ufrrj instituto de florestas programa de pÓs …r1.ufrrj.br/wp/ppgcaf/wp-content/uploads/danilo...
TRANSCRIPT

UFRRJ
INSTITUTO DE FLORESTAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
AMBIENTAIS E FLORESTAIS
DISSERTAÇÃO
FENOLOGIA REPRODUTIVA E ESTRUTURA POPULACIONAL DE
BROMELIA ANTIACANTHA EM UMA ÁREA DE RESTINGA DA ILHA
DA MARAMBAIA - MANGARATIBA / RJ.
DANILO BRAGA DE AZEVEDO
2013

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE FLORESTAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
AMBIENTAIS E FLORESTAIS
FENOLOGIA REPRODUTIVA E ESTRUTURA POPULACIONAL DE
BROMELIA ANTIACANTHA EM UMA ÁREA DE RESTINGA DA ILHA
DA MARAMBAIA - MANGARATIBA / RJ.
DANILO BRAGA DE AZEVEDO
Sob a orientação do Professor
André Felippe Nunes-Freitas
Dissertação submetida como requisito
parcial para obtenção do grau de
Mestre em Ciências, no Programa de
Pós-Graduação em Ciências Ambientais
e Florestais, Área de Concentração em
Conservação da Natureza.
Seropédica, RJ
Março de 2013

iii
UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE FLORESTAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS AMBIENTAIS E
FLORESTAIS
DANILO BRAGA DE AZEVEDO
Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Ciências,
no Programa de Pós-Graduação em Ciências Ambientais e Florestais, área de Concentração
em Conservação da Natureza.
DISSERTAÇÃO APROVADA EM /03/2013
_____________________________________________
André Felippe Nunes-Freitas. Prof. Dr. UFRRJ
(Orientador)
______________________________________________
Michelle Cristina Sampaio. Prof.ª Dr.ª UNIRIO
(Membro)
_______________________________________________
Rita de Cássia Quitete Portela. Prof.ª Dr.ª UFRJ
(Membro)

iv
Se eu pudesse deixar algum presente a você,
Deixaria aceso o sentimento de amar a vida dos seres humanos.
A consciência de aprender tudo o que foi ensinado
pelo tempo a fora. Lembraria os erros que foram cometidos para que
não mais se repetissem. A capacidade de escolher
novos rumos. Deixaria para você, se pudesse, o respeito àquilo que
é indispensável. Além do pão, o trabalho. Além do
trabalho, a ação. E, quando tudo mais faltasse, um segredo: o de buscar no
interior de si mesmo a resposta e a força para encontrar a saída.
Mahatma Gandhi

v
DEDICATÓRIA
Dedico este trabalho a minha família.
Em especial à minha Mãe e meu Pai.
Fontes de amor e confiança.

vi
AGRADECIMENTOS
A Deus, por estar sempre presente em minha vida e guiar os meus passos e minhas
escolhas.
A meus pais, pelos inúmeros ensinamentos que fizeram e fazem parte de minha
formação. Pela confiança que sempre depositaram em minhas escolhas e por estarem sempre
presentes em minha vida, pelas palavras certas nas horas certas.
A minha irmã, Geane. A todos os meus familiares por estarem presentes em minha
vida.
Ao Professor André, querido orientador. Pelos seus ensinamentos durantes estes anos
de estudos e pela amizade.
As professoras que aceitaram o convite para compor esta banca Prof.ª Alexandra Pires,
Prof.ª Dulce Mantuano, Prof.ª Michelle Sampaio e Prof.ª Rita Portela.
A Marinha do Brasil e ao Centro de Avaliação da Ilha da Marambaia (CADIM) pelo
apoio fornecido durante os anos de pesquisa na Ilha da Marambaia.
Ao Prof. Roberto de Xerez, por ser sempre gentil e atencioso e por viabilizar as
entradas na Marambaia.
A Carolina Porto, em nome da CSA - Estação Santa Cruz, por disponibilizar os dados
climáticos utilizados neste estudo.
Aos colegas do Laboratório de Ecologia Florestal e Biologia Vegetal (LEFBV), Aline,
Arthur, Bianca, Isabela, Fernanda, Juan, Kelly, Poliana, Raoni, Rafael, Tereza, Tiago, por
dividirem experiências, artigos, trabalhos de campo e a curiosa convivência no Laboratório. A
Thereza Christina, pelos conselhos sempre valiosos.
Ao Alexandre Meireles, por me convidar para minhas primeiras idas a campo para
coletar dados de fenologia e visitantes florais na Marambaia, me mostrando este mundo
fascinante.
Aos “marambaieiros” pela companhia durante as idas a campo.
A Bianca, grande amiga com quem dividi bons momentos em campo e no laboratório.
Pela amizade e conversas sempre nos momentos certos e pelas inúmeras situações engraçadas
que tornaram as idas a campo e as atividades diárias mais felizes. Ao Arthur, pelo auxílio em
campo, pela amizade e pelas idéias e conversas neste período de mestrado. A Fernanda, pela
amizade iniciada no mestrado e pelas conversas sempre animadoras sobre nossos trabalhos.
Aos moradores do Alojamento M5 - Quarto 513, a minha casa durante os anos de
2006 a 2010, por se tornarem verdadeiros amigos!
Aos moradores da República em Seropédica, pela amizade e pelos bons momentos
vividos nesta casa. Ao André Lemos pela amizade e pelos conselhos sempre precisos.
Aos grandes amigos da Turma 2006-I de Engenharia Florestal.
Aos queridos amigos de Turma do PPGCAF 2011-2012, que deixaram estes anos de
Mestrado com boas lembranças para o futuro.
Em especial, a Gabriela (Gaby) por fazer parte da minha vida! Por entender a minha
ausência quando precisei dedicar exclusividade a este trabalho (e em outras vezes também).
Por ter a certeza de que fará parte de meu futuro!

vii
RESUMO
Azevedo, Danilo Braga de. Fenologia reprodutiva e estrutura populacional de Bromelia
antiacantha em uma área de restinga da Ilha da Marambaia - Mangaratiba / RJ. 2012.
xxp Dissertação (Mestrado em Ciências Ambientais e Florestais). Instituto de Florestas,
Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2012.
Bromelia antiacantha (Bromeliaceae) é uma espécie de habito terrestre e está amplamente
distribuída nas restingas do estado do Rio de Janeiro. Neste estudo, objetivamos analisar a
estrutura populacional, o padrão fenológico, os visitantes florais e as brotações da espécie em
uma área de restinga da Ilha da Marambaia, RJ. Verificamos diferenças entre as três estações
de amostragem da população apenas para o diâmetro médio das rosetas e para a distância em
que as parcelas se encontravam da faixa de areia. Bromelia antiacantha apresenta um ciclo
fenológico anual, com floração entre os meses de outubro a dezembro e frutificação entre os
meses de dezembro a junho, estando os frutos maduros a partir do mês de março. A emissão
de brotos ocorreu ao longo de todos os meses e a maioria dos indivíduos emite apenas um
broto por estação. Houve relação significativa apenas entre a fenofase de frutificação e as
variáveis climáticas umidade relativa e temperatura, entretanto estas não explicaram
fortemente a relação. Poucos indivíduos floresceram durante os anos de observação. O
processo de formação do escapo floral ocorreu em torno de seis dias e a floração ocorreu em
três dias. O número médio de flores produzidas foi de aproximadamente 73 flores e o número
de flores abertas por dia foi de 13 flores, as quais permaneciam disponíveis para visitação das
6:00 as 18:00 horas. Dentre as espécies de visitantes florais, insetos da ordem Hymenoptera
(abelhas e vespas) e beija-flores foram os principais visitantes, com ênfase para Trigona
spinipes (85% das visitas na estação de 2011) e Amazilia fimbriata (22% das visitas na
estação de 2012). A riqueza e a abundância de visitantes florais esteve relacionada a
luminosidade durante os períodos de observação. Os brotos apresentaram maiores taxas de
crescimento (altura da roseta) nos primeiros quatro meses de acompanhamento e maiores
valores de comprimento do estolão (distância da planta-mãe) nos primeiros seis meses de
acompanhamento, os quais eram menos vigorosos após este período. A população de B.
antiacantha apresenta-se em crescimento na área de estudo, atuando como uma espécie
colonizadora. O maior número de indivíduos com brotações em relação aos indivíduos
floridos indica o predomínio da propagação vegetativa da espécie na área de estudo o que
pode ser relacionado ao grau de perturbação da área e a rápida ocupação do ambiente.
Palavras chave: dinâmica populacional, fenologia reprodutiva e propagação clonal.

viii
ABSTRACT
Azevedo, Danilo Braga de. Fenology reproductive and structure population of Bromelia
antiacantha in restinga area of Marambaia Island - Mangaratiba / RJ. xxp. Dissertation
(Master Science in Environmental and Forest Science). Instituto de Florestas, Universidade
Federal Rural do Rio de Janeiro, Seropédica, RJ, 2012.
Bromelia antiacantha (Bromeliaceae) is terrestrial species and widely distributed in restingas
of state of Rio de Janeiro. In this study, we aimed to analyze population structure,
phenological pattern, flower visitors and shoots of species in area restinga of Marambaia
Island, RJ. We observed differences among three stations sampling population just the
average diameter of rosettes and to the distance at which the parcels were in the range of sand.
Bromelia antiacantha presents an annual phenological cycle, flowering in October to
December and fruiting between December to June, the ripe fruit in March. Emission of shoots
occurred over every month and most individuals emit only a shoots for season. There was
significant relationship only between the fruiting phenology and climate variables relative
humidity and temperature, however these did not explain the relationship strongly. Few
individuals flowering during years observation. The process of formation of floral scape
occurred around six days and flowering occurred in three days. The average number of
flowers produced was approximately 73 flowers and the number of open flowers per day was
13 flowers, which remained available for visitation from 6:00a.m to 6:00p.m. Among the
species of flower visitors, insects of the order Hymenoptera (bees and wasps) and
hummingbirds were the main visitors, with emphasis on Trigona spinipes (85% of visits in
the season 2011) and Amazilia fimbriata (22% of visits in season 2012). Richness and
abundance of floral visitors was related to light during observation periods. The shoots had
higher rates of growth (height rosette) in first four months of monitoring and higher values of
stolon length (distance from the parental plant) in the first six months of monitoring, which
were less vigorous after this period. Bromelia antiacantha populations showed up in growth
in the study area, acting as a colonizing species. The largest numbers of individuals with
shoots than flowering individuals indicate predominance of vegetative propagation of the
species in the study area which may be related to the degree of disturbance of the area and the
rapid occupation of the environment.
Key words: dynamics population, reproductive phenology and growth clonal.

ix
LISTA DE FIGURAS
Figura 1. Localização Ilha da Marambaia em relação ao estado do Rio de Janeiro
(Souza et al. 2007).
11
Figura 2. Diagrama Ombrotérmico de Gaussen-Bagnouls para a Restinga da
Marambaia, RJ. Período: 1986-1998. A linha pontilhada indica a variação de
temperatura (°C) e a linha contínua indica a variação de precipitação (mm).
Modificado a partir de Mattos (2005).
12
Figura 3. Localização da restinga da Praia Grande na Ilha da Marambaia,
Mangaratiba/RJ. Círculo vermelho indica a restinga da Praia Grande. Imagem
obtida no Google Earth em 15 de janeiro de 2012.
13
Figura 4. Vista total da Praia Grande e indivíduos de Bromelia antiacantha na
área de estudo. A - Forma geral da restinga da Praia grande; B - Indivíduos de
Bromelia antiacantha na borda da faixa de areia e C - próximos a borda da faixa
de areia (Fotos: Danilo Azevedo).
14
Figura 5. Indivíduos de Bromelia antiacantha na restinga da Praia Grande na Ilha
da Marambaia, Mangaratiba/RJ. A - Indivíduo de B. antiacantha com escapo em
formação (novembro de 2012) e B - Indivíduo em floração (dezembro de 2011); C
- Indivíduo com frutos imaturos (verdes, janeiro de 2011) e D - com frutos
maduros (amarelos, abril de 2011); E - Broto de B. antiacantha em frente a
planta-mãe (abril de 2011) (Fotos: Danilo Azevedo).
17
Figura 6. Representação esquemática da alocação das parcelas de amostragens da
estrutura populacional de Bromelia antiacantha na restinga da Praia Grande na
Ilha da Marambaia, Mangaratiba/RJ.
18
Figura 7. Variação na abundância de indivíduos de Bromelia antiacantha nas três
classes de distância da faixa de areia de parcelas amostradas em uma área de
Restinga na Ilha da Marambaia, Mangaratiba/RJ. Classes de distância: 1 = borda
da faixa de areia; 2 = 20 metros de distância e 3 = 40 metros de distância.
23
Figura 8. Variação na abundância de indivíduos de Bromelia antiacantha em
relação ao grau de abertura do dossel das parcelas amostradas em uma área de
Restinga na Ilha da Marambaia, Mangaratiba/RJ.
24
Figura 9. Variação no diâmetro médio das rosetas (cm) dos indivíduos de
Bromelia antiacantha em três estações de amostragem em uma área de Restinga
na Ilha da Marambaia, Mangaratiba/RJ. Estações 1 = Outubro/2011; 2 =
Abril/2012 e 3 = Outubro/2012. A linha pontilhada indica os valores médios de
comprimento foliar dos indivíduos reprodutivos e as duas linhas cheias indicam o
desvio padrão da média.
26
Figura 10. Variação no comprimento foliar (cm) dos indivíduos de Bromelia
antiacantha em três estações de amostragem em uma área de Restinga na Ilha da
Marambaia, Mangaratiba/RJ. Estações 1 = Outubro/2011; 2 = Abril/2012 e 3 =
27

x
Outubro/2012. A linha pontilhada indica os valores médios de comprimento foliar
dos indivíduos reprodutivos e as duas linhas cheias indicam o desvio padrão da
média.
Figura 11. Variação na altura das rosetas (cm) dos indivíduos de Bromelia
antiacantha em três estações de amostragem em uma área de Restinga na Ilha da
Marambaia, Mangaratiba/RJ. Estações 1 = Outubro/2011; 2 = Abril/2012 e 3 =
Outubro/2012. A linha pontilhada indica os valores médios de comprimento foliar
dos indivíduos reprodutivos e as duas linhas cheias indicam o desvio padrão da
média.
27
Figura 12. Proporção de indivíduos de B. antiacantha emitindo brotos entre os
meses de acompanhamento das fenofases em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ.
29
Figura 13. Número de brotos de B. antiacantha mensurados durante os meses de
acompanhamento do crescimento clonal em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ.
29
Figura 14. Número de brotos emitidos por indivíduo de Bromelia antiacantha em
uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
30
Figura 15. Variação na taxa de crescimento (A) e na distância da planta-mãe (B)
dos brotos emitidos por B. antiacantha ao longo dos meses de acompanhamento
em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
31
Figura 16. Relação entre a altura e a distância da planta-mãe dos brotos emitidos
por Bromelia antiacantha durante os meses de acompanahmento em uma área de
Restinga da Ilha da Marambaia - Mangaratiba/RJ.
32
Figura 17. Variáveis climáticas analisadas durante o período de estudo na Ilha da
Marambaia, Mangaratiba/RJ. A - Variação média mensal das umidade relativa
(%); B - Variação acumulada mensal da precipitação pluviométrica (mm); C -
Variação média mensal da temperatura (°C) e D - Variação média mensal da
Radiação Solar Global (W/m2).
33
Figura 18. Proporção de indivíduos de B. antiacantha em floração (A),
frutificação (B) entre os meses de acompanhamento das fenofases em uma área de
restinga na Ilha da Marambaia, Mangaratiba/RJ.
34
Figura 19. Relação entre a porcentagem de indivíduos de B. antiacantha em
frutificação e as variáveis climáticas em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ. A - Umidade Relativa (%) e B - Temperatura (°C).
35
Figura 20. Padrão geral de visitação da guilda de visitantes florais de Bromelia
antiacantha na área de restinga da Ilha da Marambaia, Mangaratiba/RJ.
38
Figura 21. Riqueza de espécies de visitantes florais de Bromelia antiacantha por
intervalo horário em uma área de restinga da Ilha da Marambaia, Mangaratiba/RJ.
38

xi
Figura 22. Frequência e atividade ao longo do dia do visitante floral Trigona
spinipes as flores de B. antiacantha em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ.
39
Figura 23. Frequência e atividade ao longo do dia do visitante floral Halictidae
sp. as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
40
Figura 24. Frequência e atividade ao longo do dia do visitante floral Bombus sp.
as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
41
Figura 25. Frequência e atividade ao longo do dia do visitante floral Euglossa
cordata as flores de B. antiacantha em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ.
41
Figura 26. Frequência e atividade ao longo do dia do visitante floral Amazilia
fimbriata as flores de B. antiacantha em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ.
42
Figura 27. Frequência e atividade ao longo do dia do visitante floral Florisuga
fusca as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
43
Figura 28. Frequência e atividade ao longo do dia do visitante floral fêmea de
Thalurania glaucopis as flores de B. antiacantha em uma área de restinga na Ilha
da Marambaia, Mangaratiba/RJ.
43
Figura 29. Frequência e atividade ao longo do dia do visitante floral macho de
Thalurania glaucopis as flores de B. antiacantha em uma área de restinga na Ilha
da Marambaia, Mangaratiba/RJ.
44
Figura 30. Frequência e atividade ao longo do dia do visitante floral Coereba
flaveola as flores de B. antiacantha em uma área de restinga na Ilha da
Marambaia, Mangaratiba/RJ.
44
Figura 31. Frequência e atividade ao longo do dia do visitante floral Lepidoptera
sp1 as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
45
Figura 32. Frequência e atividade ao longo do dia do visitante floral Lepidoptera
sp2 as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
45
Figura 33. Variação média das variáveis microclimáticas mensuradas durante a
observação de atividades dos visitantes florais as flores de Bromelia antiacantha
em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ. A -
Luminosidade (Lux); B - Temperatura (°C) e C - Umidade relativa (%).
47
Figura 34. Relação entre a variável microclimática luminosidade (Lux) e a 48

xii
riqueza (A) e abundância (B) de visitantes florais de Bromelia antiacntha em uma
área de restinga na Ilha da Marambaia, Mangaratiba/RJ.

xiii
LISTA DE TABELAS E QUADROS
Tabela 1. Parâmetros básicos relativos ao comprimento foliar, altura e diâmetro
médio dos indivíduos de Bromelia antiacantha nas três estações de amostragem
em uma área de restinga da Ilha da Marambaia, Mangaratiba/RJ. Letras diferentes
indicam valores estatisticamente significativos.
26
Tabela 2. Valores médios (±1 desvio padrão) das variáveis morfométricas de
Bromelia antiacantha na área de restinga da Ilha da Marambaia - Mangaratiba/RJ.
(N = 13 flores).
36
Tabela 3. Visitantes florais de Bromelia antiacantha em uma área de restinga da
Ilha da Marambaia, Mangaratiba/RJ, suas respectivas taxas de visitação durante a
estação reprodutiva de 2011 e de 2012, principal recurso obtido e papel na
polinização (polinizador/pilhador).
37

xiv
SUMÁRIO
LISTA DE FIGURAS
vii
LISTA DE TABELAS
ix
1. INTRODUÇÃO
1
2. REVISÃO DE LITERATURA
2
2.1 A Família Bromeliaceae
2
2.2 Estrutura Populacional em Bromeliaceae
4
2.3 Propagação vegetativa em Bromeliaceae
6
2.4 Fenologia Reprodutiva e Influencia de Fatores Abióticos
8
2.5 Visitantes florais em Bromeliaceae
9
3. MATERIAL E MÉTODOS
11
3.1 Área de Estudo
11
3.1.1 A Ilha da Marambaia
11
3.1.2 A restinga da Praia Grande
13
3.1.3 A espécie estudada: Bromelia antiacantha Bertol.
15
3.2 Metodologia
17
3.2.1 Caracterização da estrutura populacional
17
3.2.2 Crescimento clonal
19
3.2.3 Fenologia Reprodutiva
20
3.2.4 Visitantes Florais
21
4. RESULTADOS
23
4.1 Caracterização da Estrutura Populacional
23
4.2 Crescimento clonal
28
4.3 Fenologia Reprodutiva
32
4.3.1 Variáveis Climáticas 32

xv
4.3.2 Fenologia
33
4.4 Visitantes Florais
35
4.4.1 Caracterização geral
35
4.4.2 Descrição dos visitantes florais
36
4.4.3 Variáveis microclimáticas
46
5. DISCUSSÃO
49
5.1 Caracterização da Estrutura Populacional
49
5.2 Crescimento clonal
50
5.3 Fenologia Reprodutiva
52
5.4 Visitantes Florais
53
6. CONCLUSÃO
57
7. REFERÊNCIAS BIBLIOGRÁFICAS 58

1
1 INTRODUÇÃO
A família Bromeliaceae possui 3082 espécies distribuídas em 58 gêneros (Luther,
2008), sendo predominantemente Neotropical e bem representada na Mata Atlântica do
sudeste do Brasil (BENZING, 2000; MARTINELLI et al., 2008).
Estudos sobre a demografia em populações naturais são importantes para caracterizar
a distribuição da população e o recrutamento dos indivíduos (DUARTE et al., 2007). Em
espécies vegetais os estudos fenológicos estão relacionados ao período e duração da floração,
da frutificação e da emissão foliar (ALMEIDA, 2000) e fatores climáticos como fotoperíodo,
temperatura, precipitação, umidade relativa e radiação solar podem afetar a fenologia das
plantas (BORCHERT et al., 2004). Aliado a estes fatores, o crescimento clonal também
permite o recrutamento de novos indivíduos, os quais são geneticamente idênticos a planta-
mãe e são subsidiados pela mesma durante a fase inicial de desenvolvimento (MANTUANO,
2008).
Bromelia antiacantha Bertoloni é uma espécie de hábito terrestre, com reprodução
sexuada e assexuada, nativa do Bioma Mata Atlântica, pertencente à família Bromeliaceae
(REITZ, 1983) e tipicamente encontrada nas restingas (ARAUJO & HENRIQUES, 1984). De
acordo com Pereira et al. (2001) a escolha prévia de espécies para estudos mais detalhados de
suas populações pode levar a preservação de espécies raras e ao aumento de conhecimento
sobre estratégias bem sucedidas na colonização de ambientes com solos pobres, como os de
restinga.
Estudos avaliando espécies vegetais em seu ambiente natural, abrangendo a estrutura
de sua população, comportamento fenológico, padrão de floração e as taxas de propagação
clonal são importantes para o entendimento da dinâmica destas populações, gerando
informações que poderão ser utilizadas para fomentar estratégias de conservação e de manejo
sustentável da espécie.
Este estudo possui como objetivos avaliar a estrutura populacional de Bromelia
antiacantha, descrevendo o seu padrão fenológico e relacioná-lo a variáveis climáticas,
descrever os visitantes florais da espécie para a área de estudo e verificar qual a taxa de
crescimento das brotações emitidas pela espécie. Dessa forma, visamos responder as seguintes
perguntas:
1- Qual a estrutura etária da população analisada?
2- Qual a proporção entre indivíduos jovens e adultos na população?

2
3- Qual o número de rametas e genetas na população estudada?
4- Qual a razão entre rametas e genetas?
5- Qual é o padrão fenológico exibido pela espécie na área de estudo?
6- Qual a proporção de indivíduos reprodutivos em relação aos indivíduos acompanhados?
7- Qual o grau de relação entre as fenofases e as variáveis climáticas analisadas?
8- Quais os visitantes florais registrados para a espécie? Dentre os visitantes, quais seriam
potenciais polinizadores?
9- Qual o comportamento dos visitantes durante a realização das visitas?
10- Qual é a média mensal de crescimento dos brotos?
11- Qual o tamanho de comprimento de folhas dos brotos em que estes se desligam da planta-
mãe e com quantos meses isto ocorre?
12- Qual a proporção entre brotos e indivíduos adultos da população?

3
2 REVISÃO DE LITERATURA
2.1 A Família Bromeliaceae
A família Bromeliaceae Juss. possui 3172 espécies distribuídas em 58 gêneros
(LUTHER 2008). Para a Mata Atlântica brasileira são registrados 31 gêneros, 803 espécies e
150 táxons infra-especificos (MARTINELLI et al., 2008).
As espécies de Bromeliáceas possuem um notável poder de adaptação a diferentes
condições ambientais, podendo ser encontrada em habitats muito variados (BENZING, 1980).
Sua plasticidade ecológica permite que algumas espécies ocupem os mais diversos ambientes,
podendo apresentar extensas distribuições geográficas (BENZING, 1980). Por outro lado,
algumas espécies limitam-se a áreas restritas ou isoladas devido à sua exigência de condições
ambientais particulares ou pelo isolamento geográfico, o que resultou em um considerável
endemismo na família (BENZING, 1980).
A família destaca-se como um dos principais componentes da flora e da fisionomia
dos ecossistemas brasileiros abrigando aproximadamente 36% das espécies catalogadas.
Possuem vários gêneros endêmicos, alguns deles encontrados exclusivamente na Floresta
Atlântica (MARTINELLI, 1994).
As bromélias podem possuir desde o hábito terrestre, passando pelo hábito saxícola,
ocupando acúmulos orgânicos retidos em fendas de rochas e sobre rochas nuas, até o hábito
epifítico (Benzing, 1980). Parte das espécies de bromeliáceas possui uma alta taxa de
reprodução vegetativa, o que faz com que muitas delas sejam encontradas em elevadas
abundâncias em alguns habitats (FREITAS, 2000).
A família está tradicionalmente subdividida em três subfamílias: Pitcairnioideae,
Tillandsioideae e Bromelioideae (BENZING, 2000). Estudos recentes fundamentados em
dados de filogenia molecular, com análises cladísticas baseadas na variação nucleotídica em
ndhF, um gene do genoma cloroplástico em Bromeliaceae, sugerem uma nova subdivisão
para a família (GIVNISH et al., 2007). Nesta nova subdivisão, Tillandsioideae e
Bromelioideae, que são subfamílias monofiléticas, são mantidas e Pitcairnioideae, que é
parafilética, deve ser recircunscrita, em quatro novas subfamílias (Brocchinioideae,
Lindmanioideae, Hechtioideae e Puyoideae) e duas ratificadas (Navioideae e Pitcairnioideae)
(GIVNISH et al., 2007).
A subfamília Bromeliodeae possui cerca de 30 gêneros e cerca de 430 espécies,
constituída por plantas terrestres, rupícolas e epífitas, geralmente herbáceas, variando de

4
plantas de pequeno a grande porte (LEME, 1997). Os representantes desta subfamília
apresentam caules geralmente contraídos, rizomas horizontais ou estolões, inflorescência
geralmente vistosa e folhas distribuídas em roseta, usualmente com bainha alargada na base,
propiciando a formação de um reservatório de água e nutrientes (Reitz, 1983), cujo papel
ecofisiológico é de grande importância, tanto na nutrição das bromélias, como em constituir
um micro ambiente onde habitam animais diversos.
2.2 Considerações sobre Estrutura Populacional em Bromeliaceae
Pesquisas sobre a estrutura de populações naturais se fazem necessários, pois no caso
de espécies que oferecem fontes de recursos econômicos, pode-se pensar em estratégias de
manejo sustentável, com a exploração sustentada de seus recursos e a conservação da espécie
em seu ambiente natural. Para que isto se torne possível, é necessária a existência de
pesquisas relacionadas a estas espécies que visem a contribuir e enriquecer os dados já
existentes sobre suas características (DUARTE et al., 2007).
A maior parte dos estudos de estrutura de populações em Bromeliaceae abrange a
caracterização da população em seus estágios de desenvolvimento aliados a outros
parâmetros, como por exemplo, produção de frutos (DUARTE et al., 2007), diversidade
genética (PAGGI et al. 2010), crescimento clonal (SAMPAIO et al., 2002; ROGALSKI et al.,
2007; MANTUANO & MARTINELLI, 2007).
No caso de três populações reófita Dyckia brevefolia, estudadas nas margens do Rio
Itajaí-Açu, em Santa Catarina por Rogalski et al. (2007), houve grande variação no número de
indivíduos por população e em sua distribuição em classes diamétricas, indicando diferenças
no tempo de chegada dos propágulos, tempo de colonização das áreas e disponibilidade de
substrato para colonização. Ainda segundo estes autores, o baixo número de plântulas da
espécie na área de estudo e o alto grau de agrupamento dos indivíduos indicam o predomínio
da reprodução assexuada em D. brevefolia.
Paggi et al. (2010) ao estudarem a estrutura populacional de indivíduos de Vriesea
gigantea em quatro classes de tamanho, plântulas, juvenis, adultos e adultos reprodutivos, em
um parque Estadual no Rio Grande do Sul, contabilizaram poucos indivíduos nas classes
adulto e adulto reprodutivos e a classe juvenil foi a mais numerosa, com indivíduos
provenientes de diferentes estações reprodutivas. Segundo os autores, esta espécie não possui

5
reprodução assexuada vigorosa e sua estrutura populacional é caracterizada por altas taxas de
recrutamento de sementes.
Mantuano e Martinelli (2007) ao descreverem a estrutura e dinâmica populacional de
Neoregelia cruenta em três microhabitats classificados como interior, borda e exterior de
moitas de vegetação, em uma área de restinga, identificaram que o tamanho das rosetas de N.
cruenta (comprimento foliar e área da roseta) é afetado pelos microhabitats, corroborando a
capacidade de resposta plástica da espécie a incidência de radiação luminosa, fator
caracterizado como estressante nos ambientes de restinga.
Santos et al. (2004) ao estudarem a variação no período de germinação de sementes de
Bromelia antiacantha revelaram diferenças entre as matrizes para todos os tempos de
germinação analisados, sendo mais pronunciadas após 35 dias de experimento. Segundo os
autores, estas diferenças permitem a formação de grupos germinativos distintos para Bromelia
antiacantha, permitindo que as plântulas desta espécie se adaptem a condições variadas e se
estabeleçam em ambientes contrastantes.
Segundo uma classificação proposta por Duarte et al. (2007) a partir de uma adaptação
da caracterização dos estádios de tamanho propostos por Reis et al. (1996) para uma espécie
de palmeira, indivíduos de Bromelia antiacantha com comprimento foliar menor ou igual a
20 cm foram consideradas plântulas, desde que não possuíssem vestígios que as
caracterizassem como brotações. Indivíduos que apresentaram vestígios da planta-mãe foram
denominados brotos. Os indivíduos com mais de 0,20m de comprimento de folha foram
considerados jovens quando não estavam em seu período reprodutivo e não apresentavam
características recentes de brotações, sendo subdivididos em jovem 1 e jovem 2, com 0,20 a
1,0 m e 1,00 a 2,0 m de comprimento foliar, respectivamente. Indivíduos com comprimento
foliar maior que 2,0m foram considerados adultos.
Filippon (2009) caracterizou o desenvolvimento vegetativo de Bromelia antiacantha
em uma área de Floresta Ombrófila Mista de vegetação secundária a partir de medidas de
comprimento médio foliar. Dos 88 indivíduos acompanhados pela autora, 20 foram
classificados como adultos, 22 eram jovens dois, 19 eram jovens um e 27 eram brotos. Quanto
ao recrutamento destes indivíduos, 28 foram recrutados para a classe imediatamente superior,
os quais foram provenientes, principalmente, da classe dos brotos, com maiores taxas de
incremento em comprimento médio de folhas.
Santos et al. (2004) sugerem que Bromelia antiacantha, através de uma estratégia
adaptativa, permite a entrada contínua e gradual das plântulas no ambiente. Tal fato favorece

6
seu estabelecimento em novos ambientes, pois a espécie pode continuar colonizando o
ambiente em que se encontra e ainda migrar para ambientes com alterações periódicas.
2.3 Propagação vegetativa em Bromeliaceae
As plantas podem apresentar reprodução sexuada e clonal concomitantemente e cada
uma delas pode ser mais favorável dependendo das condições ambientais existentes naquele
momento. O crescimento clonal em plantas resulta no recrutamento de novos indivíduos,
geneticamente idênticos, que tem a sua fase inicial de desenvolvimento subsidiada pelo
indivíduo parental (MANTUANO, 2008). Nas bromeliáceas a propagação clonal não é restrita
aos adultos e nem todos os adultos reprodutivos produzem clones (BENZING, 1980).
O crescimento clonal é tido como uma tendência geral nas espécies que habitam
ambientes heterogêneos (CAIN et al., 1996). A clonalidade é potencialmente ilimitada no
espaço e no tempo, sendo afetada apenas por fatores como competição, acúmulo de mutações
deletérias, fatores como clima e habilidade de dispersão (WIKBERG, 1995).
Em ambientes hostis (como as restingas), ou que sofreram algum tipo de perturbação
antrópica, como queimadas ou desmatamento, plantas que se reproduzem vegetativamente
possuem melhores chances de sobreviver, já que a reprodução assexuada garante aos
indivíduos formarem descendentes, escapando da esterilidade ou da falta de polinizadores. A
reprodução vegetativa, ou em módulos, tende, então, a resultar em um padrão de distribuição
agregado, por proporcionar um espalhamento da espécie sobre o substrato (PEREIRA &
ARAÚJO, 1995). O crescimento clonal possui papal fundamental nas restingas,
principalmente para determinadas espécies que não conseguem germinar na área nua, em
temperaturas superiores a 50°C (SAMPAIO et al., 2002).
Sendo assim, em ambientes heterogêneos, a reprodução clonal representa uma
alternativa para a persistência da população, bem como para a seleção, exploração e
colonização de novos microambientes (SAMPAIO et al., 2004).
Diferentes formas clonais representam estratégias adaptativas em habitats
heterogêneos (SONG & DONG 2002 apud GÜTSCHOW-BENTO et al., 2010). Quanto à
disponibilidade e distribuição espacial de recursos, dois tipos de crescimento clonal são
evidentes: falange, com conexões curtas entre os rametes, dispostos de forma compacta,
expandindo-se lentamente e permanecendo na área por longos períodos, e guerrilha, com
entrenós longos, de forma mais fragmentada, e com alta capacidade de penetração em novos

7
habitats, caracterizando um comportamento oportunista e agressivo (LOVETT-DOUST, 1981
apud GÜTSCHOW-BENTO et al., 2010).
A colocação seletiva de unidades vegetativas em áreas ricas em nutrientes ou
microáreas ideais pode permitir que plantas clonais escolham ativamente habitats para
crescimento futuro e reprodução (SALZMAN, 1985). De acordo com este autor, as
oportunidades para a seleção de habitat em populações naturais dependem da taxa de
crescimento do rizoma em um novo território. Uma importante questão relacionada a
ambientes com recursos distribuídos em manchas ou com flutuações sazonais é como estes
fatores afetam a estrutura e dinâmica da população (MANTUANO, 2008). Os estudos que
descrevem este fenômeno para espécies de reprodução clonal ainda são escassos,
especialmente para bromélias nos trópicos (SAMPAIO, 2004).
No caso de Bromelia antiacantha, na reprodução vegetativa (assexuada), a espécie
emite um ou mais brotos laterais logo após o termino do período floral (REITZ, 1983;
SANTOS et. al., 2004). Segundo Duarte (2007), para a maioria das bromeliáceas, incluindo a
B. antiacantha o final do ciclo reprodutivo representa também o final do ciclo de vida, onde a
espécie poderá produzir sementes e brotações.
Na vegetação Arbustiva Aberta de Clúsia na restinga de Jurubatiba, RJ, os conjuntos
de rosetas conectadas (estolão funcional) de Neoregelia cruenta podem ocorrer inteiramente
no interior das moitas de vegetação e sobre a areia sem vegetação. Nesta área, o tamanho das
rosetas e conexões em N. cruenta permite que um único conjunto conectado de rosetas possa
ocupar a borda das moitas de vegetação, local em que estão sujeitas à complementaridade das
condições ambientais do interior e exterior das moitas de vegetação (MANTUANO, 2008).
A espécie Pitcairnia geyskesii (L. B. Smith) é uma Bromeliaceae clonal que produz
novos brotos a partir do rizoma. De acordo com Sarthou et al. (2001) o florescimento desta
espécie induz a morte da roseta e leva a produção de dois brotos laterais pela reprodução
vegetativa e devido a progressiva perda da conecção física entre os rametes, a extensão do
crescimento clonal é difícil de ser documentada pela examinação direta.
Rogalski (2007) avaliou a propagação vegetativa em cinco populações de Dyckia
brevifolia Baker (Bromeliaceae), sendo verificadas as formas de emissão clonal apresentadas
pela espécie e o numero de clones axilares emitidos. Nestas populações, o recrutamento de
novas rosetas ocorreu através da entrada de plântulas (reprodução sexuada) e clones
(propagação vegetativa), e a emissão clonal ocorreu tanto em rosetas imaturas quanto
reprodutivas. A propagação clonal pode ser extremamente importante para D. brevifolia,

8
considerando que a espécie ocorre em ambiente com condições adversas (rocha exposta,
correnteza, períodos de seca e de cheia), tanto para a formação, devido ao efeito fundador
(aumentando rapidamente o tamanho das populações), como para a manutenção de suas
populações (ROGALSKI, 2007).
A bromélia epífita facultativa Aechmea nudicaulis (L.) Griseb. cresce como uma
planta terrestre clonal nas restingas, dentro e fora das ilhas de vegetação. Devido à alta
temperatura do solo (> 60°C em plena luz do sol) as sementes de A. nudicaulis não são
capazes de germinar na areia nua, sendo a germinação de sementes e o estabelecimento de
plântulas frequentemente relacionado ao interior das ilhas de vegetação, sugerindo que esta
espécie se propaga para as áreas abertas através do crescimento clonal (ZALUAR E
SCARANO, 2000). Sampaio et al. (2004) demonstraram que os fragmentos clonais de A.
nudicaulis crescendo no interior das ilhas de vegetação e na fronteira entre a areia nua e ilhas
de vegetação mostraram clara e significativa tendência de crescimento direcional para o
exterior das ilhas de vegetação.
2.4 Fenologia Reprodutiva e Influência de Fatores Abióticos
Uma parte essencial para o entendimento das relações planta-animal é o conhecimento
do padrão fenológico e do sistema de polinização da comunidade vegetal (TALORA &
MORELLATO, 2000) e de acordo com Negrele & Murato (2006) o conhecimento da
dinâmica das espécies vegetais é imprescindível para a elaboração de estratégias de
conservação e manejo.
Os estudos fenológicos analisam as fases do ciclo de vida dos vegetais a partir de
informações sobre a época e duração da floração, frutificação, dispersão e emissão foliar, as
quais podem ser relacionadas a fatores climáticos e ecológicos (ALMEIDA, 2000).
Fatores como a precipitação, o estresse hídrico, a irradiação e o fotoperíodo
influenciam o comportamento fenológico das plantas (BORCHERT et al., 2004). As
fenofases das plantas, além de estarem correlacionadas a fatores endógenos, também podem
variar em função das variáveis ambientais como, por exemplo, luz, temperatura e precipitação
(CRUZ et al., 2006).
Machado e Semir (2006) ao investigarem o padrão fenológico de floração de 14
espécies de Bromeliaceae ornitófilas da Mata Atlântica serrana de São Paulo, identificaram
períodos e picos de floração, para oito destas espécies, diferentes dos encontrados por três
outros autores em áreas também de Mata Atlântica. Segundo os autores, tais diferenças

9
poderiam indicar uma maior importância dos fatores ambientais para a fenologia destas
espécies.
A espécie epífita Vriesea incurvata estudada por Negrelle & Muraro (2006) em uma
planície litorânea paranaense, apresentou um padrão fenológico seqüencial e marcadamente
temporal, com floração iniciando em fevereiro, as primeiras brotações em maio, frutificação
em abril e dispersão a partir de setembro. Os autores evidenciaram correlação significativa
entre as fenofases floração, frutificação e emissão clonal e as variáveis fotoperíodo e
temperatura, as quais podem estar relacionadas a distribuição espacial das bromélias ser
afetada pela disponibilidade lumínica.
Meireles (2011) ao analisar a fenologia da floração e frutificação e a reprodução
vegetativa de Neoregelia cruenta e Vriesea procera na Ilha da Marambaia revelaram
correlações significativas entre as fenofases floração e frutificação e as variáveis temperatura
e precipitação para ambas as espécies, possivelmente relacionadas ao período chuvoso, em
que as condições ambientais são mais favoráveis ao desenvolvimento destas fenofases.
Rocha-Pessôa & Rocha (2008) ao analisarem a fenologia reprodutiva de Pitcairnia flammea
na floresta Atlântica da Ilha Grande e a influencia de fatores ambientais sobre os padrões de
floração e frutificação identificaram relação entre a floração e a temperatura e a frutificação e
a temperatura e fotoperíodo, os quais explicaram uma porção adicional da variação destas
fenofases.
2.5 Visitantes florais em Bromeliaceae
A família Bromeliaceae possui grande variedade de formas, estruturas e recursos
florais que durante o período de floração servem como importante fonte de alimento, atraindo
diversos tipos de polinizadores, tais como abelhas, vespas, mariposas, borboletas, beija-flores
e morcegos (ROCHA et al., 1997; ROCHA et al., 2004).
Apesar de as abelhas serem consideradas os principais agentes polinizadores nas
Angiospermas (BAWA, 1990), em Bromeliaceae, geralmente são consideradas visitantes
ocasionais e raros ou pilhadoras de recursos (SANTANA E MACHADO, 2010). No entanto,
uma maior taxa de visitação de espécies de abelhas (93,3%) às flores de Tillandsia stricta foi
verificada por Almeida et al. (2004) na restinga de Jurubatiba, RJ, evidenciando a importância
deste grupo como vetor de pólen para a espécie.

10
Segundo Machado & Semir (2006) as bromeliáceas são consideradas essenciais para
manter a fauna local de beija-flores da Mata Atlântica, beneficiando não somente estas aves,
como também a comunidade de plantas que as utilizam como vetores de pólen.

11
3. MATERIAL E MÉTODOS
3.1 Área de Estudo
3.1.1 A Ilha da Marambaia
A Ilha da Marambaia situa-se entre as coordenadas 23º 04’ S e 43º 53’ W, no litoral
Sul do estado do Rio de Janeiro, no Distrito de Itacuruçá, Município de Mangaratiba (Fig. 1).
Possui uma área de aproximadamente 42 km², ligando-se ao continente, na região de
Guaratiba, por uma faixa estreita de areia com cerca de 40 km de extensão, a Restinga da
Marambaia (PEREIRA et al., 1990). A Ilha da Marambaia propriamente dita é um trecho de
relevo montanhoso pertencente a restinga da Marambaia (MENEZES, 1996) cujo ponto
culminante é o Pico da Marambaia, com 641 m de altitude (PEREIRA et al., 1990).
Figura 1. Localização Ilha da Marambaia em relação ao estado do Rio de Janeiro (Souza et al.
2007).
A Ilha da Marambaia possui diferentes formações vegetais e sua diversidade de
ambientes pode ser relacionada a processos de sua formação, os quais originaram diferentes
feições geológicas (MENEZES E ARAÚJO, 2005). A maior parte da vegetação da Ilha é
composta por matas secundárias, com sub-bosque denso, mas também conserva uma
vegetação de restinga e de Floresta Ombrófila Densa Submontana (VELOSO et al., 1991).

12
O clima na região, segundo Köeppen, enquadra-se no macroclima Aw (tropical
chuvoso), com temperaturas do ar típicas de áreas litorâneas tropicais. As médias mensais da
temperatura do ar situam-se acima de 20,0°C e a média anual na região atinge 23,7°C, sendo
mais alta em fevereiro, com 26,8°C, e menor em julho, com 20,9°C (Menezes & Araujo,
2005; Mattos, 2005). A precipitação média anual é de 1239,7 mm, e praticamente 37% da
precipitação anual está relacionada com o período de verão, quando o número de dias
chuvosos está em torno de 40. Entre os meses de novembro e março os índices pluviométricos
médios são sempre superiores a 100 mm. O mês de março é o mais chuvoso com 140,6 mm.
Os meses de inverno são os mais secos, onde ocorrem apenas 15% da precipitação média
anual, distribuída em torno de 21 dias. Em julho e agosto a precipitação média mensal varia
de 40 a 55 mm distribuídos por um período de dois a quatro dias. Por diversos anos já foram
registrados valores abaixo de 30 mm nesta época do ano. Na primavera e outono onde o
número médio de dias de chuva em cada estação é praticamente igual e em torno de 21,
acontecem os 48% restantes do total de precipitação média anual. A relação entre a
temperatura do ar e a precipitação na área é evidenciada pelo diagrama ombrotérmico de
Gaussen-Bagnouls (Fig. 2) e caracteriza o clima da região como super-úmido, com pouco ou
nenhum déficit hídrico, em função da não existir um mês seco ao longo do ano (MENEZES &
ARAUJO, 2005; MATTOS, 2005).
Figura 2. Diagrama Ombrotérmico de Gaussen-Bagnouls para a Restinga da Marambaia, RJ.
Período: 1986-1998. A linha pontilhada indica a variação de temperatura (°C) e a linha contínua indica
a variação de precipitação (mm). Modificado a partir de Mattos (2005).

13
Desde 1614, quando se tem nota de sua primeira ocupação, e até meados de 1896, a
Ilha da Marambaia sofreu interferências de diferentes tipos e intensidades (PEREIRA et al.,
1990). Dentre suas utilizações cita-se a implantação de um entreposto negreiro (até meados de
1888), da Escola de Aprendizes de Marinheiros da Marinha do Brasil (1908), do Campo de
Provas do Exército (1933), da escola técnica de pesca, de programas de horticultura e
pecuária e de indústria para conserva de pescado (décadas de 40 e 50 do século XX) e desde
1981, a instalação do Centro de Adestramento e Instrução dos Fuzileiros Navais pela Marinha
do Brasil (ANTONINI, 2007). Na década de 1980, foi assinado um convênio entre a
Universidade Federal Rural do Rio de Janeiro (UFRRJ) e o Centro de Avaliação da Ilha da
Marambaia (CADIM), que fornece apoio logístico para a realização de pesquisas biológicas
por estudantes e professores da Universidade (ANTONINI, 2007).
3.1.2 A restinga da Praia Grande
Este estudo foi desenvolvido na área da Praia Grande, a qual está voltada para a face
oeste da Ilha da Marambaia, sofrendo influencias da Baía de Sepetiba por um lado e do
Oceano Atlântico por outro (Fig. 3).
Figura 3. Localização da restinga da Praia Grande na Ilha da Marambaia, Mangaratiba/RJ.
Círculo vermelho indica a restinga da Praia Grande. Imagem obtida no Google Earth em 15
de janeiro de 2012.

14
A Restinga da Praia Grande é recoberta por vegetação arbórea (Fig. 4a), cujo dossel
pode alcançar 13 metros e possui trechos onde as gramíneas predominam (OLIVEIRA, 2010).
O sub-bosque é pobre em plantas herbáceas, sendo dominado por espécies de Bromeliaceae
como, por exemplo, Vriesea procera, Neoregelia cruenta e N. johannis e, em menor
quantidade, por Bromelia antiacantha (MEIRELES, 2011), além de várias espécies de
Cactaceae e pela Leguminosae Sophora tomentosa L. Na Praia Grande, as espécies de
bromélias que possuem hábito terrestre formam abundantes agrupamentos (touceiras), com
poucos indivíduos ocorrendo isoladamente (MEIRELES, 2011). No trecho de vegetação
herbáceo-arbustivo (pós-praia) há o predomínio da espécie B. antiacantha, o que pode indicar
a sua tolerância, ainda que parcial, a níveis de salinização (Figs. 4b e 4c) (SANTOS et al.,
2004).
Figura 4. Vista total da Praia Grande e indivíduos de Bromelia antiacantha na área de estudo.
A - Forma geral da restinga da Praia grande; B - Indivíduos de Bromelia antiacantha na borda
da faixa de areia e C - próximos a borda da faixa de areia (Fotos: Danilo Azevedo).
A
B C

15
O solo da restinga da Praia Grande é arenoso, podendo apresentar-se exposto ou
recoberto por gramíneas (OLIVEIRA, 2010), principalmente nas áreas de clareiras, ou
recoberto por serrapilheira, que em alguns trechos forma uma densa camada sobre o solo de
aproximadamente 15 cm, como observado por Menezes & Araújo (2005) em outras áreas de
restinga com formação arbórea na Marambaia. Devido a sua localização nas proximidades do
centro urbano da Ilha da Marambaia e por fazer parte de uma área aberta a visitantes civis e
possuir algumas residências quilombolas vizinhas, a área de estudo apresenta sinais de ação
antrópica, como por exemplo, armadilhas de caça e presença de lixo doméstico (MEIRELES,
2011), sendo considerada uma área com forte influência antrópica (OLIVEIRA, 2010).
3.1.3 A espécie estudada: Bromelia antiacantha Bertol.
Bromelia antiacantha Bertol. (Fig. 5) é uma espécie de hábito terrestre pertencente à
Família Bromeliaceae, subfamília Bromeliodeae, que pode chegar a 2 metros de altura,
formando densos agrupamentos (“reboleiras”) e que apresenta características próprias de
estrutura espacial e de dinâmica populacional (Reitz, 1983; Santos, 2001). É uma espécie
nativa do Bioma Mata Atlântica, podendo habitar a Floresta Ombrófila Densa, a Floresta
Ombrófila Mista, as restingas e os afloramentos rochosos e cuja ocorrência abrange os estados
do Bahia, Minhas Gerais, Espírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina e
Rio grande do Sul (REITZ, 1983; MARTINELLI et al., 2008).
As numerosas folhas de B. antiacantha são dispostas em rosetas sem formar cisternas,
e são densamente cobertas de espinhos nas margens. Os espinhos são responsáveis pela
característica que lhe confere o nome, o qual vem do grego anti (contra) e acantha (espinhos)
em referência a disposição dos espinhos que na parte inferior da lâmina foliar estão voltados
para baixo e, ao contrário, na parte superior (REITZ, 1983).
A espécie realiza tanto reprodução sexuada quanto assexuada através da emissão de
uma ou mais brotações laterais logo após o período reprodutivo. Anteriormente ao
aparecimento da inflorescência a espécie apresenta brácteas vermelhas no centro de sua
roseta. Sua floração é anual, indo de dezembro a janeiro ou início de fevereiro (REITZ, 1983).
A frutificação ocorre de fevereiro a junho (REITZ, 1983) e seus frutos por serem comestíveis
e estarem disponíveis em grandes quantidades por um longo período do ano se tornaram um
importante recurso alimentar para a fauna (SANTOS et al., 2004).
Santos et al. (2004), baseados em seus dados, sugerem que B. antiacantha possui uma
estratégia adaptativa que permite a ocupação do ambiente através da entrada lenta e gradual

16
das plântulas. Este fato pode classificar a espécie como colonizadora, já que com essa
estratégia B. antiacantha pode ocupar habitats abertos e menos estáveis (SANTOS et al.,
2004).
Bromelia antiacantha apresenta características medicinais (com propriedades anti-
helmínticas, antitussígenas e no tratamento de cálculos renais), alimentícias, ornamentais e
industriais, como no caso do uso para fabricação de fibras para tecidos, cordoaria, o uso dos
frutos para a preparação de xaropes e para a produção de sabão. Segundo Reitz (1983) sua
utilização na medicina popular é descrita desde o ano de 1940. Cita-se ainda a sua utilização
como cercas vivas em comunidades agrícolas (BENATTO et al., s/d).
Bromelia antiacantha está amplamente distribuída nas restingas do estado do Rio de
Janeiro (ARAUJO & HENRIQUES, 1984). A ocorrência de B. antiacantha na zona arbustiva
fechada de pós-praia na restinga de Jurubatiba sugere uma maior tolerância da espécie à alta
salinidade (COGLIATTI-CARVALHO et al., 2001). Oliveira (2010) também destaca a
ocorrência de B. antiacantha exclusivamente nas unidades amostrais mais próximas à linha da
praia e, consequentemente, com os maiores teores de salinidade, chegando inclusive a sofrer a
ação das ondas em períodos de maré alta. De acordo com Scarano (2002) a comunidade de
plantas na periferia da Floresta Atlântica, incluindo as restingas, está sujeita as mais extremas
condições ambientais, tais como altas e baixas temperaturas, inundações, seca, ventos
constantes, alta salinidade e carência de nutrientes.
Nunes-Freitas et al. (2006) avaliaram a composição, abundância e similaridade da
família Bromeliaceae da restinga da Praia do Sul, na Ilha Grande. Os autores sugeriram que a
elevada salinidade, a maior ação dos ventos e a maior incidência luminosa na área de
vegetação fechada de pós- praia faz com que esta zona apresente condições menos propícias
para abrigar um elevado número de espécies, estando presentes apenas aquelas com maior
resistência a estes fatores, como por exemplo, B. antiacantha.
Bromelia antiacantha é uma planta de fácil cultivo, com alta viabilidade de sementes,
reprodução clonal e diferenças genéticas dentro e entre populações, o que torna extremamente
importante a conservação de suas populações (ZANELLA, 2009). Benatto (sem data), em seu
trabalho sobre assentamentos agroflorestais dirigidos pela sucessão natural em assentamentos
de reforma agrária, menciona a utilização da B. antiacantha em consórcio de espécies
pioneiras de hábito arbustivo e arbóreo que alcançam a maturidade e produtividade em um
período de três a sete anos. Este autor cita ainda que algumas espécies, como B. antiacantha,

17
possuem características ecológicas que as possibilitam persistir além da fase sucessional da
qual inicialmente desempenha função no sistema.
Figura 5. Indivíduos de Bromelia antiacantha na restinga da Praia Grande na Ilha da
Marambaia, Mangaratiba/RJ. A - Indivíduo de B. antiacantha com escapo em formação
(novembro de 2012) e B - Indivíduo em floração (dezembro de 2011); C - Indivíduo com
frutos imaturos (verdes, janeiro de 2011) e D - com frutos maduros (amarelos, abril de 2011);
E - Broto de B. antiacantha em frente a planta-mãe (abril de 2011) (Fotos: Danilo Azevedo).
3.2 Metodologia
3.2.1 Caracterização da estrutura populacional
Para a caracterização da estrutura populacional de B. antiacantha, estabelecemos na
área estudada um total de 50 parcelas permanentes com 100 m2 (10 x 10 metros) cada. As
parcelas situavam-se distantes entre si cerca de 10 metros, sendo amostradas três linhas com
A B
C D E

18
17, 16 e 17 parcelas, respectivamente (Fig. 6). A primeira linha localizava-se na borda da
faixa de areia, recebendo luminosidade direta. A segunda linha estava a 20 metros da borda da
faixa de areia e a terceira linha a 40 metros da borda da faixa de areia.
Mensuramos a abertura do dossel em cada parcela com a utilização de um densiômetro
de mão (%). Para os cálculos posteriores consideramos parcelas com valores inferiores a 40%
de abertura do dossel como áreas com elevada densidade e, consequentemente, com dossel
fechado, enquanto as parcelas com valores acima de 41% de abertura do dossel foram
consideradas como áreas abertas, com maior luminosidade. Verificamos se a abundância de B.
antiacantha nas três linhas de amostragem estava relacionada com o grau de abertura do
dossel e com a distância das linhas da faixa de areia através de Análise de Variância
(ANOVA) (Zar, 1999).
Contabilizamos todos os indivíduos da espécie dentro de cada parcela e tomamos as
seguintes medidas: comprimento foliar (mensuramos a maior folha periférica de cada
indivíduo, em centímetros), altura da roseta (em centímetros) e os diâmetros maiores e
menores da roseta (em centímetros). A partir dos valores de diâmetro maior e menor
calculamos o diâmetro médio para cada roseta.
Anotamos informações sobre indivíduos reprodutivos e pós-reprodutivos e a fenofase
em que se encontravam (florescendo ou frutificando, com frutos verdes ou maduros).
Também foram identificados os indivíduos que entraram na população por recrutamento ou
por brotamento e os indivíduos que saíram da população por morte. Para avaliar possíveis
mudanças na estrutura da população, as amostragens de estrutura populacional foram
realizadas ao final das estações secas (setembro/outubro de 2011 e de 2012) e da estação
chuvosa (março/abril de 2012).
Figura 6. Representação esquemática da alocação das parcelas de amostragens da estrutura
populacional de Bromelia antiacantha na restinga da Praia Grande na Ilha da Marambaia,
Mangaratiba/RJ.

19
Contabilizamos o número de indivíduos conectados por estolões a partir da presença
destas estruturas e o número de indivíduos com brotações. Visualizamos os estolões
superficiais e aqueles enterrados devido à deposição de substrato, os quais alcançamos a partir
de leve revolvimento manual do substrato próximo aos indivíduos.
Calculamos a densidade total da população, a densidade de indivíduos vegetativos,
reprodutivos e de brotações (todos os parâmetros em indivíduos/ha). Identificamos os
plântulas com base na classificação proposta por Duarte et al. (2007) e levando em
consideração o grau de isolamento destes indivíduos na área. Para identificarmos o tamanho
médio a partir dos quais os indivíduos poderiam ser considerados adultos, ou seja, que
estivessem aptos para reprodução sexuada em nossa área de estudo, calculamos a média (±
desvio padrão) dos parâmetros morfológicos dos indivíduos (comprimento foliar, altura e
diâmetro médio) que apresentavam-se reprodutivos entre as estações de amostragem.
A existência ou não de diferenças entre as estações de amostragem podem nos auxiliar
a entender se a população de B. antiacantha sofre mudanças em sua estrutura ou está em
processo de estabilização na área. Dessa forma, testamos a normalidade dos dados
morfométricos de B. antiacantha (comprimento foliar, altura e diâmetro médio) por meio do
teste de Lillifors (Zar 1999). Como os dados não apresentaram distribuição normal, utilizamos
o teste não-paramétrico de Kolmogorov-Smirnof (Zar 1999) para verificarmos possíveis
diferenças entre as três estações de amostragem para as medidas de comprimento foliar, altura
e diâmetro médio.
3.2.2 Crescimento clonal
Para verificarmos o crescimento clonal de Bromelia antiacantha na restinga da Praia
grande, marcamos 90 indivíduos no mês de maio de 2009 e o acompanhamento do
crescimento clonal ocorreu entre os meses de agosto de 2009 a dezembro de 2012.
Registramos mensalmente a emissão de brotos em cada indivíduo e aqueles que
apresentassem brotações tiveram seus brotos contados e acompanhados mensalmente,
simultaneamente ao período descrito para o acompanhamento da fenologia reprodutiva (ver
seção 3.2.3), sendo o número de brotos acompanhados dependente do número de brotos
emitidos.
Para cada broto emitido e acompanhado, medimos o comprimento da maior folha
periférica e a distância em que este broto se encontrava da planta-mãe a cada mês. Realizamos

20
as medidas até o momento (mês) em que os brotos enraizavam, contabilizamos o tempo de
enraizamento de cada broto. Casos de mortalidade dos brotos também foram registrados.
Identificamos o momento (em meses) em que os brotos encerram o seu crescimento
horizontal (crescimento do broto em paralelo ao solo) e iniciam o crescimento vertical
(crescimentos das folhas e formação da roseta). Calculamos a taxa de crescimento dos brotos
através da fórmula:
TCni = CFi - CFi-1
onde Tc é a taxa de crescimento do n-ésimo broto no i-ésimo mês; CFi é o comprimento foliar
no i-ésimo mês e CFi-1 é o comprimento foliar no mês anterior ao i-ésimo mês. Identificamos
a distância em que estes brotos se encontravam da planta-mãe através das medições do
comprimento do estolão, sendo considerada a distancia entre os centros das rosetas da planta-
mãe e do indivíduo clonal.
Verificamos a existência de relação entre a altura dos brotos e a distância em que estes
se encontravam da planta-mãe através da análise de regressão linear simples (Zar, 1999),
sendo possível identificarmos se maiores taxas de crescimento estão ou não relacionadas a
maiores distâncias de dispersão dos brotos.
3.2.3 Fenologia Reprodutiva
Para avaliarmos o padrão da fenologia reprodutiva de B. antiacantha na restinga da
Praia Grande, entre os meses de junho de 2009 a dezembro de 2012, acompanhamos os 90
indivíduos marcados para a análise do crescimento clonal.
Registramos mensalmente o estado fenológico, observando se os indivíduos estavam:
não reprodutivos, florindo ou frutificando. No caso de indivíduos reprodutivos, observamos a
presença de escapo em formação, de inflorescência, a quantidade de flores por inflorescência
e de frutos. Quando frutificando, contamos a quantidade de frutos imaturos (verdes), maduros
(amarelos) e com sinais de frugivoria (marcas de mordidas ou de bicadas, por exemplo). Para
cada uma das fenofases calculamos a frequência mensal dentro da população amostrada
através da seguinte equação:
Ff (%) = nf
100 N

21
onde Ff é a frequência da fenofase, nf é o número de indivíduos em uma determinada fenofase
e N é o número total de indivíduos amostrados no mês.
Para avaliar uma possível relação entre a proporção mensal de cada uma das fenofases
e as varáveis climáticas umidade relativa (%), precipitação pluviométrica (mm), temperatura
(°C) e radiação solar (W/m2) utilizamos a análise de regressão linear simples (Zar 1999).
Verificamos as relações utilizando os valores médios mensais de umidade relativa (%),
temperatura (°C) e radiação solar (W/m2) e os valores acumulados mensais de precipitação
pluviométrica (mm). Os dados meteorológicos foram cedidos pela ThyssenKrupp Companhia
Siderúrgica do Atlântico, EMTKCSA 4 - Estação Meteorológica CSA para o período de
junho de 2009 a dezembro de 2012.
3.2.4 Visitantes florais
Durante as estações de floração de 2011 e 2012 realizamos observações para
identificar os visitantes florais de B. antiacantha. Iniciamos as observações as 05:00 horas, a
fim de registrar o horário de abertura das flores. Caso neste horário as flores já estivessem
abertas, as observações começavam mais cedo. O término das observações ocorreu as 19:00
horas, horário em que as flores encontravam-se completamente fechadas.
Realizamos seções de observação naturalísticas, utilizando o método do indivíduo-
focal, para determinar as espécies de visitantes florais de B. antiacantha. Fizemos as
observações em intervalos de 15 minutos, com 15 minutos posteriores de descanso,
totalizando 30 minutos de observação a cada 1 hora. Eventualmente, indivíduos vizinhos em
floração também foram avistados a fim de registrar observações complementares.
Durante o período de observação registramos: as espécies de visitantes florais
(inicialmente descritas como morfoespécies), o recurso utilizado por elas (néctar ou pólen), o
tempo de duração de cada visita, a quantidade de flores visitadas por cada visitante e se estes
também visitaram inflorescências vizinhas àquelas observadas. Os visitantes foram
fotografados e/ou coletados, quando possível, para posterior identificação. O comportamento
de cada visitante também foi descrito durante as observações, verificando a presença de
diferentes espécies de visitantes em um mesmo horário no mesmo indivíduo-focal e se há
algum tipo de interação entre estas espécies. Consideramos as visitas completas quando o
visitante efetuou todas as etapas de obtenção de recurso floral, que inclui a chegada do

22
polinizador à inflorescência, inserção do bico ou aparelho bucal na flor para a obtenção do
néctar ou raspagem das anteras para obtenção de pólen, o consumo desses recursos e a saída
por completo daquela inflorescência (Rocha-Pessôa, 2004).
Registramos também, a cada intervalo de observação, as variáveis climáticas
temperatura (°C), umidade relativa (%) e luminosidade (lux).
Para a análise da morfometria floral, coletamos cinco flores de três inflorescências (n
= 15 flores) nas quais tomamos as seguintes medidas com auxílio de um paquímetro (com
precisão de 0,1 mm): comprimento da flor e das sépalas, profundidade e diâmetro da corola,
comprimento do gineceu (ovário + estilete), comprimento do ovário e do estilete e altura de
inserção dos estames. As medidas lineares destes parâmetros foram expressas como valor
médio ±1 desvio padrão.
Calculamos as taxas de visitação por intervalo horário para a guilda de visitantes
florais de B. antiacantha como um todo e para cada espécie de visitante separadamente a fim
de verificarmos o predomínio de algum visitante ou grupo funcional de visitantes em um
determinado período do dia. Verificamos também uma possível relação entre a temperatura
(°C), umidade relativa (%) e luminosidade (lux) e a abundância (N) e riqueza de visitantes
florais (S) por intervalo horário através da análise de regressão linear simples.

23
4 RESULTADOS
4.1. Caracterização da estrutura populacional
Nas três estações de amostragem da população de B. antiacantha contabilizamos um
total de 414 (outubro/2011), 533 (abril/2012) e 565 indivíduos (outubro/2012).
O número de indivíduos da população de B. antiacantha diferiu nas três distâncias da
faixa de areia (ANOVA: N = 50; F = 4,023; p = 0,024), com as faixas distantes em 20 e em 40
metros diferindo entre si (teste de Tukey: p = 0,028) (Fig. 7). Com relação ao grau de abertura
do dossel, das 50 parcelas amostradas, 28 parcelas possuíam valores superiores a 40%, as
quais consideramos como abertas, e 22 parcelas possuíam valores abaixo de 40%, sendo
fechadas. A abundância de indivíduos de B. antiacantha nas parcelas amostradas em parcelas
com dossel aberto e fechado não diferiu significativamente (p = 0,807) (Fig. 8).
Figura 7. Variação na abundância de indivíduos de Bromelia antiacantha nas três classes de
distância da faixa de areia de parcelas amostradas em uma área de Restinga na Ilha da
Marambaia, Mangaratiba/RJ. Classes de distância: 1 = borda da faixa de areia; 2 = 20 metros
de distância e 3 = 40 metros de distância.

24
Figura 8. Variação na abundância de indivíduos de Bromelia antiacantha em relação ao grau
de abertura do dossel das parcelas amostradas em uma área de Restinga na Ilha da
Marambaia, Mangaratiba/RJ.
Não observamos grandes variações entre os valores médios, mínimo e máximo do
comprimento foliar, altura e diâmetro médio dos indivíduos de B. antiacantha para as três
estações de amostragens da estrutura populacional (Tab. 1).
O comprimento foliar médio da população (agrupando as três estações de
amostragem) é igual a 126,6 (± 42,6) cm. O indivíduo com menor comprimento foliar
mensurado possuía 16,0 cm e o indivíduo com maior comprimento foliar 270,0 cm. Do
mesmo modo, a altura média da população é 120,3 (±41,8) cm, com 26,0 cm de altura para o
menor indivíduo e 310,0 cm de altura para o maior indivíduo e o diâmetro médio da
população é 148,8 (± 53,3) cm, com o diâmetro médio menor e maior igual a 29,5 e 318,0 cm,
respectivamente.
No caso dos indivíduos reprodutivos, estes parâmetros foram semelhantes entre as três
estações de amostragem, de modo que calculamos a média geral dos indivíduos reprodutivos:
comprimento foliar = 123,0 (± 38,7; mín = 48,0; máx =179,0), altura = 103,4 (± 42,1; mín =
30,0; máx = 180,0) e diâmetro médio = 169,9 (± 82,6; mín = 89,0; máx = 284,5). Na primeira
estação de amostragem todos os indivíduos reprodutivos amostrados (26 indivíduos, 6,3% do
total) estavam em fase final de frutificação ou pós-reprodutivos. Na segunda estação de
amostragem todos os indivíduos reprodutivos (38 indivíduos, 7,1% do total) estavam em

25
frutificação (com frutos verdes ou amarelo-esverdeados) e na terceira estação, dos indivíduos
reprodutivos (43 indivíduos, 7,6% do total), dois estavam com escapo floral em formação e os
demais estavam em fase final de frutificação ou pós-reprodutivos. Desse modo, indivíduos
com comprimento foliar acima de 48 cm podem ser considerados aptos para reprodução
sexuada.
Em geral, entre as três estações de amostragem, 67 indivíduos (11,9% do total)
estavam conectados a outro indivíduo via estolão. Destes, seis indivíduos (1,1% do total)
possuíam ligações com dois indivíduos.
Com relação aos brotos, contabilizamos 20 (4,8% do total de indivíduos), 42 (7,9%) e
39 brotos (6,9%) dos indivíduos amostrados reproduzindo vegetativamente nas três estações
de amostragem. O comprimento foliar médio dos brotos mensurados em cada uma das três
estações de amostragem foi de 39,4cm (± 25,5; mín = 4,0; máx = 80,0), 48,2cm (± 31,2; mín
= 7,0; máx = 115,0) e 50,5 (± 24,4; mín = 12,0; máx = 103,0). O comprimento foliar médio
dos indivíduos que produziram brotações foi de 133,6cm (± 46,5; mín = 74,0; máx = 250,0),
125,2cm (±31,0; mín = 85,0; máx = 233,0) e 135,2 (± 41,3; mín = 54,0; máx = 246,0) para a
primeira, segunda e terceira amostragem, respectivamente.
A densidade de indivíduos de Bromelia antiacantha aumentou entre as três estações de
amostragem, havendo 828 indivíduos/ha, 1066 indivíduos/há e 1130 indivíduos/há, na
primeira, segunda e terceira estação de amostragem, respectivamente.
Indivíduos de B. antiacantha classificados como plântulas foram raros. Tomando
como base a disposição destes indivíduos dentro das parcelas, isolados dos demais indivíduos
e sem vestígios de conexão com outros indivíduos, estabelecemos que indivíduos com
comprimento foliar menor que 30 cm podem ser considerados oriundos de reprodução
sexuada. Estes representam apenas um indivíduo na primeira estação (0,2% do total) e dois
indivíduos (0,4% do total) na segunda estação de amostragem. Não registramos plântulas na
terceira estação de amostragem.
A população de B. antiacantha estudada na restinga da Praia Grande diferiu
significativamente entre estações apenas em termos do diâmetro (Fig. 9), com a terceira
estação de amostragem apresentando indivíduos significativamente maiores que as outras
duas estações (primeira e terceira amostragem: KS = 0,123, p = 0,002; segunda e terceira
amostragem: KS = 0,149, p < 0,001) (Tab. 1; Fig. 8). Todos os demais parâmetros
morfométricos não apresentaram diferenças significativas entre estações (p > 0,05) (Tab. 1;
Figs. 10 e 11).

26
Tabela 1. Parâmetros básicos relativos ao comprimento foliar, altura e diâmetro médio dos
indivíduos de Bromelia antiacantha nas três estações de amostragem em uma área de restinga
da Ilha da Marambaia, Mangaratiba/RJ. Letras diferentes indicam valores estatisticamente
significativos.
Parâmetros (cm) Estação 1 Estação 2 Estação 3
Comprimento foliar
Média (± DP) 129,5 (± 46,2)a 124,3 (± 39,4)a 127,5 (± 41,6)a
Mínimo 23,0 16,0 32,0
Máximo 262,0 270,0 252,0
Altura
Média (± DP) 120,6 (± 42,5)a 119,9 (± 41,3)a 118,8 (± 42,1)a
Mínimo 29,0 26,0 26,0
Máximo 260,0 310,0 251,0
Diâmetro médio
Média (± DP) 149,7 (± 57,8)a 148,1 (± 49,5)a 162,4 (± 58,9)b
Mínimo 29,5 36,0 34,0
Máximo 318,0 290,0 379,5
Figura 9. Variação no diâmetro médio das rosetas (cm) dos indivíduos de Bromelia antiacantha em
três estações de amostragem em uma área de Restinga na Ilha da Marambaia, Mangaratiba/RJ.
Estações 1 = Outubro/2011; 2 = Abril/2012 e 3 = Outubro/2012. A linha pontilhada indica os valores
médios de comprimento foliar dos indivíduos reprodutivos e as duas linhas cheias indicam o desvio
padrão da média.

27
Figura 10. Variação no comprimento foliar (cm) dos indivíduos de Bromelia antiacantha em três
estações de amostragem em uma área de Restinga na Ilha da Marambaia, Mangaratiba/RJ. Estações 1
= Outubro/2011; 2 = Abril/2012 e 3 = Outubro/2012. A linha pontilhada indica os valores médios de
comprimento foliar dos indivíduos reprodutivos e as duas linhas cheias indicam o desvio padrão da
média.
Figura 11. Variação na altura das rosetas (cm) dos indivíduos de Bromelia antiacantha em três
estações de amostragem em uma área de Restinga na Ilha da Marambaia, Mangaratiba/RJ. Estações 1
= Outubro/2011; 2 = Abril/2012 e 3 = Outubro/2012. A linha pontilhada indica os valores médios de
comprimento foliar dos indivíduos reprodutivos e as duas linhas cheias indicam o desvio padrão da
média.

28
4.2 Crescimento clonal
A emissão de brotos pela população de Bromelia antiacantha ocorreu ao longo de
todos os meses. Em geral, houve emissão de um ou dois brotos na maioria dos meses (média
= 1,3 ± 1,8). No mês de janeiro, registramos os maiores números de brotos emitidos pela
população, com até oito brotos em 2010 e sete em 2012 (Fig. 12).
Ao longo do período de acompanhamento registramos um total de 52 brotos sendo
emitidos pelos indivíduos marcados ao longo dos meses de amostragem, sendo o mês de
setembro de 2011, com seis brotos, o mês com o menor número de brotos disponíveis para
amostragem e os meses de janeiro, fevereiro e março de 2010, com 26 brotos cada, os meses
com o maior número de brotos registrados para amostragem (Fig. 13).
As plantas-mãe podem emitir de um a quatro brotos, sendo que 49 dos indivíduos
emitiram apenas um broto, cinco indivíduos emitiram dois brotos, um indivíduo emitiu três
brotos e outro emitiu quatro brotos, com a população produzindo uma média de 1 broto (± 1)
por indivíduo (Fig. 14).
Os brotos tendem a permanecer apenas com o crescimento horizontal, em média, até o
terceiro mês (3,7 ± 2,0 meses; mín = 1; máx = 7) e o estolão cresce em comprimento em
média 15 cm (± 10 cm) por mês. O crescimento horizontal encerra-se com cerca de 8 meses
(± 3 meses). Ao enraizarem, os brotos apresentaram comprimento de estolão médio de 71,8
cm (± 29,3; mín = 30,0; máx = 162,0 cm).
Em média, os brotos crescem em comprimento foliar cerca de 8,6 cm (± 5,6 cm; mín =
4,0; máx = 22 cm) ao mês. No momento de enraizarem os brotos apresentam, em média,
comprimento foliar (altura) de 79,2 (± 15,3; mín = 58; máx = 112 cm).

29
Figura 12. Proporção de indivíduos de B. antiacantha emitindo brotos entre os meses de
acompanhamento das fenofases em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Figura 13. Número de brotos de B. antiacantha mensurados durante os meses de acompanhamento do
crescimento clonal em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.

30
Figura 14. Número de brotos emitidos por indivíduo de Bromelia antiacantha em uma área de
restinga na Ilha da Marambaia, Mangaratiba/RJ.
Os brotos tenderam a apresentar maior taxa de crescimento nos meses iniciais de
desenvolvimento (Fig. 15a). Com relação a variação na distância em que os brotos emitidos se
encontravam da planta-mãe, em geral, identificamos maiores distâncias nos primeiros meses
de acompanhamento, diminuindo a partir do quinto mês (Fig. 15b).
O tamanho dos brotos esteve significativamente relacionado com a distância da planta-
mãe (N = 408; R = 0,321 e p < 0,001) (Fig. 16).

31
Figura 15. Variação na taxa de crescimento (A) e na distância da planta-mãe (B) dos brotos
emitidos por B. antiacantha ao longo dos meses de acompanhamento em uma área de restinga
na Ilha da Marambaia, Mangaratiba/RJ.
B
A

32
Figura 16. Relação entre a altura e a distância da planta-mãe dos brotos emitidos por
Bromelia antiacantha durante os meses de acompanahmento em uma área de Restinga da Ilha
da Marambaia - Mangaratiba/RJ.
4.3 Fenologia Reprodutiva
4.3.1 Variáveis climáticas
Durante o período de estudo, a umidade relativa variou de 64,0 a 85,1%, com os
menores valores ocorrendo nos meses de fevereiro de 2010 e 2012 e maiores valores nos
meses de março de 2011 e junho de 2012 (Fig. 17a).
Os valores de precipitação pluviométrica acumulada mensal variaram ao longo dos
anos de estudo, geralmente com os menores valores nos meses de abril a maio e agosto a
setembro e com os maiores valores nos meses de outubro e dezembro a janeiro (Fig. 17b).
A temperatura média mensal variou de 18,8 a 30,5°C durante os meses de estudo, com
os maiores valores, de forma geral, entre os meses de novembro a abril e os menores valores
entre os meses de maio e setembro (Fig. 17c).
A média mensal da Radiação Solar Global (W/m2) variou de 67,7 a 297,15 W/m2, com
os maiores valores entre os meses de outubro a fevereiro e os menores valores entre os meses
de abril e julho (Fig. 17d).

33
Figura 17. Variáveis climáticas analisadas durante o período de estudo na Ilha da Marambaia,
Mangaratiba/RJ. A - Variação média mensal das umidade relativa (%); B - Variação
acumulada mensal da precipitação pluviométrica (mm); C - Variação média mensal da
temperatura (°C) e D - Variação média mensal da Radiação Solar Global (W/m2).
4.3.2 Fenologia
Bromelia antiacantha floresceu entre os meses de outubro e dezembro, ocorrendo, em
geral, em uma pequena parcela da população (11% dos indivíduos amostrados) (Fig. 18a). Já
a frutificação ocorreu entre os meses de dezembro e junho, com um pico em torno dos meses
de março e abril, quando observamos um grande número de frutos maduros (Fig. 18b).
D
A
C
B

34
Figura 18. Proporção de indivíduos de B. antiacantha em floração (A), frutificação (B) entre
os meses de acompanhamento das fenofases em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
Dentre os estados fenológicos, apenas a frutificação foi explicada pelas variáveis
climáticas umidade relativa (N = 41; p = 0,011; R2 = 0,154 ) e temperatura (N = 41; p =
0,036; R2 = 0,108) (Figs. 19a e 19b). Todos os demais estados fenológicos não responderam
as variáveis climáticas analisadas neste estudo (p > 0,05).
A
B

35
60 70 80 90
Umidade Relativa (%)
0
5
10
15F
rutifica
çã
o (
%)
15 20 25 30 35
Temperatura (°C)
0
5
10
15
Fru
tifica
çã
o (
%)
Figura 19. Relação entre a porcentagem de indivíduos de B. antiacantha em frutificação e as
variáveis climáticas em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ. A -
Umidade Relativa (%) e B - Temperatura (°C).
4.4 Visitantes florais
4.4.1 Caracterização geral
A observação dos visitantes florais de B. antiacantha ocorreu ao longo de dois dias
durante a estação reprodutiva de 2011 e ao longo de seis dias durante a estação reprodutiva de
2012, totalizando 69 horas de observação.
As flores de B. antiacantha iniciaram a antese por volta das 05:30h, estando
totalmente abertas por volta das 06:00h. As flores são diurnas e duram apenas um dia,
permanecendo abertas e disponíveis aos visitantes até aproximadamente as 18:00h,
começando a fechar as 17:30h. Tanto as anteras quanto o estigma permanecem inclusos na
corola. As pétalas apresentam coloração lilás e as sépalas são verde claro, as quais contrastam
com a coloração vermelho intensa das brácteas que circundam as inflorescencias. A Tabela 2
apresenta os valores médios (±1 desvio padrão) das variáveis morfométricas das flores de B.
antiacantha mensuradas em 13 flores de três inflorescências distintas.
A B

36
O número médio de flores produzidas por inflorescência foi de 72,8 ± 12,9 (N = 10;
mín = 45; máx = 89) e o número de flores abertas por indivíduo/dia foi de 13,0 ± 8,13 (N =
10; mín = 1; máx = 26).
Tabela 2. Valores médios (±1 desvio padrão) das variáveis morfométricas de Bromelia
antiacantha na área de restinga da Ilha da Marambaia - Mangaratiba/RJ. (N = 13 flores).
Variáveis Morfométricas Média ±1 Desvio Padrão (mm)
Comprimento da flor 49,7 ± 4,9
Profundidade da corola 28,8 ± 2,8
Diâmetro médio da corola 10,5 ± 1,9
Comprimento da sépala 12,1 ± 2,1
Altura dos estames 19,9 ± 4,6
Altura do Gineceu 22,2 ± 3,7
Comprimento do estilete 13,8 ± 1,9
Altura do ovário 11,6 ± 2,1
4.4.2 Descrição dos visitantes florais
Durante o período de observação registramos 11 espécies de visitantes florais, que
realizaram um total de 272 visitas as flores de B. antiacantha (Tab. 3). Consideramos quatro
destas espécies (36,4% do total) como pilhadoras de recursos, principalmente néctar e pólen.
As demais espécies (63,6% do total) realizaram visitas legítimas as flores de B. antiacantha,
atuando como polinizadoras.
As visitas, de uma forma geral, ocorreram durante todo o dia, desde o início até o final
da antese (Figura 20). Para a riqueza de espécies de visitantes florais por intervalo horários,
encontramos os maiores valores no intervalo horário de 12:00 as 13:00 horas, com nove
espécies de visitantes, seguido dos intervalos horários de 7:00 as 8:00 e 11:00 as 12:00 horas,
com oito espécies de visitantes (Fig. 21). Registramos os menores valores de riqueza nos

37
intervalos horários extremos, de 4:00 as 5:00 e 17:00 as 18:00 horas, com zero e duas espécies
de visitantes, respectivamente (Fig. 21).
Tabela 3. Visitantes florais de Bromelia antiacantha em uma área de restinga da Ilha da
Marambaia, Mangaratiba/RJ, suas respectivas taxas de visitação durante a estação reprodutiva
de 2011 e de 2012, principal recurso obtido e papel na polinização (polinizador/pilhador).
2011 2012
Total
Recurso
Papel Visitantes N % N %
Hymenoptera: Apoidae
Bombus sp. 14 6,4 1 0,7 15 Pólen Polinizador
Euglossa cordata 9 4,5 - - 9 Néctar Pilhador
Trigona spinipes 183 84,6 46 32,2 229 Pólen Polinizador
Halictidae sp. - - 15 10,4 15 Pólen Polinizador
Aves: Trochilidae
Amazilia fimbriata 7 3,2 31 21,7 38 Néctar Polinizador
Florisuga fusca - - 13 9,1 13 Néctar Polinizador
Thalurania glaucopis ♀ - - 21 14,7 21 Néctar Polinizador
Thalurania glaucopis ♂ - - 2 1,4 2 Néctar Polinizador
Passeriformes: Thraupidae
Coereba flaveola - - 8 5,6 8 Néctar Pilhador
Lepidoptera
Lepidoptera sp1 3 1,3 - - 3 Néctar Pilhador
Lepidoptera sp2 - - 6 4,2 6 Néctar Pilhador
TOTAL 216 100 143 100 359

38
Figura 20. Padrão geral de visitação da guilda de visitantes florais de Bromelia antiacantha
na área de restinga da Ilha da Marambaia, Mangaratiba/RJ.
Figura 21. Riqueza de espécies de visitantes florais de Bromelia antiacantha por intervalo
horário em uma área de restinga da Ilha da Marambaia, Mangaratiba/RJ.
Dentre os visitantes florais de B. antiacantha, Trigona spinipes (Fabricius, 1793) foi
responsável pela maior frequência de visitação as flores de Bromelia antiacantha em ambas
as estações reprodutivas, com 84,72% na estação reprodutiva de 2011 e 32,17% em 2012
(Tab. 3). As visitas desta espécie ocorreram ao longo do dia, principalmente entre as 07:00 e
17:00h (Fig. 22). Os indivíduos desta espécie permaneceram por um longo período de tempo
na mesma inflorescência coletando pólen e armazenando nas corbículas (chegando a ocupar

39
todos os quinze minutos de um período de observação, na maioria das vezes). Esta espécie
esteve nas duas estações reprodutivas, sendo mais frequente na estação reprodutiva de 2011
(50,97%) do que na estação reprodutiva de 2012 (12,81%), sendo considerada polinizadora.
Figura 22. Frequência e atividade ao longo do dia do visitante floral Trigona spinipes as
flores de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
As outras três espécies de Hymenopteras apresentaram taxas de visitação inferiores a
11,0%. Halictidae sp. foi responsável por 10,49% das visitas as flores de B. antiacantha e
esteve presente apenas na segunda estação reprodutiva estudada. Os indivíduos desta espécie
iniciaram suas visitas a partir das 07:00h e atingiram o pico de visitação as 11:00h,
diminuindo as taxas de visitação após este horário (Fig. 23). Durante o período de observação
registramos sempre dois indivíduos da espécie por inflorescência, os quais exibiram
comportamento similar aquele observado para T. spinipes.

40
Figura 23. Frequência e atividade ao longo do dia do visitante floral Halictidae sp. as flores
de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Bombus sp. e ocorreu nas duas estações reprodutivas, sendo responsável por 6,48%
das visitas as flores de B. antiacantha na primeira estação reprodutiva e 0,70% em 2012, com
apenas um registro de visita nesta estação reprodutiva (Tab. 3). Os indivíduos desta espécie
visitaram três a cinco flores por inflorescência, geralmente ao mesmo tempo em que vários
indivíduos de T. spinipes também buscavam recursos, com maiores frequencias entre os
horários de 8:00 as 9:00 e 11:00 as 12:00 (Fig. 24). A espécie toca os órgãos reprodutivos da
flor ao inserir sua cabeça na flor para coletar pólen, realizando a polinização.
Indivíduos de Euglossa cordata (Linnaeus, 1758), presentes apenas na estação
reprodutiva de 2011, totalizaram 4,17% das visitas às flores de B. antiacantha (Tab. 3). O fato
de possuir uma longa probóscide permite a estes indivíduos que pousem nas flores e a
introduzam no orifício da corola para a coleta de néctar, tocando eventualmente as partes
reprodutivas da flor. Posteriormente a coleta do néctar, ao se deslocar para outras flores ou
outras inflorescências, E. cordata realiza a limpeza da probóscide, que permanece estendida,
provavelmente eliminando pólen aderido a estrutura. As visitas desta espécie ocorreram entre
o período de 07:00 as 16:00h (Fig. 25).

41
Figura 24. Frequência e atividade ao longo do dia do visitante floral Bombus sp. as flores de
B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Figura 25. Frequência e atividade ao longo do dia do visitante floral Euglossa cordata as
flores de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Todas as espécies de beija-flores realizaram visitas legítimas às flores de B.
antiacantha. O comportamento destas espécies foi semelhante, geralmente realizando poucas
e espaçadas visitas as flores de B. antiacantha, não ultrapassando sete flores visitadas por
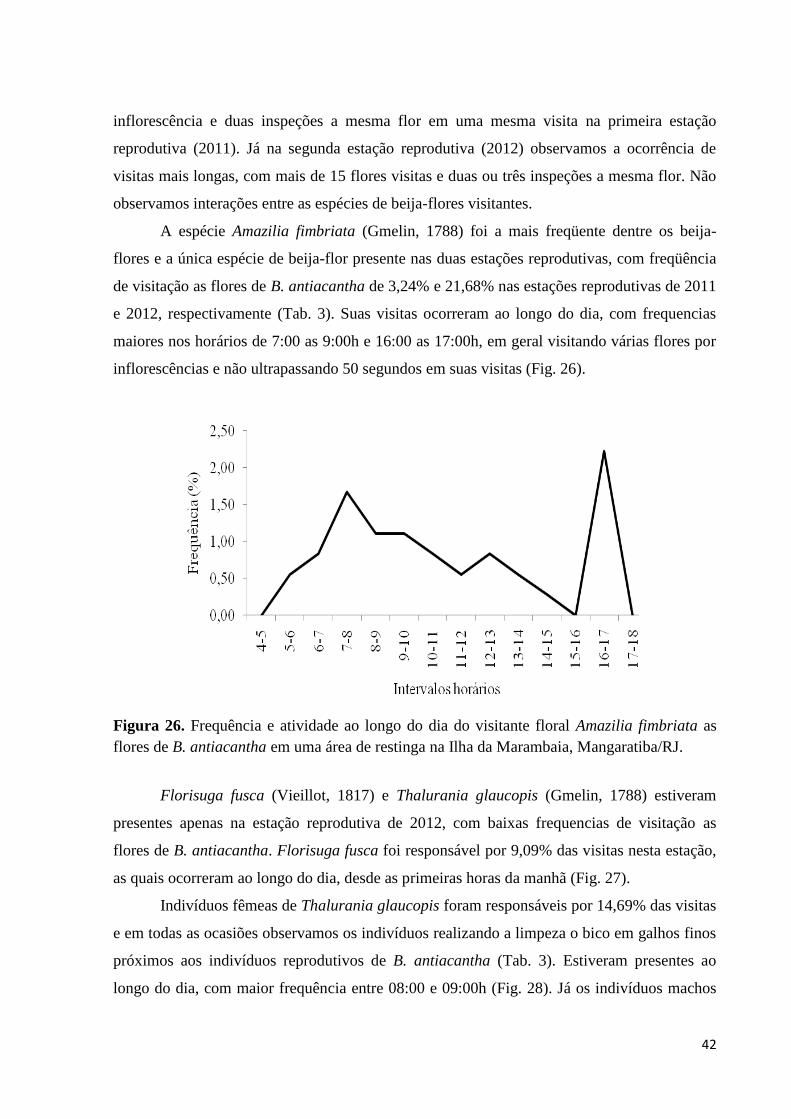
42
inflorescência e duas inspeções a mesma flor em uma mesma visita na primeira estação
reprodutiva (2011). Já na segunda estação reprodutiva (2012) observamos a ocorrência de
visitas mais longas, com mais de 15 flores visitas e duas ou três inspeções a mesma flor. Não
observamos interações entre as espécies de beija-flores visitantes.
A espécie Amazilia fimbriata (Gmelin, 1788) foi a mais freqüente dentre os beija-
flores e a única espécie de beija-flor presente nas duas estações reprodutivas, com freqüência
de visitação as flores de B. antiacantha de 3,24% e 21,68% nas estações reprodutivas de 2011
e 2012, respectivamente (Tab. 3). Suas visitas ocorreram ao longo do dia, com frequencias
maiores nos horários de 7:00 as 9:00h e 16:00 as 17:00h, em geral visitando várias flores por
inflorescências e não ultrapassando 50 segundos em suas visitas (Fig. 26).
Figura 26. Frequência e atividade ao longo do dia do visitante floral Amazilia fimbriata as
flores de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Florisuga fusca (Vieillot, 1817) e Thalurania glaucopis (Gmelin, 1788) estiveram
presentes apenas na estação reprodutiva de 2012, com baixas frequencias de visitação as
flores de B. antiacantha. Florisuga fusca foi responsável por 9,09% das visitas nesta estação,
as quais ocorreram ao longo do dia, desde as primeiras horas da manhã (Fig. 27).
Indivíduos fêmeas de Thalurania glaucopis foram responsáveis por 14,69% das visitas
e em todas as ocasiões observamos os indivíduos realizando a limpeza o bico em galhos finos
próximos aos indivíduos reprodutivos de B. antiacantha (Tab. 3). Estiveram presentes ao
longo do dia, com maior frequência entre 08:00 e 09:00h (Fig. 28). Já os indivíduos machos

43
desta espécie foram menos frequentes (1,40%) com apenas duas visitas registradas após as
13:00h (Fig. 29).
Figura 27. Frequência e atividade ao longo do dia do visitante floral Florisuga fusca as flores
de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Figura 28. Frequência e atividade ao longo do dia do visitante floral fêmea de Thalurania
glaucopis as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.

44
Figura 29. Frequência e atividade ao longo do dia do visitante floral macho de Thalurania
glaucopis as flores de B. antiacantha em uma área de restinga na Ilha da Marambaia,
Mangaratiba/RJ.
Coereba flaveola (Linnaeus, 1758) realizou um total de quatro visitas (5,59%) as
flores de B. antiacantha na estação reprodutiva de 2012 (Tab. 3). Durante as visitas,
realizadas principalmente entre as 07:00 e 14:00h (Fig. 30), a espécie pousa em botões florais
da parte superior da inflorescência e visita uma ou duas flores apenas uma vez, não sendo
observada visitando flores da parte inferior.
Figura 30. Frequência e atividade ao longo do dia do visitante floral Coereba flaveola as
flores de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.

45
As duas espécies de Lepidoptera realizaram poucas visitas as flores de B. antiacantha
(Tab. 3), geralmente pousando em botões florais próximos a flor em que iam obter o néctar.
Ambas as espécies realizaram visitas rápidas, em geral permanecendo menos de 20 segundos
em cada inflorescencia e visitaram de uma a três flores. Lepidoptera sp1 (1,39%) e
Lepidoptera sp2 (4,20%) estiveram presentes apenas na estação reprodutiva de 2011 e 2012,
respectivamente. (Fig. 31 e 32).
Figura 31. Frequência e atividade ao longo do dia do visitante floral Lepidoptera sp1 as flores
de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.
Figura 32. Frequência e atividade ao longo do dia do visitante floral Lepidoptera sp2 as flores
de B. antiacantha em uma área de restinga na Ilha da Marambaia, Mangaratiba/RJ.

46
4.4.3 Variáveis microclimáticas
Os valores médios das variáveis climáticas luminosidade (em lux), temperatura (em
°C) e umidade relativa (em %) utilizadas para caracterizar o microambiente ao redor dos
indivíduos floridos de B. antiacantha variaram ao longo do dia (Figuras 33a-c).
A luminosidade variou de 161,0 Lux nas primeiras horas da manhã a 16090,0 as 15:00
horas (Fig. 28a), diminuindo após este horário. A temperatura variou entre 25,4 e 31,7°C, com
as menores temperaturas até as 8:00 horas e as maiores de 10:00 as 16:00 horas (Fig. 28b). A
umidade relativa, em geral, é maior das 6:00 as 8:00 horas e menor das 15:00 as 16:00 horas
(Fig. 28c).
Dentre as três variáveis microclimáticas analisadas, apenas a luminosidade (Lux)
relacionou-se com a riqueza de espécies (N = 14; R2 = 0,65; p = 0,012) e a abundância (N =
14; R2 = 0,783; p = 0,001) de visitantes florais (Fig. 34a-b).

47
Figura 33. Variação média das variáveis microclimáticas mensuradas durante a observação
de atividades dos visitantes florais as flores de Bromelia antiacantha em uma área de restinga
na Ilha da Marambaia, Mangaratiba/RJ. A - Luminosidade (Lux); B - Temperatura (°C) e C -
Umidade relativa (%).
A
B
C

48
Figura 34. Relação entre a variável microclimática luminosidade (Lux) e a riqueza (A) e
abundância (B) de visitantes florais de Bromelia antiacntha em uma área de restinga na Ilha
da Marambaia, Mangaratiba/RJ.
B A

49
5 DISCUSSÃO
5.1. Caracterização da estrutura populacional
A densidade de bromélias em uma área depende da organização estrutural da
vegetação, a qual varia em função do estádio sucessional da área, sendo mais abundante em
áreas preservadas (MATOS, 2000). Assim como na população de Bromelia antiacantha na
restinga da Ilha da Marambaia, Duarte et al. (2007) também registraram um aumento no
número de indivíduos por hectare na população de B. antiacantha estudada e número médio
de indivíduos vegetativos por hectare superior a quinze vezes o número médio de indivíduos
reprodutivos para os cinco anos avaliados. Este aumento reflete o padrão de crescimento da
população na área, com altas taxas de reprodução vegetativa.
É possível que esteja ocorrendo um aumento da população em direção as áreas de
interior da restinga, uma vez que as plântulas de B. antiacantha foram encontradas na terceira
faixa de amostragem (com maior distância da faixa de areia). A instabilidade do substrato
arenoso devido a ação da erosão ou deposição de sedimentos advindos das ondas e ventos,
podem remover as plantas ou promover seu soterramento por areia (BEDUSCHI &
CASTELLANI, 2008), indicando maiores desafios para B. antiacantha neste ambiente, apesar
de um grande número de brotações serem registradas nos indivíduos próximos a faixa de
areia.
O comprimento médio das folhas dos indivíduos reprodutivos registrados por Duarte
et al. (2007) foi de 257,0 cm. Este valor é maior do que o registrado neste estudo (123,0 cm),
indicando que os indivíduos de B. antiacantha na restinga da ilha da Marambaia reproduzem-
se em menor período de tempo, provavelmente devido as condições ambientais adversas
proporcionadas pela restinga (SCARANO, 2002) .
O número médio de brotações por hectare variou entre as estações de amostragem. tal
fato também foi observado por Duarte et al. (2007), com as brotações aumentando de um ano
para o outro, totalizando 367 indivíduos por hectare e as plântulas somaram cinco indivíduos
por hectare. No caso das plântulas e brotações, os valores registrados no presente estudo
foram menores, mas com mantendo a maioria de brotos em relação às plântulas. Outras
espécies de bromélias também exibem resultados semelhantes, dos 1028 indivíduos de C.
microps contabilizados por Nunes-Freitas & Rocha (2011) para estimar o tipo de
recrutamento, 80,4% eram originários de crescimento clonal e 19,6% de sementes,

50
evidenciando o predomínio da reprodução assexuada sobre a sexuada. Rogalski et al. (2007)
registraram baixo número de plântulas de Dyckia brevifolia, com apenas 18, 7% da população
reproduzindo sexuadamente, e alta porcentagem de indivíduos agrupados, indicando
acentuada reprodução assexuada. A espécie Neoregelia cruenta na restinga de Jurubatiba, não
foi capaz de recrutar novos indivíduos através da reprodução sexuada, mesmo com a
formação de frutos e sementes ocorrendo anualmente (MANTUANO, 2008).
Sampaio et al. (2005) sugerem que apesar de a reprodução sexual de plantas em
restingas ser vista de forma contrastante, a floração destas espécies pode desempenhar um
importante papel ecológico em uma escala espacial e temporal maior, permitindo manter o
fluxo genético entre as populações nestes ambientes.
Observamos diferenças significativas, entre as três estações de amostragem da
população, apenas para o diâmetro médio da roseta, indicando que apesar da população estar
aumentando, sua dinâmica não sofre grandes modificações ou no caso de haverem
modificações estas precisam de maiores períodos de tempo para ser identificadas.
5.2 Crescimento clonal
A propagação clonal possui fundamental importância para espécies que ocorrem em
ambientes adversos, em relação à formação e manutenção de suas populações e por diminuir a
perda de diversidade genética dentro destas populações (MURAWSKI & HAMRICK, 1990) e
as populações de espécies de bromélias ocorrentes em ambientes marginais da Mata Atlântica
realizam o recrutamento de novos indivíduos majoritariamente por meio de crescimento
clonal (SCARANO, 2002). De acordo com Mantuano e Martinelli (2007) em indivíduos
conectados, fatores como a escala da variação ambiental, o tamanho dos segmentos clonais e
o comprimento do estolão são determinantes para a percepção da heterogeneidade ambiental.
A reprodução vegetativa e a propagação, além do sistema reprodutivo e dos
mecanismos de dispersão desenvolvidos pelas espécies, poderiam afetar significativamente a
composição genética e a estrutura das populações. O crescimento clonal atuaria retardando a
perda de diversidade genética dentro das populações (MURAWSKI & HAMRICK, 1990).
Sampaio et al. (2005) caracterizaram o ciclo de vida de Aechmea nudicaulis como
taxas altas de sobrevivência para rametas jovens e vegetativas, com as rametas jovens
apresentando altas taxas de crescimento. No nosso caso as rametas de B. antiacantha também

51
apresentaram altas taxas de crescimento e sobrevivência. Para Neoregelia cruenta, seu
crescimento vegetativo é uma das características que confere o seu papel facilitador na
restinga aberta de Clúsia ao amenizar o ambiente para a germinação e estabelecimento de
outras espécies, sendo o seu recrutamento majoritariamente associado à reprodução assexuada
(MANTUANO, 2008).
Bromelia antiacantha emitiu brotos ao longo de todo o ano, fato também registrado
para Neoregelia cruenta, que emite brotos laterais (de um a quatro brotos) ao longo de todo o
ano (MEIRELES, 2011) e A. nudicaulis que produz até três rametas por planta-mãe, com
média de 1,09 rametas (SAMPAIO et al., 2002). A emissão média de um broto por indivíduos
de B. antiacantha registrado no presente estudo também foi evidenciada por Duarte et al.
(2007) e Filippon (2009) que registraram a emissão de um (poucos indivíduos emitindo dois
ou mais) e 1,53 (variando de um a três) brotos por planta, respectivamente. Esses resultados
indicam que a população de B. antiacantha na Ilha da Marambaia foge dos padrões
reprodutivos esperados para a família Bromeliaceae, já que emite brotos mesmo fora do
período de reprodução sexuada, possivelmente como um mecanismo de ocupação rápida do
ambiente. O comprimento do rizoma (conexão) destes indivíduos variou de 4 a 18 cm, com a
maioria entre 10 e 14 cm (SAMPAIO et al., 2002).
As baixas taxas de mortalidade dos brotos podem estar relacionadas ao suporte
fornecido pelo indivíduo parental, uma vez que no crescimento clonal, o indivíduo oriundo de
propagação clonal possui auxílio do indivíduo parental durante a fase e estabelecimento, o
que contribui para a diminuição da mortalidade nas fases mais jovens, apesar de limitá-lo a
ocorrer no raio do comprimento do estolão (MANTUANO E MARTINELLI, 2007). Devido a
suas características de desenvolvimento, tais como a conexão entre as rametas proporcionada
pelo estolão, permitindo o transporte de água e nutrientes entre as unidades ligadas, os brotos
(unidades clonais) geralmente possuem maior tamanho e menores taxas de mortalidade
quando comparados as plântulas (ANACLETO & NEGRELE, 2009).
De acordo com Rogalski et al. (2007) as populações que apresentaram poucas rosetas
nas classes iniciais provavelmente são constituídas por poucos indivíduos gerados
sexuadamente, sendo a propagação clonal a principal forma de recrutamento. Por outro lado,
nas outras populações as duas formas (sexuada e clonal) parecem ter importância na
constituição das populações.
A produção de brotos por B. antiacantha não variou entre indivíduos jovens ou
vegetativos na população. Para Aechmea nudicaulis a produção de rametas foi maior e mais

52
variável entre os microhabitats para as rametas jovens do que para as rametas vegetativas
(SAMPAIO et al., 2005).
A variação no comprimento do estolão verificada para B. antiacantha pode estar
relacionada ao microhabitat onde estes brotos se desenvolvem, havendo a necessidade de
maiores ou menores valores de comprimento do estolão. Por outro lado, Mantuano e
Martinelli (2007) ao quantificarem os efeitos das variações ambientais na taxa de crescimento
clonal de Neoregelia cruenta na restinga de Jurubatiba constataram que o comprimento do
estolão não foi afetado pelo ambiente, sendo geneticamente determinado. Em relação a
duração da conexão entre as rametas, esta parece ocorrer até o momento em que a
sobrevivência independente dos brotos está garantida, sendo assim, quanto mais
desenvolvidos os brotos, maior será o sucesso destes indivíduos ao se tornarem independentes
(ANACLETO & NEGRELE, 2009).
As altas taxas de crescimento dos brotos de B. antiacantha estão em conformidade
com outros estudos (DUARTE et al., 2007; FILIPPON, 2009). Os brotos geralmente
apresentam comportamento diferenciado dos demais indivíduos da população, com
crescimento foliar acelerado, classes de desenvolvimento específicas e maior número de
brotos nas menores classes de tamanho, devido ao fato dos brotos maiores geralmente
perderem sua conexão com as plantas-mãe, tornando-se indivíduos adultos ou adultos
reprodutivos (DUARTE et al., 2007). Maior vigor para os brotos em relação aos demais
indivíduos e desenvolvimento rápido foram relatados por Filippon (2009).
O conjunto de características apresentado por B. antiacantha sugere que esta apresenta
uma tática de crescimento do tipo guerrilha (SAMPAIO et al., 2002), que forma um grande e
complexo sistema de rametas. Essa tática de crescimento permite que a espécie ocupe
rapidamente o ambiente, obtendo recursos de diferentes microsítios (SAMPAIO et al., 2002),
e parece ter um papel importante na dinâmica de clareiras dos ambientes de restinga, já que ao
colonizar uma determinada área, B. antiacantha pode criar sítios com condições favoráveis
para a ocupação de outras espécies.
5.3. Fenologia Reprodutiva
As baixas taxas de floração de B. antiacantha registradas neste estudo parecem ser
frequentes nos ambientes de restinga (SAMPAIO et al. 2005; MANTUANO, 2008). Nunes-
Freitas & Rocha (2011) também verificaram baixas e variadas taxas de rosetas em floração de

53
C. microps ao longo do ano. A frequência de floração desta espécie foi explicada
principalmente pelo fotoperíodo e precipitação média, enquanto a frutificação foi fracamente
relacionada às variáveis climáticas (NUNES-FREITAS & ROCHA, 2011). Meireles (2011)
registrou poucas variações para as fenofases de Neoregelia cruenta e V. procera entre as
estações reprodutivas analisadas. Para N. cruenta a floração coincidiu com o período chuvoso
e a frutificação ocorreu no final da estação chuvosa/início da estação seca. Já para V. procera,
a floração ocorreu entre os meses finais da estação chuvosa e o inicio da estação seca e a
frutificação possui maior extensão na estação seca.
A sincronia da floração das bromélias na estação chuvosa atua diversificando a oferta
de alimento neste período, principalmente para os vetores de pólen (MARQUES & LEMOS
FILHO, 2008). O ajuste da floração nesta época do ano pode estar relacionado a parte da
guilda de polinizadores da espécie, pois os insetos, em geral, possuem, altas abundâncias nos
períodos mais úmidos do ano, favorecendo sua atividade (JANZEN, 1973). Por outro lado, a
variação no período fenológico poderia ser influência das condições ambientais locais sobre
as fenofases da população (CRUZ, MELLO & VAN SULYS, 2005).
5.4 Visitantes florais
Bromelia antiacantha apresenta um padrão de floração anual, do tipo curta, com
muitas flores sendo disponibilizadas em poucos dias. Suas flores possuem características
ornitófilas e melitófilas. Suas brácteas com coloração vermelho intensa e o formato da corola
tubular com profundidade média e coloração clara (aprox. 30 mm) são características atrativas
às espécies de beija-flores e a coloração lilás e verde claro das pétalas e sépalas são
perceptíveis para os insetos, principalmente os himenópteros (abelhas e vespas) e lepidópteros
(borboletas).
Santos (2001) cita que Bromelia antiacantha aloca grande parte de seus recursos em
reprodução, pois permanece durante meses em floração e frutificação. De acordo com o
referido autor B. antiacantha é considerada uma espécie ornitófila, porém sem especialização
para um polinizador específico, pois atrai uma ampla gama de visitantes florais. Canela &
Sazima (2005) demonstraram que a espécie possui características ornitófilas e melitófilas
sendo polinizada principalmente por estes dois agentes, beija-flores e abelhas.

54
Em áreas abertas os recursos florais podem ser distribuídos mais espaçadamente e
consequentemente os beija-flores são mais oportunistas na utilização dos recursos em
contraste com os habitats florestais (GOTTSBERGER E SILBERBAUER-GOTTSBERGER,
2006). Tal fato pode ser estendido à restinga da Praia Grande, onde B. antiacantha apresenta
poucos indivíduos florescendo por estação.
A relação entre o comportamento territorial de beija-flores a disponibilidade de
recursos florais da espécie Palicourea rigida Kunth (Rubiaceae) em uma área de savana
arbustiva no cerrado mineiro observaram pode ser comprovada apenas em manchas com alta
abundância floral. Este comportamento também foi restrito a poucas espécies de beija-flores,
sendo Eupetonema macroura e Amazilia fimbriata espécies altamente dominantes e
territorialistas, defendendo as manchas de vegetação que exploram (JUSTINO et al. 2002).
Eupetonema macroura apesar de ter sido registrada por Meireles (2011) visitando as
flores de Vriesea procera na Praia Grande - Ilha da Marambaia e por Rodrigues (2008)
visitando as flores de Erythrina speciosa Andrews (Leguminosae - Faboideae), S. tomentosa e
Malvaviscus arboreus Cav. (Malvaceae) em diferentes áreas da Ilha da Marambaia, não foi
observada visitando as flores de B. antiacantha, indicando que nem todas as espécies de beija-
flores presentes na área utilizam B. antiacantha como fonte de recursos.
Já Amazilia fimbriata não apresentou comportamento territorialista em minha área de
estudo, provavelmente devido a escassez de recursos fornecidos por B. antiacantha, o que não
resulta em uma relação custo benefício favorável à defesa territorial. Este beija-flor foi
observado por Rodrigues (2008) visitando as flores de E. speciosa e Pseudananas sagenarius
(Arruda) Camargo e Vriesea neoglutinosa Mez (Bromeliaceae) em diferentes áreas da Ilha da
Marambaia e classificado como territorialista, pois defendeu manchas de P. sagenarius,
expulsando borboletas e abelhas.
Dentre as seis espécies de beija-flores registradas por Rodrigues (2008), apenas
Thalurania glaucopis ocorreu durante todos os meses do ano, sendo a espécie mais freqüente
e realizando visitas a todas as espécies de plantas estudadas pelo autor. Segundo o autor, esta
espécie apresentou comportamentos diferenciados de acordo com a espécie de planta visitada.
Em E. speciosa e P. sagenarius as visitas ocorreram em intervalos regulares e ao longo do
dia, com um padrão de forrageamento “trapliner” ou rotas de captura, também observado em
B. antiacantha. Porém, em M. arboreus, T. glaucopis defendeu o território contra outras
espécies de beija-flores, C. flaveola e borboletas (Rodrigues, 2008). O beija-flor Florisuga
fusca também foi observado visitando esporadicamente as flores de S. tomentosa e V.

55
neoglutinosa. Estas duas espécies apresentaram baixas taxas de visitação as flores de B.
antiacantha.
Dentre os insetos, as abelhas constituem o mais importante grupo em número e
diversidade de polinizadores das espécies de plantas (BAWA, 1990). A espécie Trigona
spinipes, principal abelha registrada em nosso estudo, apesar de ser considerada polinizadora,
pode ser favorecedora de autopolinização, devido ao fato de raramente mudar de
inflorescência, fato observado também por Canela e Sazima (2005). Esta espécie realizou
visitas ilegítimas as flores de Heliconia laneana var. flava ao longo do dia. Foi o principal
visitante ilegítimo (97% das visitas ilegítimas) roubando pólen e néctar (CRUZ, MELLO E
VAN SLUYS 2005).
Uma das espécies de bromélias estudadas por Santana e Machado (2010),
Hohenbergia ramageana mostrou-se a mais generalista, com corola curta e flores azuladas
atrativas para os insetos, tais como as abelhas. Apesar das visitas de insetos as flores desta
bromélia ser mínimas quando comparadas as visitas de beija-flores, os autores consideraram
as abelhas como polinizadoras, visto que as mesmas contataram os órgãos reprodutivos
durante as visitas e permanecem mais tempo nas flores do que as aves.
Nunes-Freitas & Rocha (2011) registraram, durante os anos de estudos, quase que
exclusivamente visitas de nove espécies de insetos da ordem Hymenoptera (três subfamílias:
Bombinae, Meliponinae e Euglossinae) as flores de C. microps, havendo em apenas uma das
estações reprodutivas, a visita da espécie de beija-flor Thalurania glaucopis. Ainda segundo
os autores, a composição da guilda de visitantes florais variou entre as quatro estações
reprodutivas.
Canela & Sazima (2005) classificam B. antiacantha como não autogâmica e
dependente de polinizadores. Ainda segundo estas autoras as flores de B. antiacantha
apresentam características ornitófilas e melitófilas, sendo polinizadas por estes dois tipos de
agentes e a contribuição de cada agente polinizador para o seu sucesso reprodutivo pode ser
avaliada por meio de estudos sobre a produção de sementes.
Os beija-flores são considerados vetores de pólen para aproximadamente 85% das
espécies de Bromeliaceae, com as demais espécies da família sendo visitadas por morcegos e
abelhas, em menor quantidade (CANELA & SAZIMA, 2003). Estas aves são apontadas como
os principais polinizadores de Bromelia antiacantha, porém alguns estudos também revelam a
ocorrência de outros visitantes florais, tais como abelhas e, menos freqüentemente, borboletas
e moscas (SANTOS, 2001; CANELA & SAZIMA, 2005). No presente estudo, ambos os

56
grupos, beija-flores e himenópteros, formaram a guilda de visitantes florais de B. antiacantha,
sendo Trigona spinipes a espécie de abelha mais frequente e Amazilia fimbriata a espécies de
beija-flor. Apesar da grande porcentagem de indivíduos de Trigona spinipes visitando as
flores, estes raramente mudavam de inflorescência, muitas vezes sendo contabilizados ao
longo de todo o dia no mesmo indivíduo ou em indivíduos próximos. Já as visitas de Amazilia
fimbriata ocorreram mais espaçadamente.

57
6 CONCLUSÃO
A distribuição de Bromelia antiacantha ao longo da restinga da Praia Grande
apresenta-se em processo de expansão na área, indicado pelo aumento do número de
indivíduos entre as estações de amostragem. No entanto estudos abrangendo maiores períodos
de tempo são necessários para verificar possíveis padrões de estabilização ou declínio da
população em determinadas épocas.
O crescimento clonal da espécie possui importantes funções para a manutenção de sua
população e para o seu processo de estabelecimento na área. A espécie exibe um padrão de
floração anual, ocorrente no período das chuvas e a frutificação ocorre entre o final da estação
chuvosa e o inicio da estação seca, estando relacionada, mesmo que fracamente, de
determinadas variáveis climáticas.
Os principais visitantes florais de B. antiacantha são beija-flores e abelhas. A
contribuição de cada visitante para o sucesso reprodutivo da espécie, porém, deve ser
comprovada a partir de testes de germinação de suas sementes.

58
7 REFERENCIAS BIBLIOGRÁFICAS
ALMEIDA, E.M. 2000. Fenologia, distribuição espacial e polinização de duas espécies de
Psychotria (Rubiaceae) em área de Floresta Atlântica, Rio de Janeiro. Dissertação.
Universidade Federal do Rio de Janeiro, RJ.
ALMEIDA, E.M.; STORNI, A.; RITTER, P.D.; ALVES, M.A.S. Floral visitors of Tillandsia
stricta Sol. (Bromeliaceae) at Restinga of Jurubatiba, Macaé, Rio de Janeiro, Brazil. Vidalia,
V. 2, n. 1, p. 30-35, 2004.
ANACLETO, A.; NEGRELLE, R.R.B. Extrativismo de rametes e propagação vegetativa de
Aechmea nudicaulis (L.) Griseb. (Bromeliaceae). Scientia Agraria, Curitiba, v.10, n.1,
p.085-088, 2009.
ANTONINI, R.D. Frugivoria e dispersão de sementes por aves em duas espécies de
Miconia (Melastomataceae) em uma área de Mata Atlântica na Ilha da Marambaia, RJ.
Dissertação. Universidade Federal Rural do Rio de Janeiro. Instituto de Biologia. Programa
de Pós-graduação em Biologia Animal. 78p. 2007.
ARAUJO, D.S.D.; HENRIQUES, R.P.B. Análise florística das restingas do Estado do Rio
de Janeiro. In Restingas: origem, estrutura, processos (L.D. Lacerda, D.S.D. Araujo,
R.Cerqueira & B.Turcq, orgs.).CEUFF, Niterói, p.159-193. 1984.
ARAUJO, D.S.D.; OLIVEIRA, R.R. Reserva Biológica Estadual da Praia do Sul (Ilha
Grande, estado do Rio de Janeiro): Lista preliminar da flora. Acta bot. bras. 1(2):83-94 supl.
1988.
ARAUJO, D.S.D., SCARANO, F.R., SÁ, C.F.C., KURTZ, B.C., ZALUAR, H.L.T.,
MONTEZUMA, R.C.M.; OLIVEIRA, R.C. Comunidades vegetais do Parque Nacional da
Restinga de Jurubatiba.In Ecologia das lagoas costeiras do Parque Nacional da Restinga
de Jurubatiba e do Município de Macaé (RJ) (F.A. Esteves, ed.). Universidade Federal do
Rio de Janeiro, Rio de Janeiro, p.39-62. 1998.
BAWA, S.K. Plant-pollinator interactions in tropical rain forest. Annu. Rev. Ecol. Syst., Palo
Alto, V. 21, p. 399-422, 1990.
BAZZAZ, F.A. Habitat selection in plants. Am. Nat. 137, S116-S130. 1991.

59
BEDUSCHI, T. & CASTELLANI, T.T. Estrutura populacional de Clusia criuva Cambess.
(Clusiaceae) e relação espacial com espécies de bromélias no Parque Municipal das Dunas da
lagoa da Conceição, Florianópolis, SC. Biotemas, 21(2): 41-50. 2008.
BENATTO, L. et al. Construção de sistemas agroflorestais dirigidos pela sucessão
natural em um assentamento da reforma agrária, RS – Brasil. Projeto: Diretrizes para
Revitalização do Espaço Sócio-Produtivo do Assentamento 19 de Setembro em Guaíba - RS –
Programa Universidade Solidária - Pro-Reitoria de Extensão – Universidade Federal do Rio
Grande do Sul.
BENZING, D.H. Biology of Bromeliads. Mad River Press, California, 305pp. 1980.
BENZING, D.H. Bromeliaceae: Profile and Adaptative Radiation. Cambridge, Cambridge,
University Press. 2000.
CAIN, M. L.; DUDLE, D. A.; EVANS, J. P. Spatial models of foraging in clonal plant
species. American Journal of Botany, 83 (1): 76-85, 1996.
COGLIATTI-CARVALHO, L., NUNES-FREITAS, A.F., ROCHA, C.F.D.; VAN SLUYS,
M. 2001. Variação na estrutura e na composição de Bromeliaceae em cinco zonas de restinga
no Parque Nacional da Restinga de Jurubatiba, Macaé, RJ. Revta brasil. Bot., São Paulo,
V.24, n.1, p.1-9.
CORREIA, C.M.B., DIAS, A.T.C., & SCARANO, F.R. Plant-plant associations and
population structure of four woody plant species in a patchy coastal vegetation of
Southeastern Brazil Revista Brasil. Bot. V.33, n.4, p.607-613, out.-dec. 2010.
CANELA, M.B.F. & SAZIMA, M. Aechmea pectinata: a hummingbird-dependent Bromeliad
with inconspicuous flowers from the rainforest in Southeastern Brazil. Annals of Botany 92:
731-737. 2003.
CANELA, M.B.F. & SAZIMA, M. The Pollination of Bromelia antiacantha (Bromeliaceae)
in Southeastern Brazil: Ornithophilous versus Melittophilous Features. Plant Biol. 7 411–
416. 2005.
CRUZ, D.D.; MELLO, A.R.; VAN SLUYS, M. Phenology and floral vistors of two
sympatric Heliconia species in the Brazilian Atlantic Forest. Flora, n. 201, p. 519 -527, 2006.
DUARTE, A. S. et al. Bromelia antiacantha – uma espécie nativa de múltiplo uso. NPFT –
Núcleo de Pesquisa em Florestas Tropicais / UFSC - Universidade Federal de Santa Catarina.

60
DUARTE, A.S. et al. Estrutura demográfica e produção de frutos de Bromelia antiacantha
Bertol. Revista Brasileira de Plantas Medicinais, Botucatu, v.9, n.3, p.106-112. 2007.
FILIPPON, S. Aspectos da demografia, fenologia e uso tradicional do Caraguatá
(Bromelia antiacantha Bertol.) no Planalto Norte Catarinense. [dissertação] Universidade
Federal de Santa Catarina, Centro de Ciências Agrárias, Departamento de Fitotecnia,
Programa de Pós-graduação em Recursos Genéticos Vegetais. Florianópolis, SC. 116p. 2009.
GIVNISH, T.J.; MILLAM, K.C.; BERRY, P.E.; SYTSMA, K.J. Phylogeny, adaptive
radiation, and historical biogeography of Bromeliaceae inferred from ndhF sequence data.
Aliso 23, pp. 3-26. Rancho Santa Ana Botanic Garden, Claremont, CA. 2007.
GOTTSBERGER, G.; SILBERBAUER-GOTTSBERGER, I. Life in the Cerrado: a South
American tropical seasonal ecosystem. Reta, Ulm. 2006.
GÜTSCHOW-BENTO, L. H.; CASTELLANI, T. T.; LOPES, B. C.; GODINHO, P. S.
Estratégia de crescimento clonal e fenologia de Syngonanthus chrysanthus Ruhland
(Eriocaulaceae) nas baixadas entre dunas da Praia da Joaquina, Florianópolis, SC, Brasil.
Acta bot. bras. 24(1): 205-213. 2010.
HARPER, J.L. Population Biology of Plants. Academic Press, London. 1977.
HUTCHINGS, M.J. & WIJESINGHE, D.K. Patchy habitats, division of labour and growth
dividends in clonal plants. Trends in Ecology & Evolution, 12: 390-394. 1997.
JANZEN, D.H. Sweep samples of tropical foliage insects: effects of seasons, vegetation
types, elevation, time of day, and insularity. Ecology 54(3): 687–708. 1973.
JUSTINO, D.G.; MARUYAMA, P.K. & OLIVEIRA, P.E. Floral resource availability and
hummingbird territorial behavior on a Neotropical savanna shrub. J Ornithol 153:189-197.
2012.
LOH, R.K.T.M. 2010. Estrutura clonal e estrutura genética espacial de Aechmea
nudicaulis em restinga no Sudeste brasileiro. Tese de doutorado. Universidade Federal do
Rio de Janeiro.
LUTHER, H.E. 2008. An alphabetical list of Bromeliad binomials. 11th ed. The Bromeliad
Society International, Sarasota. 114p.

61
MACHADO, C.G.; SEMIR, J. Fenologia da floração e biologia floral de bromeliáceas
ornitófilas de uma área da Mata Atlântica do Sudeste brasileiro. Revista Brasil. Bot., V.29,
n.1, p.163-174, jan-mar. 2006.
MANTUANO, D. G. Crescimento clonal em Neoregelia cruenta na Restinga de
Jurubatiba: estrutura populacional, plasticidade morfo-anatômica e integração
fisiológica. [tese] Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Escola Nacional
de Botânica Tropical. Programa de Pós-graduação em Botânica. 113p. 2008.
MANTUANO, D.G.; MARTINELLI, G. Estrutura Populacional e Crescimento da bromélia
clonal Neoregelia cruenta na Restinga de Jurubatiba. Revista Brasileira de Biociências,
Porto Alegre, v. 5, supl. 1, p. 876-878, jul. 2007.
MARQUES, A.R. & LEMOS FILHO, J.P. Fenologia reprodutiva de espécies de bromélias na
Serra da Piedade, MG, Brasil. Acta bot. bras. 22(2): 417-424. 2008.
MARTINELLI, G.; VIEIRA, C.M.; GONZALEZ, M.; LEITMAN, P.; PIRATININGA, A.;
COSTA, A.F. & FORZZA, R.C. Bromeliaceae da Mata Atlântica brasileira: lista de espécies,
distribuição e conservação. Rodriguésia 59(1): 209-258. 2008.
MATTOS, C.C.L.V. Caracterização Climática da Restinga da Marambaia, RJ. In: História
Natural da Marambaia. Editora da Universidade Federal Rural do Rio de Janeiro,
Seropédica. p. 55-66, 2005.
MEIRELES, A.S. Fenologia e ecologia da polinização de duas espécies de Bromeliaceae
na Ilha da Marambaia, RJ. [dissertação] Universidade Federal Rural do Rio de Janeiro.
Instituto de Florestas. Programa de Pós-graduação em Ciências Ambientais e Florestais. 2011.
MENEZES, L.F.T. Caracterização de comunidades vegetais praianas da Restinga da
Marambaia, R.J. [Dissertação]. Universidade Federal Rural do Rio de Janeiro. 1996, 89p.
MENEZES, L.F.T. & ARAUJO, D.S.D. Formações vegetais da Ilha da Marambaia. In:
História Natural da Marambaia. Editora da Universidade Federal Rural do Rio de Janeiro,
Seropédica. p. 67-120. 2005.
MOREIRA, M. J. S. Conservação in vitro de bromeliáceas. [Dissertação] Universidade
Federal do Recôncavo da Bahia. Centro de ciências agrárias, ambientais e biológicas, Cruz
das almas - Bahia. 2008.

62
MURAWSKI, D.A.; HAMRICK, J.L. Local genetic and clonal structure in the tropical
terrestrial bromeliad, Aechmea magdalenae. American Journal of Botany, 77(9): 1201-
1208. 1990.
NEGRELLE, R.R.B.; MURATO, D. Aspectos fenológicos e reprodutivos de Vriesea
incurvata Gaudich (Bromeliaceae). Acta Sci. Biol. Sci. Maringá, v. 28, n. 2. p. 95-102,
abril/junho, 2006.
NOGUEIRA, C.A.; CÔRTES, I.M.R. & VERÇOZA, F.C. A família Bromeliaceae na Área de
Proteção Ambiental de Grumari, Rio de Janeiro, RJ, Brasil. Natureza on line 9 (2): 91-95.
NUNES-FREITAS, A.F.; ROCHA-PESSOÔA, T.C.; COGLIATTI-CARVALHO, L.;
ROCHA, C.F.D. Bromeliaceae da restinga da Reserva Biológica Estadual da Praia do Sul:
composição, abundância e similaridade da comunidade. Acta bot. bras. 20(3): 709-717.
2006.
NUNES-FREITAS, A.F.; ROCHA, C.F.D. 'Reproductive phenology and flower visitors'
guild of Canistropsis microps (Bromeliaceae) in an Atlantic Rainforest of southeastern Brazil.
Journal of Natural History, 45: 17, 1021-1034. 2011.
OLIVEIRA, W. F. Estrutura da taxocenose de Bromeliaceae em dois fragmentos de
restinga da Ilha da Marambaia, Baía de Sepetiba, RJ. (Dissertação de mestrado).
Universidade Federal Rural do Rio de Janeiro. 2010.
PAGGI, G.M.; SAMPAIO, J.A.T.; BRUXEL, M.; ZANELLA, C.M.; GOETZE, M.;
BUTTOW, M.V.; PALMA-SILVA, C.; BERED, F. Seed dispersal and population structure in
Vriesea gigantea, a bromeliad from the Brazilian Atlantic Rainforest. Botanical Journal of
the Linnean Society, 2010, 164, 317-325.
PEREIRA, L.A.; XEREZ, R.; PEREIRA, A.J. Ilha da Marambaia (baía de Sepetiba, RJ):
resumo fisiográfico, histórico e importância ecológica atual. Revista da Sociedade
Brasileira para o Progresso da Ciência [S.I] 42(5/6):384-389. 1990.
PEREIRA, M.C.A.; ARAUJO, D.S.D.; PEREIRA, O.J. Estrutura de uma comunidade
arbustiva da restinga de Barra de Maricá - RJ. Revta Brasil. Bot., São Paulo, V.24, n.3,
p.273-281. 2001.
PEREIRA, O.J.; ARAÚJO, D.S.D. Estrutura da vegetação de entre moitas da formação aberta
de Ericaceae no Parque Estadual de Setiba, ES. Oecologia Brasiliensis 1: 245-257; 1995.
Rocha, C.F.D. et al. Bromélias: Ampliadoras da Biodiversidade. Bromélia, V. 4, n. 4, p. 7-10,
1997.

63
REITZ, R. Bromeliaceas e a malária – Bromélia endêmica. In: Flora Ilustrada Catarinense.
Itajaí: Herbário Barbosa Rodrigues, 1983. 856p.
ROCHA, C.F.D. et al. Bromélias: Amplificadoras de Biodiversidade. Bromelia, v.4, n.4, p.7-
10, 1997.
ROCHA, C.F.D.; COGLIATTI-CARVALHO, L.; FREITAS, A.F.N.; PESSÔA, T.C.R.;
DIAS, A.S.; ARIANI, C.V.; MORGADO, L.N. Conservando uma larga porção da
diversidade biológica através da conservação de Bromeliaceae. Vidália, v.2, n.1, p.52-68,
2004.
ROCHA-PESSÔA, T.C. Ecologia de Pitcairnia flammea Lindley (Bromeliaceae:
Piticairnioidea) em uma área insular de Mata Atlântica no Estado do Rio e Janeiro. [Dissertação]. Universidade Estadual do Rio de Janeiro. Instituto de Biologia Roberto
Alcântara Gomes. Programa de Pós-graduação em Biologia, Área de Concentração em
Ecologia. 2004. 69 p.
ROCHA-PESSOÔA, T.C. & ROCHA, C.F.D. Ciclo reprodutivo de Pitcairnia flammea Lindl.
(Bromeliaceae / Pitcairnioidae) em uma área de floresta tropical atlântica insular no sudeste
do Brasil. Flora 203: 229 – 233. 2008.
ROCHA-PESSÔA, TC.; NUNES-FREITAS, AF.; COGLIATTI-CARVALHO, L.; ROCHA,
CFD. Species composition of Bromeliaceae and their distribution at the Massambaba restinga
in Arraial do Cabo, Rio de Janeiro, Brazil. Braz. J. Biol., 68(2): 251-257. 2008.
ROGALSKI, J.M.; REIS, A.; REIS, M.S.; NETO, C.D. Estrutura demográfica da bromélia
clonal Dyckia brevefolia Baker, Rio Itajaí-Açu, SC. Revista Brasileira de Biociências, Porto
Alegre, v.5, supl. 1, p.264-266, jul. 2007.
RODRIGUES, M.S. Partilha de recursos florais por beija-flores em uma área de Mata
Atlântica na Ilha da Marambaia, RJ. [dissertação] Universidade Federal Rural do Rio de
Janeiro. Instituto de Biologia. Programa de Pós-graduação em Biologia Animal. 2008.
ROGALSKI, J.M. Biologia da Conservação da reófita Dyckia brevifolia Baker
(Bromeliaceae), Rio Itajaí-Açu, SC. Dissertação. Universidade Federal de Santa Catarina.
Programa de Pós-Graduação em Recursos Genéticos Vegetais, 2007.
SALZMAN, A.G. (1985) Habitat selection in a clonal plant. Science 228, 603–604.

64
SAMPAIO, M.C.; PERISSÉ, L.E.; OLIVEIRA, G.A.; RIOS, R.I. The contrasting clonal
architecture of two bromeliads from sandy coastal plains in Brazil. Flora 197, 443–451. 2002.
SAMPAIO, M. C.; ARAÚJO, T. F.; SCARANO, F. R.; STUEFER, J. F. Directional growth
of a clonal bromeliad species in response to spatial habitat heterogeneity. Evolutionary
Ecology 18: 429–442. 2004.
SAMPAIO, M.C.; PICÓ, X.; SCARANO, F.R. Ramet demography of a nurse bromeliad in
Brazilian restingas. American Journal of Botany, 92(4): 674-681. 2005.
SANTANA, C.S.; MACHADO, C.G. Fenologia de floração e polinização de espécies
ornitófilas de bromeliáceas em uma área de campo rupestre da Chapada Diamantina, BA,
Brasil. Revista Brasil. Bot. Vol.33, n.3, p.469-477, jul-set. 2010.
SANTOS, D.S. 2001. Biologia reprodutiva de Bromelia antiacantha Bertol.
(Bromeliaceae) em uma população natural sob cobertura de floresta ombrófila mista. Dissertação. Universidade Federal de Santa Catarina. Mestrado em Recursos Genéticos
Vegetais, Florianópolis. 96p.
SANTOS, D.S. et al. 2004.Variação no período de geminação de sementes em uma população
natural de Bromelia antiacantha (Bertol). Revista Brasileira de Plantas Medicinais, v.6,
n.3, p.35-41.
SARTHOU, C., SAMADI, S.; BOISSELIER-DUBAYLE, M. Genetic structure of the
saxicole Pitcairnia geyskesii (Bromeliaceae) on inselbergs in French Guiana. American
Journal of Botany 88(5): 861–868. 2001.
SCARANO, F.R. Structure, Function and Floristic Relationships of Plant Communities in
Stressful Habitats Marginal to the Brazilian Atlantic Rainforest. Annals of Botany 90: 517-
524, 2002.
SCARANO, F.R.; DUARTE, H.M.; RÔÇAS, G.; BARRETO, M.B.; AMADO, E.F.;
REINERT, F.; WENDT, T.; MANTOVANI, A.; LIMA, H.R.P.; BARROS, C.F. Acclimation
or stress sympton? An integrated study of intraspecific variation in the clonal plant Aechmea
bromelifolia, a widespread CAM tank-bromeliad. Botanical Journal of the Linnean Society,
140: 391-401, 2002.
SILVA, J.G.; OLIVEIRA, A.S. 1989. A vegetação de restinga no município de Maricá - RJ.
Acta bot. bras. 3(2):253-272.

65
SILVA, A.L.G.; PINHEIRO M.C.B. 2007. Biologia floral e polinização de quatro espécies de
Eugenia L. (Myrtaceae). Acta Botanica Brasilica 21: 235-247.
SOUZA, M.C.; MORIM, M.P.; CONDE, M.M.S.; MENEZES, L.F.T. Subtribo Myrciinae O.
Berg (Myrtaceae) na Restinga da Marambaia, RJ, Brasil. Acta bot. bras. 21(1): 49-63. 2007.
SMITH, L.B.; DOWNS, R.J. (1979) Bromelioideae (Bromeliaceae). In Flora Neotropica 14,
Part 3. New York: The New York Botanical Garden, Hafner Press, pp.1658–1660, citado por
Canela, M. B. F. & Sazima, M. 2005 The Pollination of Bromelia antiacantha (Bromeliaceae)
in Southeastern Brazil: Ornithophilous versus Melittophilous Features. Plant Biol. 7 411–416.
TALORA, D.C.; MORELLATO, P.C. Fenologia de espécies arbóreas em floresta de planície
litorânea do sudeste do Brasil. Revista Brasileira de Botânica 23:13-26. 2000.
WIKBERG, S. Fitness in clonal plants. Oikos, 72 (2): 293-297, 1995.
ZALUAR, H.L.T.; SCARANO, F.R. Facilitação em restingas de moitas: um século de
buscas por espécies focais. In F.A. Esteves and L.D. Lacerda (eds.), Ecologia de Restingas e
Lagoas Costeiras NUPEM-UFRJ, Rio de Janeiro, pp.3–23. 2000.
ZANELLA, C.M. 2009. Caracterização genética, morfológica e fitoquímica de
populações de Bromelia antiacantha (Bertol.) do Rio grande do Sul. Dissertação
(Mestrado em Genética e Biologia Molecular) - Universidade Federal do Rio Grande do Sul.
98p.