anÁlise de sobrevivÊncia nÃo-paramÉtrica da...
TRANSCRIPT

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 305
ANÁLISE DE SOBREVIVÊNCIA NÃO-PARAMÉTRICA DA IDADE AO PRIMEIRO PARTO EM FÊMEAS NELORE:
UM ESTUDO DE SIMULAÇÃO
Elizângela Emídio CUNHA1 Thaise Pinto de MELO1
� RESUMO: Foram simulados registros de tempo (em dias) até o primeiro parto de novilhas Nelore, incorporando censura à direita, a fim de estimar probabilidades de sobrevivência e taxas de risco associadas com tal evento por meio do método não-paramétrico de Kaplan-Meier de análise de sobrevivência. A função de risco de Weibull de uma fêmea no tempo t, enquanto variável dependente contínua, foi modelada incluindo o efeito fixo de rebanho (cinco níveis) e o efeito aleatório de touro (10 ou 25 níveis) com variância entre touros de 0,02 ou 0,08, que combinados originaram quatro cenários distintos: I (10_0,02); II (10_0,08); III (25_0,02) e IV (25_0,08). Os percentuais de censura por cenário foram, respectivamente, de: 26,44 e 28,32% em I e II; e 18,40 e 19,32% em III e IV. Houve diferenças significativas entre as curvas de sobrevivência dos rebanhos e dos touros (pais) de cada cenário, o que significa que eles diferiram estatisticamente entre si quanto à precocidade reprodutiva de suas fêmeas (filhas). No melhoramento genético animal, o touro cuja curva de sobrevivência decresce mais rapidamente no tempo produz filhas que parem pela primeira vez mais cedo, o que justifica seu uso como reprodutor visando reduzir a idade ao primeiro parto das matrizes nos rebanhos.
� PALAVRAS-CHAVE: Bovinocultura de corte; censura; falha; método de Kaplan-Meier
1 Introdução
As características reprodutivas são economicamente importantes para a bovinocultura de corte e vêm sendo incluídas nos programas de melhoramento genético das raças criadas no Brasil, como nos da raça zebuína Nelore que é, atualmente, a mais numerosa no país. Dentre as características femininas, a idade ao primeiro parto (IPP) possui a vantagem de ser de fácil mensuração (Pereira et al., 2000; Gunski et al., 2001; Pereira et al., 2007; Grossi et al., 2009) e não ter custo adicional para os sistemas de produção (Dias et al., 2004), servindo como indicadora da precocidade sexual e fertilidade das fêmeas, pois marca o início da vida reprodutiva das matrizes de um rebanho. É notório que fêmeas mais precoces terão maior oportunidade de deixar mais progênies em sua vida útil, e, assim, contribuir para a redução do custo de recria das novilhas, com impacto positivo sobre a lucratividade e eficiência da pecuária (Rosa et al., 2007).
No entanto, na maioria dos rebanhos, é comum algumas fêmeas não possuírem o registro de primeiro parto no momento da avaliação, o que pode acontecer por motivos
1 Universidade Federal do Rio Grande do Norte – UFRN, Departamento de Genética, Centro de Biociências,
CEP: 59.072-910, Natal, RN, Brasil. E-mail: [email protected] / [email protected]

306 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
diversos como por doença, imaturidade fisiológica, morte ou mesmo pela venda de animais, denominando-se essas observações de dados censurados.
Com uso da metodologia dos modelos lineares mistos, tradicionalmente empregados nas avaliações genéticas, são analisados apenas os registros das fêmeas que pariram (dados não-censurados), os quais podem não seguir a distribuição normal. Neste contexto, as amostras estariam viesadas o que contribuiria para mascarar a variabilidade real da característica e as diferenças genéticas entre os animais (Dias et al., 2004; Boligon et al., 2007; Boligon et al., 2008a). Em decorrência disso, as estimativas de herdabilidade para a IPP na raça Nelore têm tido, pela literatura recente (Mercadante et al., 2000; Pereira et al., 2001; Dias et al., 2004; Azevêdo et al., 2006; Boligon et al., 2008a, 2008b; Boligon e Albuquerque, 2010), de baixa a moderada magnitude (0,09 a 0,27), o que acarretaria em ganho genético por seleção apenas de médio a longo prazo. Para contornar essa situação, alguns autores têm atribuído às vacas que não pariram valores projetados com base na informação disponível para seu grupo de contemporâneos. Porém, isso compromete a análise, quando a proporção de dados censurados é alta (Donoghue et al., 2004).
A análise de sobrevivência, por sua vez, é considerada a metodologia estatística mais adequada para lidar com dados de tempo até a ocorrência de um evento de interesse (tempo de falha), na presença de censura (Colosimo e Giolo, 2006) que é sua principal característica. Como evento entende-se uma mudança qualitativa, ou seja, de um estado discreto para outro, que pode ser situada no tempo (Allison, 2010). Já a censura (observação incompleta) é definida quando os indivíduos não expressam o evento de interesse até o limite do tempo estabelecido no estudo, o que equivale a dizer que o seu tempo de falha verdadeiro é desconhecido e maior do que aquele que foi registrado. Tendo em vista que a análise de sobrevivência considera como resposta os tempos de falha (não-censurados) e de censura juntos (Colosimo e Giolo, 2006), então, a informação contida nos dados censurados também é incorporada na análise estatística. Logo, ela parece apropriada para a avaliação da IPP.
Muitos métodos estatísticos diferentes, entre paramétricos e não-paramétricos, podem ser empregados para o tratamento da censura (Allison, 2010). Dentre eles, o método não-paramétrico de Kaplan-Meier ou estimador Limite-Produto (Kaplan e Meier, 1958) não requer nenhuma suposição acerca da distribuição de probabilidade dos tempos de falha; inclusive sua vantagem está na flexibilidade para lidar com qualquer distribuição (Wienke, 2011). Colosimo e Giolo (2006) ressaltam, ainda, que este método é não-viciado para amostras grandes; converge assintoticamente para um processo gaussiano; sendo também um estimador de máxima verossimilhança da função de sobrevivência.
Na raça Nelore, algumas características de interesse econômico já foram avaliadas por meio de modelos paramétricos de análise de sobrevivência, como: o tempo em dias do nascimento à desmama que um animal leva para ganhar 160 kg de peso (Giolo et al., 2003); a idade à primeira concepção para novilhas expostas aos touros pela primeira vez em torno de 14 meses de idade (entre 11 e 16 meses) (Pereira et al., 2006; Pereira et al., 2007); a reconcepção de novilhas primíparas expostas em torno de 16 e 24 meses de idade (Pereira, 2008); e a idade da vaca ao último parto como medida de longevidade (Caetano, 2011). Contudo, não foram encontrados trabalhos relativos à idade ao primeiro parto.
O objetivo neste estudo foi simular a IPP de fêmeas Nelore, incluindo censura à direita, para avaliá-la por meio do método não-paramétrico de Kaplan-Meier, utilizado em análise de sobrevivência, a fim de estimar probabilidades (e curvas) de sobrevivência e taxas de risco (falha) associadas com o primeiro parto, em diferentes cenários propostos.

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 307
2 Material e métodos
Foram simulados 10.000 registros fenotípicos correspondentes à idade ao primeiro parto (IPP) de fêmeas Nelore, pertencentes a rebanhos bovinos de corte com estrutura semelhante ao que se observa para animais criados a pasto no Brasil, utilizando o programa simul.f do software “The Survival Kit v6.0” (Ducrocq et al., 2010), escrito na linguagem Fortran.
2.1 Definição dos cenários e obtenção do banco de dados
Os registros foram alocados equitativamente em quatro cenários distintos pela combinação de dois totais de reprodutores (10 e 25) com duas estimativas de variância entre reprodutores (���= 0,02 e 0,08). Cada cenário continha 2.500 fêmeas distribuídas por cinco rebanhos, resultando em média 500 fêmeas/rebanho/cenário. Nos cenários I (10_0,02) e II (10_0,08) foram utilizados 10 touros, que geraram exatamente 250 filhas/cenário cada, combinados às variâncias de 0,02 e 0,08, respectivamente. Nos cenários III (25_0,02) e IV (25_0,08) tais variâncias (0,02 e 0,08) foram combinadas ao uso de 25 touros, que produziram, em esquema balanceado, 100 filhas/cenário cada. Foram feitas 10 repetições/cenário, mantendo-se constantes os valores dos parâmetros combinados por cenário a fim de garantir a presença de censura nos dados. Foi constatada a igualdade entre as repetições dentro de cenário; logo, considerou-se apenas a primeira repetição/cenário. Em todas as situações simuladas, foi pressuposto o uso da inseminação artificial, processo que aumenta o número de filhas por touro nos rebanhos, de modo a garantir pelo menos cinco registros não-censurados (falhas) por touro/rebanho/cenário.
Para delimitar aquela extensão (0,09 a 0,27) das estimativas de herdabilidade para a IPP na raça Nelore, sob modelos lineares mistos, divulgadas nos estudos anteriormente mencionados, foram escolhidas as herdabilidades de 0,08 e 0,30. Em correspondência a estes valores, foram calculados os limites inferior e superior de σ�� (0,02 e 0,08) utilizando
a fórmula da herdabilidade efetiva (ℎ���� = �� �� ��) de Yazdi et al. (2002), proposta para o
modelo de riscos proporcionais de Weibull, sob modelo de reprodutor (touro), e compatível com a análise clássica por modelos lineares mistos.
Os tempos, em dias, do nascimento até a ocorrência do evento de interesse, primeiro parto (IPP), foram obtidos assumindo-se: inexistência de estação de monta controlada, garantindo parições ao longo de todo o ano e tornando contínua a distribuição da variável dependente tempo; número irrestrito de serviços por fêmea até a concepção; duração da gestação de 290 dias (9,5 meses); e novilhas começando a vida sexual a partir de 530 dias (17,4 meses) de idade, de modo que nenhuma fêmea estivesse prenhe antes disso nem tivesse parido antes de completar 820 dias.
Foram estabelecidos: o início do estudo (origem do tempo) em 820 dias (≅27 meses), pois com esta idade uma novilha Nelore é considerada apta a ter sua primeira cria; o tempo mediano de falha de 1.217 dias (≅ 40 meses); e o tempo máximo de falha de 1.460 dias (≅ 48 meses), inclusive, pois fêmeas que apresentam o primeiro parto após esta idade são consideradas tardias e não deveriam permanecer no rebanho, já que aumentam os custos de produção. A escolha dos tempos inicial e máximo foi baseada no estudo de Mercadante et al. (2000) que, na fase de edição dos dados, limitaram entre 26 e 47 meses

308 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
a idade ao primeiro parto das fêmeas Nelore de um rebanho do estado de São Paulo participante do “Programa de Melhoramento Genético da Raça Nelore” (PMGRN), para as quais registraram média de IPP de 38,3 meses. Já o tempo mediano foi escolhido mais próximo da média nacional da raça, de 1.206,22 dias (39,55 meses), divulgada pela EMBRAPA/ABCZ/MAPA (2011-2012).
Para descontar o longo período inicial (820 dias) sem ocorrência do evento e comum a todas as novilhas, a escala de tempo da IPP foi simulada no intervalo de 0 dia (820 dias) a 640 (1.460) dias, com mediana de 397 (1.217) dias. Logo, deve-se acrescentar 820 dias na interpretação dos resultados.
Na etapa de simulação, foi criada a variável qualificadora “status” que recebeu o valor de “1” para os tempos entre 0 e 640 dias, inclusive, significando que o tempo de ocorrência do evento era conhecido (tempos de falha); ou o valor de “0” para os tempos de 640 dias (limite para censura), significando que o evento teria ocorrido em qualquer tempo desconhecido após (à direita de) 640 dias (tempos censurados).
Segundo Kachman (1999), ocorre censura à direita quando a falha acontece após o final do período de estudo; observando-se, para dados censurados à direita, o tempo de censura ao invés do tempo de falha. Colosimo e Giolo (2006) classificam como censura do tipo I aquela em que o estudo é terminado após um período pré-estabelecido de tempo e até este tempo final alguns indivíduos não experimentam o evento. Assim, em cada cenário, a censura foi à direita e do tipo I, porque até o tempo de censura (640 dias) algumas fêmeas não tiveram registro do evento.
A variável aleatória contínua IPP (� ≥ 0)seguiu a distribuição paramétrica de Weibull com os parâmetros positivos λ de escala e � de forma. Segundo Colosimo e Giolo (2006), o parâmetro λ tem a mesma unidade de medida de t e � não tem unidade; além do que, a função de risco (ou de taxa de falha) desta distribuição é monótona, isto é, ela é estritamente crescente para � > 1; estritamente decrescente para � < 1; ou constante para � = 1, em qual caso tem-se a distribuição exponencial. Neste contexto, o parâmetro � da função de risco de Weibull foi simulado com o valor de 2,0, indicando um risco crescente para a fêmea de apresentar o evento de interesse com o passar do tempo. Segundo Yazdi et al. (2002), diversos autores têm utilizado tal valor de � para a análise da longevidade produtiva de bovinos.
Assumindo que ��t) representa a função densidade de probabilidade; ℎ�t), a função de risco; e ��t), a função de sobrevivência; Colosimo e Giolo (2006) descreveram tais funções, sob a distribuição de Weibull, respectivamente, por:
��t) = �λ� ���� !" #−%&
λ'�( ; h�t) = �
λ� ����;e ��t) = !" #−%&
λ'�(
As quais se relacionam da seguinte forma: ��t) = h�t)S�t). Assim, a função de risco hij�t) de uma novilha, filha do touro “j” e pertencente ao
rebanho “i”, na dependência do tempo �, foi modelada segundo a equação:
hij��) = ./h0��) !"0!1/2 3345 ou equivalentemente:
hij��) = h0��) !"0!1/2 334 + 7/8 Em que: h0�t) é a função de risco de base de Weibull descrita por ℎ9��) =:��:�)��� = �����exp��>?:), com �>?: = -12,33 como o intercepto ou média geral

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 309
correspondente ao tempo mediano de falha de 397 (1.217) dias; !1/2 é o vetor da covariável fixa rebanho “i” (i = 1 a 5) que está relacionado ao vetor β de coeficientes de regressão desconhecidos; e zB é o valor da fragilidade para o touro “j” (j = 1 a 10; ou j = 1 a 25) tal
que zB = exp0sB8, com 7/ representando o efeito doD�é�1FG touro como covariável aleatória.
A covariável fixa rebanho foi simulada sob distribuição uniforme com intervalo de riscos relativos de 0,5 a 1,5. Já a covariável aleatória touro seguiu a distribuição normal com média zero e variância entre touros ��� = H(Colosimo e Giolo, 2006), os quais eram não-aparentados entre si. Nessa função de risco, a fragilidade representa um efeito aleatório que descreve o risco comum às filhas de um mesmo touro, justificando uma maior associação entre seus tempos de sobrevivência em função de compartilharem genes.
2.2 Método de análise
O estimador de Kaplan-Meier ou limite-produto, Ŝ��), também chamado de estimador não-paramétrico de máxima verossimilhança da ���), foi obtido nos diferentes cenários por meio do procedimento LIFETEST do SAS 9.2 (SAS Institute Inc., 2009), segundo a fórmula apresentada por Allison (2010):
�J��) = K L?/ − M/?/ N/:&PQ&
= K L1 − M/?/N/:&PQ&
Para t1 ≤ t ≤ tk, supondo que existam k tempos de evento distintos com t1 < t2 ...< tk; sendo dj o número de indivíduos que falharam em tj, com j = 1, ..., k; e nj o número de indivíduos sob risco do evento em tj, ou seja, eles não falharam e não foram censurados antes do tempo tj. Se houver censura exatamente em tj, os indivíduos também estão sob risco em tj.
Em cada cenário, a função de sobrevivência foi estratificada para as covariáveis rebanho e touro a fim de comparar as curvas de sobrevivência dos diferentes rebanhos e touros entre si, empregando-se o teste não-paramétrico Log-Rank descrito por Allison (2010). A hipótese nula versa sobre a igualdade entre as curvas de sobrevivência das fêmeas dos diferentes rebanhos ou das filhas dos diferentes touros, obedecendo a distribuição Qui-quadrado (S�).
A estatística Log-Rank permite comparar a distribuição da ocorrência dos eventos observados em cada grupo com a distribuição que seria esperada, se a incidência fosse igual em todos os grupos. Se a distribuição observada for equivalente à distribuição esperada, dizemos que a função de sobrevivência dos indivíduos pertencentes ao grupo coincide com a função de sobrevivência dos indivíduos em geral, ou seja, a covariável não exerce influência sobre a sobrevivência (Bastos e Rocha, 2007).
Foram obtidas ainda, por meio do PROC LIFETEST do SAS, as seguintes curvas por cenário: da probabilidade de sobrevivência com 95% de limite de confiança; da probabilidade de sobrevivência em cada rebanho; da taxa de risco suavizada com 95% de limite de confiança; e do >?T− lnTŜ��)WW versus >?��).

310
Finalmente, usando o PROC CORR do SAS, foram estimadas correlações de Pearson e Spearman entre percentual (%) de censura, IPP média e IPP mediana, em cada cenário, considerando a análise estratificada por touro.
3 Resultados
3.1 Análise geral dos cenários
O modelo de Weibull (Figura 1) foi empregado adem cada cenário, pois os pontos do gráfico de a uma linha reta para a maior cocorrespondente ao tempo de 54,59 (tipo de curva deve ser aproximadamente linear quando o modelo de Weibull é apropriado para explicar os dados.
Figura 1 - Adequação do modelo de sobrevivência de Weibull quatro cenários: C I (10_0,02); C II (10_0,08); C III (25_0,02) e C IV (25_0,08).
O percentual de fêmeas que não tiveram o primeiro parto até a idade limite de 48
meses (fêmeas censuradas(Tabela 1), atingindo o valor médio global por cenário de: 26,44% (661 fêmeas) no I; 28,32% (708 fêmeas) no II; Percentuais mais próximos foram obtidos entre cenários que utilizaram o mesmo número de touros como pais e não entre aqueles de mesma variância entre touros (
Rev. Bras. Biom., São Paulo, v.30, n.3, p.305
usando o PROC CORR do SAS, foram estimadas correlações de Pearson e Spearman entre percentual (%) de censura, IPP média e IPP mediana, em cada cenário, considerando a análise estratificada por touro.
dos cenários: curvas e estatísticas
O modelo de Weibull (Figura 1) foi empregado adequadamente na simulação da IPPem cada cenário, pois os pontos do gráfico de lnT− lnTŜ�t�WW versus ln�t� foram próximos a uma linha reta para a maior concentração dos dados, ocorrida a partir de correspondente ao tempo de 54,59 (≅875) dias. Segundo Colosimo e Giolo (2006), esse tipo de curva deve ser aproximadamente linear quando o modelo de Weibull é apropriado para explicar os dados.
Adequação do modelo de sobrevivência de Weibull à simulação da IPP dos quatro cenários: C I (10_0,02); C II (10_0,08); C III (25_0,02) e C IV
O percentual de fêmeas que não tiveram o primeiro parto até a idade limite de 48 censuradas ou sobreviventes) variou entre rebanhos dentro de cenário
(Tabela 1), atingindo o valor médio global por cenário de: 26,44% (661 fêmeas) no I; 28,32% (708 fêmeas) no II; 18,40% (460 fêmeas) no III; e 19,32% (483 fêmeas) no IV.
róximos foram obtidos entre cenários que utilizaram o mesmo número de touros como pais e não entre aqueles de mesma variância entre touros (σ��
.3, p.305-325, 2012
usando o PROC CORR do SAS, foram estimadas correlações de Pearson e Spearman entre percentual (%) de censura, IPP média e IPP mediana, em cada
equadamente na simulação da IPP � � foram próximos
ncentração dos dados, ocorrida a partir de ln�t� = 4 875) dias. Segundo Colosimo e Giolo (2006), esse
tipo de curva deve ser aproximadamente linear quando o modelo de Weibull é apropriado
simulação da IPP dos
quatro cenários: C I (10_0,02); C II (10_0,08); C III (25_0,02) e C IV
O percentual de fêmeas que não tiveram o primeiro parto até a idade limite de 48 variou entre rebanhos dentro de cenário
(Tabela 1), atingindo o valor médio global por cenário de: 26,44% (661 fêmeas) no I; 18,40% (460 fêmeas) no III; e 19,32% (483 fêmeas) no IV.
róximos foram obtidos entre cenários que utilizaram o mesmo número
���. Em relação
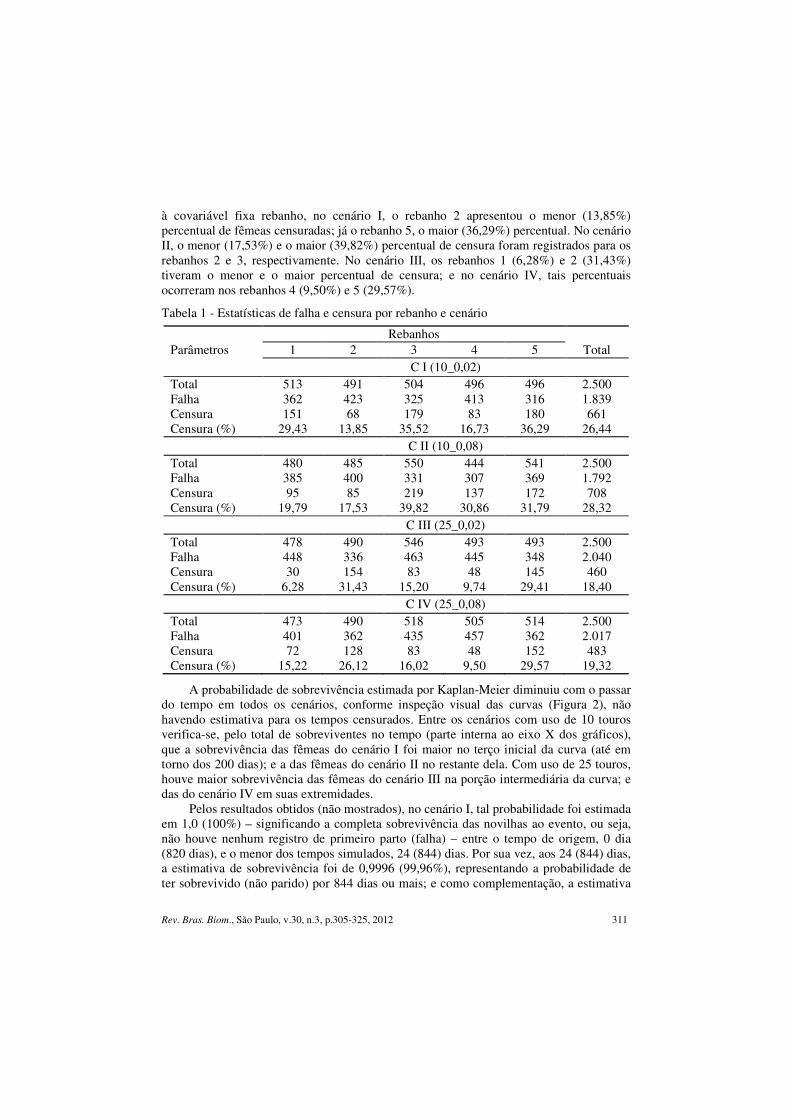
Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 311
à covariável fixa rebanho, no cenário I, o rebanho 2 apresentou o menor (13,85%) percentual de fêmeas censuradas; já o rebanho 5, o maior (36,29%) percentual. No cenário II, o menor (17,53%) e o maior (39,82%) percentual de censura foram registrados para os rebanhos 2 e 3, respectivamente. No cenário III, os rebanhos 1 (6,28%) e 2 (31,43%) tiveram o menor e o maior percentual de censura; e no cenário IV, tais percentuais ocorreram nos rebanhos 4 (9,50%) e 5 (29,57%).
Tabela 1 - Estatísticas de falha e censura por rebanho e cenário
Rebanhos Parâmetros 1 2 3 4 5 Total C I (10_0,02) Total 513 491 504 496 496 2.500 Falha 362 423 325 413 316 1.839 Censura 151 68 179 83 180 661 Censura (%) 29,43 13,85 35,52 16,73 36,29 26,44 C II (10_0,08) Total 480 485 550 444 541 2.500 Falha 385 400 331 307 369 1.792 Censura 95 85 219 137 172 708 Censura (%) 19,79 17,53 39,82 30,86 31,79 28,32 C III (25_0,02) Total 478 490 546 493 493 2.500 Falha 448 336 463 445 348 2.040 Censura 30 154 83 48 145 460 Censura (%) 6,28 31,43 15,20 9,74 29,41 18,40 C IV (25_0,08) Total 473 490 518 505 514 2.500 Falha 401 362 435 457 362 2.017 Censura 72 128 83 48 152 483 Censura (%) 15,22 26,12 16,02 9,50 29,57 19,32
A probabilidade de sobrevivência estimada por Kaplan-Meier diminuiu com o passar do tempo em todos os cenários, conforme inspeção visual das curvas (Figura 2), não havendo estimativa para os tempos censurados. Entre os cenários com uso de 10 touros verifica-se, pelo total de sobreviventes no tempo (parte interna ao eixo X dos gráficos), que a sobrevivência das fêmeas do cenário I foi maior no terço inicial da curva (até em torno dos 200 dias); e a das fêmeas do cenário II no restante dela. Com uso de 25 touros, houve maior sobrevivência das fêmeas do cenário III na porção intermediária da curva; e das do cenário IV em suas extremidades.
Pelos resultados obtidos (não mostrados), no cenário I, tal probabilidade foi estimada em 1,0 (100%) – significando a completa sobrevivência das novilhas ao evento, ou seja, não houve nenhum registro de primeiro parto (falha) – entre o tempo de origem, 0 dia (820 dias), e o menor dos tempos simulados, 24 (844) dias. Por sua vez, aos 24 (844) dias, a estimativa de sobrevivência foi de 0,9996 (99,96%), representando a probabilidade de ter sobrevivido (não parido) por 844 dias ou mais; e como complementação, a estimativa

312
de falha (parto) aos 844 dias foi deparida). Mais adiante, como se observa probabilidade de sobrevivência foi estimada em 58,08%, tinham apresentado o evento antes dos 1.220 dias e estavam sob risco; e outras 1.048 (41,92%) já tinham falhado (parido). Até (73,56%) fêmeas já tinham tido informação de IPP; restando 661 (26,44%) com registros censurados, ou seja, fêmeas para as quais o verdadeiro tempo de falha não era conhecido.
Figura 2 - Curva de sobrevivência incluindo o número de sobreviventes no limite de confiança (LC) de 95% de probabilidade para todos os cenários.
Para o mesmo número de touros, e de forma análoga, no cenário II não houve falha
(parto) no intervalo de 0 dia (820 dias) a 3 (823) dias; aos 3 (823) dias, a probabilidade de falha foi de 0,0004 (0,04%) indicando que uma fêmea havia falhado (parido) e 2.49(99,96%) estavam sob risco de falhar; aos 400 (1.220) dias, infereprobabilidade de falha foi estimada em 41,04% (1.026 fêmeas), porque a de sobrevivência o foi em 58,96% (1.474 fêmeas). Até o(71,68%) fêmeas falharam (pariram), enquanto
Os resultados para o cenário III indicam que não houve partos no intervalo de 0 dia (820 dias) a 10 (830) dias, o primeiro dos tempos após a origem. Aos 10 (830) dias, a probabilidade de falha, ou seja, de (0,04%), o que significou uma falha. Aos 400 (1.220) dias, infere(49,56%) fêmeas já tinham falhado (parido), feito, estando sob risco. Logo depois, aos 403 (1.223
Rev. Bras. Biom., São Paulo, v.30, n.3, p.305
de falha (parto) aos 844 dias foi de 0,0004 (0,04%), o que significou uma falha (fêmea parida). Mais adiante, como se observa (Figura 2) para os 400 (1.220) dias, a
bilidade de sobrevivência foi estimada em 58,08%, isto é, 1.452 fêmeas ainda não tinham apresentado o evento antes dos 1.220 dias e estavam sob risco; e outras 1.048 (41,92%) já tinham falhado (parido). Até os 640 (1.460) dias, inclusive, um total de 1.839
fêmeas já tinham tido informação de IPP; restando 661 (26,44%) com registros fêmeas para as quais o verdadeiro tempo de falha não era conhecido.
Curva de sobrevivência incluindo o número de sobreviventes no limite de confiança (LC) de 95% de probabilidade para todos os cenários.
Para o mesmo número de touros, e de forma análoga, no cenário II não houve falha (parto) no intervalo de 0 dia (820 dias) a 3 (823) dias; aos 3 (823) dias, a probabilidade de
0,0004 (0,04%) indicando que uma fêmea havia falhado (parido) e 2.49(99,96%) estavam sob risco de falhar; aos 400 (1.220) dias, infere-se, pela Figura 2, que probabilidade de falha foi estimada em 41,04% (1.026 fêmeas), porque a de sobrevivência o foi em 58,96% (1.474 fêmeas). Até o final do estudo, aos 640 (1.460) dia(71,68%) fêmeas falharam (pariram), enquanto 708 (28,32%) foram censuradas.
Os resultados para o cenário III indicam que não houve partos no intervalo de 0 dia (820 dias) a 10 (830) dias, o primeiro dos tempos após a origem. Aos 10 (830) dias, a probabilidade de falha, ou seja, de ter ocorrido o primeiro parto foi estimada em
, o que significou uma falha. Aos 400 (1.220) dias, infere-se (Figura 2) que 1.239 (49,56%) fêmeas já tinham falhado (parido), dado que 1.261 (50,44%) ainda nãofeito, estando sob risco. Logo depois, aos 403 (1.223) dias, registrou-se o tempo mediano
.3, p.305-325, 2012
0,0004 (0,04%), o que significou uma falha (fêmea 400 (1.220) dias, a
, 1.452 fêmeas ainda não tinham apresentado o evento antes dos 1.220 dias e estavam sob risco; e outras 1.048
os 640 (1.460) dias, inclusive, um total de 1.839 fêmeas já tinham tido informação de IPP; restando 661 (26,44%) com registros
fêmeas para as quais o verdadeiro tempo de falha não era conhecido.
Curva de sobrevivência incluindo o número de sobreviventes no tempo com limite de confiança (LC) de 95% de probabilidade para todos os cenários.
Para o mesmo número de touros, e de forma análoga, no cenário II não houve falha (parto) no intervalo de 0 dia (820 dias) a 3 (823) dias; aos 3 (823) dias, a probabilidade de
0,0004 (0,04%) indicando que uma fêmea havia falhado (parido) e 2.499 Figura 2, que a
probabilidade de falha foi estimada em 41,04% (1.026 fêmeas), porque a de sobrevivência final do estudo, aos 640 (1.460) dias, 1.792
foram censuradas. Os resultados para o cenário III indicam que não houve partos no intervalo de 0 dia
(820 dias) a 10 (830) dias, o primeiro dos tempos após a origem. Aos 10 (830) dias, a o primeiro parto foi estimada em 0,0004
se (Figura 2) que 1.239 que 1.261 (50,44%) ainda não o tinham
se o tempo mediano

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 313
de falha (50% de fêmeas paridas). Até os 640 (1.460) dias, inclusive, um total de 2.040 (81,60%) fêmeas tinham parido; restando 460 (18,40%) censuradas.
De modo similar, no cenário IV, não houve partos do tempo de origem, 0 dia (820 dias), até 4 (824) dias, significando uma probabilidade de sobrevivência de 1,0 (100%) neste intervalo; aos 4 (824) dias, essa probabilidade caiu para 0,9996 (99,96%) e a estimativa de falha subiu para 0,0004 (0,04%). Aos 400 (1.220) dias, 1.265 (50,60%) fêmeas já tinham parido, e outras 1.235 (49,40%) estavam sob risco. O tempo mediano de falha ocorreu um pouco antes, aos 395 (1.215) dias. Até os 640 (1.460) dias, inclusive, 2.017 (80,68%) fêmeas tinham falhado e outras 483 (19,32%) estavam censuradas.
A taxa de risco (Figura 3) estimada entre as novilhas sobreviventes (que estão sob risco) aumentou com o tempo, em todos os cenários, assim como a amplitude de seus valores (área sombreada correspondente ao limite de confiança de 95% de probabilidade). Nos tempos mais próximos do limite de 640 (1.460) dias, o risco instantâneo de o evento ocorrer esteve abaixo de 0,004 nos cenário II e IV, e acima disso nos cenários I e III. Com o aumento na taxa de risco (falha) restaram, óbvio, menos novilhas sem o registro de primeiro parto (Figura 2). Biologicamente, é o que se espera com o envelhecimento das fêmeas.
Figura 3 - Curvas da função de risco suavizada-kernel de Epanechnikov, incluindo o limite de confiança de 95% de probabilidade, para os quatro cenários.

314 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
No cenário I, a IPP apresentou média (erro-padrão) de 435,9 (3,6) dias e mediana de 451 dias com amplitude de 439 a 466 dias para o intervalo de confiança (IC) a 95% de probabilidade. Nos demais cenários, os valores destes parâmetros foram, respectivamente, de: 441 (3,7) e 464 dias com IC (a 95%) de 449 a 481 dias no cenário II; 401,9 (3,6) e 403 dias com IC (a 95%) de 391 a 413 dias no cenário III; e, finalmente, 400,7 (3,6) e 395 dias com IC (a 95%) de 382 a 409 dias no cenário IV.
Segundo Colosimo e Giolo (2006), um dos interesses nos estudos de sobrevivência é o tempo médio de vida. Porém, utilizando estimadores não-paramétricos, a estimativa do tempo médio fica subestimada quando o maior tempo observado é uma censura, tornando a mediana preferível à média, ainda que ambas sejam medidas de tendência central. Neste estudo, a média superou a mediana apenas no cenário IV.
Corrigida a escala da média e da mediana da IPP em todos os cenários, esses valores acima obtidos, corresponderam, naquela ordem a: 1.255,9 e 1.271 dias no cenário I; 1.261 e 1.284 dias no cenário II; 1.221,9 e 1.223 dias no cenário III; e 1.220,7 e 1.215 dias no cenário IV. Tempos próximos entre cenários que tiveram o mesmo número de touros como pais; tendo-se registrado a maior precocidade reprodutiva para as fêmeas do cenário IV. Contudo, tais médias estão muito acima da IPP média de 1.168,2 dias relatada por Mercadante et al. (2000) para fêmeas Nelore.
3.2 Estratificação pela covariável rebanho
Pela inspeção visual das curvas de sobrevivência dos rebanhos (Figura 4) por cenário, constatou-se, de início, homogeneidade entre elas, significando que as fêmeas dos diferentes rebanhos tiveram o primeiro parto em tempos relativamente próximos. Exemplificando, a redução de 1,0 (100%) para 0,99 (99%) na probabilidade de sobrevivência das fêmeas de cada cenário implicou na ocorrência de 25 partos. Para este pequeno percentual (1%) de redução, não houve diferença entre as curvas de sobrevivência dos rebanhos até ocorrerem, em média, os cinco primeiros partos em cada rebanho, o que levou um tempo médio/rebanho de: 65,4 (885,4) dias no cenário I; 45,6 (865,6) dias no II; 48,0 (868,0) dias no III; e 49,2 (869,2) dias no IV. De modo geral, a homogeneidade entre as curvas se estendeu por mais tempo no cenário II, em torno de 136 (956) dias; e, na maior parte do período avaliado, as curvas diferiram entre os rebanhos de um mesmo cenário e também de um cenário para o outro.
Todavia, Colosimo e Giolo (2006) ressaltam que uma vez constatada a presença de diferenças entre os estratos, torna-se necessário identificar quais curvas de sobrevivência diferem entre si aplicando-se testes de hipótese. Diferenças significativas (P<0,0001) foram detectadas entre os rebanhos de cada cenário pelos valores das estatísticas S� (I: 152,70; II: 92,75; III: 231,52; e IV: 136,84) para o teste Log-Rank, com quatro graus de liberdade. Procedeu-se, então, aos ajustamentos por comparações múltiplas usando o método de Tukey-Kramer, ao nível de 5% de probabilidade, que revelaram diferenças significativas entre os seguintes pares de rebanhos: a) 1 e 2; 1 e 4; 2 e 3; 2 e 5; 3 e 4; e 4 e 5 no cenário I; b) entre 1 e 3; 1 e 4; 1 e 5; 2 e 3; 2 e 4; 2 e 5; 3 e 4; e 3 e 5 no cenário II; c) entre 1 e 2; 1 e 3; 1 e 5; 2 e 3; 2 e 4; 3 e 5; e 4 e 5 no cenário III; e d) entre 1 e 2; 1 e 4; 1 e 5; 2 e 3; 2 e 4; 3 e 4; 3 e 5; e 4 e 5 no cenário IV.
Nos quatro cenários, a IPP média (Tabela 2) entre rebanhos variou de 1.160,9 a 1.298,8 dias; enquanto a mediana (Tabela 3), de 1.137 a 1.362,5 dias.

Rev. Bras. Biom., São Paulo, v.30
Nos cenários I e II, o rebanho 2 foi aquele cuja curva de sobrevivência decresceu mais rapidamente (vide os menores percentuais de censura)verificado média e mediana da IPP das vacas de 1.197,7 e 1.195 dias no cenário I; e de 1.218,7 e 1.207 dias no cenário II. Mercadante et al. (2000). Maior precocidade reprodutivarebanho 1 do cenário III, que exibiu as menores média e mediana, 1.165,4 e 1.153 dias, respectivamente; e pelo rebanho 4 do cenário IV cuja média foi de 1.160,9 dias, e a mediana, de 1.137 dias. muito as IPPs médias (1.033,15; 1.035,61 e 1.047,21 dias) para fêmeas Nelore relatadas por Dias et al. (2004).
Figura 4 - Curvas da probabilidade de sobrevivência dos rebanhos ao primeiro parto, em
cada cenário,
Notoriamente no cenário II, o rebanho 3 foi o que apresentou a curva de sobrevivência com decréscimos mais suaves no decorrer do tempo, ou seja, suas fêmeas foram as mais tardias respectivamente; e percentual de censura de 39,82% (vide todas as fêmeas avaliadas. No cenário I, não houve diferença significativa entre as curvas dos rebanhos 3 e 5, os mais tardios (respectivamente: 35,52 média de 1.297,3 vs. similar, no cenário III, os rebanhos 2 e 5 foram indistintamente os mais tardios (respectivamente: 31,43
., São Paulo, v.30, n.3, p.305-325, 2012
Nos cenários I e II, o rebanho 2 foi aquele cuja curva de sobrevivência decresceu mais rapidamente (vide os menores percentuais de censura) com o tempo
média e mediana da IPP das vacas de 1.197,7 e 1.195 dias no cenário I; e de 1.218,7 e 1.207 dias no cenário II. Tais valores estão muito acima da IPP média obtida por
. (2000). Maior precocidade reprodutiva, porém, foi alcançada pelo o 1 do cenário III, que exibiu as menores média e mediana, 1.165,4 e 1.153 dias,
respectivamente; e pelo rebanho 4 do cenário IV cuja média foi de 1.160,9 dias, e a mediana, de 1.137 dias. Todavia, esses resultados, nos cenários III e IV, superam em
as IPPs médias (1.033,15; 1.035,61 e 1.047,21 dias) para fêmeas Nelore relatadas . (2004).
Curvas da probabilidade de sobrevivência dos rebanhos ao primeiro parto, em cada cenário, estatisticamente diferentes entre si pelo teste Log-Rank
Notoriamente no cenário II, o rebanho 3 foi o que apresentou a curva de sobrevivência com decréscimos mais suaves no decorrer do tempo, ou seja, suas fêmeas foram as mais tardias – com IPP média e mediana de 1.298,8 e 1.362,5
ivamente; e percentual de censura de 39,82% (vide Tabela 1) – comparadas com todas as fêmeas avaliadas. No cenário I, não houve diferença significativa entre as curvas dos rebanhos 3 e 5, os mais tardios (respectivamente: 35,52 vs. 36,29% de censura; IPP
1.291,3 dias; e IPP mediana de 1.343 vs. 1.348 dias). De modo similar, no cenário III, os rebanhos 2 e 5 foram indistintamente os mais tardios (respectivamente: 31,43 vs. 29,41% de censura; IPP média de 1.277,5 vs. 1.283,4 dias; e
315
Nos cenários I e II, o rebanho 2 foi aquele cuja curva de sobrevivência decresceu com o tempo, tendo-se
média e mediana da IPP das vacas de 1.197,7 e 1.195 dias no cenário I; e de Tais valores estão muito acima da IPP média obtida por
foi alcançada pelo o 1 do cenário III, que exibiu as menores média e mediana, 1.165,4 e 1.153 dias,
respectivamente; e pelo rebanho 4 do cenário IV cuja média foi de 1.160,9 dias, e a , esses resultados, nos cenários III e IV, superam em
as IPPs médias (1.033,15; 1.035,61 e 1.047,21 dias) para fêmeas Nelore relatadas
Curvas da probabilidade de sobrevivência dos rebanhos ao primeiro parto, em Rank.
Notoriamente no cenário II, o rebanho 3 foi o que apresentou a curva de sobrevivência com decréscimos mais suaves no decorrer do tempo, ou seja, suas fêmeas
1.298,8 e 1.362,5 dias, comparadas com
todas as fêmeas avaliadas. No cenário I, não houve diferença significativa entre as curvas . 36,29% de censura; IPP
. 1.348 dias). De modo similar, no cenário III, os rebanhos 2 e 5 foram indistintamente os mais tardios
1.283,4 dias; e

316 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
IPP mediana de 1.316 vs. 1.332 dias). No cenário IV, o rebanho 5 teve as fêmeas mais tardias (IPP com média de 1.280,7 dias; e mediana de 1.315 dias). Entretanto, todas essas médias e medianas tiveram magnitude inferior a 1.376,8 dias (45,14 meses), média da IPP obtida por Azevêdo et al. (2006) para rebanhos Nelore do Norte e Nordeste do país, avaliados previamente à implementação de estação de monta pelo PMGRN.
Tabela 2 - IPP média¹ ± erro padrão (EP) nos diferentes rebanhos por cenário
Estratos i C I (10_0,02) C II (10_0,08) C III (25_0,02) C IV (25_0,08)
XY ± [\ XY ± [\ XY ± [\ XY ± [\
Rebanho
1 449,7 ± 7,9 420,8 ± 8,0 345,4 ± 7,5 380,6 ± 8,0 2 377,7 ± 7,8 398,7 ± 8,0 457,5 ± 8,1 427,0 ± 8,4 3 477,3 ± 7,6 478,8 ± 7,7 380,3 ± 7,5 391,1 ± 7,7 4 398,6 ± 8,0 454,1 ± 8,9 360,5 ± 7,7 340,9 ± 7,8 5 471,3 ± 7,8 447,2 ± 8,0 463,4 ± 7,8 460,7 ± 7,7
i – identificação; ¹escala não-corrigida.
Tabela 3 - IPP mediana¹ (dias) com intervalo de confiança (IC) a 95% de probabilidade para as fêmeas por rebanho e cenário
Cenário Rebanho Mediana IC 95%
Limite Inferior Limite Superior 1 477,0 460 498 2 375,0 344 395 I 3 523,0 492 558 4 386,5 370 425 5 528,0 491 555 1 425,5 395 459 2 387,0 360 412
II 3 542,5 513 579 4 496,5 457 552 5 477,0 449 509 1 333,0 314 359 2 496,0 459 535
III 3 365,5 343 394 4 348,0 320 370 5 512,0 486 538 1 375,0 356 396 2 430,0 394 477
IV 3 385,5 362 409 4 317,0 298 334 5 495,0 470 522
¹escala não-corrigida.

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 317
3.3 Estratificação pela covariável touro
Houve variação entre os touros de cada cenário quanto ao número de filhas que tiveram o primeiro parto (falha) até o limite do tempo de censura; e ao número de filhas censuradas (sem informação de parto) (Figuras 5 e 6). Esta variação também ocorreu entre cenários que fizeram uso da mesma quantidade de touros. Porém, o total de filhas por touro não variou: 250 filhas nos cenários I e II e 100 filhas nos cenários III e IV.
No geral, a quantidade média de filhas paridas (falhas) por touro foi maior no cenário I que no II (183,90 vs. 179,20) e ligeiramente maior no cenário III que no IV (81,6 vs. 80,68). Em contrapartida, a média de filhas censuradas por touro foi menor no cenário I que no II (66,1 vs. 70,8) e menor no III que no IV (18,4 vs. 19,32). Logo, o percentual médio de censura foi maior no cenário II que no I (28,32 vs. 26,44%) e maior no cenário IV que no III (19,32 vs. 18,40%).
Figura 5 - Percentual de fêmeas falhas (paridas: F_) por touro à esquerda; e de fêmeas censuradas (C_) à direita, nos cenários I e II (_CX) com uso de 10 touros.
Figura 6 - Percentual de fêmeas falhas (paridas: F_) por touro à esquerda; e de fêmeas censuradas (C_) à direita, nos cenários III e IV (_CX) com uso de 25 touros.
Além disso, foi rejeitada a hipótese de igualdade entre as curvas de sobrevivência
dos touros dentro de cada cenário em função da significância (P<0,0001) das estatísticas S�: 45,45; 125,92; 89,21; e 138,06 para o teste Log-Rank nos cenários I, II, III e IV, respectivamente. Isto significa que houve diferenças entre os touros quanto ao tempo de ocorrência do primeiro parto de suas filhas em cada cenário, constatando-se maiores valores de S� nos cenários em que a herdabilidade da IPP foi mais elevada (0,30).

318 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
Nem sempre foi observado para o touro com o mais baixo (ou mais alto) percentual de censura que suas filhas tiveram a menor (ou maior) idade média e/ou mediana ao evento. Mas, de modo geral, ambas as correlações de Pearson e Spearman (Tabela 4) entre essas variáveis apresentaram magnitudes muito elevadas (>0,90), nos cenários avaliados, sugerindo uma alta correspondência entre elas.
Tabela 4 - Correlações de Pearson (Spearman) entre percentual (%) de censura, IPP média e IPP mediana na estratificação por touro, em cada cenário
Variáveis IPP média IPP mediana
C I (10_0,02) % de censura 0,955 (0,948) 0,942 (0,888) IPP média 1,0 0,972 (0,952) C II (10_0,08) % de censura 0,987 (0,976) 0,992 (0,988) IPP média 1,0 0,984 (0,964) C III (25_0,02) % de censura 0,829 (0,843) 0,796 (0,783) IPP média 1,0 0,942 (0,964) C IV (25_0,08) % de censura 0,929 (0,940) 0,891 (0,875) IPP média 1,0 0,973 (0,959)
Selecionando-se, por exemplo, quatro touros por cenário usando como critério o
percentual de censura, de modo que dois deles tivessem tido os percentuais mais baixos, e outros dois, os mais altos (Tabela 5), constatamos a diferença entre eles quanto aos tempos médio e mediano ao primeiro parto de suas filhas.
No cenário I, o menor percentual de censura foi registrado para o touro 8 (18,4%) e o maior para o touro 9 (39,2%). Em conformidade, a menor IPP média, 1.234,15 dias, e a maior, 1.295,42 dias, foram constatadas para as filhas dos touros 8 e 9, respectivamente. Contudo, a menor IPP mediana, 1.237 dias, foi das filhas do touro 5; enquanto que as do touro 9 também tiveram a maior mediana, aliás bastante acima (59,58 dias) de sua média.
No cenário II, o touro 5 se destacou pelo menor percentual de filhas censuradas (12,8%) e pela maior precocidade das filhas ao primeiro parto: média de 1.191,78 dias e mediana de 1.176,50 dias. O maior (42,80%) percentual de censura foi para o touro 2, cujas filhas apresentaram o maior tempo mediano, 1.395 dias, e o segundo maior tempo médio, 1.306,10 dias.
No cenário III, o touro 22 teve o menor (6%) percentual de filhas censuradas; enquanto no cenário IV, esta posição coube ao touro 17 (3%). Mas, no cenário III (resultado não mostrado), foram as filhas do touro 2, com 14% de censura, que apresentaram tanto a menor idade média como a menor idade mediana ao primeiro parto: 1.158,08 e 1.144 dias, respectivamente. Isto se justifica pelo fato de que as correlações (Tabela 4) entre % de censura e IPP média e entre % de censura e IPP mediana do cenário III foram as mais baixas dentre todas. Já, no cenário IV, as filhas do touro 17 foram, entre todas as fêmeas, as mais precoces; ao passo que as filhas do touro 3 foram as mais tardias.

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 319
A maior variância entre touros originou uma maior diferença entre eles quanto à precocidade reprodutiva de suas filhas. Como exemplo, a diferença entre a IPP mediana das filhas dos dois touros extremos (mais tardio e mais precoce) foi de 118 vs. 218,5 dias no cenário I vs. cenário II; e de 189,5 vs. 239 dias no III vs. IV.
Tabela 5 - Estatística descritiva (% de censura; média¹; erro-padrão (EP) da média e mediana¹) para a IPP das filhas de quatro touros selecionados pelo percentual de censura em cada cenário
Touros % de censura IPP média EP da média IPP mediana
C I(10_0,02) 8 18,4 414,15 11,18 429,00 5 18,8 417,03 10,85 417,00 7 34,4 462,07 11,21 485,50 9 39,2 475,42 11,37 535,00
C II(10_0,08) 5 12,8 371,78 11,16 356,50 6 19,2 409,16 11,27 406,00 7 40,8 486,76 11,11 573,50 2 42,8 486,10 10,51 575,00
C III(25_0,02) 22 6,0 376,75 16,89 389,50 8 9,0 341,39 17,67 333,00
16* 32,0 459,45 18,12 513,50 15 35,0 427,46 18,62 437,50
C IV(25_0,08) 17 3,0 330,48 16,06 291,50 19 6,0 339,63 16,48 334,00 21 33,0 461,60 17,10 491,00 3 34,0 485,20 16,53 530,50
* % comum também ao touro 6, porém as filhas do touro 16 tiveram a maior IPP mediana; ¹escala não-corrigida.
4 Discussão
Para o objetivo de antecipar a IPP, foi constatado, utilizando o método de Kaplan-Meier, que o interesse deve recair sobre o rebanho cuja curva de sobrevivência decresce mais rapidamente com o tempo, o que significa que suas fêmeas sobrevivem ao primeiro parto (evento) por menos tempo, ou seja, apresentam-no em idades mais precoces. As diferenças entre rebanhos são, muito provavelmente, um reflexo de condições ambientais diferentes que afetam positiva ou negativamente a expressão do potencial genético das novilhas para a precocidade reprodutiva, já que a IPP é uma característica quantitativa de baixa a moderada herdabilidade. De modo semelhante, visando a melhoria genética dos rebanhos para a IPP, deve-se selecionar o touro cujas filhas expressam o primeiro parto mais cedo, isto é, aquele cuja curva de sobrevivência exibe decréscimos mais acentuados

320 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
no tempo, em função de ele possuir genes que conferem maior precocidade reprodutiva à sua progênie.
Neste estudo, os cenários que possibilitaram a análise da IPP foram obtidos sob condições mais simplificadas em termos de variáveis que, potencialmente, podem influenciar na avaliação dessa característica em rebanhos comerciais da raça Nelore. Entre as simplificações feitas, vale comentar sobre: 1) o uso de número balanceado de filhas por touro; e 2) a não implementação de estação de monta durante alguns meses do ano. De fato, a desconsideração desses dois itens proporcionou maior simplicidade na geração do banco de dados, como também na interpretação dos resultados. Antes, porém, nos preocupamos com a viabilidade dos cenários propostos a fim de garantir a validação e sustentação dos resultados obtidos, para o que nos apoiamos em trabalhos científicos.
Assim, após vasta revisão de literatura, para o primeiro item acima apontado, constatamos que outros autores também já se utilizaram de esquemas com número igual de filhas por touro em estudos de simulação, para os mais diversos propósitos, conduzidos com bovinos de corte (Cunha et al., 2006), bovinos de leite (Ducrocq e Casella, 1996; Yazdi et al., 2002) e com bovinos em geral (Carneiro et al., 2006; Carvalheiro et al., 2007; Neves et al., 2009), enquanto buscavam mimetizar a realidade de rebanhos comerciais, nos quais se espera, por obviedade, que o número de filhas por touro não seja o mesmo.
Quanto ao segundo item apontado, reconhece-se que a estação de monta é uma prática de manejo reprodutivo que vem sendo adotada pela maioria das propriedades, na bovinocultura de corte do país, a fim de melhorar os índices reprodutivos e, por conseguinte, os índices produtivos e econômicos dos rebanhos. De acordo com Boligon et
al. (2008a), esta prática é essencial para a avaliação genética dos rebanhos. Todavia, autores como Dias et al. (2004) e Boligon et al. (2008a; 2008b)
enfatizaram, em seus estudos, que a curta duração da estação de monta fixa, em torno de 60 dias, permitida aos animais dos bancos de dados que analisaram, pode ter influenciado nas estimativas de baixa magnitude da herdabilidade da IPP. Na opinião deles, tal prática, possivelmente, limitou a expressão de variabilidade genética desta característica. Segundo esses autores, se tivesse sido permitido às fêmeas entrar em reprodução durante todo o ano, as diferenças entre os animais, provavelmente, teriam sido notadas. Partilhando da mesma opinião, Pereira et al. (2007) reforçam que o manejo usado pode interferir de maneira decisiva com a expressão da IPP, especialmente o uso da estação de monta, de forma que uma análise por modelos lineares mistos apenas teria sentido se todas as fêmeas tivessem a oportunidade contínua de emprenhar.
Baseamo-nos, pois, nos argumentos apresentados por esses autores para simular os tempos até o primeiro parto desprezando a variável estação de monta, ainda que nosso objetivo aqui não tenha sido estimar parâmetros genéticos nem predizer valores genéticos supostamente afetados por ela, e nem utilizar modelos lineares mistos. Aliás, esclarece-se que o método de Kaplan-Meier pelo proc LIFETEST não produz estimativas de parâmetros (Allison, 2010). Porém, fica evidente que, quando essa prática de manejo incide nos rebanhos reais, ela deve ser considerada na análise, pois os dados coletados já foram influenciados por ela. Em razão do viés provocado pela estação de monta, alguns pesquisadores (Pereira et al., 2000; Boligon et al., 2007), analisando dados provenientes de sistemas de produção de gado de corte com estação de monta fixa, geralmente de curta duração, passaram a utilizar a característica dias para o primeiro parto (DPP) ao invés da IPP, por considerá-la mais adequada. Enquanto a IPP, em dias, corresponde à diferença

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 321
entre a data do primeiro parto e a data de nascimento da fêmea (como considerado neste estudo), a DPP, em dias, corresponde ao número de dias entre o primeiro dia da estação de monta e a data do parto subsequente (Boligon et al., 2007). A avaliação da DPP permite que se identifiquem as fêmeas com maior fertilidade, ou seja, aquelas que emprenham mais cedo dentro da estação de monta e touros cujas filhas apresentem menor número de dias para o parto (Pereira et al., 2000).
Por outro lado, a simulação é uma ferramenta que possibilita gerar bancos de dados para a análise da IPP (ou outra característica) conforme a estrutura de interesse. Logo, como não estabelecemos estação de monta, em nossos cenários, então, também não tivemos que estabelecer censura após cada ano subsequente a ela, o que seria esperado de ocorrer se acaso uma novilha não tivesse parido em função de não ter emprenhado na estação anterior. Somado a isso, o número de serviços por fêmea até a concepção era irrestrito, o que não nos exigiu impor regras para o descarte de fêmeas vazias. Por este raciocínio, todas as fêmeas tiveram oportunidade contínua de emprenhar; e as que o fizeram a menos de 9,5 meses (duração da gestação) do tempo final do estudo (48 meses), foram, logicamente, as fêmeas censuradas. Mesmo que isto pareça algo distante da realidade, foi o suficiente para atender o nosso propósito de simular a IPP, como variável aleatória contínua, tomando por base as suposições daqueles autores.
Outro fato a ser comentado, em análise de sobrevivência, diz respeito aos tempos do estudo. Neste contexto, segundo Allison (2010), a escolha da origem do tempo é um ponto problemático e uma das razões disso é que, em muitas situações, duas ou mais origens podem ser escolhidas e não há nenhum critério livre de ambiguidade para decidir entre elas. Como sugestão do autor, deve-se escolher o tempo de origem que tem um efeito mais forte sobre o risco. No caso de novilhas Nelore, estudos (Dias et al., 2004; Grossi et
al., 2009; Boligon e Albuquerque, 2010) apontam a existência de dificuldade na identificação das fêmeas mais precoces sexualmente, porque alguns produtores atrasam a entrada das novilhas na reprodução determinando uma idade ou um peso para que estas iniciem sua vida reprodutiva. Boligon et al. (2008a), por exemplo, avaliaram 28 rebanhos Nelore em que as novilhas foram submetidas à reprodução com aproximadamente 27 meses e peso médio de 330 kg. Para Laureano et al. (2011), esse atraso na exposição de novilhas Nelore à reprodução também contribui para a estimação de variâncias genéticas aditivas de pequena magnitude para a IPP. Mas, tal fato, segundo os autores, não ocorreu no rebanho comercial que eles avaliaram, porque as novilhas com idade próxima aos 17 meses participavam da estação de monta.
Diante de tais considerações, valemo-nos dos estudos acima citados, em complementação ao de Mercadante et al. (2000) e ao sumário da raça Nelore (EMBRAPA/ABCZ/MAPA, 2011-2012), para fixar os tempos de origem (820 dias), mediano (1.217 dias) e máximo (1.460 dias) para a ocorrência da IPP, a fim de que nossos cenários tivessem a maior verossimilhança possível. Sendo assim, qualquer arbitrariedade cometida na escolha desses tempos foi provavelmente mínima, pois visamos, acima de tudo, concordar com a realidade. Posto isto, limitamo-nos a efetuar poucas comparações, que julgamos pertinentes, entre nossos resultados e aqueles de outros autores.
Por meio dessas simulações – e tendo em vista a escassez de trabalhos, na literatura nacional, nos quais a idade ao primeiro parto de novilhas Nelore, na presença de censura, tenha sido avaliada por meio de método(s) classificado(s) como de análise de sobrevivência –, acreditamos que um estudo como este possa ter impacto positivo ao

322 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
apresentar uma abordagem alternativa e apropriada para avaliação de dados desta natureza, posto que ela permite revelar as diferenças entre estratos quanto ao tempo de ocorrência do evento de interesse, e vem sendo pouco explorada, quanto mais de modo pormenorizado. Por último, esclarecemos que os dados foram simulados sob a função de risco paramétrica de Weibull, que consistiu de um modelo misto de sobrevivência (ou modelo de fragilidade) de Weibull com as covariáveis rebanho como fixa e touro como aleatória, de modo a garantir a não-normalidade dos tempos; porém, toda a análise de sobrevivência, em si, foi conduzida com uso do método não-paramétrico de Kaplan-Meier, consoante nosso objetivo aqui proposto. No entanto, haja vista que a finalidade principal de uma avaliação genética é a estimação dos efeitos fixos e a predição do valor genético dos animais, abordaremos, num próximo trabalho, tais estimativas a partir do ajuste do modelo paramétrico de Weibull.
Conclusões
Curvas de sobrevivência e de taxas de risco obtidas por Kaplan-Meier mostram-se adequadas ao estudo descritivo inicial da idade ao primeiro parto de fêmeas Nelore, considerando a presença de censura à direita e a não-normalidade dos dados. Por meio dessa estatística não-paramétrica é possível detectar diferenças entre os estratos (rebanhos e touros) quanto ao tempo de ocorrência do primeiro parto.
No contexto do melhoramento genético animal, o touro com a maior proporção de fêmeas paridas (falhas) até um determinado período de tempo e, principalmente, aquele cuja curva de sobrevivência ao evento tiver decréscimos mais acentuados no tempo, em razão de o mesmo possuir genes para uma maior precocidade reprodutiva de suas filhas, devem ser selecionados para o objetivo de antecipar a idade ao primeiro parto das futuras matrizes dos rebanhos.
Agradecimentos
Ao CNPq pela concessão de uma bolsa de iniciação científica à segunda autora, durante a execução desse projeto de pesquisa, sob a coordenação da primeira autora.
CUNHA, E. E.; MELO, T. P. Nonparametric survival analysis of the age at first calving in Nellore females: a simulation study. Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012.
� ABSTRACT: Time records (in days) until the first calving from Nellore females were simulated,
incorporating right censoring, with the aim at estimating survival probabilities and hazard rates
associated with such event through the nonparametric method of Kaplan-Meier of survival
analysis. The risk function of Weibull from a female in the time t, as continuous dependent
variable, was modeled including the fixed effect of herd (five levels) and the random effect of sire
(10 or 25 levels) with sire variance of 0.02 or 0.08, that combined gave origin to distinct four
scenarios: I (10_0.02); II (10_0.08); III (25_0.02) and IV (25_0.08). The percentages of
censoring per scenario were, respectively, of: 26.44 and 28.32% at I and II; and 18.40 and
19.32% at III and IV. There were significant differences among the survival curves of the herds

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 323
and sires (fathers) of each scenario, what means that they were statistically different between
themselves for the reproductive precocity of their females (daughters). In the animal genetic
improvement, the sire whose survival curve decreases faster in the time produces daughters that
have the first calving earlier, what justifies its use as breeder aiming at reducing the age at first
calving of the females into the herds.
� KEYWORDS: Beef cattle; censoring; failure; Kaplan-Meir method.
Referências
ALLISON, P. D. Survival analysis using SAS: a practical guide. 2.ed. Cary: SAS Institute Inc. 2010. 324p.
AZEVÊDO, D. M. M. R.; MARTIN FILHO, R.; LÔBO, R. N. B.; MALHADO, C. H. M.; LÔBO, R. B.; MOURA, A. A. A.; PIMENTA FILHO, E. C. Desempenho reprodutivo de vacas Nelore no Norte e Nordeste do Brasil. Rev. Bras. Zootec., Viçosa, v.35, n.3, p.988-996, 2006 (Supl.).
BASTOS, J.; ROCHA, C. Análise de sobrevivência: métodos não-paramétricos. Arq.
Med., Belo Horizonte, v.21, n.3/4, p.111-114, 2007.
BOLIGON, A. A.; ALBUQUERQUE, L. G. Correlações genéticas entre escores visuais e características reprodutivas em bovinos Nelore usando inferência bayesiana. Pesq.
agropec. bras., Brasília, v.45, n.12, p.1412-1418, 2010.
BOLIGON, A. A.; ALBUQUERQUE, L. G.; RORATO, P. R. N. Associações genéticas entre pesos e características reprodutivas em rebanhos da raça Nelore. Rev. Bras. Zootec., Viçosa, v.37, n.4, p.596-601, 2008a.
BOLIGON, A. A.; RORATO, P. R. N.; ALBUQUERQUE, L. G. Correlações genéticas entre medidas de perímetro escrotal e características produtivas e reprodutivas de fêmeas da raça Nelore. Rev. Bras. Zootec., Viçosa, v.36, n.3, p.565-571, 2007.
BOLIGON, A. A.; VOZZI, P. A.; NOMELINI, J.; RORATO, P. R. N.; BEZERRA, L. A. F.; LÔBO, R. B. Parâmetros genéticos para a idade ao primeiro parto estimados por diferentes modelos para rebanhos da raça Nelore. Ciênc. Rur., Santa Maria, v.38, n.2, p.432-436, 2008b.
CAETANO, S. L. Estudo da idade da vaca ao último parto para avaliar longevidade em
rebanhos da raça Nelore por análise de sobrevivência. 2011. 111f. Tese (Doutorado em Genética e Melhoramento Animal) – Universidade Estadual Paulista, Jaboticabal, SP, 2011.
CARNEIRO, A. P. S.; TORRES, R. A.; LOPES, P. S.; EUCLYDES, R. F.; CARNEIRO, P. L. S.; CUNHA, E. E. Erros na classificação de touros, vacas e touros jovens geneticamente superiores avaliados na presença de heterogeneidade. Rev. Bras. Zootec., Viçosa, v.35, n.5, p.1907-1914, 2006.
CARVALHEIRO, R.; SCHENKEL, F. S.; FRIES, L. A.; QUEIROZ, S. A.; MILLER, S. P. Empirical properties of solutions from Henderson’s mixed model equations when sires are non-randomly used across contemporary groups. Rev. Bras. Biom., São Paulo, v.25, n.3. p.7-21, 2007.

324 Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012
COLOSIMO, E. A.; GIOLO, S. R. Análise de sobrevivência aplicada. São Paulo: Edgard Blücher, 2006. 370p.
CUNHA, E. E.; EUCLYDES, R. F.; TORRES, R. A.; CARNEIRO, P. L. S. Simulação de dados para avaliação genética de rebanhos de gado de corte. Arq. Bras. Med. Vet. Zootec., Belo Horizonte, v.58, n.3, p.381-387, 2006.
DIAS, L. T.; EL FARO, L.; ALBUQUERQUE, L. G. Estimativas de herdabilidade para idade ao primeiro parto de novilhas da raça Nelore. Rev. Bras. Zootec., Viçosa, v.33, n.1, p.97-102, 2004.
DONOGHUE, K. A.; REKAYA, R.; BERTRAND, J. K. Comparison of methods for handling censored records in beef fertility data: simulation study. J. An. Sci., Champaign, v.82, n.2, p.351-356, 2004.
DUCROCQ, V.; CASELLA, G. A Bayesian analysis of mixed survival models. Genet.
Sel. Evol., London, v.28, n.2, p.505-529, 1996.
DUCROCQ, V.; SÖLKNER, J.; MÉSZÁROS, G. The Survival Kit v6.0: User's manual. [S.I.: s.n.], 2010. 83p.
EMBRAPA/ABCZ/MAPA (2011-2012). Sumário nacional de touros das raças zebuínas -
Nelore. Disponível em: <http://www.cnpgc.embrapa.br/~locs/sumario/sumzebu/ nel_ index.php? tp=nel_resultado>. Acesso em: abr.2012.
GIOLO, S. R.; HENDERSON, R.; DEMÉTRIO, C.G.B. Um critério para a seleção de touros nelore usando modelos de sobrevivência. Rev. Mat. Estat., São Paulo, v.21, n.3, p.115-123, 2003.
GROSSI, D. A.; VENTURINI, G. C.; PAZ, C. C. P.; BEZERRA, L. A. F.; LÔBO, R. B.; OLIVEIRA, J. A.; MUNARI, D. P. Genetic associations between age at first calving and heifer body weight and scrotal circumference in Nellore cattle. J. Anim. Breed. Genet., Berlin, v.126, p.387-393, 2009.
GUNSKI, R. J.; GARNERO, A. V.; BEZERRA, L. A. F.; CORRADO, M. P.; LÔBO, R. B. Idade ao primeiro parto, período de gestação e peso ao nascimento na raça Nelore. Ciênc. Agron., Fortaleza, v.32, n.1/2, p.46-52, 2001.
KACHMAN, S. D. Applications in survival analysis. J. An. Sci., Champaign, v.77, p.147-153, 1999.
KAPLAN, E. L.; MEIER, P. Nonparametric estimation from incomplete observations. J.
Am. Stat. Assoc., Schaumburg, v.53, n. 282, p.457-481, 1958.
LAUREANO, M. M. M.; BOLIGON, A. A.; COSTA, R. B.; FORNI, S.; SEVERO, J. L. P.; ALBUQUERQUE, L. G. Estimativas de herdabilidade e tendências genéticas para características de crescimento e reprodutivas em bovinos da raça Nelore. Arq. Bras. Med.
Vet. Zootec., Belo Horizonte, v.63, n.1, p.143-152, 2011.
MERCADANTE, M. E. Z.; LÔBO, R. B.; OLIVEIRA, H. N. Estimativas de (co)variâncias entre características de reprodução e de crescimento em fêmeas de um rebanho Nelore. Rev. Bras. Zootec., Viçosa, v.29, n.4, p.997-1004, 2000.
NEVES, H. H. R.; CARVALHEIRO, R.; CARDOSO, V.; FRIES, L. A.; QUEIROZ, S. A. Acasalamento dirigido para aumentar a produção de animais geneticamente superiores

Rev. Bras. Biom., São Paulo, v.30, n.3, p.305-325, 2012 325
e reduzir a variabilidade da progênie em bovinos. Rev. Bras. Zootec., Viçosa, v.38, p.1201-1204, 2009.
PEREIRA, E.; ELER, J. P.; COSTA, F. A. A.; FERRAZ, J. B. S. Análise genética da idade ao primeiro parto e do perímetro escrotal em bovinos da raça Nelore. Arq. Bras.
Med. Vet. Zootec., Belo Horizonte, v.53, n.1, p.116-121, 2001.
PEREIRA, E.; ELER, J. P.; FERRAZ, J. B. S. Correlação genética entre perímetro escrotal e algumas características reprodutivas na raça Nelore. Rev. Bras. Zootec., Viçosa, v.29, n.6, p.1676-1683, 2000.
PEREIRA, E.; OLIVEIRA, H. N.; ELER, J. P.; SILVA, J. A. II V.; Van MELIS, M. H. Use of survival analysis as a tool for the genetic improvement of age at first conception in Nellore cattle. J. Anim. Breed. Genet., Berlin, v.123, n.1, p.64-71, 2006.
PEREIRA, E.; OLIVEIRA, H. N.; ELER, J. P.; SILVA, J. A. II V.; Van MELIS, M. H. Comparison among three approaches for evaluation of sexual precocity in Nellore cattle. Animal, Cambridge, v.1, n.3, p.411-418, 2007.
PEREIRA, M. C. Avaliação genética da reconcepção de fêmeas primíparas da raça
Nelore. 2008. 86f. Tese (Doutorado em Zootecnia) – Universidade Estadual Paulista, Jaboticabal, SP. 2008. ROSA, A. J. M.; BIJMA, P.; OLIVEIRA, H. N.; LÔBO, R. B.; van ARENDONK, J. A. M. A deterministic simulation study of embryo marker-assited selection for age at first calving in Nellore (Bos indicus) beef cattle. Genet. Mol. Biol., Ribeirão Preto, v.30, p.552-559, 2007.
SAS INSTITUTE INC. SAS/STAT® 9.2 User’s guide. 2.ed. Cary: SAS Institute Inc., 2009. 7869p.
WIENKE, A. Frailty models in survival analysis. Boca Raton: Chapman & Hall/CRC, 2011. 298p. (Biostatistics Series).
YAZDI, M. H.; VISSCHER, P. M.; DUCROCQ, V.; THOMPSON, R. Heritability, reliability of genetic evaluations and response to selection in proportional hazard models. J. Dairy Sci., New York, v.85, n.6, p.1563–1577, 2002.
Recebido em 10.06.2012
Aprovado após revisão em 07.12.2012