos. As maiores elevações se situam ao noroeste da sub-região e estão constituídas pelos Andes equatorianos e colombianos e as serras do norte da Venezuela. Na parte oriental se encontra o planalto do Mato Grosso. Seu limite meridional segue uma linha mais ou menos oblíqua, do noroeste ao sudeste, que parte do norte do Peru e através do leste da Bolívia e o norte da Argentina alcança o delta do Rio Paraná.
Compreende o restante do continente ao sul da sub-região Guiano-brasileira, abrangendo a maior parte do Peru, Bolívia, Argentina e toda a região do Chile. Esta unidade foi denominada por Sclater15 e Wallace16 como sub-região Chilena, mas Cabrera e Yepes3 preferem a denominação de sub-região Patagônica.
Cabrera e Yepes3 também determinaram divisões de menor hierarquia, as quais designaram distritos (considerados pelos autores como equivalentes a províncias), com base em características fisiográficas e a presença de certas espécies de mamíferos. De acordo com esses autores, sua proposta é muito similar à de Mello-Leitão 17, fundamentada nos aracnídeos. Os 1 1 distritos reconhecidos para América do Sul são os seguintes (Fig. 2. 1 ) :
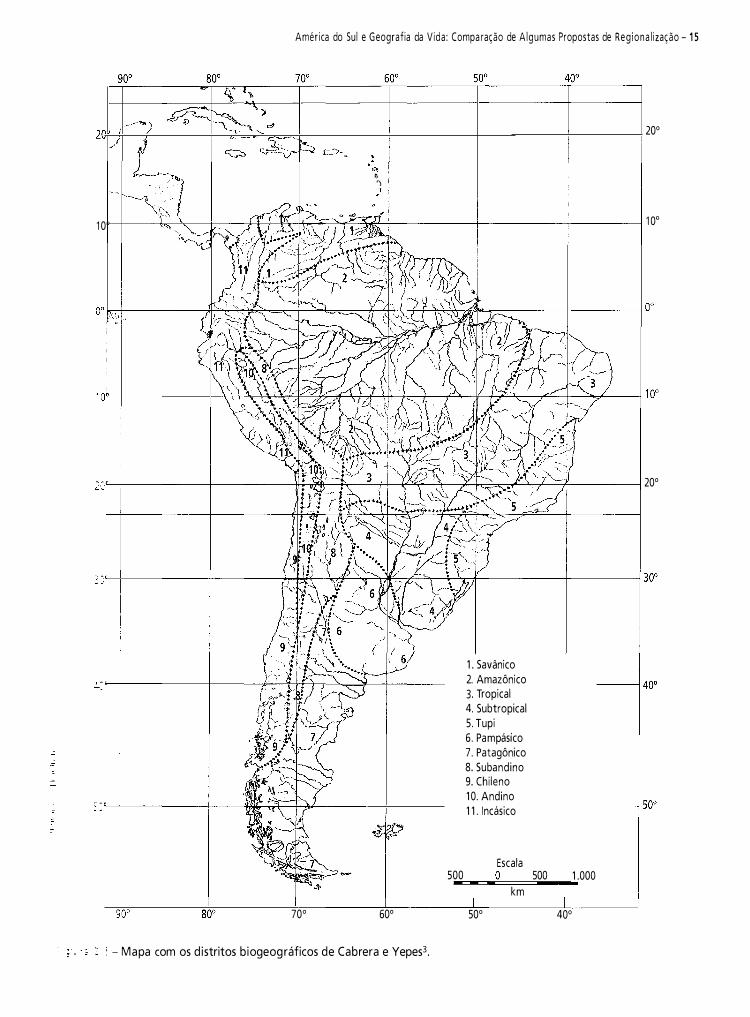
Amér ica do Sul e Geografia da Vida : Compa ração de Algumas Propostas de Reg iona l i zação - 1 5
20°
� '" n ') }
1 0 1 0°
: J'<---t--------t--------lt-i>--riJ''ft--t'I:--r--t----k'-�---'ir______;l_+c=,.._;;'I''-___r______;fj__---- 20°
1 . Savânico , -�------+---��HH�T-���4_---- 2. Amazônico
3. Tropical 4. Subtropical 5. Tupi 6. Pampásico 7 . Patagônico 8. Subandino 9. Chileno 1 0. Andino 1 1 . Incásico - '--------+-� ___ --1 50°
. .,� 3"i$.
Escala 500 O 500 1 .000
km 70° 60° 50° 40°
. - Mapa com os distritos biogeográficos de Cabrera e Yepes3.

16 - H i stórico e Conce itos
• Distrito Savânico: norte e centro da Venezuela e leste da Colômbia, em toda a zona de influência do Rio Orinoco, onde predomina a savana.
• Distrito Amazônico: centro do Brasil, nas bacias dos rios Amazonas e Madeira, desde as Guianas e a costa atlântica até o leste da Bolívia, Peru, Equador e Colômbia. Corresponde à maior parte da zona Guiano-Amazoniana de Mello-LeitãoJ7.
• Distrito Tropical: leste do Brasil, incluindo o Mato Grosso e o Chaco Boreal do Paraguai e Bolívia.
• Distrito Subtropical: sul do Paraguai, Uruguai, parte limitante do Brasil e a maior parte da bacia do Plata até o noroeste da Argentina dentro da formação chaquenha.
e Distrito Tupi: sudeste do Brasil. Corresponde à porção oriental da zona Tupi-Guaraniana de Mello-Leitão17 .
• Distrito Pampásico: litoral e centro da Argentina, através das pampas de Buenos Aires, Santa Fé e o leste de Córdoba e La Pampa. Corresponde à porção ocidental da área Tupi-Guaraniana de Mello-Leitão17 •
• Distrito Patagônico: alcança o limite mais austral de todos os distritos, compreendendo toda a estepe patagônica e os planaltos ocidentais até as proximidades da cordilheira meridional, continuando até o interior da Argentina, com forma de cunha entre os distritos Pampásico e Subandino.
• Distrito Subandino: ocupa a maior parte dos vales interiores e zonas de elevação média na Argentina, desde 45° S até o altiplano da Bolívia e Peru. Na Argentina corresponde em sua maior parte ao Monte ocidental. Pode dividir-se em uma zona norte e outra ao sul.
• Distrito Chileno: ocupa a maior parte dos vales e altitudes menores do Chile, exceto o extremo sul, que é uma continuação do distrito Patagônico, e a parte mais setentrional, que pertence ao Andino ou ao lncásico, segundo as zonas de altitude.
• Distrito Andino: grandes altitudes da Cordiheira dos Andes, desde o oeste da província de Mendoza (Argentina) até o sul do Equador através dos cumes entre Argentina, Chile, Bolívia e várias cadeias montanhosas do Peru.
o Distrito Incásico: extremo norte do Chile, na zona de menor altitude, junto com a parte pacífica do Peru, Equador e Colômbia, e adentrando até o noroeste na porção montanhosa da Venezuela.
Fittkau4 analisou as propostas de Mello-Leitão1 7 e Cabrera e Yepes3, encontrando que, mesmo que os grupos
animais sobre os quais as suas divisões se baseiam sejam diferentes (aracnídeos e mamíferos, respectivamente), as zonas delimitadas são muito similares entre si, correspondendo às principais formações de paisagens do continente. Apesar de aceitar as sub-regiões Guiano-Brasileira e Andino-Patagônica, ele considerou que estas eram difíceis de serem delimitadas com segurança, em decorrência de áreas de transição entre elas. Fittkau4 apresentou 1 3 províncias zoogeográficas, considerando que elas são muito similares aos distritos de Cabrera e Yepes3, mas levemente modificadas (Fig. 2.2) :
• Província da América Central: parte da província Centro-americana da sub-região Guiano-Brasileira. Área de florestas chuvosas tropicais da costa do Pacífico no noroeste da Colômbia.
• Província de Caquetío: bacia do Orinoco na Venezuela e oeste da Colômbia, onde predominam as savanas. Corresponde ao distrito Savânico de Cabrera e Yepes3.
• Província da Hileia: região de florestas chuvosas tropicais dentro da área dos rios Amazonas e Orinoco. Corresponde ao distrito Amazônico de Cabrera e Yepes3.
• Província Bororo: savanas e estepes desde o norte do Chaco até a cabeceira do Rio Tocantins e o Rio Paraguai. Corresponde a parte do distrito Tropical de Cabrera e Yepes3.
• Província Cariri: nordeste do Brasil, distrito semiárido na região dos rios Paranaíba e São Francisco. Corresponde a parte do distrito Tropical de Cabrera e Yepes3.
• Província Tupi: florestas costeiras do sul do Brasil, desde o sul da Bahia até Santa Catarina.
• Província Guarani: sul do Paraguai, sul do Brasil, Uruguai e nordeste da Argentina. É uma área de transição entre as sub-regiões Guiano-Brasileira e Andino-Patagônica, que inclui formações florestadas, savanas e estepes.
• Província lncásica: áreas de transição entre as regiões Guiano-Brasileira e Andino-Patagônica, e a província da América Central. Estende-se desde as áreas costeiras baixas no norte do Chile, através dos vales alinhados na Bolívia e no Peru, até as regiões montanhosas da Colômbia e o oeste da Venezuela.
• Província da Pampa: estepes das áreas centrais e costeiras da Argentina.
• Província da Patagônia: estepe patagônica e Monte setentrional.
• Província Subandina: lado oriental e áreas de altitude moderada na Argentina, desde 45° S até o altiplano da Bolívia e do Peru.
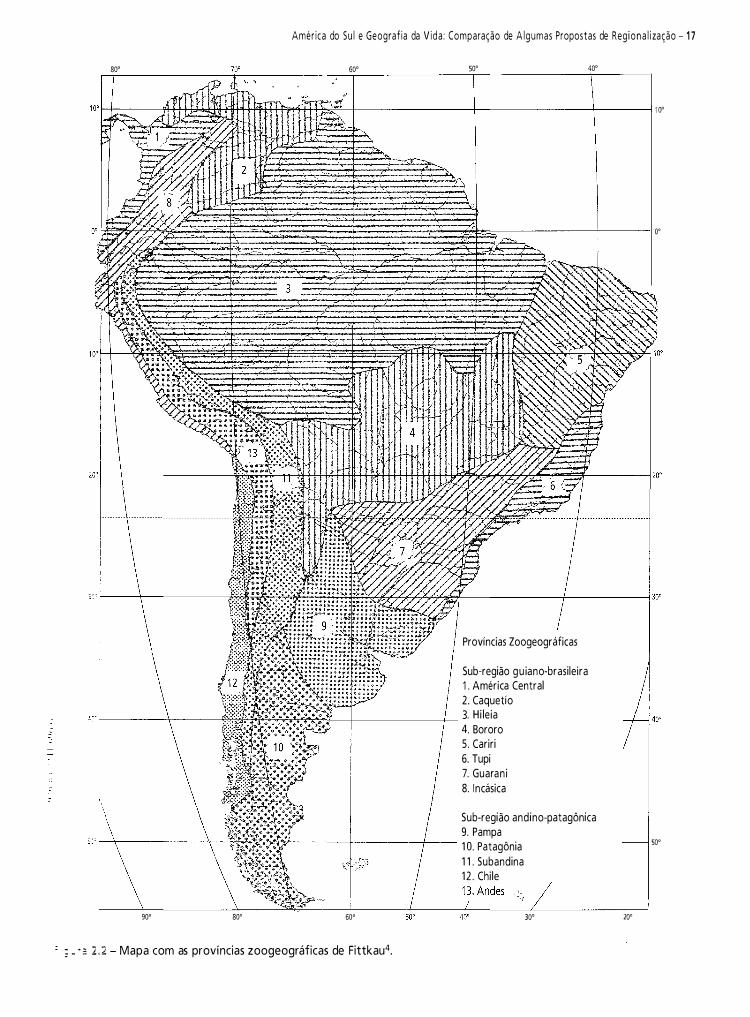
80'
lO'
20' 1--+-----------\i;*
, - o __ -+ _____ �
90' 80'
América do Su l e Geografia da V ida : Comparação de Algumas Propostas de Reg iona l i zação - 1 7
60' 50' 40' "
��_+_--------+_-------_+_---___j 1 0'
O'
:."r.,.<.A��f_------_I_-----.....j 30'
Províncias Zoogeográficas
Sub-região gu iano-brasileira 1 . América Central 2 . Caquetío
�WW�';:'--I---------J...- 3. Hi leia 4. Bororo 5. Cariri 6. Tupi 7. Guarani 8. Incásica
Sub-região andino-patagônica 9. Pampa
----+------+--- 1 0. Patagônia 1 1 . Subandina 1 2 . Chile
60' 30'
50'
20'
; _ . � 2 . 2 - Mapa com as províncias zoogeográficas de Fittkau4.

1 8 - H istórico e Conceitos
• Província do Chile: vales e áreas de altitude moderada no Chile, exceto nos extremos norte e sul.
• Província dos Andes: áreas altas dos Andes desde o norte da Argentina até o Equador.
KuscheP reconheceu duas sub-regiões, baseando-se na distribuição de insetos da ordem Coleoptera. A sua proposta é muito similar à de Sclaterl5 e de Wallace 1 6. As sub-regiões propostas pelo autor são (Fig. 2.3):
Alcança o sul do continente em Antofagasta, Chile, a oeste dos Andes, e pelo leste até o norte da Argentina, incluindo a formação do Chaco e Misiones, até Rio Grande do Sul no Brasil. Exclui os Andes e inclui as ilhas Galápagos. Nessa área se distribuem numerosas famílias e subfamílias de Coleoptera.
Compreende o restante do continente, incluindo as ilhas Desventuradas, Juan Fernández, Malvinas e Tristán da Cunha (Gough) . Esta sub-região coincide com a província ou sub-região Chilena de outros autores . Compreende os Andes acima dos 3 .000 a 3 .500m. Os táxons de Coleoptera desta sub-região estão nos níveis de subfamília e tribo.
Sick6, baseando-se em táxons animais, estabeleceu uma divisão primária entre Leste não Andino e Oeste Andino, subdividindo depois em unidades menores (Fig. 2.4).
Esta parte do continente inclui áreas tropicais e subtropicais, assim como áreas de montanha e terras baixas. São reconhecidas as seguintes unidades:
• Bacia do Amazonas: compreende a zona central na América do Sul tropical. O seu clima é conhecido por abundantes chuvas anuais (mais de 2.000mm), breves períodos secos e pelas altas temperaturas (temperatura média de 24 a 26°C). A vegetação característica é a floresta chuvosa tropical, com escassas áreas abertas (campos).
• Terras Altas da Guiana e Brasil: altitudes de 1 .000m,
alcançando altitude máxima de 3 .000m.
• Leste não Tropical: desde a bacia do Paraná-Paraguai
até os Andes, sem exceder os 200m de altitude. Tem
peratura média no inverno de 1 0 a 20°C e 1 .000 a
1 .500mm anuais de chuva. A vegetação característica
é floresta seca, com cactos e arbustos (Chaco).
Inclui as áreas andinas. Dentro desta unidade são reco
nhecidas:
• Andes Setentrionais : Venezuela, Colômbia e Equador,
alcançando pelo sul até 4° S e localizando-se comple
tamente na zona tropical.
• Andes Centrais: Peru e NOlte do Chile, entre 4 e 27° S.
• Andes Austrais: Chile e Argentina, ao sul dos 27° S ,
se localizando na zona temperada. A altitude varia de
6.000m no norte até 2.000m na Terra do Fogo. A
vegetação característica é floresta chuvosa temperada.
Cabrera e Willink7 apresentaram uma série de subdivi
sões (regiões, domínios e províncias) para América
Latina, fundamentadas na integração de dados de distri
buição de táxons animais e vegetais (para uma revisão
dos precedentes fito geográficos na obra de Cabrera, ver
Ribichich2). Na América do Sul, reconheceram seis
domínios (cinco assinalados à região Neotropical e um
à região Antártica) e 26 províncias (Fig. 2 .5) .
É o domínio mais setentrional da região. Estende-se
por quase todo México, parte da América Central,
Antilhas, Ilhas Galápagos e pequena parte da costa
atlântica da América do Sul. Inclui duas províncias na
América do Sul:
• Província Guajira: litorais do Caribe no norte da
Colômbia e parte ocidental da costa da Venezuela.
• Província das Ilhas Galápagos: Ilhas Galápagos no
Pacífico, a 1 .000km da costa do Equador, com fauna
e flora muito características e abundantes em ende
mismos.
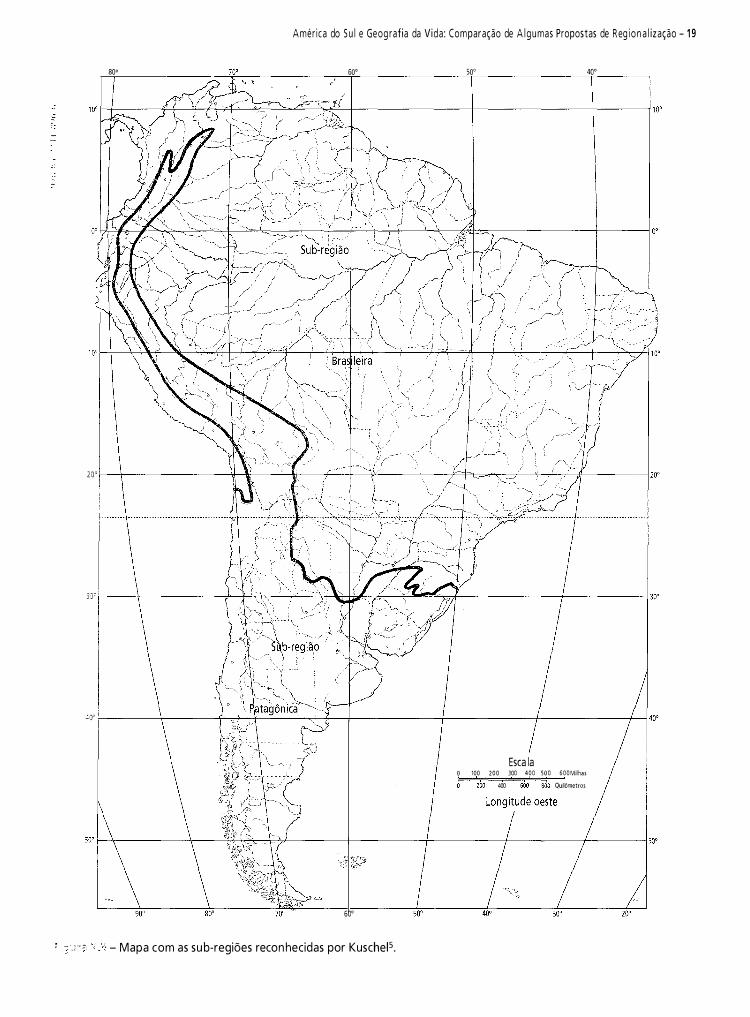
América do Su l e Geog rafia da Vida : Comparação de A lgumas Propostas de Reg iona l ização - 1 9
80' 60' 50' 40'
�---��-����-+-��-��_+---------+_--------_+---____1 1D'
20' r---\------------\\-�----'--+_-___i'-----_+-+---_:_-_f=r"t_;=-�____;_--4--+-o-__;;II_----____1 20'
H f----1----------;-i'---+'-":-:--'-----+-----'T+1!-b1T.-zic:--f:'-"'-��r----------+_-----___1 30'
':0' r----------'\-------f'---,,-'-+---=--�____;_-'----'--'---+-------I_------+_------__+____1
Esca la o 100 200 300 400 500 600 Milhas
200 400 600 800 Quilômetros
,0'r--+------+---�_f+_��-----+-----_f-----____;I_-----+_---____1
60'
r� - Mapa com as sub-regiões recon hecidas por Kuschels.

20 - H i stórico e Conce itos
80' 60' 50' 40' "
M=-----J��w(l/í�ez�t;:I���I------I------I--l l0°
40'r--------t------��7_��--��_t--t_------------r_----------_+------------7_�
Escala o 100 200 300 400 500 600 Milhas
200 400 600 800 Quilômetros
r-�--------_i----�����--------t_--------_i----------�--------_;�------J 50'
90' 80' 70' 60' 2 ,�, - Mapa biogeográfico proposto por Sick6. I = Leste não Andino; 1 . 1 = Bacia do Amazonas e á reas adjacentes; 1 . 1 a
= bacia do Amazonas; 1 . 1 b = bacia do Orinoco; 1 . 1 c = p lanícies do Mamoré; 1 .2 = terras a ltas setentrionais; 1 . 2a = terras a ltas da Gu iana; 1 .2b = terras a ltas do Brasi l ; 1.3 = Leste não Tropical ; l .3a = bacia do Paraná-Paraguai; 1 .3b = Grande Chaco; I .3c = Pampas; I .3d = Patagônia Orienta l ; 1 1 = Leste Andino; 1 1 . 1 = Andes Setentriona is; 1 1 .2 = Andes Centrais; 1 1 .3 = Andes Austra is .
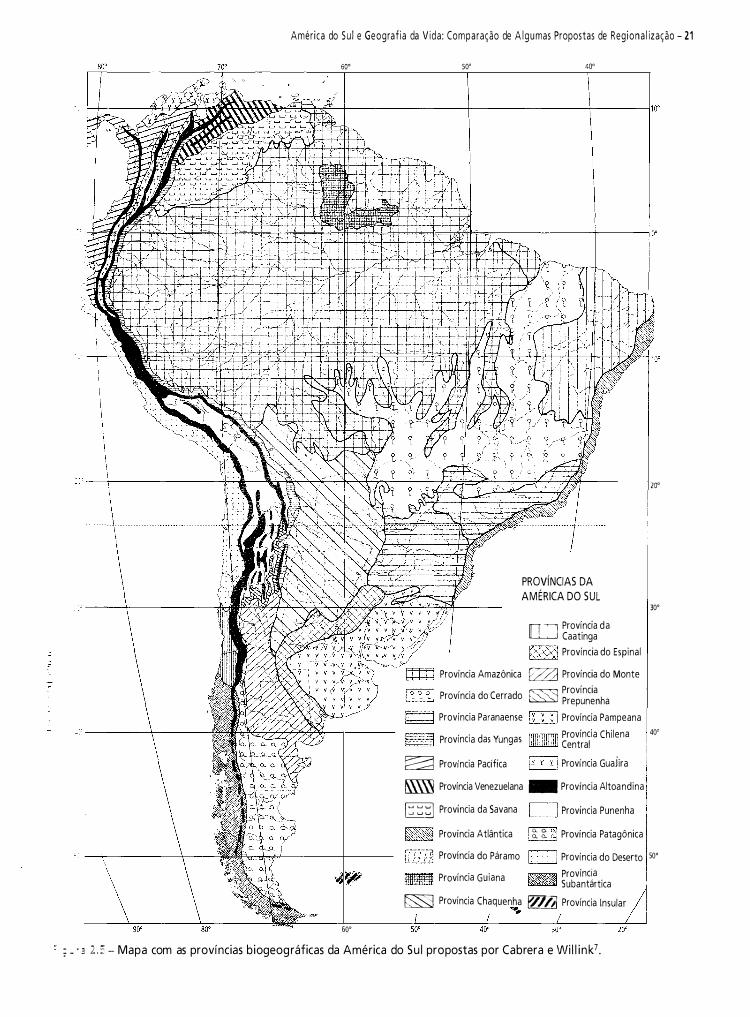
-.' -----+------4
América do Su l e Geograf ia da Vida : Comparação de A lgumas Propostas de Reg iona l ização - 21
600
600
500 400
PRovíNCIAS DA AMÉRICA DO SUL
[]]] Província da . Caatinga
Província do Espinal
Província Amazônica E2Zl Província do Monte
� Província Província do Cerrado Prepunenha
Província Paranaense Província Pampeana
Província das Yungas mmmmn Província Chilena illllilliWJj Central
� Província Pacífica Província Guajira
� Província Venezuelana _ Província Altoandina
�:;;] Província da Savana Província Punenha
Província Atlântica 1 � :5: :5:1 Província Patagônica
200
300
400
Província do Páramo [3 Província do Deserto 500 Província Guiana � Província
� Subantártica
� Província Chaquenha 1117/11 Província Insular .....
; - ' 3 ; L � - Mapa com as províncias biogeográficas da América do Su l propostas por Cabrera e Wi l l i nk7 ,

22 - H istórico e Conce itos
Cobre a maior parte da América do Sul e parte da Amé
rica Central. De acordo com o ponto de vista zoológico
é o mais abundante em formas e endemismos. Distinguem-se nove províncias:
• Província Amazônica ou Hileia: norte do Brasil, incluindo os estados do Amazonas, Pará, Acre, Amapá, noroeste de Maranhão, norte do Mato Grosso, norte de Guaparé, sul do Rio Branco, grande parte das
Guianas e da Venezuela, e o leste da Colômbia, Equador, Peru e Bolívia. Caracteriza-se pela riqueza e endemismos da sua fauna.
o Província Pacífica: compreende os lados ocidentais dos Andes equatorianos e colombianos, os vales interandinos e se estende pelas regiões baixas e os litorais da América Central.
o Província das Yungas: compreende as cordilheiras orientais dos Andes formando uma faixa estreita desde Venezuela até o noroeste da Argentina. Ao norte faz fronteira com a província Amazônica, pelas planícies da Colômbia e Venezuela, e ao Sul e ao leste, com a província Chaquenha.
o Província Venezuelana: situada entre a província da Savana da Venezuela e Colômbia, e os primeiros contrafortes dos Andes.
• Província do Cerrado: ocupa grande parte do Brasil, nos estados de Minas Gerais, Mato Grosso, Goiás, São Paulo, Paraná, Maranhão e Piauí, e atravessa até o noroeste do Paraguai.
o Província Paranaense: compreende o extremo sul do
Brasil, ao oeste da Serra do Mar, até o centro do Rio Grande do Sul, o extremo nordeste da Argentina e o leste do Paraguai. Pode subdividir-se em quatro distritos: ( 1 ) Florestas, ocupado por florestas sempre verdes; (2) Pinheirais, nas zonas elevadas do sul do Brasil , entre 600 e 1 .800m; (3) Campos, savanas entre as florestas; e (4) Serrano, nas partes mais elevadas das serras do sudeste do Brasil, acima de 1 .800m.
o Província da Savana: compreende as planícies da Venezuela e leste da Colômbia, no extremo norte da América do Sul.
• Província Atlântica: compreende os lados orientais das montanhas da costa do Brasil, desde 7° até 30° S .
o Província do Páramo: compreende as altas montanhas da Venezuela, Colômbia e Equador, aproximadamente entre 3. 800 e 4.000m. Na sua parte superior faz
fronteira com a província Altoandina.
Apresenta uma única província homônima. Ocupa su
perfície relativamente pequena no norte da América do
Sul, nos planaltos do Escudo Guiano, entre Venezuela,
Guiana e Brasil. Este domínio, também denominado
Pantepui, possui número alto de endemismos.
Ocupa uma área disjunta: a província da Caatinga no Bra
sil, e as cinco restantes ao sul.
o Província da Caatinga: compreende o nordeste do
Brasil nos estados da Bahia, Sergipe, Alagoas, Pernambuco, Paraíba, Rio Grande do Norte, Ceará e
porções do Piauí e Minas Gerais. • Província Chaquenha: compreende o sul da Bolívia,
oeste do Paraguai e norte da Argentina, até Córdoba,
San Luis e Santa Fé, desde quase a base da cordilhei
ra dos Andes até as ribeiras do Rio Paraná e um
pouco mais para o leste no noroeste de Corrientes.
o Província do Espinal: se estende pela Argentina, em
forma de arco, desde o centro de Corrientes e o norte
de Entre Ríos, pelo centro de Santa Fé e Córdoba,
grande parte de San Luis, centro de La Pampa, até o
sul de Buenos Aires. Distinguem-se três distritos: ( 1 ) Nandubay, entre Corrientes e Entre Ríos; (2) Caldén,
compreende o centro e sul de San Luis, centro de La
Pampa e sul de Buenos Aires; e (3) Algarrobo, no
centro de Santa Fe, Córdoba e norte de San Luis.
• Província Prepunenha: compreende os riachos e as
regiões secas do noroeste da Argentina, desde Jujuy
até La Rioja, entre 1 .000 e 3 .400m.
• Província do Monte: ocupa grande extensão das re
giões mais áridas da Argentina, desde 27° até 40° S , desde a província de Salta até o nordeste de Chubut.
o Província Pampeana: abrange as planícies ao leste da
Argentina, entre 30° e 39° S , Uruguai e a metade
austral do estado de Rio Grande do Sul, Brasil.
Podem-se diferenciar quatro distritos: ( 1 ) Uruguaien
se, no sul do Brasil, Uruguai, Entre Ríos e Santa Fé
na Argentina; (2) Pampeano Oriental, no norte e leste
da província de Buenos Aires, até as serranias de
Tandil e Barcarce; (3) Pampeano Ocidental, no sul
de Córdoba, nordeste de La Pampa e noroeste
de Buenos Aires; e (4) Pampeano Austral, no sul de
Buenos Aires, desde as serras de Olavarría, Tandil e
Balcarce, até próximo de Bahia Blanca.

América do Su l e Geog raf ia da V ida : Comparação de Algumas Propostas de Reg iona l ização - 23
Estende-se desde as altas cordilheiras da Venezuela e Colômbia, ao longo das cordilheiras e punas do Equador,
Peru, Bolívia e Argentina, até a Terra do Fogo, incluindo os desertos costeiros do Peru e Chile e a estepe patagônica desde Neuquén, oeste do Río Negro, Chubut e Santa Cruz. Nas regiões tropicais está limitado a altitudes superiores a 1 . 200m. A fauna deste domínio se distingue pelas suas adaptações a condições extremas da vida. Distinguem-se cinco províncias:
• ProvínciaAltoandina: compreende as altas montanhas
da cordilheira dos Andes, desde Venezuela até a Terra do Fogo. Nas zonas tropicais começa a partir de 4.200m e atinge a linha das neves perpétuas, para o sul encontra-se em altitudes menores, até 500m na Terra do Fogo.
• Província Punenha: cobre a Puna, altiplano que se
estende entre os dois braços que se abre a Cordilheira dos Andes, entre os paralelos I SO e 27° S . A altitude varia entre 3 .200 e 4.400m. Está abaixo da província Altoandina.
• Província do Deserto: compreende a costa do Pacífi
co, entre 5° e 30° S . Esta vasta província pode ser dividida em três distritos : ( 1 ) Deserto Costeiro, ocupa a zona mais seca da província; (2) Cardonales, nas vertentes ocidentais dos Andes, entre 1 . 500 e 1 .800m, por cima do distrito anterior; e (3) Coquimbano, pela costa do Chile desde a costa de Buenos Aires, ao norte de La Serena, até a altitude de Ovalle.
• Província Chilena Central: compreende a região cen
tral do Chile (exceto a alta Cordilheira), entre 32° e 38° S , aproximadamente.
• Província Patagônica: está localizada ao sul da Argentina, desde o centro da pré-cordilheira de Mendoza, se alargando até ocupar a parte ocidental de Neuquén e Río Negro, grande parte do Chubut, quase toda Santa Cruz e o norte da Terra do Fogo. Adentra no Chile na província de Aysén e no estreito de Magalhães. Distinguem-se seis distritos: ( 1 ) Payu
nia, no extremo setentrional da província; (2) Patagônico Ocidental compreende uma estreita faixa na parte ocidental da província; (3) Patagônico Central, no centro do Río Negro e Chubut e em quase toda Santa Cruz; (4) de San Jorge, nos arredores da Bahia
do mesmo nome; (5) Patagônico Subandino, estreita faixa ao longo da cordilheira austral, ao sul do paralelo 5 1 ° S, e alargando-se no extremo sul do continente; e (6) Fueguino, no nordeste da Terra do Fogo.
� ... ��� -..... - .... , � ....... -. ,-' �' . .. . �
Único domínio sul-americano designado à região Antártica. Tem por limite setentrional o Chile central, mais ou menos no paralelo 35° S, formando faixas estreitas sobre a Cordilheira da Costa e os primeiros contrafortes dos Andes. Estende-se pelo Chile até o Estreito de Magalhães e o lado leste dos Andes . Inclui a porção sudoeste da Terra do Fogo, a Ilha dos Estados, as Malvinas e as Ilhas Juan Femández. Cabrera e Willink7 descrevem três províncias:
• Província Subantártica: compreende o território inteiro do domínio, exceto as Ilhas Malvinas, Geórgias do Sul e Juan Femández. Distinguem-se cinco distritos : ( 1 ) Maulino, entre 34° 5 5 ' e 37° 20' S ; (2) Valdiviano, desde os limites do anterior até o sul, alcançando os 47° S, cobrindo todo o território chileno e estentendo-se em alguns pontos na Argentina; (3) Magalhânico, desde 47° S até o Cabo de Romos, cobre todo o sul do Chile e na Argentina compreende pequenas porções de Santa Cruz e Terra do Fogo; (4) do Pehuén, área descontínua, na cordilheira de Nahuel Butá no Chile, entre 37° 30' e 38° 40' S, e a cordilheira dos Andes, entre 37° 40' e 40° 10 ' S ; e (5) do Bosque Caducifólio, em toda a borda oriental da província Subantártica.
• Província Insular: compreende as Ilhas Malvinas e Ilhas Geórgias do Sul .
• Província de Juan Femández: compreende as ilhas do mesmo nome, localizadas no Oceano Pacífico, a 360 milhas ao oeste de Valparaíso.
Müller8 realizou uma análise da distribuição dos vertebrados na região Neotropical, propondo 40 centros de dispersão, com base na sobreposição das distribuições de espécies de anfíbios, répteis, aves e mamíferos. Destes, 33 são encontrados na América do Sul (Fig. 2.6):
• Centro Barranquilla: norte da Venezuela. • Centro Santa Marta: em florestas chuvosas de terras
baixas, localizadas na base da Serra de Santa Marta, Colômbia. Está separado do subcentro Nechí e do Centro Pacífico Colombiano pelas áreas secas do sul da Serra de Santa Marta.
• Centro Serra Nevada: acima de 1 .500m, nas florestas de montanha da Sierra Nevada, Colômbia.
• Centro Madalena: florestas chuvosas na bacia do Madalena.
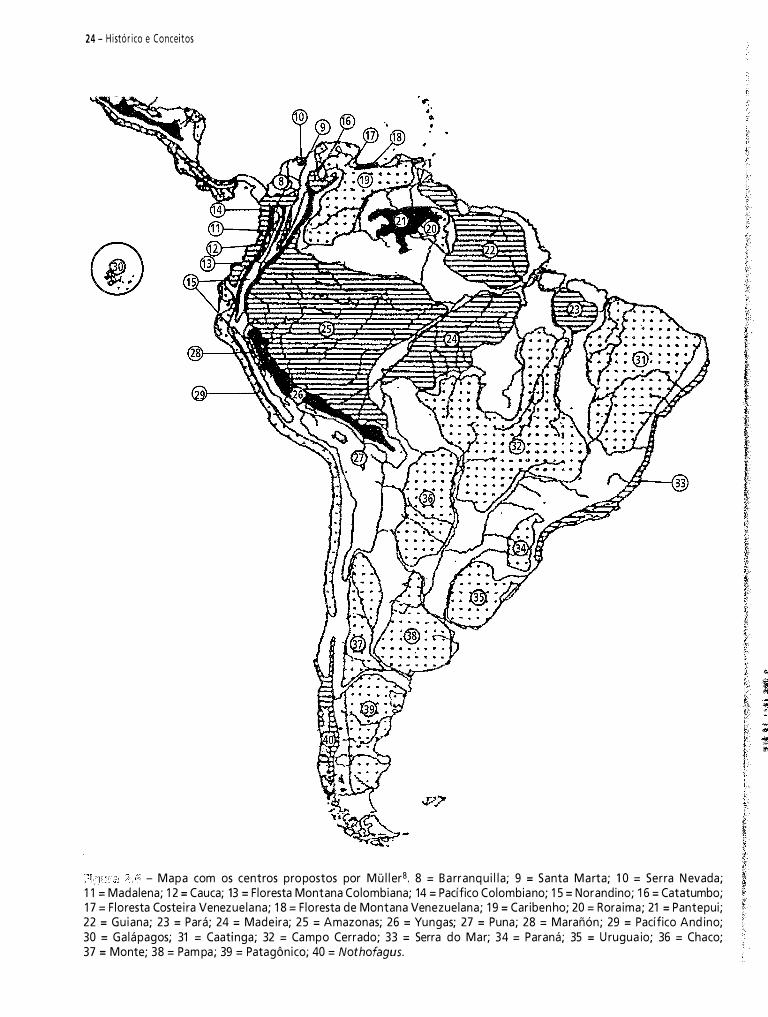
24 - H istór i co e Conceitos
- Mapa com os centros propostos por M ü l l er8. 8 = B a rranq u i l l a; 9 = Santa Marta; 1 0 = Serra N evada; 1 1 = Madalena; 1 2 = Cauca; 13 = Floresta Montana Colombiana; 14 = Pacífico Colombiano; 1 5 = Norandino; 1 6 = Catatumbo; 1 7 = F loresta Costei ra Venezuelana; 1 8 = F loresta de Montana Venezuelana; 1 9 = Caribenho; 20 = Rora ima; 21 = Pantepui; 22 = Guiana; 23 = Pará; 24 = Madeira; 25 = Amazonas; 26 = Yungas; 27 = Puna; 28 = Marafión; 29 = Pacífico And ino; 30 = Ga lápagos; 31 = Caatinga; 32 = Campo Cerrado; 33 = Serra do Mar; 34 = Paraná; 35 = U rugua io; 36 = Chaco; 37 = Monte; 38 = Pam pa; 39 = Patagônico; 40 = Nothofagus.

América do Su l e Geog rafia da Vida : Comparação de Algumas Propostas de Reg iona l ização - 25
• Centro Cauca: entre as cordilheiras oeste e central da Colômbia, incluindo o vale do Cauca e do Patía os quais se juntam ao sul, estando separados pelo altiplano de Popayán, a 1 .7S0m.
• Centro de Floresta de Montanha Colombiana: Colômbia. Podem-se distinguir dois subcentros : Andino Oeste (inclui os Andes Centrais) e Andino Leste.
• Centro Pacífico Colombiano: os limites dos elementos faunísticos deste centro estão correlacionados aos limites das florestas chuvosas baixas, que se estendem desde o oeste do Equador ao sul, até o extremo sul das montanhas do Darién ao norte; em direção ao leste, os componentes faunísticos entram ao norte na floresta do Cauca e do Madalena, mas estão ausentes dos centros Cauca e Madalena. Podem-se distinguir dois subcentros: ( 1 ) Nechí, entre os rios Sinú e Cauca; e (2) Chocó, a oeste dos Andes.
• Centro Norandino: com dois subcentros: ( 1 ) dos Andes Peruanos, páramo do noroeste do Peru; e (2) Bogotá, nos Altos Andes do Equador, Colômbia e Venezuela.
• Centro Catatumbo: Venezuela, entre sudeste da Serra de Perij á (na cabeceira do Rio Catatumbo) e o noroeste da Cordilheira de Mérida.
• Centro da Floresta da Costa Venezuelana: no noroeste das montanhas costeiras, entre Caracas ao leste e Tucacas ao noroeste; é predominantemente significativo nas terras baixas. Os componentes faunísticos típicos existem somente abaixo de I .SOOm.
• Centro da Floresta de Montanha Venezuelana: florestas de montanha da cordilheira da costa venezuelana. Os componentes faunísticos deste centro existem somente acima de 1 .400m.
• Centro Caribenho: com dois subcentros: ( 1 ) Maracaibo, que compreende as áreas secas da Península da Guajira e Falcón no Golfo de Maracaibo; e (2) Venezuelano, na região costeira venezuelana, ao leste da Península de Paraguaná, incluindo a ilha Margarita. Centro Roraima: localizado entre 600 e 1 .600m, inclui as savanas isoladas de Roraima, Ueitepui, Cuquenán, Gran Savana, monte Twekquay, Aparamán-tepui, Auyán-tepui, Uaipá-tepui, Aprada-tepui, Chimantá-tepui, Acopán-tepui e Upuigma-tepui.
• Centro Pantepui: oeste da Venezuela. • Centro Guiano: estende-se pelas Guianas, exceto as
regiões de florestas de montanha e savanas, e no sudeste da Venezuela (estado do Delta Amacuro e norte do estado de Bolívar); ao norte está limitado por uma faixa estreita de savanas ao longo da costa Atlântica; e no sul está conectado do leste ao oeste com os campos altos da Serra Tumuc-Humac, Avarai, Maquera, Pacaraima incluindo Roraima e Tapirapecó.
• Centro Pará: limitado ao oeste e norte pelo Rio Tocantins, se estende desde o norte do Rio Araguaia até Matheus; ao sul faz fronteira com a Serra do Gurupi, a porção norte de Maranhão e o Rio Grajau, ainda assim algumas espécies se estendem até o Rio Parnaíba; e ao leste está limitado pelo Rio Guaná e pela restinga da costa Atlântica.
• Centro Madeira: limitado ao norte pelo Rio Amazonas, ao oeste pelo Rio Madeira e Beni, ao leste pelo Rio Xingú, e no sul pela vertente leste da Cordilheira Boliviana.
• Centro Amazonas : limitado ao oeste pelo bioma Floresta de Montanha dos Andes, ao norte pelas áreas secas das planícies venezuelanas, a nordeste pelo Rio Negro e a Serra de Urutani, e a sudeste pelo Rio Madeira. As subespécies do norte pertencem ao subcentro Napo, com o Río Negro como limite nordeste e o Rio Amazonas como limite sul, enquanto as subespécies do sul pertencem ao subcentro Ucayali, com o Rio Amazonas como limite sul.
• Centro Yungas: os limites de distribuição dessas espécies estão correlacionados às florestas montanas andinas do leste, entre o alto Rio San Francisco e o Rio Bermejo ao sul, e o Rio Maranón ao norte.
• Centro Puna: representa a maior parte da Cordilera Andina.
• Centro Maranón: interandino. • Centro Pacífico Andino: costa do sul do Equador, Peru
e norte do Chile. • Centro Galápagos : arquipélago de Galápagos. • Centro Caatinga: nordeste do Brasil. • Centro Campo Cerrado: diagonal árida do nordeste
ao sudoeste do Brasil. • Centro Serra do Mar: compreende as florestas chuvo
sas do limite costeiro brasileiro (Serra do Mar no seu conceito mais amplo), desde o estado de Santa Catatina ao sul do estado de Pernambuco, ao norte, incluindo a linha das ilhas continentais desses estados. Seus limites são muito mais amplos que a Serra do Mar no sentido político-geográfico. No centro apresenta o maior número de gêneros de aves e anfíbios monotÍpicos. Podem-se reconhecer três subcentros : ( 1 ) Pernambuco, o limite mais ao norte, entre Salvador (Bahia) e Recife; (2) Bahia, ao sul do subcentro anterior, em floresta chuvosa; e (3) Paulista, na parte sudeste do limite costeiro brasileiro, desde Florianópolis ao sul até Cabo Frio ao norte.
• Centro Paraná: sudeste do Brasil. • Centro Uruguaio: o Rio Paraná forma seus limites
oeste e sul, e o bioma floresta chuvosa do Rio Grande do Sul e Santa Catarina constituem o limite norte.

26 - H i stórico e Conceitos
o Centro Chaco: centro norte da Argentina, Bolívia e Paraguai.
o Centro do Monte: está delimitado pelas Serras de Córdoba ao leste, os Andes a oeste, o Rio Salado ao norte e o Rio Colorado ao sul.
o Centro da Pampa: os rios Salado e Paraná constituem o limite norte desta unidade, as Serras de Córdoba formam o limite oeste, e o Rio Negro o limite sul.
o Centro Patagônico: os componentes faunísticos desse centro também alcançam as Ilhas Malvinas.
o Centro Nothofagus: florestas do sul do Chile e da Argentina.
Ringuelet9 fez uma análise global das áreas sul-americanas de peixes de água doce, analisando hipóteses paleogeográficas e estudos biogeográficos de autores prévios, e propondo genocentros. Como parte da sua revisão, ele propôs um sistema de 20 províncias ictiogeográficas (Fig. 2.7), as quais foram agrupadas em duas sub-regiões e sete domínios:
Parte tropical da América do Sul. Segue em direção ao leste até a cordilheira dos Andes e em direção ao sul, compreendendo Argentina até uma linha irregular que vai desde a bacia endorreica do Chasicó e Bahia Blanca ao sul de Córdoba, sul de San Luis e laguna Diamante em Mendoza. As principais bacias e com maior diversidade são as da Orinoquia, Amazônia, Paranaense e as vertentes do atlântico que são mais ou menos independentes . Compreende sete domínios : Madaleniense (província de Madalena), Orinoco-Venezuelano (províncias de Maracaibo, costa do Caribe, Orinoco e Trinidad), do Pacífico ou Transandino (províncias Norte-Pacífica e Guayas), Andino (províncias Norte-Andina, Titicaca e Sul-Andino Cuyana), Guiano-Amazônica (províncias de Guiana e Amazonas), Paranaense (províncias do Alto Paraguai, Alto Paraná e Párano-Platense) e do Leste do Brasil (províncias do Nordeste do Brasil, Rio São Francisco e dos Rios Costeiros do Sudeste do Brasil).
Parte temperada da América do Sul, compreendendo principalmente Chile central e austral, junto com a Patagônia andina e extra-andina. Compreende duas províncias : Chilena e Patagônica.
Ab'Sáber10 propôs uma série de domínios morfoclimáticos para América do Sul, utilizando critérios climáticos, geomorfológicos, fitogeográficos e ecológicos. O autor destacou que o mapa de vegetação de Hueck1 8 foi base t._
importante para o seu trabalho. Os domínios reconheci- -:c
dos são os seguintes (Fig. 2.8) : �
o Domínio Equatorial Amazônico: compreende quatro grupos principais de padrões: florestas em altiplanos baixos tabuliformes ; grandes planícies florestadas; sub setores mamelonizados florestados, que incluem pequenas serras com vertentes cobertas por floresta amazônica; e montanhas florestadas no oriente dos Andes, no Peru, Bolívia, Equador e Colômbia.
o Domínio Equatorial Pacífico: com florestas pluviais densas, desde o litoral até as vertentes médias dos Andes colombianos e o istmo de Panamá.
• Domínio Tropical Atlântico: reconhecido como o domínio dos "mares de morros" florestados, desenvolvido na faixa atlântica tropical do Brasil.
o Domínio dos Cerrados: nos planaltos centrais do Brasil, em áreas onde predominam climas tropicais úmidos com duas estações definidas.
o Domínio Roraima-Guianense: em posição equatorial, em terras amazônicas do hemisfério norte, na fronteira entre o território de Roraima com Venezuela e as Guianas.
o Domínio Intermontano Subequatorial: planícies da Venezuela e Colômbia, no vale do Orinoco, submetido a climas tropicais com duas estações.
o Domínio dos Andes Equatoriais: subsetor andino, com florestas pluviais densas, nas duas vertentes dos Andes (amazônica e colombiana).
o Domínio das Caatingas : no nordeste brasileiro, em áreas de depressões interplanálticas semiáridas, com drenagens intermitentes estacionárias.
o Domínio Subequatorial Caribe-Guajira: área semiárida quente, litoral e sublitoral, no Atlântico da Colômbia e Venezuela.
o Domínio do Chaco Central: área de planícies centrais sul-americanas, com florestas secas, com climas tropicais subúmidos e subtropicais semiáridos rústicos, na Bolívia, Paraguai e Argentina.
o Domínio dos Desertos Costeiros Pacíficos : faixa costeira azonal de desertos hiperáridos e nublados, que se estendem desde Tumbes, no Equador, até Chile Central (3 a 30° S) .
o Domínio das Punas e Desertos de Cordilheira dos Andes Centrais : desertos intermontanos de altitude, localizados em antigas depressões endorreicas em zonas montanhosas áridas dos Andes.
-
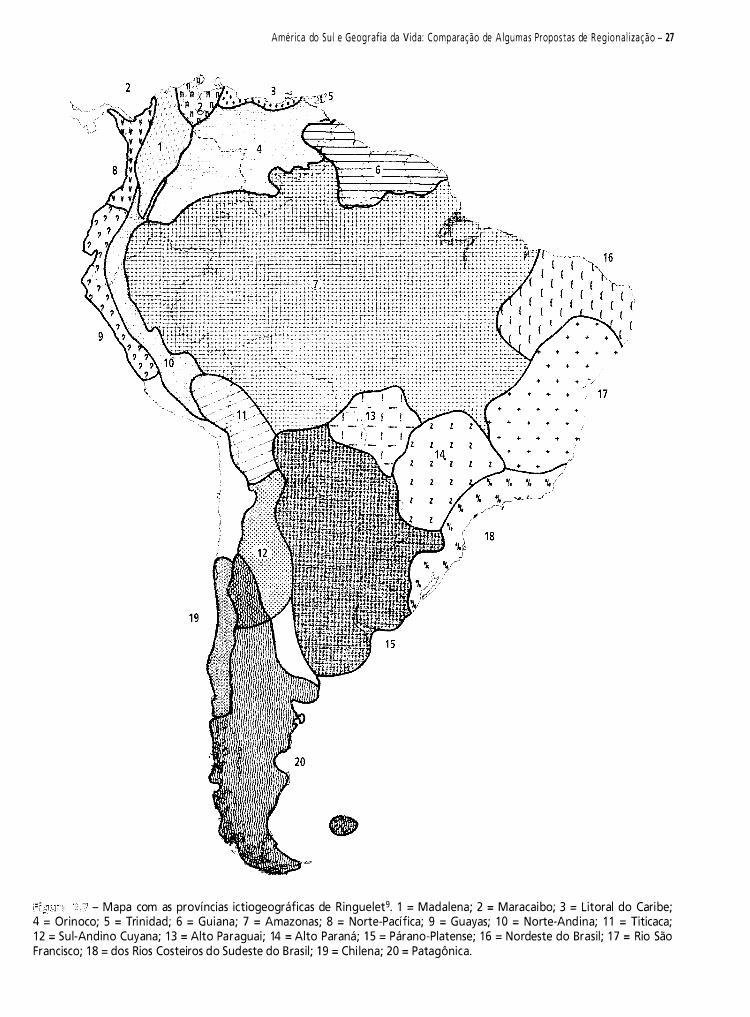
América do Su l e Geog rafia da Vida: Comparação de Algumas Propostas de Reg iona l ização - 27
- Mapa com as províncias ictiogeográficas de Ringuelet9. 1 = Madalena; 2 = Maracaibo; 3 = Litoral do Caribe; 4 = Orinoco; 5 = Trinidad; 6 = Guiana; 7 = Amazonas; 8 = Norte-Pacífica; 9 = Guayas; 1 0 = Norte-And ina; 1 1 = Titicaca; 1 2 = Su l-Andino Cuyana; 1 3 = Alto Paraguai; 14 = Alto Paraná; 1 5 = Párano-Platense; 1 6 = Nordeste do Brasi l ; 1 7 = Rio São Francisco; 1 8 = dos Rios Costeiros do Sudeste do Brasi l ; 1 9 = Chi lena; 20 = Patagônica.
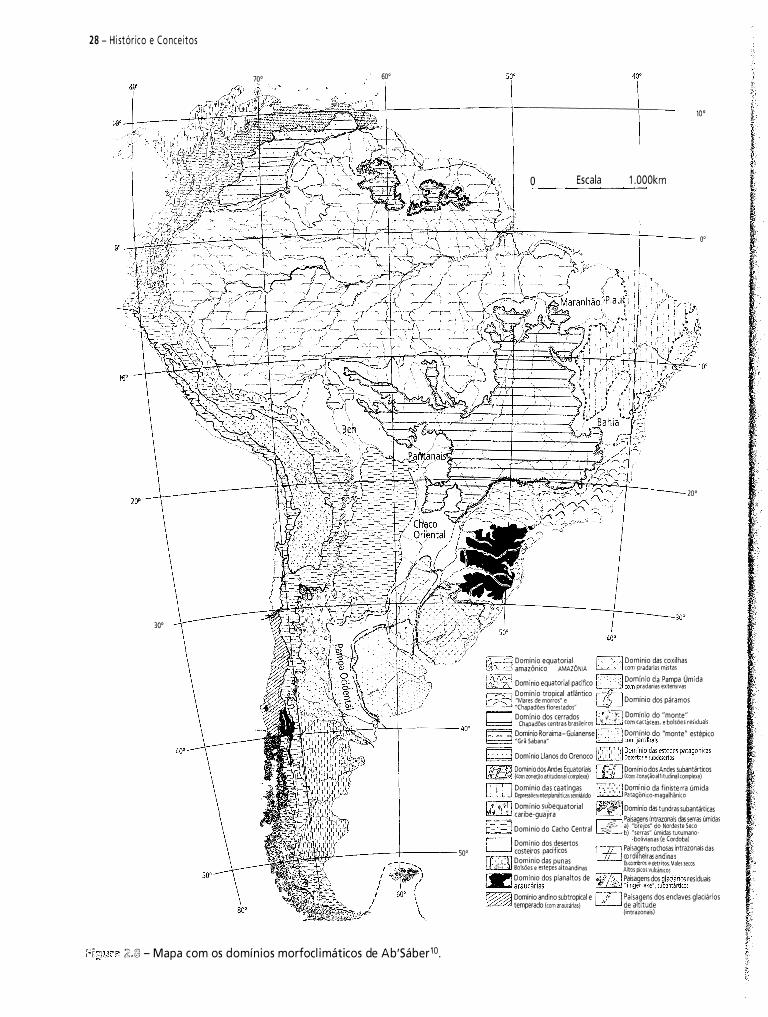
28 - H i stórico e Conceitos
70' 60'
3D'
, ___ --+----- 50'
;;US - Mapa com os domínios morfocl imáticos de Ab'Sáber10.
lO'
o Escala 1 .000km ------------------
O'
20'
40'
Domínio equatorial � Domínio das coxilhas amazônico AMAZÔNIA � com pradarias mistas
Domínio equatorial pacífico �.' Domínio �a Pampa Úmida � (om pradanas extenslvas
� Domínio tropical atlântico rI?à'i � "Mares de morros" e U ' U Domínio dos pá ramos "Chapadões florestados" Domínio dos cerrados !;4 y,: :r \ Domíni� do "mon_te" .
Chapadé5es centraiS brasileiros 't' . . • com eactaceas, e bolsoes residuais
Domínio Roraima -Guianense � Don:ín�o �o "monte" estépico "Grã Sabana" � com Jarrllhals
Domínio Llanos do Orenoco � Domínio dos �nd.es Equatoriais r-r-n Dom ínio dos Andes subantárticos � (com zOrlação atrtudlonal complexa) Ll::lLJ (com zonação altltudinal complexa)
Domínio das c�atingas 1':'� .:-.:-:1 Dom�ni,o da finis!e,rra úmida Depressõeslnterplanalticas semiáfldo •• . • . Patagonlco-magalhanlco 1,;�I4I1 tf{ I Do�ínio s��equatorial �" Domínio das tundras subantárticas canbe-guaJlra
__ Paisagens intrazonais das serras úmidas Domínio do Cacho Central I -:,::.:::;� ! a) :brejos" ?o .Nordeste Seco
b) serras" umldas tucumano-
Domínio dos desertos -bolivianas (e Cordobal
costeiros pacíficos � Pais�ge�s rocho�as intrazonais das
1 f :h:'; : �! Do�ínio das punas . UL-J ��c��b���r����i���t�le5 secos <.,.' • Bolsoes e estepes altoandmas Altos picos vulcânicos
� Domí�i? dos planaltos de fF�!� I Paisagens dos qlaciári0s residuais � araucanas . t·:"'. "finger lake", suoantártlcos
� Domínio andino subtropical e � Paisagens dos enclaves glaciários � temperado (com araucárias) L..::::-J de altitude
(intrazonaisl

América do Su l e Geograf ia da Vida : Compa ração de Algumas Propostas de Reg iona l i zação - 29
\O -O a, 00
Domínio dos Planaltos Sub-brasileiros com Araucárias: planaltos subtropicais atlânticos tapizados por
um velho núcleo de araucárias.
• Domínio Andino Subtropical e Temperado com Arau
cárias : montanhas de altitude média, com remanes
centes de araucárias, no setor meridional dos Andes.
• Domínio das Pradarias Mistas Subtropicais: prados localizados na metade sul do Rio Grande do Sul e a maior parte do Uruguai, com presença de florestas em
galerias subtropicais. , . Domínio da Pampa Úmida: grandes planícies pam
peanas com estrutura superficial e compartimentagens
complexas, cobertas por pradarias extensas . Domínio dos Páramos: paisagens residuais no setor equatorial dos Andes, em Colômbia e Venezuela, constituídas por pradarias de altitude.
� t-J, 00 00 t-a, •
• Domínio do Monte: planícies e ondulações áridas e
semiáridas, com florestas pouco densas e cactáceas,
condições arreicas generalizadas, raros rios alóctones
e riachos intermitentes, tributários de depressões lacustres (relativo a lago) residuais (tipo bolsões), no noroeste da Argentina.
• Domínio do Monte Estético: do oeste ao sudeste, na
retaguarda da pampa ocidental, com climas tempera
dos secos continentais, poucos rios alóctones e "oásis"
de piemonte. • Domínio Patagônico: com estepes subúmidas passan
do a áridas e semiáridas, do oeste para o leste da área de planaltos e ondulações baixas do sul da Argentina.
• Domínio dos Andes Subantárticos : submetidos a
climas temperados-frios e frios, com estratificação
altitudinal complexa e diferentes faixas de florestas boreais.
• Domínio de Finisterra Subúmida das Terras Patagô
nicas e Magalhânicas: coberta pela faixa mais boreal das florestas sul-americanas.
• Domínio das Tundras Subantárticas: paisagens de
tundras, na costa sul do arquipélago de Magalhães e áreas insulares (Ilhas Malvinas) .
• Paisagens das Serras Úmidas e Piemontes da Faixa Tu
cumano-Boliviana: área de climas úmidos nas vertentes
orientais dos Andes, depois da interrupção causada pela
"sombra de chuva" dos Andes meridionais. • Paisagens Rochosas: vales secos, altos picos de acu
mulação vulcânica e periglaciais, ao longo dos Andes,
em posição intrazonal. • Paisagens de Glaciais Residuais: sul dos Andes.
• Paisagens de Encraves Glaciários de Altura: picos altos
dos Andes, por cima do limite das "neves eternas".
Rivas-Martínez e Tovar1 1 propuseram que as terras elevadas dos Andes, desde Colômbia e Venezuela, os territórios da vertente pacífica entre os paralelos 5 e 38° de latitude sul e a Patagônia constituem uma unidade biogeográfica dentro do reino Neotropical, denominada sub-reino Andino. As outras unidades biogeográficas sul-americanas reconhecidas pelos autores são o sub-reino Caribenho-Amazônico, grupo de regiões chaquenhas, a região da Caatinga e a região Sub antártica. Dentro do sub-reino Andino, Rivas-Martínez e Tovarll reconheceram cinco regiões, caracterizadas pela presença de gêneros endêmicos de plantas (Fig. 2.9):
Terras andinas altas da Venezuela, Colômbia, Equador e norte do Peru; separadas na Costa Rica. Esta região possui os limites propostos para a formação do páramo, incluindo florestas e matagais perenifólios de folhas pequenas e duras.
Terras altas andinas do Peru, Bolívia, norte do Chile e norte da Argentina. Neste ponto os autores tratam como uma unidade apenas os níveis bioclimáticos mesoandinos e altoandinos.
Do norte do Peru até o paralelo 30° de latitude sul no Chile e desde o oceano até 2.500m. Possui clima extremamente árido, em razão da influência da corrente de Humboldt.
Chile entre os paralelos 30° e 38°.
Terras áridas atlânticas ao sul do paralelo 38°,
Rivas-Martínez e Navarro 12 apresentaram um mapa biogeográfico da América do Sul (Fig. 2 . 10). Os autores delimitam 44 províncias sul-americanas ao reino Neotropical-Austro-americano, dentro do qual são agrupadas em dois sub-reinos, 1 0 regiões e oito sub-regiões:
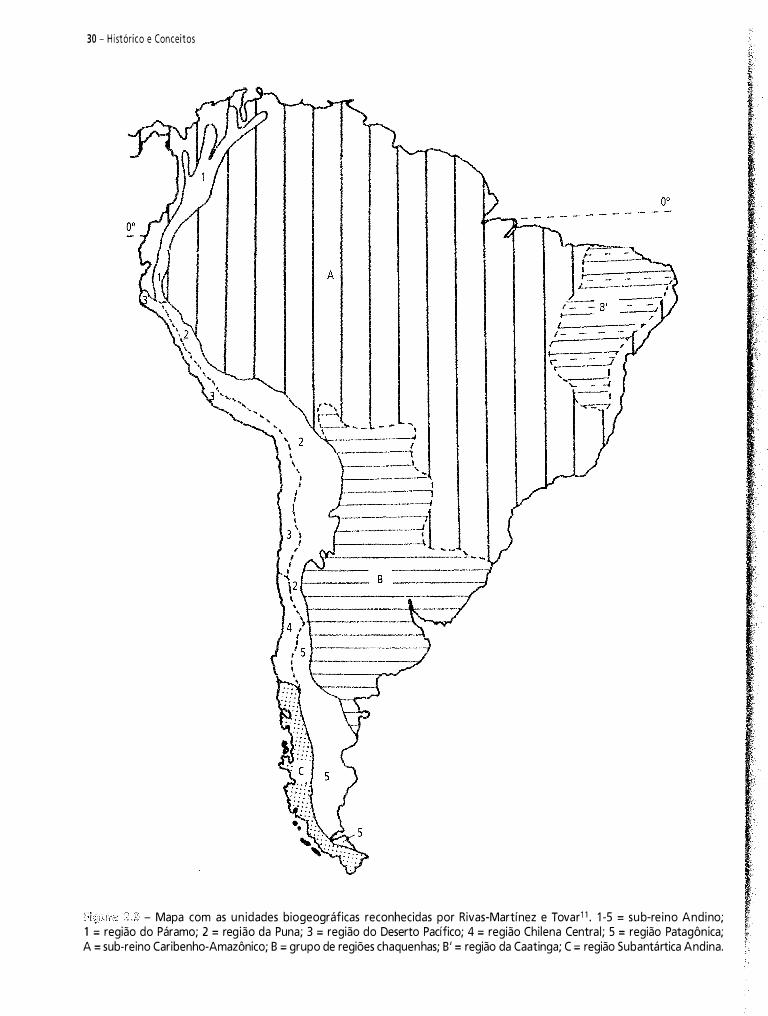
30 - H i stórico e Conceitos
- Mapa com as un idades biogeográficas reconhecidas por Rivas-Martínez e Tovar1 1 . 1 -5 = sub-reino Andino; 1 = região do Páramo; 2 = região da Puna; 3 = região do Deserto Pacífico; 4 = região Chilena Central; 5 = região Patagônica; A = sub-reino Caribenho-Amazônico; B = grupo de regiões chaquenhas; B' = região da Caatinga; C = região Subantártica Andina.
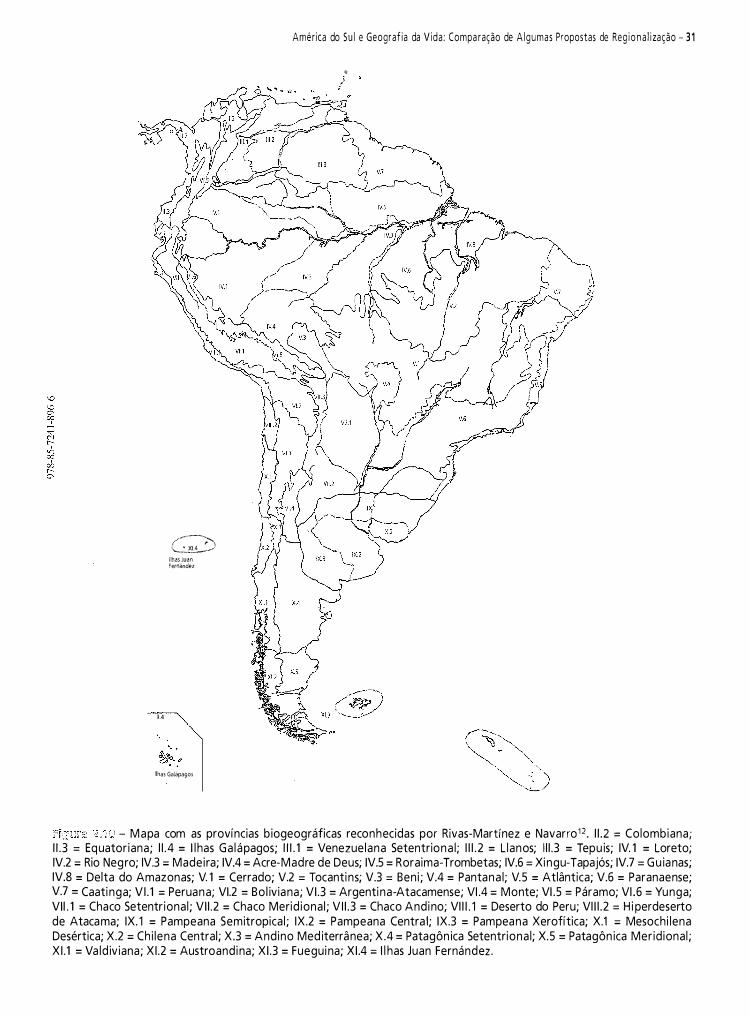
11.4
./ ........ _---........ (_ ... :.�!:..�:.-)
Ilhas Juan Fernández
Ilhas Galápago5
América do Su l e Geograf ia da Vida : Compa ração de Algumas Propostas de Reg iona l i zação - 31
Q
,:
- Mapa com as províncias biogeográficas reconhecidas por Rivas-Martínez e Navarro12 . 1 1 . 2 = Colombiana; 1 1 . 3 = Equatoriana; 1 1 .4 = I l has Ga lápagos; 1 1 1 . 1 = Venezuelana Setentriona l; 1 1 1 . 2 = L lanos; 1 1 1 .3 = Tepu is; IV. 1 = Loreto; 1V.2 = Rio Negro; IV.3 = Madeira; IV.4 = Acre-Madre de Deus; IV.5 = Roraima-Trombetas; IV.6 = Xingu-Tapajós; IV.7 = G u ianas; 1V.8 = Delta do Amazonas; V. 1 = Cerrado; V.2 = Tocantins; V.3 = Beni ; V.4 = Pantana l; V.5 = Atlântica; V.6 = Paranaense; V.7 = Caatinga; V I . 1 = Peruana; VI.2 = Bol iviana; VI.3 = Argentina-Atacamense; VI .4 = Monte; VI .5 = Páramo; VI .6 = Yunga; VI1 . 1 = Chaco Setentr ional ; VI 1 .2 = Chaco Merid ional ; V I I . 3 = Chaco Andino; VI I 1 . 1 = Deserto do Peru; VI I 1 . 2 = Hi perdeserto de Atacama; IX. 1 = Pampeana Semitropical ; IX.2 = Pampeana Centra l; IX.3 = Pampeana Xerofítica; X.1 = Mesochi lena Desértica; X.2 = Chi lena Central; X.3 = Andino Med iterrânea; X.4 = Patagôn ica Setentriona l; X.5 = Patagôn ica Meridional ; XI . 1 = Va ld iviana; XI .2 = Austroandina; XI .3 = Fueguina; XI .4 = I l has Juan Fernández.

32 - H istórico e Conceitos
Compreende sete regiões: Colombiano-Mesoamericana (províncias Colombiana, Equatoriana e das Ilhas Galápagos), Venezuelana (províncias Venezuelana Setentrional, Llanos e Tepuis), Amazônica (sub-região Ocidental: províncias Loreto, Rio Negro, Madeira e Acre-Madre de Deus; e sub-região Oriental: províncias Roraima-Trombetas, Xingu-Tapajós, Guianas e Delta do Amazonas), Brasileiro-Paranaense (sub-região Cerradense: províncias do Cerrado, Tocantins, Beni e Pantanal; e sub-região Atlântico-Paranaense: províncias Atlântica, Paranaense e Caatinga), Andina (sub-região Punenha: províncias Peruana, Boliviana, Argentino-Atacamense e Monte; e sub-região Paramuno-Yunguenha: províncias do Páramo e Yunga); Chaquenha (províncias do Chaco Setentrional, Chaco Meridional e Chaco Andino); e do Deserto Pacífico-Peruano (províncias do Deserto do Peru e do Hiperdeserto de Atacama).
Compreende três regiões: Pampeana (províncias Pampeana Semitropical, Pampeana Central e Pampeana Xerofítica), Mesochilena-Patagônica (sub-região Chilena: províncias Mesochilena Desértica e Chilena Central; e sub-região Andino-Patagônica (províncias Andino Mediterrânea, Patagônica Setentrional e Patagônica Meridional) e Valdiviano-Magalhânica (províncias Valdiviana, Austroandina, Fueguina e Ilhas Juan Femández).
Dinerstein et al. 13 propuseram um sistema de ecorregiões para América Latina e o Caribe. As 98 ecorregiões que correspondem à América do Sul foram classificadas conforme o seu tipo de hábitat principal e atribuídas a seis biorregiões (Fig. 2. 1 1 ) .
Na Colômbia, Venezuela, Equador, Peru e Bolívia. Compreende florestas tropicais úmidas de folha larga (ecorregiões das Florestas Úmidas do Chocó/Darién, Florestas Montanas do Oriente do Panamá, Florestas Montanas dos Andes Norte ocidentais, Florestas Úmidas Ocidentais Equatorianas, Florestas Montanas do Vale do Cauca, Florestas Montanas do Vale do Madalena, Florestas Úmidas do Madalena/Urabá, Florestas Montanas da Cordilheira Oriental, Florestas Montanas da Cordilheira
Real Oriental, Florestas Montanas de Santa Marta, Florestas de Montanha dos Andes Venezuelanos e Florestas Úmidas Montanas do Catatumbo), Florestas secas tropicais de folha larga (ecorregiões das Florestas Secas do Vale do Cauca, Florestas Secas do Vale do Madalena, Florestas Secas do Vale do Patía, Florestas Secas do Vale do Sinú, Florestas Secas do Equador, Florestas Secas de Tumbes/Piura, Florestas Secas de Marafion, Florestas Secas de Maracaibo e Florestas Secas de LaralFalcón), matagais alagados (ecorregião dos Matagais Alagados do
i._
Guayaquil), matagais de montanha (ecorregiões do Pára- -f mo da S erra Nevada de S anta Marta, Páramo da
Cordilheira de Mérida e Páramo do Norte dos Andes), desertos e matagais xéricos (ecorregiões do Matagal Xérico das Ilhas Galápagos, Matagal Xérico de Barranquilla e Guajira e Matagal Xérico de Paraguaná) e restingas (ecorregião das Restingas de Paraguaná).
Na Colômbia, Venezuela, Trinidad e Tobago, Guiana, Suriname e Brasil. Compreende florestas úmidas tropicais de folha larga (ecorregiões das Florestas de montanhas da Cordilheira da Costa, Florestas inundáveis do Delta do Orinoco, Florestas Úmidas de Trinidad e Tobago, Florestas Úmidas da Alta Montanha da Guiana e Tepuis), Florestas secas tropicais de folha larga (ecorregiões das Florestas Secas dos Llanos), planícies e Florestas Secas de Trinidad e Tobago, matagais, savanas e matagais (ecorregião dos Llanos), matagais inundáveis (ecorregião dos Manguezais do Orinoco), e desertos e
matagais xéricos (ecorregiões do Matagal de Cáctos de Araba, Curaçao e Bonaire, Matagal Xérico da Costa e Matagal Xérico das Penínsulas de Araya e Paría) .
Na Colômbia, Suriname, Guiana Francesa, Equador, Peru, Brasil e Bolívia. Compreende florestas úmidas tropicais de folhas largas (ecorregiões das Florestas Úmidas do Rio Napo, Florestas Montanas da Serra da Macarena, Florestas Úmidas da Região do Rio Japurá/ Río Negro, Florestas Úmidas de Uatama, Florestas Úmidas de Amapá, Florestas Úmidas da Guiana, Florestas inundáveis de Paramaribo, Florestas Úmidas do Rio Ucayali, Florestas Alagáveis da Amazônia Ocidental, Florestas Úmidas da Amazônia Sul-ocidental, Florestas Úmidas do Rio Juruá, Florestas de Várzea, Florestas Úmidas da Região do Rio Purus/Rio Madeira, Florestas Úmidas de Rondônia/Mato Grosso, Florestas
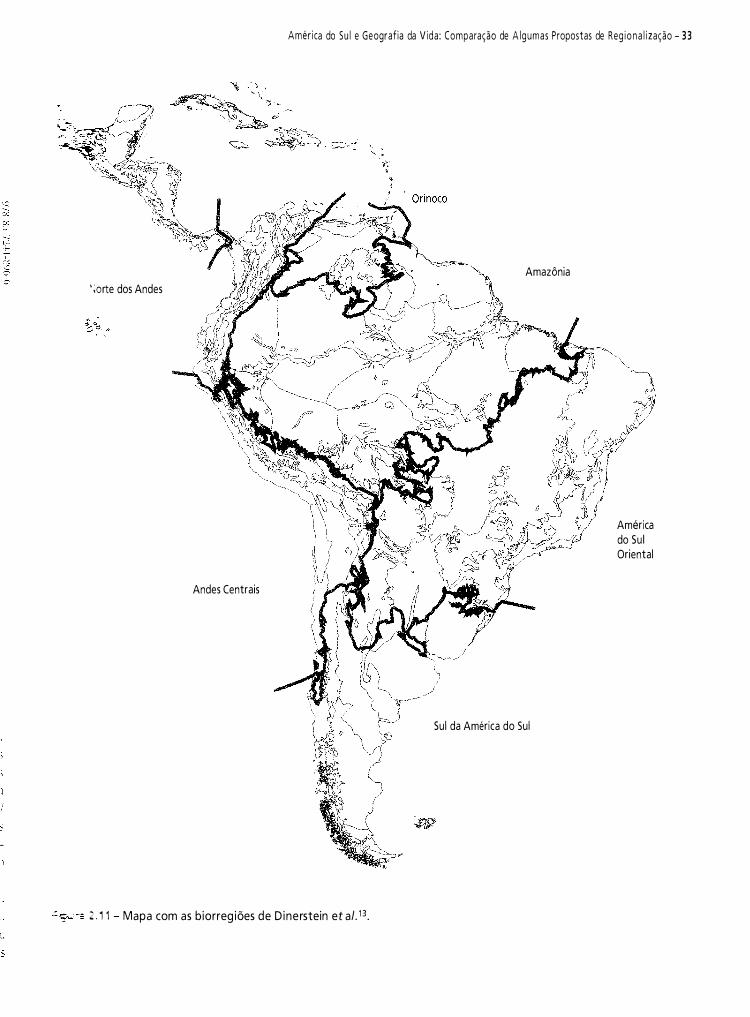
to-
L.
América do Su l e Geografia da Vida : Comparação de Algumas Propostas de Reg iona l i zação - 3 3
';orte dos Andes
Andes Centrais
2 1 1 - Mapa com as biorreg iões de Dinerstein et ai. 13 .
Amazônia
Sul da América do Sul
América do Sul Oriental

34 - H istórico e Conceitos
de Galeria e Alagáveis do Beni, Florestas Úmidas da Região do Rio TapajóslRio Xingu e Florestas Úmidas do Rio Tocantins), Florestas secas tropicais de folhas largas (ecorregião das Florestas Secas das Terras B aixas da Bolívia), matagais, savanas e matorrais (ecorregiões das Savanas da Guiana, Savanas da Amazônia e Savanas do Beni) e matagais inundáveis (ecorregiões dos Matagais Inundáveis da Amazônia Ocidental, Matagais Inundáveis da Amazônia Oriental e Matagais Inundavéis de São Luis).
No Peru, Bolívia e Argentina. Compreende Florestas úmidas tropicais de folha larga (ecorregiões das Yungas Peruanas, Yungas Bolivianas e Yungas Andinas), Florestas secas tropicais de folha larga (ecorregião das Florestas Secas Montanas da Bolívia), matagais de montanha (ecorregiões do Páramo da Cordilheira Central, Puna dos Andes Centrais, Puna úmida dos Andes Centrais e Puna Árida dos Andes Centrais), matagais mediterrâneos (ecorregião do Matagal do Chile), e desertos e matagais xéricos (ecorregiões do Deserto de Sechura e Deserto de Atacama).
No Brasil, Bolívia, Paraguai, Argentina e Uruguai. Compreende Florestas úmidas tropicais de folha larga (ecorregiões das Florestas Atânticas da costa Brasileira e Florestas Atlânticas do Interior Brasileiro), Florestas tropicais e subtropicais de coníferas (ecorregião das Florestas de Araucaria do Brasil), matagais, savanas e matorrais (ecorregiões do Cerrado, Savanas do Chaco, Chaco Úmido e Savanas Montanas de Córdoba), matagais inundáveis (ecorregiões do Pantanal e Savanas Inundáveis do Paraná), desertos e matorrais xéricos (ecorregião da Caatinga) e restingas (ecorregiões das Restingas do Nordeste do Brasil e Restingas da costa Atlântica do Brasil).
No Chile, Argentina e Uruguai. Compreende florestas temperadas (ecorregiões das Florestas de Chuva Invernal do Chile, Florestas Temperadas de Valdívia e Florestas subpolares de Nothofagus), Pastiçais, savanas e matagais (ecorregiões do Monte da Argentina, Espinheiral da Argentina, Pampas e Savanas do Uruguai) e Pastiçais de montanha (ecorregiões da Estepe do Sul dos Andes, Estepe da Patagônia e Pastiçais da Patagônia).
� I�, ir:\ <" . ' '4
Morrone14 propôs um esquema biogeográfico paraAmérica Latina e o Caribe (ver também Morrone1 9-21 ) , fundamentado em análises pan-biogeográficas e biogeográfico-cladística de animais e plantas, pelo qual _
reconheceu 70 províncias (Fig. 2 . 1 2) . As 46 províncias -sul-americanas se agrupam nas regiões N eotropical (com _
quatro sub-regiões) e Andina (com três sub-regiões), e a Zona de Transição Sul-americana (Fig. 2. 1 3) .
Corresponde aos trópicos do Novo Mundo, na maior parte da América do Sul e América Central, sul do México, Índias Ocidentais e sul da península da Flórida. Nos tempos pré-quaternários, a biota sul-americana se expandiu em direção ao norte, para América Central e
México. A região Neotropical está inserida no reino Holotropical, que inclui as áreas tropicais entre 30° S e 30° N. Assim mesmo, compreende as regiões Etiópica ou Afrotropical (África central, península Arábica, Madagascar e as ilhas do Oceano Índico Ocidental), Oriental (Índia, Burma, Malásia, Indonésia, Filipinas e as ilhas do Oceano Pacífico) e Australiana Tropical (noroeste da Austrália) . Posssui quatro sub-regiões: Caribenha, Amazônica, Chaquenha e Paranaense.
Sub-região Caribenha
Estende-se pelo sul do México, América Central, Antillas e ao noroeste da América do Sul. Possui história geo
biótica complexa, a qual é refletida nas suas múltiplas relações com outras áreas. Inclui 24 províncias, das quais as seguintes se encontram na América do Sul:
• Província do Chocó: costa do pacífico do norte do Equador, Colômbia e Panamá.
• Província de Maracaibo : norte da Colômbia e noroeste da Venezuela.
• Província do Costa Venezuelana: norte da Venezuela e Colômbia, incluindo as ilhas de Aruba, Curaçao e Donaire.
• Província de Trinidad e Tobago: ilhas de Trinidad e Tobago.
• Província de Madalena: oeste da Venezuela e noroeste da Colômbia.
• Província dos Llanos Venezuelanos : planícies de grande parte da Venezuela e o noroeste da Colômbia.
• Província de Cauca: oeste de Colômbia e Equador.
=-
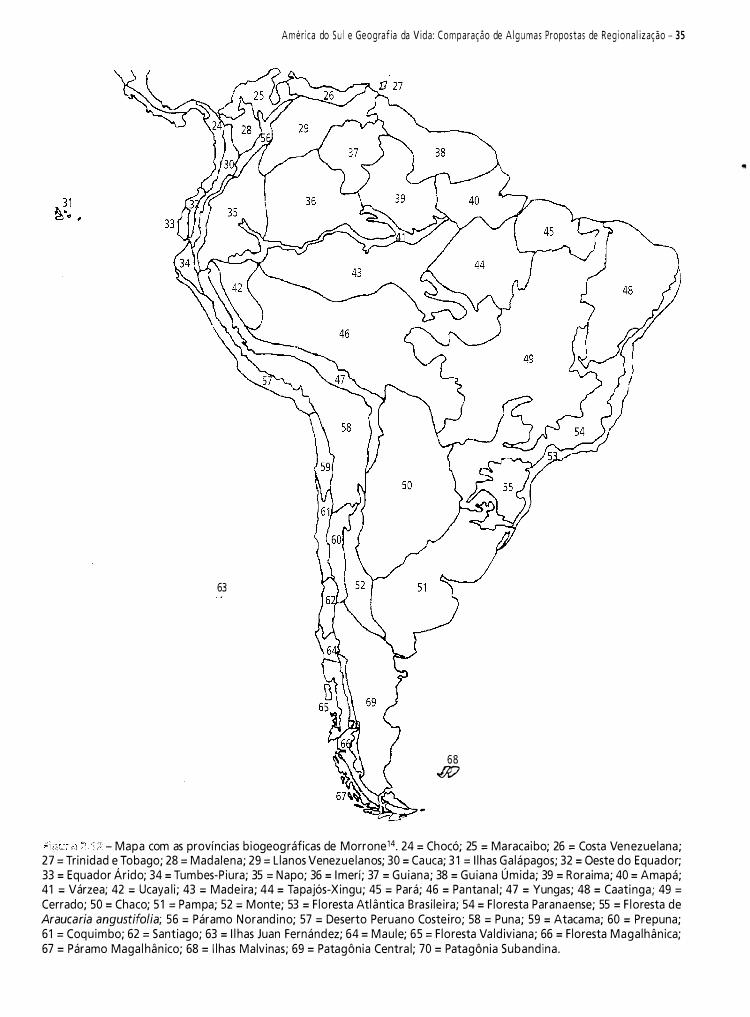
63
América do Su l e Geograf ia da V ida : Comparação de Algumas Propostas de Reg iona l ização - 35
68 ..fl{?
- Mapa com as províncias biogeográficas de Morrone14. 24 = Chocó; 25 = Maracaibo; 26 = Costa Venezuelana; 27 = Trin idad e Tobago; 28 = Madalena; 29 = Llanos Venezuelanos; 30 = Cauca; 3 1 = I l has Ga lápagos; 32 = Oeste do Equador; 33 = Equador Árido; 34 = Tumbes-Piura; 35 = Napo; 36 = I merí; 37 = Guiana; 38 = Guiana Úmida; 39 = Roraima; 40 = Amapá; 41 = Várzea; 42 = Ucaya l i ; 43 = Madeira; 44 = Tapajós-Xingu; 45 = Pará; 46 = Pantana l ; 47 = Yungas; 48 = Caatinga; 49 =
Cerrado; 50 = Chaco; 5 1 = Pampa; 52 = Monte; 53 = F loresta Atl ântica Brasi le ira; 54 = F loresta Paranaense; 55 = F loresta de Araucaria angustifolia; 56 = Páramo Norand ino; 57 = Deserto Peruano Costeiro; 58 = Puna; 59 = Atacama; 60 = Prepuna; 61 = Coquimbo; 62 = Santiago; 63 = I l has Juan Fernández; 64 = Maule; 65 = F loresta Va ld iviana; 66 = Floresta Maga lhân ica; 67 = Páramo Maga l hân ico; 68 = I l has Malvinas; 69 = Patagônia Centra l; 70 = Patagônia Subandina .
OI
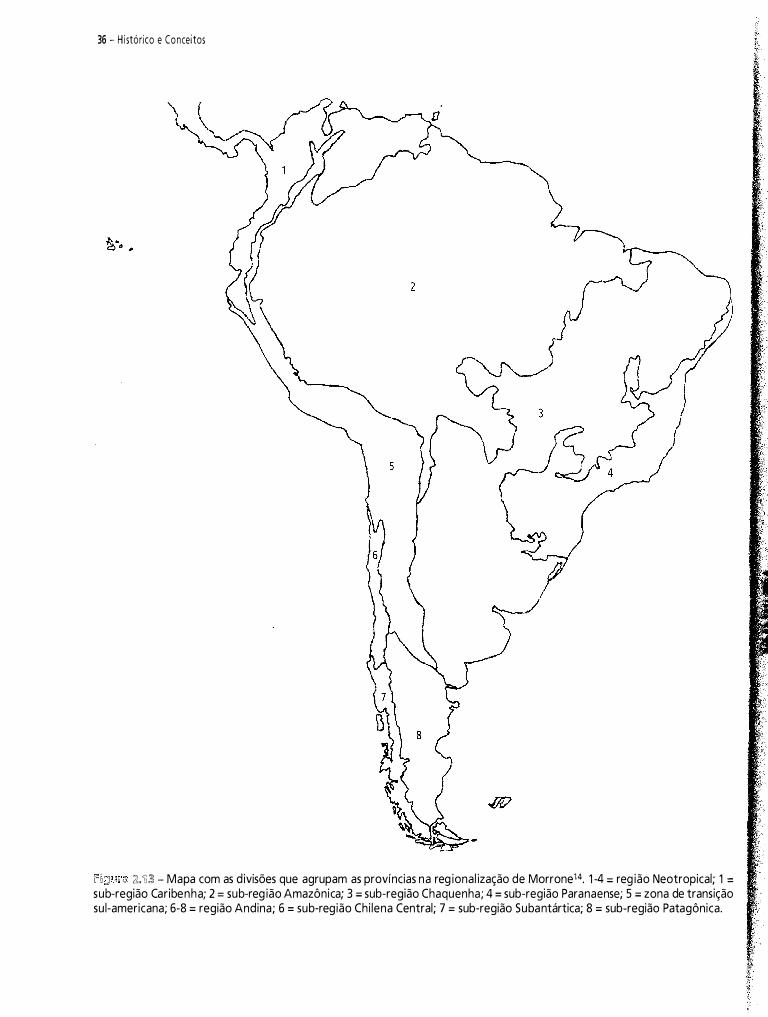
36 - H i stórico e Conceitos
2(,U - Mapa com as divisões que agrupam as províncias na reg ional ização de Morrone14. 1 -4 = reg ião Neotropical; 1 =
sub-região Caribenha; 2 = sub-reg ião Amazônica; 3 = sub-região Chaquenha; 4 = sub-região Paranaense; 5 = zona de transição sul-americana; 6-8 = região Andina; 6 = sub-região Chi lena Central; 7 = sub-região Subantártica; 8 = sub-região Patagônica.

Amér ica do Su l e Geograf ia da Vida : Comparação de Algu mas Propostas de Reg iona l i zação - 37
o Província das Ilhas Galápagos: Arquipélago de Colón, no Oceano Pacífico, a 950km da costa do Equador, que compreende 1 5 ilhas maiores e numerosas ilhas
menores.
• Província do Oeste do Equador: oeste do Equador e
sudoeste da Colômbia. • Província do Equador Árido: oeste do Equador. o Província de Tumbes-Piura: sul do Equador e norte
do Peru .
Sub-região Amazônica
É a sub-região de maior extensão da região Neotropical, se estendendo pelo Brasil, Guianas, Venezuela, Colômbia, Equador, Peru, Bolívia, Paraguai e Argentina. Compreende 13 províncias:
o Província de Napo: norte do Peru, sudoeste da Colômbia e leste do Equador.
o Província de lmerí: sul da Venezuela, sudoeste da Colômbia, nordeste do Peru e norte do Brasil.
• Província da Guiana: noroeste da América do Sul, na
plataforma Guiana, entre Venezuela, Colômbia,
Guiana, Suriname e norte do Brasil, onde ocorrem
planaltos arenosos, conhecidos como tepuis, de mais de 2.000m de altitude.
• Província da Guiana Úmida: sudoeste da Venezuela,
'D norte do Brasil, Suriname e Guiana. � • Província de Roraima: norte do Brasil, sudeste da "?
Venezuela, Suriname e Guiana.
Província de Amapá: Suriname e nordeste do Brasil.
Província da Várzea: noroeste do Brasil e noroeste do Peru.
• Província de Ucayali: leste do Peru, norte da Bolívia e oeste do Brasil.
• Província de Madeira: noroeste do Brasil, fazendo fronteira ao norte com o Rio Amazonas, ao oeste com
os rios Madeira e Beni, ao leste com o Rio Xingu e
ao oeste com a cordilheira oriental da Bolívia. o Província de Tapajós-Xingu: noroeste do Brasil.
• Província de Pará: noroeste do Brasil; faz fronteira
ao norte e oeste com os rios Tocantins e Araguaia, ao
sul com a Serra do Gurupi e o Rio Graj au, e ao leste com o Rio Guana.
• Província do Pantanal : B rasil austral e central,
noroeste da Bolívia e norte do Paraguai. o Província das Yungas: margens ocidentais dos Andes,
entre 300 e 3 .500m de altitude, desde o norte do Peru até o noroeste da Argentina.
Sub-região Chaquenha
Compreende o norte e centro da Argentina, sul da Bolívia, oeste e centro do Paraguai, Uruguai, e centro e nordeste do Brasil. Está relacionada às sub-regiões Amazônica e Paranaense. O desenvolvimento de um "corredor de savana" durante o Terciário atuou como o evento vicariante dinâmico que separou a floresta contínua Amazônico-Paranaense. Compreende quatro províncias:
o Província da Caatinga: nordeste do Brasil, nos estados da Bahia, Sergipe, Alagoas, Pernambuco, Paraiba, Rio Grande do Norte, Ceará, Piauí e Minas Gerais.
• Província do Cerrado: centro e sul do Brasil (estados de Minas Gerais, Mato Grosso, Goiás, São Paulo, Paraná, Maranhão e PiauD, nordeste do Paraguai e Bolívia.
o Província do Chaco: sul da Bolívia, oeste do Paraguai, sul do Brasil e centro norte da Argentina.
• Província da Pampa: centro oeste da Argentina entre 30 e 39° S, Uruguai, e sul do estado do Rio Grande do Sul .
Su b· região Paranaense
Localiza-se no nordeste da Argentina, leste do Paraguai, sul do Brasil (oeste da Serra do Mar em direção ao centro do Rio Grande do Sul) e leste do Brasil, entre 7 e 32° S. Compreende três províncias:
Província da Floresta Atlântica Brasileira: faixa estreita ao longo da costa atlântica do Brasil, ao leste das cordilheiras da costa, entre 7 e 32° S.
o Província da Floresta Paranaense: sudeste do Brasil, nordeste da Argentina e leste do Paraguai.
o Província da Floresta de Araucaria angustifolia: sul do Brasil e nordeste da Argentina, entre 600 e 1 .800m de altitude.
Considerada anteriormente como a sub-região Páramo-Punenha da região Andina 19,20. Estende-se ao longo dos Andes entre o oeste da Venezuela e o norte do Chile, e o centro e oeste da Argentina e compreende seis províncias:
• Província do Páramo Norandino: altas cordilheiras da Venezuela, Colômbia, Equador e Peru, acima de 3.000m de altitude.
o Província do Deserto Peruano Costeiro: faixa estreita ao longo da costa do Pacífico, desde o norte do Peru até o norte do Chile.

38 - Histórico e Conceitos
• Província da Puna: Bolívia, norte da Argentina e Chile, e sul do Peru.
• Província de Atacama: norte do Chile, entre 1 8 e 28° S. • Província da Prepuna: centro e noroeste da Argentina,
desde Jujuy até o norte de Mendoza. • Província do Monte: centro da Argentina, entre 24 e
43° S, desde Salta até o nordeste de Chubut.
Estende-se pelo Chile central e a Patagônia. Parte da sua biota evoluiu originalmente na Patagônia e entendeu-se gradualmente em direção ao norte durante o Terciário e o Pleistoceno, até a Zona de Transição Sul-americana, junto à conversão de florestas tropicais em comunidades temperadas e áridas . A região Andina designa-se ao reino Austral, o qual compreende igualmente as regiões Antártica, do Cabo ou Afrotemperada (África do Sul), Neoguineana (Nova Guinea e Nova Caledônia), Australiana Temperada (sudeste da Austrália) e Neozelandesa (Nova Zelândia). Compreende três sub-regiões: Chilena Central, Sub antártica e Patagônica.
Sub-região Chilena Central
Chile central, entre 28 e 36° S; inclui duas províncias:
• Província de Coquimbo: centro norte do Chile, entre 28 e 32° S .
• Província do Santiago: centro sul do Chile, entre 32 e 36° S.
Sub-região Sub antártica
Compreende os Andes austrais, desde 36° S até o Cabo de Romos, incluindo o arquipélago do sul do Chile e Argentina, e as Ilhas Malvinas, Geórgias do Sul e Juan Femández. Possui seis províncias:
• Província das Ilhas Juan Femández: ilhas chilenas de Masatierra ou Robinson Cru soe, Masafuera ou Alejandro Selkirk, e Santa Clara, localizadas no Oceano Pacífico, 600km ao oeste de Valparaíso, a 33° S .
• Província do Maule: sul do Chile e Argentina, entre 36 e 39° S .
• Província da Floresta Valdiviana: sul do Chile e Argentina, ao sul da província da Maule, alcançando os 47° S.
• Província da Floresta Magalhânica: sul do Chile, desde 47° S ao Cabo de Romos e sul da Argentina, em pequenas partes do oeste das províncias de Santa Cruz e Terra do Fogo.
• Província do Páramo Magalhânico: sul do Chile e Argentina, desde o Golfo de Penas (48° S) até o Cabo de Romos, fazendo fronteira a oeste com a província da Floresta Magalhânica.
• Província das Ilhas Malvinas: arquipélago argentino das Malvinas e Geórgias do sul, localizadas no Oceano Atlântico Sul, a 550km de Terra do Fogo.
Sub-região Patagônica
Sul da Argentina, desde o centro de Mendoza, alargando-se em Neuquén, Rio Negro, Chubut e Santa Cruz, até o norte da Terra do Fogo; alcança Chile em Aisén e Magalhães. Compreende duas províncias:
• Província da Patagônia Central: sudoeste da Argentina, desde o centro de Mendoza ao norte de Santa Cruz: alcançando Chile em Aisén e Magalhães.
• Província da Patagônia Subandina: sudoeste da Argentina constitui uma faixa estreita ao longo dos Andes austrais, ao sul dos 5 1 ° S, alargando-se em direção ao sul até Santa Cruz.
As regionalizações aqui analisadas são usualmente narrativas, sem aprofundar nas justificativas metodológicas. com exceção das duas mais recentes 13, 14, em que são detalhados os métodos utilizados para formulá-las. Com frequência, as regionalizações apresentam algumas deficiências na sua formulação (por exemplo, definiçõe, das regiões, províncias e distritos; detalhe dos seus limites; mapas), que toma difícil seu teste, é possível reconhecer em elas modelos gerais de referência2. Um aspecto interessante da evolução desses modelos resulta na análise da influência que tiveram os diferentes autores entre si, inferida a partir das suas citações nas referências bibliográficas (Fig. 2 . 14).
As revisões dos esquemas de regionalização biogeográfica2.22-24 permitem descobrir áreas biogeográficas robustas, sustentadas pelos diferentes grupos taxonômicos ou obtidas a partir de enfoques metodológicos diferentes. Por exemplo, a maior parte dos esquemas analisados reconhece uma divisão primária da América do Sul em uma porção tropical versus outra temperada22: as sub-regiões Guaiano-Brasileira (ou Brasílica ou Brasileira) e Patagônica (ou Austral) de Cabrera e Yepes3, Fittkau4. KuscheP; e Ringuelet9; o Leste não Andino e Oeste Andino de Sick6 e as regiões Neotropical e Antártica de Cabrera e Willink 7. Diversos autores tentaram definir o
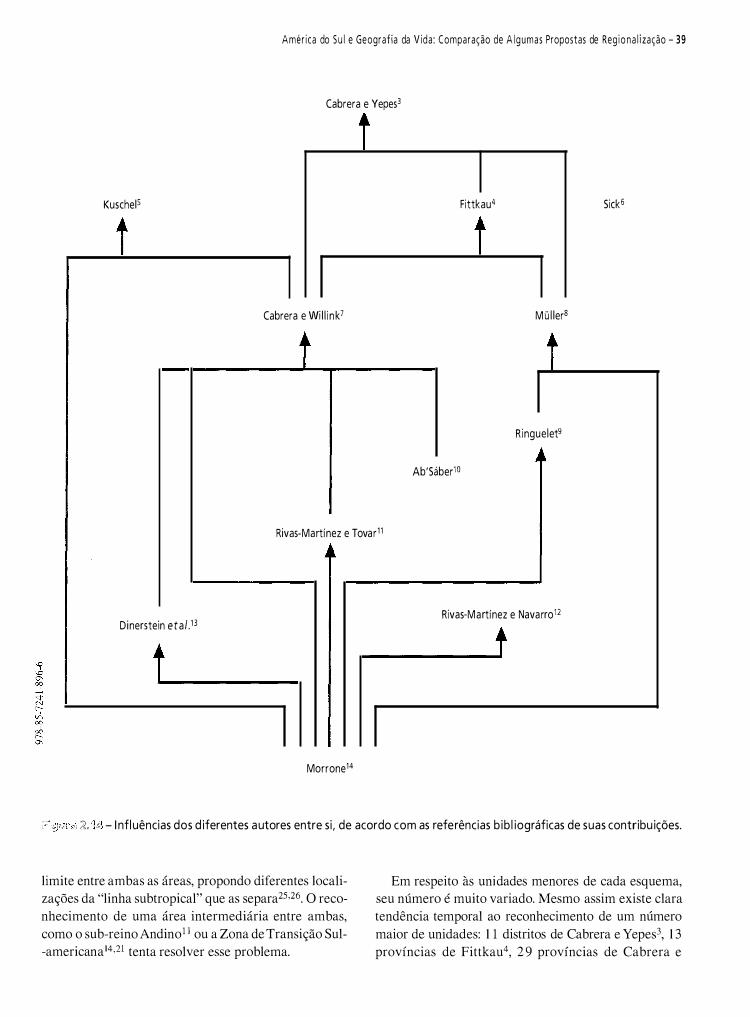
Amér ica do Su l e Geograf ia da V ida : Comparação de Algumas Propostas de Reg iona l i zação - 39
Cabrera e Yepes3 j�
KuschelS Fittkau4 Sick6
Cabrera e Wi l l i nk7 Mül ler8
R inguelet9
Ab'Sáber10
Rivas-Martinez e Tovar 1 1
D inerstein et ai . 1 3
Morrone14
Rivas-Martinez e Navarro 12
t
(2, - I nfluências dos d iferentes autores entre si, de acordo com as referências b ib l iográficas de suas contribu ições.
limite entre ambas as áreas, propondo diferentes localizações da "linha subtropical" que as separa25,26. O reconhecimento de uma área intermediária entre ambas, como o sub-reino Andinol l ou a Zona de Transição Sul-americana14.21 tenta resolver esse problema.
Em respeito às unidades menores de cada esquema, seu número é muito variado. Mesmo assim existe clara tendência temporal ao reconhecimento de um número maior de unidades: 1 1 distritos de Cabrera e Yepes3, 1 3 províncias de Fittkau4, 2 9 províncias de Cabrera e

40 - H istórico e Conceitos
Willink7, 33 centros de Müller8, 20 províncias de Ringuelet9, 23 domínios de Ab' SáberlO, 44 províncias de Rivas-Martínez e Navarro12, 98 ecorregiões de Dinerstein et al. 13 e 46 províncias de Morrone14. Uma comparação exaustiva (para algumas equivalências, ver Morrone20) permitiria deduzir semelhanças e diferenças decorrentes dos métodos utilizados e dos táxons analisados.
Os modelos de regionalização não são definitivos, pois se modificam na medida em que a nossa visão dos padrões geobióticos também se modifica14. Os esquemas analisados apresentam diferentes visões da regionalização da América do Sul. Inicialmente seu marco de referência implícito ou explícito foi dispersalista; mais recentemente foram utilizados critérios ecológicos 1 3 ou pan-biogeográficos e c1adísticos1 4. A partir dos diferentes enfoques em que se baseiam, as semelhanças entre as diversas regionalizações certamente refletem padrões naturais . Análises futuras, fundamentadas em maior número e diversidade de táxons, permitirão contrastar essas regionalizações.
REFERÊNCIAS BIBLIOGRÁFICAS 1 . DARWIN, C. R. The Origin of Species by Means of Natural
Selection, or, the Preservation ofFavoured Races in the Struggle for Life. Londres: John Murray, 1 859. 5 1 3p.
2. RlBICHICH, A. M. EI modelo clásico de la fitogeografía de Argentina: un análisis crítico. Interciencia, v. 27, p. 669-675, 2002.
3 . CABRERA, A.; YEPES, J. Mamiferos Sud-americanos (Vida, Costumbres y Descripción). Buenos Aires: Historia Natural Ediar, Compafiía Argentina de Editores, 1940. 370p.
4. FITTKAU, E. J. The fauna of South America. In: FITTKAU, E. J . ; ILLIES, 1 . ; KLINGE, H. et aI. Biogeography and Ecology in South America. La Haya: Df. W. Junk B. V. Publishers, 1 969. p. 624-658.
5 . KUSCHEL, G. Biogeography and ecology of South American Coleoptera. In: FITTKAU, E. J. ; ILLIES, J. ; KLINGE, H. et aI. Biogeography and Ecology in South America. La Haya: Df. W. Junk B . V. Publishers, 1 969. p. 709-722.
6. SICK, W. D. Geographical substance. Monographiae Biologicae, v. 1 9, p. 449-474, 1 969.
7 . CABRERA, A. L. ; WILLINK, A. Biogeografía de América Latina. Monografía nro. 13. Washington: Secretaría General de la Organización de Estados Americanos, 1973. 120p.
8 . MÜLLER, P. The dispersaI centres of terrestial vertebrales in the Neotropical realm: a study in the evolution of the NeotropicaI biota and its native landscapes. Biogeographica. La Haya: Df. W. Junk B. V. Publishers, 1973. v. 2, 244p.
9. RINGUELET, R. A. Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de Arnérica deI Sur. Ecosur, v. 2, p. 1 -122, 1975.
10. AB' SÁBER, A. N . Os domínios morfoclimáticos na América do Sul. Primeira aproximação. Geomoifología, v. 52, p. 1 -22. 1 977.
1 l . RIVAS-MARTÍNEZ, S.; TOVAR, O. Síntesis biogeográfica de los Andes. Collectanea Botanica (Barcelona), v. 14, p. 5 15-521 . 1 983.
12 . RIVAS-MARTÍNEZ, S. ; NAVARRO, G. Mapa Biogeográfico de Suramérica. Madrid: publicado por los autores, 1994.
1 3 . DINERSTEIN, E. ; OLSON, D . M.; GRAHAM, D. 1. et al. Una Evaluación dei Estado de Conservación de las Eco-regiones Terrestres de América Latina y el Caribe. Washington: B anco Mundial, 1995. 1 35p.
14. MORRONE, 1. J. Biogeographic areas and transition zones of Latin America and the Caribbean Islands based on panbiogeographic and cladistic analyses of the entomofauna. Annual Review of Entomology, v. 5 1 , p. 467-494, 2006.
1 5 . SCLATER, P. L. On the general geographic distribution of the members of the class Aves. foumal of the Linnean Societ)': Zoology, v. 2, p. 1 30- 145, 1858.
16. WALLACE, A. R. The Geographical Distribution ofAnimals. Londres: McMillan, 1 876.
17. MELLO-LEITÃO, C. Zoogeografia do Brasil. Série 5", Brasiliana, v. 77, 1937. 4 1 6p.
1 8 . HUECK, K. As Florestas da América do Sul - Ecologia, Composição e Importância Econômica. São Paulo: Editora d2 Universidade de Brasília e Editora Polígono, 1 972. 466p.
1 9. MORRONE, J. J . Presentación preliminar de un nuevo esquem� biogeográfico de América deI Sur. Biogeographica, v. 75, p. 1 -16, 1 999.
20. MORRONE, 1. 1. Biogeografia de América Latina y el Caribe. Zaragoza: M&T-Manuales & Tesis SEA, 200 1 . v. 3, 148p.
2 1 . MORRONE, J. J . Panbiogeografía, componentes bióticos : zonas de transición. Revista Brasileira de Entomologia, v. 48. p. 149-1 62, 2004.
22. SÁNCHEZ OSÉS, C . ; PÉREZ-HERNÁNDEZ, R. Revisiór. histórica de las subdivisiones biogeográficas de la región Neotropical, con especial énfasis en Suramérica. Montalbán, v. 3 1 . p . 1 69-210, 1998.
23. SÁNCHEZ OSÉS, C.; PÉREZ-HERNÁNDEZ, R. Historia \ tabla de equivalencias de las propuestas de subdivisiones bicgeográficas de la región Neotrop ical . In: LLORENTE BOUSQUETS, J.; MORRONE, J. 1. Regionalización Biogergráfica en Iberoamérica y Tópicos Afines - Primeras JornadG. Biogeográficas de la Red Iberoamericana de Biogeografía .. Entomología Sistemática (RIBES XI1.I-CYTED). México: Lo.Prensas de Ciencias, UNAM, 2005. p. 495-508.
24. MOREIRA-MUNOZ, A. The Austral f10ristic realm revisitec Joumal of Biogeography, v. 34, p. 1 649- 1660, 2007.
25. RAPOPORT, E. H. Algunos problemas biogeográficos do nuevo mundo con especial referencia a la región Neotropica: In: DELAMARE DEBOUTEVILLE, C . ; RAPOPORT, E. H Biologie de l 'Amerique Australe, París : CNRS, 1 968. v. 4. � 55- 1 10.
26. RUGGIERO, A . ; EZCURRA, C. Regiones y transicione, biogeográficas: Complementariedad de los análisis en biogec grafía histórica y ecológica. In: MORRONE, J. J . ; LLORENTE J. Una Perspectiva Latinoamericana de la Biogeograf'ía. l\1t. xico: Las Prensas de Ciencias, UNAM, 2003. p. 1 41 - 1 54.

3
Claudio J. B. de Carvalho
Fazer ciência é a procura por padrões repetidos, não simplesmente a acumulação de fatos. E para a ciência de ecologia
geográfica é, mais precisamente, a procura por padrões da vida de plantas e animais que podem ser colocados em um mapa.
Poucos conceitos na biogeografia histórica causam tantas controvérsias como o de área de endemismo. Uma área de endemismo é geralmente entendida como uma região geográfica indicada a partir da combinação de áreas de distribuição de espécies ou táxons endêmicos, isto é, espécies ou grupos de espécies com ocorrência exclusiva em uma região particular.
A área de endemismo possui claramente o componente espacial, pois é delimitada e entendida a partir das áreas de distribuição congruentes de diferentes espécies. Por outro lado, a área de distribuição é a aquela ocupada por uma espécie em determinado momento e pode ser representada pela disposição dos dados em um mapa. Esta representação indica o estado de conhecimento atual e não, necessariamente, a distribuição real das espécies! .
O s padrões de distribuição das espécies não são gerados ao acaso, pois processo(s) mais geral(is) atuou(aram) na formação do padrão de endemismo encontrado. Entretanto, o maior problema metodológico encontrado é saber qual o nível de congruência espacial, isto é, qual o grau de simpatria entre as áreas de distribuição das espécies que pode ser aceito para a delimitação da área de endemismo.
Deve-se ter em mente que o espaço na biogeografia é um conceito relativo, pois não existe espaço absoluto, mas uma área que pode apenas ser compreendida e entendida junto com os organismos que ocorrem neste local em detenninado temp02. Assim, para entendimento e definição corretos da área de endemismo devem ser avaliados também quais processos podem dar origem a esses padrões.
Robert H. MacArthur
Neste capítulo será feito um breve relato da evolução do conceito de área de endemismo enfatizando a sua origem no período pré-evolutivo. Em seguida serão apresentadas suas principais definições, mostrando que a condição de coincidência espacial total ou parcial das distribuições das espécies é a mais utilizada para reconhecimento da área. Será delineado depois o conceito de área de endemismo como área histórica e como hipótese. Após, serão indicados os principais métodos para reconhecimento dessas áreas e os possíveis problemas com a utilização de cada um deles. Por fim, o caráter de unidade histórica da área de endemismo para biogeografia será estabelecido, o que é indispensável para o correto entendimento da área para análises biogeográficas.
Hoje em dia é consenso imaginar e entender que áreas diferentes do planeta têm composições de espécies diferentes, resultantes do processo evolutivo que atinge toda a biota. Mas, antigamente, quando se descobriu os novos mundos, não era bem assim. Em meados do século XIX, Philip Lutley Sclater indicava, em seus estudos da distribuição geográfica da classe Aves, que era bem conhecido e universalmente aceito o fato de que se escolhermos duas áreas do globo, com fauna e flora diferentes, não deveria ser errado imaginar que elas foram resultados de distintas criações3.
A elaboração inicial do conceito da existência de área endêmica, com grupos específicos e característicos desta região, foi construída no período pré-evolutivo. Augustin Pyramus de Candolle, no início do século XIX, possivel-

42 - H istórico e Conceitos
mente foi o primeiro a definir o que se pode considerar hoje o que é uma área de endemismo em sua proposta para delimitação das regiões botânicas (Quadro 3. 1) . De Candolle afirmava: "De todos esses fatos, podemos deduzir que existem regiões botânicas; e este termo é definido para qualquer área que, com exceção das espécies introduzidas, contém certo número de plantas peculiares e que podem ser chamadas de verdadeiramente aborígines"4.
Dentro do paradigma evolutivo, a partir da segunda metade do século XIX, as ideias mais bem estabelecidas sobre áreas com composição particular e única de distribuição de espécies (endemismo) foram desenvolvidas por Alfred Russel Wallace, iniciadas pela sua viagem à Amazônia. Wallace constatou a importância do conhecimento do local exato da distribuição das espécies. A partir desses estudos, principalmente com macacos, foram definidas quatro grandes áreas de endemismo na Amazônia - Guiana, Equador, Peru e Brasil, delimitadas pelos grandes rioss (Fig. 3 . 1 ) . Charles Robert Darwin incluiu indicações sobre distribuição geográfica de animais e plantas no seu clássico livro The Origin the Species by Means of Natural Selection6, que foram interpretadas como oriundas dos centros de origem que podiam se dispersar por mecanismos passivos ou ativos. Esse esquema ficou mais bem estabelecido posteriormente na publicação do livro sobre a distribuição geográfica dos animais, em que começava a ser entendida a hierarquia das áreas de endemismo com a definição das regiões biogeográficas mais abrangentes a partir de reunião de áreas de nível menor de abrangência7.
Mesmo com o reconhecimento das áreas, pelo compartilhamento geográfico dos locais de ocorrência, pouco se tinha
e L " - Vinte reg iões botânicas de August in Pyramus de Candol le
1 . Ásia boreal, E u ropa e América 2. Europa. sul da região boreal e norte do Mediterrâneo 3. Sibéria 4. Região Mediterrân ica 5. Leste da E u ropa até os mares Negro e Cáspio 6. índ ia 7. China, I ndochina e Japão 8. Austrá l i a 9. Sul da África 10 . Leste da África 1 1 . Oeste da África tropical 1 2 . I lhas Canárias 1 3 . Norte dos Estados U n i dos 1 4. Costa noroeste da América do Norte 1 5 . Ant i l has 1 6. México 17. América tropical 1 8. Chi le 1 9. Sul do Bras i l e Argentina 20. Terra do Fogo
ideia sobre o porquê da relação de semelhança, pela composição das espécies, entre diferentes áreas. Sclater chamou a atenção de uma aparente incongruência nos seus estudos de Aves que "pouca ou nenhuma atenção tem sido dada ao fato de que duas ou mais regiões biogeográficas são mais relacionadas uma a outra do que a uma terceira"3. Sabe-se. obviamente, que a observação de Sclater foi realizada dentro do paradigma pré-evolutivo. No entanto, o paradigma evolutivo e a aceitação da tectônica de placas alteraram radicalmente a interpretação desta questão atualmente.
Durante o período dispersionista da biogeografia, a _
concepção de centro de origem e dispersão a partir de um local possuía grau explanatório pequeno em função do caráter pouco comparável da formação dos padrões de distribuição por processos dispersionistas. Assim mesmo, naquela época, era possível correlacionar a maioria das regiões pela proximidade geográfica (e, em alguns casos, pela proximidade política), separadas por barreiras físicas ou climáticas.
Não é de se estranhar que o reconhecimento de áreas de endemismo (de menor ou maior escala) ficou contaminado pelo reconhecimento apenas da distribuição espacial coincidente (parcial, em diferentes níveis de congruência das distribuições das espécies) dos táxons de certa região. Não era ainda perfeitamente entendida a importância da relação evolutiva de formação da área e da relação evolutiva para explicação da similaridade entre as áreas. Em outras palavras, não se sabia qual a relação entre os processos causadores e o padrão originado, ou seja, as áreas de endemismo.
Ainda no século XIX, o botânico Joseph Dalton Hooker
nas suas primeiras viagens indicou claramente que a conexão estreita entre os padrões de distribuição repetidos dos continentes do hemisfério sul devia-se ao fato de eles terem sido formados em uma época na qual esses continente< estariam unidos8. Isto ocorreu em uma época que não havÍ:: indícios consistentes da deriva continental que foi apena' entendida em seus detalhes quase 100 anos depois, pele' estabelecimento da teoria da tectônica de placas.
Existem duas razões principais para a importância de reconhecimento de áreas de endemismo: (a) elas são a< menores unidades de análise da biogeografia histórica e . como tal, são a base para construção de hipóteses sobrt os processos responsáveis pela formação da biota de um� região; (b) áreas de endemismo possuem uma reunião dt espécies únicas que devem ser priorizadas em programa' de conservaçãos (ver Capo 1 0) . Apesar de ser uma condição básica para estudos de biogeografia histórica.

Áreas de Endemismo - 43
Equador
2� , ' - As quatro grandes áreas de endemismo da Amazônia de Alfred R. Wa l lace - Gu iana, Equador, Peru e B rasi l , de l im itadas pelos grandes r ios Amazonas-Sol imões, Negro e Madeiras .
existem, disponíveis na literatura, diversos critérios para definição de uma área de endemismo. O maior número de definições está concentrado no reconhecimento dos diversos graus de congruências de distribuição de espécies ou táxons de uma área. Dentro deste tipo de definição, que utiliza apenas a simpatria, parcial ou total dos organismos, os conceitos foram resumidos por Morrone9:
• Áreas razoavelmente pequenas com um significativo número de espécies que ocorrem apenas nessas áreas; áreas delimitadas por distribuições coincidentes de táxons que não aconteceram em outras áreas4.
• Áreas definidas pelos limites de distribuição congruente de duas ou mais espécies 10.
• Áreas definidas pela distribuição de táxons endêmicos que ocorrem nessas áreas l l .
• Áreas delimitadas pela distribuição congruente de, no mínimo, duas espécies de distribuição restrita, com as áreas de distribuição das espécies fortemente congruentes 1 2 .
• Áreas com muitas espécies encontradas apenas nessas áreas e em nenhuma outra 13 .
A delimitação das áreas de endemismo indicadas anteriormente inclui apenas o critério da coincidência espacial das distribuições de duas ou mais espécies. Este é o princípio mais comumente utilizado e diversos métodos foram desenvolvidos para atender a essa indicação, dependendo do grau de simpatria e da escala utilizada do mapa. Entretanto, como se viu, outros critérios devem ser empregados conjuntamente à congruência distributiva para interpretações mais robustas de áreas de endemismo.
Axelius 14 chamou atenção que a indicação não arbitrária das áreas de endemismo está correlacionada à fidelidade e às premissas do método a ser utilizado para a sua definição. Qualquer análise de padrão biogeográfico, dentro do contexto histórico, deve estar inserida no escopo da história dos organismos, pois áreas de endemismos são fundamentalmente entidades históricas e não de distribuição15 , 1 6.
Neste contexto, claramente, devem ser usadas informações sobre a filogenia dos grupos das espécies ou

44 - Histórico e Conceitos
táxons que ocorrem em determinadas áreas, sugeridas como áreas de distribuição restrita desses táxons. Santos e Amorim17 reforçaram a ideia de que a delimitação das áreas de endemismos depende da precisa reconstrução das informações filogenéticas dos táxons que sucedem nessas áreas.
A melhor definição de área de endemismo é aquela que incorpora a história dos táxons 1 5 : área de endemismo é uma região geográfica com a distribuição de dois ou mais táxons monofiléticos que exibem congruência filo genética e distribucional, e que possuem seus respectivos grupos próximos acontecendo em outras regiões definidas do mesmo modo (Fig. 3 .2) .
Segundo Harold e Mooi1 5 existem duas fases no reconhecimento de uma área de endemismo. A primeira é a delimitação do nível de análise, em que os táxons são definidos, suas distribuições delimitadas e suas relações filogenéticas hipotetizadas. A segunda é a fase de teste em que táxons adicionais são incorporados na análise para identificar as possíveis áreas de endemismos e testar a sua realidade histórica. Essa definição de área de endemismo é derivada explicitamente do modelo vicariante de evolução de área9.
Dentro deste enfoque, uma área de endemismo é uma unidade histórica, uma vez que utiliza, na sua definição, elementos da composição de uma biota como um todo, nos seus mais variados níveis: os componentes de área, forma (a variação dos caracteres entre os táxons) e temp018. Áreas de endemismo são naturalmente hierárquicas, pois áreas menores podem estar agrupadas em uma área maior1 9, mas a delimitação dessas áreas, sem levar em consideração a filogenia das espécies que formam a área, pode resultar em errônea relação histórica entre as áreas20.
Áreas de endemismo são hipóteses que podem ser testadas e modificadas com a inclusão de novos dados de distribuição. Possuem a mesma interpretação quando táxons não monofiléticos são redefinidos4. Podem-se considerar táxons que delimitam áreas como caracteres que definem áreas. As relações entre táxons em três ou mais áreas são caracteres ou homologias que sugerem o relacionamento entre as áreasl l . Essas relações podem ser testadas à luz de novos dados disponíveis, podendo indicar a relação histórica entre as áreas de endemismos. A inclusão desses novos dados poderá dar suporte à corroboração da concepção de uma área de endemismo. Por outro lado, os novos dados também podem falsear a hipótese da área de endemismo, fornecendo ajuste metodológico com a proposição das novas hipóteses.
Têm-se empregado diversos métodos para o reconhecimento de áreas de endemismo. Talvez um dos métodos mais utilizados anteriormente foi proposto por Mül1er2 1 , Primeiramente, as áreas de distribuição das espécies eram indicadas no mapa e, em seguida, procurava-se a coincidência entre essas áreas. A extensão das áreas de endemismo deveria ser menor que os limites das distribuições das espécies que formavam a área endêmica. A principal dificuldade desse tipo de abordagem é a faltz de um critério estrito para a inclusão ou não de determinada espécie em uma área endêmica22• Dentro deste. mesma linha de raciocínio, ressalta-se que este tipo de protocolo usa métodos manuais para estimar os limites das áreas de distribuição das espécies 1, podendo causar erros na interpretação da área de distribuição das espécies,
Dentro de todas as abordagens da biogeografia histórica, a pan-biogeografia idealizada por Léon Croizat, em meados do século XX (ver Capo 5), possui metodologi", apropriada e adequada para delimitação de unidades biogeográficas naturais, sendo, por isso, elemento primordial para o entendimento da evolução do espaço-temp02�, Os métodos mais empregados (ver revisão em Morrone9 para reconhecimento das áreas de endemismo podem se� divididos em dois tipos de abordagem: aqueles que não utilizam e aqueles que utilizam algoritmos para a delimitação das áreas. O método do traço insere-se na primeirc:o abordagem e a análise parcimoniosa de endemicidade (PAE, parsimony analysis of endemicity) e o método desenvolvido por Szumik et aI. 13, usando o programc:o eNDeMismo e Visualizador de eNDeMismo (NDrvI-VNDM) , inserem-se na segunda.
Craw et aI. 2 enfatizaram que áreas de endemismo sãc unidades artificiais, dentro do enfoque da pan-biogeografia e que suportam apenas a utilização de traço, generalizados. Segundo Morrone9, a área de endemismc e o traço generalizado são diferentes representações do' componentes bióticos; a área como unidade básica de. biogeografia cladística e o traço como elemento primário da pan-biogeografia. No entanto, o traço generalizado possui um grau de equivalência com a área dê endemism09 (Fig. 3 .3) .
Consiste basicamente no estabelecimento do padrã,,' da distribuição dos grupos a partir da união de ponto ' de distribuição da espécie (ou táxon) através de linha� ,
.. -
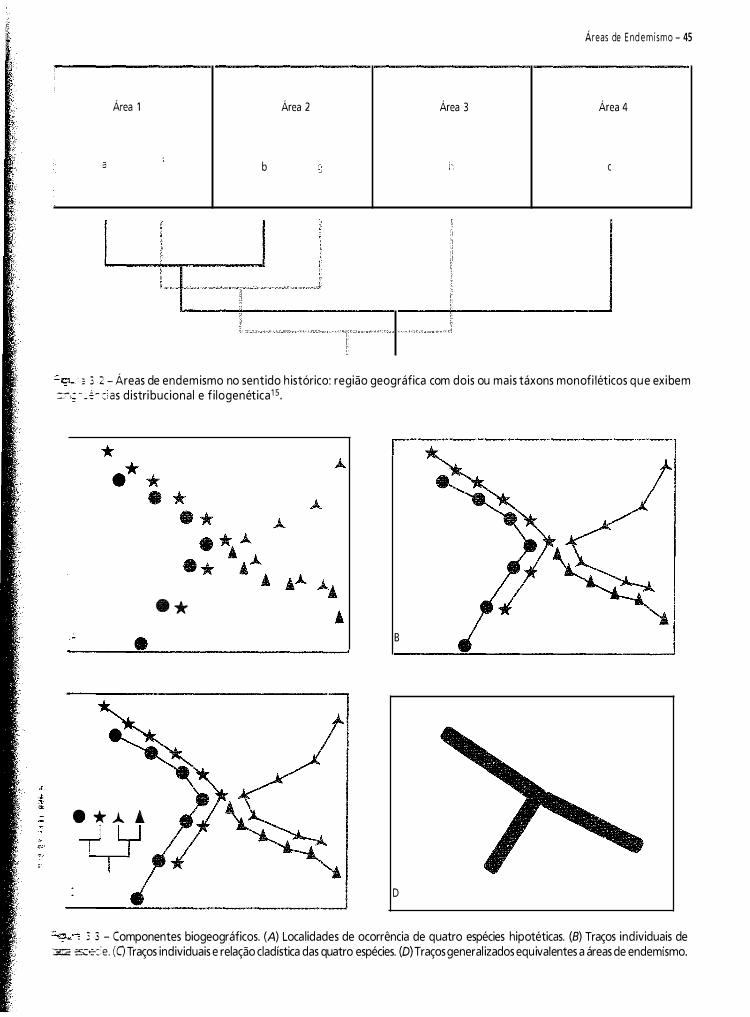
Áreas de E nd emismo - 45
Área 1 Área 2 Área 3 Área 4
a b c
I
f: :: 3 2 - Áreas de endemismo no sentido h istórico: região geográfica com dois ou mais táxons monofi léticos que exibem
:::"'1; - _�- c as distribucional e f i logenética 1 5.
- *
- B
• * Á !
Y D
3 3 - Componentes biogeográficos. (A) Localidades de ocorrência de quatro espécies hipotéticas. (B) Traços individuais de :::::a =:':>2-::e. (C) Traços individuais e relação cladística das quatro espécies. (D) Traços general izados equivalentes a áreas de endemismo.

46 - H istórico e Conceitos
respeitando-se o critério da menor distância geográfica para a ligação dos pontos. Denomina-se traço individual o conjunto de linhas que une os pontos de distribuição de um determinado táxon. Quando dois ou mais traços se sobrepõem é originado um traço generalizado. Este pode indicar áreas endêmicas segundo o critério da ocorrência de duas ou mais espécies em determinada área (Fig. 3 .3) .
Nihei e Carvalh024 sugeriram que os traços generalizados devem ser reconhecidos apenas com suporte de evidência filogenética, isto é, se eles compõem grupos irmãos (ver Capo 17). O procedimento é o seguinte:
• Indicar em um mapa, de preferência georeferenciado, as localidades onde ocorrem as espécies que estão sendo analisadas.
• Ligar as localidades de determinada espécie por um traço, respeitando, na ligação dos pontos, o critério da menor distância entre eles.
• Fazer o mesmo com todas as espécies no nível de análise do seu trabalho. Esses são os traços individuais das suas espécies.
• Comparar, dentro da escala utilizada, os traços individuais para encontrar os traços generalizados.
• Comparar nos grupos que possuem análise cladística os clados irmãos na procura de possíveis áreas de diversificação. Podem também ser incluídas na análise possíveis disjunções geológicas que ajudem no melhor entendimento da formação das áreas encontradas.
Segundo Craw et aI. 2, pode-se entender que os traços generalizados representem padrões de distribuição atuais de biotas ancestrais que foram fragmentadas, o que é suportado pela presença de grupos irmãos. Os traços generalizados equivalem a componentes bióticos que podem ser ordenados hierarquicamente em um sistema de classificação biogeográfica23.
A PAE foi criada e desenvolvida por Brian R. Rosen com o objetivo de incluir maior rigor em análises da biogeografia histórica. O método é trivial em seu uso e talvez por isso tenha sido aplicado com tanta frequência nos últimos anos. Utiliza-se a informação de presença e ausência das espécies ou táxons que ocorrem em uma região para a construção da matriz de dados. Em termos operacionais, o procedimento é análogo à análise de caráter em análises cladísticas em sistemática25. A delimitação das áreas de endemismo é oriunda da análise de uma matJiz de dados construída por presença/ausência das espécies na unidade de estudo e rodada em um pro-
grama computadorizado com algoritmo de parcimônia. Esse método permite a geração de hipóteses (cladogramas) falseáveis, maximizando a congruência de distribuição de tantos táxons quanto possível26.
A ampla utilização do método veio com a metodologia desenvolvida por Juan 1. Morrone, que empregou quadrículas como a unidade operacional da PAE27 (Fig. 3 .4). A sequência de passos é:
• Indicar as quadrículas na região do mapa a ser analisada, considerando apenas aquelas onde existe, no mínimo, uma localidade conhecida das espécies analisadas.
• Construir uma matriz de dados onde as colunas representam as espécies e as linhas representam as 1._
quadrículas. Reproduzir a presença da espécie por um � e a ausência por zero. Incluir uma quadrícula hipoté- ::
tica (área raiz) com apenas zeros para todas as colunas para servir como raiz da árvore.
• Executar uma análise da matriz de dados em um programa de computador com algoritmo de análise de parcimônia.
• Delimitar os grupos de quadrículas definidas por, no mínimo, duas espécies.
• Sobrepor os grupos delimitados no cladograma nas quadrículas e mapear as espécies endêmicas de cada grupo de quadrículas para delinear os limites de cada área.
As maiores críticas ao método apareceram quando foi salientado que a PAE basicamente unia as áreas a partir do seu compartilhamento de espécies e propondo uma história comum que podia explicar os agrupamentos encontrados. Em outras palavras, o cladograma, que é naturalmente hierárquico, mostraria as relações históricas entre as áreas de endemismo.
O método não deve ser utilizado como ferramenta da biogeografia cladística com o objetivo de descobrir as relações entre as áreas de endemism028.29. Morrone30 apontou claramente que a PAE é uma ferramenta da pan-biogeografia com o objetivo de encontrar as áreas endêmicas e que pode ser empregado para definir as homologias biogeográficas primárias da relação entre as áreas endêmicas. Este tipo de abordagem é o primeiro passo na descoberta das relações históricas entre as áreas de endemismo, mas que devem ser testadas por métodos da biogeografia cladística (ver Capo 6).
Existem outras variantes da PAE que podem ser utilizadas no sentido dinâmico - comparação e relação de áreas endêmicas em diferentes estratos geológicos de tempo - ou estático, quando a comparação de áreas

Áreas de Endemismo - 47
.------------------ Área raiz
� 8 10 1 2
2 5 6 1 1 14 1 5 � E
1 3 1 1 1 1 1 1 1
1 I 3 4 I • H Lf9-U- L 1 1
o
,-------------- C
�------------- D 1 1 1 5
Q 1 1
7 A
'--------- B A B
- Anál ise parcimon iosa de endemicidade (PAE). (A) Grade do sul da América do Su l d ividida em 22 quadrícu las de 5 graus de latitude por 5 graus de longitude (A-V). (B) Cladograma de área resultante da util ização de espécies de Palpibracus Rondani ( l nsecta, Muscidae): 1 = P. albuquerquei Carva lho; 2 = P. apicalis (Ma l loch); 3 = P. chilensis (Bigot); 4 = P. confusus (Ma l loch); 5 = P. fasciculatus (Ma l loch); 6 = P. lancifer (Ma l loch); 7 = P. nigriventris (Ma l loch); 8 = P. peruvianus (Mal loch); 9 = P. pilosus (Macquart); 1 0 = P. separatus (Ma l loch); 1 1 = P. similis (Ma l loch); 1 2 = P. spicatus (Ma l loch); 1 3 = P. trivittatus (Mal loch); 14 = P. univittatus (B igot), 1 5 = P. veneris (Bigot). Quadrículas: A, B, C, D, E, F, G, H, I, J, K, L, M, N, 0, P, Q, R, S, T, U, V. Bol inhas pretas: apomorfias (1 = presença). Bol inhas brancas: homoplasias (O = reversões). Mod ificado de Carva lho et aI. 20.
ocorre no mesmo estrat025•3 1 . Uma análise da literatura mostrou que a maioria dos trabalhos incluiu a abordagem estática na descoberta das áreas de endemism03 1 . Recentemente, foi apresentada uma proposta de análise de áreas de endemismo em diferentes estratos, mas ainda está faltando uma abordagem analítica desse tipo de abordagem32 .
Szumik et al. 1 3 propuseram um método para identificação de áreas de endemismo que usa algoritmos desenvolvidos posteriormente no programa NDM-VNDM33. O cálculo do valor de endemicidade deste algoritmo para delimitação de uma área de endemismo não utiliza nenhum modelo de evolução da área9. Este método de definição de área se baseia principalmente no conceito de área de endemismo desenvolvido por PlatnicklO.
O método, explicado por Szumik et alo 33 é bem trivial: em uma grade com quadrículas definidas, um grupo de quadrículas terá um valor de endemicidade calculado de acordo com o ajuste das localidades das espécies
endêmicas localizadas nesta área. O maior valor será indicado para as espécies que ocorrem em todas as quadrículas, diminuindo o valor para as espécies que sucedem em um número menor de quadrículas e terminando com aquelas espécies que acontecem também fora da área endêmica proposta anteriormente (Fig. 3 .5) .
Morrone9 apresentou a sequência de passos do método:
• Indicar as localidades de ocorrências das espécies no mapa com uma grade, podendo utilizar quadrículas definidas por latitude e longitude.
• Designar os valores de endemicidade de todos os dados das quadrículas, com a contagem das espécies que podem ser consideradas endêmicas a elas, de acordo com os critérios já indicadosl3.
• Escolher as quadrículas com os maiores índices de endemicidade.
• Desenhar os grupos de quadrículas no mapa mostrando as áreas de endemismo.
A comparação entre o método de endemicidade pelo programa NDM-VNDM e PAE não mostra diferenças significativas entre as áreas encontradas, mas difere principalmente no número de áreas, no número de

48 - H istórico e Conceitos
•
],:',5 - Quadrícu las com as loca l idades de ocorrência de três espécies h ipotéticas ( •• *). Quadrículas sombreadas: á rea endêmica . Cálculo do va lor de endemicidade através do Programa NDM-VNDM para determinação da área de endemismo. Mod ificado de Szumik et a/.33.
espécies endêmicas encontradas e maior sensibilidade nas áreas endêmicas limítrofes34. Como no PAE, o método implementado no NDM-VNDM não inclui a filogenia das espécies e também não sugere as relações entre as áreas, por não produzir cladogramas, que são intrinsecamente hierárquicos.
Como mencionado anteriormente, no sentido histórico devem ser incluídas e avaliadas as relações filogenéticas entre as espécies que compõem as áreas de endemismo para estabelecer as relações históricas entre as áreas.
o conceito de área de endemismo, no sentido histórico, foi desenvolvido ainda no período pré-evolutivo, quando Augustin Pyramus de Candolle caracterizou áreas a partir da singularidade exclusiva de espécies ocorrentes em uma
região4. Dentro do paradigma evolutivo, cerca de 1 50 anos depois, Léon Croizat utilizou a premissa de que Terra e
vida evoluem juntas, com a publicação do seu clássico livro Space, Time and Fonn: the Biological Synthesisl 8,
colocando a ciência da biogeografia firmemente com um dos pilares para se entender a evolução das espécies. Mesmo que área de endemismo seja usada no contexto da biogeografia ecológica, a sua singularidade pode ser apenas entendida pela distribuição congruente espacial e filo genética de táxons monofiléticos que delimitam a área.
Assim, o entendimento pode ser apenas histórico.
Como área de endemismo é a unidade histórica da biogeografia deve-se esperar uma coincidência abrangente de táxons de diferentes origens nessas áreas, em qualquer região da Terra, suportando uma história comum em resposta a eventos vicariantesl9. Em outras
palavras, a área de endemismo será singular para qual
quer táxon que ocorra na área, em razão do potencial
evolutivo da área35 .

A congruência, espacial e filogenética, deve suceder em diferentes graus e em diferentes níveis de escala analisadas. Deste modo, é razoável entender que as áreas de endemismo podem ser agrupadas e relacionadas desde um nível menor até um nível maior de abrangência, com a formação de áreas endêmicas maiores. Essas áreas podem ser indicadas como regiões biogeográficas naturais, mostrando que não podem ser definidas por táxons específicos, por exemplo, plantas e animais, isto é, regiões fitogeográficas e zoogeográficas, respectivamente. Como os processos de formação das áreas são os mesmos, deve-se utilizar a totalidade dos táxons de uma regiã036.
Áreas de Endemismo - 49
Na América do Sul, grandes áreas de endernismo são reconhecidas apesar da enorme diversidade taxonômica e de hábitat, oriunda de uma complexa história geológica (ver Capo 1 ) . É um continente biogeograficamente híbrido com distintas regiões biogeográficas e uma área de transição formada por elementos biogeográficos de regiões de diferentes origens23. O entendimento dos padrões de distribuição dos táxons da região tem sido feito geralmente com grupos específicos que podem ser extrapolados para outros táxons (Fig. 3 .6), porém é mais sensível e eficiente se, na análise, forem incluídos táxons não proximamente relacionados entre si37.
3l.i3i - Áreas de endemismo da América do Sul (estão ind icadas apenas á reas do leste do continente) com base em Aves: 1 = Napo; 2 = Imeri; 3a = Pantepui (Duida subcentro); 3b = Pantepui (Grã-Sabana subcentro); 4 = Gu iana; 5 = Inambari; 6 = Rondônia; 7 = Pará; 8 = Belém (Maranhão); 9 = Caatinga; 1 0 = Campo Cerrado; 1 1 = Chaco; 1 2 = Serra do Mar; 1 3 =
Paraná . Mod ificado de Cracraft19.

50 - H i stórico e Conceitos
Áreas de endemismo são hipóteses que podem ser corroboradas ou falseadas pela proposição de novos padrões de distribuição. As análises não podem ser apenas distribucionais procurando encontrar a maior congruência entre as áreas de distribuição das espécies, mas devem incluir também as relações filogenéticas das espécies em questão. Como áreas de endemismo são áreas naturais, pode-se inferir, a partir do relacionamento entre elas, uma visão geral da evolução espacial de uma região.
A pan-biogeografia possui métodos robustos, objetivos e empiricamente testáveis para o reconhecimento e delimitação das áreas de endemismo, no sentido histórico. Entretanto, todos esses procedimentos devem ser testados com cladogramas construídos com procedimentos da biogeografia cladística para corroborar o padrão histórico das áreas de endemismos delimitadas.
Com os dados e métodos disponíveis, ainda falta muito para compreender melhor a evolução das áreas de endemismo da América do Sul. No entanto, uma questão salta aos olhos dos interessados nesta área: é necessário que mais e mais padrões de distribuição sejam postulados para que a comparação entre eles possa ajudar no melhor entendimento do cenário evolutivo espacial da América do Sul, utilizando para isso as congruências das áreas de distribuição e o padrão filo genético dos táxons que ocorrem nessas áreas.
A Márcia Souto Couri (Museu Nacional do Rio de Janeiro) , Kirstern Lica F. Haseyama (Universidade Federal do Paraná), Eduardo A. B. Almeida (Universidade Federal do ABC) e dois revisores anônimos pela leitura crítica e sugestões. Ao Sionei Ricardo Bonatto pela confecção das figuras. Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela bolsa e auxílio concedidos (processo 300873/2008-5).
REFERÊNCIAS BIBLIOGRÁFICAS 1 . ROIG-JUNENT, S. ; CRISCI, J. v.; POSADAS, P. et a!. Áreas de
distribución y endemismo en zonas continentais. In: COSTA, c.; VANIN, S . A; LOBO, J. M. et al. Proyecto de Red lberoamericana de Biogeografia y Entomología Sistemática. Pribes. Zaragoza: Sociedad Entomológica Aragonesa, 2002. v. 2, p. 247-266.
2. CRAW, R. c. ; GREHAN, J. R.; HEADS, M. J. Panbiogeography. Tracking the History of Life. New York : Oxford University Press, 1 999. 229p.
3. SCLATER, P. L. On the general geographical distribution of the members of Class Aves. Reproduzido em LOMOLINO, M. Y.; SAX, D. F. ; BROWN, J. H. Foundations ofBiogeography. Classic Papers with Commentaries. Chicago: The University of Chicago Press, [ 1 858] 2004. parte 9, p. 1 1 8- 1 33 .
4. NELSON, G. J . ; PLATNICK, N. I. Systematics and Biogeography - Cladistics and Vicariance. New York: Columbia University Press, 198 1 . 567p.
5. SILVA, J. M. c.; RYLANDS, A B.; SILVA JR, 1. S. et aI. Primate diversity patterns and their conservation in Amazonia. In: PURVIS, A; GITTLEMAN, J. L.: BROOKS, T. Phylogeny and Conservation. Cambridge: Cambridge University Press, 2005. capo 15, p. 337-364.
6. DARWIN, C. On the Origin of Species by Means ofNatural Selection, OI; the Preservation of Favoured Races in the Strugglefor Life. London: John Munay, 1 859. 5 1 3p.
7 . WALLACE, A R . The Geographical Distribution of Animais. 2 vols. London: Macmillan, 1 876. 607p.
8 . HUMPHRIES, C. J. From dispersaI to geographic congruence: comments on cladistic biogeography in the twentieth century. In: WILLIANS, D. M . ; FOREY, P. L. Milestones in Systematics. London: CRC Press, 2004. capo 1 0, p. 225-260.
9. MORRONE, J. J. EvoIutionary Biogeography: an integrative approach with case studies. New York: Columbia University Press, 2009. 3 1 1p.
1 0. PLATNICK, N. 1. On areas of endemism. Aust. Syst. Bot. , V. 4, p. xi-xii, 1 99 1 .
1 1 . HUMPHRlES, C . J. ; PARENTI, L . R. Cladistic Biogeography: intefpreting patterns of plant and animal distributions. 2. ed. New York: Oxford University Press, 1 999. 1 87p.
J 2. LINDER, H. P. On areas of endemism, with an example fram the Ati'ican Restionaceae. Syst. Bio!. , V. 50, p. 892-9 1 2, 2001 .
1 3 . SZUMIK, C . A.; CUEZZO, F.; GOLOBOFF, P. A . et aI . An optimality criterion to determine areas of endemism. Syst. Biol. , v. S I , p. 806-816, 2002.
14. AXELIUS, B. Areas of distributions and areas of endemism. Cladistics, V. 7, p. 197-199, 1 99 1 .
1 5 . HAROLD, A S . ; MOOI, R. D . Areas o f endemism: definition and recognition criteria. Syst. Biol., V. 43, p. 2 6 1 -266, J 994.
1 6. MORRONE, J. J. Homology and areas of endemism. Divers. Distrib. , V. 7, p. 297-300, 200 1 .
1 7 . SANTOS, C. M. D . ; AMORIM, D. S. Why biogeographical hypotheses need a well supported phylogenetic framework: a conceptual evaluation. Pap. Avulsos Zool., V. 47, p. 63-73, 2007.
1 8 . CROIZAT, L. Space. Time. Form: the b iological synthesis. Caracas: published by the author, J 964. 881p .
1 9 . CRACRAFT, J. Historical biogeography and patterns of differentiation within the South American avifauna: areas of endemism. Omith Monog. , V. 36, p. 49-84, 1 985.
20. CARVALHO, C. J. B.; BORTOLANZA, M.; SILVA, M . C. C. et aI. Distributional patterns of the N eotropical Muscidae (Diptera). In: MORRONE, J. 1 . ; LLORENTE, J. Una Perspectiva Latinoamericana de la Biogeografía. México: Las Prensas de Ciencia, FacuItad de Ciencias, Universidad Autónoma de México, 2003. capo 30, p. 263-274.
2 1 . MÜLLER, P. The DispersaI Centers ofTerrestrial Vertebrates in the Neotropical Realm. The Hague: Df. W. Junk B . V. Publishers, 1 973. 244p.
22. PETERSON, A T.; WATSON, D. Problems with areal definitions of endemism: the effects of spatial scaling. Divers. Distrib . . V . 4 , p . 1 89- 1 94, 1 998.
23. MORRONE, J . J . Panbiogeografia, componentes bióticos y zonas de transición. Rev. Bras. Entomol. , V. 48, p. 1 49-162. 2004.
24. NIHEI, S. S . ; CARVALHO, C. J. B. Distributional patterns of lhe Neotropical fty genus Polietina Schnabl & Dziedzicki (Diptera, Muscidae): a phylogeny-supported analysis using Panbiogeographic tools. Papo Avulsos de Zool. (São Paulo), v. 45, p . 3 1 3-326, 2005.
25. ROSEN, B. R.; SMITH, A B. Tectonics from fossils? AnaIysis of reef-coral and sea-urchin distributions from late Cretaceous to Recent, using a new method. In: AUDLEY-CHARLES, M. G. ; HALLAM, A. Gondwana and Tethys. Geol. Soe. SP, V. 37. p. 275-306, 1988.

POSADAS, P.; MIRANDA-ESQUIVEL, D. R. El PAE (Parsi�ony Analysis of Endemicity) como una herramienta en la :,'. ",luación de la biodiversidad. Rev. Chi/. Nist. Nat. , vol. 72, p. � "9-546, 1 999. '.lORRONE, J. J. On the identification of areas of endemism. � 5t. Bio/. , v. 43, p. 438-44 1 , 1994. S �"TOS, C. M. D. Parsimony analysis of endemicity: time for ",_1 epitaph? J Biogeogr., v. 32, p. 1 284-1286, 2005. ::: -\RZÓN-ORDUNA, I. J.; MIRANDA-ESQUIVEL, D. R. ; =10:'\ATO, M. Parsimony analysis of endemicity describes but : :'os not explain: an illustrated critique. J. Biogeogr. , v. 35, p. - 3-913. '. rORRONE, J. J. Cladistic Biogeography: identity and place.
. Biogeogr., v. 32, p. 1 28 1 - 1 286, 2005. '':IHEI, S. S. Misconceptions about parsimony analysis of en:e:nicity. J. Biogeogr., v. 33 , p. 2099-2106, 2006. " IHEI, S. S. Dynamic endemism and 'general ' biogeographic � ",:tems. Biogeografía, v. 3, p. 1-6, 2008.
Áreas de Endemismo - 51
33. SZUMIK, C . ; CASAORANDA, D.; ROlO, S . Manual d e NDM/ VNDM: programas para la identificación de áreas de endemismo. Instituto Argentino de Estudios Filogenéticos, ano 5, v. 3 , p . 1 -26, 2006. Disponível em: http://www.zmuc.dk/public/ Phylogeny/Endemism/Manual_ VNDM.pdf. Acesso em: 23/09/2009.
34. MOLINE, P. M. ; LINDER, H. P. Input data, analytical methods and biogeography ofElegia (Restionaceae). J Biogeogr. , v. 33 , p. 47-62, 2006.
35. BROOKS, D. R.; MAYDEN, R. L. ; MCLENNAN, D. Phylogeny and biodiversity: Conserving our evolutionary Jegacy. Trends Eco/. Evo/. , v. 7, p. 55-59, 1992.
36. MORRONE, J. J . B iogeographical regions under track and cladistic scrutiny. J. Biogeogr. , v. 29, p. 149- 1 52, 2002.
37. SIORIST, M. S . ; CARVALHO, C. J. B. Detection of areas of endemism on two spatial scales using Parsimony Analysis of Endemicity (PAE): the Neotropical region and the Atlantic Forest. Biota Neotropica, v. 8, p. 33-42, 2008.

_____________________ 4
li �ÇI': 11<3 li ,-y LL
Eduardo A. B. Almeida
Charles Morphy D. Santos
Biogeografia é um conceito aparentemente simples. O termo exprime de maneira direta a ideia de um tratamento conjunto de informações biológicas e geográficas. A biogeografia tem como objetivos descrever a distribuição dos organismos no planeta e dar explicações para a história que teria levado a tais configurações espaciais 1-5. Trata-se de um campo de investigação interdisciplinar, que se dedica à difícil tarefa de compreender por que os táxons são encontrados em determinadas áreas e não em outras. Em sua maior escala, a biogeografia fornece a perspectiva histórica necessária tanto para a compreensão da evolução das biotas como também da evolução geológica do planeta6.
O desenvolvimento da biogeografia como ciência, especialmente ao longo dos últimos três séculos, teve forte relação com o desenvolvimento e o refinamento de sistemas cJassificatórios estáveis - que permitiram a documentação crescente da diversidade biológica - em paralelo ao aumento do conhecimento da distribuição geográfica das espécies e da geologia do planeta. A reunião de informações sobre a distribuição de um número cada vez maior de táxons permitiu a formulação de questões cruciais, como a descoberta de padrões biogeográficos e a regionalização do espaço (ver Capo 2).
O estudo de situações resultantes de eventos históricos é desafiador. Há uma multiplicidade de possíveis fatores, pretéritos e atuais responsáveis pela presença de um grupo biológico em determinada área. É exatamente o acúmulo desses diversos eventos ao longo do tempo que
8
TI (0 TiD ;{ \C '"'
0 .li L� r
torna a reconstrução da história da distribuição espacial das linhagens de organismos uma atividade complexa e que exige bem mais do que um simples método analítico. Nesse sentido, a abordagem científica da biogeografia difere daquela das ciências ditas experimentais, em que experimentos controlados permitem compreender as causas de um evento, fornecendo evidências empíricas para prever eventos futuros de natureza semelhante. Por ser uma ciência histórica, a Biogeografia, ao contrário, volta-se para o passado na tentativa de estabelecer conexões entre cadeias de eventos responsáveis por situações que não podem ser repetidas experimentalmente. Essa reconstrução do passado é uma tarefa árdua, não apenas em decorrência da complexidade dos temas ou por algum tipo de insuficiência teórica, mas da característica intrínseca dos problemas tratados. Sober7 denominou "processos destruidores da informação" aqueles que causam o desaparecimento das evidências processuais de como um padrão veio a adquirir suas características recentes. Sabe-se que as próprias histórias biológicas e geológicas destroem boa parte dos indícios que permitiriam exame direto dos padrões biogeográficos e a consequente compreensão de pelo menos parte dos processos que os originaram. Isso exige que abordagem indireta seja adotada na biogeografia, fundamentada na comparação de relações entre táxons e as áreas em que eles ocorrem.
N os seus aproximadamente 4,5 bilhões de anos, o planeta tem sofrido uma série de alterações, mais ou menos drásticas, que influenciaram, e ainda influenciam, a evolução das espécies. Movimentos das placas tectônicas, orogêneses, aparecimentos e desaparecimentos de rios e outros corpos d' água, erosões, mudanças climáticas profundas, ocorrendo em escalas de milhões, dezenas

de milhões ou centenas de milhões de anos, são alguns dos eventos responsáveis pelas contínuas modificações das feições da Terra. Esses eventos afetam as distribuições de organismos : uma população anteriormente contínua divide-se em subpopulações; uma espécie tem sua distribuição aumentada pela expansão para áreas anteriormente isoladas; grupos inteiros podem se extinguir local ou globalmente.
A evolução de um táxon por vezes segue caminhos tortuosos. No entanto, alguns desses caminhos são compartilhados por outros táxons, próxima ou distantemente relacionados. Isso acontece quando uma alteração física em dada área (Fig. 4. 1 , A) atinge mais do que uma única espécie que habita aquela região. O aparecimento de uma barreira, que divide uma área anteriormente contínua, pode separar a população ancestral de uma espécie em duas subpopulações, as quais, uma vez isoladas, podem responder independentemente às pressões do meio e se diferenciar, resultando em formação de duas novas espécies. Esse é o processo denominado vicariância (Fig. 4. 1 , B). Muitas vezes, a barreira é efetiva para espécies distintas, dividindo suas populações de maneira semelhante. Esse padrão compartilhado de disjunções, quando as linhagens irmãs de vários táxons são separadas por eventos de vicariância comuns, pode vir a ser reconhecido atualmente como um padrão biogeográfico (Fig. 4. 1 , C). Ao se assumir a possibilidade da existência de eventos comuns que alteram de modo similar as distribuições de mais de um táxon, pode-se reconstruir um panorama da história biogeográfica desses táxons com distribuição mais ou menos coincidente, a despeito do sem-número de particularidades presentes e passadas que podem ter afetado a evolução de cada grupo.
Quando apenas eventos vicariantes moldaram a distribuição dos táxons, o relacionamento filo genético entre eles estava totalmente atrelado ao relacionamento entre as áreas por eles ocupadas - as relações filogenéticas são, portanto, biogeograficamente informativas (e vice-versa). N a Figura 4. 1 , A, as áreas x e y são mais próximas entre si
J: em relação à área z, pois x e y teriam se isolado posterior-0\ Cf -'Í" '" t-tA Cf 00 t-0\
mente à separação de z. Da mesma forma, os táxons presentes em x e y (espécies 1 e 2) seriam mais proximamente relacionados entre si em relação ao táxon presente em z (espécie 3) (Fig. 4 . 1 , B). Pode-se afirmar ainda que o evento de vicariância responsável pela separação das áreas x e y (e pela origem das espécies 1 e 2) é mais recente que o evento relacionado à separação de z (que originou a espécie 3). Uma hipótese de relação entre áreas é corroborada quando filogenias de outros grupos de organismos distribuídos nessas mesmas áreas coincidem com o padrão esperado, como mostrado na Figura 4 . 1 , C.
Lógica da B iogeografia de Vica r iânc ia - 53
Como visto anteriormente, evidências de padrões biogeográficos podem ser extraídas de filogenias e das distribuições dos organismos5. O entendimento e a documentação das distribuições taxonômicas são evi
dências primárias de qualquer estudo biogeográfico, uma vez que são indicativas de possível associação histórica entre os organismos e as áreas que eles ocupam. Quando táxons diferentes encontram-se na mesma área, pode-se inferir uma hipótese inicial de que essas distribuições resultam dos mesmos eventos históricos5. Distribuições coincidentes sugerem que a área sob escrutínio constitui uma unidade biogeográfica ou área de endemismo4.
A despeito de toda controvérsia associada ao conceito de áreas de endemismo, que se estende por quase dois séculos, a sua definição invariavelmente parte do reconhecimento da repetição de configurações espaciais entre táxons distintos. Essa premissa está no cerne das diversas propostas para definir áreas de endemismo publicadas nos últimos 30 anos (ver Capo 3) . Assim como as espécies são as unidades básicas da sistemática, as áreas de endemismo são as unidades básicas da biogeografia. No entanto, ainda tem se definido endemismo de forma vaga em grande parte dos estudos biogeográficos. O reconhecimento de áreas de endemismo não depende apenas da representação de registros de ocorrências taxonômicas em um mapa, não havendo, portanto, equivalência satisfatória com áreas de distribuição. Área de endemismo é uma unidade histórica verdadeira, uma região na qual estão distribuídos dois ou mais grupos monofiléticos que exibem congruência filo genética e distributiva (ver Capo 3). Padrões de distribuição fornecem informação biogeográfica para definir áreas de endemismo apenas no contexto das relações filogenéticas dos organismos8.
Por si só, áreas de endemismo são hipóteses frágeis, já que muitos fatores isolados podem influenciar uma dada configuração espacial. A formação de padrões resultantes da repetição de conformações espaciais e de relações entre as áreas em questão são, por outro lado, candidatos menos prováveis de serem resultantes do acaso.
Neste capítulo, será explorada uma base conceitual para a compreensão da história das distribuições dos seres vivos. Mais especificamente, será tratada a Biogeografia de Vicariância (ou Biogeografia Vicariante) e o exame de padrões biogeográficos possivelmente resultantes da vicariância. A documentação do registro geográfico e a existência de hipóteses de relações filogenéticas proveem o embasamento empírico funda-

54 - H istórico e Conce itos
ti
(A)
sp . l sp . 2
t3 - - - dlllilflllll"-
t2 -
ti
(8)
t2
sp. 3
sp. 7 sp. 4
1
t3
sp . 8 sp. 5
sp . 2
(C)
sp. 9 sp. 6
sp. 3
- (A) H istória de uma área ancestral 'xyz' que ao longo da h istória geológ ica foi fragmentada, in ic ia lmente em duas á reas menores, xy e z, e posteriormente em três á reas (após separação de x e y) . (B) O grupo monofi l ético formado pelas espécies 1, 2 e 3 distribu i-se exc lusivamente nas á reas x, y e z. A h istória de d iversificação desta l i nhagem transcorreu dura nte a história geológica de separação das á reas, conforme o esquema. (C) Dois outros táxons, um com posto pelas espécies 4, 5 e 6 e o outro composto pelas espécies 7, 8 e 9, também ocupam as mesmas áreas e tiveram h istórias mu ito s imi lares à da l i nhagem que inc lu i 1 , 2 e 3. A congruência entre as três h ipóteses propostas para as três l i n hagens reforça as h ipóteses biogeográficas ind ividuais por formarem um padrão. Três momentos históricos d istintos, entre os q uais eventos de vicariância ocorreram, são denotados por ti, t2 e t3.
mental para a grande parte das inferências analíticas em biogeografia histórica por vicariância9. O propósito de uma discussão sobre a lógica de uma disciplina científica é avançar na discussão sobre os fundamentos conceituais desta última, assim como identificar paradigmas científicos cruciais para a sua sustentação.
Neste capítulo, será explorada uma base conceitual para a compreensão da história das distribuições dos seres vivos (para uma discussão sobre as duas principais abordagens da pesquisa biogeográfica, consultar Quadro 4. 1 ) . Mais especificamente, será tratada a Biogeografia de Vicariância (ou Biogeografia Vicariante) e o exame de padrões biogeográficos possivelmente resultantes da vicariância. A documentação do registro geográfico e a existência de hipóteses de relações filo genéticas proveem o embasamento empírico fundamental para a grande parte das inferências analíticas em biogeografia histórica por vicariância9. O propósito de uma discussão sobre a lógica de uma disciplina científica é avançar na discussão sobre os fundamentos conceituais desta última, assim como identificar paradigmas científicos cruciais para a sua sustentação.
���J. - Escolas da B iogeografia
Tradicionalmente, a biogeografia é dividida em biogeogra
fia ecológica e biogeografia histórica 1 -5.8,9, As diferenças
entre as duas escolas d izem respeito a ti pos de questões
levantadas, metodolog ias específicas e esca l a temporal
dos eventos.
A biogeografia ecológica l ida com processos com duração
mais restrita, cujos efeitos podem afetar a distr ibu ição de
organ ismos de uma determ inada espécie ao longo de
uma ou poucas gerações. O estudo de q u a isquer fatores
ecológicos afetando distri buição de táxons é considerado
como do escopo da biogeografia eco lógica.
A biogeografia histórica investiga as m uda nças biogeo
gráficas ocorridas ao longo do tempo evolutivo e exa mina
as causas h istóricas da d istribuição dos organismos. Esta
discip l i n a abarca a biogeografia de vicariância, q ua ndo se
fundamenta no reconhecimento de padrões vicariantes
para a investigação da evolução espaci a l9,
Apesar d e ser conveniente, por vezes, traçar uma separa
ção entre os fatores históricos e ecológicos que afetam a
d istr ibu ição dos organ ismos sobre o p l aneta, a i nteração
desses fatores para a h i stória da d istr ibuição de q u a l q uer
táxon não deve ser ig norada3.
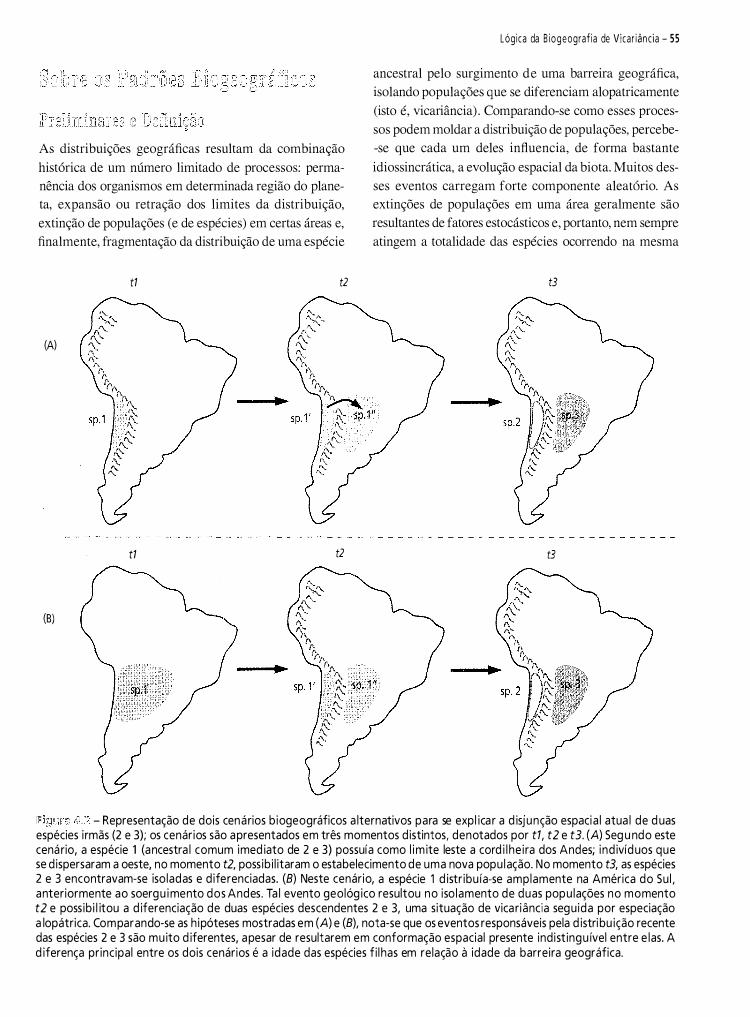
:;:;
As distribuições geográficas resultam da combinação
histórica de um número limitado de processos: perma
nência dos organismos em determinada região do planeta, expansão ou retração dos limites da distribuição,
extinção de populações (e de espécies) em certas áreas e,
finalmente, fragmentação da distribuição de uma espécie
ti t2
(A)
..
ti t2
(B)
..
Lógica da B iogeografia de Vicar iância - 55
ancestral pelo surgimento de uma barreira geográfica,
isolando populações que se diferenciam alopatricamente
(isto é, vicariância) . Comparando-se como esses proces
sos podem moldar a distribuição de populações, percebe
-se que cada um deles influencia, de forma bastante
idiossincrática, a evolução espacial da biota. Muitos des
ses eventos carregam forte componente aleatório. As
extinções de populações em uma área geralmente são
resultantes de fatores estocásticos e, portanto, nem sempre
atingem a totalidade das espécies ocorrendo na mesma
t3
t3
- Representação de dois cenários biogeográficos a lternativos para se expl ica r a d isjunção espacia l atua l de d uas espécies irmãs (2 e 3); os cenários são apresentados em três momentos distintos, denotados por ti, t2 e t3. (A) Seg u ndo este cenário, a espécie 1 (ancestra l comum imed iato de 2 e 3) possuía como l im ite leste a cord i l heira dos Andes; ind ivíduos que se dispersaram a oeste, no momento t2, possibi l itaram o estabelecimento de uma nova popu lação. No momento t3, as espécies 2 e 3 encontravam-se isoladas e diferenciadas. (B) Neste cenário, a espécie 1 d istr ibu ía-se amplamente na América do Su l , anteriormente ao soergu imento dos Andes. Ta l evento geológ ico resultou no iso lamento de duas popu lações no momento t2 e possibi l itou a d iferenciação de duas espécies descendentes 2 e 3, uma situação de vicariância seguida por especiação a lopátrica. Comparando-se as h ipóteses mostradas em (A) e (B), nota-se que os eventos responsáveis pela d istribu ição recente das espécies 2 e 3 são muito d iferentes, apesar de resultarem em conformação espacial presente indistingu ível entre e las. A d iferença principal entre os dois cenários é a idade das espécies fi l has em relação à idade da barreira geográfica.

56 - H istórico e Conceitos
t1 t2 t3
- Representação da d istr ibu ição de uma espécie (área cinza) em três momentos de sua h istória (t1, t2 e t3). A
expansão dos l im ites da d istribu ição desta espécie não envolveu a transposição de barrei ras geográficas e, por isso, não é considerada um evento de d ispersão.
região. Dispersões envolvem a transposição de barreiras geográficas preexistentes (Fig. 4.2, A), sendo eventos dependentes do acaso e de características espécie-específicas que determinam o que consiste uma barreira para os organismos de cada táxon. Expansão de distribuição geográfica (Fig. 4.3) relaciona-se à vagilidade dos organismos e à relação entre preferências ambientais e disponibilidade de hábitats ecologicamente apropriados.
O surgimento de barreiras geográficas e a consequente fragmentação da área de distribuição ancestral de espécies, resultando na formação de espécies descendentes alopátricas (Fig. 4.2, B), podem ser considerados a classe de eventos mais independente de atributos dos próprios organismos. Justamente por isso, espera-se que vários táxons habitando a área anterior à sua fragmentação respondam de maneira similar à vicariância (Fig. 4. 1 , C).
Historicamente, os cenários biogeográficos propostos para explicar distribuições disjuntas de táxons proximamente relacionados favoreceram duas classes de eventos: a expansão de distribuição pela transposição de uma
barreira geográfica preexistente (cenário dispersalista) ou a fragmentação de uma distribuição anteriormente mais ampla (cenário vicariante). De forma isolada, dados de distribuição de um táxon não são suficientes para demonstrar, de maneira inequívoca, as raízes históricas da sua atual distribuição disjunta. A melhor forma de se
abordar um problema deste tipo é a busca por padrões similares em grupos de organismos distintos2,5.
Assim como é pouco sensato pensar que a repetição dos padrões de distribuição de caracteres em um cladograma resulta do acaso, não parece razoável que padrões semelhantes de distribuição entre áreas refiram-se às histórias independentes de dispersão. É mais parcimonioso assumir que esses padrões atuais de distribuição sejam, pelo menos em parte, resultado de histórias compartilhadas causadas por respostas similares às modificações da superfície do planeta2. No entanto, um raciocínio fundamentado na ideia de que a melhor explicação inicial para uma distribuição disjunta é a fragmentação de uma população ancestral, per
se, não garante reconstrução biogeográfica confiáveL A perda de informação sobre a sequência pretérita de eventos, que acarretou distribuição atual, é o maior empecilho para que explicações biogeográficas sejam propostas. O exemplo da Figura 4.2 ilustra tal dificuldade. Quando se comparam as distribuições atuais de dois pares de espécies irmãs nos cenários distintos propostos (Fig. 4.2), percebe-se que, em ambos os casos, uma barreira geográfica separa uma espécie de sua espécie irmã. A questão fundamental é determinar o momento do surgimento da barreira em relação ao evento de especiação que causou divergência da espécie 1 em relação às espécies 2 e 3, que difere marcadamente nos dois casos.
Em suma, padrões biogeográficos são coincidências não arbitrárias (congruências) de distribuições geográficas de espécies ou grupos monofiléticos e de relacionamentos
históricos entre as áreas em que esses táxons ocorrem2,3.

Biogeógrafos são historiadores que almejam compreender
a evolução da diversidade biológica no espaço geográfico.
Infelizmente, as evidências que se mantêm como resquícios
dessa história são muitas vezes obscurecidas por outros
Área x Área y Área z
sp. 1 sp. 2 sp. 3
(A)
x y z
(C)
x w y z z
(D)
x w y w z
(E)
Lóg i ca da B iogeografia de Vicar i ânc i a - 57
sinais históricos conflitantes àqueles sugeridos pela vica
riância. Os estudos investigados do ponto de vista biogeo
gráfico normalmente incluem congruência apenas parcial
entre histórias dos táxons e das áreas em que eles são en
contrados. Parte da confusão é resultante da concomitância
entre as histórias de vicariância e as histórias de dispersões
e extinções de táxons de uma linhagem (Fig. 4.4).
x
(B)
x x y z
x w y z z
x w y w z
y
x x
w y
x w y
z
z z
z
w
- (A) Relações fi logenéticas entre as espécies 1 , 2 e 3, mostrando as áreas em que estas se d istr ibuem e sua h istória biogeográfica de vicariância . (B) Cladograma de áreas i nferido a partir de (A) . Neste caso, as relações entre as á reas correspondem à real h istória geológica da região, pois a h istória de c ladogênese do táxon composto pelas espécies 1 , 2 e 3 foi perfeitamente congruente aos eventos de vicar iância . (C a E) Três cladogramas de á reas i nferidos a partir de relações de outros táxons, cujas h istórias foram afetadas pela mesma h istória geológ ica, mas com resu ltados divergentes entre si e em relação ao caso a nterior: (C) a espécie d istribu ída na área x especiou-se simpatricamente e a espécie d istribu ída na área y se ext ingu iu; (O) a lguns ind ivíduos da espécie ocupando a á rea x d ispersaram-se para uma á rea w, dando origem a uma nova espécie e a espécie d istr ibuída na área x posteriormente se ext ingu iu; (E) i nd ivíduos das espécies ocupando as á reas y e z d ispersaram-se i ndependentemente para a á rea w e a espécie d istribuída na área z se extingu iu . L inha ponti l hada =
extinção de espécie; círcu lo preench ido (e) = especiação simpátrica .

58 - Histórico e Conce itos
Os métodos analíticos da biogeografia propõem-se jus
tamente a lidar com as "imperfeições" do registro histórico deixado pela evolução biológica e espacial dos táxons6. Essas ferramentas buscam extrair o máximo de
informação biogeográfica, até mesmo de situações ambíguas, nas quais os padrões de relação entre as áreas
estão longe de ser inequívocos. Nesse ínterim, o reco
nhecimento das relações filogenéticas entre os táxons e da sua associação ao componente espacial criam condições desejáveis para a compreensão tanto da história
biogeográfica de um grupo em particular como também
para a identificação de padrões biogeográficos gerais.
Craw et al.1 O referiram-se à pesquisa biogeográfica como o estudo da "vida em camadas geológicas". A cada estágio da história geológica do planeta, uma enormidade de fatores interagiu para resultar em conformação espacial dos organismos no estágio seguinte e, daí por diante, até o momento presente. Essa profusão de fatores influenciando as distribuições aumenta o "ruído" na análise biogeográfica, pois em grande quantidade de casos os padrões individuais não são completamente congruentes entre si, desviando do padrão biogeográfico geral, que é o objetivo dos estudos sobre o componente espacial da evolução . Page i i notou que mesmo a obtenção de congruência na sequência dos eventos de divergência em dado cenário pode acarretar erros de interpretação quanto à existência de uma história comum. Em situação ideal, deveria ser possível avaliar também a congruência temporal entre os eventos, pois os padrões observados nem sempre levam em conta que táxons com distribuições compartilhadas podem ter origens biogeográficas distintas ("pseudocongruências": os Caps. 6 e 7 discutem este tópico em mais profundidade), não passando, necessariamente, por toda a sequência de eventos que resultou na configuração atual, como é o caso dos táxons alocrônicos * .
A proliferação de métodos de estudo das relações biogeográficas foi tão grande que, nos últimos anos, classificações foram propostas para organizar essas ferramentas e abordagens, de modo a facilitar a escolha de uma ou mais
* O termo " alocrônico" foi proposto por Amorim et al. 1 2 para fazer referência a táxons que atualmente ocupam as mesmas áreas, mas cujas histórias biogeográficas teriam ocorrido em períodos geológicos distintos. Apesar da coincidência observada entre suas distribuições, não há, em casos deste tipo, congruência biogeográfica verdadeira.
delas para determinada pesquisa 1 ,3,6. Crisci et aI. 1 viram, na rápida diversificação de opções analíticas, um sinal de revolução científica da área da biogeografia.
Em uma perspectiva histórica, são identificadas três grandes fases da pesquisa biogeográfica13 : biogeografia descritiva (ou empírica), narrativa e analítica. O primeiro estágio de estudos biogeográficos foi o descritivo, em que se realizaram documentação das localidades de ocorrência das espécies e as primeiras tentativas de reconhecimento de regiões de co-ocorrência de táxons. Esse tipo de abordagem permitiu, em algum grau, indução e predição, na medida em que o registro da presença de uma espécie em determinada área permitia prever o encontro de organismos desta espécie na mesma área em momento posterior13. A biogeografia descritiva não se propõe, contudo, a gerar hipóteses sobre como as populações alcançaram as distribuições observadas.
A biogeografia narrativa inclui, sob sua alcunha, as tentativas de explicação para a origem dos padrões de distribuição13. Nela, levantam-se hipóteses - considerando os registros de ocorrência dos táxons, as relações filo genéticas presumidas entre as espécies desses grupos e eventos geológicos relevantes - que constituem cenários plausíveis para descrever a história biogeográfica. Essas narrativas procuraram descrever histórias individuais, na maior parte das vezes apoiadas em eventos de dispersão válidos apenas para o grupo taxonômico particular estudado. A validade de pesquisas narrativas é questionável, na medida em que são empiricamente fracas, em razão de sua falta de generalidade e testabilidade.
A partir da metade do século XX, a biogeografia se tomou mais analítica e, seus resultados, por conseguinte, mais testáveis do ponto de vista científico. Ano a ano, tem havido aumento no leque de opções para estudos biogeográficos analíticos, mas os pilares conceituais dessa área de pesquisa foram erguidos entre as décadas de 1950 e 1970. Três são os pilares mais fundamentais dos desenvolvimentos analíticos e conceituais da biogeografia de vicariância9: ( 1 ) a aceitação do modelo da tectônica de placas, que remonta à deriva continental proposta décadas antes por Alfred Wegener e que teve implicações profundas para a noção de vicariância como evento biogeográfico
relativamente comum; (2) o desenvolvimento da pan-biogeografia por Léon Croizat como estratégia para estudo de padrões de distribuição iluminados pela congruência espacial proveniente de vários táxons e (3) o desenvolvimento da sistemática filo genética de Willi Hennig e com o consequente estímulo à biologia comparada de modo geral, reiterando a importância da hierarquia
natural para estudos de biodiversidade.

Em termos práticos, os princípios e objetivos da
biogeografia de vicariância se delinearam com clareza
cada vez maior a partir dos anos 1970, permitindo a
formalização de hipóteses para a história espacial dos
grupos biológicos. A consequência mais notável do in
teresse por essa perspectiva histórica foi o refinamento
metodológico, com o estabelecimento de "escolas" de
pensamento biogeográfico como a pan-biogeografia (ver
Capo 5), biogeografia cladística (ver Capo 6) e de eventos
(ver Capo 7), e a filogeografia (ver Capo 8).
Alguns estudos biogeográficos são notáveis pelo seu
amplo escopo e quantidade de táxons comparados. O
trabalho de Sanmartín e Ronquistl4, apresentando uma
cuidadosa reconstrução da hierarquia da fragmentação
do supercontinente Gondwana, é um valioso exemplo
de análise que aponta para padrões gerais congruentes
com relacionamentos entre áreas derivados de estudos mais específicos 12.15.
Pesquisas biogeográficas são consideradas aceitáveis
apenas quando lidam com grande quantidade de dados?
Certamente não. Contudo, os tópicos anteriores deste
capítulo buscaram esclarecer por que a concordância
entre, por exemplo, três resultados torna uma hipótese
mais robusta que entre dois resultados. Padrões derivados de estudos com dezenas de táxons são ainda mais
convincentes, o que não deve ser tomado como sugestão
de que estudos com quantidade menor de táxons não
'D servem como evidência empírica para as conclusões -O i1 alcançadas. Os estudos biogeográficos de táxons indivi-
� duais - mesmo aqueles fundamentados em apenas um r-.;, táxon - frequentemente possuem enorme valor empírico. "? s; Relações entre áreas indicadas pelo estudo de apenas um
táxon podem ser comparadas a outros padrões, na busca
pela congruência entre os eventos vicariantes sugeridos
por eles. Da mesma forma, a sequência de eventos indi
cada por um estudo pode ser vista à luz de reconstruções
da história geológica da região estudada, também à
procura de coincidências na sequência cronológica de
eventos em ambos os casos. Além disso, pesquisas en
volvendo inferências sobre datas de origem dos táxons,
a partir da idade mínima sugerida por fósseis ou pela
aplicação de técnicas como a do relógio molecular, são
de grande interesse biogeográfico e cruciais para dar
suporte a padrões gerais de relações entre as áreas.
Lógica da B iogeografia de Vicar iânc ia - 59
Por ser uma ciência histórica, a biogeografia vicariante não se presta a especulações sobre as possíveis distribuições futuras dos táxons. A sequência de eventos que levou a qualquer padrão de distribuição espacial corrente é particular - apesar de muitas vezes compartilhada por grupos biológicos distintos - e o seu caminho futuro é único e imprevisível. O tipo de predição válido para a ciência biogeográfica parte da premissa de que há explicações comuns, ao menos para parte dos padrões encontrados. Esta condição pode indicar a existência de outros táxons, diferentes daquele em análise, com distribuições congruentes àquelas estudadas, os quais, ao serem incluídos em estudo amplo, acabam por aumentar o poder explanatório da hipótese biogeográfica4.
A biogeografia é uma ciência antiga, com longa herança intelectual, remontando às primeiras tentativas de se compreender o mundo natural2.4. l6. Somente a partir do século XVIII, porém, foi iniciada uma tradição de pesquisa sistematizada na área, com naturalistas buscando interpretar as histórias dos táxons de seu interesse, ligadas a crescente esforço de organização sistemática da natureza. No início do século XIX, destacam-se as obras de B uffon, Hooker e Augustin de Candolle3.4. 16. O ponto fundamental desse período, que se tornou a linha mestra de toda a biogeografia seguinte, foi a busca pelo reconhecimento de padrões e de generalizações acerca das possíveis causas responsáveis por tais padrões. A comparação com o estudo biogeográfico atual é um exercício interessante, pois ainda hoje os padrões são as bases para a inferência dos processos históricos que os causaram. As diferenças encontram-se especialmente em quais os tipos de conclusões derivam desses padrões observados, envoltas em formalismo metodológico obviamente inexistente na aurora da prática biogeográfica.
No século XIX, de Candolle 17 foi um dos principais responsáveis pela consolidação da biogeografia como a ciência que procura explicar quais fatores levariam os táxons a se distribuírem em determinados locais e não em outros, por meio dos seus conceitos de "habitações" ou "estações". Eles expressavam a noção de co-ocorrência de uma série de condições ecológicas que favoreceriam a presença de um dado grupo de organismos em determinadas partes do globo. Para de Candolle, as estações seriam as causas físicas que influenciariam a distribuição e que ainda estavam em operação; as habitações poderiam ser tomadas como a expressão das

60 - H istórico e Conce itos
causas geológicas não mais atuantes - essas diferenças estão no cerne da divisão tradicional entre biogeografia ecológica ("das estações") e histórica ("das habitações"). A partir da análise de semelhanças e diferenças entre áreas coincidentes, foi possível o reconhecimento de regiões botânicas, unidades históricas com uma história longa e independente.
Na segunda metade do século XIX, a biogeografia deixou de ser uma atividade meramente especulativa e narrativa e se tornou uma ciência, em que hipóteses alternativas começaram a ser comparadas e avaliadas com embasamento empírico. A maior parte desta mudança coincidiu com o aumento expressivo da formalização da biologia durante os séculos XIX e XX, em parte resultantes da expansão do conhecimento taxonômico iniciado no período após a Idade Média. No entanto, as explicações disponíveis para descrever as distribuições de plantas e animais eram, de modo geral, simplistas e apresentavam problemas práticos em consequência de seu baixo poder explicativo. A partir da aceitação da visão evolutiva, após a publicação do Origem das Espéciesl8, a disjunção entre floras e faunas passou a ser explicada unicamente a partir da ocorrência no passado de eventos de dispersão. Em linhas gerais, a distribuição orgânica resultaria de migrações seguidas de modificações subsequentes e da multiplicação das novas formas nas áreas ocupadas, ou seja, assumia-se a dispersão sobre uma geografia estável, com subsequente isolamento e posterior diferenciação das espécies. As áreas de endemismo seriam compostas por diversos elementos com distintas histórias de dispersão, isto é, por espécies provenientes de lugares e tempos diferentes. Para Nelson 16, a biogeografia dispersalista é a ciência do raro e do miraculoso, pois se concentra em improváveis eventos de dispersão para explicar porque táxons diferentes ocupam diferentes áreas.
Uma hipótese biogeográfica fundamentada em eventos de dispersão não se presta a gerar explicações gerais para a história de uma biota, apenas a história de um grupo específico. A biogeografia dispersalista é uma disciplina com pouco ou nenhum poder preditivo. Não há, tampouco, como falsear qualquer hipótese com base apenas em dispersões, já que ela não é repetível nem comparável com hipóteses alternativas. Tais reconstruções não fornecem teoria geral da história da Terra e, sim, narrativas individuais válidas para cada táxon. Hipóteses dispersalistas são, portanto, divorciadas da história geológica do planeta. Essas narrativas podem incluir o número de eventos de dispersão, ou de quaisquer outras naturezas, que parecer conveniente ao autor da hipótese. Quando um cenário dispersalista é propos-
to para a história biogeográfica de um táxon, ele não pode ser aplicado para outros estudos de caso, pois é único para determinado exemplo. Possíveis generalidades dos eventos presentes nessas situações são coinci
dências não explicadas. Em meados do século XX, Léon Croizat levou a :s
busca por padrões resultantes da sobreposição de estudos :k u.
de casos ao seu extremo mais notável. Pesquisas exten- i3 -í'-
sivas sobre grupos de animais e plantas foram registradas � e resumidas em sua obra Space, Time, Forml9 . Suas in- '/5 O-vestigações indicaram que a existência de padrões repetidos na natureza era evidência de eventos de vicariância comuns e fonte importante para reconstruir a história da área estudada, a despeito de desvios individuais do cenário geral apresentados por um ou outro táxon. Croizat identificou padrões terrestres e transoceânicos de conexões entre localidades e regiões geográficas que se repetiam em vários grupos estudados. O resultado dessas observações foi a obtenção de enormes padrões gerais (= generalizados) conectados uns aos outros. A partir do desenvolvimento das ideias centrais de Croizat, estudos dos padrões de distribuição dos táxons (sob a forma de traços ou áreas de endemismo) passaram a permitir generalizações sobre a similaridade dos padrões espaciais.
Seguindo a premissa de que a Terra e as suas biotas evoluem em conjunto 19 , a inclusão da informação sobre as relações de parentesco entre os táxons resultaria em maior entendimento sobre as relações entre as áreas. Nesse sentido, relações filogenéticas são biogeograficamente informativas, o que constitui o fundamento da biogeografia cladística (ver Capo 6) . Essa escola de pensamento biogeográfico combina o objetivo da sistemática filo genética Hennigiana de reconstrução dos padrões hierárquicos da natureza aos padrões de distribuição, de modo que se pode perguntar: as relações entre as áreas de endemismo fazem alguma referência às relações de parentesco entre as espécies que habitam essas áreas? Para três áreas definidas pela presença de táxons endêmicos, pode-se perguntar se duas delas são mais próximas entre si em relação à terceira delas. Esta indagação é parte da tradição histórica da biogeografia, ampliada por Croizat, que enfatiza as explicações de padrões observáveis por meio de um cenário que considere todas as observações2• Com relação à explicação dos padrões compartilhados, a biogeografia cladística não requer que eles tenham sido causados por vicariância. Hipóteses vicariantes supõem apenas que a congruência de padrões de distribuição originou-se de eventos vicariantes comuns aos táxons estudados.

Hipóteses científicas desejáveis são aquelas que oferecem o maior poder de indução e de explicação sobre o mundo naturaFo, o que se relaciona, obviamente, à noção de
predição científica. Na pesquisa biogeográfica, isso significa a busca por padrões de relações entre áreas inferidas pela análise simultânea de estudos de grupos taxonômicos individuais. Os padrões constituem, desse modo, hipóteses biogeográficas para o relacionamento histórico entre as áreas consideradas e, por conseguinte, das histórias biogeográficas de vicariância dos táxons associados a elas. Quanto maior o número de evidências sustentando o padrão, maior o poder empírico daquela hipótese.
O conhecimento geológico disponível sobre uma região é de grande valor para interpretações biogeográficas. E ventos geológicos concordantes com um dado padrão podem ser compreendidos como evidências complementares à hipótese biogeográfica proposta. O inverso também ocorre, pois o reconhecimento de padrões biogeográficos pode ter importância para a reconstrução da história geológica da área, muitas vezes desconhecida antes de ser conduzido o estudo com os organismos. As investigações de padrões biogeográficos são, dessa forma, atividades científicas úteis inclusive para embasar nossas interpretações acerca da história geológica do planeta6. Tanto a geologia quanto a biologia propõem hipóteses sobre a história física e biótica da Terra, com embasa
mento empírico. As duas ciências funcionam melhor conjuntamente, podendo gerar hipóteses mais robustas do que quando tomadas em separad06.
Estudos biogeográficos buscam explicar, por meio de hipóteses históricas, as distribuições dos organismos. Na menor escala possível de abrangência deste tipo de investigação, a pesquisa se restringe a um grupo (monofilético) de organismos. A sobreposição de várias histórias/ hipóteses por vezes revela padrões compartilhados que possuem maior importância empírica em decorrência de sua generalidade. Assim, a pesquisa biogeográfica deixa de ser específica para um determinado táxon ou mesmo para a biologia em si e passa à escala de padrões que descrevem uma história envolvendo tanto grupos de organismos quanto a geologia. A busca por padrões comuns
(= vicariantes), entretanto, não nega a existência passada ou a possibilidade futura de dispersões. Como dito anteriormente, defende-se aqui que a melhor explicação inicial para um padrão de distribuições disjuntas seja a divisão de uma população ancestral e que, quanto maior o número de casos congruentes, mais corroborada se torna a primeira proposição.
Lóg ica da B iogeograf ia de Vica r i ânc ia - 61
A lógica subjacente às explicações compartilhadas para distribuições coincidentes é similar àquela aplicada à análise filogenética21 : procura-se a explicação mais parcimoniosa para a distribuição dos caracteres para dada topologia22. A parcimônia em biogeografia histórica é expressa como a procura por uma causa comum para uma coincidência de distribuição espacial e relacionamento entre áreas, os quais podem se basear em processos de vicariância. A repetição de relações espaciais e a consequente formação de padrões gerais pedem explicação única, uma vez que eles provavelmente resultam de processos comuns. A vicariância é o processo capaz de moldar, ao mesmo tempo, histórias biogeográficas de linhagens distintas de organismos distribuídos por uma mesma área.
A simplicidade da biogeografia de vicariância fundamenta-se sobre apenas quatro premissas2: ( 1) as áreas se modificam com o tempo a partir de uma conformação ancestral; (2) as espécies podem ocorrer em algumas áreas, mas não em outras; (3) espécies (linhagens) se modificam com o tempo, o que se reflete em padrões históricos de relacionamentos entre linhagens descendentes; (4) mudanças nas áreas podem acarretar alterações aproximadamente sincrônicas das biotas a elas associadas, que se refletem nos padrões genealógicos entre as linhagens biológicas.
Não há, contudo, como esperar que sempre haj a cenários de vicariância para todo e qualquer grupo de organismos estudado. O que se espera é que congruências no relacionamento entre áreas sugerido pelo estudo de táxons individuais sejam mais facilmente explicadas pelo compartilhamento de histórias de vicariância que por outras explicações. Dispersões, por outro lado, são importantes como interpretações para os desvios em relação a um padrão geral, mas não podem ser utilizadas para fundamentar um panorama de pesquisas em biogeografia histórica. Discordâncias de padrões mais gerais são explicadas a posteriori, por exemplo, pela ocorrência de dispersões, expansão da área de distribuição ou extinções.
A despeito da diversificação metodológica, a unidade da biogeografia histórica como ciência tem se mantido pelo paradigma de que histórias compartilhadas de vicariância constituem a explicação mais parcimoniosa para a existência de padrões de distribuição espacial. Segundo Ruse23, uma boa teoria científica deve ser consiliente. Na biogeografia, hipóteses consilientes são aquelas que

62 - H istórico e Conceitos
explicam fenômenos não contemplados durante a sua construção, como a distribuição de outros grupos taxonômÍcos, a existência (ou ausência) de fósseis em certas camadas geológicas e as relações filo genéticas entre outros táxons distribuídos nas mesmas áreas24. Essa possibilidade de predição, ainda que não idêntica às previsões feitas por hipóteses de ciências mais simples, como a física ou a química, só é possível a partir da premissa de que há padrões biogeográficos gerais que se ajustam a mais do que apenas um táxon.
A perspectiva apresentada neste capítulo é quase consensual entre adeptos da biogeografia histórica analítica, com vozes discordantes levantando-se de tempos em tempos. Para esses críticos, a biogeografia ganharia ao adotar práticas mais quantitativas, nas quais testes estatísticos seriam árbitros da significância de hipóteses concorrentes25,26. Alguns métodos vêm incorporando, pelo menos em parte, tais sugestões - aspectos analíticos inovadores têm sido desenvolvidos continuamente nas últimas décadas (ver Caps. 5 a 8). Os amplos desenvolvimentos metodológicos recentes1 ,3 exemplificam a criatividade dos teóricos ao explorar novos modos de lidar com as dificuldades recorrentes de estudos em biogeografia histórica.
Segundo Platnick e Nelson5, a principal questão para biogeógrafos não é "o padrão é resultado de vicariância ou dispersão?", e sim "este padrão corresponde a um padrão geral de interconexões entre áreas (e, portanto, reflete a história dessas áreas) ou não?". Para a biogeografia, é necessário um método de análise que nos permita testar a hipótese se o padrão de distribuição válido para determinado grupo é na verdade um padrão geral. Como a biota não é apenas vida, mas vida ordenada e diversificada no tempo e no espaço27, a busca por padrões gerais é a única maneira robusta de compreender a história dos organismos em ambiente cambiante, como propôs Léon Croizat há mais de 40 anos.
Somos gratos à leitura crítica e comentários oferecidos por Júlia C. Almeida, Claudio J. B . de Carvalho, Jéssica P. Gillung, Rodrigo B . Gonçalves e dois revisores anônimos. Agradecemos Júlia Almeida também pela assistência na preparação de ilustrações.
REFERÊNCIAS BIBLIOGRÁFICAS I . CRISCI, J. V; KANTINAS, L.; POSADAS, P. Historical Bio
geography. An Introduction. Cambridge: Harvard University Press, 2003. 250p.
2. HUMPHRIES, C. J.; PARENTI, L. R. Cladistic Biogeography - Interpreting Patterns of Plant and Animal Distributions. 2. ed. Oxford: Oxford University Press, 1 999. 1 87p.
3 . MORRONE, J. 1. Evolutionary Biogeography -An Integrative Approach with Case Studies. New York: Columbia University Press, 2009. 301p.
4. NELSON, G. l.; PLATNICK, N. I . Systematics and Biogeography - Cladistics and Vicariance. New York: Columbia University Press, 1 98 1 . 567p.
5. PLATNICK, N. I. ; NELSON, G. A method for historie aI biogeography. Syst. Zool. , v. 27, p. 1 - 1 6, 1978.
6. EBACH, M. c.; HUMPHRIES, C. J. Cladistie biogeography and the art of diseovery. J. Biogeogr., v. 29, p. 427-444, 2002.
7. SOBER, E. Reconstructing the Pasto Parsimony, Evolution, and Inference. Cambridge: MIT Press, 1988. 265p.
8 . SANTOS, C. M. D.; AMORIM, D. S. Why biogeographical hypotheses need a well supported phylogenetic framework: a conceptual evaluation. Pap Avulsos Zoo!. , v. 47, p. 63-73, 2007.
9. WILEY, E. O. Vicariance biogeography. Annu Rev. Ecol. Syst. , v. 19, p. 5 1 3-542, 1988.
lO . CRAW, R. c.; GREHAM, 1. R.; HEADS, M. J. Panbiogeography - Tracking the History ofLife. Oxford: Oxford University Press, 1 999. 229p.
1 1 . PAGE, R. D. M. Temporal congruence and cladistic analysis of biogeography and cospeciation. Syst. Zool. , v. 39, p. 205-226, 1990.
12. AMORIM, D. S . ; SANTOS, C. M. D.; OLIVEIRA, S. S. Allochronic taxa as an altemative model to explain circumantarctic disjunetions. Syst. Entomo!. , v. 34, p. 2-9, 2009.
1 3 . BALL, I. R. Nature and forrnulation ofbiogeographical hypotheses. Syst. Zool. , v. 24, p. 407-430, 1976.
14. SANMARTÍN, I . ; RONQUIST, F. Southem hemisphere biogeography inferred by event-based models: plant versus animal pattems. Syst. Biol. , V. 53, p. 2 1 6-243, 2004.
1 5 . UPCHURCH, P. Gondwana break-up: legacies of a lost world? Trends Ecol. Evo!., v. 23, p. 229-236, 2008.
16. NELSON, G. J. From Candolle to Croizat: comments on the history of biogeography. 1. Hist. Biol. , v. l i , p. 269-330, 1 978.
1 7 . DE CANDOLLE, A. P. Essai é1émentaire de géographie botanique. Reproduzido em: LOMOLINO, M. V; SAX, D. F. ; BROWN, J. H . Foundations ofBiogeography - Classic Papers with Commentaries. Chicago: The University ofChicago Press, [ 1 820) 2004. parte 4, p. 28-48.
1 8. DARWIN, C. R. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Strugglefor Life. London: John Murray, 1 859. 5 1 3p.
1 9 . CROIZAT, L. Space, Time, Form - The Biological Synthesis. Caracas: Publicado pejo autor, (Venezuela), 1 964. 881p.
20. B OYD, R. Scientific Realism. In: ZALTA, E. N. The Stanford Encyclopedia of Philosophy (Fali 2008 ed.). 2008. Disponível em: http://plato. stanford. edu/ archives/faIl2008/entries/scientific-rea1ism!. Acesso em: 1 5/0812009.
2 1 . SOBER, E. The conceptual relationship of cladistic phylogenetics and vicanance biogeography. Syst. Zool. , v. 37, p. 245-253, 1988.
22. FARRIS, J. S . The logical basis of phylogenetic analysis. In: PLATNICK, N. L; FUNK, V A. Advances in Cladistics. New York: Columbia University Press, 1983. v. 2, p. 7-36.
23. RUSE, M. Falsifiability, consilience, and systematics. Syst. Zool., v. 28, p. 530-536, 1 979.
24. SANTOS, C. M. D. ; CAPELLARI, R. S. On Reciprocal illumination and consilience in biogeography. Evo/. Bio/., v. 36, p. 407-41 5 , 2009.
25. SIMBERLOFF, D.; HECK, K. L.; MCCOY, E. D.; CONNOR, E. F. There have been no statistical tests of cladistic biogeographical hypotheses. In: NELSON, G.; ROSEN, D. E. Vicariance Biogeography. A Critique (Symposium of the Systematics Discussion Group oftheAmericanMusewl1 ofNaturalHistory May 2-4, 1979). Nova York: Columbia University Press, 198 1 . capo 2, p. 40-63.
26. PAGE, R. D. M. Quantitative c1adistic biogeography: constructing and comparing area cladograms. Syst. Zool., V. 37, p. 254-270, 1 988.
27. WILLIAMS, D. M.; EBACH, M. C. The reforrn of palaeontology and the rise of biogeography - 25 years after 'ontogeny, phylogeny, paleontology and the biogenetic law' (Nelson, 1978). J. Biogeogr., V. 3 1 , p. 685-7 1 2, 2004.



__________________ � 5
I (� o ({b n ,iQ] li � i ! � 'Y ;;;j
John R. Grehan
_.\ diferenciação biológica é o processo espacial e temporal ao qual se dedica a pesquisa da biogeografia evolutiva. Esta área de pesquisa, ao mesmo tempo fascinante e desafiadora, une pesquisadores de várias especialidades e interesses. Para muitos evolucionistas, a biogeografia pode até mesmo parecer uma disciplina enigmática no contexto da biologia evolutiva, que se dedica predominantemente ao registro fóssil e à ação da seleção natural ao longo do tempo. Mesmo assim, a biogeografia é uma disciplina central para a teoria evolutiva, já que a evolução se dá no tempo e no espaço, assim como na forma.
A biogeografia evolutiva inclui um conjunto aparentemente diverso de técnicas, métodos e teorias ! , o que pode passar a impressão de confusão ou incerteza. Entretanto, em meio a esta complexidade, a pan-biogeografia* é distinta por ser o único método de análise e síntese que aceita dados de localidades geográficas como fonte primária de informações. Trata-se de uma abordagem distinta também porque não depende da invenção de centros de origem e rotas de migração, nem a criação de unidades biogeográficas geograficamente definidas, que exemplificam a biogeografia darwiniana2•
A pan-biogeografia representa o desafio à falta de conexão histórica entre biogeografia e teoria evolutiva. Esta disciplina foi desenvolvida e inicialmente implementada por Léon Croizat3-5 opondo-se a um contexto de conceitos críticos enraizados na história da ciência6. É a única síntese
• Segundo o novo acordo ortográfico da língua portuguesa, "Panbiogeografia" deveria ser grafada como "Pambiogeografia". Contudo, os editores optaram pelo uso de "Pan-biogeografia" pelos procedentes históricos dessa forma em português e pela proximidade ortográfica do termo original, "Panbiogeografía", como utilizado por Léon Croizat.
y' �<) () ,!f:I G /01 8 �! I� ,;;;; ',y o .i....\. \�, [0 Vy'
biológica desde a proposta da teoria da evolução de Darwin que abarcou e integrou explicitamente todas as principais dimensões e inter-relações da evolução (espaço, tempo e forma), além de gerar modelos teóricos de como a evolução biológica ocorre em seus aspectos histórico, temporal, espacial e ecológic07,8 . Embora a pan-biogeografia seja geralmente caracterizada como o método de traços (ver a seguir), esta disciplina também busca reconhecer padrões vicariantes de distribuição, em que táxons relacionados substituem-se no espaço geográfico, bem como ao longo do temp03,4. Distribuições vicariantes podem ser evidentes em qualquer nível taxonômico e aplicam-se tanto aos "bons" quanto aos "maus" dispersores . Se houver um princípio fundamental da evolução, este será a prevalência da vicariância (evolução alopátrica) na diferenciação espacial e biológica.
A pan-biogeografia não é para qualquer um, mas é essencial para todos aqueles que aceitem a localização geográfica como uma realidade evolutiva e como uma fonte de informação empírica sobre a história e o da evoluçã09. A pan-biogeografia não é um programa ultrapassado. Trata-se de metodologia e síntese que não podem ser tratadas como meras receitas de bolo ou utilizadas de forma eficaz sem sua devida compreensão. A pan-biogeografia precisa ser aprendida. Isso significa aprender sobre os lugares e a geografia, e como qualquer lugar específico no espaço está biológica e espacialmente conectado a qualquer outro lugar no mundo. A implementação de muitos "avanços técnicos importantes" para realização de análises filogenéticas tem sido confundida com progresso conceitual, enquanto a abordagem mais comum para a biogeografia ainda fundamenta-se na leitura literal do registro fóssil. Combinado com a teoria de centros de origem e dispersão, essa abordagem ignora a avaliação comparativa dos padrões biogeográficos 10, 1 1 .
A pan-biogeografia potencialmente engloba todos os aspectos da biologia evolutiva, por isso está além do

66 - Métodos e Apl i cações
escopo deste curto capítulo cobrir todos os temas pertinentes ou todas as contribuições importantes presentes na literatura. Em vez disso, este capítulo se limitará relativamente a poucos exemplos que ilustram alguns dos principais princípios e conceitos, servindo, pois, como introdução à pan-biogeografia. Possivelmente, este trabalho será demasiado simplista para o biogeógrafo avançado, mas pode contribuir de modo útil e informativo para estudantes de graduação, bem como aos biólogos que busquem uma introdução à biogeografia evolutiva. Aplicações e avanços potenciais de técnicas quantitativas podem ser encontrados em outros trabalhos1 .7, 1 2.
A maioria dos pesquisadores em biogeografia evolutiva é sistemata, cujo enfoque principal é o da Biologia Comparada à diferenciação da forma. A disposição geográfica dos táxons é de interesse apenas secundário, quase sempre em contexto explicativo para gerar interesse no que pode ser um estudo obscuro ou especializado, pelo qual poucos se interessariam em ler. O grande interesse pela biologia comparada pode explicar por que a geografia comparada é negligenciada na ciência da evolução. A biogeografia darwiniana reduz o espaço geográfico de um subproduto de dispersões a partir de centros de origem hipotéticos. Nas últimas décadas, avanços técnicos com o uso do relógio molecular passaram a impressão de que dispersões têm grande importância para a biogeografia. Contudo, as divergências moleculares continuam sendo calibradas por meio do registro fóssil, o que pode subestimar e confundir idades e origens biogeográficas 1 1 .
Se expressos como cladogramas ou qualquer outra árvore filo genética, os padrões de relacionamento puramente biológico não contêm qualquer informação espacial. O mundo é uma esfera e as relações espaciais possíveis entre quaisquer dois pontos são teoricamente infinitas. Para interpretar a origem da disjunção espacial em biogeografia, é necessário especificar qual é a conexão espacial mais provável e relevante para a explicação da origem e da evolução de uma distribuição. Somente para a pan-biogeografia este requisito é um requisito explícito para a análise7. Assim como a sistemática pode ser a expressão máxima da biologia comparada, a pan-biogeografia pode ser a expressão máxima da biogeografia comparada à síntese biológica de espaço, tempo e formas .
Para a biogeografia dispersalista (isto é, fundamentada em dispersão e em centros de origem), o significado espacial é atribuído a uma filogenia pela teorização de centros de
origem e rotas de dispersão. Assume-se, neste caso, que a sequência de diferenciação é um reflexo da sequência de dispersões. Esta atribuição de significado espacial é necessária também aos métodos que pressupõem uma conexão histórica entre padrões congruentes de relacionamento biológico e uma hipótese de sequência de eventos geológicos ou climáticos. A conexão histórica entre uma distribuição e a história geológica é muitas vezes feita pela proximidade geográfica. Por exemplo, se um mesmo táxon �
IX ocorre no Brasil e na Nigéria, assume-se que o Oceano � Atlântico tem mais relação com a origem da disjunção que ,�
os oceanos Índico e Pacífico, independentemente de hipó- .f.. D: teses de dispersão ou vicariância invocadas como explica- ? ções. A maioria dos métodos biogeográficos não inclui declaração explícita sobre as razões para a escolha de determinado relacionamento geográfico.
Os tópicos a seguir ilustrarão aspectos do método pan-biogeográfico, destacando os principais conceitos utilizados na análise pan-biogeográfica e incluindo exemplos relativos à biogeografia sul-americana. Começando com um resumo das principais ferramentas metodológicas, a sua aplicação mais ampla será, então, ilustrada com a análise vicariante (análise da diferenciação de um táxon) e análise biótica (análise dos traços concordantes e diferenciação vicariante de biotas) . Esses tópicos serão seguidos por considerações quanto às implicações para a ecologia evolutiva, e análises bióticas e da biodiversidade.
Há quatro principais ferramentas metodológicas inter-relacionadas em pan-biogeografia:
Traços, que mapeiam explicitamente as conexões espaciais entre localidades disjuntas.
• Linhas de base, que identificam características geomorfológicas atravessada por um ou mais traços.
• Nós biogeográficos, que marcam a interseção de traços diferentes (apesar de cada localidade, dentro de um traço também poder ser considerada como um nó de diferenciação).
• Centros de massa, representando a maior concentração espacial da diversidade dentro de um traço (normalmente expressada pela densidade de uma categoria taxonômica, mas podendo também incluir qualquer aspecto da diversidade biológica, inserindo diversidade molecular).
Cada um dos quatro conceitos pode ser considerado individualmente, mas abordagens integradas exigem sua utilização combinada.
c

A forma mais frequente de construção do traço formaliza o traço como um grafo linear chamado "árvore de extensão mínima" (minimum spanning tree), o que significa buscar a linha mais curta entre localidades disjuntas7 . O traço individual é a representação espacial do espaço evolutivo de um táxon em particular. O critério da extensão mínima é aplicado na ausência de outras informações, como a relações filogenéticas, em cujo caso o traço poderia conectar primeiramente os pontos de táxons mais proximamente relacionados. Em referência ao gênero de escorpiões Opisthacanthus (Ischnuridae) 13, a construção dos traços inicia-se plotando os pontos de distribuição dos táxons, incluindo os representantes fósseis, quando conhecidos (Fig. 5 . 1 , A). Localidades compartilhadas por cada uma das espécies são inicialmente ligadas pelas distâncias mínimas (Fig. 5. 1 , B), em seguida pela conexão dos táxons entre si (Fig. 5 . 1 , C) e, por fim, o traço pode ser orientado por uma linha de base Fig. 5 . 1 , D) (ver também o tópico "Linha de Base").
Traços individuais que compartilham as mesmas localidades ou um subconjunto de localidades podem ser considerados como traços generalizados (em inglês, _'flleralized tracks ou standard tracks). Na prática, a 'jentificação de traços generalizados depende da preci,ão das definições das localidades que compõem uma
Introdução à Pan·b iog eograf ia : Método e S íntese - 67
distribuição. Quando uma localidade refere-se a uma região geográfica restrita, como uma ilha, sua localização é relativamente clara. Contudo, dentro de grandes áreas contínuas de terra (ou mar), os limites geográficos terão de ser especificados. Há duas abordagens principais para avaliar a sobreposição de traços individuais. Traços que compartilham as mesmas localidades, bem como aqueles que podem aninhar-se dentro de um subconjunto dessas localidades e apresentar as mesmas conexões de distância mínima, podem ser combinados em um único traço generalizado 7. Alternativamente, apenas as porções dos traços individuais que se sobrepõem seriam designados como traços generalizadosl4.
A existência de traços generalizados derivados de traços individuais de organismos com capacidades de dispersão diferentes sugere que habilidade de dispersão não é o mecanismo comum para a origem de traços generalizados. A falta de correspondência entre os desenhos dos traços e as capacidades de dispersão levou Croizat a propor um processo de diferenciação vicariante em que cada localidade representa a diferenciação local de táxons aparentados ao longo da distribuição anteriormente ocupada por um ancestral amplamente distribuíd03. A congruência de traços individuais num traço generalizado é esperada como consequência de suas respectivas distribuições ancestrais estabelecidas através de uma história geológica compartilhada.
I �
���-� . . .. ) " y [ \
\
=[ �'.;j"� �5" - Etapas da construção do traço. (A) Loca l idades onde os táxons são encontrados. (B) Traços l igando as loca l idades :: ísjuntas de cada táxon . (C) Traços l igando todos os táxons. (O) I nd icação da l i nha de base (quadrado).

68 - Métodos e Ap l i cações
Traços individuais e generalizados podem ser identificados como espacialmente distintos, mesmo que a sobreposição geográfica entre eles seja substancial. A
distinção biogeográfica de traços ou conjunto de traços pode ser reconhecida, considerando-se características geomorfológicas, tais como falhas transformantes (do inglês, transform faults), processos orogênicos, soerguimento de cadeias de montanhas e bacias tectônicas. As características tectônicas mais extensas da Terra são as bacias oceânicas e estas representam, portanto,
as principais linhas de base para a evolução da biota moderna7. As principais linhas de base para muitos dos táxons amplamente distribuídos incluem as bacias
do Pacífico, Índico, Atlântico-Índico e lndo-PacífiC01S- 19 (Fig. 5 .2) .
Linhas de base oceânicas são relevantes para a compreensão da natureza geográfica ou geopolítica de áreas distintas (por exemplo, "Nova Zelândia", "América do Sul", "Galápagos", "África" ou qualquer subdivisão geográfica dessas áreas), que são frequentemente interpretadas como unidades biogeográficas ou áreas de endemismo2o. Entretanto, a interseção entre traços e linhas de base sugere que essas áreas são entidades biogeográfica e geologicamente híbridas. Esta situação é clara no caso de afinidades de táxons transpacíficos e transatlânticos, que acarretou predição de origens tectônicas híbridas para as Américas do Norte e do Sul, como posteriormente corroborado por evidências geológicas 7 .
- Linhas de base contrastantes e d ispersão i ncongruente. (A) Morcegos do gênero Pteropus7. (B) Lagartos do gênero Cryptoblepharus15 . (C) Gêneros irmãos de Rimanella (Odonata) (Pantepui) e Pentaphlebia (N igéria/Camarões)1 6. (O) Bromel iaceae7. (E) Peta l u ridae (Odonata)17 . (F) Heterochordeumatoidea (mi l ípedes) 1 7 . (G) Anfíbios do gênero Gymnophíona (Amphibia)18. (H) Cycada les19.

Nothofagus o gênero Nothofagus é bem conhecido como um problema clássico da biogeografia do hemisfério suI7,2 1 , Espécies recentes ocorrem em ambos os lados do Pacífico Sul e, embora muitas vezes caracterizado como um " grupo gondwânico", Nothofagus não possui registros ( recentes ou fósseis) nos continentes da África ou da Índia, Em vez disso, sua distribuição atual constitui-se de manchas nas margens de Gondwana ao longo do Pacífico, Há alguns registros de pólen fóssil na porção leste do Oceano Índico e na Antártica, mas seu centro de massa (ver "Centro de Massa") localiza-se às margens do sudeste e oeste do Pacífico Sul (Fig, 5 ,3, A), Este padrão transpacífico (inferido pelas distâncias mínimas e localização do centro de massa) também se aplica aos grupos relacionados Trigonobalanus (Fig, 5 ,3 , B), Li:hocarpus e Castanopsis (Fig, 5 ,3 , C) e Fagus (Fig, 5 .3 ,
D ) , Cada um desses táxons apresenta padrão latitudinal de relacionamentos transpacíficos que são, em grande ;>arte ou totalmente, vicariantes entre si, sugerindo que J distribuição ancestral de cada táxon foi criada ao longo je uma clina norte-sul, antes que continuidade de cada jistribuição fosse interrompida pela formação de da jacia do Pacífico,
Outro exemplo de traços transpacíficos formando ::,aralelos latitudinais é encontrado nas mariposas conhe:idas como stem-boring ghost moths (Hepialidae)22, Esses insetos formam um grupo distribuído basicamen:e nas margens do Pacífico Sul e inclui os gêneros
A
c
I ntrodução à Pan,b iogeografia: Método e Sintese - 69
Aenetus, Endoclita e Phassus, que perfuram as árvores e arbustos na Australásia, Ásia oriental e América Latina, respectivamente (Fig, 5 ,4, A), Este grupo possui padrão vicariante em relação aos gêneros Sthenopus (Fig, 5 ,4, B) e Phymatopus-Paraphymatopus (Fig, 5 ,4, C) e que ocorrem na América do Norte e partes da Eurásia, Esses exemplos demonstram que gradientes latitudinais transoceânicos não são fenômenos isolados na biogeografia de vicariância, mas são histórias compartilhadas por grupos de organismos com ecologias e meios de dispersão muito diferentes,
A maioria dos métodos e técnicas biogeográficas divide as paisagens em unidades distintas e geograficamente definidas, como "áreas", "regiões" ou "áreas de endemismo", que aparentam ter existência independente. Na verdade, essas áreas são artefatos da geografia e da ecologia modernas, que são permeadas e desconstruídas pela rede global de traços e nós biogeográficos (Fig. 5 .5). Os nós biogeográficos são os pontos ou os locais onde diferentes traços se cruzam ou se intersectam. Podem ser de importância local ou global, dependendo do âmbito geográfico e abrangência filogenética dos táxons envolvidos. Os nós biogeográficos globalmente significativos são os seguintes: Guatemala - sul do México; Distrito de Chocó da Colômbia; planalto da Guiana; o planalto Fouta Djallon na Guiné; sudoeste da Província do Cabo; Madagascar; a região em tomo da linha de Wallace e a Nova
B
D
;,;:,�; - Biogeografia do Pacífico do gênero Nothofagus e táxons relacionados aos fósseis i nd icados como círcu los (e) . A) Nothofagus. (B) Trigonobalanus. (C) Lithocarpus e Castanopsis. (O) Fagus7.

70 - Métodos e Apl i cações
B
:5),61, - Biogeografia de gêneros relacionados de Hepia l idae. (A) Endoclita (Ásia Or iental), Aenetus (Austra lásia) e Phassus (região centra l do México e América do Sul) . (B) Sthenopis (América do Norte, reg ião centra l da Ásia e Cáucaso). (C) Paraphymatopus (América do Norte) e Phymatopus (Eurásia)22.
Caledônia. Exemplos de nós biogeográficos de importância local incluem as montanhas John Crow da Jamaica, a cadeia de montanhas Atewa em Gana, as montanhas Chimanimani no Zimbabué; Aceh, na Indonésia; porção vale Watut, na Nova Guiné; noroeste de Nelson e Fiordland, na Nova Zelândia; Ilhas Fiji23.
Na medida em que traços se cruzam ocorre também a intersecção de ecologias, filogenias e distribuições diferentes. Características evolutivas podem incluir (em especial para nós biogeográficos com significado global) endemismo biológico, diversidade filo genética, limites de distribuição, ausência anômala dos táxons, disjunções, incongruência taxonômica e paralelismos (ver "Conservação Evolutiva" para aplicações à conservação). Os
pesquisadores costumam caracterizar os nós taxonômicos como casos de "engano", "enigma", "ironia" e "espanto", já que as complexidades biogeográfica, filogenética, ecológica e biológica resultam na quebra de distinções taxonômicas e filogenéticas com as quais se sentem familiarizados. Os caracteres taxonomicamente distintos em outras circunstâncias podem não ser diferentes num nó biogeográfico. Uma biota nodal pode incluir diferentes níveis taxonômicos em razão das diferenças nas taxas de diferenciação evolutiva durante uma mesma fase de modernização evolutiva. Alguns organismos podem não apresentar qualquer sinal de diferenciação taxonômica ou apenas a diferenciação de caracteres23,24.

I ntrodução à Pan-biogeografia : Método e S íntese - 7 1
;),;]í - Resumo da rede g loba l de traços e nós biogeográficos3 . Note que todas as reg iões do mundo são conectadas pela rede. Todas as á reas são conectadas e não existem áreas geográficas ú n icas que definem a estrutura biogeográfica g loba l . O Novo M undo, em particu la r, é biogeograficamente h íbrido, formado pela m istura de traços do Pacífico e do Atlântico. Porém, mesmo dentro das Américas, todas as á reas b iogeográficas locais são partes dessa rede e, deste modo, não existe uma existência evol utiva separada.
Nós biogeográficos na biogeografia darwiniana são geralmente tratados como refúgios das mudanças cliTfláticas recentes . Isso pode "explicar" endemismos, Tflas não explica as disjunções e os relacionamentos geográficos e filogenéticos para além desses nós. A :naior importância evolutiva pode estar associada às correlações com os processos tectônicos, como acreção de terrenos (terrane accretion), subducção, metamorjsmo regional, granitização, vulcanismo, falhamentos e dobramentos, soerguimento, subsidência e regressão dos mares epicontinentais . Essas correlações sugerem que as características biológicas estão ligadas em sua 'Jrigem ao processo geológico que formam as biotas e afetam seus padrões locais de diferenciação biológica e ecológica23,24.
A.brotanella As plantas da subtribo Blennospermatinae (Asteraceae) dividem-se em quatro gêneros: Crocidium, no oeste da América do Norte; Blennosperma, no oeste das Améri..::as do Norte e do Sul; Ischnea, na Nova Guiné; e _-1.brotanella, na Nova Zelândia, sudeste da Austrália, ::\ova Guiné e sul da América do Sul (Fig. 5 .6, A). Nova Guiné é um nó biogeográfico para Abrotanella e o gênero endêmico Ischnea, sendo este último o mais
proximamente relacionado a Blennosperma das Américas do Norte e do Sul (Fig. 5 .6, B).
No nó biogeográfico da Nova Guiné, a ausência de Abrotanella ao norte do cráton australiano (Fig. 5.7, A) contrasta com os endernismos de Ischnea no Mt. Wilhelm e de sua distribuição a nordeste do cráton, que inclui Jirni-Bena Bena, Finisterre e os terrenos Owen Stanley (Fig. 5.7, B). Este contraste indica que as histórias biogeográficas dos dois gêneros na região foram bastante distintas 20.
Contrastes nodais também podem ocorrer nas bacias oceânicas. As espécies sul-americanas Abrotanella diemii e A. emarginata são mais proximamente relacionadas a A. forsterioides do leste da Tasmânia (Fig. 5 . 8, A a B), enquanto A. scapigera do oeste da Tasmânia é mais próxima das espécies distribuídas no sudoeste da Nova Zelândia e ilhas Auckland e Campbell (Fig. 5 .8 , A a C). As afinidades filo genéticas diferentes das espécies orientais e ocidentais na Tasmânia são correlacionadas espacialmente à localização do terreno Jubilee oriental e do terreno Tyennan ocidental (Fig. 5 .8 ,A) . Essas relações filogenéticas e biogeográficas contrastantes mostram que existe uma unidade biogeográfica de qualquer área geográfica definida e que as origens biogeográficas de táxons não estão relacionadas às unidades geográficas como "Nova Guiné" , "Tasmânia" e "sul da América do Sul"20. Esta é a biogeografia sem áreas25 .

72 - Métodos e Apl i cações
- B iogeografia da subtribo B lennospermat inae (p lantas) . (A) Distribu ição dos gêneros: Crocidium (linhas horizontais), Blennosperma (pontilhado), Abrotanella (linha sólida) e Ischnea (linhas verticais) . (B) Nova G u iné como nó biogeográfico (círculo) para o c lado Ischnea-Blennosperma (linha tracejada) e o c lado Abrotanella (linha sólida)Zo.
50? - Mapa da Nova Gu iné i l ustrando: (A) a correlação tectôn ica contrastante de Abrotanella (e) e (B) Ischnea (.) em relação à margem do cráton Austra l iano (linha sólida) e os terrenos a nordeste (áreas pontilhadas).

I ntrodução à Pan-b iogeograf ia : Método e S íntese - 73
5,(8 - Relacionamento biogeográfico da Abrotanella da Tasmânia . (A) Correlação tectônica do A. forsterioides (leste) e A. scapigera (oeste) com distribu ição em terrenos (linhas finas). (B) A. forsterioides mais relacionada proximamente às espécies da América do Su l . (C) A. scapigera mais proximamente relacionada às espécies da Nova Zelândia/Auckland/llhas Campbel l2o.
Polietina ema análise completa do gênero Neotropical de moscas Polietina (Diptera: Muscidae) 14 resultou em 1 7 traços generalizados para a sobreposição de 1 2 localidades de espécies e 1 0 nós biogeográficos, que representam as interseções entre os traços generalizados de duas ou mais espécies (Fig. 5.9, A). Dois nós biogeográficos de Polierina (a, c) marcam os limites das distribuições de táxons irmãos (Fig. 5 .9, B), indicando que esses nós biogeográ-
.: ficos foram importantes na diferenciação evolutiva, tanto .- genérica quanto específica. O nó biogeográfico amazôni
co dos clados P. prima + P. nigra e P. flavithorax + P. major (Fig. 5.9, C) não se correlacionou com os nós biogeográficos associados aos traços generalizados (pois se restringiram às localidades conectadas compartilhadas), mas não correspondem a um nó encontrado para o gênero Cyrtoneuropsis, novamente indicando a importância de nós biogeográficos em vários níveis taxonômicos.
Por si só, traços não fornecem resolução inequívoca acerca de relações biogeográficas nem permitem a distinção biogeográfica de grupos que apresentam razoável sobreposição geográfica. Centros de massa representam a localização geográfica da diversidade, tanto taxonômica quanto biológica (morfológica, genética), e não necessariamente correspondem a uma área geográfica. O gênero Exacum (Gentianaceae) inclui 38 espécies em Madagascar e ilhas próximas, em contraste com as 24
espécies distribuídas em áreas continentais26 (Fig. 5 . 1 0, A). Neste exemplo, a extensão geográfica do centro de massa é arbitrária, embora sua localização seja inequívoca, em consequência da amplitude geográfica restrita. A padronização da abordagem pode ser alcançada pela análise com um grid de quadrículas27 (Fig. 5 . 10 , B), em que áreas iguais (isto é, quadrículas) são comparáveis7. Informações sobre a localização dos centros de massa
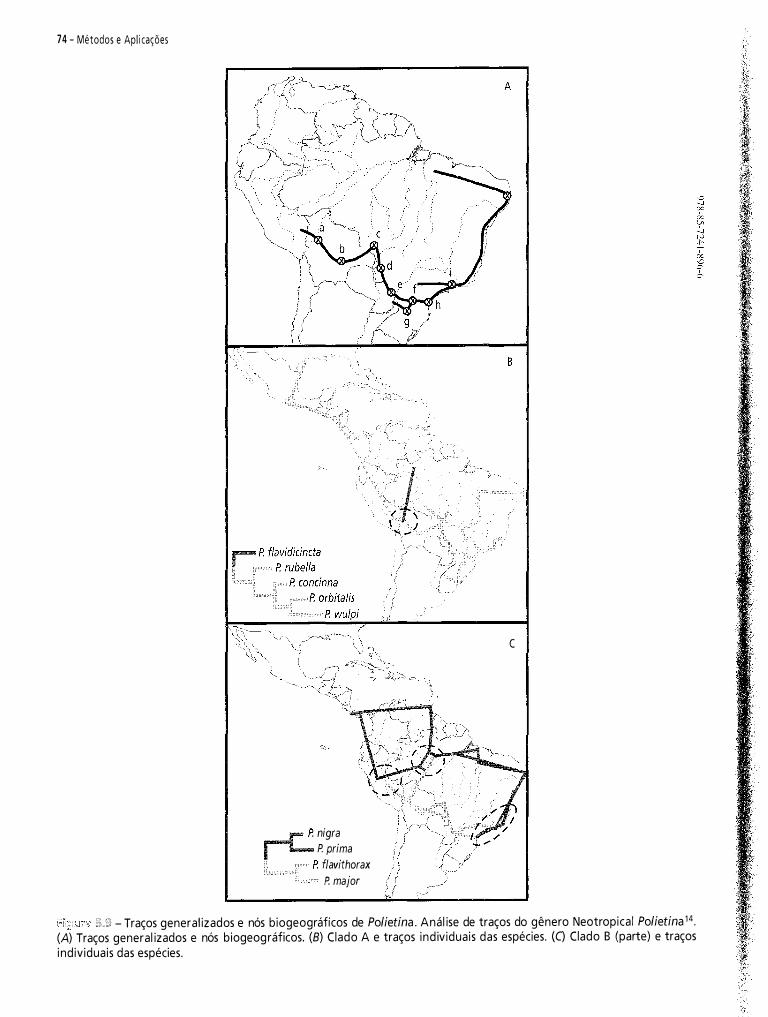
74 - Métodos e Apl i cações
nigra P. prima flavithorax P. major
c
:5D - Traços general izados e nós biogeográficos de Políetína . Anál ise de traços do gênero Neotropical Políetína 14 . (A) Traços general izados e nós biogeográficos. (B) (lado A e traços ind ividuais das espécies. (C) (lado B (parte) e traços individuais das espécies.

Introdução à Pan-b i ogeografia : Método e S íntese - 75
c , => ; �=� ;��,�'(1:· y
A
'}ijí - Centro de massa. (A) Distribuição e centro de massa no Oceano índico (Madagascar) de Exacum (Gentianaceae)26, Centro de massa representado por 38 espécies em Madagascar e em i l has próximas. Centro de massa secundário para índ ia e sudeste da Ásia em uma área maior. (B) Exemplo da aná l ise de grid de quadrícu las para identificar os centros de massa compa rando áreas de tamanhos igua is. A densidade do centro de massa pr incipal das espécies e subespécies de Hebe (Scrophu lar iaceae) é mostrada em dois pol ígonos com as l i nhas mais espessas. A densidade aqu i é três vezes maior que a densidade encontrada na Nova Zelând ia . O centro de massa está centra l i zado em Nelson e norte de Marlborough e através da I l ha Su l e então por todo o restante da Nova Zelând ia27.
podem auxiliar na orientação ou associação de traços para determinado setor geológico da Terra, onde o traço pode sugerir mais que apenas uma relação geológica.
Nothofagus Hipóteses embasadas em centros de origem e dispersão para a origem de Nothofagus afirmam que a história biogeográfica do grupo teria se dado em período demasiado tardio para que eventos geológicos transoceânicos possam ter tido importância. Essa visão ignora o fato de que não só o gênero Nothofagus encontrava-se no Cretáceo Superior, mas também seus subgêneros modernos. Os quatro subgêneros mostram uma considerável sobreposição geográfica (Fig. 5 . 1 1 ), mas isso não oblitera inteiramente seus respectivos centros de massa, os quais são incongruentes com a origem de cada subgênero relacionada às dispersões de longa distância ocorridas ao acaso21 .
O subgênero Lophozonia tem três espécies recentes no sul do Chile, uma na Nova Zelândia e uma no sudeste da Austrália. O centro de massa das espécies recentes em ambos os lados do Pacífico são iguais, mas o centro do Pacífico oriental também inclui duas espécies fósseis (pólen), e outras três espécies fósseis (pólen) foram encontradas na Antártica. Este padrão favorece o Pacífico oriental como o centro de massa para Lophozonia. O traço para o subgênero Nothofagus S.str. é muito semelhante àquele de Lophozonia, no que diz respeito à Austrália, Nova Zelândia e América do Sul, mas as espécies recentes limitam-se ao sul da América do Sul,
juntamente com quatro espécies fósseis (três delas conhecidas por pólen e uma por madeira), enquanto a Nova Zelândia e a Austrália são representados por três e duas espécies de pólen, respectivamente. Nothofagus S.str. também possui seu centro de massa localizado no Pacífico oriental, mas vicariante quando comparado a Lophozonia. O traço de Fuscospora também se estende através do Pacífico, com táxons recentes no sudeste da Austrália, Nova Zelândia e sul do Chile, mas o centro de massa das três espécies atuais e duas espécies conhecidas por pólen fossilizado é a Nova Zelândia. O subgênero Brassospora é representado por táxons fósseis na Antártica e na América do Sul, bem como sudeste da Austrália (Tasmânia) e Nova Zelândia, mas o seu centro de massa inclui 1 4 espécies recentes da Nova Guiné e cinco espécies da Nova Caledônia. Existem três espécies de fósseis de pólen na Nova Zelândia e no sudeste da Austrália, onde também há três espécies fósseis conhecidas por folhas e cinco espécies conhecidas por cúpulas, em comparação com apenas uma espécie de pólen fóssil na Antártica e outra no sul da América do Sul. O centro de massa das espécies Brassospora localiza-se no sul do Pacífico ocidental, com o centro da massa principal na Nova Guiné.
Os traços subgenéricos coletivamente compartilham uma linha de base do Pacífico e, juntos, compõem um traço generalizado no Pacífico sul. Os traços grandemente representam as mesmas localidades e, deste modo, são bastante indistinguíveis. Os seus respectivos centros de massa são claramente vicariantes em relação a cada centro de massa localizado em regiões geográficas

76 - Métodos e Apl icações
3).',]'] - Centro de massa principal dos subgêneros de Nothofagus21 : l i nhas verticais - subgênero Brassospora ( 1 4 espécies atuais), l i n has horizontais - subgênero Fuscospora (três espécies atua is), l i n has d iagonais - subgênero Lophozonia (três espécies atua is), po l ígono aberto - subgênero Nothofagus (c inco espécies atuais).
diferentes: Brassospora na Nova Guiné, Fuscospora na Nova Zelândia, Nothofagus S.str. ao sul da latitude 41°
no Chile e Lophozonia ao norte da latitude 42° no Chile (Fig. 5 . 1 1 ) . Extrapolações do registro fóssil utilizando análises de relógio molecular resultaram em hipóteses de dispersões recentes entre algumas das localidades da distribuição do gênero. No entanto, estas hipóteses são desmentidas pelos traços generalizados, pelo centro de massa vicariantes dos sub gêneros e pelo registro fóssil de cada subgênero no Cretáceo. Esses fatos sugerem que cada sub gênero seria o resultado do processo de diferenciação por vicariância do gênero Nothofagus antes do final do Cretáceo, ao longo da extensão da distribuição geográfica onde atualmente localiza-se o Pacífico SU12 1 .
o conceito de correlação geológica encontra-se implicitamente na aplicação das linhas de base, centros de massa e nós biogeográficos. As histórias geológica e climatológica são quase sempre invocadas para "explicar" relacionamentos biogeográficos . O problema desta abordagem é a remoção do conteúdo empírico do estudo biogeográfico - a biogeografia passa a não ter condições para contribuir com ideias históricas sobre a evolução. Em vez disso, a biogeografia é reduzida à invenção de estórias (teorias), em geral sobre centros de origem e dispersão fundamentadas em uma estória (teoria) geológica. Já que uma teoria é interpretação de evidências empíricas, teorias biogeográficas tomam-se metateorias, que não têm validade maior do que das teorias geológicas ou moleculares em que se
baseiam. A pan-biogeografia escapa deste paradoxo metodológico ao correlacionar espacialmente padrões biogeográficos com informações geológicas e tectônicas, que são produtos diretos da história da Terra.
A correlação geológica pode ser aplicada a traços, linhas de base, centros de massa e nós biogeográficos. Em cada caso, uma relação empírica direta entre biogeografia e geologia é identificada. Esta correlação provê predições sobre o setor geográfico mais provavelmente envolvido com a origem de uma distribuição específica ou um conjunto de distribuições (biota). Correlação geológica permite ao biogeógrafo identificar as reconstruções geo-históricas possivelmente pertinentes à interpretação histórica dos padrões. Por exemplo, a correlação entre os traços biogeográficos de efemérides (Insecta: Ephemeroptera) do gênero Choroterpes com o geossinclinal do Tétis e da falha (geológica) africana fornece uma base para avaliar as implicações geo-históricas para este grupo considerando-se os modelos de placas tectônicas28.
A correlação geológica contrapõe-se à visão de que reconstruções geológicas representam a única fonte de evidência histórica. Reconstruções geológicas são abertas à avaliação biogeográfica e à falsificação por meio de predições geológicas biogeograficamente obtidas. Por esta razão, não importava a Croizat se teorias geológicas específicas contrapunham-se às evidências biogeográficaso Teorias geológicas são teorias, independentemente de quão fortes elas pareçam ser suportadas. Não compreender este princípio significa não compreender a pan-bi o geografia.

Falhas Transformantes (Transform Faults) Disjunções laterais por meio de uma falha transformante podem estar correlacionadas aos deslocamentos laterais ao longo da falha. Um caso clássico ocorre na Nova Zelândia, em que mais de 80 táxons terrestres e marinhos incluindo subespécies, espécies, grupos de espécies, gêneros e farm1ias apresentam uma disjunção ao longo da falha Alpina (Fig. 5 . 1 2) . A explicação tradicional de geleiras exterminando populações centrais e resultando em disjunções negligencia dois fatos: ( 1 ) a frequente ocupação da região intermediária por táxons proximamente relacionados, como também (2) a existência de táxons alpinos, muitos dos quais não teriam sido afetados pela extinção por glaciação na região ao longo da disjunção. Os níveis de diferenciação taxonômica diferentes podem resultar do mesmo intervalo de tempo, neste caso, possivelmente há cerca de 20 Ma (milhões de anos). Disjunções individuais variam de acordo com diferenças na distribuição ancestral. Táxons amplamente distribuídos antes do movimento continuarão apresentando distribuição ampla após o deslocamento, ao passo que endemismos locais, limitados a um lado da falha, não sofrem disjunções. Os resultados desses processos geológicos não são apenas sobre os táxons isoladamente, mas podem afetar comunidades inteiras que tenham sido separadas. Outros testes
Introdução à Pan-b iogeografia: Método e S íntese - 77
de relacionamento de áreas podem vir de análises ecológicas para determinar se há diferenças claras entre as comunidades entre ambos os lados da falha29,3o.
Análise Integrada de Falhas
A maioria dos estudos biogeográficos é geograficamente limitada a uma área ou região, o que pressupõe limites espaço-temporais a qualquer explicação histórica. Mas, se algo pode ser dito com certeza quanto à panbiogeografia, é que a origem dos táxons numa localidade não pode ignorar sua presença ou ausência de qualquer outra localidade, pois toda a vida é biogeograficamente conectada por um conjunto traços generalizados e nós biogeográficos3. A hipótese de isolamento geográfico inerente à criação de áreas distintas muitas vezes se reflete na visão geológica de interpretar os padrões biogeográficos apenas como consequências da história geológica local. No entanto, os processos tectônicos também desafiam o isolamento geográfico na medida em que eventos locais estão interligados com os processos globais. Isso foi evidente mesmo para Charles Darwin3 1 , que observou a aparente coincidência da atividade vulcânica numa localidade ao longo da Cordi-1heira dos Andes com a atividade vulcânica a milhares de quilômetros de distância.
;;:,. 4 2 - Separação entre as espécies relacionadas Celmisia prorepens (su l ) (-) e C. sinclairii (norte) (o) (Asteraceae) ao longo da fa l ha transformante (transform fault), a fa lha Alpina da Nova Zelândia (Heads)24.

78 - Métodos e Apl icações
\ :
\�;� ��:
\ .
\ Nova 'Caledôn ia
:'), �3 - Corre lação tectônica das d isjunções da fa lha transformante (transform fault) da Nova Gu iné, Nova Caledônia e Nova Zelândia (Heads)32 .
Disjunções decorrentes de falhas transformantes nas regiões geograficamente separadas de Nova Guiné, Nova Caledônia e Nova Zelândia podem ser entendidas localmente em termos da história geológica das respectivas zonas de falha, mas uma nova hipótese biogeográfica32
sugere que todas elas compartilham um processo tectônico comum de deslocamento geológico ao longo de um eixo durante a oro gênese do Mesozoico Superior - Paleógeno (Fig. 5 . 13). Assim, um único processo tectônico pode afetar a biogeografia local de comunidades ecológicas geograficamente isoladas. Tentativas de interpretar a origem das disjunções locais como o resultado da ecologia local e dispersões não são capazes de detectar congruências tectônicas em nível regional dessas disjunções consequentes de falhas. A síntese regional e global de locais das correlações geológicas padrões biogeográficos locais representa um desenvolvimento significativo para a análise biogeográfica futura das Américas e de outras regiões.
A biogeografia de centro de origem e dispersão trata sequência filogenética como prova da dispersão sequencial a partir de um centro de origem presumível, quase sempre representado pela localização geográfica do tá-
xon mais basal. Este procedimento também é conhecido como Regra de Progressão de Hennig, mas seria mais precisamente referida como Premissa de Progressão de Hennig. Atualmente, esta premissa tem sido aplicada de modo universal em interpretações dispersão, quando se possui dados provenientes do relógio molecular. Contradições encontradas entre a hipótese de dispersão e da distribuição resultante são consideradas substituições vicariantes de táxons relacionados. Se a dispersão é tão difundida a ponto de ser responsável pelas expansões das distribuições de táxons diferenciados de forma sequencial, este processo é contrariado pela não simpatria entre descendentes. Dispersão sequencial também exige uma "barreira", que é teoricamente permeável à passagem com determinada probabilidade em determinado momento, mas posteriormente, torna-se impermeável. Esse processo estranho é necessário para explicar as hipóteses de dispersão como processos eficazes em determinados momentos e não em outros33. Essas contradições não ocorrem nos casos de distribuições vicariantes em que a sequência filo genética é representada como uma sequência de eventos de diferenciação a partir de um ancestral generalizado. Eventos sequenciais de vicariância podem ser analisados no que diz respeito às relações filo genéticas e geográficas por meio de análises de vicariância. Distribuições ancestrais são

mapeadas sequencialmente sobre a hipótese filogenética. Isso pode ser realizado, no início, de modo esquemático, sem se considerar as distribuições reais; em seguida, as distribuições são mapeadas considerando-se a geografia moderna. Anomalias geográficas resultantes podem, então, ser interpretadas no contexto de correlações geológicas e modelos geo-históricos, bem como o potencial grau de contração ou expansão da distribuição geográfica do grupo. Essa análise também pode resultar em novas avaliações da hipótese filo genética inicial fundamentada em relacionamentos biológicos, um procedimento chamado de iluminação recíproca7.
A distribuição geográfica mundial deste gênero (Fig. 5. 14, A) aparentemente é incongruente com os modelos geológicos tradicionais, que exigem que uma grande parte da sua distribuição (bacia do Pacífico) constitua-se de um oceano permanente. Migrações de longa distância extensas costumam ser invocadas para conectar localidades meridionais e setentrionais através do Oceano Pacífico. Uma análise filogenética molecular de Coriaria realizada por Yokoyama et al.34 aplicou a premissa de progressão de Hennig para propor um centro de origem asiática há cerca de 60 Ma com base em análise de relógio molecular calibrada com fósseis. Migrações sequenciais foram propostas para que o gênero alcançasse a Europa, América do Norte, América do Sul e, por fim, o Pacífico, seguido pela dispersão de volta à América do Sul. Esse dispersalismo molecular não tem fundamento biogeográfico empírico. É simplesmente o produto de centros de origem imaginados e dispersões, fundamentado ainda num relógio molecular que trata idades incorretamente como datas de divergência reais ou máximas.
Na análise de vicariância que se segue, os limites ancestrais comuns entre as distribuições são conceituais, pois as informações atuais da distribuição de táxons recentes e fósseis são incompletas (como é sempre o caso para distribuições disjuntas). As distribuições são estabelecidas com base no mapa moderno, mas a natureza disjunta da distribuição existente é indicativa das distribuições ancestrais de terem evoluído em uma paleogeografia distinta da atual. A distribuição ancestral de Coriaria (Fig. 5 . 1 4, B)
inclui regiões das Américas do Norte e do Sul, algumas ilhas do Pacífico, no sudoeste do Pacífico, e ao longo ou ao norte do cinturão de Tétis na Eurásia (uma distribuição biogeográfica clássica incluindo as bacias do Pacífico e da Europa). A diferenciação da distribuição ancestral resultou em distribuições vicariantes para o Grupo 1 : Eurásia e o Grupo 2: Pacífico (Fig. 5 . 14, C).
Introdução à Pan·b iogeografia : Método e S intese - 79
O Grupo 1 diferenciou-se, em seguida, num ancestral mediterrânico-asiático e um ancestral no Pacífico ocidental (Fig. 5 . 14, D). O ancestral mediterrânico-asiático diferenciou-se, então, em C. nepalensis (Ásia Central) e na distribuição ancestral de C. myrtifolia no Mediterrâneo e C. terminalis também na Ásia Central (Fig. 5 . 14,
E). O ancestral distribuído no Pacífico ocidental também se diferencia em C. japonica ao norte e C. intermedia ao sul (Fig. 5 . 14, E). As distribuições de C. nepalensis e C. tenninalis depois sobrepõem por meio de expansão geográfica. Por fim, eventos de extinção resultam nas disjunções modernas. A diferenciação do Grupo 2 começa com Coriaria microphylla (Fig. 5 . 15 , A), seguida por C. papuana e Coriaria sp. no sudoeste do Pacífico (Fig. 5 . 1 5, B), C. arborea na Nova Zelândia (Fig. 5 . 1 5 , C) e o ancestral de C. ruscifolia no sul da América do Sul, e C. sarmentosa e C. lurida na Nova Zelândia (Fig. 5 . 15 , D). Esta sequência filogenética e geográfica da diferenciação vicariante não cria anomalias em decorrência da dispersão e mostra que a distribuição das espécies modernas e suas relações filogenéticas são congruentes com a sua diferenciação de um ancestral amplamente distribuído no Pacífico e na Ásia Central.
Os hominídeos compreendem os seres humanos e táxons fósseis mais proximamente relacionados aos humanos que a qualquer espécie recente de primata. Uma análise cladística morfológica incluindo a espécie humana, os grandes primatas e fósseis de hominídeos (Homo, Australopithecus) e vários outros primatas extintos (fósseis do Mioceno e Plioceno) suportou a monofilia hominídeos, orangotangos e diversas espécies fósseis proximamente relacionadas ao orangotango e distribuídas entre a Europa e a Ásia35. A distribuição, em grande parte vicariante de hominídeos, orangotangos e outros grupos fósseis, sugere que esses táxons diferenciaram-se localmente de um ancestral amplamente distribuído entre a África Oriental, Mediterrâneo e Ásia central e oriental (Fig. 5 . 1 6, A). Se hominídeos constituem-se do grupo irmão de todos os fósseis e outros membros vivos do clado, como sugerido por evidências atuais, a diferenciação vicariante resultou em hominídeos na África oriental, e um clado composto por orangotangos e espécies relacionadas, distribuídos entre Espanha e Indonésia (Fig. 5 . 1 6, B).
A linhagem do orangotango deu origem ao orangotango e vários táxons fósseis completamente ou em grande parte de modo vicariante (Fig. 5 . 1 6, C), que se extinguiram há cerca de 7 Ma, à exceção dos orangotangos e de Gigantopithecus no leste da Ásia (Fig. 5 . 1 6, D).

80 - Métodos e Apl icações
A
C. termínalis (Ásia Central)
C. myrtífolía (Mediterrâneo)
'----- C. nepalíensís (Ásia Central) C. japoníca (Ásia I nsular)
C. íntermedía (Ásia Insular)
o Q. 2
\..?
C. microphy/la (América Centra l)
C. papuana (Papua Nova Guiné)
C. sp. (ilhas do sudoeste do Pacífico)
C. arborea (Nova Zelândia)
C. ruscifolía (América do Sul)
C. sarmentosa (Nova Zelândia)
C. furída (Nova Zelândia)
N o Q. 2
\..?
t_ �
����=�=�����=9=�=-�·�·=·�����=. ����: �' ���======���==�� �
B
�b_ �
:3" 'ub], - Diferenciação de Coriaria sobreposta na paisagem atua l . (A) F i logenia molecu lar do gênero . (B) Área a ncestral h ipotetizada por inc lu i r todos os fósseis e loca l i zação dos descendentes. (C) Diferenciação vicar iante dos grupos 1 e 2, com os l im ites de d iferenciação h ipotetizados no Pacífico oeste. (O) Diferenciação i n ic ia l do grupo 1 . (E) Diferenciação das espécies existentes .
. � - Diferenciação do subclado 2 de Coriaria . (A) Basa l C. mícrophy/la na América Central e norte da América do Su l . (B) C. papuana/Coriaria sp. (grupo) na Nova Guiné e oeste do Pacífico. (C) C. arborea na Nova Zelândia. (O) C. sarmentosa/C. /urida na Nova Zelândia e C. ruscífo/ia (su l da América do Sul , Ch i l e) .
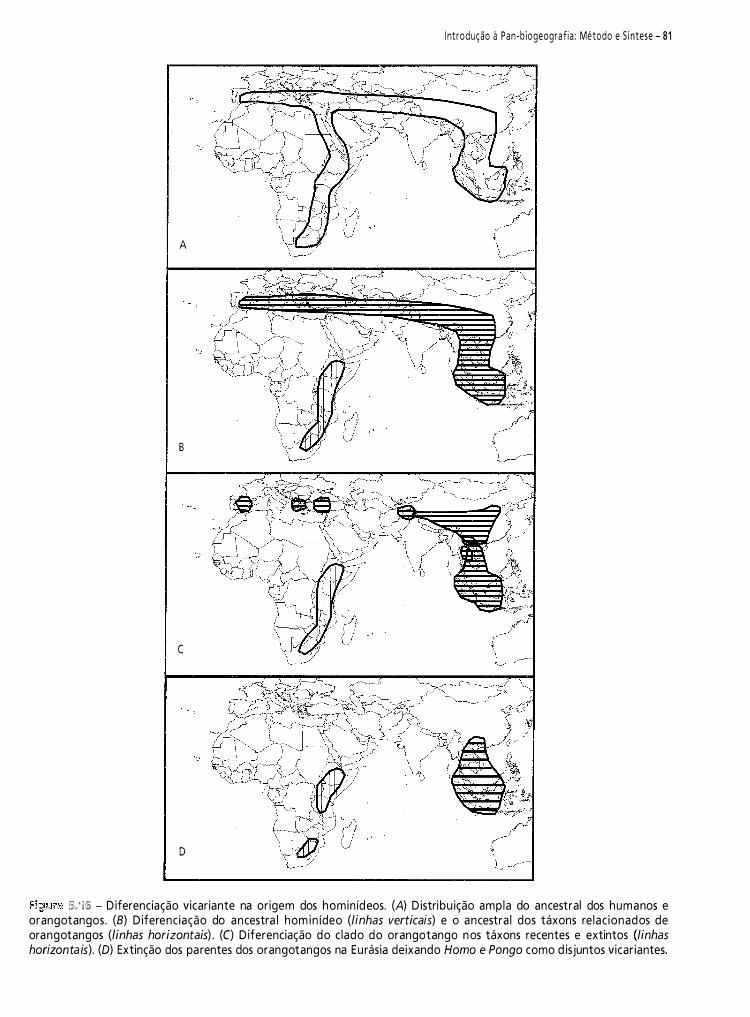
I ntrodução à Pan-biogeograf ia : Método e Síntese - 81
A
B
c
D
5o'il:E - Diferenciação vicariante na origem dos hominídeos. (A) Distribu ição ampla do ancestral dos humanos e orangotangos. (B) Diferenciação do ancestra l hom in ídeo (linhas verticais) e o ancestra l dos táxons relac ionados de orangotangos (linhas horizontais) . (e) Diferenciação do c lado do orangotango nos táxons recentes e extintos (linhas horizontais). (O) Extinção dos parentes dos orangotangos na Eurásia deixando Homo e Pongo como d isjuntos vicar iantes.

82 - Métodos e Ap l i cações
Esse padrão de extinções resultou na disjunção dos primeiros homi nídeos na África oriental e orangotangos no sudeste da Ásia (Gigantopithecus se extinguiu há cerca de 300.000 anos). Outros registros fósseis indicam
a possibilidade de orangotangos terem tido uma linhagem africana não relacionada aos hominídeos como seus parentes mais próximos, fósseis estes representados apenas por peças gastas da maxila e da dentição.
Os representantes mais antigos fósseis do clado de hominídeos no nordeste da Índia (Sivapithecus) e Tailândia (Khoratpithecus) com 1 2 a 1 3 Ma demonstram a ampla distribuição ancestral. Embora o registro fóssil de hominídeos remonte a apenas 6ma, a relação de grupo irmão com o clado dos orangotangos, que inclui fósseis de até 1 3 Ma, sugere que a linhagem dos hominídeos já havia divergido nesta época. O traço do clado dos hominídeos e orangotangos é espacialmente correlacionado à falha (geológica) da África Oriental e ao cinturão do Tétis. Se essa correlação não for um artefato de fossilização, essas formações geológicas contribuíram para a expansão do último ancestral comum antes do soerguimento tectônico do platô tibetano, cujo início estimado teria sido durante o Mioceno, há cerca de 1 5 Ma.
Opiliões da Subfamília Bourguyiinae (Arachnida)
Uma área de endemismo encontrada nas análises de Yamaguti e Pinto-da-Rocha36 só permitiu concluir que
a área de endemismo da Serra da Mantiqueira relaciona-se mais proximamente à Serra do Mar e à porção leste de São Paulo que a um conjunto de áreas que inclui a Serra dos Órgãos, o litoral do Rio de Janeiro e a Serra da Bocaina. A área de endemismo "Serra da Mantiqueira" também foi considerada como parte da Serra da Mantiqueira, ao passo que a área "Serra do Mar" foi considerada como pertencente à Serra do Mar, sendo essas duas áreas de endemismo separadas pelo vale do Rio Paraíba do Sul, caracterizado pelos autores como barreira vicariante "importante para os opiliões". A comparação do cladograma de área de Bourguyiinae com um cladograma geral de área evidenciou diferenças que foram interpretadas como resultantes de dispersão de Bourguyia trochanteralis para a área de endemismo "Sul de São Paulo e Paraná", a partir de um centro de origem imaginado na Serra da Bocaina - Serra do Mar.
O gênero Asarcus foi interpretado como tendo tido um centro de origem na Serra dos Órgãos-Serra do Mar, seguido por uma história de dispersões para a Serra do
Espinhaço, resultando emA. putunaberaba. A separação da Serra da Mantiqueira e da Serra do Mar de outras áreas foi considerada o único evento de vicariância e mesmo assim, este evento "aparentemente" não teria tido efeito sobre as espécies de Bourguyiinae.
Os traços em grande parte vicariantes dos gêneros Asarcus e Bourguyia e de suas espécies (Fig. 5 . 17 , A) são incongruentes com os centros de origem e rotas de dispersão imaginados e propostos para Bourguyiinae. A distribuição das espécies e táxons superiores mostra correspondência clara com processo de diferenciação por vicariância a partir de um ancestral amplamente distribuído (Fig. 5 . 17 , B), que se diferenciou de modo vicariante em Bourguyia e Asarcus, respectivamente (Fig. 5 . 17 , C). Bourguyia diferenciou-se em B. laevibunus e dois clados ancestrais, em que se posicionam as demais espécies do gênero (Fig. 5 . 1 7, D). O subclado do norte diferenciou-se localmente em B. mandata, ao norte, e B. bocaina, ao Sul (Fig. 5 . 17 , E). O subclado do sul também se diferenciou em B. albiomata, seguido por B. trochanteralis e B. vinosa (Fig. 5 . 17, F). O gênero Asarcus diferenciou-se em A. passarellii no Rio de Janeiro (Fig. 5 . 17 , G), e A. ingenuus na Serra da Mantiqueira e, por fim, no ancestral comum de A. longipes (Serra do Espinhaço) e A. putunaberaba (Minas Gerais) (não ilustrado) (Fig. 5 . 17 , H).
A análise de Bourguyiinae mostra que a criação de áreas de endemismo, em especial quando combinada com a especulação acerca de centros de origem, elimina as evidências de diferenciação vicariante. Isso também resulta na perda de informações biogeográficas, como a
presença de nós biogeográficos que, no caso do Bou/"guyia e Asarcus marca importante linha de diferenciação ao longo da Serra dos Órgãos e Serra do Bocaina ao sul para São Paulo.
Rhodniini (Hemiptera)
Pode parecer que a análise de vicariância só é aplicável quando vicariância é relativamente óbvia, como nos exemplos citados anteriormente. Contudo, o método pode ser aplicado em qualquer situação biogeográfica e pode ser usado para identificar os padrões de simpatria. bem como de vicariância sem que a análise de distribuição dos organismos seja subordinada à criação de áreas geograficamente definidas. No gênero Rhodnius, por exemplo, a sobreposição geográfica entre os táxons que incluem os três primeiros nós filogenéticos37 implica na dispersão como processo de expansão da distribuição dos ancestrais de cada um desses clados. Por outro lado. dentro de cada grupo a vicariância é o processo predo-
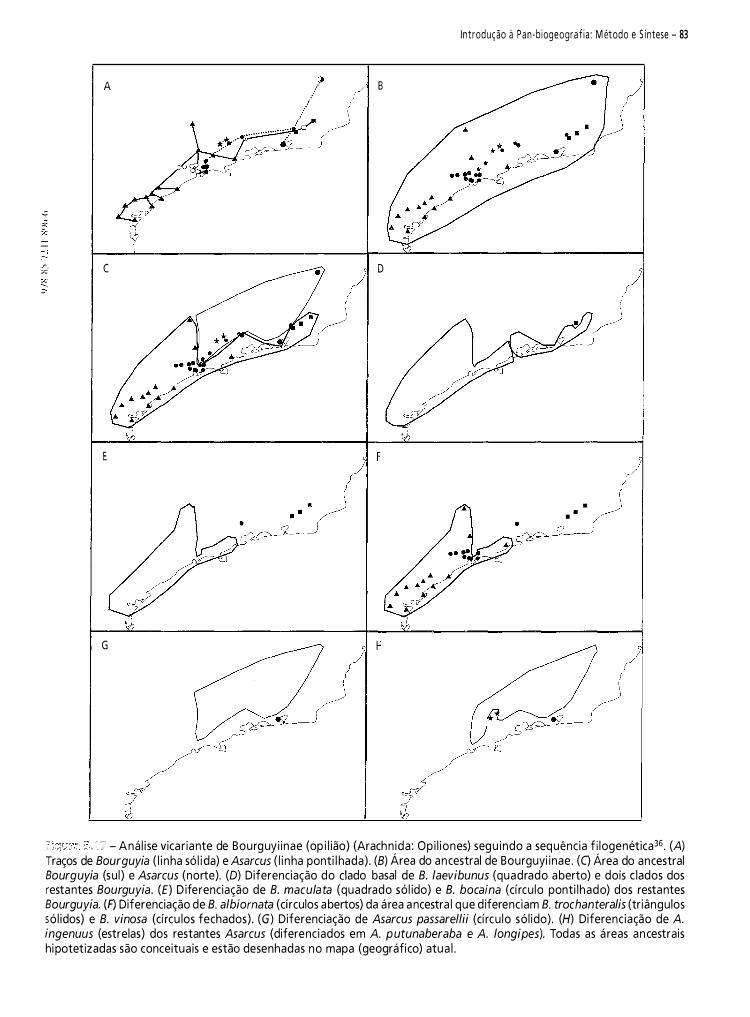
A
.=
:;;
--.: r, C x
:c
E
G
B
D
•
H
Introdução à Pan-b iogeograf ia : M étodo e S íntese - 83
• • •
' ;; - Anál ise vicariante de Bourguyi inae (op i l ião) (Arachnida: Opi l iones) segu indo a sequência f i logenética36. (A) Traços de Bourguyia ( l i nha sól ida) e Asarcus ( l i nha ponti l hada), (B) Área do ancestral de Bourguyi inae. (C) Área do ancestral Bourguyia (sul) e Asarcus (norte). (O) Diferenciação do dado basal de B. laevibunus (quadrado aberto) e dois dados dos restantes Bourguyia. (E) Diferenciação de B. maculata (quadrado sól ido) e B. bocaina (círculo ponti lhado) dos restantes Bourguyia. (F) Diferenciação de B. albiornata (círculos abertos) da área a ncestra l que diferenciam B. trochanteralis (triângulos só l idos) e B. vinosa (círcu los fechados). (G) Diferenciação de Asarcus passarellii (círcu lo sól ido), (H) Diferenciação de A. ingenuus (estrelas) dos restantes Asarcus (diferenciados em A. putunaberaba e A. longipes). Todas as á reas ancestrais hipotetizadas são conceituais e estão desenhadas no mapa (geográfico) atua l .

84 - Métodos e Apl icações
minante e a simpatria parcial está limitada a apenas algumas espécies. Uma análise de vicariância sugeriria, portanto, que a extensão da distribuição foi mais relevante para os grupos ancestrais iniciais, ao passo que as distribuições de espécies recentes têm se expandido pouco, de modo que o padrão vicariante não é ofuscado.
Este cenário vicariante para Rhodniini pode ser contrastado com a análise de dispersão-vicariância (DIVA) com áreas de endemismo predefinidas, que não resultou em padrão vicariante inequívoco; também neste caso, a sobreposição das áreas de endemismo impediu que uma explicação robusta da história biogeográfica fosse encontrada, embora as áreas sejam hipotéticas37. Uma abordagem alternativa utilizando correlação geológica conduz a possíveis explicações, que depois são confundidas se impostas sobre as áreas de endemismo. Por exemplo, o desmembramento do Andes em três cordilheiras separadas é visto como possivelmente responsável pela evolução do clado R. colombiensis/R. brethesilR. neivai. Entretanto, na análise de áreas, a espécie R. colombiensis é atribuída à vicariância na área do Noroeste Amazônico (NWAm), enquanto R. ecuadorensis na área Andina Mesoamericana (AnMa) é pensada como tendo também ocupado NW Am anteriormente, e R. pallescens no Panamá e Guatemala (área AnMa) teria supostamente dispersado a partir NWAm para AnMa, ou diferenciado quando R. ecuadoriensis desapareceu dessas áreas37. Em contraste com essa incerteza, a análise de vicariância sugere que a diferenciação de um ancestral amplamente distribuído teria resultado primeiro em R. colombiensis, seguido por R. pallescens na América Central e R. ecuadoriensis no Equador (Fig. 5 . 1 8). A distribuição ancestral para além da cordilheira andina sugere a relação de processos geológicos responsáveis pela configuração geográfica atual da América Central, bem como a elevação dos Andes.
As relações filogenéticas de um grupo de táxons vicariantes podem representar uma sequência de especiações ou pode representar agrupamentos de caracteres semelhantes que não reflete a sequência de especiações. Espécies vicariantes podem se diferenciar ao mesmo tempo, embora a distribuição dos caracteres ancestrais leve alguns descendentes a serem mais similares entre si que outros. No processo de diferenciação vicariante, as similaridades compartilhadas por quaisquer táxons são consequências da herança de caracteres ancestrais (sendo exclusivamente compartilhados ou não) por um ou mais descendentes (ver também o tópico "Ancestrais").
A análise de vicariância de caracteres biológicos pode discriminar padrões de similaridade que podem não ser aparentes na sistemática biológicaS e metodologias sistemáticas e formais para o estudo de relacionamento biológico nem sempre fornecem a representação mais precisa da evolução filogenética6,38 . Mesmo quando os grupos compartilham caracteres similares que podem ser considerados filogeneticamente não informativos, sua ocorrência compartilhada por diferentes táxons presentes em um traço generalizado sugere que eles são biogeograficamente informativos. Por exemplo, o traço que conecta grupos "não relacionados" de plantas divaricatas em Madagascar e na Nova Zelândia coincide com um traço generalizad039. A análise geográfica de cinco caracteres do gênero de libélula Tepuibasis De Marmels38 mostrou que, apesar de os diferentes estados não poderem ser interpretados como primitivos ou derivados, cada um deles identificou conjuntos diferentes de traços de vicariantes para diversas combinações de espécies (Fig. 5 . 1 9).
A análise filo genética das similaridades biológicas pode ser considerada incompleta até que sej am analisados tanto seus aspectos distributivos quanto biológicos . Isso inclui a avaliação da congruência entre proximidade espacial e similaridade biológica, já que cladogramas ou qualquer outro tipo de representação de relacionamento biológico não contêm informação espacial. Por meio de processo de iluminação recíproca. uma filogenia biológica pode fornecer informações às análises biogeográficas, mas a biogeografia, por sua vez, pode também fornecer informações à biologia e até mesmo apontar hipóteses alternativas7. A geografia de caracteres também tem implicações para a morfologia comparada. Características filogeneticamente incongruentes que possam ser julgadas como resultantes de fenômenos secundários podem, na verdade, resultar de processos morfogenéticos comuns (ver também o tópico "Ancestrais"). Androcônios, produtores de feromônios nas asas de lepidópteros, por exemplo, podem ter origem homóloga das glândulas axilares e domácias de folhas de arbustos. Em vez de subordinar a morfologia comparada à sistemática, talvez fosse melhor subordinar a sistemática a uma análise comparativa detalhada de cada característica morfológica6.
A biogeografia das comunidades ecológicas ou biotas pode ser analisada pela identificação e comparação dos traços generalizados que fornecem informações sobre a estrutura espacial de uma biota e suas origens
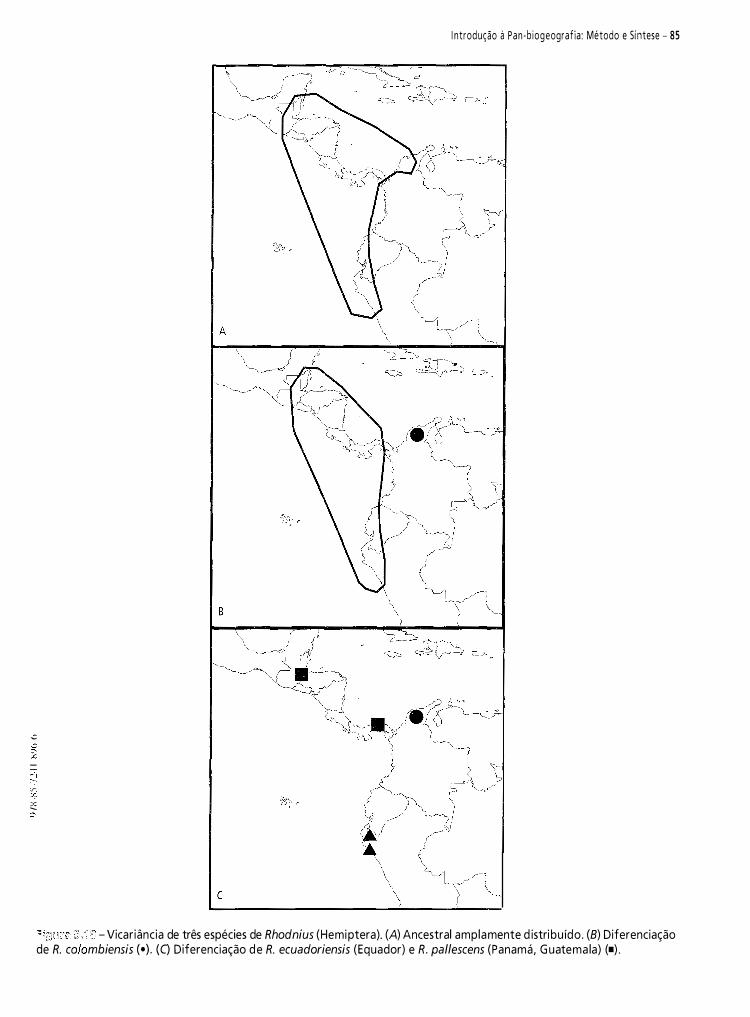
-.:; � ---, r, � 2:
I ntrodução à Pan-b iogeografia: Método e Sintese - 85
A
B
\ �� �)
(� C
yl - Vicariância de três espécies de Rhodnius (Hemiptera), (A) Ancestral amplamente d istribuído. (B) Diferenciação
de R. c% mbiensis (e) . (C) Diferenciação de R. ecuadoriensis (Equador) e R. pallescens (Panamá, G uatema la) (-) .

86 - Métodos e Apl icações
- Aná l ise de caráter do gênero Tepuibasis (Odonata). Traços vicar iantes. (A) Circus do macho arqueado para ba ixo (linha sólida) versus circus do macho reto (linha pontilhada). (B) Fronte redonda (linha sólida) versus fronte a ngu lada (linha pontilhada).
biogeográficas. O mundo é uma rede de traços interligados e de nós biogeográficos que demonstram as origens interconectadas não só de continentes inteiros, mas também do nível local de organização das comunidades ecológicas individuais24. A ecologia evolutiva de comunidades biológicas locais requer biogeografia evolutiva que identifica sua significância biogeográfica global.
As Américas Central e do Sul são comumente tratadas como uma unidade biogeográfica denominada Região Neotropical2. Após o amplo reconhecimento da estrutura biogeográfica e tectônica híbrida das Américas, hoje é reconhecida natureza artificial de uma "área NeotropicaI". Por esta razão, os esforços têm se concentrado em subdividir a região em áreas menores, que sejam consideradas unidades biogeográficas naturais4o. Essas áreas não fornecem, contudo, qualquer informação sobre a rede de traços e nós biogeográficos que as conecta com as outras áreas, e que se opõem à ideia de existência separada41 . Em pan-biogeografia, questões sobre a origem de um táxon dentro de uma área geográfica são as mesmas que as questões sobre a origem externa àquela área. Por exemplo, a questão de por que Coriaria ocorre no Chile é o mesmo que questionar por que Coriaria distribui -se na Nova Zelândia e Europa, mas aparentemente (na ausência de representantes fósseis) é ausente no Brasil, Índia e América do Norte. Nesta perspectiva global, a pan-biogeografia talvez ofereça uma alternativa mais eficaz à subdivisão da paisagem em províncias biogeográficas estanques, uma prática que remonta à época romana.
A importância global da biogeografia das Américas foi recentemente ilustrada numa análise de centros de endemismo globalmente basais42, que mostrou que clados basais de animais e plantas não são arbitrariamente espalhados pelo globo. Estes se concentram em determinados setores geográficos onde clados basais de grupos filogeneticamente independentes co-ocorrem. Em biogeografia darwiniana, regiões como estas indicam a localização dos centros de origem, enquanto a pan-biogeografia enxerga os grupos basais como os grupos irmãos menores ou menos especiosos de táxons mais diversos, que podem ser distribuídos de forma global. A localização dos grupos basais representa o local de diferenciação inicial de um ancestral amplamente distribuído (podendo até ser globalmente distribuído) . Grandes centros de diferenciação basal no Velho Mundo incluem a os mares de Coral e da Tasmânia, sudoeste da Austrália Ocidental, Sumatra/Borneo, sul da China-Taiwan-Japão, África do Sul, Madagascar, Tanzânia-sudeste de Camarões, oeste do Mediterrâneo. No Novo Mundo (Fig. 5 .20), centros importantes localizam-se em terrenos acrescidos42 (México, oeste da ColômbiaJEquador/Antilhas) e crátons da América do Sul (Norte da Colômbia/Venezuela ocidental, escudo das Guianas, Minas Gerais, sudeste do Brasil-Argentina-Nordeste) , Chile central e Ilhas Juan Fernández. Além disso, três dos grupos basais na América do Sul têm distribuições transcontinentais: Thurniaceae (grupo irmão dos grupos cosmopolitas Cyperaceae e Juncaceae) na África do Sul e do Escudo das Guianas e Welwitschia (grupo irmão do gênero pantropical Gnetum) no nordeste do Brasil (fósseis) e sudoeste da África.
A coincidência geográfica de linhagens basais mostra que suas origens não podem ser desvendadas pelo estudo

I ntrodução à Pan-biogeograf ia : Método e Síntese - 87
o
o
- Loca l ização das l i nhagens basais na América Centra l e América do Su 142.
dos efeitos locais de probabilidade de sobrevivência e de fatores ecológicos em cada área endemismo geograficamente definida. Essa necessidade é demonstrada pelo fato de as principais tendências das relações biogeográficas para as Américas serem orientadas ao longo do eixo leste-oeste e não norte-sul, como exigido pelas teorias de dispersã04. A previsão necessária de o Novo Mundo também ser uma área geologicamente híbrida e orientada ao longo do eixo leste-oeste foi corroborada por evidências geológicas para o acréscimo ocidental de terrenos do Pacífico para os blocos geológicos orientais que separavam a África da Europa7. Esta natureza biogeográfica e tectônica híbrida continua a ser ignorada na maioria dos estudos biogeográficos, particularmente aqueles que se limitam a traçar subdivisões geográficas dentro de cada um dos continentes.
A estrutura tectônica híbrida das Américas é também evidente no sudoeste do Pacífico, onde se pode observar
que Nova Zelândia, Nova Caledônia e da Nova Guiné são geologicamente polifiléticas. Todas essas áreas incluem vários terrenos orientais de origem pacífica e com afinidades diversas. Ao leste das Ilhas Salomão, localiza-se o planalto Ontong Java, originado no início do Cretáceo. Trata-se de um grande planalto, quase inteiramente submerso, que inclui sucessão espessa de rochas vulcânicas com origem subaérea, conforme evidenciado por fragmentos de madeira fossilizada ou carbonizada. Nas Américas, vários grupos basais são endêmicos do terreno Guerrero. Esta formação foi parte do arco de ilhas do Pacífico, antes de ser acrescida ao México durante o Jurássico-Cretáceo. Setores geológicos de terrenos do Pacífico acrescidos à Colômbia a oeste da zona de falha Romeral também são muito significativos para a biogeografia, como os arcos de ilha do Jurássico - Cretáceo Inferior que atualmente compõem a serra costeira na região central do Chile.

88 - Métodos e Apl icações
As afinidades tectônicas e biogeográficas das Américas com o Pacífico são congruentes com os modelos tectônicos que identificam uma história geológica complexa para a bacia do Pacífico após a formação da placa do Pacífico, no meio do Jurássico (Fig. 5 .21 ) . As formações geológicas conhecidas como Java Ontong, Manihiki e Hikurangi são três grandes planaltos ígneos atualmente localizados no Pacífico ocidental, posicio
nados nesta área no início do Cretáceo e que podem representar fragmentos de um grande planalto ancestral. O planalto Ontong Java também pode ter se formado na região do Pacífico central, próximo ao planalto Gorgona, que se encontra atualmente acrescido ao oeste do Equador e Colômbia, bem como ao Caribe. Além destas províncias ígneas, grandes terrenos-arco de ilhas no Pacífico oriental foram acrescidos às Américas do Norte, Central e do Su142.
A confiança depositada em técnicas filo genéticas complexas quando não há clareza conceitual pode confundir a compreensão biogeográfica 1 1 , como demonstrado por uma análise filogeográfica de 52 camaleões, compreendendo seis clados, dos quais três distribuem-se em Madagascar, Seychelles, África, Índia e Madagascar-Comores-Reunião (Fig. 5 . 22) . Uma abordagem utilizando o conceito de centro de origem e dispersão acarretou interpretação dos resultados filogeográficos a ser caracterizada como incompatível com a separação Gondwana. Um modelo de dispersão a partir de Madagascar foi proposto como hipótese, assumindo eventos de dispersão ao acaso sobre a água em vez de analisar os padrões e investigar a sobreposição local de distribuições de Madagascar com mais detalhes.
A ocorrência de camaleões endêmicos em Comores (formadas há 0, 13 a 5 ,4 Ma) foi tratada como corroborando a hipótese de dispersão, pois se pensava que esta origem geológica recente exigisse que os lagartos também tivessem origem recente na ilha. Esta suposição baseia-se em outra hipótese, de que não houve nenhuma massa de terra prévia na área. Embora a análise filogeográfica considere dispersão de longa distância importante, a ocorrência de camaleões, peixes de água doce e de mamíferos terrestres em Madagascar também foi interpretada como paradoxal, porque todos estes grupos são considerados como tendo baixa capacidade de dispersão. Madagascar demonstra como a redução da biogeografia a um apanhado de hipóteses de migrações reduz os problemas biogeográficos a um emaranhado de mistérios e paradoxosl l .
Atualmente, compreende-se que a pan-biogeografia de Croizat demonstra que a terra "e a vida evoluem juntos"1 .7 . Do mesmo modo, este princípio fundamental é grandemente ignorado quando a maioria dos biogeógrafos busca hipóteses de dispersão para explicar a origem das distribuições vicariantes, ignorando também as implicações ecológicas de uma história compartilhada entre a Terra e a vida. Durante a viagem do Beagle3 1 , Darwin tomou conhecimento do impacto do soerguimento de cordilheiras e revelação da distribuição altitudinal de camadas geológicas e dos fósseis nelas contidos. A taxa de elevação evidente foi, às vezes, muito impressionante e a atividade vulcânica na América do Sul indicou uma causa comum para eventos geológicos separados por milhares de quilômetros. Darwin especulou sobre os vales e planícies da América do Sul tendo se localizado anteriormente sob o mar e sobre os penhascos da Austrália tendo marcado os níveis das costas e baías. Ele também previu o surgimento de barreiras geográficas impedindo a expansão da distribuição da biota, tais como entre as Américas do Norte e do Sul, mas ele não imaginava a possibilidade de que as causas responsáveis pela distribuição vertical de estratos geológicos também poderiam afetar a vida sobre os estratos.
A pan-biogeografia integra o que Darwin não integrou: as consequências ecológicas, bem como biológicas de uma história compartilhada entre a Terra e a vida. Uma das contribuições mais importantes de pan-biogeografia à teoria da evolução é a introdução de uma nova compreensão evolutiva da dispersão. Na biogeografia de Darwin, os "meios de dispersão" presumivelmente responsáveis pela origem de táxons vicariantes são, na realidade, nada mais do que seus simples "meios de sobrevivência". Em todos os táxons, algum tipo de 10- � comoção é responsável pela distribuição da prole num ;;; hábitat, porém apenas em circunstâncias especiais que t._
estes meios de sobrevivência resultam na expansão ou � alteração de uma área geográfica. Os meios de sobrevi- 2
vência permitem a colonização de novos ambientes e é justamente este processo que permite aos organismos persistirem numa região geologicamente instável. Isso pode ocorrer em regiões extensas, onde transgressões marinhas, atividade vulcânica ou orogêneses podem destruir a vida em algumas localidades ao mesmo tempo em que criam novos ambientes, mais tarde colonizados pelos organismos sobreviventes. O mesmo processo também se aplica às ilhas oceânicas, onde os organismos persistem por sua capacidade de movimentação entre ilhas individualmente efêmeras24.42.
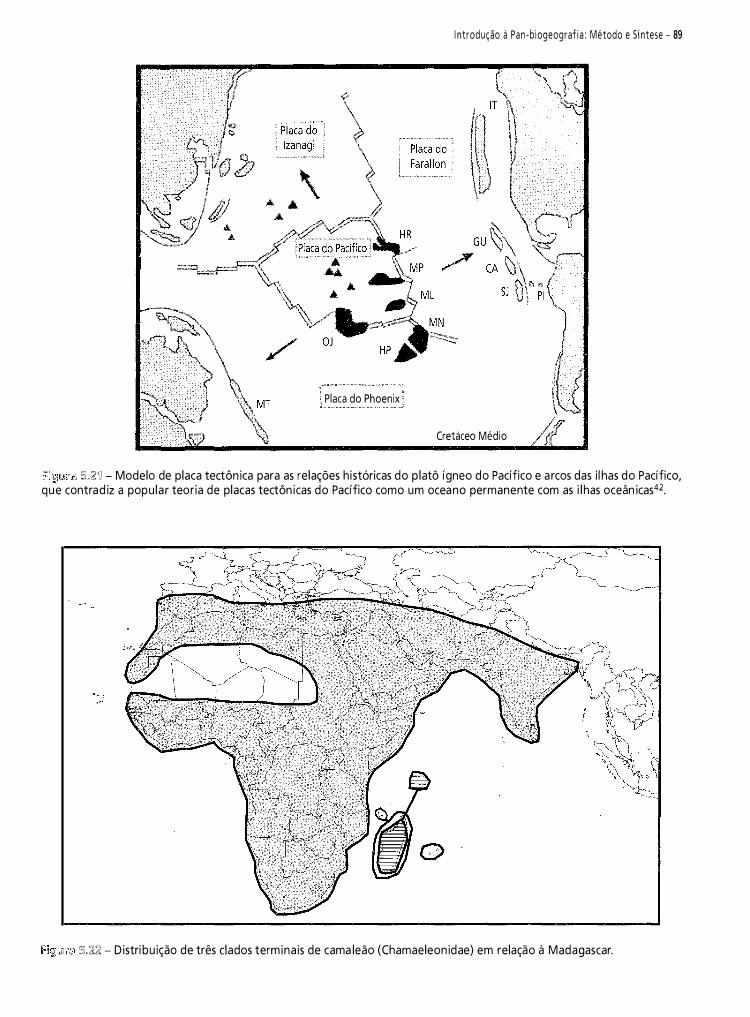
I ntrodução à Pan-biogeograf i a : Método e Síntese - 89
Placa do Phoenix
Cretáceo Médio
5.::2'] - Modelo de placa tectôn ica para as relações históricas do p latô ígneo do Pacífico e a rcos das i l has do Pacífico, que contrad iz a popular teoria de placas tectôn icas do Pacífico como um oceano permanente com as i l has oceân icas42.
3).::22 - Distribu ição de três dados terminais de cama leão (Chamaeleon idae) em relação à Madagascar.
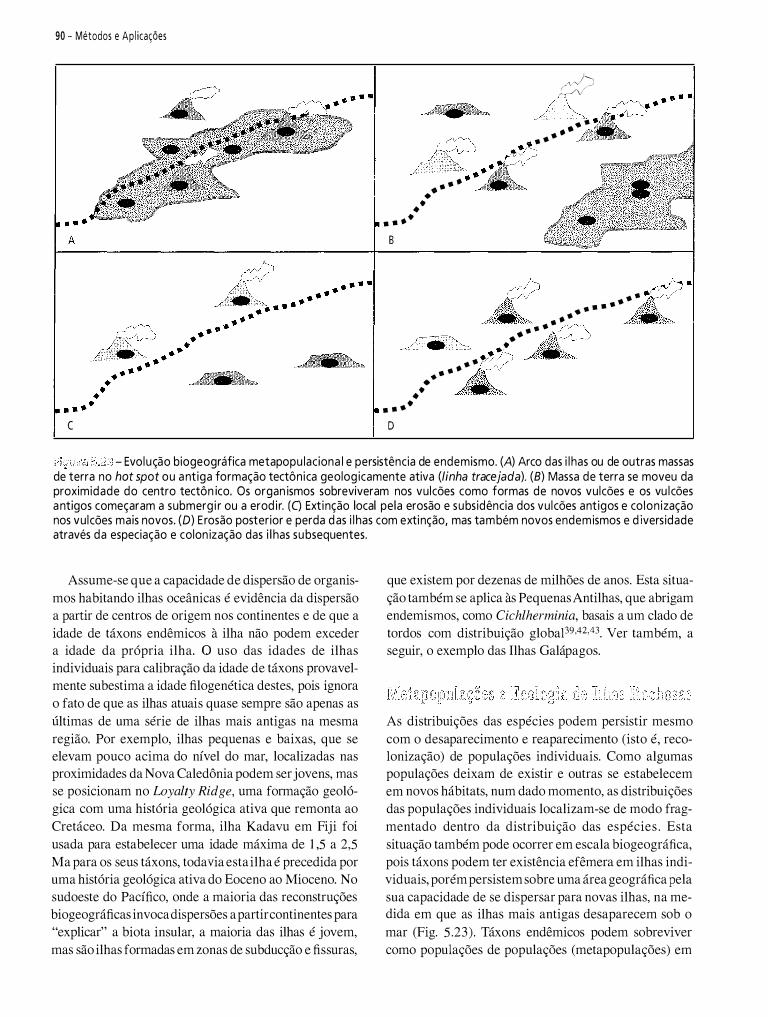
90 - Métodos e Apl icações
c
. . -.1l
.+
• • - +
B
.+
.+
• • • +
D
• •
. . -
+ 1l 1l
�') •.. �
/ � • • •
••
_ .
. -• • •
• • • • • •
• • •
- Evolução biogeográfica metapopu laciona l e persistência de endemismo. (A) Arco das i l has ou de outras massas de terra no hot spot ou a ntiga formação tectônica geolog icamente ativa (linha tracejada). (B) Massa de terra se moveu da proximidade do centro tectôn ico. Os organ ismos sobreviveram nos vulcões como formas de novos vulcões e os vu lcões a ntigos começaram a submerg i r ou a erod ir. (e) Extinção local pela erosão e subsidência dos vulcões antigos e colon ização nos vulcões mais novos. (O) Erosão posterior e perda das i l has com extinção, mas também novos endemismos e d iversidade através da especiação e colon ização das i l has subsequentes.
Assume-se que a capacidade de dispersão de organismos habitando ilhas oceânicas é evidência da dispersão a partir de centros de origem nos continentes e de que a idade de táxons endêmicos à ilha não podem exceder a idade da própria ilha. O uso das idades de ilhas individuais para calibração da idade de táxons provavelmente subestima a idade filogenética destes, pois ignora o fato de que as ilhas atuais quase sempre são apenas as últimas de uma série de ilhas mais antigas na mesma região. Por exemplo, ilhas pequenas e baixas, que se elevam pouco acima do nível do mar, localizadas nas proximidades da Nova Caledônia podem ser jovens, mas se posicionam no Loyalty Ridge, uma formação geológica com uma história geológica ativa que remonta ao Cretáceo. Da mesma forma, ilha Kadavu em Fiji foi usada para estabelecer uma idade máxima de 1 ,5 a 2,5
Ma para os seus táxons, todavia esta ilha é precedida por uma história geológica ativa do Eoceno ao Mioceno. No sudoeste do Pacífico, onde a maioria das reconstruções biogeográficas invoca dispersões a partir continentes para "explicar" a biota insular, a maioria das ilhas é jovem, mas são ilhas formadas em zonas de subducção e fissuras,
que existem por dezenas de milhões de anos. Esta situação também se aplica às Pequenas Antilhas, que abrigam endemismos, como Cichlherminia, basais a um clado de tordos com distribuição globaP9,42,43. Ver também, a seguir, o exemplo das Ilhas Galápagos.
As distribuições das espécies podem persistir mesmo com o desaparecimento e reaparecimento (isto é, recoIonização) de populações individuais. Como algumas populações deixam de existir e outras se estabelecem em novos hábitats, num dado momento, as distribuições das populações individuais localizam-se de modo fragmentado dentro da distribuição das espécies . Esta situação também pode ocorrer em escala biogeográfica, pois táxons podem ter existência efêmera em ilhas individuais, porém persistem sobre uma área geográfica pela sua capacidade de se dispersar para novas ilhas, na medida em que as ilhas mais antigas desaparecem sob o mar (Fig. 5 .23). Táxons endêmicos podem sobreviver como populações de populações (metapopulações) em

ilhas efêmeras formadas em tomo de zonas de subducção tectônica de falha geológica, para as quais a idade e a história desses processos tectônicos, não as ilhas individuais, são relevantes para a compreensão biogeográfica de sua idade e origem. O mesmo modelo pode se aplicar
::6 a outros continentes ou a grandes massas de terra, em O'> "? que táxons endêmicos sobrevivem ao se dispersarem � r-;: para novos hábitats resultantes de transgressões marinhas � ou de atividade vulcânica. Táxons alpinos ou de altitude 00 S; podem colonizar sequencialmente novas montanhas ou
vulcões, na medida em que formações mais antigas são erodidas, permitindo que táxons endêmicos antigos sobrevivam montanhas mais jovens. Táxons originados durante o Mesozoico podem acabar sendo endêmicos de sedimentos mais jovens! I ,24.
Um táxon endêmico-insular deve ser capaz de sobreviver em pequenas ilhas com variação climática ou ecológica restrita e também ser capaz de colonizar novas ilhas. Isso significa que os únicos táxons presentes em ilhas serão aqueles com habilidade de dispersão suficiente para sobreviver nessas condições. Organismos terrestres com pouca ou nenhuma tolerância à água salgada, tais como anfíbios, geralmente não são encontrados em ilhas efêmeras. Pode haver alguns mamíferos, mas comumente são limitados a roedores, morcegos e espécies marinhas. Caso contrário, a fauna será dominada por invertebrados, lagartos e aves. Este tipo de comunidade pode ser referido como tendo uma "ecologia de ilhas rochosas", típico de pilhas pequenas e pedregosas ao longo de uma costa em erosão. Se esse tipo de ecologia é posteriormente incorporado numa massa de terra maior, a biota subsequente continuará apresentando este tipo de ecologia. Um exemplo disso é a Nova Zelândia, onde invertebrados, aves e répteis são dominantes, enquanto outros vertebrados terrestres são limitados a poucos morcegos e rãs24.
O arquipélago de Galápagos localiza-se num hot spot vulcânico gerado a partir do movimento a leste das placas Cocos e Nazca. Como as ilhas são tectonicamente transportadas a partir do hot spot inicial, estas se erodem e, eventualmente, submergem como uma série de ilhas encobertas pelo oceano e que se entendem a nordeste na direção do Panamá e a leste em direção ao Equador. Na ausência de evidências de ligações continentais diretas, a maioria dos biogeógrafos atribuiu a origem de todas as espécies terrestres às dispersões sobre o oceano ocorridas ao acaso a partir do continente americano. Essa história de dispersão foi limitada no
I ntrodução à Pan-b iogeograf ia : Método e Síntese - 91
tempo à idade das ilhas atuais, cerca de 5 Ma, até que uma série mais antiga de ilhas foi descoberta, remontando a 40 Ma, no ponto onde elas estão submersas. A partir daí, passou-se a aceitar que uma dispersão inicial tenha permitido a colonização das Ilhas Galápagos mais antigas, ao que se seguiu à colonização sequencial das ilhas mais jovens. Embora este conceito de metapopulação tenha sido aplicado às Ilhas Galápagos, a premissa subjacente manteve-se como a dispersão a partir de um centro americano de origem44.
O modelo de dispersão de Galápagos foi questionado por Croizat3, pois há relacionamentos distintos mostrados pelos traços de vicariância entre Galápagos e as Américas4s (Fig. 5 .24). Por exemplo, as serpentes do gênero AIsophis são encontradas apenas em Cuba e Hispaniola e nas Ilhas Galápagos , Em contraste, os tentilhões não voadores do gênero Galapaganus ocorrem nas Ilhas Galápagos, Equador e Peru (Fig. 5 .24, A), um padrão compartilhado com outros organismos, tais como vespas aladas do gênero Tachysphex (Fig. 5 .24, B).
Outras distribuições, como escorpiões (Iuridae) ocorrem nas Américas do Norte e do Sul e as Ilhas Galápagos, mas não a América Central (Fig. 5 .24, C). Traços alternativos podem ainda ser representados por diferentes linhagens dentro de um determinado táxon, como as plantas do gênero Darwiniothamnus (Fig. 5 .24, D).
Croizat3 argumentou que as Ilhas Galápagos tinham as mesmas características biogeográficas de um nó biogeográfico localizado no interior de um continente, indicando que a biota das ilhas era "continental" em vez de simplesmente "oceânica" em sua origem. Uma predição para a costa pacífica das Américas, anterior à tectônica de placas, foi de que esta teria se estendido a oeste, como um complexo de conjuntos de ilhas, e que a Galápagos teria sido associada a uma formação tectônica principal, até então desconhecida. Essas previsões geológicas foram feitas num momento em que o fundo do mar não havia ainda sido mapeado e sua tectônica compreendida, e antes da aceitação ampla entre os geólogos da teoria de Wegener de mobilidade dos continentes. Biogeógrafos dispersalistas rejeitaram o modelo geológico proposto por Croizat, alegando não haver provas geológicas, como se a teoria geológica determinasse a história biogeográfica.
O mapeamento do fundo dos oceanos na década de 1 970 identificou o centro de expansão em que se localizam as Ilhas Galápagos e os geólogos desenvolveram modelos históricos que predizem a origem da América Central e das Antilhas na bacia do Pacífico a oeste de Galápagos. O movimento destes arcos insulares a leste, durante o Cretáceo, teria possibi litado sua interseção com

92 - Métodos e Apl icações
'3,21:) - (A) Ga/apaganus. (8) Tachysphex: espécie irmã. (C) Escorpiões ( luridae) (Grehan)43 . (O) Oarwiniothamnus: táxons i rmãos: Ga lápagos (O. /ancifo/iusIO. tenufo/ius) e Cuba (Erigeron bellidiastroides), Ga lápagos (O. a/ternifo/ius) e Chi le (E. fascicu/atuslE. /uxurians)45.
Galápagos e a redistribuição da biota do arco insular sobre o hot spot vulcânico. Alguns organismos foram capazes de persistir até o presente, por meio da colonização de novas ilhas que se formaram, enquanto as ilhas mais antigas se erodiram e foram cobertas pelo Pacífico oriental. Os arcos de ilha continuaram se movimentando rumo ao leste e foram, eventualmente, acrescidos às Américas do Norte, Central e do Sul, junto com suas biotas, incluindo organismos aparentados àqueles ocupando o hot spot de Galápagos44. O relacionamento biogeográfico entre Galápagos e as Américas inclui, portanto, uma realocação (decorrente das mudanças geológicas) de táxons para as Américas e não a partir das Américas (Fig. 5.25).
A pan-biogeografia é o único método biogeográfico capaz de gerar uma síntese evolutiva integrada da diferenciação biológica no espaço e no tempo. Um dos avanços obtidos com a pan-biogeografia foi o desenvolvimento de mode
los evolutivos que conftitam com a teoria popular da
teoria da seleção natural agindo sobre a variação "aleatória", modelos estes que se mostraram concordantes, em muitos aspectos, com teorias biológicas geradas a partir de estudos moleculares e de desenvolvimento.
O modelo de especiação por seleção natural busca mutações individuais surgidas ao acaso em determinado momento da história e se espalhando para a população em razão da seleção natural, como se a origem da nova espécie não tivesse relação com outras espécies. Com a diferenciação vicariante, a origem de qualquer "espécie" (ou qualquer outro táxon), os descendentes são necessariamente encontrados na área de distribuição do ancestral anterior, que é amplamente distribuído (em relação aos descendentes diretos) e também é ancestral aos outros descendentes vicariantes. Ao examinar a origem das espécies, é necessário estudar os mecanismos de diferenciação biológica que operam sobre uma área geográfica extensa, em vez de pontos individuais isolados46. No "Modelo de Recombinação de Caracteres", proposto por

Croizat3, a diferenciação é uma consequência de diferentes combinações de caracteres ancestrais resultando em modificações evolutivas, seja como uma consequência de novas combinações de estruturas, produzindo inovações fenotípicas, ou como um viés mutacional.
Caracteres ancestrais representam um tipo de organização que restringe os resultados possíveis de mudanças evolutivas futuras. Um camundongo, por exemplo, tem organização típica de um "roedor". Os roedores, por sua vez, representam uma recombinação especializada do que seria a organização típica de mamíferos. Isso não exclui a origem de mutações novas, mas sugere que a origem da mutação é dependente da estrutura biológica molecular preexistente do ancestral, uma possibilidade que também fora reconhecida por Charles Darwin, que desenvolveu suas leis da teoria de crescimento da teoria da mudança evolutiva sem seleção natural, fato normalmente ignorado pelos especialistas darwinianos47,48. Este
A
Arco insular
I ntrodução à Pan-b iogeografia : Método e Síntese - 93
modelo de diferenciação pan-biogeográfica sofreu oposição por parte dos evolucionistas darwinianos, porém os campos da evolução molecular e de desenvolvimento têm gerado modelos biológicos semelhantes. Mecanismos moleculares permitem que ocorram vieses na conversão de genes ou que elementos transponíveis se espalhem por uma população sem a necessidade de aumento da aptidão (jitness) reprodutiva7. A genética do desenvolvimento tem reconhecido o papel dos genes reguladores que podem rearranjar aspectos da organização biológica das espécies, sem a necessidade de mudança gradual pelo acúmulo de modificações em genes estruturais elementares49. Estes modelos de regulação podem ser comparáveis ao conceito de pan-biogeográfico de morfogenia4, em que atributos individuais são vistos como morfologias alternativas geradas a partir de um processo ancestral comum de desenvolvimento (Fig. 5 .26) .
*' Hot spot de Ga lápagos
*' Mesozoico
B
*'
Galápagos
*' Presente
:3,2:.5 - Modelo de arco insu lar de Ga lápagos. (A) Arco insu lar i n icia l deriva e se choca com o hot spot de Ga lá pagos durante o Mesozoico. Os endemismos do arco insu la r colonizam o hot spot de Ga lápagos. (B) Acreção das porções do arco insu lar com o cráton da América do Su l . Os endemismos pr imários do arco insu la r ficam isolados no hot spot de Ga lá pagos ou são transportados para a América do Su l, onde podem ser loca lmente d ispersos7,44.

94 - Métodos e Aplicações
- Exemplo de substituição morfológica. =
"normal"; 2 = com apêndices (leg-like)5.
o modelo morfogenético elimina a necessidade de se explicar a origem da estrutura biológica como consequência da função, uma forma de raciocínio que exige a teleologia ou propósito como uma explicação à Oligem47. Em vez disso, é a evolução estrutural que afeta os resultados funcionais48. Pernas, antenas, apêndices e genitália de um inseto, por exemplo, não evoluem para atender algum objetivo funcional, mas representam diferentes subprodutos morfogenéticos de um apêndice ancestral generalizado em posições diferentes no corpo do insetos. Esta relação é geneticamente representada pela expressão diferencial de genes homeobo.09. Um modelo morfogenético de evolução das fi ores foi proposto por Croizat4, indicando que as origens comuns foram ofuscadas e que definições particulares para morfologias distintas de determinados táxons individuais haviam sido favorecidas erroneamente por outros autores50.
A pan-biogeografia fornece os fundamentos para a conservação evolutiva porque documenta a estrutura evolutiva da biodivcrsidadc no espaço e no tempo. Os nós biogeográficos proveem informações sobre os centros de diversidade, ao passo que traços identificam suas inter-relações espaciais. Centros de massa representam gradientes de diversidade para os táxons e, juntamente com a linha de base, fornecem informações sobre as origens tectônicas e a significância de táxons individuais e biotas inteiras. As análises pan-biogeográficas são sinônimos de análises de biodiversidade, que identificam a estrutura evolutiva e a importância das distribuições de plantas e animais.
Uma ferramenta fundamental para a conservação evo
lutiva é o desenvolvimento de um Atlas da Biodiversidade,
fornecendo informações biogeográficas empíricas sobre táxons individuais e sobre a biota. sem a necessidade da aceitação de unidades artificiais na fOlwa de áreas ecoló
gicas ou biogeográficas geograficamente delimitadas. Este
Atlas enfatizaria a vida como uma rede global de traços e nós biogeográficos, na qual a origem da biodiversidade dentro de uma região local está conectada ao desenvolvimento da vida planetária em geral. As aplicações particulares do conceito de um Atlas da Biodiversidade já foram consideradas por vários biogeógrafos51.
Um método que combine traços e informações filoge
néticas para criação de ranking de prioridades para a biodiversidade em diferentes áreas de endemismo foi proposto por Morrone51. Neste método, os nós biogeográficos são considerados como prioridades primárias e são classificados dentro de cada traço generalizado nas áreas de endemismo de acordo com seu valor filogenético (o número de grupos incluindo cada espécie). Uma aplicação desta abordagem foi feita para áreas de florestas de neblina no estado de Hidalgo (México), utilizando uma análise de parcimônia de presença e ausência de espécies para gerar um cladograma53. As espécies definidoras de clados foram sequencialmente removidas e os dados foram reanalisados para identificar outros clados. As localidades em cada clado foram mapeadas como árvores de distâncias mínimas (traços) e as áreas de floresta foram classificadas
de acordo com o número de interseções de traços (Fig. 5.27). A área mais bem posicionada no ranking, Chapulhuacán, apresentou relacionamentos de seis traços generalizados e, pOltanto, alto valor potencial para conservação, embora nenhuma das áreas de floresta de neblina de Hidalgo estivesse sob proteção oficial.
A utilização de vários táxons por Prevedello e CarvalhoS! para os biomas brasileiros do Cerrado e Pantanal resultaram em traços para 55 espécies de plantas, mamíferos, aves e insetos. Na região do Cerrado, 48 nós biogeográficos foram reconhecidos como áreas prioritárias para conservação da biodiversidade. Os nós biogeográficos foram classificados de acordo com o número de faixas generalizadas aos quais estes se conectam, com 22 nós biogeográficos sustentando dois traços generalizados, 15 nós sustentando três, sete nós sustentando quatro e quatro sustentando cinco traços generalizados. Os nós biogeográficos das duas últimas categorias foram considerados como tendo a mais alta prioridade para conservação. Constatou-se que apenas 2 1 % dos nós biogeográficos localizavam-se total ou parcialmente dentro de áreas protegidas e apenas 13%
estavam totalmente protegidas; por outro lado, 77% dos nós localizavam-se em áreas identificadas como priori
tárias para conservação (Fig. 5.28). Observou-se ainda
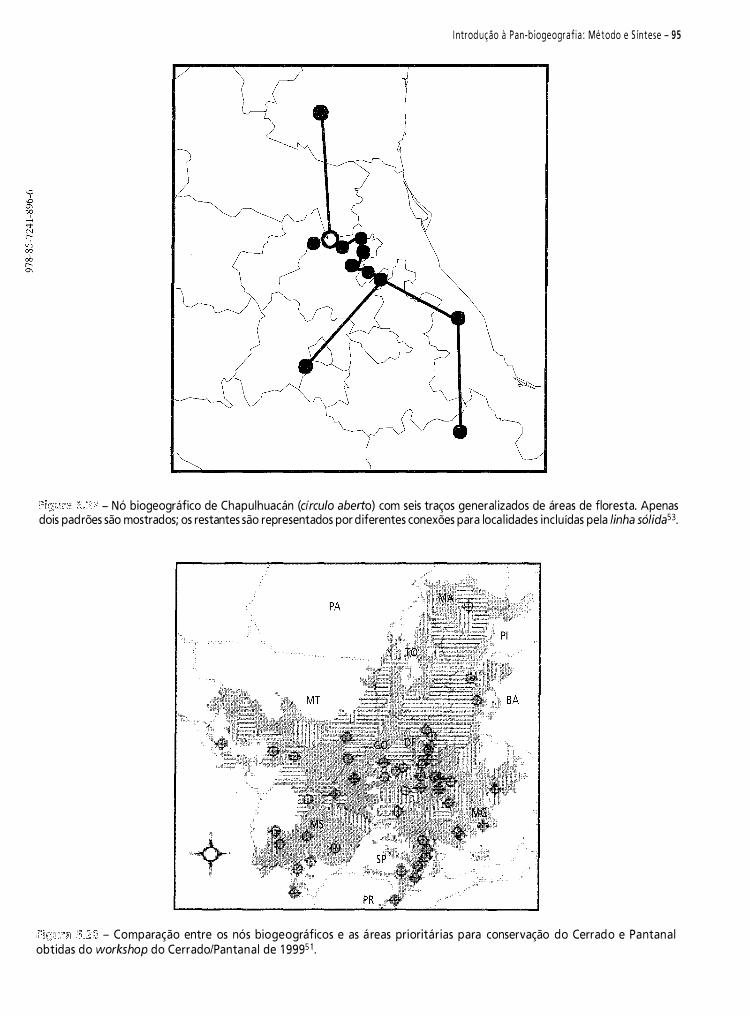
I ntrodução à Pan-biogeografia : Método e S íntese - 95
- Nó biogeográfico de Chapulhuacán (círculo aberto) com seis traços general izados de áreas de floresta. Apenas dois padrões são mostrados; os restantes são representados por d iferentes conexões para loca l idades inc luídas pela linha sólida53.
- Comparação entre os nós biogeográficos e as á reas prioritá rias para conservação do Cerrado e Pantana l obtidas do workshop do Cerrado/Pantana l de 1 9995 1 .

96 - Métodos e Ap l i cações
que 10 nós biogeográficos encontravam-se num nível avançado de degradação ecológica, indicando que suas localidades-chave estavam ameaçadas ou que já haviam sido perdidas. Prevedello e Carvalh05l concluíram que as análises revelaram deficiências atuais no sistema de áreas protegidas, especialmente no que diz respeito à sua história e biogeografia histórica, bem como corroboraram a importância das áreas protegidas que abrangem os nós biogeográficos.
A comparação de presenças e ausências de espécies de áreas disjuntas representa um passo importante no desenvolvimento de um arcabouço biogeográfico para análise da biodiversidade. Para capturar o significado espaço-temporal de traços de espécies e de nós biogeográficos, também será necessário integrar as relações filogenéticas dos táxons, tanto dentro das áreas sendo consideradas como externamente a estas, com relação ao mundo como um todo.
A ciência da biogeografia é fortemente influenciada por programas de pesquisa darwinista desenvolvidos nos Estados Unidos e no Reino Unido. Ironicamente, Darwin tinha, de modo relativo, pouco a contribuir para a ciência da biogeografia, embora sua premissa fundamental da exigência de centros de origem e capacidade de dispersão para a evolução biológica talvez seja mais difundida hoje do que no seu tempo. Sob a égide aparentemente protetora da teoria do relógio molecular, biogeógrafos darwinianos se apressaram ao assumir várias hipóteses de dispersão com base no falso pressuposto de que as datas são precisas ou representam idades máximas. Esta popularidade foi erroneamente representada como um progresso em decorrência da aplicação de "poderosas" ferramentas de novas. Contudo, as teorias do relógio molecular de dispersão são tão fictícias quanto as suas antecessoras que usaram dados fósseis e exigem que as evidências de incongruências sejam refutadas ou ignoradas (em geral o segundo caso).
Pode ser um pouco desconfortável pensar que a pan-biogeografia discorde da escola biogeográfica dispersalista de centro de origem. Entretanto, não é menos verdade que a pan-biogeografia abraça algumas das chamadas "verdades" mais comumente aceitas da biogeografia e da evolução. A pan-biogeografia, por conseguinte, foi inicialmente declarada morta a partir de uma série de perspectivas biogeográficas, incluindo do teórico darwiniano Ernst Mayr54, do cladista de vicariância ale Seberg55, do dispersalista John Briggs56, e do "dispersalista molecular" Mat-
thew McGlone57. Essas afirmações repetidas exigem saber
por que é necessário continuar fazendo tais declarações.
Por meio dos esforços pioneiros de Robin Craw e
Michael Heads (enquanto ainda estudantes de pós-gra
duação), a pan-biogeografia começou a florescer na Nova
Zelândia durante a década de 198058 . Naquele tempo,
os pesquisadores quase sempre gozavam do privilégio
de poderem escolher seus tópicos de pesquisa e adotarem :5 ex
suas próprias perspectivas, em vez de serem subordina- 3; -'-'
dos às prioridades de um complexo científico-industrial. 'l;: Aquele também era um momento em que o reducionismo � molecular ainda não havia ofuscado as ciências naturais & e o modelo de financiamento estudantil não drenava o
tempo e os recursos para longe do pensamento inde
pendente. a predomínio crescente da evolução e bio
geografia darwiniana posteriormente acabou com essas
vantagens e pan-biogeografia foi marginalizada na Nova
Zelândia6. Entretanto, como uma erva daninha per
sistente, a pan-biogeografia tem crescido em meio à
diáspora intelectual global, particularmente entre pes
quisadores do Chile, Argentina, Brasil, Venezuela, Co
lômbia, México, França e Itália59.6o•
Se a literatura recente for uma boa indicação dessa
tendência, há mais interesse em pan-biogeografia naAmé
rica Latina do que em qualquer outra região do mundo.
Este interesse foi manifestado pelo apoio explícito ao
programa de pesquisa pan-biogeográfico, enquanto outros
métodos se esforçam para incorporar aspectos de pan
-biogeografia, quando esses se mostram problemáticos. a
entusiasmo intelectual da biogeografia latino-americana
foi evidente no XXVII Congresso Brasileiro de Zoologia,
em 2008, durante o Simpósio de Biogeografia Neotropical,
organizado por Claudio J. B . de Carvalho e Dalton S .
Amorim6l . As palestras foram variadas e muito procuradas
pelos participantes (em especial por estudantes), tendo
abrangido todos os aspectos de teorias e métodos biogeo
gráficos. Juntamente com o predomínio da biogeografia
em apresentações de pôsteres sobre a biodiversidade bra
sileira, isso mostra que a ciência da biogeografia no Brasil
é dinâmica e progressiva. Se há alguma lição da história
da biogeografia, é a persistência de alternativas. Não exis
te um imperativo histórico exigindo que a biogeografia
futura da América Latina continue a se subordinar à teoria
da evolução de Darwin. as estudantes de hoje serão os
líderes de amanhã. Há cinco décadas, Léon Croizat deu o
primeiro grande passo além do darwinismo. Duas décadas
atrás, os neozelandeses iniciaram uma segunda etapa.
Quem será o pioneiro da terceira?

Sou grato à minha esposa, Claudia Violette, que permitiu minha participação permanente na exploração e na aplicação da pan-biogeografia por meio de seu apoio, incentivo e pensamento crítico. Tenho especial gratidão ao Claudio J. B . de Carvalho, pela oportunidade de participação deste livro. Agradeço ainda ao Michael Heads pela edição e comentários a uma versão preliminar deste manuscrito, bem como comentários de Carlos Mielke e Juan Morrone. Serei sempre grato aos meus colegas Robin Craw e Michael Heads, que em todo momento se mostraram gentis e pacientes perante minha ingenuidade sobre a maioria dos aspectos da teoria da evolução, quando me interessei inicialmente pela pan-biogeografia.
A formatação final das figuras foi realizada graças ao apoio de Thorn Furtado e Ashlei Jachimovicz (Exhibits Department, Buffalo Museum of Science).
REFERÊNCIAS BIBLIOGRÁFICAS 1 . MORRONE, J. J. Evolutionary Biogeography - an lntegrative
Approach with Case Studies. Nova York: Colombia University Press, 2009. 301 p.
2. WALLACE, A R. The Geographical Distribution of Animais.
London: Macmillan, 1 876. 638p. 3 . CROIZAT, L. Panbiogeography. Caracas: published by the
author, 1958. 2755p. 4. CROIZAT, L. Principia Botanica. Caracas: published by the
author, 1 96 1 . 1 821p. 5 . CROIZAT, L . Space, Time, Form, The Biological Synthesis.
Caracas: published by the author, 1964. 881p. 6. HEADS, M. The history and philosophy of panbiogeography.
In: LLORENTE, J.; MORRONE, 1. 1. Regionalización Bio
geográfica en lberoamérica y Tópicos Afines. México : Universidad Nacional Autónoma d e México, 2005. p. 67- 123.
7. CRAW, R. c.; GREHAN, 1. R.; HEADS, M. J. Panbiogeography: tracking the history of life. Nova York: Oxford University Press, 1 999. 229p.
8. MORRONE, J. J. El tiempo de Darwin y el espacio de Croizat: rupturas epistémicas en los estudios evolutivos. Ciencia, v. 5 1 , p . 39-46, 2000.
9. GREHAN, J. R. Panbiogeography y biogeografia de la vida. In: LLORENTE, J.; MORRONE, 1. J. lntroduccion a la Biogeografia: Teorias, conceptos, metodos y aplicaciones. México: Universidad Nacional Autónoma, 2001 . p. 1 8 1 - 1 96.
10. HEADS, M. A review of "R.T. Pennington, Q.C.B. Cronk, and J.A. Richardson (eds .), 2004. Plant phylogeny and the origin of major biomes". The Systematist, v. 24, p. 19-22, 2005.
J 1. HEADS, M. Dating nodes on molecular phylogenies: a critique of molecular biogeography. Cladistics, v. 2 1 , p. 62-78, 2005.
12. ROJAS-PARRA, C. A Una herramienta automatizada para realizar análisis panbiogeográficos. Biogeografia, v. 1 , p. 3 J -33, 2007.
1 3 . LOURENÇO, W. R. Nouvelles considerations sur la classification et la biogeographie des Opisthacanthus neotropicaux
(Scorpiones, Ischnuridae). Biogeographica, v. 7, p. 75-82, J 995. 14. NIHEI, S . S . ; CARVALHO, C. J. B. Distributional patterns of
the Neotropical fly genus Polietina Schnabl & Dziedzicki (Diptera, Muscidae): a phylogeny-supported analysis using panbiogeographic tools. Pap Avulsos Zool. (Sao Paulo), v. 45, p. 3 1 3-326, 2005.
I ntrodução à Pan-biogeografia: M étodo e Síntese - 97
15. ROCHA, S.; CARRETERO, M.A; VENCES, M. et aI. Deciphering pattems of transoceanic dispersai: the evolutionary origin and biogeography of coastal lizards (Cl)ptoblepharus) in the western Indian Ocean region. 1. Biogeogr., v. 33, p. 1 3-32, 2006.
1 6 . DE MARMELS, J . A new species of Xylophanes tersa (Sphingidae) from Venezuela. J. Lep. Soe., v. 50, p. 303-308, 1 996.
17. GREHAN, J. R. A brieflook at Pacific biogeography: the transoceanic travels of Microseris (Angiosperms: Asteraceae). In: EBACH, M. c.; TANGNEY, R S. Biogeography in a Changing World. Boca Raton: CRC Press, 2007. p. 83-94.
1 8 . PILLAI, R. S . ; RAVICHANDRAN, M. S. Gymnophiona (Amphibia) of India: a taxonomic study. Ree. Zool. Surv. India, Occ Pap, n. 1 72, p. 1 - 1 26, 2005.
19. HILL, K. D. Cycads in the Pacifico In: KEAST, A . ; MILLER, S. E. The Origin and Evolution of Pacijic Island Biotas, New Guinea to Eastem Polynesia: patterns and processes. Amsterdam: SPB Academic Publishing, 1996. p. 267-274.
20. HEADS, M. Vicariance biogeography and terrane tectonics in the South Pacific: an analysis of the genus Abrotanella (Compositae). Biol. J. Linn. Soe. , v. 67, p. 391-432, 1 999.
2 1 . HEADS, M. Panbiogeography of Nothofagus (Nothofagaceae): analysis of the main species massings. J. Biogeogr. , v. 33, p . 1 066- 1 075, 2006.
22. GREHAN, 1. R; RAWLINS, J. E. Larval description of a New World ghost moth, Phassus sp., and the evolutionary biogeography of wood-boring Hepialidae (Lepidoptera: Exoporia: Hepialoidea). Proc. Entomol. Soe. Wash, v. 1 05, p. 733-755, 2003.
23. HEADS, M. What is a node? 1. Biogeog/:, v. 3 1 , p. 1 883-1891, 2004. 24. HEADS, M. Integrating earth and life sciences in New Zealand
natural history: the parallel arcs model. NZJ Zool. , v. 1 6, p. 549-585, 1 990.
25. HENDERSON, I. M. Biogeography without area? Aust. Syst. Bot. , V. 4, p. 59-7 1 , 1 99 1 .
26. YUAN, Y. M . ; WOHLHAUSSER, S . ; MOLLER, M . , e t aI. Phylogeny and biogeography of Exacum (Gentianaceae): a disjunctive distribution in the Indian Ocean basin resulted from long distance dispersaI and extensive radiation. Syst. Biol. , V. 54, p. 2 1 -34, 2005.
27. HEADS, M. Biogeography and biodiversity in Hebe, a South Pacific genus of Scrophulariaceae. Candollea, v. 48, p. 19-60, 1993.
28. FLOWERS, R. W. Life was a beach: a panbiogeographic analysis of the cosmopolitan mayfly genus Choroterpes (Ephemeroptera: Leptophlebeiidae: Atalophlebiinae. Aquat. lnsects, v. 3 1 , p. 585-593, 2009.
29. HEADS, M. 1. Biogeographic disjunction along theAlpine fault, New Zealand. Biol. J. Linn. Soe. , V. 63, p . l 6 1 - 176, 1998.
30. HEADS, M. J.; CRAW, R. C. The Alpine Fault biogeographic hypothesis revisited. Cladistics, v. 20, p . 1 84-190, 2004.
3 1 . DARWIN, C. The Voyage ofthe Beagle. Nova York: P. F. Collier & Son, 1 845 ( 1 909 edition). 547p.
32. HEADS, M. 1. Biological disjunctions along the West Caledonian fault, New Caledonia: a synthesis of molecular phylogenetics and panbiogeography. Bot. 1. Linn. Soe., V. 1 58, p. 470-488, 2008a.
33. HEADS, M. J. Vicariance. In: GILLESPIE, R.; CLAGUE, D. Encyclopedia of Islands. B erkeley: University of California Press, 2009. p. 947-950.
34. YOKOYAMA, J.; SUZUKI, M.; IWATSUKI, K. et aI. Molecular phylogeny of Coriaria, with special emphasis on the disjunct distribution. Moi. Phylogenet. Evol. , v. 14, p. 1 1 - 19, 2000.
35. GREHAN, J. R.; SCHWARTZ, J. H. Evolution 01' the second orangutan: phylogeny and biogeography of hominid origins. 1. Biogeog r. , v. 36, p. 1 823-1 844, 2009.
36. YAMAGUTI, H. Y.; PINTO-DA-ROCHA, R Taxonomic review of Bourguyiinae, cladistic analysis, and a new hypothesis of biogeographic re1ationships of the Brazilian Atlantic rainforest (Arachnida: OpiJiones, Gonyleptidae). Zool. J. Linn. Soe. , v. 1 56, p. 3 1 9-362, 2009.
37. SILVA DE PAULA, A; DIOTAIUTI, L.; GALVÃO, C. Systematics and biogeography ofRhodniini (Heteroptera: Reduviidae: Triatominae) based on 1 6S mitochondrial rDNA sequences. J. Biogeogr., v. 34, p. 699-7 12, 2007.

98 - Métodos e Apl icações
38. DE MARMELS, J. Tepuibasis gen.nov. from the Pantepui region of Venezuela, with descriptions of four new species, and with biogeographic, phylogenetic and taxonomic considerations on the Teinobasinae (Zygoptera: Coenagrionidae). Odonatologica, v. 36, p. 1 17-146, 2007.
39. HEADS , M. Mesozoic tectonics and the deconstruction of biogeography: a new model of Australasian biology. J. Biogeog/:, v. 17, p. 223-225, 1990.
40. MORRONE, J. J. Biogeografía de América Latina y el Caribe. Manuales & Tesis SEA, v. 3, p. 1 - 1 48, 200 1 .
4 1 . HENDERSON,1. M . Quantitative biogeography: an investigation into concepts and methods. NZJ Zool., v. 1 6, p. 495-5 1 0, 1 99 1 .
42. HEADS, M. Globally basal centres o f endemism: the Tasman-Coral Sea region (south-west Pacific), Latin America and Madagascar/South Africa. Bio!' J. Linn. Soe. , v. 96, p. 222- 245, 2009.
43. HEADS, M. Seed plants of Fiji: an ecological analysis. Biol. J. Linn. Soe. , v. 89, p. 407 -431.
44. GREHAN, J. R. Biogeography and evolution of the Galapagos: integration of the biological and geological evidence. Bio!' 1. Linn. Soe. , v. 74, p. 267-287, 200 1 .
45. ANDRUS, N . ; TYE, A . ; NESOM, G . e t aI. Phylogenetics of Darwiniothamnus (Asleraceae: Astereae) - molecular evidence for multiple origins in the endemic flora of the Galápagos 1slands. J Biogeogr., v. 36, p. 1 055- 1 069, 2009.
46. HEADS, M. On the nature of ancestors. Syst. Zool. , v. 34, p . 205-2 1 5, 1985.
47. GREHAN, J. R. Evolution, issues in. In: BIRX, H. J. Encyclopedia ofTime. Beverly Hills: SAGE Publications, 2009. v. 1 , p. 469-48 1 .
48. HEAD S , M . Darwin's changing views o n evolution : from ccntres of origin and teleology to vicariance and incomplete lineage sorting. J. Biogeogr., v. 36, p. 1 0 1 8-1026, 2009c.
49. SCHWARTZ. J. H. Sudden Origins: Fossils, genes, and the emergence of species. Nova York: John Wiley and Sons, 1999. 42Op.
50. HEADS. M. Principia Botanica: Croizat's contribution to botany. Tuatara, v. 27, p. 26-48, 1984.
5 1 . PREVEDELLO, J. A.; CARVALHO, C. J. B. The Brazilian Cerrado conservation: the panbiogeographic method as a tool for selecting priority areas. Nat. Conservação, v. 4, p. 1 27-145, 2006.
52. MORRONE, J. J. How can biogeography and cladistics interact for the selection of areas for biodiversity conservation? A view from Andean weevils (Coleoptera: Curculionidae). Biogeographica, v. 75, p. 89-96, 1999.
53. LUNA, 1. ; ALCÁNTRA, O.; MORRONE, J. J. et aI. Track analysis and conservation priorities in the cloud forests of Hidalgo, Mexico. Divers Distrib. , v. 6, p. 1 37- 143, 2000.
54. CROIZAT, L. Mayr vs. Croizat: Croizat vs. Mayr - an enquiry. Tuatara, v. 27, p. 49-66, 1 984.
55. SEBERG, O. A critique of the theory and methods of panbiogeography. Syst. Zoo!. , v. 35, p. 369-380, 1 986.
56. BRIGGS, J. C. Panbiogeography: its origins, metamorphosis and decline. Rus. J. Marine Bio!', v. 33, p. 273-277, 2007.
57. MCGLONE, M. Goodbye Gondwana. J. Biogeog/: , v. 32, p. 739-749, 2005.
58. MATTHEWS, C. Panbiogeography special issue. New Zeal J Zool. , v. 16, p. 1-8 1 5, 1990.
59. MORRONE, J. J . ; LLORENTE-BOUSQUETS, J. Una Perspectiva Latinoamericana de la Biogeografía. Méxic o : Universidad Nacional Autônoma de México, 2003 . 307p.
60. MORRONE, J. J.; LLORENTE-B OUSQUETS, J. Componentes Bióticos Principales de la Entomofauna Mexicana. México: Universidad Nacional Autônoma de México, 2006. 1025p.
6 1 . LOWENBERG-NETO, P.; HASEYAMA, K. L. F. B razil as the stage for some inspiring discussions on biogeography. Biogeografía, v. 4, p. 3-4, 2009.

__________________ � 6
Silvio Shigueo Nihei
Deve-se fazer menção a Gareth Nelson, Norman Platnick e Donn Rosen como os principais responsáveis pelo desenvolvimento da biogeografia cladística ao longo da década de 1970 e início da de 1 980 (por exemplo, Nelsonl , Rosen2, Platnick e Nelson3, Nelson e Platnick4). A publicação de Systematics and Biogeography: Cladistics and Vicariance por Nelson e Platnick4 não representou um marco somente para a sistemática, mas também o foi para a biogeografia histórica, sobretudo a biogeografia cladística, para a qual, sem dúvida alguma, figura como uma das principais referências, mesmo passados quase 30 anos.
A biogeografia histórica está alicerçada sobre três pilares teóricos, a partir da integração dos conhecimentos da tectônica de placas (fundamentada, sobretudo, na deriva continental de Alfred Wegener), da sistemática filo genética de Willi Hennig e do processo de vicariância de Léon Croizat. A associação entre a história do planeta e a história dos organismos tomava-se cada vez mais evidente e factual aos biogeógrafos, como foi notoriamente enfatizado por Croizat5. Entretanto, foi apenas com a integração destes três pilares que se tomou possível sustentar e analisar de forma concreta e objetiva a correspondência entre o relacionamento filogenético dos táxons, seu padrão de distribuição e a história da terra.
Não é nada surpreendente que qualquer explanação biogeográfica antes da década de 1 960 fora baseada sob uma abordagem estritamente dispersalista, uma vez que se acreditava firmemente que as posições dos continentes eram estáticas. A teoria da tectônica de placas emergiu, assim, como o novo paradigma das geociências (para melhor entendimento dos processos de dispersão e vicariância, ver Capo 4). Vale mencionar que, apesar de hipóteses e modelos geológicos serem extremamente importantes para explicar o que determinou o padrão de
distribuição dos organismos, não se devem assumir evidências geológicas como definitivas e irrefutáveis. Neste sentido, é sempre fundamental procurar por congruência entre hipóteses biogeográficas e hipóteses geológicas6.
A sistemática filo genética trouxe-nos a possibilidade de inferir as relações filo genéticas dos táxons sob uma metodologia rigorosa e objetiva. A noção de ancestralidade comum dos membros de um grupo e, também, de compartilhamento de caracteres entre estes membros como resultado desta ancestralidade, permitiu que a história dos organismos fosse estudada empiricamente. Sua história genealógica é representada por diagramas ramificados (cladogramas) que denotam hipóteses de relacionamento entre os ramos terminais.
Até que Hennig tivesse esclarecido o conceito de relacionamento entre táxons, a questão sobre a relação entre áreas não poderia ser estudada e elucidada7, pois, além da suma importância que a filogenia dos organismos tem para a biogeografia, de certa forma, diagramas ramificados também são úteis para representar sequências de eventos de disjunção espacial, neste caso, evidenciando dicotomias entre áreas (= cladograma de áreas). Assim, os nós de um cladograma são potencialmente informativos não somente acerca da história distribucional dos organismos, mas também sobre o relacionamento entre as áreas ocupadas8. A fragmentação na distribuição de um táxon ancestral amplamente distribuído (cosmopolitismo primitivo) em duas porções, uma de cada lado do evento vicariante (por exemplo, evento geológico/abiótico), conduziria, com o passar do tempo, à diferenciação dos táxons descendentes isolados pelo evento vicariante. Se diferentes grupos de organismos apresentarem semelhanças distribucional (isto é, mesmo padrão de distribuição na área) e filo genética (táxons relacionados como grupos-irmãos), pode-se inferir que estas duas áreas são proximamente relacionadas (Fig. 6. 1 ) .

100 - Métodos e Ap l i cações
Fragmentação da área
I A I B I C I t
o
I I BC o.. A E �
t ABC
A B C
BC
ABC
Diversificação biológica
A sp.1
B sp. 2
C
sp. 3
Cenário geológico Cladograma geológ ico
Cladograma taxonômico
� - Correspondência entre as h istórias geológica e b iológica: cenário geológ ico com eventos de fragmentação da área, c ladograma geológico denotando a sequência de eventos de fragmentação da área e um cladograma de táxons endêmicos para a á rea.
Apesar da clara inspiração da biogeografia cladística na pan-biogeografia de Croizat e na sistemática filogenética de Hennig4,9, a leitura de um e de outro notabiliza a sua incompatibilidade, Croizat foi bastante enfático em rejeitar qualquer relação conceituaj5,1O e Hennig, por sua vez, assim como vários outros sistematas e biogeógrafos daquela (e desta) época, não fez questão alguma de referenciar Croizat4,
Mesmo com a comprovada relação de parentesco entre a biogeografia cladística e a pan-biogeografia, é bastante aparente a dicotomia metodológica existente entre ambas. Os traços, construídos na pan-biogeografia, representam as coordenadas primárias de um táxon no espaço, e é a análise destas coordenadas primárias que abre margem a questionamentos mais particulares sobre a interação entre forma, tempo e espaços, Entretanto, pelo lado da biogeografia cladística, critica-se, por exemplo, o significado e o conteúdo explanatório dos traços. Muitas das críticas direcionadas à pan-biogeografia devem-se ao fato de as relações filogenéticas dos táxons não serem levadas em conta e, ainda, pela falta de critérios precisos para determinar relacionamentos entre áreas 7. Mesmo que traços possam ser interpretados como cladogramas de áreas pouco informativos7, l l , apenas dados de distribuição geográfica seriam insuficientes para procurar padrões espaciais históricos e para reconhecer os fatores causais (por exemplo, vicariância, dispersão) dos padrões de disjunção distribucional12. De fato, Hennig foi o pioneiro no emprego direto de hipóteses filogenéticas para explicar o padrão de distribuição dos organismos, mas sua análise e interpretação eram limitadas com relação
aos processos causais implicados. Seu tipo de abordagem caracteriza o programa de pesquisa que, mais tarde, foi denominado de biogeografia filogenética (Quadro 6 . 1 ) .
A biogeografia cladística fundamenta-se principalmente na premissa de que existe uma correspondência entre o relacionamento filogenético dos táxons e seu padrão de distribuição e a história geológica da Terra (Fig. 6. 1 ) . Se a forma como os organismos estão distribuídos é entendida como o resultado da sua história ao longo do espaço e do tempo, então é possível encontrar uma associação entre a história dos organismos e a história do planeta. Nas palavras de Donn Rosen (em Nelson e Rosen 1S) seria uma "história natural integrada dos sistemas geológico e biológico". Em síntese, a biogeografia cladística tem como objetivo central a procura por padrões gerais de relacionamento entre áreas e, após o padrão geral ser descoberto, seus processos causais podem ser inferidos e explicados3,4,7 , 16 . A comparação entre cladogramas de áreas derivados de diferentes táxons que habitam uma mesma região espacial possibilita que padrões gerais sejam reconhecidos. Os padrões descobertos podem ser explicados primariamente por eventos vicariantes, uma vez que dispersões aleatórias não poderiam explicar padrões recorrentes, nem seriam falseáveis2,6. No entanto, à parte da disputa que possa existir em determinados setores da biogeografia sobre qual evento seria mais determinante na formação de padrões, tanto eventos de vicariância quanto de dispersão são interpretações subjetivas para os padrões de distribuição, e tais interpretações devem ser consideradas meramente hipóteses e não observações factuaisl7 .

B iogeografia C lad ística - 1 01
0:t,,;;:(;;CO� 5" - A Biogeografia F i logenética de Wi l l i Hennig
Apesar de pensar e desenvolver o seu "método parasitológico", no qual cladogramas de parasitas e de hospedeiros estariam fortemente associados entre si em relação a seus eventos cladogenéticos, o d i pterista alemão Wi l l i Hennig13 não foi capaz de pensar numa mesma associação entre organismos e espaço. Esta associação foi, sim, pensada de forma pioneira pelo botânico franco-ita l iano Léon Croizat, formal izada na célebre sentença "terra e vida evoluem juntas"5. Mesmo assim, Hennig13 avançou significativamente ao interpretar a história espacial dos organ ismos com base em sua fi logenia. De fato, foi o primeiro a praticar ta l abordagem7,9, a pesar de as i nterpretações derivadas terem sido fortemente ancoradas à noção de centro de origem e à diversificação progressiva do grupo a partir deste. Sua metodologia analítica e i nterpretativa ficou conhecida como Biogeografia F i logenética e, ao contrário do que poderia se supor, existe uma clara d istinção entre B iogeografia C ladística e B iogeografia F i logenética. E n q u a nto a p r i m e i ra assume pr imar iamente o processo de vicariância em sua interpretação dos padrões de d istri b u ição, a segunda é basicamente d ispersa l i sta . Hennig13 (ver também Brundin 14) postulou o que e l e próprio chamou de " regra de progressão" ou " l e i de para l e l ismo entre progressão morfológica e corológ ica " . Para um determi nado grupo (A (B (C (D, E)))), no q u a l se observa uma série de transformação (caráter) 0 - 1 - 2 - 3 - 4, com cada estado presente em um ún ico táxon na mesma sequência do cladograma, se for encontrada a l g u ma correspondência (d i recional e não sobreposta) entre a d istr ibuição dos estados e a d i str ibuição dos táxons A-E no espaço, então, pode-se afirmar que existe uma certa tendência e associação da h istória evolutiva deste caráter com a d istribuição do grupo no espaço. Sob esta i nterpretação, os táxons portadores de estados mais prim itivos seriam mais a ntigos e, por isso, esta riam m a i s próximos do centro de origem, enquanto os táxons mais derivados seriam mais recentes e estar iam na periferia, mais d i stantes deste centro. Portanto, a série de transformação ordenada i ndicaria, a lém da sequência de evolução do ca ráter, tam bém a sequência de evolução dos táxons no espaço. Seria isto u m parale l ismo entre progressão morfológica do táxon e a progressão do táxon no espaço. Uma segunda regra postulada por Hennig13 foi a regra de derivação ou divergência ("deviation rufe") . Nos eventos de especiação, a espécie ancestral dá origem a duas espécies descendentes, e sempre a espécie periférica a presenta mais atributos derivados em relação à espécie-irmã, que possui maior área de d istribu ição e é mais pr imitiva em seus atributos.
Algumas das questões implicadas pela biogeografia cladística sã03,4,12 :
l . Por que os táxons estão distribuídos onde estão hoje? 2. Como os organismos estão distribuídos? 3. Existem padrões de endemicidade? 4. É possível reconhecer padrões de relacionamentos
entre áreas? S . O padrão de relacionamento apresenta correlação com
a história geológica?
Embora sejam questões que preocupam os biogeógrafos em diferentes níveis, não necessariamente todas elas podem (ou devem) ser respondidas pelos métodos analíticos preconizados atualmente na biogeografia cladística.
Cladogramas de áreas (Fig. 6. 1 ) representam hipóteses de relacionamentos entre unidades de áreas com base em informações históricas e distribucionais dos táxons. As dicotomias presentes nos cladogramas de áreas possivelmente representam eventos de fragmentação, digamos, de uma área original A nas novas áreas B e C (Fig. 6. 1 ) .
O efeito destes eventos de fragmentação sobre a biota pode resultar em processos de vicariância, caso ocorra isolamento geográfico e diferenciação dos táxons.
Áreas de endemismo são as unidades espaciais básicas que estão sendo relacionadas num cladograma de
áreas. Posto isto, fica bastante claro que um estudo de identificação de áreas de endemismo consiste em etapa prévia obrigatória na biogeografia cladística. Esta etapa é considerada fundamental e, de modo análogo, é tão importante quanto a correta delimitação das espécies em uma análise cladística. As áreas de endemismo serão os ramos terminais do cladograma de áreas e, obviamente, qualquer equívoco em sua delimitação resultará em drásticas consequências nas etapas posteriores da análise. A questão das áreas de endemismo, sua importância, identificação e problematização são discutidas apropriadamente no Capítulo 3 .
N a biogeografia cladística assume-se a possibilidade de haver uma correspondência entre relacionamentos entre espécies e relacionamentos entre áreas (Fig. 6. 1 ) . Assim, comparações de cladogramas de vários grupos taxonômicos que habitam uma mesma região geográfica podem revelar a existência de padrões gerais contendo hipóteses sobre a história da biota e da área (Fig. 6.2).
A descoberta de um padrão comum de relacionamento entre áreas é evidência de que os táxons analisados possuem uma história comum7. 17 .
A seguinte sequência de etapas resume a execução de uma análise generalizada na biogeografia cladística (Figs. 6.2 e 6.3):
1 . Reconstrução de cladogramas para os táxons em estudo. 2 . Obtenção de cladogramas taxonômicos de áreas (TAC,
taxon-area cladogram), pela substituição do nome do táxon pela sua área de endemismo.

102 - Métodos e Apl icações
Amphibia A1 A2 A3 A4 AUS ORI AS AF
8ryophyta 81 82 83 84 AUS ORI AS AF
Crustacea C1 C2 C3 C4 AUS ORI AS AF
Cladogramas taxonômicos
Cladogramas taxonômicos
de áreas
AUS IND AS
Cladograma geral de áreas
AF
- Em busca de congruência: reconstrução e comparação de cladogramas taxonômicos de áreas de d iferentes táxons para reconhecimento de um cladograma geral de á reas.
Amphibia
\y�W �\)YF
Crustacea
(\��/5 �'wF :'\Sl Cladogramas taxonômicos
Cladogramas taxonômicos
de áreas
Cladogramas resolvidos de áreas
AUS IND AS AF
W Cladograma
geral
- Em busca de congruência: durante a comparação de cladogramas taxonômicos de áreas de d iferentes táxons, podem existir i nformações distribucionais ambíguas que necessitam de uma resolução. Neste caso, cladogramas taxonômicos de áreas são convertidos em cladogramas resolvidos de áreas, que então são comparados e se reconhece um cladograma geral de áreas.

B i ogeograf ia C l ad ística - 1 03
Diptera Táxon ampl i locado
Distribuições redundantes E1 E2 E3 E4 E5 AUS ORI AS ORI AF
Fabaceae
Áreas ausentes F1 F2 F3 AUS AS AF
V- V - Cenários prob lemáticos: táxons amp l i locados, d istr ibu ições redundantes e á reas ausentes.
3. Se necessário, conversão dos cladogramas taxonômicos de áreas em cladogramas resolvidos de áreas (RAC, resolved areas cladogram) (também denominados cladogramas fundamentais de áreas) .
4. Procura por congruência no padrão de relacionamento entre áreas e reconhecimento de um cladograma geral de áreas (ou areagrama).
Os métodos analíticos da biogeografia cladística requerem previamente um cladograma taxonômico (ou um conjunto de cladogramas) a partir do qual se constrói um cladograma de áreas. Este é o ponto de partida para as etapas subsequentes da análise. Para a obtenção de
� um cladograma de áreas, os nomes dos táxons simples-� '? mente são substituídos pelas suas respectivas áreas de � endemismo (Fig. 6 .2) . Este primeiro cladograma é tA '? 00 r�
denominado cladograma taxonômico de áreas ou cladograma de táxon-área (TAC) . Como se pode observar, a construção de um TAC em que todos seus táxons são endêmicos, ou seja, exclusivos de uma única área, é um procedimento bastante simples (Fig. 6.2).
Entretanto, ao se estudarem casos reais podemos nos deparar com algumas complicações (Fig. 6.3) que, na
biogeografia cladística, estão convencionados como sendo de três tipos (Fig. 6.4) :
1 . Táxons amplamente distribuídos ou amplilocados* (táxon que ocorre em mais de uma área de endemismo).
2. Distribuições redundantes (área onde dois ou mais táxons co-ocorrem e, assim, a área aparece repetidamente no cladograma) .
3. Áreas ausentes (áreas que estão ausentes em um determinado cladograma de um conjunto de TAC).
Estes cenários são considerados problemáticos, pois são fontes de ambiguidade e podem trazer menos resolução à análise. Para contornar estes problemas foram implementados três pressupostos (= assumptions): AO, AI e A2. Dessa forma, os TAC contendo estes casos problemáticos são manipulados e tratados com a aplicação dos pressupostos para a obtenção dos chamados
* O termo "táxon ampl i locado" foi cr iado, bastante oportunamente, por Márc io Bernard ino DaS i lva em sua tese de Doutorado (2008) em referência ao termo widespread taxon, cuja tradução d i reta ser ia um termo relativamente longo e cansativo (táxon amplamente d istribuído). O Capítu lo 1 4 emprega o termo pela primei r a vez e m uma pub l i cação.

1 04 - Métodos e Ap l i cações
cladogramas resolvidos de áreas (RAC). O "ruído biogeográfico" resultante desta resolução artificial é relativamente pequeno e não seria suficiente para mascarar o padrão de relacionamento entre áreas4,6.
Com a finalidade de se entender cada um desses pressupostos, considere-se o seguinte cladograma de áreas não resolvido contendo um táxon amplilocado, conforme mostrado na Figura 6.5. Sob o pressuposto A07,1 8-22, a espécie amplilocada é considerada sinapomórfica para as áreas A e B que compõem, assim, um grupo monofilético, o clado A + B, em um único cladograma resolvido. Aqui, assume-se que os relacionamentos entre áreas são resultantes exclusivamente de eventos de vicariância23 e que, neste caso, não teria afetado a espécie amplilocada em A e B . Sob o pressuposto AI4,7, 18, 19,23, as áreas A e B compõem um grupo monofilético (espécie amplilocada é sinapomórfica) ou, como alternativa, parafilético (espécie amplilocada é homoplástica), em que as áreas A ou B estariam mais proximamente relacionadas ao clado C + D. As resoluções parafiléticas baseiam-se na hipótese da espécie amplilocada não ter respondido a suposto evento vicariante que teria frag-
Táxon ampl i locado
AO
A-\)!
A1
mentado A de BCD (ou B de ACD), ou ainda, que algumas espécies teriam se extinguido e, ausentes na análise, não indicariam de forma direta um relacionamento de A ou B com o clado C + D. Sob o pressuposto A24,7, 18 , 19,23, as áreas A e B são consideradas monofiléticas, parafiléticas ou polifiléticas (espécie amplilocada é homoplástica). Dessa forma, as resoluções indicariam um clado A + B, ou as áreas A ou B estariam mais proximamente relacionadas ao clado C + D, ou ainda que as áreas A ou B estariam relacionadas às áreas C ou D, desfazendo-se então da informação do clado C + D. As resoluções polifiléticas consideram a hipótese da espécie amplilocada ter assumido a distribuição atual por dispersão, :s
cc tenha a espécie se originado em A e dispersado poste-riormente para B, ou o inverso.
No caso anterior, a utilização dos pressupostos AO, AI e A2 resultaria em um, três e sete cladogramas resolvidos, respectivamente (Fig. 6 .5) . Se em um extremo o pressuposto AO restringe a resolução do problema para um relacionamento monofilético das áreas A e B, no outro extremo, o pressuposto A2 permite todas as resoluções possíveis, inclusive possibilitando o descarte dos componentes originalmente informativos (clado C + D). Em geral, os pressupostos possuem uma relação de inclusão, na qual as resoluções de AO estariam incluídas
B D A C
W A2
00 li> .:., � , CIO \C
o-&
- Táxons ampl i locados: resolução pela implementação de pressupostos AO, A 1 e A2.

B iogeografia C ladística - 1 05
A B C D A
A
Distribuições red u nda ntes
B C D
AO
A A B C D
Al
A B C D
A VI B C D A
VI A2
'0,6 - Distr ibuições redundantes: resolução pela imp lementação de pressupostos AO, A 1 e A2.
em AI que estariam incluídas em A223. A aplicação dos
pressupostos para um cenário simplificado com três
táxons, quatro áreas e um táxon amplilocado resultou
entre um a sete cladogramas resolvidos. Logicamente,
o número de cladogramas resolvidos será maior de
acordo com o número de táxons, de áreas e de casos
problemáticos4.
Muitos autores preferem a utilização do pressuposto
A2 para o tratamento de táxons amplilocados e os mo
tivos explicitados geralmente levam em conta que este
pressuposto relativiza a existência de equívocos não
conhecidos e/ou não atestados:
1 . Trabalhos adicionais podem revelar que a espécie
amplilocada é, na realidade, constituída de várias
espécies distintas e endêmicas de uma única área4 ("taxonomia imatura"18) .
2. Uma espécie tornou-se amplilocada por simples au
mento de sua área de distribuiçã04.
3 . Uma espécie pode ser amplilocada porque não res
pondeu ao evento vicariante3.4.24. 4. Uma espécie pode ser amplilocada porque não teve
tempo suficiente para sofrer especiaçãol8 .
5 . Além de erros taxonômicos, as áreas podem ter sido
mal delimitadas2s .
No caso de distribuições redundantes (ou "área de simpatria"18 ou "paralogia geográfica"26), a simpatria não interfere na conversão do TAC para RAC4.18 quando os táxons que ocupam uma mesma área formam um grupo monofilético. Entretanto, embora este seja um padrão quase sempre encontrado, situações em que estes táxons simpátricos não formam um grupo monofilético também são bastante frequentes (Fig. 6.6). Sob AI , as ocorrências nas áreas redundantes são consideradas igualmente válidas e considera-se que são resultado de evento de duplicação com subsequentes extinções nas demais áreas de um dos ramos duplicadosI 8.23•27. Sob A2, as ocorrências são consideradas individualmente, com um RAC para cada uma delas 18.27. Neste, a interpretação é que as distribuições redundantes resultam de eventos de duplicação e dispersão. O pressuposto AO, assim como o A I , considera as ocorrências como igualmente válidas, apesar de não considerar nenhuma interpretação específica para o casol8. As possíveis causas para a origem de redundâncias ou paralogias geográficas são muito variadas: ação tectônica, dispersão, especiação simpátrica, relacionamentos equívocos entre espécies, má delimitação das áreas de endemismo etc.26.
Para a resolução de áreas ausentes (Fig. 6 .7) , os pressupostos A 1 e A2 consideram tais áreas como não inforrnativasl9. Neste caso, a área ausente é codificada

1 06 - Métodos e Aplicações
como "?" e assim os cladogramas resolvidos assumirão todas as posições possíveis para esta área. Esta implementação permite recuperar história de áreas nas quais aconteceram eventos de extinção. Outra alternativa é utilizar o pressuposto AO, que considera a área ausente como primitivamente ausente, sendo codificada como
zero19. Isto permitiria que uma análise com vários táxons pudesse indicar as reversões como ausências secundárias ou extinçã028.29. Qualquer que seja o pressuposto utilizado, nenhum assegura logicamente a informação fornecida pela área ausente, isto é, de que, de fato, não há informação (nem de ausência nem de presença). Conforme já notado por Page27, se uma área está ausente, não há nenhuma informação sobre o relacionamento desta área. Qualquer uma das codificações usualmente empregadas ("O" ou "?") consiste numa implementação inadequada.
Nas decisões sobre qual pressuposto aplicar, as escolhas não são mutuamente excludentes, é possível (e até aconselhado por alguns autores) implementar os pressupostos de forma combinada - por exemplo, tratar os táxons amplilocados sob AO, enquanto as distribuições
redundantes sob A218. O uso do pressuposto AO é o mais amplamente criticado e controverso. Enquanto o pres-
A C D Áreas
V B Ausente
ausentes
A C
AO
D B A C D
suposto )\.: rêlatiyíza a existência de possíveis equívocos em alguma êtap a prévia (taxonomia, filogenia, distribuição. áre as de endemismo) , AO é completamente inflexÍ\ el no trato das informações distribucionais. Sob AO. assume-se que a hipótese filo genética reconstruída é a melhor estimatin possível da verdadeira filogenia22. Também. se uma espécie amplilocada for confirmada como sendo formada por diferentes espécies endêmicas, sob AO assume-se que estas são espécies irmãs.
Alguns dos estudos sobre a implementação dos pressupostos AO, A I e A2 com descrições detalhadas, testes comparativos e/ou discussões na resolução de casos particulares foram publicados por Ebach et ai. 30, Enghoff18, Humphries e Parenti7, Morrone23, Nelson e Ladiges31 , Nelson e Platnick4, Page24.27, Van Veller et
ai. 20, Wiley21 .32, Zandee e Ross22.
De fato, o primeiro método analítico proposto para se lidar com padrões comuns na biogeografia cladística foi
A B C
A1/A2
D A C B D
- Áreas a usentes: resolução pela imp lementação de pressupostos AO, A 1 e A2.

o protocolo de redução de cladograma de áreas de Rosen33. Neste protocolo, aplicado por Rosen a dois gêneros de peixes dulcícolas da Mesoamérica, os cladogramas dos táxons em estudo são convertidos em cladogramas taxonômicos de áreas, estes são comparados entre si e todas as informações ambíguas (isto é, relações incongruentes contidas nos cladogramas) são simplesmente excluídas para, enfim, obter-se um cladograma geral de áreas. Áreas com distribuições redundantes e áreas ausentes seriam alguns exemplos de informação descartada. Esse método não tem sido utilizado já há bastante tempo. Críticas apontam como falhas a análise não permitir qualquer explicação sobre as informações ambíguas (que são simplesmente eliminadas), além do fato de que o cladograma reduzido de áreas representa apenas uma explicação parcial para a história da biota na área32.
Como exposto anteriormente, o TAC representa o ponto de partida para todos os métodos analíticos da biogeografia cladística. Entretanto, como será visto adiante, as etapas subsequentes da análise variam significativamente de um método para outro, por exemplo, na conversão de cladograma taxonômico de áreas em cladograma resolvido, a aplicação dos pressupostos AO, A I e A2, a forma como dados de distribuição e relacionamentos de espécies são implementados e "lidos", entre outros .
Uma recente proposta de categorização dos métodos em biogeografia cladística34 divide-os em "métodos baseados em padrão" e "métodos baseados em eventos" (Quadro 6.2). Métodos baseados em padrão procuram
B iogeografia C lad ística - 107
reconhecer padrões gerais de relacionamentos entre áreas e, somente então, tentam inferir os processos (basicamente, vicariância, dispersão e extinção) que teriam afetado de forma comum a história da biota destas áreas. Por outro lado, métodos baseados em eventos assumem modelos explícitos para os processos que teriam afetado a história de um ou mais táxons. Tai s modelos compreendem34 a filogenia do táxon associada aos seus dados de distribuição geográfica para a obtenção de um c1adograma geral de áreas e à derivação final de um modelo biogeográfico especificando os tipos de eventos que teriam produzido as distribuições, assim como os custos requeridos para cada evento (ver Capo 7).
Este capítulo aborda apenas os métodos baseados em padrão, em particular: (1) análise de componentes; (2) análise de parcimônia de Brooks; (3) análise de enunciados de 3-áreas; e (4) análise de subárvores livres de paralogias. O Quadro 6.3 apresenta uma sinopse dos programas computacionais implementados para estes métodos analíticos.
Este método analítico foi desenvolvido por Gareth Nelson e Norman Platnick em 19783,4. Nele, c1adogramas taxonômicos de áreas são convertidos em cladogramas resolvidos de áreas após tratamento (se necessário) com os pressupostos AO, A l ou A2. No passo seguinte, procura-se por congruência no padrão de relacionamento entre áreas, percorrendo-se os c1adogramas de áreas em busca do maior número de componentes em comum. Um
Ç:CJ;,2d�'�� 5):: - Biogeografia vicar iante, biogeografia c ladística etc.
Em 1 978, Norman Platn ick e Ga reth Nelson3 propuseram um novo método de b iogeografia h istórica. Este novo método abr iu caminho para o surgimento de uma nova escola biogeográfica, passando a ser denominada biogeografia vicariante (por exemplo, N elson e Platnick4, Nelson e Rosen 1 5, W i l ey32, entre outros). Mas esta denomi nação era um tanto descabida37, uma vez que a Síntese (Pa n)biogeográfica de Croizat foi a pioneira tanto na idea l ização do processo de vicariância quanto na exe m p l ificação e va loração da vica riância como fenômeno gerador dos padrões de d istribu ição. Como era de se esperar, o recon hecimento do e q u ívoco veio pouco tempo depois. A denom i n ação biogeografia cladística foi empregada "del iberadamente" no l ivro Cladistic Biogeography de Christopher H umph ries e Lynne Parenti publ icado em 1 9867,14. Apesar do claro entendimento quanto à fundamentação da b iogeografia cladística, existem debates na l iteratura sobre o escopo metodológ ico da b iogeog rafia c l a d ística. Além da controvérsi a de quais e q u a ntos métodos a n a l íticos compreender iam a b iogeog rafia c ladística, tam bém há controvérsias de quais e q u a ntas a bordagens ou progra mas de pesq u isa existi r iam na
:6 Biogeografia H istórica. Jorge CrisciB (ver também Crisci et aJ.9) reconheceu nove a bordagens gerais: centros de origem e dispersão,
:3? pan-biogeografia, biogeografia f i logenética, biogeografia c ladística, f i logeografia, a n á l ise de parci mônia de endem icidade, ,..., métodos baseados em eventos, áreas ancestrais e b iogeografia experimenta l . Em uma classificação mais lumper, procurando � t- agrupar os métodos de acordo com suas s imi laridades e complementaridades, Juan Morrone36 reconheceu somente duas abordagens:
� Dispersal ismo e B iogeografia Vicariante (Tabela 6 . 1 ) . Nesta classificação, coerentemente, a Biogeografia Vicar iante abrange a � Pan-biogeografia e a B iogeografia C ladística. Esta última i nc l u i, lado a lado, métodos considerados opostos por a lguns a utores. 0'\ Tam bém, têm-se proposto a lgumas tentativas de c lassificação no i ntu ito de categorizar e d iscrim i n a r os i n ú m eros métodos de
biogeografia cladística de acordo com a lguns critérios operacionais, tipos de a bordagem, modo de m a n ip u lação dos dados or ig ina is, enfoque em modelos predefin idos, entre outros. Por exemplo, temos as seguintes pola rizações: métodos a posteriori versus métodos a priori20,35, métodos fundamentados em padrão versus métodos baseados em eventos20, b iogeografia de táxons versus biogeografia de áreasB,9 etc.

1 08 - Métodos e Apl icações
• COM PONENT versão 1 . 5: Autoria: Roderic page38 Apl icação: a n á l ise de com ponentes
• COMPON ENT versão 2.0: Autoria: Roderic page39 Apl icação: a n á l ise de reconcil iação de árvores, a n á l ise de componentes
• TAS: Autoria: Gareth Nelson e Pau l i ne Ladiges40 Apl icação: anál ise de enunciados de 3-áreas
• TASS: Autoria: Gareth Nelson e Pau l ine Ladiges41 Apl icação: a n á l ise de subárvores l ivres de paralogias
• N E LSON05: Autoria: Jacques Ducasse, Nathanaél Cao e René Zaragüeta-Bagi ls42 Apl icação: a n á l ise de enu nciados de 3-á reas, Anál ise de subárvores l ivres de paralogias
• NONA, PAU P * e TNT: Autoria: Pablo Goloboff43 (NONA); David Swofford44 (PAUP*); Pablo Goloboff, Steve Farris e Kevin Nixon4s (TNT) Apl icação: anál ise de parcimônia de Brooks
componente é basicamente um conjunto de áreas relacionadas (conectadas a um nó) ou um grupo monofilético de áreas. Alguns autores equiparam o componente ao traço pan-biogeográfico 1 J ,32. Sendo assim, um componente formado por América do Sul + África + Austrália seria equivalente a um traço conectando três táxons endêmicos de América do Sul, África e Austrália, respectivamente. Existe uma noção de hierarquia subjacente aos componentes, no sentido de que um componente inclui componentes menores, conforme indicado por relacionamentos internos entre suas áreas. Por exemplo, temos um componente formado por AS + AF + AUS, que inclui um outro formado por AF + AS.
Procedimentos operacionais4,7,9,23 (Fig. 6.8) :
1 . Reconstruçã% btenção dos cladogramas dos táxons em estudo.
2. Obtenção de TAC pela substituição do nome do táxon pela sua área de endemismo.
3. Se necessário, conversão de cladogramas taxonômicos de áreas em RAC pela implementação dos pressupostos AO, A 1 e A2 para resolução de casos problemáticos (táxons amplilocados, áreas ausentes e distribuições redundantes ) .
4. Intersecção dos conjuntos de cladogramas resolvidos de áreas de cada grupo taxonômico para encontrar um cladograma comum a todos os conjuntos, este é denominado cladograma geral de áreas.
5 . Se mais de um cladograma geral for encontrado, construção de uma árvore de consenso para sumarizar os relacionamentos consistentes. Caso nenhum cladograma geral de áreas seja encontrado na intersecção, deve-se verificar se há algum cladograma comum a alguns conjuntos.
Como alternativa, quando dois ou mais cladogramas gerais de áreas são encontrados, em lugar do consenso, os cladogramas gerais obtidos podem ser considerados igualmente válidos, uma vez que o padrão de relacionamento entre áreas não necessariamente deve ser um único para explicar a história da área ou do táxon9. Além disso, se nenhum cladograma geral de áreas for encontrado (ou sej a, se nenhum dos cladogramas resolvidos de áreas for comum a todos os conjuntos), pode-se construir uma árvore de consenso com base nos cladogramas resolvidos de áreas9, a fim de preservar alguma informação de relacionamento.
Uma das críticas feitas à análise de componentes é exatamente o emprego de consenso21,22,32. Para Wiley32, esta é sua maior fraqueza. Árvores de consenso são informativas e representativas quando os cladogramas fundamentais apresentam informações logicamente consistentes. Mas, de forma geral, cladogramas fundamentais são inconsistentes em seus relacionamentos e o resultado são árvores de consenso pouco resolvidas (com politomias). Uma modificação sugerida à análise de componentes seria não aplicar técnicas de consenso, mas implementar uma análise de parcimônia dos componentes21, algo ligeiramente parecido com o que pode ser praticado na análise de subárvores livres de paralogias. Por outro lado, alguns autores 18,24 distinguem a análise de componentes como sendo composta por dois métodos distintos: ( 1 ) a resolução de um TAC em um RAC ou em vários RAC igualmente parcimoniosos e (2) o exame comparativo de RAC derivados de diferentes TAC para obter um cladograma geral de áreas.
Este método analítico foi desenvolvido por Daniel Brooks em 1981 28 para estudar a associação histórica

Gastropoda
G\%�� \ AUS OR! AS AF i
- \0/ .
. . . / /©
.. ... .. C
B iogeografia C ladística - 1 09
Oip lopoda . , 0\;i1 02 03 AUS-\XOR! AS AF AU\jS OR! AS AF AUwS OR! AS AF/ ORw! AUS AS AF ORw! AS AUS AF
" : � � ..
. i
® ® ............... ..
.. ... . i i
OR! AF AUS AS AUS AS OR! AF AUS AF OR! AS
W W W© Palmae
P 1 P2 P3 P4 P5 P6 AUS OR! AS OR! AF AF ÂÚS OR! AS AF AUS AS OR! AF :AU�
\X-\X- w/ w© Cladograma geral de
áreas
- Anál ise de componentes: procedimentos operaciona is. A = C1adograma taxonômico. B = Cladograma taxonômico de áreas. C = Cladogramas resolvidos de áreas que são i nterseccionados para se obter o D = c1adograma geral de áreas.
parasita-hospedeiro, com o propósito de solucionar um suposto problema do "método parasitológico" de Hennig13 . De acordo com Brooks28, por não ser um parasitologista, Hennig olhava a questão apenas sob o ponto de vista do hospedeiro. Para Hennig1 3 existiria um forte paralelo entre a história do parasita e a de seu hospedeiro, estando a evolução e diversificação do parasita fortemente associada (e, de certa forma, dependente) à de seu hospedeiro. Sendo assim, seria possível explicar eventos de especiação ou extinção ao se estudar parasitas quando conjugados à filogenia dos hospedeiros. Alternativamente, Brooks28 propõe um método baseado em análise de parcimônia com o objetivo de reconstruir a filogenia do hospedeiro a partir das relações filogenéticas conhecidas dos parasitas. Neste procedimento, uma matriz de dados é construída com hospedeiros como táxons e a presença/ausência de parasitas como caracteres. Contudo, em casos com parasitas altamente específicos (um parasita para um hospedeiro), a total ausência de compartilhamento de caracteres (parasitas) conduz à obtenção de um cladograma sem qualquer
resolução (Fig. 6.9, A a D). Como solução, Brooks28 considerou a inclusão dos nós ancestrais dos parasitas (Fig. 6.9, E a H) . Posteriormente, Wiley21,32 reconsiderou o método proposto por Brooks, mas com vistas para sua aplicação em biogeografia histórica. Wiley batizou-o como análise de parcimônia de Brooks (BPA, de Brooks parsimony analysis) e disseminou seu uso para o estudo da associação táxon-área.
Brooks46 distinguiu o método em duas etapas : BPA primária e BPA secundária. A BPA primária consiste no método originalmente proposto por Brooks28, mas caracterizado e formulado para biogeografia principalmente por Wiley21 ,32. A BPA secundária incorporou algumas modificações na análise com a implementação de medidas adicionais21 ,32,46, tendo sido caracterizada primeiramente por Brooks46. Em razão das supostas confusões na literatura, passados 1 1 anos, Brooks et aI. 47 tentaram novamente, de forma mais clara, descrever as duas etapas da BPA.
Procedimentos operacionais da B PA primá
ria9,21 ,23,28,32,46 (Fig. 6. 10):

1 1 0 - Métodos e Apl icações
2 3 4 A B C D
®
A B C D 1 2 3 4
A 1 O O O B O 1 O O C O O 1 O D O O O 1
©
2 3 4 A B C D
A B C D 1 2 3 4 5 6 7 4
A 1 O O O O O 1 B O 1 O O O 1 1 C O O 1 O 1 1 1 D O O O 1 1 1 1
® ®
- Aná l ise de parcimônia de Brooks (BPA). (A a O) Quando os parasitas são a ltamente específicos ao seu hospedeiro, não há comparti l hamento de caracteres (= parasitas). (E a H) Como solução, B rooks28 inc lu iu informação dos nós a ncestra is. Mod ificado de Brooks28.
1 . Reconstruçã% btenção dos cladogramas dos táxons em estudo.
2. Obtenção de TAC pela substituição do nome do táxon pelo nome de sua área de endemismo.
3. Confecção de uma matriz binária de áreas X táxons (inclusive nós ancestrais), com ausência (O) e presença ( 1 ), com uma área hipotética tudo-zero para enraizamento dos cladogramas.
4. Aplicação de análise de parcimônia e obtenção de cladograma de áreas. O cladograma de áreas resultante de análise de um único táxon representa um cladograma individual de áreas, enquanto um cladograma geral de áreas resulta da análise de vários táxons.
Na BPA primária e secundária, os cladogramas taxonômicos de áreas não precisam ser convertidos em cladogramas resolvidos, uma vez que a análise implementa unicamente a aplicação do pressuposto AO para resolver os casos de táxons amplilocados. As áreas ausentes são consideradas não informativas e codificadas simplesmente como "7". Para distribuições redundantes, a BPA primária não aplica nenhum tratamento específico (as informações são consideradas tais quais), enquanto a BPA secundária pode implementar alguma manipulação a posteriori (após obter a obtenção do cladograma geral de áreas da BPA primária).
Segundo Brooks et aI. 47 , a função da BPA primária é verificar se há um padrão geral de relacionamento entre
-o --J 00 00 U1 c.., N � � , 00 -o o-'"
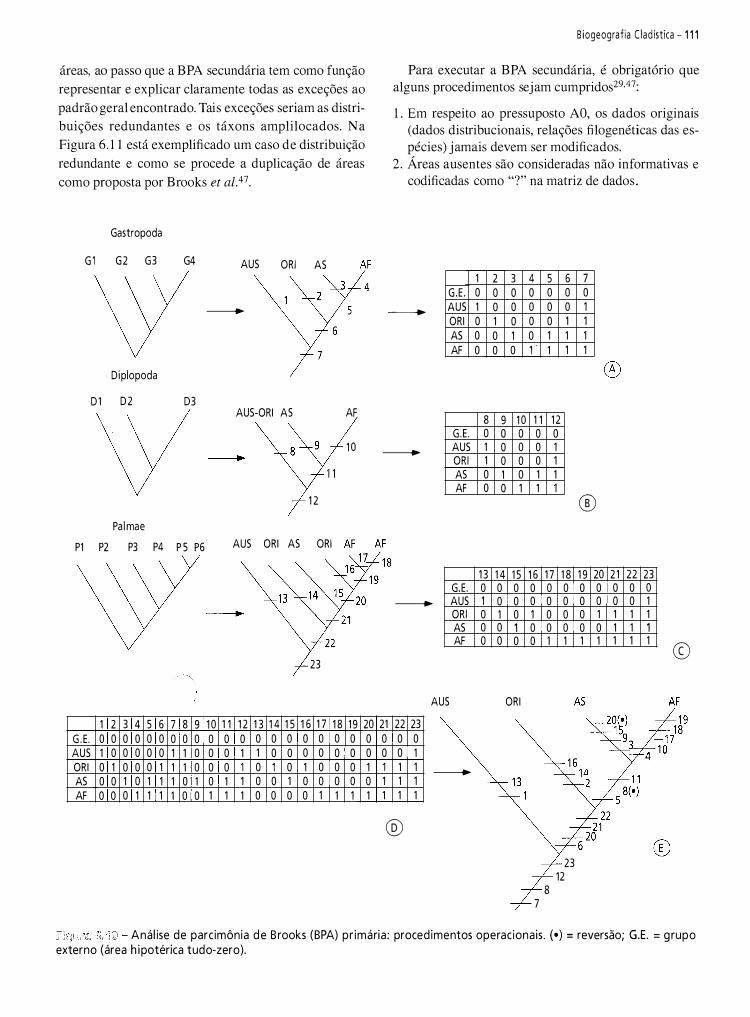
áreas, ao passo que a BPA secundária tem como função
representar e explicar claramente todas as exceções ao
padrão geral encontrado. Tais exceções seriam as distri
buições redundantes e os táxons amplilocados. Na
Figura 6. 1 1 está exemplificado um caso de distribuição
redundante e como se procede a duplicação de áreas
como proposta por Brooks et al.47.
Pl
Gastropoda
G l G2 G3
Diplopoda
D l D2
Pa lmae P2 P3 P4
1 2 3 4 5 6 G . E. O O O O O O
G4 AUS ORI AS
-----.
D3 AUS-ORI AS
1 2
P 5 P6 AUS ORI AS ORI
23
7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 O O O O O O O O O O O O
AF
1 0
1 9 20 O O
B iogeografia C ladística - 1 1 1
Para executar a BPA secundária, é obrigatório que alguns procedimentos sejam cumpridos29,47:
1 . Em respeito ao pressuposto AO, os dados originais (dados distribucionais, relações filo genéticas das espécies) jamais devem ser modificados.
2. Áreas ausentes são consideradas não informativas e codificadas como "7" na matriz de dados.
1 2 3 4 5 6 7 G .E . O O O O O O O
-----. AUS 1 O O O O O 1 ORI O 1 O O O 1 1 AS O O 1 O 1 1 1 AF O O O , . 1 1 1
8 9 1 0 1 1 1 2 G .E . O O O O O
-----. AUS 1 O O O 1 ORI 1 O O O 1 AS O 1 O 1 1 AF O O 1 1 1
®
1 3 1 4 1 5 1 6 1 7 1 8 1 9 20 2 1 22 23 G.E. O O O O O O O O O O O
-----. AUS 1 O O O O O O O O O 1 ORI O 1 O 1 O O O 1 1 1 1 AS O O 1 O O O O O 1 1 1 AF O O O O 1 1 1 1 1 1 1 ©
AUS ORI
21 22 23 O O O
AUS 1 O O O O O 1 1 O O O 1 1 O O O O O O O O O 1 ORI O 1 O O O 1 1 1 O O O 1 O 1
AS O O 1 O 1 1 1 O 1 O 1 1 O O
AF O O O 1 1 1 1 O O 1 1 1 O O
O 1 O O O
1 O O O O
O O 1 1 1
1 1 1 O 1 1 1 1 1
®
1 -----. 1 1
8 7
23 12
- Anál ise de parcimônia de Brooks (BPA) pr imária: procedimentos operacionais . (e) = reversão; G.E. = grupo externo (área h ipotérica tudo-zero).

1 1 2 - Métodos e Apl icações
5 1
1 G.E. O AU5 1 ORI O A5 O AF O
5quamata
52 S3
2 3 4 O O O
O O O 1 O O
O 1 O
O O 1
5 O O
1 O
O
S4 55
® 6 7 8 9 O O O O
O O O O 1 1 1 1 O O O 1 O O 1 1
56 AU5
1 0 1 1 AU5 O O O 1 1 1 1 1 1 1
©
AU5 ORI 1
ORI A5
1 1
A5
A5 AF
1 1
AF ORI
ORI
l ORI2
ORI 6
® AF
4
®
ORI3
6
� '.i - Anál ise de parcimônia de Brooks (BPA) secundária : exemplo de dup l icação de áreas para caso de d istr ibu ições redundantes. G .E . = grupo externo (área hipotética tudo-zero). Modificado de Brooks et a/.47.
3. Após obter o cladograma geral de áreas pela BPA primária é permitida a duplicação de áreas para se resolverem eventuais ambiguidades que possam falsear a análise (isto é, áreas com histórias reticuladas, que são evidenciadas na análise por distribuições redundantes e espécies amplilocadas) (Fig. 6. 1 1 ) .
4. Três ou mais táxons devem ser analisados para que o cladograma geral de áreas encontrado baseie-se presumivelmente em padrões gerais e não em casos particulares e ruidosos.
Os procedimentos 3 e 4, de fato, representam novas implementações à análise; o procedimento 2 já era normalmente implementado durante a execução da BPA
primária21 ; e a medida 1 impõe, agora enfaticamente, a implementação do pressuposto AO. Existe uma notável contradição entre os procedimentos, pois enquanto o procedimento 1 respeita a integridade dos dados originais (em obediência ao pressuposto AO), o procedimento 3
permite que os dados originais sejam modificados conforme surjam necessidades (para explicar as "exceções" ao padrão geral) . De forma paradoxal, a duplicação é um procedimento que satisfaz AO, de acordo com Brooks29. É exatamente o respeito ao pressuposto AO e a não modificação dos dados originais a maior qualidade da BPA apontada pelos seus usuários e defensores20,35,46,47. É também este o critério utilizado20,35 para categorizar os métodos de biogeografia cladística em
'D -l 00 00 u. � N +--
, 00 'D O> o,

dois grupos, os métodos a priori (permitem manipular TAC para resolver os casos problemáticos previamente à análise - análise de componentes, análise de subárvores livres de paralogia, análise de enunciados de 3-áreas) e métodos a posteriori (não manipulam as TAC antes da análise - BPA primária, embora permitam manipular em momento posterior - BPA secundária) . Por outro lado, na visão de Ebach e Humphries 1 7 , existiriam dois paradigmas metodológicos: "paradigma de geração" e "paradigma de descoberta". Métodos a priori estariam sob o paradigma de geração, pois, com base em premissas dogmáticas de evolução e biogeografia, conseguem reconhecer congruência em dados ambíguos. Estes métodos não descobrem e, sim, geram congruência.
A duplicação de áreas visa recuperar a história de áreas supostamente reticuladas, pois a priori a BPA primária é proibitiva à ocorrência de áreas com histórias reticuladas47. Assim, a BPA secundária permitiria que relacionamentos espúrios pudessem ser evitados. Em teoria, esta medida representa um aperfeiçoamento efetivo para a BPA e, até para a biogeografia cladística, em termos de melhor entendimento da história da área, uma vez que é amplamente sabido que as áreas possuem, de fato, uma história reticulada (por exemplo, Platnick e Nelson3, Brooks29). Porém, se em teoria a duplicação de áreas for uma implementação justificável, na prática, a realidade será muito mais complexa, e tal implementação pode resultar em uma série de decisões arbitrárias (por exemplo, quais áreas duplicar?). Os exemplos utilizados por Brooks et al.47 para demonstrar como implementar a BPA secundária e executar a duplicação de áreas são simplificados (e didáticos), pois, em casos reais, o cladograma de áreas resultante da análise de três ou mais táxons usualmente não envolve somente um ou dois casos problemáticos (redundâncias e distribuição ampla). Embora o princípio da parcimônia seja clamado durante as decisões de duplicação de áreas, o que se verifica em alguns dos exemplos de Brooks46 são decisões aplicadas "caprichosamente" e alienadas à parcimônia48.
Originalmente proposta para estudos sistemáticos, a análise de enunciados de 3-itens consiste em um método para inferência de relacionamento de parentesco entre táxons, tendo como base a reinterpretação das observações (caracteres) em unidades mínimas de relacionamento, por exemplo, enunciados compostos por três táxons49•5o. Para três táxons A, B e C, sempre dois deles estão mais proximamente relacionados entre si que com o terceiro, assim, por exemplo: (A (BC)). Caracte-
B iogeograf ia C lad ística - 1 1 3
res binários incorporam elementos de "identidade" (estado 1) e de "diferença" (estado O). Se B e C apresentam o atributo observado (estado 1) e A não (estado O), logo, temos o enunciado (A(BC)). Para quatro táxons A-D, se B , C e D apresentam o estado 1 e A, o estado 0, este caráter binário expressa o relacionamento (A(BCD» , que produz três enunciados de 3-itens: (A(BC» , (A(BD» e (A(CD»). Para cinco táxons A-E, se um caráter binário expressa (AB(CDE» , logo, temos seis enunciados de 3-itens: (A(CD)), (A(CE»), (A(DE) , (B(CD»), (B(CE» e (B(DE)). O número total de enunciados possíveis depende do número de táxons com o estado informativo (n) e do número total de táxons (t), conforme a equação: s = (t-n).n. (n- l )/2. Em seguida, uma nova matriz de dados é construída somente com enunciados de 3-itens, onde cada enunciado é considerado um novo caráter. O enunciado (A(CD)) é codificado 071 1 ? para A-E, respectivamente; o enunciado (B(CE)) é codificado?Ol 7 1 , e assim por diante. A matriz é submetida à análise de parcimônia, e o resultado são cladogramas mais "parcimoniosos" (mais ajustados) que acomodam a maior quantidade de enunciados.
Concomitantemente, este método foi também empregado para análise biogeográfica por Gareth Nelson e Pauline Ladiges em 1 991 3 1 , apesar de despretensiosamente como uma simples apresentação de novas implementações para os pressupostos AO, AI e A2 com base em enunciados de 3-itens. A implementação para estudo de relacionamento entre áreas é semelhante àquela descrita anteriormente para estudos sistemáticos, com a diferença de que as observações distribucionais primárias são representadas somente por caracteres binários, a partir dos quais são elaborados os enunciados de 3-áreas. Outra diferença é que, em consequência da presença eventual de casos problemáticos (táxons amplilocados e distribuições redundantes), os enunciados são definidos como informativos e não informativos. Por exemplo, para o cladograma (A(B(C,D))) temos o TAC (AS(AF(AUS-AF,ORI), que produz sete enunciados de 3-áreas: (AS(AF,AUS)), (AS(AF,AF)), (AS(AF,ORI)), (AS (AUS,ORI)) , (AS (AF,ORI)), (AF(AUS,ORI)) e (AF(AF, ORI) ) . Destes , cinco são i nformativos : (AS (AF,AUS) ) , (AS (AF,ORI») , (AS (AU S ,ORI)) , (AS(AF,ORI») e (AF(AUS,ORI)) . Somente o s enunciados informativos são codificados como caracteres na matriz de dados.
Métodos como a análise de componentes ou a BPA obtêm informações primárias de um cladograma taxonômico de áreas pela derivação de componentes (grupo de áreas conectadas a um nó). Por exemplo: para o cladograma « A(B ,C))(D,E)) temos os componentes

1 1 4 - Métodos e Apl icações
ABCDE, ABC, BC e DE. Embora estes componentes relacionem áreas sob a forma de agrupamento, não há qualquer informação de relacionamento interno. Assim, um dado componente ABC, de forma independente, não fornece qualquer informação de relacionamento entre as áreas A, B e C. Por outro lado, de forma dependente, o componente ABC denota relacionamento entre suas áreas quando combinado ao componente BC; o componente ABCDE denota relacionamento se combinado com os componentes ABC e DE. Como alternativa a esta noção de componentes como agrupamentos, componentes podem ser concebidos como relacionamentos se forem minimamente traduzidos em enunciados de 3-áreas31 . Dessa forma, em vez de ABC temos A(BC), com inferência de agrupamento e de relacionamento; portanto, um componente na análise de 3-áreas é independente e contém mais informação.
A implementação dos pressupostos AO e A I é considerada superior quando aplicada para enunciados de 3-áreas3 J , visto que, em um enunciado qualquer, A(BC), seu nó informativo (BC) corresponde ao nó informativo do cladograma taxonômico. Desse modo, a informação distribucional de táxons amplilocados é dissolúvel e nula (não informativa), ao passo que a informação dos nós é efetivamente incorporada. Para melhor compreensão, compare as resoluções apresentadas aqui com aquelas apresentadas anteriormente nas implementações dos pressupostos AO, AI e A2, sob as quais a informação do táxon amplilocado é indissolúvel e preservada sempre (AO) ou eventualmente (AI e A2) . Por maximizar a incorporação de informações biogeograficamente relevantes, Nelson e Ladiges3 1 advogam a superioridade da análise de enunciados de 3-áreas.
Procedimentos operacionais9,23,3 1 (Fig. 6. 1 2) :
1 . Reconstruçã% btenção dos cladogramas dos táxons em estudo.
2. Obtenção de TAC pela substituição do nome do táxon pela sua área de endemismo.
3. Confecção de uma matriz binária de áreas X componentes, na qual todas as informações distribucionais contidas nos TAC são codificadas sob a forma de enunciados de 3-áreas, com ausência (O), presença ( I ) e dados inaplicáveis (7), e adiciona-se uma área hipotética apenas com zeros para enraizamento dos cladogramas.
4. Aplicação de análise de parcimônia ou análise de compatibilidade para obtenção de cladograma geral de áreas.
As críticas à análise de enunciados de 3-áreas são reflexo daquelas feitas à análise de enunciados de 3-itens
aplicada a estudos sistemáticos 19,5 1 . De acordo com Humphries e ParentF, a aplicação de análise de parcimônia em estudos biogeográficos não é apropriada, pois seria equivocada a analogia entre caracteres X táxons e táxons X áreas. Como os próprios advogam, uma alternativa seria a aplicação do critério de parcimônia, mas sob a forma descrita na análise de 3-itens.
Este método analítico foi desenvolvido por Gareth Nelson e Pauline Ladiges em 1 99626. O propósito da análise de subárvores é especificar os dados relevantes à biogeografia cladística. TAC são decompostos em subárvores para excluir ou minimizar informação redundante. Como visto antes, redundância consiste na sobreposição espacial de dois táxons em uma área qualquer. Redundância foi definida e caracterizada com mais precisão e num contexto evolutivo por Gareth Nelson, Pauline Ladiges e Roderic Page26,3 1 que a denominaram de paralogia geográfica, que é evidenciada pela duplicação ou sobreposição na distribuição geográfica de táxons relacionados. Em um cladograma, um nó é considerado parálogo quando relaciona organismos às distribuições que se sobrepõem em algum grau e tais distribuições são consideradas parálogas26.
É evidente que em cladogramas contendo muitos táxons a ocorrência de paralogias aumenta quanto mais próximo da base do cladograma; assim, a partir de determinado ponto, todos os nós basais são provavelmente parálogos26.
Procedimentos operacionais 7,23,26 (Figs. 6. 1 3 e 6. 1 4):
1 . Reconstruçã% btenção dos cladogramas dos táxons em estudo.
2. Obtenção de TAC pela substituição do nome do táxon pela sua área de endemismo.
3. Extração e produção das sub árvores livres de paralogias a partir dos TAC. As subárvores representam componentes não parálogos e são, por isso, geograficamente informativas.
4. Construção de uma matriz binária de áreas X componentes, ou áreas X enunciados de 3-itens.
5. Análise de parcimônia para obtenção de um cladograma geral de áreas.
Para extração e produção de sub árvores livres de paralogias é identificado cada nó terminal não parálogo (e geograficamente informativo), progressivamente em direção à base (Fig. 6. 1 3) . Este procedimento é repetido várias vezes até a exaustão, de nó em nó, sempre em

Gastropoda G1 G2 G3 G4 AUS ORI AS AF
v-Diplopoda
D 1 D2 D3 AUS-ORI AS AF
v-Fabaceae
F 1 F2 F3 AUS AS AF
\/- \/ �
P1 P2 P3 P4 P5 P6 AUS ORI AS ORI AF AF
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 G.E. O O O O O O O O O O O O O
AUS O O O ? O ? 1 1 O O O O ?
ORI 1 1 ? O ? O 1 1 ? 1 1 ? O
AS 1 ? 1 1 1 1 O ? 1 ? 1 1 1 AF ? 1 1 1 1 1 ? O 1 1 ? 1 1
AUS (ORI, AS)
AUS (ORI, AF)
AUS (AS, AF) �
ORI (AS, AF)
AUS (AS, AF)
ORI (AS, AF) �
AS (AUS, ORI)
AF (AUS, ORI)
AUS (AS, AF) �
AUS (ORI, AF)
AUS (AS, ORI)
AUS (AS, AF) �
ORI (AS, AF)
AS (ORI, AF)
AUS 1 4 O
?
1 O
1
co
7
G .E . AUS ORI AS AF
G.E. AUS ORI AS AF
G .E . AUS ORI AS AF
G.E. AUS ORI AS AF
ORI
14 8
B iogeografia Cladística - 1 1 5
1 2 3 4 O O O O
O O O ?
1 1 ? O
1 ? 1 1 ? 1 1 1 ® 5 6 7 8 O O O O
O ? 1 1 ? O 1 1 1 1 O ?
1 1 ? O ® 9 O
O
?
1 1 ©
1 0 1 1 1 2 1 3 1 4 O O O O O
O O O ? ?
1 1 ? O 1
? 1 1 1 O
1 ? 1 1 1 ®
AS
'j2 - Anál ise de enunciados de 3-áreas: procedimentos operacionais. (A a D) Cladograma taxonômico convertido para c1adogramas taxonômicos de áreas (TAC), enunciados informativos e sua codificação na matriz, para cada um dos táxons. (E) Matriz de dados concatenada. (F) Cladograma geral de áreas. (e) = reversão; G.E . = grupo externo (área h ipotética tudo-zero).

1 1 6 - Métodos e Ap l i cações
NG NZ AS 25 24 23
AS 1 6
AUS AS 22 2 1
NZ NG 1 5 1 4
Subárvore 1
NC 1 3
NZ 20
AS
AUS AS 1 9 1 8
2
NZ 2 1 20
AUS AS
AUS 1 9
1 7 1 6
4
Y Subárvore 2
NZ NG 1 5 1 4
®
NG NZ AS 25 24 23
6 3
Subárvore 3
NC 1 3
1 0
AUS 22
® - Aná l ise de subárvores l ivres de paralog ias. Procedimentos operaciona is, parte 1 . (A) Cladograma taxonômico
de áreas (TAC). (B) Extração de subárvores. Mod ificado de Nelson e Lad iges26.
Nós das subárvores 7 1 1 5 3 6
G.E . O O O O O
AUS ? ? 1 1 1 NC 1 1 ? ? ?
NG 1 1 ? O O
AS O O O 1 1 NZ 1 O 1 1 O ®
Su posição/su bá rvore 1/1 2/1 3/1 4/1 5/2 6/3 7/3
G . E . O O O O O O O
AUS ? ? ? ? 1 1 1 NC 1 1 ? 1 ? ? ?
NG 1 ? 1 1 ? O O
AS O O O 7 O 1 7 NZ 7 1 1 O 1 7 1
NC
8/3 9/3 O O
? 1 ? ?
O ?
1 1 1 O
©
NC
NG NZ
7 5
AUS
NG NZ
5 2 3
AS
5(0)
AUS AS
5(0)
® - Aná l ise de subárvores l ivres de paralogias. Proced imentos operacionais, parte 2. (A) Matriz de componentes.
(B) Um dos seis cladogramas mais parcimoniosos obtidos. (C) Matriz de enunciados de 3-itens. (O) Um dos quatro cl adogramas mais parcimoniosos obtidos. (o) = reversão; G .E . = grupo externo (área h ipotética tudo-zero).

direção à base, até que todo o cladograma seja averiguado. As informações geográficas não parálogas são mantidas e incorporadas à análise, enquanto as "informações" parálogas são descartadas. Apesar de simples, a produção de subárvores pode ser algo bastante complexo, em especial quando nos deparamos com táxons amplilocados. Sua presença pode confundir nós não parálogos com nós parálogos, o que pode ocasionar perda de informação geográfica. Assim, excepcionalmente, se um nó resulta diretamente a um ou mais táxons terminais que são amplilocados e parte dessa distribuição se sobrepõe com a de outro(s) táxon(s), então a distribuição ampla é reduzida ao elemento geográfico não sobreposto. Seguindo a exemplificação de Nelson e Ladiges26, para cinco táxons com distribuição A, AB, B, BC e D, cujo TAC denota o relacionamento (D((A,AB)(B,BC» ), uma primeira conclusão seria de que todos os nós são parálogos, mas, se resolvermos os táxons amplilocados pela implementação do pressuposto A2, teremos o TAC parcialmente resolvido como (D((A,B)(B ,C))) . E, em seguida, as redundâncias são eliminadas com a produção das seguintes subárvores: (D(A,B» e (D(B,C» . Neste exemplo, coincidentemente as sub árvores correspondem a enunciados de 3-áreas.
Ebach25 (ver também Ebach e Humphries 17) denominou como "Cladística de Áreas" um novo método que combina alguns procedimentos operacionais específicos: implementação pelo pressuposto A2, produção de sub árvores livres (ou com redução) de paralogias e análise com enunciados de 3-itens. Assim, em lugar de análise de parcimônia, a matriz de dados é submetida à análise de 3-itens.
Antes da execução de qualquer dos métodos analíticos algumas precauções preliminares devem ser tomadas. As seguintes etapas são consideradas primárias e fundamentais para a execução de uma análise biogeográfica de modo adequado. Não é preciso dizer o quanto a falta de cuidados nestas etapas terá influência direta nos resultados finais de uma análise.
É imprescindível que o conhecimento taxonômico de cada espécie incluída na análise biogeográfica estej a apurado e determinado de forma inequívoca, sem dúvidas quanto à validade de seu status como espécie nominal. A inclusão de espécies duvidosas (por exemplo,
B iogeografia C lad ística - 1 1 7
que possa consistir num complexo de espécies ou que possa ser parte de outra) incorpora falsas informações distribucionais à análise, como distribuição ampla e distribuição sobreposta (redundância)4, 18.
Se, há 25 anos, Nelson52 reconheceu a escassez de clado gramas para níveis taxonômicos inferiores como um dos principais problemas daquela época, hoje este problema inverteu-se por completo. Respeitando-se as diferenças quanto ao nível de conhecimento sistemático de cada grupo taxonômico, ainda assim, existe abundância de cladogramas para níveis inferiores e um problema atualmente enfrentado é quanto à acurácia e à confiabilidade das informações contidas nesses cladogramas. O esforço dos estudos sistemáticos deve ter como finalidade última a obtenção de cladogramas completamente resolvidos cujos nós são confiáveis (a confiabilidade dos nós depende de estudos adicionais que forneçam corroboração/refutação )26 .
Tão importante quanto a delimitação taxonômica e hipóteses filogenéticas acuradas. Esta é uma das maiores fontes de erro na análise, no que concerne à origem de problemas de táxons amplilocados, redundâncias e áreas ausentes. Áreas de endemismo representam as unidades básicas de relacionamento nos cladogramas de áreas, sejam TAC, RAC ou cladogramas gerais de áreas. Um pequeno erro inicial tende a gerar um enorme erro ao final, pois falsos relacionamentos entre áreas produzirão erros nos eventos de vicariância e dispersão inferidos, nos eventos geológicos associados, na interpretação e na explicação da história da biota e da área, entre outros (para áreas de endemismo, ver Capo 3).
São várias as fontes disponíveis para a compilação de dados de distribuição como observação e/ou captura no campo, rótulos de exemplares em coleções, dados de literatura (revisões taxonômicas, monografias, relatos de ocorrência, levantamentos de diversidade, estudos de comunidade etc.). E, assim, são também muito diversos os tipos de erros: amostragem insuficiente ou deficiente, erros de identificação do táxon, erros de rotulação dos exemplares (rótulos trocados, rótulos com informações imprecisas etc.), registros imprecisos (informações deficientes, localidades homônimas, localidades antigas renomeadas) etc.

1 1 8 - Métodos e Apl icações
A relação e a comparação de cladogramas de diferentes grupos somente são possíveis quando há um denominador comum52 . A própria teoria evolutiva, com a mudança de forma no espaço e ao longo do tempo, representa este elemento comum.
A informação temporal na filogenia possibilita inferir
quando se deram a origem e a diversificação das linhagens e, por conseguinte, possibilita corroborar ou refutar hipóteses de associação causa-efeito entre eventos biogeográficos e eventos cladogenéticos . Entretanto, quase nunca a informação temporal é formalmente apresentada e, com frequência, estudos comparativos ou estudos de busca por padrões não assumem qualquer informação de tempo sobre os táxons em estudo. Parece óbvio dizer, mas não faz sentido algum comparar-se um cladograma de áreas de um táxon com diversificação durante o período Jurássico com outro de um táxon com diversi-
ficação no Paleoceno. Os eventos causais que interferiram na história destes dois táxons não contemporâneos foram completamente diferentes, embora possam ter ocorrido numa mesma área (isto é, eventos coincidentes no espaço).
Há muito tempo tem-se salientado a importância da informação temporal na congruência biogeográfica (por exemplo, Nelson e Platnick4) . Mais recentemente, Donoghue e Moore53 criticaram ostensivamente os estudos de busca por padrões da biogeografia cladística, nos quais a marginalização da informação temporal vinha (ou vem) sendo realizada de forma indiscriminada e despreocupada. Estes autores reconheceram e caracterizaram o que seria uma autêntica congruência e algumas modalidades de não congruências3 (Fig. 6 . 1 5) .
Reconhece-se a existência de "congruência" quando cladogramas de áreas apresentam congruência topológica e coincidência temporal. Dentre os casos de não congruência, uma "incongruência" é reconhecida quando cladogramas de áreas não apresentam congruência topológica apesar de sua coincidência temporal.
Coincidência temporal
'" u '0, 'o o D.. o +-' co
'u c <QJ ::> o, c o
u
S IM
NÃO
S IM
Congruência
A B C A B C
2
A B C A B C
2
Incongruência
NÃO
Pseudocongruência
A B C
A B C
A B C
2
A B C
2
Pseudoincongruência
3i - Congruência autêntica e modal idades de não congruência. Mod ificado de Donoghue e MooreS3.
'O -...J 00 00 V> c.., N ::: 00 \C 0\ �

"Pseudo congruência" é reconhecida quando cladogramas de áreas apresentam congruência topológica, mas não são temporalmente coincidentes. E, por fim, a "pseudoincongruência" consiste no caso de cladogramas sem congruência topológica e sem coincidência temporal. Nos casos de "congruência" e "incongruência", os táxons tiveram sua diversificação numa mesma faixa temporal e estiveram sujeitos aos mesmos eventos geológicos, conquanto suas respostas tenham sido diferentes a estes eventos. Por outro lado, nos casos de "pseudocongruência" e "pseudoincongruência", a diversificação dos táxons não teria ocorrido num mesmo intervalo de tempo e, então, não poderiam ter sofrido efeitos dos mesmos eventos geológicos.
Embora se releve a importância da informação temporal das linhagens em análises biogeográficas, infelizmente os métodos de estimativas de datação hoje disponíveis são altamente questionáveis. Existem três abordagens para se estimar a idade de um táxons4: ( 1 ) com base na idade do fóssil mais antigo conhecido; (2) fundamentada na idade da camada estratigráfica da qual o táxon é endêmico; e (3) baseada na idade de eventos paleogeográficos supostamente associados à história do táxon. Dentre estas, o registro fóssil é o mais amplamente utilizado nos protocolos moleculares de estimativas de idades para calibrar as árvores e é, por isso, o mais criticado. Além do problema inerente de escassez de registros fósseis para virtualmente todos os grupos de organismos, a informação temporal fornecida por fósseis é limitada. Registros fósseis fornecem a idade mínima de um táxon e não idade absoluta como equivocadamente assumida e interpretada nas filogenias e biogeografias moleculares. Um dado fóssil de 35 milhões de anos atrás nos informa que o táxon ao qual pertence estava representado naquele exato ponto da história geológica, embora não nos informe simplesmente nada sobre a existência do táxon nas épocas anteriores. Exemplos de quão efêmero pode ser o status de "mais antigo" para um registro fóssil são apresentados por Headss4.
É inegável o revigoramento trazido pelas técnicas moleculares aos estudos sistemáticos e biogeográficos, mas, por outro lado, as estimativas de idades de linhagens inacreditavelmente recentes e anunciadas quase como verdades incontestes têm sido extremamente prejudiciais, bem como o ressurgimento de hipóteses biogeográficas fundamentadas em centros de origem e dispersalismo (ver Parenti 16, Nelson e LadigesSS, HeadsS4). A noção viciada de que táxons "must-be-too-young"55 é usual nestes estudos em que mais fácil é assumir uma postura C' est la vie e explicar padrões de distribuição com as informações disponíveis (sejam boas
B i ogeografia C lad ística - 1 1 9
ou não, sejam suficientes ou não) do que simplesmente assumir o desconhecimento dos inúmeros fatos ora obscurecidosss. Muitos dos equívocos contidos nas interpretações (dispersalistas) de biogeografias moleculares (encabeçada pela filogeografia) devem ser consequência direta dos prováveis erros nas estimativas de datação.
Lê-se no trecho a seguir, extraído de Nelson e Platnick4, um importante questionamento: "Geralmente desconhece-se, mesmo hoje, se relacionamento de áreas é algo simples ou complexo: se um único cladograma de áreas inclui todos os relacionamentos de espécies, ou se dois ou mais cladogramas são necessários para explicar todos os relacionamentos de espécies dos táxons endêmicos para uma área qualquer". O trecho revela uma preocupação antiga e atual da biogeografia - se os padrões de distribuição da biota de uma área poderiam ser explicados por um único cladograma de áreas, por um único padrão de relacionamento entre áreas. Desde sempre, a procura por padrões gerais para cada região espacial, nos quais se "enquadrariam" todos ou grande parte dos organismos que compõem a biota da região, representa uma obsessão da biogeografia cladística, cujo objetivo maior é mesmo o de procurar por padrões gerais. A consciente noção de que a resposta biológica a eventos geográficos e geológicos é diferenciada conforme propriedades intrínsecas de cada grupo de organismos e que as áreas possuem histórias reticuladas trouxeram importantes elucidações para a biogeografias6. Como já exemplificado em alguns estudos empíricos57,S8, uma biota particular é composta por organismos com diferentes histórias na área, com diferentes padrões biogeográficos, sendo assim, é ingênuo pensar em um único padrão geral. Platnick e Nelson3 já alertavam que "não é necessário, ou mesmo esperado, que encontremos que a biota de uma área apresente somente um único padrão geral, ou que, caso encontremos mais de um, que apenas um padrão contribua com informação sobre a história das áreas envolvidas".
Para Crisci8 (ver também Crisci et az.9 e Enghoff18) estudos de biogeografia histórica podem apresentar diferentes perspectivas, dentre elas, a "biogeografia de táxon" (análise da história espacial de táxons individuais) e "biogeografia de áreas" (análise da história da área, busca pelo padrão geral de relacionamento entre áreas com base na congruência de diferentes táxons). A biogeografia cladística é predominantemente um programa

1 20 - Métodos e Apl icações
So - Classificação dos métodos de B iogeografia H istórica
Abordagens
Dispersalismo
Biogeografia vicariante
Pan-biogeografia
B iogeografia cladística
Modificada de Morrone35.
de pesquisa da "biogeografia de áreas", pois grande parte de seus métodos analíticos tem como objetivo a busca por padrões gerais de relacionamento entre áreas. Entretanto, a biogeografia cladística, no sentido amplo dado por Morrone36 (ver Quadro 6.2 e Tabela 6. 1 ), também compreende métodos que buscam explicar a história espacial de táxons individuais.
É consensual que o objetivo central da biogeografia cladística seja a procura por padrões gerais de relacionamento entre áreas e, descoberto o padrão, seus processos causais podem, então, ser inferidos e interpretados3.4,7, 16 . Usualmente, eventos vicariantes são considerados os processos explicativos primários, pois dispersões aleatórias não poderiam explicar padrões recorrentes6. No entanto, tanto eventos de vicariância quanto de dispersão são interpretações subjetivas para os padrões de distribuição, e como tais representam unicamente hipóteses e não observações factuais 17 . Nenhum método analítico da biogeografia cladística é capaz de determinar objetivamente quais eventos, se vicariância ou dispersão, foram responsáveis por um padrão biogeográfico, já que ambos podem sim gerar padrões semelhantes 17. Deve-se ter, portanto, a noção clara de que o padrão descoberto é basicamente uma hipótese de relacionamento entre áreas fundamentada na congruência dos dados analisados (filogenias de vários táxons, dados distribucionais) e que não implica diretamente sobre qualquer inferência processual ou causal. A inferência dos processos causais dependerá de evidências independentes da história da terra3 (isto é,
Métodos analíticos
Reconstrução m a n u a l Biogeografia f i l ogenética Áreas ancestrais F i l ogeografia
Reconstrução m a n u a l Pan-biogeografia q u a ntitativa Compatib i l idade de traços Anál ise de parci mônia de endemicidade
Cladograma de áreas reduzido B iogeografia f i l ogenética quantitativa Aná l ise de componentes Anál ise de parcimônia de Brooks Compatibi l idade de componentes A n á l ise de enu nciados de 3-áreas Anál ise de reconci l iação de árvores Aná l ise de subárvores l ivres de paralog ias Anál ise de eventos vicariantes A n á l ise de d ispersão-vica riância F i l ogeografia comparativa
evidências geográficas e geológicas) . Para alguns biogeógrafos, hipóteses dispersalistas não seriam falseáveis (por exemplo, Rosen2); para outros, seriam difíceis de se testar, pois a aleatoriedade dos eventos de dispersão \C
-J torna a sequência de eventos particular para cada grupo, Cf'
00 u. suas histórias sendo, por conseguinte, incomparáveis (por exemplo, Humphries e Parente). Por outro lado, é importante destacar que qualquer teste dependerá da disponibilidade de informações temporais tanto dos organismos quanto dos eventos geológicos supostamente associados à sua história na área.
A Ricardo Pinto da Rocha, Márcio Bernardino DaSilva e J éssica P. Gillung pela leitura crítica e sugestões ao manuscrito. Ricardo Pinto da Rocha, Márcio Bernardino DaSilva, Eduardo Almeida e Antônio Carlos Marques auxiliaram na escolha de alguns termos mais adequados em língua portuguesa. Aos editores, Claudio 1. B. de Carvalho e Eduardo A. B . Almeida, que foram bastante cuidadosos durante a revisão final. Também, um revisor anônimo forneceu importantíssimas sugestões e críticas ao conteúdo e à redação do manuscrito. À equipe técnica da Editora Roca pela preparação final das figuras. Auxílio financeiro recebido da FAPESP (proc. n . 2007/50836-7) e CNPq (proc. n. 477 1 86/2008-4). Bolsista de Produtividade em Pesquisa do Conselho Nacional de Desenvolvimento Científico e Tecnológico (proc. n. 303897/2008-2).
.:., � 00 \C � '"

REFERÊNCIAS BIBLIOGRÁFICAS 1 . NELSON, G. Historical biogeography: an altemative forrnaliza
tion. Syst. Zool. , v. 23, p. 555-558, 1 974. 2 . ROSEN, D. E. A vicariance model of Caribbean biogeography.
Syst. Zool. , v. 24, p. 43 1 -464, 1976. 3 . PLATNICK, N. L; NELSON, G. A method of analysis for
historical biogeography. Syst. Zool. , v. 27, p . 1 - 1 6, 1 978. 4. NELSON, G.; PLATNICK, N. !. Systematics and Biogeography:
cladistics and vicariance. New York: Columbia University Press, 198 1 . 567p.
5. CROIZAT, L. Space, Time, Form: the hiological synthesis. Caracas: publicado pelo autor, 1 964. 8 8 1 p.
6. LLORENTE, J. ; PAPAVERO, N. ; BUENO, A. Síntesis histórica de la biogeografía. In: LLORENTE, 1 . ; MORRONE, J. J. (orgs.). Introducción a la Biogeografía en la Latinoamérica: teorías, conceptos, métodos y aplicaciones. Mexico: Las Prensas de Ciencias/UNAM, 200 1 . p. 1 -1 4.
7. HUMPHRIES, C. J . ; PARENTI, L. R. Cladistic Biogeography. 2. ed. Oxford: Clarendon Press, 1 999. 1 87p.
8 . CRISCI, J. V. The voice of historical biogeography. 1. Biogeogr. , v. 28, p. 1 57- 1 68, 2001 .
9. CRISCI, J. v.; KATINAS, L . ; POSADAS, P Historical Biogeography. Cambridge: Harvard University Press, 2003. 250p.
1 0. PLATNICK, N. L; NELSON, G. Spanning-tree biogeography: shortcut, detour, or dead-end? Syst. Zool. , v. 37, p. 4 1 0-4 1 9, 1 988.
1 1 . PLATNICK, N. L; NELSON, G. Composite areas in vicariance biogeography. Syst. Zool. , v. 33, p. 328-335, 1984.
12. NELSON, G.; PLATNICK, N. L The perils of plesiomorphy: widespread taxa, dispersaI, and phenetic biogeography. Syst. Zool., v. 27, p. 474-477, 1 978.
13. HENNIG, W. Phylogenetic Systematics. Urbana: University of Illinois Press, 1 966. 263p.
14. BRUNDIN, L. Z. Phylogenetic biogeography. In: MYERS, A. A.; GILLER, P S. (orgs.). Analytical Biogeography: an integrated approach to the study of animal and plant distributions. London: Chapman & Hal!, 199 1 . p. 343-369.
15. NELSON, G.; ROSEN, D. E. (orgs.). Vicariance Biogeography:
a critique. New York: Columbia University Press, 1 98 1 . 593p. 16. PARENTI, L. R. Common cause and historical biogeography.
In: EBACH, M. c.; TANGNEY, R. S. (orgs.). Biogeography in a Changing World. Boca Raton: CRC Press, 2007. p. 61 -82.
17. EBACH, M . c.; HUMPHRIES, C. J. Cladistic biogeography and the art of discovery. J. Biogeogr., v. 29, p. 427-444, 2002.
1 8 . ENGHOFF, H. Widespread taxa, sympatry, dispersai, and an algorithm for resolved area cladograrns. Cladistics, v. 1 2, p. 349-364, 1 996.
1 9. MORRONE, 1. J.; CRISCI, 1. V. Historical biogeography: introduction to methods. Ann. Rev. Ecol. Syst., v. 26, p. 373-401 , 1995.
20. VAN VELLER, M . G. P; KORNET, D. 1.; ZANDEE, M. Methods in vicariance b i ogeograph y : assessment of the implementations of assumptions zero, 1 and 2. Cladistics, v. 16 , p. 3 1 9-345, 2000.
2 1 . WILEY, E. O. Parsimony analysis and vicariance biogeography. Syst. Zool. , v. 37, p. 27 1 -290, 1988.
22. ZANDEE, M.; ROSS, M . C. Component-compatibility in historical biogeography. Cladistics, v. 3 , p. 305-332, 1 987.
23. MORRONE, J. J. Evolutionary Biogeography: an integrative approach with case studies. New York: Columbia University Press, 2009. 301 p.
24. PAGE, R. D. Component analysis: a valiant failure? Cladistics, � 6, p. 1 1 9- 1 36, 1 990.
25. EBACH, M. C. Paralogy and the centre of origin concept. Cladistics, v. 15 , p. 3 87-39 1 , 1 999.
B iogeografia C lad ística - 12 1
2 6 . NELSON, G. ; LADIGES, P Y. Paralogy in cladistic biogeography and analysis of paralogy-free subtrees. Am. Mus. Novit. , v. 3 167, p. 1 -58, 1 996.
27. PAGE, R. D. Quantitative cladistic biogeography: constructing and comparing area cJadograrns. Syst. Zool. , v. 37, p. 254-270, 1988.
28. BROOKS, D. R. Hennig's parasitological method: a proposed solution. Syst. Zool. , v. 30, p. 229-249, 1 9 8 1 .
29. BROOKS, D. R . Reticulations i n historical biogeography: the triumph of time over space in evolution. In: LOMOLINO, M.; HEANEY, L. (orgs.). Frontiers ojBiogeography: new directions in the geography ofnature. Sunderland: Sinauer, 2004. 436p.
30. EBACH, M. C . ; HUMPHRIES, C. J . ; NEWMAN, R. A . ; WILLIAMS, D. M.; WALSH, S . A . Assumption 2: opaque to intuition? J. Biogeog/: , v. 32, p . 7 8 1 -787, 2005.
3 1 . NELSON, G.; LADIGES, P Y. Three-area statements: standard assumptions for biogeographic analysis. Syst. Zool. , v. 40, p. 470-485, 1 99 1 .
32. WILEY, E . O . Vicariance biogeography. Ann. Rev. Ecol. Syst. , v. 19, p. 5 13-542, 1 988.
3 3 . ROSEN, D. E. Vicariant pattems and historical explanation in biogeography. Syst. Zool. , v. 27, p. 1 59- 1 88, 1 978.
34. SANMARTÍN, L Event-based biogeography: integrating pattems, processes, and time. In: EBACH, M. c.; TANGNEY, R. S. (orgs.). Biogeography in a Changing World. Boca Raton: CRC Press, 2007. p. 1 35-159.
35. VAN VELLER, M. G. P ; BROOKS, D. R. When simplicity is not parsimonious: a priori and a posteriori methods in historical biogeography. J. Biogeogr., v. 28, p. 1 - 1 1 , 2001 .
36. MORRONE, J . 1 . Cladistic biogeography: identity and place. J. Biogeogr. , v. 32, p. 1 28 1 - 1 286, 2005.
37. CROIZAT, L. Vicariance/vicariism, panbiogeography, "vicariance biogeography", etc. : a clarification. Syst. Zool. , v. 3 1 , p . 29 1 -304, 1 982.
38. PAGE, R. D. Component User's Manual. Version 1 .5. Auckland: Publicado pelo autor, 1989.
39. PAGE, R. D. Component User's Manual. Version 2.0. London: The Natural History Museum, 1 993. Disponível em: http://taxonomy.zoology.gla.ac.uk/rod/cpw.html.
40. NELSON, G.; LADIGES, P. Y. TAS. New York: Publicado pelos autores, 1 99 1 .
4 1 . NELSON, G.; LADIGES, P Y. TASS. New York: Publicado pelos autores, 1995.
42. DUCASSE, 1.; CAO, N.; ZARAGÜETA-BAGILS, R. NELSON05: Three- item Analysis Software Package. Paris : Université Pierre e t Marie Curie, 2008.
43. GOLOBOFF, P. A. NONA. Version 2.0. Publicado pelo autor, 1 993. Disponível em http://www.zmuc.dk/public/phylogeny/ Nona-PeeWee/.
44. SWOFFORD, D. L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0betalO. Sunderland: Sinauer, 2001 .
4 5 . GOLOBOFF, P. ; FARRIS, S . ; NIXON, K. TNT (Tree analysis using New Technology). Version 1 . 1. Tucumán: Publicado pelos autores, 2007. Disponível em http://www.cladistics.org/tnt.html.
46. BROOKS, D. R. Parsimony analysis in historical biogeography and coevolution: Methodological and theoretiacal update. Syst. Zool. , v. 39, p. 14-30, 1 990.
47. BROOKS, D. R.; VAN VELLER, M. G. P; MCLENNAN, D. A. How to do BPA real1y. J. Biogeogr. , v. 28, p. 345-358, 200 1 .
4 8 . SIDALL, M . E. Bracing for arrother decade o f deception: the promise of secondary brooks parsimony analysis. Cladistics, v. 2 1 , p. 90-99, 2005.
49. NELSON, G.; PLATNICK, N. L Three-taxon statements: a more precise use of parsimony? Cladistics, v. 7, p. 3 5 1 -366, 199 1 .

1 22 - Métodos e Aplicações
50. KITCHING, I. J.; FOREY, P. L.; HUMPHRIES, C. J. ; WILLIAMS, D. M. Cladistics: the theory and p ractice of parsimony analysis. New York: Oxford University Press, 1998. 228p.
5 1 . KLUGE, A. G. Three-taxon transformation in phlogenetic inference: ambiguity and distortion as regards explanatory power. Cladistics, v. 9, p. 246-259, 1993.
52. NELSON, G. C1adistics and biogeography. In: DUNCAN, T; STUESSY, T F. (orgs.). Cladistics: perspectives on the reconstruction of evolutionary history. New York: Columbia University Press, 1984. p. 273-293.
53. DONOGHUE, M. J.; MOORE, B. R. Toward an Integrative Historical Biogeography. Integ/: Comp. Biol. , v. 43, p. 261-270, 2003.
54. HEADS, M. Dating nodes on molecular phylogenies: a critique of molecular biogeography. Cladistics, v. 2 1 , p. 62-78, 2005.
5 5 . NELSON, G . ; LADIGES, P. Y. Biogeography and the molecular dating game: a futile revival of phenetics? Buli. Soco Geol.
Fr. , v. 1 80, p. 39-43, 2009. 56. NIHEI, S. S. Dynamic endemism and 'general' biogeographic
pattems. Biogeografía: Buli. Syst. Evo!' Biogeog/: Assoe., v. 3 , p. 2-6, 2008.
57. ESCALANTE, T; RODRÍGUEZ, G.; CAO, N.; EBACH, M. C.; MORRONE, J. J . Cladistic biogeographic analysis suggests an early Caribbean diversification in Mexico. Naturwissensehaften, v. 94, p. 561-565, 2007.
5 8. NIHEI, S. S . ; CARVALHO, C. J. B. Systematics and biogeography of Polietina Schnabl & Dziedzicki (Diptera: Muscida� ):_ Neotropical areas relationships and Amazonia as composite area. Syst. Entomol. , v. 32, p. 477-5 0 1 , 2007 .

__________________ �_ 7
, l'Çl :§ �0 \�J
Eduardo A. B. Almeida
o desenvolvimento teórico e metodológico em biogeografia ao longo das últimas décadas reflete o interesse crescente na compreensão da distribuição geográfica dos organismos e sua relação com a história geológica da Terra. O reconhecimento da vicariância como um tipo de fenômeno gerador de padrões norteou o desenvolvimento teórico e as pesquisas empíricas, que se embasaram na busca por congruência entre relacionamentos filogenéticos e a história do planeta! . Longe de ter sido uma história de simples refinamento de métodos previamente propostos, a biogeografia analítica foi muitas vezes modificada por propostas de métodos com características muito diferentes das de outros já disponíveis. Este foi justamente o caso do desenvolvimento dos métodos de eventos.
Tanto a biogeografia cladística quanto a pan-biogeografia (ver Caps. 5 e 6) partem do reconhecimento de evidências de histórias de vicariância, a despeito das peculiaridades que distinguem essas escolas da biogeografia histórica. Neste contexto, distribuições de táxons que não se encaixam em padrões explicáveis como resultantes de eventos de vicariância comuns são então relegados à classe de "ruído biogeográfico", cuja origem seria em eventos como dispersão ou extinções de linhagens. A análise é realizada, portanto, em duas etapas : na primeira buscam-se evidências de relacionamentos filogenéticos afetados por uma causa comum (por exemplo, vicariância), ao passo que uma avaliação a posteriori indica possíveis hipóteses de como processos distintos da vicariância teriam afetado as histórias dos táxons. O problema mais conhecido em biogeografia analítica é a perda do sinal histórico da vicariância quando sobrepujado por outros processos, como dispersões. Nessas situações, os padrões históricos são tão transfigurados
'J /?), 6'� (S� -1 in �y '\.� W \J
que nenhum método fundamentado em padrões seria capaz de reconstruir a história biogeográfica de modo satisfatório.
As abordagens coletivamente denominadas "métodos de eventos" ou "métodos baseados em eventos" buscam estabelecer uma relação direta entre o procedimento analítico e um modelo que considera a vicariância conjuntamente com outros eventos históricos relevantes à inferência biogeográfica2-s . Com os métodos de eventos propõe-se, assim, a avaliação da influência de vários eventos simultaneamente por meio de um modelo explícito. Este modelo de eventos nada mais é que um conjunto de regras relacionadas aos processos que podem ter influenciado as associações entre as histórias dos organismos e das áreas. Deste modo, os métodos de eventos permitem uma avaliação comparativa de hipóteses biogeográficas concorrentes.
Os métodos de eventos aplicáveis para estudos biogeográficos são também utilizados por duas outras disciplinas da biologia de modo bastante similar: coespeciação de parasitas e hospedeiros, e evolução gênica3•6. De modo geral, os métodos apresentados neste capítulo foram desenvolvidos em paralelo por teóricos das disciplinas e posteriormente aplicados às demais6-9. Essas três áreas de estudo investigam associações históricas entre uma entidade denominada "associado" e outra, "hospedeiro": em biogeografia, associações entre táxons e áreas; em coespeciação, entre parasitas e hospedeiros; em evolução gênica, entre genes e organismos (Tabela 7 . 1 ) . Em todos os casos, espera-se que a evolução dos associados seja influenciada pela evolução dos hospedeiros. Em biogeografia, isso significa que os eventos de vicariância das áreas refletem-se em cladogênese dos táxons nelas distribuídosI .3,7, o que resultaria em congruência espacial e temporal (ver Capo 4). Apesar de biogeografia, coespeciação e evolução lidarem com

1 24 - Métodos e Apl icações
:IG1i))0,�2 , ': - Comparação dos ti pos de eventos, equ iva lências e termino logias próprias das três discip l inas q ue tratam de associações h istóricas entre duas entidades, uma chamada de "associado" e a outra de " hospedeiro " 1 ,3,4,6
Disciplina
Entidades associadas
I . Biogeografia
"Associado" Táxons
11. Associação de 111. Evolução gênica parasitismo
Parasitas Genes
" H ospedeiro " Áreas Hospedei ros Organismos ------------�------------------------------------�------------�------------------------- ,�
Codivergência Vicariância: especiação a lopátrica Coespeciação Herança de a le lo gên ico por a m bas as ;3;3 Eventos relacionada ao surgimento de uma l in hagens resultantes de cladogênese; e;; barreira à d ispersão, que pode ortologia e.., afetar vários táxons � s i m u ltaneamente �
0\ Dupl icação Especiação na área; por exemplo, Especiação do Dupl icação gênica; paralogia Q,
especiação a lopátrica re lacionada à parasita sem que formação de barreira temporá ria; ocorra especiação especiação s impátrica do hospedeiro
Transferência horizontal
Dispersão (= transposição de I nfecção de novo Transferência horizonta l de gene (por exemplo, por meio de i ntrogressão por h i bridização); xenologia
barreira geográfica), seguida por hospedeiro
Extinção
especiação
Extinção de um táxon de á rea em que este seria esperado ocorrer
problemas ontologicamente muito distintos, todos convergem para padrões que podem frequentemente ser representados por cladogramas e suas resoluções podem partir de princípios muito similares 10. Os paralelos que podem ser traçados entre coespeciação, evolução gênica e biogeografia são ilustrados na Figura 7 . 1 .
Se as histórias dos táxons e das áreas forem representadas como cladogramas, a congruência entre elas permite que uma árvore seja sobreposta à outra, ou como se refere a isso comumente, uma história pode ser mapeada sobre a outra4. No entanto, o resultado de uma tentativa de se mapear a história de um táxon sobre uma hipótese de história geológica nem sempre é completamente congruente, o que gera uma série de questionamentos. Por exemplo, quando há congruência parcial entre as histórias, deve-se descartar ou aceitar a hipótese vicariante? E o que as similaridades ou diferenças entre as histórias de táxons e de áreas dizem sobre a história evolutiva da associação? Essas questões constituem alguns dos desafios metodológicos da biogeografialO- 1 2• Para PageJO, respostas a esse tipo de questão são inerentemente estatísticas. Os métodos de eventos surgiram como uma reação à insatisfação quanto às abordagens que consideram suficiente a busca por padrões de vicariância e, em parte, uma reação também à falta de rigor estatístico na comparação de hipóteses alternativas no caso da maioria dos métodos tradicionais.
A adoção de modelos constituídos por eventos explícitos permite, em muitos casos, a variação de parâmetros da análise, de modo a avaliar a credibilidade de diferentes cenários biogeográficos5. Pode-se, por exemplo,
Extinção do parasita
Deleção gênica ou extinção a lél ica
estimar qual a frequência relativa de dispersões e vicariância para a história de um grupo taxonômic05. De maneira geral, os eventos de dispersão e de extinção são minimizados nessas análises, ao passo que explicações embasadas em vicariância são favorecidas. A definição dos parâmetros analíticos pode ser expressa por um modelo em que cada uma das classes de eventos considerados possui um custo. Neste contexto, a análise biogeográfica consistirá na busca pela explicação com o menor custo total, de modo análogo à análise cladística, que seleciona a topologia mais parcimoniosa para um conjunto de dados.
O modelo é mais realista na medida em que os custos dos eventos são inversamente proporcionais às chances de ocorrerem2,4,5 . Assim sendo, o custo para explicações de vicariância deve ser baixo relativamente ao custo de dispersões, de modo a favorecer cenários históricos nos quais pressupõe-se que disjunções sejam consequências naturais de eventos de vicariância e que a transposições de barreiras geográficas por dispersão possam ocorrer, embora sejam raras4, 1 2 . Os custos dos eventos dos modelos nem sempre podem ser facilmente determinados, o que gera as principais críticas a esta classe de métodos analíticos 13 .
Além das justificativas apresentadas anteriormente para a utilização de métodos de eventos, o estudo da congruência temporal dos eventos também pode ser facilitado pelo uso de modelos sobre um arcabouço filogenético. As filogenias dos táxons representam, neste caso, os sistemas de referência sobre os quais as inferências biogeográficas podem ser feitas3. As árvores

Modelos de Eventos para Reconstrução B iogeográfica - 125
A B Coespeciação Evolução gênica
Hospedeiro A --1 Parasita
B --2
2
C --3
B iogeog rafia histórica
A B 2
C áreas 3 táxons
r _ H istórias b iogeográficas, de coespeciação e de l i nhagens gênicas podem ser extremamente s imi la res na forma como as associações e ntre duas entidades podem ser representadas graficamente: (A) associação coevolutiva entre três espécies de parasita e três de hospedeiro. (B) A evol ução de um gene ( l i nha preta), cuja h istória de d ivergência coincide com a h istória fi logenética dos organ ismos (ci l i ndros acinzentados) que possuem este gene. (C) Área ABC progressivamente se fragmentou em três á reas menores: A, B e C. Relações fi logenéticas de táxons cujos ancestrais habitavam a área ABC podem ter sido afetadas pela história de vicariância, como mostrado para o clado formado pelas espécies 1 , 2 e 3.
auxiliam a compreensão de padrões históricos porque a divergência entre os ramos de um cladograma representa um esquema natural de organização de tempos relativos de divergência filogenética, que podem ser traduzidos como ordenação temporal dos eventos que moldaram a história de um clad03,7, 14 . O tempo relativo é de enorme importância, mas o tempo absoluto também tem sido apontado como uma informação muito desejáveI ls . Mesmo havendo congruência entre a ordem dos eventos de cladogênese e de vicariância, os primeiros não poderiam ocorrer anteriormente aos últimos, pois a hipótese de relação de causa e efeito seria quebrada (Fig. 7.2) . Idealmente, a congruência temporal poderia se manifestar pela detecção dos eventos de cladogênese como concomitantes ou seguidos aos eventos geológicos 7,8, 14.
Nas seções seguintes deste capítulo, serão abordados os seis principais métodos de eventos disponíveis hoje: vicariância máxima (VM)16, Jungles 17 , análise de dispersão-vicariância (DIVA, dispersal-vicariance analysis)1 8,19, TreeFitting4,20, análise de verossimilhança de evolução de distribuição geográfica (Modelo DEC)2 1 ,22 e métodos frequentistas e bayesianos para
inferências estatísticas de associação entre histórias 1 1 , 12.
Todos esses métodos, exceto DIVA e DEC, buscam conciliação de estudos de vários grupos ou de um táxon individual e uma hipótese de área em busca de padrões biogeográficos mais gerais. A compreensão da maioria dos métodos de eventos é dificultada por requerer não somente o entendimento das premissas teóricas da biogeografia histórica, mas também uma base matemática e estatística. Nas apresentações feitas a seguir, será priorizada a descrição das características principais dos métodos em detrimento do formalismo matemático.
Todos os métodos discutidos neste capítulo dependem da utilização de hipóteses filogenéticas para os táxons sendo estudados, como também de conhecimento sobre as áreas onde se distribuem cada terminal das filogenias. Essas áreas de distribuição poderiam ser delimitadas segundo os princípios para delimitação de áreas de endemismo (ver Capo 3), apesar de não haver preocupação explícita por parte da maioria dos autores de métodos

126 - Métodos e Apl icações
A .---- 3 B .---- C C .------- 3
1-.....- 2 '---- B '------- 2
i------ A
3 2 o 3 2 o 3 2 o Tempo Tempo Tempo
- Congruência re lativa e a bsol uta entre h istórias de táxons e á reas. As espécies 1 , 2 e 3 d istr ibuem-se, respectivamente, nas á reas A, B e C. Caso os tempos de divergência re lativos entre espécies e á reas sejam s imi lares à situação mostrada em (A) e (B), há congruência tempora l entre os eventos e é possível supor que a h istória de vicariância das á reas pode se relacionar às relações de parentesco entre os táxons. Como a lternativa, se a d ivergência entre os táxons (C) tiver ocorrido anteriormente às mudanças geológ icas das á reas (B), não há como supor q ue exista uma associação entre as duas.
de eventos quanto ao formalismo e à rigidez desta etapa. A despeito disso, a escolha do modo como se realiza a delimitação de áreas tem influência crucial no resultado de análises biogeográficas.
As abordagens de eventos necessitam de dois procedimentos principais3:
1. Identificação dos processos históricos relevantes à reconciliação das histórias (isto é, filogenia dos táxons, hipóteses para a história das áreas e distribuição dos táxons nas áreas).
2. Formulação de um modelo que considere o relacionamento filogenético entre táxons e áreas, e os eventos possivelmente relacionados a esta associação, em que os eventos recebem custos relacionados à probabilidade de ocorrência.
No caso de métodos que não operem visando reconciliar histórias de áreas e de táxons (ver a seguir: DIVA e DEC), apenas a parte 2 é necessária para propor hipóteses sobre um cenário que teria levado os terminais de uma filogenia a adquirirem a distribuição que possuem.
No caso de métodos que se fundamentam em reconciliação de cladogramas de áreas aos cladogramas de táxons, pode-se generalizar que há três passos principais na análise biogeográfica:
1 . Os cladogramas de táxons e de áreas são posicionados frente a frente, um como a imagem espelhada do outro, e os terminais são associados, de tal maneira
que os táxons são conectados às áreas em que se distribuem. O conjunto dos dois cladogramas com terminais associados é denominado "conectograma"* (Fig. 7 .3 , A).
2 . Os nós dos dois cladogramas são associados . Táxons irmãos associados a áreas irmãs são considerados associados, assumindo-se que isso se deve à vicariância. A representação gráfica da associação é normalmente feita por meio de um trilhograma **, após a sobreposição dos cladogramas de áreas e de táxons (Fig. 7 .3 , B).
3 . Os nós não associados no conectograma podem ser explicados por eventos não vicariantes: dispersão, duplicação e extinção, de modo que o número desses eventos seja o menor possível, minimizando o custo total da reconstrução.
A associação dos cladogramas do conecto grama da Figura 7 .4, A exige que eventos diferentes da vicariância sejam invocados para construção de trilho gramas (Fig. 7 .4, B).
* "Conectograma" é a tradução adotada aqui para o termo orig inal em ing lês tanglegram4. Dois c1adogramas sendo associados são posicionados frente a frente, como imagens espel hadas que permitem a avaliação da congruência topológica entre eles.
** "Tri lhograma" é a tradução adotada aqui para o termo original em inglês trackogram4.

A A---1
B---2
(---3
D---4
Modelos de Eventos para Reconstrução B iogeog ráfica - 1 27
B A 1
B 2
( 3
D 4
- (A) O conectograma apresenta a associação entre os táxons termina is 1 -4 e as á reas A-D onde se distribuem, a lém de h ipóteses de relacionamento para ambos. Como se observa uma congruência perfeita dos eventos de d ivergência entre os cladogramas de á reas e de táxons, o tr i l hograma (B) para a associação necessita apenas de eventos de vicariância para permitir a sobreposição das duas h istórias. • = cladogênese em resposta à vicariância.
A A--- 1
B---2
(---3
D---4
( ( 3 3
D D 4 4
A A 1 1
B B 2 2
/! - (A) O conectograma apresenta a associação entre os táxons term inais 1 -4 e as á reas A-D onde se d istribuem (ver F ig. 7.3 para mais detalhes). Os eventos de d ivergência entre os cladogramas de áreas e de táxons não são completamente congruentes, de modo que se faz necessário invocar eventos além de vicariância para sobreposição das á rvores e formação dos tri l hogramas. Na h ipótese representada pelo primeiro tr i l hograma (B), foi representado um evento de dispersão; na segunda h ipótese de reconci l iação (C), foram representadas uma dup l icação a ncestral e três extinções . • = c ladogênese em resposta à vicar iância; � = cladogênese e dispersão de uma l i nhagem f i l ha; D = dup l icação; ___ = cladogênese e extinção de uma l i nhagem fi l ha.

128 - Métodos e Apl icações
o primeiro método de eventos formalmente proposto foi o de coespeciação máxima 16, que no caso da biogeografia histórica pode ser referido como VMl8. De modo geral, o método pode ser descrito como uma tentativa de conciliação de duas histórias hierárquicas em que eventos de vicariância (= codivergência) são maximizados, minimizando-se a ocorrência de outros tipos de eventos. Matematicamente, isso significa dar custo nulo a todos os eventos que afetariam a história biogeográfica de um táxon, exceto à vicariância, que recebe custo negativo. As hipóteses biogeográficas são comparadas e prefere-se aquela com menor custo. VM15 foi pioneiro em realizar uma análise de reconciliação de duas árvores buscando um cenário que considere simultaneamente vicariância, dispersão, duplicação e extinção. Ao contrário de outros métodos considerados adiante (por exemplo, Jungles e TreeFitting), VM possui a propriedade atraente de não depender de uma escolha dos custos do modelo, pois a distinção entre vicariância e todos os demais eventos toma esta etapa desnecessária. Por outro lado, o método mostra-se insatisfatório em certas situações por não ser capaz de distinguir os casos em que o sinal de vicariância é pujante daqueles em que este sinal é marginal, quando explicações alternativas deveriam também ser consideradas4• 17.
Quando se faz uma reconstrução como em VM, ou em outros métodos de associação, inevitavelmente uma associação entre os cladogramas de áreas e de táxons será alcançada4,1O. Isso não significa, contudo, que a associação histórica seja estatisticamente mais robusta que uma associação proposta entre duas topologias arbitrárias, pois a associação, por si só, não constitui um teste de significância. VM oferece uma forma útil de se investigar se o número de casos de vicariância observados como resultado de uma análise é maior que o número esperado ao acas03,16. Para tanto, é necessário realizar um teste de hipóteses, em que seja calculada a distribuição do número de eventos de associação por codivergência por meio de aleatoriedade de um dos cladogramas que compõem as associações. As relações entre os táxons (hipótese filogenética para os táxons) ou das áreas (cladograma de áreas) são alteradas e, em cada passo deste procedimento, o número de eventos de codivergência é calculado. Para isso, são gerados vários cladogramas com o mesmo número de terminais que o cladograma de táxons ou que o cladograma de áreas por meio de um processo de Markov (ou de Yule) e cada um é associado ao cladograma complementar4, 16. Após
o cálculo do número de eventos de vicariância para as replicações ter sido realizado por um número estatisticamente suficiente de vezes, obtém-se uma distribuição das frequências deste evento. Essa distribuição estende-se do menor ao maior número possível de eventos desta natureza, servindo como distribuição esperada das frequências de vicariância para o estudo sendo feito. O número de eventos de vicariância observado é então comparado à distribuição esperada para que se verifique 'D se este se situa, digamos, na faixa dos 95% com maior �
00 probabilidade. Se isso ocorrer, a hipótese nula de que a � congruência entre as relações das áreas e dos táxons � resulta somente de uma associação por acaso pode ser �
a, rejeitada ao nível de ex = 0,054, 16. o,
Anteriormente à proposta do método de VM, uma das formas propostas para reconciliação de cladogramas de áreas e táxons foi a análise de componentesl (ver Capo 6). Page 7,8, l O, 14 realizou testes de associação de cladogramas com esse tipo de ferramenta, realizando também testes de hipótese para medir a significância estatística da congruência entre as árvores comparadas. As associações, denominadas "reconciliações de árvores"8, buscam reduzir o número de eventos de extinções e duplicações em relação às co divergências (dispersões não são um tipo de evento considerado explicitamente neste caso). As diferenças entre duas hipóteses são quantificadas por "itens de erro", uma medida indireta (e abstrata) da incongruência entre dois cladogramas. Itens de erro não podem ser diretamente atribuídos à dispersão, à extinção, ou outra classe específica de evento, mas são pensados como entidades abstratas com valor heurístico para descrição da incongruência entre cladogramas. Após o cálculo do número de itens de erro, pode-se realizar um teste estatístico similar àquele descrito anteriormente para VM, em que se investiga haver ou não evidências suficientes para descartar a hipótese nula (de que a congruência observada entre os as árvores deve-se somente ao acaso). O cálculo de itens de erro e o teste estatístico de significância pela aleatoriedade de topologias podem ser realizados com o auxílio do programa TreeMap v. 1 23.
Em comparação ao cálculo de itens de erro para reconciliação de cladogramas, a análise de VM possibilita uma análise mais direta e com possibilidade de incorporação de dispersões à explicação das possíveis incongruências entre as relações entre áreas e táxons.
Métodos de padrão, como análise de parcimônia de Brooks (BPA, Brooks parsimony analysis) (ver Capo 6), não lidam diretamente com dispersões, pois operam de modo a minimizar o número de hipóteses ad hoc em relação ao padrão

vicariante. No caso de VMI6, os eventos de dispersão são considerados, mas, segundo Charleston 17, de modo insatisfatório. Hipóteses de dispersão que seriam classificadas como marginalmente prováveis são excluídas dos cenários possíveis gerados por análises de VM4,17. Jungles17,24 foi concebida como uma alternativa às propostas de reconciliação de árvores, mais flexível que qualquer proposta anterior, pois permitiria a avaliação quantitativa de um número maior de hipóteses, que, por sua vez, poderiam ser modeladas em termos dos custos de eventos considerados mais apropriados. Jungles foi definido como uma estrutura conceitual (e gráfica) em que se listam todas as soluções potencialmente ótimas para um problema de associação de árvores ao longo do tempo17,24.
Durante esta busca pelos cenários mais parcimoniosos, o número total de eventos é minimizado e o número de hipóteses de associação decorrente da vicariância é maximizado. Para tanto, as análises levam em consideração topologia, associação entre áreas e táxons, e, se possível, tempos de divergência. Em Jungles, não há custo para vicariância (custo zero), mas custos de duplicações, extinções e dispersões podem ser determinados como qualquer valor positiv04. Podem-se buscar todas as soluções plausíveis para um mesmo problema de reconciliação de árvores, testando-se uma gama de combinações de custos1 7,24. Com Jungles, buscam-se todas as soluções igualmente parcimoniosas para a reconciliação, considerando-se não apenas os custos dos eventos presentes no modelo, como também a coincidência temporal de ocorrência desses eventos.
Jungles foi originalmente proposto para estudos de coespeciação e possivelmente em razão de sua complexidade e dificuldade de estabelecimento dos custos dos eventos, foi pouco empregado em estudos empíricos, em particular de biogeografia. A implementação do método em programas de computador, realizada por M. Charleston, encontra-se disponível em duas versões: v.2�, em coautoria com R. D. M. Page, e a versão 3 .0, de autoria de Charleston. Ambas podem ser encontradas em http:// www.it.usyd.edu.au/-mcharles/software/software.html.
Os dois trilhogramas da Figura 7.4 foram obtidos com uma análise de Jungles, aplicando custo = O para vicariância e duplicação, custo = 1 para extinção e custo = 2 para dispersão.
Quando foram propostos, os métodos de Huelsenbeck et af. 1 1 , 12 para estudos de associações históricas representaram um avanço em rigor estatístico das pesquisas
Modelos de Eventos para Reconstrução B iogeográfica - 129
deste tipo. Essas metodologias apoiam-se na máxima verossimilhança como critério de otimização e tratam-se, portanto, de abordagens paramétricas que, no caso dos estudos de associações históricas, tomam as filogenias fornecidas pelo pesquisador, incluindo os comprimentos de ramos, como parte de um modelo proposto para compreender a associação. Para Huelsenbeck et al. 1 1 , 12, a frequência de eventos de dispersão em relação à ocorrência de vicariância e a congruência topológica e temporal poderiam ser inferi das por meio da verossimilhança estatística ou por probabilidades posteriores bayesianas, no contexto de um modelo estocástico.
A associação entre dois cladogramas não necessita ser completamente congruente para que se aceite as similaridades filogenéticas como resultantes de uma associação histórica. Não há, contudo, uma medida absoluta do que vem a ser "suficientemente congruente" nessas situações. Para avaliar a significância estatística das associações, Huelsenbeck et al. l l , 1 2 propuseram um procedimento consistindo em dois testes de hipótese principais: no primeiro, congruência verdadeira é diferenciada de mera similaridade entre duas histórias. Caso a associação entre cladogramas seja confirmada como congruente, o próximo passo seria a comparação das idades dos eventos de divergência entre as histórias sendo comparadas . Quando há evidências favoráveis à aceitação da congruência em ambos os casos, esta é interpretada como resultante de processos históricos comuns.
A congruência topológica pode ser testada comparando-se o valor de verossimilhança ass ociado à filogenia das espécies à verossimilhança da filogenia das mesmas espécies esperada no caso em que haja congruência perfeita entre as histórias das áreas e dos táxons. A diferença entre os dois valores pode é facilmente testável quanto à significância estatística. De modo semelhante, a congruência temporal entre as árvores de táxons e as áreas poderia ser testada. Como se pode perceber, a avaliação da significância estatística da associação histórica não depende de comparações de parte dos dados originais a um conjunto de árvores aleatórias, como em VMI6. Os métodos probabilísticos descritos aqui realizam os testes estatísticos de significância diretamente a partir das informações observadas (por exemplo, árvores sendo reconciliadas), o que é uma propriedade atraente, pois simplifica a análise.
Apesar dos desenvolvimentos teóricos de Huelsenbeck et ai . representarem avanços metodológicos importantes para o estudo de associações históricas, ao menos quatro modificações poderiam ser incorporadas aos modelos para torná-los mais realistas 1 1 , 12:

1 30 - Métodos e Apl i cações
1 . Duplicação. 2. Extinção. 3 . Distribuição redundante, isto é, mais de uma espécie
em uma mesma área. 4. Taxas de dispersão variáveis ao longo dos ramos da
filogenia dos táxons.
Esses métodos 1 1 . 12 foram inicialmente pensados para estudos de coespeciação entre parasitas e hospedeiros, apesar de ter sido feita menção à possibilidade da aplicabilidade dos mesmos métodos à pesquisa biogeográfica l I . A grande diferença prática entre um caso e outro é que na pesquisa envolvendo a reconciliação entre filogenias de dois grupos de espécies as árvores são obtidas por métodos similares (isto é, árvores obtidas por verossimilhança e incluindo comprimentos de ramos a partir de dados moleculares, uma para o hospedeiro e outra para o parasita) . Neste caso, as filogenias são extremamente comparáveis em muitos aspectos, até mesmo na taxa estimada de substituição dos nucleotídeos ao longo do tempo. Quando se representam histórias de áreas para estudos biogeográficos, pode-se utilizar um cladograma para a representação desta história, mas sua comparabilidade à filogenia dos organismos distribuídos nas áreas é mais limitada.
Incerteza Topológica
A qualidade das conclusões de qualquer estudo biogeográfico que se baseie em hipóteses filogenéticas é dependente da qualidade destas hipóteses25, mas, em geral, é difícil a expressão da confiança a elas associada. Na quase totalidade dos casos, busca-se a melhor hipótese de relacionamento filo genético para um conjunto de táxons e esta será assumida como correta para a realização das análises biogeográficas subsequentes. No caso da inferência bayesianal l , admite-se a impossibilidade da certeza quanto às relações representadas por qualquer hipótese filogenética. Por outro lado, o valor de verossimilhança atribuído a cada hipótese permite a definição de probabilidades distintas para qualquer topologia (e comprimentos de ramos a ela associados).
A inferência bayesiana de associações 10 foi o primeiro método capaz de lidar com incerteza acerca das topologias das árvores utilizadas para representar as relações entre os táxons e entre as áreas. Segundo esta proposta, todas as hipóteses filogenéticas para as relações entre os táxons podem ser testadas, pois estas são consideradas conjuntamente e recebem um peso relativo proporcional à probabilidade de estarem corretas diante da evidência empírica disponível 10. 1 1 ,25. Desse modo,
vários cenários biogeográficos podem ser considerados conjuntamente, porém com atribuição de probabilidades relativas que se associam às probabilidades das hipóteses filogenéticas a que se referem.
Como o próprio nome sugere, o princípio deste método é o de reconciliação de árvores, de modo similar aos métodos descritos anteriormente : VM, Jungles e abordagens probabilísticas. Contudo, a estrutura dos modelos de eventos e os cálculos dos parâmetros são radicalmente diferentes entre esses métodos4. No caso de ajuste parcimonioso de árvores (do original em inglês, parsimony-based tree fitting), há um modelo explicitamente constituído pelos quatro eventos normalmente considerados em análises deste tipo: vicariância, duplicação, dispersão e extinçã04.2o. Este método possibilita a inferência da significância estatística da frequência estimada de cada um dos eventos por meio de permutações do cladograma de táxons, após a reconciliação das árvores4,26. Além disso, as análises permitem grande flexibilidade na determinação dos custos dos diferentes eventos do modelo. O cenário biogeográfico mais parcimonioso pode ser calculado buscando-se a reconstrução histórica mais parcimoniosa (menor custo total). As relações entre táxons individuais podem ser combinadas para elaboração de cladogramas gerais de áreas ou pode-se verificar a congruência entre as histórias dos táxons e um cladograma de áreas obtido, por exemplo, a partir de reconstruções geológicas1 3·26.
Na implementação do método, no programa TreeFit
ter20, podem ser reconciliados cladogramas de um ou vários táxons à árvore representando a história das áreas de distribuição desses táxons. Segundo os custos predefinidos (default) do modelo do programa, à dispersão é atribuído custo 2 , à extinção custo 1 e custo nulo para vicariância e duplicaçã04•2o• Esta combinação foi demonstrada como adequada para o reconhecimento eficiente do sinal histórico na maioria dos estudos biogeográficos em que a vicariância teve papel relevante na evolução espacial das linhagens de organismos4,26. A lógica dos custos reduzidos de vicariância e duplicação, em relação às dispersões e extinções, é que esses eventos geram padrões de distribuição congruentes com relações filogenéticas4•27. Por outro lado, extinções e dispersões são eventos que tendem a gerar distribuições discordantes da história das áreas e, por conseguinte, incongruentes com padrões biogeográficos.
Qualquer outra combinação de custos pode ser utilizada nas análises, assim como a realização de testes que

Pranchas Coloridas
Figura 12.1 - Áreas de endemismo identificadas para vertebrados terrestres na Amazônia
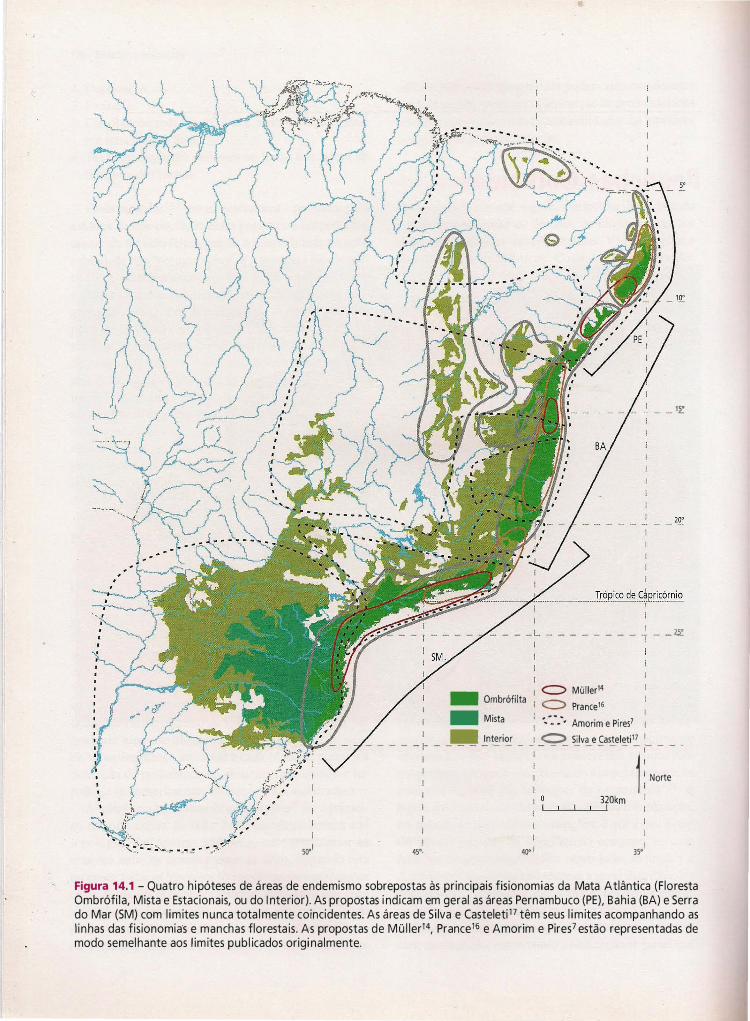
50,1
_ _ 12:'
1 ___ L ________ { __ �
1 c:::> Müller14 _ Ombrófilta 1 _ 1 c:::> Prance16 1 I Mista I • .-::. Amorim e Pires) I I I _ Interior 1 c:::::::> Silva e Casteleti17 1
---- - r ------- - -------- ----- -
, ' j ! Nort.
I 45'· 40' 1
320km 1 I
1 I
35'
Figura 14.1 - Quatro h ipóteses de áreas de endemismo sobrepostas às principais fisionomias da Mata Atlântica (Floresta Ombrófila, Mista e Estacionais, ou do I nterior). As propostas i nd icam em geral as áreas Pernambuco (PE), Bah ia (BA) e Serra do Mar (SM) com lim ites nunca tota lmente coincidentes. As áreas de Silva e Casteleti17 têm seus lim ites acompanhando as l i nhas das fisionomias e manchas floresta is. As propostas de Mü l ler14, Prance16 e Amorim e Pires7 estão representadas de modo semelhante aos l im ites publ icados originalmente.

Figura 14.2 - Espécies de opil iões das subfamílias endêmicas da Mata Atlântica. (A) Thereza poranga - Caelopyginae; (8) Goniosoma carum - Goniosomatinae; (C) Heliella singularis - Progonyleptoidell inae; (D) Sodreana sodreana - Sodrean inae; (E) Tribunosoma disciêpans - Heteropachylinae; (F) Asarcus longipes - Bourguyiinae; (G) Piassagera brieni - Hernandari inae.
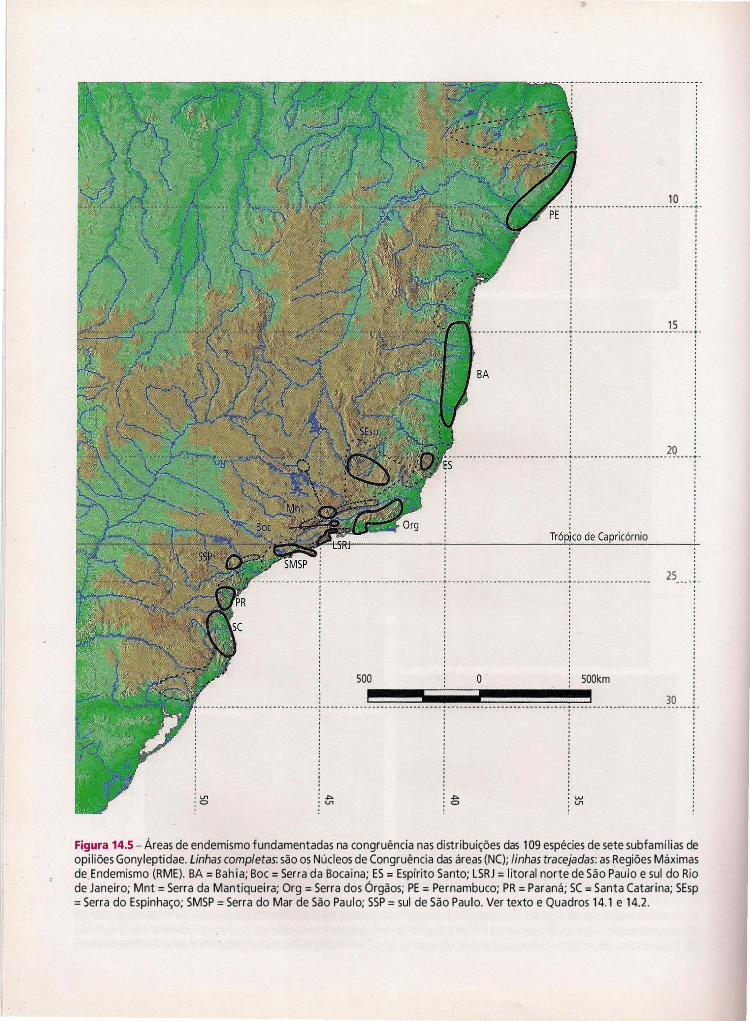
1 0
15 - - � - _ .. � _R. ____ - _ .......... + ____ .. __ .. ___ ...... _______ ............ __ ..
. ___________________________ • _______________________ ?9 ____ •. o o o o o o o o o o o o
icórnio
___ _ __ _ _ _ _ ._. ; __ _ ' _ _ . __ . ___________________ � _____________________________ � ______ _ _ _ _ _ ____ _ _ _ _ 25
___ -l-
500 o 500km
. - .. - - - - - - --. -. - - - - - -- - - - ---- f -- -- - - - . - - - - ----------- - - - - ---: - - - - -- -- -------.... --. -- - --. -I -.---------.. --- --.-.--�9·--·t·
\J1 .j::> .j::> o \J1 o
o o
w \J1
Figura 14.5 - Áreas de endemismo fundamentadas na congruência nas distribuições das 109 espécies de sete subfamí lias de opil iões Gonyleptidae. Unhas completas: são os Núcleos de Congruência das áreas (NC); linhas tracejadas: as Regiões Máximas deJndemismo (RME). BA = Bahia; Boc = Serra da Boca ina; ES = Espírito Santo; LSRJ = l itoral norte de São Paulo e sul do Rio de Janeiro; Mnt = Serra da Mantiqueira; Org = Serra dos Órgãos; PE = Pernambuco; PR = Paraná; SC = Santa Catarina; SEsp = Serra do Espinhaço; SMSP = Serra do Mar de São Paulo; SSP = sul de São Paulo. Ver texto e Quadros 14.1 e 14.2.
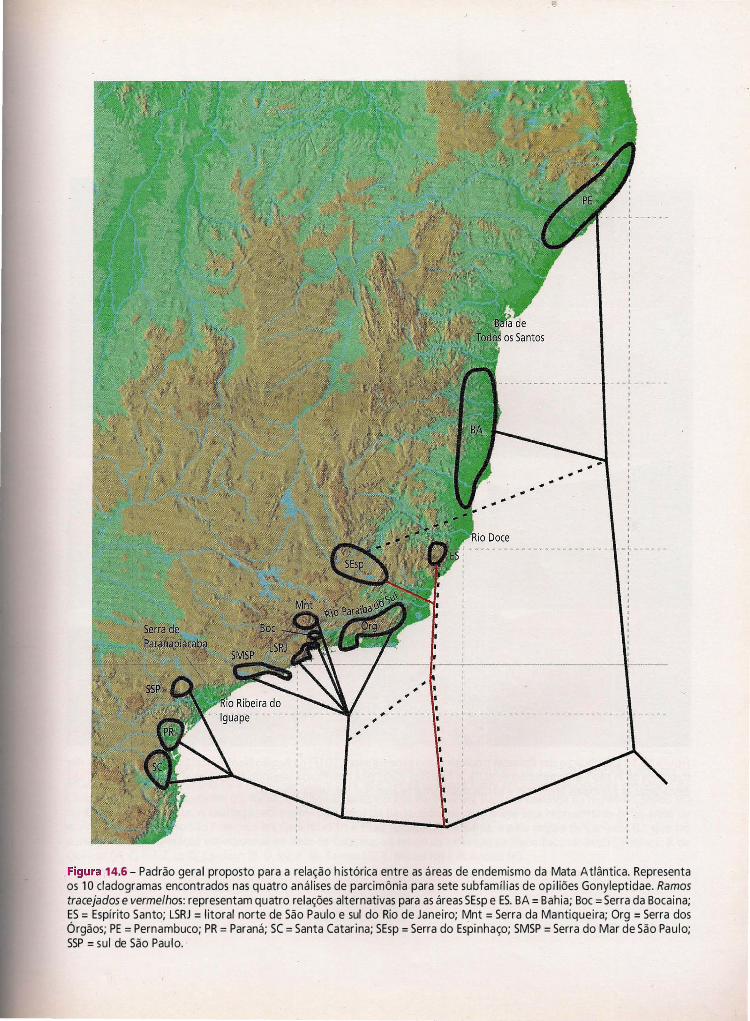
.. ..
.. .. ....
_ N _l. _____ �
, _L ____ N _ ,
, F igura 14.6 - Padrão geral proposto para a relação h istórica entre as á reas de endemismo da Mata Atlântica. Representa os 10 cladogramas encontrados nas quatro aná l ises de parcimônia para sete subfamíl ias de opiliões Gonyleptidae. Ramos tracejados e vermelhos: representam quatro relações alternativas para as áreas SEsp e ES. BA = Bahia; Boc = Serra da Bocaina; ES = Espírito Santo; LSRJ = litoral norte de São Paulo e sul do Rio de Janeiro; Mnt = Serra da Mantiqueira; Org = Serra dos Órgãos; PE = Pernambuco; PR = Paraná; SC = Santa Catarina; SEsp = Serra do Espinhaço; SMSP = Serra do Mar de São Pau lo; SSP = su l de São Paulo.

S
• SDTF
Caatinga arenosa
Savana
• Floresta úmida
• Chaco
Figura 15.1 - Distribuição das florestas tropicais sazonalmente secas (SDTF) na Região Neotropica l (modificado a partir dos trabalhos de Prado 10 e Penn ington et al. 11). A caatinga (Caa) está del im itada segundo Vel loso et a/.12, dentro da qua l são destacadas as principais á reas sobre bacias sedimentares a renosas (caatinga arenosa).

Figura 15.2 - Diferentes tipos vegetacionais de caatinga sobre embasamento cristalino (A a I), representados durante as estações seca (A a F) e úmida (G a I), com destaque para as áreas de caatinga sobre afloramentos calcários em Morro do Chapéu (D) e Serra do Ramalho (E), Bahia; caatinga arbórea em Ruy Barbosa (G), Bah ia; e caatinga da região do Seridó (H) no Rio G rande do Norte. A caatinga sobre solos derivados da bacia sedi mentar arenosa está representada pelas á reas de dunas interiores do Rio São Francisco em Casa Nova (1) e Pi lão Arcado (K) na Bahia, e pela região de Canudos (L), que faz parte da mesma formação vegetacional do Raso da Catarina, ao longo da extensão da Bacia Tucano-Jatobá. Fotos: L. P. de Queiroz (A, C, F, I, J, L); D. Cardoso (B, D, H); R . M. Santos (E); E. Matos (G) e A. Rapini (K).

Figura 15.3 - Exemplos da d iversidade florística de leguminosas de caatinga: A = Libidibia ferrea; B = Amburana cearensis; C = Mimosa modesta; D = Pterocarpus vi/losus; E = Mimosa verrucosa; F = Erythrina ve/utina; G = Apu/eia /eiocarpa; H =
Craty/ia mo/lis; I = Coursetia rostrata; J = Mimosa /epidophora; K = Pterocarpus monophy/lus; L = Aeschynomene martii; M = Poinciane/la pyramida/is; N = Luetze/burgia bahiensis; O = Goniorrhachis marginata; P = Pithece/lobium diversifo/ium; Q = Senna bracteosa; R = Trischidium mo/le; 5 = Mysanthus u/eanus; T = Chaetoca/yx scandens. Fotos: D. Cardoso (A a J, L a Ti e A. P. Prata (K).

permitam estimar os valores mais apropriados para um conjunto de dados26. Com Tree Fitting, pode-se buscar aplicar o princípio de que os custos de cada evento, de modo ideal, seriam inversamente proporcionais à sua probabilidade de ocorrência2 e a hipótese escolhida é aquela que minimiza o custo total dos eventos assumidos. O próprio TreeFitter pode ser usado para este tipo de avaliação de custos dos eventos do modelo, como demonstrado por Ronquist4.
De maneira análoga à análise de significância estatística de VM, com TreeFitter as árvores utilizadas nas análises podem ser aleatorizadas por meio de permutações4•26. Entretanto, em vez de se modificar completamente a topologia de um dos cladogramas, como em VM, com o TreeFitter, a posição dos terminais é rearranjada para realização dos testes de significância3.4. O custo da associação entre os cladogramas originais é comparado à distribuição de custos de associação gerada por meio da permutação para se avaliar a significância do custo de associação para os táxons e áreas estudados. O procedimento de permutações permite, dependendo de como for realizado, compreensão mais detalhada da associação histórica, buscando-se estimar, por exemplo, a significância de uma classe de eventos específica (por exemplo, dispersão), ou a avaliação da importância da vicariância para um conjunto de áreas em comparação com outras áreas.
Quando uma hipótese geral para o padrão de relacionamento entre áreas não estiver disponível, pode-se ainda assim realizar análises de Tree Fitting, porém os resultados não seriam muito diferentes daqueles obtidos com DIVA (ver a seguir)26.
Análises com TreeFitter dependem de um cladograma dicotômico representando as relações entre os táxons. Caso o cladograma não seja completamente resolvido, o programa resolve as politomias de modo arbitrário, o que não é desejável nem justificável. Nesses casos, é preferível que as análises de todos os cladogramas compatíveis com as politomias originais sejam fornecidas ao programa e posterior interpretação do impacto deste ato sobre os resultados obtidos.
Muitas das possibilidades metodológicas de Tree -= Fitting foram exploradas por Sanmartín e Ronquist26. x Naquele trabalho, foi realizada uma síntese dos padrões � biogeográficos gondwânicos. Um conjunto de 73 estudos � de caso de animais e plantas que se distribuem primax riamente pelos continentes austrais foi analisado con'"
juntamente em análises de Tree Fitting, de modo a elucidar padrões comuns, além de terem sido comparados a padrões gerais entre áreas inferidos a partir de dados geológicos.
Modelos de Eventos para Reconstrução B iogeográfica - 13 1
Tratamento de Táxons Amplamente Distribuídos Em análises de biogeografia cladística, táxons amplamente distribuídos necessitam de um tratamento especial, pois podem mascarar o sinal histórico de vicariância de um conjunto de dados, visto que são dados ambíguos. No caso das análises de componentes, a escolha dos pressupostos AO, A 1 e A2 reflete interpretações distintas das informações provenientes desses táxons (ver Capo 6), mas não são facilmente aplicáveis em análises de eventos. Na tentativa de solucionar este problema e propor uma maneira de tratar táxons amplamente distribuídos de modo satisfatório, Sanmartín et al.28 propuseram uma interpretação alternativa aos pressupostos, porém também incluindo três opções, denominadas "Livre", "Recente" e "Ancestral", aplicáveis em Tree Fitting26. Segundo a opção Ancestral, táxons amplamente distribuídos seriam resultantes de eventos de duplicações (mas extinções também são permitidas) e todas as áreas do terminal são, então, interpretadas como ocupadas por seu ancestral (similar a uma combinação dos pressupostos AO e A 1 )26.27.' Com a opção Livre, entende-se que uma combinação de eventos é necessária para explicar sua distribuição ampla (similar ao pressuposto A2). Os terminais amplamente distribuídos são interpretados como um conjunto politômico de terminais, cada um destes distribuído em apenas uma das áreas em que o terminal verdadeiro ocorre26.27. Por fim, com a opção Recente, assume-se que dispersão(ões) recente(s) é(são) responsável(is) pela distribuição ampla e que apenas uma das áreas ocupadas pelo táxon terminal constituiria a área do seu ancestral (não há um paralelo entre está opção e os pressupostos da análise de componentes )26.27 .
A DIVA é um dos poucos métodos de análise biogeográfica que permite a formulação de um cenário para a história biogeográfica de um grupo sem a necessidade de qualquer hipótese prévia para história geológica das áreas, um cladograma geral de áreas. Tudo quanto é necessário para a realização da análise é uma hipótese filogenética para um grupo de organismos e a distribuição geográfica de cada terminal em uma ou mais áreas. A partir desse conjunto de dados, DIVA reconstrói as distribuições ancestrais e especifica os eventos biogeográficos que priorizam hipóteses de vicariância em detrimento de hipóteses de dispersão. Possivelmente em decorrência dessas características, DIVA se tornou um dos métodos biogeográficos mais amplamente empregados nos últimos anos29.

1 32 - Métodos e Apl icações
Com DIVA, áreas ancestrais são estimadas por meio de um procedimento de otimização, similar à otimização de caracteres em um cladograma18, 19,30. Com isso, pode-se inferir a prevalência de eventos vicariantes para explicação da história dos táxons e a ordem (ou até mesmo a idade) desses eventos de vicariância pode ser comparada a padrões biogeográficos gerais para as áreas às quais os organismos estudados se associam.
Da mesma forma que em uma otimização de caracteres convencional, a análise DIVA calcula a distribuição de cada nó considerando ao mesmo tempo as distribuições dos nós imediatamente anterior e posterior. Ronquist18 ,19,30 representou o problema da inferência das distribuições ancestrais na forma de matriz tridimensional de custos, em referência às três variáveis analisadas ao se inferir uma distribuição ancestral (pensado como estado de um caráter) :
1 . Estado imediatamente ancestral ao nó sendo analisado. 2. Estado do primeiro descendente. 3 . Estado do segundo descendente.
A otimização compõe-se de duas passagens pela árvore30: a primeira, dos terminais em direção à raiz (passagem preliminar) e, uma segunda, em direção aos terminais (passagem final), para determinar as distribuições ancestrais dos ancestrais e eventos necessários para se chegar a tais distribuições.
Quanto maior for a restrição imposta ao número máximo de áreas das reconstruções dos nós, maior é a semelhança entre otimização de caracteres de Fitch e DIVA (quando o número máximo é 1, os dois procedimentos são equivalentes). As reconstruções são realizadas utilizando-se uma matriz tridimensional de custos de eventos, de acordo com as seguintes regras1 8 :
1 . Não há custo associado à vicariância. Neste caso, assume-se que a especiação ocorreu após a divisão da área ancestral em dois conjuntos de áreas mutuamente exclusivas.
2. Duplicação também recebe custo zero. 3 . O custo de eventos de dispersão é um, por unidade de
área adicionada à distribuição. 4. Eventos de extinção recebem custo um, por unidade
de área eliminada da distribuição,
O uso de custos relativos para mudanças dos estados de caráter considerados, no caso, áreas de distribuição, torna o cálculo de DIVA similar à otimização de Sankoff, que também se baseia em matriz de passos para estimar a reconstrução mais plausível para um nó30. A
possibilidade de reconstrução com custo zero permite que vicariância seja preferencialmente assumida para explicação de determinado caso; custos positivos para os eventos de dispersão e/ou extinção fazem com que estes eventos sejam assumidos apenas em condições particulares.
Para o cálculo das matrizes de custo tridimensionais, uma das preocupações matemáticas é a possibilidade de �
-J violação do princípio matemático da metricidade do ;k
u. model03o. Para evitar violações à metricidade da matriz .:., N e garantir que dispersões e extinções sej am penalizados : em relação às explicações vicariantes, é necessário que �
"" duplicação tenha custo nul03o. Além disso, nenhum evento deve receber custo negativo, de modo que vicariância também recebe custo zero, ao passo que dispersões e extinções têm custo um, por evento.
Em razão da simplicidade geral do método e de seu modelo, e principalmente do fato de as conclusões poderem se derivar de apenas um estudo de caso, os resultados de DIVA devem ser tratados com extremo cuidadoI8 ,29,3 1 . A reconstrução fornecida para os nós mais antigos merecem, de modo amplo, o maior ceticismo na interpretação, o que é uma recomendação para procedimentos de otimização em geral18. O nó mais antigo de uma árvore normalmente é o mais afetado por esse tipo de incerteza e, segundo Ronquist 1 8,19, a reconstrução deste nó só pode ser realizada com alguma confiança quando o grupo irmão do clado de interesse e, se possível, outras linhagens próximas forem amostradas para o estudo.
É comum que o resultado da análise contenha incerteza quanto às reconstruções propostas, refletindo o grande número de hipóteses igualmente parcimoniosas por vezes disponíveis para um determinado problema. Além da possibilidade de várias reconstruções do mesmo modo plausíveis, pode-se também obter nós antigos reconstruídos como tendo sido amplamente distribuídos. Neste caso, pode-se artificialmente restringir o número máximo de áreas incluídas nas reconstruções, de modo a verificar o efeito deste procedimento sobre o resultado da análiseI8,19. A análise depende de um cladograma dicotômico por completo representando as relações entre os táxons, além de informações sobre as áreas de ocorrência de cada táxon. Caso o cladograma não seja completamente resolvido, pode-se realizar a análise com cada um dos cladogramas compatíveis com as politomias originais e fazer avaliação a posteriori dos efeitos das várias análises sobre as conclusões biogeográficas finais que se pode obter do conjunto de resultados de DIVAI8.
A implementação original de DIVA não permite lidar com a incerteza topológica do relacionamento entre

táxons. Nylander et al.25 propuseram uma análise denominada Bayes-DIVA e Yu et af32 desenvolveram outro método denominado S-DIVA. Esses métodos são modificações do método DIVA original, capaz de explorar o efeito da incerteza filogenética sobre os resultados biogeográficos. Em vez de realizarem a inferência sobre as áreas ancestrais (e eventos biogeográficos responsáveis por tais distribuições hipotéticas) considerando apenas uma hipótese filogenética, Bayes-DIVA e S-DIVA podem analisar simultaneamente uma amostra de árvores grande o suficiente para representar as incertezas topológicas25,32.
Hierarquia e Reticulação
Quando a história de um conjunto de áreas atuais pode ser explicada por uma série de eventos de vicariância que acarretam quebra de uma área ancestral em áreas cada vez menores, esta história pode ser representada por um diagrama ramificado (isto é, cladograma) de áreas, pois é perfeitamente hierárquica. Apesar de cladogramas serem
Austrál ia
I '�- Nova Guiné
Modelos de Eventos para Reconstrução B iogeog ráfica - 1 33
representações muito úteis para as relações de parentesco entre grande parte dos organismos vivos, a evolução de áreas não necessariamente se dá de fonna hierárquica. Não se sabe se histórias de áreas geográficas exclusivamente divergentes, como em hierarquias, são raras. A hipótese mais conhecida de uma história geológica hierárquica é de Gondwana, em que se supõe que quebras sucessivas desse supercontinente ao longo dos últimos 160 Ma (Fig. 7.5, A) teriam dado origem aos continentes austrais atuais: Antártica, África, América do Sul, Austrália, além de Madagascar e a placa Índia26. Por outro lado, a história do supercontinente Laurásia não pode, de maneira alguma, ser considerada hierárquica (Fig. 7.5, B)28. O aspecto dinâmico da história do planeta tanto contribui para a formação de barreiras e novas configurações espaciais, como, em alguns casos, resulta na aproximação de regiões anteriormente separadas (disjuntas) e desaparecimento de barreiras geográficas. Isso sendo verdade, os padrões formados são reticulados e representações hierárquicas nem sempre constituem o modelo mais fiel da evolução espacial.
---Sul da América do Sul
B
Nova Caledônia
Nova Zelândia
índia
Madagascar
África
Norte da América do Sul
- Histórias dos supercontinentes Gondwana e Laurásia. (A) A maior parte das h ipóteses para a sequência de eventos geológicos que ter ia l evado à fragmentação de Gondwana ao longo dos ú lt imos 1 60 m i lhões de anos26. (B) A reconstrução da h istória geológ ica do supercont inente Lau rásia i nc lu iu disj unções geradas pelos movimentos tectônicos e reconexões entre as á reas, resu ltando em h istória essenc ia lmente ret icu lada entre as reg iões Neártica e Paleárt i ca28: houve períodos quando a porção oeste da Reg ião Paleártica conectava-se à porção leste do Neártico por pontes transatlânticas ao mesmo tempo em que a porção leste do Paleárt ico conectava-se ao oeste do Neárt ico através do estreito de Bering; houve períodos em que as quatro á reas tiveram pouca conexão; e períodos, como o atual , em que a região Neártica forma um continente (América do Norte) e a Reg ião Pa leártica forma outro (Eurásia), separados um do outro.

134 - Métodos e Ap l i cações
DIVA seria particulannente útil para casos chamados de reticulados, em que há uma história de vicariância, mas com alguns relacionamentos entre áreas do mesmo estudo tendo sido gerados por vários outros processos. DIVA ajuda a apresentar uma explicação histórica que pode ser comparável a padrões mais gerais, ao menos no que diz respeito aos eventos vicariantes reconhecíveis, já que hipóteses não estritamente hierárquicas (isto é, reticuladas) são tão aceitáveis quanto as hierárquicas. Isso depende apenas do peso das evidências a favor de cenários de vicariância ou da inclusão de novos eventos.
Este método foi originalmente proposto para biogeografia, em vez de se tratar de uma adaptação de métodos inicialmente pensados para estudos de coespeciação ou de evolução gênica. O método compartilha com DIVA a característica de não depender de uma hipótese prévia para a história das áreas ocupadas pelos táxons de interesse. O estudo da evolução da distribuição por meio de uma análise de verossimilhança é um dos métodos de eventos aplicado à biogeografia mais recentemente propostos2J.22. Segundo os autores do método, não são buscadas histórias gerais de vicariância, mas procura-se compreender como a distribuição de uma linhagem de organismos modificou-se ao longo da evolução, resultando nas áreas de ocorrência de seus descendentes . Trata-se de uma proposta que reduz a ênfase na procura de cladogramas gerais de áreas e enfoca, de modo específico, o estudo de casos individuais.
De forma simplificada, o modelo biogeográfico necessita de três componentes2 1 : hipótese filogenética para o grupo de estudo, áreas de distribuição dos grupos (assumidas como unidades estáveis ao longo do tempo) e informações paleogeográficas, paleogeológicas e/ou quaisquer dados históricos que possam esclarecer possíveis rotas de dispersão e conexão pretérita entre as áreas. Apesar de os autores do método tratarem qualquer evento que resulte em alteração da área ancestral como sendo um caso de dispersão, vale ressaltar que se trata de uma simplificação analítica, pois incluem-se aqui tanto casos de transposição de barreiras geográficas (=
dispersão no sentido mais restrito) , como de expansão de área de distribuição.
A reconstrução desta história é realizada por meio de uma abordagem paramétrica em que ocorrência de dispersão, extinção e resposta à vicariância se dão segundo
um modelo estocástico de evoluçãos. A utilização de modelos estocásticos é uma característica compartilhada com as abordagens probabilísticas de Huelsenbeck et al .ll, 12 descritas anteriormente. O comprimento de ramos (ou idade absoluta estimada das linhagens) é de crucial importância, uma vez que o tempo transcorrido é proporcional à verossimilhança de ocorrência dos eventos representados no modelo. Este método propõe-se a integrar infonnações de várias fontes e fornecer um cenário biogeográfico que busca elucidar a variação histórica das distribuições dos ancestrais hipotéticos dos táxons estudados, de modo similar à DIVA, mas com maior sofisticação analítica. As áreas em que os táxons se distribuem são tratadas como estados de caráter otimizados sobre a hipótese filogenética para esses táxons. As distribuições geográficas dos ancestrais são estimadas considerando-se três fontes de infonnação:
1 . Probabilidades de dispersão e extinção aparecem no modelo para o estudo de caso.
2. Comprimentos dos ramos adjacentes. 3. Distribuição geográfica de nós e/ou de tenninais ad
jacentes.
Contudo, diferentemente dos métodos usuais de otimização de estados de caracteres que preveem a herança da condição ancestral pelos descendentes, no caso desta análise21 ,22 os estados (= áreas de distribuição) das espécies resultantes de uma especiação podem ser distintos.
As hipóteses produzidas por esta análise são apresentadas em tennos de probabilidades e não como a única hipótese plausível (ou conjunto de hipóteses igualmente plausíveis); neste último caso sem que haja avaliação quantitativa da verossimilhança dos eventos incluídos na hipótese. Na versão mais recente do método, Ree e Smith22 passaram a chamar o modelo de DEC, em referência aos três elementos que o compõem: dispersão, extinção e cladogênese. Este modelo foi implementado em linguagem Python no programa LAGRANGE (do original em inglês, Likelihood Analysis of Geographic Range Evolution; disponibilizado em: http://code.google. comlp/lagrange/).
Trata-se, sem dúvida, de uma proposta ambiciosa, que certamente será ainda testada e aperfeiçoada nos próximos anos. No futuro breve, poderá revelar se tratar de um método importante para iluminar histórias de táxons e áreass,29. Em comparação à DIVA, por exemplo, uma das inovações atraentes do modelo DEC é a possibilidade da avaliação da variação das taxas de dispersão para as áreas de análise e incorporação desta variação ao modelo de eventoss.

..:;
Todos os métodos de padrão abordados neste capítulo oferecem a possibilidade de incorporação das idades das linhagens e de cada um dos clados de uma filogenia na análise. No caso de se possuir informações confiáveis sobre as idades dos grupos, isso representa uma fonte de evidências espetacular para análise e discussão de cenários biogeográficos. Além da congruência espacial, pode-se estudar a congruência temporal das histórias de grupos ocupando as mesmas regiões do planeta concomitantemente5,13-15 . Isso é relevante não só para eventos de vicariância, que permitem buscar evidências geológicas para um determinado padrão, mas para extinção (por exemplo, desaparecimento de várias linhagens após um evento físico-químico) ou dispersão (por exemplo, favorecimento da dispersão de uma área para outra durante alguns milhões de anos, quando as correntes de \'ento facilitaram transposição de uma barreira).
Havendo hipóteses sobre as idade dos clados, tem-se também conhecimento sobre as idades do eventos apontados por exemplo, pela análise de dispersão-vicariância. Partindo-se de evidências para um evento de vicariância promovendo separação de linhagens distribuídas em duas áreas anteriormente conectadas, podem-se buscar outros grupos cujas histórias indiquem um cenário similar, em
� especial quando se têm ideias sobre os períodos quando :::: se passaram essas histórias3-15. Pode-se, portanto, passar x de um estudo de caso e buscar-se histórias congruentes x
entre si, ou hipóteses para determinado táxon, que seja congruente à hipótese geológica para uma área15,25,27.
Por outro lado, há histórias biogeográficas de grupos para os quais não se espera congruência com padrões mais gerais. Este é o caso das histórias de dispersões e colonização de ilhas oceânicas não originadas por vicariância. Para lidar com esse tipo de situação, Sanmartín et al.33 desenvolveram uma abordagem bayesiana em que as taxas de dispersão são os parâmetros mais relevantes do modelo biogeográfico, em vez da vicariância. É bem possível que este e outros métodos de eventos sej am cada vez mais utilizados para iluminar inferências biogeográficas em que a biogeografia de vicariância tradicional tem pouco a contribuir33.
Apesar das diferenças, à primeira vista, muito acentuadas entre métodos de padrão (ver Caps. 5 e 6) e de eventos, os objetivos de qualquer análise, em última
Modelos de Eventos para Reconstrução B iogeográfica - 135
instância, residem em se encontrar evidências para padrões de vicariância (ver Capo 4). A diferença entre métodos de eventos e outras classes de métodos analíticos de biogeografia histórica é principalmente epistemológica. Ademais, Ronquist4 demonstrou ser matematicamente possível converter um método de padrão da biogeografia cladística, como BPA, num modelo de como dos métodos de eventos.
Uma questão relevante, contudo, diz respeito à escolha da análise mais apropriada para uma pesquisa. Em meio à tamanha diversidade de opções para análises biogeográficas, qual o melhor método para um determinado estudo? Embora a frustração que isso possa causar, atualmente não há uma resposta para essa questão. Desde o primeiro contato com os dados de distribuição de um grupo de espécies, há muitas opções metodológicas a se considerar. O melhor modo de se começar a compreender as diferenças e as vantagens relativas dos vários métodos é testando-os aos dados disponíveis e por meio do estudo da literatura primária para compreensão mais aprofundada do assunto. A separação entre métodos de eventos e métodos de padrão, métodos que dependem do uso de hipóteses filogenéticas para os táxons e aqueles não dependem disso ajudam a categorizar essas formas de analisar os dados, mas a decisão das melhores opções em situações diversas é algo que realmente se amadurece com a familiaridade teórica e empírica . Deve-se lembrar ainda que a comparação entre resultados de mais de uma abordagem pode ser muito útil para que evidências históricas sejam iluminadas e elucidadas5,29,31. Finalmente, e não menos importante, a integração de várias metodologias é possível e recomendada, até porque os métodos são complementares, mais que exclusivos ou concorrentes9.
A única previsão segura sobre o futuro da biogeografia analítica é de que novos métodos e abordagens empíricas continuarão sendo propostos. Os métodos tratados neste capítulo não existiam formalmente há poucas décadas, mas o número de novas publicações recentes de cunho teórico-metodológic05,13,22,25,32,33 sobre o assunto indica o vigor desta escola da biogeografia.
Sou profundamente grato aos responsáveis por minha iniciação à biogeografia histórica, Jim Liebherr e Melissa Luckow, dois brilhantes cientistas e educadores. Agradeço aos comentários e sugestões oferecidos por Claudio J. B . de Carvalho, Júlia C. Almeida e dois revisores anônimos.

1 36 - Métodos e Ap l i cações
REFERÊNCIAS BIBLIOGRÁFICAS 1. NELSON, G. 1.; PLATNICK, N. L Systematics andBiogeogra
phy: Cladistics and Vica rianee. New York: Columbia University Press, 1981. 567p.
2. RONQUIST, F; NYLIN, S. Process and pattern in the evolution of species associations. Syst. Zool., v. 39, p. 323-344, 1 990.
3. RONQUIST, F Phylogenetic approaches in coevolution and biogeography. Zool. Ser., v. 26, p. 313-322, 1997.
4. RONQUIST, F Parsimony analysis of coevolving species associations. In: PAGE, R. D. M. Tangled Trees: Phylogeny, Cospeeiation, and Coevolution. Chicago: Chicago University Press, 2003. capo 2, p. 22-64.
5. REE, H. R.; SANMARTÍN, I. Prospects and challenges for parametric models in historical biogeographical inference. 1. Biogeogr., V. 36, p. 1 211-1220, 2009.
6. PAGE, R.; CHARLESTON, M. A. Trees within trees: Phylogeny and historical associations. Trends Ecol. Evol., V. 13, p. 356-359, 1998.
7 . PAGE, R. D. M. Genes, organisms, and areas: The problem of multiple lineages. Syst. Biol., V. 42, p. 77-84, 1993.
8 . PAGE, R. D. M. Maps between trees and cladistic analysis of historical associations among genes, organisms, and areas. Syst. Biol., v. 43, p. 58-77,1994.
9. CRISCI, J. V; KANTINAS, L.; POSADAS, P. Historieal Biogeography. An Introduction. Cambridge: Harvard University Press, 2003 . 250p.
10. PAGE, R. D. M. Quantitative c1adistic biogeography: constructing and comparing area cladograms. Syst Zool., V. 37, p. 254-270, 1988.
11. HUELSENBECK, J. P; RANNALA, B . ; LARGET, B.A Bayesian framework for the analysis of coespeciation. Evolutian, V. 54, p. 352-364, 2000.
12. HUELSENBECK, J. P.; RANNALA, B . ; LARGET, B . A statistical perspective for reconstructing the history of host-parasite associations. In: PAGE, R. D. M. Tangled Trees: Phylogeny, Cospeciation, and Coevalution. Chicago: Chicago University Press, 2003. capo 4, p. 93-119.
13. SANMARTÍN, L Event-based biogeography: Integrating pattems, processes, and time. In: EBACH, M. C.; TANGNEY, R. S. Biogeography in a Changing World. Boca Rataon, CRC Press, 2007. cap. 6, p. 135-159.
1 4. PAGE, R. D. M. Temporal congruence and cladistic analysis of biogeography and cospeciation. Syst. Zool., V. 39, p. 205-226, 1990.
1 5 . UPCHURCH, P. Gondwana break-up: legacies of a 10st world? Trends Eca!. Eval., V. 23, p. 229-236, 2008.
J 6. PAGE, R. D. M. Paralle1 phylogenies: Reconstructing the history ofhost-parasite assemblages. Cladistics, V. 10, p. 1 55-173, 1994.
17. CHARLESTON, M. A. Jungles: A new solution to the host/ parasite phylogeny reconciliation problem. Math B/usei., v. 149, p. 191-223, 1998.
18. RONQUIST, F. Dispersal-Viearianee Analysis (DN A). Versão 1.1. Programa para computador e manual, 1996. Disponível em: http://www.ebc.uu.se/systzoo/research/diva/diva.html. Acesso em 20/ago/2009.
19. RONQUIST, F. Dispersal-vicariance analysis: A new biogeographic approach to the quantification ofhistorical biogeography. Syst. Biol., V. 46, p. 195-203, 1997.
20. RONQUIST, F TreeFitter. Versão 1.0. Programa para computador e manual, 200 1 . Disponível em: http://www.ebc.uu.se/ systzoo/research/treefitter/treefitter.htm1. Acesso em 20/ ago/2009.
21. REE, R. H.; MOORE, B. R.; WEBB, C. O. ; DONOGHUE, M. J. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution, v. 59, p . 2299-2311, 2005.
22. REE, R. H. ; SMITH, S. A Maximum likelihood inference of geographic range evolution by dispersaI, local extinction, and cladogenesis. Syst. Biol., V. 57, p. 4-14, 2008.
23. PAGE, R. D. M. TreeMap. Versão 1.0. Programa para computador e manual, 1995. Disponível em: http://taxonomy.zoology. gla.ac.uk/rod/treemap.html. Acesso em 20/ago/2009.
24. CHARLESTON, M. A; PERKINS, S. L. Lizards, malaria, and jungles in the Caribbean. In: PAGE, R. D. M. Tangled Trees: Phylogeny, Cospeciation, and Coevolution. Chicago: Chicago University Press, 2003. Capo 3, p. 65-92.
25. NYLANDER,J.AA.; OLSON, U.;ALSTROM,P; SANMARTÍN, I. Accounting for phylogenetic uncertainty in biogeography: A Bayesian approach to Dispersal-Vicariance Analysis of the thrushes (Aves: Turdus). Syst. Biol., V. 57, p. 257-268, 2008.
26. SANMARTÍN, I.; RONQUIST, F Southem hemisphere biogeography inferred by event-based models: plant versus animal patterns. Syst. Riol., V. 53, p. 2 J 6-243, 2004.
27. SANMARTÍN, I.; RONQUIST, F New solutions to old problems: widespread taxa, redundant distributions and missing areas in event-based biogeography. Anim. Biodivers. Conserv., V. 25, p. 75-93, 2002.
28. SANMARTÍN, L; ENGHOFF, H. ; RONQUIST, F. Pattems of animal dispersaI, vicariance and diversification in the Holarctic. Biol. J. Linn. Soe., V. 73, p. 345-390, 200 1 .
29. KODANDARAMAIAH, U . Use of dispersal-vicariance analysis in biogeography - a critique. J. Biogeogr., V. 37, p. 3-11, 2010.
30. RONQUIST, F Three-dimensional cost-matrix optimization and maximum coespeciation. Cladistics, V. 1 4, p. 167-1 72, 1998.
31. BROOKS, D. R.; MCLENNAN, D. A. A comparison of a discovery-based and an event-based method of historical biogeography. J. Biogeogr., V. 28, p. 757-767, 2001.
32. YU, Y.; HARRIS, A J. ; HE, X. S-DIVA (Statistical DispersalVicariance Analysis) : A to 01 for inferring biogeographic histories. Molec. Phylog. Eval., v. 56, p. 848-850, 2010.
33. SANMARTÍN, 1.;VAN DER MARK, P; RONQUIST, F. Inferring dispersaI: a Bayesian approach to phylogeny-based island biography with specia1 refcrcncc to the Canary Islands. J. Biogeogr., V. 35, p. 428-449, 2008.

Felipe de Mello Martins
M arcus Vinicius Domingues
A filogeografia é definida como o estudo dos princípios e processos que determinam a distribuição geográfica de linhagens genealógicas!. Como o próprio nome indica, a filo geografia lida não somente com as relações filogenéticas entre os táxons estudados, mas com os fatores históricos responsáveis pela distribuição espacial das linhagens. A análise e a interpretação da distribuição de linhagens requerem o processamento conjunto de informações de uma série de disciplinas, incluindo sistemática filogenética, genética de populações, etologia, demografia, paleontologia, geologia e modelos paleogeográficos e paleoclimáticos. Dessa forma, o caráter multidisciplinar da filogeografia faz com que esta disciplina crie uma ponte entre processos micro e macroevolutivos.
Apesar de os processos micro e macroevolutivos serem historicamente tratados de forma separada, estes processos sempre estiveram intimamente ligados, um como extrapolação do outro. Os ramos de uma árvore filogenética, que considera linhagens em escala macroevolutiva que possuem uma sub estruturação hierárquica, dividem-se em ramos cada vez menores, resultando, em última análise, numa relação de pedigree de uma geração para outra (Fig. 8 .1) . Tanto a genética de populações quanto a sistemática filogenética lidam com relações de ancestralidade, pois é por meio desses pedigrees que os genes são transmitidos, traçando o caminho de hereditariedade (conceito da genética de populações) que, por sua vez, estarão impressos na filogenia (conceito da sistemática).
A filo geografia é considerada uma subdisciplina da biogeografia. Estudos filo geográficos incluem preceitos
8
da ecogeografia em uma perspectiva histórica e multidisciplinar, tendo em vista que:
1 . Pressões ecológicas contemporâneas não são as únicas responsáveis pela distribuição espacial de características presentes nos organismos.
2. Seleção natural não é o único mecanismo capaz de gerar padrões geográficos em atributos genéticos, pois para populações delimitadas historicamente e que experimentaram pouco ou nenhum fluxo gênico por longos períodos de tempo, a divergência genética ocorrerá inexoravelmente tanto em genes neutros como em genes sob seleção.
Em sua essência, a filogeografia lida com a distribuição espacial de alelos, cuja relação filogenética pode ser inferida. Porém, ao contrário da percepção comum, esta disciplina não precisa basear-se exclusivamente em dados moleculares. Pesquisas filogeográficas podem ser realizadas com dados morfológicos, comportamentais ou outros quaisquer desde que analisados por uma perspectiva filogenética ao longo de uma distribuição geográfica 1 .
A história da filo geografia está intimamente ligada à história das pesquisas com ácido desoxirribonucleico (DNA, desoxyribonucleic acid) mitocondrial (DNAmt) (Quadro 8 .1) . Foi a partir da descoberta das enzimas de restrição (enzimas que reconhecem sequências específicas de DNA e fragmentam o DNA em sítios específicos) e da pesquisa em DNAmt no início da década de 1970 que se estabeleceram as bases que permitiriam o desenvolvimento da filogeografia. No fim da mesma década, foram desenvolvidos algoritmos que permitiam não só o mapeamento de genomas mitocondriais por meio das enzimas de restrição, mas também o cálculo de distâncias

138 - Métodos e Aplicações
Catarrhini Hy/obates
pongo
Hominoidea Homo
Platyrrh in i Pan
5trepsirh in i Gorilla
Macroevolução ......-: ........................................................................................... Microevolução
Figura 8.1- Processos micro e macroevolutivos estão intimamente ligados, um como extrapolação do outro. Ao submeter uma árvore filogenética (por exemplo, filogenia da Ordem Primata) a a umento de resolução, em última análise chega-se a uma relação de pedígree (heredograma de indivíduos da espécie Homo sapíens).
genéticas que culminaram com a implementação de redes de haplótipos. A constatação do ritmo acelerado de evolução do DNAmt instigou os pesquisadores a utilizarem este marcador em estudos de populações naturais. Os primeiros trabalhos com uma abordagem verdadeiramente filogeográfica surgiram no ano de 1 9792. Após uma década de desenvolvimento epistemológico nesta área da biologia comparada, Avise eta!. 3 formularam os alicerces defuma nova disciplina que denominaram filogeografia.
Esses trabalhos pioneiros e o surgimento da filogeografia como disciplina trouxeram o aspecto revolucionário de utilizarem conceitos filogenéticos em nível intraespecífico, introduzindo-se a noção não ortodoxa de considerar os indivíduos de uma espécie como unidades taxonômicas operacionais (OTU, operational
taxonomic units) em análises de genética de populações. Nos trabalhos anteriores de genética de populações usando principalmente alozimas, as unidades analíticas eram sempre populações definidas a priori, em geral por sua localidade geográfica. Após a análise dos dados de loei nucleares, tais unidades poderiam ou não ser confirmadas pela comparação de frequências alélicas empregando-se abordagens estatísticas, extremamente sensíveis a erros de amostragem. A_ partir do momento em que é possível recuperar a história evolutiva de determinados marcadores em indivíduos amostrados, não há necessidade de elaboração de premissas sobre a história da população, bem como podem ser descartadas preocupações exageradas quanto aos erros de amostragem tão comuns em estudos anteriores.
Quadro 8.1 - Ácido desoxirribonucleico mitocondrial (DNAmt)
o DNAmt é uma molécula dupla fita circular, comumente contendo de 15 a 17 quilobases. Essa molécula contém, em sua maioria, sequências não repetitivas: 13 genes que codificam proteínas, dois genes de ácido ribonucleico (RNA) ribossômico, 22 de RNA transportador e uma região controle não codificadora que contém sítios para replicação e início de transcrição. O DNAmt de metazoários é estruturalmente muito estável- há pouca variação de tamanho genômico e arranjo gênico entre as espécies1. O DNAmt de vegetais, por outro lado, possui características inversas: tem grande variação na ordem dos genes, mas sua taxa de substituição nucleotídica é muito lenta. Muitas hipóteses foram geradas para explicar a alta taxa de mutação nesse genoma; em alguns casos, ordens de grandeza maior
que as do DNA nuclear. Entre elas estão o fato do DNAmt não possuir histonas, estar exposto a radicais livres decorrentes da fosforilação oxidativa que ocorre nesta organela, não possuir mecanismos de reparo do DNA e ser duplicado muitas e muitas vezes nas linhagens celulares. A questão de evolução neutra envolvendo as mitocôndrias muitas vezes é pouco explorada e pouco clara em muitos livros. A descoberta do fato de que o DNAmt apresentava uma taxa de mutação maior que o DNA nuclear foi uma surpresa para a época, pois a mitocôndria apresenta algo próximo ao que se acredita ser um "genoma mínimo" e por ter função tão importante no organismo. O DNAmt não está livre de pressões seletivas, uma vez que quase todo seu genoma codifica proteínas fundamentais para a fosforilação oxidativa. Dessa forma, está sob severa restrição funcional. A grande maioria das mutações encontradas neste marcador, em regiões codificadoras, ocorre na terceira posição do códon, ou seja, são, em geral, sinônimas. Como uma mutação misense ou nonsense em um gene fundamental (por exemplo, o do citocromci b) tornaria o indivíduo inviável, não é possível afirmar que o DNAmt pode evoluir livremente como esperado de um marcador neutro. Esta mesma característica, porém, lhe confere sua maior vantagem: com restrição funcional tão forte, é certo que todas as mutações encontradas em um estudo, mesmo que não sinônimas, são seletivamente neutras. Enquanto a literatura lista alguns raros casos de recombinação, heteroplasmia (cenário onde "populações" de mitocôndrias geneticamente diferentes habitam o mesmo indivíduo) e de herança paterna, nunca foi publicado um estudo que demonstrasse de forma consistente a atuação de seleção neste marcador.
\O --J 'l" 00 LIl .c., N .j>. -
, 00 \O 0\ O-

Filogeografia - 1 39
- Conceitos básicos em genética de popu lações
Para compreender a l iteratura em fi logeografia é necessár io dominar os conceitos e a term i nologia da genética de populações. A segui r, a lguns dos termos uti l izados neste capítu lo. O primeiro é o modelo de Hardy-Weinberg (HW), no qual se assume que os a le ios nos loei uti l i zados no estudo são seletivamente neutros, que não há mutação ou fluxo gênico, que a população é pan-mítica (ou seja, não há acasa lamento preferencial ; os acasalamentos ocorrem ao acaso em relação ao loeus de i nteresse) e que a população pode ser considerada i nfi nita mente grande. Dentro destas cond ições, a frequência dos genótipos na popu lação é s implesmente o produto das freq uências gênicas, que se mantém estável ao longo das gerações (eq u i l íbrio de HW). Em populações dentro do modelo de HW, mas com tamanho fin ito, as frequências gênicas variam de uma geração para outra ao acaso e a probabi l idade de um ale lo ser transmitido para a próxima geração corresponde à sua frequência relativa. O processo pelo qual essas frequências variam a leatoriamente ao longo do tempo é denomi nado deriva genética. Por ser um processo estocástico, a deriva não possui d ireção e seus efeitos são acumulativos. Esse processo causa a perda de variabi l idade genética dentro da população e aumenta as d iferenças entre popu lações. Em marcadores seletivamente neutros, como o ácido desoxirribonucleico m itocondrial (DNAmt), a deriva é o principal processo responsável pela d iferenciação populaciona l . Como a maior pa rte das populações naturais v io la um ou mais princípios do modelo de HW, é i nteressante saber qua l o N que em um modelo ideal geraria as frequências a lé l icas que estão sendo observadas. A resposta para esta pergunta é o tamanho efetivo da população (Ne). O Ne não está relacionado ao número de censo da espécie. Estatísticas F são referentes às a bordagens que procuram med i r ou ava l i ar o desvio das frequências gênicas em popu lação(ões) do eq u i l íbrio esperado pelo modelo de HW. O mais uti l izado em estudos populacionais é o Fst, que pode ser defi n ido como o desvio corrigido da heterozigosidade em uma amostra.
o DNAmt possui uma série de características que o tomam um marcador particularmente apropriado para estudos filo geográficos. Em decorrência do grande número de mitocôndrias presentes nas células dos tecidos de animais, o DNAmt é extraído e amplificado com facilidade. Além disso, o DNAmt é encontrado em qualquer organismo eucarioto, de forma a possibilitar comparações homólogas entre espécies. O DNAmt possui um padrão de herança simples (predominante ou exclusivamente materna) e, ao contrário do genoma nuclear, o DNAmt animal possui uma estrutura simples, sem elementos móveis ou repetitivos, pseudogenes ou íntrons, além, de maneira geral, não estar sujeito à recombinação. Estes atributos permitem que a relação filogenética entre os alelos possa ser facilmente reconstruída considerando eventos mutacionais como evidência de relação de ancestralidade e descendência entre haplótipos.
As características já descritas conferem ao genoma mitocondrial outras propriedades importantes em estudos filogeográficos. Como o DNAmt é, em geral, herdado uniparentalmente e é haploide, seu tamanho populacional efetivo (Ne) é um quarto do Ne do genoma nuclear (Quadro 8 .2). Isto significa também, para fins filogeográficos, que o DNAmt atinge a monofilia recíproca entre populações com baixo ou nenhum fluxo gênico em cerca de um quarto do tempo que o DNA nuclear4. Após a separação das populações (ou espécies), a deriva genética e o sorteio de linhagens atuam nas entidades em evolução.
De acordo com Moore4, se um pesquisador estuda um grupo cuj a divergência entre linhagens ocorreu no Pleistoceno, é muito provável que uma filogenia baseada em DNAmt apresentará monofilia recíproca entre as
populações, enquanto em média os Zoei nucleares permanecerão sem monofilia recíprocaS (Fig. 8 .2). De acordo com Moore4, o número médio de Zoei nucleares necessário para recuperar um evento de divergência entre linhagens que o DNAmt recupera com 95% de confiança seria de 16 . Entretanto este número pode chegar a 40, dependendo do método utilizado para realizar esta estimativa. Consequentemente, seriam necessárias 40 árvores de genes nucleares para recuperar a mesma história de uma única sequência de DNAmt.
Diante das propriedades distintas do número efetivo entre estes genomas, os primeiros esforços contemplando marcadores nucleares em estudos filo geográficos utilizaram marcadores microssatélites (Quadro 8.3). Os estudos utilizando este marcador foram de grande serventia para a disciplina, ao demonstrar empiricamente algumas limitações do DNAmt diante de cenários demográficos como filopatria (permanência no local de nascimento e/ou reprodução) de fêmeas e dispersão mediada por machos. Como o DNAmt é herdado maternalmente, o que se recupera ao realizar uma filogenia deste marcador é a história da linhagem materna, que nem sempre pode ser generalizada para a espécie como um todo.
Por outro lado, os micros satélites apresentam como limitação o fato de que não é possível extrair informações filogenéticas de seus dados pelo fato de seu mecanismo de mutação ser pouco conhecido. Portanto, não é possível inferir relações de ancestralidade entre os alelos encontrados. Sendo assim, comparações entre populações ficam limitadas à demografia histórica, concentradas principalmente em estatísticas que descrevem estrutura populacional (por exemplo, estatísticas F, dependentes de frequência) .
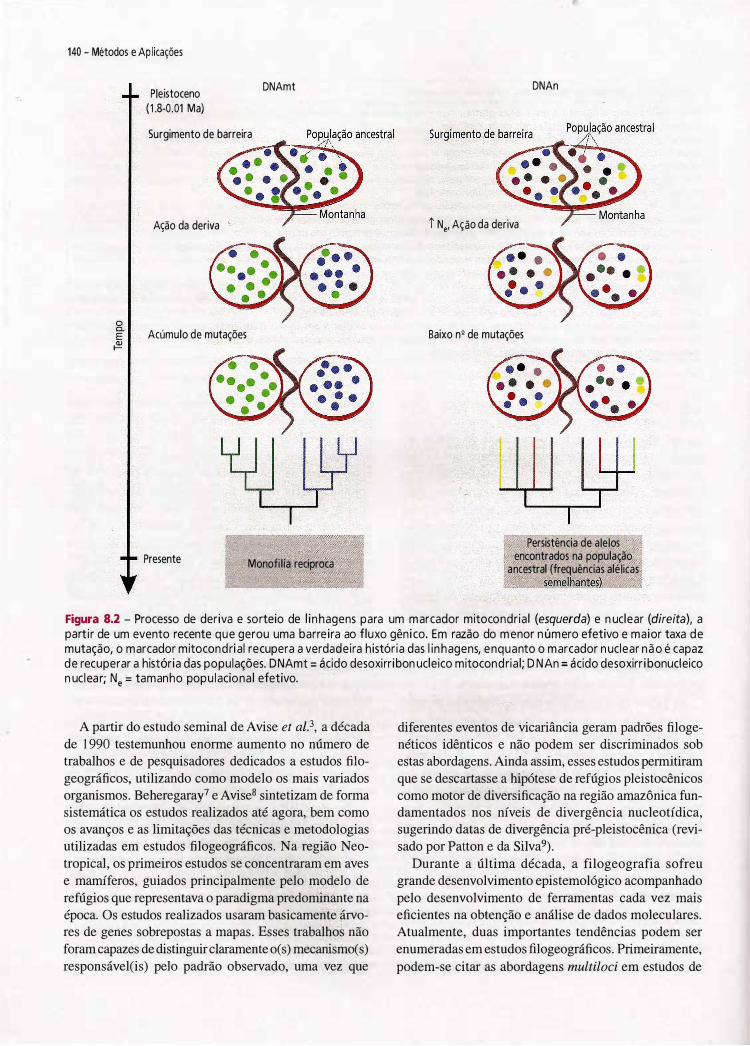
140 - Métodos e Aplicações
Pleistoceno ( 1 .8-0.01 Ma)
DNAmt
Surgimento de barreira
o o.. E �
Ação da deriva
Acúmulo de mutações
Presente Monofi lia recíproca
DNAn
i Ne, Ação da deriva
Baixo nº de mutações
Persistência de alelos encontrados na população
ancestral (frequências alélicas semelhantesl __ _
Figura 8.2 - Processo de deriva e sorteio de linhagens para um marcador mitocondria l (esquerda) e n uclear (direita), a part ir de um evento recente que gerou uma barreira ao fluxo gênico. Em razão do menor número efetivo e maior taxa de mutação, o marcador mitocondrial recupera a verdadeira histór ia das l inhagens, enquanto o marcador nuclear não é capaz de recuperar a história das populações. DNAmt = ácido desoxirribon ucleico mitocondria l; DNAn = ácido desoxirribonucleico n uclear; Ne = tamanho popu lacional efetivo.
A partir do estudo seminal de Avise et az.3, a década de 1 990 testemunhou enorme aumento no número de trabalhos e de pesquisadores dedicados a estudos filogeográficos, utilizando como modelo os mais variados organismos. Beheregaray 7 e Avise8 sintetizam de forma sistemática os estudos realizados até agora, bem como os avanços e as limitações das técnicas e metodologias utilizadas em estudos filogeográficos. Na região Neotropical, os primeiros estudos se concentraram em aves
. e mamíferos, guiados principalmente pelo modelo de refúgios que representava o paradigma predominante na época. Os estudos realizados usaram basicamente árvores de genes sobrepostas a mapas. Esses trabalhos não foram capazes de distinguir claramente o( s) mecanismo( s) responsável(is) pelo padrão observado, uma vez que
diferentes eventos de vicariância geram padrões filogenéticos idênticos e não podem ser discriminados sob estas abordagens. Ainda assim, esses estudos permitiram que se descartasse a hipótese de refúgios pleistocênicos como motor de diversificação na região amazônica fundamentados nos níveis de divergência nucleotídica, sugerindo datas de divergência pré-pleistocênica (revisado por Patton e da Silva9).
Durante a última década, a filogeografia sofreu grande desenvolvimento epistemológico acompanhado pelo desenvolvimento de ferramentas cada vez mais eficientes na obtenção e análise de dados moleculares. Atualmente, duas importantes tendências podem ser enumeradas em estudos filogeográficos. Primeiramente, podem-se citar as abordagens multiloci em estudos de

F i logeografia - 141
Quadro 8.3 - Microssatélites
Microssatél ites são caracterizados por pequenos motivos (por exemplo, TAT) de DNA repetidos em sequência, diversas vezes. Esses motivos podem ter de um a seis pares de base. M icrossatélites existem em grande número no genoma de animais e vegetais, espalhados por diversas regiões do genoma. Essas regiões do genoma são não codificadoras e apresentam alta taxa de mutação. Desta forma, são marcadores altamente pol imórficos e informativos em estudos inter e i ntrapopulacionais. As mutações que ocorrem internamente nos microssatélites são mudanças no n úmero das unidades de repetição. O mecanismo mais provável para explicar as altas taxas de mutação seria a ocorrência de erros de pareamento, ocasionados pelo deslizamento das fitas de DNA no momento da repl icação (DNA replication slippage). Além do alto grau de polimorfismo, os m icrossatélites são marcadores que apresentam a lelos-codominantes e são, de modo geral, seletivamente neutros. Os microssatélites são marcadores efetivos de fluxo gênico e h istória populacional, mesmo em espécies com variação gênica l imitada, além de serem muito versáteis e ideais para a identificação das relações entre populações naturais de determinada espécieS.
populações naturais. Em segundo, o emprego de abordagens explicitamente estatísticas para a inferência de processos históricos responsáveis pelo padrão observado. Essas duas abordagens serão tratadas a seguir com mais detalhes.
Diversos estudos mostram que a análise de sequências de diferentes Zoei nucleares se faz necessária para melhor compreensão da história natural de espécies, bem como do seu status taxonôrnico. Genealogias reconstruídas a partir de um único gene podem ser problemáticas porque cada reconstrução é somente um ponto no espaço de genealogias possíveis. Dependendo da história genealógica da população, esta abordagem pode causar sérios erros quando se iguala a árvores de genes à da espécielO. Dessa forma, a árvore do gene pode ter sido inferida corretamente, mas não corresponde à árvore da espécie (Fig. 8 .3).
Ao estudar distintas regiões do genoma é possível se obter uma história evolutiva mais completa, pois reduz o risco de não se identificar um evento ou processo em
A B
razão da ausência de uma mutação apropriada no tempo e espaço em uma sequência de DNA em particularl l . Para uma abordagem multiloei são necessárias análises de Zoei nucleares, uma vez que o DNA rnitocondrial não sofre recombinação e é herdado como um único Zoeus, com vários genes com diferentes taxas de mutação. Como o DNAmt é herdado de forma uniparental, o padrão encontrado para estes marcadores pode não ser representativo da espécie caso haja dispersão e/ou seleção ligada ao sexo12. A seleção age localmente no genoma, enquanto espera-se que a demografia afete de forma uniforme todos os Zoei neutros13•
As abordagens muZtiloei em filogeografia foram fundamentais para demonstrar não somente eventos de dispersão diferencial entre machos e fêmeas, mas também que eventos de introgressão (hibridização entre espécies ou populações) são mais comuns no DNAmt que no DNA nuclear. Além disso, verificou-se ser possível recuperar mais de um evento demográfico em
c D
- Organismo
"-J Gene
Figura 8.3 - Incongruência entre história filogenética de populações e de um marcador molecular hipotético decorrente da estocasticidade dos processos de mutação e deriva genética. Note que a população D não é monofilética para o gene estudado.

1 42 - Métodos e Ap l i cações
- Teoria neutra
A teoria neutra de evolução postula q ue a maior parte da variação a lé l ica e substituições no DNA são neutras16. Isto i m p l ica que a maior parte da variabi l idade observada em espécies é fruto da deriva genética e não da seleção. Vários pesquisadores durante a década de 1 960 chegaram a esta ideia de forma independente, por ser uma ideia muito s imples. Alelos neutros são definidos como a le los que não possuem nenhuma vantagem adaptativa com relação ao seu ale lo a ncestra l, ou seja, são equiva lentes em termos de chance de serem repl icados e transmitidos à próxima geração. Marcadores neutros são, portanto, marcadores em que todos os a le los possuem o mesmo valor adaptativo. A teoria neutra surg i u na década de 1 960 a partir da observação d ireta de dados de variabi l idade genética em populações naturais proporcionado pelo surgimento das técnicas de e letroforese e seq uenciamento de proteínas. Estas técnicas permiti ram observar a ltos índices de variab i l idade genética que não contribuem para o fenótipo, a lém de demonstrar que a quantidade de diferenças acumu ladas entre espécies no n ível de proteínas está relacionada ao tempo de divergência entre e las (que deu origem à ideia de u m relógio molecular) e não do seu g ra u de parentesco ou tamanho efetivo. Em termos matemáticos, isso s ign ifica que a d istância genética de dois organismos depende somente da taxa de m utação para aquele marcador. O equi l íbrio entre m utação e deriva é responsável pela manutenção dos pol imorfismos em espécies ou populações. Desta forma a Teoria Neutra de K imura conci l ia as ide ias sobre relógio molecu lar e os a ltos níveis de pol imorfismo intraespecífico observadas por Dobzhansky e Sturtevant na década de 1 93017. A teoria neutra faz uma série de previsões de como devem se comportar parâmetros como o nú mero de sítios pol i mórficos e proporção entre substituições s inônimas e não sinônimas em popu lações de ta manho constante. Portanto, é possível a p l icar testes de neutra l idade em um conju nto de dados para testar a h ipótese de seleção atua ndo em determ inado !oeus ou de h istória demográfica. Dois dos mais uti l i zados são os testes de Taj ima e H KA.
diferentes escalas temporais de acordo com a característica de cada loeus utilizado. Apesar de serem muito comuns em estudos de evolução em organismos modelos (como humanos e camundongos), estudos multiloei em organismos não modelo não são a regra - em especial no caso de espécies Neotropicais. A falta de dados sobre o conteúdo informativo de certos marcadores aliados a dificuldade em desenvolver e aperfeiçoar marcadores nucleares para espécies não modelo pode ser citada como causas para este fenômeno.
O surgimento e o emprego de abordagens explicitamente estatísticas em filogeografia representaram importantes passos para a disciplina. Com este advento, pode-se abandonar o caráter exploratório dos seus primeiros esforços para realizar testes explícitos de hipóteses biogeográficas. Pela utilização de testes de neutralidade e relógio molecular (Quadro 8.4), por exemplo, é possível distinguir entre eventos vicariantes gerados por barreira física segregando uma população contínua e estável e fenômeno como refúgios pleistocênicos, uma vez que o último pressupõe um gargalo populacional e divergência nos últimos 1 ,8 milhão de anos. Estas hipóteses podem ser testadas estatisticamente. O termo filogeografia estatística foi introduzido por Knowles e Maddisonl4. Os autores fizeram uso de simulações coalescentes para gerar distribuições nulas e testar, com confiança estatística, diferentes cenários históricos para um conjunto de dados . Porém, a abordagem pioneira no uso de testes estatísticos em filo geografia é a análise de clados agrupados (NCA, nested clade analysis), desenvolvido por Templetonl5. A primeira etapa desta abordagem é converter uma árvore haplotípica em uma série hierárquica de ramos ou
clados utilizando-se um algoritmo de parcimônia estatística 15 . O resultado desta análise não é uma árvore com padrão de bifurcações, mas sim uma rede de haplótipos não enraizada que reflete, de forma mais fiel , o verdadeiro processo coalescente atribuído à linhagem, segundo os autores (Fig. 8 .4). Os haplótipos são então hierarquicamente agrupados em clados de acordo com o número de passos mutacionais entre eles e classificados como clados de interior ou de ponta, conforme sua localização na rede de haplótipos. Clados interiores e que possuam maior frequência relativa de espécimes são considerados linhagens mais antigas. Na próxima etapa desta abordagem, calcula-se a correlação estatística entre haplótipos e coordenadas geográficas. Tal análise revela índices para serem utilizados em uma chave de inferência que, a partir dos resultados obtidos, pode discriminar o processo que deu origem ao padrão observado entre fragmentação alopátrica, expansão da área de ocorrência (contígua ou não), isolamento por distância, fluxo gênico (restrito ou irrestrito) e colonização de longas distâncias 15 .
Engloba uma coleção de tratamentos matemáticos formais às relações de ancestralidade e descendência. Esta teoria é considerada o maior avanço em genética de populações dos últimos 20 anos e é particularmente conveniente para dados moleculares, como sequências de DNA - que contêm ricas informações sobre relações de ancestralidade. Com o grande acúmulo de informações de dados moleculares a partir da década de 1980,

F i l ogeografia - 1 43
o 1 - 1
2-1
1 -2 (
2-2
1 -3
1 -4 - ""'--_-I _/
C: '�- ',c," - Representação de uma rede de haplótipos não enra izada. Círculos: representam estados intermed iários entre os hap lótipos amostrados. Estruturas ovais e quadradas: representam os dados observados. O tamanho relativo dessas estruturas é proporcional à frequência relativa dos a lelos. Os números apresentados representam o grau h ierárqu ico de cada agrupamento.
esta teoria tomou-se ferramenta importante para estudos genéticos, sobretudo em genética evolutiva humana13 . Enquanto a genética de populações clássica preocupa-se com o futuro das frequências gênicas nas populações a partir de determinado ponto, a teoria de coalescência parte do presente para inferir o passado utilizando-se uma abordagem probabilística13. A teoria de coalescência, porém, apresenta-se como um campo bastante árido, de difícil compreensão para não matemáticos. Em consequência, pesquisadores mais voltados para a coleta de dados mostraram certa resistência ao uso dessas técnicas em seus programas de pesquisa, mesmo diante do reconhecimento de que a integração entre a filo geografia comparativa e a teoria de coalescência traria grandes benefícios para a primeira18 .
A implementação e o uso comum de novos métodos não dependem apenas da aceitação de seus preceitos epistemológicos, quase sempre associados ao desenvolvimento teórico de uma disciplina, mas também de ferramentas que viabilizem sua aplicação. No caso da teoria da coalescência, sua aplicação foi rapidamente viabilizada pelo fato de seu desenvolvimento coincidir com a disponibilidade de computadores cada vez mais eficientes e acessíveis. A integração entre a teoria de coalescência e filogeografia tomou-se possível a partir do trabalho desenvolvido por Kuhner et al. 19 na década de 1990, empregando-se uma abordagem estatística em conjunção com um algoritmo que já havia sido descrito na literatura (Metropolis-Hastings) para calcular o parâmetro 8, ou seja, a taxa de mutação escalonada (2pNell, em que p representa a ploidia do

144 - Métodos e Ap l i cações
marcador, Ne o número efetivo da população e I.l a taxa de mutação). Este parâmetro é também denominado taxa de mutação populacional e pode ser interpretado como o número esperado de mutações entre duas sequências amostradas ao acaso. De maneira análoga, outros parâmetros populacionais podem ser calculados a partir de e, com base nesta abordagem. O surgimento de métodos computacionais baseados no algoritmo de Metropolis-Hastings foi fundamental para o desenvolvimento do aparato necessário para a análise de dados empíricos tendo a teoria de coalescência como base teórica.
Na última década observou-se grande aumento no número de programas computacionais que implementaram metodologia coalescente, bem como no número de trabalhos que utilizam estes métodos. A teoria de coalescência é usada não somente para a estimativa de parâmetros populacionais de interesse para a filogeografia - como Ne, taxas de migração (M) e tempo de divergência (T) - mas também para estimar taxas de recombinação, diferentes tipos de seleção natural atuando em sequências de DNA (em genes ou regiões adjacentes) e taxas de mutação. Ao contrário do que ocorreu na década de 1990 com programas para construção de filogenias, todos os programas computadorizados que utilizam o coalescente são de domínio público; muitos também disponibilizam o souree eode, isto é, o código original que permite àqueles que dominam linguagens de programação modificar o programa. Adicionalmente, grande parte desses programas foge do padrão comum daqueles que utilizam interfaces gráficas, reduzindo o caráter amigável do software. Entre os programas mais usados neste tipo de análise podem-se citar o IM e o IMa2o. Esses programas estimam os seguintes parâmetros entre pares de populações: e (das populações um e dois e ancestral), duas taxas de migração (da população um para dois e vice-versa) e o tempo de separação entre suas populações. É possível testar também modelos de isolamento total e fluxo gênico.
Mais rccentemente foram desenvolvidos programas de simulações coalescentes. Tais programas simulam sequências de DNA segundo determinados parâmetros fornecidos pelo usuário. Em alguns casos, é possível realizar simulações com cenários demográficos extremamente complexos, como a colonização das Américas pelo Homo sapiens. Estas simulações permitem estimar qual a probabilidade associada a diferentes cenários históricos originarem o padrão filo geográfico observado.
Avise et ai. 1 ,3 , com base nos estudos publicados até então, sumarizaram os possíveis padrões filogeográficos que podem ser descritos ao se estudar organismos. São
basicamente cinco padrões resultantes da combinação de dois fatores: a magnitude da divergência genética encontrada entre as linhagens e o grau de localização (ou circunscrição) geográfica delas. Os diferentes padrões estão representados na Figura 8 .5 :
• Padrão tipo I - alta divergência genética, linhagens alopátricas: este padrão é caracterizado por clados geograficamente circunscritos separados por grande número de mutações. O cenário histórico responsável por este padrão é provavelmente a presença de uma conspícua barreira ao fluxo gênico separando as populações por longos períodos.
• Padrão tipo II - alta divergência genética, linhagens simpátricas : duas explicações são possíveis para este padrão. A primeira é de que a deriva em populações grandes possa fazer com que linhagens divergentes surjam e persistam mesmo na presença de fluxo gênico. A atuação de seleção balanceadora, com vantagem seletiva para heterozigotos de alelos divergentes, poderia influenciar a dinâmica dos genes levando um determinado loeus a apresentar este padrão. Não existe, porém nenhum caso na literatura onde isto foi sugerido para marcadores neutros. O cenário mais provável para o surgimento deste padrão é o de contato secundário entre linhagens que divergiram alopatricamente, criando as condições para o surgimento das linhagens com alta divergência genética. O desaparecimento da barreira ao fluxo gênico torna as linhagens simpátricas outra vez.
• Padrão tipo lU -baixa divergência genética, linhagens alopátricas: neste padrão os alelos são filo geneticamente próximos, mas separados espacialmente. Neste caso, as populações estão, com grande probabilidade, intimamente ligadas do ponto de vista histórico, com uma história de baixo ou nulo fluxo gênico mais ou menos recente. Também pode ser fruto de vicariância em espécies com baixa taxa de evolução nucleotídica, como as aves ou os anfíbios.
• Padrão tipo IV - baixa divergência genética, linhagens simpátricas : este padrão é atribuído a espécies com grande capacidade de dispersão, ou seja, em sua distribuição não há barreiras ao fluxo gênico entre os indivíduos provenientes de diferentes localidades. A livre movimentação dos indivíduos ao longo da distribuição e o efeito homogeneizante do fluxo gênico impedem o surgimento de linhagens muito divergentes.
• Padrão tipo V - baixa divergência genética, linhagens parcialmente separadas: este padrão pode ser considerado intermediário entre os padrões III e IV e envolve linhagens com ampla distribuição geográfica

\O ..o a.. O? -v "" r;-or> co 00 t-a..
o .", -O &.
o .", -O '" c...
-o .� -c '" c...
::: o .� -c '" c...
> o .� -c '" "-
Área 1
@0{Q
0{D-(D
0
[D
0
CD0
Distância genética
Área 2
@0{Q �
0{D-(D
0
CD
0
0
Área 3
@0{Q
0{D-(D
0
[D
0
0{D
Grande � pequena �
Fi logeografia - 145
Figura 8.5 - Representação dos cinco padrões filogeográficos descritos por Avise 1 demonstrando a relação entre distância genética e distribuição geográfica das linhagens genéticas.

146 - Métodos e Apl icações
e algumas linhagens confinadas a determinadas localidades. Acredita-se que este padrão seja fruto de populações intimamente ligadas do ponto de vista histórico, que apresentam limitado ou nenhum fluxo gênico contemporâneo. As linhagens de distribuição ampla são as ancestrais, enquanto as raras e de distribuição restrita são as mais derivadas.
Historicamente, novas abordagens e interpretações para fenômenos naturais nascem da descoberta de descontinuidades. Por isso o padrão que mostra máxima descontinuidade intraespecífica é o tipo 1. Os estudos pioneiros em filogeografia - tanto nos Neotrópicos quanto no resto do mundo - tinham como objeto de estudo pequenos roedores e apresentaram como resultados este padrão. Pequenos roedores são animais que dispersam muito pouco em relação ao local onde nasceram. Além disso, barreiras aparentemente banais para outros animais podem ser intransponíveis para estes animais, como rios, fragmentação de hábitat ou barreiras geológicas como montanhas, resultando em alto grau de diferenciação entre populações de uma mesma espécie. Exemplos de animais terrestres que exibem este padrão de localização geográfica de suas linhagens incluem uma coleção de pequenos mamíferos não voadores (marsupiais e roedores) da região amazônica estudados pelo grupo do professor James Patton9. O Rio Doce, no Espírito Santo, mostrou-se uma barreira importante para limitar a dispersão de lagartos gekonídeos21 .
Outros organismos que comumente apresentam este padrão são organismos dulcícolas de ampla distribuição, uma vez que esses organismos são incapazes de transpor as barreiras físicas (por exemplo, bacias hidrográficas que não se comunicam) ou físico-químicas (por exemplo, composição química da água) que os separam. Corpos de águas continentais oferecem oportunidade ímpar para estudar a diversificação e a distribuição de biotas. Esses ambientes podem apresentar padrões replicados de subdivisão de táxons em subpopulações discretas e possibilitam a caracterização da dinâmica histórica de uma região e sua influência na diferenciação geográfica e formação de novas populações ou espécies.
Dois exemplos ilustrativos das premissas já apresentadas envolvem espécies de peixes sulamericanos dos gêneros Prochilodus (Characiformes) e Potamorrhaphis (Beloniformes). No primeiro caso, Sivasundar et az. 22
encontraram um clado mitocondrial de espécies de Prochilodus para cada bacia hidrográfica da América do Sul amostrada (isto é, Madalena, Orinoco, Amazonas e Paraná) (Fig. 8 .6, A), a partir da análise de genes ATPase 6 e 8. No segundo caso, espécies de Potamorrhaphis
manifestam diferentes clados mitocondriais geograficamente localizados para espécies que ocorrem em uma mesma bacia. Para este grupo de organismos pode-se verificar que a barreira é ecológica e não física. Pota- :s
'f' morrhaphis exibe preferência por ambientes lacustres e 3;
.:.., pequenos cursos d' água, sendo limitada a dispersão por � -
meio de grandes corpos d' água. Sendo assim, grandes i!2: rios aparentemente representam uma barreira para fluxo � gênico para estes organismos23.
O que distingue os padrões I e III é basicamente a profundidade da divergência entre populações amostradas. Se esta divergência é baixa (tipo III), isto pode ser resultado de separação recente ou de baixa taxa de mutação para um marcador. O estudo realizado por Carnaval et al.24, com anuros do gênero Hypsiboas, é bom exemplo de como um organismo pode apresentar mais de um padrão. Para três espécies deste gênero há uma profunda divergência entre linhagens ao Norte e ao Sul do Rio São Francisco, acompanhadas de divergências menores, mas ainda geograficamente localizadas dentro das linhagens j á citadas.
No caso do padrão tipo II - linhagens simpátricas altamente divergentes, provavelmente fruto de contato secundário entre populações que divergiram de modo alopátrico - poucos exemplos são descritos na literatura e destes nenhum abordando a fauna N eotropical. Todavia, um exemplo interessante foi descrito por Worthington-Wilmer et al.25 para o morcego australiano Macroderma gigas. Duas linhagens distintas ocupam uma mesma caverna. No entanto, dados geológicos indicam que esta caverna é fruto da união de duas cavernas distintas por um evento tectônico que fez com que duas populações com longa história de alopatria se unissem recentemente. No caso do morcego Carollia perspicillata, Hoffman e Baker26 descreveram três clados mitocondriais sem correlação com localidade geográfica para esta espécie. Esta espécie é muito comum e certamente possui alto número efetivo. Este provavelmente é um caso em que a deriva genética resultou em surgimento de linhagens monofiléticas em população de alto número efetivo, mimetizando um padrão como o descrito para M. gigas.
No caso do padrão IV - ausência de diferenciação geográfica e baixa variabilidade, vários exemplos que incluem animais de alto poder de dispersão podem ser encontrados na literatura. Um deles é o próprio ser humano: nossa espécie apresenta baixa variabilidade genética quando comparada, por exemplo, aos chimpanzés, e a análise de marcadores moleculares demonstra que existiu e existe fluxo gênico ao longo de nossa distribuição suficiente para "homogeneizar" a distribuição
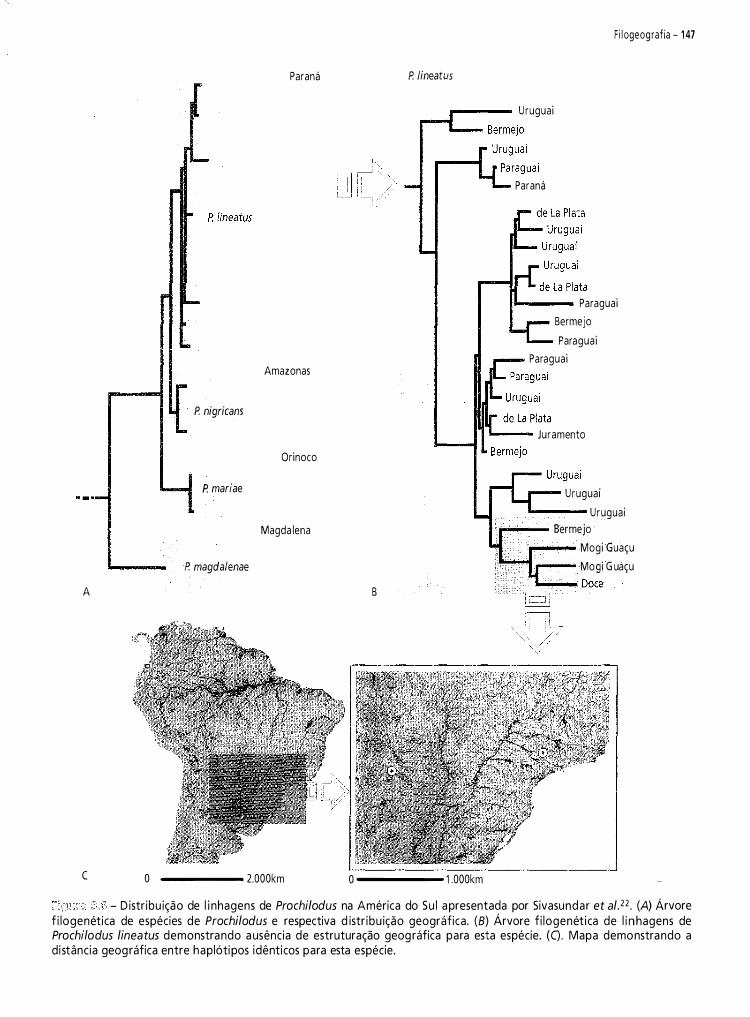
Paraná
Amazonas
p. nigricans
Orinoco
P. maríae
Magda lena
P. magdalenae
A
c o ----- 2.000km
F i l ogeografia - 1 47
P. lineatus
.----- Uruguai
B
o ------ 1 .000km
Paraná
. ... _-- Paraguai
Bermejo
Paraguai
Paraguai
---- Juramento
Uruguai
Uruguai
_--- Bermejo
.---- Mogi Guaçu
.... -- Mogi G uaçu
"'-- Doce
- D istr ibu ição de l inhagens de Prochílodus na América do Su l apresentada por Sivasundar et al.22 . (A) Árvore f i logenética de espécies de Prochilodus e respectiva d istr ibu ição geográfica. (B) Árvore f i logenét ica de l i nhagens de Prochílodus líneatus demonstrando ausência de estruturação geográfica para esta espécie. (C). Mapa demonstrando a d istância geográfica entre haplótipos idênticos para esta espécie.

148 - Métodos e Apl i cações
de alelos, apesar de historicamente existirem tentativas de dividir esta espécie em raças ou outras múltiplas divisões. Este padrão também está exemplificado na espécie de morcego Artibeus lituratus27, em que o mesmo haplótipo de DNAmt ocorre em localidades com até 3 .200km de distância como Foz de Iguaçu, Bahia e México. Este tipo de padrão não se restringe apenas a animais terrestres. Outro exemplo é descrito por Sivasundar et al.22 para o peixe dulcícola Prochilodus lineatus. Esses autores encontraram haplótipos de DNAmt compartilhados entre os rios Mogi Guaçu e Bermejo separados por uma distância de 2.600km (Fig. 8.6, B e C). Todos os exemplos supracitados tratam de organismos com alta vagilidade, ou seja, uma alta capacidade de dispersão.
No caso do padrão filogeográfico tipo V, o morcego vampiro comum Desmodus rotundus apresenta ao mesmo tempo este e outro padrão. Para o marcador mitocondrial esta espécie exibe um padrão tipo I, enquanto para um marcador nuclear esta mesma espécie possui aleI os amplamente distribuídos acompanhados de aleI os em menor frequência restritos a determinadas localidades, demonstrando que para este marcador o tempo de separação não foi suficiente para gerar monofilia recíproca em um marcador nuclear (enquanto, para um marcador mitocondrial, gerou um alto grau de diferenciação), mas sim uma diferenciação incipiente2s.
É possível notar que a maior parte dos estudos citados foi realizada com animais de pequeno porte. Esses modelos são indicados para estudos filogeográficos, que necessitam de amostragem considerável para estimar a vmiabilidade genética das populações, por serem animais cuja captura é relativamente simples e existirem em grandes números em seu hábitat. Animais de grande porte apresentam como principal dificuldade para estudos filogeográficos a amostragem - animais maiores costumam exibir menor densidade populacional e, portanto, uma dificuldade inerente de amostragem significativa da variabilidade populacional.
Como pode ser notada pelos exemplos enumerados, grande importância é dada à vagilidade das espécies sendo estudadas em filogeografia, como fator preponderante em determinar qual dos padrões de I a V determinada espécie deveria apresentar. Com o intuito de testar diferentes hipóteses sobre a disparidade entre padrões filogeográficos manifestados por aves e pequenos mamíferos não voadores, Ditchfield27 realizou estudo aprofundado em quatro espécies de morcegos e preliminar em outras 13 (todas espécies Neotropicais) e constatou que os morcegos possuíam taxa de substituição maior que a das aves e dos pequenos mamíferos não voadores, mas, ainda assim, um padrão filogeográfico
tipo IV. O trabalho de Ditchfield27 concluiu que é a capacidade de dispersão que determina qual o padrão esperado para um determinado táxon e não outras características como proximidade filo genética. Após a publicação desse estudo, porém, várias outras publicações mostraram padrões de alta localização geográfica de linhagens genealógicas para morcegos e aves passeriformes. No caso das aves, algumas habitam somente 'D
o interior de florestas, inclusive possuindo fotofobia. 0;3 00 Estas espécies não são capazes, a princípio, de atravessar � áreas abertas ou até mesmo de cruzar rios, sendo assim suscetíveis à fragmentação histórica. Recentemente foram descritos padrões filogeográficos dos tipos I e/ou III para espécies de aves na Mata Atlântica, do gênero Xyphorhynchus29. Em outros casos, fatores comportamentais como filopatria podem ser preponderantes sobre a capacidade de dispersão. Assim, o pesquisador deve procurar compreender todos os aspectos relacionados à história natural de uma espécie para ser capaz de interpretar seu padrão filogeográfico e não apenas seu potencial para dispersão que, como discutido aqui, é potencial, mas não necessariamente real.
Mesmo que o termo filogeografia tenha sido cunhado apenas em 19873, o trabalho abordando polimorfismo de DNAmt de roedores do sudeste norte-americano publicado por Avise et af.2 em 1979 é considerado um marco em estudos filogeográficos. De 1 987 a 2009, conforme dados disponíveis no ISI Web of ScienceSM, 4.376 artigos que apresentam a palavra filogeografia (ou sua derivação, filogeográfico) em seu título, resumo ou palavras-chave foram publicados (Fig. 8.7), em que virtualmente, representantes dos cinco reinos conhecidos e incluindo também os vírus, ao menos uma vez já foram abordados filo geograficamente em alguma publicação. Constatou-se também que houve aumento exponencial de publicações na última década, na qual o número de publicações saltou de 97 (em 1999) para 1 . 1 26 (em 2008 e primeiro semestre de 2009). Este recente progresso na área é fruto de melhor compreensão de processos e padrões de distribuição dos organismos aliada a novos métodos, técnicas e computadores mais potentes (revisado por Beheregaray7 e AviseS).
Todavia, o número de publicações abordando aspectos filogeográficos de organismos Neotropicais quando comparados aos dados já citados é pouco expressivo (Fig. 8 .7) . Paradoxalmente, a região Neotropical é conhecida por sua megadiversidade, uma vez que
� 00 'D (]'; '"'
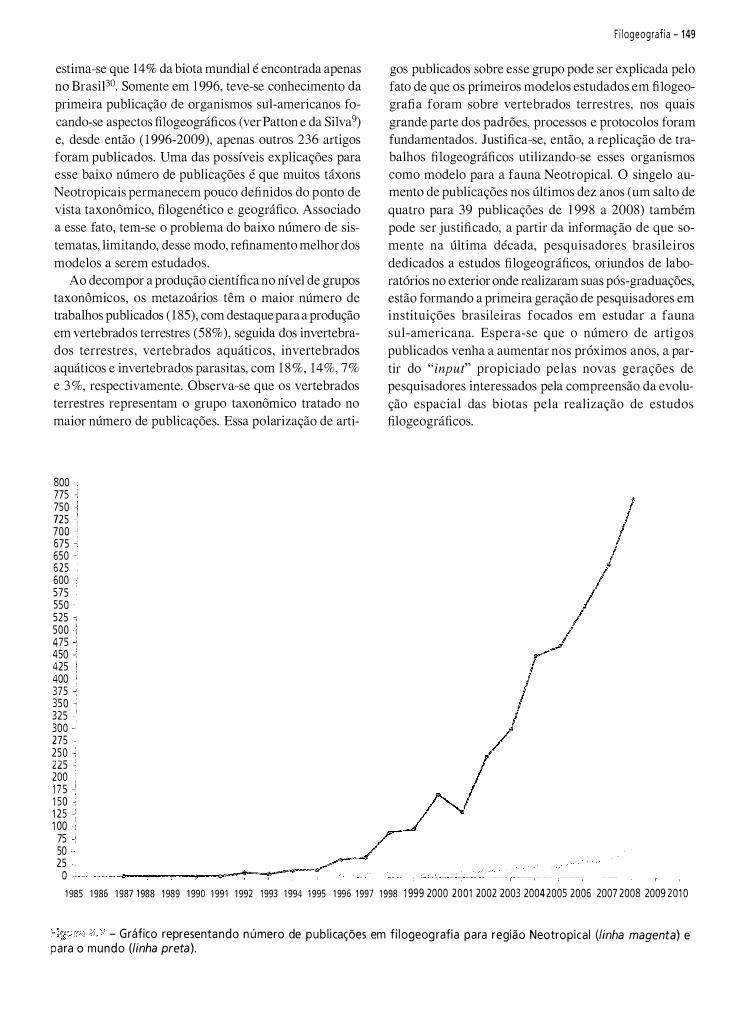
estima-se que 14% da biota mundial é encontrada apenas no BrasiPo. Somente em 1996, teve-se conhecimento da primeira publicação de organismos sul-americanos focando-se aspectos filogeográficos (ver Patton e da Si1va9) e, desde então ( 1996-2009), apenas outros 236 artigos foram publicados. Uma das possíveis explicações para esse baixo número de publicações é que muitos táxons Neotropicais permanecem pouco definidos do ponto de vista taxonômico, filogenético e geográfico. Associado a esse fato, tem-se o problema do baixo número de sistematas, limitando, desse modo, refinamento melhor dos modelos a serem estudados.
Ao decompor a produção científica no nível de grupos taxonômicos, os metazoários têm o maior número de trabalhos publicados ( 1 85), com destaque para a produção em vertebrados terrestres (58%), seguida dos invertebrados terrestres, vertebrados aquáticos, invertebrados aquáticos e invertebrados parasitas, com 18%, 14%, 7% e 3%, respectivamente. Observa-se que os vertebrados terrestres representam o grupo taxonômico tratado no maior número de publicações. Essa polarização de arti-
800 775 -i
750 , 725 700 675 650 625 600 575 550 " 525 500 475 450 -425 400 375 350 325 300 , 27S 250 225 200 1 7 5 -1 50 1 25 -1 00
75 50 25
O
F i logeografia - 149
gos publicados sobre esse grupo pode ser explicada pelo fato de que os primeiros modelos estudados em fi10geografia foram sobre vertebrados terrestres, nos quais grande parte dos padrões, processos e protocolos foram fundamentados. Justifica-se, então, a replicação de trabalhos fi10geográficos utilizando-se esses organismos como modelo para a fauna Neotropical. O singelo aumento de publicações nos últimos dez anos (um salto de quatro para 39 publicações de 1 998 a 2008) também pode ser justificado, a partir da informação de que somente na última década, pesquisadores brasileiros dedicados a estudos filo geográficos, oriundos de laboratórios no exterior onde realizaram suas pós-graduações, estão formando a primeira geração de pesquisadores em instituições brasileiras focados em estudar a fauna sul-americana. Espera-se que o número de artigos publicados venha a aumentar nos próximos anos, a partir do "input" propiciado pelas novas gerações de pesquisadores interessados pela compreensão da evolução espacial das biotas pela realização de estudos filogeográficos.
1 985 1 986 1 987 1 988 1 989 1 990 1 991 1 992 1993 1 994 1 995 1 996 1997 1 998 1 999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 201 0
- Gráfico representando número de pub l i cações em fi logeografia para região Neotropical (linha magenta) e para o mundo (linha preta).

1 50 - Métodos e Apl icações
Ao Prof. Dr. Fernando Portella de Luna Marques, da Universidade de São Paulo, pela revisão e sugestões que contribuíram para a conclusão deste capítulo.
REFERÊNCIAS BIBLIOGRÁFICAS 1. AVISE, J. C. Phylogeography: the History and Formation of
Species. Cambridge, Massachusetts, London, England: Harvard University Press, 2000. 447p.
2. AVISE, J. C. ; GIBLIN-DAVIDSON, C. ; LAERM, J. et aI. Mitochondrial DNA evolution clones and matriarchal phylogeny within and among geographic populations of the pocket gopher, Geomys pinetis. Prac. Natl. Aead. Sei. USA., v. 76, p . 6694-6698, 1 979.
3 . AVISE, J. C.; ARNOLD, 1.; BALL, R. M., et a!. Intraspecific phylogeography: The mitochondrial DNA bridge between Population Genetics and Systematics. Ann. Rev. Ecol. Syst., v. 1 8 , p. 489-522, 1987.
4. MOORE, W. S . Inferring phylogenies from mtDNA variation: mitochondrial-gene versus nuclear-gene trees. Evolution, v. 49, p. 7 1 8-26, 1 995.
5 . HARE, M. P. Prospects for nuclear gene phylogeography. Trends Eco!. Evol., v. 1 6, p. 700-706, 200 1 .
6 . SCHLÓTTERER, C . 1 998. Microsatellites. In: HOELZEL, A. R. Molecular Genetic Analysis of Populations. 2 ed. USA: Oxford University Press, 1 998. capo 7, p. 237-261 .
7 . BEHEREGARAY, L. B . Twenty years of phylogeography: the state ofthe field and the challenges for the Southern Hemisphere. Mo!. Ecol., V. 17, p. 3754-3774, 2008.
8. AVISE, 1. C. Phylogeography: retrospect and prospect. J. Biogeogr., V. 36, p . 3 - 1 5 , 2009.
9. PATTON, J. L . ; DA SILVA, M. N. F. Rivers, refuges and ridges: the geography of speciation of Amazonian mammals. In: BERLOCHER, S., HOWARD, D. Endless Forms: species and speciation. NewYork: Oxford University Press, 1 998. Capo 1 5 , p. 202-21 3 .
1 0. WAKELEY, J. Coalescent Theory: a n introduction. Greenwood Village, Colorado: Roberts and Company Publishers, 2008. 432p.
1 1. TEMPLETON, A. R. Using haplotype trees for phylogeographic and species inference in fish populations. Enviran. Bio!. Fish., V. 69, p. 7-20, 2004.
1 2. HOELZER, G. A. Inferring phylogenies from mtDNA variation: mitochondrial-gene trees versus nuclear-gene trees revisited. Evolution, V. 5 1, p. 622-626, 1997.
13. FU, Y-X.; LI, W.-H. Coalescing into the 2 1 st century: an overview and prospects of coalescent theory. Theor. Pop. Riol., v. 56, p. 1 - 1 0, 1 999.
1 4. KNOWLES, L. L.; MADDISON, W. P. Statistical phylogeography. Moi. Ecol. , V. 1 1 , p. 2623-2635, 2002.
1 5 . TEMPLETON, A. R. Nested clade analyses of phylogeographic data: testing hypotheses about gene fiow and population history. MoI. Ecol., V. 7, p. 3 8 1 -397, 1 998.
16. KIMURA, M. Genetic variability maintained in a finite population due mutation production of neutral and nearly neutral isoalleles. Genet. Res., V. 1 1 , p. 247-270, 1968.
1 7 . TEMPLETON, A. R . Population Genetics and Microevolutionary Theory. Hoboken, New Jersey: John Wiley & Sons, 2006. 705p.
1 8 . BERMINGHAM, E.; MORITZ, C. Comparative phylogeography: concepts and applications. Mal. Ecol., V. 7, p. 367-369, 1998.
19 . KUHNER, M . K. ; YAMATO, J . ; FELSESTEIN, J. Estimating effective population size and mutation rate from sequence data using Metropolis-Hastings sampling. Genetics, v. 1 40, p. 1 42 1 -1430, 1995.
20. HEY, J . ; NIELSEN, R. Multilocus methods for estimating populations sizes, migration rates and divergence times, with application to the divergence of Drosophila pseudoobscura and D. persimilis. Geneties, V. 1 67, p. 747-760, 2004.
2 1 . PELLEGRINO, K. C. M.; RODRIGUES, M. T.; WAITE, A. N. et a! . Phylogeography and species limits in the Gymnodaetylus darwinii complex (Gekkonidae, Squamata): genetic structure coincides with river systems in the Brazilian Atlantic Forest. Biol. J. Linn. Soe. , V. 85, p. 1 3-26, 2005.
22. SIVASUNDAR,A.; BERMINGHAM, E.; ORTÍ, G. Population structure and biogeography of migratory freshwater fishes (Prochilodus: Characiformes) in major South America rivers. Mal. Ecol., V. 10, p. 407-417, 200 1 .
2 3 . LOVEJOY, N. R . ; DE ARAÚJO, M. L. G . Molecular systematics, biogeography and population structure of Neotropical freshwater needlefishes of the genus Potamorrhaphis. Mo!. Eco!' , v. 9, p. 259-268, 2000.
24. CARNAVAL, A. c. ; HlCKERSON, M. J. ; HADDAD, C. F. B.; RODRIGUES, M. T. ; MORITZ, C. Atlantic Forest Hotspot Stability Predicts Genetic Diversity in the Brazilian Atlantic Forest Hotspot. Science, v. 323, p. 785-789, 2009.
25. WORTHlNGTON-WILMER, 1 . ; MORITZ, c.; HALL, L. et a!. Extreme population structuring in the threatened ghost bat, Macrodenna gigas: evidence from mitochondrial DNA. P Roy. SOCo Lond. B. Bio., V. 257, p. 1 93- 198, 1994.
26. HOFFMAN, F. G.; BAKER, R. 1. Comparative phylogeography of short-tailed bats (Carollia: Phyllostomidae). MoI. Eco!' , V. 12, p. 3403-3414, 2003.
27. DITCHFIELD, A. D. The comparative phylogeography of Neotropical mammals: patterns of intraespecific mitochondrial DNA variation among bats contrasted to nonvolant small mammais. Moi. Eco!' , V. 9, p. 1 307-1 3 1 8, 2000.
28. MARTINS, F. M.; TEMPLETON, A. R.; PAVAN, A. C. O. ; KOHLBACH, B . C. ; MORGANTE, J. S . Phylogeography of the common vampire bat (Desmodus rotundus): mark population structure, Pleistocene vicarience and incongruence between nuclear and mtDNA markers. BMC Evo!' Biol., (no prelo).
29. CABANNE, G. S . ; SANTOS, F. R.; MIYAKI, C. Y Phylogeography of Xip h o rhynchus fuscus (Pas seriformes , Dendrocolaptidae): vicariance and recent demographic expansion in southern Atlantic forest. Biol. J. Linn. Soc. , V. 9 1 , p. 73-84, 2007.
30. LEWINSOHN, T. M. , PRADO, P. L Quantas espécies há no Brasil? Belo Horizonte: Megadiversidade, V. 1 , p . 36-42, 2005.

__________________ �_�o _ 9
, J i " -1 I L
José Alexandre Felizola Diniz Filho
Miguel Bastos Araújo
A macroecologia pode ser compreendida como uma síntese entre diferentes campos do conhecimento biológico, incluindo Ecologia, Biogeografia, Fisiologia, Genética e Evolução, e que procura avaliar empiricamente os padrões de diversidade em grandes escalas de tempo e espaçol . Seu objetivo, portanto, é utilizar os conhecimentos desses diferentes campos a fim de alcançar compreensão integrada dos processos ecológicos e evolutivos envolvidos na origem e manutenção dos padrões de diversidade. De fato, segundo a proposta original de J. H. Brownl , a macroecologia pode ser definida como:
" . . . uma investigação estatística e não experimental da relação entre a dinâmica e interação entre populações das espécies que têm sido estudadas em escalas locais por ecólogos, e processos de especiação, extinção e expansão e contração de distribuições geográficas que têm sido investigadas em escalas mais amplas por biogeógrafos, paleontólogos e macroevolucionistas".
Embora a macroecologia fosse originalmente baseada em análises comparativas (cross-species) dos padrões de distribuição estatística e das relações entre tamanho do corpo, densidade populacional local e distribuição geográfica, o programa de pesquisa se consolidou pela incorporação de muitas questões gerais que já estavam sendo investigadas há bastante tempo por ecólogos e biogeógrafos desde o século XIX, incluindo a análise dos padrões latitudinais de riqueza de espécies, das regras "eco geográficas" (por exemplo, Bergmann, Rapoport, Allen), das relações espécie-área e das distri-
' 1 " ! - � "� , I�, ..l '-, ,-'
buições espécie-abundância, dentre outras . A análise inicial de todas essas questões está, em grande parte, centrada na reconstrução das distribuições geográficas das espécies em diferentes escalas espaciais e de sua dinâmica ao longo do tempo. Nos últimos 10 a 20 anos, a compreensão dos padrões e processos associados às distribuições geográficas das espécies passou a ter grande importância para otimizar as estratégias para conservação da biodiversidade, que se tomaram críticas diante de crescentes ameaças causadas pela ocupação cada vez mais intensa do planeta pela população humana.
Restam poucas dúvidas de que a crescente intensidade com que as sociedades humanas ocupam o planeta está causando, direta ou indiretamente, impacto significativo sobre os seus ecossistemas naturais2. A destruição de hábitats e a conversão das áreas naturais para uso em atividades antrópicas são certamente os mecanismos que possuem maiores impactos diretos sobre a diversidade. Existe, do mesmo modo, evidência que as mudanças climáticas acarretam impactos indiretos assinaláveis sobre a biodiversidade nomeadamente alterações na fenologia, distribuição das espécies e, em certos casos, a sua extinçã02,3. As mudanças climáticas atuais podem ser encaradas, de fato, como oportunidade de realizar experimentos ecológicos em escala global, já que mani" pulações do clima estão causando respostas nas espécies que podem ser observadas, modeladas e validadas. Esta é, portanto, oportunidade ímpar para rever o conhecimento que existe sobre os mecanismos envolvidos na distribuição geográfica das espécies.
Estima-se que as mudanças climáticas globais atuais sejam consequências do aumento da concentração de gases de efeito de estufa na atmosfera terrestre, incluindo o CO
2• Este aumento de concentração de gases de efeito

1 52 - M étodos e Ap l i cações
de estufa deve-se essencialmente ao crescimento da população humana, que foi acompanhado de elevação progressiva no uso de combustíveis fósseis, em especial a partir do século XIX. Diversas atividades humanas, tais como desmatamento e a conversão de hábitats para agricultura intensiva, também contribuem para liberar grande quantidade de CO
2 na atmosfera. Estes fatores tendem,
por meio de uma série de sistemas complexos de retroalimentação, a gerar mudanças climáticas em escalas locais, regionais e globais3. ° aumento da temperatura média é apenas uma das consequências destas mudanças e os padrões observados ao nível planetário são bem mais complexos e envolvem mudanças nas médias, variâncias e picos tanto de temperatura quanto de umidade. Nos últimos anos progrediu-se de forma considerável no conhecimento dos mecanismos que geram os padrões atmosféricos, permitindo, assim, a construção dos chamados modelos de circulação global (global circulation models - GCM), ou os mais complexos modelos acoplados oceânico-atmosféricos de circulação global (AOGCM, coupledAtmosphere-Ocean Global Circulation Models)3.
Apesar de os efeitos das mudanças climáticas ocorrerem em escalas espaciais amplas, uma parte importante dos trabalhos publicados sobre os impactos climáticos nos padrões e processos biológicos tem sido realizada em escalas locais, utilizando-se abordagem experimental4. Sem dúvida, a avaliação de processos por meio de técnicas experimentais possui maior potencial preditivo por permitir avaliação causal mais bem determinada. Entretanto, essa abordagem oferece algumas limitações por não permitir generalizações para escalas geográficas mais amplas. Assim, como alternativa aos procedimentos experimentais, pode-se pensar em abordagens macroecológicas mais gerais e fundamentadas na análise estatística dos padrões geográficos de diversidade, com base tanto nas espécies consideradas de forma individual, como na agregação de espécies em padrões de riqueza e outras métricas de diversidade biológica.
Na verdade, o exemplo dado no primeiro livro sobre macroecologia para descrever a lógica da investigação em macroecologia 1 refere-se exatamente a uma análise da extinção de pequenos mamíferos, no topo de montanhas, sob efeito de mudanças climáticas. Este é um exemplo interessante, mas pontual, e de fato ainda é necessário avaliar de forma mais genérica como as dificuldades que existem na compreensão das relações padrão-processo, a partir de dados observacionais pode ser compensada pela possibilidade de compreender e predizer os efeitos das mudanças climáticas em escalas geográficas mais amplas4,s. De fato, após fase inicial na qual os padrões macroecológicos foram caracterizados,
passou a haver recentemente uma ênfase cada vez maior na compreensão e análise dos processos ecológicos e evolutivos subjacentes a esses padrões, utilizando procedimentos de simulação, pelo estabelecimento mais firme das bases teóricas e das predições únicas que podem ser associadas aos mecanismos responsáveis pela
'-O origem dos padrõess,6. Assim, apesar das dificuldades ?l inferenciais, espera-se que abordagens macroecológicas
00 U1 c.., N t passem a ser cada vez mais comuns para avaliar os
efeitos das mudanças globais sobre a biodiversidade em ::jg 0\ grandes escalas. Em certo sentido, a utilização de mo- '" delos de distribuição geográfica com base em nicho ecológico (ver a seguir), que cresceu consideravelmen-te nos últimos 10 anos, mostra que essas aplicações já estão, de fato, aumentando.
Apesar de dificuldades operacionais, a distribuição geográfica das espécies (range) pode ser pensada como a unidade fundamental de análise biogeográfica7. Não obstante, a compreensão dos padrões biogeográficos a partir das distribuições das espécies é controversa, principalmente pelas dificuldades em desdobrar os mecanismos envolvidos na origem e manutenção dessas distribuições e pelas dificuldades em compreender os mecanismos que operam em diferentes escalas espaciais e evolutivas.
Em relação aos aspectos teóricos e conceituais, é importante destacar que há certo consenso de que as espécies não se distribuem aleatoriamente no espaço (tanto no espaço geográfico quanto no espaço ambiental) . A maior parte das espécies possui distribuição geográfica restrita, com grande número de espécies ocorrendo em áreas pequenas e poucas espécies ocorrendo em muitas regiões (Fig. 9. 1 , A) . Por outro lado, a maioria das espécies concentra-se nas regiões tropicais do globo (Fig. 9 . 1 , B). Esses dois padrões são comuns à grande parte dos grupos de organismos, ainda que existam algumas exceções em níveis taxonômicos mais baixos (por exemplo, as abelhas mamangavas do gênero Bombus, que possuem maiores concentrações de riqueza fora da região tropical). A discussão sobre os mecanismos envolvidos na origem e manutenção desses padrões centra-se no balanço entre processos atuais (ligados ao impacto dos fatores ambientais atuais, designadamente o clima, sobre
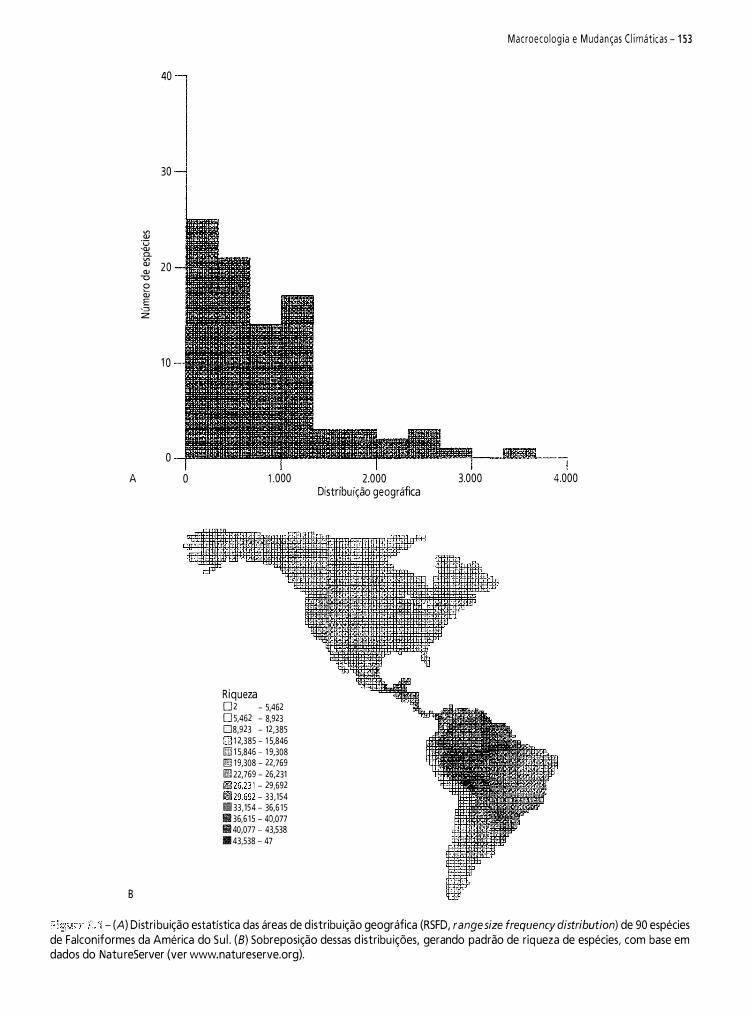
40
30
'" <lJ 'u '<lJ � <lJ 20 <lJ
""O o W E ':J
Z
1 0
o
A
B
o 1 .000
Riqueza 02 - 5,462 05,462 - 8,923 08,923 - 12,385 D 1 2,385 - 1 5,846 ll1Il 1 5,846 - 1 9,308 1!!lll 1 9,308 - 22,769 11!!1 22,769 - 26,231
- 29,692 - 33,154
1133,154 - 36,6 1 5 1136,6 1 5 - 40,077 1140,077 - 43,538 II1II43,538 - 47
2.000 Distribuição geográfica
3.000
Macroecologia e Mudanças C l i máticas - 1 53
4.000
- (A) Distribuição estatística das áreas de distribu ição geográfica (RSFD, range size frequency distribution) de 90 espécies de Falconiformes da América do Su l . (B) Sobreposição dessas d istribu ições, gerando padrão de riqueza de espécies, com base em dados do NatureServer (ver www.natureserve.org).

1 54 - Métodos e Apl i cações
as distribuições das espécies) e históricos (fatores históricos ligados aos padrões geográficos de especiação e extinção e inércia filogenética, ou não equilíbrio) afetando a distribuição geográfica das espécies e propriedades derivadas de sua sobreposição, tais como riqueza. Entender os mecanismos ecológicos e evolutivos associados diretamente aos padrões macroecológicos nas distribuições geográficas e na riqueza de espécies é, com certeza, fundamental para prever a resposta da diversidade biológica às mudanças climáticas previstas para os próximos anos como consequência da ação humana.
Na realidade, a distribuição geográfica de uma espécie, tal e qual considerada nos estudos de macroecologia, é uma abstração. A distribuição geográfica das espécies é, de fato, o resultado de um conjunto de populações locais (ou mesmo indivíduos) distribuídos no espaço geográfico e ligados entre si por dispersão e migração, o que garante a "coesão" genética (e, portanto, microevolutiva) da espécie num dado instante de tempo. Em um primeiro momento, poder-se-ia pensar em modelo integrador no qual os limites de ocorrência das populações seriam determinados pela tolerância a variações ambientais. Na verdade, esses limites são usualmente difusos e caracterizados por redução gradual da abundância da espécie8. Ao mesmo tempo, processos estocásticos e históricos (barreiras) estariam envolvidos na ocupação de regiões de ocupação potencial, considerando-se os limites de tolerância ambiental. Haveria, assim, forte componente demográfico na estrutura da abundância da espécie dentro da sua distribuição geográfica9, determinado simultaneamente por probabilidades de extinção local, conectividade espacial entre indivíduos e populações e adaptações genéticas às condições ambientais locais, que, por sua vez, determinaria as taxas de crescimento da população local. Além disso, a alocação da espécie ao longo do gradiente ecológico estaria fortemente ligada aos processos evolutivos e às contingências históricas.
Dado esse modelo integrador da distribuição geográfica de uma espécie, é possível avançar algumas questões operacionais que permitiam, ao mesmo tempo, avaliar os processos e fazer predições sobre as mudanças na distribuição geográfica em resposta a diferentes eventos . Certamente, as primeiras fontes de informação sobre a distribuição geográfica de uma espécie são os locais de ocorrência (Fig. 9.2,A), que são, na verdade, uma amostra (em geral, enviesada) das localidades onde a espécie ocorre ou já ocorreu. A partir dessa informação, os biogeógrafos constroem mapas de ocorrência (ou mais precisamente mapas de extensão de ocorrência), que estão, de fato, procurando estabelecer limites geográficos em escalas amplas, dados por extrapolação geral dos pontos com base no
conhecimento empírico ou assumindo-se certos limites ambientais ou geográficos (Fig. 9.2, B). Modelos mais sofisticados que levam em consideração características ecológicas das espécies têm sido muito atualizados (ver a seguir) e geram mapas bem mais complexos (Fig. 9.2, C). De qualquer modo, a partir desses mapas podem ser estabelecidas diversas "propriedades" da distribuição geográfica das espécies, tais como sua extensão latitudinal, sua extensão longitudinal ou seu centroide geográfico (o ponto médio de ocorrência, que pode ser dado simplesmente pela média de latitude e longitude ou por métodos geométricos mais complexos). A análise, quase sempre comparativa, dessas propriedades foi denominada nos anos 1970 como areografia, a partir dos trabalhos do biogeógrafo argentino Eduardo Rapoport7,8.
Considerando-se esse modelo teórico subjacente à distribuição geográfica das espécies, pode-se extrair dos dados empíricos alguns aspectos importantes. Em primeiro lugar, fica claro que a área de ocorrência da espécie é uma métrica de difícil avaliação operacional e extremamente dependente da escala, mas que a maior parte dos processos ecológicos e evolutivos poderia ser avaliada a partir da extensão de ocorrência da espécie, estabelecida para dada escala. Em segundo lugar, as distribuições geográficas não são constantes no tempo, de modo que a extensão de ocorrência pode refletir processos passados e não estar em "equilíbrio" com o clima atual 10. Além disso, é comum hoje contrastar a distribuição atual e a distribuição história, sendo esta última definida (usualmente de forma imprecisa) pela distribuição da espécie antes dos efeitos antrópicos recentes (por exemplo, após o último máximo glacial). Dessa forma, a extensão de ocorrência possui erros inerentes à sua construção empírica. Os mais conhecidos seriam os erros de comissão (ou seja, atribuir a ocorrência da espécie a uma região onde ela não sucede) e os erros de omissão (omitir a ocorrência onde a espécie ocorre). Esses dois erros têm tanto origem metodológica, associada ao baixo conhecimento sobre os processos que determinam a distribuição geográfica da espécie, quanto origem mais conceitual, dada pela dificuldade em estabelecer corretamente o balanço entre processos determinísticos (ambiente) e estocásticos (história ou "não equilíbrio").
De qualquer modo, o ponto importante da definição da extensão de ocorrência é que ela é, de fato, um modelo da distribuição geográfica da espécie, sendo, desse modo, uma simplificação da realidade com fins operacionais. Com base nessa concepção, pode-se tentar estabelecer, de maneira mais formal, modelos matemáticos/estatísticos que, ao incorporar os processos ecológicos e evolutivos que estão possivelmente ligados

à origem e manutenção das distribuições geográficas, sejam capazes de delinear, de forma mais objetiva, a extensão de ocorrência de uma espécie. Os modelos mais utilizados para esse propósito atualmente são os modelos de nicho.
O conceito de nicho tem sido amplamente discutido desde a emergência da ciência ecológica e existem várias definições conceituais e metodológicas (operacionais) de nicho que se aplicam à modelagem da distribuição
Macroecologia e Mudanças C l imáticas - 1 55
- Diferentes maneiras de apresentar distribuição geográfica, usando como exemplo uma espécie de Falconiformes (Harpya haruja) . (A) Dados de ocorrência. (B) Mapa de extensão de ocorrência. (C) Modelo de n icho obtido usando o a lgoritmo MAXENT.
das espécies1 1 ,12. Sob um ponto de vista operacional, o aspecto central do conceito de nicho, que se liga aos modelos de distribuição geográfica das espécies, é a ideia de que existe um nicho multi dimensional. Esta foi uma das ideias pioneiras propostas por Hutchinson, que definiu o nicho ecológico como sendo o conjunto de condições ambientais que permitem com que uma espécie sobreviva indefinidamente. Este conceito de nicho, também conhecido como nicho abiótico, foi definido por Hutchinson como o nicho fundamental da espécie que seria, na prática, reduzido em razão dos processos de interação (competição) decorrentes da sobreposição dos

1 56 - Métodos e Apl icações
nichos fundamentais de diferentes espécies que interagem, gerando, assim, o chamado nicho realizado da espécie. Operacionalmente, o aspecto importante dessa definição de nicho é que permite estabelecer um modelo de distribuição de espécies com base na caracterização do espaço multidimensional formado pelas variáveis ambientais que definem o nicho.
Assumindo-se que os limites de tolerância de uma espécie em relação a uma variável ambiental, como a temperatura, são conhecidos (por exemplo, a espécie sobrevive entre 1 0 e 20°C), seria possível abstrair inicialmente que a espécie poderia ocorrer em qualquer lugar no qual a temperatura se encontra dentro desta faixa de variação. Com base nesta premissa simples constrói-se um modelo simplificado da distribuição potencial da espécie ou, se quisermos ser mais precisos, um modelo do efeito da variável ambiental temperatura na distribuição da espécie considerada. Esse modelo pode ser ampliado para incorporar mais variáveis, por exemplo, construindo-se um espaço bidimensional que expresse o efeito combinado de temperatura e precipitação. Na teoria, esta abordagem poderia ser expandida indefinidamente a fim de incorporar todas as dimensões do nicho hutchinsoniano, delimitando-se, assim, um envelope ambiental ou bioclimáticolO. Existem, porém, alguns problemas tanto conceituais quanto metodológicos nesse processo de delimitação e estabelecimento do nicho que merecem ser discutidos.
Em primeiro lugar, é fundamental reconhecer que, mesmo considerando que as variáveis que determinam, ou compõem, o nicho ecológico da espécie sejam conhecidas e incorporadas no modelo; é importante lembrar que este seria um modelo do nicho realizado da espécie e que este determinaria apenas a sua distribuição ou extensão geográfica potencial, e não, necessariamente, sua distribuição real (ou atual). Seria preciso incorporar nos modelos de nicho outros componentes, que descrevam processos de dispersão, interação biótica, dispersão e de inércia na ocupação do ambiente potencial, a fim de transformar efetivamente os modelos de nicho em modelos de distribuição geográfica.
Outro ponto essencial está na definição das variáveis que compõe o nicho de uma espécie. Na definição original de Hutchinson, há uma separação entre os fatores de tolerância ambiental e os processos de interação, e, de fato, é possível definir os chamados nichos grinneliano (condições e fatores ambientais) e o nicho eltoniano (processos de interação e utilização de recursos), embora nem sempre seja simples separar efetivamente esses componentes12. Por exemplo, a radiação solar está diretamente correlacionada à temperatura e como tal tem
sido utilizada como indicador de variável hutchinsonia-na para definir os limites da tolerância dos animais e plantas à temperatura. Para as plantas, porém, a radiação é uma variável que se relaciona diretamente à obtenção de recursos por via da fotos síntese e assim é passível de ser sujeita a processos de interação (por exemplo, competição por luz) que caracteriza o nicho eltoniano. Esses dois conceitos de nicho podem ser compreendidos também no espaço multi dimensional e a sua sobreposição 'D
-.J gera os nichos fundamentais e realizados de Hutchinson. â::
UI De qualquer modo, na grande maioria dos casos, o com- .:.,
� ponente grinneliano do nicho seria a base dos modelos , 00
utilizados atualmente e necessários para fazer predições 2í': o,
sobre os padrões de distribuição em grandes escalas, havendo, ainda, muitas dificuldades operacionais para incorporar componentes eltonianos nos modelos.
No contexto do nicho grinneliano, outro ponto importante no processo de delimitação do envelope climático é que seus limites são, em geral, estabelecidos a partir de dados de ocorrência obtidos em coleções zoológicas ou botânicas, de modo que o próprio processo de construção das regras (o modelo) pode ser enviesado pelos processos não ambientais subjacentes à distribuição das espécies. Seria muito interessante que mesmo o componente grinneliano do nicho fosse determinado a partir de dados fisiológicos que permitissem estabelecer de modo "experimental" os limites ecofisiológicos de tolerância da espécie. É óbvio que essa análise também teria suas próprias limitações, visto que não é tão simples realizar experimentos para grande número de espécies, ou mesmo conhecer que variáveis climáticas seriam importantes fisiologicamente para estabelecer os limites de ocorrência (embora disponibilidade de energia e água sej a determinante fundamental para a vida no planeta).
Por fim, existem diversas técnicas estatísticas e computacionais para estabelecer um modelo de nicho ecológico e projetar sua distribuição geográfica como base neste1 3, 14. Essas técnicas, que vão desde simples processos de construção de regras bioclimáticas como as usadas anteriormente até técnicas complexas de inteligência artificiaIl4, são amplamente utilizadas para avaliar as distribuições geográficas das espécies (Fig. 9.2, C) e, a partir daí, discutir padrões e processos biogeográficos ou estabelecer estratégias de conservação. Os detalhes sobre cada um desses métodos e sua avalia-ção estão além do alcance desse texto, mas é suficiente entender, no presente momento, que existem várias técnicas, com diferentes propriedades estatísticas e diferentes capacidades de predição. Não obstante, ainda há dúvidas sobre quais delas são mais adequadas para modelar a distribuição das espécies.

Dadas as muitas incertezas existentes, tanto ecológicas como algorítmicas, uma estratégia possível para diminuir os erros nas diversas fases do processo de modelagem é empregar técnicas de consenso para realizar as projeções (ensembleforecasting) 1 5 . A ideia subjacente a essa estratégia é que, assumindo-se que as idiossincrasias do processo de modelagem e suas incertezas estão aleatoriamente distribuídas no espaço geográfico, é mais conservador sobrepor as diversas soluções alternativas obtidas nos processos de modelagem e utilizar fazer previsões sobre a ocorrência de espécie apenas às regiões de consenso entre essas alternativas. Essas incertezas viriam de condições iniciais (por exemplo, vieses nas coleções zoológicas e botânicas), classes de modelos (por exemplo, diferentes técnicas estatísticas utilizadas para modelar o nicho a partir da correspondência entre ocorrências e variação ambiental) , parametrização dos modelos (por exemplo, definição de variáveis ambientais, restrições ou não à dispersão) e condições de contorno (por exemplo, diferentes GCM e cenários de emissão).
Sem dúvida, as técnicas de consenso não são soluções finais para alguns dos problemas já discutidos, tais como a questão do não equilíbrio e do pressuposto de conservação de nicho. Esses problemas são, na realidade, mais associados à própria natureza da inferência de processos causais relacionando ocorrência à variação ambiental com base em método correlativo. No entanto, de qualquer modo, as técnicas de consenso podem ser muito úteis no sentido de minimizar a variabilidade inerente ao processo de modelagem. É possível ainda avaliar os componentes de variação associados a cada uma das fontes de incerteza, de modo a investir em melhor conhecimento das fontes mais importantes .
Uma vez que são construídos modelos de nicho ecológico e se tenta estabelecer uma relação explícita entre nicho e distribuição geográfica, pode-se avaliar como as espécies seriam afetadas por mudanças nas condições ambientais que definem seu nicho grinneliano. Como já discutido, existem dificuldades inerentes ao processo de modelagem e à própria relação nicho-distribuição, mas mesmo que essas dificuldades sejam minimizadas, ainda é preciso considerar alguns aspectos a fim de fazer predições sobre a distribuição geográfica futura das espécies como resposta às mudanças climáticas.
Em primeiro lugar, é preciso considerar que as mudanças climáticas fazem parte da própria dinâmica
Macroecologia e Mudanças C l imáticas - 157
evolutiva dos nichos das espécies e que, na verdade, as distribuições são dinâmicas e mudam com o passar do tempo em função dessas flutuações (os melhores exemplos conhecidos são as mudanças de distribuições de espécies temperadas respondendo às variações climáticas no Pleistoceno tardio, acompanhando os ciclos glaciais) 16. Entretanto, no caso das mudanças atuais (ou futuras), é preciso considerar que as mudanças ocorrem muito rapidamente e que, por conseguinte, pode haver problemas relacionados principalmente aos processos de dispersão.
É notoriamente difícil incorporar dispersão de forma realista nos modelos de nicho, embora sej a possível estabelecer funções probabilísticas da ocorrência da espécie no futuro a partir de acontecimentos atuais. Há, contudo, pouco conhecimento da biologia das espécies que permita parametrizar esses modelos e, mais importante, há interação entre dispersão e conversão de hábitats por ocupação humana que se torna difícil de modelar, pois não encontra paralelo na história do planeta. Isso implica que modelos bem mais complexos, que incorporem a estrutura espacial de conectividade entre hábitats e a capacidade de dispersão das espécies, sejam necessários (ver a seguir) .
Outro ponto fundamental é que, ao utilizar modelos de nicho para fazer predições, está assumindo-se que o modelo permanece o mesmo no momento atual e no futuro, o que quer dizer, ecologicamente, que o nicho da espécie se mantém, ou seja, há conservação do nicho. Em resumo, a capacidade de predição com base no nicho ecológico depende basicamente de dois processos básicos:
1 . Há conservação do nicho, de modo que processos adaptativos gerando evolução do nicho não ocorrem e a espécie mantém seu nicho atual e muda sua distribuição atual.
2. Há possibilidade total de dispersão, não existindo barreiras naturais ou antrópicas para a espécie se deslocar para os ambientes adequados (que obviamente precisam continuar a existir), ou não há qualquer capacidade de dispersão pelo que qualquer modificação na distribuição geográfica do nicho resulta em extinções 10caisl , lO.
Se essas duas condições não acontecerem, os modelos atuais tendem a representar, de forma imprecisa, a distribuição futura da espécie.
Embora os dois processos provoquem falhas nas predições dos modelos de nicho, suas implicações para a conservação da diversidade biológica são diferentes.

1 58 - Métodos e Apl icações
No caso do primeiro processo, ou sej a, se não há conservação de nicho, a espécie ou população pode continuar a existir onde ocorre hoje, mas com características biológicas diferentes da espécie ou população ancestral (decorrentes do processo de adaptação local). Ela pode continuar a ocorrer mesmo em condições subótimas, caso o processo de adaptação não seja tão eficientel . Neste caso, a perda se dá em termos de características biológicas que existiam no tempo anterior, porém isso pode ser encarado como parte do processo de evolução e adaptação da própria linhagem às mudanças. Por outro lado, a incapacidade de modelar a dispersão de forma explícita implica incapacidade de prever extinções que resultem de modos de dispersão situados entre os dois extremos do gradiente: dispersão ilimitada e nula.
A macroecologia busca estabelecer padrões gerais e tenta encontrar os processos ecológicos e evolutivos associados a esses padrões. Esses processos são gerais e não devem ser únicos na história da Terra, de modo que não se devem considerar características únicas ou particulares de espécies ou taxa. De certa forma, assume-se, portanto, que as espécies sejam de fato apenas replicações e o mais interessante nesse contexto seja avaliar simultaneamente o comportamento de grande número de espécies e avaliar suas respostas comuns.
Para tal, é possível então calcular algumas métricas a partir das distribuições modeladas das espécies, a mais óbvia destas sendo o número de espécies numa dada área (riqueza). Por exemplo, modelando-se simultaneamente a distribuição de 90 espécies de aves da ordem Falconiformes (Acciptridae e Falconidae) nas Américas é possível perceber que aparece redução na riqueza de espécies predita para 2080-2 1 00 no Centro da América do Sul, na região Amazônica, o que se tem quase sempre atribuído a uma redução de precipitação nessa região e conversão de hábitats florestais e áreas mais abertas de Cerrado (Fig. 9 .3, A).
Em um contexto de mudanças climáticas globais é interessante também avaliar o número de espécies que entram ou desaparecem destas unidades, usando uma métrica de "turnover" de espécies, que pode ser calculado por (G + L)/(S + G), em que S é a riqueza
inicial da unidade espacial, enquanto G e L são o número de espécies que aparecem e desaparecem, respectivamente, de cada unidade espacial 1 7 . No caso dos falconiformes, as análises sugerem maior substituição de espécies na região dos Andes e do Leste da América do Norte e pouca redução nas áreas de Floresta Atlântica (Fig. 9 .3 , B).
Discussão interessante sob um ponto de vista macroe- :s 't"
cológico é avaliar como as predições de modelo ambiental f:j; .:., da riqueza de espécies estão correlacionadas às predições � de riqueza obtidas a partir da sobreposição dos modelos �
o, das espécies obtidos individualmente18• A ideia é que se a "" riqueza é gerada apenas pela sobreposição das distribuições das espécies (que por sua vez já incorporaria processos de não equilíbrio), sem nenhum processo de interação ligando essas espécies e gerando padrões emergentes, os dois modelos estariam altamente correlacionados.
No caso dos falconiformes americanos, a riqueza de espécies predita para 2080-2100 a partir de um modelo linear usando as mesmas variáveis empregadas no processo de modelagem mostra que, até dado valor de riqueza, há boa convergência dos padrões, mas os modelos de sobreposição de distribuição subestimam as riquezas mais elevadas preditas pelo modelo de riqueza (Fig. 9.4). Ademais, um aspecto importante a ser considerado é que diferentes métodos produzem correlações diferentes, de certo modo ligadas à complexidade dos métodos. Por exemplo, a correlação obtida a partir de métodos simples de envelope, como método de envelopes bioclimáticos (BIOCLIM), e Distâncias Mahalanobis é relativamente baixa (r = 0,488 e r = 0,733, respectivamente), ao passo que a correlação entre a riqueza predita e a gerada pelo método de máxima entropia (MAXENT) é bem mais elevada (r = 0,882). É difícil estabelecer, em um primeiro momento, qual das duas ideias é mais adequada e essa avaliação "multi escala" pode ser uma das principais linhas de pesquisa na área.
As implicações das mudanças climáticas para a biodiversidade são óbvias, já que podem forçar mudanças substanciais em termos de ganho e perda de espécies (Fig. 9 .4). Na realidade, isso traz consequências importantes que obrigam uma revisão dos princípios inerentes aos processos de planejamento do território. Por exemplo, seria essencial verificar se o planejamento de áreas prioritárias para conservação leva esses deslocamentos em consideração, ou se os sistemas de reservas (a princípio planejados de forma ótima) perderão eficácia no futuro19. Na verdade, o principal "problema" em relação
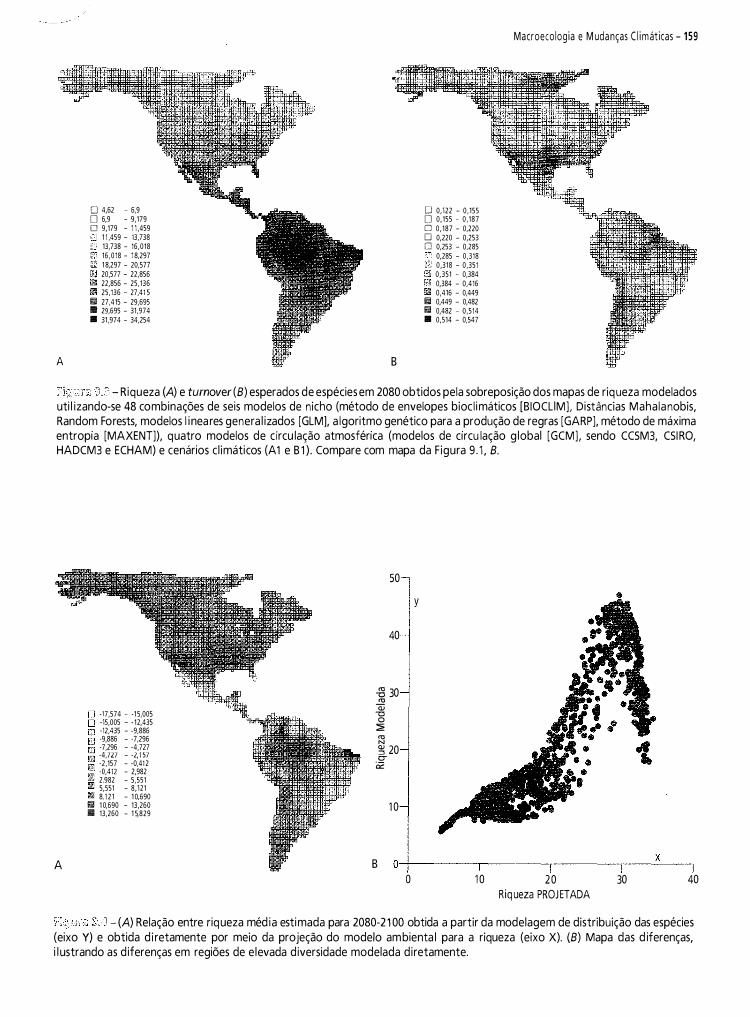
A
o 4,62 - 6,9 O 6,9 - 9,179 O 9,179 - 1 1 ,459
1 1 ,459 - 13,738 13,738 - 16,018 16,018 - 1 8,297 1 8,297 - 20,577 20,577 - 22,856 22,856 - 25,136 25,136 - 27,41 5
11 27,415 - 29,695 !li 29,695 - 31,974 li 31,974 - 34,254
B
0 0,122 - 0,155 O 0,155 - 0,187 o 0,187 - 0,220 o 0,220 - 0,253 o 0,253 - 0,285
0,285 - 0,318 0,318 - 0,351 0,351 - 0,384 0,384 - 0,416 0,416 - 0,449
11 0,449 - 0,482 11 0,482 - 0,514 li 0,514 - 0,547
Macroecologia e M udanças C l imáticas - 159
- Riqueza (A) e turnover (B) esperados de espécies em 2080 obtidos pela sobreposição dos mapas de riqueza modelados uti l izando-se 48 combinações de seis modelos de nicho (método de envelopes bioc l imáticos [BIOCLlM], Distâncias Mahalanobis, Random Forests, modelos l i neares genera l izados [GLM], a lgoritmo genético para a produção de regras [GARP], método de máxima entropia [MAXENT]), quatro modelos de circu lação atmosférica (modelos de circu lação g lobal [GCM], sendo CC5M3, C5IRO, HADCM3 e ECHAM) e cenários cl imáticos (A1 e B 1 ) , Compare com mapa da Figura 9 . 1 , B,
A
·17,574 - ·1 5,005 ·15,005 - -12,435 -12,435 - -9,886 -9,886 - -7,296 -7,296 - -4,727 -4,727 - -2,1 57 -2,157 - -0,41 2 -0,412 - 2,982 2.982 - 5,551 5,551 - 8,121 8, 121 - 1 0,690
11 1 0,690 - 13,260 11 13,260 - 1 5,829
50
40
� 30 '" (jj "O o � '" N � 20 o
o::
1 0
y
B O�--------'--------'---------r------� O 1 0 2 0
Riqueza PROJ ETADA 30 40
- (A) Relação entre riqueza méd ia estimada para 2080-2 1 00 obtida a partir da modelagem de distribuição das espécies (eixo Y) e obtida di retamente por meio da projeção do modelo ambienta l para a riqueza (eixo X). (B) Mapa das d iferenças, i l ustrando as diferenças em regiões de elevada diversidade modelada diretamente.

1 60 - Métodos e Apl i cações
à biodiversidade é exatamente que, como os nichos tendem a ser conservados em escalas de tempo reduzidas, as mudanças climáticas deverão desencadear deslocamentos nas distribuições (e não adaptações ou persistência em condições subótimas, que manteriam as reservas com a mesma eficácia), forçando as espécies a se deslocarem e eventualmente encontrarem refúgio fora das áreas que são atualmente reservadas para a conservação.
Não obstante, é possível incorporar os efeitos das mudanças climáticas em programas de planejamento para conservação da biodiversidade. Um primeiro passo será a modelagem das distribuições no presente e no futuro para avaliar mudanças nos padrões de distribuição, riqueza ou composição de espécies e mudanças fundamentais nas interações das distribuições das espécies com os sistemas de unidades de conservação, uso do solo, ocupação humana, etc. Um segundo passo será integrar a informação obtida pelos processos de modelagem das distribuições em um sistema de apoio à tomada de decisão em conservação. De fato, alguns progressos têm sido feitos neste contexto. De concreto foram desenvolvidos métodos que permitem integrar metodologias "ótimas" (mas estáticas) de seleção de reservas com modelagem de distribuições e identificação de corredores ecológicos para dispersão em contexto de modificação do clima. Os primeiros exemplos práticos desta nova metodologia foram desenvolvidos para um conjunto de plantas na província do Cabo na África do Sul, tendo dado resultados promissores2o.
A questão essencial é que existem muitas incertezas nos processos de modelagem das distribuições das espécies e estas podem prejudicar as aplicações práticas ao nível de políticas de conservação. Uma alternativa aos modelos de nicho, que começa a ser explorada, é usar aspectos de modelagem dinâmica que considerem cenários e custos que mudam com o tempo. Neste caso, é importante levar em consideração também outros componentes, como dispersão, que, por sua vez, podem ser afetados pelos padrões de ocupação humana, conforme já discutido. Análises de deslocamentos de distribuição geográfica em tempos relativamente recentes (por exemplo, Pleistoceno) sugerem que mesmo modelos de dispersão com alta assimetria (long tails) são factíveis e podem efetivamente permitir deslocamentos a grandes distâncias espaciais, dado que o número de gerações seria alto.
Entretanto, no caso das mudanças climáticas atuais, há pelo menos dois problemas que poderão condicionar o uso desses padrões para extrapolar a resposta das espécies às mudanças climáticas:
1 . As mudanças atuais devem ocorrer de forma muito mais rápida, de modo que o balanço entre deslocamento de distribuição sob conservação de nicho e dispersão não permitirá que as espécies persistam (ainda que exista atualmente um debate sobre a velocidade das mudanças climáticas do passado e se discuta a possibilidade de que as mudanças atuais não sejam, afinal, excepcionais no que respeita a velocidade).
2. As trilhas de dispersão não estão necessariamente "livres", pois a fragmentação e a perda de hábitats fazem com que, mesmo que as distâncias não sejam excessivamente grandes, a falta de conectividade pode impedir uma dispersão gradual e exigir que a nova distribuição seja alcançada por eventos raros de dispersão de longa distância (diminuindo, assim, a probabilidade de persistência).
Dessa forma, mesmo que um dado ambiente adequado para o estabelecimento da espécie no futuro esteja "ao alcance" de processos de dispersão (ou seja, a espécie possa alcançar o novo local com alta probabilidade), pode não haver mais conectividade entre o ambiente atual e o futuro que permita essa dispersão. Seria preciso incorporar ao mesmo tempo em um modelo o nicho ecológico das espécies (com todas as dificuldades e incertezas já discutidas), o processo de dispersão estruturado por hábitats e os padrões socioeconômicos de ocupação humana, a fim de fazer predições mais realistas sobre alterações nas distribuições das espécies em resposta às mudanças climáticas.
Por fim, é interessante notar que a maioria dos trabalhos mostrando efeitos das mudanças climáticas na biodiversidade tem sido feita nas regiões temperadas do hemisfério norte, onde os gradientes climáticos, sendo mais intensos, devem (a princípio) gerar maiores impactos sobre as distribuições e as abundâncias das espécies. Entretanto, é preciso lembrar que a resposta das espécies depende também (e talvez principalmente) das suas pró- :s prias características ecológicas e fisiológicas que, em última instância, definem o seu nicho. Como colocaram Bush e Hooghiesmtra21 , " . . . uma vez que os sistemas tropicais úmidos possuem tanta diversidade, e porque acredita-se que a maioria das espécies possua nichos restritos . . . , há potencial para que mesmo pequenas perturbações climáticas tenham um efeito profundo". Mais estudos na região Neotropical são necessários para avaliar, de forma efetiva, os impactos das mudanças climáticas, mas de qualquer modo é importante levar em consideração que mesmo pequenas mudanças podem gerar grandes impactos se os nichos das espécies são mais estreitos.
00 00 UI .:., �
, 00 'D 0\ b,

Agradecemos a L. M. Bini, Paulo de Marco Jr. e aos estudantes e investigadores do BIOCHANGE Lab de Madrid pelas discussões, a Thiago F. L. V. B . Rangel pelo desenvolvimento do software Bioensembles e a João Carlos Nabout e Guilherme de Oliveira pela ajuda na confecção dos mapas e processamento dos dados utilizados aqui. Nosso trabalho em modelos de distribuição potencial e impactos das mudanças climáticas na região N eotropical tem sido apoiado pela Fundação BB VA, por meio do projeto BIOIMPACTO. J. A. F. Diniz-Filho tem sido continuamente apoiado por uma bolsa de Produtividade do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
REFERÊNCIAS BIBLIOGRÁFICAS 1 . BROWN, 1. H. Macroecology. Chicago: Chicago University
Press, 1995. 269p. 2. PARMESAN, C. Ecological and evolutionary responses to
recent c1imate change. Ann. Rev. Eco!, Evo!' Syst., v. 37, p. 637-669, 2006.
3. LOVEJOY, T. E.; HANNAH, L. Climate Change and Biodiversity. New Haven: Yale University Press, 2005. 4 1 8p.
4. KERR, J. T.; KHAROUBA, H. M . ; CURRIE, D. 1. 2007. The macroecological contribution to global change solutions. Science, v. 3 1 6, p. 1 5 8 1 - 1 584.
5 . CURRIE, D. 1.; MITTELBACH, G. G.; CORNELL, H. V. et a!. Predictions and tests of c1imate-based hypotheses of broadscale variation in taxonomic richness. Ecol. Lett., v. 7, p . 1 12 1 - 1 134, 2004.
6. ARAÚJO, M. B . ; NOGUÉS-BRAVO, D . ; DINIZ-FILHO, 1. A. F. et a!. Quaternary c1imate changes explain diversity among reptiles and amphibians. Ecography, v. 3 1 , p. 8-15, 2008.
7. GASTON, K. J. The Structure and Dynamics of Geographic Ranges. Oxford: Oxford University Press, 2003. 266p.
Macroeco log ia e M udanças C l imáticas - 1 61
8. MAC ARTHUR, R. Geographical Ecology. Princeton, Princeton University Press, 1972. 269p.
9. MAURER, B. A. Untangling Ecological Complexity. Chicago: Chicago University Press, 2000. 251p.
10. ARAÚJO, M . B . ; PEARSO�, R. G. Equilibrium of species' distributions with c1imate. Ecography, v. 28, p. 693-695, 2005.
1 1 . ARAÚJO, M. B . ; GUISAN, A. Five (or so) challenges for species distribution modelling. J Biogeogl:, v. 33, p. 1677-1688, 2006.
1 2 . SOBERÓN, 1. Grinnellian and Eltonian niches and geographic distributions of species. Eco!, Lett. , v. 10, p. 1 1 1 5 - 1 1 23 , 2007.
1 3 . SEGURADO, P.; ARAÚJO, M. B. An evaluation of methods for modelling species distributions. 1. Biogeogr. , v. 3 1 , p. 1 555-1 568, 2004.
14. ELITH, J . ; GRAHAM, C. H. ; ANDERSON, R. P. et a!. Novel methods improve prediction of species' distributions from occurrence data. Ecography, v. 29, p. 1 29- 1 5 1 , 2006.
1 5 . ARAÚJO, M. B . ; NEW, M. Ensemble forecasting of species distributions. Trends Eco/. Evol. , v. 22, p. 42-47, 2007.
16 . MARTINEZ-MEYER, E . ; PETERSON, A. T. Ecological niches as stable distributiona1 constraints on mammal species, with implications for Pleistocene extinctions and climate change projections for biodiversity. Global Eco!, Biogeogr., v. 1 3 , p. 305-3 14, 2004.
1 7 . THUILLER, W. Patterns and uncertainties of species' range shifts under climate change. Global Change Biol. , v. 10, p. 2020-2027, 2004.
1 8. ALGAR, A. c. ; KHAROUBA, M. ; YOUNG, E. c.; KERR, 1. T. Predicting the future of species diversity: macroecological theory, climate change. Ecography, v. 32, p. 22-33, 2009.
1 9. ARAÚJO, M. B . ; CABEZA, M . ; THUILLER, W. et a!. Would c1imate change drive species out of reserves? An assessment of existing reserve-selection methods. Global Change Biol. , v. 10, p. 1 6 1 8 - 1 626, 2004.
20. WILLIAMS, P. ; HANNAH, L. ; ANDELMAN, S . ; MIDGLEY, G. ; ARAÚJO, M. B . ; HUGHES, G. ; MANNE, L. L. ; MARTINEZ-MEYER, E.; PEARSON, R. P1anning for c1imate change: Identifying minimum dispersaI corridors for the Cape Proteaceae. Consel". Biol. , v. 19, p. 1060- 1 073, 2005.
2 1 . BUSH, M. A. ; HOOGHIEMSTRA, H. Tropical biotic responses to c1imate change. In: LOVEJOY, T. E . ; HANNAH, L. Climate Change and Biodiversity. New Haven & London: Yale University Press, 2005. capo 9, p . 1 25 - 1 56.

� 1 0 ---------------------------------------------------
.J
]
Peter Lowenberg Neto
É uma pergunta fundamental para planejar: é necessário conservar a biodiversidade. Simples? Não ! Biodiversidade parece ser um daqueles termos de múltiplas interpretações que cada um, de forma intuitiva, adota o significado que quer e no final ninguém sabe exatamente do que estão falando. Dificilmente há um consenso do que é biodiversidade, em especial nos meios políticos, sociais e científicos, e isso dificulta a comunicação e ações em conservação. Portanto, o primeiro passo para uma abordagem analítica será delimitar o que conservar com precisão.
O termo "biodiversidade" é a contração de "diversidade biológica" e foi criado em 1 985 durante o planejamento de um evento sobre a preservação da natureza, o Fórum Nacional de BioDiversidade. O termo surgiu em um momento que a comunidade científica admitiu que a diversidade da vida estivesse sob ameaça. A Biologia da Conservação começou a emergir como ciência e, gradativamente, a atenção política, pública e científica se voltou para este assunto.
Depois da criação do termo, a definição de biodiversidade teve várias interpretações. Edward O. Wilson, um dos cientistas mais conhecidos na divulgação da biodiversidade e sua conservação, afirmou que a biodiversidade encerra toda a manifestação e variedade biológica da Terra, ou seja, de certo modo, biodiversidade é tudo ! . A ideia de que biodiversidade é tudo satisfaz o desejo comunal de se preservar toda natureza. Entretanto, a ideia de totalidade associada ao termo
dificulta a sua clareza analítica e a objetividade de ações em relação a ela. Considerar que biodiversidade é tudo o que se quer conservar não é eficiente em um contexto de planejamento e manejo. Aspectos relacionados às múltiplas interpretações de biodiversidade exigiram que o termo tivesse uma definição formal. Biodiversidade foi definida na Convenção sobre Diversidade Biológica, assinada no Rio de Janeiro em 1992, como sendo a variabilidade entre os organismos vivos de todas as origens incluindo, entre outras coisas, ecossistemas terrestres, marinhos e aquáticos e os complexos ecológicos nos quais eles fazem parte: isso inclui diversidade intraespecífica, entre espécies e de ecossistemas.
Uma ideia bastante comum que permeia a interpretação de alguns cientistas é que biodiversidade é sinônimo de diversidade de espécies e suas abundâncias relativas. Esta perspectiva está diretamente vinculada com a definição de "diversidade" em ecologia que antecede o termo biodiversidade. No entanto, assumir que biodiversidade é diversidade de espécies significa ignorar a diversidade nos demais níveis hierárquicos de organização biológica e inutilizar o neologismo. Biodiversidade é a diversidade de espécies e, sobretudo, diversidade de genes e diversidade de ecossistemas arranjadas hierarquicamente seguindo os níveis de organização biológica.
A biodiversidade pode ser contextualizada em diferentes abordagens filosóficas2. Por exemplo, a distinção temporal em biodiversidade contemporânea, quando tratada em relação à atual biodiversidade e biodiversidade histórica (por exemplo, linhagens antecessoras, fósseis); a distinção espacial em biodiversidade local e global, remetendo às diversidades alfa, beta e gama em ecologia; a distinção em biodiversidade natural e biodiversidade gerada pelo homem etc.

Há uma perspectiva filosófica que é importante para definir o que efetivamente se deve conservar: trata-se da abordagem processual da biodiversidade. A biodiversidade processual é oposta à biodiversidade de entidades ou estática. A perspectiva estática assume que biodiversidade é, semanticamente, apenas a diversidade de genes, espécies e ecossistemas sensu stricto. Noss3 afirmou que muitas definições de biodiversidade falharam na inclusão de processos ecológicos, tais como interações interespecíficas, distúrbios naturais e ciclos de nutrientes. Ele afirmou que tais processos são essenciais para a manutenção da biodiversidade e, por esta razão, devem ser contempladas no contexto da biodiversidade. Entretanto, DeLong4 argumentou que a inclusão de processos à biodiversidade confunde a definição com relações funcionais . Seria o mesmo que incluir ar e água como componentes do corpo humano porque eles são necessários para a sobrevivência humana.
Um segundo ponto que sustenta a biodiversidade estática é que a ideia de diversidade é inaplicável para processos biológicos porque eles são, teoricamente, os mesmos para todos os organismos, por exemplo, especiação, predação, fotos síntese. Sob esta perspectiva, a diversidade está relacionada à variedade de sujeitos e circunstâncias de um dado processo e não aos processos em si. Além disso, alguns processos, como ciclos de nutrientes e água, são processos abióticos e semanticamente não seriam contemplados em um termo com prefixo bi04.
Na perspectiva processual, os mecanismos e processos que mantêm e geram a diversidade têm lugar de destaque. Biodiversidade, neste contexto, consiste na diversidade de genes, espécies e ecossistemas e nos processos ecológicos e evolutivos que geram e sustentam a biodiversidades. Em O Desenvolvimento do Pensamento Biológico, Ernst Mayr6 apresentou o lugar da biologia na ciência. Motivado a argumentar contra o senso de que a física era o paradigma da ciência e que todos os fenômenos biológicos poderiam ser reduzidos
-= às leis físico-químicas, Mayr sumarizou propriedades "" :: únicas da biologia. Algumas das propriedades estão :::2 relacionadas à composição química de diversas macro=;; moléculas biológicas como o ácido nucleico e enzimas, ?:: à singularidade e variabilidade observadas em indiví-
duos, que ainda sendo da mesma população, são únicos, o porte de um programa genético que permite a herança e modificação de informações biológicas e outros aspectos. Uma propriedade retrata bem a essência processual das ciências biológicas: não existe nenhum fenômeno que seja absoluto em biologia. Em outras palavras, os elementos biológicos carregam, de maneira intrínseca,
Conservação da B i od iversidade e B i ogeografia H i stór ica - 163
um fardo histórico de milhões de anos que resultou em sistemas diferentes dos sistemas não vivos e que são quase incomparavelmente mais complicados. "Tudo é temporalmente e espacialmente delimitado. O animal ou a planta ou o micro-organismo com o qual ele está trabalhando nada mais é do que um elo da cadeia evolutiva da mudança na forma, nenhum dos quais tem alguma validade permanente" 7. Processo é uma condição sine qua non da biologia e, portanto, da biodiversidade. É importante reconhecer que a biodiversidade é a diversidade de elementos em seus níveis de organização biológica, desde genes até ecossistemas, e os processos que promovem e mantêm a diversidade biológica. Além de ser coerente com a ontologia das ciências biológicas6, a perspectiva processual valoriza o lastro evolutivo da biodiversidade que contemplamos hoje.
-, --�--;-T .-, -'] - ' ;"-- '
�, - ;-" � - -, : �
A manutenção da diversidade biológica pode ser exercida sob duas estratégias de conservação : a conservação ex situ e a conservação in situo A estratégia de conservação ex situ preserva genes e espécies fora da natureza ou em lugares especializados para este fim que não o seu local de origem. Como exemplos, a preservação de genes e espécies em zoológicos, jardins botânicos, bancos de sementes etc. É uma das mais antigas atividades de preservação e pode cooperar com a conservação in situ da natureza.
Entretanto, como delineado anteriormente, é necessário conservar a diversidade biológica em todos os seus níveis de organização biológica e, em particular, os processos ecológicos e evolutivos que a sustenta. Isso só é possível em conservação in situ, estratégia que busca assegurar que genes, espécies e ecossistemas sejam preservados em seu local de ocorrência na natureza. A conservação in situ possibilita que os processos naturais de transformação, fluxo e transferência de matéria e energia possam ser mantidos assim como processos genealógicos de herança, conservação e modificação da informação biológica.
A conservação in situ da biodiversidade é feita pelo estabelecimento de áreas protegidas ou unidades de conservação como são conhecidas no Brasil. As unidades de conservação são áreas estabelecidas pela lei e regidas sob diretrizes que assegurem a conservação da natureza. Conforme o Sistema Nacional de Unidades de Conservaçã08, no Brasil existem dois grupos de categorias: ( 1 ) unidades de conservação de proteção integral que têm o objetivo principal de resguardar a natureza em seu estado natural. Neste grupo estão as categorias de Estação

1 64 - Métodos e Ap l i cações
Ecológica, Reserva Biológica, Parque Nacional, Monumento Natural e Refúgio de Vida Silvestre; e (2) unidades de conservação de uso sustentável que buscam compatibilizar a conservação com uso de recursos naturais . Neste grupo estão as categorias de Área de Proteção Ambiental, Área de Relevante Interesse Ecológico, Floresta N acionaI, Reserva Extrativista, Reserva de Fauna, Reserva de Desenvolvimento Sustentável e Reserva Particular do Patrimônio Natural.
Em âmbito internacional, as áreas protegidas estão organizadas em seis categorias9: I - Reserva Natural/ Área Selvagem; H - Parque Nacional; IH - Monumento Natural; IV - Área de Manejo de Espécies/Hábitats ; V - Paisagem Terrestre ou Marítima Protegida; e VI - Área Protegida para Manejo de Recursos. Elas foram discutidas no Congresso Mundial de Parques Nacionais e Áreas Protegidas, realizado na Venezuela em 1992, e são mediadas e atualizadas pela União Internacional para a Conservação da Natureza e dos Recursos Naturais (UICN). Em um paralelo com o sistema brasileiro, as categorias I, H e III apresentam objetivos e diretrizes compatíveis com as unidades de conservação de proteção integral (uso indireto) e as categorias IV, V e VI são compatíveis com as unidades de uso sustentável (uso direto).
Unidades de conservação de proteção integral encerram categorias que apresentam maior restrição ao uso dos recursos naturais e não admitem a ocupação humana. Apesar de haver discussões em relação a sua efetividade, a unidade de conservação de proteção integral é atualmente a melhor opção para a preservação in situ da biodiversidade.
As unidades de conservação existem porque os ambientes em seus estados naturais estão sob ameaça. Ações em conservação da natureza competem com outras modalidades do uso das áreas, como o extrativismo e a ocupação humana. Além disso, os recursos financeiros destinados à conservação são sempre limitados. A soma de todos estes fatores torna a escolha de áreas uma etapa estratégica para a preservação da biodiversidade.
Diversos critérios j á foram utilizados para priorizar áreas para a conservação. Na Amazônia, por exemplo, as primeiras áreas foram escolhidas com base em amostras de regiões fito geográficas, refúgios do Pleistoceno, distintas formações vegetais, formações geológicas, hidrológicas e de SOlOlO. Outras feições, como beleza cênica, sítios naturais raros ou singulares, são critérios
condicionais para o estabelecimento de Parques e Monumentos Naturais. Os famosos "hot spots de biodiversidade" prioritários para conservação foram apontados em áreas que apresentaram excepcional concentração de \O
espécies endêmicas de plantas e altas taxas de conversão � de hábitatsl l . Espécies e ecossistemas ameaçados, lacu- � nas espaciais, complementaridade de espécies também t
00 são informações utilizadas para priorizar áreas. \O 9'
A maioria dos critérios já mencionados, no entanto, 0\
não é de critérios que favorecem a conservação da biodiversidade processual. Em outras palavras, os critérios j á citados asseguram a proteção de amostras espaciais da biodiversidade, mas em consequência da natureza quantitativa e estática dos critérios, não há evidências de que os processos serão conservados ao longo do tempo. Neste contexto, a biogeografia histórica é a melhor alternativa para a conservação da biodiversidade, fornecendo um arcabouço teórico e analítico para o reconhecimento de padrões espaciais da biodiversidade processual.
Busca interpretar os padrões espaciais dos organismos por processos históricos que ocorreram há muito temp012. Ela é, por natureza, uma ciência multidisciplinar e esta característica promove que, em um contexto espacial e temporal, o conhecimento biológico se concilie com o conhecimento abiótico e geológico para entender os padrões e os processos da natureza13 . Em biogeografia, a busca pelo "padrão" e pelo "processo" é importante, pois eles estruturam a sua ontologia em duas práticas 14: ( 1 ) a prática descritiva, que busca reconhecer e descrever o arranjo espacial dos organismos; e (2) a prática interpretativa, que busca interpretar as causas que geraram o padrão descrito.
Na prática interpretativa é onde reside a dicotomia da biogeografia: a biogeografia histórica e a biogeografia ecológica 14. A diferença fundamental entre a biogeografia histórica e a ecológica é a escala espacial e temporal de análise e, como consequência disso, a natureza dos processos envolvidos. Entender em escala global as causas do padrão espacial de espécies de um táxon monofilético, por exemplo, exige que eventos geológicos remotos sejam invocados (por exemplo, deriva continental, mudança climática) e interpretados conjuntamente com mecanismos evolutivos (vicariância, dispersão e extinção). Já entender as causas espaciais de padrões locais, como a de uma comunidade em um fragmento florestal, exige o entendimento da influência de distúr-

bios ambientais e conversão de hábitats, eventos em escala local e muitíssimo recentes na escala geológica.
É importante salientar que a biogeografia histórica foi consolidada como disciplina da Biologia Comparada 14 e tradicionalmente teve acentuada atuação de cientistas que atuavam em sistemática e taxonomia. Motivados a entender a história espacial dos táxons de interesse e, sobretudo, da biodiversidade, estes cientistas promoveram que ideias da biologia comparada permeassem constantemente teorias e métodos da biogeografia histórica. Muitas das práticas consistem em observar e descrever os padrões espaciais, construir narrativas históricas e comparar os padrões de diversos táxons. A influência da biologia comparada na biogeografia continua presente e um exemplo disso é o Código Internacional de Nomenclatura de Área (CINA), proposto recentemente para sistematizar o conhecimento biogeográfico.
A biogeografia histórica busca reconstruir e interpretar a trajetória espaço-temporal dos organismos, com o respaldo do conhecimento de eventos antecedentes, e promover o entendimento do arranjo espacial da biodiversidade na atualidade. Para a conservação da biodiversidade ela contribui de duas maneiras : ( 1 ) métodos que reconhecem padrões espaciais de distribuição geográfica dos organismos15 e (2) princípios de sistematização do conhecimento espacial da biodiversidade.
A biogeografia histórica é marcada pela grande variedade de métodos e técnicas. É possível sumarizar mais de 30 métodos e abordagens em biogeografial3, tais como centro de origem e dispersão, biogeografia filo genética, áreas ancestrais, pan-biogeografia, biogeografia cladística, análise parcimoniosa de endemismo, métodos baseados em eventos, filo geografia e biogeografia experimental. Alguns cientistas criticam que a pluralidade de métodos da biogeografia histórica gera ou reflete uma crise de identidade da ciência em questão. Entretanto, outros cientistas argumentam que a diversidade de técnicas apenas reflete a complexidade que permeia a arena biogeográfica.
Em biogeografia há três abordagens básicas 1 3 : biogeografia do táxon - que busca reconstruir a narrativa histórica espacial de um grupo de espécies ; biogeografia de área - que busca reconstruir o relacionamento entre as áreas de endemismo; e homologia espacial - que busca reconstruir a historia espacial das biotas. Depen-
Conservação da B iod ivers idade e B iogeografia H istórica - 1 65
dendo do perfil da pesquisa e da pergunta científica há um ou mais métodos que podem ser utilizados e, até mesmo, o emprego de métodos em conjunto.
A aplicação de métodos para a conservação da biodiversidade não é diferente. É essencial que de antemão se saiba o que é importante para a conservação e o perfil da abordagem para que a escolha do método seja a mais apropriada. As três abordagens apontadas anteriormente (táxon, área e homologia espacial) geram informações para a conservação. A abordagem de táxon pode ser usada para investigar a biogeografia de uma espécie ou gênero que é chave em um ecossistema ou indicador de biodiversidade; a abordagem de área pode ser usada para o planejamento de uma rede de unidades de conservação que leva em conta o relacionamento histórico das áreas ; e o estudo da homologia espacial fornece informações sobre a disposição espacial e historicidade das áreas.
Neste capítulo serão enfatizados os métodos da abordagem de homologia espacial, pois é, atualmente, a abordagem que mais pode contribuir para o reconhecimento de áreas prioritárias para se conservar. Dentre os elementos geográficos utilizados para o apontamento de áreas para se conservar estão as áreas de endemismo e os complexos biogeológicos.
Área de Endemismo e Métodos para o seu Reconhecimento
As áreas de endemismo são áreas que contêm dois ou mais táxons exclusivos e de distribuição restrita àquela área16. Elas são as unidades fundamentais de análise na biogeografia cladística e são importantes no entendimento dos padrões de distribuição de uma região (ver Capo 3) . No contexto da conservação, as áreas de endemismo são elementos biogeográficos empregados para priorizar áreas para se conservar, pois enfatizam espacialmente feições singulares da biodiversidade.
A natureza da área de endemismo não é a priori evidente. Não há como saber de antemão se uma área de endemismo resultou de processos ecológicos ou históricos. Por isso, o estudo das áreas de endemismos, em especial na reconstrução da história espacial das biotas, segue duas etapas 17 : ( 1 ) delineamento de áreas de endemismo que apresentarem simpatria de táxons exclusivos. Nesta etapa, assume-se que as áreas de endemismo encontradas são hipóteses de homologia biogeográfica primária, ou seja, unidades biogeográficas naturais que apresentam táxons exclusivos pela associação histórica entre si e com a área geográfica; e (2)

1 66 - Métodos e Apl icações
teste da homologia biogeográfica primária com informações filo genéticas dos táxons. Nesta etapa as áreas são testadas quanto à congruência topológica dos relacionamentos entre os táxons.
A primeira etapa da análise, que consiste no delineamento das áreas de endemismo (homologia biogeográfica primária), pode ser feita por várias técnicas. A seguir serão apresentados um método manual e três métodos numéricos (agrupamento fenético e a análise parcimoniosa de endemicidade [PAE, parsimony analysis of endemicity] e eNDeMism [NDM]) para o reconhecimento das áreas.
O método manual consiste na delimitação das áreas de endemismo pela inspeção visual da sobreposição espacial das áreas de distribuição18 • As áreas de distribuição dos táxons em estudo são manualmente contornadas, agregadas em um mapa e as áreas de endemismo são visualmente delimitadas onde há simpatria total ou parcial de táxons com distribuição restrita. Este método foi muito utilizado antes do surgimento de métodos computacionais. As desvantagens deste método são a delimitação subjetiva e a dificuldade em analisar uma grande quantidade de dados simultaneamente.
O agrupamento fenético e a PAE são métodos numéricos e hierárquicos. Numéricos porque a implementação destes métodos segue rotinas de transformação de dados espaciais em matrizes binárias, o que permitiu que pudessem ser analisados em programas computacionais; e hierárquicos porque o resultado são dendrogramas de áreas agrupados ou caracterizados pelas espécies que elas compartilham.
O procedimento de análise do agrupamento fenético e da PAE19 segue basicamente os mesmos passos: ( 1 ) estabelecimento de unidades geográficas operacionais de análise. Em geral, são utilizadas localidades ou quadrículas que podem variar de tamanho dependendo da área. A análise por quadrículas, ou qualquer outro polígono, é interessante porque gera um cenário de reconhecimento das áreas sem interferência de delimitações bióticas prévias; (2) transformação de dados espaciais em matriz binária. As matrizes são codificadas O para ausência e 1 para presença de cada táxon em cada unidade geográfica operacional. No caso da PAE, a matriz tem que apresentar uma área hipotética codificada com ausência de todos os táxons; (3) análise da matriz. Neste passo é que a análise fenética e a PAE se diferenciam. Na análise fenética utilizam-se coeficientes de similaridade para agregar as áreas e na PAE a matriz é submetida ao algoritmo de parcimônia. O resultado de ambas as análises são dendrogramas de áreas agrupadas pelas espécies. Os dendrogramas não representam o relacionamento histórico das áreas e a topologia é interpretada apenas quanto ao aninhamento hierárquico das áreas.
O NDM16 consiste em um método numérico que implementa e busca as áreas de endemismo por meio de índices de endemismo. Em relação a uma dada área inicial, cada táxon tem seu índice de endemismo calculado. O índice é influenciado pela distribuição do táxon pelas quadrículas da área: táxons que ocupam todas as quadrículas da área têm índice um ( 1 ) enquanto táxons que também ocupam áreas externas ou ocupam parcialmente a área em questão têm índices inferiores. A área de endemismo é delimitada pela soma dos índices de endemismo dos táxons presentes na área.
Estudos usando o NDM têm mostrado que o método é capaz de encontrar mais áreas de endemismo, principalmente áreas em simpatria, que os demais métodos numéricos. Isto ocorre porque o NDM é um método que delimita as áreas de endemismo conforme um índice e não necessita resolver o arranjo das áreas em um contexto hierárquico e limitado. Ele gera o número necessário de áreas de endemismo para incluir todos os arranjos de espécies exclusivas existentes indiferentemente ao aninhamento das áreas. Atualmente é método numérico mais robusto e detalhado de reconhecimento de áreas de endemismo.
Os métodos para o reconhecimento de endemismo geram valiosas informações para a conservação. O simples fato de identificar as áreas que albergam táxons exclusivos j á fornece informações importantes para priorizar áreas para se conservar. No Brasil, um estudo buscou delimitar áreas de endemismo na região sul do Brasil e compará-las com a disposição das unidades de conservação de proteção integral2o. O estudo levou em conta a distribuição de espécies de diversos grupos como angiospermas, plantas vasculares sem sementes, líquens, invertebrados e vertebrados e delimitou as áreas de endemismo empregando o método de parcimônia. As áreas de endemismos delimitadas na região sul estavam parcialmente sob proteção integral e, por isso, foram sugeridas como prioritárias e, como alternativa, foi proposta a recategorização quanto ao manejo das unidades de conservação de uso sustentável que estavam em coincidência com as áreas delimitadas. Uma ressalva particular quanto a este estudo é a delimitação da área estudada (região sul do Brasil) : o delineamento político deve ser evitado em estudos biogeográficos por não compreender formação natural.
O passo seguinte da análise em duas etapas pode gerar informações ainda mais relevantes para a conservação. O segundo passo trata-se do teste da homologia biogeográfica da área e busca entender se os táxons são endêmicos pela associação histórica entre si e a área geográfica. Utiliza-se a informação filogenética dos

táxons e verifica-se a congruência topológica entre os táxons não relacionados. Caso haja congruência espacial e topológica dos relacionamentos, a área pode ser considerada uma área de endemismo sob os critérios de Harold e MooFl. A vantagem desta perspectiva é que a homologia, ou seja, o vínculo espacial e a correspondência evolutiva são certificados e a área de endemismo é considerada como uma unidade histórica natural (ver Capo 3) . A conservação destas áreas implica que não somente táxons exclusivos serão preservados, mas principalmente a herança de todos os processos evolutivos e paleoecológicos que moldaram aquela biota.
Área de Endemismo e Complexo Biogeológico : Pan-biogeografia Área de endemismo e complexo biogeológico podem ser estimados pela Pan-biogeografia (ver Capo 5) . A Pan-biogeografia não se trata apenas de um método; ela pode ser considerada um programa de pesquisa em biogeografia histórica que apresenta suas próprias premissas e interpretações. Apesar de bastante criticada até os dias atuais, a Pan-biogeografia conquistou seu espaço científico e hoje é largamente utilizada para o entendimento espacial dos organismos.
A Pan-biogeografia é um conjunto de métodos que busca enfatizar a dimensão espacial da diversidade da vida. Ela reconhece a importância das localidades como fonte primária e fundamental de informação para a análise biogeográfica permeada por mudanças evolutivas. As localidades geográficas de ocorrência dos táxons têm posição de destaque no procedimento metodológico e processos ecológicos e evolutivos são interpretados em um contexto geográfico. A ontologia pan-biogeográfica assume que a evolução dos organismos é estruturada por três sustentáculos - espaço, tempo e forma, e que a Terra e vida evoluem juntas22.
Na análise de traços, que é a modalidade mais difundida da pan-biogeografia, os pontos de ocorrência de um táxon são conectados por traços seguindo a regra da distância mínima geográfica entre os pontos. Cada táxon
� tem sua hipótese de distribuição, chamada de traço in-dividual, que consiste no traço desenhado sobre os
� r. pontos de ocorrência. Se, ao juntar diversos traços indi-x: :!: viduais em um mesmo mapa, houver coincidência '='
espacial de dois ou mais traços, então a área em coincidência é chamada de traço generalizado (ver Capo 5).
o traço generalizado é interpretado como uma hipótese de biotas ancestrais que foram fragmentadas por eventos de mudança climática ou tectônica. Por isso, os
Conservação da B iod ivers idade e Bi ogeografia H istórica - 167
traços generalizados também são considerados como homologia biogeográfica e inseridos no contexto da análise em dois estágios 17 . Podem ser usados como áreas prioritárias para se conservar devido ao possível endemismo das espécies envolvidas.
Quando há a intersecção de dois ou mais traços generalizados , um nó biogeográfico é estabelecido. A natureza biogeográfica do nó pode, no início, não ser clara. Fundamentalmente, o nó biogeográfico representa uma área onde vários eventos vicariantes ocorreram ocasionados por diferentes atividades geológicas. Por conta disso, o nó pode ser biologicamente interpretado de diversas maneiras, como endemismo, alta diversidade, limites distribucionais, ausências anômalas, populações disjuntas, entre outras e isso depende da informação disponível e do modelo de especiação adotad023.
Independente das interpretações do nó biogeográfico, este elemento representa essencialmente um complexo biogeológico, ou seja, uma área onde houve uma história geológica dinâmica que ocasionou que feições biológicas extraordinárias surgissem. Nenhum outro elemento biogeográfico é tão singular quanto a sua interpretação e potencialidade em caracterizar uma nova feição da biodiversidade. Por isso, ao se proteger um nó biogeográfico, não apenas a herança de uma narrativa evolutiva marcada por eventos geológicos é conservada, mas todos os elementos e características biológicas envolvidas com esta história.
No Brasil, a abordagem da análise de traços foi usada para priorizar áreas para a conservação no Cerrad024. A análise de traços foi realizada com várias espécies, incluindo insetos, aves, mamíferos e plantas e os nós biogeográficos delimitados em análise de todas as espécies em conjunto. Os nós biogeográficos foram comparados à atual disposição das unidades de conservação e às áreas prioritárias para conservação definidas pelo workshop do Ministério do Meio Ambiente de 1 998 e atualizadas em 2004. Verificou-se que a maioria dos nós biogeográficos (80%) não apresentou coincidência espacial com unidades de conservação; o restante dos nós (20%) estava protegido, entretanto, em grande parte, em unidades de conservação de uso sustentável; e 77% dos nós foram delimitados em áreas prioritárias. Concluiu-se que a biodiversidade do Cerrado ainda está desprotegida e sugeriu-se que os nós biogeográficos encontrados em áreas ameaçadas sejam incorporados à política de priorização de áreas para conservação.
Outro exemplo de complexo biogeológico são as áreas de diversificação. As áreas de diversificação são hipóteses de locais onde ocorrera especiação devido a um ou vários eventos de vicariância25 . O reconhecimento

1 68 - Métodos e Ap l icações
de tais áreas é feito pela associação da análise de traços com informações filo genética dos táxons de interesse. Esta abordagem consiste em dois passos:
1. Desenham-se apenas traços generalizados suportados por espécies irmãs. Considerando que os traços generalizados são hipóteses de biotas ancestrais que foram fragmentadas por eventos de mudança climática ou tectônica, um traço suportado por espécies irmãs permite inferir, com respaldo temporal, uma hipótese de local onde a especiação ocorreu.
2. Desenham-se os traços generalizados suportados por táxons irmãos e buscam-se os nós biogeográficos. A área que compreende os traços e os nós é delineada manualmente e denominada área de diversificação.
Esta metodologia foi empregada com 1 1 gêneros de Muscidae (Insecla: Diptera) para os quais hipóteses filo genéticas estão disponíveis e que ocorriam na região Neotropical. Por meio da metodologia já descrita foi possível reconhecer quatro áreas de diversificação: Panamá, Amazônia, Floresta Atlântica e Chile central. Surpreendentemente, os traços e os nós coincidiram com regiões do continente onde houvera diversas atividades geológicas25 . Acredita-se que estas áreas fornecem um conhecimento fundamental da trajetória espacial dos táxons e que deve ser considerada em um contexto conservacionista.
As premissas da Pan-biogeografia diferem da perspectiva da ecologia espacial de que os nichos são os principais responsáveis pela ocupação e arranjo espacial dos organismos. Sabe-se que os seres vivos são influenciados por aporte e demanda de matéria e energia ; contudo, a perspectiva pan-biogeográfica busca enfatizar e resgatar os elementos geográficos responsáveis pela ocupação espacial. Esta perspectiva proporciona que elementos biogeográficos como complexos biogeológicos sejam reconhecidos e empregados como critério para priorizar áreas para conservação.
Em qualquer ramo da ciência, a organização do conhecimento é fundamental para se estabelecer o estado da arte e estruturar seus avanços. A relação ente os métodos e a sistematização do conhecimento consiste em um ciclo virtuoso: os métodos são importantes ferramentas para transformar informações em conhecimento e a organização do conhecimento é fundamental para a
comunicação e o avanço científico. Em biogeografia histórica a sistematização do conhecimento é realizada por duas práticas: classificação biogeográfica e o CINA.
Classificação Biogeográfica
Uma classificação biogeográfica é a compilação do conhecimento espacial da biodiversidade. Exemplos de classificações são as regiões fitogeográficas, reinos e regiões zoogeográficas e mapas de ecorregiões. Eles são interessantes para a conservação, pois fornecem um panorama geral da organização espacial da vida, auxiliam na priorização das áreas e geram subsídios para a construção de uma rede de unidades de conservação biogeograficamente consciente.
As classificações biogeográficas têm a característica de compilar e delimitar as áreas seguindo determinado critério. As regiões zoogeográficas propostas por Sclater, por exemplo, foram delimitadas pelo endemismo de aves, enquanto as regiões de Wallace foram delimitadas pelo endemismo de mamíferos. Elas apresentam áreas em comum, mas alguns detalhes particulares de determinado grupo são somente vistos em um dos esquemas.
As regiões fito geográficas propostas por de Candolle são diferentes das ecorregiões, por exemplo. As regiões fitogeográficas foram construídas considerando os padrões de endemismo das plantas, ao passo que as ecorregiões consideram a organização funcional e paisagística. Os diferentes critérios utilizados para a construção do esquema biogeográfico não são mutuamente exclusivos; de fato, eles salientam diferentes aspectos da biodiversidade.
No âmbito da biogeografia histórica, os esquemas biogeográficos são construídos sob o critério da homologia das unidades geográficas, ou seja, as áreas de endemismo e todos os níveis superiores da hierarquia biogeográfica (províncias, sub-regiões, regiões, reinos) são elementos naturais e históricos. Um esquema que bem ilustra o critério da homologia são os reinos biogeográficos mundiais apresentados por Morrone26. Este esquema baseou-se no pressuposto de que todas as regiões reconhecidas hoje derivaram do último supercontinente Pangea. No estudo, foram compilados vários estudos biogeográficos de plantas e animais e verificou-se que a relação e delimitação das regiões biogeográficas encontradas hoje são equivalentes às áreas que no passado estiveram juntas. Neste esquema, o reino Holártico equivale ao paleocontinente da Laurásia, o reino Holotropical equivale à porção leste do Gondwana e o reino Austral equivale à porção oeste do Gonduana. O esquema é impressionantemente compatível com hipóteses

prévias apresentadas para plantas e animais, com a vantagem de ser conoborada pela homologia biogeográfica (Fig. 10. 1 , A).
As classificações em econegiões enfatizam atributos da biodiversidade ligados à funcionalidade. As paisagens são delimitadas levando-se em conta formações fitológicas, clima, solo etc. A relação entre econegiões e as áreas de endemismo é facilmente entendida quando se pensa na Floresta Amazônica. A Floresta Amazônica, sob os critérios de ecorregião, apresenta feições paisagísticas uniformes que permitem delinear toda a floresta tropical como uma única unidade funcional (Fig. 10. 1 , B). Entretanto, ao explorar a composição de espécies, verifica-se que a Amazônia é um grande mosaico de áreas de endemism028, cada uma delas com sua identidade histórica e composição de espécies. Isto já foi observado em estudos na Floresta Atlântica e, certamente, será encontrado em todos os estudos que assumem uma formação paisagística e comparam com a composição espacial das espécies. Concomitantemente, entende-se que as diferentes espécies, que constituem as áreas de endemismo, apresentam redundância em suas funções, já que é possível delinear uma unidade paisagística que inclui todas as áreas de endemismo.
Em estratégias de conservação que buscam a representatividade, ou seja, conservar amostras substanciais da biodiversidade, os esquemas biogeográficos históricos têm vantagens sobre esquemas em ecorregiões . Isto principalmente porque as áreas de endemismos enfatizam feições singulares da biodiversidade resultante de processos pretéritos e com o respaldo da homologia biogeográfica. As econegiões enfatizam a paisagem e não a composição da biodiversidade. Isso ocasiona que
� as formações paisagísticas sejam delimitadas em mais =-- de uma unidade de homologia. Em outras palavras, há x: - ecossistemas semelhantes em continentes de distintas :: >r. histórias biogeográficas. Este padrão é também conheci-� do como a primeira lei da biogeografia ou lei de B uffon29. =- Para a conservação da representatividade biológica
sugere-se que esquemas que salientem feições singulares da biodiversidade sejam valorizados na escolha de áreas e na montagem de uma rede de áreas protegidas. Encoraja-se que a avaliação seja feita com econegiões e regiões biogeográficas em conjunto, idealmente com esquemas que considerem os dois critérios, favorecendo as econegiões sob estratégia de persistência funcional e as regiões biogeográficas sob a estratégia da representatividade.
Recentemente, foi elaborado um esquema biogeográfico para a América Latina e Caribe que analisou a distribuição geográfica de centenas de espécies de insetos por intermédio da análise de traços30. Os traços genera-
Conservação da B iod ivers idade e B iogeograf ia H i stór ica - 169
lizados foram interpretados como áreas de endemismo ou homologia primária biogeográfica e foram compilados em um esquema geral para a região (Fig. 10.2).
Para verificar a representatividade das feições singulares da biodiversidade que estão sob proteção, as províncias da América Latina e Caribe foram sobrepostas por áreas protegidas. Apesar de as províncias apresentam áreas muito mais amplas em relação às áreas protegidas, o cenário é válido para se avaliar a disposição espacial das unidades de conservação e verificar se todas as províncias têm frações sob proteção. Em uma inspeção visual é possível observar que, embora a maioria das províncias contenha pelo menos uma área protegida, há várias províncias que não estão sob proteção (Fig. 10.2).
Código Internacional de Nomenclatura de Área
Em estudos biogeográficos, é comum que ao delimitar uma área de endemismo se atribua um nome a ela. Normalmente os nomes são relacionados a formações vegetais (por exemplo, Pampa, Floresta Valdiviana), delineamento político (por exemplo, Guiana, Oeste do Equador), rios (por exemplo, Madeira, Xingu-Tapajós) ou qualquer outro nome que identifique e forneça uma noção da localização geográfica da área. Entretanto, com o aumento de técnicas de identificação de áreas de endemismo e, sobretudo, o crescente interesse pela biogeografia, muita informação sobre áreas de endemismo foram geradas e estas áreas nomeadas sem critérios formais. A consequência disso foi que algumas áreas eram conhecidas por mais de um nome e um nome era associado a mais de uma área.
Em 2007, membros da Associação de Biogeografia Sistemática e Evolutiva discutiram e formalizaram o CINA, que promove um sistema universal de nomes das áreas de endemismo. O objetivo do código é padronizar o nome das áreas utilizadas em biogeografia e outras disciplinas com a finalidade de melhorar a comunicação e construir classificações3 l .
A ideia geral que sustenta o CINA é análoga aos demais códigos de nomenclatura biológica. Uma vez que um nome válido é estabelecido, ele deve ser acompanhado do nome do autor e ano (por exemplo, Região Neotropical Sclater, 1 858). Caso seja proposto um novo nome de área, o nome deve estar vinculado a uma localidade tipo acompanhado de descrição e diagnose. A diagnose deve ser detalhada quanto a sua delimitação espacial, táxons endêmicos e acompanhado de um mapa biogeográfico. Os nomes das áreas são ananjados sob nomes mais inclusivos para representar a hierarquia biogeográfica (distritos, províncias, regiões, reinos).
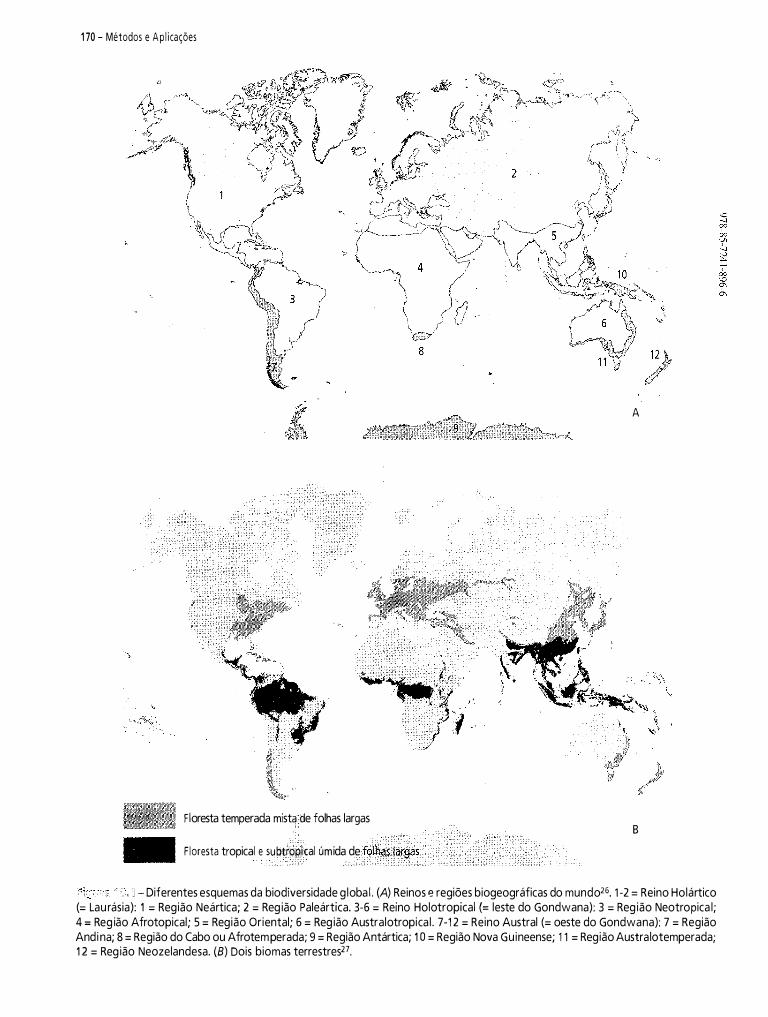
1 70 - Métodos e Ap l i cações
A
F loresta temperada mista de folhas largas B
F loresta tropical e subt�º�i(al úmida de tO'lI1ª�ja�lJas
J - Diferentes esquemas da biodiversidade g loba l . (A) Reinos e regiões biogeográficas do mundo26. 1 -2 = Reino Holártico (= Laurásia): 1 = Região Neártica; 2 = Reg ião Paleártica. 3-6 = Reino Holotropical (= leste do Gondwana): 3 = Reg ião Neotropica l; 4 = Reg ião Afrotopical; 5 = Região Oriental; 6 = Reg ião Australotropica l . 7-1 2 = Reino Austral (= oeste do Gondwana): 7 = Região Andi na; 8 = Região do Cabo ou Afrotemperada; 9 = Região Antártica; 1 0 = Região Nova Gu ineense; 1 1 = Região Austra lotemperada; 1 2 = Reg ião Neozelandesa. (B) Dois biomas terrestres27.

Conservação da B i od ivers idade e B i ogeograf ia H i stór ica - 1 71
c _ Províncias da América Latina e Caribe del i neadas pelo padrão de distribuição de insetos3o• Os polígonos pretos são as áreas proteg idas categorias I, II e 1 1 1 e os polígonos cinza são as áreas protegidas categorias IV, V e VI da Un ião Internacional para a Conservação da Natureza e dos Recursos Naturais (UICN). Fonte: www.wdpa.org - World Database on Protected Areas.
Um exemplo da utilização do CINA foi o estudo desenvolvido por López et al. 32, que delimitou áreas de endemismo na Argentina. O estudo analisou dados de distribuição de peixes de água doce e utilizou PAE, similaridade e análise de agrupamento como metodologia para o delineamento das áreas de endemismo. As áreas foram descritas e organizadas conforme o código de áreas. A seguir um exemplo: Reino Holotropical (Rapoport, 1 968); Região Neotropical (Selater, 1 858); Província Great Rivers (López et al. 32) :
• Localidade tipo: médio Rio Paraná (27° 1 8 ' S , 58°36' S a 32°07 ' S, 60°37 ' W) .
• Diagnose: esta província corresponde à grande região subtropical definida como a Mesopotamia argentina (área entre os rios Paraná e Uruguai) e áreas adjacen-
tes o No sentido usado aqui, a província é mais restrita que a província Párano-Platense de Ringuelet (ver Capo 2). Esta diferença resultou da extensão da província Pampeana de nossa análise.
• Descrição: existem 394 espécies de peixes na província Great Rivers, na qual é a mais rica da Argentina. Noventa e nove espécies (23, 1 %) são compartilhadas com a província Pampean. Na província Great Rivers, Salminus brasiliensis, Prochilodus lineatus e Synbranchus marmoratus ocorrem em todas as localidades; Eigenmannia virescens, Roeboides bonariensis, Loricariicthys melanocheilus, Astyanax abramis, Hoplias malabaricus, Pimelodus maculatus, Pimelodella gracilis, Astyanax bimaculatus, A. fasciatus, Odontostilbe pequira, Pimelodus albicans e Pygocentrus nattereri ocorrem na maioria das localidades.

172 - Métodos e Ap l i cações
o CINA representa um avanço para a biogeografia em relação à sistematização do conhecimento espacial da biodiversidade. Isso certamente contribuirá para a melhoria do entendimento biogeográfico e, por consequência, favorecerá a conservação da natureza.
A utilização da biogeografia histórica tem muito a contribuir para a conservação da biodiversidade33 por se tratar de uma ciência de base que gera conhecimentos para o pleno entendimento da espacialidade da biodiversidade. Um ponto importante desta ciência, e que certamente a distingue das demais envolvidas com a geografia dos seres vivos, é a construção do entendimento espacial da biodiversidade pela reconstrução da trajetória histórico-evolutiva de organismos e biotas. O passado é o prólogo e a vocação histórica da biogeografia favorece que, em um contexto de análise espacial, os mecanismos evolutivos e ecológicos que geraram e mantiveram a diversidade biológica sejam valorizados.
REFERÊNCIAS BIBLIOGRÁFICAS 1. WILSON, E. O. Introduction. In: REAKA-KUDLA, M . J. ;
WILSON, D. E.; WILSON, E. O. Biodiversity 11: understanding and protecting our biological resources. Washington: Joseph Henry Press, 1 997. capo 1 , p. 1 -3.
2. KORlCHEVA, J.; SUPI, H. The phenomenon ofbiodiversity. In: OKSANEN, M.; PIETARINEN, J. Philosophy and Biodiversity. New York: Cambridge University Press, 2004. capo 1 , p. 27-53.
3 . N OSS, R F. Indicators for monitoring biodiversity: a hierarchical approach. Conser Biol., v. 4, p. 355-364, 1 990.
4. DELONG, D. C. Defining Biodiversity. Wildlife Soe. B., V. 24, p. 738-749, 1 996.
5 . GASTON, K. J. Biodiversity: a biology ofnumbers and difference. Oxford: Blackwell Science, 1 996. 396p.
6. MAYR, E. O Desenvolvimento do Pensamento Biológico - Diversidade, Evolução e Herança. Brasília: Editora UnE, 1998. 1 107p.
7. DELBRÜCK, M. A physicist look at biology. Trans Conn Acad Arts Sei, v. 38, p. 173- 1 90, 1 949.
8. BRASIL. Lei n0 9.985. SNUC - Sistema Nacional de Unidades de Conservação, 1 8 jul. 2000.
9. DUDLEY, N. GuidelinesforApplying ProtectedArea Management Categories. Gland, Switzerland: UICN, 2008. 86p.
10. RYLANDS, A. B . ; BRANDON, K. Brazilian protected areas. Conser. Biol. , v. 1 9, p. 6 1 2-618, 2005.
1 1 . MYERS, N.; MITTERMEIER, R A; MITTERMEIER, C. G. et aI. Biodiversity hotspots for conservation priorities. Nature, v. 403, p. 853-858. 2000.
12 . CRISCI, J. J.; SALA, O. E.; KATINAS, L. et aI. Bridging historical and ecological approaches in biogeography. Austral. Syst. Bot. , v. 19 , p. 1 - 1 0, 2006.
1 3 . CRISCl, J. V; KATINAS, L.; POSADAS, P Historieal Biogeography: an introduetion. Cambridge: Harvard Unviersity Press, 2003. 25 p.
14. POSADAS, P.; CRISCI, J. V; KATINAS, L. Historical biogeography: a review of its basic concepts and criticaI issues. J. Arid. Environ. , v. 66, 389-403, 2006.
1 5 . CARVALHO, C. J. B. Ferramentas atuais da biogeografia histórica para utilização em conservação. In: MILANO, M. S . ; TAKAHASHI, L. Y.; NUNES, M. L. Unidades d e Conservação: atualidades e tendências. Curitiba: FBPN, 2004. p. 92- 1 03.
1 6. SZUMICK, C.; GOLOBOFF, P Areas of endemism: An improved optimality criterion. Syst. Biol. , v. 53, p. 968-977, 2004.
1 7 . MORRONE, J. J. Homology, biogeography and areas of endemism. Diversity Distrib. , v. 7, p. 297-300, 200 1 .
1 8 . MULLER, P The dispersaI centres of terrestrial vertebrates in the Neotropical realm. Biogeographica, v. 2, p. 1-244, 1973.
1 9. MORRONE, J. J. On the identification of areas of endemism. Syst. Biol., v. 43, p. 438-441 , 1 994.
20. LGWENBERG-NETO, P.; CARVALHO, C. J. B. Análise Parcimoniosa de Endemicidade (PAE) na delimitação de áreas de endemismos: inferências para conservação da biodiversidade na Região Sul do Brasil. Nat. Conserv. , v. 2, p. 58-65, 2004.
2 1 . HAROLD, A S . ; MOOI, R. D. Areas of endemism: definition and recognition criteria. Syst. Siol. , v. 43, p. 261 -266, 1994.
22. CRAW, R. C. ; GREHAN, J. R; HEADS, M. J . Panbiogeography: tracking the history of life. New York: Oxford University Press, 1 999. 229p.
23. HEADS, M. What is anode? 1. Biogeogr., v. 3 1 , p. 1 883-1 89 1 , 2004. 24. PREVEDELLO, J. A; CARVALHO, C. J. B. Conservação do
cerrado brasileiro: o método pan-biogeográfico como ferramenta para a seleção de áreas prioritárias. Nat. Conserv., v. 4, p. 39-57, 2006.
25. LGWENBERG-NETO, P; CARVALHO. C. J. B. Areas of endemism and spatial diversification of the Muscidae (Insecta: Diptera) in the Andean and Neotropical regions. J. Biogeogr. , v. 36, p. 1 750- 1759, 2009.
26. MORRONE, J. J. Biogeographical regions under track and cladistic scrutiny. 1. Biogeogl: , v. 29, p. 149- 1 52, 2002.
27. OLSON, D. M.; DINERSTEIN, E. The Global 200: Priority ecoregions for global conservation. Ann. Missouri Bot. Gard. , v. 89, p. 199-224, 2002.
28. SILVA, J. M. C. ; RYLANDS, A. B . ; DA FONSECA, G. A B . O destino das áreas de endemismo d a Amazônia. Megadiversidade, v. 1 , p. 124- 1 3 1 . 2005.
29. BUFFON, G. L. L. Histoire Naturelle Générale et Particuliere. Paris: Imprimerie Royale, 1 76 1 .
30. MORRONE, J . J . Biogeographic areas and transition zones of Latin America and the Caribbean islands based on panbiogeographic and cladistic analyses of the entomofauna. Annu Rev. Entomol., v. 5 1 , p. 467-494, 2006.
3 1 . EBACH, M. c.; MORRONE, J. J . ; PARENTI, L. R. et a!. Intemational Code of Area Nomenclature. 1. Biogeogr., v. 35, p. 1 153-1 1 57, 2008.
32. LÓPEZ, H. L.; MENNI, R. c.; DONATO, M. et aI. Biogeographical revision of Argentina (Andean and Neotropical Regions): an analysis using freshwater fishes. J. Biogeogr. , v. 35, p. 1 564-1 579, 2008.
33. WHITTAKER, R J.; ARAÚJO, M. B . ; JEPSON, P. et aI. Conservation Biogeography: assessment and prospect. Diversity Distrib., v. 1 1 , p. 3-23, 2005.

,�/
. :. - \


Paula Posadas
Edgardo Ortiz-Jaureguizar
A diversidade biológica atual da América do Sul é o resultado de extenso e dinâmico processo histórico. Este foi particularmente marcado por mudanças importantes nas condições climáticas e ambientais, longos períodos de isolamento geográfico, quebra desse isolamento e amplas flutuações no nível do mar, que isolaram, por completo, partes do continente. A conjunção das mudanças tectônicas e do nível do mar dirigiu a evolução climática e da paisagem e, por conseguinte, as mudanças de composição da biota, resultando nas atuais condições biogeográficas do continente, como é evidenciado, por exemplo, pela sua fauna de manúferos 1 .
Postula-se que, como consequência desses processos de isolamento e de conexão com massas continentais diferentes, a América do Sul apresenta uma biota de origem híbrida2• Nesta biota se diferencia um componente austral relacionado a outras áreas meridionais e um componente tropical relacionado às biotas da América do Norte e África.
Além da sua relação com a biota de outras áreas, essas duas biotas sul-americanas têm características que claramente as diferenciam. Assim, a maioria das regionalizações biogeográficas da América do Sul evidencia essa diferenciação biótica a partir do século XIX, nos esquemas propostos por autores como Philip Lutley Sclater e Alfred Russel Wallace. Estes autores reconhecem duas grandes áreas biogeográficas em nosso continente, que representam os dois componentes já mencionados. A maioria dos esquemas mais recentes também reconhece essa divisão, mas os seus limites variam de um autor para outro e de acordo com os táxons e os critérios que foram levados em conta em cada caso (ver Capo 2).
1 1
Uma das divisões biogeográficas mais recentes da América do Sul foi proposta por Morrone3. Ele postulou a divisão do continente em duas regiões (Andina e Neotropical) e uma zona de transição sul-americana, que apresenta uma sobreposição de elementos bióticos neotropicais e andinos. Neste esquema, a região Andina (Fig. 1 1 . 1 ) estende-se desde o Chile Central em direção ao sul, incluindo a Patagônia argentino-chilena. É esta definição de região Andina que será considerada neste capítulo.
Os objetivos deste capítulo são: ( 1 ) descrever as principais mudanças ocorridas na América do Sul austral, que resultaram na atual diferenciação da região Andina e seus componentes bióticos; e (2) exemplificar como a biogeografia histórica permite compreender estes fenômenos a partir da informação dos padrões de distribuição dos táxons atuais.
A região Andina (sensu Morrone3) está atualmente caracterizada pela presença de três biotas (a chilena central, a subantártica e a erêmica ou da estepe patagônica) . Biotas notavelmente diferentes umas das outras, tanto na sua composição taxonômica como de ecossistemas. Entretanto, como se verá mais adiante, essas biotas não são distinguíveis se voltarmos no tempo geológico. Por exemplo, as biotas subantártica e erêmica começaram a se diferenciar, durante o Mioceno, como consequência da ação da fase Quechua da orogenia andina5.
Em razão de suas peculiaridades geológico-geográficas (por exemplo, escassa variação latitudinal durante o Cenozoico, proximidade da Antártica, influência oceâ-
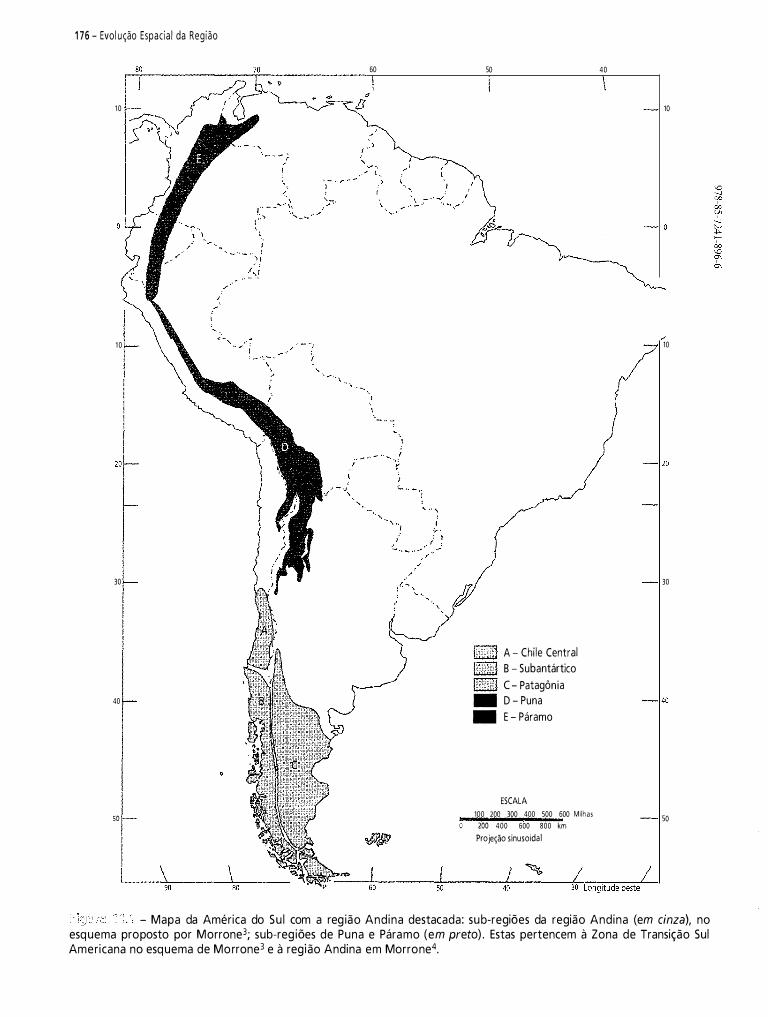
1 76 - Evol ução Espacial da Região
60
1 0
1 0
zo
30
40
50
50
liiii :,m:m A - Chile Central IU!l!!mml B - Subantártico
ImEEHil'. C Patago' n l'a ;;;;;1::w; -
_ D - Puna _ E - Páramo
ESCALA 100 ZOO 300 400 500 600 Milhas
ZOO 400 600 800 km Projeção sinusoidal
40
10
10
30
50
- Mapa da América do Su l com a reg ião Andina destacada: sub-regiões da região Andina (em cinza), no esquema proposto por Morrone3; sub-reg iões de Puna e Páramo (em preto) , Estas pertencem à Zona de Transição Sul Americana no esquema de Morrone3 e à região Andina em Morrone4,

nica), a biota da região Andina tem estado mais exposta às mudanças climáticas-ambientais ocorridas durante o Cenozoico, do que a biota das regiões localizadas ao norte dessa região.
A biota chilena central se localiza no território chileno entre 30 e 37°S, exceto pela Alta Cordilheira. O seu principal tipo de vegetação é uma formação xérica de tipo matorral mediterrâneo. Esta biota caracteriza-se pela riqueza de espécies e endemismos em diferentes níveis taxonômicos. O matorral alterna-se com florestas de pouca altura.
A biota subantártica compreende as florestas temperadas dos lados leste e oeste do sul da cadeia andina, as Ilhas Juan Fernández (Oceano Pacífico) e Ilhas Malvinas (Atlântico Sul). Uma das comunidades mais evidentes pela qual é caracterizada consiste em florestas de Nothofagus. De acordo com a altitude e a latitude, estas florestas são dominadas por espécies diferentes, que se estendem ao longo dos Andes ao sul de 35°S na Argentina e o Chile. Em áreas com maior umidade, a sua distribuição é descontínua, albergando endemismos em diferentes níveis taxonômicos.
Por outro lado, a biota erêmica é representada na sub-região Patagônica, que compreende o sul de Mendoza, Neuquén ocidental, Rio Negro, grande parte do Chubut, Santa Cruz e norte da Terra do Fogo. A vegetação dominante é o estepe arbustivo, com predomínio de plantas em "cojín". Do ponto de vista florístico, a família mais importante e com maior número de endemismos é Asteraceae (Nassauvia, Perezia, Chuquiraga etc .) . Em relação à fauna, existem muitas espécies adaptadas a este tipo de vegetação (por exemplo, coleópteros como Cylydrorhinus, Acrostomus, Cnemalobus e Barypus).
No entanto, como já mencionado, a distinção destas três biotas que atualmente representam a região Andina, no sentido de Morrone3, não é tão clara no passado geológico.
Antes de começar desenvolver o esquema de mudanças paleogeográficas e bióticas que resultou no que hoje se conhece como região andina, salienta-se que não é tarefa simples reconstruir a história de determinada região biogeográfica. A este respeito, deve-se lembrar que uma unidade biogeográfica (por exemplo, Reino, Região, Província) é uma área geográfica que é reconhecida pela presença de determinados táxons e as suas características fisionômicas (tipo de vegetação), que, por sua vez, são
Evo lução da Reg ião And ina da Amér ica do Su l - 177
influenciadas pelas condições geográficas, edáficas, climáticas etc. Como pode ser visto, nenhum dos componentes que permitem reconhecer determinada unidade biogeográfica, permanece estável ao longo do tempo geológico. Além disso, cada um tem sua própria taxa de mudança, de modo que à medida que caminhamos no sentido oposto ao da seta do tempo, inexoravelmente deixaremos de reconhecer as características que nos permitiam identificar a unidade biogeográfica, até um ponto em que nenhuma delas é reconhecível.
Levando-se em conta as considerações já citadas, este tópico descreverá a modificação da região Andina ao longo do tempo, tomando como unidade de análise a área geográfica que esta região ocupa hoje. Tentar-se-á definir o tempo que se estabeleceram as condições climáticas-ambientais que permitem o seu reconhecimento e das unidades biogeográficas que a constituem. Então serão usadas as informações geológicas e paleontológicas, especialmente plantas vasculares e mamíferos, dois dos grupos com melhor e mais completo registro do Cenozoico sul-americano. A limitação ao Cenozoico (isto é, os últimos 65 milhões de anos da história do nosso planeta) não é arbitrária, porque durante a sua história ocorreu o soerguimento da cordilheira dos Andes. No Paleoceno foram registrados os primeiros mamíferos "modernos" (placentários e marsupiais) da América do Sul. As plantas com flores se tornaram dominantes nos ambientes continentais. As referências citadas no texto correspondem a trabalhos da paleontologia, nos quais pode ser encontrada bibliografia mais detalhada, tanto paleontológica quanto geológica.
Ao longo do Mesozoico e do Cenozoico, o desenvolvimento tectônico do sul da América do Sul era controlado por um sistema complexo de subducção na sua margem ocidental e a expansão da crosta oceânica ao longo da Cordilheira Centro Atlântica, na sua margem oriental. A evolução tectônica, em conjunto com outras grandes forças geodinâmicas como a mudança no nível do mar, glaciações e temperaturas do oceano, mudou a paleogeografia do continente. Estes processos ocorreram principalmente na sua porção sul, direcionando a evolução das paisagens e clima, assim como as mudanças na composição da biota. Por conseguinte, neste trabalho comentamos, a partir da literatura, os principais eventos na tectônica de placas, vulcanismo, mudanças no nível do mar, as temperaturas marinhas e as glaciações, a fim de caracterizar as principais mudanças que aconteceram na paleogeografia do continente, nas áreas em que hoje se desenvolve a biota andina.
A fim de comparar o padrão de mudanças nas condições climático-ambientais do Cenozoico derivado de informação geológica, foram utilizadas informações do registro fóssil de plantas vasculares e os mamíferos continentais.

1 78 - Evo l ução Espacial da Reg ião
Ambos os registros são os mais completos (geográfica e estratigraficamente) e têm sido muito empregados para inferir condições climático-ambientais (como exemplo, ver Ortiz-Jaureguizar e Cladera5 e literatura citada). No caso da informação paleobotânica, usamos as últimas sínteses paleoflorísticas, que, por sua vez, se baseiam principalmente em informações de pólen e foliares6-8. No caso dos mamíferos, foram utilizados os trabalhos de Pascual et al. 1 e Ortiz-Jaureguizar e Claderas.
Durante a parte mais tardia do Cretáceo Superior e o Paleoceno Inferior, a ausência de grandes barreiras topográficas permitiu extensa transgressão marinha do Atlântico. Essa transgressão cobriu a maior parte do sul da América do Sul, desde o extremo mais austral da Patagônia até os atuais territórios da Bolívia e do Peru (Fig. 1 1 .2). Essa transgressão, conhecida como "Mar Salamanquense", dividiu o continente em duas regiões principais, a Norte Gondwânica e a Sul Gondwânica; esta última, por sua vez, subdividindo-se em vários territórios insulares relativamente extensos I. A partir do PaJeoceno Médio a maior parte deste vasto mar epicontinental regrediu, passando a cobrir apenas o extremo
mais austral da Patagônia e o atual território da Terra do Fogo. Os territórios anteriormente cobertos pelo "Mar Salamanquense" se transformaram em extensas planícies aluviais e grandes bacias de lagos. Os sedimentos predominantes são piroclásticos, derivados dos cinturões vulcânicos correspondentes aos Andes norte-patagônicos e o Maciço Somuncurá I .
Ao longo do Paleoceno, a América do Sul permaneceu unida à Antártica, que, por sua vez, conectava-se à Austrália (Fig. 1 1 .2). Esta conexão Gondwânica teve grande influência na circulação oceânica e atmosférica e, por conseguinte, nas temperaturas marinha e continental, além de ter facilitado o intercâmbio de fauna e flora (ver, por exemplo, Pascual e Ortiz-Jaureguizar9). As temperaturas do oceano durante o Paleoceno foram superiores aos valores atuais, aproximando-se de 20°C nas águas das latitudes médias e 1 2°C nas latitudes polares. Da mesma forma, no final do Paleoceno, registrou-se o chamado "Máximo Térmico do Paleoceno" (Fig. 1 1 .3) .
Em concordância com as altas temperaturas nos oceanos, na Patagônia registrou-se predomínio de selvas e florestas tropicais, assim como de manguezais em áreas próximas à costa (Fig. 1 1 .3) . Estimativas climáticas derivadas do estudo desta flora são consistentes com as temperaturas do oceano, uma vez que indicam a
'1 '! ,/.; - Diferenciação do Norte Gondwânico (província) e Su l Gondwân ico (província) (cerca de 85-63 Ma). Em cinza, as á reas emergidas durante a transgressão do "Mar Sa lamanquense" na América do Su l austral que a inda está l igada à Antártica e à Austrá l i a . Modificado de Pascual e Ortiz-Jaureguizar9.
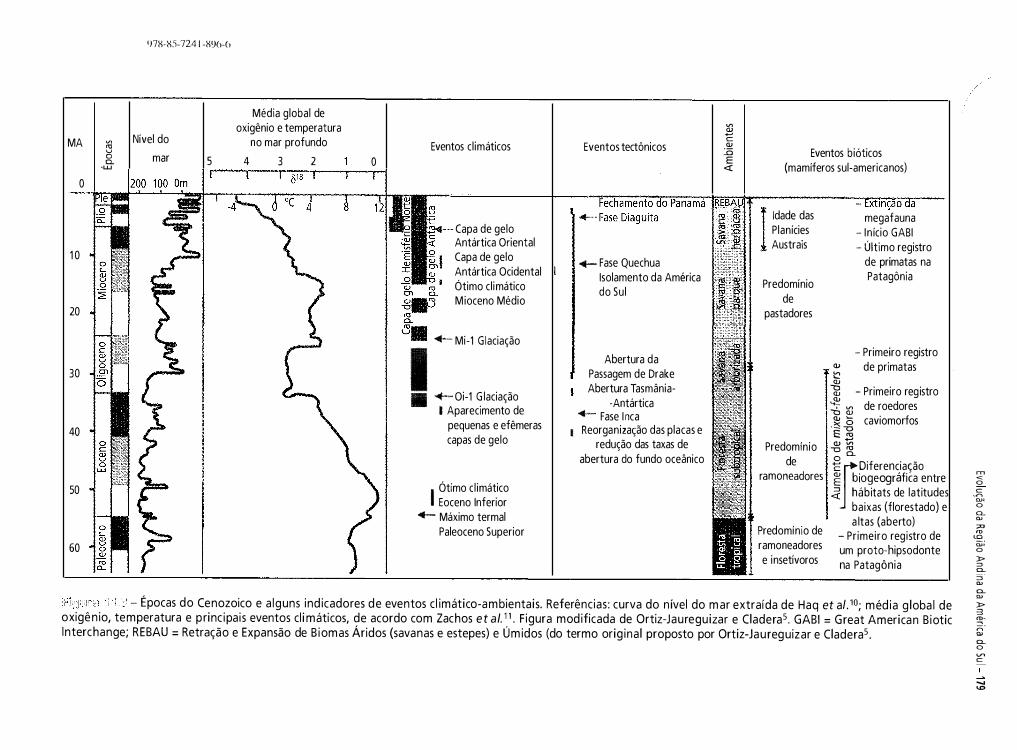
MA
o
1 0
20
30
40
50
60
V> n:l v o o.. 'LLl
lJ7X-X5-724 1 -X%-tl
Nível do
mar 5
Média global de oxigênio e temperatura
no mar profundo
4 3 2 o
I •
Eventos cl imáticos
Capa de gelo Antártica Oriental Capa de gelo Antártica Ocidental Ótimo cl imático Mioceno Médio
"iIf- Mi-1 G laciação
"iIf- Oi-1 G laciação I Aparecimento de
pequenas e efêmeras capas de gelo
I Ótimo c l imático Eoceno Inferior
"iIf- Máximo termal Paleoceno Superior
Eventos tectônicos
"iIf- Fase Quechua Isolamento da América do Sul
Abertura da Passagem de Drake Abertura Tasmânia
-Antártica "iIf- Fase I nca
I Reorganização das placas e redução das taxas de
abertura do fundo oceânico
V> <lJ ..... C <lJ :o E
<t: Eventos bióticos
(mamíferos sul-americanos)
Idade das megafauna Planícies - In ício GABI Austrais - Último registro
de primatas na
Predomínio Patagônia
de pastadores
- Primeiro registro <lJ de primatas � (IJ
-o - Primeiro registro (IJ � de roedores -6 <D � o caviomorfos ' - -0 E m .....
Predomínio I � ti de � Diferenciação
"moo"doco, � fl ;09eog'áfka "'CO J. hábitats de latitudes
baixas (florestado) e
Predomínio de a ltas (aberto)
- Primeiro registro de ramoneadores um proto-hipsodonte e insetívoros na Patagônia
- Épocas do Cenozoico e a lguns ind icadores de eventos c l imático-ambientais. Referências : curva do n ível do mar extraída de Haq et ai. 10; média g loba l de oxigênio, temperatura e princ ipa is eventos climáticos, de acordo com Zachos et al. 1 1 . Figura mod ificada de Ortiz-Jaureguizar e Cladera5. GABI = Great American B iotic Interchange; REBAU = Retração e Expansão de B iomas Áridos (savanas e estepes) e Úmidos (do termo orig ina l proposto por Ortiz-Jaureguizar e Cladera5.
rn < o c
,� "n o CL cu ou ro LO "n o » :::> CL :::> cu CL ru » '3 �� � cu CL o <.r> c
..... \O

1 80 - Evolução Espaci a l da Reg ião
existência de temperaturas médias anuais de 25°C e precipitações superiores a l .SOOmm por ano. Isto permite a inferência de condições climáticas tropicais e sem sazonalidade, pelo menos até 46°S7,8.
Em relação aos dados de mamíferos, também são consistentes a indicação do predomínio de clima quente e úmido e os ambientes de floresta (Fig. 1 1 .3) . No Paleoceno Inferior, prevaleceram na Patagônia os insetívoros-frugívoros (muitos deles provavelmente com hábitos escansoriais e os ramoneadores-onívoros* [herbívoros])5. Durante o restante do Paleoceno, a fauna de mamíferos esteve dominada pelos ramoneadores, embora os insetívoros mantivessem diversidade relativamente alta. Um fato marcante é o registro dos primeiros mamíferos "ungulados" na Patagônia, com dentes molares de coroas altas (proto-hipsodontes). Como classicamente este tipo de molar foi associado a hábitos pastadores ** e, por conseguinte, ao desenvolvimento das pastagens, este registro pode ser interpretado como a primeira evidência deste tipo de ambiente na América do Sul. No entanto, não há registro de pólen de gramíneas ou fitólitos na Patagônia naquela época, ao passo que se registram sedimentos piroclásticos, ricos em vidro vulcânico e, em consequência, altamente abras i -vos. Assim, a presença de herbívoros com molares proto-hipsodontes poderia ser interpretada como resposta à tais "poluentes" na dieta5 .
No Eoceno Inferior o nível do mar permaneceu elevado e as temperaturas continuaram altas, coerente com o chamado "Ótimo Climático do Eoceno Inferior" (Fig. 1 1 .3). Ao longo do Eoceno Médio, registraram-se na Antártica Ocidental os primeiros glaciais do tipo Alpino. Finalmente, no Eoceno Superior registrou-se deformação tectônica na zona ocidental do continente, que corresponderam às ações da chamada "Fase Inca" da orogenia andinal2. Sob a ação desta fase diastrófica, os Andes austrais ainda incipientes reativam-se na Patagônia Central. Enquanto isso, na Antártica registrou-se a abertura do Mar da Tasmânia, no final do Eoceno e começo do Oligoceno. Isto se traduz em mudança no fluxo das águas da Antártica e queda significativa da temperatura e do nível do mar13. Correspondendo a estas mudanças,
* Ramoneadores: designação dada a an imais que se a l imentam de fol has de á rvores e arbustos.
** Pastadores: designação dada a an ima is com o hábito de pastejar a relva ou plantas rasteiras.
foram registrados eventos glaciais efêmeros na Antártica Oriental (Fig. 1 l .3) .
Durante o Eoceno Inferior se manteve o predomínio de elementos florísticos indicadores de climas quentes e úmidos (Fig. 1 1 .3) . As reconstruções climáticas resultan- \O
tes do estudo desta flora indicam temperaturas subtropicais (oscilando em tomo de 1 6 a 1 9°C) e precipitações biesta-
-.) 00 00 Y' -.) N ... cionais, com valores de até l .SOOmm e provável contri-00 buição da umidade que vinha tanto do leste como do '&-
oeste7. No entanto, em certas áreas restritas do interior da o, Patagônia teriam prevalecido condições quentes, embora mais secas, a julgar pelo registro de táxons indicadores dessas condições como as anacardiáceas8. Do Eoceno Médio ao Oligoceno Inferior, registrou-se expansão das florestas de Nothofagus, documentando-se substituição progressiva da flora subtropical por uma mais temperada, dominada por Nothofagus e Podocarpaceae, e o aparecimento de alguns táxons indicadores de ambientes xéricos, como Schinopsis6,8 . Também ao longo do Eoceno Superior preservaram-se nos paleossolos da Patagônia os primeiros fitólitos de gramíneass.
Em relação aos mamíferos, as inferências paleoambientais concordam com aquelas obtidas a partir da flora. Durante o Eoceno Inferior e Médio, as comunidades foram dominadas pelos tipos ramoneadores, registrando baixa diversidade de mixed-feeders e pastadoress (Fig. 1 1 .3) . Os ramoneadores, juntamente com os tipos de insetívoros e onívoro-insetívoros, permitem inferir a existência de ambientes de floresta subtropical, desenvolvidos sob climas quentes e úmidoss. Por fim, durante o Eoceno Superior os grupos dominantes continuaram sendo os ramoneadores, mas aumentou a diversidade de mixed-feeders e de pastadores (Fig. 1 1 .3) . A maior diversidade dos dois grupos sugere elevação similar na aridez e maior porcentagem de gramíneas nas comunidades vegetais5.
A maior parte do Oligoceno transcorreu com relativa calma tectônica. Ao longo do Oligoceno Superior, as modificações na direção da convergência das placas de Nazca e Sul-americana provocaram importantes mudanças no arco andino, reativando os principais cinturões vulcânicos. A atividade ígnea ocorreu em vastas áreas da zona central e ocidental da Argentina, Bolívia e Peru, mas não no sul da Argentina e no Chile, onde a atividade magmática foi escassa. No centro da Patagônia, desenvolveu-se significativo vulcanismo basáltico de intraplaca, com características petrológicas e geoquímicas semelhantes ao vulcanismo da intraplaca havaiano-oceânica. No entanto,

a partir do Oligoceno Superior começou a tomar forma a configuração atual morfoestrutural dos Andes do Chile central. Ao mesmo tempo, uma nova transgressão marinha do Atlântico, conhecida como "Mar Patagônico", ocorreu em tomo das mesmas áreas cobertas durante o Cretáceo Superior e Paleoceno Inferior pelo "Mar Salamanquense" (ver Fig. 1 1 .2). Durante as últimas fases dessa transgressão marinha, registrou-se generalizada transgressão pacífica na margem ocidental do sul da América do Sul.
Como já mencionado, a Austrália separou-se da Antártica no limite entre Eoceno e Oligoceno. Essa ruptura deu origem a uma corrente de água fria em tomo da Antártica que, por sua vez, iniciou a glaciação no leste desse continente. A temperatura da superfície das águas do oceano caiu entre 7 e 8°C em relação ao registrado antes da abertura do Mar da Tasmânia, atingindo temperaturas de 4 a soe. Além disso, a contribuição das águas frias da Antártida fez diminuir numa escala semelhante à temperatura da água em regiões tropicais, que chegaram a ter temperatura próxima de 20°C, em vez de 28°C registrados anteriormente. Em meados do Oligoceno (ver Fig. 1 1 .3), a Passagem de Drake abriu-se de modo permanente e, ao mesmo tempo, houve queda acentuada nas temperaturas marinhas (ver Fig. 1 1 .3). Em paralelo, houve uma era do gelo importante em toda a Antártica (ver Fig. 1 1 .3) .
A flora do Oligoceno Inferior da América do Sul foi semelhante à flora do Eoceno Superior, com substituição gradual da flora quente para uma mais temperada, dominada pelos Nothofagus e as Podocarpaceae, com poucos indicadores de ambientes xéricos. A partir do Oligoceno Superior, as comunidades de plantas começaram a ter aparência similar à de hoje, com a presença marcante de grupos com hábitos herbáceo-arbustivos. Formações xerófiticas teriam ocupado ambientes costeiros e manchas de vegetação aberta em áreas interioranas, apesar de ainda se notar a presença de florestas notofagáceas, araucárias e podocarpáceas no território atual da sub-região da Patagônia. Isto permite inferir o estabelecimento de contraste florístico bem marcado entre ambientes continentais e costeiros8.
A flora estabelecida a partir do Eoceno Superior nas latitudes médias e altas da América do Sul caracteriza-se por ser formada por uma mistura peculiar de elementos com distribuição atual austral-antártica junto com outros de distribuição neotropical e pantropical, e outros de ampla distribuição geográfica. Esse arranjo de elementos peculiares temperados-frios e quentes levou a chamá-lo de "Paleoflora Mista"6. Estimativas climáticas derivadas dessa flora permitem inferir temperatura média anual próxima de 15°C e precipitação média anual muito inferior ao período anterior, atingindo valores que variam entre 570 e 880mm7.
Evolução da Reg ião And ina da América do Su l - 181
As comunidades de mamíferos continentais do Oligoceno inferior foram dominadas por ramoneadores, mas os mixed-feeders e os pastadores foram mais diversos que durante o Eocenos. Isso é consistente com a existência de savanas arborizadas (ver Fig. 1 1 .3) , estabelecidas sob regime de precipitação pluvial anual baixa. No Oligoceno Superior (e pela primeira vez durante o Cenozoico) , as comunidades de mamíferos passaram a ser dominadas pelos pastadores, apesar de os ramoneadores e frugívoros ainda apresentarem alta diversidades. Esta distribuição dos tipos tróficos permite inferir que as comunidades também foram desenvolvidas em uma vegetação de savana arborizada (ver Fig. 1 1 .3), embora um pouco mais aberta do que no Oligoceno Inferiorl . A dominância dos tipos pastadores , portadores de molares de coroa alta (ambos proto-hipsodontes como eu-hipsodontes) é de grande relevância, já que permite sugerir que os matagais não só estiveram presentes, mas que tiveram ampla distribuição geográfica na Patagônia central durante o Oligoceno Superior. Essa inferência é corroborada pelo registro abundante de fitólitos opalizados nos mesmos sedimentos que incluem os mamíferos5.
A partir do Mioceno, a atividade tectônica se tomou mais importante em razão do aumento da taxa de subducção entre as placas de Nazca e Sul-americana. As mudanças oro gráficas associadas a este aumento da atividade tectônica tiveram notável impacto sobre o clima e a biota, ao resultarem em contraste entre a biota ocidental e oriental do continente. O resultado claramente foi a diferenciação da região andinas.7.9.
Ao longo do Mioceno Inferior, as características geológicas e geográficas do continente permaneceram praticamente inalteradas em relação Oligoceno Superior. Entretanto, no final desta época, o vulcanismo explosivo registrado da Patagônia central até o Estreito de Magalhães, injetou na atmosfera grande quantidade de cinzas vulcânicas, que deram origem aos espessos depósitos de toba conhecidos para esse tempo na Patagônia. Também durante este período, houve nova glaciação Antártica, conhecida como "Mil " (ver Fig. 1 1 .3) e sazonalidade climática bem definida desenvolveu-se na Patagônias. Os níveis do mar encontravam-se relativamente baixos e as temperaturas mantiveram-se mais ou menos constantes até o final do Mioceno Inferiorl3.
Durante o período que vai do Mioceno Inferior ao Médio, o clima tomou-se relativamente mais quente e chuvoso do que no período anterior, em concordância

1 82 - Evolução Espac ia l da Reg ião
com um evento de aquecimento global conhecido como o "Ótimo Climático do Mioceno Médio" (ver Fig. 1 1 .3). Ao longo deste evento, a temperatura do mar aumentou cerca de 2°C em relação às temperaturas do período anterior. No Mioceno Médio, as paisagens do sul da América do Sul começaram a ser dominadas pelos processos que levaram à configuração atual do cinturão tectônico e magmático dos Andes. Também durante o Mioceno Médio e Superior foram registradas três sucessivas transgressões marinhas, conjuntamente referidas pelo nome "Mar Paranaense" l . Este mar cobriu, no sul da América do Sul, aproximadamente o mesmo território abrangido pelo "Mar Salamanquense" no Cretáceo Superior-Paleoceno1 .5 (ver Fig. 1 1 .2).
Depois do "Ótimo Climático do Mioceno Médio" (ver Fig. 1 1 . 3 ) , houve redução rápida e sustentada da temperatura e da precipitação em nível mundial. Estas mudanças climáticas parecem ter sido associadas a eventos como a conclusão da formação da camada de gelo permanente na Antártica (ver Fig. 1 1 .3) e o concomitante aumento do gradiente térmico entre o Equador e os polos. Este gradiente térmico afetou a circulação atmosférica e oceânica, elevando a influência da Corrente Fria do Peru ao longo da costa pacífica do continente e o Anticiclone Subtropical do Pacífico sue.
Além disso, o aumento sustentado da taxa de elevação dos Andes, do Mioceno Superior Médio ao Superior elevou o efeito de "Zona de Sombra de Chuva" aos ventos úmidos, tanto do leste quanto do oeste. Nesse sentido, deve-se salientar que durante o Mioceno Superior os Andes centrais alcançaram altura aproximadamente igual à metade de sua altura atual, ou seja, cerca de 2.000 a 2.S00m.
Esses fatores, conjuntamente, favoreceram o desenvolvimento progressivo da "Diagonal de Formações Abertas Secas" sul-americana (ver Capo 1 3), que, por sua vez, teve efeitos diversos e significativos sobre a evolução e a biogeografia da biota continenta17.
Durante o Mioceno Superior-Plioceno Inferior o "Mar Paranaense" retraiu-se, desenvolvendo-se no seu lugar planícies igualmente extensas, que cobriram a área do norte da Patagônia até o Uruguai, o centro e norte da Argentina, e ao longo das margens orientais dos Andes que se desenvolviam na Bolívia, Peru e Venezuela, assim como a parte superior da bacia amazônica 1. Por esse motivo, denomina-se este período de "a Época das Planícies Austrais". A regressão do "Mar Paranaense" e o subsequente desenvolvimento das planícies estiveram associados à chamada "Fase Quechua" do diastrofismo andino, iniciada durante o Mioceno Médio (ver Fig. 1 l .3). Sob sua influência, as cordilheiras Patagônica e Principal
foram sucessivamente elevadas, tomando-se a já mencionada barreira para os ventos úmidos do oeste e do Amazonas. Este efeito de "Zona de Sombra de Chuva" resultou na diferenciação do que hoje se conhece como região Andina1 . Por fim, no período do Mioceno Superior-Plioceno Inferior foi registrada a primeira evidência de um avanço glacial no extremo austral da Argentina. Assim, durante a "Idade das Planícies Austrais" os climas foram mais frios e a sazonalidade mais pronunciada do que no Mioceno Médio, com maior vmiedade de subdivisão ambiental 1 .
As comunidades vegetais do Mioceno Inferior foram similares às do Oligoceno Superior. Durante o Mioceno Médio se registrou aumento significativo na diversidade e abundância de táxons xerofíticos (por exemplo, asteráceas, quenopodiáceas, convolvuláceas). Este aumento, somado à extinção dos táxons indicadores de climas subtropicais não estacionais, concorda com elevação da aridez e da sazonalidade. No Mioceno Superior, as floras áridas eram muito semelhantes às atuais, observando-se a partir desse momento a expansão da estepe na Patagônia extra-andina (atual sub-região Patagônica) e a retração das florestas à faixa ocidental dos Andes (atual sub-região Sub antártica, mais úmida)8.
A fauna de mamíferos do Mioceno Inferior foi semelhante à do Oligoceno Superior, dominada pelos pastadores, embora com alta diversidade de ramoneadores e frugívoros. A principal diferença entre elas é o número maior de mixed-feeders no Mioceno Inferior, indicando que nesse tempo houve equilíbrio entre as pastagens e as florestas, provavelmente representadas por savanas-parque (Fig. 1 1 . 3 ; ver também Ortiz-Jaureguizar e Cladera5). O registro de primatas e outros vertebrados climático-sensitivos em latitudes tão elevadas como Terra do Fogo permite inferir que os climas quentes e úmidos, assim como as formações f1orestadas, foram bem desenvolvidos na Patagônia. Junto com estes registros também se encontraram mamíferos fossoriais, que permitem inferir a existência de eventos climáticos de maior aridez, ou de ambientes complexos, representados por dunas que rodeiam áreas de f1orestas 1,5.
A fauna de mamíferos do Mioceno Médio se desenvolveu sob influência do referido "Mar Paranaense". As comunidades estiveram dominadas por pastadores (que foram muito mais diversos do que no Mioceno Inferior), seguido por mixed-feeders, frugívoros e ramoneadores. Esses dois últimos tipos tróficos foram menos diversos do que no Mioceno Inferior5. Essa mudança de composição tem sido interpretada como uma resposta à passagem de savanas-parque a pastagens e estepes1 . Mudança relacionada, por sua vez, à ação das primeiras

subfases da fase diastrófica Quechua da orogenia andina 1 . De qualquer maneira, a persistência de primatas no Mioceno Médio da Patagônia indica que os climas quentes e os ambientes florestados foram desenvolvidos, mesmo nas latitudes mais austrais dessa região1 (ver Fig. 1 1 .3) .
Ao longo do Mioceno Superior, a maioria dos mamíferos associados a ambientes florestais se extinguiu, tomaram-se escassos ou desapareceram da Patagônia (por exemplo, para primatas, consulte Pascual et al. 1). A fauna desse período foi dominada por pastadores, seguido por mixedjeeders, enquanto os ramoneadores e frugívoros apresentaram diversidade muito baixas. Essa alta diversidade de pastadores, juntamente com a redução de ramoneadores, corresponde à expansão das planícies austrais e à expansão simultânea das savanas herbáceas e das pastagens1.5 (ver Fig. 1 1 .3).
Como indicado anteriormente no Mioceno Superior, o Plioceno caracterizou-se pelo auge das planícies austrais. Assim como seu início está ligado à "Fase Quechua" da orogenia Andina, a seu término relacionou-se à ação da "Fase Diaguita" da mesma orogenia1,5 (Fig. 1 1 .3) . Esta é a causa da elevação da Cordilheira Central da Argentina e do Chile, bem como dos sistemas orográficos do leste da Argentina (por exemplo, Puna, Sierras Pampeanas). A elevação final das Serras Pampeanas e dos Andes Centrais teve consequências marcantes para a biota da América do Sul, uma vez que o efeito da "Zona de Sombra de Chuva" se intensificou, culminando com as condições climáticas extremamente áridas observadas em áreas situadas entre elas1 ,5 .
Durante o Plioceno Superior, a comunicação entre o Atlântico e o Pacífico foi fechada em razão do surgimento do Istmo do Panamá (ver Fig. 1 1 .3) . Este fechamento alterou a circulação em ambos os oceanos, facilitando, entre outras coisas, a instalação do gelo no Ártico e no hemisfério sul, a criação da corrente circum-Antártica e a formação de uma espessa camada de gelo na Antártica Ocidental. Na Patagônia, registrou-se grande glaciação há cerca de 2,3 Ma. Depois disso, o clima do Plioceno foi relativamente mais frio que do Mioceno Superior,
-o com subdivisão ambiental mais marcada 1 . '" � Ao longo do Pleistoceno, os ciclos de avanço e recuo ;:;; dos glaciares produziram expansões e contrações de r-J-. ambientes áridos e úmidos. Este padrão possibilitou a "? � alternância de oportunidades para as espécies de ambos '"
os ambientes. Assim, durante a expansão do clima frio e seco, as áreas ocupadas por biota tropical e subtropical foram reduzidas, resultando na expansão simultânea e interligação da biota de ambientes abertos. Em conse-
Evo l u ção da Região And ina da América do S u l - 1 83
quência, ao longo dos lados orientais dos Andes formou-se um corredor de savanas que permitiu a conexão das estepes e matagais do sul da Argentina às pradarias colombianas. Inversamente, as florestas e selvas se expandiram quando ambientes abertos se retraíram, durante os períodos interglaciais quentes e úmidos. Assim, por exemplo, durante a expansão das geleiras as espécies típicas da Patagônia extra-andina, como a pequena doninha marsupial, Lestodelphis halli, expandiu a sua distribuição até a província de Buenos Aires, ao passo que se retraiu novamente para a estepe patagônica, durante os períodos interglaciais.
Infelizmente, o registro de mamíferos da Patagônia no período Plioceno-Pleistoceno é muito baixo (exceto para o Pleistoceno Superior). Isso provavelmente se deve à predominância de agentes da erosão sobre os depósitos, em resposta à elevação dos Andes e da elevação epeirogênica do território patagônico1. Ainda assim, as condições físicas são compatíveis com as poucas evidências biológicas disponíveis, indicando que os diferentes componentes bióticos que compõem a região andina podiam expandir ou retrair suas áreas de distribuição de acordo com a expansão/contração das massas de gelo.
A biogeografia é dividida em duas grandes subdisciplinas que tradicionalmente se desenvolveram de formas diferentes, mas têm um objetivo comum: a explicação causal das distribuições dos organismos. Uma dessas subdisciplinas, a biogeografia histórica, enfoca os processos que afetaram a distribuição dos organismos em grandes períodos (por exemplo, a evolução, tectônica de placas). A outra, a biogeografia ecológica, enfoca os processos que afetam a distribuição dos organismos em uma escala de tempo curta14 (ver Capo 4).
Ambas as disciplinas inicialmente estudam a distribuição geográfica das espécies, ou seja, sua área de distribuição. Do ponto de vista da biogeografia histórica, a congruência de distribuição de duas ou mais espécies é considerada não aleatória e define a unidade de estudo: a área de endemismo14 (ver Capo 3). A sobreposição e a nidificação de áreas de endemismo permitem definir unidades maiores: as unidades biogeográficas (o conceito de unidades biogeográficas refere-se às unidades das regionalizações, independentemente da sua categoria, ou seja, aplica-se a ambos os distritos, províncias, como sub-regiões etc.). Estas três categorias de áreas ou arranjos espaciais (áreas de distribuição, áreas de endemismo e unidades biogeográficas) são moldadas e/ou modifi-

1 84 - Evo lução Espac ia l da Reg ião
cadas por processos espaço-temporais, tais como a dispersão, a vicariância e a extinção, que são eles mesmos o resultado da história da Terra.
A biogeografia histórica é uma disciplina que se tornou cada vez mais importante nas últimas décadas1-U5. Segundo Craw et aI. 16, a geografia é o substrato sobre o qual a história da vida tem lugar, havendo uma ligação inevitável entre história e espaço. Portanto, não há processos evolutivos e ecológicos que não estejam ligados de forma essencial ao local e ao momento histórico particular em que eles ocorreram. Disto, pode-se concluir que um dos requisitos necessários para o desenvolvimento de quadro coerente e abrangente dentro da biogeografia histórica é uma interação maior com a paleontologia e com a geologia.
Uma revisão abrangente dos princípios, fundamentos e métodos utilizados na biogeografia histórica pode ser encontrada em Crisci et aI. 14 e Morrone1S e nos Capítulos 4 a 7. Em resumo, pode-se dizer que, a partir das ide ias de Croizat1 7 começou na biogeografia histórica uma "revolução científica", no sentido do filósofo Thomas Kuhn. Croizat propôs que a Terra e a vida evoluem juntas, de maneira que os padrões de distribuição da biodiversidade que se vê hoje representam o resultado dessa evolução conjunta. A partir desta hipótese, Croizat15 postulou uma teoria chamada pan-biogeografia. Com a base teórica proposta por Croizat, desenvolveram-se duas diferentes escolas biogeográficas : a pan-biogeografial6J7 (ver Capo 5) e a biogeografia cladística1 8 (ver Capo 6). A partir daqui, tem-se associado o desenvolvimento posterior da biogeografia histórica ao uso de filogenias como uma fonte de informação histórica, que se reflete na expressão "phylogeny-based biogeography" usada por Wiens e Donoghuel9, para se referir à biogeografia histórica.
A necessidade de considerar o fator tempo nos estudos de biogeografia histórica tem sido reconhecida por vários autores, j á que ignorar a informação temporal obscurece a conexão entre os padrões biogeográficos e suas possíveis causas subjacentes (ver Crisci et aI. 14 e literatura citada). Uma forma possível de comparar os resultados biogeográficos com informação temporal é pelo uso de elementos fornecidos pelos fósseisl,5,20. Além disso, e como discutido na seção anterior, os fósseis, junto com a evidência geológica (por exemplo, tafonomia, sedimentologia), fornecem informações fundamentais sobre os paleoambientes, permitindo contextualizar os resultados dos métodos biogeográficos históricos.
A América do Sul em geral, e a região andina em particular, têm sido objeto de vários estudos biogeográficos históricos nas últimas décadas. Os trabalhos que
consideram a região andina (ou América do Sul austral), do ponto de vista biogeográfico histórico, poderiam se dividir em duas categorias principais : ( 1 ) aqueles que consideram a relação da biota da região andina com a biota de outras áreas; e (2) aqueles que enfocam a formulação de hipóteses dentro das unidades biogeográficas menores que compõem essa região. Entre os primeiros poderiam ser mencionados, a título de exemplo, os tra- 'Ú
-..l balhos de Brundin21, Crisci et aI. 2, Roig-Jufient22, Katinas �
U\ et ai. 23, Gilibert e Edgecombe24. Dentre os trabalhos que .:.,
� investigam as relações biogeográficas entre as áreas
(Xl
austrais (incluindo a região Andina), destacam-se os re- '5-, '"
feridos ao gênero Nothofagus. Este gênero de árvores tem sido um táxon-chave na biogeografia do hemisfério sul, em decorrência de sua ampla distribuição geográfica e temporal, o seu adequado registro fóssil, os seus frutos não dispersos pelo vento e por ter constituído e ainda constituir um elemento dominante nas florestas temperadas do hemisfério sul. Crisci et alo 14 apresentaram um capítulo sobre Nothofagus como estudo de caso no qual se discutem as principais abordagens que foram aplicadas ao estudo biogeográfico deste gênero. Entre os trabalhos que postulam ou discutem hipóteses das relações entre as unidades biogeográficas menores que constituem a região andina poder-se-iam mencionar: Morrone25, Morrone et az. 26 ; Posadas et aI. 27, Posadas e Morrone28, Dominguez et az.29, Roig-Jufient et al. 3o e Lowemberg-Neto et al. 3 1 . Todos esses trabalhos são alguns exemplos de abordagens da problemática biogeográfica histórica da região Andina em diferentes escalas espaciais, por meio da utilização de diferentes métodos e com base em diferentes táxons. Serão utilizados dois desses trabalhos para ilustrar o uso da biogeografia histórica para o estudo da região andina.
Como mencionado anteriormente neste capítulo, a maioria dos esquemas biogeográficos propostos para a América do Sul reconheceu uma grande subdivisão do continente em duas áreas : a biota temperada Andina-Patagônica que ocupa o sul e oeste do continente (ver Fig. 1 1 . 1 ) e

uma biota neotropical que ocupa o centro, norte e leste da América do Sul. Ademais, pelo estudo das semelhanças entre estas duas biotas com as de outras áreas, foram geradas hipóteses sobre a origem híbrida da biota da América do Sul. Crisci et al. 2 propuseram-se testar esta hipótese utilizando ferramentas de biogeografia cladística (ver Capo 6). Para fazer isso, esses autores consideraram a América do Sul dividida em duas unidades : América do Sul austral e norte da América do Sul.
Material e Métodos
Seleção de áreas. Com base na observação dos padrões de distribuição de vários táxons animais e vegetais foram estabelecidas as áreas que tinham afinidade com América do Sul austral. As unidades de área analisadas foram: América do Sul austral (SSA), Norte da América do Sul (NSA), Austrália (AUS), Nova Guiné (NG), Nova Caledônia (NC), Tasmânia (TAS), Nova Zelândia (NZ), África do Sul (AF) e América do Norte (NA) .
Seleção dos táxons. Foram selecionados 1 7 táxons, os quais tinham informação filo genética disponível. Estes 17 táxons corresponderam a oito filogenias de insetos, oito plantas vasculares e um fungo. Entre as primeiras, os táxons envolvidos corresponderam às
Evolução da Região And ina da América do S u l - 1 85
famílias Silphidae, Carabidae e Staphylinidae (Coleoptera), Siphlonuridae (Ephemeroptera), Nannochoristidae (Mecoptera) , Ericoccidae (Hemiptera) e Chironomidae (Diptera) . Dentre as filogenias de plantas, encontraram-se representados táxons das fanu1ias : Elaeocarpaceae, Proteaceae, Fagaceae e Nothofagaceae, Gesneriaceae, Apiaceae e Winteraceae. Os fungos foram representados pela família Cyttareaceae.
Análise dos dados: foram utilizadas três técnicas de biogeografia cladística: análise de parcimônia de Brooks (BPA, Brooks parsimony analysis), análise de componentes e quantificação da análise de componentes. Para discussão sobre os métodos mencionados aqui ver Crisci et alo 14 e literatura citada.
Resultados
A anábse dos dados resultou em mais de uma hipótese de relações entre as áreas consideradas . No entanto, elementos comuns foram encontrados em todos os clado gramas gerais de áreas obtidos: América do Sul Austral (SSA) está ligada a outras áreas austrais (exceto África do Sul), ao passo que o norte da América do Sul (NSA) relaciona-se à América do Norte ou forma uma tricotomia com América do Norte e África do Sul (Fig. 1 1 .4) .
, I - Representação dos padrões comuns dos c ladogramas gerais de área obtidos por Crisci et ai. 2 . AF = África; AN = América do Norte; AUS = Austrá l ia; NC = Nova Caledônia; NG = Nova Gu iné; NSA = Norte da América do Sul; NZ =
Nova Zelândia; SSA = Su l da América do Su l; TAS = Tasmân ia .

186 - Evolução Espacia l da Região
Conclusões
Os resultados obtidos por Crisci et aI. 2 são consistentes com uma origem híbrida da biota da América do Sul, já que nenhum dos cladogramas de área resultantes mostra SSA e NSA como áreas irmãs, mas que estão em clados separados. Além disso, relações conflituosas entre SSA e outras áreas austrais, sugerem que esta área tem, por sua vez, uma origem composta.
Os Listroderina (Curculionidae: Cyclominae: Rhythirrhinini) são uma subtribo monofilética amplamente distribuída em regiões temperadas-frias da América do Sul e do Norte, Austrália e Nova Zelândia. Esta tribo é um elemento característico da curculiofauna da região andina, na qual o grupo é extremamente diversificado. Os objetivos de Donato et al. 2o foram: (1) analisar os padrões de distribuição da subtribo com aplicação da análise de dispersão-vicariância (DNA, dispersal-vicariance analysis), proposto por Ronquist32; e (2) interpretar a história biogeográfica do grupo no contexto da evolução geobiótica da América do Sul.
DIVA é um método para reconstruir as distribuições ancestrais de um táxon, maximizando os eventos de vicariância e minimizando os eventos de dispersão e extinção com base em informações filo genéticas e de distribuição do táxon (ver Capo 7). Este método, ao contrário dos métodos biogeográficos cladísticos, que possibilitam apenas estabelecer relações hierárquicas entre as áreas envolvidas, permite o estabelecimento de relações reticuladas entre elas.
Material e Métodos
A história biogeográfica dos Listroderina foi analisada usando-se a filogenia do grupo proposta por Morrone33. Como os terminais dessa filogenia foram gêneros, a distribuição de cada terminal foi determinada como a área ancestral resultante de aplicar DIVA à filogenia de cada gênero que tivesse mais de três espécies; no caso de não haver filogenias de gêneros ou de se tratar de uma ou duas espécies, foi considerada como distribuição de gênero a soma das áreas habitadas pelas espécies. As áreas consideradas pelos autores foram as sub-regiões da região andina (sensu Morroné) como mostrado na Figura 1 1 . 1 , mas também foram incluídas outras áreas nas que estão os Listroderina analisados: Neotrópico e Neártico. Os dados foram analisados usando-se DIVA 1 . 1 32.
Resultados
Os resultados da DIVA estão resumidos na Figura 1 1 .5 . Os autores postularam uma possível origem dos Listroderina no sul da América do Sul, em razão da grande diversificação do grupo na área e do fato de que o mais antigo registro fóssil da faIlll1ia Curculionidae vem de sedimentos do Cretáceo no sul do Chile. De acordo com 'D
-J os resultados obtidos se propõe a hipótese de que os � listroderina tinham distribuição ancestral confinada às �
\:,: áreas ocupadas pelo Chile Central, Páramo, Puna e região , 00
sub antártica. Posteriormente, a distribuição do grupo se a:: '" restringiu às áreas hoje ocupadas pelo Páramo e a Puna.
Isso teria ocorrido em consequência de eventos de extinção local que seriam decorrentes das mudanças climáticas-ambientais que sucederam no sul da América do Sul durante o período Eoceno Superior-Oligoceno Inferior (ver anteriormente). Os listroderinos teriam sido limitados neste período às áreas do norte da sua distribuição onde as condições climáticas teriam sido mais favoráveis do que no sul do continente. Esta mudança climática se reflete em mamíferos herbívoros em fósseis, que na Patagônia mostram aumento progressivo da altura dos molares durante o Paleoceno Superior e o Eoceno inferior. Ao mesmo tempo, as formas extrapatagônicas formavam comunidades de mamíferos dominadas por herbívoros de molares baixos tipo ramoneadores. Além disso, a flora Antártica e da Patagônia para esse momento foi denominada paleoflora mista (ver anteriormente), ao passo que as comunidades subtropicais e tropicais típicas ocupavam o centro e norte do continente sul-americano.
Depois, os Listroderina sofreram um evento de dispersão que ampliou novamente sua área de ocorrência, alcançando o subantártico. Este evento pode ter acontecido por causa da relativa quiescência tectônica que caracterizou o Oligoceno, refletida na escassa atividade magmática na América do Sul austral.
Um evento vicariante fragmentou a ampla distribuição dos Listroderina, gerando um clado subantártico (que resultou nos gêneros Listroderes, Listronotus, Falklandius e Antarctobius) e outro páramo-punefío (que deu origem ao grupo genérico Macrostyphlus). Posteriormente, ambos os dados se desenvolveram de forma independente. Esse evento de vicariância que separou o clado subantártico do páramo-punefío poderia ser explicado pela transgressão marinha Paranaense e uma nova etapa de diastrofismo andino, que poderia ter gerado uma barreira responsável pela fragmentação da distribuição do grupo durante o Mioceno Médio. Em seguida, parte do dado páramo-punefío, representado pelo grupo

Evo lução da Região And ina da América do Su l - 1 87
--------------- Rhígopsidius
.---------------- Philippius
..._---------------- Trachodema
,.....---------------- Lamiarhinus
r----------------- Acrorius
...----------------Acroriellus ...---------- Hyperoides
Acrostomus Listroderes
.---- Falklandiellus
Grupo Listroderes
Subantártico
Falklandiopsis Telurus
Falklandius Lanteriella
Grupo Falklandius
Vicariância ..... ---1 Dispersão
Páramo + Puna + Subantártico ,/
Haversiella Listronotus Neopachytychius
..._--- Germainiellus 10-_____ -1 Nacodius
Grupo Listronotus
Grupo Antarctobius
Páramo + Puna
Antarctobíus ..._----- Rupanius
.---Adíorístídíus L_-t---Amathynetoídes
Puna Puranius Macrostyphlus Andesianellus
Grupo Macrostyphlus
,S - Cladograma de Lístroderína mostrando os pr incipais eventos da história b iogeográfica, de acordo com os resu ltados da aná l ise de dispersão-vicar iância . Modificado de Donato et aI. 20.
genérico Macrostyphlus, ficou restrito à Puna. Isto se deu durante a fase Diaguita da Orogenia Andina, terminando com o soerguimento da Puna, criando um ambiente isolado de condições climáticas extremas, apesar de alguns de seus gêneros apresentarem, como nos outros clados, dispersão recente para outras áreas.
As amplas distribuições que exibem alguns dos táxons terminais dos clados subantártico e páramo-punefio, por exemplo, Listroderes, são atribuídos a fenômenos de dispersão recentes . No caso do clado sub antártico, é possível que a ampla diversificação estivesse associada aos fenômenos de isolamento e subsequente expansão das populações que teriam sua origem nas mudanças climáticas do Plio-Pleistoceno.
A evidência mostrada de várias fontes (por exemplo, geologia, paleontologia, biogeografia histórica) converge, permitindo postular hipóteses sobre os processos, que resultaram na configuração atual da diversidade biológica da América do Sul austral. A evolução da região
Andina foi marcada por processos geológicos importantes, que resultaram em mudanças climáticas e ambientais e que devem ser consideradas quando se postula hipóteses sobre padrões de distribuição dos táxons que nele habitam.
No caso da biogeografia histórica, a história geobiótica das áreas envolvidas fornece um quadro explicativo temporário que permite interpretar os resultados obtidos por meio de estudos de padrões de distribuição dos atuais táxons. Para isto é importante levar em conta a antiguidade do táxon na área de estudo pela interação com a paleontologia.
Os autores agradecem a Claudio J. B . de Carvalho e Eduardo A. B . Almeida pelo seu generoso convite para participar deste livro. Agradecemos também aos Drs. Rosendo Pascual e Jorge V. Crisci pelas frutíferas discussões e que enriqueceram os temas discutidos neste capítulo. Ambos os autores são membros do Consejo Nacional de Investigaciones Científicas y Técnicas de

188 - Evo l ução Espacia l da Região
Argentina (CONICET) e Universidad Nacional de La Plata, Argentina (UNLP), cujo apoio continuado agradecemos. Este trabalho está no marco dos projectos PICT 26298 (ANPCyT, Argentina) e PIP 1 12-200801 -00729 (CONICET, Argentina).
REFERÊNCIAS BIBLIOGRÁFICAS 1. PASCUAL, R. ; ORTIZ-JAUREGUIZAR, E.; PRADO, J. L.
Land mammals: paradigm for Cenozoic South American geobiotic evolution. In: ARRATIA, G. Contribution of Southem South America to Vertebrate Paleontology. Miincher Geowiss Abh (A), v. 30, p. 265-3 1 9, 1996.
2. CRISCI, J. V; CIGLIANO, M. M.; MORRONE, J. J. et a!. Historical biogeography of southern South America. Sys Zoo/., v. 40, p. 1 5 2- 1 7 1 , 1 99 1 .
3 . MORRONE, J . J . Biogeographic areas and transition zones of Latin America and the Caribbean Islands based on panbiogeographic and cladistic analyses of the entomofauna. Ann. Rev. Entomol., v. 5 1 , p. 467-494, 2006.
4. MORRONE, J. J. Presentación preliminar de un nuevo esquema biogeográfico de América deI Sur. Biogeographica, v. 75, p. 1 - 16, 1 999.
5 . ORTIZ-JAUREGUIZAR, E.; CLADERA, G. Paleoenvironmental evolution of southem South America during the Cenozoic. J. Arid. Environ. , v. 66, p. 489-532, 2006.
6. TRONCOSO, A.; ROMERO, J. Evolución de las comunidades florísticas en el extremo sur de Sudamérica durante el Cenofítico. Monog. Syst. Bot. Missouri Botanical Garden, v. 68, p. 149-1 72, 1 998.
7 . VILLAGRÁN, C.; HINOJOSA, L. F. Esquema biogeográfico de Chile. In: LLORENTE-BOUSQUETS, J.; MORRONE, J. J. Regionalización Biogeográfica en Iberoamírica y Tópicos Afines. México: Universidad Nacional Autónoma de México, Jiménez Editores, 2005. p. 5 5 1 -577.
8. BARREDA, V; PALAZZESI, F. Patagonian vegetation tumovers during the Paleogene-Early Neogene: origin of arid-adapted floras. Bot. Rev., v. 73, p. 3 1 -50, 2007.
9. PASCUAL, R.; ORTIZ-JAUREGUIZAR, E. The Gondwanan and South American Episodes: Two major and unrelated moments in the history of the South American mammals. J. Mammal. Evol. , v. 14, p. 75- 1 37, 2007.
10. HAQ, B. U.; HARDENBOL, J.; VAIL, P. R. Chronology of fluctuating sea leveIs since the Triassic. Scienee, v. 235, p . 1 156- 1 1 67, 1 987.
1 1 . ZACHOS , J . ; PAGANI, M . ; SLOAN, L . ; THOMAS, E . ; BILLUPS, K. Trends, rhythms, and aberrations i n global climate 65 Ma to present. Scienee, v. 292, p. 686-693, 200 1 .
1 2. MARSHALL, L. G . ; SEMPERE, T. Evolution o f the NeotropicaI Cenozoic land mammal fauna in its geochronologic, stratigraphic, and tectonic context. In: GOLDBLATT, P Biologieal Relationships Belween Africa and South America. New Haven: Yale University Press, 1 993. p. 329-392.
1 3 . MARKGRAF, V; ROMERO, E.; VILLAGRAN, C. History and paleoecology of SouthAmerican Nothofagus forests. In: VEBLEN, T. T.; HILL, R. S. The Ecology and Biogeography of Nothofagus Forests. New Haven: Yale University Press, 1996. p. 354-386.
1 4. CRISCI, J. V; KATINAS, L . ; POSADAS, P. Historical Biogeography: an introduc/ion. Cambridge: Harvard University Press, 2003. 250p.
1 5 . MORRONE, J. J. Evolutionary Biogeography: an intergrative approach with case studies. New York: Columbia University Press, 2008. 301p.
16 . CRAW, R. C. ; GREHAN, J. R.; HEADS, M. J. Panbiogeography: tracking the histOl)' of lzfe. New York: Oxford University Press, 1 999. 229p.
1 7 . CROIZAT, L. Panbiogeography. v. 1 , 2a. and 2b. Caracas: publicado por el autor, 1 958, 1 73 1p.
18. NELSON, G.; PLATNICK, N. I. Systematics andBiogeography - Cladistics and Vicariance. Nova York: Columbia University Press, 1 98 1 . 587p.
19. WIENS, J. J.; DONOGHUE. M. J. Historical biogeography, ecology and species richness. Trends Eco/. Evo/. , v. 1 9, p. 639--644, 2004.
20. DONATO, M . ; POSADAS, P; MIRANDA-ESQUIVEL, D. R. et aI. Historical biogeography of the Andean subregion: evidence from Listroderina (Coleoptera: CurcuJionidae: Rhytirrhinini) in the context of the South American geobiotic scenario. Bio/. J. Linnean Soe. , v. 80, p. 339-352, 2003.
2 1 . BRUNDIN, L. Transantarctic relationships and their significance. Kungl Svens Vetenskapakad Handl, v. I ! , p. 1 -472, 1966.
22. ROIG-JUNENT, S . Historia biogeográfica de América deI Sur austral. Multequina, v. 3, p. 1 67-203, 1 994.
23. KATINAS, L . ; MORRONE, J. J.; CRISCI, J. V Track analysis reveals the composite nature of the Andean biota. Austr. J. Bot., v. 47, p. 1 1 1 - 1 30, 1999.
24. GIRIBET, G.; EDGECOMBE, G. D. The importance of looking at small-scale pattems when inferring Gondwanan biogography: a case study of the centipede Paralamyctes (Chilopoda, Lithobyomorpha, Henicopidae). Bio/. J. Linnean Soe., v. 89, p. 65-78, 2006.
25. MORRONE, J. J. Distributional patterns of species of Rhytirrhinini (Coleoptera: Curculionidae) and the historical relationships of the Andean provinces. Glob. Ecol. Biogeog. LeU. , v. 4, p. 1 88- 1 94, 1994.
26. MORRONE, J. J. ; KATINAS, L.; CRISCI, J. V A cladistic biogeographic analysis of Central Chile. J. Comparat. Bio/., v. 2, p. 25-42, 1997.
27. POSADAS, P.; ESTÉVEZ, J. M.; MORRONE, J . J. Distributional patterns and endemism areas of vascular plants in the Andean subregion. Fontqueria, v. 48, p. 1 - 10, 1 997.
28. POSADAS, P; MORRONE, J. J. Biogeografía histórica de la familia Curculionidae (Insecta: Coleoptera) en las subregiones Subantártica y Chilena Central. Rev. Soe. Entomol. Argentina, v. 62, n. 1 -2, p. 75-84, 2003.
29. DOMÍNGUEZ, M. C.; ROIG-JUNENT, S . ; TASSIN, J . J. et al. Areas of endemism of the Patagonian steppe: an approach based on insect distributional patterns using endemicity analysis. J. Biogeog. , v. 33, p. 1 5 27 - 1 537, 2006.
30. ROIG-JUNENT, S . ; DOMÍNGUEZ, M. c.; FLORES, G. E. et aI. Biogeographic history of South American arid lands: A view from its arthropods using TASS analysis. J. Arid. Environ., v. 66, p. 404-420, 2006.
3 1 . LOWEMBERG-NETO, P; DE CARVALHO, C. J. B . ; DINIZ-FILHO, J. A. F. Spatial congruence between biotic history and species richness of Muscidae (Diptera, Insecta) in the Andean and Neotropical regions. J. Zoo/. Syst. Evo/. Res. , v. 46, p. 374-380, 2008.
32. RONQUIST, F. Dispersal-Vicariance Analysis (DIVA). Programa para computador e manual, 1 996 - versão 1 . 1 . Disponível em: http://www.ebc.uu.se/systzoo/research/diva/diva.html. Acesso em: 20/08/2009.
33. MORRONE, J. J. Cladistics of the New World genera of Listroderina (Coleoptera: Curculionidae: Rhytirrhinini). Cladisties, v. 1 3 , p. 247-266, 1997.

.::, I 0 I V ''-.�
José Maria Cardoso da Silva
Adrian Antonio Garda
o trabalho da biogeografia consiste em três etapas : ( 1 ) documentar a distribuição dos organismos (vivos ou fósseis) ; (2) identificar padrões biogeográficos; e (3) determinar os processos biogeográficos que explicam os padrões observados. Assume-se, portanto, que a ocorrência de um padrão na natureza foi causada por um processo ou um conjunto de processos gerais . Esta relação de causalidade é elemento essencial em todo o trabalho de um biogeógrafo.
Como em outras ciências, a biogeografia possui hipóteses sobre padrões e hipóteses sobre processos. Um padrão é um enunciado sobre relações regulares e repetitivas entre várias observações da natureza. O padrão, uma vez estabelecido, representa algo que deve ser explicado. A explicação, por sua vez, requer a compreensão do processo que causa um padrão ser o que ele é. O processo é, portanto, sinônimo com causa, a operação de um ou mais fatores que produzem uma relação particular entre observações.
Há hipóteses sobre padrões e hipóteses sobre processos. É importante não confundi-las. De modo geral, as hipóteses sobre padrões precisam ser rigorosamente testadas antes de se propor uma hipótese de processo para explicar o padrão detectado. Em ambos os casos, estas hipóteses são testadas de forma rigorosa até que sejam consideradas como necessárias ou suficientes para tornarem-se amplamente aceitas. Imaginemos que determinada área possui várias espécies endêmicas a ela. Então, podemos construir uma hipótese de que esta área representa uma área de endemismo. Esta é uma hipótese de padrão. Para testarmos esta hipótese, podemos verificar se o número de espécies restritas à área é maior
1 2
._��
do que seria esperado ao acaso. Se este for o caso, então a hipótese de padrão é apoiada e ela passa a exigir uma explicação. A explicação para a concentração de espécies endêmicas naquela área específica passa a ser uma hipótese de processo, cuj a avaliação requer métodos apropriados. Como é comum na ciência moderna, a ampla aceitação de uma hipótese não implica, de forma alguma, que ela esteja protegida de questionamentos, pois novas informações e novas interpretações podem resultar em derrubada de hipóteses tradicionalmente aceitas e surgimento de novas hipóteses com maior poder explicativo.
A Amazônia é a região de maior biodiversidade do planeta. Cobrindo mais de 6 milhões de km2 em nove países do norte da América do Sul, a região abriga pelo menos 40.000 espécies de plantas, 427 de mamíferos, 1 . 294 de aves, 378 de répteis, 427 de anfíbios e mais de 3 .000 espécies de peixes1 . Em geral, estima-se que a Amazônia abrigue cerca de 10% das espécies do planeta. Explicar a origem da extraordinária riqueza de espécies na Amazônia sempre foi um grande desafio para gerações de cientistas 1 ,2. No processo de investigação é muito importante distinguir entre os processos que deram origem à grande diversidade de espécies na região e os processos que atuam de forma integrada para manter esta biodiversidade. Compreender a origem das espécies na região é um tema que pode ser investigado de forma adequada com os métodos da biogeografia, enquanto entender como a biodiversidade é mantida atualmente é um tema que requer os métodos da ecologia.
Este capítulo trata da origem e evolução da moderna biota amazônica. Iniciar-se-á discutindo os esforços que foram feitos para identificar quais são as unidades biogeográficas básicas da Amazônia. Depois, será feita uma

1 90 - Evol ução Espacia l da Região
síntese sobre quais as relações históricas entre estas unidades biogeográficas. Em seguida, será analisada qual a contribuição dos diferentes processos biogeográficos na formação das biotas modernas das áreas de endemismo amazônicas. Por fim, será discutido o problema do tempo de origem das espécies na Amazônia.
As unidades biogeográficas básicas para a compreensão da história das biotas regionais modernas são as áreas de endemismo, que são definidas aqui como espaços geográficos determinados pela congmência nas distribuições de duas ou mais espécies que não ocorrem em nenhum outro lugar (ver Capo 3). Há vários métodos propostos para identificar áreas de endemismo, mas serão contrastados apenas dois métodos: método tradicional e método proposto por Morrone3. O método tradicional sobrepõe os mapas com as distribuições de espécies de distribuição restrita para identificar os lugares com altas concentrações de tais espécies. Este método é limitado porque faltam critérios claros para determinar se uma espécie possui ou não distribuição restrita. Além disso, o método não revela a natureza hierárquica das áreas de endemismo. O método proposto por Morrone3 analisa a distribuição das espécies utilizando o critério da parcimônia para identificar subconjuntos de "unidades geográficas operacionais" (geralmente localidades ou quadrados de 0,5° ou 1°), que são definidos de forma não ambígua por, pelo menos, duas espécies. O método de Morrone é considerado mais objetivo e completo que o método tradicional, pois não requer nenhuma premissa sobre a extensão das distribuições das espécies e revela, de forma clara, a hierarquia das áreas de endemismo.
O estudo comparativo da distribuição das espécies na Amazônia indica dois padrões biogeográficos básicos. O primeiro padrão é que as espécies não estão distribuídas de forma homogênea na região: a maioria das espécies possui distribuição restrita, definindo várias áreas de endemismo. O segundo padrão é o da substituição das espécies endêmicas a uma área de endemismo por espécies aparentadas nas áreas de endemismo adjacentes. Em muitos casos, estas substituições de espécies estão associadas à presença de barreiras físicas bem definidas, tais como rios, planaltos ou manchas de vegetação aberta. Entretanto, em alguns casos, essa substituição se dá em lugares sem nenhuma barreira atual visível conhecida. Os dois padrões não passaram
despercebidos pelos primeiros naturalistas que visitaram a região1 .2 e todos os esforços feitos pelos biogeógrafos até hoje visaram basicamente identificar os processos biogeográficos que causaram estes padrões.
Wallace2 dividiu a Amazônia em quatro áreas de endemismo (que ele denominou de "distritos"), com base no estudo da distribuição das espécies de primatas:
'D Guiana, Equador, Pem e Brasil. As bordas destas áreas Cf de endemismo correspondiam aos rios Amazonas-Soli- e;; .:., mões, Negro e Madeira. A hipótese de Wallace foi t apoiada por vários estudos de diversos gmpos de verte- �
o-, brados como também pela reanálise das informações o, atualizadas sobre os primatas amazônicos.
Examinando as distribuições das espécies de aves e usando o método tradicional de identificar áreas de endemismo, Haffer propôs seis áreas de endemismo para aves, modificando as áreas identificadas por Wallace. Assim, a área Guiana permaneceu como uma área de endemismo distinta, a área Equador foi dividida em duas áreas de endemismo (Imeri e Napo), a área Pem foi renomeada Inambari e a área Brasil foi dividida em duas áreas (Rondônia e Belém). Cracraft5 fez uma análise mais detalhada das distribuições das espécies e subespécies de aves na América do Sul e chegou quase à mesma conclusão que Haffer, no que diz respeito ao número de áreas de endemismo na Amazônia. A única diferença foi que Cracraft reconheceu mais uma área de endemismo, que foi denominada de Pará, para toda a região entre os rios Tocantins e Tapajós.
A partir da publicação do trabalho de Morrone3, vários estudos foram feitos para avaliar a proposta destas sete áreas de endemismo utilizando a nova metodologia. Em todos os estudos, a divisão da Amazônia em sete áreas de endemismo foi apoiada. Mais recentemente, Silva et a!. 6 analisaram novas informações sobre a distribuição e taxonomia de aves e sugeriram que a área de endemismo Pará é, de fato, composta por duas áreas bem distintas, cada qual com o seu próprio conjunto de espécies endêmicas, que foram nomeadas como Xingu e Tapajós. Além disso, eles redefiniram os limites da área de endemismo Imeri, fazendo-a concordar com o interflúvio Negro-Japurá/Solimões e estendendo-a até o baixo Rio Negro.
Desta forma, oito grandes áreas de endemismo podem ser reconhecidas para vertebrados terrestres na Amazônia (Fig. 12 . 1 ) . As áreas de endemismo reconhecidas para subespécies de borboletas florestais e para espécies de plantas vasculares são geralmente coincidentes ou estão contidas dentro das oito áreas de endemismo de vertebrados terrestres, indicando, assim, uma boa congmência espacial entre os padrões de endemismo de diferentes gmpos taxonômicos 7.

Padrões e Processos B iogeográficos na Amazôn ia - 1 91
'J 2, - Áreas de endemismo identificadas para vertebrados terrestres na Amazônia . Ver Prancha Colorida.
As áreas de endemismo na Amazônia variam consideravelmente em extensão, desde a pequena Belém (201 .54Ikm2) até a enorme Guiana ( 1 .700.532km2). As outras possuem as seguintes extensões : Imeri (679 . 8 67km2) , Napo ( 5 08 . 1 04km2) , Inambari ( l . 326 .684km2), Rondônia (675 .454km2) , Tapajós (648. 862km2) e Xingu (392.468km2).
O número e os limites das áreas de endemismo na Amazônia devem ser vistos como hipóteses de trabalho, exigindo constante reavaliação quando novos dados taxonômicos e biogeográficos de diferentes grupos de organismos tornarem-se formalmente disponíveis . É
possível predizer, por exemplo, que algumas áreas de endemismo, como Guiana, Imeri e lnambari, serão subdivididas em uma ou mais áreas de acordo com o aumento do conhecimento sobre suas biotas.
Para determinar os padrões de relações históricas entre áreas de endemismo é necessário reconstruir primeiro as
relações históricas entre as espécies que caracterizam as áreas de endemismo. Para estabelecer as relações históricas entre as espécies, utilizam-se métodos de análise filogenética. Esses métodos usam caracteres morfológicos, comportamentais ou moleculares para produzir um diagrama no formato de árvore, que agrupa as espécies somente com base nas sinapomorfias. Este diagrama de relações é denominado cladograma. Em um cladograma, cada ramo representa uma linhagem. A linhagem se divide, dando origem a duas ou mais espécies descendentes. Espécies que se originaram de um ancestral comum imediato são denominadas de espécies irmãs. Quando o pesquisador utiliza dados moleculares é possível também estimar a idade dos eventos que deram origem a cada uma das espécies. Isso é feito a partir da calibração das distâncias genéticas entre as espécies irmãs e a idade de fósseis ou barreiras geológicas de datas conhecidas. Apesar das críticas a este procedimento, estas estimativas sobre a origem das espécies são bastante úteis para estudos biogeográficos.
Os cladogramas de grupos de espécies podem ser combinados para reconstruir a história das áreas de endemismo na Amazônia. Os métodos da biogeografia cladística (ver Capo 6) geram o que se denomina

1 92 - Evo lução Espac ia l da Reg ião
"cladograma de área ou areagrama", ou seja, uma representação gráfica das relações históricas entre as áreas de endemismo estimadas a partir das relações de parentesco das espécies endêmicas.
Prum8 foi o primeiro a realizar um estudo para avaliar as relações históricas entre as áreas de endemismo da Amazônia empregando a biogeografia cladística. Ele comparou os cladogramas de 1 3 grupos de aves que ocorrem nas várias áreas de endernismo reconhecidas para as florestas tropicais de terras baixas sul-americanas. Em vez de um diagrama simples de relações entre as áreas de endemismo, este autor propôs um areagrama complexo, no qual algumas áreas de endemismo aparecem mais de uma vez (Guiana e Rondônia), indicando que estas áreas foram separadas e reagrupadas com outras áreas de endemismo várias vezes durante a formação de suas biotas (Fig. 1 2.2). De grande interesse é a sugestão de que as áreas de endemismo amazônicas não formam uma unidade biogeográfica natural, pois partes da Floresta Atlântica são mais próximas do conjunto formado pelas áreas de endemismo Belém, Xingu,
Tapajós e Rondônia, do que de outros setores da Floresta Amazônica.
Amorim9 combinou vários cladogramas de distintos grupos de insetos e primatas e indicou também que a Amazônia não é uma unidade natural, mas, sim, composta por dois grupos de áreas . O primeiro grupo é composto pelas áreas de endemismo Belém, Xingu, Guiana, Imeri, N apo e lnambari e está mais associado ao Escudo das Guianas, ao norte da América do Sul e à América Central. O segundo grupo é composto pelas áreas de endernismo Rondônia e Tapajós e é mais relacionado ao Escudo Brasileiro, incluindo aí a Floresta Atlântica (Fig. 1 2.3) . O único resultado em comum entre os estudos de Prum e Amorin é a evidência de que a Amazônia não forma uma unidade biogeográfica, pois as sequências de divisão das áreas amazônicas entre as duas propostas são completamente discordantes.
Cracraft10 usou filogenias de espécies de aves amazônicas para demonstrar que os métodos da biogeografia cladística não funcionam adequadamente quando empregados em biotas continentais marcadas por histórias
... ----------------- Floresta Atlântica
.. -------------- Chocá
... ----- Chocá
.. --- Nechi
América Central Pacífica
América Central Caribenha
Guiana
Guiana
Belém
Xingu-Tapajás
.. --- Rondônia
.. ----- Floresta Atlântica
.------ Rondônia
.. --- Imeri
Napo
I nambari
- Relações h istóricas entre áreas de endemismo nas florestas Neotropicais conforme Prums. Note que a lgumas áreas de endemismo aparecem no cladograma de área mais de uma vez, ind icando complexas relações h istóricas entre as á reas.

Pad rões e Processos B iogeográficos na Amazôn ia - 193
.. _______________ Andes + América Central
.. --- Inambari
... _- Napo
.. --------- Imeri
... ----- Guiana
.--- Belém
"' __ Xingu
.-__ Tapajós
... __ Rondônia
.. _____ Floresta Atlântica
- Relações históricas entre áreas de endemismo nas florestas Neotropicais conforme Amorim9.
geológicas muito complexas. Ele argumentou que processos como junção de áreas em consequência de desaparecimento de barreiras, extinções e eventos vicariantes de diferentes idades localizados nas mesmas áreas tendem a apagar os resultados dos eventos vicariantes anteriores, limitando a possibilidade de reconstruir a história das relações entre as áreas de endemismo, de forma adequada, utilizando métodos reducionistas.
Bates ! ! reconheceu que a história das áreas de endemismo da Amazônia é, de fato, muito complexa, dada a enorme variedade de fatores geológicos que afetaram a região nos últimos 20 milhões de anos. Entretanto, ele sugeriu uma divergência primária e instantânea, do ponto de vista evolutivo da Amazônia, em três grupos basais de áreas de endemismo: Nordeste (área de endemismo Guiana), Oeste (áreas de endemismo Napo e Inambari) e Sudeste (incluindo as áreas de endemismo Tapajós, Xingu e Belém). Ele também sugeriu que as duas outras áreas de endemismo (Rondônia e Imeri) possuem uma biota derivada a partir de espécies ancestrais que colonizaram estas regiões a partir das três áreas basais. Bates! ! testou as predições deste modelo utilizando todas as filogenias de grupos de aves amazônicas existentes e encontrou um grande apoio.
Camargo e Pedro! 2 sumarizaram o conhecimento sobre sistemática, taxonomia e biogeografia de abelhas sem ferrão (Meliponini) neotropicais. A filogenia produzida para o gênero Partamona permitiu a delimitação de áreas de endemismo para o grupo nas Américas e o cladograma de áreas resultante corroborou a divisão proposta por Bates! ! . Três componentes amazônicos são claramente distinguíveis nas porções oeste, nordeste e sudeste do bioma. Algumas espécies de Partamona mais relacionadas aos táxons de um componente biogeográfico podem ocorrer em outros componentes ou mesmo estar restritas a áreas onde suas espécies irmãs não acontecem. Esses resultados sugerem um padrão biogeográfico complexo, no qual as concordâncias gerais entre táxons são permeadas por casos específicos. Essas exceções podem contribuir para o entendimento de eventos de menor escala que certamente influenciaram na distribuição das espécies e subespécies de animais e plantas na região.
Em resumo, os estudos recentes indicam dois resultados interessantes e provocativos. Primeiro, a Amazônia, tal como definida de forma ecológica, possivelmente não é uma unidade biogeográfica natural, pois há evidências de que algumas de suas áreas de endemismo são historicamente mais relacionadas às áreas de endemismo de outras

1 94 - Evol ução Espac i a l da Região
regiões sul-americanas do que às demais áreas de endemismo amazônicas. Segundo, os métodos tradicionais de biogeografia cladística têm muitas limitações práticas quando são aplicados a biotas continentais complexas e dinâmicas 10. Isto indica que o uso de relações dicotômicas, que podem ou não ser congruentes temporalmente, talvez não seja a melhor forma de representar as complexas relações existentes entre áreas de endemismo continentais. Forçar uma história complexa em um aerograma dicotômico é estratégia reducionista e acarreta perda de informações importantes para a compreensão da evolução de biotas regionais. Talvez o uso de redes de relações para representar relações biogeográficas, tendo como base teórica a teoria dos grafos, seja um caminho metodológico a ser mais explorado pelos biogeógrafos históricos.
Quais os processos que determinaram os padrões atuais de riqueza e endemismo das biotas das áreas de endemismo que compõem a Amazônia? Para responder esta pergunta é necessário primeiro reconhecer que nem sempre diversidade de espécies e a porcentagem do número de espécies endêmicas a uma biota são relacionadas . Há vários casos em que áreas ricas em espécies são pobres em espécies endêmicas e áreas ricas em espécies endêmicas que são pobres de espécies. Esta relação precisa ser mais estudada por biogeógrafos.
Ricklefs e Schluter1 3 discutiram as relações entre diversidade regional e diversidade local de espécies e propuseram que a diversidade regional (número de espécies em determinada região) é um produto basicamente de três processos biogeográficos básicos: produção de espécies, intercâmbio biótico e extinção em massa. O grande desafio teórico da biogeografia moderna é o de desenvolver métodos para estimar a contribuição relativa de cada um desses processos na formação das modernas biotas regionais.
As espécies são produzidas quando uma espécie ancestral dá origem a duas ou mais espécies descendentes via um processo denominado de especiação. Há vários modelos de especiação. Entretanto, em vertebrados terrestres, o modelo mais comumente aceito é da especiação alopátrica por vicariância. Neste modelo, uma espécie ancestral tem sua distribuição fragmentada por
fatores geológicos ou ecológicos. Assim, as populações tornam-se isoladas e passam por processo de diferenciação, dando origem a duas ou mais espécies descendentes. A produção de espécies quase sempre aumenta a diversidade de espécies em uma região, mas nem sempre este 'D
é o caso. Dessa forma, toma-se necessário fazer a distinção entre produção intrarregional de espécies (a
...., 00 00 V> .c., � divisão de uma espécie ancestral em duas ou mais espé-00
cies descendentes dentro de uma região biogeográfica) � da produção inter-regional de espécies (a divisão de uma ""
espécie ancestral em duas ou mais espécies descendentes ao longo dos limites de duas ou mais regiões biogeográficas). Esta separação é importante, pois apenas a primeira aumenta a diversidade regional, mas os dois tipos, desde que não sejam seguidos imediatamen-te por eventos de dispersão, aumentam o número de espécies endêmicas em dada região.
O intercâmbio biótico é o fluxo natural de espécies entre regiões adjacentes. A diversidade de espécies aumenta quando uma área é colonizada via dispersão, mas a porcentagem de espécies endêmicas reduz-se. Enquanto a dispersão por saltos pode ser importante para explicar a formação de biotas em ilhas oceânicas, a difusão e a dispersão secular são os tipos mais prováveis de dispersão responsáveis pela formação de biotas re-gionais dentro de continentes.
Os eventos de extinção em massa diminuem tanto a diversidade regional como a porcentagem de espécies endêmicas e podem ser causados tanto por fatores bióticos como por fatores abióticos . No entanto, eles costumam estar associados às mudanças ambientais drásticas. Seus efeitos quantitativos sobre as biotas modernas não podem ser facilmente estimados pelos métodos de análise tradicionais, pois isso requer fósseis abundantes e bem preservados.
Silva14 comparou as biotas dos cinco grandes biomas brasileiros e sugeriu que a produção de espécies (especiação intrarregional) contribuiu mais para a formação da moderna biota na Amazônia e da Floresta Atlântica do que o intercâmbio biótico. Em contraste, o intercâmbio biótico é mais importante do que a produção de espécies no Pantanal. A Caatinga e o Cerrado localizam-se entre estes dois extremos, com posições dependendo do grupo estudado. Para aves, o intercâmbio biótico parece ter desempenhado o papel mais fundamental, enquanto para plantas a produção de espécies é, de longe, o processo biogeográfico mais relevante. Silva14 sugeriu que a formação de áreas de endemismo na Amazônia e na Floresta Atlântica poderia ser consequência de um ou vários eventos de especiação, que atingiram, ao mesmo tempo, um conjunto grande de linhagens

ancestrais que possuíam ampla distribuição na região. Esta conclusão geral não é nova. Na verdade, ela apenas sintetiza a principal conclusão de todos os principais estudos j á produzidos sobre a biogeografia neotropical. Neste momento, não é possível avaliar, com as informações atualmente disponíveis, a contribuição do processo de extinção em massa para explicar a atual diversidade de espécies das áreas de endemismo amazônicas; contudo, este processo pode ter sido muito mais importante na determinação de alguns padrões biogeográficos atuais do que se imagina, pois há fortes evidências indicando que a diversidade de plantas na região, por exemplo, foi mais alta no Mioceno do que é atualmente1S.
Nos últimos 30 anos, tem-se discutido bastante o papel das flutuações climáticas do Quaternário - os últimos dois milhões e meio de anos - como a principal causa da origem das espécies na Amazônia e, por consequência, da formação das áreas de endemismo na Amazônia. Esta ideia compõe o núcleo da "teoria dos refúgios" e foi proposta inicialmente por Haffer. A hipótese de Haffer é simples e elegante, apesar de gerar poucas predições testáveis. Ele sugeriu que durante os vários períodos glaciais do Quaternário, a floresta amazônica foi dividida em vários pequenos pedaços de florestas que foram isolados uns dos outros por extensas áreas de vegetação aberta, não florestal. Os remanescentes de floresta serviram como "refúgios" para numerosas populações de animais e plantas florestais, que se diferenciaram umas das outras via seleção natural e oportunidade durante períodos de isolamento. Os pequenos blocos de floresta foram novamente reunidos nos períodos interglaciais, quando as áreas antes recobertas por vegetações abertas foram outra vez dominadas por florestas, permitindo que as populações dos "refúgios" expandissem de novo as suas distribuições. Durante este processo de expansão, as populações entraram novamente em contato com suas populações irmãs que tinham ficado isoladas em outros "refúgios". O resultado deste encontro pode ter sido variável, dependendo de quanto estas populações tinham se diferenciado em isolamento e desenvolvido ou não incompatibilidade reprodutiva e/ou ecológica. Se as populações isoladas desenvolveram tanto incompatibilidade reprodutiva como ecológica, elas tinham se transformado em espécies distintas e tenderiam a se excluir geograficamente ao longo de uma zona de contato. Se as populações desenvolveram incompatibilidade reprodutiva, mas não incompatibilidade ecológica, então
Padrões e Processos B iogeográfi cos na Amazôn ia - 1 95
as espécies resultantes poderiam estabelecer áreas de sobreposição (ou simpatria) ao longo das zonas de contato. Se as populações não desenvolveram nem incompatibilidade reprodutiva e nem incompatibilidade ecológica, então as populações poderiam estabelecer faixas de hibridização, cujas larguras poderiam variar bastante, ou mesmo se fundirem completamente e, assim, eliminar toda a diferenciação acumulada durante o período de diferenciação. Este processo de separação e contato de vários blocos de floresta na Amazônia foi provavelmente repetido várias vezes durante o Quaternário e resultou em diferenciação da biota florestal da região em tempo geológico relativamente recente. Esta hipótese teve grande aceitação durante as décadas de 1970 e 1980, mas foi lentamente abandonada nas últimas duas décadas. Algumas razões para isso são:
• Os estudos geológicos recentes na região indicam que a Amazônia foi muito mais dinâmica do que anteriormente tinha sido imaginado. Mesmo o proponente da "teoria dos refúgios", um geólogo por profissão, imaginava que a Amazônia tinha sido relativamente estável durante o Terciári04. Entretanto, os dados atuais1S.l6 demonstram a importância de eventos ocorridos durante o Terciário, como as transgressões marinhas, mudanças no nível do mar, neotectonismo e dinâmica fluvial para o processo de formação da paisagem amazônica moderna.
• As evidências paleoecológicas coletadas até o momento 1 7 não demonstram, de forma inequívoca, que grande parte da floresta amazônica tenha sido, em algum momento, substituída tão extensamente por formações abertas e não florestais, tal como cerrados e caatingas abertas, como foi sugerido pelos proponentes da teoria dos refúgios. Os poucos estudos paleopalinológicos existentes que apóiam a substituição da floresta por vegetações abertas1 S estão localizadas na periferia da Amazônia (Katira e Georgetown) ou em regiões dominadas por mosaicos formados por savanas e florestas no interior da Amazônia (Carajás).
• Os estudos moleculares sobre espécies de vertebrados amazônicos continuam a indicar que estas são, em grande parte, muito mais antigas que o Quaternário 1 1 . 1 8-20 , Um estudo molecular envolvendo vários grupos de plantas21 concluiu que mudanças climáticas do Quaternário não foram a maior força responsável pelas taxas de especiação de plantas sul-americanas.
• A conclusão de que a "teoria dos refúgios"4 não pode ser testada por métodos filogenéticos, pois ela não especifica qualquer sequência de fragmentação dos refúgios para ser avaliada pela sequência de eventos de separação de táxons1 8. Ainda assim, o papel das

1 96 - Evolução Espac i a l da Reg ião
flutuações climáticas durante o Pleistoceno sobre distribuição e diversidade de alguns elementos da fauna Amazônica não pode ser descartado.
A descoberta de que a paisagem da Amazônia foi muito mais dinâmica no passado do que se imaginava e que as espécies são mais antigas do que se previa serviu de pano de fundo para que outras questões interessantes pudessem ser abordadas. Isso implicou em reestruturar as velhas hipóteses em bases mais sólidas e gerar predições quantitativas que possam ser testadas no campo com o uso de novas ferramentas da moderna biologia evolutiva (ver Capo 8).
Uma das abordagens mais promissoras é a da filogeografia. Diferentemente das análises filogenéticas tradicionais, que enfocam relações no nível de espécies para cima, as análises filogeográficas estão centradas nas relações históricas entre populações de uma mesma espécie. Para tanto, sequências de ácido desoxirribonucleico (DNA, desoxyribonucleic acid) de indivíduos de diversas populações são analisadas tanto por meio de técnicas de reconstrução filogenética como por métodos quantitativos de genética de populações e as frequências gênicas são usadas para avaliar a existência ou não de fluxo gênico entre as populações em questão. A filogeografia funciona, portanto, como uma ponte entre disciplinas microevolutivas (demografia e genética de populações) e disciplinas macroevolutivas (sistemática filogenética, paleontologia e biogeografia histórica)22. Assim, com amostragem adequada, pode-se empregar cada uma dessas metodologias para testar diferentes tipos de hipóteses, por exemplo:
1 . Se diversas populações correspondem realmente a uma mesma espécie (filogeografia).
2. Se existe fluxo gênico entre elas (genética de populações).
3. Na ausência de fluxo gênico, há quanto tempo esse fluxo cessou (demografia e genética de populações).
4. Se existe concordância entre o padrão de estruturação hierárquico entre as populações e as barreiras geográficas que as separam (sistemática filogenética e biogeografia).
5. Se existem evidências para expansões populacionais ou contrações recentes (genética de populações) .
Expansões e contrações populacionais, concordância entre a estruturação hierárquica e filogenética de populações e o tempo de separação entre elas podem rigorosamente testar predições de hipóteses que visem explicar os padrões biogeográficos atuais na Amazônia.
Aleix023 reformulou três hipóteses principais utilizadas geralmente para explicar a diversificação de aves na
Amazônia, extraindo delas predições testáveis. Então, ele estudou a filogeografia de um grupo de espécies de aves (Xiphorhynchus) que habitam as florestas de terra firme para testar estas predições. Ele encontrou que a hipótese dos rios como barreiras, que sugere que a formação dos rios foi o evento principal para formação das áreas de endemismo, foi apoiada apenas para os rios de água escura. A hipótese dos refúgios foi refutada em todas as ocasiões, pois as populações de áreas instáveis durante o Pleistoceno não apresentaram sinais de redução populacional seguido de expansão, conforme seria esperado. A hipótese de que as incursões marinhas do Mioceno provocaram muitos eventos de especiação quando o aumento do nível do mar causou a entrada da água do mar ao longo da planície do Rio Amazonas, inundando vastas áreas na região, foi comprovada, em parte, pois a data desse evento coincide bem com a separação de algumas populações de Xiphorhynchus.
Santos et al. 20 utilizaram uma análise filogeográfica de 406 indivíduos pertencentes a 137 espécies descritas e 34-89 espécies (estimativa de mínimo e máximo) não descritas de rãs venenosas (família Dendrobatidae) amazônicas e andinas para testar, com um relógio molecular calibrado, as origens e a diversificação dos táxons amazônicos. Os autores encontraram uma história complexa. Primeiro, as espécies amazônicas destas rãs foram formadas a partir de 14 linhagens que se dispersaram para a região a partir dos Andes, principalmente após o término das incursões marinhas do Mioceno (cerca de 10 milhões de anos atrás). Segundo, essas linhagens passaram por um processo de rápida diversificação in situ dentro da Amazônia durante a transição Mioceno-Plioceno, dando origem ao atual conjunto de espécies endêmicas existentes atualmente na Amazônia.
A crescente evidência de que eventos vicariantes mais antigos foram os mais importantes no processo de formação das espécies que vivem hoje na Amazônia não implica que as mudanças climáticas globais que ocorreram no Quaternário não tiveram impOliância alguma em todas as regiões do planeta. Na Amazônia, com topografia relativamente mais suave, as espécies possuem áreas de distribuição grandes quando comparadas com espécies vivendo em outras regiões tropicais. Como consequência, as populações são também grandes e, portanto, menos propensas a se diferenciarem em resposta a barreiras relativamente instáveis e de curta duração, tal como as associadas às mudanças climáticas do Quaternário. Em contraste, as espécies que habitam regiões biogeográficas com topografia complexa e gradientes altitudinais marcantes quase sempre possuem distribuição geográfica restrita e populações pequenas. Nestas regiões de topo-

grafia complexa, como os Andes, as espécies são muito mais propensas a se diferenciarem como consequência
'D das mudanças climático-vegetacionais que ocorreram no � � Quaternário. Um estudo recente por Weir24 apoia essa ;'!; ideia. Ele apresentou uma das poucas análises compara-t""-tA tivas entre taxas de diversificação de um grupo de '1 00 t""-0\
organismos (aves) entre terras baixas e terras altas na região neotropical. Weir24 encontrou que há importantes diferenças no acúmulo de espécies ao longo do tempo. Nas terras altas, a taxa de diversificação aumentou no Plioceno e no Pleistoceno e culminou com uma taxa de diversificação no final do Pleistoceno duas vezes mais alta do que nos períodos anteriores. Este padrão é consistente com a ideia de que mudanças climáticas no Quaternário tiveram um impacto na diversificação das espécies andinas. Em contraste, as taxas de diversificação nas terras baixas atingiram o seu ápice no Mioceno e foram reduzindo ao longo do tempo até taxas muito baixas durante o final do Pleistocen024. Entretanto, essa abordagem tem sido muito criticada, em particular pela amostragem deficiente e pela adoção da taxonomia vigente, que se vale de grande número de subespécies e muito provavelmente subestima a diversidade de aves na Amazônia25.
As espécies que vivem na Amazônia possuem complexa história evolutiva. As suas linhagens ancestrais foram influenciadas por vários eventos de vicariância associados aos grandes processos de reorganização da paisagem regional como consequência dos vários períodos de tectonismo intenso dos Andes. Todas as evidências atuais indicam papel limitado das mudanças climático-vegetacionais do Quaternário para explicar a origem das espécies amazônicas. Dada a intensa dinâmica geomorfológica da Amazônia, nenhuma hipótese fundamentada em um único processo geológico ou paleoecológico será suficiente para explicar a origem dos padrões biogeográficos observados atualmente na Amazônia.
REFERÊNCIAS BIBLIOGRÁFICAS 1 . MITTERMEIER, R. A. ; MITTERMEIER. C. G. ; GIL, P. R. et
a!. Wildemess - Earth 's Last Wild Places. Mexico City: CEMEX, 2002. 576p.
2. WALLACE, A. R. On the monkeys of the Amazon. Prac. Zoo/. Soco Lond. , v. 20, p. 107- 1 10, 1 852.
3 . MORRONE, J. J. On the identification of areas of endemism. Syst. Biol., v. 43, p. 438-44 1 , 1 994.
4. HAFFER, J. Speciation in Amazonian forest birds. Science, v. 1 65, p . 1 3 1 - 1 37, 1 969.
5 . CRACRAFT, J. Historical biogeography and patterns of differentiation within the South American avifauna: areas of endemism. Omithol. Monogr. , v. 36, p. 49-84, 1 985.
6. SILVA, J. M. c.; NOVAES, F. c. ; OREN, D. C. Differentiation of Xiphocolaptes (Dendroco1aptidae) across the river Xingu, Brazilian Amazonia: recognition of a new phylogenetic species and biogeographic implications. Bul!. Brit. Omith. Cluh, v. 1 22, p. 1 85 - 194, 2002.
Padrões e Processos B iogeográficos na Amazônia - 1 97
7. HALL. J. P. w.; HARVEY, D. 1. The phylogeography of Amazonia revisited: new evidence from riodinid butterflies. Evolution, v. 56, p. 1 489- 1497, 2002.
8. PRUM, R. Historical relationships among avian forest areas of endemism in the neotropics. Acta x/x Congressus Intemationalis Ornithologici, v. 19 , p. 2562-2572, 1 988.
9. AMORIM, D. S. Dos Amazonias. In: BOUSQUETS, 1. L.; MORRONEJ. J. (ed.). Introducción a la Biogeografia em LatinoAmérica - Teorias, Conceptos, Métodos yAplicaciones. Mexico: Las Prensas de Ciencias, Facultad de Ciencia" UNAM. 2001 . p. 245-255.
10. CRACRAFT, J. Deep-history biogeography: retrieving the historical pattern of evolving continental biotas. Sys. Zool. , v. 37, p. 221-236, 1 988 .
1 1 . BATES, J. Avian diversification in Amazonia: evidence for historical complexity and a vicariance mo deI for basic diversification pattern. In: VIEIRA, L C. G.: SILVA, 1. M. C.; OREN, D. C. et a!. (ed.). Diversidade Biológica e Cultural da Amazônia. Belém: Museu Paraense Emílio Goeldi. 200 1 . p. 1 19- 137 .
12 . CAMARGO, J. M. F.; PEDRO, S . R. M. Meliponini neotropicais: o gênero Partamona Schwarz, 1 939 (Hymenoptera, Apidae, Apinae) -bionornia e biogeografia. Rev. Bras. Entomol. , v. 47, p. 3 1 1 -372, 2003.
13 . RICKLEFS, R. E.; SCHLUTER, D. Species diversity: regional and historical infiuences. In: RICKLEFS, R. E.; SCHLUTER, D. (ed.). Species Diversity in Ecological Communities. Chicago: The University of Chicago Press, 1993. p. 350-363.
14. SILVA, J. M. C. La importancia relativa de los processos que determinan la diversidad regional de las aves en las grandes regiones ecológicas brasilefias. In: SILVA, H. G. D. ; ITA, A. O. D. (ed.). Conservación de Aves: Experiencias en México. Ciudad deI México: CIPAMEX, 2004. p. 63-67.
15. VAN DER HAMMEN, T. Paleoecology of Amazonia. In: VIEIRA, L C. G.; SILVA, J. M. C.; OREN, D. C. et a!. (ed.). Diversidade Biológica e Cultural da Amazônia. Belém: Museu Paraense Emílio Goeldi, 2001 . p. 1 9-44.
16 . PETRI, S . ; FÚLFARO, V. J. Geologia do Brasil. São Paulo: Editora da Universidade de São Paulo (EDUSP), 1 983. 63 1p.
1 7. KASTNER, T. P.; GONI, M. A. Constancy in the vegetation of the Amazon Basin during the late Pleistocene: Evidence from the organic matter composition of Amazon deep sea fan sediments. Geology, v. 3 1 , p. 291-294, 2003.
1 8 . PATTON, J. L.; SILVA, M. N. F. Molecular phylogenetics and the diversification of Amazonian mammals. In: VIEIRA, L C. G.; SILVA, J. M. C. ; OREN, D. C. et a!. (ed.). Diversidade Biológica e Cultural da Amazônia. Belém: Museu Paraense Emílio Goeldi, 2001 . p. 138- 1 64.
19. GIUGLIANO, L. G.; COLLEVATTI, R. G. ; COLLI, G. R. Molecular dating and phylogenetic relationships among Teiidae (Squamata) inferred by molecular and morphological data. Moi. Phylogenet. Evo/. , v. 45, p. 168- 1 79, 2007.
20. SANTOS, 1. C. ; COLOMA, L. A.; SUMMERS, K. et aI. Amazonian amphibian diversity is primarily derived from late Miocene andean 1ineages. PIos. Biol. , v. 7, p. 448-461 , 2009.
2 1 . PENNINGTON, R. T.; LAVIN, M.; PRADO, D. E. et a!. Historical climate change and speeiation: neotropical seasonally dry forest plants show patterns of both Tertiary and Quaternary diversification. Philos. T. Roy. Soe. B. , v. 359, p. 5 1 5-537, 2004.
22. AVISE, J. C. Molecular Markers, Natural Histo/); and Evolution. Sunderland: Sinauer, 2004. 5 1 1 p.
23. ALEIXO, A. Historieal diversification of a Terra Firme forest bird superspecies: a phylogeographic perspective on the role of different hypotheses of Amazonian diversification. Evolution, v. 58 , p. 1 303- 1 3 1 7, 2004.
24. WEIR, J. T. Divergent timing and patterns of species accumulation in lowland and highland neotropical birds. Evolution, v. 60, p. 842-855, 2006.
25. TOBIAS, J. A.; BATES J. M.; HACKETT, S. 1. et a!. Comment on the latitudinal gradient in recent speciation and extinction rates ofbirds and mammals. Science, v. 3 19, n. 5865, p. 90 1e , 2008.

Fernando César Vieira Zanella
A porção tropical/subtropical da América do Sul, situada a leste dos Andes, compreende extensos biomas cujos limites são determinados principalmente por diferenças nos níveis pluviométricos. Em um esquema simples, pode-se separá-los em dois grupos fisionômicos: florestas tropicais pluviais, em regiões mais pluviosas sem período bem definido de deficiência hídrica, e florestas e formações abertas tropicais estacionais secas, que apresentam estação seca bem definida. As primeiras incluem a Floresta Amazônica e a Floresta Atlântica, esta próxima à costa do Brasil. As outras formam uma ampla faixa que se estende do Nordeste do Brasil ao Noroeste da Argentina, incluindo as províncias biogeográficas da Caatinga, do Cerrado e do Chaco1•2 (Fig. 1 3 . 1 ) . Essa ampla faixa de clima sazonal e restrição hídrica em parte do ano apresentam uma variedade de formações vegetacionais, sendo as de vegetação aberta, na qual grande parte da luminosidade incidente atinge o solo ou a vegetação herbácea, aquelas com maior contraste ecológico e de composição de sua biota em relação às de vegetação floresta densa e fechada, típicas das florestas tropicais úmidas. A ocorrência de um conjunto florístico e faunístico cuja distribuição se estende por essas três províncias, resultou em denominar esse espaço geográfico como diagonal de formações abertas ou diagonal de áreas secas.
Caatinga, Cerrado e Chaco são três províncias biogeográficas que exibem componentes bióticos endêmicos, indicando histórias evolutivas de suas floras e faunas com certo grau de independência. A compreensão dos fatores atuais e passados, que determinam os padrões de
1 3
fi
distribuição, demanda uma variedade de abordagens, o que inclui o estudo da diversidade atual de espécies e seus padrões de distribuição, relações evolutivas entre
elas e características ecológicas dos ecossistemas onde ocorrem, complementado pelas informações sobre alterações geológicas e climáticas do passado, e de mudanças nas características das comunidades locais por meio do estudo de fósseis, especialmente do pólen fóssil de camadas geológicas.
Neste capítulo é apresentada uma síntese do conhecimento atual sobre a evolução da biota da diagonal de formações abertas secas, sendo ressaltadas interpretações aparentemente contraditórias e resultados recentes obtidos em estudos de biogeografia histórica com abelhas. São citados no texto apenas os trabalhos de revisão mais amplos e acessíveis, porém em razão da abrangência do tema é necessário mencionar que o relato manifestado só foi possível pela contribuição de centenas de pesquisadores e que uma relação mais completa das fontes deve ser procurada nas referências citadas.
Morrone3 revisou os padrões de distribuição de grupos de animais e plantas que sucederam na sub-região Chacoana (ou Chaquenha), que, em sua definição, inclui como províncias, além da Caatinga, Cerrado e Chaco, o deserto do Monte e o Pampa. Um total de 1 6 1 táxons com traços biogeográficos representativos dessa sub-região foi relacionado por esse autor, além daqueles representativos de cada província (Tabela 13 . 1) . Como exemplo de um traçado típico da sub-região, ele menciona o da ema, Rhea americana (Linnaeus) (Fig. 1 3 .2).
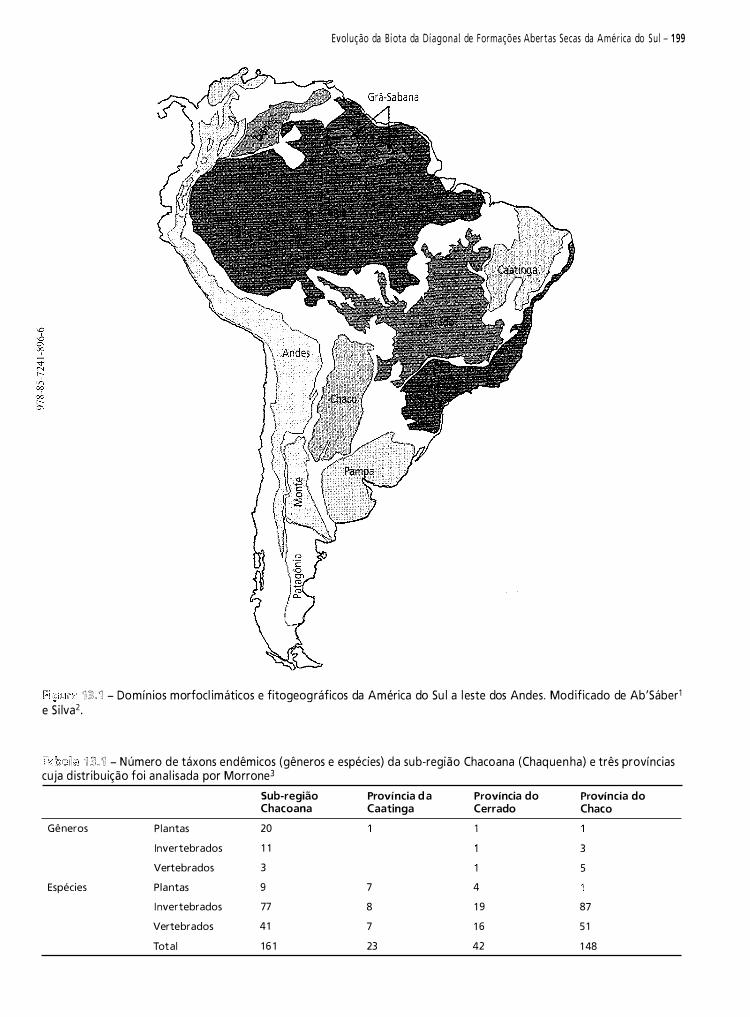
Evol ução da B iota da D iagona l de Formações Abertas Secas da Amér ica do Su l - 1 99
1;31,'] - Domín ios morfocl imáticos e fitogeográficos da América do Su l a l este dos Andes. Mod ificado de Ab'Sáber1 e S i lva2•
'EU - Número de táxons endêmicos (gêneros e espécies) da sub-região Chacoana (Chaquen ha) e três províncias cuja d istr ibu ição foi ana l isada por Morrone3
Gêneros P lantas
Invertebrados
Vertebrados
Espécies P lantas
Invertebrados
Vertebrados
Total
Sub-região Chacoana
20
1 1
3
9
77
41
1 6 1
Província d a Caatinga
7
8
7
23
Província do Cerrado
4
1 9
1 6
42
Província do Chaco
3
5
87
5 1
1 48
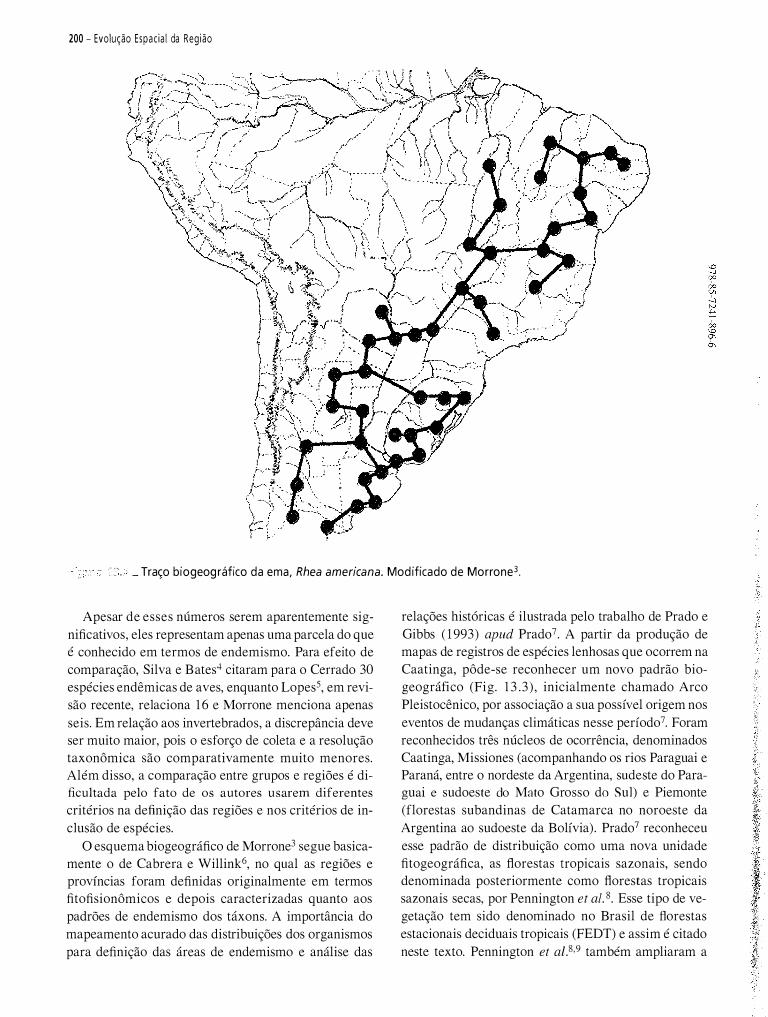
200 - Evolução Espacial da Região
� = , - Traço biogeográfico da ema, Rhea americana. Modificado de Morrone3.
Apesar de esses números serem aparentemente significativos, eles representam apenas uma parcela do que é conhecido em termos de endemismo. Para efeito de comparação, Silva e Bates4 citaram para o Cerrado 30 espécies endêmicas de aves, enquanto Lopes5, em revisão recente, relaciona 16 e Morrone menciona apenas seis. Em relação aos invertebrados, a discrepância deve ser muito maior, pois o esforço de coleta e a resolução taxonômica são comparativamente muito menores. Além disso, a comparação entre grupos e regiões é dificultada pelo fato de os autores usarem diferentes
critérios na definição das regiões e nos critérios de inclusão de espécies.
O esquema biogeográfico de Morrone3 segue basicamente o de Cabrera e Willink6, no qual as regiões e províncias foram definidas originalmente em termos fitofisionômicos e depois caracterizadas quanto aos padrões de endemismo dos táxons. A importância do mapeamento acurado das distribuições dos organismos para definição das áreas de endemismo e análise das
relações históricas é ilustrada pelo trabalho de Prado e Gibbs ( 1 993) apud Prad07. A partir da produção de mapas de registros de espécies lenhosas que ocorrem na Caatinga, pôde-se reconhecer um novo padrão biogeográfico (Fig. 1 3 .3) , inicialmente chamado Arco Pleistocênico, por associação a sua possível origem nos eventos de mudanças climáticas nesse períod07. Foram reconhecidos três núcleos de ocorrência, denominados Caatinga, Missiones (acompanhando os rios Paraguai e Paraná, entre o nordeste da Argentina, sudeste do Paraguai e sudoeste do Mato Grosso do Sul) e Piemonte (florestas subandinas de Catamarca no noroeste da Argentina ao sudoeste da Bolívia). Prad07 reconheceu esse padrão de distribuição como uma nova unidade fito geográfica, as florestas tropicais sazonais, sendo denominada posteriormente como florestas tropicais sazonais secas, por Pennington et a!. 8. Esse tipo de vegetação tem sido denominado no Brasil de florestas estacionais deciduais tropicais (FEDT) e assim é citado neste texto. Pennington et al.8,9 também ampliaram a
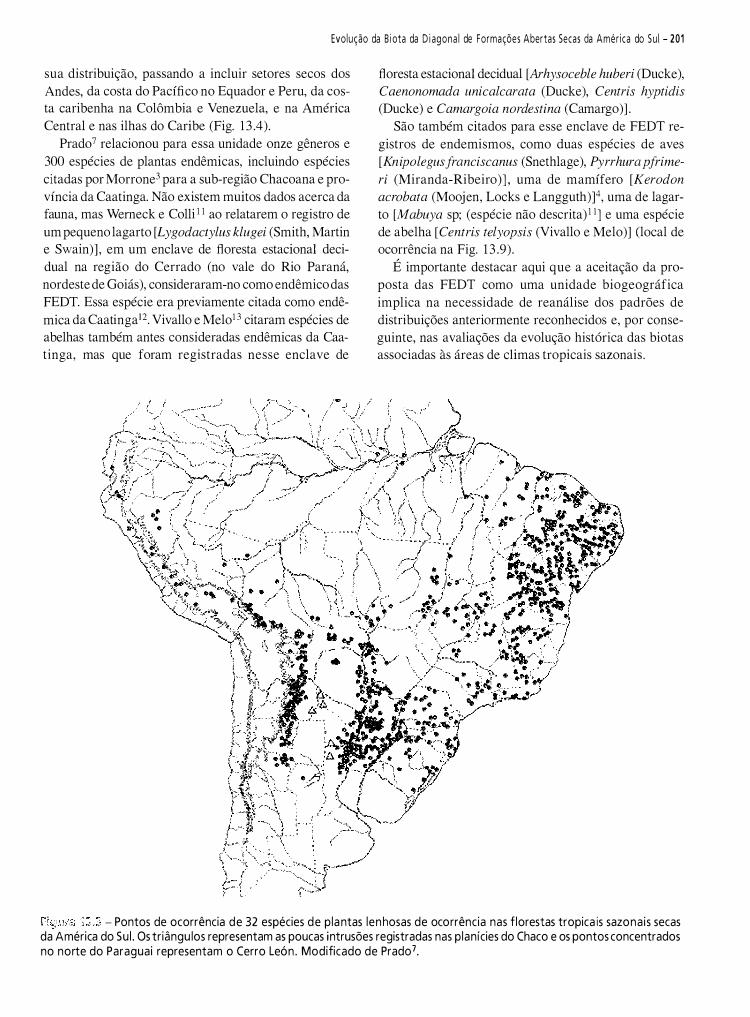
Evolução da B i ota da D i agona l de Formações Abertas Secas da América do Su l - 201
sua distribuição, passando a incluir setores secos dos Andes, da costa do Pacífico no Equador e Peru, da costa caribenha na Colômbia e Venezuela, e na América Central e nas ilhas do Caribe (Fig. 13 .4).
Prad07 relacionou para essa unidade onze gêneros e 300 espécies de plantas endêmicas, incluindo espécies citadas por Morrone3 para a sub-região Chacoana e província da Caatinga. Não existem muitos dados acerca da fauna, mas Werneck e Colli 1 1 ao relatarem o registro de um pequeno lagarto [Lygodactylus klugei (Smith, Martin e Swain)] , em um enclave de floresta estacionaI decidual na região do Cerrado (no vale do Rio Paraná, nordeste de Goiás), consideraram-no como endêmico das FEDT. Essa espécie era previamente citada como endêmica da Caatinga12. Vivallo e Melo1 3 citaram espécies de abelhas também antes consideradas endêmicas da Caatinga, mas que foram registradas nesse enclave de
floresta estacionaI decidual [Arhysoceble huberi (Ducke), Caenonomada unicalcarata (Ducke), Centris hyptidis (Ducke) e Camargoia nordestina (Camargo)] .
São também citados para esse enclave de FEDT registros de endemismos, como duas espécies de aves [Knipolegus franciscanus (Snethlage), Pyrrhura pfrimeri (Miranda-Ribeiro)] , uma de mamífero [Kerodon acrobata (Moojen, Locks e Langguth)]4, uma de lagarto [Mabuya sp; (espécie não descrita) l 1] e uma espécie de abelha [Centris telyopsis (Vivallo e Melo)] (local de ocorrência na Fig. 1 3 .9).
É importante destacar aqui que a aceitação da proposta das FEDT como uma unidade biogeográfica implica na necessidade de reanálise dos padrões de distribuições anteriormente reconhecidos e, por conseguinte, nas avaliações da evolução histórica das biotas associadas às áreas de climas tropicais sazonais.
- Pontos de ocorrência de 32 espécies de plantas lenhosas de ocorrência nas f lorestas tropica is sazona is secas da América do Su l . Os triângu los representam as poucas intrusões registradas nas planícies do Chaco e os pontos concentrados no norte do Paraguai representam o Cerro León . Modificado de Prad07.

202 - Evol ução Espac i a l da Região
A descrição das características predominantes nessas três províncias biogeográficas pode parecer tarefa fácil e formal, resultado simplesmente da compilação dos principais aspectos físicos e da ecologia de suas biotas. No entanto, como será evidenciado a seguir, a literatura, que têm por referência o reconhecimento da diagonal de áreas de formações abertas, a qual se pode referir aqui como tradicional, destaca características opostas às das florestas tropicais úmidas, em especial as diferenças nos
8
níveis pluviométricos e à vegetação aberta. Por outro lado, no novo esquema biogeográfico, que considera as FEDT, a Caatinga é levada em conta como formação eminentemente florestal, em oposição ao Cerrado, de vegetação aberta, com estrato graminoso característico, determinado por particularidades do solo e ocorrência do fog09.
Dentre as três províncias biogeográficas, o Cerrado apresenta maiores valores pluviométricos . Resultam assim, na diagonal de formações abertas, dois polos de maior aridez: a Caatinga, isolada entre áreas mais chuvosas, e o Chaco, vizinho a áreas mais úmidas a leste e oeste, mas representando o ponto extremo a nordeste de
- Áreas de endemismo das f lorestas tropicais sazona is secas da região Neotropical (cinza escuro). 1 = Núcleo Caatinga; 2 = F lorestas Sazonais do Sudeste do Brazi l ; 3 = Núcleo M issiones; 4 = Florestas Ch iqu itanas do norte da Bo l ívia; 5 = Núcleo Piemonte; 6 = Va l es interand inos da Bo l ívia; 7 = Va les interandinos do Peru e Equador; 8 = Costa do Equador e Peru; 9 = Va les interandinos da Colômbia e Venezuela; 1 0 = Costa caribenha da Colômbia e Venezuela; 1 1 = América Centra l e México; 1 2 = I l has do Caribe; 1 3 = Flórida; B = Bol ívia; Ce = Cerrado; Ch = Chaco (cinza intermediário); Co = Costal (Amapá, B R-Rupununi); LI = L1anos; Rr = Rio B ranco-Rupununi; Savanas (cinza claro). Modificado de Pennington et al.9, 1 0.

Evo lução da B i ota da D iagona l de Formações Abertas Secas da América do Su l - 203
um conjunto de áreas secas denominado de Diagonal Arreica, e sendo contíguo ao deserto do Monte, na Argentina. Deve-se chamar a atenção que a área do Chaco considerada aqui (e representada na Fig. 1 3 .4) corresponde ao que se pode chamar de Chaco seco, pois o Chaco em sentido amplo inclui áreas mais úmidas ao leste do Paraguai e província de Missiones, Argentina (redefinição dos limites do Chaco em Prad07).
Se a pluviosidade for o fator ecológico-chave para a determinação da distribuição dos táxons restritos a cada região, é evidente que alterações nessa variável no passado, como o suposto aumento da aridez nos períodos glaciais do Pleistoceno deve ter resultado em modificações no seu padrão de distribuição. É deste modo que têm sido interpretados muitos casos de prováveis disjunções entre a Caatinga e o Chaco, com ausência ou poucos registros dentro do Cerrado.
No entanto, características do solo são também importantes para a ocorrência de vegetação de cerrado. Isto é demonstrado pela existência desse tipo de ecossistema em locais com padrões climáticos muito distintos, incluindo áreas de pluviosidade acima de 2.000mm anuais e áreas próximas à caatinga, com menos de 650mm anuais. Ocorre também em locais próximos vegetações de cerrado e florestas secas, com sutis variações de textura de solo ou com diferentes níveis de acúmulo de água. O Cerrado se encontra majoritariamente sobre o Planalto Central Brasileiro, em locais com 500 a 1 .200m de altitude e os solos quase sempre mais intensamente lixiviados, ácidos e pobres em nutrientes em comparação aos da Caatinga e Chaco, além de apresentarem elevada concentração de alumínio, que é tóxico para plantas J4.
As paisagens típicas de Caatinga e Chaco correspondem a terras baixas, sendo na primeira, em geral, depressões interplanálticas de solos rasos e pedregosos, sem ou com lençol freático de distribuição restrita, mas com setores de solo sedimentar. O Chaco se encontra em ampla planície muito mais uniforme, que corresponde a
::% uma bacia sedimentar de solos de sedimentos finos trans-a, o;' portados pelo vento (loess), profundos e compactada, � quase sem rochas, o que dificulta a infiltração e deixa o .;, o;' lençol freático usualmente fora do alcance das raízes das 00 s:; plantas. A Caatinga e o Cerrado são tipicamente tropicais,
com temperaturas relativamente altas o ano todo. O Chaco, em posição subtropical, apresenta grande variação sazonal, com elevadas temperaturas no verão, que corresponde ao período chuvoso. O verão no Chaco exibe as temperaturas mais altas do continente, chegando a 49°C, mas no inverno ocorrem geadas frequentes8.
Pennington et al.8 , proponentes da unidade fitogeográfica FEDT, reconhecem basicamente dois tipos de
vegetação florística e ecologicamente distintas nas regiões tropicais sazonais, as florestas sazonais secas e as savanas, onde as primeiras apresentam basicamente um porte florestal, com um esparso sub-bosque herbáceo, enquanto as savanas têm um componente herbáceo-graminoso representativo, adaptado a suportar o efeito do fogo. As FEDT são descritas como possuindo porte, área basal e produtividade menores que as florestas tropicais úmidas, com copas relativamente fechadas (no período chuvoso), espécies espinhosas frequentes e estrato herbáceo esparso. Como são florestas decíduas, a luminosidade atinge o solo durante a estação seca, reduzindo a decomposição em momento em que há aumento na deposição do folhiço. Diferentemente do Cerrado, os solos onde ocorrem são descritos como férteis, de pH moderado a alto. No mesmo trabalh08, os autores destacam a existência de um gradiente de formações vegetacionais nas FEDT, incluindo florestas de porte relativamente grande nos locais mais úmidos e formações arbustivas com abundância de cacto nos locais mais secos, resultando em uma variedade de nomes locais (florestas secas tropicais e subtropicais, caatinga, florestas mesotróficas, mesófilas, mesofíticas, semidecíduas ou decíduas, bosque caducifólio , bosque espinhoso). Mais especificamente para a região da Caatinga uma variedade de classificações para a vegetação também foi proposta (caatinga alta, caatinga baixa, Cariri, Curimataú, Sertão, Agreste, Seridó) . O caso do Seridó é interessante por se tratar de uma região de caatinga mais aberta que apresenta um estrato graminoso típico, o que contrasta com a caracterização de Pennington et alo 8. Entretanto, esse aspecto aberto j á foi considerado como resultado da degradação por ação humana, podendo, assim, não ser uma formação natural.
Em relação ao Chaco, Pennington et al. 8 destacaram que se trata da única região do mundo onde a transição entre as zonas tropicais e temperadas não é um deserto, mas uma região semiárida com florestas. Há um estrato arbustivo descontínuo e o herbáceo é esparso. A composição florÍstica é considerada distinta da presente nas FEDT, sendo mais relacionada à do deserto do Monte e a de formações andinas de pré-Puna, possivelmente em decorrência do efeito das frequentes geadas. Desse modo, em termos biogeográficos, é descrita como uma extensão subtropical de formações temperadas. Deve-se destacar aqui que essa interpretação aparentemente contradiz os esquemas biogeográficos que consideram a diagonal de formações abertas.
Para o Cerrado (em sentido estrito), ressaltam a intolerância de sua vegetação ao encharcamento do solo e o importante papel ecológico do fogo como determinante

204 - Evolução Espacia l da Região
da presença de um estrato graminoso, muitas vezes denso, e de vegetação lenhosa com adaptações a esse fator, como casca espessa, raízes subterrâneas e gemas laterais que podem rebrotar quando as apicais são queimadas.
É fundamental ressaltar que todas as províncias da diagonal de formações abertas apresentam vegetação polimórfica, compreendendo fácies mais abertas a formações florestais, além da presença de enclaves de vegetação diversa e de matas de galeria ao longo do curso dos rios. Como a disponibilidade de água no solo é um dos fatores principais para determinação do tipo de vegetação que pode ocorrer em cada local, o relevo, a profundidade e textura do solo, além das variações locais na precipitação, estão relacionados à intensidade de estresse hídrico no período seco e, em consequência, com o tipo de vegetação. No caso da Caatinga, Pennington et al. 8 enfatizam a ausência de um estrato herbáceo significativo. Entretanto, apesar de ainda não existir avaliação abrangente da importância desse componente herbáceo, Costa et al. I5, ao estudarem uma área de caatinga arbórea e caatinga aberta no Ceará, encontraram predominância de espécies herbáceas, em particular terófitas, em relação às espécies arbóreas.
Além disso, há uma paisagem de exceção na Caatinga, que apresenta feição savaniforme, com predominância de gramíneas e outras plantas herbáceas, chamada localmente de arisco. Essa formação ocorre em solos do tipo solonetz solodizado/planossolo nátrico, cuja ocorrência é determinada por condições de encharcamento temporário. A existência de setores savaniformes também é relatada para o ChacoJ 6•
Quando se analisam dados que possam fornecer informações sobre a origem de uma biota é necessário estudar a história evolutiva dos táxons endêmicos e os que apresentam populações disjuntas (no caso de táxons amplamente distribuídos, a abordagem filogeográfica pode ser utilizada para analisar a estruturação genética geográfica, o que seria equivalente ao endemismo na presente discussão). Os grupos que não exibem distribuição restrita à região em análise, seja como espécie ou populações isoladas, não permitem a construção de um relato da sua evolução na biota. Por exemplo, a análise da história evolutiva do grupo das aves ratitas, ao qual pertencem a ema, o avestruz e o kiwi, não fornecerá elementos para apreciação da história do Chaco,
do Cerrado ou da Caatinga, em particular, pois a ema ocorre amplamente em regiões de vegetação aberta, incluindo as três j á citadas e outras mais ao sul. No entanto, pode corroborar a hipótese da existência de formações abertas na América do Sul, ao menos desde a quebra da Gondwana (especialmente a porção mais ao sul, que ocorreu no Cretáceo inferior).
Ademais, interpretações acerca da existência prévia das formações vegetacionais na configuração espacial atual podem não representar exatamente a origem das biotas, uma vez que os diferentes componentes podem ter se originado em condições distintas. Por exemplo, quando a vegetação encontrava-se espalhada em setores específicos da paisagem e não amplamente distribuída, como observado na atualidade.
No caso do Cerrado, os extensos planaltos de terrenos sedimentares onde ocorre devem ter se formado durante todo Terciário, em regiões com vegetação aberta sob climas secos. Do final do Cretáceo à metade do Oligoceno (cerca de 65 milhões de anos atrás [maa] a 28maa), houve uma fase mais úmida, que deve ter promovido a intensa intemperização das rochas e solos. A partir da metade do Oligoceno o clima tornou-se mais frio e seco, quando deve ter se intensificado a formação desses solos sedimentares profundosl4. Para esse momento, é interpretado também que tenha havido expansão de formações vegetacionais abertas, com abundância de ervas adaptadas a condições secas, similares a savanas. Gottsberger e Silberbauer-Gottsbergerl4 assumiram que o desenvolvimento inicial do Cerrado se deu entre 28 e 25maa, no Oligoceno superior. Entretanto, como já colocado, esse pode ter sido o momento de expansão de uma biota representativa de Cerrado para uma configuração espacial próxima à atual, e a origem de seus vários componentes pode ser mais antiga. Esses autoresI4 mencionaram, por exemplo, que, segundo Van der Hammen, algumas gramíneas de savana têm registros históricos até o Eoceno médio, cerca de 50maa.
A origem da vegetação savânica na América do Sul é associada à Província Microfloral Mista CRomero, 1993 apud Gottsberger e Silberbauer-Gottsberger1 4 ; Colli 12), conjunto paleovegetacional predominantemente de vegetação aberta e amplamente distribuído na porção sul do continente desde o final do Cretáceo, que ocorria em condições temperadas e mais secas do que as documentadas para a paleoflora do norte do continente.
A origem do Chaco, em sua configuração espacial atual, é correlacionada à subsidência da extensa planície sedimentar onde acontece, a qual está associada ao intenso soerguimento dos Andes que sucedeu na transição entre o Plioceno e o Pleistoceno, cerca de 3 a 2maa. Esse
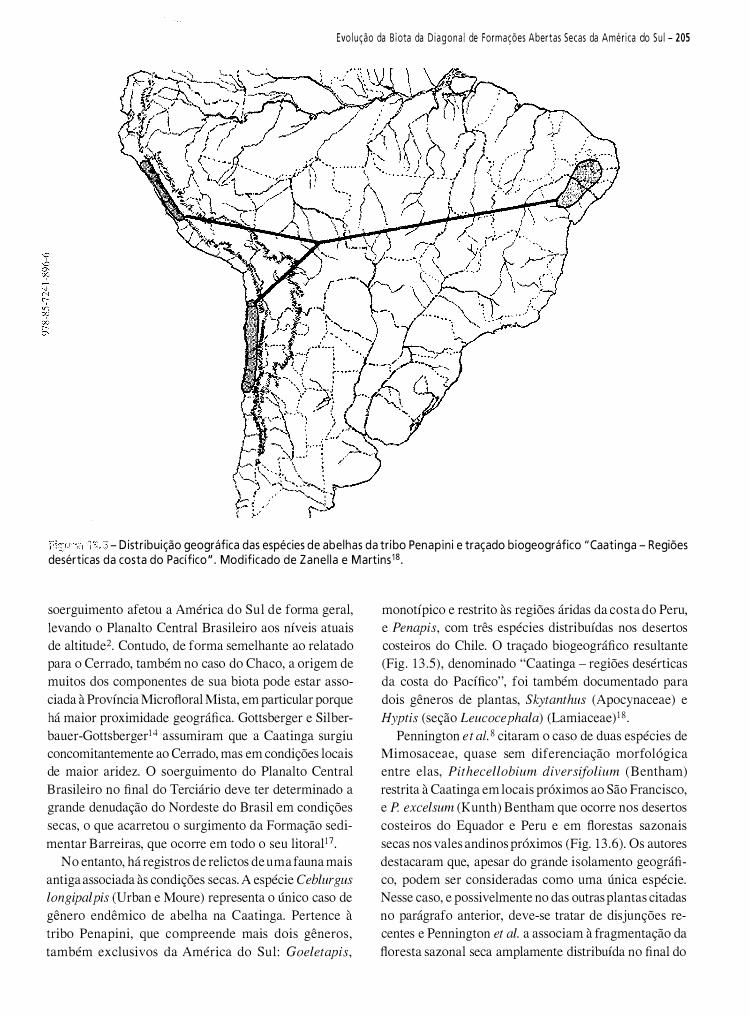
Evo l ução da B iota da D iagona l de Formações Abertas Secas da Amér ica do Su l - 205
- Distribuição geográfica das espécies de abelhas da tribo Penapin i e traçado biogeográfico "Caatinga - Regiões desérticas da costa do Pacífico" . Mod ificado de Zane l la e Martins18 .
soerguimento afetou a América do Sul de forma geral, levando o Planalto Central Brasileiro aos níveis atuais de altitude2. Contudo, de forma semelhante ao relatado para o Cerrado, também no caso do Chaco, a origem de muitos dos componentes de sua biota pode estar associada à Província Microfloral Mista, em particular porque há maior proximidade geográfica. Gottsberger e Silberbauer-Gottsberger14 assumiram que a Caatinga surgiu concomitantemente ao Cerrado, mas em condições locais de maior aridez. O soerguimento do Planalto Central Brasileiro no final do Terciário deve ter determinado a grande denudação do Nordeste do Brasil em condições secas, o que acarretou o surgimento da Formação sedimentar Barreiras, que ocorre em todo o seu litoral17•
No entanto, há registros de relictos de uma fauna mais antiga associada às condições secas. A espécie Ceblurgus longipalpis (U rban e Moure) representa o único caso de gênero endêmico de abelha na Caatinga. Pertence à tribo Penapini, que compreende mais dois gêneros, também exclusivos da América do Sul: Goeletapis,
monotípico e restrito às regiões áridas da costa do Peru, e Penapis, com três espécies distribuídas nos desertos costeiros do Chile. O traçado biogeográfico resultante (Fig. 1 3 .5), denominado "Caatinga - regiões desérticas da costa do Pacífico", foi também documentado para dois gêneros de plantas, Skytanthus (Apocynaceae) e Hyptis (seção Leucocephala) (Lamiaceae) 1 8 .
Pennington et aI. 8 citaram o caso de duas espécies de Mimosaceae, quase sem diferenciação morfológica entre elas, Pithecellobium diversifolium (Bentham) restrita à Caatinga em locais próximos ao São Francisco, e P excelsum (Kunth) Bentham que ocorre nos desertos costeiros do Equador e Peru e em florestas sazonais secas nos vales andinos próximos (Fig. 13 .6). Os autores destacaram que, apesar do grande isolamento geográfico, podem ser consideradas como uma única espécie. Nesse caso, e possivelmente no das outras plantas citadas no parágrafo anterior, deve-se tratar de disjunções recentes e Pennington et al. a associam à fragmentação da floresta sazonal seca amplamente distribuída no final do

206 - Evo lução Espac ia l da Região
Pleistoceno. No caso das abelhas, a diferenciação observada entre os gêneros implica em fragmentação mais antiga. A hipótese de um antigo evento de vicariância em grupos associados às regiões secas anteriores à formação dos Andes, com representantes na Caatinga e em desertos secos da costa do Pacífico, foi corroborada recentemente por Danforth et al. 19 , que, utilizando marcadores moleculares para estudarem as relações históricas entre espécies de abelhas Halictidae, chegaram à estimativa de divergência entre Goeletapis e Penapis anterior a 50maa. Apesar de não ter sido incluída na análise a espécie de Ceblurgus, é provável que a separação deste ramo em relação ao ancestral dos Penapini da América do Sul tenha sido anterior ao de Goeletapis + Penapis, tanto em razão da maior divergência morfológica, como do maior isolamento geográfico. Dessa forma, esse resultado amplia as evidências acerca da existência de uma fauna antiga na América do Sul adaptada às condições xéricas. Também considerados como parte dessa fauna, estão os representantes do componente erêmico, assim denominado por Kuschel ( 1964, in
Roig-Jufíent et al. 20), que exclui grupos sem parentes próximos em outros continentes gondwânicos. No caso dos Penapini, o possível grupo irmão ocorre na África.
Um possível representante do componente erêmico com presença na Caatinga é o do gênero de escorpiões Bothriurus Peters. Segundo Roig-Jufíent et aI. 20, a separação desse ramo evolutivo em relação ao das outras espécies que compõem o gênero, que se distribuem nas áreas secas do cone sul, teria acontecido antes do evento inicial de formação dos Andes, que se deu cerca de 37 a 25maa. Esses autores analisaram as relações históricas entre regiões secas da Amética do Sul a partir dos cladogramas biológicos de área de táxons pertencentes aos elementos do componente erêmico da fauna de artrópodes da América do Sul . A metodologia usada, denominada Análise de enunciados de 3-áreas (TASS), é implementada por meio de um programa de computador que permite extrair informações não ambíguas de relacionamento histórico entre áreas a partir de cladogramas complexos (ver Capo 6). Dentre os grupos analisados somente um possuía representantes na Caatinga, o gênero
- Distr ibuição de PítheceJlobíum díversífolíum e P. excelsum. Mod ificado de Penn ington et al.s.

Evo l ução da B iota da D iagonal de Formações Abertas Secas da América do Su l - 207
de escorpiões Bothriurus Peters. O consenso estrito obtido, que apresentou a Caatinga como grupo irmão do conjunto das outras regiões, só foi obtido após a eliminação das áreas de Cerrado e Llanos Chaquenhos. Esses registros de artrópodes ressaltam a natureza relictual de alguns componentes da biota da Caatinga e a antiguidade de uma fauna adaptada às condições xéricas.
Outro caso em que se pode interpretar como semelhante é o do gênero de abelhas Protomeliturga. Esse grupo é o único representante da tribo Protomeliturgini, que constitui o grupo irmão dos Calliopsini, grupo também restrito ao Novo Mundo, mas bem diversificado, apresentando 1 5 gêneros e pelo menos 93 espécies distribuídas entre o Canadá e o Chile e Argentina2! . Há somente duas espécies de Protomeliturga: P turnerae Ducke, endêmica do Nordeste do Brasil, mas com registros em setores mais úmidos do litoral do Nordeste do Brasil além da Caatinga, e P catimbaui Schlindwein e Moure, conhecida apenas da localidade-tipo, no vale do C atimbau, setor de altitude relativamente alta no interior da Caatinga, Estado de Pernambuco. Apesar de P turnerae não ser restrita à Caatinga, ela ocorre apenas em locais de vegetação aberta, sendo especializada na coleta de pólen de plantas herbáceas da família Turneraceae. Desse modo, apesar de não ser restrita aos ambientes xéricos, a sua ocorrência marginal nos domínio de Floresta Atlântica e no Maranhão, pode ter sido resultado de expansão posterior, relacionada à derrubada da floresta pela ocupação humana. Assim, poderia também ser interpretada como um registro da ocorrência antiga de vegetação aberta no Nordeste do Brasil.
Neste item, são relatados os achados que podem ser associados aos eventos de vicariância da biota da diagonal de formações abertas anteriores aos eventos climáticos do Quaternário. São descritas as interpretações disponíveis na literatura, sendo destacadas possíveis inconsistências, relacionando os padrões que originaram essas interpretações aos padrões correlatos observados em outros trabalhos, mesmo que não tenham sido originalmente comparados. Deve-se destacar aqui que as evidências para as inferências acerca do momento de ocorrência dos eventos ainda são poucas e dados independentes, como metodologias com datação molecular, podem servir para corroborar ou não as interpretações apresentadas.
Silva2 reconheceu o soerguimento do Planalto Central Brasileiro e a subsidência do Chaco e áreas próximas, ocorrido próximo à transição entre Plioceno e Pleistoceno, como o evento determinante para a vicariância de grupos de aves que hoje se encontram com distribuição disjunta, com espécies no Cerrado e/ou Cadeia do Espinhaço, e em áreas de vegetação aberta mais ao sul, como Chaco, Pampa, Patagônia e setores subandinos. Ele avaliou as espécies de aves consideradas endêmicas do domínio do Cerrado (incluindo as terras altas do Espinhaço), de ocorrência restrita aos ambientes de vegetação aberta, e os dados disponíveis sobre a distribuição de grupos próximos filo geneticamente. Das seis espécies que apresentam parentes em outras regiões, de forma disjunta, três são restritas ao Espinhaço [Asthenes luizae Vielliard, Embernagra longicauda Strickland, Augastes scutatus (Temminck)] e as outras ocorrem amplamente no Cerrado, incluindo o setor do Espinhaço [Geobates poecilopterus (Wied), Melanopareia torquata (Wied-Neuwied) e Poospiza cinerea Bonapartel Os parentes próximos de cinco dessas espécies acontecem em áreas de vegetação aberta do cone sul, incluindo Chaco, Pampa, Patagônia e setores subandinos.
No entanto, sua análise é comprometida, pois, segundo Vasconcelos22, a biota das serras do leste do Brasil, incluindo a cadeia do Espinhaço, apresenta uma história independente da do Cerrado e deveria ser analisada separadamente. Além disso, Silva e Bates4 classificaram duas das três espécies de ampla ocorrência no Cerrado como neoendemismos, assumindo, a partir de inferências de estudos moleculares, que a divergência com suas possíveis espécies irmãs teria ocorrido após 1 ,8maa, posteriormente, portanto, ao momento assumido da ocorrência do evento de subsidência do Chaco.
Por sua vez, Colli 12 relacionou a divergência entre grupos da herpetofauna que sucedeu no Cerrado com parentes próximos nas áreas mais ao sul do continente a uma grande transgressão marinha ocorrida no Mioceno, citando como exemplo, os lagartos Tupinambis merianae (Duméril e Bibron) e T. duseni Lonnberg versus T. rufescens GÜnther. Essa transgressão se deu de forma cíclica e na sua maior extensão cobriu a planície do Chaco, parte do Pampa e o norte da Patagônia23 (Fig. 1 3 .7).
Porzecanski e Cracraft24, em análise de dados de distribuição das espécies de aves de regiões secas da América do Sul, na qual enfocaram especialmente as relações históricas entre as áreas de endemismo da diagonal de formações abertas, chegaram à hipótese de relacionamento histórico mais recente entre as avifaunas do Cerrado e do Chaco, com a Caatinga representando
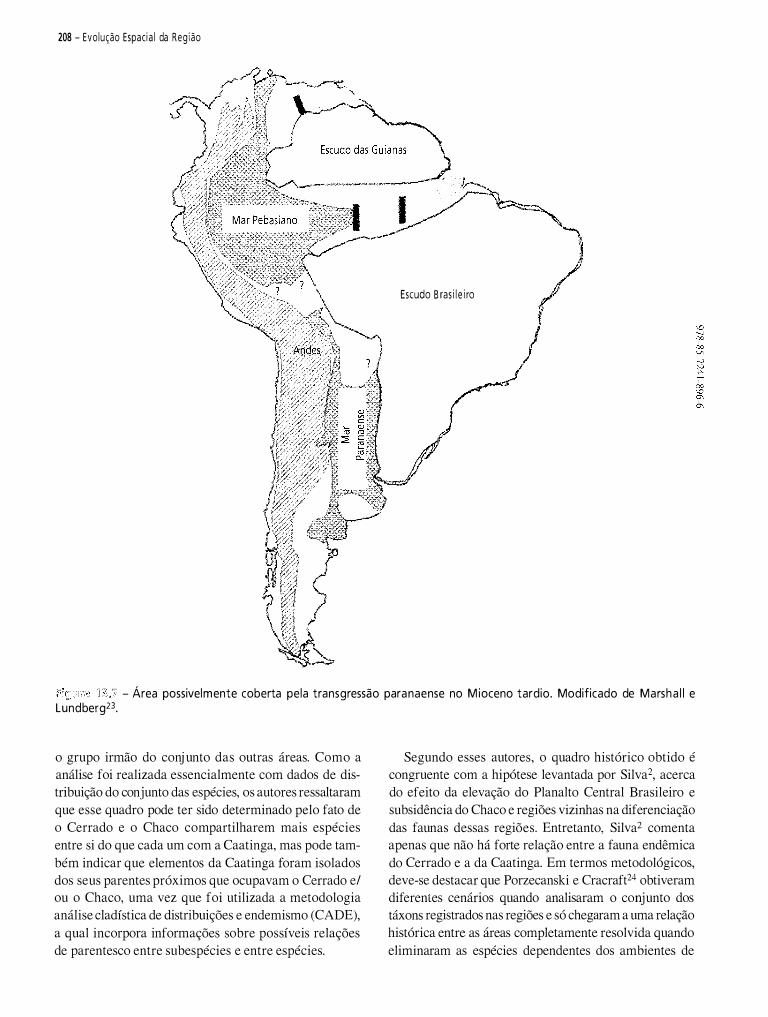
208 - Evolução Espacia l da Região
Escudo B ras i l e i ro
� - Área possivelmente coberta pela transgressão paranaense no M ioceno tardio. Modificado de Marsha l l e Lundberg23.
o grupo irmão do conjunto das outras áreas. Como a análise foi realizada essencialmente com dados de distribuição do conjunto das espécies, os autores ressaltaram que esse quadro pode ter sido determinado pelo fato de o Cerrado e o Chaco compartilharem mais espécies entre si do que cada um com a Caatinga, mas pode também indicar que elementos da Caatinga foram isolados dos seus parentes próximos que ocupavam o Cerrado e/ ou o Chaco, uma vez que foi utilizada a metodologia análise cladística de distribuições e endemismo (CADE), a qual incorpora informações sobre possíveis relações de parentesco entre subespécies e entre espécies.
Segundo esses autores, o quadro histórico obtido é congruente com a hipótese levantada por Silva2, acerca do efeito da elevação do Planalto Central Brasileiro e subsidência do Chaco e regiões vizinhas na diferenciação das faunas dessas regiões. Entretanto, Silva2 comenta apenas que não há forte relação entre a fauna endêmica do Cerrado e a da Caatinga. Em termos metodológicos, deve-se destacar que Porzecanski e Cracraft24 obtiveram diferentes cenários quando analisaram o conjunto dos táxons registrados nas regiões e só chegaram a uma relação histórica entre as áreas completamente resolvida quando eliminaram as espécies dependentes dos ambientes de

Evolução da B iota da D iagona l de Formações Abertas Secas da Amér ica do Su l - 209
floresta que ocorrem no Cerrado. Essa interferência é justificada, pois a avifauna associada aos ambientes florestais deve ter história diferente da de ambientes abertos e analisá-las conjuntamente não permite reconhecer a história de cada grupo.
Grupos de abelhas recentemente estudados, que são exclusivos da América do Sul e com espécies distribuídas ao longo da diagonal de formações abertas, apresentaram de forma consistente, o par de espécies presentes no Cerrado e Chaco compartilhando uma história evolutiva mais recente, em relação à espécie presente na Caatinga (ver a seguir), de forma consistente ao encontrado por Porzecanski e Cracraft24.
No caso de Caenonomada, são reconhecidas três espécies: C. bruneri Ashmead, de distribuição mais meridional, sobre todo o Chaco e o Pampa; C. labrata Zanella, do Brasil central e C. unicalcarata, que previamente era considerada endêmica da Caatinga, mas, após revisão, sendo registradas populações meridionais, entre o Mato Grosso e o norte da Argentina25 (Fig. 13.8). Recentemente, Aguiar e Mel026 fizeram novos registros de locais intermediários, na Bolívia e Mato Grosso do Sul, e interpretaram que essas populações do sul, entre o Mato Grosso e o norte da Argentina, constituiriam uma outra espécie, próxima a C. unicalcarata.
A decisão de considerá-las ou não como uma espécie distinta não muda a interpretação histórica apresentada por Zanella25 , de que um antigo evento de vicariância separou o ancestral de Caenonomada em uma população na Caatinga e outra correspondendo ao ancestral de C. bruneri + C. labrata, mais ao sul. Posteriormente, um outro evento separou a população ancestral destas duas espécies. A presença de uma população de C. unicalcarata mais ao sul, em simpatria parcial com as outras duas espécies foi interpretada como resultado da expansão das formações abertas xerófilas ocorridas nos períodos glaciais do Quaternário.
Não foi indicado nesse trabalho nenhum possível evento geológico associado aos dois eventos de vicariância. Caso se assuma a proposição de Silva2 do soerguimento do Planalto Central Brasileiro como evento importante na diferenciação das biotas do Cerrado e Chaco, fica em aberto a questão do evento que determinou a vicariância anterior, separando uma população no interior do Nordeste do Brasil. Não há citações de um outro evento geológico ou climático anterior, mas há a possibilidade desse processo de soerguimento ter determinado as duas divisões, uma vez que esse amplo soerguimento das superfícies da América do Sul também
- Distribuição das espécies de Caenonomada e relação histórica entre e las . Mod ificado de Zanel la25.

210 - Evo l ução Espacia l da Reg ião
afetou a região hoje ocupada pela Caatinga, pois determinou uma intensa denudação dos terrenos.
O grupo hyptidis do gênero de abelhas Centris compreende três espécies, em que C. hyptidis é considerada endêmica na Caatinga, com expansões marginais para o leste e sudoeste; C. h)ptidoides Roig-Alsina ocorre no Chaco, e C. thelyopsis Vivallo e Melo, recentemente descrita do nordeste de Goiás, um local com um enclave de FEDT no interior do domínio do Cerrado 13 (Fig. 13 .9). Nesse trabalho não foi fornecida uma hipótese de relação histórica entre as espécies, porém, de acordo com a filogenia apresentada por Vivallo-Martinez27, o estado de caráter citado como autapomorfia de C. hyptidoides, mancha supraclipeal de forma retangular, está também presente em C. te lyopsis13 . Dessa forma, esse estado pode ser considerado uma sinapomorfia do dado formado por essas duas espécies. Assim, se confirmada essa interpretação, tem-se hipotetizado um quadro histórico semelhante ao descrito para Caenonomada.
A distribuição do gênero Tapinotaspoides foi também associada por Aguiar e Melo26 à diagonal de formações
abertas. Com sete possíveis espécies, o dado formado por T rufescens (Friese), T aff. nifescens, T tucumana (Vachal) e T serraticornis (Friese), apresenta distribuição e hipótese de relacionamento histórico semelhantes às observadas para Caenonomada, com a primeira espécie de distribuição similar a C. unicalcarata, e as duas últimas, com distribuição e relacionamento histórico comparável a C. bruneri e C. labrata. Tapinotaspoides aff. rufescens corresponde às populações meridionais de T nifescens, como também ocorre com às populações meridionais de Caenonomada unicalcarata, interpretadas como uma espécie distinta por Aguiar e Mel026. Um aspecto distintivo da biogeografia desse grupo é a presença de uma espécie em áreas de vegetação aberta no norte da América do Sul. A posição basal desse ramo evolutivo na história do grupo evidencia que ocorreu um antigo intercâmbio de fauna entre essa região e as áreas similares do nordeste e sul do continente26.
Como exemplos de possível relacionamento histórico entre táxons de ocorrência no Cerrado e no Chaco podem-se mencionar ainda os pares: Geotrigona mombuca
- Registros de locais de ocorrência de espécies de abe lhas Centris g rupo hyptidis. Mod ificado de Viva l l o e

Evo l u ção da B iota da D iagona l de Formações Abertas Secas da América do Su l - 2 1 1
(Smith) - Geotrigona argentina Camargo e Moure e Paratrigona lineata lineata (Lepeletier) - Paratrigona lineata glabella Camargo e Moure. Esses grupos de abelhas sociais de ninhos subterrâneos foram estudados por Camargo e Moure28, que levantaram a possibilidade de um evento de vicariância como fator para a diferenciação. É interessante ressaltar aqui que em decorrência da grande similaridade entre elas, o que torna difícil o reconhecimento como entidades distintas, o evento deve ter sido mais recente que o assumido para Caenonomada bruneri + C. labrata.
Deve ser ainda mencionado que após a formação do Istmo do Panamá, a partir de 3 a 3,5maa, que conectou por via terrestre a América do S uI a América Central e do Norte (ver Capo 1), ocorreu um intercâmbio de elementos bióticos, com vários grupos de origem setentrional tendo ocupado espaços da diagonal de formações abertas e até mesmo se diversificado. São citados como exemplos desses grupos: cervos, felinos, canídeos, porcos-da-mato (Tayassuidae) e serpente cascavel (Crotalus durissus L.), abelhas mamangavas sociais (Bombus) e abelhas solitárias do gênero Anthidium, bem como grupos atualmente extintos, como tigre de dentes de sabre (Smilodon), cavalos, ursos, elefantídeos e camelídeos, que faziam parte da megafauna pleistocênica29.3o, Os fatores que resultaram na diversificação de alguns desses grupos na América do Sul ainda não estão bem estudados. Admite-se que houve duas rotas para a invasão, uma pelas terras altas dos Andes e outra pelas terras baixas do leste do continente, neste caso sendo invocado o papel dos períodos glaciais na ampliação das formações vegetais abertas na bacia amazônica para permitir expansão para o sul dos organismos adaptados a esses ambientes. É interessante mencionar que a rota andina não permitiu a passagem de répteis, por não se adaptarem às baixas temperaturas, sendo utilizada sem tanta restrição por animais de sangue quente31 .
São relatados quatro períodos glaciais no Quaternário, quando sucedem diminuições generalizadas nas temperaturas, nos níveis pluviométricos e, em consequência, alterações nas paisagens, com a expansão das vegetações sazonalmente secas (do tipo savana, vegetação xerófila, florestas estacionais decíduas ou semidecíduas) e retração
das florestas pluviais32. Mais recentemente, tem-se destacado o papel da redução dos níveis de CO
2 nos períodos
glaciais na determinação dos tipos de vegetação, por ampliar o déficit hídrico por meio da lirni tação de carbon033,34. ° último período glacial, considerado o mais intenso, terminou há cerca de 10 mil anos (final do Pleistocen035; mas ver também Wuster et az. 36, que apresenta hipótese divergente de que o último ou os dois últimos foram menos intensos). Nos períodos interglaciais, como este em que nos encontramos atualmente, houve uma expansão das florestas tropicais pluviais e retração das vegetações sazonalmente secas. As avaliações das mudanças ocorridas nos períodos glaciais no conjunto do espaço geográfico da América do Sul têm se restringido ao último pico glacial, mesmo assim ainda há muita divergência, por exemplo, sobre a natureza das formações vegetais que ocuparam a Amazônia, em quais setores sucedeu essa substituição da floresta pluvial, se houve um cOlTedor de vegetação xerófila na parte central do Cerrado possibilitando trocas bióticas entre a Caatinga e o Chaco, e quanto ao papel dessas mudanças na diversificação da biota8,14,37,38.
Tem-se aventado a hipótese de uma ampla coalescência entre as biotas xéricas da Caatinga e do Chaco, inclusive com propostas de mapeamento da sua distribuição no último pico glacial do Pleistoceno (Fig. ] 3. 10) . No esquema biogeográfico de Cabrera e Willink6, essa relação biótica é expressa pela inclusão da Caatinga na região Chaquenha, como uma província disjunta, enquanto o Cerrado foi incluído na região Amazônica.
No entanto, ao mapear a distribuição de espécies lenhos as da Caatinga, Prado e Gibbs ( 1993, in Prad07) não encontraram suporte para essa suposta conexão biogeográfica, pois as espécies de plantas lenhosas citadas como exemplos de uma disjunção Caatinga-Chaco em realidade não ocorrem na área seca do Chaco, sua distribuição está limitada a setores mais úmidos periféricos ou somente acontecem na área seca nas zonas de influência de cursos d'água (Fig. 13 .3). Segundo a distribuição atual das espécies de FEDT há um setor de maior proximidade entre os núcleos da Caatinga e de Missiones, via sudeste do Brasil (Fig. 13.4). Prad07 ressaltou a possibilidade de uma via alternativa de contato através do Cerrado, mais especificamente, do oeste da Bahia, aos afloramentos de calcáreo do Distrito Federal e áreas próximas, depois a leste de Goiânia, no Mato Grosso de Goiás, e áreas de Xavantina/ Cachimbo e Ilha do Bananal até o norte de Cuiabá e, por fim, nas elevações calcáreas de Corumbá e Urucum no Mato Grosso do Sul, aíjá próximo das florestas Chiquitanas na Bolívia (Figs. 13 .3 e 13 . 10).
Em relação às florestas do Chaco seco, Pennington et al.8 ressaltam que apresentam maior afinidade florística

212 - Evolução Espacial da Região
lorestas pluvia is tropicais
orestas de araucária
Florestas temperadas d e a lt itude
� �� , ',' "'; - Distr ibu ição h ipotética dos domín ios fitomorfocl i máticos da América do Su l ao l este dos Andes no ú lt imo p ico g lac ia l , entre 18 e 13 mi l a nos atrás. Poucos rios estão representados, a penas para i l u strar seu possível papel como corredor para e lementos da biota. Mod ificado de Ab'Sáber35.
com a vegetação do deserto temperado de Monte na Argentina e as regiões altoandinas e não com a Caatinga, o que seria determinado climaticamente pelas frequentes ocorrências de geadas. Segundo esses autores a assumida ligação entre Caatinga e Chaco, em sentido estrito, seria um erro de interpretação determinado pelas similaridades fitofisionômicas e na falta de análises florísticas.
Ao rever as possíveis relações históricas entre a Caatinga e o Chaco, Prado e Gibbs ( 1993) apud Prad07 analisaram somente táxons de plantas lenhosas. Não foram encontradas análises biogeográficas de plantas não lenhosas, mas dados de distribuição de abelhas solitárias, que dependem de plantas herbáceas e apresentam um traço biogeográfico semelhante, têm resultado em mapeamento um pouco distinto do observado para as plantas lenhosas. Nesses casos, as espécies ou populações disjuntas de distribuição meridional dentro da diagonal de
formações abertas aparentemente ocorrem nas planícies do Chaco secol6. Esse é o caso de Caenonomada bruneri (ver Fig. 1 3 .9), Centris hyptidoides (ver Fig. 1 3 .8) e Tapinotaspoides aft. rufescens. Como essas abelhas são especializadas na sua relação com as fontes de recursos florais, em especial no caso de pólen e óleo, é provável que entre as plantas hospedeiras apresentem um padrão de distribuição similar. Assim, deve-se encontrar no Chaco seco elementos tropicais misturados a elementos temperados e o traço biogeográfico Caatinga-Chaco resulta sobreposto, em parte ao traço que une os núcleos Caatinga-Missiones dentro do mapeamento das FEDT.
De modo distinto, Gottsberger e Silberbauer-Gottsberger14 postularam que a porção central do Brasil se manteve coberta predominantemente por cerrado. Essa interpretação baseou-se nos estudos da distribuição da megafauna pleistocênica da América do Sul realizados

Evo l ução da B iota da Diagona l de Formações Abertas Secas da América do Su l - 2 1 3
por Ochsenius ( 1 982) apud Gottsberger e Silberbauer-Gottsberger14. No entanto, a interpretação da distribuição dos paleoambientes apresentada por esse autor39 ressalta apenas que a área coberta pelo Cerrado no Pleistoceno tardio deve ter sido muito maior do que ocupada pela Caatinga, o que seria evidenciado pelo registro de animais pastadores em áreas atualmente ocupadas por florestas pluviais tropicais.
A ausência de camelídeos fósseis no Brasil centraP9 poderia ser interpretada como indicadora da não coalescência de biotas de ambientes semiáridos nessa região durante o pico glacial, já que esse grupo de animais pastadores é considerado indicador de ambientes áridos e semiáridos (ver Capo 1 1 ) . Todavia, é possível que a ausência de fósseis seja apenas decorrente de impedimentos para o processo de fossilização, uma vez que não são conhecidos sítios fossilíferos de vertebrados no Brasil central (Báez e Gasparini, 1 979 e Estes e Báez, 1 985 apud Colli 12). Além disso, o registro de fósseis do camelídeo Macrauchenia em áreas disjuntas como Argentina, Sul e Sudeste do Brasil, no Nordeste do Brasil e na região de Guajira (Venezuela e Colômbia) permite sugerir que deve ter ocorrido em algum momento nas áreas intermediárias, mesmo que não haja registro fóssil.
Prado e Gibbs ( 1 993) apud Prad07 assumiram que o padrão de disjunção na distribuição das espécies observados nas FEDT é resultante da retração na distribuição das espécies após o último período glacial. Posteriormente, Pennington et al. 10 avaliaram a hipótese de que gêneros de plantas com espécies distribuídas em setores das FEDT pudessem ter se diversificado por alopatria resultante de eventos de vicariância durante o Pleistoceno. Para isso, selecionaram vários táxons de diferentes famílias, de acordo com o seu padrão de distribuição, e realizaram análises moleculares de parentesco calibradas temporalmente por meio de registros fósseis e/ou de eventos geológicos. O resultado obtido, porém, não corroborou a hipótese de diversificação recente (anterior a 1 ,8maa), especialmente na América do Sul, sendo postulado para quatro gêneros que o início da diversificação se deu entre 20 e l l maa, muito anterior, portanto, ao Pleistoceno e seus ciclos glaciais.
Distintamente do resultado para a América do Sul, foram reconhecidas espécies de origem pleistocênica na América Central, bem como de espécies de florestas úmidas derivadas de grupos associados às florestas estacionais deciduais tropicais; contudo, como ressaltado por Pennington et ai. lO, a ausência de registros de diversificação pleistocênica nas FEDT na América do Sul não implica necessariamente na inexistência de alterações na distribuição dos táxons e de mudanças nas paisagens
durante os ciclos climáticos do Pleistoceno, mas que essas mudanças podem simplesmente não ter resultado em especiação. De qualquer forma, a ocorrência de populações isoladas de muitas espécies nas várias áreas de FEDT na América do Sul e Central é relacionada como evidência de distribuição prévia contínua.
Em relação à bacia amazônica, tem havido intenso debate acerca do tipo de vegetação que a ocupou nos períodos glaciais. Enquanto alguns autores assumem que a floresta pluvial tropical teria sido substituída predominantemente por vegetação aberta de savana (Ochsenius39; Webb e Rancy, 1 986 apud Colinvaux et a!. 33; Gottsberger e Silberbauer-Gottsberger14), outros sustentam que a área central da bacia manteve cobertura florestal, tendo as savanas ocupado apenas áreas marginais, ao norte e ao SUP3.34,38. E essa cobertura florestal pode ter sido, em boa pmte, de florestas estacionais deciduais ou sernideciduaiss,34.
Colinvaux et al. 33 (e trabalhos anteriores) relataram dois estudos de paleopalinologia interpretados como evidências fortes para a hipótese de que a região se manteve amplamente florestada durante os períodos glaciais: um de sedimentos da Lagoa Pata, na Amazônia ocidental, e outro da foz do Rio Amazonas, depositados em período glacial. O fato de a manutenção da cobertura florestal, porém, não significa que a floresta se manteve nas mesmas condições, pois eles registraram a ocorrência de espécies de árvores intolerantes ao calor, como Podocarpus, que hoje somente são encontradas em locais de maior altitude. Além disso, eles revisaram criticamente as supostas evidências apresentadas antes como suporte à hipótese de uma ampla distribuição prévia de vegetação de savana na Amazônia: ( 1 ) a ocorrência de campos de dunas em regiões fora da bacia amazônica supostamente ativos em períodos glaciais, o que permitiria supor, de forma generalizada, o acontecimento de um período mais seco na América do Sul tropical; (2) a existência de grandes depósitos de areia arcósica nos sedimentos da foz do Amazonas, um tipo de material que teria se degradado rapidamente em condições tropicais úmidas, cuja origem foi datada para o período glacial; (3) as linhas de pedra no solo, como supostos paleopavimentos formados em condições xéricas; (4) as extensas áreas de areias brancas no interior da Amazônia, interpretadas como de origem eólica; (5) a ocorrência de descontinuidades na distribuição geográfica de táxons ocorrentes na bacia e associados à floresta pluvial; (6) o registro de uma megafauna pleisto cênica no oeste da Amazônia, composta por animais pastadores adaptados a uma vegetação de savana, incluindo preguiças gigantes, tatus gigantes (gliptodontes), toxodontes, mastodontes e camelídeos.

214 - Evolução Espac i a l da Reg ião
Em síntese, os argumentos dos autores são: (1) não há evidências substanciais de que os campos de dunas efetivamente documentados, na região dos Llanos (Venezuela) e no nordeste do Brasil, próximo à porção média do Rio São Francisco, tenham estado ativos no último período glacial. E no caso do nordeste do Brasil, dados paleopalinológicos permitem interpretar que houve uma época mais úmida no final do Pleistoceno; (2) os depósitos de areia arcósica teriam origem fora da bacia, a partir do transporte de sedimentos dos Andes, ou da calha do Rio Amazonas, quando ela teria sido escavada profundamente por ocasião do rebaixamento do nível dos oceanos no período glacial; (3) o material de linhas de pedra observadas na bacia examinado foi interpretado como sendo concreções formadas no interior do próprio solo e, portanto, sem relação com paleossolos de ambientes xéricos, e também não há evidências de que tenham se formado no Pleistoceno; (4) as areias brancas teriam se formado em condições úmidas durante o Terciário; (5) no caso das descontinuidades biogeográficas, ressaltam que a suposta substituição da floresta por vegetação de savana é apenas uma proposta de explicação para os padrões encontrados. Desse modo, esses padrões não podem ser utilizados como evidência da expansão dessa vegetação aberta na região, uma vez que eles podem ter sido produzidos de outra forma; (6) em relação ao registro de animais fósseis pastadores, afirmam que a sua ocorrência não implica no acontecimento de vegetação de savana de forma ampla, podendo eles ter apresentado uma distribuição limitada aos vales dos rios, onde ocorreriam setores de vegetação baixa.
Posteriomente, Pennington et al. 8 reinterpretaram os dados palinológicos exibidos por Colinvaux et ai. 33, sugerindo que 32 dos 40 gêneros listados para o período glacial ocorrem também nas florestas estacionais deciduais tropicais e que os poucos representantes exclusivos de florestas úmidas poderiam ter sucedido apenas em florestas de galeria. Desse modo, os dados não permitiriam desconsiderar a hipótese de que no pico glacial a Floresta Amazônica tenha sido substituída em boa parte por florestas estacionais secas , seja deciduais ou semideciduais. Mayle et al. 34 avaliaram a possibilidade de se distinguir palinologicamente esses dois tipos de florestas, mas descartaram o uso da abundância de pólen de árvores da família Moraceae e de gramíneas como indicadores, sugerindo que a presença de pólen de árvores do gênero Anadenanthera (Mimosaceae) poderia ser interpretada como sinal da ocorrência de florestas estacionais. Reavaliando os dados da Lagoa Pata, os autores34 concluíram que, em razão da ausência desse tipo de pólen, não teria acontecido florestas estacionais na região durante o
período glacial. De modo complementar, por meio de modelagem da dinâmica da vegetação sob diferentes condições climáticas e de concentração de CO2, May le et al.34 obtiveram um quadro que corrobora a hipótese de que a bacia amazônica se manteve amplamente florestada no último período glacial. Em apoio à sugestão de Pennington et al. 8, porém, grande parte da metade sul da bacia teria sido ocupada por florestas decíduas.
A existência de táxons associados às formações vegetacionais estacionais com distribuição disjunta separada pela floresta amazônica também tem sido utilizada como evidência de prévia expansão dessas formações sobre a bacia amazônica. No entanto, as interpretações têm de ser feitas com cautela, buscando caracterizar se realmente há associação a um tipo determinado de vegetação e, mesmo que haja essa relação, a ocorrência de distribuição disjunta não implica em ampla alteração da paisagem, podendo ter sido resultado do surgimento de apenas um corredor por onde as espécies expandiram a distribuição de suas populações em certo momento. Além disso, há a possibilidade de ter acontecido dispersão de longa distância, como sugerido por Mayle (2006) apud Furley e Metcalfe38, para explicar as ocorrências disjuntas das FEDT.
Silva2 avaliou as possíveis conexões pleistocências entre savanas ao sul e ao norte da Amazônia, com base nas distribuições atuais de aves (Fig. 1 3 . 1 1 ) , encontrando suporte apenas para o corredor do Andes e o da costa do Atlântico. Segundo o autor, esses corredores seriam de savana; entretanto, pelo mapeamento da distribuição das espécies analisadas, elas quase sempre não ocorrem apenas em áreas de Cerrado (Fig. 1 3 . 12). Assim, é possível que a vegetação aberta que ocupou a área desses corredores, permitindo a expansão na distribuição dessas espécies, não tenha sido de Cerrado.
Lourenço e Silva40 sugeriram que houve uma conexão entre áreas de vegetação aberta na bacia amazônica, com base na ocorrência de espécies de escorpião associadas aos ambientes de vegetação aberta. São citadas Rhopalurus amazonicus Lourenço, registradas unicamente nas savanas de Alter do Chão (Pará), cuja possível espécie irmã sucede no Brasil central, e Tytius sylviae Lourenço e Silva, descrita de manchas de vegetação aberta, tipo campinarana, próximas ao Rio Negro na Amazônia ocidental, que é relacionada a T. marajoensis, da Ilha de Marajó, e T. blaseri Mello-Leitão, do norte de Goiás e T. confluens Borelli, que ocorre principalmente no Chaco, estendendo-se, em parte, do Pantanal e Cerrado.
Wuster et al. 36, ao estudarem por meio de marcadores moleculares a divergência entre populações de cascavel (Crotalus durrissus), estimaram que a separação de
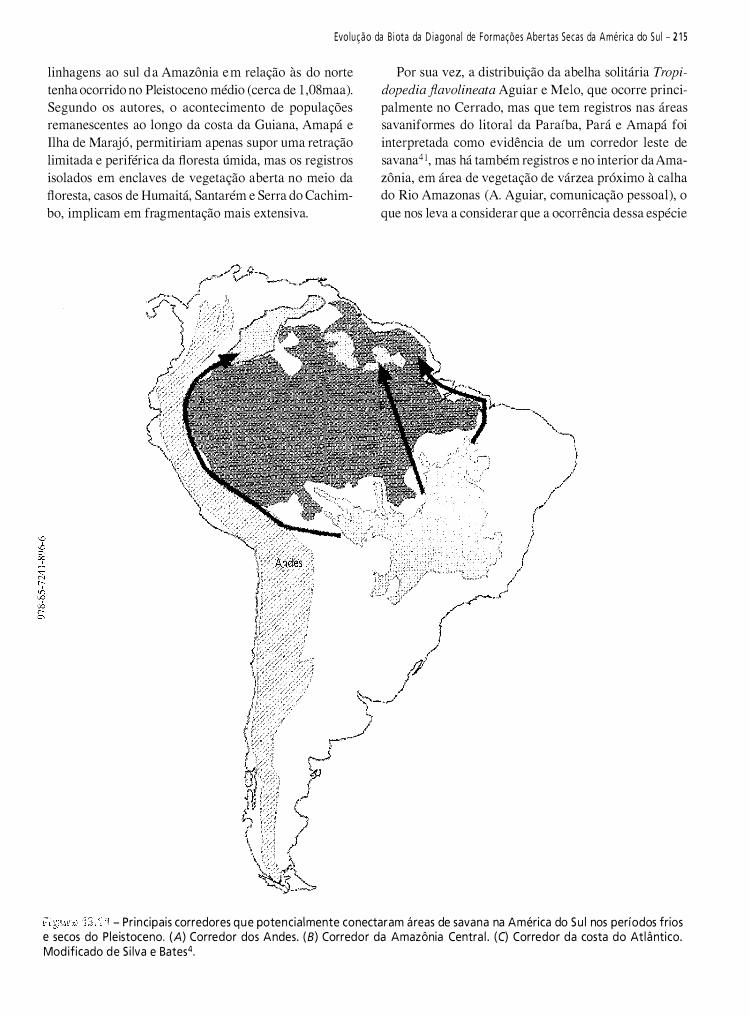
Evo lução da B iota da D iagonal de Formações Abertas Secas da América do Su l - 2 1 5
linhagens ao sul da Amazônia em relação às do norte tenha ocorrido no Pleistoceno médio (cerca de 1 ,08maa). Segundo os autores, o acontecimento de populações remanescentes ao longo da costa da Guiana, Amapá e Ilha de Marajó, permitiriam apenas supor uma retração limitada e periférica da floresta úmida, mas os registros isolados em enclaves de vegetação aberta no meio da floresta, casos de Humaitá, Santarém e Serra do Cachimbo, implicam em fragmentação mais extensiva.
Por sua vez, a distribuição da abelha solitária Tropidopedia flavolineata Aguiar e Melo, que ocorre principalmente no Cerrado, mas que tem registros nas áreas savaniformes do litoral da Paraíba, Pará e Amapá foi interpretada como evidência de um corredor leste de savana41, mas há também registros e no interior da Amazônia, em área de vegetação de várzea próximo à calha do Rio Amazonas (A. Aguiar, comunicação pessoal), o que nos leva a considerar que a ocorrência dessa espécie
- Principais corredores que potencialmente conectaram áreas de savana na América do Su l nos períodos frios e secos do P le istoceno. (A) Corredor dos Andes. (B) Corredor da Amazônia Centra l . (C) Corredor da costa do Atlântico. Mod ificado de S i lva e Bates4.
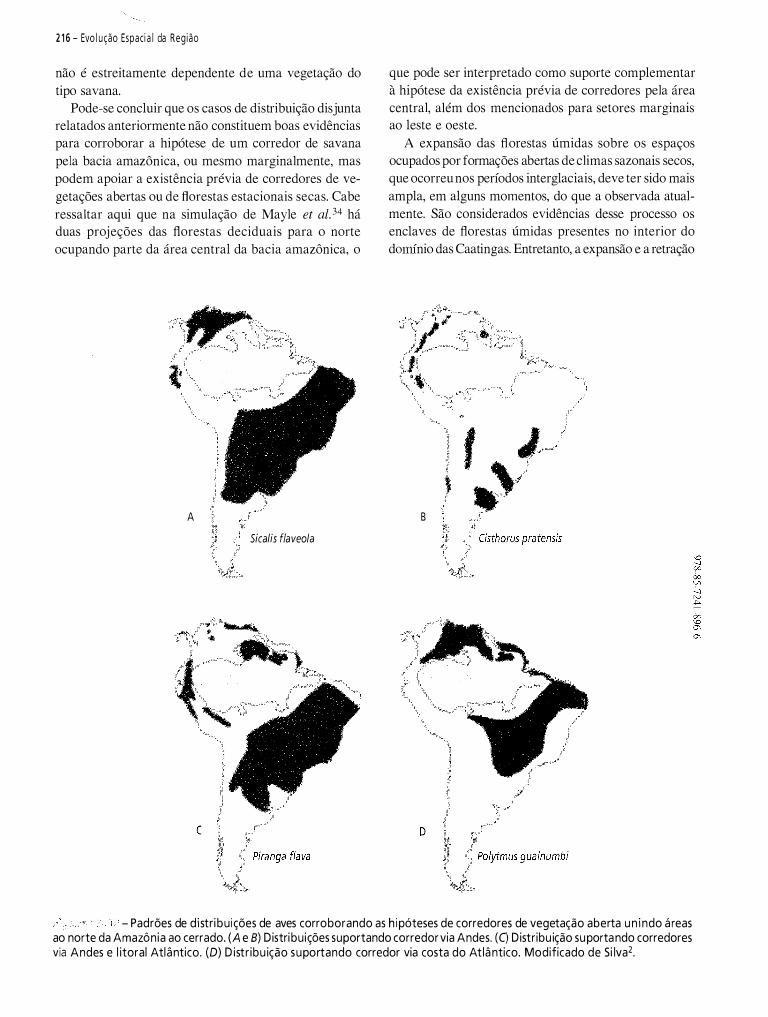
2 1 6 - Evo l ução Espaci a l da Reg ião
não é estreitamente dependente de uma vegetação do tipo savana.
Pode-se concluir que os casos de distribuição disjunta relatados anteriormente não constituem boas evidências para corroborar a hipótese de um corredor de savana pela bacia amazônica, ou mesmo marginalmente, mas podem apoiar a existência prévia de corredores de vegetações abertas ou de florestas estacionais secas. Cabe ressaltar aqui que na simulação de Mayle et aI. 34 há duas projeções das florestas deciduais para o norte ocupando parte da área central da bacia amazônica, o
A
Sicalis flaveola
que pode ser interpretado como suporte complementar à hipótese da existência prévia de corredores pela área central, além dos mencionados para setores marginais ao leste e oeste.
A expansão das florestas úmidas sobre os espaços ocupados por formações abertas de climas sazonais secos, que ocorreu nos períodos interglaciais, deve ter sido mais ampla, em alguns momentos, do que a observada atualmente. São considerados evidências desse processo os enclaves de florestas úmidas presentes no interior do dOllÚnio das Caatingas. Entretanto, a expansão e a retração
B
- Padrões de d istribu ições de aves corroborando as hipóteses de corredores de vegetação aberta un indo áreas ao norte da Amazônia ao cerrado. (A e B) Distribuições suportando corredor via Andes. (C) Distribu ição suportando corredores via Andes e l itoral Atlântico. (O) Distr ibu ição suportando corredor via costa do Atlântico. Modificado de Si lva2.

Evolução da B iota da D iagona l de Formações Abertas Secas da América do Su l - 2 1 7
Oi - Distribu ição das espécies Geotrigona mattogrossensis, G . aequinoctialis e G. xanthopoda que formam um possível dado do componente Sudeste. Mod ificado de Camargo e Moure28.
das florestas úmidas também contribuíram para diversidade da biota das áreas secas. O traçado biogeográfico formado pelas espécies de abelhas Geotrigona xanthopoda Camargo e Moure + G. aequinoctialis (Ducke) + G. mattogrossensis (Ducke) foi denominado Caatinga-Amazônia SE16. Essas três espécies formam um clado segundo Camargo e Moure28 e enquanto a primeira foi registrada em setores localizados de caatinga, as outras acontecem em áreas de floresta úmida, no caso de G. aequinoctialis, incluindo enclaves no Ceará (Fig. 13 . 13). De modo semelhante, Aguiar e Me1041 interpretaram que pelo menos três espécies de abelhas do gênero Tropidopedia, grupo punctifrons, que sucedem no Brasil central, em áreas de cerrado e florestas secas, são derivadas de grupos associados aos ambientes de florestas úmidas.
Outras fontes de espécies documentadas para o Cerrado, neste caso possivelmente associada aos períodos glaciais, são as regiões mais frias localizadas ao sul. Um caso foi observado por Almeida e Silveira42, ao estudarem a biogeografia de abelhas do subgênero Exomalopsis (Phanomalopsis) Michener e Moure, em que a espécie E. holotricha Silveira e Almeida, que ocorre no
Cerrado, se restringe a setores mais elevados e também acontece em grande parte do sul do Brasil, na sub-região paranaense (no sentido de Cabrera e Willink6). No entanto, conforme ressaltado por Vasconcelos22, devem-se distinguir as espécies que sucedem realmente em hábitats de vegetação de Cerrado daquelas que OCOlTem apenas em locais de maior altitude, em particular quando restritas às áreas de campos rupestres, pois a biota desta região parece ter história evolutiva independente.
A história da biota da diagonal de formações abertas secas da América do Sul é complexa e ainda pouco conhecida. A origem de parte significativa de sua biota é quase sempre associada à Província Microfloral Mista, originalmente distribuída mais ao sul e depois se expandindo durante o Cenozoico, especial após a metade do Oligoceno. Possivelmente, os biomas da Caatinga, Cerrado e Chaco chegaram a uma configuração espacial próxima a atual com o soerguimento do Planalto Central

218 - Evolução Espacia l da Região
Brasileiro, que levou os terrenos onde se encontra a maior parte do Cerrado aos níveis altitudinais atuais, e a subsidência da planície do Chaco e denudação da maior parte do nordeste brasileiro, onde se encontra a Caatinga. No entanto, registros de distribuição relictual no Nordeste do Brasil, como o de Ceblurgus e de Bothriurus, permitem sugerir que houve elementos xéricos na região desde a metade inicial do Cenozoico.
Os dois eventos do Terciário citados na literatura que possivelmente afetaram a biota da diagonal de formações abertas secas: o soerguimento do Planalto Central Brasileiro e o mar paranaense, ainda carecem de dados mais consistentes para corroborar seu papel na diversificação biológica. Além disso, o efeito das mudanças climáticas do Pleistoceno precisa ser reavaliado considerando-se a resposta de cada fisionomia de vegetação nas diferentes fases e de acordo com os vários condicionantes ecológicos. O esquema básico inicialmente proposto, no qual a Caatinga e o Chaco, mais secos do que o Cerrado, coalesceram nas fases secas e frias dos períodos glaciais, tem se demonstrado limitado, por exemplo, pela interpretação de momentos de climas úmidos e mais frios que o atual no Maranhão e em Minas Gerais32. Também, pela interpretação de que as florestas tropicais sazonais secas podem ser distintas da vegetação do Chaco e das formações abertas da Caatinga. E, no caso do Cerrado, como além do fator climático as características edáficas são determinantes da sua distribuição, há a necessidade da compreensão de como se deram as alterações nos solos durante as mudanças climáticas. Furley e Metcalfe38 sugerem a integração de abordagens distintas, incluindo dados de paleolimnologia, microrremanescentes orgânicos, minerais e isótopos estáveis . A paleomodelagem climática também é promissora, mas deve contemplar um detalhamento envolvendo as várias fisionomias de vegetaçã034.43.
Em todo o caso, a distribuição de Caenonomada unicalcatara e C. aif. unicalcarata e de Tapinotaspoides rufescens e T afr rufescens parece ter sido resultado de uma prévia expansão de formações abertas. Registros como os de Geotrigona xanthopoda e de espécies de Tropidopedia, grupo punctifrons, servem de exemplos do papel da expansão das florestas úmidas sobre o espaço geográfico da Caatinga e do Cerrado, respectivamente, e sua contribuição na diversificação da biota desses biomas.
Em termos da distribuição da biota atual, apesar da limitação dos dados, deve-se destacar que diferentes grupos analisados têm fornecido informações distintas acerca das alterações passadas na paisagem. É possível que, em vez de serem ruídos na análise, as divergências observadas representem respostas diferentes a fatores
ambientais. Assim, o fato de a vegetação lenhosa do Chaco seco ter influência eminentemente temperada, segundo Pennington et a!. 8 e a co-ocorrência de grupos de abelhas tropicais associados à vegetação herbácea, como Caenonomada, pode ter resultado de uma mistura de elementos de origens diferentes.
A ocorrência de uma espécie endêmica de escorpião em área de vegetação aberta no interior da bacia amazônica pode ser um indício de que a expansão da :s 00 vegetação aberta na região tenha permitido ao menos a t;
.:.., invasão por elementos da fauna de artrópodes e posterior � diversificação. Essas são questões em aberto, que preci- 00 '" sam ser investigadas com maior quantidade de dados, 6: com levantamentos intensivos das biotas dessas áreas, buscando o acúmulo de evidências para cada padrão encontrado.
Quanto aos esquemas biogeográficos propostos, deve-se entender que eles são reduções da história, que servem como ponto de partida para as análises biogeográficas, mas podem limitá-las, se não for compreendida a complexidade da história das biotas. Pode-se admitir que há somente uma história para cada região, porém cada componente da biota de uma região tem sua história, que pode ser independente, em certo grau da história, dos outros componentes. A complexidade da história de suas biotas tem sido reconhecida para muitas regiões estudadas como resultado da presença de elementos de origens distintas.
Tome-se o caso do Cerrado como exemplo. Essa província foi incluída na região [sic, sub-região] Amazônica por Cabrera e Willink6, deixando a Caatinga como uma província disjunta da região Chacoana. Morrone3 transferiu o Cerrado para a sub-região Chacoana.
Como há na região do Cerrado uma sobreposição de traçados de elementos da biota associados às florestas úmidas com elementos associados às formações abertas secas, a decisão de associá-la à região Amazônica ou à região Chacoana significa privilegiar o registro de uma informação histórica em detrimento de outra.
Nesse sentido, também se pode levar em conta a limitação das análises redutoras, que obrigatoriamente resulta em apenas uma hipótese de relação histórica entre áreas (ou, de forma mais apropriada, em somente uma hipótese entre biotas atuais de diferentes áreas), como se houvesse uma média, ou consenso, entre as histórias dos diferentes grupos que habitam uma região.
Pode-se, assim, interpretar a melhor resolução da hipótese de relação histórica entre as áreas de endemismo da diagonal de formações abertas secas obtida por Porzecanski e Cracraft24 quando eliminam da análise os táxons associados aos ambientes florestais, pois se trata

Evo lução da B iota da D iagona l de Formações Abertas Secas da América do S u l - 2 1 9
certamente de elementos com uma história diferente dos táxons associados aos ambientes abertos. Em todo caso, isso não elimina a possibilidade de haver táxons de ambientes xéricos com diferentes histórias, mas o seu reconhecimento só será possível com análises filogenéticas e de distribuição de um amplo conjunto de grupos.
Morrone e Crisci44 propuseram, seguindo Croizat, que o passo inicial de análises de biogeografia histórica seria o reconhecimento da homologia espacial por meio do achado de traçados generalizados. Cabe aqui ressaltar que deve ser considerada também a homologia temporal, pois traçados aparentemente equivalentes em termos espaciais são, de modo claro, representativos de eventos históricos distintos e, portanto, não são equivalentes. Servem de exemplo os traçados já citados de Penapini e de Pithecellobium excelsum-P. diversifolium, bem como os traçados de Caenonomada e o de Tapinotaspoides. Lembremos ainda de que há fatores potencialmente causadores de mudanças da distribuição das espécies e determinantes de vicaliância nas populações que ocorreram de forma cíclica, como as glaciações no Pleistoceno e a transgressão paranaense do Mioceno.
Aguiar e Mel026 sugeriram, como meio de avançar no conhecimento da história da biota das formações abertas secas, que as análises considerassem as distintas áreas de endemismo presentes na Caatinga e no Cerrado. Isso certamente representará um avanço, de modo semelhante ao reconhecimento por Prad07, de setores do Chaco com diferentes biotas e de áreas de endemismo de distribuição restrita no meio do domínio do Cerrado, por Silva e Bates4. No entanto, cabe lembrar ao final, o provocativo trabalho de Henderson45, entitulado Biogeography without areas ?, no qual ressalta que o objetivo da biogeografia deve ser a história das biotas e não a história das áreas. De acordo com ele, a segunda opção limitou o escopo dessa área da ciência a um apêndice da geologia e levou a se desconsiderar aspectos ecológicos nas análises de biogeografia histórica. Sugere-se, assim, que o reconhecimento das várias biotas ancestrais, das quais a diversidade biológica atual de cada região descende, caracterizadas para diferentes momentos históricos e condições ecológicas, seja parte essencial do processo de conhecimento da história biogeográfica das regiões.
A Fundação de Apoio à Pesquisa da Paraíba (FAPESQ), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e Ministério do Meio Ambiente pelo apoio recebido aos projetos de pesquisa do Laboratório
de Ecologia e Biogeografia de Insetos da Caatinga! UFCG. Ao Dr. Antonio Aguiar (UnB) e Dra. Francisca Soares de Araújo (UFC) e dois consultores anônimos pela revisão do manuscrito, sugestões e referências indicadas. Aos editores Dr. Claudio J. B. de Carvalho e Dr. Eduardo A. B. Almeida, pelo convite para participar deste projeto. Este trabalho é oferecido em memória do Dr. João Maria Franco de Camargo, recentemente falecido, amigo, orientador e reconhecido pesquisador da biogeografia das abelhas sem ferrão da América do Sul.
REFERÊNCIAS BIBLIOGRÁFICAS 1. AB' SÁBER, A. N. Os dOTIÚnios morfoclimáticos na América do
Sul. Primeira aproximação. GeomOlfologia, v. 52, p. 1 -24, 1 977. 2 . SILVA, 1. M. C. Biogeographic analysis ofthe South American
avifauna. Steenstrupia, v. 2 1 , p. 49-67, 1 995. 3. MORRONE, 1. What is the Chacoan subregion? Neotropica, v.
46, p. 5 1 -68, 2000. 4. SILVA, 1. M. c.; BATES, J. M. Biogeographic patterns and
conservation in the South American Cerrado: a tropical savanna hotspot. Bioscience, v. 52, p. 225-233, 2002.
5. LOPES, L. E. Taxonomia, Biogeograjia e Conservação de Aves Endêmicas do Cerrado. Belo Horizonte: UFMG, 2009, 26 1p. Tese (Doutorado) - Pós-graduação em Ecologia, Conservação e Manejo da Vida Silvestre -Universidade Federal de Minas Gerais.
6. CABRERA, A. L.; WILLINK, A. Biogeograjía de América Latina. 1 . ed. Washington: Secretaría General de la Organización de los Estados Americanos, 1 973. 1 26p.
7. PRADO, D. E. Seasonally dry forests of tropical SouthAmerica: ±fom forgotten ecosystems to a new phytogeographic unit. Edinb J. Bot. , v. 57, p. 437-461 , 2000.
8. PENN1NGTON, R. T.; PRADO, D. A.; PENDRY, C. Neotropical seasonally dry forests and P1eistocene vegetation changes. J. Biogeogr. , v. 27, p. 26J -273, 2000.
9. PENNINGTON, R. T.; LEWIS, G. P.; RATTER, J. A. An overview of the plant diversity, biogeography and conservation of neotropical savannas and seasonaJly dry forests. In: Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography and Conservation. Boca Raton: CRC Press Taylor & Francis Group, 2006. capo I , p. 1 -30.
10. PENNINGTON, R. T.; LAVIN, M. ; PRADO, D. A. et aI. Historical climate change and speciation: neotropical seasonally dry forest plants show patterns of both Tertiary and Quaternary diversification. Phil Trans Royal Soe. London Biol. Sei. , V. 359, p. 5 1 5-537, 2004.
1 1 . WERNECK, F. P; COLLI, G. R. The lizard assemblage from Seasonally Dry Tropical Forest enclaves in the Cerrado biome, Brazil, and its association with the Pleistocenic Are. 1. Biogeop:, V. 33, p. 1 983- 1 992, 2006.
12 . COLLI, G. R. As origens e a diversificação da herpetofauna do Cerrado. In: SCARIOT, A.; SOUZA-SILVA, 1. c.; FELFILI, J. M. (eds.). Cerrado: ecologia, biodiversidade e conservação. Brasília: Ministério do Meio Ambiente, 2005. capo 14, p. 249-264.
1 3 . VIVALLO, F; MELO, G. A. R. Taxonomy and geographic distribution of the species of Centris of the hyptidis group (Hymenoptera: Apidae: Centridini), with description of a new species from central BraziJ. Zootaxa, V. 2075, p. 33-44, 2009.
1 4. GOTTSBERGER, G.; SlLBERBAUER-GOTTSBERGER, 1. Life in the Cerrado - a South American Tropical Seasonal Ecossystem. Origin, strueture anel plant use. Ulm: Reta Verlag, 2006. V. 1. 277p.
1 5 . COSTA, R. c.; ARAÚJO, F. c. ; LIMA-VERDE, L. W. Flora and life-form spectrum in an area of deciduous thom woodland (caatinga) in Northeastem Brazi1. J. Arid Environ., v. 68, p . 237-247 , 2007.

220 - Evolução Espaci a l da Reg ião
1 6. STRAUBE, F. C. ; DI-GIÁCOMO, A. A avifauna das regiões subtropical e temperada do Neotrópico: desafios biogeográficos. Ciência e Ambiente, v. 35, p. 1 37- 166, 2007. Disponível em: http://w3.ufsm.brlreciaml
1 7. SUGUIO, K. ; ANGULO, R. J.; CARVALHO, A. M. et aI. Paleoníveis do mar e paleolinhas da costa. In: SOUZA, C. R. G.; SUGUIO, K. ; OLIVEIRA, A. M. S . et aI. Quaternário do Brasil. 1. ed. Ribeirão Preto: Editora Holos, 2005. capo 6, p. 1 14-1 29.
1 8 . ZANELLA, F. C. v.; MARTINS, C. F. Abelhas da Caatinga: Biogeografia, ecologia e conservação. In: LEAL, I. R. ; TABARELLI, M. ; SILVA, J. M. C. (eds.). Ecologia e Conservação da Caatinga. Recife:Editora UFPE, 2003. capo 2, p. 75- 134.
19. DANFORTH, B. N . ; BRADY, S . G.; SIPES, S . D. et ai. Singlecopy nuclear genes recover Cretaceus-age divergences in bees. Syst. Biol. , V. 53, p . 309-326, 2004.
20. ROIG-JUNENT, S . ; DOMÍNGUEZ, M. C.; FLOREZ, G. E. et aI. Biogeographic history of South American arid lands: A view from its arthropods using TASS analysis. 1. Arid Environ. , V. 66, p. 404-420, 2006.
2 1 . RUZ, L. Classification and phylogenetic relationships of the panurginae bees: Calliopsini and allies (Hymenoptera, Andrenidae). Kans Univ. Sci. Buli., v. 57, p. 209-253, 199 1 .
22. VASCONCELOS, M . F. Mountaintop endemism in eastern Brazil : why some bird species from campos rupestres of the Espinhaço Range are not endemic to the Cerrado region? Rev. Bras. Omitol. , V. 16, p. 348-362, 2008.
23. MARSHALL, L.; LUNDBERG, J. Technical comments. Science, V. 273, p. 1 24, 1 996.
24. PORZECANSKI, A. L. ; CRACRAFT, 1. Cladistic analysis of distributions and endemism (CADE): using raw distributions of birds to unravel the biogeography of the South American Aridlands. J. Biogeogr. , V. 32, p. 261-275, 2005.
25. ZANELLA, F. C. V. Systematics and biogeography of the bee genus Caenonomada Ashmead, 1 899 (Hymenoptera:Apidae: Tapinotaspidini) . Stud. Neotrop. Fauna Environ. , V. 37, p. 249-261 , 2002.
26. AGUIAR, A. 1. C. ; MELO, G. A. R. Biogeografia das áreas abertas da América do Sul com ênfase em abelhas da tribo Tapinotaspidini. In: VIII ENCONTRO SOBRE ABELHAS, 2008. Ribeirão Preto. Anais do VIII Encontro sobre Abelhas, 2008, p. 322-326.
27. VIVALLO-MARTlNEZ, F. Sistemática, filogenia )' distribuición geográfica de las especies dei gênero Centris Fabricius, con ênfasis en los subgêneros Wagenknechtia Moure, 1 950, Ptilocentris Snelling, 7984 y AphemisiaAyala, 2002, incluyendo un análiis .filogenêtico dei grupo Melacentris. Valparaiso: Universidad Catolica de Valparaiso, 2004, 221p. Dissertação (Mestrado) - Posgraduacion en Ecologia y Sistematica - Universidad Catolica de Valparaiso.
28. CAMARGO, J. M. F.; MOURE, J. S. Meliponini neotropicais: o gênero Geotrigona Moure, 1 943 (Apinae, Apidae, Hymenoptera). Arq. Zool. , V. 33, p. 95- 1 6 1 , 1 996.
29. SIMPSON, B. B . ; NEFF, 1. L. Plants, their pollinating bees and the GreatAmerican Interchange. In: STEHLI; F. G.; WEBB, S . D. The GreatAmerican Biotic Interchange. Nova York: Plenum, 1 985. cap. 1 3 , p. 427-452.
30. BURNHAM, R. J. ; GRAHAM, A. The history of neotropical vegetation: new developments and status. Ann. Missouri Bot. Gard. , V. 86, p. 546-589, 1 999.
3 1 . WEBB, S . D. A history of savanna verlebrates in lhe New World. Par! II: South America and the Great Interchange. Ann. Rev. Ecol. Syst., V. 9, p. 393-426, 1 978 .
32 . OLIVEIRA, P. E.; BEHLING, H. ; LEDRU, M. P. et aI. Paleovegetação e paleoclimas do Quaternário do Brasil. In: SOUZA, C. R. G. ; SUGUIO, K.; OLIVEIRA, A. M. S. el al. Quaternário do Brasil. Ribeirão Preto: Holos, 2005. capo 3, p. 52-74.
33. COLINVAUX, P. A.; DE OLIVEIRA, P. E . ; BUSH, M. B . Amazonian and neotropical plant communities o n glacial timescales: the failure ofthe aridity and refuge hypotheses. Quatem. Sci. Rev. , V. 19 , p. 1 4 1 - 169, 2000.
34. MAYLE, F. E. ; BEERLING, D. J . ; BUSH, M. B. el aI. Responses of Amazonian ecosystems to climatic and atmospheric carbon dioxide changes since the last glacial maximum. Phil. Trans. R. Soe. B. , v. 359, p. 499-5 14, 2004.
35. AB 'SÁBER, A. N. Paleoclimas quaternários e pré-história da América Tropical. I . Rev. Brasil Biol. , V. 50, p. 805-820, 1 990.
36. WUSTER, w.; FERGUSON, J. E.; QUIJADA-MASCARENAS, J. A. et aI. Tracing an invasion: landbridges, refugia, and the phylogeography of the NeotropicaJ rattlesnake (Serpentes: Viperidae: erotalus durissus). MoI. Ecol. , v. 1 4, p. 1 095-1 1 08, 2005.
37. KIPNIS, R.; SCHEEL-YBERT, R. Arqueologia e paJeoambienteso In: SOUZA, C. R. G. ; SUGUIO, K.; OLIVEIRA, A. M. S . e t aI. Quaternário do Brasil. Ribeirão Preto: Holos, 2005. capo 16 , p. 343-362.
38. FURLEY, P. A.; METCALFE, S . E. Dynamic changes in savanna and seasonally dry vegetation through time. Prog. Phys. Geog. , V. 3 1 , p. 633-642, 2007.
39. OCHSENIUS, C. Pleniglacial desertization, large-animal mass extinction and Pleistocene-HoJocene boundary in South America. Rev. de Geog., Norte Grande, V. 12, p. 35-47, 1985.
40. LOURENÇO, W. R. ; SILVA, E. A. New evidence for a disrupted distribution pattern ofthe Tytius confluens' complex, with the description of a new species from the State of Pará, Brazil (Scorpiones, Buthidae). Amazoniana, V. 19, p. 77-86, 2007.
4 1 . AGUIAR, A. J. C. ; MELO, G. A. R. Taxonomic revision, phylogenetic analysis , and biogeography of the bee genus Tropidopedia (Hymenoptera, Apidae, Tapinotaspidini). Zool. J. Linll. Soc., V. 1 5 1 , p. 5 1 1-554, 2007.
42. ALMEIDA, E. A. B . ; SILVEIRA, F. A. Biogeografia histórica de Exomalopsis (Phanomalopsis) Michener e Moure 1957 (Apidae: Exomalopsini). In : VIII ENCONTRO SOBRE ABELHAS, 2008. Ribeirão Preto. Anais do VIII Encontro sobre Abelhas, 2008, p. 327-3 3 1 .
43. CARNAVAL, A . c . ; MORITZ, C. Historical climate modelling predicts patterns of currenl biodiversity in the Brazilian Atlantic forest. J. Biogeogr. , V. 35, p. 1 1 87-120 1 , 2008.
44. MORRONE, 1.; CRISCI, J. V. Historical biogeography: introduction to methods. Ann. Rev. Ecol. Syst. , V. 26, p. 373-401 , 1 995.
45. HENDERSON, I. M. Biogeography without area? Austral. Syst. Bot., V. 4, p. 59-7 1 , 1 99 1 .

/t d �, ',Y ;. '
T
Marcio Bernardino DaSilva
Ricardo Pinto-da-Rocha
Antes do início da devastação e consequente fragmentação da Mata Atlântica, iniciada há cinco séculos com a chegada dos colonizadores europeus, esse bioma cobria quase toda a costa leste do Brasil, desde o Rio Grande do Norte até o Rio Grande do Sul . Na porção meridional, alcança a bacia do Rio Paraná e do centro ao norte limita-se pelo Cerrado e Caatinga. Essa grande extensão determina ampla variação nas suas características, desde uma floresta muito úmida próxima ao litoral, a uma mais seca no interior, de sempre quente nas menores latitudes a com invernos rigorosos nos seus limites ao sul. Toda essa variação geográfica faz com que a Mata Atlântica apresente "muitas florestas", ou seja, fisionomias muito diferentes entre porções desse bioma. Na porção leste, próxima ao litoral, está a Floresta Ombrófila Densa, caracterizada pela ausência de estação seca, isto é, umidade alta durante o ano inteiro. As encostas do Planalto Atlântico voltadas para o mar, conhecidas genericamente na parte sul como Serra do Mar, e a Serra da Mantiqueira, mais ao interior, retêm boa parte da umidade vinda do oceano e provocam as chuvas orográficas, principais mantenedoras das condições climáticas para essa floresta mais desenvolvida e exuberantel . Ainda, adjacentes à Floresta Ombrófila, existem ecossistemas mais relacionados às caracterís� ticas físicas típicas de ambientes costeiros, como os manguezais e as restingas.
1 i
1 4
Em direção ao interior, observa-se a redução da umidade dos ecossistemas da Mata Atlântica, e existe um amplo gradiente fisionômico da Floresta EstacionaI Semidecidual que aos poucos se torna a zona de contato com o domínio do Cerrado. Essa floresta é caracterizada pelo inverno bem mais seco, que se apresenta como uma floresta sazonal e menos desenvolvida. Mais ao sul, principalmente no Paraná, a floresta do interior (conhecida como Floresta Mista) também exibe características distintas, pela presença marcante do pinheiro-do-Paraná, Araucaria angustifolia, espécie cujo gênero sempre está em regiões mais frias ou temperadas da América do Sul. Essa formação, marcada pela ocorrência da Araucaria, também ocorre mais ao norte em grandes altitudes (acima de 1 .200m) até o sul de Minas Gerais, onde o frio do inverno determina as condições para o desenvolvimento dessa fisionomia diferenciadal .
Mesmo nas encostas voltadas para o mar, a zonação altitudinal das serras também é refletida em fisionomias distintas, desde as florestas de planícies, passando pela Submontana e a Montana; esta última acima de 300m de altitude, e que em muitos pontos transformam-se em vegetações abertas, como campos de altitudes. No caso das florestas de planície da Mata Atlântica, elas atingem maior extensão no nordeste brasileiro, onde são conhecidas como Florestas de Tabuleiros e apresentam características físicas diferenciadas pela sua origem e desenvolvimento sobre terreno sedimentarmarinho em grande extensão na regiãol .
Toda esta zonação ecológica reflete processos físicos atuantes cotidianamente ou, pelo menos, anualmente na floresta. É evidente que esses processos determinam

222 - Evolução Espac i a l da Região
diferenças na composição das espécies entre uma e outra fisionomia. Por exemplo, ao se traçar um transecto das florestas mais úmidas da costa em direção a uma floresta mais seca no interior, observa-se grande variação entre as espécies vegetais e animais (esta última menor) de um local a outro, em particular entre os extremos. O mesmo acontece ao se comparar a diferença entre a composição de espécies de qualquer grupo taxonômico entre as matas de planície em direção às matas de altitude. Sobre o primeiro exemplo, Oliveira-Filho e Fontes2 demonstraram que existe um continuum na substituição de espécies de árvores entre as duas fisionomias, do litoral ao interior, e que a Floresta Semidecidual é formada majoritariamente por uma fração das espécies encontradas na Floresta Ombrófila, as mais resistentes à estação seca. No caso da transição para a altitude, os padrões de substituição de espécies também são graduais e, em geral, nas áreas mais altas ocorrem as espécies mais generalistas (também presentes nas partes baixas) a condições físicas diferentes, obedecendo aos padrões gerais de distribuição de espécies em gradientes climáticos3.
Entretanto, existem outros padrões de distribuição das espécies que vivem na Mata Atlântica. As espécies que se distribuem em todo o bioma ou em alguma das fisionomias já citadas apresentam limitações de distribuição provavelmente relacionadas às condições físicas que são observáveis com facilidade nos dias de hoje, por exemplo, a alta umidade em contraste à maior aridez da diagonal seca formada por caatinga e cerrado. Por outro lado, existem milhares de espécies endêmicas de áreas relativamente menores, limitadas por processos históricos, ainda não esclarecidos por completo.
Desde sua origem, quais teriam sido os grandes acontecimentos que resultaram nos endemismos e a diversidade da Mata Atlântica? Por meio de distribuições não relacionadas apenas ao clima atual, é possível inferir outros processos que determinaram a distribuição das espécies e pode-se propor hipóteses sobre a história do bioma.
Este capítulo pretende contar o pouco do que se sabe dessa história. Plimeiramente, será apresentada uma hipótese de sua origem e como ela se isolou de outros biomas florestais e tomou-se uma unidade biogeográfica. Em seguida, será sumarizado o conhecimento sobre o seu endemismo, sobre as suas subáreas e as divisões históricas mais importantes. Discutem-se ainda os principais processos que podem ter influenciado mais significativamente na alta diversificação de espécies e endemismos comuns em florestas tropicais. Por último, demonstram-se como estudos sobre opiliões estão ajudando a compreender o passado de um dos mais importantes hot spots para a conservação da biodiversidade do mundo, a Mata Atlântica.
Morley4 exibiu uma síntese de como teriam surgido e se desenvolvido as florestas tropicais, que está resumida a seguir, com foco na Mata Atlântica. Até o fim do Cretáceo
-o (mais de 65 milhões de anos atrás [maaJ), não havia con- cf dições para a existência de florestas tropicais como a Mata f7: .:., Atlântica em nenhum lugar do mundo. Isso porque, àque- :;:: la época, o clima era bem mais seco do que o atual e do �
O' que se seguiu durante o Terciário (a partir de 65maa). Além "" disso, as angiospermas ainda não haviam se diversificado e dominado ecossistemas florestais tropicais. A partir de 80maa, houve rápida diversificação das angiospermas, com primeiro aparecimento no registro fóssil de farm1ias de árvores tropicais importantes como Palmae, Bombacaceae, Caesalpinaceae, Euphorbiaceae, Meliaceae e Rutaceae, e de sementes relativamente grandes com muito endosperma, próprias de florestas com dos sei fechado. Essa diversificação coincidiu com o estabelecimento de um clima bastante úmido e a zonação latitudinal, que caracteriza diferentes províncias ftorísticas, estabelecidas com paleopalinologia (estudo de pólens fósseis). Naquela época, o continente sul-americano já se encontrava bastante distanciado da África, em sua deriva em direção oeste e o Oceano Atlântico já tinha características semelhantes às atuais. Estas evidências indicam a provável existência de uma antiga floresta tropical na costa leste da América do Sul, como uma Mata Atlântica ancestral4.
O Cretáceo terminou com grande evento de extinção da biota terrestre, provavelmente causado pelo choque de um meteorito no atual México há 65maa. Esse evento resultou na extinção da megafauna dos dinossauros e de cerca de 40% das espécies de plantas. Entretanto, no final do Paleoceno e início do Eoceno (pouco mais de 50maa), o clima predominantemente quente e úmido, favoreceu a recuperação da diversidade das angiospermas. Muitas novas espécies de plantas surgiram em decorrência do período de grande diversificação e, assim, foi garantida a manutenção de florestas úmidas na América do Su14. Provavelmente nessa época essas florestas eram contínuas do Pacífico ao Atlântico e ainda eram mais extensas que as atuais durante o Mioceno Médio ( 1 6 a l Omaa)4. Contudo, a partir do Plioceno (-5maa), com o esfriamento e ressecamento global e os ciclos glaciais-interglaciais que permanecem até hoje, houve expansão de áreas abertas savânicas, representadas majoritariamente na América do Sul pela "Diagonal de formações abertas", formada pelos biomas da Caatinga, Cerrado e Chaco (ver Capo 13) . O aparecimento dessa diagonal seca separou

H i stór ia B iogeográfica da Mata At l ânt ica : Opi l iões (Arachn ida) como Mode lo para sua I nferênc i a - 223
definitivamente a Mata Atlântica das outras florestas sul-americanas e deu início à evolução isolada do bioma. Esse isolamento resultou em alto nível de endemismo, chegando a 54% das espécies de árvores5, 60% de anfíbios5 e 97,5% de opiliões6, demonstrando a alta diversificação que ocorreu dentro de seus limites.
A biota da Mata Atlântica é composta por táxons de diferentes origens biogeográficas. Grande parte dos estudos que analisaram as relações históricas da Mata Atlântica aponta para uma relação biogeográfica mais próxima destes com a biota da Floresta Amazônica, em particular a porção sudeste deste bioma7-9. Alguns outros estudos mostram relação mais próxima com o componente florestal do cerrado, por meio de florestas de galerialO-12, ou da parte meridional da Mata Atlântica com áreas dos Andes, em especial as áreas mais altas, relacionadas à presença de Araucaria angustifolia, ou mesmo com o sudoeste da Amazônia 1 .
Essas relações com a Floresta Amazônica e florestas residuais no Cerrado são explicadas pela possível ligação passada dos dois blocos de floresta em épocas mais úmidas, principalmente no Mioceno Médio ( l6maa) . Além disso, após a separação dessas florestas e a expansão da vegetação aberta, provavelmente continuou acontecendo intercâmbio de espécies por dispersão pelas manchas florestais. No caso das florestas com araucária e na relação com os Andes, pode-se explicar pelo avanço dessa biota, originada nas áreas temperadas, para o norte em épocas mais frias, como durante as glaciações cíclicas ocorridas desde o final do Terciário (por volta de l Omaa) .
A partir desse contexto de isolamento da floresta e a diferenciação da sua biota, quais teriam sido os principais processos que resultaram na alta diversificação observada nos dias de hoje e onde eles teriam ocorrido na história do bioma?
Uma das formas para se inferir a história biogeográfica da Mata Atlântica como um todo é procurar por padrões distribucionais atuais que se repetem em diversos táxons (ver Capo 3). Analisando as distribuições da maioria das espécies animais e vegetais da Mata Atlântica, observa-se que os táxons são restritos a alguma porção menor do bioma. Morrone13 buscou padronizar uma classificação biogeográfica fundamentada em padrões de endemismo gerados pela análise conjunta de vários grupos biológicos. Nessa classificação, a Mata Atlântica está inserida na sub-região Paranaense, dividida em
três províncias 1 3 : "Bosque Atlântico Brasilefio", que inclui as florestas de encosta e planície do Rio Grande do Norte a Santa Catarina, "Bosque Paranaense", compreendendo as florestas de planalto e relacionada à Bacia do Rio Paraná, e "Bosque de Araucaria angustifolia", formado pelas florestas mistas de planalto do Paraná e Santa Catarina. Essa classificação13 resgata as principais fisionomias do bioma, que se reproduz na distribuição de grupos taxonômicos diferentes . Entretanto, existem padrões de endemismo para a Mata Atlântica diferentes desta proposta, mais restritos e/ou discordantes, discutidos a seguir.
As propostas de divisão em áreas de endemismo convergem para a presença de quatro áreas, mas os limites não são os mesmos de estudo para estudo (Fig. 14 . 1 ) . Três delas são subdivisões da província "Bosque Atlântico Brasilefio" de Morrone13 : uma área tem centro em Pernambuco (por exemplo, Müller14, com anfíbios, répteis, aves e marrúferos, Costa et al. lS, com marrúferos, Amorim e Pires 7, com primatas e dípteros), uma com centro no sul da Bahia (por exemplo, Prance16, com plantas lenhosas, Silva e Casteleti 17, com aves, primatas e borboletas) e uma relacionada à Serra do Mar14. 16. 17. A quarta área relaciona-se às florestas de araucária, podendo alcançar áreas a oeste do Rio Paraná7• 13 (Fig. 14. 1 ).
Para entender as implicações históricas desses padrões de endemismo, ou como foram criadas essas áreas de endemismo, é de suma importância analisar a história filogenética dos grupos biológicos que vivem na Mata Atlântica. Ou seja, caso se assuma que duas espécies possuem um ancestral mais recente que aquele compartilhado com uma terceira espécie, pode-se inferir que as áreas em que as duas primeiras habitam poderiam ter uma história comum mais recente. Caso se observe repetição nesses padrões de distribuição e filogenia, podem-se inferir padrões gerais, que, em última instância, podem ajudar a reconstruir a história da floresta como um todo. Amorim e Pires7 fizeram uma análise biogeográfica usando distribuições e filogenias de primatas (Callitrichidae) e dípteros (Sciaridae e Ditomyiidae) e tiveram como resultado uma divisão principal do bioma, a mais antiga divisão detectada, no vale do Rio Doce (Espírito Santo). Seis áreas de endemismo foram delimitadas7: três ao norte e três ao sul da divisão principal, congruentes em seus limites mais amplos com as delimitações citadas anteriormente (Fig. 14. 1 ) . Outros trabalhos também têm apontado esta região do Espírito Santo como importante divisor de áreas biogeográficas, como os já citados14. l6 e os trabalhos de filogeografia de Pellegrino et al. 1 S e Cabanne et al. 19 . A porção média da Bahia, nas regiões do vale do Rio
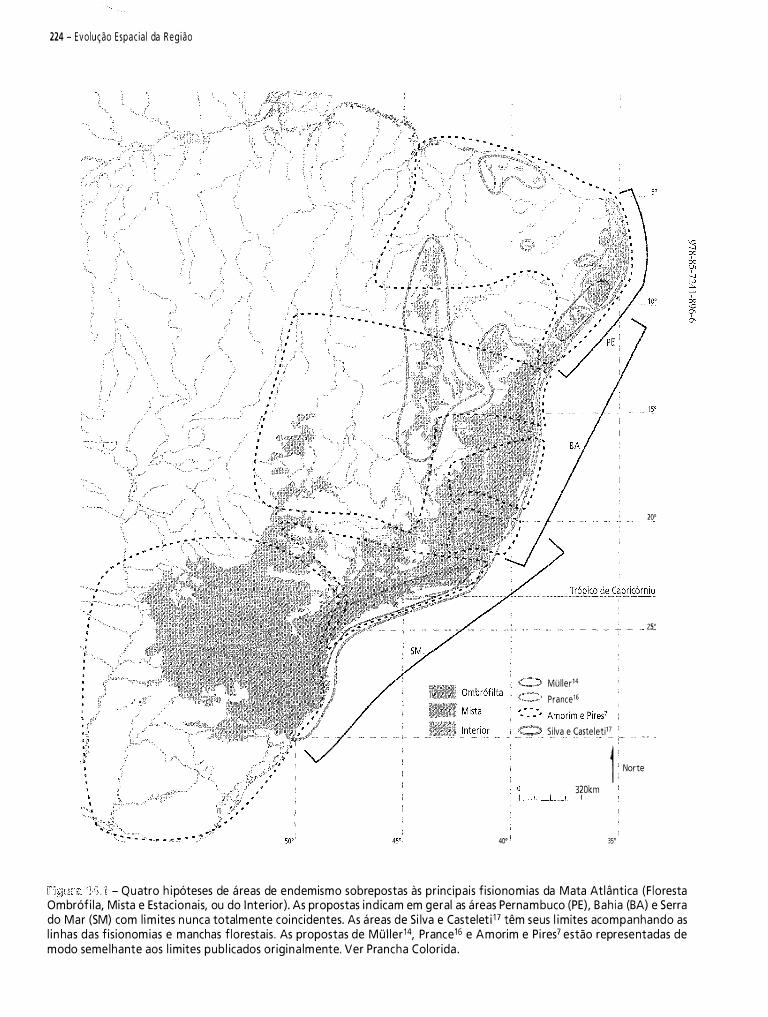
224 - Evolução Espacia l da Região
40'
Müller14 Prance16
Si lva e Casteleti17
1 ;
Norte
320km '---"----'--'----11
35'
20'
25�
- Quatro h ipóteses de áreas de endemismo sobrepostas às pr incipais f is ionomias da Mata Atlântica (Floresta Ombróf i la, Mista e Estacionais, ou do I nterior). As propostas i nd icam em geral as á reas Pernambuco (PE), Bah ia (BA) e Serra do Mar (SM) com l imites n unca totalmente coincidentes. As á reas de Si lva e Castelet i 1 7 têm seus l imites acompanhando as l i n has das fis ionomias e manchas f lorestais . As propostas de Mü l ler14, Prance16 e Amorim e Pi res7 estão representadas de modo semelhante aos l imites pub l icados orig ina lmente. Ver Prancha Colorida .

H i stória B iogeográfica da Mata Atlânt ica : Op i l iões (Arachn ida) como Modelo para sua I nferênc ia - 225
Jequitinhonha à Baía de Todos os Santos, também tem aparecido em alguns trabalhos como importante divisor de áreas8, 14, 1 6, 1 8 ,
A divisão de biotas norte-sul do bioma também é marcada pela presença de fisionomias xeromórficas chegando à costa, o que resulta na interrupção das florestas úmidas no litoral, no norte do Rio de Janeiro e na porção de baixa altitude do Espírito Santo, do Rio Paraíba do Sul ao Rio Doce1 ,
Além dos trabalhos citados7,8 , não se conhecem outras hipóteses biogeográficas gerais para a diversificação da Mata Atlântica, Cabe destacar que, apesar disso, os processos que teriam sido mais importantes para a especiação nessa floresta e em florestas tropicais como um todo têm sido muito debatidos2o,21 ,
Têm-se desenvolvido diversos modelos de especiação e diversificação das florestas neotropicais que buscam explicar a notável diversidade desses ecossistemas e, com mais detalhes, os padrões de distribuição e endemismo de sua biota, O modelo dos refúgios florestais pleistocênicos foi amplamente utilizado no final do século XX22,23. De acordo com esse modelo, as oscilações climáticas e a redução das florestas durante as glaciações ocorridas principalmente durante o Pleistoceno (começou há 2maa) teriam criado refúgios onde as populações poderiam especiar e, em períodos interglaciais, hibridizar ou se misturar a espécies provenientes de diferentes refúgios. Whitmore e Prance23, utilizando as distribuições de aves, borboletas e plantas, empregaram este modelo para propor que centros de endemismo encontrados na região Neotropical teriam resultado de especiação nestes refúgios. A maioria dos estudos que aceitam e empregam o modelo dos refúgios na porção centro-leste da América do Sul, porém, concentrou-se na região amazônica. Em muitos desses trabalhos, o uso acrítico e monótono do modelo gerou explicações inconsistentes, como o pouco tempo para gerar tantas especiações desde o Pleistoceno ou a existência de padrões biogeográficos diferentes daqueles apontados como resultado de especiação em refúgios.
Atualmente há uma tendência de se buscar a interação de diversos modelos que se complementam na formulação de hipóteses sobre a história destas biotas, incluindo mudanças geológicas, climáticas e vegetacionais, que teriam gerado relações bastante complexas entre as áreas2o. Lara et aI. 21 sintetizaram as principais hipóteses ou modelos de diversificação propostos para as florestas
tropicais. Além da Hipótese dos Refúgios, os autores discutiram o papel de mais dois modelos que também se baseiam em especiação alopátrica: a "Hipótese de Isolamento em Montanhas", em que as florestas teriam seus centros de diversificação em áreas montanhosas, e a "Hipótese de Rios como Barreiras", em que o aparecimento ou a presença dos grandes rios serviriam como barreiras que favorecem as especiações. Uma quarta hipótese baseia-se em especiação parapátrica, a "Hipótese dos Gradientes Ecológicos", na qual nos gradientes ambientais, como em ecótonos ou diferenças altitudinais ao longo do mesmo bioma, ocorreria seleção divergente e consequente especiação.
Entretanto, a procura por padrões repetitivos, seja nas distribuições das espécies, delimitando áreas de endemismo, ou em hipóteses de relacionamento entre as áreas sobre uma base filo genética, é essencial para a proposição de modelos de diversificação para as florestas neotropicais como um todo. Na próxima parte do capítulo, será mostrado como o estudo desses aspectos da biogeografia de opiliões pode ajudar a evidenciar as principais áreas históricas da Mata Atlântica e propor processos que tenham sido importantes para a diversificação da biota nas florestas tropicais.
A ordem de aracnídeos Opiliones tem-se demonstrado como bom objeto para o estudo da biogeografia histórica, especialmente na Mata Atlântica, onde existe a maior diversidade de espécies do grupo no mundo. São mais de 600 espécies descritas para esse bioma, dentre as 900 registradas no Brasi16 e muitas novas que têm sido descobertas nos últimos anos. As espécies de opiliões em geral distribuem-se em áreas bastante restritas, normalmente limitadas a uma única cadeia de montanhas, e mostram um alto grau de endemismo, maior que observado para a maioria dos grupos biológicos6.
Algumas características dos opiliões provavelmente favoreceram esse alto grau de isolamento e diversificação . Em geral, esses animais são, em especial, higrófilos, tendo dificuldades fisiológicas para evitar perda de água e apresentando comportamento críptico para compensá-la. Os opiliões têm relativamente baixa capacidade de dispersão em ambientes com vegetação aberta, onde a perda de água os impede de viver. Na Mata Atlântica, tais características restringem estes animais a hábitats com alta umidade, de preferência a uma floresta mais madura e pouco alterada24. Alguns trabalhos também têm demonstrado alta filopatria de
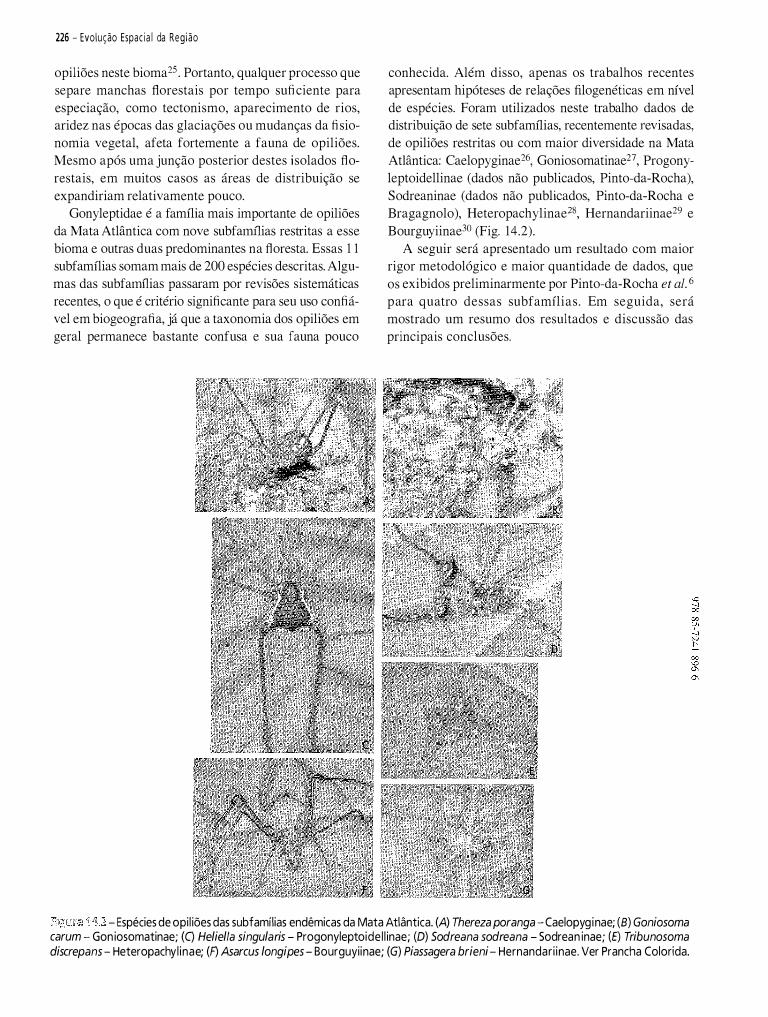
226 - Evolução Espac i a l da Região
opiliões neste bioma2s . Portanto, qualquer processo que separe manchas florestais por tempo suficiente para especiação, como tectonismo, aparecimento de rios, aridez nas épocas das glaciações ou mudanças da fisionomia vegetal, afeta fortemente a fauna de opiliões. Mesmo após uma junção posterior destes isolados florestais, em muitos casos as áreas de distribuição se expandiriam relativamente pouco.
Gonyleptidae é a família mais importante de opiliões da Mata Atlântica com nove subfaffillias restritas a esse bioma e outras duas predominantes na floresta. Essas 1 1 subfamílias somam mais de 200 espécies descritas. Algumas das subfaffillias passaram por revisões sistemáticas recentes, o que é critério significante para seu uso confiável em biogeografia, já que a taxonomia dos opiliões em geral permanece bastante confusa e sua fauna pouco
conhecida. Além disso, apenas os trabalhos recentes apresentam hipóteses de relações filogenéticas em nível de espécies. Foram utilizados neste trabalho dados de distribuição de sete subfamílias, recentemente revisadas, de opiliões restritas ou com maior diversidade na Mata Atlântica: Caelopyginae26, Goniosomatinae27, Progonyleptoidellinae (dados não publicados, Pinto-da-Rocha), Sodreaninae (dados não publicados, Pinto-da-Rocha e Bragagnolo), Heteropachylinae28, Hernandariinae29 e Bourguyiinae30 (Fig. 14.2).
A seguir será apresentado um resultado com maior rigor metodológico e maior quantidade de dados, que os exibidos preliminarmente por Pinto-da-Rocha et a!. 6 para quatro dessas subfamílias. Em seguida, será mostrado um resumo dos resultados e discussão das principais conclusões.
- Espécies de opil iões das subfamílias endêmicas da Mata Atlântica. (A) Thereza poranga - Caelopyginae; (B) Goniosoma carum - Gon iosomatinae; (C) Heliel/a singularis - Progonyleptoidel l inae; (O) Sodreana sodreana - Sodrean inae; (f) Tribunosoma discrepans - Heteropachyl inae; (F) Asarcus longipes - Bourguyiinae; (G) Piassagera brieni - Hernandari inae. Ver Prancha Colorida.

História B iogeográfica da Mata Atl ânt ica : Op i l iões (Arachnidal como Modelo para sua I nferênc i a - 227
A primeira parte da análise utiliza as distribuições de cada uma das espécies para, em seguida, buscar coincidência entre elas (ver Capo 3). Se duas ou mais espécies apresentam distribuições sobrepostas, ou são extensivamente simpátricas, podem-se criar hipóteses de áreas de endemismo. Se existe essa congruência, tem-se alguma evidência de que houve algum processo histórico, seja ele climático, geomorfológico ou ecológico, que teria influenciado e restringido boa parte da biota que vive
ali, gerando estas áreas. Procurou-se, assim, por padrões comuns de distribuição da fauna opiliológica, o que pode indicar eventos históricos.
As sete subfamílias citadas somaram 109 espécies com mais de um registro na Mata Atlântica. Todas essas espécies têm distribuições sobrepostas a de alguma outra e apenas uma espécie de Hemandariinae (Hemandaria scabricula
Sorensen, 1884) e uma de Heteropachylinae (Pseudopu
crolia rugosa [Roewer, 1 930]) têm ocorrências exclusivas e que se estendem além dos limites do bioma (Fig. 14.3).
Trópico de Capricórnio
o Pseudopucrolia rugosa
..&. Hernandaria scabricula
• 109 espêcies
O'
5'
lO'
1 5'
20'
25'
lO'
500 o 500km
'" o
,....--;.;;.;.;;;;;;,...--
- D istr ibu ição das 1 09 espécies usadas no presente estudo e duas espécies, H. scabricula e P. rugosa, exclu ídas das aná l ises por não se sobreporem às demais . A grade de 1/2° x 1/2° foi usada nos métodos numéricos aná l ise parcimon iosa de endemicidade (PAE) e eNDeMismo (NDM). No detalhe, mostra-se a grade ma is deta l hada, de %0 x %0. Foram ana l isadas 48x39 célu l as na g rade menos deta lhada e 97x79, na mais deta l hada.
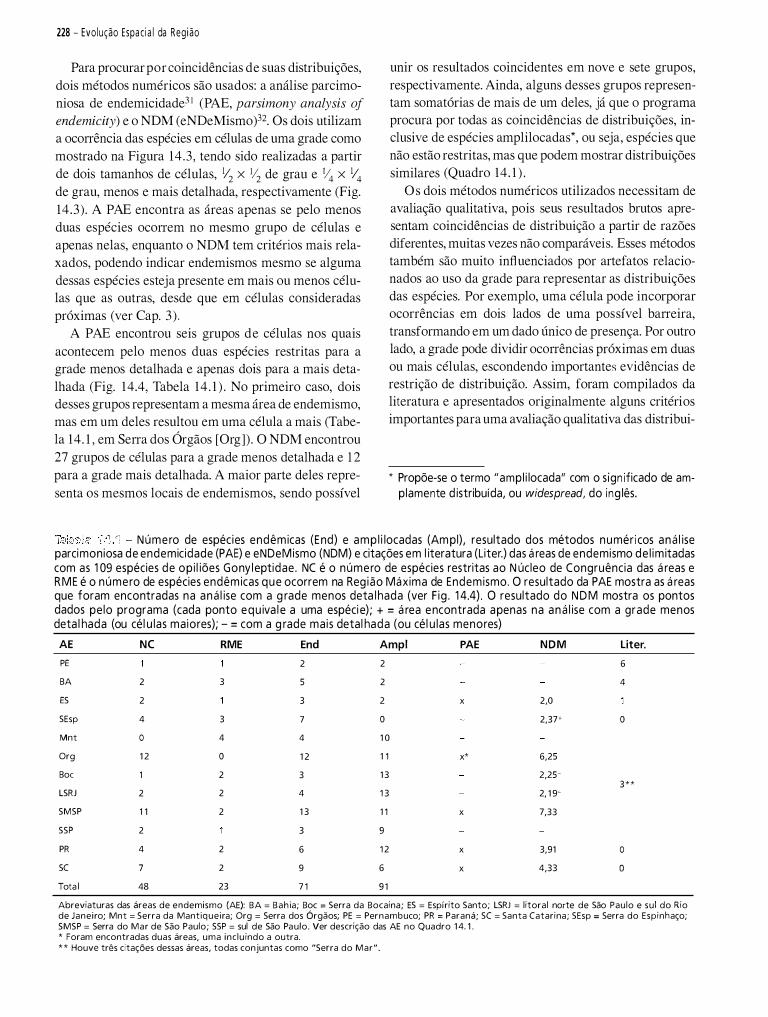
228 - Evolução Espac i a l da Região
Para procurar por coincidências de suas distribuições, dois métodos numéricos são usados: a análise parcimoniosa de endemicidade3 ! (PAE, parsimony analysis of endemicity) e o NDM (eNDeMismo)32. Os dois utilizam a oconência das espécies em células de uma grade como mostrado na Figura 14.3 , tendo sido realizadas a partir de dois tamanhos de células, V
2 x 1/
2 de grau e 1/4 x V
4 de grau, menos e mais detalhada, respectivamente (Fig. 14.3) . A PAE encontra as áreas apenas se pelo menos duas espécies oconem no mesmo grupo de células e apenas nelas, enquanto o NDM tem critérios mais relaxados, podendo indicar endemismos mesmo se alguma dessas espécies esteja presente em mais ou menos células que as outras, desde que em células consideradas próximas (ver Capo 3) .
A PAE encontrou seis grupos de células nos quais acontecem pelo menos duas espécies restritas para a grade menos detalhada e apenas dois para a mais detalhada (Fig. 14.4, Tabela 14. 1 ) . No primeiro caso, dois desses grupos representam a mesma área de endemismo, mas em um deles resultou em uma célula a mais (Tabela 14. 1 , em Sena dos Órgãos [Org]). O NDM encontrou 27 grupos de células para a grade menos detalhada e 1 2 para a grade mais detalhada. A maior parte deles representa os mesmos locais de endemismos, sendo possível
unir os resultados coincidentes em nove e sete grupos, respectivamente. Ainda, alguns desses grupos representam somatórias de mais de um deles, já que o programa procura por todas as coincidências de distribuições, inclusive de espécies amplilocadas*, ou seja, espécies que não estão restritas, mas que podem mostrar distribuições similares (Quadro 14. 1 ) .
Os dois métodos numéricos utilizados necessitam de avaliação qualitativa, pois seus resultados brutos apresentam coincidências de distribuição a partir de razões diferentes, muitas vezes não comparáveis. Esses métodos também são muito influenciados por artefatos relacionados ao uso da grade para representar as distribuições das espécies. Por exemplo, uma célula pode incorporar ocorrências em dois lados de uma possível baneira, transformando em um dado único de presença. Por outro lado, a grade pode dividir oconências próximas em duas ou mais células, escondendo importantes evidências de restrição de distribuição. Assim, foram compilados da literatura e apresentados originalmente alguns critérios importantes para uma avaliação qualitativa das distribui-
* Propõe-se o termo "ampl i locada" com o sign ificado de amplamente d istribuída, ou widespread, do ing lês.
" - Número de espécies endêmicas (End) e amp l i locadas (Ampl), resultado dos métodos numéri cos aná l ise parcimoniosa de endemicidade (PAE) e eNDeMismo (NDM) e citações em l iteratura (Liter.) das á reas de endemismo de l im itadas com as 1 09 espécies de op i l iões Gony leptidae. NC é o número de espécies restritas ao N úcleo de Congruência das á reas e RME é o número de espécies endêmicas que ocorrem na Reg ião Máxima de Endemismo. O resultado da PAE mostra as á reas que foram encontradas na aná l ise com a g rade menos deta lhada (ver F ig . 1 4.4) . O resultado do N D M mostra os pontos dados pelo programa (cada ponto equ iva l e a uma espéci e); + = á rea encontrada apenas na aná l ise com a grade menos deta l hada (ou cél u las ma iores); - = com a g rade mais detal hada (ou cél u las menores)
AE NC RME End Ampl PAE N D M Liter.
PE
BA 2 3
ES 2
SEsp 4 3
Mnt o 4
Org 1 2 o Boe 2
LSRJ 2 2
SMSP 1 1 2
SSP 2
PR 4 2
SC 7 2
Tota l 48 23
2 2
5 2
3 2
7 o 4 1 0
1 2 1 1
3 1 3
4 1 3
1 3 1 1
3 9
6 1 2
9 6
7 1 9 1
x
x*
x
x
x
2,0
2,37+
6,25
2,25-
2, 1 9-
7,33
3,91
4,33
6
4
o
3 * *
o
O
Abreviaturas das áreas de endemismo (AE): BA = Bah ia; Boc = Serra da Bocaina; ES = Espírito Santo; LSRJ = l itoral norte de São Paulo e sul do Rio de Janeiro; Mnt = Serra da Mantiq ueira; Org = Serra dos Órgãos; PE = Pernambuco; PR = Paraná; SC = Santa Catarina; SEsp = Serra do Espinhaço; SMSP = Serra do M a r de São Paulo; SSP = sul de São Pa ulo. Ver descrição das AE no Quadro 1 4. 1 . * Foram encontradas duas áreas, uma inc lu indo a outra. * * Houve três citações dessas áreas, todas conjuntas como "Serra do Mar".

3 7 40 58 8084 a34-19
H istória B iogeográfica da Mata At lâ ntica: Op i l iões (Arachn i da) como Mode lo para sua I nferênc ia - 229
7 62 8 103 r"'� a66-35 ü·--U�a66_36
, 10 , a85-24 �-La86-24
a35- 18 41
1 1 4752 101
98 106 49 5085 92 a37- 12
101 a38- 12
1 9 25 31 99 a27-27 a28-27
7 18 77 79
44 90 91 101
a 17-30 yt---t--<o--- a 1 8-29
a 1 8-30 a46-9
1 1 14
97
a38- 1 1 PR a39-1 1
ES a32-22
'--0-00-.-0-- a32 -2 3
se
a35-16
Org
yt-oo-oo-o.o-- a3 5-17 SMSP
a36-14 '-0-00-0.0-- a 3 7-1 1
86 a86-22 :-::-C a86-23
, a77-16 1 1 33 I
I ' a79-11 L a84-2
Org SMSP
,,' - Cladogramas resu ltantes da aná l ise parcimoniosa de endemicidade (PAE), para a grade menos deta lhada (à esquerda) e mais deta lhada (à direita), com as á reas de endemismo reconhecidas pelo agrupamento das cél u las com duas ou mais espécies exclusivas. É mostrada a parte dos c ladogramas de consenso onde há agrupamento de cél u l as. Círculos pretos: representam as presenças exclusivas das espécies. " * ,, e " * * ,, são as duas possibi l i dades de áreas inclusivas na área Org (Serra dos Órgãos). ES = Espírito Santo; PR = Paraná; SC = Santa Catar ina; SMSP = Serra do Mar de São Pau lo .

230 - Evolução Espacia l da Região
Ç;rv;�cbJ '1 - Critérios combinados de ava l iação dos endemismos
1 . Coincidência na distr ibuição de pelo menos duas espécies: critério básico para a procura por áreas de endem ismo, como ind icado por aná l ise parci moniosa de endem icidade (PAE) e eNDe M ismo (NDM)
2. As espécies devem ser classificadas em três n íveis de endem ismo: ava l iando as distri buições das espécies, percebe-se que existem aquelas cujas d istribuições são restritas e congruentes com a de outras espécies, como encontrado pelos métodos numéricos. Chama-se essa pequena área de " N úcleo de Congruêncía" (NC). Além dessas, existem vá rias espécies que ocorrem a penas em um NC, ou seja, são endêmicas, mas têm sua d istr ibu ição m a ior que essa restrição: chama-se essa área maior de "Reg ião Máxima de Endem ismo " . Ainda há outro g rupo de espécies, aquelas d istribu ídas em mais de um NC, ou espécies a m p l i locadas, e que não podem servir de evidência de áreas de endemismo
3. A congruência geral de d istri buições é diferente dos endem ismos específicos: é comum ocorrer congruência entre espécies ampl i locadas tam bém, o que não pode ser usado para de l imitar áreas de endemismo. Assim, é importante d iferenciar os dois padrões. Como exemplo, temos a lguns gru pos de células encontrados pelo NDM que devem ser descartados por representarem esse tipo de congruência geral e somarem mais de um núc leo de congruência
4. Áreas de endemismo devem ser exclusivas: como h ipóteses de iso lamento e restrição histórica de u m g rupo de espécies, as á reas devem ser exclusivas. Alguns resultados do N D M tam bém devem ser ava l iados por a presentarem á reas contraditórias
5. Espécies d istribuídas fora de qualquer NC podem representar outras áreas de endemismo: essas espécies podem se sobrepor a outras, mas não podem coincidir espacia lmente para esta belecer um NC. Apesar disso, elas podem indicar um possível endemismo, o que pode ser evi denciado posteriormente quando de posse de dados de ocorrência adicionais
6. Evidências geográficas: possíveis barrei ras ou unidades geog ráficas podem ind icar áreas de endemismo, como gra ndes rios, a ltos de serras, i nterrupções de floresta, va les etc. Essas evidências em geral são mu ito mais precisas do que os l imites im postos pelas cél u las das grades
ções dos opiliões. São chamados Critérios Combinados (Quadro 14. 1 ), pois devem ser utilizados juntos, de forma dinâmica, e confrontados com os resultados dos métodos numéricos.
Empregando os métodos numéricos e a avaliação qualitativa por meio dos Critérios Combinados com as 1 09 espécies de opiliões, foram encontradas 12 áreas de endemismo para a Mata Atlântica (Fig. 14.5, Tabela 14. 1) , que estão descritas no Quadro 1 4.2.
Como esperado, os opiliões forneceram evidências de endemismo em áreas muito menores que as apresentadas por outros grupos. As áreas Pernambuco (PE) e Bahia (BA) são as mais corroboradas pelos dados da literatura, tendo sido citadas seis e quatro vezes, respectivamente, nos trabalhos reunidos por nós, conforme indicações derivadas de estudos de outros organismos (ver seção "Endemismos e Relações Internas", anteriormente). Espírito Santo (ES) havia sido citada uma vez, enquanto as outras não haviam sido propostas antes. Grande parte das áreas concentra-se na porção sul da Mata Atlântica e algumas delas haviam sido citadas unidas à grande área "Serra do Mar" por três trabalhos (Tabela 14 .1) .
Cinco delas foram encontradas pela PAE com a grade menos detalhada, seis pelo NDM com a grade menos detalhada e sete pela grade mais detalhada (Tabela 14. 1) . As áreas mais ao norte, PE e BA, não foram encontradas pelos métodos numéricos porque são as menos representadas em número de ocorrências e são muito extensas, deixando os registros muito esparsos (Fig. 14.3). Provavelmente, elas seriam encontradas com esses dados usando-se uma grade com células maiores ou com a
intensificação de estudos na região. O mesmo aconteceu com a área Serra do Espinhaço (SEsp), que foi encontrada apenas com a grade menos detalhada pelo NDM. Três áreas, Serra da Bocaina (Boc), litoral norte de São Paulo e sul do Rio de Janeiro (LSRJ) e sul de São Paulo (SSP), sofreram o problema de artefato do uso da grade já citado. Nos dois primeiros casos, como são áreas adjacentes, elas foram incluídas numa mesma célula na grade menos detalhada; por isso, só foram encontradas com a grade mais detalhada pelo NDM. No caso de SSP, algumas células incluíam ocorrências nela e ocorrências na área ao sul, Paraná (PR), do outro lado de um grande rio, o Ribeira do Iguape.
As áreas Serra da Mantiqueira (Mnt) e SSP foram as menos corroboradas pelos dados de ocorrência, apesar de haver evidências para as suas delimitações como áreas de endemismo (Tabela 14. 1) , como duas espécies endêmicas e formarem unidades geográficas com barreiras as limitando. Por outro lado, algumas áreas são hipóteses bastante robustas, como Serra dos Órgãos (Org), Serra do Mar de São Paulo (SMSP), PR e Santa Catarina (SC).
As áreas de endemismo definidas na Figura 14.5 foram delimitadas com um Núcleo de Congruência, ou a área propriamente dita, e uma Região Máxima de Endemismo (RME), conforme discutido nos Critérios Combinados (Quadro 14 .1) . A RME pode ser interpretada de duas formas : pode significar uma região de transição entre áreas de endemismo, como de recente colonização por espécies de maior dispersão, por exemplo. A RME também pode ser interpretada do ponto de vista metodológico, sendo pensada como uma região

H istór ia B iogeográfica da Mata At lâ ntica: Op i l iões (Arachn ida) como Mode lo para sua I nferência - 231
As áreas aqui a presentadas são resultados da presente aná l ise. Algumas delas corroboram traba l hos a nteriores, i l ustrados na F igura 14.1 e Tabela 1 4. 1 : • Pernambuco (PE) (7°50'-1 0°50' S 34°50'-37°40' W). Estreita faixa n a costa d e Pernambuco a Sergipe, formada por p la n ície costeira ampla na foz dos rios, ou substituídas por fa lésias e Tabuleiros Costeiros baixos (até 1 00m de altura), formando um relevo ondu lado cortado pelo Rio São Francisco. C l ima dominado por ventos a l ísios de sudeste, o que permite estab i l idade à reg ião: chove muito durante todo o ano (até 2 .200mm anua is), média anua l de 25°C. Ao sul da área, a floresta é considerada Estaciona i Semidecidua e ao norte, Ombrófi la Densa e Aberta. O núcleo de congruência (NC) se estende do Rio I pojuca, ao sul do Rio São Francisco, quase chegando ao Rio Vaza-Barris • Bahia (BA) (1 4°30'- 1 8°50' S 38°50'-39°40' W). Estreita faixa l itorânea formada pela P lan ície Costeira i rreg u lar, ampla nos deltas, principal mente no Rio Doce, ou com falésias, e Tabule iros Costeiros (até 1 00m de a ltitude e 1 20km de largu ra) . Restrita a oeste pelos maciços crista l i nos, mais a ltos e acidentados. Possui a lta umidade, ao norte do NC a p l uvios idade média é su perior a 1 .500mm anuais (concentrada no outono), enquanto na porção S, próxima do va le do Rio Doce, de cerca de 1 .200mm anuais (concentrada na primavera). A temperatura va ria pouco, média em torno de 25°C. Formada pelas Florestas dos Ta buleiros, dominada pela Floresta Ombrófi la Densa, sendo substituída por formações pioneiras nas p lan icies costeiras. O NC se estende do Rio de Contas (Bahia) até o Rio Doce no Espírito Santo • Espírito Santo (ES) (1 9°50'-20°20' S 40°30'-40°40' W). Abra nge serras próximas à costa com patamares/degraus de l imitados por frentes escarpadas com cai mento topográfico para SE. Essas serras atingem de 800 a 1 .000m de a ltitude, formando pontões rochosos no a lto e va les profundos q ue drenam a porção sul da bacia do Rio Doce. A pl uviosidade varia de 1 .400 a 1 .800mm anua is: possui estação mais seca no inverno. A temperatura média anua l varia de 20 a 22°C, no p lana lto e l itora l, respectivamente. A vegetação dominante é a Floresta Ombrófila Densa, de Terras Ba ixas, Submonta na e Montana, e F loresta Ombrófi la Aberta ao su l do N C. O NC está no centro do estado de Espírito Santo, l i m itado ao norte pelo va le do Rio Doce • Serra do Espinhaço (SEsp) (20°-20°50' S 42°20'-43°40' W). Composta por três un idades de relevo: ao noroeste o Quadri látero Ferrífero, i n ício sul da Serra do Espinhaço, marcado por cristas escarpadas e grandes paredões das serras de Ouro Branco e do Caraça, a ltitude média de 1 .000m; a SE as serras com cristas e a longadas, como a Serra do Brigadeiro (altitude superior a 1 .000m); e entre as duas a Depressão Interplanática do Médio Rio Doce com formas col inosas a longadas de topos convexados (altitude média de 550m). Pluviosidade anual na porção norte superior a 1 .700mm anua is, na depressão de cerca de 1 . 1 00mm, com uma estação mais seca com quatro meses no inverno. Temperatura méd ia da porção mais baixa é de cerca de 20°C. Coberta pela F loresta Estacionai Semidecidual M ontana, sendo substituida por campos de a ltitude acima de 1 . 500m. O NC é l i m itado ao sul e ao oeste pelo f inal nordeste da Serra da Mantiqueira, ao leste pela Serra do Caparaó e ao norte pela Serra do Espinhaço em M i nas Gerais • Serra dos Órgãos (Org) (2 1 °50'-22°50' S 41 °40'-43°30' W). Composta pela Ba ixada F luminense e seus Maciços Costeiros, com pães-de-açúcar e serras isoladas, e a Serra dos Órgãos nas suas escarpas íngremes e festonadas voltadas para o mar. Caracterizada por inúmeros va les su lcados divid indo-as em espigões, que descem até a baixada, e e levações rochosas de 2.000m de a l titude. Ao leste perde a forma de paredão e red uz de tamanho, inf luenciada pelo amplo vale do Rio Macaé. A pl uviosidade na Serra dos Órgãos tem média de 2 . 1 OOmm anua is, reduzi ndo na p lanície para 1 .300mm, sem estação seca. Temperatura média anua l de 1 8°C no alto da serra e 23°C na baixada. Coberta por F loresta Ombrófi la Densa, com domín io de F loresta Submontana. Del imitada ao norte pelo a lto da Serra dos Órgãos, ao leste pela interrupção da mata no va l e do Rio Paraíba do Sul e ao su l pelo fim dos maciços da Ba ixada F luminense • Serra da Mantiqueira (Mnt) (22°1 0'-22°30' S 44°30'-44°50' W). Abrange o Maciço de Itat ia ia e sua encosta até o vale do Rio Paraíba do Sul , fazendo pa rte da porção su l da Serra da Mantiqueira. O Maciço é caracterizado por relevo montanhoso de a ltitude superior a 2.000m e íngreme, com vales profundos. Os topos das montanhas são desnivel ados e estreitos, formando cristas e p icos, muitas vezes rochosos. A precipitação média anua l é de 1 . 500mm no reverso da serra, para o interior, e de 2 .300 mm no a lto da escarpa sul , com dois meses secos no a no, e a temperatura média geral é de 22°C, no a lto do p lana lto é de 1 1 °C. A vegetação é a Floresta Ombrófi la Densa Montana e Alto-montana, com manchas de Araucaria angustifolia e Podocarpus lambertii na ú ltima, e Cam pos de Altitude (acima de 1 .600m). O NC é pequeno, a penas nas matas do Pa rque Nacional de Itatia ia • Serra da Bocaina (Boc) (22°40'-22°50' S 44°30'-44°40' W). Está no planalto da Serra da Bocaina, b loco montanhoso da Serra do Mar, com a ltitudes de cerca de 1 .000 a 2 . 132m, entre o vale do Rio Paraíba do Su l e o litoral. O planalto apresenta topos convexos e cristas aguçadas, com relevo dissecado em formas alongadas, apresentando vales de até 1 92m de profundidade. A precipitação média anual é de 1 .700mm com dois a três meses secos e a temperatura média é 1 4°C. A vegetação é dominada pela Floresta Ombrófi la Densa Alto-montana e F loresta Ombrófi la Mista (com Araucaria angustifolia e Podocarpus Jambertit), com uma mancha de campos de a ltitude • Litoral norte de São Pau lo e sul do Rio de Janeiro (LSRJ) (22°50'-23°40' 5 44°20'-45° 1 0' W). Formada pelas encostas e l itora l adjacente da Serra da Bocaina e do Plana lto Paraitinga-Para ibuna da Serra do Mar. As escarpas são vertentes íngremes e festonadas com cerca de 1 .000m de a ltitude, com vales curtos e profundos, sendo ma is irregu lar com grandes reentrâncias próxima à Serra da Bocaina. A plan ície l itorânea é bastante estreita, com muitas enseadas e baías decorrentes das sa l i ências da escarpa, com seus esporões mu itas vezes chegando no mar. A precipitação é superior a 2 .000mm mensais, sem período seco, com temperaturas médias a nuais de 21 a 23°C. Coberta pela F loresta Ombrófila Densa, variando de Terras Ba ixas, Su bmontana e Montana. O NC se estende do n ível do mar a 1 .300m de a ltitude na divisa entre Rio de Janeiro e São Paulo, de Angra dos Reis à U batuba • Serra do Mar de São Paulo (SMSP) (23°30'-24°S 45°1 0'-46°50'W). Porção da Serra do Mar no Plana lto Paulistano e l itora l . As escarpas formam cristas e espigões (de 1 .200m em média) subpara lelos à l in ha da costa na porção nordeste, reduzindo a menos de 1 .000m, com vertentes íngremes na porção sudoeste. Na primeira porção, o l itoral é estreito e os rios que descem da escarpa são curtos e torrenciais; na segunda, a serra é separada do mar por extensas p lanícies flúvio-marinhas. A precipitação e a temperatura anua l médias são de 2.000mm e 1 7°C no plana lto a mais de 3.000mm na escarpa e litoral, com cerca de 22°C de temperatura, sem estação seca. Dominada pela Floresta Ombrófi la Densa, variando entre Floresta de Terras Baixas, Submontana e Montana, com campos de a ltitude e cerrado no planalto. No planalto, o NC é l imitado ao norte pelo va le do Rio Tietê, situando-se em volta da bacia de São Paulo nas col inas crista l inas. No litora l e escarpa da Serra do Mar, ele se estende da I lha Bela ao norte à Ba ixada Sa ntista ao su l .
(continua)

232 - Evolução Espacia l da Região
... ; c, ] ::: . , c - Áreas de endemismo (continuação)
• Sul de São Paulo (55P) (24°1 0'-25°30' 5 48° 1 0'-48°40' W). Situa-se nas escarpas da Serra de Paranapiacaba, entre o p lana lto e o va le do Rio Ribeira do I guape que é contínuo à p lan ície l itorânea. O relevo é montanhoso com serras a longadas, vales profundos, escarpas festonadas e espigões d ig itados. A base da escarpa na p lan ície está a a proximadamente 1 00m de a ltitude e o seu topo passa dos 1 .000m. O c l ima é subtropical úmido, sem estação seca, com precipitação anua l de cerca de 1 . 500mm e temperatura méd ia de 18 a 1 9°C. Coberta pela Floresta Ombrófi l a Densa Submontana e Montana. O NC está restrito aos Pa rques Estaduais Turístico do Alto do Ribe i ra (PETAR) e I ntervales, em São Paulo • Paraná (PR) (24°50'-26°5 48°20'-49°20' W). Formada pela Serra do Mar do Paraná e o Pr imeiro P lanalto Paranaense. A serra é dividida em d iversos maciços e leva ndo-se de 500 a 1 .000m acima do plana lto, ati ngindo quase 2.000m (a noroeste). A serra é bastante i rregu lar, apresentando escarpas íngremes, monta nhas isoladas, cones regu lares, cumes rochosos e vales profundos. Ao sudoeste, a escarpa forma a penas um degrau íngreme do p lana lto e ocorrem serras iso ladas mais próximas da costa. O primeiro p lanalto é u n iforme de pa isagem ondulada e a ltitude de cerca de 900m. C l ima é temperado úmido e as médias anuais de precipitação e temperatura var iam do planalto (1 .400mm e cerca de 1 6°C) para o litora l (3.000mm e 2 1 °C). Dominada pela F loresta Ombrófi la Densa Submontana, Montana e Alto-montana, com Araucaria angustifolia acima de 500m . O NC ocupa a porção central e sudoeste da Serra do M a r e i n ício do Plana lto de Curiti ba . O l i m ite da área ao sul coincide com o fim da Serra do Mar como escarpa próxima à costa, que se d i reciona a sudoeste até o Va le do Itajaí e o i n ício da Serra Geral • Santa Catarina (5e) (24°50'-26°5 48°20'-49°20' W). Situa-se no entorno do vale do Rio Itajaí, na sua pa rte baixa e porções montan hosas ao norte e su l . Ao norte está um relevo acidentado com encostas íngremes, cu lminando em largos cha padões, e va les profundos e estreitos. Ao s u l o vale é interrompido pelo G rupo B rusque, complexo rochoso com falhas intensas que formam os vales do Rio Itaj aí-Mir im e Rio Tijucas. Mais ao sudoeste, na costa, estão as Serras do Tabule iro e a I l ha de Santa Catarina, onde se formam cristas montanhosas, com a ltitudes superiores a 1 .000m na pr imeira e 500m na segunda. Cl ima influenciado pelas massas de ar oceânicas, precipitação de 1 .400 a 1 .600mm anuais, sem estação seca, e temperatura média de 2 1 °C, podendo cair para a ba ixo de zero no inverno. Vegetação domi nada pela Floresta Ombrófi l a Densa de Terras Baixas, Submontana e M ontana, com Araucaria angustifolia. O NC incl u i as encostas do fim da Serra do Mar e das serras do leste catarinense, as p lan ícies costei ra e fluviais das bacias do Itaja í e do Tijucas e a I lha de Santa Catar ina
com carência de estudos que possam complementar as ocorrências, o que serviria como fonte de falseabilidade da hipótese de delimitação.
A partir dessa divisão histórica em áreas, tem-se uma hipótese inicial sobre uma divisão da Mata Atlântica em unidades históricas. Isso só tem significado, porém, além de uma proposta de regionalização biogeográfica, caso se some o vetor temporal à análise, pelo emprego das filogenias dos opiliões, como descrito a seguir.
Ao comparar filogenias de organismos que vivem em um mesmo conjunto de áreas, procura-se por padrões gerais de repetição de uma mesma história. Se existe repetição entre essas filogenias, pode-se concluir que resultam de eventos comuns que teriam afetado grande parte da biota. Apesar do padrão geral, porém, cada táxon tem também o seu grau de história individualizada, pois as espécies podem se dispersar, extinguir, ou se especiar por motivos únicos. Por isso, esta comparação não é trivial e exige aprofundamento metodológico dentro da disciplina da Biogeografia Cladística (ver Capo 6).
No presente estudo de caso, utilizaram-se sete hipóteses filo genéticas de grupos monofiléticos, a maior contendo 36 espécies terminais (Goniosomatinae) e a menor apenas seis (Sodreaninae). Essas espécies estão distribuídas nas 12 áreas de endemismo descritas anteriormente. Noventa e quatro dessas espécies sucedem em
apenas uma área de endemismo, enquanto as outras 34 acontecem em mais de uma, ou seja, são amplilocadas. Nem todas essas filogenias apresentam espécies presentes nas 12 áreas de endemismo. Heteropachylinae, por exemplo, está presente em apenas três delas. Ao se observarem as relações entre as espécies e as áreas em que elas vivem, nota-se que relacionar as áreas é muito difícil, pois existem muitas redundâncias. Para procurar um padrão geral, um cladograma de área generalizado, utilizaram-se análises de parcimônia, considerando as áreas de endemismo como terminais e os nós e terminais das hipóteses filogenéticas dos opiliões como caracteres. Foram feitas quatro análises : ( 1 ) com todos os nós e terminais (ou análise de parcimônia de Brooks [BPA, Brooks parsimony analysis]) ; (2) apenas com os nós; (3) utilizando os nós que não apresentavam redundância; (4) apenas com os terminais. Empregando esse protocolo, podem-se representar camadas temporais diferentes nas análises, ilustradas pelos vários níveis de inclusão das filogenias, desde os nós mais antigos das histórias dos grupos, até as especiações mais recentes, representadas pela análise apenas com os terminais.
Essas quatro análises resultaram em 10 hipóteses diferentes relacionando as 12 áreas de endemismo. Esses 10 cladogramas de áreas são sumarizados na Figura 14.6, que visa apresentar o padrão geral das relações entre as áreas. Nele, podem-se ver as áreas de endemismo reunidas em três blocos principais: norte, formado por PE e BA, central, formado por Mnt, Org, Boc, LSRJ e SMSP,

H istória B i ogeográf ica da Mata Atl â ntica: Op i l iões (Arachn ida) como Modelo para sua iderE i c , ê - 233
e sul, formado por SSP, PR e se. As áreas ES e SEsp apresentam relações conflitantes, podendo estar mais relacionadas cada uma delas com blocos diferentes. Dentro de cada bloco, as relações não estão claramente resolvidas, representadas pelas politomias. No atual estado do conhecimento acredita-se que relações entre áreas continentais, sem barreiras muito marcantes, sejam
mais difíceis de serem detectadas. Esse resultado representa com maior realidade uma história bastante complexa dessas áreas, com reticulações e trocas bióticas entre elas. Mas que tipo de inferências pode ser tirado de um resultado como esse? Quais foram os eventos e/ou processos mais importantes para a diversificação dos opiliões e como se pode interpretá-los para toda a biota?
';tL5 - Áreas de endemismo fundamentadas na congruência nas distribuições das 1 09 espécies de sete subfamíl ias de opil iões Gonyleptidae. Linhas completas: são os Núcleos de Congruência das á reas (NC); linhas tracejadas: as Regiões Máximas de Endemismo (RME). BA = Bahia; Boc = Serra da Bocaina; ES = Espírito Santo; LSRJ = l itoral norte de São Paulo e sul do Rio de Janeiro; Mnt = Serra da Mantiqueira; Org = Serra dos Órgãos; PE = Pernambuco; PR = Paraná; se = Santa Catarina; SEsp = Serra do Espin haço; SMSP = Serra do Mar de São Pau lo; SSP = sul de São Paulo. Ver texto e Quadros 1 4. 1 e 1 4.2. Ver Prancha Colorida.
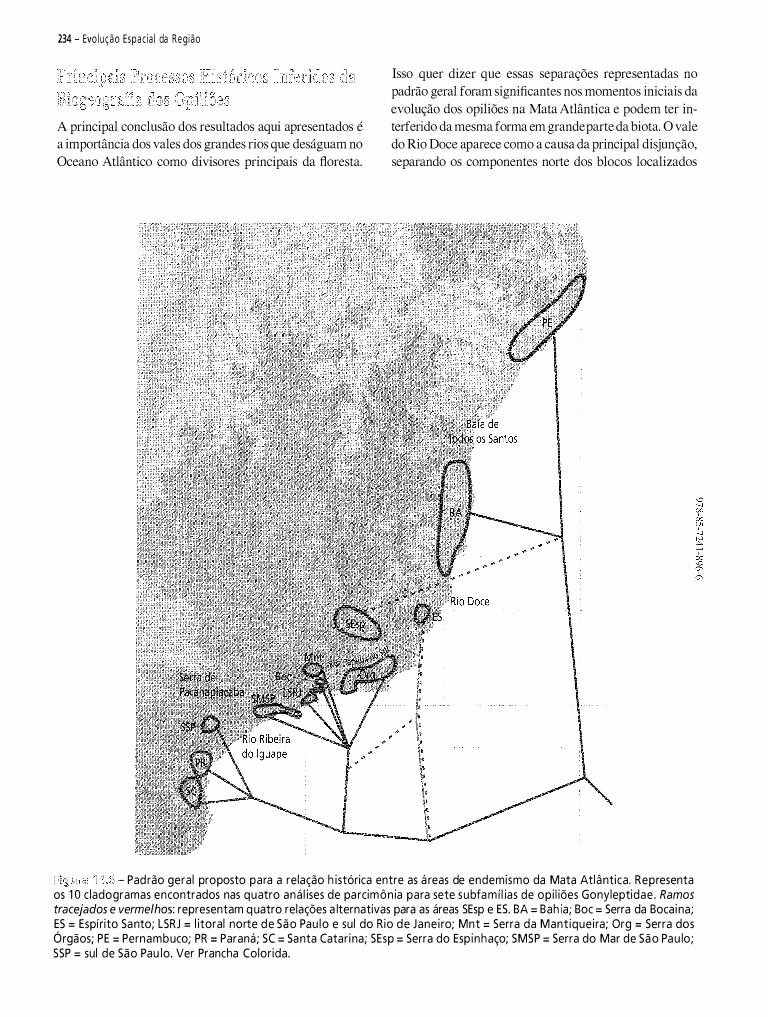
234 - Evo lução Espac ia l da Região
A principal conclusão dos resultados aqui apresentados é a importância dos vales dos grandes rios que deságuam no Oceano Atlântico como divisores principais da floresta.
Isso quer dizer que essas separações representadas no padrão geral foram significantes nos momentos iniciais da evolução dos opiliões na Mata Atlântica e podem ter interferido da mesma forma em grande parte da biota. O vale do Rio Doce aparece como a causa da principal disjunção, separando os componentes norte dos blocos localizados
- Padrão geral proposto para a relação h istórica entre as á reas de endemismo da Mata Atlântica. Representa os 1 0 cladogramas encontrados nas quatro aná l ises de parcimôn ia para sete subfam íl ias de opi l iões Gonyleptidae. Ramos tracejados e vermelhos: representam quatro relações a lternativas para as áreas SEsp e ES. BA = Bah ia; Boc = Serra da Boca ina; ES = Espírito Santo; LSRJ = l itoral norte de São Pau lo e sul do Rio de Jane i ro; Mnt = Serra da Mant iqueira; Org = Serra dos Órgãos; PE = Pernambuco; PR = Paraná; SC = Santa Catar ina; SEsp = Serra do Esp inhaço; SMSP = Serra do Mar de São Pau lo; SSP = sul de São Pau lo. Ver Prancha Colorida.

H i stória B iogeográfica da M ata Atl ântica: Opi l iões (Arachnida) como Mode lo para sua I nferência - 235
no centro e sul, como também apontado por trabalhos anteriores 7,14,16, 1 8, 19, Pela sua relativa antiguidade, pode-se inferir que os processos tectônicos responsáveis pelo surgimento do vale e a origem do rio podem já ter sido de grande importância para essa disjunção, Infelizmente, existe pouco conhecimento geológico sobre esse processo, em especial datações desses eventos, Outros eventos que podem ter separado essa biota ancestral relacionam-se às transgressões marinhas ocorridas nos períodos interglaciais, desde o Pleistoceno pelo menos33. Atualmente, podem-se observar nesse grande vale sistemas lacustres e de sedimentação marinha a até 50km para dentro do continente, o que indica que tais transgressões teriam afetado bastante a distribuição de populações de espécies florestais.
Outro fator a ser considerado é a diferenciação geomorfológica entre as porções norte e sul da Mata Atlântica, e que ocorre por volta dessa região do Rio Doce. Grande parte das áreas de endernismo situa-se nas encostas do planalto atlântico ao sul, que estão bastante próximas do litoral, do Rio de Janeiro a Santa Catarina, enquanto no Espírito Santo e Serra do Espinhaço situam-se em serras afastadas da costa e na Bahia e em Pernambuco, em extensas planícies litorâneas.
De acordo com o padrão geral, a divisão subsequente da floresta estaria relacionada ao vale do Rio Paraíba do Sul, esse sim com boa documentação do seu passado. Esse vale acompanha a principal falha do sudeste brasileiro que se abriu desde o Oligoceno a partir de pulsos tectônicos ocorridos desde o começo do Cenozoico, causados pela deriva do continente sul-americano para oeste34. Além do seu surgimento, as transgressões marinhas, como no caso do Rio Doce, também parecem ter sido importantes para a configuração atual, além do que, se observam ainda hoje em dia nesse vale a interrupção natural das florestas e a existência de vegetação xeromórfica.
Outra vicariância importante parece ter ocorrido na região da Serra de Paranapiacaba, onde a Serra do Mar de São Paulo se afasta da costa e forma o vale do Rio Ribeira do Iguape, separando o bloco central do bloco sul . O surgimento do vale do Rio Ribeira teve grandes consequências na morfologia da Serra do Mar, fazendo com que a escarpa do planalto se situe mais ao interior, formando a Serra de Paranapiacaba. Existem evidências de eventos tectônicos causadores da abertura do vale desde o início do Terciário, o que modificou bastante a estrutura escarpada da Serra do Mar que chega ao sul até a Serra de Itatins33. Os grandes eventos j á citados podem ter determinado esta separação mais antiga, apesar de não ser o próprio vale o principal fator de disjunção nesse caso, pois a área SSP, mais relacionada com o bloco sul, encontra-se ao norte do rio.
As disjunções dentro de cada um dos três blocos, apesar de não apresentarem resolução quando observado o padrão geral da Figura 14.6, são inferidas pelo alto grau de endemismo das espécies de opiliões e pela delimitação das áreas pela congruência de suas distribuições. Assim, pode-se inferir que cada uma delas se manteve isolada por grande período depois das principais vicariâncias que separaram os três blocos j á citadas. Os processos que isolaram essas áreas podem não ter afetado, da mesma forma, grande parte da biota e parece ter havido muitas trocas entre elas por meio de dispersão, o que explica a falta de congruência nas suas relações.
No bloco norte, a principal disjunção parece ter ocorrido na região da Baía de Todos os Santos, como também mostrada em outros trabalhosS.14,16, 1S . Nos casos dos blocos central e sul, observam-se a importância das serras e suas escarpas na formação desses endemismos, como a Serra do Mar e a Serra da Mantiqueira. Pode-se relacioná-los à própria formação dessas serras e seus vales, de origem nos pulsos tectônicos ocorridos principalmente no Terciári033. Entretanto, eventos mais recentes podem ter influenciado mais nas últimas especiações, na parte final das filogenias. As reduções das florestas durante os períodos glaciais podem explicar esse alto endemismo, posto que, nessa época, elas estariam concentradas nas escarpas das serras voltadas para o mar, locais onde as áreas de endemismo estão delimitadas e ocorrem as maiores precipitações do leste brasileiro, causadas pelas chuvas orográficas. Mesmo com o aumento na extensão das florestas nos períodos interglaciais, como hoje, muitos opiliões mantiveram restrição nas suas distribuições pela sua baixa vagilidade. Pode-se verificar esse fenômeno entre áreas como SMSP e LSRJ, onde se encontra continuidade entre as matas, mas grandes diferenças entre as faunas de opiliões. Em outros casos, como entre Boc e LSRJ, ou Mnt e Org, ainda ocorre interrupção da floresta, seja pela altitude das serras, como no primeiro caso, seja pela sombra de chuva nos reversos dessas serras, como no segundo caso. O vale do Rio Paraíba do Sul, localizado em um desses reversos em grande parte da sua extensão, ainda divide as áreas Org, Boc e LSRJ, de um lado, e SEsp e Mut, de outro.
Outra inferência importante que se deriva dos padrões filogenéticos dos opiliões é que várias destas barreiras, em particular as mais novas, teriam que ser cíclicas. Isto é, apareceriam e davam tempo à especiação, e desapareciam ou reduziam sua importância, dando tempo a muitas dessas espécies expandirem suas áreas de distribuição, voltando a aparecer novamente em locais onde haviam sido extintas localmente, e assim por diante. Como exemplo, pode-se observar a filogenia do gênero Heteromitobates, com seis espécies que vivem nas áreas

236 - Evolução Espacial da Região
de endemismo Boc, LSRJ e SMSP (Fig. 14.7). As redundâncias nas relações entre as áreas podem ser explicadas nesse modelo por dois ciclos, admitindo-se que as espécies ampliariam suas distribuições para as três áreas nos ciclos de ampliação da floresta e sofreriam três especiações em cada período vicariante, com duas extinções em Boc e LSRJ. Claro que, neste caso, admite-se também que não houve extinções de linhagens mais antigas dentro do gênero e que os processos são simétricos. Mas é justamente o caráter dinâmico desses processos cíclicos, agindo em unidades isoladas, que são
as espécies, e permitindo dispersões, isolamentos e extinções desiguais, que criam um padrão politômico no padrão biogeográfico geral e um alto endemismo e diversidade na floresta. Esses tipos de processos, que Dalton Amorim (comunicação pessoal) chamou de "barreiras reiterativas", podem ser relacionados aos refúgios de florestas em épocas secas, às transgressões marinhas em épocas úmidas e até mesmo à destruição ambiental causada pelos pulsos tectônicos e movimentos geológicos de massa, ocorridos na constituição do relevo acidentado da costa oriental brasileira.
Expansão geográfica
Vicariância e especiação
Expansão geográfica
Vicariância e especiação
Boe - H. a/ienus
SMSP - H. har/equin
LSRJ - H. disco/ar
SMSP - H. inscriptus
SMSP - H. a/biscriptus
Boc/LSRJ - H. anarchus
- Modelo de padrão redundante da filogenia das espécies de Heteromitobates (Goniosomatinae) em relação às á reas de ocorrência. A partir da espécie ancestral que vivia em uma única á rea ancestral, diversificam-se seis espécies atuais (Soc = Serra da Socaina; LSRJ = litoral norte de São Pau lo e SMSP = Serra do Mar de São Paulo) . A redundância nas relações entre essas á reas pode ser explicada por dois ciclos em que barreiras reiterativas agem separando as á reas (elipses cinzas) e causando especiação (coluna bege), a lternando com épocas sem barreiras, quando ocorre a expansão das espécies sobre as três á reas (coluna verde). Para completar o quadro, duas extinções recentes são hipotetizadas (ramos pontilhados) e a não especiação da espécie H. anarchus, mesmo após a separação das á reas Soc e LSRJ. Exemplos de processos que coincidem com esse modelo são os gerados pelas f l utuações climáticas, criando refúgios ou manchas f lorestais separadas em épocas secas (coluna bege) e florestas contínuas entre as á reas em épocas mais úmidas (co/una verde). t = tempo.

H istória B i ogeográfica da Mata Atl â ntica: Op i l iões (Arachn ida) como Mode lo para sua Inferência - 237
A Mata Atlântica é caracterizada por grandes variações em sua biota, acompanhando as fisionomias diferentes, atualmente marcadas por regimes climáticos diferentes, e endemismos mesmo dentro de uma mesma fisionomia, causados por processos históricos variados.
Os opiliões têm características importantes para o estudo de biogeografia histórica como o seu alto grau de endemismo, principalmente em florestas tropicais como a Mata Atlântica. Seus padrões biogeográficos refletem a história da porção mais úmida desse grande domínio formado pelas diversas fisionomias. Espera-se que os seus padrões possam se repetir em muitos organismos que tenham as restrições climáticas e ecológicas semelhantes aos opiliões, como grilos e pererecas. Mesmo em organismos com gerações mais longas ou menores restrições, como vertebrados, os mesmos eventos podem ter causado diferenciação em nível populacional.
Após o isolamento e a formação da Floresta Atlântica como unidade biogeográfica, as principais disjunções da floresta teriam se relacionado aos vales do Rio Doce e Rio Paraíba do Sul, seja pelo tectonismo responsável pelos seus surgimentos ou os próprios rios servindo como barreiras, principalmente em períodos de transgressão marinha. Os vales do Rio Ribeira do Iguape e a Baía de Todos os Santos podem ter sido disjunções secundárias e teriam contribuído para a delimitação das 1 2 áreas de endemismo encontradas. O soerguimento das serras teria influenciado mais recentemente também o desenvolvimento do padrão de endemismo encontrado, sendo mais importante na formação das escarpas onde ocorrem as chuvas orográficas, e reversos com sombra de chuva e ambientes altos, onde se dá a interrupção da floresta. Os refúgios florestais em períodos glaciais, que aconteceram pelo menos desde o Plioceno, também parecem ter influenciado bastante na alta diversidade e endemismo, além de terem criado um padrão biogeográfico bastante reticulado.
A diversificação na floresta, pelo menos para opiliões, parece ser impulsionada por processos cíclicos de restrição e vicariância alternados com simpatria e dispersão, com barreiras reiterativas como áreas não florestadas entre refúgios e transgressões marinhas. Entretanto, deve-se levar em conta que processos descritos por modelos diferentes se complementam para explicar especiação e diversidade, ainda mais em áreas continentais. Assim, o tectonismo que abre o vale e forma a serra teria influenciado a formação de refúgios florestais em momentos diferentes e alternados, que, por sua vez, teriam sido influenciados por escarpas e vales, alternados
por transgressões marinhas e influência sedimentar-marinha. Esses processos funcionam de forma dinâmica e conjunta e não impedem a formação de um padrão geral que possa ser revelado como o aqui apresentado pelos opiliões. Pelo contrário, ele sugere que o entendimento detalhado das relações biogeográficas entre as áreas da Mata Atlântica e de seu próprio passado depende da compreensão de uma história geomorfológica e climática bastante complexa.
REFERÊNCIAS BIBLIOGRÁFICAS 1 . POR, F. D. Sooretama - The Atlantic Rain Forest of Brazil. The
Hague: SPB Academic Publishing, 1 992. 1 30p. 2. OLIVEIRA-FILHO, A. T; FONTES, M. A. L. Patterns of
fioristic differentiation among Atlantic Forests in Southeastern Brazil and the infiuence of c\imate. Biotropica, v. 32, p. 793-8 10, 2000.
3. ALMEIDA-NETO, M.; MACHADO, G.; PINTO-DA-ROCHA, R. et aI. Harvestman (Arachnida: Opiliones) species distribution along three Neotropical elevational gradients: an alternative rescue effect to explain Rapoport's rule? J. Biogeogr. , v. 33, p. 361 -375, 2006.
4. MORLEY, R. J. Origin and Evolution ofTropical Rain Forests. John Wiley & Sons, 2000. 378p.
5 . FONSECA, G. A. B . ; RYLANDS, A; PAGLIA, A et a\. Atlantic Forest. In : MITTERMEIER, R. A . ; GIL, P. B . ; HOFFMANN, M . et aI. Hotspots revisited. Earth 's biologically richest and most endangered terrestrial ecoregions. Mexico: CEMEX & Agrupación Sierra Madre, 2004. p. 84-9 l .
6 . PINTO-DA-ROCHA, R.; DASILVA, M. B . ; BRAGAGNOLO, C. Faunistic similarity and historical biogeography of the harvestmen of southern and southeastern Atlantic Rain Forest of BraziI. 1. Arachnol. , v. 33, p. 290-299, 2005.
7. AMORIM, D. S . ; PIRES, M. R. S. Neotropical biogeography and a method for a maximum biodiversity estimation. In: BICUDO, C. E. M.; MENEZES, N. A. Biodiversity in Brazil - A First Approach. São Paulo: Conselho Nacional de Desenvolvimento Científico e Tecnológico, 1 996. p. 1 83-219 .
8 . BATES, J . M.; HACKETT, S. J . ; CRACRAFT, J. Area-relationships in the Neotropical lowlands: an hyphothesis based on raw distributions of Passerine birds. J. Biogeogr. , v. 25, p. 783-793, 1 998.
9 . EBERHARD, J. R. ; BERMINGHAM, E. Phylogeny and comparative biogeography of Pionopsitta parrots and Pteroglossus toucans. Mal. Phylogenet. Eva!' , v. 36, p. 288-304, 2005.
10 . ROIG-JUNENT, S. R.; COSCARÓN, S. Biogeographical history of the Neotropical and Neantarctic Simuliidae (Diptera). Rev. Mus. Argentino Ciene. Nat. , v. 3, p. 1 19- 1 34, 2001 .
1 1 . COSTA, L . P. The historical bridge between the Amazon and the Atlantic Forest of Brazil: a study of molecular phylogeography with small mammals . J. Biogeog r. , v. 30, p. 7 1 -86, 2003.
1 2 . PORZECANSKI, A. L.; CRACRAFT J. Cladistic Analysis of Distributions and Endemism (CADE): Using raw distributions of birds to unravel the biogeography of the South American aridlands. 1. Biogeog r. , v. 32, p. 261 -275, 2005.
1 3 . MORRONE, J. J. Biogeografía de América Latina y el Caribe. Zaragoza: Manuales y Tesis, SEA, v. 3, 200 l . 1 48p.
1 4. MÜLLER, P The Dispersai Centers ofTerrestrial Vertebrates in the Neotropical Realm. The Hague: dr. W. Junk B . V. Publishers, 1973. 244p.
1 5 . COSTA, L. P; FONSECA, G. A B.; FONSECA, M. T Biogeography of South America forest mammals: endemism and diversity in theAtlantic Forest. Biotropica, v. 32, p. 872-881 , 2000.
16 . PRANCE, G. T Forest refuges: evidence from woody angiosperms. In: Biological Diversijication in the Tropics. Nova York: Columbia University Press, 1 982. p. 1 37- 158 .

238 - Evolução Espac ia l da Reg ião
17 . SILVA, J. M. c.; CASTELETI, C. H. M. Estado da biodiversidade da Mata Atlântica brasileira. In: GALINDO-LEAL, C. ; CÂMARA, I. G. Mata Atlântica: Biodiversidade, ameaças e perspectivas. São Paulo: Fundação SOS Mata Atlântica; Belo Horizonte: Conservação Internacional, 2005. p. 43-59.
18. PELLEGRINO, K. C. M.; RODRIGUES, M. T; WAITE, A. N. et aI. Phylogeography and species limits in the Gymnodactylus darwinii complex (Gekkonidae, Squamata): genetic structure coincides with river systems in the Brazilian Atlantic Forest. Biol. J. Linn. Soc. , v. 85, p. 1 3-26, 2005.
19. CABANNE, G. S.; SANTOS, F. R.; MIYAKI, C. Y Phylogeography ofXiphorhynchus fuscus (passeriformes, Dendrocolaptidae): vicariance and recent demographic expansion in southern Atlantic forest. Biol. J Linn. Soc. , v. 9 1 , p. 73-84, 2007.
20. TUOMISTO, H. Interpreting the biogeography of South America. J Biogeogr. , v. 34, p. 1 294- 1 295, 2007.
2 1 . LARA, M. c.; GEISE, L. ; SCHNEIDER C. J. Diversification of small mammals in the Atlantic forest of Brazil: testing the alternatives. In: LACEY, E. A. ; MYERS P. Mammalian Diversification:from chromossomes to phylogeography (a celebration of the carrer of James L. Patton). Berkeley, Los Angeles, and London: University of California Press, 2005. p. 3 1 1 -333.
22. HAFFER, J. Speciation in Amazonian forest birds. Science, v, 1 65 , p. 1 3 1 -1 37, 1 969.
23. WHITMORE, T c. ; PRANCE G. T Biogeography and Quatemary History in Tropical America. Oxford: Clarendon Press, 1 987. 2 14p.
24. BRAGAGNOLO, c.; NOGUEIRA, A. A.; PINTO-DA-ROCHA, R. et aI. Harvestmen in an Atlantic forest fragmented landscape: Evaluating assemblage response to habitat quality and quantity. Biol. Conserv. , v. 1 39, p. 389-400, 2007.
25. WILLEMART, R. H.; GNASPINI, P. Spatial distribution, mobility, gregariousness and defensive behaviour in a Brazilian cave harvestman Goniosoma albiscriptum (Arachnida, Opiliones, Gonyleptidae). Anim. Biol. , v. 54, p. 221 -235, 2004.
26. PINTO-DA-ROCHA, R. Systematic review and cladistic analysis of the Brazilian subfamily Caelopyginae (Opiliones: Gonyleptidae) . Arq. Zoo/. , v. 36, p. 357-464, 2002.
27. DASILVA, M. B . ; GNASPINI, P. A systematic revision of Goniosomatinae (Arachnida: Opiliones: Gonyleptidae), with a cladistic analysis and biogeographical notes. Invertebr. Syst. , v. 23, p. 530-624, 2009.
28. MENDES, A. C. Revisão Sistemática e Análise Filogenética de Heteropachy/inae KUI}� 1994 (Arachnida, Opiliones, Gonyleptidae) . Rio de Janeiro: UFRJ, 2005 , 1 6 1p . Dissertação (Mestrado) - Museu Nacional da Universidade Federal do Rio de Janeiro, 2005.
29. DASILVA, M. B . ; PINTO-DA-ROCHA, R. Systematic review and cladistic analysis of the Hernandariinae (Opiliones: Gonyleptidae). Zoologia, v. 27, p. 577-642, 2010.
30. YAMAGUTI, H. Y; PINTO-DA-ROCHA, R. Taxonomic review of Bourguyiinae, c1adistic analysis, and a new hypothesis of biogeographic relationships of the Brazilian Atlantic Rainforest (Arachnida: Opiliones, Gony1eptidae). Zool. J. Linn. Soc., v. 1 66, p . 3 19-362, 2009.
3 1 . MORRONE, J. J. On the identification of Areas of Endemism. Syst. Biol. , v. 43, p. 438-441 , 1994.
32. SZUMIK, C. A.; GOLOBOFF, P. Areas of endemism: improved optimality criteria. Syst. Biol. , v. 53, p. 968-977, 2004.
33. SUGUIO, K.; ANGULO, R. 1 . ; CARVALHO, A. M. et aI. Paleoníveis do mar e paleo1inhas de costa. In: SOUZA, C. R. G. ; SUGUIO, K. ; OLIVEIRA, A. M. S . et aI . Quaternário do Brasil. Associação Brasileira de Estudos do Quaternário. Ribeirão Preto : Ho1os, 2005. p. 1 14- 1 29.
34. SAADI, A.; BEZERRA, F. H. R. ; COSTA, R. D. et aI. Neotectônica da plataforma brasileira. In: SOUZA, C. R. G. ; SUGUIO, K.; OLIVEIRA, A. M. S. et aI. Quaternário do Brasil. Associação Brasileira de Estudos do Quaternário. Ribeirão Preto: Ho1os, 2005. p. 2 1 1 -234.


j j j j j j j j j j j j j j j j j j j j j j j j j j J
j j j j j j j j j j j j j
j j j j j j j
j j j
j j
j j j j

Domingos Benício Oliveira Silva Cardoso
Luciano Paganucci de Queiroz
o bioma caatinga estende-se por uma área de mais de 800.000km2, cobrindo a maior parte dos estados brasileiros do Piauí, Ceará, Rio Grande do Norte, Paraíba, Pernambuco, Alagoas, Sergipe, Bahia e o norte de Minas Gerais. Este bioma está entre os tipos vegetacionais deciduais mais ameaçados da Região Neotropical, sendo alvo de destruição de grandes áreas naturais, o que tem resultado em crescente processo de fragmentaçãol . Apesar disso, a caatinga é desvalorizada nos estudos de biodiversidade e tem recebido poucos esforços conservacionistas, possuindo a menor extensão protegida dentre todos os biomas brasileiros2.
As paisagens desoladoras da vegetação da caatinga, quase completamente desfolhada durante a estação mais seca, transmitem a ideia de que elas possuem uma diversidade muito baixa de espécies e que seriam apenas vegetações secundárias derivadas da flora de outros biomas, como a Mata Atlântica ou o Chaco. Nos últimos anos, vários estudos 1 ,3.4 têm contestado este conceito, mostrando que a caatinga possui alta diversidade de espécies, com sua composição florística única e rica em táxons endêmicos.
Leguminosae (= Fabaceae) é a família de plantas mais diversa na caatinga. Um estudo recente mostra que a família está representada por 320 espécies e 86 gêneros, dentre os quais 1 24 spp. e os gêneros Blanchetiodendron, Goniorrhachis e Tabaroa são endêmicos5, este último recentemente descoberto para o estado da Bahia. Esta
?�-:
�,-, E 1 5
riqueza advém, muito provavelmente, do fato de as leguminosas possuírem longa história de diversificação em vegetações secas6•7, associada ainda à estabilidade florística destas áreas8. Além disso, as leguminosas apresentam várias adaptações morfológicas que permitem às suas espécies colonizarem as áreas secas, onde recursos hídricos são escassos e as plantas estão sujeitas a uma série de restrições ambientais, como altas temperaturas e baixa umidade9.
Com base no padrão de distribuição de espécies arbóreas, Prado 1 0 e Pennington et alo 1 1 propuseram que a caatinga faz parte de uma unidade fitogeográfica maior, a Província das Florestas Tropicais Sazonalmente Secas (SDTF, Seasonally Dry Tropical Forests), que deve ter apresentado sua expansão máxima durante as fases mais secas do Pleistoceno. Atualmente, as SDTF distribuem-se disjuntamente na caatinga, nas florestas estacionais do sudeste e centro-oeste do Brasil e nordeste da Argentina (núcleo Misiones), no Piemonte dos Andes, noroeste da Argentina e sul da Bolívia (núcleo Piemonte), seguindo para o norte através de vales secos encravados nos Andes, formando, assim, o chamado Arco PleistocênicolO, l l (Fig. 1 5 . 1 ). Várias espécies de leguminosas exibem este padrão mais amplo de distribuição, como Anadenanthera colubrina, Amburana cearensis, Enterolobium contortisiliquum, Machaerium acutifolium, Piptadenia viridifiora, Pterogyne nitens e Senna spectabilis, ou ocorrem como populações disjuntas entre a caatinga e em algumas outras áreas de SDTF, a exemplo de Mimosa tenuifiora e M. hexandra.

242 - Padrões e Processos - Estudos de Casos
W
N
E
S
• SDTF
Caatinga arenosa
Savana
Floresta úmida
Chaco
': - Distribuição das f lorestas tropica is sazona lmente secas (SDTF) na Reg ião Neotropical (mod ificado a part i r dos traba lhos de Prado1o e Penn ington et al. 1 1 ) . A caatinga (Caa) está de l im itada segundo Vel l oso et al. 1 2, dentro da qua l são destacadas as pr incipais áreas sobre bacias sedimentares a renosas (caatinga arenosa). Ver Prancha Colorida.
Grandes mudanças geológicas (como tectonismos) e flutuações climáticas durante o Pleistoceno foram propostas como as principais hipóteses para explicar a origem da diversidade e os padrões de distribuição de plantas nas florestas secas neotropicais 10, I I . De acordo com as hipóteses influenciadas pela escola da biogeografia de vicariância, a distribuição disjunta de leguminosas nas diferentes áreas de SDTF teria se originado a partir da fragmentação de populações que apresentavam distribuição mais ampla durante o Pleistoceno, quando as SDTF atingiram o máximo de sua expansão; ao mesmo tempo, a origem da diversidade de espécies endêmicas nestas áreas, a exemplo da caatinga, teria resultado do isolamento dessas populações ancestrais (origem por vicariância).
Por manifestarem grande diversidade de táxons, diferentes padrões de distribuição na caatinga e diversificação antiga em áreas secas6,7,9, as leguminosas são
consideradas um grupo-chave para o entendimento das relações históricas entre as SDTF e da evolução de sua flora. Estudos recentes6,8,9, J 3, 14 têm mostrado uma explicação alternativa para os padrões filo genéticos e biogeográficos em leguminosas das SDTF, utilizando a teoria neutra unificada de biodiversidade e biogeografia 1 5 em uma visão de metacomunidades. Essa perspectiva, no entanto, ainda tem sido pouco explorada com os outros grupos de plantas destas áreas . Neste capítulo, portanto, serão discutidas a biogeografia e a diversidade da caatinga dentro do contexto geral da teoria de metacomunidades 1 5 . Dados de distribuição geográfica, endemismos, padrões de estrutura filogenética e abundância de espécies em leguminosas serão usados como evidência de que a origem e a manutenção da diversidade, bem como os padrões biogeográficos da caatinga podem ser mais bem explicados por meio de processos de metacomunidades.
'C -J 00 00 lh .co N :: 00 'C o, o,

Caati nga no Contexto de uma Metacom u n idade : Evidências da B iogeografia, Padrões F i logenéticos e . . . - 243
Muitos autores1 ,5 j á reconheceram a diversidade de fatores físicos no bioma caatinga ao longo da sua distribuição, além de fisionomias da vegetação (Fig. 1 5 .2), determinando a existência de oito ecorregiões 12: Complexo de Campo Maior, Complexo Ibiapaba-Araripe, Depressão Sertaneja Setentrional, Planalto da Borborema, Depressão Sertaneja Meridional, Dunas do São Francisco, Complexo da Chapada Diamantina e Raso da Catarina. Dentre estas, a Depressão Sertaneja Meridional ocupa a maior área (622.000km2 ou 73% do total da área do bioma caatinga) e apresenta o maior número de leguminosas
endêmicas: 57 táxons (39% do total de leguminosas de caatinga), incluindo os gêneros monoespecíficos Blanchetiodendron, Goniorrhachis e Tabaroa1 •
No bioma caatinga, as temperaturas médias anuais estão entre as mais elevadas do BrasiJl6, variando entre 26 e 28°C, embora as médias das temperaturas máximas raramente ultrapassem 40°C. As precipitações pluviais são baixas e irregulares, em geral limitadas a curto período do ano. Cerca de metade da área do bioma recebe menos de 750mm/ano, havendo núcleos onde essas taxas são inferiores a 500mmlano. Em quase toda a caatinga, 50 a 70% da precipitação anual estão concentradas em três meses consecutivos, caracterizando um clima marca-
- D iferentes t ipos vegetacionais de caatinga sobre embasamento crista l ino (A a 1), representados durante as estações seca (A a F) e úmida (G a I), com destaque para as á reas de caatinga sobre afloramentos calcários em M orro do Chapéu (O) e Serra do Ramalho (E), Bah ia; caat inga arbórea em Ruy Barbosa (G), Bah ia; e caatinga da reg ião do Seridó (H) no Rio Grande do Norte. A caatinga sobre solos derivados da bacia sed imentar a renosa está representada pelas á reas de dunas i nteriores do Rio São Francisco em Casa Nova (J) e P i lão Arcado (K) na Bahia, e pe la reg ião de Canudos (L), que faz parte da mesma formação vegetacional do Raso da Catarina, ao longo da extensão da Bacia Tucano-Jatobá. Fotos: L. P. de Queiroz (A, C, F, /, J, L); D. Cardoso (B, D, H); R. M. Santos (E); E. Matos (G) e A. Rap in i (K). Ver Prancha Colorida .

244 - Padrões e Processos - Estudos de Casos
damente sazonal com estação seca longa, variando de seis a nove meses, mas podendo chegar, em alguns núcleos mais secos, a dez ou onze meses16. Assim, mais do que a precipitação total, é a concentração das chuvas em curtos períodos intercalados por longos períodos secos que exerce efeito mais acentuado sobre as características morfofuncionais das plantas. O clima da caatinga, além de apresentar uma estação chuvosa muito curta, é marcado pela grande inegularidade das chuvas de um ano para outro, de modo que alguns anos ficam praticamente sem chuvas, caracterizando secas ainda mais prolongadas.
Há ainda encraves de outros tipos vegetacionais dentro dos limites do bioma caatinga, como os campos rupestres e cenados (tipos de savanas inseridos no Complexo da Chapada Diamantina) e as florestas senanas (tipos de florestas úmidas encontrados no interior da Depressão Sertaneja Setentrional e na Chapada Diamantina) . Tais tipos vegetacionais exibem composição florística distinta da vegetação de caatinga e não serão discutidos neste capítulo.
A chamada vegetação de caatinga é um tipo de floresta estacionaI decidual caracterizada pelo seu porte baixo, formada por árvores e arbustos caducifólios, que muitas vezes apresentam espinhos ou acúleos e microfilia. Possui estrato herbáceo presente apenas durante a curta estação chuvosa, composto principalmente por espécies de Acanthaceae, Asteraceae, Convolvulaceae, Rubiaceae e Leguminosae. Plantas suculentas das famílias Cactaceae, Euphorbiaceae e Bromeliaceae, além de trepadeiras das famílias Bignoniaceae e Sapindaceae, são comuns. Algumas plantas exibem órgãos especializados para acúmulo de água, às vezes subtenâneos (como no umbuzeiro, Spondias tuberosa Anuda, Anacardiaceae), às vezes em dilatações do tronco (como nas banigudas dos gêneros Ceiba e Cavanillesia, Malvaceae). Leguminosae é uma das famílias mais importantes, por representar a maior parte da diversidade florística (Fig. 1 5 .3), tendo os gêneros Bauhinia, Calliandra, Chamaecrista, Luetzelburgia, Mimosa, Poincianella, Senegalia e Senna como os mais conspícuos e representativos em número de espécies, compondo, juntamente com as cactáceas, a paisagem característica da caatinga 1 (Fig. 1 5.2).
A proporção de espécies que mantêm as folhas na estação seca varia de 26% a quase 0% em áreas onde somente Ziziphus joazeiro Mart. (Rhamnaceae), Capparis yco Mart. (Brassicaceae) ou Licania rigida Benth. (Chrysobalanaceae) podem ser consideradas perenifólias l , 17 . Machado et al. 17 demonstraram que a fenologia da floração da maioria das espécies lenhosas é sincronizada com o início da estação chuvosa, um padrão também observado em outras áreas de SDTF neotropicais 1 8 .
Diferentemente da vegetação de cenado, a caatinga é compostas por plantas não tolerantes ao fogo e ocupa faixas onde o solo costuma ser rico em nutrientes.
Na maior parte dos estudos biogeográficos sobre as florestas secas neotropicais, o bioma caatinga tem sido tratado como uma unidade vegetacionaP,7, 10, 1 1 , 19 . No entanto, alguns autores têm argumentado que ela pode ser composta por dois conjuntos florísticos distintos1,20.2 1 , De fato, em estudo realizado por Queiroz1 , utilizando padrões d e distribuição das leguminosas, verificou-se que, sob a denominação de caatinga, pode ser diferenciada uma biota associada aos afloramentos do embasamento cristalino Pré-cambriano, de uma outra que ocupa solos arenosos das superfícies sedimentares. Cardoso e Queiroz2 1 também observaram nítida separação entre estas duas biotas, mesmo em escala local, ao analisarem a similaridade entre as leguminosas de uma região no nordeste da Bahia,
A flora do embasamento c1istalino conesponde à fisionomia mais típica de caatinga, como já descrita, Ela abrange dois grandes centros de endemismo, definidos com base nos padrões de distribuição das leguminosas I : ( 1 ) a Depressão Sertaneja Setentrional, que vai em direção ao limite norte do semiárido a partir da borda dos estados de Pernambuco e Ceará, incluindo a econegião do Planalto da Borborema; e (2) a Depressão Sertaneja Meridional, que se alonga em direção ao sul a partir dos limites da Depressão Sertaneja Setentrional até o norte de Minas Gerais. Queirozl demonstrou que, na verdade, a flora do embasamento cristalino, e não a da caatinga como um todo, é relacionada à flora geral das SDTF De fato, todos os gêneros endêmicos das SDTF que também oconem na caatinga 10 são encontrados apenas nestes dois centros de endemismo, Os gêneros de leguminosas Amburana, Apuleia, Holocalyx e Pterogyne, juntamente com Myracrodruon (Anacardiaceae), Patagonula (Boraginaceae), Brasiliopuntia (Cactaceae), Quiabentia (Cactaceae) e Balfourodendron (Rutaceae) demonstram ligação histórica entre a flora do cristalino e as demais áreas de SDTFlO. É importante ainda destacar que quase todas as espécies de Leguminosae que sucedem disjuntamente na caatinga e em alguma outra SDTF pertencem apenas às áreas de solos derivados do embasamento cristalino1 .
A vegetação do bioma caatinga que ocupa os solos arenosos das bacias sedimentares cobre mais de

Caat inga no Contexto de uma Metacom u n i dade : Ev idênc ias da B iogeograf ia , Padrões F i l ogenét icos e , , , - 245
� §)":1 - Exemplos da diversidade florística de leguminosas de caatinga: A = Líbidibia ferrea; B = Amburana cearensis; C = Mimosa modesta; D = Pterocarpus vil/osus; E = Mimosa verrucosa; F = Erythrina ve/utina; G = Apu/eia /eiocarpa; H =
Craty/ia mol/is; I = Coursetia rostrata; J = Mimosa /epidophora; K = Pterocarpus monophy/lus; L = Aeschynomene martii; M = Poincianel/a pyramida/is; N = Luetze/burgia bahiensis; O = Goniorrhachis marginata; P = Pithecel/obium diversifo/ium; Q = Senna bracteosa; R = Trischidium moI/e; S = Mysanthus u/eanus; T = Chaetoca/yx scandens. Fotos: D. Cardoso (A a J, L a n e A. P. Prata (K). Ver Prancha Colorida .
136.000km2 (cerca de 1 6% da área total do bioma) e inclui quatro principais áreas que se encontram atualmente disjuntas (ver Fig. 15 . 1 ) : ( 1 ) Chapada do Ibiapaba, entre os estados do Ceará e do Piauí, e Chapada do Ara-
::6 ripe, na borda dos estados do Ceará e Pernambuco; (2) � Raso da Catarina, que inclui toda a extensão da Bacia � Tucano-Jatobá, desde o nordeste da Bahia até o centro-sul ::2 de Pernambuco; (3) dunas interiores do Rio São Francis-00 :;; co, que são depósitos eólicos continentais que alcançam
até 100m de altura; e (4) depressões sazo-nalmente inundadas do sudeste do Piauí, oeste de Pernambuco e norte da Bahia. Outras pequenas áreas arenosas ao longo do bioma caatinga acontecem no nordeste do Rio Grande do Norte, Chapada Diamantina e em locais associados a
elevações residuais. Esta unidade fiorística se dá em grandes áreas descontínuas, sendo definida pela presença de espécies disjuntas, o que sugere uma ligação pretérita entre as fioras destas áreasl .2o. Dentre as Leguminosae que caracterizam este padrão disjunto, podem ser citadas: Aeschynomene martii, Calliandra depauperata, C. macrocalyx, C. squarrosa, Cenostigma macrophyllum, Copaifera arenicola, Cratylia mollis, Dioclea marginata, Galactia remansoana, Hymenaea eriogyne, H. velutina, Lonchocarpus araripensis, Mimosa verrucosa, Poincianella microphylla, Senna gardneri, Trischidium molle e Zornia echinocarpa. Plantas de outras famílias também apresentam esse padrão de distribuição, tais como: Harpochilus neesianus

246 - Padrões e Processos - Estudos de Casos
Mart. ex Nees (Acanthaceae), Godmania dardanoi (J.c.Gomes) A.H.Gentry (Bignoniaceae), Pilosocereus tuberculatus (Werderm.) Byles & G.D.Rowley, Tacinga inamoena (Cactaceae), Pavonia glazioviana Gürke (Malvaceae) e Jatropha mutabilis (Pohl) Baill. (Euphorbiaceae) 1 .20.
Outras evidências confirmam que a flora da vegetação sobre as bacias sedimentares arenosas deve ser tratada como uma unidade fito geográfica à parte. Por exemplo, os dados fenológicos apresentados por Rocha et al. 2o para uma área de dunas continentais no Vale Médio do Rio São Francisco mostraram que os ciclos vegetativos e reprodutivos não são governados pela distribuição das chuvas. Os autores verificaram que o brotamento e a queda foliares, desenvolvimento floral e antese, bem como a produção de fmtos e dispersão das sementes não foram sincronizados entre as espécies, e que pelo menos 50% dos indivíduos produziam folhas ao longo de todo o an020. Essas características são contrastantes com o padrão marcadamente decíduo das folhas e com a acentuada sincronia fenológica encontrados nas florestas secas neotropicais, incluindo a caatinga sobre embasamento cristalino. Esses autores relataram ainda que a zoocoria é a síndrome de dispersão mais frequente entre as fanerófitas (3 1 ,3% das espécies, 78,7% da frequência total), ao contrário do padrão predominantemente anemocórico ou autocórico nas áreas do embasamento cristalin022.
Esses dois conjuntos florísticos das áreas do cristalino e das bacias sedimentares arenosas, além de serem diferenciados por composição, estmtura e ritmos fenológicos, podem ter tido origem fito geográfica distinta. Queirozl propôs uma hipótese de vicariância para a vegetação das superfícies arenosas. Segundo ele, a distribuição disjunta desta vegetação teria surgido a partir de intenso processo de pediplanaçã023 durante o Terciário Superior, quando ela ainda possuía distribuição mais ampla e contínua. Este processo resultou no isolamento da biota nas superfícies sedimentares residuais e, ao mesmo tempo, teria aberto caminho para a invasão da flora das SDTF, que passou a dominar as superfícies do cristalino. De acordo com esta hipótese, a vegetação original da caatinga seria a das bacias sedimentares arenosas, por ser a mais antiga. Esta hipótese também traz implicações ao entendimento dos processos de especiação que deram origem à diversidade e ao endemismo na caatinga. Ao sugerir uma idade mais antiga para os eventos de especiação, Queirozl contesta a ideia de que grande parte da diversidade da caatinga foi originada durante as mudanças climáticas do Pleistocen03,l0. E, de fato, estudos recentes6,7 sobre a
diversificação das leguminosas encontradas em diferentes florestas secas têm indicado que as plantas da caatinga teriam surgido antes do Pleistoceno.
Embora a caatinga sobre embasamento cristalino seja historicamente mais relacionada às SDTF, neste capítu- v:;
lo, o termo "caatinga" será utilizado também para as fisionomias vegetacionais que ocorrem sobre as bacias
-J 9" 00 VI :.,
sedimentares arenosas. No entanto, serão excluídas as � fisionomias que claramente representam disjunções de � outros biomas, apesar de estarem inseridas na delimita- &. ção geográfica do bioma caatinga. É o caso do cerrado, relacionado à flora do Brasil Central, e dos campos rupestres da Chapada Diamantina, que ocorrem em altitudes acima de 900m e apresentam composição florística muito distinta da caatinga.
Como exibido anteriormente, os padrões de distribuição geográfica das espécies de caatinga foram utilizados por Prado 1 0 e Pennington et aI. 1 1 para propor a província fitogeográfica das SDTF. Para eles, a origem da diversidade e os padrões biogeográficos encontrados ao longo desta província teriam sido influenciados pelas flutuações climáticas do Pleistoceno que atingiram a região N eotropical. As SDTF teriam passado por fases de expansão, permitindo a dispersão de espécies entre as áreas secas atualmente disjuntas, e retração, ocasionando especiação por vicariância. Contudo, estudos realizados nos últimos anos em filogenia de diferentes grupos de leguminosas de áreas secas6,7 . 13,14 têm apontado uma explicação alternativa para o entendimento da biogeografia das SDTF, tendo em vista que a diversificação destes grupos é anterior ao Pleistoceno. As ideias que estão sendo discutidas a esse respeito têm como base a teoria neutra unificada de biodiversidade e biogeografia em um contexto de metacomunidades 15 .
A teoria neutra unificada, proposta por Hubbell 15 , assemelha-se à teoria de biogeografia de ilhas. Ambas tratam do papel da dispersão na formação de comunidades ecológicas, de modo que as diferentes ilhas ou comunidades locais mantêm ligação pelo intercâmbio de espécies. Na teoria original de biogeografia de ilhas, a riqueza de espécies em ilhas ou em comunidades locais representa um equilíbrio dinâmico entre as taxas de imigração de espécies e a taxa de extinção local sub sequente destas mesmas espécies. A teoria da biogeografia

Caatinga no Contexto de uma Metacomun idade : Ev idênc ias da B iogeograf ia, Padrões F i logenét icos e . . . - 247
de ilhas também se fundamenta na neutralidade porque assume que as espécies são idênticas (simétricas) em suas possibilidades de chegada (imigração) e sobrevivência em determinada comunidade local15 . Esta é uma visão contrária aos fundamentos da ecologia contemporânea, que predizem a natureza assimétrica das espécies, ou seja, que as comunidades são formadas por espécies diferenciadas pelos seus nichos, sendo cada uma delas um melhor competidor em seu próprio nicho ecológico. De forma interessante, a teoria neutra unificada construiu uma ponte de fundamentação teórica entre estas duas perspectivas divergentes, por meio da incorporação da dinâmica de abundância relativa de espécies na teoria de biogeografia de ilhasJ 5 . Além disso, ela substitui o conceito de "área fonte" de espécies (mainland source area) pela metacomunidade.
De modo geral, a teoria neutra unificada favoreceu a proposta de hipóteses quantitativas para origem, manutenção e perda de biodiversidade em contexto biogeográfic024. Na formulação original de Hubbelll5, a dinâmica de metacomunidades é controlada por dois princípios básicos. O primeiro é da neutralidade, equivalência, ou simetria25, o qual também inclui especiação neutra. Este pressuposto fundamenta-se na observação de que os indivíduos de diferentes espécies em uma mesma comunidade ecológica parecem ser controlados por taxas similares de nascimento, morte e migração. No entanto, ao contrário da teoria de biogeografia de ilhas, a teoria de Hubbell 15 pressupõe a neutralidade no nível de indivíduo, e não no de espécie. Isso permite que as espécies de uma comunidade local mudem a abundância relativa ao longo do tempo através da deriva ecológica (isto é, estocasticidade demográfica)l5 . Neste caso, os tempos de persistência das espécies sob deriva são, portanto, influenciados pelas suas abundâncias, de modo que a taxa de extinção pode ser também prevista pela teorials. O segundo princípio baseia-se na observação de que os sistemas ecológicos são saturados, resultando no que Hubbellls chama "dinâmica de soma zero" (zero-sum dynamics). O pressuposto de soma zero significa que o número total de indivíduos em uma comunidade local é fixo e, por conseguinte, um indivíduo só consegue ser introduzido na mesma comunidade quando outro é eliminado (isto é, quando algum indivíduo morre)24.
A abordagem de metacomunidades é bastante conveniente para discutir a biogeografia e a diversidade da caatinga, tendo em vista a distribuição atual das SDTF de forma fragmentada (isto é, como ilhas de vegetação dentro do continente). Nos tópicos seguintes serão discutidas algumas evidências que demonstram que a
caatinga está inserida em contexto biogeográfico e ecológico que se chama metacomunidade das SDTF6, o qual, por sua vez, faz parte, em escala global, do bioma rico em suculentas9. Para isso, se buscará suporte em dados filogenéticos (estrutura das filogenias), biogeográficos (distribuição e endemismos) e ecológicos (abundância de espécies), reunidos na ampla literatura disponível sobre as leguminosas.
As Leguminosae apresentam forte predileção pelas SDTF, o que pode ser evidenciado pela sua alta diversidade e endemismos, bem como pelo sucesso adaptativo de suas espécies nestes locais marcados por restrições ambientais9. Padrões de distribuição disjunta de Leguminosae ao longo dos diferentes fragmentos das SDTF, assim como aqueles analisados inicialmente por Prado e Gibbs3, Pradolo e Pennington et al. ll para várias famílias de plantas neotropicais, permitem testar a hipótese de que a caatinga exibe uma ligação histórica com as demais SDTF. As disjunções das leguminosas de caatinga com outras SDTF podem ser exemplificadas por espécies com ampla distribuição nas SDTF, como AI11-burana cearensis, Anadenanthera colubrina, Apuleia leiocarpa, Chloroleuconfoliolosum, Erythrina velutina. Geoffroea spinosa, Machaerium acutifolium, Platypodium elegans, Poeppigia procera, Poincianella pluviosa, Pterogyne nitens, Senna spectabilis e Vachellia farnesiana, e por espécies que ocorrem em poucas áreas de SDTF, como Copaifera duckei (sudeste brasileiro) , Luetzelburgia andrade-limae (áreas de afloramento calcário no Brasil Central), Mimosa arenosa (costa da Venezuela), M. hexandra (descontinuamente do sul do México ao norte da Venezuela e ao longo do Rio Paraguai, no Mato Grosso do Sul [Brasil] , Paraguai e Argentina), M. tenuifiora (norte da Venezuela e Colômbia e vales secos no sul do México, Honduras e EI Salvador), Phaneraflexuosa (leste do Brasil), Piptadenia viridifiora (região noroeste da Argentina e norte do Paraguai), Pityrocarpa moniliformis (região de Sucre, Venezuela), Pseudopiptadenia brenanii (florestas secas da Bahia e Minas Gerais) e Trischidium molle (bosque seco Chiquitano, Bolívia) (Tabela 1 5 . 1 ) . No total, pelo menos 45 espécies, cerca de 14% da diversidade da família encontrada na caatingaS, manifestam disjunções com alguma floresta seca neotropical (Tabela 1 5 . 1 ) . Vale lembrar que quase todas as espécies de Leguminosae que acontecem disjuntamente na caatinga e em alguma outra área de SDTF pertencem somente às áreas de solos derivados do embasamento cristalinol .

248 - Padrões e Processos - Estudos d e Casos
'" 3 '1 - Lista das espécies de Legum inosae endêmicas da caatinga e/ou com d istr ibu ição d isjunta entre a caatinga e em pelo menos u ma outra á rea da metacomun idade de f lorestas secas neotropicais (SDTF)
Táxons
Subfamília Caesalpinioideae Apuleia grazielanae Afr. Fern.
*Apuleia leiocarpa (Vogel) Macbride
Bauhinia aculeata L.
Bauhinia acuruana Moric.
Bauhinia cheilantha (Bong.) Steud.
Bauhinia harleyi Wu nderl i n sp. novo ined.
Bauhinia forficata Link
Bauhinia pentandra (Bong .) Vogel ex Steud.
Bauhinia subclavata Benth.
Chamaecrista amiciella (H.S . lrwin e Barneby) H .S . l rwin e Barneby
Chamaecrista belemii (H.S . l rwin e Barneby) H .S . lrwin e Barneby
Chamaecrista brevicalyx (Benth.) H .S . l rwin e Ba rneby
Chamaecrista carobinha (H.S . l rwin e Barneby) H .S . l rwin e Barneby
Chamaecrista cuprea H.S . l rwin e Barneby
Chamaecrista duckeana (P.Bezerra e Afr. Fern.) H .S . l rwin e Barneby
Chamaecrista tenuisepala (Benth.) H .S . l rwin e Barneby
Copaifera arenicola (Ducke) J .A.S.Costa e L.P.Queiroz
Copaifera coriacea Mart.
Erythrostemon ca/yeina (Benth.) L.P.Queiroz
*Goniorrhachis marginata Taub .
Hymenaea eriogyne Benth.
Hymenaea velutina D ucke
*Libidibia ferrea (Mart. ex Tu l . ) L.P.Queiroz
Martiodendron mediterraneum (Benth.) K6eppen
Parkinsonia aculeata L. peltogyne pauciflora Benth.
Peltophorum dubium (Spreng.) Taub.
Phanera flexuosa (Moric.) L.P.Queiroz
Phanera trichosepala L.P.Queiroz
Poeppigia procera C.Presl
Poineianel/a gardneriana (Benth.) L .P.Quei roz
Poincianel/a laxiflora (Tu ! .) L .P.Queiroz
Poincianella microphyl/a (Mart. ex G .Don) L.P.Queiroz
Poinciane/la pluviosa (DC.) L.P.Queiroz
*Poincianella pyramidalis (Tu ! .) L.P.Queiroz
Pterogyne nitens Tul .
Senna aversiflora (Herbert) H .S . l rwin e Ba rneby
*Senna bracteosa D.B .O.S .Cardoso e L.P.Queiroz
Senna catingae (Ha rms) L.P.Queiroz
Senna cearensis Afr. Fern.
Hábito Distribuição na caatinga e/ou disjunção com as SDTF
Arv
Arv
Arb
Arb
Arb
Arb
Arv
Arb
Arb
Erv
Arb
Arb
Erv
Subarb
Subarb
Subarb
Arv
Arv
Arb
Arv
Arv
Arv
Arv
Arv
Arv
Arv
Arv
Trep
Trep
Arv
Arv
Arb
Arb
Arv
Arb
Arv
Arb
Arb
Arb
Arb
Endêmica
Ampla em SDTF
Endêmica
Endêmica
Disjunta com o Mato Grosso e Parag uai
Endêmica
Disjunta com outras SDTF do leste d o Brasi l
Disj unta com outras SDTF do leste e reg ião central do Brasi l
Endêmica
Endêm ica
Endêm ica
Endêmica
Endêm ica
Endêmica
Endêm ica
Endêmica
Endêm ica
Endêmica
Endêmica
Endêmica
Endêmica
Endêm ica
Disjunta com outras SDTF do nordeste do Brasi l
Endêmica
Ampla em SDTF
Endêmica
Ampla em SDTF
Disj unta com outras SDTF do leste do Brasi l
Endêmica
Ampla em SDTF
Endêmica
Endêmica
E ndêm ica
Ampla em SDTF
Endêmica
Ampla em SDTF
Endêmica
Endêmica
Endêmica
Endêm ica

Caati nga no Contexto de uma Metacom u n idade: Ev idênc ias da B iogeograf ia, Padrões F i l ogenét icos e . . . - 249
;$iitl@üGJ 'J §,'TI - Lista das espécies de Leguminosae endêmicas da caatinga e/ou com d istr ibu ição d isj unta entre a caat inga e em pelo menos uma outra á rea da metacomun idade de florestas secas neotropicais (SDTF) (continuação)
Táxons
Senna gardneri (Benth.) H .S . l rwin e Barneby
Senna harleyi H.S . l rwin e Barneby
Senna lechriosperma H.S . l rwin e Barneby
*Senna martiana (Benth.) H .S . l rwin e Barneby
Senna rizzinii H .S . l rwin e Barneby
Senna spectabilis (DC.) H .S . l rwin e Barneby
Senna trachypus (Benth.) H . S. l rwin e Barneby
Subfamilia Mimosoideae Anadenanthera colubrina (Ve l l .) B renan
81anchetiodendron blanchetii (Benth.) Barneby e J .W.G rimes
CaJliandra aeschynomenoides Benth.
CaJliandra depauperata Benth.
Calliandra duckei Barneby
Calliandra fernandesii Barneby
Calliandra imperialis Barneby
Calliandra leptopoda Benth.
Calliandra macrocalyx Harms
Calliandra pilgeriana Harms
Calliandra spinosa Ducke
Calliandra squarrosa Benth.
Calliandra ulei H a rms
Calliandra umbeJlifera Benth.
Chloroleucon extortum Ba rneby e J .W.G rimes
Chloroleucon foliolosum (Benth.) G . P. Lewis
Enterolobium timboüva Mart.
Leucochloron limae Barneby e J .W.Gr imes
Mimosa arenosa (Wi l ld.) Poir.
Mimosa borboremae Harms
Mimosa brevipinna Benth.
Mimosa caesalpiniifolia Benth.
Mimosa coruscocaesia Barneby
Mimosa exalbescens Barneby
Mimosa glaucula Barneby
Mimosa hexandra Michel i
Mimosa hirsuticaulis Harms
Mimosa hortensis Barneby
Mimosa irrigua Barneby
*Mimosa lepidophora Rizz in i
Hábito Distribuição na caatinga e/ou disjunção com as SDTF
Arb
Arb
Arb
Arb
Arb
Arb/ Arv
Arb
Arv
Arv
Arb
Arb
Arb
Arb
Arb
Arb
Arb
Arb
Arb
Arb
Arb
Arb
Arv
Arb/ Arv
Arv
Arv
Arb
Erv
Subarb
Arv
Arb
Arb
Subarb
Arb/ Arv
Subarb
Subarb
Arb
Arb/ Arv
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Ampla em SDTF
Endêm ica
Ampla em SDTF
Endêm ica
E ndêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Endêmica
Ampla nas SDTF
Ampla nas SDTF
Endêm ica
D isjunta com outras SDTF da costa da Venezuela
Endêm ica
Endêmica
Disjunta com outras SDTF do Maranhão
Endêmica
D isjunta com outras SDTF do Maranhão
Endêm ica
D isjunta com outras SDTF do sul do México ao norte da Venezuela e ao longo do Rio Paraguai
E ndêmica
Endêmica
Endêmica
Endêmica
(continua)

250 - Pad rões e Processos - Estudos de Casos
' 3)" - Lista das espécies de Legum inosae endêm icas da caatinga e/ou com d istribu ição d isju nta entre a caat inga e em pelo menos uma outra á rea da metacomun idade de f lorestas secas neotropicais (SDTF) (continuação)
Táxons
*Mimosa modesta Mart.
Mimosa morroensis Barneby
Mimosa nothopteris Barneby
Mimosa ophthalmocentra Mart.ex Benth.
Mimosa paraibana Barneby
Mimosa poculata Ba rneby
Mimosa pseudosepiaria Harms
Mimosa setuligera Harms
Mimosa tenuiflora (Wi l ld . ) Poir.
*Mimosa verrucosa Benth.
Mimosa xiquexiquensis Barneby
Parapiptadenia blanchetii (Benth.) Vaz e M . P. Lima
Parapiptadenia zehntneri (Harms) M . P.L ima e H.C .L ima
Piptadenia irwinii G . P. Lewis
Piptadenia stipulacea (Benth.) Ducke
Piptadenia viridiflora (Kunth) Benth.
*Pithecellobium diversifolium Benth.
Pityrocarpa moniliformis (Benth.) Luckow e Jobson
Pityrocarpa obliqua (Pers.) Brenan
Pseudopiptadenia bahiana G . P. Lewis e M . P. Lima
Pseudopiptadenia brenanii G . P. Lewis e M . P. Lima
Samanea inopinata (Harms) Barneby e J.W.Grimes
Senegalia bahiensis (Benth.) Seigler e Ebi nger
Senegalia globosa (Bocage e M iotto) L. P.Queiroz
Senegalia kallunkiae (J .W.Grimes e Barneby) Seigler e Ebinger
Senegalia langsdorffii (Benth.) Seigler e Ebinger
Senegalia limae (Bocage e M iotto) L.P.Queiroz
Senegalia monacantha (Wi l ld.) Seigler e Ebinger
Senegalia piauhiensis (Benth.) Seig ler e E binger
Senegalia ricoae (Bocage e Miotto) L.P.Queiroz**
Senegalia santosii (G . P.Lewis) Seig ler e E binger
Vachellia farnesiana (L.) Wight e Arn.
Hábito
Erv
Subarb
Arb
Arb/ Arv
Arb/ Arv
Arb
Arb/ Arv
Subarb
Arb/ Arv
Arb
Subarb
Arv
Arv
Trep
Arb
Arv
Arb
Arv
Arb
Arv
Arv
Arv
Arb/ Arv
Arb
Arv
Arb
Arb
Arb
Arv
Arb
Arv
Arb
D istribuição na caatinga e/ou disjunção com as SDTF
Endêm ica
Endêmica
Endêmica
Endêm ica
Endêmica
Endêmica
Endêmica
Endêmica
Disjunta com outras SDTF do nordeste do Brasi l , norte da Venezuela e Colôm bia, e vales secos no sul do México, Honduras e EI Salvador
Endêmica
Endêmica
Endêmica
Disjunta com outras SDTF do nordeste do Brasil
Disjunta com outras SDTF da Bahia
Endêm ica
D isjunta com outras SDTF do nordeste do Brasi l e noroeste da Argentina e norte do Paraguai
Endêm ica
D isjunta com outras SDTF da reg ião de Sucre (Venezuela)
Disjunta com outras SDTF do México a EI Sa lvador e Bahia
Disjunta com outras SDTF da Bahia
Disjunta com outras SDTF da Bahia e M inas Gerais
Disjunta com outras SDTF do norte da América do Sul e nordeste do Brasi l
Endêm ica
Endêm ica
Endêmica
Disjunta com outras SDTF do nordeste do Brasi l e M i nas Gerais
Endêm ica
Disjunta com outras SDTF do nordeste do Brasi l e Minas Gerais
Endêm ica
Endêm ica
Endêm ica
Ampla nas SDTF
'D -..J � 00 UI e., N :: , 00 'D 0\ '"

Caatinga no Contexto de uma Metacomun idade: Ev idênc ias da B i ogeografia, Pad rões F i logenéticos e" . - 251
c� 3 ,1 - Lista das espécies de Legum inosae endêmicas da caatinga e/ou com d istribu ição d isj unta entre a caatinga e em pelo menos uma outra á rea da metacomun idade de florestas secas neotropicais (SDTF) (continuação)
Táxons
Subfamília Papilionoideae Acosmium diffusissimum (Mohlenbr.) Yakovlev
*Aeschynomene martii Benth.
Aeschynomene matosii Afr. Fern.
Aeschynomene monteiroi Afr. Fern. e P. Bezerra
Aeschynomene sabu/ico/a L.P.Queiroz e D.B .O.S. Ca rdoso
Aeschynomene soniae G . P. Lewis
*Amburana cearensis (Al lemão) A.C.Sm.
Arachis dardani Krapovickas e W.c.Gregory
Arachis pusilla Benth.
Arachis triseminata Krapovickas e W.c.Gregory
Canava/ia brasi/iensis Mart.ex Benth.
Centrosema rotundifo/ium Mart. ex Benth.
Chaetoca/yx b/anchetiana (Benth.) Rudd
*Chaetoca/yx scandens (L.) U rb.
*Coursetia rostrata Benth.
Coursetia vicio ides (Nees e Mart.) Benth.
Coursetia sp. novo
Craty/ia bahiensis L .P.Queiroz
*Craty/ia mollis Mart. ex Benth.
Crota/aria bahiensis Windler e S .G .Sk i nner
Crota/aria brachycarpa Benth.
Crota/aria h% sericea Nees e Mart.
Da/bergia catingico/a Harms
Da/bergia cearensis Ducke
Da/bergia decipu/aris Rizzin i e A. M attos
Dioc/ea grandif/ora Mart. ex Benth.
Dioc/ea marginata Benth.
*Erythrina ve/utina Wil ld .
Ga/actia remansoana H a rms
Geoffroea spinosa Jacq.
H% ca/yx ba/ansae M iche l i
/ndigofera b/anchetiana Benth.
Lonchocarpus araripensis Benth.
Lonchocarpus castaneifo/ius M.J .S i lva e A.M.G .Azevedo
Luetzelburgia andrade-/imae H . C. Lima
Luetze/burgia auricu/ata (Al lemão) Ducke
Hábito Distribuição na caatinga e/ou disjunção com as SDTF
Arv
Arb
Arb
Arb
Subarb
Arb
Arv
E rv
E rv
E rv
Trep
Trep
Trep
Trep
Arb
Arb
Arb
Arb
Arb
Subarb
Subarb
Subarb
Arv
Arv
Arv
Trep
Trep
Arv
Erv
Arv
Arv
Arb
Arv
Arv
Arv
Arbl Arv
Endêm ica
Endêm ica
Endêm ica
Endêm ica
E ndêm ica
Endêm ica
Disjunta com outras SDTF do B ras i l Centra l (Goiás e Mato G rosso do Sul) e norte do Paraguai, Argentina e Bol ívia
Endêm ica
Endêm ica
E ndêm ica
Endêm ica
Endêm ica
Endêm ica
D isj unta com outras SDTF do leste do B rasi l e México, Colômbia e Venezuela
Endêm ica
Endêm ica
Endêm ica
Endêm ica
E ndêm ica
Endêm ica
E ndêm ica
E ndêm ica
E ndêm ica
Endêm ica
Disj u nta com outras SDTF da Bahia e M inas Gerais
Endêmica
E ndêmica
Disju nta com outras SDTF do oeste da América do Sul, no Peru e Equador, e no Caribe
Endêmica
Ampla em SDTF
Ampla em SDTF
Endêm ica
D isj unta com outras SDTF da Bol ívia e M ato G rosso do Su l
Endêm ica
Disj u nta com outras SDTF do B ras i l Central
E ndêm ica
(continua)

252 - Padrões e Processos - Estudos de Casos
"fu�0®Ücl �,1 - Lista das espécies de Legum inosae endêmicas da caatinga e/ou com distribu ição d isju nta entre a caatinga e em pelo menos uma outra á rea da metacomun idade de f lorestas secas neotropicais (SDTF) (continuação)
Táxons Hábito Distribuição na caatinga e/ou disjunção com as SDTF
Arb/ Endêm ica
Arv *Luetzelburgia bahiensis Yakovlev
Luetzelburgia harleyi D . B.a.S.Cardoso, L.P.Queiroz e H .CLima
Luetzelburgia neurocarpa D.B .a .S .Cardoso, L.P.Queiroz e H.CLima
Luetzelburgia purpurea D. B.a.S .Cardoso, L .P.Queiroz e H.CLima
Machaerium acutifolium Vogel
Arv
Arb
Arv
Arv
Endêm ica
Endêmica
Endêm ica
Ampla em SDTF
Machaerium hirtum (Ve l l .) Ste l lfeld
Machaerium ovalifolium G laziou ex Rudd
Machaerium punctatum (Poir.) Pers.
*Mysanthus uleanus (Harms) G . P. Lewis e A.Delgado-Sa l i nas
Platypodium elegans Vogel
Poecilanthe ulei (Harms) Arroyo e Rudd
Pterocarpus monophyllus Kl itgaard, L.P.Queiroz e G . P. Lewis
*Pterocarpus villosus (Mart. ex Benth.) Benth.
Pterocarpus zehntneri Harms
Rhynchosia sp. novo
Riedeliella magalhaesii (Rizzi ni) H .CLima e Vaz
Sty/osanthes bahiensis 't Man netje e G . P.Lewis
Stylosanthes seabrana B . L.Maass e 't Mannetje
Tabaroa sp. novo (gen. et sp. novo ined .)
Trischidium decipiens (R.S.Cowan) H .E . l re land
*Trischidium moi/e (Benth.) H . E . l re land
Zornia afranioi R.vann i
Zornia harmsiana Sta ndl .
Zornia ulei Harms
* Refere-se às espécies q u e estão representadas na F igura 1 5 .3.
Arv Ampla em SDTF
Arv Endêmica
Arv Disjunta com outras SDTF do nordeste e região central do Brasi l e Bol ívia
Trep Disjunta com outras SDTF do Brasi l Centra l
Arv Ampla em SDTF
Arv Endêm ica
Arv Endêm ica
Arv Endêmica
Arv Endêmica
Trep Endêmica
Arb Endêm ica
Erv Endêm ica
Erv Endêmica
Arv Endêmica
Arb Endêmica
Arb Disjunta com as SDTF da reg ião da Chiquitania boliviana
Erv Endêmica
Erv Endêmica
Subarb Endêm ica
Arb = arbusto; Arv = á rvore; Erv = erva; Subarb = subarbusto; Trep = trepadeiras lenhosas ou herbáceas.
Por outro lado, estas disjunções podem ser tomadas como prova da ligação entre as áreas descontínuas de SDTP por meio de dispersão aleatória a longa distância, como prediz a teoria de metacomunidades. Diferentemente do que pressupõe a biogeografia de vicariância, as disjunções seriam resultados de vários eventos de dispersão. Estudos filo geográficos permitem testar as duas hipóteses, uma vez que possibilitam traçar a história evolutiva das populações (e/ou das espécies próximas com distribuição disjunta) por datação dos eventos de diversificação. Outra abordagem interessante utiliza genética de populações, correlacionando a estrutura genética atual das populações aos eventos históricos de isolamento ou dispersão. A utilização destas duas técnicas moleculares para a reconstrução histórica da
distribuição das SDTP pode ser promissora nos seguintes grupos de Leguminosae com ocorrência marcante nessas florestas: Amburana cearensis, Anadenanthera colubrina e o gênero Luetzelburgia. A variabilidade genética de populações de Astronium urundeuva Engl. (Anacardiaceae), uma espécie característica das SDTP foi analisada em um estudo pioneiro feito por Caetano et az. 19 , empregando-se dados moleculares de ácido desoxirribonucleico (DNA, desoxyribonucleic acid) de cloroplasto e microssatélites. Estes autores verificaram que a estrutura genética encontrada favorece a existência pretérita de uma formação contínua de SDTP no leste da América do Sul. Dessa forma, a variabilidade genética encontrada seria resultado do isolamento das populações com distribuição mais ampla no passado.
� -J oc 00 U1 c.., N ..,. , 00 � a-o.,

Caati nga no Contexto de uma Metacomunidade : Ev idênc ias da B iogeograf ia, Padrões F i logenéticos e . . . - 253
Dentre os núcleos das SDTF, a caatinga é o que apresenta maior extensão e maior riqueza de espécies e de endemismos. Giulietti et aI. 4 listaram 3 1 8 espécies e 1 8 gêneros endêmicos da caatinga. As Leguminosae figuram como a família mais rica em número de espécies e endemismos. A análise da distribuição de 274 táxons de Leguminosae de caatinga realizada por Queiroz! revelou 1 44 táxons endêmicos, o que representa pouco mais da metade (52,55%) da diversidade da família no bioma. Apesar de este número incluir também os táxons infraespecíficos, ainda assim a quantidade de espécies endêmicas ( 1 24) é bem significativa, de acordo com a listagem mais atualizada, apresentada na Tabela 1 5 . 1 .
Exemplos de leguminosas endêmicas da caatinga com distribuição mais ampla no bioma são: Aeschynomene martii, Calliandra depauperata, Dalbergia cearensis, Dioclea grandiflora, Luetzelburgia auriculata, Piptadenia stipulacea, Senna aversiflora, S. catingae e S. martiana. Várias outras espécies mostram distribuição mais restrita, às vezes ocorrendo em apenas uma ou duas localidades, tais como: Aeschynomene sabulicola, A. soniae, Calliandra ulei, Dioclea marginata, Luetzelburgia harleyi, L. neurocarpa, Mimosa hortensis, M. lepidophora, M. nothopteris, M. xiquexiquensis, Pterocarpus monophyllus, Senegalia kallunkiae, Senna bracteosa e Trischidium decipiens. Algumas espécies de Bauhinia, Coursetia e Rhynchosia foram descobertas recentemente e ainda não estão descritas, e um novo gênero monoespecífico afim a Harpalyce só é conhecido em uma área restrita de caatinga ao sul da Chapada Diamantina.
Esse alto número de espécies endêmicas na caatinga pode ser explicado em termos dos processos de metacomunidades. O isolamento da caatinga em relação a outros fragmentos de SDTF neotropicais favorece a evolução de uma flora autóctone, assim como geralmente observado em ilhas oceânicas. De acordo com a teoria de metacomunidades, o fato de a migração entre os diferentes fragmentos da metacomunidade ser estocástica e em taxa muito baixa, quando comparada ao observado nas florestas úmidas, favorece o estabelecimento das espécies residentes. Ademais, espécies imigrantes de outros biomas dificilmente conseguiriam vantagem competitiva em relação às espécies j á adaptadas às áreas secas, marcadas por restrições ambientais. A vegetação de caatinga sucede, de modo geral, em áreas sujeitas à acentuada escassez hídrica, com chuvas irregulares de menos de l .OOOmml ano concentradas em somente 3 a 5 meses do ano. Assim, os diferentes fragmentos da metacomunidade estarão isolados suficientemente para permitir que a deriva
ecológica atue na sua divergência26. Além disso, as populações de cada espécie estarão geneticamente isoladas, de modo que a deriva genética também tenderá a causar divergência e formação de novas espécies26. A combinação de baixas taxas de imigração e competição, isolamento vegetacional e sucesso reprodutivo das populações residentes garante, portanto, a evolução de uma flora altamente endêmica.
O sucesso em diversidade de espécies e endemismos das Leguminosae em florestas secas também está intrinsecamente relacionado às adaptações morfológicas que permitem o estabelecimento dessas plantas nestes ambientes, onde os recursos são escassos. Tais adaptações incluem: folhas compostas com grande capacidade fotos sintética em períodos favoráveis e diminuição da perda de água por meio da queda dos folíolos em períodos desfavoráveis; alto metabolismo de nitrogênio a partir de relações simbióticas com bactérias, conferindo maior vantagem competitiva na colonização de ambientes sazonalmente secos e oligotróficos; além de adaptações da semente, por exemplo, testa resistente, alta dormência, alta viabilidade e habilidade para armazenar nitrogênio, promovendo rápido desenvolvimento de plântulas9.
A teoria de metacomunidades também pode explicar por que a abundância das espécies de ecossistemas formados por vegetação descontínua, a exemplo das SDTF, quase sempre é superior a das espécies de florestas úmidas. Em áreas descontínuas é comum encontrar espécies localmente endêmicas e abundantes, enquanto nas florestas úmidas elas tendem a ser raras e com ampla distribuiçã026. Segundo a teoria de metacomunidades, a elevada abundância relativa observada em florestas secas está relacionada à baixa migração de espécies, em razão do isolamento dos fragmentos . Restrições ambientais impostas aos novos indivíduos, que tentam colonizar as áreas secas, também desfavorecem a imigração. Desta forma, a competição local diminui, possibilitando o crescimento populacional das espécies residentes.
Ainda não foram realizados estudos discutindo a abundância relativa de espécies de florestas secas sob a perspectiva ecológica neutra de metacomunidades, não obstante vários autores6•8•27 tenham sugerido que as SDTF neotropicais apresentem todas as características de um sistema de metacomunidades em nível continental. Na Tabela 1 5 .2 são exibidos dados comparando a abundância de espécies de Leguminosae de caatinga e

254 - Padrões e Processos - Estudos de Casos
de florestas úmidas dos domínios Atlântico e Amazônico. A abundância é representada pela densidade absoluta (DA), medida em indiv./ha, uma vez que muitos trabalhos de fitossociologia não forneciam os valores de abundância relativa (AR) . Os dados da Tabela 15 . 2 foram sumarizados na Figura 15 .4. Entre as Leguminosae de caatinga, Poincianella gardneriana apresentou o maior valor de densidade absoluta (mais de 1 .000 indiv./ha), enquanto o maior valor para as florestas úmidas foi de apenas 7 1 indiv./ha, observado em Chamaecrista ensiformis. Em conjunto, os dados da Tabela 15 .2 sugerem que a abundância de espécies de Leguminosae de caatinga se comporta exatamente como prediz a teoria de metacomunidades, ou seja, as espécies tendem a ser muito comuns (Fig. 15 .4), porém com distribuição restrita a determinado fragmento da metacomunidade. Já o padrão de abundância de Leguminosae nas florestas úmidas analisadas é consideravelmente diferente, visto que as espécies tendem a ser raras (Fig. 15 .4) e com ampla distribuição na América do Sul (Tabela 15 .2).
Esses diferentes padrões de abundância nas SDTF e nas florestas úmidas poderiam ser testados de maneira indireta pela análise coleções de herbári027. Caso se considere que a dispersão nas SDTF permite que as espécies endêmicas de um fragmento (por exemplo, caatinga) aumentem a sua frequência por meio de deriva ecológica, então tais espécies deveriam estar bem
25
20
V> CIJ 'u 1 5 'CIJ o. � CIJ "O e CIJ
1 0 E ':::> z:
5
o
representadas em materiais de herbário, uma vez que elas seriam muito comuns na área e, por isso, mais prováveis de serem coletadas. Por outro lado, se a alta taxa de imigração nas florestas úmidas mantém as espécies a uma taxa de abundância muito baixa, então, tais espécies, mesmo sendo amplamente distribuídas, não seriam bem representadas nos herbários, tendo em vista que elas são localmente raras27•
O padrão de estrutura filogenética dos clados confinados predominantemente às florestas secas neotropicais também é consistente com a teoria de metacomunidades6,8, indicando ainda que existe grande limitação na dispersão entre os fragmentos isolados das SDTF, diferentemente do observado dentro e entre as grandes áreas de florestas úmidas. De acordo com a teoria ecológica neutra, as taxas de especiação e o grau de limitação da dispersão entre os fragmentos moldam a filogenia das linhagens que constituem a metacomunidade. Estudos filogenéticos em vários gêneros que possuem espécies endêmicas dos diferentes fragmentos de SDTF neotropicais têm mostrado que as filogenias possuem alta estruturação geográfica, ou seja, as espécies que ocorrem em determinada comunidade local são também filo geneticamente mais próximas e, com frequência, formam grupos monofiléticos6,8 , 13 , 14 . Isso
Florestas úmi das
F lorestas secas (caatinga)
\O -...l 00 00 Vl .:.,
- - N -l"-, 00 \O c;,
b --- -
<5 6 - 1 5 1 6 - 30 3 1 - 45 46 - 60 61 - 75 76 - 1 00 10 1 - 200 >200
Densidade absoluta ( indivíduos/hectare)
- D istr ibu ição da abundância de espécies de leguminosas em florestas secas (caatinga) e úmidas (Amazônia e Mata Atlântica). O gráfico foi obtido a part ir dos dados da Tabela 1 5 .2 .

Caati nga no Contexto de uma M etacom un idade : Ev idênc ias da B iogeograf ia, Padrões F i l ogenéticos e . . . - 255
Ti;;; - Abundância (medida a part i r da densidade absoluta [DA] defin ida como o número de ind ivíduos de cada espécie por hectare) de espécies lenhosas de Leguminosae em florestas secas (caatinga) e úmidas. Os dados fora m obtidos a part i r de vários estudos fitossociológ icos disponíveis na l iteratura, sendo 1 1 á reas de caati nga e 1 1 de florestas úmidas (domínio Atlântico e Amazônico)
Número de espécies DA** Referência Vegetação de leguminosae Táxons* (indivíduos! Distribuição
amostradas hectare)
Florestas secas Poincianella pyramidalis 700 Endêm ica
Caatinga Mimosa tenuiflora 2 3 1 , 7 Ampla
28 (Rio G rande do 9 Piptadenia stipulacea 1 86,7 Endêmica Norte) Anadenanthera colubrina 1 50 Ampla
Libidibia ferrea 1 03,3 Endêmica
Chloroleucon foliolosum 562 Ampla
Caatinga Mimosa acutistipula 287 Ampla
29 (Rio G rande do 6 Anadenanthera colubrina 1 23 Ampla Norte) Poincianella pyramidalis 202 Endêmica
8auhinia cheilantha 1 5 Disj u nta
Senegalia sp. 330
Piptadenia stipulacea 253 Endêmica
30 Caatinga
1 2 8auhinia cheilantha 43 Disj u nta (Paraíba)
Senegalia polyphylla 27 Ampla
Amburana cearensis 20 Ampla
Poincianella pyramidalis 1 97,5 Endêmica
8auhinia cheilantha 1 32,5 Disjunta
3 1 Caati nga
6 Mimosa sp. 52,5 (Paraíba)
Piptadenia stipulacea 52,5 Endêm ica
Erythrina velutina 2,5 Ampla
Poincianella pyramidalis 775 Endêmica
Mimosa tenuiflora 62,5 Ampla
32 Caatinga 6 Piptadenia stipulacea 37,5 Endêmica
(Paraíba) Mimosa adenophylla 32,5 Ampla
Anadenanthera colubrina 1 0 Ampla
8auhinia pulchella 537 Ampla
Pityrocarpa moniliformis 1 22 Disjunta
33 Caatinga
1 4 Swartzia f1aemingii 99 Ampla (Caatinga) Copaifera martii 79 Ampla
Senegalia langsdorffii 74 Disj unta
Poincianella gardneriana 1 .099 Endêm ica
Mimosa ophthalmocentra 40 Endêmica
34 Caatinga
8 Piptadenia stipulacea 1 0 Endêmica (Pernambuco) Anadenanthera colubrina 5 Ampla
Amburana cearensis 4 Ampla
Poincianella pyramidalis 5 1 3,3 Endêmica
Bauhinia cheilantha 408,3 Disj u nta
3 5 Caatinga
1 0 Mimosa arenosa 233,3 Ampla (Pernambuco)
Piptadenia stipulacea 1 36,7 Endêmica
Anadenanthera colubrina 8 1 ,7 Ampla
(continua)

256 - Pad rões e Processos - Estudos de Casos
'1 - Abundância (medida a part ir da densidade absoluta [DA] def in ida como o número de ind ivíduos de cada espécie por hectare) de espécies lenhosas de Leguminosae em florestas secas (caatinga) e úmidas. Os dados foram obtidos a partir de vários estudos fitossociológicos disponíveis na l i teratura, sendo 1 1 áreas de caatinga e 1 1 de f lorestas úmidas (domínio Atlântico e Amazônico) (continuação)
Número de espécies DA** Referência Vegetação de leguminosae Táxons* (indivíduos! Distribuição
amostradas hectare)
Senegalia langsdorffii 1 .057 Disjunta
Cratylia mollis 3 1 0 Endêmica
36 Caatinga
20 Bauhinia acuruana 2 5 1 , 7 Endêmica (Piau í)
Platypodium elegans 2 1 3, 3 Ampla
Cenostigma macrophyllum 1 06, 1 Ampla '" -..J 00 Senegalia martii 387,5 Ampla 00 Ul
Poincianella pluviosa 1 40 Ampla .:., N +-
37 Caatinga
1 4 Anadenanthera colubrina 2 5 Ampla 00 (Minas Gerais) '" a-
Mimosa tenuiflora 1 7, 5 Ampla o, Leucochloron limae 7, 5 Endêm ica
Copaifera coriacea 1 60 Endêm ica
Chamaecrista belemii 62 Endêm ica
20 Caati nga
7 Trischidium molle 29 Disjunta (Bahia)
Peltogyne pauciflora 5 Endêm ica
Cratylia mollis 3 Endêm ica
Florestas úmidas
Balizia pedicellaris 1 3,3 Ampla
Copaifera trapezifolia 8,9 Ampla
38 Mata Atlântica
1 4 Inga capitata 8,9 Ampla (Rio de Jane iro)
Swartzia apetala 8,9 Ampla
Copaifera langsdorffii 4,4 Ampla
Inga brevialata 6 Ampla
Macrolobium latifolium 3,8 Ampla
39 Amazônia
41 Inga paraensis 3,5 Ampla (Pará)
Abarema jupunba 1 ,6 Ampla
Eperua bijuga 0,8 Ampla
Machaerium paraguariense 3,9 Ampla
Lonchocarpus subglaucescens 1 ,9 Endêmica
40 Mata Atlântica
6 Dalbergia frutescens 1 ,9 Ampla (Paraná) Machaerium brasiliense 1 , 9 Ampla
Lonchocarpus cultratus 1 , 1 Ampla
Dalbergia frutescens 23 Ampla
Mata Atlântica Machaerium paraguariense 1 5 Ampla
41 (Rio G rande do 6 Albizia niopoides 4 Ampla Sul)
Parapiptadenia rigida 3 Ampla
Inga vera Ampla
Diplotropis incexis 23 Endêmica
Inga edulis 1 6 Ampla
42 Mata Atlântica
1 7 Copaifera langsdorffii 9 Ampla (Pernambuco) Inga thibaudiana 9 Ampla
Balizia pedicellaris 6 Ampla

Caat i nga no Contexto de uma Metacomun idade : Ev idências da B iogeografia, Padrões F i logenéticos e . . . - 257
�Oiél1i):"Jiél - Abundância (medida a part i r da densidade absoluta [DA] defin ida como o número de i nd ivíduos de cada espécie por hectare) de espécies lenhosas de Legum inosae em f lorestas secas (caatinga) e úm idas. Os dados foram obtidos a part i r de vários estudos fitossociológ icos d ispon íveis na l iteratura, sendo 1 1 á reas de caat inga e 1 1 de f lorestas úmidas (domín io Atl ântico e Amazôn ico) (continuação)
N úmero de espécies DA** Referência Vegetação de Leguminosae Táxons* (indivíduos/ Distribuição
amostradas hectare)
Inga heterophyl/a 1 3,6 Ampla
Inga alba 7,8 Ampla
43 Amazôn ia
36 Tachigali me/inonii 6,6 Ampla (Pará)
Tachiga/i me/anocarpa 6,4 Ampla
Chamaecrista sp. 6,3
Chamaecrista ensiformis 7 1 Ampla
Andira nitida 9,7 Ampla
44 Mata Atlântica
7 /nga capitata 8,6 Ampla (Espírito Santo)
Ormosia arborea 5,4 Endêm ica
Andira fraxinifo/ia 3,2 Ampla
Swartzia reticu/ata 48 Ampla
Macr% bium /atifo/ium 28 Ampla
45 Mata Atlântica 42 Har/eyodendron unifo/io/atum 24 Endêmica
(Bahia) Arapatiel/a emarginata 1 6 Endêmica
P/atymiscium speciosum 1 6 Endêmica
Poeppigia procera 25 Ampla
Tachiga/i rugosa 25 Ampla
46 Mata Atlântica
1 6 Da/bergia vil/osa 20,2 Ampla (Minas Gerais)
Senna macranthera 1 5,4 Ampla
Senna mu/tijuga 9,6 Ampla
Eperua duckeana 7 Endêmica
Eperua g/abrif/ora 4 Endêmica
47 Amazônia
23 Macr% bium suaveo/ens 4 Ampla (Amazonas)
/nga a/ba 3 Ampla
Parkia mu/tijuga 3 Ampla
Pseudopiptadenia contorta 36 Ampla
Senega/ia po/yphyl/a 22 Ampla
48 Mata Atlântica
1 7 Parapiptadenia pterosperma 9 Ampla (Rio de Jane i ro)
Machaeríum íncorruptíbí/e 7 Endêmica
Myrocarpus frondosus 5 Ampla
* Os nomes dos táxons que apareciam nas publ icações originais foram atualizados. ** Os valores são apresentados apenas para os cinco primeiros táxons de Leg uminosae com maior densidade absoluta.
acontece porque as espécies endêmicas originadas ao longo do tempo em cada uma das áreas isoladas das SDTF não são substituídas por espécies imigrantes, resultando em pequenos dados de espécies confinadas a cada fragment027. Esses dados contrastam com os resultados obtidos para leguminosas com diversificação predominante em áreas úmidas, a exemplo dos gêneros Inga8,49 e
Swartzia50, os quais apresentam estruturação filogenética geográfica muito baixa, ou seja, mesmo as espécies simpátricas não são proximamente relacionadas. Neste caso, a distribuição geográfica não prediz a relação filogenética entre as espécies.
Ao analisar a estrutura filo genética de alguns gêneros de leguminosas que possuem espécies endêmicas da

258 - Padrões e Processos - Estudos de Casos
caatinga, verificou-se que em geral elas formam um clado irmão de espécies de outras florestas secas. No gênero Coursetia, por exemplo, as únicas espécies que ocorrem na caatinga (C. rostrata, C. vicioides e Coursetia sp. nov.) são endêmicas e formam um grupo monofilético irmão de um outro composto por espécies de florestas secas da América do NorteS (Matt Lavin, comunicação pessoal). lndigofera blanchetiana, uma espécie endêmica da caatinga, aparece relacionada a três outras espécies de lndigofera (1. thibaudiana, 1. cuerna
vacana e I. platycarpa) distribuídas em florestas secas do Méxic014. Este padrão de distribuição disjunta em gêneros característicos de florestas secas neotropicais está sendo também estudado em Luetzelburgia (dados não publicados, Cardoso et a!. ), um gênero pequeno (circa 14 spp.), cuja maioria das espécies é endêmica da caatinga, duas ocorrendo em florestas semidecíduas do sudeste brasileiro, outras duas nas florestas secas do Brasil Central e duas espécies ainda inéditas sucedem na região da Chiquitania boliviana e dos vales secos interandinos. Isso torna Luetzelburgia um grupo muito promissor para estudar a influência dos processos de metacomunidades na estrutura filogenética.
O padrão de estruturação filo genética na metacomunidade das SDTF também tem sido muito similar ao encontrado em vários grupos de plantas e animais que habitam ilhas oceânicas6.7. Essa é mais uma evidência de que a distribuição fragmentada das SDTF funciona como um sistema de "ilhas", em que a distância geográfica e o tamanho dos fragmentos influenciam nas taxas de dispersão, endemismos, distribuição e manutenção da diversidade de espécies.
O padrão de diversificação nas filogenias das leguminosas de florestas secas é outro aspecto que deve ser ressaltado. Filogenias que incorporam informação temporal obtidas por técnicas de datação utilizando calibração fóssil têm indicado uma diversificação antiga, quase sempre há mais de 1 5 Ma, associada à taxa de especiação muito lenta6-8 . Tem-se observado um padrão distinto nas leguminosas de florestas úmidas (por exemplo, lnga), que costumam apresentar diversificação bem mais recente, às vezes há menos de 2 Ma, e taxa de evolução rápida, o que resulta em filogenias menos resolvidasS.49. A manutenção de uma flora antiga nas SDTF pode ser entendida também do ponto de vista de metacomunidades. A baixa taxa de imigração em determinada comunidade local de floresta seca diminui a competição, evitando grande perda de espécies (isto é, baixa taxa de extinção), o que resulta na continuidade da maioria das linhagens ao longo do tempo.
Neste capítulo buscou-se entender a biogeografia da caatinga dentro do contexto da metacomunidade das SDTF neotropicais, fundamentada nos avanços recentes dos estudos filogenéticos em Leguminosae, uma família que tem longa história de diversificação nas florestas :g
00 secas de todo o mundo. A teoria de metacomunidades, B; para isso, tem se mostrado desafiadora, trazendo nova � abordagem ecológica e histórica para o entendimento 00 '-O do funcionamento destas florestas. Viu-se, por exemplo, � que a elevada abundância de espécies de Leguminosae da caatinga se comporta como prediz a teoria, enquanto as espécies de florestas úmidas tendem a ser raras em determinada comunidade local.
A datação de filogenias de leguminosas confinadas primariamente às SDTF tem revelado que a diversificação destas florestas é muito antiga. Isso contradiz a ideia de que a riqueza de espécies e endemismos nos fragmentos de SDTF tenham sido originados apenas pelos processos de vicariância ocorridos na Região Neotropical durante o Pleistoceno. Entretanto, não deve ser excluída a hipótese de que estes processos também contribuíram, de alguma forma, para a evolução da flora das SDTF e, sobretudo, da caatinga.
Os resultados obtidos com as leguminosas podem ser, portanto, mais bem interpretados de outra forma. Eles mostram que eventos contínuos, como os processos ecológicos de metacomunidades, podem estar atuando há milhares de anos na manutenção da diversidade e endemismos das florestas secas, ao contrário de pressupor apenas eventos históricos únicos, como deriva continental ou mudanças climáticas. Desta forma, diferentemente das hipóteses de vicariância, a teoria neutra de metacomunidades traz uma visão mais ecológica e dinâmica para a biogeografia das SDTF, ou sej a, mostra que os processos ainda estão acontecendo.
A biogeografia da caatinga, como discutida aqui, segundo uma visão de metacomunidades, foi desenvolvida a partir da combinação de vários estudos que vêm sendo realizados apenas com as leguminosas. Seria importante verificar se a teoria também consegue suporte de padrões biogeográficos e filogenéticos de outras famílias de plantas ou mesmo de animais . Para isso, seria imprescindível selecionar grupos de organismos que possuem compreensão filo genética robusta e que apresentam distribuição relacionada à caatinga e aos demais fragmentos da metacomunidade de SDTF neotropicais.

Caati nga no Contexto de uma Metacomun idade : Ev idênc ias da B i ogeograf ia , Padrões F i logenét icos e . . . - 259
Agradecemos ao Dr. Alessandro Rapini e dois revisores anônimos pelas críticas e sugestões feitas ao manuscrito; Quézia C. dos Santos pelas sugestões na redação final do texto; José Marcos da Silva pela atualização da distribuição das espécies de Lonchocarpus de caatinga; e Dr. Ramiro Camacho (UERN) e Diego Nathan N. Souza pelo apoio em coletas na caatinga da região do Seridó, Rio Grande do Norte. Trabalhos de campo foram parcialmente financiados pelos projetos IMSEAR (CNPq) e PPBio Semiárido (MCT). Agradecemos ainda ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela concessão de bolsa de doutorado ao primeiro autor, através do Programa de Pós-graduação em Botânica da Universidade Estadual de Feira de Santana (UEFS) e de produtividade em pesquisa ao segundo (processos 143 1 1 6/2008-8 e 301 308/2007-1) .
REFERÊNCIAS BIBLIOGRÁFICAS
1. QUEIROZ, L. P. The Brazilian Caatinga: phytogeographical patterns inferred from distribution data of the Leguminosae. In: PENNINGTON, R. T; LEWIS, G. P; RATTER, J. A. Neo
tropical Savannas and Seasonally D ry Forests - Plant
Diversity, Biogeography, and Conservation. Boca Raton: CRC Press, 2006. capo 6, p. 1 2 1 - 1 57 .
2. LEAL, L R ; SILVA, J. M. c. ; TABARELLI, M. et a!. Changing the course of biodiversity conservation in the Caatinga of Northeastern Brazil. Consel". Bio/. , v. 1 9, p. 701-706, 2005.
3. PRADO, D. E . ; GIBBS, P E. Patterns of species distributions in the dry seasonal forests of South America. Ann. Missouri
Bot. Gard. , v. 80, p. 902-927, 1 993. 4. GIULIETTI, A. M. ; HARLEY, R. M.; QUEIROZ, L. P et aI.
Espécies endêmicas da caatinga. In: SAMPAIO, E. V. S. B . ; GIULIETTI A. M. ; VIRGÍNIO, J . ; GAMARRA-ROJAS, C. F. L. Vegetação e Flora da Caatinga. Recife: APNE/CNIP, 2002. capo 7, p. 103-1 1 8 .
5 . QUEIROZ, L. P Leguminosas da Caatinga. Feira de Santana: Universidade Estadual de Feira de Santana, 2009. 467p.
6. LAVIN, M.; SCHRlRE, B. D. ; LEWIS, G. P et aI. Metacommunity process rather than continental tectonic history better explains geographically structured phylogenies in legumes. Phil. TraiU. R. Soc. Lond., B. Biological Sciences, v. 359, p. 1 509- 1 522, 2004.
7. PENNINGTON, R. T; LAVIN, M.; PRADO, D. E. et aI. Historical climate change and speciation: neotropical seasonally dry forest plants show patterns ofboth Tertiary and Quaternary diversification. Phil. Trans. R. Soco Lond., B. Biological Sci
ences, v. 359, p. 5 15-537, 2004. 8. LAVIN, M. Floristic and geographical stability of discontinuous
seasonally dry forests explains patterns of plant phylogeny and endemism. In: PENNINGTON, R T; LEWIS, G. P; RATTER, 1. A. Neotropical Savannas and Seasonally Dry Forests - Plant Diversity, Biogeography, and Conservation. Boca Raton, Florida: CRC Press, 2006. capo 19 , p. 433-447. -
9. SCHRlRE. B. D.; LAVIN, M.; LEWIS, G. P Global distribution patterns of the Leguminosae: insights from recent phylogenies.
In: FRIlS, L; BALSLEV, H. Plant diversity and complexity patterns: local, regional and global dimensions. Bio/. Skr., v. 55, p . 375-422, 2005.
1 0. PRADO, D. E. Seasonally dry forests of tropical South America: from forgotten ecosystems to a new phytogeographic unit. Edinb. J. Bot. , v. 57, p. 437-461 , 2000.
1 1 . PENNINGTON, R T.; PRADO, D. E.; PENDRY, C. A. Neotropical seasonally dry forests and Quaternary vegetation changes. 1. Biogeogr. , V. 27, p. 261 -273, 2000.
12 . VELLOSO, A. L.; SAMPAIO, E. V. S. B . ; GIULIETTI, A. M. et aI. Ecorregiões - Propostas para o Bioma Caatinga. Recife: APNE, The Nature Conservancy do Brasil, 2002. 76p.
13 . LAVIN, M.; BEYRA-MATOS, A. The impact of ecology and biogeography on legume diversity, endemism, and phylogeny in the Caribbean Region: a new direction in historical biogeography. Bot. Rev., v. 74, p. 178-196, 2008.
14. SCHRlRE, B. D.; LAVIN, M.; BARKER, N. P et a!. Phylogeny of the tribe Indigofereae (Leguminosae - Papilionoideae): geographically structured more in succulent-rich and ternperate settings than in grass-rich environments. Amer. 1. Bot., V. 96, p. 8 16-852, 2009.
1 5 . HUBBELL, S. P The Unified Neutral Theory of Biodiversity andBiogeography. Princeton: Princeton University Press, 2001 . 448p.
16. NIMER, E. Climatologia da região Nordeste do Brasil. Introdução à climatologia dinâmica. Rev. Bras. Geogr. , v. 34, p. 3-5 1 , 1 972.
17. MACHADO, L C. S . ; BARROS, L. M.; SAMPAIO, E. V. S. B . Phenology of the caatinga species at Serra Talhada, PE, Northeastern BraziI. Biotropica, v. 29, p. 57-68, 1 997.
1 8. BULLOCK, S. H. Plant reproduction in neotropical dry forests. In: BULLOCK, S . ; MOONEY, H.; MEDINA, E. Seasollalh
Dry Tropical Forests. Cambridge: Cambridge University Press. 1 995. capo 1 1 , p. 277-297.
19 . CAETANO, S . ; PRADO, D.; PENNINGTON, R T et aI. The history of seasonally dry tropical forests in eastern South America: inferences from the genetic structure of the tree As
tronium urundeuva (Anacardiaceae) . MoI. Eco/. , v. 1 7 , p. 3 147-3 1 59, 2008.
20. ROCHA, P L. B . ; QUEIROZ, L. P. ; PIRANI, 1. R Plant species and habitat structure in a sand dune field in the Brazilian Caatinga: a homogenous habitat harbouring an endemic biota. Rev.
Bras. Bot. , v. 27, p. 739-755, 2004. 2 1 . CARDOSO, D. B. O. S . ; QUEIROZ, L. P Diversidade de Le
guminosae nas caatingas de Tucano, Bahia: implicações para a fitogeografia do semi-árido do Nordeste do Brasil. Rodriguésia,
v. 58, p. 379-391 , 2007. 22. GRIZ, L. M. S.; MACHADO, L C. S . Fruiting phenology and
seed dispersaI syndromes in caatinga, a tropical dry forest in the northeast of Brazil. J. Trop. Eco/. , v. 1 7, p . 303-321 , 200 1 .
23. AB'SABER, A . N. O doilÚnio morfoclimático semi-árido das caatingas brasileiras. Geomoifologia, v. 43, p. 1 -39, 1 974.
24. ALONSO, D. ; ETIENNE, R S.; McKANE, A. 1. The merits of neutral theory. Trends Eco/. Eval. , v. 2 1 , p. 45 1 -457 , 2006.
25. HUBBELL, S. P Neutral theory in community eco10gy and the hypothesis of functional equivalence. Funct. Eco/. , V. 1 9, p. 166-172, 2005.
26. LATIMER, A. M.; SILANDER, J. A.; COWLING, R. M. NeutraI ecological theory reveals isolation and rapid speciation in a biodiversity hot spot. Science, v. 309, p. 1 722- 1 725, 2005.
27. PENNINGTON, R T; RICHARDSON, 1. E.; LAVIN, M. Insights into the historical construction of species-rich biomes fiom dated p1ant phy1ogenies, neutral ecological theory and phylogenetic community structure. New Phyto/. , v. 172, p. 605-61 6, 2006.

260 - Padrões e Processos - Estudos de Casos
28. SANTANA, 1. A. S. Estrutura .fitossociológica, produção de serapilheira e ciclagem de nutrientes em uma área de Caatinga no Seridó do Rio Grande do Norte. Areia, 2005, 1 84p. Tese (Doutorado) - Universidade Federal da Paraíba.
29. AMo.RIM, r. L.; SAMPAIO., E. V. S. B . ; ARAÚJO, E. L. Flora e estrutura da vegetação arbustivo-arbórea de uma área de caatinga do Seridó, RN, Brasil. Acta Bot. Bras. , v. 19, p. 615-623, 2005.
30. PEREIRA, L M.; ANDRADE, L. A.; BARBo.SA, M. R. V. et aI. Composição florística e análise fitos sociológica do componente arbustivo-arbóreo de um remanescente florestal no agreste paraibano. Acta Bot. Bras. , v. 1 6, p. 357-369, 2002.
3 1 . QUEIRo.Z, J. A.; TRo.VÃo., D. M. B. M.; o.LIVEIRA, A. B. et aI. Análise da estrutura fitossociológica da Serra do Monte, Boqueirão, Paraíba. Rev. Biol. e Ciênc.Terra, v. 6, p. 25 1 -259, 2006.
32. FABRICANTE, J. R.; ANDRADE, L. A. Análise estrutural de um remanescente de caatinga no Seridó paraibano. Gecol. Bras., v. ! l , p. 341 -349, 2007.
33. LIMA, J. R.; SAMPAIO., E. V. S. B . ; Ro.DAL, M. J. N. et aI. Estrutura da floresta estacionai decidual montana (mata seca) da RPPN Serra das Almas, Ceará. Rev. Bras. Biociências, v. 5 , p. 438-440, 2007.
34. Ro.DAL, M. J. N; Co.STA, K. C. c.; SILVA, A. C. B. L. Estrutura da vegetação caducifólia espinhosa (caatinga) de uma área do sertão central de Pernambuco. Hoehnea, v. 35, p. 209-217, 2008.
35. ALCo.Fo.RADo.-FILHo., F. G.; SAMPAIO., E. V. S. B . ; Ro.DAL, M. 1. N. Florística e fitos sociologia de um remanescente de vegetação caducifólia espinhosa arbórea em Caruaru, Pernambnco. Acta Bot. Bras. , v. 1 7, p. 287-303, 2003.
36. LEMo.S, J. R. ; Ro.DAL, M. 1. N. Fitossociologia do componente lenhoso de nm trecho da vegetação de caatinga no Parqne Nacional Serra da Capivara, Piauí, Brasil. Acta Bot. Bras. , v. 16 , p. 23-42, 2002.
37. SANTo.S, R. M.; VIEIRA, F. A.; SANTo.S, P. F. et aI. Estrutura e florÍstica de um remanescente florestal na fazenda Ribeirão, mnnicípio de Juvenília, MG, Brasil. Rev. Caatinga, v.21 , p. 1 54- 1 62, 2008.
38. PESSo.A, S. V. A.; o.LIVEIRA, R. R. Análise estrutural da vegetação arbórea em três fragmentos florestais na Reserva Biológica de Poço das Antas, Rio de Janeiro, Brasil. Rodriguésia, v. 57, p. 39 1 -4 1 1 , 2006.
39. PINHEIRO., K. A. 0.. ; CARVALHO., J. o.. P. ; QUANZ, B. et aI. Fitossociologia de uma área de preservação permanente no leste da Amazônia: indicação de espécies para recuperação de áreas alteradas. Floresta, v. 37, p. 175- 1 87, 2007.
40. Ko.ZERA, C.; DITTRICH, V. A. 0..; SILVA, S. M. Fitossociologia do componente arbóreo de um fragmento de floresta ombrófila mista montana, Curitiba, PR, BR. Floresta, v. 36, p. 225-237, 2006.
4 1 . BUDKE, J. C . ; GIEHL, E. L. H.; ATHAYDE, E. A. et aI. Florística e fitos sociologia do componente arbóreo de uma floresta ribeirinha, arroio Passo das Tropas, Santa Maria, RS, Brasil. Acta Bot. Bras. , v. 1 8 , p. 58 1 -589, 2004.
42. FERRAZ, E. M. N.; Ro.DAL, M. J. N. Caracterização fisionômica-estrntural de um remanescente de floresta ombrófila montana de Pernambuco, Brasil. Acta Bot. Bras. , v. 20, p. 9 1 1 -926, 2006.
43. ESPÍRITo.-SANTo., F. B . ; SHIMABUKURo., Y. E.; ARAGÃO., L. E. o.. C. et aI. Análise da composição florística e fitossociológica da floresta nacional do Tapajós com o apoio geográfico de imagens de satélites. ActaAmazonica, v. 35, p. 1 55- J 73, 2005.
44. SIMONELLI, M.; So.UZA, A. L.; PEIXo.To., A. L. et aI. Floristic composi tion and strncture of the tree component of a muçunnnga forest in the Linhares Forest Reserve, Espírito Santo, Brazil. Mem. New York Bot. Gard., v. 100, p. 35 1 -370, 2008.
45. THo.MAS, W W; CARVALHO., A. M.; AMo.RIM, A. M. A. et aI. Diversity of wood plants in the Atlantic Coastal Forest of Southern Bahia, Brazil. Mem. New York Bot. Gard. , v. 100, p. 2 1 -66, 2008.
46. CARVALHO., D. A.; o.LIVEIRA-FILHo., A. T; VAN DEN BERG, E. et aI. Variações florÍsticas e esttuturais do componente arbóreo de uma floresta ombrófila alto-montana às margens do rio Grande, Bocaina de Minas, MG, Brasil. Acta Bot. Bras. , v. 1 9, p . 9 1 - 109, 2005.
47. SILVA, K. E.; MATo.S, F. D. A.; FERREIRA, M. M. Composição florística e fitossociologia de espécies arbóreas do Parque Fenológico da Embrapa Amazônia o.cidental. ActaAmazonica, v. 38, p. 2 13-222, 2008.
48. SILVA, G. C.; NASCIMENTO., M. T Fitossocio10gia de um remanescente de mata sobre tabuleiros no norte do estado do Rio de Janeiro (Mata do Carvão). Rev. Brasil. Bot., v. 24, p . 5 1 -62, 200 1 .
49. RICHARDSo.N, 1. E.; PENNINGTo.N, R. T; PENNINGTo.N, T D. et aI. Recent and rapid diversification of a species-rich genus ofneotropical trees. Science, v. 293, p. 2242-2245, 200 I .
50. To.RKE, B . M. ; SCHAAL, B . A. Molecular phylogenetics of the species-rich neotropical genus Swartzia (Leguminosae, Papilionoideae) and related genera of the swartzioid c1ade. Amer. 1. Bot. , v. 95, p. 2 15-228, 2008.

J l
Alexandre Cunha Ribeiro
Flávio César Thadeo de Lima
Naércio Aquino Menezes
As águas continentais da região Neotropical abrigam a mais diversificada fauna de peixes de água doce do planeta. O número de espécies conhecidas se aproxima de 5.000 e estimativas quanto ao número real, considerando aquelas que ainda estão por serem descritas, giram em tomo de 6 .000 a 8 .000 espéciesl .
A composição desta diversidade é resultado de longo processo de irradiação de diversas linhagens ao longo de milhões de anos, mas também da extinção de grupos outrora diversificados. Esta riqueza resulta, em última instância, de processos ecológicos e históricos complexos, contínuos através do tempo geológico. A história de nossa fauna moderna, cujas raízes remontam principalmente ao período Mesozoico, ainda não é compreendida na sua totalidade. Desde as primeiras ideias dos autores precursores das atuais hipóteses fundamentadas no arcabouço conceitual de métodos modernos de reconstrução filogenética e em teorias unificadoras, por exemplo, a Tectônica de Placas, houve avanços significativos. É
notório, entretanto, que a história de nossa fauna aquática ainda contém lacunas e grandes questões que poderão ocupar a mente dos biogeógrafos por muitas gerações.
Neste capítulo pretende-se fornecer uma visão geral do conhecimento da biogeografia dos peixes de água doce da América do Sul tentando correlacionar a evolução da fauna com a evolução geográfica do continente, porém não se esgotará o assunto. Tentar-se-á demonstrar
1 6
aqui, com base em exemplos da nossa rica fauna de peixes que, conforme já declarado por um dos mais importantes autores em biogeografia, Léon Croizat. "Terra e vida evoluem juntas".
É imprescindível, em uma discussão acerca da biogeografia dos peixes de água doce Neotropicais, vislumbrar a megabiodiversidade da ictiofauna, abordando seus componentes principais. Em sua composição ocorrem representantes de grupos antigos, outrora diversificados em Gondwana, mas hoje escassos, grupos Gondwânicos remanescentes e diversos ainda hoje e grupos resultantes de irradiações mais recentes, presumivelmente posteriores a sua ruptura final, ou seja, grupos endêmicos de um continente sul-americano j á isolado. Somam-se a estes componentes grupos de ancestrais marinhos relativamente recentes, invasores secundários da água doce e, em menor escala, grupos que transitam entre os ambientes marinho e dulciaquícola.
Uma caracterização resumida nos obriga a analisar a composição da ictiofauna Neotropical em nível de agrupamentos taxonômicos mais inclusivos, o que pode ser feito tendo como pano de fundo a filogenia simplificada dos grandes grupos modernos de peixes (Fig. 16 . 1 ) . Detalhes sobre esta composição podem ser vistos em Reis et aI. 1 .

262 - Padrões e Processos - Estudos de Casos
A ictiofauna dominante pertence a um grande grupo denominado Ostariophysi, que inclui todos os peixes que possuem um órgão de audição, o aparelho de Weber, por meio do qual ondas sonoras que chegam à bexiga natatória são transmitidas ao ouvido interno por meio de uma série de ossículos modificados na parte anterior
da coluna vertebral e transformadas em impulsos elétricos transmitidos ao cérebro. Desse grupo faz parte a maioria dos "peixes de escama" (ordem Characiformes), os "peixes de couro", vulgarmente conhecidos como bagres e cascudos (ordem Siluriformes) e os "peixes elétricos" sul-americanos (ordem Gymnotiformes).
Myxiniformes (peixes feiticeira)
Petromyzontiformes ( Iampreias)
Holocephal i (qu imeras)
Hybodontidae t Galeomorphi (tubarões comuns)
Batoidea (raias marinhas e de água doce)
Squal iformes (cações-anjo e cia.)
Polypteriformes
Chondrostei (esturjões, peixes-espátu la e cia.)
Lapisosteidae (gars)
Ami iformes
Osteoglossomorpha (pirarucu, aruanãs e ( ia .)
E lopomorpha (tarpões, enguias, morei as e (ia.)
C lupeomorpha (sardinhas, anchovas e c ia .)
Cyprin iformes (carpas e cia.) 1fr � Characiformes (traíras, piranhas, lambaris e cia.) � �
ãJ
Gonorynchiformes ,10 Q
-__ ----Siluriformes (bagres) �.
Gymnotiformes (poraquê, tuviras)
Protacanthopteryg i i (salmões e cia.)
Esociformes
Stenopterygi i
Scopelomorpha
Lam pridiomorpha
Paracanthopterygi i (baca l haus e c ia .)
Atherinomorpha (peixes-rei, barrigudinhos, peixes-agulha e c ia .) Gasterosteiformes
Scorpaeniformes (peixes-escorpião)
Tetraodontiformes (baiacus e peixe-cofre)
Pleuronectiformes ( l inguados)
Perciformes (corvinas, carás e cia.)
"tl ro ;:; o 3 o
-O ::Y '"
J
rn c ,-+ � ro o ::q. �.
- Fi logenia dos g ra ndes grupos de vertebrados i l ustrando as relações entre os componentes da fauna de peixes da América do Su l discutidos neste capítu lo.

Dentre os Characiformes, a farm1ia Characidae (lambaris, piranhas, pacus, matrinxãs e outros) inclui 1 .057 espécies válidas reconhecidas atualmente (esta e as demais estimativas de espécies válidas por grupo aqui apresentadas, segundo Eschmeyer e Fricke2). Algumas famílias, como a dos corimbatás (Prochilodontidae) e a dos piaus (Anostomidae), incluem os peixes mais importantes na pesca de subsistência, em razão de grande abundância e biomassa. Outras famílias, como a das traíras (Erythrinidae), possuem menor diversidade, mas são bem conhecidas popularmente. Além da América do Sul, os Characiformes ocorrem também na África (quatro famílias) e América do Norte, no sul do Texas e Novo México [apenas uma espécie, Astyanax mexicanus (De Filippi, 1853)] .
Na Região Neotropica1, os Siluriformes são representados por 15 famílias, a maior parte delas com espécies na América do Sul. Sua diversidade, em número de espécies, ultrapassa a dos Characiformes. Só a família Loricariidae (popularmente conhecidos como cascudos, caris ou bodós) inclui cerca de 792 espécies conhecidas. Coridoras e tamboatás (família Callichthyidae) somam 197 espécies. Os grandes bagres como jaús, pintados e cacharas, assim como os mandis da farm1ia Pimelodidae, são representados por aproximadamente 102 espécies. Outras famílias importantes, menos conhecidas por serem representadas por formas diminutas, crípticas e de hábitos noturnos, são os bagrinhos da família Heptapteridae ( 199 espécies) e as cambevas e candirus da família Trichomycteridae (235 espécies). Duas famílias, Ariidae e Plotosidae, incluem espécies que invadiram secundariamente o hábitat marinho. Além da América do Sul, os Siluriformes ocorrem em todos os continentes, inclusive a Antártica (como fósseis).
Os Gymnotiformes, ou peixes elétricos sul-americanos, são endêmicos da região Neotropical, porém com diversidade menor (algo em torno de 150 espécies), que os dois grupos anteriores. Todas as espécies das cinco famílias da ordem possuem a capacidade de gerar campo elétrico a partir de sua musculatura especializada, utilizado para eletropercepção e comunicação. Uma espécie, Electrophorus electricus (Linnaeus, 1766), conhecida como poraquê, produz descarga elétrica capaz de atordoar e mesmo matar presas ou predadores.
Os peixes ósseos (Osteichthyes) incluem também alguns grupos antigos, mais abundantes como fósseis, mas atualmente representados por poucas espécies, como a piramboia (Lepidosiren paradoxa [Fitzinger, 1 837]) "peixe pulmonado" da ordem Lepidosireniformes, encontrada na Amazônia e bacias do Paraguai e do Prata, o pirarucu (Arapaima gigas [Schinz, 1 822]) e os aruanãs
B iogeografia dos Peixes de Água Doce da América do Su l - 263
(Osteoglossum bicirrhosum [Cuvier, 1829]) e O. ferreirai (Kanazawa, 1966) da ordem Osteoglossiformes, restritos à bacia Amazônica e outras drenagens ao norte da América do Sul .
O tarpão (Megalops atlanticus [Valenciennes, 1847]) da ordem Elopiformes ocorre em águas marinhas e estuarinas, mas eventualmente é encontrado em água doce. Entre os Anguillliformes apenas duas espécies, a enguia (Anguillidae) e Stictorhinus potamius (Bi:ihlke e McCosker, 1 975) da família Ophichthidae, são registradas em águas doces sul-americanas, a primeira no norte da América do Sul (Colômbia e Venezuela) e a segunda na bacia do Rio Tocantins, Brasil.
Representantes da ordem Clupeiformes (manjubas, apapás e sardinhas), um grupo tipicamente marinho, possuem, no entanto, moderada representação em água doce. Nas águas continentais da América do Sul ocorrem três gêneros da família Clupeidae (Platanichthys e Ramnogaster na parte inferior da bacia do Prata e Rhinosardinia nos cursos inferiores de rios do norte e nordeste do continente) e espécies dos gêneros Anchoa, Anchovia, Amazonsprattus, Anchoviella, Jurengaulis.
Pterengraulis e Lycengraulis, da família Engraulidae. na Amazônia, bacia do Orinoco, Rio São Francisco e rios das Guianas. Em Pristigasteridae são conhecidos representantes dos gêneros Ilisha e Pristigaster na bacia Amazônia e Pellona nas bacias Amazônica e Paraná-Paraguai e do Orinoco.
No grande grupo que inclui os teleósteos mais derivados, Acanthopterygii, predominantes no ambiente marinho, algumas ordens são representadas por grupos que invadiram secundariamente a água doce. Em Atheriniformes são encontrados alguns poucos representantes de peixes-agulha (Belonidae e Hemiramphidae) e os peixesrei (Atherinopsidae). Uma ordem primariamente de água doce e com grande diversidade é Cyprinodontiformes, que inclui os barrigudinhos (Poeciliidae), os peixes anuais (Rivulidae), os tralhotos (Anablepidae), além de Cyprinodontidae em sistemas costeiros do norte da América do Sul e os representantes de Orestiidae do altiplano andino.
O grupo com a maior diversidade de teleósteos, coletivamente chamados Percomorpha, é representado em águas continentais sul-americanas, por famílias com poucos representantes, tais como Polycentridae (peixes-folha, duas espécies), Synbranchidae (muçum, quatro espécies), Gobiidae (amborés, três espécies exclusivas de água doce e pouco mais de uma dezena de espécies anfídromas), Sciaenidae (corvinas e pescadas, 19 espécies exclusivas de água doce), Percichthyidae (cinco espécies patagônicas/chilenas), Percilidae (duas espécies no Chile) e outras de

264 - Padrões e Processos - Estudos de Casos
grande diversidade, tais como Cichlidae (acarás, jacundás e tucunarés), que abrigam mais de 330 espécies conhecidas em águas doces sul-americanas.
Grupos ainda mais derivados em função dos caracteres morfológicos exclusivos que possuem, também são representados por um número mais reduzido de espécies em água doce, como os baiacus (Colomesus asellus [Müller e Troschel, 1 849] [Tetraodontidae: Tetraodontiformes]), e alguns gêneros de linguados (Achiridae: Pleuronectiformes) .
Os demais não Osteichthyes têm representatividade muito reduzida. Entre os peixes cartilaginosos (tubarões e raias) apenas uma espécie de tubarão, Carcharhinus leucas (Müller e Renle, 1 839) e duas espécies de peixes-serra, Pristis pristis (Linnaeus, 1758) e Pristis pectinata (Latham, 1794) penetram em água doce, mas no grupo das raias existe toda uma farm1ia, Potamotrygonidae, exclusiva de água doce e representada por três gêneros (Potamotrygon, Paratrygon e Plesiotrygon) e pelo menos 1 8 espécies.
Neste tópico, são comentadas algumas obras consideradas de referência para o estudo da biogeografia de peixes de água doce da América do Sul. São incluídos trabalhos que, por seu escopo amplo e marcada influência, trouxeram grandes avanços no entendimento da biogeografia de peixes de água doce neotropicais. Estudos mais pontuais, embora não menos dotados de importância intrínseca, não serão mencionados por questões práticas.
O primeiro autor a realizar uma ampla análise de padrões biogeográficos em peixes sul-americanos foi Eigenmann3. Este autor dividiu a América do Sul em três regiões "ictiogeográficas" : "Brasileira", "Andina" e "Patagônica". A Região Patagônica foi definida como possuindo uma ictiofauna muito pobre, composta por elementos "migrantes do mar", como os Atherinopsidae e os Percichthyidae, "imigrantes" da Região Brasileira (Characidae e Trichomycteridae), formas "autóctones" ou de "origem duvidosa" (Nematogenyidae e Diplomystidae) e grupos de relação biogeográfica transcontinental (Galaxiidae, Aplochitonidae, atualmente incluídos em Galaxiidae e Petromyzontidae, hoje incluídos em Mordaciidae e Geotriidae). A Região Andina, definida
como compreendendo rios drenando ambas as vertentes andinas e sistemas endorreicos da Bolívia e Peru (isto é, Lago Titicaca), também bastante pobre ictiofaunisticamente, sendo caracterizada pela presença de alguns poucos grupos endêmicos (por exemplo, Astroblebidae e Orestiidae). A Região Brasileira, a mais vasta e diversa região ictiogeográfica, inclui, na concepção de Eigenmann3, além de quase toda a América do Sul, toda a América Central, até o sul do México. Eigenmann3 subdivide essa região em dez "províncias", das quais duas ("pacífica" e "Magdalena") estão parcialmente e seis ("Amazônica", "Guiana", 'Trinidad", "São Francisco", "costeira" e "La Plata") completamente situadas na América do Sul. A divisão em regiões e províncias ictiofaunísticas de Eigenmann3 obedece tanto a critérios de composição faunística como relações de grandes grupos; por exemplo, a ocorrência de Galaxiidae e lampreias na região patagônica foi por ele considerada como indício da relação desta área com outras massas continentais austrais que contêm esses mesmos elementos (isto é, Austrália e Nova Zelândia), assim como a ocorrência de Orestiidae, uma família endêmica de Cyprinodontiformes do altiplano andino, que provavelmente teria se originado, nas palavras de Eigenmann3, "when the lake was still an arm of the sea". É notável que Eigenmann3A j á percebera a importância da história geológica na distribuição dos peixes de água doce sul-americanos, ao verificar que as terras altas dos escudos, geologicamente mais antigas, apresentavam ictiofauna bastante distinta daquela das terras baixas das planícies, e que esta relação poderia ser mais significativa biogeograficamente do que os limites dos sistemas hidrográficos. Por exemplo, a região guianense de Eigenmann3 inclui não só os rios guianenses, como também os tributários da margem norte do Rio Amazonas que drenam o escudo guianense, bem como o Rio Branco. Em sentido amplo, a província amazônica incluiria, de fato, as terras baixas de todos os grandes sistemas hidrográficos sul-americanos, ou nas palavras de Eigenmann3 : "East of the Co rdilleras, and therefore east of the Magdalena basin, is found the most extensive and intricate fresh water system in the world . . . a network of rivers practically ininterrumpted, extending from the mouth of the Orinoco through the Cassiquiare, Rio Branco, Rio Negro, Rio Madeira, Rio Guaporé, Rio Paraguay, Parana and La Plata to Buenos Aires".
Somente 60 anos depois, com Géry5, a biogeografia de peixes sul-americanos de água doce voltou a ser abordada de forma ampla. Géry5 identificou oito grandes regiões ictiogeográficas na América do Sul, que coincidem, em parte, com as regiões e províncias identificadas

por Eigenmann3. As regiões ictiofaunísticas identificadas por Géry5 foram "Orinoco-Venezuelana", abrangendo a bacia do Rio Orinoco, em conjunto com a bacia do lago Maracaibo, a ilha de Trinidade e alguns pequenos sistemas hidrográficos do Mar do Caribe; "Magdalena", abrangendo apenas o sistema do Rio Madalena na Colômbia; "Transandino", compreendendo os sistemas hidrográficos a oeste dos Andes, situados entre o norte da Colômbia e Panamá até o norte do Peru; "Andina", compreendendo a porção alta dos Andes, da Colômbia ao Chile; "Paranaense", abrangendo a bacia platina e alguns sistemas do leste do Brasil; "Patagônica", abarcando as drenagens do sul da Argentina e Chile; "Guiano-Amazônica", abrangendo os rios guianenses e a bacia amazônica, e "Leste do Brasil", incluindo os sistemas costeiros do nordeste, leste e sul do Brasil, incluindo o Rio São Francisco. Ao contrário de Eigenmann3, Géry5 não reconheceu uma divisão de terras baixas e terras altas, mas as suas regiões são bastante similares às "províncias" de Eigenmann3, tendo como principal diferença o reconhecimento da região do Orinoco-Venezuela, uma área virtualmente desconhecida de modo ictiológico quando Eigenmann3 escreveu sua monografia.
Ringuelet6 apresentou uma nova classificação das áreas de endemismo de peixes na América do Sul. Ele reconheceu vinte "áreas ictiogeográficas" na América do Sul, a maioria delas seguindo a proposta de subdivisão de áreas maiores que haviam sido reconhecidas por Eigenmann3 e Géry5. Por exemplo, a área "Orinoco-Venezuelana" foi por ele dividida em três áreas distintas: sistema do Lago de Maracaibo, drenagens costeiras do Caribe venezuelano e bacia do Rio Orinoco propriamente. A bacia platina foi dividida em três áreas: alto Paraná, alto Paraguai e sistema Párano-platense. Embora o reconhecimento de maior número de áreas por Ringuelet6 reflita os avanços no conhecimento ictiofaunístico da América do Sul desde Eigenmann3, a desvantagem em reconhecer muitas regiões de endemismo reside em que as relações faunísticas entre as áreas são obscurecidas no processo, uma vez que as semelhanças (= táxons
� compartilhados) são preteridas ante as diferenças (= 0\ "? táxons endêmicos). � Weitzman e Weitzman 7 apresentaram uma revisão � crítica das hipóteses biogeográficas sobre peixes sul-co s; -americanos formuladas até aquela data. Eles notaram a
falta de embasamento em hipóteses filo genéticas dos esquemas biogeográficos previamente propostos e a necessidade de conhecimento filogenético, taxonômico e distribucional muito mais detalhado da ictiofauna sul-americana para que hipóteses biogeográficas mais
B iogeografia dos Peixes de Água Doce da Amér ica do Sul - 265
específicas pudessem ser formuladas. Weitzman et al.8 exemplificaram o tipo de abordagem necessário, ao exibir uma discussão biogeográfica sobre os representantes da tribo Glandulocaudini (Characidae) no leste da América do Sul.
Fundamentado em ampla revisão sistemática da família Curimatidae, Vari9 apresentou revisão das regiões de endemismo em peixes de água doce sul-americanos. Como Weitzman e Weitzman7, Vari9 também enfatizou a necessidade de incorporar hipóteses filo genéticas e, assim, entender o componente histórico por trás das presentes associações faunísticas. Vari9 identificou oito áreas de endemismo habitadas por curimatídeos na América do Sul: "westem" (incluindo os sistemas hidrográficos transandinos, do lago de Maracaibo ao Rio Chira no norte do Peru), "Orinoco" (correspondendo à bacia do Rio Orinoco); "Guianas" (sistemas hidrográficos da Guiana, Suriname e Guiana Francesa) ; "Amazon" (bacia amazônica, incluindo a bacia do Rio Tocantins); "northeast" (drenagens nordestinas ao norte do Rio São Francisco); "São Francisco" (correspondendo à bacia homônima) ; "coastal" (sistemas hidrográficos entre o sul do Rio São Francisco até o Rio Paraíba do Sul) : "upper Paraná" (correspondendo à bacia do Rio Paraná. acima do salto de Sete Quedas, hoje afogado sob o lago de Itaipu) e "Paraguay" (incluindo a toda a bacia platina. com exceção do alto Paraná, e também os sistemas costeiros do Uruguai ao sul do estado de São Paulo). Apesar de repetir autores precedentes em estabelecer áreas de endemismo para peixes de água doce sul-americanos com base na presença de espécies endêmicas nos diferentes sistemas, a grande importância do trabalho de Vari9 consiste em ser o primeiro a discutir relações históricas entre diferentes bacias hidrográficas, fundamentado em filogenias de alguns gêneros de Curimatidae. Assim, Vari9 aponta relação entre o lago de Maracaibo com as bacias amazônica, do Orinoco e platina, baseada na filogenia do gênero Potamorhina, da bacia do Rio Madalena com a bacia do Rio Orinoco e a bacia amazônica, embasado na filogenia de um grupo de espécies do gênero Curimata9, bem como a origem "híbrida" da fauna do Rio São Francisc09.
Ao longo da década de 1 990 houve grande incremento no número de estudos de revisão e filogenia de grupos de peixes Neotropicais com base em metodologias cladísticas modernas. Alguns estudos importantes trouxeram visão nova das afinidades zoogeográficas do continente sul-americano. Muito significante é a contribuição de LundberglO, que reviu criticamente as evidências das supostas relações das faunas neotropical e africana, demonstrando que as evidências fósseis e filogenéticas

266 - Padrões e Processos - Estudos de Casos
sugerem padrões muito mais complexos que não podem, na maioria das vezes, serem identificados simplesmente como relações de grupos irmãos associados a um evento vicariante simples, por exemplo, a ruptura de Gondwana.
Com relação aos grupos neotropicais, o ápice desta tendência moderna foi a publicação, em 1998, dos resultados de um simpósio sobre filogenia e classificação de peixes neotropicais na obra Phylogeny and Classification of Neotropical Fishes. Nesta importante obra foi publicado um dos trabalhos de maior impacto para a biogeografia de peixes Neotropicais. Trata-se do artigo de Lundberg et al. 1 1 , o primeiro a associar, de forma muito clara, a biogeografia de peixes de água doce aos processos tectônicos em escala continental a remodelar o continente sul-americano desde sua ruptura com a África. Naquele artigo ficou claro o papel dos processos tectônicos na remodelagem da paisagem em escala continental e de sua influência nos proccssos vicariantes a afetar a distribuição geográfica da fauna de peixes de água doce na América do Sul. A contribuição de Lundberg et al. 1 1 , entretanto, enfatiza eventos históricos desencadeados em porções amazônicas e ao longo do sopé andino, áreas de marcada influência do atual processo de soerguimento da cordilheira dos Andes ao longo principalmente do Cenozoico. Mais recentemente outros estudos trouxeram contribuições relevantes à compreensão da biogeografia de peixes Neotropicais. Ribeiro12 analisou a biogeografia dos peixes das drenagens costeiras do Brasil, na margem leste do continente sul-americano, abordando detalhadamente a história tectônica e sua significância na configuração de padrões de distribuição ao longo da margem atlântica em evolução a partir do Mesozoico. Hubert e Renno13 identificaram e propuseram relações entre 1 1 grandes áreas de endemismo com base em análise parcimoniosa de endemicidade (PAE, parsimony analysis of endemicity) (ver Capo 3).
Atualmente sabe-se que a evolução da paisagem de um continente deve ser compreendida no contexto da tectônica global. A Teoria da Tectônica de Placas de Alfred Wegener possui aceitação global atingida apenas por poucas teorias científicas (ver Capo 1) , o que lhe confere um status em Geociências comparável àquela atingida pela Teoria da Evolução nas Biociências ou Teoria da Relatividade na Física.
Sabe-se hoje que as posições relativas dos continentes não são fixas, mas, por possuírem constituição rochosa majoritariamente granítica e, portanto, pouco densa, flutuam em equilíbrio isostático sobre o assoalho oceânico, de composição majoritariamente basáltica e mais 'C
--..) densa. As placas tectônicas, como são denominados 00
De estes gigantescos blocos continentais, têm estado em � movimento, por conseguinte, continuamente ao longo � das eras geológicas, impelidos por forças advindas das � correntes de convecção do manto. O resultado é que os continentes estão em constante "deriva", colidindo uns com os outros e também se fraturando e se fragmentando em continentes menores, continuamente extinguindo e originando bacias oceânicas ao longo das eras geológicas. Este ciclo contínuo de amalgamação e ruptura de paleocontinentes é chamado "Ciclo de Wilson", ou "abertura e fechamento de bacias oceânicas" ou ainda "Ciclos Oro genéticos" . Este último nome vem do fato de que, quando colidem, as placas tectônicas deformam-se originando cadeias de montanhas como o Himalaia e os Andes (oros = montanha, genesis = origem).
O que hoje é identificado como continente sul-americano, com suas grandes bacias hidrográficas (Fig. 16.2), é resultante de pelo menos quatro grandes ciclos orogenéticos, ou seja, quatro grandes fusões e outras quatro grandes rupturas continentais, além do atual processo orogenético andinol4. Durante estes ciclos, processos de deformação continental, magmatismo e falhamentos originaram grande parte da atual estrutura geológica do continente sul-americano.
Dentre os ciclos oro genéticos que afetaram e resultaram na atual estrutura geológica do continente sul-americano, dois possuem significado especial para o entendimento da evolução atual da paisagem sul-americana e, consequentemente, de sua biogeografia. São eles: o terceiro episódio de fusão continental e os quatro eventos de fissão, ou ruptura continental. Tais eventos são conhecidos na literatura especializada como ciclo Brasiliano/Pan-Africano e Ruptura do supercontinente Pangeia.
O ciclo Brasiliano/Pan-Africano constitui, no conjunto, uma série de eventos complexos ocorridos entre 900 e 540 milhões de anosl4. A esta altura alguém pode estar se perguntando como eventos geológicos tão antigos (já que os primeiros peixes aos quais se tem notícia não possuem muito mais do que 550 milhões de anos) podem ser de relevância no entendimento da biogeografia da fauna atual? Pois foi durante este período em que antigos continentes se amalgamaram e originaram a maior parte das províncias estruturais hoje presentes no continente sul-americano. Foi durante o ciclo Brasiliano/Pan-
""

B i ogeograf ia dos Peixes de Água Doce da América do Su l - 267
- G randes bacias h idrográficas su l-americanas. Modificado de Lundberg et al. 1 1 •
-Africano que antigos continentes (sua porções são hoje denominados crátons) colidiram entre si formando extensas áreas de dobramentos em seu entorno (hoje denominados cinturões orogenéticos antigos). Juntas, estas diferentes províncias constituem-se na maioria dos chamados Escudos Cristalinos, ou seja, a maior porção do atual continente sul-americano. Destas antigas cadeias montanhosas, hoje totalmente erodidas, restam expostas apenas suas raízes mais profundas, contíguas aos crátons adjacentes. Os crátons, centrais nos escudos, são mais estáveis tectonica-
mente. Já os antigos cinturões orogenéticos circundantes guardam heranças de sua origem enquanto resultado de colisões continentais, entre elas, um intrincado sistema de antigas falhas geológicas. Ao longo dos eventos tectônicos que se desenrolaram posteriormente, muitos deles ativos até hoje, estes sistemas de falhas pré-cambrianas foram e continuam sendo reativados, originando importantes elementos da paisagem sul-americana, tais como bacias tectônicas modernas como o Pantanal e a depressão do Araguaia-Tocantins, entre outras 15,16. Além

268 - Pad rões e Processos - Estudos de Casos
disso, é a reativação destas antigas falhas por forças tectônicas modernas que gera processos de capturas de rios entre bacias adjacentes 1 2, 1 7 , A compreensão acerca da geologia estrutural do continente sul-americano é, portanto, de crucial importância àqueles que pretendem estudar a biogeografia de peixes ou quaisquer outros organismos,
O outro grande evento tectônico global de crucial relevância é a ruptura da Pangeia, mais especificamente, os momentos finais de fragmentação de um de seus últimos grandes segmentos, Gondwana, que culminou com a ruptura da África e América do SuL A ruptura de Gondwana não é um processo simples, ou mesmo sincronizado. Este evento tectônico magno, conhecido como "Evento Sul-Atlântico" tem suas origens ainda no Triássico, com indícios geológicos associados ao processo de ruptura identificados nas margens equatoriais da América do Sul, da Guiana Francesa ao delta do Amazonas entre 230 e 1 70 milhões de anos. Depois, entre 1 70 e 120 milhões de anos surgem indícios de atividade associada ao processo de ruptura mais ao sul, desde as margens costeiras da Argentina, Uruguai e Brasil, atingindo a região da atual costa do Estado do Espírito Santo. Há cerca de 90 milhões de anos desfez-se definitivamente a conexão existente entre a África e a América do Sul na atual região do Nordeste Brasileiro e a costa da Nigéria (ver referências citadas em Ribeiro 12).
A certa altura do processo de ruptura continental evoluiu, entre as margens divergentes dos futuros continentes Africano e Sul-americano, uma grande bacia fluvial alongada em sentido norte-sul, estendendo-se entre zero e 20° de latitude sul, a chamada "Depressão Afro-brasileira". Este sistema fluvial interconectado, nos quais estavam presentes sistemas fluviais e lacustres, persistiu por milhões de anos, subdividindo-se em sistemas fluviais menores, ao longo do tempo que antecedeu a fase de influência marinha entre as margens continentais 18 .
Dentre as principais heranças da ruptura de Gondwana para a atual paisagem sul-americana está o sistema de drenagem do planalto cristalino brasileiro, pelo menos em sua porção mais ao leste, marcadamente influenciada pelos processos de ruptura continentaL De acordo com alguns modelos, plumas ascendentes do manto causariam deformações em abóboda com centenas de quilômetros de diâmetro entre os continentes em separação, causando um padrão de drenagem característico, no qual os rios passam a drenar do centro soerguido (localizado na região central do rifte) para suas margens (localizadas tanto para as vertentes africanas como sul-americanas) 19. Tal padrão de drenagem é característico e visível ainda hoje em
diversos sistemas fluviais sul-americanos, tais como os dos rios Paraná, São Francisco, Doce e Uruguai, os quais parecem nascer e "fugir" de centros de abóbodas ou megadomos soerguidos em direção ao interior dos continentes, ou diretamente ao ocean020. Concomitantemente à abertura do Oceano Atlântico inicia-se, por conseguinte, a evolução das drenagens costeiras sul-americanas. Muitas delas evoluindo e encontrando seu caminho ao longo dos sistemas de falhas geológicas pré-cambrianas, tais como o Rio Paraíba do Sul, que segue retilíneo, encaixado em antigo sistema de falhas, outros, erodindo o embasamento e aproveitando também os sistemas de falha, mais suscetíveis à erosão, como no caso do vale do Rio Ribeira de Iguape2 1 .
No lado oposto do continente, em sua margem convergente, a Cordilheira dos Andes segue sua evolução concomitante, decorrente da subducção entre a placa continental sul-americana e a placa oceânica de Nazca. Embora a história andina ultrapasse no passado os limites do Mesozoico, é a partir deste período que sua evolução moderna se institui. Esta possui íntima correlação com o atual processo de deriva continental, com a placa continental sul-americana convergindo contra a placa oceânica de Nazca, cuja deformação gera, em última instância, o cinturão orogenético Andinol4. De acordo com McQuarrie et alo 22, o início do soerguimento andino remonta a pelo menos 70 milhões de anos.
Margens convergentes, independentemente de sua extensão, idade ou estágio de desenvolvimento, demonstram similaridades topográficas que permitem a distinção de uma série de fisiografias. Ao sopé do cinturão dobrado propriamente dito (jold-thrust belt) desenvolve-se uma área deprimida denominada bacia de antepaís (joreland basin) . Tipicamente, ocorre uma flexura da crosta nos flancos da bacia de antepaís denominada forebulge1 1 . Como a deformação do orógeno andino progride para o leste, os sedimentos da bacia de antepaís vão sendo incorporados ao complexo montanhoso e a bacia de antepaís, assim como o arco flexural migram para leste juntamente com o orógeno. Áreas soerguidas dos planaltos adjacentes podem ser incorporadas ao conjunto de terras baixas da bacia de antepaís, por exemplo, o Pantanal, uma bacia tectônica interconectada as terras baixas do Chacol 6. As bacias de antepaís formam um conjunto de terras baixas ao sopé andino que podem sofrer transgressões marinhas esporádicas. Isto pode se desenvolver em consequência do aumento do nível dos oceanos em determinado momento, ou por afundamento (subsidência) da própria bacia de antepaís, impulsionada, por exemplo, por maior taxa de compressão entre as margens convergentes em interação.

Diversos representantes da atual fauna de peixes de água doce da América do Sul claramente pertencem às linhagens de ampla distribuição em Gondwana, antecedendo a separação entre os continentes africano e sul-americano23. A piramboia sul-americana (Lepidosiren paradoxa: Lepidosirenidae) corresponde ao grupo irmão dos peixes pulmonados africanos do gênero Protopterus (Protopteridae) e ambos correspondem ao grupo irmão do peixe pulmonado australiano (Neoceratodus forsteri [Krefft, 1 870] : Ceratodontidae). Fósseis de Lepidosiren cf. paradoxa são conhecidos do Cretáceo Superior ao Paleoceno no Peru e Bolívia e de Lepidosiren paradoxa no Eoceno da Argentina I 0. Uma outra família, Ceratodontidae, cujo único representante vivente é o peixe pulmonado australiano, ocorre na América do Sul apenas como fósseis. Estudos moleculares têm sugerido que a divergência entre Ceratodontidae e o clado formado por Lepidosirenidae e Protopteridae tenha ocorrido ainda no final do Paleozoico, há pelo menos 277 milhões de anos, enquanto a divergência entre os peixes pulmonados sul-americanos e africanos gira em torno de 1 20 milhões de anos24. É possível, portanto, que neste último caso a separação entre a África e a América do sul corresponda ao evento cladogenético responsável pela divergência entre as linhagens africanas e sul-americana modernas.
A superordem Osteoglossomorpha (que na América do Sul inclui o pirarucu e os aruanãs) também se enquadra no grupo de peixes cujas linhagens antecedem a ruptura afrobrasileira. Os Osteoglossomorpha estão entre as linhagens mais basais dentre os peixes atuais (Fig. 1 6. 1 ) e com exceção da irradiação moderna da família africana Mormyridae (com mais de 200 espécies), esta superordem é representada por relativamente poucos grupos recentes, e seu registro fóssil é extensolO. Arapaima gigas (Schinz, 1 822), o pirarucu é considerado o grupo-irmão de Heterotis niloticus (Cuvier, 1829) da África, ambos pertencentes à farmlia Arapaimatidae. Já os aruanãs sul-americanos (Osteoglossum) estão mais proximamente
'D aparentados com Scleropages, os "Aruanãs" da Ásia e '" � Austrália e são todos inclusos na farm1ia OsteoglossidaelO. -,r '" t-.n o;' 00 ta-,
Por fim, dentre os representantes sul-americanos, cujas raízes filéticas comprovadamente remontam ao período que antecede a ruptura afrobrasileira, estão os Characiformes e Siluriformes. Além da América do Sul, os Characiformes também estão na África, embora lá o grupo apresente irradiação menor, tanto em número de farmlias (quatro) quanto em espécies (pouco mais de 250).
B iogeograf ia dos Peixes de Água Doce da América do Su l - 269
Alguns grupos sul-americanos estão mais proximamente relacionados aos grupos africanos do que com os demais representantes Neotropicais. Estudos filogenéticos25 identificam, por exemplo, que as bicudas sul-americanas (gêneros Boulengerella e Ctenolucius) e as traíras (Erythrinidae) estão mais intimamente relacionadas ao gênero Hepsetus da áfrica do que aos demais grupos sul-americanos. O mesmo ocorre com o gênero Neotropical Chalceus, cujas afinidades de parentesco se dão com demais membros da farmlia africana Alestidae26 . É possível que, com o avanço dos estudos filogenéticos, vários outros grupos neotropicais venham a ser considerados como pertencentes aos clados transoceânicos, ou seja, cujas raízes evolutivas antecedem a ruptura de Gondwana. Enquanto fósseis de Characifonnes são conhecidos já do Terciário da Europa23 e de vários depósitos da América do Sul e África27, a identidade de fósseis de Characiformes mais antigos é um assunto debatido. Possíveis dentes de traíras (Erythrinidae) são encontrados em depósitos do Cretáceo Superior e início do Paleocen027, constituindo-se evidências importantes da idade mínima de alguns subgrupos dentre os Characiformes.
Dentre os Siluriformes, alguns estudos sugerem que diversas linhagens sul-americanas estão mais proximamente relacionadas às linhagens africanas, tais como 0 5 Doradoidea neotropicais e Mochokidae da África. L o ricarioidea (Região Neotropical) e Amphiliidae (África I . assim como Aspredinidae (Região Neotropical) e Sisoroidea (Ásia)28.
Um exemplo da contínua evolução faunística em íntima associação aos eventos geológicos que governam a evolução das bacias hidrográficas ao longo do tempo, pode ser visto na fauna das drenagens costeiras do leste brasileiro. Ribeiro1 2 revisou os padrões de diversidade envolvendo os rios costeiros e as drenagens do escudo adjacentes, e os relacionou à história geológica da região.
Quando entendida do ponto de vista de suas relações filogenéticas, conclui-se que a grande diversidade de peixes endêmicos dos rios costeiros é também compartilhada com as áreas adjacentes do escudo brasileiro, embora em diferentes níveis hierárquicos. Tratam-se de faunas irmãs, desde níveis muito inclusivos (relações entre subfamílias) passando por níveis intermediários (relações entre gêneros) aos níveis menos inclusivos onde se compartilham populações da mesma espécie (padrões designados por Ribeiro12 como padrões A, B e C, respectivamente).

270 - Padrões e Processos - Estudos de Casos
Os diferentes níveis de compartilhamento faunístico entre estas regiões sugerem que os eventos cladogenéticos entre as grandes bacias do escudo cristalino brasileiro e os rios costeiros são contínuos ao longo do tempo. Eventos antigos são responsáveis pelo reconhecimento dos níveis hierárquicos mais inclusivos, enquanto eventos recentes promovem o compartilhamento de populações da mesma espécie nas diferentes áreas.
Desde o final da ruptura do supercontinente Gondwana aos dias atuais, as drenagens costeiras vêm evoluindo na margem leste do continente. Tal evolução geomorfológica está intimamente relacionada aos eventos tectônicos antigos e recentes ocorridos na região. Diferentemente do que se acreditava há algumas décadas. a margem leste da América do Sul apresenta intensa atividade tectônica29-3 1 •
o que promove intenso realTanjo entre drenagens entre o escudo cristalino e os tributários diretos do Oceano Atlântico com consequente miscigenação faunística12.
Ribeiro et alY, ao analisar a modesta ictiofauna da bacia do Guaratuba, uma cabeceira situada no alto da SelTa do Mar paulista, identificaram, de forma muito clara, a maneira pela qual diferentes bacias hidrográficas permutam sua ictiofauna. A SelTa do Mar cOlTesponde a uma das inúmeras feições geomorfológicas que atuam como divisores de água entre grandes bacias hidrográficas. Tal qual a SelTa do Mar, a maior parte das regiões selTanas divisoras de água entre grandes bacias hidrográficas do escudo brasileiro originou-se exatamente nas zonas de dobramentos e falhamentos antigos, caracterizadas por intensa atividade tectônica ressurgente (Fig. 1 6.2). Ainda que drenando diretamente para o Oceano Atlântico, a ictiofauna do trecho superior da bacia do Rio Guaratuba, o qual está isolado de seu trecho inferior pelos contrafortes escarpados da SelTa do Mar, possui a mesma fauna de peixes de sua bacia vizinha, a do Rio Claro. Esta cOlTesponde a um dos inúmeros tributários do alto Rio Tietê, cuja fauna é muito distinta daquela típica da província costeira. O fato de a fauna do Guaratuba ser idêntica daquela do Tietê levantou a hipótese de que o trecho superior do Guaratuba fosse um fragmento capturado do Rio Claro para a drenagem do Guaratuba, que deságua no Oceano Atlântico. Uma análise morfotectônica, efetuada pelos autores demonstrou que a reativação de falhas geológicas presentes na região, fora a responsável pela captura do trecho superior do Rio Guaratuba l evando consigo sua antiga ictiofauna para seu novo curso, como tributário direto do Oceano Atlântico e não mais do alto Rio Tietê 17•
Talvez o grupo de peixes de água doce da América do Sul mais bem estudado do ponto de vista biogeográfico seja Glandulocaudinae, uma subfamília de Characidae. Weitzman
et aI. 8 delinearam o padrão de distribuição do grupo (então considerado como tlibo Glandulocaudini), com base no estudo de relações filogenéticas conhecido na época. Foi invocada uma combinação dos fenômenos de capturas e cabeceiras entre rios de drenagens distintas e abaixamento do nível do mar durante o fim do Pleistoceno como determinante nos processos de dispersão e vicariância. O conhecimento mais recente da filogenia de Glandulocaudinae ::3 e de aspectos da evolução tectônica da parte sul Cisandina � da América do Suj32 possibilitaram uma explicação algo � N diferente. Foi reconhecida a importância das flutuações do �
:;;c nível do mar na distribuição geográfica do grupo. Entretan- � to. a di\'ersificação atual de Glandulocaudinae foi admitida a-
como resultante principalmente de reativações tectônicas ao longo da margem oeste da bacia do Paraná supelior, da história tectônica da margem leste da mesma bacia e da atividade tectônica do arco de Ponta Grossa. uma área sujeita a levantamento tectônico recente que provavelmente proporcionou contato entre rios costeiros e afluentes da pm1e alta adjacente a leste do Rio Paraná supe110r.
As áreas dos antigos cinturões orogenéticos, dos quais nascem cabeceiras de diversos quadrantes hidrográficos, são sabidamente áreas preferenciais das reativações tectônicas antigas e recentes 1 5 . Diversos divisores de água são transecionados por extensas zonas de falha pré-cambrianas, muitas delas com atividade tectônica comprovada de menos de 1 ,6 milhão de anos33. Mecanismos análogos aos oconidos na bacia do Guaratuba são potencialmente passíveis de acontecer ao longo de extensas áreas que representam diYÍsores de água entre grandes bacias, por exemplo, entre as bacias do Paraná. Tocantins, Araguaia. São Francisco. PaI"aíba do Sul, Rio Doce, entre outras . As defonnações tectônicas nestas regiões ocasionam rearranjos de drenagem de diversas magnitudes. Desde capturas de cursos pequenos. como o Guaratuba, a grandes eventos, como a famosa captura do trecho superior do PaI"aíba do SuL antigo tl1butálio do Rio Tietê3o. Outros eventos de grande magnitude também são associados aos movimentos tectônicos recentes, por exemplo, a origem da planície alagada do Pantanal Mato-grossense, e as depressões do Araguaia e Tocantins, todas associadas às reativações de grandiosas zonas de fratura continentais pré-camblianas, cuja importância no intercâmbio faunístico entre bacias não pode ser negligenciado.
A paisagem sul-americana possui dois grandes componentes que podem ser prontamente identificados . Seu amplo território se subdivide em um conjunto de terras

baixas, cujos limites podem ser arbitrariamente definidos como aqueles com cotas abaixo de 250m acima do nível do mar34 e um conjunto de terras altas, excetuando-se os Andes, cujas altitudes normalmente ultrapassam a cota dos 500m (Fig. 1 6.3). Tal divisão, embora arbitrária, é de grande significado biogeográfico e ecológico para a fauna de peixes da América do Sul.
Nas regiões planálticas sul-americanas predominam rios com elevado gradiente e, por conseguinte, muita energia, o que se traduz em rios encachoeirados, encaixados em seus vales. Nestes sistemas, planícies de inundação são relativamente reduzidas, restritas às porções baixas das grandes bacias planálticas. Uma das características de muitos sistemas hidrográficos planálticos é que estes possuem rios denominados "superimpostos", o que significa que seu curso ignora ou se sobrepõe às estruturas litológicas. O motivo para isso é que tais drenagens são antigas e já deveriam estar estabelecidas em períodos em que as estruturas por elas cortadas ainda não afloravam. Com a evolução do processo erosivo acarretado pela drenagem, esta, j á estabelecida, simplesmente "passou por cima" de estruturas que, de outra forma, condicionariam seu curso. De fato, muitas das drenagens planálticas sul-americanas tiveram seus cursos gerais determinados pelos processos de soerguimento que antecederam a ruptura de Gondwana19 .
B iogeografia dos Peixes de Água Doce da Amér ica d o Su l - 271
Muitas espécies de peixes endêmicos de regiões de grande altitude nos escudos cristalinos parecem representar grupos antigos, com distribuições relictuais. Dentre os Glandulocaudinae já citados, por exemplo, os grupos mais basais na filogenia possuem distribuição restrita às regiões serranas do Brasil Central. Lophiobrycon weitzmanni, grupo irmão dos demais gêneros da subfamília, é conhecido apenas da Serra da Canastra em Minas Gerais. O grupo irmão subsequente, gênero Glandulocauda, também está restrito ao alto da Serra do Mar. Apenas o gênero Mimagoniates, mais diversificado, possui distribuição em áreas de terras baixas17·32. Ribeiro et al. 35, ao incluírem uma espécie recém-descoberta de lambari do gênero Creagrutus na filogenia do gênero, evidenciaram que aquela espécie, ocorrente na bacia do alto Paranaíba em Minas Gerais e Goiás, correspondia ao grupo irmão de todas as demais espécies do gênero, com ampla distribuição Trans e Cisandina. As antigas terras altas do planalto brasileiro abrigam, portanto, representantes relativamente antigos da irradiação moderna da fauna de peixes de água doce da América do Sul.
O fato de grandes bacias hidrográficas serem imensas áreas isoladas umas das outras levou a inevitável ideia, muito difundida, de que cada bacia deveria constituir grande área de endemismo. Realmente, muitas espécies
- (A) Topografia do conti nente su l-americano visto a part ir das i magens de Shuttle Radar Topography M ission (SRTM) da Nat iona l Aeronautics and Space Adm i n istrat ion (NASA) obtidas a part i r de i nterferometria de radar. (B) Compart imenta l i zação do continente em terras a ltas e baixas (abaixo de 250m acima do n ível do mar) .

272 - Padrões e Processos - Estudos de Casos
são endêmicas de detenllinadas bacias hidrográficas, mas a verdade é que muitas outras possuem distribuições que ultrapassam os limites dos divisores de água entre bacias. Certos grupos com distribuição tipicamente planáltica podem, por exemplo, ocorrer em diversas drenagens, transpondo os limites de seus divisores, porém sempre associadas às suas cabeceiras, ou seja, nas porções mais altas da bacia. Diversos exemplos são dados por Lima e Ribeiro34 para as drenagens dos rios Tapajós, Madeira e Tocantins-Araguaia. Tal ocorrência demonstra que, tal qual se dá na margem divergente da América do Sul, capturas entre bacias adjacentes impulsionada por reativações tectônicas certamente acontecem em muitas outras regiões, incluindo o Brasil central. Grandes feições da paisagem atual são resultantes destes processos tectônicos, tais como a depressão do Araguaia-Tocantins e a depressão do Pantanal mato-grossense. Todos resultantes de reativações tectônicas relativamente recentes que aproveitaram o sistema de falhas geológicas antigas15 .
Outro padrão biogeográfico bastante recorrente se refere às espécies comuns, porém disjuntas entre o Escudo das Guianas e o Escudo Brasileir034. Tal padrão poderia constituir um típico padrão relictual . Dados geológicos apontam, de fato, que as cabeceiras do que hoje são os tributários das margens sul e norte da Amazônia Oriental poderiam constituir cabeceiras de uma grande drenagem tipicamente planáltica2o, hoje marcadamente influenciada pela atual foz do amazonas, atuante como uma barreira ecológica atual para estes grupos planálticos34.
O soerguimento da cordilheira dos Andes é um evento magno, de grandes consequências para a evolução da paisagem sul-americana e, por conseguinte, de sua ictiofauna. Tradicionalmente, a ictiofauna sul-americana se divide em um componente Cis (a leste) e outro Trans (oeste) andino, dada a grande distinção que se reconhece entre as faunas separadas entre os dois lados desta cordilheira. Entretanto, grupos transandinos possuem relações filogenéticas com grupos cisandinos em diferentes níveis. Com certeza, a evolução da cordilheira dos Andes, que se iniciou ainda no Cretáceo, não corresponde a um evento de vicariâncias simples, mas assincrônico e regionalmente diferenciado.
A porção transandina da América do Sul pode ser dividida em três regiões: uma região meridional, que corresponde ao Chile e que apresenta uma ictiofauna tipicamente austral, a região situada entre o norte do
Chile e norte do Peru, semi desértica e com pouquíssimos peixes de água doce, e uma região setentrional, do norte do Peru às bacias do lago de Maracaibo na Venezuela, com uma fauna de peixes relativamente bem diversificada. A afinidade dessa fauna com àquela ocorrendo na América do Sul cisandina já havia sido apontada por Eigenmann36, que escreveu: "the fauna [dos rios transandinos} is largely a part ofthe general SouthAmerican fauna which has been pinched oif by the formation of the Andes, and has gane its own way since the Andes have become high enough to form an eifective barrier against the ready intermigration between the cisandean and transandean parts of the continent". A conectividade das drenagens do noroeste da América do Sul com àquelas do resto do continente começou a ser interrompida no meio do Mioceno, com o começo do soerguimento da cordilheira oriental na Colômbia, que determinou o atual curso do Rio Madalena ( 1 2 a 1 1 ,8 Ma) e culminou com a mudança da foz do Rio Orinoco das bacias do Maracaibo/Falcon para uma posição mais ao leste, no final do Mioceno (8 Ma) l l . Esse grande evento vicariante pode ser traçado tanto pela ocorrência de diversos fósseis , tanto de peixes como de outros vertebrados aquáticos, hoje restritos a drenagens do norte da América do Sul cisandina, como grande bagres, pirarucus, tambaquis, tartarugas mata-matas, jacaré-açu e botos, em formações miocênicas nas bacias dos rios Madalena e do Lago de Maracaibol l ,37,38, como pela ocorrência de múltiplas relações de grupos irmãos encontrados entre táxons trans e cisandinos9,34.
Dentre os padrões biogeográficos envolvendo os peixes da América do Sul, alguns são muito peculiares. Por exemplo, alguns grupos endêmicos dos altiplanos andinos possuem afinidades inesperadas. O gênero Orestias, um grupo de aproximadamente 40 espécies de peixes da ordem Cyprinodontiformes são endêmicos de rios e lagos de altitude, entre a região central do Peru e o norte do Chile, sendo mais diversificados no Lago Titicaca, onde se dá mais de 60% da diversidade do grup039. É um grupo cujas afinidades filogenéticas são controversas, tendo sido já considerados como relacionados aos subgrupos de Cyprinodontiformes norte-americanos e, como alternativa, a grupos sul-americanos, ou ainda aos membros da famI1ia Cyprinodontidae do velho mundo, habitantes de água doce ao redor dos mares Mediterrâneo, Negro, Vermelho e da Arábia. Costa39, entretanto, identificou que Orestias consiste no grupo irmão de um clado mais inclusivo que abrange os membros da América Central e

ao redor do Mediterrâneo. A constatação de que a fauna de Orestias do altiplano andino possui afinidades com outras regiões biogeográficas é surpreendente, uma vez que para a maior parte dos grupos Neotropicais de água doce, as afinidades se dão com outros componentes da região Neotropical ou da Gondwana. Outros grupos de ocorrência nos altiplanos possuem suas relações com outras áreas da região NeotropicaP9.
A fanu1ia Galaxiidae (ordem Salmoniforrnes) é um grupo que compreende cerca de 50 espécies, que apresenta uma distribuição austral, ocorrendo no sudeste da Austrália, Tasmânia, Nova Zelândia, Nova Caledônia, sul da América do Sul (Chile e Patagônia argentina), sul da África do Sul, além de diversas ilhas neozelandesas, a ilha Lord Howe (Austrália) e as ilhas Falk:lands (Malvinas) (Argentina)4o. Sete espécies ocorrem na América
'CO do Sul, no Chile, Argentina e ilhas Falk:lands4o. Há grande � controvérsia sobre a biogeografia da família. Para alguns, "? � ela constitui um dos melhores exemplos de táxons apre-:A sentando uma distribuição ancestral gondwânica4 1 . � Contudo, a presença de diadromia no grupo, a pouca r--0\ diferenciação das linhagens de Galaxiidae entre os dife-
rentes continentes (por exemplo, a ocorrência de uma mesma espécie, Galaxias maculatus, uma espécie diádroma, no sul da América do Sul, Nova Zelândia, Austrália, em diversas ilhas neozelandesas e australianas, bem como nas ilhas Falklands) e a não relação entre espécies ocorrentes em determinada área com a história geológica (por exemplo, o acontecimento de espécies com afinidades sul-americanas, Aplochiton zebra e Galaxias maculatus nas ilhas Falk:lands/Malvinas, um bloco cflistal que se desprendeu do sul da África) indicam que dispersão transoceânica deve ter tido papel preponderante no estabelecimento do atual padrão de distribuição geográfica do grup042. O mesmo é verdadeiro para as lampreias sul-americanas, Geotriidae e Mordaciidae, distribuídas através do sul da Austrália, Tasmânia, Nova Zelândia, Chile e Argentina, cujos juvenis, após longo período larval em água doce, vivem por três a quatro anos crescendo e se alimentando no mar2.
Abaixo de 250m de altitude estendem-se, no continente sul-americano, milhões de quilômetros quadrados de planícies, muitas delas extensas áreas alagáveis, tais como a planície pantaneira e as terras baixas amazônicas. Caso se considere que a atual plataforma continental esteve emersa durante períodos glaciais do Pleistoceno,
B iogeograf ia dos Peixes de Água Doce da América do Su l - 273
onde o nível dos oceanos foi mais abaixo do que o atual em até 100m, aumenta-se consideravelmente a extensão das terras baixas sul-americanas. Em muitas das terras baixas sul-americanas, a diversidade de peixes de água doce atinge seu auge no que se refere ao número de espécies em coexistência.
Diferentemente dos planaltos adjacentes, as terras baixas sul-americanas (as quais se constituem em grandes bacias sedimentares mesozoicas e cenozoicas) possuem sistemas fluviais caracterizados por extensas planícies de inundação de grandes rios meandrantes. Enquanto nos planaltos predominam rios encaixados, nas terras baixas seus leitos oscilam ao longo de toda sua bacia de sedimentação durante milhares de anos. Desenvolve-se ainda, em áreas onde rios Andinos atingem as terras baixas adjacentes, a evolução de megacones fluviais: testemunhos dos graus de oscilação que os canais perfazem ao longo das planícies em sua longa evolução gcomorfológica43. Dependendo de onde se localizam suas cabeceiras, grandes rios das terras baixas sul-americanas podem ser caracterizados como de água preta (carregados de ácidos húmicos, resultado da decomposição de matéria vegetal, lixiviado dos solos arenosos), tais como aqueles que descem as vertentes do Escudo das Guianas; de água branca, carregados de sedimentos proveniente dos Andes, ou de água clara, quando descem das áreas lavadas do Escudo Brasileiro. Tal distinção acarreta grandes consequências ecológicas nas comunidades aquáticas sul-americanas34.
As terras baixas sul-americanas podem ser divididas em dois grandes conjuntos quanto sua origem geológica: depressões tectônicas encravadas nos escudos e bacias de antepaís. As grandes depressões, tal qual a Depressão do Araguaia-Tocantins são oriundas de reativações tectônicas de falhas geológica antigas dos escudos, ao passo que as bacias de Antepaís constituem-se terras rebaixadas que se estabelecem ao sopé andino como um arco flexural consequente do mesmo processo que gera a deformação da cadeia montanhosa vizinha, num processo típico de margens convergentes.
As bacias de antepaís, tais como a do Chaco, que se interconecta com a depressão do Pantanal, são áreas rebaixadas que recebem esporadicamente transgressões marinhas, quer seja em decorrência do aumento geral dos níveis dos oceanos, quer seja pelo seu rebaixamento tectônico eventual. Embora a extensão e a frequência das transgressões marinhas ao longo das terras baixas sul-americanas sejam assunto de grande debate44, acredita-se que a maior e mais recente delas tenha inundado todo o conjunto de terras baixas do Chaco-Pantanal, assim como o sopé andino ao norte, criando uma conexão marinha entre o Oceano Atlântico Sul e o Caribe

274 - Padrões e Processos - Estudos de Casos
através de seu braço nOIte, e estendendo-se a leste pelas terras baixas amazônicas até a atual foz do Rio Amazonas aproximadamente entre 15 e 1 2 milhões de anos45.
Dados de distribuição de espécies de peixes de água doce demonstram que as terras baixas são áreas propicias à expansão da distribuição geográfica da fauna aquática. Diversas espécies de peixes e outros grupos de água doce possuem distribuição que ultrapassa os limites dos atuais divisores de água entre as grandes bacias de antepaís ao sopé andino, estendendo-se também por outras áreas baixas sul-americanas, tais como a extensa bacia do Prata, ao sul, a maior parte das terras baixas amazônicas e atingindo as terras baixas tectonicamente desenvolvidas nos escudos cristalinos, tais como a depressão do Araguaia-Tocantins34. O padrão também se repete para muitos táxons em que já existem disponíveis filogenias, onde se percebe a relação de grupos irmãos entre bacias de antepaís adjacentes34. O dinamismo tectônico e geomorfológico associado às terras baixas sul-americanas certamente promove a expansão da distribuição geográfica de diversos táxons, porém também promove vicariância, o que se reflete na relação de grupos irmãos ao longo das bacias de antepaís. Os diferentes níveis de inclusão destes padrões de grupos irmãos demonstram que a dinâmica destas terras baixas é constante, culminando em padrões semelhantes, mas que podem estar distantes no tempo.
As grandes transgressões marinhas que invadem a plataforma sul-americana pelas extensas baixadas representam a porta de entrada de táxons de origem marinha, podendo explicar muitos dos grupos secundariamente de água doce hoje encontrados nos sistemas fluviais sul-americanos. Entretanto, esta óbvia correlação deve ser vista com cautela. A distribuição secundariamente marinha de muitos dos atuais grupos, tais como as raias de água doce da família Potamotrygonidae, tem sido explicada a partir destes eventos de transgressão. Lovejoy46 propôs, com base em relações filo genéticas e relógios moleculares, que as raias de água doce neotropicais teriam invadido o continente sul-americano através de transgressões marinhas a partir da região do Caribe entre 15 e 23 milhões de anos. No entanto, a inclusão de diversos grupos fósseis em análise filogenética mais abrangente indicou que a linhagem de Potamotrygonidae seria ainda muito mais antiga, com idade mínima de 50 milhões de anos47. Tal hipótese não relaciona, portanto, a invasão das águas continentais da América do Sul pelo ancestral comum de Potamotrygonidae aos eventos de transgressão marinhos mais recentes associados à evolução cenozoica da Cordilheira dos Andes, mas a prováveis eventos mesozoicos.
;, • ..:....J.
Neste capítulo são abordados alguns aspectos considerados relevantes acerca da biogeografia de peixes de água doce da América do Sul. Contudo, como já salientado em seu início, não se pretende aqui esgotar o assunto, o qual abarca muitos detalhes complexos, cuja abordagem não cabe nesta pequena introdução. O crescente avanço nas técnicas utilizadas em estudos de biologia comparativa, dentre eles aqueles com possibilidades de obtenção de filogenias e datações cada vez mais precisas, aliado ao aumento da compreensão dos processos em \O
-J geologia histórica e tectônica conduzem a um terreno �
V1 fértil de exploração científica. Se por um lado muitos � padrões antigos parecem já quase indistinguíveis, dada : a intensa dinâmica de paisagem sul-americana, e apenas �
bpodem ser acessados com base em estudos comparativos abrangentes (incluindo grupos fósseis), por outro lado abordagens em nível molecular poderão contribuir para a identificação de padrões de irradiação modernos a partir de estudos filogeográficos. Entretanto, não podemos esquecer que estes avanços só serão possíveis com o continuo aumento da informação disponível sobre a verdadeira identidade taxonômica e a real conhecimen-to acerca da distribuição geográfica das espécies de peixes de água doce. Embora se tenha progredido consideravelmente neste assunto, ainda se está muito longe de vislumbrar a verdadeira grandeza da diversidade de peixes da América do Sul, no que se refere aos seus padrões e processos. Sem tais estudos taxonômicos básicos, métodos analíticos avançados e caros correm o risco de tornarem-se apenas desperdícios frustrantes.
REFERÊNCIAS BIBLIOGRÁFICAS 1 . REIS, R. E. ; KULLANDER, S . O. ; FERRARIS, C. 1. Check
List Df the Freshwater Fishes Df South and Central America. Porto Alegre: Edipucrs, 2003. 729p.
2. ESCHMEYER, W. N. ; FONG, R. Catalog ofFishes electronic version 2009. Disponível em: http://research.calacademy.org/ ichthyology/catalog/fishcatmain.asp. Acesso em: 12/0 1120 10.
3 . EIGENMANN, C. H. The freshwater fishes of Patagonia and an examination of the Archiplata-Archhelenis theory. In: SCOTT, W. B . Reports ofthe Princenton University Expeditions to Patagonia, 1 896-1899. Princenton, NJ: Princeton University Press, 1909. p. 225-374. v. 3 (Zoology)
4. EIGENMANN, C. H. The freshwater fishes oí' British Guiana, inc1uding a study of the ecological grouping of species and the relation ofthe fauna ofthe plateau to that ofthe lowlands. Mem. Carnegie Mus. , v. 5, n. 1 , p. i-xx, 1 9 12.
5. GÉRY, J. The freshwater fishes of SouthAmerica. In: FITTKAU, E. J . ; ILLIES, 1 . ; KLINGE, H. et a!. Biogeography and Ecology in South America. DI. W. 1. Junk Publishers: The Hague, 1 969. p. 828-848.
6. RINGUELET, R. A. Zoogeografia y ecología de los peces de águas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América de Sur. Ecosur., v. 2, n. 3, p. 1-1 5 1 , 1 975.

7. WEITZMAN, S. H . ; WEITZMAN, M. Biogeography and evolutionary diversification in Neotropical freshwater fishes with comments on the refuge theory. In: PRANCE, G. T Biological Diversijication in the Tropics. New York: Columbia University Press, 1 982. p. 403-422.
8. WEITZMAN, S. H.; MENEZES, N. A; WEITZMAN, M. 1. Phylogenetic biogeography of the Glandulocaudini (Teleostei: Characiformes, Characidae) with comments on the distribution of other freshwater fishes in eastern and southeastern Brazil. In: VANZOLINI, P. E. : HEYER W. R. WORKSHOP ON NEOTROPICAL DISTRIBUTION PATTERNS, 1988 . Rio de Janeiro. Proceedings of a Workshop on Neotropical Distribution Patterns. Rio de Janeiro: Academia Brasileira de Ciências, 1 988. p. 379-427.
9. VARI, R. P. The Curimatidae, a lowland Neotropical fish famiIy (Pisces: Characiformes) ; distribution, endemism, and phylogenetic biogeography. In: VANZOLINI, P. E.; HEYER W. R. WORKSHOP ON NEOTROPICAL DISTRIBUTION PATTERNS, 1988 . Rio de Janeiro. Proceedings of a Workshop on Neotropical Distribution Patterns. Rio de Janeiro: Academia Brasileira de Ciências, 1988 . p. 343-377.
10. LUNDBERG, J. G. African-South American freshwater fish clades and continental drift: Problems with a paradigm. In: GOLDBLATT, P. Biological Relatiunships Between Africa and SouthAmerica. New Haven: Yale Univ. Press, 1 993. p. 156- 1 99.
1 1 . LUNDBERG, J. G.; MARSHALL, L. G.; GUERRERO, J. et a!. The stage for Neotropical fish diversification: a history of Tropical South American rivers. In: MALABARBA, L. R. ; REIS, R. E . ; VARI, R. P. et a! . Phylogeny and Classijication of Neotropical Fishes. Porto Alegre: Edipucrs, 1 998. p. 13 -48.
12. RIBEIRO, A C. Tectonic history and the biogeography of the freshwater fishes from the coastal drainages of eastern Brazil: an example of faunal evolution associated with a divergent continental margin. Neotrop. Ichthyo/., v. 4, p. 225-246, 2006.
13 . HUBERT, N.; RENNO, 1. F. Historical biogeography of South American freshwater fishes. J Biogeogr., v. 33, p. 1414-1436, 2006.
1 -1. BRITO-NEVES, B . B. América do Sul: quatro fusões, quatro fissões e o processo acrescionário andino. Revista Brasileira de Geociências, v. 29, p. 379-392, 1 999.
15. SAADI, A. Neotectônica da plataforma brasileira: esboço e interpretação preliminares. Geonomos, v. 1, p. 1 -15 , 1 993.
16. ASSINE, M. L. A bacia sedimentar do Pantanal Mato-Grossense . I n : MANTESS O-NETO, Y. ; BARTORELLI, A . ; CARNEIRO, C . D . R.; BRITO-NEVES, B . B . Geologia do Continente Sul-Americano - Evolução da obra de Fernando
Flávio Marques de Almeida. São Paulo: Beca, 2004. p. 6 1 -74. 1 7 . RIBEIRO, A. C . ; LIMA, F. C. T. ; RICCOMINI, C. et aI.
Fishes of the Atlantic Rainforest ofBoraceia: testimonies ofthe Quaternary fault reactivation within a Neoproterozoic tectonic province in Southeastern Brazil. Ichthyol. Explor. Fres. , v. 17 , p . 157-1 64, 2006.
18 . DA ROSA, Á. A. S . ; GARCIA, A J. V. Paleobiogeographic aspects of northeast Brazilian basins during the Berriasian before the break up of Gondwana. Cretaceous Res., v. 2 1 , p 221-339, 2000.
19. COX, K G. The role of mande plumes in the development of continental drainage patterns. Nature, v. 342, p. 873-876, 1 989.
20. POTTER, P. E. The Mesozoic and Cenozoic paleodrainage of South America: a natural history. J. S. Am. Eartl? Sei. , v. 1 0, p. 331 -344, 1997.
2 1 . ALMEIDA, F. F. M.; CARNEIRO, C. D. R. Origem e evolução da Serra do Mar. Rev. Bras. Geoc. , v. 28, p. 1 35 - 150, 1 998.
22. MCQUARRIE, N.; HORTON, B . K ; ZANDT, G. et aI . Lithospheric evolution of the Andean fold-thrust belt, Bolivia, and the origin of the central Andean plateau. Teetonuphysics, v. 399, p. 1 5-37, 2005.
23. BRITO, P. M.; MEUNIER, F. J.; LEAL, M. E. C. Origine et diversification de l' ichtyofaune néotropicale: une revue. Cybium, v. 3 1 , p. 149- 163, 2007.
B iogeograf ia dos Peixes de Água Doce da América do Su l - 275
24. HEINICKE, M. P.; SANDER. 1. M.; HEDGES, S. B. Lungfishes (Dipinoi) In: HEDGES, S. B. ; WATSON, 1. D. The Timetree of Life. Disponível em: http://www.timetree.orglbook.php.
25. BUCKUP, P. A. Relationships of the Characidiinae and Phylogeny of the Characiform fishes (Teleostei: Ostariophysi). In: MALABARBA, L. R.; REIS, R. E.; VARI, R. P. et aI. Phylugeny and Classijication of Neotropical Fishes. Porto Alegre: Edipucrs, 1 998. p. 1 23-144.
26. ZANATA, A M.; VARI, R. P. The family Alestidae (Ostariophysi, Characiformes): a phylogenetic analysis of a trans-Atlantic c1ade. Zool. J. Linn. Soe. , v. 145, p. 1 - 144, 2005.
27. LUNDBERG, J. G. The Temporal Context for Diversification of Neotropical Fishes. In: MALABARBA, L. R.; REIS, R. E. ; VARI, R. P. et aI. Phylogen} and Classification of Neotropical Fishes. Porto Alegre: Edipucrs, 1998. p. 49-68.
28. PINNA, M. C. C. Phylogenetic relationships of Neotropical Siluriformes (Teleostei: Ostariophysi): Historical overview and synthesis of hypothesis. In: MALABARBA, L. R. ; REIS, R. E . ; VARI, R. P. et aI. Phylogeny and Classification of Neotropical Fishes. Porto Alegre: Edipucrs, 1 998. p. 279-330.
29. SAADI, A.; BEZERRA, F. H. R.; COSTA, R. D. et aI. Neotectônica da Plataforma Brasileira. In: SOUZA, C. R. G.; SUGUIO, K ; OLIVEIRA, A M. S. et al. Quaternário do Brasil. Ribeirão Preto: Halos, 2005. p. 2 1 1 -234.
30. COBBOLD, P. R.; MEISLING, K. E.; MOUNT, Y. S. Reactivation of an obliquely rifted margin, Campos and Santos basins, southeastern Brazil. Am. Assoe. Petl: Geo/. B. , v. 85, p. 1 925-1 944, 200 1 .
3 1 . RICCOMINI, C . ; SANT' ANNA, L . G . ; FERRARI, A . L . Evolução geológica do rift continental do sudeste do Brasil. In: MANTESSO-NETO, Y. ; BARTORELLI, A; CARNEIRO, C. D. R. et aI. Geologia do Continente Sul-americano - Evolução da Obra de Fernando Flávio Marques de Almeida. São Paulo: Beca, 2004. p. 383-405.
32. MENEZES, N. A; RIBEIRO, A C.; WEITZMAN, S. et aI. Biogeography of Glandulocaudinae (TeJeostei: Characiformes: Characidae) revisited: phylogenetic patterns, hist0l1cal geology and genetic connectivity. Zootaxa, v. 1726, p. 33-48, 2008.
33. SAADI, A; MACHETTE, M. N.; HALLER, K M. et aJ. Map and database of Quaternary faults and 1ineaments in BraziJ. U.S. Geological Survey, Open-File Report 02-230. Disponível em: http://pubs.usgs.gov/of/2002/ofr-02-230. Acesso em: 1 7/01/2010.
34. LIMA, F. C. T; RIBEIRO, A. C. Continental-scale tectonic controls ofbiogeography and ecology In: ALBERT, J. S . ; REIS R. E. Historical Biogeography of Neotropical Freshwater Fishes. Califórnia: University of California Press (no prelo).
35. RIBEIRO, A C.; BENINE, R. C.; FIGUEIREDO, C. A. A new species of Creagrutus Günther (Teleostei: Characiformes), from the upper Rio Paraná basin, central Brazil. J. Fisl? Bio/., v. 64, p. 597-6 \ 1 , 2005.
36. EIGENMANN, C. H. The origin and distriblltion of the genera of the fishes of South America west of the Maracaibo, Orinoco, Amazon, and Titicaca basins. Proc. Am. Philos. Soe. , v. LX, A, p. 1 -6, 1 9 1 2.
37. LUNDBERG, 1. G.; MACHADO-ALLISON, A ; KAY, R. F. Miocene characid fishes from Colombia: evolutionary stasis and extirpation. Science, v. 234, p. 208-209, 1 986.
38 . LUNDBERG, J. G.; CHERNOFF, B. A Miocene fossil of the Amazonian fish Arapaima (Teleostei, Arapaimidae) from the Magdalena River region of Colombia - biogeographic and evolutionary implications. Biotropica, v. 24, p. 2-14, 1 992.
39. COSTA, W. 1. E. M. PhyJogeny and classification of the Cyprinodontidae revisited (Teleostei: Cyprinodontiformes): Are Andean and Anatolian killifishes sister taxa? 1. Comp. Eio/. , v. 2, p. 1 - 17 , 1 997.
40. MCDOWALL, R. M. Crying wolf, crying foul, or crying shame: alien salmonids and a biodiversity crisis in the southern cooltemperate galaxioid fishes? Rev. Fish. Biol. Fish, v. 16, p. 233-422, 2006.

276 - Pad rões e Processos - Estudos de Casos
41 . CAMPOS, H. Godwana and neotropical galaxioid fish biogeography. In: ZARET, T. M. Evolutionary Eeology ofNeotropical
Freshwater Fishes. Boston: Df. Junk, 1 984. p. 1 13 - 125 . 42. MCDOWALL, R. M. Accumulating evidence for a dispersaI
biogeography of southern cool temperate freshwater fishes. J.
Biogeogr., v. 29, p. 207-2 19 , 2002. 43. HORTON. B. K.; DECELLES, P. G. Modern and ancient flu
vial megafans in the foreland basin system of the central Andes, southern Bolivia: implications for drainage network evolution in fold-thrust belts. Basin. Res. , v. 1 3, p. 43-63, 200 1 .
44. LOVEJOY, N . R ; ALBERT, J. S . ; CRAMPTON, W. G . R Miocene marine incursions and marine/freshwater transitions: evidence from neotropical fishes. J. S. Am. Earth. Sei., v. 2 1 , p. 5- 1 3 , 2006.
45. RAMOS, V; ALEMAN, A. Tectonic of the Andes. In: CORDANI, U. G.; MILANI, E. J.; THOMAZ-FILHO, A.; CAMPOS, D. A. Tectonic Evolution of SouthAmerica. XXXI INTERNATIONAL GEOLOGICAL CONGRESS, 2000. Rio de Janeiro. Proeeedings of the XXXI International Geologieal Congresso Rio de Janeiro: Academia Brasileira de Ciências e Departamento Nacional da Produção Mineral (DNPM), 2000. p. 635-685.
46. LOVEJOY, N. R. Systematics of myliobatoid elasmobranches with emphasis on the phylogeny and historical biogeography of Neotropical freshwater stingrays (Potamotrygonidae: Rajiformes). Zool. J. Linn. Soe. , V. 1 17, p . 207-257, 1 996.
47. CARVALHO, M. R; MAISEY, J. G.; GRANDE, L. Freshwater stingrays of the Green River Formation of Wyoming (Early Eocene), with the description of a new genus and species and an analysis of its phylogenetic relationships (Chondrichthyes: Myliobatiformes). Bull. Am. Mus. Nat. Hist., V. 284, p. 1 - 136, 2004.

/1\
Claudio J. B. de Carvalho
Márcia Souto Couri
Por mais de 100 anos, biologistas têm proposto diferentes teorias para explicar a origem e a relação evolutiva da biota da América do Sul com outras regiões. Desde os estudos primordiais de Augustin Pyramus de Candolle no início do século XIX, diferentes metodologias têm sido utilizadas para entender a composição e a evolução da fauna e flora do continente. Esta multiplicidade de hipóteses e teorias para o entendimento da biota é decorrente, em particular, da formação geológica e biogeograficamente híbrida do continente! . Esta condição pode ser visualizada com facilidade pelo reconhecimento das duas regiões biogeográficas, Neotropical e Andina, encontradas no continente e de uma zona de transição sul-americana, recentemente caracterizada2.
A ocorrência de qualquer espécie em determinado local é explicada, no mínimo, a partir de elementos históricos, por três processos : vicariância, dispersão e extinção. Esses processos devem estar na mente de qualquer biogeógrafo, quando da indicação de explicação de qualquer padrão biogeográfico. Os táxons mais adequados para análise biogeográfica são aqueles que não têm problemas no reconhecimento da precisa localização da distribuição geográfica de suas espécies e aqueles que possuam taxonomia bem resolvida, sem dúvidas na identidade e identificação das espécies. Táxons com grande riqueza de espécies são também bem interessantes, pois têm como pressupostos a ocupação de grande variedade de locais e ambientes.
Entre os insetos, os dípteros são umas das ordens mais bem catalogadas, tanto taxonomicamente como geogra-
1 7
.. ," ; :' c�
ficamente em todas as regiões biogeográficas . Esta condição faz com que, de maneira geral, a ordem seja um bom modelo para análise biogeográfica. Muscidae é uma das maiores famílias com cerca de 5.000 espécies descritas no mundo em 1 80 gêneros. As espécies estão naturalmente distribuídas em todas as regiões biogeográficas, exceto nas regiões polares. A família é bem conhecida pela presença de Musca domestica Linnaeus, espécie comum e altamente sinantrópica e por outras espécies de interesse médico e veterinário que são vetores de doenças ao homem e aos seus animais domésticos. Entretanto, a maioria das espécies nunca entra em contato com o homem, pois seus hábitats não incluem o ambiente antropofílico.
Os Muscidae estão distribuídos em quase todos os ambientes da América do Sul, desde o nível do mar, com poucas espécies encontradas em rochas umedecidas pelo mar no litoral brasileiro e até mais de 3 .000m de altitude nos Andes. São mais encontradas em regiões de florestas e áreas com alta pluviosidade. Os muscídeos não ocorrem em regiões muito áridas ou regiões extremamente frias, como os picos gelados de altas montanhas, mas são relativamente comuns em regiões de altitude. São menos comuns em áreas abertas ou expostas e em solos arenosos ou ácidos.
Na América do Sul existem áreas ainda amostradas com deficiência (Fig. 17 . 1 ) . Na região ocorrem cerca de 700 espécies, 15% de todas as espécies descritas no mundo, agrupadas em 80 gêneros3.
O principal objetivo deste capítulo é descrever os padrões de distribuição das espécies de Muscidae e indicar possíveis explicações da formação desses padrões. Serão

278 - Padrões e Processos - Estudos de Casos
e
'-� e e e'e e •
. .. e e • •
e
, •
•
'; - Loca l idades de coleta das espécies de Muscidae na América do Su l .
• e e e •
e •

analisados e discutidos os gêneros que possuem hipótese de relação filogenética para suas espécies. Por fim, serão discutidos os possíveis caminhos para o melhor conhecimento da evolução espacial dos Muscidae na região.
A primeira explicação da presença de espécies na América do Sul foi feita por Willi Hennig. Em uma época propícia, houve dispersão de espécies por meio de ligações terrestres entre as massas continentais do Norte e Sul. Hennig4 indicou que através dessas conexões ocorreu a dispersão, iniciada no final do Cretáceo e início do Terciário, há mais ou menos 65 milhões de anos. Esta visão essencialmente dispersalista para a explicação da ocorrência das espécies, utiliza o conceito de centro de origem, localizado, neste caso, no Hemisfério Norte. Segundo Hennig4, esse intercâmbio faunístico se deu em diversas ocasiões. A América do Sul ficou isolada dos continentes do norte desde o início do Terciário até o Plioceno, quando foi reconstruída uma ligação terrestre estável entre os continentes do norte e sul, pela elevação do istmo do Panamá.
Para Hennig4, as grandes diferenças taxonômicas entre as espécies da América do Norte e América do Sul seriam resultados de grandes extinções das linhagens do norte e maior diferenciação das linhagens do sul. A grande diferenciação dos grupos de Muscidae encontrados no sul da América do Sul seria resultado de uma evolução de longo tempo realizada in situo Por outro lado, os grupos que se dispersaram mais tardiamente seriam mais próximos das espécies da América do Norte4.
Similar às ideias apresentadas por Hennig4 para os Muscidae, Michelsens também propôs a ocorrência primeiramente de espécies de Anthomyiidae, grupo irmão de Muscidae, na América do Sul por dispersão. O clado basal de Anthomyiidae formado por Coenosopsia Malloch + Phaonantho Albuquerque teria se dispersado para o sul por conexões terrestres, a partir da América do Norte. Coenosopsia é um gênero monofilético com espécies distribuídas desde o sul dos Estados Unidos ao sul do Brasil6. Bortolanza et af.7 indicaram que as diferentes espécies encontradas em áreas abertas não mostram relação filogenética próxima entre elas, sugerindo que diferentes processos podem ter dado origem a essas espécies.
A visão dispersalista indicada por Hennig4 e Michelsens, que assume um centro de origem e dispersão
B iogeograf ia de M usc idae (I nsecta, D iptera) da América do S u l - 279
deste local para a periferia, tem sido gradualmente substituída nos últimos anos por uma visão da formação de padrões por processos primariamente vicariantes8. Segundo Croizat9, a Terra e a vida evoluem juntas, pois evolução é resultado da interação entre espaço, tempo e forma (ver Capo 5).
A seguir são apresentadas descrições do padrão de distribuição das espécies de 1 1 gêneros de Muscidae e a relação cladística de suas espécies. Primeiramente é exibida a descrição do padrão de distribuição das espécies de gêneros de ocorrência essencialmente N eotropical e posteriormente aqueles com espécies de ocorrência na região Andina e na região de transição sul-americana.
Gênero monofilético com cinco espécies distribuídas exclusivamente nas regiões de florestas úmidas daAmérica do Sul. Não foram encontradas espécies nas Antilhas nem nos países da América Central. As espécies estão distribuídas na região Amazônica e Floresta Atlântica. Na Figura 17 .2 estão apresentados os traços individuais de quatro espécies e na Figura 17 .3 a relação cladística das cinco espécies8. Não foi mostrado o traço individual de Souzalopesmyia sulina Carvalho por ser conhecida apenas da localidade-tipo (Reserva Natural deI BosquelMbaracyu, Paraguai).
Gênero monofilético com 1 1 espécies distribuídas do México a Argentina, com algumas espécies tendo grande área de distribuição em ambientes abertos e de florestas. Na Figura 17.4 estão apresentados os traços de nove espécies e, na Figura 17.5, a relação cladística das espécies 10. Bithoracochaeta nigricornis Malloch que ocorre na Costa Rica e B. pacifera (Giglio-Tos) distribuída apenas no México não foram incluídas nos mapas por ter apenas uma ou duas localidades de distribuição conhecidas.
Gênero com 10 espécies distribuídas em áreas de floresta e não encontrado no sul da América do Sul. Pamplona 1 1 indicou a monofilia do gênero e a relação de grupo irmão com Cyrtoneuropsis Malloch. Na Figura 17.6 estão exibidos os traços individuais de sete espécies e na Figura 17.7 a relação cladística das espéciesll . 12 . Não foram apresentados os dados de distribuição de C. arleriopsis

280 - Padrões e Processos - Estudos de Casos
':1 7.!( - Traços i nd ividua is de Souzalopesmyia Albuquerque. (A) S. amazonica Albuquerque. (B) S. carioca Albuquerque. (C) S. paraensis Carva l ho. (D) S. singularis 5tein .

B i ogeograf ia de Musc idae (I nsecta, D i ptera) d a América do S u l - 281
..-------- Souza/opesmyía amazonica
r---- Souzalopesmyia paraensis
Souzalopesmyia singularis
Souzalopesmyia sulina
....... ---- Souzalopesmyia carioca
,':: - Cladograma de Souza/opesmyia Albuquerque8.
- Traços i nd ividuais de Bithoracochaeta 5te in . (A) B. annu/ata 5tein . (B) B. (a/opus (Bigot). (C) B. equatoria/is (Couri e Marques) . (O) B. f/avicoxa (Mal loch). (E) B. /eucoprocta (Wiedemann), onde a l inha tracejada l im ita o traço norte norte americano. (F) B. maricaensis (Couri e Motta). (G) B. nigricoxa Couri . (H) B. p/uma ta Albuquerque. (I) B. varicornis (Coqu i l lett) .

282 - Padrões e Processos - Estudos de Casos
Bithoracochaeta flavicoxa
,-----------l-- Bithoracochaeta nigricornis
Bithoracochaeta pacifera
,----------- Bithoracochaeta leucoprocta
r-------- Bithoracochaeta atricornis
,.------ Bithoracochaeta annulata
f----- Bithoracochaeta varicornis
- Cladograma de Bithoracochaeta Stein 1 0 .
Bithoracochaeta maricaensis
Bithoracochaeta plumata
- Traços ind iv idua is de Cyrtoneurina Gig l io-Tos. (A) C. alifusca Couri . (B) C. biseta Snyder. (C) C. confusa Snyder. (D) C. costalis (Wa lker). (E) C. crispaseta Snyder. (F) C. geminata (Ste in) . (G) C. uber Gig l io-Tos.

B iogeograf ia de M usc idae ( lnsecta, D i ptera) da Amér ica do Su l - 283
,.------------ Cyrtoneurina uber
,--------- Cyrtoneurina confusa
Cyrtoneurina arleriopsis
Cyrtoneurina biseta
,------ Cyrtoneurina geminata
.. - Cladograma de Cyrtoneurina Gig l io-ToSl l , 1 2 .
Couri (Sinop, Brasil), C. cylindrica (Macquart) (Brasil) e C. monstrata (Wulp) (Xucumanatlán, México) por serem conhecidas apenas da localidade-tipo.
Gênero monofilético composto por 33 espécies distribuídas em áreas de florestas úmidas e, como Cyrtoneurina, não é encontrado no sul da América do Sul. Na Figura 17 .8 estão exibidos os traços individuais de 30 espécies e na Figura 17 .9 a relação cladística das espécies 1 1 , 12. Não foram mostrados os dados de distribuição de C. fuscisquama (Snyder) (Rio de Janeiro, Brasil), C. pallipes (Stein) (Paraguai), e C. similata (Couri) (Santarém, Brasil) por serem conhecidas apenas da localidade-tipo.
Gênero monofilético com 10 espécies distribuídas da Nicarágua ao sul do Brasil. Não ocorrem espécies no sul da América do Sul. Na Figura 17 . 10 estão expostos os traços individuais de nove espécies e na Figura 17. 1 1 a relação cladística das espécies13• Não foram apresentados os dados de distribuição de P. elbida Schuehli e Carvalho (Reserva Natural deI Bosque/Mbaracyu, Paraguai) por ser apenas conhecida da localidade-tipo.
Gênero monofilético com a maioria das 17 espécies distribuídas apenas em áreas de florestas úmidas. Poucas espécies são encontradas em áreas abertas, tal como o Cerrado brasileiro. Nas Figuras l 7 . 1 2 e l 7 . 1 3 estão apresentados os traços individuais das espécies e cladogramas. A área de conexão entre P. flavidicincta (Stein) e o clado restante [P. rubella (Wulp) + P. concinna
(Wulp) + P. orbitalis (Stein) + P. wulpi (Couri e Carva-
Cyrtoneurina costalis
Cyrtoneurina crispaseta
lho)] está indicada por um círculo (Fig. l 7 . 12) . Na Figura 17. 1 3 também estão indicadas por círculos as áreas de conexão entre os clados [P. nigra Couri e Carvalho + P. prima (Couri e Machado)] + (P. flavithorax + P. major Albuquerque) 14. Na Figura l 7 . l4 está representada a relação cladística de todas as espécies15 .
Apsil é um gênero monofilético com todas as 10 espécies endêmicas da região Andina, distribuídas na Patagôni2 e sul do Chile. As espécies estão distribuídas do nÍ\ el do mar até as altas altitudes nos Andes. Apsil e ReYlloldsú;
Malloch são grupos irmãos e possuem semelhantes áreas de distribuição das espécies 16. Na Figura 17 . 1 5 estão indicados os traços individuais de sete espécies de Apsil e na Figura 17. 1 6 a relação cladística das espécies 1 6. Não foram apresentados os dados de distribuição deA. diminuta Couri (Parque Nacional Puyehue, Chile), A. flavipalpis (Malloch) (Bariloche, Argentina) e A.
maculiventris Malloch (Perales, Chile) por serem conhecidas apenas da localidade-tipo.
Reynoldsia é um gênero monofilético com nove espécies, endêmicas na região Andina. As espécies são encontradas do nível do mar a altas altitudes nos Andes. Reynoldsia é considerado grupo irmão de Apsil e possui espécies distribuídas nos mesmos tipos de ambientes das espécies de Apsil. Na Figura l 7 . l 7 estão exibidos os traços individuais e na Figura 17 . 1 8 a relação cladística das espécies 16. Não foram apresentados os dados de distribuição de R. brevitarsis Malloch, R. robusta (Stein) e R. trochanterata Malloch por serem conhecidas apenas de uma a duas localidades.

284 - Padrões e Processos - Estudos de Casos
- Traços i nd iv iduais de Cyrtoneuropsis Mal loch. (A) C. armipes (Stein) . (8) C. beebei (Curran) . (C) C. brunnea (Hough). (O) C. conspersa (Ste in) . (E) C. dubia (Snyder). (F) C. flaviantennata (Couri) . (G) c. fuscicosta Curran . (H) C. gemina (Wiedemann) . (I) C. gluta (G ig l io-Tos). (1) C. immunda (Ste in) . (K) C. incognita. (L) C. inuber (G ig l io-Tos). (M) C. maculipennis (Macquart) . (N) C. mellina (Ste in) . (O) C. mimica (Snyder) . (P) C. multomaculata (Ste in) . (Q) C. neotrita (Snyder) . (R) C. ocasionalis (Couri) . (5) C. pararescita (Couri) . m c. polystigma (Wu lp) . (U) C. praenubila (Snyder) . (V) C. protosetosa (Snyder) . (W) C. seriata (Stein) . (X) C. rescita (Wa lker). (Y) C. spiloptera (Wiedemann) . (Z) C. steíni (Snyder) . (A 1) C. varicolor (Hough). (81 ) C. veniseta (Ste in) . (O) C. walkeri (Pont) . (0 1) C. wulpi (Snyder).

B iogeog rafia de Musc idae ( Insecta, D i ptera) da Amér ica do S u l - 285
,..---------------------------------- Cyrtoneuropsis armipes Cyrtoneuropsis conspersa Cyrtoneuropsis flaviantennata
,..-____________________________ -1,..---- Cyrtoneuropsis g/uta Cyrtoneuropsis wa/keri
,..------ Cyrtoneuropsis fuseicosta I-------------------------r------ Cyrtoneuropsis gemina
,..---- Cyrtoneuropsis steini C yrtoneuropsis seriata Cyrtoneuropsis immunda
,..----------------------------- Cyrtoneuropsis dubia ,..-------------------------- Cyrtoneuropsís fuscíquama .------------------------ Cyrtoneuropsis íncogníta ...--------------------- Cyrtoneuropsís po/ystigma
,.-------- Cyrtoneuropsis varíc% r ,-------------1 Cyrtoneuropsís me/lína
Cyrtoneuropsís inuber Cyrtoneuropsis veniseta
r---------------- Cyrtoneuropsis wu/pi
- Cladograma de Cyrtoneuropsís Ma l loch1 1 , 1 2 .
Gênero monofilético com 19 espécies de distribuição exclusiva na Patagônia e sul do Chile (região Andina). As espécies são encontradas desde o nível do mar até os Andes. Na Figura 17. 1 9 estão apresentados os traços individuais de 1 2 espécies e na Figura 17 .20 a relação cladística de 15 espécies17• Não foram mostrados os dados de distribuição de sete espécies: P. albuquerquei Carvalho, P. carvalho i Lopes e Khouri, P. chilensis Bigot, P. darwini Soares e Carvalho, P. flavithorax Couri e Penny, P. schlingeri Couri e Penny e P. vittatus Couri e Penny.
Gênero monofilético com 10 espécies, até recentemente conhecido apenas da região Andina, porém hoje com espécies identificadas e descritas da região norte da
,..---- Cyrtoneuropsís sími/ata .----------1 Cyrtoneuropsis mimica
Cyrtoneuropsis mu/tomacu/ata ,..----------- Cyrtoneuropsis macu/ipennis
Cyrtoneuropsís praenubi/a Cyrtoneuropsis spi/optera
,.- Cyrtoneuropsis protosetosa .-------j-- Cyrtoneuropsís beebeí
Cyrtoneuropsis neotríta ,------- Cyrtoneuropsis ocasionalís
Cyrtoneuropsís parareseita Cyrtoneuropsis rescita
(Port Famine, Chile), B. maculata Couri, Caf\�alho e Pont (próximo de Maipu, Chile), B. valdiviensis (Pamplona) (Valdivia, Chile).
Drepanocnemis é um gênero monofilético com três espécies distribuídas na zona de transição sul-americana. As espécies de Drepanocnemis ocorrem também em altas altitudes, pois exemplares de D. hirticeps Stein foram coletados a 5 .000m nos Andes. Na Figura 17.23 estão mostrados os traços individuais e na Figura 17.24, a relação cladística das espécies 19.
'O América do Sul, na zona de transição sul-americana2. '.Ó
Exceto Apsil, Reynoldsia, Palpibracus e Brachygaste
rina, com espécies de ocorrência na região Andina e Drepanocnemis com espécies acontecendo na zona de transição sul-americana, os outros gêneros de Muscidae possuem espécies distribuídas apenas na área NeotropicaI da América do Sul. Essas espécies mostram grande dependência com as regiões de florestas tropicais, não ocorrendo exclusivamente em áreas não florestadas. O traço generalizado de Polietina na América do Sul indica também este padrão (Fig. 17.25).
:3? As espécies estão distribuídas desde o nível do mar em -('!; regiões de latitudes altas e em altas altitudes nos Andes, c-;2 Equador e Colômbia. Na Figura 17 .21 estão apresen-r tados os traços individuais e na Figura 17 .22, a relação O"-
cladística das espécies18 . Não foram exibidos os dados de distribuição de B. andina Carvalho e Pont (Pichincha, Equador), B. bochica Soares e Carvalho (Chinacota, Colômbia), B. fulvohumeralis (Malloch)

286 - Padrões e Processos - Estudos de Casos
- Traços ind iv idua is de Pseudopti/o/epis Snyder. (A) P. centra/is Schueh l i e Carva lho . (B) P. chrysella Schueh l i e Carva lho . (C) P. confusa Snyder. (O) P. crocina Schueh l i e Carva lho. (E) P. f/uminensis Albuquerque. (F) P. fu/vapoda Snyder. (G) P. /atipa/pis (Ste in) . (H) P. nigripoda Snyder. (I) P. nudap/eura Snyder.

B iogeografia de M usc idae ( Insecta, Di ptera) da América do S u l - 287
,----------- Pseudoptí/o/epís centra/ís
.----- Pseudoptí/o/epís chrysella
Pseudoptí/o/epís f/uminensis
Pseudopti/o/epís fu/vapoda
,.------- Pseudoptí/o/epís nudap/eura
,.------ Pseudopti/o/epis e/bida
- Cladograma de Pseudopti/o/epis Snyder13 .
.. ----------- Po/íetina f/avícincta
--------- Po/ietina rubella
------ Po/ietina concínna
__ �--IH Po/ietina orbita/is
.. __ Po/ietina wu/pi
Pseudopti/o/epís nigripoda
Pseudopti/o/epís crocína
') (;; - Traços ind ividua is de Po/ietina Schnab l e Dziedzicki e cladograma - I . O círcu lo tracejado (em preto) i nd ica á rea de vicariância antiga entre os clados14.

288 - Padrões e Processos - Estudos de Casos
,...-- Polietina nigra _--t
--- Polietína prima
2-'-'0''''' Polietina flavithorax íY/'_W_dV,,=Y�
lyhh"m_,yy�, Polietína major
" :? " � :s - Traços i nd ividuais de Polietina Schnab l e Dziedzicki e cladograma - 1 1 . Os círcu los e a e l ipse tracejados (em preto) i nd icam áreas de vicariância antiga entre os c lados14.
Políetína flavícíncta
Polietina rubella
Políetina concinna
t Políetina orbitalis I Polietina wulpi '" --J 00
Polietina steíni 00 U>
Polietina nigra c.., N ...
Polietina prima 00 '" 0\ Polietina flavithorax b,
Polietina major
Polietina univíttata
Polietina bicolor
Polietina minor
- Cladograma de Polietina Schnabl e Dziedzicki 1 5 .

B i ogeografia de Musc idae (I nsecta, D i ptera) da América do Su l - 289
B
E
,�D(')'1;cTC' " - Traços ind iv idua is de Apsil Mal loch. (A) A. apicata Ma l loch. (B) A. atripes Ma l loch. (e) A. biseta Ma l loch. (D) A. dilata Mal loch. (E) A. maculipennis Ma l loch. (F) A. pennata Mal loch . (G) A. spatulata Mal loch.

290 - Padrões e Processos - Estudos de Casos
': -, 1 ' - Cladograma de Apsil Mal loch 16 .
A
C
E
,---- Apsil pennata
Apsíl dílata
Apsíl spatulata
r-- Apsíl maculíventrís ,----1
Apsíl apícata
,--- Apsíl maculípennís
Apsíl biseta
L-________ Apsíl atrípes
B
D
< ,' , - Traços i nd iv iduais de Reynoldsía Mal loch . (A) R. aurífera (B igot). (B) R. coxata (Mal loch). (C) R. pectínata Mal loch. (O) R. pteropleuralís Mal loch . (E) R. rufoapicata Mal loch. (F) R. scutelfata Mal loch .

B iogeograf ia de M usc idae ( Insecta, D i ptera) d a América do Sul - 291
..---- Reynoldsia pectinata
Reynoldsia pteropleuralis
Reynoldsia rufoapicata
,--------- Reynoldsia coxata
,------- Reynoldsia brevitarsis
,----- Reynoldsia aurifera
L--__ Reynoldsia scutel/ata
0- , ] ' _ Cladograma de Reynoldsia Mal loch16 .
K L
0;' - Traços ind iv iduais de Palpibracus Rondan i . (A) P. confusus (Mal loch). (B) P. fasciculatus (Mal loch). (e) P. lancifer (Ma l loch). (O) P. nigriventris (Mal loch). (E) P. peruvianus (Mal loch). (F) P. pilosus (Macquart) . (G) P. separatus (Mal loch). (H) P. similis (Ma l loch). (I) P. spicatus (Mal loch). (J) P. trivittatus (Mal loch). (K) P. univittatus (Bigot) . (L) P. veneris (Bigot) .

292 - Padrões e Processos - Estudos de Casos
r--------------------------- Palpíbracus venerís
.------ Palpíbracus fascículatus
I
- Cladograma de Palpibracus Rondan i 1 7 .
,.----------{ '--- Palpíbracus nígríventrís
,------- Palpíbracus albuquerqueí
l..-----iL Palpíbracus peruvíanus
Palplbracus tnvlttatus
I I
Palpíbracus spícatus
Palpíbracus unívíttatus
Palpíbracus lancífer
Palpíbracus darwíní
Palpíbracus chílensís
Palpíbracus confusus
Palpíbracus separatus
'---- Palpíbracus símílís
'-------- Palpíbracus pílosus
- Traços ind iv iduais de Brachygasterina. (A) B. humboldtí Soares e Carva lho. (B) B. major Mal loeh. (C) B. muísca Soares e Carva lho . (O) B. stuebeli (R6der). (E) B. violaceiventrís Maeq uart.

B iogeografia de M uscidae ( I nsecta, D iptera) da América d o Su l - 293
Brachygasterina andina
Brachygasterina stuebeli
Brachygasterina major
Brachygasterina valdiviensis
Brachygasterina violaceiventris
- Cladograma de Brachygasterina Macquart18.
- Traços individuais de Drepanocnemis 5tein . (A) D. dorae 5tein. (B) D. griseovirens Mal loch. (C) D. hirticeps Stein.

294 - Padrões e Processos - Estudos de Casos
.. __________ Drepanocnemis dorae
.. _____ Drepanocnemis griseovirens
.. ---- Drepanocnemis hirticeps
CJ: _ Cladograma de Drepanocnemis Stei n 19 ,
1] 70((S - Traços genera l izados de Polietina Schnab l e Dziedzicki na América do SU1 14 ,

Os padrões de distribuição de Muscidae12 são fortemente coincidentes com as sub-regiões e províncias delimitadas por Morrone20. Comparando os padrões de distribuição das espécies dos gêneros anteriores pode-se encontrar explicação partindo de uma história biogeográfica comum para alguns gêneros . É possível, a partir da pan-biogeografia e dos métodos biogeográficos cladísticos, fazer uma correspondência e conexão entre as relações cladísticas das espécies e o relacionamento das áreas onde as espécies são encontradas2 1 .
A evolução geológica da América do Sul influenciou a evolução dos organismos na região. Amorim e Pires22 apresentaram um cladograma geral de área que, desde então, tem sido uma hipótese de trabalho de como teria ocorrido a evolução da área. Tem-se comparado este cladograma geral aos cladogramas particulares de grupos mostrando uma possível explicação da evolução de grupos, em especial desde a última décadas. Na Figura 17 .26 são exibidos os principais eventos vicariantes da América do Sul. A reconstrução é iniciada no Cretáceo com a separação da placa Caribenha e, posteriormente, pela elevação dos Andes e a formação, mais recente, dos mares epicontinentais durante o final do Terciári06.
A comparação dos padrões de distribuição das espécies de Pseudoptilolepis e Bithoracochaeta com os padrões de distribuição dos outros gêneros estudados sugere que outros processos, como dispersão e extinção, foram importantes para a evolução desses gêneros. Isto indica que a evolução desses táxons não é plenamente compatível com a história ligada à evolução de toda a área 12. Boa estratégia de análise biogeográfica é verificar o padrão de distribuição de grupos irmãos. A área de distribuição de Cyrtoneuropsis e Cyrtoneurina sugere que a área de distribuição da espécie ancestral ocupava áreas tropicais das Américas Central e do Suj12, pois as espécies atuais desses gêneros estão distribuídas em toda a região Neotropical.
Aceitando que o padrão de distribuição das espécies encontrado em alguns gêneros de Muscidae (por exemplo, Souzalopesmyia) foi ocasionado por eventos acontecidos no Cretáce022 poderia levar a idade de alguns táxons para o Cretáceo com distribuição no Gondwana. Essa é uma hipótese alternativa para a ocorrência de espécies na América do Su123.
O padrão de distribuição de Polietina mostra, por exemplo, a importância da evolução da região amazônica
Biogeog raf ia de M usc idae ( l n secta, Diptera) da Amér ica d o Su l - 295
na diferenciação das espécies do gênero (ver Figs. 17. 1 2 e 17. 1 3) . Entretanto, as idades desses eventos cladogenéticos ainda não são entendidas e impedem discussão mais pormenorizada de como esses grupos evoluíram na região.
Eventos como a elevação dos Andes são importantes para a evolução das espécies que ocorrem na região, por exemplo, para Palpibracus e Brachygasterina, grupos irmãosl8 . A diversificação de Palpibracus no Chile. abaixo da latitude 30° Sul, pode ser explicada pela história geológica e eventos na área. Introgressões marinhas ocorridas entre 26 e 20 milhões de anos atrás, durante o Oligoceno Superior ao Mioceno Inferior, sugerem a importância desses eventos na evolução das espécies do gênero. Os poucos registros de Palpibracus ao sul do paralelo 42° Sul sugerem que os padrões de distribuição dessas espécies foram influenciados por eventos de glaciação do Quaternário, pois essa região ficou congelada em diversos períodos recentes. Assim, espécies encontradas hoje em dia na Patagônia e no extremo sul do Chile provavelmente atingiram essas áreas por dispers3.li. em período recente com temperatura mais amena 1 - .
Diferente das espécies de Palpibracus, a área de distribuição das espécies de Brachygasterina se estende da extremidade sul do continente até as altas altitudes dos Andes, na Colômbia. Isto sugere um cenário biogeográfico mais elaborado fundamentado na ocorrência da área ancestral dos gêneros irmãos. É provável que a espécie ancestral de Brachygasterina, com distribuição restrita ao sul do continente, tenha expandido sua área de distribuição para o norte. Após, esta área de distribuição ancestral foi fragmentada pelos movimentos tectônicos da elevação e formação dos Andesl 8 .
Por outro lado, o padrão de distribuição das espécies de Apsil e Reynoldsia é mais restrito, se assemelhando à área de distribuição das espécies de Palpibracus. Reynoldsia pectinata ocorre na Terra do Fogo, extremo sul do continente, e a análise do padrão de distribuição das espécies do gênero, comparado à reconstrução biogeográfica do sul do Chile sugere que possam ser encontradas espécies de Reynoldsia nas ilhas Malvinas l6. Essas ilhas fazem parte da região Subantártica, região entendida como monofilética22, indicando padrão de distribuição ancestral das espécies do gênero.
Dentre todos os gêneros estudados, apenas Drepanocnemis possui espécies distribuídas integralmente

296 - Pad rões e Processos - Estudos de Casos
A
c
Separação da placa caribenha
Formação de um mar epicontinental restrito
Formação de um mar epicontinental maior
B
D
Separação do noroeste e sudeste neotropical
Conexão entre as bacias do Parnaíba e Paraná
Soergu imento final dos Andes
- Mapas com a reconstrução de eventos da América do Su l . (A) Separação da p laca caribenha do continente com i n ício no f ina l do Cretáceo e f ina l izando no Mioceno-Pl ioceno Médio. (B) Separação dos componentes noroeste e sudoeste neotropical no f ina l do Cretáceo em razão da formação do l ago ao longo dos rios Amazonas, Madeira e Mamoré; i n ício da e levação dos Andes no Cretáceo Médio. (C) Formação do mar epicont inenta l pelo portal oceân ico do norte (Macaraibo) e l este (Amazonas) (a l i nha ponti lhada demarca os l im ites do mar). (O) Conexão entre as bacias do Parna íba e Paraná que existiam no Cretáceo Médio e Superior. (E) Formação do mar epiconti nental pela i nvasão através do norte, leste e su l (bacia do Paraná) no M ioceno; a e levação dos Andes atinge um estág io avançado. (F) E levação dos Andes no estágio f ina l , ating indo a conformação atua l no Mioceno-Pl ioceno. Mod ificado de N ihei e Carva l h06.

dentro da zona de transição, sugerindo que os eventos de especiação se deram apenas nessa área. Segundo Morrone2, zonas de transição merecem atenção especial, pois são áreas limítrofes entre regiões biogeográficas, indicando intensa interação biótica com diversidade bastante alta.
A descoberta de outros padrões biogeográficos semelhantes aos conhecidos poderá sugerir homologia biogeográfica primária se referindo à hipótese de uma história biogeográfica comum24. Como se sabe, um padrão geral biogeográfico de qualquer região será mais informativo e detalhado quando mais padrões individuais forem propostos e testados6. Por exemplo, as áreas de endemismo encontradas em Muscidae na América do Sul sugerem que processos vicariantes foram importantes na formação desses padrões, sugerindo um padrão mais geral para a evolução dos grupos das regiões Neotropical e Andina25.
Na biogeografia histórica existem problemas metodológicos detectados recentemente. Donoghue e Moore26 indicaram que alguns padrões biogeográficos podem não ser comparáveis, isto é, as congruências topológicas encontradas nos cladogramas de área não foram causadas pelos mesmos eventos de disjunções, se esses eventos ocorreram em épocas diferentes (ver Capo 6). Assim, não é possível comparar esses padrões, pois foram gerados a partir de eventos de diferentes idades.
Para solucionar essa questão é necessário ter outra estimativa da datação dos eventos. Eventos geológicos conhecidos, registros fósseis e também análise de dados moleculares pode nos auxiliar na estimativa do tempo de divergência das linhagens. Bortolanza et al. 7 chamaram atenção da pseudocongruência e a necessidade da introdução de componentes explícitos temporais na análise do padrão de distribuição das espécies de Coenosopsia (Anthomyiidae). Esses componentes temporais devem ser incluídos em qualquer análise biogeográfica de organismos na América do Sul.
É provável que os eventos vicariantes, sugeridos como possíveis causadores de disjunção anteriormente para o Cretáceo Médio ou Superior22, tenham sido gerados durante o Cenozoic027. A América do Sul foi coberta por mares epicontinentais em épocas diferentes (por exemplo, Cretáceo e Mioceno, Fig. 17 .26) e os padrões originados a partir dessas disjunções não são comparáveis.
B iogeograf ia de M usc idae ( Insecta, D i ptera) da América do Su l - 297
o entendimento detalhado da época que ocorreram as disjunções, que podem ter causado os padrões, nos auxiliará a entender melhor o quebra-cabeça que é a evolução biogeográfica da América do Sul. Isto auxiliará não apenas o entendimento da evolução biogeográfica de táxons de Muscidae, mas de todos os organismos que estão distribuídos na região.
/. �
Ao Peter Lowenberg-Neto (Universidade Federal da Integração Latino-americana) por ceder seus dados adicionais da distribuição geográfica de Muscidae e pela confecção da Figura 17. 1 . A Sílvio S. Nihei (Universidade de São Paulo) , Kirstern Lica F. Haseyama (Universidade Federal do Paraná) e Eduardo A. B . Almeida (Universidade Federal do ABC) pela leitura crítica e sugestões do manuscrito; Kirstern Lica F. Haseyama também confeccionou as demais figuras. Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela bolsa e auxílio concedidos, processos 300873/2008-5 (CJBC) e 30130112007-7 (MSC).
REFERÊNCIAS B IBLIOGRÁFICAS 1. CRISCl, 1. Y.; CIGLIANO. M. M.; MORRONE. 1. J . ç: .,
Historical biogeographical of southern South America. S .
Zool. , v. 42, p. 1 52- 1 7 1 , 1 99 1 . 2 . MORRONE, J . 1 . Panbiogeografia, componentes biótico, :
zonas de transición. Rev. Bras. Entomol. , v. 48, p. 149-162, 20ü�. 3. CARVALHO, C. J. B . ; COURI, M. S.; PONT, A. C. et al. . .1,.
Catalogue of the Muscidae (Diptera) of the Neotropical Region. Zootaxa, v. 860, p. 1 -282, 2005.
4. HENNIG, W. Vorarbeiten zu einem phylogenetischen System der Muscidae (Diptera: Cyc1orrhapa). Stllttg Beitr Natllrk, \ . 1 4 1 , p . 1 - 1 00, 1 965.
5. MICHELSEN, Y. Revision of the aberrant New World genus Coenosopsia (Diptera: Anthomyiidae), with a discussion of anthomyiid relationships. Syst. Entomol., v. 1 6, p. 85- 1 04, 1 99 1 .
6 . NIHEI, S . S . ; CARVALHO, C . 1 . B. Taxonomy, c1adistics and biogeography of Coenosopsia Malloch (Diptera, Anthomyiidae) and its significance to the evolution of anthomyiids in the Neotropics. Syst. Entomol., v. 29, p. 260-275, 2004.
7. BORTOLANZA, M. ; CARVALHO, C. J. B . ; LARA, A. P. C. A new species of Coenosopsia Malloch (Diptera, Anthomyiidae) from southern Brazil and a reappraisal of the cladistic analysis of the genus. Zootaxa, v. 1 242, p. 37-52, 2006.
8. CARVALHO, C. 1. B. Revision, Cladistics and Biogeography of the Neotropical Genus Souzalopesmyia Albuquerque (Diptera: Muscidae). P Entomol. Soe. Wash, V. 101 , p. 1 23- 1 37, 1 999.
9 . CROIZAT, L. Spaee, Time, Forrn - The Biologieal Synthesis. Caracas : pub1ished by the author, 1 964. 8 8 1 p.
lO. COURI, M. S . ; MOTTA, H. C. G. Análise cladística de Bithoraeochaeta Stein (Diptera, Muscidae). Rev. Bras. Entornol. , v. 44, p. l 05- 108, 2000.
1 1 . PAMPLONA, D. M. Nova caracterização de Cyrtoneurina Giglio-Tos, 1 893 e descrição de Paracyrtoneurina gen. novo (Diptera, Muscidae). Rev. Bras. Entornol., V. 43, p. 9-24, 1 999.
12. CARVALHO, C. 1. B.; BORTOLANZA, M.; SILVA, M. C. C. et a!. Distributional patterns of the Neotropical Muscidae (Dip-

298 - Padrões e Processos - Estudos de Casos
tera). In: MORRONE, J. J . ; LLORENTE, J. Una Perspectiva Latinoamericana de la Biogeograjra. México: Las Prensas de Ciencia, Facultad de Ciencias, Universidad Autônoma de México, 2003. capo 30, p. 263-274.
1 3 . SCHUEHLI, G. S . ; CARVALHO, C. J. B. Revision and cladistics of the Neotropical genus Pseudoptilolepis Snyder, 1 949 (Diptera: Muscidae). Rev. Bras. Zool. , V. 22, p. 23-34, 2005.
14. NIHEI, S . S . ; CARVALHO, C. J. B. Distributional patterns of the Neotropical fiy genus Polietina Schnabl e Dziedzicki (Diptera, Muscidae) : a phylogeny-supported analysis using Panbiogeographic tools. Papo Avulsos de Zool. (São Paulo), V. 45, p. 3 1 3-326, 2005.
1 5 . NIHEI, S . S.; CARVALHO, C. J. B. Systematics and biogeography of Po/ietina Schnabl & Dziedzicki (Diptera, Muscidae): evidences on Neotropical area relationships toward the hypothesis of Amazonia as composite area. Syst. Entomol. , v. 32, p. 477-501, 2007.
16. CARVALHO, C. J. B. ; COURI, M. S . A Cladistic and Biogeographic analysis of Apsil Malloch and Reynoldsia Malloch (Diptera, Muscidae) of southern South America. P Entomol. Soe. Wash, V. 1 04, p. 309-3 17 , 2002.
1 7 . SOARES, E. D. G. ; CARVALHO, C. J. B. Biogeography of Palpibracus (Diptera: Muscidae): an integrative study using panbiogeography, parsimony analysis of endemicity and component analysis . In : LLORENTE, J . ; MORRONE, J. J . Regionalización Biogeográfica en Iberoamérica y Tópicos Afines. México: Universidad Autonoma de Mexico, 2005. capo 30, p. 485-494.
1 8 . CARVALHO, C. J. B . ; PONT, A. C. Revision, cladistics and biogeography of the South American genus Brachygasterina (Diptera: Muscidae). Zootaxa, V. 1 1 5 1 , p. 1 -26, 2006.
19 . COURI, M. S . ; CARVALHO, C. 1. B. A review of the Neotropical genus Drepanocnemis Stein (Diptera, Muscidae), with phylogenetic and biogeographic analysis of its species. J. Nat. Rist. , V. 42, p. 2659-2678, 2008.
20. MORRONE, J. J. Biogeografía de América Latina y el Caribe. M & T - Manuales & Tesis SEA, v. 3, Zaragoza, 200 1 . 148p.
2 1 . NELSON, G.; PLATNICK, N. Systematics and Biogeography, Cladistics and Vicariance. Nova York: Columbia University Press, 1 9 8 1 . 567p.
22. AMORIM, D. S.; PIRES, M. R. S . Neotropical Biogeography and a method for maximum biodiversity estimation. In: BICUDO, C. E. M.; MENEZES, N. A. Biodiversity in Brazil, a First Approach. São Paulo: CNPq, 1996. p. 1 83-2 19.
23. COURI, M. S . ; CARVALHO, C. J. B . Systematic relations among Philornis Meinert, Passeromyia Rodhain & Villeneuve and allied genera (Diptera, Muscidae). Braz. J. Biol., V. 63, p . 223-232, 2003.
24. MORRONE, J. J. Homology and areas of endemism. Divers. Distrib. , V. 7, p. 297-300, 2001 .
25. LOWENBERG-NETO, P. ; CARVALHO, C . J . B . Areas of endemism and spatial diversification of the Muscidae (Insecta: Diptera) in the Andean and Neotropical regions. J. Biogeogr., V. 36, p. 1 750-1759. 2009.
26. DONOGHUE, M. J . ; MOORE, B. R. Toward an Integrative Historical Biogeography. Integr. Comp. Biol. , V. 43, p. 261 -270, 2003.
27. AMORIM, D. S . Neotropical Diptera diversity: richness, patterns, and perspectives. In: PAPE, T; BlCKEL, D. ; MEIER, R. Diptera Diversit)' - Status, Challenges and Tools. Nova York: Koninklijke Brill, 2009. capo 3, p. 7 1 -97.

Algoritmo : sequência de regras para execução de uma tarefa específica.
Alopatria (adj.: alopátrico): ocorrência em áreas distintas; disjunção; distribuições mutuamente exclusivas.
Amplilocado: amplamente distribuído.
Ancestral: Espécie, indivíduo ou área que dá origem a espécies, indivíduos ou áreas descendentes.
Apomorfia (adj.: apomórfico): estado derivado de um caráter.
Área de endemismo : unidade fundamental do estudo biogeográfico, em sua definição mais tradicional, reconhecida pela distribuição congruente de dois ou mais táxons.
Autapomorfia (adj.: autapomórfico): apomorfia presente em um táxon terminal, mas não compartilhada com seu grupo irmão.
Barreira: característica geográfica ou biótica de uma região que impede (parcial ou totalmente) a dispersão de indivíduos entre as porções do espaço isoladas por este elemento. Alguns exemplos de barreira são corpos d' água, cadeias de montanhas, descontinuidades vegetacionais (para organismos terrestres), diferenças de salinidade e separação física de bacias hidrográficas (para organismos aquáticos) , entre muitos outros . Características que constituem uma barreira para os indivíduos de uma espécie podem não impedir o movimento de indivíduos de outras espécies; o conceito de barreira não pode ser considerado absoluto.
Biodiversidade: variação ou variabilidade da vida em todos os níveis de sua organização.
Biogeografia: estudo da distribuição dos organismos no espaço através do tempo.
Biogeografia ecológica: estudo dos efeitos das condições ambientais e das interações bióticas sobre a distribuição dos organismos.
Biogeografia histórica: estudo da evolução espacial das biotas e dos seus táxons.
Biogeografia vicariante (de vicariância): ramo da biogeografia histórica fundamentada na busca por padrões
biogeográficos, os quais, quando encontrados, são interpretados como resultantes de eventos de vicariância. Pan-biogeografia, Biogeografia cladística e parte dos métodos de eventos são incluídas nesta escola da biogeografia.
Biota: conjunto de espécies encontradas em determinada região geográfica.
Centro de origem: segundo o paradigma dispersionista, trata-se da região onde ocorreu a evolução de uma espécie ancestral e de onde indivíduos desta espécie se dispersaram e colonizaram outras áreas.
Cladística: metodologia aplicada à inferência de relacionamento filogenético entre táxons fundamentada na organização mais parcimoniosa das informações de caracteres homólogos compartilhados; busca de padrões de agrupamento pelo reconhecimento de sinapomorfias e aplicação do critério da parcimônia.
Clado: ver grupo monofilético.
Cladograma: Representação das relações de parentesco entre três ou mais táxons ou áreas ; diagrama ramificado com o qual são representadas as relações hierárquicas entre táxons ou áreas . Nos cladogramas, a maior proximidade de dois elementos em comparação a um terceiro é interpretada como um reflexo da história evolutiva ou geológica.
Congruência: coincidência gerada por histórias compartilhadas; concordância.
ConservaçãolBiologia da conservação: campo de pesquisa multidisciplinar voltado para a preservação da biodiversidade.
Datação: análise que visa estimar idades de origem de espécies ou clados, podendo incluir informações do registro fóssil, da evolução molecular dos táxons recentes, assim como também de outras informações relevantes às hipóteses filo genéticas e/ou biogeográficas.
Disjunção (adj.: disjunto): distribuição descontínua de uma espécie ou táxon no espaço.
Dispersão: transposição de uma barreira por um ou mais indivíduos de uma espécie, sucedida pelo estabelecimento

300 - G lossár io
de nova população disjunta (ver também expansão da distribuição ) .
Dispersionismo (adj. : dispersionista, dispersalista): paradigma biogeográfico segundo o qual é possível propor explicações históricas para a distribuição de um táxon pelo uso de eventos de dispersão a partir de um centro de origem.
Divergência genética: medida das diferenças observadas ou estimadas entre aleIos, populações ou linhagens.
Diversidade genética: número relativo de linhagens gênicas distintas encontradas em uma população ou espécie, em que não é contabilizada a relação entre os alelos ou a divergência genética entre estes.
Endêmico: relativo a um táxon cuja distribuição geográfica pode ser considerada restrita; a definição do que se considera "restrito" não é absoluta e depende do contexto do estudo em questão.
Endemismo: ver área de endemismo.
Especiação: surgimento de duas ou mais espécies descendentes a partir de uma espécie ancestral; cladogênese.
Especiação alopátrica: especiação em decorrência do isolamento geográfico de populações de uma espécie com distribuição mais ampla.
Espécies irmãs: duas espécies são ditas "irmãs" quando forem consideradas mais próximas entre si que em relação a qualquer outra espécie. A mesma definição pode ser aplicada a táxons, clados ou áreas (isto é, táxons irmãos, clados irmãos, áreas irmãs).
Evento biogeográfico: qualquer processo capaz de modificar as distribuições de táxons, por exemplo, vicariância, dispersão, extinção, desaparecimento de barreira.
Expansão da distribuição: aumento da área de ocorrência de uma população ou táxon; normalmente, utilizado para situações que não envolvem transposição de barreira e, portanto, sem ocorrência de dispersão.
Extinção: desaparecimento de um táxon ou população de uma área.
Filogenia: Relações evolutivas entre táxons.
Grupo monofilético: grupo formado exclusivamente por uma espécie ancestral e todos os seus descendentes; grupo diagnosticável pela presença de sinapomorfia(s); grupo natural; clado.
Grupo parafilético: grupo formado por uma espécie ancestral e parte de suas espécies descendentes, mas não todas; conjunto de táxons que compartilha uma ou mais plesiomorfias, mas nenhuma sinapomorfia; grupo artificial.
Grupo polifilético: agrupamento reconhecido por caracteres convergentes (homoplasias); grupo formado por espécies descendentes de vários ancestrais; grupo artificial.
Grupos externos: táxons não pertencentes ao grupo de estudo, mas incluídos em análises filogenéticas para enraizamento dos cladogramas resultantes.
Haplótipo: combinação distinta de nucleotídeos em um único cromossomo em um loeus.
Homologia: Similaridade compartilhada por dois ou mais organismos em razão da herança de um ancestral comum. Em biogeografia, áreas homólogas são aquelas :s
'?" que compartilham uma história. �
c.., Íntrons: sequências que fazem parte de um gene, porém � não são codificadas, mas descartadas durante a síntese �
0\ proteica. b
Migração: expansão da distribuição geográfica de uma espécie com ou sem transposição de uma barreira.
Nós: pontos de ramificação de um cladograma.
Otimização de Fitch: algoritmo de parcimônia propos-to por Walter Fitch no início da década de 1 970 para estudo de mudanças de estado de caracteres discretos sobre cladogramas.
Padrão biogeográfico: congruência distribucional e de relacionamento histórico entre as áreas.
Plesiomorfia (adj. : plesiomórfico): estado ancestral de um caráter.
Processo: causa ou conjunto de causas responsáveis pela distribuição geográfica de um táxon; ver evento biogeográfico.
Pseudogenes: genes não funcionais espalhados pelo genoma. Normalmente surgem por duplicação de genes funcionais e acumulam mutações que os tomam inativos.
Recombinação: processo que ocorre durante a meiose e consiste na troca de fragmentos cromossômicos entre cromátides não irmãs. Este processo forma novas combinações e consequentemente novos haplótipos na ausência de mutação.
Redundância (adj.: redundante): em biogeografia, diz respeito à distribuição de duas ou mais espécies não irmãs numa mesma área de endernismo.
Simpatria: distribuição de dois ou mais táxons em áreas parcial ou completamente coincidentes.
Sinapomorfia (adj. : sinapomórfico) : Apomorfia compartilhada pelos táxons terminais que compõem um grupo monofilético.
Táxon (pl. táxons): grupo de organismos ou de táxons em qualquer nível da hierarquia sistemática. Um táxon pode possuir um nome formal ou não.

Teoria dos refúgios: teoria originalmente elaborada por Jürgen Haffer em 1969. A teoria propõe que as oscilações climáticas ocorridas durante o Pleistoceno teriam causado a aridificação da região amazônica acarretando refúgios florestais mais úmidos, separados por áreas de vegetação aberta. Nesses refúgios, as populações evoluiriam em alopatria. Com a expansão das florestas durante o período interglacial, as populações isoladas da espécie ancestral teriam se diferenciado em espécies distintas e voltariam a entrar em contato. Para Haffer, esse seria o mecanismo responsável pela alta diversidade encontrada na Amazônia.
G lossár i o - 301
Traço biogeográfico: um grafo ou linha que conecta dois ou mais registros de distribuição (ou áreas de endemismo) num mapa.
Vagilidade: qualidade de organismos de uma determi
nada espécie de deslocamento geográfico; capacidade de organismos de mudança de localização ou de distribuição.
Variabilidade genética: ver diversidade genética.
Vicariância (= evento de vicariância) : divisão da área de distribuição de um táxon ancestral em duas ou mais áreas pelo surgimento de barreira(s) natural(is) entre
elas, seguida por especiação alopátrica.


A Abordagem multiloci, 141 Abrotanella, 71 Ab' Sáber, domínios morfodimáticos, 26 Agrupamento fenético, 1 66 Algoritmo de Metropolis-Hastings, 144 Alozimas, 138 Amazônia
áreas de endemismo, 1 90 idade das espécies, 195 origem e evolução da moderna biota, 1 89 unidades biogeográficas básicas, 1 90
América do Sul biogeografia
da ictiofauna, 269 dos peixes de água doce, 261
distribuiçôes disjuntas, 270 singulares, 272
escudos cristalinos antigos, 270 estrutura geológica, 266 grupos gondwânicos, 269 história tectônica, 266 ictiofauna, 261 margem atlântica em evolução, 269 padrões biogeográficos, 269 ruptura afro-brasileira, 269
Análise biótica, 86 cladística de distribuições e endemismo, 208 comparativas, 1 5 1 de dados agrupados, 1 42 de componentes, 1 07 de dispersão-vicariância, 84, 1 3 1 , 186 de enunciados
de 3-áreas, 1 1 3 , 206 de 3-itens, 1 1 3
de parcimônia de Brooks, 108, 1 09, 1 28, 232 de subárvores livres de paralogia, 1 14 de traços, 1 67 de verossimilhança de evolução de distribuição geográfica,
1 34 integrada de falhas, 77 parcimoniosa de endemicidade, 1 66, 228, 266 vicariante, 78
geografia de caracteres, 84 Antártica, 3 Arco pleistocênico, 200
Área de diversificação, 1 67 de endemismo, 44
amazônicas formação das biotas, 1 94 relações históricas. 1 9 1
análise parcimoniosa. 46 complexo biogeológico, 167 conceitos, 4 1 definição, 42 delimitação, 42, 1 17 método
dos traços. 44 para reconhecimento, 44, 1 65
pan-biogeografia, 167 unidade histórica. 43
de manejo de espécies/hábitats, 1 64 de simpatria, 105 protegida para manejo de recursos, 1 64 unidade histórica da biogeografia, 48
Árvores, ajuste parcimonioso, 130 Associação de biogeografia sistemática e evolutiva, 1 69 Aulacógenos, 7
B Barreiras
geográficas, 54 reiterativas, 236
Biodiversidade atlas, 94 conhecimento espacial, 168 estática, 1 63 processual, 163
Biogeografia dadística, 99, 1 00, 123, 1 35 , 1 84, 1 9 1 , 232
informação temporal, 1 1 8 métodos analíticos, 106
de área, 165 de eventos, 135 de vicariância, 53, 252 do táxon. 165 ecológica, 62, 1 64 evolutiva, 65 filogenética, 1 0 1 q histórica, 62, 99, 1 64, 1 83, 2 1 9
aplicações, 1 84 exemplos, 1 84
Biologia da conservação, 1 62 Biota sul-americana, origem híbrida, 1 84
As letrasfe q que se seguem aos números de páginas correspondem, respectivamente, afiguras e quadros.

304 - í nd ice Remissivo
Bosque atlântico brasileiro, 223 de Araucaria angustifolia, 223 paranaense, 223
c Caatinga, 24 1 , 243
biogeografia, 247 características, 202 clima, 244 endemismos, 253 espécies, 253 fiora relictual, 244 padrões filo genéticos, 254 vegetação, 244, 253
Cabrera e Willink, domínio
amazônico, 22 andino-patagônico, 23 Caribe, 1 8 chaquenho, 22 guiano, 22 subantártico, 23
e Yepes, 1 4 Carbonífero, 4 , 6 Cenozoico, 6 Cerrado, características, 202 Chaco, características, 202 Ciclo
de Wilson, 266 orogenéticos, 266
Cladograma de áreas, 99, 1 0 1 , 1 03, 1 27, 1 92 Classificação biogeográfica, 168 Código Internacional de Nomenclatura de Área, 165, 169 Conectograma, 1 26 Congresso Mundial de Parques Nacionais e Áreas Protegidas, 164 Congruência biogeográfica, 1 1 8 Conservação
da biodiversidade, 1 62 ex situ, 163 in situ, 1 63
Convenção sobre Diversidade Biológica, 1 62 Cordilheira dos Andes
consequências biogeográficas, 272 relevos, 8 soerguimento, 8, 272
Coriaria, vicariância, 79 Cosmopolitismo primitivo, 99 Crátons, 6 Cretáceo, 4, 8, 9
D Datação molecular, 207 Densidade absoluta, 254 Depressão afro-brasileira, 268 Deserto de Botucatu, 8 Diagonal
arreica, 203 de formações abertas, 222
secas, 1 82 evolução, 1 9 8 história, 2 1 7 origem, 204 padrões de endemismo, 1 9 8 relações históricas entre regiões, 207 variações climáticas do quaternário, 2 1 1
Diferenciação biológica, 65 Dinâmica de soma zero, 247 Dinossauros, 1 0 Dípteros, 277 Disjunções, 252, 297 Dispersão, 1 00, 1 35
de longa distância, 2 1 4 ilimitada e nula, 1 5 8
Distribuições geográficas, 1 55
das espécies em contexto macroecológico, 1 52 propriedades, 152 respostas às mudanças climáticas, 157
redundantes, 1 05
E Ecologia
de ilhas rochosas, 90 espacial, 168 evolutiva, 89
Efeito de sombra de barreira, 9 estufa, 4
concentração de gases, 1 5 1 Eoceno, 1 1 Escudo
brasileiro, 272 cristalinos, 267 das Guianas, 272
Espécies amplilocadas, 228 delimitação taxonômica, 1 1 7
Estocasticidade demográfica, 247 Estudos filogeográficos na região neotropical, 148 Evento
de extinção em massa, 10 sul-atlântico, 268 vicariantes, 142, 297
Evolução alopátrica, 65 Extinção, 1 35
em massa, 1 0
F Falhas transformantes, 77 Fase
diaguita, 1 8 3 inca, 1 80 quechua, 1 75, 1 82, 1 83
Filogeografia, 1 96 conceitos, 1 37 estatística, 142 histórico, 1 37
Fittkau, províncias zoogeográficas, 1 6 Florestas
estacionais deciduais tropicais, 200 tropicais sazonalmente secas, 241
metacomunidade, 246 Fórum Nacional de Biodiversidade, 162 Fotossíntese, 3
G Genética de populações, 1 37, 1 96 Glaciação, 1 1 Gondwana, 4, 7, 8
história geológica hierárquica, 1 3 3

H Hipótese
de isolamento em montanhas, 225 de rios como barreiras, 225 dos gradientes ecológicos, 225 dos refúgios, 225 filogenética dos táxons, 1 27
Hominídeos, vicariância, 80 Homologia
biogeográfica primária, 166 espacial, 1 65 , 2 1 9 temporal , 2 1 9
Hot spots, 7 de biodiversidade, 164
I Ictiofauna sul-americana, 261 Ilhas rochosas, 89 Incerteza topológica, 1 30 Inferência
bayesiana, 1 30 probabilísticas sobre associação de árvores, 1 29
Introgressão, 1 4 1
J Jungles, 1 28 Jurássico, 4, 7, 8
K Kuschel, 1 8
L Laurásia, 4, 6, 1 33 Listriderinos andino-patagônicos, biogeografia histórica, 1 86
M Macroecologia, 1 5 1 Mar
paranaense, 1 82 patagônico, 1 8 1 salamanquense, 178, 1 8 1 , 1 82
Mata Atlântica biota, 223 de tabuleiros, 221 endemismos, 223 floresta
estacionai semidecidual, 221 mista, 221 ombrófila densa, 221
história biogeográfica, 223 hot spots, 222 opiliões, 225 origens, 222 processos
de diversificação, 225 tectônicos, 235
regionalização ecológica, 221 relações
históricas com outras áreas, 222 internas, 223
Mesozoico, 4, 6, 8, 1 1
Metacomunidades, 242, 247, 252, 253 , 258 Metapopulações, 89 Métodos
índ ice Rem iss ivo - 305
aplicados à conservação d a biodiversidade, 1 65 de eventos, 123 , 125
Microssatélites, 1 39 , 141 Mioceno, 9, 1 1 Modelo
de circulação global, 1 5 2 d e dispersão
com alta assimetria, 1 60 de Galápagos, 9 1 -extinção-cladogênese, 134
de eventos, 1 23 de Hardy-Weinberg, 1 39q de recombinação de caracteres, 93 estocástico de evolução, 1 34
Monumento natural, 1 64 Morrone
região andina, 38 neotropical, 34
zona de transição sul-americana, 37 Mudanças climáticas, 1 5 1 Müller, centros de dispersão, 23 Muscidae
Apsil Malloch, 283 Bithoracochaeta Stein, 279 Brachygasterina Macquart, 285 Cyrtoneurina Giglio-Tos, 279 Cyrtoneuropsis Malloch, 283 Drepanocnemis Stein, 285 espécies
localidades de coleta, 278f padrões de distribuição, 277, 279 relação evolutiva, 279
evolução, 295 Palpibracus Rondani, 285 Po/ietina Schnabl e Dziedzicki, 283 problemas, 297 províncias, 295 Pseudoptilolepis Snyder, 283 regiões, 295 Reynoldsia Malloch, 283 soluções biogeográficas, 297 Souzalopesmyia Albuquerque, 279
Mutação populacional, taxa, 144
N Nicho
ecológico, ISS eltoniano, 1 5 6 grinneliano, 1 56 multidimensional, ISS
Norte gondwânica, 178 Nós biogeográficos, 94 Nothofagus, 69, 75, 76, 1 77 , 1 80, 1 8 1 , 1 84
o Oceano Atlântico, abertura, 7 Oligoceno, 8, I I Opiliões
biogeografia, 225, 234 endemismo, 235, 237 padrões biogeográficos, 237 principais processos históricos inferidos, 234

306 - índ ice Remiss ivo
p Padrões
biogeográficos, 54, 58 de distribuição das espécies, 41 filogeográficos descritos na literatura, 144 macroecológicos e mudanças climáticas, 158
Paleoceno, 178 Paleofiora mista, 1 8 1 Paleolatitude, 3 Paleopalinologia, 2 1 3 , 222 Paleozoico, 4, 6 Pan-biogeografia, 65, 1 23 , 1 84
centro de massa, 73 conceitos metodológicos, 66 correlação geológica, 76 linha de base, 68 nó biogeográfico, 69 traço, 67
Pangeia, 3, 4, 6-8, 1 0 Panthalassa, 6 Paralogia geográfica, 1 05 , 1 14 Parque nacional, 1 64 Patagonides, 8 Peixes de água doce sul-americanos, 264 Permiano, 4, 6 Placa de Nazca, 8 Planícies austrais, idade, 1 82 Pleistoceno, 1 1 Pluma mantélica, 9 Pré-cambriano, 4 Processo
de interação (competição), 1 5 5 d e vicariância, 99
Província rnicrofioral mista, 204, 205, 2 1 7 Pseudo congruência, 57, 1 1 9, 297 Pseudoincongruência, 1 1 9
R Recolonização, 89 Região
andina, 175, 177 botânicas, 42 fitogeográficas, 1 68 neotropical, 86 zoogeográficas, 1 68
Regionalizações biogeográficas, 1 4 Regra d e progressão de Hennig, 7 8 Re1ictos, 205 Reserva natural/área selvagem, 164 Ringuelet, 26 Rivas-Martínez
e Navarro, 29, 32 e Tovar, 29
Rodínia, 3, 4 Ruído biogeográfico, 1 23
s Sick, 1 8 Síntese biológica
ancestrais, 92 conservação evolutiva, 94
Sistema Nacional de Unidades de Conservação, 163, 1 64 Sistemática
conceito, 1 37 filo genética, 99, 196
Subducção, 8 Sul gondwânica, 1 7 8
T Tanglegram, 1 26 Taxonomia imatura, 1 05 Tectônica de placas, 99, 261 Teoria
da coalescência, 142 da deriva dos continentes, 4 da tectônica de placas, 266 dos refúgios, 1 95 neutra
de evolução, 142 unificada, 247
Terras baixas sul-americanas bacias de antepaís, 273 depressões tectônicas encravadas nos escudos, 273 transgressões marinhas, 273
Terrenos, acreção, 7 1 Trackogram, 1 26 Transgressões marinhas, 274 Triássico, 4, 6, 7, 10 Trilhograma, 1 26, 1 29 Turnover, 1 5 8
u União Internacional para a Conservação da Natureza e dos
Recursos Naturais, 1 64 Unidades
geográficas operacionais, 1 90 taxonômicas operacionais, 1 3 8
v Vagilidade, 148 Vicariância, 53, 54, 65, 99, 100, 123 , 242
áreas de endernismo, 8 1 máxima, 1 27
Visão dispersalista, 279 Vulcanismo, 3, 4, 9
z Zona de sombra de chuva, 1 82, 1 83

