autoecologia alimentar e reprodutiva de cnemidophorus ... · pelo auxílio de campo e ... incluído...
TRANSCRIPT

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
DEPARTAMENTO DE SISTEMÁTICA E ECOLOGIA
Autoecologia alimentar e reprodutiva de Cnemidophorus
ocellifer (Squamata, Teiidae) em uma região savânica do
Nordeste brasileiro
LUCAS BARBOSA DE QUEIROGA CAVALCANTI
Monografia apresentada à Coordenação do curso
de Ciências Biológicas da Universidade Federal
da Paraíba Campus I, para a obtenção do título de
Bacharel em Ciências Biológicas.
João Pessoa - PB, Novembro de 2011

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
DEPARTAMENTO DE SISTEMÁTICA E ECOLOGIA
Autoecologia alimentar e reprodutiva de Cnemidophorus
ocellifer (Squamata, Teiidae) em uma região savânica do
Nordeste brasileiro
LUCAS BARBOSA DE QUEIROGA CAVALCANTI
ORIENTADOR: DR. DANIEL OLIVEIRA MESQUITA
Monografia apresentada à Coordenação do curso
de Ciências Biológicas da Universidade Federal
da Paraíba Campus I, para a obtenção do título de
Bacharel em Ciências Biológicas.
João Pessoa - PB, Novembro de 2011

Agradecimentos
Ao meu orientador Prof. Daniel Oliveira Mesquita, pela excelente orientação,
oportunidades, apoio e amizade;
Ao CNPq e à FAPESQ pelo suporte financeiro deste trabalho e ao IBAMA e
ICMBIO/REBIO GUARIBAS (e todos seus funcionários) pela licença de coleta e liberação
do trabalho junto à logística e alojamento, dos quais foram essenciais para o bom desempenho
deste estudo;
Aos professores Frederico França e Gustavo Vieira, os quais considero co-
orientadores, pela amizade, ajuda e contribuições sempre construtivas para minha formação e
pela presença na banca examinadora;
Ao Prof. Alan Loures Ribeiro, pela ajuda no decorrer do curso e pela presença como
suplente nesta banca examinadora;
À todos os professores que me lecionaram ou me auxiliaram neste curso;
À minha namorada Talita, que deu e ainda dá apoio em todas as etapas e também à sua
família;
Aos companheiros e companheiras da Herpetologia: Daniel Orsi, Taís, Ralph, Isabella,
Edinaldo, “Felipes” (Luiz, Boca e Xulipa), Laura, Diego, Camila, Cynthia, Samuel, Ricardo,
Rachid e principalmente ao Linccon e Carmem pela parceria e coletas, e ao Arielson e Fagner
pelo auxílio de campo e ensinamentos herpetológicos;
Aos familiares que desde pequeno me auxiliaram no sonho de me tornar biólogo;
Aos meus colegas: Elis, Juliana, Talita, Caetano, Altielys, Ariadne, Janaína, Everton,
Maíra, Rodrigo, Anne e Sara, e outros de demais períodos;

Na Caatinga, no Cerrado,
Ocelos para todo lado,
Com uma ampla distribuição,
Forrageiam ativamente vasculhando todo chão...
Que calango é esse? ...
Amazonia, Mata Atlântica, possui alguns parentes...
America do Norte já não é mais, Aspidoscelis tanto faz...
Cnemidophorus gr. ocellifer, Mesquita sabe o que é...
Coletamos mais de dez, para evitar o viés...
E pseudoreplicação...
Que calango é esse? ...
Espécie nova se for...
Publica no exterior...
O gênero vai ficar rico...
Com muito mais descrição...
É só contarmos, todas as escamas...
Dos bichos da coleção...
Que calango é esse?
Cavalcanti, 2011; adaptado de Russo, 1987.

ÍNDICE:
Introdução.............................................................................................................................. 01
Materiais e Métodos.............................................................................................................. 03
Resultados.............................................................................................................................. 08
Discussão................................................................................................................................ 10
Bibliografia............................................................................................................................ 17
Figuras.................................................................................................................................... 24
Tabelas................................................................................................................................... 33

1
Introdução:
A utilização de répteis, em especial lagartos, como modelos para estudos ecológicos
foi e vem sendo extremamente relevante em consideração a hipóteses, teorias ecológicas, em
diversas áreas da ecologia, como nos estudos de populações e comunidades, biologia
comparativa, teorias de história de vida, entre outros (Huey et al., 1983; Pianka e Vitt, 2003;
Vitt e Pianka, 1994). Esta escolha pela utilização de lagartos como modelos pode se dar ao
fato destes organismos serem razoavelmente abundantes, além da relativa facilidade de se
coletar (Vitt et al., 2007).
Como todo e qualquer organismo, lagartos são afetados em seus padrões de
comportamento e história natural por diversos fatores, podendo estes serem de origem
histórica (origem filogenética, uma herança genética que molda as características ecológicas)
e/ou ecológica (origem recente, está diretamente relacionada com o ambiente em que o
organismo está inserido atualmente, por exemplo competição e predação).
Fatores de origem histórica podem influenciar significantemente os hábitos
alimentares de algumas famílias de lagartos (Vitt e Pianka, 2005). Por exemplo, dentro da
família Teiidae, pode-se ressaltar o gênero Cnemidophorus, um grupo de ampla distribuição e
que apresenta pouca variação nos seus itens alimentares (em grande maioria, cupins),
caracterizando assim um forte conservatismo filogenético (Mesquita e Colli, 2003b; Teixeira-
Filho et al., 2003; Vitt, 1991b). Por outro lado, a influência de fatores ecológicos pode ser
evidenciada quando se observa as características reprodutivas de algumas espécies de lagartos
tropicais (Mesquita e Colli, 2010). Em regiões onde a pluviosidade é bem distribuída ao longo
do ano ou é um fenômeno imprevisível, algumas espécies de lagartos tendem a se reproduzir
durante o ano todo, apresentando várias ninhadas de poucos ovos (Mesquita e Colli, 2003b;
Mesquita e Colli, 2010; Vitt e Goldberg, 1983), enquanto em regiões de maior sazonalidade

2
eles tendem a ser reproduzir ciclicamente apresentando basicamente uma ou poucas ninhadas
maiores (Mesquita e Colli, 2003b; Mesquita e Colli, 2010; Vitt, 1992).
O gênero Cnemidophorus representa um táxon de lagartos da família Teiidae, que era
distribuído por quase todo o Novo Mundo dentro da faixa tropical e subtropical. Porém, uma
revisão taxonômica dividiu o gênero em Cnemidophorus e Aspidoscelis (Reeder et al., 2002),
sendo atualmente Cnemidophorus restrito à América do Sul e parte da América Central. Estes
animais geralmente habitam regiões que possuem vegetação aberta com incidência direta do
sol e solo arenoso, como praias, bordas de florestas, formações desérticas e savanas (Pianka e
Vitt, 2003; Wright e Vitt, 1993), podendo até mesmo serem encontrados em regiões bastante
antropizadas. Cnemidophorus ocellifer representa um complexo de espécies distribuído por
quase todo o território brasileiro, ao leste/sudeste da Bacia Amazônica. Diversos estudos
taxonômicos das últimas duas décadas vem separando populações antes reconhecidas como C.
ocellifer em novas espécies, como: C. nativo, um lagarto partenogênico do bioma costeiro
Restinga (Rocha et al., 1997), C. littoralis, endêmico de restingas no sudeste do Brasil (Rocha
et al., 2000), C. mumbuca, da região do Jalapão no Cerrado do Brasil Central (Colli et al.,
2003a), C. parecis, de isolados de Cerrado no sudoeste da Amazônia (Colli et al., 2003b), C.
jalapensis, da região do Jalapão no Cerrado do Brasil Central (Colli et al., 2009), C.
confusionibus e C. venetacaudus, duas espécies recém descritas para o bioma semi-árido da
Caatinga (Arias et al., 2011a), e C. cyanurus e C. nigrigula, também espécies recém descritas
do Complexo de Dunas de Areia do Rio São Francisco, incluído no domínio da Caatinga
(Arias et al., 2011b).
Dentro deste âmbito, é notável a necessidade de esforços nos estudos dos aspectos
ecológicos destas populações, pois o domínio do conhecimento a cerca dos padrões de
história natural pode ser uma peça chave no clareamento das dúvidas sobre este complexo.
Além do mais, estudos recentes apontam Cnemidophorus como um gênero bastante

3
importante em estudos de história de vida (Mesquita e Colli, 2010). Estes apresentam uma
alta inércia filogenética em suas características reprodutivas, um padrão significantemente
peculiar para espécies de lagartos tropicais (Mesquita e Colli, 2010). Vale também salientar
que nenhuma informação sobre aspectos ecológicos de C. ocellifer que habitam savanas de
Mata Atlântica foi publicada até então.
O presente trabalho teve como objetivo geral entender os aspectos ecológicos da dieta
e reprodução de C. ocellifer em uma região de Tabuleiro nordestino, respondendo as
seguintes perguntas: Quais são os principais itens predados por C. ocellifer? Como são as
características reprodutivas em C. ocellifer? Existe correlação entre as dimensões da cabeça
dos lagartos e as dimensões das presas ingeridas? Existe variação sazonal nos principais itens
predados por C. ocellifer? Existe variação na proporção de indivíduos reprodutivos e não
reprodutivos ao longo do ano, ou seja, existe sazonalidade na reprodução? Existe correlação
entre o tamanho da ninhada e o comprimento rostro-cloacal das fêmeas reprodutivas?
Materiais e Métodos:
Área de estudo - O estudo foi desenvolvido na Reserva Biológica Guaribas. Esta
reserva é uma unidade de conservação coordenada pelo Instituto Chico Mendes de
Biodiversidade (ICMBIO). Sua vegetação é típica de Mata Atlântica, com mata ombrófila
semi decídua. Ela é localizada entre e nas proximidades dos municípios de Mamanguape (6�
50�20�S, 35�7�33�W) e Rio Tinto (6�48�10�S, 35�4�51�W), ambos dentro do estado da Paraíba,
Nordeste do Brasil. A reserva é dividida em três áreas separadas fisicamente, chamadas
SEMAS (Fig. 1): SEMA I (616,4 ha), SEMA II (3.378,2 ha) e SEMA III (327 ha). O estudo
em questão foi desenvolvido somente na SEMA II (6°41’ e 6°45’S; 35°7’ e 35°12’W), a qual

4
possui inserido dentro da Mata Atlântica, fragmentos de Tabuleiro. O Tabuleiro é uma
vegetação savânica do nordeste brasileiro, muito parecida com o bioma do Cerrado, pois
apresenta um estrato herbáceo que forma um tapete de gramíneas, apresentando árvores e
arbustos espaçados de troncos retorcidos e suberosos (Oliveira Filho e Carvalho, 1993) (Fig.
2). Atualmente, devido à expansão agrícola, os Tabuleiros estão sofrendo grande ameaça em
termos conservacionistas (Oliveira Filho e Carvalho, 1993).
Coleta de dados - Foram coletados um total de 278 C. ocellifer (Fig. 3). Os animais
foram coletados durante o final do ano de 2008, e durante os anos de 2009 e 2010. Foram
utilizados dois métodos de captura: Armadilhas de interceptação e queda (pitfall traps) e
busca ativa. A proporção do número de animais capturados em armadilhas em relação aos
animais coletados ativamente foi respectivamente de 3:2.
Durante todo o período de coleta, as armadilhas permaneceram montadas, porém os
animais só foram coletados nas mesmas até o dezembro de 2009. Foram realizadas vistorias
duas vezes por semana das armadilhas. Foram dispostos numa trilha de aproximadamente 2
km pela região de Tabuleiro, 25 conjuntos de armadilhas. Cada conjunto é formado por 4
baldes de 60 litros enterrados até o nível do solo, dispostos de maneira que formem um Y,
sendo um balde central e 3 nos vértices. Entre o balde central e cada balde das pontas, foram
colocadas chapas galvanizadas de 5 metros de comprimento, formando cercas guia,
totalizando então 100 baldes (Fig. 4).
As buscas ativas foram realizadas mensalmente por equipes de dois integrantes em
dois dias de campo, até atingir o número mínimo de aproximadamente 15 animais/mês. Estas
buscas ocorreram principalmente nos anos de 2009 e 2010, pelo período de atividade do sol,
das 8:00 h até as 18:00 h, totalizando aproximadamente 480 horas/homem. Os animais

5
coletados por busca ativa foram capturados com auxílio de tubos de látex/garrotes formando
elásticos de coleta (“rubber bands”).
Os animais que ainda estavam vivos, foram mortos em laboratório pela injeção de
cloridrato de lidocaína 2%, foram mensurados e em seguida fixados em formalina 4% por 48
horas. Após a fixação, todos os animais foram acondicionados em recipientes de vidro com
álcool etílico 70%. Cada lagarto coletado foi depositado e tombado na Coleção Herpetológica
da Universidade Federal da Paraíba (CHUFPB).
Variáveis morfométricas - Antes de fixados, os animais foram pesados e mensuradas
as seguintes variáveis: comprimento rostro-cloacal (CRC), comprimento da cauda (CCAU),
largura do corpo (LCO), altura do corpo (ACO), largura da cabeça (LCA), altura da cabeça
(ACA), comprimento da cabeça (CCA), comprimento do membro anterior (CMA) e
comprimento do membro posterior (CMP). Todas estas variáveis foram mensuradas em
milímetros e com auxílio de paquímetros digitais Mitutoyo® de precisão 0,01mm.
Dieta - A análise da dieta foi realizada através da observação dos itens alimentares
presentes no estômago de cada animal. Os animais foram dissecados, os estômagos foram
removidos e a observação foi feita com o auxílio de um estereomicroscópio de aumento
máximo de 40x. As presas foram identificadas e categorizadas ao nível taxonômico de ordem,
com exceção de algumas, que foram identificadas ao nível de família, e outras que foram
incluídas em categorias artificiais (e.g., larvas de inseto).
Utilizou-se neste estudo tanto a apresentação e tratamento dos dados de dieta na forma
de estômagos individuais (na forma de média e desvio padrão, que demonstra a informação
do que C. ocellifer está predando em média) e na forma de estômagos agrupados (na forma de
somatório, que considera todos os estômagos como um só indivíduo, e demonstra o que a
população como um todo está predando e as implicações na teia alimentar).

6
Para cada categoria de presa, foram calculados os valores de ocorrência (frequência
absoluta e relativa), os valores de número (bruto e percentual) e volume (bruto e percentual),
em mm³. Para o cálculo do volume, foi mensurado a largura e o comprimento de cada presa
encontrada relativamente intacta, com auxílio de um paquímetro digital. Foi utilizada a
fórmula de um elipsóide a seguir:
V =43
πl2
2 c2
Onde c é o comprimento da presa e l é a largura da presa. Utilizando as variáveis percentuais
volumétricas e numéricas, calculou-se a largura de nicho (B) para cada lagarto utilizando o
inverso do índice de Simpson (Simpson, 1949):
B =1
pi2
i=1
n
∑
Finalmente, para determinar a contribuição de cada categoria de presa numa escala relativa,
utilizou-se um índice de importância (I) para os estômagos individuais e para os estômagos
agrupados, utilizando a seguinte fórmula:
,3
%%% VNFI
++=
Onde F% é a frequência relativa, N% é o valor numérico percentual e V% é o valor
volumétrico percentual. Foram então ranqueadas as três primeiras categorias de cada valor
(frequência, número, volume e importância).
Para testar se existe relação entre as dimensões da cabeça dos animais em relação as
dimensões da presa, realizou-se uma correlação canônica com dois conjuntos de variáveis:
comprimento máximo e largura máxima de presa (por estômago) versus comprimento, largura

7
e altura da cabeça (por lagarto). Para observar a variação sazonal na dieta de C. ocellifer,
foram observados os valores percentuais e a largura do nicho de volume e número para a
estação seca e para a estação chuvosa, baseando-se nas médias mensais de pluviosidade (Fig.
5)
Reprodução - Foi determinado o sexo e o estágio reprodutivo dos lagartos. A
metodologia utilizada para determinação de estágio reprodutivo foi a observação direta das
gônadas com auxílio de um estereomicroscópio. Machos foram considerados reprodutivos
quando apresentaram testículos dilatados/alargados junto a epidídimos enovelados. O
testículo esquerdo de cada macho foi mensurado baseando-se na fórmula elipsoide, descrita
acima.
As fêmeas foram consideradas reprodutivamente ativas quando apresentaram folículos
vitelogênicos, corpo lúteo e/ou ovos no oviduto. O tamanho da ninhada foi determinado pela
soma total de ovos e/ou folículos. A presença concomitante de folículos vitelogênicos junto à
presença de ovos no oviduto ou corpo lúteo foi considerada um indício para a presença de
mais de uma ninhada por ano. Também foram mensurados o comprimento, a largura e o
volume (igualmente pela fórmula elipsoide, descrita acima) de cada ovo. O recrutamento foi
determinado baseando-se nas maiores amplitudes de CRC, por mês, observadas.
Também foi estimado o tamanho de maturidade sexual baseando-se no menor macho
reprodutivo e na menor fêmea reprodutiva. Para verificar se existe relação do tamanho do
corpo das fêmeas com o tamanho da ninhada, foi realizada uma regressão linear simples.
Análises estatísticas- Todos os testes estatísticos foram realizados no SYSTAT 12 e no
SAS para Windows. O nível de significância para as hipóteses testadas foi de 5%. Os valores
de média estão no formato �� ± DP, onde �� é a média e DP o desvio padrão.

8
Resultados:
Dieta - Do total de estômagos dissecados, 37 estavam vazios (16%). Foram
encontradas 19 categorias de presa (Tabela 1), dentre as quais foram mais frequentes
ortópteras, larvas de inseto e aranhas/hemípteras. Considerando os estômagos individuais,
numericamente as principais categorias de presa foram ortópteras, larvas de inseto e
artrópodes não identificados. Volumetricamente, ortópteras, larvas de inseto e aranhas foram
as principais categorias de presa. Finalmente, o índice de importância para estômagos
individuais mostram que ortópteras, seguido de larvas de inseto e aranhas são os itens que
mais contribuem nos hábitos alimentares de C. ocellifer do Tabuleiro (Tabela 1). A largura de
nicho numérica foi de 1,91 ± 1,03 e a largura de nicho volumétrica foi de 1,38 ± 0,61, o que
indica que C. ocellifer do Tabuleiro se alimenta de aproximadamente 1-2 tipos de presas.
Levando-se em conta os valores para os estômagos agrupados, numericamente, as
principais presas foram isópteras, ortópteras e larvas de inseto. Volumetricamente, ortópteras,
larvas de inseto e material vegetal foram os principais itens alimentares. O índice de
importância para estômagos agrupados foi diferente dos estômagos individuais, apontando
isópteras como o terceiro item mais importante no lugar de aranhas. O primeiro e o segundo
item permaneceram ortópteras e larvas de inseto (Tabela 1).
Observando os resultados numa perspectiva sazonal, na estação chuvosa (fevereiro a
julho, Tabela 2), levando em consideração as médias por estômago, a dieta foi composta
numericamente por ortópteras, larvas de inseto e isópteras. Volumetricamente, a dieta foi
composta de ortópteras, larvas de inseto e aranhas. Levando-se em conta os estômagos
agrupados, a dieta foi composta numericamente por isópteras, ortópteras e larvas de inseto.
Volumetricamente, a dieta foi composta por ortópteras, material vegetal e larvas de inseto. Os
resultados do índice de importância para as médias por estômago foram semelhantes aos da

9
população durante o ano todo (Ver Tabela 1 e 2), porém para estômagos agrupados os
isópteras vem em segundo lugar na lista, sendo ortópteras ainda em primeiro e larvas de
inseto em terceiro (Tabela 2). Na estação seca, as médias por estômago apresentaram
resultados semelhantes aos da população como um todo (ver Tabela 1 e 3). Para estômagos
agrupados, a dieta é composta numericamente por isópteras, ortópteras e larvas de inseto e
volumetricamente por ortópteras, larvas de inseto e aranhas. Os índices de importância
apresentaram resultados semelhantes entre si, demonstrando que ortópteras, seguido de larvas
de inseto e aranhas são os itens mais importantes (Tabela 3). A largura de nicho numérica e
volumétrica para a chuva foi de 1,97 ± 1,07 e 1,12 ± 0,84. A largura de nicho numérica e
volumétrica da estação seca apresentou resultados semelhantes, sendo 1,86 ± 1,00 e 1,33 ±
0,55.
A correlação canônica apresentou uma associação positiva (Wilk’s Lambda = 0,89, P
= 0,004) entre as variáveis de tamanho da presa e das variáveis de cabeça, onde a primeira
variável canônica explicou 67% desta variação (Tabela 4).
Reprodução - O menor macho reprodutivo apresentou 45 mm de CRC, enquanto a
menor fêmea reprodutiva apresentou 44 mm de CRC, o que mostra que provavelmente ambos
os sexos atingem a maturidade com a mesmo tamanho/idade (Fig. 6). A média do tamanho da
ninhada foi de 1,89 ± 0,75 (amplitude 1 - 4, N = 35). As médias do comprimento e largura dos
ovos foram de 14,44 ± 1,58 mm e 7,48 ± 1,10 mm (N = 9), respectivamente. A média do
volume dos ovos foi de 288,97 ± 17,67 mm³ (N = 9).
A época reprodutiva ocorreu no início da estação seca (meses de agosto a dezembro,
com pico em setembro), porém apresentando ainda fêmeas reprodutivas no final da estação
seca e início da chuvosa (Fig. 7). Durante todos os meses do ano, foram encontrados machos
reprodutivos (Fig. 8). Foi encontrada apenas uma fêmea possuindo folículos vitelogênicos e

10
corpo lúteo simultaneamente, assim como uma fêmea reprodutiva no mês de março. Portanto,
C. ocellifer do Tabuleiro possui provavelmente apenas uma ninhada por ano, porém podendo
ocasionalmente apresentar indivíduos exibindo mais de uma ninhada por estação reprodutiva.
O recrutamento se inicia principalmente no final da estação seca e início da chuvosa, porém
parece existir um pico no final de estação chuvosa, no mês de julho (Fig. 9).
A regressão linear simples demonstrou que não existe absolutamente nenhuma relação
entre o tamanho da ninhada e o tamanho do corpo das fêmeas (r < 0,001 ; P = 1,00).
Discussão:
Dieta - Usualmente, Cnemidophorus é considerado um táxon que se alimenta de
insetos com frequência, onde os cupins seriam as presas teoricamente mais importantes
(Menezes et al., 2006; Menezes et al., 2008; Mesquita e Colli, 2003b). No entanto, diferente
das demais populações de C. ocellifer que habitam savanas, a população do Tabuleiro não
apresentou um valor tão alto de cupins em seus hábitos alimentares. Ao invés disto, ortópteras
e larvas de inseto foram observados em grande quantidade e frequência. Recentemente,
alguns trabalhos em regiões de Restinga no Nordeste brasileiro com uma população de C.
ocellifer (Santana et al., 2010), e outra com a congenérica C. abaetensis (Dias & Rocha,
2007), também apresentaram resultados semelhantes. Ambos demonstraram alta influência de
gafanhotos, grilos e larvas de inseto na dieta de Cnemidophorus. Uma das hipóteses para
explicar a baixa incidência de isópteras da dieta de C. ocellifer do Tabuleiro é a baixa
densidade e diversidade de cupins terrícolas na área de estudo e outras áreas relacionadas à
mesma (Vasconcellos et al., 2008). Esta baixa diversidade de isópteras pode estar relacionada
com pertubações antrópicas do passado, o que modificou as características físico-químicas do
solo, deixando o mesmo mais pobre em nutrientes e assim resultando numa baixa

11
disponibilidade de recursos para os cupins (Vasconcellos et al., 2008). Este fenômeno da
substituição de cupins na dieta por ortópteras e larvas de inseto, possivelmente associado à
baixa disponibilidade no ambiente, foi também sugerido em um estudo com C. ocellifer numa
área de Restinga nordestina (Santana et al., 2010). Junto a isto, muitos ortópteras apresentam
alta densidade e diversidade em áreas de Mata Atlântica, especialmente nas bordas de mata
(Barbosa et al., 2005). Como já conhecido, áreas de Restinga e de Tabuleiro são associadas
com regiões de Mata Atlântica, portanto a artropofauna presente nas bordas da mata pode
exercer um importante papel na disponibilidade de alimento para as populações de C. ocellifer
que habitam estas regiões. Resultados semelhantes foram relatados para outras populações de
Cnemidophorus, as quais também vivem em fragmentos de savanas dentro de regiões
florestais, onde estes tenderam a se alimentar de larvas de inseto, ortópteras e até mesmo
frutos ao invés de cupins (Mesquita e Colli, 2003b; Vitt, 1973; Vitt e Carvalho, 1995; Vitt et
al., 1997). Também foram observados em estudos de ecologia e segregação de nicho entre
espécies simpátricas de Cnemidophorus, onde C. ocellifer ingeriu altos índices de cupim em
detrimento a C. abaetensis que praticamente não ingeriu cupim, mas ingeriu altos índices de
larvas de inseto (Dias e Rocha, 2007). Ainda, vale salientar também que em um trabalho
experimental com algumas populações do gênero Aspidoscelis, foi encontrado uma correlação
negativa entre a ingestão de ortópteras e tamanho do corpo, sugerindo que essa frequência de
ortópteras pode estar relacionado com o tamanho do corpo (Eifler e Passek, 2000). Entretanto,
estudos de variação geográfica na ecologia são necessários para testar essa hipótese.
Apesar da relativa baixa frequência de isópteras dos hábitos alimentares de C. ocellifer
do Tabuleiro, os valores de importância das presas secundárias, como aranhas e larvas de
inseto foi similar ao encontrado em outras populações de outras regiões, como Cerrado e
Caatinga (Mesquita e Colli, 2003a; Vitt, 1991b; Vitt, 1995). Estas categorias de presa são
também importantes componentes dos hábitos alimentares de um significante número de

12
espécies congenéricas, como C. cryptus, C. gramivagus, C. parecis, C. mumbuca (Mesquita e
Colli, 2003b; Mesquita et al., 2006b; Mesquita et al., 2006a), C. lemniscatus (Magnusson e
Silva, 1993; Mesquita e Colli, 2003b), C. nativo (Menezes et al., 2008), C. littoralis (Teixeira-
Filho et al., 2003) e até mesmo outros teídeos (e.g., Vitt, 1991a; Vitt e Colli, 1994; Vitt e Zani,
1996), o que parecer ser consequência do modo de forrageamento, uma importante
característica dos lagartos da família Teiidae (Pianka e Vitt, 2003; Schoener, 1991). Como a
maioria dos forrageadores ativos aparentemente utilizam discriminação química para buscar
seu alimento (Cooper Jr. et al., 2000), eles podem acessar estes tipos de presas que geralmente
possuem baixa atividade durante o dia (como ortópteras e aranhas) e que podem ser
encontrados em locais mais escondidos, como serrapilheira, tocas e troncos caídos.
Numa perspectiva sazonal, não foram observadas grandes variações nos itens mais
importantes na dieta nem na largura do nicho, o que ainda confirma a tendência de escolha de
certos tipos de presa por C. ocellifer. A constante presença destes itens pode estar relacionada
também ao fato de que ambas as categorias de presa são relativamente abundantes o ano todo
(Barbosa et al., 2005).
Um outro resultado interessante foram as altas taxas de material vegetal ingerido pela
população do Tabuleiro. Algumas espécies de Cnemidophorus de isolados, como C. murinus
de Bonaire, nas Antilhas, que é estritamente herbívoro (Dearing, 1993), e outras espécies,
como C. lemniscatus, de isolados de Cerrado na Amazônia, podem utilizar grandes
quantidades de frutos na sua dieta (Vitt et al., 1997).
Outro resultado peculiar foi a correlação positiva entre as dimensões da presa e as
dimensões da cabeça dos lagartos, não observada na população do Cerrado (Mesquita e Colli,
2003a). Um estudo realizado com C. littoralis (Teixeira-Filho et al., 2003) mostrou que a
especialização dos hábitos alimentares desses animais em cupins pode provocar uma inércia

13
ontogenética nas dimensões das presas e na diversidade destas também. A ausência desta
inércia, explicada pela correlação positiva na população do Tabuleiro está provavelmente
relacionada com a composição da dieta.
Os resultados observados neste trabalho ainda sugerem uma significante influência
filogenética no nicho alimentar de C. ocellifer, o que parece ser uma característica comum em
lagartos da família Teiidae (Vitt e Pianka, 2005). Porém, mesmo com estes fatores históricos
constringindo os hábitos alimentares de C. ocellifer, pode-se observar a partir deste estudo
que este táxon possui uma plástica tolerância à baixa disponibilidade de presas importantes
como cupins ou uma variação de nicho por possíveis competidores, o que sugere um papel
significativo de fatores ecológicos nos aspectos alimentares de C. ocellifer do Tabuleiro.
Reprodução - No Tabuleiro, C. ocellifer se reproduz sazonalmente, concentrando seu
ciclo reprodutivo durante toda a estação seca, com o recrutamento ocorrendo principalmente
durante o final da estação seca e início da estação chuvosa, com um pequeno pico de
recrutamento no meio da estação chuvosa (julho). Primariamente, estudos clássicos de
história de vida apontam que a grande maioria dos lagartos das regiões neotropicais tende a se
reproduzir continuamente durante todo o ano e apresentando várias ninhadas ao longo do
mesmo (Fitch, 1970, 1982). Porém um estudo um pouco mais recente indicou que na verdade
existem muitas formas e tipos de estratégias reprodutivas em lagartos neotropicais (Vitt,
1992).
No geral, se observa que espécies de lagartos neotropicais que habitam regiões onde os
fatores ambientais e as condições climáticas são estáveis (e.g., Amazônia) ou que as
condições fossem relativamente imprevisíveis (e.g., Caatinga), os lagartos tenderiam a se
reproduzir continuamente durante todo o ano apresentando várias ninhadas de poucos ovos
(Colli, 1991; Mesquita e Colli, 2010; Vitt e Colli, 1994). Por outro lado, em regiões onde as

14
condições ambientais são sazonais (e.g., Cerrado), os lagartos tenderiam a se reproduzir
ciclicamente, apresentando poucas ou apenas uma ninhada durante o ano, porém com
ninhadas maiores (Colli, 1991; Mesquita e Colli, 2010; Vitt e Colli, 1994).
Originalmente, a maioria dos estudos consideravam que os fatores ecológicos,
principalmente as condições ambientais, seriam os fatores mais influentes dos padrões de
história de vida dos lagartos (Tinkle et al., 1970), apesar de que alguns já passaram a
considerar influências históricas nestes padrões (Dunham e Miles, 1985; Dunham et al.,
1988). Mais recentemente, um estudo sobre padrões de história de vida de lagartos
neotropicais, relata que Cnemidophorus é um táxon que apresenta uma forte influência
histórica nos padrões e estratégias reprodutivas (Mesquita e Colli, 2010), o que é corroborado
no presente estudo, pois as populações de C. ocellifer do Cerrado (Mesquita e Colli, 2003a;
Mesquita e Colli, 2003b), se reproduzem também sazonalmente (durante a estação seca),
similar à população do Tabuleiro. Como o Tabuleiro recebe forte influência da região de Mata
Atlântica, a reprodução poderia ser relativamente contínua, similar a regiões de mata
ombrófila, onde os lagartos tendem a se reproduzir continuamente (Colli, 1991; Mesquita e
Colli, 2010; Vitt e Colli, 1994). Entretanto, observando a distribuição do tamanho dos animais
ao longo do ano, é visível a existência de uma maior amplitude no tamanho dos animais
durante o mês de julho, o que pode caracterizar um segundo recrutamento. Além disto, foi
encontrada uma fêmea com folículos vitelogênicos e corpo lúteo simultaneamente no mês de
dezembro, e uma fêmea reprodutiva no mês de março, o que sugere que mesmo que C.
ocellifer aparentemente se reproduza sazonalmente, o ciclo parece ser mais contínuo do que
em outras regiões, como o Cerrado (Mesquita e Colli, 2003a), o que seria um indício de
influência ecológica nos padrões reprodutivos desta população de C. ocellifer. Entretanto, o
número amostral de fêmeas reprodutivas pode não ter sido suficiente para se visualizar o
padrão claramente.

15
Mesmo com estes resultados, existem dificuldades em assumir que a filogenia é o fator
chave influenciando o ciclo reprodutivo de C. ocellifer do Tabuleiro. Apesar de ser mais
úmido e de está diretamente relacionado com uma área de mata ombrófila, o Tabuleiro (e a
Mata Atlântica nordestina como um todo) também apresenta uma sazonalidade bem definida
dividida em estação seca e chuvosa. Levando-se em consideração as temperaturas médias, não
existem grandes diferenças entre os biomas durante o ano, porém as temperaturas mínimas do
Cerrado são consideravelmente menores e mais variáveis ao longo do ano que no Tabuleiro
(Nimer, 1989b; Nimer, 1989a). Um estudo com Anolis carolinensis demonstrou a influência
da temperatura e do fotoperiodismo na recrudescência testicular, sendo que temperaturas mais
altas e constantes durante o dia e a noite aceleram a espermatogênese (Licht, 1966). Desta
forma, um fenômeno como este poderia promover uma reprodução mais contínua no
Tabuleiro. Além disto, na Caatinga, C. ocellifer se reproduz continuamente durante o ano
apresentando diversas ninhadas ao longo do mesmo (Mesquita e Colli, 2003b; Vitt, 1983), o
que segundo trabalhos de história de vida, seria um resultado esperado, pois assim como as
matas, em regiões imprevisíveis como a Caatinga, os lagartos também tendem a se reproduzir
continuamente (Colli, 1991; Mesquita e Colli, 2010; Vitt e Colli, 1994). Visto a proximidade
geográfica entre as matas de tabuleiro e a Caatinga, não seria difícil de se esperar alguma
influencia.
Outra hipótese para explicar a sazonalidade da reprodução, é a disponibilidade de
alimentos. Como a densidade de artrópodes é influenciada pela sazonalidade em algumas
regiões tropicais (Levings e Windsor, 1982; Smythe, 1982), isto pode afetar diretamente o
ciclo reprodutivo de seus predadores. Porém, muitos estudos mostram que lagartos acumulam
corpos adiposos para a reprodução durante a estação seca (Colli et al., 1997), portanto a
densidade sazonal de alimentos pode não ser um modelador significante nos padrões de ciclos
reprodutivos em lagartos.

16
Outra característica reprodutiva, como o tamanho da ninhada também foi similar a
outras populações de C. ocellifer que habitam savanas (Mesquita e Colli, 2003a; Mesquita e
Colli, 2003b), apesar de ser numericamente menor que a população do Cerrado. Entretanto, a
sazonalidade na reprodução não é o único fator que influencia o tamanho da ninhada.
Forrageadores ativos possuem certas características (sinapomorfias, como alongamento do
corpo) que restringem o número de ovos por ninhada (Vitt e Congdon, 1978; Vitt e Price,
1982). Os resultados da regressão mostraram que não existe nenhuma relação entre o tamanho
da ninhada e o tamanho do corpo das fêmeas. Em geral, o tamanho do corpo é correlacionado
positivamente com o tamanho da ninhada (Dunham et al., 1988), inclusive em C. ocellifer
(Mesquita e Colli, 2003a; Mesquita e Colli, 2003b). Um estudo com populações de
Cnemidophorus de savanas amazônicas apresentou um resultado semelhante, porém mesmo
que baixa, foi encontrada uma correlação positiva (Vitt et al., 1997). Existem diversos casos
de ninhadas fixas em lagartos (principamente em geckonídeos e anolíneos). Além do que esta
característica parecer ter evoluído diversas vezes (Dunham et al., 1988; Mesquita e Colli,
2010; Tinkle et al., 1970). Ainda, o tamanho e a quantidade dos ovos pode ser determinado
pela espaço da cavidade corporal da fêmea (Shine e Schwarzkopf, 1992), tendo então a
morfologia um papel importante na determinação do tamanho da ninhada. Portanto as
adaptações morfológicas ao forrageamento ativo observadas em Cnemidophorus poderiam
explicar a falta de relação entre tamanho do corpo e tamanho da ninhada (Vitt e Congdon,
1978; Vitt e Price, 1982). Além disto, o número de fêmeas reprodutivas foi relativamente
baixo, dificultando assim conclusões sobre a ausência de correlação, ou se a mesma não
estaria simplesmente relacionada com o tamanho amostral. Neste contexto, ambos fatores,
históricos e ecológicos, exercem influência nos padrões reprodutivos de C. ocellifer do
Tabuleiro.

17
Referências
ARIAS, F., C. M. CARVALHO, M. T. RODRIGUES, e H. ZAHER. 2011a. Two new species of
Cnemidophorus (Squamata: Teiidae) from the Caatinga, Northwest Brazil. Zootaxa.
2787:37-54.
ARIAS, F., C. M. CARVALHO, M. T. RODRIGUES, e H. ZAHER. 2011b. Two new species of
Cnemidophorus (Squamata: Teiidae) of the C. ocellifer group, from Bahia, Brazil.
Zootaxa. 3022:1-21.
BARBOSA, V. S., I. R. LEAL, L. IANNUZZI, e J. ALMEIDA-CORTEZ. 2005. Distribution Pattern
of Herbivorous Insects in a Remnant of Brazilian Atlantic Forest. Neotropical
Entomology. 34:701-711.
COLLI, G. R. 1991. Reproductive ecology of Ameiva ameiva (Sauria: Teiidae) in the cerrado
of central Brazil. Copeia. 1991:1002-1012.
COLLI, G. R., J. P. CALDWELL, G. C. COSTA, A. M. GAINSBURY, A. A. GARDA, D. O.
MESQUITA, C. M. M. R. FILHO, A. H. B. SOARES, V. N. SILVA, P. H. VALDUJO, G. H.
C. VIEIRA, L. J. VITT, F. P. WERNECK, H. C. WIEDERHECKER, e M. G. ZATZ. 2003a. A
new species of Cnemidophorus (Squamata, Teiidae) from the Cerrado biome in central
Brazil. Occasional Papers of the Oklahoma Museum of Natural History. 14:1-14.
COLLI, G. R., G. C. COSTA, A. A. GARDA, K. A. KOPP, D. O. MESQUITA, A. K. PÉRES JR, P.
H. VALDUJO, G. H. C. VIEIRA, e H. C. WIEDERHECKER. 2003b. A critically
endangered new species of Cnemidophorus (Squamata, Teiidae) from a Cerrado
enclave in southwestern Amazonia, Brazil. Herpetologica. 59:76-88.
COLLI, G. R., L. G. GIUGLIANO, D. O. MESQUITA, e F. G. R. FRANÇA. 2009. A new species of
Cnemidophorus from the Jalapão region, in the Central Brazilian Cerrado.
Herpetologica 65:311-327.

18
COLLI, G. R., A. K. PÉRES, JR., e M. G. ZATZ. 1997. Foraging mode and reproductive
seasonality in tropical lizards. Journal of Herpetology. 31:490-499.
COOPER JR., W. E., M. A. PAULISSEN, e J. J. HABEGGER. 2000. Discrimination of prey, but not
plant, chemicals by actively foraging, insectivorous lizards, the lacertid Takydromus
sexlineatus and the teiid Cnemidophorus gularis. Journal of Chemical Ecology.
26:1623-1634.
DEARING, M. D. 1993. An alimentary specialization for herbivory in the tropical whiptail
lizard Cnemidophorus murinus. Journal of Herpetology. 27:111-114.
DIAS, E. J. R., e C. F. D. ROCHA. 2007. Niche differences between two sympatric whiptail
lizards (Cnemidophorus abaetensis and C. ocellifer, Teiidae) in the restinga habitat of
northeastern Brazil. Brazilian Journal of Biology. 67:41-46.
DUNHAM, A. E., e D. B. MILES. 1985. Patterns of covariation in life history traits of squamate
reptiles: the effects of size and phylogeny reconsidered. The American Naturalist.
126:231-257.
DUNHAM, A. E., D. B. MILES, e D. N. REZNICK. 1988. Life history patterns in squamate
reptiles, p. 441-522. In: Biology of the Reptilia. Vol. 16, Ecology B. Defense and Life
History. C. Gans e R. B. Huey (eds.). Alan R. Liss, Inc., New York.
EIFLER, D. A., e K. M. PASSEK. 2000. Body size effects on pursuit sucess and interspecific
diet differences in Cnemidophorus lizards. Amphibia-Reptilia. 21:477-484.
HUEY, R. B., E. R. PIANKA, e A. SCHOENER. 1983. Lizard Ecology: Studies of a Model
Organism. Harvard University Press, Cambridge, Mass.
LEVINGS, S. C., e D. M. WINDSOR. 1982. Seasonal and annual variation in litter arthropod
populations, p. 355-387. In: Advances in Herpetology and Evolutionary Biology:
Essays in Honor of Ernest E. Williams. A. G. Rhodin e K. Miyata (eds.). Smithsonian
Institution Press, Washington, D.C.

19
LICHT, P. 1966. Reproduction in lizards: influence of temperature on photoperiodism in
testicular recrudescence. Science. 154:1668-1670.
MAGNUSSON, W. E., e E. V. SILVA. 1993. Relative effects of size, season and species on the
diets of some amazonian Savanna lizards. Journal of Herpetology. 27:380-385.
MENEZES, V. A., V. C. AMARAL, M. VAN SLUYS, e C. F. D. ROCHA. 2006. Diet and foraging
of the endemic lizard Cnemidophorus littoralis (Squamata, Teiidae) in the Restinga de
Jurubatiba, Macaé, RJ. Brazilian Journal of Biology. 66:803-807.
MENEZES, V. A., G. F. DUTRA, e C. F. D. ROCHA. 2008. Feeding habits of the endemic
tropical parthenogenetic lizard Cnemidophorus nativo (Teiidae) in a restinga area of
northeastern Brazil. Journal of Natural History. 42:2575-2583.
MESQUITA, D. O., e G. R. COLLI. 2003a. The ecology of Cnemidophorus ocellifer (Squamata,
Teiidae) in a neotropical savanna. Journal of Herpetology. 37:498-509.
MESQUITA, D. O., e G. R. COLLI. 2003b. Geographical variation in the ecology of populations
of some Brazilian species of Cnemidophorus (Squamata, Teiidae). Copeia. 2003:285-
298.
MESQUITA, D. O., e G. R. COLLI. 2010. Life history patterns in South American tropical
lizards, p. 45-71. In: Reproducción en Reptiles: Morfología, Ecología y Evolución. O.
H. Gallegos, F. R. M. Cruz, e J. F. M. Sánchez (eds.). Universidad Autónoma del
Estado de México, México.
MESQUITA, D. O., G. R. COLLI, F. G. R. FRANÇA, e L. J. VITT. 2006a. Ecology of a Cerrado
lizard assemblage in the Jalapão region of Brazil. Copeia. 2006:460-471.
MESQUITA, D. O., G. C. COSTA, e G. R. COLLI. 2006b. Ecology of an amazonian savanna
lizard assemblage in Monte Alegre, Pará State, Brazil. South American Journal of
Herpetology. 1:61-71.

20
NIMER, E. 1989a. Climatologia da região centro-oeste, p. 393-421. In: Climatologia do Brasil.
Fundação Instituto Brasileiro de Geografia e Estatística - IBGE, Rio de Janeiro,
Brazil.
NIMER, E. 1989b. Climatologia da região nordeste, p. 315-361. In: Climatologia do Brasil. F.
I. B. d. G. e. Estatística (ed.). Fundação Instituto Brasileiro de Geografia e Estatística -
IBGE, Rio de Janeiro, Brazil.
OLIVEIRA FILHO, A. T., e D. A. CARVALHO. 1993. Florística e fisionomia da vegetação no
extremo norte do litoral da Paraíba. Revista Brasileira de Botânica. 16:115-130.
PIANKA, E. R., e L. J. VITT. 2003. Lizards: Windows of the Evolution of Diversity. University
of California Press, Los Angeles, CA.
REEDER, T. W., C. J. COLE, e H. C. DESSAUER. 2002. Phylogenetic relationships of Whiptail
lizards of the genus Cnemidophorus (Squamata: Teiidae): a test of monophyly,
reevaluation of karyotypic evolution, and review of hybrid origins. American Museum
Novitates. 3365:1-61.
ROCHA, C. F. D., A. F. B. ARAÚJO, D. VRCIBRADIC, e E. M. M. COSTA. 2000. New
Cnemidophorus (Squamata; Teidae) from coastal Rio de Janeiro State, Southeastern
Brazil. Copeia. 2000:501-509.
ROCHA, C. F. D., H. G. BERGALLO, e D. PECCININI-SEALE. 1997. Evidence of an unisexual
population of the Brazilian whiptail lizard genus Cnemidophorus (Teiidae), with
description of a new species. Herpetologica. 53:374-382.
SANTANA, G. G., A. VASCONCELLOS, Y. E. A. GADELHA, W. L. S. VIEIRA, W. O. ALMEIDA,
R. P. NÓBREGA, e R. R. N. ALVES. 2010. Feeding habits, sexual dimorphism and size
at maturity of the lizard Cnemidophorus ocellifer (Spix, 1825) (Teiidae) in a
reforested resting habitat in Northeastern Brazil. Brazilian Journal of Biology. 70:409-
416.

21
SCHOENER, T. W. 1991. Theory of Feeding Strategies. Annual Review of Ecology and
Systematics. 2:369-404.
SHINE, R., e L. SCHWARZKOPF. 1992. The evolution of reproductive effort in lizards and
snakes. Evolution. 46:62-75.
SIMPSON, E. H. 1949. Measurement of diversity. Nature. 163:688.
SMYTHE, N. 1982. The seasonal abundance of night-flying insects in a Neotropical forest, p.
309-318. In: The Ecology of a Tropical Forest: Seasonal Rhythms and Long-term
Changes. E. G. Leigh, Jr., A. S. Rand, e D. M. Windsor (eds.). Smithsonian Institution
Press, Washington, D.C.
TEIXEIRA-FILHO, P. F., C. F. D. ROCHA, e S. C. RIBAS. 2003. Relative feeding specialization
may depress ontogenetic, seasonal, and sexual variations in diet: the endemic lizard
Cnemidophorus littoralis (Teiidae). Brazilian Journal of Biology. 63:321-328.
TINKLE, D. W., H. M. WILBUR, e S. G. TILLEY. 1970. Evolutionary strategies in lizard
reproduction. Evolution. 24:55-74.
VASCONCELLOS, A., A. G. BANDEIRA, W. O. ALMEIDA, e F. M. S. MOURA. 2008. Térmitas
construtores de ninhos conspícuos em duas áreas de Mata Atlântica com diferentes
níveis de perturbação antrópica. Neotropical Entomology. 37:15-19.
VITT, L. J. 1973. Reproductive biology of the anguid lizard, Gerrhonotus coeruleus principis.
Copeia. 1973:176-184.
VITT, L. J. 1983. Reproduction and sexual dimorphism in the tropical teiid lizard
Cnemidophorus ocellifer. Copeia. 1983:359-366.
VITT, L. J. 1991a. Ecology and life history of the wide-foraging lizard Kentropyx calcarata
(Teiidae) in Amazonian Brazil. Canadian Journal of Zoology. 69:2791-2799.
VITT, L. J. 1991b. An introduction to the ecology of Cerrado lizards. Journal of Herpetology.
25:79-90.

22
VITT, L. J. 1992. Diversity of reproductive strategies among brazilian lizards and snakes: the
significance of lineage and adaptation, p. 135-149. In: Reproductive Biology of South
American Vertebrates. W. C. Hamlett (ed.). Springer-Verlag, New York.
VITT, L. J. 1995. The ecology of tropical lizards in the Caatinga of northeast Brazil.
Occasional Papers of the Oklahoma Museum of Natural History. 1:1-29.
VITT, L. J., e C. M. CARVALHO. 1995. Niche partitioning in a tropical wet season: lizards in
the Lavrado area of Northern Brazil. Copeia. 1995:305-329.
VITT, L. J., e G. R. COLLI. 1994. Geographical ecology of a neotropical lizard: Ameiva ameiva
(Teiidae) in Brazil. Canadian Journal of Zoology. 72:1986-2008.
VITT, L. J., G. R. COLLI, J. P. CALDWELL, D. O. MESQUITA, A. A. GARDA, e F. G. R.
FRANÇA. 2007. Detecting variation in microhabitat use in low diversity lizard
assemblages across small-scale habitat gradients. Journal of Herpetology. 41:653-662.
VITT, L. J., e J. D. CONGDON. 1978. Body shape, reproductive effort, and relative clutch mass
in lizards: resolution of a paradox. The American Naturalist. 112:595-608.
VITT, L. J., e S. R. GOLDBERG. 1983. Reproductive ecology of two tropical iguanid lizards:
Tropidurus torquatus and Platynotus semitaeniatus. Copeia. 1983:131-141.
VITT, L. J., e E. R. PIANKA. 1994. Lizard Ecology. Princeton University Press, Princeton, NJ.
VITT, L. J., e E. R. PIANKA. 2005. Deep history impacts present-day ecology and biodiversity.
Proceedings of the National Academy of Science. 102:7877-7881.
VITT, L. J., e H. J. PRICE. 1982. Ecological and evolutionary determinants of relative clutch
mass in lizards. Herpetologica. 38:237-255.
VITT, L. J., e P. A. ZANI. 1996. Ecology of the lizard Ameiva festiva (Teiidae) in Southeastern
Nicaragua. Journal of Herpetology. 30:110-117.

23
VITT, L. J., P. A. ZANI, J. P. CALDWELL, M. C. D. ARAUJO, e W. E. MAGNUSSON. 1997.
Ecology of whiptail lizards (Cnemidophorus) in the Amazon region of Brazil. Copeia.
1997:745-757.
WRIGHT, J. W., e L. J. VITT. 1993. Biology of Whiptail Lizards (Genus Cnemidophorus). The
Oklahoma Museum of Natural History, Norman, Oklahoma.
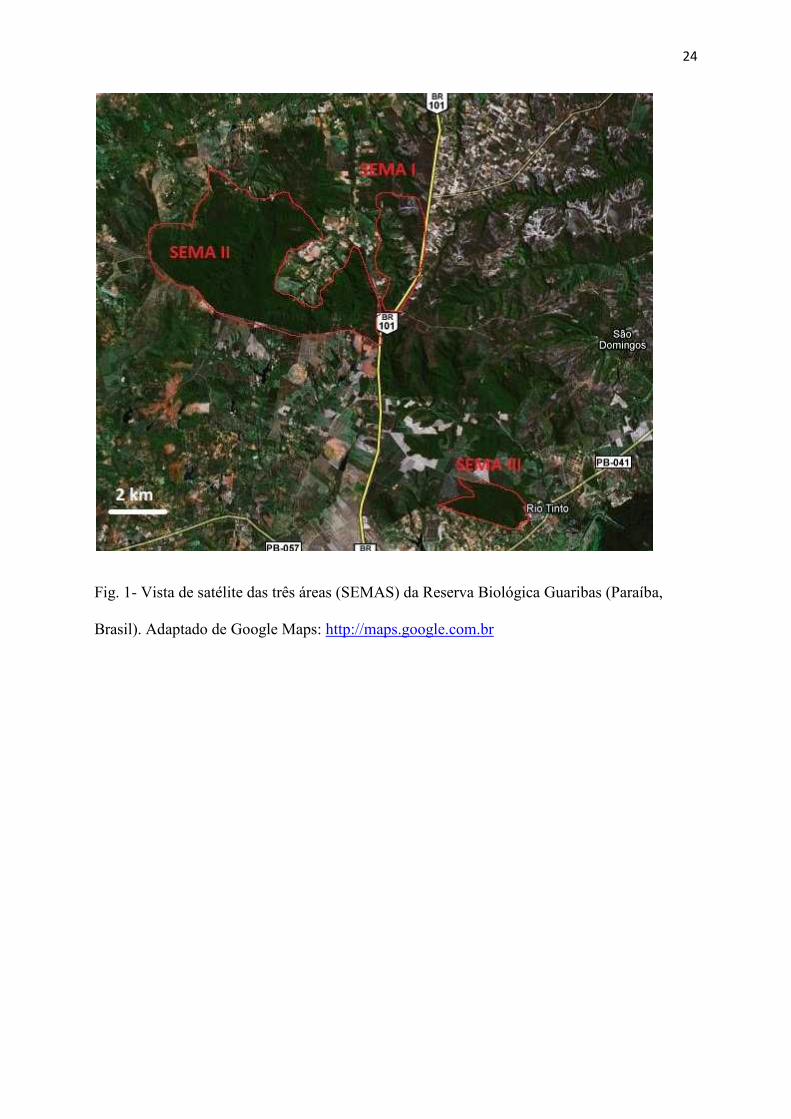
24
Fig. 1- Vista de satélite das três áreas (SEMAS) da Reserva Biológica Guaribas (Paraíba,
Brasil). Adaptado de Google Maps: http://maps.google.com.br

25
Fig. 2- Região de Tabuleiro da Reserva Biológicas Guaribas SEMA II (Paraíba, Brasil). Foto:
Carmem Pedro.

26
Fig. 3- Macho (acima) e fêmea (abaixo) de Cnemidophorus ocellifer da Reserva Biológica
Guaribas (Paraíba, Brasil). Foto: Daniel Mesquita
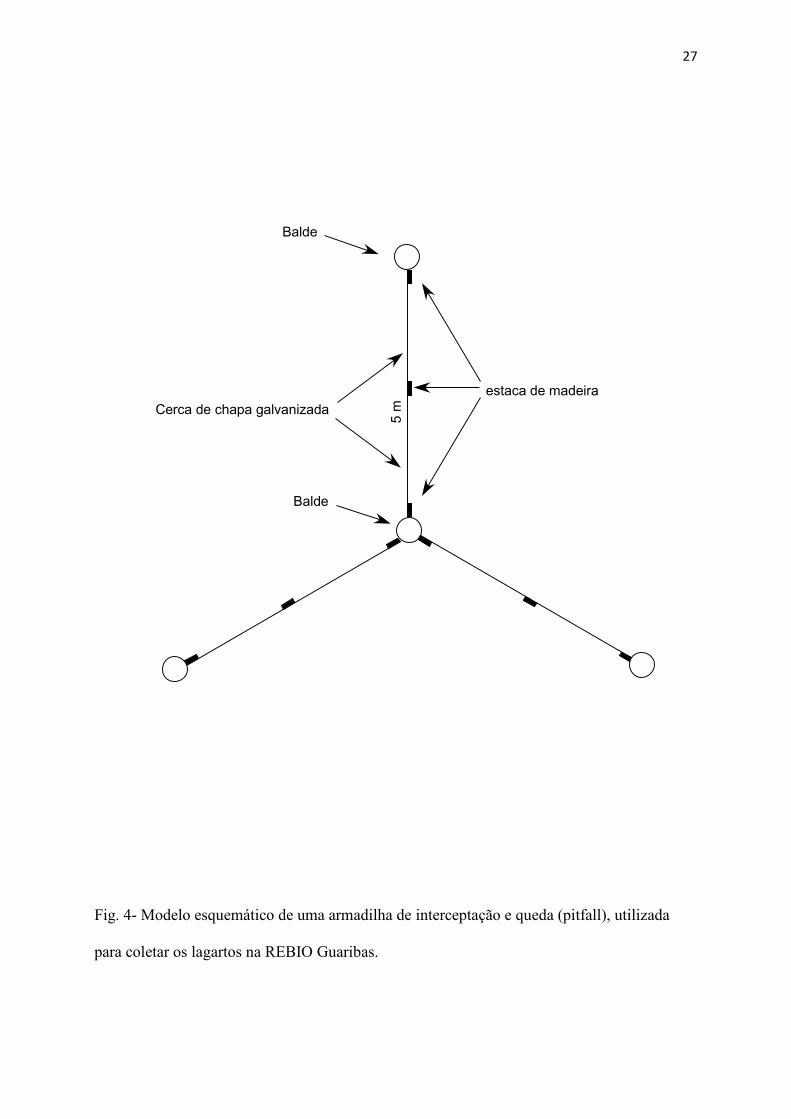
27
Fig. 4- Modelo esquemático de uma armadilha de interceptação e queda (pitfall), utilizada
para coletar os lagartos na REBIO Guaribas.
5 m
Balde
Cerca de chapa galvanizadaestaca de madeira
Balde

28
Fig. 5- Pluviosidade média mensal dos últimos cinco anos (2006-2011) na REBIO Guaribas,
no município de Mamanguape-PB. Fonte: PROCLIMA (www.cptec.inpe.br/proclima).

29
Fig. 6- Distribuição de frequência baseando-se no tamanho do corpo e condição reprodutiva
de machos e fêmeas de Cnemidophorus ocellifer do Tabuleiro na Rebio Guaribas - PB.
Barras em cinza escuro representam indivíduos não reprodutivos e as barras em cinza claro
representam indivíduos reprodutivos.

30
Fig. 7 – Distribuição mensal de machos e fêmeas de Cnemidophorus ocellifer do Tabuleiro
na Rebio Guaribas – PB de acordo com a condição reprodutiva. Números nos topo das barras
representam o tamanho amostral.
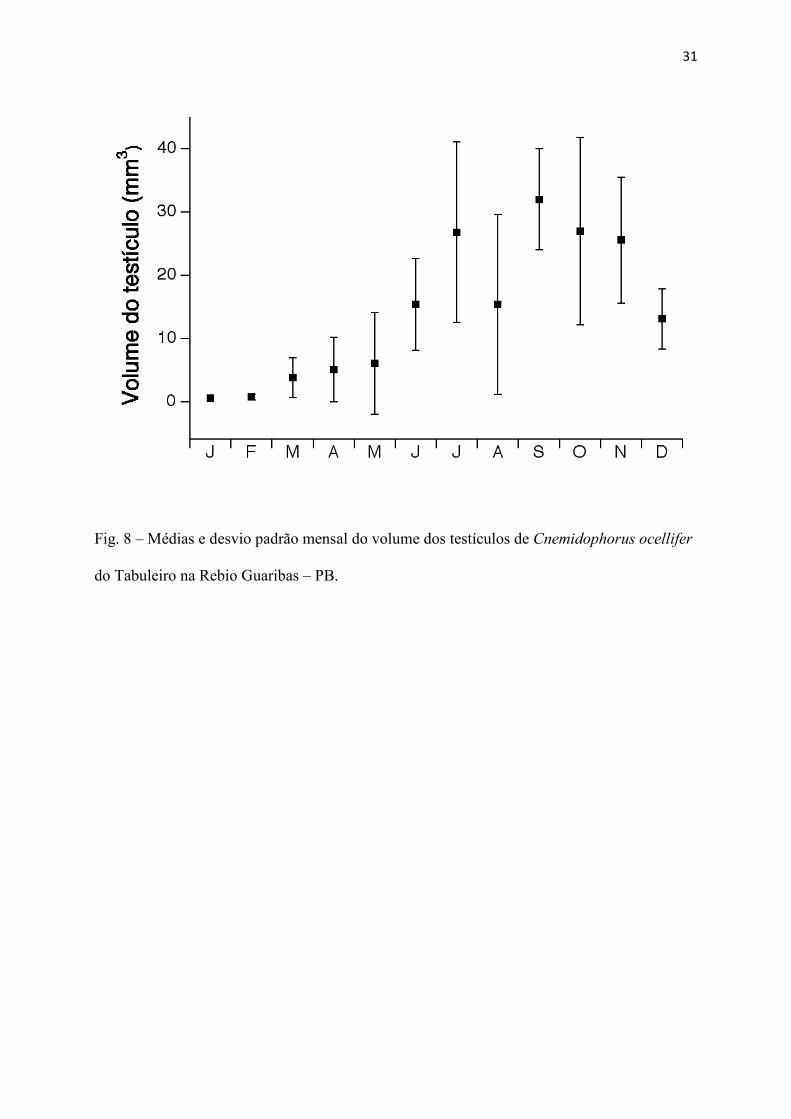
31
Fig. 8 – Médias e desvio padrão mensal do volume dos testículos de Cnemidophorus ocellifer
do Tabuleiro na Rebio Guaribas – PB.

32
Fig. 9 – Distribuição mensal de indivíduos do Cnemidophorus ocellifer no Tabuleiro da Rebio
Guaribas – PB, de acordo com o comprimento rostro-cloacal.

33
Fig. 10- Relação entre o tamanho da ninhada e comprimento rostro cloacal de Cnemidophorus
ocellifer, no Tabuleiro da Rebio Guaribas – PB.

34
Tabela 1: Composição da dieta de Cnemidophorus ocellifer (N=233) da Reserva Biológica Guaribas (Mamanguape, Paraíba, Brasil). a: Número de
estômagos contendo aquela categoria de presa, b : Índice de importância baseado nas médias por estômago, c : Índice de importância baseado nos
estômagos agrupados. Valores brutos de volumes estão em milímetros cúbicos (mm³). As três categorias de maior valor para cada coluna estão em
negrito.
Categorias
Ocorrência
Média por estômago
Estômagos agrupados
Importância
fa
f%
N
N%
Vol.
Vol.%
N
N%
Vol.
Vol.% IEIb IEAc
Aranae
63
27,04 0,40±0,82
8,24±18,04
5,62±31,39
7,51
±22,29 93 8,39 1310,32
7,05 14
,26 14,16
Blattaria
18 7,73 0,08±0,27
3,94±17,03
2,00±18,15
2,45±13,91 18 1,62 466,35
2,51 4,70 3,95
Chilopoda
2
0,86 0,01±0,09
0,64±7,31
0,32±3,44
0,84±9,06
2
0,18
74,16
0,40 0,78 0,48
Coleoptera
49 21,03 0,36±0,94
7,84±19,42
3.56±18,78
3.43±15,37 84 7,57 829,02
4,46 10,78 11,02
Diplopoda
1
0,43 <0,01±0,07 0,04±0,66
0,01±0,18
0,01±0,10
1
0,09
2,81
0,02 0,16 0,18
Diptera
3
1,29 0,01±0,11
0,59±6,79
0,05±0,75
0,04±0,55
3
0,27
11,42
0,06 0,64 0,54
Hemiptera
24 10,30 0,14±0,46
2,79±9,88
1,15±6,28
3,09±14,87 33 2,98 268,11
1,44 5,39 4,91
Hymenoptera
- Formicidae
- Outros
37
15
15,88
6,44
0,34±1,00
0,07±0,31
6,24±17,83
3,95±16,83
0,42±2,38
0,06±0,60
2,28±13,26
0,47±6,56
79
17
7,12
1,53
97,49
14,37
0,52
0,08
8,13
3,62
7,84
2,68
Isoptera
34 14,59
1,71
±7,40
8,80±23,36
1,32±5,42
3,80±14,84 39
8 25
,89
306,69
1,65 9,06
17,38
Larva de inseto
72
30,90
0,57
±1,47
11,13±
20,90
14,44,47
,37
15,03±
31,07 13
3 11
,99
3365
,14
18,11
19
,02
20,33
Lepidoptera
4
1,72 0,02±0,13
0,31±2,81
0,28±2,80
0,15±1,47
4
0,36
65,27
0,35 0,73 0,81

35
Mantodea
7
3,00 0,03±0,17
0,62±4,30
0,25±3,45
0,24±3,36
7
0,63
57,37
0,31 1,29 1,31
Mat. vegetal
16 6,87 0,11±0,47
3,52±14,81
11,47±
102,96 2,95±16,01 25 2,25
2672
,47
14,38
4,44 7,83
Não ident.
33 14,16 0,14±0,35
9,67
±27,53
1,66±19,91
1,47±11,20 33 2,98 386,22
2,08 8,44 6,41
Odonata
4
1,72 0,02±0,13
0,68±6,96
0,56±5,52
0,66±6,88
4
0,36 131,05
0,71 1,02 0,93
Orthoptera
11
6 49,79
0,65
±0,79
25,87±
35,87
27,69±
69,83
24,63±
39,57 15
1 13
,62
6452
,89
34,73
33
,43
32,71
Ovo de inseto
1
0,43 0,01±0,13
0,17±2,62
0,01±0,20
0,23±3,48
2
0,18
3,00
0,02 0,28 0,21
Ovo de réptil
1
0,43 <0,01±0,07 0,43±6,55
3,23±49,34
0,43±6,55
1
0,09 753,20
4,05 0,43 1,52
Pupa de inseto 3
1,29 0,01±0,11
0,28±2,46
1,22±14,43
0,55±6,65
3
0,27 283,22
1,52 0,70 1,03
Scorpionida
16 6,87 0,08±0,31
4,22±18,72
4,42±29,36
3,54±17,70 18 1,62 1030,48
5,55 4,88 4,68

36
Tabela 2: Composição da dieta de Cnemidophorus ocellifer (N=109) da Reserva Biológica Guaribas (Mamanguape, Paraíba, Brasil) durante a estação
chuvosa (Fevereiro até Julho). a : Número de estômagos contendo aquela categoria de presa em porcentagem, b: Índice de importância baseado nas
médias por estômago, c : Índice de importância baseado nos estômagos agrupados. As três categorias de maior valor para cada coluna estão em negrito.
Categoria
Ocorrência
Média por estômago
Estômagos agrupados
Importância
fa %
N%
Vol.%
N%
Vol.%
IEIb
IEAc
Aranae
26
,61
7,55±17,84
9,27
±22,39
7,64
4,60
14
,48
12,95
Blattaria
6,42
3,42±16,80
2,18±12,57
1,16
0,86
4,01
2,81
Chilopoda
-
-
-
- -
-
-
Coleoptera
22,02
8,33±21,40
2,39±11,49
5,98
2,42
10,91
10,14
Diplopoda
0,92
0,09±0,96
0,02±0,16
0,17
0,04
0,34
0,38
Diptera
1,83
1,03±9,64
0,10±0,92
0,33
0,16
0,99
0,77
Hemiptera
17,43
4,83±12,70
8,04±23,71
4,49
3,40
10,10
8,44
Hymenoptera
- Formicidae
- Outros
-
16,51
4,59
-
6,22±17,38
2,94±15,05
-
2,06±11,36
1,20±10,98
-
5,81
0,83
-
0,53
0,08
-
8,26
2,91
-
7,62
1,83
Isoptera
17,43
10
,69±
25,72
7,28±20,19
44
,02
3,06
11,80
21,50
Larva de inseto
34
,86
11
,20±
20,17
22,16±
35,77
10
,30
11,14
22
,74
18,77
Lepidoptera
2,75
0,44±3,35
0,38±2,43
0,50
0,86
1,19
1,37
Mantodea
3,67
0,92±5, 66
0,01±0,14
0,66
0,04
1,53
1,46

37
Mat. vegetal
8,26
3,83±14,73
4,30±18,86
2,33
29,01
5,46
13,20
Não ident.
13,76
8,41±25,63
1,26±9,88
2,49
4,23
7,81
6,83
Odonata
1,83
0,46±3,37
1,41±10,76
0,33
1,61
1,23
1,26
Orthoptera
51
,38
27
,86±
37,89
36,10±
43,57
11
,63
35,60
38
,45
32,87
Ovo de inseto
0,92
0,37±3,83
0,64±5,83
0,33
0,04
0,64
0,43
Ovo de réptil
-
-
-
- -
-
-
Pupa de inseto
0,92
0,18±1,92
0,36±3,29
0,17
1,09
0,49
0,73
Scorpionida
4,59
1,22±6,30
0,85±6,61
0,83
1,23
2,22
2,22
38
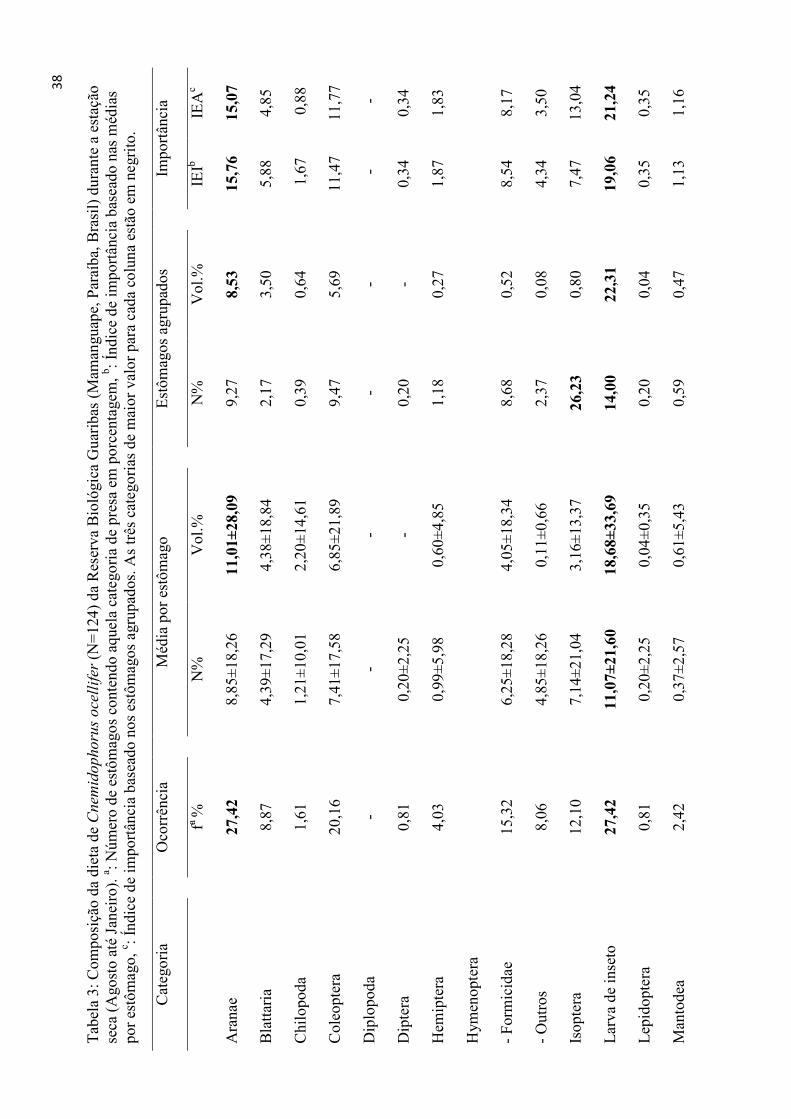
Tabela 3: Composição da dieta de Cnemidophorus ocellifer (N=124) da Reserva Biológica Guaribas (Mamanguape, Paraíba, Brasil) durante a estação
seca (Agosto até Janeiro). a: Número de estômagos contendo aquela categoria de presa em porcentagem, b: Índice de importância baseado nas médias
por estômago, c : Índice de importância baseado nos estômagos agrupados. As três categorias de maior valor para cada coluna estão em negrito.
Categoria
Ocorrência
Média por estômago
Estômagos agrupados
Importância
fa %
N%
Vol.%
N%
Vol.%
IEIb
IEAc
Aranae
27
,42
8,85±18,26
11,01±
28,09
9,27
8,53
15
,76
15,07
Blattaria
8,87
4,39±17,29
4,38±18,84
2,17
3,50
5,88
4,85
Chilopoda
1,61
1,21±10,01
2,20±14,61
0,39
0,64
1,67
0,88
Coleoptera
20,16
7,41±17,58
6,85±21,89
9,47
5,69
11,47
11,77
Diplopoda
-
-
-
- -
-
-
Diptera
0,81
0,20±2,25
-
0,20
-
0,34
0,34
Hemiptera
4,03
0,99±5,98
0,60±4,85
1,18
0,27
1,87
1,83
Hymenoptera
- Formicidae
- Outros
15,32
8,06
6,25±18,28
4,85±18,26
4,05±18,34
0,11±0,66
8,68
2,37
0,52
0,08
8,54
4,34
8,17
3,50
Isoptera
12,10
7,14±21,04
3,16±13,37
26
,23
0,80
7,47
13,04
Larva de inseto
27
,42
11
,07±
21,60
18,68±
33,69
14
,00
22,31
19
,06
21,24
Lepidoptera
0,81
0,20±2,25
0,04±0,35
0,20
0,04
0,35
0,35
Mantodea
2,42
0,37±2,57
0,61±5,43
0,59
0,47
1,13
1,16

39
Mat. vegetal
5,65
3,25±14,94
3,71±18,33
2,17
5,58
4,20
4,47
Não ident.
14,52
10
,78±
29,16
2,68±15,38
3,55
0,78
9,33
6,28
Odonata
1,61
0,88±9,01
0,42±3,98
0,39
0,16
0,97
0,72
Orthoptera
48
,39
24
,12±
34,05
30,81±
42,13
15
,98
34,21
34
,44
32,86
Ovo de inseto
-
-
-
- -
-
-
Ovo de réptil
0,81
0,81±8,98
1,12±10,60
0,20
6,49
0,91
2,50
Pupa de inseto
1,61
0,36±2,86
1,09±10,29
0,39
1,78
1,02
1,26
Scorpionida
8,87
6,86±24,72
8,48±27,29
2,56
8,14
8,07
6,52

40
Tabela 4. - Análise de correlação canônica e análise de redundância canônica das dimensões de
presa versus dimensões de cabeça em C. ocellifer das savanas de Tabuleiro.
Coeficientes canônicos ajustados para as dimensões de cabeça
1ª variável canônica 2ª variável canônica
Largura da cabeça 0,53 -2,17
Altura de cabeça 0,45 0,92
Comprimento da cabeça 0,06 1,44
Coeficientes canônicos ajustados para as dimensões de presa
1ª variável canônica 2ª variável canônica
Largura max. de presa 0,97 -0,57
Comprimento max. de presa 0,06 1,13
Variáveis Canônicas Correlação
Canônica
Correlação Canônica
Ajustada
F (apróx.) P
I 0,31 0,27 3,26 0,004
II 0,25 0,14 1,81 0,17
Wilk’s Lambda = 0,89, P = 0,004
Variação ajustada das dimensões de presa explicadas pelas variáveis canônicas das dimensões de
cabeça
Variável canônica Proporção Proporção acumulada
I 0,63 0,63
II 0,37 1,00