universidade federal do rio grande do norte … · “que eu continue a acreditar no outro mesmo...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
DEPARTAMENTO DE BIOQUÍMICA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
ANA CELLY BEZERRA CRUZ
PURIFICAÇÃO, CARACTERIZAÇÃO E ANÁLISE DA ATIVIDADE
BIOINSETICIDA, DE UM INIBIDOR DE TRIPSINA EM SEMENTES
DE CATANDUVA (Piptadenia moniliformis)
NATAL/RN 2008

ANA CELLY BEZERRA CRUZ
PURIFICAÇÃO, CARACTERIZAÇÃO E ANÁLISE DA ATIVIDADE
BIOINSETICIDA, DE UM INIBIDOR DE TRIPSINA EM SEMENTES
DE CATANDUVA (Piptadenia moniliformis).
Dissertação apresentada ao Departamento de
Bioquímica da Universidade Federal do Rio
Grande do Norte como requisito parcial para
obtenção do título de Mestre em Bioquímica.
Orientador: Prof. Dr. Maurício Pereira de Sales.
NATAL 2008

Divisão de Serviços Técnicos Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial Leopoldo Nelson.
Cruz, Ana Celly Bezerra. Purificação, caracterização e análise da atividade bioinseticida, de um inibidor de tripsina em sementes de catanduva (Piptadenia moniliformis) / Ana Celly Bezerra Cruz. - Natal(RN), 2008. 97 f.
Orientador: Mauricio Pereira de Sales.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Departamento de Bioquímica. Programa de Pós-Graduação em Bioquímica.
1. Bioquímica - Dissertação. 2. Inibidor de tripsina - Dissertação. 3. Piptadenia moniliformis - Dissertação. 4. Insetos praga - Dissertação. 5. Inibidor Kunitz – Dissertação. I. Sales, Mauricio Pereira. II. Universidade Federal do Rio Grande do Norte. III. Título. RN/UF/BC CDU 577.1(043.3)

ANA CELLY BEZERRA CRUZ
PURIFICAÇÃO, CARACTERIZAÇÃO E ANÁLISE DA ATIVIDADE BIOSINSETICIDA DE UM INIBIDOR DE TRIPSINA EM SEMENTES DE
CATANDUVA (Piptadenia moniliformis)
Dissertação apresentada ao Departamento de Bioquímica da Universidade Federal do Rio Grande do Norte como requisito parcial para obtenção do título de Mestre em Bioquímica.
Aprovado em: 20/10/2008
BANCA EXAMINADORA
___________________________________________________Prof. Dr. Elizeu Antunes dos Santos
Departamento de Bioquímica - UFRN Coordenador
____________________________________________________ Profª. Drª. Antônia Elenir Amâncio Oliveira
Centro de Biociências- UENF 1º Examinador
____________________________________________________ Profª. Drª. Luciana Duarte Martins da Matta
Departamento de Bioquímica – UFRN 2º Examinador

Dedico
Aos meus pais, Edilson e Gorett que de diferentes formas me ajudaram a finalizar mais uma etapa da minha vida.
À Marcos, presente de Deus, pela paciência e dedicação ao nosso amor.
A Adeliana, mulher maravilhosa, que nunca mediu esforços para me ajudar na hora ou no dia que fosse necessário. Esse trabalho também é seu!
Ao meu orientador, professor Maurício, obrigada pela oportunidade de fazer parte do LQFPB, pela paciência e ensinamentos.

AGRADECIMENTOS
A DEUS por me dar tanta força quando achei que não conseguiria concluir
essa etapa da minha vida e não deixou que o desespero tomasse conta do meu
coração e colocou as pessoas certas nas horas certas para que eu conseguisse
chegar até aqui.
À minha família (alicerce), meus pais e meu marido, por tudo que eles
representam em minha vida. O meu amor é de vocês.
A Marcos, que me ajudou e esteve sempre comigo em todos os momentos
difíceis. Não tenho palavras para agradecer. Amo você.
À Pureza e Macau (in memorian), meus sogros, que de diferentes formas
me ajudaram a alcançar mais uma meta em minha vida. Obrigada pelas orações. E
valeu Tio Macau pelo exemplo de força...!
A Tia Fátima pelas palavras de apoio e força, a atenção nos momentos
difíceis.
Ao professor Maurício pela força nos momentos mais difíceis que enfrentei
durante o mestrado sem sua amizade e carinho não teria conseguido chegar até
aqui. MUITO OBRIGADA!

Ao professor Elizeu pela serenidade e sabedoria. Suas palavras são muito
acalentadoras. Obrigada!
Aos professores que fazem o Departamento de Bioquímica: Edda Lisboa,
Hugo, Luiz, Roberto, Fernanda, Suely, Jacira, Selma e, em especial, à professora
Dilma Ferreira Lima (uma mãe), pelo apoio e confiança transmitidos nos anos de
iniciação cientifica.
À Creusa, que me ensinou muito quando precisei. Nunca esquecerei de
você!
A grande amiga Virginia, que passou a ser a irmã que não tive neste período de
convivência, compartilhando os momentos difíceis e principalmente as vitórias.
Nossos momentos de alegria foram muito importantes. Obrigada por tudo!
Aos amigos da turma do mestrado, Vi, Mi, Vide, Dani, Lí, Adri, Pablito,
Ludo, Rho e Sérgio. Valeu a todos!
Aos companheiros de laboratório, Sheyla (Negona, a amiga que Deus
mandou para mim), Adeliana (Mãezona), Hugo (Gaguinho), Norba (Norbinha),
Jannison (Jan), Dayse (Daysoca, caladinha, mas muito especial para mim), Ludovico
(Ludinho, amigo do coração mesmo que às vezes meio maluquinho), Rodrigo (Rho)
e Fabinho (Binho, pelo inicio desse trabalho!) e a todos aqueles que foram
importantes e não estão no laboratório, mas estarão sempre no meu coração, Ibson,
Léo Caicó (meu braço direito por muitos meses), Danielle (companheira e ouvinte

para muitos desabafos), Ticiana (pela ajuda incondicional), Gioconda (pelas palavras
de amizade), Léo Pepino, Kátya e Joelma (pela alegria e carinho).
Às professoras da Pré-qualificação, Adriana Uchôa e Kátia pelas
sugestões. Obrigada!
A Adriana Uchoa e Adeliana pelas sugestões dadas e pelo tempo a mim
concedido para as correções... Muito obrigada!
A todos os funcionários e amigos do Departamento de Bioquímica, em especial
a Ana Katarina e Eliene que de alguma forma contribuíram para a realização
deste trabalho.

“Que eu continue a acreditar no outro mesmo sabendo de alguns valores tão
esquisitos que permeiam o mundo; Que eu continue otimista, mesmo sabendo que o futuro que nos espera nem
sempre é tão alegre; Que eu continue com a vontade de viver, mesmo sabendo que a vida é, em muitos
momentos, uma lição difícil de ser aprendida; Que eu permaneça com a vontade de ter grandes amigos (as), mesmo sabendo que
com as voltas do mundo, eles (as) vão indo embora de nossas vidas; Que eu realimente sempre a vontade de ajudar as pessoas, mesmo sabendo que
muitas delas são incapazes de ver, sentir, entender ou utilizar esta ajuda.... E, acima de tudo...
Que eu lembre sempre que todos nós fazemos parte desta maravilhosa teia chamada vida, criada por alguém bem superior a todos nós!
E que as grandes mudanças não ocorrem por grandes feitos de alguns e, sim, nas pequenas parcelas cotidianas de todos nós!“
CHICO XAVIER

RESUMO
Um inibidor de tripsina da família Kunitz (PmTI) foi purificado de sementes de Piptadenia moniliformis, uma árvore da sub-família Mimosoideae, através da precipitação com ácido tricloroacético (TCA), cromatografia de afinidade com tripsina acoplada em sepharose, coluna DEAE- celulose(troca iônica) e Superose 12 (exclusão molecular) em sistema FPLC/AKTA. O inibidor possui massa molecular de 25 kDa como confirmado através de SDS-PAGE e cromatografia de exclusão molecular. A seqüência do N-terminal deste inibidor mostrou alta homologia com outros inibidores da família Kunitz. Este também é estável as variações de temperatura e pH, e apresentou um pequeno decréscimo na sua atividade quando incubado com DTT na concentração de 100 mM por 120 minutos. A inibição da tripsina foi do tipo competitiva com Ki de 1,57x10-11 mM. A atividade da tripsina foi inibida efetivamente com percentual de inibição de 100%, entre as outras enzimas testadas não foi detectada inibição para a bromelaína, foi fracamente inibidor da elastase pancreática (3,17% de inibição), inibiu em 76,42% elastase de neutrófilos, inibiu de forma moderada quimotripsina e papaína com percentual de inibição de 42,96% e 23,10%, respectivamente. Ensaios in vitro foram realizados com as proteinases digestivas de Lepidóptera, Coleóptera e Díptera. Vários graus de inibição foram encontrados. Para Anthonomus grandis e Ceratitis capitata a inibição foi de 89,93% e 70,52%, respectivamente, e as enzimas de Zabrotes subfasciatus e Callosobruchus maculatus foram inibidas com percentuais de 5,96% e 9,41% respectivamente, e as enzimas de Plodia interpunctella e Castnia licus foram inibidas com percentuais de 59,94% e 23,67%, respectivamente. No ensaio in vivo, foi observada redução no desenvolvimento de larvas em 4º ínstar de C. capitata, quando PmTI foi adicionado à dieta artificial, obtendo WD50 de 0,30% e LD50 0,33% . Estes resultados sugerem que este inibidor possa ser um forte candidato para programas de melhoramentos de plantas via transgenia.
Palavras-chave: Inibidor de tripsina, Piptadenia moniliformis, Insetos praga, Bioinseticida, Inibidor Kunitz.

ABSTRACT
One Kunitz-type trypsin inhibitors (PmTI) was purified from Piptadenia moniliformisseeds, a tree of the sub-family Mimosoideae, by TCA precipitation, affinity chromatography on immobilized trypsin-Sepharose, DEAE – cellulose (ion exchange) and Superose 12 (molecular exclusion) column FPLC/AKTA. The inhibitor has Mr of 25 kDa by SDS-PAGE and chromatography molecular exclusion. The N-terminal sequence of this inhibitor showed high homology with other family Kunitz inhibitors. This also stable variations in temperature and pH and showed a small decrease in its activity when incubated with DDT in the concentration of 100mM for 120 minutes. The inhibition of trypsin by PmTI was competitive, with Ki of 1.57 x10-11 M. The activity of trypsin was effectively inhibited by percentage of inhibition of 100%, among enzymes tested, was not detected inhibition for the bromelain, was weak inhibitor of pancreatic elastase (3.17% of inhibition) and inhibited by 76.42% elastase of neutrophils, and inhibited in a moderate, chymotrypsin and papain with percentage of inhibition of 42.96% and 23.10% respectively. In vitro assays against digestive proteinases from Lepidoptera, Diptera and Coleoptera pests were carried out. Several degrees of inhibition were found. For Anthonomus grandis and Ceratitis capitata the inhibition was 89.93% and 70.52%, respectively, and the enzymes of Zabrotes subfasciatus and Callosobruchus maculatus were inhibited by 5.96% and 9.41%, respectively, and the enzymes of Plodia. interpunctella and Castnia licuswere inhibited by 59.94% and 23.67, respectively. In vivo assays, was observed reduction in the development of larvae in 4rd instar of C. capitata, when PmTI was added to the artificial diet, getting WD50 and LD50 of 0.30% and 0.33%, respectively. These results suggest that this inhibitor could be a strong candidate to plant management programs cross transgenic.
Keywords: Trypsin inhibitor, Piptadenia moniliformis, Insect pests, Bioinsecticide, Kunitz inhibitor.

LISTA DE FIGURAS
FIGURA 1 Estrutura primária de inibidor de soja da família Bowman-Birk.
23
FIGURA 2 Adulto de Callosobruchus maculatus 27
FIGURA 3 Efeito de incorporação de PDTI em dieta artificial 29
FIGURA 4 Árvore de Catanduva (Piptadenia moniliformis) 31
FIGURA 5 Larvas e insetos adultos 32
FIGURA 6 Esquema de hidrólise do BApNA 37
FIGURA 7 Esquema da tripsina acoplada a Sepharose 39
FIGURA 8 Esquema da metodologia empregada no trabalho 51
FIGURA 9 Atividade anti-tríptica de frações protéicas de sementes de Catanduva
52
FIGURA 10 Análise cromatográfica de CT1% em Tripsina-Sepharose 53
FIGURA 11 Perfil cromatográfico de PmTI em coluna DEAE- celulose FPLC/AKTA
54
FIGURA 12 Perfil da PmTI purificada em FPLC – AKTA, coluna Superose 12
55
FIGURA 13 Eletroforese em Gel de Poliacrilamida (SDS-PAGE 20%) 56
FIGURA 14 Determinação da massa molecular de PmTI por cromatografia Superose-12 em sistema FPLC-AKTA
58
FIGURA 15 Seqüência parcial e alinhamento comparativo de PmTI 59
FIGURA 16 Estabilidade a diferentes pHs 60
FIGURA 17 Estabilidade térmica de PmTI 61
FIGURA 18 Efeito do desnaturante do DTT na atividade de PmTI 62
FIGURA 20 Mecanismo de inibição de PmTI sobre a atividade catalítica
64
FIGURA 21 Determinação do valor de Ki de PmTI contra tripsina 65
FIGURA 22 Peso médio das larvas de 4º ínstar de C. capitatata 69
FIGURA 23 Sobrevivência das larvas de 4º ínstar de C. capitatata 70

LISTA DE TABELAS
TABELA 1 Família de inibidores de proteinase de plantas 21
TABELA 2 Tabela de purificação 56
TABELA 3 Especificidade de PmTI para proteinases serínicas e cisteínicas
66
TABELA 4 Especificidade de inibição de PmTI para enzimas de diferentes insetos
67

LISTA DE ABREVIATURAS
AcTKI- Inibidor de Acacia confusa
ApTKI- Inibidor de Adenanthera pavonina
[S] - Concentração de substrato
μg - Micrograma
μL - Microlitro
BApNA - α- benzoil-DL-arginina-ρ-nitroanilida
BBI – Bowman- Birk
CT1% - Fração de Catanduva obtida com 1% de TCA
CT2% - Fração de Catanduva obtida com 2% de TCA
CT3% - Fração de Catanduva obtida com 3% de TCA
CT4% - Fração de Catanduva obtida com 4% de TCA
DMSO - Dimetilsulfóxido
DTT- Dithiothreitol
EB- Extrato Bruto de Piptadenia moniliformis
EcTKI- Inibidor de Enterelobium contortisiliquum
EDTA- Ácido etilenodiamino-tetra-acético
kDa- Quilodaltons
Ki - Constante de inibição
LD50 – Dose letal
LiTKI- Inibidor de Leucena leucocephala
M - Molar
mg - Miligrama
Min - Minuto
mL- Mililitro
mM - Milimolar
NaCl – Cloreto de sódio
NCBI- National Center for Biotechnology Information
ND - Não detectado
nm - Nanômetro
ºC - Graus Celcius
PBS - Tampão fosfato

PjTKI- Inibidor de Prosopis juliflora
PmTI - Inibidor de tripsina de Piptadenia moniliformis
SDS - Dodecil sulfato de sódio
SDS-PAGE-Eletroforese em gel de poliacrilamida contendo SDS
TCA - Ácido tricloroacético
TEMED – N, N,N’,N’- tetrametiletilenodiamino
Tris - Tris hidroximetil aminometano
UA/mg prot - Unidades de atividade por proteína
UI - Unidade de inibição
UR - Umidade relativa
Vmax - Velocidade máxima
v/v – Relação volume / volume
WD50 – Quantidade de inibidor que reduz em 50% o peso das larvas

SUMÁRIO
1 INTRODUÇÃO 16
1.1 Considerações gerais 16
1.2 INIBIDORES: distribuição, ocorrência e características. 18
1.3 Inibidores da família Bowman Birk 22
1.4 Inibidores da família Kunitz. 23
1.5 Ação de inibidores sobre insetos 25
2 OBJETIVOS 30
3 MATERIAIS 31
3.1 Sementes 31
3.2 Insetos 32
3.3 Enzimas adquiridas comercialmente 33
3.4 Substratos 33
3.5 Reagentes 34
3.6 Equipamentos 34
4 MÉTODOS 35
4.1 Preparação do extrato protéico de sementes de Catanduva 35
4.2 Fracionamento protéico com ácido tricloroacético (TCA) 35
4.3 Determinação de proteínas 36
4.4 Preparo das soluções usadas como substrato 36
4.5 Determinação da atividade anti-tríptica 36
4.6 Isolamento e Purificação 38
4.6.1 Cromatografia de afinidade em Tripsina-Sepharose 38
4.6.2 Cromatografia de troca iônica em coluna DEAE-celulose em
sistema FPLC/AKTA
38
4.6.3 Determinação da Massa Molecular por Cromatografia de
Exclusão Molecular em Superose 12 10 300 GL em Sistema
FPLC-AKTA.
39
4.7 Determinação da seqüência de aminoácidos N- terminal da
proteína purificada
40
4.8 Eletroforese em gel de poliacrilamida (SDS-PAGE) 41

4.8.1 Em condições desnaturantes e não redutoras 41
4.8.2 Em condições desnaturantes e redutoras 42
4.8.3 Coloramento com Comassie Blue. 42
4.8.4 Revelação com Nitrato de Prata 42
4.9 Caracterização do inibidor 43
4.9.1 Estabilidade do inibidor em diferentes pHs 43
4.9.2 Estabilidade térmica do inibidor 44
4.8.3 Efeito do DTT na atividade do inibidor 44
4.8.4 Especificidade do inibidor para proteinases serínicas e
cisteínicas
45
4.8.4.1 Inibição da elastase pancreática 45
4.8.4.2 Inibição da elastase de neutrófilos 45
4.8.4.3 Inibição da quimotripsina 46
4.8.4.4 Inibição da bromelaína 46
4.8.4.5 Inibição da papaína 47
4.9 Determinação do mecanismo e da constante de inibição do
inibidor
47
4.10 Preparo das enzimas digestórias de insetos 48
4.10.1 Ensaios in vitro sobre extrato intestinal de larvas de insetos 49
4.10.1.1 Ensaio in vitro para C. maculatus, A grandis e Z. subfaciatus 49
4.10.1.2 Ensaio in vitro para C. capitata, C.licus, e P. interpunctela 49
4.11 Bioensaio para C. capitata 50
5 RESULTADOS 52
5.1 Isolamento e purificação 52
5.1.1 Detecção da atividade inibitória para tripsina em extratos
protéicos de Catanduva
52
5.1.2 Análise cromatográfica de CT1% em Tripsina-Sepharose 53
5.1.3 Isolamento do inibidor de tripsina em cromatografia DEAE-
celulose/AKTA
54
5.1.4 Análise cromatográfica em Superose - 12 AKTA 55
5.1.5 Análise das etapas de purificação 56
5.1.5.1 Eletroforese em gel de poliacrilamida na presença de SDS
(SDS-PAGE) da fração protéica de Catanduva
57

5.2 Caracterização bioquímica dos inibidores de sementes de
Catanduva
58
5.2.1 Determinação da massa molecular relativa 58
5.3 Alinhamento e análise da seqüência N-terminal 59
5.4 Estabilidade em diferentes pHs 60
5.5 Estabilidade térmica 61
5.6 Efeito do DTT na atividade do inibidor 62
5.7 Mecanismo de inibição e valor de Ki 63
5.7.1 Mecanismo de inibição sobre a tripsina 63
5.8 Especificidade para proteinases serínicas e cisteínicas 66
5.9 Especificidade dos inibidores para enzimas digestórias de
insetos
67
5. 10 Bioensaio para C. capitata 68
6 DISCUSSÃO 71
7 CONCLUSÕES 80
REFERÊNCIAS BIBLIOGRÁFICAS 81

16 Introdução
1 INTRODUÇÃO
1.1 Considerações gerais.
O aumento da população mundial demanda concomitante incremento da
produção de alimentos em escala global. A expansão da área disponível para o
cultivo poderá causar um impacto negativo ao meio ambiente e aos recursos
naturais, desta forma faz-se necessário o desenvolvimento de novas tecnologias que
promovam aumento da produção, sem aumento da área plantada, além de
proporcionar alimentos de melhor qualidade nutricional, resistentes a estresses
abióticos (ex., salinidade, seca, etc) (KRATTIGER, 1997) e bióticos (pragas e
patógenos) (KOGAN, 1986).
As perdas na produção mundial de grãos sem o uso de pesticidas e de outras
estratégias de controles não-químicas são calculadas em cerca de 70%, chegando a
um prejuízo de 400 bilhões de dólares. As perdas da pré-colheita devido à ação de
insetos, apesar da utilização de inseticidas, são estimadas em 15% da produção
total, representando um valor de 100 bilhões de dólares (Ferry et al., 2004). O custo
anual do controle de insetos equivale a 8 bilhões de dólares, justificando assim o
emprego de medidas mais econômicas e urgentes para o controle (LAWRENCE;
KOUNDAL, 2002). As pragas e os patógenos (fungos, bactérias, vírus e nematóides)
são responsáveis por grandes prejuízos na agricultura mundial por se alimentarem
dos tecidos das plantas. As perdas chegam a 37%, sendo 13% destas causadas por
insetos e doenças (GATEHOUSE; BOULTER; HILDER, 1992).
Segundo LAURENCE; KOUNDAL, (2002), a busca e o desenvolvimento de
técnicas ambientalmente favoráveis tornam-se a solução para diminuir, a utilização
de produtos químicos, gastos em energia e, portanto a produção de materiais

17 Introdução
prejudiciais, como resíduos de pesticidas. O uso exclusivo de pesticidas químicos,
além de resultar num rápido desenvolvimento de resistência nos insetos, é
responsável por alterar o balanço entre as pragas e seus predadores naturais,
geralmente em favor das primeiras. Sendo assim, um programa de manejo integrado
proporcionaria a melhor opção, pois ele seria baseado na combinação de práticas
onde ocorra o uso criterioso de pesticidas, rotação de culturas e acima de tudo a
exploração de variedades de plantas resistentes.
As sementes de leguminosas são fontes ricas de proteínas, dentre as
diversas classes de proteínas de sementes, algumas são consideradas fatores anti-
nutricionais, como os inibidores de proteinases, quitinases, as lectinas, proteínas
alergênicas e outras toxinas protéicas (XAVIER-FILHO; VENTURA, 1988; PUZSTAI,
1989; CARLINI; Sales et al., 1996; GROSSI-DE-SÁ, 2002). Estas substâncias tem a
função de proteger ou defender a semente e a planta do ataque de patógenos e de
pragas, e dentre estas os inibidores de leguminosas vêm sendo intensamente
estudados (NEURATH, 1984; Macedo et al., 2000; Pando et al., 2001; Cursino-
Santos et al., 2003; Santos et al., 2004).
A primeira transferência de gene bem sucedida para outra espécie vegetal foi
realizada por Hilder et al., (1987), onde genes de feijão de corda (Vigna unguiculata),
que codifica um inibidor de tripsina, foram transferidos para plantas de tabaco
(Nicotiana tabacum). Nesse contexto, a busca de genes e produtos gênicos de
proteínas de defesa, tais como lectinas, inibidores de enzimas entre outras
moléculas que possam ser utilizados em biotecnologias têm sido amplamente
estimuladas, como, por exemplo, na formulação de bioinseticidas e para modificação
genética de plantas cultivadas.

18 Introdução
1.2 - INIBIDORES: distribuição, ocorrência e características.
No final do século XIX, as substâncias inibidoras de proteinases foram
encontradas em nematóides e no soro humano (RICHARDSON, 1991). Entretanto,
especulações sobre inibidores de plantas iniciaram-se a partir da década de 40
quando KNEEN; SANDSTEDT (1943) encontraram um inibidor de α-amilase em
grãos de vários cereais, e posteriormente, KUNITZ (1945) purificou uma proteína
termoestável da soja com capacidade de inibir a tripsina.
Os inibidores de proteinases são proteínas ou peptídeos, amplamente
encontrados em vegetais, animais e microorganismos, com propriedades de inibir a
ação catalítica das enzimas, ao interagir em diversos graus de afinidade com os
sítios reativos das enzimas, formando complexos estequiométricos inativos (RYAN,
1973; RICHARDSON, 1977; WEDER, 1981; RICHARDSON, 1991). Nas plantas os
inibidores vão se concentrar principalmente nos órgãos reprodutivos e de reserva,
tais como, sementes e tubérculos (RYAN, 1981; BRZIN; KIDRIC, 1995). No entanto,
já foram encontrados em polpa de frutas (Araújo et al., 2004), folhas, frutos, raízes e
vagens (XAVIER-FILHO, 1992; CUNNINGHAM; VOLENEC, 1996; XU et al., 2001;
SIN; CHYE, 2004) e, podem apresentar diversas funções como: proteínas de
reserva, agentes reguladores de proteases endógenas, importantes para proteger
fluidos e tecidos da degradação pela atividade proteolítica (NEURATH, 1984; RYAN,
1990; RICHARDSON, 1991; Walker et al.,1997) além de serem parte do sistema
natural de defesa das plantas mostrando especificidade para a maioria das
proteases digestivas dos insetos herbívoros (BODE; HUBER, 2000) e patógenos
(BRODWAY; DUFFEY, 1986; Johnson et al.,1989; BRODWAY, 1995; TORNERO et
al., 1997; Schuler et al., 1998; TIFFIN; GAUT, 2001).

19 Introdução
A expressão dos inibidores pode ser constitutiva ou induzida (Tornero et al.,
1997; VAN DER HOORN; JONES, 2004). O mecanismo de defesa das plantas são
constitutivas, se sua ação faz-se dentro do programa de desenvolvimento normal da
planta nos diferentes tecidos vegetais; ou induzidas quando estão envolvidas
diretamente na resposta a infecção ou estímulos ambientais (CHESIN; ZIPF, 1990).
Essa resposta induzida pode resultar em efetivos mecanismos de resistência a
doenças quando ela é expressa pela planta, sistemicamente. Nesse caso, os
agentes envolvidos induzem uma resposta do hospedeiro, não apenas em torno das
partes atingidas, como também em partes da planta distante da área onde ocorreu à
injúria, sendo este processo (DEAN; KUC, 1986; GOTTSTEIN; KUC, 1989). As
defesas induzidas são mais importantes para as defesas de partes vegetativas das
plantas, enquanto que as defesas constitutivas são mais importantes para as
defesas das sementes (XAVIER-FILHO, 1993, Uchôa et al., 2002).
Em relação à distribuição dos inibidores em plantas, sabe-se que as
angiospermas dicotiledôneas englobam o maior número de espécies contendo
essas proteínas, merecendo destaque as famílias Leguminosae (ou Fabaceae)
sendo representada pelos feijões, soja e ervilha (XAVIER-FILHO, 1992; Macedo et
al., 2000), e a família Solanaceae representada pela batata, tomate e tabaco.
Enquanto que entre as monocotiledôneas os inibidores estão mais amplamente
distribuídos na família Gramineae (atualmente denominada Poaceae) tendo como
representantes, arroz, milho, trigo e centeio (OLIVEIRA, 2007). No entanto, a
quantidade de inibidores é extremamente variável, dependendo de vários fatores
como o estágio de maturação da planta, sua localização nos tecidos, tempo de
colheita e de armazenamento (SHEWRY; LUCAS, 1997).

20 Introdução
A especificidade é uma característica marcante no estudo das interações
entre enzima-inibidor, sendo determinada pela dinâmica das interações envolvidas e
pela estrutura nativa do inibidor e da enzima (MACEDO; XAVIER - FILHO, 1993). A
inibição exibida por estas proteínas é altamente variável e muitos dos inibidores de
tripsina também inibem quimotripsina. Em alguns casos, o sítio reativo é o mesmo
para ambas as enzimas ou a inibição ocorre através de sítios reativos
independentes (BIRK, 1994).
Os inibidores de proteinases são caracterizados de acordo com sua
especificidade e seu mecanismo de ação. Baseado nessa observação quatro
classes de inibidores foram estabelecidas: inibidores de proteinases serínicas; de
proteinases cisteínicas; aspárticas e de metalo-proteinases (RYAN, 1990;
RICHARDSON, 1991) (Tabela 1). Os mais estudados e caracterizados são os
inibidores de proteinases serínicas, (Koiwa et al., 1997). Entretanto, alguns
inibidores de proteinases cisteínicas foram caracterizados e seus efeitos observados
contra insetos pragas (BODE; HUBER, 1992; 2000; Oliveira et al., 2003; Macedo et
al., 2004).
Os inibidores de proteinases serínicas presentes em plantas são classificados
em sete famílias (Tabela 1) através de características da homologia da estrutura
primária, massa molecular, conteúdo de cisteína e de pontes dissulfetos
(RICHARDSON, 1991; Koiwa et al., 1997). Os demais inibidores compreendem
apenas uma família (BODE; HUBER, 1992; 2000; Koiwa et al., 1997; MARGIS;
REIS; VILLERET, 1998). Os inibidores de plantas são tipicamente pobres nos
aminoácidos metionina, histidina e triptofano; porém, são ricos em ácido aspártico,
ácido glutâmico, serina, lisina e cisteína (BOWLES, 1990). Dentre estes, os
inibidores de proteinases serínicas da família Kunitz e Bowman-Birk têm sido

21 Introdução
bastante estudados e diferem principalmente em massa molecular, conteúdo de
cisteína e números de sítios reativos (RICHARDSON, 1977).
Tabela 1 - Família de inibidores de proteinases de plantas
Classes de inibidores Famílias de inibidores
Inibidores de proteinases serínicas Kunitz
Bowman-Birk
Batata I
Batata II
Superfamilia de cereais
Taumatina
Ragi I-2/milho
Inibidores de proteinases cisteínicas Cistatinas de planta (Fitocistatinas)
Inibidores de proteinases aspárticas Inibidor de proteinase aspártica
Inibidores de metalo-proteinases Inibidores de carboxipeptidases A e B
Existem relatos sobre inibidores de proteinases serínicas que também são ativos
contra enzimas de outras classes, como por exemplo, inibidores de tripsina que
também inibem papaína (uma proteinase cisteínica) (Macedo et al., 2004; Oliveira
et al., 2007), e também aqueles que apresentam uma atividade inibitória sobre α-
amilase (Koiwa et al., 1997, BELITZ; GROSCH, 1997).

22 Introdução
Os inibidores de proteinases mais abundantes e estudados são aqueles capazes
de inibir as proteinases serínicas, grupo da tripsina e quimotripsina, enzimas
digestivas encontradas em insetos das ordens Lepidoptera, como em Spodoptera
frugiperda (TERRA; FERREIRA, 1994) e Diptera, como em Ceratitis capitata
(Gomes et al., 2005; Silva et al., 2006). Inibidores de proteinases cisteínicas e
aspárticas, enzimas que são predominantes nas ordens Coleoptera e Hemiptera,
como no Calosobruchus maculatus, Zabrotes subfasciatus e Dysdercus
peruvianus (SILVA; XAVIER-FILHO, 1991; Campos et al., 1989; Haq et al., 2004;
StanisçuaskI et al., 2005) são menos conhecidos.
1.3 - Inibidores da família Bowman-Birk
Os inibidores da família Bowman-Birk (BBI) são proteínas globulares solúveis,
caracterizadas por suas pequenas massas moleculares que variam de 8 a 10
kDa, com alto conteúdo de cisteína podendo apresentar 7 pontes dissulfeto, que
conferem grande estabilidade à sua estrutura, distribuídas em cerca de 70 a 90
aminoácidos (LASKOWSKI; KATO, 1980; RICHARDSON, 1991; BIRK, 1994).
O primeiro inibidor pertencente à família Bowman-Birk (Figura 1), que deu
origem a esta família, foi purificado de sementes de soja (Glycine max), constituído
de uma cadeia polipeptídica com dois sítios reativos, um para a tripsina e o outro
para quimotripsina, e formado de 71 resíduos de aminoácidos (ODANI; IKENAKA,
1973a, b).

23 Introdução
1.4 - Inibidores da família Kunitz
Os inibidores da família Kunitz são proteínas que apresentam uma ou duas
cadeias polipeptídicas, em geral, com apenas um sítio reativo, e massa molecular
entre 18 e 24 kDa, correspondendo a aproximadamente 180 resíduos de
aminoácidos, geralmente com quatro resíduos de cisteína que formam duas
pontes dissulfeto, que proporcionam estabilidade a estrutura protéica (Svendnsen
et al., 1984; KIM; LAMBOOY, 1985; RICHARDSON, 1991; Batista et al., 1996).
Os inibidores que apresentam um sítio reativo são conhecidos como inibidores de
Figura 1: Estrutura primária de inibidor de soja da família Bowman-
Birk. A estrutura primária do inibidor de soja da família Bowman-Birk
mostra a organização das sete pontes dissulfetos. Os dois sítios
reativos localizados nos loops e a posição do resíduo P1 estão
indicados: O sítio reativo para tripsina (Lisina16-Serina-17), e
quimotripsina (Leucina44-Serina45) (ODANI; IKENAKA, 1973 a, b).

24 Introdução
uma cabeça “single headed”. Entretanto, alguns representantes dessa família
foram caracterizados como inibidores que possuem dois sítios reativos “double
headed” (RICHARDSON, 1991). A interação direta do resíduo do sítio reativo P1
do inibidor no sítio ativo da enzima caracteriza um mecanismo de inibição
competitivo, comum para muitos inibidores de tripsina da família Kunitz (BODE;
HUBER, 1992, 2000; Franco et al., 2002; Mandal et al., 2002, Bhattacharyya et
al., 2006).
Os inibidores de proteinases serínicas da família Kunitz foram purificados de
vários tecidos e órgãos de plantas incluindo látex do mamão (OdanI et al., 1996;
Azarkan et al., 2006), folhas de azolla (MAITY; PATRA, 2003), rizomas tuberosos
de várias aráceas (OLIVEIRA, 2001; SUMATHI; PATTABIRAMAN, 1979),
tubérculos de solanáceas (Valueva et al., 1998; VALUEVA; REVINA; MOSOLOV,
1997, 1999), sementes de jaca (BHAT; PATTABIRAMN, 1989), sementes de
mustarda (Mandal et al., 2002), e particularmente de sementes de leguminosas
(RICHARDSON, 1991).
Esses inibidores podem ser encontrados em quantidades relativamente grandes
em sementes das três subfamílias de leguminosas: Mimosoideae,
Caesalpinoideae e Papilionoideae (Faboideae). Sendo as Mimosoideae as mais
primitivas, as Papilionoideae as mais evoluídas e as Caesalpinoideae
pertencentes a um estágio intermediário de evolução (CRONQUIST, 1988).
Os pertencentes à subfamília Mimosoideae são formados por duas cadeias
polipeptídicas após redução das pontes dissulfeto, como é o caso do inibidor de
tripsina isolado da Leucaena leucocephala (Oliva et al., 1999), do inibidor de
tripsina da batata, denominado PSPI-21 (Valueva, et al., 1998), do inibidor de
tripsina de Prosopis juliflora (Negreiros et al., 1991), do inibidor de tripsina de

25 Introdução
Archidendron ellipticum (Bhattacharyya et al., 2006), do inibidor de tripsina de
Dimorphandra mollis (Macedo et al., 2000).
2- Ação de inibidores sobre insetos
A planta, durante todo o seu ciclo de vida, está exposta ao ataque de uma
grande variedade de insetos, microorganismos e outros agentes que afetam sua
integridade. A presença de um mecanismo de defesa no sistema vegetal torna-se
então, fundamental para alcançar o sucesso na manutenção do seu crescimento e
desenvolvimento (Mello et al., 2001). Este mecanismo abrange os mais variados
tipos de sistemas de defesa, desde a ocorrência de barreiras físicas ou de
modificações estruturais na morfologia vegetal, até a presença de compostos
químicos tais como alcalóides, toxinas, inibidores enzimáticos e outras substâncias
biologicamente ativas contra os agentes predadores (BROADWAY, 1996; SHEWRY;
LUCAS, 1997; HARBORNE, 1997; Tamhane et al., 2005; Dunaevsky et al., 2005).
As plantas desenvolveram características extraordinárias contra as proteinases
dos insetos tais como o aumento da atividade do inibidor em seus tecidos (Rakwal
et al., 2001), produção de um conjunto heterogêneo de inibidores ativos contra
amilases e proteinases (ROY; GUPTA, 2000), além de inibidores de enzimas de
insetos altamente específicos (Falco et al., 2001), resistentes à proteólise e ativos
sob condições diversas de pH (Christeller et al., 1990, CHRISTELLER, 2005).

26 Introdução
Para a obtenção de aminoácidos essenciais utilizados nos processos
fisiológicos, os insetos utilizam uma combinação de proteases em seus processos
digestivos (MCFARLANE, 1985; Terra et al., 1990). Essas proteases digestivas
catalisam a liberação de peptídeos e aminoácidos das proteínas da dieta e são
encontradas abundantemente na região do intestino médio do trato digestivo de
insetos (JONGSMA; BOLTER, 1977). Por um longo tempo pensava-se que,
semelhante aos vertebrados, os insetos continham apenas proteinases serínicas do
tipo tripsina e quimotripsina, e aspárticas do tipo pepsina, pois a maioria das
pesquisas foi realizada com insetos da ordem Lepidoptera e Diptera, que geralmente
usam proteinases serínicas para a digestão das proteínas, possuindo em seus
intestinos médio um pH alcalino variando entre 8 a 11,5, ideal para atividade dessa
classe de enzimas (APLEBAUM, 1964). Entretanto, estudos mostraram que outras
classes de insetos utilizam proteinases cisteínicas e aspárticas para degradar as
proteínas da dieta. Dentre esses estão os insetos da ordem Coleoptera, com pH
ótimo do intestino entre 5 e 7 para proteinases cisteínicas e 4,5 para proteinases
aspárticas (BARRET, 1986; TURK; BODE, 1991; BODE; HUBER, 1992). Diante
disso, existe a hipótese de que o uso de proteinases cisteínicas pode ter sido uma
adaptação evolucionária dos insetos que se alimentam de sementes de leguminosas
e outros tecidos de plantas que possuem, naturalmente, altas concentrações de
inibidores de proteinases serínicas (JONGSMA; BOLTER, 1997). Como exemplo
temos o bruquídeo Callosobruchus maculatus, pertencente à classe Coleóptera que
infesta variedades de Vigna unguiculata (feijão-de-corda) que apresentam elevados
níveis de inibidores de tripsina em suas sementes (Gatehouse et al., 1979
GATEHOUSE; BOULTER, 1983) (Figura 2).

27 Introdução
A presença de inibidores em sementes ou em tubérculos também facilita o
acúmulo e armazenamento de proteínas pela diminuição da atividade das
proteinases endógenas (Koiwa et al., 1997). O estudo dessas proteínas de defesa
tem mostrado que a introdução de genes de inibidores de proteinases em plantas
transgênicas pode conferir resistência a insetos, comprovando seu envolvimento nas
defesas de vegetais (Hilder et al., 1987; Johnson et al., 1989; Boulter et al., 1990;
Mcmanaus et al., 1994). Foi observado que a indução de inibidores de proteinases
em folhas de tomate reduz o crescimento de larvas do cartucho da beterraba
(Spodoptera exigua), quando comparada com as larvas controle alimentadas com
Figura 2: Adulto de Callosobruchus maculatus em grãos
apresentando ovos e orifício para saída dos insetos
(Fonte: EMBRAPA MEIO-AMBIENTE, 2003).

28 Introdução
folhas sem inibidor (Brodway et al., 1986). Vários outros trabalhos têm demonstrado
que inibidores de proteinases podem inibir a atividade de enzimas digestivas in vitro.
Como exemplos, temos o inibidor de tripsina de soja que é efetivo contra enzimas do
besouro da farinha, Tenebrio molitor (Applebaum et al., 1985), a tripsina de Manduca
sexta (MILLER, KRAMER, LAW, 1974) e a elastase de Teleogryllus commodus
(Christeller et al., 1990). A presença de inibidores também tem sido relatada por
retardar o desenvolvimento larval quando esses são incorporados em ensaios in vivo
utilizando dietas artificiais (Macedo et al., 2000).
Inúmeros trabalhos demonstraram que os inibidores de proteinases,
principalmente, serínicas e cisteínicas presentes em dietas artificiais ou em plantas
transgênicas, são capazes de interferir no crescimento e desenvolvimento de
algumas espécies de insetos principalmente aquelas pertencentes às ordens
Lepidóptera e Coleóptera (Jouanin et al., 1998). Ensaios biológicos, nos quais
inibidores são incorporados em dietas artificiais de insetos (Macedo et al., 2003)
(Figura 03), fornecem uma valiosa informação sobre os tipos de inibidores com
potencial para serem usados como fatores de resistência na manipulação genética e
programas de melhoramento de plantas economicamente importantes (McManus et
al., 1995; BROADWAY, 1997).

29 Introdução
Diante do exposto, a purificação e caracterização de inibidores de proteinases
presentes em sementes e a determinação da especificidade de inibição é um passo
importante para a indicação desse inibidor para os programas de melhoramento de
plantas cultivadas e também no seu uso como bioinseticida. Dentro desse contexto,
nenhum estudo foi desenvolvido sobre proteínas de defesa de sementes de
Catanduva (Piptadenia moniliformis) e sua ação bioinseticida sobre os insetos
pragas.
Figura 03: Efeito de incorporação de PDTI em dieta artificial
para Anagasta kuenhiella (Lepidoptera: Pyralidae).
Variações no tamanho de larvas de quarto instar de A.
kuenhiella alimentada sobre dieta controle (A), e dieta
contendo 0,4%(B); 0,8%(C) e 16% de PDTI (D) (Macedo et
al., 2003).
A B C D

30 Objetivos
2 OBJETIVOS
Esse trabalho teve como objetivos:
• Identificar a presença de inibidores de enzimas do tipo serínicas em sementes
de catanduva;
• Purificar um inibidor de tripsina presente em sementes de Catanduva
(Piptadenia moniliformis);
• Caracterizar o inibidor de tripsina purificado e determinar sua especificidade
para diferentes enzimas;
• Determinar a atividade inibitória in vitro sobre enzimas digestórias de
Lepidópteras (Plodia interpuncttela, Castnia licus), Coleópteras
(Callosobruchus maculatus e Zabrotes subfasciatus, Anthonomus grandis) e
Díptera (Ceratitis capitata);
• Avaliar a ação do inibidor purificado in vivo para o díptera C. capitata (mosca-
das-frutas), através da determinação da sua WD50 e LD50.

_________________________________________________________________ Materiais e Métodos
31
3 MATERIAIS
3.1 Sementes
As sementes de Catanduva (Piptadenia moniliformis) (Benth.) (Figura 4) foram
gentilmente fornecidas pela Divisão Técnica do Setor de Sementeiras do Instituto
Brasileiro do Meio ambiente (IBAMA), Natal-RN.
P. moniliformis (Benth.), pertence à familia Mimosaceae (leguminosae), é uma
árvore que pode atingir até 20 metros de altura, encontrada em áreas de caatinga e
clima semi-árido em quase todo Brasil, sendo amplamente distribuída nos Estados
da Bahia, Ceará, Pernambuco, Piauí e Rio Grande do Norte. A planta P. moniliformis
é conhecida popularmente como Angico, Angico-de-bezerro, Carrasco, Catanduba,
Jurema-preta, Marmeleiro, Quipembe, Rama-de-bezerro, além de Catanduva
(LEWIS, 2003).
Figura 4: Árvore e sementes de Catanduva (P. moniliformis)
Fonte: http://www.cnip.org.br/bdpn/fotosdb/32.jpg (acesso em 05 de maio de 2008).

_________________________________________________________________ Materiais e Métodos
32
3.2 Insetos
Espécimes de Callosobruchus maculatus (Coleoptera: Bruquideo; gorgulho do feijão-
de-corda), Zabrotes subfasciatus (Coleoptera: Bruquideo; gorgulho do feijão-de-
corda e feijão comum) e Plodia interpunctela (Lepidoptera: Pyralidae; praga
generalista) foram coletados da criação mantida no Laboratório de Química e
Função de Proteínas Bioativas (LQFPB), do Departamento de Bioquímica da UFRN.
Ceratitis capitata (Diptera: Tephritidae; mosca-das-frutas,) foram gentilmente cedidos
pelo Prof. Dr. Francisco Pepino de Macedo do Laboratório de Mosca-das-frutas, no
Departamento de Biologia Celular e Genética da UFRN. As larvas de Anthonomus
grandis (Coleoptera: Curculionidae; bicudo do algodoeiro) foram gentilmente cedidas
pela Dra. Márcia Vidal, do Centro Nacional de Pesquisa do Algodão, de Campina
Grande, Paraíba. E as larvas de Castnia licus (Lepidoptera: Castniidae; broca
gigante) foram gentilmente cedidas pela Dra. Fátima Grossi de Sá, do CENARGEN,
de Brasília.
Figura 5: Larvas e insetos adultos utilizados nos ensaios com enzimas digestórias. Larvas de: A - C.
licus; e B - A. grandis. Insetos adultos de: C- C. maculatus; D- C. capitata; E- P. interpunctella; e F- Z.
subfasciatus.
A B C
D E F

_________________________________________________________________ Materiais e Métodos
33
3.3 Enzimas adquiridas comercialmente
Proteinase serínica: Tripsina – pâncreas bovino (Sigma)
Proteinase serínica: Quimotripsina - pâncreas bovino (Sigma)
Proteinase serínica: Elastase - pâncreas bovino (Sigma)
Proteinase serínica: Elastase – neutrófilo humano (Sigma)
Proteinase cisteínica: Papaína – látex de mamão (Sigma)
Proteinase cisteínica: Bromelaína do talo de abacaxi (Sigma)
3.4 Substratos
Azocaseína- (Sigma)
BApNA- (Sigma): α- benzoil-DL-arginina-p- nitroanilida
SAAvNA (Bachem Feinchemikalien AG): N-succinil-L-alanil-L-valina-p-nitroanilida
3.5 Reagentes
BSA (albumina sérica bovina) – Sigma
Comassie blue G-250 e R- 250 – Sigma
Dimetilformamida (N’N’ – Dimetilformamida) - Sigma
DMSO (dimetilsulfóxido) - VTEC Química Fina LTDA
DTT (ditiotreitol) - Sigma
EDTA (ácido etileno dianimotetracético) – Sigma
Mercaptoetanol – VETEC Química Fina LTDA
Padrão de massa molecular -Fermentas life sciences:

_________________________________________________________________ Materiais e Métodos
34
Beta-amilase (200 kDa)
Álcool desidrogenase (150 kDa)
β-galactosidase (116 kDa);
Albumina sérica bovina (66,2 kDa);
Ovalbumina (45 kDa);
Lactato desidrogenase (35 kDa);
Endonuclease de restrição (25 kDa);
β-lactoglobulina (18,4 kDa);
Lisozima (14,4 kDa).
SDS (dodecil sulfato de sódio) – Sigma
Sepharose-4B – Sigma
TCA (ácido tricloroacético) - VETEC Química Fina LTDA
TEMED (N, N, N-tetrametiletilinodiamino) – Sigma
3.6 Equipamentos
Agitador Magnético Tecnal TE-081
Balança analítica eletrônica Tecnal classe II
Banho-Maria -Tecnal Te 056
Centrífuga HITACHI CR 21
Espectrofotômetro Femto 700 plus
FPLC AKTA Purifier – GE Healthcare Bio-Sciences Corp.
Microcentrífuga Eppendorf 5410

_________________________________________________________________ Materiais e Métodos
35
4 MÉTODOS
4.1 Preparação do extrato protéico de sementes de Catanduva
As sementes de Catanduva (P. moniliformis) foram moídas em um processador até a
formação de um pó de granulação fina, em torno de 40 mesh. As proteínas totais
desta farinha foram extraídas em tampão tetraborato de sódio 50 mM, pH 7,5, na
proporção de 1:10 (p/v). O extrato protéico foi submetido à agitação constante, por 3
horas em temperatura ambiente. Decorrido esse tempo, o extrato protéico foi
centrifugada a 12.000 xg por 30 minutos a 4 0C e o sobrenadante obtido foi
denominado extrato bruto (EB).
4.2 Fracionamento protéico com ácido tricloroacético (TCA)
O extrato bruto obtido foi submetido à precipitação com solução aquosa de TCA
20% para remoção de proteínas de alta massa molecular. Alíquotas de 2 ml de
extrato bruto foram precipitadas com solução aquosa de TCA 20%, em banho de
gelo, para obter frações protéicas com concentrações finais de 1, 2, 3 e 4% de TCA.
Decorridos 30 minutos da adição lenta de TCA em agitação constante, as frações
protéicas foram centrifugadas a 12.000 xg por 30 minutos a 4 0C e os sobrenadantes
obtidos foram dialisados contra tampão tetraborato de sódio 50 mM, pH 7,5 por 16
horas, e em seguida foram submetidos a ensaio de atividade inibitória para tripsina.
Após essa primeira análise, a fração protéica que apresentou maior atividade
inibitória foi utilizada nos passos subseqüentes. O restante do extrato bruto foi

_________________________________________________________________ Materiais e Métodos
36
submetido ao mesmo tratamento e a fração protéica foi utilizada nas etapas
subseqüentes de isolamento e purificação protéica.
4.3 Determinação de proteínas
As proteínas foram quantificadas pelo método de BRADFORD (1976), utilizando-se
albumina sérica bovina (BSA) como padrão. Medidas de densidade óptica de 280
nm foram utilizadas para acompanhamento das corridas cromatográficas.
4.4 Preparo das soluções usadas como substrato
BApNA (1,25 mM) foi dissolvido em DMSO 1% (v/v) e o volume completado com
tampão Tris-HCl 50 mM, pH 7,5. Para os ensaios cinéticos também foram
preparados BApNA a 0,5; 0,75; 1,0; 1,25; 2,0; e 2,5 mM, utilizando o mesmo
procedimento descrito anteriormente.
Azocaseína a 1% foi preparada em tampão Tris-HCl 50 mM, pH 7,5 e fervida por
cerca de 15 minutos, após resfriamento o volume foi completado com água
destilada. A solução foi reservada em congelador até sua utilização.
SAAvNA (300 mM ) foi dissolvido em N’N’ – Dimetilformamida 100%.
4.5 Determinação da atividade anti-tríptica
A atividade anti-tríptica das frações foi determinada utilizando a seguinte
metodologia (ERLANGER; KOLOWSKY; COHEN, 1961): alíquota de 30 μL da
solução de tripsina bovina (0,3 mg/ mL tampão Tris-HCl 50 mM, pH 7,5) foi pré-

_________________________________________________________________ Materiais e Métodos
37
incubada com 390 μL de tampão Tris-HCl 50 mM, pH 7,5 e 50 μL do inibidor por 10
minutos a 37 0C. Após esse período a reação foi iniciada adicionando-se 500 μL de
solução de substrato (BApNA 1,25 mM). A reação processou-se por mais 15 minutos
nas mesmas condições de incubação, e em seguida foi paralisada adicionando-se
120 μL de solução de ácido acético 30%. Todos os ensaios foram feitos em
triplicatas e provas em branco foram realizadas. O p-nitroanilida hidrolisado foi
monitorado em espectrofotômetro a 410 nm, como mostrado na Figura 6.
Figura 6: Esquema de hidrólise do BApNA (Adaptado de OLIVEIRA, 2007).
p-nitroanilida Positivo= amarelo
Tripsina H2O

_________________________________________________________________ Materiais e Métodos
38
4.6 Isolamento e Purificação
4.6.1 Cromatografia de afinidade em Tripsina-Sepharose 4B
Para a cromatografia de afinidade foi feito a acoplagem de tripsina bovina a uma
resina de Sepharose 4B ativada com brometo de cianogênio, seguindo instruções do
fabricante. Para seu preparo, pesou-se 1g da Sepharose para cada 3,5 mL final de
gel. O pó foi ressuspendido em 50 mL de HCl 1 mM e lavado por 15 min sendo
filtrado várias vezes com auxílio de bomba a vácuo. O gel foi retirado do filtro,
adicionou-se 15 mL de HCl 1 mM e deixou-se em repouso por 15 min.
A tripsina para o acoplamento a resina Sepharose 4B ativada (Figura 7) foi
preparada dissolvendo-se 10 mg de enzima, por mL de gel, em tampão bicarbonato
de sódio 100 mM, pH 8,3 contendo NaCl 500 mM. A solução de tripsina foi então
adicionada ao gel e incubou-se por 16h a 4°C. Decorrido o tempo, lavou-se o
excesso de ligante (tripsina) com 5 volumes de tampão bicarbonato de sódio 100
mM contendo NaCl 500 mM. Em seguida o gel foi transferido para um recipiente
contendo tampão Tris-HCl 100 mM, pH 8,0 por duas horas para bloquear quaisquer
grupos ativos remanescentes na resina. Decorrido esse tempo, o gel foi lavado com
5 volumes de tampão acetato de sódio 100 mM contendo NaCl 500 mM, pH 4,0
seguido por 5 volumes de Tris-HCl 100 mM contendo NaCl 500 mM, pH 8,0. Após
esse procedimento o gel foi transferido para uma coluna (10 cm x 1,5 cm) e
equilibrada com tampão tetraborato de sódio 50 mM, pH 7,5.

_________________________________________________________________ Materiais e Métodos
39
Figura 7: Esquema da tripsina acoplada a Sepharose (Adaptado de OLIVEIRA, 2007).
A fração mais enriquecida com atividade inibitória foi dialisada contra tampão
tetraborato de sódio 50 mM, pH 7,5, proveniente da precipitação com solução
aquosa de TCA 20%, denominada CT1%, foi submetida a cromatografia de
afinidade em Tripsina-Sepharose (10 cm X 1,5 cm) equilibrada com tampão
tetraborato de sódio 50 mM, pH 7,5. As proteínas adsorvidas na matriz foram eluídas
com solução de HCl 5 mM. Frações de 2 mL foram coletadas em fluxo constante de
2 mL/min e o perfil protéico foi acompanhado por espectrofotometria a 280 nm. O
pico retido da matriz de afinidade, denominado CT1Af foi dialisado em tampão
tetraborato de sódio 50 mM, pH 7,5 e submetido a ensaio de atividade inibitória para
tripsina.
4.6.2 - Cromatografia de troca iônica em coluna DEAE-celulose em sistema
FPLC/AKTA
O pico com atividade antitríptica (CT1Af), detectado na coluna de afinidade Tripsina
–Sepharose, foi dialisado e aplicado em uma coluna de troca iônica DEAE- celulose
equilibrada previamente com tampão tetraborato de sódio 50 mM, pH 7,5. Foi
OH
NH2
O C NH Tripsina Sepharose

_________________________________________________________________ Materiais e Métodos
40
utilizado um gradiente salino de NaCl de 0-1,0 M, para eluição das proteínas
absorvidas, sendo coletadas frações de 0,5 mL por tubo sob um fluxo de 0,5 mL/min
e absorbância medida a 280 nm .
4.6.3-Determinação da Massa Molecular por Cromatografia de Exclusão Molecular
em Superose 12 10 300 GL em Sistema FPLC-AKTA.
Para confirmação da purificação e determinação da massa molecular de
PmTI, cerca de 0,034 mg de proteína foi submetido a cromatografia de exclusão
molecular Superose 12 10 300 GL, em sistema FPLC/AKTA em tampão Tris-HCl 50
mM, pH 7,5. Esta coluna foi previamente calibrada com os seguintes padrões de
proteínas: Beta-amilase (200 kDa); álcool desidrogenase (150 kDa); anidrase
carbônica (29 kDa) e tripsina (24 kDa). Frações de 0,5 mL foram coletadas sob um
fluxo de 0,5 mL/min e absorbância medida a 280 nm. A massa molecular aparente
do inibidor foi estimada a partir da interpolação logarítmica entre massas
moleculares dos diferentes marcadores protéicos utilizados e os seus respectivos
volumes de eluição.
4.7 Determinação da seqüência de aminoácidos N- terminal da proteína
purificada
O método de degradação automática de Edman foi utilizado para determinar
a seqüência da porção N-terminal do inibidor de PmTI, que foi realizada no
Departamento de Bioquímica e Biologia Molecular, ICB, na Universidade Federal do

_________________________________________________________________ Materiais e Métodos
41
Ceará. O peptídeo de interesse foi submetido à degradação de Edman em
seqüenciador automático da Shimadzu PPSQ23.
4.8 Eletroforese em gel de poliacrilamida (SDS-PAGE)
4.8.1 Em condições desnaturante e não redutoras
A eletroforese das amostras foi feita segundo o método desenvolvido por Laemmli
(1970). Foram utilizadas placas de vidro de dimensões 10 x 14 cm, espaçadores de
0,75 mm e solução estoque de acrilamida/bisacrilamida (30%/ 0,8%, p/v) dissolvidas
em água destilada em volume para 100 mL. Esta solução foi filtrada em papel de
filtro Whatman n01 e estocada em frascos escuros a 50C. O gel de separação foi
preparado numa concentração de 20% contendo: 1,2 mL de água destilada, 1,3 mL
de tampão Tris-HCl 1500 mM, pH 8,8, 50 μL de SDS 10 %, 2,5 mL de solução
estoque de acrilamida/bisacrilamida, 25 μL de persulfato de amônio e 2,5 μL de
Temed. O gel de concentração foi preparado com 1,5 mL de água destilada, 0,625
mL de tampão Tris-HCl 1500 mM, pH 6,8, 25 μL de SDS 10%, 0,330 mL de solução
estoque de acrilamida/bisacrilamida, 12,5 μL de persulfato de amônio e 2,5 μL
Temed. A fração protéica contendo 15 μg de proteína, foi adicionada tampão de
amostra constituído de Tris-HCl 62,5 mM, pH 6,8, SDS 2 %, glicerol 10 % v/v, e 0,01
% de azul de bromofenol. O tampão de corrida continha trizma base 25 mM, glicina
192 mM e SDS 10 %. A eletroforese foi realizada sob corrente constante de 30 mA
por aproximadamente 2 horas e após o marcador de corrida (azul de bromofenol)
atingir o final do gel, a corrida foi parada.

_________________________________________________________________ Materiais e Métodos
42
4.7.2 Em condições desnaturantes e redutoras
Essa eletroforese foi realizada com amostras tratadas com β-mercaptoetanol 100
mM a 100 °C por 10 min. Após esse procedimento as amostras foram submetidas à
migração eletroforética como descrito no item anterior.
4.7.3 Coramento com Comassie Blue
Após a eletroforese o gel foi corado segundo procedimento descrito por Weber e
Osborne (1969). A solução corante foi preparada usando-se Comassie Blue R-250 a
1%, metanol 40%, ácido acético 10% em água destilada. O descoloramento foi feito
com uma solução contendo ácido acético 10 % e etanol 30 %.
4.7.4 Revelação com Nitrato de Prata
Para melhor visualização das bandas protéicas o gel foi submetido a revelação com
nitrato de prata, o qual foi desidratado com uma solução de etanol 50%,
submetendo-se a três lavagens de 20 minutos cada. Em seguida, foi adicionada uma
solução de tiossulfato de sódio (20 mg/100 mL H20) e mantida por 1 minuto sob leve
agitação. Decorrido esse tempo, foram feitas três lavagens rápidas em água
destilada, sendo então adicionada a solução de nitrato de prata (200 mg + 74 μL de
formaldeído em 100 mL de H20) mantendo-se por 20 minutos sob leve agitação.
Após esse tempo, o gel foi submetido a três lavagens rápidas em água destilada e

_________________________________________________________________ Materiais e Métodos
43
adicionou-se a solução reveladora (carbonato de cálcio 6 g + 50 μL de formaldeído +
2 mL de tiossulfato de sódio em 100 mL de H20). Para cessar a revelação adicionou-
se uma solução de ácido acético 13 % (BLUM; BEIER; GROSS, 1987).
4.8 Caracterização do inibidor purificado
4.8 1 Estabilidade do inibidor em diferentes pHs
Para avaliar a estabilidade do inibidor à variação de pHs, seguiu-se a metodologia
como descrita em Gomes et al., (2005), com pequenas modificações, na qual
alíquotas de 1 mL do inibidor foram dialisadas contra o tampão com o pH desejado
por 16 horas. Estes tampões foram, glicina-HCl pH 2-3; fosfato de sódio pH 6 e 8; e
glicina-NaOH pH 11-12, nos quais a concentração de soluto foi de 100 mM. Após
esse período as alíquotas foram incubadas por 30 min a 37 0C nos referidos
tampões, em seguida as amostras foram dialisadas por cerca de 4 horas em Tris-
HCl 50 mM, pH 7,5, com a finalidade de ajustar ao pH do ensaio. Os ensaios de
atividade inibitória para tripsina foram feitos em triplicata utilizando-se 30μL do
inibidor (6,9 μg). Os ensaios foram realizados em triplicata e provas em branco
foram realizadas conforme descrito no item 4.5.
4.8.2 Estabilidade térmica do inibidor

_________________________________________________________________ Materiais e Métodos
44
Para determinar a estabilidade térmica dos inibidores seguiu-se a metodologia como
descrita por Gomes et al., (2005), na qual alíquotas de 1 mL do inibidor foram
incubadas por 30 minutos a 37, 40, 60, 70, 90 e 100 0C. Decorrido o tempo de
incubação, as amostras foram resfriadas a 40C e após esse procedimento alíquotas
de 30μL (6,9 μg) dos inibidores foram utilizadas no ensaio de atividade inibitória
sobre a tripsina. Provas em branco foram realizadas e os ensaios foram feitos em
triplicata.
4.8.3 Efeito do DTT na atividade do inibidor
Para avaliar a atividade do PmTI, na presença de agente redutor, seguiu-se a
metodologia descrita por Mello (2005), onde amostras foram incubadas com
soluções de DTT em concentrações finais de 1,10 e 100 mM. A reação foi encerrada
pela adição de iodoacetamida, na quantidade de duas vezes a concentração final de
DTT. Durante a incubação a 37°C, alíquotas foram coletadas em intervalos de tempo
de 15, 30, 60 e 120 minutos. Para todos esses intervalos, um controle da amostra
sem DTT foi feito. Após o período de incubação, essas alíquotas foram submetidas a
ensaios antitrípticos para verificar a atividade do inibidor. Os resultados mostrados
foram obtidos a partir da média de triplicatas.
4.8.4 Especificidade do inibidor para proteinases serínicas e cisteínicas

_________________________________________________________________ Materiais e Métodos
45
4.8.4.1 Inibição da elastase pancreática
A inibição da atividade da elastase pancreática, foi avaliada usando 30 μL da enzima
(0,1mg/mL em Tris-HCl 50 mM, pH 7,5) foram incubados com 310 μL de tampão
Tris-HCl 50 mM, pH 7,5 e 50 μL (11,5 μg ) do inibidor PmTI por 15 min a 37 °C, após
esse tempo foram acrescentados 500μL de azocaseína a 1%. Após 30 min a reação
foi parada pela adição de 150 μL de TCA a 20%. A mistura de reação foi
centrifugada por 10 min a 12000 xg. Alíquotas de 700 μL dos sobrenadantes foram
adicionados a 700 μL de NaOH 2 N, e a absorbância foi medida a 440 nm. Provas
em branco foram realizadas e os ensaios foram feitos em triplicata.
4.8.4.2 Inibição da elastase de neutrófilos
Para determinar a atividade inibitória sobre a elastase de neutrófilos, 50 μL da
enzima (PBS 0,1 M, pH 7,4) foi pré-incubada com 450 μL de tampão PBS 0,1 M pH
7,4 e 50 μL do inibidor PmTI (11,5 μg) por 15 min. A reação foi iniciada após adição
de 2,5 μL de SAAvNA 300 mM. O tempo de reação do ensaio foi de 1 hora, a
temperatura de 37 ºC. A reação foi interrompida com a adição de 250 μL de solução
de ácido cítrico 2,0%. Os produtos da reação foram lidos a 405 nm. Os ensaios
foram feitos em triplicata e provas em branco foram realizadas.
4.8.4.3 Inibição da quimotripsina

_________________________________________________________________ Materiais e Métodos
46
Para verificar a atividade inibitória sobre a atividade da quimotripsina, 30 μL da
enzima (0,1mg/mL de Tris-HCl 50 mM, pH 7,5) foram incubados com 320 μL de
tampão Tris-HCl 50 mM, pH 7,5 e 50 μL do inibidor PmTI (11,5 μg) por 15 min a 37
°C, após esse tempo foram acrescentados 500 μl de azocaseína a 1%. Decorridos
30 min de incubação a 37 °C a reação foi parada pela adição de 150 μL de TCA a 20
%. A mistura de reação foi centrifugada por 10 min a 12000 x g. Alíquotas de 700 μL
dos sobrenadantes foram adicionados a 700 μL de NaOH 2N, e a absorbância foi
medida a 440 nm. Provas em branco foram realizadas e os ensaios foram feitos em
triplicata.
4.8.4.3 Inibição da bromelaína
Para determinação da atividade inibitória para a bromelaína, 10 μL de solução de
bromaleína (1mg/mL de tampão acetato de sódio 300 mM, pH 5,5) foi pré-incubada
com 330 μL de tampão Tris-HCl 50 mM, pH 7,5, e 50 μL do inibidor PmTI (11,5 μg)
por um período de 20 min a 45 0C. Após esse tempo foram acrescentados 500 μL de
azocaseína a 1%. Decorridos 30 min de incubação a reação foi parada pela adição
de 150 μL de TCA a 20%. A mistura de reação foi centrifugada por 10 min a 12000 x
g. Alíquotas de 700 μL dos sobrenadantes foram adicionados a 700 μL de NaOH 2
N, e a absorbância foi medida a 440 nm. Provas em branco foram realizadas e os
ensaios foram feitos em triplicata.
4.8.4.4 Inibição da papaína

_________________________________________________________________ Materiais e Métodos
47
Para determinação da atividade anti-papainásica, 10 μL de solução de papaína
(1mg/mL Tris-HCl 50 mM, pH 7,5) foi pré-incubada com 40 μl de solução ativadora
contendo L-cisteína 50 mM e EDTA 20 mM pH 8,0, 400 μL tampão Tris-HCl 50 mM,
pH 7,5 e 50 μL do inibidor (11,5 μg) por um período de 15 minutos a 37 0C. Após
este período foi acrescentado 200 μL de solução de azocaseína 1%. Passados mais
30 minutos a reação foi paralisada com 300 μL de TCA 20%. A mistura de reação foi
centrifugada por 10 min a 12000 x g. Alíquotas de 700 μL dos sobrenadantes foram
adicionados a 700 μL de NaOH 2 N, e a absorbância foi medida a 440 nm. Provas
em branco foram realizadas e os ensaios foram feitos em triplicata.
4.9 Determinação do mecanismo e da constante de inibição
Para a determinação do mecanismo de inibição os ensaios foram realizados como
descrito em 4.5. Para o controle positivo, alíquotas de 10 μL de solução de tripsina
(0,3 mg/mL tampão Tris-HCl 50 mM, pH 7,5 foram incubadas por 15 minutos a 370 C
com concentrações crescentes de BApNA (0,5; 0,66; 0,8; 1; 1,25; 1,5 e 2,0 mM) em
volumes de 250 μL livres de inibidor. Para análise da cinética de PmTI, alíquotas de
20, 40, 60 e 80 μL de PmTI, foram incubadas com 10 μL de solução de tripsina e
250 μL das referidas solução de BApNA com leitura de absorbância a 410 nm.
O valor de Ki foi determinado de acordo com Dixon et al., 1979. O ensaio enzimático
com tripsina para a determinação da constante de inibição (Ki) foi realizado na
presença de concentrações crescentes de PmTI (0,209; 0,418 e 0,836 ug) e duas
diferentes concentrações de BApNA (0,5 mM e 1 mM) e o valor de Ki foi obtido da
intercessão de duas linhas correspondentes as concentrações do substrato.

_________________________________________________________________ Materiais e Métodos
48
4.10 Preparo das enzimas digestórias de insetos
Larvas de C. maculatus, Z. subfaciatus e A. grandis foram coletadas entre 17 e 20
dias de vida pós-postura (3° ínstar), larvas de P. interpunctella do 5o ínstar, e larvas
de C. capitata de 5 dias de idade foram mergulhadas em solução de NaCl a 150 mM
e dissecadas com auxílio de pinças anti-magnéticas número 4 e lupa estereoscópica
para retirada de todos os tecidos estranhos as paredes intestinais. Cerca de 25
intestinos médios de P. interpunctella assim obtidos foram imediatamente colocados
em tubos para microcentrífuga contendo 100 μL de tampão Tris-HCl, 50 mM, pH 9,5.
Cerca de 90 intestinos de C. maculatus, e 50 de Z. subfasciatus e A. grandis,
respectivamente, foram colocados em tampão Tris-HCl, 50 mM, pH 7,5 e 5 intestinos
de C. licus foram colocados em tampão Tris-HCl, 50 mM, pH 7,5, e 80 intestinos de
C. capitata foram colocados em tampão Tris-HCl, 50 mM, pH 8,5, e estocados a -20
°C. Os homogenatos intestinais das larvas foram preparados seguindo a
metodologia estabelecida por Terra, Ferreira e Bianchi (1977). Os intestinos das
larvas dos insetos foram homogeneizados com auxílio de um “potter” em banho de
gelo por aproximadamente 10 minutos, usando-se como extrator 1 mL dos tampões
descritos a cima. Em seguida, os homogenatos foram centrifugados a 12.000 x g por
10 min a 4 °C e os sobrenadantes obtidos foram imediatamente utilizados para
ensaios de atividade enzimática e inibitória como descrito a seguir.
4.10.1 Ensaio de inibição contra enzimas dos extratos intestinais de larvas de
insetos (in vitro).
4.10.1.1 Ensaio para enzimas digestórias de C. maculatus, A. grandis e Z.
subfaciatus

_________________________________________________________________ Materiais e Métodos
49
Nos ensaios in vitro, alíquotas de 100 μL de extrato intestinal de Z. subfaciatus, 50
μL de A. grandis e 20 μL de C. maculatus foram incubados com 150, 200 e 230 μL
de tampão Tris-HCl, 50 mM, pH 7,5, respectivamente, e 10 μL (0,418 μg) do inibidor
PmTI por 10 min a 37 °C. Após este período de incubação, foram adicionados 250
μL de BapNA e a incubação procedeu-se por mais 15 minutos na mesma
temperatura. Decorrido este tempo, a reação foi parada pela adição de 60 μL de
ácido acético 30 %, e a absorbância foi medida a 410 nm. Todos os ensaios foram
realizados em triplicata e com provas em branco.
4.10.1.2 Ensaio para enzimas digestórias de C. capitata, C.licus, e P. interpunctela
(in vitro).
Para determinar a atividade inibitória in vitro para o extrato intestinal de C. capitata,
C. licus e P. interpunctela, alíquotas de 50, 10 e 5 μL dos extratos intestinais,
respectivamente, foram pré-incubados com 320, 370 e 375 μL de tampão Tris-HCl,
50 mM, pH 8,5, 7,5 e 9,5, respectivamente, e 10 μL (0,418 μg) do inibidor isolado por
10 min a 37 °C. Após este período de incubação, foram adicionados 250 μL de
BapNA e a incubação procedeu-se por mais 15 minutos na mesma temperatura.
Decorrido esse tempo, a reação foi parada pela adição de 60 μL de ácido acético 30
%, e a absorbância foi medida a 410 nm. Todos os ensaios foram realizados em
triplicata e com provas em branco.
4.11 Bioensaio para C. capitata

_________________________________________________________________ Materiais e Métodos
50
As larvas de C. capitata foram submetidas a ensaio in vivo para avaliação do
efeito da ingestão de PmTI no crescimento e desenvolvimento do inseto (MACEDO
et al., 2008). Para tal, dietas artificiais foram preparadas contendo concentrações
crescentes de PmTI: 0,05, 0,1, 0,2, 0,5 e 1,0 %, e dieta controle sem inibidor. Foram
usadas larvas de 1° dia de vida pós-eclosão acondicionadas em tubos de ensaio
contendo 500 mg de dieta, 8 larvas/tubo, sendo 3 tubos por tratamento. Os tubos
foram então fechados com filme plástico e incubados por 5 dias sobre condições
controladas (28°C e 90% U.R). No 5° dia, as larvas foram retiradas das dietas,
lavadas e pesadas individualmente em balança de precisão. Foram calculadas as
médias das massas e da sobrevivência e os desvios padrões para cada tratamento.
• Dieta artificial para C. capitata:
Para o preparo de 500 g da dieta, segundo Walder, (2002), foi adicionando gérmen
de trigo (15 g), farinha de trigo (32,5 g), açúcar cristal (60 g), extrato de levedura
(49,5 g), benzoato de sódio (1,5 g), HCl (4,5 g), H2O (285 g) e farinha de bagaço da
cana-de-açúcar (52 g). O material foi misturado até a formação de uma pasta
homogênea. Para os bioensaios com larvas de C. capitata diferentes quantidades do
inibidor liofilizado foram adicionadas e os valores subtraídos da massa de água.

Materiais e Métodos 51
Figura 8: Esquema da metodologia empregada no trabalho
EXTRATO BRUTO
Precipitação com TCA (1%, 2%, 3%, e 4%).
CT1% CT2% CT3% CT4%
Cromatografia Tripsina-Sepharose
Cromatografia DEAE-celulose
Cromatografia Superose 12
Tampão Bórax 50 mM, pH 7, 5, 1:10, agitação 3h Centrifugação: 12.000 x g por 30 min a 4°C
Farinha de sementes de Piptadenia moniliformis
Especificidade
Enzimas serínicas e cisteínicas
Atividade in vitro paraenzimas digestorias de
insetos
Bioensaio
Larvas
SDS – PAGE
Determinação da massa molecular
Caracterização bioquimica
Determinação do mecanismo e da
constante de inibição
pH, temperatura e DTT
WD50
LD50

Resultados 52
5 RESULTADOS
5.1 - Isolamento e purificação
5.1.1 - Detecção da atividade inibitória para tripsina em extratos protéicos de
Catanduva
A figura 9 apresenta as atividades inibitórias sobre a tripsina presente no extrato
bruto (EB) e nas frações protéicas de sementes de Catanduva obtidas após
tratamento do EB com ácido tricloroacético (TCA 20 %) em concentrações finais de
1, 2, 3 e 4%, denominadas de CT1%, CT2%, CT3% e CT4%. O EB inibiu em torno
de 70% a atividade tríptica enquanto que as frações inibiram em 71,0, 42,5, 10,7 e
13,6%, respectivamente. A fração CT1% apresentando maior atividade anti-tríptica
foi usada nos passos subseqüentes de purificação.
0
10
20
30
40
50
60
70
80
1 2 3 4 5
% In
ibiç
ão
Figura 9: Atividade anti-tríptica de frações protéicas de sementes de Catanduva: No ensaio para
determinar a atividade inibitória das frações protéicas sobre a atividade da tripsina foi utilizado 50
μL de cada fração, tendo BApNA 1,25 mM como substrato.
CT1% CT2%EB CT4%

Resultados 53
5.1.2 - Análise de CT1% em cromatografia de afinidade Tripsina-Sepharose
A fração protéica de sementes de Catanduva denominada de CT1% proveniente do
fracionamento do extrato bruto de sementes com TCA, apresentando cerca de 71%
de inibição para a tripsina, foi submetida à cromatografia de afinidade em Tripsina-
Sepharose. As frações não adsorvidas na matriz foram eluídas com tampão
tetraborato de sódio 50 mM, pH 7,5, e as frações protéicas adsorvidas foram eluídas
com HCl 5 mM em fluxo constante de 2 mL/min, e a densidade óptica foi monitorada
a 280 nm. As frações protéicas foram dialisadas contra tampão tetraborato de sódio
50 mM, pH 7,5 e submetidas a ensaio para determinação da atividade inibitória
sobre a atividade catalítica da tripsina, no qual foi detectado aproximadamente 100
% de inibição nas frações adsorvidas que foram reunidas e denominadas CT1Af
(Figura 10).
Figura 10: Perfil de eluição de CT1% (0,52 μg/mL de proteína) em Tripsina-Sepharose: A coluna
foi previamente equilibrada com tampão tetraborato de sódio 50 mM, pH 7,5. As frações
protéicas foram eluídas em fluxo constante de 2 mL/min., a densidade óptica medida a (�) 280
nm e (ο) atividade anti-tríptica foi avaliada usando 50 μL de cada fração, tendo BApNA 1,25 mM
como substrato.
0
500
1000
1500
2000
2500
3000
3500
1 11 21 31 41 51
Frações
D.O
280
nm
0
20
40
60
80
100
120%
In
ibiç
ão
HCl 5 mM
(mL)

Resultados 54
5.1.3 - Isolamento do inibidor em cromatografia DEAE-celulose/AKTA
A fração (CT1Af) com atividade inibitória foi dialisada contra tampão tetraborato de
sódio 50 mM, pH 7,5 e submetida a cromatografia de troca iônica em coluna DEAE-
celulose em sistema de purificação FPLC/AKTA. A Figura 11 ilustra o perfil
cromatográfico de CT1Af em DEAE-celulose que apresentou um pico protéico
predominante com atividade inibitória sobre a atividade catalítica da tripsina o qual
foi eluído com gradiente salino, em torno de 0,7 M de NaCl, o qual foi dialisado para
dar seqüência aos passos de purificação.
Figura 11: Perfil cromatográfico de PmTI em coluna DEAE-celulose em sistema FPLC/AKTA. A
coluna foi previamente equilibrada com tampão tetraborato de sódio 50 mM, pH 7,5. As frações
protéicas foram eluídas em fluxo constante de 0,5 mL/min e a densidade óptica medida a 280 nm. A
atividade anti-tríptica foi avaliada usando 50 μL de cada fração, tendo como substrato BApNA 1,25
mM. ( ) frações com atividade inibitória.
Frações
M

Resultados 55
5.1.4 - Análise cromatográfica de Catanduva em SUPEROSE-12 AKTA
Para confirmar o grau de purificação e determinar a massa molecular do
inibidor de tripsina de sementes de Catanduva, a fração protéica obtida em coluna
DEAE-Celulose em sistema FPLC/AKTA, apresentando atividade inibitória sobre a
atividade catalítica da tripsina foi submetida à cromatografia de exclusão molecular
Superose 12 10 300 GL em sistema FPLC/AKTA. O perfil cromatográfico (Figura 12)
confirmou a purificação do inibidor de tripsina de sementes de Catanduva, P.
moniliformis que foi denominado de PmTI.
Figura 12: Perfil cromatográfico de PmTI purificada na coluna Superose 12 FPLC – AKTA. A
coluna foi previamente equilibrada com tampão Tris-HCl 50 mM, pH 7,5. As frações protéicas
foram eluídas em fluxo constante de 0,5 mL/min, a densidade óptica medida a 280 nm e atividade
anti-tríptica foi avaliada usando 50 μL de cada fração, tendo como substrato BApNA 1,25 mM. ( )
as frações com atividade inibitória.
Frações (mL)

Resultados 56
5.1.5 - Análise das etapas de purificação
As etapas utilizadas no processo de purificação do inibidor de sementes de
Catanduva estão sumarizadas na Tabela 2. O extrato bruto (EB), obtido na primeira
etapa do processo de purificação foi utilizado como parâmetro para as demais
frações protéicas. O valor de purificação 1 e um rendimento de 100% foram
atribuídos a essa fração. O grau de purificação das demais frações foi calculado
como sendo a razão entre sua atividade específica e a do extrato bruto. A
percentagem de rendimento das demais frações foi calculada com base na razão
entre a sua atividade inibitória e a atividade inibitória do extrato bruto multiplicado
por 100. A atividade especifica (UI) foi calculada com base na razão entre a
atividade inibitória da fração obtida e proteína total da mesma fração. A tabela de
purificação mostra que PmTI foi purificado 21,3 vezes em Superose-12/AKTA com
recuperação de 0,02%.
Tabela 2 - Tabela de purificação
Uma unidade de inibidor foi definida como a quantidade de inibidor que diminui a atividade da enzima
em 0,01 da absorbância a 410 nm.
Etapas Proteína Total (mg)
UI Total
Atividade Específica (UI/mgP)
Purificação (X)
Rendimento(%)
EB 207,0 16284 78,7 1,0 100 CT1% 146,9 21775 148,2 1,9 71 CT1Af 2,24 473,2 211,3 2,7 1,1
DEAE-Celulose
0,6 169,0 281,7 3,6 0,3
PmTI 0,034 57,0 1676,5 21,3 0,02

Resultados 57
5.1.5.1 - Eletroforese em gel de poliacrilamida na presença de SDS (SDS-PAGE)
A análise por eletroforese da fração protéica obtida da Superose 12/AKTA em gel de
poliacrilamida (SDS-PAGE) na ausência de agente redutor revelou a presença de
uma única banda protéica com massa molecular aparente de 25 kDa (Figura 13). A
análise de PmTI em SDS-PAGE tratado com agente redutor (β - mercaptoetanol)
revelou a presença de uma única banda protéica sugerindo que o inibidor é
composto por uma única cadeia polipeptídica de 25 kDa.
Figura 13: Eletroforese em Gel de Poliacrilamida (SDS-PAGE 20%)
M – Padrão de massa molecular: Álcool desidrogenase (150 kDa), galactosidase (116
kDa), albumina sérica bovina (66 kDa), ovalbumina (45 kDa), lactato desidrogenase (35
kDa), endonuclease de restrição (25 kDa), lactoalbumina (18,4 kDa); PmTI, fração
protéica com atividade inibitória (15 μg); PmTI, tratada com β- mercaptoetanol. A seta
indica a banda de PmTI com massa molecular de aproximadamente 25 kDa.
PmTI PmTI + β
kDa

Resultados 58
5.2 - Caracterização bioquímica dos inibidores de sementes de Catanduva
5.2.1 - Determinação da massa molecular relativa em cromatografia de exclusão
molecular Superose 12 10 300GL em sistema FPLC-AKTA
Através da curva de calibração com proteínas de massas moleculares
conhecidas, foi possível determinar a massa molecular relativa de PmTI. Apos
determinação dos volumes de eluição das proteínas marcadoras e do inibidor PmTI
foi construída uma curva em escala semi-log (Figura 14), e calculada a massa
molecular de PMTI. A determinação da massa relativa foi realizada através do
gráfico de log Mr x Ve/Vo. Assim encontramos uma massa molecular para PmTI de
25 kDa confirmando o observado em SDS-PAGE.
3
4
5
6
1 1,5 2
Ve/Vo
Lo
g (P
eso
Mo
lecu
lar)
Figura 14: Determinação da massa molecular de PmTI por cromatografia Superose-12 em
sistema FPLC-AKTA. Curva de calibração dos pesos moleculares: 1, Beta-amilase (200 kDa);
2, Álcool desidrogenase (150 kDa); 3, Anidrase carbônica (29 kDa); 4, PmTI (25 kDa) e 5,
Tripsina (24 kDa). Ve/Vo - relação entre o volume de eluição da proteína e o volume total da
coluna.
12
3 4 5

Resultados 59
5.3 – Alinhamento e análise da seqüência N-terminal
A Figura 15 mostra o alinhamento comparativo de PmTI com outros inibidores de
tripsina da família Kunitz de sementes de leguminosas da subfamília Mimosoideae.
A seqüência N-terminal de PmTI mostrou forte identidade com inibidor de tripsina de
sementes de Acacia confusa (AcTKI), Enterelobium contortisiliquum (EcTKI),
Prosopis juliflora (PjTKI), Adenanthera pavonina (ApTKI) e Leucena leucocephala
(LlTKI). De acordo com a Figura 15, foi observada a presença de resíduos de
aminoácidos fortemente conservados em todas as seqüências analisadas, como:
glicina (G8, G14, G27, G28, e G29), asparagina (N13), tirosina (Y17 e Y18), leucina
(L3 ,L11 e L30), Isoleucina (I19) e ácido aspártico (D5 e D7).
Os resultados obtidos a partir do alinhamento de PmTI com inibidores de
tripsina de sementes de leguminosas da família Kunitz confirmam que o inibidor de
tripsina purificado de sementes de Catanduva é um novo membro de inibidor de
tripsina da família Kunitz.
10 20 30 % identidade
AcTKI_m KELLDADGDILRNGGAYYILPALRGKGGGL 86,67%EcTKI_m KELLDSDGDILRNGGTYYILPALRGKGGGL 86,67%PmTKI_m KELLDADGDLLRNGGTYYILPVFRGKGGGLPjTKI_m QELLDVDGEILRNGGSYYILPAFRGKGGGL 80% ApTKI_m RELLDVDGNFLRNGGSYYIVPAFRGKGGGL 76,67%LlTKI_m QVLVDLDGDPLYNGMSYYILPVARGKGGGL 70% : *:* **: * ** :***:*. *******
Figura 15: Seqüência parcial e alinhamento comparativo de PmTI com outros inibidores de tripsina da
família Kunitz de sementes de leguminosas: A. confusa (AcTKI, gi: 166234,NCBI), E. contortisiliquum
(EcTKI, Batista el al., 1996), P. juliflora (PjTKI, gi: 243386, NCBI), A. pavonina (ApTKI, gi: 225058,
NCBI), L. leucocephala (LiTKI, gi, 18202442,NCBI). Os resíduos de aminoácidos em vermelho são
idênticos e conservados, em verde, forte similaridade, e em azul, indicam fraca similaridade.
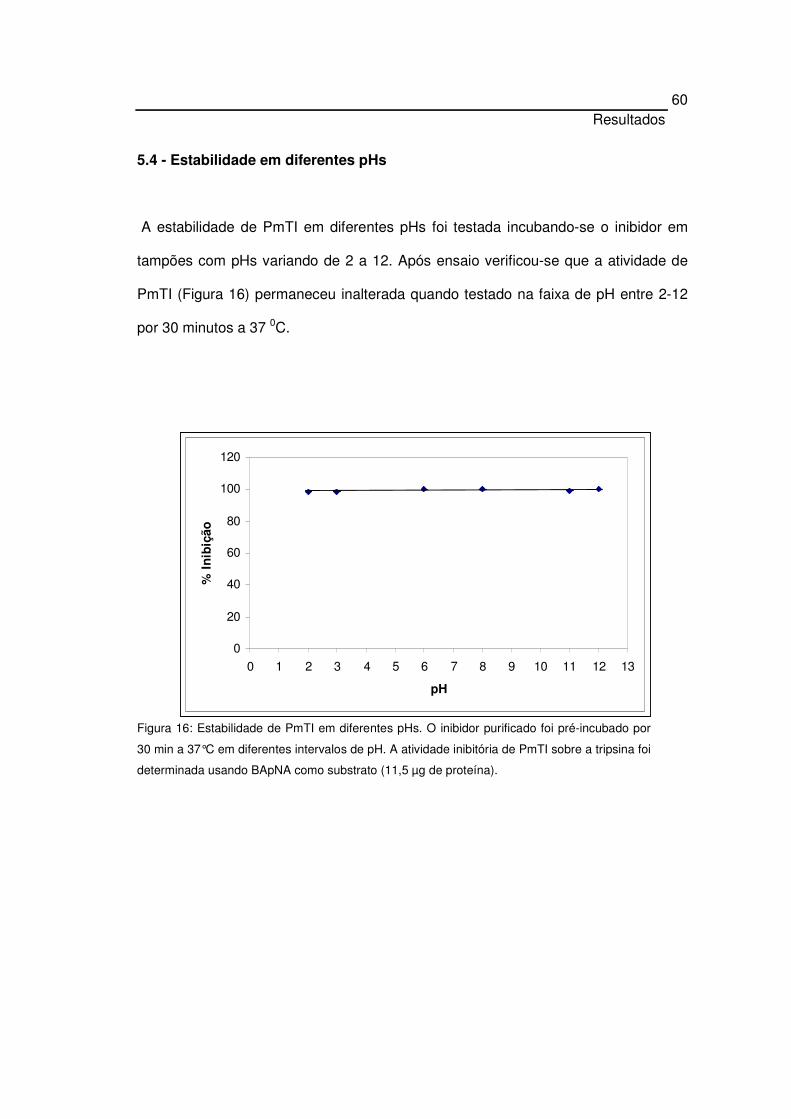
Resultados 60
5.4 - Estabilidade em diferentes pHs
A estabilidade de PmTI em diferentes pHs foi testada incubando-se o inibidor em
tampões com pHs variando de 2 a 12. Após ensaio verificou-se que a atividade de
PmTI (Figura 16) permaneceu inalterada quando testado na faixa de pH entre 2-12
por 30 minutos a 37 0C.
Figura 16: Estabilidade de PmTI em diferentes pHs. O inibidor purificado foi pré-incubado por
30 min a 37°C em diferentes intervalos de pH. A atividade inibitória de PmTI sobre a tripsina foi
determinada usando BApNA como substrato (11,5 μg de proteína).
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7 8 9 10 11 12 13
pH
% I
nib
ição

Resultados 61
5.5 - Estabilidade térmica
O inibidor purificado de sementes de Catanduva também foi avaliado quanto a sua
resistência em diferentes temperaturas (37 a 100 °C). Na Figura 17 observa-se que
PmTI manteve sua atividade máxima de inibição sobre a atividade catalítica da
tripsina quando testado em temperaturas de 37, 40 e 60 °C, 10% dessa atividade foi
perdida em temperatura 70 °C. A pré-incubação desse inibidor em temperaturas de
90 e 100 °C por 30 minutos reduziu sua capacidade inibitória em 20%.
0
20
40
60
80
100
120
30 40 50 60 70 80 90 100 110
Temperatura (°C)
% I
nib
ição
Figura 17: Estabilidade térmica de PmTI. O inibidor purificado foi pré-incubado por 30 min
em diferentes temperaturas. A atividade inibitória de PmTI sobre a tripsina foi determinada
usando BApNA como substrato (11,5 μg de proteína).

Resultados 62
5.6 - Efeito do DTT na atividade do inibidor
PmTI foi incubadas com DTT, nas concentrações finais de 1, 10 e 100 mM,
em intervalos de tempo de 15, 30, 60 e 120 minutos para determinar sua
estabilidade funcional. A Figura 18 mostra os resultados obtidos após ensaio
enzimático. PmTI mostrou-se estável nas concentrações de 1 e 10 mM. Observou-se
pouca redução quando o mesmo se encontrava na concentração de 100 mM, após
duas horas de incubação com o agente desnaturante foi observada perda de 20 %
de sua atividade inibitória.
0
10
20
30
40
50
60
70
80
90
100
110
120
1 2 3 4
Tempo (minutos)
% d
e A
tivi
dad
e R
esid
ual
Controle 1 mM 10 mM 100 mM
Figura 18: Efeito de desnaturante DTT na atividade de PmTI (11,5 μg de proteína). A atividade
foi determinada como descrito 4.5 em diferentes concentrações finais de DTT e por intervalo de
tempos específicos.
1 3 6 120

Resultados 63
5.7– Mecanismo de inibição e valor de Ki
PmTI purificado de sementes de Catanduva foi avaliado quanto ao seu mecanismo
de inibição sobre a atividade catalítica da tripsina e o valor de Ki foi determinado.
5.7.1– Mecanismo de inibição sobre a tripsina
Após a realização de ensaios utilizando concentrações variáveis de inibidor e
substrato (BApNA) foi construído um gráfico duplo recíproco de Lineweaver-Burk,
para determinação do tipo de mecanismo de inibição de PmTI para Tripsina. A
Figura 19 mostra que PmTI suprimiu a atividade catalítica da tripsina de modo
competitivo, caracterizado pela variação do valor de Km (constante de Michaelis-
Menten) e inalteração da velocidade máxima de reação.
A Figura 20, mostra a inibição da tripsina em pH 7,5 por PmTI (concentrações
crescentes) sobre a hidrolise de BApNA como substrato nas concentrações de 0,5 e 1
mM. A análise do gráfico de Dixon et al., 1979 confirmou que PmTI inibiu a tripsina de
modo competitivo similar ao observado no gráfico duplo recíproco de Lineweaver-Burk
e a constante de inibição (Ki) calculada foi de 1,5 x 10-11 mM ou 0,416 μg para PmTI.

Resultados 64
Figura 19: Mecanismo de inibição de PmTI sobre a atividade catalítica da tripsina. A atividade
inibitória de PmTI foi determinada usando várias concentrações de BApNA (0,5; 0,66; 0,8; 1;
1,25; 1,5 e 2,0 mM), e ilustrados através do gráfico do duplo recíproco de LIneweaver-Burk. �,
controle positivo, �, 0,209 ug; �, 0,418 ug; �, 0,836 ug de PmTI.

Resultados 65
0
2
4
6
8
10
12
14
-10 -5 0 5 10 15
PmTI (ug)
1/v
(DO
410/
h/m
L) -1
BApNA [1 mM] BApNA [0,5 mM]
Figura 20: Determinação do valor de Ki de PmTI contra tripsina. A constante de inibição
(Ki) de PmTI sobre a tripsina foi obtido pelo método de Dixon et al., 1979. O valor de Ki foi
obtido dos interceptos de duas retas das duas concentrações de substrato utilizada.

Resultados 66
5.8 - Especificidade para proteinases serínicas e cisteínicas
Para determinar a especificidade de inibição do PmTI foram realizados ensaios
utilizando-se proteinases serínicas e cisteínicas. Como se pode observar na Tabela
3, com relação a proteinases serínicas, PmTI foi potente na inibição da atividade
catalítica da tripsina suprimindo esta atividade em torno de 99,76 % também foi
observada inibições para elastase de neutrófilos, quimotripsina e elastase
pancreática em 76,42, 42,96 e 20,77 %, respectivamente. E com relação às
proteinases cisteínicas foi observada uma inibição de 23,10 % para papaína e de
5,28 % para bromelaína.
Tabela – 3: Especificidade de inibição de PmTI para enzimas do tipo serínicas e cisteínicas.
Enzimas Atividade
enzimática Atividade residual
da enzima Percentual de
inibição
Tripsinaa 28,13 ± 1,75 0,06 ± 0,12 99,76 %
Elastase dec
neutrófilos 31,96 ± 6,31 7,20 ± 2,12 76,42 %
Elastase pancreática b
17,0 ± 1,76 13,4 ± 0.65 3,17%
Quimotripsinab 17,33 ± 0,77 9,86 ± 0,57 42,96 %
Papaínab 36,90 ± 5,2 28,13 ± 2,28 23,10 %
Bromelaínab 27,33 ± 1,95 25,53 ± 0,51 NDd
a Atividade inibitória usando BApNA como substrato. b Atividade inibitória usando azocaseína
como substrato. c Atividade inibitória usando SAAvNA como substrato. d não detectado

Resultados 67
5.9 - Especificidade do inibidor para enzimas digestórias de insetos
A atividade in vitro de PmTI (inibidor purificado de sementes de Catanduva) para
enzimas digestórias de diversos insetos-praga foi testada (Tabela 4). PmTI inibiu de
forma seletiva a atividade das enzimas digestórias de Coleópteras como
Anthonomus grandis em 89,93 %, Zabrotes subfasciatus em 5,96 % e
Callosobruchus maculatus em 9,41 % de inibição, quando testadas as enzimas dos
Lepidópteras como Plodia interpunctella houve inibição de 59,94 % e para Castnia
licus de 23,67 % e para Ceratitis capitata (Diptera) houve 70,52 % de inibição.
A partir deste resultado larvas de C. capitata foram escolhidas para serem
submetidas a ensaio in vivo com dieta preparada com PmTI.
Tabela 4 - Especificidade de inibição de PmTI para enzimas de diferentes insetos.
Os ensaios inibitórios sobre a atividade catalítica das enzimas digestórias de insetos praga foram
realizadas em triplicata usando azocaseína como substrato e os valores na tabela correspondem à
media do percentual de inibição e seus respectivos desvios padrões.
Insetos Atividade
enzimática Atividade residual
da enzima Percentual de
inibição
A. grandis 15,90 ± 1,01 1,60 ± 0,81 89,93%
Z.subfasciatus 12,33 ± 0,98 11,56 ± 0,40 5,96%
C. maculatus 22,96 ± 0,28 20,83 ± 0,64 9,41%
P. interpunctela 32,73 ± 1,86 13,1 ± 0,86 59,94%
C. licus 29,86 ± 1,25 22,7 ± 0,57 23,67%
C. capitata 38,96 ± 0,28 11,50 ± 0,17 70,52%

Resultados 68
5.10- Bioensaio para C. capitata
Para o bioensaio concentrações crescentes de PmTI (liofilizado) foram utilizados e
dois parâmetros foram avaliados, a quantidade de PmTI que reduzia a massa das
larvas em 50% (WD50) e a quantidade que reduzia a população das larvas em 50%
(LD50).
Em relação à massa das larvas de quarto ínstar foram observadas alterações
significativas, quando as larvas foram alimentadas com PmTI na concentração de
1% não foi observado ganho de massa. Enquanto que a massa média das larvas
alimentadas em dieta controle foi de 10 mg as larvas alimentadas em dietas
contendo PmTI em diferentes concentrações variou de 8,0 mg a 3 mg,
aproximadamente (Figura 21).
A sobrevivência das larvas de quarto ínstar de C. capitata alimentadas com dietas
artificiais contendo concentrações crescentes de PmTI (0,05 a 1%) apresentou um
decréscimo linear na taxa de sobrevivência quando comparada com as larvas do
grupo controle, quando submetidas a dieta contendo 1 % de PmTI foi observada
aproximadamente 90% de mortalidade larval (Figura 22).
A massa das larvas foi afetada com uma WD50 de 0,30% e sua sobrevivência com
uma LD50 de 0,33%.

Resultados 69
Figura 21: Peso médio das larvas de 4º ínstar de C. capitata alimentadas com dieta controle sem inibidor e dietas artificiais contendo PmTI em concentrações crescentes de 0,05 % a 1 %. Cada valor representa à média ± SD.
y = -2.5218x + 12.925R2 = 0.9979
0
2
4
6
8
10
12
Controle 0, 05% 0, 20% 0, 50% 1%
Concentração
Mas
sa L
arva
l (m
g)

Resultados 70
Figura 22: Sobrevivência das larvas de 4º ínstar de C. capitata alimentadas com dieta controle
sem inibidor e dietas artificiais contendo PmTI em concentrações crescentes de 0,05 % a 1 %.
Cada valor representa à média ± SD.
y = 18.889x - 12.222
R2 = 0.9633
0
20
40
60
80
100
Contole (mg) 0, 05% 0, 20% 0, 50% 1%Concentração
Controle
% M
ort
alid
ade

_________________________________________________________________ Discussão
71
6 DISCUSSÃO
Em plantas, os inibidores de proteinases atuam como proteínas de reserva
(MOSOLOV, 1995; VALUEVA; MOSOLOV, 1999) ou podem estar envolvidos no
mecanismo de defesa vegetal contra pragas e doenças causadas por patógenos
(RYAN, 1990; Macêdo et al., 2003; Haq et al., 2004; Tamhane et al., 2005;
Dunaevsky et al., 2005). Os inibidores podem ser também utilizados como
ferramentas alternativas no tratamento de patologias humanas tais como
hemorragias, inflamação e câncer (Oliva et al., 2000, CHEN; SHAW, 2003; Fook et
al., 2005; Mello et al., 2006).
Considerando o conhecimento de que as plantas utilizam substâncias
químicas como proteínas para se defender da agressão por herbívoros e
fitopatógenos, muitas pesquisas estão sendo realizadas com o intuito de verificar a
eficácia de proteínas de defesa de plantas e trazer assim contribuições futuras para
a agricultura (Jouanin et al., 1998).
A ação bioinseticida de um inibidor de Piptadenia moniliformis foi avaliada
para viabilizar um potencial emprego na aplicação desta molécula como ferramenta
biotecnológica.
Muitos inibidores presentes nas sementes das mais variadas espécies de
plantas são purificados e isolados com a utilização de técnicas cromatográficas.
Assim cromatografias em colunas de troca iônica e de exclusão molecular são
amplamente utilizados como passos obrigatórios durante o processo de purificação
de inibidores (RICHARDSON, 1991). A fração CT1% da Catanduva foi fracionada
numa coluna de afinidade Tripsina-Sepharose, cujo perfil cromatográfico está
ilustrado na Figura 10. O "pool" obtido, detectado através de ensaio anti-tríptico, foi
denominado de CT1Af e apresentou um rendimento de 1,1% (Tabela 2), os passos

_________________________________________________________________ Discussão
72
seguintes foram a cromatografia DEAE - celulose e após submetido a Superose 12
mostrando um rendimento final de 0,02%, resultado esse inferior aos encontrados
por outros autores (Mello et al., 2001; Gomes et al., 2005; Bhattacharyya et al.,
2006; Macedo et al., 2007; BHATTACHARYYA, LEIGHTON, BABU; 2007).
A eletroforese em gel de poliacrilamida (20%), na presença de SDS, permitiu
verificar que o PmTI, (Figuras 13 e 14), apresentou uma massa molecular de
aproximadamente, 25 kDa, uma única cadeia polipeptídica e exibiu potente
capacidade inibitória sobre a atividade catalítica da tripsina, sugerindo ser um
inibidor da família Kunitz como relatado na literatura por Richardson, 1991; e
Oliveira, 2001. Um inibidor de tripsina da família Kunitz encontrado em
Dimorphandra mollis apresentou massa molecular similar com 23 kDa (Macedo et
al., 2002), que semelhante ao PmTI é composto de apenas uma cadeia
polipeptídica, em contrapartida, um inibidor encontrado em Crotalaria pallida
(GOMES, 2004) apresentou massa molecular de 32,5 kDa, sendo este composto por
duas subunidades,uma de 27,7 kDa e uma menor de 5,6 kDa. No entanto, nenhum
inibidor da família Kunitz, até o momento, foi relatado ter uma massa molecular de
25 kDa.
O sequenciamento da estrutura N-terminal de PmTI indicou um alto grau de
homologia com inibidores da família Kunitz, conforme é mostrado na figura 15. Na
seqüência de PmTI foi observado a presença do resíduo de asparagina (N13), que é
conservado em vários inibidores de tripsina da família Kunitz relatados na literatura
(Batista et al., 1996, Macêdo et al., 2003, Iwanaga et al., 2005, Bhattacharyya et al.,
2006), resíduo este que está envolvido na interação entre tripsina- inibidor (Iwanaga
et al., 2005).

_________________________________________________________________ Discussão
73
Estudos para avaliar a estabilidade do inibidor foram realizados com o
objetivo de verificar os efeitos causados por diferentes condições de pH,
temperatura e concentrações do agente desnaturante (DTT).
As variações de pH alteram o estado iônico das cadeias laterais de
aminoácidos, alterando, portanto, a distribuição de cargas e a necessidade da
existência de pontes de hidrogênio, que contribuem para a estabilidade das
proteínas. A atividade inibitória de PmTI não sofreu variação considerável quando
submetida à diferentes condições de pH (2- 12); revelando que o inibidor é
notavelmente estável a uma ampla faixa de pH, inclusive nas condições extremas
(Figura 16). A estabilidade dos inibidores da família Kunitz a diferentes condições de
pH já é fato registrado na literatura (RICHARDSON, 1991). Nossos resultados foram
similares ao observado para outros inibidores da família Kunitz (GRUEN; TAO;
KORTT, 1984; Mello et al., 2002; HAQ; KLAN, 2003; Bhattacharyya et al., 2006), no
entanto, outros inibidores mostraram maior estabilidade à variação de pH, como
observado para o inibidor de tripsina purificado de tubérculo de C. esculenta que
manteve sua atividade inibitória máxima sobre a atividade catalítica da tripsina em
pH ácido (1-4) e básico (12-13) após pré-incubação por aproximadamente 17 horas
(OLIVEIRA, 2001).
Resultados de termoestabilidade de PmTI (Figura 17) mostraram que este
inibidor não apresentou diminuição considerável de sua atividade inibitória até 60°C,
por 30 minutos. Assim, PmTI é uma proteína relativamente termoestável apesar de
ser da família de Kunitz e possivelmente apresentar poucas pontes dissulfeto. Este
resultado é compatível com alguns inibidores da família Kunitz, como foi relatado por
Souza et al., 1995, Osman, et al., 2002 e Oliveira et al., 2007, no entanto, Garcia et
al., 2004, caracterizaram um inibidor presente em sementes de Poecilanthe

_________________________________________________________________ Discussão
74
parviflora, o PPTI, com uma estabilidade térmica superior quando submetido à pré-
incubação do inibidor a temperatura de 100 ºC por 30 minutos.
A ação de um agente redutor, como o DTT, sobre as pontes dissulfeto
presentes nos inibidores de proteinases pode causar danos a sua integridade, e
provavelmente, afetar a atividade antitríptica dos mesmos, dependendo da
localização destas pontes em relação aos seus sítios reativos (Macedo, et al., 2007).
Assim, foi realizado um estudo para verificar a ação do DTT sobre a atividade
inibitória de PmTI. Observamos que o DTT a 100 mM provocou uma redução de 20
% da capacidade inibitória de PmTI (Figura 18). Lehle et al., 1996, também
relataram que ETI, um inibidor de tripsina da família Kunitz presente em sementes
de Erythrina caffra, conservou sua atividade inibitória após redução com DTT.
Macedo et al., 2003, observaram que quando o inibidor PDTI foi incubado, por 120
minutos, com 10 mM de DTT, a atividade inibitória sofreu um decréscimo de 60% e
quando incubado com 100 mM do agente redutor, o mesmo perdeu 90% de sua
atividade. Fato semelhante foi demonstrado por Garcia et al., 2004, cujo inibidor
PPTI perdeu 85% de sua atividade quando incubado com a mesma concentração de
DTT por 120 minutos. Aparentemente, a estabilidade do inibidor de P. moniliformis
não está relacionada à presença de pontes dissulfeto. Os resultados mostrados na
Figura 18 sugere que a estabilidade da atividade inibitória de PmTI pode estar
relacionada com a presença de pontes dissulfetos intra-cadeias ou que as pontes
afetadas pela redução estariam relativamente distantes do sítio reativo e que
somente uma redução e alquilação das mesmas ocasionaria a perda total da
atividade inibitória.
A cinética de PmTI é do tipo competitiva (Figura 19) corroborando com outros
inibidores de tripsina da família Kunitz (LAKOWSKI; KATO,1980; FUSHIKI; KAZUO,

_________________________________________________________________ Discussão
75
1989). Nossos resultados, no entanto, contrastam com os resultados relatados para
inibidores que interagiram com a tripsina de modo não competitivo como observado
para um inibidor de tripsina purificado de tubérculo de C. esculenta (OLIVEIRA,
2001) e C. antiquorum (SUMATHI; PATTABIRAMAN, 1979), e de sementes de
leguminosas como, CapTI (Gomes et al., 2005a), ApTI de sementes de A. pavonina
(Macedo et al., 2004), TTI de sementes de Tamarindus indica (Araújo et al., 2005a),
e uma fitocistatina purificada de Castanea sativa, denominada CsC, que inibiu não
competitivamente a tripsina com valor de Ki de 3,49 x 10-6 M (Pernas et al., 1998).
Em adição, também foi descrito que a atividade da tripsina foi inibida de modo
incompetitivo pelo inibidor purificado de sementes de Caesalpinia pulcherima
(HASE; KOYAMA; DAIYASU, 1986) e Artocarpus integrifolia (BHAT;
PATTABIRAMAN, 1989).
O valor de Ki (constante de inibição) de 1,57 x 10-11 mM, calculado utilizando o
modelo proposto por Dixon (1979),(Figura 20) demonstra que PmTI tem uma alta
afinidade pela tripsina, semelhante ao encontrado para outros inibidores de tripsina
vegetais, purificados e já caracterizados (BIRK, 1994). Comparando-se os valores
de Ki de PmTI com outros inibidores da família Kunitz foi observado que esse
inibidor é mais potente inibidor de tripsina do que os encontrados em C. esculenta
(OLIVEIRA, 2001), ETIa de sementes de Erythrina variegata (Kouzuma et al., 1992;
Iwanaga et al., 2005) que apresentaram valor de Ki na ordem de 10-6 M, CapTI
(Gomes et al., 2005), PPTI (Garcia et al., 2004), PPI de sementes de Carica papaya
(Odani et al., 1996), e COTI de sementes de Cassia obtusifolia (Liao et al., 2007)
cujos valores de Ki foram determinados na ordem de 10-7 M. No entanto, o inibidor
de Catanduva apresentou valores de Ki similares em ordem de grandeza àqueles
determinados para os inibidores de TTI (Araujo et al., 2005a), BrTI de sementes de

_________________________________________________________________ Discussão
76
Bauhinia rufa (Nakahata et al., 2006), SWTI de sementes de Swartizia pickelli
(Cavalcanti et al., 2002), ECTI (Batista et al., 1996), DMTI-II de sementes de
Dimorphandra mollis (Mello et al., 2002), DMTI (Macedo et al., 2000), AeTI de
sementes de Archidrendon ellipticum (Bhattacharyya et al., 2006), BbKI de sementes
de Bauhinia bauhinoides (Oliva et al., 2001), PDTI de sementes de Peltophorum
dubium (Macedo et al., 2003), ILTI de sementes de Inga laurina (Macedo et al.,
2007), CbTI-2 de sementes de Caesalpinia bonduc (BHATTACHARYYA, RAI; BABU,
2007), e de sementes de Murraya koenigii (SHEE; SHARMA, 2007) que
apresentaram valores de Ki na ordem de 10-9 e 10-10 M. confirmando que o valor de
Ki determinado para o inibidor de Catanduva encontra-se dentro do padrão do valor
de Ki determinado para outros inibidores da família Kunitz.
Diante das características e propriedades apresentadas por ele no decorrer
do estudo, concluímos que o inibidor presente em sementes de P. moniliformis
pertence à família Kunitz.
Análises da inibição in vitro, de enzimas obtidas comercialmente ou das
enzimas presentes no trato digestório dos insetos alvos, foram feitos para determinar
a especificidade do inibidor sobre estas proteases (BROADWAY, 1997). Através
destas análises, o potencial de proteínas em inibir o desenvolvimento de insetos
quando ingeridas pode ser avaliado.
Na ordem Lepidóptera e Díptera são encontradas proteinases serínicas, como
tripsina, quimotripsina e elastase (TERRA; FERREIRA, 1994, HAQ; ATIF; KHAN,
2004, Gatehouse et al., 1999, Silva et al., 2006), e sabe-se que C. capitata, expressa
proteinases serínicas em seus intestinos médios (Applebaum et al., 1985, SILVA,
2003; Gatehouse et al., 1999; Zeng et al., 2001). Enquanto da ordem coleóptera

_________________________________________________________________ Discussão
77
apresentam proteinases do tipo cisteínicas (Lemos et al., 1990, Lecardonnet et al.,
1999),
Os inibidores de proteinases serínicas são considerados como agentes
naturais de controle contra insetos herbívoros, devido à capacidade de suprimir a
atividade dessas enzimas proteolíticas in vitro (Harsulkar et al., 1999; REED;
CHANDLER; SANDEMAN, 1999; De Leo et al., 2001; Macedo et al., 2003; Telang et
al., 2003, 2005; Tamhane et al., 2005; Garcia et al., 2004; Gomes et al., 2005b;
Srinivansan et al., 2005; Liao et al., 2007), retardar o crescimento e desenvolvimento
larval (Gatehouse et al., 1999), diminuir a fertilidade e fecundidade de fêmeas (DE
LEO; GALLERANI, 2002; Telang et al., 2003; TAMHANE et al., 2005), causar
deformidades (Franco et al., 2004; Gomes et al., 2005b) e aumentar a taxa de
mortalidade de insetos pragas (Macedo et al., 2002; Gomes et al., 2005a; Araujo et
al., 2005; Moura et al., 2006).
A atividade de PmTI foi testada, in vitro, contra extratos enzimáticos de
intestinos de 6 espécies de insetos-praga, pertencentes a diferentes ordens,
Lepidópteras (P. interpunctella e C. licus), Coleópteras (C. maculatus , Z.
subfasciatus e A. grandis) e Díptera (C. capitata) (Tabela 4). PmTI foi capaz de inibir
em 89,93% as enzimas de A. grandis, apresentando uma inibição de 59,94% para
as enzimas de P. interpunctella, este resultado foi similar ao observado para o
inibidor de tripsina de sementes de T. indica, denominado TTI, este inibidor suprimiu
a atividade de proteinases serínicas de Alabama argillacea (Lepidóptera) em
aproximadamente 54% (Araujo et al., 2005a). As enzimas de C. capitata, foram
inibidas em 70,52% resultado este que corrobora com o encontrado por Oliveira,
(2007), quando utilizou o inibidor de Pithecellobium dumosum, denominado JB2, que

_________________________________________________________________ Discussão
78
apresentou uma inibição de 70%. PmTI apresentou inibição de 5,96% para Z.
subfasciatus, de 9,41% para C. maculatus e de 23,67% para C. licus.
Os resultados obtidos nos experimentos in vitro conduziram a uma segunda
etapa do estudo da atividade inseticida do inibidor PmTI, o ensaio in vivo, no qual ele
foi oferecido como fonte de alimento em várias concentrações em dieta artificial com
a finalidade de analisarmos os parâmetros de crescimento e sobrevivência (WD50 %
e LD50 %) de C. capitata que é uma importante praga para o agronegócio potiguar.
Para verificar a aplicabilidade da ação inseticida dos inibidores e ou de
qualquer outra proteína, são utilizados métodos que envolvem ensaios de inibição in
vitro das proteases presentes no trato digestivo do inseto pelos inibidores alvo de
estudo, testes de alimentação de insetos contendo a proteína de estudo e numa fase
mais avançada a produção de plantas transgênicas, onde o aumento de sua
resistência seria decorrente da transferência de genes codificantes de inibidores de
proteinases (Macedo et al., 2001).
A presença de PmTI na dieta artificial reduziu significativamente a
sobrevivência (WD50 de 0,33) e o peso médio (LD50 de 0,30) das larvas de C.
capitata quando comparado ao controle (Figura 21 e 22), sugerindo um efeito
subseqüente no desenvolvimento do inseto, Macedo et al., (2000), demonstraram
que um inibidor de Dimorphandra mollis apresentou um WD50 de 0,53%, para larvas
de C. maculatus, resultado esse superior ao encontrado para PmTI. No entanto, o
nosso resultado é bem superior ao inibidor de tripsina presente nas sementes de
Peltophorum dubium que reduziu em 50% o ganho de peso de Anagasta kuehniella
(Lepidoptera) quando presente a 1% em dieta artificial, (Macedo et al., 2003). Mello
et al., (2004) quando alimentaram larvas de A. kuehniella com DMTI-II em
concentrações crescentes de 0,5 a 2,0% não observou efeitos significativos sobre a

_________________________________________________________________ Discussão
79
sobrevivência das larvas, resultado esse inferior ao encontrado para PmTI quando
testado contra as larvas de C. capitata, que na concentração de 1% não foi
observado crescimento e em relação a sobrevivência observou-se uma taxa de
mortalidade crescente nas diferentes concentrações testadas. Franco et al., (2004)
demonstraram que a presença de 500 μM de SKTI na dieta oferecida a A. grandis
causou uma diminuição no peso médio larval de 75%. O resultado obtido com PmTI
foi superior ao encontrado por GOMES, (2004) que alimentou C. capitata com o
inibidor de C. pallida e não observou mudança significativa em relação a nenhum
dos parâmetros analisados, o que explicaria neste caso uma adaptação do inseto ao
inibidor fato este já relatado na literatura (DE LEO; GALLERANI, 2002; Franco et al.,
2004).

Conclusões 80
7 CONCLUSÕES
A massa molecular, a potente atividade inibitória sobre a atividade catalítica
da tripsina, bem como, a similaridade da seqüência de aminoácidos do inibidor de
tripsina purificado de sementes de Catanduva (P.moniliformis), denominado PmTI,
com outros inibidores de sementes de leguminosas da família Kunitz são indicativos
de que este é o mais novo membro de inibidor de tripsina da família Kunitz;
A estabilidade do inibidor purificado de sementes de Catanduva às variações
de temperaturas, pHs e agente redutor (DTT) pode sugerir a presença de pontes
dissulfeto intramoleculares ou a presença de inúmeras pontes de hidrogênio;
PmTI mostrou especificidade de inibição, in vitro, para enzimas proteolíticas
pertencentes às classes mecanísticas das proteinases serínicas e cisteínicas;
PmTI purificado de sementes de Catanduva exibiu capacidade de inibir
diferencialmente, in vitro, as enzimas digestórias de insetos pragas pertencentes a
diferentes ordens e apresentou uma atividade tóxica para larvas de mosca-das-
frutas (C. capitata);
A perspectiva que se abre em relação aos resultados dos ensaios in vitro e in
vivo obtidos com PmTI é que este inibidor possa ser um forte candidato aos
programas de melhoramento via transgenia.

Referências 81
REFERÊNCIAS
APPLEBAUM, S. W. Biochemistry of digestion. In: Kerkut, g. A.; Gilbert, L. I. (eds.) Comprehensive Insect Physiology, Biochemistry and Pharmacology, Pergamon Press, Oxford, p. 270-311,1985.
ARAÚJO, C. L.; BEZERRA, I. W. L.; DANTAS, I. C.; LIMA, T. V. S.; OLIVEIRA, A. S.; MIRANDA, M. R. A., LEITE, E. L.; SALES, M. P. Biological activity of proteins from pupls of tropical fruits. Food Chemistry. 5, 107-110, 2004.
ARAÚJO, C. L.; BEZERRA, I. W. L.; OLIVEIRA, A. S.; MOURA, F. T.; MACEDO, L. L. P.; GOMES, C. E. M.; BARBOSA, A. E. A. D.; MACEDO, F. P.; SOUZA, T. M. S.; FRANCO, O. L.; BLOCH-JR, C.; SALES, M. P. In vivo bioinsecticidal activity toward Ceratitis capitata (fruitfly) and Callosobruchus maculatus (cowpea weevil) and in vitro bioinsecticidal activity toward different orders of insect pests of a typsin inhibitor purified from tamarind tree (Tamarindus indica) seeds. Journal Agricultural and Food Chemistry. 53: 4381-4387, 2005.
AZARKAN, M.; DIBIANI, R.; GOORMAGHTIGH, E.; RAUSSENS, V.; BAEYENS-VOLANT, D. The papaya Kunitz-Type trypsin inhibitor is a highly stable β-sheet glycoprotein. Biochimica et Biophysica Acta. 1764, 1063-1072, 2006.
BATISTA, I. F. C.; OLIVA, M. L. V.; ARAUJO, M. S.; SAMPAIO, U. M.; RICHARDSON, M.; FRITZ, H.; SAMPAIO, C. A. M. Primary structure of a kunitz-type trypsin inhibitor from Enterolobium contortisiliquum seeds. Phytochemistry. 41, 1017-1022, 1996.
BARRET, A. J. The cystatins: a new class of peptidase inhibitor. Trends in Biochemistry Science, 12,193-196, 1987.
BELITZ,H.D.;GROSCH, W. Quimica de los alimentos. Espanha: Acribia, 1087, 1997.
BELITZ, H.D.; WEDER, J.K.P. Protein inhibitors of hydrolases in plants foodstuffs.Food Reviews International, 6, 151-211, 1990.
BHAT, A. V.; PATTABIRAMAN, T. N. Protease inhibitors from jackfruit seed (Artocarpus integrifolia). Journal of Bioscience. 14(4), 351-365, 1989.

Referências 82
BHATTACHARYYA, A.; RAI, S.; BABU, C. R. A trypsin and chymotrypsin inhibitor from Caesalpinia bonduc seeds: Isolation, partical characterization and insecticidal properties. Plant Physiology and Biochemistry. 45, 169-177, 2007.
BHATTACHARYYA, A.; MZUMDAR, S.; LEIGHTON, S. M.; BABU, C. R. A Kunitz proteinase inhibitor from Archidendron ellipticum seeds: Purification, characterization, and kinetic properties. Phytochemistry. 67, 232-241, 2006.
BIRK, Y., Protein proteinase inhibitors in legume seeds- overview. Archivos Latinoamericanos de Nutricion.44, 4, 26-30, 1994
BLUM, H., BEIER, H., GROSS, H. J. Improved silver staining of plant-proteins, Rna and DNA in polyacrilamide gels. Electrophoresis. 8 (2), 93-99, 1997.
BODE, W.; HUBER, R. Natural protein proteinase inhibitors and their interaction with proteinases. European Journal of Biochemistry. 204, 433-451, 1992.
BODE, W.; HUBER, R. Structural basis of the endoproteinases-protein inhibitor interaction. Biochimica et Biophysica Acta. 1477, 241-252, 2000.
BOULTER, D.; EDWARDS, G. A.; GATEHOUSE, A. M. R.; GATEHOUSE, J.A.,HILDER, V. A. Additive protective effects of incorporating two different higher plant derived insect resistance genes in transgenic tobacco plants. Crop protection. 9,351-354, 1990.
BOWLES, J. D. Defense-related proteins in higher plants. Annual Revision Biochemistry. 59: 873-875, 1990.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of die-binding. Analytical Biochemistry, 72, 218-254, 1976.
BROADWAY, R. M. Are insects resistant to plant proteinase inhibitors? Journal of Insect Physiology. 41(2): 107-116, 1995.
BROADWAY, R. M. Plant dietary proteinase inhibitors alter complement of midgut proteases. Archives of Insect Biochemistry and Physiology. 32, 39–53, 1996.

Referências 83
BROADWAY, R. M. Dietary regulation of serine proteinases that are resistant to serine proteinase inhibitors. Journal of Insect Physiology. 43, 855–874, 1997.
BROADWAY, R. M & DUFFEY, S.S. Plant Proteinases Inhibitors: Mechanism of Action and effect on the growth and digestive physiology of larval Heliothis zea and Spodoptera exigua. Journal of Insect Physiology. 32, 827-833, 1986.
BRZIN, J.; KIDRIC, M. Proteinases and their inhibitors in plants: role in normal growth and in response to various stress conditions. Biotechnology and Genetic Engineering Reviews. 13, 421-467, 1995.
CAMPOS, F. A. P.; XAVIER-FILHO, J.; SILVA, C. P.; ARY, M. B. Resolution and partial characterization of proteinases and α-amylases from midguts of larvae of the bruchid beetle Callosobruchus maculatus (F.). Comparative Biochemistry and Physiology, part B: Biochemistry & Molecular Biology. 92, 51-57, 1989.
CARLINI, C.R.; GROSSI-DE-SÁ, M.F. Plant toxic proteins with insecticidal properties. A review on their potentialities as bioinseticides. Toxicon, 40, 1515-1539. 2002.
CAVALCANTI, M. S. M., OLIVA, M. L.V., FRITZ, H., JACHUM, M., MENTELE, R., SAMPAIO, M., COELHO, L. C. B. B., BATISTA, I. F.C., SAMPAIO, C. A. M. Characterization of a kunitz trypsin inhibitor with one disulfide bridge purified from Swartzia pickellii. Biochemical and Biophysical Research Communications. 29, 635-639, 2002.
CHEN, T.B.; SHAW, C. Identification and molecular cloning of novel trypsin inhibitor analogs from the dermal venom of the Oriental firebellied toad (Bombina orientalis) and the European yellow-bellied toad (Bombina variegata). Peptides (6), 873–880. 2003.
CHESIN, M.; ZIPF, A. E. Alam systems in highterplants. The Botanical Review. 56, 193-235, 1990
CHRISTELLER, J. T., LAING, W.A.; SHAW, B.P.; BURGUESS, E.P.J., Characterization and partial purification of the digestive protease of the black field cricket Teleogryflus commodus (Walker): elastase is a major component. Insect Biochemistry. 20,157-164, 1990.
CHRISTELLER, J. T. Evolutionary mechanism acting on proteinase inhibitor variability. FEBS Journal. 272, 5710-5722, 2005.

Referências 84
CRONQUIST, A. The evolution and classification of flowering plants. New York: The New York Botanical Garden, 555,1988.
CUNNINGHAM, S.M., VOLENEC, J.J. Purification and caracterization of vegetative storage proteins from alfafa(Medicago sativa L.) taproots. Journal Plant Physiology, 147: 625-632, 1996.
CURSINO-SANTOS, J.R., DEFINA, T.P.A., MARTINEZ-ROSSI, N.M. Os segredos das plantas e de seus patogenos na era molecular. Ribeirão Preto: Sociedade Brasileira de Genética, 1,41. 2003.
DEAN, R.; KUC, J. Induction systemic protection in curcumber. The source of the signal. Physiology and Molecular Plant Pathology, 28, 227-233, 1986.
DE LEO, F.; BONADE-BOTINO, M.; CECI,R. L.; GALERANI, R.; JOUANIN, L. Effects of mustard trypsin inhibitors expressed in different plants on three different lepidopteran pests. Insect Biochemistry and Molecular Biology. 31, 593-602, 2001
DE LEO, F.; GALLERANI, R. The mustard trypsin inhibitor 2 affects the fertility of Spodoptera littoralis larvae fed on transgenic plants. Insect Biochemistry and Molecular Biology. 32, 489–496, 2002.
DIXON, M., WEBBER, R.C., THERONE, C.J.R., TRIPTON, K. F. Inhibition and activation in enzymes. Academic Press, New York, 332-381, 1979.
DUNAEVSKY, Y.E.; ELPIDINA, E.N.; VINOKUROV, K.S.; BELOZERSKY, M.A. Protease inhibitors in improvement of plant resistance to pathogens and insects. Molecular Biology 39, 608–613. 2005.
EMBRAPA MEIO – NORTE. Cultivo de Feijão – caupi. Disponível em: http://sistemasdeprodução.cnptia.embrapa.br/FontesHTML/Feijão/Feijão Caupi/pragas.htm. Acesso em: 10 de maio de 2008.
ERLANGER, B.F.; KOKOWSKY, N.; COHEN, W. The preparation and properties of two chromogenic substrates of trypsin. Archives of Biochemistry and Biophysics, 95, 271-278. 1961.

Referências 85
FALCO, M,C., MARBACH, P.A.S., POMPERMAYER, P., LOPES, F.C.C, SILVA-FILHO, M.C. Mechanisms of sugarcane response to herbivory. Genetic. Molecular. Biology. 24:113-122. 2001
FERRY, N., EDWARDS, M. G., GATEHOUSE, J.A., GATEHOUSE, A.M.R. Plant - insect interactions: molecular approaches to insect resistance. Current Opinion on Biotechnology, 15:1-7.2004.
FOOK, J.M.S.L.L.; MACEDO, L.L.P.; MOURA, G.E.D.D.; TEIXEIRA, F.M.; OLIVEIRA, A.S.; QUEIROZ, A.F.S.; SALES, M.P. A serine proteinase inhibitor isolated from Tamarindus indica seeds and its effects on the release of human neutrophil elastase. Life Science. 76, 2881–2891. 2005.
FRANCO, O. L.; RIGDEN, D. J.; FRANCISLETE R. MELO, F. R; GROSSI-DE-AS, M. F. Plant a-amylase inhibitors and their interaction with insect a-amylases: Structure, function and potential for crop protection. European Journal of Biochemistry. 269, 397-412, 2002.
FRANCO, O. L.; DIAS, S. C.; MAGALHÃES, C. P.; MONTEIRO, A. C. S.; BLOCH-JR, C.; MELO, F. R.; OLIVEIRA-NETO, O. B.; MONNERAT, R. G.; GROSSI-DE-SÁ, M. F. Effects of soybean kunitz trypsin inhibitor on the cotton boll weevil (Anthonomus grandis). Phytochemistry. 65, 81-89, 2004.
FUSHIKI, T., KAZUO, I. Two hypothesis on the feedback regulation of pancreatic enzyme secretion. FASEB J., 3: 121-126. 1989
GARCIA, V. L; FREIRE, G. M; NOVELLO, J. C; MARANGONI, S; MACEDO, M. L. R. Trypsin inhibitor from Poecilantha parviflora seeds: Purification, characterization, and activity against pest protease. The Protein Journal. 23, 343-350, 2004.
GATEHOUSE, A. M. R., BOULTER, D, HILDER, V. A. Potential of plant-derived genes in the genetic manipulation of crops for insect resistance. In: Biotechnology in Agriculture, Plant Genetic Manipulation for Crop Protection, CAB Internacional, 7, 155-181, 1992.
GATEHOUSE, A. M.; NORTON, E.; DAVISON, G. M.; BARBE, S. M.; NEWELL, C. A.; GATEHOUSE, J. A. Digestive proteolytic activity in larvae of tomato moth, Lacanobia oleracea; effects of plant protease inhibitors in vitro and in vivo. Journal of Insect Physiology. 45, 545-558, 1999.

Referências 86
GATEHOUSE, A.M.R., GATEHOUSE, J.A., DOBIE, P., KILMINSTER, A.M.,BOULTER, D. Biochemical basis of insect resistance in Vigna unguiculata.Journal Science Food Agriculture.30,948-958, 1979
GOMES, C. E. M. Purificação, caracterização e atividade Bioinseticida, de um inibidor de tripsina de sementes de Crotalaria pallida. 2004. 74p. Dissertação (Mestrado em Bioquímica) - Universidade Federal do Rio Grande do Norte, Natal, 2004.
GOMES, C. E. M; BARBOSA, E. A. D; MACEDO, L. L. P; PITANGA, J. C. M; MOURA, F. T; OLIVEIRA, A. S; MOURA, R. M; QUEIROZ, A. F. S; MACEDO, F. P; ANDRADE, L. B. S; VIDAL, M. S; SALES, M. P. Effect of trypsin inhibitor from Crotalaria pallida seeds on Callosobruchus maculates (cowpea weevil) and Ceratitis capitata (fruit fly). Plant Physiology and Biochememistry. 46, 1095-1102, 2005a.
GOMES, A. P. G.; SIMONI C. DIAS, S. C.; CARLOS BLOCH JR., C.; MELO, F. R; FURTADO JR, J. R.; MONNERAT, R. G.; MARIA F. GROSSI-DE-SA, M. F.; FRANCO, O. L. Toxicity to cotton boll weevil Anthonomus grandis of a trypsin inhibitor from chickpea seeds. Comparative Biochemistry and Physiology, part B: Biochemistry & Molecular Biology. 140, 313–319, 2005b.
GOTTSTEIN, H. D.; KUC, J. Induction of systemic resistance in cuccumber by phosphates. Phytopathology, 79, 176-179, 1989.
GRUEN, L. C., TAO, Z. J., KORTT, A.A. Stability and physicochemical properties of a trypsin inhibitor from winged bean seed (Psophocarpus tetragonolobus (L) DC). Biochemica et Biophysica Acta. 791 (3), 285-293,1984
HARBONE, J. B. Biochemical plant ecology. Plant Biochemistry. 14, 503-515,1997.
HARSULKAR, A.M.; GIRI, A.P.; GUPTA, V.S.; PATANKAR, A.G.; SAINANI, M.N.; RANJEKAR, P.K.; DESHPANDE, V.V. Successive use of non-host plant proteinase inhibitors required for effective inhibition of Helicoverpa armigera gut proteinases and larval growth. Plant Physiology. 121: 497-506. 1999.
HASE, S., KOYAMA, S.,DAIYASU , H., TAKEMOTO, H., HARA, S. , KOBAYASHI Y., KYOGOKU, Y., IKENAKA, T. Structure of a sugar chain of a protease inhibitor isolated from Barbados pride (Caesalpinia pulcherrima) seeds. Journal of Biochemistry 100(1): 1-10.1986.

Referências 87
HAQ, S. K.; KHAN, R. H. Characterization of a proteinase inhibitor from Cajanus cajan (L.). Journal of Protein Chemistry. 22 (6), 543-554, 2003.
HAQ, S. K.; ATIF, S. M.; KHAN, R. H. Protein proteinase inhibitor genes in combat against insects, pests, and pathogens: natural and engineered phytoprotection. Archives of Biochemistry and Biophysics, 431, 145-159, 2004.
HILDER, V. A.; GATEHOUSE, A. M. R.; SHEERMAN, S.E.; BAKER, R. F.; BOULTER, D. A novel mechanism of insect resistance engineered into tobacco. Nature, 330, 160-163, 1987.
IWANAGA, S.; YAMASAKI, N.; KIMURA, M.; KOUZUMA, Y. Contribution of conserved Asn residues to the inhibitory activities of Kunitz-type protease inhibitors from plants. Bioscience, Biotechnology, and Biochemistry. 69 (1), 220-223, 2005.
JOUANIN, L. BONADÉ-BOTTINO, M., GIRARD, C., MORROT, G., GIBAND, M. Transgenic plants for insect resistance. Plant Science. 131(1):1-11, 1998.
JONGSMA, M.A.; BOULTER, C. The adaptation of insects to plant proteaseinhibitors. Journal Insect Physiology. 43: 885-895. 1997.
JOHNSON, R.,NARVAEZ, J., AN, G., RYAN, C.A. Expression of proteinase inhibitors I and II in transgenic tobacco plants: effects on natural defense against Manduca sexta larvae. Proc. Natl. Academy Science.86: 9871-9875, 1989.
KIM, Y.S., LAMBOOY, J.P. Covalent bonding of riboflavin homologs by hepatics mitochondrial mmbrane enzymes. International Journal Biochemistry.17, 9, 975-981, 1985.
KNEEN, E.; SANDSTEDT, R. M. An amylase inhibitor from certain cereals. Journal American Chemical Society. 65, 1247,1943.
KRATIGER, A.F. Insect resistence in Crops: a case study of Bacillus thuntingiensis (bt) and its transfer to developing countries. ISAAA Briefs. 2: 42, 1997.
KOGAN, M. Natural chemicals in plant resistance to insects. Iowa State Journal Research. 60, 501-527, 1986.

Referências 88
KOIWA, H.; SUBRAMANIAN, L.; SHADE, R. E.; ZHU, K. Y.; NIELSEN, S. S.; MURDOCK, L. L.; BRESSAN, R. A.; HASEGAWA, P. M. Insecticidal activities of soybean cysteine proteinase inhibitors are correlated with their papain-binding affinities as phage-displayed proteins. Plant Physiology. 114, 1115-1120, 1997.
KOUZUMA, Y., SUETAKE, M., KIMURA, M., YAMASAKI, N. Isolation and primary structure of proteinase inhibitors from Erythrina variegate (Linn.) var. Orientalis seeds. Bioscience Biotechnology Biochemistry.56, 1819-1824, 1992.
KUNITZ, M. Crystalline soybean trypsin inhibitor. Journal Genetic Physiology. 29, 149-154, 1945.
LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature.,227: 680-685. 1970.
LASKOWISKI, M. JR.; E KATO, I. Protein inhibitors of proteinases. Annual Review Biochemistry. 49:593-626. 1980.
LAWRENCE, P. L.; KOUNDAL, K. R. Plant protease inhibitors in control of phytophagous insects. Electronic Journal of Biotechnology. 5 (1), 93-109, 2002.
LECARDONNEL, A. CHAUVIN, I. JOUANIN, L. BEAUJEAN, A., PREVOST, G. SANGSWAN-NORREEL,B. Effects of rice cystatin I expression in transgenic potato on Colorado potato beetle larvae. Plant Science. 140, 7179,1999.
LEHLE, K., KOHNERT, U. STERN, A. POPP, F., JAENICKE, R. Effect of dissulfide bonds on the structure and stability of the trypsin/tPA inhibitor from Erythrina caffra: site-directed mutagenesis, expression, and physiochemical characterization. Nature Biotechnology. 14:476-480, 1996.
LEMOS, F.J.A., CAMPOS, F.A.P., SILVA, C.P., XAVIER – FILHO, J. Proteinases and amylases of larval midgut of Zabrotes subfasciatus reared on cowpea (Vigna unguiculata) seeds. Entomology Experiment Applicad. 56:219-227, 1990.
LEWIS, G.P. - Revisao Manuscrita da Familia Mimosaceae APNE-CNIP, 2003
LIAO, H., REN, W., KANG, Z., JIANG, J-H., ZHAO, X-J., DU, L-F. A trypsin inhibitor from Cassia obtusifolia seeds: isolation, characterization and activity against Pieris rapae. Biotechnology letters. 29 (4), 653-658, 2007.

Referências 89
MACEDO, M. L. R., XAVIER, J. Purification and Partial Characterization of Trypsin-Inhibitors from Seeds of Clitoria-Ternatea. Journal of the Science of Food and Agriculture. 58, 1, 55-58, 1992.
MACEDO, M. L. R.; ANDRADE, L. B. S.; MORAES, R. A.; XAVIER-FILHO, J. Vicilin variants and the resistance of cowpea (Vigna unguiculata) seeds to the cowpea weevil (Callosobruchus maculatus). Comparative Biochemistry and Physiology. Toxicology & Pharmacology. 105, 89-94, 1993.
MACEDO, M. L. R; FREIRE, M. G. M; CABRINI, E. C; TOYAMA, M. H; NOVELLO, J. C; MARANGONI, S. A trypsin inhibitor from Peltophorum dubium seeds active against pest proteases and its effect on the survival of Anagasta Kuehniella. Biochimica et Biophysica Acta. 1621, 170-182, 2003.
MACEDO, M. L. R.; MATOS, D. G. G.; MACHADO, O. L. T.; MARANGONI, S.; NOVELLO, J. C. Trypsin inhibitor from Dimorphandra mollis seeds: purification and properties. Phytochemistry. 54, 553-558, 2000.
MACEDO, M.L.R.; MELLO, G.C.; FREIRE, M.G.M.; NOVELLO, J.C.; MARANGONI, S.; MATOS, D.G.G. Effect of a trypsin inhibitor from Dimorphandra mollis seeds on the development of Callosobruchus maculatus. Plant Physiology and Biochemistry. 40, 891-898, 2002.
MACEDO, M. L. R.; SA, C. M.; FREIRE, M. G. M.; PARRA, J. R. P. A kunitz-type inhibitor of Coleopteran proteases, isolated from Adenanthera pavonina L. seeds and its effect on Callosobruchus maculatus. Journal of Agricultural and Food Chemistry. 52, 2533-2540, 2004.
MACEDO, M. L. R.; GARCIA, V. A.; FREIRE, M. G. M.; RICHARDSON, M. Characterization of a kunitz trypsin inhibitor with a single disulfide bridge from seeds of Ingra laurina (SW.) Willd. Phytochemistry. 68, 1104-111, 2007.
MACEDO, L. L. P.; AMORIM, T. M. L.; OLIVEIRA, A. S.; Uchoa, A. F.; MACEDO, F. P; RIBEIRO, J. K. C; SANTOS, E. A.; SALES, M. P.. Larvicidal effects of a chitin-binding vicilin from Erythrina velutina seeds on the mediterranean fruit fly Ceratitis capitata. Journal of Agricultural and Food Chemistry, 56, 802-808, 2008.
MAITY, J.; PATRA, B. C. Isolation and characterization of trypsin Inhibitor from the water fern, Azolla pinnata r. Br. Journal of Food Biochemistry. 27, 281-294, 2003.

Referências 90
MANDAL, S.; KUNDU, P.; ROY, B.; MANDAL, R. K. Precursor of the inactive 2S seed storage protein from the indian mustard Brassica juncea is a novel trypsin inhibitor. The Journal of Biological Chemistry. 277, 37161-37168, 2002.
MARGIS, R., REIS, .E.M., VILLERET, V. Structural and phylogenetic relationships among plant and animal cystatins. Archives Biochem Biophysic. 359, 1, 24-30, 1998.
MCFARLANE, J. E. Nutrition and digestive organs. Fundamentals of Insect Physiology. M. Blum. New York, John Wiley & Sons: 59-89.1985.
MCMANUS, M. T.; BURGESS, E. P. J.; Effects of the Soybean (Kunitz) Trypsin-Inhibitor on Growth and Digestive Proteases of Larvae of Spodoptera-Litura. Journal of Insect Physiology. 41, 9, 731-738, 1995.
MELLO, G. C., OLIVA, V.L.M., SUMIKAWA, J. T., MACHADO, O.L.T., MARANGONI, S., NOVELLO, J., C., MACEDO, M.L.R. Purification and a characterization of a new trypsin inhibitor from Dimorphandra mollis seeds Journal of Protein Chemistry, 20,8, 625-632, 2001
MELLO, M.O, SILVA-FILHO, M.C. Plant-insect interactions: an evolutionary arms race between two distinct defense mechanisms. Brazilian Journal Plant Physiology, 14: 71-81, 2002.
MELLO, G.C.; DESOUZA, I.A.; MARANGONI, S.; NOVELLO, J.C.; ANTUNES, E.; MACEDO, M.L.R. Oedematogenic activity induced by Kunitz type inhibitors from Dimorphandra mollis seeds. Toxicon 47, 150–155. 2006.
MILLER, J.W., KRAMER, K. J., LAW, J. H. Isolation and partial caractherization of the larval midgut tripsyn from the tobacco hornworm Manduca sexta (Lepdoptera: Sphin gidae). Comportament Biochemistry and Physiology. 48,117-129, 1974.
MOSOLOV, V.V. Protein inhibitors of proteinases and alfa-amylases in plants(a review). Applicad Biochemistry Microbiology, 31: 3-7, 1995.
MOURA, F. T.; OLIVEIRA, A. S.; MACEDO, L. L. P.; VIANNA, A. L. B. R; ANDRADE, L. B. S.; MARTINS-MIRANDA, A. S.; OLIVEIRA, J. T. A.; SANTOS, E. A. ; SALES, M. P. Effects of a Chitin-Binding Vicilin from Enterolobium contortisiliquum Seeds on Bean Bruchid Pests (Callosobruchus maculatus and Zabrotes subfasciatus) and

Referências 91
Phytopathogenic Fungi (Fusarium solani and Colletrichum lindemuntianum). Journal Agricultural and Food Chemistry. 55, 260-266, 2006.
NAKAHATA, A. M.; BUENO, N. R.; ROCHA, H. A. O., FRANCO, C. R. C.; CHAMMAS, R.; NAKAIE, C. R.; JOSIUTIONIS, M. G.; NADER, H. B.; SANTANA, L. A.; SAMPAIO, U. M.; OLIVA, M. L. V.; Structural and inhibitory properties of a plant proteinase inhibitor containing the RGD motif. International Journal of Biological Macromolecules. 40(1), 22-29, 2006.
NEGREIROS, A. N. M.; CARVALHO, M. M.; XAVIER-FILHO, J.; BLANCO-LABRA, A.; SHEWRY, P. R.; RICHARDSON, M. Phytochemistry. 30 (9), 2829-2833, 1991.
NORIOKA, N., HARA, S., IKENAKA, T., ABE, J. Distribution of the kunitz and Bowman-Birk family proteinase inhibitor in leguminosae seeds. Agricultural and Biological Chemistry. 52, 1245-1252, 1988.
ODANI, S.; YOKOKAWA, Y.; TAKEDA, H.; A. B. E, S.; ODANI, S. The primary structure and characterization of carbohydrate chains of the extracellular glycoproteinase inhibitor from látex of Carica papaya. European Journal of Biochemistry. 241,70-82, 1996.
ODANI, S.; IKENAKA, T. Studies on soybean trypsin inhibitor. (VII)-Dissulfide bridges in soybean Bowman-Birk proteinase inhibitor. Journal Biochemistry. 74(4), 697-715, 1973a.
ODANI, S.; IKENAKA, T. Scission of soybean Bowman-Birk proteinase inhibitor into two small fragments having either trypsin or chymotrypsin inhibitory activity. Journal Biochemistry. 74(4), 857-860, 1973b.
OLIVA, M.L., MENDES, C.R.,. JULIANO, M.A,. CHAGAS J.R, ROSA, J.C., GREENE L.J., SAMPAIO M.U., SAMPAIO C.A.M. Characterization of a tissue kallikrein inhibitor isolated from Bauhinia bauhinioides seeds: inhibition of the hydrolysis of kininogen related substrates, Immunopharmacology 45 ,163–169. 1999.
OLIVA, M. L.; MENDES, C. R.; SANTOMAURO-VAZ, E. M.; JULIANO, M. A.; MENTELE, R.; AUERSALD, E. A.; SAMPAIO, U. M.; SAMAPAIO, C. A. Bauhinia bauhinoides plasma kallikrein inhibitor: interaction with synthetic peptides and

Referências 92
fluorogenic peptide substrates related to the reactive site sequence. Current Medicinal Chemistry. 8, 977-984, 2001.
OLIVA, M. L. V; SOUZA-PINTO, J. C; BATISTA, I. F. C; ARAÚJO, M. S; SILVEIRA, V. F; AUERSWALD, E. A; MENTELE, R; ECKERSKORN, C; SAMPAIO, U. M; SAMPAIO, C. A. M. Leucaena leucocephala serine proteinase inhibitor: prymary structure and action on blood coagulation, kinin release and paw edema. Biochimica et Biophysica Acta. 1477, 64-74, 2000.
OLIVEIRA, C. M. Isolamento e caracterização parcial de inibidores de tripsina e quimotripsina de tubérculos de Colocasia esculenta. 2001. 100f. Dissertação (Mestrado em Bioquímica) - Universidade Federal do Rio Grande do Norte, Natal, 2001.
OLIVEIRA, A. S. Purificação e caracterização de inibidores de tripsina da família Kunitz de sementes de Pithecellobium dumosum e seus efeitos In vitro sobre insetos pragas. 2007. 157f. Tese (Doutorado em Bioquímica) - Universidade Federal do Ceará, Ceará, 2007.
OLIVEIRA, A. S.MIGLIOLO, L.AQUINO, R. O.RIBEIRO, J. K. C.MACEDO, L. L. P.ANDRADE,L. B. S.BEMQUERER, M. P.SANTOS, E. A.KIYOTA, S.SALES, M. P. Identification of a Kunitz-type proteinase inhibitor from Pithecellobium dumosum seeds with insecticidal properties and double activity. Journal of Agricultural and Food Chemistry. 55, 18, 7342-7349, 2007.
OLIVEIRA, A.S., XAVIER-FILHO, J., SALES, M.P. Cysteine proteinase and cystatins. Brazilian Archives Biology and Technology.46 (1):91-104.2003.
OSMAN, M. A.; REID, P. M.; WEBER, C. W. Thermal inactivation of tepary bean (Phaseolus acutifolius), soybean and lima bean protease inhibitors: effect of acidic and basic pH. Food Chemistry. v. 78, n. 4, p. 419-423, 2002.
PANDO, S. C; OLIVA, M. L. V; SAMPAIO, C. A. M; DI CIERO, L; NOVELLO, J. C; MARANGONI, S. Prymary sequence determination of a Kunitz inhibitor siolated from Delonix regia seeds. Phytochemistry. 57, 625-631, 2001.
PERNAS, M.; SANCHEZ-MONGE, R.; GÓMEZ, L.; SALCEDO, G. A. Chestnut seed cystatin differentially effective against cysteine proteinases from closely related pests. Plant Molecular Biology. 38, 1235–1242, 1998.

Referências 93
RAKWAL, R.,AGRAWAL, G.K.,JWA, N.S. Characterization of a rice (Oriza sativa L.) Bowman-Birk proteinase inhibitor: tightly light regulated induction in response to cut, jasmonic acid, ethylene and protein phosphatase 2A inhibitors. Gene,263: 189 198.2001.
REED, B.J., CHANDLER, D.S., SANDEMAN, R.M. Expression of the insecticidal bean alpha-amylase inhibitor transgene has minimal detrimental effect on the nutritional value of peas fed to rats at 30% of the diet. International Journal Parasitology. 29, 839– 850.1999.
RYAN, C. A. Protease inhibitor in plants: Genes for Improving defenses against insects and pathogens. Annual Review of Phytopathology. 28, 425-449, 1990.
RYAN, C. A. Proteolytic enzymes and their inhibitor in plants. Annual Version Plant Physiology. 24,173-196, 1973.
RYAN, C. A. Plant proteinases. The biochemistry of plants.Academic Press, New York, 1981.
RICHARDSON, M. The proteinase inhibitors of plants and microorganisms. Phytochemistry. 16, 1959-169, 1977.
RICHARDSON, M. Seed storage proteins: The enzyme inhibitor. In: Methods in plants Biochemistry. New York, Academic Press. 5, 259-305. 1991.
ROY, I., GUPTA, M.N. Purification of a double-headed inhibitor of alpha-anylase/proteinase K from wheat germ by expanded bed chromatography.Bioseparation, 9;239-245, 2000.
SALES, M. P.; GOMES, V. M.; FERNANDES, K. V. S.; XAVIER-FILHO, J. Chitin binding proteins from cowpea (Vigna unguiculata) seeds. Brazilian Journal of Medical and Biological Research. 29, 319-326, 1996.
SANTOS, I. S.; DA CUNHA, M.; MACHADO, O. L. T.; GOMES, V. M. A chitinase from Adenanthera pavonina L. seeds: purification, characterization and immunolocalization. Plant Science. 167, 6, 1203-1210, 2004.
SCHULER, T. H.; POPPY, G. M.; KERRY, B. R; DENHOLM, I. Insect-resistant transgenic plants. Trends in Biotechnology, 16, 168-175, 1998.

Referências 94
SHEE, C; SHARMA, A. K. Purification and characterization of a trypsin inhibitor from seeds of Murraya koenigii. Journal of Enzyme Inhibition and Medicinal Chemistry. 22(1), 115-120, 2007.
SHEWRY, P. R., LUCAS, J.A. Plant proteins that confer resistance to pest in pathogens. Advances in Botanical Research, 26: 135-192, 1997.
SILVA, C. P.; XAVIER-FILHO, J. Comparasion between the levels of aspartic and cysteine proteinases of the larval midguts of Callosobruchus maculatus (F.) and Zabrotes subfasciatus (Boh.)(Coleoptera: Bruchidae). Comparative Biochemistry and Physiology, part B: Biochemistry & Molecular Biology. 99, 529-533, 1991.
SILVA, C. B. L. F.; ALCAZER, A. A.; MACEDO, L. L. P.; OLIVEIRA, A. S.; MACEDO, F. P.; ABREU, L. R. D.; SANTOS, E. A.; SALES, M. P. Digestive enzymes during development of Ceratitis capitata (Diptera:Tephritidae) and effects of SBTI on its digestive serine proteinase targets. Insect Biochemistry and Molecular Biology, 36, 561-569, 2006.
SILVA, C. B. L. F. Identificação e Caracterização Parcial das enzimas digestivas de Ceratitis capitata (Díptera: Tephritidae) durante o desenvolvimento larval e no inseto adulto, 2003. 91f. Dissertação (Mestrado em Bioquímica) - Universidade Federal do Rio Grande do Norte, Natal, 2003.
SIN, S.F. & CHYE, M.L. Expression of proteinase inhibitor II proteins during floral development in Solanum americanum. Plant 219: 1010-1022. 2004.
SOUZA, E. M. T.; MIZUTA, K.; SAMPAIO, M. U.; SAMPAIO, C. M. Purification and partial characterization of a Schizolobium parahyba chymotrypsin inhibitor. Phytochemistry. 39, 521-525, 1995.
SRINIVASAN, A.; CHOUGULE, N. P.; ASHOK P. GIRI, A. P.; Gatehouse, J. A.; Gupta, V. S. Podborer (Helicoverpa armigera Hubn.) does not show specific adaptations in gut proteinases to dietary Cicer arietinum Kunitz proteinase inhibitor. Journal of Insect Physiology. 51, 1268–1276, 2005.
STANISÇUASKI,F. FERREIRA-DASILVA,C.T. MULINARI,F. . PIRES-ALVES M, CARLINI, C.R. Insecticidal effects of canatoxin on the cotton stainer bug Dysdercus peruvianus (Hemiptera: Pyrrhocoridae).Toxicon.45(6): 753-760.2005.

Referências 95
SUMATHI, S., PATTABIRAMAN, T. N. Natural plant enzyme inhibitors IV. Studies on trypsin inhibitors of Colocasia antiquorum tubers. Biochimica et Biophysica Acta. 566, 115-127, 1979.
SVENDSEN I., HANSEN S.I., HOLM J., LYNGBYE, J. The complete amino acid sequence of the folate-binding protein from cow's milk. Carlsberg Research Communications 49,123–131.1984.
TAMHANE, V. A.; CHOUGULE, N. P.; GIRI, A. P.; DIXIT, A. R.; SAINANI, M. N.; GUPTA, V. S. In vitro and in vivo effects of Capsicum annum proteinase inhibitors on Helicoverpa armigera gut proteinases. Biochimica et Biophysica Acta. 1722, 155–167, 2005.
TELANG, M. A.; SRINIVASAN, A.; PATANKAR, A. G.; HARSULKAR, A. M.; JOSHI, V.; DAMLE, A.; DESHPANDE, V. V.; SAINANI, M. N.; RANJEKAR, P. K.; GUPTA, G. P.; BIRAH, A.; RANI, S.; KACHOLE, M.; GIRI, A. P.; GUPTA, V. S. Bitter gourd proteinase inhibitors: potential growth inhibitors of Helicoverpa armigera and Spodoptera litura. Phytochemistry. 63, 643–652, 2003.
TELANG, M. A.; GIRI, A. P.; SAINANI, M. N.; GUPTA, V. S. Elastase-like proteinase of Helicoverpa armigera is responsible for inactivation of a proteinase inhibitor from chickpea. Journal of Insect Physiology. 51, 513–522, 2005.
TERRA, W.R., Evolution of digestive systems of insects. Annual Version Entomology. 35, 181-200.1990.
TERRA, W. R.; FERREIRA, C. Insect digestive enzymes: properties, compartmentalization and function. Comparative Biochemistry and Physiology, part B: Biochemistry & Molecular Biology. 109, 1-62, 1994.
TIFFIN, P., GAUT, B.S. Molecular evolution of the wound-induced serine protease inhibitor wip1 in Zea and related genera. Molecular Biology Evolucion. 18: 2092-2101. 2001.
TORNERO, P. CONEJERO, V. VERA, P. Identification of a new pathogen-induced member of the subtilisin-like processing proteases family from plants. Journal Biological Chemistry 2729(22), 14412-14419, 1997.
TURK, V., BODE, W. The cystatins: protein inhibitors of cysteine proteinases. FEBS Letters, 285, 2,213-219.1991.

Referências 96
UCHÔA, A. F., CARVALHO, C. D. S. C. OLIVEIRA, A E A, XAVIER-FILHO, J. Sementes e suas defesas contra insetos, Disponível em: http://www.jxavierfilho.hpg.ig.com.br/semente.htm (Acesso em 05 de julho de 2007).
VALUEVA, T. A.; REVINA, T. A; MOSOLOV, V. V. Potato tuber protein proteinase inhibitor belonging to the Kunitz soybean inhibitor family. Biochemistry. 62, 1367-1600, 1997.
VALUEVA, T. A.; REVINA, T. A.; KLADNITSKAYA, G. V.; MOSLOV, V. V. Kunitz-type proteinase inhibitors from intact and Phytophthora-infected potato tubers. FEBS Letters. 426, 131-134, 1998.
VALUEVA, T.A.; MOSOLOV, V.V. Reactive sites of the 21-kD protein inhibitor of serine proteinases from potato tubers. Biochemistry (Mosc), 64, 9, 1074-1078, 1999.
VAN DER HOORN, R.A.L; JONES, J.D.G.The plant proteolytic machinery and its role in defense. Current Opinion In Plant Biology 7: 400-4007,2004.
WALDER, J. M.M. Produção de moscas-de-frutas e seus inimigos naturais: associação de moscas estéreis e controle biológico. In: Controle biológico no Brasil: Parasitóides e predadores. José Roberto Pastolli Parra. Ed. Manole, São Paulo, 2002.
WALKER, A.J., FORD, L., MAJERUS, M. E.N., GEOGHEGAN, A.E., BIRCH,N., GATEHOUSE, J.A, GATEHOUSE A.M.R. Characterization of the mid gut digestive proteinase activity of the two-spot ladybird(Adalia bipunctata) and its sensitivity to proteinase inhibitors. Insect Biochemistry and Molecular Biology, 28,173-180, 1997.
WEBER, K., OSBORNE, M. The reability of molecular weight determination by sodium dodecil sulfate polyacrilamide gel eletroforesis. Journal of Biological Chemistry, 244, 4406-4412.1969.
WEDER, J. K. P. Protease inhibitors in the leguminosae. In: Advances in Legumes Systematics (Pholhil, R. M. & Roven, P.H. eds.). Royal Botanic Gardens, Kew, Richmond, Surrey. TW9 3AE, England.1981.
XAVIER-FILHO, J. The Biological roles of serine and cysteine proteinase inhibitors in plants. Revista Brasileira de Fisiologia Vegetal. 4, 1-6, 1992.

Referências 97
XAVIER-FILHO, J. Sementes e suas defesas contra insetos. Projeto Multinacional de Biotecnologia e Alimentos. Organização dos Estados Americanos-OEA. 1-31. 1993.
XAVIER-FILHO, J.; VENTURA, M.M. Trypsin inhibitors in cowpea: A review. Journal Agriculture and food chemistry, 5: 239-264, 1988
XU, Z.F.; QI, W.Q.; OUYANG, X.Z.; YEUNG, E.; CHYE, M.L. A proteinase inhibitor II of Solanum americanum is expressed in phloem. Plant Molecular Biology. 47: 727-738. 2001.
ZENG, F., ZHU, Y. C., COHEN, A. C. Molecular cloning and partial characterization of a trypsin-like protein in salivary glands of Lygus Hesperus (Hemiptera:Miridae). Insect Biochem Molecular Biology. 32(4): 455-464, 2001.