variação diária e anual do comportamento do sotalia … · a deus, “inteligência...
TRANSCRIPT

i
FERNANDO ROBERTO GUILHERME SILVEIRA
VARIAÇÃO DIÁRIA E ANUAL DE FREQÜÊNCIA E ATIVIDADE DO
BOTO CINZA Sotalia guianensis, EM PIPA, NORDESTE DO BRASIL
Natal/RN
2006

ii
FERNANDO ROBERTO GUILHERME SILVEIRA
VARIAÇÃO DIÁRIA E ANUAL DE FREQÜÊNCIA E ATIVIDADE DO
BOTO CINZA Sotalia guianensis, EM PIPA, NORDESTE DO BRASIL
Dissertação apresentada ao Programa de
Pós-Graduação em Psicobiologia do
Departamento de Fisiologia da Universidade
Federal do Rio Grande do Norte, Como parte dos
requisitos para obtenção do título de Mestre.
Orientador: Prof. Dr. Flávio José de Lima Silva
Natal/RN
2006

iii
Divisão de Serviços Técnicos
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede
Silveira, Fernando Roberto Guilherme. Variação diária e anual de freqüência e atividade do Boto Cinza Sotalia guianensis em Pipa, Nordeste do Brasil
/ Fernando Roberto Guilherme Silveira. – Natal, RN, 2006.
69 f
Orientador : Flávio José de Lima Silva.
Dissertação (Mestrado) - Universidade Federal do Rio Grande do Norte. Departamento de Fisiologia.
Programa de Pós-Graduação em Psicobiologia.
1. Comportamento animal – Dissertação. 2. Boto Cinza (Sotalia guianensis) -
Dissertação. 3. Variação diária - Dissertação. 4. Sazonalidade – Dissertação. I. Silva, Flávio
José de Lima. II. Título.
RN/UF/BCZM CDU 591.5 (043.3)

v
A minha filha Rafaela, razão pela
qual eu tenho perseverado.

vi
AGRADECIMENTOS
Todo mundo tem um modo diferente e particular de ver a vida. Entre os
vários modos como poderia defini-la, diria que é uma espiral, cujo círculo
superior é sempre maior que o inferior. Cada um deles representa uma
experiência, um relacionamento, uma etapa, um momento em nossas vidas que
marca de forma positiva ou não para o resto da eternidade. Assim vejo estes
últimos dois anos. Como etapas de uma espiral, que compõe o que sou. Ao
longo do período que passei no Centro de Biociências, adquiri um pouco mais.
Conheci colegas, professores e funcionários, uns mais afins que outros. Passei
bons momentos e outros nem tanto. O conhecimento e a experiência foram
conquistados com sacrifício, determinação, amor, responsabilidade,
simplicidade, humildade, admiração, respeito e confiança. Alguns destes valores
nasceram comigo. Outros, adquiri convivendo. Essa espiral não se expandiria,
não fosse a colaboração de todos com quem convivi, sejam de forma agradável
ou não. Desta forma o que sou devo a muitos.
A DEUS, “inteligência suprema, causa primeira de todas as coisas”, onde fui
sorver amor, caridade, simplicidade e humildade e que me abençoou com outras
tantas.
A minha filha Rafaela, razão pela qual eu tenho perseverado.
A minha Família, que mesmo sem saber é meu sustentáculo emocional.
As Famílias Vianna e Soares, cujo apoio emocional, financeiro e fraterno,
permitiu-me chegar até aqui, mas que antes de tudo, são pessoas com quem
contei e poderei contar sempre. Amigos de todas as horas, cujo valor é
inestimável.
As Famílias Guilherme, Custódio, Menezes.
Ao meu Orientador Profº. Dr. Flávio José de Lima Silva, que na prática,
discutiu estes dados e orientando-me ajudou-me a delinear este trabalho, mas
acima de tudo mostrou-se um amigo também inestimável
A todos os Professores que contribuíram para com a minha formação como
biólogo e mestre e aqueles que de forma indireta também o fizeram.

vii
A todo aquele funcionário que se dispôs a realizar suas dignas funções e com
isto facilitaram a minha vida e de todos aqueles que ainda se encontram dentro
do processo.
A todos os colegas da Base de Pesquisa com Cetáceos, que me ensinaram a
vê-los e compreendê-los em seu ambiente natural; e cederam os dados que
culminaram neste trabalho.
A todos os camaradas que conheci nas salas de aula e nos corredores do
Biociências, ao longo destes anos de mestrado.
A todos os camaradas de outras Bases de Pesquisa e Laboratórios que
contribuíram com este trabalho.
Ao amigo e Professor Gilson Volpato. O tenho na mais alta estima, pois me
mostrou o caminho de uma boa redação.
Certamente é difícil nomear à todos com quem convivemos até porque
alguns convívios são passageiros e sutis, mas não menos importantes, uma vez
que as experiências ficam ao passo que as lembranças, por vezes se perdem na
memória. De qualquer forma, sou graças a eles.
Não poderia deixar de mencionar ainda, o apoio financeiro dado pela CAPES,
cujos resultados são as duas publicações contidas neste trabalho; ao Programa
de Pós-Graduação do Departamento de Fisiologia do Centro de Biociências e a
Universidade Federal do Rio Grande do Norte – UFRN na forma da Instituição.

viii
RESUMO
Neste estudo avaliamos a variação diária e anual na freqüência de adultos e
imaturos e nos comportamentos de socialização, atividade aérea e forrageio
numa população de boto cinza, Sotalia guianensis, na Enseada do Curral
(6º13’00”S; 35º3’36”W), no nordeste brasileiro. Após detectarmos algumas
variações procuramos associá-las a parâmetros ambientais. Associamos as
variações diárias ao período do dia, às variações da maré e ao período do ano;
e as sazonais, a variação do fotoperíodo e da precipitação. De Outubro de 1999
a Setembro de 2003, através de coletas quatro vezes ao mês, efetuaram-se
registros instantâneos a cada 5min, em seis horas de observação. As horas de
observação estiveram inseridas dentro da fase clara das 24 horas, ou seja, no
intervalo das 06:00 h as 17:00 h. Essas observações eram realizadas
simultaneamente por dois observadores, num ponto fixo no alto da falésia a
partir do qual visualizava-se toda a enseada. Observamos que os animais são
mais ativos e freqüentes pela manhã. Não apresentaram associação com a
maré, apesar de apresentarem maior atividade na maré baixa e menor atividade
na maré alta. Quanto à sazonalidade, observamos ainda, que no período de
junho a novembro (inverno e primavera) houve maior freqüência dos
comportamentos de socialização, atividade aérea e forrageio embora o número
de indivíduos (adultos e imaturos) não tenha variado significativamente ao longo
do ano. A maior concentração desses comportamentos não foi associada ao
fotoperíodo e nem à precipitação na região. Desses resultados, mostramos a
existência de variação diária e sazonalidade comportamental nesta espécie,
mesmo nas regiões equatoriais.

ix
ABSTRACT
The aim of this study was to investigate the daily and annual variation in
frequency and in activity of an estuarine dolphin population, Sotalia guianensis,
in Brazilian northeastern. Thus we estimated the daily variation and seasonality
in frequency of adult and immature and in behavior of socialization, aerial activity,
foraging of a boto equatorial population in the Curral inlet (6º13’00”S; 35º3’36”W).
We detect some variation. Thus we associate the daily variation with daily period,
tide fluctuation and annual season. And the seasonality variation we associate
with the fotoperíodo and precipitation variation. From October 1999 to September
2003, through collect four times at month, we made instantaneous record at each
five minute in six hours of survey. The surveys hours were ranged into light time
of 24 hours, that is, between 06:00 and 17:00 o clock. These observations were
developed simultaneous for two observers in a land-based survey at coastline
cliff top of which observer have a fully visualization of whole inlet. We found that
animals were more active and frequent during the morning. They were more
active during low tide than high tide. However they did not show association with
tide. We found yet that in a period from June to November (winter-spring) there
was the highest socialization, aerial activity and foraging frequency. However
adult and immature individual number did not have significance variation along
the year. The larger concentration of this behaviors were not associate with
photoperiod neither precipitation in the region. These results showed the
existence of behavioral daily variation and seasonality in a species that live in
equatorial region.

iv
Título: Variação diária e anual de freqüência e atividade do boto cinza
Sotalia guianensis, em pipa, nordeste do brasil.
Autor: Fernando Roberto Guilherme Silveira
Data da Defesa: 22 de maio de 2006.
Banca Examinadora:

1
SUMÁRIO
INTRODUÇÃO GERAL..................................................................................... 2
ARTIGO 1.......................................................................................................... 5
ARTIGO 2........................................................................................................ 29
DISCUSSÃO GERAL...................................................................................... 57
CONCLUSÃO GERAL.................................................................................... 60
LITERATURA CITADA (INTRODUÇÃO E DISCUSSÃO GERAIS) .............. 60
ANEXOS.......................................................................................................... 63
ANEXO 1: Normas de Publicação – Canadian Journal Zoology .............................. 64

2
INTRODUÇÃO GERAL
Os organismos vivos, em geral, apresentam variações no seu repertório
comportamental ao longo do dia e em épocas distintas do ano como forma de
ajuste às oscilações das condições ambientais e climáticas dos locais onde
habitam. Desta maneira, essas oscilações climáticas interferem na expressão de
comportamentos como forrageio, socialização, reprodução, repouso entre
outros.
É possível observar, que mamíferos aquáticos geralmente, mantêm uma
estreita associação com algum fator ambiental (disponibilidade de recursos,
estabilidade ambiental, temperatura da água, estação do ano, etc.), quando se
trata de adquirir seu alimento (Similä et al. 1996; Urian et al. 1996; Baird 2000;
Claphan 2000; Iñíguez 2001).
Mamíferos aquáticos como os misticetos da ordem cetácea possuem uma
área de uso extensa e antecipam-se ao inverno. Estes animais migram das
áreas de alimentação (altas latitudes), para as áreas de acasalamento e
procriação (baixas latitudes) (Claphan et al. 1993; Claphan 2000).
Os odontocetos, animais menores do que os misticetos possuem uma
notável plasticidade, ajustando seus comportamentos aos ciclos geofísicos. As
orcas, Orcinus orca, por exemplo, podem pertencer tanto a grupos que migram
(transeuntes), quanto a grupos que não migram (as residentes) (Baird 2000).
Populações diferentes desse mesmo animal deslocam-se sazonalmente entre
norte e sul, seguindo suas presas, seja na Noruega na migração do arenque
(Similä et al. 1996), seja na Patagônia no período reprodutivo dos leões e
elefantes marinhos (Iñíguez 2001).
O golfinho nariz de garrafa, Turciops truncatus, um dos mamíferos
aquáticos mais versáteis a quaisquer tipos de ambientes e adversidades, possui
uma grande capacidade de sincronização às condições ambientais locais. Essa
adaptação exerce uma forte influência sazonal no ciclo reprodutivo em
populações no Golfo do México (Urian et al. 1996).
Este ajuste de comportamentos devido às variações diárias e anuais do
ambiente está presente como importante ferramenta de adaptação seletiva, em
todas as espécies, inclusive mamíferos aquáticos entre eles os cetáceos.

3
Esta flutuação comportamental é mais observada geralmente em populações
que habitam as altas latitudes (Read 1990; Similä et al. 1996; Wilson et al. 1997;
Larkman e Veit 1998; Wood 1998; Hindell et al. 2003; Thayer et al. 2003).
Embora seja freqüente considerar que as condições climáticas ou fatores
ambientais sejam sutis ou irrelevantes para influenciar a flutuação dos
comportamentos de populações nas regiões tropicais e equatoriais, estudos
anteriores têm indicado possíveis alterações do comportamento reprodutivo em
cetáceos decorrentes de variações de fotoperíodo e precipitação entre outros
fatores (Urian et al. 1996; Santos et al. 2001; Rosas e Monteiro-Filho 2002;
Pastor e Aguilar 2003).
Para o Sotalia guianensis, segundo Geise et al. (1999), os animais em
Cananéia, São Paulo, apresentam uma flutuação diária, sendo o número de
indivíduos associado ao momento do dia cuja disponibilidade de alimento é
maior; e o tamanho dos grupos varia conforme a atividade realizada. Segundo
as autoras, não foi verificado sazonalidade no período reprodutivo e no
deslocamento da população, levando a crer que apresentam um padrão difuso
de ocorrência e reproduzem-se durante o ano todo. No estudo de Lodi (2003)
em Paraty, Rio de Janeiro, ela verificou a permanência destes animais em
determinado habitat o ano todo devido à disponibilidade de recursos; proteção
contra predadores; diversidade de nichos; águas rasas, protegidas e quentes.
O presente trabalho teve como objetivo verificar a flutuação diária e a
sazonalidade no comportamento de uma população equatorial de cetáceo.
Deste modo investigamos a freqüência dos indivíduos (adultos e imaturos) e os
comportamentos de socialização, atividade aérea e forrageio em uma população
de boto cinza, S. guianensis, no nordeste do Brasil. Também verificamos o efeito
de parâmetros ambientais, particularmente o fotoperíodo a precipitação e a
variação da maré, sobre a expressão dos comportamentos desses animais.

4
Objetivos Específicos:
1º) Identificar uma possível sazonalidade na freqüência de indivíduos e nos
comportamentos de socialização, atividade aérea e forrageio em uma população
de boto cinza, Sotalia guianensis, no nordeste do Brasil (Artigo 1);
2º) Verificar o efeito dos parâmetros ambientais fotoperíodo e precipitação,
na expressão dos comportamentos desses animais (Artigo 1);
3º) Investigar a existência de variação diária nos comportamentos de
socialização, atividade aérea, forrageio e na freqüência de indivíduos de uma
população de boto cinza, Sotalia guianensis, no nordeste Brasileiro (Artigo 2);
4º) Investigar a influência dos parâmetros ambientais, período do dia e
variação da maré na expressão desses comportamentos (Artigo 2).

5
ARTIGO 1
BEHAVIORAL SEASONALITY OF THE ESTUARINE DOLPHIN, Sotalia
guianensis, IN NORTHEASTERN BRAZIL
Silveira, F.R.G.1 and Silva, F.J.L.2
1 Programa de Pós-Graduação em Psicobiologia, Departamento de Fisiologia.
Universidade Federal do Rio Grande do Norte. Campus Central – Natal, RN,
Brazil. E-mail: [email protected] 2 Departamento de Ciências Biológicas, Universidade do Estado do Rio Grande
do Norte, Avenida Antônio de Campos, s/n. CEP: 59600-970. Mossoró, RN,
Brazil.
e-mail:[email protected]. Fax: ++ 55 84 3315-2235.
www.golfinhorotador.org.br
A SER SUBMETIDO AO
CANADIAN JOURNAL OF ZOOLOGY
(INSTRUÇÕES PARA AUTORES – ANEXO)

1
Behavioral seasonality of the estuarine dolphin, Sotalia guianensis, in
Northeastern Brazil
Silveira, F.R.G.
Programa de Pós-Graduação em Psicobiologia, Departamento de Fisiologia.
Universidade Federal do Rio Grande do Norte. Campus Central – Natal, RN, Brazil. E-
mail: [email protected]
Silva, F.J.L.
Departamento de Ciências Biológicas, Universidade do Estado do Rio Grande do
Norte, Avenida Antônio de Campos, s/n. CEP: 59600-970. Mossoró, RN, Brazil. e-
mail:[email protected]. Fax: ++ 55 84 3315-2235.
www.golfinhorotador.org.br
Editor: please send correspondence to Silveira F.R.G.
Key words: Delphinidae, boto, estuarine dolphin, Sotalia guianensis, seasonal,
behavior fluctuation, circannual.
"This paper has not been submitted elsewhere in identical or similar form, nor
will it be during the first three months after its submission."

2
RESUMO
Neste estudo avaliamos a sazonalidade na freqüência de indivíduos e nos
comportamentos de socialização, atividade aérea e forrageio numa população de boto
cinza, Sotalia guianensis, na Enseada do Curral (6º13’00”S; 35º3’36”W), no nordeste
brasileiro. Após detectarmos algumas variações sazonais, procuramos associá-las com
variações de fotoperíodo e precipitação. De Outubro de 1999 a Setembro de 2003, por
meio de coletas mensais (4 por mês) com registros instantâneos realizada a cada 5 min
ao longo de seis horas, no período claro do dia, das 06:00 h às 17:00 h. As
observações eram realizadas simultaneamente por dois observadores, num ponto fixo
no alto da falésia a partir do qual visualizava-se toda a enseada. Observamos que no
período de junho a novembro (inverno e primavera) houve maior freqüência dos
comportamentos de socialização, atividade aérea e forrageio embora o número de
indivíduos não tenha variado significativamente ao longo do ano. A maior concentração
desses comportamentos não foi associada ao fotoperíodo e nem à precipitação na
região. Desses resultados, mostramos a existência de sazonalidade comportamental
nesta espécie, mesmo nas regiões equatoriais.

3
INTRODUÇÃO
Os organismos vivos, em geral, apresentam variações de comportamento ao longo
do ano como forma de ajuste às oscilações sazonais das condições ambientais e
climáticas dos locais onde habitavam. Desta maneira, as condições ambientais e
climáticas em épocas distintas do ano interferem na expressão de diferentes categorias
de comportamentos. Este ajuste de comportamentos a determinada época do ano
devido às condições climáticas que se estabelecem em cada estação, está presente
como importante ferramenta de adaptação seletiva, em todas as espécies, inclusive
mamíferos aquáticos e em especial entre os cetáceos.
Entre os cetáceos a sazonalidade nos comportamentos é observada em populações
que habitam as altas latitudes (Read 1990; Boyd et al. 1994; Similä et al. 1996; Wilson
et al. 1997; Larkman and Veit 1998; Wood 1998; Hindell et al. 2003; Thayer et al.
2003). Embora seja freqüente considerar que as condições climáticas ou fatores
ambientais são sutis ou irrelevantes para influenciar a flutuação dos comportamentos
de populações nas regiões tropicais e equatoriais, estudos anteriores indicam possíveis
alterações do comportamento reprodutivo em cetáceos decorrentes de variações de
fotoperíodo e pluviosidade (Urian et al. 1996; Santos et al. 2001; Rosas and Monteiro-
Filho 2002; Pastor and Aguilar 2003).
O presente trabalho teve como objetivo verificar a sazonalidade no
comportamento de indivíduos de uma população equatorial de cetáceo. Deste
modo investigamos a freqüência dos indivíduos e dos comportamentos de
socialização, atividade aérea e forrageio em uma população de boto cinza, S.
guianensis, na enseada do Curral, distrito de Pipa, nordeste do Brasil. Também
verificamos o efeito de parâmetros ambientais, particularmente o fotoperíodo e a
precipitação, sobre a expressão dos comportamentos desses animais. Usamos o

4
Fotoperíodo e a Precipitação, por considerarmos estes dois fatores ambientais de
extrema importância na expressão dos comportamentos nesta latitude.
MATERIAL E MÉTODOS
Espécie e área de estudo
Investigamos o boto cinza, que na nomenclatura de Monteiro–Filho et al. (2002) e
Cunha et al. (2005) chama-se Sotalia guianensis van Bénéden 1864 (Cetacea,
Delphinidae). Essa espécie possui distribuição restrita entre a Lagoa de Waunte,
Nicarágua ao norte e a Baía Norte, no Estado de Santa Catarina, Brasil ao sul (da Silva
and Best 1996).
Este estudo restringiu-se aos indivíduos que ocorreram na enseada do Curral
(6º13’00”S; 35º3’36”W), no distrito de Pipa, RN, Brasil. A enseada possui
aproximadamente um quilômetro de extensão. A área de estudo ficou compreendida
entre a praia e uma linha reta imaginária traçada ligando as extremidades da enseada
(Fig. 1). Essa linha tem cerca de 1000 m e a maior distância da praia é na região
central (cerca de 500 m). As extremidades dessa enseada apresentam formações
rochosas (arenito sedimentar) que se estendem do andar infralitoral para o supralitoral
até unir-se à falésia, cuja altura atinge cerca de 30 m em sua porção mais elevada
(local usado para as observações). O fundo na enseada é constituído exclusivamente
de arenito, com profundidade máxima de cinco metros. É uma região abrigada de
ventos (< 1,0 na escala de Beaufort) e das correntes costeiras. O clima na região varia
ente semi-árido mediano e tropical úmido com temperaturas variando de 24° a 30°C,
sendo uma região sem variações sazonais evidentes.

5
Delineamento Observacional
As observações foram realizadas entre outubro de 1999 a setembro de 2003, por
meio de coletas mensais (4 por mês) com registros instantâneos realizada a cada 5 min
ao longo de seis horas, no período claro do dia, das 06:00 h às 17:00 h. As
observações foram realizadas simultaneamente por dois observadores, com auxílio de
binóculos (10x50mm), posicionados em um ponto fixo no alto da falésia, com condições
de visualização de toda a enseada. Registrava-se o número de indivíduos visíveis por
emergirem ao respirar, discriminando os comportamentos realizados.
Para quantificação do número de indivíduos, a observação era feita continuamente,
sendo o registro do número de animais realizado ao som de um sinal sonoro disparado
a cada 5 min. Com isso, a contagem era continuamente avaliada pelos observadores e
o registro era pontual, evitando assim a contagem repetida de um mesmo indivíduo.
As freqüências dos seguintes comportamentos foram registradas (Connor et al.
2000):
Forrageio: o boto deslocava-se em nado rápido, geralmente solitário, com o ventre
voltado para cima (identificado pela coloração clara) e perseguindo a presa (peixe) que
saltava sucessivamente, podendo ser capturada por meio de um salto ou bote, já na
região próxima à praia.
Atividade aérea: saltos em que os animais saem total ou parcialmente da água,
independente da posição do animal.
Socialização: toda e qualquer atividade envolvendo contato físico entre os animais,
incluindo contextos agonísticos ou afiliativos.
Análise dos dados
Consideramos a taxa máxima de indivíduos ocorrida ao longo do dia para cada dia
de observação. Em relação aos comportamentos, calculamos o valor diário através da

6
razão entre a soma de cada comportamento observado ao longo do dia pelas seis
horas observadas. Em seguida, para essas taxas diárias foram então calculadas as
medianas mensais.
Para verificar a variação sazonal do número de indivíduos e comportamentos
observamos se havia um período do ano de maior atividade. Esse se mostrou entre os
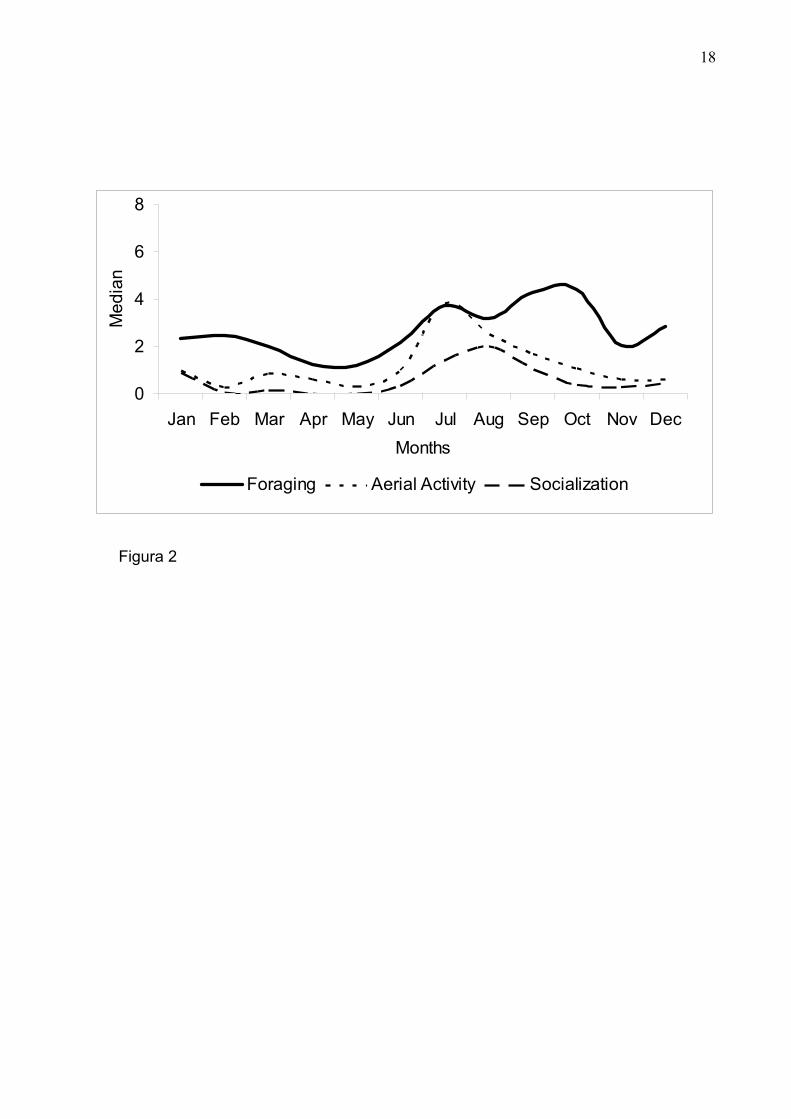
meses de junho e novembro (inverno – primavera) e o de menor atividade entre
dezembro e maio (verão – outono) (Fig. 2).
Em relação ao fotoperíodo analisamos os dados de acordo com dias longos e dias
curtos. Baseado no equinócio, tanto outonal (20/03) como primaveril (23/09), que é de
12:06 h., o período de dias longos correspondeu aos meses de outubro a março com
mais de 12:06 h de luz e o período de dias curtos entre abril e setembro com menos de
12:06 h de luz. Obtivemos os dados de fotoperíodo do Observatório Nacional
(coordenadas locais: 4º48’00”S; 35º3’36”W; fuso: -3).
Consideramos 0.50 mm3 como a quantidade de precipitação que determina um dia
de chuva conforme dados do Instituto Nacional de Meteorologia – INMET datado de
1999. Assim a estação chuvosa foi entre março e agosto, cuja mediana pluviometria foi
acima de 0,50 mm3 e a estação seca, entre setembro e fevereiro, cuja mediana
pluviometria foi abaixo de 0.50 mm3. Obtivemos os dados pluviométricos na Empresa
de Pesquisa Agropecuária do Estado do Rio Grande do Norte S/A – EMPARN.
Compararam-se estatisticamente as medianas de cada conjunto de dados por meio
do teste de Wilcoxon (medidas dependentes), adotando o nível de significância de 5%.
RESULTADOS
Número de Indivíduos
A presença de indivíduos na área foi registrada durante todos os dias de estudo (n
= 192) e a mediana variou entre 3 e 11 animais (Fig. 3).

7
Quando comparamos a freqüência de indivíduos entre os dois períodos (junho -
novembro e dezembro - maio), não verificamos diferenças significativas (MdJN = 5.000;
MdDM = 5.000; Z = -1.161; p = 0.246). Também não verificamos diferença significativa
na freqüência de indivíduos entre os períodos de dias curtos e dias longos (MdDC =
5.000; MdDL = 5.000; Z = -0.467; p = 0.640) e na freqüência de indivíduos entre as
estações chuvosa e seca (MdEC = 5.000; MdES = 5.250; Z = -1.353; p = 0.1769) (Fig. 4).
Freqüência de Comportamentos
A análise da freqüência dos comportamentos expressos pelos botos, mostrou que o
comportamento de socialização representou 12.6% de toda a atividade realizada
durante o período de estudo. A atividade aérea representou 30.7% e forrageio 56.7%.
A freqüência de socialização foi significativamente menor que a atividade aérea (Z = -
3.037; p = 0.002) e o forrageio (Z = - 6.462; p = 0.0001). A atividade de forrageio ainda
se mostro significativamente maior do que a atividade aérea (Z = -4.089; p = 0.0001)
(Fig. 5).
Comparamos a freqüência dos comportamentos entre os períodos junho-novembro
(inverno-primavera) e dezembro-maio (verão-outono) e verificarmos que houve
diferença significativa para todas as três atividades estudadas: socialização (MdJN =
0.542; MdDM = 0.099; z = -2.329; p = 0.020); atividade aérea (MdJN = 1.268; MdDM =
0.521; z = -2.224; p = 0.026) e forrageio (MdJN = 3.268; MdDM = 1.762; z = -2.906; p =
0.004) (Fig. 4).
Por outro lado, entre os períodos de dias curtos e dias longos, não houve diferença
significativa na freqüência das atividades estudadas: socialização (MdDC = 0.339; MdDL
= 0.183; z = -1.738; p = 0.082); atividade aérea (MdDC = 1.295; MdDL = 0.625; z = -1.607;
p = 0.108) e forrageio (MdDC = 2.417; MdDL = 2.583; z = -0.438; p = 0.661) (Fig. 4).

8
De forma semelhante, entre a estação seca e estação chuvosa, também não houve
diferença significativa da freqüência das atividades: socialização (MdES = 0.229; MdEC =
0.225; z = -0.828; p = 0.408); atividade aérea (MdES = 0.625; MdEC = 1.000; z = -1.656;
p = 0.098) e forrageio (MdES = 3.017; MdEC = 2.292; z = -0.730; p = 0.465) (Fig. 4).
DISCUSSÃO
Freqüência de adultos e imaturos
A freqüência dos botos em todos os dias de observação, assim como a ausência de
variação ao longo do ano, nos permite indicar a existência de um padrão difuso da
ocorrência dos indivíduos dessa população na área. Assim como no presente trabalho,
outros estudos evidenciaram um padrão difuso de ocorrência tanto de outras espécies
de golfinhos, como do boto cinza, S. guianensis em diferentes áreas.
Barros e Wells (1998) verificaram que a população de golfinho Tursiops truncatus
em Sarasota (Florida) não realiza migração e possui um padrão difuso de freqüência na
área. Defran e Weller (1999) observaram o mesmo padrão, também para T. truncatus,
em San Diego (Califórnia). Wilson et al. (1997) estudando a população de T. truncatus
em Moray Firth (Escócia), também verificaram a presença dos golfinhos na área ao
longo de todo o ano, com pouca variação do número de indivíduos entre as estações.
Esses autores consideraram que este padrão de freqüência difusa deve-se às
condições ambientais favoráveis ao longo do ano na região, principalmente no que se
refere à elevação da temperatura da água no verão e aumento da disponibilidade de
recursos alimentares no inverno.
Em se tratando do boto cinza, S. guianensis, assim como em nosso estudo, Geise
et al. (1999) verificaram que não há variação da freqüência do número de indivíduos ao
longo do ano no complexo estuarino de Cananéia, estado de São Paulo (litoral sudeste
do Brasil). Nessa mesma área, Santos et al. (2001) demonstraram que esses animais

9
possuem um padrão de freqüência difuso e apontaram a existência de áreas abrigadas,
pouca variação de parâmetros climáticos e grande disponibilidade de recursos
alimentares como fatores que influenciam na freqüência constante dos animais na
região.
Um considerável número de estudos não publicados (PhD e MSc thesis) ou
publicados em português indicam situação semelhante para outras populações de S.
guianensis ao longo de sua distribuição no litoral do nordeste do Brasil. Até onde
sabemos, este é o primeiro estudo caracterizando esse padrão difuso de freqüência de
ocorrência da espécie nessa região.
Lodi (2003) verificou a presença de indivíduos durante todo o ano em Paraty,
estado do Rio de Janeiro, litoral sudeste do Brasil. Em consonância com nosso
trabalho, esses estudos confirmam um padrão difuso de reprodução do boto cinza,
S.guianensis.
Essa condição também é relatada para outras espécies de cetáceos que habitam
regiões de latitudes intermediárias. Urian et al. (1996) indicam padrões reprodutivos
distintos do T. truncatus para latitudes diferentes (de 20º a 60º), sendo sazonal em
altas latitudes e difuso em zonas de latitudes mais baixas. Segundo este estudo o
comportamento reprodutivo reflete adaptações que variam conforme a espécie, local e
população. Além disso, considera que fatores ambientais por si só, como fotoperíodo
ou temperatura da água, não interferem na sazonalidade do comportamento
reprodutivo.
Read (1990) afirma ser esperado um padrão difuso na freqüência de populações de
odontocetos em zonas tropicais devido a pouca variação das condições ambientais
nessas áreas facilitarem a reprodução ao longo de todo o ano. Por outro lado,
populações de cetáceos que ocorrem em zonas de altas latitudes, onde as alterações
ambientais são severas, apresentam comportamento migratório, deslocando-se

10
sazonalmente entre diferentes regiões em busca de recursos alimentares, a exemplo
de orcas Orcinus orca (Similä et al. 1996; Iñíguez 2001) e do golfinho Phocoena
phocoena (Neave and Wright 1968).
Sazonalidade dos Comportamentos
É interessante notar que enquanto o número de botos não se altera ao longo do
ano, ocorre uma oscilação da freqüência dos comportamentos, constituindo um padrão
sazonal de variação.
Verificamos que socialização, atividade aérea e forrageio se mostraram
significativamente mais expressivos no período entre junho e novembro (inverno –
primavera). Apesar da sazonalidade desses comportamentos, não foi possível verificar
associação com fotoperíodo e precipitação.
É esperado que nas regiões tropicais e equatoriais, a sincronização de qualquer
comportamento com o fotoperíodo seja bastante discreta. Diferente das altas latitudes,
os fatores ambientais nas baixas latitudes são mais sutis e flexíveis para influenciar a
expressão dos comportamentos. Por outro lado, as chuvas nas regiões tropicais e
equatoriais marcam as estações seca e chuvosa, influenciando o comportamento dos
organismos vivos. A despeito desse padrão de influência sazonal da pluviometria sobre
a expressão dos comportamentos, não verificamos diferenças significativas nos
comportamentos analisados na população de botos em Pipa em relação a esse
parâmetro.
Consideramos que outros parâmetros ambientais e fatores biológicos podem estar
influenciando a expressão desses comportamentos. Além disso, é provável que a
expressão de um determinado comportamento possa influenciar a freqüência de outros
estados comportamentais. Nesse caso, em nosso estudo, o aumento da atividade de
forrageio pode estar influenciando a elevação da atividade de socialização, bem como

11
da atividade aérea. Como essa elevação da freqüência de forrageio pode representar
aumento da eficácia de captura, os animais podem redirecionar o tempo e esforço
dedicado à procura de presas para outras atividades, particularmente a socialização.
De forma semelhante ao presente trabalho, Neumann (2001) estudando o
orçamento de atividades de Delphinus delphis na Nova Zelândia, analisou entre outros
padrões comportamentais, a socialização. Nesse estudo verificou-se que o tempo em
que os golfinhos permaneciam envolvidos em socialização estava relacionado com a
disponibilidade alimentar ou com a satisfação dos animais após o forrageio. Nesse
caso os animais permaneciam engajados por mais tempo em atividades sociais quanto
maior a disponibilidade de alimento em determinadas épocas do ano.
Também é interessante destacar que semelhante ao nosso estudo, o boto tucuxi S.
fluviatilis na bacia do Amazonas apresenta variação anual dos comportamentos,
principalmente o forrageio que é mais intenso no período seco devido ao aumento da
densidade das presas com o baixo nível das águas represadas em lagos naturais (da
Silva and Best 1996). Esse mesmo padrão foi verificado para o boto vermelho, Inia
geoffrensis, na bacia do Orinoco, Venezuela (McGuire and Winerniller 1998). Da
mesma maneira, os registros do comportamento de forrageio de boto cinza, S.
guianensis, em Paraty (sudeste do Brasil) foi mais elevado durante o verão, período em
que as Águas Centrais do Atlântico Sul (ACAS) e a precipitação das chuvas provocam
o fenômeno da ressurgência na área (Lodi 2003). Em Cananéia (sudeste do Brasil) o
comportamento de forrageio do boto cinza foi mais intenso durante o verão e outono
(Geise et al. 1999).
A freqüência do S. guianensis foi verificada durante todos os dias de estudo,
indicando um importante papel desse sistema marinho para os botos na região. Além
disso, os resultados aqui encontrados mostraram que não existe variação sazonal tanto
no número de adultos, como de imaturos, constituindo um padrão difuso de freqüência

12
dos animais na região. Por outro lado, verificamos que a freqüência dos
comportamentos de socialização, atividade aérea e forrageio variam ao longo do ano,
apresentando um padrão sazonal. Este padrão sazonal está relacionado com fatores
ambientais, como por exemplo a disponibilidade alimentar.
AGRADECIMENTOS
Somos extremamente gratos ao prof. Gilson Volpato pela participação nas
sugestões e correções do manuscrito, aos amigos Érico dos Santos Junior; Kelly
Cristina de Araújo Pansard; Lídio França do Nascimento; Mariana Alves Gondim; Luiza;
Priscila Izabel Alves Pereira de Medeiros; Sandra Mara de Araújo Ananias; Rose Emília
Macedo de Queiroz pela ajuda nas atividades de campo, sem as quais este trabalho
não teria sido realizado.
REFERÊNCIAS
Barros, N.B., and Wells R.S. 1998. Prey and feeding patterns of resident bottlenose
dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Journal of Mammology.
79(3):1045-1059;
Boyd, I.L., Arnould, J.P.Y., Barton, T., and Croxall, J.P. 1994. Foraging behaviour of
antarctic fur seals during periods of contrasting prey abundance. Journal of Animal
Ecology. 63(3):703-713;
Connor, R.C., Wells, R., Mann, J., and Read, J. 2000. The bottlenose dolphin In
Cetacean Societies: Field Studies of Dolphins and Whale. Edited by Mann, J.,
Connor, R.C., Tyack, P.L., and Whitehead, H. University of Chicago press,
Chicago. pp.91-126;
Cunha, H.A., da Silva, V.M.F., Lailson-BritoJr, J., Santos, M.C.O., Flores, P.A.C.,
Martin, A.R., Azevedo, A.F., Fragoso, A.B.L., Zanelatto, R.C., and Sole-Cava, A.M.

13
2005. Riverine and marine ecotypes of Sotalia dolphins are different species.
Marine Biology ;
da Silva, V.M.F., and Best, R.C. 1996. Sotalia fluviatilis. Mammalian Species. Americam
Society of Mammologists. 527:1-7;
Defran, R.H., and Wellemr, D.W. 1999. Occurrence, distribution, site fidelity and school
size of bottlenose dolphins (Tursiops truncatus) off San Diego, Califórnia. Marine
Mammal Science. 15(2):366-380;
Geise, L., Gomes, N., and Cerqueira, R. 1999. Behavior, habitat use and population
size of Sotalia fluviatilis (Gervais, 1853) (Cetácea, Delphinidae) in the Cananéia
estuary region. São Paulo, Brasil. Revista Brasileira de Zoologia. 59:2;
Hindell, M.A., Bradshaw, C.J.A., Summer, M.D., Michael, K.J., and Burton, H.R. 2003.
Dispersal of female southern elephant seals and their prey consumption during the
austral summer relevance to management and oceanographic zones. Journal of
Applied Ecology. 40:703-715;
Iñíguez, M.A. 2001. Seasonal distribuition of Killer Whale (Orcinus orca) in northen
Patagonia, Argentina. Aquatic Mammals. 27(2):154-161;
Larkman, V., and Veit, R. 1998. Seasonality and abundance of blue whales off southern
California. CalCOFI Rep. 39:236-239;
Lodi, L.F. 2003. Tamanho e composição de grupo dos botos-cinza, Sotalia guianensis
(van Bénéden 1864) (CETACEA, DEPHINIDAE), na Baía de Paraty, Rio de
Janeiro, Brasil. Atlântica, Rio Grande. 25(2): 135-146;
McGuire, T.L., and Winemiller, K.O. 1998. Occurrence patterns, habitat associations,
and potential prey of the river dolphins Inia geoffrensis, in the Cinaruco river,
Venezuela. Biotropica. 30(4):625-638;

14
Monteiro-Filho, E.L.A., Monteiro, L.R., and dos Reis, S.F. 2002. Skull shape and size
divergence in the dolphins of the genus Sotalia: a tridimensional morphometric
analysis. Journal Mammology. 83(1):125-134;
Muller, M., Boutière, H., Weaver, A., and Candelon, N. 1998. Ethogram of the
bottlenose dolphin (Tursiops truncatus) with special reference to solitary and
sociable dolphins. English Translation of Vie Millieu. 48(2):89-104;
Neave, D.J., and Wright, B.S. 1968. Seasonal migrations of the harbor porpoise
(Phocoena phocoena) and other cetacea in the Bay of Fundy. Journal
Mammology. 49(2):259-264;
Neumann, D.R. 2001. Activity budget of free-ranging common dophins (Delphinus
delphis) in the northwestern Bay of Plenty, New Zealand. Aquatic Mammals.
27(2):121-136;
Norris, K.S., Würsig, B., and Wells, R.S. 1994. Aerial Behavior In The Hawaiian Spinner
Dolphin. Edited by Norris, K.S., Würsig, B., Wells, R.S. e Würsig M. University of
California Press, Berkeley. pp. 103-120.
Pastor, T., and Aguilar, A. 2003. Reproductive cycle of the female mediterraneam monk
seal in the western Sahara. Marine Mammal Science. 19(2):318-330;
Read, A.J. 1990. Reprodutive seasonality in Harbor Porpoise, Phocoena phocoena,
from the Bay of Fundy, Can. J. Zool. 68:284-288;
Rosas, F.C.W., and Monteiro-Filho, E.L.A. 2002. Reproduction of the estuarine dolphin
(Sotalia guianensis) on the coast of Paraná, solthern Brazil. Journal of
Mammology. 83(2):507-515;
Santos, M.C.O., Açuña, L.B., and Rosso, S. 2001. Insights on site fidelity and calving
intervals f the marine tucuxi dophin (Sotalia fluviatilis) in south-eastern Brasil, J.
Mar. Biol. Ass. U.K. 81:1049-1052;

15
Similä, T., Holst, J.C., and Christensen, I. 1996. Occurrence and diet of Killer Whale in
northern Norway: Seasonal patterns relative to the distribution and abundance of
Norwegian Spring-Spawning Herring. Can. J. Fish. Aquat. Sci. 53:769-779;
Smolker, R.A., Richards, A.F., Connor, R.C., and Pepper, J.W. 1992. Sex difference in
patterns of association among indian ocean bottlenose dolphins. Behaviour. 123:1-
2;
Urian, K.W., Duffield, D.A., Read, A.J., Well, R.S., and Shell, E.D. 1996. Sasonality of
reproduction in bottlenose dolphins, Tursiops truncatus. Jornal of Mammalogy.
77(2):394-403;
Whiteread, H., and Mann, J. 2000. Female reproductive strategies of cetaceans In
Cetacean Societies: Field Studies of Dolphins and Whale. Edited by Mann, J.,
Connor, R.C., Tyack, P.L., and Whitehead, H. University of Chicago press,
Chicago. pp.219-246;
Wilson, B., Thomposon, P.M., and Hammond, P.S. 1997. Habit use by bottlenose
dophins: seasonal distribution and stratified movement patterns in Moray Firth,
Scotland. Journal of Applied Ecology. 34(6):1365-1374.

16
LEGENDAS DE FIGURAS
Figura 1: Localização da área de estudo (Enseada do Curral, Praia de Pipa, estado
do Rio Grande do Norte, Nordeste do Brasil). Fonte: IDEMA.
Figura 2: Freqüência das atividades de Forrageio (Foraging); Atividade Aérea (Aerial
Activity) e Socialização (Socialization) dos estuarine dolphins ao longo do ano na
enseada do Curral, nordeste do Brasil.
Figura 3: A freqüência de ocorrência dos estuarine dolphins ao longo do ano na
Enseada do Curral, nordeste do Brasil.
Figura 4: Distribuição das medianas das variáveis dependentes freqüência de
indivíduos, forrageio, atividade aérea e socialização, conforme o período do ano de
maior atividade verificada, fotoperíodo e precipitação. As letras maiúsculas diferentes
indicam a existência de diferença significativa a 5% dentro da variável independente.
Figura 5: Freqüência relativa das atividades de Forrageio (Foraging); Atividade Aérea
(Aerial Activity) e Socialização (Socialization) dos estuarine dolphins na Enseada do
Curra, nordeste do Brasil. As letras minúsculas diferentes indicam a existência de
diferença significativa a 5% entre as variáveis independentes.

17
Figura 1
Pipa Beach
MadeiroBeach
Imaginary Line
Concentration Area
N
18
Figura 2
0
2
4
6
8
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Months
Me
dia
n
Foraging Aerial Activity Socialization

19
Figura 3:
0
2
4
6
8
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Months
Me
dia
n
INDIVIDUAL FREQUENCY

20
Figura 4:
SOCIALIZATIONAERIAL ACTIVITYFORAGINGINDIVIDUAL
PR
EC
IPIT
AT
ION
PH
OT
OP
ER
IOD
AC
TIV
ITY
10
5
0
ME
DIA
N
10
5
0
ME
DIA
N
10
5
0
ME
DIA
N
HIGH LOW HIGH LOW HIGH LOW HIGH LOW
LONG DAY SHORT DAY LONG DAY SHORT DAY LONG DAY SHORT DAY LONG DAY SHORT DAY
RAINY DRY RAINY DRY RAINY DRY RAINY DRY
A
BA
B
A
B

21
Figura 5:
FORAGINGAERIAL ACTIVITYSOCIALIZATION
69,330,7%
28,612,6%
128,056,7%
FREQUENCY ACTIVITY
A
B
C

29
ARTIGO 2
BEHAVIORAL DAILY VARIATION OF THE ESTUARINE DOLPHIN, Sotalia
guianensis, IN NORTHEASTERN BRAZIL
Silveira, F.R.G.1 and Silva, F.J.L.2
1 Programa de Pós-Graduação em Psicobiologia, Departamento de Fisiologia.
Universidade Federal do Rio Grande do Norte. Campus Central – Natal, RN,
Brazil. E-mail: [email protected] 2 Departamento de Ciências Biológicas, Universidade do Estado do Rio Grande
do Norte, Avenida Antônio de Campos, s/n. CEP: 59600-970. Mossoró, RN,
Brazil.
e-mail:[email protected]. Fax: ++ 55 84 3315-2235.
www.golfinhorotador.org.br
A SER SUBMETIDO AO
CANADIAN JOURNAL OF ZOOLOGY
(INSTRUÇÕES PARA AUTORES – ANEXO)

1
Behavioral daily variation of the estuarine dolphin, Sotalia guianensis, in
Northeastern Brazil
Silveira, F.R.G.
Programa de Pós-Graduação em Psicobiologia, Departamento de Fisiologia.
Universidade Federal do Rio Grande do Norte. Campus Central – Natal, RN, Brazil. E-
mail: [email protected]
Silva, F.J.L.
Departamento de Ciências Biológicas, Universidade do Estado do Rio Grande do
Norte, Avenida Antônio de Campos, s/n. CEP: 59600-970. Mossoró, RN, Brazil. e-
mail:[email protected]. Fax: ++ 55 84 3315-2235.
www.golfinhorotador.org.br
Editor: please send correspondence to Silva F.J.L.
Key words: Delphinidae, boto, estuarine dolphin, Sotalia guianensis, daily behavior,
diurnal, circatidal.
"This paper has not been submitted elsewhere in identical or similar form, nor
will it be during the first three months after its submission."

2
RESUMO
Neste estudo investigamos a variação diária na freqüência de adultos e imaturos e
nos comportamentos de socialização, atividade aérea e forrageio numa população de
Boto Cinza (Sotalia guianensis) na Enseada do Curral (6º13’00”S 35º3’36”W;), no
nordeste brasileiro. Após detectarmos algumas variações diárias, procuramos associá-
las ao período do dia, às variações da maré e período do ano. De Outubro de 1999 a
Setembro de 2003, através de coletas quatro vezes ao mês, efetuaram-se registros
instantâneos a cada 5 min, em seis horas de observação. As horas de observação
estiveram inseridas dentro da fase clara das 24 horas, ou seja, no intervalo das 06:00 h
as 17:00 h. Essas observações eram realizadas simultaneamente por dois
observadores, num ponto fixo no alto da falésia a partir do qual visualizava-se toda a
enseada. Observamos que os animais são mais ativos e freqüentes pela manhã. Não
apresentaram associação com a maré, apesar de apresentarem maior atividade na
maré baixa e menor atividade na maré alta. E quando comparado com as estações do
ano verificamos que existe uma variação significativa da variação diária dos
comportamentos nas estações inverno e primavera (de junho a novembro). Já a
freqüência de indivíduos não apresentou esta associação. Desses resultados,
mostramos a existência de uma variação comportamental diária nesta espécie, mesmo
nas regiões equatoriais.

3
INTRODUÇÃO
Há um crescente aumento no número de estudos sobre o comportamento dos
odontocetos na natureza. Desses estudos, observa-se que os comportamentos são
afetados por variáveis ambientais, como latitude, temperatura da água, maré,
disponibilidade alimentar, profundidade, tipo de fundo e relevo. Deste modo,
características topográficas de determinada região afetam o comportamento desses
animais (Saayman et al. 1973; Geise et al. 1999; Karczmarski et al. 2000; Neumann,
2001). Contudo existe uma carência de estudos realizados na baixa latitude e em
ambiente como a enseada, cuja topografia é diferente daquelas onde se concentram a
maioria dos trabalhos.
A espécie Sotalia guianensis, tem ampla distribuição geográfica, tanto no que se
refere à latitude (de 15º N a 27º S) quando a regiões topográficas (enseadas, baias,
estuários e rios). Sua importância em termos de estudos é estar presente em habitats
onde seus comportamentos têm sido pouco estudados. Nesse sentido, é uma espécie
que ocorre em baixas latitudes (nordeste do Brazil), ao contrário da maioria das
espécies até aqui estudadas, que mostraram alterações comportamentais,
influenciadas por parâmetros ambientais marcados pelas altas latitudes (Norris et al.
1994; Wilson et al. 1997; Rogan et al. 2000; Nichols et al. 2001).
Além disso, os estudos geralmente investigam espécies em ambientes como ilhas
oceânicas (Norris et al. 1994; Rogan et al. 2000; Neumann, 2001), baías (Karczmarski
et al. 2000) e estuários (Wilson et al. 1997; Geise et al. 1999). Os estudos de golfinhos
em enseadas são mais restritos. A enseada oferece condições ambientais peculiares,
dada às condições topográficas apresentadas. Ela estaria entre as condições de baía
(proteção oferecida) e as condições oceânicas (mar aberto), o que pode interferir no
comportamento desses animais.

4
Assim, os comportamentos dessa espécie foram enfocados no presente estudo,
que se restringiu a investigar a variação comportamental em função do horário no
período claro do dia. Bem como, a verificar a influência das marés nesses
comportamentos, uma vez que esses efeitos são geralmente observados em
odontocetos que se alimentam de presas que vivem na região intertidal.
Desta forma, este estudo teve como objetivo mostrar a flutuação diária no
comportamento de uma população equatorial de golfinhos. Investigou-se a freqüência
dos indivíduos (adultos e imaturos) e os comportamentos de socialização, atividade
aérea e forrageio em indivíduos de uma população de boto cinza, S. guianensis, na
enseada do Curral, distrito de Pipa, no nordeste do Brasil. Em seguida, verificou-se a
influência dos parâmetros ambientais, período do dia e variação da maré sobre essa
atividade comportamental e freqüência de indivíduos.
MATERIAL E MÉTODOS
Espécie e área de estudo
Investigamos o boto cinza que na nomenclatura de Monteiro–Filho et al. (2002) e
Cunha et al. (2005) é Sotalia guianensis van Bénéden 1864 (Cetacea, Delphinidae).
Essa espécie possui distribuição restrita entre o Norte da Lagoa de Waunte, Nicarágua
e o sul da Baía Norte, no Estado de Santa Catarina, Brasil (da Silva and Best 1996).
Este estudo restringiu-se aos indivíduos que ocorreram na enseada do Curral
(6º13’00”S; 35º3’36”W), no distrito de Pipa, RN, Brazil. A enseada possui
aproximadamente um quilômetro de extensão. A área de estudo ficou compreendida
entre a praia e uma linha reta imaginária traçada ligando as extremidades da enseada
(Fig. 1). Essa linha tem cerca de 800 m e a maior distância da praia é na região central
(cerca de 500 m). As extremidades dessa enseada apresentam formações rochosas
(arenito sedimentar) que se estendem do andar infralitoral para o supralitoral até unir-

5
se à falésia, cuja altura atinge cerca de 30 m em sua porção mais elevada (local usado
para as observações). O fundo na enseada é constituído exclusivamente de arenito,
com profundidade máxima de cinco metros. É uma região abrigada de ventos (< 1,0 na
escala de Beaufort) e das correntes costeiras. O clima na região varia ente semi-árido
mediano e tropical úmido com temperaturas variando de 24° a 30°C, sendo uma região
sem variações sazonais evidentes.
Delineamento Observacional
As observações foram realizadas entre outubro de 1999 a setembro de 2003, por
meio de coletas mensais (4 por mês) com registros instantâneos realizada a cada 5 min
ao longo de seis horas, no período claro do dia, das 06:00 h às 17:00 h.
As observações foram realizadas com auxílio de binóculos (10x50 mm)
simultaneamente por dois observadores, posicionados em um ponto fixo no alto da
falésia, com condições de visualização de toda a enseada. Registrava-se o número de
indivíduos visíveis por emergirem total ou parcialmente, discriminando-se a faixa etária
(adulto e imaturo) e os comportamentos emitidos.
Para quantificação do número de indivíduos, a observação era feita continuamente,
sendo o registro do número de animais realizado ao som de uma campainha disparado
a cada 5 min. Com isso, a contagem era continuamente avaliada pelo observador e o
registro era pontual, evitando assim a contagem repetida de um mesmo indivíduo.
As freqüências dos seguintes comportamentos foram registradas (baseado em
Connor et al. 2000):
Forrageio: o boto deslocava-se em nado rápido, geralmente solitário, com o ventre
voltado para cima (identificado pela coloração clara) e perseguindo a presa (peixe) que
saltava sucessivamente, podendo ser capturada por meio de um salto ou bote, já na
região próxima à praia.

6
Atividade aérea: saltos em que os animais saem total ou parcialmente da água,
independente da posição do animal.
Socialização: toda e qualquer atividade envolvendo contato físico entre os animais,
incluindo contextos agonísticos ou afiliativos.
Análise dos dados
Como o número total de animais variava a cada dia de observação, corrigiu-se tal
efeito calculando-se a taxa percentual da freqüência de indivíduos para cada dia de
observação. Essas taxas, bem como as freqüências dos itens comportamentais, foram
então calculadas por hora de observação e esses valores comparados entre os
períodos do dia ou fases de maré.
Os períodos do dia foram considerados das 06:00 h e 09:00 h, das 09:00 h às 12:00
h, das 12:00 h às 15:00 h e das 15:00 h às 18:00 h. Com relação às marés,
estabeleceu-se 4 períodos de 2 h cada, variando em torno da maré baixa e alta. Com
isso, discriminou-se os seguintes períodos: maré subindo, maré alta, maré descendo e
maré baixa.
Comparou-se estatisticamente as medianas de cada conjunto de dados por meio do
teste de Wilcoxon (medidas dependentes), adotando o nível de significância a 5%.
RESULTADOS
Freqüência de Indivíduos
Durante todos os dias de observação, verificamos a presença de indivíduos. O
número variou de um a dezesseis indivíduos na enseada. A freqüência de indivíduos
variou significativamente durante os períodos do dia, com maior freqüência entre as
09:00 h e 12:00 h (Md9h–12h = 1.00). Esse período do dia apresentou diferença
significativa em relação ao primeiro período (Md6h–9h = 0.50) (Z = -5.341; p = 0.0001);

7
terceiro (Md12h–15h = 0.67) (Z = -3.957; p = 0.0001) e quarto período (Md15h–18h = 0.60)
(Z = -2.338; p = 0.019) (figura 2a). Foi verificada ainda, variação da freqüência de
indivíduos entre os estados de maré. A maré alta (MdHigh = 0.57), apresentou diferença
significativa entre a maré baixa (MdLow = 0.75) (Z = -1.964; p = 0.049); maré subindo
(MdFood = 0.80) (Z = -2.310; p = 0.021) e maré baixando (MdEbb = 0.83) (Z = -2.192; p =
0.028) (figura 2b).
Freqüência de Comportamentos
O comportamento de socialização representou 12.6% de toda a atividade realizada
durante o período de estudo. A atividade aérea representou 30.7% e forrageio 56.7%.
A freqüência de socialização foi significativamente menor que a atividade aérea (Z = -
3.037; p = 0.002) e o forrageio (Z = - 6.462; p = 0.0001). A atividade de forrageio ainda
se mostro significativamente maior do que a atividade aérea (Z = -4.089; p = 0.0001)
(figura 3).
A variação da freqüência da atividade de socialização não apresentou diferença
significativa entre os períodos do dia (figura 4a) e entre os intervalos de maré. A
mediana tanto para os períodos do dia como as maré foram zero (figura 4b).
A variação da freqüência da atividade aérea também não apresentou diferença
significativa entre os períodos do dia (figura 4a), porém apresentou diferença
significativa entre maré baixa (MdLow = 0.850) e subindo (MdFlood = 0.001) (Z = -2.544; p
= 0.011) e entre maré subindo (MdFlood = 0.001) e baixando (MdEbb = 0.500) (Z = -2.572;
p = 0.010) (figura 4b).
A variação da freqüência de forrageio no primeiro período do dia (Md6h–9h = 1.580)
foi significantemente diferente do segundo (Md9h–12h = 3.000) (Z = -2.084; p = 0.037) e
terceiro (Md12h–15h = 3.000) (Z = -2.299; p = 0.022), sendo esses dois últimos onde se
concentrou a maior freqüência de atividade (figura 4a). Houve diferença significativa na

8
freqüência de forrageio entre a maré alta (MdHigh = 1.500) e baixando (MdEbb = 2.670)
(Z = -2.554; p = 0.011), com maior atividade durante a maré baixando (figura 4b).
DISCUSSÃO
Os resultados aqui encontrados nos permitem indicar que o Boto Cinza (Sotalia
guianensis) é mais ativo e freqüente na Praia do Curral no período da manhã. A
diferença foi significativa para freqüência de indivíduos e forrageio, não sendo para
atividade aérea e socialização.
Semelhante ao que verificamos no presente estudo, a população de Boto Cinza no
estuário de Cananéia também apresentou uma maior freqüência de indivíduos (adulto,
juvenil e filhote) no período da manhã, mais precisamente entre as 09:00 h e 10:00 h.
Nesse estudo os autores descrevem técnicas diferentes de forrageio, cada qual em
horários distintos do dia. A estratégia de pesca casual, na superfície, a mais comum e
semelhante ao que ocorre em Pipa, se deu pela manhã entre as 06:00 h as 07:00 h.
Por outro lado, a população de Cananéia possui um pico de atividade aérea no final da
tarde e após o meio dia (Geise et al. 1999). Consideramos que em Pipa, diferente do
que ocorre em Cananéia, a atividade aérea e socialização estejam associadas com o
forrageio.
Neumann (2001) verificou em uma população de Delphinus delphis, na baia de
Plenty, Nova Zelândia, que após a atividade de forrageio, deu-se início a socialização e
que o tempo de socialização esteve relacionado com a disponibilidade alimentar ou
com o quanto os indivíduos estariam satisfeitos após o forrageio. O golfinho Tursiops
truncatus, nos estuários da costa do Texas apresenta um padrão, cujo pico de
atividade é durante a fase clara do dia com uma queda de atividade durante a fase
escura (Lynn 1995). Segundo esse autor, o golfinho nariz de garrafa apresentou um
pico de atividade principalmente no início da manhã diminuindo ao longo do dia, sendo

9
o forrageio a atividade mais freqüente. Ainda segundo ele, após a atividade de
forrageio, os animais envolveram-se em atividades de socialização. Comportamento
semelhante apresentou o golfinho nariz de garrafa do oceano Indico (Tursiops aduncus
ehrenburg) na Baía de Algoa, África do Sul (Saayman et al. 1973) e o golfinho
corcunda (Souza chinensis) também em Algoa, África do Sul (Karczmarski et al. 2000).
Em todos esses estúdios os animais apresentaram um padrão, cujo pico de atividade
foi durante a fase clara do dia decrescendo durante a fase escura, com atividades
como a socialização ocorrendo após o forrageio. De forma semelhante, verificamos no
presente estudo que a população de boto cinza além de ter apresentado uma maior
freqüência das três atividades comportamentais estudadas pela manhã, tem mostrado
que após a atividade de forrageio, segue-se a atividade de socialização.
Corroborando com o que verificamos para o boto cinza em Pipa, Norris et al. (1994),
no Havaí, afirmaram que a atividade diária do golfinho rotador (Stenella longirostris) na
baía de kealake`akua tem pelo menos duas finalidades. A primeira e primordial foi o
descanso; a segunda foi entre outras, a socialização. Após a entrada dos animais na
baía nas primeiras horas do dia, houve um período de repouso de quatro a cinco horas.
Em seguida, o tempo restante entre o repouso e o início do forrageio (que ocorre no
início da noite) foi gasto de varias maneiras como a atividade aérea e socialização.
Semelhante ao Havaí, o golfinho de Hector (Cephalorhynchus hectori) no porto de
Akaroa, Nova Zelândia, também usa aquela área durante o dia para repouso,
socialização e forrageio (Nichols et al. 2001). A maioria dos mamíferos aquáticos
apresenta um padrão de atividade, que oscila dentro das 24 h. É o que ocorre em
relação ao forrageio e repouso, com as focas (Siniff et al. 1971; Thompon et al. 1989;
Orthmann 1998); leões marinhos (Sepúlveda et al. 2001); lontras (Estes et al. 1982) e
morças (Salter 1979). No caso da Foca de pêlo e Leões marinhos do Arquipélago das
Galápagos, ocorreu uma distribuição temporal onde as focas são diurnas e os leões

10
noturnos. Tal situação foi indicada como adaptação à pressão imposta pela competição
interespecífica por alimento, separando-os em nichos diferentes (Dellinger and Trillmich
1999). O equilíbrio entre presa e predador, em dada região, pode refletir-se na variação
da atividade de forrageio ao longo do dia com picos no início e final dele e um grande
custo de energia. Já o desequilíbrio entre ambos, onde haja abundância alimentar (de
presa), pode refletir um padrão homogêneo da atividade de forrageio ao longo do dia
com um baixo custo energético nessa atividade, dado a grande disponibilidade de
presas (Estes et al. 1982). Os resultados aqui apresentados para o boto de Pipa,
apontam um padrão de equilíbrio semelhante.
Alguns mamíferos aquáticos sincronizam suas atividades, como repouso ou
forrageio, não só com o ciclo claro-escuro, mas também com o ciclo de maré
(Thompson et al. 1989; Thompson and Miller 1990; Bjørge et al. 2002). A maré pode
ainda, influenciar o forrageio devido à sincronia que determinadas presas tem com sua
flutuação. É o caso da Phoca vitulina no estuário de Moray (Thompson and Miller 1990)
e Orkney (Thompson et al. 1989), ambas na Escócia. O golfinho T. truncatus no
estuário de Shannon, Irlanda, repousa no estofo da maré e socializa e forrageia
durante a subida e descida (Rogan et al. 2000).
O forrageio durante a subida e descida da maré pode estar relacionada à estratégia
de captura, considerando o aumento da densidade de presas devido à diminuição da
lâmina de água (Karczmarski et al. 2000). Pode estar ainda, relacionado à facilidade de
emboscar, arrebanhar, perseguir as prezas ou ainda minimizar os gastos energéticos
(Wilson et al. 1997).
Em nosso estudo, verificamos que durante a maré baixando, baixa e subindo,
freqüência de indivíduos; atividade aérea e forrageio foram maiores do que na maré
alta. Isso indica que esta população de Pipa ajusta-se às condições ambientais como

11
disponibilidade de alimento, densidade de presa, profundidade, respondendo a pressão
das condições locais.
Os resultados apresentados indicam que o boto cinza é mais ativo e mais freqüente
durante o período da manhã. Verificamos ainda, que os comportamentos do boto
sofrem influências do estado da maré, com maior atividade na fase de maré baixa.
Além disso, a variação diária da freqüência e atividade variam ao longo do ano
respondendo a variação ambiental.
AGRADECIMENTOS
Somos extremamente gratos ao prof. Gilson Volpato pela participação nas
sugestões e correções do manuscrito, aos amigos Érico dos Santos Junior; Kelly
Cristina de Araújo Pansard; Lídio França do Nascimento; Mariana Alves Gondim; Luiza;
Priscila Izabel Alves Pereira de Medeiros; Sandra Mara de Araújo Ananias; Rose Emília
Macedo de Queiroz pela ajuda nas atividades de campo, sem as quais este trabalho
não teria sido realizado.
REFERÊNCIAS
Bjørge, A., Bekkby, T., and Bryant, E.B. 2002. Summer home range and habitat
selection of Harbor Seal (Phoca vitulina) pups. Marine Mammal Science
18(2):438-454;
Connor, R.C., Wells, R., Mann, J., and Read, J. 2000. The bottlenose dolphin. In:
Cetacean Societies: Field Studies of Dolphins and Whale. Edited by Mann, J.,
Connor, R.C., Tyack, P.L., and Whitehead, H. University of Chicago press.
Chicago. pp.173-196;
Cunha, H.A., da Silva, V.M.F., Lailson-BritoJr, J., Santos, M.C.O., Flores, P.A.C.,
Martin, A.R., Azevedo, A.F., Fragoso, A.B.L., Zanelatto, R.C., and Sole-Cava, A.M.

12
2005. Riverine and marine ecotypes of Sotalia dolphins are different species.
Marine Biology ;
da Silva, V.M.F., and Best, R.C. 1996. Sotalia fluviatilis. Mammalian Species. Americam
Society of Mammologists 527:1-7;
Dellinger, T., and Trillmich, F. 1999. Fish prey of the synpatric Galapagos fur seals and
sea lions seasonal variation and niche separation. Canadian Journal of Zoology
77:1204-1216;
Estes, J.A., Ronald, J., and Rhode, E.B. 1982. Activity and prey election in the sea otter
influence of population status on community structure. Americam Naturalist
120(2):242-258;
Geise, L., Gomes, N., and Cerqueira, R. 1999. Behavior, habitat use and population
size of Sotalia fluviatilis (Gervais, 1853) (Cetácea, Delphinidae) in the Cananéia
estuary region. São Paulo, Brasil. Revista Brasileira de Zoologia 59:2;
Karczmarski, L., Thornton, M., and Cockcroft, V.G. 2000. Daylight occurrence of
humpback dolphins, Sousa chinensis, in Algoa Bay, South Africa. African Journal
Ecology 38:86-90;
Lammers, M.O., Albinsson, L., Bird, K.B., and Davis, L. 2001. The occurrence and
behavior of whales and dolphins near kalaeloa barbers point harbor: a study to
assess the potential interactions with proposed harbor modification activities.
Hawaii State Department of Transportation – Harbor Division. 27pp. Hawaii;
Lynn, S.K. 1995. Movements, site fidelity and surfacing patterns of Bottlenose Dolphins
on the central Texas coast. Texas A &M University. [Thesis of Master]. 139pp.
Texas;
Monteiro-Filho, E.L.A., Monteiro, L.R., and dos Reis, S.F. 2002. Skull shape and size
divergence in the dolphins of the genus Sotalia: a tridimensional morphometric
analysis. Journal Mammology 83(1):125-134;

13
Muller, M., Boutière, H., Weaver, A., and Candelon, N. 1998. Ethogram of the
bottlenose dolphin (Tursiops truncatus) with special reference to solitary and
sociable dolphins. English Translation of Vie Millieu 48(2):89-104;
Neumann, D.R. 2001. Activity budget of free-ranging common dophins (Delphinus
delphis) in the northwestern Bay of Plenty, New Zealand. Aquatic Mammals
27(2):121-136;
Nichols, C., Stone, G., Hutt, A., Brown, J., and Yoshinaga A. 2001. Observations of
interactions between hector dolphin and boat and people at Akaroa Harbour. New
Zealand Science for Concervation 178. 49p;
Norris, K.S., Würsig, B., and Wells, R.S. 1994. Aerial Behavior. In: The Hawaiian
Spinner Dolphin. Edited by Norris, K.S., Würsig, B., Wells, R.S., and Würsig M.
University of California Press. Berkeley. pp.103-120;
Orthmann, T. 1998. Tracking harbour seals (Phoca vitulina richardsi) to determine dive
behavior, foraging activity, and haulout site use. Marine Mammal Science
14(2):361-372;
Rogan, E., Ingram, S., Holmes, B., and O´Flanagan, C. 2000. A survey of Bottlenose
dolphins (Tursiops truncatus) in the Shannon estuary. The Marine Research
Measure. [Project]. 49pp. Irlanda;
Saayman, G.S., Tayler, C.K., and Bower, D. 1973. Diurnal activity cycles in captive and
free-ranging Indian ocean bottlenose dolphins (Tursiops aduncus ehrenburg).
Behaviour 44(3-4):212-233;
Salter, R.E. 1979. Site utilization, activity budget, and disturbance responses of Atlantic
walruses during terrestrial haulout. Canadian Journal of Zoology 57(6):1169-1180;
Siniff, D.B., Tester, J.R., and Kuechle, V.B. 1971. Some observations on the activity
patterns of weddell seal as recorded by telemetry. Antarctic Research Series
18:173-180;

14
Smolker, R.A., Richards, A.F., Connor, R.C., and Pepper, J.W. 1992. Sex difference in
patterns of association among indian ocean bottlenose dolphins. Behaviour 123:1-
2;
Thompson, P.M., Fedak, M.A., McConnell, B.J., and Nicholas, K.S. 1989. Seasonal and
sex-related variation in the activity patterns of common seals (Phoca vitulina).
Journal of Applied Ecology 26(2):521-535;
Thompson P.M., and Miller, D. 1990. Summer foraging activity and movements of radio-
tagged common seals (Phoca vitulina. L.) in the Moray firth Scotland. Journal of
Applied Ecology 27(2):492-501;
Whiteread, H., and Mann, J. 2000. Female reproductive strategies of cetaceans In
Cetacean Societies: Field Studies of Dolphins and Whale. Edited by Mann, J.,
Connor, R.C., Tyack, P.L., and Whitehead, H. University of Chicago press,
Chicago. pp.219-246;
Wilson, B., Thompson, P.M., and Hammond, P.S. 1997. Habit use by bottlenose
dophins: seasonal distribution and stratified movement patterns in Moray Firth,
Scotland. Journal of Applied Ecology 34(6):1365-1374;

15
LEGENDAS DAS FIGURAS
Figura 1: Localização geográfica da Enseada do Curral, distrito de Pipa. Fonte:
IDEMA
Figura 2: Freqüência de indivíduos (em medianas) na enseada do Curral nos
períodos do dia e sob a influência da maré (b). As letras maiúscula diferentes indicam
uma diferença significativa (5%).
Figura 3: Freqüência dos comportamentos de Forrageio (Foraging), Atividade Aérea
(Aerial Activity) e Socialização (Socialization) durante o período de estudo. As letras
maiúsculas diferentes indicam uma diferença significativa (5%).
Figura 4: Freqüência (em mediana) do Forrageio (Foraging); Atividade Aérea (Aerial
Activity) e Socialização (Socialization) nos períodos do dia e sob a influência da maré;
As letras maiúsculas diferentes indicam uma diferença significativa (5%).

16
Figura 1
Pipa Beach
MadeiroBeach
Imaginary Line
Concentration Area
N

17
Figura 2
TIDEDAY PERIOD
1,0
0,5
0,0
ME
DIA
N
6h-9h 9h-12h 12h-15h 15h-18h LOW FLOOD HIGH EBB
AB A A A A B A

18
Figura 3:
FORAGINGAERIAL ACTIVITYSOCIALIZATION
69,330,7%
28,612,6%
128,056,7%
FREQUENCY ACTIVITY
A
B
C

19
Figura 4
SOCIALIZATIONAERIAL ACTIVITYFORAGING
TID
ED
AY
PE
RIO
D
15h-18h12h-15h9h-12h6h-9h 15h-18h12h-15h9h-12h6h-9h 15h-18h12h-15h9h-12h6h-9h
15
10
5
0
15
10
5
0LOW FLOOD HIGH EBB LOW FLOOD HIGH EBB LOW FLOOD HIGH EBB
B
A
B
A
A
A
B A

57
DISCUSSÃO GERAL
Freqüência de Indivíduos
Foi possível verificar a presença de adulto e imaturo na enseada do Curral
durante todos os dias de observação, indicando a importância deste sistema
marinho para a população em estudo. Também foi possível verificar que não
houve variação do número de indivíduos ao longo do ano, caracterizando um
padrão difuso de ocorrência da população de boto cinza em Pipa.
Tais fatos associados podem indicar fidelidade ao habitat, semelhante ao que
ocorre na região sudeste do Brasil, onde populações apresentam um elevado
grau de fidelidade (Geise et al. 1999; Santos et al. 2001; Rosas e Monteiro-Filho
2002; Lodi 2003). A região de Pipa apesar de pouco influenciada pelo ambiente
estuarino, se compararmos com Cananéia ou Sepetiba, apresenta em suas
enseadas um ambiente estável às correntes costeiras, ventos e estabilidade
oceanográfica. Desta forma, apresentando condições satisfatórias para
reprodução, alimentação repouso e socialização.
Estudos realizados com a mesma espécie, Sotalia guianensis, tem
apresentado os mesmos resultados, apontando a estabilidade do habitat como
fator que influenciaria a disponibilidade alimentar, menor risco de predação e
possibilidade de investimento parental (Geise et al. 1999; Santos et al. 2001;
Rosas e Monteiro-Filho 2002; Lodi 2003), maximizando os benefícios e
minimizando os custos da permanência difusa na área.
Outros estudos com o golfinho nariz de garrafa, Tursiops truncatus, em
Sarasota, Florida (Barros e Wells 1998), San Diego, Califórnia (Defran e Weller
1999) e Moray Firth na Escócia (Wilson et al. 1997) evidenciaram o mesmo
padrão difuso de residência aqui verificado, apesar de serem regiões de altas
latitudes. Esses autores associaram o padrão difuso a fatores bióticos e
abióticos como o fotoperíodo e a temperatura da água. Urian et al. (1996),
estudando o padrão de reprodução, na mesma espécie, em localidades
diferentes (de 20º a 60º de latitude) também encontrou um padrão difuso de
reprodução. Entretanto, esses autores afirmam que uma variável apenas não
seria suficiente para prever períodos reprodutivos, muito embora um somatório

58
delas o fizesse. Essa proposição corrobora com os resultados que encontramos
para a população de Pipa, mostrando o mesmo padrão difuso de freqüência na
área.
Além disso, os resultados obtidos podem estar associados ao ajuste da
população a um conjunto de fatores ambientais locais que possibilitariam a esta
população permanecer na área o ano todo, dadas as melhores condições. Read
(1990) afirma ser previsível este padrão difuso de residência, tendo em vista a
pouca variação das condições ambientais nas baixas latitudes.
Análises da variação diária nos permitiu verificar que esta população é mais
freqüente durante o período da manhã, mais precisamente entre as 09:00h e
12:00h. Quanto à variação da maré, verificamos que não houve associação
entre a freqüência de indivíduos e a mesma.
Poucos são os trabalhos que abordam a variação diária de indivíduos. Geise
et al. (1999) verificaram essa variação, sendo sua maior freqüência durante a
manhã entre as 09:00h e 10:00h. Esta elevação da freqüência de indivíduos em
um período do dia pode manter relações com fatores ambientais, tais como
hábito diário das presas; facilidade de arrebanhar, encurralar e capturar a
presas; condições de luminosidade; temperatura da água, ventos, correntes e
outras condições oceanográficas, que neste trabalho não foram analisados ou
identificados por nós.
Freqüência de Comportamentos
É interessante notar que apesar de não ter havido variação da freqüência de
indivíduos ao longo do ano, houve variação comportamental, sendo o período de
junho a novembro (primavera-inverno) o período de maior freqüência de
atividade. Contudo, esta sazonalidade não apresentou associação com os
fatores ambientais aqui analisados, fotoperíodo e precipitação.
Embora não tenhamos verificado tal associação, a variação comportamental
ao longo do ano está associada a outros fatores ambientais dada a sazonalidade
apresentada. Estudos com S. fluviatilis na bacia do Amazonas (Da Silva e Best
1996) e Inia geoffrensis na bacia do Orenoco (McGuire e Winerniller 1998),

59
mostraram um padrão anual da atividade de forrageio associado ao período de
seca, quando a densidade dos peixes é maior dado ao baixo volume das águas.
Em Sarasota, Florida o T. truncatus é residente, porém com variações
sazonais de comportamento. Forrageia só ou em pequenos grupos e em águas
rasas (entre 2 e 3m). Alimenta-se de espécies demersal sedentária vivendo
entre campos de algas marinhas ou de espécies que formam pequenos
cardumes fora desses campos. Semelhante à Sarasota, a população de Pipa
apresenta a mesma técnica de forrageio, sendo possível que a disponibilidade
de presas na área seja um dos fatores da variação da expressão dos
comportamentos dos botos.
É interessante destacar que observamos uma relação entre os
comportamentos de forrageio, atividade aérea e socialização expressos pelos
botos em Pipa e que a freqüência da atividade de forrageio nitidamente
influencia a expressão das demais atividades. Neumann (2001) estudando o
orçamento de atividades para o Delphinus delphis na Nova Zelândia, verificou
que após o forrageio os indivíduos se envolviam em atividades de socialização,
atribuindo essa dinâmica comportamental ao fato dos animais estarem saciados.
Afirmou ainda que quanto maior a disponibilidade alimentar, maior era o tempo
de socialização dos animais. De maneira semelhante, observamos que em Pipa
as maiores freqüências comportamentais se deram na mesma época, (de junho
a novembro). O que nos leva a considerar que após a atividade de forrageio os
animais se envolveriam em outras atividades como a socialização.
Quanto à variação diária dos comportamentos, verificamos que os botos são
mais ativos durante a manhã, entre as 09:00h e 12:00h. Geise et al. (1999)
também verificaram que o boto cinza forrageia mais pela manhã entre as 06:00h
e 07:00h. Segundo as autoras o forrageio ocorreria nesse horário devido à
disponibilidade alimentar.
Os comportamentos aqui estudados apesar de não apresentarem diferença
significativa entre os estados da maré, foram mais freqüentes durante a maré
baixa. Consideramos que tal fato se dê devido ao aumento da densidade dos
peixes com a diminuição da lâmina de água e a facilidade de arrebanhar,
encurralar e capturar as presas nessas condições. Da Silva e Best (1996) e

60
McGuire e Winerniller (1998), observaram o mesmo para o tucuxi e o boto rosa
respectivamente.
CONCLUSÃO GERAL
Durante todo o período de estudo foi observado a presença do Sotalia
guianensis na enseada do Curral distrito de Pipa, indicando um importante papel
desse sistema marinho para essa população.
Também verificamos que a freqüência de animais adultos e imaturos não
variou ao longo do ano mostrando que esta população apresenta um padrão
difuso de ocorrência. Contudo, a freqüência dos comportamentos de
socialização, atividade aérea e forrageio apresentaram um padrão sazonal.
Os resultados indicam ainda, que o boto cinza foi mais ativo e mais freqüente
pela manhã; sofreu influência da maré baixa e que, a variação diária da
freqüência dos indivíduos e atividades comportamentais variaram ao longo do
ano como resposta às variações ambientais.
LITERATURA CITADA (INTRODUÇÃO E DISCUSSÃO GERAIS)
Baird, R.W. (2000). The Killer Whale: Foraging specializations and group hunting
In: Mann, J., Connor, R.C., Tyack, P.L. e Whitehead, H., Cetacean Societies:
Field Studies of Dolphins and Whale, 1ª Ed. pp.127-153, Chicago, University
of Chicago press;
Barros, N.B., & Wells R.S. (1998). Prey and feeding patterns of resident
bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Journal of
Mammology 79(3):1045-1059;
Claphan, P.J. (2000). The Humpback Whale: Seasonal feeding and breeding in a
Baleen Whale In: Mann, J., Connor, R.C., Tyack, P.L. e Whitehead, H.,
Cetacean Societies: Field Studies of Dolphins and Whale, 1ª Ed. pp.173-196,
Chicago, University of Chicago press;
Claphan, P.J., Baraff, L.S., Carlson, C.A., Christian M.A., Mattila, D.K., Mayo,
C.A., Murthy, M.A., & Pittman,S. (1993). Seasonal occurrence and annual

61
return of Humpback Whale, Megaptera novaengiae, in the southern Gulf of
Maine, Can. J. Zool. 71:440-443;
da Silva, V.M.F., & Best, R.C. (1996). Sotalia fluviatilis. Mammalian Species.
Americam Society of Mammologists. 527:1-7;
Defran, R.H., & Wellemr, D.W. (1999). Occurrence, distribution, site fidelity and
school size of bottlenose dolphins (Tursiops truncatus) off San Diego,
Califórnia. Marine Mammal Science. 15(2):366-380;
Geise, L., Gomes, N., & Cerqueira, R. (1999). Behavior, habitat use and
population size of Sotalia fluviatilis (Gervais, 1853) (Cetácea, Delphinidae)
in the Cananéia estuary region. São Paulo, Brasil. Revista Brasileira de
Zoologia. 59:2;
Iñíguez, M.A. (2001). Seasonal distribuition of Killer Whale (Orcinus orca) in
northen Patagonia, Argentina. Aquatic Mammals. 27(2):154-161;
Karczmarski, L., Thornton, M., & Cockcroft, V.G. (2000). Daylight occurrence of
humpback dolphins, Sousa chinensis, in Algoa Bay, South Africa. African
Journal Ecology 38:86-90;
Larkman, V., & Veit, R. (1998). Seasonality and abundance of blue whales off
southern California. CalCOFI Rep. 39:236-239;
Lodi, L.F. (2003). Tamanho e composição de grupo dos botos-cinza, Sotalia
guianensis (van Bénéden 1864) (CETACEA, DEPHINIDAE), na Baía de Paraty,
Rio de Janeiro, Brasil. Atlântica. 25(2): 135-146;
McGuire, T.L., & Winemiller, K.O. (1998). Occurrence patterns, habitat
associations, and potential prey of the river dolphins Inia geoffrensis, in the
Cinaruco river, Venezuela. Biotropica. 30(4):625-638;
Neumann, D.R. (2001). Activity budget of free-ranging common dophins
(Delphinus delphis) in the northwestern Bay of Plenty, New Zealand. Aquatic
Mammals. 27(2):121-136;
Pastor, T., & Aguilar, A. (2003). Reproductive cycle of the female mediterraneam
monk seal in the western Sahara. Marine Mammal Science. 19(2):318-330;
Read, A.J. (1990). Reprodutive seasonality in Harbor Porpoise, Phocoena
phocoena, from the Bay of Fundy, Can. J. Zool. 68:284-288;

62
Rosas, F.C.W., & Monteiro-Filho, E.L.A. (2002). Reproduction of the estuarine
dolphin (Sotalia guianensis) on the coast of Paraná, solthern Brazil. Journal
of Mammology. 83(2):507-515;
Saayman, G.S., Tayler, C.K., & Bower, D. (1973). Diurnal activity cycles in
captive and free-ranging Indian ocean bottlenose dolphins (Tursiops
aduncus ehrenburg). Behaviour 44(3-4):212-233;
Santos, M.C.O., Açuña, L.B., & Rosso, S. (2001). Insights on site fidelity and
calving intervals f the marine tucuxi dophin (Sotalia fluviatilis) in south-
eastern Brasil, J. Mar. Biol. Ass. 81:1049-1052;
Similä, T., Holst, J.C., & Christensen, I. (1996). Occurrence and diet of Killer
Whale in northern Norway: Seasonal patterns relative to the distribution and
abundance of Norwegian Spring-Spawning Herring. Can. J. Fish. Aquat. Sci.
53:769-779;
Thayer, V.G., Read, A.J., Colby, D.R., Horn, A.A., McLellan, W.A., Pabst, D.A.,
Dearolf, J.L., Bowles, N.I., Russell, J.R., & Rittmaster, K.A. (2003).
Reproductive seasoanlity of western atlantic bottlenose dolphins off North
Caroline, USA. Marine Mammal Science 19(4):617-629;
Urian, K.W., Duffield, D.A., Read, A.J., Well, R.S., & Shell, E.D. (1996).
Sasonality of reproduction in bottlenose dolphins, Tursiops truncatus. Jornal
of Mammalogy. 77(2):394-403;
Wilson, B., Thomposon, P.M., and Hammond, P.S. 1997. Habit use by bottlenose
dophins: seasonal distribution and stratified movement patterns in Moray Firth,
Scotland. Journal of Applied Ecology. 34(6):1365-1374.
Wood, C.J. (1998). Movement of bottlenose dolphins arond the sothwest coast of
Britain. J.Zool. 246:155-163;

63
ANEXOS

64
Normas de Publicação – Canadian Journal Zoology
Language
Papers must be clearly and concisely written in good English or French.
Format and organization
The manuscript should be typewritten, double-spaced, on paper 21.5 × 28 cm
(8.5 × 11 in. or ISO A4). Typing should be on one side of the page only. Each
page should be numbered, beginning with the title page. For material that is to be
set in italics, use an italic font; do not underline. Use capital letters only when the
letters or words should appear in capitals. All articles should contain a title page
(p. 1), an abstract (p. 2), followed by Introduction (p. 3), Materials and
methods, Results, Discussion, and Acknowledgements sections, plus
references, tables, figure captions, and appendices, in that order. (See
descriptions of each part of the manuscript, below.) Tables and captions for
illustrations should be on separate pages
Title page
The title page should contain the following. ( i) The full title of the paper. ( ii)
Authors’ names (with initials only) listed in the order in which they are to appear
at the head of the printed article. ( iii) Affiliation and address (including e-mail
address) for each author. This should reflect the affiliation and address at the
time of the study. Indicate current affiliations and addresses (including e-mail
addresses) that differ from those in the by-line in a footnote. ( iv) Name, address,
telephone number, fax number, and e-mail address of the author responsible for
correspondence.
Abstract
An abstract is required for every contribution and should contain accurate
descriptive words that will draw the reader to the content. This is particularly
important because contemporary alerting services and search engines will
search this text. It should not be more than 200 words and should appear on a
separate page. All authors’ names and initials, as well as the complete title of
the paper, must be typewritten at the top of the page (a copy of this page is sent
separately to the translator). The concise abstract should present the paper
content accurately and should supplement, not duplicate, the title in this respect.

65
Authors able to submit abstracts in both fluent English and French are
encouraged to do so. Abstracts submitted in one language will be translated into
the other official language by the Journal translator. References should not be
cited in the abstract unless they are absolutely essential, in which case full
bibliographic information must be provided.
References
General form
The author is responsible for verifying each reference against the original article.
Each reference must be cited in the text using the surnames of the authors and
the year, for example, (Walpole 1985) or Green and Brown (1990). Depending on
the sentence construction, the names may or may not be in parentheses, but the
year always is. If there are three or more authors, the citation should give the
name of the first author followed by et al. (e.g., Green et al. 1991). References
must be listed in alphabetical order according to the name of the first author and
not numbered.
Journal article with DOI:
Sterling, J.T., and Ream, R.R. 2004. At-sea behavior of juvenile male northern
fur seals ( Callorhinus ursinus). Can. J. Zool. 82(10): 1621–1637.
doi:10.1139/Z04-136.
Book:
Williams, R.A. 1987. Communication systems analysis and design. Prentice-Hall,
Inc., Englewood Cliffs, N.J.
Part of book:
Healey, M.C. 1980. The ecology of juvenile salmon in Georgia Strait, Britsh
Columbia. In Salmonid ecosystems of the North Pacific. Edited by W.J. McNeil
and D.C. Himsworth. Oregon State University Press, Corvallis, Oreg. pp. 203–
229.
Figure captions
Figure captions should be listed on a separate page and placed after the tables.
The caption should informatively describe the content of the figure, without need
for detailed reference to the text. Experimental conditions should not be included,
but should be adequately covered in the Methods. For graphs, captions should

66
not repeat axis labels, but should describe what the data show. A single caption
can be provided for multipart (composite) figures, with necessary details on the
separate parts, identified by their individual labels. If the separate parts require
enough information to warrant separate captions, then the composite should be
separated into individual figures.
Maps
Maps must have very clear, bold patterns and must show longitudes and
latitudes (or UTM coordinates) and a scale, to ensure proper identification of
study locations.
Nomenclature
Authors are required to follow the International Code of Zoological Nomenclature
(4th ed., 1999), published by The International Trust for Zoological
Nomenclature, c/o The Natural History Museum, Cromwell Road, London SW7
5BD, UK. In subsequent mentions of the same species, the genus is abbreviated
to the first letter (e.g., S. plicata). Genera are always written in full at the
beginning of a sentence.
Writing numbers
In writing long numbers the digits should be separated into groups of three,
counted from the decimal marker to the left and right. The separator should be a
space and not a comma, period, or any other mark, for example, 25 562 987 and
not 25,562,987. In English text, the decimal marker should be a point, for
example, 0.1 mL and not 0,1 mL. The decimal point in all numbers between 1
and –1, except 0, must be preceded by a 0. The sign × should be used to
indicate multiplication, e.g., 3 × 106 and not 3·106.