universidade federal do rio grande do sul …livros01.livrosgratis.com.br/cp066495.pdf ·...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
DINÂMICA FOLICULAR EM ÉGUAS MINIPÔNEI
BIANCA DO PRADO LIMA PETRUCCI
Porto Alegre, Agosto de 2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
AVALIAÇÃO DA DINÂMICA FOLICULAR EM ÉGUAS MINIPÔNEI
BIANCA DO PRADO LIMA PETRUCCI
Dissertação apresentada como um dos requisitos
ao grau de mestre em Medicina Veterinária, área
de Reprodução Animal.
Orientador: Prof. Dr. Rodrigo Costa Mattos
Porto Alegre, Agosto de 2006

T787p Petrucci, Bianca do Prado Lima Dinâmica folicular em éguas Minipônei/ Bianca do Prado Lima Petrucci. –
Porto Alegre: UFRGS, 2006.
52 f.; il. – Dissertação (Mestrado) – Universidade Federal do Rio Grande do Sul, Faculdade de Veterinária, Programa de Pós-graduação em Ciências Veterinárias, Porto Alegre, RS-BR, 2006. Rodrigo Costa Mattos, Orient.
1. Minipônei: eqüinos 2. Ciclo estral
3. Dinâmica folicular I. Mattos, Rodrigo Costa, Orient. II. Título.
CDD 619.38
Catalogação na fonte: Biblioteca da Faculdade de Veterinária da UFRGS

ii
Bianca do Prado Lima Petrucci
DINÂMICA FOLICULAR EM ÉGUAS MINIPÔNEI
APROVADO EM 31 DE AGOSTO DE 2006.
Prof. Dr. Rodrigo Costa Mattos Orientador e Presidente da Comissão Profa. Dra. Sandra Mara Fiala Membro da Comissão
Prof. Dr. Maria Inês Mascarenhas Jobim Membro da Comissão
Prof. Dr. Ricardo Macedo Gregory Membro da Comissão

iii
RESUMO
Dissertação de Mestrado Programa de Pós-graduação em Medicina Veterinária Área de Concentração Fisiopatologia da Reprodução
Universidade Federal do Rio Grande do Sul
DINÂMICA FOLICULAR EM ÉGUAS MINIPÔNEI
Autor: Bianca do Prado Lima Petrucci
Orientador: Rodrigo Costa Mattos
O presente estudo teve como objetivo conhecer a dinâmica folicular das éguas minipônei
durante um intervalo interovulatório. Durante a estação reprodutiva foram utilizadas 12
éguas minipôneis com histórico reprodutivo desconhecido, clinicamente sadias, cíclicas e
não lactantes, com idades variando entre 3 e 7 anos. As éguas foram avaliadas diariamente,
durante um ciclo estral, por um único operador, através de palpação e ultra-sonografia
trans-retal. Não foi realizado nenhum tipo de terapia hormonal nestes animais durante o
experimento. Todos os folículos visíveis à ultra-sonografia tiveram o antro medido e os
dados, de cada ovário, transferidos para ficha especial, identificando-se a exata posição de
cada folículo. Considerou-se como emergência o momento em que os folículos atingiram
6mm, com subseqüente crescimento. A divergência foi determinada como o dia em que as
taxas de crescimento entre os dois maiores folículos tornaram-se discrepantes. A altura,
largura e área do corpo lúteo foram medidos. A ecotextura uterina foi avaliada em escore de
1 a 4 durante todo o ciclo. As minipôneis apresentam um ciclo estral de 23,3 ± 3,2 dias com
uma onda folicular maior, não ocorrendo ondas secundárias na maioria dos casos. O
número de folículos durante o ciclo é estável e somente um, acima de 25 mm, foi observado
em cada ciclo, em todas as éguas. A emergência ocorre a partir do primeiro dia do ciclo e a
divergência, no 14o dia, junto com o desaparecimento do corpo lúteo. O maior diâmetro
folicular foi observado dois dias antes da ovulação. Finalmente, foi observado que os
escores das ecotexturas uterinas elevaram-se a partir do 9o dia do ciclo.
Palavras-chave: Minipôneis, ciclo estral, dinâmica folicular.

iv
ABSTRACT
Dissertação de Mestrado Programa de Pós-graduação em Medicina Veterinária Área de Concentração Fisiopatologia da Reprodução
Universidade Federal do Rio Grande do Sul
FOLLICULAR DYNAMICS IN MINI-PONEY MARES
Autor: Bianca do Prado Lima Petrucci
Orientador: Rodrigo Costa Mattos
This study aimed to verify the follicular dynamics from mini-pony mares during an
interovulatory interval. During the breeding season 12 mini-pony mares with unknown
reproductive history, clinically normal, cyclic, non lactating with age ranging between three
to seven years old were used. Mares were examined daily, during one estrus cycle, by a
single operator using transrectal ultrasonography. The mares were not submitted to any
hormonal therapy during the experiment. All the visible follicles were monitored daily,
when the antrum was measured and the position was taken. The data was then recorded in a
formulary. Follicle emergence was considered when it reached 6 mm diameter and
subsequent growth. Deviation was defined as the beginning of a marked difference in
growth rates between the two largest follicles. The height, width and area of the corpus
luteum were measured. The uterine ecotexture was also daily taken during the estrus cycle.
The data obtained suggest that the mini-pony mares present an estrus cycle of 23.3 ± 3.2
days, with one major follicular wave. Minor follicular waves were observed in a few cases.
The total number of follicles was similar throughout the cycle and only one follicle greater
than 25 mm was detected per cycle. The emergence was observed at the first day of the
cycle and the divergence at the 14th day together with the last observation of the corpus
luteum. The maximum diameter of the largest follicle was observed two days before
ovulation. Finally, it was observed that the uterine ecotexture grade raised until the 9th day
of the cycle.
Keywords: Mini-pony, estrus cycle, follicular dynamics.

v
AGRADECIMENTOS
Em primeiro lugar, agradeço ao meu prezado Orientador e amigo Rodrigo Costa
Mattos que fez possível a conquista deste sonho, através de seus ensinamentos e dedicação
à Medicina Veterinária e à instituição (UFRGS).
Ao amigo e Co-orientador Carlos Afonso de Castro Beck, agradeço por ter me aberto
a “primeira porta” desta instituição, que me acolheu de braços abertos desde o primeiro dia
em que aqui estive e que tantas outras “portas” me abriu no decorrer desta jornada.
A Melba e Eduardo Gastal por terem proporcionado a metodologia e auxiliado à
distância no desenvolvimento do projeto.
Agradeço também aos meus pais, Carmen Lúcia e Carlos Eugênio, pelo amor
incondicional e incentivo em todos os momentos de minha vida.
A minha amada irmã, Ana Carolina, meu muito obrigada pela amizade e pelo carinho
que sempre se fazem presentes entre nós.
Ao meu namorado, Dimas Rocha, agradeço pela tranqüilidade e bom humor
inabaláveis, pela alegria sempre presente, pelo carinho e pela paciência, que por muitas
vezes se fez necessária nos passos finais deste caminho.
À Médica Veterinária Adriana Neves, agradeço pela parceria e disponibilidade,
durante toda a realização deste experimento, se fazendo sempre presente, e aos
ensinamentos durante toda esta trajetória.
Agradeço também à aluna de graduação em Medicina Veterinária e bolsista do
Reprolab, Fabiana Castro, por ter sido de importância fundamental no êxito deste trabalho,
prestando auxílio na totalidade de sua realização.
E, por fim, ao amigo e aluno de graduação em Medicina Veterinária Émerson Rocha,
pela alegria que sempre o acompanha, pela seriedade e auxílio desde os primeiros
momentos do experimento.

6
SUMÁRIO
RESUMO....................................................................................................................... iii
ABSTRACT................................................................................................................... iv
AGRADECIMENTOS................................................................................................. v
SUMÁRIO..................................................................................................................... 6
LISTA DE FIGURAS.................................................................................................. 7
LISTA DE TABELAS.................................................................................................. 8
1. INTRODUÇÃO......................................................................................................... 9
2. REVISÃO BIBLIOGRÁFICA................................................................................ 11
2.1 Ciclo Reprodutivo..................................................................................................... 11
2.2 Dinâmica Folicular................................................................................................... 14
2.3 Ultra-sonografia e Dinâmica Folicular..................................................................... 19
3. ARTIGO - Dinâmica Folicular Em Éguas Minipônei........................ 23
4. CONCLUSÕES......................................................................................................... 46
5. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................... 47
ANEXO.......................................................................................................................... 52

7
LISTA DE FIGURAS
Figura 1 – Desenho experimental da coleta de dados realizada em 12 éguas
minipôneis.....................................................................................................
29
Figura 2 – Diâmetro médio e desvio padrão do diâmetro médio do folículo
dominante e do subordinado pelo método identificado. A – Dia 0 foi
considerado o dia da emergência. B – Dia 0 foi considerado o dia da
divergência....................................................................................................
35
Figura 3 – Média dos seis maiores folículos observados pelo método sem
identificação, durante o ciclo estral de 12 minpôneis. .................................
36
Figura 4 - Média e desvio padrão do número de folículos agrupados por diâmetro (A)
e totais (B), pelo método identificado, durante o ciclo estral de 12 éguas
minipôneis.....................................................................................................
36
Figura 5 – Média e desvio padrão (A) do diâmetro (mm) e (B) da área (cm2) do
corpo lúteo e (C) da ecotextura do útero (1-4) do ciclo estral de 12 éguas
minipôneis, considerando-se 0 o dia da primeira ovulação..........................
38

8
LISTA DE TABELAS
Tabela 1 - Médias (± desvio padrão) do dia da ocorrência e dos intervalos
observados, pelo método não identificado, durante o ciclo estral de 12
éguas minipôneis..........................................................................................
32
Tabela 2 - Médias (± desvio padrão) dos diâmetros foliculares e taxas de crescimento
observadas, pelo método não identificado, durante o ciclo estral de 12
éguas minipôneis..........................................................................................
34

9
1. INTRODUÇÃO
Atualmente, as raças de cavalos miniatura tem se tornado popular. No Brasil é criada
a Raça Brasileira de Minipôneis originada do cruzamento entre Shetland e o Falabella e,
mais recentemente, a introdução do American Miniature Horse (BERGMANN et al.,
1997). É um eqüino eumétrico, ágil, de bom temperamento para o serviço, dócil, com
proporções equilibradas entre a altura da cernelha e o comprimento do corpo. A altura
máxima aos 36 meses deverá ser de 1,00 m para os machos e de 1,10 m para as fêmeas.
Pouco é conhecido sobre os parâmetros reprodutivos desta raça. Os poucos artigos
existentes tratam de características seminais e concluem que os mesmos parâmetros
existentes nos animais de maior porte não podem ser utilizados para os minipôneis
(PACCAMONTI et al., 1999; NEVES et al., 2005).
Compreender os mecanismos do desenvolvimento folicular das diferentes espécies é
de grande importância para o estudo da reprodução, uma vez que é o folículo que
proporciona um ambiente ótimo para a maturação do oócito, preparando-o para a
subseqüente fertilização após a ovulação. O conhecimento pleno destes mecanismos
fisiológicos é fundamental para que ocorra o aperfeiçoamento ou desenvolvimento de
novas técnicas para manipular a função reprodutiva dos animais domésticos (ADASHI &
ROBAN, 1992).
A dinâmica folicular pode ser precisamente analisada através da ultra-sonografia
transretal. Esta técnica permite a visualização freqüente e de forma não invasiva da
dinâmica da população de folículos, do processo ovulatório e do desenvolvimento,
manutenção e regressão do corpo lúteo (GINTHER & PIERSON, 1984a, 1984b;
GINTHER, 1986; GINTHER, 1998).
Estudos em éguas e em vacas são realizados desde a década de 80 e serviram de
modelo para o estudo em diversas espécies, como em ovelhas, cabras, porcas, lhamas
(BARTLEWSKI et al, 2000; DE CASTRO et al, 1999; COX, 1997; ADAMS et al, 1990).
Entretanto, devido à similaridade em diversas características da dinâmica folicular, a égua
demonstrou ser um possível modelo experimental para o estudo da foliculogênese da
mulher, com a vantagem de apresentar foliculos maiores (GINTHER et al., 2004).

10
A hipótese deste trabalho é de que a dinâmica folicular das éguas minipôneis seja
similar a dos eqüinos de outras raças já estudadas, apesar de seus respectivos portes.
O presente estudo teve o objetivo de verificar a dinâmica folicular das éguas
minipôneis durante um intervalo interovulatório e comparar os dados obtidos com
resultados já conhecidos de éguas de maior porte.

11
2. REVISÃO BIBLIOGRÁFICA
2.1 Ciclo Reprodutivo
O ciclo reprodutivo das éguas pode ser definido em duas estações distintas: a
ovulatória e a anovulatória, interligadas por períodos denominados transicionais (BAKER
& KENNEY, 1980).
A estação anovulatória geralmente compreende os meses de inverno e início da
primavera e apresenta uma duração variável de 40 dias a oito meses (HUGHES et al.,
1972). Durante este período menos de 25% das éguas chegam à ovulação, menos de 10%
permitem a monta do garanhão e cerca de 19% ficam indiferentes a sua presença (SHARP,
1980).
Observa-se uma fase transicional da atividade ovariana durante a primavera, quando
estros anovulatórios normalmente ocorrem (OSBORNE, 1966). Um estudo realizado com
éguas demonstrou um nítido aumento de peso dos ovários nos meses de julho e agosto, mas
a taxa ovulatória ainda encontrava-se em declínio e sofreu um acréscimo somente no mês
de setembro. Este período foi caracterizado como transicional de primavera (TAROUCO et
al., 1995). Durante a estação ovulatória, que corresponde ao final da primavera e verão, a
maioria das éguas ciclam em intervalos regulares e desenvolvem estros ovulatórios
(OSBORNE, 1966).
Em pôneis estas estações costumam ser mais rigidamente definidas quando
comparadas com as raças de maior porte, que apresentam estações ovulatórias mais longas
e um número maior de fêmeas ciclando e ovulando durante todo o ano (WESSON &
GINTHER, 1981). Da mesma forma, a duração do ciclo estral das éguas pôneis é, em
média, dois dias mais longa. Respectivamente, o estro, diestro e ciclo estral apresentam 7,
15 e 22 dias nas éguas maiores e 8, 16 e 24 dias em pôneis (GINTHER, 1992).
O ciclo estral é definido como o período entre duas ovulações consecutivas,
acompanhadas por sinais de estro (HUGHES et al., 1975; 1980). Pode ser dividido em duas
fases: uma folicular ou de estro (que compreende apenas 20% do ciclo), caracterizada pela
receptividade sexual, crescimento folicular, maior secreção de estrógeno e concentrações
plasmáticas de progesterona abaixo de 1ng/ml, e outra luteal ou de diestro, que inicia logo

12
após a ovulação e caracteriza-se por uma ativa resistência ao garanhão, formação do corpo
lúteo e maior secreção de progesterona (HUGHES et al., 1972).
O intervalo interovulatório inicia com uma ovulação, associada ao estro, e termina
com a subseqüente. É a melhor referência para as pesquisas relacionadas a ondas foliculares
e tem duração média de 21 a 22 dias em éguas e de 24 dias em pôneis (GASTAL, 1999).
Entretanto, pode sofrer variações de acordo com a época do ano em que é observado, bem
como o próprio estro, que costuma ser significativamente mais curto nos meses de verão
em relação aos de outono, inverno e início da primavera (GINTHER, 1992).
Segundo Hansen (1985), a duração diária de horas de luz tem importante papel na
reprodução dos mamíferos. Os fotoperíodos longos estimulam e os curtos inibem a
espermatogênese e ocorrência de estro nas espécies cujo acasalamento ocorre na primavera
e verão. Isto explica o acréscimo das taxas ovulatórias observado durante a primavera,
atingindo incidências máximas no verão, e a redução do número de ovulações observada
nos meses de outono e inverno (GINTHER, 1992).
As estações do ano também interferem no tamanho do folículo pré-ovulatório que,
nos meses de verão, quando a duração do estro normalmente é menor, costuma apresentar
diâmetros menores do que os encontrados no início e final da estação reprodutiva
(GRAVES et al., 1995).
Com o aumento do fotoperíodo, a atividade antigonadal diminui e a liberação das
gonadotrofinas se inicia proporcionalmente. Essa mudança gradual resulta em
manifestações “transicionais” caracterizadas pela formação de múltiplos e pequenos
folículos e pela ocorrência de ciclos prolongados, nos quais o grau de receptividade sexual
varia até o estro ser evidente (HUGHES et al., 1980).
Os raios solares incidem sobre a retina ocular estimulando seus receptores que,
através das fibras simpáticas do nervo óptico e conexões na base do cérebro, transmitem
mensagens para o gânglio cervical superior e então para a glândula pineal, iniciando os
eventos cerebrais, onde estão envolvidos também o hipotálamo e a hipófise anterior, e
finalmente para os ovários (SENGER, 2003).
A glândula pineal é responsável pela síntese e secreção da melatonina (N-acetil 5-
metoxitriptofano) a partir do aminoácido triptofano presente na circulação (LINCOLN,
1987) que, nos animais que acasalam durante fotoperíodo positivo, exerce um efeito

13
depressor ou antigonadal nos ovários, inibindo a liberação do Hormônio Liberador de
Gonadotropinas (GnRH) pelo hipotálamo (GINTHER, 1992). A taxa de síntese deste
hormônio é inversamente proporcional à duração da luminosidade ambiental diária. Assim,
durante o estímulo luminoso, a síntese e secreção da melatonina são inibidas promovendo a
liberação do GnRH pelo hipotálamo que, por sua vez, estimula a liberação dos Hormônios
Folículo Estimulante (FSH) e Luteinizante (LH), a ovulação e a formação do corpo lúteo
(SHARP, 1980; GINTHER, 1992).
A fase folicular do ciclo estral, que tem início logo após a luteólise, é controlado pelo
hipotálamo, lobo anterior da hipófise e ovários através da produção de estrógeno na
ausência de progesterona. Este momento envolve quatro principais eventos: liberação
maciça de gonadotropinas pelo lobo anterior da glândula pituitária, crescimento folicular,
receptividade sexual e ovulação (ADASHI & ROBAN, 1992).
Na ausência do corpo lúteo, observa-se um grande decréscimo nas concentrações
séricas de progesterona. A interrupção do feedback negativo exercido por ela no
hipotálamo tem como resultado a liberação de GnRH em altas amplitudes e freqüências até
o final da fase folicular, no momento da ovulação e formação de um novo corpo lúteo
(GINTHER et al., 1997).
A secreção de GnRH, na fêmea, é controlada por duas áreas distintas do hipotálamo:
o centro tônico e o centro pré-ovulatório. O primeiro é responsável pela secreção basal de
GnRH, liberando-o em pequenos e múltiplos pulsos, com variadas frequências e
amplitudes, durante toda a extensão do ciclo estral. O segundo, por sua vez, realiza a
liberação pré-ovulatória de GnRH, estimulando o desenvolvimento da onda de LH que
antecede a ovulação. Entretanto, este centro também libera níveis basais de GnRH até que
ocorra a secreção maciça de estrógeno na ausência de progesterona, o que caracteriza o
período de estro (GINTHER, 1992).
Inicialmente, a elevação dos níveis de GnRH promove a liberação de grandes
concentrações de FSH e LH pelo lobo anterior da glândula pituitária e, por conseguinte, o
desenvolvimento de folículos e produção de estrógeno (esta seqüência de eventos
caracteriza o período de diestro). O LH, em particular, além de estimular a produção de
estrógeno, dá início a um processo de feedback positivo com o centro pré-ovulatório do

14
hipotálamo, ocasionando a liberação de concentrações ainda maiores de GnRH (FAY &
DOUGLAS, 1987; GINTHER, 1992).
Quando os folículos recrutados começam a desenvolver dominância, nos momentos
finais da fase folicular, secretam estrógeno e inibina, ambos envolvidos no feedback
negativo com o lobo anterior da hipófise que tem como desfecho a supressão da liberação
de FSH que, juntamente com a elevada secreção de estrógeno verificada neste momento,
dará início a um pico pré-ovulatório de LH, responsável pela ovulação (BERGFELT et al.,
1991; GINTHER, 1992; GINTHER & BERGFELT, 1993; GINTHER et al., 1997).
2.2 Dinâmica Folicular
Como onda folicular entende-se a emergência de diversos folículos, que inicialmente
crescem e se desenvolvem em sincronia, até o momento em que apenas um (ou o número
específico de cada espécie) continua crescendo até a ovulação, enquanto os demais tendem
a sofrer atresia e regredir (GINTHER et al., 2004; GINTHER & PIERSON, 1989).
O número de ondas foliculares observadas durante o ciclo estral varia entre as
diferentes espécies (GINTHER & BERGFELT, 1992). Em éguas, as ondas foliculares
foram divididas em menores, nas quais não ocorre divergência e o maior folículo não se
torna dominante, e maiores, quando ocorre divergência e desenvolvimento de dominância
(GINTHER & BERGFELT, 1992; GINTHER, 1993).
As ondas maiores são ainda subdivididas em primárias, quando dão origem ao
folículo que irá ovular durante o período de estro, e secundárias, quando existe dominância,
mas a ovulação não ocorre ou se dá após a onda primária ou estro, caracterizando uma
ovulação secundária ou de diestro. As ondas menores, bem como as secundárias, ocorrem
com maior freqüência durante a primavera (GINTHER, 1993).
A foliculogênese ovariana é um processo dinâmico caracterizado por uma marcante
proliferação e diferenciação de componentes e células, no qual o folículo adquire,
progressivamente, algumas propriedades essenciais para seu futuro desenvolvimento. Deve
ocorrer no tempo certo e numa seqüência exata. Do contrário, acarretará prejuízos no
desenvolvimento com posterior atresia (CAMPBELL et al., 1995).

15
Pesquisas recentes sugerem que a dinâmica folicular pode ser dividida em dois
momentos: o primeiro denomina-se fase de recrutamento inicial, envolvendo o
recrutamento contínuo de folículos que terminarão por sofrer atresia, e o segundo é
caracterizado pelo recrutamento cíclico, que inicia após a puberdade e é resultado de níveis
elevados de FSH observados durante o ciclo estral (GASTAL, 1999).
A dinâmica folicular envolve quatro processos distintos: a emergência, a seleção, a
dominância e a atresia. Este último caracteriza-se pelo encerramento ou desaparecimento
do antro folicular, seguido de degeneração. Em qualquer ponto durante o período
reprodutivo pós-púbere a proporção de folículos em algum estágio de atresia é bastante alta
(SENGER, 2003).
Ao primeiro dia em que é possível observar o crescimento dos folículos de uma onda,
através da ultra-sonogrfia, dá-se o nome de emergência folicular, freqüentemente utilizado
como sinônimo de recrutamento folicular (BERGFELT & GINTHER, 1993; GINTHER et
al., 1996). Pode também ser definida como o momento em que os folículos em crescimento
tornam-se suscetíveis à ação das gonadotropinas e iniciam a produção de pequenas
concentrações de estrógeno (DRIANCOURT, 2001).
A seleção é o processo que resulta na redução do número de folículos de uma onda e
varia de acordo com o número específico de ovulações apresentado por cada espécie
animal. Seu término se dá no momento em que as taxas de crescimento entre os dois
maiores folículos de uma onda tornam-se discrepantes, caracterizando o início do processo
de divergência (GINTHER et al., 1996; 1997).
O processo de dominância caracteriza-se por um ou mais folículos maiores exercendo
efeito inibitório nos demais presentes nos grupos de recruta e seleção. O dominante é o
único que continua a se desenvolver enquanto os demais, chamados de subordinados têm
seu crescimento inibido (GINTHER, 1993). Seu número é específico em cada espécie e
está diretamente relacionado com o tamanho da progênie (GINTHER, 1992; 1995).
É possível ainda classificar os folículos de uma onda de acordo com os diâmetros por
eles apresentados. São considerados pequenos, médios e grandes aqueles com menos de
10mm, entre 10 e 20mm e acima de 20mm de diâmetro, respectivamente (SENGER, 2003).
Os ovários das éguas, durante o ciclo estral, contêm cerca de 5 a 12 folículos medindo
aproximadamente 10mm de diâmetro (PIERSON & GINTHER, 1987) e o maior chega a

16
medir entre 35 e 55mm (SIROIS et al., 1991; GINTHER & BERGFELT, 1992; GINTHER,
1993). O folículo ovulatório de uma onda primária maior emerge cerca de 6 ou 7 dias após
a última ovulação e ovula em média 15 dias depois, com aproximadamente 45mm de
diâmetro (GINTHER, 1993; GINTHER & BERGFELT, 1993).
O desenvolvimento de ondas foliculares não ocorre exclusivamente durante o ciclo
estral, mas também antes da puberdade e durante a gestação, anestro e puerpério.
Entretanto, não costumam produzir folículos dominantes e, futuramente, são responsáveis
por uma redução significativa do número de folículos para as ovulações seguintes
(SENGER, 2003).
As ondas foliculares foram caracterizadas em vacas (EVANS et al., 1994a, b) e éguas
(NOGUEIRA & GINTHER, 2000) pré-púberes. Aparentemente, durante este período,
existe um aumento gradual do diâmetro máximo dos folículos, que culmina na puberdade e
ciclo estral. Foram também identificadas em períodos de anestro em éguas (GINTHER,
1990; DONADEU & GINTHER, 2002), vacas (STAGG et al., 1995; BOSSIS et al., 2000)
e ovelhas (BARTLEWINSKI et al., 1998; EVANS et al., 2001), demonstrando que o
fenômeno do desenvolvimento folicular ocorre durante muitos estágios fisiológicos.
A preparação do folículo para a ovulação ocorre sob uma seqüência de eventos
endócrinos, característicos da onda folicular pré-ovulatória, no momento em que as
concentrações de FSH e LH encontram-se elevadas, quando não mais se observa a inibição
da secreção de GnRH pela presença da progesterona (GINTHER et al., 1998).
O acréscimo nas concentrações de FSH é pré-requisito para que ocorra o
desenvolvimento de uma onda folicular (GINTHER et al., 1996; DRIANCOURT, 2001;
FORTUNE et al., 2001) e, segundo Gastal et al.(1997), em éguas tem início quando o
maior folículo apresenta um tamanho médio de 6mm e atinge seu pico quando ele encontra-
se com cerca de 13mm, com subseqüente declínio. Próximo à luteólise tem-se o início do
aumento das concentrações de estradiol e LH. As concentrações de LH aumentam de forma
bifásica, mais lentamente na primeira fase do que na seguinte. O término da primeira e
início da segunda fase ocorre no dia em que o estradiol atinge suas concentrações séricas
máximas (GINTHER et al., 2004).
Uma vez recrutados os folículos, inicia-se a produção e secreção de concentrações
reduzidas de estrógeno e inibina que, a partir do momento da seleção, acabam elevando-se.

17
Desta forma, acentua-se o feedback negativo com o lobo anterior da hipófise e os níveis de
FSH começam a decrescer e, quando atingem valores mínimos, o recrutamento chega ao
fim. Neste momento (aproximadamente no 16º dia do ciclo), o LH torna-se mais importante
para o crescimento dos folículos maiores e os demais folículos até então recrutados
começam a sofrer o processo de atresia (GINTHER et al., 1998).
Durante a seleção folicular o futuro folículo dominante é o primeiro a se tornar apto a
elevar a concentração intrafolicular e sistêmica de estradiol, reduzir os níveis sistêmicos de
FSH, impedindo que novos folículos sejam recrutados, e responder ao aumento dos níveis
de LH e, assim, estabelecer dominância (GASTAL, 1999).
A seleção do folículo pré-ovulatório, durante a fase folicular, envolve um decréscimo
nas concentrações de FSH e um acréscimo nas freqüências dos pulsos de LH. Este aumento
da razão LH:FSH acentua o crescimento do folículo dominante, que apresenta receptores
para o LH nas células da teca e da granulosa, e induz a atresia dos pequenos, que são
desprovidos desse recurso (SENGER, 2003).
Ginther et al., (1996) sugeriram que um determinado nível de LH altera a
sensibilidade do maior folículo frente às gonadotropinas (FSH e LH), próximo ao momento
da divergência, e parece ser necessário para o crescimento folicular após este momento. Os
fatores que influenciam a sensibilidade dos folículos selecionados ao decréscimo das
concentrações de FSH podem envolver o aumento da vascularização desses folículos e do
número de células da granulosa ou de receptores para o FSH presentes nas mesmas.
Aparentemente, a supressão final da onda de FSH, juntamente com o acréscimo dos
níveis de LH, tem importante papel na divergência das taxas de crescimento entre o futuro
folículo dominante e os demais (GINTHER et al., 1996; 1998). Um dos grandes
questionamentos sobre a relação entre o declínio do FSH e a divergência consiste em como
o folículo selecionado para ser o dominante continua a crescer frente ao decréscimo das
concentrações deste hormônio (GASTAL, 1999).
Segundo Acosta et al. (2004) e Bollwein et al. (2002), a combinação entre a produção
de grandes concentrações de estrógeno e inibina pelo folículo dominante é responsável pela
redução do aporte sangüíneo aos demais folículos que, juntamente com as baixas taxas de
FSH circulante, acabam por sofrer atresia. Somente aqueles que mantiverem um adequado
suprimento circulatório, e por conseqüência estiverem sujeitos a altos níveis de

18
gonadotropinas, continuarão a crescer até a ovulação. A diferença entre o aporte sangüíneo
ao folículo dominante e ao seu maior subordinado inicia cerca de dois dias antes da
divergência entre suas taxas de crescimento. Fatores angiogênicos, de crescimento
endotelial, foram encontrados em grande quantidade no fluído do folículo dominante,
demonstrando a capacidade de controlar seu próprio aporte sanguíneo (ACOSTA et al.,
2004; BOLLWEIN et al., 2002).
Gastal (1999), em estudos realizados em éguas, defendeu a hipótese de que o futuro
folículo dominante apresenta um desenvolvimento mais avançado no momento da
divergência e é o primeiro a atingir um estágio decisivo, medindo cerca de 21-23mm. Neste
momento, mecanismos que fazem parte dos eventos da divergência inibem o maior
subordinado antes que ele atinja o mesmo estágio, em média um dia depois. Observou
também, que o início da divergência entre as curvas de crescimento desses folículos ocorre,
em média, no dia 17-18 do ciclo ou seis dias após a emergência do maior e sete dias antes
da ovulação.
Em suma, dois aspectos parecem ser importantes para que ocorra o mecanismo da
divergência: a supressão do FSH pelo futuro folículo dominante, resultando em prejuízos
para os subordinados, e a sensibilização ao LH desenvolvida por ele (GASTAL, 1999).
Baseados nos resultados de um estudo realizado em éguas, sobre a liberação pulsátil
de LH pela hipófise, Silvia et al. (1995) concluíram que a progesterona endógena resulta na
redução da freqüência de liberação de GnRH e causa um decréscimo na concentração de
LH circulante sem interferir nos níveis de FSH. Após definir a dominância o folículo
continua a crescer, ainda que os níveis de FSH encontrem-se reduzidos, atinge seu tamanho
máximo e passa a secretar níveis elevados de estradiol. A partir deste momento, o LH pré-
ovulatório surge causando profundas alterações nas funções deste folículo, reduzindo de
forma abrupta a produção de estradiol (GINTHER & BERGFELT, 1993).
A ovulação é um complexo processo que resulta na reorgenização do tecido folicular.
O fluxo sanguíneo no ovário é controlado pelos níveis teciduais de histamina e PGE2, e um
grande aporte sanguíneo já está previamente estabelecido em decorrência da presença do
folículo dominante. O aumento da permeabilidade vascular causado pela histamina torna a
teca interna bastante edematosa e, por conseguinte, um aumento da pressão hidrostática ao

19
redor do folículo fará com que ele se torne mais frágil e suscetível à ruptura (SENGER,
2003).
A partir da liberação da onda de LH pré-ovulatória as células ovarianas da teca
interna passam a produzir progesterona. A elevação das concentrações deste hormônio
estimula a síntese local da colagenase, enzima que degrada o colágeno, maior componente
do tecido conjuntivo entre o folículo e o ovário. Neste mesmo momento, ocorre a síntese e
secreção local de PGF2α e PGE2. A primeira é responsável por contrações da musculatura
lisa ovariana, ocasionando uma superficialização ainda maior do folículo, e, a segunda, tem
a função de reorganizar o tecido folicular em corpo lúteo após a ovulação, ativando uma
enzima denominada de plasminogênio, responsável pela formação de um coágulo após a
ruptura do folículo e pela reorganização celular (SENGER, 2003).
O corpo lúteo (CL) é uma estrutura que produz progesterona e sua presença implica
na supressão do comportamento de estro, mantendo a égua não receptiva ao garanhão. A
progesterona prepara o trato reprodutivo para receber o embrião, mas se a prenhez não
estiver presente, o CL tende a regredir e um novo estro deverá ocorrer. Em caso de
gestação, o CL se mantém e continua a produzir progesterona enquanto se fizer necessário
(GINTHER, 1995).
O local da ovulação pode ser diagnosticado facilmente, através da palpação retal,
somente no primeiro dia após a ovulação, quando se percebe uma grande massa macia no
local. A partir das 24 horas subseqüentes, a cavidade preenche-se de sangue, recebendo o
nome de corpo hemorrágico, facilmente confundido com um grande folículo à palpação. A
partir deste momento, o diagnóstico definitivo da ovulação se dá somente através da ultra-
sonografia, observando-se a presença do CL (GINTHER, 1995).
2.3 Ultra-sonografia e Dinâmica Folicular
A dinâmica folicular pode ser precisamente analisada através da ultra-sonografia
transretal. Esta técnica permite a visualização freqüente e de forma não invasiva da
dinâmica da população de folículos, do processo ovulatório e do desenvolvimento,

20
manutenção e regressão do corpo lúteo (GINTHER & PIERSON, 1984a, 1984b;
GINTHER, 1986; GINTHER, 1998), além de tornar viável a pesquisa sobre os folículos
menores, com diâmetro entre 2 e 10mm (GINTHER e BERGFELT, 1993).
O fenômeno da dinâmica folicular foi descoberto e descrito através da ultra-
sonografia, observando-se as mudanças de tamanho e número dos folículos. Esta técnica
está entre as mais importantes utilizadas nas pesquisas reprodutivas e tem grande
importância como meio de diagnóstico. É minimamente invasiva, não requer a utilização de
técnicas cirúrgicas e pode ser utilizada para o diagnóstico de gestação, idade e crescimento
embrionário e fetal e para detectar mudanças das estruturas ovarianas, anormalidades fetais
e casos de gestações gemelares em vacas e éguas (GINTHER, 1995).
Estudando o fenômeno da seleção do folículo dominante em éguas pôneis, através da
ultra-sonografia trans-retal, Gastal (1999) verificou que ele emerge cerca de um dia antes
do maior subordinado. Estes dois folículos crescem paralelamente, mas o maior deles
mantém uma vantagem em tamanho, equivalente a um dia, por cerca de seis dias ou até
estar com cerca de 23mm, quando inicia o processo da divergência.
Redmer & Reynolds (1996), em estudos realizados em éguas através da ultra-
sonografia, observaram um crescimento folicular médio de 3mm por dia, sete dias antes da
ovulação, e um diâmetro médio de 45mm, um dia antes de ela ocorrer. Relataram também
que em 82% dos casos analisados o folículo pré-ovulatório tornou-se o maior cerca de seis
dias antes da ovulação e 85% deles sofreram alterações de suas formas esféricas para
cônicas devido ao aumento da flutuação e ação da PGF-2α.
Mudanças na vascularização dos folículos pré-ovulatórios têm sido estudadas em
várias espécies. A alteração morfológica mais marcante ocorre com o sistema
microcirculatório da teca interna que, durante a onda pré-ovulatória de gonadotropinas é
seguida de hiperemia, edema e extravasamento de eritrócitos para o seu interior
(CAVENDER & MURDOCH, 1998).
Em decorrência das elevadas concentrações de estrógeno próximo ao momento da
ovulação, o folículo pré-ovulatório torna-se mais vascularizado (CAVENDER &
MURDOCH, 1998; KERBAN et al., 1997). Ecograficamente, observa-se uma camada
anecóica na granulosa folicular, atribuída à hipervascularização e edema da teca interna
(RITCHIE, 1986). Estas alterações podem ser utilizadas para determinar o momento prévio

21
à ovulação em mulheres (JAFFE et al., 1987) e éguas (GASTAL, 1997). Em éguas, nota-se
também, o aumento da espessura (PIERSON & GINTHER, 1985) e ecogenicidade
(PIERSON & GINTHER, 1985; CARNEVALE, 1998) da parede do folículo.
Segundo Acosta et al. (2004), em estudos realizados com éguas, a divergência é
morfologicamente identificada através do uso do ultra-som, pelo aumento da espessura da
camada anecóica que circunda a teca interna do folículo dominante. Esta alteração da
ecotextura folicular permite a distinção entre o futuro dominante e seu maior subordinado
cerca de um dia antes de iniciar o processo de divergência, e também é atribuída ao
aumento da vascularização (GASTAL et al., 1999).
De acordo com Ginther (1995), a ecotextura uterina também sofre alterações em
relação aos diferentes momentos durante o ciclo estral das éguas. Durante o estro, as
imagens uterinas obtidas através do ultra-som caracterizam-se por áreas alternadas de hipo
e hiperecogenicidade. As porções hiperecogênicas correspondem ao endométrio e, as
hipoecogênicas, ao edema uterino verificado durante este período. Entretanto, durante o
diestro, a ecotextura uterina costuma ser mais homogênea e verifica-se a predominância de
áreas hiperecogênicas.
A ecotextura uterina pode ser classificada, segundo o grau de edema, em três escores:
1 ( ausência de edema e imagem homogênea, caracterizando o período de diestro), 2 (nível
intermediário de edema e imagem heterogênea, sem evidenciar as camadas uterinas) e 3
(edema máximo, imagem bastante heterogênea e identificação das camadas do endométrio,
caracterizando o período do estro). Pode ainda ser utilizado um escore subdividido em
quatro graus, variando de 1 a 4 de acordo com o edema uterino, sendo 1 o menor grau e 4 o
máximo (GINTHER, 1995).
Observando os ovários das éguas através da utilização de um transdutor de 5.0 MHz,
durante o intervalo inter-ovulatório, percebe-se que o CL pode ser detectado, em média, até
o Dia 17, para um ciclo de 22 dias e é facilmente visualizado neste período. O diâmetro do
maior folículo no dia em que o CL é identificado pela última vez pode auxiliar a distinção
entre as éguas cíclicas e as que se encontram durante a estação anovulatória. Se o maior
folículo medir 20mm ou menos e o CL não estiver presente, considera-se a égua não
cíclica. Na maioria delas o folículo dominante apresenta diâmetros maiores do que 20mm
no momento em que o CL não pode mais ser identificado, caracterizando a estação

22
ovulatória. A presença de um folículo com diâmetro igual ou superior a 25mm de diâmetro,
durante mais de 10 dias consecutivos, demonstra uma possível fase transicional do ciclo
reprodutivo das éguas (GINTHER, 1995).
O CL pode apresentar duas morfologias distintas: um tecido luteal com ecogenicidade
uniforme ou um coágulo sangüíneo central, caracterizado como uma porção anecóica,
circunscrita por tecido ecogênico, denominado de corpo hemorrágico (GINTHER, 1986).
Esta estrutura é resultado do preenchimento do centro do CL com sangue, iniciando, em sua
maioria, 24 horas após a ovulação. Entretanto, em alguns casos, pode ser observado
imediatamente após a ruptura do folículo pré-ovulatório. O diâmetro do CL aumenta entre
os Dias 1 e 3, quando então tende a regredir até o desaparecimento completo (GINTHER,
1995).

23
3 ARTIGO
DINÂMICA FOLICULAR EM ÉGUAS MINIPÔNEI
Bianca P. L. Petrucci1*, Adriana P.Neves1,4, Fabiana S. Castro1, Ricardo M. Gregory1,4. Melba O. Gastal2, Eduardo L. Gastal3, Rodrigo C. Mattos1,4
1 – Reprolab, Departamento de Medicina Animal, Universidade Federal do Rio Grande do Sul,
Porto Alegre, Brasil. 2 – Animal Health and Biomedical Sciences, University of Wisconsin, Madison, WI, EUA.
3 – Eutheria Foundation, Cross Plains, WI, EUA. 4 – Bolsista do CNPq
Resumo O presente estudo teve como objetivo conhecer a dinâmica folicular das éguas minipônei
durante um intervalo interovulatório. Durante a estação reprodutiva foram utilizadas 12
éguas minipôneis com histórico reprodutivo desconhecido, clinicamente sadias, cíclicas e
não lactantes, com idades variando entre 3 e 7 anos. As éguas foram avaliadas diariamente,
durante um ciclo estral, por um único operador, através de palpação e ultra-sonografia
trans-retal. Não foi realizado nenhum tipo de terapia hormonal nestes animais durante o
experimento. Todos os folículos visíveis à ultra-sonografia tiveram o antro medido e os
dados, de cada ovário, transferidos para ficha especial, identificando-se a exata posição de
cada folículo. Considerou-se como emergência o momento em que os folículos atingiram
6mm, com subseqüente crescimento. A divergência foi determinada como o dia em que as
taxas de crescimento entre os dois maiores folículos tornaram-se discrepantes. A altura,
largura e área do corpo lúteo foram medidos. A ecotextura uterina foi avaliada em escore de
1 a 4 durante todo o ciclo. As minipôneis apresentam um ciclo estral de 23,3 ± 3,2 dias com
uma onda folicular maior, não ocorrendo ondas secundárias na maioria dos casos. O
número de folículos durante o ciclo é estável e somente um, acima de 25 mm, foi observado
em cada ciclo, em todas as éguas. A emergência ocorre a partir do primeiro dia do ciclo e a
divergência, no 14o dia, junto com o desaparecimento do corpo lúteo. O maior diâmetro
folicular foi observado dois dias antes da ovulação. Finalmente, foi observado que os
escores das ecotexturas uterinas elevaram-se a partir do 9o dia do ciclo.
Palavras-chave: Minipôneis, ciclo estral, dinâmica folicular.
* Endereço para correspondência: Bento Gonçalves 9090, 91540-000, Porto Alegre, Brasil. [email protected].

24
Abstract This study aimed to verify the follicular dynamics from mini-pony mares during an
interovulatory interval. During the breeding season 12 mini-pony mares with unknown
reproductive history, clinically normal, cyclic, non lactating with age ranging between three
to seven years old were used. Mares were examined daily, during one estrus cycle, by a
single operator using transrectal ultrasonography. The mares were not submitted to any
hormonal therapy during the experiment. All the visible follicles were monitored daily,
when the antrum was measured and the position was taken. The data was then recorded in a
formulary. Follicle emergence was considered when it reached 6 mm diameter and
subsequent growth. Deviation was defined as the beginning of a marked difference in
growth rates between the two largest follicles. The height, width and area of the corpus
luteum were measured. The uterine ecotexture was also daily taken during the estrus cycle.
The data obtained suggest that the mini-pony mares present an estrus cycle of 23.3 ± 3.2
days, with one major follicular wave. Minor follicular waves were observed in a few cases.
The total number of follicles was similar throughout the cycle and only one follicle greater
than 25 mm was detected per cycle. The emergence was observed at the first day of the
cycle and the divergence at the 14th day together with the last observation of the corpus
luteum. The maximum diameter of the largest follicle was observed two days before
ovulation. Finally, it was observed that the uterine ecotexture grade raised until the 9th day
of the cycle.
Keywords: Mini-pony, estrus cycle, follicular dynamics.

25
Introdução
O intervalo interovulatório, que inicia com uma ovulação associada ao estro e termina
com a subseqüente, é a melhor referência para as pesquisas relacionadas a ondas foliculares
e tem duração média de 21 a 22 dias em éguas e de 24 dias em pôneis (GASTAL, 1999). O
número de ondas foliculares observadas durante o ciclo estral varia entre as diferentes
espécies (GINTHER & BERGFELT, 1992). Em éguas, as ondas foliculares foram
divididas em menores, nas quais não ocorre divergência e o maior folículo não se torna
dominante, e maiores, quando ocorre divergência e desenvolvimento de dominância
(GINTHER & BERGFELT, 1992; GINTHER, 1993).
A dinâmica folicular envolve quatro processos distintos: a emergência, a seleção, a
dominância e a atresia. Ao primeiro dia em que é possível observar o crescimento dos
folículos de uma onda, através da ultra-sonografia (BERGFELT & GINTHER, 1993;
GINTHER et al., 1996), ou ainda ao momento em que os folículos em crescimento tornam-
se suscetíveis à ação das gonadotropinas e iniciam a produção de pequenas concentrações
de estrógeno (DRIANCOURT, 2001), dá-se o nome de emergência folicular.
A seleção é o processo que resulta na redução do número de folículos de uma onda e
varia de acordo com o número específico de ovulações apresentado por cada espécie. Seu
término se dá no momento em que as taxas de crescimento entre os dois maiores folículos
de uma onda tornam-se discrepantes, caracterizando o início do processo de divergência
(GINTHER et al., 1996; 1997). Durante a seleção folicular o futuro folículo dominante é o
primeiro a se tornar apto a elevar a concentração intrafolicular e sistêmica de estradiol,
reduzir os níveis sistêmicos de FSH, impedindo que novos folículos sejam recrutados, e
responder ao aumento dos níveis de LH e, assim, estabelecer dominância (GASTAL, 1999).
O processo de dominância, por sua vez, caracteriza-se por um ou mais folículos
maiores exercendo efeito inibitório nos demais presentes nos grupos de recrutamento e
seleção. O dominante é o único que continua a se desenvolver enquanto os demais,
chamados subordinados, têm seu crescimento inibido (GINTHER, 1993). Seu número é
específico de cada espécie e está diretamente relacionado com o tamanho da progênie
(GINTHER, 1992; 1995). Aparentemente, a supressão final da onda de FSH, juntamente

26
com o acréscimo dos níveis de LH, tem importante papel na divergência das taxas de
crescimento entre o futuro folículo dominante e os demais (GINTHER et al., 1996; 1998).
A produção de grandes concentrações de estrógeno e inibina pelo folículo dominante
é responsável pela redução do aporte sangüíneo aos demais folículos que, juntamente com
as baixas taxas de FSH circulante, acabam por sofrer atresia. Somente aqueles que
mantiverem um adequado suprimento circulatório e, por conseqüência, estiverem sujeitos a
altos níveis de gonadotropinas continuarão a crescer até a ovulação (ACOSTA et al. 2004,
e BOLLWEIN et al. 2002).
A preparação do folículo para a ovulação ocorre sob uma seqüência de eventos
endócrinos, característicos da onda folicular pré-ovulatória, no momento em que as
concentrações de FSH e LH encontram-se elevadas, quando não mais se observa a inibição
da secreção de GnRH pela presença da progesterona (GINTHER et al., 1998).
A partir da liberação da onda de LH pré-ovulatória as células ovarianas da teca
interna passam a produzir progesterona, que estimula a síntese local da colagenase, enzima
que degrada o colágeno, maior componente do tecido conectivo entre o folículo e o ovário.
Neste mesmo momento, a produção local de PGF2α e PGE2 ocasiona contrações da
musculatura lisa ovariana, superficializando o folículo, e reorganiza o tecido folicular na
forma de corpo lúteo após a ovulação, através da ativação da enzima plasminogênio,
responsável pela formação de um coágulo após a ruptura do folículo (SENGER, 2003).
Compreender os mecanismos do desenvolvimento folicular das diferentes espécies é
de grande importância para o estudo da reprodução, uma vez que é o folículo que
proporciona um ambiente ótimo para a maturação do oócito, preparando-o para a
subseqüente fertilização após a ovulação. O conhecimento pleno destes mecanismos
fisiológicos é fundamental para que ocorra o aperfeiçoamento ou desenvolvimento de
novas técnicas para manipular a função reprodutiva dos animais domésticos (ADASHI &
ROBAN, 1992).
A dinâmica folicular pode ser precisamente analisada através da ultra-sonografia
transretal. Esta técnica permite a visualização freqüente e de forma não invasiva da
dinâmica da população de folículos, do processo ovulatório e do desenvolvimento,

27
manutenção e regressão do corpo lúteo (GINTHER & PIERSON, 1984a, 1984b;
GINTHER, 1986; GINTHER, 1998).
Estudos em éguas e em vacas são realizados desde a década de 80 e serviram de
modelo para o estudo em diversas espécies, como em ovelhas, cabras, porcas, lhamas
(BARTLEWSKI et al, 2000; DE CASTRO et al, 1999; COX, 1997; ADAMS et al, 1990).
Entretanto, devido à similaridade em diversas características da dinâmica folicular a égua
demonstrou ser um possível modelo experimental para o estudo da foliculogênese da
mulher com a vantagem de apresentar tamanhos foliculares maiores (GINTHER et al.,
2004).
Este trabalho tem como hipótese que a dinâmica folicular das minipôneis deverá ser
similar aos eqüinos até então estudados, apesar destes animais apresentarem um porte
bastante reduzido (acredita-se que as minipôneis devem apresentar diversos tipos de ondas
foliculares e que o diâmetro folicular na emergência e na divergência sejam semelhantes ao
dos eqüinos de maior porte).
O presente estudo teve como objetivo conhecer a dinâmica folicular das éguas
minipôneis durante um intervalo interovulatório e comparar os dados obtidos com
resultados já conhecidos de éguas de maior porte.
Material e Métodos
Animais Experimentais
Foram utilizadas 12 éguas minipôneis com histórico reprodutivo desconhecido nos
meses de janeiro a março, durante a estação reprodutiva. Antes de iniciar o experimento os
animais foram examinadas quanto à aptidão reprodutiva por meio de exame da
conformação perineal, exame retal e ultra-sonografia. Todos foram submetidos, também, a
um exame clínico geral, avaliando-se principalmente alterações locomotoras, respiratórias e
dentárias.

28
Somente foram utilizadas éguas clinicamente sadias, cíclicas e não lactantes com
idade média de cinco anos (3 a 7 anos), altura máxima de 90cm e peso vivo médio de 117
Kg (110 a 156 kg).
O experimento foi realizado no REPROLAB, localizado na Faculdade de Veterinária
da Universidade Federal do Rio Grande do Sul, Porto Alegre, Brasil (30º 04’ S, 51º 07’ W)
durante os meses de janeiro, fevereiro e março. Os animais foram everminados no início do
experimento e a cada 45 dias, alternando-se a utilização de uma associação de ivermectina
e pamoato de pirantel com febendazol. Durante o período que compreendeu o experimento,
as minipôneis foram mantidas a campo e receberam suplementação alimentar de
concentrado (com 12% de proteína bruta) e feno de alfafa, duas vezes por dia, e água à
vontade.
Ultra-sonografia
Para melhor compreender a dinâmica folicular das éguas minipôneis, 12 destes
animais foram submetidos a avaliações diárias através de ultra-sonografia, utilizando-se um
modelo de identificação individual de folículos. Buscando minimizar a porcentagem de
erros, as éguas eram examinadas calmamente por cerca de 15 minutos cada. Observaram-se
alguns eventos durante o desenvolvimento de uma onda folicular, tais como a emergência e
a divergência dos folículos, o máximo diâmetro atingido pelo folículo pré-ovulatório, o dia
da ovulação e a última observação do CL, bem como alguns intervalos entre estes eventos e
suas respectivas taxas de crescimento folicular, além de achados sobre ecotextura uterina.
As éguas foram avaliadas diariamente através de palpação trans-retal e ultra-
sonografia utilizando aparelho de ultra-som† e transdutor linear de 5.0 MHz até a
identificação da primeira ovulação (D 0). Quinze dias após a detecção da primeira
ovulação, utilizando a técnica da ultra-sonografia, o útero e os ovários das éguas foram
diariamente examinados, através do uso do ultra-som, e os dados anotados em fichas de
acompanhamento (Anexo 1), até 6 dias após a 3ª ovulação (Fig. 1). As fichas continham
† Aloka SSD 500V, Berger, São Paulo, Brazil

29
uma representação gráfica de cada ovário para cada dia de exame, com a localização e
tamanho das estruturas ovarianas.
As éguas foram avaliadas durante todo o experimento por um único operador. Não foi
realizado nenhum tipo de terapia hormonal nestes animais, nem para fins de sincronização
de estros, tampouco para promover a regressão precoce do CL ou indução da ovulação.
Figura 1 – Desenho experimental da coleta de dados realizada em 12 éguas minipôneis.
1ªOV 2ªOV 3ªOV
Dia 0 Dia 15 Dia 0 Dia 0 Dia 6
Coleta de dados diária por ultra-sonografia
Início do Experimento Fim do
Experimento
Folículos
Todos os folículos visíveis à ultra-sonografia tiveram o antro medido, duas vezes por
exame, no eixo de sua maior altura e largura, tomando-se como base a interface existente
entre a parede interna do folículo e o líquido folicular. Os dados de cada ovário foram
transferidos para a ficha especial, tomando-se o cuidado de identificar a exata posição de
cada folículo em relação à fossa ovulatória e aos principais eixos do ovário.
Os diâmetros eram medidos diariamente com a finalidade de acompanhar as
respectivas curvas de crescimento, o momento da divergência, o estabelecimento da
dominância e os tamanhos pré-ovulatórios destes folículos.
Considerou-se como emergência o momento em que os folículos atingiram 6mm com
subseqüente crescimento. A divergência foi determinada como o dia em que as taxas de
crescimento do maior e do segundo maior folículos tornaram-se discrepantes.

30
Corpo Lúteo (CL)
Quando existente, a maior altura e largura do corpo lúteo foram medidos. Utilizando a
ferramenta de desenho, demarcaram-se e mediram-se as áreas totais. Os diâmetros, as áreas
e suas posições foram devidamente anotados nas fichas de acompanhamento.
Útero
A ecotextura uterina foi avaliada, durante todo o período de exames, e classificada de
acordo com o grau de edema numa escala crescente de 1 a 4, equivalente a à ausência de
edema e grau máximo, respectivamente.
A ecotextura uterina pode ser classificada, segundo o grau de edema, em três escores:
1 ( ausência de edema e imagem homogênea, caracterizando o período de diestro), 2 (nível
intermediário de edema e imagem heterogênea, sem evidenciar as camadas uterinas) e 3
(edema máximo, imagem bastante heterogênea e identificação das camadas do endométrio,
caracterizando o período do estro). Pode ainda ser utilizado um escore subdividido em
quatro graus, variando de 1 a 4 de acordo com o edema uterino, sendo 1 o menor grau e 4 o
máximo (GINTHER, 1995).
Análise Estatística
Com a finalidade de conhecer a dinâmica folicular das éguas minipôneis observou-se,
através da ultra-sonografia trans-retal, os ovários e úteros destes animais, diariamente,
durante o período já previamente citado. Uma série de pontos foi analisada e registrada
durante este experimento, no que diz respeito ao comportamento folicular, luteal e uterino
durante duas ovulações consecutivas. Após o término do experimento, os dados foram
organizados dividindo-os em dia da ocorrência dos eventos de uma onda folicular,
intervalos (em dias) entre estes eventos, as respectivas taxas de crescimento folicular e o
diâmetro dos maiores folículos em cada um desses momentos. Utilizaram-se os métodos
com e sem identificação (GINTHER, 1995) na mensuração dos folículos. O método

31
identificado permitiu observar o folículo dominante e o maior subordinado, enquanto que o
não identificado permitiu mensurar o maior e segundo maior folículo.
Observaram-se as médias e desvios padrões do dia da ocorrência da emergência e da
divergência dos dois maiores folículos, da última observação do CL, do diâmetro máximo
(em mm) dos dois maiores folículos e da ovulação. Também foram determinados os
intervalos, em dias, entre a emergência e a divergência, entre a divergência e o máximo
diâmetro atingido pelos dois maiores folículos, entre a divergência e a ovulação e entre o
diâmetro máximo e a ovulação. Os diâmetros destes dois folículos foram mensurados no
dia da emergência, no início da divergência, ao atingir o diâmetro máximo e no dia anterior
à ovulação. E, suas respectivas taxas de crescimento foram avaliadas entre a emergência e a
divergência, entre a divergência e o maior diâmetro, entre a divergência e a ovulação e
entre o Dia -4 e a ovulação.
As diferenças entre as médias do dia da emergência, do diâmetro máximo, do
intervalo em dias entre a divergência e o máximo diâmetro e entre o maior diâmetro e a
ovulação, bem como, o diâmetro folicular as taxas de crescimento na emergência, na
divergência, no maior diâmetro e no dia anterior à ovulação, entre o maior e o segundo
maior folículo foram avaliadas através do teste T, considerando-se significativa uma
diferença P < 0,05.
Resultados
Na Tabela 1 estão descritos os dias de ocorrência da emergência dos dois maiores folículos,
da divergência, da última observação do corpo lúteo, do maior diâmetro folicular observado
pelos dois maiores folículos e o dia da ovulação, bem como os intervalos em dias da
emergência até a divergência, da divergência até o dia do maior diâmetro, da divergência
até a ovulação e do maior diâmetro até a ovulação. Ao se avaliar a Tabela 1 verifica-se que
o ciclo estral médio das 12 éguas minipôneis foi de 23 dias. Verifica-se, também, que os
dois maiores folículos emergem até o oitavo dia do ciclo estral e a divergência ocorre no
mesmo dia da última observação do corpo lúteo. O maior folículo atinge o máximo
diâmetro sete dias após a divergência, enquanto o segundo maior folículo chega ao maior

32
diâmetro imediatamente após ela ocorrer (cerca de um dia), quando então inicia um
processo de atresia. Após atingir o máximo diâmetro o maior folículo demorou em média
dois dias até a ocorrência da ovulação.
Tabela 1. Médias (± desvio padrão) do dia da ocorrência e dos intervalos observados, pelo
método não identificado, durante o ciclo estral de 12 éguas minipôneis.
Observação Média ± DP P Número de Éguas 12 Dia da Ocorrência
Emergência1 com 6mm Folículo Dominante 7,8 ± 5,9 Maior Folículo Subordinado 6,3 ± 4,9
= 0,50
Divergência2
14,4 ± 3,1
Última observação do CL
14,8 ± 2,3
Diâmetro máximo
Maior Folículo 21,5 ± 2,9 Segundo Maior Folículo 15,2 ± 3,1
< 0,01
Ovulação
23,3 ± 3,2
Intervalos (dias)
Emergência até Divergência
6,6 ± 6,2
Divergência até Máximo Diâmetro
Maior Folículo 6,2 ± 2,4 Segundo Maior Folículo 0,5 ± 1,0
< 0,01
Divergência até Ovulação
7,1 ± 2,2
Diâmetro Máximo até Ovulação
Maior Folículo 2,1 ± 1,3 Segundo Maior Folículo 8,3 ± 3,0
<0,01
1 A emergência de um folículo foi definida quando este atingia 6mm 2 Divergência = dia do início da maior diferença na taxa de crescimento entre os dois maiores folículos

33
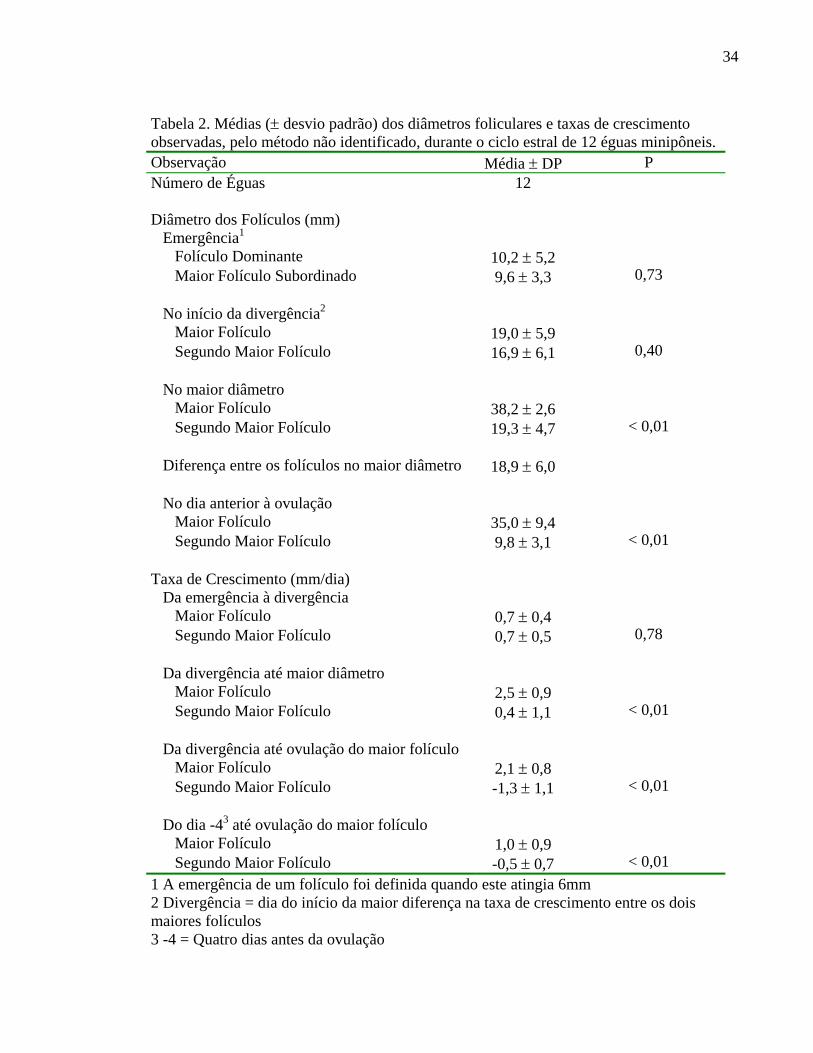
Na Tabela 2 estão descritos os diâmetros observados do maior e do segundo maior
folículo no dia da emegência, no início da divergência, no dia do maior diâmetro, no dia
anterior à ovulação bem como, as taxas de crescimento observadas nos diferentes
momentos do ciclo estral. Verifica-se que os diâmetros dos dois maiores folículos são
semelhantes na emergência até o início da divergência. Entretanto, há uma diferença
significativa (P < 0,01) entre o tamanho observado do maior e do segundo maior folículo
quando estes atingem o maior diâmetro, diferença esta superior a 18 mm. Os dois maiores
folículos diferem significativamente (P <0,01) também, no dia anterior à ovulação. Da
mesma forma, observa-se que as taxas de crescimento dos dois maiores folículos são
semelhantes (P > 0,05) entre a emergência e a divergência, porém diferem a partir da
divergência até a ovulação. A taxa de crescimento observada do maior folículo da
emergência à divergência é significativamente menor (P < 0,05) que as taxa observadas
entre a divergência e o maior diâmetro observado. Entretanto, a taxa de crescimento nos
quatro dias que precedem a ovulação do maior folículo foi semelhante (P = 0,38) à
observada entre a emergência e a divergência.
34
Tabela 2. Médias (± desvio padrão) dos diâmetros foliculares e taxas de crescimento observadas, pelo método não identificado, durante o ciclo estral de 12 éguas minipôneis. Observação Média ± DP P Número de Éguas 12 Diâmetro dos Folículos (mm)
Emergência1 Folículo Dominante 10,2 ± 5,2 Maior Folículo Subordinado 9,6 ± 3,3
0,73
No início da divergência2
Maior Folículo 19,0 ± 5,9 Segundo Maior Folículo 16,9 ± 6,1
0,40
No maior diâmetro
Maior Folículo 38,2 ± 2,6 Segundo Maior Folículo 19,3 ± 4,7
< 0,01
Diferença entre os folículos no maior diâmetro
18,9 ± 6,0
No dia anterior à ovulação
Maior Folículo 35,0 ± 9,4 Segundo Maior Folículo 9,8 ± 3,1
< 0,01
Taxa de Crescimento (mm/dia)
Da emergência à divergência Maior Folículo 0,7 ± 0,4 Segundo Maior Folículo 0,7 ± 0,5
0,78
Da divergência até maior diâmetro
Maior Folículo 2,5 ± 0,9 Segundo Maior Folículo 0,4 ± 1,1
< 0,01
Da divergência até ovulação do maior folículo
Maior Folículo 2,1 ± 0,8 Segundo Maior Folículo -1,3 ± 1,1
< 0,01
Do dia -43 até ovulação do maior folículo
Maior Folículo 1,0 ± 0,9 Segundo Maior Folículo -0,5 ± 0,7
< 0,01
1 A emergência de um folículo foi definida quando este atingia 6mm 2 Divergência = dia do início da maior diferença na taxa de crescimento entre os dois maiores folículos 3 -4 = Quatro dias antes da ovulação
35
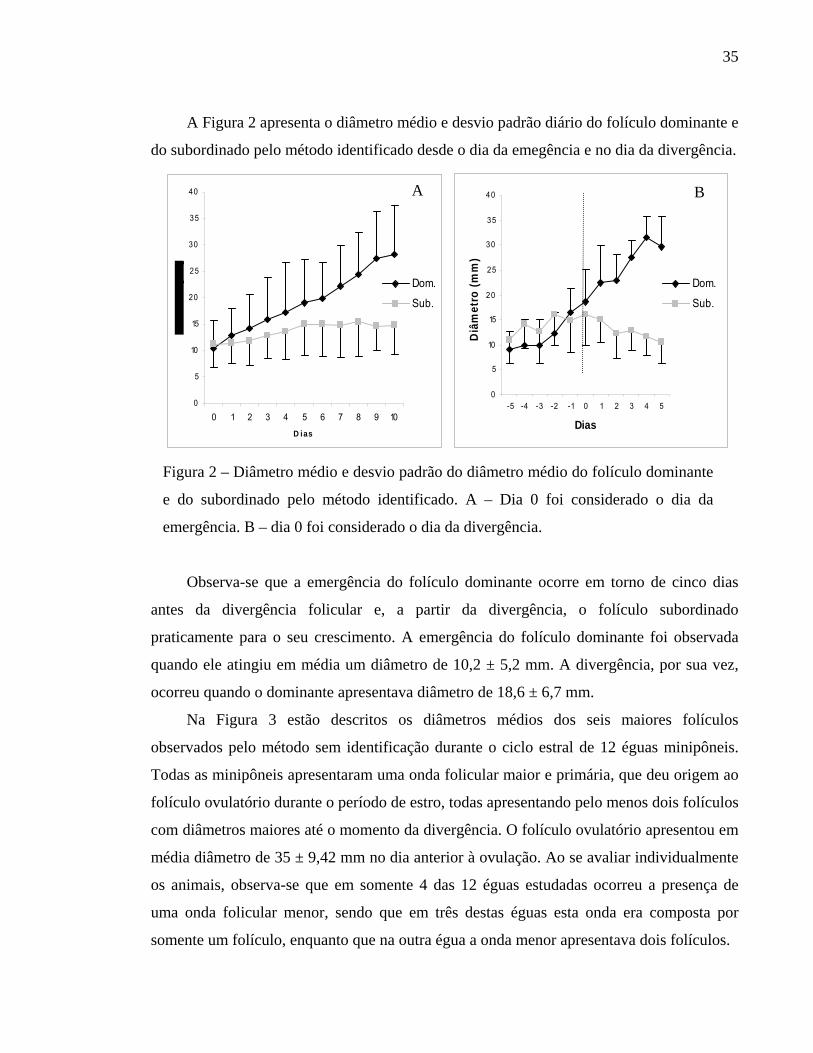
A Figura 2 apresenta o diâmetro médio e desvio padrão diário do folículo dominante e
do subordinado pelo método identificado desde o dia da emegência e no dia da divergência.
Figura 2 – Diâmetro médio e desvio padrão do diâmetro médio do folículo dominante
e do subordinado pelo método identificado. A – Dia 0 foi considerado o dia da
emergência. B – dia 0 foi considerado o dia da divergência.
Observa-se que a emergência do folículo dominante ocorre em torno de cinco dias
antes da divergência folicular e, a partir da divergência, o folículo subordinado
praticamente para o seu crescimento. A emergência do folículo dominante foi observada
quando ele atingiu em média um diâmetro de 10,2 ± 5,2 mm. A divergência, por sua vez,
ocorreu quando o dominante apresentava diâmetro de 18,6 ± 6,7 mm.
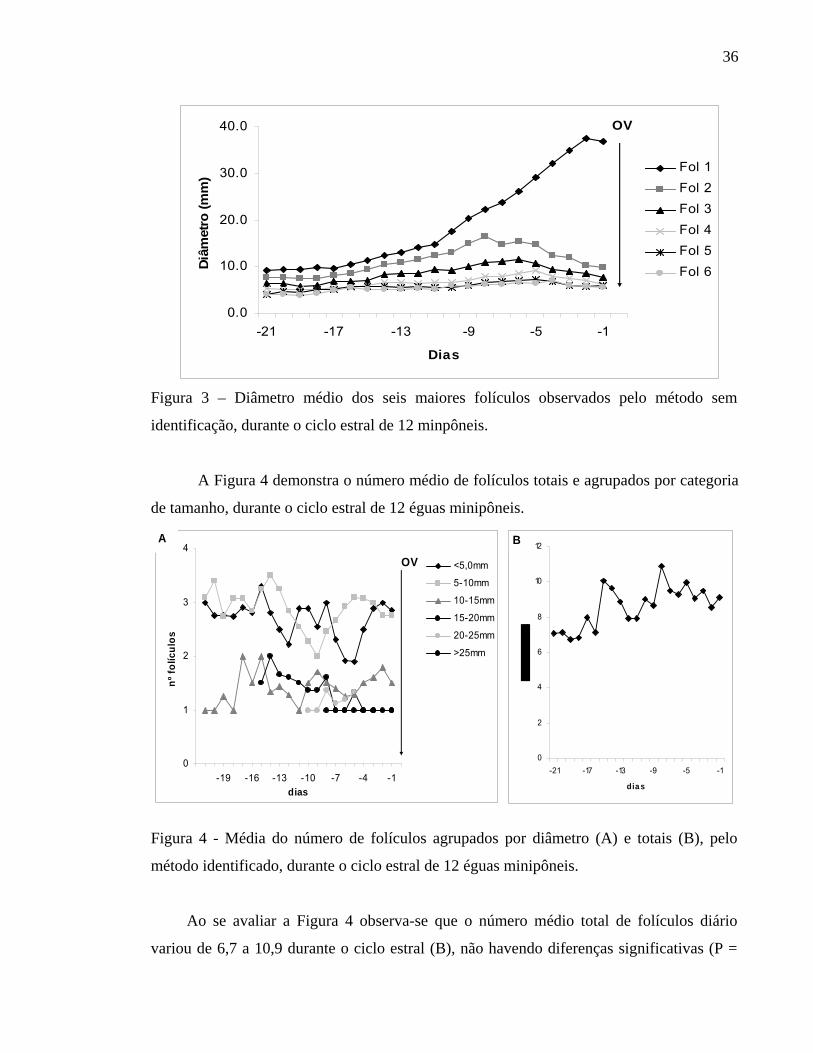
Na Figura 3 estão descritos os diâmetros médios dos seis maiores folículos
observados pelo método sem identificação durante o ciclo estral de 12 éguas minipôneis.
Todas as minipôneis apresentaram uma onda folicular maior e primária, que deu origem ao
folículo ovulatório durante o período de estro, todas apresentando pelo menos dois folículos
com diâmetros maiores até o momento da divergência. O folículo ovulatório apresentou em
média diâmetro de 35 ± 9,42 mm no dia anterior à ovulação. Ao se avaliar individualmente
os animais, observa-se que em somente 4 das 12 éguas estudadas ocorreu a presença de
uma onda folicular menor, sendo que em três destas éguas esta onda era composta por
somente um folículo, enquanto que na outra égua a onda menor apresentava dois folículos.
0
5
10
15
20
25
30
35
40
-5 -4 -3 -2 -1 0 1 2 3 4 5
Dias
Diâ
met
ro (m
m)
Dom.
Sub.
0
5
10
15
20
25
30
35
40
0 1 2 3 4 5 6 7 8 9 10
D ias
Dom.
Sub.
A B
36
0.0
10.0
20.0
30.0
40.0
-21 -17 -13 -9 -5 -1
Dias
Diâm
etro
(mm
)
Fol 1Fol 2Fol 3Fol 4Fol 5Fol 6
OV
Figura 3 – Diâmetro médio dos seis maiores folículos observados pelo método sem
identificação, durante o ciclo estral de 12 minpôneis.
A Figura 4 demonstra o número médio de folículos totais e agrupados por categoria
de tamanho, durante o ciclo estral de 12 éguas minipôneis.
0
1
2
3
4
-19 -16 -13 -10 -7 -4 -1dias
nº fo
lícul
os
<5,0mm
5-10mm
10-15mm
15-20mm
20-25mm
>25mm
OV
0
2
4
6
8
10
12
-21 -17 -13 -9 -5 -1
dia s
A B
Figura 4 - Média do número de folículos agrupados por diâmetro (A) e totais (B), pelo
método identificado, durante o ciclo estral de 12 éguas minipôneis.
Ao se avaliar a Figura 4 observa-se que o número médio total de folículos diário
variou de 6,7 a 10,9 durante o ciclo estral (B), não havendo diferenças significativas (P =

37
0,88). Observa-se deste total uma predominância (A) durante o ciclo estral de folículos
menores do que 10mm que apresentaram uma variação diária de 4,8 a 6,6 folículos.
Nenhuma égua apresentou mais do que um folículo superior a 25 mm.
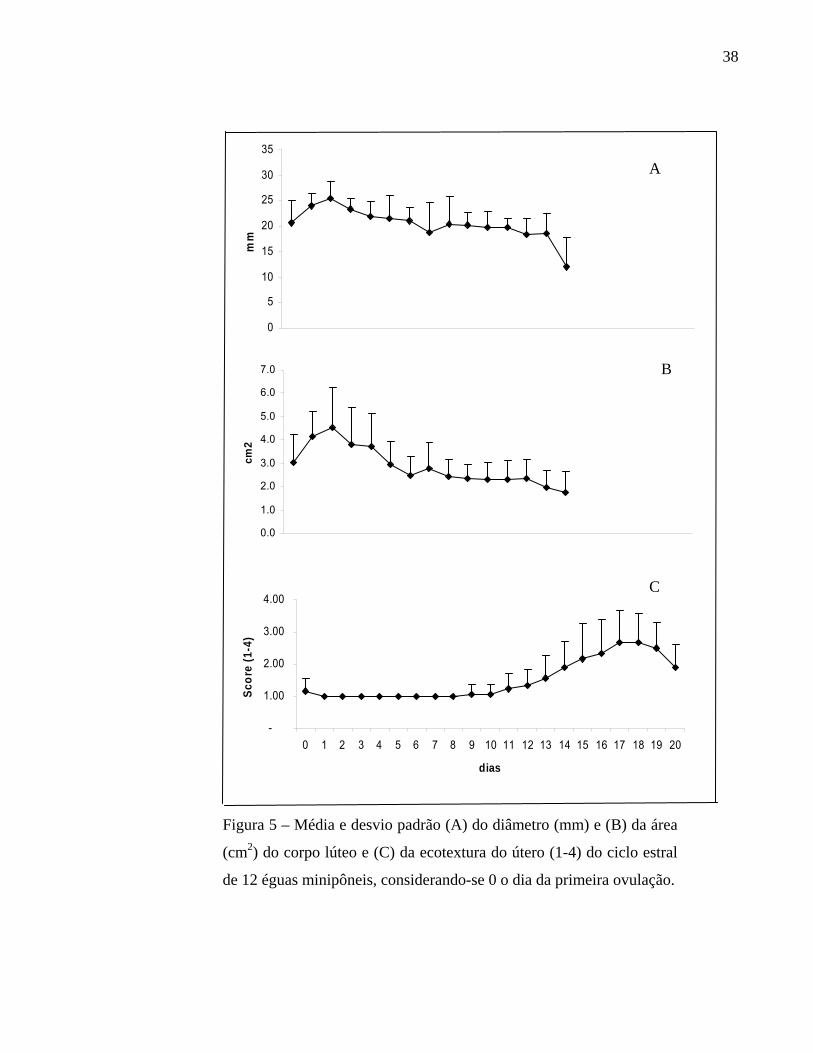
Na figura 5 estão descritas as médias e desvios padrões do diâmetro e da área do
corpo lúteo e a ecotextura uterina média observada durante um ciclo estral de éguas
minipôneis. O CL apresentou seu maior diâmetro (25,5 ± 3,3 mm) e sua maior área (4,5 ±
1,8 cm2) no segundo dia após a ovulação. Tanto área quanto diâmetro decaíram levemente
até o 14º dia não sendo mais possível a visualização no 15º dia. Por sua vez a ecotextura
manteve-se no grau 1 até o 8º dia do ciclo, quando iniciou um leve aumento que tornou-se
ainda maior a partir do 12º dia do ciclo e atingiu seu ápice no 18° dia. Em 8 éguas foi
observada a ecotextura 4, entretanto este alto grau de edemaciação ocorreu entre o 15º e o
19º dia do ciclo e teve duração máxima de um dia em 6 éguas e de dois dias em duas éguas.
38
0
5
10
15
20
25
30
35
mm
A
B
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
cm2
-
1.00
2.00
3.00
4.00
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
dias
Scor
e (1
-4)
C
Figura 5 – Média e desvio padrão (A) do diâmetro (mm) e (B) da área
(cm2) do corpo lúteo e (C) da ecotextura do útero (1-4) do ciclo estral
de 12 éguas minipôneis, considerando-se 0 o dia da primeira ovulação.

39
Discussão
Como onda folicular, segundo Ginther et al. (2004), entende-se a emergência de
diversos folículos que inicialmente crescem em sincronia até o momento em que apenas
um, ou o número específico de cada espécie, continua crescendo até a ovulação enquanto os
demais tendem a regredir. O acréscimo nas concentrações de FSH é pré-requisito para que
uma onda folicular seja desencadeada (GINTHER et al., 1996; ROCHE, 1996;
DRIANCOURT, 2001; FORTUNE et al., 2001). Isto inicia quando o maior folículo
apresenta cerca de 6mm de diâmetro e tem seu pico quando ele atinge aproximadamente
13mm (GASTAL, 1997).
Em éguas, as ondas foliculares classificam-se como menores, quando não se observa
divergência e o maior folículo não se torna dominante, e maiores, onde ambos estes
fenômenos ocorrem (GINTHER & BERGFELT, 1992; GINTHER, 1993). Estas últimas
podem ainda ser subdivididas em primárias, quando dão origem a um folículo que irá
ovular durante o estro, ou secundárias, quando existe dominância mas a ovulação não
ocorre ou se dá após a onda primária ou estro, caracterizando uma ovulação secundária ou
de diestro. As ondas menores, bem como as secundárias, ocorrem com maior freqüência
durante a primavera (GINTHER, 1993). No decorrer deste experimento foram observadas,
em 8 das 12 éguas (cerca de 67%), somente ondas foliculares maiores e primárias,
provavelmente pelo fato de que o acompanhamento dos ciclos se deu principalmente
durante os meses de verão e pelo reduzido número de folículos presentes nos ovários das
minipôneis, quando comparados com éguas de médio a grande porte, em qualquer fase do
ciclo estral. As 4 restantes (33%) apresentaram ondas menores no decorrer de seus ciclos
estrais, aproximadamente um dia após o desaparecimento do CL, sendo que 3 delas eram
compostas por apenas um folículo.
Segundo Ginther et al. (1996; 1997) o processo de seleção resulta na redução do
número de folículos de uma onda, fenômeno não observado no presente trabalho.
Observou-se que as minipôneis apresentam um menor número de folículos durante toda a
extensão do ciclo estral, possivelmente explicada pelas reduzidas dimensões ovarianas.
Pierson & Ginther (1987) relatam a presença de até 12 folículos medindo cerca de 10mm

40
de diâmetro em ovários de éguas. Neste experimento encontrou-se um número máximo de
6,6 folículos com diâmetro inferior a 10mm.
Como emergência folicular entende-se o primeiro dia em que é possível observar o
crescimento dos folículos de uma onda (BERGFELT & GINTHER, 1993; GINTHER et al.,
1996), ou ainda o momento em que eles se tornam suscetíveis à ação das gonadotropinas e
iniciam a produção de pequenas concentrações de estrógeno (GINTHER et al., 1996;
1997). No presente trabalho, devido à indisponibilidade de mensuração hormonal,
estipulou-se a emergência como o momento em que os folículos atingiram diâmetros
mínimos de 6mm, com subseqüente crescimento. Este estágio foi observado já no Dia 7,8
do ciclo para o maior folículo e no Dia 6,3 para o segundo maior , entretanto, não se
observou diferença em diâmetro entre o maior e o segundo maior folículo no dia da
emergência conforme relatado por Gastal et al. (1997) para éguas de maior porte.
Ginther et al. (1996; 1997) descrevem a divergência como o momento em que os dois
maiores folículos de uma onda passam a apresentar taxas de crescimento discrepantes.
Gastal (1999) cita ainda que este ponto ocorre aproximadamente no Dia 17 do ciclo e que o
folículo dominante, em éguas, costuma medir entre 21 e 23mm de diâmetro. Entretanto, nas
minipôneis observaram-se folículos com cerca de 19mm, diferença já esperada pelo
reduzido tamanho ovariano e diâmetro folicular apresentados por estes animais quando
comparados com éguas maiores. Observou-se também que o diâmetro máximo alcançado
pelos folículos pré-ovulatórios das minipôneis foi de 38mm em média e, segundo Ginther
et al. (2004), em éguas costumam medir cerca de 45mm.
O dia de início da divergência verificado nas minipôneis (Dia 14) foi verificado cerca
de três dias antes quando comparado aos achados de Gastal et al. (1997) em estudos
realizados em éguas maiores e, o ciclo estral, apresentou um dia a menos (23). Verificou-se
que o intervalo entre a divergência e a ovulação das minipôneis foi, em média, de 7 dias,
similar ao descrito por Gastal et al. (1997) em éguas de maior porte. O início da
divergência parece estar relacionado com a regressão e desaparecimento do CL, verificado,
em média, também no Dia 14. Provavelmente o decréscimo nas concentrações de
progesterona, a partir da ausência do CL, e o conseqüente aumento das taxas de LH
circulante, sejam responsáveis pelo desenvolvimento do folículo pré-ovulatório.

41
O decréscimo das concentrações de progesterona a partir da ausência do CL e o
conseguinte aumento das taxas de LH circulante são responsáveis pelo desenvolvimento
final do folículo pré-ovulatório. O início da onda de LH, responsável pela ovulação, é
ocasionado pela redução da progesterona circulante e pelo aumento das concentrações de
estrógenos. A produção de inibina pelo folículo dominante, a partir da divergência, levará à
diminuição da produção de FSH e a conseqüente regressão dos demais folículos
(BERGFELT et al., 1991; GINTHER, 1992; GINTHER & BERGFELT, 1993; GINTHER
et al., 1997).
Acosta et al. (2004) e Bollwein et al. (2002) sugerem a ocorrência de um acréscimo
do aporte sangüíneo ao folículo dominante e uma redução do suprimento sangüíneo ao
maior subordinado cerca de dois dias antes da divergência. Conseqüentemente, o
dominante será exposto a grandes quantidades de gondotropinas e continuará crescendo até
a ovulação. Entretanto, os demais folículos, submetidos a um reduzido aporte sangüíneo,
juntamente com o decréscimo das concentrações de FSH verificadas no momento da
divergência, acabam por sofrer atresia.
As taxas de crescimento do maior e do segundo maior folículo, verificadas entre a
emergência e a divergência, entre a divergência e o máximo diâmetro alcançado pelo
folículo dominante, entre a divergência e a ovulação e entre o Dia -4, quatro dias antes da
ovulação, até ela ocorrer foram semelhantes com as encontradas por Gastal et al. (1997). A
partir do início da divergência, o segundo maior folículo sofre um brusco decréscimo em
suas taxas de crescimento. O futuro folículo ovulatório apresenta uma elevada taxa de
crescimento até o quarto dia antes da ovulação, quando o crescimento torna-se mais lento.
Em 66,6% das éguas minipôneis avaliadas o folículo dominante atingiu seu diâmetro
máximo no dia anterior à ovulação, concordando com os achados de Ginther et al. (2004),
em trabalho realizado com éguas de maior porte. Os autores descreveram ainda que, em
27,6% dos casos, o tamanho folicular máximo foi alcançado dois dias antes da ovulação,
6,9% três dias e 3,4% quatro dias antes dela ocorrer. Em 16% das éguas minipôneis
observou-se o maior diâmetro do folículo pré-ovulatório 2 e 3 dias antes da ovulação.
Em acordo com os dados descritos por Pierson & Ginther (1985) e Redmer &
Reynolds (1996), observou-se neste trabalho uma pronunciada alteração na forma esférica

42
do folículo pré-ovulatório para cônica ou de pêra, em cerca de 85% das éguas avaliadas,
possivelmente relacionada com um acréscimo da flutuação que ocorre próximo à ovulação.
A ecotextura uterina também sofre alterações durante o ciclo estral das éguas
(GINTHER, 1995). Durante o estro, as imagens uterinas obtidas através do ultra-som
caracterizam-se por áreas alternadas de hipoecogenicidade e hiperecogenicidade. As
porções hiperecogênicas correspondem ao endométrio e, as hipoecogênicas, ao edema
uterino verificado durante este período. Entretanto, durante o diestro, a ecotextura uterina
costuma ser mais homogênea e verifica-se a predominância de áreas hiperecogênicas. Estas
alterações foram também observadas no útero das minipôneis.
Com relação à ecotextura uterina, Ginther (1995) descreve ainda que 61% das éguas
atingem os escores máximos no período pré-ovulatório e 65% apresentam uma redução do
edema uterino nos Dias -3 ao -1 do ciclo, possivelmente relacionado com o declínio da
concentração de estrógeno circulante, bem como observado nas minipôneis durante este
experimento. Esta observação pode ser válida para a identificação de uma ovulação
iminente. Os escores tendem a diminuir entre os Dias -15 e -9 e então passam a aumentar
progressivamente até o dia -3, quando decrescem e atingem valores mínimos no Dia +2,
caracterizando o período de diestro.
Observando os ovários das éguas através da utilização de um transdutor de 5.0 MHz,
durante o intervalo inter-ovulatório, observa-se que o CL pode ser detectado, em média, até
o Dia 17, para um ciclo de 22 dias e é facilmente visualizado diariamente dentro deste
período (GINTHER, 1986). Entretanto, nas minipôneis a última visualização do CL ocorreu
em média no Dia 14, para um ciclo com extensão semelhante ao das éguas de maior porte.
Esta diferença pode estar relacionada com a menor dimensão de todas as estruturas
ovarianas das minipôneis, em relação às éguas maiores.
O diâmetro do maior folículo no dia em que o CL é observado pela última vez pode
auxiliar à distinção entre as éguas cíclicas e as que se encontram-na fase anovulatória. Se o
maior folículo medir 20mm ou menos e o CL não estiver presente, considera-se a égua não-
cíclica (GINTHER, 1995). Em 67% das minipôneis o folículo dominante apresentou
diâmetros maiores do que 20mm no momento em que o CL não pode mais ser identificado,
caracterizando a estação ovulatória. Entretanto, em 4 minipôneis que ovularam, o maior
folículo não atingiu 20 mm no dia da divergência, variando entre 14,5 e 19,4 mm, porém

43
não se observou a presença de folículos medindo mais de 25mm por períodos superiores a
10 dias nos ovários destes animais o que, segundo Ginther (1995), também caracterizaria
uma fase trasnsicional.
No presente experimento observou-se um aumento do diâmetro e da área do CL entre
o primeiro e terceiro dia do ciclo, quando, então iniciou uma lenta diminuição até sua
completa regressão. Estes dados são semelhantes aos observados por Ginther (1995) em
éguas de maior porte. Entretanto, nas minipôneis a ecotextura uterina iniciou um aumento
nos escores no nono dia do ciclo, seis dias antes do completo desaparecimento do CL.
Referências
ACOSTA, T. J. et al. Differential blood flow changes between the future dominant and subordinate follicles precede diameter changes during follicle selection in mares. Biology of Reproduction, v. 71, p. 502-507, 2004. ADAMS, G. P.; SUMAR, J. GINTHER, O. J. Effects of lactational and reproductive status on ovarian follicular wave in llamas (Lama Glama). Journal of Reproduction and Fertility, v. 90, p. 535-545, 1990. ADASHI, E. Y.; ROBAN, R. M. Intraovarian regulation. Peptidergic signalling systems. Trens in Endocrinology nd Metabolism, v. 3, p. 139-146, 1992. BARTLEWSKI, P. M. et al. Ovarian antral follicular dynamics and their associations with peripheral concentrations of gonadotropins and ovarian steroids in anoestrous Finnish Landrace ewe. Animal Reproduction Science, v. 58, p. 273-291, 2000. BERGFELT, D. R. et al. Circulating concentrations of immunoreactive inhibin and FSH during the estrous cycle of mares. Journal of Equine Veterinarian Science, v. 11, p. 319-322, 1991. BERGFELT, D. R.; GINTHER, O. J. Relationships between FSH surges and follicular waves during the estrous cycle in mares. Theriogenology, v. 39, n. 4, p. 781-796, 1993. BOLLWEIN, H. et al. Uterine and ovarian blood flow during the estrous cycle in mares. Theriogenology, v. 57, p. 2129-2138, 2002. de CASTRO, T. et al. Ovarian dynamics, serum estradiol and progesterone concentrations during the interovulatory interval in goats. Theriogenology, v. 52, p. 399-411, 1999. COX, N. M. Control of follicular development and ovulation rate in pigs. Journal of Reproduction and Fertility, v. 52, p. 31-46, 1997. Supplement.

44
DRIANCOURT, M. A. Follicular dynamics in farm animals. Implicaion for manipulation of reproduction. Theriogenology, v. 55, p. 1211-1239, 2001. FORTUNE, J. E. et al. Differentiation of dominant versus subordinate follicles in cattle. Biology of Reproduction, v. 65, p. 648-654, 2001. GINTHER, O. J. Ultrasonic Imaging and Reproductive Events in Mare. Cross Plains, WI: Equiservices, 1986. GINTHER, O. J. Major and Minor follicular waves during the equine estrous cycle. Journal of Equine Veterinarian Science, v. 13, p. 18-25, 1993. GINTHER, O. J. Reproductive Biology of the Mare, Basic and Applied Aspects, 2nd ed. Cross Plains, WI: Equiservices Publishing, 1992. GINTHER, O. J. Ultrasonic Imaging and Animal Reproduction: Horses. Book 2. Cross Plains, WI: Equiservices Publishing, 1995. GINTHER, O. J. Ultrasonic Imaging and Animal Reproduction: Fundamentals. Book 1. Cross Plains, WI: Equiservices Publishing, 1995. GINTHER, O. J. et al. Comparative study of the dynamics of follicular waves in mares and woman. Biology of Reproduction, v. 71, p. 1195-1201, 2004. GINTHER, O. J et al. Pulsatility of systemic FSH and LH concentrations during follicular wave development in catle. Theriogenology, v. 50, P. 507-519, 1998. GINTHER, O. J. et al. Selection of the dominant follicle in cattle. Biology of Reproduction, v. 55, p. 1187-11194, 1996. GINTHER, O. J et al. Sampling follicular fluid without altering follicular status in catle: oestradiol concentrations early in a follicular wave. Journal of Reproduction and Fertility , v. 109, p. 181-186, 1997 GINTHER, O. J.; BERGFELT, D. R. Growth of small follicles and concentrations of FSH during the equine oestrous cycle. Journal of Reproduction and Fertility , v. 99, p. 105-111, 1993. GINTHER, O. J.; BERGFELT, D. R. Ultrasonic characterization of follicular wave in mares without maintaining identity of individual follicles. Journal of Equine Veterinarian Science, v. 12, p. 349-354, 1992. GINTHER, O. J.; PIERSON, R.A. Ultrasonic anatomy and pathology of the equine uterus. Theriogenology, v. 21, p. 471-483, 1984a.

45
GINTHER, O. J.; PIERSON, R.A. Ultrasonic evaluation of the reproductive tract of the mare: ovaries. Journal of Equine Veterinarian Science, v. 4, p. 11-16, 1984b. GASTAL, E. L. Selection of the dominant follicle in mares: role of follicle-diameter differences, gonadotropins, and estradiol. 1999. 258 f. Tese (Doctor of Phisiology) – University of Wisconsin-Madison, 1999. GASTAL, E. L. et al. Echotextural changes in the follicular wall during follicle deviation in mares. Theriogenology, v.52, p. 803-814, 1999. GASTAL, E. L. et al. Role of diameter diferences among follicle in selection of a future dominant follicle in mares. Biology of Reproduction, v. 57, n. 6, p. 1320-1327, 1997. PIERSON, R. A.; GINTHER, O. J. Follicular populations during the estrous cycle of the mare. Animal Reproduction Science, v. 14, p. 219-231, 1987. PIERSON, R. A.; GINTHER, O. J. Ultrasonic evaluation of the preovulatory follicle in the mare. Theriogenology, v.24, p.359-368, 1985. REDMER, D. A.; REYNOLDS, L. P. Angiogeneses in the ovary. Reviews of Reproduction, v.1, p. 182-192, 1996. SENGER, P. L. Pregnancy and Parturition, 2nd ed. Pullman: Current Conceptions, Inc, 2003.

46
4 CONCLUSÕES
1 – O ciclo estral das éguas minipôneis é de 23,3 ± 3,2 dias.
2 - As éguas minipôneis apresentam uma onda folicular maior primária, não ocorrendo
ondas secundárias na maioria dos ciclos.
3 – O número de folículos durante o ciclo estral das minipôneis é estável e menor do que o
número de folículos encontrados nos ovários de éguas de maior porte. Somente um folículo
acima de 25 mm foi observado por ciclo.
4 – A emergência folicular ocorre a partir dos primeiros dias do ciclo, bem como observado
em éguas de portes maiores.
5 – A divergência folicular das minipôneis ocorre no 14o dia, juntamente com o
desaparecimento do corpo lúteo. Diferindo de dados descritos em éguas maiores, quando a
divergência é observada no 11° dia do ciclo, em média.
6 – O maior diâmetro folicular foi observado cerca de dois dias antes da ovulação.
7 – O útero das éguas Minipônei aumenta sua ecotextura a partir do oitavo dia do ciclo,
atingindo seu grau máximo no 18° dia.

47
5 REFERÊNCIAS BIBLIOGRÁFICAS
ACOSTA, T. J. et al. Differential blood flow changes between the future dominant and subordinate follicles precede diameter changes during follicle selection in mares. Biology of Reproduction, v. 71, p. 502-507, 2004. ADAMS, G. P.; SUMAR, J.; GINTHER, O. J. Effects of lactational and reproductive status on ovarian follicular wave in llamas (Lama Glama). Journal of Reproduction and Fertility, v. 90, p. 535-545, 1990. ADAMS, G. P. et al. Association between surges of follicle-stimulating hormane and the emergence of follicukar wave in heifers. Journal of Reproduction nd Fertility, v. 94, p. 177-188, 1992. ADASHI, E. Y.; ROBAN, R. M. Intraovarian regulation. Peptidergic signalling systems. Trens in Endocrinology nd Metabolism, v. 3, p. 139-146, 1992. BAKER, C. B.; KENNEY, R. M. Systemathic approach to the diagnosis of the intertile and subfertil mare. In: MORROW (ed) Current therapy in teriogenology. Phyladelphia: W. B. Saunders Co., 1980. p. 721-736. BARTLEWSKI, P. M. et al. Ovarian follicular dynamics during anoestrus in ewes. Journal of Reproduction and Fertility, v. 113, p. 275-285, 1998. BARTLEWSKI, P. M. et al. Ovarian antral follicular dynamics and their associations with peripheral concentrations of gonadotropins and ovarian steroids in anoestrous Finnish Landrace ewe. Animal Reproduction Science, v. 58, p. 273-291, 2000. BERGFELT, D. R. et al. Circulating concentrations of immunoreactive inhibin and FSH during the estrous cycle of mares. Journal of Equine Veterinarian Science, v. 11, p. 319-322, 1991. BERGFELT, D. R.; GINTHER, O. J. Relationships between FSH surges and follicular waves during the estrous cycle in mares. Theriogenology, v. 39, n. 4, p. 781-796, 1993. BERGMANN, J. A. G. et al. Formação e estrutura da raça pônei Brasileira. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 49, n.2, p. 251-259, 1997. BOLLWEIN, H. et al. Uterine and ovarian blood flow during the estrous cycle in mares. Theriogenology, v. 57, p. 2129-2138, 2002. BOSSIS, I. et al. Nutritionally induced anovulation in beef heifers: ovarian and endocrine function during realimentation and resumption of ovulation. Biology of Reproduction, v. 62, p. 1436-1444, 2000.

48
CAMPBELL, B. K.; SCARAMUZZI, R. J.; WEBB, R. Control of antral follicle development and selection in sheep and cattle. Journal of Reproduction nd Fertility, v. 49, p. 335-350, 1995. Supplement. de CASTRO, T. et al. Ovarian dynamics, serum estradiol and progesterone concentrations during the interovulatory interval in goats. Theriogenology, v. 52, p. 399-411, 1999. CAVENDER, J. L.; MURDOCH, W. J. Morphological studies of the circulatory system of preovulatory ovine follicles. Biology of Reproduction, v. 39, p. 989-997, 1998. COX, N. M. Control of follicular development and ovulation rate in pigs. Journal of Reproduction and Fertility, v. 52, p. 31-46, 1997. Supplement. DONADEU, F. X.; GINTHER, O. J. Follicular waves and circulating concentrations of gonadotrophins, inhibin and oestradiol during the anovulatory season in mares. Reproduction, v. 124, p. 875-885, 2002. DRIANCOURT, M. A. Follicular dynamics in farm animals. Implicaion for manipulation of reproduction. Theriogenology, v. 55, p. 1211-1239, 2001. EVANS, A. C. O.; ADAMS, G. P.; RAWLINGS, N. C. Endocrine and ovarian follicular changes leading up to first ovulation in prepubertal heifers. Journal of Reproduction and Fertility, v. 100, p. 187-194, 1994a. EVANS, A. C. O.; ADAMS, G. P.; RAWLINGS, N. C. Follicular and hormonal development in prepubertal heifers from 2 to 36 weeks of age. Journal of Reproduction and Fertility, v. 102, p. 463-470, 1994b. EVANS, A. C. O. et al. Follicular waves are associated with transient fluctuations in FSH but not oestradiol or inhibin-A concentrations in anoestrous ewes. Animal Science, v. 72, p. 547-554, 2001. FAY, J. E.; DOUGLAS, R. H. Changes in thecal and granulosa cell LH and FSH receptor content associated with follicular fluid and peripheral plasma gonadotrophin and steroid hormone concentrations in preovulatory follicle of mares. Journal of Reproduction and Fertility, v. 35, p. 169-181, 1987. Supplement. FORTUNE, J. E. et al. Differentiation of dominant versus subordinate follicles in cattle. Biology of Reproduction, v. 65, p. 648-654, 2001. GASTAL, E. L. Selection of the dominant follicle in mares: role of follicle-diameter differences, gonadotropins, and estradiol. 1999. 258 f. Tese (Doctor of Phisiology) – University of Wisconsin-Madison, 1999. GASTAL, E. L. et al. Role of diameter diferences among follicle in selection of a future dominant follicle in mares. Biology of Reproduction, v. 57, n. 6, p. 1320-1327, 1997.

49
GASTAL, E. L. et al. Echotextural changes in the follicular wall during follicle deviation in mares. Theriogenology, v.52, p. 803-814, 1999. GASTAL, E. L. et al. Follicle deviation and intrafolicular and systemic estradiol concentrations in mares. Biology of Reproduction, v. 61, n. 1, p. 31-39, 1999. GRAVES, P. E. et al. Cloning of a receptor for prostaglandin F2-alfa from ovine corpus luteum. Endocrinology, v. 136, p. 3430-3436, 1995. GINTHER, O. J. Folliculogenesis during the transitional period and early ovulatory season in mares. Journal of Reproduction and Fertility , v. 90, p. 311-320, 1990. GINTHER, O. J. Major and Minor follicular waves during the equine estrous cycle. Journal of Equine Veterinarian Science, v. 13, p. 18-25, 1993. GINTHER, O. J. Ultrasonic Imaging and Reproductive Events in Mare. Cross Plains, WI: Equiservices, 1986. GINTHER, O. J. Reproductive Biology of the Mare, Basic and Applied Aspects, 2nd ed. Cross Plains, WI: Equiservices Publishing, 1992. GINTHER, O. J. Ultrasonic Imaging and Animal Reproduction: Horses. Book 2. Cross Plains, WI: Equiservices Publishing, 1995. GINTHER, O. J. Ultrasonic Imaging and Animal Reproduction: Fundamentals. Book 1. Cross Plains, WI: Equiservices Publishing, 1995. GINTHER, O. J. et al. Selection of the dominant follicle in cattle. Biology of Reproduction, v. 55, p. 1187-11194, 1996. GINTHER, O. J. et al. Comparative study of the dynamics of follicular waves in mares and woman. Biology of Reproduction, v. 71, p. 1195-1201, 2004. GINTHER, O. J et al. Pulsatility of systemic FSH and LH concentrations during follicular wave development in catle. Theriogenology, v. 50, P. 507-519, 1998. GINTHER, O. J et al. Sampling follicular fluid without altering follicular status in catle: oestradiol concentrations early in a follicular wave. Journal of Reproduction and Fertility , v. 109, p. 181-186, 1997. GINTHER, O. J.; BERGFELT, D. R. Growth of small follicles and concentrations of FSH during the equine oestrous cycle. Journal of Reproduction and Fertility , v. 99, p. 105-111, 1993.

50
GINTHER, O. J.; BERGFELT, D. R. Ultrasonic characterization of follicular wave in mares without maintaining identity of individual follicles. Journal of Equine Veterinarian Science, v. 12, p. 349-354, 1992. GINTHER, O. J.; PIERSON, R.A. Regular and irregular characteristics of ovulation and the interovulatory interval in mares. Journal of Equine Veterinarian Science, v. 9, p. 4-12, 1989. GINTHER, O. J.; PIERSON, R.A. Ultrasonic anatomy and pathology of the equine uterus. Theriogenology, v. 21, p. 471-483, 1984a. GINTHER, O. J.; PIERSON, R.A. Ultrasonic evaluation of the reproductive tract of the mare: ovaries. Journal of Equine Veterinarian Science, v. 4, p. 11-16, 1984b. HANSEN, P. J. Photoperiodic regulation of reproduction in mammals breeding during long days versus mammal breeding during short days. Animal Reproduction Science, v. 9, p. 301-315, 1985. HUGHES, J. P. et al. The estrous cycle of the mare and its uterine control. Australian Veterinary Journal, v. 53, p. 415-419, 1975. HUGHES, J. P. et al. The estrous cycle and selected functional and pathologic ovarian abnormalities in the mare. Veterinary Clinics of North American (Large Animal Practice), v. 2, n. 22, p. 225-239, 1980. HUGHES, J. P. et al. Estrous cycle and ovulation in the mare. Journal of American Veterinary Medicine Association, v. 161, p. 1367-1374, 1972. JAFFE, R.; ABRAMOWICS, J.; BEN-ADERET, N. Correlation between the endocrine profile of ovulation and the ultrasonically detected ‘double contour’ of the preovulatory follicle. Gynecological Obstetric Investigation, v. 24, p. 119-124, 1987. KERBAN, A.; DORE, M.; SIROIS, J. Cellular and vascular changes in equine follicles prior to ovulation. Biology of Reproduction, v. 56 (suppl 1), p. 117 abstr., 1997. KINDER, J. E. et al. Progestin and estrogen regulation of pulsatile LH release and development of persistent ovarian follicles in cattle. Journal of Animal Science, v. 74, p. 1424-1440, 1996. LINCOLN, G. A. The pineal gland. In: AUSTIN, G. H.; SHORT, F. R. S. (eds). Reproduction in Mammals, Hormonal control of Reproduction. 2. ed. New York: Cambridge University press, 1987. p. 52-75. NEVES, A. P. et al. Reproductive parameters of mini-Shetland stallions in north Germany proceedings of the 4th International Symposium on Stallion Reproduction. Animal Reproduction Science, v. 89, p.267-270, 2005.

51
NOGUEIRA, G. P.; GINTHER, O. J. Dynamics of follicle populations and gonadotropin concentrations in fillies age two to ten months. Equine Veterinary Journal, v. 32, p. 482-488, 2000. OSBORNE, V. E. Analysis of the pattern of ovulation as it occurs in the annual reproductive cycle of the mare in Australia. Australian Veterinary Journal, v. 42, p. 149-154, 1966. PACCAMONTI, D. L. et al. Reproductive parameters of miniatures stallions. Theriogenology, v. 51, p. 1343-1349, 1999. PIERSON, R. A.; GINTHER, O. J. Follicular populations during the estrous cycle of the mare. Animal Reproduction Science, v. 14, p. 219-231, 1987. PIERSON, R. A.; GINTHER, O. J. Ultrasonic evaluation of the preovulatory follicle in the mare. Theriogenology, v.24, p.359-368, 1985. REDMER, D. A.; REYNOLDS, L. P. Angiogeneses in the ovary. Reviews of Reproduction, v.1, p. 182-192, 1996. SENGER, P. L. Reproductive Ciclicity- The Follicular Phase- In: Pathways to Pregnancy and Parturition, 2nd ed. Pullman: Current Conceptions, Inc, 2003. p. 164-187. SHARP, D. C. Enviromental influences on reproduction in horses. Veterinary Clinics of North America, v. 2, p. 207-223, 1980. SILVIA, P. J.; MEYER, S. L.; FITZGERALD, B. P. Pulsatile gonadotropin secretion determined by frequent sampling from the intercavernous sinus of the mare: possible modulatory role of peogesterone during luteolysis. Biology of Reproduction, v. 53, p. 438-446, 1995. SIROIS, J.; KIMMICH, T. L.; FORTUNE, J. E. Steroidogenesis by equine preovulatory follicles relatives roles of theca interna and granulosa cells. Endocrinology, v. 128, p. 1159-1166, 1991. STAGG, K. et al. Follicular development in long-term anoestrous suckler beef cows fed two levels of energy postpartum. Animal Reproduction Science, v. 38, p. 49-61, 1995. TAROUCO, A. K. et al. Estacionalidade reprodutiva de éguas abatidas em Pelotas-RS. Arquivo da Faculdade de Veterinária da UFRGS, v. 23, p. 26-42. WESSON, J. A.; GINTHER, O. J. Influence of season and age on reproductive activity in pony mares on the basis of a slaughterhouse study. Journal of American Science, v. 52, p. 119-129, 1981.
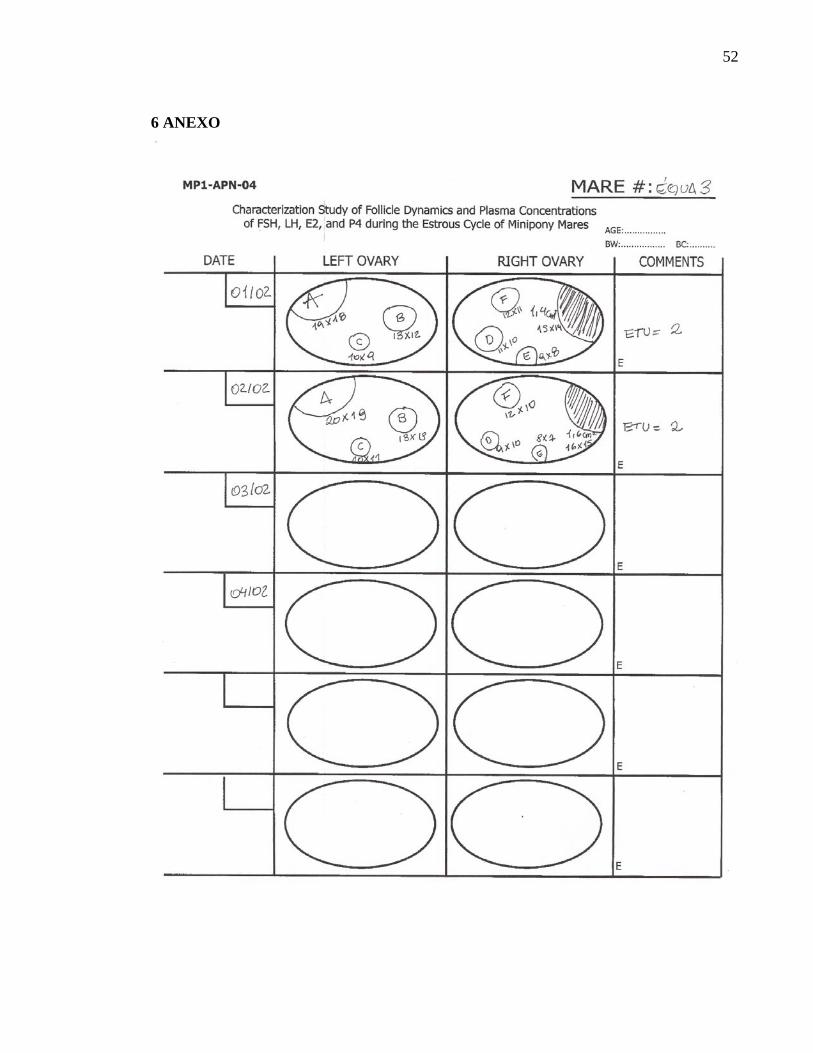
52
6 ANEXO
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo