universidade de sÃo paulo escola de engenharia...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO ESCOLA DE ENGENHARIA DE LORENA
AMANDA MONTEIRO ELIAS
Ação das enzimas de Ceriporiopsis subvermispora no biobranqueamento de polpas kraft de Eucalyptus grandis
Lorena 2007

2
AMANDA MONTEIRO ELIAS
Ação das enzimas de Ceriporiopsis subvermispora no biobranqueamento de polpas kraft de Eucalyptus grandis
Dissertação apresentada à Escola de Engenharia de Lorena da Universidade de São Paulo para obtenção do título de mestre em Biotecnologia Industrial. Área de concentração: Bioquímica e Microbiologia Orientadora: Profa Dra. Adriane Maria Ferreira Milagres
Lorena 2007

3
AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE TRABALHO, POR QUALQUER MEIO CONVENCIONAL OU ELETRÔNICO, PARA FINS DE ESTUDO E PESQUISA, DESDE QUE CITADA A FONTE.
Catalogação na Publicação Biblioteca Universitária
Escola de Engenharia de Lorena da Universidade de São Paulo
Elias, Amanda Monteiro
Ação das enzimas de Ceriporiopsis subvermispora no biobranqueamento de polpas kraft de Eucalyptus grandis / Amanda Monteiro Elias; orientadora Adriane Maria Ferreira Milagres - 2007
106 f: fig.
Dissertação (Mestrado – Programa de Pós-Graduação em Biotecnologia Industrial. Área de Concentração: Bioquímica e Microbiologia) – Escola de Engenharia de Lorena da Universidade de São Paulo, 2007
1. Biotecnologia 2. Xilanase 3. Ceriporiopsis subvermispora 4. Polpas kraft. I. Título.
574.6 - CDU

4
DEDICATÓRIA
Em primeiro lugar a Deus que escreve nossos caminhos. Aos meus pais, César e Fátima, com amor, admiração e gratidão por sua compreensão, carinho, incentivo e incansável apoio ao longo do período de elaboração deste trabalho.

5
AGRADECIMENTOS
À profa
Dra. Adriane Maria Ferreira Milagres que muito me ensinou, pela paciência e ensinamentos, contribuindo para meu crescimento intelectual e científico. Aos professores, funcionários e técnicos do Departamento de Biotecnologia (DEBIQ). Aos membros da banca. À CAPES pela concessão da bolsa de mestrado e pelo apoio financeiro para a realização deste trabalho. Aos meus irmãos, Luciane e Vinicius que mesmo distantes me incentivaram e trocaram idéias, enriquecendo este trabalho. Ao Luiz Homero e à Dona Dora pelo apoio, carinho e suporte durante a elaboração e conclusão deste trabalho. Às meninas companheiras, aos amigos que já tinha, e aos que ganhei neste caminho. Aos companheiros de trabalho, principalmente ao Valdeir, Joseana, Luís Ricardo e Michel, que me ensinaram muitas coisas importantes e sobretudo colaboraram no meu aprendizado.
Obrigada!

6
“Já percorri esse longo caminho da liberdade. Procurei não vacilar e dei
muitos passos em falso no percurso. No entanto, descobri que depois de subir um
monte bem alto a gente apenas verifica que há muitos outros montes a escalar. Tirei
um instante para descansar, para dar uma olhadela no panorama glorioso que me
cerca, para olhar para trás e ver a distância que percorri. Porém só posso descansar
um instante, pois com a liberdade vêm as responsabilidades e eu não ouso demorar-
me, minha longa caminhada ainda não terminou.”
Nelson Mandela

7
RESUMO
ELIAS, A. M. Ação das enzimas de Ceriporiopsis subvermispora no biobranqueamento de polpas kraft de Eucalyptus grandis. 2007. 106 f. Dissertação (Mestrado em Biotecnologia Industrial) – Escola de Engenharia de Lorena da Universidade de São Paulo, Lorena, 2007.
Enzimas extraídas de madeiras biotratadas por Ceriporiopsis subvermispora foram utilizadas no processo de biobranqueamento de polpas kraft de Eucalyptus grandis, seguida da seqüência de branqueamento químico EQP (extração alcalina, quelação e peroxidação). A caracterização dos extratos foi feita através da determinação das atividades enzimáticas do complexo xilanolítico: β-xilanase (346 UI/kg), β-xilosidase (62,6 UI/kg), β-glicosidase (87 UI/kg), β-mananase (231 UI/kg), endoglucanase (133 UI/kg) e da enzima manganês peroxidase (765 UI/kg). Xilanas de bétula, de fibra de trigo, de carvalho e de bagaço de cana-de-açúcar foram tratadas com xilanases de C. subvermispora e analisada a liberação de açúcares redutores que foi crescente por até 96 h. As xilanas de bétula, de fibra de trigo e de carvalho liberaram principalmente xilotetraose (0,63, 0,76, 0 g/L), xilotriose (0,24, 0,38, 0,19 g/L), xilobiose (0,52, 0,70, 0,39 g/L) e xilose (0, 0, 0,37 g/L), respectivamente. Ensaios com o extrato bruto de C. subvermispora e um extrato comercial sobre as polpas kraft de Eucalyptus grandis mostraram uma atuação semelhante quanto à liberação de açúcares redutores dos dois extratos nas polpas. Às polpas lavadas contendo 78% de celulose, 12,8% de polioses e 5,3% de lignina em sua constituição química e kappa e viscosidade iniciais de 12,9 e 24,8 cP, respectivamente, foram aplicados os extratos enzimáticos em função da atividade de xilanase nas concentrações de 0,5, 1, 1,5 e 2 UI/g, em diferentes tempos de tratamento (0,5, 1, 2, 3 e 5 h) a 40ºC e 10% de consistência. Foram feitas as medidas de absorbância a 237, 254, 280 e 465 nm e a quantificação dos açúcares redutores. Houve liberação de cromóforos a 254, 280 e 465 nm apenas para as cargas de 1,5 e 2 UI/g, enquanto que a 237 nm houve liberação de quantidades crescentes de cromóforos com o aumento da carga enzimática. A concentração máxima de açúcares redutores (23,6 g/L) foi obtida após 3 h de tratamento com 2 UI/g. O número kappa e a viscosidade das polpas foram medidos após o tratamento enzimático e após o branqueamento químico e o cálculo da eficiência da deslignificação e a seletividade foram medidos após a etapa de branqueamento químico. A diminuição do número kappa foi de 7,8% no tratamento enzimático, enquanto que no químico foi de 11,4%, ambos com a carga de 2 UI/g com 3 h de reação. As viscosidades de todas as amostras tratadas enzimaticamente foram maiores do que os controles. Após o branqueamento químico a carga de 2 UI/g foi a que apresentou os maiores valores de seletividade. A carga que propiciou a máxima redução no número kappa, aliados à maior redução de cromóforos a 237 e 465 nm e à maior liberação de açúcares redutores foi a de 2 UI/g, com tempos variando de 2 a 3 h de tratamento.
Palavras-chave: Biotecnologia Xilanase Ceriporiopsis subvermispora Polpa kraft

8
ABSTRACT
ELIAS, A. M. Action of enzymes from Ceriporiopsis subvermispora in the biobleaching of Eucalyptus grandis kraft pulps. 2007. 106 f. Dissertation (Master of Science in Industrial Biotechnology) – Escola de Engenharia de Lorena, Universidade de São Paulo, Lorena, 2007.
Enzymes obtained from biopulping of wood by Ceriporiopsis subvermispora were used in the biobleaching process of Eucalyptus grandis kraft pulps, followed by an EQP bleaching sequence (alkaline extraction, chelating and peroxidation). Characterization of crude extracts were performed by determination of enzymatic activities of xylanolitic complex: β-xylanase (346 UI/kg), β-xylosidase (62,6 UI/kg), β-glucosidase (87 UI/kg), β-mannanase (231 UI/kg), endoglucanase (133 UI/kg) and for manganese peroxidase (765 UI/kg). Xylans from birchwood, oat spelt, beechwood and sugarcane bagasse were treated with xylanases from C. subvermispora and reducing sugars arose until 96 h. Birchwood xylan, oat-spelt xylan, beechwood xylan and sugarcane bagasse xylan released different quantities of xylotetraose (0,63, 0,76, 0 g/L), xylotriose (0,24, 0,38, 0,19 g/L), xylobiose (0,52, 0,70, 0,39 g/L) and xylose (0, 0, 0,37 g/L), respectively. Comparisons of kraft pulps assayed with a commercial extract or enzymatic extract of C. subvermispora showed similar results with respect to reducing sugars released. Pulps containing 78% of cellulose, 12,8% of hemicelluloses and 5,3% of lignin in its chemical composition, kappa number of 12,9 and viscosity of 24,8 cP, were submitted to the enzymatic treatment based on xylanase activity, at the concentrations of 0,5, 1, 1,5 and 2 UI/g for 0,5, 1, 2, 3 and 5 h at 40ºC and 10% consistency. It were measured the release of chromophores at 237, 254, 208 and 465 nm and reducing sugars. The chomophores were released at 254, 280 and 465 nm with xylanase dose of 1,5 and 2 UI/g, while the absorbance at 237 nm arose out of the xylanase dose increased. The maximum reducing sugars (23,6 g/L) was achieved after 3 h of treatment with 2 UI/g. Kappa number and viscosity of pulps were measured after enzymatic step and after chemical bleaching sequence and delignification efficiency and selectivity were determined after chemical bleaching sequence. The kappa number reduction was 7,8% after enzymatic pretreatment and 11,4% after the chemical bleaching sequence, both with xylanase dose of 2 UI/g for 3 h. Viscosity of all pulps enzymatically treated was higher than those without enzymatic treatment. The highest value of selectivity was achieved with 2 UI/g of xylanase. The highest kappa number reduction joined with the highest release of chomophores at 237 and 465 nm and reducing sugars were achieved with 2 UI/g of xylanase for 2 to 3 h of treatment.
Key-words: Biotechnology Xylanase Ceriporiopsis subvermispora kraft pulp

9
LISTA DE FIGURAS
Figura 2.1 - Estrutura da celulose, parte central da cadeia molecular ......................................21
Figura 2.2 - Estrutura proposta para a macromolécula de lignina de Eucalyptus grandis .......22
Figura 2.3 - Estrutura dos monossacarídeos que formam as polioses ......................................23
Figura 2.4 - (A) Enzimas xilanolíticas envolvidas na degradação da xilana. Ac: grupo acetil;
α-Araf: α-arabinofuranose. (B) hidrólise de xilooligossacarídeos pela β-xilosidade...............26
Figura 2.5 - Cromóforos incorporados na lignina ....................................................................32
Figura 2.6 - Grupos cromógenos da lignina .............................................................................32
Figura 5.1 - Porcentagem de hidrólise da xilana de polpas kraft pelo extrato enzimático de C.
subvermispora e pelo extrato comercial. A carga enzimática aplicada foi de 2 UI/g nos dois
ensaios realizados .....................................................................................................................66
Figura 5.2 - Perfil da liberação de açúcares das xilanas por enzimas de C. subvermispora ....68
Figura 5.3 - Liberação de cromóforos a 237 nm em função do tempo de tratamento..............71
Figura 5.4 - Liberação de cromóforos a 254 nm em função do tempo de tratamento.............72
Figura 5.5 - Liberação de cromóforos a 280 nm em função do tempo de tratamento..............72
Figura 5.6 - Liberação de cromóforos a 465 nm em função do tempo de tratamento..............73
Figura 5.7 - Liberação de cromóforos pelo tratamento enzimático com 2 UI/g de xilanase nos
diferentes tempos de tratamento ...............................................................................................74
Figura 5.8 - Liberação de cromóforos pelo tratamento enzimático com 1,5 UI/g de xilanase
nos diferentes tempos de tratamento ........................................................................................74
Figura 5.9 - Liberação de cromóforos pelo tratamento enzimático com 1 (a) e 0,5 (b) UI/g de
xilanase nos diferentes tempos de tratamento ..........................................................................75
Figura 5.10 - Açúcares redutores liberados pelo tratamento enzimático nas polpas kraft em
função da carga enzimática referente à xilanase aplicada ........................................................76
Figura 5.11 - Hidrólise das polioses da polpa em diferentes cargas enzimáticas.....................77
Figura 5.12 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga
enzimática de 2 UI/g.................................................................................................................80
Figura 5.13 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga
enzimática de 1,5 UI/g..............................................................................................................81
Figura 5.14 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga
enzimática de 1 UI/g.................................................................................................................81
Figura 5.15 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga
enzimática de 0,5 UI/g..............................................................................................................82

10
Figura 5.16 - Viscosidade da polpa tratada com 2 UI/g de xilanase após o branqueamento
químico .....................................................................................................................................83
Figura 5.17 - Viscosidade da polpa tratada com 1,5 UI/g de xilanase após o branqueamento
químico .....................................................................................................................................83
Figura 5.18 - Viscosidade da polpa tratada com 1 UI/g de xilanase após o branqueamento
químico .....................................................................................................................................84
Figura 5.19 - Viscosidade da polpa tratada com 0,5 UI/g de xilanase após o branqueamento
químico .....................................................................................................................................84
Figura 5.20 - Número kappa das polpas resultantes do tratamento enzimático utilizando a
carga de xilanase de 2 UI/g e das polpas resultantes do branqueamento químico ...................86
Figura 5.21 - Número kappa das polpas resultantes do tratamento enzimático utilizando a
carga de xilanase de 1,5 UI/g e das polpas resultantes do branqueamento químico ................87
Figura 5.22 - Número kappa das polpas resultantes do tratamento enzimático utilizando a
carga de xilanase de 1 UI/g e das polpas resultantes do branqueamento químico ...................88
Figura 5.23 - Número kappa das polpas resultantes do tratamento enzimático utilizando 0,5
UI/g e das polpas resultantes do branqueamento químico .......................................................89
Figura 5.24 - Viscosidade e número kappa em função da concentração de xilanase para os
tempos de tratamento enzimático de 1, 2 e 3 horas ..................................................................90
Figura 5.25 - Viscosidade e número kappa em função da concentração de xilanase para os
tempo de tratamento de 1, 2 e 3 horas para o branqueamento químico ...................................91
Figura 5.26 - Cromóforos (237 e 465 nm) e açúcares redutores liberados e número kappa em
função do tempo de tratamento (2 UI/g de xilanase)................................................................93

11
LISTA DE TABELAS
Tabela 2.1 - Composição química das madeiras macias e duras..............................................21
Tabela 2.2 - Quadro resumido dos diferentes processos de separação dos componentes
lignocelulósicos ........................................................................................................................28
Tabela 2.3 - Características da celulose industrial obtida em diferentes processos de polpação
e seus usos ................................................................................................................................29
Tabela 2.4 - Notação dos Estágios de Branqueamento ............................................................33
Tabela 4.1 - Caracterização da polpa kraft de eucalipto...........................................................52
Tabela 4.2 - Constituição química do E. grandis utilizado neste trabalho ...............................52
Tabela 4.3 - Condições gerais de branqueamento ....................................................................60
Tabela 5.1 -.Produção enzimática de C. subvermispora em cavacos de E. grandis .................63
Tabela 5.2 - Concentração de açúcares redutores liberados dos tratamentos com os extratos
enzimáticos sobre polpas kraft. A carga enzimática aplicada foi de 2 UI/g nos dois ensaios
realizados ..................................................................................................................................65
Tabela 5.3 - Quantificação dos oligômeros detectados por CLAE ..........................................67
Tabela 5.4 - Quadro comparativo do grau de hidrólise entre as diferentes xilanas..................68
Tabela 5.5 - Condições utilizadas no branqueamento enzimático............................................69
Tabela 5.6 - Atividades aplicadas sobre a polpa referente às demais enzimas contidas no
extrato bruto de C. subvermispora............................................................................................70
Tabela 5.7 - Dados obtidos dos filtrados das polpas kraft após o tratamento com o extrato
bruto de C. subvermispora em diferente concentrações (0,5, 1, 1,5, e 2 UI/g) pela
cromatografia líquida de alta eficiência (CLAE) .....................................................................79
Tabela 5.8 - Resultados quanto à redução da viscosidade (%) e seletividade após o
branqueamento químico ...........................................................................................................91

12
LISTA DE EQUAÇÕES
Equação 4.1 - Relação entre massa seca, massa úmida, volume de enzima e volume de água a
ser acrescentada à mistura de reação para ajuste da consistência da polpa ..............................53
Equação 4.2 - Cálculo da lignina Klason .................................................................................54
Equação 4.3 - Cálculo da lignina solúvel .................................................................................55
Equação 4.4 - Cálculo da lignina solúvel .................................................................................55
Equação 4.5 - Cálculo do número kappa das polpas kraft........................................................57
Equação 4.6 - Cálculo do número kappa das polpas kraft........................................................57
Equação 4.7 - Cálculo do número kappa das polpas kraft........................................................57
Equação 4.8 - Cálculo da constante do viscosímetro ...............................................................58
Equação 4.9 - Cálculo da viscosidade ......................................................................................58
Equação 4.10 - Cálculo da redução da viscosidade (%)...........................................................59
Equação 4.11 - Cálculo da eficiência de deslignificação e seletividade ..................................59
Equação 4.12 - Cálculo da eficiência de deslignificação e seletividade ..................................59
Equação 4.13 - Taxa de hidrólise da xilana..............................................................................60

13
LISTA DE ABREVIATURAS E SIGLAS
AOX – Adsorbable Organic Halogen (‘Compostos Halogenados Absorvíveis’);
BRACELPA – Associação Brasileira de Papel e Celulose;
CMCase – Carboximetilcelulase ou Endo-1-4-β-glicanase;
CLAE – Cromatografia Líquida de Alta Eficiência;
DNS – Ácido 3,5-dinitrosalicílico;
DP – Degree of Polymerization (‘Grau de Polimerização’);
DTPA – Diethylene Triamine Penta Acetate (‘Dietileno-Triamino-Penta-Acetato’);
EC – Enzyme Comission (‘Comissão que delibera sobre a nomenclatura das enzimas’);
ECF – Elemental Chlorine Free (‘Totalmente Livre de Cloro Elementar’);
EDTA – Ethylene Diamine Tetra Acetic Acid (‘Ácido Etileno-Diamino-Tetra-Acético’);
HexA’s – Hexenuronic Acids (‘Ácidos Hexenurônicos’);
HPLC – High Performance Liquid Chromatography (‘Cromatografia Líquida de Alta
Eficiência’);
IPT – Instituto de Pesquisas Tecnológicas;
LCC – Lignin Carbohydrate Complex (‘Complexo Lignina-carboidrato’);
LiP – Lignina Peroxidase;
MnP – Manganês Peroxidase;
P&D – Pesquisa e Desenvolvimento;
PIB – Produto Interno Bruto;
pNP – P-nitrofenol;
pNPG – P-nitro-fenil-β-D-glicopiranosídeo;
pNPX – P-nitrofenil-β-D-xilopiranosídeo;
RI – Refraction Index (‘Índice de Refração’);
TAPPI - Technical Association of Pulp and Paper Industry (‘Associação Técnica da Indústria
do Papel e da Celulose’);
TCDF – 2,3,7,8-tetracloro-dibenzeno-furano;
TCF – Totally Chlorine Free (‘Totalmente Livre de Cloro’);
TCDD – 2,3,7,8-tetracloro-dibenzeno-p-dioxina.

14
SUMÁRIO
1. INTRODUÇÃO....................................................................................................................16
2. REVISÃO BIBLIOGRÁFICA .............................................................................................20
2.1. COMPOSIÇÃO QUÍMICA DA MADEIRA.................................................................20
2.2. ENZIMAS ENVOLVIDAS NA DEGRADAÇÃO DA MADEIRA .............................25
2.3. PROCESSOS DE POLPAÇÃO .....................................................................................28
2.4. PROCESSO KRAFT......................................................................................................29
2.5. PROCESSOS DE BRANQUEAMENTO......................................................................30
2.6. BRANQUEAMENTO ENZIMÁTICO..........................................................................37
2.7. HISTÓRICO DO USO DE XILANASES NA INDÚSTRIA DE PAPEL E CELULOSE
..................................................................................................................................................38
2.8. UTILIZAÇÃO DA ENZIMA MANGANÊS PEROXIDASE .......................................41
2.9. Ceriporiopsis subvermispora .........................................................................................42
3. OBJETIVOS.........................................................................................................................44
4. MATERIAIS E MÉTODOS.................................................................................................45
4.1. PREPARO DOS EXTRATOS ENZIMÁTICOS ...........................................................45
4.1.1. Extrato de Ceriporiopsis subvermispora ..................................................................45
4.1.2. Extrato comercial......................................................................................................46
4.1.3. Caracterização dos extratos ......................................................................................46
4.1.3.1. β-Xilanase...........................................................................................................46
4.1.3.2. β-Xilosidase........................................................................................................47
4.1.3.3. β-Glicosidase ......................................................................................................48
4.1.3.4. β-Mananase ........................................................................................................49
4.1.3.5. Endoglucanase ....................................................................................................50
4.1.3.6. Manganês Peroxidase (MnP)..............................................................................51
4.2. PREPARO DAS XILANAS...........................................................................................51
4.3. POLPA............................................................................................................................52
4.3.1. Consistência das polpas (%) .....................................................................................53
4.3.2. Tratamento Enzimático das Polpas...........................................................................53
4.4. CARACTERIZAÇÃO FÍSICO-QUÍMICA DAS POLPAS ..........................................54
4.4.1. Determinação da lignina insolúvel (Lignina Klason)...............................................54
4.4.2. Determinação da lignina solúvel ..............................................................................55

15
4.4.3. Determinação da Lignina Total ................................................................................56
4.4.4. Determinação dos carboidratos ................................................................................56
4.4.5. Determinação do número kappa ...............................................................................56
4.4.6. Determinação da Viscosidade ..................................................................................58
4.4.7. Redução da Viscosidade...........................................................................................59
4.4.8. Cálculo da Eficiência de Deslignificação e Seletividade .........................................59
4.5. TAXA DE HIDRÓLISE DA XILANA..........................................................................60
4.6. BRANQUEAMENTO DAS POLPAS...........................................................................60
4.6.1. Determinação do pH das polpas ...............................................................................61
4.6.2. Extração alcalina (E) ................................................................................................61
4.6.3. Quelação (Q).............................................................................................................61
4.6.4. Peroxidação (P).........................................................................................................62
4.7. REPRODUTIBILIDADE DOS RESULTADOS...........................................................62
5. RESULTADOS E DISCUSSÃO..........................................................................................63
5.1. DETERMINAÇÃO DAS ATIVIDADES ENZIMÁTICAS NOS EXTRATOS DE C.
SUBVERMISPORA...................................................................................................................63
5.2. EFEITO DAS ENZIMAS DE C. SUBVERMISPORA E DO EXTRATO COMERCIAL
NA HIDRÓLISE DAS POLPAS .............................................................................................64
5.3. ANÁLISE DOS PRODUTOS DAS XILANAS DE DIFERENTES ORIGENS APÓS
AÇÃO DA XILANASE ...........................................................................................................66
5.4. BRANQUEAMENTO DAS POLPAS...........................................................................69
5.4.1. Análise dos produtos das polpas kraft após a ação do complexo enzimático de C.
subvermispora ..........................................................................................................................70
5.4.2. Características da polpa tratada com diferentes concentrações de enzimas e
branqueada................................................................................................................................79
5.5. CORRELAÇÃO ENTRE CROMÓFOROS, NÚMERO KAPPA E LIBERAÇÃO DE
AÇÚCARES REDUTORES ....................................................................................................92
6. CONCLUSÕES....................................................................................................................94
REFERÊNCIAS .......................................................................................................................96
APÊNDICES ..........................................................................................................................105

16
1. INTRODUÇÃO
A indústria de papel e celulose representa um dos mais expressivos setores industriais do
mundo. O mercado mundial de celulose atingiu cerca de 28 milhões de toneladas em 1995,
movimentando US$ 25 bilhões, resultado do aumento de preços ocorrido a partir de 1994. No
período de 1995/2005 as taxas anuais médias ficaram ao redor de 3,3%, percentual elevado a 4,6%
quando se trata das fibras curtas de eucalipto, a especialidade brasileira. Entre 1993 e 2003, foram
investidos US$ 12 bilhões e para o período de 2003 até 2012, a meta é ultrapassar os US$ 14
bilhões. Nos últimos dez anos, o crescimento do Produto Interno Bruto (PIB) do país foi de 25%,
mas a indústria de celulose e papel registrou, no mesmo período, crescimento na produção de
celulose de 70% e de 50% na produção de papel. Segundo estimativas da Bracelpa, a produção de
celulose em 2006 alcançou 10,5 milhões de toneladas, 5% a mais em relação a 2005. Desse total, 6
milhões de toneladas foram destinadas à exportação, volume 16% superior ao ano anterior. A
produção nacional de celulose de eucalipto (celulose de fibra curta) responde pela metade da
produção mundial desse tipo de fibra, com uma produção ao redor de 6 milhões de toneladas de
polpa por ano, dos quais 98% são branqueadas.
A indústria de papel e celulose é importante na economia da América do Sul devido à
grande disponibilidade de recursos florestais, sendo o Brasil e o Chile os maiores produtores de
celulose da região. Os grandes avanços nos campos da silvicultura e da biotecnologia permitiram
que o Brasil se tornasse o 7º maior produtor mundial de celulose (incluindo fibras curtas e longas),
o maior produtor mundial de celulose branqueada de eucalipto, líder nas exportações de celulose
de fibra curta e 11º na produção de papel. Na década de 90 havia no Brasil mais de 90 empresas
papeleiras, como por exemplo as grandes Aracruz, Suzano, Champion e Rio Cell que obtiveram
um faturamento líquido em torno de R$ 940 milhões em 2005 (CABALLERO, 2005). Hoje, são
220 empresas de papel e celulose, que empregam cerca de 100 mil pessoas em 255 unidades
industriais. O setor exporta 90% da sua celulose de mercado (a não integrada com a produção do
papel). Com capacidade atual instalada de 7,8 milhões de toneladas/ano de celulose de fibra curta,
das quais 5,5 milhões de toneladas destinadas ao mercado externo, notadamente Europa (48%),
Ásia (27%), América do Norte (22%) e demais países (3%), o Brasil assumiu a liderança mundial
na exportação de celulose de fibra curta branqueada e deverá notabilizar-se cada vez mais nesse
segmento como provedor do mercado internacional (FURTADO, 2006).

17
A principal vantagem comparativa do Brasil para o aumento de suas exportações foi o
desenvolvimento da celulose de fibra curta de eucalipto e a sua aceitação no mercado
internacional. Esse tipo de celulose significou uma redução do tempo de corte da madeira, o que
representa em torno de 25% do custo da celulose. Em 2000, a produção de papel atingiu 7,2
milhões de toneladas e a de celulose 7,6 milhões. E a previsão é de que o consumo de celulose
aumente nessa década sendo previsto um incremento na produção de 4 milhões de toneladas de
celulose por volta de 2017-2018. O superávit da balança comercial em 2000 foi de US$ 1,8 bilhão,
enquanto o faturamento total do setor alcançou em torno de R$ 13,7 bilhões.
O potencial de crescimento do setor de papel no mercado brasileiro é infinitamente superior
em relação aos níveis atuais, apesar do consumo per capita ser considerado um dos menores do
mundo, havendo, portanto, grande necessidade de se reduzir a carga tributária sobre os
investimentos, como forma de contribuir para a instalação de novas fábricas de papel.
Enquanto no Brasil são consumidos apenas 40 quilos de papel por habitante ao ano, em países
desenvolvidos são encontrados níveis de consumo bem mais elevados, como é o caso dos Estados
Unidos (312 kg per capita/ano), Japão (247 kg per capita/ano), Alemanha (236 kg per capita/ano),
Canadá (223 kg per capita/ano), Reino Unido (210 kg per capita/ano) e Itália (195 kg per
capita/ano), ou mesmo nos demais países latino-americanos como Chile (67 kg per capita/ano),
México (58 kg per capita/ano) e Argentina (50 kg per capita/ano). Isso porque o consumo está
vinculado a dois fatores muito importantes: renda e escolaridade.
Como a produção de celulose e a produção de papel estão integradas, os investimentos se
direcionam para a integração da cadeia produtiva, ou seja, desde o plantio das árvores até o
acabamento do produto final, por exemplo, papel de impressão, tornando as empresas auto-
suficientes em matérias primas. Isso faz com que os investimentos dessa indústria tenham um
longo prazo de retorno, dado que os investimentos são da ordem de US$ 1.400 por tonelada de
papel; as escalas mínimas de investimentos atualmente estão acima de 100 mil toneladas. Além
disso, há a necessidade de investimentos contínuos em torno de US$ 100 milhões por parte dos
grandes produtores para a manutenção da capacidade produtiva. Outros investimentos são
direcionados para a atividade de reflorestamento, principal fonte de matéria-prima, e para o
desenvolvimento tecnológico. Os investimentos em pesquisa e desenvolvimento da indústria
brasileira representam apenas 1% do seu faturamento. Há também o impacto sócio-ambiental do
setor. O segmento de celulose e papel planeja investir no Brasil US$ 14,4 bilhões, no período de
2003 a 2012, para ampliar a capacidade produtiva e abrir 60 mil novos empregos. Nos últimos dez
anos, foram aplicados US$ 12 bilhões na ampliação da capacidade (BRACELPA, 2006).

18
A competição nesta indústria se dá por preço e qualidade. Nesse setor é necessário conjugar
qualidade, baixo custo e, principalmente, escala de produção. A diferenciação dos produtos torna-
se cada vez mais relevante, havendo a valorização do produto final. Nos últimos anos, acelerou-se
o processo de criação de produtos, ocorrendo algumas inovações na tecnologia de processos,
possibilitando ganhos expressivos de produtividade, e conseqüentemente, incentivando os gastos
com P&D. Pressões ambientalistas têm levado as empresas a investir no desenvolvimento e
implantação de novas tecnologias de processos, com destaque para a área de branqueamento, além
de pesados gastos com controle ambiental. Essa pressão é exercida, principalmente, em três
vetores: a exigência de produtos que não agridam o meio ambiente, o uso de tecnologias limpas e
o deslocamento de matéria-prima de origem florestal por papel reciclado. O principal objetivo das
indústrias tem sido o desenvolvimento de novas tecnologias que possam reduzir ou suprimir a
presença de poluentes, principalmente compostos organoclorados, nos efluentes das plantas de
branqueamento. Uma das escolhas para esse objetivo é a tecnologia baseada no tratamento
enzimático. Uma classe de enzimas já utilizada em escala industrial são as xilanases,
principalmente devido ao seu baixo custo de produção. Em 1995 o custo da enzima foi de US$
3,50 por tonelada de polpa (FARREL et al., 1996) e há relatos de que esse preço atualmente é mais
baixo. O tratamento com xilanase é muito versátil, associando as vantagens do pré-tratamento
enzimático, como redução de reagentes químicos, com modificações mínimas nos processos e
conseqüentemente um capital de investimento mínimo. É realizado em plantas que tratam tanto
madeiras macias, quanto madeiras duras que produzem polpas de mercado, plantas integradas,
plantas com deslignificação e plantas com 3-, 4- e 5- seqüências de branqueamento. Sabe-se que a
enzima não branqueia ou deslignifica a polpa, mas torna a polpa mais fácil para branquear nas
fases subseqüentes do branqueamento químico. Muitas xilanases têm sido isoladas e
caracterizadas, contudo pouco se sabe da atuação do complexo em substratos insolúveis, como em
polpas kraft. Para se tirar o máximo proveito do uso das enzimas, há necessidade de conhecer seus
mecanismos de reação com a lignina residual. A partir desse conhecimento, é possível combinar
de maneira mais efetiva os vários reagentes de branqueamento numa seqüência que seja de alta
eficiência e baixo impacto ambiental. O tratamento com xilanase também diminui o conteúdo de
ácidos hexenurônicos (HexA’s) das polpas, diminuindo o consumo de agentes branqueadores.
Existem 20 fábricas de celulose na América do Norte utilizando xilanase, sendo 7 no
Canadá e 13 nos Estados Unidos. Acima de 3,9 milhões de toneladas de polpa kraft foi tratada em
2001 e 6 milhões de toneladas em 2002. No Canadá, 4,2 toneladas de polpa são tratadas
diariamente, enquanto nos Estados Unidos são 6,9 toneladas. A utilização da xilanase em fábricas

19
americanas tem sido de aproximadamente 67% em madeiras duras e 37% em madeiras macias.
Uma razão para o aumento na utilização de xilanase é que o desempenho da xilanase tem
melhorado significativamente desde sua primeira implementação em fábricas anos atrás (VAN
DER BURGT et al., 2002). A efetividade do tratamento enzimático depende das propriedades da
enzima utilizada e da procedência desta, do tipo da madeira, do processo de cozimento e da
seqüência branqueadora que será utilizada em conjunto com a incubação com xilanase. Por estas
razões, estudos preliminares são requeridos havendo a necessidade de uma caracterização
detalhada da polpa e do papel resultantes através da utilização de xilanases de diferentes origens,
para um melhor entendimento dos efeitos enzimáticos nas fibras, primeiramente em escala
laboratorial para que seja possível então a subseqüente utilização desta tecnologia em escala
industrial.
Estudos feitos em laboratório mostram que não são caros os investimentos necessários para
adaptar o tratamento enzimático às plantas industriais existentes e o tratamento enzimático tem se
mostrado completamente compatível com os equipamentos instalados. Além disso, linhagens de
microrganismos que apresentam maior eficiência na produção de xilanase aliada à tecnologia,
oferecem baixos investimentos em xilanases branqueadoras de polpa, que são ambientalmente e
economicamente vantajosas. Indústrias de papel e celulose iniciaram pesquisas que relacionam a
produção de xilanases livres de celulase por microrganismos.
Neste estudo foi utilizado o extrato enzimático produzido pelo fungo de decomposição
branca Ceriporiopsis subvermispora para a avaliação do efeito biobranqueador de polpas kraft de
Eucaliptus grandis. Em seguida a polpa tratada enzimaticamente foi submetida a uma seqüência de
branqueamento químico. Ceriporiopsis subvermispora tem sido um dos fungos mais estudados nos
últimos anos devido ao complexo enzimático produzido e pelo seu potencial de deslignificar
seletivamente materiais lignocelulósicos, sendo por essa razão escolhido para este estudo.

20
2. REVISÃO BIBLIOGRÁFICA
A biodegradação da madeira é um processo natural de reciclagem da matéria orgânica e
ocorre em todos os ambientes naturais (terrestres e aquáticos) quando as condições ambientais são
favoráveis. Do ponto de vista tecnológico, a biodegradação ou biopolpação, da madeira pode ser
utilizada em diversos processos industriais de transformação destinados à produção de celulose e
papel, além de outros.
Durante o processo de biopolpação um grande volume de enzimas pode ser recuperado.
Manganês-peroxidase é a principal enzima oxidativa produzida por Ceriporiopsis subvermispora,
enquanto xilanase e mananase são as principais enzimas hidrolíticas. Tais enzimas são
potencialmente importantes no biobranqueamento de polpas celulósicas, em que a lignina é
removida quase completamente após o processo de polpação. Porém uma parte dessa lignina
permanece na polpa interferindo na qualidade do produto final (CHRISTOV e PRIOR, 1997).
Estudos têm mostrado que a aplicação de enzimas hemicelulolíticas como a xilanase e a mananase
nesse processo, é capaz de solubilizar as polioses facilitando indiretamente a retirada de lignina.
Porém, tem-se descrito que a extensão da hidrólise das polioses residuais na polpa é dependente
entre outros fatores, das propriedades das enzimas, a massa molar, as cargas líquidas, o ponto
isoelétrico, a estabilidade térmica, a estabilidade ao pH e a interferência de inibidores da sua
atividade (MAGALHÃES, 2005).
2.1. COMPOSIÇÃO QUÍMICA DA MADEIRA
Sob o ponto de vista químico, a madeira é constituída principalmente de celulose, lignina e
polioses, em diferentes proporções. Extrativos, minerais, amido e pectina também estão presentes
na madeira num baixo teor (Tabela 2.1).

21
Tabela 2.1 - Composição química das madeiras macias e duras
Componentes Madeira dura (%) Madeira macia (%)
Celulose 40-50 45-50
Galactoglucomanana 2-5 20-25
Xilana 15-30 5-10
Lignina 18-25 25-35
Extrativos 1-5 3-8
(BIERMANN, 1996)
- Celulose
É o componente mais abundante nos lignocelulósicos, sendo um polímero linear (parte
amorfo e parte cristalino) formado exclusivamente por moléculas de anidro-glicose unidas por
meio de ligações β-(1-4)-glicosídicas. Estritamente a celulose é composta por unidades
monoméricas de celobiose (Figura 2.1) que se repetem sempre pela junção de duas moléculas de
glicose seguida da eliminação de água através das hidroxilas ligadas ao carbono 1 e 4 (FENGEL e
WEGENER, 1989). As pontes de hidrogênio inter e intramolecular são responsáveis pela
manutenção das regiões cristalinas, e tornam a celulose altamente resistente à hidrólise ácida,
alcalina ou enzimática (WOOD e SADDLER, 1988; CONVERSE e WARE, 1994). O tamanho da
cadeia de celulose é normalmente especificado pelo grau de polimerização (GP), que corresponde
ao número médio de unidades de glicose presentes na molécula de celulose.
Figura 2.1 - Estrutura da celulose, parte central da cadeia molecular (FENGEL e WEGENER, 1989)

22
- Lignina
A lignina tem um papel significante na proteção natural da madeira (DANIEL, 2003). É
composta basicamente de unidades fenilpropano formando uma macromolécula tridimensional e
amorfa, representando de 20 a 30% do total de lignocelulósicos (Figura 2.2). O acoplamento das
unidades fenilpropano não ocorre de forma regular e repetitiva, o que é atribuído ao mecanismo de
biossíntese da lignina, que se processa por via radicalar a partir da reação de três diferentes álcoois
cinamílicos precursores (sinapílico, coniferílico e para-cumarílico).
Os diferentes tipos de acoplamento entre os precursores dão origem a vários tipos de
ligação entre as unidades fenilpropano. As mais abundantes são: β-O-4 e α-O-4 (50 a 65%), β-5 (6
a 15%), β-1 (9 a 15%), 5-5 (2 a 9%) e β-β (2 a 5%). Esses vários tipos de ligações formadas
originam uma estrutura tridimensional complexa (KANTELINEN, 1992).
Figura 2.2 - Estrutura proposta para a macromolécula de lignina de Eucalyptus grandis (BRASILEIRO et al., 2001)

23
- Polioses
As hemiceluloses ou polioses são oligômeros de baixa massa molar formadas por xilana,
galactana e glucomanana como componentes principais, podendo ainda apresentar quantidades
variáveis de ácidos urônicos e desoxi-hexoses em alguns tipos de madeira. As polioses estão
intimamente associadas à celulose na parede da célula vegetal e são compostas por diferentes
unidades de açúcares formando cadeias ramificadas (FENGEL e WEGENER, 1989). A estrutura
dos monossacarídeos que formam as polioses está mostrada na Figura 2.3.
Figura 2.3 - Estrutura dos monossacarídeos que formam as polioses (FENGEL e WEGENER, 1989)
A classificação é feita geralmente de acordo com o resíduo de açúcar presente na cadeia
principal. A cadeia principal pode conter quantidades variadas de diversas ramificações como
acetil, metil-glucuronil e arabinofuranosil. Os grupamentos arabinofuranosil podem ser
esterificados por ácidos aromáticos tais como ácido ferúlico e p-cumárico, que podem participar na
ligação lignina-hemicelulose (JEFFRIES, 1990; COUGHLAN e HAZLEWOOD, 1993). O teor de
polioses em diferentes tipos de lignocelulósicos é bastante variável, mas pode-se admitir um valor
médio de cerca de 20% (FERRAZ et al., 2002). Nas madeiras duras, tipo eucalipto, cerca de 25 a
40% da massa é constituída por polioses do tipo xilana, O-acetil-4-O-metil glucoronoxilana (20 a

24
35%) e glucomanana (3 a 5%) (TIMELL, 1965; JANES, 1969; COWLING e KIRK, 1976;
FENGEL e WEGENER, 1989; HON e SHIRAISHI, 2001).
Em seu estado natural as polioses não são cristalinas em razão da heterogeneidade de seus
constituintes, da presença de pequenos grupos laterais e em alguns casos ramificações,
diferentemente da estrutura altamente linear das moléculas de celulose (JANES, 1969).
- Extrativos
Os denominados extrativos da madeira são terpenos, resinas e polifenóis localizados
principalmente no cerne, e são responsáveis pela cor, cheiro e resistência da madeira (HARTLEY
e CLIVE, 1989; DESCHAMPS, 1989). As leucoantocianinas são os polifenóis mais abundantes
nas plantas depois da lignina, e são os precursores dos flavonóides e taninos presentes na madeira
(BARISKA e PIZZI, 1986). Os extrativos na madeira podem representar até 10% de sua
composição (FENGEL e WEGENER, 1989).
- Elementos inorgânicos
Na madeira os elementos inorgânicos encontram-se em baixas concentrações e
representam de 0,2 a 0,3% de seu peso seco (KOLLMAN e COTÊ, 1984). Os elementos
inorgânicos mais comuns são Ca (500-800 ppm), K (60-100 ppm) e Mg (50-80 ppm) (ELLIS,
1965). Também tem sido detectado P (20 ppm), Mn (4-100 ppm), B (2-15 ppm), Ba (2-15 ppm),
Fe (1-3 ppm), Sr (1-6 ppm), Al (0,5-6 ppm), Cu (0,5-1 ppm) e Zn (0,3-0,8 ppm) (GUYETTE et al.,
1992). Os íons metálicos na madeira podem existir na forma de compostos inorgânicos insolúveis
ou unidos à lignina e pectinas (CUTTER e GUYETTE, 1993). A presença dos elementos Mn, Fe,
Ca, Cr e Zn em maiores concentrações tem sido detectadas nas células do parênquima e na lamela
média (GUYETTE et al., 1992; HEIJNESSON et al., 1995).

25
2.2. ENZIMAS ENVOLVIDAS NA DEGRADAÇÃO DA MADEIRA
A biodegradação da celulose ocorre pela ação de três grupos de enzimas que atuam
sinergicamente. Esses grupos de enzimas compreendem as endo-1,4-β-glicanases, as exo-1,4-β-
glicanases e as 1,4-β-glicosidases. As endoglucanases rompem a molécula de celulose ao acaso e
liberam fragmentos menores que servem de substrato para as exo-glucanases. As exo-glucanases
hidrolisam, pelas pontas, os fragmentos de menor massa molecular, liberando moléculas de
celobiose. As β-glicosidases hidrolisam a celobiose até glicose (BHAT e BHAT, 1997).
A biodegradação da lignina ocorre através de pelo menos duas classes distintas de
enzimas: as fenoloxidases e as enzimas que produzem peróxido de hidrogênio. Entre as
fenoloxidases, ainda se podem descrever dois grupos: um contém as enzimas dependentes de
peróxido ou peroxidases. As peroxidases que estão envolvidas na biodegradação da lignina são
lignina peroxidase (LiP, EC 1.11.1.14) e peroxidase dependente de manganês (MnP, EC
1.11.1.13). As lacases (EC 1.10.3.2, benzenodiol oxigênio oxidoredutase), o outro subgrupo, são
cuproproteínas que não dependem de peróxido para atuar (FERRAZ, 2001; HAKALA et al.,
2005). As enzimas que produzem peróxido de hidrogênio são acessórias às peroxidases, gerando
peróxido de hidrogênio ‘in situ’ possibilitando que as peroxidases atuem.
A biodegradação das polioses ocorre de forma semelhante à da celulose, porém requer
um conjunto de enzimas extracelulares mais complexo. Isso ocorre devido a sua estrutura de
heteropolissacarídeo ramificado. São classificadas de acordo com o substrato em que atuam, sendo
também agrupadas sob o nome genérico de glicano-hidrolases (SUNNA e ANTRANIKIAN,
1997).
Por exemplo, a degradação completa de xilana requer a ação combinada de diferentes
enzimas (Figura 2.4). Assim, as endo-β-1,4-D xilanases rompem ligações glicosídicas ao acaso
produzindo grandes quantidades de xilo-oligossacarídeos substituídos e não-substituídos de
diversos tamanhos. As exo-β-1,4-D-xilanases removem somente unidades de xilose a partir das
extremidades da cadeia de xilana. As β-xilosidases hidrolisam sacarídeos como xilobiose e xilo-
oligossacarídeos maiores até xilose. Entre as enzimas que participam da quebra das cadeias laterais
destacam-se as α-L-arabinofuranosidases e as α-D-glucuronidases, além das esterases que
participam na liberação dos substituintes acetil, cumaril e feruloil, que atuam sinergicamente com
as endo-xilanases e as β-xilosidases (ERIKSSON, 1990; COUGHLAN e HAZLEWOOD, 1993).

26
As mananases atuam sobre ligações glicosídicas entre moléculas de manose e as glucuronidases
sobre ligações de ácidos urônicos com moléculas de açúcares.
Figura 2.4 - (A) Enzimas xilanolíticas envolvidas na degradação da xilana. Ac: grupo acetil; α-Araf: α-arabinofuranose. (B) hidrólise de xilooligossacarídeos pela β-xilosidade (SUNNA e ANTRANIKIAN, 1997)
A disponibilidade de informações sobre as propriedades das diferentes xilanases vem do
estudo das enzimas de fungos e bactérias. Uma maneira de classificar as xilanases as dividem em
quatro grupos. Dois capazes de hidrolisar pontos de ramificação, diferindo nos produtos finais
formados. Assim, o primeiro produz xilobiose e xilose e o segundo produz xilo-oligossacarídeos
de tamanho intermediário. Os outros grupos não são capazes de hidrolisar ramificações. Um deles
produz xilo-oligossacarídeos maiores que a xilobiose e o outro produz principalmente xilobiose e
xilose (KULKARNI et al., 1999).
Xilanases de diferentes organismos são geralmente estáveis em uma ampla faixa de pH
(3-10) e apresentam uma faixa ótima que pode variar de 4-7. As xilanases produzidas pelo fungo
Aspergillus kawachii, por exemplo, e Penicillium herque exibem pH ótimo na faixa ácida (pH 2-
6). Endo-xilanases de fungos têm massas molares na faixa de 7 a 60 kDa e são geralmente mais
ativas a pH 3,5-6,0 e 40-60ºC (TAN et al., 1987; BISWAS et al., 1990; BAILEY et al., 1992).

27
Xilanases microbianas são formadas por proteínas de apenas uma cadeia com massas moleculares
por volta de 8 a 145 kDa (KULKARNI et al., 1999).
As β-xilosidases purificadas de fungos, bactérias e leveduras hidrolisam pequenos xilo-
oligossacarídeos a partir do terminal não redutor liberando xilopiranoses. Além da atividade
hidrolítica muitas β-xilosidases tem uma alta atividade de transferase, o que provoca a formação de
oligossacarídeos de massa molar maior que o substrato original (REILLY, 1981). Em comparação
às endo-xilanases, β-xilosidases apresentam uma massa molar mais elevada. Elas são mais ativas
na faixa de pH de 4,0-7,0 e a temperaturas entre 40-80ºC (KITPREECHAVANICH et al, 1986;
JOHN e SCHMIDT, 1988). A afinidade de β-xilosidases em degradar xilana e xilo-
oligossacarídeos é fundamental na conversão total da biomassa a monossacarídeos. Pequenos xilo-
oligossacarídeos lineares podem ser convertidos completamente a xilose por β-xilosidases,
contudo a afinidade em relação aos xilo-oligossacarídeos diminui com o aumento da cadeia
(TAKENISHI e TSUJISAKA, 1973; REILLY, 1981).
As α-L-arabinofuranosidases são enzimas que atuam em L-arabinosídeos de baixa massa
molar e seus oligossacarídeos. Este grupo de enzimas atua juntamente com as endo-xilanases e β-
xilosidases para hidrólise de arabino(glucorono)xilanas. As α-L-arabinofuranosidases tem sido
purificadas de fungos, bactérias e leveduras. Em comparação às endo-xilanases, apresentam massa
molar relativamente alta (400 kDa) e pH ótimo entre 2,5-7,0 (BRILLOUET et al., 1985;
POUTANEN, 1988). A utilização desta enzima para hidrólise de arabinoxilanas reduz a
solubilidade do substrato devido à agregação intermolecular. Um pré-tratamento de arabinoxilana
com endo-xilanases que não apresentem atividade desramificadora, seguido de um tratamento com
arabinofuranosidase, seria a melhor alternativa para prevenir a agregação de longas cadeias e a
obtenção de um rendimento máximo em monossacarídeos (ANDREWARTHA et al., 1979).
Um grupo de enzimas de grande importância na hidrólise da xilana proveniente de
madeiras duras são as acetil esterases. Estas enzimas hidrolisam ligações éster de cadeias alifáticas
e aromáticas e removem grupos O-acetil de xilanas ou oligossacarídeos. As esterases apresentam
pesos moleculares na faixa de 30-70 kDa, com poucas exceções. Elas são mais ativas na faixa de
pH de 5,0-8,0 e a temperaturas variando de 40-80ºC (POUTANEN e SUNDBERG, 1988;
SUNDBERG et al., 1990). Acetil esterases são importantes na degradação de xilanas acetiladas
porque em geral os substituintes acetil impedem a ação de endo-xilanases. Tem sido demonstrado
que a acetil esterase e endo-xilanase atuam cooperativamente na hidrólise de acetil xilana (BIELY,
1985). Dependendo do substrato e da especificidade da acetil esterase, uma distinção pode ser feita
entre acetil xilana esterase e acetil esterase. Acetil xilana esterase é capaz de liberar ácido acético

28
de xilana acetilada sem a cooperação de endo-xilanases, enquanto acetil esterase necessita da
cooperação de uma endo-xilanase para liberar ácido acético (POUTANEN e SUNDBERG, 1988;
POUTANEN et al., 1990; SUNDBERG e POUTANEN, 1991).
As α-glucuronidases são enzimas que hidrolisam resíduos (4-O-metil)glucurônico, e que
auxiliam na conversão total de (arabino)glucuronoxilanas a monossacarídeos. As análises
estruturais da parede celular mostraram que grupos carboxílicos do ácido metilglucurônico estão
ligados a grupos hidroxilas da lignina (SMITH e FORSBERG, 1991). Possuem massas molares
elevadas e são mais ativas a altas temperaturas e condições ácidas.
2.3. PROCESSOS DE POLPAÇÃO
O termo polpação se refere a diferentes processos de separação das fibras de materiais
lignocelulósicos, principalmente da madeira. Assim, de acordo com o processo, a pasta resultante
pode ser química, semiquímica, quimimecânica, quimiotermomêcanica e mecânica.
(CARVALHO, 2003).
Os processos de polpação em geral se dividem em dois grandes grupos, polpação de alto
rendimento (mecânico) e polpação química. Mais recentemente processos biotecnológicos vêm
sendo desenvolvidos (Tabela 2.2).
A polpação química apresenta rendimento médio de 50%, já que a maior parte da lignina
e das polioses são removidas durante o processo de cozimento. Entre os processos químicos de
polpação, o mais utilizado é o sulfato ou kraft, correspondendo a mais de 80% da produção anual
mundial de polpas celulósicas.
Tabela 2.2 - Quadro resumido dos diferentes processos de separação dos componentes lignocelulósicos Tipo de processo Descrição sucinta do processo Observações
Físico Utilizam apenas energia mecânica, não envolvendo emprego de reagente químicos. a) Irradiação de raios gama; b) Tratamento a vapor (tratamento térmico) c) Steam-explosion (aquecimento e rápida descompressão do material).
Elevado custo de energia, tanto elétrica, quanto mecânica. a)Pouco eficaz. b)Pouco eficaz, podendo provocar reações entre os produtos secundários oriundos da fração hemicelulósica e o complexo lignina-celulose. c)Efetivo, promove a separação integral dos três componentes poliméricos.
Continua

29
Conclusão
Tabela 2.2 - Quadro resumido dos diferentes processos de separação dos componentes lignocelulósicos Químico Utiliza agentes químicos
específicos para cozinhar o material sob pressão. Os processos podem ser: ácidos (sulfito) ou alcalinos (sulfato e soda).
Efetivo. Amplamente empregado na indústria de celulose e papel, tendo como desvantagem a formação de resíduos altamente poluentes.
Biotecnológico a)Utiliza a ação de microrganismos selecionados capazes de promover a deslignificação dos materiais lignocelulósicos. b)Utilização de enzimas provenientes de microrganismos.
Parcialmente efetivo. Necessita de processos complementares de deslignificação, devido à baixa velocidade de degradação do material (período de meses). Processo ainda em fase experimental. Efetivo no pré-tratamento complementar aos processos industriais convencionais. Necessita aprimoramento.
(CASTRO, 2001)
Assim, as propriedades do papel e da celulose dependem do processo industrial utilizado
(Tabela 2.3).
Tabela 2.3 - Características da celulose industrial obtida em diferentes processos de polpação e seus usos
Tipos Características Usos
Pasta mecânica Resistência física reduzida,
baixo custo, boa capacidade de impressão, alta opacidade.
Papel de jornal, catálogos, revistas, papéis de parede, papéis absorventes e papelão.
Celulose semiquímica
Características bem variáveis de processo para processo.
Papelão corrugado, papel de jornal, papel de impressão, escrita e desenho.
Celulose sulfato/
kraft
Escura, opaca e bastante resistente.
Não-branqueada: papéis, papelões e cartões para embalagem e revestimentos.
Branqueada: papéis de para embalagens, impressão (livros).
(CASTRO, 2001)
2.4. PROCESSO KRAFT
Entre os processos químicos de polpação, o mais bem estabelecido e utilizado
mundialmente em larga escala é o kraft. Na polpação kraft, os cavacos de madeira são submetidos
à reação com solução de hidróxido de sódio (NaOH) e sulfato de sódio (Na2SO4): o “licor

30
branco”, formando então sulfeto de sódio (Na2S). Isso ocorre dentro do digestor, mantido a altas
pressões e temperaturas. Os produtos químicos fragmentam a lignina, quebrando-a em substâncias
de baixa massa molar, solúveis na solução alcalina, podendo então ser removidas das fibras por
inúmeras etapas de lavagem. A polpa ou pasta celulósica resultante da polpação (polpa marrom)
ainda não é adequada para a produção de determinados tipos de papel, exatamente pela sua
coloração escura. Essa coloração é devida, principalmente, a pequenas quantidades de lignina que
não foram removidas das fibras, chamada agora de lignina residual. A transformação tecnológica
da madeira em polpa celulósica, pelos processos químicos convencionais resulta na degradação e
perda de 50% do peso da madeira. Além da clivagem e solubilização da lignina, parte dos
produtos químicos é utilizada para neutralização de compostos ácidos provenientes da degradação
dos carboidratos (ácido acético e ácidos urônicos removidos das polioses) e pelos produtos de
degradação da lignina (FENGEL e WEGENER, 1989; DYER, 2004). A presença de sulfeto de
sódio (Na2S) produz íons hidrossulfito (HS-) que aceleram a remoção da lignina, resultando em
uma polpa de melhores propriedades mecânicas. O cozimento é controlado até atingir-se um
número kappa pré-estabelecido o que indica a quantidade de lignina residual na polpa (MIMMS et
al., 1993). A reação é realizada a temperaturas entre 160-180°C, com tempo de cozimento entre
0,5-6 horas, dependendo das propriedades desejadas para a polpa (FENGEL e WEGENER, 1989;
BIERMANN, 1993).
Em madeiras duras, como o eucalipto, o principal constituinte das polioses é uma O-
acetil-2-O-(4-O-metil-α-D-glucurono)-β-(1,4)-D-xilana. Durante a polpação kraft, a estrutura desta
poliose é extensivamente modificada.
2.5. PROCESSOS DE BRANQUEAMENTO
No branqueamento das pastas químicas, em que a maior parte da lignina foi removida
previamente pelo processo de polpação, devem ser removidos derivados de lignina ainda
remanescentes na pasta. O branqueamento reduz a quantidade de certas impurezas na polpa, tais
como feixes de fibras contendo um teor de lignina mais alto do que a média da polpa (shives) e
fragmentos de casca (flecks). O valor comercial de certos tipos de polpa é altamente dependente da
quantidade dessas impurezas. O branqueamento também ajuda a reduzir o conteúdo de extrativos
resinosos (pitch) na polpa, que pode gerar pintas e depósitos de resina durante o processo de
fabricação de papel, além do conteúdo de "pitch" influenciar as propriedades de envelhecimento

31
do papel. Essas são as mudanças das características da polpa mais importantes que ocorrem
durante o branqueamento. Outras incluem um aumento da capacidade de absorção de água,
redução do grau de polimerização, retenção de corante e resinas e a alteração da composição dos
carboidratos (SINGH, 1979). Para obter uma polpa com elevada alvura e estabilidade de alvura, a
lignina tem que ser removida. Isto não pode ser feito no digestor porque se o cozimento for muito
prolongado ocorrerá degradação e dissolução dos carboidratos.
Os grupos funcionais que conferem cor às substâncias são conhecidos como cromóforos.
A maioria deles possui ligações insaturadas como C =O, C = C ou N = N, em sistemas conjugados.
Alguns grupos funcionais como as hidroxilas (-OH), as aminas (-NH) e os halogêneos (Cl, Br),
não conferem cor às substâncias, porém conseguem aumentar a absorção de um cromóforo e
deslocar seu comprimento máximo de absorção, sendo denominados ‘auxôcromos’. Denomina-se
cromógena uma estrutura básica, capaz de absorver radiação ultravioleta ou visível e que tem essa
absorção deslocada em comprimento de onda pela ação de um grupo funcional. Inicialmente,
assumiu-se que a fonte primária de cor na madeira era a estrutura da lignina. Mais tarde, por meio
de análises espectrais, constatou-se que outras estruturas possuindo propriedades cromóforas
específicas precisam também estar presentes para justificar a cor apresentada pelas pastas, já que
nenhuma das estruturas básicas da lignina, ou seja, guaiacil propano e siringil propano, absorve
radiação de comprimento de onda na região visível. A madeira possui vários cromóforos
incorporados em sua estrutura, indicadas na Figura 2.5 (NAVARRO, 2004).

32
Figura 2.5 - Cromóforos incorporados na lignina (RAPSON, 1969)
Os mais importantes grupos cromógenos entre os radicais das moléculas da lignina são
as carbonilas conjugadas, as duplas ligações e combinações de ambas (Figura 2.6) (SILVA, 1986).
Figura 2.6 - Grupos cromógenos da lignina (RAPSON, 1969)

33
Embora os polissacarídeos, celulose e polioses não absorvam na região visível, quando
são formados grupos carbonila nos carboidratos de uma pasta branqueada, é observado um
amarelamento, pois o envelhecimento pela luz do dia, temperatura ou um ataque mais acentuado
do álcali durante o cozimento resultam na degradação, especialmente das polioses, associada à
reversão de alvura (PERISSOTO, 2000).
Tendo observado a natureza da cor na pasta celulósica, verifica-se que para torná-la mais
clara, ou seja, branqueá-la, é necessário remover ou alterar quimicamente as substâncias coloridas,
interrompendo as conjugações de duplas ligações por oxidação, redução ou hidrólise dos grupos
saturados (REDKO, 1978; SJÖSTRÖM, 1993).
A remoção da lignina residual de polpas normalmente é conduzida em uma seqüência
multiestágios (Tabela 2.4), variando de acordo com o grau de alvura desejado, para melhor
preservação das propriedades mecânicas das fibras de celulose (FENGEL e WEGENER, 1989). A
seleção de um processo de branqueamento depende da matéria-prima tratada (madeira, gramíneas,
resíduos agrícolas e outros), do tipo de polpação (mecânico, sulfato, semiquímica e outras) e da
finalidade a qual se destina o produto (impressão, embalagem, uso industrial, higiênico, dissolução
ou outras).
Tabela 2.4 - Notação dos Estágios de Branqueamento Estágio Notação Reagente Cloração C Cloro gasoso ou água de cloro Dioxidação D Dióxido de cloro (ClO2) Dióxido de cloro / Cloro DC Dióxido de cloro (ClO2) seguido de Cloro (Cl2) sem
lavagem intermediária Extração alcalina E Hidróxido de sódio (NaOH) Extração oxidativa (E+H) Inclusão do hipoclorito no estágio de extração Extração oxidativa (E+P) Inclusão do peróxido no estágio de extração Hipocloração H Hipoclorito de sódio (NaClO) ou de Cálcio (Ca(ClO)2) Peroxidação P Peróxido de hidrogênio (H2O2) Quelação Q Tratamento da polpa com quelante (EDTA, DTPA, etc) Oxigenação (Pré- branqueamento) O Oxigênio (O2) e Hidróxido de sódio (NaOH) Branqueamento enzimático X Xilanase, lacase e/ou outras
(SENAI CETCEP, 2001)
Modificações no processo kraft de polpação tem permitido obter polpas com menor teor
de lignina residual, possibilitando o desenvolvimento de técnicas de branqueamento livre de cloro
elementar (ECF) e totalmente livre de cloro (TCF). No processo ECF o dióxido de cloro (ClO2) e o
hipoclorito de sódio (NaClO) são usados nas seqüências de branqueamento e no processo TCF

34
usam-se oxidantes não clorados como o oxigênio, ozônio e peróxido de hidrogênio. No entanto,
esses reagentes são menos seletivos e eficientes quando comparados ao cloro elementar e ao
dióxido de cloro (ERIKSSON, 1997; WONG et al., 1997a).
Um dos maiores inconvenientes do uso da cloração, processo convencional e mais
utilizado no branqueamento de polpas, é a geração de produtos organoclorados (mais de 200), cuja
toxicologia de mistura é muito complexa. Uma das maneiras de se quantificar esses compostos é
através da medida do conteúdo em organoclorados que podem ser adsorvidos em carvão ativado
(AOX). Desses AOX, de 1-3% são solúveis em solventes orgânicos não polares e são os de maior
significância ambiental, já que são bioacumulativos. Os compostos de maior toxicidade são as
dioxinas e os compostos dibenzeno furanos, dentre os quais o 2,3,7,8-tetracloro-dibenzeno-p-
dioxina (TCDD) e o 2,3,7,8-tetracloro-dibenzeno-furano (TCDF) que são os mais tóxicos (CPRH,
1998).
As operações de polpação e branqueamento têm forte efeito na química dos carboidratos
de polpas kraft. Essencialmente, a estrutura química das xilanas é modificada devido à conversão
dos grupos de ácidos 4-O-metilglucurônicos em grupos de ácidos hexenurônicos (HexA’s).
Durante esta conversão ácidos 4-O-metilglucorônicos são primeiro isomerizados para ácidos 4-O-
metilidurônicos e, posteriormente, convertidos via β-eliminação em ácidos hexenurônicos. Estes
contêm uma dupla ligação, que pode reagir com vários agentes químicos de branqueamento, como
ozônio, dióxido de cloro ou perácidos. A eficiência do branqueamento pode ser melhorada
removendo-se os ácidos hexenurônicos da polpa através de uma hidrólise ácida, resultando na
economia de reagentes de branqueamento. Os ácidos hexenurônicos também reagem com
permanganato e contribuem para o aumento do número kappa das polpas kraft.
Durante a hidrólise ácida, os ácidos hexenurônicos são convertidos principalmente em
ácido 2-furanocarboxílico, ácido fórmico e 5-carboxi-2-furaldeído. Sob condições ácidas brandas,
os HexAs são hidrolisados muito mais rapidamente do que outras estruturas de carboidratos.
Simultaneamente, o número kappa da polpa é decrescido. Os ácidos urônicos presentes na polpa
participam da reversão de alvura de polpas kraft, então parece que os ácidos hexenurônicos,
devido ao fato de conterem na sua estrutura uma dupla ligação, têm maior significância no papel
de reversão de alvura do que os ácidos metilglucurônicos (JEFFRIES, 2001).
A remoção seletiva de ácidos hexenurônicos tem aplicação no branqueamento ECF e
TCF de polpas kraft. As principais vantagens são um menor consumo de reagentes de
branqueamento, maior estabilidade na alvura da polpa branqueada e decréscimo na formação de
depósitos nos equipamentos de branqueamento, que é devido à redução na formação de ácido

35
oxálico. Adicionalmente, o controle de metais no branqueamento TCF é mais fácil porque a
hidrólise seletiva dos hexenurônicos remove, parcialmente, os sítios quelantes das polioses.
- Extração alcalina
A extração alcalina da polpa, também conhecida como pré-deslignificação, pode ser
considerada uma parte integrante de uma seqüência de branqueamento de múltiplos estágios. Seu
objetivo é remover os componentes coloridos da polpa, parcialmente branqueada, que se tornam
solúveis em soluções alcalinas diluídas mornas, pela ação de reagentes químicos usados no pré-
branqueamento (SENAI CETCEP, 2001). No estágio de extração alcalina ocorre uma remoção
extensiva de lignina clorada e oxidada e, conseqüentemente, o grau de alvura da polpa no
branqueamento subseqüente aumenta, sendo necessário menores quantidades de reagente de
branqueamento para atingir o grau de alvura desejado. Portanto, a economia do branqueamento e a
manutenção das propriedades de resistência da polpa são favorecidas. Nesse sentido, a extração
alcalina pode ser considerada um estágio de branqueamento sem significativa degradação
oxidativa das fibras. Entretanto, pode ocorrer a solubilização das polioses, sendo necessário um
controle de temperatura e concentração da soda cáustica, de forma a limitar o problema. Outros
benefícios provenientes dos processos de extração alcalina são relacionados à estabilidade da
alvura, opacidade, maciez e algumas propriedades mecânicas da polpa. Os principais objetivos nos
estágios que precedem a extração são a remoção da maioria da lignina e a solubilização da lignina
residual em meio alcalino. O uso de hidróxido de sódio para lavar a polpa após a cloração tem se
tornado uma etapa vital em plantas de branqueamento (NAVARRO, 2004).
- Peroxidação
Os peróxidos usados no branqueamento são o peróxido de hidrogênio e o peróxido de
sódio. O branqueamento com H2O2 remove pouca lignina em comparação com os processos
convencionais que utilizam oxigênio e compostos clorados. Esta característica é extremamente
importante quando se trata de pastas de alto rendimento. Para estes tipos de pasta, o estágio de

36
branqueamento visa apenas a modificação da estrutura das substâncias que estão presentes na
massa, sem solubilizá-las. Entretanto, sua utilização no branqueamento de polpas químicas vem se
destacando em função de fortes pressões ambientais contra os compostos clorados, responsáveis
pelos elevados níveis de AOX (compostos organoclorados) presentes nos efluentes do processo de
branqueamento. O íon hidroperóxido (OOH-) é a espécie ativa do branqueamento com peróxido.
Por esta razão, o branqueamento ocorre em meio alcalino, o que favorece o aparecimento do
hidroperóxido. A decomposição do peróxido é controlada pela adição de materiais tais como,
silicato de sódio e sulfato de magnésio, que além de formar um complexo com as formas oxidadas
das glucopiranoses, estabilizam as ligações glicosídicas da molécula de celulose, evitando sua
despolimerização (HORTAL e LLUCIÁ, 1984); agentes quelantes, tais como: EDTA (ácido
etileno-diamino-tetra-acético), DTPA (dietileno-triamino-penta-acetato) e tratamento ácido com
HCl, H2SO4, SO2-. O efeito benéfico de agentes quelantes é geralmente creditado às suas
habilidades em desativar metais de transição que catalisam a decomposição do peróxido. É muito
importante uma lavagem perfeita da pasta após o pré-tratamento com ácido ou agentes quelantes
para remoção dos metais seqüestrados (SENAI CETCEP, 2001).
O peróxido de hidrogênio é um ácido fraco, com constante de ionização 1,72 a 20ºC e,
em solução, forma o íon hidroperóxido ao qual se atribui a ação branqueadora. Embora a formação
de íons hidroperóxido e a razão do branqueamento aumentem com a alcalinidade, a decomposição
do peróxido de hidrogênio em oxigênio e água é acelerada em meio fortemente alcalino. A
decomposição do peróxido é catalisada ainda por um grande número de impurezas metálicas,
como prata, platina, cobre, cromo, manganês, cobalto, molibdênio e tungstênio, que estão
presentes no licor alcalino na forma de hidróxidos. A eficiência dos branqueadores à base de
peróxido depende da inativação desses catalisadores de decomposição. Durante a peroxidação, a
adição de sais de magnésio tem comprovado ser eficiente na diminuição da degradação dos
carboidratos. Diversas teorias explicam essa função de proteção dos carboidratos (SINGH e
DILLNER, 1979; NUNN e LINDE, 1980), mas de acordo com COLLODETE et al. (1990), a
função do magnésio seria a de prevenir a propagação das reações em cadeia dos radicais livres
gerados no processo. Esses radicais livres, e em particular, o radical hidroxil (HO*) podem causar
a clivagem das cadeias de celulose. A perda de viscosidade resultante do uso de peróxido de
hidrogênio na extração não traduz em resistências inferiores da polpa branqueada, e que de fato
seu uso facilita o refino da polpa, além de reduzir a reversão da alvura (NAVARRO, 2004).

37
2.6. BRANQUEAMENTO ENZIMÁTICO
O uso de enzima como auxiliar no branqueamento passou de escala laboratorial no final
dos anos 80 (VIIKARI et al., 1987) para aplicações industriais atualmente, com a vantagem de não
requerer grandes modificações no processo tradicional (SUURNAKKI, 1997; FERRAZ, 1999;
DUARTE et al., 2003).
Na indústria de papel e celulose, as enzimas de maior interesse e aplicabilidade são as
hemicelulases, poligalacturonases, pectinases, lacases, celulases, lignina peroxidases, manganês
peroxidase e glicose oxidases (MONTEIRO, 1997). Os compostos convencionais de
branqueamento com reagentes à base de cloro vêm sendo substituídos alternativamente por
oxigênio, peróxido de hidrogênio, ozônio e perácidos, com boa aceitação comercial. Mais
recentemente, o tratamento enzimático da polpa com hemicelulases, principalmente xilanases,
resultou na diminuição dos branqueadores químicos utilizados no processo, diminuindo a poluição
ambiental (SUBRAMANIYAN e PREMA, 2002).
Nos processos de branqueamento enzimático das polpas a dose ótima de enzima está entre
2 e 5 UI/g de polpa seca e a consistência da polpa entre 5 a 10% (BEG et al., 2001). Muitos dos
efeitos benéficos do pré-branqueamento da xilanase pode ser obtido de 1 a 2 h de tratamento (BEG
et al., 2000; GUPTA et al., 2000).
O objetivo principal do branqueamento químico é remover a lignina residual das polpas,
retendo os carboidratos. Há várias hipóteses propostas para explicar o efeito que o pré-tratamento
com hemicelulases tem no branqueamento das polpas. A primeira é a remoção de cromóforos
derivados de xilana, já que no cozimento alcalino as moléculas de xilose e xilana podem sofrer
modificações, formando estruturas parcialmente aromáticas e coloridas. Esses cromóforos
derivados da degradação de xilana podem ser removidos mais facilmente pelo uso de xilanase do
que por reagentes químicos (WONG et al., 1997a). Outra hipótese seria a de que há uma ligação
entre lignina e polioses na polpa que restringe a remoção da lignina residual. O rompimento das
cadeias de xilana pela xilanase separa as ligações de lignina-carboidrato, que ocorrem após o
período de cozimento, melhorando o acesso dos reagentes de branqueamento e facilitando a
remoção da lignina em subseqüentes seqüências químicas de branqueamento (BAJPAI e BAJPAI,
1996; YOUNG e AKHTAR, 1998; ERIKSSON, 1997; WONG et al., 1997a). As interações entre
a celulose e a xilana podem contribuir para a integridade das fibras da polpa dificultando a
remoção da lignina presente no interior dessa estrutura. O uso de xilanase pode romper essa
estrutura facilitando a subseqüente remoção da lignina residual. Ocorre também o inchamento das

38
fibras que gera poros maiores do que as macromoléculas de xilanas removidas (WONG et al.,
1997a). Dentre os mecanismos citados, os mais aceitos são a clivagem da porção de carboidrato do
complexo lignina-carboidrato e a remoção das xilanas redepositadas sobre as fibras. Também há
evidências de que a presença de ácidos hexenurônicos derivados de xilana que escurecem as
polpas e levam a um maior consumo de reagentes químicos de branqueamento (BUCHERT et al.,
1996; SUURNÄKKI et al., 1996; DYER, 2004).
2.7. HISTÓRICO DO USO DE XILANASES NA INDÚSTRIA DE PAPEL E CELULOSE
O primeiro registro sobre a possibilidade da xilanase aumentar o branqueamento de
polpa kraft foi apresentado em Estocolmo em 1986, durante a Terceira Conferência Internacional
sobre Biotecnologia na Indústria de Polpa e Papel (VIIKARI et al., 1986). O pré-tratamento da
polpa com hemicelulases brutas, preparadas a partir de um fungo Ascomycete, melhorou a
deslignificação por peróxido de polpas de bétula e pinus, reduzindo a quantidade de lignina
remanescente na polpa, medida pelo número kappa. A deslignificação por uso de dióxido de cloro
e cloro com subseqüente extração alcalina também aumentou, permitindo uma redução na
quantidade de cloro usado no estágio de branqueamento. Trabalhos subseqüentes usando enzimas
parcialmente purificadas sugeriram que xilanases, e não outras enzimas hemicelulolíticas ou
desramificantes, são responsáveis pelo aumento na deslignificação da polpa bem como pelo
branqueamento (KANTELINEN et al., 1988). Embora alguns trabalhos tenham mostrado que
mananase também contribui para o branqueamento de polpas de madeira macia, a habilidade de
xilanases para aumentar o branqueamento de polpa kraft tem sido verificado usando xilanases
clonadas geneticamente, que não apresentam outra atividade enzimática conhecida (WONG et al.,
1997b).
PAICE et al. (1992) encontraram que os efeitos de xilanases no branqueamento da polpa
estão associados a uma marcante redução no grau de polimerização (DP) da xilana. Essas
observações foram consistentes com a hipótese de que os efeitos do pré-branqueamento resultam
primariamente da despolimerização, mas não necessariamente na solubilização das polioses
derivadas da xilana.
Os fungos são excelentes produtores de xilanase, mas freqüentemente secretam
celulases, que pode afetar negativamente a qualidade das polpas. Muitos trabalhos tem estudado a
produção de xilanases livres de celulases utilizando diversas fontes de carbono, principalmente

39
resíduos, como palha de cana-de-açúcar, palha de milho e outros. Alternativamente, organismos
geneticamente modificados tem sido utilizados para produzir exclusivamente xilanases.
Entretanto, aplicando-se métodos e meios de crescimento apropriados é possível isolar e
selecionar microrganismos que produzem xilanases totalmente livres de celulases ou que
contenham quantidades negligenciáveis. Aspergillus foetidus (ATCC 14916); Aspergillus oryzae
(NRRL 1808); Aspergillus niger (ATCC 10864); Aspergillus niger (NRRL 3536); Aspergillus
phoenicis (ATCC 15555); Gliocladium viride (CBS 658.70) e Aureobasidium pullulans (NRRL
Y-2311) são excelentes produtores de xilanase apresentando menos de 0,1 UI/mL de atividade de
enzimas celulolíticas (CHRISTOV at al., 1999).
Características desejáveis das xilanases para o branqueamento de polpas kraft incluem:
baixo peso molecular (assegurando o acesso à matriz da polpa), atividade em pH alcalino (para
funcionar bem nas polpas kraft), especificidade para xilana (para evitar a degradação da celulose)
(BARAZNENOK et al., 1999).
Preparações de xilanases de Trichoderma spp. foram usadas em branqueamento. Tais
pré-tratamentos foram realizados utilizando-se de 1 a 5 kg de enzima comercial para cada tonelada
de polpa seca, com tempos de incubação de 0,5 a 3 horas. A comparação de duas xilanases de T.
reesei Rut C30, sugeriu que a enzima que mais solubilizou a xilana de polpa de pinus também
forneceu um melhor efeito no aumento do branqueamento (TENKANEN et al., 1992).
As modificações nas propriedades do papel após pré-tratamento da polpa com xilanase
do actinomicete termofílico, Saccharomonospora viridis, também foram estudadas. Este
microrganismo produz xilanase extracelular sem apresentar atividade para celulose. As
propriedades foram exploradas pela aplicação da enzima bruta em polpa de bétula e resultou na
remoção seletiva de aproximadamente 20% do total de xilana presente. Os resultados, juntamente
com aqueles do pré-tratamento da polpa de algodão com a xilanase de S. viridis e uma celulose
comercial, sugeriram que a remoção específica de xilana reduz o grau de ligação entre as fibras
mas não as rompe (BARAZNENOK et al., 1999).
Dentre as preparações comerciais de xilanases indicadas para tratamento da polpa de
madeira, três são produzidas a partir de: Trichoderma ssp.: Pulpzyme HA (Novo Laboratórios
Ltda), de T. reesei: Albazyme (Cultor Ltd.) e de T. longibrachiatum: Ecopulp (Alko Ltd.)
(WONG e SADDLER, 1992). As faixas de pH e temperatura usuais de algumas enzimas
comerciais são 5-7 e 40-55ºC para Cartazyme, pH 7-8 e 50-60ºC para Irgazyme, pH 7-8 e 50ºC
para Ecopulp e pH 7 e 45-50ºC para Resinase, respectivamente. Bleachzyme, Amano 90 e

40
Xilanase GS35 são outras enzimas comerciais com temperatura de aplicação de 30-40ºC e pH 4,5-
7 (TECHAPUM et al., 2003).
Em um estudo comparativo realizado com duas xilanases comerciais e um extrato bruto
de xilanase isolado de Streptomyces thermoviolaceus para aplicação em seqüência de
branqueamento CEDED de polpas de bétula, foi obtida uma redução no consumo de ClO2 de cerca
de 30 a 35%. Como decorrência da redução na quantidade de cloro utilizado no branqueamento a
carga de compostos organoclorados produzidos e descartados nos efluentes também é reduzida.
Por exemplo, a quantidade de matéria orgânica clorada (AOX) produzida no branqueamento de
uma polpa kraft de 91% de alvura, é reduzido de 4,3 para 3,5 kg de AOX/tonelada métrica de
polpa (cerca de 19%), quando xilanases são aplicadas antes do branqueamento com Cl2 e ClO2
(FERRAZ et al., 2001). Os estudos de aplicação de novas xilanases em seqüências de
branqueamento são necessários porque cada tipo de polpa e processo pode ter uma resposta frente
a um tratamento enzimático específico, ou seja, os preparados comerciais não atuam com a mesma
eficiência sobre qualquer tipo de polpa.
O desenvolvimento de seqüências de branqueamento ECF que utilizam agentes
oxidantes não clorados em substituição a um dos estágios com dióxido de cloro também tem se
mostrado promissor se aplicado em conjunto com xilanases. No estudo de uma seqüência de
branqueamento (XDEP) foi obtida uma polpa final com maior índice de alvura e viscosidade, com
redução de 26 a 42% no consumo de dióxido de cloro em relação à polpa controle. As
características do efluente também foram melhores para a seqüência com utilização de xilanases,
com redução na cor e no conteúdo de compostos AOX (ELEGIR et al., 1995).
Estudos detalhados feitos em laboratório mostram que não são caros os investimentos
necessários para adaptar o tratamento enzimático às plantas industriais, exceto em relação ao pH.
O tratamento enzimático tem se mostrado completamente compatível com os equipamentos
industriais existentes. Desde 1991, o biobranqueamento, juntamente com os processos ECF e TCF,
tem sido continuamente usado em escala industrial na Finlândia. A quantidade de cloro requerida
no branqueamento é reduzida de 20-30% e como resultado tem-se a redução da carga de AOX nos
efluentes de branqueamento de 15-20%. Maior número de testes industriais tem sido feitos na
Europa, principalmente na Escandinávia, onde a maioria da polpa kraft é produzida (KULKARNI
et al., 1999).

41
2.8. UTILIZAÇÃO DA ENZIMA MANGANÊS PEROXIDASE
A enzima manganês peroxidase é secretada por muitos fungos degradadores de lignina,
incluindo Phanerochaete chrysosporium, T. versicolor, P. sordida, e outros (HARAZONO et al.,
1994).
O íon Mn(II) presente nos cavacos de madeira pode ser oxidado durante o cozimento
kraft (sob condições alcalinas) (HIRAI et al., 2002). Embora o Mn(II) necessário para o sistema
catalítico da MnP esteja presente nas polpas kraft, a polpa não é substancialmente branqueada a
menos que MnSO4 seja adicionado durante o branqueamento ‘in vitro’ contendo tampão malonato
50 nM (pH 4,5) (HARAZONO et al., 1994). A presença de surfactantes também é um fator
importante no branqueamento de polpas com MnP. O surfactante pode dispersar a lignina
hidrofóbica degradada em soluções aquosas impedindo a repolimerização da lignina degradada.
Foi mostrado que MnP exerce um importante papel no branqueamento de polpas kraft de
madeiras não branqueadas por fungos de decomposição branca e a enzima tem se mostrado
eficiente no pré-tratamento de polpas kraft de madeiras duras. Harazono et al. (1994) reportaram a
deslignificação da lignina residual em polpas kraft com MnP isolada. MnP proporcionou um
aumento na alvura de cerca de 10 pontos e reduziu o número kappa em cerca de 6 pontos na
presença de MnSO4 e H2O2. Quando os tratamentos com MnP foram repetidos seis vezes em
combinação com a extração alcalina, a alvura da polpa aumentou 43 pontos. Com polpas kraft de
eucalipto foi obtida uma redução de 14% do valor kappa (MILAGRES et al., 1992; DURÁN et al.,
1995). Resultados similares foram obtidos com polpa organosolv (ácido fórmico). Reduções do
valor kappa e aumentos da alvura em solventes orgânicos também foram observados (DURÁN,
1996). A enzima MnP do fungo Phanerochaete sordida foi isolada e testada ‘in vitro’ com polpas
kraft de eucalipto deslignificadas com oxigênio, e a redução no número kappa foi utilizada como
uma indicação da oxidação da lignina. A preparação enzimática contendo MnP aplicada na dose de
60 UI/g de polpa por 6 horas (com agitação de 150 rpm a 27°C) causou um significante
decréscimo de 13% no número kappa na polpa em relação aos controles. O tratamento foi iniciado
com a adição de peróxido de hidrogênio (H2O2) em pulsos a cada 30 min, dando uma concentração
final de 0,5 mM a cada adição. Esta redução é comparável àquelas obtidas para várias polpas
utilizando MnP de outras fontes fúngicas, que obtiveram uma redução variando de 9 a 32%. Neste
estudo foi observado que doses maiores do que as utilizadas não trouxeram benefícios adicionais
às polpas. Esta observação pode sugerir limitações quanto à acessibilidade ou a susceptibilidade à

42
oxidação da lignina residual nas polpas (HIRAI et al., 2002). Foi observado uma correlação
positiva entre os níveis da enzima MnP produzida pelo fungo Bjerkandera sp. linhagem BOS55 e
o branqueamento de polpas quando a atividade de MnP variou entre 0 a 40 UI/ml, entretanto não
houve um grande aumento no branqueamento quando foram adicionados atividades maiores. É
possível que a solubilização da lignina e dos cromóforos da polpa seja limitante ao processo de
branqueamento. Esse processo é bem regulado na presença do microrganismo, gerando um bom
branqueamento, o que não acontece quando a enzima é isolada (MOREIRA et al., 2001).
2.9. Ceriporiopsis subvermispora
O gênero Ceriporiopsis Dom. é um gênero poróide que pertence à família Polyporaceae
(HAROLD, 1998). C. subvermispora vem sendo utilizado há alguns anos em pesquisas realizadas
por um consórcio formado por indústrias papeleiras e a Universidade de Wisconsin, em Madison,
EUA. Resultados bastante animadores já foram reportados envolvendo a utilização de C.
subvermispora no pré-tratamento de madeira anterior a processos mecânicos e termomecânicos
(SETLIFF et al., 1992; AKHTAR et al., 1992, 1993).
Muitas publicações têm mostrado que o crescimento fúngico em substrato sólido e líquido,
apresentou baixa atividade celulolítica, mas uma alta atividade hemicelulolítica e ligninolítica
(SETHURAMAN et al., 1998, SOUZA-CRUZ et al, 2004; HEIDORNE et al., 2006).
Considerando a atividade celulolítica, detectou-se baixa atividade para as endo-celulases e
quantidades não significantes de atividade para exo-celulases. Em cultivos em cavacos de
eucalipto e pinus, em condições de biopolpação, a xilanase foi a principal enzima hidrolítica
produzida. O perfil eletroforético do extrato foi obtido em gel desnaturante de poliacrilamida
revelando cinco bandas protéicas com massa molecular de 85,7 kDa, 60,2 kDa, 56 kDa, 52,1 kDa
e 50,9 kDa. A temperatura ótima das enzimas dos extratos enzimáticos foi 60oC e o valor de pH
ótimo para as enzimas foi 4,0 para a β-glicosidase, 4,5 para a β-xilosidase, 5,0 para a xilanase, 4,5
para a mananase e 3,5 para a CMCase (Carboximetilcelulase). A estabilidade ao pH foi
determinada em pH 3,5, 5,0 e 6,5 em tampão fosfato-citrato 50 mM e incubadas a 50oC por
períodos de até 6 horas, separadamente dos substratos. Verificou-se que as enzimas apresentaram
maior estabilidade em pH 5,0 (HEIDORNE et al, 2006). A estabilidade térmica das enzimas foi
determinada a 40, 50 e 60oC e incubadas nas respectivas temperaturas por períodos de 3 a 8 horas,

43
separadamente dos substratos. Observou-se que as enzimas apresentaram maior estabilidade à
30oC, porém a 50oC a estabilidade foi alta por 2 horas.
C. subvermispora degrada lignina de um material lignocelulósico através da ação da
manganês peroxidase e lacase. (RUTTIMANN-JOHNSON et al., 1993). Entretanto, lacases não
foram detectadas em condições de cultivo onde o único substrato orgânico é o próprio composto
lignocelulósico (FERRAZ et al., 2002; SOUZA-CRUZ, 2002). Com isso, a degradação de lignina
por esse fungo deveria, a princípio, ser atribuída principalmente a ação da MnP.

44
3. OBJETIVOS
O objetivo principal deste trabalho foi avaliar a ação do extrato enzimático extraído de
madeiras biodegradadas por Ceriporiopsis subvermispora no biobranqueamento de polpas kraft de
Eucalyptus grandis.
Para alcançar este objetivo foram realizadas as seguintes etapas:
- Análise dos produtos de xilanas de bétula, carvalho, fibra de trigo e bagaço de cana-de-
açúcar após ação das xilanases;
- Aplicações do extrato enzimático de C. subvermispora e de um extrato comercial em
polpas kraft para avaliação da taxa de hidrólise das polpas;
- Aplicações do extrato enzimático em polpas kraft em diferentes concentrações e diferentes
tempos de tratamento;
- Identificação dos monômeros e oligômeros liberados das polpas;
- Branqueamento químico das polpas pré-tratadas enzimaticamente pela seqüência EQP;
- Avaliação da ação enzimática nas polpas kraft por viscosidade, número kappa e liberação
de cromóforos.

45
4. MATERIAIS E MÉTODOS
4.1. PREPARO DOS EXTRATOS ENZIMÁTICOS
4.1.1. Extrato de Ceriporiopsis subvermispora
Os extratos enzimáticos produzidos pelo fungo Ceriporiopsis subvermispora foram
produzidos no laboratório de Microbiologia e Bioquímica do Departamento de Biotecnologia da
EEL-USP. C. subvermispora SS-3, mantido em agar extrato de malte (2%) a 4 ºC, foi repicado (20
discos com diâmetro de 8 mm) em frascos Erlenmeyer de 2 L contendo 200 mL de meio à base de
extrato de batata-dextrose (2,4%) e de extrato de levedura (0,7%). Após 12 dias de incubação
estática a 27ºC, o micélio obtido foi filtrado, lavado e macerado com água esterilizada.
Biorreatores de polietileno com capacidade de 20 L foram preenchidos com 2 kg de cavacos de E.
grandis (base seca) e mantidos imersos em água por um período de 12 h. Após drenagem e
esterilização a 121ºC por 15 min, os biorreatores foram inoculados com uma alíquota da suspensão
de micélio, de forma a se obter uma concentração inicial de micélio igual a 500 mg/kg de madeira
(base seca). Os biorreatores inoculados foram então agitados para homogeneização de seu
conteúdo e incubados a 27ºC, utilizando-se fluxo de ar igual a 23 L/h. Foram inoculados três
biorreatores e cada um deles forneceu os cavacos biodegradados para o preparo dos extratos
enzimáticos no tempo de 28 dias.
Para a obtenção dos extratos enzimáticos, o conteúdo do biorreator (micélio e cavacos)
foi transferido, em frações de 200 g, para frascos Erlenmeyer de 2 L contendo 500 mL do tampão
de extração (acetato de sódio 50 mM, pH 5,5, adicionado de 0,01% de Tween 60) e mantido sob
agitação de 120 rpm a 10ºC por um período de 4 h. Os extratos enzimáticos assim obtidos foram
centrifugados a 3400 g por 15 min, para a remoção de materiais particulados, e armazenados à
temperatura de - 80°C.
As atividades enzimáticas relacionadas à degradação da lignina, celulose e polioses
foram determinadas no momento do uso, utilizando-se o Espectrofotômetro U-2001 (Hitachi) e
expressas como unidades internacionais (UI/ml).

46
4.1.2. Extrato comercial
O extrato comercial cedido para este trabalho continha 107 UI/mL de atividade de
xilanase e 7 UI/mL de atividade de endoglucanase. As atividades enzimáticas relacionadas à
degradação da lignina, celulose e polioses foram determinadas no momento do uso, utilizando-se o
Espectrofotômetro U-2001 (Hitachi) e expressas como unidades internacionais (UI/ml).
4.1.3. Caracterização dos extratos
A caracterização dos extratos foi realizada através da determinação das atividades
enzimáticas das enzimas β-xilanase, β-xilosidase, β-glicosidase, β-mananase,
Carboximetilcelulase e Manganês peroxidase, além da quantificação de açúcares redutores. Os
procedimentos estão descritos abaixo.
4.1.3.1. β-Xilanase
O método baseia-se na liberação de açúcares redutores a partir da xilana 1% como
substrato (BAILEY et al., 1992), seguido da quantificação de açúcares redutores liberados pelo
método de DNS (MILLER, 1959).
Uma alíquota de 0,9 mL de xilana foi colocada em tubos de ensaio e incubada em banho
termostatizado à temperatura de 50oC durante 2 minutos. Após incubação, foram adicionados
100 μL da enzima diluída apropriadamente no tampão, e a amostra incubada por 5, 15 e 30
minutos. A reação foi interrompida pela adição de 1,5 mL do reagente do DNS, sendo a mistura
fervida durante 5 minutos. Os tubos foram resfriados em banho de gelo, e a absorbância lida a 540
nm em espectrofotômetro. Para calibração do aparelho foi feito um controle onde se substituiu o
volume da enzima pelo tampão acetato de sódio 50 mM, pH 5,4.
Foram feitos controles para cada amostra, adicionando-se o reagente DNS antes do
extrato enzimático. Nessas condições é possível dosar compostos redutores que não são
provenientes da reação enzimática.

47
A curva padrão foi construída com solução de xilose (Merck), nas concentrações entre 2
e 20 μmol/mL. Uma unidade de atividade enzimática corresponde à quantidade desta capaz de
catalisar a liberação de 1 μmol de açúcar redutor por minuto.
- Preparo da solução de xilana 1%
A 80 mL de tampão acetato de sódio 50 mM, pH 5,4 adicionou-se 1 g de xilana
(Bichwood-Sigma-St. Louis, USA). A solução foi aquecida até ebulição em forno de microondas,
e após retornar à temperatura ambiente o volume foi ajustado para 100 mL com o referido tampão.
A solução permaneceu em agitação mecânica até completa solubilização da xilana na solução.
- Preparo do reagente DNS
O reagente foi preparado dissolvendo-se 10,6 g de ácido 3,5-dinitrosalicílico, 19,8 g de
hidróxido de sódio, 306 g de tartarato de sódio e potássio e 8,3 g de bissulfito de sódio em 1 L de
água destilada. Após agitação para total dissolução dos reagentes, foram adicionados 7,6 mL de
fenol, e o volume completado para 1,4 L com água destilada (MILLER, 1959).
4.1.3.2. β-Xilosidase
A atividade de β-Xilosidase foi determinada segundo TAN et al., (1987), através da
estimativa do p-nitrofenol (pNP) liberado a partir do p-nitrofenil-β-D-xilopiranosídeo (pNPX).
Uma alíquota de 0,2 mL de extrato enzimático foi adicionada a 0,8 mL de solução 0,1%
de pNPX em tampão acetato de sódio 50 mM e pH 4,8. A reação foi conduzida incubando os tubos
durante 15, 30 e 60 minutos a 50oC. Após incubação, a reação foi interrompida pela adição de 2
mL de solução de bicarbonato de sódio 10% e a absorbância lida a 410 nm. Foram feitos controles
para cada amostra, adicionando-se a solução de bicarbonato de sódio 10% antes do extrato

48
enzimático. Para calibração do aparelho foi feito um controle onde se substituiu o volume da
enzima pelo tampão acetato de sódio 50 mM, pH 4,8.
O pNP liberado foi determinado através de curva de calibração com padrão de p-
nitrofenol nas concentrações entre 50 e 350 μmol/mL. Uma unidade de atividade enzimática é
definida como a quantidade de enzima capaz de catalisar a liberação de 1 μmol de pNP por minuto
a 50oC.
- Preparo da solução de para-nitro-fenil-β-D-xilopiranosídeo (pNPX)
A solução de pNPX foi preparada dissolvendo-se 0,1 g de pNPX (Sigma-St. Louis,
USA) em 90 mL de tampão acetato de sódio 50 mM, pH 4,8, sob agitação mecânica e o volume
ajustado para 100 mL com o referido tampão.
- Preparo do bicarbonato de sódio 10%
A solução de bicarbonato de sódio 10% foi preparada dissolvendo-se 10 g de bicarbonato
de sódio em 100 mL de água destilada, sob agitação e aquecimento em agitador mecânico.
4.1.3.3. β-Glicosidase
A atividade de β-Glicosidase foi determinada segundo TAN et al., (1987), através da
estimativa do p-nitrofenol (pNP) liberado a partir do p-nitrofenil-β-D-glicopiranosídeo (pNPG).
Uma alíquota de 0,2 mL de extrato enzimático foi adicionada a 0,8 mL de solução 0,1%
de pNPG em tampão acetato de sódio 50 mM, pH 4,8. A reação foi conduzida durante 15, 30 e 60
minutos a 50oC. Após incubação, a reação foi interrompida pela adição de 2 mL de solução de
bicarbonato de sódio 10% e a absorbância lida a 410 nm. Foram feitos controles para cada
amostra, adicionando-se a solução de bicarbonato de sódio 10% antes da adição do extrato

49
enzimático. Para calibração do aparelho foi feito um controle em que se substituiu o volume da
enzima pelo tampão acetato de sódio 50 mM, pH 4,8.
O pNP liberado foi determinado através de curva de calibração com padrão nas
concentrações entre 50 e 350 μmol/mL. Uma unidade de atividade enzimática é definida como a
quantidade de enzima capaz de catalisar a liberação de 1 μmol de pNP por minuto a 50oC.
- Preparo da solução de para-nitro-fenil-β-D-glicopiranosídeo (pNPG)
A solução de pNPG foi preparada dissolvendo-se 0,1 g de pNPG (Sigma-St. Louis,
USA) em 90 mL de tampão acetato de sódio 50 mM, pH 4,8, sob agitação mecânica e o volume
ajustado para 100 mL com o referido tampão.
4.1.3.4. β-Mananase
O método baseia-se na liberação de açúcares redutores a partir de galactoglucomanana,
0,5%, seguido da quantificação de açúcares redutores liberados pelo método de DNS (RATTO e
POUTANEN, 1988).
Uma alíquota de 0,1 mL de extrato enzimático foi adicionada a 0,9 mL do substrato em
tubos de ensaio durante 5, 15 e 30 minutos à temperatura de 50oC. Após incubação, a reação foi
interrompida pela adição de 1,5 mL do reagente de DNS. Para calibração do aparelho foi feito um
controle em que se substituiu o volume da enzima por tampão acetato de sódio 50 mM, pH 5,4.
Foram feitos controles para cada amostra, adicionando-se o reagente DNS antes do
extrato enzimático para a determinação de compostos redutores que não são provenientes da
reação enzimática.
A curva padrão foi construída com solução de manose (Merck), nas concentrações entre
1 e 15 μmol/mL. Uma unidade de atividade enzimática é definida como sendo a quantidade de
enzima capaz de catalisar a liberação de 1 μmol de manose por minuto a 50oC.

50
- Preparo da solução de manana
A solução de manana foi preparada dissolvendo-se 0,5 g de galactoglucomanana
("locust bean gum"-Sigma-St. Louis, USA) em 80 mL de tampão acetato de sódio 50 mM, pH 5,4.
A solução foi submetida a aquecimento até aproximadamente 80ºC, em forno de microondas, e
após retornar a temperatura ambiente o volume foi ajustado para 100 mL com o referido tampão.
A solução permaneceu em agitação mecânica até completa dissolução da manana em solução.
4.1.3.5. Endoglucanase
A atividade de endo-1-4-β-glucanase ou carboximetilcelulase (CMCase) foi
determinada através da hidrólise de uma solução de carboximetilcelulose a 0,44% em tampão
acetato de sódio 50 mM e pH 5,0 (TANAKA et al., 1981). A quantidade de açúcares redutores foi
determinada pelo método do DNS (MILLER, 1959). Os valores de absorbância lidos a 540 nm
foram transformados em μmol de glicose pela comparação da curva padrão de glicose.
Uma alíquota de 0,1 mL de extrato enzimático, devidamente diluído em tampão acetato
de sódio, foi adicionada a 0,9 mL do substrato durante 30, 60 e 90 minutos à temperatura de
50oC. Após incubação, a reação foi interrompida pela adição de 1,5 mL do reagente DNS, e os
tubos fervidos por 5 minutos. Foram feitos controles para cada amostra adicionando-se o reagente
DNS antes do extrato enzimático. Para calibração do aparelho foi feito um controle em que se
substituiu o volume da enzima pelo tampão acetato de sódio 50 mM, pH 5,4.
A curva padrão foi construída com solução de glicose (Merck), nas concentrações entre
1 e 12 μmol/mL. Uma unidade de atividade enzimática é definida como sendo a quantidade de
enzima capaz de catalisar a liberação de 1 μmol de glicose por minuto a 50oC.
- Preparo da solução de carboximetilcelulose
A 80 mL de tampão acetato de sódio 50 mM, pH 5,4 adicionou-se 0,44 g de
carboximetilcelulose média viscosidade (Sigma-St. Louis, USA). A solução foi fervida em forno

51
de microondas, e após retornar a temperatura ambiente o volume foi ajustado para 100 mL com o
referido tampão. A solução permaneceu em agitação mecânica até completa dissolução da
carboximetilcelulose na solução.
4.1.3.6. Manganês Peroxidase (MnP)
A atividade de manganês peroxidase foi determinada acompanhando-se a oxidação do
vermelho de fenol (ε610 = 22.000 M-1.cm-1) (LUNDELL et al., 1990). A mistura reacional foi
constituída por 1,75 mL de tampão succinato de sódio 50 mM (pH 3,2), 1,5 mL de lactato de sódio
50 mM, 0,5 mL de vermelho de fenol 0,1%, 0,5 mL de sulfato de manganês 1 mM, 0,25 mL de
albumina bovina 1,8%, 0,25 mL de peróxido de hidrogênio 2 mM e 0,25 mL de extrato
enzimático.
Após o início da reação, frações de 1 mL foram retiradas do tubo contendo a mistura
reacional a cada 1 min e transferidas para cubetas contendo 65 μL de hidróxido de sódio 6,5 M,
previamente à leitura da absorbância. Uma unidade de atividade enzimática foi considerada como
a quantidade de enzima capaz de catalisar a formação de 1 μmol de vermelho de fenol oxidado por
minuto.
4.2. PREPARO DAS XILANAS
Xilanas de bétula, fibra de trigo e carvalho foram obtidas da Sigma Chemical Co., St.
Louis, USA. As xilanas (1%) preparadas em água deionizada, foram mantida a 4ºC por 24 h e
então centrifugadas a 5000 g por 20 min. O sobrenadante foi filtrado em papel de filtro Watman nº
1 (LI et al., 2000). A solução foi concentrada com uma membrana Millipore (de corte molecular
5000) de 100 para 10 mL. Água deionizada foi utilizada para restaurar o volume do ultrafiltrado. A
xilana de bagaço-de-cana foi preparada através de hidrólise com NaOH (FENGEL e WEGENER,
1989). Em seguida foi seca em estufa e lavada com água deionizada em abundância. Alíquotas das
soluções das xilanas (10 mL) foram secas em estufa a 102ºC por 24 h e então determinadas as
massas secas. As massas obtidas foram convertidas para mg/mL. Cada ensaio foi realizado em

52
duplicata. As massas secas obtidas foram: xilana de bétula- 8,78 mg/mL; xilana de carvalho- 4,78
mg/mL; xilana de fibra de trigo- 8,77 mg/mL e xilana de bagaço-de-cana- 4,63 mg/mL.
Para a hidrólise enzimática das xilanas, a mistura de reação consistia de 19 mL de cada
xilana em 0,8 mL de tampão citrato de sódio 50 mM (pH 6,0) e 2 UI/g de xilanase. Essa mistura
foi incubada a 40ºC por até 96 h. Alíquotas de 3,5 mL foram retiradas nos tempos 0, 24, 48, 72 e
96 h durante a hidrólise. Cada alíquota foi fervida durante 5 min a 100ºC para inativação das
enzimas. As amostras foram concentradas em liofilizador e então filtradas em filtro Sep-Pak
Cartridges C18 (Waters), para remoção de impurezas e compostos fenólicos. As liberações de
xilose e oligômeros nos filtrados foram analisadas em cromatógrafo líquido WATERS, modelo
515 HPLC, coluna Aminex HPX 42A de troca iônica (300 x 7,8 mm, Bio-Rad Laboratories Ltd),
sendo o eluente água, fluxo de 0,6 mL/min e temperatura de 45°C. Os compostos foram
monitorados em um detector de índice de refração (RI).
4.3. POLPA
Foi utilizada uma polpa kraft industrial de eucalipto. A amostra de polpa foi
exaustivamente lavada com água deionizada e o pH foi ajustado para 5,0 pela adição de H2SO4
4M. A polpa foi deixada em contato com o ácido durante uma noite, o pH foi verificado pelo
contato com uma fita de pH e posteriormente caracterizada em termos de número kappa e
viscosidade (Tabela 4.1) e composição química (Tabela 4.2).
Tabela 4.1 - Caracterização da polpa kraft de eucalipto
Parâmetros ResultadosNúmero kappa 12,9
Viscosidade (cP) 24,8
Tabela 4.2 - Constituição química do E. grandis utilizado neste trabalho
Componentes Componentes na madeira (%) Componentes na polpa kraft (%)Celulose 47,8±0,9 78,1±0,3 Polioses 23,1±0,9 12,8±0,8 Lignina 28,6±0,4 5,3±0,5 Extrativos 1,5±0,4/0,9±0,8 ND

53
4.3.1. Consistência das polpas (%)
A consistência foi determinada pela relação massa seca/massa úmida, sendo expressa em
porcentagem. A polpa foi pesada determinando-se a massa úmida (m.u.). Aproximadamente 1,0 g
de polpa úmida foi colocada em uma balança Mettler Toledo moisture analyser, com aquecimento
por infra-vermelho. Quando a massa da polpa estabilizou, foi determinada a massa seca (m.s.) e a
consistência foi calculada.
4.3.2. Tratamento Enzimático das Polpas
Foi utilizado 25 g de polpa seca para as seqüências de branqueamento. Foi adicionado o
extrato enzimático às polpa em diferentes concentrações (0,5, 1, 1,5 e 2 UI/g de xilanase) e
acrescentada água até a consistência de 10%. Este ajuste foi realizado segundo a Equação 4.1. A
mistura de enzima e polpa foi colocada em sacos de polietileno, vedados com seladora e
devidamente incubados em banho com temperatura de 40ºC.
Equação 4.1 - Relação entre massa seca, massa úmida, volume de enzima e volume de água a ser acrescentada à mistura de reação para ajuste da consistência da polpa
massa seca (g) – massa úmida (g) – vol. enzima (mL) = volume de água a ser (4.1) consistência adicionado
O tratamento foi avaliado durante 0,5, 1, 2, 3 e 5 horas. Após os tratamentos as polpas
foram filtradas em funil de Buchner e lavadas com 500 ml de água destilada. A polpa foi usada
para determinação dos parâmetros número kappa e viscosidade. O filtrado foi tratado
termicamente (100ºC/5 min) e armazenado para a determinação da absorbância a 237, 254, 280 e
465 nm relativas à liberação de compostos fenólicos, produtos de degradação das polioses e
hidrofóbicos da polpa, respectivamente (PATEL e PAVITT, 1993; GARG et al., 1996; WONG et
al., 1997b; GUPTA et al, 2000; LI et al., 2005; SALLES et al., 2005). As liberações de xilose,
xilobiose e xilotriose foram avaliadas nos filtrados das polpas após os períodos de incubação e
então identificados por cromatografia líquida de alta eficiência (CLAE) em um cromatógrafo

54
líquido WATERS, modelo 515 HPLC. As soluções foram filtradas em filtro Sep-Pak C18 (Waters)
e injetadas diretamente em uma coluna Aminex HPX 42A (300 x 7,8 mm, Bio-Rad Laboratories
Ltd), sendo o eluente água deionizada, fluxo de 0,6 mL/min e temperatura de 45ºC. Os compostos
foram monitorados em um detector de índice de refração (RI).
A identificação dos oligômeros de xilose presentes nos filtrados foram determinadas a
partir de curvas de calibração externas obtidas com padrões analíticos de xilose, xilobiose,
xilotriose e xilotetraose (Megazyme-Ireland).
4.4. CARACTERIZAÇÃO FÍSICO-QUÍMICA DAS POLPAS
4.4.1. Determinação da lignina insolúvel (Lignina Klason)
A quantidade de lignina Klason insolúvel em meio ácido foi determinada de acordo com
o método ASTM (ASTM Methods, 1966). Uma amostra de 1,2 g de polpa, previamente seca a
80°C durante uma noite, foi transferida para um becher de 100 ml e tratada com 10 mL de ácido
sulfúrico 72% (66,5% v/v) durante 1 h a 30°C e misturada continuamente com o auxílio de um
bastão de vidro. A reação foi interrompida com a adição de 50 ml de água destilada e a mistura foi
transferida quantitativamente para Erlenmeyer de 500 mL usando 270 mL de água e a seguir
autoclavada a 125°C por 1 h. Após esfriar a temperatura ambiente, a mistura foi filtrada em funil
de vidro sinterizado (previamente seco e pesado) sendo então adicionada água destilada até que
fosse completado um volume de 500 mL. A mistura assim obtida foi filtrada e o material retido no
funil foi lavado com aproximadamente 1,8 L de água destilada e seco em estufa a 110°C até massa
constante e a massa de lignina determinada por diferença.
O percentual de lignina insolúvel foi determinado no material insolúvel retido no
cadinho sendo calculado em relação à massa seca das amostras, conforme Equação 4.2.
Equação 4.2 - Cálculo da lignina Klason
% Lignina Klason = massa de lignina Klason . 100 (4.2) massa de polpa seca

55
Sendo:
% Lignina Klason = lignina insolúvel (% em relação à polpa);
Massa de lignina Klason = massa de lignina Klason seca (g);
Massa de polpa seca = massa de polpa base seca (g).
4.4.2. Determinação da lignina solúvel
A quantidade de lignina solúvel em meio ácido foi determinada pela medida da
absorbância a 205 nm de uma diluição 1:2 do hidrolisado obtido no item anterior. Para o cálculo
da concentração de lignina solúvel foi utilizada uma absortividade de 105 g-1cm-1L no
comprimento de onda de 205 nm através das Equações 4.3 e 4.4 (FERRAZ et al., 2000).
Equações 4.3 e 4.4 - Cálculo da lignina solúvel
[Lig] = (Abs 205 nm / A . B) . D (4.3)
% Lignina solúvel = [Lig] . V (4.4)
massa de polpa seca
Sendo:
[Lig] = concentração de lignina no hidrolisado (g.L-1);
Abs 205 nm = absorbância do hidrolisado a 205 nm;
A = absortividade (105 L/g.cm);
B = caminho óptico (1 cm);
D = fator de diluição;
% Lignina solúvel = lignina solúvel (% em relação à polpa);
V = volume da amostra no hidrolisado;
Massa de polpa seca = massa de polpa base seca (g).

56
4.4.3. Determinação da Lignina Total
A lignina total foi determinada pela soma da lignina insolúvel (lignina Klason) com a
lignina solúvel.
4.4.4. Determinação dos carboidratos
Os carboidratos presentes no hidrolisado foram identificados em cromatógrafo líquido
WATERS, modelo 515 HPLC. O hidrolisado foi extraído em cartuchos Sep-Pak C18 (waters) para
remoção de compostos aromáticos, e então injetado em uma coluna Aminex BIORAD HPX 87H
(Bio-Rad Laboratories Ltd), acoplada a uma pré-coluna trocadora de cátions (Bio-Rad
Laboratories Ltd) usando-se ácido sulfúrico 0,005 M como fase móvel a um fluxo de 0,6 mL/min a
45°C. Os compostos foram monitorados por um detector de índice de refração. Nestas condições a
glicose e celobiose é separada dos demais açúcares provenientes da hemicelulose (xilose, manose,
galactose e arabinose). O teor de celulose foi calculado como o teor de glicose x 0,9 e teor de
celobiose x 0,95 (fator de hidrólise). De maneira similar, xilose x 0,88 foi convertida em xilana
(ROCHA, 2000).
As concentrações de celobiose, glicose, xilose, arabinose e ácido acético foram
determinadas a partir de curvas de calibração externas obtidas com padrões analíticos.
4.4.5. Determinação do Número Kappa
O número kappa é a medida indireta da lignina residual na pasta tratada. Determina
também o grau de deslignificação ou a branqueabilidade da celulose e corresponde aos mililitros
de uma solução padrão de permanganato de potássio 0,1 N consumida por 1,0 g de celulose, nas
condições especificadas nas normas TAPPI (Technical Association of Pulp and Paper Industry,
1985).

57
Para avaliação do número kappa foram utilizados entre 0,5 a 1,0 g de polpa seca. A
amostra foi desintegrada com a mão em 10 mL de água destilada até ficar livre dos grumos e dos
feixes de fibras não dispersas. Este procedimento teve por objetivo separar as fibras evitando seu
corte excessivo. Depois de desintegradas, as polpas foram transferidas para um Erlenmeyer de 300
mL, seguidas de sucessivas lavagens com 140 mL de água destilada e colocadas em banho de
temperatura constante (25 ± 0,1°C). Separadamente em um Erlenmeyer de 125 mL foram
pipetados 25 mL de KMnO4 0,1 N + 0,0005 e 25 mL de H2SO4 4 M. O Erlenmeyer contendo a
solução foi colocado em um banho de temperatura constante (25 ± 0,1°C). Esta solução foi
transferida para o Erlenmeyer com a polpa. O Erlenmeyer de 125 ml foi lavado utilizando–se 50
ml de água destilada, que foi adicionada à polpa. A solução contendo a polpa foi deixada reagir
por 10 min. Para interromper a reação foram adicionados 5 mL de solução de KI 1,0 M. O iodo
livre na suspensão foi titulado com tiossulfato de sódio 0,1 N + 0,0005 até a solução (com agitação
constante) ficar com uma coloração amarelo claro. Foi então acrescentado 0,5 ml de solução de
amido 1%, continuando-se a titulação até a viragem da cor azul para o branco.
Foi realizada uma determinação do branco sem a polpa, utilizando 150 ml de água
destilada (ao invés de 140 ml), adotando-se o mesmo procedimento descrito anteriormente. O
permanganato oxida a lignina residual, e assim, a titulação foi realizada para se determinar quanto
de permanganato foi consumido. Quanto maior o conteúdo de lignina da polpa, maior o volume de
permanganato de potássio consumido por esta e conseqüentemente menor o volume de tiossulfato
de sódio utilizado para titular o iodo formado. Para o cálculo do número kappa foram utilizadas as
Equações 4.5, 4.6 e 4.7.
Equações 4.5, 4.6 e 4.7 - Cálculo do número kappa das polpas kraft
Sendo:
P = volume de KMnO4 que reagiu (mL);
a = volume de Na2SO4 gasto na titulação da amostra (mL);
b = volume de Na2SO4 gasto na titulação do branco (mL);
f = fator de correção para um consumo de 50% de permanganato de potássio,
determinado para cada P;
P = {(b – a) x N}/ 0,1 (4.5) f = 0,0084 P + 0,895 (4.6)
Nº kappa = (P x f)/w x [1 + 0,013 (25 – T)] (4.7)

58
N = normalidade da solução de Na2SO4;
T = temperatura de análise (ºC);
w = massa da polpa seca (g).
4.4.6. Determinação da Viscosidade
Exatamente 0,125 g de polpa seca foi colocada em um frasco com tampa sendo em
seguida adicionados 25 mL de etilenodiamina cúprica, solução 0,5 M em cobre. A mistura foi
agitada por 10 minutos com auxílio de uma barra magnética. Após esse tempo, a mistura foi
filtrada em cadinho de vidro sinterizado (nº 3) e transferida para a pipeta de viscosidade Fenske-
Otswald imersa em banho termostatizado a 25,0±0,1°C, previamente aferida com H2SO4
concentrado e utilizada dentro dos limites de viscosidade apropriados. Após equilíbrio térmico, a
solução foi aspirada e o tempo de escoamento, cronometrado. O tempo necessário para o menisco
do líquido passar entre as 2 marcas foi determinado em triplicata (TAPPI, 1985). Assim, com os
valores de densidade e viscosidade do H2SO4 concentrado na temperatura de medida, é feito o
cálculo da constante do viscosímetro pela Equação 4.8. E conhecida a constante do viscosímetro é
possível calcular o valor da viscosidade pela Equação 4.9.
Equação 4.8 - Cálculo da constante do viscosímetro
Sendo:
kv = constante do viscosímetro (cP.s-1.g-1.cm3);
V = viscosidade do H2SO4 a 25°C - 19,25 cP (TAPPI, 1982);
d = densidade do H2SO4 a 25°C - 1,84 g.cm-3 (WEAST, 1984);
t = tempo de escoamento (s).
Equação 4.9 - Cálculo da viscosidade
(4.8)
(4.9)

59
Sendo:
V = viscosidade da solução (cP);
kv = constante do viscosímetro (cP.s-1.g-1.cm3);
t = tempo de escoamento (s);
d = densidade da solução de celulose a 25ºC - 1,052 g.cm-3 (TAPPI, 1982).
4.4.7. Redução da Viscosidade
O percentual de redução da viscosidade, dado pelo grau de polimerização da celulose,
durante os tratamentos foi calculado conforme a Equação 4.10.
Equação 4.10 - Cálculo da redução da viscosidade (%)
5.3.4. Determinação da Alvura
4.4.8. Cálculo da Eficiência de Deslignificação e Seletividade
Para a análise dos resultados obtidos no branqueamento da polpa outros cálculos foram
realizados, conforme as Equações 4.11 e 4.12.
Equações 4.11 e 4.12 - Cálculo da eficiência de deslignificação e seletividade
Redução da Viscosidade (%) = (viscosidade inicial – viscosidade final) x 100 (4.10) viscosidade inicial
% Eficiência de deslignificação = Nº kappa inicial - Nº kappa final . 100 (4.11) Nº kappa inicial Seletividade = Eficiência de deslignificação (4.12) % de redução da viscosidade da polpa

60
4.5. TAXA DE HIDRÓLISE DA XILANA
Para a determinação da taxa de hidrólise das polpas com relação à xilana foi utilizada a
Equação 4.13.
Equação 4.13 - Taxa de hidrólise da xilana
Sendo:
[ALX] = Açúcares redutores liberados pela ação da xilanase;
[ALC] = Açúcares redutores liberados pelos controles;
Fator de hidrólise da xilana = 0,88.
4.6. BRANQUEAMENTO DAS POLPAS
O branqueamento foi realizado utilizando-se uma seqüência curta de branqueamento,
constituída de três estágios: extração alcalina com hidróxido de sódio (E), etapa de quelação com
EDTA (Q) e tratamento com peróxido de hidrogênio (P) para posterior avaliação das polpas. As
condições utilizadas estão mostradas na Tabela 4.3.
Tabela 4.3 - Condições gerais de branqueamento
Condições E Q P Consistência (%) 10 10 10
NaOH (%) 3 - - EDTA (%) - 0,4 -
H2O2 (%)/MgSO4* - - 3/0,05 pHfinal 10-11 5,5-7,0 10
Tempo (min) 60 30 60 Temperatura (ºC) 60±3 60±3 60±3
*0,05 g de MgSO4/100 g de polpa.
% Hidrólise = ([ALX] – [ALC]) x fator de hidrólise x 100 (4.13) Massa seca da amostra

61
4.6.1. Determinação do pH das polpas
Após lavagem com água, o pH das polpas obtidas medido de forma aproximada pelo
contato de uma tira de papel indicador universal com a polpa úmida.
O pH inicial foi determinado antes do branqueamento, ou seja, após a mistura da polpa
com o reagente de branqueamento (enzimático ou químico) e o pH final foi determinado após o
branqueamento das amostras para controle do processo.
4.6.2. Extração alcalina (E)
A seqüência de branqueamento químico foi iniciada pelo tratamento das polpas com
hidróxido de sódio. Os ensaios foram realizados em sacos de polietileno com quantidade suficiente
de polpa para as análises posteriores e conforme as condições da Tabela 4.3. Após mistura da
polpa nos sacos de polietileno e pré-aquecimento em forno de microondas, as polpas foram
mantidas na temperatura desejada em banho termostatizado. Após equilíbrio térmico foi
adicionado hidróxido de sódio (3% base polpa seca), sendo mantidas sob agitação por 60 min.
Após esse tempo, as polpas tratadas foram filtradas em funil de Buchner, lavadas com água
destilada até pH próximo ao neutro, secas à temperatura ambiente e guardadas adequadamente
para a etapa seguinte.
4.6.3. Quelação (Q)
A etapa de quelação foi usada visando a remoção de possíveis metais presentes nas
polpas, pois a presença desses metais pode consumir o peróxido de hidrogênio usado na posterior
etapa de branqueamento. Após a adição da dosagem adequada de EDTA (0,4% base polpa seca),
adicionou-se água à polpa, para ajuste de consistência, e suficiente quantidade de H2SO4 4 N, para
ajuste de pH (Tabela 4.3). A temperatura de reação foi ajustada pelo aquecimento da suspensão em
banho termostatizado. Terminada a reação, após 30 min, as polpas tratadas foram filtradas em funil

62
de Buchner, lavadas com água destilada até pH próximo ao neutro, secas à temperatura ambiente e
guardadas adequadamente para a etapa seguinte.
4.6.4. Peroxidação (P)
Após mistura da polpa com água e sulfato de magnésio (0,05 g MgSO4/100 g de polpa)
em sacos de polietileno, a temperatura do material foi ajustada pelo seu aquecimento em forno de
microondas e mantida no valor em banho termostatizado. O sulfato de magnésio aqui adicionado
teve a função de proteger a celulose da degradação, pois o peróxido, além de reagir com a lignina,
pode também reagir com a celulose o que ocasionaria uma possível despolimerização, causando
danos à polpa (HORTAL e LLUCIÁ, 1984).
Após equilíbrio térmico foi adicionado H2O2 (3% base polpa seca), corrigindo o pH para
10 com adição de hidróxido de sódio e os frascos mantidos sob agitação por 60 min (Tabela 4.3).
Após esse tempo, as polpas tratadas foram filtradas em funil de Buchner, lavadas com água
destilada até pH próximo ao neutro, secas à temperatura ambiente e guardadas adequadamente
para as análises seguintes.
4.7. REPRODUTIBILIDADE DOS RESULTADOS
Todos os experimentos foram realizados em triplicata para a confirmação dos resultados.

63
5. RESULTADOS E DISCUSSÃO
5.1. DETERMINAÇÃO DAS ATIVIDADES ENZIMÁTICAS NOS EXTRATOS DE C. subvermispora
C. subvermispora foi utilizado para a biopolpação de cavacos de E. grandis e os extratos
obtidos a partir desses cavacos biodegradados foram avaliados quanto à atividade de algumas
enzimas oxidativas e hidrolíticas (Tabela 5.1). As atividades totais foram relacionadas com a
quantidade de cavacos de madeira utilizados durante o processo de biopolpação e expressas como
UI/kg (μmol/min x kg de cavaco).
Tabela 5.1 -.Produção enzimática de C. subvermispora em cavacos de E. grandis
Característica Xilanase Mananase Endoglucanase Β-glicosidase Β-xilosidase Mn- peroxidase
Atividade (UI/kg) 376 213 133 87 62,6 765
Ph ótimo 5,0 4,0 3,5 4,5 4,0 3,2
Temperatura ótima 60 60 70 60 60 40
C. subvermispora produziu baixas atividades de celulases, mas altas atividades de
hemicelulases e Mn-peroxidases, as quais são de interesse em aplicações tecnológicas, por
exemplo em processos de biobranqueamento em fábricas de celulose e papel. A produção de
xilanases e Mn-peroxidases livres de celulases pode ser realizada em grande escala durante a
biopolpação de cavacos e empregadas na remoção da lignina residual de polpas celulósicas. As
enzimas produzidas dessa maneira podem ser recuperadas e utilizadas como produtos finais sendo
úteis em seqüências de branqueamento nas próprias plantas que utilizam esse processo. Por
exemplo, se pelo menos 5% das enzimas produzidas puderem ser recuperadas da madeira
biopolpada através de algum procedimento de lavagem, mesmo em uma pequena planta de
polpação termo-mecânica, que produza 200 toneladas de polpa por dia, poderão ser produzidas 3,7
x 106 UI de xilanase e 7,6 x 106 UI de Mn-peroxidase por dia. Isto é mais do que a produção usual
de culturas líquidas submersas ou agitadas de Bacillus spp., Trichoderma spp., Phanerochaete
chrysosporium ou outros microrganismos (HARAZONO et al., 1996; SUBRAMANIYAN et al.,
2002; HEIDORNE et al., 2006).
A ação de MnP com o intuito de oxidar estruturas fenólicas da lignina ocorre na presença

64
de MnSO4, porém o conteúdo Mn na polpa kraft é muito baixo (4,8 ppm) (SILVA, 1997). Assim,
as polpas não são branqueadas sem a adição externa de MnSO4 (KONDO et al., 1994). Para uma
ação eficiente de MnP é necessário ainda que H2O2 e ácidos orgânicos, como o málico ou oxálico,
sejam adicionados continuamente durante a etapa de branqueamento, o que torna o processo mais
dispendioso e de difícil controle. Portanto, mesmo a MnP estando em maior proporção no extrato
de C. subvermispora do que outras enzimas, ela não é considerada um eficiente agente de redução
da lignina em polpas nas condições utilizadas. Além disso, elevadas concentrações de H2O2 podem
inativar irreversivelmente a enzima através da formação de um intermediário da enzima,
denominado composto III (WARIISHI et al., 1988).
As hemicelulases produzidas por C. subvermispora apresentam uma faixa de pH e
temperatura ótima de acordo com as comumente observadas, variando entre 4,0–6,0 e 60–70ºC,
respectivamente. As xilanases são estáveis em pH entre 5,0–6,0 e são mais ativas a 40–50ºC. As
atividades enzimáticas decaem durante longos períodos de incubação e em temperaturas maiores
que 50ºC. As enzimas são, geralmente, inativadas acima de 80ºC. Com relação às MnPs, verificou-
se que as atividades foram mais altas com o uso de tampão com menor pH (3,2) e em temperatura
de 40oC.
As enzimas produzidas por C. subvermispora apresentam algumas propriedades atípicas
quando comparadas a outras enzimas produzidas por fungos e podem ser úteis na indústria devido
a essas características. Talvez, a propriedade mais notável seja a alta especificidade (HEIDORNE
et al, 2006).
5.2. EFEITO DAS ENZIMAS DE C. subvermispora E DO EXTRATO COMERCIAL NA HIDRÓLISE DAS POLPAS
Foram realizados alguns ensaios para a comparação da ação do extrato bruto de C.
subvermispora e do extrato comercial em relação à extensão da liberação de açúcares redutores
(Tabela 5.2).
Observou-se uma atuação muito semelhante dos dois extratos na polpa. Os açúcares
redutores foram detectados somente a partir de 2 horas de incubação. Com o prosseguimento da
reação foram detectadas quantidades crescentes de açúcares redutores nos extratos até 12 horas de
tratamento. Após este tempo, a ação hidrolítica de ambos os extratos se estabiliza. A análise dos
dados mostra que tempos de tratamento muito longos (acima de 8 horas) não aumentam a extensão

65
de hidrólise significativamente nas polpas, confirmando alguns resultados encontrados na literatura
(LI et al., 2002). Esse fato poderia ser atribuído à inativação ou perda de função das enzimas em
tempos mais prolongados de tratamento. Entretanto, resultados anteriores mostraram que as
hemicelulases de C. subvermispora são estáveis durante mais de 6 h a 40oC, descartando a
possibilidade de redução da hidrólise da polpa por inativação térmica da enzima.
Tabela 5.2 - Concentração de açúcares redutores liberados dos tratamentos com os extratos enzimáticos sobre polpas kraft. A carga enzimática aplicada foi de 2 UI/g nos dois ensaios realizados
Açúcares redutores (g/L) Tempo de tratamento (h) Extrato de C. subvermispora Extrato comercial
0 2,21 2,21 2 6,64 7,74 4 12,17 12,21 8 16,59 17,53 12 24,53 22,32 24 24,62 21,16
Outros fatores a serem considerados quanto à diminuição da velocidade de hidrólise das
polioses é a redução da concentração e da disponibilidade dos substratos na polpa. Em tempos
mais prolongados de ação da enzima ocorrem alterações na estrutura das polioses. Para uma
hidrólise total, as enzimas têm que agir liberando substratos para que outras enzimas completem a
hidrólise. Esse processo na presença do fungo é bem regulado, no entanto quando se trabalha com
extrato livre de células, as enzimas podem sofrer inibição pelos produtos, o que restringiria a sua
ação (TECHAPUN et al., 2003).
A Figura 5.1 relaciona os resultados obtidos quanto à porcentagem de hidrólise de xilana
da polpa. Até 5 horas de tratamento, as enzimas de C. subvermispora e a comercial hidrolisaram
cerca de 45% das xilanas existentes na polpa. Acompanhando o perfil de liberação de açúcares
redutores, até 12 horas de reação a taxa de hidrólise de ambos os extratos é alta sendo que acima
desse tempo de tratamento a ação hidrolítica de ambos os extratos diminui. Em tempos mais
prolongados as enzimas de C. subvermispora tiveram uma ação mais eficiente na hidrólise das
xilanas em comparação ao extrato comercial. As diferenças percentuais obtidas foram de 14%
para 2 horas de tratamento, 0,33% para 4 horas, 5,4% para 8 horas, 9% para 12 horas e finalmente,
14% para 24 horas de tratamento com relação à enzima comercial.. As enzimas, principalmente as
xilanases, agem com sucesso na polpa sem perda de sua estabilidade, que permanecem adequadas

66
ao processo durante 6 horas em temperaturas de 40ºC e pH 5,0 (HEIDORNE et al., 2006). Dessa
maneira foi possível estabelecer um tempo de tratamento das polpas de até 5 horas, o que é um
tempo apropriado aos processos industriais.
0
10
20
30
40
50
60
70
0 3 6 9 12 15 18 21 24
Tempo de tratamento (h)
Hid
rólis
e (%
)
C. subvermisporaExt. comercial
Figura 5.1 - Porcentagem de hidrólise da xilana de polpas kraft pelos extratos enzimáticos de C. subvermispora e pelo extrato comercial. A carga enzimática aplicada foi de 2 UI/g nos dois ensaios realizados
5.3. ANÁLISE DOS PRODUTOS DE XILANAS DE DIFERENTES ORIGENS APÓS AÇÃO DA XILANASE
Para o estudo dos produtos de hidrólise das xilanases de C. subvermispora utilizou-se
xilanas de bétula, de fibra de trigo, de carvalho e de bagaço de cana-de-açúcar. Cerca de 94% da
estrutura da xilana de bétula corresponde a xilose, o que a torna um substrato ideal para
padronizar as atividades de várias xilanases. Além da xilose, há pequenas quantidades de glicose e
galactose. A xilana de fibra de trigo contém pequenas quantidade de ácidos glucurônicos,
enquanto que na xilana de carvalho, o carboidrato mais abundante é a xilose (52%), seguida da
arabinose (22%) (LI et al., 2000). A xilana de cana de açúcar contém principalmente açúcares
facilmente hidrolisados por enzimas (ADSUL et al, 2005), principalmente xilose (75-80%),
manose, arabinose e galactose (GOMES et al., 2005).
Amostras de xilana de fibra de trigo liberaram xilopentaose, xilotetraose, xilotriose e
uma pequena quantidade de xilobiose. Em tempos de tratamento mais longos (96 horas), toda a

67
xilopentaose desapareceu, sendo convertida a xilobiose. A xilose, embora tenha sido detectada,
não foi quantificada, pois sua concentração ficou abaixo da curva de calibração. Nas xilanas de
bétula, a enzima liberou xilobiose, xilotriose e xilotetraose. Após 72 horas também foi detectada
xilose, permanecendo o mesmo perfil até 96 horas. A xilana de carvalho liberou principalmente
xilotriose e xilobiose a partir de 48 horas de reação. Xilose foi liberada com 72 horas, sendo
verificada redução na quantidade dos xilooligômeros liberados. Além desse tempo de reação
houve pouca redução na quantidade dos produtos liberados. A Tabela 5.3 traz os valores das
concentrações dos xilooligômeros detectados por CLAE.
Tabela 5.3 - Quantificação dos oligômeros detectados por CLAE
Xilanas Xilooligômeros Fibra de trigo xilopentaose Xilotetraose xilotriose xilobiose xilose
72 h 0,590 0,760 0,387 0,157 - 96 h - 0,680 0,235 0,697 -
Bétula 24h - 0,672 0,244 0,515 - 48h - 0,672 0,244 0,334 - 72h - 0,672 0,244 0,334 0,221 96h - 0,419 0,129 0,105 0,221
Carvalho 48h - - 0,192 0,390 - 72h - - 0,182 0,232 0,366 96h - - 0,182 0,220 0,366
Unidades em g/L; O símbolo (-) indica níveis abaixo do limite de detecção.
Por tratarem-se de substratos que se degradam com mais facilidade, os controles (sem
adição de enzima) apresentaram glicose e xilose em pequenas quantidades. Os produtos obtidos da
hidrólise da xilana mostraram que as enzimas de C. subvermispora apresentaram um padrão de
endo-xilanase, atacando a cadeia de xilana de uma maneira aleatória. Esses resultados estão em
acordo com os dados da literatura (LI et al., 2005). Endo-1,4-β-xilanases (EC 3.2.1.8) produzem
oligossacarídeos derivados da quebra aleatória da xilana enquanto que as 1,4-β-xilosidases (EC
3.2.1.37) atuam principalmente nos oligossacarídeos de xilana produzindo xilose. Algumas endo-
xilanases parecem ter grande especificidade por substratos de cadeia linear, e outras, por cadeias
mais ramificadas. A presença de ramificações em alguns desses substratos na cadeia de xilana
parece prevenir sua hidrólise completa pela xilanase. Enzimas capazes de hidrolisar as ligações α-
1,3 e α-1,2 podem ser necessárias para a hidrólise completa da xilana (ANAND et al., 1996).

68
A ação da xilanase nas xilanas foi analisada pelo teor de açúcares redutores e os
resultados mostraram que para as xilanas de bétula, fibra de trigo e carvalho já existia produto
suficiente para detecção antes de 24 horas de hidrólise. A hidrólise da xilana de bagaço de cana foi
lenta, mas com 48 horas de hidrólise havia produtos (Figura 5.2). No final da reação, a xilana de
bagaço de cana foi a que apresentou maior grau de hidrólise seguida da xilana de fibra de trigo,
em relação ao tempo total de hidrólise.
0
5
10
15
20
25
0 24 48 72 96
Tempo (horas)
Açú
care
s re
duto
res
(g/L
)
Bagaço de cana Carvalho Bétula Fibra de trigo
Figura 5.2 - Perfil da liberação de açúcares redutores das xilanas de diferentes origens
Calculando-se a relação entre a concentração de açúcares redutores e a massa seca de
cada xilana solúvel, foi possível obter o grau máximo de hidrólise com o tratamento enzimático
(Tabela 5.4). Esses valores são relativos ao tempo de 96 horas de tratamento para as xilanas de
bétula, carvalho, bagaço de cana e 72 horas para a xilana de fibra de trigo.
Tabela 5.4 - Quadro comparativo do grau de hidrólise entre as diferentes xilanas
Xilanas Grau de hidrólise das xilanas (%)
Bétula 22,9
Carvalho 18,3
Fibra de trigo 34,0
Bagaço de Cana 61,6

69
A xilana de carvalho foi a que apresentou o menor grau de hidrólise entre as xilanas
estudadas. Aparentemente isto se deve ao fato desta xilana apresentar mais ramificações com
resíduos arabinosil.
5.4. BRANQUEAMENTO DAS POLPAS
As amostras de polpa kraft foram submetidas a um tratamento enzimático seguido de
uma seqüência de branqueamento químico. Foram adicionadas diferentes concentrações de
enzima, tendo como base os valores normalmente empregados no branqueamento de polpas com
xilanases.
A carga enzimática e as condições dos ensaios estão mostrados na Tabela 5.5. Para
estimar o potencial branqueador do extrato enzimático nas polpas, as seguintes análises foram
realizadas: medidas de absorbância a 237, 254, 280 e 465 nm, número kappa, viscosidade e
liberação de açúcares redutores.
Tabela 5.5 - Condições utilizadas no branqueamento enzimático
Condições Xilanase
Enzima (UI/g) 0,5 1 1,5 2
Tempo de reação (horas) 0,5 1 2 3 5
Temperatura/Consistência/pH 40ºC/10%/5,0
Quando a polpa kraft é pré-tratada com xilanase, a xilose e outros açúcares redutores
liberados resultam em um aumento no conteúdo de açúcar livre em meio à polpa. A xilana, como
uma parte das polioses, que está entre a barreira de lignina e celulose, quando hidrolisada pela
xilanase, em adição à xilose, também libera lignina e outros compostos fenólicos presentes nas
fibras da polpa. São esses compostos fenólicos que causam um aumento na absorbância a 237, 254
e 280 nm das amostras tratadas com xilanase comparadas ao controle. Assim, a correlação entre a
liberação dos cromóforos (principalmente a 237 nm), a liberação de compostos hidrofóbicos (465
nm) e a redução no número kappa associados com a liberação de açúcares redutores sugerem a
dissociação do complexo lignina-carboidrato das fibras da polpa (BEG et al., 2000). Esses
fenômenos propiciam uma redução na quantidade de reagentes de branqueamento, pois as fibras

70
mais livres desses complexos facilitam a penetração desses reagentes, branqueando melhor a polpa
em relação aos controles.
De acordo com as cargas de xilanase aplicadas e conhecendo-se as atividades das outras
enzimas estudadas (Tabela 5.1) foi possível calcular as cargas enzimáticas de cada enzima
presente no extrato utilizado. A Tabela 5.6 relaciona os resultados obtidos.
Tabela 5.6 - Atividades aplicadas sobre a polpa referente às demais enzimas contidas no extrato bruto de C. subvermispora
Enzima Atividade (UI/g)
B-xilanase 2,0 1,5 1,0 0,5
B-mananase 1,0 0,75 0,5 0,25
Endoglucanase 0,06 0,05 0,03 0,01
B-glicosidase 0,04 0,03 0,02 0,007
B-xilosidase 0,04 0,03 0,02 0,007
Mn-peroxidase 4,0 3,0 2,0 1,0
5.4.1. Análise dos produtos das polpas kraft após a ação do complexo enzimático de C. subvermispora
Foi analisada a ação do extrato enzimático de C. subvermispora nas polpas kraft. O
processo kraft remove a maior parte da lignina e dos carboidratos. Durante o período inicial de
aquecimento, a xilana é parcialmente despolimerizada e são removidos seus grupos acetil e
arabinosil. Após a polpação, parte dessa xilana pode estar recristalizada na superfície da fibra,
constituindo uma barreira física ao fluxo dos reagentes nas posteriores etapas de tratamento das
fibras. É nessa polpa rica em celulose (78%), com baixo teor de lignina (5,3%) e parcialmente
recoberta por xilana (12,8%) que as enzimas de C. subvermispora vão agir.
- Cromóforos liberados da polpa em função da concentração de enzima
Os cromóforos são estruturas insaturadas atribuídas principalmente a produtos derivados
da degradação da lignina e das polioses, por exemplo, grupos carbonila e ácidos urônicos

71
formados durante a polpação kraft. Foram feitas as determinações das absorbâncias a 237, 254 e
280 nm relativos à liberação de compostos fenólicos e produtos de degradação das polioses e 465
nm relativo à liberação de compostos hidrofóbicos da polpa.
Pode-se observar pela Figura 5.3 que para o comprimento de onda de 237 nm, as retas
correspondentes a todas as cargas enzimáticas apresentaram um aumento na inclinação até o
ponto correspondente a 3 horas de reação. Em seguida houve uma diminuição na liberação de
substâncias que absorvem nesse comprimento de onda, à exceção da reta correspondente a 0,5
UI/g, que apresentou um aumento da taxa a partir do ponto correspondente a 3 horas de reação.
0
0,1
0,2
0,3
0,4
0 1 2 3 4 5
Tempo (h)
Abs
2 UI/g
1,5 UI/g
1 UI/g
0,5 UI/g
Figura 5.3 - Liberação de cromóforos a 237 nm em função do tempo de tratamento
As Figuras 5.4, 5.5 e 5.6 mostram a liberação de cromóforos nos comprimentos de onda
de 254, 280 e 465 nm. As cargas enzimáticas de 0,5 e 1 UI/g não apresentaram liberação de
quantidades significantes de compostos que absorvem em nenhum desses comprimentos de onda.
Para o comprimento de onda de 254 nm é possível observar que as retas correspondentes
às cargas enzimáticas de 1,5 e 2 UI/g apresentaram uma inclinação constante e gradual, embora
pequena, até o ponto correspondente a 2 horas de reação, apresentando em seguida uma inclinação
maior e mais acentuada entre 2 e 3 horas de reação e um pouco menor entre 3 e 5 horas de reação.

72
0
0,1
0,2
0,3
0,4
0,5
0 1 2 3 4 5Tempo (h)
Abs
2 UI/g
1,5 UI/g
1UI/g
0,5 UI/g
Figura 5.4 - Liberação de cromóforos a 254 nm em função do tempo de tratamento
Para o comprimento de onda de 280 nm é possível observar que as retas correspondentes
às cargas enzimáticas de 1,5 e 2 UI/g apresentaram comportamentos diferenciados. Enquanto a
reta correspondente a 1,5 UI/g apresentou uma inclinação acentuada e constante para os tempos de
reação de 2 a 5 horas, a reta correspondente a 2 UI/g apresentou inclinações diferentes a cada
intervalo de tempo.
0
0,1
0,2
0,3
0,4
0,5
0 1 2 3 4 5
Tempo (h)
Abs
2 UI/g
1,5 UI/g
1 UI/g
0,5 UI/g
Figura 5.5 - Liberação de cromóforos a 280 nm em função do tempo de tratamento
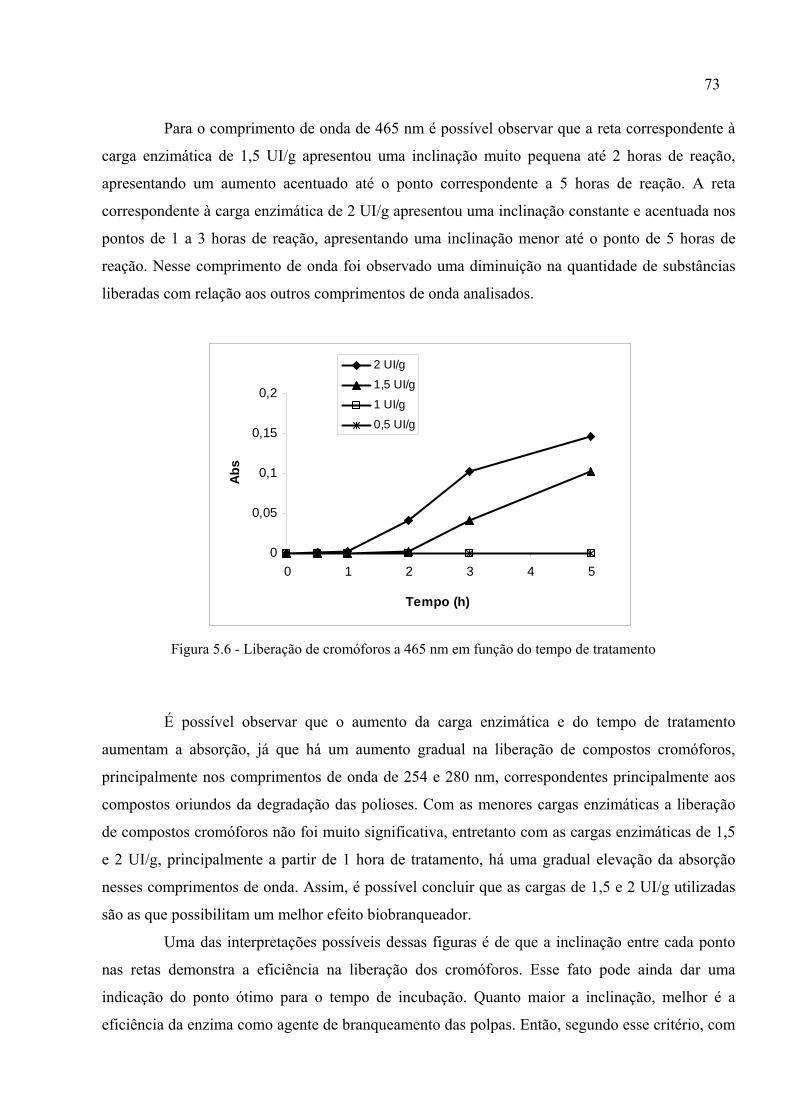
73
Para o comprimento de onda de 465 nm é possível observar que a reta correspondente à
carga enzimática de 1,5 UI/g apresentou uma inclinação muito pequena até 2 horas de reação,
apresentando um aumento acentuado até o ponto correspondente a 5 horas de reação. A reta
correspondente à carga enzimática de 2 UI/g apresentou uma inclinação constante e acentuada nos
pontos de 1 a 3 horas de reação, apresentando uma inclinação menor até o ponto de 5 horas de
reação. Nesse comprimento de onda foi observado uma diminuição na quantidade de substâncias
liberadas com relação aos outros comprimentos de onda analisados.
0
0,05
0,1
0,15
0,2
0 1 2 3 4 5
Tempo (h)
Abs
2 UI/g
1,5 UI/g
1 UI/g
0,5 UI/g
Figura 5.6 - Liberação de cromóforos a 465 nm em função do tempo de tratamento
É possível observar que o aumento da carga enzimática e do tempo de tratamento
aumentam a absorção, já que há um aumento gradual na liberação de compostos cromóforos,
principalmente nos comprimentos de onda de 254 e 280 nm, correspondentes principalmente aos
compostos oriundos da degradação das polioses. Com as menores cargas enzimáticas a liberação
de compostos cromóforos não foi muito significativa, entretanto com as cargas enzimáticas de 1,5
e 2 UI/g, principalmente a partir de 1 hora de tratamento, há uma gradual elevação da absorção
nesses comprimentos de onda. Assim, é possível concluir que as cargas de 1,5 e 2 UI/g utilizadas
são as que possibilitam um melhor efeito biobranqueador.
Uma das interpretações possíveis dessas figuras é de que a inclinação entre cada ponto
nas retas demonstra a eficiência na liberação dos cromóforos. Esse fato pode ainda dar uma
indicação do ponto ótimo para o tempo de incubação. Quanto maior a inclinação, melhor é a
eficiência da enzima como agente de branqueamento das polpas. Então, segundo esse critério, com

74
a aplicação da carga enzimática de 0,5 UI/g, o melhor tempo de tratamento seria de 5 horas e para
cargas maiores, o tempo ótimo de tratamento poderia ser reduzido a 3 horas.
As Figuras 5.7, 5.8 e 5.9 apresentam os resultados referentes às liberações de cromóforos
nos comprimentos de onda estudados (237, 254, 280 e 465 nm) em função do tempo de tratamento
para cada carga enzimática aplicada.
2 UI/g
0
0,1
0,2
0,3
0,4
0,5
0,6
0 1 2 3 4 5
Tempo (h)
Abso
rção
237 nm
254 nm
280 nm
465 nm
Figura 5.7 - Liberação de cromóforos pelo tratamento enzimático com 2 UI/g de xilanase nos diferentes tempos de tratamento
1,5 UI/g
0
0,1
0,2
0,3
0,4
0,5
0,6
0 1 2 3 4 5
Tempo (h)
Abso
rção
237 nm
254 nm
280 nm
465 nm
Figura 5.8 - Liberação de cromóforos pelo tratamento enzimático com 1,5 UI/g de xilanase nos diferentes tempos de tratamento

75
1 UI/g 0,5 UI/g
0
0,05
0,1
0,15
0,2
0 1 2 3 4 5
Tempo (h)
Abso
rção
237 nm a
237 nm b
Figura 5.9 - Liberação de cromóforos pelo tratamento enzimático com 1 (a) e 0,5 (b) UI/g de xilanase nos diferentes tempos de tratamento
Os materiais cromóforos liberados a 237, 254 e 280 nm mostram que houve decréscimo
na aromaticidade da lignina residual quando foram aplicadas as cargas de 1,5 e 2 UI/g. Além de
compostos fenólicos, esses comprimentos de onda referem-se também a produtos de degradação
das polioses que contribuem em grande parte na coloração dessas polpas kraft. Entretanto, mesmo
havendo um aumento na absorbância dos filtrados não ocorreu uma concomitante diminuição na
cor das polpas.
- Açúcares redutores liberados da polpa em função da concentração de enzima
O extrato enzimático de C. subvermispora não teve efeito significante na polpa quando
utilizado em baixas atividades. Com a carga enzimática de 0,5 UI/g de xilanase houve pequena
liberação de açúcares redutores. O controle e o ensaio apresentaram concentrações muito
próximas, em torno de 0,30 g/L. Com a aplicação de 1 UI/g de xilanase a concentração de açúcares
aumentou discretamente, e a partir de 2 horas de tratamento a liberação ficou mais ou menos
estável, em 0,96 g/L. A aplicação de 1,5 UI/g de xilanase aumentou consideravelmente a
concentração de açúcares redutores liberados, chegando a 2,7 g/L em 30 minutos de reação. A
maior liberação de açúcares redutores foi observada quando foi aplicada a carga enzimática de 2

76
UI/g. Em 30 minutos foram produzidos 17 g/L de açúcares redutores e a máxima concentração
observada foi de 23,6 g/L no tempo de 3 horas de tratamento (Figura 5.10).
0
5
10
15
20
25
0,5 1 1,5 2
Carga enzimática (UI/g)
Açú
care
s re
duto
res
(g/L
) 0,5 hora1 hora2 horas3 horas5 horas
Figura 5.10 - Açúcares redutores liberados pelo tratamento enzimático nas polpas kraft em função da carga enzimática referente à xilanase aplicada
A Figura 5.11 mostra a taxa de hidrólise das polioses nas diferentes cargas enzimáticas. A
retirada das polioses é uma condição necessária para alterar a interface entre a celulose e a lignina,
facilitando a remoção da fração associada à lignina com o mínimo de dano à polpa (TECHAPUN
et al., 2003). O pré-tratamento com xilanase pode liberar uma maior superfície das fibras, expondo
a lignina e facilitando assim a posterior etapa de branqueamento da polpa, possibilitando também
uma economia de reagentes, além de preservar e até melhorar algumas propriedades da polpa
resultante, já que não retira carboidratos excessivamente.

77
0
10
20
30
40
50
60
70
0 1 2 3 4 5Tempo de tratamento (h)
Hid
rólis
e (%
)2 UI/g1,5 UI/g1 UI/g0,5 UI/g
Figura 5.11 - Hidrólise das polioses da polpa em diferentes cargas enzimáticas
O tempo de tratamento, juntamente com a carga enzimática aplicada, é um dos principais
fatores que influenciam a quantidade de açúcares redutores liberados. Durante o tratamento
enzimático é necessário levar em consideração também a consistência da polpa e a agitação do
meio reacional, que além de diluir, permite a difusão dos produtos finais dispersando os produtos
que poderiam inibir a ação das enzimas.
A liberação de açúcares pela aplicação de cargas enzimáticas entre 0,5 e 1,5 UI/g de
xilanase às polpas é praticamente idêntica em todos os tempos estudados. Quando foi aplicado 2
UI/g é possível notar um aumento progressivo na liberação de açúcares redutores que se torna mais
acentuado a partir de 3 horas e depois se estabiliza.
É importante ressaltar que no extrato estavam presentes outras enzimas que também
atuaram nas polioses presentes na polpa, portanto também responsáveis por essa ação hidrolítica.
Vários mecanismos podem estar envolvidos na dissolução e hidrólise dessas polioses facilitando o
passo subseqüente de extração da lignina. As xilanas recristalizadas sobre as fibras de celulose e
aquelas associadas à lignina (LCC) podem ser parcialmente ou totalmente removidas e
quantificadas como açúcares redutores. Essa liberação de açúcares redutores é uma indicação de
que o uso de enzimas do complexo xilanolítico facilita o posterior branqueamento com reagentes
químicos.
A liberação de açúcares redutores e a liberação de lignina e compostos fenólicos são
fenômenos inter-relacionados. Quando a polpa kraft é pré-tratada com xilanase, a xilose e outros
açúcares redutores são liberados das polioses, resultando em um aumento na quantidade de

78
açúcares redutores da amostra. A xilana é uma parte das polioses que está localizada entre a
lignina e a celulose. Quando a xilana é degradada pela xilanase, não só xilose é liberada das fibras,
mas também pequenas quantidades de lignina e compostos fenólicos (KHANDEPARKAR et al.,
2006).
- Identificação dos produtos da hidrólise dos carboidratos da polpa
Para estudo dos monossacarídeos e oligossacarídeos obtidos da hidrólise das polpas kraft
pelas enzimas de C. subvermispora foi utilizada cromatografia líquida de alta eficiência (CLAE).
As liberações de celobiose, glicose, arabinose, xilose, xilobiose, xilotriose e outros oligômeros
foram avaliadas nos filtrados das polpas após os períodos de incubação.
Os ensaios com xilanase não liberaram xilooligômeros em quantidades detectáveis em
nenhuma carga enzimática utilizada (Tabela 5.7). Os ensaios com xilanase na concentração de 1
UI/g liberaram apenas celobiose em quantidades detectáveis pela CLAE nos tempos de tratamento
utilizados. Na concentração de 1,5 UI/g foram detectados xilose e celobiose. Pelos resultados
obtidos é possível verificar que o tempo de 2 e 3 horas de tratamento foram os que liberaram as
maiores quantidades de xilose. Os ensaios com xilanase na concentração de 2 UI/g liberaram as
maiores quantidades de açúcares redutores. Arabinose foi detectada com apenas 0,5 hora de
tratamento, o que não ocorreu com a aplicação das menores cargas enzimáticas. Isso pode ter
ocorrido devido à presença de outras enzimas no extrato, como por exemplo a enzima
arabinofuranosidase. O aumento na concentração de xilanase aumenta, conseqüentemente, a
concentração dessas outras enzimas.

79
Tabela 5.7 - Dados obtidos dos filtrados das polpas kraft após o tratamento com o extrato bruto de C. subvermispora em diferentes concentrações (0,5, 1, 1,5 e 2 UI/g) pela cromatografia líquida de alta eficiência (CLAE)
0,5 UI/g Tempo de tratamento (h) 0,5 1 2 3 5
Celobiose, Glicose, Xilose, Arabinose, Xilooligômeros*
-
1 UI/g Tempo de tratamento (h) 0,5 1 2 3 5
Celobiose - - - - - Glicose - - 0,211 0,212 0,197 Xilose - - - - -
Arabinose - - - - - Xilooligômeros* - - - - -
1,5 UI/g Tempos de tratamento (h) 0,5 1 2 3 5
Celobiose - 0,569 0,870 0,922 0,685 Glicose - - - - - Xilose - 0,180 0,260 0,277 0,237
Arabinose - - - - - Xilooligômeros* - - - - -
2 UI/g Tempos de tratamento (h) 0,5 1 2 3 5
Celobiose 0,224 0,406 0,804 1,032 0,560 Glicose - 0,206 0,254 0,272 0,242 Xilose - - 0,224 0,260 0,242
Arabinose 0,330 0,333 0,346 0,866 0,351 Xilooligômeros* - - - - -
Valores de açúcares redutores (g/L); * Foram usados como padrões xilobiose, xilotriose, xilotetraose e xilopentaose; O símbolo (-) indica níveis abaixo do limite de detecção.
5.4.2. Características da polpa tratada com diferentes concentrações de enzimas e branqueada
As polpas tratadas com diferentes concentrações de enzima foram branqueadas com a
seqüência X/E/Q/P e foram realizadas as análises de número kappa e viscosidade. Analisando os
valores da viscosidade após a etapa de tratamento enzimático (Apêndice A) é possível verificar
que houve um pequeno aumento da viscosidade das polpas branqueadas em relação ao controle.
As concentrações de enzima empregadas resultaram em viscosidades muito próximas, ou seja, a
faixa escolhida não afetou essa propriedade da polpa (Figuras 5.12, 5.13, 5.14 e 5.15). Tal fato
pode estar relacionado à ação da xilanase, que promove mudanças morfológicas na superfície das
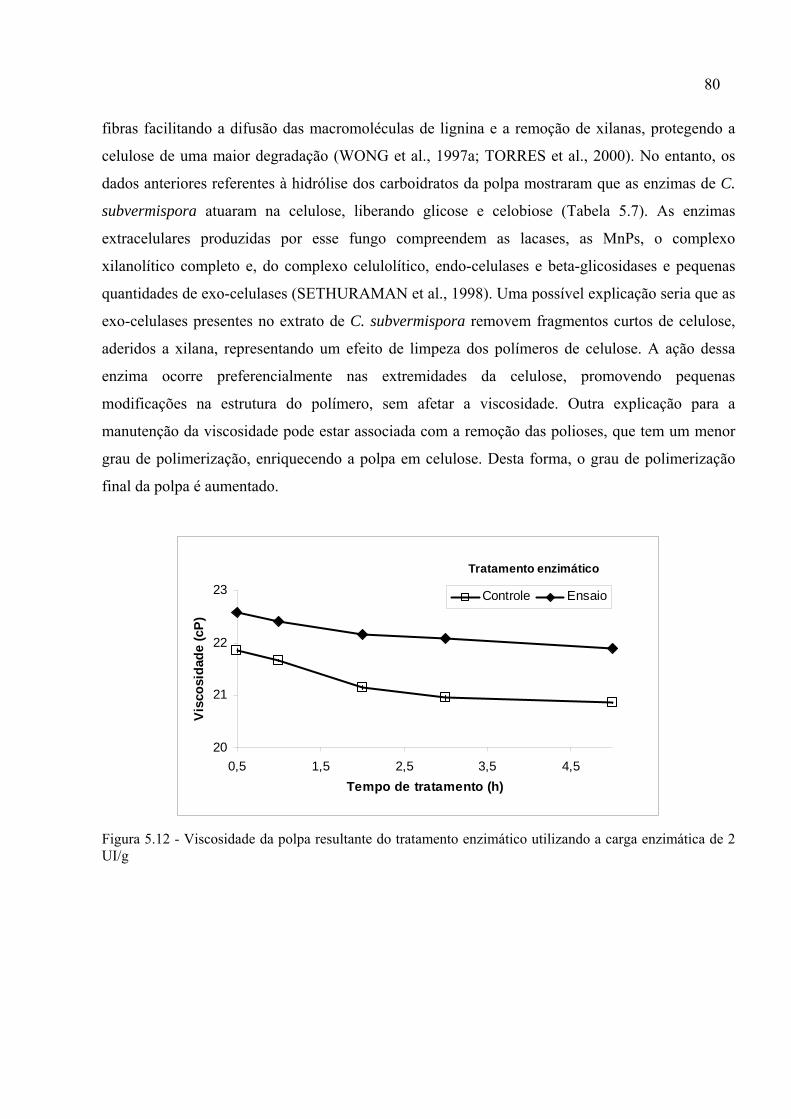
80
fibras facilitando a difusão das macromoléculas de lignina e a remoção de xilanas, protegendo a
celulose de uma maior degradação (WONG et al., 1997a; TORRES et al., 2000). No entanto, os
dados anteriores referentes à hidrólise dos carboidratos da polpa mostraram que as enzimas de C.
subvermispora atuaram na celulose, liberando glicose e celobiose (Tabela 5.7). As enzimas
extracelulares produzidas por esse fungo compreendem as lacases, as MnPs, o complexo
xilanolítico completo e, do complexo celulolítico, endo-celulases e beta-glicosidases e pequenas
quantidades de exo-celulases (SETHURAMAN et al., 1998). Uma possível explicação seria que as
exo-celulases presentes no extrato de C. subvermispora removem fragmentos curtos de celulose,
aderidos a xilana, representando um efeito de limpeza dos polímeros de celulose. A ação dessa
enzima ocorre preferencialmente nas extremidades da celulose, promovendo pequenas
modificações na estrutura do polímero, sem afetar a viscosidade. Outra explicação para a
manutenção da viscosidade pode estar associada com a remoção das polioses, que tem um menor
grau de polimerização, enriquecendo a polpa em celulose. Desta forma, o grau de polimerização
final da polpa é aumentado.
Tratamento enzimático
20
21
22
23
0,5 1,5 2,5 3,5 4,5Tempo de tratamento (h)
Visc
osid
ade
(cP)
Controle Ensaio
Figura 5.12 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga enzimática de 2 UI/g

81
Tratamento enzimático
20
21
22
23
0,5 1,5 2,5 3,5 4,5Tempo de tratamento (h)
Visc
osid
ade
(cP)
Controle Ensaio
Figura 5.13 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga enzimática de 1,5 UI/g
Tratamento enzimático
20
21
22
23
0,5 1,5 2,5 3,5 4,5
Tempo de tratamento (h)
Vis
cosi
dade
(cP)
Controle Ensaio
Figura 5.14 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga enzimática de 1 UI/g
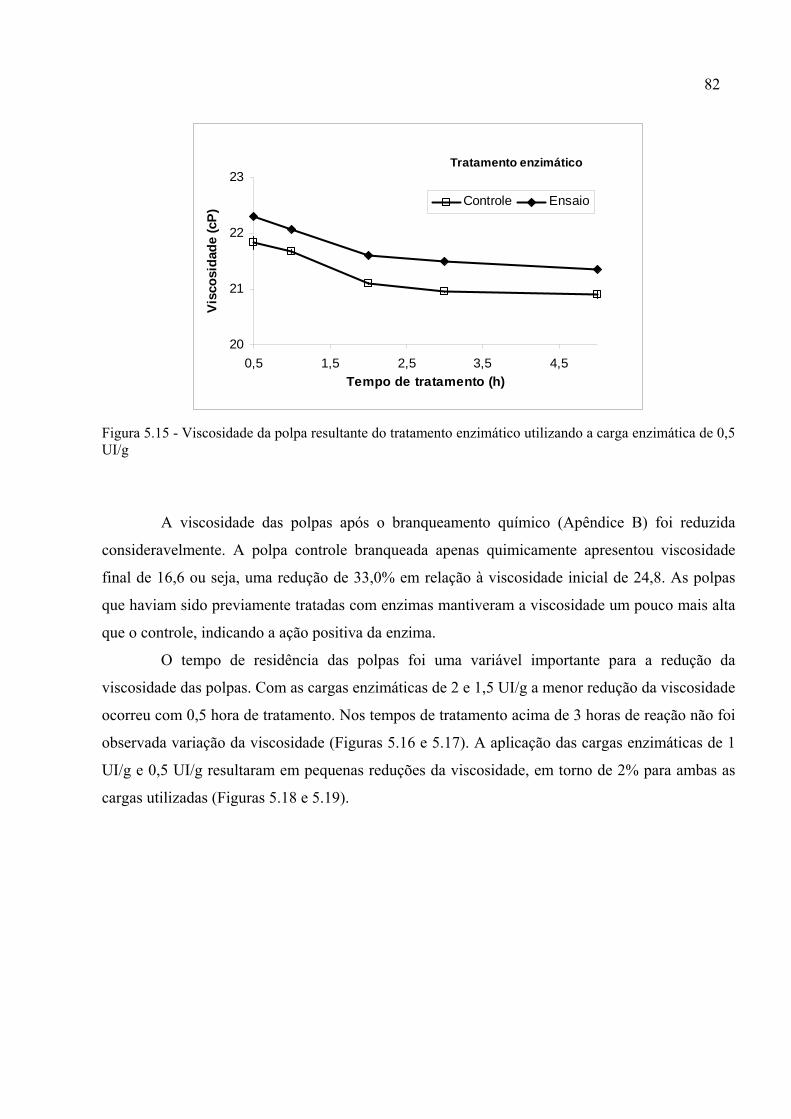
82
Tratamento enzimático
20
21
22
23
0,5 1,5 2,5 3,5 4,5Tempo de tratamento (h)
Vis
cosi
dade
(cP
) Controle Ensaio
Figura 5.15 - Viscosidade da polpa resultante do tratamento enzimático utilizando a carga enzimática de 0,5 UI/g
A viscosidade das polpas após o branqueamento químico (Apêndice B) foi reduzida
consideravelmente. A polpa controle branqueada apenas quimicamente apresentou viscosidade
final de 16,6 ou seja, uma redução de 33,0% em relação à viscosidade inicial de 24,8. As polpas
que haviam sido previamente tratadas com enzimas mantiveram a viscosidade um pouco mais alta
que o controle, indicando a ação positiva da enzima.
O tempo de residência das polpas foi uma variável importante para a redução da
viscosidade das polpas. Com as cargas enzimáticas de 2 e 1,5 UI/g a menor redução da viscosidade
ocorreu com 0,5 hora de tratamento. Nos tempos de tratamento acima de 3 horas de reação não foi
observada variação da viscosidade (Figuras 5.16 e 5.17). A aplicação das cargas enzimáticas de 1
UI/g e 0,5 UI/g resultaram em pequenas reduções da viscosidade, em torno de 2% para ambas as
cargas utilizadas (Figuras 5.18 e 5.19).

83
Branqueamento químico
10
15
20
25
0 0,5 1 2 3 5
Tempo de tratamento (h)
Visc
osid
ade
(cP
)
0
2
4
6
8
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.16 - Viscosidade da polpa tratada com 2 UI/g de xilanase após o branqueamento químico
Branqueamento químico
10
15
20
25
0 0,5 1 2 3 5
Tempo de tratamento (h)
Visc
osid
ade
(cP
)
0
2
4
6
8
Red
ução
(%)
Controle Ensaio % de Redução
Figura 5.17 - Viscosidade da polpa tratada com 1,5 UI/g de xilanase após o branqueamento químico

84
Branqueamento químico
10
15
20
25
0 0,5 1 2 3 5
Tempo de tratamento (h)
Visc
osid
ade
(cP)
0
2
4
6
8
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.18 - Viscosidade da polpa tratada com 1 UI/g de xilanase após o branqueamento químico
Branqueamento químico
10
15
20
25
0 0,5 1 2 3 5
Tempo de tratamento (h)
Visc
osid
ade
(cP)
0
2
4
6
8
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.19 - Viscosidade da polpa tratada com 0,5 UI/g de xilanase após o branqueamento químico
A redução da viscosidade é indesejável, pois esta propriedade se relaciona com o grau de
polimerização da celulose e, indiretamente, com a resistência do papel. A redução após o
branqueamento químico pode ter ocorrido pela ação do peróxido de hidrogênio na polpa, apesar
da adição do sulfato de magnésio. O peróxido, além de reagir com a lignina, pode também reagir
com a celulose o que pode ocasionar uma possível despolimerização e conseqüente redução da
viscosidade, interferindo na qualidade final da polpa produzida. A adição do sulfato de magnésio à

85
etapa de branqueamento com peróxido de hidrogênio teve a função de proteger a celulose,
minimizando a sua degradação.
Após a etapa de tratamento enzimático observa-se que o número kappa (Apêndice C) das
polpas ficou menor do que na polpa controle, sendo a redução máxima atingida em 3 horas de
tratamento. Essa redução ocorre provavelmente pela retirada de parte da lignina durante a lavagem
da polpa, realizada antes da análise do número kappa (Figuras 5.20, 5.21, 5.22 e 5.23).
O efeito da redução do número kappa em função da concentração de enzima e do tempo
de reação fica mais evidente depois do branqueamento químico (Apêndice D). A maior carga de
xilanase utilizada (2 UI/g) foi eficaz na redução do número kappa em tempos curtos de reação. O
número kappa das polpas foi reduzido em 7,7% após o tratamento com 2 UI/g de xilanase e em
10,9% após o branqueamento químico, com 3 horas de reação (Figura 5.20). Nos ensaios em que
se utilizou uma menor carga de enzima verificou-se uma menor redução do kappa após o
tratamento enzimático e conseqüentemente o kappa final foi maior (Figura 5.21). O tratamento das
polpas com 1 UI/g e 0,5 UI/g não proporcionou uma ação eficiente nas fibras, quando se avalia o
efeito de redução no número kappa. Este resultado foi comprovado após o tratamento químico das
polpas, em que o kappa praticamente não foi alterado (Figuras 5.22 e 5.23).
A redução do número kappa tem sido atribuída à remoção da lignina, porém há
evidências de que a remoção de ácidos hexenurônicos (HexAs) formados durante a polpação kraft,
derivados dos ácidos 4-O-metil-D-glucurônico das xilanas, também são responsáveis pelo
mecanismo básico de redução do número kappa. Esses HexAs formados possuem ligações C4-C5
insaturadas, resultando em um aumento de absorbância na região de 230-265 nm. Foi
demonstrado também que esses HexAs interferem positivamente na determinação do número
kappa de madeiras duras da ordem de 30-40%. Outro dado importante é que a liberação de
material que absorve nesses comprimentos de onda, principalmente a 237 nm, fornece uma boa
correlação com o aumento na alvura nas polpas branqueadas (JEFFRIES, 2001).

86
Tratamento enzimático
11,5
12
12,5
13
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
Redu
ção
(%)
Controle Ensaio % de Redução
Branqueamento químico
7
8
9
10
11
12
13
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
12
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.20 - Número kappa das polpas resultantes do tratamento enzimático utilizando a carga de xilanase de 2 UI/g e das polpas resultantes do branqueamento químico

87
Tratamento enzimático
11,5
12,0
12,5
13,0
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
Redu
ção
(%)
Branqueamento químico
7
8
9
10
11
12
13
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
12
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.21 - Número kappa das polpas resultantes do tratamento enzimático utilizando a carga de xilanase de 1,5 UI/g e das polpas resultantes do branqueamento químico

88
Tratamento enzimático
11,5
12,0
12,5
13,0
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
Red
ução
(%)
Branqueamento químico
7
8
9
10
11
12
13
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
12
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.22 - Número kappa das polpas resultantes do tratamento enzimático utilizando a carga de xilanase de 1 UI/g e das polpas resultantes do branqueamento químico

89
Tratamento enzimático
11,5
12,0
12,5
13,0
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
Redu
ção
(%)
Branqueamento químico
7
8
9
10
11
12
13
0 0,5 1 2 3 5Tempo de tratamento (h)
Nº k
appa
0
2
4
6
8
10
12
Redu
ção
(%)
Controle Ensaio % de Redução
Figura 5.23 - Número kappa das polpas resultantes do tratamento enzimático utilizando 0,5 UI/g e das polpas resultantes do branqueamento químico
A perda da lignina após o tratamento enzimático, tem sido atribuída a redução dos
complexos lignina-carboidrato. As xilanases, catalisando a despolimerização das xilanas na polpa,
permitem que a lignina que estava ligada às polioses se difunda mais facilmente para fora da fibra.
Ocorrem também mudanças morfológicas na superfície das fibras como fendas, filamentos e
desfibramento devido ao tratamento enzimático. Estas fendas facilitam a difusão das
macromoléculas de lignina degradada da parede das fibras, além da remoção das xilanas,
proporcionando uma diminuição mais acentuada do número kappa das polpas branqueadas
quimicamente (WONG et al., 1997a; TORRES et al., 2000).

90
Comparando-se os resultados da viscosidade e do número kappa em função da
concentração de xilanase para os tempos de 1, 2 e 3 horas é possível verificar que a viscosidade é
melhor preservada com as maiores cargas enzimáticas embora, com maiores tempos de tratamento
ela seja reduzida. Em relação ao número kappa, os maiores tempos de tratamento permitem uma
maior redução, o que é desejável (Figura 5.24).
Tratamento enzimático
21,4
21,8
22,2
22,6
0,5 1 1,5 2Carga enzimática (UI/g)
Vis
cosi
dade
(cP
)
11,5
12
12,5
13
Nº k
appa
1 h viscosidade 2 h viscosidade 3 h viscosidade1 h kappa 2 h kappa 3 h kappa
Figura 5.24 - Viscosidade e número kappa em função da concentração de xilanase para os tempos de tratamento enzimático de 1, 2 e 3 horas
Após o branqueamento químico verifica-se que a viscosidade das polpas tratadas com
xilanase nos diferentes tempos foi pouco alterada. Quanto ao número kappa, o tempo de 3 horas
foi o que proporcionou os menores valores, em todas as cargas enzimáticas utilizadas (Figura
5.25).

91
Branqueamento químico
15,5
16
16,5
17
17,5
0,5 1 1,5 2Carga enzimática (UI/g)
Vis
cosi
dade
(cP)
7,5
8
8,5
9
9,5
10
Nº k
appa
1 h viscosidade 2 h viscosidade 3 h viscosidade1 h kappa 2 h kappa 3 h kappa
Figura 5.25 - Viscosidade e número kappa em função da concentração de xilanase para os tempo de tratamento de 1, 2 e 3 horas para o branqueamento químico
Os resultados quanto à redução da viscosidade e seletividade após o branqueamento
químico nas diferentes cargas enzimáticas utilizadas estão apresentados na Tabela 5.8.
Tabela 5.8 - Resultados quanto à redução da viscosidade (%) e seletividade após o branqueamento químico
Redução da viscosidade (%)*
Tempo de reação (h) 2 UI/g 1,5 UI/g 1 UI/g 0,5 UI/g 0 4,15 3,32 2,83 1,84
0,5 2,96 3,57 2,31 2,38 1 4,88 4,59 2,04 2,28 2 5,48 5,86 2,35 2,48 3 6,79 6,73 3,00 3,02 5 6,46 6,58 3,83 3,67
Seletividade Tempo de reação (h) 2 UI/g 1,5 UI/g 1 UI/g 0,5 UI/g
0 0,40 0,37 0,12 0,06 0,5 1,35 1,17 1,06 1,03 1 1,50 1,34 1,10 1,02 2 1,61 1,32 1,13 1,08 3 1,74 1,53 1,12 1,06 5 1,43 1,30 1,09 1,05
*Valores referentes à diferença entre os ensaios e os controles.

92
Pelos resultados obtidos é possível observar que a seletividade foi maior quando foram
utilizadas as maiores cargas enzimáticas antes do branqueamento químico. Isso porque a xilanase
branqueia a polpa de uma forma indireta, agindo apenas sobre as polioses, embora tenha sido
observada uma pequena diminuição na viscosidade das polpas. Esses valores indicam que o pré-
tratamento com xilanase aumenta a deslignificação com uma melhor proteção dos carboidratos
(RONCERO et al., 2000). Assim ela apenas provoca um efeito indireto que auxilia o posterior
branqueamento com reagentes químicos. Esses resultados confirmam dados da literatura sobre a
xilanase (RONCERO et al., 2003; SALLES et al., 2005) e mostram que o extrato enzimático de C.
subvermispora apresenta um efeito biobranqueador significativo.
5.5. CORRELAÇÃO ENTRE CROMÓFOROS, NÚMERO KAPPA E LIBERAÇÃO DE AÇÚCARES REDUTORES
A correlação entre a liberação de cromóforos, principalmente a 237 nm, e compostos
hidrofóbicos (465 nm) e a redução no número kappa aliados a liberação de açúcares redutores
sugerem a dissociação do complexo lignina-carboidrato (LCC) das fibras da polpa (Figura 5.26).

93
Figura 5.26 - Cromóforos (237 e 465 nm) e açúcares redutores liberados e número kappa em função do tempo de tratamento (2 UI/g de xilanase)
Priorizando a redução do número kappa e a máxima liberação de compostos coloridos da
polpa e açúcares redutores é possível verificar que com a carga de xilanase de 2 UI/g, o tempo de
tratamento de 3 horas foi o que melhor aliou essas características, portanto, sendo o tempo de
tratamento mais eficiente. Maiores períodos de incubação não melhoram as propriedades da polpa.
0 1 2 3 4 50,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
18
19
20
21
22
23
24
11,6
11,8
12,0
12,2
12,4
12,6 465 nm 237 nm Açúcares redutores Nº kappa
237
nm, 4
65 n
m
Tempo de tratamento (h)
Açú
care
s re
duto
res
(g/L
)
Nº k
appa

94
6. CONCLUSÕES
Através dos resultados obtidos neste trabalho foi possível concluir que:
- Houve ação efetiva das hemicelulases presentes no extrato de C. subvermispora. Esse
fato pode ser evidenciado pelos dados referentes à liberação de açúcares redutores, à liberação de
substâncias cromóforas nos comprimentos de onda estudados e à diminuição do número kappa nas
polpas tratadas enzimaticamente, principalmente nas maiores cargas utilizadas.
- Acima de 12 horas de tratamento o extrato enzimático de C. subvermispora sofre uma
diminuição na sua ação hidrolítica sobre as polpas devido a uma possível inibição das enzimas
pelos produtos formados. Além disso, a redução da concentração e da disponibilidade das polioses
diminui a velocidade de hidrólise das polpas, portanto tempos de tratamento acima de 8 horas não
aumentam a extensão da hidrólise dos carboidratos da polpa.
- Os produtos obtidos da hidrólise das xilanas mostraram que as xilanases de C.
subvermispora apresentaram um padrão de endo-xilanase, estando esses resultados de acordo com
os dados encontrados na literatura.
- O fato das polioses presentes nas polpas kraft estarem mais condensadas e ligadas
quimicamente à lignina e à celulose proporcionou uma boa atuação tanto das xilanases, quanto das
enzimas acessórias presentes no extrato, permitindo altos graus de hidrólise quando utilizada a
carga de 2 UI/g.
- Comparando-se os resultados da viscosidade em função da concentração de xilanase é
possível verificar que a viscosidade é melhor preservada com as maiores cargas enzimáticas
utilizadas, ou seja, 1,5 e 2 UI/g.
- A carga enzimática que proporcionou as maiores reduções do número kappa foi a de 2
UI/g, sendo a diminuição máxima de 7,7% após o tratamento enzimático e de 10,9% após o
branqueamento químico, ambos com 3 h de tratamento. Após o branqueamento químico o efeito
na redução do número kappa em função da carga enzimática utilizada fica mais evidente,
confirmando as hipóteses do mecanismo de biobranqueamento da xilanase.
- Pelos resultados obtidos é possível observar que a seletividade foi maior quando foram
utilizadas as maiores cargas enzimáticas antes do branqueamento químico.
- A carga enzimática que permitiu uma melhor combinação entre a máxima redução no
número kappa, uma maior redução de cromóforos a 237 e 465 nm e uma maior liberação de

95
açúcares redutores foi a de 2 UI/g de xilanase com um tempo variando de 2 a 3 horas de
tratamento.
- A ação da enzima manganês peroxidase sobre polpas kraft não foi eficiente, mesmo
havendo uma alta atividade dessa enzima no extrato de C. subvermipora. Para que a enzima possa
atuar seria necessário o acréscimo de MnSO4 e pequenas quantidades de peróxido de hidrogênio, o
que poderia inativar outras enzimas presentes no extrato.

96
REFERÊNCIAS ANDREWARTHA, K. A., PHILIPS, D. R., STONE, B. A. Solution properties of wheat-flour arabinoxylans and enzymically modified arabinoxylans. Carbohydrate Research. v. 77, p. 191-204, 1979. AKTHAR M.; ATTRIDGE, M. C.; MYERS, G. C.; KIRK, T. K.; BLANCHETTE, R. A. Biomechanical pulping of loblolly pine chips with different strains of the whit-rot fungus Ceriporiopsis subvermispora. Tappi Journal. v. 75, n. 2, p. 105-109, 1992. AKTHAR M., ATTRIDGE, M. C., MYERS, G. C., BLANCHETTE, R. A. Biomechanical pulping of loblolly pine chips with selected white-rot fungi. Holzforschung. v.47, p. 36-40, 1993. ASSOCIAÇÃO BRASILEIRA DE CELULOSE E PAPEL (Bracelpa). Disponível em: http://www.bracelpa.org.br/br/estudantes/floresta/socioeconomico.htm. Acessado em: 07/08/2006. BAILEY, M. J.; BIELY, P.; POUTANEN, K. Inter-laboratory testing of methods for assay of xilanases activity. Journal of Biotechnology. v. 23, p. 257-270, 1992. BAJPAI, P., BAJPAI, P. K. Application of xylanases in prebleaching of bamboo kraft pulp. Tappi Journal. v. 79, n. 4, p. 225-230, 1996. BARAZNENOK, V. A., BECKER, E. G., ANKUDIMOVA, N. V., OKUNEV, N. N. Characterization of neutral xylanases from Chaetomium cellulolyticum and their biobleaching effect on eucalyptus pulp. Enzyme and Microbial Technology. n. 25, p. 651–659, 1999. BARISKA, M., PIZZI, A. The interaction of polyflavonoid tannins with wood cell walls. Holzforschung. v. 40, p. 299-302, 1986. BEG, Q. K.; KAPPOR, M.; MAHAJAN, L.; HOONDAL, G. S. Microbial xylanase and their industrial applications: a review. Applied Microbial Biotechnology. v. 56, p. 326-338, 2001. BEG, Q. K.; BHUSHAN, B.; KAPPOR, M.; HOONDAL, G. S. Enhanced production of a thermostable xylanase from Streptomyces sp. QG-11-3 and its application in biobleaching of eucalyptus kraft pulp. Enzyme Microbiology Technology. v.27, p. 459-466, 2000. BIELY, P. Microbial xylanolitic systems. Trends in Biotechnology. v. 3: 286-290, 1985. BIERMANN, C. J. Handbook of Pulping and Papermaking, San Diego: Academic Press, 1996. 754 p. BIERMANN, C. J. Handbook of Pulping and Papermaking, San Diego: Academic Press, 1993. p. 123-136.

97
BISWAS, S. R.; JANA, S. C.; MISHA, A. K.; NANDA, G. Production, purification and characterization of xylanase from a hyperxylanolytic mutant of Aspergillus ochraceus. Biotechnology and Bioengineering. v. 35: 244-251, 1990. BRASILEIRO, L. B.; COLODETTE, J. L.; PILÓ-VELOSO, D. A utilização de perácidos na deslignificação e no branqueamento de polpas celulósicas. Quim. Nova. v. 24, n. 6, p. 819-829, 2001. BRILLOUET, J. M.; MOULIN, J. C.; AGOSIN, E. Production, purification and properties of a α-L-arabinofuranosidase from Dichomitus squalens. Carbohydrate Research. v. 144, p. 113-126, 1985. BUCHERT, J.; CARLSSON, G.; VIIKARI, L.; STRÖM, G. Holzforschung. v. 50, p. 69-74, 1996. CABALLERO, N. E. D. Notícias do Instituto de Química, Unicamp, Campinas, 2005. CARVALHO, M. E. A. Produção de Lignina Peroxidase e sua Aplicação no Biobranqueamento de Pasta de Celulose. 2003. p. 29-36. Tese (Doutorado) - IQ/UFRJ Rio de Janeiro, 2003. CASTRO, H. F. Processos químicos industriais II (apostila 4). EEL-Usp, Lorena, 2004. Disponível em: http://dqisrv.dequi.eel.usp.br/heizir/. Acesso em: 30/06/2006. COLODETTE, J. L.; GOMIDE, J. L.; BATISTA, J. C. Efeito de aditivos na deslignificação com oxigênio de polpa kraft de eucalipto. O Papel, p. 25-35, Maio, 1990. CONVERSE, A. O.; WARE, W. On the reactivity of cellulosic substrates in enzymatic hydrolysis. In: IEA/BIOFOR WORKSHOP ON APPLICATIONS OF BIOTECHNOLOGY IN BIOENERGY SYSTEMS. 18-20 Out., 1994, Canada. COUGHLAN, M. P.; HAZLEWOOD, G. P. β-1,4-D-Xylan- degrading enzyme systems: biochemistry molecular biology and applications. Biotechnology Applied Biochemistry. v. 17, p. 259-289, 1993. COWLING, E.; KIRK, K. T. Properties of cellulose and lignocellulosics materials as substrate for enzymatic conversion processes. Biotechnol. Bioeng. Symp. v. 6, p. 95-123, 1976. CPRH. Roteiro Complementar de Licenciamento e Fiscalização: Indústria de Papel e Celulose. Recife, 1998. 95p. CHRISTOV, L. P.; PRIOR, B. A. Bleaching response of sulfite pulps to pretreatment with xylanase. Biotech. Progress. v. 13, p. 695-698, 1997. CUTTER, B.; GUYETTE, R. Anatomical, chemical and ecological factors affecting tree species choice in dendrochemistry studies. J Environ. Qual. v. 22, p. 611-619, 1993. DANIEL, G. Microview of under degradation by bacteria and fungi. In: Wood deterioration and preservation: advances in our changing world, Washington, 2003. v. 845, p. 34-72. ACS Symposium Series.

98
D’ALMEIDA, M. L. O. Composição química dos materiais lignocelulósicos. In: D’ALMEIDA, M. L. O. Papel e Celulose I: Tecnologia de Fabricação de Pastas Celulósicas. São Paulo: Instituto de Pesquisas Tecnológicas do Estado de São Paulo, 1978. p. 169-312. DANILAS, R. M. Branqueamento de pastas celulósicas. In: D’AMEIDA, M. L. O. Papel e Celulose I: Tecnologia de Fabricação de Pastas Celulósicas. São Paulo: Instituto de Pesquisas Tecnológicas do Estado de São Paulo, 1998. p. 169-312. DESCHAMPS, A. M. Microbial degradation of tannins and related compounds. Plant cell wall polymers. American Chemical Society, 1989. p. 559-567. DEKKER, R. F. H. The enzymatic hydrolysis of biomass. IN: SEMINÁRIO DE HIDRÓLISE ENZIMÁTICA DE BIOMASSA, 2. 1985, Maringá. DURÁN, N. Recentes Desenvolvimentos da Utilização de Enzimas na Indústria de Papel e Celulose. Campinas: Instituto de Química, Laboratório de Química Biológica, Universidade Estadual de Campinas, 1996. DURÁN, N. Proc. 5th. Braz. Symp. Chem. Lignins and Other Wood Comp. n. 5, p. 164, 1995. DYER, T. J. Elucidating the Formation and Chemistry of Chromophores during Kraft Pulping. 2004. University of Wisconsin-Stevens Point, Institute of Paper Science and Technology, Dissertation (Mastership), 2004. ELEGIR, G.; SYKES, M.; JEFFRIES, T. W. – Differential and synergistic action of Streptomyces endoxylanases in prebleaching of kraft pulps. Enzyme Microb. Technol. v. 17, p. 954, 1995. ELLIS, E. L. Cellular Ultra structure of Wood Plants. Ed. Syracuse, 1965. p. 181-189. ERIKSSON K. L. E. Biotechnology in the pulp and paper industry. Wood Sci. Technol. v. 24, p. 79-101, 1990. ERIKSSON, K. L. E. Biotechnology in Pulp and Paper Industry (Ed.). Advances in Biochemical Engineering Biotechnology. Berlin: Springer, 1997. v. 57. FARREL, R. L.; VIIKARI, L.; SENIOR, D. Enzyme treatments of pulp. In: Pulp Bleaching- Principles and Practice. Eds. Dence, C. W., Reeve, D. W. Atlanta, USA: Tappi Press, 1996. p. 363. FENGEL, D.; WEGENER, G. Wood Chemistry, Ultra structure, Reactions. New York: Walter de Gryuter, USA, 1989. 613p. FERRAZ, A.; GUERRA, A.; MENDONÇA, R. Molecular weight distribution of wood components extracted from Pinus taeda biotreated by Ceriporiopsis subvermispora. Enzyme and Microbial Technology. n. 33, p. 12-18, 2002. FERRAZ, A. Aplicações da Biotecnologia na produção de papel e celulose. In: LIMA, U. A.; AQUARONE, E.; BORZANI, W.; SCHMIDELL, W. (Eds.). Biotecnologia Industrial. São Paulo: Edgar Blucher, 2001. v.3, p. 465-484.

99
FERRAZ, A.; RODRIGUEZ, J.; FREER, J.; BAEZA, J. Estimating chemical composition of biodegraded pine and eucalyptus by DRIF spectroscopy and multivariate analysis. Bioresouce Tecnhology. v. 74, p.201-212, 2000. FERRAZ, A. Aplicações da biotecnologia na produção de papel e celulose. In: BORZANI, W.; ALMEIDA, U.; AQUARONE, E. (eds). Processos Enzimáticos e Fermentativos. São Paulo: Edgard Blücher, 1999. FURTADO, M. FIBRA CURTA EM ALTA FAZ SETOR DESENGAVETAR GRANDES PROJETOS. Disponível em: http://www.quimica.com.br/revista/qd386/papel_celulose1.htm. Acessada em: 11/08/2006. GARG, A. P.; McCARTHY, A. J.; ROBERTS, J. C. Biobleaching effect of Streptomyces thermoviolaceus xylanase preparations on birchwood kraft pulp. Enzyme and Microbial Technology. n. 18, p.261-267, 1996. GUPTA R.; BEG, Q, K.; LORENZ, P. Bacterial alkaline proteases: molecular approaches and industrial applications. Appl. Micobiol. Biotechnol. v. 59, p. 15-32, 2000. GUYETTE, R. P., HENDERSON, G. S., CUTTER, B. E. Reconstructing soil pH from manganese concentrations in tree-rings. Forest Sci. v. 38, n. 4, p. 727-737, 1992. HAKALA, T. K.; LUNDELL, T.; GALKIN, S.; MAIJALA, P.; KALKKINEN, N.; HATAKKA, A. Manganese peroxidases, laccases and oxalic acid from the selective white-rot fungus Physisporinus rivulosus grown on spruce wood chips. Enzyme and Microbial Technology. v. 36, n. 4, p. 461-468, 2005. HARAZONO, K .; KONDO, R.; SAKAI, K. Bleaching of Hardwood Kraft Pulp with Manganese Peroxidase secreted from Phanerochaete sordida YK-624. Applied and Environmental Microbiology. v. 60, n. 3, p. 4359–4363, 1994. HAROLD, H. B. J. An overview of biomechanical pulping research. In: Environmentally Friendly Technologies for the Pulp and Paper Industry. New York: John Wiley, 1998. p. 259-272. HARTLEY, R. D.; CLIVE, W. F. Phenolic constituents of plant cell walls and wall biodegradability. Plant cell wall polymers. American Chemical Society, 1989. p. 137-145. HEIDORNE F.; MAGALHÃES, P.; FERRAZ, A.; MILAGRES, A. F. Characterization of hemicellulases and cellulases produced by Ceriporiopsis subvermispora grown on wood under biopulping conditions. Enzyme and Microbial Technology. n.38, p. 436-442, 2006. HEIJNESSON, A.; SIMNSON, U.; WESTERMARK, U. Metal ion content of material removed from the surface of unbleached kraft fibres. Holzforschung. v. 49, n. 1, p. 75-80, 1995. HON, D. N. S., SHIRAISHI, N. Wood and Cellulosic Chemistry. New York: Marcel Deckker, 2001. 914p. HORTAL, J. A. G.; LLUCIÁ, T. V. Branqueamento de pastas na indústria papeleira. Rio de Janeiro: Tecnografic, 1984.

100
INSTITUTO DE PESQUISAS TECNOLÓGICAS DO ESTADO DE SÃO PAULO, IPT. Celulose e Papel. Tecnologia de Fabricação da Pasta Celulósica. São Paulo, 1988. JANES, R. L. The chemistry of wood and fibers. In: MACDONALD, R. G.; FRANKLIN, J. N. The Pulping of Wood. New York: McGraw-Hill, 1969, p 33. JEFFRIES, T. W. Conserved Motifs in Xylanases for Pulp Bleaching. USDA: Forest service, 2001. JEFFRIES, T. W.; CHOL, S.; KIRK, T. K. Nutritional regulation of lignin degradation by Phanerochaete chrysosporium. Appl. Environ. Microbiol. v.42, n. 2, p. 290-296, 1990. JOHN, M.; SCHMIDT, J. Xylanase and β-xylosidase of Trichoderma. Methods in Enzymology. v. 160, p. 662-671, 1988. KANTELINEN, A. Enzymes in bleaching of kraft pulp. 1992. 86 p. Dissertation (Doctor of Technology) - Helsinki University of Technology, Espoo, Finland. 1992. KANTELINEN, A.; RANUA, M.; RATTO, M.; VIIKARI, L.; SUNDSQUIST, J.; LINKO, M. Hemicellulases and Their Potential Role in Bleaching. In: TAPPI INTERNATIONAL PULP BLEACHING CONFERENCE. Proceedings. Tappi Press, Atlanta, 1988. p. 1. KITPREECHAVANICH, V.; HAYASHI, M.; NAGAI, S. Purification and characterization of extracellular β-xilosidase and β-glicosidase from Aspergillus fumigatus. Agricultural and Biological Chemistry. v. 50, n. 7, p. 1703-1711, 1986. KOLLMAN, F. P.; COTÊ, W. A. Jr. Principles of Wood Sciences and Technology VY: Solid Wood. Springer-Verlag, 1984. 563p. KULKARNI, N.; SHENDYE, A.; RAO, M. Molecular and biotechnological aspects of xylanases. FEMS Microbiology Reviews. n. 23, p. 411-456, 1999. MAGALHÃES, P. O. Purificação de hemicelulases e de uma β-glicosidase de Ceriporiopsis subvermispora produzidas em condições de biopolpação. 2005. 113 p. Tese (Doutorado) - Faculdade de Engenharia Química de Lorena, 2005. MAGEE, R. J. E KOSARIC, N. Bioconversion of hemicellulosics: Advances in Biochemical Engineering and Biotechnology. n. 32, p. 61-93, 1985. MILLER, G. L., Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal Chem. v. 31, p. 426-428, 1959. MIMMS, A.; KOCUREK, M. J.; PYATTE, J. A.; WRIGHT, E. E. (Eds.). Kraft pulping: a compilation of notes. Atlanta: Tappi Press, 1993. MONTEIRO, M. C. Produção e utilização de complexos enzimáticos de diferentes fungos no biobranqueamento de polpa kraft. 1997. Dissertação (Mestrado) - Faculdade de Engenharia Química de Lorena, 1997.

101
MOREIRA, M. T.; SIERRA-ALVAREZ, R.; LEMA, J. M.; FEIJOO, G.; FIELD, J. A. Oxidation of lignin in eucalyptus kraft pulp by manganese peroxidase from Bjerkandera sp. strain BOS55. Bioresource Technology. n. 78, p. 71-79, 2001. NAVARRO, R. M. S. Estudo dos diferentes tipos de processos de branqueamento de celulose objetivando a comparação entre seus métodos e a geração do potencial de poluentes em seus respectivos efluentes. 2004. Dissertação (Mestrado) – Unicamp, Campinas, 2004. NUNN, J. R.; LINDE, J. V. D. The protective action of magnesium in oxygen bleaching of pulp. In: GRATZL, J. S., NAKANO, J., SINGH, R. P. Eds. Chemistry of delignification with oxygen, ozone and peroxides. Tokyo: Uni. Publ., 1980. p. 79-88. PATEL, P.; PAVITT, K. A technological competency in the world’s largest firms: characteristics, constraints and scope for managerial choice. University of Sussex. STEEP Discussion Paper, 1993. n. 13, SRPU. PAICE M. G.; GURNAGUL, N.; PAGE, D. H.; JURASEK, L. Mechanism of hemicellulose directed prebleaching of kraft pulps. Enzyme and Microbial Technology. v. 14 (4), p. 272-276, 1992. PERISSOTTO, D. O. Estudo da composição química do extrato em dioxano:HCl da polpa kraft de Eucalyptus grandis, após reações de branqueamento. 2000. Dissertação (Mestrado) - Universidade Federal de Viçosa, 2000. POUTANEN, K.; SUNDBERG, M.; KORTE, H.; PULS, J. Deacetylation of xylans by acetyl esterases of Trichoderma reesei. Applied Microbiology and Biotechnology. v. 33, p. 506-510, 1990. POUTANEN, K.; BAILEY, M.; SUNDBERG, M.; RATTO, M.; PULS, J. Enzymatic solubilization of xylans. In: RALEIGH (Ed). Biotechnology in pulp and paper industry. 1989. p. 559-565. POUTANEN, K., SUNDBERG, M. An acetyl esterase of Trichoderma reesei and its role in the hydrolysis of acetyl xylans. Applied Microbiology and Biotechnology. v. 28, p. 419-424, 1988. RAPSON, W. H. Mechanisms of Groundwood Bleaching. Appita. Set, 1969. REDKO, B. V. P. Aspectos do Comportamento do Eucalipto e da Bétula frente aos Agentes de Branqueamento. In: CONGRESSO ANUAL DA ABCP, 1978, São Paulo. REILLY, P. J. Xylanases: structure and function. In: Alexander Hollaender. Trends in the Biology of Fermentations for Fuels and Chemicals. p. 111-129, 1981. ROCHA, G., J. M. Deslignificação do bagaço de cana-de-açúcar assistida por oxigênio. 2000. Tese (Doutorado). Universidade de São Paulo/Instituto de Química de São Carlos, São Carlos, 2000. p. 40-41. RONCERO, M. B.; PASTOR, F. I. J.; VIDAL, T.; GALLARDO, O.; DIAZ, P.; COLOM, J. F.; TORRES, A. L. Novel xylanases for ECF bleaching of oxygen delignified Eucalyptus kraft pulps.

102
Influence on permanence and on refining properties. In: CONFERENCE ON PAPER AND PULP INDUSTRY, 9º. 2000. p. 186. RUTTIMANN-JOHNSON, C.; SALAS, L.; VICUÑA, R.; KIRK, K. Extracellular enzyme production and synthetic lignin mineralization by Ceriporiopsis subvermispora. Applied and Environmental Microbiology. v. 59, p. 1792-1797, 1993. SETLIFF E. C.; MARTON, R.; GRANZOW, S. G.; ERIKSSON, K. L., Biomechanical pulping with white-rot fungi. Tappi Journal. v.73, n. 8, p. 141-147, 1992. SETHURAMAN, A.; AKIN, D. E.; ERIKSSON, K. E. L. Plant cell wall degrading enzymes produced by the white-rot fungus Ceriporiopsis subvermispora. Biotechnology Applied Biochemistry. v. 27, p. 37-47, 1998. SJÖSTRÖM, E. Wood chemistry, fundamentals and applications. San Diego: Academic Press, 1993. 293 p. SILVA, O. F. Propriedades Ópticas de Pastas Celulósicas. São Paulo, 1986. Apostila do Curso de Especialização em Celulose e Papel. SINGH, R. P. The Bleaching of Pulp. 3ed. Atlanta: Tappi Press, 1979. SINGH, R. P.; DILLNER, B. C. Oxygen bleaching. In: SINGH, R. P. Ed. The bleaching of pulp. 3ed. Atlanta: Tappi Press, 1979. p. 159-210. SMITH, D. C.; FORSBERG, C. W. α-glucuronidase and other hemicellulase activies of Fibrobacter succinogenes S85 grown on crystalline cellulose or ball-milled barley straw. Applied and Environmental Microbiological. v. 57, p. 3552-3557, 1991. SOUZA-CRUZ, P. B. Estudo das enzimas produzidas por Ceriporiopsis subvermispora (Pilát) Gilbn. e Ryv. Durante a Biodegradação de Pinus taeda L. 2002. Dissertação (Mestrado) - Faculdade de Engenharia Química de Lorena, 2002. SUBRAMANIYAN, S.; PREMA, P. Biotechnology of microbial xylanase enzymology, molecular biology and application. Critical Reviews in Biotechnology. v. 22, n. 1, p. 33-64, 2002. SUNNA, A. E.; ANTRANIKIAN, G. Xylanolitic enzymes from fungi and bacteria. Cr. Rev. Biotechnol. v. 17, n. 1, p. 39-67, 1997. SUNDBERG, M.; POUTANEN, K.; MARKKANEN, P.; LINKO, M. An extracellular esterase of Aspergillus awamori. Biotechnology and Applied Biochemistry. v. 12, p. 670-680, 1990. SUNDBERG, M.; POUTANEN, K. Purification and properties of two acetylxylan esterases of Trichoderma reesei. Biotechnology and Applied Biochemistry. v. 13, p. 1-11, 1991. SUURNÄKKI, A.; TENKANEN, M.; BUCHET, J.; VIIKARI, L. Hemicellulases in the bleaching of chemical pulps. In: ADVANCES IN BIOCHEMICAL ENGINEERING BIOTECHNOLOGY. Scheper, T. (Ed.), 1997. p. 262-290.

103
SUURNÄKKI, A.; CLARK, T. A.; ALLISON, R. W.; VIIKARI, L.; BUCHERT, J. Journal Tappi. v. 79, n. 7, p. 111-117, 1996. TAN, L. U. L.; MYERS, P.; SADDLER, J. N. Purification and characterization of a thermostable xylanase from a thermophilic fungus Thermoascus aurantiacus. Canadian Journal of Microbiology. v. 33, n. 8, p. 689-692, 1987. TAKENISHI, S.; TSUJISAKA, Y. Purification and some properties of three xylanases from Penicillium janthinellum. Journal of Fermentation Technology. v. 51, n. 7, p. 458-468, 1973. TAPPI -Technical Association of the Pulp and Paper Industry. TAPPI Standard Methods. T 230 om-82. 1982. TAPPI - Technical Association of the Pulp and Paper Industry TAPPI Standard Methods. T 236 cm-85. 1985. TECHAPUN, C.; POOSARAN, N.; WATANABE, M.; SASAKI, K. Thermostable and alkaline-tolerant microbial cellulose free xylanases produced from agricultural wastes and the properties required for use in pulp bleaching processes: a review. Process Biochem. n.38, p. 1237-1340, 2003. TIMELL, T. E. Wood hemicelluloses: Part II. Advances in Carbohydrate Chemistry and Biochemistry. n. 19, p. 409-483, 1965. TENKANEN, M.; PULS, J.; POUTANEN, K. Two major xylanases of Trichoderma reesei. Enzyme Microbial Technology. v. 14, p. 566-574, 1992. THOMPSON, N. S. Hemicellulose as a biomass resource. In: Soltes, J. (ed) Journal Wood and Agricultural Residues: research on use for feed, fuels and chemicals. New York: Academic Press, 1983. p. 101-119. VAN DER BURGT, T.; TOLAN, J. S.; THIBAULT, L. C. US kraft mills lead in xylanase implementation. Tappi Fall Conference & Trade Fair. LOGEN Corporation. Ontario: 2002. VIIKARI, L.; RANUA, M.; KANTELINEN, A.; LINKO, M.; SUNDQUIST, J. In: INTERNATIONAL SYMPOSIUM ON WOOD AND PULPING CHEMISTRY NOTES, ATIP, 1987, Paris. n. 3, p.151. VIIKARI, L.; RANUA, M.; KANTELINEN, A.; SUNDQUIST, J.; LINKO, M. Bleaching with enzymes. In: INTERNATIONAL CONFERENCE ON BIOTECHNOLOGY IN THE PULP AND PAPER INDUSTRY, 16-19 Jun., 1986, Stocolmo. WONG, K. K.; JONG, E. D.; SADDLER, J. N.; ALLISON, R. W. (a) Mechanisms of xylanase aided bleaching of kraft pulp. Part 1: Process parameters. Appita Journal. v. 50, n. 5, 1997. WONG, K. K.; JONG, E. D.; SADDLER, J. N.; ALLISON, R. W. (b) Mechanisms of xylanase aided bleaching of kraft pulp. Part 2: Target substrates. Appita Journal. v. 50, n. 5, 1997. WONG, K. K. Y.; SADDLER, J. N. Trichoderma xylanase, their properties and applications. Critical Reviews in Biotechnology. v. 12, n. 5/6, p. 413-435, 1992.

104
WOOD, T. M.; SADDLER, J. N. Increasing the availability of cellulose in biomass materials. Methods Enzymol. v. 160, p. 3-11, 1988. WOODWARD, J. Xylanases: Functions, properties and applications. In: WISEMAN, A. Topics in Enzyme and Fermentation Biotechnology. p. 9-30, 1984. YOUNG, R. A.; AKHTAR, M. In: Developments in organosolv pulping- an overview. In: ENVIRONMENTALLY FRIENDLY TECHNOLOGIES FOR THE PULP AND PAPER INDUSTRY. New York: John Wiley, 1998. p. 5-69.

105
APÊNDICES
Tabela A - Viscosidade das polpas após o tratamento enzimático com as diferentes cargas enzimáticas utilizadas
Tempo de reação (h) Controle 2 UI/g 1,5 UI/g 1 UI/g 0,5 UI/g
0 24,8±0,20 24,8±0,80 24,8±0,80 24,8±0,80 24,8±0,80
0,5 21,8±0,10 22,6±0,04 22,6±0,02 22,4±0,04 22,3±0,02
1 21,7±0,03 22,4±0,04 22,4±0,01 22,2±0,03 22,1±0,01
2 21,1±0,01 22,2±0,01 22,0±0,01 21,8±0,02 21,6±0,01
3 21,0±0,01 22,1±0,02 21,9±0,01 21,8±0,03 21,5±0,02
5 20,9±0,07 21,9±0,05 21,7±0,02 21,6±0,04 21,4±0,02
Tabela B - Viscosidade das polpas após o branqueamento químico com as diferentes cargas enzimáticas utilizadas
Tempo de reação (h) Controle 2 UI/g 1,5 UI/g 1 UI/g 0,5 UI/g
0 20,9±0,07 21,9±0,05 21,7±0,02 21,6±0,04 21,4±0,02
0,5 15,3±0,05 16,7±0,02 16,7±0,03 16,3±0,01 16,1±0,02
1 15,2±0,03 17,0±0,01 16,8±0,01 16,2±0,01 16,0±0,02
2 15,1±0,02 17,1±0,06 17,0±0,02 16,1±0,01 16,0±0,01
3 14,9±0,03 17,1±0,04 17,0±0,01 16,1±0,01 15,9±0,02
5 14,7±0,07 16,8±0,03 16,7±0,01 16,0±0,03 15,8±0,01

106
Tabela C - Número kappa das polpas após o tratamento enzimático com as diferentes cargas enzimáticas utilizadas
Tempo de reação (h) Controle 2 UI/g 1,5 UI/g 1 UI/g 0,5 UI/g
0 12,9±0,01 12,9±0,20 12,9±0,20 12,9±0,10 12,9±0,20
0,5 12,9±0,05 12,5±0,30 12,6±0,40 12,8±0,50 12,9±0,25
1 12,8±0,30 12,3±0,20 12,5±0,10 12,8±0,01 12,8±0,50
2 12,8±0,10 11,9±0,20 12,2±0,50 12,7±0,40 12,6±0,25
3 12,8±0,20 11,8±0,20 12,0±0,40 12,5±0,30 12,5±0,40
5 12,8±0,20 12,1±0,30 12,3±0,20 12,6±0,40 12,8±0,20
Tabela D - Número kappa das polpas após o branqueamento químico com as diferentes cargas enzimáticas utilizadas
Tempo de reação (h) Controle 2 UI/g 1,5 UI/g 1 UI/g 0,5 UI/g
0 12,8±0,20 12,3±0,20 12,5±0,20 12,5±0,05 12,7±0,20
0,5 9,6±0,30 8,8±0,05 9,4±0,03 9,5±0,20 9,6±0,20
1 9,5±0,10 8,6±0,10 9,0±0,10 9,3±0,10 9,6±0,08
2 9,6±0,10 8,3±0,10 9,2±0,10 9,2±0,10 9,4±0,09
3 9,4±0,05 8,0±0,10 8,6±0,10 9,2±0,05 9,4±0,20
5 9,4±0,10 8,6±0,10 9,0±0,10 9,2±0,20 9,4±0,10