universidade federal de sÃo carlos · la criptococosis es una micosis oportunista, adquirida por...
TRANSCRIPT

I
CARACTERIZACIÓN GENÓMICA Y PROTEÓMICA DE AISLADOS
AMBIENTALES Y CLÍNICOS DE Cryptococcus gattii y C. neoformans EN CÚCUTA,
NORTE DE SANTANDER, COLOMBIA.
ASBLEIDE KARINA ANGARITA SANCHEZ
17861018
UNIVERSIDAD DE SANTANDER
FACULTAD DE CIENCIAS DE LA SALUD
MAESTRÍA EN INVESTIGACIÓN EN ENFERMEDADES INFECCIOSAS
BUCARAMANGA
2019

II
CARACTERIZACIÓN GENÓMICA Y PROTEÓMICA DE AISLADOS
AMBIENTALES Y CLÍNICOS DE Cryptococcus gattii y C. neoformans EN CÚCUTA,
NORTE DE SANTANDER, COLOMBIA.
Proyecto de grado presentado como parte de los requisitos para la obtención del título de
magister en investigación en enfermedades infecciosas
Tutor: PhD. Claudia Marcela Parra Giraldo
Co-Tutor: MSc. Denny Miley Cárdenas Sierra
UNIVERSIDAD DE SANTANDER
FACULTAD DE CIENCIAS DE LA SALUD
MAESTRÍA EN INVESTIGACIÓN EN ENFERMEDADES INFECCIOSAS
2019

III

IV
___________________________________
Tutor
DRA. CLAUDIA MARCELA PARRA GIRALDO

V
Dedicatoria:
A mis hijos Julián y Esteban.
A mis padres, por sus esfuerzos, trabajo y lucha constante.

VI
AGRADECIMENTOS
A Dios por la vida y fuerza para seguir siempre adelante.
A mis padres por su apoyo constante y ejemplo de perseverancia.
A Julián y Esteban por sus risas, amor y energía.
A mi tutora Claudia Marcela Parra, a mi asesora Científica Patricia Escandón y cotutora
Denny Cárdenas por su confianza y tiempo.
A Alba Rojas y Carlos Moncada por su apoyo y porque como abuela y padre cuidaron
de mis hijos durante mis extensas jornadas de estudio y viajes constantes en mi proceso
de formación.
A la Universidad de Santander y a cada uno de quienes hacen parte de ella que siempre
abrieron sus puertas durante estos dos años.
Al cuerpo docente, especialmente a la Dra. Liliana Torcoroma por su excelente labor y
constante apoyo.

VII
TABLA DE CONTENIDO
INTRODUCCIÓN 1
2.MARCO TEÓRICO 4
2.1 CICLO BIOLOGICO Cryptococcus neoformans/C. gattii 4
2.2 FACTORES DE VIRULENCIA Y EVASIÓN DEL SISTEMA INMUNE 5
2.3 TAXONOMIA Cryptococcus neoformans/C. gattii 6
2.4 CRIPTOCOCCOSIS 8
2.5 IDENTIFICACION MOLECULAR 10
2.5.1 PCR para la identificación de hongos patógenos a nivel de especie 11
2.5.2 Identificación genotípica de tipos moleculares de C. neoformans y C. gattii 13
2.5.3 Identificación proteómica de tipos moleculares de C. neoformans y C. gattii 15
3.OBJETIVOS 18
3.1 OBJETIVO GENERAL 18
3.2 OBJETIVOS ESPECÍFICOS 18
4.MATERIALES Y MÉTODOS 19
4.1. POBLACIÓN 19
4.2 PROCESAMIENTO DE MUESTRAS AMBIENTALES 20
4.3 PROCESAMIMENTOS DE AISLADOS CLÍNICOS 23
4.4 EXTRACCIÓN DE ADN 23
4.5 DETERMINACIÓN DE PATRÓN MOLECULAR PCR-HUELLA DIGITAL 24
4.6 DETERMINACIÓN DEL PATRÓN MOLECULAR POR RFLP- GEN URA5 25
4.7 CARACTERIZACIÓN PROTEOMICA POR MALDI TOF 26
4.8 ANÁLISIS MOLECULAR: 28

VIII
5. RESULTADOS 30
5.1 CARACTERIZACIÓN DE MUESTRAS AMBIENTALES 30
5.2 CARACTERIZACIÓN DE MUESTRAS CLÍNICAS 33
5.3 RELACIÓN GENÉTICA ENTRE LOS AISLADOS DE c. neoformans 35
6. DISCUSIÓN 37
7. CONCLUSIONES 44
8. RECOMENDACIONES 45
BIBLIOGRAFIA 46
ANEXOS 49

IX
LISTA DE FIGURAS
Figura 1 Modelo del ciclo de vida de Cryptococcus neoformans (Voelz, 2010). .............. 4
Figura 2 Preparación en tinta china 100X. Realizada en el Laboratorio LIIBAAM de la
Universidad de Santander. 2a C. neoformans aislamiento clínico. 2b tamaño
capsular alcanzado post-inoculación en Galleria mellonella.(Angarita A 2019) ...... 5
Figura 3 Principales componentes implicados en la virulencia de Cryptococcus
neoformans (Patcharin 2018) ..................................................................................... 6
Figura 4 Estructura de la región del gen rRNA, mostrando las posiciones de los
iniciadores de PCR. ................................................................................................. 12
Figura 5: Posición del exón estudiado de la Gen URA5. Un fragmento de 345 pb del
exón. (posiciones 1145–1492) ................................................................................ 15
Figura 6 Transcripción, traducción y modificación postraduccional y/o proteólisis de las
proteínas (Cazzulo). ................................................................................................. 16
Figura 7. Ubicación geográfica de la ciudad de Cúcuta en el Departamento Norte de
Santander, en donde se obtuvieron los aislamientos ............................................... 20
Figura 8. Cryptococcus spp. Agar semilla de girasol (Producción de melanina).
Universidad de Santander, Cúcuta. .......................................................................... 22
Figura 9 a. Pruebas de Urea positiva para Cryptococcus spp. b. Control positivo y
control negativo. Universidad de Santander, Cúcuta. .............................................. 22
Figura 10 Prueba de CGB para identificación diferencial de C. neoformans y C. gattii.
Universidad de Santander, Cúcuta. .......................................................................... 23

X
Figura 11Cryptococcus spp muestra de líquido cefalorraquídeo en tinta china. Instituto
Nacional de Salud, Colombia). ................................................................................ 23
Figura 12. Flujograma de identificación por PCR- Huella digital. Universidad de
Santander, Cúcuta. ................................................................................................... 25
Figura 13 Descripción de la técnica de MALDI TOF MS.(Croxatto et al., 2012). ......... 27
Figura 14. Prevalencia para Cryptococcus sp en áreas públicas de Cúcuta. .................... 32
Figura15. PCR huella digital de aislados clínicos (CL) y ambientales de Cryptococcus
neoformans var. grubii usando el cebador (GTG), con marcador de peso molecular
1kb y cepas de control. ............................................................................................. 34
Figura 16. Espectros de masas obtenidos en el MALDI TOF para Cryptococcus
neoformans VNI y Cryptococcus gattii VGII. ......................................................... 34
Figura 17. Dendrograma de aislamientos clínicos y ambientales. BioGalaxy module of
the Biolomics program, versión 3.0 ......................................................................... 36
Figura 18. Dendograma de análisis proteómico de aislamientos clínicos y ambientales.37

XI
LISTA DE TABLAS
Tabla 1 Clasificación grupos moleculares 7
Tabla 2 Descripción de muestras positivas para C.neoformans y C.gattii 31
Tabla 3 Descripción de muestras clínicas positivas para C,neoformans VNI 34

XII
LISTA DE ABREVIATURAS
ADN: Ácido desoxirribonucleico.
AFLP: “Amplified Fragment Length Polymorphism” Fragmento amplificado de longitud
polimórfica
AP-PCR: Reacción en cadena de la polimerasa con oligonucleótidos arbitrarios.
CGB: “L-Canavanina-Glicina- Azul de bromotimol”
dNTP: “Deoxynucleoside triphosphates” desoxirribonucleótidos trifosfato “Matrix-
Assisted Laser Desorption/Ionization” desorción/ionización láser asistida por matriz.
MLST: “Multilocus séquenme tilinga” Tipificación Secuencia Mutilocus
pb: acrónimo de pares de bases.
PCR: “Polimerice Chin Recatico” Reacción en cadena de la polimerasa.
RAPD: amplificación de ADN polimórfico al azar
RFPL: “Restrictos Fragment Length Polymorphism” Fragmentos de restricción de
longitud polimórfica.
SDA: “Saboureaud Dextrose Agar” Medio de cultivo semisintético para levaduras
SIDA: Síndrome de la inmunodeficiencia adquirida.
TOF: “Time-Of-Flight” tiempo de vuelo.
URA 5: Gen de levaduras que codifica para la orotidina monofosfato pirofosforilasa.
Var: Variedad.
VIH: Virus de la inmunodeficiencia humana.

XIII
RESUMEN
Título: CARACTERIZACIÓN GENÓMICA Y PROTEÓMICA DE AISLADOS
AMBIENTALES Y CLÍNICOS DE Cryptococcus gattii y C. neoformans EN CÚCUTA,
NORTE DE SANTANDER, COLOMBIA
Autor: Asbleide Karina Angarita Sánchez
Palabras claves: Criptococosis, genómica, proteómica, prevalencia, hábitat.
Descripción
La Criptococosis es una micosis oportunista, adquirida por la inhalación de
propágulos fúngicos presentes en el ambiente, siendo una infección potencialmente fatal
que afecta los pulmones y el sistema nervioso central, tanto en personas inmunosuprimidas
como inmunocompetentes, causada por dos especies patógenas para humanos. El Grupo
Colombiano para el estudio de la Criptococosis ha reportado aislamientos de
Cryptococcus neoformans y C. gattii tanto a partir de muestras clínicas como ambientales
en la ciudad de Cúcuta, considerado un municipio relevante respecto al reporte de casos
de ambas especies, en encuestas nacionales. El objetivo de este estudio fue establecer la
concordancia entre relación genética y proteómica de aislados clínicos y ambientales de
C. neoformans y C. gattii en la ciudad de Cúcuta, se colectaron un total 1300 muestras de
446 árboles de 10 especies diferentes a partir de 10 zonas públicas; adicionalmente, se
obtuvieron 6 aislamientos clínicos de C. neoformans durante junio de 2016 a junio de
2017, provenientes de la Empresa Social del Estado Hospital Universitario Erasmo Meoz,
los cuales fueron cultivados en agar semillas de Guizottia abysinica para identificación
fenotípica por pruebas bioquímicas y se realizó tipificación genética por PCR-huella
digital, RFLP-URA5 y caracterización proteómica empleando MALDI TOF MS. Se
obtuvieron 19/446 (4,3%) árboles positivos para aislamiento de C. neoformans y 1/446
(0,2%) para C. gatti en la población arbórea evaluada para un total de 21 aislamientos (dos
de un mismo individuo), de los cuales 10/21 (47,6%), correspondientes a C. neoformans,
provenían del Parque Santander y el único aislado de C. gattii provenía del parque
Mercedes Ábrego. Con relación a los aislamientos clínicos 6/6(100%) fueron
identificados como C. neoformans var. grubii VNI.

XIV
ABSTRACT
Title: GENOMIC AND PROTEOMIC CHARACTERIZATION OF
ENVIRONMENTAL AND CLINICAL ISOLATES OF Cryptococcus gattii and C.
neoformans IN CÚCUTA, NORTH OF SANTANDER, COLOMBIA
Author: Asbleide Karina Angarita Sánchez
Key words: Cryptococosis, genomics, proteomics, prevalence, habitat.
Description
Cryptococosis is an opportunistic mycosis, acquired by the inhalation of fungal propagules
present in the environment, being a potentially fatal infection that affects the lungs and
the central nervous system, is that affects immunocompetent and immunodeficient
individuals, caused by two pathogenic species for humans. The Colombian Group for the
study of Criptococosis has reported isolates of Cryptococcus neoformans and C.gattii in
both clinical and environmental samples in the city of Cúcuta. considered a relevant city
regarding the reporting of cases of both species, in national surveys. The objective of this
study las to establish the concordance between the genetic and proteomic relationship of
clinical and environmental isolates of Cryptococcus neoformans and Cryptococcus gattii
in the city of Cúcuta. A total of 1300 samples were collected from 446 trees of 10 different
species from 10 public areas; additionally, they were obtained 6 clinical isolates of
Cryptococcus neoformans. were obtained during June 2016 to June 2017, from Hospital
Universitario Erasmo Meoz were cultivated in Guizottia abysinica medium and were then
biochemically identified and genetic typing by Fingerprinting PCR, RFLP of the URA5
gene and proteomics by MALDI TOF MS. This yielded 19/446 (4.3%) positive trees for
isolation of C. neoformans and 1/446 (0.2%) for C. gatti, the tree population evaluated for
a total of 21 isolates (two from the same individual), of which 10/21 (47.6%),
corresponding to C. neoformans, came from the Santander Park and the only one isolated
from C. gattii came from the Mercedes Ábrego Park. In relation to clinical isolates 6/6
(100%) were identified as C. neoformans var. grubii VNI.

1
INTRODUCCIÓN
La Criptococosis es una micosis oportunista, adquirida por la inhalación de
propágulos fúngicos presentes en el ambiente. Constituye una infección
potencialmente fatal que afecta a los pulmones y el sistema nervioso central, tanto en
individuos inmunosuprimidos como inmunocompetentes (Lin & Heitman, 2006;
Perfect, 2007; Perfect et al., 2010; Ye et al., 2012), siendo su agente causal la levadura
capsulada Cryptococcus sp., con sus dos especies patógenas C. neoformans y C. gattii.
la primera, se ha descrito la variedad grubii (correspondiente al serotipo A, patrones
moleculares VNI y VNII), la variedad neoformans (correspondiente al serotipo D,
patrón VNIV) y el híbrido serotipo AD con el patrón molecular VNIII. Por su parte,
Cryptococcus gattii comprende los serotipos B y C (patrones moleculares VGI-VGIV)
(Kwon-Chung, 2014).
En el año 2015 se publicó una revisión de la taxonomía de ambos complejos,
con una nueva propuesta incluyendo siete especies: C. neoformans (C.neoformans var
grubii) C. deneoformans(C.noeformans var neoformans), C.gattii(C.gattii VGI),
C.bacillisporus(C.gattii VGIII),C. deuterogattii (C.gattii VGII), C. tetragattii(C.gattii
VGIV) y decagattii (C.gattii VGIVc) ( (Hagen et al., 2015).
La mayor frecuencia de reportes corresponde a VNI y VNII este último con
menor proporción en el número de aislamientos(Cattana et al., 2014; Mak, Vélez,
Castañeda, & Escandón, 2015; Nweze, Kechia, Dibua, Eze, & Onoja, 2015), en
Estados Unidos y Europa, la variedad molecular VNIV es mayor que en otras áreas
geográficas, en el noreste pacífico de Estados Unidos y en Vancouver, la mayor
frecuencia reportada se relaciona con VGII, este tipo molecular también reportado en

2
Brasil, Colombia, Australia y algunos casos en Europa(Campbell, Fraser, et al., 2005;
Cogliati, 2013) Por su parte, el tipo molecular VGIII ha sido recuperado en México,
Colombia y Estados Unidos. La mayor frecuencia de VGI se ha descrito en Oceanía
seguida por Asia, América y Europa y el tipo Molecular VGIV ha sido aislado en
África, India Colombia y México(Campbell, Currie, et al., 2005; Firacative, Trilles, &
Meyer, 2017; Meyer et al., 2011).
C. neoformans var. grubii patrón molecular VNI es el principal agente
etiológico de la Criptococosis en Colombia; sin embargo, se ha descrito que Cúcuta
presenta una alta prevalencia para C. gattii patrón molecular VGII, el cual se asocia
con alta virulencia y su potencial para generar brotes (Lizarazo, Escandón, Agudelo,
Firacative, et al., 2014) , así, aunque la infección por C. gattii es minoritaria en
Colombia respecto a C. neoformans (serotipos A y D), resulta preocupante la
prevalencia en la ciudad de Cúcuta, Norte de Santander, la cual llega al 60% en
pacientes VIH negativos(Lizarazo, Escandón, Agudelo, & Castañeda, 2014)
La incidencia de la Criptococosis en la población colombiana es de 0,23 casos
por cada 100.000 habitantes, siendo Norte de Santander el departamento con mayor
tasa de incidencia con 0,56 casos por cada 100,000 habitantes, lo que justifica la
importancia de estudios con aislamientos clínicos y ambientales de Cryptococcus, en
el departamento.(P. Escandón, J. Lizarazo, C. Agudelo, & E. Castañeda, 2018a)
En la ciudad de Cúcuta, C.gattii serotipo C fue aislado por primera vez a partir
del ambiente en el año 1998, este aislamiento es el primer reporte de C.gattii serotipo
C en el ambiente (Callejas, Ordonez, Rodriguez, & Castaneda, 1998) y
posteriormente, en el año 2011 se obtuvo el primer aislamiento ambiental de C. gattii
serotipo B(VGIa) en esta misma ciudad, Este informe confirmó la presencia de C.

3
gattii en un ecosistema terrestre, y aunque el aislado recuperado era molecular tipo
VGI, en contraposición a los aislados clínicos que se han recuperado en Cúcuta, que
son el tipo molecular mayoritario VGII , este hallazgo fue de gran importancia en el
estudio de la ecoepidemiología de C.gattii (Lizarazo et al., 2012). Los aislamientos de
Cryptococcus neoformans y Cryptococcus gattii a partir de muestras ambientales han
tomado importancia a nivel mundial debido a la identificación de nuevos nichos
ecológicos, y así mismo el reconocimiento a diversos focos de infección siendo estos
de gran impacto para la población, es importante identificar la relación entre los
asilamientos clínicos y los ambientales para intervenciones que permitan contribuir en
las medidas de pronostico control y manejo de los focos de infección.
Teniendo en cuenta la alta incidencia referida de Criptococosis en la ciudad de
Cúcuta, el objetivo del presente fue caracterizar genética y proteomicamente aislados
ambientales y clínicos de Cryptococcus gatti, C. neoformans en Cúcuta, Norte de
Santander, Colombia, buscando aportar información que adicionalmente permita en
un futuro a las autoridades sanitarias y demás entidades competentes, tomar medidas
preventivas en cuanto a las normas de bioseguridad adoptadas por comunidad
susceptible en zonas de riesgo y de alta concurrencia, las cuales sean consideradas
focos ambientales del hongo en el municipio de San José de Cúcuta, ante la posible
relación entre los aislados de procedencia clínica y ambiental.

4
2. MARCO TEORICO
2.1 CICLO BIOLÓGICO Cryptococcus neoformans/C. gattii
Cryptococcus spp es una levadura capsulada cuya fase teleomorfa se denomina
Filobasidiella (K. Kwon-Chung, 1975), Fillobasidiella neoformans var neoformans
se deriva de Cryptococcus neoformans y Fillobasidiella neoformans var bacillispora,
se deriva de Cryptococcus gattii(K. J. Kwon-Chung et al., 2014). Se encuentran en el
ambiente, principalmente en árboles y suelos contaminados con heces de aves, entre
las cuales sobresale la paloma urbana (Columba livia) es la más importante como
reservorio del hongo.(Timarán et al., 2016)
El ciclo de vida inicia cuando se fusiona una blastoconidia (α) y una blastocodia
(a) dando como resultado un filamento dicariótico, seguido de división y formación de
una blastospora, de donde puede germinar la hifa que se divide mitóticamente en la
forma de levadura; algunas hifas pueden formar clamidiosporas. En el estado basidio
los dos núcleos se someten a meiosis y producen cuatro células meióticas que forman
cadenas de basiodiosporas por mitosis, brotando de la superficie de la basidia (Lin &
Heitman, 2006). Figura 1.
Figura 1 Modelo del ciclo de vida de Cryptococcus neoformans (Voelz, 2010).

5
2.2 FACTORES DE VIRULENCIA Y EVASIÓN DEL SISTEMA
INMUNE
Cryptococcus spp mide 5-7µm de diámetro, pero cuando está en el hospedero
su tamaño puede aumentar drásticamente hasta llegar a medir >25 µm (Dykstra,
Friedman, & Murphy, 1977) debido a su cápsula, la cual tiene una composición de
glucuronoxilomanano, galactoxilomanano, y manoproteínas, estudios recientes
muestran como las células titanic (tamaño capsular hasta 25 micras) presentan
elevados niveles de quitina que son reconocidas por las quitinasas del huésped lo que
induce un disminución de la respuesta inmune adaptativa; adicionalmente, las células
más viejas presentes en la infección inicial denominadas células fundadoras, tienen
mayor capacidad de resistir tanto la fagocitosis como a los fármacos. (Zaragoza et al.,
2009)Figura 2 .
Figura 2 Preparación en tinta china 100X. Realizada en el Laboratorio LIIBAAM de
la Universidad de Santander. 2a C. neoformans aislamiento clínico. 2b tamaño
capsular alcanzado post-inoculación en Galleria mellonella.(Angarita A 2019)
Además de la cápsula, los principales factores de patogenicidad los constituyen
la presencia de melanina, y enzimas lacasas, ureasas y fosfolipasa B, puesto que están
implicados en la adaptación y en el proceso infectivo del hongo, además de estar
a b

6
implicados en la evasión del sistema inmunológico del hospedero (Alspaugh, 2015).
Figura 3.
Estudios comparativos entre aislamientos clínicos y ambientales han permitido
evidenciar la mayor producción de enzima fosfolipasa entre los aislamientos clínicos
lo cual permite evidenciar que esta enzima puede estar implicada en la patogénesis de
la Criptococosis. (Pini, Faggi, & Campisi, 2017)
Otra característica importante de Cryptococcus es la producción de melanina
la cual para su formación requiere compuestos exógenos difenólicos como la L
DOPA, la síntesis de la melanina depende de la enzima difeniloxidasa la cual se
codifica por los genes LAC1 y LAC2(Zaragoza, 2019),
Cryptococcus sintetiza grandes cantidades de ureasas, compuesto importante
por su capacidad de catalizar la hidrolisis de la urea a amoniaco, la actividad de la
ureasa se ha relacionado con la patogénesis de la Criptococosis (figura 3).
Polisacáridos
Melanina
Ureasas
Figura 3 Principales componentes implicados en la virulencia de Cryptococcus
neoformans (Patcharin 2018)
2.3 TAXONOMIA Cryptococcus neoformans/C. gattii
El género Cryptococcus abarca alrededor de 37 especies. Filogenéticamente
presenta la siguiente clasificación taxonómica: Organismo celular Eukaryota,

7
opiskonta, reino Fungi, filum: Basidiomycota, subfilum: Agaricomycotica, clase:
Tremellomycetes, orden; Tremellales. familia en su fase anamorfo Cryptococcacea y
en la fase teleomorfo Filobasidiaceae de las cuales las especies C. neoformans, C.gattii
han mostrado ser patógenos para el ser humano(Kurtzman, Fell, Boekhout, & Robert,
2011; Tello et al., 2013). El complejo de especies C. neoformans-C. gattii, incluye
dos especies anamórficas (K. J. Kwon-Chung et al., 2014).
La primera presenta dos variedades: Cryptococcus neoformans var. grubii
(comprendiendo el serotipo A, con subgrupos genéticos VNI y VNII) y Cryptococcus
neoformans var. neoformans (comprendiendo por el serotipo D, con subgrupo genético
VNIV); éstas presentan un híbrido, correspondiente al serotipo AD y subgrupo
genético VNIII. Por su parte, Cryptococcus gattii comprende los serotipos B
(subgrupos genéticos VGI y VGII) y serotipo C (subgrupos genéticos VGIII y VGIV)
(K. J. Kwon-Chung et al., 2014; Rohatgi & Pirofski, 2015).
Estudios recientes proponen reconocer a C. neoformans var. grubii como
deneoformans y C. neoformans var neoformans como C. neoformans y cinco especies
dentro C. gattii las cuales corresponden así: C.gatti a VGI, C.bacillisporus a VGIII,
C.deuterogattii a VGII, C.tetragattii VGIV y C.decagattii (Hagen et al., 2015)
Tabla 1. Clasificación grupos moleculares (Escandón, 2010)

8
C.neoformans y C. gattii habitan naturalmente en suelo, aire y árboles, siendo
mucho mayor la prevalencia ambiental de C. neoformans, especialmente C.
neoformans var grubii (serotipo A), cuyo habitat se extiende también a excretas de
aves.(Chowdhary, Randhawa, Prakash, & Meis, 2012). C. gattii estaba clasificado
como un microorganismo restringido a zonas tropicales y subtropicales, sin embargo,
en el año 2000 se reportó la primera epidemia de Criptococosis en el mundo por C.
gattii, en la isla de Vancouver, Canadá, la cual afectó a humanos y animales
inmunocompetentes, generando un incremento inusual en la incidencia de
casos(Stephen, Lester, Black, Fyfe, & Raverty, 2002)
2.4 CRIPTOCOCCOSIS
La vía de entrada del agente es mediante inhalación de esporas, causando
enfermedad pulmonar, aunque el agente puede diseminarse mediante vía sanguínea y
alojarse en el Sistema Nervioso Central; además se ha referido infección en piel, tejidos
blandos y tracto genitourinario (P. Escandón, J. Lizarazo, C. I. Agudelo, & E.
Castañeda, 2018b; Firacative et al., 2017; Perfect & Bicanic, 2015). Los casos de
Criptococosis en las diferentes presentaciones clínicas (Bese 1987) incrementan
paralelamente al aumento de casos de pacientes con SIDA (Nakanishi, Almeida, &
Romiti, 2006).
La meningitis criptococócica corresponde a la micosis más común causada por
el género Cryptococcus en pacientes VIH positivos, a modo de meningoencefalitis
subaguda (recuento de linfocitos T CD4 inferior a 100 células/µl) (Williamson et al.,
2017). La incidencia global de la Criptococosis meníngea es de 223,100 casos anuales.
(IC 95% 150600–282400), lo que resultó en 181,100 (IC 95% 119400-234300)

9
muertes anuales en el año 2014, con elevada mortalidad asociada en países en vía de
desarrollo (entre 35% y 65% de los casos), o incluso hasta el 70%, particularmente en
África sub Sahariana (Lessells, Mutevedzi, Heller, & Newell, 2011; Rajasingham et
al., 2017).
La Criptococosis pulmonar: es la presentación más común de la Criptococosis
puede afectar a personas tanto inmunocompetentes como inmunocomprometidos
siendo poco frecuentes en los primeros, las manifestaciones clínicas pulmonares
abarcan desde hallazgos radiológicos asintomáticos hasta trastornos agudos. Pueden
presentarse nódulos e infiltrados.(Gal, Koss, Hawkins, Evans, & Einstein, 1986;
Guilarde, Andrade, De Sousa, De Oliveira, & Sugita, 2019).
La Criptococosis cutánea: puede ser de tipo primaria o secundaria en el caso de
la primera solo se presentan manifestaciones en piel y en ausencia de enfermedad
diseminada este tipo de Criptococosis se asocia con personas expuestas a heces de
palomas (Beatson, Harwood, Reese, & Robinson-Bostom, 2019; Valenzuela-Oñate et
al., 2019) en el caso de la Criptococosis secundaria se presenta diseminación
hematógena del hongo a l piel(Noguchi et al., 2016)
Según la encuesta nacional de Criptococosis, la presentación clínica de ésta
micosis en Colombia muestra como principal síntoma clínico el dolor de cabeza en el
73,3% de los casos, fiebre en el 53,3%, náuseas y vomito en el 48,6% y confusión en
el 40,3% de los casos. El 46,1% de los pacientes se reporta manifestaciones
neurológicas (rigidez de cuello, hidrocefalia, hipertensión intracraneal y
convulsiones), con una tasa de mortalidad alta en 290/610 casos (47,5%), mortalidad

10
temprana, en el 44,3%, de la población entre 26 y 40 años de edad y con el mayor
registro de letalidad para los pacientes con SIDA (48,1%) (Escandón et al., 2018b).
2.5 IDENTIFICACION MOLECULAR
Los métodos de identificación molecular han permitido grandes avances en el
estudio de las enfermedades infecciosas mediante el conocimiento del genoma de los
microrganismos aportando al estudio a nivel clínico de las enfermedades, sin
embargo, es claro que el estudio completo de las células requiere además del estudio
genotípico, el estudio fenotípico celular y su proteóma entendiéndose este término
como el conjunto de proteínas que un organismo expresa es así como el ADN
mantiene la información genética de generación en generación mediante la replicación
y esta información se expresa al transcribirse al ARN mensajero el cual se traduce a
proteínas (Wilkins et al., 1996) de ahí la importancia de la integración de estas dos
disciplinas como avance en la identificación molecular de los microrganismos.
La PCR es hoy en una herramienta fundamental en el abordaje de la
Criptococosis y es el método más ampliamente utilizados en los laboratorios de
biología molecular y en el diagnóstico clínico. La aplicación de esta tecnología ha
aumentado, este tipo de técnicas han aportado grandes avances en el estudio de la
epidemiologia molecular de la Criptococosis.
Una gran variedad de genes se utilizan actualmente para el estudio filogenético
en hongos siendo el grupo (cluster) de genes ADN ribosomal (ADNr) el que se usa
más comúnmente para identificación de taxonomía y posterior identificación de
microrganismos llegando a la especie, existen otras técnicas como lo es la PCR
modificada, conocida amplificación de ADN polimórfico (RAPD) PCR anillamiento

11
arbitrario (AP-PCR) o PCR Huella digital, en donde los perfiles de ADN multilocus
generados permiten la identificación de la especie así como relación de las diferentes
cepas de una especie(Fernández-Cuenca, 2004).
Por su parte la proteómica como herramienta basada en la identificación y
análisis de proteínas permite evaluar el fenotipo, generando identificaciones
confiables, además, permite entender los múltiples procesos celulares por los que pasa
la célula en un momento determinado. Procesos que son en su mayoría regulados por
genes, los cuales son estudiados desde la genómica. En este estudio hicimos uso del
MALDI TOF MS para la identificación de las levaduras y para llevar a cabo análisis
de los espectros obtenidos.
2.5.1 PCR para la identificación de hongos patógenos a nivel de especie
Aunque ambas especies de Cryptococcus se pueden diferenciar por métodos
fenotípicos, Existen métodos moleculares que pueden contribuir a la identifación de
estas y sobre todo a diferenciar con otros hongos con mayor sensibilidad y
especificidad.
Amplificación de ADN ribosomal (ADNr) Son varios los genes utilizados
para estudios filogenéticos en hongos, el usado con mayor frecuencia es el cluster de
genes ADN ribosomal (ADNr) por las siguientes características:
- Es un gen multicopias en la mayoría de los casos, por lo cual se necesitan
pequeñas cantidades de ADN templado.
- Contiene genes altamente conservados de ADNr que codifican la
subunidad grande y pequeña del ARN ribosomal.

12
- Contienen regiones altamente variables llamadas Internal Transcribed
Sparcers (ITS) que a diferencia del ADNr estas regiones pueden acumular
cambios entre organismos estrechamente relacionados.(Meyer, 2014).
- Los genes de Arna 17s, 5.8s y 25s, son en esencia idénticos en todas las
especies. Sin embargo, la longitud de la región ITS (internal transcribed
spacer, o espaciador interno del transcripto) depende de la especie. Las ITS
se localizan entre los genes de ARNr 17s y 25s, y está dividida en la región
ITS1 entre los genes 17s y 5.8s; y la región ITS2, entre los genes ARNr
5.8s y 25s. Por esta razón, son secuencias que se han utilizado como
marcadores moleculares para la identificación de géneros y especies de
hongos(Vilgalys & Hester, 1990).Figura 4.
Figura 4 Estructura de la región del gen rRNA, mostrando las posiciones de
los iniciadores de PCR.

13
2.5.2 Identificación genotípica de tipos moleculares de C. neoformans y
C. gattii
La identificación de los tipos moleculares es un gran aporte a la epidemiologia
molecular de la Criptococosis que ha permitido identificar los factores de riesgo
ambientales y distribución entre poblaciones representando importantes
contribuciones a la salud pública.
Para la identificación a nivel intraespecífico se han utilizados varias pruebas
genotípicas incluyendo PCR huella digital con iniciadores como M13, (GACA)₄ y
(GTG)₅, polimorfismo de los fragmentos de restricción(RFLP), polimorfismo de
longitud de los fragmentos amplificados (AFLP) de los genes PBLₗ y URA₅,
tipificación de multilocus microsatelite (MLMT) y tipificación de secuencias
moltilocus (MLST) en la cual se emplean los genes: CAP59, GPD1, LAC1, PLB1,
SOD1, URA5 y la región /GS1 todos estos métodos de tipificación han identificados
los principales 8 tipos moleculares.(Tabla 1)
PCR Huella digital para la identificación de especies y caracterización de
cepas: esta técnica PCR huella digital se utilizan oligonucleótidos que son
específicos minisatelites o secuencias microsatelites y que fueron utilizados
inicialmente como sondas de hibridación en la huella digital de ADN convencional,
debido a la alta especificidad (temperatura de anillamiento 50°C) del iniciador
utilizado la técnica se combinan las técnicas ADN huella digital con la PCR en donde
el polimorfismo de la banda obtenida puedes usarse para estimar la relación de
diferentes cepas de una especie(Meyer, Mitchell, Freedman, & Vilgalys, 1993)
La amplificación de ADN polimórfico al azar ( RAPD) y PCR con
oligonucleótidos arbitrarios (AP-PCR) son métodos utilizados para amplificar

14
fragmentos de ADN desconocidos y polimórficos, en ambas técnicas se emplean
iniciadores cortos a temperatura de anillamiento (36°C) generalmente con 40 ciclos
de amplificación, estas técnicas permite detectar la secuencia DNA repetida e
hipervariable, la baja especificidad del iniciador no discrimina a numerosos sitios en
cualquier genoma, por lo cual se hizo necesario la utilización de método Southern con
una categoría de polimorfismo de repetición debido a diferencias en el número de
copias de secuencias repetidas o tándem estos son los minisatélites o microsatélites
se han empleado con éxito para amplificar secuencias hipervariables interrepetidas en
varios genomas de hongos. Los cebadores que se han utilizado para caracterización de
Cryptococcus han sido un que detecta secuencia minisatelite, un oligonucleótido
(GAGGGTGGCGGTTCT) de la secuencia de fago M13 y cebadores que detectan
secuencias de ADN microsatélite: (GTG)₅ y (GACA) ₄(Meyer et al., 1993).
Polimorfismo de longitudes de los fragmentos de restricción (RFPL)
URA5: Los RFLP (Restriction Fragment Length Polimorphism) se define como
variantes individuales de las secuencia de ADN y pueden presentar sobre secuencias
de genes o extragenicas para su evidencia se utilizan enzima de restricción, estas
provienen de bacterias y su función en este método es unirse a sitios específicos de la
cadena de ADN para así realizar cortes, son muchas las enzimas de restricción
estudiadas a la fecha. En esté trabajo fue utilizado el gen URA5, se utilizan primers
derivados del exón sucesivos."Intrón B" del gen URA5 (Fig. 5), y con el posterior
análisis discriminatorio de RFLP desarrollado para ayudar a un diagnóstico rápido y
la caracterización de los ocho tipos moleculares de C.neoformans y C. gattii.
(Velegraki et al., 2001)

15
A“intron A” B:“intron A”
Figura 5: Posición del exón estudiado de la Gen URA5. Un fragmento de 345
pb del exón. (posiciones 1145–1492) (GenBank, accesión no. M34606) (Velegraki
2001)
Tipificación Secuencia Multilocus (MLST): Dado los avances de la biología
molecular ha sido posible implementar dentro de las técnicas de tipificación de
Cryptococcus la metodología MLST propuesta por el Grupo de Genotipificación de
Cryptococcus neoformans/ Cryptococcus gattii de la Internanational Society for
Human and Animal Mycoses(ISHAM) como método estándar universal para la
genotipiciación molecular, mediante la utilización de ocho loci polimórficos: CAP59,
GPD1, LAC1, PLB1, SOD1, URA5 y IGS1, aunque este mismo ente reconoce las
diversas técnicas utilizadas en epidemiologia molecular para el estudio de la
Criptococosis que han permitido de la nomenclatura de los ocho patronoes moleculares
productos de la utilización de las técnicas PCR huella digital utilizando iniciadores
microsatelites o minisatelites y análisis mediante RFLP(Escandón & Montilla, 2011)
2.5.3 Identificación proteómica de tipos moleculares de C. neoformans y C.
gattii
El termino proteoma proviene de la analogía entre proteína-genoma y
corresponde al número de proteínas que una célula u organismo expresa (Wilkins et
al., 1996) las metodología aplicable del proteoma es denominado proteómica y sus
inicios datan del año 1964. Es importante aclarar cuando se establecen relaciones entre
genoma y proteoma, que un gen da origen a seis o más proteínas diferentes (figura 6),

16
la producción de proteínas es un proceso dinámico de las células que puede variar por
factores externos siendo la heterogeneidad de las proteínas es mucho mayor que la de
los genes (Scherl et al., 2002), es así como en el área de investigación de las
enfermedades infecciosas los estudios de los microorganismo se deben hacer acorde
con los avances de las ciencias biológicas con la utilización de diversas herramientas
moleculares como lo son además de la genómica y proteómica, la interactomica donde
confluyen la bioinformática, transcriptómica y metabólomica.
Figura 6 Transcripción, traducción y modificación postraduccional y/o
proteólisis de las proteínas (Cazzulo).
Espectrofotometría de masas: La espectrometría de masas es una tecnológica
básica en el estudio de la proteómica por la alta capacidad de análisis, su sensibilidad
y precisión en la identificación de masas moleculares proteómicas. Los
espectrofotómetros de masas se dividen en dos partes: fuente de iones y detector.
- Fuente de iones: la cual genera e introduce los iones analitos en el
instrumento, se utilizan principalmente dos tipos de técnicas de ionización,
Una en la cual se utiliza electrospray también llamada ionización por
electronebulización y la otra técnica fue la utilizada en nuestro trabajo
desorción/ionización mediante laser asistida por matriz llamada ionización

17
por MALDI, esta última por lo general suele asociarse a un analizador de
tiempo de vuelo TOF (Time-of- Fligth) en los que los iones se separan en
su relación masa-carga después de ser acelerados en un campo eléctrico.
- Detector: para medir las cargas de los iones generados.(Dingle & Butler-
Wu, 2013)

18
3.OBJETIVOS
3.1 OBJETIVO GENERAL
Caracterizar genética y proteomicamente aislados ambientales y clínicos de
Cryptococcus gatti, C. neoformans en Cúcuta, Norte de Santander, Colombia.
2.2 OBJETIVOS ESPECÍFICOS
Evaluar la prevalencia de C. gattii y C. neoformans en árboles de parques
públicos de la ciudad de Cúcuta.
Caracterizar genéticamente los aislamientos ambientales y clínicos de C.gattii
y C. neoformans, usando técnicas moleculares.
Caracterizar proteómicamente los aislamientos ambientales y clínicos de
C.gattii y C.neoformans, usando MALDI TOF.

Discusión
4. MATERIALES Y MÉTODOS
4.1 POBLACIÓN
La población corresponde a los arboles de parques públicos de la ciudad de
Cúcuta, los criterios para selección de la muestra fueron: ubicación en zona de alto
tránsito de personas y/o antecedentes de reportes de aislamientos de Cryptococcus sp.
Se colectaron 1300 muestras de 446 árboles distribuidas así: suelo (442), corteza (434),
frutos secos (40) y hojas (384); en el momento de recolección de muestras se tuvo en
cuenta principalmente la presencia heces de palomas en su superficie. El primer
muestreo se realizó en el mes octubre de 2016 en los parques Mercedes Abrego, Simón
Bolívar, Arcoíris (La Libertad) y zona periférica al estadio General Santander. El
segundo muestreo se llevó a cabo en el mes de enero del 2017 en los Parques Antonia
Santos, La Victoria, Juana Rangel de Cuellar y Parque Santander; el tercer, muestreo
se realizó en el mes de abril del 2017, en los parques Nacional y fuentes de Leones.
(Figura 7). Las variables meteorológicas relacionadas fueron en el primer muestreo
una precipitación (mm) de 5,3, en el segundo muestreo de 0,0 y en el tercer muestreo
de 51,1; con relación a la temperatura(°C) promedio en el primer muestreo de 29,05,
en el segundo muestreo de 25,1 y en el tercer muestreo 26,08 .

20
Figura 7. Ubicación geográfica de la ciudad de Cúcuta en el Departamento
Norte de Santander, en donde se obtuvieron los aislamientos en árboles de Parques
Públicos.
Por otra parte, se obtuvieron 6 aislados de C. neoformans de líquido
cefalorraquídeo (LCR) de pacientes VIH positivos con Cryptococosis meníngea,
procedentes del Hospital Universitario Erasmo Meoz de Cúcuta entre, los meses de
junio de 2016 y junio del 2017. Estas muestras correspondieron al 4,3% de la totalidad
de las reportadas a través de la vigilancia pasiva de la Cryptococosis que se realiza en
el país mediante la encuesta epidemiológica sobre la Cryptococosis en Colombia.
4.2 PROCESAMIENTO DE MUESTRAS AMBIENTALES
Se realizó un mapeo de los lugares de muestreo donde se identificó a cada
individuo (árbol) para la recolección de muestras en cada una de las zonas
seleccionadas del área metropolitana de Cúcuta, empleando bolsas plásticas (Anexo
1).
Estas fueron procesadas mediante la técnica de extracción de buffer fosfato
salino (PBS), suplementando con antibiótico, como se describió previamente

21
(Escandón et al., 2005; Paliwal & Randhawa, 1978). Después, 5g de muestra fueron
resuspendidos en 25 ml de PBS 1X para su homogenización y reposo durante 60
minutos. Posteriormente, se filtró cada homogenizado con gasa estéril, se adicionó el
antibiótico y se dejó actuar por 60 minutos. Un total de 100µl de cada preparación
fueron sembrados en agar semilla de girasol para posterior incubación a 27°C durante
20 días, con observación periódica semanal, como ha sido referido (Staib, Seibold,
Antweiler, & Fröhlich, 1989).
Las colonias cremosas, elevadas, con bordes regulares y pigmento café
(producción de melanina) (Figura 8) obtenidas, fueron observadas al microscopio
empleando tinta china. Toda colonia correspondiente a levaduras grandes, redondas y
con presencia de cápsula fue sometida a pruebas de degradación de urea, prueba que
se basa en la capacidad de producir la enzima ureasa, la cual desdobla la urea en
dióxido de carbono y amonio, incrementando el pH del medio y produciendo un
cambio de color rojo – púrpura usando el indicador rojo de fenol. La prueba se
considera positiva cuando se alcaliniza el medio lo que produce un cambio de color
original (amarillo) a rosa o rojo. (K. Kwon-Chung, Wickes, Booth, Vishniac, &
Bennett, 1987) Se utilizaron como controles positivos cepas ATCC de C.neorformans
y como control negativo cepas ATCC de Candida albicans. (Figura 9). Finalmente,
para el análisis bioquímico se determinó la especie mediante crecimiento en agar CGB
(canavanina glicina azul de bromotimol sódico) (Min & Kwon-Chung, 1986).
Colonias con crecimiento y viraje a azul del medio corresponden a C. gattii y colonias
sin cambio del medio a C. neoformans (Figura 10).

22
Figura 8. Cryptococcus spp. Agar semilla de girasol (Producción de melanina).
Universidad de Santander, Cúcuta.
a. b.
Figura 9 a. Pruebas de Urea positiva para Cryptococcus spp. b. Control
positivo y control negativo. Universidad de Santander, Cúcuta.

23
Figura 10 Prueba de CGB para identificación diferencial de C. neoformans y
C. gattii. Universidad de Santander, Cúcuta.
4.3 PROCESAMIMENTOS DE AISLADOS CLINICOS
A los aislados de procedencia clínica (Figura 11) igualmente se les realizó
cultivo en agar semilla de girasol, pruebas de degradación de urea(K. Kwon-Chung et
al., 1987) y determinación de la especie en agar CGB (canavanina glicina azul de
bromotimol sódico) (Min & Kwon-Chung, 1986).
Figura 11Cryptococcus spp muestra de líquido cefalorraquídeo en tinta china.
Instituto Nacional de Salud, Colombia).
4.4 EXTRACCION DE ADN
El ADN de alto peso molecular fue extraído, utilizando el método
fenol:cloroformo:alcohol isoamílico según Casali et al (Casali et al., 2003) Este
procedimiento se llevó a cabo utilizando biomasa obtenida de las cepas clínicas y
ambientales una vez cultivadas en agar saboureaud a 27°C por 48 horas.
Posteriormente, se transfirieron colonias del cultivo a tubos de microcentrífuga de 1,5
ml y se almacenaron a -20°C durante una noche. Seguidamente, las colonias se
suspendieron en 500μl de buffer de extracción (Tris - HCl 50 mM, EDTA 50 mM,

24
dodecil sulfato sódico al 3%, 2 - mercaptoetanol 1%), agitando vigorosamente con
vórtex y se incubaron a 65°C durante 1 hora. El lisado se extrajo con fenol-cloroformo-
alcohol isoamílico. El ADN se recuperó mediante precipitación con isopropanol a -
20°C durante una noche; se lavó con etanol al 70 % (vol / vol) y finalmente, se
resuspendió en buffer TRIS – EDTA (TE). La concentración se midió mediante
fluorescencia (Quibit 3.0 fluorómetro)
4.5 DETERMINACION DE PATRON MOLECULAR POR PCR-
HUELLA DIGITAL
La caracterización molecular de las especies de Cryptococcus spp, se realizó
mediante PCR - huella digital, utilizando como cebador único de secuencia especifica
microsatélite (GTG)5 (5’-GTGGTGGTGGTGGTG-3’) descrito por Escandón y
colaboradores en 2006 (Escandón, Sánchez, Martínez, Meyer, & Castañeda, 2006).
La mezcla de PCR de 50 µL contenía; 31,5 µL de agua desionizada estéril, 25
ng de ADN, 5 µl de tampón de PCR 10X (Tris / HCl 10 mM, pH 8,3, 50 mM KCl, 1,5
mM MgCl ) (Invitrogen, Carlsbad, CA), 0,02 mM de cada dNTP (dATP, dCTP, dGTP
y dTTP) (Promega, Madison, WI), acetato de sodio 3 mM (Sigma, Atlanta, GA), O,8
ng de cebador (para GTG5 la concentración fue 10 ng), 2 mM MgCl, 1 ml de albúmina
de suero bovino (BSA, 200 mg mL-1 \ Delta 1) y 0,05 U Amplitaq. La PCR se realizó
durante 35 ciclos en un SimpliAmp™ Thermal Cycler utilizando, las siguientes
condiciones: desnaturalización a 94ºC durante 20 segundos, hibridación a 50ºC
durante 1 minuto, extensión a 72ºC durante 20 segundos y un ciclo de extensión final
a 72ºC durante 6 minutos. Los productos de la amplificación se separaron por

25
electroforesis en un gel de agarosa al 1,4% en tampón Tris-borato EDTA (TBE) 1X a
100 V durante 2 horas.
Los productos de la amplificación GTG5 se tiñeron con Gel red Nucleid Acid
Gel Stain Biotium a 0,3 mg mL-1, durante 30 minutos. Las bandas se visualizaron bajo
luz UV utilizando marcador de tamaño molecular de 1 kb en tres pozos para permitir
la normalización de los geles. Los tipos moleculares (VNI-VNIV y VGI-VGIV) se
asignaron para comparar las cepas de referencia de los ocho tipos moleculares
principales (Patrones moleculares William Meyer) cargados en cada gel (Escandón et
al., 2012). (Figura 12).
Figura 12. Flujograma de identificación por PCR- Huella digital. Universidad de
Santander, Cúcuta.
4.6 DETERMINACIÓN DEL PATRÓN MOLECULAR POR RFLP-
GEN URA5 Y ANÁLISIS DE ASOCIACIÓN.
La identificación de los tipos moleculares de Cryptococcus spp. Se realizó tanto
a las cepas clínicas como a las ambientales, empleando los dos cebadores: URA5

26
(5’ATGTCCTCCCAAGCCCTCGACTCCG3’) y SJ01
(5’TTAAGACCTCTGAACACCGTACTC3’). La reacción de la PCR se llevó a cabo
según lo descrito por Meyer et al. 2003 (Meyer et al., 2003).
Los productos de la PCR se sometieron a una doble digestión enzimática con
Sau 96I y Hha I a 37 °C durante 3 horas. Los fragmentos de restricción fueron
separados por electroforesis en gel de agarosa al 3% en tampón Tris-borato EDTA
(TBE) 1X a 100 V durante 5 horas.
4.7 CARACTERIZACIÓN PROTEOMICA POR MALDI TOF
Las pruebas proteómicas fueron realizadas en el Laboratorio de Micosis
humana y proteómica de la Pontificia Universidad Javeriana.
La tecnología MALDI-TOF-MS está siendo aplicada a la identificación de
microorganismos y ha surgido en los laboratorios como una herramienta de
diagnóstico en todo el mundo; debido a la precisión de la técnica, se compara
favorablemente con la secuenciación genómica y se obtiene a un costo
significativamente menor a ésta última(Lau, Drake, Calhoun, Henderson, & Zelazny,
2013).
Principio de MATDI TOF MS: La muestra se mezcla con una matriz sobre una
placa metálica conductora. Después de la cristalización de la matriz y del material
microbiano, la placa de metal se introduce en el espectrómetro de masas y se
bombardea con breves pulsos de láser. Las moléculas ionizadas se aceleran a través de
un campo electrostático y se expulsan a través de un tubo de vuelo de metal, sometidas
a vacío hasta que alcanzan un detector. Por lo tanto, los bioanalitos separados generan
un espectro de masas compuesto por picos de relación masa / carga (m / z) con
intensidades variables. Un espectro es por lo tanto una huella microbiana que se

27
compara con una base de datos para la identificación a nivel de especie o
género(Croxatto, Prod'hom, & Greub, 2012) (Figura 13).
Figura 13 Descripción de la técnica de MALDI TOF MS.(Croxatto et al.,
2012).
La extracción con ácido fórmico : Para el análisis de espectros en el MALDI
TOF MS Biotyper, es necesaria para romper la pared celular y separar las
proteínas.(Clark, Kaleta, Arora, & Wolk, 2013).
Como preparación de la muestra para el análisis proteómico, los aislamientos
de Cryptococcus spp fueron incubados en agar saboureaud 24 horas a 37°C; 1 colonia
grande fue tomada con el uso de un asa estéril y a ésta se le agregaron 300 ul de agua
grado HPLC, luego se adicionaron 900 ul de etanol grado HPLC seguido de
homogenización y centrifugación a máxima velocidad (13000 rpm) por 2 minutos; el
sobrenadante se decantó y al pellet restante se le agregaron 50 ul de ácido fórmico al

28
100%. Se dio nuevamente vórtex y se dejó reposar durante aproximadamente 5
minutos, seguido de lo cual se añadieron 50 ul de acetonitrilo al 100%; se homogenizó
y centrifugó a máxima velocidad (13000 rpm) por 2 minutos se transfirió 1ul de
sobrenadante en la placa de acero, se superpuso 1 ul del matrix, y se dejó secar.
Finalmente, se introdujo el montaje en el equipo MALDI-TOF MS para su análisis.
4.8 ANALISIS MOLECULAR:
Los perfiles de restricción productos de la pruebas RFLP de las cepas clínicas
y ambientales fueron comparados con los patrones de las cepas de referencia(William
Meyer donados por Instituto Nacional de Salud), utilizando análisis en módulo
BioGalaxy del programa Biolomics, versión 3.0, el cual emplea para la detección
automática de carriles y bandas, alogaritmos incorporados en un software que para una
identificación rápida y precisa, esta tecnología permitió la lectura de los geles
electroforéticos analizando las fotos originales para producir una lista de pesos
moleculares que fueron comparadas con una base de datos de referencia, además, una
vez realizada las lecturas los módulos de identificación permitieron mostrar los
resultados de la técnica de similitud mediante la agrupación de los resultados en un
dendograma (Robert et al., 2011)BioloMICS es un sistema que permite crear base de
datos personalizadas con solo seguimientos aun proceso sencillo y rápido el cual no
requiere conocimientos especializados en bioinformática.
Identificación de especies de Cryptococcus spp por espectrometría de masa
MALDI Biotyper (BRUKER)
Una vez se obtuvo el crecimiento en fase exponencial de los aislamientos, se
preparó una hoja de trabajo de la placa MALDI Biotyper con las identificaciones de
las muestras, dejando las dos posiciones iniciales de la placa para el Bacterial test

29
Standar (BTS) y la cepa H99 de Cryptococcus; para estas y para los aislamientos se
realizó una extracción de proteínas citoplasmáticas, mediante lisis con solventes
orgánicos, del cual ce coloco 1ul en la placa de acero con 96 pozos (las muestras se
procesaron por duplicado), se adiciono 1 µl de ácido fórmico al 100%, se dejó secar
y luego se le adiciono 1 µl de Bruker Matrix HCCA a cada posición de la placa con
las muestras, se dejó secar a temperatura ambiente y se analizó en el equipo, el cual
detecta proteínas ribosomales y citoplasmáticas presentes en la muestra, en donde se
genera una huella espectral que permite determinar la identificación del
microorganismo mediante el Software Biotyper 3.1, el cual hace una comparación con
la librería de espectros, con un nivel de confianza que se calcula con base al porcentaje
de probabilidad (0.000-1.699 no identificación, 1.700-1.999 identificación a nivel de
género, 2.000-3.000 identificación a nivel de especie); este módulo también permitió
determinar la relación de los diferentes aislamientos y llevar a cabo una agrupación
jerárquica de los mismos, representando los resultados en dendogramas, por el método
de correlación y agrupamiento completo(Bellido & de Buitrago Arriero, 2015; Zhou
et al., 2017). Para elaborar el dendograma primero se creó una base de datos con los
espectros de los aislados en estudio, posteriormente se realizó un análisis de la
similitud de los espectros mediante el software (ClinProTools, flex Analysis), este
análisis dio la información sobre la hetero/homogeneidad del juego de datos
(espectros) representados en el dendograma.(Rojo, 2014)

30
5. RESULTADOS
5.1 CARACTERIZACIÓN DE MUESTRAS AMBIENTALES
El 75% de los árboles analizados provenían de 6 parques: parque Simón
Bolívar en primer lugar con 80 individuos, seguido de los parques Santander, La
Victoria, Nacional, y Mercedes Ábrego (con 70, 68, 67 y 50 individuos,
respectivamente), siendo esta última cantidad igual para la zona circundante al Estadio
General Santander. El restante 25% de los arboles analizados correspondían a los
parques Arcoíris, Antonia Santos, Juana Rangel de Cuellar y Fuente de Leones con
(20, 14, 14 y 13 individuos respectivamente).
A su vez, la diversidad de los individuos (árboles) analizados en el presente
estudio comprendió 10 especies, siendo oití (Licania tomentosa) la más ampliamente
distribuida y predominante con el 66,4% del total de individuos, seguida del almendro
(Terminalia catappa) con 13,2%, abarcando éstas dos especies aproximadamente el
80 % de la muestra total. El 20% restante se distribuyó entre mamoncillo (Melicoccus
bijugatus) 7,2%, ficus (Ficus benjamina) 5,4%, tamarindo (Tamarindus indica) con
2,9%, chiminango y totumo (Pithecellobium dulce) con 1,8% cada uno; samán
(Samanea saman), limón swingle (Swinglea glutinosa) y mango (Mangifera indica),
0,4% cada uno.
El cultivo inicial reveló la presencia de 21 aislados compatibles con
Cryptococcus spp, 20 de los cuales correspondían a C. neoformans (95,2%) y 1 a C.
gattii (4,8%), obtenidos a partir de 20 de los 446 individuos, ya que a partir de un
mismo árbol se obtuvo doble positividad para C. neoformans (Tabla 2). Por lo anterior,
31
se obtuvieron prevalencias de 4,3% y 0,2%, para C. neoformans y C. gattii,
respectivamente, tomando en cuenta la totalidad de los individuos analizados.
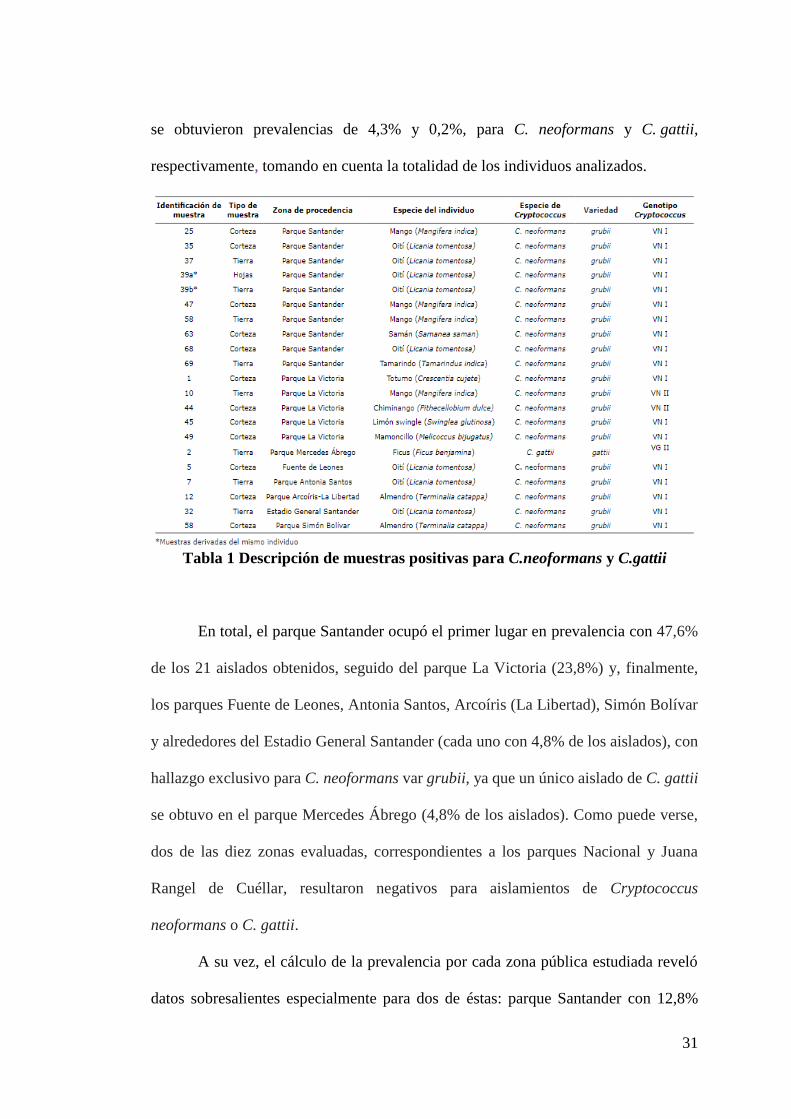
Tabla 1 Descripción de muestras positivas para C.neoformans y C.gattii
En total, el parque Santander ocupó el primer lugar en prevalencia con 47,6%
de los 21 aislados obtenidos, seguido del parque La Victoria (23,8%) y, finalmente,
los parques Fuente de Leones, Antonia Santos, Arcoíris (La Libertad), Simón Bolívar
y alrededores del Estadio General Santander (cada uno con 4,8% de los aislados), con
hallazgo exclusivo para C. neoformans var grubii, ya que un único aislado de C. gattii
se obtuvo en el parque Mercedes Ábrego (4,8% de los aislados). Como puede verse,
dos de las diez zonas evaluadas, correspondientes a los parques Nacional y Juana
Rangel de Cuéllar, resultaron negativos para aislamientos de Cryptococcus
neoformans o C. gattii.
A su vez, el cálculo de la prevalencia por cada zona pública estudiada reveló
datos sobresalientes especialmente para dos de éstas: parque Santander con 12,8%
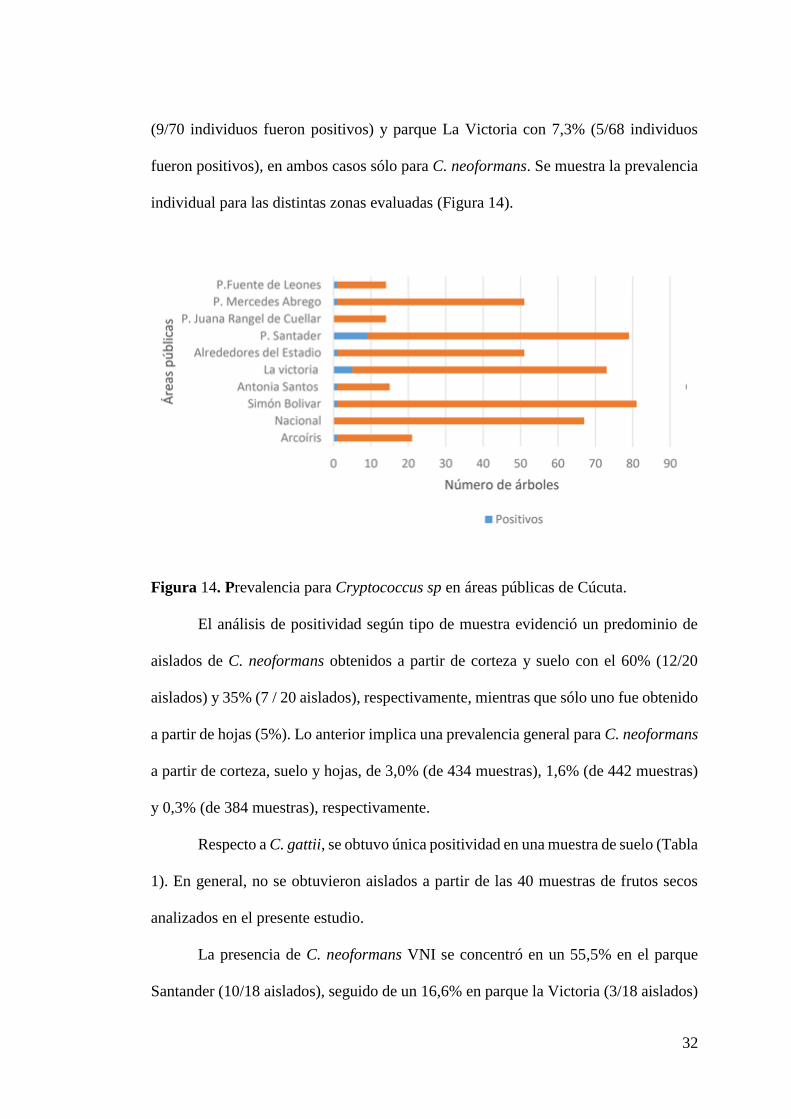
32
(9/70 individuos fueron positivos) y parque La Victoria con 7,3% (5/68 individuos
fueron positivos), en ambos casos sólo para C. neoformans. Se muestra la prevalencia
individual para las distintas zonas evaluadas (Figura 14).
Figura 14. Prevalencia para Cryptococcus sp en áreas públicas de Cúcuta.
El análisis de positividad según tipo de muestra evidenció un predominio de
aislados de C. neoformans obtenidos a partir de corteza y suelo con el 60% (12/20
aislados) y 35% (7 / 20 aislados), respectivamente, mientras que sólo uno fue obtenido
a partir de hojas (5%). Lo anterior implica una prevalencia general para C. neoformans
a partir de corteza, suelo y hojas, de 3,0% (de 434 muestras), 1,6% (de 442 muestras)
y 0,3% (de 384 muestras), respectivamente.
Respecto a C. gattii, se obtuvo única positividad en una muestra de suelo (Tabla
1). En general, no se obtuvieron aislados a partir de las 40 muestras de frutos secos
analizados en el presente estudio.
La presencia de C. neoformans VNI se concentró en un 55,5% en el parque
Santander (10/18 aislados), seguido de un 16,6% en parque la Victoria (3/18 aislados)

33
y una minoría se distribuyó en los parques Fuente de Leones, Antonia Santos, Arcoíris
(La Libertad), Simón Bolívar y alrededores del Estadio General Santander, con 5,3%
de los aislados para cada zona (1/18 aislados), mientras que el tipo molecular VNII se
halló exclusivamente en el parque la Victoria, aislado a partir de suelo y corteza de las
especies arbóreas mango (Mangifera indica) y chiminango (Pithecellobium dulce),
respectivamente.Por otra parte, el único aislado obtenido de C. gattii correspondió al
tipo molecular VGII, siendo a su vez el único obtenido a partir del parque Mercedes
Abrego, a partir de suelo asociado a un individuo de ficus (Ficus benjamina) (Tabla
2).
5.2 CARACTERIZACIÓN DE AISLAMIENTOS A PARTIR DE L.C.R.
Para la identificación del tipo molecular se realizó la caracterización por
métodos genotípicos y por espectometria de masas, a partir de aislados obtenidos de
liquidos cefaloraquídeos (LCR), donde en la identificación a nivel de genotipo, el
100% mostraron patrones del grupo VNI., Mientras que, en los aislamientos
ambientales, dentro de las levaduras identificadas bioquímicamente como del
complejo Cryptococcus neoformans, el 85,7% correspondieron al genotipo molecular
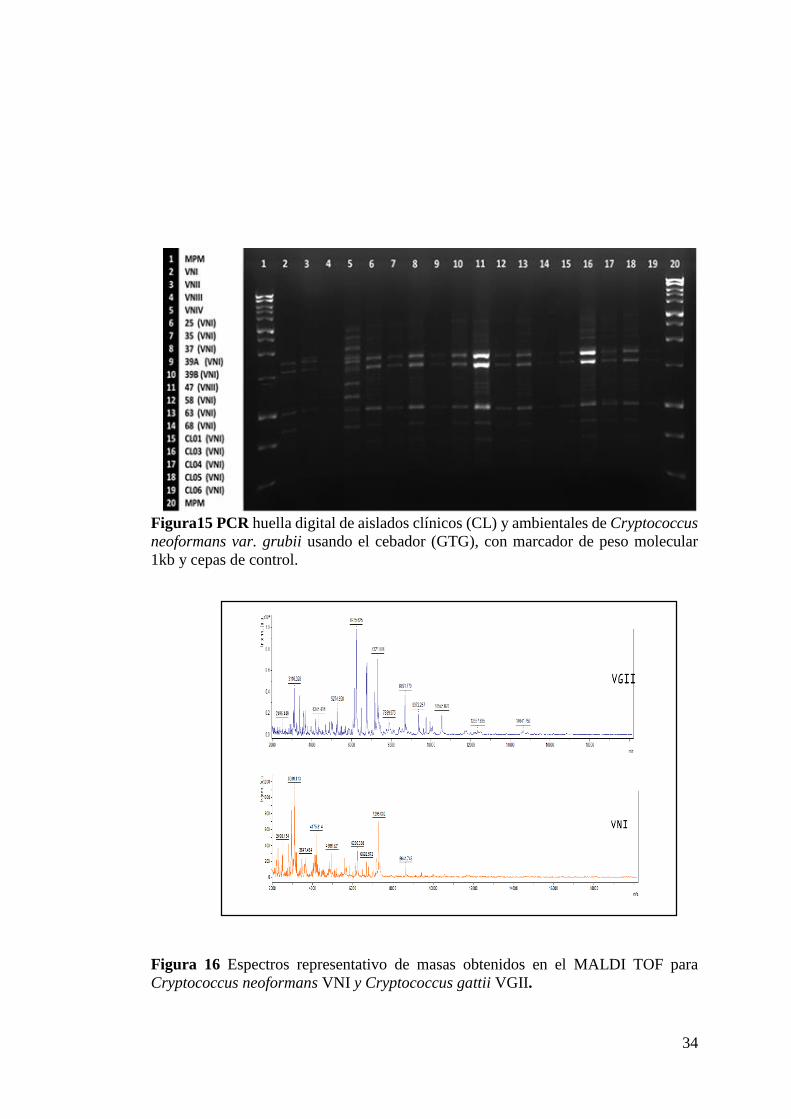
VNI y el 9.5% al tipo molecular VNII (Figura 15 y 16).
Tabla 3 Descripción de muestras Clínicas para C.neoformans VNI por técnicas
genómicas y proteómicas
34
Figura15 PCR huella digital de aislados clínicos (CL) y ambientales de Cryptococcus
neoformans var. grubii usando el cebador (GTG), con marcador de peso molecular
1kb y cepas de control.
Figura 16 Espectros representativo de masas obtenidos en el MALDI TOF para
Cryptococcus neoformans VNI y Cryptococcus gattii VGII.
a
b

35
5.3 RELACIÓN GENÉTICA ENTRE LOS AISLADOS DE
C. neoformans DE PROCEDENCIA CLÍNICA Y AMBIENTAL.
Como se describió anteriormente, mientras que a nivel ambiental se obtuvieron
aislados tanto de C. neoformans como de C. gattii, el primero incluso exponiendo los
tipos moleculares VNI (mayoritariamente) y VNII, a nivel clínico se identificó
únicamente la variante VNI en la totalidad de los casos. Teniendo en cuenta lo anterior,
se procedió a analizar la relación filogenética entre los aislados de C. neoformans var
grubii con tipo molecular VNI de procedencias tanto ambiental como clínica
La caracterización genética de los aislamientos clínicos en el patrón molecular
VNI mediante PCR huella digital y RFLP del gen URA5 reveló que el patrón genético
encontrado en aquellos aislamientos provenientes de pacientes está igualmente
presente en la mayoría de los aislamientos ambientales.
Se observó una correlación entre estos dos tipos de aislamientos a nivel genético.
(Figura 17).
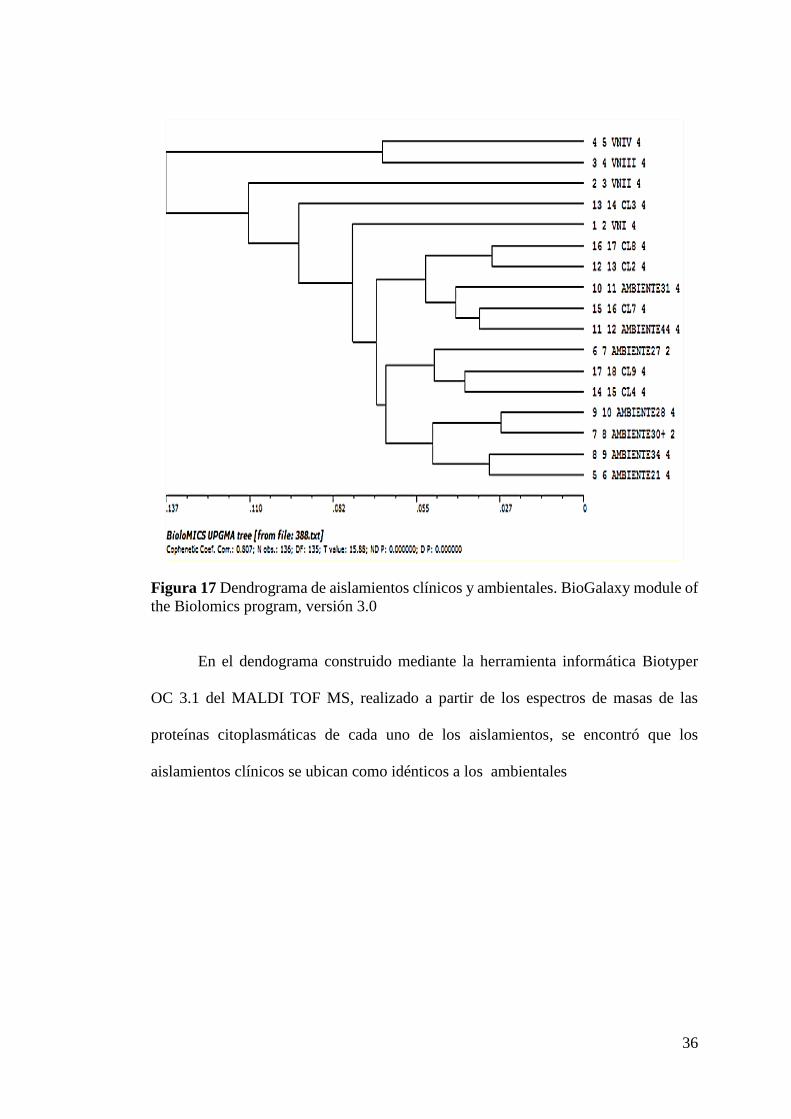
36
Figura 17 Dendrograma de aislamientos clínicos y ambientales. BioGalaxy module of
the Biolomics program, versión 3.0
En el dendograma construido mediante la herramienta informática Biotyper
OC 3.1 del MALDI TOF MS, realizado a partir de los espectros de masas de las
proteínas citoplasmáticas de cada uno de los aislamientos, se encontró que los
aislamientos clínicos se ubican como idénticos a los ambientales
37
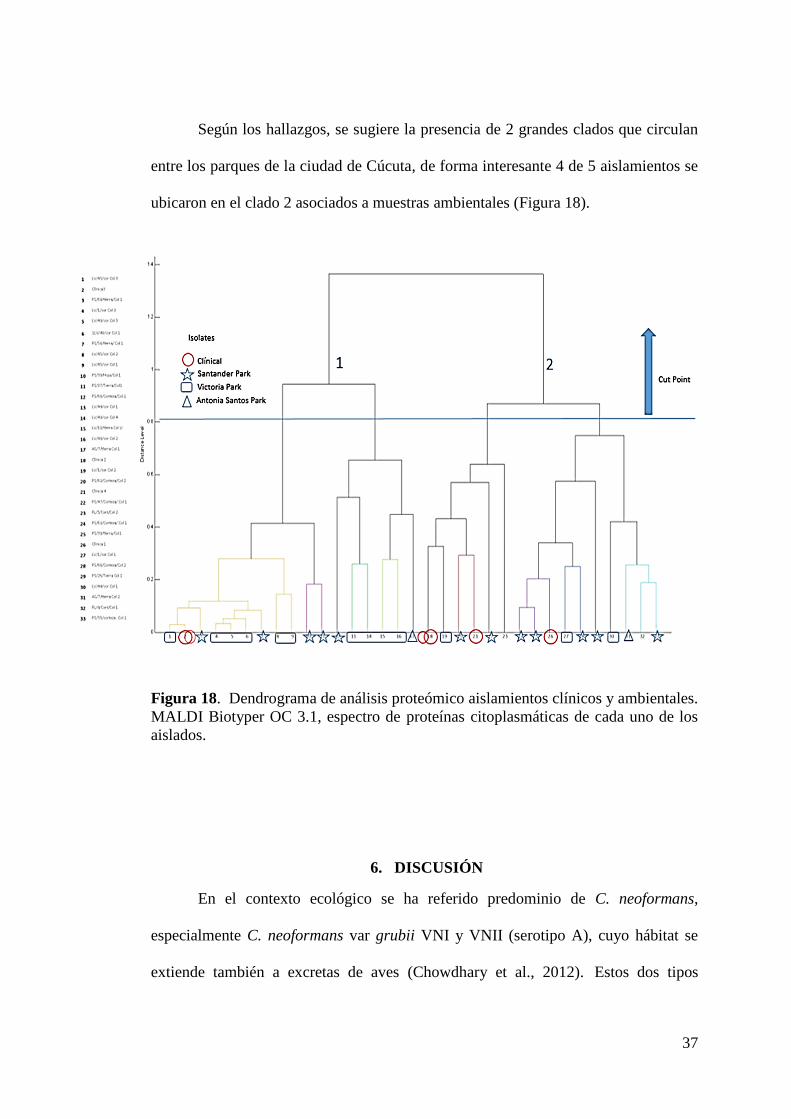
Según los hallazgos, se sugiere la presencia de 2 grandes clados que circulan
entre los parques de la ciudad de Cúcuta, de forma interesante 4 de 5 aislamientos se
ubicaron en el clado 2 asociados a muestras ambientales (Figura 18).
Figura 18. Dendrograma de análisis proteómico aislamientos clínicos y ambientales.
MALDI Biotyper OC 3.1, espectro de proteínas citoplasmáticas de cada uno de los
aislados.
6. DISCUSIÓN
En el contexto ecológico se ha referido predominio de C. neoformans,
especialmente C. neoformans var grubii VNI y VNII (serotipo A), cuyo hábitat se
extiende también a excretas de aves (Chowdhary et al., 2012). Estos dos tipos

38
moleculares son igualmente los más prevalentes en casos de Criptococosis (97%) en
individuos inmunosuprimidos en Colombia (Escandón et al., 2012; Lizarazo et al.,
2007) y otros países incluso europeos (Cogliati, 2013). Sin embargo, la presencia
ambiental de C. gattii reviste especial relevancia en torno a su capacidad de generar
enfermedad en individuos inmunocompetentes y su potencial de adaptación a distintas
condiciones geográficas, dejando de ser exclusivo de zonas tropicales y subtropicales,
para pasar a hallarse también en zonas templadas y lluviosas(Mak, Vélez, Castañeda,
Escandón, & Group, 2015).
El presente estudio abarcó 10 zonas públicas altamente concurridas de la ciudad
de Cúcuta, en las cuales en su mayoría se reveló la presencia de patógenos del género
Cryptococcus sp, exceptuando los parques Nacional y Juana Rangel de Cuéllar. La
prevalencia hallada fue C. neoformans (4,3%) respecto a C. gattii (0,2%) a razón de
21.5:1, similar a lo referido previamente (Chowdhary et al., 2012), por otra parte,
contrastante con datos previos reportados para la ciudad de Cúcuta en particular (2008
y 2009), habiéndose referido como de 0,07% de manera similar para ambas especies
del hongo, incluso con un tamizaje significativamente mayor de individuos (3634)
procedentes de zonas La Libertad, San Eduardo y alrededores del Estadio General
Santander (Firacative, Torres, Rodríguez, & Escandón, 2011).
Según nuestros hallazgos, la positividad para C. neoformans mostró
predominio en el parque Santander (47,6% de los 21 aislados), seguido del parque La
Victoria (23,8%) y finalmente los parques Fuente de Leones, Antonia Santos, Arcoíris
(La Libertad), Simón Bolívar y alrededores del Estadio General Santander (cada uno
con 4,8% de los aislados), exclusivamente para C. neoformans variedad grubii.
Tomando en cuenta lo anterior, podemos concluir inicialmente que los parques

39
Santander (especialmente y con prevalencia individual superior al 10%) y La Victoria,
constituyen nichos ecológicos relevantes para C. neoformans, aportando éste último
además variabilidad genética al hallarse tanto el tipo molecular VNI como el VNII
(única zona con hallazgo de éste último).
Se contó con una variedad de especies arbóreas que evidenciaron ser nichos del
hongo, especialmente oití (Licania tomentosa), ampliamente distribuida en las zonas
evaluadas, a partir de la cual se identificó el 38% del total de aislados (8 / 21 aislados),
todos correspondientes a C.neoformans VNI, además de hallarse en diversidad de
fuentes (corteza, tierra y hojas), seguida del mango (Mangifera indica), a partir del
cual se obtuvieron 4 aislados (19%), tanto en tierra como en corteza, correspondientes
a C. neoformans variedad grubbii (VNI y VNII) y el almendro (Terminalia catappa),
con dos aislados (9,5%), identificados como C. neoformans VNI, en contraste con el
reporte previo de éste último como hábitat tanto de C. neoformans como de C. gattii
(Cogliati, 2013).
En el parque Santander de manera interesante, los cinco aislados obtenidos de
la especie arbórea oití (Licania tomentosa) correspondieron a cuatro individuos, puesto
que uno de ellos fue doble positivo, revelando la presencia de C.neoformans VNI
simultáneamente en suelo o tierra adyacente al mismo y en sus hojas. Este caso único
de doble positividad para Cryptococcus neoformans, registrado en el presente trabajo
no tiene antecedente en la ciudad, en lo que respecta al tipo de árbol (Licania
tomentosa).
Según lo publicado por Firacative et al (Firacative et al., 2011) el rastreo de
más de 4300 muestras procedentes de árboles de diversas zonas de Cúcuta y áreas

40
cercanas a la vivienda de pacientes infectados por C. gattii serotipo B (VGI/a), arrojó
sólo un aislamiento de éste patógeno, dos aislamientos de C. gattii serotipo C (VGIII/a)
y tres de C.neoformans serotipo A (VNI/a), donde uno de éstos últimos, junto con el
único aislado de C.gattii (VGI/a) se obtuvieron de un mismo árbol pero en ese caso de
especie Ficus (Ficus benjamina) en el sector San Eduardo.
De igual manera, en los alrededores del Estadio General Santander, también en
suelo de un mismo árbol Ficus (Ficus benjamina) se hallaron simultáneamente C.
gattii (VGIII/a) y C.neoformans serotipo A (VNI/a).
En relación a lo anterior, el presente trabajo reveló positividad para ésta última
especie de árbol, pero correspondiente al aislamiento de C. gattii VGII a partir de
muestra de tierra en el parque Mercedes Ábrego (sin antecedente), en plena zona
céntrica de la ciudad de Cúcuta. Ésta fue la cuarta zona con mayor cantidad de
individuos tamizados, sin embargo, sólo uno de sus 50 árboles resultó positivo,
tratándose de un individuo Ficus (Ficus benjamina).
El hallazgo de que Ficus benjamina alberga tanto a C. gattii como a C.
neoformans (Firacative et al., 2011) reviste importancia si se considera la presencia
de los genotipos VGI, VGII y VGIII tanto en muestras ambientales como en pacientes
con enfermedad criptocóccica (Escandón et al., 2012; Lizarazo, Escandón, Agudelo,
Firacative, et al., 2014; Lizarazo et al., 2007) Estos tres tipos moleculares también han
sido asociados a casos de Criptococosis humana y animal en el occidente de Estados
Unidos. Particularmente los genotipos VGII y VGIII de origen ambiental se han
identificado como el posible agente etiológico de la micosis en individuos VIH
positivo (Springer et al., 2014).

41
Los hallazgos en la zona circundante al Estadio General Santander son
consistentes con un reporte previo, el cual mostró la presencia de C. neoformans
(Firacative et al., 2011), permitiendo concluir sobre su persistencia en el área. Sin
embargo, esta positividad se observó por primera vez en árboles de la especie Licania
tomentosa, reflejando una variación tanto de los tipos de hábitat del hongo, como de
las especies del patógeno que circulan en una zona determinada.
Dada la amplia distribución ambiental de C. neoformans y C. gattii, y su
asociación a patología en individuos inmunosuprimidos e incluso inmunocompetentes,
resulta relevante el rastreo permanente de zonas comunes de alta confluencia
poblacional (Mak, Vélez, Castañeda, Escandón, et al., 2015). A esto se suma que la
incidencia de Criptococosis afecta también a menores de edad en Colombia (menores
de 16 años), donde la infección por VIH constituye un factor de riesgo reconocible en
apenas la mitad de los casos, quedando por establecer factores adicionales. Resulta
preocupante, sin embargo, que esta incidencia es siete veces mayor para el mismo
grupo poblacional en Norte de Santander (0,122 casos/100,000 habitantes), respecto a
la media nacional (0,017 casos/100,000 habitantes) (Lizarazo, Escandón, Agudelo,
Firacative, et al., 2014).
Teniendo en cuenta todo lo anterior y el propósito del presente estudio de
determinar la relación existente entre los diversos aislados de procedencia ambiental,
con aquellos obtenidos a partir del tamizaje ambiental, el reconocimiento de la
identidad entre dichos aislados de Cryptococcus sp mediante la comparación de
genotipos circulantes cobra una enorme importancia teniendo en cuenta además el
aumento de los casos de Criptococosis en individuos VIH positivos, al comparar los
lapsos de tiempo 1997-2005 con el periodo 2006-2010 (3000 casos/millón de

42
habitantes y 3300 casos/millón de habitantes, respectivamente)(Escandón et al., 2012;
Lizarazo et al., 2007), a su vez que la determinación en el presente caso de ambas
especies patógenas del hongo en zona públicas ampliamente concurridas, incluso
mostrando una variedad de tipos moleculares. Sin embargo, resultó particularmente,
de interés en esta investigación el contraste entre los aislados del C. neoformans var
grubbi, VNI, común a todas las muestras procedentes de pacientes VIH positivos, con
aquellos del mismo tipo aislados de siete de las ocho zonas públicas de la ciudad de
Cúcuta, que, según los hallazgos anteriormente referidos, evidenciaron la presencia de
éste mismo tipo molecular VNI, accediendo a importantes herramientas de análisis
tanto genómico como proteómico.
Este estudio se evidencia la concordancia entre relación genética y proteómica
de aislados ambientales y clínicos de Cryptococcus neoformans de tipo molecular VNI
en Cúcuta, es necesario realizar estudios de MLST a los aislados obtenidos para poder
obtener información de la relación entre secuencias ST como se observa en estudios
como el realizado en Apulia, Italia donde se compararon los tipos moleculares y los
resultados ST los resultados de ST obtenidos de 21 aislamientos clínicos recolectados
en Apulia mostró que un aislado clínico de VNI de C. neoformans compartió un tipo
de secuencia idéntico de un aislado arbóreo (ST61) y que un aislado clínico de VGI de
C. gattii emparejado con el ST principal (ST156) presente en el medio ambiente en
este estudio se da a conocer los asilamientos clínicos circulantes y su relación con los
ambientales como contribución a los programas de Salud pública de esta
región(Montagna et al., 2018)
Adicionalmente, el análisis genotípico de las especies patógenas procedentes
de 6 muestras clínicas en el presente trabajo reveló exclusivamente la presencia de C.

43
neoformans var grubii VNI (100% de los casos) , en consistencia con el amplio
predominio registrado en estudios como el de González y Colaboradores en México,
durante 12 años, a partir de 166 muestras clínicas (no se especifica si expresamente de
pacientes VIH+), quienes refirieron prevalencia para C. neoformans de 92,2%,
respecto a C. gattii (7,8%) y correspondientes a VNI (74,6%), VNII (9%), VNIII
(4,8%), VNIV (3,6%), VGI (3,6%), VGII (1,8%), VGIII y VGIV (1.2% cada uno)
(González et al., 2016), dejando entrever una vez más, la alta prevalencia registrada
para el tipo molecular VNI respecto a los demás en la presentación clínica de la
enfermedad en población muy susceptible, pero a lo que de acuerdo con los hallazgos
revelados por el presente estudio, se suma la identidad genética y proteómica
evidenciada entre las cepas circulantes libremente en el ambiente, especialmente en
zonas de alta circulación de población humana en nuestra región y aquellas causantes
de una de las formas más agresivas de la Criptococosis.

7. CONCLUSIONES
La alta prevalencia de Criptococosis en la ciudad de Cúcuta, el constante
reporte de casos tanto en la región como a nivel mundial, el aumento de las
enfermedades predisponente como lo es en Síndrome de Inmunodeficiencia Adquirida
y la de resistencia a los tratamientos antifúngicos evidencia la importancia de estudios
de nichos ecológicos de hongos, especialmente en las zonas relacionadas con el reporte
de la enfermedad.
El muestreo longitudinal de nichos ecológicos donde el hongo se ha recuperado
previamente revela su persistencia en nichos donde Cryptococcus encuentra
condiciones favorables para sobrevivir, este tipo de estudios es obligatorio para
realizarse en áreas donde la enfermedad afecta continuamente a una gran proporción
de la población y debe ser realizado de forma rutinaria por las autoridades de salud
pública.
Los estudios ecológicos son relevantes en el estudio epidemiológico de este
patógeno; por lo tanto, la continuidad de la investigación es importante para identificar
áreas donde el hongo puede encontrar condiciones favorables para desarrollarse en
Colombia.
La utilización de pruebas diagnósticas moleculares que permitan identificar la
frecuencia de casos relacionados con las especies y los patrones moleculares son de
gran aporte en el manejo de los tratamientos terapéuticos y la patogenia de esta
micosis.

8. RECOMENDACIONES
Es necesario ampliar el número de aislamientos clínicos que permitan
determinar asociaciones con los aislamientos ambientales del patrón VNII, así como
C.gattii presente en el ambiente.
Es de resaltar que Cúcuta se caracteriza por sectores ampliamente arborizados
y la presencia de gran cantidad de aves destacándose principalmente la Paloma de la
especie Columbia livia en los parques estudiados, siendo más representativa la
presencia de estas en el Parque Santander en donde este trabajo permite confirmar el
nicho de la levadura, otros estudios evidencian la relación existente entre los nichos
ecológicos y estas aves(Montagna et al., 2018); de ahí la importancia de una vigilancia
zoonótica y epidemiológica debido a la cercanía con los humanos y el hecho de que
pueden trasladarse de un parque al otro.
Dada la presencia de aislamientos ambientales y el alto reporte de
Criptococosis en pacientes humanos en la ciudad de Cúcuta es importante continuar
estudios que permitan búsqueda activa de aislamientos en el área veterinaria
especialmente en caninos y felinos.

46
REFERENCIAS BIBLIOGRÁFICAS
Alspaugh, J. A. (2015). Virulence Mechanisms and Cryptococcus neoformans pathogenesis. Fungal
genetics and biology: FG & B, 78, 55.
Beatson, M., Harwood, M., Reese, V., & Robinson-Bostom, L. (2019). Primary cutaneous
cryptococcosis in an elderly pigeon breeder. JAAD case reports, 5(5), 433-435.
Bellido, J. L. M., & de Buitrago Arriero, J. M. G. (2015). Espectrometría de masas MALDI-TOF en
microbiología clínica. Situación actual y perspectivas futuras. Enfermedades infecciosas y
microbiologia clinica, 33(6), 369-371.
Callejas, A., Ordonez, N., Rodriguez, M., & Castaneda, E. (1998). First isolation of Cryptococcus
neoformans var. gattii, serotype C, from the environment in Colombia. Medical mycology,
36(5), 341-344.
Campbell, L. T., Currie, B. J., Krockenberger, M., Malik, R., Meyer, W., Heitman, J., & Carter, D.
(2005). Clonality and recombination in genetically differentiated subgroups of Cryptococcus
gattii. Eukaryotic cell, 4(8), 1403-1409.
Campbell, L. T., Fraser, J. A., Nichols, C. B., Dietrich, F. S., Carter, D., & Heitman, J. (2005). Clinical
and environmental isolates of Cryptococcus gattii from Australia that retain sexual fecundity.
Eukaryotic cell, 4(8), 1410-1419.
Casali, A. K., Goulart, L., Rosa e Silva, L. K., Ribeiro, Â. M., Amaral, A. A., Alves, S. H., . . . Vainstein,
M. H. (2003). Molecular typing of clinical and environmental Cryptococcus neoformans
isolates in the Brazilian state Rio Grande do Sul. FEMS yeast research, 3(4), 405-415.
Cattana, M. E., de los Ángeles Sosa, M., Fernández, M., Rojas, F., Mangiaterra, M., & Giusiano, G.
(2014). Native trees of the Northeast Argentine: Natural hosts of the Cryptococcus
neoformans–Cryptococcus gattii species complex. Revista iberoamericana de micologia,
31(3), 188-192.
Clark, A. E., Kaleta, E. J., Arora, A., & Wolk, D. M. (2013). Matrix-assisted laser desorption ionization–
time of flight mass spectrometry: a fundamental shift in the routine practice of clinical
microbiology. Clinical Microbiology Reviews, 26(3), 547-603.
Cogliati, M. (2013). Global molecular epidemiology of Cryptococcus neoformans and Cryptococcus
gattii: an atlas of the molecular types. Scientifica, 2013.
Croxatto, A., Prod'hom, G., & Greub, G. (2012). Applications of MALDI-TOF mass spectrometry in
clinical diagnostic microbiology. FEMS microbiology reviews, 36(2), 380-407.
Chowdhary, A., Randhawa, H. S., Prakash, A., & Meis, J. F. (2012). Environmental prevalence of
Cryptococcus neoformans and Cryptococcus gattii in India: an update. Critical reviews in
microbiology, 38(1), 1-16.
Dingle, T. C., & Butler-Wu, S. M. (2013). MALDI-TOF mass spectrometry for microorganism
identification. Clinics in laboratory medicine, 33(3), 589-609.
Dykstra, M. A., Friedman, L., & Murphy, J. (1977). Capsule size of Cryptococcus neoformans: control
and relationship to virulence. Infection and immunity, 16(1), 129-135.
Escandón, P., de Bedout, C., Lizarazo, J., Agudelo, C. I., Tobón, Á., Bello, S., . . . Castañeda, E. (2012).
Cryptococcosis in Colombia: Results of the national surveillance program for the years 2006-
2010. Biomédica, 32(3), 386-398.
Escandón, P., Lizarazo, J., Agudelo, C., & Castañeda, E. (2018a). Cryptococcosis in Colombia:
Compilation and analysis of data from laboratory-based surveillance. Journal of Fungi, 4(1),
32.
Escandón, P., Lizarazo, J., Agudelo, C. I., & Castañeda, E. (2018b). Cryptococcosis in Colombia:
Compilation and Analysis of Data from Laboratory-Based Surveillance. Journal of Fungi,
4(1), 32.
Escandón, P., & Montilla, A. (2011). Tipificación molecular de aislamientos del complejo Cryptococcus
neoformans/Cryptococcus gattii. Infectio, 14(2S).
Escandón, P., Quintero, E., Granados, D., Huérfano, S., Ruiz, A., & Castañeda, E. (2005). Isolation of
Cryptococcus gattii serotype B from detritus of Eucalyptus trees in Colombia. Biomédica,
25(3), 390-397.
Escandón, P., Sánchez, A., Martínez, M., Meyer, W., & Castañeda, E. (2006). Molecular epidemiology
of clinical and environmental isolates of the Cryptococcus neoformans species complex reveals
a high genetic diversity and the presence of the molecular type VGII mating type a in
Colombia. FEMS yeast research, 6(4), 625-635.

47
Fernández-Cuenca, F. (2004). Aplicaciones de las técnicas de PCR a la epidemiología molecular de las
enfermedades infecciosas. Enfermedades infecciosas y microbiologia clinica, 22(6), 355-360.
Firacative, C., Torres, G., Rodríguez, M. C., & Escandón, P. (2011). First environmental isolation of
Cryptococcus gattii serotype B, from Cúcuta, Colombia. Biomédica, 31(1), 118-123.
Firacative, C., Trilles, L., & Meyer, W. (2017). Advances in the understanding of the Cryptococcus
neoformans and C. gattii species complexes and cryptococcosis. Microbiology Australia,
38(3), 106-111.
Gal, A., Koss, M., Hawkins, J., Evans, S., & Einstein, H. (1986). The pathology of pulmonary
cryptococcal infections in the acquired immunodeficiency syndrome. Archives of pathology &
laboratory medicine, 110(6), 502-507.
González, G. M., Casillas-Vega, N., Garza-González, E., Hernández-Bello, R., Rivera, G., Rodríguez,
J. A., & Bocanegra-Garcia, V. (2016). Molecular typing of clinical isolates of Cryptococcus
neoformans/Cryptococcus gattii species complex from Northeast Mexico. Folia
microbiologica, 61(1), 51-56.
Guilarde, A. O., Andrade, A. C. A., De Sousa, M. A., De Oliveira, A. M., & Sugita, D. M. (2019).
Placental and pulmonary cryptococcosis associated with fungemia in patient with acquired
immunodeficiency syndrome. Le infezioni in medicina: rivista periodica di eziologia,
epidemiologia, diagnostica, clinica e terapia delle patologie infettive, 27(2), 187-189.
Hagen, F., Khayhan, K., Theelen, B., Kolecka, A., Polacheck, I., Sionov, E., . . . Boekhout, T. (2015).
Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species
complex. Fungal genetics and biology, 78, 16-48.
Kurtzman, C. P., Fell, J. W., Boekhout, T., & Robert, V. (2011). Methods for isolation, phenotypic
characterization and maintenance of yeasts. The yeasts, a taxonomic study, 5th edn. Elsevier,
Amsterdam, 87-110.
Kwon-Chung, K. (1975). A new genus, Filobasidiella, the perfect state of Cryptococcus neoformans.
Mycologia, 67(6), 1197-1200.
Kwon-Chung, K., Wickes, B. L., Booth, J., Vishniac, H. S., & Bennett, J. E. (1987). Urease inhibition
by EDTA in the two varieties of Cryptococcus neoformans. Infection and immunity, 55(8),
1751-1754.
Kwon-Chung, K. J., Fraser, J. A., Doering, T. L., Wang, Z., Janbon, G., Idnurm, A., & Bahn, Y.-S.
(2014). Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of
cryptococcosis. Cold Spring Harb Perspect Med, 4(7), a019760.
Lau, A. F., Drake, S. K., Calhoun, L. B., Henderson, C. M., & Zelazny, A. M. (2013). Development of
a clinically comprehensive database and a simple procedure for identification of molds from
solid media by matrix-assisted laser desorption ionization–time of flight mass spectrometry.
Journal of clinical microbiology, 51(3), 828-834.
Lessells, R. J., Mutevedzi, P. C., Heller, T., & Newell, M.-L. (2011). Poor long-term outcomes for
cryptococcal meningitis in rural South Africa. SAMJ: South African Medical Journal, 101(4),
251-252.
Lin, X., & Heitman, J. (2006). The biology of the Cryptococcus neoformans species complex. Annu.
Rev. Microbiol., 60, 69-105.
Lizarazo, J., Chaves, Ó., Peña, Y., Escandón, P., Agudelo, C. I., & Castañeda, E. (2012). Comparación
de los hallazgos clínicos y de supervivencia entre pacientes VIH positivos y VIH negativos
con criptococosis meníngea en un hospital de tercer nivel. Acta Médica Colombiana, 37(2),
49-61.
Lizarazo, J., Escandón, P., Agudelo, C. I., & Castañeda, E. (2014). Cryptococcosis in Colombian
children and literature review. Memórias do Instituto Oswaldo Cruz, 109(6), 797-804.
Lizarazo, J., Escandón, P., Agudelo, C. I., Firacative, C., Meyer, W., & Castañeda, E. (2014).
Retrospective study of the epidemiology and clinical manifestations of Cryptococcus gattii
infections in Colombia from 1997–2011. PLoS neglected tropical diseases, 8(11), e3272.
Lizarazo, J., Linares, M., de Bedout, C., Restrepo, Á., Agudelo, C. I., & Castañeda, E. (2007). Estudio
clínico y epidemiológico de la criptococosis en Colombia: resultados de nueve años de la
encuesta nacional, 1997-2005. Biomédica, 27(1), 94-109.
Mak, S., Vélez, N., Castañeda, E., & Escandón, P. (2015). The fungus among us: Cryptococcus
neoformans and Cryptococcus gattii ecological modeling for Colombia. Journal of Fungi, 1(3),
332-344.

48
Mak, S., Vélez, N., Castañeda, E., Escandón, P., & Group, C. (2015). The Fungus among Us:
Cryptococcus neoformans and Cryptococcus gattii Ecological Modeling for Colombia. Journal
of Fungi, 1(3), 332.
Meyer, W. (2014). Primer Curso Lationamericano en diagnostico, identificación molecular y
epidemiologia de hongos patógenos humanos Grupo de Microbiologia Instituto Naciona de
Salud
Meyer, W., Castañeda, A., Jackson, S., Huynh, M., Castañeda, E., & Group, I. C. S. (2003). Molecular
typing of IberoAmerican Cryptococcus neoformans isolates. Emerging infectious diseases,
9(2), 189.
Meyer, W., Gilgado, F., Ngamskulrungroj, P., Trilles, L., Hagen, F., Castañeda, E., & Boekhout, T.
(2011). Molecular typing of the Cryptococcus neoformans/Cryptococcus gattii species
complex Cryptococcus (pp. 327-357): American Society of Microbiology.
Meyer, W., Mitchell, T. G., Freedman, E., & Vilgalys, R. (1993). Hybridization probes for conventional
DNA fingerprinting used as single primers in the polymerase chain reaction to distinguish
strains of Cryptococcus neoformans. Journal of Clinical Microbiology, 31(9), 2274-2280.
Min, K., & Kwon-Chung, K. (1986). The biochemical basis for the distinction between the two
Cryptococcus neoformans varieties with CGB medium. Zentralblatt für Bakteriologie,
Mikrobiologie und Hygiene. Series A: Medical Microbiology, Infectious Diseases, Virology,
Parasitology, 261(4), 471-480.
Montagna, M. T., De Donno, A., Caggiano, G., Serio, F., De Giglio, O., Bagordo, F., . . . Cogliati, M.
(2018). Molecular characterization of Cryptococcus neoformans and Cryptococcus gattii from
environmental sources and genetic comparison with clinical isolates in Apulia, Italy.
Environmental research, 160, 347-352.
Noguchi, H., Hiruma, M., Maruo, K., Jono, M., Miyata, K., Tanaka, H., . . . Ihn, H. (2016). Localized
Cutaneous Cryptococcosis: summary of reported cases in Japan. Medical mycology journal,
57(3), E35-E39.
Nweze, E. I., Kechia, F. A., Dibua, U. E., Eze, C., & Onoja, U. S. (2015). Isolation of Cryptococcus
neoformans from environmental samples collected in Southeastern Nigeria. Revista do
Instituto de Medicina Tropical de São Paulo, 57(4), 295-298.
Paliwal, D., & Randhawa, H. (1978). Evaluation of a simplified Guizotia abyssinica seed medium for
differentiation of Cryptococcus neoformans. Journal of clinical microbiology, 7(4), 346-348.
Perfect, J. R. (2007). Management of cryptococcosis: how are we doing? PLoS Medicine, 4(2), e47.
Perfect, J. R., & Bicanic, T. (2015). Cryptococcosis diagnosis and treatment: what do we know now.
Fungal genetics and biology, 78, 49-54.
Perfect, J. R., Dismukes, W. E., Dromer, F., Goldman, D. L., Graybill, J. R., Hamill, R. J., . . . Nguyen,
M.-H. (2010). Clinical practice guidelines for the management of cryptococcal disease: 2010
update by the Infectious Diseases Society of America. Clinical Infectious Diseases, 50(3), 291-
322.
Pini, G., Faggi, E., & Campisi, E. (2017). Enzymatic characterization of clinical and environmental
Cryptococcus neoformans strains isolated in Italy. Revista iberoamericana de micologia,
34(2), 77-82.
Rajasingham, R., Smith, R. M., Park, B. J., Jarvis, J. N., Govender, N. P., Chiller, T. M., . . . Boulware,
D. R. (2017). Global burden of disease of HIV-associated cryptococcal meningitis: an updated
analysis. The Lancet infectious diseases, 17(8), 873-881.
Robert, V., Szoke, S., Jabas, B., Vu, D., Chouchen, O., Blom, E., & Cardinali, G. (2011). BioloMICS
software: Biological data management, identification, classification and statistics. Open
Applied Informatics Journal, 5, 87-98.
Rohatgi, S., & Pirofski, L.-a. (2015). Host immunity to Cryptococcus neoformans. Future microbiology,
10(4), 565-581.
Rojo, M. D. (2014). MALDI-TOF MS en la detección de resistencias bacterianas y estudios
epidemiologicos.
Scherl, A., Couté, Y., Déon, C., Callé, A., Kindbeiter, K., Sanchez, J.-C., . . . Diaz, J.-J. (2002).
Functional proteomic analysis of human nucleolus. Molecular biology of the cell, 13(11),
4100-4109.
Springer, D. J., Billmyre, R. B., Filler, E. E., Voelz, K., Pursall, R., Mieczkowski, P. A., . . . Filler, S.
G. (2014). Cryptococcus gattii VGIII isolates causing infections in HIV/AIDS patients in

49
Southern California: identification of the local environmental source as arboreal. PLoS Pathog,
10(8), e1004285.
Staib, F., Seibold, M., Antweiler, E., & Fröhlich, B. (1989). Staib Agar Supplemented with a Triple
Antibiotic Combination for the Detection of Cryptococcus neoformtans in Clinical Specimens:
Staib‐Agar mit einer Dreifach‐Antibiotika‐Kombination für den Nachweis von Cryptococcus
neoformans in klinischem Untersuchungsmaterial. Mycoses, 32(9), 448-454.
Stephen, C., Lester, S., Black, W., Fyfe, M., & Raverty, S. (2002). British Columbia: Multispecies
outbreak of cryptococcosis on southern Vancouver Island, British Columbia. The Canadian
Veterinary Journal, 43(10), 792.
Tello, M., Gutiérrez, E., Béjar, V., Galarza, C., Ramos, W., & Ortega-Loayza, A. G. (2013).
Cryptococcosis. Revista Médica de Risaralda, 19(2), 147-153.
Timarán, V., Antonio, D., Benavides Melo, C. J., Chaves Velásquez, C. A., Morillo Caicedo, M. I., &
Castillo Ceballos, A. M. (2016). Aislamiento de Cryptococcus neoformans en heces de
palomas (Columba livia) en el casco urbano del municipio de Pasto, Colombia. Biosalud,
15(1), 62-71.
Valenzuela-Oñate, C., Magdaleno-Tapial, J., von der Weth, M. G., Perez-Ferriols, A., Carazo, J. L. S.,
Ricart-Olmos, C., . . . de Miquel, V. A. (2019). Disseminated skin nodules in a migrant patient.
JAAD case reports, 5(5), 430-432.
Velegraki, A., Kiosses, V., Kansouzidou, A., Smilakou, S., Mitroussia-Ziouva, A., & Legakis, N.
(2001). Prospective use of RFLP analysis on amplified Cryptococcus neoformans URA 5 gene
sequences for rapid identification of varieties and serotypes in clinical samples. Medical
mycology, 39(5), 409-417.
Vilgalys, R., & Hester, M. (1990). Rapid genetic identification and mapping of enzymatically amplified
ribosomal DNA from several Cryptococcus species. Journal of bacteriology, 172(8), 4238-
4246.
Wilkins, M. R., Pasquali, C., Appel, R. D., Ou, K., Golaz, O., Sanchez, J.-C., . . . Humphery-Smith, I.
(1996). From proteins to proteomes: large scale protein identification by two-dimensional
electrophoresis and arnino acid analysis. Bio/technology, 14(1), 61.
Williamson, P. R., Jarvis, J. N., Panackal, A. A., Fisher, M. C., Molloy, S. F., Loyse, A., & Harrison,
T. S. (2017). Cryptococcal meningitis: epidemiology, immunology, diagnosis and therapy.
Nature Reviews Neurology, 13(1), 13.
Ye, F., Xie, J.-x., Zeng, Q.-s., Chen, G.-q., Zhong, S.-q., & Zhong, N.-s. (2012). Retrospective analysis
of 76 immunocompetent patients with primary pulmonary cryptococcosis. Lung, 190(3), 339-
346.
Zaragoza, O. (2019). Basic principles of the virulence of Cryptococcus. Virulence, 10(1), 490-501.
Zaragoza, O., Rodrigues, M. L., De Jesus, M., Frases, S., Dadachova, E., & Casadevall, A. (2009). The
capsule of the fungal pathogen Cryptococcus neoformans. Advances in applied microbiology,
68, 133-216.
Zhou, Y., Shen, N., Hou, H.-Y., Lu, Y., Yu, J., Mao, L., & Sun, Z. (2017). Identification accuracy of
matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF
MS) for clinical pathogenic bacteria and fungi diagnosis: a meta-analysis. Int. J. Clin. Exp.
Med, 10(2), 4057-4076.
ANEXOS
Anexo 1: Mapas de los sitios de muestreo.
50
51

52
Anexo 2: Fotos de individuos positivos para Cryptococcus neoformans

53
Anexo 3. Respuesta Minambiente contrato de Acceso a Recursos Genéticos y sus
Productos Derivados

54
Anexo 4. Divulgación de resultados. Ponencia Internacional: 10th International
conference Cryptococcus and Cryptococosis, Foz do Iquacu, Brazil from 26 -30th
March 2017.

55

56
Anexo 5. Divulgación de resultados. Ponencia nacional: "I Simposio Regional de Enfermedades Infecciosas y Buenas Prácticas clínicas" Bucaramanga noviembre
2017(certificado)

57

58
Anexo 6. Divulgación de resultados. Libro de memorias del "I Simposio Regional de
Enfermedades Infecciosas y Buenas Prácticas clínicas" Bucaramanga noviembre 2017

59
Anexo 7. Divulgación de resultados. Reconocimiento primer puesto modalidad
de poster "I Simposio Regional de Enfermedades Infecciosas y Buenas Prácticas
clínicas" Bucaramanga noviembre 2017
60
Anexo 8. Divulgación de resultados. Libro memorias del 10 International conference
Cryptococcus and Cryptococosis. Foz do Iquacu, Brazil

61
Anexo 9. Divulgación de resultados. Ponencia Oral, VII Encuentro Académico del
Programa de Bacteriología y Laboratorio Clínico, Tunja abril 2018

62
Anexo 10. Divulgación de resultados. Ponencia modalidad poster "II Simposio
Regional de Enfermedades Infecciosas y Buenas Prácticas clínicas" Bucaramanga
octubre 2019.

63

64
Anexo 11. Divulgación de resultados. Ponencia modalidad oral. 5th International week of science
Tecnology and innovation. November 2018.

65
Anexo 12. Artículo publicado: Revista MVZ Córdoba 2019: 24(1).
Link: http://revistas.unicordoba.edu.co/index.php/revistamvz/article/view/1601

66

67

68

69

70