universidade de sÃo paulo faculdade de ciÊncias … · 2007-10-10 · universidade de sÃo paulo...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO FACULDADE DE CIÊNCIAS FARMACÊUTICAS
Programa de Pós-Graduação em Ciência dos Alimentos
Área de Bromatologia
Enterococos em amostras de alimentos e águas: avaliação da virulência e do desempenho como indicadores de higiene
Bruna Carrer Gomes
Tese para obtenção do grau de
DOUTOR
Orientadora: Profa. Dra. Bernadette D. G. M. Franco Co-Orientadora: Profa. Dra. Elaine Cristina Pereira De Martinis
São Paulo 2007

Bruna Carrer Gomes
Enterococos em amostras de alimentos e águas: avaliação da virulência e do desempenho como indicadores de higiene
Tese para obtenção do grau de
DOUTOR
Orientadora: Profa. Dra. Bernadette D. G. M. Franco Co-Orientadora: Profa. Dra. Elaine Cristina Pereira De Martinis
São Paulo 2007

Ficha Catalográfica Elaborada pela Divisão de Biblioteca e
Documentação do Conjunto das Químicas da USP.
Gomes, Bruna Carrer G633e Enterococos em amostras de alimentos e águas: avaliação da
virulência e do desempenho como indicadores de higiene / Bruna Carrer Gomes. -- São Paulo, 2007.
151p. Tese (doutorado) - Faculdade de Ciências Farmacêuticas da
Universidade de São Paulo. Departamento de Alimentos e Nutrição Experimental
Orientador: Franco, Bernadette Dora Gombossy de Melo Co-orientador: Martinis, Elaine Cristina Pereira de 1. Alimento: Análise : Ciência dos alimentos 2. Microbiologia de
alimentos 3. Alimento: Segurança: Saúde pública 4. Bacteriologia I. T. II. Franco, Bernadette Dora Gombossy de Melo, orientador. III. Mart in is , Elaine Cris t ina Pereira de, co-orientador .
641 CDD

BRUNA CARRER GOMES
Enterococos em amostras de alimentos e águas: avaliação da virulência e do desempenho como indicadores de higiene
Comissão Julgadora da
Tese para obtenção do título de Doutor
________________________________________
Profa. Dra. Bernadette D. G. M. Franco Orientadora/Presidente
________________________________________ 1º Examinador
________________________________________ 2º Examinador
________________________________________ 3º Examinador
________________________________________ 4º Examinador
São Paulo, ________________ de 2007.

Ao meu marido, Iran, pela compreensão e apoio. Muito obrigada por sua cumplicidade,
carinho e paciência.
Aos meus pais, Elias e Nilvia e minhas irmãs, Carin e Vivian pelo constante apoio e
carinho fundamentais na minha vida.

Agradecimentos
À Profa. Dra. Bernadette D. G. M. Franco, pela orientação, incentivo
e sugestões sempre tão oportunas.
À Profa. Dra. Elaine C. P. De Martinis, pela co-orientação, incentivo
e disponibilização do laboratório para realização dos experimentos.
À Profa. Dra. Ana Lúcia da Costa Darini, da FCFRP-USP, pelo
auxílio durante realização de parte dos experimentos.
À Carolina Tonani Esteves, pela ajuda indispensável e pelos
momentos de descontração.
À Izabel Cristina V. Palazzo, especialista em laboratório da FCFRP-
USP, pela disponibilidade e auxílio na identificação dos isolados de
enterococos.
Aos amigos do laboratório de Microbiologia de Alimentos da
FCFRP-USP Rafael, Cláudia, Carlos Eduardo, Regiane, Virgínia,

Maria Aparecida, Lizziane, Fabrício e Natália pelos alegres
momentos de descontração e pela convivência.
À técnica Vanessa Maciel, da FCFRP-USP, pelas dicas de informática
durante a elaboração da tese.
Aos colegas do laboratório de Microbiologia de Alimentos da FCF-
USP Janine e Eb e às técnicas, Kátia e Lúcia, pelas dicas durante
minha permanência em São Paulo.
Aos funcionários da Secretaria do departamento de Alimentos da
FCF-USP, Mônica e Edilson, e da Secretaria de Pós-Graduação,
Elaine e Jorge, por terem sempre colaborado nos momentos que
necessitei.
Ao Prof. Dr. Leonardo Sechi da Università di Sassari que permitiu a
realização do doutorado sanduíche na Itália, pela convivência
agradável e confiança. Grazie mille per la disponibilità e oportunità.

Alle persone che ho conosciuto nel laboratorio dell’ Università di
Sassari: Antonella, Silvia, Giovanna, Donatella, Paola, Sara, Daniela,
Domenico, Signore Manca, Monica, Adriana, Alessandra, Sara, Luca
e Domenico, perché sono stati sempre disponibili. Grazie delle
parole di stimolo, dei tanti fini di settimana con le vostre famiglia
che tante volte me hanno fatto sentire a casa, del cambiamento di
cultura, dell’ amicizia e di tutte belle ricordazione...Un
ringraziamento speciale a Giovanna Felis dell’ aiuto professionale e
della disponibilità. Grazie Mille.
À Università di Sassari - Itália pela oportunidade de estudo.
À Giovanna Felis e Anna Castioni - Laboratorio di Microbiologia
Alimentare, Dipartamento Scientifico e Tecnologico della Università
di Verona que colaboraram com a análise dos dados de RAPD.
Ao Departamento de Água e Esgotos de Ribeirão Preto - DAERP,
especialmente ao Sr. Carlos Farjani Neto, pela colaboração na
obtenção das amostras de água analisadas neste trabalho.

À 3M do Brasil pela doação de placas Petrifilm® EC utilizadas neste
trabalho.
Ao Centro de Zootecnia - Pólo Regional Leste de Ribeirão Preto, da
Secretaria Estadual de Agricultura e Abastecimento (SAA) do
Estado de São Paulo, pela doação das amostras de leite cru
analisadas neste trabalho.
À Dra. V. Vankerckhoven da Universidade Antwerp na Bélgica pela
doação das cepas de E. faecalis MMH594 e E. faecium C68.
Ao CNPq pela bolsa de doutorado e à CAPES pela bolsa de
doutorado sanduíche.
À FAPESP, pelo financiamento do trabalho (processo n°
2003/11986-2).
À todos que de alguma forma colaboraram durante todos esses anos
para realização do meu trabalho.

GOMES, B. C. Enterococos em amostras de alimentos e águas: avaliação da virulência e do desempenho como indicadores de higiene. 2007. 151f. Tese (Doutorado) – Faculdade de Ciências Farmacêuticas, Universidade de São Paulo, São Paulo, 2007.
RESUMO
Enterococcus spp. pertencem ao grupo das bactérias láticas e estão presentes em solos, águas, plantas, microbiota autóctone de vários alimentos e como membros da microbiota intestinal de humanos e animais. Esses microrganismos foram considerados por muito tempo como comensais, mas o aumento da severidade das infecções nosocomiais causadas por enterococos mutirresistentes a antimicrobianos e, a falta de conhecimento sobre seus fatores de virulência geram insegurança na utilização de cepas deste gênero na produção de alimentos como culturas fermentadoras e/ou probióticas. A diferença entre uma cepa de enterococos com potencial patogênico e outra aparentemente segura para uso em processamento de alimentos não é clara, e a probabilidade de que esta última adquira fatores de virulência merece investigação. O objetivo do presente projeto foi determinar características fenotípicas e genotípicas de Enterococcus spp. isolados de amostras de alimentos e águas correlacionando sua presença com indicadores clássicos de higiene e contaminação fecal. De 812 colônias indicativas do gênero enterococos obtidas a partir de 120 amostras de alimentos, 299 isolados (37%) foram presuntivamente caracterizados como Enterococcus spp. Após identificação por PCR, 139 (46,5%) E. faecium, 80 (26,8%) E. faecalis, 36 (12%) E. casseliflavus e 8 (2,7%) E. gallinarum. Produção de gelatinase foi detectada apenas em isolados de E. faecalis (60%). Um isolado de E. faecium (0,7%) e 31 isolados de E. faecalis (38,7%) apresentaram perfil β-hemolítico. Produção de bacteriocina contra Lactobacillus sakei e/ou Listeria monocytogenes foi observada para 10% dos isolados de E. faecalis e 23% dos isolados de E. faecium. Hidrólise de sais biliares foi observada para 100% dos isolados de E. gallinarum, 86% E. casseliflavus, 65% E. faecalis e 62,6% de E. faecium. Alguns isolados de E. faecium apresentaram resistência à vancomicina, eritromicina e tetraciclina. Entre os isolados de E. faecalis não houve resistência à vancomicina, mas foi observada resistência à tetraciclina, eritromicina e alta concentração de gentamicina. Houve uma maior prevalência dos genes de virulência (esp, gel, ace, as, efaA e cylA) entre os isolados de E. faecalis quando comparado a E. faecium. Além disso, os isolados de E. faecalis, resistentes a antibióticos, mostraram forte adesão a células Caco-2 e capacidade de formação de biofilme em superfície abiótica. RAPD-PCR individualizou 14 cepas de E. faecium e 17 cepas de E. faecalis dentre os 52 isolados Enterococcus spp. resistentes a antibióticos. A variabilidade dos resultados impediu o estabelecimento de uma correlação entre a presença ou contagem de coliformes, E. coli e enterococos nas amostras analisadas. Os dados deste trabalho sobre marcadores fenotípicos e genotípicos de virulência, e a presença de cepas resistentes a antibióticos evidenciam a necessidade da avaliação cuidadosa de linhagens de enterococos para aplicações em alimentos. Palavras-chave: Probióticos, bacteriocinas, enterococos, fatores de virulência, indicadores de higiene.

GOMES, B. C. Enterococci in samples of food and water: evaluation of their virulence markers and their suitability as hygiene indicators. 2007. 151f. Thesis (Doctoral) – Faculdade de Ciências Farmacêuticas, Universidade de São Paulo, São Paulo, 2007.
ABSTRACT
Enterococcus spp. belong to the group of lactic acid bacteria widely distributed in soil, plants, foods, animals and humans. In the past, these microorganisms were considered commensals but the increase of antibiotic-resistant enterococci and the lack of knowledge about their virulence markers, had raised concerns regarding the safety of using strains of this genus in the food production as fermentative or probiotic cultures. Besides this, literature data suggests the use of enterococci as sanitary indicator for foods. Differences between enterococci strains with pathogenic potential and an apparently safe ones is unclear and there is a concern about virulence markers transfer. The aim of this work was to determine phenotypic and genotypic characteristics of Enterococcus spp. isolated from foods and water and also to correlate their presence with classical indicators of sanitary quality. Out of 812 presumptive enterococci colonies obtained from 120 food samples, 299 isolates (37%) were presuntively characterized as Enterococcus spp. Isolates were identified by PCR: 139 (46.5%) E. faecium, 80 (26.8%) E. faecalis, 36 (12.0%) E. casseliflavus and 8 (2.7%) E. gallinarum. Only E. faecalis isolates (60%) produced gelatinase. One E. faecium (0.7%) and 31 E. faecalis (38.7%) were β-haemolytic. Bacteriocin activity against Lactobacillus sakei and/or Listeria monocytogenes was observed for 10% of the E. faecalis and for 23% of the E. faecium isolates. All E. gallinarum isolates, 86% of the E. casseliflavus, 65% of the E. faecalis and 62.6% of the E. faecium isolates showed bile salt hydrolysis activity. Some E. faecium isolates were resistant to vancomycin, erythromycin and tetracycline. Vancomycin resistance was absent among the E. faecalis but, resistance to tetracycline, erythromycin and high-level gentamicin was observed. There was a higher prevalence of virulence genes (esp, gel, ace, as, efaA e cylA) among the E. faecalis isolates when compared to the E. faecium. Antibiotic resistant E. faecalis isolates strongly adhered to Caco-2 cells and formed biofilm on abiotic surface. Using RAPD-PCR 14 E. faecium and 17 E. faecalis strains could be individualized from the 52 antibiotic resistant enterococci. It was not possible to correlate the presence of total coliforms, E. coli and Enterococcus spp. in the samples of food and water analysed due to results variability. Data obtained regarding phenotypic and genotypic virulence markers and the presence of antibiotic resistant enterococci raise the needs of a carefully evaluation of the Enterococcus spp. strains before future applications in foods. Keywords: Probiotics, enterococci, bacteriocins, virulence factors, hygiene indicators.

LISTA DE FIGURAS
Figura 1- Riscos e benefícios associados à presença de Enterococcus
spp. em alimentos, amostras ambientais e clínicas..................
16
Figura 2- Foto ilustrativa de géis de agarose com os produtos de PCR
para identificação das espécies de Enterococcus obtidos a
partir de amostras de alimentos...............................................
52
Figura 3- Perfil de resistência de Enterococcus faecium a antibióticos
de uso clínico isolados a partir de amostras de queijo (a),
produtos cárneos (b) e leite (c)...............................................
62
Figura 4- Perfil de resistência de Enterococcus faecalis a antibióticos
de uso clínico isolados a partir de amostras de queijo (a),
produtos cárneos (b), leite (c) e vegetais (d)...........................
63
Figura 5- Gel de agarose contendo produto de PCR dos genes de
virulência de enterococos.......................................................
68
Figura 6- Formação de biofilme em superfície abiótica por E. faecalis
e E faecium resistentes a antibióticos (RA) e não
resistentes...............................................................................
74
Figura 7- Fotomicrografia (Olympus BX51) mostrando isolado de E.
faecalis 591 fortemente adesivo (A e B) e isolado 568 (C e
D) com ausência de adesão às células da linhagem Caco-2,
coradas com May Grunwald – Giemsa...................... ............
75
Figura 8- Perfis de RAPD dos isolados de Enterococcus spp. com o
primer M-13.............................................................................
77

LISTA DE TABELAS
Tabela 1- Sequência dos primers utilizados e tamanho dos
fragmentos obtidos na identificação de E. faecalis, E.
faecium, E. casseliflavus e E. gallinarum.............................
24
Tabela 2- Sequência dos primers utilizados e tamanho dos
fragmentos obtidos na pesquisa dos genes de virulência
de Enterococcus spp........................................................... 29
Tabela 3- Caracterização fenotípica e molecular de Enterococcus
spp. obtidos a partir das amostras de alimentos
semeadas nos meios KEA e BE.......................................... 36
Tabela 4- Resultados presuntivos e finais da população de
Enterococcus spp. nas amostras de carnes, queijos,
vegetais, leite e águas a partir dos meios Bile Esculina
(BE) e Kanamicina Esculina Azida (KEA)............................ 45
Tabela 5- Número de isolados obtidos nos meios KEA e BE
identificados como Enterococcus spp. por testes
fenotípicos e PCR............................................................... 50
Tabela 6- Distribuição das espécies dos 263 isolados de
Enterococcus spp. obtidos a partir de amostras de
alimentos e identificados por PCR....................................... 53
Tabela 7- Comparação da identificação de Enterococcus spp. por
PCR e API 20 STREP (Biomérieux, França)....................... 54
Tabela 8- Perfil de Enterococcus spp. quanto à capacidade de
hidrolisar sais biliares, presença fenotípica dos
marcadores de virulência (β-hemólise e gelatinase) e
atividade inibitória frente a L. sakei ATCC 15521 e/ou L.
monocytogenes ATCC 19115.............................................. 57
Tabela 9- Perfil de resistência de E. faecium isolados de amostras
de queijo, produtos cárneos e leite adquiridas no comércio
de Ribeirão Preto – SP, Brasil..............................................
60

Tabela 10- Perfil de resistência de E. faecalis isolados de amostras de
queijos, produtos cárneos, leite e vegetais adquiridas no
comércio de Ribeirão Preto – SP, Brasil................................ 61
Tabela 11- Perfil de resistência a antibióticos apresentado por E.
faecalis e E. faecium obtidos de amostras de queijos,
produtos cárneos e vegetais adquiridas em Ribeirão Preto
– SP, Brasil........................................................................... 64
Tabela 12- Resultados da Concentração Inibitória Mínima (CIM) para
isolados de E. faecalis e E. faecium obtidos de amostras
de queijo, produtos cárneos, leite e vegetais adquiridas na
cidade de Ribeirão Preto – SP, Brasil.................................. 65
Tabela 13- Distribuição dos genes de virulência em E. faecium e E.
faecalis obtidos de amostras de alimentos.......................... 69
Tabela 14- Genes de virulência, formação de biofilme e adesão a
células Caco-2 em E. faecium e E. faecalis resistentes a
antibióticos de uso clínico.................................................... 72
Tabela 15- Comparação da prevalência de genes de virulência entre
enterococos resistentes a antibióticos (RA) e enterococos
não resistentes..................................................................... 73
Tabela 16- Enumeração de E. coli e coliformes totais (log UFC/g ou
mL) das amostras de carne, leite, queijo, vegetais e águas
em Petrifilm EC (3M) e membrana filtrante.......................... 79
Tabela 17- Contagem de E. coli, coliformes totais e Enterococcus
spp. (log UFC g-1 ou ml-1) nas amostras de alimentos e
águas................................................................................... 83
Tabela 18- Comparação da população de Enterococcus spp.,
coliformes totais e E. coli nas amostras de alimentos e
águas.................................................................................... 84

SUMÁRIO
1. INTRODUÇÃO ....................................................................................... 1
1.1 Histórico.............................................................................................. 2
1.2 Características fenotípicas, identificação e isolamento ...................... 3
1.3 Patogenicidade de Enterococcus spp................................................. 6
1.4 Enterococos como indicadores de contaminação fecal ...................... 10
1.5 Importância de Enterococcus em alimentos ....................................... 12
2. OBJETIVOS........................................................................................... 17
2.1 Objetivo geral...................................................................................... 18
2.2 Objetivos específicos.......................................................................... 18
3. MATERIAS E MÉTODOS ...................................................................... 19
3.1 Coleta das amostras de alimentos e águas........................................ 20
3.2 Preparo das amostras ........................................................................ 20
3.3 Isolamento e enumeração de Enterococcus spp ................................ 21
3.4 Identificação presuntiva de Enterococcus spp.................................... 23
3.5 Identificação das espécies de Enterococcus spp. por testes
bioquímicos............................................................................................... 23
3.6 Identificação das espécies de enterococcus empregando-se PCR.... 24
3.7 Caracterização fenotípica das cepas de Enterococcus spp................ 24
3.7.1 Hidrólise de gelatina ................................................................... 24
3.7.2 Atividade hemolítica ................................................................... 25
3.7.3 Determinação da produção de bacteriocina ............................... 25
3.7.4 Avaliação da hidrólise de sais biliares (BSH) ............................. 26
3.8 Determinação da resistência a antibióticos......................................... 27
3.9 Detecção de genes de virulência em isolados de E. faecium e E.
faecalis ..................................................................................................... 28
3.10 Formação de biofilme em superfície abiótica.................................... 31
3.11 Adesão em células da linhagem Caco-2........................................... 31
3.12 Análise de fingerprinting por meio de RAPD-PCR............................ 32
3.13 Enumeração de coliformes totais e E. coli ....................................... 32

4. RESULTADOS....................................................................................... 34
4.1 Isolamento de Enterococcus spp........................................................ 35
4.2 Identificação de espécies de Enterococcus spp ................................. 51
4.3 Caracterização fenotípica de Enterococcus spp................................. 55
4.4 Determinação da resistência a antibióticos......................................... 58
4.5 Detecção de genes de virulência em isolados de E. faecium e E.
faecalis ..................................................................................................... 67
4.6 Formação de biofilme em superfície abiótica e adesão em células
da linhagem Caco-2.................................................................................. 73
4.7 Análise de fingerprinting por meio de RAPD-PCR.............................. 76
4.8 Enumeração de coliformes totais e E. coli nas amostras de
alimentos .................................................................................................. 78
5. DISCUSSÃO .......................................................................................... 88
6. CONCLUSÕES ...................................................................................... 104
7. REFERÊNCIAS BIBLIOGRÁFICAS ...................................................... 107
APÊNDICE ............................................................................................... 128

_________________1. Introdução

Introdução________________________________________________________________________ 2
1.1 Histórico
A classificação taxonômica dos membros do gênero Streptococcus e
Enterococcus passou por alterações significativas nos últimos anos. A partir do
início do século vinte Thiercelin e Jouhaud propuseram, pela primeira vez, a
criação do gênero Enterococcus para designar diplococos Gram-positivos de
origem intestinal (Morrison et al., 1997). Entretanto, estes microrganismos
foram agrupados, por similaridade, no gênero Streptococcus, sendo
denominados S. faecalis. Posteriormente, em função das suas características
antigênicas foram classificados como Streptococcus do grupo D de Lancefield
(Hardie e Whiley, 1997).
Em 1937, Sherman propôs a criação de quatro subgrupos neste gênero:
“piogênico”, ”viridans”, ”láctico” e ”enterococos”. Além disso, este autor afirmou
que o grupo dos enterococos apresentava características notoriamente
distintas em relação aos outros subgrupos (Hardie e Whiley, 1997).
Posteriormente, estudos detalhados de caracterização molecular baseados em
critérios como, seqüência nucleotídica do DNA ribossômico 16S e hibridização
de DNA:DNA e DNA:RNA de diferentes cepas, forneceram bases para
reorganização do gênero Streptococcus em três grupos geneticamente
distintos: Streptococcus sensu stricto, Enterococcus e Lactococcus (Stiles e
Holzapfel, 1997; Facklam et al., 1999).
A criação do gênero Enterococcus para agrupar as espécies de S.
faecalis e S. faecium foi então proposto por Kalina (1970), baseando-se em
diferenças morfológicas, biológicas e sorológicas. Entretanto, essa
classificação só foi aceita a partir das evidências genéticas fornecidas por
estudos moleculares e quimiotaxonômicos, conduzidos por Schleifer e Kilpper-

Introdução________________________________________________________________________ 3
Bälz (1984). Estes autores confirmaram que as espécies de Streptococcus
faecalis e Streptococcus faecium, agrupadas no grupo D de Lancefield,
deveriam ser transferidas para o gênero Enterococcus por apresentarem
características marcadamente distintas dos demais Streptococcus.
Segundo Facklam et al. (1999), o gênero Enterococcus compreende 16
espécies: E. avium, E. malodoratus, E. raffinosus, E. pseudoavium, E.
saccharolyticus, E. faecalis, E. faecium, E. casseliflavus, E. mundtii, E.
gallinarum, E. durans, E. hirae, E. dispar, E. sulfureus, E. cecorum e E.
columbae. Entretanto, alguns pesquisadores aceitam a existência de até 28
espécies de enterococos (Foulquié-Moreno et al. 2006). Dentre essas
espécies, E. faecalis e E. faecium são as mais freqüentemente isoladas de
amostras clínicas (Moellering, 1992; Vancanneyt et al., 2002).
1.2 Características fenotípicas, identificação e isolamento
Os enterococos estão amplamente distribuídos na natureza, estando
presentes em solos, águas, plantas, vegetais, e microbiota autóctone de vários
alimentos (Hardie e Whiley, 1997; Eaton e Gasson, 2001; Giraffa, 2002; Iversen
et al., 2002). Este gênero compreende cocos Gram-positivos que ocorrem
isolados, aos pares ou em pequenas cadeias. São anaeróbios facultativos,
alguns são móveis e a temperatura ótima de crescimento é de 35 a 37°C,
sendo que muitas cepas apresentam capacidade de multiplicação entre 10 e
45°C. Estes microrganismos não requerem atmosfera contendo elevada
concentração de CO2 para sua multiplicação embora, algumas cepas o façam
melhor sob esta condição. O metabolismo fermentativo resulta em L (+) ácido

Introdução________________________________________________________________________ 4
lático como produto principal da fermentação da glicose (Hardie e Whiley,
1997; Domig et al., 2003).
Os meios ágar soja tripticase ou ágar infusão cérebro coração
adicionados de 5% de sangue de carneiro, ou qualquer ágar base contendo 5%
de sangue de outro animal, são suficientes para o crescimento destes
microrganismos. Alguns autores utilizam caldo de Man Rogosa, Sharpe (MRS)
ou o caldo Todd – Hewitt para o isolamento de enterococos (Ike et al., 1987;
Franz et al., 2001a; Franz et al., 2001b; Vancanneyt et al., 2002).
O isolamento seletivo - diferencial de enterococos a partir de amostras
clínicas ou de alimentos pode ser realizado em meio contendo como inibidores
seletivos, azida sódica, antibióticos (kanamicina ou gentamicina) e sais biliares
e, como indicadores, esculina ou tetrazólio (Ike et al., 1987; Devriese et al.,
1995; Morrison et al., 1997; Facklam et al., 1999).
Domig et al. (2003) afirmaram que o ágar Bile Esculina (BE) foi muito
empregado para a diferenciação e identificação presuntiva de enterococos, já
que a tolerância aos sais biliares e hidrólise da esculina é indicativa da
presença de membros deste gênero. O ágar Kanamicina Esculina Azida (KEA)
é um meio amplamente usado para isolamento e enumeração de enterococos
a partir de alimentos (Devriese et al., 1995; Calicioglu et al., 1999; Ingham et
al., 2000; Andrighetto et al., 2001; Gelsomino et al., 2001; Gelsomino et al.,
2002) e contém azida sódica (inibidor enzimático da cadeia de transporte de
elétrons) em combinação com o antibiótico kanamicina. O produto da hidrólise
da esculina reage com íons de ferro presente em ambos os meios e leva à
formação de halos escuros ao redor das colônias de enterococos (Domig et al.,
2003). Embora existam mais de 100 meios seletivos de cultura diferentes para

Introdução________________________________________________________________________ 5
o isolamento de enterococos, Morrison et al. (1997) afirmaram que nenhum é
suficientemente seletivo.
Segundo Facklam et al. (1999) e Domig et al. (2003), a identificação
presuntiva do gênero Enterococcus, dentre cocos Gram-positivos e catalase
negativos, pode ser confirmada com resultados positivos para hidrólise de
pirrolidonil –β – naftilamida (PYR), leucina– β – naftilamida (LAP), e
crescimento na presença de 6,5% de cloreto de sódio e a 45ºC.
Considerando-se que o gênero Enterococcus não é filogeneticamente
homogêneo, é comum o isolamento de linhagens de Enterococcus atípicas
(Franz et al., 1999). Sendo assim, para uma identificação adequada é
necessária a separação deste gênero em cinco grupos, baseando-se na
formação de ácido em caldo manitol e sorbose e, na hidrólise da arginina. Em
seguida, as espécies de cada grupo são identificadas por reações mais
específicas (Facklam et al., 1999).
Embora, atualmente, venha se tornando comum o uso de métodos
moleculares para a identificação de espécies de Enterococcus, ainda é comum
o emprego de características fenotípicas e de testes bioquímicos para a
classificação das espécies deste gênero (Facklam e Collins, 1989; Devriese et
al., 1993; Devriese et al., 1995; Franz et al., 2001b; Chingwaru et al., 2003).
Existem alguns testes rápidos, comercialmente disponíveis, para a
identificação de enterococos. Contudo, segundo Facklam et al. (1999) esses
testes devem ser utilizados com ressalvas, já que muitos identificam
corretamente apenas E. faecalis. Dentre os sistemas de identificação rápida
destacam-se: ID 32 STREP, API 20 STREP, API 50CH, Api zym (BioMérieux,
França) e Phene Plate PhP Plate System (PhPlate Microplate Techniques,

Introdução________________________________________________________________________ 6
Suécia) (Ike et al., 1987; Devriese et al., 1995; Borgen et al., 2001; Franz et al.,
2001b; Vancanneyt et al., 2002).
1.3 Patogenicidade de Enterococcus spp.
Por muito tempo as espécies do gênero Enterococcus foram
consideradas comensais. Entretanto, nos últimos anos, estes microrganismos
vêm se destacando como importantes patógenos nosocomiais, sendo
reconhecidos hoje como a segunda causa de infecções do trato urinário e a
terceira nos casos de bacteremia nosocomial (Aguirre e Collins, 1993; Franz et
al., 2001a). Foulquié-Moreno et al. (2006) afirmaram que dentre as bactérias
láticas, o gênero Enterococcus é o que causa mais controvérsia.
O aumento da severidade das infecções causadas por enterococos pode
estar relacionado ao uso irracional de antimicrobianos, permitindo sua
sobrevivência e disseminação no ambiente hospitalar. Além disso, bactérias
desse gênero apresentam uma alta capacidade de adquirir e transferir genes
de resistência a antibióticos de uso clínico (Vancanneyt et al., 2002).
Enterococcus spp. apresentam resistência intrínseca a vários agentes
antimicrobianos como penicilinas, cefalosporinas, sulfonamidas e um perfil de
baixa resistência a aminoglicosídeos e lincosamidas. A resistência adquirida
baseada na aquisição de plasmídeos e transposons é relevante para
cloranfenicol, eritromicina e para altas concentrações de clindamicina,
aminoglicosídeos, tetraciclinas, β-lactâmicos, fluoroquinolonas e glicopeptídeos
(Moellering, 1992; Facklam et al., 1999; Franz et al., 1999). Outro fato relevante
é a emergência de vários perfis fenotípicos de E. faecalis e E. faecium
resistentes à vancomicina (em inglês, VRE – “vancomycin resistant

Introdução________________________________________________________________________ 7
enterococci”). Palazzo et al. (2006) analisaram 51 cepas de enterococos
resistentes à vancomicina, que continham o elemento de inserção Tn1546,
isoladas de diferentes hospitais no Brasil. Estes autores observaram que na
maior parte das cepas analisadas os genes responsáveis pela resistência à
vancomicina estavam localizados em plasmídeos conjugativos de 70Kb,
facilitando a transferência de resistência.
Todas essas características vêm contribuindo para que estes
microrganismos se destaquem dentre os patógenos causadores de infecções
em humanos. Entretanto, a patogenicidade dos enterococos não pode ser
explicada apenas pelo seu perfil de resistência aos antimicrobianos, devendo-
se levar em consideração também, outros fatores de virulência (Franz et al.,
1999).
Atualmente, procura-se relacionar os fatores de virulência apresentados
por Enterococcus spp. com a severidade das infecções por eles causadas, bem
como sua origem (Ike et al., 1987; Morrison et al., 1997; Franz et al., 1999;
Eaton e Gasson, 2001; Franz et al., 2001a; Vancanneyt et al., 2002; Duprè et
al., 2003; Franz et al., 2003).
A hemolisina, um dos marcadores fenotípicos que possivelmente atua
como um fator de virulência para algumas cepas de E. faecalis, é capaz de lisar
eritrócitos humanos, de cavalos e de coelhos. Cepas de enterococos que
apresentam perfil β-hemolítico também podem produzir um peptídeo
antimicrobiano (bacteriocina) codificado por plasmídeos conjugativos com
atividade inibitória contra muitas bactérias Gram-positivas (Coque et al., 1995;
Hickey et al., 2003). Para alguns membros do gênero, quando hemolisina e

Introdução________________________________________________________________________ 8
bacteriocina são expressas pelo mesmo determinante genético, recebem a
denominação genérica de citolisina (Booth et al., 1996).
A citolisina está presente em cerca de 60% dos E. faecalis envolvidos
em infecções e causa reação do tipo β-hemólise em ágar contendo sangue
humano ou de cavalo, mas não é capaz de hemolisar sangue de carneiro, que
é rotineiramente empregado em laboratórios clínicos (Mundy et al., 2000). Além
disso, especula-se que essa proteína confira às cepas que a produzem um
mecanismo de auto-proteção.
Ike et al. (1987) procuraram relacionar a produção de hemolisina por
cepas de E. faecalis isolados de amostras de pacientes com infecções e fezes
de pacientes sadios. Aproximadamente 60% das cepas de E. faecalis isoladas
de pacientes com infecção apresentavam perfil hemolítico, enquanto que a
análise de material fecal de indivíduos saudáveis revelou uma prevalência de
17% deste tipo de enterococos. Estes autores também observaram que o perfil
hemolítico poderia ser transferido a alta freqüência por meio de plasmídeos
conjugativos.
A produção de uma adesina denominada substância de agregação (AS)
é considerada outro fator de virulência de enterococos e, contribui para a
formação de agregados celulares, facilitando a troca de material genético entre
as células bacterianas (Franz et al., 2001a). A proteína AS é codificada por
plasmídeos que são incorporados pelas células receptoras por meio de
conjugação bacteriana mediada por substâncias quimioatraentes (feromônios).
Estas substâncias induzem a expressão de AS na superfície de células
doadoras, facilitando a formação de agregados celulares e conseqüente troca
de material genético (Wells et al., 2000).

Introdução________________________________________________________________________ 9
A proteína de superfície “Esp” (do inglês, "Enterococcus surface protein")
também se relaciona à patogenicidade de algumas cepas de enterococos, e
parece estar envolvida no processo de adesão célula-célula. Esta proteína
apresenta várias regiões, características de proteínas de superfície,
reconhecidamente envolvidas no processo de adesão às células eucarióticas e
evasão da resposta imune (Franz et al., 1999; Shankar et al., 1999; Duprè et
al., 2003).
Toledo-Arana et al. (2001) analisaram 93 cepas de E. faecalis que
possuíam o gene esp, e mostraram que destas cepas, 87 foram capazes de
formar biofilmes. Todas as 59 cepas deficientes de esp não apresentaram essa
mesma capacidade. Estes autores sugeriram que a formação de biofilmes
aumentaria a resistência inata de alguns microrganismos frente a antibióticos e,
conseqüentemente, sua patogenicidade.
Um outro fator de virulência que vem sendo amplamente pesquisado
para Enterococcus spp. é a gelatinase (Gel), uma protease responsável pela
hidrólise de gelatina, colágeno, hemoglobina e outros peptídeos bioativos.
Alguns pesquisadores relacionam a produção de gelatinase com a indução de
processos inflamatórios (Coque et al., 1995; Hickey et al., 2003).
Hialuronidase (Hyl) é um outro fator de virulência descrito para E.
faecium que apresenta um certo grau de similaridade à hialuronidase
previamente descrita para Streptococcus pyogenes, Staphylococcus aureus e
Streptococcus pneumoniae. Embora ainda não esteja completamente
estabelecido, acredita-se que esta enzima seja responsável por injúria tecidual,
aumento do poder invasivo do microrganismo e até mesmo pela progressão da
pneumonia pneumocócica (Rice et al., 2003; Kayaoglu e Orstavik, 2004;

Introdução________________________________________________________________________ 10
Vankerckhoven et al., 2004).
De acordo com Kayaoglu e Orstavik (2004) a proteína EfaA, antígeno A
de E. faecalis, possui cerca de 55 a 60% de homologia com um grupo de
proteínas de estreptococos conhecidas como adesinas e em enterococos
parece funcionar como uma adesina de endocardite. A presença do gene efaA
que codifica esta proteína foi relatada em vários isolados clínicos de E. faecalis
e também em vários isolados obtidos a partir de amostras de alimentos (Eaton
e Gasson, 2001).
Uma outra proteína que também parece estar relacionada com o
aumento de patogenicidade de E. faecalis é a adesina de colágeno, codificada
pelo gene ace. Esta proteína é capaz de aderir aos colágenos tipo I e IV,
similar à proteína Cna de Staphylococcus aureus (Nallapareddy et al., 2000;
Mannu et al., 2003).
1.4 Enterococos como indicadores de contaminação fecal
O interesse do uso de Enterococcus spp. como indicador de higiene e
contaminação fecal pode ser justificado pelo fato que este gênero é geralmente
mais tolerante à refrigeração quando comparado com E. coli e outros
coliformes e, parece sobreviver por mais tempo durante a estocagem
refrigerada de carcaças de carnes (Ingham e Schmidt, 2000).
Entretanto, embora os enterococos façam parte da microbiota intestinal
de humanos e de diferentes espécies animais, o uso deste gênero como
indicador de contaminação fecal pode ser limitado devido ao fato destes
microrganismos apresentarem uma alta resistência ao calor, serem capazes de
sobreviver no ambiente extra-enteral, e poderem sobrepujar facilmente a

Introdução________________________________________________________________________ 11
microbiota de um alimento termicamente processado (Smoot e Pierson, 1997;
Borgen et al., 2001). Ao contrário do grupo de coliformes totais e fecais, não há
um nível estabelecido para relacionar a presença de Enterococcus spp. e a
qualidade higiênica de um determinado produto (Giraffa et al., 1997).
Atualmente, as populações de coliformes totais, coliformes fecais e
Escherichia coli são freqüentemente pesquisadas em amostras de água e em
alimentos como indicadores da presença de patógenos humanos associados à
contaminação fecal (BAM, 2001; Vail et al., 2003).
Segundo o "Health Products and Food Branch" (Canadá, 2001) dentre
as metodologias disponíveis para a enumeração de microrganismos do grupo
dos coliformes e de E. coli em amostras de água e alimentos, destaca-se o uso
de placas PetrifilmTM (3M). Estas placas são formadas por filmes de papel
quadriculado revestidos com polietileno, nutrientes desidratados e gel
hidrossolúvel a frio. São comercializadas prontas para o uso, permitindo a
análise de um grande número de amostras em pouco tempo.
A viabilidade de se utilizar coliformes totais, fecais e enterococos como
microrganismos indicadores também foi avaliada por Noble et al. (2003). Estes
autores monitoraram a qualidade higiênica de amostras de água provenientes
da costa da Califórnia (EUA) e os resultados obtidos não permitiram concluir
quais os microrganismos indicadores mais adequados para o monitoramento
da qualidade de água naquela região.
Calicioglu et al. (1999) compararam a recuperação de Enterococcus spp.
e E. coli a partir de carcaças bovinas artificialmente inoculadas com material
fecal. Utilizando o Ágar Kanamicina Esculina Azida e placas de PetrifilmTM (3M)
para contagem de, respectivamente, Enterococcus spp. e E. coli, estes autores

Introdução________________________________________________________________________ 12
concluíram que a contagem de enterococos poderia ser útil como indicador de
contaminação fecal de carcaças bovinas.
Quando comparados aos coliformes fecais, os enterococos são capazes
de sobreviver por mais tempo no ambiente marinho e são mais resistentes aos
processos de tratamento de efluentes, havendo boa correlação deste
microrganismo com a presença de patógenos de interesse. Por este motivo, o
estado da Califórnia (EUA) incorporou a enumeração de enterococos aos
métodos tradicionais para determinação da qualidade de água recreacional
(Choi et al., 2003).
Existem, contudo, algumas limitações para o uso de enterococos como
indicadores de contaminação fecal recente: (i) são microrganismos
encontrados não apenas no trato intestinal de humanos mas também de outros
animais de sangue quente e não apresentam, dessa forma, especificidade de
hospedeiro; (ii) não é bem conhecida a distribuição das espécies de
enterococos em águas residuais e, (iii) a presença de enterococos é desejável
em muitos alimentos. Sendo assim, a interpretação da presença de bactérias
desse gênero, deve ser feita com cautela (Giraffa, 2002; Blanch et al., 2003;
Choi et al., 2003).
1.5 Importância de Enterococcus em alimentos
A natureza ubíqua dos enterococos faz com que seja comum o seu
isolamento de uma ampla variedade de alimentos (Klein et al., 1998; Robredo
et al., 2000; Chingwaru et al., 2003). Em vegetais, E. casseliflavus é a espécie
mais comum (Devriese et al., 1995; Gelsomino, 2002). Além disso,
processamentos tecnológicos de alguns alimentos como, salga e defumação,

Introdução________________________________________________________________________ 13
expõem os microrganismos a extremos de temperatura, pH e salinidade.
Nestas condições, os enterococos, que são altamente resistentes, podem se
multiplicar e até provocar a deterioração do produto (Giraffa, 2002; Foulquié-
Moreno et al., 2006).
Enterococcus spp. também podem contaminar produtos acabados
durante o próprio processamento. De acordo com Gelsomino et al. (2002) os
enterococos podem sobreviver e até se multiplicar em equipamentos e tanques
de estocagem de laticínios, favorecendo assim, a contaminação direta dos
produtos. Entretanto, enquanto alguns estudos revelam a relação entre
enterococos e a deterioração de alguns produtos, outros defendem o papel
deste microrganismo no desenvolvimento de características sensoriais em
queijos e produtos fermentados (Foulquié-Moreno et al., 2006).
A presença de enterococos no trato gastrintestinal de animais pode levar
à contaminação da carne durante o abate. E. faecalis e E. faecium são as
espécies predominantes em diversos tipos de produtos cárneos fermentados
ou não (Franz et al., 2003; Foulquié-Moreno et al., 2006). A alta resistência
térmica apresentada por enterococos também favorece a persistência destes
microrganismos em produtos cárneos processados (Franz et al., 1999;
Foulquié-Moreno et al., 2006) e pode contribuir para o desenvolvimento de
aromas pela ação glicolítica, proteolítica e lipolítica (Hugas et al., 2003).
Algumas propriedades bioquímicas dos enterococos como, alta
tolerância a sal e ácido, são desejáveis no processamento de alguns queijos
(Andrighetto et al., 2001; Giraffa, 2002; Jurkovič et al., 2006). Em particular, E.
faecium, é freqüentemente utilizado na fermentação de vários queijos de
produção artesanal no sul da Europa (Stiles e Holzapfel, 1997). A população de

Introdução________________________________________________________________________ 14
enterococos em diversos queijos pode variar de 104 UFC g-1 a 107 UFC g-1
dependendo do tipo de queijo, da fase da maturação e da contaminação inicial
do leite utilizado no processamento (Franz et al., 1999).
A avaliação da microbiota de queijos da região mediterrânea, produzidos a
partir de leite cru de ovelha e cabra, revelou que o metabolismo de Enterococcus
spp. é importante na maturação e no desenvolvimento de características
sensoriais desses queijos, quer seja por proteólise, lipólise ou até mesmo
metabolismo do citrato (Foulquié-Moreno et al., 2006). Os enterococos, ao
contrário de outras bactérias láticas, são capazes de metabolizar o citrato com
conseqüente produção de diacetil, acetaldeído, acetoína, 2-3-butanodiol e dióxido
de carbono, que conferem características e textura particulares em queijos e
outros produtos lácteos (Foulquié-Moreno et al., 2006).
Além do emprego tecnológico, a utilização de Enterococcus spp. como
probióticos na diminuição dos riscos de doenças intestinais, em humanos e
animais também é possível (Stiles e Holzapfel, 1997). Zeyner e Boldt (2005)
demonstraram que a administração oral diária de E. faecium CF68 foi eficaz na
redução de casos de diarréia em porcos. Adicionalmente, E. faecium CF68
pareceu estar envolvido no aumento da concentração sérica de IgG contra
Giardia e IgA específica e total, em fezes de ratos (Benyacoub et al., 2005).
De acordo com Klingberg et al. (2005) para que um microrganismo seja
considerado probiótico, deve-se garantir que cerca de 108 células cheguem
viáveis ao trato gastrintestinal e que estes microrganismos resistam a baixo pH
e sais biliares, garantindo a sobrevivência à passagem gastrintestinal. Além
disso, é fundamental garantir que as cepas probióticas sejam seguras para
utilização na produção de alimentos (Saavedra et al., 2003; De Vuyst et al.,

Introdução________________________________________________________________________ 15
2003). De acordo com Salminen et al. (1996) algumas características devem
ser verificadas para que um microrganismo seja classificado como probiótico:
capacidade de aderir e de multiplicar na mucosa intestinal, impedir ou reduzir a
adesão de patógenos, ser seguro, não invasivo, não carcinogênico, produzir
ácidos, peróxido de hidrogênio e bacteriocinas.
De acordo com Strompfova et al. (2006) as bacteriocinas de
enterococos, conhecidas como enterocinas, pertencem ao grupo IIa que são
peptídeos pequenos, catiônicos, hidrofóbicos, termicamente estáveis e com
atividade contra uma ampla variedade de microrganismos. Estas bacteriocinas
vêm despertando muito interesse para o uso na indústria de alimentos como
antimicrobianos naturais (Cleveland et al., 2001; Drider et al. 2006)
Essas bacteriocinas podem apresentar atividade inibitória frente a alguns
patógenos. Particularmente, E. faecalis e E. faecium foram capazes de
controlar a multiplicação de Listeria sp. durante o processamento de alguns
queijos (Sulzer e Busse, 1991; Giraffa et al., 1997).
Dentre as características probióticas de Enterococcus spp. cabe-se
destacar a capacidade de redução de colesterol sérico pela atividade de
hidrólise de sais biliares (BSH). De acordo com Franz et al. (2001b) a
desconjugação de sais biliares no lúmen intestinal contribui para a redução de
nivéis séricos de colesterol, pois os ácidos biliares livres são mais facilmente
excretados pelo trato gastrointestinal.
Por outro lado, o aumento da severidade das infecções nosocomiais
causadas por cepas de Enterococcus mutirresistentes a antimicrobianos e a falta
de conhecimento sobre seus fatores de virulência geram insegurança na utilização
de cepas deste gênero na produção de alimentos, como culturas fermentadoras e

Introdução________________________________________________________________________ 16
probióticas (Giraffa et al., 1997; Franz et al., 1999; Giraffa 2002; Franz et al.,
2003). As várias aplicações de Enterococcus spp. em alimentos, bem como os
riscos associados com sua presença, estão ilustrados na Figura 1 .
Figura 1: Riscos e benefícios associados à presença de Enterococcus spp. em alimentos, amostras ambientais e clínicas. Adaptado de Domig et al. (2003).
Eaton e Gasson (2001) afirmaram que a diferença entre uma cepa de
enterococos com potencial patogênico e outra aparentemente segura para uso
em processamento de alimentos não é clara, e a probabilidade de que esta
última adquira fatores de virulência merece maior investigação.
Diante do exposto e, considerando-se a escassez de dados sobre
enterococos em alimentos consumidos no Brasil, este trabalho foi realizado
para investigar isolados obtidos a partir de amostras de águas e alimentos,
bem como para determinar perfis de resistência a antibióticos e fatores de
virulência de Enterococcus spp. Ainda, foi estudada a viabilidade de sua
utilização como indicador de contaminação fecal.

___________________2. Objetivos

Objetivos________________________________________________________________________ 18
2.1 Objetivo geral
Determinar características fenotípicas e moleculares de Enterococcus spp.
isolados de amostras de alimentos e águas e correlacionar a presença destes
microrganismos com indicadores clássicos de higiene e contaminação fecal.
2.2 Objetivos específicos
• Fornecer dados da ocorrência de Enterococcus spp. em alimentos
comercializados na cidade de Ribeirão Preto-SP;
• Avaliar a eficácia do sistema API20 Strep na identificação das espécies
dos isolados obtidos;
• Caracterizar os isolados obtidos quanto à produção de hemolisina,
bacteriocina, gelatinase e capacidade de hidrólise de sais biliares (BSH);
• Verificar a presença de marcadores de virulência (as, ace, efaA, gelE,
esp, cyl e hyl) nos isolados de E. faecium e E. faecalis;
• Determinar o perfil de resistência de E. faecalis e E. faecium aos
antibióticos de uso clínico: ampicilina, tetraciclina, vancomicina,
eritromicina e gentamicina;
• Analisar os isolados de E. faecalis e E. faecium quanto à capacidade de
formação de biofilme em superfície abiótica;
• Analisar os isolados de E. faecalis e E. faecium, resistentes a antibióticos,
quanto à capacidade de adesão em células da linhagem Caco-2;
• Avaliar a similaridade dos isolados de E. faecalis e E. faecium,
resistentes a antibióticos, aplicando-se a técnica de RAPD-PCR;
• Correlacionar a presença de Enterococcus spp. com indicadores
clássicos de higiene e contaminação fecal, E. coli e coliformes totais.

___________3. Materiais e Métodos

Materiais e Métodos_________________________________________________________________ 20
3.1 Coleta das amostras de alimentos e águas
Foram analisadas 120 amostras de alimentos, sendo: 30 de queijo
(mussarela, queijo Minas, ricota, gouda, cheddar, parmesão, provolone,
chancliche e gorgonzola), 30 de produtos cárneos (carne bovina e suína crua,
embutidos, miúdos, carne de frango e de peixe), 30 de vegetais (verduras e
vegetais frescos e minimamente processados, grãos cozidos, conservas e
ervas) e 11 amostras de leite pasteurizado adquiridas no comércio varejista de
Ribeirão Preto – SP e 19 de leite cru coletadas na fazenda de Zootecnia do
Pólo Regional Leste de Ribeirão Preto-SP. Também foram analisadas 30
amostras de água destinada ao consumo humano, sendo 25 amostras
representativas do sistema de distribuição municipal de Ribeirão Preto, cedidas
pelo Departamento de água e Esgotos de Ribeirão Preto – DAERP e 5
amostras de água mineral comercializadas em Ribeirão Preto – SP. As
amostras foram transportadas para o laboratório em caixas isotérmicas com
gelo reciclável e mantidas sob refrigeração até o momento da análise.
3.2 Preparo das amostras
As amostras de produtos cárneos, queijos, e vegetais foram fracionadas
assepticamente em porções de 25 g, acondicionadas em embalagens plásticas
esterilizadas e adicionadas de 225 mL de peptona bacteriológica (Oxoid –
Reino Unido) a 0,1% (p/v). Alíquotas de 10 mL de leite foram diluídas em 90
mL de peptona a 0,1% (p/v). Essas amostras foram homogeneizadas por um
minuto em homogeneizador de amostras para microbiologia (“Bag Mixer”,
Interscience, França). As amostras de água foram homogeneizadas no próprio
recipiente da coleta. A partir destas suspensões foram preparadas diluições

Materiais e Métodos_________________________________________________________________ 21
decimais seriadas (10-2, 10-3 , 10-4 ,10-5) para enumeração de enterococos,
coliformes totais e E. coli.
3.3 Isolamento e enumeração de Enterococcus spp.
Para o isolamento de Enterococcus spp. a partir de amostras de
alimentos foram utilizados os ágares "Bile Esculina" (BE) e "Kanamicina
Esculina Azida" base suplementado com 20mg/L kanamicina (KEA), ambos da
Oxoid, Reino Unido. Estes meios foram escolhidos devido a relatos de bons
resultados para o isolamento de enterococos a partir de amostras contendo
uma densa população microbiana (Ike et al., 1987; Devriese et al., 1995;
Morrison et al., 1997; Facklam et al., 1999; Andrighetto et al., 2001).
Alíquotas de 0,1 mL das diluições das amostras (10-1, 10-2, 10-3, 10-4,10-5)
foram semeadas em superfície nos meios BE e KEA. Os meios inoculados
foram incubados por 24 horas a 37°C. Para a enumeração de Enterococcus
spp. foram selecionadas as placas contendo entre 25 e 250 UFC (Unidade
Formadora de Colônia), representativas de cada amostra. Utilizou-se a raiz
quadrada do número de UFC para definir o número de colônias a serem
submetidas aos testes posteriores de identificação presuntiva de Enterococcus
spp. Para as amostras sólidas e líquidas o resultado foi expresso,
respectivamente, em UFC/ g da amostra e UFC/ mL da amostra.
A enumeração de Enterococcus spp. em amostras de água foi feita
empregando-se a técnica da membrana filtrante. Alíquotas de 100 mL foram
filtradas utilizando filtros com poro de 0,45 µm (Millipore, EUA). A membrana foi
transferida para a superfície de ágar m-Enterococcus (Becton Dickinson, EUA)
e incubada a 37°C por 24-48 horas. A presença de colônias com coloração

Materiais e Métodos_________________________________________________________________ 22
roxa, devido a redução do indicador trifenil tetrazólio (TTC) era considerada
indicativa de Enterococcus spp.
3.4 Identificação presuntiva de Enterococcus spp.
Após o período de incubação, as colônias características do gênero
Enterococcus foram purificadas por inoculação em ágar soja tripticase (TSA,
Oxoid) suplementado com 0,6% de extrato de levedura (Oxoid) e incubadas por
24 horas a 37°C. Em seguida, uma colônia isolada foi transferida para um tubo
inclinado contendo TSA suplementado com 0,6% de extrato de levedura e
incubada por 24 horas a 37°C. A partir desta cultura foi realizado o teste de
catalase. As colônias catalase negativa foram submetidas à coloração de Gram
e os isolados que se apresentaram como cocos Gram-positivos aos pares ou
isolados foram mantidos para testes posteriores.
A identificação presuntiva foi feita baseando-se na capacidade de
multiplicação dos enterocos em caldo de Man Rogosa Sharpe – MRS (Oxoid)Q
contendo (i) 6,5% (p=-21/v) de cloreto de sódio, (ii) caldo MRS ajustado
para pH 9,6 com solução de hidróxido de sódio (10N), (iii) multiplicação em
caldo MRS a 10 e 45°C e (iv) teste do L-pirrolidonil-β-naftilamida - PYR
(DrySlide PYR Kit - Becton, Dickinson and Company, EUA) de acordo com a
metodologia proposta por Schillinger e Lücke (1987), Franz et al. (1999) e
Andrighetto et al. (2001). Para a realização destes testes, alíquotas de 100 µL
das culturas provenientes do crescimento de 24 horas em caldo MRS foram
inoculadas nos respectivos meios de cultura. A turvação dos meios, após
incubação por 24 horas, era indicativa de teste positivo. Para o teste do PYR,
eram utilizadas colônias isoladas de enterococos provenientes do crescimento

Materiais e Métodos_________________________________________________________________ 23
de 24 horas em TSA adicionado de 5% de sangue de cavalo. Estas colônias
eram transferidas para a área de reação do kit, impregnadas com uma solução
de revelação e o desenvolvimento de coloração rosa/roxa, devido à reação
entre β-naftilamina e p-dimetilaminocinamaldeído, era indicativo de teste
positivo, conforme instruções do fabricante.
3.5 Identificação das espécies de Enterococcus spp. por testes
bioquímicos
Isolados que apresentaram resultados presuntivos para o gênero
enterococos foram identificados pelo teste API 20 STREP (BioMérieux,
França), conforme especificações do fabricante.
3.6 Identificação das espécies de Enterococcus empregando-se PCR
Os isolados indicativos do gênero enterococos foram cultivados em
placas de TSA suplementado com 0,6% de extrato de levedura por 24 horas.
As culturas de E. faecalis NCTC 775, E. faecium NCTC 7171, E. gallinarum
NCTC 12359 e E. casseliflavus NCTC 12361 foram utilizadas como controles
positivos. As colônias foram raspadas com auxílio de uma alça esterilizada e
transferidas para um tubo Eppendorf contendo 100 µL de água para PCR (w-
3500, Sigma). Esta suspensão foi homogeneizada e em seguida centrifugada
por aproximadamente 15 segundos. O sobrenadante obtido foi utilizado na
PCR para identificação dos genes específicos das espécies de E. faecium, E.
faecalis, E. casseliflavus e E. gallinarum (Dutka-Malen et al., 1995). Os primers
utilizados bem como o tamanho dos fragmentos obtidos estão descritos na
Tabela 1.

Materiais e Métodos_________________________________________________________________ 24
Tabela 1: Seqüência dos primers utilizados e tamanho dos fragmentos obtidos
na identificação de E. faecalis, E. faecium, E. casseliflavus e E. gallinarum
Espécie Gene amplificado Seqüência (5 ’- 3’) Tamanho produto (pb)
E. faecalis ddlE. faecalis + ATCAAGTACAGTTAGTCT - ACGATTCAAAGCTAACTG
941
E. faecium ddlE. faecium + GCAAGGCTTCTTAGAGA - CATCGTGTAAGCTAACTTC 550
E. gallinarum vanC-1 + GGTATCAAGGAAACCTC - CTTCCGCCATCATAGCT 822
E. casseliflavus vanC-2, vanC-3 + CTCCTACGATTCTCTTG - CGAGCAAGACCTTTAAG 439
Fonte: Dutka-Malen et al.(1995)
A PCR foi feita em termociclador (Mastercycler – Eppendorf) com
volume final de 25 µL que consistiu de 60 ng de DNA, 25 pmol de cada primer,
0,2 mM de dATP, dCTP, dTTP, e dGTP, 2 mM de tampão contendo MgCl2 e
0,625 U de Taq DNA polimerase. Foi utilizado um ciclo de 94°C por dois
minutos seguido de 30 ciclos de 94°C por um minuto, 54°C por um minuto,
72°C por um minuto e um ciclo final de 72°C por dez minutos. Os produtos da
PCR foram submetidos a eletroforese em um gel de 1% (p/v) de agarose em
tampão Tris-borato-EDTA e corados com brometo de etídeo (Dutka-Malen et
al., 1995).
3.7 Caracterização fenotípica das cepas de Enterococcus spp.
3.7.1 Hidrólise de gelatina
A produção de gelatinase foi avaliada no ágar Todd – Hewitt
suplementado com 30 g/L de gelatina bacteriológica, ambos da Oxoid. A
cultura proveniente do crescimento de 24 horas a 37°C no caldo MRS foi
transferida para a superfície deste meio com auxílio de uma alça esterilizada e

Materiais e Métodos_________________________________________________________________ 25
a placa foi incubada a 37°C por 18 horas e em seguida resfriada a 4°C por 5
horas. A hidrólise da gelatina foi visualizada pela formação de um halo turvo ao
redor da colônia (Ike et al., 1987; Eaton e Gasson, 2001).
3.7.2 Atividade hemolítica
Para investigação da produção de hemolisina foi utilizado o ágar
tripticase de soja (TSA) suplementado com 5% de sangue de cavalo pois
segundo Domig et al. (2003), para este gênero bacteriano a reação β-
hemolítica baseada na atividade da citolisina é favorecida neste meio.
Culturas puras provenientes do crescimento de 24 horas em caldo MRS
foram semeadas nas placas contendo este meio e incubadas a 37°C por 24-48
horas. A formação de zonas claras ao redor da colônia foi indicativa da
produção de β-hemólise (Eaton e Gasson, 2001).
3.7.3 Determinação da produção de bacteriocina
Para avaliação da atividade inibitória, culturas puras de Enterococcus
spp. foram submetidas ao teste “spot-on-the-lawn” com Lactobacillus sakei
ATCC 15221 e Listeria monocytogenes ATCC 19115 como microrganismos
indicadores. Resumidamente, alíquotas de 2µL das culturas de Enterococcus
spp. reativadas em caldo MRS a 37°C durante 24 horas foram inoculadas em
placas de Petri contendo ágar TSA adicionado de 0,6% de extrato de levedura
(TSAYE) e incubadas a 37°C por 24 horas em jarras contendo envelopes
geradores de anaerobiose (Anaerogen, Oxoid). Em seguida, adicionou-se
nestas placas uma sobrecamada de caldo infusão cérebro-coração (BHI) semi-
sólido contendo 1% do microrganismo indicador. As placas foram incubadas

Materiais e Métodos_________________________________________________________________ 26
mais uma vez em anaerobiose a 37°C por 24 horas e, após esse tempo, a
presença de um halo de inibição ao redor das colônias de Enterococcus spp.
foi indicativa da presença de substância inibitória (Lewus et al., 1991)
Empregando a metodologia descrita por Lewus et al. (1991) foi excluída
a possibilidade de inibição devido à produção de ácidos orgânicos, peróxido de
hidrogênio e atividade de bacteriófagos líticos. Além disso, para a
caracterização da substância inibitória como bacteriocina foi avaliada sua
natureza protéica utilizando as enzimas Proteinase K (P-8044) e Protease (P-
5147) de Streptomyces griseus (ambas da Sigma, Alemanha) de acordo com a
metodologia proposta por Lewus et al. (1991).
3.7.4 Avaliação da hidrólise de sais biliares
Os isolados de enterococos foram avaliados quanto à capacidade de
hidrolisar sais biliares (BSH). Para isso, alíquotas de 10 µL das culturas
proveniente do crescimento de 24 horas em caldo Man, Rogosa, and Sharpe –
MRS (Oxoid) foram transferidas para placas de ágar MRS suplementado com
0,5% (p/v) de sal sódico do ácido taurodeoxicólico e 0,37 g/L de cloreto de
cálcio (CaCl2). As placas foram incubadas a 37°C por 72 horas em jarras
contendo envelopes geradores de anaerobiose (Anaerogen, Oxoid). Baseando-
se no diâmetro da zona de precipitação ao redor das colônias, após o período
de incubação, a atividade de hidrólise de sais biliares foi classificada como:
ausente (ausência de zona de precipitação), baixa (zona de precipitação de até
10mm), média (zona de precipitação entre 10 e 15mm) e alta (zona de
precipitação > 15mm) (Franz et al., 2001 b).

Materiais e Métodos_________________________________________________________________ 27
Experimentos realizados durante estágio de doutorado no exterior
Esta parte do trabalho (itens 3.8 a 3.12) foi realizada durante Estágio de
Doutorando no Exterior com bolsa da CAPES, desenvolvido no período de
05/2005 a 05/2006 com a colaboração do Prof. Dr. Leonardo A. Sechi, da
Università degli Studi di Sassari – Dipartamento di Scienze Biomediche, Itália.
3.8 Determinação da resistência a antibióticos
A presença de enterococos resistentes a cinco antibióticos de uso clínico
foi avaliada através de uma triagem em ágar BHI com as seguintes
concentrações de antibióticos: 32 µg/mL vancomicina, 16 µg/mL tetraciclina, 8
µg/mL eritromicina, 16 µg/mL ampicilina e 500 µg/mL gentamicina (Willey et al.,
1992, CLSI, 2005). Para tanto, os isolados de E. faecium e E. faecalis
provenientes do crescimento de 24 horas em TSA foram transferidas para tubo
contendo solução salina esterilizada, para obtenção de uma suspensão
correspondente à turbidez 0,5 da escala de MacFarland. Dois microlitros desta
suspensão foram transferidos para as placas contendo ágar BHI com as
concentrações adequadas de antibióticos. Os meios inoculados foram
incubados a 37°C e a leitura foi realizada após 24 e 48 horas de incubação.
Em seguida, os isolados que apresentaram resistência aos antibióticos
testados foram submetidos à determinação da concentração inibitória mínima
(CIM). A determinação da CIM foi realizada pelo método de diluição em ágar
(Yousif et al., 2005; CLSI, 2005) com concentrações de vancomicina,
tetraciclina e eritromicina variando de 1 µg/mL a 256 µg/mL enquanto que a
concentração de gentamicina variou de de 1 µg/mL a 500 µg/mL.

Materiais e Métodos_________________________________________________________________ 28
3.9 Detecção de genes de virulência em isolados de E. faecium e E.
faecalis O DNA genômico dos isolados foi extraído conforme descrito por Sechi
et al. (1998 a, b) com algumas modificações e usado para detecção dos
seguintes genes de virulência: esp, as, ace, efaA, gelE, cylA e hyl.
Em todas as reações as seguintes cepas foram usadas como controle
positivo: E. faecalis JH2-2 para ace e gelE, E. faecalis ATCC 47077 para efaA,
E. faecalis 740 (nossa coleção) para o gene as, E. faecalis MMH594 para cylA
e E. faecium C68 para hyl (Vankerckhoven et al., 2004) e E. faecium 18 R320
para esp (Mannu et al., 2003). As seqüências dos primers utilizados para
amplificação dos genes de interesse estão apresentadas na Tabela 2.

29M
ateriais e Métodos________________________________________________________________________
Tabela 2: Sequência dos primers utilizados e tamanho dos fragmentos obtidos na pesquisa dos genes de virulência de
Enterococcus spp.
Gene Marcador de virulência Primers Seqüência (5’ - 3’) Tamanho
produto (pb) Referência
as substância de agregação AS 1 CCAGTAATCAGTCCAGAAACAACC 406 Mannu et al. 2003
AS 2 TAGCTTTTTTCATTCTTGTGTTTGTT
ace adesina de colágeno de E. faecalis ACE 1 AAAGTAGAATTAGATCCACAC 320 Mannu et al. 2003
ACE 2 TCTATCACATTCGGTTGCG
gelE Gelatinase gel E1 AGTTCATGTCTATTTTCTTCAC 402 Mannu et al. 2003
gel E2 CTTCATTATTTACACGTTTG
efaA antígeno A de E. faecalis efaA1 CGTGAGAAAGAAATGGAGGA 499 Mannu et al. 2003
efaA2 CTACTAACACGTCACGAATG
esp proteína de superfície ESP46 TTACCAAGATGGTTCTGTAGGCAC 913 Shankar et al. 1999
ESP47 CCAAGTATACTTAGCATCTTTTGG
cylA Citolisina CYT I ACTCGGGGATTGATAGGC 688 Vankerckhoven et al. 2004
CYT IIb GCTGCTAAAGCTGCGCTT
hyl Hialuronidase HYL n1 ACAGAAGAGCTGCAGGAAATG 276 Vankerckhoven et al. 2004
HYL n2 GACTGACGTCCAAGTTTCCAA

Materiais e Métodos_________________________________________________________________
30
Para os genes esp, as, ace, efaA e gelE as reações foram feitas em um
total de 30 µl, usando 5 µl de DNA como molde, 0,5 µM de cada primer, 1x de
tampão PCR, 4,0 mM de MgCl2, 200 µM de cada dNTPs, 1U de Taq platinum
DNA polymerase (Invitrogen Life Technologies, EUA). As reações de PCR
consistiam de uma etapa inicial de 94°C por 3 minutos, seguida de 30 ciclos de
desnaturação a 94°C por 1 minuto, anelamento a 56°C por 1 minuto, extensão
a 72°C por 1 minuto e uma etapa final de 72°C por 5 minutos, para os genes
ace, efaA and gelE.
O gene as foi amplificado utilizando-se 30 ciclos de PCR com
desnaturação a 94°C por 1 minuto, anelamento a 54°C por 1 minuto, extensão
a 72°C por 1 minuto, enquanto que para o gene esp a desnaturação foi feita a
94°C for 1 minuto, anelamento a 58°C por 1 minuto, extensão a 72°C por 1
minuto, por 30 ciclos (Mannu et al., 2003).
Os genes cylA e hyl foram amplificados por meio de uma multiplex PCR,
de acordo com a metodologia proposta por Vankerckhoven et al. (2004) com
algumas modificações. Resumidamente, as reações foram feitas utilizando-se
um volume final de 30 µl com 5 µl de DNA molde, 0,1 µM de cada primer de hyl
e 0,2 µM de cada primer de cylA, 1x de tampão PCR, 2,5 mM de MgCl2, 200
µM de cada dNTP, 1U of Taq platinum DNA polymerase (Invitrogen Life
Technologies). As reações de PCR foram feitas com uma etapa inicial de 94°C
por 3 minutos, seguida de 30 ciclos de desnaturação a 94°C por 1 minuto,
anelamento a 56°C por 1 minuto, extensão a 72°C por 1 minuto e uma etapa
final de 72°C por 10 minutos.
Os produtos de PCR foram separados por eletroforese utilizando-se gel
de agarose a 1,5% corado com brometo de etídeo e visualizado sob luz UV.

Materiais e Métodos_________________________________________________________________
31
3.10 Formação de biofilme em superfície abiótica
A formação de biofilme em superfície abiótica foi quantificada de acordo
com a metodologia descrita por Toledo-Arana et al. (2001) com algumas
modificações. Resumidamente, os 80 isolados de E. faecalis e 139 de E.
faecium foram reativados em BHI a 37°C por 24 horas. As culturas foram
diluídas 1:20 em BHI e 200 µl desta suspensão foram transferidas para placas
de microtitulação de 96 poços (Becton Dickinson, EUA). As placas foram
incubadas a 37°C por 24 horas, e então os poços foram lavados com solução
de tampão fosfato (PBS), secos na posição invertida e corados com solução de
1% de cristal violeta por 15 minutos. Os poços foram enxagüados mais uma
vez e o corante remanescente foi solubilizado com 200 µl de uma solução de
etanol-acetona (80:20, v/v). A densidade ótica foi determinada a 595 nm (A595)
usando um leitor de microplacas (Versa max - Molecular Devices, EUA). Cada
teste foi feito em triplicata e os valores obtidos classificados como a seguir: A595
≤ 1 não formador de biofilme (-); 1 <A595 ≤ 2 fraco (+); 2 < A595 ≤ 3 moderado
(++); A595 > 3 forte (+++), conforme descrito por Duprè et al. (2003).
3.11 Adesão em células da linhagem Caco-2
Células da linhagem Caco-2 foram obtidas da American Type Culture
Collection - ATCC (Manassas, EUA), cultivadas em RPMI 1640 - Glutamax I
(Gibco, EUA) suplementado com 10% de soro fetal bovino (SFB) e 100
unidades/mL de estreptomicina e penicilina. As células foram incubadas a 37°C
em 5% CO2 e quando apresentavam crescimento confluente foram transferidas
para placas de 24 poços e incubadas por mais 24 horas a 37°C em 5% de CO2.
As culturas de E. faecalis e E. faecium (resistentes a antibióticos) foram

Materiais e Métodos_________________________________________________________________
32
cultivadas em caldo BHI e diluídas no próprio meio de preparo das células sem
adição de antibióticos. Dez microlitros desta suspensão contendo cerca de 106
bactérias, foram transferidos para cada poço contendo as células Caco-2 e a
placa foi incubada a 37°C por 3 horas em 5% CO2. Após fixação e coloração
com May–Grunwald–Giemsa a adesão foi estimada pela contagem em
microscópio (aumento 1.000 vezes) do número de células de enterococos
aderidas às células Caco-2 (Duprè et al., 2003).
3.12 Análise de fingerprinting por meio de RAPD-PCR
A similaridade dos isolados de E. faecalis e E. faecium, resistentes a
antibióticos, foi analisada empregando-se a técnica de RAPD-PCR (do inglês,
Random Amplified Polymorphic DNA – Polymerase Chain Reaction) utilizando-
se o primer aleatório M13: 5’-GAGGGTGGCGGTTCT-3’ (Huey e Hall 1989;
Andrighetto et al. 2001; Vancanneyt et al., 2002; Cosentino et al., 2004;
Rossetti e Giraffa, 2005; Yousif et al., 2005).
Os produtos de PCR foram separados por eletroforese utilizando-se gel
de agarose a 1,5% (p/v), corados com brometo de etídeo e visualizados sob luz
UV. A imagem foi transferida para o computador e a análise dos padrões
obtidos foi feita utilizando-se o software Gelcompar e o algoritmo UPGMA
(Applied Maths, Bélgica).
3.13 Enumeração de coliformes totais e E. coli
Para as amostras de produtos cárneos, queijos, vegetais e leite a
enumeração dos coliformes totais e E. coli foi realizada pela semeadura em
placas PetrifilmTM EC (3M, EUA). Para isso, alíquotas de 1 mL das suspensões
preparadas conforme descrito no item 3.2 foram semeadas no centro das

Materiais e Métodos_________________________________________________________________
33
placas. Após geleificação do meio, as placas foram incubadas a 35°C por 24
horas para enumeração de coliformes e a 35°C por 48 horas para enumeração
de E. coli, conforme especificações do fabricante (BAM, 2001; Canadá, 2001).
A técnica da membrana filtrante foi aplicada para a pesquisa de
coliformes totais e E. coli nas amostras de água (Celico et al., 2004; Juárez-
Figueroa et al., 2003). Alíquotas de 100 mL de amostra foram filtradas em
duplicata utilizando filtros com poro de 0,45 µm (Millipore, EUA). Uma das
membranas foi pré-incubada em uma placa sobre uma esponja umedecida com
1,8 mL de caldo lauril triptose (Difco, EUA) a 35°C por 2 horas. Em seguida,
esta membrana foi transferida para a superfície do ágar m-Endo LES (Becton
Dickinson, EUA) e incubada a 35°C por 24 horas. A presença de colônias com
coloração vermelho escuro (devido à reação do acetaldeído, resultante do
metabolismo das bactérias que utilizam a lactose, com o sulfato de sódio e
fucsina, presentes no meio) e brilho verde-metálico (devido à rápida
metabolização da lactose com produção de aldeído) foram consideradas
coliformes totais. A outra membrana, utilizada para a enumeração de
coliformes fecais, foi incubada em ágar m-FC a 44,5°C por 24 horas. Colônias
apresentando coloração azul (na presença dos indicadores ácido rosólico e
azul anilina) foram submetidas a testes para confirmação de E. coli.

___________________4. Resultados

Resultados________________________________________________________________________
35
4.1 Isolamento de Enterococcus spp.
De janeiro de 2004 a março de 2005 foram analisadas 120 amostras de
alimentos e 30 de águas. Um total de 812 colônias com características
indicativas do gênero Enterococcus spp. foram selecionadas nos meios BE e
KEA (Apêndice A, Tabela 3). A tabela 3 apresenta somente as amostras que
tiveram isolados identificados como Enterococcus spp.
Os resultados presuntivos da enumeração de Enterococcus spp. a partir
das amostras de alimentos, obtidos nos meios BE e KEA, estão apresentados
na Tabela 4. Observa-se que a média da contagem de enterococos no meio
BE variou de 3,0 a 5,8 log de UFC por grama ou mililitro das amostras de
carne, queijo, vegetais e leite. No meio KEA a contagem de enterococos ficou
entre 2,5 a 5,2 log de UFC por grama ou mililitro das amostras citadas
anteriormente. Não foi obtida nenhuma colônia característica de Enterococcus
spp. a partir das amostras de água analisadas, sendo que a contagem em
todas as amostras ficou abaixo do limite de detecção do método que é de 1
UFC/ 100 mL.
Após a identificação das espécies dos enterococos por PCR, a média
da contagem de Enterococcus spp. nos alimentos analisados foi de 3,0 log
UFC/g para as amostras de produtos cárneos, de 4,8 log UFC/g para as
amostras de queijos, de 4,4 log UFC/g para as amostras de vegetais e 2,4 log
UFC/mL para as amostras de leite (Tabela 4).

Continuação
36Resultados_______________________________________________________________________________
Tabela 3: Caracterização fenotípica e molecular de Enterococcus spp. obtidos a partir das amostras de alimentos semeadas
nos meios KEA e BE
Multiplicação
Amostra Alimento Isolado Meio isolamentoa
Cocos Gram + Catalase
10°C 45°C 6,5% NaCl pH 9,6 PYRb API 20 STREP PCR
1 Mussarela 5 BE + - + + + + + E. durans ** 8 BE + - + + + + + Leuconostoc sp. E. faecium 4 KEA + - + + + + + Leuconostoc sp. E. faecium
2 Queijo minas 13 KEA + - + + + + + E. faecalis E. faecalis 10 BE + - + + + + + E. faecalis E. faecalis 12 KEA + - + + + + + E. faecalis E. faecalis
4 Queijo minas 20 KEA + - + + + + + Aerococcus viridans E. gallinarum 6 Carne moída 21 KEA + - + + + + + E. avium E. faecalis 22 KEA + - + + + + + result. não conclusivo E. faecalis
8 Mussarela 36 BE + - + + + + + E. durans E. faecium 37 BE + - + + + + + baixa discriminação E. faecium 38 BE + - + + + + + n.a E. faecium 39 KEA + - + + + + + n.a E. faecium 40 KEA + - + + + + + n.a E. faecium 41 KEA + - + + + + + n.a E. faecium
9 Presunto 30 BE + - + + + + + baixa discriminação E. faecium 32 BE + - + + + + + Leuconostoc sp. E. faecium 34 KEA + - + + + + + E. durans E. faecium 35 KEA + - + + + + + result. não conclusivo E. faecium
10 Apresuntado 43 KEA + - + + + + + E. durans E. faecium 44 KEA + - + + + + + baixa discriminação E. faecium
11 Ricota 48 KEA + - + + + + + baixa discriminação E. faecium 12 Lingüiça frango 53 KEA + - + + + + + baixa discriminação **
57 BE + - + + + + + Enterococcus sp. E. faecalis 14 Salsicha 60 KEA + - + + + + + E. faecalis E. faecalis
62 BE + - + + + + + Enterococcus sp. E. faecalis 63 BE + - + + + + + E. avium E. faecalis
15 Queijo prato 65 BE + - + + + + + result. não conclusivo E. faecium 66 BE + - + + + + + Enterococcus sp. E. faecium 67 KEA + - + + + + + n.a E. faecalis 68 KEA + - + + + + + n.a E. faecium

Continuação
37Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
69 KEA + - + + + + + n.a E. faecium 16 Queijo Minas 70 BE + - + + + + + E.durans E. faecium
71 BE + - + + + + + Aerococcus viridans E. gallinarum 73 KEA + - + + + + + n.a E. faecalis 74 KEA + - + + + + + n.a E. gallinarum 75 KEA + - + + + + + n.a E. faecium
17 Queijo gouda 76 KEA + - + + + + + E. durans E. faecium 77 KEA + - + + + + + Enterococcus sp. E. faecium
18 Mussarela 100 KEA + - + + + + + E. faecalis E. faecalis 101 KEA + - + + + + + n.a E. faecium 102 KEA + - + + + + + n.a E. faecium 103 KEA + - + + + + + n.a E. faecium 104 KEA + - + + + + + n.a E. faecium 105 KEA + - + + + + + n.a E. faecium 112 BE + - + + + + + n.a E. faecium 113 BE + - + + + + + n.a E. faecium 114 BE + - + + + + + n.a E. faecium 115 BE + - + + + + + n.a ** 116 BE + - + + + + + n.a ** 117 BE + - + + + + + n.a E. faecium 118 BE + - + + + + + n.a E. faecium 119 BE + - + + + + + n.a E. faecium 120 BE + - + + + + + n.a E. faecium
20 lagarto bov. def. 82 BE + - + + + + + E. faecalis E. faecalis 83 BE + - + + + + + Enterococcus sp. E. faecalis
21 Rosbife 87 KEA + - + + + + + E. faecium E. faecium 23 Salsicha + picles 97 KEA + - + + + + + E. faecium E. faecium
98 KEA + - + + + + + n.a E. faecalis 99 KEA + - + + + + + n.a E. faecium
26 Mussarela 129 KEA + - + + + + + Enterococcus sp. E. faecium 130 KEA + - + + + + + n.a E. faecium 131 KEA + - + + + + + n.a E. faecium 132 KEA + - + + + + + n.a E. faecium 137 BE + - + + + + + E. durans ** 138 BE + - + + + + + E. durans E. faecium 139 BE + - + + + + + n.a ** 140 BE + - + + + + + n.a E. faecalis

Continuação
38Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
141 BE + - + + + + + n.a ** 142 BE + - + + + + + n.a E. faecium
27 Salame 143 KEA + - + + + + + Enterococcus sp. E. faecium 144 KEA + - + + + + + n.a E. faecium 146 KEA + - + + + + + n.a E. faecium 147 KEA + - + + + + + n.a ** 154 BE + - + + + + + E. durans **
28 Mortadela 156 KEA + - + + + + + n.a ** 157 KEA + - + + + + + result. não conclusivo E. faecium 158 KEA + - + + + + + n.a E. faecium 163 BE + - + + + + + n.a E. faecium
29 Queijo prato 170 KEA + - + + + + + E. faecium E. faecium 171 KEA + - + + + + + n.a E. faecium 172 KEA + - + + + + + n.a ** 173 KEA + - + + + + + n.a E. faecium 174 KEA + - + + + + + n.a E. faecium
30 Mussarela 175 BE + - + + + + + E. faecium E. faecium 176 BE + - + + + + + n.a ** 177 BE + - + + + + + n.a ** 178 BE + - + + + + + n.a ** 179 BE + - + + + + + E. durans E. faecium 180 BE + - + + + + + E. faecium E. faecium 181 BE + - + + + + + n.a E. faecium 182 BE + - + + + + + n.a E. faecium 183 BE + - + + + + + n.a E. faecium 184 BE + - + + + + + n.a E. faecium 185 BE + - + + + + + n.a E. faecalis 186 BE + - + + + + + n.a E. faecium 187 BE + - + + + + + n.a E. faecium 188 BE + - + + + + + n.a E. faecium 189 KEA + - + + + + + E. durans E. faecium 190 KEA + - + + + + + n.a E. faecium 191 KEA + - + + + + + n.a E. faecium 192 KEA + - + + + + + n.a E. faecium 193 KEA + - + + + + + n.a E. faecium 194 KEA + - + + + + + n.a E. faecium 195 KEA + - + + + + + n.a E. faecium

Continuação
39Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
196 KEA + - + + + + + n.a E. faecium 197 KEA + - + + + + + n.a E. faecium 198 KEA + - + + + + + n.a E. faecium 199 KEA + - + + + + + n.a E. faecium 200 KEA + - + + + + + n.a E. faecium
32 Presunto peru 213 BE + - + + + + + Leuconostoc sp. E. faecium 214 BE + - + + + + + n.a E. faecium 215 BE + - + + + + + n.a E. faecium 217 BE + - + + + + + n.a E. faecium 218 KEA + - + + + + + Leuconostoc sp. E. faecium 219 KEA + - + + + + + n.a E. faecium 225 KEA + - + + + + + n.a E. faecium 227 KEA + - + + + + + n.a E. faecium 228 KEA + - + + + + + n.a E. faecium 232 KEA + - + + + + + n.a E. faecium
33 Lingüiça frango 233 KEA + - + + + + + E. avium E. faecium 234 KEA + - + + + + + baixa discriminação E. casseliflavus 237 KEA + - + + + + + n.a E. faecalis 238 KEA + - + + + + + n.a E faecium
34 Lingüiça de porco 246 KEA + - + + + + + E. avium E. casseliflavus 247 KEA + - + + + + + n.a E. casseliflavus 248 KEA + - + + + + + n.a E. casseliflavus 249 KEA + - + + + + + n.a E. casseliflavus 250 KEA + - + + + + + n.a E. casseliflavus 251 KEA + - + + + + + n.a E. casseliflavus 253 BE + - + + + + + n.a E. casseliflavus 254 BE + - + + + + + n.a E. casseliflavus 256 BE + - + + + + + n.a E. casseliflavus 259 BE + - + + + + + n.a E. casseliflavus
36 Salada repolho/beterraba 272 KEA + - + + + + + Aerococcus viridans E. casseliflavus
38 Moela frango 288 KEA + - + + + + + E. avium E. faecalis 42 Queijo parmesão 309 KEA + - + + + + + n.a E. faecium
310 KEA + - + + + + + n.a E. faecalis 311 BE + - + + + + + n.a E. faecium
43 Presunto 316 KEA + - + + + + + n.a E. faecium 317 KEA + - + + + + + n.a E. faecium

Continuação
40Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
318 KEA + - + + + + + n.a E. faecium 44 Queijo gorgonzola 329 KEA + - + + + + + n.a E. gallinarum 46 Queijo minas 340 KEA + - + + + + + n.a E. faecalis
341 KEA + - + + + + + n.a E. faecalis 49 Bacon 349 BE + - + + + + + n.a E. faecium
352 BE + - + + + + + n.a E. faecium 354 KEA + - + + + + + n.a E. faecium 355 KEA + - + + + + + n.a E. faecium 356 KEA + - + + + + + n.a E. faecium 357 KEA + - + + + + + n.a E. faecium 358 KEA + - + + + + + n.a E. faecium
52 Coxão mole 377 KEA + - + + + + + n.a E. faecalis 55 Queijo Provolone 395 KEA + - + + + + + n.a E. faecalis
396 KEA + - + + + + + n.a E. faecium 397 KEA + - + + + + + n.a E.faecium 398 KEA + - + + + + + n.a ** 399 BE + - + + + + + n.a E. faecalis 400 BE + - + + + + + n.a E. faecalis 401 BE + - + + + + + n.a E. faecium 402 BE + - + + + + + n.a E. faecalis 403 BE + - + + + + + n.a E. faecalis
58 Broto de Feijão 425 KEA + - + + + + + n.a E. casseliflavus 426 KEA + - + + + + + n.a E. casseliflavus 427 KEA + - + + + + + n.a E. casseliflavus 428 KEA + - + + + + + n.a E. casseliflavus 429 KEA + - + + + + + n.a E. casseliflavus 431 KEA + - + + + + + n.a E. casseliflavus 432 KEA + - + + + + + n.a E. casseliflavus 433 KEA + - + + + + + n.a E. casseliflavus 434 KEA + - + + + + + n.a E. casseliflavus 435 KEA + - + + + + + n.a E. casseliflavus 437 KEA + - + + + + + n.a E. casseliflavus 438 KEA + - + + + + + n.a E. casseliflavus
59 Couve manteiga 439 KEA + - + + + + + n.a E. casseliflavus 440 KEA + - + + + + + n.a E. casseliflavus 442 KEA + - + + + + + n.a E. casseliflavus 443 KEA + - + + + + + n.a E. casseliflavus
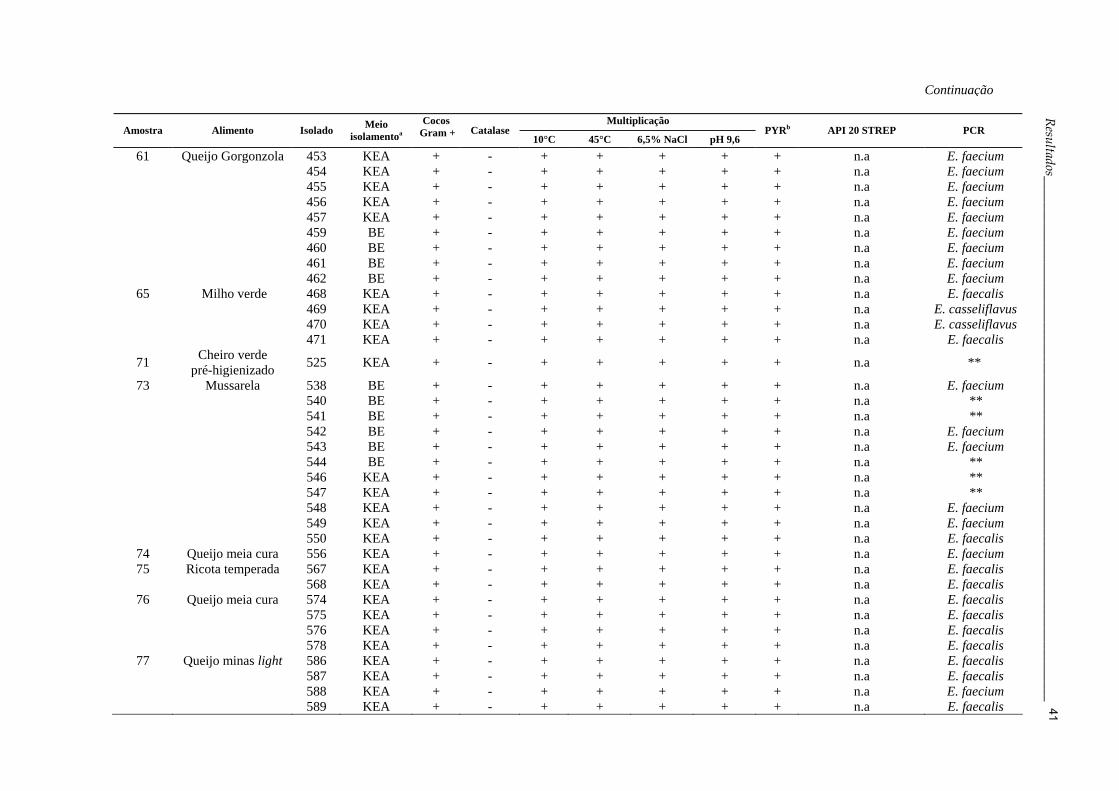
Continuação
41Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
61 Queijo Gorgonzola 453 KEA + - + + + + + n.a E. faecium 454 KEA + - + + + + + n.a E. faecium 455 KEA + - + + + + + n.a E. faecium 456 KEA + - + + + + + n.a E. faecium 457 KEA + - + + + + + n.a E. faecium 459 BE + - + + + + + n.a E. faecium 460 BE + - + + + + + n.a E. faecium 461 BE + - + + + + + n.a E. faecium 462 BE + - + + + + + n.a E. faecium
65 Milho verde 468 KEA + - + + + + + n.a E. faecalis 469 KEA + - + + + + + n.a E. casseliflavus 470 KEA + - + + + + + n.a E. casseliflavus 471 KEA + - + + + + + n.a E. faecalis
71 Cheiro verde pré-higienizado 525 KEA + - + + + + + n.a **
73 Mussarela 538 BE + - + + + + + n.a E. faecium 540 BE + - + + + + + n.a ** 541 BE + - + + + + + n.a ** 542 BE + - + + + + + n.a E. faecium 543 BE + - + + + + + n.a E. faecium 544 BE + - + + + + + n.a ** 546 KEA + - + + + + + n.a ** 547 KEA + - + + + + + n.a ** 548 KEA + - + + + + + n.a E. faecium 549 KEA + - + + + + + n.a E. faecium 550 KEA + - + + + + + n.a E. faecalis
74 Queijo meia cura 556 KEA + - + + + + + n.a E. faecium 75 Ricota temperada 567 KEA + - + + + + + n.a E. faecalis
568 KEA + - + + + + + n.a E. faecalis 76 Queijo meia cura 574 KEA + - + + + + + n.a E. faecalis
575 KEA + - + + + + + n.a E. faecalis 576 KEA + - + + + + + n.a E. faecalis 578 KEA + - + + + + + n.a E. faecalis
77 Queijo minas light 586 KEA + - + + + + + n.a E. faecalis 587 KEA + - + + + + + n.a E. faecalis 588 KEA + - + + + + + n.a E. faecium 589 KEA + - + + + + + n.a E. faecalis

Continuação
42Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
590 KEA + - + + + + + n.a E. faecalis 591 KEA + - + + + + + n.a E. faecalis 595 BE + - + + + + + n.a E. faecalis
78 Mussarela 598 KEA + - + + + + + n.a E. faecalis 599 KEA + - + + + + + n.a E. faecalis 600 KEA + - + + + + + n.a E. faecalis 601 KEA + - + + + + + n.a E.gallinarum 602 KEA + - + + + + + n.a E. faecalis 603 BE + - + + + + + n.a E. faecalis 604 BE + - + + + + + n.a E. faecalis 605 BE + - + + + + + n.a E. faecalis 606 BE + - + + + + + n.a E. faecalis 607 BE + - + + + + + n.a E. faecalis 608 BE + - + + + + + n.a ** 609 BE + - + + + + + n.a E. faecalis
79 Queijo prato 610 KEA + - + + + + + n.a E. faecalis 611 KEA + - + + + + + n.a E. faecalis
80 Queijo meia cura 612 BE + - + + + + + n.a E. faecium 613 BE + - + + + + + n.a E. faecium 614 BE + - + + + + + n.a E. faecium 615 BE + - + + + + + n.a E. faecium 616 BE + - + + + + + n.a E. faecium 617 BE + - + + + + + n.a E. faecium 618 BE + - + + + + + n.a E. faecium 619 BE + - + + + + + n.a E. faecium 620 BE + - + + + + + n.a E. faecium 622 KEA + - + + + + + n.a E. faecium 623 KEA + - + + + + + n.a E. faecium 624 KEA + - + + + + + n.a E. faecium 625 KEA + - + + + + + n.a E. faecium 626 KEA + - + + + + + n.a E. faecium 627 KEA + - + + + + + n.a E. faecium 628 KEA + - + + + + + n.a E. faecium 629 KEA + - + + + + + n.a E. faecium
82 Salsinha 649 BE + - + + + + + n.a E. casseliflavus 88 alface crespa 679 BE + - + + + + + n.a E. casseliflavus
692 KEA + - + + + + + n.a E. casseliflavus

Continuação
43Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
693 KEA + - + + + + + n.a E. casseliflavus 95 Leite tipo A 716 KEA + - + + + + + n.a **
717 KEA + - + + + + + n.a E. casseliflavus 718 KEA + - + + + + + n.a ** 719 KEA + - + + + + + n.a ** 720 KEA + - + + + + + n.a **
96 Leite tipo C 726 KEA + - + + + + + n.a ** 728 KEA + - + + + + + n.a E. faecium 729 KEA + - + + + + + n.a E. faecium
97 Leite cru de vaca 730 BE + - + + + + + n.a E. faecalis 732 KEA + - + + + + + n.a E. faecalis
98 Leite cru de vaca 733 KEA + - + + + + + n.a E. faecalis 734 KEA + - + + + + + n.a E. faecalis 735 KEA + - + + + + + n.a E. faecalis
101 Leite cru de vaca 738 KEA + - + + + + + n.a E. faecalis 739 KEA + - + + + + + n.a E. faecalis 740 KEA + - + + + + + n.a E. faecalis
102 Leite cru de vaca 741 KEA + - + + + + + n.a E. faecalis 742 KEA + - + + + + + n.a E. faecalis 743 KEA + - + + + + + n.a E. faecalis 744 KEA + - + + + + + n.a E. faecalis 745 KEA + - + + + + + n.a E. faecalis 746 KEA + - + + + + + n.a E. casseliflavus
103 Leite cru de vaca 750 KEA + - + + + + + n.a E. faecalis 105 Leite cru de vaca 755 KEA + - + + + + + n.a E. faecalis 106 Leite cru de vaca 756 KEA + - + + + + + n.a E. faecalis
757 KEA + - + + + + + n.a E. faecalis 758 KEA + - + + + + + n.a E. faecalis 759 KEA + - + + + + + n.a E. faecalis 760 KEA + - + + + + + n.a E. faecalis 761 KEA + - + + + + + n.a E. faecalis
107 Leite cru de vaca 763 KEA + - + + + + + n.a E. faecalis 764 KEA + - + + + + + n.a ** 765 KEA + - + + + + + n.a **
108 Leite cru de vaca 768 BE + - + + + + + n.a ** 769 KEA + - + + + + + n.a ** 771 KEA + - + + + + + n.a **

Continuação
44Resultados_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
109 Leite cru de vaca 778 KEA + - + + + + + n.a ** 779 KEA + - + + + + + n.a ** 780 KEA + - + + + + + n.a ** 782 KEA + - + + + + + n.a E. faecalis
112 Leite de cabra 792 KEA + - + + + + + n.a E. gallinarun 794 KEA + - + + + + + n.a E. gallinarum 798 KEA + - + + + + + n.a E. gallinarum
115 Leite cru de vaca 806 KEA + - + + + + + n.a ** 116 Leite cru de vaca 808 KEA + - + + + + + n.a E. faecalis 120 Leite cru de vaca 812 KEA + - + + + + + n.a E. faecium
813 KEA + - + + + + + n.a E. faecalis (+) positivo, (-) negativo. a BE: ágar Bile Esculina, KEA: Kanamicina Esculina Azida n.a: não avaliado até o momento b hidrólise de pirrolidonil –ß – naftilamida (PYR) ** PCR não conclusivo com os primers para E. faecalis, E. faecium, E. gallinarum e E. casseliflavus
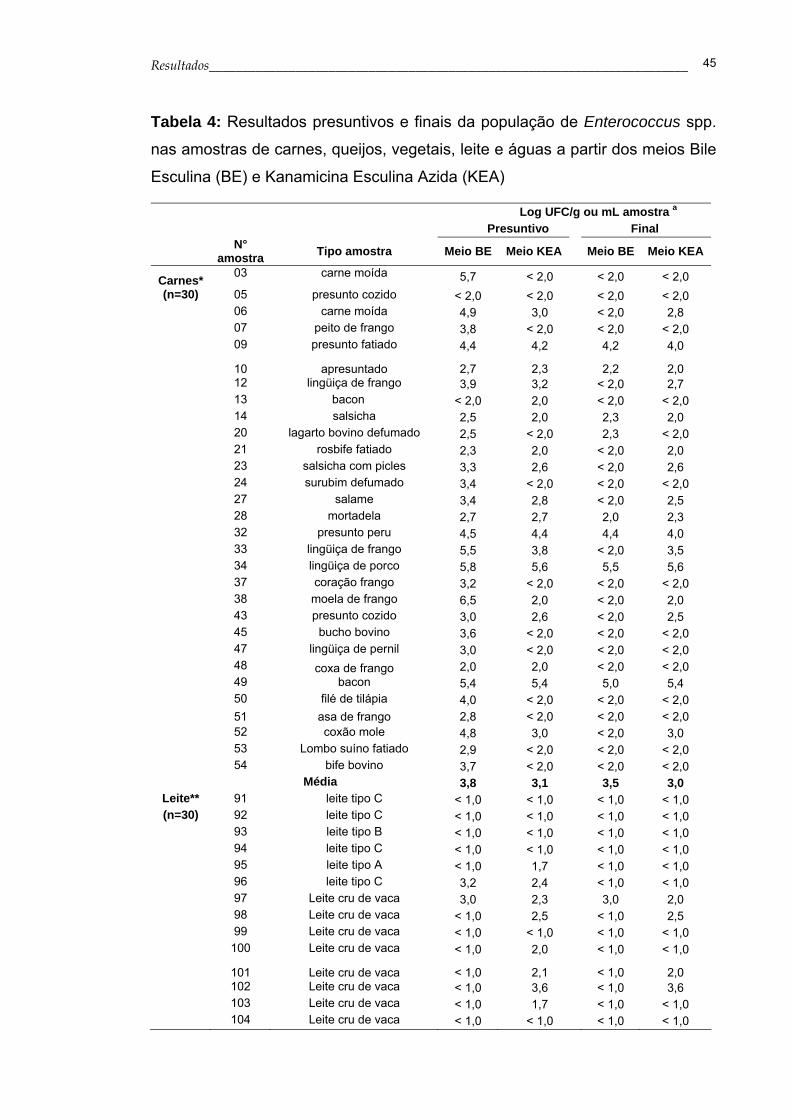
Resultados________________________________________________________________________
45
Tabela 4: Resultados presuntivos e finais da população de Enterococcus spp.
nas amostras de carnes, queijos, vegetais, leite e águas a partir dos meios Bile
Esculina (BE) e Kanamicina Esculina Azida (KEA)
Log UFC/g ou mL amostra a Presuntivo Final
N° amostra Tipo amostra Meio BE Meio KEA Meio BE Meio KEA
Carnes* 03 carne moída 5,7 < 2,0 < 2,0 < 2,0 (n=30) 05 presunto cozido < 2,0 < 2,0 < 2,0 < 2,0
06 carne moída 4,9 3,0 < 2,0 2,8 07 peito de frango 3,8 < 2,0 < 2,0 < 2,0 09 presunto fatiado 4,4 4,2 4,2 4,0
10 apresuntado 2,7 2,3 2,2 2,0 12 lingüiça de frango 3,9 3,2 < 2,0 2,7 13 bacon < 2,0 2,0 < 2,0 < 2,0 14 salsicha 2,5 2,0 2,3 2,0 20 lagarto bovino defumado 2,5 < 2,0 2,3 < 2,0 21 rosbife fatiado 2,3 2,0 < 2,0 2,0 23 salsicha com picles 3,3 2,6 < 2,0 2,6 24 surubim defumado 3,4 < 2,0 < 2,0 < 2,0 27 salame 3,4 2,8 < 2,0 2,5 28 mortadela 2,7 2,7 2,0 2,3 32 presunto peru 4,5 4,4 4,4 4,0 33 lingüiça de frango 5,5 3,8 < 2,0 3,5 34 lingüiça de porco 5,8 5,6 5,5 5,6 37 coração frango 3,2 < 2,0 < 2,0 < 2,0 38 moela de frango 6,5 2,0 < 2,0 2,0 43 presunto cozido 3,0 2,6 < 2,0 2,5 45 bucho bovino 3,6 < 2,0 < 2,0 < 2,0 47 lingüiça de pernil 3,0 < 2,0 < 2,0 < 2,0 48 coxa de frango 2,0 2,0 < 2,0 < 2,0 49 bacon 5,4 5,4 5,0 5,4 50 filé de tilápia 4,0 < 2,0 < 2,0 < 2,0 51 asa de frango 2,8 < 2,0 < 2,0 < 2,0 52 coxão mole 4,8 3,0 < 2,0 3,0 53 Lombo suíno fatiado 2,9 < 2,0 < 2,0 < 2,0 54 bife bovino 3,7 < 2,0 < 2,0 < 2,0
Média 3,8 3,1 3,5 3,0 Leite** 91 leite tipo C < 1,0 < 1,0 < 1,0 < 1,0 (n=30) 92 leite tipo C < 1,0 < 1,0 < 1,0 < 1,0
93 leite tipo B < 1,0 < 1,0 < 1,0 < 1,0 94 leite tipo C < 1,0 < 1,0 < 1,0 < 1,0 95 leite tipo A < 1,0 1,7 < 1,0 < 1,0 96 leite tipo C 3,2 2,4 < 1,0 < 1,0 97 Leite cru de vaca 3,0 2,3 3,0 2,0 98 Leite cru de vaca < 1,0 2,5 < 1,0 2,5 99 Leite cru de vaca < 1,0 < 1,0 < 1,0 < 1,0 100 Leite cru de vaca < 1,0 2,0 < 1,0 < 1,0
101 Leite cru de vaca < 1,0 2,1 < 1,0 2,0 102 Leite cru de vaca < 1,0 3,6 < 1,0 3,6 103 Leite cru de vaca < 1,0 1,7 < 1,0 < 1,0 104 Leite cru de vaca < 1,0 < 1,0 < 1,0 < 1,0

Resultados________________________________________________________________________
46
Log UFC/g ou mL amostra a Presuntivo Final
N° amostra Tipo amostra Meio BE Meio KEA Meio BE Meio KEA
105 Leite cru de vaca < 1,0 2,2 < 1,0 < 1,0 106 Leite cru de vaca < 1,0 2,7 < 1,0 2,3 107 Leite cru de vaca < 1,0 2,7 < 1,0 < 1,0 108 Leite cru de vaca 4,0 1,8 < 1,0 < 1,0 109 Leite cru de vaca 5,0 3,4 < 1,0 2,7 110 leite tipo B 1,0 < 1,0 < 1,0 < 1,0 111 leite tipo C 2,4 < 1,0 < 1,0 < 1,0 112 leite cabra 2,5 2,1 < 1,0 1,5 113 leite tipo C < 1,0 < 1,0 < 1,0 < 1,0 114 leite tipo C < 1,0 < 1,0 < 1,0 < 1,0 115 Leite cru de vaca < 1,0 3,3 < 1,0 < 1,0 116 Leite cru de vaca 3,0 2,0 < 1,0 2,0 117 Leite cru de vaca < 1,0 < 1,0 < 1,0 < 1,0 118 Leite cru de vaca < 1,0 3,3 < 1,0 < 1,0 119 Leite cru de vaca < 1,0 3,0 < 1,0 < 1,0 120 Leite cru de vaca < 1,0 3,3 < 1,0 3,3
Média 3,0 2,5 3,0 2,4 Queijos* 01 mussarela 6,7 6,6 6,0 6,0 (n=30) 02 queijo minas 8,4 7,8 7,9 7,8
04 queijo minas 8,5 6,8 < 2,0 6,4 08 mussarela 5,8 5,7 5,8 5,7 11 ricota 7,2 6,0 6,7 5,5 15 queijo prato 4,6 4,2 4,4 4,2 16 queijo minas frescal 6,8 6,3 6,6 6,3 17 queijo gouda 5,5 2,6 <2,0 2,4 18 mussarela 5,9 5,9 5,8 5,6 25 queijo árabe 2,5 < 2,0 < 2,0 < 2,0 26 mussarela 5,5 5,6 5,2 5,3 29 queijo prato 4,5 3,3 < 2,0 3,2 30 mussarela 6,3 6,2 6,2 6,2 31 queijo prato 7,5 7,5 < 2,0 < 2,0 42 queijo parmesão 2,0 2,3 2,0 2,3 44 queijo gorgonzola 4,5 3,1 < 2,0 2,5 46 queijo minas 6,2 5,3 < 2,0 5,0 55 queijo provolone 2,7 2,6 2,7 2,5 56 ricota 5,7 7,0 < 2,0 < 2,0 57 queijo cheedar < 2,0 < 2,0 < 2,0 < 2,0 61 queijo gorgonzola 5,4 4,4 4,3 4,4 62 mussarela < 2,0 < 2,0 < 2,0 < 2,0 73 mussarela 5,9 5,4 5,4 5,4 74 queijo meia cura 6,4 4,4 < 2,0 3,7 75 ricota temperada 7,3 6,3 < 2,0 5,8 76 queijo meia cura 6,2 4,9 < 2,0 4,6 77 queijo minas frescal light 6,8 5,6 6,0 5,5 78 mussarela 4,7 3,5 4,6 3,5 79 queijo prato < 2,0 2,3 < 2,0 2,3 80 queijo meia cura 8,0 7,8 7,9 7,8
Média 5,8 5,2 5,5 4,8 Vegetais*
(n=30) 19 seleta de legumes < 2,0 < 2,0 < 2,0 < 2,0
22 cenoura ralada 6,7 6,0 < 2,0 < 2,0 35 repolho/cenoura/escarola 5,6 4,4 < 2,0 < 2,0
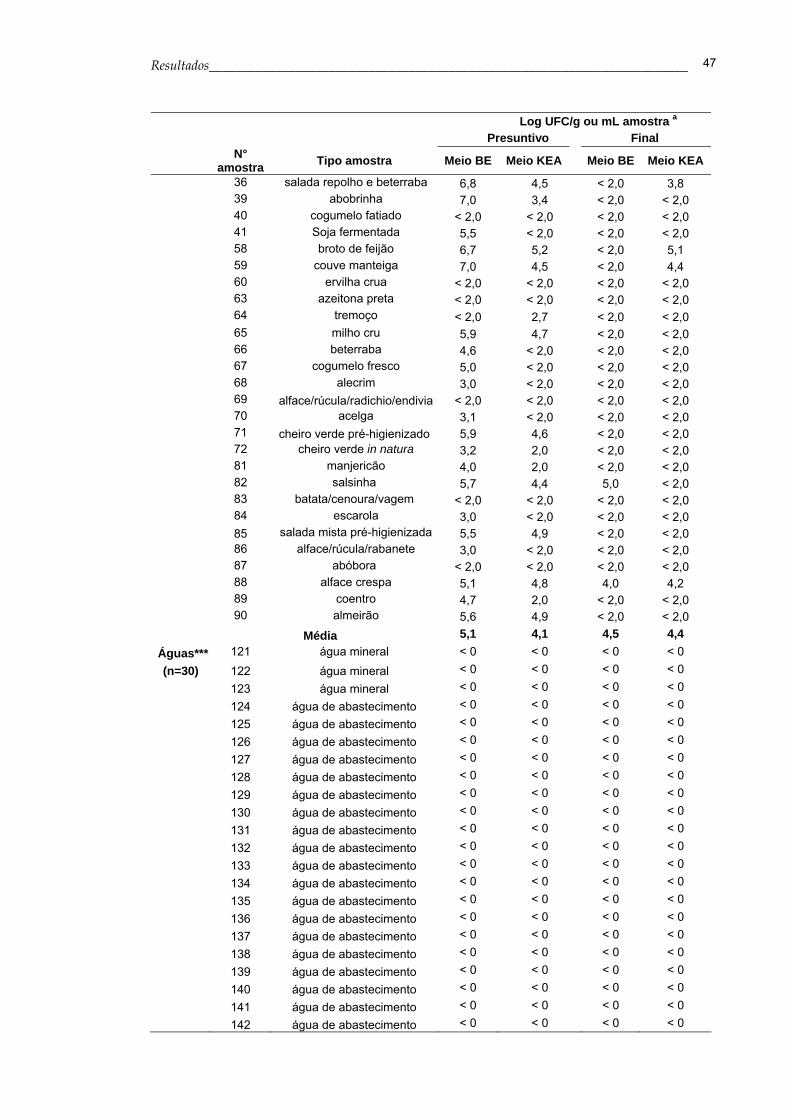
Resultados________________________________________________________________________
47
Log UFC/g ou mL amostra a Presuntivo Final
N° amostra Tipo amostra Meio BE Meio KEA Meio BE Meio KEA
36 salada repolho e beterraba 6,8 4,5 < 2,0 3,8 39 abobrinha 7,0 3,4 < 2,0 < 2,0 40 cogumelo fatiado < 2,0 < 2,0 < 2,0 < 2,0 41 Soja fermentada 5,5 < 2,0 < 2,0 < 2,0 58 broto de feijão 6,7 5,2 < 2,0 5,1 59 couve manteiga 7,0 4,5 < 2,0 4,4 60 ervilha crua < 2,0 < 2,0 < 2,0 < 2,0 63 azeitona preta < 2,0 < 2,0 < 2,0 < 2,0 64 tremoço < 2,0 2,7 < 2,0 < 2,0 65 milho cru 5,9 4,7 < 2,0 < 2,0 66 beterraba 4,6 < 2,0 < 2,0 < 2,0 67 cogumelo fresco 5,0 < 2,0 < 2,0 < 2,0 68 alecrim 3,0 < 2,0 < 2,0 < 2,0 69 alface/rúcula/radichio/endivia < 2,0 < 2,0 < 2,0 < 2,0 70 acelga 3,1 < 2,0 < 2,0 < 2,0 71 cheiro verde pré-higienizado 5,9 4,6 < 2,0 < 2,0 72 cheiro verde in natura 3,2 2,0 < 2,0 < 2,0 81 manjericão 4,0 2,0 < 2,0 < 2,0 82 salsinha 5,7 4,4 5,0 < 2,0 83 batata/cenoura/vagem < 2,0 < 2,0 < 2,0 < 2,0 84 escarola 3,0 < 2,0 < 2,0 < 2,0 85 salada mista pré-higienizada 5,5 4,9 < 2,0 < 2,0 86 alface/rúcula/rabanete 3,0 < 2,0 < 2,0 < 2,0 87 abóbora < 2,0 < 2,0 < 2,0 < 2,0 88 alface crespa 5,1 4,8 4,0 4,2 89 coentro 4,7 2,0 < 2,0 < 2,0 90 almeirão 5,6 4,9 < 2,0 < 2,0
Média 5,1 4,1 4,5 4,4 Águas*** 121 água mineral < 0 < 0 < 0 < 0 (n=30) 122 água mineral < 0 < 0 < 0 < 0
123 água mineral < 0 < 0 < 0 < 0 124 água de abastecimento < 0 < 0 < 0 < 0 125 água de abastecimento < 0 < 0 < 0 < 0 126 água de abastecimento < 0 < 0 < 0 < 0 127 água de abastecimento < 0 < 0 < 0 < 0 128 água de abastecimento < 0 < 0 < 0 < 0 129 água de abastecimento < 0 < 0 < 0 < 0 130 água de abastecimento < 0 < 0 < 0 < 0 131 água de abastecimento < 0 < 0 < 0 < 0 132 água de abastecimento < 0 < 0 < 0 < 0 133 água de abastecimento < 0 < 0 < 0 < 0 134 água de abastecimento < 0 < 0 < 0 < 0 135 água de abastecimento < 0 < 0 < 0 < 0 136 água de abastecimento < 0 < 0 < 0 < 0 137 água de abastecimento < 0 < 0 < 0 < 0 138 água de abastecimento < 0 < 0 < 0 < 0 139 água de abastecimento < 0 < 0 < 0 < 0 140 água de abastecimento < 0 < 0 < 0 < 0 141 água de abastecimento < 0 < 0 < 0 < 0 142 água de abastecimento < 0 < 0 < 0 < 0

Resultados________________________________________________________________________
48
Log UFC/g ou mL amostra a Presuntivo Final
N° amostra Tipo amostra Meio BE Meio KEA Meio BE Meio KEA
143 água de abastecimento < 0 < 0 < 0 < 0 144 água de abastecimento < 0 < 0 < 0 < 0 145 água de abastecimento < 0 < 0 < 0 < 0 146 água de abastecimento < 0 < 0 < 0 < 0 147 água mineral < 0 < 0 < 0 < 0 148 água mineral < 0 < 0 < 0 < 0 149 água de torneira < 0 < 0 < 0 < 0 150 água de bebedouro < 0 < 0 < 0 < 0
Médiaa n.d n.d n.d n.d
*Os valores referentes às contagens de colônias indicativas do gênero Enterococcus nas amostras de produtos cárneos, queijos e vegetais, representadas pelo valor <2 (inferior a 2 log UFC/g amostra) são valores estimados já que encontravam-se abaixo do limite detecção do método que é de 100 UFC/g de amostra ou 2 log UFC/g. **Os valores referente às contagens de colônias indicativas do gênero Enterococcus nas amostras de leite, que encontravam-se abaixo do limite detecção do método (10 UFC/mL de amostra ou 1 log UFC/mL), são valores estimados e estão representadas por <1 (inferior a 1log UFC/mL amostra). ***Os valores referentes às contagens de colônias indicativas do gênero Enterococcus nas amostras de águas que se encontravam abaixo do limite detecção do método que é de 1 UFC/ 100 mL de amostra ou 0 log UFC/ 100 mL, são valores etimados e estão representadas por <0. a n.d: não determinado pois valores estimados não foram utilizados para obtenção da média.
A avaliação dos dados obtidos (Tabela 5) revela que, de forma geral,
apesar de terem sido isoladas mais colônias características do gênero
Enterococcus no meio BE, a especificidade não foi muito alta, pois muitos
isolados não foram posteriormente confirmados como Enterococcus spp. A
taxa de confirmação de Enterococcus spp. a partir da identificação dos
isolados provenientes do meio KEA foi maior quando comparado ao BE
(Tabela 5).
Para as amostras de produtos cárneos apenas 14,8% dos 128 isolados
obtidos no meio BE foram presuntivamente classificados como membros do
gênero Enterococcus, enquanto que 64,3% dos 70 isolados obtidos no meio
KEA foram presuntivamente classificados como Enterococcus spp. Em relação
ao número de amostras positivas observa-se que em 90% das 30 amostras de
queijo analisadas houve crescimento característico de colônias de enterococos
nos dois meios empregados. Da mesma forma que no caso anterior, a

Resultados________________________________________________________________________
49
confirmação presuntiva dos isolados inicialmente suspeitos de Enterococcus
spp. a partir do ágar BE (51,4%) foi inferior ao número confirmado a partir do
ágar KEA (63,4%).
Para os leites analisados, de forma geral, foi obtido um menor número
de amostras positivas nos dois meio de cultura em comparação com as
amostras anteriores. Das 30 amostras deste alimento, 19 (63,3%) forneceram
colônias indicativas do gênero Enterococcus spp. no meio KEA, enquanto que
no meio BE apenas 8 (26,6%) foram positivas. Das colônias confirmadas como
Enterococcus spp. foram obtidos 45 isolados (57,7%) a partir de colônias
características no meio KEA e 2 isolados (10%) dos 20 obtidos no meio BE.
Para as amostras de vegetais nota-se que nos meios KEA e BE foram
obtidas 53,3% e 73,3%, respectivamente, de amostras com colônias
características de enterococos, sendo que 24,2% e 1,4% dos isolados obtidos
a partir de KEA e BE foram confirmados como Enterococcus spp.
Com a identificação dos isolados por PCR, a quantidade de amostras
positivas obtidas a partir da utilização do ágar KEA foi reduzida a 16 (53,3%)
para as amostras de produtos cárneos, 25 (83,3%) das amostras de queijos, 4
(13,3%) das amostras de vegetais, e 9 (30,0%) das amostras de leite. Quando
as amostras foram semeadas em ágar BE estes valores foram reduzidos a 8
(26,6%), 16 (53,3%), 2(6,6%) e 2(6,6%), respectivamente (Tabela 4).
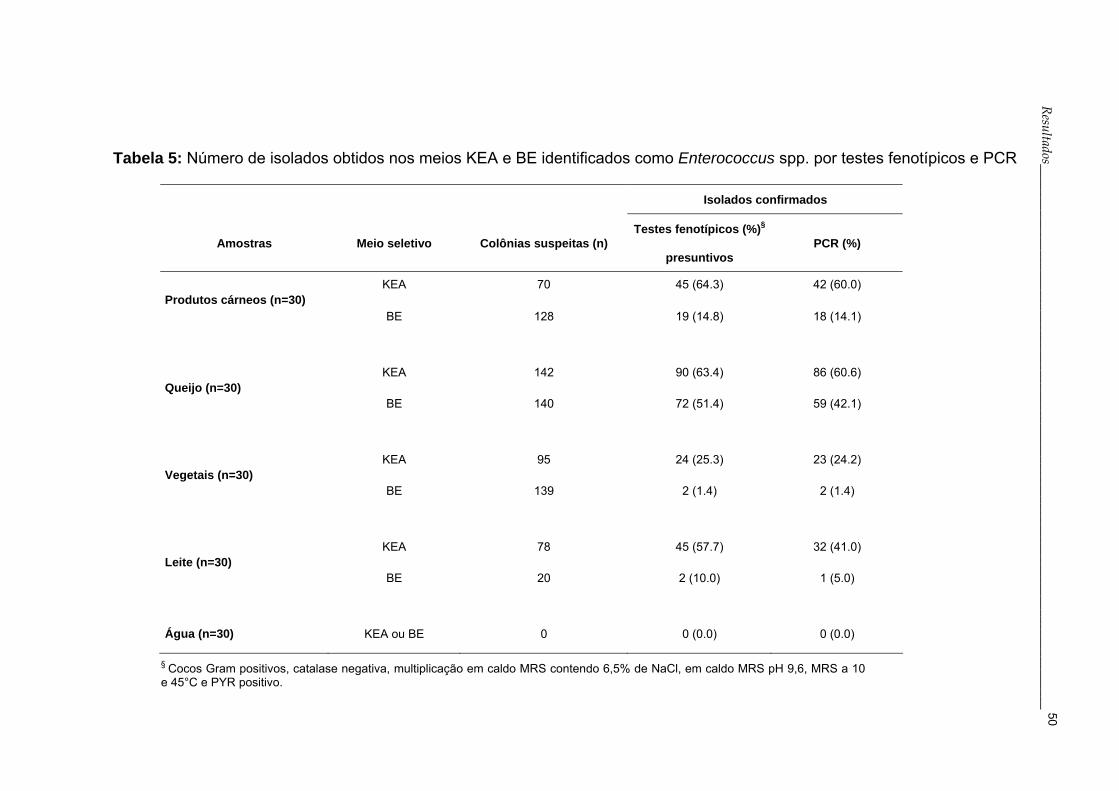
50Resultados__________________________________________________________________________________
Tabela 5: Número de isolados obtidos nos meios KEA e BE identificados como Enterococcus spp. por testes fenotípicos e PCR
Isolados confirmados
Amostras Meio seletivo Colônias suspeitas (n) Testes fenotípicos (%)§
presuntivos PCR (%)
KEA 70 45 (64.3) 42 (60.0) Produtos cárneos (n=30)
BE 128 19 (14.8) 18 (14.1)
KEA 142 90 (63.4) 86 (60.6) Queijo (n=30)
BE 140 72 (51.4) 59 (42.1)
KEA 95 24 (25.3) 23 (24.2) Vegetais (n=30)
BE 139 2 (1.4) 2 (1.4)
KEA 78 45 (57.7) 32 (41.0) Leite (n=30)
BE 20 2 (10.0) 1 (5.0)
Água (n=30) KEA ou BE 0 0 (0.0) 0 (0.0)
§ Cocos Gram positivos, catalase negativa, multiplicação em caldo MRS contendo 6,5% de NaCl, em caldo MRS pH 9,6, MRS a 10 e 45°C e PYR positivo.

Resultados________________________________________________________________________
51
4.2 Identificação de espécies de Enterococcus spp.
Das 812 colônias com morfologia indicativa do gênero enterococos nos
meios Bile Esculina (BE) e Kanamicina Esculina Azida (KEA), foram obtidos
299 isolados (37%) caracterizados bioquimicamente como pertencentes ao
gênero Enterococcus spp. Dentre estes isolados foram obtidos, após
identificação por PCR, 139 E. faecium (46,5%), 80 E. faecalis (26,8%), 36 E.
casseliflavus (12%) e 8 E. gallinarum (2,7%), sendo que 36 isolados (12%) não
foram identificados com os primers disponíveis para as espécies citadas. A
figura 2 ilustra o perfil obtido com os primers específicos para as espécies de
E. faecalis, E. faecium, E. casseliflavus e E. gallinarum.
Quanto à distribuição das espécies de enterococos nos grupos de
alimentos analisados, foi observado que dos 139 isolados de E. faecium, 71,2%
foram obtidos a partir de amostras de queijo, 26,6% a partir de produtos
cárneos e 2,2% a partir das amostras de leite. Em relação à origem dos 80
isolados de E. faecalis a distribuição obtida foi de 51,3% provenientes de
amostras de queijos, 31,2% de leite, 15% de produtos cárneos e 2,5% de
amostras de vegetais. Os isolados de E. casseliflavus foram obtidos na grande
maioria (64%) a partir de amostras de vegetais, 30,5% de amostras de
produtos cárneos e 5,5% a partir de amostras de queijo. Cinco isolados de E.
gallinarum (62,5%) foram obtidos de queijos e os três restantes (37,5%) de
amostras de leite (Tabela 6).

Resultados________________________________________________________________________
52
A
B
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
500
100
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
500 _
Figura 2: Foto ilustrativa de géis de agarose com os produtos de PCR para identificação das espécies de Enterococcus obtidos a partir de amostras de alimentos. A) Canaleta M: marcador de 100 pb; 1: controle positivo de E. faecium NCTC 7171 (550pb), 2: controle positivo de E. faecalis NCTC 775 (941pb), 3: controle positivo de E. gallinarum NCTC 12359 (822pb), 4: controle positivo de E. casseliflavus NCTC 12361 (439pb), 5 a 15: cepas provenientes da amostra 30 de mussarela identificadas como E. faecium. B) Canaleta M: marcador de 100 pb; 1: controle positivo de E. gallinarum NCTC 12359 (822pb), 2: controle positivo de E. casseliflavus NCTC 12361 (439pb), 3: controle positivo de E. faecium NCTC 7171 (550pb), 4: controle positivo de E. faecalis NCTC 775 (941pb), 5, 7, 13, 14 e 15: E. faecium isolado de amostras de mussarela e presunto (amostras 1, 8 e 9), 6: cepa não identificada com os primers disponíveis, 8, 9, 10 e 12: E. faecalis proveniente de amostras de queijo Minas e carne moída (amostras 2 e 6), 11: E. gallinarum isolado a partir da amostra 4 de queijo Minas.

Resultados________________________________________________________________________
53
Tabela 6: Distribuição das espécies dos 263 isolados de Enterococcus spp.
obtidos a partir de amostras de alimentos e identificados por PCR
Espécies Produtos Cárneos*
Queijos* Vegetais* Leites* Total
E. faecium 37 (26,6) 99 (71,2) 0 (0) 3 (2,2) 139
E. faecalis 12 (15) 41 (51,3) 2 (2,5) 25 (31,2) 80
E. gallinarum 0 (0) 5 (62,5) 0 (0) 3 (37,5) 8
E. casseliflavus 11 (30,5) 0 (0) 23 (64) 2 (5,5) 36
* O número entre parênteses indica a porcentagem da espécie isolada naquele grupo de alimento.
A Tabela 7 mostra o perfil das espécies que seria obtido, considerando a
identificação dos isolados com o sistema API 20 STREP comparando-a com
resultados de identificação por PCR. Dos 52 isolados avaliados, seis
bioquimicamente identificados como E. faecalis e 5 como E. faecium tiveram
sua identificação confirmada por PCR.
Foi observado que um total de 41 isolados (78,8%) foram erroneamente
identificados pelo API 20 STREP. Dentre esses isolados, 10 não tiveram o
perfil identificado, e posteriormente foram classificados por PCR como E.
faecalis (3), E. faecium (5) e E. casseliflavus (1).
Além disso, sete isolados que foram classificados pelo API 20 STREP
como pertencentes ao gênero Enterococcus, sem resultado conclusivo para
identificação de espécie, foram re-agrupados dentro das espécies de E.
faecium (6) e E. faecalis (1).

54Resultados_________________________________________________________________________________
Tabela 7: Comparação da identificação de Enterococcus spp. por PCR e API 20 STREP (Biomérieux, França)
API 20 STREP
PCR n E. faecalis E. faecium E. durans E. avium Enterococcus spp. Aerococcus viridans Leuconostoc sp. Não identificado
E. faecalis 13 6 3 1 3
E. faecium 30 5 8 1 6 5 5
E. gallinarum 2 2
E. casseliflavus 3 1 1 1
Não identificado 4 3 1
Total 52 6 5 11 5 7 3 5 10

Resultados________________________________________________________________________
55
4.3 Caracterização fenotípica de Enterococcus spp.
Na Tabela 8 estão apresentados os resultados do perfil fenotípico de 80
isolados de E. faecalis, 139 isolados de E. faecium, 8 isolados de E. gallinarum
e 36 isolados de E. casseliflavus, obtidos de amostras de produtos cárneos,
queijos, vegetais e leite. Observa-se que dos 80 isolados de E. faecalis
analisados, 48 (60%) produziram gelatinase. As demais espécies não
apresentaram resultado positivo para avaliação fenotípica de produção de
gelatinase.
Quanto à produção de bacteriocina, foi observado que dos 80 isolados
de E. faecalis analisados, oito (10%) mostraram atividade inibitória frente a L.
sakei ATCC 15521 e/ou frente a L. monocytogenes ATCC 19115. Seis isolados
de E. faecalis que produziram bacteriocina também apresentaram resultados
positivos para produção de gelatinase.
Entre os isolados de E. faecium analisados não foi obtido resultado
positivo para produção de gelatinase. Entretanto, 32 (23%) mostraram
atividade inibitória frente a L. sakei e/ou L. monocytogenes. Para os isolados de
E. faecium não foi observada uma relação entre a produção de bacteriocina e
de gelatinase. Da mesma forma dentre os isolados de E. gallinarum e E.
casseliflavus não foi detectada atividade bacteriocinogênica e produção de
gelatinase.
A pesquisa da atividade hemolítica revelou que dos 263 isolados de
Enterococcus spp. (80 E. faecalis, 139 E. faecium, 8 E. gallinarum e 36 E.
casseliflavus) 81% apresentaram hemólise parcial (tipo alfa), 12%
apresentaram atividade hemolítica total (tipo beta) e 7% não apresentaram

Resultados________________________________________________________________________
56
atividade hemolítica (tipo gama). O perfil β-hemolítico foi observado em um
isolado de E. faecium (0,7%) e em 31 (38,7%) isolados de E. faecalis. Dentre
os isolados de E. faecalis, um apresentou o perfil β-hemolítico/bateriocina
indicativo da presença de citolisina.
Foram obtidos 178 isolados (67,7%) de Enterococcus spp. (52 E.
faecalis, 87 E. faecium, 31 E. casseliflavus e 8 E. gallinarum) com capacidade
de hidrolisar sais biliares. De acordo com a classificação proposta por Franz et
al. (2001b) 60,9% dos isolados acima demonstraram uma baixa capacidade de
hidrolisar sais biliares (BSH), 6,0% uma média capacidade de hidrolisar sais
biliares, 0,8% uma alta capacidade de hidrolisar sais biliares e 32,3% não
demonstraram atividade de hidrólise de sais biliares. A produção de BSH,
analisada segundo a espécie de enterococos, foi de 65% para E. faecalis,
62,6% para E. faecium, 86% para E. casseliflavus e 100% para E. gallinarum.
Em alguns casos, isolados obtidos de uma mesma amostra de alimento
apresentaram perfil fenotípico diverso. Como exemplo, podem ser citados os
isolados de E. faecalis 598, 602, 603 e o isolado de E. gallinarum 601, obtidos
da mesma amostra de queijo tipo mussarela, que apresentaram resultados
diversos em relação à presença de alguns marcadores fenotípicos de
virulência.

57Resultados_________________________________________________________________________________
Tabela 8: Perfil de Enterococcus spp. quanto à capacidade de hidrolisar sais biliares, presença fenotípica dos marcadores
de virulência (β-hemólise e gelatinase) e atividade inibitória frente a L. sakei ATCC 15521 e/ou L. monocytogenes ATCC
19115
Hemólise§ Hidrólise de sais biliares Espécies Alimentos
Nº de
isolados Bac+ * Gel+*
α β γ Ausente Baixa Média Alta
E. faecalis Produtos
cárneos 12 3 9 9 0 3 10 2 0 0
Queijo 41 4 19 32 7 2 14 27 0 0
Vegetais 2 0 2 0 2 0 2 0 0 0
Leite 25 1 18 3 22 0 2 23 0 0
Total 80 8 48 44 31 5 28 52 0 0
E. faecium Produtos
cárneos 37 6 0 30 0 7 14 23 0 0
Queijo 99 26 0 93 0 6 36 62 1 0
Leite 3 0 0 2 1 0 2 1 0 0
Total 139 32 0 125 1 13 52 86 1 0
E. casseliflavus Produtos
cárneos 11 0 0 10 0 1 3 6 2 0
Vegetais 23 0 0 23 0 0 1 9 11 2
Leite 2 0 0 2 0 0 1 1 0 0
Total 36 0 0 35 0 1 5 16 13 2
E. gallinarum Queijo 5 0 0 5 0 0 0 5 0 0
Leite 3 0 0 3 0 0 0 1 2 0
Total 8 0 0 8 0 0 0 6 2 0 ∗ Bac+: atividade bacteriocinogênica, Gel+: producão gelatinase § α= hemólise parcial, β= hemólise total, γ= ausência de hemólise

Resultados________________________________________________________________________
58
4.4 Determinação da resistência a antibióticos
O perfil de resistência dos 139 isolados de E. faecium e 80 de E. faecalis
foi analisado frente a cinco antibióticos de uso clínico: vancomicina, ampicilina,
tetraciclina, eritromicina e gentamicina. De acordo com os dados apresentados
nas Tabelas 9, 10 e 11 nenhum dos isolados de E. faecium e E. faecalis
mostraram resistência à ampicilina. O mesmo resultado foi obtido frente à alta
concentração de gentamicina, exceto para os isolados de E. faecalis obtidos a
partir de amostras de leite. Neste caso, foram obtidos 72% de isolados
resistentes a alta concentração de gentamicina (500µg/mL). Dentre os isolados
de E. faecalis não foi observada resistência à vancomicina enquanto que para
E. faecium obtidos a partir de amostras de queijos e produtos cárneos foram
obtidos, respectivamente, 2% e 3% dos isolados resistentes a este antibiótico.
Não foi observada resistência à vancomicina para E. faecium obtido de
amostras de leite (Figuras 3 e 4).
Para os isolados de E. faecium obtidos a partir de amostras de produtos
cárneos e leite foram obtidas, respectivamente, 22% e 33% de cepas
resistentes à tetraciclina. Entretanto, dentre os isolados de E. faecium obtidos a
partir de queijo não foi observada resistência a este antibiótico. Para E. faecalis
obtidos a partir de amostras de queijo, produtos cárneos e leite, a resistência a
este antibiótico foi de, respectivamente, 15%, 8% e 72%. Dentre os isolados de
E. faecalis obtidos a partir de amostras de vegetais não foi encontrada
resistência à tetraciclina. Todas os isolados de E. faecalis resistentes à
gentamicina também apresentaram resistência à tetraciclina.
A resistência à eritromicina dentre os isolados de E. faecium obtidos a
partir de amostras de queijo, produtos cárneos e leite foi de respectivamente,

Resultados________________________________________________________________________
59
10%, 8% e 0%. Para os isolados de E. faecalis obtidos de amostras de queijo e
leite foi obtido o mesmo número de resistentes à eritromicina (12%). Não foi
observada resistência a este antibiótico para os isolados de E. faecalis obtidos
a partir de produtos cárneos e vegetais.
Nenhum isolado de E. faecalis e E. faecium apresentou resistência a
todos os antibióticos testados, mas 36,2% de E. faecalis e 16,5% de E. faecium
apresentaram resistência a pelo menos um dos antibióticos testados.
Os isolados que apresentaram perfil de resistência aos antibióticos
anteriores foram submetidos à determinação da Concentração Inibitória Mínima
(CIM) e os resultados obtidos estão apresentados na Tabela 12.
Os resultados obtidos de CIM confirmaram a triagem em ágar BHI
adicionado de antibióticos. Todos os isolados resistentes à vancomicina
apresentaram uma CIM ≥ 256 µg/mL e eram sensíveis aos outros antibióticos.
A CIM apresentada por todos os isolados resistentes à gentamicina foi superior
a 500 µg/mL, caracterizando o perfil de elevada resistência. A CIM para
eritromicina variou de 16 µg/mL a valores superiores a 256 µg/mL enquanto
que a CIM para tetraciclina variou de 32 µg/mL a 256 µg/mL (Tabela 12).
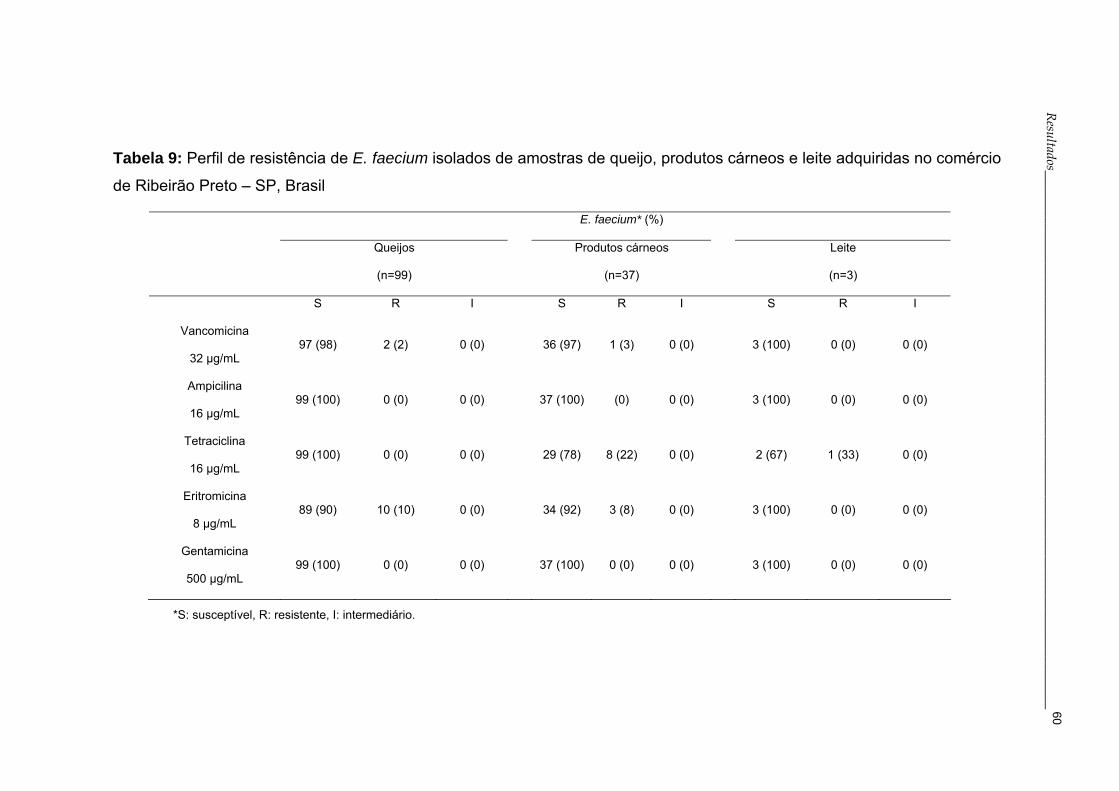
60Resultados_________________________________________________________________________________
Tabela 9: Perfil de resistência de E. faecium isolados de amostras de queijo, produtos cárneos e leite adquiridas no comércio
de Ribeirão Preto – SP, Brasil
E. faecium* (%)
Queijos
(n=99)
Produtos cárneos
(n=37)
Leite
(n=3)
S R I S R I S R I
Vancomicina
32 µg/mL 97 (98) 2 (2) 0 (0)
36 (97) 1 (3) 0 (0)
3 (100) 0 (0) 0 (0)
Ampicilina
16 µg/mL 99 (100) 0 (0) 0 (0)
37 (100) (0) 0 (0)
3 (100) 0 (0) 0 (0)
Tetraciclina
16 µg/mL 99 (100) 0 (0) 0 (0)
29 (78) 8 (22) 0 (0)
2 (67) 1 (33) 0 (0)
Eritromicina
8 µg/mL 89 (90) 10 (10) 0 (0)
34 (92) 3 (8) 0 (0)
3 (100) 0 (0) 0 (0)
Gentamicina
500 µg/mL 99 (100) 0 (0) 0 (0)
37 (100) 0 (0) 0 (0)
3 (100) 0 (0) 0 (0)
*S: susceptível, R: resistente, I: intermediário.

61Resultados_________________________________________________________________________________
Tabela 10: Perfil de resistência de E. faecalis isolados de amostras de queijos, produtos cárneos, leite e vegetais adquiridas
no comércio de Ribeirão Preto – SP, Brasil
E. faecalis* (%)
Queijos
(n=41)
Produtos cárneos
(n=12)
Leite
(n=25)
Vegetais
(n=2)
S R I S R I S R I S R I
Vancomicina
32 µg/mL 41 (100) 0 (0) 0 (0) 12 (100) 0 (0) 0 (0) 25 (100) 0 (0) 0 (0) 2 (100) 0 (0) 0 (0)
Ampicilina
16 µg/mL 41 (100) 0 (0) 0 (0) 12 (100) 0 (0) 0 (0) 25 (100) 0 (0) 0 (0) 2 (100) 0 (0) 0 (0)
Tetraciclina
16 µg/mL 35 (85) 6 (15) 0 (0) 11 (92) 1 (8) 0 (0) 7 (28) 18 (72) 0 (0) 2 (100) 0 (0) 0 (0)
Eritromicina
8 µg/mL 36 (88) 5 (12) 0 (0) 12 (100) 0 (0) 0 (0) 22 (88) 3 (12) 0 (0) 2 (100) 0 (0) 0 (0)
Gentamicina
500 µg/mL 41 (100) 0 (0) 0 (0) 12 (100) 0 (0) 0 (0) 7 (28) 18 (72) 0 (0) 2 (100) 0 (0) 0 (0)
*S: susceptível, R: resistente, I: intermediário.

Resultados________________________________________________________________________
62
020406080
100
vanc
omic
ina
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário
020406080
100
vanc
omic
ina
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário
a
b c Figura 3: Perfil de resistência de Enterococcus faecium a antibióticos de uso clínico isolados a partir de amostras de queijo (a), produtos cárneos (b) e leite (c).
020406080
100va
ncom
icin
a
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário

Resultados________________________________________________________________________
63
020406080
100
vanc
omic
ina
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário
020406080
100
vanc
omic
ina
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário
020406080
100
vanc
omic
ina
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário
020406080
100
vanc
omic
ina
ampi
cilin
a
tetra
cicl
ina
eritr
omic
ina
gent
amic
ina
%
resistentesusceptívelintermediário
a b c d Figura 4: Perfil de resistência de Enterococcus faecalis a antibióticos de uso clínico isolados a partir de amostras de queijo (a), produtos cárneos (b), leite (c) e vegetais (d).

Resultados________________________________________________________________________
64
Tabela 11: Perfil de resistência a antibióticos apresentado por E. faecalis e E.
faecium obtidos de amostras de queijos, produtos cárneos e vegetais
adquiridas em Ribeirão Preto – SP, Brasil
N°. de isolados resistentes Fonte
Nº de isolados
Perfil de resistência* Produtos
cárneos Queijo Leite
E. faecalis 29 Tc 1 5
Er 4
Tc, Er 1
Ge, Tc 15
Ge, Er, Tc 3
E. faecium 23 Van 2
Tc 6 1
Er 2 10
Tc, Er 1
Van, Tc 1
*Resistência definida após triagem em ágar BHI com as seguintes concentrações de antibióticos: 32 µg/mL vancomicina, 16 µg/mL tetraciclina, 8 µg/ml, eritromicina, 500 µg/mL gentamicina e 16 µg/mL ampicilina. Tc: tetraciclina, Van: vancomicina, Er: eritromicina, Ge: gentamicina.

Resultados________________________________________________________________________
65
Tabela 12: Resultados da Concentração Inibitória Mínima (CIM) para isolados
de E. faecalis e E. faecium obtidos de amostras de queijo, produtos cárneos,
leite e vegetais adquiridas na cidade de Ribeirão Preto – SP, Brasil
Concentração inibitória mínima CIM
(µg/mL)
Alimento Isolado
Tc Van Er Ge
Perfil de resistência *
E. faecalis Queijos 340 128 > 256 Tc, Er
(n=29) 341 64 Tc
568 32 Tc
591 128 Tc
598 128 Tc
599 128 Tc
600 > 256 Er
604 > 256 Er
605 > 256 Er
606 > 256 Er
Produtos
cárneos 57 128 Tc
Leite 733 256 > 500 Tc, Ge
734 256 > 256 > 500 Tc, Er, Ge
735 256 > 256 > 500 Tc, Er, Ge
738 256 > 500 Tc, Ge
739 256 > 500 Tc, Ge
740 256 > 256 > 500 Tc, Er, Ge
742 256 > 500 Tc, Ge
743 256 > 500 Tc, Ge
744 256 > 500 Tc, Ge
756 256 > 500 Tc, Ge
757 256 > 500 Tc, Ge
758 256 > 500 Tc, Ge
759 256 > 500 Tc, Ge
760 256 > 500 Tc, Ge
761 256 > 500 Tc, Ge
763 256 > 500 Tc, Ge
782 256 > 500 Tc, Ge
813 256 > 500 Tc, Ge
E. faecium Queijos 77 > 256 Van
(n=23) 397 > 256 Van
401 32 Er
612 16 Er
613 16 Er
614 16 Er
615 64 Er

Resultados________________________________________________________________________
66
Concentração inibitória mínima CIM
(µg/mL)
Alimento Isolado
Tc Van Er Ge
Perfil de resistência *
620 64 Er
623 16 Er
624 16 Er
625 32 Er
627 32 Er
Produtos
cárneos 87 32 Er
97 16 Er
233 64 Tc
349 256 > 256 Tc, Er
352 256 256 Van, Tc
354 256 Tc
355 256 Tc
356 256 Tc
357 256 Tc
358 256 Tc
Leite 812 256 Tc
* Tc: tetraciclina, Van: vancomicina, Er: eritromicina, Ge: gentamicina.

Resultados________________________________________________________________________
67
4.5 Detecção de genes de virulência em isolados de E. faecium e E.
faecalis
Os 139 isolados de E. faecium e 80 de E. faecalis foram analisados por
PCR para verificar a presença dos determinantes de virulência: gelatinase
(gel), proteína de superfície (esp), substância de agregação (as), adesina de
colágeno de E. faecalis (ace), antígeno A de E. faecalis (efaA), citolisina (cylA)
e hialuronidase (hylA). A Figura 5 nos mostra os perfis das bandas obtidas para
os genes avaliados.
De uma forma geral, a Tabela 13 mostra que a distribuição destes genes
de virulência foi mais uniforme para o grupo de E. faecalis quando comparado
a E. faecium. Nenhum dos 139 isolados de E. faecium obtidos a partir de
amostras de queijo, produtos cárneos e leite apresentaram resultado positivo
para o gene esp, as, hyl e cylA. Apenas 3 isolados de E. faecium (2,1%) foram
positivos para o gene ace, sendo que dois isolados foram obtidos a partir de
amostras de produtos cárneos e um de queijo. O gene responsável pela
produção de gelatinase (gel) foi encontrado em 33,8% dos isolados de E.
faecium com uma proporção maior para os isolados obtidos de produtos
cárneos (56,7%) seguido daqueles obtidos a partir de amostras de queijos
(26,3%). De uma forma geral, o gene efaA foi o mais prevalente entre E.
faecium, sendo que 72,7% dos isolados obtidos de amostras de queijos
apresentaram este gene, seguido de 56,7% dos isolados obtidos de produtos
cárneos e 33,3% de leite.
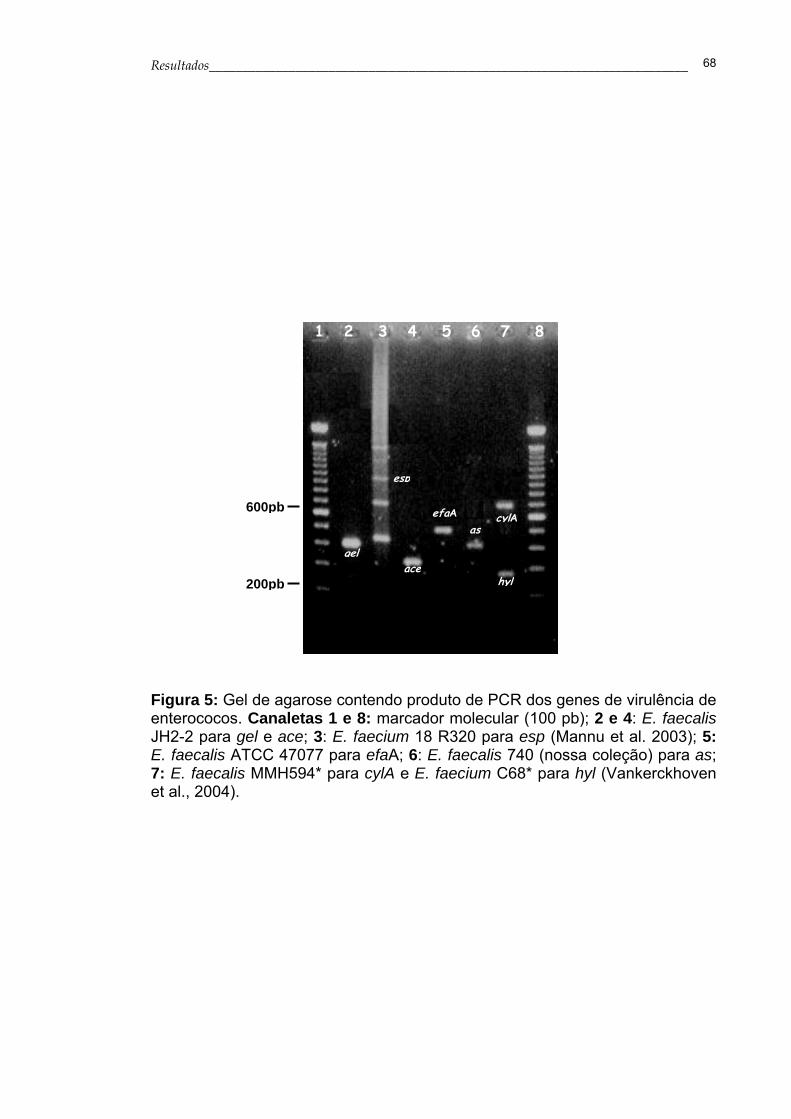
Resultados________________________________________________________________________
68
Figura 5: Gel de agarose contendo produto de PCR dos genes de virulência de enterococos. Canaletas 1 e 8: marcador molecular (100 pb); 2 e 4: E. faecalis JH2-2 para gel e ace; 3: E. faecium 18 R320 para esp (Mannu et al. 2003); 5: E. faecalis ATCC 47077 para efaA; 6: E. faecalis 740 (nossa coleção) para as; 7: E. faecalis MMH594* para cylA e E. faecium C68* para hyl (Vankerckhoven et al., 2004).
gel
esp
ace
efaA
1 2 3 4 5
as
hyl
cylA
6 7 8
600pb
200pb

Resultados________________________________________________________________________
69
Tabela 13: Distribuição dos genes de virulência em E. faecium e E. faecalis
obtidos de amostras de alimentos
Positividade para os genes (%)
N° de
isolados esp gel ace as efaA hyl cylA
E. faecium Produtos
cárneos
37 0
(0)
21
(56,7)
2
(5,4)
0
(0)
21
(56,7)
0
(0)
0
(0)
Queijos 99 0
(0)
26
(26,3)
1
(1,0)
0
(0)
72
(72,7)
0
(0)
0
(0)
Leite 3 0
(0)
0
(0)
0
(0)
0
(0)
1
(33,3)
0
(0)
0
(0)
Total 139 0
(0)
47
(33,8)
3
(2,1)
0
(0)
94
(67,6)
0
(0)
0
(0)
E. faecalis Produtos
cárneos
12 0
(0)
12
(100,0)
10
(83,3)
1
(8,3)
12
(100,0)
0
(0)
0
(0)
Queijos 41 3
(7,3)
39
(95,1)
41
(100,0)
11
(26,8)
40
(97,5)
0
(0)
0
(0)
Leite 25 0
(0)
24
(96,0)
25
(100,0)
21
(84,0)
25
(100,0)
0
(0)
22
(88,0)
Vegetais 2 0
(0)
2
(100,0)
2
(100,0)
2
(100,0)
2
(100,0)
0
(0)
2
(100,0)
Total 80 3
(3,7)
77
(96,2)
78
(97,5)
35
(43,7)
79
(98,7)
0
(0)
24
(30,0)

Resultados________________________________________________________________________
70
Dentre os isolados de E. faecalis (Tabela 13) obtidos a partir de
amostras de produtos cárneos, queijos, leites e vegetais também foi
encontrado uma ampla distribuição do gene efaA (100%, 97,5%, 100% e 100%,
respectivamente). Em seguida, foi observado que 100% dos isolados de E.
faecalis obtidos de amostras de queijo, leite e vegetais e 83,3% daqueles
obtidos de produtos cárneos apresentaram resultados positivos para o gene
ace. Todas os isolados de E. faecalis obtidos de produtos cárneos e vegetais
foram positivos para o gene gel sendo que, para aqueles obtidos de amostras
de queijo e leite este valor foi de, respectivamente, 95,1% e 96%. Já para os
isolados de E. faecium obtidos de amostras de produtos cárneos e queijo foi
encontrado, respectivamente, 56,7% e 26,3% de positividade.
Foi observada uma distribuição diversa do gene as entre os isolados de
E. faecalis obtidos de diferentes grupos de alimentos, sendo que para aqueles
provenientes das amostras de vegetais foi obtido 100% de resultado positivo
seguido de 84% para os obtidos de leite. Já para os isolados provenientes de
queijos e produtos cárneos esse valor foi de, respectivamente, 26,8% e 8,3%.
Uma elevada porcentagem de E. faecalis obtidos de amostras de leite (88%) e
de vegetais (100%) foram positivos para o gene cylA. Por outro lado, este gene
não foi detectado em nenhum isolado de E. faecalis obtido a partir de produtos
cárneos e queijo. O gene esp foi detectado em apenas três isolados (7,3%) de
E. faecalis obtidos a partir de amostras de leite. Dentre os genes de virulência
pesquisados neste trabalho, o gene hyl foi o único que apresentou resultado
negativo para todos os isolados de E. faecalis analisados (Tabela 13).
A prevalência dos genes de virulência para os 23 isolados de E. faecium
e 29 isolados de E. faecalis resistentes a antibióticos está apresentada na

Resultados________________________________________________________________________
71
Tabela 14. De acordo com esta tabela, todos os isolados de E. faecalis e 8
(35%) E. faecium foram positivos para um ou mais dos seguintes genes de
virulência: esp, gel, ace, as, efaA, hyl e cylA. Foi observado que 26 (89,6%)
isolados de E. faecalis resistentes a antibióticos e 6 (26%) isolados de E.
faecium resistentes a antibióticos apresentaram resultado positivo para o gene
gel. Quanto à produção fenotípica de gelatinase, foi observado que 17 E.
faecalis (58,6%) foram capazes de hidrolisar a gelatina enquanto que, nenhum
E. faecium apresentou esta característica.
Dos 21 isolados de E. faecalis que produziram biofilme em superfície
abiótica, 19 (90,5%) apresentaram o genótipo gel+. Para os 3 isolados de E.
faecium que produziram biofilme, 2 (66,7%) possuíam o gene gel. Dos 13 E.
faecalis que apresentaram o gene cylA, 12 (92,3%) apresentaram reação de
beta-hemólise, revelando uma correlação positiva entre o teste fenotípico e o
genotípico para este grupo de microrganismo. Por outro lado, entre os 26
isolados de E. faecalis gel+, apenas 17 (65,4%) produziram gelatinase no teste
fenotípico.
Comparando a prevalência de genes de virulência entre os enterococos
resistentes a antibióticos e os não resistentes, a Tabela 15 evidencia que não
havia maior prevalência dos genes de virulência para o grupo dos enterococos
resistentes a antibióticos.

Resultados________________________________________________________________________
72
Tabela 14: Genes de virulência, formação de biofilme e adesão a células Caco-2
em E. faecium e E. faecalis resistentes a antibióticos de uso clínico
Genes virulência* Espécie/origem Isolado Biofilme§ Adesão† Resistência
a antibiótico esp gel ace as efaA hyl cylA
E. faecalis Queijos 340 + + Tc, Er - + + - - - -
341 + +++ Tc - + + - + - - 568 - - Tc - + + - + - - 591 - +++ Tc - - + - + - - 598 + +++ Tc - - + - + - - 599 + +++ Tc - + + - + - - 600 + - Er - + + - + - - 604 - - Er - + + + + - - 605 + - Er - + + - + - - 606 + - Er - + + - + - - Produtos cárneos 57 + - Tc - + + + + - -
Leite 733 + - Tc, Ge - + + + + - - 734 + - Tc, Er, Ge - + + - + - - 735 + - Tc, Er, Ge - + + - + - - 738 + - Tc, Ge - + + + + - - 739 + - Tc, Ge - + + + + - - 740 + - Tc, Er, Ge - + + - + - + 742 - - Tc, Ge - + + + + - + 743 + - Tc, Ge - + + + + - + 744 + - Tc, Ge - + + + + - + 756 - - Tc, Ge - + + - + - + 757 + - Tc, Ge - + + + + - + 758 - - Tc, Ge - + + + + - + 759 + + Tc, Ge - + + + + - + 760 - - Tc, Ge - + + + + - + 761 + - Tc, Ge - + + + + - + 763 - - Tc, Ge - + + + + - + 782 + - Tc, Ge - - + + + - + 813 + - Tc, Ge - + + + + - + E. faecium Queijos 77 - - Van - + - - + - -
397 - - Van - - - - + - - 401 - - Er - - - - - - - 612 - - Er - - - - - - - 613 - - Er - - - - - - - 614 - - Er - - - - - - - 615 - - Er - - - - - - - 620 - - Er - - - - - - - 623 - - Er - - - - - - - 624 - - Er - - - - - - - 625 - - Er - - - - - - - 627 - - Er - - - - - - - Produtos cárneos 87 + - Er - + - - - - -
97 - - Er - + + - + - - 233 + - Tc - - - - - - - 349 + - Tc, Er - + + - - - - 352 - - Van, Tc - - - - - - - 354 - - Tc - - - - - - - 355 - - Tc - - - - - - - 356 - - Tc - - - - - - - 357 - - Tc - + - - - - - 358 - - Tc - + - - - - -
Leite 812 - - Tc - - - - + - - § Biofilme ( Duprè et al. 2003) : A595 ≤ 1 não formador (-);1 <A595 ≤ 2 fraco (+);2 < A595 ≤ 3 moderado (++); A595 > 3 forte (+++).† Adesão: < 10 bactérias por célula: (-) não aderente, 10-16 bactérias por célula: (+) fraca adesão, 16-60 bactérias or célula: (++) adesão moderada, > 60 bactérias por célula: (+++) forte adesão.*(+): positivo (-): negativo para os genes pesquisados.

Resultados________________________________________________________________________
73
Tabela 15: Comparação da prevalência de genes de virulência entre
enterococos resistentes a antibióticos (RA) e enterococos não resistentes
No. de isolados (%)
esp gel ace as efaA hyl cylA
E. faecium
(n=139)
0
(0,0)
47
(33,8)
3
(2,1)
0
(0,0)
94
(67,6)
0
(0,0)
0
(0,0)
E. faecium RA
(n=23)
0
(0,0)
6
(26)
2
(8,7)
0
(0,0)
4
(17,4)
0
(0,0)
0
(0,0)
E. faecalis
(n=80)
3
(3,7)
77
(96,2)
78
(97,5)
35
(43,7)
79
(98,7)
0
(0,0)
24
(30)
E. faecalis RA
(n=29)
0
(0,0)
26
(89,6)
29
(100,0)
16
(55,2)
28
(96,5)
0
(0,0)
13
(44,8)
4.6 Formação de biofilme em superfície abiótica e adesão em células da
linhagem Caco-2
De acordo com a classificação proposta de formação de biofilme em
superfície abiótica, foram obtidos 48 isolados de E. faecalis (60,0%) e 57
isolados de E. faecium (41,0%) com uma fraca capacidade de formação de
biofilme e 6 (7,5%) E. faecalis e 16 (11,5%) E. faecium com uma moderada
capacidade de formação de biofilme (Figura 6 e Tabela 14). Se for considerado
apenas o grupo de E. faecalis e E. faecium resistentes a antibióticos, observa-
se que 72,4% dos isolados de E. faecalis e 13% de E. faecium (Tabela 14 e
Figura 6) demonstraram uma fraca capacidade de formação de biofilme.
Quanto ao teste de adesão às células da linhagem Caco-2, que foi realizado
somente com os isolados de E. faecium e E. faecalis resistentes a antibióticos,
foi observado que 4 E. faecalis (14%) mostraram um perfil altamente adesivo a
estas células (Tabela 14 e Figura 7), enquanto que para E. faecium, este
resultado não foi observado.

Resultados________________________________________________________________________
74
0102030405060708090
100
- + ++ +++
Classificação *
% fo
rmaç
ão b
iofil
me E. faecium
E. faecalis
E. faecium (RA)
E. faecalis (RA)
Figura 6: Formação de biofilme em superfície abiótica por E. faecalis e E faecium resistentes a antibióticos (RA) e não resistentes. *Biofilme classificado de acordo com Duprè et al. (2003): A595 ≤ 1 não formador (-); 1 <A595 ≤ 2 fraco (+); 2 < A595 ≤ 3 moderado (++); A595 > 3 forte (+++).

Resultados________________________________________________________________________
75
Figura 7: Fotomicrografia (Olympus BX51) mostrando isolado de E. faecalis 341 fortemente adesivo (A e B) e E. faecalis 568 (C e D) com ausência de adesão às células da linhagem Caco-2, coradas com May Grunwald – Giemsa (aumento 1.000 vezes).

Resultados________________________________________________________________________
76
4.7 Análise de fingerprinting por meio de RAPD-PCR
Os 52 isolados de Enterococcus spp. resistentes a antibióticos foram
agrupados em 7 diferentes clusters com um nível de similaridade de 80%
(Figura 8). Os isolados de E. faecium e E. faecalis foram agrupados em 4 e 3
diferentes clusters, respectivamente. A análise do dendrograma mostrou que
os perfis de resistência a antibióticos estavam bem distribuídos (Figura 8). No
geral, não foi encontrada uma correlação entre os tipos de alimentos e a
localização dos isolados nos clusters formados, exceto para o cluster 3 (a e b)
que inclui somente isolados obtidos de diferentes amostras de leite.
O nível de similaridade dos isolados de E. faecium resistentes à
tetraciclina e distribuídos nos clusters 1, 4 e 6d foi aproximadamente 70%. Os
isolados de E. faecalis com perfil de resistência múltipla à tetraciclina,
eritromicina e alta concentração de gentamicina foram agrupados nos clusters
2 e 3a com um nível de similaridade de 77%. É importante notar que os dois
isolados de E. faecium resistentes à vancomicina foram obtidos de diferentes
amostras de queijo. Estes isolados foram distribuídos nos clusters 1 e 6a com
um nível de similaridade de 69%. O perfil de resistência à eritromicina foi o
mais distribuído nos clusters, enquanto que o perfil de resistência à alta
concentração de gentamicina esteve presente nos clusters 2, 3a e 3b. Este
experimento foi feito para verificar se os isolados de enterococos resistentes a
antibióticos representavam diferentes cepas ou se poderiam ser considerados
múltiplos isolados de uma mesma cepa. A análise dos resultados de RAPD
aliados ao perfil de resistência a antibióticos e características fenotípicas
sugerem a existência de 14 cepas diferentes de E. faecium e 17 de E. faecalis
(marcadas em cinza no dendrograma).

Resultados________________________________________________________________________
77
Cluster Resistência a antibióticos
fm 352 fm 354 fm 355 fm 349 FM LMG 11423T fm 233 fm 620 fm 623 fm 615 fm 625 fm 624
10090 80 7060
Tc
Tc
Tc
Er
Van
Tc, Er, Ge
Tc, Ge
Tc, Er,Ge
Tc, Ge
Tc, Er
Er Tc
Er
Van
Er
Tc, Er
Tc
Er
Van, Tc
1
2
3a
5
6a, 6b, 6c
7
3b
3c
3d
4
6d
fm 357 fm 358 fm 356 fm 401 fm 613 fm 614 fm 612 fm 397 fs 734 fs 735 fs 759 fs 760 fs 761 fs 813 fs 740 fs 763 fs 782 fs 733 fs 757 fs 758 fs 756 fs 743 fs 744 fs 742 fs 738 fs 739 fs 340 fs 591 fs 598 fs 599 fs 57 fs 341 fs 568 fm 627 fm 812 fs 605 fs 606 fs 604 fs 600 fm 77 fm 87 fm 97
Figura 8: Perfis de RAPD dos isolados de Enterococcus spp. com o primer M-13. fs: E. faecalis, fm: E. faecium, FM LMG 11423: cepa de referência

Resultados________________________________________________________________________
78
4.8 Enumeração de coliformes totais e E. coli nas amostras de alimentos
Na Tabela 16 observa-se que entre os quatro grupos de alimentos a
população de E. coli variou de 0 a valores superiores a 6,4 log UFC/g ou mL, e
a de coliformes totais variou de 0 a 7,8 log UFC/g ou mL. Nas amostras de
água não foram detectados coliformes totais e E. coli, ou seja, estes
microrganismos estavam abaixo dos limite de detecção do método que era de
1 UFC/100 mL.
As amostras de leite cru apresentaram o maior número de amostras
positivas para E. coli (18 amostras), seguido das amostras de queijo (12
amostras), carne (7 amostras) e vegetais (3 amostras). Para a contagem de
coliformes totais foi observado que no grupo de produtos cárneos foi obtido o
maior número de amostras positivas (23) seguido das amostras de queijo, leite
e vegetais (22 amostras) (Tabela 16).
Para as 11 amostras de leite pasteurizado (tipos A, B, C e de cabra) as
contagens de E. coli ficaram abaixo do limite de detecção do método
empregado (<1 UFC/mL ou < 0 log UFC/mL). Em apenas três amostras de leite
pasteurizado a população de coliformes totais estava acima do limite de
detecção da técnica. Por outro lado, todas as amostras de leite cru
apresentaram contagem de coliformes totais.

Resultados________________________________________________________________________
79
Tabela 16: Enumeração de E. coli e coliformes totais (log UFC/g ou mL) das
amostras de carne, leite, queijo, vegetais e águas em Petrifilm EC (3M) e
membrana filtrante
Log UFC/g ou mL amostra
Amostra N° amostra E. coli a,b Coliformes totais a,b
Produtos carne moída 03 3 3,9 cárneos a presunto cozido 05 <1 <1
(n=30) carne moída 06 1 3,5 peito de frango 07 <1 2,4 presunto fatiado 09 <1 2,2 apresuntado 10 <1 2,1 lingüiça frango 12 <1 1 bacon 13 <1 <1 salsicha 14 <1 1,8 lagarto bovino defumado 20 <1 1 rosbife fatiado 21 <1 <1 salsicha com picles 23 <1 2,4 surubim defumado 24 <1 <1 salame 27 <1 <1 mortadela 28 <1 <1 presunto peru 32 <1 1,8 lingüiça frango 33 2,5 3,7 lingüiça de porco 34 2,3 3,9 coração frango 37 1,3 2,3 moela frango 38 <1 4 presunto cozido 43 <1 1,8 bucho bovino 45 <1 3,5 lingüiça pernil 47 1 1,5 coxa de frango 48 <1 2,5 bacon 49 <1 <1 filé tilápia 50 <1 1,3 asa de frango 51 <1 2,2 coxão mole 52 1,6 2,6 lombo suíno fatiado 53 <1 2,1 bife bovino 54 <1 2,7
Queijos a mussarela 01 3,5 3,6 (n=30) queijo minas 02 >6,4 >6,4
queijo minas 04 6,8 7,8 mussarela fatiada 08 <1 1,5 ricota 11 <1 5,7 queijo prato 15 <1 2 queijo minas 16 6,8 7,4 queijo gouda 17 <1 2,1 mussarela 18 <1 <1 queijo árabe 25 <1 <1 mussarela 26 <1 <1 queijo prato fatiado 29 3,6 4,4 mussarela 30 <1 1,7 queijo prato 31 2,2 3 queijo parmesão 42 <1 1,7 queijo gorgonzola 44 <1 1,6 queijo minas 46 5,5 6 queijo provolone 55 <1 <1 ricota 56 <1 7 queijo cheedar 57 <1 <1 queijo gorgonzola 61 2,2 2,6 mussarela 62 <1 <1 mussarela 73 1 1,5 queijo meia cura 74 3,9 4,2 ricota temperada 75 5,8 5,9 queijo meia cura 76 <1 4,2

Resultados________________________________________________________________________
80
Log UFC/g ou mL amostra
Amostra N° amostra E. coli a,b Coliformes totais a,b
queijo minas light 77 5 7,4 mussarela fatiada 78 <1 <1 queijo prato 79 <1 2,4 queijo meia cura 80 3,0 3,5
Vegetais A seleta legumes 19 <1 <1 (n=30) cenoura ralada 22 <1 6,1
salada mista 35 <1 4,6 salada repolho e beterraba 36 <1 4,5 abobrinha 39 <1 2,6 cogumelo 40 <1 <1 soja fermentada 41 <1 2,9 broto de feijão 58 <1 7,4 couve manteiga 59 2 3,7 ervilha crua 60 1,7 1,9 azeitona preta 63 <1 <1 tremoço 64 <1 2,4 milho cru 65 <1 5,0 beterraba 66 <1 <1 cogumelo 67 <1 4,7 Alecrim 68 <1 3,1 salada tricolor 69 <1 1,6 Acelga 70 <1 3,6 cheiro verde pré-higienizado 71 <1 6,6 cheiro verde in natura 72 <1 <1 manjericão 81 <1 2,4 salsinha 82 <1 4,4 batata/cenoura/vagem 83 <1 <1 escarola 84 <1 <1 salada mista 85 <1 5,0 alface/ rúcula/ rabanete 86 <1 1,7 abóbora cozida 87 <1 <1 alface crespa 88 <1 3,4 coentro 89 1 1,8 almeirão 90 <1 5
Leite b leite tipo c 91 <0 <0 (n=30) leite tipo c 92 <0 <0
leite tipo b 93 <0 <0 leite tipo c 94 <0 <0 leite tipo a 95 <0 <0 leite tipo c 96 <0 2,4 leite cru 97 2,5 3 leite cru 98 3,5 3,6 leite cru 99 3,1 3,2 leite cru 100 4,1 4,1 leite cru 101 2,9 3,1 leite cru 102 3,4 3,7 leite cru 103 2,8 3 leite cru 104 4 4,1 leite cru 105 3 3,2 leite cru 106 2,8 3,2 leite cru 107 2,9 3,2 leite cru 108 2,5 3,0 leite cru 109 2,9 3,3 leite tipo b 110 <0 <0 leite tipo c 111 <0 2,2 leite cabra 112 <0 2,6 leite tipo c 113 <0 <0 leite tipo c 114 <0 <0 leite cru 115 4,6 5,1 leite cru 116 3,3 4 leite cru 117 3,9 4,2 leite cru 118 4,3 5,3

Resultados________________________________________________________________________
81
Log UFC/g ou mL amostra
Amostra N° amostra E. coli a,b Coliformes totais a,b
leite cru 119 3,8 4,2 leite cru 120 <0 4,3
Águas c água mineral 121 < 0 < 0 (n=30) água mineral 122 < 0 < 0
água mineral 123 < 0 < 0 água abastecimento 124 < 0 < 0 água abastecimento 125 < 0 < 0 água abastecimento 126 < 0 < 0 água abastecimento 127 < 0 < 0 água abastecimento 128 < 0 < 0 água abastecimento 129 < 0 < 0 água abastecimento 130 < 0 < 0 água abastecimento 131 < 0 < 0 água abastecimento 132 < 0 < 0 água abastecimento 133 < 0 < 0 água abastecimento 134 < 0 < 0 água abastecimento 135 < 0 < 0 água abastecimento 136 < 0 < 0 água abastecimento 137 < 0 < 0 água abastecimento 138 < 0 < 0 água abastecimento 139 < 0 < 0 água abastecimento 140 < 0 < 0 água abastecimento 141 < 0 < 0 água abastecimento 142 < 0 < 0 água abastecimento 143 < 0 < 0 água abastecimento 144 < 0 < 0 água abastecimento 145 < 0 < 0 água abastecimento 146 < 0 < 0 água mineral 147 < 0 < 0 água mineral 148 < 0 < 0 água torneira 149 < 0 < 0 água bebedouro 150 < 0 < 0
a: as contagens de E. coli e coliformes totais presentes nas amostras de carnes, queijos e vegetais que encontravam-se abaixo do limite detecção do método (10 UFC/g), são valores estimados e estão representadas por <1 (inferior a 1 Log UFC/g amostra). b: contagens de E. coli e coliformes totais presentes nas amostras de leites que se encontravam abaixo do limite detecção do método (1 UFC/mL) de amostra, são valores estimados e estão representadas por < 0 (inferior a 0 Log UFC/mL amostra). c: contagens de E. coli e coliformes totais presentes nas amostras de águas que se encontravam abaixo do limite detecção do método (1 UFC/100mL) de amostra, são valores estimados e estão representadas por < 0 (inferior a 0 Log UFC/100 mL amostra).

Resultados________________________________________________________________________
82
A Tabela 17 agrupa as contagens de coliformes totais, E. coli e enterococos
por grupo de alimento. Foi considerada a contagem de Enterococcus spp. obtida no
meio KEA por ter sido mais adequado para enumeração deste microrganismo. De
uma forma geral, não houve correlação entre a contagem de enterococos,
coliformes totais e E. coli. Analisando casos isolados (Tabela 18), a amostra 34 de
lingüiça apresentou uma contagem de E. coli de 2,3 log UFC/g enquanto que a de
Enterococcus foi de 5,6 log UFC/g. Por outro lado, a amostra de carne moída nº 3
apresentou uma contagem de 3,0 log UFC/g amostra de E. coli enquanto que a
contagem de Enterococcus estava abaixo do limite de detecção do método.
Para as amostras de leite cru nº 99 e 100 observou-se que enquanto a
contagem de E. coli foi de 3,1 e 4,1, respectivamente, a contagem de
Enterococcus spp. ficou abaixo do limite de detecção do método. Resultado
semelhante foi obtido para a amostra 31 (queijo prato) que apresentou
contagem de E. coli de 2,2 log UFC/g enquanto que a contagem de
enterococos estava abaixo do limite de detecção do método. Já a amostra 61
de queijo gorgonzola apresentou uma contagem de E. coli de 2,2 e uma
contagem de Enterococcus spp. de 4,4 log UFC/g amostra. As amostras 29
(queijo tipo prato) e 2 (queijo minas) apresentaram valores similares para a
contagem destes dois grupos de microrganismos.
É importante ressaltar que as amostras nº 8, 18, 26 e 30 de queijo tipo
mussarela apresentaram uma contagem de Enterococcus spp. que variou de
5,3 a 6,2 log UFC/g enquanto que a contagem de E. coli foi de < 1,0 log UFC/g
e a de coliformes totais variou de < 1,0 a 1,7 UFC/g (Tabela 18).
Com as amostras de águas foi observado que a contagem de todos os
microrganismos (E. coli, coliformes totais e Enterococcus spp.) ficou abaixo do
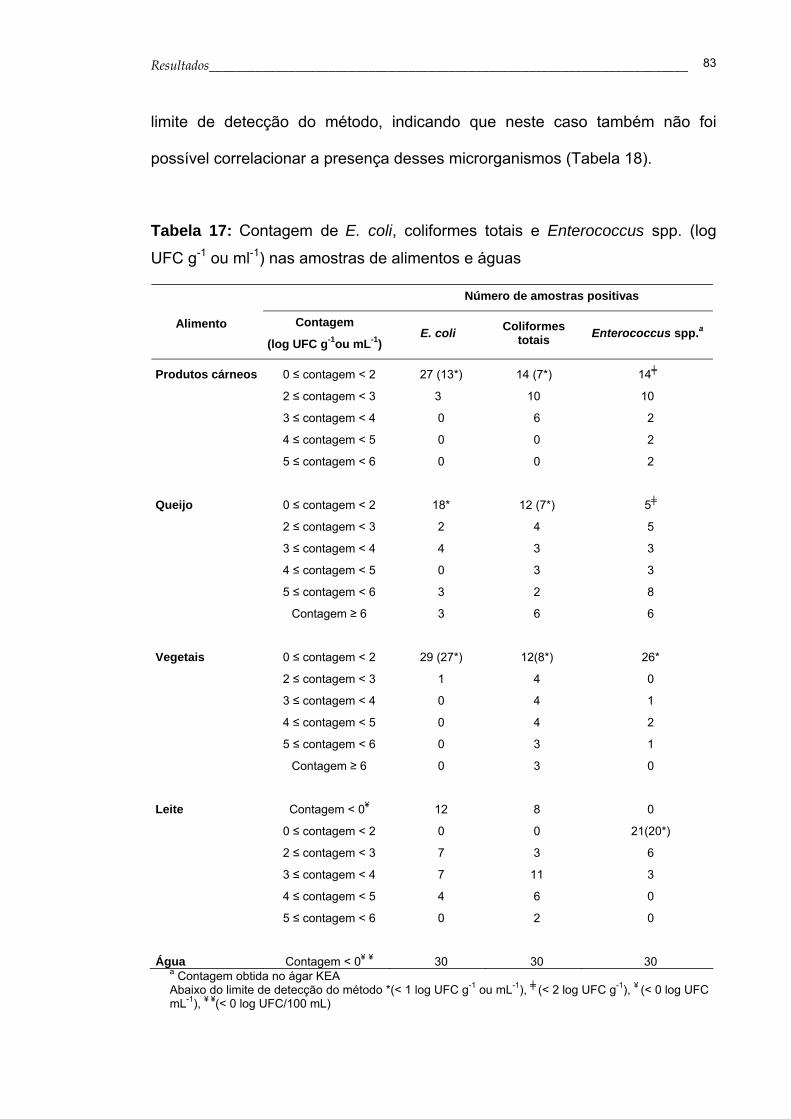
Resultados________________________________________________________________________
83
limite de detecção do método, indicando que neste caso também não foi
possível correlacionar a presença desses microrganismos (Tabela 18).
Tabela 17: Contagem de E. coli, coliformes totais e Enterococcus spp. (log
UFC g-1 ou ml-1) nas amostras de alimentos e águas
Número de amostras positivas
Alimento Contagem
(log UFC g-1ou mL-1) E. coli Coliformes
totais Enterococcus spp.a
Produtos cárneos 0 ≤ contagem < 2 27 (13*) 14 (7*) 14╪
2 ≤ contagem < 3 3 10 10
3 ≤ contagem < 4 0 6 2
4 ≤ contagem < 5 0 0 2
5 ≤ contagem < 6 0 0 2
Queijo 0 ≤ contagem < 2 18* 12 (7*) 5╪
2 ≤ contagem < 3 2 4 5
3 ≤ contagem < 4 4 3 3
4 ≤ contagem < 5 0 3 3
5 ≤ contagem < 6 3 2 8
Contagem ≥ 6 3 6 6
Vegetais 0 ≤ contagem < 2 29 (27*) 12(8*) 26*
2 ≤ contagem < 3 1 4 0
3 ≤ contagem < 4 0 4 1
4 ≤ contagem < 5 0 4 2
5 ≤ contagem < 6 0 3 1
Contagem ≥ 6 0 3 0
Leite Contagem < 0¥ 12 8 0
0 ≤ contagem < 2 0 0 21(20*)
2 ≤ contagem < 3 7 3 6
3 ≤ contagem < 4 7 11 3
4 ≤ contagem < 5 4 6 0
5 ≤ contagem < 6 0 2 0
Água Contagem < 0¥ ¥ 30 30 30 a Contagem obtida no ágar KEA Abaixo do limite de detecção do método *(< 1 log UFC g-1 ou mL-1), ╪ (< 2 log UFC g-1), ¥ (< 0 log UFC mL-1), ¥ ¥(< 0 log UFC/100 mL)

Resultados________________________________________________________________________
84
Tabela 18: Comparação da população de Enterococcus spp., coliformes totais
e E. coli nas amostras de alimentos e águas
Log UFC/g ou mL amostra
Enterococcus spp. Indicadores clássicos
N° amostra Meio BE Meio KEA
Coliformes totais
E. coli
Produtos cárneos
3 < 2,0 < 2,0
3,9 3
(n=30) 5 < 2,0 < 2,0 <1 1
6 < 2,0 2,8 3,5 1
7 < 2,0 < 2,0 2,4 1
9 4,2 4,0 2,2 1
10 2,2 2,0
2,1 <1
12 < 2,0 2,7 1 <1
13 < 2,0 < 2,0 < 1 <1
14 2,3 2,0 1,8 1
20 2,3 < 2,0 1 <1
21 < 2,0 2,0 <1 1
23 < 2,0 2,6 2,4 1
24 < 2,0 < 2,0 <1 <1
27 < 2,0 2,5 <1 1
28 2,0 2,3 < 1 1
32 4,4 4,0 1,8 1
33 < 2,0 3,5 3,7 2,5
34 5,5 5,6 3,9 2,3
37 < 2,0 < 2,0 2,3 1,3
38 < 2,0 2,0 4 <1
43 < 2,0 2,5 1,8 1
45 < 2,0 < 2,0 3,5 <1
47 < 2,0 < 2,0 1,5 1
48 < 2,0 < 2,0 2,5 <1
49 5,0 5,4 < 1 <1
50 < 2,0 < 2,0 1,3 <1
51 < 2,0 < 2,0 2,2 <1
52 < 2,0 3,0 2,6 1,6
53 < 2,0 < 2,0 2,1 <1
54 < 2,0 < 2,0 2,7 <1
Leite 91 < 1,0 < 1,0 <0 <0
(n=30) 92 < 1,0 < 1,0 <0 <0
93 < 1,0 < 1,0 <0 <0
94 < 1,0 < 1,0 <0 <0
95 < 1,0 < 1,0 <0 <0
96 < 1,0 < 1,0 2,4 <0

Resultados________________________________________________________________________
85
Log UFC/g ou mL amostra
Enterococcus spp. Indicadores clássicos
N° amostra Meio BE Meio KEA
Coliformes totais
E. coli
97 3,0 2,0 3 2,5
98 < 1,0 2,5 3,6 3,5
99 < 1,0 < 1,0 3,2 3,1
100 < 1,0 < 1,0 4,1 4,1
101 < 1,0 2,0
3,1 2,9
102 < 1,0 3,6 3,7 3,4
103 < 1,0 < 1,0 3 2,8
104 < 1,0 < 1,0 4,1 4
105 < 1,0 < 1,0 3,2 3
106 < 1,0 2,3 3,2 2,8
107 < 1,0 < 1,0 3,2 2,9
108 < 1,0 < 1,0 3 2,5
109 < 1,0 2,7 3,3 2,9
110 < 1,0 < 1,0 <0 <0
111 < 1,0 < 1,0 2,2 <0
112 < 1,0 1,5 2,6 <0
113 < 1,0 < 1,0 <0 <0
114 < 1,0 < 1,0 <0 <0
115 < 1,0 < 1,0 5,1 4,6
116 < 1,0 2,0 4 3,3
117 < 1,0 < 1,0 4,2 3,9
118 < 1,0 < 1,0 5,3 4,3
119 < 1,0 < 1,0 4,2 3,8
120 < 1,0 3,3 4,3 <0
Queijo 1 6,0 6,0 3,6 3,5
(n=30) 2 7,9 7,8 >6,4 >6,4
4 < 2,0 6,4 7,8 > 6,4
8 5,8 5,7 1,5 <1
11 6,7 5,5 5,7 <1
15 4,4 4,2 2 <1
16 6,6 6,3 7,4 6,8
17 <2,0 2,4 2,1 <1
18 5,8 5,6 <1 <1
25 < 2,0 < 2,0 <1 <1
26 5,2 5,3 <1 <1
29 < 2,0 3,2 4,4 3,6
30 6,2 6,2 1,7 <1
31 < 2,0 < 2,0 3 2,2
42 2,0 2,3 1,7 <1
44 < 2,0 2,5 1,6 <1

Resultados________________________________________________________________________
86
Log UFC/g ou mL amostra
Enterococcus spp. Indicadores clássicos
N° amostra Meio BE Meio KEA
Coliformes totais
E. coli
46 < 2,0 5,0 6 5,5
55 2,7 2,5 <1 <1
56 < 2,0 < 2,0 7 <1
57 < 2,0 < 2,0 <1 <1
61 4,3 4,4 2,6 2,2
62 < 2,0 < 2,0 <1 <1
73 5,4 5,4 1,5 1
74 < 2,0 3,7 4,2 3,9
75 < 2,0 5,8 5,9 5,8
76 < 2,0 4,6 4,2 <1
77 6,0 5,5 7,4 5
78 4,6 3,5 <1 <1
79 < 2,0 2,3 2,4 <1
80 7,9 7,8 3,5 3
Vegetais 19 < 2,0 < 2,0 <1 <1
(n=30) 22 < 2,0 < 2,0 6,1 <1
35 < 2,0 < 2,0 4,6 <1
36 < 2,0 3,8 4,5 <1
39 < 2,0 < 2,0 2,6 <1
40 < 2,0 < 2,0 <1 <1
41 < 2,0 < 2,0 2,9 <1
58 < 2,0 5,1 7,4 <1
59 < 2,0 4,4 3,7 2
60 < 2,0 < 2,0 1,9 1,7
63 < 2,0 < 2,0 <1 <1
64 < 2,0 < 2,0 2,4 <1
65 < 2,0 < 2,0 5 <1
66 < 2,0 < 2,0 <1 <1
67 < 2,0 < 2,0 4,7 <1
68 < 2,0 < 2,0 3,1 <1
69 < 2,0 < 2,0 1,6 <1
70 < 2,0 < 2,0 3,6 <1
71 < 2,0 < 2,0 6,6 <1
72 < 2,0 < 2,0 <1 <1
81 < 2,0 < 2,0 2,4 <1
82 5,0 < 2,0 4,4 <1
83 < 2,0 < 2,0 <1 <1
84 < 2,0 < 2,0 <1 <1
85 < 2,0 < 2,0 5 <1
86 < 2,0 < 2,0 1,7 <1

Resultados________________________________________________________________________
87
Log UFC/g ou mL amostra
Enterococcus spp. Indicadores clássicos
N° amostra Meio BE Meio KEA
Coliformes totais
E. coli
87 < 2,0 < 2,0 <1 <1
88 4,0 4,2 3,4 <1
89 < 2,0 < 2,0 1,8 1
90 < 2,0 < 2,0 5 <1
Água* 121 < 0 < 0 < 0 < 0
(n=30) 122 < 0 < 0 < 0 < 0
123 < 0 < 0 < 0 < 0
124 < 0 < 0 < 0 < 0
125 < 0 < 0 < 0 < 0
126 < 0 < 0 < 0 < 0
127 < 0 < 0 < 0 < 0
128 < 0 < 0 < 0 < 0
129 < 0 < 0 < 0 < 0
130 < 0 < 0 < 0 < 0
131 < 0 < 0 < 0 < 0
132 < 0 < 0 < 0 < 0
133 < 0 < 0 < 0 < 0
134 < 0 < 0 < 0 < 0
135 < 0 < 0 < 0 < 0
136 < 0 < 0 < 0 < 0
137 < 0 < 0 < 0 < 0
138 < 0 < 0 < 0 < 0
139 < 0 < 0 < 0 < 0
140 < 0 < 0 < 0 < 0
141 < 0 < 0 < 0 < 0
142 < 0 < 0 < 0 < 0
143 < 0 < 0 < 0 < 0
144 < 0 < 0 < 0 < 0
145 < 0 < 0 < 0 < 0
146 < 0 < 0 < 0 < 0
147 < 0 < 0 < 0 < 0
148 < 0 < 0 < 0 < 0
149 < 0 < 0 < 0 < 0
150 < 0 < 0 < 0 < 0
*Valor estimado; abaixo do limite de detecção do método <1UFC/100mL

____________________5. Discussão

Discussão________________________________________________________________________
89
Estudos sobre caracterização de Enterococcus spp. isolados de
diferentes amostras de alimentos consumidos no Brasil, são muito escassos e
de acordo com Devriese et al. (1995) a identificação de enterococos presentes
nos alimentos pode ajudar a elucidar sua origem de contaminação.
Para o isolamento de Enterococcus spp. a partir das amostras de
alimentos e águas o ágar KEA apresentou melhores resultados quando
comparado ao ágar BE. Resultado semelhante foi obtido por García et al.
(2002) com o isolamento de enterococos a partir de diversas etapas de
fabricação do queijo San-Simón, fabricado na Espanha. Estes autores
observaram que 79,6% das culturas provenientes do ágar KEA puderam ser
confirmadas como pertencentes ao gênero Enterococcus por meio de testes
bioquímicos.
Domig et al. (2003) revisaram os diferentes meios e métodos
empregados para o isolamento de enterococos e afirmaram que nenhum foi
suficientemente seletivo para o isolamento deste microrganismo.
Com o emprego de testes fenotípicos para a caracterização presuntiva
de enterococos, foram obtidos muitos isolados pertencentes a outros gêneros
como Lactococcus, Pediococcus e Leuconostoc que apresentaram perfil
semelhante a Enterococcus spp. em alguns testes propostos (Devriese et al.,
1993).
Pavia et al. (2000) analisaram 100 amostras de carne comercializadas
em Catanzaro – Itália. Estes autores observaram que 45% das amostras de
alimentos eram positivas para enterococos, sendo que as espécies mais
freqüentes foram E. faecium (35,6%) e E. faecalis (33,3%). Estes autores

Discussão________________________________________________________________________
90
relataram ainda que a população deste microrganismo variou de 2,0 x 101 a
8,0 x 103 UFC/g.
Enterococcus spp. são comumente isolados de uma ampla variedade de
alimentos, já que são capazes de resistir ao processamento térmico, a
processos de salga, cura e maturação de alguns produtos fermentados
(Giraffa, 2002). Além disso, a ocorrência de Enterococcus spp. em leite cru e
em queijos produzidos com leite cru é indicativa de contaminação fecal e
práticas higiênicas inadequadas durante a ordenha (Mannu et al., 2003).
Santos et al. (2004) analisaram a ocorrência de Enterococcus spp. em
amostras de alimentos enterais administrados em ambiente hospitalar e
observaram que 16,6% das 30 amostras industrializadas e artesanais estavam
contaminadas com Enterococcus spp. identificados somente com base em
características bioquímicas. Segundo estes autores, a presença deste
microrganismo indicava práticas inadequadas de manipulação.
E. faecium é a espécie de enterococos mais freqüentemente encontrada
em amostras de alimentos (Moellering 1992, Vancanneyt et al., 2002). Da
mesma forma, no presente trabalho, foram obtidos 139 isolados (46,5%) de E.
faecium, 80 (26,8%) de E. faecalis, 36 (12%) de E. casseliflavus e 8 (2,7%) de
E. gallinarum. Estes resultados também estão de acordo com os dados
relatados por alguns autores de que a espécie de enterococos mais comum em
frutas e vegetais é E. casseliflavus (Devriese et al., 1995; Gelsomino 2002;
McGowan et al., 2006).
Foi observado freqüentemente o isolamento de mais de uma espécie de
enterococos a partir da mesma amostra de alimento. Na prática, isto significa
que testar apenas uma colônia de cada amostra subestimaria a distribuição

Discussão________________________________________________________________________
91
real de Enterococcus spp. nas amostras analisadas e diminuiria as
possibilidades de avaliação de diferenças fenotípicas entre os membros da
mesma espécie.
Neste trabalho, inicialmente, a identificação das espécies de
Enterococcus spp. foi feita com base no perfil fenotípico obtido com reações
bioquímicas e enzimáticas fornecido pelo sistema de identificação API 20
STREP. Entretanto, foram obtidos resultados insatisfatórios para alguns
isolados e a técnica de PCR foi empregada para um estudo comparativo de
identificação. Considerando-se o custo-benefício, a identificação da totalidade
dos isolados foi realizada posteriormente, empregando-se somente a técnica
de PCR.
Nesta técnica, a identificação das espécies de enterococos foi feita por
amplificação de fragmentos característicos que codificam as ligases D-
alanina:D-alanina e as enzimas relacionadas à resistência a glicopeptídeos
específica para algumas espécies deste gênero (Dutka-Malen et al., 1995).
De acordo com os resultados obtidos para a identificação de
Enterococcus spp., observamos que dos isolados inicialmente identificados
pelo sistema API 20 STREP, apenas aqueles identificados como E. faecalis e
E. faecium tiveram a identificação confirmada com os primers específicos para
as espécies. Este resultado está de acordo com o encontrado por Velasco et al.
(2004) e Devriese et al. (1995) que afirmaram que o sistema API 20 STREP foi
mais preciso para identificação de E. faecalis e pouco adequado para
identificação das demais espécies. Segundo estes autores, este sistema não
possui todos os testes necessários para uma correta diferenciação das
espécies de enterococos.

Discussão________________________________________________________________________
92
Da mesma forma, Robredo et al. (2000) utilizaram o sistema API 20
STREP e identificação molecular por hibridização de colônias para a
identificação das espécies de enterococos resistentes à vancomicina em 101
amostras de alimentos de origem animal. Para as cepas identificadas como E.
faecalis pelo API 20 STREP, não foi observada discrepância de resultados.
Entretanto, estes autores relataram que oito cepas previamente identificadas
como E. durans e E. casseliflavus pelo sistema bioquímico não tiveram o perfil
confirmado pelo método molecular, que identificou essas cepas como E.
faecium.
Facklam et al. (1999) em uma revisão sobre Enterococcus afirmaram
que os sistemas comerciais destinados à identificação das espécies deste
microrganismo são falhos e muitas vezes apenas identificam corretamente E.
faecalis.
Em relação à caracterização fenotípica observamos que, ao contrário de
E. faecalis, os isolados de E. faecium não produziram gelatinase. Este
resultado está de acordo com o relatado por outros autores que observaram
uma ausência de produção de gelatinase dentre as cepas de E. faecium
(Coque et al., 1995; Elsner et al., 2000; Eaton e Gasson 2001; Franz et al.,
2001a; Mannu et al., 2003).
A gelatinase é uma protease responsável pela hidrólise de gelatina,
colágeno, hemoglobina e outros peptídeos bioativos. Alguns pesquisadores
relacionam a produção de gelatinase com a indução de processos inflamatórios
(Coque et al., 1995; Hickey et al., 2003).
Segundo Ike et al. (1987), aproximadamente 60% das cepas de E.
faecalis isoladas de pacientes com infecção apresentavam perfil hemolítico,

Discussão________________________________________________________________________
93
enquanto que a análise de material fecal de indivíduos saudáveis revelou uma
prevalência de 17% deste tipo de enterococos. Estes autores também
observaram que, os genes responsáveis pelo perfil hemolítico poderiam ser
transferidos facilmente por meio de plasmídeos conjugativos.
Para a pesquisa da atividade β-hemolítica (citolisina) em cepas de
Enterococcus spp. isoladas a partir de amostras clínicas, Elsner et al. (2000)
utilizaram a mesma metodologia que foi empregada neste trabalho e
observaram que 16% dos E. faecalis apresentavam atividade enquanto que
entre os isolados de E. faecium este perfil não foi observado.
Os resultados de hemólise observados na presente pesquisa estão de
acordo com os resultados obtidos por Fifadara et al. (2003) que analisaram o
perfil hemolítico de cepas de E. faecalis resistentes à vancomicina, isoladas de
150 amostras de carne provenientes da Malásia. Estes autores observaram
que doze das 22 (54,5%) cepas isoladas apresentavam o perfil β-hemolítico.
Miguel et al. (1995) obtiveram, após a análise de 13 cepas de E. faecalis
e 5 de E. faecium isoladas de 55 amostras de alimentos, apenas uma cepa de
E. faecalis com perfil beta-hemolítico enquanto que as demais produziam
hemólise parcial do tipo alfa.
Psoni et al. (2006) também verificaram a presença de atividade β-
hemolítica em 34 isolados de E. durans obtidos de queijo de cabra e
encontraram 29,4% destes isolados positivos para esta característica.
O isolamento de enterococos hemolíticos a partir de alimentos pode ter
implicações sanitárias (Eaton e Gasson 2001; Franz et al., 2001a). Por este
motivo, a ausência de atividade hemolítica deve ser considerada um importante

Discussão________________________________________________________________________
94
critério para a seleção de culturas iniciadoras úteis na produção de alimentos
fermentados (Fifadara et al., 2003).
Por outro lado, algumas cepas de enterococos podem ter características
que justifiquem seu uso como probióticas e/ou como conservadores em
alimentos. Além disso, E. faecalis e E. faecium são capazes de controlar a
multiplicação de Listeria sp. durante o processamento de alguns queijos
através da produção de bacteriocinas (Sulzer e Busse, 1991; Giraffa et al.,
1997).
De acordo com Strompfova et al. (2006) as bacteriocinas de
enterococos, conhecidas como enterocinas, pertencem ao grupo IIa que são
peptídeos pequenos, catiônicos, hidrofóbicos, termicamente estáveis e com
atividade contra uma ampla variedade de microrganismos. Estas bacteriocinas
vêm despertando muito interesse para o uso na indústria de alimentos como
antimicrobianos naturais (Cleveland et al., 2001, Drider et al., 2006). Muitas
pesquisas procuram avaliar esta capacidade e ao mesmo tempo garantir que
as cepas de enterococos que apresentam esta característica sejam seguras
para serem utilizadas na produção de alimentos como probióticas, starters ou
conservadoras.
A prevalência encontrada para produção de bacteriocina por E. faecalis
e E. faecium em nosso trabalho, está de acordo com relatos da literatura.
Yousif et al. (2005) analisaram as propriedades tecnológicas de enterococos
isolados a partir de um produto africano fermentado e dentre as 22 cepas de E.
faecium obtidas, seis (27,3%) foram capazes de produzir bacteriocinas contra
L. monocytogenes ATCC 7644, L. innocua WS 2258 e S. aureus DSM 6732.
De Vuyst et al. (2003) analisaram a atividade inibitória de 426

Discussão________________________________________________________________________
95
enterococos de diferentes origens contra diversas bactérias indicadoras
incluindo espécies relacionadas, bactérias láticas (BAL), bactérias patogênicas
e deteriorantes de alimentos. Dentre as 426 cepas, 122 pertencentes
principalmente às espécies E. faecalis e E. faecium mostraram atividade
bacteriocinogênica.
Saavedra et al. (2003) selecionaram 11 cepas de E. faecium obtidas de
um queijo artesanal, Tafí, característico de Tucumán - Argentina para avaliação
das propriedades tecnológicas. Das cepas analisadas, duas (18,2%)
produziram bacteriocina com atividade inibitória frente à L. innocua e L.
monocytogenes. Essas cepas apresentaram ausência de fatores de virulência
e poderiam desta forma ser aplicadas como culturas iniciadoras garantindo a
segurança na fabricação deste tipo de queijo e mantendo as características
sensoriais desejadas.
Ben Omar et al. (2004) analisaram algumas características de
enterococos obtidos de diversos alimentos da Espanha. Dos 50 isolados de
enterococos obtidos, três E. faecalis (21,4%) e 14 E. faecium (38,8%) foram
capazes de inibir o crescimento de E. faecalis A-48-32, evidenciando a possível
aplicação destas cepas na conservação de alimentos, como probióticas ou
como culturas iniciadoras.
Uma outra característica de enterococos e algumas bactérias láticas que
vem despertando interesse é a capacidade de hidrolisar sais biliares
contribuindo, conseqüentemente, com a diminuição da concentração de
colesterol na circulação sanguínea, quando utilizados como probióticos (Franz
et al., 2001b; Pereira et al., 2003). Franz et al. (2001b) analisaram 117 cepas
de enterococos previamente isoladas de alimentos quanto à capacidade de

Discussão________________________________________________________________________
96
hidrolisar sais biliares em MRS suplementado com ácido taurodeoxicólico e
cloreto de cálcio. Dentre as cepas analisadas, houve uma maior prevalência de
hidrólise de sais biliares para E. faecalis (81%) seguido por E. faecium (50%) e
E. durans (44%).
Rossi et al. (1999) avaliaram as características probióticas de E. faecium
CRL183 que seria utilizado na formulação de um leite de soja fermentado e
observaram que este microrganismo na presença de Lactobacillus jugurti foi a
melhor combinação em termos sensoriais e tecnológicos e além disso,
promoveu uma redução de 43% dos níveis de colesterol, in vitro.
Em relação à ocorrência de determinantes fenotípicos de virulência em
Enterococcus spp., nossos dados revelam uma maior prevalência destes
fatores em E. faecalis quando comparado à E. faecium. Entretanto,
considerando-se que ainda não foram totalmente esclarecidos quais fatores
são realmente responsáveis pela patogenicidade de enterococos, a ausência
fenotípica de alguns dos fatores de virulência não implica necessariamente que
se trata de uma cepa não patogênica.
Por este motivo, Franz et al. (2001a) analisaram a prevalência fenotípica
e molecular dos marcadores de virulência entre isolados de enterococos
obtidos de amostras de alimentos e que poderiam ser utilizados como culturas
iniciadoras. Das 48 cepas de E. faecium analisadas, apenas 10% foram
positivas para um ou mais determinantes de virulência enquanto que, para as
47 cepas de E. faecalis analisadas, esse valor foi de 78,7%. Neste trabalho, foi
observada uma maior prevalência de marcadores genotípicos de virulência
para E. faecalis quando comparado a E. faecium.
De uma forma geral, 33,8% dos isolados de E. faecium e 96,2% de E.

Discussão________________________________________________________________________
97
faecalis foram gel+. Eaton e Gasson (2001) mostraram que 78,0% das cepas
de E. faecalis obtidas de alimentos possuíam o gene gel enquanto que todas
as cepas de E. faecium foram negativas para o mesmo gene. Resultados
semelhantes foram obtidos por Franz et al. (2001a). Estes autores relataram
que 23 (48,9%) cepas de E. faecalis e nenhuma de E. faecium isoladas de
alimentos produziam gelatinase.
Analisando cepas de enterococos obtidas de amostras clínicas, Duprè et
al. (2003) observaram que 73,3% das cepas de E. faecalis foram gel+,
enquanto que Vankerckhoven et al. (2004) não encontraram este gene em 271
cepas de E. faecium. Da mesma forma, Camargo et al. (2006) não obtiveram
isolados positivos para gel e cyl, quando analisaram a ocorrência de
marcadores de virulência em 56 isolados de E. faecium resistentes ou não à
vancomicina obtidos de amostras fecais e de infecções em pacientes no Brasil.
A prevalência de efaA e cyl encontrada em nosso trabalho está de
acordo com os resultados relatados na literatura (Eaton e Gasson, 2001; Yousif
et al. 2005). Além disso, também foi observado que o grupo de alimento que
apresentou a maior prevalência de E. faecium positivo para efaA foi o grupo de
queijos. Mannu et al. (2003) também registraram uma maior prevalência
(47,5%) de gene efaA em cepas de E. faecium isoladas de produtos lácteos.
Os genes esp e as não foram encontrados em nenhum isolado de E.
faecium analisado neste trabalho. Estes resultados estão de acordo com os
resultados previamente relatados por Eaton e Gasson (2001), Franz et al.
(2001a) e Mannu et al. (2003). Ao contrário dos dados obtidos em nosso
trabalho, Camargo et al. (2006) observaram que 13 (56%) isolados de E.
faecium resistentes à vancomicina possuíam o gene esp. No trabalho de

Discussão________________________________________________________________________
98
Duprè et al. (2003), o gene ace foi detectado em 9 das 15 cepas de E. faecalis
enquanto que entre E. faecium este gene não foi encontrado. No presente
trabalho, todas os isolados de E. faecalis possuíam o gene ace.
De acordo com Di Rosa et al. (2006) produção de biofilme foi
recentemente reconhecida como um importante fator de patogenicidade de
Enterococcus spp. e sua produção pode ser afetada por condições ambientais,
atividade de gelatinase e presença do gene esp (Toledo-Arana et al., 2001; Di
Rosa et al., 2006). Para os isolados de E. faecalis, obtidos neste trabalho,
produtores de biofilme em superfície abiótica foi observada uma maior
ocorrência do gene gel.
Toledo - Arana et al. (2001) avaliaram a possível relação entre a
capacidade de cepas de E. faecalis produzirem biofilme e a presença do gene
esp. Eles observaram que 87 das 93 cepas de E. faecalis (93,5%) que
possuíam o gene esp, foram capazes de produzir biofilme in vitro. A mesma
associação não foi observada para as 59 cepas que não possuíam este gene.
Por outro lado, Mohamed e Murray (2005) mostraram uma ausência de
correlação entre formação de biofilme e produção de gelatinase para
enterococos isolados de amostras fecais e clínicas.
De acordo com os resultados obtidos, foi obtido um maior número de
isolados de E. faecalis adesivos às células da linhagem Caco-2 quando
comparados a E. faecium. Duprè et al. (2003) avaliaram a prevalência de
genes de virulência em cepas clínicas de E. faecium e E. faecalis obtidos na
Sardenha – Itália. Estes autores observaram que 6 das 15 cepas de E. faecalis
(40%) foram capazes de aderir às células da linhagem Caco-2, enquanto que
apenas 3 cepas de E. faecium (9,4%) apresentaram o mesmo resultado.

Discussão________________________________________________________________________
99
Além dos fatores de virulência já discutidos, é reconhecido que a
presença de enterococos em alimentos é controversa devido ao aumento da
ocorrência de cepas resistentes a antibióticos de uso clínico e relacionadas a
infecções nosocomias (Ben Omar et al., 2004). No Brasil, desde o isolamento
da primeira cepa de E. faecium resistente à vancomicina a partir de amostra de
sangue de uma criança com anemia aplástica (Dalla Costa et al., 1998), várias
cepas resistentes a este antibiótico foram isoladas em diferentes hospitais em
São Paulo e outras regiões (Zanella et al., 2003; Camargo et al., 2004).
Neste estudo, 16,5% dos isolados de E. faecium e 36,2% dos isolados
de E. faecalis mostraram perfil de resistência a um ou mais dos antibióticos
testados. Foram obtidos E. faecium com perfil de resistência à vancomicina,
tetraciclina e eritromicina, enquanto que as cepas de E. faecalis apresentaram
resistência à alta concentração de gentamicina, tetraciclina e eritromicina.
Busani et al. (2004) também relataram a existência de cepas de E.
faecium e E. faecalis resistentes à eritromicina e tetraciclina, obtidas de carne
crua de frango e porco. Neste trabalho, foi observado o perfil de resistência
múltipla à gentamicina, eritromicina e tetraciclina para três cepas de E.
faecalis.
De acordo com Franz et al. (1999) a presença de resistência baseada
na aquisição de plasmídeos e transposons é relevante para eritromicina, altas
concentrações de aminoglicosídeos (gentamicina), tetraciclinas, beta-
lactâmicos (ampicilina) e glicopeptídeos (vancomicina). Da mesma forma, a
ocorrência concomitante de resistência à tetraciclina e gentamicina é comum
entre cepas de enterococos.
Vários trabalhos relatam a existência de cepas de enterococos isoladas

Discussão________________________________________________________________________
100
de alimentos resistentes à vancomicina. Araya et al. (2005) encontraram 8%
das cepas de Enterococcus spp., isoladas de leite não pasteurizado,
resistentes à vancomicina. Já, Çitak et al. (2004) analisaram 101 cepas de
Enterococcus spp. obtidas de amostras de queijo turco e relataram que
resistência à estreptomicina, eritromicina, oxacilina e vancomicina foi
freqüentemente encontrada entre os isolados sendo que, 96,8% das cepas de
E. faecalis e 76% das cepas de E. faecium eram resistentes à vancomicina. De
acordo com estes autores, a presença de enterococos resistentes a
antibióticos em alimentos, reforça o possível papel dos alimentos na
persistência e transmissão de cepas resistentes a antimicrobianos.
Harada et al. (2004) analisaram 137 cepas de Enterococcus spp.
isoladas de carne de frango, proveniente de diversas regiões (Brasil, China e
Japão) e observaram resistência à alta concentração de gentamicina. Este tipo
de resistência apresentada por enterococos, segundo estes autores, pode ser
explicada pela ação bifuncional da enzima modificadora de aminoglicosídeo
AAC(6’)-APH(2’’). Esta enzima anula o efeito bactericida sinérgico quando
gentamicina é usada em combinação com ampicilina ou vancomicina. Dos
isolados de E. faecalis obtidos a partir de amostras de leite neste estudo, 72%
foram resistentes à gentamicina, reforçando as observações descritas por
Harada et al. (2004).
Franz et al. (2001a) analisaram o perfil de resistência de 48 cepas de E.
faecium e 47 cepas de E. faecalis a antibióticos de uso clínico. Os autores
encontraram uma cepa de E. faecium (2,1%) resistente à vancomicina, 27,1%
resistentes à eritromicina, 6,3% resistentes à tetraciclina e 2,1% resistentes à
gentamicina e nenhuma cepa resistente à ampicilina. Para E. faecalis

Discussão________________________________________________________________________
101
encontraram 44,7% das cepas resistentes à tetraciclina, 31,9% à eritromicina,
25,5% à gentamicina e nenhuma resistente à vancomicina. Estes mesmos
autores afirmam ainda que é difícil prever o impacto da resistência a
antibióticos apresentada por enterococos isolados de alimentos em relação à
sua patogenicidade.
Por este motivo, além da caracterização fenotípica e molecular dos
fatores de virulência apresentados por Enterococcus sp, vários autores
buscam correlacionar estes resultados com a origem dos isolados. Para tanto,
a técnica de análise por RAPD-PCR utilizando o primer M13 vem sendo
empregada com sucesso para avaliação da diversidade genética de isolados
de enterococos (Andrighetto et al., 2001; Vancanneyt et al., 2002; Cosentino et
al., 2004; Yousif et al., 2005). A aplicação desta metodologia para análise dos
52 isolados de enterococos resistentes a antibióticos obtidos neste estudo,
mostrou que isolados com diferentes perfis fenotípico e genotípico podiam
apresentar o mesmo perfil de RAPD.
Gelsomino et al. (2001) relataram uma baixa sensibilidade deste método
para distinguir isolados de enterococos, obtidos de amostras de queijo tipo
Cheddar e do local de produção, e relacionar uma possível fonte de
contaminação.
Ao contrário do que ocorre com o grupo de coliformes totais e fecais,
não há um nível estabelecido para relacionar a presença de Enterococcus spp.
e a qualidade higiênica de um determinado produto (Giraffa et al., 1997). A
legislação brasileira não preconiza a contagem de enterococos em alimentos,
mas devido à importância deste grupo de microrganismo e de relatos da
literatura sobre a utilidade de pesquisar enterococos em águas, foi comparada

Discussão________________________________________________________________________
102
a ocorrência de enterococos e indicadores clássicos de higiene. Além disso,
Ingham e Schmidt (2000) afirmam que Enterococcus spp. são geralmente mais
tolerantes à refrigeração quando comparados com E. coli e outros coliformes e
parecem sobreviver por mais tempo durante a estocagem refrigerada de
carcaças de carnes.
Não houve neste trabalho consenso em relação à presença destes
microrganismos. Em alguns casos, embora para algumas amostras de
produtos cárneos (9, 32, 34, 49 e 52), queijos (8, 11, 15, 18, 26, 30, 73 e 76) e
vegetais (36, 58, 59 e 88) a contagem de E. coli estivesse próxima dos valores
de coliformes fecais, contemplados pela legislação (Brasil, RDC nº 12, de 02 de
janeiro de 2001), a contagem de Enterococcus spp. superava a contagem
deste indicador.
Por outro lado, em algumas das amostras de alimentos, principalmente
amostras de leite cru, a contagem de E. coli estava alta, enquanto que a
contagem de enterococos permaneceu abaixo do limite de detecção do
método. Este resultado, em particular evidencia a necessidade de estudos mais
detalhados estabelecendo critérios mais adequados para uma possível
aplicação de enterococos como indicador de contaminação fecal.
Para as amostras de água mineral também não foi possível correlacionar
a presença de coliformes totais, E. coli e enterococos, já que em todas as
amostras a contagem destes microrganismos ficou abaixo do limite de
detecção dos métodos empregados. Entretanto, em todos os casos estes
microrganismos estavam dentro do limite exigido pela legislação que é de < 1
UFC em 100 mL para coliformes totais e enterococos e ausência em 100 mL
para E. coli (Brasil - RDC nº 54, de 15 de junho de 2000).

Discussão________________________________________________________________________
103
Alguns pontos devem ser apontados em relação ao uso de enterococos
como indicadores de contaminação fecal. O primeiro problema é que nem
todos os isolados presuntivos de enterococos foram identificados como
Enterococcus spp. Desta forma, a existência de um método mais específico
(Petrifilm) para detecção de E. coli faz com que o uso deste microrganismo
como indicador de higiene seja mais adequado quando comparado a
Enterococcus spp. (Ingham e Schmidt 2000). Além disso, a presença de
enterococos é desejável em muitos alimentos, sendo assim, a interpretação de
sua presença nesses casos deve ser feita com cautela (Giraffa, 2002; Blanch et
al., 2003; Choi et al., 2003).

__________________6.Conclusões

Conclusões________________________________________________________________________105
Com base nos resultados obtidos e na discussão apresentada, pode-se
concluir que:
• Enterococcus spp. foram isolados a partir de diversos alimentos
comercializados na cidade de Ribeirão Preto – SP, e foi freqüente a
ocorrência de diferentes espécies de enterococos na mesma amostra de
alimento, evidenciando a ampla distribuição deste microrganismo;
• O meio de cultura Kanamicina Esculina Azida foi mais adequado para a
enumeração de Enterococcus spp. a partir de amostras de alimentos,
quando comparado ao meio Bile Esculina;
• Foi detectada a ocorrência de Enterococcus spp. resistentes a antibióticos,
indicando que os alimentos podem desempenhar um papel importante na
disseminação de cepas resistentes a antibióticos de uso clínico;
• A ocorrência de diversas espécies, além de E. faecalis e E. faecium, limitou
o emprego do API 20 Strep para a identificação dos isolados obtidos a partir
de amostras de alimentos;
• Houve uma maior prevalência de marcadores de virulência (fenotípicos e
genotípicos) entre os isolados de E. faecalis quando comparado à E.
faecium, independentemente da resistência a antibióticos, evidenciando o
risco associado à presença destes microrganismos em alimentos;

Conclusões________________________________________________________________________106
• Isolados de E. casseliflavus e E. gallinarum mostraram-se capazes de
hidrolisar sais biliares, justificando a necessidade de estudos mais
detalhados com estes microrganismos para futura aplicação como
probióticos;
• A análise de 52 isolados por meio da técnica de RAPD-PCR, perfil de
resistência a antibióticos e características fenotípicas permitiu a
individualização de 17 cepas de E. faecalis e 14 cepas de E. faecium;
• Para as amostras de água, não foi possível estabelecer uma correlação
entre a enumeração de coliformes, E. coli e enterococos, pois todas as
contagens desses indicadores estavam abaixo do limite de detecção do
método. Para as amostras de vegetais, carnes, leite e queijos, a
variabilidade dos resultados impediu o estabelecimento de uma correlação
entre a presença ou contagem de coliformes, E. coli e enterococos;
• Embora algumas das cepas analisadas apresentassem características
tecnológicas que justificassem o seu uso como probióticas e até como
conservadoras, a identificação de marcadores fenotípicos de virulência e a
presença de cepas com perfil de resistência a antibióticos de uso clínico
evidenciam a necessidade de aprofundamento dos estudos com estas
cepas.

______7. Referências Bibliográficas*
* De acordo com: Universidade de São Paulo. Sistema Integrado de Bibliotecas / Grupo diTeses. Diretrizes para apresentação de dissertações e teses da USP: documento eletrônico e impresso. Vânia M. B. de Oliveira Funaro, coord. Et al. – São Paulo: SIBi/USP, 2004.

Referências Bibliográficas____________________________________________________________108
AGUIRRE, M.; COLLINS, M.D. Lactic acid bacteria and human clinical infection.
Journal of Applied Bacteriology, v. 75, p. 95-107, 1993.
ANDRIGHETTO, C., KNIJFF, E., LOMBARDI, A., TORRIANI, S.,
VANCANNEYT, M., KERSTERS, K., SWINGS, J., DELLAGLIO, F. Phenotypic
and genetic diversity of enterococci isolated from Italian cheeses. Journal of
Dairy Research, v. 68, p. 303-316, 2001.
ARAYA, M., DAVIDOVICH, G., ARIAS, M.L., CHAVES, C. Identification of
Enterococcus spp. isolated from raw milk samples coming from the metropolitan
area of Costa Rica and evaluation of its antibiotic sensibility pattern. Archivos
Latinoamericanos Nutrición, v. 55, p. 161-166, 2005. (Resumo)
BAM - Bacteriological Analytical Manual Online. FENG, WEAGANT, S.D.,
GRANT, M.A. Enumeration of Escherichia coli and the coliform bacteria. In:
Food and Drug Administration. Washington: FDA, 2001, chapter 4. Disponível
em: http://www.cfsan.fda.gov/~ebam/bam-toc.html. Acesso: 28/07/03.
BENYACOUB, J., PÉREZ, P. F., ROCHAT, F., SAUDAN, K. Y., REUTELER,
G., ANTILLE, N. HUMEN, M., DE ANTONI, G. L., CAVADINI, C., BLUM, S.,
SCHIFFRIN, E. J. Enterococcus faecium SF68 enhances the immune response
to Giardia intestinalis in mice. Nutritional Immunology. v.135, p.1171–1176,
2005.

Referências Bibliográficas____________________________________________________________109
BEN OMAR, N., CASTRO, A., LUCAS, ABRIOUEL, H, YOUSIF, N.M.K.,
FRANZ, C.M.A.P., HOLZAPFEL, W.H., PÉREZ-PULIDO, R., MARTÍNEZ-
CANAMERO, M., GÁLVEZ, A. Functional and safety aspects of enterococci
isolated from different Spanish foods. Systematic and Applied Microbiology,
v. 27, p.118-130, 2004.
BLANCH, A.R., CAPLIN, J.L., IVERSEN, A., KÜHN, I., MANERO, A., TAYLOR,
H.D., VILANOVA, X. Comparison of enterococcal populations related to urban
and hospital wastewater in various climatic and geographic European regions.
Journal of Applied Microbiology, v. 94, p.994-1002, 2003.
BOOTH, M.C., BOGIE, C.P., SAHL, H.G., SIEZEN, R.J., HATTER, K.L.,
GILMORE, M.S. Structural analysis and proteolytic activation of Enterococcus
faecalis cytolysin, a novel lantibiotic. Molecular Microbiology, v. 21, p.1175-
1184, 1996.
BORGEN, K., SORUM, M., WASTESON, Y., KRUSE, H. VanA-type
vancomycin-resistant enterococci (VRE) remain prevalent in poultry carcasses
3 year after avoparcin was banned. International Journal of Food
Microbiology, v. 64, p. 89-94, 2001.
BRASIL. Resolução -RDC nº. 54, de 15 de junho de 2000. Dispõe sobre o
Regulamento Técnico para Fixação de Identidade e Qualidade de Água Mineral
Natural e Água Natural. Diário Oficial da União, Brasília, 19 de junho de 2000.

Referências Bibliográficas____________________________________________________________110
BRASIL. Resolução - RDC nº 12, de 02 de janeiro de 2001. Aprova o
Regulamento Técnico sobre padrões microbiológicos para alimentos. Diário
Oficial da União, Brasília, 10 de janeiro de 2001.
BUSANI, L., GROSSO, M. PALADINI, C., GRAZIANI, C., PANTOSTI, A.,
BIAVASCO, F. CAPRIOLI, A. Antimicrobial susceptibility of vancomycin-
susceptible and -resistant enterococci isolated in Italy from raw meat products,
farm animals, and human infections. International Journal of Food
Microbiology, v.97, p. 17-22, 2004.
CALICIOGLU, M., BUEGE, D.R., INGHAM, S.C., LUCHANSKY, J.B. Recovery
of Escherichia coli biotype I and Enterococcus spp. during refrigerated storage
of beef carcasses inoculated with a fecal slurry. Journal of Food Protection,
v.62, p.944-947, 1999.
CAMARGO, I. L. B. C., DEL PELOSO, P. F., DA COSTA LEITE, C. F.,
GOLDMAN, G. H., DARINI, A. L. C. Identification of an unusual vanA element in
glycopeptide-resistant Enterococcus faecium in Brazil following international
transfer of a bone marrow transplant patient. Canadian Journal of
Microbiology, v. 50, p. 767-770, 2004.
CAMARGO, I. L. B. C., GILMORE, M. S., DARINI, A. L. C. Multilocus sequence
typing and analysis of putative virulence factors in vancomycin-resistant and
vancomycin-sensitive Enterococcus faecium isolates from Brazil. Clinical
Microbiology and Infection, v. 12, p. 1123-1130, 2006.

Referências Bibliográficas____________________________________________________________111
CANADÁ. Health Products and Food Branch. Enumeration of E.coli and
coliforms in food products and food ingredients using 3MTM PetrifilmTM E. coli
count plates. 2001. Disponível em: http://www.hc-sc.gc.ca/food-aliment. Acesso
em 28/07/03.
CELICO, F., VARCAMONTI, M., GUIDA, M., NACLERIO, G. Influence of
precipitation and soil on transport of fecal enterococci in fractured limestone
aquifers. Applied and Environmental Microbiology, v.70, p. 2843–2847,
2004.
CHINGWARU, W., MPUCHANE, S.F., GASHE, B.A. Enterococcus faecalis and
Enterococcus faecium isolates from milk, beef, and chicken and their antibiotic
resistance. Journal of Food Protection, v. 66 n. 6, p. 931-936, 2003.
CHOI, S., CHU, W., BROWN, J., BECKER, S.J., HARWOOD, V.J., JIANG, S.C.
Application of enterococci antibiotic resistance patterns for contamination
source identification at Huntington Beach, California. Marine Pollution
Bulletin, v. 46, p. 748-755, 2003.
ÇITAK, S., YUCEL, N. ORHAN, S. Antibiotic resistance and incidence of
Enterococcus species in turkish white cheese. International Journal of Dairy
Technology, v. 57, p. 27-31, 2004.

Referências Bibliográficas____________________________________________________________112
CLEVELAND, J., MONTVILLE, T. J., NES, I. F., CHIKINDAS, M. L.
Bacteriocins: safe, natural, antimicrobials for food preservation. International
Journal of Food Microbiology, v. 71, p. 1-20, 2001.
CLSI – Clinical and Laboratory Standards Institute. Methods for dilution
antimicrobial susceptibility tests for bacteria that grow aerobically. 5th ed., 20,
approved standard M7-A5. Wayne, Pennsylvania, 2005.
COQUE, T.M., PATTERSON, J.E., STECKELBERG, J.M., MURRAY, B.E.
Incidence of hemolysin, gelatinase, and aggregation substance among
enterococci isolated from patients with endocarditis and other infections and
from feces of hospitalized and community-based persons. Journal of
Infectious Diseases, v. 171, p.1223-1229, 1995.
COSENTINO, S., PISANO, M.B., CORDA, A., FADDA, M.E., PIRAS, C.
Genotypic and technological characterization of enterococci isolated from
artisanal Fiore Sardo cheese. Journal of Dairy Research, v. 71, p.444-450,
2004.
DALLA COSTA, L. M., SOUZA, D. C., MARTINS, L. T., ZANELLA, R. C.,
BRANDILEONE, M. C., BOKERMANN, S., SADER, H. S., SOUZA, H. A.
Vancomycin-resistant Enterococcus faecium: first case in Brazil. Brazilian
Journal of Infectious Disease, v. 2, p. 160-163, 1998 (Resumo).

Referências Bibliográficas____________________________________________________________113
DE VUYST, L., FOULQUIE MORENO, M.R., REVETS, H. Screening for
enterocins and detection of hemolysin and vancomycin resistance in
enterococci of different origins. International Journal of Food Microbiology,
v. 84, p.299-318, 2003.
DEVRIESE, L.A., POT, B., COLLINS, M.D. Phenotypic identification of the
genus Enterococcus and differentiation of phylogenetically distinct enterococcal
species and species groups. Journal of Applied Bacteriology, v. 75, p. 399-
408, 1993.
DEVRIESE, L.A., POT, B., DAMME, L.V., KERSTERS, K., HAESEBROUCK, F.
Identification of Enterococcus species isolated from foods of animal origin.
International Journal of Food Microbiology, v. 26, p. 187-197, 1995.
DI ROSA, R, CRETI, R., VENDITTI, M., D’AMELIO, R., ARCIOLA, C.R.,
MONTANARO, L, BALDASSARRI, L. Relationship between biofilm formation,
the enterococcal surface protein (Esp), and gelatinase in clinical isolates of
Enterococcus faecalis and Enterococcus faecium. FEMS Microbiology
Letters, v. 256, p.145-150, 2006.
DOMIG, K.J., MAYER H.K., KNEIFEL, W. Methods used for the isolation,
enumeration, characterisation and identification of Enterococcus spp. 1. Media
for isolation and enumeration. International Journal of Food Microbiology, v.
88, p. 147-164, 2003.

Referências Bibliográficas____________________________________________________________114
DRIDER, D., FIMLAND, G., HÉCHARD, Y., McMULLEN, L. M., PRÉVOST, H.
The continuing story of class IIa bacteriocins. Microbiology and Molecular
Biology Reviews, v. 70, p. 564-582, 2006.
DUPRÈ, I., ZANETTI, S., SCHITO, A.M., FADDA, G., SECHI, L.A. Incidence of
virulence determinants in clinical Enterococcus faecium and Enterococcus
faecalis isolates collected in Sardinia (Italy). Journal of Medical Microbiology,
v.52, p.491-498, 2003.
DUTKA-MALLEN, S., EVERS, S., COURVALIN, P. Detection of glycopeptide
resistance genotypes and identification to the species level of clinically relevant
enterococci by PCR. Journal of Clinical Microbiology, v. 33, p.24-27, 1995.
EATON , T.J., GASSON M.J. Molecular screening of Enterococcus virulence
determinants and potential for genetic exchange between food and medical
isolates. Applied and Environmental Microbiology, v. 67, p.1628-1635, 2001.
ELSNER, H.A, SOBOTTKA, I., MACK, D., CLAUSSEN, M., LAUFS, R., WIRTH,
R. Virulence factors of Enterococcus faecalis and Enterococcus faecium blood
culture isolates. European Journal of Clinical Microbiology and Infectious
Diseases, v. 19, p.39-42, 2000.
FACKLAM, R.R., COLLINS, M.D. Identification of Enterococcus species
isolated from human infections by a conventional test scheme. Journal of
Clinical Microbiology, v. 27, p. 731-734, 1989.

Referências Bibliográficas____________________________________________________________115
FACKLAM, R., SAHM, D.F., TEIXEIRA, L.M. Enterococcus. In: MURRAY, P.R.,
BARON, E.J., PFALLER, M.A, TENOVER, F.C., YOLKEN, R.H. Manual of
Clinical Microbiology. 7 ed. Washington D. C.: ASM Press, 1999, p.297-305.
FIFADARA, N., RADU, S., HASSAN, Z., BEUCHAT, L.R., RUSUL, G.
Hemolytic and nonhemolytic vancomycin-resistant Enterococcus faecalis
isolated from beef imported to Malaysia. Journal of Food Protection, v.66,
p.1845-1850, 2003.
FOULQUIÉ-MORENO, M.R., SARANTINOPOULOS, P., TSAKALIDOU, E., DE
VUYST, L. The role and application of enterococci in food and health.
International Journal of Food Microbiology, v. 106, p. 1-24, 2006.
FRANZ, C.M.A.P., HOLZAPFEL, W.H., STILES, M.E. Enterococci at the
crossroads of food safety? International Journal of Food Microbiology, v. 47,
p. 1-24, 1999.
FRANZ, C.M.A.P., MUSCHOLL-SILBERHORN, A.B., YOUSIF, M.K.,
VANCANNEYT, M., SWINGS, J., HOLZAPFEL, W. Incidence of virulence
factors and antibiotic resistance among enterococci isolated from food. Applied
and Environmental Microbiology, v. 67, p. 4385-4389, 2001(a).
FRANZ, C.M.A.P., SPECHT, I. HABERER, P., HOLZAPFEL, W.H. Bile salt
hydrolase activity of enterococci isolated from food: screening and quantitative
determination. Journal of Food Protection, v. 64, p. 725-729, 2001(b).

Referências Bibliográficas____________________________________________________________116
FRANZ, C.M.A.P., STILES, M.E., SCHLEIFER, K.H., HOLZAPFEL, W.H.
Enterococci in foods – a conundrum for food safety. International Journal of
Food Microbiology, v. 88, p.105-122, 2003.
GARCÍA, M.C., RODRÍGUEZ, M.J. BERNARDO, A TORNADIJO, M.E.,
CARBALLO, J. Study of enterococci and micrococci isolated throughout
manufacture and ripening of SAN Simón cheese. Food Microbiology, v. 19,
p.23-33, 2002.
GELSOMINO, R., VANCANNEYT, M., COGAN, T.M., CONDON, S., SWINGS.
Source of enterococci in a farmhouse raw-milk cheese. Applied and
Environmental Microbiology, v. 68, p. 3560-3565, 2002.
GELSOMINO, R., VANCANNEYT, M., CONDON, S., SWINGS, J., COGAN,
T.M. Enterococcal diversity in the environment of an Irish cheddar-type
cheesemaking factory. International Journal of Food Microbiology, v. 71,
p.177-188, 2001.
GIRAFFA, G. Enterococci from foods. FEMS Microbiology Reviews, v.26,
p.163-171, 2002.
GIRAFFA, G., CARMINATI, D., NEVIANI, E. Enterococci isolated from dairy
products: a review of risks and potential technological use. Journal of Food
Protection, v. 60, p. 732-738, 1997.

Referências Bibliográficas____________________________________________________________117
HARADA, T., MITO, Y., OTSUKI, K e MURASE, T. Resistance to gentamicin
and vancomycin in enterococcal strains isolated from retail broilre chickens in
Japan. Journal of Food Protection, v. 67, p. 2292-2295, 2004.
HARDIE, J.M., WHILEY, R.A. Classification and overview of the genera
Streptococcus and Enterococcus. Journal of Applied Microbiology, v. 83, p.
1S-11S, 1997.
HICKEY, R.M., TWOMEY, D.P., ROSS, R.P., HILL, C. Production of enterolysin
A by raw milk enterococcal isolate exhibiting multiple virulence factors.
Microbiology, v. 149, p.655-664, 2003.
HUEY B., HALL, J. Hypervariable DNA fingerprinting in Escherichia coli:
minisatellite probe from bacteriophage M13. Journal of Bacteriology, v.171,
p.2528-2532, 1989.
HUGAS, M., GARRIGA, M., AYMERICH., M. T. Functionalty of enterococci in
meat products. International Journal of Food Microbiology, v. 88, p. 223-
233, 2003.
IKE, Y., HASHIMOTO, H., CLEWELL, D.B. High incidence of hemolysin
production by Enterococcus (Streptococcus) faecalis strains associated with
human parenteral infections. Journal of Clinical Microbiology, v. 25, p.1524-
1528, 1987.

Referências Bibliográficas____________________________________________________________118
INGHAM, S.C. e SCHMIDT, D. Alternative indicator bacteria analyses for
evaluating the sanitary condition of beef carcasses. Journal of Food
Protection, v. 63, p. 51-55, 2000.
IVERSEN, A., KÜHN, I., FRANKLIN, A., MÖLLBY, R. High prevalence of
vancomycin-resistant enterococci in Swedish sewage. Applied and
Environmental Microbiology, v.68, p. 2838-2842, 2002.
JUÁREZ-FIGUEROA, L.A., SILVA-SÁNCHEZ, J., URIBE-SALAS, F.J.,
CIFUENTES-GARCÍA, E. Microbiological indicators of water quality in the
Xochomilco canals, Mexico City. Salud Pública de México, v.45, p. 389-395,
2003.
JURKOVIČ, D., KRIZKOVÁ, L., DUSINSKÝ, R., BELICOVÁ, A., SOJKA, M.,
KRAJČOVIČ, J., EBRINGER, L. Identification and characterization of
enterococci from bryndza cheese. Letters of Applied Microbiology, v.42, p.
553-559, 2006.
KALINA, A.P. The taxonomy and nomenclature of enterococcus. International
Journal of Systematic Bacteriology, v.20, p.185-189, 1970 apud
MORRISON, D., WOODFORD, N., COOKSON, B. Enterococci as emerging
pathogens of humans. Journal of Applied Microbiology, v. 83, p. 89S-99S,
1997.

Referências Bibliográficas____________________________________________________________119
KAYAOGLU, G. ORSTAVIK, D. Virulence factors of Enterococcus faecalis:
relationship to endodontic disease. Critical Reviews in Oral Biology and
Medicine, v. 15, p. 308-320, 2004.
KLEIN, G., PACK, A., REUTER, G. Antibiotic resistance patterns of enterococci
and occurrence of vancomycin-resistant enterococci in raw minced beef and
pork in Germany. Applied and Environmental Microbiology, v. 64, p.1825-
1830, 1998.
KLINGBERG, T.D., AXELSSON, L., NATERSTAD, K., ELSSER, D., BUDDE,
B.B. Identification of potential probiotic starter cultures for Scandinavian-type
fermented sausages. International Journal of Food Microbiology, v. 105, p.
419-431, 2005.
LEWUS, C.B., KAISER, A., MONTVILLE, T.J. Inhibition of foodborne bacterial
pathogens by bacteriocins from lactic acid bacteria. Applied and
Environmental Microbiology, v. 57, p.1683-1688, 1991.
MANNU, L., PABA, A., DAGA, E., COMUNIAN, R., ZANETTI, S., DUPRÈ, I.,
SECHI, L.A. Comparison of the incidence of virulence determinants and
antibiotic resistance between Enterococcus faecium strains of dairy, animal and
clinical origin. International Journal of Food Microbiology, v. 88, p.291-304,
2003.

Referências Bibliográficas____________________________________________________________120
McGOWAN, L.L., JACKSON, C. R., BARRETT, J. H. HIOTT, L. M., FEDORKA-
CRAY, P. J. Prevalence and antimicrobial resistance of enterococci isolated
from retail fruits, vegetables and meats. Journal of Food Protection, v. 69, p.
2976-2982, 2006.
MIGUEL, M.A.L., TEIXEIRA, L.M., NOLETO, A.L.S. Characterization of
Enterococcus strains isolated from ready-to-eat foods. Revista de
Microbiologia, v. 26, p. 121-124, 1995.
MOELLERING Jr., R.C. Emergence of Enterococcus as a significant pathogen.
Clinical Infectious Diseases, v. 14, p.1173-1178, 1992.
MOHAMED, J.A., MURRAY, B.E. Lack of correlation of gelatinase production
and biofilm formation in a large collection of Enterococcus faecalis isolates.
Journal of Clinical Microbiology, v. 43, p. 5405-5407, 2005.
MORRISON, D., WOODFORD, N., COOKSON, B. Enterococci as emerging
pathogens of humans. Journal of Applied Microbiology, v. 83, p. 89S-99S,
1997.
MUNDY, L.M., SAHM, D.F. and GILMORE, M. Relationships between
enterococcal virulence and antimicrobial resistance. Clinical Microbiology
Reviews, v. 13, p.513-522, 2000.

Referências Bibliográficas____________________________________________________________121
NALLAPAREDDY, S.R., QIN, X., WEINSTOCK, G.M., HOOK, M. AND
MURRAY, B.E. Enterococcus faecalis adhesin, ace, mediates attachment to
extracellular matrix proteins collagen type IV and laminin as well as collagen
type I. Infection and Immunity, v.68, p.5218-5224, 2000.
NOBLE, R.T., MOORE, D.F., LEECASTER, M.K., McGEE, C.D., WEISBERG,
S.B. Comparison of total coliform, fecal coliform, and enterococcus bacterial
indicator response for ocean recreational water quality testing. Water
Research, v. 37, p. 1637-1643, 2003.
PALAZZO, I. C. V., CAMARGO, I. L. B. C., ZANELLA, R. C., DARINI, A. L. C.
Evaluation of clonality in enterococci isolated in Brazil carrying Tn1546-like
elements associated with vanA plasmids. FEMS Microbiology Letters, v. 258,
p. 29-36, 2006.
PAVIA, M., NOBILE, C., G.A., SALPIETRO, L., ANGELILLO, I. Vancomycin
resistance and antibiotic susceptibility of enterococci in raw meat. Journal of
Food Protection, v. 63, p.912-915, 2000.
PEREIRA, D.I.A., McCARTNEY, A.L., GIBSON, G.R. An in vitro study of the
probiotic potential of a bile salt hydrolyzing Lactobacillus fermentum strain, and
determination of its cholesterol lowering properties. Applied and
Environmental Microbiology, v.60, p. 4723-4752, 2003.

Referências Bibliográficas____________________________________________________________122
PSONI, L., KOTZAMANIDES, C., ANDRIGHETTO, C., LOMBARDI, A.,
TZANETAKIS, N., LITOPOULOU-TZANETAKI, E. Genotypic and phenotypic
heterogeneity in Enterococcus isolates from Batzos, a raw goat milk cheese.
International Journal of Food Microbiology, v. 109, p. 109-120, 2006.
RICE, L.B., CARIAS, L., RUDIN, S., VAEL, C., GOOSSENS, H., KONSTABEL,
C., KLARE, I., NALLAPAREDDY, S.R., HUANG, W., MURRAY, B.E. A potential
virulence gene, hylefm, predominates in Enterococcus faecium of clinical origin.
Journal of Infectious Diseases, v.187, p.508-512, 2003.
ROBREDO, B., SINGH, K.V., BAQUERO, F., MURRAY, B.E., TORRES, C.
Vancomycin-resistant enterococci isolated from animals and food. International
Journal of Food Microbiology, v. 54, p. 197-204, 2000.
ROSSETTI, L. e GIRAFFA, G. Rapid identification of dairy lactic acid bacteria
by M13-generated, RAPD-PCR fingerprint databases. Journal of
Microbiological Methods v.63, p.135-144, 2005.
ROSSI, E. A., VENDRAMINI, R. C., CARLOS, I. Z., PEI, Y. C., VALDEZ, G. F.
Development of a novel fermented soymilk product with potential probiotic
properties. European Food Research and Technology, v. 209, p. 305-307,
1999.

Referências Bibliográficas____________________________________________________________123
SAAVEDRA, L., TARANTO, M.P., SESMA, F., VALDEZ, G.F. Homemade
traditional cheeses for the isolation of probiotic Enterococcus faecium strains.
International Journal of Food Microbiology, v.88, p.241-245, 2003.
SALMINEN, S.; DEIGHTON, M. A.; BENNO, Y.; GORBACH, S. L. Lactic acid
bacteria in health and disease. In: Lactic acid bacteria: microbiology and
functional aspects. 2nd ed. New York: Marcel Dekker, p. 211-253, 1998.
SANTOS, B.H.C., SOUZA, E.L., ODDI, I.C. Presença de Enterococcus sp. em
alimento enteral e perfil de resistência a antimicrobianos. Revista Brasileira de
Análises Clínicas, v. 36, p.47-49, 2004.
SCHILLINGER U. e LÜCKE, F-K. Identification of lactobacilli from meat and
meat products. Food Microbiology., v. 4, p. 199-208, 1987.
SCHLEIFER, K.H., KILPPER-BALZ, R. Transfer of Streptococcus faecalis and
Streptococcus faecium to the genus Enterococcus nom. rev. as Enterococcus
faecalis comb. nov. and Enterococcus faecium comb. nov. International
Journal of Systematic Bacteriology, v. 34, p. 31-34, 1984.
SECHI, L.A., FRANKLIN, R., DUPRE, I., ZANETTI, S., FADDA, G., DANEO-
MOORE, L. Characterization of new insertion-like sequences of Enterococcus
hirae and their dissemination among clinical Enterococcus faecium isolates.
FEMS Microbiology Letters, v.161, p.165-172, 1998a.

Referências Bibliográficas____________________________________________________________124
SECHI, L.A., ZANETTI, S., DUPRE, I., CAPIELLO, M.G., DELOGU, G.,
MORTENSEN, J.E., DANEO-MOORE, L., FADDA, G. Molecular epidemiology
by ribotyping and PCR-ribotyping of Enterococcus faecium strains isolated from
intercontinental areas. New Microbiologica., v. 21, p.113-122, 1998b.
SHANKAR, V., BAGHDAYAN, A.S., HUYCKE, M.M., LINDAHL, G., GILMORE,
M.S. Infection-derived Enterococcus faecalis strains are enriched in esp, a gene
encoding a novel surface protein. Infection and Immunity, v. 67, p.193-200,
1999.
SMOOT, L.M., PIERSON, M.D. Indicator microorganisms and microbiological
criteria. In: DOYLE, M.P., BEUCHAT, L.R., MONTVILLE, T.J. Food
Microbiology: fundamentals and frontiers. Washington D.C.: ASM Press,
1997. p.76.
STILES, M.E., HOLZAPFEL, W.H. Lactic acid bacteria of foods and their
current taxonomy. International Journal of Food Microbiology, v. 36, p. 1-29,
1997.
STROMPFOVA, V., MARCINAKOVA, M., SIMONOVA, M., GANCARCIKOVA,
S., JONECOVA, Z., SCIRANKOVA, L., KOSCOVA, J., BULECA, V.,
COBANOVA, K., LAUKOVA, A. Enterococcus faecium EK13-an enterocin
producing strain with probiotic character and its effect in piglets. Anaerobe, v.
12, p.242-248, 2006.

Referências Bibliográficas____________________________________________________________125
SULZER, G., BUSSE, M. Growth inhibition of Listeria sp. on camembert cheese
by bacteria producing inhibitory substances. International Journal of Food
Microbiology, v. 14, p.287-296, 1991.(Resumo)
TOLEDO-ARANA, A., VALLE, J., SOLANO, C., ARRIZUBIETA, M.J.,
CUCARELLA, C., LAMATA, M., AMORENA, B., LEIVA, J., PENADÉS, J.R.,
LASA, I. The enterococcal surface protein, Esp, is involved in Enterococcus
faecalis biofilm formation. Applied and Environmental Microbiology, v. 67, p.
4538-4545, 2001.
VAIL, J.H., MORGAN, R., MERINO, C.R., GONZALES, F., MILLER, R., RAM,
J.L. Enumeration of waterborne Escherichia coli with petrifilm plates:
comparison to standard methods. Journal of Environmental Quality, v. 32, n.
1, p. 368-373, 2003.
VANCANNEYT, M., LOMBARDI, A., ANDRIGHETTO, C., KNIJFF, E.,
TORRIANI, S., BJÖRKROTH, K.J., FRANZ, C.M.A.P., MORENO, M.R.F.,
REVETS, H., VUYST, L. SWINGS, J., KERSTERS, K., DELLAGLIO, F.,
HOLZAPFEL, W.H. Intraspecies genomic groups in Enterococcus faecium and
their carrelation with origin and pathogenicity. Applied and Environmental
Microbiology, v. 68, p. 1381-1391, 2002.
VANKERCKHOVEN, V., AUTGAERDEN, T.V., VAEL, C., LAMMENS, C.,
CHAPELLE, S., ROSSI, R., JABES, D. AND GOOSSENS, H. Development of a
multiplex PCR for the detection of asa1, gelE, cylA, esp, and hyl genes in

Referências Bibliográficas____________________________________________________________126
enterococci and survey for virulence determinants among european hospital
isolates of Enterococcus faecium. Journal of Clinical Microbiology, v. 42,
p.4473-4479, 2004.
VELASCO, D.; PEREZ, S.; PEÑA, F.; DOMINGUEZ, M.A.; CARTELLE, M.;
MOLINA, F.; MOURE, R.; VILLANUEVA, R.; BOU, G. Lack of correlation
between phenotypic techniques and PCR-based genotypic methods for
identification of Enterococcus spp. Diagnostic Microbiology and Infectious
Disease, v. 49, p. 151-156, 2004.
ZANELLA, R.C., BRANDILEONE, M. C. C., BOKERMANN, S., ALMEIDA, S. C.
G., VALDETARO, F., VITÓRIO, F., MOREIRA, M. F. A., VILLINS, M.,
SALOMÃO, R., PIGNATARI, A.C. C. Phenotypic and genotypic characterization
of VanA Enterococcus isolated during the first nosocomial outbreak in Brazil.
Microbial Drug Resistance, v. 9, p. 283-291, 2003 (Resumo).
ZEINER, A., BOLDT, E. Effects of a probiotic Enterococcus faecium strain
supplemented from birth to weaning on diarrhoea patterns and performance of
piglets. Journal of Animal Physiology and Animal Nutrition, v. 90, p. 25–31,
2006.
WELLS, C.L., MOORE, E.A., HOAG, J.A., HIRT, H., DUNNY, G.M.,
ERLANDSEN, S.L. Inducible expression of Enterococcus faecalis aggregation
substance surface protein facilitates bacterial internalization by cultured
enterocytes. Infection and Immunity, v. 68, p. 7190-7194, 2000.

Referências Bibliográficas____________________________________________________________127
WILLEY, B.M., KREISWIRTH, B.N., SIMOR, A.E., WILLIAMS, G., SCRIVER,
S.R., PHILLIPS, A., LOW, D.E. Detection of vancomycin resistance in
Enterococcus species. Journal of Clinical Microbiology, v. 30, p.1621-1624,
1992.
YOUSIF, N.M.K., DAWYNDT, P., ABRIOUEL, H., WIJAYA, A., SCHILLINGER,
U., VANCANNEYT, M., SWINGS, J., DIRAR, H.A, HOLZAPFEL, W.H., FRANZ,
C.M.A.P. Molecular characterization, technological properties and safety
aspects of enterococci from Hussuwa, an African fermented sorghum product.
Journal of Applied Microbiology, v.98, p.216-228, 2005.

Referências Bibliográficas____________________________________________________________128
_________________Apêndice

Continuação
129A
pêndice_______________________________________________________________________________
APÊNDICE A - Caracterização fenotípica e molecular dos isolados de enterococos obtidos a partir das amostras de
alimentos semeadas nos meios KEA e BE
Multiplicação
Amostra Alimento Isolado Meio isolamentoa
Cocos Gram + Catalase
10°C 45°C 6,5% NaCl pH 9,6 PYRb API 20 STREP PCR
1 Mussarela 5 BE + - + + + + + E. durans ** 6 BE + - + + + - 7 BE + - - + + - 8 BE + - + + + + + Leuconostoc sp. E. faecium 1 KEA + - - + + + 2 KEA + - - + + + 3 KEA + - - + + - 9 KEA + - - + + + 4 KEA + - + + + + + Leuconostoc sp. E. faecium
2 Queijo minas 11 BE + - - + + + 13 KEA + - + + + + + E. faecalis E. faecalis 10 BE + - + + + + + E. faecalis E. faecalis 12 KEA + - + + + + + E. faecalis E. faecalis
3 Carne moída 16 BE - - + - - + 14 BE + 15 BE +
4 Queijo minas 17 BE + 18 BE + 19 KEA + 20 KEA + - + + + + + Aerococcus viridans E. gallinarum
5 Presunto cozido nenhuma colônia característica 6 Carne moída 21 KEA + - + + + + + E. avium E. faecalis 22 KEA + - + + + + + result. não conclusivo E. faecalis 23 KEA + - + + - + 24 BE + 25 BE + 26 BE +
7 Peito frango 27 BE + - - - - - 28 BE + 29 BE +
8 Mussarela 36 BE + - + + + + + E. durans E. faecium

Continuação
130A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
37 BE + - + + + + + baixa discriminação E. faecium 38 BE + - + + + + + n.a E. faecium 39 KEA + - + + + + + n.a E. faecium 40 KEA + - + + + + + n.a E. faecium 41 KEA + - + + + + + n.a E. faecium
9 Presunto 30 BE + - + + + + + baixa discriminação E. faecium 31 BE + 32 BE + - + + + + + Leuconostoc sp. E. faecium 33 KEA + 34 KEA + - + + + + + E. durans E. faecium 35 KEA + - + + + + + result. não conclusivo E. faecium
10 Apresuntado 42 KEA + - - + - - 43 KEA + - + + + + + E. durans E. faecium 44 KEA + - + + + + + baixa discriminação E. faecium 45 BE + 46 BE +
11 Ricota 47 KEA + - - - + + 48 KEA + - + + + + + baixa discriminação E. faecium 49 KEA * 50 BE + - - + - - 51 BE + 52 BE *
12 Lingüiça frango 53 KEA + - + + + + + baixa discriminação ** 54 KEA * 55 KEA + - - + + - 56 BE * 57 BE + - + + + + + Enterococcus sp. E. faecalis 58 BE + - + - - -
13 Bacon 59 KEA + - - + - + 14 Salsicha 60 KEA + - + + + + + E. faecalis E. faecalis
61 BE + 62 BE + - + + + + + Enterococcus sp. E. faecalis 63 BE + - + + + + + E. avium E. faecalis
15 Queijo prato 64 BE + 65 BE + - + + + + + result. não conclusivo E. faecium 66 BE + - + + + + + Enterococcus sp. E. faecium 67 KEA + - + + + + + n.a E. faecalis

Continuação
131A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
68 KEA + - + + + + + n.a E. faecium 69 KEA + - + + + + + n.a E. faecium
16 Queijo Minas 70 BE + - + + + + + E.durans E. faecium 71 BE + - + + + + + Aerococcus viridans E. gallinarum 72 BE + 73 KEA + - + + + + + n.a E. faecalis 74 KEA + - + + + + + n.a E. gallinarum 75 KEA + - + + + + + n.a E. faecium
17 Queijo gouda 76 KEA + - + + + + + E. durans E. faecium 77 KEA + - + + + + + Enterococcus sp. E. faecium 78 KEA * 79 BE * 80 BE * 81 BE *
18 Mussarela 100 KEA + - + + + + + E. faecalis E. faecalis 101 KEA + - + + + + + n.a E. faecium 102 KEA + - + + + + + n.a E. faecium 103 KEA + - + + + + + n.a E. faecium 104 KEA + - + + + + + n.a E. faecium 105 KEA + - + + + + + n.a E. faecium 106 KEA * 107 KEA + - - + - - 108 KEA * 109 KEA * 110 KEA + - (+) + - - 111 KEA + - (+) + - - 112 BE + - + + + + + n.a E. faecium 113 BE + - + + + + + n.a E. faecium 114 BE + - + + + + + n.a E. faecium 115 BE + - + + + + + n.a ** 116 BE + - + + + + + n.a ** 117 BE + - + + + + + n.a E. faecium 118 BE + - + + + + + n.a E. faecium 119 BE + - + + + + + n.a E. faecium 120 BE + - + + + + + n.a E. faecium
19 Seleta legumes pré-cozida nenhuma colônia característica

Continuação
132A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
20 lagarto bov. def. 82 BE + - + + + + + E. faecalis E. faecalis 83 BE + - + + + + + Enterococcus sp. E. faecalis 84 BE *
21 Rosbife 85 BE + 86 BE + - + - - - 87 KEA + - + + + + + E. faecium E. faecium
22 Cenoura ralada 88 KEA + 89 KEA - - 90 KEA - + 91 BE + 92 BE + 93 BE + - - - - -
23 Salsicha + picles 94 BE + 95 BE + 96 BE + 97 KEA + - + + + + + E. faecium E. faecium 98 KEA + - + + + + + n.a E. faecalis 99 KEA + - + + + + + n.a E. faecium
24 Surubim defumado 121 BE + 122 BE + 123 BE + 124 BE + 125 BE +
25 Queijo árabe 126 BE + 127 BE + 128 BE +
26 Mussarela 129 KEA + - + + + + + Enterococcus sp. E. faecium 130 KEA + - + + + + + n.a E. faecium 131 KEA + - + + + + + n.a E. faecium 132 KEA + - + + + + + n.a E. faecium 133 KEA + - + + - - 134 KEA + - + + + - 135 KEA + - + + + - 136 KEA + - + + - - 137 BE + - + + + + + E. durans ** 138 BE + - + + + + + E. durans E. faecium 139 BE + - + + + + + n.a **

Continuação
133A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
140 BE + - + + + + + n.a E. faecalis 141 BE + - + + + + + n.a ** 142 BE + - + + + + + n.a E. faecium
27 Salame 143 KEA + - + + + + + Enterococcus sp. E. faecium 144 KEA + - + + + + + n.a E. faecium 145 KEA * 146 KEA + - + + + + + n.a E. faecium 147 KEA + - + + + + + n.a ** 148 KEA * 149 KEA + - + + + - 150 BE + - + - + + 151 BE + - + + + + - 152 BE + 153 BE - - 154 BE + - + + + + + E. durans **
28 Mortadela 155 KEA + - - + - + 156 KEA + - + + + + + n.a ** 157 KEA + - + + + + + result. não conclusivo E. faecium 158 KEA + - + + + + + n.a E. faecium 159 KEA + - + + + - 160 BE + 161 BE + 162 BE + 163 BE + - + + + + + n.a E. faecium 164 BE +
29 Queijo prato 165 BE + 166 BE + 167 BE + 168 BE + 169 BE + 170 KEA + - + + + + + E. faecium E. faecium 171 KEA + - + + + + + n.a E. faecium 172 KEA + - + + + + + n.a ** 173 KEA + - + + + + + n.a E. faecium 174 KEA + - + + + + + n.a E. faecium
30 Mussarela 175 BE + - + + + + + E. faecium E. faecium 176 BE + - + + + + + n.a **

Continuação
134A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
177 BE + - + + + + + n.a ** 178 BE + - + + + + + n.a ** 179 BE + - + + + + + E. durans E. faecium 180 BE + - + + + + + E. faecium E. faecium 181 BE + - + + + + + n.a E. faecium 182 BE + - + + + + + n.a E. faecium 183 BE + - + + + + + n.a E. faecium 184 BE + - + + + + + n.a E. faecium 185 BE + - + + + + + n.a E. faecalis 186 BE + - + + + + + n.a E. faecium 187 BE + - + + + + + n.a E. faecium 188 BE + - + + + + + n.a E. faecium 189 KEA + - + + + + + E. durans E. faecium 190 KEA + - + + + + + n.a E. faecium 191 KEA + - + + + + + n.a E. faecium 192 KEA + - + + + + + n.a E. faecium 193 KEA + - + + + + + n.a E. faecium 194 KEA + - + + + + + n.a E. faecium 195 KEA + - + + + + + n.a E. faecium 196 KEA + - + + + + + n.a E. faecium 197 KEA + - + + + + + n.a E. faecium 198 KEA + - + + + + + n.a E. faecium 199 KEA + - + + + + + n.a E. faecium 200 KEA + - + + + + + n.a E. faecium
31 Queijo prato 201 KEA + - - + - - 202 KEA + - - + - - 203 KEA + - - + - - 204 KEA + - - + + + 205 KEA + - - + - - 206 KEA + 207 BE + - - + - - 208 BE + - - + - - 209 BE + - - + - - 210 BE + - - + - - 211 BE + - - + - - 212 BE Perdeu viabilidade
32 Presunto peru 213 BE + - + + + + + Leuconostoc sp. E. faecium

Continuação
135A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
214 BE + - + + + + + n.a E. faecium 215 BE + - + + + + + n.a E. faecium 216 BE + - - + - - 217 BE + - + + + + + n.a E. faecium 218 KEA + - + + + + + Leuconostoc sp. E. faecium 219 KEA + - + + + + + n.a E. faecium 220 KEA + - - + - - 221 KEA + - - + - - 222 KEA + - - + - - 223 KEA + - - + - - 224 KEA + - - + - + 225 KEA + - + + + + + n.a E. faecium 226 KEA + - - + - - 227 KEA + - + + + + + n.a E. faecium 228 KEA + - + + + + + n.a E. faecium 229 KEA + - - + + - 230 KEA Perdeu viabilidade 231 KEA Perdeu viabilidade 232 KEA + - + + + + + n.a E. faecium
33 Lingüiça frango 233 KEA + - + + + + + E. avium E. faecium 234 KEA + - + + + + + baixa discriminação E. casseliflavus 235 KEA + 236 KEA + 237 KEA + - + + + + + n.a E. faecalis 238 KEA + - + + + + + n.a E faecium 239 KEA + 240 KEA + 241 BE + - + - + + 242 BE Perdeu viabilidadade 243 BE + - + - + + 244 BE + - + - + + 245 BE +
34 Lingüiça de porco 246 KEA + - + + + + + E. avium E. casseliflavus 247 KEA + - + + + + + n.a E. casseliflavus 248 KEA + - + + + + + n.a E. casseliflavus 249 KEA + - + + + + + n.a E. casseliflavus 250 KEA + - + + + + + n.a E. casseliflavus

Continuação
136A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
251 KEA + - + + + + + n.a E. casseliflavus 252 BE + 253 BE + - + + + + + n.a E. casseliflavus 254 BE + - + + + + + n.a E. casseliflavus 255 BE + 256 BE + - + + + + + n.a E. casseliflavus 257 BE + 258 BE + 259 BE + - + + + + + n.a E. casseliflavus
35 Salada mista 260 KEA + 261 KEA + 262 KEA + 263 KEA + 264 KEA + 265 BE + 266 BE + 267 BE + 268 BE +
36 Salada repolho/beterraba
269 KEA +
270 KEA - - 271 KEA - - 272 KEA + - + + + + + Aerococcus viridans E. casseliflavus 273 KEA + 274 BE - - 275 BE + - + - - + 276 BE + - + + + + - 277 BE Perdeu viabilidade 278 BE - - 279 BE + - + - - + 280 BE + - + - - - 281 BE + - - - - + 282 BE + - - - - -
37 Coração frango 283 BE + 284 BE + 285 BE - - 286 BE +

Continuação
137A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
287 BE + 38 Moela frango 288 KEA + - + + + + + E. avium E. faecalis
289 BE + 290 BE - - 291 BE * 292 BE * 293 BE * 294 BE + 295 BE - -
39 Abobrinha 296 KEA + - + - - - 297 KEA + - + - - + 298 KEA + 299 KEA + 300 KEA + 301 KEA + 302 KEA + - + - + + 303 KEA + - - + + + 304 KEA + - + + + + 305 BE + - + - + + 306 BE + - + - + +
40 Cogumelo nenhuma colônia característica 41 Soja fermentada 307 BE +
308 BE + 42 Queijo parmesão 309 KEA + - + + + + + n.a E. faecium
310 KEA + - + + + + + n.a E. faecalis 311 BE + - + + + + + n.a E. faecium
43 Presunto 312 BE + - + - - + 313 BE - - 314 BE - - 315 BE - - 316 KEA + - + + + + + n.a E. faecium 317 KEA + - + + + + + n.a E. faecium 318 KEA + - + + + + + n.a E. faecium 319 KEA +
44 Queijo gorgonzola 320 BE + - - + - - 321 BE - - 322 BE - -

Continuação
138A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
323 BE - - 324 BE + - - + - + 325 BE - - 326 KEA + - - + - + 327 KEA - - 328 KEA - - 329 KEA + - + + + + + n.a E. gallinarum
45 Bucho bovino 330 BE + 331 BE + 332 BE + 333 BE +
46 Queijo minas 334 BE + 335 BE + 336 BE + 337 BE + 338 KEA + - - + - - 339 KEA + - + + + + 340 KEA + - + + + + + n.a E. faecalis 341 KEA + - + + + + + n.a E. faecalis
47 Lingüiça de pernil 342 BE + 343 BE + 344 BE * 345 BE + - + - + + 346 BE - -
48 Frango 347 BE - - 348 KEA + -
49 Bacon 349 BE + - + + + + + n.a E. faecium 350 BE * 351 BE - - 352 BE + - + + + + + n.a E. faecium 353 BE - - 354 KEA + - + + + + + n.a E. faecium 355 KEA + - + + + + + n.a E. faecium 356 KEA + - + + + + + n.a E. faecium 357 KEA + - + + + + + n.a E. faecium 358 KEA + - + + + + + n.a E. faecium
50 Filé tilápia 359 BE +

Continuação
139A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
360 BE - - 361 BE - - 362 BE - - 363 BE + 364 BE +
51 Asa frango 365 BE - - 366 BE - - 367 BE + 368 BE + 369 BE + 370 BE +
52 Coxão mole 371 BE - - 372 BE - - 373 BE - - 374 BE - - 375 BE - - 376 BE - - 377 KEA + - + + + + + n.a E. faecalis
53 Lombo suíno fatiado
378 BE +
379 BE + 380 BE + 381 BE + 382 BE + 383 BE + 384 BE +
54 Bife bovino 385 BE + 386 BE + 387 BE + 388 BE + 389 BE + 390 BE + 391 BE + 392 BE + 393 BE + 394 BE +
55 Queijo Provolone 395 KEA + - + + + + + n.a E. faecalis

Continuação
140A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
396 KEA + - + + + + + n.a E. faecium 397 KEA + - + + + + + n.a E.faecium 398 KEA + - + + + + + n.a ** 399 BE + - + + + + + n.a E. faecalis 400 BE + - + + + + + n.a E. faecalis 401 BE + - + + + + + n.a E. faecium 402 BE + - + + + + + n.a E. faecalis 403 BE + - + + + + + n.a E. faecalis
56 Ricota 404 BE - - 405 BE - - 406 BE + - + - - - 407 BE + - + - - + 408 BE + 409 KEA + 410 KEA + 411 KEA + 412 KEA + 413 KEA + 414 KEA + 415 KEA + 416 KEA + 417 KEA + 418 KEA + 419 KEA +
57 Queijo cheddar Nenhuma colônia característica 58 Broto de Feijão 420 BE + - + - - -
421 BE + - + - - - 422 BE + - + - - - 423 BE + - + - - + 424 BE + - + - - + 425 KEA + - + + + + + n.a E. casseliflavus 426 KEA + - + + + + + n.a E. casseliflavus 427 KEA + - + + + + + n.a E. casseliflavus 428 KEA + - + + + + + n.a E. casseliflavus 429 KEA + - + + + + + n.a E. casseliflavus 430 KEA + - - - - + 431 KEA + - + + + + + n.a E. casseliflavus

Continuação
141A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
432 KEA + - + + + + + n.a E. casseliflavus 433 KEA + - + + + + + n.a E. casseliflavus 434 KEA + - + + + + + n.a E. casseliflavus 435 KEA + - + + + + + n.a E. casseliflavus 436 KEA + - - + + + 437 KEA + - + + + + + n.a E. casseliflavus 438 KEA + - + + + + + n.a E. casseliflavus
59 Couve manteiga 439 KEA + - + + + + + n.a E. casseliflavus 440 KEA + - + + + + + n.a E. casseliflavus 441 KEA - 442 KEA + - + + + + + n.a E. casseliflavus 443 KEA + - + + + + + n.a E. casseliflavus 444 BE + 445 BE + 446 BE + 447 BE + 448 BE + 449 BE + 450 BE + 451 BE + 452 BE +
60 Ervilha Crua nenhuma colônia característica 61 Queijo Gorgonzola 453 KEA + - + + + + + n.a E. faecium
454 KEA + - + + + + + n.a E. faecium 455 KEA + - + + + + + n.a E. faecium 456 KEA + - + + + + + n.a E. faecium 457 KEA + - + + + + + n.a E. faecium 458 BE + - - + - + 459 BE + - + + + + + n.a E. faecium 460 BE + - + + + + + n.a E. faecium 461 BE + - + + + + + n.a E. faecium 462 BE + - + + + + + n.a E. faecium
62 Mussarela nenhuma colônia característica 63 Azeitona Preta nenhuma colônia característica 64 Tremoço 463 KEA + - + + + + -
464 KEA + - + + + - 465 KEA + - + + + + -

Continuação
142A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
466 KEA + - + + + + - 467 KEA + - + + + -
65 Milho Verde 468 KEA + - + + + + + n.a E. faecalis 469 KEA + - + + + + + n.a E. casseliflavus 470 KEA + - + + + + + n.a E. casseliflavus 471 KEA + - + + + + + n.a E. faecalis 472 KEA + - - - + + 473 KEA + - + - + - 474 KEA + - + - + - 475 KEA + - + - + - 476 KEA + - - - + - 477 KEA + - + - + - 478 BE + - + - + - 479 BE + - + - + - 480 BE + - - - + - 481 BE + - + + + - 482 BE + - + - + - 483 BE + - + - + - 484 BE + - + - + - 485 BE + - + - + -
66 Beterraba 486 BE + 487 BE + 488 BE + 489 BE + 490 BE + 491 BE +
67 Cogumelo 492 BE + 493 BE + 494 BE + 495 BE + 496 BE + 497 BE + 498 BE + 499 BE + 500 BE + 501 BE +
68 Alecrim 502 BE +

Continuação
143A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
69 Salada tricolor nenhuma colônia característica 70 Acelga 503 BE +
504 BE + 505 BE + 506 BE + 507 BE + 508 BE + 509 BE + 510 BE + 511 BE + 512 BE + 513 BE + 514 BE + 515 BE +
71 Cheiro Verde 516 BE + pré-higienizado 517 BE + 518 BE + 519 BE + 520 BE + 521 BE + 522 BE + 523 BE + 524 BE + 525 KEA + - + + + + + n.a ** 526 KEA + 527 KEA + 528 KEA + 529 KEA + 530 KEA + 531 KEA - -
72 Cheiro verde in natura
532 BE +
533 BE + 534 BE + 535 BE + 536 KEA + - - - - +
73 Mussarela 537 BE - -

Continuação
144A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
538 BE + - + + + + + n.a E. faecium 539 BE + - + - + + 540 BE + - + + + + + n.a ** 541 BE + - + + + + + n.a ** 542 BE + - + + + + + n.a E. faecium 543 BE + - + + + + + n.a E. faecium 544 BE + - + + + + + n.a ** 545 BE - - 546 KEA + - + + + + + n.a ** 547 KEA + - + + + + + n.a ** 548 KEA + - + + + + + n.a E. faecium 549 KEA + - + + + + + n.a E. faecium 550 KEA + - + + + + + n.a E. faecalis
74 Queijo meia cura 551 BE 552 BE + - - - - - 553 BE + - - - - - 554 BE + - - - - - 555 BE - - 556 KEA + - + + + + + n.a E. faecium 557 Cepa inexistente devido a erro na numeração 558 KEA + 559 KEA + 560 KEA + 561 KEA +
75 Ricota temperada 562 BE + - + + - + 563 BE + - + + - + 564 BE - - 565 BE + - + - - + 566 BE + - + - - + 567 KEA + - + + + + + n.a E. faecalis 568 KEA + - + + + + + n.a E. faecalis 569 KEA + - - + - + 570 KEA + - - + - + 571 KEA + - - + - - 572 KEA + - - + - -
76 Queijo meia cura 573 KEA + - + - - + 574 KEA + - + + + + + n.a E. faecalis
Continuação
145A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
575 KEA + - + + + + + n.a E. faecalis 576 KEA + - + + + + + n.a E. faecalis 577 KEA + - + + - + 578 KEA + - + + + + + n.a E. faecalis 579 KEA + - + + - + 580 KEA + - + + - + 581 KEA + - + - - + 582 BE + - + - - + 583 BE + - + - - + 584 BE + - + - - + 585 BE + - + - - +
77 Queijo minas light 586 KEA + - + + + + + n.a E. faecalis 587 KEA + - + + + + + n.a E. faecalis 588 KEA + - + + + + + n.a E. faecium 589 KEA + - + + + + + n.a E. faecalis 590 KEA + - + + + + + n.a E. faecalis 591 KEA + - + + + + + n.a E. faecalis 592 BE + - + - - + 593 BE + - + + - - 594 BE + - + - - + 595 BE + - + + + + + n.a E. faecalis 596 BE + - + + - + - 597 BE + - + - - +
78 Mussarela 598 KEA + - + + + + + n.a E. faecalis 599 KEA + - + + + + + E. faecalis 600 KEA + - + + + + + n.a E. faecalis 601 KEA + - + + + + + n.a E.gallinarum 602 KEA + - + + + + + n.a E. faecalis 603 BE + - + + + + + n.a E. faecalis 604 BE + - + + + + + n.a E. faecalis 605 BE + - + + + + + n.a E. faecalis 606 BE + - + + + + + n.a E. faecalis 607 BE + - + + + + + n.a E. faecalis 608 BE + - + + + + + n.a ** 609 BE + - + + + + + n.a E. faecalis
79 Queijo prato 610 KEA + - + + + + + n.a E. faecalis 611 KEA + - + + + + + n.a E. faecalis

Continuação
146A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
80 Queijo meia cura 612 BE + - + + + + + n.a E. faecium 613 BE + - + + + + + n.a E. faecium 614 BE + - + + + + + n.a E. faecium 615 BE + - + + + + + n.a E. faecium 616 BE + - + + + + + n.a E. faecium 617 BE + - + + + + + n.a E. faecium 618 BE + - + + + + + n.a E. faecium 619 BE + - + + + + + n.a E. faecium 620 BE + - + + + + + n.a E. faecium 621 BE + - + - - - 622 KEA + - + + + + + n.a E. faecium 623 KEA + - + + + + + n.a E. faecium 624 KEA + - + + + + + n.a E. faecium 625 KEA + - + + + + + n.a E. faecium 626 KEA + - + + + + + n.a E. faecium 627 KEA + - + + + + + n.a E. faecium 628 KEA + - + + + + + n.a E. faecium 629 KEA + - + + + + + n.a E. faecium
81 Manjericão 630 BE + 631 BE + 632 BE - - 633 BE + 634 BE - - 635 BE - - 636 BE + 637 BE + 638 BE - - 639 BE + 640 BE + 641 KEA +
82 Salsinha 642 KEA - - 643 KEA + - + + - + 644 KEA + - + + - + 645 KEA + - + - - + 646 KEA - - 647 BE + 648 BE +

Continuação
147A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
649 BE + - + + + + + E. casseliflavus 650 BE + 651 BE +
83 Batata/cenoura/ vagem nenhuma colônia característica
84 Escarola lavada 652 BE + 653 BE + 654 BE + 655 BE + 656 BE + 657 BE + 658 BE + 659 BE + 660 BE + 661 BE + 662 BE +
85 Salada de vegetais 663 KEA + 664 KEA + 665 KEA + 666 KEA + 667 KEA + 668 KEA + 669 KEA + 670 KEA + 671 BE + 672 BE + 673 BE + 674 BE + 675 BE + 676 BE +
86 Salada alface/rúcula/
rabanete
677 BE +
87 abóbora cabocha nenhuma colônia característica 88 alface crespa 678 BE +
679 BE + - + + + + + n.a E. casseliflavus 680 BE + - + - + +

Continuação
148A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
681 BE + - + - + + 682 BE + 683 BE + - + - + + 684 BE + - + - - + 685 BE + 686 BE + - - - + + 687 BE + 688 BE + 689 BE + 690 KEA + - + - + + 691 KEA + - + - + + 692 KEA + - + + + + + n.a E. casseliflavus 693 KEA + - + + + + + n.a E. casseliflavus 694 KEA + - - - - + 695 KEA + - + - + + 696 KEA + - - - - + 697 KEA + - + - + +
89 coentro 698 BE + 699 BE + 700 BE + 701 BE + 702 BE + 703 KEA +
90 almeirão 704 BE + - + - - + 705 BE + 706 BE + - + - - - 707 BE + 708 KEA + - - + - + 709 KEA + - - + - + 710 KEA + - - - - + 711 KEA + - - - - + 712 KEA + - - - - + 713 KEA + - - - - + 714 KEA + - - - - + 715 KEA + - + - - +
91 Leite tipo C nenhuma colônia característica 92 Leite tipo C nenhuma colônia característica

Continuação
149A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
93 Leite tipo B nenhuma colônia característica 94 Leite tipo C nenhuma colônia característica 95 Leite tipo A 716 KEA + - + + + + + n.a **
717 KEA + - + + + + + n.a E. casseliflavus 718 KEA + - + + + + + n.a ** 719 KEA + - + + + + + n.a ** 720 KEA + - + + + + + n.a **
96 Leite tipo C 721 BE + - - - - + 722 BE + - - + + + 723 BE + - - + + + 724 BE + - - + + + 725 KEA + - - + + + 726 KEA + - + + + + + n.a ** 727 KEA + - - + + + 728 KEA + - + + + + + n.a E. faecium 729 KEA + - + + + + + n.a E. faecium
97 Leite cru de vaca 730 BE + - + + + + + n.a E. faecalis 731 KEA + - - + - - 732 KEA + - + + + + + n.a E. faecalis
98 Leite cru de vaca 733 KEA + - + + + + + n.a E. faecalis 734 KEA + - + + + + + n.a E. faecalis 735 KEA + - + + + + + n.a E. faecalis
99 Leite cru de vaca nenhuma colônia característica 100 Leite cru de vaca 736 KEA - - 101 Leite cru de vaca 737 KEA *
738 KEA + - + + + + + n.a E. faecalis 739 KEA + - + + + + + n.a E. faecalis 740 KEA + - + + + + + n.a E. faecalis
102 Leite cru de vaca 741 KEA + - + + + + + n.a E. faecalis 742 KEA + - + + + + + n.a E. faecalis 743 KEA + - + + + + + n.a E. faecalis 744 KEA + - + + + + + n.a E. faecalis 745 KEA + - + + + + + n.a E. faecalis 746 KEA + - + + + + + n.a E. casseliflavus
103 Leite cru de vaca 747 KEA * 748 KEA * 749 KEA *
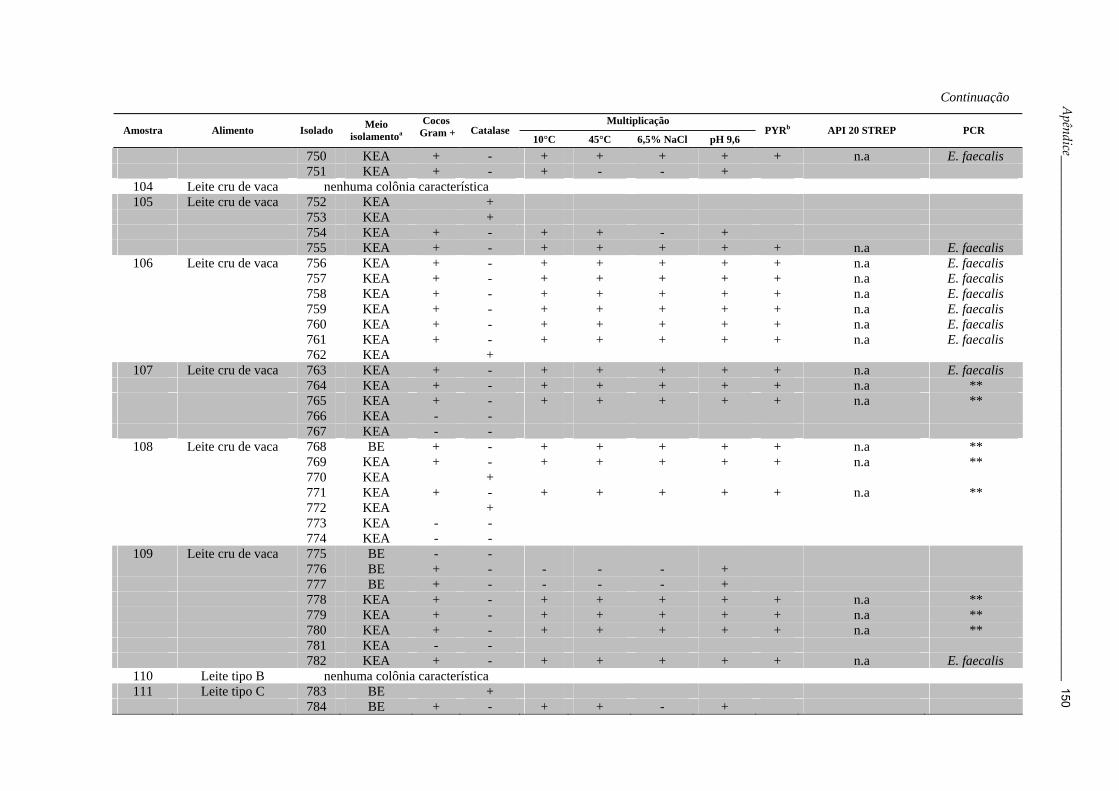
Continuação
150A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
750 KEA + - + + + + + n.a E. faecalis 751 KEA + - + - - +
104 Leite cru de vaca nenhuma colônia característica 105 Leite cru de vaca 752 KEA +
753 KEA + 754 KEA + - + + - + 755 KEA + - + + + + + n.a E. faecalis
106 Leite cru de vaca 756 KEA + - + + + + + n.a E. faecalis 757 KEA + - + + + + + n.a E. faecalis 758 KEA + - + + + + + n.a E. faecalis 759 KEA + - + + + + + n.a E. faecalis 760 KEA + - + + + + + n.a E. faecalis 761 KEA + - + + + + + n.a E. faecalis 762 KEA +
107 Leite cru de vaca 763 KEA + - + + + + + n.a E. faecalis 764 KEA + - + + + + + n.a ** 765 KEA + - + + + + + n.a ** 766 KEA - - 767 KEA - -
108 Leite cru de vaca 768 BE + - + + + + + n.a ** 769 KEA + - + + + + + n.a ** 770 KEA + 771 KEA + - + + + + + n.a ** 772 KEA + 773 KEA - - 774 KEA - -
109 Leite cru de vaca 775 BE - - 776 BE + - - - - + 777 BE + - - - - + 778 KEA + - + + + + + n.a ** 779 KEA + - + + + + + n.a ** 780 KEA + - + + + + + n.a ** 781 KEA - - 782 KEA + - + + + + + n.a E. faecalis
110 Leite tipo B nenhuma colônia característica 111 Leite tipo C 783 BE +
784 BE + - + + - +

Continuação
151A
pêndice_______________________________________________________________________________
Multiplicação Amostra Alimento Isolado Meio
isolamentoa Cocos
Gram + Catalase 10°C 45°C 6,5% NaCl pH 9,6
PYRb API 20 STREP PCR
785 BE + - + + - + 786 BE + - + + - + 787 BE + - + + - + 788 BE + - + + - + 789 BE - - + + - +
112 Leite de cabra 790 KEA + - + + - + 791 KEA + - + + - + 792 KEA + - + + + + + n.a E. gallinarun 793 KEA + - + + - + 794 KEA + - + + + + + n.a E. gallinarum 795 KEA + - + + - + 796 KEA + - + + - + 797 KEA + - + + - + 798 KEA + - + + + + + n.a E. gallinarum 799 KEA + - + + - + 800 KEA + - + + - + 801 KEA + - + + - + 802 BE + - + + - + 803 BE + - + + - + 804 BE + - + + - +
113 Leite tipo C nenhuma colônia característica 114 Leite tipo C nenhuma colônia característica 115 Leite cru 805 KEA +
806 KEA + - + + + + + n.a ** 116 Leite cru 807 BE +
808 KEA + - + + + + + n.a E. faecalis 117 Leite cru nenhuma colônia característica 118 Leite cru 809 KEA + - + - - -
810 KEA + 119 Leite cru 811 KEA + 120 Leite cru 812 KEA + - + + + + + n.a E. faecium
813 KEA + - + + + + + n.a E. faecalis Os espaços em branco indicam que o teste não foi realizado já que pelo menos um teste não estava de acordo com o esperado. (+) positivo, (-) negativo. * cepas que não se multiplicaram no TSAYE a BE: ágar Bile Esculina, KEA: Kanamicina Esculina Azida n.a: não avaliado até o momento b hidrólise de pirrolidonil –ß – naftilamida (PYR) ** PCR não conclusivo com os primers para E. faecalis, E. faecium, E. gallinarum e E. casseliflavus