aspectos da microsporogênese em acessos poliplóides de ... · kellen regina boldrini aspectos da...
TRANSCRIPT

KELLEN REGINA BOLDRINI
Aspectos da microsporogênese em acessos
poliplóides de Brachiaria (Poaceae: Paniceae)
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Biológicas (área de concentração – Biologia Celular), da Universidade Estadual de Maringá para obtenção do grau de Mestre em Ciências Biológicas.
MARINGÁ- PR
2006

2
KELLEN REGINA BOLDRINI
Aspectos da microsporogênese em acessos
poliplóides de Brachiaria (Poaceae: Paniceae)
Orientadora
Dra Maria Suely Pagliarini
Co-Orientadora
Dra Cacilda Borges do Valle
MARINGÁ- PR
2006

3
Dados Internacionais de Catalogação-na-Publicação (CIP) (Biblioteca Central - UEM, Maringá – PR., Brasil)
Boldrini, Kellen Regina B687a Aspectos da microsporogênese em acessos poliplóides
de Brachiaria (Poaceae: Paniceae) / Kellen Regina Boldrini. -- Maringá : [s.n.], 2006.
51 f. : il. Orientador : Prof. Dr. Maria Suely Pagli arini; Co-
orientadora: Cacilda Borges do Valle Dissertação (mestrado) - Universidade Es tadual de
Maringá. Programa de Pós-graduação em Ciências Biológicas, 2006.
1. Biologia celular. 2.Citogenética vege tal. 2.
Melhoramento genético. 3.Meiose. 4. Poliploidia. 5. Apomixia. 6. Citomixia. 7. Fusão celular. 8. Gameta 2n. 9. Brachiaria humidicola. 10. Brachiaria decumbens. 11. Brachiaria dura. 12. Gramineae. Universidade Estadual de Maringá. Programa de Pós-graduação em Ciências Biológicas.
CDD 21.ed. 571.6845

4
Dedico
Para meus pais... Meus mentores e heróis.
AGRADECIMENTOS
A Deus, luz na minha vida.

5
À professora Dra Maria Suely Pagliarini, pela determinação, dedicação, ensinamentos e
profissionalismo, a quem admiro muito.
À Dra Cacilda Borges do Valle, pesquisadora do Centro Nacional de Pesquisa de Gado
de Corte/Embrapa (Campo-Grande- MS), pela co-orientação e concessão do material
analisado.
À Coordenação e demais docentes do Programa de Pós-Graduação em Ciências
Biológicas/UEM, pela oportunidade de realização desse curso.
À Capes, pela bolsa de estudo concedida.
Às amigas, Andréa, Eleniza e Neide, pelo carinho e presença constante em todos os
momentos e, principalmente, pela amizade.
A todos os colegas de curso e de laboratório, pela amizade e convivência.
Em especial, agradeço à minha família, cuja força e apoio foram determinantes para a
realização deste trabalho e pelo carinho demonstrado nas horas mais difíceis.

6
SUMÁRIO
Apresentação..................................................................................................... 1
Resumo.............................................................................................................. 2
Abstract………………………………………………………………...…...… 3
Artigos: 1. A new meiotic mechanism for 2n gamete formation in Brachiaria (Poaceae: Paniceae)………………………………………………………..…. 4 2. Cell fusion and cytomixis during microsporogenesis in Brachiaria humidicola (Poaceae)……………………………………………………….... 17 3. Abnormal timing of cytokinesis in microsporogenesis of Brachiaria humidicola (Poaceae: Paniceae)…………………………………….………… 30 Apêndice……………………………………………………………..….......... 40

7
APRESENTAÇÃO
Esta dissertação é composta por três artigos originados a partir da análise do
comportamento meiótico em acessos de três espécies de Brachiaria (B. humidicola, B.
decumbens e B. dura), coletados junto à coleção de germoplasma desta gramínea,
alocada na Embrapa Gado de Corte (Campo Grande, MS).
Os artigos estão apresentados de acordo com as normas estabelecidas pelas
revistas a que foram submetidos.
1. Kellen Regina Boldrini, Patrícia Helena Gallo, Pamela Lonardoni Micheletti, Claudicéia Risso-Pascotto, Maria Suely Pagliarini, and Cacilda Borges do Valle. A new meiotic mechanism for 2n gamete formation in Brachiaria (Poaceae: Paniceae). Sexual Plant Reproduction (submetido)
2. Kellen Regina Boldrini, Maria Suely Pagliarini, and Cacilda Borges do Valle. Cell fusion and cytomixis during microsporogenesis in Brachiaria humidicola (Poaceae). South African Journal of Botany (submetido)
3. Kellen Regina Boldrini, Maria Suely Pagliarini, and Cacilda Borges do Valle. Abnormal timing of cytokinesis in microsporogenesis of Brachiaria humidicola (Poaceae: Paniceae). Journal of Genetics (aceito)
Aspectos da microsporogênese em acessos poliplóides de Brachiaria (Trin.) Griseb. (Poaceae: Paniceae)
Kellen Regina Boldrini Maria Suely Pagliarini
Cacilda Borges do Valle
RESUMO. A coleção de germoplasma de Brachiaria spp da Embrapa Gado de Corte
conta com 475 acessos de 15 espécies. Algumas espécies encontram-se sob estudos
citológicos a fim de subsidiar o programa de melhoramento desta forrageira.
Anormalidades meióticas têm sido descritas entre acessos já analisados. Acessos
poliplóides de três espécies de Brachiaria, incluindo B. humidicola (2n = 4x = 36), B.
decumbens (2n = 4x = 36) e B. dura (2n = 6x = 54) apresentaram citocinese anormal. O
primeiro sinal de citocinese apareceu somente em metáfase II e não dividiu o meiócito

8
em díade. Ausência total de citocinese também foi detectada entre meiócitos durante a
segunda divisão. Em ambos os casos, a proximidade entre as duas placas metafásicas
facilitou a reunião dos cromossomos após a anáfase II, formando um núcleo de
restituição em telófase II. Na maioria dos meiócitos, a segunda citocinese ocorreu após
a telófase II. Mônades, díades e tríades com núcleos n ou 2n foram observados entre os
produtos meióticos. Entre 28 acessos poliplóides de B. humidicola analisados, fusões
celulares foram encontradas em dois acessos e transferência de cromossomos entre
meiócitos em um deles. Nesta espécie, anormalidades meióticas e pós-meióticas
relacionadas à citocinese também foram encontradas em um acesso. A primeira
citocinese ocorreu após a telófase II, enquanto a segunda citocinese ocorreu por
invaginação em micrósporos binucleados após a dissolução da parede de calose. Esta
citocinese tardia não afetou a viabilidade do pólen uma vez que o produto meiótico,
embora formado tardiamente, foi caracterizado por quatro micrósporos haplóides. A
meiose é controlada por um grande número de genes, geralmente dominantes, que são
estágio, local e tempo-específicos. As anormalidades meióticas causadas por mutações
afetam a viabilidade do pólen e comprometem a produção de sementes. No gênero
Brachiaria, acessos poliplóides são, em geral, apomíticos, embora pseudógamos.
Conseqüentemente, pólen fértil é essencial para fertilizar o núcleo central do saco
embrionário e garantir a produção de sementes viáveis. Assim, acessos com alta
freqüência de anormalidades meióticas devem ser eliminados dos programas de
melhoramento.
Aspects of microsporogenesis in polyploid accessions of Brachiaria (Trin.) Griseb. (Poaceae: Paniceae)
Kellen Regina Boldrini Maria Suely Pagliarini
Cacilda Borges do Valle
ABSTRACT. The Brazilian Brachiaria spp collection at Embrapa Beef Cattle
encompasses 475 accessions of 15 species. Some species are under detailed analysis of
microsporogenesis. Meiotic abnormalities have been recorded among accessions.
Polyploid accessions of three Brachiaria species, including B. humidicola (2n = 4x =
36), B. decumbens (2n = 4x = 36), and B. dura (2n = 6x = 54) presented abnormal
cytokinesis. The first sign of cytokinesis appeared only in metaphase II and it did not

9
divid the meiocyte into a dyad. Total absence of cytokinesis was also detected among
meiocytes in the second division. In both cases, the two metaphase plates were very
close facilitating the rejoining of chromosome sets after anaphase II, forming a
restitutional nucleus in telophase II. In the majority of meiocytes, the second cytokinesis
occurred after telophase II. Monads, dyads, and triads with n or 2n nuclei were recorded
among meiotic products. Among 28 polyploid accessions of B. humidicola analyzed,
cell fusions were recorded in two accessions and chromosome transfer among
meiocytes in one of them. In this species, meiotic and post-meiotic abnormalities related
to cytokinesis were observed in one accession. The first cytokinesis occurred after
telophase II, and the second cytokinesis occured by invagination in binucleated
microspores only after callose wall dissolution. These late cytokinesis did not affect
pollen viability since the meiotic product, although formed late, was characterized by
four reduced normal microspores. Meiosis is controlled by a large number of genes,
generally dominant, which are stage, site- and time-specific. Meiotic abnormalities
caused by mutations compromise pollen viability and impair seed production. In the
Brachiaria genus, polyploid accessions are, in general, apomictic, albeit pseudogamous.
Consequently, fertile pollen is essential to fertilize the central nucleus of the embryo-sac
and ensure viable seed production. Thus, accessions with high frequencies of meiotic
abnormalities should be eliminated early from the breeding program.

10
A new meiotic mechanism for 2n gamete formation in Brachiaria (Poaceae: Paniceae)
Kellen Regina Boldrini1,Patrícia Helena Gallo1, Pamela Lonardoni Micheletti1, Claudicéia Risso-Pascotto1, Maria Suely Pagliarini1, and Cacilda Borges do Valle2.
1. Department of Cell Biology and Genetics, State University of Maringá, 87020-900 Maringá PR Brazil. 2. Embrapa Beef Cattle, P.O. Box 154, 79002-970 Campo Grande MS Brazil.
* Author for correspondence: Maria Suely Pagliarini (E-mail: [email protected]).
Abstract. Microsporogenesis of several Brachiaria species of the Brazilian collection at
Embrapa Beef Cattle was analysed in detail. Accessions of three species (B. humidicola,
A new meiotic mechanism for 2n gamete formation in Brachiaria
(Poaceae: Paniceae)

11
2n=4x=36, B. decumbens, 2n=4x=36, and B. dura, 2n=6x=54) presented abnormal
cytokinesis. Chromosomes paired in bi-, tri-, and quadrivalents in these accessions,
whereas chromosome segregation at meiosis I was characterized by exclusion of
laggards as micronuclei. In a high number of meiocytes, the first sign of cytokinesis
appeared only in metaphase II and did not divide the meiocyte into a dyad. Total
absence of cytokinesis was also detected among meiocytes in the second division. Since
in both cases the two metaphase plates were very close, they favored the rejoining of
chromosome sets after anaphase II and formed a restitutional nucleus in telophase II.
Second cytokinesis occurred after telophase II in most meiocytes. Monads, dyads, and
triads with n or 2n nuclei were observed among meiotic products. The 2n gametes
observed correspond to the first division restitution (FDR). The number of affected cells
in each accession was variable, but the number of microspores with restitutional
nucleus, including those scored in tetrads and the released ones, did not exceed 9%.
Although polyploidy is common in the genus Brachiaria, its origin is not well known.
Current results suggest that 2n gametes may have contributed to the evolutionary
history of the genus.
Key words: abnormal cytokinesis, Brachiaria, 2n gamete, microsporogenesis,
restitutional nucleus, polyploidization.
Introduction
Polyploidy is a key element in the evolution of higher plants and leads toward the
formation of new species. Most flowering species have evolved through one or more
rounds of polyploidization, either by the doubling of their chromosome number
(autopolyploidy) or by combining chromosome sets from distinct related species
(allopolyploidy). Although this behavior suggests that polyploidy must confer some
selective advantage (Osborn, 2004), the evolutionary success of polyploidy relies on the
ability of polyploid individuals to reproduce and transmit their genes to subsequent
generations (Pannell et al., 2004). For nearly 90 years since the discovery of polyploidy,
it has not been known what percentage of the angiosperm is polyploid. Estimates of

12
polyploidy among angiosperms range from a liberal 70 – 80% to a conservative 30%
(Bennett, 2004).
Polyploidization may be asexual through somatic chromosome doubling and
sexual through the formation of 2n gametes. The first assumption was seriously
questioned by Harlan and De Wet (1975) since it has been shown that 2n gametes are
widespread in plants (Harlan and De Wet, 1975; Veilleux, 1985; Bretagnolle and
Thompson, 1995). Actually, 2n gametes are considered to be the dominant process
involved in the origin of polyploidy in plants. 2n gametes result by modified meiosis
affecting specific stages of micro- and megasporogenesis, which lead to the formation
of restitutional nucleus. Several mechanisms involved in meiotic nuclear restitution
have been reported in several plant species (see Ramanna, 1979; Veilleux, 1985;
Bretagnolle and Thompson, 1995).
During an extensive study on cytogenetic behavior in several Brachiaria species
from a Brazilian collection at Embrapa Beef Cattle (Campo Grande, MS), a new meiotic
mechanism for 2n gamete formation was recorded in three different species (B.
humidicola, B. decumbens, and B. dura). This mechanism, leading to first division
restitution (FDR), is currently discussed.
Materials and Methods
Accessions of several Brachiaria species from the Embrapa Beef Cattle collection
(Campo Grande MS Brazil), comprising B. humidicola (23 accessions), B. decumbens
(15 accessions), and B. dura (2 accessions), were cytologically analyzed. All accessions
were previously collected in the wild African savannas in the mid-1980s by the Centro
Nacional de Agricultura Tropical (CIAT, Colombia) and introduced in Brazil between

13
1986 and 1990. Site’s characteristics in Brazil are: climate type Aw: tropical humid
savanna; average annual precipitation = 1526 mm; average temperature = 22°C; altitude
520 m; latitude = 20° 28’ S; longitude = 55° 40’ W; poor Dark Red Latossol (59% sand;
8% silt; 33% clay; pH = 4.2).
Inflorescences from several plants representing each accession were collected
for meiotic study and fixed in a mixture of ethanol 95%, chloroform and propionic acid
(6:3:2) for 24 hours, transferred to 70% alcohol and stored under refrigeration until use.
Microsporocytes were prepared by squashing and staining with 0.5% propionic
carmine. Photomicrographs were made with a Wild Leitz microscope using Kodak
Imagelink – HQ, ISO 25 black and white film.
Results and Discussion
The Brazilian Brachiaria collection comprises of 475 accessions of 15 species, of which
nearly half has been cytologically analyzed. Among these, three of them belonging to
distinct species, B. humidicola (H047, 2n=4x=36), B. decumbens (D076, 2n=4x=36)
and B. dura (Du001, 2n=6x=54), revealed total absence or an abnormal type of
cytokinesis following the first or the second meiotic division. As all accessions were
polyploid, characteristica polyploidy abnormalities related to irregular chromosome
segregation were also found among meiocytes. However, meiosis proceeded normally
in its course till telophase I (Fig. 1 a to c).
Brachiaria is a genus of African origin widely used as forage grass in the humid
and sub-humid tropical regions, especially in Brazil. As a monocotyledonous species, it
shows successive cytokinesis, or rather, the first cytokinesis occurs after telophase I,
dividing the cell into a dyad, followed by the second cytokinesis after telophase II,

14
giving rise to microspore tetrads. In the accessions under study, a large number of
meiocytes (Table 1) displayed the first signal of cytokinesis in metaphase II (Fig. 1 d),
albeit partial, and, it did not divide the meiocyte into a dyad. During prophase II,
cytokinesis was not evident in most cells. In metaphase II, anaphase II, and telophase II,
the number of meiocytes with partial cytokinesis was much higher than those lacking it.
In both cases, the two metaphase plates were very close (Fig. 1 d) favoring the rejoining
of chromosome sets after anaphase II (Fig. 1 e, f). Consequently, a restitutional nucleus
in telophase II was formed (fig. 1 g, h). The above phenomenon was prevalent and
restitutional nucleus formation was increased by moderate convergence of spindles, i.e.,
tripolar spindles (Fig. 1 e). Depending on its position, the restitutional nucleus occupied
either one cell (Fig. 1 p) or shared the cytoplasm with another n nucleus (Fig. 1 h). After
telophase II, the start of the second cytokinesis occurred although the first cytokinesis
was either not yet complete (Fig. 1 h and j) or totally absent (Fig. 1 k, l). As a
consequence, the number of tetrads with incomplete cytokinesis was high, and monads,
dyads, and triads with n or 2n nuclei were recorded among meiotic products. Abnormal
microspores with different combinations of n and 2n nuclei were observed (Fig. 1 m to
p).
Table 1. Frequency of meiotic abnormalities in the three accessions analyzed. Phase Abnormalities Accession H047 D076 Du001 Analyzed
cells No. of cells
affected (%)
Analyzed cells
No. of cells affected
(%)
Analyzed cells
No. of cells affected
(%) Prophase II Total absence of cytokinesis 366 227 (62.0) 150 85 (56.7) 146 1 (0.7) Partial cytokinesis 36 (9,8) 0 80 (54.8)

15
Metaphase II Total absence of cytokinesis 721 22 (3.1) 199 21 (10.6) 244 6 (2.5) Partial cytokinesis 83 (11.5) 23 (11.6) 134 (54.9) Anaphase II Total absence of cytokinesis 177 2 (1.1) 140 11 (7.9) 147 3 (2.0) Partial cytokinesis 87 (49.2) 34 (24.3) 69 (46.9) Tripolar spindle 21 (11.9) 22 (15.7) 0 Telophase II Total absence of cytokinesis 564 19 (3.4) 215 13 (6.1) 206 3 (1.46) Partial cytokinesis 210 (37.2) 35 (16.3) 124 (60.2) Restitutional nucleus 43 (7.6) 36 (16.7) 12 (5.8) Tetrad Partial cytokinesis 519 132 (25.4) 1235 146 (11.8) 166 24 (14.5) Monad trinucleated 15 (2.9) 23 (1.9) 0 Dyad binucleated 13 (2.5) 81 (6.6) 0 Triad 86 (6.6) 0 4 (2.4) Restitutional nucleus in dyad or triad 45 (8.7) 0 7 (4.2) Microspores Trinuclead 676 22 (3.3) 620 14 (2.3) 539 0 Restitutional nucleus 3 (0.4) 13 (2.1) 7 (1.3) Binucleated 104 (15.4) 60 (9.7) 60 (11.1) Tetranucleated 0 7 (1.3 ) 0

16
Figure 1. Aspects of abnormal cytokinesis and restitutional nucleus formation in Brachiaria. a to c) Regular meiosis I: metaphase I (a), anaphase I (b), and telophase I (c). d) Metaphase II with incomplete cytokinesis. The proximity of the two metaphase plates may be observed. e) Anaphase II with convergent spindles in a cell with total absence of cytokinesis. Chromosomes rejoining in one pole is shown. f) Early telophase II with a restitutional nucleus. g) Telophase II with the formation of a restitutional nucleus. h) Telophase II with the restitutional nucleus sharing the cytoplasm with one n nucleus. i, j) Different aspects of telophase II in relation to cytokinesis. k, l) Telophase II with total absence of cytokinesis. m, n) Triads with one restitutional nucleus in one microspore. o) Tetranucleated monad and two normal n microspores. p) Binucleated microspore and a normal one. (Magnification 400x)

17
Similar mechanism was reported in Hierochloë odorata, a rhizomatous perennial grass
(Ferris et al., 1992). The number of affected cells in each accession was variable, but the
number of microspores with restitutional nuclei, including those scored in tetrads and
the released ones, did not exceed 9%. The trend to form 2n gametes in plants is highly
variablea and it varies among individuals within a single taxonomic group or even
among flowers of an individual plant (Bretagnolle and Thompson, 1995).
Meiotic nuclear restitution may be caused by different mechanisms, including
semiheterotypic division, pseudohomotypic division, mitotised meiosis, first division
restitution (FDR), second division restitution (SDR), premature cytokinensis 1 and 2,
and pre- and post-meiotic doubling of chromosomes (Ramanna, 1979; Veilleux, 1985,
Bretagnolle and Thompson, 1995). Most mechanisms have been described in
dicotyledons with scanty studies and revealed 2n gamete formation in monocots
(Pagliarini et al., 1999; Lim et al., 2001; Barba-Gonzalez et al., 2004). In monocots it is
very important to determine how events of chromosomal division and cytokinesis occur
to yield restitutional nucleus. Although 2n gametes may result from several different
meiotic abnormalities, two types of 2n gametes are the product of one out of two basic
processes and depend on the mode of nuclear restitution: FDR and SDR. The 2n
gametes in the present accessions resulted from FDR. In spite of correct chromosome
pairing and homologous segregation at anaphase I, the homologous chromosomes were
rejoined in the second division. Heterozygosis level in 2n gametes is influenced by the
timing of nuclear restitution. Although FDR generally conserves heterozygosity of
proximal segments, it reduces that of distal segments by half (Veilleux, 1985;
Bretagnolle and Thompson, 1995). According to Lim et al. (2001), an important feature
of restitutional meiosis with successive cytokinesis, such as that observed in the present
Brachiaria accessions, is the manner in wich chromosomal division and cytokinesis

18
events occur. Cytological observations in these accessions clearly indicated that meiosis
had been modified: chromosomes paired normally in prophase I, formed bi-, tri-, and
quadrivalents and then segregated simultaneously in anaphase I, in spite of certain
abnormal segregation due to their polyploid condition. As genetic recombination
occurred, FDR nucleus gave rise to 2n gametes with recombinant chromosomes.
Recombination is one of the most important events for introgression. Most of the
evidence for in using 2n gametes in breeding programs has been focused on their use in
autopolyploids or their polysomic polyploids (Mariani and Tavoletti, 1992; Carputo et
al., 2000). In these cases, the importance of FDR gametes lies in the transference of
heterosis and intact parental gene combinations to sexual polyploids. The 2n gametes
allow breeders to broaden the genetic bases of cultivated species.
Although the genetic determination of 2n pollen production has been studied in
detail, the genetic control of 2n egg formation is still poorly understood. Bretagnolle
and Thompson (1995) presented an extensive list of genes responsible for this feature in
several plant species, in which monogenic recessive status of mutant alleles is largely
predominant. Although the genetic base of abnormal cytokinesis found in these
accessions is not known, the fact that accessions of three different Brachiaria species
have precisely the same abnormality, has led the authors to hypothesize that the
characteristic is genetically controlled. 2n gametes resulting from total absence either of
the first or the second cytokinesis have been reported in B. brizantha (Risso-Pascotto et
al., 2003), but in much lower frequency. Some exhaustive reviews have shown the
influence of seasonal and environmental factors, such as high and low temperature, on
2n gamete production (see Bretagnolle and Thompson, 1995).
In the genus Brachiaria, polyploidy is of common occurrence (Valle and
Savidan, 1996). Studies performed by flow cytometry on 435 accessions, belonging to

19
13 species, revealed that only 13% are diploids. The ploidy level ranged from 2n = 4x to
2n = 7x, with predominance (58.13%) of tetraploidy (Penteado et al., 2000). The origin
of polyploidy in the genus is not well known. Taking into account the meiotic pairing
and the meiotic behavior of several accessions of different Brachiaria species studied,
there is evidence of autopolyploidy and segmental allopolyploidy (Mendes-Bonato et
al., 2002, 2006; Utsunomiya et al., 2005), and even of true allopolyploidy (Mendes et
al., 2006). At any rate, 2n gametes may have contributed to the evolutionary history of
the genus. In fact, recent studies of natural polyploidy complexes have shown that the
production of 2n gametes may have played a role in the creation of new polyploids by
unilateral or bilateral sexual polyploidization (see Bretagnolle and Thompson, 1995).
The forage potential of Brachiaria species was acknowledged in Brazil about 40
years ago. However, the importance of the genus was felt only in the past three decades
when two to three Brachiaria cultivars, were extensively sown in tropical America
(Miles et al., 1996, 2004). Brachiaria decumbens cv. Basilisk (D) and B. brizantha cv.
Marandu (B) undoubtedly make up the most extensively planted cultivars, covering
over 50 million hectares of poor and acid soils of central Brazil and Latin America. To
increase genetic variability in the genus, a dynamic breeding program based on intra-
and interspecies hybridization is underway at the Embrapa Beef Cattle Center since
1988. Hybridization in the genus Brachiaria is rather difficult mainly owing to ploidy
differences among accessions and species, and to reproduction by apomixis (Valle and
Savidan, 1996). Most accessions in promising species are tetraploid and apomictic.
Thus, several tetraploid interspecific hybrids involving different accessions of (D) and
(B) were synthesized by the use of artificially tetraploidized sexual accessions of B.
ruziziensis as female genitors. The success of such program depends heavily on the high
pollen fertility of the apomictic accessions used as the male parent. These crosses

20
obviously involve 2n gametes generated by regular chromosome segregation of both
tetraploid genitors. Since 2n gametes from restitutional nucleus have never been
explored in the genus for breeding purposes, current results open new perspectives,
possibilities and further exploration for Brachiaria breeding.
References Barba-Gonzalez R, Lokker AC, Lim KB, Ramanna MS, Tuyl JM (2004) Use of 2n
gametes for the production of sexual polyploids from sterile Oriental x Asiatic
hybrids of lilies (Lilium). Theor Appl Genet 109:1125-1132
Bennett MD (2004) Perspectives on polyploidy in plants – ancient and neo. Biol J Linn
Soc 82:411–423
Bretagnolle F, Thompson JD (1995) Gametes with the somatic chromosome number:
mechanisms of their formation and role in the evolution of autopolyploid plants.
New Phytol 129:1-22
Carputo D, Barone A, Frusciante L (2000) 2n gametes in the potato: essential
ingredients for breeding and germplasm transfer. Theor Appl Genet 101:805-813
Ferris C, Callow RS, Gray AJ (1992) Mixed first and second division restitution in male
meiosis of Hierochloë odorata (L.) Beauv (Holy grass). Heredity 69:21-31
Harlan JR, De Wet JMJ. 1975. On Ö Winge and a prayer: the origins of polyploidy. Bot
Rev 41:361-390
Lim KB, Ramanna MS, Jong JH, Jacobsen E, Tuyl JM (2001) Indeterminate meiotic
restitution (IMR): a novel type of meiotic nuclear restitution detected in interspecific
lily hybrids by GISH. Theor Appl Genet 103:219-230
Mariani A, Tavoletti S (1992) Alfalfa evolution and breeding through 2n gametes. In:
Veronesi et al. (eds). Gametes with somatic chromosome number in the evolution
and breeding of polyploidy polysomic species: achievements and perspectives. Proc.
Workshop, Tipografia Forziuncula, Assisi, Italy, pp 73-81

21
Mendes DV, Boldrini KR, Mendes-Bonato AB, Pagliarini MS, Valle CB (2006)
Cytological evidence of natural hybridization in Brachiaria brizantha Stapf
(Gramineae). Bot J Linn Soc (in press).
Mendes-Bonato AB, Pagliarini MS, Forli F, Valle CB, Penteado MIO (2002)
Chromosome number and microsporogenesis in Brachiaria brizantha (Gramineae).
Euphytica 125:419-425
Mendes-Bonato AB, Pagliarini MS, Risso-Pascotto C, Valle CB (2006) Chromosome
number and meiotic behavior in Brachiaria jubata (Gramineae). J Genet (in press).
Miles JW, Maass BL, Valle CB (1996) Brachiaria: Biology, Agronomy, and
Improvement. CIAT/Embrapa, 288 p
Miles JW, Valle CB, Rao IM, Euclides VPB (2004) Brachiaria-grasses. In: L.
Sollenberger (ed) Warm-Season (C4) Grasses. ASA, CSSA, SSSA, Madison.
Agronomy Monograph, n.45. pp 745-760
Osborn TC (2004) The contribution of polyploidy to variation in Brassica species.
Physiol Plantarum 121:531-536
Pagliarini MS, Takayama SY, Freitas PM, Carraro LR, Adamowski EV, Silva N (1999)
Failure of cytokinesis and 2n gamete formation in Brazilian accessions of
Paspalum. Euphytica 108:129-135
Pannell JR, Obbard DJ, Buggs RJA (2004) Polyploidy and the sexual system: what can
we learn from Mercurialis annua? Biol J Linn Soc 82:547-560
Penteado MIO, Santos ACM, Rodrigues IF, Valle CB, Seixas MAC, Esteves A (2000)
Determinação de poliploidia e avaliação da quantidade de DNA total em diferentes
espécies de gênero Brachiaria. Boletim de Pesquisa, 11. Campo Grande-MS,
Embrapa Gado de Corte. 19p.
Ramanna MS (1979) A re-examination of the mechanisms of 2n gametes formation in
potato and its implications for breeding. Euphytica 28:537-561

22
Risso-Pascotto C., Pagliarini MS, Valle CB, Mendes-Bonato AB (2003) Chromosome
number and microsporogenesis in pentaploid accession of Brachiaria brizantha
(Gramineae). Plant Breed 122:136-140
Utsunomiya KS., Pagliarini MS, Valle CB (2005) Microsporogenesis in tetraploid
accessions of Brachiaria nigropedata (Ficalho & Hiern) Stapf (Gramineae). Biocell,
(in press).
Valle CB, Savidan YH (1996) Genetics, cytogenetics, and reproductive biology of
Brachiaria. In: Miles JW, Maass BL, Valle CB (eds) Brachiaria: Biology,
Agronomy, and Improvement. Colombia: CIAT, pp 147-163
Veilleux R (1985) Diploid and polyploid gametes in crop plants: mechanisms of
formation and utilization in plant breeding. Plant Breed Rev 3:253-288

23
Cell fusion and cytomixis during microsporogenesis in Brachiaria
humidicola (Poaceae)

24
Cell fusion and cytomixis during microsporogenesis in
Brachiaria humidicola (Poaceae)
Kellen Regina Boldrini1, Maria Suely Pagliarini1, and Cacilda Borges
do Valle2
1 Department of Cell Biology and Genetics, State University of Maringá, 87020-900 Maringá PR, Brazil; 2 Embrapa Beef Cattle, P.O. Box 154, 79002-970 Campo Grande MS, Brazil.
ABSTRACT
During cytological analysis for breeding purposes of microsporogenesis in 28 polyploid
accessions of Brachiaria humidicola (Poaceae) from the Embrapa Beef Cattle
germplasm collection assembled, cell fusions were recorded in two accessions and
chromosome transfer among meiocytes in one of them. Cell fusion between two to more
than ten cells was recorded from prophase I to telophase II. In the syncyte, each nucleus
maintained its integrity. In one of these accessions, cytomixis with characteristics never
reported in any other plant species was recorded. It occurred among very small
meiocytes that transferred the entire genome or part of it to normal meiocytes.
Chromosome transfer occurred preferentially during telophase I and, during migration,
chromatin showed structural alteration. Both abnormalities compromise pollen fertility.
In the Brachiaria genus, polyploid accessions are, in general, apomictic, albeit
pseudogamous. Consequently, fertile pollen is essential to fertilize the central nucleus of
the embryo-sac and ensure viable seed production. Thus, accessions with high
frequencies of meiotic abnormalities should be eliminated early from the breeding
program.
Key words: Brachiaria humidicola, cell fusion, cytomixis, forage grass,
microsporogenesis.

25
INTRODUCTION
Meiosis is a continuous process involving several cytological events that result in the
reduction of chromosome number by half, thus ensuring the constancy of ploidy in the
species after fertilization. There is ample evidence that meiosis is controlled by a large
number of genes (Gottschalk and Kaul, 1974; Baker et al., 1976; Golubovskaya, 1979,
1989). Disruption in any step of meiosis, due to environmental or genetic factors can
affect gametic fertility. Depending on the severity of the abnormality, total sterility can
be expected.
Cytological analyses performed on several Brachiaria species of the Embrapa
Beef Cattle collection revealed a large amount of different meiotic (Mendes-Bonato et
al., 2001 a, b; 2002 a, b, 2003; Risso-Pascotto et al., 2002, 2003 a; Utsunomiya et al.,
2004, 2005) and post-meiotic abnormalities (Junqueira Filho et al., 2003; Mendes-
Bonato et al.; 2004, Risso-Pascotto et al., 2005 a) which compromise pollen viability. In
spite of the prevalent asexual reproduction by apospoy of the Panicum type (Valle and
Savidan, 1996; Araújo et al., 2000), these polyploid accessions in the genus Brachiaria
are pseudogamous, i.e. fertile pollen grains are necessary to fertilize the two secondary
nuclei of the embryo sac to ensure endosperm development (Alves et al., 2001).
B. humidicola is a species natural to Africa and widely used for pastures in the
tropics, especially under poorly drained conditions, where its is about the only option
available for pasture establishment. New cultivars are urgently needed to minimize the
risk of extensive contiguous areas being planted to the only apomictic cultivar available
commercially. New varieties are being sought to explore either the natural genetic
variability among accessions or to generate novel genetic variability by intraspecific
hybridization, since tetraploid sexual accessions have been identified (Valle and

26
Glienke, 1991; Valle and Savidan, 1996). A new cultivar, cv Tupi, is scheduled to be
released in Brazil in 2007 and was selected from the germplasm collection. There is an
increase in interest in this species by breeders and producers thus justifying the effort to
analyze the microsporogenesis of all the accessions of this species and related ones in
the Embrapa Beef Cattle collection. During cytological characterization of 28
accessions of B. humidicola, two showed cell fusion and one of these also cytomixis
among meiocytes. These abnormalities are reported here.
MATERIAL AND METHODS
Twenty eight of about 60 accessions of Brachiaria humidicola (Rendle) Schweick from
the Embrapa Beef Cattle germplasm collection collected in wild East African savannas
in the 1980s were analyzed cytologically. Site characteristics of accessions cultivated at
the Embrapa Beef Cattle Research Center at Campo Grande, Mato Grosso do Sul,
Brazil were: climate type Aw: tropical humid savanna; average annual precipitation =
1526 mm; average temperature = 22°C; altitude 520 m; latitude = 20° 28’ S; longitude =
55° 40’ W; poor Dark Red Latossol soil composed of 59% sand; 8% silt and 33% clay;
pH = 4.2).
Inflorescences of each accessions were collected for the meiotic study in plots
to represent each accession and fixed in a mixture of 95% ethanol, chloroform and
propionic acid (6:3:2) for 24 hours, transferred to 70% alcohol and stored under
refrigeration until use. Microsporocytes were prepared by squashing and staining with
0.5% propionic carmine. Photomicrographs were taken in a Wild Leitz microscope
using Kodak Imagelink – HQ, ISO 25 black and white film.

27
RESULTS AND DISCUSSION
In addition to the expected meiotic abnormalities typical of polyploidy and affecting
pollen viability, such as irregular chromosome segregation in the first and the second
meiotic divisions, cell fusions were recorded in two accessions (H012, BRA004979 and
H003, BRA004812) and chromosome transfer among meiocytes in one of them (H003,
BRA004812). Cytological details during microsporogenesis classifies H012 as a
hexaploid (2n = 6x = 54), with a basic chromosome number x = 9 and suggests that
H003 is a heptaploid (2n = 7x = 42), with a basic chromosome number x = 6. Recent
cytological analyses in Brachiaria revealed that the majority of species are polyploid
(Penteado et al., 2000), derived from the predominant basic chromosome number in the
genus x = 9, followed by x = 7 (Mendes-Bonato et al., 2002 a; Utsunomiya et al., 2005).
A new basic chromosome number x = 6 has recently been described in B. dictyoneura
(Risso-Pascotto et al., 2006).
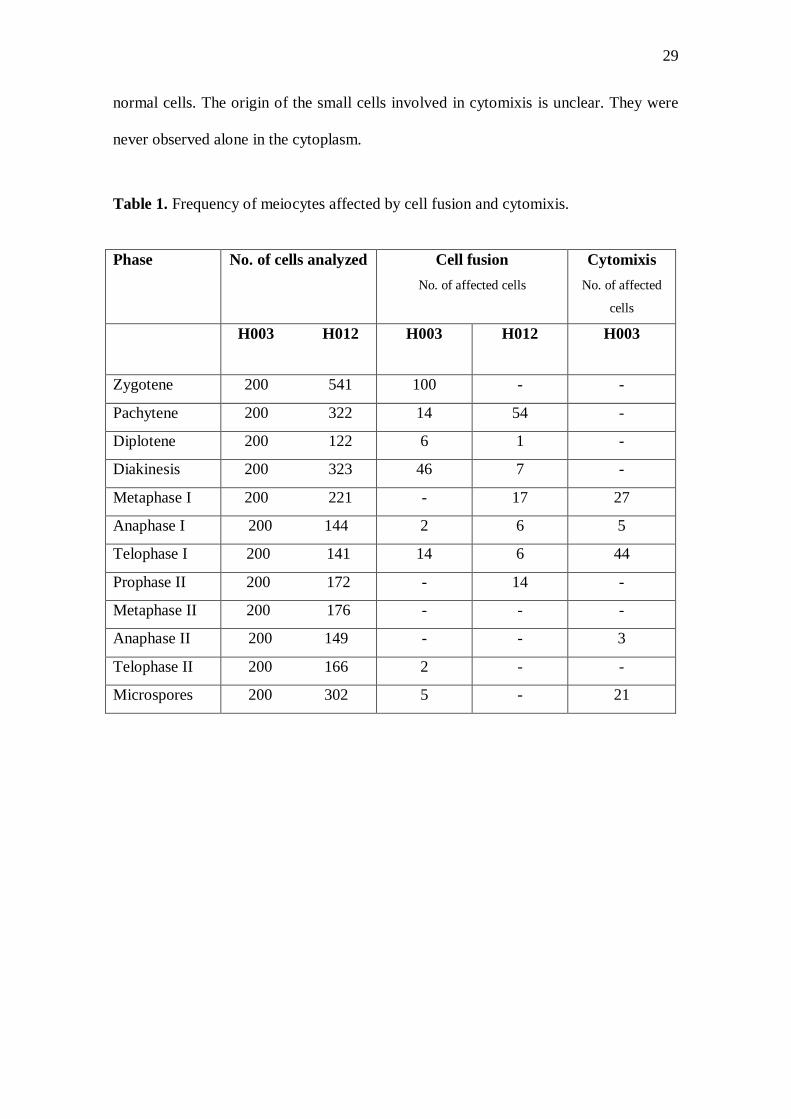
Table 1 shows the frequency of cells involved in these abnormalities. Cell fusion
was detected among some meiocytes in several anthers. The fusions involved from 2 to
more than 10 cells (Fig. 1). The majority of fused cells occurred in prophase I (Fig. 1 a,
b), but fusions were recorded until telophase I (Fig. 1 e) giving rise to abnormal meiotic
products (Figs. 1 f). There was no nuclear fusion in the syncytes. Each genome
maintained its integrity. Another interesting aspect observed in H003 was the difference
in size of fused cells. Very small cells with an apparently normal genome were found
fused with normal cells (Fig. 1 c). Cell fusion had been reported in some Brachiaria
species. In some B. brizantha genotypes, this phenomenon was restricted to male
flowers of the raceme (Mendes-Bonato et al., 2001 c); however, in accessions of other

28
species they occurred in the hermaphrodite flowers (Mendes-Bonato et al., 2001 a;
Risso-Pascotto et al., 2003a; Utsunomyia et al., 2005). In 5 of 22 accessions of B.
jubata, cell fusion was found to occur among two cells and, after normal cytokinesis, it
produced eight normal microspores (Mendes-Bonato et al., 2003).
Cell fusion was also reported in several other plant species (Nirmala and Rao,
1996), and may result from suppression of cell wall formation during premeiotic
mitoses. In general, cell fusion leads to abnormal formation of pollen grains. In the
present accessions that was also the case. According to Nirmala and Rao (1996), several
factors may cause cell fusion such as exposure to chemicals, temperature, culture
conditions, and genetic factors. Considering that the present accessions of B. humidicola
were cultivated under similar environmental conditions, the results suggest genetic
control of cell fusion.
Chromosome transfer among meiocytes of H003 was recorded in low
frequencies, but showing a pattern never reported before in other plant species. The
transfer of chromosomes always occurred between cells of different sizes. Normal
microsporocytes received the entire genome, or part of it, from very small cells through
large inter-cytomictic channels (Fig. 1 g, h). In general, the genomes of the small cells
involved showed chromosome stickiness. A similar process of structural alteration of
migrating chromatin was also recorded in B. nigropedata (Utsunomiya et al., 2004).
According to Feijó and Pais (1989) such agglutination eases the passage for migrating
chromatin. Hyperploid cells involved in cytomixis were also observed in our accession
(Fig. 1 h). The result of cytomixis was the increase of the genome in the cells (Fig. 1 i).
The phenomenon was found to occur from metaphase I to microspore stage, but most
frequently at telophase I. A normal meiocyte could receive chromosomes from two cells
in different meiotic stages or one small cell could transfer part of its genome to two
29
normal cells. The origin of the small cells involved in cytomixis is unclear. They were
never observed alone in the cytoplasm.
Table 1. Frequency of meiocytes affected by cell fusion and cytomixis.
Phase No. of cells analyzed Cell fusion
No. of affected cells
Cytomixis
No. of affected
cells
H003 H012 H003
H012 H003
Zygotene 200 541 100 - -
Pachytene 200 322 14 54 -
Diplotene 200 122 6 1 -
Diakinesis 200 323 46 7 -
Metaphase I 200 221 - 17 27
Anaphase I 200 144 2 6 5
Telophase I 200 141 14 6 44
Prophase II 200 172 - 14 -
Metaphase II 200 176 - - -
Anaphase II 200 149 - - 3
Telophase II 200 166 2 - -
Microspores 200 302 5 - 21

30
Figure 1. Aspects of cell fusion and chromosome transfer among meiocytes in B.
humidicola H003. a) Fusion of two cells in pachytene. b) Fusion of three cells in
diakinesis. c) Fusion of a normal cell and two small cells. d) Fusion of three cells in
anaphase I. e) Fusion between two cells in telophase I. f) Abnormal meiotic products
resulting from cell fusions. g, h) Chromosome transfer among meiocytes in telophase I
(g) and metaphase (h). Observe that in g the normal cell is receiving chromosomes from
the smallest cells, and that the receptive cell is hyperploid. i) Telophase II with an extra
nucleus resulting from cytomixis. (Scale bar = 1 µm)

31
Cytomixis is commonly reported in meiocytes especially during prophase I,
when cytoplasmic channels exist among cells. Heslop-Harrison (1966a, b) demonstrated
that cytoplasmic channels initiated in the preleptotene stage, persisted throughout the
meiotic prophase and disappeared before meiosis II, when each meiocyte became totally
isolated within the enclosing callose wall. Cytomixis has been reported to occur
preferentially between genetically unbalanced types such as polyploids, hybrids, and
apomictics (Gottschalk 1970; Bahl and Tyagl, 1988). Perhaps the polyploid and the
apomictic condition of the accession H003 (Valle, unpublished data) predispose it to
chromosome transfer among meiocytes. However, we cannot exclude the possibility of
some genetic factor interfering with this phenomenon, because among the 25 accessions
of B. humidicola analyzed, only this one was affected.
Despite the number of species in which cytomixis has been reported, its origin
and significance are still unknown. Its role in the evolutionary process is contradictory,
because it results in the formation of hyperploid and hypoploid cells, compromising
pollen fertility. The influence of cytomixis on the generation of polyploid gametes can
be expected in Brachiaria, a genus where polyploidy is predominant (Valle and
Savidan, 1996; Penteado et al., 2000). However, when only a part of the genome is
transferred, unbalanced and sterile gametes are formed. In the present accessions not
only cell fusion and cytomixis contributed to pollen sterility, but also many other
meiotic abnormalities typical of the polyploid condition were recorded. Were it not for
apomixis, in wich meiosis is by passed in megagametogenesis, and this genotype evoult
probably not be preserved.
The Brachiaria breeding program depends on hybridization to produce novel
genetic variability using sexual genotypes and the pollen of selected apomictic
accessions or hybrids. The hybrids that are produced are then selected, among other

32
traits, for good seed production in order for this technology to be widely adopted.
Therefore accessions with high frequencies of meiotic abnormalities such as the ones
observed for H003 and H012 present serious problems and should be eliminated early
from the breeding program, thus enhancing breeding efficiency and success.
References
Alves ER, Carneiro VTC, Araújo ACG (2001) Direct evidence of pseudogamy in
apomictic Brachiaria brizantha (Poaceae). Sexual Plant Reproduction, 14: 207-
212.
Araújo ACG, Mukhambetzhanov S, Pozzobon MT, Santana EF, Carneiro VTC (2000)
Female gametophyte development in apomictic and sexual Brachiaria brizantha
(Poaceae). Revue de Cytologie et Biologie Végétales – Le Botaniste, 23: 13-26.
Bahl JR, Tyagl BR (1988) Cytomixis in pollen mother cells of Papaver dubium L.
Cytologia, 53: 771-775.
Baker BS, Carpenter ATC, Esposito MS, Esposito RE, Sandler L (1976) The genetic
control of meiosis. Annual Review of Genetics 10: 53-134.
Feijó JA, Pais MSS (1989) Cytomixis in meiosis during the microsporogenesis in
Ophrys lutea: an ultrastructural study. Caryologia 42: 37-48.
Golubovskaya IN (1979) Genetic control of meiosis. International Review of Cytology
58: 247-290.
Golubovskaya IN (1989). Meiosis in maize: mei genes and conception of genetic
control of meiosis. Advances in Genetics 26: 149-192.
Gottschalk W (1970) Chromosome and nucleus migration during microsporogenesis of
Pisum sativum. The Nucleus 13: 1-9.

33
Gottschalk W, Kaul MLH (1974) The genetic control of microsporogenesis in higher
plants. The Nucleus 17: 133-166.
Heslop-Harrison J. (1966 a) Cytoplasmic connections between angiosperm meiocytes.
Annals of Botany 30: 221-230.
Heslop-Harrison J. (1966 b) Cytoplasmic continuities during spore formation of
flowering plants. Endeavour 25: 67-72.
Junqueira Filho R G, Mendes-Bonato AB, Pagliarini MS, Bione NCP, Valle CB,
Penteado MI.O (2003) Absence of microspore polarity, symmetric divisions and
pollen cell fate in Brachiaria decumbens (Gramineae). Genome 46: 83-88.
Mendes-Bonato AB, Pagliarini MS, Silva N, Valle CB (2001 a) Meiotic instability in
invader plants of signal grass Brachiaria decumbens Stapf (Gramineae). Acta
Scientiarum 23: 619-625.
Mendes-Bonato AB, Pagliarini MS, Valle CB, Penteado MIO (2001 b) A severe case of
chromosome stickiness in pollen mother cells of Brachiaria brizantha (Hochst)
Staph (Gramineae). Cytologia 66: 287-291
Mendes-Bonato AB, Pagliarini MS, Valle CB, Penteado MIO (2001 c) Archesporial
syncytes restricted to male flowers in a hexaploid accession of Brachiaria brizantha
(Hochst) Stapf (Gramineae). The Nucleus 44: 137-140.
Mendes-Bonato AB, Pagliarini MS, Forli F, Valle CB, Penteado MIO (2002 a)
Chromosome number and microsporogenesis in Brachiaria brizantha (Gramineae).
Euphytica 125: 419-425.
Mendes-Bonato AB, Junqueira Filho RG, Pagliarini MS, Valle CB, Penteado MIO
(2002 b) Unusual cytological patterns of microsporogenesis in Brachiaria
decumbens: abnormalities in spindle and defective cytokinesis causing precocious
cellularization. Cell Biology International 26: 641-646.

34
Mendes-Bonato A.B., Pagliarini MS, Risso-Pascotto C, Valle CB (2004) Abnormal
pollen mitoses (PM I and PM II) in an interspecific hybrid from Brachiaria
ruziziensis x Brachiaria decumbens (Gramineae). Journal of Genetics 83: 279-283.
Mendes-Bonato AB, Risso-Pascotto C, Pagliarini MS, Valle CB (2003) Normal
microspore production after cell fusion in Brachiaria jubata (Gramineae). Genetics
and Molecular Biology, 26: 517-520.
Nirmala A, Rao PN (1996) Genesis of chromosome numerical mosaicism in higher
plants. The Nucleus 39: 151-175.
Penteado MIO, Santos ACM, Rodrigues IF, Valle CB, Seixas MAC, Esteves A. (2000)
Determinação de poliploidia e avaliação da quantidade de DNA total em diferentes
espécies de gênero Brachiaria. Boletim de Pesquisa, 11. Campo Grande-MS,
Embrapa Gado de Corte. 19p.
Risso-Pascotto C., Pagliarini MS, Valle CB (2002) Abnormal nucleolar cycle in
microsporogenesis of Brachiaria decumbens (Gramineae). Cytologia 67: 355-360.
Risso-Pascotto C., Pagliarini MS, Valle CB, Mendes-Bonato AB (2003 a) Chromosome
number and microsporogenesis in a pentaploid accession of Brachiaria brizantha
(Gramineae). Plant Breeding 122:136-140.
Risso-Pascotto C, Pagliarini MS, Valle CB (2003 b) A mutation in the spindle
checkpoint arresting meiosis in Brachiaria ruziziensis. Genome 46: 724-728.
Risso-Pascotto C, Pagliarini, MS, Valle CB (2005 a) Symmetric pollen mitosis I and
suppression of pollen mitosis II prevent pollen development in Brachiaria jubata
(Gramineae). Brazilian Journal of Biological and Medical Research 38: 1603-1608.
Risso-Pascotto C, Pagliarini MS, Valle CB (2006) A new basic chromosome number
for the genus Brachiaria (Trin.) Griseb. (Poaceae: Panicoideae: Paniceae). Genetic
Research and Crop Evolution (in press).

35
Utsunomiya KS, Pagliarini MS, Valle CB (2004) Chromosome transfer among
meiocytes in Brachiaria nigropedata (Ficalho & Hiern) Stapf (Gramineae).
Cytologia 69:395-398.
Utsunomiya KS, Pagliarini MS, Valle CB (2005) Microsporogenesis in tetraploid
accessions of Brachiaria nigropedata (Ficalho & Hiern) Stapf (Gramineae). Biocell
(in press).
Valle CB, Glienke C (1991) New sexual accessions in Brachiaria. Apomixis Newsletter
3: 11-13.
Valle CB, Savidan Y (1996) Genetics, cytogenetics, and reproductive biology of
Brachiaria. In: Miles JW, Maass BL, Valle CB (eds). Brachiaria: Biology,
Agronomy, and Improvement Centro Internacional de Agricultura Tropical –
CIAT/Empresa Brasileira de Pesquisa Agropecuária –CIAT pp-147-163.

36
ABNORMAL TIMING OF CYTOKINESIS IN MICROSPOROGENESIS OF Brachiaria
humidicola (Poaceae: Paniceae)

37
ABNORMAL TIMING OF CYTOKINESIS IN
MICROSPOROGENESIS OF BRACHIARIA HUMIDICOLA
(POACEAE: PANICEAE)
Running title: Cytokinesis in Brachiaria humidicola
Key words: Brachiaria humidicola, microsporogenesis, cytokinesis, 2n gamete, forage
grass.
Kellen Regina Boldrini1, Maria Suely Pagliarini1, and Cacilda Borges
do Valle2
1 Department of Cell Biology and Genetics, State University of Maringá, 87020-900
Maringá PR, Brazil; 2 Embrapa Beef Cattle, P.O. Box 154, 79002-970 Campo Grande
MS, Brazil. ([email protected])
Meiosis is controlled by a large number of genes, generally dominant, which are stage,
site- and time-specific (Gottschalk and Kaul, 1974, 1980 a, b; Baker et al., 1976;
Golubovskaya 1979, 1989). Among genes acting in the meiotic process, those
responsible for the partitioning of the cytoplasm after nuclear division play a very
important role in the formation of viable gametes. After two rounds of chromosome
segregation (karyokinesis) and one simultaneous or two successive cytoplasmic
divisions (cytokinesis), the final product of male meiosis in flowering plants emerges as
a tetrad of haploid microspores enclosed in a callose wall. The timing of cytokinesis
varies among angiosperms. In most monocotyledons, cytokinesis is successive, i.e, one

38
partitioning of the cytoplasm occurs after telophase I and another after telophase II, so
that there is a distinct dyad stage; however, in most dicots, it is simultaneous and occurs
after telophase II (Peirson et al., 1996).
Many meiotic mutants have been reported in higher plants affecting the pattern
of cytokinesis (see Peirson et al., 1996). In Brachiaria, a promising African genus of
forage grass for the Brazilian savannas, absence of cytokinesis leading to 2n
microspores and binucleated or tetranucleated microspores have been reported in B.
brizantha (Risso-Pascotto et al., 2003) and B. nigropedata (Utsunomiya et al., 2005).
The present study details meiotic and post-meiotic abnormalities related to cytokinesis
observed in one accession of B. humidicola. In this accession the first cytokinesis
occurred after telophase II, and the second cytokinesis occured by invagination in
binucleated microspores only after callose wall dissolution. These late cytokinesis did
not affect pollen viability since the meiotic product, although formed lately, was
characterized by four reduced normal microspores.
Twenty five accessions of Brachiaria humidicola from the Embrapa Beef Cattle
germplasm collection (Campo Grande, state of Mato Grosso do Sul, Brazil) originally
collected in the wild African savannas in the mid 1980s by CIAT (Colombia) were
cytologically analyzed. Inflorescences for meiotic study were fixed in a mixture of
ethanol 95%, chloroform and propionic acid (6:3:2) for 24 hours, transferred to 70%
alcohol and stored under refrigeration until use. Microsporocytes were prepared by
squashing and staining with 0.5% propionic carmine. Photomicrographs were made
with a Wild Leitz microscope using Kodak Imagelink – HQ, ISO 25 black and white
film.
Cytological characterization revealed one accession (H003) presenting several
meiotic abnormalities, mainly due to its polyploid condition. Chromosome number in

39
this accession was defined as 2n = 42. Among abnormalities recorded, those related to
abnormal timing of cytokinesis are worth discussing. As a monocotyledonous species,
B. humidicola was expected to present successive cytokinesis. However, a large number
of meiocytes did not show the first cytokinesis after telophase I (Table 1), and the
second division occurred in a common cytoplasm (Fig. 1 a to c). The first cytokinesis
occurred after telophase II, giving rise to a dyad with two binucleated microspores (Fig.
1 d). Following callose wall dissolution, the binucleated microspores were released
(Fig. 1 e) and the second cytokinesis begun to occur discreetly by invagination (Fig. 1 f
to h). The furrow of invagination progressed up to the opposite side (Fig. 1 i) dividing
the binucleated microspores in two normal uninucleated microspores (Fig. 1 j). A
similar pattern of cytokinesis was reported in an intergeneric hybrid between Zea mays
x Tripsacum dactyloides (Kindiger, 1993), but affected only a few microspores and was
one among several types of abnormal microspore behavior observed. This pattern of
cytokinesis is being reported for the first time in Brachiaria. In other Brachiaria species
where cytokinesis was absent (Risso-Pascotto et al., 2003; Utsunomiya et al., 2005), the
failure of cytokinesis occurred after telophase I or telophase II, but never in released
microspores, or by invagination. For this accession of B. humidicola, the first
cytokinesis occurred after telophase II, and the second cytokinesis, also programmed by
the cell, occurred beyond that time. It is suggested that the genetic control for
cytokinesis exists in these meiocytes and was activated, but it was not synchronous with
karyokinesis.
The absence of the first and/or the second cytokinesis in H003 also predispose it
to the occurrence of other abnormalities in the second division. Meiocytes that
underwent only one cytokinesis gave rise to binucleated dyads (Fig. 1 k). Some

40
meiocytes that did not suffer the first cytokinesis regrouped the two chromosome sets
(Fig. 1 m) originating a restitutional nucleus.
Table 1. Percentage of cells with abnormal cytokinesis in the accession H003 of B.
humidicola.
Phase No. of cells analyzed
/No. of abnormal cells
Abnormalities (%)
Prophase II 104 / 91 Absence of cytokinesis 91 (87.5)
Metaphase II 274 / 164 Absence of cytokinesis 164 (58.8)
Anaphase II 184 / 132 Absence of cytokinesis 100 (54.3)
Tripolar spindle 32 (17.8)
Telophase II 564 / 268 Absence of cytokinesis 184 (32.6)
Restitutional nucleus 84 (14.9)
Meiotic products 450 / 438 Absence of cytokinesis 16 (3.6)
Uninucleated dyad 10 (2.2)
Binucleated dyad 373 (82.9)
Triads 30 (6.8)
Monads 9 (2.1)
Microspores 2234 / 1403 Binucleated 557 (24.9)
Initial cytokinesis 222 (9.9)
Advanced cytokinesis 607 (27.2)
Trinucleated 10 (0.4)
Tetranucleated 17 (0.8)

41
Figure 1. Aspects of abnormal cytokinesis. a to c) Phases of meiosis II with absence of the first
cytokinesis: metaphase II (a), anaphase II (b), and telophase II (c). d) Telophase II presenting the first
cytokinesis. e) Binucleated microspore released from the dyad. f to j) Progressive stages of the second
cytokinesis by invagination in binucleated microspores. Observe that in j one microspore is initiating the
cytokinesis whereas the other just completed it. k) Dyad with two binucleated microspores. l)
Mononucleated n microspore resulted from the second cytokinesis. m) Meiocytes in the second division
without the first cytokinesis (prophase II and metaphase II). Observe that in one meiocyte in metaphase
II, the two chromosome sets were rejoined due to absence of cytokinesis, resulting in a restitutional
nucleus (arrowhead). n) Trinucleated telophase II with a restitutional nucleus (arrowhead) resulting from
tripolar spindle orientation. o) Products of meiosis with different genetic constitution: a binucleated dyad,
a triad with a restitutional nucleus in one microspore (arrow), and a future dyad with restitutional nucleus
in both microspores (arrowhead). p) Tetranucleated microspore resulted from the absence of both
cytokinesis. (Scale bar = 1 µm)

42
Tripolar spindles in anaphase II were detected in a considerable number of meiocytes
and gave rise again to restitutional nucleus in telophase II (Fig. 1 n). When these cells
underwent cytokinesis, they gave rise to triads with one uninucleated (2n) and two
binucleated microspores (Fig. 1 o). However, if these cells did not suffer any
cytokinesis, they originated trinucleated microspores with one 2n nucleus. On the other
hand, in those meiocytes that lack both cytokinesis, a tetranucleated microspore was
formed (Fig. 1 p). Among a large number of microspores analyzed, more than 60% was
involved in some event related to irregular cytokinesis, including late but programmed
cytokinesis, and total absence of one or both cytokinesis.
2n microspores resulting from abnormal cytokinesis have been reported in B.
brizantha (Risso-Pascotto et al., 2003) and B. nigropedata (Utsunomiya et al., 2005).
2n gametes might have contributed to the evolutionary history of the genus Brachiaria.
In this genus, the majority of species are polyploid, mainly tetraploid (Valle and
Savidan, 1996; Penteado et al., 2000; Mendes-Bonato et al., 2002, 2006; Utsunomiya et
al., 2006). The origin of polyploidy is not yet well known. However, evidences from
conventional cytological studies point to the hypothesis that some polyploid accessions
originated by autotetraploidy, segmental allotetraploidy (Mendes-Bonato et al., 2002,
2006; Risso-Pascotto et al., 2003; Utsunomiya et al., 2005) and, in some cases, by
allopolyploidy (Mendes et al., 2006). According to Veilleux (1985) and Bretagnolle and
Thompson (1995), 2n gametes play an important role in evolution of higher plants, and
even more so in grasses, where polyploidy is widely reported (Hunziker and Stebbins,
1986; De Wet, 1986).
Among Brachiaria species available at Embrapa Beef Cattle Research Center,
the B. humidicola germplasm collection is represented by 60 accessions (Valle and
Savidan, 1996). Determination of ploidy level by flow cytometry showed that all of

43
them are polyploids, with the ploidy level ranging from 4n to 7n (Penteado et al., 2000).
Due to their adaptation to poorly drained and infertile acid soils (Keller-Grein et al.,
1996), such as those found in the Brazilian “Pantanal” region, some promising
apomictic accessions of this species are under careful agronomic and grazing evaluation
in hopes of selecting new cultivars. Other accessions, however, may be used in intra- or
interspecific hybridization as pollen donors. In this genus polyploidy is correlated with
apomixis, but for seed development, the secondary nuclei of the embryo sac need to be
fertilized by a male gamete - pseudogamy. Thus, accessions with meiotic abnormalities
which severely impair pollen viability need to be discarded from the breeding program.
The occurrence of late cytokinesis, per se, did not affect pollen viability because the
meiotic product, although lately formed, was characterized by four n normal
microspores. However, many other meiotic abnormalities due to polyploidy were
recorded in this accession and compromised pollen fertility by generating unbalanced
gametes.
REFERENCES
Baker B. S., Carpenter A.T.C., Esposito M.S., Esposito R.E. and Sandler L. 1976 The genetic control of meiosis. Annu. Rev. Genet. 10, 53-134.
Bretagnolle F. and Thompson J. D. 1995 Gametes with the somatic chromosome number: mechanisms of their formation and role in the evolution of autopolyploid plants. New Phytol. 129, 1-22.
de Wet J.M.J. 1986 Hybridization and polyploidy in the Poaceae. In Grass: Systematics and Evolution. (ed. Soderstrom TR, Hilu WH, Campbell CS, and Barkworth ME), pp 179-187. Smithsonian Institution Press, Washington DC.
Golubovskaya I. N. 1979 Genetic control of meiosis. Int. Rev. Cytol. 58, 247-290.
Golubovskaya I. N. 1989 Meiosis in maize: mei genes and conception of genetic control of meiosis. Adv. Genet. 26, 149-192.
Gottschalk W. and Kaul M. L. H. 1974 The genetic control of microsporogenesis in higher plants. Nucleus 17, 133-166.

44
Gottschalk W. and Kaul M. L. H. 1980 a Asynapsis and desynapsis in flowering plants. I. Asynapsis. Nucleus 23, 1-15.
Gottschalk W. and Kaul M.L.H. 1980 b Asynapsis and desynapsis in flowering plants. II. Desynapsis. Nucleus 23, 97-120.
Hunziker JH, Stebbins LG. 1986 Chromosomal evolution in the Gramineae. In Grass: Systematics and Evolution (ed. Soderstrom TR, Hilu WH, Campbell CS, and Barkworth ME), pp 179-187. Smithsonian Institution Press, Washington DC.
Keller-Grein G., Maass, B. L., and Hanson, J. 1996 Natural variation in Brachiaria and existing germoplasma collections. In Brachiaria: Biology, Agronomy, and Improvement (ed. Miles J. W., Maass, B. L. and Valle, C. B.) CIAT, Colombia.
Kindiger B. 1993 Aberrant microspore development in hybrids of maize x Tripsacum dactyloides. Genome 36, 987-997.
Mendes D. V., Boldrini K. R., Mendes-Bonato A. B., Pagliarini M. S. and Valle C. B. 2006 Cytological evidence of natural hybridization in Brachiaria brizantha Stapf (Gramineae). Bot. J. Linn. Soc. (in press).
Mendes-Bonato A. B., Pagliarini M. S., Forli F., Valle C. B. and Penteado M. I. O. 2002 Chromosome number and microsporogenesis in Brachiaria brizantha (Gramineae). Euphytica 125, 419-425.
Mendes-Bonato A. B., Pagliarini M. S., Risso-Pascotto C. and Valle C. B. 2006 Chromosome number and meiotic behavior in Brachiaria jubata (Gramineae). J. Genet. (in press).
Peirson B. N., Owen H. A., Feldmann K. A. and Makaroff C. A. 1996 Characterization of three male-sterile mutants of Arabidopsis thaliana exhibiting alterations in meiosis. Sex. Plant Reprod. 9, 1-16.
Penteado M. I. O, Santos A. C. M, Rodrigues I. F, Valle C. B, Seixas M. A. C. and Esteves A. 2000 Determinação de poliploidia e avaliação da quantidade de DNA total em diferentes espécies de gênero Brachiaria. Boletim de Pesquisa, 11. Campo Grande-MS, Embrapa Gado de Corte. 19p.
Risso-Pascotto C., Pagliarini M. S., Valle C. B. and Mendes-Bonato A. B. 2003 Chromosome number and microsporogenesis in pentaploid accession of Brachiaria brizantha (Gramineae). Plant Breed 122., 136-140.
Utsunomiya K. S., Pagliarini M. S. and Valle C. B. 2005 Microsporogenesis in tetraploid accessions of Brachiaria nigropedata (Ficalho & Hiern) Stapf (Gramineae). Biocell (in press).
Valle C. B. and Savidan Y. 1996 Genetics, cytogenetics, and reproductive biology of Brachiaria. In Brachiaria: Biology, Agronomy, and Improvment (ed. Miles J. W., Maass, B. L. and Valle, C. B.) CIAT, Colombia.

45
Veilleux R. 1985 Diploid and polyploid gametes in crop plants: mechanisms of formation and utilization in plant breeding. Plant Breed. Rev. 3, 253-288.

46
APÊNDICE