universidade federal de goiÁs instituto de …1).pdfassigned to different t. cucumeris ags (ag1-ia,...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
IDENTIFICAÇÃO MOLECULAR E CARACTERIZAÇÃO DE ISOLADOS
DE FUNGOS DO GÊNERO RHIZOCTONIA RECUPERADOS DO
CERRADO AGRÍCOLA DO BRASIL
ANGEL JOSÉ VIEIRA BLANCO
GOIÂNIA-GO
2016

ANGEL JOSÉ VIEIRA BLANCO
IDENTIFICAÇÃO MOLECULAR E CARACTERIZAÇÃO DE ISOLADOS
DE FUNGOS DO GÊNERO RHIZOCTONIA RECUPERADOS DO
CERRADO AGRÍCOLA DO BRASIL
Tese apresentada ao Programa de Pós-
Graduação em Ciências Biológicas do
Instituto de Ciências Biológicas da
Universidade Federal de Goiás, como
requisito parcial para obtenção do título de
Doutor em Ciências Biológicas.
Área de Concentração: Bioquímica e Genética
Orientador: Prof. Dr. Cirano José Ulhoa Co-orientador: Dr. Murillo Lobo Jr.
GOIÂNIA-GO
2016

ANGEL JOSÉ VIEIRA BLANCO
IDENTIFICAÇÃO MOLECULAR E CARACTERIZAÇÃO DE ISOLADOS
DE FUNGOS DO GÊNERO RHIZOCTONIA RECUPERADOS DO
CERRADO AGRÍCOLA DO BRASIL
BANCA EXAMINADORA
_____________________________________________
Orientador: Prof. Dr. Cirano José Ulhoa Departamento de Bioquímica e Biologia Molecular– ICB-II
Universidade Federal de Goiás
_____________________________________________ Co-Orientador: Dr. Murillo Lobo Júnior
Embrapa Arroz e Feijão
_____________________________________________
Prof. Dr. Adalberto Café Filho Departamento de Fitopatologia – Instituto de Biologia
Universidade de Brasília
_____________________________________________
Profa. Dra Silvana Petrofeza da Silva Departamento de Bioquímica e Biologia Molecular– ICB-II
Universidade Federal de Goiás
_____________________________________________
Prof. Dr Marcos Gomes da Cunha Departamento de Fitopatologia – Escola de Agronomia e de Engenharia de Alimentos
Universidade Federal de Goiás
_____________________________________________
Dra. Marta Cristina Corsi de Felippi Embrapa Arroz e feijão
Aprovada em: ____/____/________

SUMÁRIO
RESUMO ................................................................................................................................................. i
ABSTRACT ............................................................................................................................................ ii
Introdução .............................................................................................................................................. 9
Objetivos .............................................................................................................................................. 17
Metodologia ......................................................................................................................................... 18
Coleta de amostras e isolamento dos Fungos .......................................................................... 18
Isolamento e quantificação do DNA Genômico ......................................................................... 22
Amplificação da região ITS e sequenciamento ......................................................................... 23
Morfologia de Colônia e Condição nuclear ................................................................................ 23
Determinação de AGs e inferência filogenética dos isolados ................................................. 23
Testes de patogenicidade em feijão e milho .............................................................................. 24
Resultados ........................................................................................................................................... 26
Identificação molecular, morfologia de colônias e condição nuclear ..................................... 26
Determinação de AG e Análises filogenéticas ........................................................................... 26
Testes de patogenicidade em Feijão e Milho ............................................................................ 32
Discussão ............................................................................................................................................ 35
Conclusões .......................................................................................................................................... 44
Referências Bibliográficas ................................................................................................................. 45
Anexos ................................................................................................................................................. 55
Artigo em formatado para submissão à revista Plant Disease. .................................................. 56

LISTA DE FIGURAS
Fig. 1. Distribuição espacial dos locais de amostragem dos isolados de Rhizoctonia
identificados neste estudo. Esses locais de amostragem abrangem as cinco principais
regiões brasileiras sob domínio do bioma Cerrado (área destacada em cinza). ............. 18
Fig. 2. Padrões culturais das colônias de AGs/variedades identificados neste trabalho. A-F:
Ceratobasidium (AGA, AGF, AGFa, AGG, AGP e AGR); G-L: T. cucumeris (AG1-IA,
AG1-IB, AG1-ID, AG1-IE, AG4-HGII e AG4-HGIII); M-O: W. circinata (var. circinata, var.
oryzae, var. zeae). ...................................................................................................................... 27
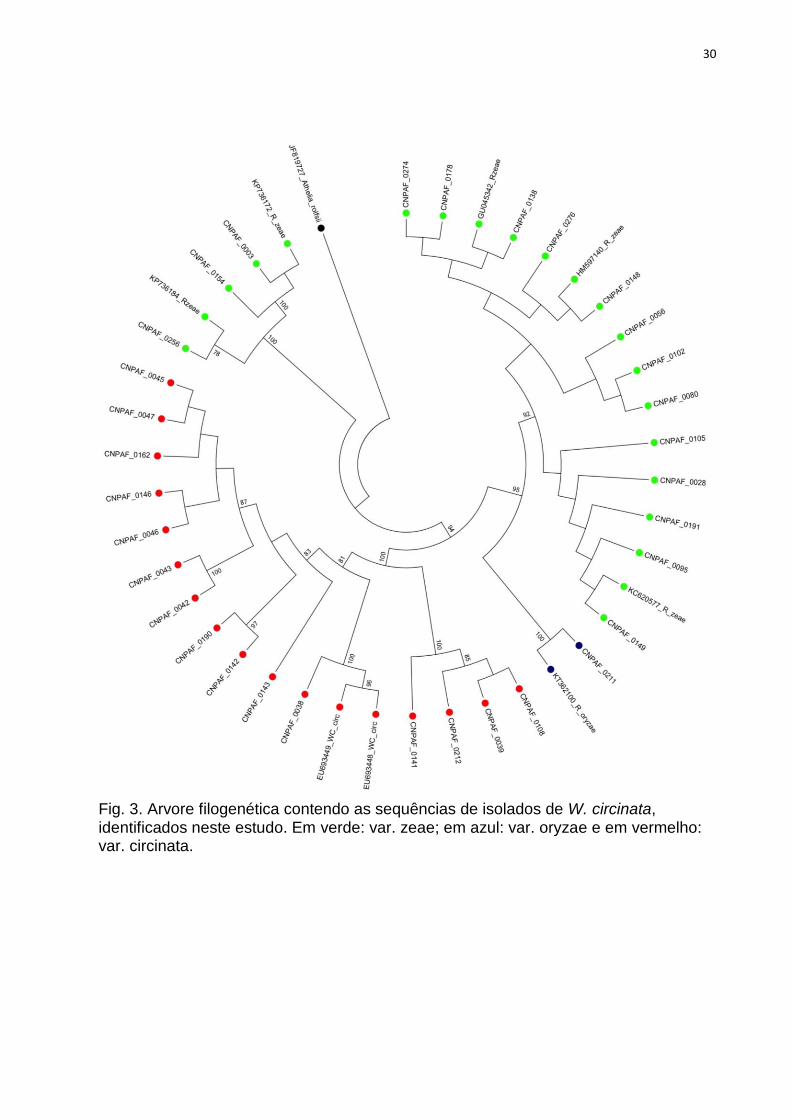
Fig. 3. Arvore filogenética contendo as sequências de isolados de W. circinata, identificados
neste estudo. Em verde: var. zeae; em azul: var. oryzae e em vermelho: var. circinata.30
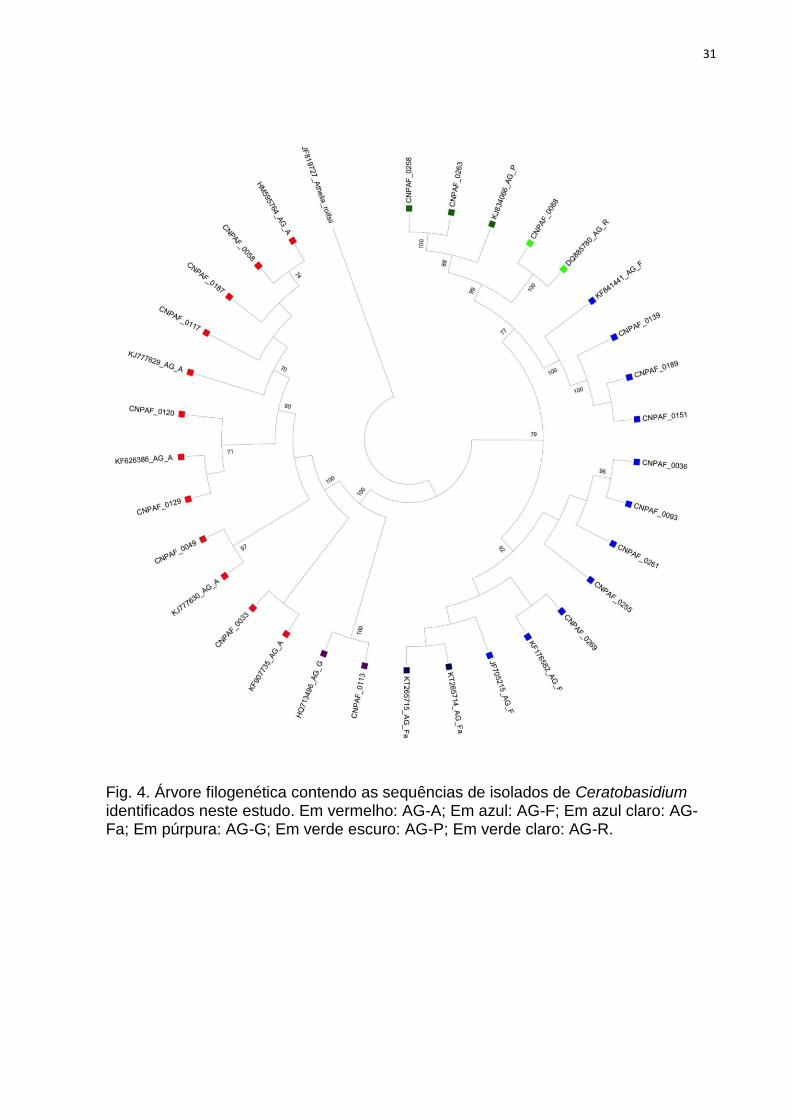
Fig. 4. Árvore filogenética contendo as sequências de isolados de Ceratobasidium
identificados neste estudo. Em vermelho: AG-A; Em azul: AG-F; Em azul claro: AG-Fa;
Em púrpura: AG-G; Em verde escuro: AG-P; Em verde claro: AG-R. ............................... 31
Fig. 5. Árvore filogenética contendo as sequências de isolados de T. Cucumeris identificados
neste estudo. Em vermelho: isolados de AG1 (ISGs IA, IB, ID e IE); em azul: isolados
AG4 (ISGs HGI e HGIII). ........................................................................................................... 32
Fig. 6. Severidade média de podridão radicular em plantas de feijão e de milho, causada por
isolados dos gêneros Ceratobasidium, Thanatephorus e Waitea. Severidade da doença
estimada através da escala proposta por Schoonhoven e Pastor-Corrales (1987). ....... 33
Fig. 7. Severidade média de podridão radicular em plantas de feijão e de milho, causada por
diferentes Grupos de Anastomose identificados neste estudo. Severidade da doença
estimada através da escala proposta por Schoonhoven e Pastor-Corrales (1987). ....... 34
Fig. 8. Distribuição geográfica da severidade da doença (podridão de raiz) no feijão,
considerando os diferentes AGs/variedades identificados. AGs não patogênicos não são
mostrados. A área da savana brasileira é destacada em cinza. ........................................ 42
Fig. 9. Distribuição geográfica da severidade da doença (podridão de raiz) no milho,
considerando os diferentes AGs/variedades identificados. AGs não patogênicos não são
mostrados. A área da savana brasileira é destacada em cinza. ........................................ 43

LISTA DE TABELAS
Tabela 1. Relação de isolados de Rhizoctonia spp. caracterizados neste trabalho com seus
respectivos números de identificação, condição nuclear, sequência mais similar (best hit
Blast para cada sequência), número de registro das sequências depositadas no
Genbank, substrato ou histórico recente de cultivo e região de coleta. ............................ 19
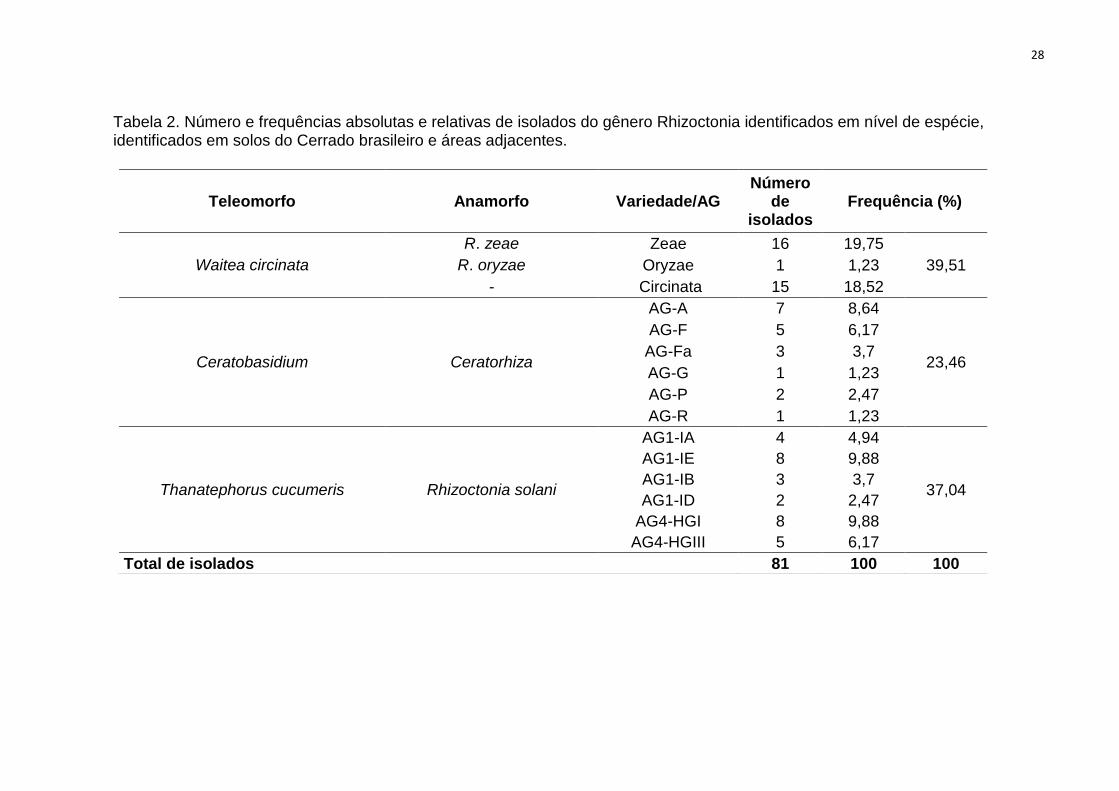
Tabela 2. Número e frequências absolutas e relativas de isolados do gênero Rhizoctonia
identificados em nível de espécie, identificados em solos do Cerrado brasileiro e áreas
adjacentes. .................................................................................................................................. 28

i
RESUMO
Fungos do gênero Rhizoctonia constituem um complexo grupo taxonômico de
fitopatógenos. Espécies multinucleadas como Thanatephorus cucumeris
(Rhizoctonia solani), Waitea circinata var. oryzae (Rhizoctonia oryzae), Waitea
circinata var. zeae (Rhizoctonia zeae), Waitea circinata var. circinata (anamorfo não
definido) e espécies de Rhizoctonia binucleadas estão associados a muitas doenças
em diversas plantas economicamente importantes, incluindo as principais espécies
utilizadas na alimentação humana, como feijão, arroz, milho e trigo. Com o objetivo
principal de avaliar a distribuição de espécies e grupos intraespecíficos do gênero
Rhizoctonia no Cerrado agrícola brasileiro e áreas de transição, 81 isolados foram
identificados através de métodos moleculares e de coloração de núcleos após serem
recuperados de amostras de solo. Além da caracterização morfológica e cultural dos
isolados, análises filogenéticas utilizando a região ITS1-5.8S-ITS2 confirmaram a
identificação de quase todos os isolados realizada através do algoritmo Blast do
Genebank – NCBI. Aproximadamente 40 % dos isolados recuperados pertencem a
anamorfos de W. circinata (variedades zeae, oryzae e circinata); 37,04% à grupos
de anastomose (AG) do teleomorfo T. cucumeris (AG1-IA, AG1-IB, AG1-ID, AG1-IE,
AG4-HGI e AG4-HGIII) e 23,46% a AGs de Ceratobasidium (AG-A, AG-F, AG-Fa,
AG-G, AG-P e AG-R). Aparentemente é o primeiro relato sobre a ocorrência da
variedade circinata no Brasil. AG1 foi identificado entre o Centro e o Norte do Brasil;
AG3 entre o Centro e o Sul; R. zeae em regiões de grande produção de Milho e AG-
F e AG-Fa em áreas de produção de Feijão Os resultados sugerem que a cultura
adotada e as características ambientais podem ter sido determinantes na
distribuição geográfica dos AGs/variedades encontrados. Testes de patogenicidade
mostraram que parte significativa dos isolados estudados são patogênicos a milho e
feijão e que os AGs/variedades identificados apresentaram diferenças significativas
na agressividade a essas duas plantas.
Palavras-Chave: Genética de Microrganismos; Rhizoctonia; Cerrado agrícola;
Fitopatógen

ii
ABSTRACT
Fungi from Rhizoctonia genus constitute a complex taxonomic group of
phytopathogens. Multinucleated species such as Thanatephorus cucumeris
(Rhizoctonia solani), Waitea circinata var. oryzae (Rhizoctonia oryzae), Waitea
circinata var. Zeae (Rhizoctonia zeae), Waitea circinata var. Circinata (anamorph not
defined) and binucleate Rhizoctonia species, are associated with many diseases in
several economically important crops, including species used in human nutrition,
such as beans, rice, corn and wheat. In order to evaluate the species distribution and
intraspecific groups of Rhizoctonia in Brazilian agricultural Cerrado and transitional
areas, 81 isolates were identified through molecular methods and nucleate coloring
after being recovered from soil samples. In addition to the morphological and cultural
characterization of the isolates, phylogenetic analyzes using the ITS1-5.8S-ITS2
region confirmed the identification of almost all isolates using the Genebank-NCBI
Blast algorithm. Approximately 40% of the recovered isolates belong to W. circinata
anamorphs (zeae, oryzae and circinata varieties); 37.04% of total isolates were
assigned to different T. cucumeris AGs (AG1-IA, AG1-IB, AG1-ID, AG1-IE, AG4-HGI
and AG4-HGIII) and 23.46% to different Ceratobasidium AGs (AG-A, AG-F, AG-Fa,
AG-G, AG-P and AG-R). Apparently, it is the first report of the circinata variety in
Brazil. AG1 was identified between the Center and the North of Brazil; AG3 between
Central and South; R. zeae in regions of high production of maize and AG-F and AG-
Fa in Bean production areas. The results suggests that cultivation mechanisms and
environmental characteristics are determinant on geographical distribution of AGs
varieties used in this study. Pathogenicity tests showed that a significant part of the
isolates studied are pathogenic and presented a significant variation of
aggressiveness in maize and beans.
Keywords: Genetics of Microorganisms; Rhizoctonia; agricultural Cerrado;
Phytopathogens

9
Introdução
Os fungos são considerados importantes constituintes de ecossistemas
naturais e podem interagir de modo positivo ou negativo com outros organismos
(Sha et al. 2008), exercendo várias atividades ecológicas funcionais, sob influência
de diferentes parâmetros ambientais (Branco et al. 2013). Embora sejam vistos por
grande parte da população leiga como organismos prejudiciais, pois, de fato, alguns
deles são perigosos à saúde de plantas e animais, incluindo a espécie humana, a
maioria é benéfica, podendo-se admitir que sem os fungos dificilmente outras
espécies poderiam existir (Azevedo e Pizzirani-Kleiner 2002).
Bruns et al. (1991) destacam uma série de razões que enfatizam a grande
importância dos fungos: a) são decompositores primários em todos os ecossistemas
terrestres, b) formam importantes associações simbióticas, envolvendo mutualismo
ou parasitismo, com plantas vasculares, c) constituem a esmagadora maioria de
patógenos de plantas e por isso provocam impacto econômico significativo (vários
patógenos humanos importantes também são fungos), d) oferecem vários modelos
genéticos para biologia molecular (ex: Saccharomyces cerevisiae, Neurospora
crassa, Aspergillus nidulans), e e) são cruciais na indústria de fermentação e
biotecnologia.
De fato, a literatura especializada mostra que os fungos possuem aplicações
diversas, quase sempre associadas de maneira estreita ao cotidiano dos seres
humanos, como por exemplo, a confecção de produtos e serviços. Alimentos e
bebidas, ácidos orgânicos, enzimas, aromas e fragrâncias, agentes cardiovasculares
e esteroides (Melo 2002), representam exemplos de produtos oriundos da ação
direta ou indireta de fungos. Além disso, estes microrganismos encontram-se
também relacionados a segmentos considerados estrategicamente importantes para
o desenvolvimento de qualquer país, como meio ambiente e energia (Azevedo e
Pizzirani-Kleiner 2002).
Diversas indústrias têm utilizado a capacidade bioquímica dos fungos para
produzir substâncias orgânicas: há alguns anos leveduras começaram a ser
utilizadas na produção de proteínas para enriquecimento de ração animal (Raven e
Johnson 2002); e espécies do gênero Aspergillus na produção em larga-escala de
ácido cítrico e enzimas de aplicação industrial como amilases, proteases e lipases
(May e Adams 1997). Muitos antibióticos, inclusive o primeiro utilizado em larga

10
escala (penicilina) são derivados de fungos. Há fungos também sendo utilizados na
recuperação de ambientes contaminados, onde atuam convertendo moléculas
orgânicas complexas em outras, desintoxicando o meio (Raven e Johnson 2002) e
diminuindo a emissão de gases de efeito estufa (Lange 2014).
Alguns fungos estão direta ou indiretamente relacionados com a ocorrência e
o controle de doenças em lavouras. Avanços atuais neste campo têm permitido o
trabalho conjunto entre melhoristas, microbiologistas e produtores na busca de
plantas mais resistentes e de soluções alternativas para minimizar as perdas
recorrentes no setor agrícola. Nesse sentido, é uma premissa básica conhecer a
variabilidade dos fungos fitopatógenos, principalmente aqueles de maior importância
econômica e social, como é o caso do gênero Rhizoctonia.
As Rhizoctonias são um importante gênero de fitopatógenos que podem se
associar a diversos órgãos de plantas (Basseto et al. 2008; González 2013;
Kuramae et al. 2007; Youssef et al. 2012). São organismos cosmopolitas, que
podem ser encontradas em solos utilizados para a agricultura ou mantidos sob
vegetação nativa (González-Garcia et al. 2006). Suas espécies se destacam pela
capacidade de parasitar e causar doenças importantes em diversas espécies de
plantas (Sneh et al. 1991). As espécies do gênero Rhizoctonia são altamente
diversificadas, apresentando ciclo de vida com dois estágios sexuais (sexuado ou
teleomórfico e assexuado ou anamórfico).
Além de parasitas de plantas economicamente importantes, o gênero
Rhizoctonia apresenta ainda espécies saprófitos e simbiontes (González-Garcia et
al. 2006). Há também no grupo espécies mutualísticas (Otero et al. 2002) e com
potencial para ser explorado no controle biológico (Sneh et al. 1991). Dessa forma,
observa-se que estes fungos assumem importância em diversos contextos, sendo
organismos particularmente interessantes do ponto de vista agronômico, econômico
e também ecológico.
A primeira descrição de espécies de Rhizoctonia foi realizada por De Candolle
em 1815 (Sneh et al. 1991) e, desde então, muitas espécies foram identificadas e
incluídas no gênero conforme revisado por González-Garcia et al. (2006). No
passado, isolados de Rhizoctonia eram classificados de acordo com características
culturais (morfologia de colônia e coloração de micélio) e quanto ao número de
núcleos, (Sneh et al. 1991). Entretanto, a taxonomia de Rhizoctonia utilizando
apenas análise morfológica de colônias sempre foi incerta, uma vez que a ausência

11
de conídios e condição imperfeita do fungo complicam o processo de identificação.
Além disso, Rhizoctonia representa uma mistura complexa de fungos filamentosos
que compartilham características em seu estado anamórfico (González-Garcia et al.
2006), o que também dificulta a escolha de caracteres úteis para descrever e
diferenciar espécies dentro do gênero.
Atualmente, além da análise morfológica, coloração de núcleos e grupos de
anastomose (AG), o sequenciamento da região ITS1-5.8S-ITS2 do rDNA é um dos
principais métodos utilizados para organizar (e reorganizar também) a taxonomia de
Rhizoctonia. Dados sobre a ultraestrutura do fungo, patogenicidade e distribuição
geográfica, são levantados para se caracterizar isolados e definir sua posição
sistemática dentro do gênero (Carling et al. 2002a; Fang et al. 2013; González 2013;
González et al. 2006; Kuninaga et al. 2000; Kuramae et al. 2007; Mathew et al. 2012;
Sharon et al. 2008; 2006; Vilgays e Cubeta 1994), Além disso, o avanço dos
métodos de bioestatística tem permitido também a caracterização de genes e da
diversidade genética das espécies e populações de Rhizoctonia (Ceresini et al.
2003; Ciampi et al. 2005), disponibilizando assim informações cada vez mais
refinadas para identificar e classificar de maneira acurada, isolados coletados nos
mais diferentes tipos de solo ou hospedeiros.
Muitos estudos têm descrito a diversidade genética de Rhizoctonia em uma
série de plantas cultivadas, tais como: alface (Youssef et al. 2012); arroz
(Chaijuckam e Davis 2010; Ciampi et al. 2009; Taheri et al. 2007); batata (Ceresini et
al. 2003; Kuninaga et al. 2000); couve (Kuramae et al. 2007); ervilha (Mathew et al.
2012), feijão (Godoy-Lutz et al. 2003); melancia (Aiello et al. 2012); morango (Fang
et al. 2013); seringueira (Ceresini et al. 2010; Youssef et al. 2012); soja (Ciampi et al.
2009; Ciampi et al. 2005; Fenille et al. 2003; González 2013); tabaco (Kuninaga et al.
2000) e milho (Summer e Minton 1989).
Também já foram observados ataques de Rhizoctonia spp. a espécies
florestais como a teca (Poltronieri et al. 2008), e o ipê (Costa et al. 2013); em plantas
ornamentais como azaleia (Rinehart et al. 2007) e bastão-do-imperador (Verginassi
et al. 2008) e, até mesmo, em gramíneas invasoras (Chen et al. 2009).
Existem três principais grupos de espécies de Rhizoctonia associados a
doenças de plantas: Thanatephorus cucumeris, espécie multinucleada (anamorfo =
Rhizoctonia solani); Ceratobasidium sp, (compreende espécies binucleadas e são
ocasionalmente classificadas no gênero anamorfo Ceratorhiza), e Waitea circinata,

12
espécie multinucleada que tem cinco anamorfos: Rhizoctonia zeae, Rhizoctonia
oryzae, Waitea circinata var. circinata (Gunnell 1986; Leiner e Carling 1994; Roberts
1999, Vilgays e Cubeta 1994), além de dois outros anamorfos recentemente
relacionados a W. circinata: var. prodigus e var. agrostis (Kammerer et al. 2011;
Toda et al. 2007), cujo epíteto “Rhizoctonia” ainda não foi adotado.
Das espécies anteriormente descritas, T.cucumeris é reconhecidamente a
mais importante, causando doenças em mais de 200 espécies de plantas, incluindo
Arroz, Milho, Trigo, Soja, Algodão, além de espécies florestais, frutíferas, gramíneas
e plantas ornamentais (Gonzalez et al. 2006). Talvez por isso haja tanto interesse
em estudar este fungo, que foi inicialmente descrito por Julius Kühn, em batata, em
1858 (Kareen e Hassan, 2013). Estudos já realizados no Brasil relataram a
ocorrência de R. Solani em culturas importantes como: feijão (Ceresini e Souza
1997; Godoy-Lutz et al. 2003; Kuramae et al. 2002); arroz (Souza et al. 2007); e soja
(Yorinori 1998; Meyer e Yorinori 1999).
De acordo com os critérios utilizados tradicionalmente para reações de
anastomose de hifas, isolados de T. cucumeris são divididos em 14 grupos de
anastomose (AG): AG 1-13, além AG-BI (Sneh et al. 1991; Carling et al. 2002b;
Guillemaut et al., 2014). Além disso, muitos AGs agrupam indivíduos que diferem
por uma ou mais características, sendo por isso subdivididos em subgrupos (grupos
intraespecíficos – ISG). Essa subdivisão considera variações em características
genéticas, bioquímicas, morfológicas e de patogenicidade, entre outras, observadas
entre isolados dentro de um mesmo AG (Ogoshi 1987), permitindo a diferenciação
de 23 ISGs, dentro dos 14 AGs de R. solani (Carling 2000).
O acúmulo de informações geradas pelos diferentes métodos de
caracterização de isolados, incluindo as mais diversas técnicas e modelos
matemáticos utilizados nas análises de agrupamentos, faz com que o sistema de
classificação de T. cucumeris esteja em constante atualização. Exemplo disso são
as evidências filogenéticas obtidas por Carling et al. (2002-b), nas quais observou-se
o agrupamento de isolados de AG-BI num cluster de AG-2, o que fez com que estes
autores propusessem a inclusão de AG-BI como um subgrupo de AG-2, com a
denominação de AG2-BI, o que reduziria o número de AGs de T. Cucumeris para
13, (Lübeck, 2004). Informações detalhadas sobre os hospedeiros e a
sintomatologia provocada por cada um dos AGs e seus respectivos ISGs em T.

13
cucumeris podem ser encontradas em diversas revisões sobre a espécie (González-
Garcia et al. 2006; Nakatani 2006; Genhua e Chengyum 2012).
O gênero teleomorfo Ceratobasidium é bastante diversificado,
compreendendo patógenos, saprófitas e formadores de micorrizas (Ogoshi 1987).
Como patógeno, Ceratobasidium spp podem causar o tombamento (damping off em
inglês), cujos sintomas são podridão das raízes, talos e sementes. Outras
consequências da ação desses patógenos incluem a queima da bainha em milho,
deterioração dos frutos e a mela em uma significativa variedade de plantas
importantes para agricultura (Li 1998; Nechet e Half-Vieira, 2006; Sneh et al. 1991;
Yan, 1984). Entre seus hospedeiros estão o feijão (Cardoso e Echandi, 1987); a
batata (Escandi e Echandi, 1991); o pepino (Villajuan-Abgona et al. 1996); o repolho
(Ross et al., 1998); e o Arroz (Taheri, et al., 2007).
Assim como acontece com T. cucumeris, a classificação das formas de
Ceratobasidium, leva em consideração a anastomose de hifas. O sistema de
classificação dos anamorfos de Ceratobasidium também é muito dinâmico e como
discutido por Sharon et al. (2008), o número de grupos de anastomose, que chegou
a 21, já foi reduzido para 16. Na realidade, na década de 1980 existiam dois
sistemas de classificação para as formas de Ceratobasidium, que acabaram se
fundindo, prevalecendo a nomenclatura do sistema japonês (Anastomosis Groups -
AGs) em detrimento ao sistema americano (Ceratobasidium Anastomosis Groups -
CAGs). Como era menos abrangente e todos os isolados de seus diferentes CAGs
estavam também presentes no sistema japonês, o sistema americano, proposto por
Burpee et al. (1980), deixou de ser utilizado e os seus sete CAGs foram
subsequentemente incorporados ao sistema japonês de AGs desenvolvido por
Ogoshi et al. (1983).
Na década de 80, a espécie W. circinata era classificada em três variedades,
circinata, oryzae e zeae, com base na morfologia de colônias no estado anamorfo,
incluindo coloração e diâmetro de escleródios (Gunnell 1986). Posteriormente,
Leiner e Carling (1994) assumiram W. circinata var. oryzae e Waitea circinata var.
zeae, como os teleomorfos de Rhizoctonia zeae e Rhizoctonia oryzae,
respectivamente, embora, tivessem afirmado que a nomenclatura para o anamorfo
de W. circinata var. circinata ainda não estava plenamente estabelecida.
Atualmente, existem designações para cinco variedades dentro de Waitea
circinata, com base na biologia molecular e na análise de diferenças genéticas:

14
oryzae, zeae, circinata, agrostis e prodigus (Leiner e Carling 1994; Sharon et al.
2006; De la Cerda et al. 2007; Toda et al. 2007), entretanto, não é comum encontrar
na literatura, designações correspondentes para os anamorfos das variedades
circinata, agrostis e prodigus como acontece em R. oryzae e R. zeae.
As variedades de Waitea circinata anteriormente descritas causam uma série
de diferentes doenças em gramíneas (Kammerer 2011). Nesse sentido, esse mesmo
autor propôs o reconhecimento da variedade prodigus, com base em características
morfológicas e análise de dados de sequências de DNA, após isolar o fungo de
grama em campos de golfe nos EUA. Trabalhos envolvendo a variedade prodigus
ainda são escassos na literatura, assim como dados moleculares no GenBank.
A variedade zeae parece ser a mais importante dentro do gênero Waitea, não
só pela quantidade de diferentes hospedeiros: arroz (Oniki et al. 1985); milho
(Sumner e Bell 1982); cebola (Erper et al. 2006); beterraba (Kuznia e Windels 1994);
trigo e cevada (Ogoshi et al. 1990), mas também pela quantidade de estudos
disponíveis na literatura. Esta espécie é mundialmente distribuída e já foi isolada em
países como China (Li et al. 1998); Brasil (Poltronieri et al. 2008); Argentina
(Gutiérrez et al. 2007) e Irã (Telmadarrehei et al. 2011), entre outros.
O método de identificação de Rhizoctonia baseado na anastomose de hifas
foi proposto por Matsumoto e colaboradores há aproximadamente 90 anos, e implica
que isolados que possuem a habilidade de se fundir são geneticamente relacionados
(González 2013). Embora ainda seja válido e de nos últimos anos ter sido
corroborado por dados genéticos (Carling et al. 2002b; González-Garcia et al. 2006;
Sharon et al. 2008), as reações de anastomose nem sempre são bem definidas, pois
há isolados que geneticamente se posicionam de maneira intermediária entre dois
AGs, podendo assim fundir suas hifas com ambos. Além disso, alguns isolados
podem, inclusive, perder a capacidade relacionada à fusão de suas hifas (Sharon et
al. 2008), e assim dificultar a definição de sua posição sistemática.
A distinção de espécies em meio de cultura é difícil na maioria das vezes,
principalmente devido à ausência de características morfológicas estáveis para
suportar a classificação do gênero Rhizoctonia (Nakatani, 2006). Nesse sentido, nos
últimos anos, métodos moleculares vêm sendo amplamente utilizados para confirmar
a identidade de espécies, variedades e AGs de Rhizoctonia, com destaque para a
análise da região intergênica do DNA ribossomal (ITS-rDNA), que vem se

15
consolidando como uma importante ferramenta na elucidação da sistemática do
gênero (Kuninaga et al. 1997; Sharon et al. 2006, 2008; White et al. 1990;).
Identificar isolados de Rhizoctonia é uma ação importante para fins de manejo
dos sistemas de produção agrícola, pois a gama de hospedeiras, a capacidade
saprofítica e a ausência de cultivares resistentes dificultam o controle desses fungos
através de resistência genética e práticas culturais. Isto porque muitos AGs são
patógenos de diferentes hospedeiros, normalmente utilizados em sistemas de
rotação de cultura por critérios econômicos ou sociais, contribuindo assim para o
aumento anual do inóculo no solo, que pode levar a perdas consideráveis de
produtividade (Lanoiselet 2005; Paulitz e Okubara 2006). Em especial, é importante
considerar que a aplicação de fungicidas pode ser uma estratégia de controle pouco
efetiva uma vez que a eficácia desses compostos em Rhizoctonia varia entre
diferentes espécies e AGs (Amaradasa et al. 2014; Carling et al. 1990; Kataria et al.
1991).
Apesar de já haver uma grande quantidade de estudos sobre a diversidade de
Rhizoctonia no Brasil (Ceresini et al. 2010; Ciampi et al. 2009; 2005) pouco se sabe
sobre a distribuição de suas espécies no Cerrado agrícola e em áreas de transição
com outros biomas, uma vez que a maioria dos estudos sobre a diversidade de
Rhizoctonia no Brasil é geralmente direcionada para uma cultura específica ou para
algum grupo de anastomose relacionado a uma cultura de interesse (Basseto et al.
2008; Ceresini et al. 2010; Ciampi et al. 2005, 2009; Fenille et al. 2003; Goulart et al.
2011; Kuramae et al. 2007; Souza et al. 2007).
Com exceção do trabalho realizado por Bolkan e Ribeiro (1985), pouco se
sabe sobre a distribuição de espécies de Rhizoctonia em maior escala geográfica. A
escassez de informações sobre a distribuição e diversidade de espécies de
Rhizoctonia limita a compreensão sobre os mecanismos que favorecem a dispersão
e o estabelecimento destes fitopatógenos em regiões produtivas do Cerrado agrícola
e em áreas de transição com outros biomas.
O Cerrado é o segundo maior Bioma brasileiro, distribuindo-se por
aproximadamente dois milhões de Km2, equivalentes a 23,92% do território nacional,
(IBGE, 2004). Considerando a extensão territorial deste bioma, a possibilidade de
obtenção de duas a três safras anuais e a preferência de produtores por poucas
espécies cultivadas (ex: milho, soja, feijão comum e hortaliças, hospedeiras de
Rhizoctonias), é provável que as práticas culturais prevalentes e os fatores

16
ambientais ajudam a regular a distribuição de espécies e AGs de Rhizoctonia nos
solos do Cerrado agrícola e áreas de transição.

17
Objetivos
Este trabalho teve como objetivos:
i. Identificar espécies e grupos intraespecíficos do gênero Rhizoctonia,
recuperados de solos de áreas agrícolas do Cerrado brasileiro e áreas
de transição;
ii. Estimar possíveis relações entre a distribuição de espécies, práticas
agrícolas e a diversidade climática na região-alvo;
iii. Verificar a patogenicidade dos isolados em milho e feijão, devido à
importância econômica e social destas espécies.

18
Metodologia
Coleta de amostras e isolamento dos Fungos
Os isolados utilizados neste estudo foram obtidos em diferentes áreas
agrícolas e vegetação nativa do Cerrado e áreas de transição com este bioma,
distribuídas pelas cinco regiões brasileiras. Assim, a distribuição espacial das
amostras se deu da seguinte forma: Norte (Roraima e Tocantins); Nordeste (Bahia);
Centro-Oeste (Goiás e Mato Grosso); Sudeste (Minas Gerais e São Paulo); Sul
(Paraná). As amostras compostas de solos foram coletadas na camada 0-10 cm,
armazenadas em sacos plásticos, sendo posteriormente armazenadas em câmara
fria a 4 + 1ºC. A distribuição espacial dos locais de coleta no território brasileiro está
apresentada na Figura 1. A relação contendo os municípios sede, as quais
pertencem as áreas de coleta e suas coordenadas geográficas podem ser
observadas na Tabela 1.
Fig. 1. Distribuição espacial dos locais de amostragem dos isolados de Rhizoctonia identificados neste estudo. Esses locais de amostragem abrangem as cinco principais regiões brasileiras sob domínio do bioma Cerrado (área destacada em cinza).
19
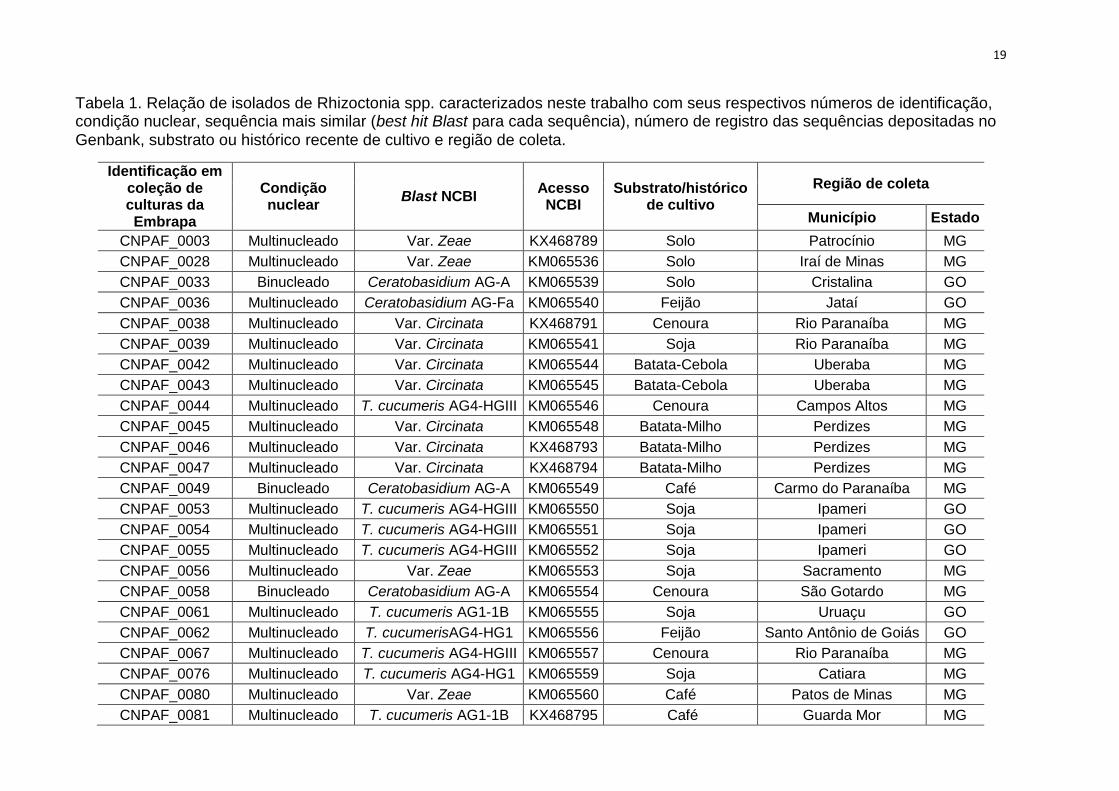
Tabela 1. Relação de isolados de Rhizoctonia spp. caracterizados neste trabalho com seus respectivos números de identificação, condição nuclear, sequência mais similar (best hit Blast para cada sequência), número de registro das sequências depositadas no Genbank, substrato ou histórico recente de cultivo e região de coleta.
Identificação em coleção de culturas da Embrapa
Condição nuclear
Blast NCBI Acesso NCBI
Substrato/histórico de cultivo
Região de coleta
Município Estado
CNPAF_0003 Multinucleado Var. Zeae KX468789 Solo Patrocínio MG
CNPAF_0028 Multinucleado Var. Zeae KM065536 Solo Iraí de Minas MG
CNPAF_0033 Binucleado Ceratobasidium AG-A KM065539 Solo Cristalina GO
CNPAF_0036 Multinucleado Ceratobasidium AG-Fa KM065540 Feijão Jataí GO
CNPAF_0038 Multinucleado Var. Circinata KX468791 Cenoura Rio Paranaíba MG
CNPAF_0039 Multinucleado Var. Circinata KM065541 Soja Rio Paranaíba MG
CNPAF_0042 Multinucleado Var. Circinata KM065544 Batata-Cebola Uberaba MG
CNPAF_0043 Multinucleado Var. Circinata KM065545 Batata-Cebola Uberaba MG
CNPAF_0044 Multinucleado T. cucumeris AG4-HGIII KM065546 Cenoura Campos Altos MG
CNPAF_0045 Multinucleado Var. Circinata KM065548 Batata-Milho Perdizes MG
CNPAF_0046 Multinucleado Var. Circinata KX468793 Batata-Milho Perdizes MG
CNPAF_0047 Multinucleado Var. Circinata KX468794 Batata-Milho Perdizes MG
CNPAF_0049 Binucleado Ceratobasidium AG-A KM065549 Café Carmo do Paranaíba MG
CNPAF_0053 Multinucleado T. cucumeris AG4-HGIII KM065550 Soja Ipameri GO
CNPAF_0054 Multinucleado T. cucumeris AG4-HGIII KM065551 Soja Ipameri GO
CNPAF_0055 Multinucleado T. cucumeris AG4-HGIII KM065552 Soja Ipameri GO
CNPAF_0056 Multinucleado Var. Zeae KM065553 Soja Sacramento MG
CNPAF_0058 Binucleado Ceratobasidium AG-A KM065554 Cenoura São Gotardo MG
CNPAF_0061 Multinucleado T. cucumeris AG1-1B KM065555 Soja Uruaçu GO
CNPAF_0062 Multinucleado T. cucumerisAG4-HG1 KM065556 Feijão Santo Antônio de Goiás GO
CNPAF_0067 Multinucleado T. cucumeris AG4-HGIII KM065557 Cenoura Rio Paranaíba MG
CNPAF_0076 Multinucleado T. cucumeris AG4-HG1 KM065559 Soja Catiara MG
CNPAF_0080 Multinucleado Var. Zeae KM065560 Café Patos de Minas MG
CNPAF_0081 Multinucleado T. cucumeris AG1-1B KX468795 Café Guarda Mor MG
20
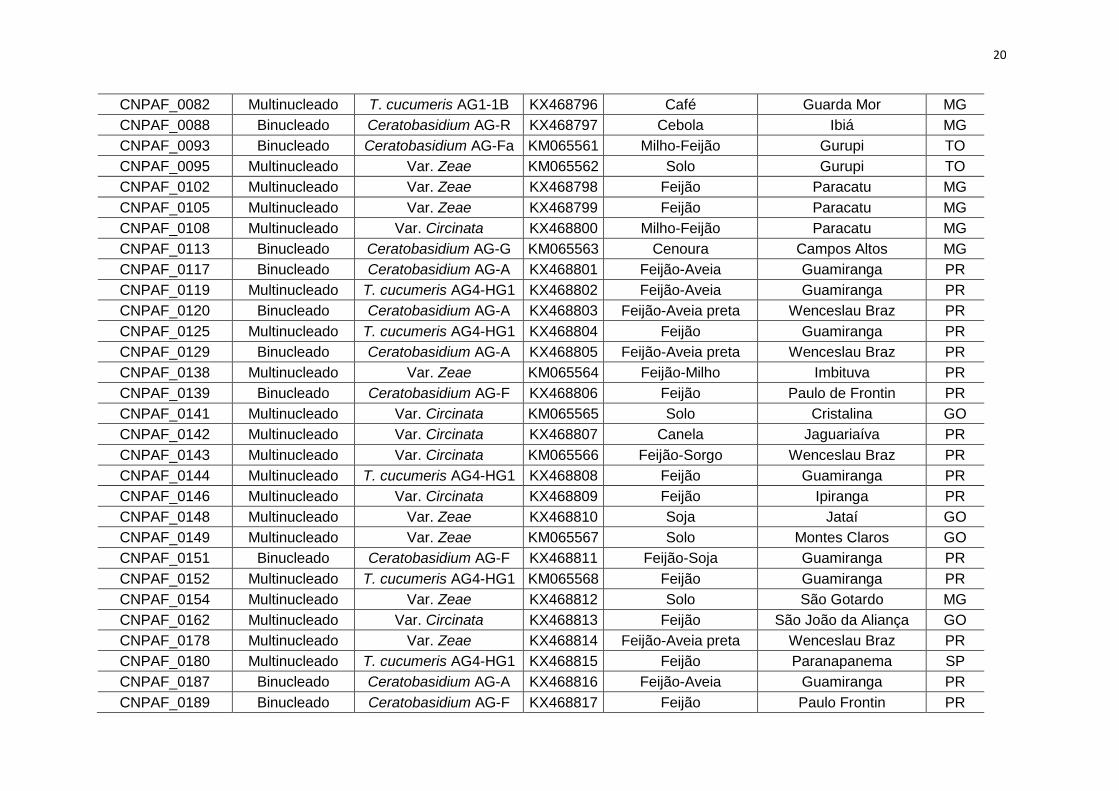
CNPAF_0082 Multinucleado T. cucumeris AG1-1B KX468796 Café Guarda Mor MG
CNPAF_0088 Binucleado Ceratobasidium AG-R KX468797 Cebola Ibiá MG
CNPAF_0093 Binucleado Ceratobasidium AG-Fa KM065561 Milho-Feijão Gurupi TO
CNPAF_0095 Multinucleado Var. Zeae KM065562 Solo Gurupi TO
CNPAF_0102 Multinucleado Var. Zeae KX468798 Feijão Paracatu MG
CNPAF_0105 Multinucleado Var. Zeae KX468799 Feijão Paracatu MG
CNPAF_0108 Multinucleado Var. Circinata KX468800 Milho-Feijão Paracatu MG
CNPAF_0113 Binucleado Ceratobasidium AG-G KM065563 Cenoura Campos Altos MG
CNPAF_0117 Binucleado Ceratobasidium AG-A KX468801 Feijão-Aveia Guamiranga PR
CNPAF_0119 Multinucleado T. cucumeris AG4-HG1 KX468802 Feijão-Aveia Guamiranga PR
CNPAF_0120 Binucleado Ceratobasidium AG-A KX468803 Feijão-Aveia preta Wenceslau Braz PR
CNPAF_0125 Multinucleado T. cucumeris AG4-HG1 KX468804 Feijão Guamiranga PR
CNPAF_0129 Binucleado Ceratobasidium AG-A KX468805 Feijão-Aveia preta Wenceslau Braz PR
CNPAF_0138 Multinucleado Var. Zeae KM065564 Feijão-Milho Imbituva PR
CNPAF_0139 Binucleado Ceratobasidium AG-F KX468806 Feijão Paulo de Frontin PR
CNPAF_0141 Multinucleado Var. Circinata KM065565 Solo Cristalina GO
CNPAF_0142 Multinucleado Var. Circinata KX468807 Canela Jaguariaíva PR
CNPAF_0143 Multinucleado Var. Circinata KM065566 Feijão-Sorgo Wenceslau Braz PR
CNPAF_0144 Multinucleado T. cucumeris AG4-HG1 KX468808 Feijão Guamiranga PR
CNPAF_0146 Multinucleado Var. Circinata KX468809 Feijão Ipiranga PR
CNPAF_0148 Multinucleado Var. Zeae KX468810 Soja Jataí GO
CNPAF_0149 Multinucleado Var. Zeae KM065567 Solo Montes Claros GO
CNPAF_0151 Binucleado Ceratobasidium AG-F KX468811 Feijão-Soja Guamiranga PR
CNPAF_0152 Multinucleado T. cucumeris AG4-HG1 KM065568 Feijão Guamiranga PR
CNPAF_0154 Multinucleado Var. Zeae KX468812 Solo São Gotardo MG
CNPAF_0162 Multinucleado Var. Circinata KX468813 Feijão São João da Aliança GO
CNPAF_0178 Multinucleado Var. Zeae KX468814 Feijão-Aveia preta Wenceslau Braz PR
CNPAF_0180 Multinucleado T. cucumeris AG4-HG1 KX468815 Feijão Paranapanema SP
CNPAF_0187 Binucleado Ceratobasidium AG-A KX468816 Feijão-Aveia Guamiranga PR
CNPAF_0189 Binucleado Ceratobasidium AG-F KX468817 Feijão Paulo Frontin PR
21
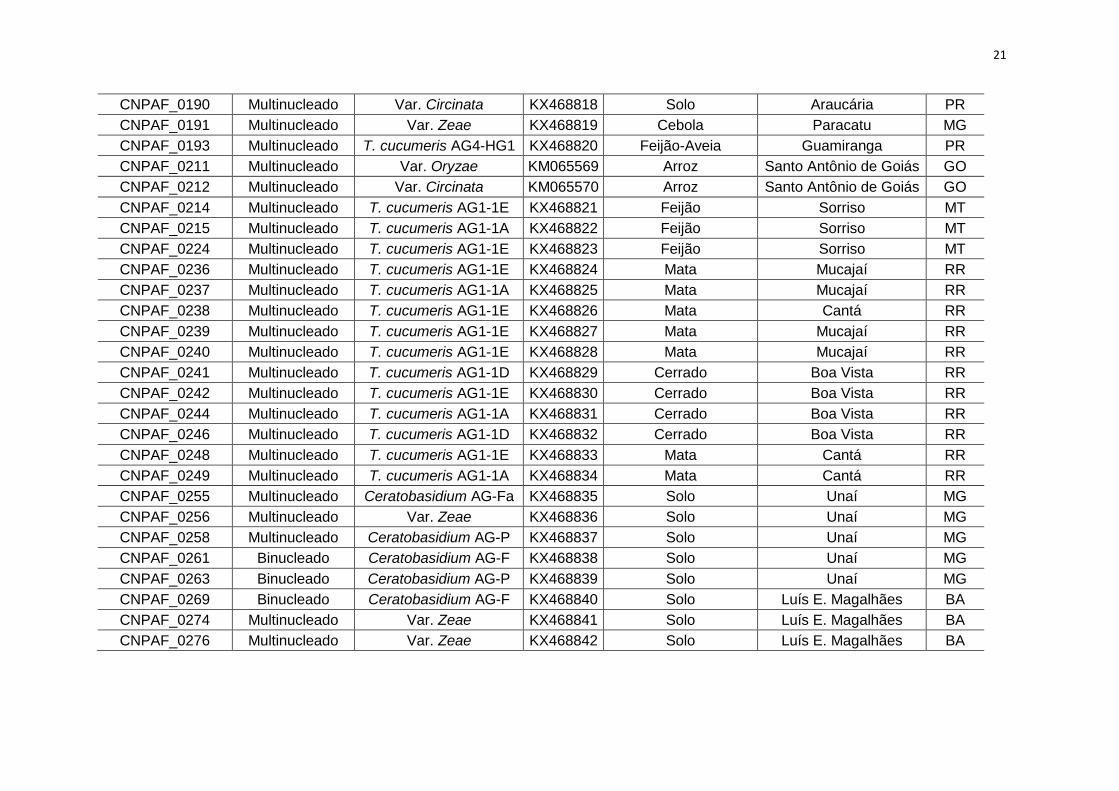
CNPAF_0190 Multinucleado Var. Circinata KX468818 Solo Araucária PR
CNPAF_0191 Multinucleado Var. Zeae KX468819 Cebola Paracatu MG
CNPAF_0193 Multinucleado T. cucumeris AG4-HG1 KX468820 Feijão-Aveia Guamiranga PR
CNPAF_0211 Multinucleado Var. Oryzae KM065569 Arroz Santo Antônio de Goiás GO
CNPAF_0212 Multinucleado Var. Circinata KM065570 Arroz Santo Antônio de Goiás GO
CNPAF_0214 Multinucleado T. cucumeris AG1-1E KX468821 Feijão Sorriso MT
CNPAF_0215 Multinucleado T. cucumeris AG1-1A KX468822 Feijão Sorriso MT
CNPAF_0224 Multinucleado T. cucumeris AG1-1E KX468823 Feijão Sorriso MT
CNPAF_0236 Multinucleado T. cucumeris AG1-1E KX468824 Mata Mucajaí RR
CNPAF_0237 Multinucleado T. cucumeris AG1-1A KX468825 Mata Mucajaí RR
CNPAF_0238 Multinucleado T. cucumeris AG1-1E KX468826 Mata Cantá RR
CNPAF_0239 Multinucleado T. cucumeris AG1-1E KX468827 Mata Mucajaí RR
CNPAF_0240 Multinucleado T. cucumeris AG1-1E KX468828 Mata Mucajaí RR
CNPAF_0241 Multinucleado T. cucumeris AG1-1D KX468829 Cerrado Boa Vista RR
CNPAF_0242 Multinucleado T. cucumeris AG1-1E KX468830 Cerrado Boa Vista RR
CNPAF_0244 Multinucleado T. cucumeris AG1-1A KX468831 Cerrado Boa Vista RR
CNPAF_0246 Multinucleado T. cucumeris AG1-1D KX468832 Cerrado Boa Vista RR
CNPAF_0248 Multinucleado T. cucumeris AG1-1E KX468833 Mata Cantá RR
CNPAF_0249 Multinucleado T. cucumeris AG1-1A KX468834 Mata Cantá RR
CNPAF_0255 Multinucleado Ceratobasidium AG-Fa KX468835 Solo Unaí MG
CNPAF_0256 Multinucleado Var. Zeae KX468836 Solo Unaí MG
CNPAF_0258 Multinucleado Ceratobasidium AG-P KX468837 Solo Unaí MG
CNPAF_0261 Binucleado Ceratobasidium AG-F KX468838 Solo Unaí MG
CNPAF_0263 Binucleado Ceratobasidium AG-P KX468839 Solo Unaí MG
CNPAF_0269 Binucleado Ceratobasidium AG-F KX468840 Solo Luís E. Magalhães BA
CNPAF_0274 Multinucleado Var. Zeae KX468841 Solo Luís E. Magalhães BA
CNPAF_0276 Multinucleado Var. Zeae KX468842 Solo Luís E. Magalhães BA

22
O isolamento das cepas foi realizado adaptando-se os protocolos de
Weinhold (1977) e de Paulitz e Schroeder (2005). Pelo método de Weinhold, obtém-
se os isolados de Rhizoctonia spp. originados de fragmentos de matéria orgânica
distribuídos em placas de Petri com meio ágar-água + cloranfenicol 200 ppm, após
incubação a 25 + 1°C por cinco dias. No protocolo de Paulitz e Schroeder (2005),
resumidamente, seis palitos de dente autoclavados foram inseridos em 200g de solo
disposto em potes de polipropileno identificados e incubados durante cinco dias a 25
+ 1°C. Após esse período, os palitos colonizados foram transferidos para placas de
Petri contendo meio de cultura ágar-ágar com cloranfenicol a 200 ppm.
Após cinco dias, as colônias com características morfológicas típicas do
gênero Rhizoctonia (hifas com septos “doliporo”, com espessura acima de 5mm e
ramificações com ângulo de 90º, além de crescimento geralmente rápido a 25º C,
sem produção de esporos nesta fase anamórfica, conforme observações ao
microscópio óptico a 100x foram repicadas para placas de Petri contendo meio BDA
+ Cloranfenicol 200 ppm, sendo incubadas também por cinco dias, a 25 + 1°C. Os
isolados obtidos passaram por crescimento em meio líquido enriquecido PD (potato,
dextrose) sendo cultivados em shaker a 180 rpm e 28°C por 72h para a obtenção de
micélio. Decorrido este tempo, o micélio foi filtrado através de um funil de “Buchner”.
Isolamento e quantificação do DNA Genômico
O micélio foi macerado em almofariz com nitrogênio líquido até se tornar pó.
Ao micélio em pó foi acrescentado 0,8 ml do tampão de extração de DNA (Tris-HCl
100mM, NaCl 1,4 M, 0,02% β-mercaptoetanol, 10µl de RNAse A e EDTA 5mM pH
8,0), pré-aquecidos a 65ºC em banho-maria (Lopes et al. 2012). Esta solução foi
mantida em banho-maria a 65 °C, sob suave agitação, durante 20 minutos e
centrifugada a 12000 x g durante 10 minutos. Após a centrifugação, foi coletado o
sobrenadante e acrescentado 0,4 ml de fenol equilibrado em tampão Tris-HCl, pH
8,0 e 0,4 ml de clorofórmio e álcool isoamílico (49:1) e centrifugado a 12000 x g por
10 minutos. Novamente 0,4 ml de clorofórmio e álcool isoamílico (49:1) foi
adicionada ao sobrenadante coletado e em seguida centrifugado a 12000 x g
durante 15 minutos. Após a extração do sobrenadante, foi adicionado 0,8 ml de
isopropanol e em seguida centrifugado a 12000 x g por 15 minutos. O pellet de DNA
foi lavado em solução de etanol 70% (v/v) e ressuspendido 100 µl de água estéril. As

23
amostras de DNA foram então quantificadas e avaliadas quanto à sua qualidade por
espectrofotometria ultravioleta nos comprimentos de onda de 260 e 280 nm.
Amplificação da região ITS e sequenciamento
A região ITS1-5.8S-ITS2 foi amplificada por reação em cadeia da polimerase
(PCR) usando a combinação dos primers ITS1 (5'-TCCGTAGGTGAACCTGCGG-3')
e ITS4 (5'- TCCTCCGCTTATTGATATGC-3') (White et al., 1990). Após amplificação,
os produtos de PCR foram submetidos à eletroforese em gel de agarose 1,2 % e
purificados utilizando o protocolo baseado em precipitação por polietileno glicol
(PEG) - 20% PEG, 2,5 M NaCl, 80% etanol e etanol puro. Os produtos da
purificação foram usados na reação de sequenciamento utilizando DYEnamicTM ET
Terminator Cycle Sequencing kit (GE Healthcare, USA) O sequenciamento foi
realizado no sequenciador ABI Prism 3100 (Applied Biosystems).
Morfologia de Colônia e Condição nuclear
Os isolados foram analisados através de inspeção visual para observação da
morfologia de colônias (coloração de micélio e presença/ausência de escleródios). O
procedimento para a avaliação da condição nuclear envolveu a retirada um disco de
5mm de diâmetro contendo meio BDA e micélio, transferindo-o para uma lâmina de
vidro para microcultura. A coloração de núcleos foi realizada utilizando-se o corante
Safranina-O, como descrito por (Bandoni 1979). A contagem de núcleos foi realizada
observando-se dez células por lâmina. Para cada isolado foram feitas dez lâminas.
Determinação de AGs e inferência filogenética dos isolados
As sequências obtidas foram identificadas através do algoritmo BLAST
(Altschul et al. 1997), comparando-as com as sequências depositadas no banco de
dados não redundante do National Center for Biotechnology Information, disponível
em (http://www.ncbi.nlm.nih.gov/). A identificação das sequências foi procedida
utilizando-se o melhor resultado BLAST (best hit) de cada comparação. O banco de
dados do NCBI para Rhizoctonia apresenta muitas sequências parcialmente
identificadas e assim, nos casos em que o best hit não permitiu a identificação direta
das sequências utilizadas neste estudo, a identificação foi procedida a partir do
melhor score, cujas informações do banco de dados permitissem chegar ao nível de

24
espécie. Nesses casos, a observação de descritores importantes, como coloração
de micélio e condição nuclear auxiliou na identificação dos isolados.
Após identificação, as sequências foram alinhadas utilizando-se o programa
Clustal X – v2.1.0.0 (Thompson et al. 1997). O alinhamento foi ajustado
manualmente, tendo as extremidades sido podadas através do programa Bioedit –
v7.1.7.0 (Hall 1999). Todas as sequências foram depositadas no Genbank, e as
culturas dos isolados em estudo foram depositadas na Coleção de Fungos e
Microrganismos Funcionais da Embrapa Arroz e Feijão (Santo Antônio de Goiás,
GO).
As sequências mais similares e que serviram para identificação dos isolados
foram utilizadas para a análise de agrupamentos filogenéticos. Objetivando uma
melhor descrição dos agrupamentos, foram construídas três árvores filogenéticas:
uma contendo as sequências dos isolados do gênero teleomorfo Thanatephorus
(R. solani), outra com as sequências de Ceratobasidium (Rhizoctonias binucleadas)
e uma última contendo sequências dos isolados pertencentes ao gênero Waitea. O
software Mega (Molecular Evolution Genetics Analyses), versão 6.06 (Tamura et al.
2013), foi utilizado para a escolha do modelo evolutivo melhor adaptado a homologia
da sequência de nucleotídeos, de cada conjunto de dados, e para produzir as
topologias de árvores filogenéticas com base no menor valor do critério de
informação Bayesiana (BIC). Para Thanatephorus e Waitea o modelo sugerido foi
GTR + I (General Time Reversible + Invariant Sites), enquanto para Ceratobasidium
o modelo sugerido foi Jukes-Cantor (JC) (Jukes e Cantor 1969). Todas as árvores
filogenéticas foram construídas utilizando-se o método estatístico máxima
verossimilhança, com base em 1000 bootstraps.
Testes de patogenicidade em feijão e milho
A avaliação da patogenicidade dos isolados foi conduzida de acordo com o
método da “camada única de inóculo” descrito por Chaudhary et al. (2006).
Recipientes de polipropileno de 500 mL foram utilizados para plantio em vermiculita
na presença dos isolados testados. Cada recipiente foi preparado na seguinte
ordem: 6cm de vermiculita autoclavada; uma colônia de 9 cm de diâmetro do isolado
cultivado em BDA, cuidadosamente removida de placa de Petri; outra camada
vermiculita, agora de 1,0; e finalmente, cinco sementes de feijão cv. Pérola

25
previamente desinfestadas em NaClO 0,5% e enxaguadas por três vezes em água
destilada autoclavada.
As sementes foram cobertas com mais uma camada de 2,0 cm de vermiculita,
e incubadas a 20+ 2ºC por 21 dias em câmara de crescimento sob fotoperíodo de 12
horas, e irrigadas três vezes por semana. O mesmo procedimento foi utilizado para
sementes de milho. Os experimentos foram montados utilizando-se o delineamento
inteiramente casualizado, com duas repetições e cinco plantas por parcela
experimental, utilizando-se também testemunhas não infestadas.
Após o período de incubação, as plantas foram lavadas em água corrente
para a retirada de resíduos de vermiculita e posterior avaliação da severidade da
doença. Para a avaliação da severidade da doença em raízes de feijão comum e de
milho, utilizou-se a escala de notas de 1 a 9, conforme protocolo desenvolvido por
Schoonhoven e Pastor-Corrales (1987), sendo considerados patogênicos os
isolados que apresentaram índice de severidade maior ou igual a 2. Os resultados
obtidos para a severidade da doença foram então submetidos ao teste de Kruskal-
Wallis (KW) com nível de significância de 5% (α= 0,05). O teste de KW é um teste
não paramétrico utilizado na comparação de três ou mais amostras independentes e
é análogo ao teste F usado na análise de variância (ANOVA). O teste KW foi
utilizado para a realização de duas análises: uma delas considerando os índices de
severidade da doença obtidos pelos isolados dentro de cada um dos gêneros e outra
considerando os índices de severidade da doença obtidos pelos isolados dentro de
seus respectivos AGs/variedade, individualmente. Todas as análises estatísticas
foram realizados no software Statistica (StatSoft Inc., 2007).
Análises de distribuição espacial associando a identificação dos isolados e os
resultados de testes de patogenicidade ao ponto geográfico em que os isolados
foram coletados foram conduzidas utilizando-se o software ArcGIS (ESRI 2011).

26
Resultados
Identificação molecular, morfologia de colônias e condição nuclear
Todos os 81 isolados avaliados pertencem ao gênero Rhizoctonia de acordo
com descritores morfológicos e sequências avaliadas com o Blast-NCBI. Os isolados
identificados como W. circinata apresentaram-se multinucleados e com micélio de
coloração alaranjada, salmão ou marrom claro. Após 40 dias de cultivo em BDA,
isolados da var. zeae e var. circinata formaram escleródios. Não foram observados
escleródios no isolado identificado como var. oryzae. Os isolados identificados como
T. cucumeris também apresentaram vários núcleos por célula, com a coloração do
micélio variando entre o marrom claro e o marrom escuro. Isolados de AG1 e AG4
apresentaram formação de escleródios, cujas dimensões, quantidades e coloração
variaram em função dos ISGs. Quase todos os isolados identificados como
Ceratobasidium apresentaram dois núcleos por célula, com exceção dos isolados
CNPAF_0036, CNPAF_0255, CNPAF_0258, que foram multinucleados. A maior
parte dos isolados de Ceratobasidium apresentaram micélio branco-amarelado, de
aspecto pulverulento, com anéis concêntricos e número variável de escleródios,
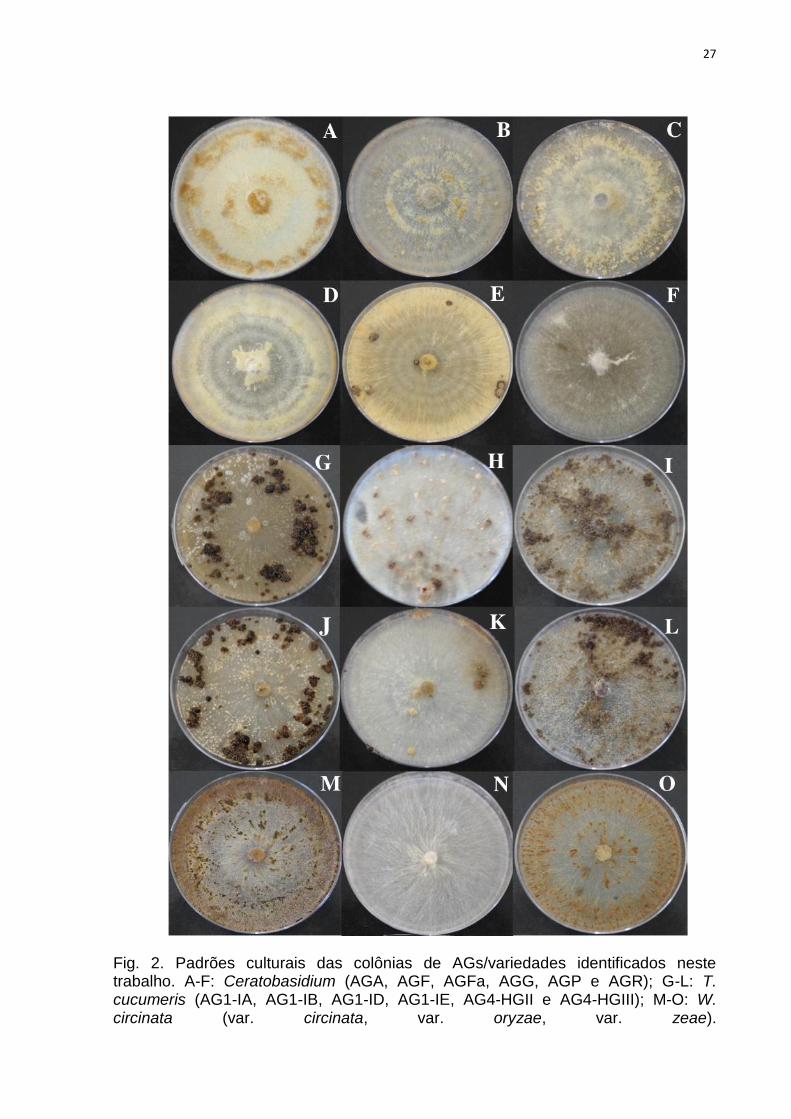
também branco-amarelados (Tabela 1) (Figura 2).
Determinação de AG e Análises filogenéticas
A maior parte dos isolados estudados foi identificada como W. circinata,
aproximadamente 40% do total, sendo var. zeae a espécie mais representativa do
gênero (var. zeae = 19,75%, var. circinata = 18,5%, var. oryzae = 1,23%). Isolados
identificados como pertencentes à AGs de T. cucumeris corresponderam a 37% do
total. AG1-1E e AG4-HGI foram os grupos mais frequentes (AG1-IA = 4,94%, AG1-
1B = 3,70%, AG1-ID = 2,47%, AG1-1E = 9,88%, AG4-HGI = 9,88% e AG4-HGIII =
6,17%). O gênero Ceratobasidium correspondeu a 23% do total de isolados, com
maior ocorrência para AG-A (AG-A = 8,64%, AG-F = 6,17%, AG-Fa = 3,70%, AG-P =
2,47 %, e AG-G e AG-R ambos com 1,23% dos isolados)
De acordo com a distribuição geográfica e a frequência dos isolados
amostrados (Tabelas 1 e 2) observa-se que isolados de W. circinata foram
concentrados em Goiás, Minas Gerais, Paraná e Tocantins. Apenas dois grupos de
anastomose de T. cucumeris foram identificados: AG1 e AG4. O grupo AG1 se
27
Fig. 2. Padrões culturais das colônias de AGs/variedades identificados neste trabalho. A-F: Ceratobasidium (AGA, AGF, AGFa, AGG, AGP e AGR); G-L: T. cucumeris (AG1-IA, AG1-IB, AG1-ID, AG1-IE, AG4-HGII e AG4-HGIII); M-O: W. circinata (var. circinata, var. oryzae, var. zeae).
28
Tabela 2. Número e frequências absolutas e relativas de isolados do gênero Rhizoctonia identificados em nível de espécie, identificados em solos do Cerrado brasileiro e áreas adjacentes.
Teleomorfo Anamorfo Variedade/AG Número
de isolados
Frequência (%)
Waitea circinata
R. zeae Zeae 16 19,75
39,51 R. oryzae Oryzae 1 1,23
- Circinata 15 18,52
Ceratobasidium Ceratorhiza
AG-A 7 8,64
23,46
AG-F 5 6,17
AG-Fa 3 3,7
AG-G 1 1,23
AG-P 2 2,47
AG-R 1 1,23
Thanatephorus cucumeris Rhizoctonia solani
AG1-IA 4 4,94
37,04
AG1-IE 8 9,88
AG1-IB 3 3,7
AG1-ID 2 2,47
AG4-HGI 8 9,88
AG4-HGIII 5 6,17
Total de isolados 81 100 100

29
apresentou em quatro ISGs: IA, IB, ID e IE, enquanto os isolados de AG4 se
distribuíram nos ISGs HGI e HGIII. Todos os isolados de AG4 foram provenientes de
áreas agrícolas. Já os isolados de AG1 foram oriundos de áreas de cultivo e também
de regiões de mata e cerrado. Isolados de AG1 foram distribuídos entre o centro e o
norte do Brasil, enquanto isolados de AG4 entre o centro e o sul do país. Em
Ceratobasidium, a maior parte dos isolados identificados como AG-A foi oriunda de
áreas de produção de feijão, assim como AG-F e AG-Fa. Os grupos AG-G e AG-R
foram isolados de solos provenientes de áreas de produção de cenoura e cebola,
respectivamente.
Nas análises filogenéticas, os agrupamentos formados entre as sequências
dos isolados estudados e aquelas obtidas no NCBI confirmaram a identificação de
quase todos os isolados, obtida através do algoritmo Blast. Em geral, os
agrupamentos ocorreram entre isolados de um mesmo AG/variedade, mas isolados
de AG1-IA e AG1-IE ocuparam um mesmo clado. Var. zeae, AG4-HGI e AG4-HGIII
dividiram-se em dois clados cada, localizados separadamente em suas respectivas
árvores. É possível observar que em alguns casos houve estruturação geográfica,
com agrupamentos de isolados ocorrendo preferencialmente por estado, como em
Roraima, Mato Grosso, Paraná e Minas Gerais (Tabela 1; Figuras 3, 4, 5).
30
Fig. 3. Arvore filogenética contendo as sequências de isolados de W. circinata, identificados neste estudo. Em verde: var. zeae; em azul: var. oryzae e em vermelho: var. circinata.
31
Fig. 4. Árvore filogenética contendo as sequências de isolados de Ceratobasidium identificados neste estudo. Em vermelho: AG-A; Em azul: AG-F; Em azul claro: AG-Fa; Em púrpura: AG-G; Em verde escuro: AG-P; Em verde claro: AG-R.

32
Fig. 5. Árvore filogenética contendo as sequências de isolados de T. Cucumeris identificados neste estudo. Em vermelho: isolados de AG1 (ISGs IA, IB, ID e IE); em azul: isolados AG4 (ISGs HGI e HGIII).
Testes de patogenicidade em Feijão e Milho
Os testes de patogenicidade em milho e feijão revelaram que 53,1% dos
isolados infectam feijão e 45,7% infectam milho. Vinte e dois isolados (27,1% do
total) atacam raízes das duas culturas. Destes, 12 pertencem a T. Cucumeris
(CNPAF - 0076, CNPAF-0119, CNPAF - 0144, CNPAF - 0152, CNPAF - 0180,
CNPAF - 0189, CNPAF - 0193, CNPAF - 0215, CNPAF - 0238, CNPAF - 0240,
CNPAF - 0246, CNPAF – 0248); seis a Ceratobasidium (CNPAF - 0088, CNPAF -
0093, CNPAF - 0117, CNPAF - 0129, CNPAF - 0255, CNPAF – 0258) e quatro a
33
variedades de W. circinata (CNPAF - 0138, CNPAF - 0148, CNPAF - 0178, CNPAF –
0211).
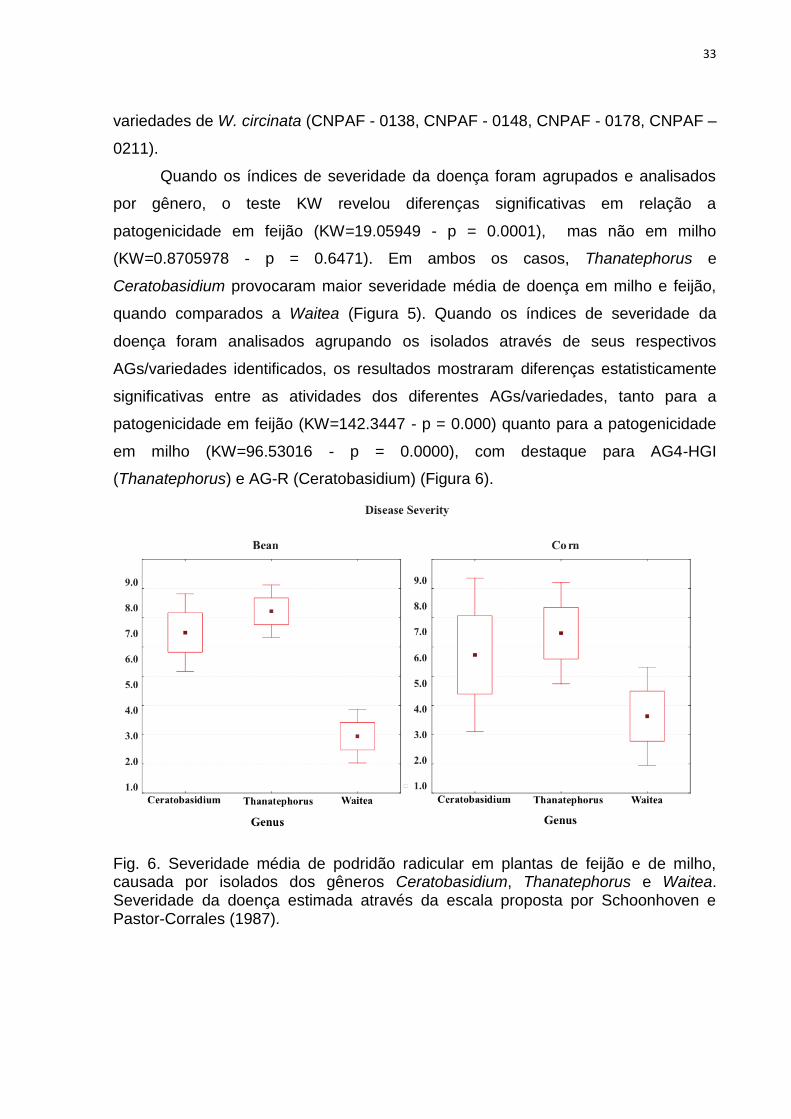
Quando os índices de severidade da doença foram agrupados e analisados
por gênero, o teste KW revelou diferenças significativas em relação a
patogenicidade em feijão (KW=19.05949 - p = 0.0001), mas não em milho
(KW=0.8705978 - p = 0.6471). Em ambos os casos, Thanatephorus e
Ceratobasidium provocaram maior severidade média de doença em milho e feijão,
quando comparados a Waitea (Figura 5). Quando os índices de severidade da
doença foram analisados agrupando os isolados através de seus respectivos
AGs/variedades identificados, os resultados mostraram diferenças estatisticamente
significativas entre as atividades dos diferentes AGs/variedades, tanto para a
patogenicidade em feijão (KW=142.3447 - p = 0.000) quanto para a patogenicidade
em milho (KW=96.53016 - p = 0.0000), com destaque para AG4-HGI
(Thanatephorus) e AG-R (Ceratobasidium) (Figura 6).
Fig. 6. Severidade média de podridão radicular em plantas de feijão e de milho, causada por isolados dos gêneros Ceratobasidium, Thanatephorus e Waitea. Severidade da doença estimada através da escala proposta por Schoonhoven e Pastor-Corrales (1987).

34
Fig. 7. Severidade média de podridão radicular em plantas de feijão e de milho, causada por diferentes Grupos de Anastomose identificados neste estudo. Severidade da doença estimada através da escala proposta por Schoonhoven e Pastor-Corrales (1987).

35
Discussão
Os padrões morfológicos observados nos isolados foram compatíveis com a
descrição na literatura atualmente válida para Waitea, Ceratobasidium e
Thanatephorus (Demirci 1998; Leiner e Carling 1994; Kammerer et al. 2011; Muzhinji
et al. 2015; Sneh 1991). Especialmente em W. Circinata, a comparação de
morfologia de colônias tem sido considerada um importante instrumento para
diferenciar variedades (Toda et al., 2007). Entretanto, algumas descrições
observadas na literatura apresentam divergências, principalmente relacionadas à
coloração do micélio (Demicri 1998; De la Cerda et al. 2007; Toda et al. 2005; 2007).
Tais discrepâncias podem ser resultado do processo de adaptação do fungo aos
diferentes hospedeiros ou aos diferentes ambientes ocupados por fungos e
hospedeiros. Por outro lado, podem também ser resultado da avaliação fenotípica
em diferentes tempos de vida dos isolados. Toda et al. (2007) enfatizam que são
necessários 30 dias, ou mais, para comparação de morfologia de colônia, já que
durante este período, isolados trabalhados por ele passaram por modificações
fenotípicas.
O conflito existente entre a condição multinuclear dos isolados de
Ceratobasidium CNPAF_0036, CNPAF_0255, CNPAF_0258 e suas respectivas
identificações no Genebank não é um fato novo, e pode ser explicado pela sua
proximidade genética com o gênero Thanatephorus (anamorfo = R.solani). Esses
gêneros parecem ainda não ter passado por separação evolutiva completa (Ceresini
2014), o que pode ocasionar instabilidade na sistemática do grupo. Segundo Lübeck
(2004), existem relatos que colocam em dúvida a posição taxonômica de
determinados AGs dentro destes dois gêneros. Ainda de acordo com esta autora, já
foi observado, por exemplo, isolados de AG-6 (Thanatephorus) fundindo hifas com
AG-F (Ceratobasidium). Análises filogenéticas envolvendo rDNA, com forte suporte
estatístico, sugerem que alguns isolados classificados como Ceratobasidium, com
base na condição nuclear e reação de anastomose de hifas, podem ser mais
corretamente classificados dentro do gênero Thanatephorus (Gonzalez et al. 2001).
A ocorrência de determinadas espécies, AGs, subgrupos ou variedades de
Rhizoctonia em regiões geográficas distintas no Brasil pode estar relacionada não só
ao tipo de planta cultivada no momento da amostragem, mas também à
susceptibilidade da cultura precedente a estes fungos. Este efeito já foi observado

36
em Rhizoctonias associadas a batatas na África do Sul (Muzhinji et al. 2015). A
rotação de culturas é uma prática comum adotada por produtores rurais e é definida
regionalmente conforme critérios econômicos, sociais e climáticos. Juntamente com
a facilidade de sobrevivência de Rhizoctonias nos solos, o perfil de plantas
hospedeiras prevalentes em cada região pode ter influenciado na incidência e na
composição de AGs/variedades de Rhizoctonia identificados no presente trabalho,
em suas respectivas áreas de origem.
O histórico de cultivo das áreas amostradas, apresentado na Tabela 1 e a
análise da relação de hospedeiros de Rhizoctonia, descritos por Genhua e
Chengyum (2012) e Lakshman e Amaradasa (2014) sugerem que a distribuição
geográfica da maior parte dos isolados identificados neste trabalho pode mesmo ter
sido influenciada pela escolha das espécies cultivadas nestas áreas. A identificação
das variedades de W. circinata em cinco dos oito estados amostrados neste estudo
(Tabela 1), ilustra bem o aspecto acima descrito. Estes organismos causam uma
série de doenças em diferentes tipos de gramíneas (Kammerer et al. 2011), inclusive
milho e arroz (Lakshman e Amaradasa, 2014). Nesse sentido, é importante destacar
que Paraná, Goiás, Bahia, Minas Gerais (32, 492 mil toneladas de milho) e
Tocantins (600 mil toneladas de arroz), são grandes produtores destas duas
importantes culturas de grãos no Brasil (BRASIL 2015), podendo assim ter
colaborado para a fixação do patógeno nessas regiões, ao longo do tempo.
A espécie var. zeae foi a mais abundante entre os 81 isolados estudados,
tendo sido identificada em áreas agrícolas da Bahia, Goiás, Minas Gerais, Paraná e
Tocantins, apresentando-se, portanto, amplamente distribuída no Brasil (Tabelas 1 e
2). Esta espécie também já foi isolada por Poltronieri et al. (2002), que a descreveu
como causadora da podridão do milho no estado do Pará, o que aumenta mais
ainda a amplitude geográfica de sua distribuição. Dessa forma, sua ocorrência
parece não estar necessariamente associada a fatores ambientais ou climáticos, o
que pode ser constatado a partir da observação de trabalhos envolvendo isolados
dessa espécie nos mais variados climas e ambientes, de países como Turquia,
Estados Unidos, Argentina, China e Irã (Demirci 1998; Gutiérrez et al. 2007; Li et al.
1998; Telmadarrehei et al. 2011).
A var. oryzae teve baixa representatividade nas áreas amostradas, sendo
identificado apenas um isolado, em amostra de Santo Antônio de Goiás (Tabela 1).
Estudos sobre a ocorrência e distribuição da var. oryzae no Brasil ainda são

37
escassos, embora Prabhu et al. (2002) tenham isolado essa espécie em lavouras de
arroz irrigado, no Estado do Tocantins. Diferentemente das variedades oryzae e
zeae, W. circinata var. circinata é uma espécie descrita há relativamente pouco
tempo e tem sido relacionada a doenças em espécies de gramas pertencentes a
família Poaceae (Chen, et al. 2009). Os isolados brasileiros desta espécie foram
identificados em sua maior parte em Minas Gerais, formando o segundo maior
percentual de isolados amostrados (Tabelas 1 e 2). Aparentemente, este é o
primeiro relato sobre a ocorrência dessa variedade no Brasil.
A composição da diversidade dentro de Ceratobasidium no Cerrado agrícola
também parece ter sofrido influência das espécies de plantas escolhidas para cultivo
nas regiões de onde os fungos foram isolados. Dos seis grupos de anastomose
identificados neste trabalho, pelo menos cinco estão diretamente relacionados ao
feijoeiro comum: AG-A, AG-F, AG-G, AG-R e AG-P (Cardenas et al. 2015; Genhua e
Chengyum 2012) e um está indiretamente associado a esta cultura, já que
evidências filogenéticas sugerem que AG-Fa é um subgrupo de AG-F (Gurkanli e
Ozkoc 2011; Sharon et al. 2008). Com exceção de Tocantins, os demais estados em
que AGs de Ceratobasidium foram identificados são grandes produtores de feijão.
Juntos, Paraná, Minas Gerais, Bahia e Goiás produziram quase dois milhões de
toneladas na safra 2014/2015 (BRASIL 2015). A hipótese de que a diversidade de
Ceratobasidium pode ter sido influenciada pelo histórico de cultivos adotados nas
diferentes regiões agrícolas amostradas pode ser melhor analisada a partir da
observação conjunta das informações apresentadas na Tabela 1. Isolados
identificados como AG-A e AG-F no Paraná foram amostrados em áreas onde
tradicionalmente há produção de feijão. Em Minas Gerais, AG-A também foi
identificado em área de produção de feijão, assim como AG-Fa em Goiás e
Tocantins. Há ainda outras situações em que se pode observar uma provável
correlação entre grupos de anastomose identificados nas amostras de solo e a
cultura implantada na área de coleta. AG-P foi encontrado em Minas Gerais e
embora não tenha sido proveniente diretamente de áreas de plantio de feijão, a
região de origem das amostras (Unaí) é produtora de diferentes variedades desta
cultura (Barbosa 2009). Também em Minas Gerais, identificou-se o grupo de
anastomose AG-R em área de produção de cebola. A associação de AG-R a
doenças em cebola já foi observada por Burpe et al. (1980).

38
A ocorrência de AG-G (isolado CNPAF - 0113) em área de produção de
Cenoura (Campos Altos – MG) não indica necessariamente que já exista uma
associação específica entre esta cultura e o respectivo AG. Entretanto, AG-G já foi
identificado no Brasil causando podridão de raiz em Yacon (Fenile et al. 2005). Além
disso, cenoura e yacon pertencem a um mesmo táxon vegetal (Asterales) e
compartilham sinapomorfias morfológicas e moleculares (Judd et al. 2007). Esses
resultados sugerem a necessidade de estudos com maior representatividade,
envolvendo amostragem maior dentro de áreas geograficamente menores, além de
testes de patogenicidade para investigar possível associação entre AG-G e cenoura.
A análise da distribuição geográfica dos AGs de T. cucumeris revelou
semelhanças com os resultados obtidos para W. circinata e Ceratobasidium sp.,
acima discutidos. Observou-se uma estreita associação entre os grupos de
anastomose identificados e o histórico recente de cultivo das áreas amostradas,
para a maioria dos isolados. Em áreas de produção de feijão foram identificados os
grupos de anastomose AG1-IA e AG1-IE (Mato Grosso), AG4-HGI (Goiás, Paraná e
São Paulo) e AG4-HGIII (Goiás). AG4-HGI e AG4-HGIII também foram identificados
em áreas de soja (Goiás e Minas Gerais), também hospedeira destes grupos. AG1-
1B foi identificado em áreas de soja (Goiás) e Café (Minas Gerais) (Tabela 1). AG1-
IA e AG1-IE já foram descritos como patógenos de feijoeiro (González et al. 2012;
Mora-Umaña et al. 2013; Youssef et al. 2012), assim como AG4-HGI e AG4-HGIII
(Çebi Kılıçoğlu e Özkoç 2013; Ceresini 2014; Genhua e Chengyum 2012; Lakshman
e Amaradasa 2014). AG4-HGI e AG4-HGIII também já foram associados a doenças
em soja (Ceresini 2014; Fenile et al. 2002, 2003). A soja também é hospedeira de
AG1-1B (Lakshman; Amaradasa 2014; Youssef et al. 2012). AG1-IB já foi descrito
por Gaino et al. (2010) como patógeno de café.
Apesar dos resultados indicarem que a maior parte dos AGs de T. cucumeris
pode ter sido influenciada pelo histórico de cultivo para se estabelecer nas regiões
produtivas analisadas, é importante considerar, entretanto, que AG1-IA, AG1-ID e
AG1-1E também foram identificados em amostras de solo de mata e Cerrado
(Tabela 1). A identificação desses ISGs de AG1 em solos não cultivados no extremo
norte do Brasil (Roraima) sugere a existência de Rhizoctonias nativas, bem
adaptadas ao clima equatorial, onde prevalecem elevadas temperaturas e alta
pluviosidade.

39
Em países da América Central e Caribe, onde as condições climáticas são
semelhantes às de Roraima, a ocorrência e a associação do grupo AG1 e seus
respectivos ISGs ao feijoeiro estão descritas na literatura (Godoy-Lutz et al. 2008;
González et al. 2012; Mora-Umaña et al. 2013). Os grupos AG1-IA e AG1-IE, por
exemplo, estão entre os causadores da “mela” do feijoeiro (Godoy-Lutz et al. 2008),
doença severa que ataca a parte aérea, diminuindo o valor comercial dos grãos,
acarretando perdas econômicas importantes, e em muitos casos causando a morte
das plantas (Godoy-Lutz et al. 1996).
Por outro lado, apesar da distribuição geográfica dos AGs de T. cucumeris
identificados neste trabalho estar aparentemente correlacionada com as
especificidades culturais estabelecidas nas áreas estudadas, deve-se considerar
que os efeitos do ambiente podem ser igualmente determinantes na ocorrência de
AG1 no Brasil. O mesmo acontece com AG4. De fato, quando se compara a
distribuição dos isolados de AG1 e AG4 neste estudo, independente de seus
respectivos ISGs, observa-se que enquanto AG1 foi encontrado em áreas de
latitudes mais ao norte, AG4 ficou restrito a latitudes mais ao sul do país. O impacto
das condições climáticas e ambientais na distribuição desses respectivos AGs já foi
anteriormente detectado por Fenille et al. (2002) em áreas de produção de soja no
Brasil. Esses autores observaram em seu trabalho que enquanto isolados
provenientes de áreas agrícolas do Mato Grosso foram identificados como AG1 e
provocaram sintomas da “mela”, isolados identificados como AG4 e provenientes de
áreas agrícolas de Minas Gerais, Paraná e São Paulo foram associados com a
podridão de hipocótilo e raiz.
Analisando-se as árvores filogenéticas dos três gêneros identificados neste
estudo, é possível observar que isolados de uma mesma variedade ou AG tenderam
a se agrupar de acordo com a proximidade de suas origens, na maioria dos ramos,
sugerindo que dentro de uma mesma variedade ou AG pode haver correlação entre
distância genética e distância geográfica e restrição ao fluxo gênico nas áreas
estudadas.
Ainda que sejam levadas em consideração as diversas possibilidades para
dispersão das Rhizoctonias (sementes, compartilhamento de máquinas e utensílios
agrícolas entre diferentes lavouras, etc), os resultados sugerem que a distribuição
geográfica dos AGs/variedades nas áreas estudadas está, muito provavelmente,
mais relacionada com o histórico recente dos cultivos e às condições climáticas

40
locais do que com o mecanismo de fluxo gênico entre diferentes populações desses
fungos. Entretanto, estudos populacionais utilizando marcadores moleculares
específicos são necessários para investigar melhor essas questões.
Isolados de AG1-IA e AG1-IE ocuparam um mesmo clado, não apresentando
padrão de agrupamentos tipicamente esperado para grupos que já se separaram
evolutivamente. É importante considerar, entretanto, que ambos ISGs pertencem ao
mesmo AG e assim, o isolamento reprodutivo pode ainda não ter acontecido
totalmente na região onde foram coletados. Alternativamente, deve-se observar a
que a existência de múltiplas cópias da região gênica analisada, decorrente da
condição multinuclear desses fungos, pode significar uma limitação na reconstrução
das relações filogenéticas destes organismos. Novos estudos que contemple a
adição de mais regiões gênicas à ITS-rDNA nas análises deve esclarecer de
maneira definitiva as relações filogenéticas entre isolados brasileiros desses dois
grupos.
Analisando-se a Figura 2, é possível observar que os isolados AG1-IA e AG1-
IE são muito semelhantes, morfologicamente. Principalmente em relação ao número,
tamanho e disposição dos escleródios. É preciso considerar também que estudos
relatando a presença de AG1-IE no Brasil não são comuns, enquanto AG1-IA já foi
identificado nos estados do Acre, Maranhão, Norte do Mato Grosso, Rondônia, Sul
do Pará e Tocantins (Ceresini 2014; Poltronieri 2008; Youssef 2012), cujos territórios
pertencem à mesma região climática dos isolados AG1-IA - AG1-IE analisados
nesse estudo. Assim, é possível que todos os isolados localizados neste clado
sejam na realidade pertencentes apenas à AG1-IA.
Não é difícil supor também que tal semelhança morfológica possa conduzir a
equívocos de descrição de isolados, no momento de assinar as sequências durante
o depósito no NCBI. Como discutido por Sharon et al. (2006, 2008), muitas
sequências de Rhizoctonia são erroneamente assinadas no Genebank, o que pode
ocasionar equívocos durante processo de identificação. Nesse caso, porém, se
houve equívoco no processo de assinatura das sequências que serviram de base
para a identificação de AG1-IA e AG1-IE, obtidas no Norte do Brasil, está claro que o
equívoco se resume ao ISG (IA ou IE) e não à AG1, que é tipicamente encontrado
em zonas equatoriais (Godoy-Lutz et al. 2008; González et al. 2012; Mora-Umaña et
al. 2013). Estudos envolvendo reações de anastomose são indicados para
solucionar esta dúvida.

41
Os resultados dos testes de patogenicidade mostraram que parte significativa
dos isolados apresentou atividade fitopatogênica, com diferentes graus de
severidade entre os diferentes gêneros, entre diferentes AGs e entre isolados dentro
de um mesmo AG, em milho e feijão (Figuras 5 e 6). Essas diferenças se devem,
provavelmente, muito mais à variabilidade genética dos isolados do que às
especificidades dos AG identificados. É possível observar nas Figuras 7 e 8, por
exemplo, que isolados de AG1-1A e AG1-IB apresentaram-se hipovirulentos às
raízes de milho e feijão, respectivamente, apesar dessas plantas serem
reconhecidamente descritas com hospedeiras dessas duas espécies de plantas
(Ceresini 2014). Nesse caso, conclui-se que a variação genética presente nesses
isolados não foi suficientemente eficaz para desenvolver sintomas de sua atividade
fitopatogênica em milho e feijão.
42
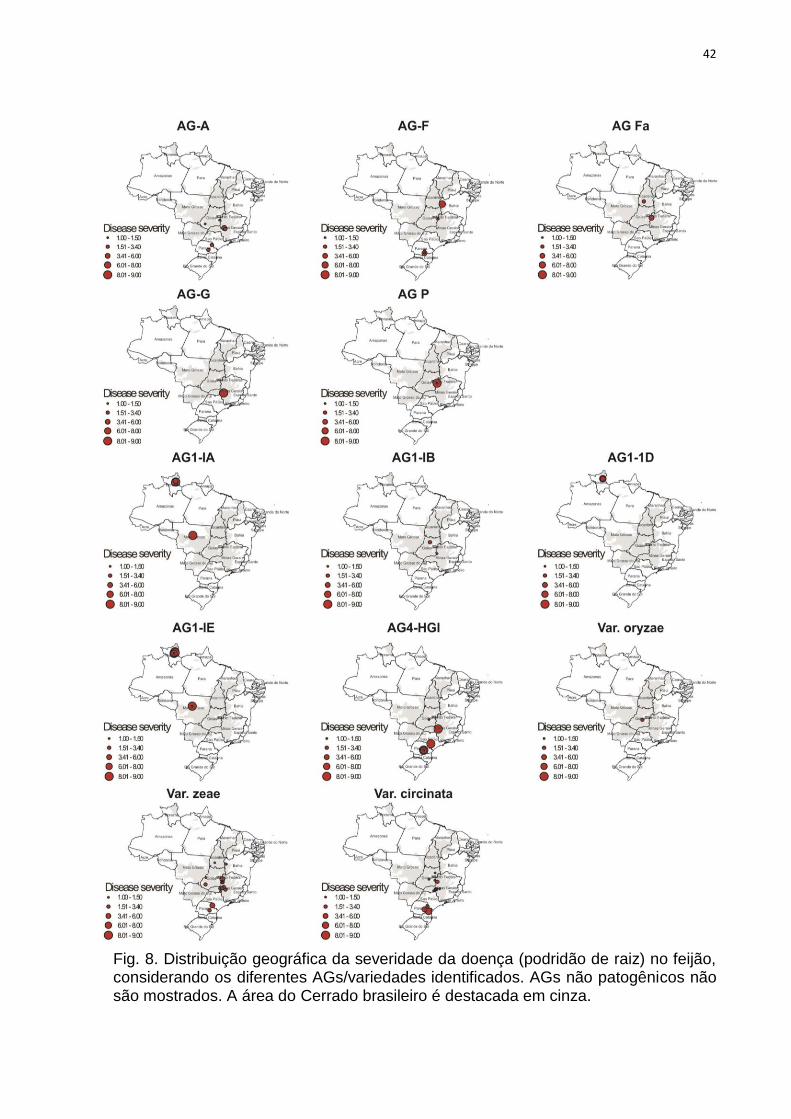
Fig. 8. Distribuição geográfica da severidade da doença (podridão de raiz) no feijão, considerando os diferentes AGs/variedades identificados. AGs não patogênicos não são mostrados. A área do Cerrado brasileiro é destacada em cinza.
43
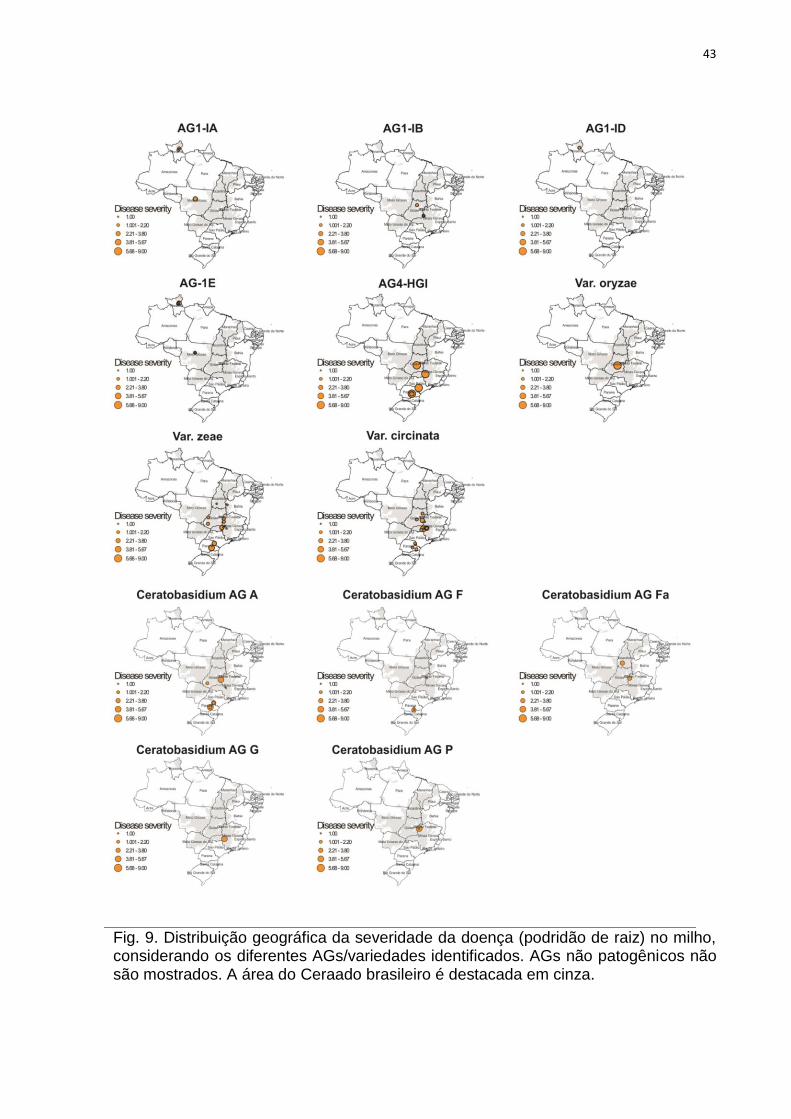
Fig. 9. Distribuição geográfica da severidade da doença (podridão de raiz) no milho, considerando os diferentes AGs/variedades identificados. AGs não patogênicos não são mostrados. A área do Ceraado brasileiro é destacada em cinza.

44
Conclusões
Nesse estudo foi possível observar que o gênero Rhizoctonia encontra-se
bastante diversificado em áreas agrícolas brasileiras, tendo sido identificados seus
principais grupos de fitopatógenos. Observou-se também que as suas espécies,
grupos de anastomose e variedades tendem a se distribuir preferencialmente
seguindo um gradiente latitudinal e de acordo com disposição geográfica das
culturas que utilizam como respectivas hospedeiras.
Os resultados aqui apresentados reafirmam a importância do gênero
Rhizoctonia no contexto da produção agrícola brasileira, sugerindo que mais estudos
com maior representatividade de amostras por área e com um conjunto mais
abrangente e diversificado de análises irão contribuir de maneira significativa para a
compreensão de aspectos importantes para o manejo e controle deste importante
grupo de fitopatógenos.

45
Referências Bibliográficas Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., Lipman, D. J., Gapped BLAST and PSI-BLAST: a new generation of protein database search programs, Nucleic Acids Res. 25 (1997) 3389-3402. Aiello, D., Vitale, A., Hyakumachi, M., Polizzi, G. 2012. Molecular characterization and pathogenicity of binucleate Rhizoctonia AG-F associated to the watermelon vine decline in Italy. European Journal of Plant Pathology 134, 1: 161-165. Amaradasa, B. S., Lakshman, D.K., McCall, D. Horvath, B. J. 2014. In Vitro Fungicide Sensitivity of Rhizoctonia and Waitea Isolates Collected from Turfgrasses. J. Environ. Hort. 32: 126-132. Azevedo, J. L., Pizzirani-Kleiner, A. A. 2002. Melhoramento de fungos de importância na agricultura. In: Melo, I. S., Valadares-Inglis, M. C., Nass, L. L., Valois, A. C. C. (orgs.) Recursos Genéticos e Melhoramento: microorganismos. Jaguariúna: Embrapa Meio Ambiente. Bandoni, R. J. Safranin O as a rapid nuclear stain for fungi. Mycologia 71:873-874. 1979. Barbosa, F. R., Silva, C. C. Da., Gonzaga, A. C. De O., Silveira, P. M. Da., Quintela, E. D., Lobo Júnior, M., Cobucci, T., Del Peloso, M. J., Junqueira, R. B. M. Sistema de produção integrada do feijoeiro comum na região central brasileira. Santo Antônio de Goiás: Embrapa Arroz e Feijão, 2009. 28 p. (Embrapa Arroz e Feijão. Circular técnica, 86). Basseto, M. A., Filho, W. V. V., Filho, W. V. V. F., Souza, E. C., Ceresini, P. C. 2008. O papel de Rhizoctonia spp. binucleadas na indução de resistência a mela da soja. Acta Scientiarum. Agronomy, 30, 2: 183-189. Bolkan, H. A., Ribeiro, W. R. C. 1985. Anastomosis groups and pathogenicity of Rhizoctonia solani isolates from Brazil. Plant Disease 69:599-601. Branco, S., Bruns, T. D., Singleton, I. 2013. Fungi at a small scale: spatial zonation of fungal assemblages around single trees. PLoS One, 8: 10, e78295. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Projeções do agronegócio: Brasil 2014/15 a 20124/25: projeções de longo prazo. 6. ed. Brasília: AGE/Mapa, 2015. Disponível em: <http://www.agricultura.gov.br/arq_editor/PROJECOES_DO_AGRONEGOCIO_2025_WEB.pdf>. Acesso em 15 jun. 2016. Bruns, T. D., White, T. J., Taylor, J.W. 1991. Fungal molecular systematics. Annual Review of Ecology and Systematics 22: 525-564. Burpee, L. L., Sunders, P. L., Cole, H. Jr., Sherwood, R. T. 1980. Anastomosis groups among isolates of Ceratobasidium cornigerum and related fungi. Mycologia, 72: 689–701

46
Cardenas, G. E. M., Galvan, M. Z., Barrera, V. A., Rodriguero, M. S., Carmona, M. A., March, G. J., Ramallo, A. C., Shew, H. D. 2015. Molecular identification and pathogenicity of Rhizoctonia spp. from tobacco growing areas in northwestern Argentina. Tropical Plant Pathology, 40(3), 160-168. Cardoso, J. E., Echandi, E. 1987. Biological control of Rhizoctonia root rot of snap bean with binucleate Rhizoctonia-like fungi. Plant Disease, 71:167-170. Carling, D. E., Helm, D. J., Leiner. R. H. 1990. In vitro sensitivity of Rhizoctonia solani and other multinucleate and binucleate Rhizoctonia to selected fungicides. Plant Disease, 74: 860–863. Carling, D. E. 2000. Anastomosis group and subets of anastomosis group of Rhizoctonia solani. In: International Symposium on Rhizoctonia 3, Taichung, Abstract Taichung, ISR, p. 14, 2000. Carling, D. E., Baird, R. E., Gitaitis, R. D., Brainard, K. A., Kuniaga, S. 2002a. Characterization of AG-13, a Newly Reported Anastomosis Group of Rhizoctonia solani. Phytopathology 92: 893-899. Carling, D. E., Kuninaga, S., Brainard, K. A. 2002b. Hyphal Anastomosis Reactions, rDNA-Internal Transcribed Spacer Sequences, and Virulence Levels Among Subsets of Rhizoctonia solani Anastomosis Group-2 (AG-2) and AG-BI. Phytopathology, 92, 1: 43-50. Çebi Kılıçoğlu, M., Özkoç, İ. 2010. Molecular characterization of Rhizoctonia solani AG4 using PCR-RFLP of the rDNA-ITS region. Turk. J. Biol, 34: 261–269. Ceresini, P. C. 2014. Rhizoctonia como fitopatógeno: biologia e diversidade de Rhizoctonia solani em agroecosistemas tropicais e perspectivas do manejo da rhizoctoniose usando resistência de plantas. Pages 177 – 190 in: Sanidade de Raízes. 1ed. NEFIT – Núcleo de Estudos em Fitopatologia, São Carlos, SP. Ceresini, P. C., Gaino, A. P. S. C., Basseto, M. A., Gasparotto, L., Poltronieri, L. S 2010. Inferência filogenética revela a complexa etiologia das manchas areolada e foliar em seringueira e em outras espécies cultivadas na Amazônia. Acta Scientiarum. Agronomy 32, 3: 385-395. Ceresini, P. C., Shew, P. C., Vilgays, R. J., Gale, L. R., Cubeta, M. A. 2003. Detecting Migrants in Populations of Rhizoctonia solani Anastamosis Group 3 from potato in North Carolina Using Multilocus Genotype Probabilities. Phytopathology 93, 5: 610-615. Ceresini, P.C., Souza, N. L. 1997. Associação de Rhizoctonia spp. binucleadas e R. solani Kühn AG4 HGI e AG 2-2 IIIB ao feijoeiro (Phaseolus vulgaris L.) no Estado de São Paulo. Summa Phytopathologica, 23: 14-24. Chaijuckam, P., Davis, R. M. 2010. Characterization of Diversity Among Isolates of Rhizoctonia oryzae-sativae from California Rice Fields. Plant Disease 946: 690-696.

47
Chaudhary, T. T., Anderson, T. T., Park, S. J., Yu, K. 2006. Comparison of screening methods for resistance to Fusarium root rot in common beans (Phaseolus vulgaris L.). Journal of Phytopathology, 154: 303- 308. Chen, C-M., de la Cerda, K. A., Kaminski, J. E., Douhan, G. W., Wong, F. P. 2009. Geographic Distribution and rDNA-ITS Region Sequence Diversity of Waitea circinata var. circinata Isolated from Annual Bluegrass in the United States. Plant Disease 93: 906-11. Ciampi, M. B., Gale, L. R., Lemos, E. G. de M., Ceresini, P. C. 2009. Distinctively variable sequence-based nuclear DNA markers for multilocus phylogeography of the soybean-and rice-infecting fungal pathogen Rhizoctonia solani AG-1 IA. Genetics and Molecular Biology. 32: 840-846. Ciampi, M. B., Kuramae, E. E., Fenille., R. C. Meyer, M. C., Souza, N. L., Ceresini, P. C. 2005. Intraspecific Evolution of Rhizoctonia solani AG-1 IA Associated with Soybean and Rice in Brazil based on Polymorphisms at the ITS-5.8S rDNA Operon. European Journal of Plant Pathology 1132: 183-196. Costa, R. C., Verzignassi, J. R., Poltronieri, T. P. S., Poltronieri, L. S., Monteiro, L. C. 2013. Novos hospedeiros de Ceratobasidium ochroleucum, agente causal da queima-do-fio no Pará. Summa Phytopathologica, 39, 1: 62. De la Cerda, K., Douhan, G., Wong, F. 2007. Discovery and characterization of Waitea circinatavar. circinataaffecting annual bluegrass from the Western United States. Plant Disease, 91: 791-797. Demirci, E. 1998. Rhizoctonia species and anastomosis groups isolated from barley and wheat in Erzurum, Turkey. Plant Pathology, 47, 1: 10-15. ESRI. 2011. ArcGIS Desktop: Release 10. Redlands, CA: Environmental Systems Research Institute. Erper, I. Karaca, G. H. Turkkan, M.; Ozkoc, I. 2006. Characterization and pathogenicity of Rhizoctonia spp. from onion in amasya. Journal of Phytopathology, 154: 75-79. Escandi, A. R., Echandi, E. 1991. Protection of potato from Rhizoctonia canker with binucleate Rhizoctoniafungi. Plant Pathology, 40: 197-202. Fang, X., Finnegan, P. M., Barbetti, M. J. 2013. Wide variation in virulence and genetic diversity of binucleate Rhizoctonia isolates associated with root rot of strawberry in Western Australia. PLoS One, 8, 2: 1-14. Fenille, R. C., Ciampi, M. B., Souza, N. L., Nakataniand, A. K., Kuramae, E. E. 2005 Binucleate Rhizoctonia sp. AG-G causing rot rot in yacon (Smallanyhussonchifolius) in Brazail. Plant Pathology, 54,325-330.

48
Fenille, R. C., Kuramae, E. E., Souza, N. L. 2003. Identification of Rhizoctonia solani Associated with Soybean in Brazil by rDNA-ITS Sequences. Fitopatologia Brasileira, 28, 4: 413-419. Fenille, R. C., Souza, N. L., Kuramae, E. E. 2002. Characterization of Rhizoctonia solani associated with soybean in Brazil. European Journal of Plant Pathology, 108, 8: 783-792. Gaino, A. P. S. C., Basseto, M. A., Gasparotto, l., Poltronieiri, l. S., Ceresini, P. C. 2010. Inferência filogenética revela a complexa etiologia das manchas areolada e foliar em seringueira e em outras espécies cultivadas na Amazônia. Acta Scientiarum. Agronomy, 32: 385-395. Genhua, Y., Chengyun, L. 2012. General Description of Rhizoctonia Species Complex, Plant Pathology, Dr. Christian Joseph Cumagun (Ed), InTech, DOI: 10.5772/39026. Disponível em: http://www.intechopen.com/books/plant-pathology/general-description-of-rhizocotonia-species-complex. Acesso: 20/12/14. Godoy-Lutz, G., Arias. J., Steadman, J. R., Eskridge, K. M. 1996. Role of natural seed infection by the web blight pathogen in common bean seed damage, seedling emergence, and early disease development. Plant Disease, 80: 887–890. Godoy – Lutz, G., Steadman, J. R., Higgins, B., Powers, K. 2003. Genetic Variation Among Isolates of the Web Blight Pathogen of common bean. Plant Disease, 87, 7: 766-771. Godoy-Lutz, G., Kuninaga, S., Steadman, J. R., Powers, K. 2008. Phylogenetic analysis of Rhizoctonia solani subgroups associated with web blight symptoms on common bean based on ITS-5.8S rDNA. Journal of General Plant Pathology, 74: 32–40. González-García, V., Portal-Onco, M. A., Rubio-Susan, V. 2006. Review: Biology and Systematics of the form genus Rhizoctonia. Span. J. Agric. Res. 4: 55-79. González, M. D. 2013. Identification, molecular characterization, and evolution of group I introns at the expansion segment D11 of 28S rDNA in Rhizoctonia species. Fungal Biology 117: 623-637. González, M. D., Carling, D. E., Kuninaga, S., Vilgalys, R., Cubeta, M. A. 2001. Ribosomal DNA systematics of Ceratobasidium and Thanatephorus with Rhizoctonia anamorphs. Mycologia, 93: 1138-1150. González, M. D., Cubeta, M. A., Vilgalys, R. 2006. Phylogenetic utility of indels within ribosomal DNA and beta-tubulin sequences from fungi in the Rhizoctonia solani species complex. Molecular Phylogenetic Evolution, 40, 2: 459-470. González, N., Godoy-Lutz, G., Steadman, J. R., Higgins, R., Eskridge, K. M. 2012. Assessing genetic diversity in the web blight pathogen Thanatephorus cucumeris (anamorph = Rhizoctonia solani) subgroups AG 1-IE and AG 1-IF with molecular markers. Journal of General Plant Pathology, 78: 85-98.

49
Goulart, A. C. P., Assis, J. B., Ciampi, M. B., Ceresini, P. C. 2011. Ocorrência de mela causada por Rhizoctonia solani AG4-HGI em plântulas de algodoeiro no brasil. Summa Phytopathologica 37, 1: 68-69. Gunnell, P. S. 1986. Characterization of the teleomorphs of Rhizoctonia oryzae-sativae, Rhizoctonia oryzae, and Rhizoctonia zeae, an the effect of cultural practices on aggregate sheath spot of rice, caused by R. oryzae sativae. Ph.D. thesis. University of California, Davis. Guillemaut, C., Edel-Herman, V., Camporota, P., Alabouvette, C., Richard-Molard, M., Steinberg, C., 2003. Typing of anastomosis groups of Rhizoctonia solani by restriction analysis of ribosomal DNA. Canadian Journal of Microbiology, 49: 556–568.
Gutiérrez, S. A., Cúndom, M. A., Barrera, V., Gasoni, L. 2007. First record of Rhizoctonia Zeae on corn in Argentina. Australasian Plant Disease Notes, 2, 137-138. Gurkanli, C. T., Ozkoc, I. 2011. First report of B. N. Rhizoctonia from Tobacco (Nicotiana tabacum L.) in Samsum, Turkey. Pakistan Journal of Botany, 43: 51–57. Hall, T. 2014. BioEdit v7.0.9: Biological sequence alignment editor for Win95/98/2K/XP/7. Disponível em: http://www.mbio.ncsu.edu/bioedit/page2.html. Acesso em: Julho de 2015. IBGE. Mapas de biomas e de Vegetação. 2004. Disponível em: http://www.ibge.gov.br/home/presidencia/noticias/21052004biomashtml.shtm. Acesso em: 29 de setembro de 2016. Judd, W. S., Campbell, C. S., Kellogg, E. A., Stevens, P. F., Donoghue, M.G. 2007. Sistemática Vegetal: um enfoque filogenético. Porto Alegre: Artmed. 3ª ed. Jukes, T. H; Cantor, C. R . Evolution of protein molecules. In: Munro, H. N. (Ed.). Mammalian Protein Metabolism, New York: Academic Press,1969, p. 21-132. Kammerer, S. J., Burpee, L. L., Harmon, P. F. 2011. Identification of a New Waitea circinata Variety Causing Basal Leaf Blight of Seashore Paspalum. Plant Disease, 95, 5: 515-522. Kareem, T. A., Hassan, M. S. 2013. Molecular Characterization of Rhizoctonia solani. Diyala Agricultural Sciences Journal, 5, 2: 45-54. Kataria, H. R., Hugelshofer, U., Gisi, U. 1991. Sensitivity of Rhizoctonia species to different fungicides. Plant Pathol. 40:2 03–211. Kuninaga, S., Carling, D. E., Takeuchi, T., Yokosawa, R. 2000. Comparison of rDNA-ITS Sequences between Potato and Tobacco Strains in Rhizoctonia solani AG-3. Journal of General Plant Pathology, 66: 2-11.

50
Kuninaga, S., Natsuaki, T., Takeuchi, T., Yokosawa, R. 1997. Sequence variation of the rDNA ITS regions within and between anastomosis groups in Rhizoctonia solani. Current Genetics, 32: 237-243. Kuramae, E. E., Nozaki, D.N., Fenille, R. C., Ceresini, P. C., Souza, N. L. 2002. Elemento extracromossomal dsRNA em Rhizoctonia AG4 HGI e AG2-2 IIIB e Rhizoctonia spp. binucleada associados às culturas de feijão e amendoim. Summa Phytopatholgica, 28: 52-57. Kuramae E. E., Buzeto, A. L., Nakatani, A. K., Souza, N. L. 2007. rDNA-based characterization of a new binucleate Rhizoctonia spp. causing root rot on kale in Brazil. European Journal of Plant Pathology, 119: 469–475. Kuznia, R. A.; Windels, C. E. 1994. Rhizoctonia zeae pathogenic to spring wheat and sugarbeet seedlings. Phytopathology, 84: 1159. Lange, L. The importance of fungi and mycology for addressing major global challenges. 2014. Ima Fungus. 5, 2: 463-471. Lanoiselet, V. M., Cother, E. J., Ash, G. J., Harper, J. D. I. 2005. Yield loss in rice caused by Rhizoctonia oryzae and R. oryzae-sativae in Australia. Australasian Plant Pathology, 4: 175-179. Lakshman, D. K., Amaradasa, B. S. 2014. The pathogen biology, identification and management of Rhizoctonia species with emphasis on isolates infecting turfgrasses. Indian Phytopathology, 67 (4): 327-345. Leiner, R. H., Carling, D. E. 1994. Characterization of Waitea circinata (Rhizoctonia) isolated from agricultural soils in Alaska. Plant Disease, 78: 385-388. Li, H. R., Wu, B. C., Yan, S. Q., 1998. Aetiology of Rhizoctonia in sheath blight of maize in Sichuan. Plant Pathology, 47: 16-21. Lübeck, M. 2004. Molecular Characterization of rhizoctonia. Applied Mycology & Biotechnology, 4. Fungal Genomes, 205-224. Mathew, F. M., Lamppa, R. S., Chittem, K., Chang, Y. W., Botschner, M., Kinzer, K., Goswami, R. S., Markell, S. G. 2012. Characterization and Pathogenicity of Rhizoctonia solani Isolates Affecting Pisum sativum in North Dakota. Plant Disease, 96, 5: 666-672. May, G. S., Adams, T. H. 1997. The Importance of Fungi to Man. Genome Research, 7 (11): 1041-1044. Melo, I. S. Recursos genéticos microbianos. 2002. In: Melo, I. S., Valadares-inglis, M. C., Nass, L. L., Valois, A. C. C. (orgs.). Recursos Genéticos e Melhoramento: Microorganismos. Jaguariúna: Embrapa Meio Ambiente.

51
Meyer, M. C., Yorinori, J. T. 1999. Incidência de doenças da soja em regiões tropicais. In: Congresso Brasileiro de Soja, Londrina. Anais. Londrina, Embrapa Soja, 1999, p. 457. (Embrapa Soja. Documentos, 124). Mora-Umaña, F., Barboza, N., Alvarado, R., Vásquez, M., Godoy-Lutz, G., Stedman, J.R., Ramírez, P. 2013. Virulence and Molecular Characterization of Costa Rican Isolates of Rhizoctonia solani from Common Bean. Tropical Plant Pathology, 38: 461-471. Muzhinji, N., Truter, M., Woodhall, J. W., van der Waals, J. E. 2015. Anastomosis Groups and Pathogenicity of Rhizoctonia solani and Binucleate Rhizoctonia from Potato in South Africa. Plant Disease, 99, 12: 1790 – 1802. Nakatani, A. K. 2006. Diversidade genética de Rhizoctonia spp. e análise de seqüências multilocos 2006. Tese. Universidade Estadual Paulista – UNESP. 98 f. Nechet, K. L., Halfeld-Vieira, B. A. 2006. Caracterização de isolados de Rhizoctonia spp., associados à mela do feijão caupi (Vigna unguiculata), coletados em Roraima. Fitopatologia Brasileira 31:505-508. Ogoshi, A., Oniki, M., Araki, T., Ui, T. 1983. Anastomosis groups of binucleate Rhizoctoniain Japan and North America and their perfect states. Trans Mycol Soc Jpn, 24: 79–87. Ogoshi, A. 1987. Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kühn. Annual Review of Phytopathology 25, 1: 125-143. Ogoshi, A.; Cook, R. J.; Bassett, E. N. 1990. Rhizoctonia species and anastomosis groups causing root rot of wheat and barley in the Pacific Northwest. Phytopathology, 80: 784-788. Oniki, M.; Ogoshi, A.; Araki, T.; Sakai, R.; Tanaka, S. 1985. The perfect state of Rhizoctonia oryzae and R. zeae and anastomosis groups of Waitea circinata. Trans. Mycol. Soc. Jpn, 26: 189-198. Otero, J. T., Ackerman, J. D., Bayman, P. 2002. Diversity and Host Specificity of Endophytic. American Journal of Botany 89, 11: 1852–1858. Paulitz, T. C., Okubara, P. A. 2006. First report of damping-off of canola caused by Rhizoctonia solani AG2–1 in Washington State. Plant Disease, 90, 6: 829.
Paulitz, T. C., Schroeder, K. L. 2005. A New Method for the Quantification of Rhizoctonia Solani and R. oryzae from Soil. Plant Disease, 89, 7: 767-772. Poltronieri, L. S., Trindade, D. R., Albuquerque, F. C. 2002. Rhizoctonia zeae Causando Podridão em Milho no Estado do Pará. Fitopatologia. Brasileira, 27, 4: 423-423 Poltronieri, L. S., Verzignassi, J. R., Benchimol, R. L. 2008. Tectonia grandis, nova hospedeira de Rhizoctonia solani no Pará. Summa Phytopathologica, 34, 2: 291.

52
Prabhu, A. S., Filippi, M. C., Silva, G. B., Santos, G. R. Resistência de cultivares de arroz a Rhizoctonia solani e Rhizoctonia oryzae. Pesquisa Agropecuária Brasileira, Brasília, 37, 5: 589-595, 2002. Raven, P. H., Johnson, G. B. Biology. 5ª (Ed), Boston, MA. McGraw-Hill, 2002. 1344pp. Rinehart, T. A., Copes, W. E., Toda, T., Cubeta, M. A. 2007. Genetic Characterization of Binucleate Rhizoctonia Species Causing Web Blight on Azalea in Mississippi and Alabama. Plant Disease, 91, 5: 616-623. Roberts, P. 1999. Rhizoctonia-forming Fungi. A Taxonomic Guide. Royal Botanic Gardens, Kew, London, UK. Ross, R. E., Keinath, A. P., Cubeta, M. A. 1998. Biological control of wirestem on cabbage using binucleate Rhizoctonia spp. Crop Prot, 17: 99-104. Schoonhoven, A. V., Pastor-Corrales, M. A. Standard system for the evaluation of bean germplasm. Cali: CIAT - Centro Internacional de Agricultura Tropical, 1987, 53p. Sha, T., Xu, J., Palanichamy, M. G., Zhang, T. L., Zhao, Z., Zhang, Y. 2008. Genetic diversity of the endemic gourmet mushroom Thelephora ganbajun from south-western China. Microbiology, 154: 3460-3468. Sharon, M., Sneh, B., Kuninaga, S., Hyakumachi, M. 2006. The advancing identification and classification of Rhizoctonia spp. using molecular and biotechnological methods compared with the classical anastomosis grouping. Mycoscience, 47: 299-316. Sharon, M., Sneh, B., Kuninaga, S., Hyakumachi, M., Naito, S. 2008. Classification of Rhizoctonia spp. using rDNA-ITS sequence analysis supports the genetic basis of the classical anastomosis grouping. Mycoscience, 49, 2: 93-114. Sneh B., Burpee L., Ogoshi, A. 1991. Identification of Rhizoctonia species. APS Press, St. Paul, Minnesota, USA. Souza, E. C., Kuramae, E. E., Nakatani, A. K., Basseto, M. A., Prabhu, A. S., Ceresini, P. C. 2007. Caracterização citomorfológica, cultural, molecular e patogênica de Rhizoctonia solani Kühn associado ao arroz em Tocantins, Brasil. Summa Phytopathologica, 33, 2: 129-136. StatSoft, Inc. 2007. STATISTICA (Data Analysis Software System), Version 8. http://www.statsoft.com. Summer, D. R., Minton, N. A. 1989. Crop Losses in Corn Induced by Rhizoctonia solani AG-2-2 and Nematodes. Phytopathology, 79: 934-931.

53
Sumner, D. R., Bell, D. K. 1982. Root diseases induced in corn by Rhizoctonia solani and Rhizoctonia zeae. Phytopathology, 72: 86-91. Tamura, K., Stecher, G., Peterson, D., Filipski., A., Kumar, S. MEGA6: molecular evolutionary genetics analysis, Version 6.0. Molecular Biology Evolution, 30, 12: 2725-2729, 2013. Taheri, P., Gnanamanickam, S., Hofte, M. 2007. Characterization, Genetic Structure, and Pathogenicity of Rhizoctonia spp. Associated with Rice Sheath Diseases in India. Phytopathology, 97: 3, 373-383. Telmadarrehei, T., Tajick Ghanbary, M. A., Rahimian, H., Rezazadeh, A., Javadi, M. A. 2011. Isolation and some pathologic properties of Rhizoctonia zeae from cultural soils of Golestan and Mazandaran Provinces, Iran. World Applied Sciences Journal, 14, 3: 374-377. Thompson, J. D., Gibson T. J., Plewniak, F., Jeanmougin, F., Higgins. D. G. 1997. The Clustal-X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 25: 4876-4882. Toda, T., Mushika, T., Hayakawa, T., Tanaka, A., Tani, T., Hyakumachi, M. 2005. Brown Ring Patch: A New Disease on Bentgrass Caused by Waitea circinata var. circinata. Plant Disease, 89, 6: 536-542. Toda, T., Hyakawa, T., Mwafaida Mghalu, J., Yaguchi, S., Hyakumachi, M. 2007. A new Rhizoctonia sp. closely related to Waitea circinata causes a new disease of creeping bentgrass. Journal of General Plant Pathology, 73: 379-387. Verignassi, J. R., Poltronieri, L. S., Benchimol, R. L. 2008. Ocorrência de Rhizoctonia solani AG1 em bastão-do-imperador no Estado do Pará. Summa Phytopathologica, 34, 3: 290. Vilgalys, R., Cubeta, M. A. 1994. Molecular systematics and population biology of Rhizoctonia, Annual Review of Phytopathology, 32: 135-155. Villajuan-Abgona, R., Kageyama, K., Hyakumachi, M. 1996. Biocontrol of Rhizoctonia damping-off of cucumber by nonpathogenic binucleate Rhizoctonia damping-off of cucumber by non pathogenic binucleate Rhizoctonia. European Journal Plant Pathology, 102, 227-235. Weinhold, A. R. 1977. Population of Rhizoctonia solani in agricultural soils determined by a screening procedure. Phytopathology, 67: 566-569. White, T. J., Bruns, S. Lee., Taylor, J. W. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Pp. 315-322 In: PCR Protocols: A Guide to A Guide to Methods and Applications, eds. Innis, M. A., D. H. Gelfand, J. J. Sninsky, and T. J. White. Academic Press, Inc., New York.

54
Yan, S. Q., Wu, B. C., Tang, X. F., Liu, Z. J., Jiang, L. 1984. Sheath blight of cereal crops: I. On the relation between sheath blight of rice, maize and wheat as well as soreshin of cotton. Acta Phytopathol Sin, 14: 25–31. (in Chinese with English abstract) Yorinori, J. T. 1998. Estratégias de controle das doenças da soja. Correio Agrícola, 2, 8-12. Youssef, D. R., Souza, G. R., Nechet, K. L., Halfed-Vieira, B. A. 2012. Caracterização de isolados de Rhizoctonia associados à queima foliar em Roraima. Revista Agro@mbiente On-line, 6, 2: 158-165.

55
Anexos

56
Artigo em formatado para submissão à revista Plant Disease.


Plant Disease

IDENTIFICATION, DIVERSITY AND PATHOGENICITY OF Rhizoctonia-1
LIKE PATHOGENS FROM THE BRAZILIAN SAVANNA 2
3
Angel José Vieira Blanco and Marília Oliveira Costa, Goiás Federal Institute 4
of Education, Science and Technology - Campus Inhúmas, Inhúmas, GO, Brazil, 5
75400-000; Roberto do Nascimento Silva, Department of Biochemistry and 6
Immunology, Ribeirão Preto Medical School, University of São Paulo, SP, 7
Brazil, 14049-900; Fábio Suzart de Albuquerque, Science and Mathematics 8
Faculty, College of Integrative Sciences and Arts, Arizona State University; 9
Arthur Tavares de Oliveira Melo, Department of Biological Sciences, College 10
of Life Sciences and Agriculture, University of New Hampshire, Durham, NH, 11
USA, 03824; Fabyano Alvares Cardoso Lopes, Institute of Biology, Federal 12
University of Rio de Janeiro, Rio de Janeiro, RJ, Brazil, 21941-599; Andrei 13
Stecca Steindorff, Department of Cell Biology, Biological Sciences Institute, 14
Brasília University Campus Darcy Ribeiro, Brasília, DF, Brazil, 70910-900; 15
Cirano José Ulhoa, Biological Sciences Institute, Biochemistry and Molecular 16
Biology Department, Goiás Federal University, Campus Samambaia, Goiânia, 17
GO, Brazil, 74690-900; Elder Tadeu Barbosa and Murillo Lobo Junior, 18
Brazilian Corporation for Agricultural Research, Embrapa Rice and Beans, 19
Santo Antônio de Goiás, GO, Brazil, 75375-000. 20
21
Corresponding author: C. J. Ulhoa; E-mail: [email protected] 22

Abstract 23
Blanco, A. J. V., Costa, M. O., Silva, R. N., Albuquerque, F. S., Melo, A. T. O., 24
Lopes, F. A. C., Steindorff, A. S., Ulhoa, C. J, Barbosa, E. T. and Lobo Junior, M. 25
Identification, diversity and pathogenicity of Rhizoctonia-like pathogens from 26
the Brazilian Savanna. 27
28
Eighty-one Rhizoctonia-like isolates from natural and agricultural soils 29
from the Brazilian Savanna were identified by genetic-based, morphology and 30
nuclei-coloring methods. The nucleotide similarity analysis of ITS2-ITS1-31
5.8S region identified 15 different taxa, with 39.5% of isolates assigned to 32
Waitea circinata (zeae, oryzae and circinata varieties), while 37.0% belonged to 33
Thanatephorus cucumeris AG1-IA, AG1-IB, AG1-ID, AG1-IE, AG4-HGI and AG4-34
HGIII. Ceratobasidium spp. AG-A, AG-F, AG-Fa, AG-P and AG-R summed 35
23.5%. Rhizoctonia zeae (19.75%), R. solani, AG1-IE (9.88%), R. solani AG1-IA 36
(8.64%) and binucleate Rhizoctonia AG-A (8.64%) were the most frequent 37
anamorphic states found. Root rot severity caused by the different taxa varied 38
from low to high on common beans, and tended to be low to average in maize. 39
The results indicate that cropping history and environmental characteristics 40
affect the geographical arrangement of AGs, with group AG1 identified in the 41
tropical zone from Center to North of Brazil while the AG4 group was found in 42
opposite direction, from Center towards the subtropical South. Waitea circinata 43
var. zeae was found predominantly in soil underneath corn production and the 44

Blanco, AJV – Plant Disease 3
AG-F and AG-Fa were found in areas of common bean production. Apparently, 45
this is the first report on the occurrence of W. circinata var. circinata in Brazil. 46
47
Keywords: Rhizoctonia diversity; ITS sequencing; Brazilian savanna; Root rot. 48
49
Introduction 50
51
The Rhizoctonias are an important gender of phytopathogenic fungi 52
associated with various plant tissues (Basseto et al. 2008; González 2013; 53
Kuramae et al. 2007; Youssef et al. 2012). They are cosmopolitan organisms 54
found in both soils used for agriculture or under native vegetation (Gonzalez-55
Garcia et al. 2006) and survive saprophytically or remain dormant as sclerotium 56
(Bruehl 1987). The species of this genus are highly diversified, showing life 57
cycle with teleomorphic or anamorphic sexual stages, which can cause 58
important diseases in several plant species (Sneh et al. 1991). 59
There are three main species of Rhizoctonias associated with plant 60
diseases: Thanatephorus cucumeris, a multinucleated species (Rhizoctonia solani 61
in anamorphic state), Ceratobasidium spp., a binucleate species often classified 62
like Ceratorhiza (anamorphic gender of Ceratobasidium) and Waitea circinata, a 63
multinucleated species with five known varieties: zeae, oryzae, circinata, 64
prodigus and agrostis (Kammerer et al. 2011; Leiner and Carling 1994; Roberts 65
1999; Toda et al. 2007; Vilgays and Cubeta 1994). 66

Blanco, AJV – Plant Disease 4
Thanatephorus cucumeris is regarded as the most important plant 67
pathogen within this group, causing diseases in over 200 hosts, including major 68
crops such as rice, maize, wheat, soybean and cotton, along with vegetables, 69
fruits, trees, weeds and ornamental species (Gonzalez et al. 2006). This species 70
causes either root or foliar diseases and, according to hyphal fusion between 71
compatible isolates, it is divided in 13 Anastomosis Groups (AG1 – AG13). 72
Some Ags may be subdivided into intraspecific groups (ISGs), based on 73
phenotypic traits like colony morphology, nutritional requirements, growth 74
temperature, host range and pathogenicity (Carling et al. 2002; Guillemaut et 75
al. 2003). 76
The teleomorphic genus Ceratobasidium is quite diverse, comprising 77
pathogens, saprophytes and mycorrhizal formers (Ogoshi 1987). 78
Ceratobasidium spp. infects common bean (Cardoso and Echandi 1987); potato 79
(Escandi and Echandi 1991); cucumber (Villajuan-Abgona et al., 1996); 80
Cabbage (Ross et al., 1998); and rice (Taheri et al., 2006). There are also some 81
reports about sheath blight (Li 1998), web blight (Nechet and Half-Vieira 2006) 82
(normally attributed to T. cucumeris) and fruit decay in a variety of hosts (Sneh 83
et al. 1991; Yan 1984). The classification system of Ceratobasidium is also 84
based on hyphal anastomosis, currently organized in 16 Gas, as discussed by 85
Sharon et al. (2008). 86
The five varieties oryzae, zeae, circinata, agrostis and prodigus of W. 87
circinata were stablished with the support of molecular biology and genetic 88

Blanco, AJV – Plant Disease 5
dissimilarity analysis (De la Cerda et al. 2007; Sharon et al. 2006; Toda et al. 89
2007). The epithet “Rhizoctonia” was not yet adopted for the agrostis and 90
prodigus varieties and, in general, and only the varieties oryzae and zeae have 91
been reported with relevancy. Waitea circinata var. zeae is the most common 92
variety due to its host range, which includes widespread crops such as maize 93
(Sumner and Bell 1982), rice (Oniki et al. 1985), onions (Erper et al. 2006), 94
sugar-beet (Kuznia and Windels 1994) and wheat and barley (Ogoshi et al. 95
1990). 96
The identification of Rhizoctonia species and their ISGs is a key measure 97
for the management of agricultural areas. The huge number of hosts, the 98
saprophytic capacity, the absence of resistant plant cultivars and the low 99
efficacy of fungicide applications (Amaradasa et al. 2014; Carling et al. 1990; 100
Kataria et al. 1991) makes disease management through genetic resistance and 101
cultural practices a challenge for growers and the rural extension service. 102
Without efficient management, there is an increase of the inoculum density on 103
soil, causing significant yield losses (Lanoiselet et al. 2005; Paulitz and 104
Okubara 2006). 105
The identification of Rhizoctonias still considers morphological, nuclei-106
staining and cultural aspects, but relies mostly in genetic-based methods to 107
confirm the identity of species, varieties and AG's. Out of these molecular-108
based methods, the genetic nucleotide analysis of the intergenic ITS region of 109
ribosomal DNA (rDNA) has been consolidated as an important tool to support 110

Blanco, AJV – Plant Disease 6
the Rhizoctonia species classification (White et al. 1990; Kuninaga et al. 1997; 111
Sharon et al. 2006, 2008). 112
The majority of the morphological and genetic diversity studies of 113
Rhizoctonias in Brazil is generally specific either for a single crop or an specific 114
AG (Basseto et al. 2008; Ceresini et al. 2010; Ciampi et al. 2005, 2009; Fenille et 115
al. 2003; Goulart et al, 2011; Kuramae et al. 2007, Souza et al. 2007), accounting 116
on lack of information about the geographic distribution and genetic diversity 117
of Rhizoctonia species across the Brazilian territory. 118
Hypothetically, cultural practices and environmental factors affect the 119
distribution of Rhizoctonia species and AGs in the country’s wide territory. 120
Thus, it’s also pertinent to associate species distribution to mechanisms linked 121
to the spread and establishment of these pathogens in relevant agricultural 122
regions such as the Cerrado (Brazilian Savanna), where intensive agriculture 123
and the achievement of 2-3 yields per year facilitate the establishment of soil 124
borne pathogens. 125
So, the objectives of this study are: i) identify species and ISGs of 126
Rhizoctonia spp. recovered from soils under native vegetation or agricultural 127
areas from the Brazilian Cerrado, ii) verify the pathogenicity of isolates in 128
maize and common beans roots and iii) estimate the relationship between 129
species distribution, farming practices and climate conditions in the studied 130
regions. 131
132

Blanco, AJV – Plant Disease 7
Material and Methods 133
Sampling sites and strain isolation 134
Eighty-one Rhizoctonia and Rhizoctonia-like isolates retrieved from 135
soils samples in different agricultural and native vegetation areas from the 136
Brazilian Savanna were investigated. The samples within this biome were 137
spatially distributed through the five Brazilian political regions (North, 138
Northeast, Midwest, Southeast and South) (Figure 1). Seventy samples come 139
from agricultural areas, with 37 under monoculture, 17 from soils that a crop 140
rotation was reported and 16 from soils where no information about the 141
cropping history was available (Table 1). The other 11 samples were recovered 142
from native vegetation areas. All soil samples were retrieved from the 0-10 cm 143
top soil, stored in plastic bags for transport and then preserved at 4°C in the 144
laboratory, until use. 145
The isolates were obtained using either the Weinhold (1977) method or 146
the Paulitz and Schroeder (2005) protocol and then, grown in agar-water with 147
chloramphenicol 200 ppm at 25ºC for five days. After checking morphological 148
traits typical of Rhizoctonia-like species at 100x, isolates were preserved in 149
sterilized paper stripes at -20ºC or -80ºC, and stored at the Fungal and 150
Functional Microorganisms Culture Collection from the Brazilian Company of 151
Agricultural Research (EMBRAPA), at the National Research Center for Rice 152
and Beans. 153

Blanco, AJV – Plant Disease 8
154
Nuclei staining and colony morphology 155
Isolates were evaluated for typical Rhizoctonia morphological traits, 156
such as 90º mycelial branches, absence of spores, lack of connection clamps, 157
number of nuclei per cell, mycelium color and presence of sclerotia in culture 158
media. For morphological analysis, a 5 mm diameter mycelial disc was 159
transferred to a glass slide for microculture. Fungal nuclei were colored with 160
Safranin-O (Bandoni 1979) and counted observing ten cells per glass slide, in 161
ten replicates. When necessary, microculture procedures were repeated three 162
times. 163
164
Genomic DNA extraction and sequencing of ribosomal DNA 165
The genomic DNA (gDNA) of each isolate was extracted according to 166
Lopes et al. (2012), with quality and amount checked with ultraviolet 167
spectrophotometry at 260 and 280 nm. Therefore, high quality gDNA samples 168
were amplified using the ITS1-5.8S-ITS2 region through the polymerase chain 169
reaction employing the pair of primers ITS1 (5'-TCCGTAGGTGAACCTGCGG-3 170
') and ITS4 (5'TCCTCCGCTTATTGATATGC- 3 ') (White et al. 1990). The PCR 171
products were submitted to agarose gel electrophoresis (1.2%) and then 172
purified using polyethylene glycol (PEG) 20%, 2.5 M NaCl and ethanol). The 173
purified products were sequenced following the DYEnamicTM ET Terminator 174

Blanco, AJV – Plant Disease 9
Cycle Sequencing kit (GE Healthcare, USA) procedures, in a ABI Prism 3100 175
(Applied Biosystems). 176
177
Species, varieties and AGs identification and phylogenetic analysis 178
To identify the 81 Rhizoctonia-like isolates at a synapormophic level, a 179
nucleotide similarity analysis through the BLASTn v.2.2.30 algorithm (Altschul 180
et al. 1997) was performed using the rDNA sequences as query sequences and 181
the non-redundant nucleotide database of NCBI as a reference. The best-hit 182
sequences with lower e-value that maximized the coverage of query sequences 183
were chosen as homologous copies, used to classify the samples at species, 184
variety and AG level. Eventually, to avoid misidentification due to presence of 185
GenBank sequences partially characterized, the next best-hits and the 186
morphological traits were used to ensure isolate classification. 187
Subsequently, the nucleotide rDNA sequences were submitted to local 188
alignment using Clustal X v.2.1 (Thompson et al. 1997) software and then 189
manually edited with Bioedit v.7.1.7 software (Hall 1999). All rDNA nucleotide 190
sequences were deposited at the GenBank database, and identified as on Table 191
1. A phylogenetic analysis was carried out with the maximum likelihood 192
method and 1,000 bootstraps to test the consistency of tree nodes. Three 193
distinct specific (Ceratobasidium spp., Thanatephorus spp. and Waitea spp.) 194
phylogenetic trees were constructed separately for group description with a 195
better resolution. The most appropriate evolutionary model for each of the 196

Blanco, AJV – Plant Disease 10
three datasets to reconstruct the phylogenetic tree topologies was chosen 197
based on the lowest value of the Bayesian information criterion (BIC) were 198
achieved using MEGA v.6.06 (Tamura et al. 2013). 199
200
Pathogenicity tests 201
Pathogenicity tests were conducted with the single inoculum layer 202
method (Chaudhary et al. 2006). Five seeds of common bean (var. Pérola) and 203
maize (MCP10 genotype), previously disinfested, were tested separately, in 500 204
ml polyethylene containers filled with vermiculite and a PDA layer with a 5-day 205
colony of the testing isolate. Pathogenicity tests were carried out at 20°C for 21 206
days in a growth chamber with 12-h photoperiod and irrigation three times a 207
week. The experiments were performed under a completely randomized design 208
with two replications and five plants per experimental unit, with non-infested 209
containers as control plots. 210
After harvest, plant roots were washed in tap water and checked for 211
pathogenicity and disease severity assessment using 1-9 scale from 212
Schoonhoven and Pastor-Corrales (1987). Isolates were compared for disease 213
severity with the Kruskal-Wallis (KW) non-parametric test at 5% significance, 214
considering severity among the different genders (inter-gender test), and in 215
intra-gender tests regarding the different species, variety and AGs. Statistical 216
procedures were performed on Statistica software (StatSoft Inc. 2007). The 217
spatial distribution analysis gathering results from isolate identification, their 218

Blanco, AJV – Plant Disease 11
geographical origin and pathogenicity tests were supported by ArcGis software 219
(ESRI 2011). 220
221
Results 222
Species, varieties and AG identification 223
All the 81 isolates collected across the Brazilian Cerrado were 224
successfully classified within the Rhizoctonia gender, in 15 different taxa. While 225
39.5% of the isolates were assigned to W.circinata varieties zeae, oryzae and 226
circinata, 37.0% belonged to T. cucumeris (AG1-IA, AG1-IB, AG1-ID, AG1-IE, 227
AG4-HGI and AG4-HGIII), while 23.5% were characterized as Ceratobasidium 228
(AG-A, AG-F, AG-Fa, AG-G, AG-P and AG-R). 229
All W. circinata isolates were multinucleated with mycelial color ranging 230
from orange to salmon or light brown (Figure 2 M-O). The W. circinata var. zeae 231
and W. circinata var. circinata isolates formed sclerotia after 40 days of growth, 232
in contrast to W. circinata var. oryzae. Waitea circinata was the most 233
representative “Rhizoctonia” species across the Brazilian Savanna, identified in 234
32 out of the 81 sampling sites. Zeae and circinata were the main W. circinata 235
varieties representing, respectively, 19.75% and 18.52% of total assignments 236
(Table 2). According to the geographical distribution and the relative and 237
absolute frequencies of the isolates, W. circinata had a higher incidence in 238
States of Goiás, Minas Gerais and Paraná (Table 1). 239

Blanco, AJV – Plant Disease 12
The T. cucumeris accounted for 37% of the total and were all 240
multinucleated, with the mycelium color ranging from light brown to dark 241
brown (Figure 2 G-L). The only two T. cucumeris AGs identified were AG1 and 242
AG4. Isolates belonging to AG1 were labeled in four ISGs: IA, IB, ID and IE, 243
while the AG4 were from the HGI and HGIII subgroups. All T. cucumeris AG4 244
isolates were sampled from agricultural areas between the center and the south 245
of Brazil, while the AG1 isolates were found in both agricultural soils or under 246
native vegetation located between the center and the north of Brazil (Table 1). 247
Isolates classified as AG1 and AG4 presented sclerotia formation, whose 248
dimensions, amounts and staining apparently varied according to the ISG. The 249
T. cucumeris isolates classified as AG1-1E and AG4-HGI were the most frequent, 250
with 9.88% each. The isolates labeled as AG1-IA, AG1-1B, AG1-ID and AG4-251
HGIII corresponded respectively to 4.94%, 3.70%, 2.47% and 6.17% of the 252
relative frequency within this group (Table 2). 253
Almost all isolates classified as Ceratobasidium presented two nuclei per 254
cell, except for isolates CNPAF_0036, CNPAF_0255 and CNPAF_0258, which 255
were multinucleate. All Ceratobasidium isolates also showed a yellowish-white 256
mycelium with powdery texture and variable number of light-colored sclerotia 257
(Figure 2 A-F). This gender also showed the lowest rate of isolates (only 23% of 258
the total), with AG-A and AG-F accounting respectively in 8.64% and 6.17% of 259
isolates within this gender. The other Ceratobasidium AGs were AG-Fa, AG-P, 260
AG-G and AG-R, and resulted together in lower than 10% of assignments (Table 261

Blanco, AJV – Plant Disease 13
2). The Ceratobasidium AG-A group showed a high occurrence in soils under 262
common bean cultivation, along with AG-F and AG-Fa. Furthermore, 263
Ceratobasidium AG-G and AG-R were isolated mainly from soils under carrot 264
and onion cultivation, respectively. 265
266
Pathogenicity and disease severity tests in common bean and maize 267
The pathogenicity tests on common bean and maize showed a different 268
response between isolates. First, in the inter-gender tests, there were no 269
differences in disease severity in maize roots (KW = 0.87; p-value = 0.6471). In 270
contrast, differences were found on root rot severity in common beans (KW = 271
19.05; p-value = 0.0001), with the highest average of disease severity caused by 272
T. cucumeris and Ceratobasidium spp. With regard to the intra-gender analysis, 273
there were significant differences between for both plant species, with KW = 274
142.34 (p-value = 0.000) for common bean and KW = 96.53 (p-value = 0.0000) 275
maize (Figure 3). 276
277
Phylogenetic analysis 278
The evolutionary model suggested to best fit the nucleotide sequence 279
homology and produce the phylogenetic tree topologies for both T. cucumeris 280
and Waitea spp. was the GTR + I (General Time Reversible + Invariant sites). 281
The Jukes-Cantor (JC) (Jukes and Cantor 1969) was the best model for 282
Ceratobasidium spp. Respectively, 81.46% and 69.43% nucleotide sites were 283

Blanco, AJV – Plant Disease 14
considered polymorphic and parsimoniously informative and, on average, 284
60.95% of bootstraps values supported monophyletic groups between the 285
isolates and their best hit sequences from NCBI. 286
In general, the clusterization occurred among isolates of the same AG 287
and/or variety, except for T. cucumeris isolates AG1-IA and AG1-IE that formed 288
a monophyletic group. The W. circinata var. zeae and the AGs from T. cucumeris 289
AG4-HGI and AG4-HGIII were clustered in two different groups each, and in 290
some cases, a geographic structure was observed with some isolates grouping 291
preferably by area, as for the Brazilian States of Roraima, Mato Grosso, Paraná 292
and Minas Gerais (Figures 4, 5, 6). 293
294
Discussion 295
296
Species, varieties and AG identification 297
The morphological patterns observed for Waitea, Ceratobasidium and 298
Thanatephorus isolates were consistent with the current descriptions for these 299
three genders by several authors (Demirci 1998; Kammerer et al. 2011; Leiner 300
and Carling 1994; Muzhinji et al. 2015; Sneh 1991). However, some 301
inconsistencies were not solved even with the support of the molecular 302
approach. Specifically, there was a conflict between the identification of the 303
multinuclear isolates CNPAF_0036, CNPAF_0255 and CNPAF_0258 that were 304
labeled as Ceratobasidium (supposedly binucleate), according to their 305

Blanco, AJV – Plant Disease 15
identification based on NCBI database. This mismatch is not a new report, and 306
can be explained by the genetic similarity between Ceratobasidium and 307
Thanatephorus, which is a multinucleated gender. 308
Probably, an evolutionary complete separation have not occurred yet for 309
these two genus (Ceresini 2014), causing a phylogenetic instability for the 310
group. In addition, according to Lubeck (2004), the taxonomic position of some 311
AGs within these two genus is not clear, because isolates from AG-6 312
(Thanatephorus) can merge hyphae with AG-F (Ceratobasidium), for example. 313
The phylogenetic analyzes also suggest that some isolates classified as 314
Ceratobasidium, based on a nuclei number and hyphae anastomosis can be 315
assigned to Thanatephorus genus (Gonzalez et al. 2001). 316
317
Disease severity evaluation for bean and maize 318
The pathogenicity tests revealed the prevalence of phytopathogenic 319
Rhizoctonia and Rhizoctonia-like isolates in a wide geographic area formed by 320
the Brazilian Savanna biome. These isolates differ in terms of disease severity 321
when compared within the same AG or between AGs, for the three species 322
found – T. cucumeris, W. circinata and Ceratobasidium spp. These differences 323
were attributed to the genetic variability of the isolates and specific traits of 324
their respective AGs. Although common bean and maize are known hosts of 325
both AG1-1A and AG1-1B (Ceresini, 2014), there were isolates causing low or 326
high disease severity in common bean, and most of them were hypovirulent on 327

Blanco, AJV – Plant Disease 16
maize roots. Therefore, their importance as root pathogens deserves a careful 328
look. 329
330
Geographic Distribution of Rhizoctonias in the Brazilian savanna. 331
The distribution of species, varieties and AGs of Rhizoctonia in such a 332
wide geographical region should not be related not only with the presence of 333
susceptible hosts at the sampling moment (Muzhinji et al. 2015). The 334
distribution of Rhizoctonia AGs on the sampling sites is also influenced by 335
climate variables, a combination of host plants sequentially cropped in the 336
field, cultural practices and the capacity of isolates to compete with other 337
microorganisms and persist in soil over sequential cropping seasons. 338
Despite the differences in time (years) of anthropic interference, not 339
assessed here, the cropping history of the sampling sites is formed by a narrow 340
range of crops, where soybean-maize-common bean is the main cropping 341
sequence. Even though some other hosts such as rice or vegetables may be 342
locally relevant, nevertheless, specialized agriculture is a trend in all 343
agricultural regions investigated, which probably favors the establishment of 344
specialized forms of Rhizoctonia. (Genhua and Chengyum 2012; Lakshman and 345
Amaradasa 2014). 346
347
The W. circinata distribution 348

Blanco, AJV – Plant Disease 17
The identification of W. circinata varieties and their distribution across 349
the five major Brazilian follows the same rationalization described above for 350
Rhizoctonias. The Waitea species attack different grass species (Kammerer et 351
al, 2011.) such as maize (Lakshman and Amaradasa, 2014), commonly found 352
and the Brazilian States of Paraná, Goiás, Bahia, Minas Gerais. The W. circinata 353
presence in the Tocantins state was reported in a relevant cropping region of 354
tropical rice (Brasil 2015) and, in both maize and rice cases, the cropping 355
history seems to contribute for to the establishment of W. circinata varieties in 356
these regions. 357
Specifically, the W. circinata var. zeae was the most abundant W. 358
circinata variety among the 81 isolate collection and was identified in 359
agricultural regions across the sampling sites. This variety was found in 360
agricultural sites and not identified in soils under native vegetation areas at the 361
Roraima State. Despite the popularity of maize crops in all sampling regions, 362
there are few reports in Brazil about root rots caused by the zeae variety of W. 363
circinata (Poltronieri et al. 2002), and it’s not clear if the pathogen represents 364
an underrated threat to maize in the country. Its occurrence seems not being 365
necessarily associated with environmental or climatic factors, which also can 366
be supported by studies that have found this variety in several climates and 367
environments from different countries around the world (Demirci 1998; 368
Gutiérrez et al. 2007; Li et al. 1998; Telmadarrehei et al. 2011). 369

Blanco, AJV – Plant Disease 18
In contrast, the W. circinata var. oryzae had a very low representation 370
across the sampling sites, found in a single location in the Goiás State. Studies 371
reporting the occurrence and distribution of this variety in Brazil are scarce, 372
although Prabhu et al. (2002) reported this species in rice fields at Tocantins 373
State (North region). 374
Contrasting with the former two W. circinata varieties discussed above, 375
the circinata variety has a recent taxonomical characterization, and it’s also 376
related to diseases Poaceae family hosts (Chen et al. 2009). Most of W. circinata 377
var. circinata isolates were found at Southeast region, mainly at Minas Gerais 378
State, forming the second largest group of isolates. Apparently, this is the first 379
report of this variety in Brazil. 380
381
The Ceratobasidium spp. distribution 382
The diversity of Ceratobasidium isolates also seems to be influenced by 383
the plant species cultivated over the Brazilian Cerrado. Five Ceratobasidium 384
AGs (AG-A, AG-F, AG-G, AG-R and AG-P) were previously related to areas 385
where common bean have been cultivated (Cardenas et al. 2015; Genhua and 386
Chengyum 2012). The “Fa” AG is not directly associated to common bean areas, 387
mainly because this AG have been considered a subgroup of AG-F (Gurkanli and 388
Ozkoc 2011; Sharon et al. 2008). With exception for the Tocantins State, our 389
results match the reports described above, with the record of Ceratobasidium 390
AGs in Paraná, Minas Gerais, Bahia and Goiás States that are important 391

Blanco, AJV – Plant Disease 19
common bean producers, with nearly two million tons of common bean yielded 392
in the 2014/2015 crop season (Brasil 2015). Although the AG-P found in Minas 393
Gerais was not isolated directly from common bean areas, the sampling site 394
(city of Unaí) is the largest producer of common bean in Brazil and an 395
important grower of maize and soybean. 396
In regard with minor crops, the identification of Ceratobasidium AG-R in 397
soils under onion cropping is consistent with the report of Burpe et al. (1980) 398
that linked AG-R with onions diseases. Also, the occurrence of AG-G in soils 399
cropped with carrot in a single site (Campos Altos - MG) does not necessarily 400
indicate a host x pathogen association, even though AG-G causes root rots in 401
yacon (Fenile et al. 2005), which shares morphologic and molecular 402
synapomorphies with carrot (Judd et al. 2007). 403
404
The T. cucumeris distribution 405
The geographic distribution of T. cucumeris followed the same 406
assumptions described for W. circinata and Ceratobasidium spp., especially for 407
the association between AGs and the recent cultivation history of their 408
respective sampling sites. In areas with common bean cultivation we found 409
AG1-IA and AG1-IE (Mato Grosso State), AG4-HGI (Goiás, Paraná and São 410
Paulo) and AG4-HGIII (Goiás). The AG4-HGI and AG4-HGIII were identified in 411
soils under soybean production in Goiás and Minas Gerais states, while The 412

Blanco, AJV – Plant Disease 20
AG1-1B was also identified in soybean areas in Goiás and in coffee-growing 413
areas in Minas Gerais (Table 1). 414
The AG1-IA and AG1-IE are known common bean pathogens, causing 415
web blight (AG1 subgroups) and root rots (AG4) (Gonzalez et al. 2012; Mora-416
Umaña et al. 2013; Youssef et al. 2012), AG4-HGI and AG4-HGIII (Çebi et al. 417
2010; Ceresini 2014; Genhua and Chengyum 2012; Lakshman and Amaradasa 418
2014). The AG4-HGI and AG4-HGIII (Ceresini 2014; Fenile et al. 2002; 2003) 419
and the AG1-1B (Lakshman and Amaradasa 2014; Youssef et al. 2012) also 420
infect soybeans. Although the results can indicate that establishment of most 421
T. cucumeris AGs was influenced by local crop management, it is also 422
significant that AG1-IA, AG1-ID and AG1-1E were present in areas under 423
native vegetation. 424
The identification of AG1 ISGs in non-agricultural soils on Northern 425
Brazil (Roraima State) confirms the existence of native Rhizoctonias, well 426
adapted to the equatorial environment, in the presence of high temperature 427
and rainfall through the year. In Central America and Caribbean countries, with 428
environmental conditions similar to Northern Brazil, AG1 and their ISGs have 429
been reported (Godoy-Lutz et al. 2008; González et al. 2012; Mora-Umaña et al. 430
2013), mainly as important common bean pathogens, causing web blight and 431
major economic losses (Godoy-Lutz et al. 1996; 2008). 432
Despite the geographical distribution of T. cucumeris AGs apparently 433
correlates with mainstream agricultural crops, the environmental traits are also 434

Blanco, AJV – Plant Disease 21
considered an important causes to explain the distribution of AG1 and AG4 in 435
the Brazilian Savanna, with AG1 found in Northern areas, while AG4 was 436
restricted to Southern regions. These results are consistent with Fenille et al. 437
(2002) who identified AG1 isolates causing web blight in soybeans grown in 438
Mato Grosso State, while AG4 was found in isolates from Minas Gerais, Paraná 439
and São Paulo, causing root rots. 440
441
Phylogenetic-based clustering 442
The phylogenetic analysis was effective to differentiate the taxa showing 443
that isolates belonging to the same variety or AGs generally are clustered, 444
based on the geographic distance among the sampling sites. It suggests that 445
within the same variety or AGs an important correlation between genetic 446
dissimilarity and geographic distance can be found. Furthermore, this 447
correlation also indicates a limited gene flow among the sampling sites. 448
Differently from what is expected for divergent groups, a monophyletic 449
grouping between AG1-IA and AG1-IE was found and, even both isolates are 450
from the same AGs, it seems that their reproductive separation may not have 451
completed. 452
The multinucleated pattern found for this AG could skew the 453
phylogenetic analysis once multiple copies of the ITS2-ITS1-5.8S region can be 454
sequenced. In addition, AG1-IA and AG1-IE are morphologically very similar, 455
especially regarding the number, size and disposition of sclerotia, not assessed 456

Blanco, AJV – Plant Disease 22
here (Figure 2 G-L). There are scarce reports of AG1-IE in Brazil, while AG1-IA 457
has been identified in several Brazilian regions (Ceresini 2014; Poltronieri, 458
2008; Youssef 2012), and so it is possible that all AG1-IE isolates could actually 459
belong to AG1-IA (Figure 5). 460
The morphological similarity among the Waitea, Ceratobasidium and 461
Thanatephorus colonies is obviously not enough for species identification, but 462
the molecular characterization of Rhizoctonias also requires attention (Sharon 463
et al. 2006; 2008), to avoid mistakes during the sequence assignment at the 464
NCBI database. Assuming that reference sequences support correctly our 465
results and that there are still some gaps to clarify all phylogenetic 466
relationships within Rhizoctonia-related genus, our results endorse previous 467
reports for AG1 distribution typically in the equatorial zones (Godoy-Lutz et al. 468
2008; Gonzalez et al. 2012; Mora-Umaña et al 2013), even tough some 469
misunderstanding is possible to differentiate closely-related ISGs (IA or IE). 470
In conclusion, the Rhizoctonias diversity in the Brazilian Cerrado is 471
formed by different species, which are divided in varieties and AGs. Their 472
geographic distribution varies in a latitudinal gradient and is influenced by 473
environmental variables and presence of host crops, forming a pattern of 474
distribution across this biome. The gene flow between distant regions seems to 475
be limited. These results reiterate the phytopathogenic importance of the 476
Rhizoctonia gender concerning the Brazilian agriculture, mostly with common 477
beans, suggesting that further studies with more representative, larger number 478

Blanco, AJV – Plant Disease 23
of samples by narrower areas can add new relevant information, to support the 479
disease management. 480
481
Acknowledgments 482
The authors would like to thanks the Federal Institute of Education, Science 483
and Technology of Goiás (IFG), the Brazilian Company for Agricultural 484
Research (EMBRAPA Rice and Beans), the Federal University of Goiás (UFG) 485
and the Brazilian National Council for Scientific and Technological 486
Development (CNPq) that have been supported the execution of this study with 487
both financial resources and infrastructure. 488
489
Literature Cited 490
491
Altschul, S. F., Madden, T. L., Scha ffer, A. A., Zhang, J., Zhang, Z., Miller, W., 492
Lipman, D. J., Gapped BLAST and PSI-BLAST: a new generation of protein 493
database search programs, Nucleic Acids Res. 25 (1997) 3389-3402. 494
495
Amaradasa, B. S., Lakshman, D. K., McCall, D. Horvath, B. J. 2014. In Vitro 496
Fungicide Sensitivity of Rhizoctonia and Waitea Isolates Collected from 497
Turfgrasses. J. Environ. Hort. 32: 126-132. 498
499
Bandoni, R. J. Safranin O as a rapid nuclear stain for fungi. Mycologia 71:873-500
874. 1979. 501
502
Basseto, M. A., Filho, W. V. V., Filho, W. V. V. F., Souza, E. C., Ceresini, P. C. 503
2008. O papel de Rhizoctonia spp. binucleadas na indução de resistência a mela 504
da soja. Acta Scientiarum. Agronomy, 30, 2: 183-189. 505

Blanco, AJV – Plant Disease 24
506
BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Projeções do 507
agronegócio: Brasil 2014/15 a 20124/25: projeções de longo prazo. 6. ed. 508
Brasília: AGE/Mapa, 2015. Disponível em: 509
<http://www.agricultura.gov.br/arq_editor/PROJECOES_DO_AGRONEGOCIO_2510
025_WEB.pdf>. Acesso em 15 jun. 2016. 511
512
Bruehl, G.W. Soilborne plant pathogens. New York NY. Mac Millan Publishing 513
Company. 1987. 514
515
Burpee, L. L., Sunders, P. L., Cole, H. Jr., Sherwood, R. T. 1980. Anastomosis 516
groups among isolates of Ceratobasidium cornigerum and related fungi. 517
Mycologia, 72: 689–701. 518
519
Cardenas, G. E. M., Galvan, M. Z., Barrera, V. A., Rodriguero, M. S., Carmona, 520
M. A., March, G. J., Ramallo, A. C., Shew, H. D. 2015. Molecular identification 521
and pathogenicity of Rhizoctonia spp. from tobacco growing areas in 522
northwestern Argentina. Tropical Plant Pathology, 40(3), 160-168. 523
524
Cardoso, J. E., Echandi, E. 1987. Biological control of Rhizoctonia root rot of 525
snap bean with binucleate Rhizoctonia-like fungi. Plant Disease, 71: 167-170. 526
527
Carling, D. E., Helm, D. J., Leiner. R. H. 1990. In vitro sensitivity of Rhizoctonia 528
solani and other multinucleate and binucleate Rhizoctonia to selected 529
fungicides. Plant Disease, 74: 860–863. 530
531
Carling, D. E., Kuninaga, S., Brainard, K. A. 2002. Hyphal Anastomosis 532
Reactions, rDNA-Internal Transcribed Spacer Sequences, and Virulence Levels 533
Among Subsets of Rhizoctonia solani Anastomosis Group-2 (AG-2) and AG-BI. 534
Phytopathology, 92, 1: 43–50. 535
536
Çebi Kılıçoğlu, M., Özkoç, İ. 2010. Molecular characterization of Rhizoctonia 537
solani AG4 using PCR-RFLP of the rDNA-ITS region. Turk. J. Biol, 34: 261–269. 538
539
Ceresini, P. C. 2014. Rhizoctonia como fitopatógeno: biologia e diversidade de 540
Rhizoctonia solani em agroecosistemas tropicais e perspectivas do manejo da 541
rhizoctoniose usando resistência de plantas. Pages 177 – 190 in: Sanidade de 542
Raízes. 1ed. NEFIT – Núcleo de Estudos em Fitopatologia, São Carlos, SP. 543
544
Ceresini, P. C., Gaino, A. P. S. C., Basseto, M. A., Gasparotto, L., Poltronieri, L. 545
S. 2010. Inferência filogenética revela a complexa etiologia das manchas 546
areolada e foliar em seringueira e em outras espécies cultivadas na Amazônia. 547
Acta Scientiarum. Agronomy, 32, 3: 385-395. 548
549

Blanco, AJV – Plant Disease 25
Chaudhary, T. T., Anderson, T. T., Park, S. J., Yu, K. 2006. Comparison of 550
screening methods for resistance to Fusarium root rot in common beans 551
(Phaseolus vulgaris L.). Journal of Phytopathology, 154: 303- 308. 552
553
Chen, C-M., de la Cerda, K. A., Kaminski, J. E., Douhan, G. W., Wong, F. P. 2009. 554
Geographic Distribution and rDNA-ITS Region Sequence Diversity of Waitea 555
circinata var. circinata Isolated from Annual Bluegrass in the United States. 556
Plant Disease, 93: 906-911. 557
558
Ciampi, M. B., Gale, L. R., Lemos, E. G. de M., Ceresini, P. C. 2009. Distinctively 559
variable sequence-based nuclear DNA markers for multilocus phylogeography 560
of the soybean-and rice-infecting fungal pathogen Rhizoctonia solani AG-1 IA. 561
Genetics and Molecular Biology, 32: 840-846. 562
563
Ciampi, M. B., Kuramae, E. E., Fenille, R. C., Meyer, M. C., Souza, N. L., 564
Ceresini, P. C. 2005. Intraspecific Evolution of Rhizoctonia solani AG-1 IA 565
Associated with Soybean and Rice in Brazil based on Polymorphisms at the ITS-566
5.8S rDNA Operon. European Journal of Plant Pathology, 1132: 183-196. 567
568
De la Cerda, K., Douhan, G., Wong, F. 2007. Discovery and characterization of 569
Waitea circinata var. circinata affecting annual bluegrass from the Western 570
United States. Plant Disease, 91: 791-797. 571
572
Demirci, E. 1998. Rhizoctonia species and anastomosis groups isolated from 573
barley and wheat in Erzurum, Turkey. Plant Pathology, 47, 1: 10-15. 574
575
Elliott, M. L. 1999. Comparison of Rhizoctonia zeae Isolates from Florida and 576
Ohio Turfgrasses. HortScience, 34: 298-300. 577
578
Erper, I. Karaca, G. H. Turkkan, M.; Ozkoc, I. 2006. Characterization and 579
pathogenicity of Rhizoctonia spp. from onion in Amasya, Turkey. Journal of 580
Phytopathology, 154: 75-79. 581
582
Escandi, A. R., Echandi, E. 1991. Protection of potato from Rhizoctonia canker 583
with binucleate Rhizoctonia fungi. Plant Pathology, 40: 197-202. 584
585
ESRI 2011. ArcGIS Desktop: Release 10. Redlands, CA: Environmental Systems 586
Research Institute. 587
588
Fenille, R. C., Ciampi, M. B., Souza, N. L., Nakataniand, A. K., Kuramae, E. E. 589
2005 Binucleate Rhizoctonia sp. AG-G causing root rot in yacon (Smallanyhus 590
sonchifolius) in Brazil. Plant Pathology, 54,325-330. 591
592

Blanco, AJV – Plant Disease 26
Fenille, R. C., Kuramae, E. E., Souza, N. L. 2003. Identification of Rhizoctonia 593
solani Associated with Soybean in Brazil by rDNA-ITS Sequences. Fitopatologia 594
Brasileira, 28, 4: 413-419. 595
596
Fenille, R. C., Souza, N. L., Kuramae, E. E. 2002. Characterization of Rhizoctonia 597
solani associated with soybean in Brazil. European Journal of Plant Pathology, 598
108, 8: 783-792. 599
600
Genhua, Y., Chengyun, L. 2012. General Description of Rhizoctonia Species 601
Complex, Plant Pathology, Dr. Christian Joseph Cumagun (Ed), InTech, DOI: 602
10.5772/39026. Disponível em: http://www.intechopen.com/books/plant-603
pathology/general-description-of-rhizocotonia-species-complex. Acesso: 604
20/12/14. 605
606
Godoy-Lutz, G., Arias. J., Steadman, J. R., Eskridge, K. M. 1996. Role of natural 607
seed infection by the web blight pathogen in common bean seed damage, 608
seedling emergence, and early disease development. Plant Disease, 80: 887–609
890. 610
611
Godoy-Lutz, G., Kuninaga, S., Steadman, J. R., Powers, K. 2008. Phylogenetic 612
analysis of Rhizoctonia solani subgroups associated with web blight symptoms 613
on common bean based on ITS-5.8S rDNA. Journal of General Plant Pathology, 614
74: 32–40. 615
616
González-García, V., Portal-Onco, M. A., Rubio-Susan, V. 2006. Review: 617
Biology and Systematics of the form genus Rhizoctonia. Spanish Journal of 618
Agricultural Research, 4: 55-79. 619
620
González, M. D. 2013. Identification, molecular characterization, and evolution 621
of group I introns at the expansion segment D11 of 28S rDNA in Rhizoctonia 622
species. Fungal Biology, 117: 623-637. 623
624
González, M. D., Carling, D. E., Kuninaga, S., Vilgalys, R., Cubeta, M. A. 2001. 625
Ribosomal DNA systematics of Ceratobasidium and Thanatephorus with 626
Rhizoctonia anamorphs. Mycologia, 93: 1138-1150. 627
628
González, M. D., Cubeta, M. A., Vilgalys, R. 2006. Phylogenetic utility of indels 629
within ribosomal DNA and beta-tubulin sequences from fungi in the 630
Rhizoctonia solani species complex. Molecular Phylogenetic Evolution, 40, 2: 631
459-470. 632
633
González, N., Godoy-Lutz, G., Steadman, J. R., Higgins, R., Eskridge, K. M. 2012. 634
Assessing genetic diversity in the web blight pathogen Thanatephorus cucumeris 635

Blanco, AJV – Plant Disease 27
(anamorph = Rhizoctonia solani) subgroups AG 1-IE and AG 1-IF with molecular 636
markers. Journal of General Plant Pathology, 78: 85-98. 637
638
Goulart, A. C. P., Assis, J. B., Ciampi, M. B., Ceresini, P. C. 2011. Ocorrência de 639
mela causada por Rhizoctonia solani AG4-HGI em plântulas de algodoeiro no 640
brasil. Summa Phytopathologica, 37, 1: 68-69. 641
642
Guillemaut, C., Edel-Herman, V., Camporota, P., Alabouvette, C., Richard-643
Molard, M., Steinberg, C., 2003. Typing of anastomosis groups of Rhizoctonia 644
solani by restriction analysis of ribosomal DNA. Canadian Journal of 645
Microbiology, 49: 556–568. 646
Gurkanli, C. T., Ozkoc, I. 2011. First report of B. N. Rhizoctonia from Tobacco 647
(Nicotiana tabacum L.) in Samsum, Turkey. Pakistan Journal of Botany, 43: 51–648
57. 649
650
Gutiérrez, S. A., Cúndom, M. A., Barrera, V., Gasoni, L. 2007. First record of 651
Rhizoctonia Zeae on corn in Argentina. Australasian Plant Disease Notes, 2: 652
137-138. 653
654
Hall, T. 2014. BioEdit v7.0.9: Biological sequence alignment editor for 655
Win95/98/2K/XP/7. Disponível em: 656
http://www.mbio.ncsu.edu/bioedit/page2.html. Acesso em: Julho de 2013. 657
658
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor 659
and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 660
41: 95-98. 661
662
Judd, W. S., Campbell, C. S., Kellogg, E. A., Stevens, P. F., Donoghue, M.G. 2007. 663
Sistemática Vegetal: um enfoque filogenético. Porto Alegre: Artmed. 3ª ed. 664
665
Jukes, T. H; Cantor, C. R . Evolution of protein molecules. In: Munro, H. N. 666
(Ed.). Mammalian Protein Metabolism, New York: Academic Press, 1969, p. 21-667
132. 668
669
Kammerer, S. J., Burpee, L. L., Harmon, P. F. 2011. Identification of a New 670
Waitea circinata Variety Causing Basal Leaf Blight of Seashore paspalum. Plant 671
Disease, 95, 5: 515-522. 672
673
Kareem, T. A., Hassan, M. S. 2013. Molecular characterization of Rhizoctonia 674
solani. Diyala Agricultural Sciences Journal, 5, 2: 45-54. 675
676
Kataria, H. R., Hugelshofer, U., Gisi, U. 1991. Sensitivity of Rhizoctonia species 677
to different fungicides. Plant Pathology, 40:2 03–211. 678

Blanco, AJV – Plant Disease 28
679
Kuninaga, S., Natsuaki, T., Takeuchi, T., Yokosawa, R. 1997. Sequence variation 680
of the rDNA ITS regions within and between anastomosis groups in Rhizoctonia 681
solani. Current Genetics, 32: 237-243. 682
683
Kuramae E. E., Buzeto, A. L., Nakatani, A. K., Souza, N. L. 2007. rDNA-based 684
characterization of a new binucleate Rhizoctonia spp. causing root rot on kale in 685
Brazil. European Journal of Plant Pathology, 119: 469–475. 686
687
Kuznia, R. A.; Windels, C. E. 1994. Rhizoctonia zeae pathogenic to spring wheat 688
and sugar beet seedlings. Phytopathology, 84: 1159. 689
690
Lanoiselet, V. M., Cother, E. J., Ash, G. J., Harper, J. D. I. 2005. Yield loss in rice 691
caused by Rhizoctonia oryzae and R. oryzae-sativae in Australia. Australasian 692
Plant Pathology, 4: 175-179. 693
694
Lakshman, D. K., Amaradasa, B. S. 2014. The pathogen biology, identification 695
and management of Rhizoctonia species with emphasis on isolates infecting 696
turfgrasses. Indian Phytopathology, 67 (4): 327-345. 697
Leiner, R. H., Carling, D. E. 1994. Characterization of Waitea circinata 698
(Rhizoctonia) isolated from agricultural soils in Alaska. Plant Disease, 78: 385-699
388. 700
701
Li, H. R., Wu, B. C., Yan, S. Q. 1998. Aetiology of Rhizoctonia in sheath blight of 702
maize in Sichuan. Plant Pathology, 47: 16-21. 703
704
Lopes, F. A. C., Steindorff, A. S., Geraldine, A. M. Brandão, R. S., Monteiro, V. 705
N., Lobo-Junior, M., Coelho, A. S. G., Ulhoa, C. J., Silva, R. N. 2012. Biochemical 706
and metabolic profiles of Trichoderma strains isolated from common bean 707
crops in the Brazilian Cerrado, and potential antagonism against Sclerotinia 708
sclerotiorum. Fungal Biology, 116: 815-824. 709
710
Lübeck, M. 2004. Molecular Characterization of Rhizoctonia. Applied Mycology 711
& Biotechnology, 4. Fungal Genomes, 205-224. 712
713
Mora-Umaña, F., Barboza, N., Alvarado, R., Vásquez, M., Godoy-Lutz, G., 714
Steadman, J.R., Ramírez, P. 2013. Virulence and Molecular Characterization of 715
Costa Rican Isolates of Rhizoctonia solani from Common Bean. Tropical Plant 716
Pathology, 38: 461-471. 717
718

Blanco, AJV – Plant Disease 29
Muzhinji, N., Truter, M., Woodhall, J. W., van der Waals, J. E. 2015. 719
Anastomosis Groups and Pathogenicity of Rhizoctonia solani and Binucleate 720
Rhizoctonia from Potato in South Africa. Plant Disease, 99, 12: 1790 – 1802. 721
722
Nechet, K. L., Halfeld-Vieira, B. A. 2006. Caracterização de isolados de 723
Rhizoctonia spp., associados à mela do feijão caupi (Vigna unguiculata), 724
coletados em Roraima. Fitopatologia Brasileira, 31: 505-508. 725
726
Ogoshi, A., Cook, R. J., Bassett, E. N. 1990. Rhizoctonia species and 727
anastomosis groups causing root rot of wheat and barley in the Pacific 728
Northwest. Phytopathology, 80: 784-788. 729
730
Ogoshi, A. 1987. Ecology and pathogenicity of anastomosis and intraspecific 731
groups of Rhizoctonia solani Kühn. Annual Review of Phytopathology, 25:125-732
143. 733
734
Oniki, M., Ogoshi, A., Araki, T., Sakai, R., Tanaka, S. 1985. The perfect state of 735
Rhizoctonia oryzae and R. zeae and anastomosis groups of Waitea circinata. 736
Transactions of the Mycological Society of Japan, 26: 189-198. 737
738
Prabhu, A. S., Filippi, M. C., Silva, G. B., Santos, G. R. Resistência de cultivares 739
de arroz a Rhizoctonia solani e Rhizoctonia oryzae. Pesquisa Agropecuária 740
Brasileira, Brasília, 37, 5: 589-595, 2002. 741
742
Paulitz, T. C., Okubara, P. A. 2006. First report of damping-off of canola caused 743
by Rhizoctonia solani AG2–1 in Washington State. Plant Disease, 90, 6: 829. 744
745
Paulitz, T. C., Schroeder, K. L. 2005. A New Method for the Quantification of 746
Rhizoctonia Solani and R. oryzae from Soil. Plant Disease, 89, 7: 767-772. 747
748
Poltronieri, L. S., Trindade, D. R., Albuquerque, F. C. 2002. Rhizoctonia zeae 749
Causando Podridão em Milho no Estado do Pará. Fitopatologia. Brasileira, 27, 750
4: 423-423 751
752
Poltronieri, L. S., Verzignassi, J. R., Benchimol, R. L. 2008. Tectonia grandis, 753
nova hospedeira de Rhizoctonia solani no Pará. Summa Phytopathologica, 34, 2: 754
291. 755
756
Roberts, P. 1999. Rhizoctonia-forming Fungi. A Taxonomic Guide. Royal 757
Botanic Gardens, Kew, London, UK. 758
759
Ross, R. E., Keinath, A. P., Cubeta, M. A. 1998. Biological control of wirestem on 760
cabbage using binucleate Rhizoctonia spp. Crop Protection, 17: 99-104. 761
762

Blanco, AJV – Plant Disease 30
Schoonhoven, A. V., Pastor-Corrales, M. A. Standard system for the evaluation 763
of bean germplasm. Cali: CIAT - Centro Internacional de Agricultura Tropical, 764
1987, 53p. 765
766
Sharon, M., Sneh, B., Kuninaga, S., Hyakumachi, M. 2006. The advancing 767
identification and classification of Rhizoctonia spp. using molecular and 768
biotechnological methods compared with the classical anastomosis grouping. 769
Mycoscience, 47: 299-316. 770
771
Sharon, M., Sneh, B., Kuninaga, S., Hyakumachi, M., Naito, S. 2008. 772
Classification of Rhizoctonia spp. using rDNA-ITS sequence analysis supports 773
the genetic basis of the classical anastomosis grouping. Mycoscience, 49, 2: 93-774
114. 775
776
Sneh B., Burpee L., Ogoshi, A. 1991. Identification of Rhizoctonia species. APS 777
Press, St. Paul, Minnesota, USA. 778
779
Souza, E. C., Kuramae, E. E., Nakatani, A. K., Basseto, M. A., Prabhu, A. S., 780
Ceresini, P. C. 2007. Caracterização citomorfológica, cultural, molecular e 781
patogênica de Rhizoctonia solani Kühn associado ao arroz em Tocantins, Brasil. 782
Summa Phytopathologica, 33, 2: 129-136. 783
784
StatSoft, Inc. 2007. STATISTICA (Data Analysis Software System), Version 8. 785
http://www.statsoft.com. 786
787
Sumner, D. R., Bell, D. K. 1982. Root diseases induced in corn by Rhizoctonia 788
solani and Rhizoctonia zeae. Phytopathology, 72: 86-91. 789
790
Taheri, P., Gnanamanickam, S., Höfte, M. 2007. Characterization, Genetic 791
Structure, and Pathogenicity of Rhizoctonia spp. Associated with rice Sheath 792
Diseases in India. Phytopathology, 97, 3: 373-383. 793
794
Tamura, K., Stecher, G., Peterson, D., Filipski, A., Kumar, S. MEGA6: molecular 795
evolutionary genetics analysis, Version 6.0. Molecular Biology Evolution, 30, 796
12: 2725-2729, 2013. 797
798
Telmadarrehei, T., Tajick Ghanbary, M. A., Rahimian, H., Rezazadeh, A., Javadi, 799
M. A. 2011. Isolation and some pathologic properties of Rhizoctonia zeae from 800
cultural soils of Golestan and Mazandaran Provinces, Iran. World Applied 801
Sciences Journal, 14, 3: 374-377. 802
803
Thompson, J. D., Gibson T. J., Plewniak, F., Jeanmougin, F., Higgins. D. G. 1997. 804
The Clustal-X windows interface: flexible strategies for multiple sequence 805

Blanco, AJV – Plant Disease 31
alignment aided by quality analysis tools. Nucleic Acids Research, 25: 4876-806
4882. 807
808
Toda, T., Hyakawa, T., Mwafaida Mghalu, J., Yaguchi, S., Hyakumachi, M. 2007. 809
A new Rhizoctonia sp. closely related to Waitea circinata causes a new disease 810
of creeping bentgrass. Journal of General Plant Pathology, 73: 379-387. 811
812
Vilgalys, R., Cubeta, M. A. 1994. Molecular systematics and population biology 813
of Rhizoctonia, Annual Review of Phytopathology, 32: 135-155. 814
815
Villajuan-Abgona, R., Kageyama, K., Hyakumachi, M. 1996. Biocontrol of 816
Rhizoctonia damping-off of cucumber by nonpathogenic binucleate 817
Rhizoctonia damping-off of cucumber by non-pathogenic binucleate 818
Rhizoctonia. European Journal Plant Pathology, 102: 227-235. 819
820
Weinhold, A. R. 1977. Population of Rhizoctonia solani in agricultural soils 821
determined by a screening procedure. Phytopathology, 67: 566-569. 822
823
White, T. J., Bruns, S. Lee., Taylor, J. W. 1990. Amplification and direct 824
sequencing of fungal ribosomal RNA genes for phylogenetics. Pp. 315-322 In: 825
PCR Protocols: A Guide to A Guide to Methods and Applications, eds. Innis, M. 826
A., D. H. Gelfand, J. J. Sninsky, and T. J. White. Academic Press, Inc., New York. 827
828
Yan, S. Q., Wu, B. C., Tang, X. F., Liu, Z. J., Jiang, L. 1984. Sheath blight of 829
cereal crops: I. On the relation between sheath blight of rice, maize and wheat 830
as well as soreshin of cotton. Acta Phytopathol Sin, 14: 25–31. (in Chinese with 831
English abstract). 832
833
Youssef, D. R., Souza, G. R., Nechet, K. L., Halfeld-Vieira, B. A. 2012. 834
Caracterização de isolados de Rhizoctonia associados à queima foliar em 835
Roraima. Revista Agro@mbiente On-line, 6, 2: 158-165. 836
837
838
839
840
841
842
843
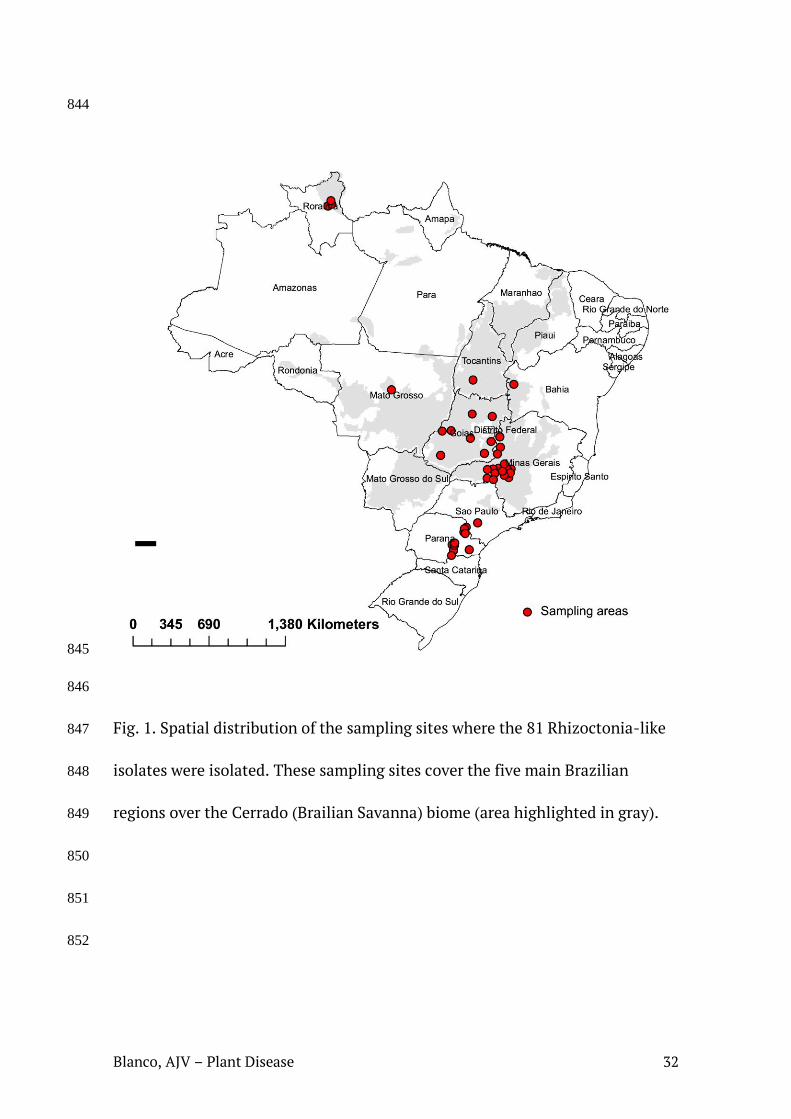
Blanco, AJV – Plant Disease 32
844
845
846
Fig. 1. Spatial distribution of the sampling sites where the 81 Rhizoctonia-like 847
isolates were isolated. These sampling sites cover the five main Brazilian 848
regions over the Cerrado (Brailian Savanna) biome (area highlighted in gray). 849
850
851
852
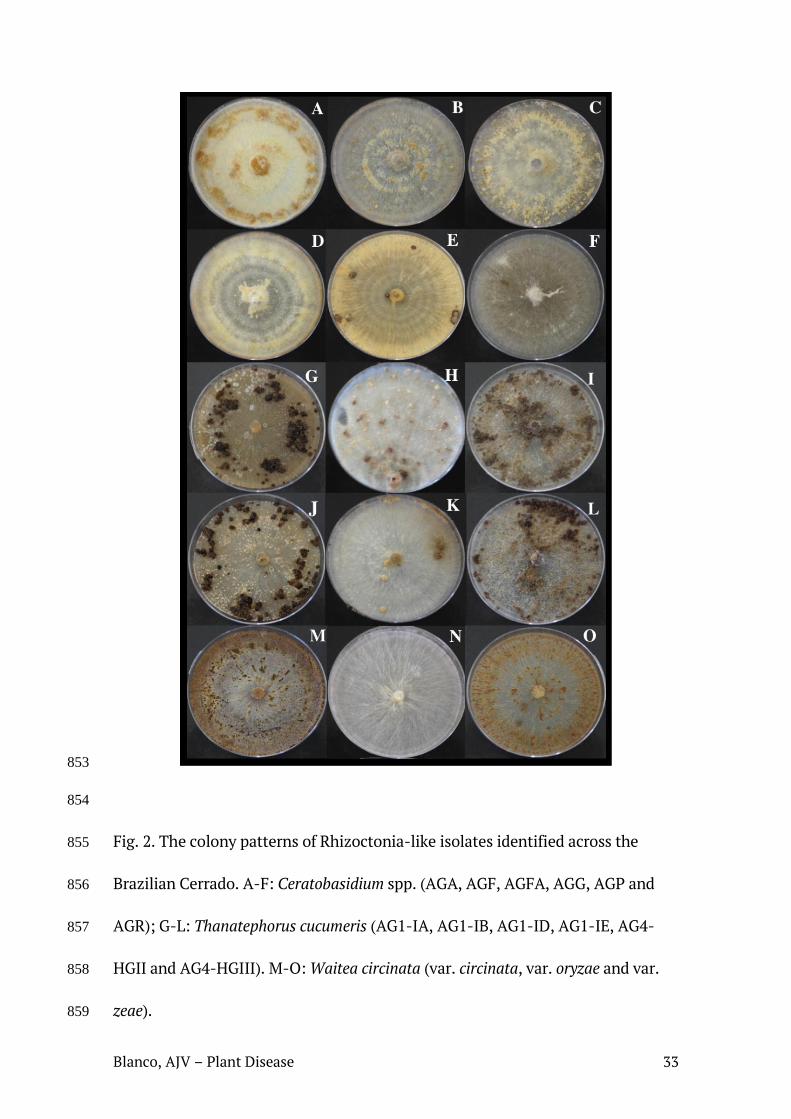
Blanco, AJV – Plant Disease 33
853
854
Fig. 2. The colony patterns of Rhizoctonia-like isolates identified across the 855
Brazilian Cerrado. A-F: Ceratobasidium spp. (AGA, AGF, AGFA, AGG, AGP and 856
AGR); G-L: Thanatephorus cucumeris (AG1-IA, AG1-IB, AG1-ID, AG1-IE, AG4-857
HGII and AG4-HGIII). M-O: Waitea circinata (var. circinata, var. oryzae and var. 858
zeae). 859
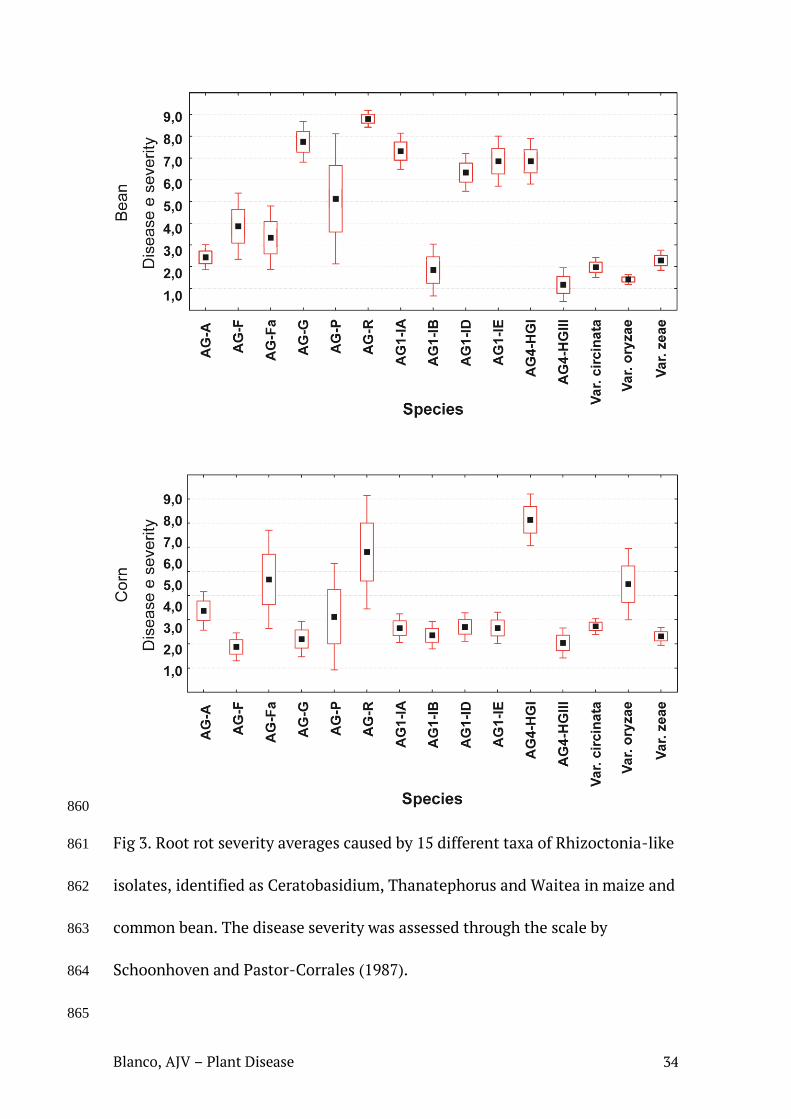
Blanco, AJV – Plant Disease 34
860
Fig 3. Root rot severity averages caused by 15 different taxa of Rhizoctonia-like 861
isolates, identified as Ceratobasidium, Thanatephorus and Waitea in maize and 862
common bean. The disease severity was assessed through the scale by 863
Schoonhoven and Pastor-Corrales (1987). 864
865
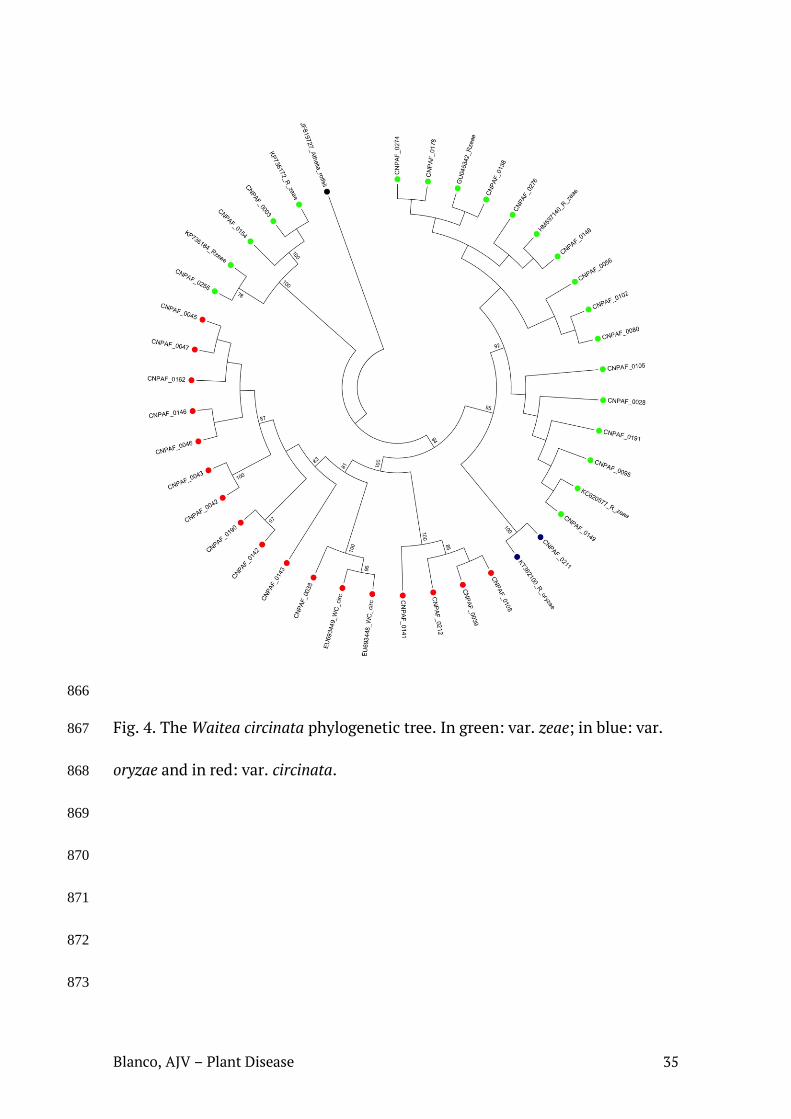
Blanco, AJV – Plant Disease 35
866
Fig. 4. The Waitea circinata phylogenetic tree. In green: var. zeae; in blue: var. 867
oryzae and in red: var. circinata. 868
869
870
871
872
873

Blanco, AJV – Plant Disease 36
874
875
876
Fig. 5. The Ceratobasidium spp. phylogenetic tree. In red: AG-A; in blue: AG-F; 877
in light blue: AG-Fa; in purple: AG-G; in dark green: AG-P; in light green: AG-878
R. 879
880
881
Blanco, AJV – Plant Disease 37
882
883
884
Fig. 6. The Thanatephorus cucumeris phylogenetic tree. In red: AG1 isolates 885
(ISGs IA, IB, ID and IE); in blue: AG4 isolates (ISGs HGI and HGIII). 886
887
888
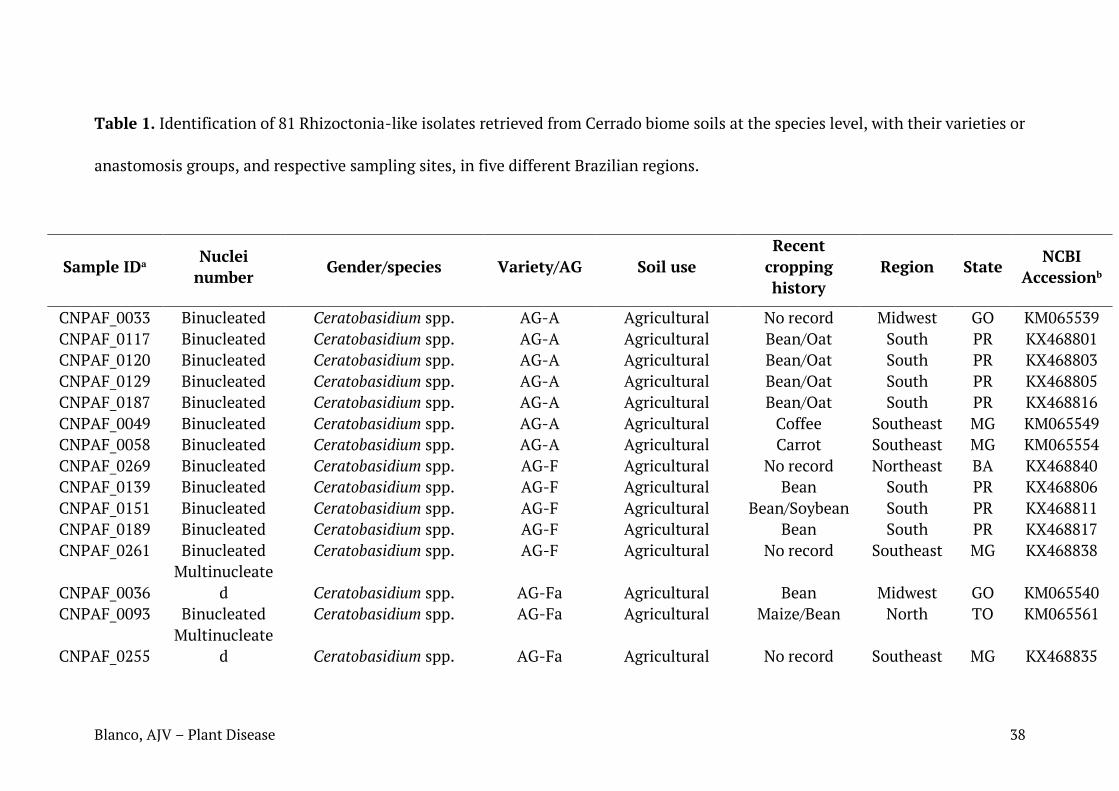
Blanco, AJV – Plant Disease 38
Table 1. Identification of 81 Rhizoctonia-like isolates retrieved from Cerrado biome soils at the species level, with their varieties or
anastomosis groups, and respective sampling sites, in five different Brazilian regions.
Sample IDa Nuclei number Gender/species Variety/AG Soil use
Recent cropping history
Region State NCBI Accessionb
CNPAF_0033 Binucleated Ceratobasidium spp. AG-A Agricultural No record Midwest GO KM065539 CNPAF_0117 Binucleated Ceratobasidium spp. AG-A Agricultural Bean/Oat South PR KX468801 CNPAF_0120 Binucleated Ceratobasidium spp. AG-A Agricultural Bean/Oat South PR KX468803 CNPAF_0129 Binucleated Ceratobasidium spp. AG-A Agricultural Bean/Oat South PR KX468805 CNPAF_0187 Binucleated Ceratobasidium spp. AG-A Agricultural Bean/Oat South PR KX468816 CNPAF_0049 Binucleated Ceratobasidium spp. AG-A Agricultural Coffee Southeast MG KM065549 CNPAF_0058 Binucleated Ceratobasidium spp. AG-A Agricultural Carrot Southeast MG KM065554 CNPAF_0269 Binucleated Ceratobasidium spp. AG-F Agricultural No record Northeast BA KX468840 CNPAF_0139 Binucleated Ceratobasidium spp. AG-F Agricultural Bean South PR KX468806 CNPAF_0151 Binucleated Ceratobasidium spp. AG-F Agricultural Bean/Soybean South PR KX468811 CNPAF_0189 Binucleated Ceratobasidium spp. AG-F Agricultural Bean South PR KX468817 CNPAF_0261 Binucleated Ceratobasidium spp. AG-F Agricultural No record Southeast MG KX468838
CNPAF_0036 Multinucleate
d Ceratobasidium spp. AG-Fa Agricultural Bean Midwest GO KM065540 CNPAF_0093 Binucleated Ceratobasidium spp. AG-Fa Agricultural Maize/Bean North TO KM065561
CNPAF_0255 Multinucleate
d Ceratobasidium spp. AG-Fa Agricultural No record Southeast MG KX468835
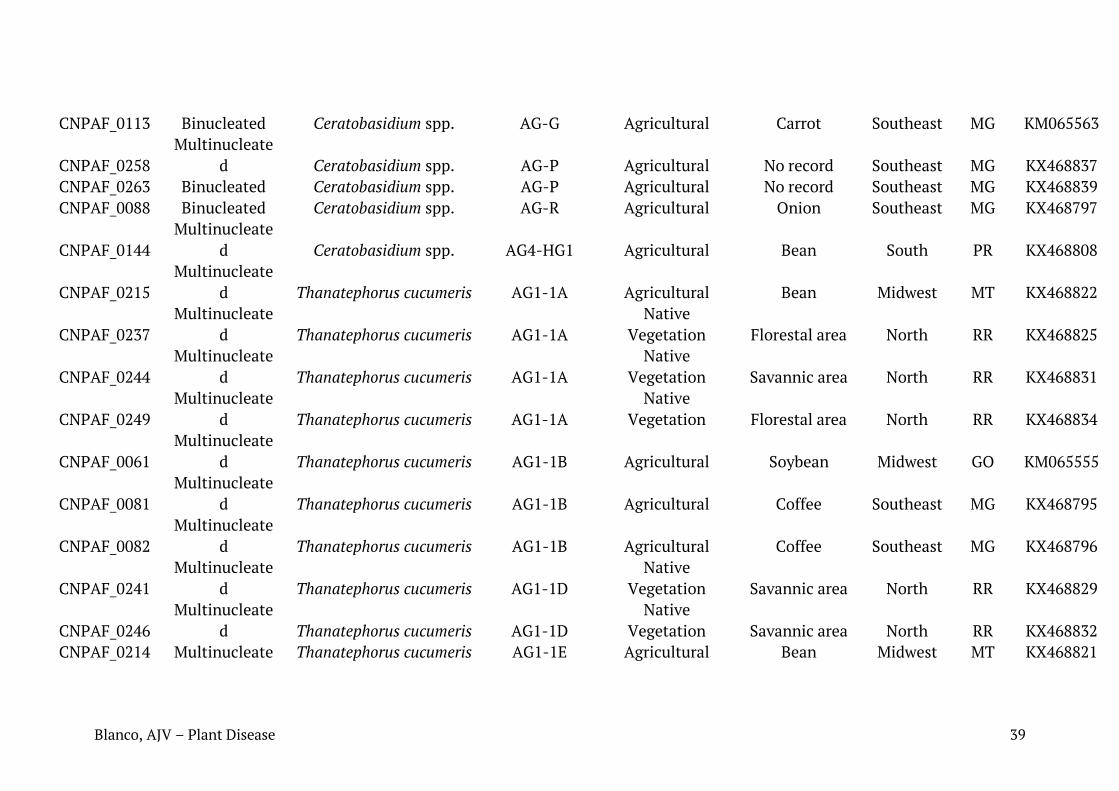
Blanco, AJV – Plant Disease 39
CNPAF_0113 Binucleated Ceratobasidium spp. AG-G Agricultural Carrot Southeast MG KM065563
CNPAF_0258 Multinucleate
d Ceratobasidium spp. AG-P Agricultural No record Southeast MG KX468837 CNPAF_0263 Binucleated Ceratobasidium spp. AG-P Agricultural No record Southeast MG KX468839 CNPAF_0088 Binucleated Ceratobasidium spp. AG-R Agricultural Onion Southeast MG KX468797
CNPAF_0144 Multinucleate
d Ceratobasidium spp. AG4-HG1 Agricultural Bean South PR KX468808
CNPAF_0215 Multinucleate
d Thanatephorus cucumeris AG1-1A Agricultural Bean Midwest MT KX468822
CNPAF_0237 Multinucleate
d Thanatephorus cucumeris AG1-1A Native
Vegetation Florestal area North RR KX468825
CNPAF_0244 Multinucleate
d Thanatephorus cucumeris AG1-1A Native
Vegetation Savannic area North RR KX468831
CNPAF_0249 Multinucleate
d Thanatephorus cucumeris AG1-1A Native
Vegetation Florestal area North RR KX468834
CNPAF_0061 Multinucleate
d Thanatephorus cucumeris AG1-1B Agricultural Soybean Midwest GO KM065555
CNPAF_0081 Multinucleate
d Thanatephorus cucumeris AG1-1B Agricultural Coffee Southeast MG KX468795
CNPAF_0082 Multinucleate
d Thanatephorus cucumeris AG1-1B Agricultural Coffee Southeast MG KX468796
CNPAF_0241 Multinucleate
d Thanatephorus cucumeris AG1-1D Native
Vegetation Savannic area North RR KX468829
CNPAF_0246 Multinucleate
d Thanatephorus cucumeris AG1-1D Native
Vegetation Savannic area North RR KX468832 CNPAF_0214 Multinucleate Thanatephorus cucumeris AG1-1E Agricultural Bean Midwest MT KX468821
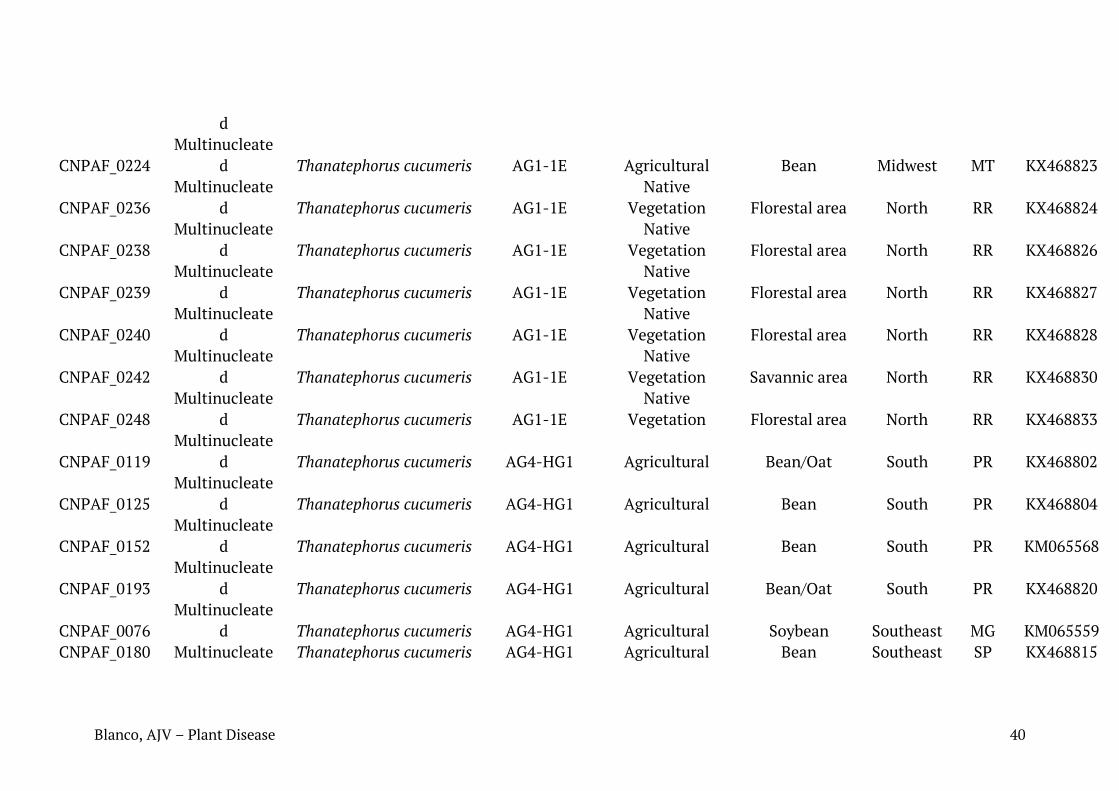
Blanco, AJV – Plant Disease 40
d
CNPAF_0224 Multinucleate
d Thanatephorus cucumeris AG1-1E Agricultural Bean Midwest MT KX468823
CNPAF_0236 Multinucleate
d Thanatephorus cucumeris AG1-1E Native
Vegetation Florestal area North RR KX468824
CNPAF_0238 Multinucleate
d Thanatephorus cucumeris AG1-1E Native
Vegetation Florestal area North RR KX468826
CNPAF_0239 Multinucleate
d Thanatephorus cucumeris AG1-1E Native
Vegetation Florestal area North RR KX468827
CNPAF_0240 Multinucleate
d Thanatephorus cucumeris AG1-1E Native
Vegetation Florestal area North RR KX468828
CNPAF_0242 Multinucleate
d Thanatephorus cucumeris AG1-1E Native
Vegetation Savannic area North RR KX468830
CNPAF_0248 Multinucleate
d Thanatephorus cucumeris AG1-1E Native
Vegetation Florestal area North RR KX468833
CNPAF_0119 Multinucleate
d Thanatephorus cucumeris AG4-HG1 Agricultural Bean/Oat South PR KX468802
CNPAF_0125 Multinucleate
d Thanatephorus cucumeris AG4-HG1 Agricultural Bean South PR KX468804
CNPAF_0152 Multinucleate
d Thanatephorus cucumeris AG4-HG1 Agricultural Bean South PR KM065568
CNPAF_0193 Multinucleate
d Thanatephorus cucumeris AG4-HG1 Agricultural Bean/Oat South PR KX468820
CNPAF_0076 Multinucleate
d Thanatephorus cucumeris AG4-HG1 Agricultural Soybean Southeast MG KM065559 CNPAF_0180 Multinucleate Thanatephorus cucumeris AG4-HG1 Agricultural Bean Southeast SP KX468815

Blanco, AJV – Plant Disease 41
d
CNPAF_0053 Multinucleate
d Thanatephorus cucumeris AG4-HGIII Agricultural Soybean Midwest GO KM065550
CNPAF_0054 Multinucleate
d Thanatephorus cucumeris AG4-HGIII Agricultural Soybean Midwest GO KM065551
CNPAF_0055 Multinucleate
d Thanatephorus cucumeris AG4-HGIII Agricultural Soybean Midwest GO KM065552
CNPAF_0044 Multinucleate
d Thanatephorus cucumeris AG4-HGIII Agricultural Carrot Southeast MG KM065546
CNPAF_0067 Multinucleate
d Thanatephorus cucumeris AG4-HGIII Agricultural Carrot Southeast MG KM065557
CNPAF_0062 Multinucleate
d Thanatephorus cucumeris AG4-HG1 Agricultural Bean Midwest GO KM065556
CNPAF_0141 Multinucleate
d Waitea circinata var. circinata Agricultural No record Midwest GO KM065565
CNPAF_0162 Multinucleate
d Waitea circinata var. circinata Agricultural Bean Midwest GO KX468813
CNPAF_0212 Multinucleate
d Waitea circinata var. circinata Agricultural Rice Midwest GO KM065570
CNPAF_0142 Multinucleate
d Waitea circinata var. circinata Agricultural Cinnamon South PR KX468807
CNPAF_0143 Multinucleate
d Waitea circinata var. circinata Agricultural Bean/Sorgo South PR KM065566
CNPAF_0146 Multinucleate
d Waitea circinata var. circinata Agricultural Bean South PR KX468809 CNPAF_0190 Multinucleate Waitea circinata var. circinata Agricultural No record South PR KX468818
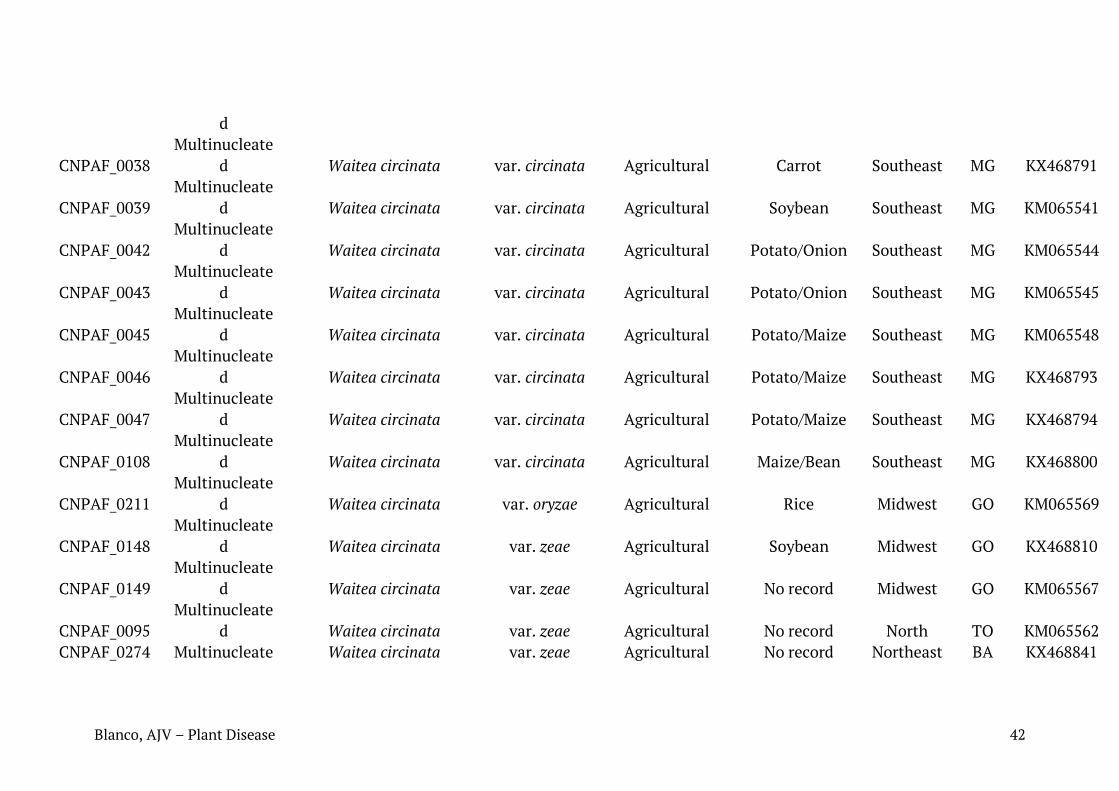
Blanco, AJV – Plant Disease 42
d
CNPAF_0038 Multinucleate
d Waitea circinata var. circinata Agricultural Carrot Southeast MG KX468791
CNPAF_0039 Multinucleate
d Waitea circinata var. circinata Agricultural Soybean Southeast MG KM065541
CNPAF_0042 Multinucleate
d Waitea circinata var. circinata Agricultural Potato/Onion Southeast MG KM065544
CNPAF_0043 Multinucleate
d Waitea circinata var. circinata Agricultural Potato/Onion Southeast MG KM065545
CNPAF_0045 Multinucleate
d Waitea circinata var. circinata Agricultural Potato/Maize Southeast MG KM065548
CNPAF_0046 Multinucleate
d Waitea circinata var. circinata Agricultural Potato/Maize Southeast MG KX468793
CNPAF_0047 Multinucleate
d Waitea circinata var. circinata Agricultural Potato/Maize Southeast MG KX468794
CNPAF_0108 Multinucleate
d Waitea circinata var. circinata Agricultural Maize/Bean Southeast MG KX468800
CNPAF_0211 Multinucleate
d Waitea circinata var. oryzae Agricultural Rice Midwest GO KM065569
CNPAF_0148 Multinucleate
d Waitea circinata var. zeae Agricultural Soybean Midwest GO KX468810
CNPAF_0149 Multinucleate
d Waitea circinata var. zeae Agricultural No record Midwest GO KM065567
CNPAF_0095 Multinucleate
d Waitea circinata var. zeae Agricultural No record North TO KM065562 CNPAF_0274 Multinucleate Waitea circinata var. zeae Agricultural No record Northeast BA KX468841
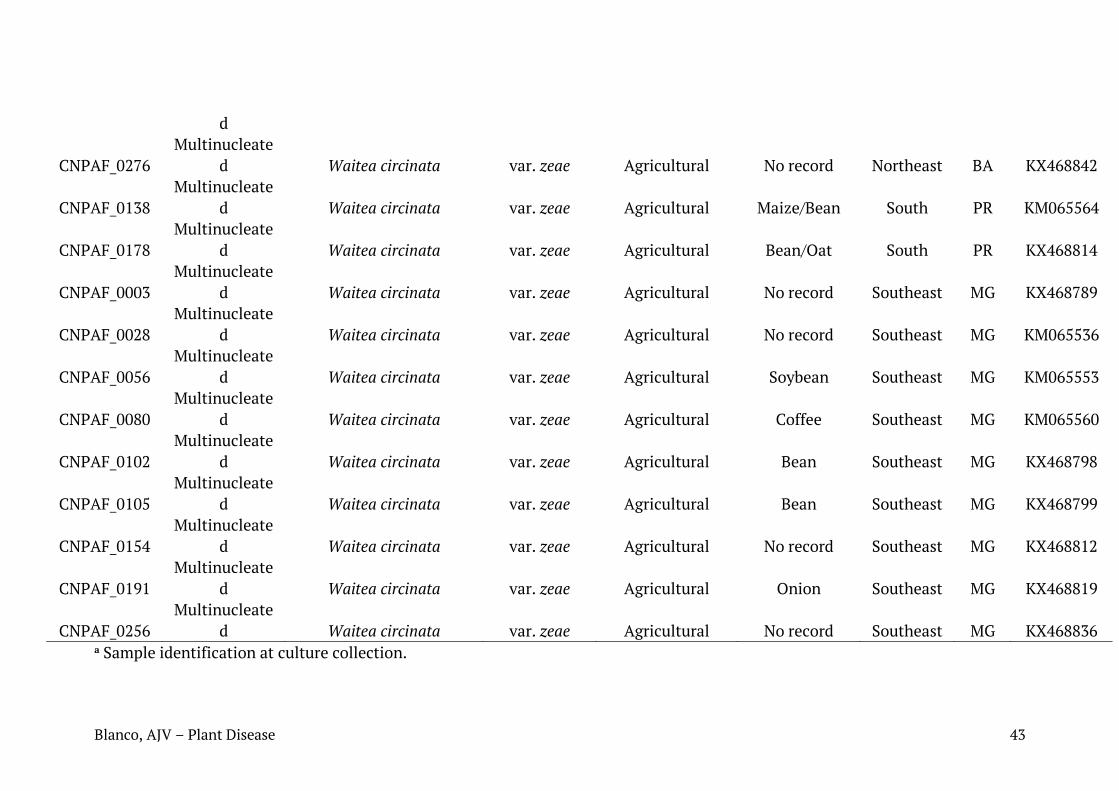
Blanco, AJV – Plant Disease 43
d
CNPAF_0276 Multinucleate
d Waitea circinata var. zeae Agricultural No record Northeast BA KX468842
CNPAF_0138 Multinucleate
d Waitea circinata var. zeae Agricultural Maize/Bean South PR KM065564
CNPAF_0178 Multinucleate
d Waitea circinata var. zeae Agricultural Bean/Oat South PR KX468814
CNPAF_0003 Multinucleate
d Waitea circinata var. zeae Agricultural No record Southeast MG KX468789
CNPAF_0028 Multinucleate
d Waitea circinata var. zeae Agricultural No record Southeast MG KM065536
CNPAF_0056 Multinucleate
d Waitea circinata var. zeae Agricultural Soybean Southeast MG KM065553
CNPAF_0080 Multinucleate
d Waitea circinata var. zeae Agricultural Coffee Southeast MG KM065560
CNPAF_0102 Multinucleate
d Waitea circinata var. zeae Agricultural Bean Southeast MG KX468798
CNPAF_0105 Multinucleate
d Waitea circinata var. zeae Agricultural Bean Southeast MG KX468799
CNPAF_0154 Multinucleate
d Waitea circinata var. zeae Agricultural No record Southeast MG KX468812
CNPAF_0191 Multinucleate
d Waitea circinata var. zeae Agricultural Onion Southeast MG KX468819
CNPAF_0256 Multinucleate
d Waitea circinata var. zeae Agricultural No record Southeast MG KX468836 a Sample identification at culture collection.

Blanco, AJV – Plant Disease 44
b The NCBI accession ID of the ribossomal DNA sequenced from those 81 isolates.
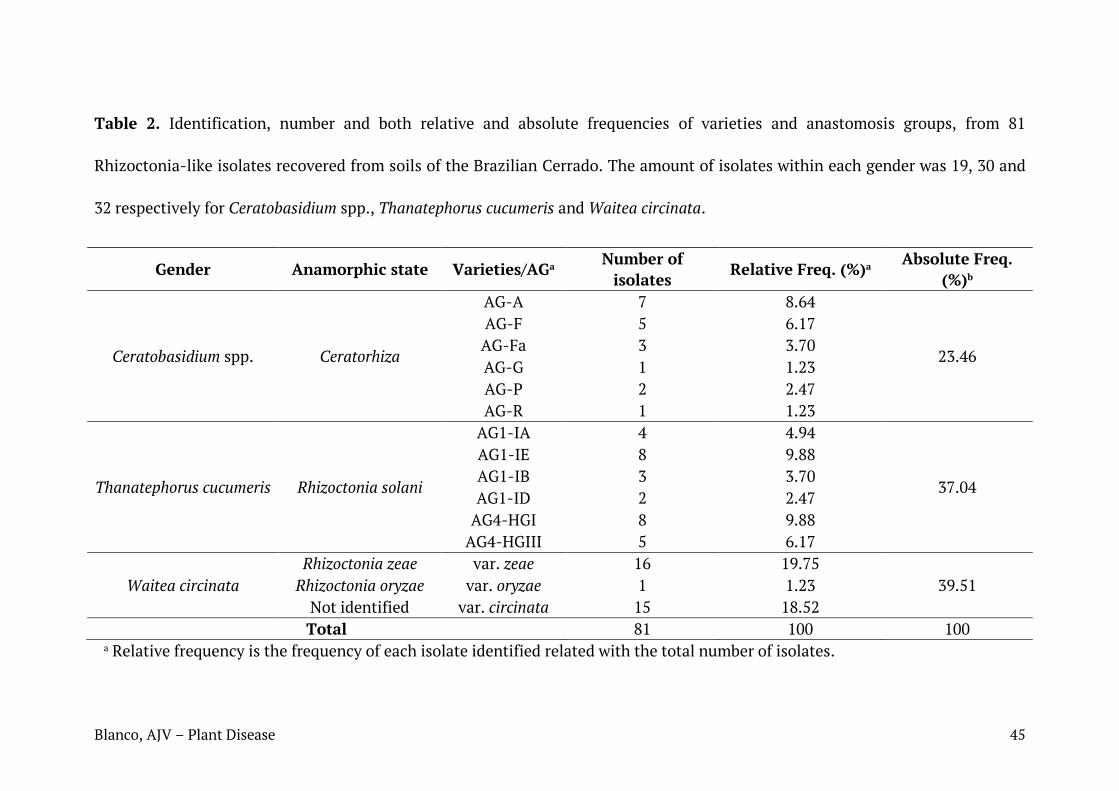
Blanco, AJV – Plant Disease 45
Table 2. Identification, number and both relative and absolute frequencies of varieties and anastomosis groups, from 81
Rhizoctonia-like isolates recovered from soils of the Brazilian Cerrado. The amount of isolates within each gender was 19, 30 and
32 respectively for Ceratobasidium spp., Thanatephorus cucumeris and Waitea circinata.
Gender Anamorphic state Varieties/AGa Number of isolates Relative Freq. (%)a Absolute Freq.
(%)b
Ceratobasidium spp. Ceratorhiza
AG-A 7 8.64
23.46
AG-F 5 6.17 AG-Fa 3 3.70 AG-G 1 1.23 AG-P 2 2.47 AG-R 1 1.23
Thanatephorus cucumeris Rhizoctonia solani
AG1-IA 4 4.94
37.04
AG1-IE 8 9.88 AG1-IB 3 3.70 AG1-ID 2 2.47
AG4-HGI 8 9.88 AG4-HGIII 5 6.17
Waitea circinata Rhizoctonia zeae var. zeae 16 19.75
39.51 Rhizoctonia oryzae var. oryzae 1 1.23 Not identified var. circinata 15 18.52 Total 81 100 100
a Relative frequency is the frequency of each isolate identified related with the total number of isolates.

Blanco, AJV – Plant Disease 46
b Absolute frequency is the frequency of each of those three genders.

Blanco, AJV – Plant Disease 47