universidade sÃo judas tadeu pÓs-graduaÇÃo … · “você faz suas escolhas, e suas escolhas...
TRANSCRIPT

UNIVERSIDADE SÃO JUDAS TADEU
PÓS-GRADUAÇÃO STRICTO SENSU EM EDUCAÇÃO FÍSICA
Janaina de Oliveira Brito
INFLUÊNCIA DA HIPERTENSÃO NOS AJUSTES INDUZIDOS PEL O
TREINAMENTO FÍSICO NA MODULAÇÃO AUTONÔMICA CARDIOVA SCULAR
E NO ESTRESSE OXIDATIVO EM UM MODELO EXPERIMENTAL
DE MENOPAUSA E DISFUNÇÃO METABÓLICA
SÃO PAULO
2008

UNIVERSIDADE SÃO JUDAS TADEU
PÓS-GRADUAÇÃO STRICTO SENSU EM EDUCAÇÃO FÍSICA
Janaina de Oliveira Brito
INFLUÊNCIA DA HIPERTENSÃO NOS AJUSTES INDUZIDOS PEL O
TREINAMENTO FÍSICO NA MODULAÇÃO AUTONÔMICA CARDIOVA SCULAR
E NO ESTRESSE OXIDATIVO EM UM MODELO EXPERIMENTAL D E
MENOPAUSA E DISFUNÇÃO METABÓLICA
Dissertação apresentada ao Programa de Mestrado em Educação Física da Universidade São Judas Tadeu como requisito parcial à obtenção do título de Mestre em Educação Física.
Área de Concentração: Bases Biodinâmicas da Atividade Física.
Orientadora: Profa. Dra. Kátia De Angelis.
SÃO PAULO
2008

Brito, Janaina de Oliveira
Influência da hipertensão nos ajustes induzidos pelo treinamento físico na modulação autonômica cardiovascular e no estresse oxidativo em um modelo experimental de menopausa e disfunção metabólica / Janaina de Oliveira Brito. - São Paulo, 2008.
103 f. ; 30 cm
Dissertação (mestrado) – Universidade São Judas Tadeu, São Paulo, 2008. Orientador: Kátia De Angelis
1. Hipertensão. 2. Menopausa. 3. Estresse Oxidativo. 4. Treinamento Físico. I.
De Angelis, Kátia. II. Título
CDD - 613.71

DEDICATÓRIA Dedico aos meus pais, José de Brito e Elza Francisca de Oliveira Brito, não somente esta
dissertação, mas todos os anos de estudo que me proporcionaram. Aproveito para agradecer a
compreensão nos momentos de correria em casa (e de ausência) e por todo o auxílio (com o qual
posso contar sempre). As próximas páginas são resultado de uma longa trajetória, repleta de
esforços e dedicação para que tudo saísse da melhor maneira possível. O conteúdo desta dissertação
é fruto do que, antes de mais ninguém, vocês plantaram!
Dedico também ao meu irmão Sebastião de Brito, meu amigo durante 23 anos da minha
vida! A pessoa que está ao meu lado em vários momentos bons e tristes, me ajudando nas horas
mais difíceis.
Dedico ao meu namorado Emerson Monzani, por compreender as minhas ausências nos
finais de semana, as minhas viagens para Congressos, a troca do “cinema em casa” pelos artigos.
Enfim, por estar ao meu lado desde o início da minha trajetória acadêmica.
Dedico esta dissertação aos meus familiares: meus avós que não estão mais presentes, meus
tios, tias, primos, primas, cunhadas, sogro, sogra, ex-professores e amigos. Todos, mesmo aqueles
que moram longe e encontro pouquíssimo, contribuem para a minha formação enquanto ser
humano.
Dedico aos meus amigos e eternos ANGELITOS : Bruno Rodrigues, Cristiano Mostarda,
Danielle Dias, Demilto Yamaguchi, Diego Figueroa, Geórgia Cândido, Henrique Marchet, Íris
Callado Sanches, Jacqueline Freire, Janaina Paulini, Juliana Francica, Karin Flues, Kátia Ponciano,
Luciana Jorge, Lucinar Flores, Marcelo Heeren, Márcia Val, Marcio Tubaldini, Michelle Sartori e
Renata Juliana. Pessoas que, às vezes, passam mais tempo ao meu lado do que meus próprios pais.
Pessoas com características ímpares, sempre ouvindo o que tenho a falar e me ensinando com suas
próprias experiências. Amigos que ajudam a transformar meus sonhos em realidade!

AGRADECIMENTOS À Professora Doutora Kátia De Angelis, por ter acreditado em mim desde o 2º ano
da graduação, orientando minha pesquisa de Iniciação Científica, meu Trabalho de
Conclusão de Curso, esta dissertação!! Enfim, o meu mais sincero muito obrigada por fazer
parte da minha vida, por passar noites em claro, por ser justa com todos os seus alunos, por
confiar na minha pessoa.
À Professora Doutora Maria Cláudia Irigoyen por estar sempre presente em nosso
grupo, nos auxiliando com o suporte técnico e acrescentando seus conhecimentos em
nossos trabalhos de maneira doce e significativa!
À minha amiga de graduação, companheira de TCC, colega de laboratório e parceira
de todas as horas, Nathalia Bernardes, por me aturar quando estou com muita fome e fico
de mau-humor, por ouvir as minhas diversas histórias com atenção e sempre rir das minhas
palhaçadas....É nada!!
Aos profissionais responsáveis pelos laboratórios e equipamentos da Universidade
São Judas Tadeu, em especial a Leide, Rosana e João Paulo, pela disposição em nos
ajudar e bom humor sempre!
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pelo
apoio financeiro.
À Universidade São Judas Tadeu (USJT) e todos os professores, coordenadores,
secretários e funcionários que, direta ou indiretamente, contribuíram para a realização deste
trabalho!

“A ciência não pode prever o que vai acontecer.
Só pode prever a probabilidade de algo acontecer.”
César Lattes
“Você faz suas escolhas, e suas escolhas fazem você.”
Steve Beckman
“Você nasceu para vencer,
mas para ser um vencedor você precisa planejar para vencer,
se preparar para vencer e
esperar para vencer.”
Zig Ziglar

SUMÁRIO
Lista de figuras
Lista de tabelas e quadros
Lista de abreviaturas
Resumo
Abstract
1. INTRODUÇÃO 1
1.1. Doença Cardiovascular e Gênero: O Impacto da Menopausa........................................... 1
1.2. Doença Cardiovascular e Síndrome Metabólica .............................................................. 4
1.3. Doença Cardiovascular e Disfunção Autonômica............................................................. 10
1.4. Doença Cardiovascular e Estresse Oxidativo ................................................................... 12
1.5. Doença Cardiovascular e Treinamento Físico .................................................................. 15
2. OBJETIVOS.......................................................................................................................... 20
2.1. Objetivo Geral................................................................................................................... 20
2.2. Objetivos Específicos........................................................................................................ 20
3. MATERIAIS E MÉTODOS................................................................................................. 21
3.1. Amostra............................................................................................................................. 21
3.2. Seqüência Experimental.................................................................................................... 22
3.3. Procedimentos .................................................................................................................. 22
3.3.1. Indução do Modelo Experimental de Síndrome Metabólica....................................... 22
3.3.2. Ooforectomia Bilateral................................................................................................ 23
3.3.3. Teste de Esforço Máximo........................................................................................... 24
3.3.4. Treinamento Físico...................................................................................................... 25

3.3.5. Medida da Glicemia e dos Triglicerídeos Plasmáticos............................................... 27
3.3.6. Canulação.................................................................................................................... 28
3.3.7. Registro de Pressão Arterial ....................................................................................... 30
3.3.8. Avaliação da Modulação Autonômica Cardiovascular............................................... 31
3.3.8.1. Análise da Variabilidade da Pressão Arterial Sistólica......................................... 31
3.3.8.2. Análise da Variabilidade do Intervalo de Pulso..................................................... 31
3.3.8.3. Análise da Sensibilidade Barorreflexa Espontânea (Índice Alfa).......................... 32
3.3.9. Teste de Resistência à Insulina.................................................................................... 32
3.3.10. Eutanásia dos Animais.............................................................................................. 32
3.3.11. Preparação dos Tecidos............................................................................................. 33
3.3.12. Dosagem de Proteínas............................................................................................... 33
3.3.13. Estresse Oxidativo e Enzimas Antioxidantes............................................................ 34
3.3.13.1. Medida de Lipoperoxidação (LPO): Quimiluminescência Iniciada por
t-BOOH (QL).........................................................................................................................
34
3.3.13.2. Razão GSH/GSSG............................................................................................ 35
3.3.13.2.1. Glutationa Total.......................................................................................... 35
3.3.13.2.2. Glutationa Oxidada.................................................................................... 36
3.3.13.3. Superóxido Dismutase (SOD).......................................................................... 36
3.3.13.4. Catalase (CAT)................................................................................................. 37
3.3.13.5. Glutationa Peroxidase (GPx)............................................................................ 38
3.4. Análise Estatística.............................................................................................................. 39
4. RESULTADOS………………….......................................................................................... 40
4.1. Avaliação da Capacidade Física……………………….……………………………..….. 40
4.2. Avaliação do Peso Corporal……………………………………………………………... 41

4.3. Avaliação do Tecido Adiposo………..……………………………………………........... 43
4.4. Avaliações Metabólicas…………………………………………………………….......... 44
4.5. Avaliações Hemodinâmicas……………………………………………………………… 47
4.6. Avaliações da Modulação Autonômica da Freqüência Cardíaca....................................... 49
4.7. Avaliação da Variabilidade da Pressão Arterial................................................................. 53
4.8. Avaliação da Sensibilidade Barorreflexa Espontânea........................................................ 56
4.9. Avaliações de Estresse Oxidativo....................................................................................... 57
4.10. Análise das Enzimas Antioxidantes.................................................................................. 60
5. DISCUSSÃO.......................................................................................................................... 62
5.1. Avaliações da Capacidade Física........................................................................................ 62
5.2. Avaliações Metabólicas e de Pesos Corporal e do Tecido Adiposo................................... 64
5.3. Avaliações Hemodinâmicas e Autonômicas....................................................................... 70
5.4. Avaliações do Perfil Oxidativo........................................................................................... 76
6. CONCLUSÃO........................................................................................................................ 84
7 REFERÊNCIAS BIBLIOGRÁFICAS................................................................................. 85

LISTA DE FIGURAS
Figura 1. Esquema geral das principais fontes celulares de produção e metabolização das
ERO. UQH• + ubisemiquinona; GSSG = glutationa oxidada; GSH = glutationa reduzida;
DH2 e D = sistemas redutores de NADP não-específicos; SOD = superóxido dismutase;
GPx = glutationa peroxidase; CAT = catalase; B e BH2 = doadores de hidrogênio........13
Figura 2. Seqüência Experimental do protocolo..............................................................22
Figura 3. Tratamento de D-frutose na água de beber.......................................................23
Figura 4. Etapas de realização da ooforectomia bilateral em ratas..................................23
Figura 5. Correlação entre o consumo de oxigênio (VO2) e a velocidade do teste de
esforço (km/h) em ratas ooforectomizadas.......................................................................24
Figura 6. Fotografia de ratos submetidos ao protocolo de treinamento físico em esteira
ergométrica a USJT...........................................................................................................25
Figura 7. Aparelhos que foram utilizados para análises das concentrações sangüíneas de
glicose e triglicerídeos......................................................................................................28
Figura 8. Esquema do local da canulação da artéria carótida e veia jugular...................29
Figura 9. Foto do animal com a cânula exteriorizada......................................................29
Figura 10. Sistema de registro de pressão arterial e conexão entre a cânula e o transdutor
eletromagnético.................................................................................................................30
Figura 11. Velocidade máxima alcançada no teste de esforço inicial, intermediário (4ª
semana) e final (8ª semana) nos grupos FOS (frutose ooforectomizado sedentário), FOT
(frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário
tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com
frutose)..............................................................................................................................41

Figura 12. Peso corporal dos grupos FOS (frutose ooforectomizado sedentário), FOT
(frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário
tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose)
ao longo das últimas semanas de protocolo......................................................................42
Figura 13. Tecido adiposo dos grupos FOS (frutose ooforectomizado sedentário), FOT
(frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário
tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com
frutose)..............................................................................................................................44
Figura 14. Glicemia nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose
ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com
frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose)..................46
Figura 15. Concentrações sanguíneas de triglicerídeos nos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)...........................................................................43
Figura 16. Constante de decaimento da gliemia (KITT) durante o teste de tolerância à
insulina nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose
ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com
frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose)..................46
Figura 17. Pressão Arterial Média nos grupos FOS (frutose ooforectomizado
sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado
hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado
tratado com frutose)..........................................................................................................47

Figura 18. Freqüência Cardíaca nos grupos FOS (frutose ooforectomizado sedentário),
FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário
tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com
frutose)..............................................................................................................................49
Figura 19. Variância do intervalo de pulso nos grupos FOS (frutose ooforectomizado
sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado
hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado
tratado com frutose)..........................................................................................................52
Figura 20. Banda de baixa freqüência do intervalo de pulso normalizada nos grupos FOS
(frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)...........................................................................52
Figura 21. Banda de alta freqüência do intervalo de pulso normalizada nos grupos FOS
(frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)..........................................................................53
Figura 22. Variância da pressão arterial sistólica nos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)...........................................................................55
Figura 23. Banda de baixa freqüência da pressão arterial sistólica nos grupos FOS
(frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)...........................................................................55

Figura 24. Índice alfa nos grupos FOS (frutose ooforectomizado sedentário), FOT
(frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário
tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com
frutose)..............................................................................................................................56
Figura 25. Quimiluminescência no tecido cardíaco nos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)...........................................................................58
Figura 26. Razão da glutationa reduzida pela glutationa oxidada (GSH/GSSG) no tecido
cardíaco nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose
ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com
frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose)..................58
Figura 27. Correlação negativa obtida entre a QL no tecido cardíaco e o índice alfa nos
grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado
treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT
(ooforectomizado hipertenso treinado tratado com frutose).............................................59
Figura 28. Correlação negativa entre a QL no tecido cardíaco e o RMSSD obtida nos
grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado
treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT
(ooforectomizado hipertenso treinado tratado com frutose).............................................60
Figura 29. Atividade da enzima catalase no tecido cardíaco nos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado
hipertenso treinado tratado com frutose)...........................................................................61

LISTA DE TABELAS E QUADROS Tabela 1. Velocidade máxima alcançada no teste de esforço inicial, intermediário (4ª
semana) e final (8ª semana) nos grupos FOS (frutose ooforectomizado sedentário),
FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso
sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado
com frutose).................................................................................................................40
Tabela 2. Peso corporal dos grupos FOS (frutose ooforectomizado sedentário), FOT
(frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário
tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com
frutose) ao início (dia ooforectomia) e ao final do protocolo.....................................43
Tabela 3. Valores sanguíneos de glicose (GLIC), triglicerídeos (TG) e constante de
decaimento da glicemia (KITT) dos grupos FOS (frutose ooforectomizado
sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado
hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso
treinado tratado com frutose) no início e ao final do protocolo..................................45
Tabela 4. Parâmetros hemodinâmicos avaliados em repouso dos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT
(ooforectomizado hipertenso treinado tratado com frutose).......................................48

Tabela 5. Variabilidade da freqüência cardíaca dos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT
(ooforectomizado hipertenso treinado tratado com frutose).......................................51
Tabela 6. Variabilidade da pressão arterial sistólica dos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT
(ooforectomizado hipertenso treinado tratado com frutose).......................................54
Tabela 7. Atividade das enzimas catalase (CAT), superóxido dismutase (SOD) e
glutationa peroxidase (GPx) no tecido cardíaco dos grupos FOS (frutose
ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS
(ooforectomizado hipertenso sedentário tratado com frutose) e FOHT
(ooforectomizado hipertenso treinado tratado com frutose).......................................61
Quadro 1. Protocolo de treinamento físico para ratas Wistar....................................26
Quadro 2. Protocolo de treinamento físico para ratas SHR.......................................27
Quadro 3. Alterações metabólicas dos grupos controle (CS), ooforectomizada (OS),
ooforectomizada hipertensa (OHS), frutose ooforectomizada (FOS) e frutose
ooforectomizada hipertensa (FOHS)...........................................................................66
Quadro 4. Alterações hemodinâmicas e autonômicas dos grupos controle (CS),
ooforectomizada (OS), ooforectomizada hipertensa (OHS), frutose ooforectomizada
(FOS) e frutose ooforectomizada hipertensa (FOHS).................................................71

LISTA DE ABREVIATURAS %AF = banda de alta freqüência normalizada
%BF = banda de baixa freqüência normalizada
AF = banda de alta freqüência
ATRAMI =Autonomic Tone and Reflexes After
Myocardial Infarction
B e BH2 = doadores de hidrogênio
BF = banda de baixa freqüência
CAT = catalase
CO3HK = bicarbonato potásico
COEP = Comitê de Ética em Pesquisa
CuSO4.= sulfato de cobre
DH2 e D = sistemas redutores de NADP não-
específicos
DM = diabetes mellitus
DNA = ácido desoxirriblonucleico
DTNB = acido ditionitrobenzóico
ERO = espécies reativas de oxigênio
FC = frequência cardíaca
FFT = Transformada Rápida de Fourrier
FOHS = frutose hipertensa ooforectomizada
sedentária
FOHT = frutose hipertensa ooforectomizada
treinada
FOS = frutose ooforectomizada sedentária
FOT = frutose ooforectomizada treinada
GPx = glutationa peroxidase
GSH = glutationa reduzida
GSH/GSSG = razão glutationa reduzida pela
glutationa oxidada
GSSG = glutationa oxidada
H2O = água
HAS = hipertensão arterial sistêmica
HDL = lipoproteína de alta densidade
i.p. = intra-peritonial
IP = intervalo de pulso
KCl = cloreto de potássio
Kitt = constante de queda da glicose plasmática
KNaC4H4O6 = tartarato de sódio e potássio
LPO = lipoperoxidação
Na2HPO4 = fosfato dissódico
NADPH = forma reduzida da nicotinamida
adenina dinucleotídeo fosfato
NaHCO3 = bicarbonato de sódio
NaOH = hidróxido de sódio
NO = óxido nítrico
PA = pressão arterial
PAD = pressão arterial diastólica
PAM = pressão arterial média
PAS = pressão arterial sistólica
PMSF = fluoreto de fenil metil sulfonila
QL = quimiluminescência
RMSSD = raiz quadrada da média dos quadrados
das diferenças entre os intervalos R-R normais
sucessivos
SHR = rato espontaneamente hipertenso
SM = síndrome metabólica
SNA = sistema nervoso autônomo
SOD = superóxido dismutase
t-BOOH = hidroperóxido de tert-butil
UQH• = ubisemiquinona
VAR = variância
VO2 = volume de oxigênio
WHI = Women´s Health Initiative

RESUMO
O objetivo do presente estudo foi verificar os efeitos do treinamento físico (TF) em parâmetros
cardiovasculares e metabólicos, na modulação autonômica cardiovascular e no estresse
oxidativo em ratas fêmeas Wistar e em ratas espontaneamente hipertensas (SHR), ambas
ooforectomizadas submetidas à sobrecarga de frutose na água de beber. Foram utilizadas 40
fêmeas Wistar e SHR divididas em 4 grupos (n=10 em cada grupo) de ratas ooforectomizadas
(remoção bilateral dos ovários) submetidas à sobrecarga de frutose (100g/L de água): sedentária
(FOS), treinada (FOT), hipertensa sedentária (FHOS), hipertensa treinada (FHOT). Os grupos
treinados foram submetidos a um programa de TF em esteira ergométrica (1 hora/dia, 5
dias/semana, 8 semanas, 40-60% da velocidade máxima no teste de esforço). A concentração
sanguínea de glicose e triglicerídeos e o teste de resistência à insulina foram utilizados para
avaliar o perfil metabólico. Ao final do protocolo, os animais foram canulados para registro
direto de pressão arterial (PA). Além disso, avaliou-se a modulação autonômica cardiovascular
no domínio do tempo e da freqüência (análise espectral). O perfil oxidativo foi verificado no
tecido cardíaco avaliando-se a quimiluminescência (QL), a relação glutationa
reduzida/glutationa oxidada (GSH/GSSG), bem como a atividade das enzimas antioxidantes:
superóxido dismutase (SOD), glutationa peroxidase (GPx) e a catalase (CAT). O grupo FOS
apresentou redução dos valores de triglicerídeos e do KITT (constante de decaimento da
glicose) no teste de tolerância à insulina quando comparado ao grupo FOT. Os grupos FOHS e
FOHT apresentaram valores de resistência à insulina e valores triglicerídeos semelhantes ao
grupo FOS. Os valores de PA foram maiores nos grupos FOHS e FOHT em relação aos grupos
FOS e FOT. Além disso, os animais do grupo FOHS apresentaram taquicardia de repouso em
relação aos demais grupos estudados. O TF induziu bradicardia de repouso nos grupos
treinados. A hipertensão induziu redução do RMSSD (raiz quadrada da média dos quadrados

das diferenças entre os intervalos R-R normais sucessivos) do intervalo de pulso (IP); e o TF
induziu aumento do RMSSD somente no grupo FOT em relação aos demais grupos estudados.
Com relação à banda de baixa freqüência do IP (BF), os grupos FOHS e FOHT demonstraram
uma redução exacerbada desses valores em relação aos grupos FOS e FOT. A banda de alta
freqüência do IP (AF) foi menor no grupo FOHT quando comparados ao grupo FOT. A
hipertensão induziu um aumento na VAR-PAS e da banda de BF da PAS. No entanto, o TF
atenuou tais disfunções nos grupos FOT e FOHT. A hipertensão provocou uma redução no
índice alfa, representativa da sensibilidade barorreflexa espontânea que foi atenuada pelo TF
(FOT e FOHT). A QL estava reduzida somente no grupo FOT em relação ao grupo FOS. Já a
relação GSH/GSSG estava maior no grupo FOHT em relação ao grupo FOHS. A atividade da
CAT foi maior nos grupos treinados, mas a atividade da GPx e da SOD foi semelhante entre os
grupos estudados. Observaram-se correlações negativas entre a QL e o RMSSD (r=-0,60) e o
índice alfa (r=-0,63), sugerindo que animais que reduziram o estresse oxidativo cardíaco
apresentavam melhora na modulação autonômica cardiovascular. Concluindo, os resultados
demonstraram prejuízos no perfil metabólico e autonômico em ratas ooforectomizadas
sedentárias normotensas tratadas com sobrecarga frutose, e que a hipertensão induziu
disfunções hemodinâmica e autonômica adicionais nesses animais. Entretanto, o achado mais
importante de nosso trabalho foi que o treinamento físico atenuou algumas dessas disfunções
decorrentes da privação dos hormônios ovarianos e do consumo crônico de frutose, pelo menos
em parte associado à redução do estresse oxidativo; todavia, a presença de hipertensão aboliu
alguns benefícios observados no grupo normotenso treinado.

ABSTRACT
The purpose of this study was to evaluate the effects of physical training on cardiovascular and
metabolic parameters, on autonomic cardiovascular modulation and on oxidative stress in
female rats Wistar and in spontaneously hypertensive rats (SHR), both ovariectomized and
submitted to fructose overload in drinking water. Ovariectomized (bilateral ovary removal)
female Wistar rats and female SHR submitted to fructose overload (100g/L) were divided into 4
groups (n=10 each): sedentary (SOF), trained (TOF), sedentary hypertensive (SHOF), trained
hypertensive (THOF). The trained groups were submitted to an exercise training protocol on a
treadmill (1 hour/day; 5 days/week; 8 weeks; 40-60% of the maximum velocity of the exercise
test). The blood glucose and triglycerides concentrations and the insulin tolerance test were
performed to evaluate the metabolic profile. At the end of the protocol all the rats were
cannulated to arterial pressure (AP) direct recording. Moreover, the cardiovascular autonomic
control was evaluated in the time and the frequency (spectral analysis) domains. The oxidative
profile was verified in the heart tissue by the chemiluminescence (CL), the glutathione
reductase/glutathione oxidase ratio (GSH/GSSG), as well as by the antioxidant enzymes
superoxide dismutase (SOD), glutathione peroxidase (GPx) and catalase (CAT) activities. The
SOF group presented reduced triglycerides values and KITT (constant of glucose reduction) in
the insulin tolerance test when compared with the TOF group. The SHOF and THOF rats
showed similar KITT and triglycerides values when compared to SOF rats. The AP values were
higher in SHOF and THOF groups when compared with SOF and TOF groups. Furthermore,
SHOF rats presented resting tachycardia in relation to the other studied groups. Exercise
training (ET) induced resting bradycardia in the trained groups. The hypertension induced to a
reduction in RMSSD (root mean square of successive differences) of the pulse interval (PI);
and the ET induced an increase in the RMSSD only in the TOF group when compared to the

other studied groups. Regarding the low frequency band (LF) of the PI, the SHOF and THOF
groups showed a exacerbated reduction as compared to SOF and TOF groups. The high
frequency band (HF) of the PI was lower in the THOF group in relation to the TOF group. The
hypertension induced an enhancement in the variance of the PA (VAR-PAS) and the LF band
of the PA. However, the ET attenuated such dysfunctions in the SOF and THOF groups. The
hypertension induced a reduction in the alpha-index (which represents the spontaneous
baroreflex sensitivity), that was attenuated by the ET (TOF e THOF). The CL was reduced in
the TOF rats as compared to the SOF rats. The GSH/GSSG ratio was increased in the SHOF
group in relation to THOF group. The CAT activity was higher in the trained groups, but the
GPx and SOD activities were similar between studied groups. Negative correlations were
obtained between CL and RMSSD (r=-0.60) and the alpha index (r=-0.63), suggesting that
animals that reduced oxidative stress showed improvement in the autonomic cardiovascular
modulation. In conclusion, the results demonstrated impairments in the metabolic and
autonomic profiles in ovariectomized sedentary normotensive rats submitted to fructose
overload, and that hypertension induced additional hemodynamic and autonomic dysfunctions
in these animals. However, the main finding of our study was that the ET had attenuated some
dysfunctions observed in female rats submitted to ovarian hormones deprivation and chronic
fructose overload, at least in part associated with oxidative stress reduction; nevertheless the
presence of hypertension abolished some benefits that were observed in the normotensive
trained group.

1
1. INTRODUÇÃO
1.1. Doença Cardiovascular e Gênero: O Impacto da Menopausa
Atualmente, as doenças cardiovasculares constituem a mais importante causa de
morte em ambos os sexos em todas as regiões do país e no mundo ocidental (CASTANHO
et al., 2001; NAHAS, 2001; BOUCHARD, 2003). A morte devido às doenças
cardiovasculares é maior em homens (39%) do que em mulheres na faixa etária dos 45 a 64
anos. Entretanto, após os 65 anos, a taxa de mortalidade das mulheres por doenças
cardiovasculares ultrapassa à dos homens em até 22% (NATIONAL CENTER FOR
HEALTH STATISTICS, 1997). Além disso, a taxa de mortalidade devido às doenças
cardiovasculares no sexo feminino elevou-se rapidamente nas últimas décadas. Isso
provavelmente ocorreu, pois atualmente, as mulheres estão mais expostas a fatores de risco
como o estresse, o fumo, os maus hábitos alimentares e o sedentarismo, sem contar com a
sua inserção no mercado de trabalho. No Brasil, a prevalência de morte por doença
cardiovascular aumentou de 10 para 25% entre os anos 60 e 70 para o sexo feminino
(CASTANHO et al., 2001).
Castanho et al. (2001) citam como fatores de risco para doença cardiovascular: o
diabetes, o excesso de peso e a obesidade, a inatividade física, o fumo, a
hipercolesterolenia e a hipertensão, sendo estes dois últimos fatores de risco mais
prevalentes em mulheres do que em homens. Neste contexto, vale destacar que a diferença
em mortalidade cardiovascular entre os sexos pode ser devida a vários fatores, como
prevalência diferenciada dos fatores de risco cardiovascular citados acima, bem como
diferenças na modulação autonômica cardiovascular. Estudos clínicos e experimentais

2
parecem concordar que o sexo feminino, antes da privação dos hormônios ovarianos, tem
maior predomínio vagal e maior sensibilidade dos pressorreceptores (o mais importante
regulador da pressão arterial (PA) à curto prazo) e, portanto, maior proteção cardiovascular,
em relação ao sexo masculino (KUO et al., 1999; HUIKURI et al., 1996; GREGOIRE et
al., 1996; LEINWAND, 2003). Entretanto, é importante enfatizar que essa proteção
autonômica cardiovascular apresentada pelo sexo feminino é atenuada após a privação dos
hormônios ovarianos (KUO et al., 1999). Além disto, um estudo demonstrou que 24% das
mulheres com mais de 40 anos de idade apresentam uma marcante diminuição na
sensibilidade dos barorreceptores em relação a mulheres jovens (LAITINEN et al., 1998).
Considerando esses achados, parece razoável supor que a disautonomia cardiovascular
possa estar relacionada à equivalência nas taxas de eventos cardiovasculares entre os sexos
após o advento da menopausa (BRENNER, 1988; NCEP, 2001).
De fato, após a menopausa (a última menstruação da mulher), observaram-se
progressivamente importantes alterações fisiológicas que podem afetar vários locais do
organismo e determinar sinais e sintomas conhecidos por síndrome climatérica e que
podem estar relacionadas a alterações no controle do sistema nervoso autônomo sobre
órgãos e sistemas (GUYTON & HALL, 2002; ANTUNES, MARCELINO & AGUIAR,
2003; DE ANGELIS et al., 2004). Vale destacar que quanto mais cedo ocorrer à parada da
produção dos hormônios femininos, (em especial induzidos cirurgicamente), mais efeitos
negativos são observados, gerando um abrupto aumento do risco de infarto agudo do
miocárdio (COLDITZ et al., 1987). Conforme Schouw et al. (1996) quanto mais cedo o
climatério, maiores os riscos, ou seja, quanto mais precoce a mulher entra no período de
restrição dos hormônios femininos maior é o risco de eventos cardíacos. Neste contexto,

3
estudos vêm demonstrando que os hormônios ovarianos podem ser responsáveis por tais
alterações no risco cardiovascular. Corroborando a importância dos hormônios ovarianos
no controle cardiovascular, estudos demonstram que a PA é mais baixa em mulheres pré-
menopausa e também que se eleva em mulheres menopausadas (STAESSEN et al., 1997;
WEIS, 1972) ou em animais submetidos à privação dos hormônios ovarianos (IRIGOYEN
et al., 2005; RECCKELHOLF et al., 2000).
Trabalhos experimentais vêm colaborando para o melhor entendimento dos
processos envolvidos no aumento do risco cardiovascular após a menopausa
(SHAWAERY et al., 1997; ADAMS et al., 1995). Estudos demonstram disfunção
autonômica relacionada a privação dos hormônios ovarianos (KUO et al., 1999,
LAITINEN et al., 1998; IRIGOYEN et al., 2005; JURCA et al., 2004). Além disto,
trabalhos sugerem que um dos vários mecanismos de cardioproteção dos estrogênios seria a
preservação da função endotelial, através, por exemplo, da inibição da proliferação da
célula muscular lisa, das ações antioxidantes e da melhora na reatividade vascular, os quais
induziriam um melhor equilíbrio na função vasodilatadora/vasoconstrictora com
conseqüente diminuição dos riscos cardiovasculares. Por fim, não se pode esquecer que em
paralelo a privação dos hormônios ovarianos começa a ocorrer mudanças desfavoráveis em
lipídios, na tolerância à glicose (AMODEO & HEIMANN, 2003), aumento do peso
corporal e da prevalência de diabetes e redução na capacidade de exercício, na força
muscular e na massa óssea (SOWERS & LA PIETRA, 1995). Essas alterações aumentam
significativamente o risco de eventos cardiovasculares nesta fase de vida da mulher e
também podem estar relacionadas à disfunção autonômica, uma vez que esses fatores

4
associados podem desencadear o aparecimento de doenças cardiovasculares (SINAGRA &
CONTI, 2007).
1.2. Doença Cardiovascular e Síndrome Metabólica
Nos últimos anos houve um aumento na incidência de doenças crônicas (obesidade,
diabetes, doenças cardiovasculares) e fatores de risco associados para essas doenças (como
tabagismo, estresse, sedentarismo e alimentação inadequada) o que ocasionou o aumento
da morbidade e da mortalidade da população adulta mundial. Estima-se que nos Estados
Unidos a atual prevalência da síndrome metabólica (SM) seja de 21,8% da população
adulta, o que representa cerca de 47 milhões de norte-americanos (GUIMARÃES &
CIOLAC, 2004).
A SM é um termo que tem sido utilizado para relacionar a doença coronariana, a
hipertensão, o diabetes Tipo II (DM2) e a obesidade da porção superior do corpo a
resistência à insulina e à hiperinsulinemia. Essa síndrome foi também denominada
síndrome X e síndrome da civilização. Não está totalmente claro quando essa síndrome
começa, mas foi observado que a obesidade da porção superior do corpo está associada à
resistência à insulina e que esta está relacionada a um maior risco de doença coronariana,
hipertensão e DM2. Essa síndrome tornou-se um importante tópico de pesquisa na década
de 1990, devendo os resultados ajudar-nos a compreender melhor a fisiopatologia dessas
doenças e suas inter-relações (WILMORE & COSTILL, 2001).
A SM foi provavelmente identificada em 1923, quando Kylin notou que a
hipertensão e a hiperglicemia se correlacionavam. Nos anos 60, surgiu uma definição mais
moderna desta síndrome, que incluía obesidade, hipertensão, diabetes, e hiperlipidemia.

5
Investigadores alemães dos anos 70 foram os primeiros a usar o termo “síndrome
metabólica“, começando a explorar a associação da síndrome com a arteriosclerose. Após
os anos 90, Ferrannini e colaboradores (1990) sugeriram que a causa subjacente da
síndrome era resistência à insulina e, por isso, sugeriram o termo “síndrome de resistência à
insulina”.
De fato, há aumento do risco cardiovascular decorrente da associação entre
obesidade, hipertensão arterial e alterações no metabolismo lipídico e glicêmico. Tem sido,
inclusive, demonstrado que a distribuição da gordura no corpo é mais importante do que o
aumento de peso (HALPEN, 1998). Além disto, o acúmulo de gordura abdominal, mesmo
em não obesos, está relacionado a doenças metabólicas comuns na meia-idade. Denominou
esse estado de "Síndrome X" que passou posteriormente a ser chamada de "Síndrome
Plurimetabólica" ou "Metabólica" (REPETTO, 1998).
Além disso, foi demonstrado que as mulheres que apresentaram obesidade
abdominal tinham maior dificuldade na redução da glicemia do que o grupo controle de
mulheres eutróficas e do que um grupo de obesas com gordura localizada perifericamente
(WARRAM et al., 1990). Este fato pode, talvez, ser explicado porque os adipócitos
abdominais são muito maiores e têm aumentada tendência em converter os lipídios em
ácidos graxos, quando comparados aos adipócitos acumulados em outros compartimentos
corporais (WARRAM et al., 1990). Ao liberar rapidamente seu conteúdo de ácido graxo na
corrente sangüínea, os adipócitos viscerais provocam um aumento dos ácidos graxos
circulantes, ocasionando aumento dos níveis séricos de glicose e triglicérides. Ainda, os
ácidos graxos livres dificultam a entrada de glicose nas células musculares, levando à
hiperglicemia com aumento do risco de desenvolvimento de DM2. Esses ácidos graxos têm

6
acesso direto ao fígado pela veia porta, interferindo no metabolismo da insulina e afetando
conseqüentemente a captação celular de glicose, por sua vez levando a resistência à
insulina. Esse estado fisiopatológico é considerado como elemento fundamental na
etiologia da SM (ISSA & FRANCISCO, 1996).
Vários estudos correlacionaram a hiperinsulinemia e resistência à insulina com risco
aumentado para aterosclerose, hipertensão arterial, cardiopatia isquêmica, dislipidemias,
intolerância à glicose, obesidade abdominal e DM2 (ROCCHINI et al.,1989; DALY &
LANDSBERG, 1991). Dessa forma, observa-se que a SM engloba variáveis que aumentam
o risco para as doenças cardiovasculares (LOPES, 2005). Trabalhos recentes têm
demonstrado que pessoas com SM baseados no critério de NCEP/ATP III apresentam
maior risco de diabetes e doenças cardiovasculares (FORD et al., 2004;
DASKALOPOULOU et al 2004; MATSUZAWA, 2005). Vale destacar que a associação
da SM com a doença cardiovascular está aumentando a mortalidade geral em cerca de 1,5
vezes e a cardiovascular em cerca de 2,5 vezes (LAKKA et al., 2002; FORD & GILES,
2003; HAFFNER & TAEGTMEYER, 2003; GANG et al.; 2004; GIRMAM et al., 2004).
As evidências da literatura levam a crer que a SM resulta da influência do meio
ambiente em indivíduos geneticamente predispostos. Tudo indica que a obesidade central, a
pressão arterial aumentada, o aumento de triglicérides, a glicemia de jejum alterada e o
baixo HDL-colesterol são os principais componentes para definir a SM (LOPES, 2005). A
resistência à insulina, e até mesmo o diabetes, podem não estar presente no paciente com
SM conforme os critérios da NCEP/ATP III (2001). Porém, de acordo com a Organização
Mundial da Saúde (OMS), o ponto de partida para definição da SM é a avaliação da
resistência à insulina ou do distúrbio do metabolismo da glicose (ALBERTI & ZIMMET,

7
1998). De fato, um estudo de Goff e colaboradores (2003) indicaram haver uma correlação
inversa entre sensibilidade à insulina e incidência de hipertensão, independente de idade,
sexo ou etnia.
A associação de disfunções como intolerância à glicose, adiposidade abdominal,
elevação de triglicérides, baixos níveis de colesterol HDL, hipertensão, dislipidemia,
hipertrigliceridemia aumentam ainda mais a doença cardiovascular (KAMITANI et al.,
2005). Contudo, acreditam que resistência à insulina é um fator chave para a patogênese da
SM. Sendo que, a resistência metabólica à insulina constitui a maior disfunção na
patogênese do DM tipo 2 e condições relacionadas, incluindo alterações do tecido
endotelial. Carneiro e colaboradores (2003) citam em seu estudo que a maior prevalência
de hipertensão tem sido atribuída a hiperinsulinemia decorrente da resistência à insulina
presente em indivíduos obesos, principalmente naqueles que apresentam excesso de
gordura na região abdominal. Neste estudo foi observado um aumento significativo na
prevalência da hipertensão relacionada com o aumento do índice de massa corpórea
(CARNEIRO et al., 2003).
A relação entre dislipidemia e doença cardiovascular na população geral está bem
estabelecida, sendo independente do sexo, idade, história de tabagismo e presença de
hipertensão arterial ou DM (BATISTA & RODRIGUES, 2004). Além da hipertensão e da
dislipidemia, o diabetes citado anteriormente representa um importante fator de risco para
desenvolvimento e morte por doença cardiovascular (EDWING et al., 1980; VINIK et al.,
2003; KASETA et al., 1999). Indivíduos com DM do tipo 2 apresentam 2 a 4 vezes mais
risco de doenças cardiovasculares do que não-diabéticos, sendo a doença cardiovascular a
causa de morte em até 80% deles (KANNEL & MCGEE, 1979; STAMLER et al.,1993).

8
O modelo experimental de diabetes por estreptozotocina tem sido utilizado por muitos
investigadores, inclusive por nosso grupo, no estudo das alterações metabólicas e
cardiovasculares, bem como na busca dos benefícios do treinamento físico nesta doença
(MAEDA et al., 1995; DE ANGELIS et al., 2000, 2002; DALL’AGO, 2007; MAEDA et
al., 2007; HARTHMANN et al., 2007). É importante destacar ainda que estudos
demonstram que o diabetes dobra o risco de desenvolvimento das doenças cardio-
circulatória no homem e triplica nas mulheres (MUIR et al., 1992; KASETA et al., 1999).
Nosso grupo recentemente demonstrou que o treinamento físico induziu melhora na função
autonômica associada à redução da mortalidade em ratas ooforectomizadas diabéticas por
estreptozotocina (SOUZA et al., 2007). Todavia, este modelo experimental não apresenta
aumento de peso corporal ou hipertensão, diferenciando-se nestes aspectos de um modelo
de SM.
Neste aspecto, novos modelos experimentais têm sido desenvolvidos para o estudo
da SM, entre eles o modelo de sobrecarga de frutose. Interessantemente, acompanhando o
aumento da epidemia de obesidade e SM, o consumo de frutose na dieta (principalmente
em alimentos industrializados), casualmente ou não, aumentou cerca de 250% nos últimos
15 anos (BASCIANO et al., 2005; BRAY et al., 2004). Dessa forma, recentemente sugeriu-
se que o aumento marcante no consumo de frutose poderia favorecer o ganho de peso e a
obesidade (BASCIANO et al., 2005; ELLIOT et al., 2002). Estudos em humanos
verificaram que o consumo de frutose pode induzir ganho de peso, redução da sensibilidade
à insulina, hipertrigliceridemia e aumento da PA (ELLIOT et al., 2002). Já estudos
experimentais em animais machos, com sobrecarga de frutose na ração ou na água de
beber, verificaram aumento dos triglicerídeos e da insulina plasmática, elevação discreta da

9
PA e resistência à insulina, alterações compatíveis com um quadro inicial de
desenvolvimento de SM (SUZUKI et al., 1997; FARAH et al., 2006; CUNHA et al., 2006;
TEFF et al., 2004; ELLIOT et al., 2002).
Recentemente, um estudo realizado por nosso grupo em camundongos machos
submetidos à sobrecarga de frutose demonstrou intolerância à glicose, além de correlação
entre disfunções renais e as alterações cardiovasculares e autonômicas (CUNHA et al.,
2007). Corroborando os resultados obtidos em machos, resultados recentes de nosso
laboratório evidenciaram que a resistência à insulina estava associada à disfunção
autonômica (redução do tônus vagal) em ratas fêmeas saudáveis submetidas à sobrecarga
de frutose na água de beber de ratas (BRITO et al., 2008). Verificamos também que o
treinamento físico neste modelo em ratas (fase não ovulatória) induziu redução da
resistência à insulina, normalização da PA e do exacerbado efeito simpático cardíaco, além
de aumento do efeito vagal (BRITO et al., 2006).
Estudos recentes do Women´s Health Initiative (WHI) têm mostrado relação das
mudanças metabólicas que ocorrem nas mulheres na transição da pré-menopausa com a
pós-menopausa, diretamente com a falência ovariana ou alternativamente como um
resultado metabólico indireto da redistribuição da gordura central decorrente da deficiência
estrogênica (CARR & BRUNZELL, 2003). Diante desses achados, a detecção da SM é
fundamental para estratificar o risco global do indivíduo, principalmente da mulher e
instituir um tratamento adequado, com o objetivo de controlar todos os distúrbios
fisiológicos presentes.

10
1.3. Doença Cardiovascular e Disfunção Autonômica
Walter Canon por volta de 1920 definiu que o sistema nervoso autônomo (SNA) era
fundamental para manutenção do equilíbrio do organismo, denominando esta situação com
o termo “homeostasia” (CANON, 1939). Atualmente, é consenso na literatura que a
regulação neural do coração ocorre através da integração da atividade nervosa do simpático
e do parassimpático, consistindo na manutenção da função cardíaca. Além disso, a atuação
dos reflexos originados pelos pressorreceptores arteriais e sua integração central, dependem
do controle cardiovascular (MANCIA et al., 1997; IRIGOYEN et al., 2005). Estes reflexos
contribuem para manter a perfusão tecidual adequada, uma vez que a PA não sofra grandes
variações. Contudo, as alterações da atividade nervosa simpática são bem mais conhecidas
e estudadas que as do parassimpático, uma vez que as doenças cardiovasculares
representam uma das mais importantes causas de morte nos países ocidentais (NAHAS,
2001; BOUCHARD, 2003), logo essas alterações constituem as mais fortes evidências da
disfunção autonômica (FRANCHINI & KRIEGER, 1989). Entretanto, vale ressaltar que a
função vagal relaciona-se com a proteção e preservação é benéfica na manutenção da
variabilidade da PA, com conseqüente proteção de lesão de órgão alvo (SU & MIAO,
2001).
Uma das formas que vem sendo muito utilizada para avaliar o controle autonômico
é o estudo da variabilidade da freqüência cardíaca (FC). Até 20 anos atrás, variações do
ritmo cardíaco (ou da PA) eram completamente ignoradas pelos fisiologistas e
cardiologistas. A variabilidade natural de parâmetros cardiovasculares como PA e FC
reflete a interação de diversos fatores que, em sua maioria, envolvem uma influência do
SNA sobre o aparelho cardiovascular (JOAQUIM et al., 2005). Hoje se sabe que

11
irregularidades na variabilidade da FC e da PA significam algum tipo de anormalidade, e
que a diminuição da variabilidade da FC é um mau prognóstico (RIBEIRO & MORAES,
2005).
A avaliação da varibilidade da FC e da PA e de seus componentes também permite
a avaliação da sensibilidade espontânea dos pressorreceptores, que são mecanorreceptores
responsáveis pelo controle da PA através da atividade simpática e parassimpática em um
curto espaço de tempo (DE ANGELIS et al., 2004; IRIGOYEN et al., 2003). Neste
aspecto, vale destacar que recentemente, o controle reflexo da circulação comandado pelos
barorreceptores tem sido reconhecido também como um importante preditor de risco após
evento cardiovascular (TASK FORCE, 1996; LA ROVERE et al., 1998).
De fato, estudos experimentais e clínicos vêm demonstrando que a disautonomia
(disfunções no SNA) está presente em uma série de patologias, tais como a hipertensão
arterial, a insuficiência cardíaca, o diabetes mellitus e outras alterações metabólicas (DE
ANGELIS et al., 2004; IRIGOYEN & KRIEGER, 1998; ZANCHETTI & MANCIA, 1991;
LA ROVERE et al., 1998; EWING et al., 1980; VINIK et al., 2003; DE ANGELIS et al.,
2004, FARAH et al., 2007). Neste contexto, estudos vêm demonstrando de forma
consistente que a hiperatividade simpática aumenta o risco cardiovascular, ao passo que
uma função vagal preservada ou aumentada tem sido considerada um fator de proteção
cardiovascular (TASK FORCE, 1996; KLEIGER et al., 1987). Mais recentemente, o
estudo ATRAMI (Autonomic Tone and Reflexes After Myocardial Infarction) forneceu
evidências clínicas do valor prognóstico da disfunção autonômica cardiovascular na
mortalidade cardíaca pós-infarto do miocárdio (LA ROVERE et al.,1998). Dessa forma,
intervenções no sentido de detectar, prevenir e/ou atenuar a disfunção autonômica

12
cardiovascular tem sido vistas como novas e/ou importantes estratégias no manejo das
doenças cardiovasculares (LA ROVERE et al., 2002). Todavia, os estudos que verificaram
disfunção autonômica foram realizados em sua grande maioria em indivíduos do sexo
masculino. Em contrapartida, é consenso na literatura que machos e fêmeas apresentam
diferenças fisiológicas importantes que merecem ser mais bem estudadas.
1.4. Doença Cardiovascular e Estresse Oxidativo
Os mecanismos pelos quais o estrogênio reduz o desenvolvimento de doenças
cardiovasculares são multifatoriais e incluem alterações no metabolismo lipídico, ações nos
componentes das paredes vasculares (endotélio, músculo liso e células adventiciais), e em
elementos do sangue (plaquetas e leucócitos), bem como alterações no controle do
autonômico cardiovascular (MILLER, 1999). De uma forma geral, a participação do
estrogênio como cardioprotetor, agindo como um antioxidante, atribui-se a sua estrutura
fenólica que pode agir como neutralizador dos radicais livres (NIKKI, 1990). O
grupamento hidrofenólico do estrogênio doa o hidrogênio para uma molécula instável,
tornando-se um radical menos lesivo; assim o anel da molécula do estrogênio se reorganiza
e se estabiliza, retirando do meio um radical livre. Dessa forma, o estrogênio tem ação
antioxidante importante agindo como “scavenger” de radicais livres, ativando enzimas
antioxidantes, aumentando a expressão da enzima superóxido dismutase (SOD),
aumentando a síntese de mediadores vasoativos derivados do endotélio e diminuindo a
expressão de enzimas pró-oxidantes (NADPH oxidase) (NIKI, 1992; KIM et al., 1996).
Considerando o papel do estrogênio como uma molécula que pode neutralizar as
espécies reativas de oxigênio (ERO), vale lembra que as ERO são substâncias que

13
apresentam alta reatividade para outras biomoléculas, principalmente lipídios e proteínas
das membranas celulares e para o DNA. Todas as células aeróbias possuem mecanismos
para combater os efeitos agressivos das ERO, que incluem as enzimas SOD citada
anteriormente, a catalase (CAT) e a glutationa peroxidase (GPx) e o sistema não
enzimático (DORMANDY, 1978; SIES, 1986). As ERO promovem estresse oxidativo
quando as defesas antioxidantes da célula são insuficientes para deter a produção pró-
oxidante (NORDMANN, 1994).
Observa-se na figura 1 as principais fontes celulares de ERO e sua metabolização.
Figura 1: Esquema geral das principais fontes celulares de produção e metabolização das ERO. UQH• + ubisemiquinona; GSSG = glutationa oxidada; GSH = glutationa reduzida; DH2 e D = sistemas redutores de NADP não-específicos; SOD = superóxido dismutase; GPx = glutationa peroxidase; CAT = catalase; B e BH2 = doadores de hidrogênio (Adaptado de BOVERIS & CHANCE, 1973).

14
Um número crescente de trabalhos tem demonstrado o papel fundamental do estresse
oxidativo na patogênese das doenças cardiovasculares (CAI & HARRISON, 2000). A
geração de ERO em maiores quantidades causa uma diminuição do (NO) biodisponível, o
que induz prejuízo na função endotelial e cardíaca (PANZA et al., 1990). O NO pode ser
destruído pelo radical superóxido e protegido por mecanismos antioxidantes como a
enzima superóxido dismutase (SOD) (RUBANYI & VANHOUTE, 1986; GRYGLEWSKI
et al., 1986). Contudo, pacientes com hipertensão, hipercolesterolemia, diabetes, fumantes,
e até mesmo no processo fisiológico do envelhecimento demonstraram relação na
disfunção da vasodilatação e do endotélio, por sua vez com o estresse oxidativo (BERRY et
al., 2001; CAI & HARRISON, 2000; ORIEN et al., 1999).
É interessante notar que em trabalhos do nosso grupo, a redução do estresse
oxidativo e o aumento das enzimas antioxidantes têm sido correlacionados com melhora
em parâmetros cardiovasculares e autonômicos, como a sensibilidade dos
pressorreceptores, em ratos machos velhos, com insuficiência cardíaca ou hipertensão e em
ratas fêmeas submetidas à privação dos hormônios ovarianos (DE ANGELIS et al., 1997;
RABELO et al., 2001; IRIGOYEN et al., 2005; BERTAGNOLI et al., 2006). Dessa forma,
o óxido nítríco, e conseqüentemente a redução de sua biodisponibilidade em situações de
aumento de estresse oxidativo, tem sido destacado como um potencial mecanismo que pode
estar envolvido na disfunção dos pressorreceptores, uma vez que é um regulador central da
função autonômica e da sensibilidade dos pressorreceptores (CHOWDHARY et al., 2000;
DE ANGELIS et al., 1999), é um fator que pode alterar a distensibilidade arterial (KASSIS
& AMTORP, 1987), bem como o efeito direto do NO ou do ânion superóxido pode

15
modificar a descarga dos pressorreceptores (LI et al., 1996; SHULTZ & USTINOVA,
1998).
Nosso grupo vem estudando há alguns anos os efeitos do treinamento físico
dinâmico aeróbio em modelos animais, ratos e camundongos machos, como uma
abordagem não farmacológica capaz de induzir alterações favoráveis na regulação
autonômica cardiovascular, bem como tem buscado os possíveis mecanismos, entre eles o
estresse oxidativo, envolvidos nos benefícios dessa abordagem (DE ANGELIS et al., 1997,
1999, 2000, 2004; PARENTE COSTA et al., 2004, BERTAGNOLLI et al., 2006,
HARTHMANN et al., 2007). Todavia, deve-se considerar que a maior parte dos trabalhos
publicados na literatura com relação aos efeitos hemodinâmicos, autonômicos e no estresse
oxidativo em animais e humanos foi realizado em amostras do sexo masculino, ficando a
dúvida se o sexo feminino, em situações fisiológicas, como o climatério, e fisiopatológicas,
como a SM, se adaptaria de forma semelhante. Além disto, é necessária a busca de
alternativas terapêuticas para atenuar e ou tratar as disfunções decorrentes da privação dos
hormônios ovarianos. Neste aspecto, vários estudos têm demonstrado o papel benéfico do
treinamento físico em situações fisiológicas e patológicas.
1.5. Doença Cardiovascular e Treinamento Físico
Estudos epidemiológicos têm demonstrado que o sedentarismo é um grande fator de
risco para o desenvolvimento da hipertensão e do diabetes não insulino dependente
(NIDDM) (HARDMANN, 1996). Os efeitos benéficos do exercício físico têm sido
demonstrados na prevenção e tratamento da HAS, na resistência à insulina, no DM, na
dislipidemia e na obesidade, no qual são fatores predominantes na síndrome metabólica

16
(NCEP, 2001; TUOMILEHTO et al., 2001; WHELTON et al., 2002; HENRISSEN, 2002;
ROSS et al., 2000; TORJESEN et al., 1997; HOUMARD et al., 2004; KNOWLER et al.,
2002; PAN et al., 1997; BACON et al., 2004; HAGBERG et al., 2000; CARROL &
DUDFIELD, 2004; GUIMARÃES & CIOLAC, 2004).
A prática regular de atividade física é considerada um tratamento não-
farmacológico para o manejo e/ou prevenção de diversas patologias. Nesse contexto, sem
dúvida o estilo de vida adotado pelas sociedades modernas pode colaborar para esses altos
índices de sedentarismo, o qual pode contribuir de forma importante para o
desenvolvimento de doenças crônicas (FRANCISCHI, 2000).
Gregoire e colaboradores (1996) observaram uma maior variabilidade da FC, um
achado associado à menor risco cardiovascular, em mulheres jovens (treinadas e não-
treinadas) e de meia-idade (treinadas e não-treinadas) quando comparadas a homens. As
jovens não-treinadas e as mulheres de meia-idade (treinadas e não-treinadas) tiveram uma
atividade simpática de repouso significativamente menor em relação aos homens nas idades
correspondentes. Além disso, as jovens não-treinadas e as de meia-idade treinadas tiveram
uma maior atividade parassimpática de repouso do que os seus correspondentes do sexo
masculino. Esses dados em conjunto, mesmo indicando melhor modulação autonômica
cardiovascular no sexo feminino do que no masculino, não permitem, entretanto, concluir
sobre os efeitos do treinamento físico na proteção cardiovascular da mulher, pois outros
fatores, como os genéticos poderiam modular esses resultados.
Considerando que a hipertensão é um achado comum em pacientes com SM, é
importante destacar a redução dos níveis pressóricos pós-treinamento observada de forma
consistente tanto em homens como em mulheres hipertensos, pré ou pós-menopausa

17
(WHELTON et al., 2002; KELLEY, 1999; SEALS et al., 1997). Um dos fatores que pode
contribuir para reduzir a PA é reversão da atenuação da disfunção barorreflexa após
treinamento físico dinâmico em indivíduos hipertensos (BRUM et al., 2000;
O´SULLIVAN & BELL, 2000).
Estudos em mulheres no climatério vêm demonstrando que o treinamento físico
também induz melhora no perfil lipídico principalmente em presença de sobrepeso ou
dislipidemia (ASIKAINEN et al., 2004). Sugawara e colaboradores (2004), em estudo em
mulheres após a menopausa, verificaram que o treinamento físico de baixa ou de moderada
intensidade melhorou a complacência arterial. Sabe-se que a redução na complacência
arterial resulta no aumento progressivo da PAS relacionado ao envelhecimento,
aumentando também a função ventricular esquerda, diminuindo a pressão diastólica, e com
isso alterando a perfusão coronariana.
O treinamento físico pode provocar alterações cardiovasculares e autonômicas
importantes tais como bradicardia de repouso (NEGRÃO et al., 1992; DE ANGELIS et al.,
1997, 1999, 2004; KATONA et al., 1982; FRICK, 1967), redução da PA em ratos
espontaneamente hipertensos (SHR) (SILVA et al., 1997; BERTAGNOLLI et al., 2006) e
melhora da sensibilidade dos pressorreceptores em sujeitos normotensos (MC`DONALD et
al., 1993; BARNEY et al., 1988; DE ANGELIS et al., 2004; NEGRÃO et al, 1992;
BEDFORD & TIPTON, 1987) e em ratos SHR e diabéticos (SILVA et al., 1997;
BERTAGNOLLI et al., 2006; HARTHMANN et al., 2007). Além disto, estudos
demonstraram adaptações das enzimas antioxidantes e redução do estresse oxidativo em
resposta ao treinamento físico (JI & FU, 1992; MARGARATIS et al., 1997; VENDITTI E

18
DI MEO, 1997; DE ANGELIS et al., 1997; IRIGOYEN et al., 2005; BERTAGNOLLI et
al., 2006).
Apesar dos vários trabalhos evidenciando os benefícios do treinamento físico, a
grande maioria desses estudos foi realizada em amostras do sexo masculino. Em estudos
mais recentes tem sido demonstrado que treinamento físico aeróbio pode induzir melhora
nos perfis metabólico e lipídico, reduzir a inflamação e as moléculas de adesão (WEGGE et
al., 2004) e aumentar a variabilidade da FC (JURCA et al., 2004) em mulheres
menopausadas (ASIKAINEN et al., 2004), bem como melhorar a resposta da insulina
estimulada pelo teste de tolerância a glicose em ratas ooforectomizadas (LATOUR et al.,
2001). Um trabalho recente de nosso laboratório evidenciou que o treinamento físico
aeróbio em um modelo experimental de menopausa em ratas induziu aumento da
capacidade aeróbia, redução do peso corporal, da PA e da FC de repouso e melhora na
sensibilidade dos pressorreceptores associada à redução no estresse oxidativo e ao aumento
nas defesas antioxidantes no tecido cardíaco (IRIGOYEN et al., 2005).
Por fim, vale destacar que a taxa de mortalidade devido a doenças cardiovasculares
aumentou de 10 para 25% nos anos 60 e 70 em mulheres (CASTANHO et al., 2001) é
eminente a necessidade da busca de alternativas terapêuticas para a prevenção e o
tratamento das doenças cardiovasculares e metabólicas na mulher. Cabe lembrar que
atualmente os efeitos de proteção cardiovascular através da terapia hormonal são altamente
controversos (WRITING GROUP FOR THE WOMEN`S INITIATIVE
INVESTIGATORS, 1996). Em contrapartida, os benefícios obtidos através da atividade
física regular têm cada vez mais reforçando a importância desta abordagem na prevenção e
no tratamento das doenças (PEDERSEN & SALTIN, 2006; MOSCA et al., 2007).

19
Considerando o importante papel da disautonomia como fator de risco de doença
cardiovascular, bem como o possível envolvimento do estresse oxidativo nesta disfunção,
intervenções que reduzam o estresse oxidativo e/ou melhorem a função autonômica tem
sido vistas como potenciais estratégias no manejo do risco cardiovascular. Neste projeto
avaliamos o efeito da sobrecarga de frutose na água de beber (um modelo de disfunção
cardiovascular e metabólico) tanto em ratas fêmeas Wistar como em fêmeas
espontaneamente hipertensas (SHR), ambas ooforectomizadas, na tentativa de
alcançarmos um modelo de SM, e, assim, pudemos avaliar os efeitos do treinamento
físico em diferentes fatores de risco muitas vezes observados de forma concomitante em
mulheres no climatério.

20
2. OBJETIVOS
2.1. Objetivo Geral
O objetivo do presente estudo foi verificar os efeitos do treinamento físico em
parâmetros cardiovasculares e metabólicos, na modulação autonômica cardiovascular e no
estresse oxidativo em ratas fêmeas ooforectomizadas normotensas e hipertensas submetidas
à sobrecarga de frutose.
2.2. Objetivos Específicos
Os objetivos específicos do presente projeto foram avaliar os efeitos do treinamento
físico em ratas fêmeas normotensas e hipertensas submetidas à privação dos hormônios
ovarianos e a ingestão crônica de frutose na água de beber nos seguintes parâmetros:
• peso corporal;
• glicose e nos triglicerídeos sanguíneos;
• resistência à insulina;
• pressão arterial e na freqüência cardíaca;
• variabilidade da frequência cardíaca (RMSSD, VAR-IP, %BF e %AF);
• variabilidade da pressão arterial (VAR-PAS, BF);
• sensibilidade barroreflexa espontânea (Índice alfa);
• estresse oxidativo (QL e GSH/GSSG);
• atividade de enzimas antioxidantes (CAT, SOD e GPx).

21
3. MATERIAIS E MÉTODOS
O presente estudo foi aprovado pelo Comitê de Ética em Pesquisa da Universidade
São Judas Tadeu (COEP-USJT) de acordo com os seguintes protocolos: 024/2005,
064/2006 e 01/2008.
3.1. Amostra
Foram utilizadas 20 ratas Wistar fêmeas e 20 ratas SHR (ratas espontaneamente
hipertensas) fêmeas, pesando entre 50-60g, provenientes do biotério da Universidade São
Judas Tadeu e do Biotério da Universidade Federal de São Paulo. Os animais foram
mantidos em gaiolas, contendo no máximo 4 animais em cada uma, em ambiente com
temperatura controlada (220 - 240C) e com luz controlada em ciclo de 12 horas (claro -
escuro, invertido). Os animais foram divididos em 4 grupos experimentais com 10 ratas em
cada grupo:
Grupo I - Frutose ooforectomizada sedentária (FOS) (n=10): ração industrial
para ratos + solução de água com frutose (10%), durante um período de 18 semanas.
Grupo II - Frutose ooforectomizada treinada (FOT) (n=10): ração industrial +
solução de água com frutose (10%), durante um período de 18 semanas, sendo que foram
submetidas à treinamento físico em esteira ergométrica rolante (Imbramed TK-01) a partir
da 9ª semana de protocolo.
Grupo III - Frutose hipertensa ooforectomizada sedentária (FOHS) (n=10):
ração industrial + solução de água com frutose (10%) durante 18 semanas.
Grupo IV - Frutose hipertensa ooforectomizada treinada (FOHT) (n=10): ração
industrial + solução de água com frutose (10%) durante 18 semanas, sendo que foram

22
submetidas à treinamento físico em esteira ergométrica rolante (Imbramed TK-01) a partir
da 9ª semana de protocolo.
3.2. Seqüência Experimental
Os grupos experimentais seguiram a seqüência experimental ilustrada abaixo:
Figura 2: Seqüência Experimental do protocolo.
3.3.PROCEDIMENTOS
3.3.1. Indução do Modelo Experimental de Síndrome Metabólica
Todos os grupos foram submetidos a ingestão de frutose na água de beber (D-
frutose, 100g/L) (SUZUKI et al., 1997). O tratamento foi iniciado após o desmame e
seguiu até o final do protocolo (Figura 3).
Ava
liaçõ
es B
ioqu
ímic
as
(FOT e FOHT)
(FOS e FOHS)
Tratamento com
Frutose
(FOS e FOHS)
Ava
liaçõ
es B
ioqu
ímic
as
(FOT e FOHT)
(FOS e FOHS)
Tratamento com
Frutose
(FOS e FOHS)

23
Figura 3: Tratamento de D-frutose na água de beber.
3.3.2. Ooforectomia Bilateral
As ratas foram anestesiadas com cloridrato de cetamina (Ketalar) e cloridrato de
xilazina (Rompum) e colocadas em decúbito dorsal para que se realize uma pequena
incisão (1cm) em paralelo com a linha do corpo na pele e na musculatura no terço inferior
na região abdominal. Os ovários foram localizados e foi realizada a ligadura dos ovidutos,
incluindo os vasos sangüíneos. Os ovidutos foram seccionados e os ovários removidos. A
musculatura e a pele foram suturadas e uma dose de antibiótico foi administrada
(Benzetacil, 40 000 U/Kg, i.m) (LATOUR et al., 2001; IRIGOYEN et al., 2005) (Figura 4).
Figura 4: Etapas de realização da ooforectomia bilateral em ratas.
24
3.3.3. Teste de Esforço Máximo
O teste de esforço constituiu em um protocolo escalonado com incrementos de
velocidade de 0,3 km/h a cada 3 minutos, até que seja atingida a velocidade máxima
suportada pelos animais. O critério utilizado para a determinação da exaustão do animal e
interrupção do teste foi o momento em que o rato não foi mais capaz de correr mediante o
incremento de velocidade da esteira (BROOKS & WHITE, 1978). Vale ressaltar que
recentemente demonstramos relação entre velocidade do atingida no teste de esforço e a
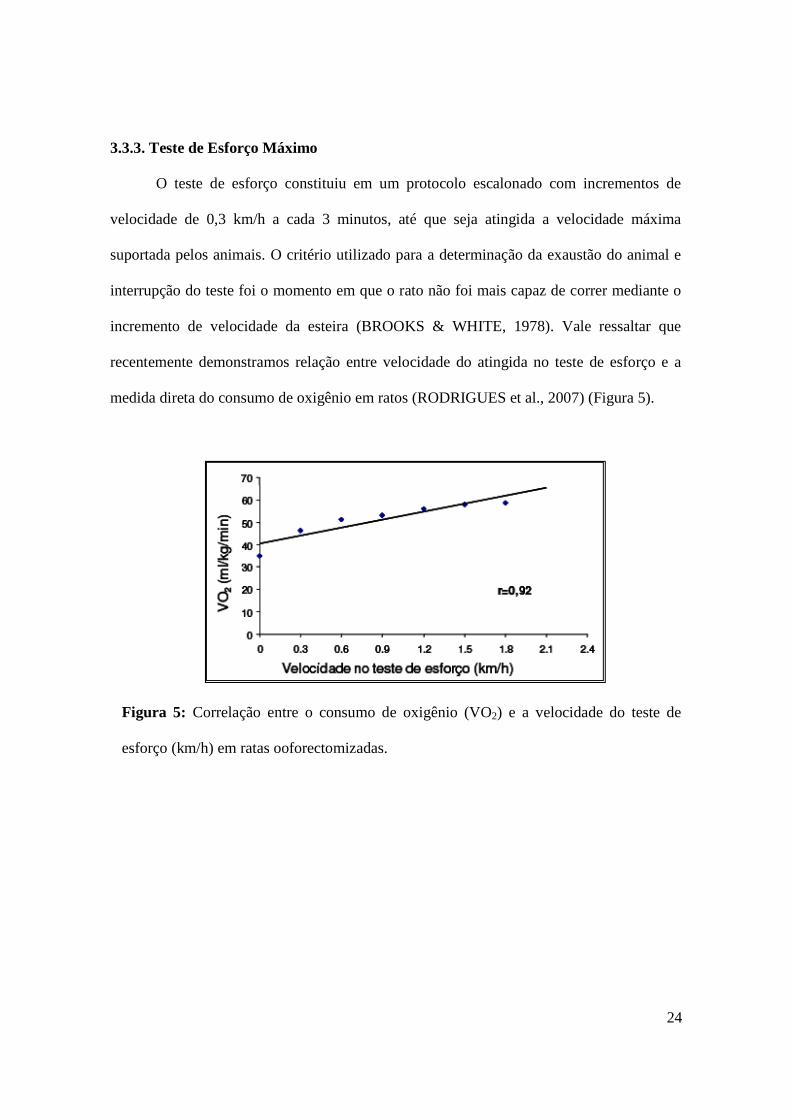
medida direta do consumo de oxigênio em ratos (RODRIGUES et al., 2007) (Figura 5).
Figura 5: Correlação entre o consumo de oxigênio (VO2) e a velocidade do teste de
esforço (km/h) em ratas ooforectomizadas.

25
3.3.4. Treinamento Físico
O grupo de ratas treinadas foi submetido a um protocolo de treinamento físico (40-
60% da velocidade máxima alcançada no teste de esforço) em esteira ergométrica com
velocidade e carga progressiva durante 8 semanas conforme descrito resumidamente abaixo
(IRIGOYEN et al., 2005, DE ANGELIS et al., 1997, 1999) (Figura 6, Quadro 1 e Quadro
2).
Figura 6: Fotografia de ratos submetidos ao protocolo de treinamento físico em esteira
ergométrica na USJT.
26
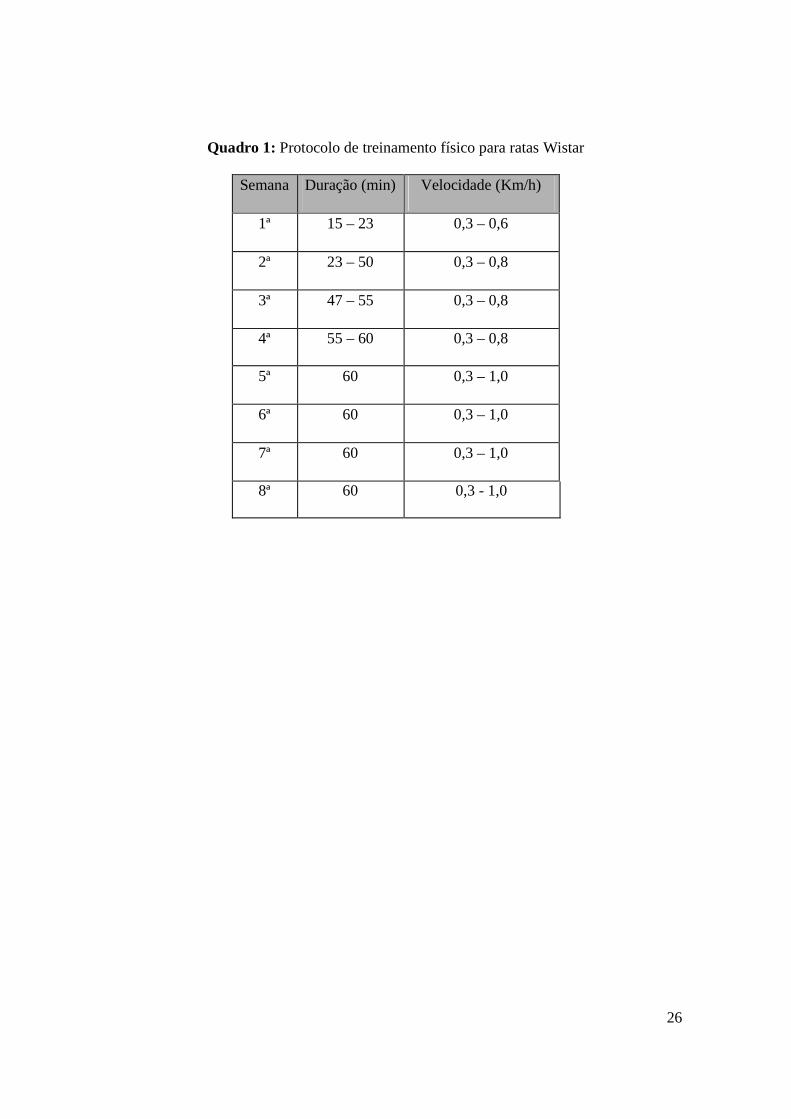
Quadro 1: Protocolo de treinamento físico para ratas Wistar
Semana Duração (min) Velocidade (Km/h)
1ª 15 – 23 0,3 – 0,6
2ª 23 – 50 0,3 – 0,8
3ª 47 – 55 0,3 – 0,8
4ª 55 – 60 0,3 – 0,8
5ª 60 0,3 – 1,0
6ª 60 0,3 – 1,0
7ª 60 0,3 – 1,0
8ª 60 0,3 - 1,0

27
Quadro 2: Protocolo de treinamento físico para ratas SHR
Semana Duração (min) Velocidade (Km/h)
1ª 15 – 23 0,3 – 0,9
2ª 23 – 50 0,3 – 1,1
3ª 47 – 55 0,3 – 1,1
4ª 55 – 60 0,3 – 1,2
5ª 60 0,3 – 1,4
6ª 60 0,3 – 1,5
7ª 60 0,3 – 1,5
8ª 60 0,3 - 1,5
3.3.5. Medida da Glicemia e dos Triglicerídeos Sangüíneos
Ao final do protocolo (18 semanas) os animais foram submetidos a jejum de 4 horas
e, após isto foi retirada uma gota de sangue da cauda para análise da glicose plasmática
pelo glicosímetro (Accucheck, Roche) e uma gota para medida dos triglicerídeos
sanguíneos do aparelho Accutrend GTC, Roche (Figura 7).

28
Figura 7: Aparelhos que foram utilizados para análises das concentrações sangüíneas de
glicose e triglicerídeos.
3.3.6. Canulação
Após 18 semanas de protocolo, as ratas foram anestesiadas (i.p.) com cloridrato de
cetamina (50mg/Kg, Ketalar, Parke-Davis) e cloridrato de xilazina (12mg/Kg, Rompum,
Bayer) e colocadas em decúbito dorsal para que se realize uma pequena incisão na região
do pescoço para implantação de uma cânula na artéria carótida em direção ao ventrículo
esquerdo, para registro direto da PA e na veia jugular para administração das drogas. Após
a correta e firme implantação das cânulas na artéria carótida e veia jugular, as extremidades
mais calibrosas das cânulas foram passadas subcutâneamente, exteriorizadas no dorso da
região cervical e fixadas com fio de algodão na pele. As cânulas foram confeccionadas com
tubos de Policloreto de Vinila (Abbott) equivalente ao polietileno PE10 e PE50. Estes
foram soldados por aquecimento e logo após, as cânulas foram preenchidas com solução
fisiológica e mantidas ocluídas com pinos de aço inoxidável (MAEDA et al., 1995; DE
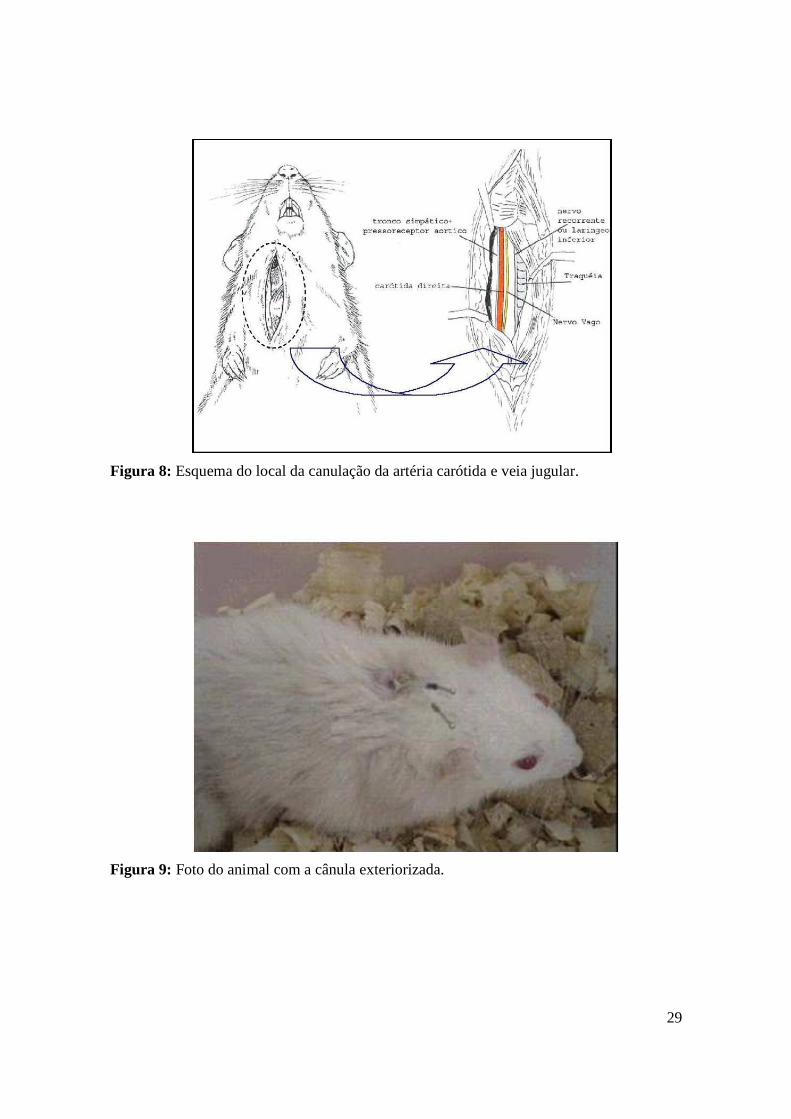
ANGELIS et al.,1999, 2000) (Figura 8 e Figura 9).
29
Figura 8: Esquema do local da canulação da artéria carótida e veia jugular.
Figura 9: Foto do animal com a cânula exteriorizada.

30
3.3.7. Registro de Pressão Arterial
No dia seguinte à canulação, com o animal acordado, a cânula arterial foi conectada
a uma extensão de 20 cm (PE-50), permitindo livre movimentação do animal pela caixa,
durante todo o período do experimento. Esta extensão foi conectada a um transdutor
eletromagnético (Blood Pressure XDCR, Kent© Scientific, Litchfield, CT, EUA)
que, por sua vez, esteve conectado a um pré-amplificador (STEMTECH BPMT-2,
Quintron Instrument© Inc, Milwaukee, EUA). Sinais de PA foram gravados durante um
período de 30 minutos em um microcomputador equipado com um sistema de aquisição de
dados (CODAS, 1Kz, DATAQ Instruments, Akron, OH, EUA), permitindo análise dos
pulsos de pressão, batimento-a-batimento, com uma freqüência de amostragem de 2000 Hz
por canal, para estudo dos valores de PAS, PA diastólica (PAD), PA média (PAM) e FC.
Os valores de FC foram derivados do sinal pulsátil da PA (Figura 10).
Figura 10: Sistema de registro de pressão arterial e conexão entre a cânula e o transdutor
eletromagnético.

31
3.3.8. Avaliação da Modulação Autonômica Cardiovascular
3.3.8.1. Análise da Variabilidade da Pressão Arterial Sistólica A partir do registro basal dos animais acordados, foi possível utilizar a ferramenta
de análise tempo-freqüência da variabilidade da PAS. Os parâmetros para análise no
domínio do tempo consistiram em calcular os valores médios da PAS, sendo a sua
variabilidade quantificada pela variância da PAS.
A análise no domínio da freqüência consistiu-se da decomposição do sistograma
pela Transformada Rápida de Fourier (FFT). Após esse remodelamento matemático, foram
obtidas as potências absolutas da banda de baixa freqüência (BF: 0,20-0,75 Hz) (SOARES
et al., 2004).
3.3.8.2 Análise da Variabilidade do Intervalo de Pulso A variabilidade do intervalo de pulso foi obtida pela análise do tacograma a partir
do registro da PAS, no qual a freqüência dos batimentos foi determinada pelo intervalo
entre dois picos sistólicos. Para essa análise foram utilizados registros estáveis, de no mínimo 5
minutos e com freqüência de amostragem de 2.000 Hz. Também dois componentes foram
obtidos na análise espectral: muito baixa freqüência (MBF: banda de muito baixa freqüência),
baixa freqüência (BF: 0,20-0,75 Hz) e alta freqüência (AF: 0,75-3,0 Hz). O componente BF foi
usado como um índice da atividade simpática. O componente AF foi usado como um índice da
atividade parassimpática. Além disso, avaliou-se também as variáveis % BF (banda de baixa
freqüência do intervalo de pulso), %AF (banda de alta freqüência do intervalo de pulso),
RMSSD (raiz quadrada da média dos quadrados das diferenças entre os intervalos intervalos R-
R normais sucessivos) e VAR-IP (variância do intervalo de pulso) (ISHISE & ASANOI et al.,
1998).

32
3.3.8.3. Análise da Sensibilidade Barorreflexa Espontânea (Índice Alfa)
Valores de batimento a batimento da pressão arterial sistólica e intervalo RR foram usados
para estimar a sensibilidade barorreflexa pelo método de análise espectral, utilizando o índice alfa
da banda BF (0,20–0,75hz). A coerência entre a variabilidade dos sinais de intervalos RR e pressão
arterial sistólica foi realizada pelo método de análise espectral cruzada. O índice alfa foi calculado
somente quando a magnitude da coerência ao quadrado entre os sinais de RR e pressão arterial
sistólica excederam 0,5 (amplitude 0 a 1) na banda BF depois do cálculo da coerência, o índice alfa
foi extraído da raiz quadrada da razão da variabilidade entre RR e PAS nas duas principais bandas
de BF.
3.3.9. Teste de Resistência à Insulina
No dia seguinte das avaliações hemodinâmicas os animais foram submetidos à
jejum de 2 horas, foram anestesiados com pentobarbital sódico (40 mg/kg), e receberam
uma injeção endovenosa de insulina (0,75 U/kg peso corporal). A glicose plasmática foi
medida a partir de amostras de sangue obtidas da veia caudal utilizando-se de um
glicosímetro (Accucheck, Roche) nos tempos 0, 4, 8, 12 e 16 min após a injeção de
insulina. Os valores de glicemia dos minutos 4 a 16 foram usados para calcular a constante
de queda da glicose plasmática (Kitt) de acordo com a descrição de Bonora et al. (1989).
3.3.10. Eutanásia dos Animais
No dia seguinte ao término das avaliações hemodinâmicas, os animais de todos os
grupos foram submetidos a eutanásia por decapitação e os tecidos foram pesados e
congelados para avaliações do estresse oxidativo e das concentrações dos nitritos e nitratos.

33
3.3.11. Preparação dos Tecidos
Após as avaliações citadas acima, os animais foram pesados e mortos através de
decapitação. O coração foi coletado e foi homogeneizado durante 30 segundos em um
homogeneizador Ultra-Turrax, com KCl 1,15% e fluoreto de fenil metil sulfonila (PMSF),
na concentração de 100mmol/L em isopropanol e na quantidade de 10µL/mL de KCl
adicionado. Em seguida, os homogeneizados foram centrifugados por 10 minutos a
3000rpm, em centrífuga refrigerada entre 0 e 4°C (Eppendorf, 5804-R), e o sobrenadante
foi congelado em freezer a -70°C para as dosagens (LLESUY et al., 1985).
3.3.12. Dosagem de Proteínas
As proteínas foram quantificadas pelo método descrito por Lowry e
colaboradores, que utiliza como padrão uma solução de albumina bovina na concentração
de 1mg/mL (LOWRY et al., 1951). Para a realização das dosagens foram utilizados os
seguintes reagentes:
Reativo de Folin Ciocaulteau diluído em água destilada na proporção de 1:3.
Reativo C, que é composto por 50mL do reativo A, 0,5mL do reativo B1 e 0,5mL
do reativo B2, cujos reativos A, B1 e B2 são respectivamente:
• NaHCO3 (bicarbonato de sódio) 2% em NaOH (hidróxido de sódio) 0,1N.
• CuSO4.5H2O (sulfato de cobre) 1%.
• KNaC4H4O6.4H2O (tartarato de sódio e potássio) 2%.

34
Em tubos de ensaio, foram adicionados 20µL de amostra (homogeneizado de
tecido) em 0,78mL de água destilada e 2mL de reativo C preparado a fresco, aguardando-se
10 minutos. Depois, adicionava-se 0,2mL do reativo de Folin Ciocaulteau, aguardando-se
mais 30 minutos. Após, a solução adquiria uma coloração azulada que foi medida em
espectrofotômetro, marca Varian modelo Cary, a 625 nm.
O cálculo foi feito utilizando-se um fator de correção médio calculado a partir da
curva de calibração construída utilizando-se a solução padrão de albumina bovina. Os
resultados foram expressos em mg de proteína.
3.3.13. Estresse oxidativo e enzimas antioxidantes
Para medidas de estresse oxidativo e enzimas antioxidantes foi utilizado um “n” de
7 animais por grupo.
3.3.13.1. Medida de Lipoperoxidação (LPO): Quimiluminescência iniciada por t-
BOOH (QL)
Este método consistiu em adicionar um hidroperóxido orgânico de origem
sintética, o hidroperóxido de tert-butil (t-BOOH), a amostra do homogeneizado de tecido,
avaliando a capacidade de resposta produzida pela amostra. A realização deste tipo de teste
consistiu no fato de que os hidroperóxidos são espécies químicas bastante instáveis,
reagindo com lipídios por um mecanismo radicalar, o qual gera produtos que emitem luz
pela amostra em estudo. Este método é um dos mais sensíveis para a medida de LPO.

35
A QL foi medida em um contador beta (TriCrab 2800TR, PerkinElmer) com o
circuito de coincidência desconectado e foi utilizado o canal de trítio. As determinações
foram realizadas em câmara escura, em frascos de vidro mantidos na penumbra para evitar
a fosforescência ativada pela luz fluorescente. O meio de reação no qual foi realizado o
ensaio consistiu em 3,5 mL de uma solução tampão de fosfatos 20 mmol/L, contendo KCl
140 mmol/L (pH 7,4), à qual foi adicionado 0,5 mL de homogeneizado. Após esse
momento, foi realizada uma leitura inicial, considerada como a emissão basal de luz pelo
homogeneizado. O hidroperóxido de tert-butila foi usado na concentração de 400 mmol/L,
dos quais foram adicionados 30 µL no meio de reação para obter-se uma concentração final
de 3 mmol/L. Foi medida a emissão de luz e desta foi descontada a emissão basal do
homogeneizado para fins de cálculo (GONZALES FLECHA et al., 1991).
3.3.13.2. Razão GSH/GSSG
3.3.13.2.1. Glutationa Total
A glutationa total mediu a reação de óxido redução entre a glutationa reduzida
(GSH) e a glutationa oxidada (GSSG). O meio de reação no qual foi realizado o ensaio
consistiu em uma solução de tampão fosfato 300 mM (Na2HPO4.1H2O), e uma solução de
DTNB (acido ditionitrobenzóico). No momento do ensaio, foi agregado á 1 mL de tampão
fosfato, 100µL de DTNB, foi zerado o espectro (Biospectro) e após foi acrescentado 250
µL amostra (BEUTLER et al., 1963).

36
3.3.13.2.2. Glutationa Oxidada (GSSG)
Foi adicionado a amostra ácido perclórico, os quais foram colocados em ependorf e
centrifugados, posteriormente foi neutralizado com bicarbonato potásico (CO3HK).
Desta foi extraída uma alíquota de 60mL + 60 mL DNTB + 60 mL GSSH e 60 mL
NADPH. A leitura foi realizada em espectrofotômetro 414 nm. Uma vez neutralizada
com CO3HK foi adicionado 5 ml de 2 vinil piridina e deixará repousar por uma hora a
temperatura ambiente. Logo se repetirá o processo anterior. A diferença entre a
primeira e a segunda leitura foi a GSSG (TIETZE, 1969).
A GSH foi calculada a partir da subtração da glutationa total pela GSSG, e em
seguida foi calculada a razão GSH/GSSG.
3.3.13.3. Superóxido Dismutase (SOD)
A enzima SOD catalisa a reação de dois ânions superóxido, com a conseqüente
formação de peróxido de hidrogênio, que é menos reativo e pode ser degradado por outras
enzimas, como a catalase (CAT) e glutationa peroxidase (GPx). A velocidade da reação
catalisada pela SOD é 104 vezes maior do que a velocidade da dismutação espontânea em
pH fisiológico (SOUTHORN & POWIS, 1988; BOVERIS, et al. 1983).
22222 2 OOHHOO +⇒++ +−•−•
A técnica utilizada foi baseada na inibição da reação do radical superóxido com o
piragalol. Uma vez que não se consegue determinar a concentração da enzima nem sua

37
atividade em termos de substrato consumido por unidade de tempo, utilizou-se a
quantificação em unidades relativas. Uma unidade de SOD foi definida como a quantidade
de enzima que inibe em 50% a velocidade de oxidação do detector. A oxidação do
pirogalol levou à formação de um produto colorido, e foi detectado
espectrofotometricamente a 420 nm (Biospectro) durante 2 minutos. A atividade da SOD
foi determinada medindo-se a velocidade de formação do pirogalol oxidado. No meio de
reação, foram utilizados 20 µL de homogeneizado, 973 µL de tampão Tris-Fosfato a 50
mmol/L (pH 8,2), 8 µL de pirogalol a 24 mmol/L, 4 µL de CAT a 30 µmol/L. Esta curva
obtida foi utilizada como branco. Foi feito também uma curva padrão utilizando três
concentrações distintas de SOD (0,25U, 0,5U e 1U), através da qual foi obtida a equação da
reta para realização dos cálculos.
3.3.13.4. Catalase (CAT)
A enzima CAT é altamente específica e possui atividade apenas para peróxido de
hidrogênio, hidroperóxidos de metila e etila (WEBSTER & NUNN, 1998). No homem, a
CAT é uma hemoproteína e catalisa a decomposição do peróxido de hidrogênio à água e
oxigênio (BOVERIS & CHANCE, 1973).
A taxa de decomposição do peróxido de hidrogênio foi diretamente proporcional à
atividade da CAT. Desta forma, o consumo de H2O2 pode ser utilizado como uma medida
de atividade da enzima CAT. O ensaio consistiu em medir a diminuição da absorbância a
240nm, comprimento de onda onde houve a maior absorção pelo peróxido de hidrogênio,
utilizando-se cubetas de quartzo. Para a realização das medidas foi usada uma solução

38
tampão constituída de fosfatos a 50 mmol/L em pH 7,4. Foram adicionados 9µL deste
tampão e 10µL de amostra de tecido na cubeta do espectrofotômetro, sendo esta mistura
descontada contra um branco de tampão fosfato. A seguir foram adicionados 35µL de
peróxido de hidrogênio (0,3 mol/L) e foi monitorada a diminuição da absorbância no
espectrofotômetro (Biospectro) (BOVERIS & CHANCE, 1973).
22222 22 OOHOH +⇒
3.3.13.5. Glutationa Peroxidase (GPx)
A enzima GPx catalisa a reação de hidroperóxidos com a glutationa reduzida (GSH)
para formar glutationa oxidada (GSSG) e o produto da redução do hidroperóxido, por isso,
a sua atividade pode ser determinada medindo-se o consumo de NADPH, na presença de
GSH e GR na reação de redução acoplada à reação da GPx. A azida sódica (N3Na) é
adicionada para inibir a catalase (WENDEL, 1981).
OHGSSGROHGSHROOH GPx22 ++→+
A atividade da GPx foi medida em um espectrofotômetro (Biospectro). Foi
monitorada a diminuição de absorbância do NADPH a 340 nm. Na cubeta do
espectrofotômetro, foram adicionados 330 µL de tampão, 50 µL do homogeneizado
(amostra), 500 µL de NADPH, 10 µL de azida sódica, 50 µL de GSH e 10 µL de GR. Foi
registrada a absorbância por um período de aproximadamente 2 minutos, para obtenção da
linha de base. Após esse momento, foram adicionados 50 µL de hidroperóxido de tert-

39
butila, e a diminuição da absorbância devida ao consumo de NADPH foi monitorada por
mais 3 minutos (FLOHÉ & GUNZLER, 1984).
3.4. Análise Estatística
Para análise dos dados foi utilizado o software STATISTICS® 6.0 (Statsoft©). O
teste de Kolmogorv-Smirnov foi utilizado para verificar a normalidade das variáveis. O
teste de análise de variância (ANOVA) two way ou multifatorial seguido do teste
complementar de Student Newman-Keuls foi devidamente aplicado para análise dos dados.
Os resultados são apresentados como média ± erro padrão. Valores de p<0,05 foram
considerados estatisticamente significantes.
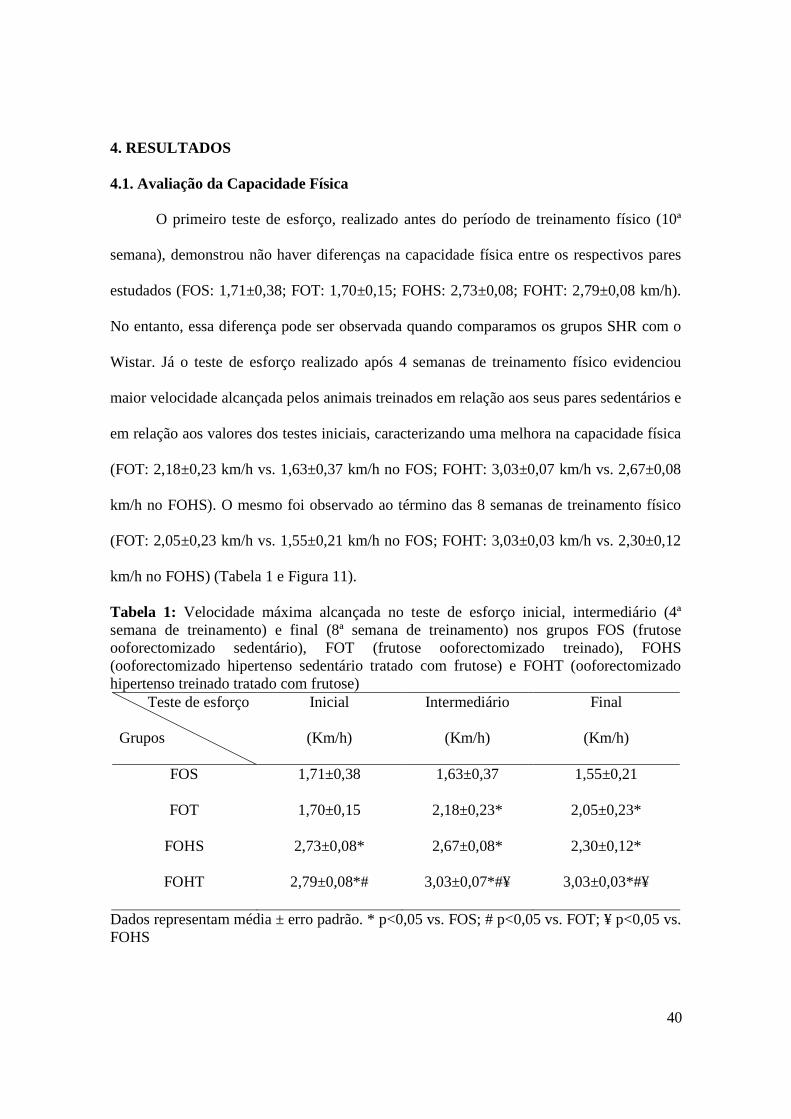
40
4. RESULTADOS 4.1. Avaliação da Capacidade Física
O primeiro teste de esforço, realizado antes do período de treinamento físico (10ª
semana), demonstrou não haver diferenças na capacidade física entre os respectivos pares
estudados (FOS: 1,71±0,38; FOT: 1,70±0,15; FOHS: 2,73±0,08; FOHT: 2,79±0,08 km/h).
No entanto, essa diferença pode ser observada quando comparamos os grupos SHR com o
Wistar. Já o teste de esforço realizado após 4 semanas de treinamento físico evidenciou
maior velocidade alcançada pelos animais treinados em relação aos seus pares sedentários e
em relação aos valores dos testes iniciais, caracterizando uma melhora na capacidade física
(FOT: 2,18±0,23 km/h vs. 1,63±0,37 km/h no FOS; FOHT: 3,03±0,07 km/h vs. 2,67±0,08
km/h no FOHS). O mesmo foi observado ao término das 8 semanas de treinamento físico
(FOT: 2,05±0,23 km/h vs. 1,55±0,21 km/h no FOS; FOHT: 3,03±0,03 km/h vs. 2,30±0,12
km/h no FOHS) (Tabela 1 e Figura 11).
Tabela 1: Velocidade máxima alcançada no teste de esforço inicial, intermediário (4ª semana de treinamento) e final (8ª semana de treinamento) nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose)
Teste de esforço
Grupos
Inicial
(Km/h)
Intermediário
(Km/h)
Final
(Km/h)
FOS 1,71±0,38 1,63±0,37 1,55±0,21
FOT 1,70±0,15 2,18±0,23* 2,05±0,23*
FOHS 2,73±0,08* 2,67±0,08* 2,30±0,12*
FOHT 2,79±0,08*# 3,03±0,07*#¥ 3,03±0,03*#¥
Dados representam média ± erro padrão. * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS
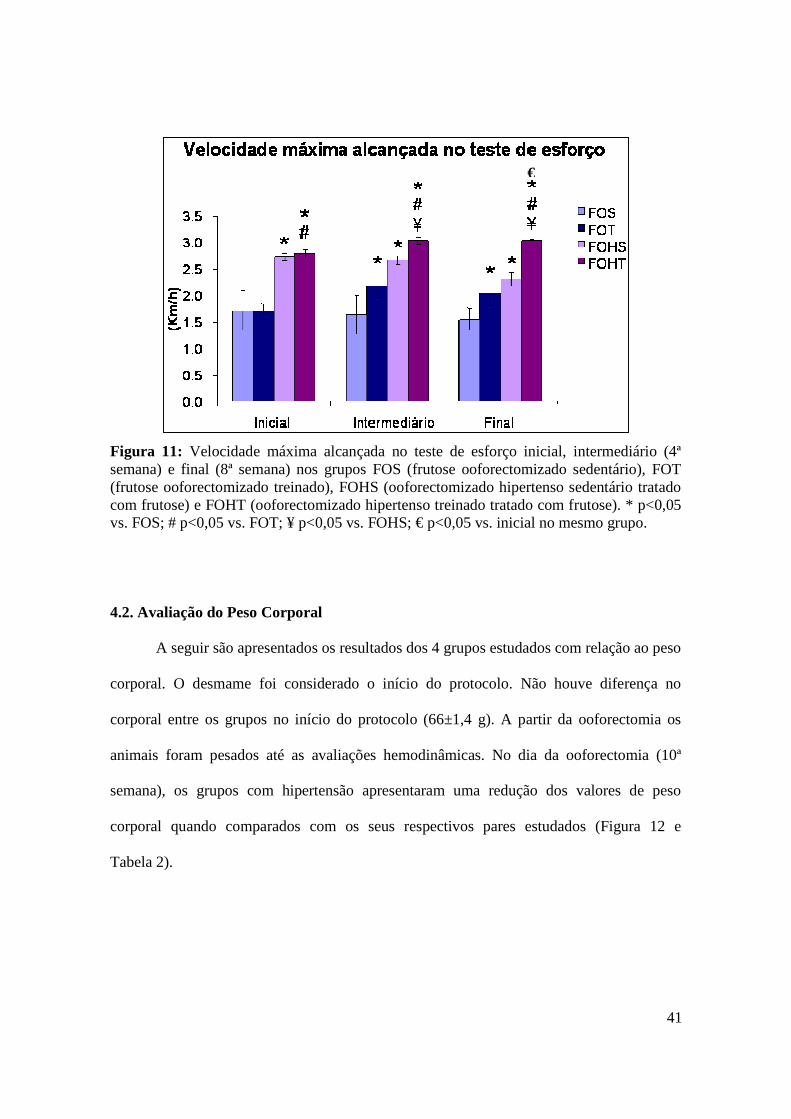
41
Figura 11: Velocidade máxima alcançada no teste de esforço inicial, intermediário (4ª semana) e final (8ª semana) nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS; € p<0,05 vs. inicial no mesmo grupo. 4.2. Avaliação do Peso Corporal A seguir são apresentados os resultados dos 4 grupos estudados com relação ao peso
corporal. O desmame foi considerado o início do protocolo. Não houve diferença no
corporal entre os grupos no início do protocolo (66±1,4 g). A partir da ooforectomia os
animais foram pesados até as avaliações hemodinâmicas. No dia da ooforectomia (10ª
semana), os grupos com hipertensão apresentaram uma redução dos valores de peso
corporal quando comparados com os seus respectivos pares estudados (Figura 12 e
Tabela 2).
€
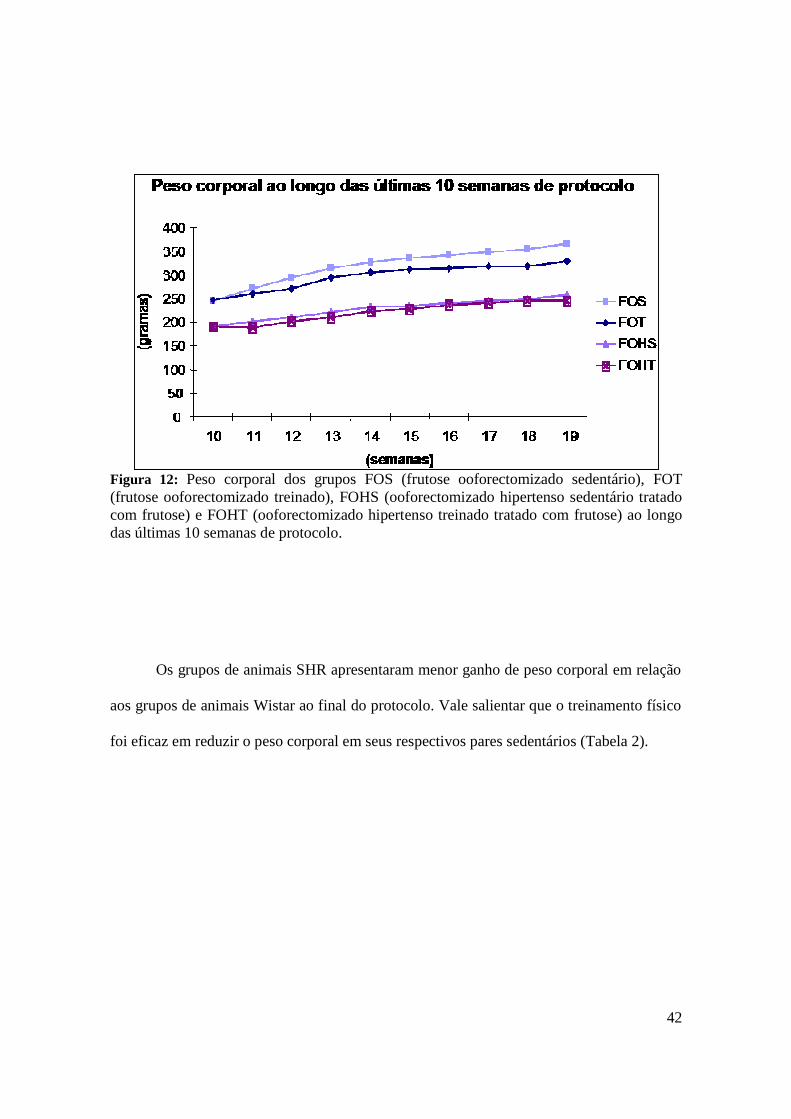
42
Figura 12: Peso corporal dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose) ao longo das últimas 10 semanas de protocolo.
Os grupos de animais SHR apresentaram menor ganho de peso corporal em relação
aos grupos de animais Wistar ao final do protocolo. Vale salientar que o treinamento físico
foi eficaz em reduzir o peso corporal em seus respectivos pares sedentários (Tabela 2).

43
Tabela 2: Peso corporal dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose) na 10ª semana (dia da ooforectomia) e no final do protocolo.
Dados representam média ± erro padrão.* p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS. 4.3. Avaliação do Tecido Adiposo
A Figura 13 apresenta os valores de tecido adiposo dos 4 grupos estudados ao final
do protocolo. Pode-se observar que o treinamento físico reduziu o tecido adiposo no grupo
FOT (5,00±0,60g) em comparação ao grupo FOS (9,93±0,70g). Interessantemente, os
grupos hipertensos (FOHS: 5,25±0.39 e FOHT: 4,32±0,54g).apresentaram menor
quantidade de tecido adiposo quando comparado ao grupo FOS.
Peso corporal
Grupos
10ª semana
(gramas)
Final
(gramas)
FOS 246±8,9 365±10,9
FOT 248±4,9 329±15,7*
FOHS 192±11,3* 257±9,6*
FOHT 191±8,9*# 245±10,8*#¥
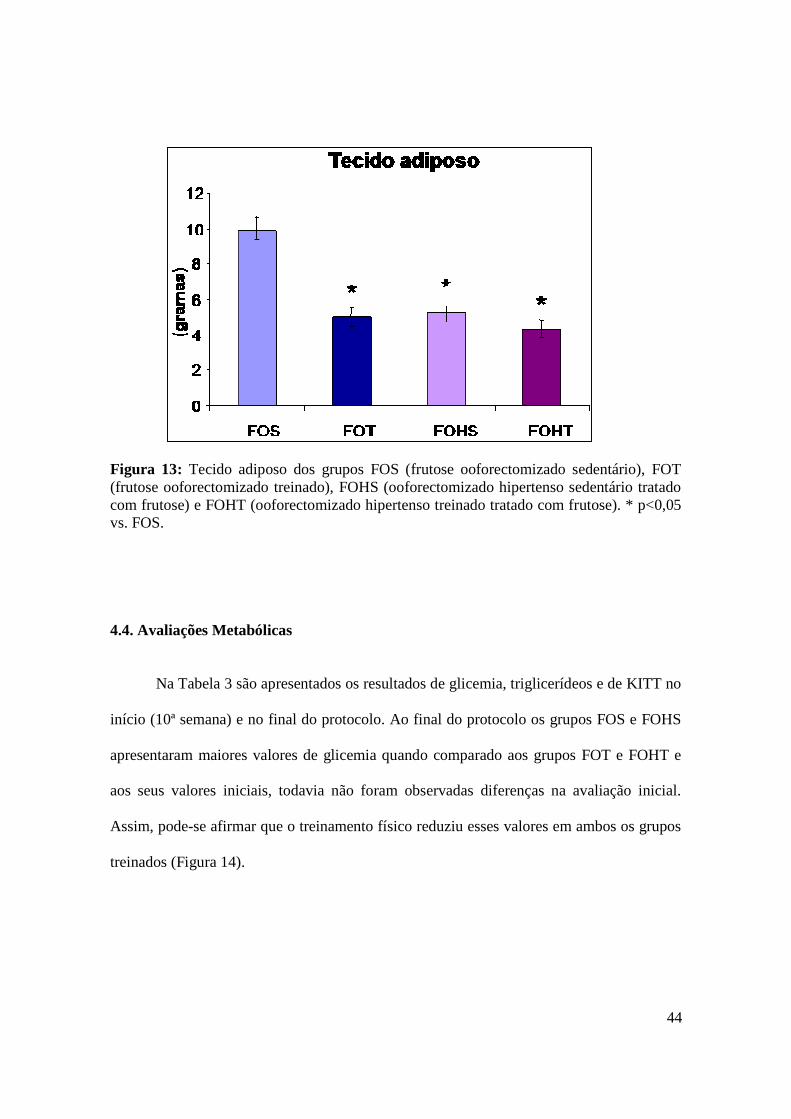
44
Figura 13: Tecido adiposo dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS. 4.4. Avaliações Metabólicas
Na Tabela 3 são apresentados os resultados de glicemia, triglicerídeos e de KITT no
início (10ª semana) e no final do protocolo. Ao final do protocolo os grupos FOS e FOHS
apresentaram maiores valores de glicemia quando comparado aos grupos FOT e FOHT e
aos seus valores iniciais, todavia não foram observadas diferenças na avaliação inicial.
Assim, pode-se afirmar que o treinamento físico reduziu esses valores em ambos os grupos
treinados (Figura 14).
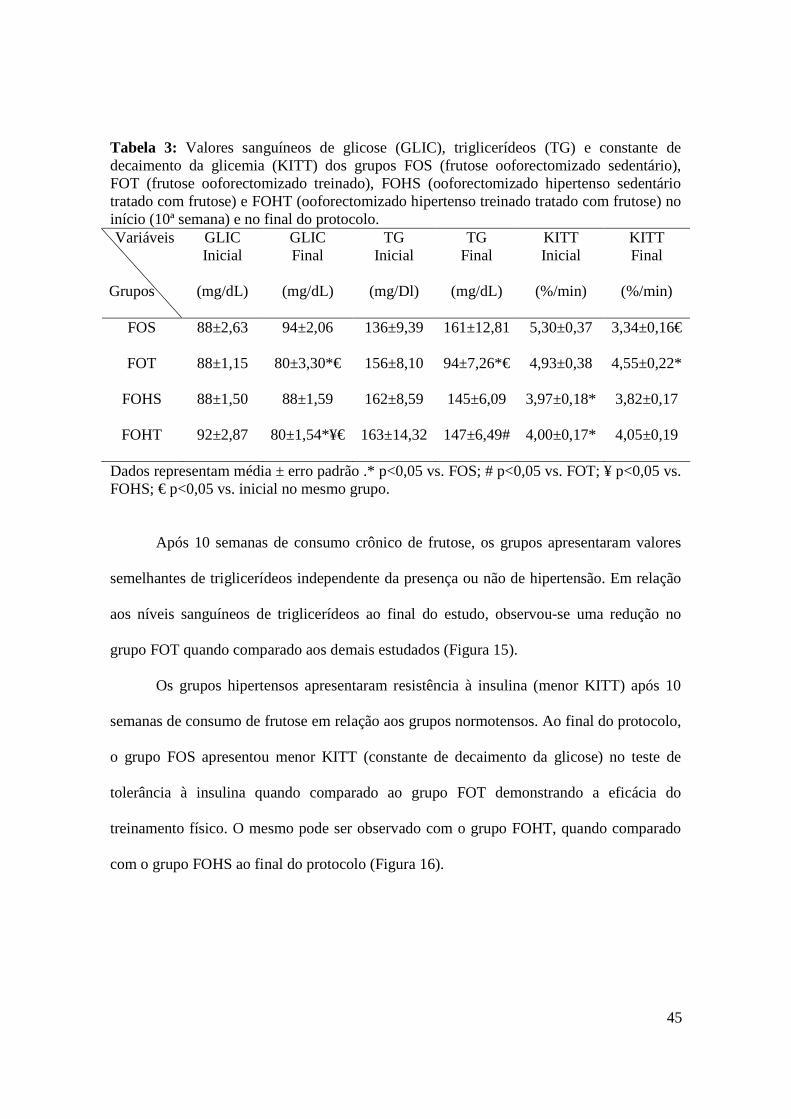
45
Tabela 3: Valores sanguíneos de glicose (GLIC), triglicerídeos (TG) e constante de decaimento da glicemia (KITT) dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose) no início (10ª semana) e no final do protocolo. Variáveis
Grupos
GLIC Inicial
(mg/dL)
GLIC Final
(mg/dL)
TG Inicial
(mg/Dl)
TG Final
(mg/dL)
KITT Inicial
(%/min)
KITT Final
(%/min)
FOS 88±2,63 94±2,06 136±9,39 161±12,81 5,30±0,37 3,34±0,16€
FOT 88±1,15 80±3,30*€ 156±8,10 94±7,26*€ 4,93±0,38 4,55±0,22*
FOHS 88±1,50 88±1,59 162±8,59 145±6,09 3,97±0,18* 3,82±0,17
FOHT 92±2,87 80±1,54*¥€ 163±14,32 147±6,49# 4,00±0,17* 4,05±0,19
Dados representam média ± erro padrão .* p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS; € p<0,05 vs. inicial no mesmo grupo.
Após 10 semanas de consumo crônico de frutose, os grupos apresentaram valores
semelhantes de triglicerídeos independente da presença ou não de hipertensão. Em relação
aos níveis sanguíneos de triglicerídeos ao final do estudo, observou-se uma redução no
grupo FOT quando comparado aos demais estudados (Figura 15).
Os grupos hipertensos apresentaram resistência à insulina (menor KITT) após 10
semanas de consumo de frutose em relação aos grupos normotensos. Ao final do protocolo,
o grupo FOS apresentou menor KITT (constante de decaimento da glicose) no teste de
tolerância à insulina quando comparado ao grupo FOT demonstrando a eficácia do
treinamento físico. O mesmo pode ser observado com o grupo FOHT, quando comparado
com o grupo FOHS ao final do protocolo (Figura 16).

46
Figura 14: Glicemia nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose) ao final do protocolo. * p<0,05 vs. FOS; ¥ p<0,05 vs. FOHS.
Figura 15: Concentrações sanguíneas de triglicerídeos nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose) ao final do protocolo. * p<0,05 vs. FOS; # p<0,05 vs. FOT.
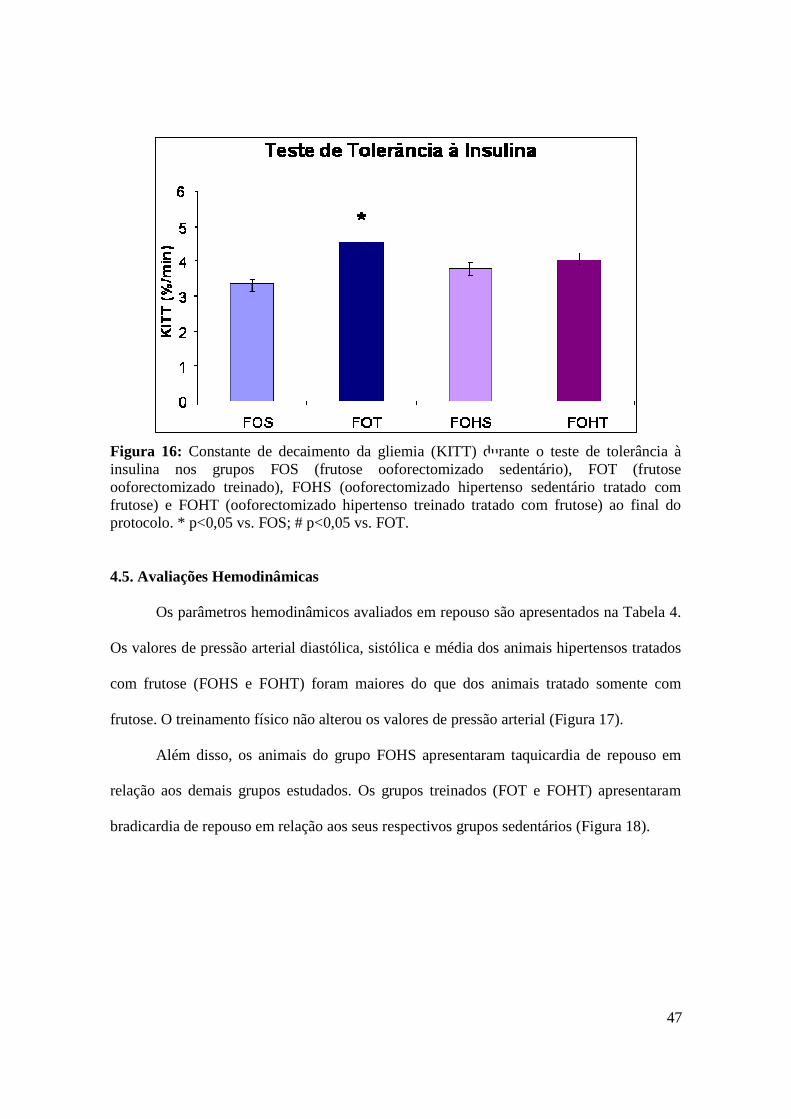
47
Figura 16: Constante de decaimento da gliemia (KITT) durante o teste de tolerância à insulina nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose) ao final do protocolo. * p<0,05 vs. FOS; # p<0,05 vs. FOT.
4.5. Avaliações Hemodinâmicas
Os parâmetros hemodinâmicos avaliados em repouso são apresentados na Tabela 4.
Os valores de pressão arterial diastólica, sistólica e média dos animais hipertensos tratados
com frutose (FOHS e FOHT) foram maiores do que dos animais tratado somente com
frutose. O treinamento físico não alterou os valores de pressão arterial (Figura 17).
Além disso, os animais do grupo FOHS apresentaram taquicardia de repouso em
relação aos demais grupos estudados. Os grupos treinados (FOT e FOHT) apresentaram
bradicardia de repouso em relação aos seus respectivos grupos sedentários (Figura 18).
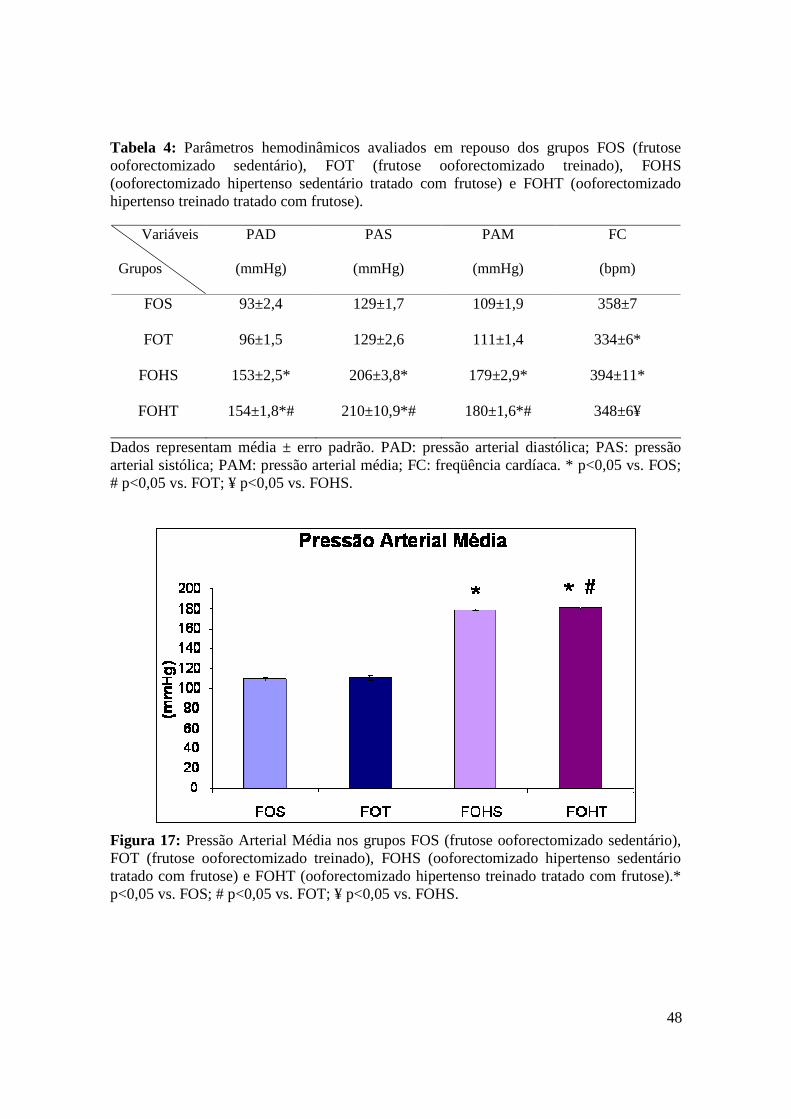
48
Tabela 4: Parâmetros hemodinâmicos avaliados em repouso dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).
Variáveis
Grupos
PAD
(mmHg)
PAS
(mmHg)
PAM
(mmHg)
FC
(bpm)
FOS 93±2,4 129±1,7 109±1,9 358±7
FOT 96±1,5 129±2,6 111±1,4 334±6*
FOHS 153±2,5* 206±3,8* 179±2,9* 394±11*
FOHT 154±1,8*# 210±10,9*# 180±1,6*# 348±6¥
Dados representam média ± erro padrão. PAD: pressão arterial diastólica; PAS: pressão arterial sistólica; PAM: pressão arterial média; FC: freqüência cardíaca. * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS. Figura 17: Pressão Arterial Média nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).* p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.
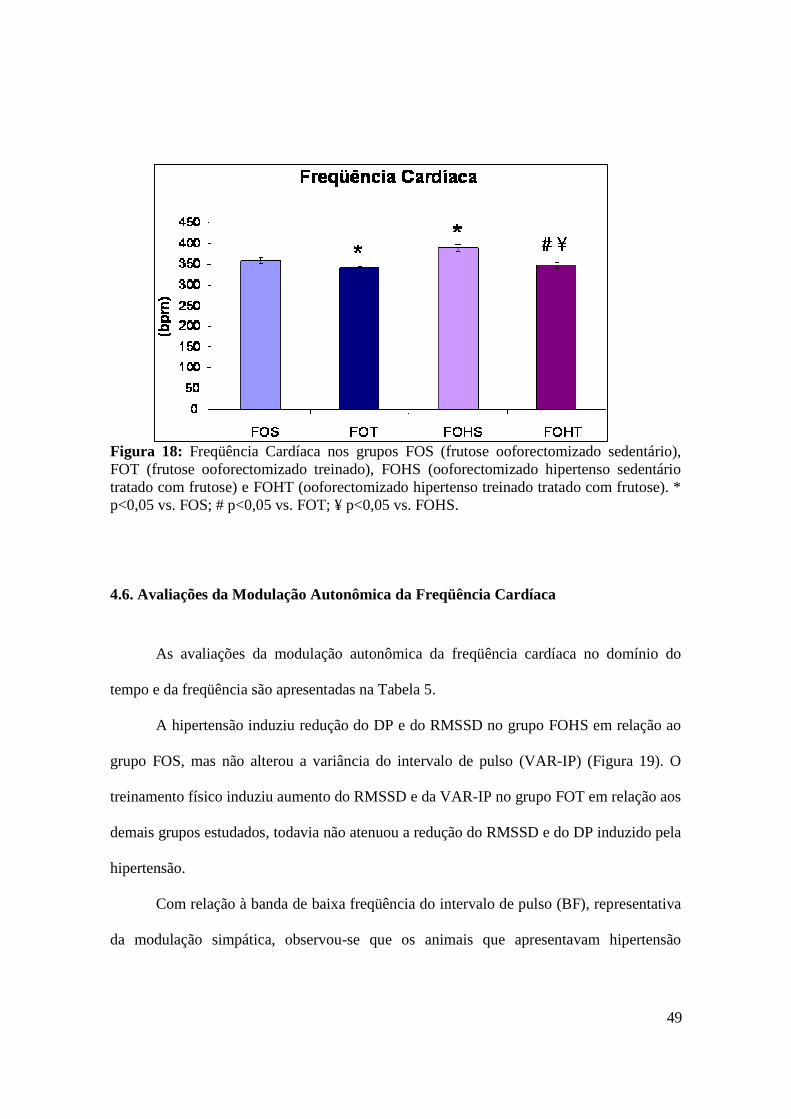
49
Figura 18: Freqüência Cardíaca nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.
4.6. Avaliações da Modulação Autonômica da Freqüência Cardíaca
As avaliações da modulação autonômica da freqüência cardíaca no domínio do
tempo e da freqüência são apresentadas na Tabela 5.
A hipertensão induziu redução do DP e do RMSSD no grupo FOHS em relação ao
grupo FOS, mas não alterou a variância do intervalo de pulso (VAR-IP) (Figura 19). O
treinamento físico induziu aumento do RMSSD e da VAR-IP no grupo FOT em relação aos
demais grupos estudados, todavia não atenuou a redução do RMSSD e do DP induzido pela
hipertensão.
Com relação à banda de baixa freqüência do intervalo de pulso (BF), representativa
da modulação simpática, observou-se que os animais que apresentavam hipertensão

50
associada ao consumo de frutose (FOHS e FOHT) apresentraram uma redução exacerbada
desses valores em relação aos grupos normotensos tratados com frutose (FOS e FOT). A
banda de alta freqüência do intervalo de pulso (AF), representativa da modulação
parassimpática, foi menor nos grupos FOHS e FOHT quando comparados ao grupo FOT.
A banda de muito baixa freqüência do intervalo de pulso (MBF) foi menor apenas no grupo
FOHT em relação ao grupo FOT. A banda de BF normalizada foi menor nos grupos
hipertensos em relação ao grupo normotenso sedentário (FOS) (Figura 20). O treinamento
físico reduziu a banda de BF normalizada no grupo FOT quando comparado com o grupo
FOS. Não foram observadas diferenças para a banda de AF normalizada (Figura 21).
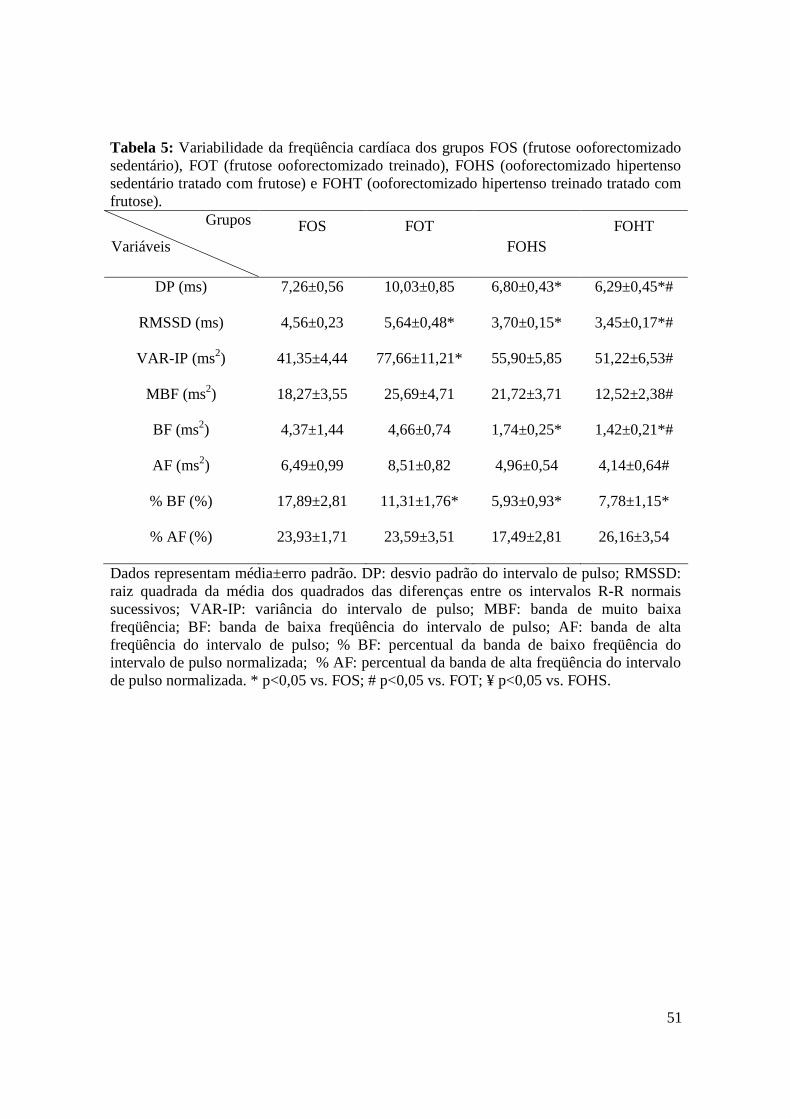
51
Tabela 5: Variabilidade da freqüência cardíaca dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).
Grupos
Variáveis FOS
FOT
FOHS FOHT
DP (ms) 7,26±0,56 10,03±0,85 6,80±0,43* 6,29±0,45*#
RMSSD (ms) 4,56±0,23 5,64±0,48* 3,70±0,15* 3,45±0,17*#
VAR-IP (ms2) 41,35±4,44 77,66±11,21* 55,90±5,85 51,22±6,53#
MBF (ms2) 18,27±3,55 25,69±4,71 21,72±3,71 12,52±2,38#
BF (ms2) 4,37±1,44 4,66±0,74 1,74±0,25* 1,42±0,21*#
AF (ms2) 6,49±0,99 8,51±0,82 4,96±0,54 4,14±0,64#
% BF (%) 17,89±2,81 11,31±1,76* 5,93±0,93* 7,78±1,15*
% AF (%) 23,93±1,71 23,59±3,51 17,49±2,81 26,16±3,54
Dados representam média±erro padrão. DP: desvio padrão do intervalo de pulso; RMSSD: raiz quadrada da média dos quadrados das diferenças entre os intervalos R-R normais sucessivos; VAR-IP: variância do intervalo de pulso; MBF: banda de muito baixa freqüência; BF: banda de baixa freqüência do intervalo de pulso; AF: banda de alta freqüência do intervalo de pulso; % BF: percentual da banda de baixo freqüência do intervalo de pulso normalizada; % AF: percentual da banda de alta freqüência do intervalo de pulso normalizada. * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.
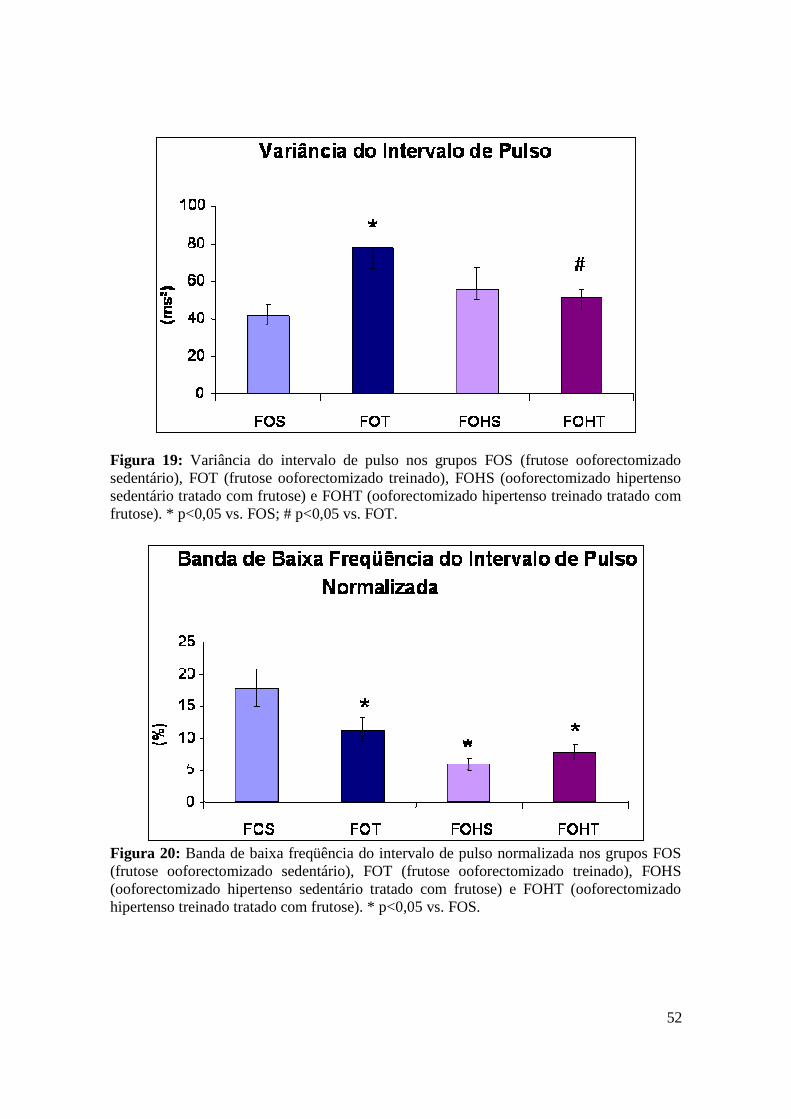
52
Figura 19: Variância do intervalo de pulso nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS; # p<0,05 vs. FOT. Figura 20: Banda de baixa freqüência do intervalo de pulso normalizada nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS.
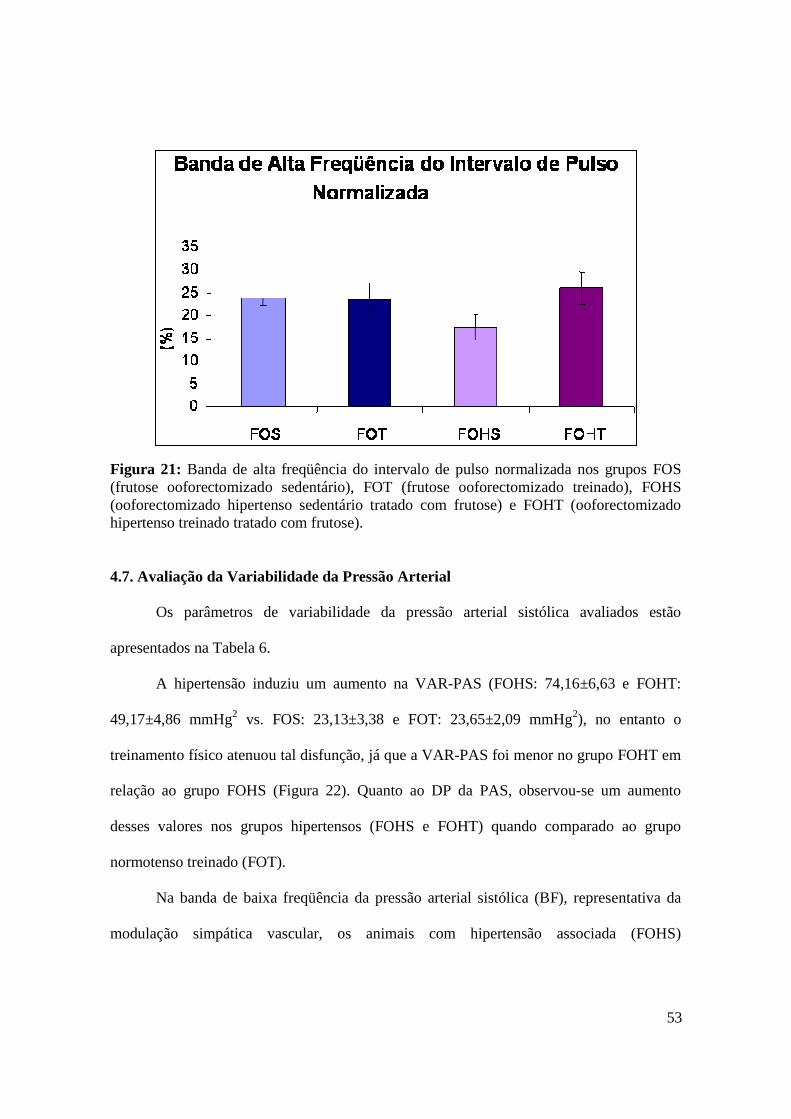
53
Figura 21: Banda de alta freqüência do intervalo de pulso normalizada nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).
4.7. Avaliação da Variabilidade da Pressão Arterial
Os parâmetros de variabilidade da pressão arterial sistólica avaliados estão
apresentados na Tabela 6.
A hipertensão induziu um aumento na VAR-PAS (FOHS: 74,16±6,63 e FOHT:
49,17±4,86 mmHg2 vs. FOS: 23,13±3,38 e FOT: 23,65±2,09 mmHg2), no entanto o
treinamento físico atenuou tal disfunção, já que a VAR-PAS foi menor no grupo FOHT em
relação ao grupo FOHS (Figura 22). Quanto ao DP da PAS, observou-se um aumento
desses valores nos grupos hipertensos (FOHS e FOHT) quando comparado ao grupo
normotenso treinado (FOT).
Na banda de baixa freqüência da pressão arterial sistólica (BF), representativa da
modulação simpática vascular, os animais com hipertensão associada (FOHS)
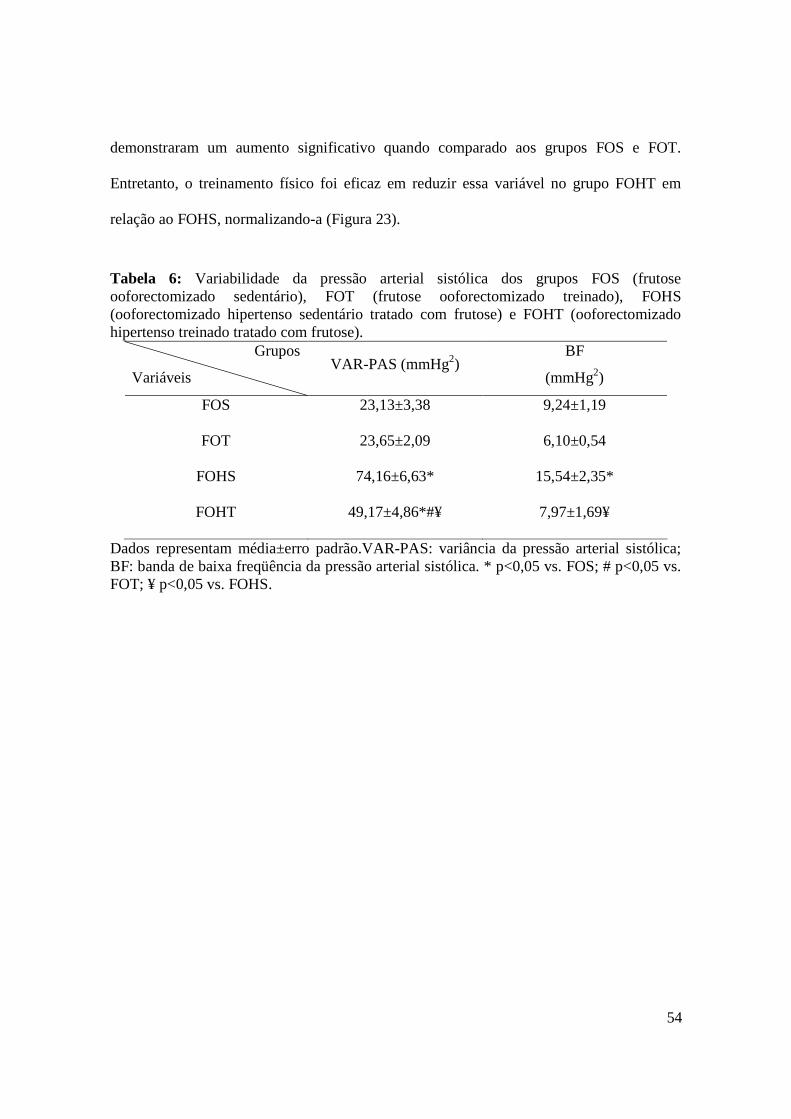
54
demonstraram um aumento significativo quando comparado aos grupos FOS e FOT.
Entretanto, o treinamento físico foi eficaz em reduzir essa variável no grupo FOHT em
relação ao FOHS, normalizando-a (Figura 23).
Tabela 6: Variabilidade da pressão arterial sistólica dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).
Grupos
Variáveis VAR-PAS (mmHg2)
BF
(mmHg2)
FOS 23,13±3,38 9,24±1,19
FOT 23,65±2,09 6,10±0,54
FOHS 74,16±6,63* 15,54±2,35*
FOHT 49,17±4,86*#¥ 7,97±1,69¥
Dados representam média±erro padrão.VAR-PAS: variância da pressão arterial sistólica; BF: banda de baixa freqüência da pressão arterial sistólica. * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.
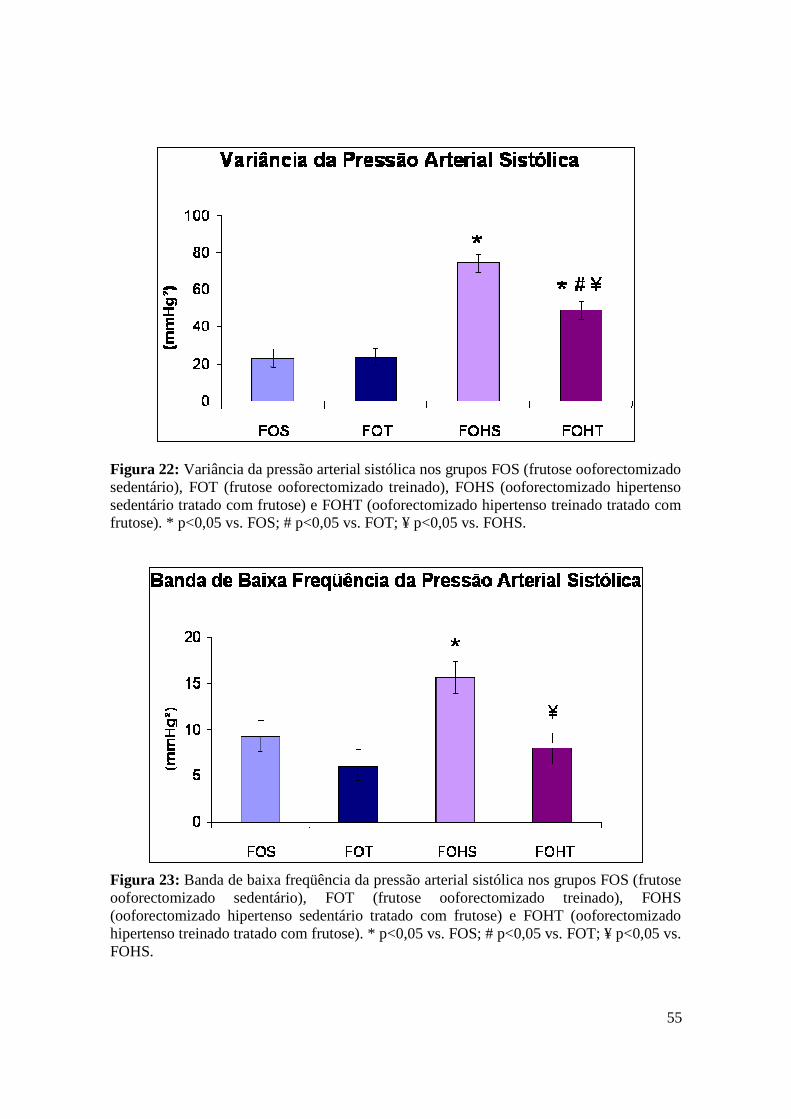
55
Figura 22: Variância da pressão arterial sistólica nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.
Figura 23: Banda de baixa freqüência da pressão arterial sistólica nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.
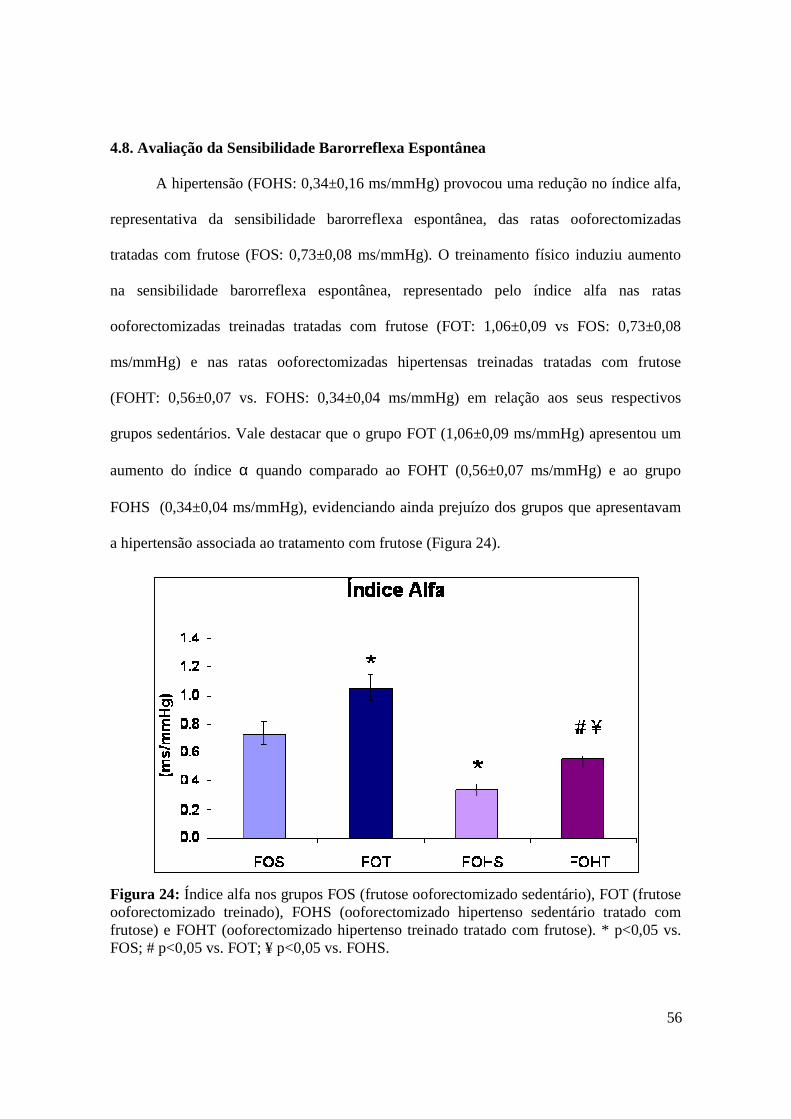
56
4.8. Avaliação da Sensibilidade Barorreflexa Espontânea
A hipertensão (FOHS: 0,34±0,16 ms/mmHg) provocou uma redução no índice alfa,
representativa da sensibilidade barorreflexa espontânea, das ratas ooforectomizadas
tratadas com frutose (FOS: 0,73±0,08 ms/mmHg). O treinamento físico induziu aumento
na sensibilidade barorreflexa espontânea, representado pelo índice alfa nas ratas
ooforectomizadas treinadas tratadas com frutose (FOT: 1,06±0,09 vs FOS: 0,73±0,08
ms/mmHg) e nas ratas ooforectomizadas hipertensas treinadas tratadas com frutose
(FOHT: 0,56±0,07 vs. FOHS: 0,34±0,04 ms/mmHg) em relação aos seus respectivos
grupos sedentários. Vale destacar que o grupo FOT (1,06±0,09 ms/mmHg) apresentou um
aumento do índice α quando comparado ao FOHT (0,56±0,07 ms/mmHg) e ao grupo
FOHS (0,34±0,04 ms/mmHg), evidenciando ainda prejuízo dos grupos que apresentavam
a hipertensão associada ao tratamento com frutose (Figura 24).
Figura 24: Índice alfa nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS; # p<0,05 vs. FOT; ¥ p<0,05 vs. FOHS.

57
4.9. Avaliações de Estresse Oxidativo
Para as avaliações do perfil oxidativo foram utilizados 7 animais por grupo.
Os parâmetros de estresse oxidativo avaliados estão apresentados nas Figuras 25 e
26. A hipertensão não induziu aumento adicional dos valores de QL no tecido cardíaco nos
animais submetidos ao consumo crônico de frutose (FOS: 15432±1010 e FOHS:
14914±2159 cps/mg proteína). O treinamento físico reduziu esses valores (FOT:
9270±1438 cps/mg proteína) somente no grupo que não apresentava a hipertensão
associada, demonstrando a eficácia desta abordagem em reduzir o estresse oxidativo. Os
valores de QL foram semelhantes entre os grupos FOHS e FOHT (14914±2159 vs
13337±1118 cps/mg proteína).
Para a razão GSH/GSSG o grupo FOHS (2,72±0,14) apresentou uma diminuição
dos valores quando comparado ao grupo FOHT (4,00±0,45), evidenciando redução do
estresse oxidativo após o treinamento físico. Vale salientar que durante a realização dessas
dosagens nos grupos não hipertensos houve problemas com as medidas, o que permitiu a
análise de apenas dois animais dos grupos FOS e FOT. Apesar do “n” reduzido, o que não
possibilitou a comparação estatística, observou-se que os valores desta relação foram
maiores no grupo FOT em comparação ao grupo FOS (FOS: 1,95±1,02 vs FOT: 7,69±2,82,
n=2 cada grupo).
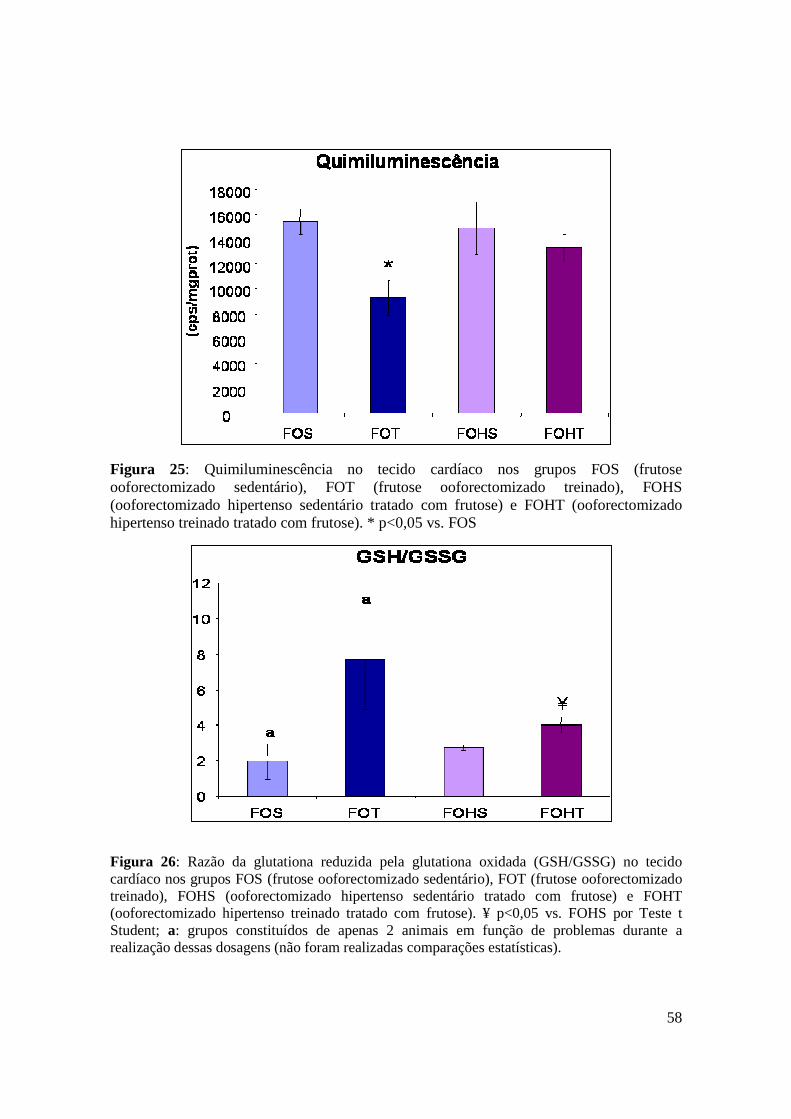
58
Figura 25: Quimiluminescência no tecido cardíaco nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS
Figura 26: Razão da glutationa reduzida pela glutationa oxidada (GSH/GSSG) no tecido cardíaco nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). ¥ p<0,05 vs. FOHS por Teste t Student; a: grupos constituídos de apenas 2 animais em função de problemas durante a realização dessas dosagens (não foram realizadas comparações estatísticas).
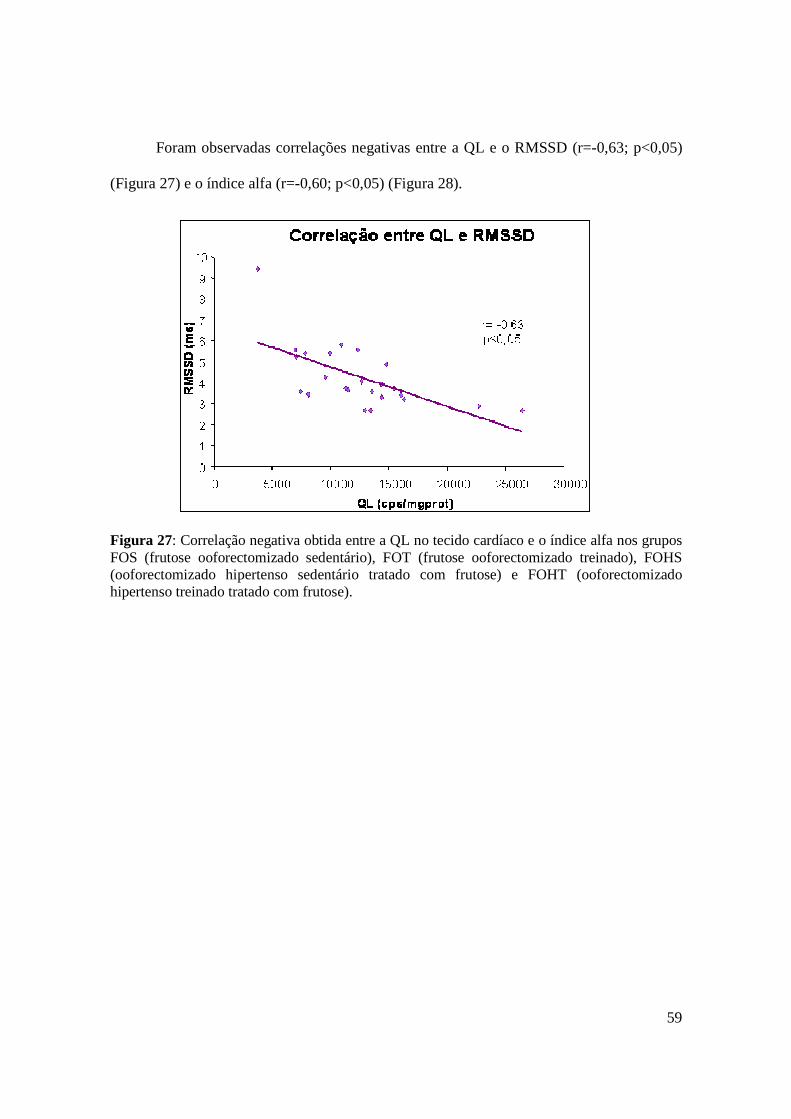
59
Foram observadas correlações negativas entre a QL e o RMSSD (r=-0,63; p<0,05)
(Figura 27) e o índice alfa (r=-0,60; p<0,05) (Figura 28).
Figura 27: Correlação negativa obtida entre a QL no tecido cardíaco e o índice alfa nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).
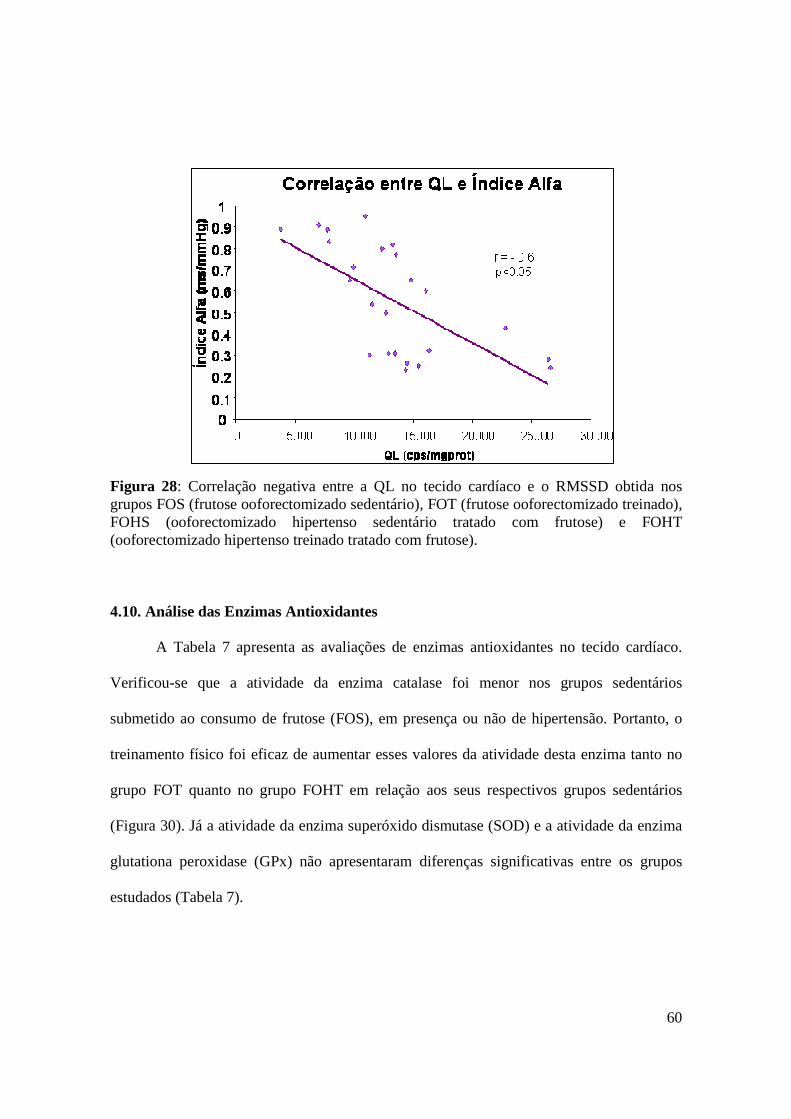
60
Figura 28: Correlação negativa entre a QL no tecido cardíaco e o RMSSD obtida nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). 4.10. Análise das Enzimas Antioxidantes
A Tabela 7 apresenta as avaliações de enzimas antioxidantes no tecido cardíaco.
Verificou-se que a atividade da enzima catalase foi menor nos grupos sedentários
submetido ao consumo de frutose (FOS), em presença ou não de hipertensão. Portanto, o
treinamento físico foi eficaz de aumentar esses valores da atividade desta enzima tanto no
grupo FOT quanto no grupo FOHT em relação aos seus respectivos grupos sedentários
(Figura 30). Já a atividade da enzima superóxido dismutase (SOD) e a atividade da enzima
glutationa peroxidase (GPx) não apresentaram diferenças significativas entre os grupos
estudados (Tabela 7).
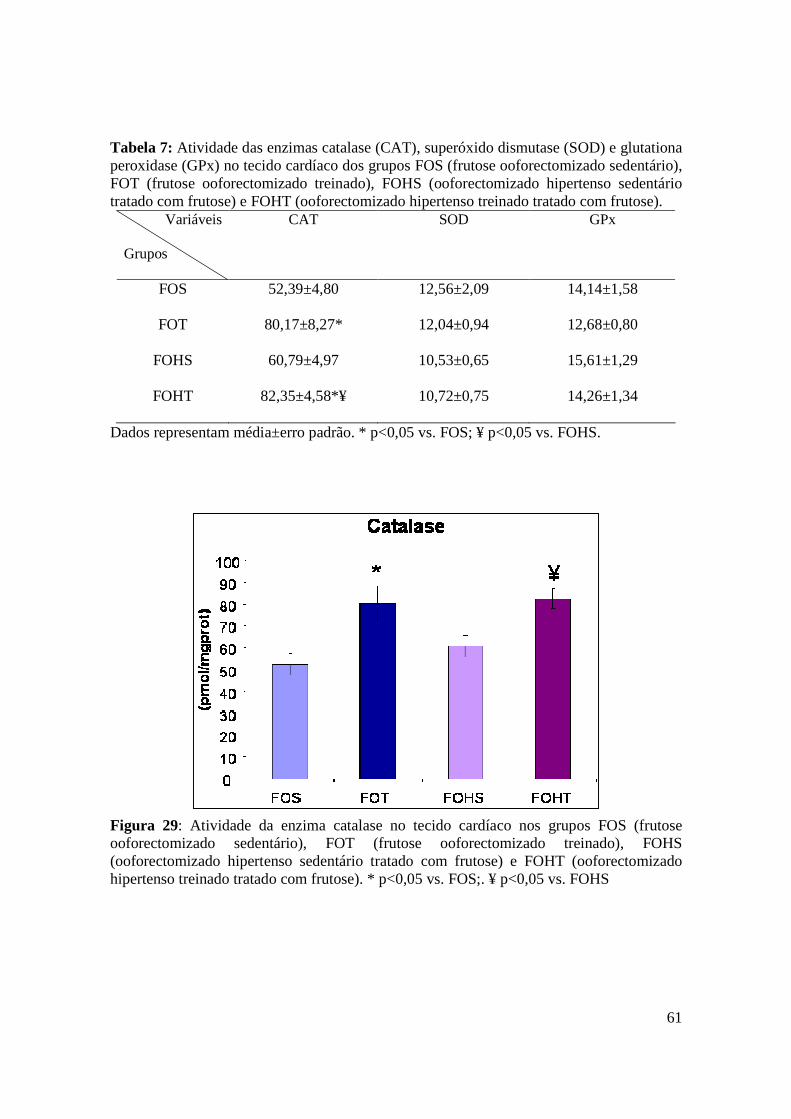
61
Tabela 7: Atividade das enzimas catalase (CAT), superóxido dismutase (SOD) e glutationa peroxidase (GPx) no tecido cardíaco dos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose).
Variáveis
Grupos
CAT SOD GPx
FOS 52,39±4,80 12,56±2,09 14,14±1,58
FOT 80,17±8,27* 12,04±0,94 12,68±0,80
FOHS 60,79±4,97 10,53±0,65 15,61±1,29
FOHT 82,35±4,58*¥ 10,72±0,75 14,26±1,34
Dados representam média±erro padrão. * p<0,05 vs. FOS; ¥ p<0,05 vs. FOHS.
Figura 29: Atividade da enzima catalase no tecido cardíaco nos grupos FOS (frutose ooforectomizado sedentário), FOT (frutose ooforectomizado treinado), FOHS (ooforectomizado hipertenso sedentário tratado com frutose) e FOHT (ooforectomizado hipertenso treinado tratado com frutose). * p<0,05 vs. FOS;. ¥ p<0,05 vs. FOHS

62
5. DISCUSSÃO
O objetivo do presente estudo foi verificar os efeitos do treinamento físico em
parâmetros cardiovasculares e metabólicos, na modulação autonômica cardiovascular e no
estresse oxidativo em ratas fêmeas ooforectomizadas normotensas e hipertensas submetidas
à sobrecarga de frutose. Nossos resultados demonstram que associação do consumo de
frutose com a hipertensão, geneticamente determinada nos SHR, possibilitou o estudo das
alterações metabólicas, cardiovasculares e autonômicas em um modelo que apresenta
disfunções semelhantes às observadas na síndrome metabólica. Considerando que essa
síndrome é mais prevalente em mulheres no climatério, neste estudo, a privação dos
hormônios ovarianos possibilitou avaliar as disfunções características da síndrome
metabólica na ausência dos hormônios ovarianos. Por fim, a aplicação do treinamento
físico possibilitou evidenciar os benefícios desta abordagem não farmacológica que, em
parte, parece ser atenuada quando são somados fatores de risco cardiovascular.
5.1. Avaliações da Capacidade Física
Nosso trabalho evidenciou aumento na capacidade física nos grupos que foram
submetidos ao treinamento físico. Recentemente, foi demonstrado por nosso grupo que se
pode estimar o consumo máximo de oxigênio (VO2máx), ou seja, o transporte, consumo e
utilização de oxigênio, a partir dos resultados do teste de esforço máximo utilizando-se a
equação de regressão linear entre VO2máx e teste de esforço. Além disto, diferenças na
capacidade de exercício podem ser detectadas pelo teste de esforço uma vez que a
velocidade máxima obtida nesse teste foi correlacionada com o VO2 máx em ratos machos
saudáveis (RODRIGUES et al., 2006). De fato, um estudo piloto do nosso grupo em ratas

63
fêmeas ooforectomizadas também demonstrou correlação entre VO2 e velocidade do teste
de esforço.
Vale destacar que os animais SHR apresentaram maior capacidade física em relação
aos animais Wistar, isso se deve à característica hiperativa da linhagem SHR
(TAKAHASHI, 2006). Os grupos FOT e FOHT alcançaram maiores velocidades nos testes
de esforço (intermediário e final) quando comparados aos seus valores no início do estudo,
e aos seus respectivos pares sedentários. Os grupos FOS e FOHS apresentaram manutenção
da velocidade alcançada ao longo do estudo em relação ao teste de esforço inicial. Em
estudo publicado recentemente, demonstramos melhora da capacidade física em ratas
ooforectomizadas após oito semanas de treinamento (IRIGOYEN et al., 2005). Resultados
semelhantes foram obtidos em mulheres pré-menopausa (GREEN et al., 2002),
menopausadas sem (GREEN et al., 2002; KIRWAN et al., 2003; IRVING et al., 2003;
AIELLO et al., 2004) e com reposição hormonal (GREEN et al., 2002; TEIXEIRA et al.,
2003). Protocolos com dieta e treinamento físico, realizando ou não a reposição hormonal,
também evidenciaram melhora de capacidade física (STEFANICK et al.,1998).
Considerando a melhora de performance nos grupos treinado no presente estudo,
vale lembrar que o teste de esforço é um dos exames não-invasivos mais utilizados para
avaliar pacientes com doença cardiovascular. O teste de esforço tem por objetivo submeter
o paciente a estresse físico, com finalidade de avaliar a resposta clínica, hemodinâmica,
eletrocardiográfica e metabólica ao esforço. Essa avaliação permite detectar isquemia
miocárdica, arritmias cardíacas, distúrbios hemodinâmicos esforço-induzidos, avaliar
capacidade funcional, avaliar diagnóstico e prognóstico das doenças cardiovasculares,
prescrever exercícios (NEGRÃO & BARRETO, 2005). Vale lembrar que o VO2máx, o qual

64
foi relacionado a velocidade no teste de esforço em ratos (RODRIGUES et al., 2007),
representa hoje não só um indicador de performance, mas um marcador prognóstico em
cardiopatas (MYERS et al., 1998; ARMOSTRONG et al., 2005).
5.2. Avaliações Metabólicas e dos Pesos Corporal e do Tecido Adiposo
Várias alterações fisiológicas têm sido associadas ao advento da menopausa, dentre
elas: a redução da massa corporal magra, da densidade óssea, da taxa metabólica de
repouso, da capacidade física e o aumento dos depósitos de gordura corporal, marcado por
grande aumento da gordura abdominal. Em conjunto, essas alterações normalmente
induzem aumento do peso corporal total e maior incidência de doenças cardiovasculares e
metabólicas, as quais têm sido relacionadas à restrição hormonal observada no climatério
(SOWERS & PIETRA, 1995; HERNÁNDEZ et al., 2000; TEIXEIRA et al., 2003;
PEIXOTO et al., 2006). Em ratas fêmeas, a retirada dos ovários leva a privação dos
hormônios ovarianos semelhante à observada em mulheres após a menopausa, e induz
aumento da ingestão de alimentos, do peso corporal e da resistência à insulina
(WATTANAPERMPOOL & REISER, 1999; HERNÁNDEZ et al., 2000; LATOUR et al.,
2001; IRIGOYEN et al., 2005). Além disto, estas mudanças podem ficar mais acentuadas
em presença de sedentarismo tanto em mulheres quanto em animais de experimentação
(HASSAGER & CHRISTIANSEN, 1989; DAWSON-HUGES & HARRIS, 1992).
Confirmando esses achados, em nosso laboratório verificamos que ratas
ooforectomizadas sedentárias apresentaram maior peso corporal do que ratas controles
(ciclo estral normal) após 9 semanas de privação dos hormônios ovarianos (FLORES et al.,
2005). Neste aspecto, vários estudos demonstraram que o treinamento físico pode ser uma

65
abordagem favorável para redução e/ou controle do aumento de peso corporal, tanto em
humanos (BOUCHARD, 2003; SHANGOLD, 1990; TEIXEIRA et al., 2003) quanto em
animais de experimentação (DE ANGELIS et al., 1997; MELO et al., 2003). Todavia,
Latour e colaboradores (2001) avaliaram o efeito do treinamento físico em ratas
ooforectomizadas e não verificaram redução do peso corporal.
Ao final do protocolo, as ratas ooforectomizadas hipertensas tratadas com frutose
apresentaram menor peso corporal do que as ratas ooforectomizadas normotensas tratadas
com frutose. Isso se deve às características da linhagem SHR, que possui um porte físico
relativamente pequeno (CICOGNA et al., 1997). Vale ressaltar que o consumo de frutose
não induziu ganho de peso corporal adicional no grupo ooforectomizado hipertenso em um
estudo realizado previamente em nosso grupo (SANCHES, 2007). De fato, estudos que
utilizaram sobrecarga de frutose não detectaram alterações no peso corporal ao final do
protocolo (KOTCHEN et al., 1997; BEZERRA et al., 2001; CATENA et al., 2003; SONG
et al., 2004; D´ANGELO et al., 2005). Todavia, existem alguns trabalhos que associam o
consumo de frutose a maior ganho de peso corporal em humanos e animais (TORDOFF &
ALLEVA, 1990; ANDERSON et al., 1989). Estudos anteriores de nosso laboratório
demonstraram que ratas Wistar saudáveis ou ooforectomizadas submetidas ao tratamento
de sobrecarga de frutose apresentam aumento do peso corporal e do tecido adiposo,
acompanhado de aumento da glicemia e triglicerídeos sanguíneos (PONCIANO et al.,
2006; BRITO et al., 2008).
No presente estudo, apesar de não termos alteração no peso corporal entre os grupos
sedentários, observamos que o grupo FOS apresentou aumento do tecido adiposo em
relação à ratas somente ooforectomizadas (PONCIANO, 2006). Novamente, o treinamento

66
físico promoveu redução do peso corporal, induzindo diminuição desta variável no grupo
FOT em relação ao grupo FOS. Interessantemente, o tecido adiposo estava reduzido nos
grupos hipertensos, o que provavelmente está relacionado ao menor peso corporal, maior
metabolismo e conseqüentemente melhor resposta no teste de esforço em relação a animais
Wistar (RICHARD et al., 1987).
O quadro 3 ilustra as alterações metabólicas induzidas pela privação dos hormônios
ovarianos (OS), em ratas saudáveis (CS), bem como as alterações adicionais induzidas pela
adição de fatores de risco como hipertensão (OHS), consumo crônico de frutose (FOS) e
associação desses 2 fatores de risco (FOHS) em fêmeas ooforectomizadas (OS) com
relação à glicemia, triglicerídeos sanguíneos e KITT.
Quadro 3: Alterações metabólicas dos grupos controle (CS), ooforectomizada (OS),
ooforectomizada hipertensa (OHS), frutose ooforectomizada (FOS) e frutose
ooforectomizada hipertensa (FOHS).
Grupos
Variáveis
CS x OS
OS x OHS
OS x FOS
OS x FOHS
Glicemia (mg/dL) ↑ 7% ↑4% ↑14% ↑8%
Triglicerídeos (mg/dL) ↑ 5% ↑10% ↑ 30% ↑22%
KITT (%/min) ↓15% ↑25% ↓ 11% ↑ 2%
Dados obtidos de Dissertações de Mestrado (SANCHES, 2007; PONCIANO, 2006; FLORES,
2005) e artigos científicos (IRIGOYEN et al., 2005; SOUZA et al., 2007; LIMA et al., 2007).

67
Observe que o consumo de frutose induziu aumento nas concentrações de
triglicerídeos desde a 10ª semana até o final do protocolo em relação à dados obtidos por
Ponciano, 2006 e Flores, 2005. Esse achado pode ser decorrente do fato da frutose ser
rapidamente absorvida e metabolizada pelo fígado, e estar diretamente relacionado à rápida
estimulação da lipogênese e ao acúmulo de triglicérides, favorecendo a redução da
sensibilidade à insulina, a resistência à insulina hepática e a intolerância à glicose
(BASCIANO et al., 2005). Apesar de existir controvérsia com relação ao consumo de
frutose e o aumento no ganho de peso, um achado consistente na literatura em ratos e
camundongos machos é que a sobrecarga de frutose induz hipertrigliceridemia, aumento da
glicemia, hiperinsulinemia e resistência à insulina (CUNHA et al., 2007; FARAH et al.,
2006; GALIPEAU et al., 2002; TAKAGAWA et al., 2002; SONG et al., 2004).
A hipertensão associada ao consumo de frutose não induziu aumento adicional nas
concentrações de triglicerídeos. Contudo, o aumento dos níveis sanguíneos de triglicerídeos
nos grupos FOS, FOHS e FOHT pode estar relacionado com a diminuição do número de
receptores insulínicos, desta forma, reduzindo a sensibilidade à insulina (BIEGER et al.,
1984 ). Por sua vez, o treinamento físico induziu redução dos triglicerídeos sanguíneos
somente no grupo não hipertenso (FOT), sugerindo que apesar da hipertensão não agravar a
disfunção no perfil lipídico impede o benefício do treinamento físico.
Além disto, discreta alteração de glicemia foi observada em função da privação dos
hormônios ovarianos, que não foi mais agravada de forma significativa pela somatória de
fatores de risco como a hipertensão ou a sobrecarga de frutose (Quadro 3). No presente
estudo, o treinamento físico promoveu redução do peso corporal, além de redução da
glicemia plasmática nas ratas FOT e FOHT quando comparadas aos grupos FOS e FOHS,

68
reforçando a importância dessa abordagem na melhora do perfil metabólico na privação
dos hormônios ovarianos com sobrecarga de frutose associado ou não a hipertensão.
A privação dos hormônios ovarianos tem sido relacionada à resistência à insulina e
maior prevalência de diabetes (SOWERS & LA PIETRA, 1995). Observe no Quadro 3 que
a ooforectomia em ratas induziu redução do KITT (15%) obtido após a sobrecarga de
insulina (ITT). O consumo de frutose induziu prejuízo adicional (↓11%) nas ratas
ooforectomizadas. Neste aspecto, apesar de agudamente a frutose não provocar aumento
nos níveis de insulina, em longo prazo esta substância pode induzir hiperinsulinemia e
obesidade através de mecanismos indiretos. Trabalhos demonstraram que a dieta rica em
frutose induziu resistência à insulina em roedores (HALLFRISCH et al., 1979; REISER &
HALLFRISCH, 1977; ZAVARONI et al., 1980) e cães (MARTINEZ et al., 1994). Em
ratos submetidos a uma dieta com 66% de frutose por 2 semanas, o RNA mensageiro para o
receptor de insulina e o próprio receptor de insulina no músculo esquelético e no fígado
estavam significativamente diminuídos quando comparados aos ratos com dieta normal
(ração padrão). Em um outro estudo, ratos tratados com frutose por 28 dias não
apresentaram alterações na concentração de receptores de insulina, mas a autofosforilação
estimulada pela insulina, um mecanismo necessário para a ação da insulina, estava reduzida
em 72% no fígado. Somado a isso, observou-se uma redução na fosforilação dos substratos
do receptor de insulina (IRS), outro passo fundamental para a ação insulínica, no fígado e
no músculo de animais submetidos a sobrecarga de frutose (UENO et al., 2000). Há ainda
fortes evidências sugerindo que o aumento dos AGLs nos modelos tratados com frutose
desempenhem um importante papel na resistência à insulina. Se os AGLs não são
removidos dos tecidos, como ocorre na resistência à insulina nos modelos tratados com

69
frutose, há um aumento no fluxo de AGL e que leva a um aumento na secreção de
triglicerídeos. A resistência à insulina também tem sido correlacionada com os estoques de
triglicerídeos celular, os quais estão envolvidos em lipotoxicidade e falha das células beta,
levando ao diabetes (ZIEGLER et al., 2001).
Além disso, há algumas diferenças importantes entre as rotas metabólicas da glicose
e da frutose. A absorção gástrica de ambas, glicose e frutose, é realizada através da veia
porta ao fígado. Acredita-se que o fígado, ao metabolizar altas doses de frutose,
rapidamente direciona esta substância para a rota glicolítica. Neste aspecto, parece de
fundamental importância à habilidade da frutose passar às etapas regulatórias da glicólise,
ou seja, esta molécula é convertida de glicose-6-fosfato em frutose 1-6-difosfato pela
fosfofrutoquinase (PFK). O fato da PFK ser um dos passos limitantes da rota glicolítica, e a
frutose ser rapidamente convertida por esta enzima explicaria, pelo menos em parte, a
rápida incorporação desta molécula ao metabolismo glicolítico. Assim, enquanto o
metabolismo de glicose é negativamente regulado pelo fosfofrutoquinase, a frutose pode
continuamente entrar na rota glicolítica. Nesse sentido, a frutose pode incontrolavelmente
produzir glicose, glicogênio, lactato e piruvato, fornecendo glicerol e acil para a formação
de moléculas de acilglicerol, promovendo uma super produção de triglicerídeos (MAYES,
1993), conforme observado no presente estudo no grupo hipertenso ooforectomizado
tratado com frutose.
Interessantemente, a presença de hipertensão associado à presença dos hormônios
não induz alteração no KITT de forma significativa, pelo contrário, até melhora. Esse fato
pode estar relacionado ao aumento do metabolismo em SHR (RICHARD et al., 1987;
TAKAHASHI, 2006). Todavia, observa-se no Quadro 3 que o consumo de frutose no

70
grupo hipertenso induziu aumento neste índice de resistência à insulina de mais de 20%.
Tais achados sugerem que a adição de fatores de risco aumenta a resistência à insulina.
Além disto, vale lembrar que o KITT já estava aumentado após 10 semanas de tratamento
nos grupos hipertensos, o que não foi observado nos grupos normotensos, demonstrando
que a hipertensão induz aparecimento precoce das disfunções metabólicas induzidas pela
sobrecarga de frutose.
No presente estudo observou-se um aumento da sensibilidade à insulina somente
no grupo FOT em relação ao grupo FOS, evidenciando o papel do treinamento físico. De
forma semelhante, Latour e colaboradores (2001) observaram melhora na resposta da
insulina após estimulação pelo teste de tolerância de glicose após treinamento físico de oito
semanas em ratas ooforectomizadas. Todavia, vale destacar que a presença de hipertensão
associada ao tratamento com a frutose e a privação dos hormônios ovarianos aboliu os
benefícios do treinamento físico na resistência à insulina, já que os grupos FOHS e FOHT
apresentaram valores semelhantes de KITT.
5.3. Avaliações Hemodinâmicas e Autonômicas
O início da equivalência nas taxas de eventos cardiovasculares entre os sexos
coincide com o advento da menopausa e, conseqüentemente, da privação estrogênica.
Estudos vêm demonstrando que mulheres menopausadas com mais de 55 anos apresentam
aumentado risco para doenças cardiovasculares, parte do qual tem sido atribuído a
disfunções do endotélio vascular, que parecem estar associadas ao aumento da pressão
arterial.
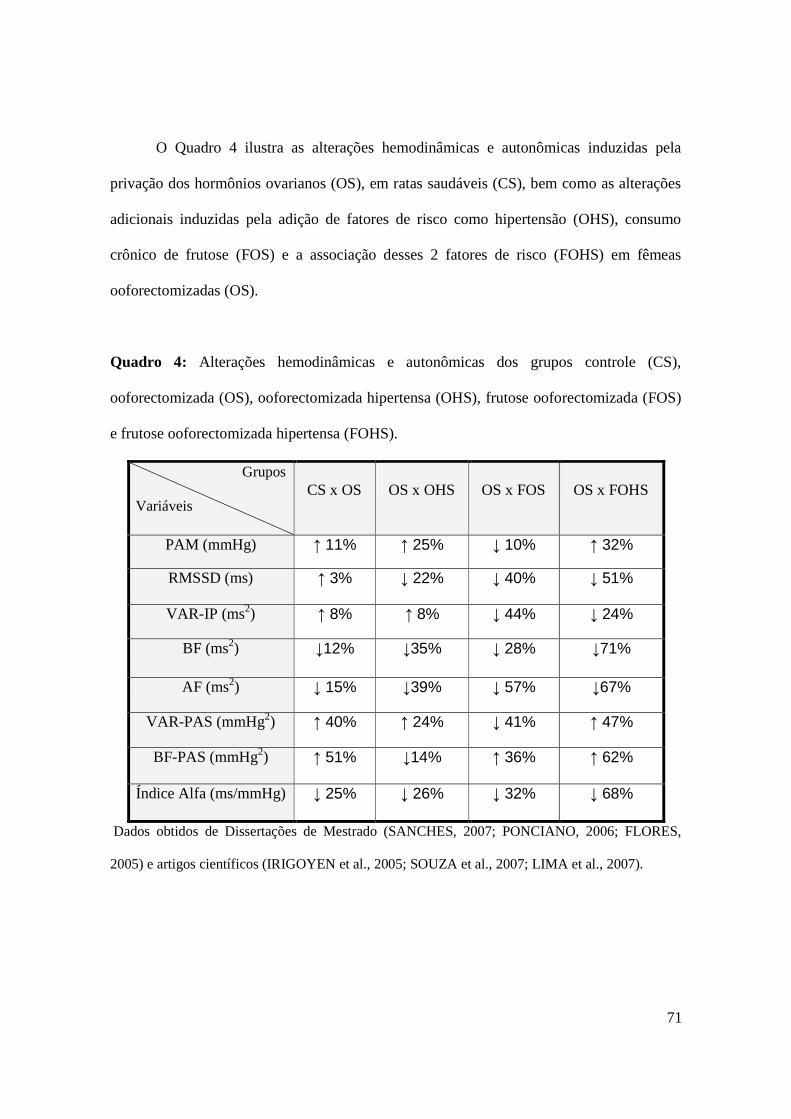
71
O Quadro 4 ilustra as alterações hemodinâmicas e autonômicas induzidas pela
privação dos hormônios ovarianos (OS), em ratas saudáveis (CS), bem como as alterações
adicionais induzidas pela adição de fatores de risco como hipertensão (OHS), consumo
crônico de frutose (FOS) e a associação desses 2 fatores de risco (FOHS) em fêmeas
ooforectomizadas (OS).
Quadro 4: Alterações hemodinâmicas e autonômicas dos grupos controle (CS),
ooforectomizada (OS), ooforectomizada hipertensa (OHS), frutose ooforectomizada (FOS)
e frutose ooforectomizada hipertensa (FOHS).
Grupos Variáveis
CS x OS
OS x OHS
OS x FOS
OS x FOHS
PAM (mmHg) ↑ 11% ↑ 25% ↓ 10% ↑ 32%
RMSSD (ms) ↑ 3% ↓ 22% ↓ 40% ↓ 51%
VAR-IP (ms2) ↑ 8% ↑ 8% ↓ 44% ↓ 24%
BF (ms2) ↓12% ↓35% ↓ 28% ↓71%
AF (ms2) ↓ 15% ↓39% ↓ 57% ↓67%
VAR-PAS (mmHg2) ↑ 40% ↑ 24% ↓ 41% ↑ 47%
BF-PAS (mmHg2)
↑ 51% ↓14% ↑ 36% ↑ 62%
Índice Alfa (ms/mmHg) ↓ 25%
↓ 26%
↓ 32%
↓ 68%
Dados obtidos de Dissertações de Mestrado (SANCHES, 2007; PONCIANO, 2006; FLORES,
2005) e artigos científicos (IRIGOYEN et al., 2005; SOUZA et al., 2007; LIMA et al., 2007).

72
Interessantemente, os grupos submetidos à sobrecarga de frutose (FOS e FOT), não
apresentaram aumento da pressão arterial no presente estudo. Existem evidências na
literatura de um papel importante do simpático e do SRA nas alterações cardiovasculares
induzidas pelo tratamento com frutose. A simpatectomia atenuou o desenvolvimento da
hipertensão em ratos tratados com frutose (VERMA et al., 1999). O tratamento com frutose
também aumentou a excreção urinária de catecolaminas e expressão de receptores
adrenérgicos (KAMIDE et al., 2002). Somado a isto, estudos demonstraram aumento na
expressão de receptores de Angiotensina na vasculatura e no efeito depressor de
antagonistas do receptor de Angiotensina em ratos (KATOVICH et al., 2001; HSIEH,
2005) e ativação do SRA em camundongos (SHINOZAKI et al., 2004). Farah e
colaboradores (2004) evidenciaram níveis aumentados de Angiotensina II em camundongos
que consumiram frutose (60 dias).
Além da ingestão de frutose induzir à ativação do sistema SRA, estudo de nosso
laboratório demonstrou que ratas submetidas à ooforectomia também apresentaram
ativação deste sistema. Assim é possível que os animais dos grupos FOS e FOT
apresentassem um aumento expressivo do sistema SRA, com conseqüente maior atividade
da ECA2 levando à maior produção da angiotensina 1-7, cujo efeito é vasodilatador. Esta
hipótese poderia explicar a não alteração da PA nos grupos FOS e FOT. Corroborando esta
hipótese Farah e colaboradores (2004) observaram atividade da ECA2 aumentada em
animais AT1 knockout submetidos à sobrecarga de frutose.
No entanto, como já era de se esperar, observamos um aumento dos valores de PAD,
PAS e PAM nos animais hipertensos tratados com frutose (FOHS e FOHT) em relação aos
animais tratados somente com frutose sem a hipertensão associada (FOS e FOT). Com isso,

73
pode-se perceber que a associação da ooforectomia à hipertensão espontânea dos animais
SHR (FOS) possibilitou a avaliação de alterações hemodinâmicas e autonômicas induzidas
pela privação dos hormônios ovarianos na presença de hipertensão estabelecida, como
muitas vezes verifica-se em mulheres. De fato, estudos demonstram que a PA de mulheres
é mais baixa do que a de homens até a faixa etária dos 50-60 anos. Após essa fase, que
coincide com o advento da menopausa, a PA (particularmente a sistólica) aumenta nas
mulheres e a hipertensão torna-se mais prevalente (STAMLER et al., 1976) ou, pelo menos,
igualmente prevalente entre homens e mulheres, sugerindo que os hormônios ovarianos
possam ser responsáveis pela PA mais baixa em mulheres pré-menopausa e a sua ausência
pelo aumento da PA em mulheres menopausadas (STAESSEN et al., 1997). Em
camundongos tratados com frutose na ração foi observado um aumento da PA, que foi
atribuído ao aumento da VAR-PAS e conseqüentemente ao aumento da banda de BF
(representativa da modulação simpática) (FARAH et al., 2006).
O aumento da PA em ratos SHR tem sido relacionado a hiperatividade simpática e ao
prejuízo na sensibilidade do barorreflexo (SILVA et al., 1997; GAVA et al.,1995).
Confirmando esses achados verificamos aumento da banda de BF da PAS, indicando um
aumento da atividade simpática vascular que poderia explicar o aumento da resistência
vascular periférica, bem como alterações estruturais e/ou funcionais, que podem estar
relacionadas à hipertensão e a atenuação do barorreflexo (IRIGOYEN et al., 2003;
IRIGOYEN et al., 2005). Adicionalmente, provavelmente associada à redução da
sensibilidade barorreflexa espontânea (índice alfa), a VAR-PAS mostrou-se aumentada no
grupo hipertenso ooforectomizado sedentário tratado com frutose no presente estudo.
Observe no Quadro 4 que o consumo de frutose em ratas hipertensas induz aumento

74
adicional da pressão arterial, provavelmente relacionado a disfunção associada da VAR-
PAS, da banda de BF da PAS e do índice alfa nesse grupo. Vale destacar que o treinamento
físico reduziu a modulação simpática vascular e atenuou a aumentada variabilidade da PAS
no grupo hipertenso submetido ao consumo crônico de frutose (FOHT).
O tratamento com frutose no grupo hipertenso ooforectomizado sedentário (FOHS)
induziu também taquicardia de repouso. A taquicardia nos animais hipertensos
provavelmente deve estar associada ao aumento da atividade simpática cardíaca,
evidenciada pela redução exacerbada do componente de BF do intervalo de pulso e do
componente de BF do intervalo de pulso normalizado, o que tem sido relacionado a
hiperatividade simpática. Apesar de em um primeiro momento parecer estranho associar a
redução da banda de BF a um aumento da modulação simpática cardíaca, estudos têm
evidenciado que o fato da atividade simpática estar extremamente exacerbada (saturada)
em determinadas condições patológicas faz com que a modulação (variação) deste
componente seja reduzida, o que tem sido relacionado inclusive à pior prognóstico (VAN
DE BORNE et al., 1997; MORTARA et al., 1994). Adicionalmente, a diminuição da banda
de AF e do índice RMSSD do intervalo de pulso possivelmente desempenham um
importante papel na taquicardia observada no grupo sedentário hipertenso tratado com
frutose (FOHS). Assim, nossos resultados demonstram que a variabilidade da freqüência
cardíaca, avaliada no domínio do tempo e da freqüência, confirma o prejuízo vagal e o
predomínio simpático no coração das ratas hipertensas ooforectomizadas tratadas com
frutose (FOHS).
Além disso, vale destacar que o treinamento físico aumentou a VAR do IP e reduziu
a banda de BF do IP na ausência de hipertensão associada (FOT vs. FOS), sendo que tal

75
benefício não foi evidenciado no grupo treinado que apresentava hipertensão (FOHT). A
redução da taquicardia no grupo FOHT em relação ao grupo FOHS provavelmente foi
decorrente de uma melhora discreta do balanço simpato/vagal (aumento da modulação
vagal sem diferença significativa) no grupo FOHT em relação ao grupo FOHS. Além disto,
a melhora do índice alfa também poderia estar relacionada a redução da atividade simpática
(BRUM et al., 2000), e conseqüente redução da freqüência cardíaca nesse grupo treinado
hipertenso.
Com relação a sensibilidade barorreflexa espontânea (índice alfa), observou-se que
esta mostrou-se adicionalmente reduzida nos grupos com hipertensão associada ao
consumo crônico de frutose (FOHS e FOHT), o que provavelmente está relacionado à
elevação da PA, ao aumento da variabilidade da PA (aumento do DP-PAS e da VAR-PAS).
Vale destacar que a piora na sensibilidade barorreflexa em SHR tem sido associada
a aumentos na variabilidade da pressão arterial e ao favorecimento de lesões em órgãos
alvos (SHAN et al., 1999). Neste aspecto, um trabalho recente de nosso grupo demonstrou
em camundongos tratados dois meses com frutose que as alterações na modulação
autonômica cardiovascular estavam associadas a prejuízos estruturais e funcionais no tecido
renal (CUNHA et al., 2007). Neste aspecto vale ressaltar que o treinamento físico foi eficaz
em melhorar a sensibilidade barorreflexa em ambos os grupos treinados (FOT e FOHT) em
relação aos seus respectivos grupos sedentários, além de reduzir os valores de VAR-PAS e
da BF-PAS no FOHT em comparação ao grupo FOHS. Todavia, mesmo após a melhora do
índice alfa, a sensibilidade barorreflexa espontânea permaneceu reduzida no grupo FOHT
em relação aos grupos normotensos (FOS e FOT).

76
5.4. Avaliações do Perfil Oxidativo
Atualmente, confere-se aos radicais livres participação em diversos processos
patológicos e também nas alterações verificadas durante o envelhecimento. Os radicais
livres de oxigênio, chamado também de espécies reativas de oxigênio (EROs), são
produzidos naturalmente em nosso organismo através dos processos metabólicos oxidativos
e, muitas vezes, são de extrema utilidade, como nas situações em que há necessidade de
ativação do sistema imunológico, na desintoxicação de drogas e nos processos que
desencadeiam o relaxamento dos vasos sanguíneos em resposta a atividade do óxido
nítrico, que é um radical livre (HALLIWELL, 1992). A produção de radicais livres e de
outras substâncias altamente reativas em decorrência do metabolismo do oxigênio (EROs) é
contrabalanceado por muitos mecanismos de defesa antioxidante para limitar os níveis e
impedir a indução de danos celulares (SIES, 1993).
O desequilíbrio entre a produção de moléculas oxidantes e a capacidade de inativação
dos mecanismos antioxidantes, que resulta na indução de danos celulares pelos radicais
livres, tem sido chamado de estresse oxidativo, ou seja, quando há um desequilíbrio entre
ações pró-oxidantes e a defesa antioxidante (EO) (SIES, 1993). Os danos oxidativos
induzidos nas células e tecidos têm sido relacionados com a etiologia de várias doenças
degenerativas tais como as cardiopatias, aterosclerose e problemas pulmonares (AMES et
al., 1993; WITZUM, 1994; STAHL & SIES, 1997). Esse importante papel no
desenvolvimento de patologias vinculado aos radicais livres se deve a razão de que todos os
componentes celulares são suscetíveis à ação das EROs. No entanto, vale salientar que o
estrogênio tem ação antioxidante e sua ausência acarreta danos no perfil oxidativo (NIKKI
et al.,1990, KIM et al., 1996). Em um estudo de Hernándes e colaboradores (2000) foi

77
avaliado se a ausência do estrogênio estava associada a aumento do estresse oxidativo e
diminuição do NO. Nesse estudo foi observado que a reposição estrogênica em ratas
ooforectomizadas preveniu a perda dos níveis de nitritos/nitratos, a redução do nível
antioxidante e o aumento de lipoperóxidos. Nossos resultados, anteriormente publicado,
realmente demonstram aumento do estresse oxidativo, o qual pode ser revertido pela
reposição com 17β- estradiol (LIMA et al., 2007). Outro trabalho demonstra ainda que o
estrogênio reduz a hipercolesterolemia em suínos e a lipoperoxidação em humanos (TANG,
1996).
Além disso, o consumo crônico de frutose tem sido associado ao aumento de estresse
oxidativo, resultado do desenvolvimento de hiperglicemia, hiperinsulinemia e resistência à
insulina (THIRUNAVUKKARASAU & ANURADHA, 2004). Assim, uma conseqüência
direta dessas anormalidades metabólicas, o estresse oxidativo, parece constituir um ponto
inicial para as complicações cardiovasculares associadas com à síndrome metabólica
(OUDOT et al., 2008). De fato, o consumo de frutose promove um aumento das EROS
reduzindo a biodisponibilidade por inativação do NO peroxinitrito (OUTDOT et al., 2008).
Estudos demonstraram que a utilização desse açúcar altera a produção de radicais livres e a
defesa antioxidante em ratos, conduzindo para um aumento suscetível de lipoperoxidação
(BUSSEROLLES et al., 2002). Dessa forma, postula-se que a membrana plasmática é uma
das estruturas mais atingidas, fato esse reconhecido como peroxidação lipídica ou
lipoperoxidação, que acarreta alterações na estrutura e na permeabilidade das membranas
celulares (MELLO FILHO et al., 1983). Sabe-se que a lipoperoxidação está associada ao
aumento da morbidade cardiovascular em indivíduos velhos (PATRICO et al., 2002) e com
diabetes (LIGUORI et al., 2001). Thirunavukkarasau e colaboradores (2004) mostraram um

78
aumento da lipoperoxidação em ratos que consumiam frutose. Tal fato pode estar
relacionado com a associação do aumento da circulação da glicose. Por sua vez, a
hiperglicemia tem efeito direto na oxidação lipídica e modificação da proteína assim como
na formação de radicais livres durante o processo da glicação.
De fato, no presente estudo observamos aumento da QL no grupo FOS se
comparado a dados anteriormente publicados por nosso grupo (IRIGOYEN et al., 2005). É
importante salientarmos que a lipoperoxidação se inicia com o seqüestro do hidrogênio do
ácido graxo poli-insaturado da membrana celular promovendo, dessa forma, a perda da
seletividade na troca iônica e liberação do conteúdo de organelas, e formação de produtos
citotóxicos culminando com a morte celular (HERSHKO, 1989).
Outro importante agente antioxidante presente na maioria das células é a glutationa
reduzida (GSH). A GSH pode ser considerada um dos agentes mais importantes do sistema
de defesa antioxidante da célula, protegendo-a contra a lesão resultante da exposição a
agentes como o íon ferro, a radiação e à luz utravioleta. Quando exposta ao agente
oxidante, ocorre sua oxidação e forma-se a glutationa oxidadada (GSSG). A recuperação da
GSH é feita através da enzima glutationa redutase, etapa essencial para manter íntegro o
sistema de proteção celular (GILBERT & MC LEAN, 1990).
Em situações em que o sistema de óxido-redução está integro, haverá recuperação
da GSH. Entretanto, sob condições de excesso de agentes oxidantes e/ou deficiência do
sistema protetor, haverá desequilíbrio entre o consumo de GSH e a produção de GSSG, o
que caracteriza o estresse oxidativo (HALLIWELL, 1993). Dessa forma, a magnitude do
estresse oxidativo pode ser monitorada pela razão GSH/GSSG. De forma semelhante ao
observado com as outras enzimas antioxidantes, estudos demonstram que o treinamento

79
físico também produz efeitos sobre a enzima glutationa reduzida. Entretanto, os dados
ainda são muito controversos, tendo o aumento e/ou a diminuição da GSH nos diferentes
tipos musculares sido encontrados em diversos estudos.
O estresse oxidativo pode ainda contribuir para o início e manutenção da
hipertensão através da inativação do NO (GIRARD et al., 2006). No presente estudo, a
razão GSH/GSSG mostrou-se reduzida no grupo FOHS em relação ao grupo FOS,
indicando maior estresse oxidativo decorrente da associação de fatores de risco. Assim,
esses resultados sugerem o aumento da produção EROS com conseqüente inativação da NO
em decorrência da hipertensão arterial (SAGAR et al., 1992; SUZUKI et al., 1998).
Todavia, o achado mais importante deste estudo foi a redução da QL cardíaca no
grupo FOT em comparação ao grupo FOS, bem como o aumento da razão GSH/GSSG nos
grupos treinados (apesar de não termos a comparação estatística entre os grupos FOS e
FOT por problemas de “n”), demonstrando que esta abordagem foi eficaz em reduzir o
estresse oxidativo. Tal achado provavelmente está associado à melhora autonômica nesse
grupo (FOT), já que inclusive observamos correlações da QL com o RMSSD e o índice
alfa. Essa correlação com a sensibilidade barorreflexa espontânea baseia-se no fato que a
redução do estresse oxidativo poderia induzir alterações na complacência arterial, alterando
a aferência do barorreflexo. Segundo o conceito mecâno-elástico aplicado sobre os
barorreceptores, quanto maior a complacência vascular sob a mesma pressão de pulso,
maior será a ativação dos pressorreceptores (KIRCHHEIM, 1976) e, portanto melhora o
controle barorreflexo arterial. Relacionado a este paradigma, o treinamento físico tem se
mostrado eficaz em aumentar a complacência vascular tanto em ratos (KINGWELL et al.,

80
1997) como em humanos saudáveis (CAMERON & DART, 1994) e, mais especificamente,
em ratos geneticamente hipertensos (ULRIKA et al., 2004). Assim, é possível sugerir que
após o treinamento físico, a complacência arterial estaria melhorada nos leitos vasculares,
incluindo as artérias aorta e carotídeas, aprimorando a transdução mecânica dos
pressorreceptores e, conseqüentemente, o controle barorreflexo arterial.
Nesse sentido, Bertagnolli e colaboradores (2006) demonstraram, em ratos
espontaneamente hipertensos, que após 10 semanas de treinamento físico ocorria uma
expressiva melhora no estresse oxidativo da artéria aorta, possivelmente, aumentando a sua
complacência. Além disso, esses autores (BERTAGNOLLI et al., 2006) observaram
associação direta entre a diminuição no estresse oxidativo e o aumento no controle
barorreflexo da freqüência cardíaca desses animais. Recentemente demonstramos também
melhora da sensibilidade barorreflexa associada a redução do estresse oxidativo e aumento
das enzimas antioxidativas (IRIGOYEN et al., 2005). Outros estudos abordam redução de
estresse oxidativo como forma de alteração benéfica da sensibilidade barorreflexa atuando
no aumento da biodisponibilidade do óxido nítrico, que em mulheres pós menopausa pode
estar comprometido devido a privação dos hormônios ovarianos (HERNANDEZ et al.,
2000; MULLAN et al., 2002).
Sabe-se, ainda, que a hipertensão arterial leva, ao longo do tempo, a importantes
alterações nas estruturas vasculares, tais como, o espessamento da parede vascular e
conseqüentemente o lúmen do vaso arterial (HEERKENS et al., 2007). Alguns autores
(WIJNEN et al., 1991; HUONKER et al., 1996) documentaram que indivíduos saudáveis
treinados possuem a espessura intima média e a razão parede/luz reduzidas quando
comparados aos seus pares sedentários. Na hipertensão arterial experimental, estudos

81
realizados em animais geneticamente hipertensos tem mostrado que 13 semanas de
treinamento físico aeróbio promovem um positivo remodelamento do leito vascular dos
pacientes hipertensos e que essas adaptações estariam envolvidas, de fato, na melhora da
complacência vascular desses pacientes (AMARAL et al., 2000; MELO et al., 2003).
Entretanto, não podemos excluir a possibilidade de que a melhora desse reflexo
esteja associada a outras alterações nos componentes centrais do ramo barorreflexo. Neste
aspecto, Pan e colaboradores (2007) verificaram que o treinamento físico preveniu a
disfunção barorreflexa provocada pela administração central de angiotensina II em ratos
previamente saudáveis. Especificamente na hipertensão arterial, Felix e colaboradores
(2007) observaram recentemente que o treinamento físico normaliza os elevados níveis
centrais de RNA mensageiro do angiotensinogênio em ratos espontaneamente hipertensos.
Esses autores sugerem que a normalização dos níveis do RNA mensageiro do
angiotensinogênio pode ser um possível mecanismo relacionado à melhora do controle
barorreflexo arterial observado na hipertensão arterial após o treinamento físico.
Adicionalmente, Kishi e colaboradores (2003) demonstraram que o aumento na produção
de óxido nítrico sintase endotelial na região do bulbo ventrolateral rostral melhorava o
controle barorreflexo da freqüência cardíaca de ratos espontaneamente hipertensos.
A diminuição do estresse oxidativo tem sido vinculada à melhora da atividade das
enzimas antioxidantes e conseqüente melhora do perfil oxidativo (SCHENEIDER &
OLIVEIRA, 2004). O treinamento físico diminuiu a peroxidação lipídica, sendo sua
melhora associada ao aumento da CAT e manutenção da concentração da enzima SOD no
músculo esquelético (ALESSIO & GOLDFARB, 1988). A enzima SOD é o principal
mecanismo de inativação do radical superóxido durante o exercício e/ou após o exercício.

82
Estudos já demonstraram de forma consistente que a atividade da SOD está aumentada
após uma sessão de exercício (JI et al., 1990; QUINTANILHA et al., 1983) bem como após
um período de treinamento físico (HIGUSHI et al., 1985; SEN et al., 1992). Essa resposta
aumentada verificada na atividade da SOD é considerada uma adaptação do sistema para a
aumentada produção de superóxido induzido pelo exercício físico (JI, 1993). No presente
estudo não encontramos diferenças entre os grupos estudados na atividade da enzima SOD.
Este fato é interessante, pois a sobrecarga de frutose (10%) pode induzir uma redução na
atividade da SOD em animais machos (BUSSEROLLES et al., 2003; BUSSEROLLES et
al., 2002; GIRARD et al., 2006)
Outra enzima atuante no balanço redox da célula é a glutationa peroxidase GPx,
considerada uma das enzimas chaves que fazem parte das defesas antioxidantes primárias
(MILLS, 1957; MILLS, 1960). A família das glutationas peroxidases (GPx) removem o
H2O2 acoplando sua redução à água com a oxidação da glutationa reduzida (GSH). A GPx
se utiliza de uma variedade de doadores de elétrons e também de glutationa reduzida
(GSH), esta por sua vez pode ser oxidada (GSSG) pelo peróxido de hidrogênio,
removendo-o e formando água. A glutationa peroxidase também catalisa a redução de
lipoperóxidos, prevenindo, desta forma, a lipoperoxidação, ou seja, impedindo assim a fase
de propagação desse processo (DEL MAESTRO, 1980; SOUTHORN & POWIS, 1988). A
GPx tem alta afinidade no fígado e nos eritrócitos, moderada atividade no coração e nos
pulmões e baixa atividade no músculo (MILLS, 1960). Estudos demonstraram que o
consumo de frutose ou a hipertensão espontânea podem promover uma redução da
atividade da GPx, associada com um aumento da TBARS e com uma diminuição da
atividade da SOD no sangue (KOJO, 2004; CHAUDIERRE & FERRARI, 1999). Nossos

83
resultados demonstraram não haver diferença significativa entre os grupos estudados na
atividade da GPx no tecido cardíaco, de acordo com resultados obtido no tecido cardíaco de
fêmeas somente ooforectomizadas (IRIGOYEN et al., 2005)
Adicionalmente, entretanto, de forma um pouco menos consistente, estudos
demonstram um aumento na atividade da enzima CAT em reposta a uma sessão aguda de
exercício (JI et al., 1992) e também ao treinamento físico (QUINTANILHA et al., 1984;
OH-TSHI et al., 1997). No presente estudo, observamos que os grupos FOT e FOHT
apresentaram um aumento na atividade da enzima CAT em relação ao grupo FOS,
reforçando o papel do treinamento físico no aumento das defesas antioxidantes com
conseqüente melhora do perfil oxidativo.

84
6. CONCLUSÃO
Concluindo, os resultados demonstraram prejuízos no perfil metabólico e
autonômico em ratas ooforectomizadas sedentárias normotensas tratadas com sobrecarga
frutose, e que a hipertensão induziu disfunções hemodinâmica e autonômica adicionais
nesses animais.
Entretanto, o achado mais importante de nosso trabalho foi que o treinamento físico
atenuou algumas dessas disfunções decorrentes da privação dos hormônios ovarianos e do
consumo crônico de frutose, pelo menos em parte associado à redução do estresse
oxidativo; todavia, a presença de hipertensão aboliu alguns benefícios observados no grupo
normotenso treinado.

85
7. REFERÊNCIAS BIBLIOGRÁFICAS ADAMS, S.; WILLIAMS, V.; VESSEY, M.P. Cardiovascular disease and hormone replacement treatment: a pilot case-control study. BMJ , v.13, p.510-516, 1995. AIELLO E.J., YASUI Y., TWOROGER S.S., ULRICH C.M., IRWIN M.L., BOWEN D., SCHWARTZ R.S., KUMAL C., POTTER J.D, McTIERNAN A. Effect of yearlong, moderate-intensity exercise intervention on the occurrence and severity of menopause symptoms in postmenopausal women. Menopause. 2004 11(4):382-388. ALBERTI, F.G.M.M.; ZIMMET, P.Z. For the WHO Consultation. Definition, diagnosis and classification of diabetes mellitus and ist complications. Part 1: Diagnosis and classification of diabetes mellitus, provisional report of a WHO consultation. Diabet Med, v.15, p.539-5539, 1998. ALESSIO H M, GOLDFARB A H. Lipid peroxidation and scavenger enzymes during exercise: adaptive response to training. J Appl Physiol n.64, p.1333-1336, 1988. AMES BN, SHIGENAGA MK, and HAGEN TM. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. PNAS; 90: 79151993. AMODEO, C. & HEIMANN, J.C. Endotélio e hipertensão arterial sistêmica: mecanismos de lesão/novo alvo terapêutico? Rev Soc Cardiol Estado de São Paulo, v.1, p.121-129, 2003. ANDERSON, J.W.; STORY, L.J.; ZETTWOCH, N.C.; GUSTAFSON, N.J.; JEFFERSON, B.S. Metabolic effects of fructose supplementation in diabetic individuals. Diabetes Care, v.12, p.337-344, 1989. ANTUNES, S.; MARCELINO, O.; AGUIAR, T. Fisiopatologia da menopausa. Port Clin Geral. v.19, p.353-357, 2003. ASIKAINEN, T.M.; KUKKONEN-HARJULA, K.; MIILUNPALO, S. Exercise for health for early postmenopausal women: a systematic review of randomized controlled trials. Sports Med, v.34, n.11, p.753-778, 2004. BACON, S.L.; SHERWOOD, A.; HINDERLITER, A.; BLUMENTHAL, J.A. Effects of exercise, diet and weight loss on high blood pressure. Sports Med, v.34, p.207-316, 2004. BARNEY, J.A.; EBERT, T.J.; GROBAN, L.; FARREL, P.A.; HUGHES, C.V.; SMITH, J.J. Carotid baroreflex responsiveness in high-fit and sedentary young men. Journal of Apllied Physiology, n.65, p.2190-2194, 1988. BARON, A.D & CLARK M.G. Role of blood flow in the regulation of muscle glucose uptake. Annu Rev Nutr. p. 487-499, 1997.

86
BASCIANO, H.; FEDERICO, L.; ADELI, K. Fructose, insulin resistance and metabolic dyslipidemia. Nutrition & Metabolism , n.5, p.1-14, 2005. BATISTA, M. & RODRIGUES, J.O. Alterações metabólicas. Journal Bras. De Nefrologia, , v.3, n. 26, p.15-19. 2004 BEDFORD, T.G. & TIPTON, C.M. Exercise training and the arterial baroreflex. Journal of Apllied Physiology, n.63, p.1926-1932, 1987. BEHR-ROUSSEL D.; CHAMIOT-CLERC P.; BERNABE, J.; MEVEL K.; ALEXANDRE L, SAFAR ME, GIULIANO F. Erectile dysfunction in spontaneously hypertensive rats: pathophysiological mechanisms.Am J Physiol Regul Integr Comp Physiol. n.284 v.3, p.682-8, 2003. BERRY, C.M.J.; BROSNAN, J; FENNELL, C.A.; HAMILTON, A.F. Oxidative stress and vascular damage in hypertension. Curr Opin Nephrol Hypertens, n.10, p.247-255, 2001. BERTAGNOLLI, M.; CAMPOS, C.; SCHENKEL, P.C.; DE OLIVEIRA, V.L.; DE ANGELIS, K.; BELLO-KLEIN, A.; RIGATTO, K.; IRIGOYEN, M.C. Baroreflex sensitivity improvement is associated with decreased oxidative stress in trained spontaneously hypertensive rat. J Hypertens, v.24, n.12, p.2437-2443, 2006. BEUTLER, E; DURAN, O; KELLY, BM. Improved method for determination of blood glutatione. J Lab Clin Med, n.61, p.802-888, 1963. BEZERRA, R.M.N.; BUENO, M.; SILVA, M.S.; TAVARES, D.Q.; CARVALHO, C.R.O.; SAAD, M.J.A.; GONTIJO, J.A.R. A high-fructose diet induces insulin resistance but not blood pressure changes in normotensive rats. Brazilian Journal of Medical and Biological Research, 34: 1155-1160, 2001. BIEGER , W.P.; MICHEL, G.;BARWICH , D.; WIRTH , A.; Diminihed insulin receptors on monocytes and erytrhrocytes in hypertriglyceridemia. Metabolism, 33; p. 982-987, 1984. BONORA, E.; MOGHETTI, P.; ZANCANARO, C.; CIGOLINI, M.; QUERENA, M.; CACCIATONI, V.; CORGNATI, A.; MUGGEO, M. Estimates of in vivo insulin action in man: comparison of insulin tolerance tests with euglycemic and hyperglycemic glucose clamp studies. Journal of Clinical Endocrinology and Metabolism, n.68, p.374-378, 1989. BRAY, G.A.; NIELSEN, S.J.; POPKIN, B.M. Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. Am J Clin Nutr , n.79, p.537-543, 2004. BOUCHARD, C. Atividade física e obesidade. Barueri-Sp: Editora Manole, 2003.

87
BOVERIS, A.; CHANCE, B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem J, n.134, p.707-716, 1973.
BOVERIS, A.; FRAGA, C.G.; VARSAVSKY, A. & KOCH, O. Arch. Biochem. Biophys. 227: 534-541, (1983). BRENNER, P.F. The menopausal Syndrome. Obstet Gynecol, v.72, n.5, p.6-11, 1988. BRITO, J.O & DE ANGELIS, K. Efeitos do treinamento físico no perfil hemodinâmico e metabólico em um modelo experimental de síndrome metabólica. In: XII Simpósio Multidisciplinar da USJT, 2006, São Paulo. XII Simpósio Multidisciplinar. São Paulo. XI Simpósio Multidisciplinar da Universidade São Judas Tadeu, 2006. BRITO, J.O.; PONCIANO, K.; FIGUEROA, D; BERNARDES, N.; SANCHES, I.C.; IRIGOYEN, MC; DE ANGELIS, K. Parasympathetic dysfunction is associated with insulin resistance in fructose-fed female rats. Brazilian Journal of Medical and Biological Research, v.41, n.9, p. 804-808, 2008. BROOKS, G.A. & WHITE, T.P. Determination of metabolic and heart rate responses of rats to treadmill exercise. J Appl Physiol, v.45, n.6, p.1009-15, 1978. BRUM, P.C.; DA SILVA, G.J.; MOREIRA, E.D.; IDA, F.; NEGRÃO, C.E.; KRIEGER, E.M. Exercise training increases baroreceptor gain sensitivity in normal and hypertensive rats. Hypertension, v.36, p.1018-22, 2000. BUEGE, J.A. & AUST, S.D. Microssomal Lipid Peroxidation. Methods Enzymology, n.52, p.302-309, 1978. BUSSEROLLES, J.; MAZUR, A.; GUEUX, E.;, ROCK, E.; RAYSSIGUIER, Y. Metabolic syndrome in the rat: females are protected against the pro-oxidant effect of a high sucrose diet. Exp Biol Med, v. 227, p.837-842, 2002. BUSSEROLLES, J.; GUEUX, E.;, ROCK, E.; DEMIGNE, C.; , MAZUR, A.; RAYSSIGUIER, Y. Oligofructose Protects against the Hypertriglyceridemic and Pro-oxidative.Effects of a High Fructose Diet in Rats. J Nutrition , p.1903-1908, 2003. CAI, H. & HARRISON, D.G. Endothelial dysfunction in cardiovascular diseases: the role of oxidant stress. Circ Res, n.87, p.840-844, 2000. CAMPOS, C.; BERTAGNOLLI, M.; EVANGELHO, J.; TUSSET, C.; BELLÓ –KLEIN, A.; IRIGOYEN, M.C, RIGATTO, K. Baroreflex sensitivity and oxidative stress in the LDL receptor knockout mice Experimental and Toxicologic Pathology, n.60 p.329–335, 2008. CANNON, W.B. The Wisdom of the Body. New York: W.W. Norton, 1939.

88
CARNEIRO, G.; FARIA, A.N.; FILHO, F.F.R.; GUIMARÃES, A.; LERÁRIO, D.; FERREIRA, S.R.G.; ZANELLA, M.T. Influência da distribuição da gordura corporal sobre a prevalência de hipertensão arterial e outros fatores de risco cardiovasculares em indivíduos obesos. Rev Assoc. Med. Brasil, v.49, n.3, p.306-311, 2003. CARR, M.C. & BRUNZELL, J.D. Abdominal obesity and dyslipidemia in the metabolic syndrome: importance of type 2 diabetes and familial combined hyperlipidemia in coronary artery disease risk. The Journal Of Clinical Endocrinology & Metabolism, v.89, n.6, p.2601–2607, 2003. CARROL, S.; DUDFIELD, M. What is the relationship between exercise and metabolic abnormalities? A review of the metabolic syndrome. Sports Med, v.34, p.371-418, 2004. CASTANHO, V.S.; OLIVEIRA, L.S.; PINHEIRO, H.P.; OLIVEIRA, H.C.F.; FARIA, E.C. Sex differences in risk factors for coronary heart disease: a study in a Brazilian population. BMC Public Health, v.1, n.3, 2001. CATENA, C.; GIACCHETTI, G.; NOVELLO, M.; COLUSSI, G.; CAVARAPE, A.; SECHI, L. Cellular mechanisms of insulin resistance in rats with fructose-induced hypertension. AJH , v.16, p.973-978, 2003. CICOGNA, A.C.; ROBINSON, K.G.; CONRAD, C.H.; SQUIRE, R.; OKOSHI, M.P.; BING, O.H.L. Participação do estado contrátil e do relaxamento miocárdico na disfunção ventricular durante a transição hipertrofia-falência cardíaca. Arq Bras Cardiol , v.69, n.6, 1997. CHOWDHARY, S.; VAILE, J.C.; FLETCHER, J.; ROSS, H.F.; COOTE, J.H.; TOWNEND, J.N. Nitric oxide and cardiac autonomic control in humans. Hypertension, n.36, p.264–269, 2000. CRAIG, B.W .The influence of fructose feeding on physical performance Am J Clin Nutr. n.58 (suppl) p.815S-819S. 1993 COLDITZ GA, WILLETT WC, STAMPFER MJ, ROSNER B, SPEIZER FE, HENNEKENS CH. Menopause and the risk of coronary heart disease in women. New Engl J Med, n.316, p.1105-1110, 1987. CUNHA, T.S.; FARAH, V.A.M.; PAULINI, J.; PAZZINE, M.; ELASED, K.; MARCONDES, F.; IRIGOYEN, M.C.; DE ANGELIS, K.; MORRIS, M. Relationship between renal and cardiovascular changes in a murine model of glucose intolerance. Regulatory Peptides, v.1, n.139, s.1-3, p.1-4, 2007. DALL'AGO, P.; SCHAAN, B.D.; SILVA, V.O.; WERNER, J.; SOARES, P.P.S.; DE ANGELIS, K.; IRIGOYEN, M.C. Parasympathetic dysfunction is associated with baroreflex and chemoreflex impairment in streptozotocin-induced diabetes in rats. Autonomic Neuroscience: Basic and Clinical, v.131, p.28-35, 2007.

89
DALY PA, LANDSBERG L. Hypertension in obesity and NIDDM. Role of insulin and sympathetic nervous system. Diabetes Care, v.14, p.240-248, 1991. D´ANGELO, G.; ELMARAKBY, A.A.; POLLOCK, D.M.; STEPP, D.W. Fructose feeding increases insulin resistance but not blood pressure in sprague-dawley rats. Hypertension, v.46, p.806-811, 2005. DASKALOPOULOU, S.S.; MIKHAILIDIS, D.P.; ELISAF, M. Prevention and treatment of the metabolic syndrome. Angiology, n.55, p.589-612, 2004. DAWSON-HUHGES, B & HARRIS, S. Regional changes in body composition by time of year in healthy postmenopausal women. Am J Clin Nutri . v.3,n.56, p.307-313,1992. DE ANGELIS, K.L.D.; OLIVEIRA, A.R.; WERNER, A.; BOCK, P.; BELLÓ-KLEIN, A.; IRIGOYEN, M.C. Exercise training in aging: hemodynamis, metabolic, and oxidative stress evaluations. Hypertension, v.30, n.3, p. 767-771, 1997. DE ANGELIS, K.; GADONSKI, G.; FANG, J.; DALL’AGO, P.; ALBUQUERQUE, V.L.; PEIXOTO, L.R.A.; FERNANDES, T.G.; IRIGOYEN, M.C. Exercise reverses peripheral insulin resistance in trained L-NAME-hypertensive rats. Hypertension, v.34, n.2, p.768-772, 1999. DE ANGELIS, K.; OLIVEIRA, A.R.; DALL’AGO, P.; PEIXOTO, L.R.A.; GADONSKI, G.; FERNANDES, T.G.; IRIGOYEN, M.C. Effects of exercise training in autonomic and myocardial dysfunction in streptozotocin-diabetic rats. Braz J Med Biol Res, n.33, p.635-641, 2000. DE ANGELIS, K.; SCHAAN, B.D.; MAEDA, C.Y.;DALL’AGO, P.; WICHI, R.B.; IRIGOYEN, M.C. Cardiovascular control in experimental diabetes. Brazilian Journal Of Medical Biological Research, v.35, n.9, p.1091-1100, 2002. DE ANGELIS, K.; SANTOS, M.S.B.; IRIGOYEN, M.C. Sistema nervoso autônomo e doença cardiovascular. Revista de da Sociedade de Cardiologia do Rio Grande do Sul, v.3, 2004. DEL MAESTRO, RF. An Approach to Free Radicals in Medicine and Biology, in: Act Physiol Scand, Suppl. 492: 153-168, 1980 DORMANDY, T.L. Free-radical oxidation and antioxidants. Lancet, 1:647-650, 1978. ELLIOT, S.S.; KEIM, N.L.; STERN, J.S.; TEFF, K.; HAVEL, P.T. Fructose, weight gain, and the insulin resistance syndrome. Am J Clin Nutrit, n.76, p. 911-922, 2002. EWING, D.; CAMPBELL, I.; CLARKE, B. The natural history of diabetic autonomic neuropathy. Quart J Med, n.49, p.95-108, 1980.

90
FARAH, V.; DE ANGELIS, K.; CANDIDO,G.; BERNARDES, N.; JOAQUIM, L.F; SCHAAN, B.D.; IRIGOYEN, M.C. Autonomic modulation of arterial pressure and heart rate variability in hypertension diabetic rats. Clinics, v.62, n.4, p.477-482, 2007. FARAH, V.; ELASED, K.; CHEN, Y.; KEY, M.; CUNHA, T.; IRIGOYEN, M.C.; MORRIS, M. Nocturnal hypertension in mice consuming a high fructose diet. Autonomic Neuro Sciense Basic and Clinical, p.1-10, 2006. FERRANNINI, E., HAFFNER, S.M.; STERN, M.P. Essential hypertension and insulin-resistant state. J. Cardiovasc. Pharmacol, 15: S18-S25, 1990.
FLOHE, L.; GUNZLER, W.A. Assays of glutathione peroxidase. Methods Enzymol, n.105, p.114-121, 1984. FLORES L.J.F., SANCHES, I., AGUIAR, J., FLUES, K., JORGE, L., PUREZA, D.Y., IRIGOYEN, M.C., DE ANGELIS, K. Privação estrogênica induz disautonomia cardiovascular em ratas. In: XX Reunião Anual da Fesbe, 2005, Águas de Lindóia. Anais da XX Reunião Anual da Fesbe. 2005 (15): 056. FORD, E.S.; GILES, W.H. A comparision of the prevalence of the metabolic syndrome using two poposed definitions. Diabetes Care, v.26, p.575-581, 2003. FORD, E.S., GILES, W.H., MOKDAD, A.H. Increasing prevalence of the metabolic syndrome among u.s. adults. Diabetes Care, n.27, p.2444–2449, 2004. FRANCHINI, K.G & KRIEGER, E.M. Bradycardic responses to vagal stimulation and methacholine injection in sino-aortic denervated rats. Brazilian Journal Of Medical Biological Research, n.22, p.757-60, 1989. FRANCISCHI, R.P. Obesidade: atualização sobre sua etiologia, morbidade e tratamento. Rev Nutr, v.13, p.17-28, 2000. FRICK, M.H. The mechanisms of bradycardia evoked by physical training. Cardiologia, n.51, p.46-54, 1967. GALIPEAU D, VERMA S, MCNEILL JH. Female rats are protected against fructose induced changes in metabolism and blood pressure. Am J Physiol Heart Circ Physiol 2002; 283: 2478-2484. GANG, H.; QIAO, Q.; TUOMLIEHTO, J.; BALKAU, B.; BORCH-JOHNSEN, K.; PYORALA, K. For the DECODE Study Group. Prevalence of the metabolic syndrome and its relation to all-cause and cardiovascular mortality in nondiabetic European men and women. Arch Intern Med , v.164, p.1066-1076, 2004.

91
GAVA, N.S.; VÉRAS-SLIVA, A.S.; NEGRÃO, C.E.; KRIEGER, E.M. Low-intensity exercise training attenuates cardiac �-adrenergic tone during exercise in spontaneously hypertensive rats. Hypertension, v.26, p.1129-33, 1995. GIRARD, A.; MADANI, S., BOUSTANI,E.S.E.; BELLEVILLE, J.; PROST. Changes in lipid metabolism and antioxidant defense status in spontaneously hypertensive rats and Wistar rats fed a diet enriched with fructose and saturated fatty acids., Nutrition v.21 p.240–248, 2005. GIRARD A, MADANI S, BOUKORTT F, CHERKAOUI-MALKI M, BELLEVILLE J, PROST J. Fructose-enriched diet modifies antioxidant status and lipid metabolism in spontaneously hypertensive rats. Nutrition . v.22 p.758-66, 2006 GIRMAM, C.J.; RHODES, T.; MERCURI, M.; PYORALA, K.; KJEKSHUS, J.; PEDERSENT, R. et al. for the 4S Group and the AFCAPS/TexCAPS Research Group. The metabolic syndrome and risk of major coronoary events in the Scandinavian Simvastatin Survival Study (4S) and the Air Force/Texas Coronary Atherosclerosis Prevention Study (AFCAPS/TexCAPS). Am J Cardiol , v.93, p.136-141, 2004. GOFF Jr., D. C.; HAFFNER, S.M.; ZACCARO, D.J.; SAAD, M.F. Insulin sensitivity and the risk of incident hypertension. Diabetes Care, v.26, n.3, p.805-809, 2003. GONZALEZ FLECHA, B.; LLESUY, S.; BOVERIS, A. Hydroperoxide-initiated chemiluminescence: an assay for oxidative stress in biopsies of heart, liver, and muscle. Free Radical Biol Med, n.10, p.1093-100, 1991. GRANGER, D.L.; ANSTEY, N.M.; MILLER, W.C.; WEINBERG, J.B. Measuring nitric oxide production in human clinical studies. Methods in Enzymology, n.301, p.58-61, 1999. GREEN J.S., STANFORTH P.R., GAGNON J., LEON A.S., RAO D.C., SKINNER J.S., BOUCHARD C., RANKINEN T. WILMORE J.H. Menopause, estrogen, and training effects on exercise hemodynamics the HERITAGE study. Med Sci Sports Exerc,v.2, n.34, p.74-82, 2002. GREGOIRE, J.; TUCK, S.; YAMAMOTO, Y.; HUGHSON, R. Heart rate variability at rest and exercise: influence of age, gender and physical training. Can J Apll Physiol, v.21, n.6, p.455-470, 1996. GRYGLEWSKI, R.J.; PALMER, R.M.; MONCADA, S. Superoxide anion is involved in the breakdown of endothelium-derived vascular relaxing factor. Nature, n.320, p.454-456, 1986. GUIMARÃES, G.V & CIOLAC, E.G. Síndrome metabólica: abordagem do educador físico. Rev Soc Cardiol de São Paulo, v.14 , pp. 659-70, 2004.

92
GUYTON, A.C & HALL, J.E. Fisiologia Médica. 10.ed. Rio de Janeiro: Guanabara Koogan, 2002. HALLFRISCH, J.; LAZAR, F.; JORGENSEN, C.; REISER, S. Insulin and glucose responses in rats fed sucrose or starch. Am J Clin Nutr , 32:787-793, 1979. HAFFNER, S.; TAEGTMEYER, H. Epidemic obesity and the metabolic syndrome. Circulation , v.108, p.1541-1545, 2003. HAGBERG, J.M.; PARK, J.J.; BROWN, M.D. The role of exercise training in the treatment of hypertension: an update. Sports Med, v.30, p.193-206, 2000. HALLIWELL B. Reactive oxygen species and the central nervous system. J Neurochem. 59:1609-23, 1992. HALPEN, A. A importância da obesidade. Diab Metab, 2: 114-115,1998. HARDMAN, A.E. Exercise in prevention of atherosclerotic, metabolic and hipertensive diseases: a review. Journal of sports Science, v.14, n.3, p.201-218, 1996. HARTHMANN, A.D.; DE ANGELIS, K.; COSTA, L.P.; SENADOR, D.; SCHAAN, B.D.; KRIEGER, E.M.; IRIGOYEN, M.C. Exercise training improves arterial baro- and chemoreflex in control and diabetic rats. Auton Neurosci, n.133, p.115-120, 2007. HASSAGER, C. & CHRISTIANSEN, C. Estrogen/gestagen therapy changes soft tissue body composition in postmenopausal women. Metabolism. v.7, n.38, p.662-665, 1989. HENRISSEN, E.J. Exercise effects of muscle insulin signaling and action. Invited review: effects of acute exercise and exercise training on insulin resistance. J Appl Physiol, v.93, p.788-796, 2002. HÉRNANDEZ, I.; DELGADO, J.L,; DIAZ, J.; QUESADA, T.; TERUEL, M.J.G.; CARMENL, M.; CARBONELL, L.F. 17�-Estradiol prevents oxidative stress and decreases blood pressure in ovariectomized rats. Am J Physiol Regulatory Integrative Comp Physiol. n.279, p.R1599-R1605, 2000. HERSHKO C. Mechanism of iron toxicity and its possible role in red cell membrane damage. Semin Hematol. 26(4):p. 277-85, 1989. HOUMARD, J.A.; TANNER, C.J.; SLENTZ, C.A. et al. The effect of the volume and intensity of exercise training on insulin sensitivity. J Appl Physiol, v.96, p.101-106, 2004. HSIEH, P.S. Reversal of fructose-induced hypertension and insulin resistance by chronic losartan treatment is independent of AT2 receptor activation in rats. J.Hypertens. 2005 23: 2209-2217.

93
HUIKURI, H.V.; PIKKUJAMSA, S.M.; AIRAKSINEN, J.; IKAHEIMO, M.J.; RANTALA, A.O.; KAUMA, H.; LILJA, M.; KESANIEMI, Y.A . Sex-related differences in autonomic modulation of heart rate in middle-aged subjects. Circulation , v.94, p.122-125, 1996. IRIGOYEN, M.C & KRIEGER, E.M. Baroreflex control of sympathetic activity in experimental hypertension. Brazilian Journal Of Medical Biological Research, n.31,p.1213-20, 1998. IRIGOYEN, M.C.; LACCHINI, S.; DE ANGELIS, K.; MICHELINI, L.C. Fisiopatologia da hipertensão: o que avançamos? Rev Soc Cardiol Estado São de Paulo, v.13, n.1, p.20-45, 2003. IRIGOYEN, M.C.; PAULINI, J.; FLUES, K.; FLORES, L.J.F.; CONSOLIM-COLOMBO, F.M.; MOREIRA, E.D.; BELLÓ-KLEIN, A.; DE ANGELIS, K. Exercise training improves baroreflex sensitivity associated with oxidative stress reduction in ovariectomized rats. Hypertension, v.46, n.2, p.998-1003, 2005. IRWING, M.L.; YASUI, Y.; ULRICH, C.M.; BOWEN, D.; RUDOLPH, R.E.; SCHWARTZ, R.S.; YUKAWA, M.; AIELLO, E.; POTTER, J.D.; e MCTIERNAN, A. Effects of exercise on total and intra-abdominal body fat in postmenopausal women: a randomized controlled trial. JAMA . n.289, p.323-330, 2003. ISHISE, HISANARI, HIDETSUGU ASANOI, SHINJI ISHIZAKA, SHUJI JOHO, TOMOKI KAMEYAMA, KATSUMI UMENO, AND HIROSHI INOUE. Time course of sympathovagal imbalance and left ventricular dysfunction in conscious dogs with heart failure. J. appl. physiol. 84(4): 1234–1241, 1998. ISSA, J.S.; FRANCISCO, Y.A. Obesidade e doenças cardiovasculares. Revista Soc Cardiol São Paulo, v.5, p.558-630, 1996. JI, L.L. & FU, R. Responses of glutathione system and antioxidant enzymes to exhaustive exercise and hidroperoxide. J Appl Physiol, n.72, p.549-54, 1992. JOAQUIM, L.F.; SALGADO, H.C.; FAZAN, R. Variabilidade da pressão arterial e da freqüência cardíaca, e sensibilidade do barorreflexo, em animais geneticamente manipulados. Revista Brasileira de Hipertensão, v. 12, p. 36-40, 2005. JURCA, R.; CHURCH, T.S.; MORSS, G.M.; JORDAN, A.N.; EARNEST, C.P. Eight weeks of moderate-intensity exercise training increases heart rate variability in sedentary postmenopausal women. Am Heart J, v.147, n.5, p.21, 2004. KAMIDE K, RAKUGI H, HIGAKI J, OKAMURA A, NAGAI M, M ORIGUSHI K et al. The renin-angiotensin and adrenergic nervous system in cardiac hypertrophy in fructose-fed rats. Am J Hypertens. 2002 (15): 66-71.

94
KAMITANI, A.; KAWAMOTO, R.; TOMITA, H.; OKA, Y.; KO DAMA, A. Metabolic syndrome amplifies de LDL –cholesterol associated increases in carotidatherosclerosis. Internal Medicine, v.44, n.12, 2005. KANNEL, W..B. & MCGEE, D.L. Diabetes and cardiovascular disease: the Framingham study. JAMA , n.241, p.2035-2038, 1979. KASETA, J.R.; SKAFAR, D.F.; RAM, J.L.; JACOBER, S.J.; SOWERS, J.R. Cardiovascular disease in the diabetic woman. J Clin Endocrinol Metab, v.84, p.1835–1838, 1999. KASSIS, E. & AMTORP, O. Cardiovascular and neurohumoral postural responses and baroreceptor abnormalities during a course of adjunctive vasodilator therapy with felodipine for congestive heart failure. Circulation , v.75, p.1204–1213, 1987. KATONA, P.G.; McLEAN, M.; DIGHTON, D.H.; GUZ, A. Sympathetic and parasympathetic cardiac cointrol in athletes and nonathletes at rest. Journal of Applied Physiology, v.52, p.1652-1657, 1982. KATOVICH,M.J., REAVES,P.Y., FRANCIS,S.C. ET AL. Gene therapy attenuates the elevated blood pressure and glucose intolerance in an insulin-resistant model of hypertension. J Hypertens. 2001 19: 1553-1558. KELLEY, G.A. Aerobic exercise and resting blood pressure among women: a meta-analysis. Prev Med, v.28, n.3, p.264-75, 1999. KIM, D. H., H. IIJIMA, K. GOTO, J. SAKAI, H. ISHII, H. J. KIM, H. SUZUKI, H. KONDO, S. SAEKI, AND T. YAMAMOTO. Human apolipoprotein E receptor 2. A novel lipoprotein receptor of the low density lipoprotein receptor family predominantly expressed in brain. J. Biol. Chem. n. 271, p.8373–8380, 1996. KIRWAN, L.D., MERTENS, D.J., KAVANAGH, T. THOMAS S.G. e GOODMAN, J.M. Exercise training in women with heart disease: influence of hormone replacement therapy. Med Sci Sports Exerc. v.2, n.35, p.185-192, 2003. KLEIGER, R.E.; MILLER, J.P., BIGGER, J.T.; MOSS, A.J. Decreased heart rate variability and its association with increased mortality after acute myocardial infarction. Am J Cardiol , v.59, n.4, p.256-62, 1987. KNOWLER, W.C.; BARRET-CONNOR, E.; FOWLER, S.E. et al. Reduction of the incidence of type II diabetes with lifestyle intervention of metformin. N Engl J Med, v.344, p.393-403, 2002. KOTCHEN, T.A.; REDDY, S.; ZHANG, H.Y. Increasing insulin sensitivity lowers blood pressure in the fructose-fed rat. AJH , 10: 1020-1026, 1997.

95
KUO, T.B.; LIN, T.; YANG, C.C.; LI, C.L.; CHEN, CF; CHOU, P. Effect of aging on gender differences in neural control of heart rate. Am J Physiol (Heart Circ Physiol), v.277, n.46, p.H2233-2239, 1999. LA ROVERE, M.T.; BERSANO, C.; GNEMMI, M.; SPECCHIA, G.; SCHWARTZ, P.J. Exercise-induced increase in baroreflex sensitivity predicts improved prognosis after myocardial infarction. Circulation, v.106, n.8, p.945-9, 2002. LA ROVERE, M.T.; BIGGER, J.T. JR; MARCUS, F.I.; MORTARA, A.; SCHWARTZ, P.J. Baroreflex sensitivity and heart-rate variability in prediction of total cardiac mortality after myocardial infarction. ATRAMI (Autonomic Tone and Reflexes After Myocardial Infarction) Investigators. Lancet, v.351, p.478-84, 1998. LAITINEN, T.; HARTIKAINEN, J.; VANNINEN, E.; NISKANEN, L.; GEELEN, G.; LANSIMIES, E. Age and gender dependency of baroreflex sensitivity in health subjects. J Appl Physiol, v.84, n.2, p.576-583, 1998. LAKKA, H.M.; LAAKSONDEN, D.E.; LAKKATA, NISKANEM, L .K.; KUMPUSALO, E.;TUOMILEHTO, J. et al. The metabolic syndrome and total cardiovascular disease mortality in middle-aged men. JAMA , v.288, p2709-2716, 2002. LATOUR, M.G.; SHINODA, M.; LAVONE, J.M. Metabolic effects of physical training in ovariectomized and hyperestrogenic rats. Am J Physiol, v.90, p.235-241, 2001. LEINWAND, L.A. Sex is a potent modifier of the cardiovascular system. The Journal of Clinical Investigation, v.112, n.3, p.302-307, 2003. LI, Z.; MAO, H.Z.; ABBOUD, F.M.; CHAPLEAU, M.W. Oxygen-derived free radicals contribute to baroreceptor dysfunction in atherosclerotic rabbits. Circ. Res, n.79, p.802–811, 1996. LIGUORI A, ABETE P, HAYDEN JM, CACCIATORE F, RENGO F, AMBROSIO G, BONADUCE D, CONDORELLI M, REAVEN PD, NAPOLI C. Effect of glycaemic control and age on low-density lipoprotein susceptibility to oxidation in diabetes mellitus type 1. Eur Heart J v.22 p.2045– 2047, 2001 LIMA, S.R.R.M.; BELÓ-KLEIN, A.; FLUES, K.;PAULINI, J.;MONTE, O.;IRIGOYEN, M.C.; DE ANGELIS, K. Efeitos da suplementação do 17β estradiol no dano oxidativo cardíaco de ratas submetidas a privação dos hormônios ovarianos. Revista Brasileira de Ginecologia e Obstetrícia, Rio de Janeiro, v.29, n. 1, p. 27-33, 2007. LLESUY, S.F.; MILEI, J.; MOLINA, H.; BOVERIS, A.; MILEI, S. Comparison of lipid peroxidation and myocardial damage induced by adriamycin and 4'-epiadriamycin in mice. Tumori , v.71, p.241-249, 1985.

96
LOPES, H. F. Hipertensão, Obesidade, Resistência À Insulina E síndrome metabólica. Rev Bras Hipertens, v.12, n.3, p.154-158, 2005. LOWRY, O.H.; ROSENBROUGH, N.T.; FARR, A.L. Protein measurements with the Folin phenol reagent. J Biol Chem, n.193, p.265-275, 1951. MAEDA, C.Y.; FERNANDES, T.G.; TIMM, H.B.; IRIGOYEN, M.C. Autonomic dysfunction in short-term experimental diabetes. Hypertension, n.26, p.1100–1104, 1995. MAEDA, C.Y.; SCHAAN, B.D.; OLIVEIRA, E.M.; OLIVEIRA, V.L.; DE ANGELIS, K.; IRIGOYEN, M.C. Chronic salt loading and cardiovascular-associated changes in experimental diabetes in rats. Clin Exp Pharmacol Physiol, v.34, n.7, p.574-80, 2007. MAGARITIS, I.; TESSIER, F.; RICHARD, M.J.; MARCONNET, P. No evidence of oxidative stress after a triathlon race in highly trained competitors. Int J Sports Med, n.18, p.186-90, 1997. MANCIA, G.; GRASSI, G.; FERRARI, A.U. Reflex control of circulation in experimental and human hypertension. In: ZANCHETTI, A.; MANCIA, G. Handbook of hypertension, Pathophysiology of hypertension. Amsterdam: Elsevier Science Publishers BV, p.568-601, 1997. MATSUZAWA, Y. Metabolic syndrome-definition and diagnostic criteria in Japan. J JPN Soc. Int. Med, (850): 188-203, 2005. MAYES, P.A. Intermediary metabolism of fructose. Am J Clin Nutr , 58:754S-765S, 1993. MELO RM, MARINHO E, MICHELLINE LC. Exercise training normalizes wall-tolumen ratio of the gracilis muscle arterioles and reduces pressure in spontaneously hypertensive rats. J Hipertenses, v.18, p.1563-1572, 2000. MCDONALD, P.M.; SANFILIPO, A.J.; SAVARD, G.K. Baroreflex function and cardiac structure with moderate endurance training in normotensive men. Journal of Apllied Physiology, n.4, p.2469-2477, 1993. MILLS GCJ. Glutathione peroxidase and the destruction of hydrogen peroxide in animal tissues. Arch. Biochem. Biophys. 86: 1-5, 1960. MILLS GCJ. Glutathione peroxidase and erytrocyte enzyme which protects hemoglobin from oxidative breakdoown. Biol. Chem. 229: 189-197, 1957 MILLER VM. Gender, Estrogen, and NOS. Cautions about Generalizations. Cir Res. 85:979-981, 1999.

97
MOSCA, L.; BANKA, C.L.; BENJAMIN, E.J.; BERRA, M. Evidence-based guidelines for cardiovascular disease prevention in women: 2007 update. Circulation, v.20, n.115, supl.11, p.1481-501, 2007. MORTARA A, LA ROVERE MT, SIGNORINI MG et al. Can power spectral analysis of heart rate variability identify a high risk subgroup of congestive heart failure patients with excessive sympathetic activation? A pilot study before and after heart transplantation. Br Heart J,n.71, p.422–430, 1994 MUIR, A.; SCHATZ, D.A.; MACLAREN, N.K.The pathogenesis, prediction, and prevention of insulin-dependent diabetes mellitus. Endocrinology Metabolism Clinic North American, v.21, n.2, p.199-219. 1992. MYERS J, GULLESTAD L, VAGELOS R, DO D, BELLIN D, ROSS H, FOWLER MB: Clinical hemodynamic and cardiopulmonary exercise test determinants of survival in patients referred for evaluation of heart failure. Ann Intern Med 1998, 129:286-293. NAHAS, M.V. Atividade física, saúde e qualidade de vida: conceitos e sugestões para um estilo de vida ativo. Londrina, Midiograf, 2001. NATIONAL CENTER FOR HEALTH STATISTICS. Statistical abstract of the United States. 117th edition. US Bureau of the Census. Washington, DC, USA, p.83, 1997. http://www.census.gov/prod/www/statistical-abstract-us.html. NCEP- Executive summary of the third report of the national cholesterol education program expert panel on detection, evaluation and treatment of high blood cholesterol in adults (adult treatment panel III) JAMA , n.285, p.2486-2497, 2001. NEGRÃO, C.E.; MOREIRA, E.D.; BRUM, P.C.; DENADAI, M.L.D.R.; KRIEGER, E.M. Vagal and simpathetic control of heart rate during exercise by sedentary and exercise-trained rats. Braz J Med Biol Res, n.25, p.1045-1052, 1992. NEGRÃO C.E., BARRETO A.C.P. Cardiologia do exercício – Do atleta ao cardiopata. Editora Manole. 2005 (1): 1-8. NIKI E, NAKANO M. Estrogens as Antioxidants. Methods in Enzymology. Vol 186, 330-333, 1990. NORDMANN, R. Alcohol and antioxidant systems. Alcohol, n.29, p.513-522, 1994. ORIEN, N.; ZIDEK W.; TEPEL, M. Reactive oxygen species in essential hypertension and non-insulin- dependent diabetes mellitus. Am J Hypertens, n.12, p.1169-1174, 1999. O'SULLIVAN, S.E.; BELL, C. The effects of exercise and training on human cardiovascular reflex control. J Auton Nerv Syst, n.81, p.16–24, 2000.

98
OUDOT, A.;BEHR-ROUSSEL, D.; COMPAGNIE, S.;CAISEY,S.;LE COZ O.; GORNY D.; ALEXANDRE, L.;GIULIANO, F. Endothelial dysfunction in insulin resistant rats is associatedwith oxidative stress and cox pathway dysregulation. Physiol Res, 2008 PAN, X.; LI, G.; HUY, H. et al. Effects of diet and exercise in preventing NIDDM in people with impaired glucose tolerance. The Da Qing IGT Diabetes Study. Diabetes Care, v.20, p.537-544, 1997. PANZA, J.A.; QUYYUMI, A.A.; BRUSH, JR, JE; EPSTEIN, E. Abnormal endothelium-dependent vascular relaxation in patients with essential hypertension. N Engl J Med, n.323, p.22-27, 1990. PARENTE COSTA, L.P.; HARTHMANN, A.; ARRUDA, P.; DE ANGELIS, K.; IRIGOYEN, M.C. Treinamento físico melhora a disfunção quimiorreflexa em ratos diabéticos por estreptozotocina. Rev Bras Educ Fís Esp, São Paulo, v.18, n.3, p.293-301. PRATICO D, CLARK CM, LUIN F, LEE VY, TROJANOWSKI JQ. Increase of brain oxidative stress in mild cognitive impairment: a possible predictor of Alzheimer disease. Arch Neurol, n.59, p.972–976, 2002. PEDERSEN, B.K.; SALTIN, B. Evidence for prescribing exercise as therapy in chronic disease. Scand J Med Sci Sports, v.16 ,supl.1, p.3–63, 2006. PEIXOTO, R.T., PEIXOTO, E.C., SENA, M.A., TEDESCHI, A.L., BORGES, I.P. e RACHID, M.B. Gender influence on the immediate and medium-term progression after primary percutaneous coronary intervention and analysis of independent risk factors for death or events. Arq Bras Cardiol ; 86(3):211-8, 2006. PONCIANO, K.R. Efeitos cardiovasculares e metabólicos do treinamento físico em ratas submetidas à sobrecarga de frutose e à privação dos hormônios ovarianos. Dissertação de mestrado, 2006. RABELO, E.; DE ANGELIS, K.; BOCK, P.; FERNANDES, T.; CERVO, F.; BELLO-KLEIN, A.; CLAUSSEL, N.; IRIGOYEN, M.C. Baroreflex sensitivity and oxidative stress in adriamycin-induced heart failure. Hypertension, EUA, v.38, n.2, p.576-580, 2001. RECKELHOFF JF, ZHANG H, SRIVASTAVA K. Gender differences in the development of hypertension in SHR: role of the renin angiotensin system. Hypertension, v.35, p.480-483, 2000. REISER, S.; HALLFRISCH, J. Insulin sensitivity and adipose tissue weight of rats fed starch or sucrose diets ad libitum or in meals. J Nutr, 107:147-155, 1977 REPETTO, G. A Síndrome Plurimetabólica, também chamada de Síndrome da "vida boa" ou dos "boas vidas". Diab Metab, 2: 56- 57, 1998.

99
RIBEIRO, J.P.; MORAES FILHO; R.S. Variabilidade da Freqüência Cardíaca como Instrumento de Investigação do Sistema Nervoso Autônomo. Revista Brasileira de Hipertensão, v. 12, p. 14-20, 2005. RICHARD D., ROCHON L. e DESHAIES Y. Effects of exercise training on energy balance of ovariectomized rats. Am J Physiol. v.253 p.740-745. MILLS GCJ. Glutathione peroxidase and the destruction of hydrogen peroxide in animal tissues. Arch. Biochem. Biophys. 86: 1-5, 1960. MILLS GCJ. Glutathione peroxidase and erytrocyte enzyme which protects hemoglobin from oxidative breakdoown. Biol. Chem, v.229 p.189-197, 1957. ROCCHINI, A.P.; KATSCH, V.; KVESELIS, D. et al. Insulin and renal sodium retention in obese adolescents. Hypertension, 14 : 367-374, 1989. RODRIGUES, B.; IRIGOYEN, M.C.; DE ANGELIS, K. Correlation between speed and oxygen consumption (VO2) in rats submitted to maximum exercise test. FIEP Bulletim , n.76, p.231-233, 2006. ROSS, R.; FREEMAN, J.A.; JANSSEN, I. Exercise alone is an effective strategy for reducting obesity and related comorbidities. Exerc Sport Sci Rev, v.28, p.165-170, 2000. RUBANYI, G. & VANHOUTE, M. The role of endothelium in cardiovascular homeostasis and diseases. J Cardiovasc Pharmacol, n.22, supl. 4, p.S1-14, 1993. SANCHES, I.C. Perfil metabólico e cardiovascular de ratas hipertensas submetidas a um modelo experimental de menopausa e síndrome metabólica: papel do treinamento físico Dissertação de mestrado, 2007. SCHULTZ, H.D & USTINOVA, E.E. Capsaicin receptors mediate free radical-induced activation of cardiac afferent endings. Cardiovasc Res, n.38, p.348–355, 1998. SEALS, D.R.; SILVERMAN, H.G.; REILING, M.J.; DAVY, K.P. Effect of regular aerobic exercise on elevated blood pressure in postmenopausal women. Am J Cardiol , v.80, n.1, p.49-55, 1997. SHAN, Z.; DAI, S., SU, D. Relationship between baroreceptor reflex function and end-organ damage in spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol, n.277, p.H1200- H1206, 1999. SHANGOLD, M.M. Exercise in the menopausal woman. Obstet Gynecol. 1990 75: 52s. SHINOZAKI K., AYAJIKI K., NISHIO Y., SUGARA T., KASHIWAGI A., OKAMURA T. Evidence for causal role of renin-angiotensin system in vascular dysfuntion associated with insulin resistance. Hypertension. 2004 (43): 255-262.

100
SIES, H. Biochemistry of oxidative stress. Angew Chem Int Ed Ingl, n.25, p.1058-71, 1986. SILVA, G.J.J.; BRUM, P.C.; NEGRÃO, C.E.; KRIEGER, E.M. Acute and chronic effect of exercise or baroreflexes in spontaneausly hypertensive rats. Hypertesion, n.30, p.714-719, 1997. SINAGRA, D & CONTI, M. Metabolic syndrome in menopause: physiopathological, clinic and therapeutic aspects for cardiovascular prevention. Recenti Prog Med, v.98, n.3:185-91, 2007. STEFANICK, M.L., MACKEY S., SHEEHAN M., ELLSWORTH N., HASKELL W., e WOOD P.D. Effects of diet and exercise in men and postmenopausal women with low levels of HDL-cholesterol and high levels of LDL-cholesterol. N Engl J Med, n.339, p.12-20. 1998 SONG, D.; ARIKAWA, E.; GALIPEAU, D.; BATTELL, M.; McNEILL, J.H. Androgens are necessary for the development of fructose-induced hypertension. Hypertension, n.43, p. 667-672, 2004. SOUTHORN P & POWIS G. Free Radicals in Medicine. I. Chemical Nature and Biologic Reactions, in: Mayio Clin Proc., 63: 381-389, 1988. SOUZA, S.B.C.; FLUES, K.; PAULINI, J.; MOSTARDA, C.; RODRIGUES, B.; SOUZA, L.E.; IRIGOYEN, M.C.; DE ANGELIS, K. Role of exercise training in cardiovascular autonomic dysfunction and mortality in diabetic ovariectomized rats. Hypertension, n.30, p.786-791, 2007. SOWERS, M.R. & LA PIETRA, M. Menopause: its epidemiology and potencial association with chronic diseases. Epidemiol Rev, n.17, p. 287-302, 1995. SCHNEIDER C. D, OLIVEIRA A. R. Radicais livres de oxigênio e exercício: mecanismos de formação e adaptação ao treinamento físico. Rev Bras Med Esporte. vol.10 n. 4, 2004 SCHOUW, Y.T., GRAAF, Y.; STEYERBERG, E.W,; EIJKEMANS, J.C; BANGA, J.D. Age at menopause as a risk factor for cardiovascular mortality. Lancet, 347: 714-718, 1996. SHAWAERY, G.; VITA, J.A. & KEANEY, J.F. Antioxidant protection of LDL by physiological concentrations of 17β estradiol. Circulation, n.95, p.1378-1385, 1997. STAESSEN, J.Á., GINNOCHIO G., THIJS L., FAGARD R. Conventional and ambulatory blood pressure and menopause ina prospective population study. Journal of Human Hypertension, v.11, p.507-514, 1997.

101
STAMLER, J.; VACCARO, O.; NEATON, J.D.; WENTWORTH, D. Diabetes, other factors, and 12-yr cardiovascular mortality for men screened in the multiple risk factor intervention trial. Diabetes Care, v.16, n. 2, p.434-442. 1993. SIES H & MURPHY ME. Role of Tocopherols in the protection of Biological Against Oxidative Damage. J. Photochem. Photobiol. B. Biol. 8: 211-224, 1991. SU, D.F & MIAO, C.Y. Blood pressure variability and organ damage. Clin Exper Pharmac Physiol, n.28, p.709-715, 2001. SUGAWARA, J.; HIRONOBU, I.; KOICHIRO, H.; TAKASHI, Y.; KONO, I. Effect of low-intensity aerobic exercise training on arterial compliance in postmenopausal women. Hypertension, v.27, p.897-901, 2004. SUZUKI, M.; NOMURA, C.; ODAKA, H.; IKEDA, H. Effect of an insulin sensitizer, pioglitazone, on hypertension in fructose-drinking rats. Jpn J Pharmacol, v.74, n.4, p.297-302, 1997. TAKAHASHI, R.N. Animais espontaneamente hipertensos (ratos SHR) como modelo experimental de TDAH. Anais da 58ª Reunião Anual da SBPC, Florianópolis, 2006. TASK FORCE OF THE EUROPEAN SOCIETY OF CARDIOLOGY AND THE NORTH AMERICAN SOCIETY OF PACING AND ELECTROPHYSIOLOGY: Heart rate variability: Standards of measurement, physiological interpretation, and clinical use. Circulation , n.93, p.1043-1065, 1996. TEIXEIRA, P.J., GOING, S.B., HOUTKOOPER, L.B., METCALFE, L.L., BLEW, R.M., FLINT-WAGNER, H.G., CUSSLER, E.C., SARDINHA, L.B. e LOHMAN, T.G. Resistance training in postmenopausal women with and without hormone therapy. Med Sci Sports Exerc, v.4, n.35, p.555-562, 2003. TEFF, K.L.; ELLIOTT, S.S.; TSCHOP, M.; KIEFFER, T.J.; RADER, D.; HEIMAN, M.; TOWNSEND, R.R.; KEIM, N.L.; D´ALESSIO, D.; HAVEL, P. Dietary fructose reduces circulating insulin and leptin, attenuates postprandial suppression of ghrelin, and increases triglycerides in women. J Clin Endocrinol Metab , n.89, p.2963-2972, 2004. THIRUNAVUKKARASU, V. & ANURADHA C. V. Influence of a-lipoic acid on lipid peroxidation and antioxidant defence system in blood of insulin-resistant rats. Diabetes, Obesity and Metabolism, v.6, p.200–207, 2004. TIETZE, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: applications to mammalian blood and other tissues. Analytical Biochemistry, v.27, n.3, p.502-522,1969.

102
TORDOFF, M.G.; ALLEVA, A.M. Effect of drinking soda sweetened with aspartame or high-fructose corn syrup on food intake and body weight. Am J Clin Nutr , n.51, p.963-969, 1990. TORJESEN, P.A.; BIRKELAND, K.I.; ANDERSSEN, S.A. et al. Lifestyle changes may reverse development of the insulin resistance syndrome. The Olso Diet and Exercise Study: a randomized trial. Diabetes Care, v.20, p.26-31, 1997. TUOMILEHTO, J.; LINDSTROM, J.; ERIKSSON, J.G.; VALLE, T.T.; HAMALAINEN, H.; ILANNE-PARIKKA, P.; KEINANEN-KIUKAANNIEMI, S.; LAAKSO, M.; LOUHERANTA, A.; RASTAS, M.; SALMINEN, V.; UUSITUPA, M. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. NEngl J Méd, v.344, n.18, p.1343-1350, 2001. BUENO, M.; BEZERRA, R.M.; SILVA, M.S.; TAVARES, D.Q.; CARVALHO, C.R.; SAAD, M.J. A high-fructose diet induces changes in pp185 phosphorylation in muscle and liver of rats. Braz J Med Biol Re, 33:I421-I427, 2000. VAN DE BORNE P, MONTANO N, PAGANI M et al. Absence of low-frequency variability of sympathetic nerve activity in severe heart failure. Circulation , n.95, p.1449–1454, 1997. VENDITTI, P. & DI MEO, S. Effect of training on antioxidant capacity, tissue damage, and endurance of adult male rats. Int J Sports Exerc, n.18, p.497-502, 1997. VERMA, S.; BHANOT, S.; YAO, L.; McNEILL, J.H. Defective endothelium-dependen relaxation in fructose-hypertensive rats. Am J Hypertens, 9, 370, 376, 1996. VINIK, A.I.; MASER, R.E.; MITCHELL, B.D.; FREEMAN, R. Diabetic autonomic neuropathy. Diabetes Care, v.26, n.5, p.1553-79, 2003. WARRAM, J.H.; MARTIN, B.C.; KROLEWSKI, A.S.; SOELDNER, J.S., KAHN, C.R. Slow glucose removal rate and hyperinsulinemia precede the development of type II diabetes in the offspring of diabetes parents. Ann Inter Méd , 113 : 909-915, 1990. WATTANAPERMPOOL, J. & REISER, P.J. Differential effects of ovariectomy on calcium activation of cardiac and soleus myofilaments. Am J Physiol, v.46, n.277, p.H467-H473, 1999. WEGGE, J.K.; ROBERTS, C.K.; NGO, T.H.; BARNARD, R.J. Effect diet and exercise intervention on inflammatory and adhesion molecules in postmenopausal women on hormone replacement therapy and at risk for coronary artery disease. Metabolism, v.53, n.3, p.377-81, 2004. WEISS, N.S. Relationship of menopause to serum cholesterol and arterial pressure. The United States Health Examination Survey of Adults. Am J Epidemiol. n.96, p.237-241, 1972.

103
WELTY, F.K. Preventing clinically evident coronary heart disease in the postmenopausal women. Menopause, v.4, n.11, p. 484-494, 2004. WENDEL, A . Glutathione peroxidase. Meth. Enzymol. 77: 325-333, 1981. WHELTON, S.P.; CHIN, A.; XIN, X.; HE, J. Effect of aerobic exercise on blood pressure: a meta-analysis of randomized, controlled trials. Ann Intern Med , v.136, p.493-503, 2002. WILMORE JH.; COSTILL DL. Fisiologia do esporte e do exercício. Trad. de Marcos Ikeda. 2.ed. São Paulo: Manole, 2001. WRITING GROUP FOR THE WOMEN`S HEALTH INITIATIVE INVESTIGATORS. Risk and benefits of estrogen plus progestin in healthy postmenopausal women. Principal results from the Women`s Healthy Initiative randomized controlled trial. JAMA , n.347, p.714-718, 1996. ZANCHETTI, A. & MANCIA, G. Cardiovascular reflexes and hypertension. Hypertension, n.18, p.III13-III21, 1991. ZIEGLER, O.; QUILLIOT, D.; GUERCI, B.; DROUIN, P. Macronutrients, fat mass, fatty acid flux and insulin sensitivity. Diabetes Metab, 27: 261-270, 2001.