bases neurais do controle postural e o envolvimento da...
TRANSCRIPT

139
Science in Health set-dez 2012; 3(3): 139-51
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
Bases neurais do controle postural e o envolvimento da suBunidade Glur1 nas vias vestiBulares em aves
NeuroNal bases of postural coNtrol aNd the iNvolvemeNt of the Glur1 subuNit iN the aviaN vestibular pathways
Maria de Fátima Passetto1
João Roberto Rocha1,2
Renato Figueiredo de Santana1 Cláudio A.B. de Toledo (in memoriam)1
1 Laboratório de Neurociência, Universidade Cidade de São Paulo, São Paulo, SP, 03071-000.2 IAMSPE, Instituto de Assistência Médica ao Servidor Público Estadual, São Paulo, SP, 04029-000
R E S U M O
introdução: Controle postural é a resposta proveniente da integração de múltiplos circuitos sensórios-motores na manutenção do equilíbrio e da postura. De todos os sistemas envolvidos destaca-se o sistema vestibular, que através de seus receptores capta os deslocamentos do corpo em relação à gravidade; e através de vias eferentes modula as respostas reflexas posturais. O glutamato é o principal neurotransmissor excitatório do sistema ves-tibular, liberado pelo nervo vestibular e captado pelos receptores de glutamato do tipo AMPA. O receptor da família AMPA é constituído por quatro subunidades nomeadas de GluR1 a GluR4, sendo que a subunidade GluR1 é fortemente relacionada com a formação dos circuitos sinápticos. Este trabalho teve como objetivos fazer uma revisão sobre esses conceitos e apresentar diferenças da expressão da subunidade GluR1 do receptor do tipo AMPA durante o desenvolvimento e mapear a terminação das fibras aferentes do canal semicircular horizontal nos neurô-nios dos núcleos vestibulares em pombos. métodos: Utilizamos a técnica de imunihistoquìmica para identificar as subunidades GluR1 e GluR4 nos núcleos do complexo nuclear vestibular, a projeção do canal semicircular horizontal foi identificada com auxílio de um traçador neural. A técnica de Giemsa serviu com guia histológico da região. resultados e conclusão: Os dados mostraram uma diminuição da expressão da subunidade GluR1 nos neurônios vestibulares durante o desenvolvimento, e que os axônios do canal semicircular horizontal terminam em todos os núcleos vestibulares, sem demonstrar nítida preferência. A discussão pretendeu discorrer sobre o papel das vias eferentes vestibulares no controle postural.Pal avr as - chave : Equilíbrio postural • Vestíbulo do labirinto • Neurotransmissores • Glutamatos
A B S T R A C T
introduction: Posture adjustments are performed with the aid of several neural circuitries that integrate sensorial stimulus with motor outputs. The vestibular system (VS) has a critical role in detecting the corporal movements by the vestibular labyrinth in the temporal bone and integrating these signals with motor responses that are needed to compensate the gravity forces. The VS sends neural information to somatic motoneurons forming the neuro-logical basis of the postural reflexes. The amino acid glutamate is the major excitatory neurotransmitter in the VS and their release from pre-synaptic structures activates glutamate receptors (GluRs) which responses promotes the final muscular activation necessary to compensate body movements. methods: The fastest responses of the GluRs are provided by the AMPA-type family, constituted by four subunits named GluR1 to GluR4. The GluR1 subunit is related with synaptic formation and we aimed today to review these concepts and to present the differences in the GluR1 expression on neurons belonging to the mesencephalic vestibular nuclei and to map the terminal of afferents fibers from the vestibular receptors of the horizontal semicircular Channel present into the labyrinth of pigeons. We identify the appearance of GluR1 in vestibular neurons and testify their innervations pattern by peripheral vestibular fibers. results and conclusion: We found a reduction of the GluR1 presence according the animal evolves and that the vestibular fibers arrive at practically all mesencephalic vestibular nuclei. We centered our discussion on the vestibular pathways and their roles on postural control.Ke y words : Postural balance • Vestibule, labyrinth • Neurotransmitter agents • Glutamates.

140
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
INTRODUÇÃO O controle postural, através da percepção
dos movimentos e da orientação espacial, é um comportamento neuromotor que depende da rápida detecção das alterações do corpo no espaço, seguida de adequada resposta mo-tora para sua manutenção (Balasubramaniam e Wing1, 2002, Skoyles2, 2006). Essa resposta é ob-tida através de ajustes posturais cuja manuten-ção depende da interação de três sistemas sen-soriais: 1) Proprioceptivo - detecta o movimento das articulações, o estado de contração e tensão muscular e a força de contato entre os pés e o chão; 2) Visual - capta a posição dos olhos em relação ao meio externo e 3) Vestibular- detec-ta a orientação e movimento da cabeça e do corpo em relação à gravidade (Diaz e Puelles3, 2002, Ali et al.4, 2003). Esse atributo é fundamen-tal na realização das tarefas do dia a dia, inclu-sive durante a locomoção, cabendo ao sistema vestibular a dupla função sensorial e motora de identificar e corrigir as alterações posturais du-rante a realização dessas tarefas.
1. ANATOMIA DO SISTEMA VESTIBULAR
O sistema vestibular atuando como um sis-tema sensorial é capaz de detectar, através dos canais semicirculares e órgãos otolíticos, pequenas acelerações angulares e lineares da cabeça resultante do deslocamento corporal. Como um sistema motor, o sistema vestibular modula as respostas reflexas posturais através das projeções dos núcleos vestibulares para a medula espinal (Horak et al.5, 1994, Gdowski e McCrea6, 1999). O sistema vestibular sensorial periférico é formado pelo labirinto ósseo, labi-rinto membranáceo e pelo nervo vestibular (VIII par craniano). O labirinto ósseo está localizado no osso temporal e protege internamente o labi-rinto membranáceo, constituído pelos recepto-res sensoriais: órgãos otolíticos (sáculo e utrículo) e os canais semicirculares. Os receptores sen-soriais possuem no seu interior células ciliadas, que convertem o deslocamento provocado pelo movimento da cabeça em descarga neural, libe-rando neurotransmissores que por sua vez se li-
gam a receptores químicos específicos localiza-dos no terminal periférico do nervo vestibular. O nervo vestibular, constituído por fibras aferentes dos neurônios bipolares localizados no gânglio vestibular (antigo gânglio de Scarpa), transmite si-nais dos receptores vestibulares para os núcleos vestibulares, cerebelo e alguns núcleos motores oculares (Büttner-Ennever7, 2000, Bonaldi8, 2004).
A porção motora do sistema vestibular com-preende o complexo nuclear vestibular (CNV) constituído por quatro núcleos principais: Nú-cleo Vestibular Superior (NVS), Núcleo Vesti-bular Medial (NVM), Núcleo Vestibular Lateral (NVL) e o Núcleo Vestibular Descendente (NVD) (Büttner-Ennever7, 2000, Bonaldi8, 2004, Hain e Hillman9, 1994).
Envolvido em uma extensa rede de conexões nervosas, o CNV recebe informações aferentes proprioceptivas, visuais, corticais e cerebelares (Büttner-Ennever7, 2000, Bonaldi8, 2004, Fanard-jian et al.10, 2001). A partir dessas informações os neurônios dos núcleos vestibulares elaboram uma representação interna tridimensional da posição e movimento da cabeça, empregada no contro-le da estabilização da mirada e do ajuste postural através dos reflexos vestíbulo-ocular e vestíbulo--espinal, respectivamente (Peusner11, 2001).
2. ASPECTOS DO DESENVOLVIMENTO DO SISTEMA VESTIBULAR
Bipedalismo é um termo que descreve o uso de duas pernas para o ortostatismo e locomo-ção, aplicado em várias espécies de animais, incluindo primatas e aves (Alexander12, 2004). Durante a evolução filogenética, a mudança da posição quadrúpede para bípede durante a locomoção impôs modificações anatômicas e neuroquímicas de todo o sistema nervoso, en-volvendo principalmente o sistema vestibular (Ciarac et al.13, 1998) que, em conjunto com a formação reticular, constituem o mais antigo sis-tema do controle motor (Fanardjian et al.10, 2001).
Estudos do desenvolvimento da atividade motora em várias espécies têm mostrado a pre-sença de um padrão cíclico no comportamento

141
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
de todos os músculos esqueléticos. Esse padrão mostra diferenças entre o período embrionário e o período fetal. No início, ele alterna salvas de atividade seguidas de longos períodos de inati-vidade no começo da ontogênese; contrastan-do-se depois com a atividade motora contínua que prevalece no período final do desenvolvi-mento fetal. Diferentes grupos musculares são ativados de forma independente pela chega-da de informações supraespinais em diferentes tempos em diferentes espécies (Deliagina et al.14, 2006). Embora as projeções encefálicas aportem os motoneurônios medulares na fase inicial da embriogênese, a atividade funcional dos moto-neurônios ocorre em um período mais tardio da fase de feto (pré-nascimento) quando as sinap-ses tornam-se finalmente funcionais pela síntese e liberação de neurotransmissores, que ocorre em fase mais tardia (Stockx et al.15, 2007).
3. ASPECTOS NEUROQUÍMICOS DO SISTEMA VESTIBULAR
Todo o complexo de informações neurais, que inicia com a captação dos estímulos senso-riais e termina com a resposta motora adequada para os ajustes posturais, depende de uma ex-tensa rede de comunicação sináptica modulada pela liberação de neurotransmissores e capta-ção pelos seus receptores. O glutamato é con-siderado um dos principais neurotransmissores excitatórios do sistema nervoso, estando alta-mente correlacionado com neurônios eferentes motores. Em relação aos núcleos vestibulares, o glutamato parece ser o mais constante agente envolvido, sendo liberado pelo nervo vestibular e também pelas aferências comissurais vestibu-lares excitatórias (Popper et al.16, 1997).
Assim, investigar seus receptores é de vital importância para desvendar as propriedades da transmissão nervosa nesse sistema. Os recepto-res glutamatérgicos são classificados em iono-trópicos, quando formam canais iônicos, e meta-botrópicos, quando atuam através de segundos mensageiros. De acordo com suas propriedades farmacológicas, os receptores do tipo ionotrópi-cos são divididos em NMDA e não NMDA, sen-
do este último ainda subdividido em receptores tipo AMPA e do tipo cainato (Chan et al.17, 2003).
Os receptores do tipo AMPA são compostos pela combinação entre quatro diferentes subuni-dades designadas de GluR1, GluR2, GluR3 e GluR4 (Popper et al.16, 1997, Chan et al.17, 2003) e essa combinação determina a condutância iônica e as propriedades cinéticas dos receptores AMPA. Este artigo mostra a distribuição temporal (pós--nascimento) das subunidades dos receptores glutamatérgicos do tipo AMPA em neurônios do complexo nuclear vestibular (CNV) de aves, correlacionando seu aparecimento com o papel funcional do próprio CNV.
MATERIAL E METODOSPombos com idade variando do dia do nas-
cimento (eclosão) e adulto foram anestesiados, tiveram o sangue substituído por solução salina, seguido de uma substância fixativa para preser-vação do tecido neural. O encéfalo foi retirado do crânio, congelado e seccionado com uma es-pessura de 40µm. Os cortes foram processados imunohistoquimicamente para identificar a subu-nidade GluR1, do receptor de glutamato do tipo AMPA, utilizando-se um anticorpo específico contra uma sequência de aminoácidos de uma proteína presente nessa subunidade (Passetto et al.18, 2008). Essa marcação foi posteriormente revelada, tornando possível identificar as célu-las que continham essa subunidade. Os cortes foram colocados em lâminas gelatinizadas, desi-dratados e cobertos com lamínula para observa-ção em microscópio.
Em alguns pombos adultos, foi injetado o neurotraçador BDA 10K no canal semicircular horizontal, que serviu para marcar o trajeto dos axônios deste canal até os núcleos vestibulares. Após uma sobrevida de dez dias os animais fo-ram submetidos ao mesmo processo de preser-vação neural, secção do encéfalo, processamen-to para identificar a marcação e montagem das lâminas. Em alguns cortes, usou-se um anticorpo específico contra a subunidade GluR4, do re-ceptor de glutamato do tipo AMPA, feito para

142
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
melhor identificar os núcleos do complexo ves-tibular. Em outra parte dos cortes, foi utilizada como guia histológico a técnica de Giemsa, que a princípio é capaz de corar células neuronais em geral.
Todos os experimentos foram realizados de acordo com os Princípios Éticos na experimentação animal, editado pelo Colégio Brasileiro em Experimentação Animal (COBEA), mediante protocolo aprovado pela Comissão de Ética em Experimentação Animal (CEEA) da UNICAMP (protocolo número 759-2, aprovado em 17/02/05).
RESULTADOS 1. Localização e características dos núcleos
vestibulares
O complexo nuclear vestibular está situado no assoalho do quarto ventrículo. O núcleo ves-tibular superior faz limite com a parede do quar-to ventrículo; o lateral limita-se com a junção bulbopontina; o medial com o quarto ventrículo e o núcleo descendente limita-se com a parte inferior do bulbo (Fig. 1).
• Núcleo vestibular superior (NVS): Forma-do por células de tamanho pequeno e médio, limita-se rostralmente com o núcleo trigêmeo, medialmente com o núcleo vestibular medial, lateralmente com o corpo restiforme e caudal-mente com o núcleo vestibular lateral.
• Núcleo vestibular lateral (NVL): Formado por células de tamanho variado, é facilmente identificado pela presença de células gigantes. Faz limite rostral com o núcleo superior, medial com o núcleo vestibular medial, lateral com o corpo restiforme e caudal com o núcleo vestibular descendente.
• Núcleo vestibular medial (NVM): Constituído por células de tamanho pequeno e médio, limitando-se lateralmente, em toda sua extensão rostro-caudal, com os núcleos vestibulares superior, lateral e descendente e medialmente com o assoalho do quarto ventrículo.
• Núcleo vestibular descendente (NVD): Constituído principalmente de células de tamanho médio, limita-se rostralmente com o núcleo vestibular lateral, medialmente com o núcleo vestibular medial, lateralmente é possível visualizar entre as fibras do nervo vestibular e caudalmente limita-se com o núcleo do trato solitário.
2. Distribuição das subunidades GluR1 do receptor de glutamato do tipo AMPA nos núcleos vestibulares durante o desenvolvimento.
No primeiro dia após a eclosão, muitos neurônios no núcleo vestibular lateral mostraram conter a subunidade GluR1. A análise do padrão da imunorreatividade contra essa subunidade mostrou que durante o desenvolvimento ocorre
Figura 1 - corte coronal do encéfalo de pombo, co-rado pela técnica de Giemsa, mostrando a posição do complexo nuclear vestibular den-tro do tronco encefálico. esse complexo é composto por quatro núcleos ocupando po-sições distintas. o núcleo vestibular superior (Nvs) ocupa a posição mais rostral, enquanto os núcleos vestibulares lateral (Nvl) e medial (Nvm) ocupam uma posição mais ventral e o núcleo vestibular descendente (Nvd) a posição caudal no tronco encefálico.
143
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
complexo nuclear vestibular mostrou que todos os núcleos vestibulares recebem aferências do canal semicircular horizontal, com distribuição desigual entre os núcleos (Fig. 4), com presença de botões terminais, supostamente indicando um terminal sináptico (Fig. 5).
DISCUSSÃOO controle postural tem como objetivos
funcionais: 1) orientação postural - envolvendo
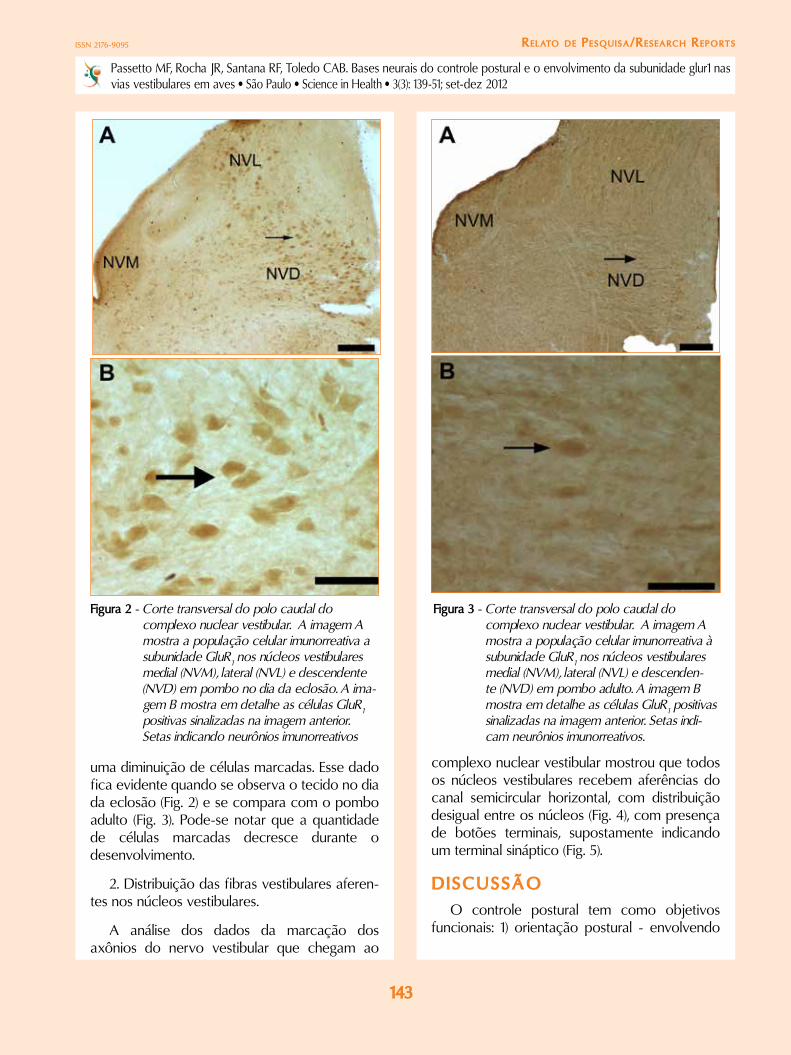
Figura 2 - corte transversal do polo caudal do complexo nuclear vestibular. a imagem a mostra a população celular imunorreativa a subunidade Glur1 nos núcleos vestibulares medial (Nvm), lateral (Nvl) e descendente (Nvd) em pombo no dia da eclosão. a ima-gem b mostra em detalhe as células Glur1 positivas sinalizadas na imagem anterior. setas indicando neurônios imunorreativos
uma diminuição de células marcadas. Esse dado fica evidente quando se observa o tecido no dia da eclosão (Fig. 2) e se compara com o pombo adulto (Fig. 3). Pode-se notar que a quantidade de células marcadas decresce durante o desenvolvimento.
2. Distribuição das fibras vestibulares aferen-tes nos núcleos vestibulares.
A análise dos dados da marcação dos axônios do nervo vestibular que chegam ao
Figura 3 - corte transversal do polo caudal do complexo nuclear vestibular. a imagem a mostra a população celular imunorreativa à subunidade Glur1 nos núcleos vestibulares medial (Nvm), lateral (Nvl) e descenden-te (Nvd) em pombo adulto. a imagem b mostra em detalhe as células Glur1 positivas sinalizadas na imagem anterior. setas indi-cam neurônios imunorreativos.

144
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
Figura 4 - corte transversal do complexo nuclear ves-tibular (cNv) mostrando as projeções afe-rentes do canal semicircular horizontal. Nas imagens a e b as setas indicam a presença das fibras aferentes chegando ao NVS (A) e aos Nvl e Nvm (b). legenda: Nvm: núcleo vestibular medial; Nvs: núcleo vestibular superior; Nvl: núcleo vestibular superior.
o alinhamento do controle ativo do corpo em relação à gravidade, superfície de suporte, am-biente visual e referências internas, baseado na interpretação da convergência das informações sensoriais dos sistemas somatossensorial, vesti-bular e visual (Ali et al.4, 2003, Gdowski e Mc-Crea6, 1999) e 2) equilíbrio postural - envolvendo a coordenação das estratégias para estabilizar o centro de massa corporal (CMC) durante as os-cilações posturais em relação à base de suporte (Deliagina et al.14, 2006). As estratégias motoras são movimentos usados para retornar o corpo a uma posição de equilíbrio estável; destacando-
Figura 5 - imagem ampliada de um neurônio imunorre-ativo a Glur4 do complexo nuclear vestibu-lar de um animal adulto. as setas indicam o contato de botões terminais das vias aferen-tes do canal semicircular horizontal com o neurônio, sugerindo uma possível sinapse.
-se as estratégias do tornozelo (o movimento é realizado na articulação do tornozelo, emprega-do na manutenção da postura durante pequenos deslocamentos com o indivíduo parado sobre uma superfície estável) e a estratégia do quadril (empregada para deslocamento rápido utilizado quando ocorre rápido deslocamento do CMC) (Gdowski e McCrea6, 1999).
As similaridades do sistema vestibular entre diferentes classes de vertebrados em relação ao padrão temporal do desenvolvimento do siste-ma vestibular (Fanardjian et al.10, 2001, Ciarac et al.13, 1998), citoarquitetura (Passetto et al.18, 2008, Wold19, 1976) e neuroquímica (Büttner-Ennever7, 2000, Passetto et al.18, 2008) permitem a utili-zação de aves como modelo experimental no estudo do envolvimento do sistema vestibular no controle postural. Na verdade, sendo animais bípedes, predominantemente visuais e de hábi-tos diurnos, as aves são um modelo bem mais fidedigno do que roedores quando os estudos compreendem aspectos de controle motor e postural.
Estudos têm mostrado que a o sistema ner-

145
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
voso altera a origem da informação sensorial para o controle do equilíbrio postural de acordo com as variações biomecânicas e do ambiente (Deliagina et al.14, 2006, Cenciarini e Peterka20, 2006). Em deslocamentos de pequena amplitude da superfície de suporte ou de movimentos do campo visual a contribuição dos sistemas pro-prioceptivo, visual e vestibular é de 50, 35 e 15 %, respectivamente (Peterka21, 2002).
As informações provenientes do sistema ves-tibular tornam-se mais relevantes para o con-trole postural em situações críticas para a ma-nutenção do equilíbrio (Day e Cole22, 2002) ou quando há comprometimento de outras fontes sensoriais, como a perda da propriocepção (Ho-rak e Hlavacka23, 2001, Blouin et al.24, 2007, Ho-rak25, 2010).
Durante o desenvolvimento ontogenético, o sistema vestibular é o primeiro sistema envolvido no controle dos ajustes posturais durante a aqui-sição motora funcional, supostamente devida à necessidade de um controle cervical que segue o padrão de desenvolvimento céfalo-caudal do sistema nervoso (Ciarac et al.13, 1998). Sua atu-ação se faz principalmente no ajuste do tônus extensor, favorecendo o desenvolvimento dos sistemas visuais e proprioceptivos no controle tônico postural (Horak e Hlavacka23, 2001), cujas respostas dependem da natureza neuroquímica de seus circuitos.
1. ASPECTOS IMUNO-HISTOQUÍMICOS DOS NÚCLEOS VESTIBULARES
Investigações eletrofisiológicas e imuno-his-toquímicas têm mostrado grande diversidade de neurotransmissores presentes no sistema vesti-bular envolvidos no controle do ajuste postural nas várias classes de vertebrados (Popper et al.16, 1997, Chan et al.17, 2003), com nítido destaque para o neurotransmissor glutamato (Popper et al.16, 1997). A transmissão de informações entre as células ciliadas vestibulares e seus neurônios aferentes se faz através de receptores glutama-térgicos do tipo AMPA e NMDA (Chan et al.17, 2003, Sans et al.26, 2000, Chen et al.27, 2009), como também na transmissão de disparos das
vias vestibulares aferentes para neurônios do complexo nuclear vestibular (Smith28, 2000). As-sim, aparentemente, a descarga dos neurônios vestibulares é dada pelo sistema glutamatérgi-co, além de outros agentes neuroquímicos no exercício de sua função nos ajustes posturais. Embora bastante sugestivos, há necessidade de experimentos eletrofisiológicos para comprovar ou rejeitar esse dado.
O padrão de expressão das subunidades dos receptores de glutamato varia durante a onto-gênese do encéfalo. De modo geral, os autores relatam que os receptores AMPA mostram um aumento na sua expressão com a idade. Essa informação sugere que os receptores do tipo AMPA têm importante papel na formação das sinapses no sistema nervoso central. É forte a noção de que essas subunidades estão envolvi-das na conversão de sinapses silentes em sinap-ses funcionais durante o desenvolvimento (Sans et al.26, 2000).
Através de estudo com neurotraçadores e métodos imuno-histoquímicos tem sido identifi-cada intensa aferentação glutamatérgica a par-tir dos sensores vestibulares (Büttner-Ennever7, 2000) para os núcleos vestibulares, com neurô-nios imunorreativos às quatro subunidades dos receptores glutamatérgicos do tipo AMPA (Po-pper et al.16, 1997, Passetto et al.18, 2008, Hirate et al.29, 2000), envolvidos principalmente na trans-missão excitatória pós-sináptica nos neurônios do CNV (Hirate et al. 29, 2000).
Os dados deste trabalho mostram que durante o desenvolvimento pós-eclosão há significativa diferença de células imunorreativas a GluR1. Imediatamente após a eclosão (Fig. 2) há uma maior distribuição de células marcadas nos núcleos vestibulares, enquanto que no adulto observam-se poucas células dispersas pelos núcleos (Fig. 3).
2. ASPECTOS FUNCIONAIS DO SISTEMA VESTIBULAR
Embora nas atividades do dia a dia todos os receptores vestibulares sejam estimulados,

146
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
situações específicas como andar de elevador ou frear um carro, por exemplo, exigem maior interferência de um determinado tipo de receptor sensorial para se obter uma resposta motora mais adequada ao ajuste postural nessa situação. Essa resposta depende de informações que chegam aos neurônios dos núcleos vestibulares e de suas projeções para outras estruturas do sistema nervoso central.
Cada um dos canais semicirculares apresenta estimulação preferencial, em seu próprio eixo, durante a rotação da cabeça, embora acelera-ções angulares ao longo de qualquer plano no espaço possam estimular ao menos dois canais pelo alinhamento não perfeito entre os canais. Os órgãos otolíticos são estimulados pela gra-vidade em aceleração linear da cabeça, embora possam estar sujeitos a forças tangenciais e cen-trífugas durante a rotação da cabeça (Bonaldi8, 2004, Naito et al.30, 1995).
Alguns neurônios vestibulares estão envolvi-dos nos ajustes posturais quando os movimentos ocorrem na horizontal (receptores utriculares) ou na vertical (receptores saculares); outros, com o movimento da cabeça e/ou do corpo (recepto-res proprioceptivos) no controle postural (Boyle et al.31, 1992, Cullen e Minor32, 2002) e ainda ou-tros neurônios do complexo vestibular parecem estar envolvidos com os movimentos dos olhos e do controle da postura ao mesmo tempo (Cul-len e Minor32, 2002). Esses achados sugerem a existência de um padrão de diferencial das afe-rências dos receptores otolíticos e dos canais nos neurônios dos núcleos vestibulares, que também estão organizados de forma funcional em relação às eferências vestibulares. Dessa forma, informações dos movimentos e posição da cabeça em qualquer direção no espaço po-dem ser precisamente interpretadas e combina-das durante os ajustes posturais (McCrea et al.33, 1987, Haque et al.34, 2008). As projeções vesti-bulares primárias terminam em todos os núcleos vestibulares, entretanto, com diferentes padrões de distribuição entre as fibras dos canais semi-circulares e otolíticos. Essa variação de aporte (a distribuição diferencial dentro de cada núcleo
vestibular), supostamente ocorre devido a dife-renças funcionais tanto dos receptores sensoriais quanto das projeções eferentes dos núcleos ves-tibulares. Regiões dos núcleos vestibulares que recebem aferentação dos canais semicirculares e dos órgãos otolíticos parecem estar organiza-das de forma que as áreas que recebem pro-jeções aferentes utriculares também recebem primariamente terminações do canal semicircu-lar horizontal, enquanto que as projeções sacu-lares aferentes terminam nas mesmas áreas de terminação das eferências do canal semicircu-lar vertical (Büttner-Ennever7, 2000, McCrea et al.33, 1987). Nos núcleos vestibulares, essas infor-mações são processadas e a eferência motora postural ocorre principalmente por três reflexos. Três dos principais reflexos vestibulares estão re-lacionados com o controle postural.
2.a Reflexo vestíbulo-ocular (RVO): Respon-sável pelo controle do movimento dos olhos conjugados com os movimentos da cabeça per-mitindo a sustentação do olhar fixo no espaço, mantendo a visualização clara do alvo durante o movimento do corpo (Gdowski e McCrea6, 1999, Bonaldi8, 2004, Peusner11, 2001, Day e Cole22, 2002).
Estudos eletrofisiológicos têm mostrado que todos os receptores vestibulares, bem como todos os núcleos vestibulares participam da modulação desse reflexo, mas diferenciando no grau de participação.
O núcleo superior é o principal núcleo do complexo vestibular na participação do reflexo vestíbulo-ocular, devido à intensa aferentação dos canais semicirculares que chegam a todo o núcleo, e ao maciço envio de eferências para os núcleos motores oculares (núcleo troclear e abducente) (McCrea et al.33, 1987). Os núcleos vestibulares lateral, medial e descendente têm uma menor participação do RVO, apresentado similaridades na localização de chegada das vias vestibulares sensoriais. A região medial destes núcleos recebe projeções utriculares e do canal horizontal, enquanto que a região mais lateral re-cebe aferências saculares e do canal posterior.

147
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
Estas regiões enviam projeções axonais que al-cançam os núcleos motores oculares (Dickman e Fang35, 1996, Kleine et al.36, 2004).
Os dados desta pesquisa com marcação das aferências que partem do canal semicircular horizontal são coincidentes com os dados acima citados, mostrando a chegada dessas fibras na região mais medial dos núcleos vestibulares, como mostra a Figura 6, em relação ao núcleo vestibular descendente.
Esse complexo circuito neuronal permite, por exemplo, que, ao andar a cavalo, seja possível fixar o olhar em um objeto a sua frente. Durante o movimento do cavalo, seu corpo movimenta--se no sentido céfalo-caudal, mantendo a esta-bilização da cabeça primariamente à custa das informações que partem do sáculo, enquanto que os olhos se movem no sentido contrário à custa das informações primariamente dos canais semicirculares que enviam informações para os núcleos motores oculares, estabilizando a ima-gem na retina.
2.b Reflexo vestíbulo-espinal (RVE): Respon-sável pelos movimentos compensatórios do cor-po em relação aos movimentos da cabeça na
manutenção da estabilidade postural (Gdowski e McCrea6, 1999, Bonaldi8, 2004, Peusner11, 2001). A modulação do RVE depende da informação dos cinco receptores vestibulares que terminam em todos os núcleos do complexo vestibular e de suas vias eferentes que terminam de forma direta ou indireta nos núcleos motores da me-dula espinal.
Na região lateral do núcleo vestibular supe-rior chegam terminações sensoriais predominan-temente otolíticas. Os neurônios desSe núcleo enviam projeções eferentes para a formação reticular, cerebelo e tálamo, envolvidos de forma indireta com os ajustes corporais (Büttner-Enne-ver7, 2000, Haines37, 1975).
No núcleo vestibular lateral somente a por-ção ventral recebe aferências primárias, que par-tem do sáculo e utrículo terminam na porção mais lateral da região ventral, sendo suas proje-ções mais densas do que as dos canais semicir-culares. Os núcleos vestibular medial e vestibular descen dente também recebem aferências otolí-ticas, embora em menor quantidade (Dickman e Fang35, 1996). Os axônios motores desses três núcleos terminam nos neurônios motores da medula espinal, participando dos ajustes para o controle postural. Essas vias eferentes alcançam a medula espinal através do trato vestíbulo-espi-nal que, segundo seu trajeto descendente, é divi-dido em duas porções: lateral e medial (Büttner--Ennever7, 2000, Fanardjian et al.10, 2001).
• Trato vestíbulo-espinal lateral - Formado por fibras descendentes do núcleo ves-tibular lateral e descendente, projeta-se ipsolateralmente para a medula espinal (Dickman e Fang35, 1996, Wold38, 1978, Tellegen et al.39, 2001) com ação facilita-tória tônica sobre os motoneurônios alfa e gama para os músculos do pescoço, do tronco e músculos extensores dos mem-bros inferiores e flexores dos membros superiores e ação inibitória sobre os mús-culos flexores dos membros inferiores, possibilitando a manutenção do corpo na postura ereta (Ghez40, 1991). Esse trato
Figura 6 - corte transversal do complexo nuclear vestibular mostrando o maior contingente de terminações do canal semicircular hori-zontal na região medial do núcleo vestibular descendente (Nvd) indicado pela seta.

148
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
apresenta uma organização somatotópi-ca, com os segmentos medulares cervi-cais supridos pela porção rostroventral, os segmentos lombares e torácicos da porção dorsocaudal (Pompeiano e Bro-dal41, 1957, Rose et al.42, 1992).
• Trato vestíbulo-espinal medial - Com-posto principalmente de fibras do núcleo vestibular medial e, em menor contingen-te, de fibras do núcleo vestibular lateral e do núcleo vestibular descendente, termi-na bilateralmente na medula espinal, tam-bém regulando a atividade dos motoneu-rônios para os músculos do pescoço e axiais (Büttner-Ennever7, 2000, Fanardjian et al.10, 2001).
2.c Reflexo vestíbulo-cólico (RVC): Respon-sável pela estabilização da cabeça no espaço durante os movimentos rotacionais iniciados pelo tronco (Gdowski e McCrea6, 1999, Peus-ner11, 2001) e tem ação compensatória entre os músculos do pescoço e dos olhos. Esse reflexo envolve a existência de projeções diretas e/ou indiretas dos núcleos vestibulares sobre os mo-toneurônios inervando os músculos do pescoço (Tellegen et al.39, 2001).
Assim, através de complexa circuitaria, os
neurônios dos núcleos vestibulares decodificam com precisão informações da posição do corpo e direção do movimento, elaborando respostas motoras para controlar de forma precisa os movimentos dos olhos e a resposta postural na manutenção da postura e do equilíbrio.
CONCLUSÕESNeste trabalho, foram ressaltadas as proprie-
dades das vias vestibulares e os princípios neu-rais que fundamentam a atividade do circuito envolvido com ajuste postural em vertebrados. Além disso, foi apresentada a importância do glutamato como neurotransmissor-chave des-se sistema e foi introduzida a subunidade GluR1 como tendo um papel crucial na formação e consolidação do circuito vestibular.
AGRADECIMENTOS Gostaríamos de agradecer a Márcia Maria
Tsuruta pelo apoio na condução dos experimen-tos e ao Prof. Dr. Cláudio A. B. Toledo (in me-moriam), idealizador e coordenador do NUPEN, assim como orientador e mentor deste trabalho. O Prof Cláudio foi exemplo de dedicação e res-peito, discutindo e semeando o pensar científi-co. Apoio Financeiro: FAPESP (processos núme-ro: 04/11039-6 e 08/51110-2

149
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
reFerÊNcIas
1- Balasubramaniam R, Wing AM. The dyna-mics of standing balance. Trends Cogn Sci 2002; 6(12):531-536.
2- Skoyles JR. Human balance, the evolution of bipedalism and dysequilibrium syndro-me. Medical Hypotheses 2006; 66:1060-1068.
3- Diaz C, Puelles L. Organización segmen-tária de las eferencias del complejo ves-tibular en el embrión de pollo: ejemplo del caso general? Rev Neurol 2002; 35(10): 922-930.
4- Ali AS, Rowen KA, Iles J.F. Vestibular ac-tions on back and lower limb muscles du-ring postural tasks in man. J Physiol 2003; 15(546):615-624.
5- Horak FB, Shupert CL, Dietz G, Horstmann G. Vestibular and somatosensory contribu-tions to responses to head and body dis-placements in stance. Exp Brain Res 1994; 10011(1):93-106.
6- Gdowski GT, McCrea, RA. Integration of Vestibular and Head Movement Signals in the Vestibular Nuclei During Whole-Body Rotation. J Neurophysiol 1999; 82(1):436-449.
7- Büttner-Ennever JA. Overview of the vesti-bular system: Anatomy. In: Beitz AJ, Ander-son JH. Neurochemistry of the vestibular system. Boca Raton: CRC Press; 2000. p. 3-45.
8- Bonaldi LV. Bases anatômicas da audição e do equilíbrio. São Paulo: Santos; 2004. p. 65-74.
9- Hain TC, Hillman MA. Anatomy and Phy-siology of Normal Vestibular. In: Herdman SJ. Vestibular Rehabilitation. Philadelphia: F.A Davis Company; 1994. p. 3-24.
10- Fanardjian VV, Manvelyan LR, Nasoyan AM. Spatial distribution of the vestibulos-pinal neurons in the frog vestibular nuclei. Neuroscience 2001; 104(3):853-862.
11- Peusner KD. Development of the gravi-ty sensing system. J Neurosci Res 2001; 63(2):103-108.
12- Alexander RMcN. Bipedal animals, and their differences from humans. J. Anat 2004:321–330.
13- Clarac F, Vinay L, Cazalets JR, Fady JC, Ja-mon M. Role of gravity in the development of posture and locomotion in the neonatal rat. Brain Res Rev 1998; 28(1-2):35-43.
14- Deliagina TG, Orlovsky GN, Zelenin PV, Beloozerova In. Neural Bases of Postural Control. Physiology 2006; 21: 216–225.
15- Stockx EM, Anderson CR, Murphy SM, Cooke IRC, Berger PJ. The development of descending projections from the brains-tem to the spinal cord in the fetal sheep. BMC Neuroscience 2000; 78(40):1-12.
16- Popper P, Rodrigo JP, Alvarez JC, Lopez I, Honrubia V. Expression of the AMPA-se-lective receptor subunits in the vestibular nuclei of the chinchilla. Brain Res Mol Brain Res 1997; 44(1):21-30.
17- Chan YS, Chen LW, Lai CH, Shum DKY, Yung KKL, Zhang FX. Receptors of glutamate and neurotrophin in vestibular neuronal functions. J Biomed Sci 2003; 10(6):577-587.
18- Passetto MF, Britto LRG, Toledo CAB. Mor-phometric analysis of the AMPA type neu-rons in the Deiters vestibular complex of the chick brain. J Chem Neuroanat 2008; 35:334–345.
19- Wold JE. The Vestibular Nuclei in the Do-mestic Hen. I-Normal Anatomy. Anat Em-bryol 1976; 149(1):29-46.
20- Cenciarini M, Peterka RJ. Stimulus-depen-dent changes in the vestibular contribution to human postural control. J Neurophysiol 2006; 95:2733-2750.
21- Peterka RJ. Sensorimotor Integration in Human Postural Control. J Neurophysiol 2002; 88:1097-1118.

150
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
22- Day BL, Cole J. Vestibular –evoked pos-tural responses in the absence of so-matosensory information. Brain 2002; 125:2081-2088.
23- Horak FB, Hlavacka F. Somatosensory loss increases vestibulospinal sensitivity. J Neurophysiol 2001; 86(2):575-585.
24- Blouin J, Teasdale N, Mouchnino L. Vesti-bular signal processing in a subject with somatosensory deafferentation: The case of sitting posture. BMC Neurology 2007; 7:25.
25- Horak FB. Postural compensation for vestibular loss and implications for rehabilitation. Restor Neurol Neurosci. 2010;28(1):57-68.
26- Sans NA, Montcouquiol ME, Raymond J. Postnatal developmental changes in AMPA and NMDA receptors in the rat vestibular nuclei. Brain Res Dev Brain Res 2000; 123(1):41-52.
27- Chen W, Prithviraj R, Mahnke AH, McGloin KE. Ampa glutamate receptor subunits 1 and 2 regulate dendrite com-plexity and spine motility in neurons of the developing neocortex. Neuroscience 2009; 159:172–182.
28- Smith PF. Pharmacology of the vestibular system. Curr Opin Neurol 2000; 13:31-7.
29- Hirate H, Kitayama A, Furuyab N. Roles f glutamate receptor subtypes in the deve-lopment of vestibular compensation after unilateral labyrinthectomy in the guinea pig. Neurosci Lett 2000; 296:158-162.
30- Naito Y, Newman A, Lee WS, Beykirch K, Honrubia V. Projections of the individual vestibular end-organs in the brain stem of the squirrel monkey. Hear Res 1995; 87:141-156.
31- Boyle R, Goldberg JM, Highstein SM. Inputs from regularly and irregularly discharging vestibular nerve afferents to seconda-ry neurons in squirrel monkey vestibular nuclei. III. Correlation with vestibulospinal and vestibuloocular output pathways. J Neurophysiol 1992; 68(2):471-84.
32- Cullen KE, Minor LB. Semicircular canal afferents similarly encode active and pas-sive head-on-body rotations:Implications for the role of vestibular efference. J Neurosci 2002; 22:1-7.
33- McCrea RA, Strassman A, Highstein SM. Anatomical and physiological characteris-tics of vestibular neurons mediating the vertical vestibulo-ocular reflexes of the squirrel monkey. J Comp Neurol 1987; 264(4):571-94.
34- Haque A, Zakir M, Dickman JD. Recovery of gaze stability during vestibular regene-ration. J Neurophysiol 2008; 99: 853–865.
35- Dickman JD, Fang Q. Differential central projections of vestibular afferents in pige-on. J Comp Neurol 1996; 367:110-131.
36- Kleine JF, Guan Y, Kipiani E, Glonti L, Hoshi M, Büttner U. Trunk position influences vestibular responses of fastigial nucleus neurons in the alert monkey. J Neurophy-siol 2004; 91:1090-2100.
37- Haines DE. Cerebellar corticovestibular fibers of the posterior lobe in a prosi-mian primate, the lesser bushbaby (Ga-lago senegalensis). J Comp Neurol 1975; 160(3):363-97.
38- Wold JE. The Vestibular Nuclei in the Do-mestic Hen. IV. The projection to the spi-nal cord. Brain Behav Evol 1978; 15(1):41-62.
39- Tellegen AJ, Arends JJ, Dubbeldam JL. The vestibular nuclei and vestibuloreti-cular connections in the mallard (Anas platyrhynchos L.). An anterograde and re-trograde tracing study. Brain Behav Evol 2001; 58(4):205-217.

151
Passetto MF, Rocha JR, Santana RF, Toledo CAB. Bases neurais do controle postural e o envolvimento da subunidade glur1 nas vias vestibulares em aves • São Paulo • Science in Health • 3(3): 139-51; set-dez 2012
ISSN 2176-9095 Relato de Pesquisa/ReseaRch RePoRts
40- Ghez C. The control of movement. In: Kandel ER, Schwartz JH, Jessel TM. Prin-ciples of neural science. 3rd ed. London: Appleton & Lange; 1991. p. 533-547.
41- Pompeiano O, Brodal A. Spinovestibular fibers in the cat; an experimental study. J Comp Neurol 1957; 108(3):353-81.
42- Rose PK, Wainwright K, Neuber-Hess M. Connections from the lateral vestibular nucleus to the upper cervical spinal cord of the cat: a study with the anterogra-de tracer PHA-L. J Comp Neurol 1992; 321(2):312-24.