trabalho de conclusão de curso - gaby cardoso de lima - orientador: milton luiz da paz lima
TRANSCRIPT

INSTITUTO FEDERAL GOIANO câmpus URUTAÍ
Curso de Ciências Biológicas
ABDIAS RODRIGUES DA MATA NETO
CARACTERIZAÇÃO BIOQUÍMICA DE ISOLADOS DE Colletotrichum gloeosporioides.
URUTAÍ, GO, 16 de dezembro de 2014
1

ABDIAS RODRIGUES DA MATA NETO
CARACTERIZAÇÃO BIOQUÍMICA DE ISOLADOS DE Colletotrichum gloeosporioides.
Trabalho de Conclusão de Curso apresentado para obtenção do grau de Licenciado em Ciências Biológicas ao Instituto Federal Goiano – Câmpus Urutaí.
Orientador: Prof. D.Sc. Milton Luiz da Paz Lima.
URUTAÍ, GO, 16 de dezembro de 2014
2

ABDIAS RODRIGUES DA MATA NETO
CARACTERIZAÇÃO BIOQUÍMICA DE ISOLADOS DE Colletotrichum gloeosporioides.
COMISSÃO EXAMINADORA
______________________________D.Sc. Milton Luiz da Paz Lima
Orientador(Instituto Federal Goiano campus Urutaí)
______________________________D.Sc. Gleina Costa Membro avaliador
(Instituto Federal Goiano campus Urutaí)
______________________________M.Sc. Gesiane Ribeiro Guimarães
Membro avaliador (Universidade Estadual de Goiás UnU Ipameri)
Urutaí, 16 de dezembro de 2014
3

4
Agradeço a Deus por todas as vitórias concebidas em minha vida.

SUMÁRIO
RESUMO .................................................................................................................................. 7
ABSTRACT .............................................................................................................................. 8
RESULTADOS E DISCUSSÃO ............................................................................................ 17
CONCLUSÕES ....................................................................................................................... 24
5

LISTA DE TABELAS E FIGURAS
Tabela 1. Informações dos hospedeiros por onde os isolados foram obtidos considerando o nome científico, família botânica e local de coleta. ________________________________15
Tabela 2. Valores F da análise de variância dos fatores hospedeiros de origem e tratamentos e os valores dos parâmetros das variáveis dependentes peso seco de micélio e área abaixo da curva de progresso do crescimento micelial (AACPCM).____________________________18
Tabela 3. Médias dos pesos secos de micélio (PS) e dos Valores de área abaixo da curva de progresso do crescimento micelial (AACPCM) dos isolados submetidos a diferentes concentrações de carbono e nitrogênio (C/N).____________________________________________________________________19
Tabela 4. Médias dos pesos secos do micélio (PS) e área abaixo da curva de progresso do crescimento micelial (AACPCM) nos diferentes tratamentos contendo fontes de carbono e nitrogênio.________________________________________________________________20
Figura 1. Efeito dos tratamentos 1 a 5 na área abaixo da curva de progresso do crescimento micelial (AACPCM) de oito isolados apresentados.________________________________21
Figura 2. Efeito dos tratamentos 6 a 9 na área abaixo da curva de progresso do crescimento micelial (AACPCM) de oito isolados apresentados.________________________________21
Figura 3. Efeito dos tratamentos 1 a 5 na taxa de crescimento de oito isolados.___________22
Figura 4. Efeito dos tratamentos 6 a 9 na taxa de crescimento de oito isolados.___________22
6

RESUMO
MATA NETO, A.R. Caracterização bioquímica de isolados de Colletotrichum gloeosporioides. Trabalho de Conclusão de Curso. 2014.
A nutrição de C/N exerce efeito nos processos fisiológicos de fungos, principalmente aqueles relacionados ao crescimento, produção de conídios, germinação, peso seco e permite também, estabelecer diferenças bioquímicas entre Colletotrichum gloeosporioides, pela sua habilidade em usar determinada fonte de carbono e nitrogênio. Vinte e quatro isolados oriundos de 24 hospedeiras distintas foram incubados em meio, contendo os seguintes tratamentos (fontes de carbono x nitrogênio): i) dextrose x asparagina, ii) dextrose x triptona, iii) dextrose x nitrato de potássio, iv) sacarose x asparagina, v) sacarose x triptona, vi) sacarose x nitrato de potássio, vii) sorbitol x asparagina, viii) sorbitol x triptona, ix) sorbitol x nitrato de potássio, x) controle [BDA] crescidos sob duas repetições. As placas foram incubadas sob 12 horas de luz, a 25 oC, num delineamento inteiramente casualisado, sendo o fator fontes CN constituídos de 10 tratamentos, fator isolados 24 tratamentos, duas repetições, totalizando 480 UE. Foi avaliado o crescimento micelial, durante o período de sete dias, sendo calculado através de regressão a taxa de crescimento micelial (mm.dia-1) e a área abaixo da curva de progresso do crescimento micelial (AACPCM). A combinação sorbitol x nitrato de potássio promoveu as maiores taxas de crescimento para os isolados de arvore da fortuna, amora e banana. Com relação às combinações C/N, a análise estatística revelou diferença significativa entre os isolados, sob efeito da interação C/N, bem como dos fatores independentes, sobre o crescimento micelial, produção de esporos e peso seco do micélio. Os isolados de iuca e mandioca foram os que resultaram maior crescimento micelial. Através desse trabalho podemos verificar que isolados como de iuca e mandioca possuem atividade metabólica mais elevada em relação aos demais, e a fonte de C originada de dextrose é a que promove maior produção de micélio. Nenhuma fonte de N mereceu destaque.
Palavras-chave: crescimento micelial, carbono, nitrogênio, peso seco, nutrição.
7

ABSTRACT
MATA NETO, A.R. Biochemical characterization of Colletotrichum gloeosporioides. Course Final Report. 2014.
The nutrition using Carbon and nitrogen has an effect in the physiological processes of fungi, especially those related to growth, conidia production, germination, dry weight, it is enabling to establish biochemical differences between Colletotrichum gloeosporioides. For their ability to use certain carbon source and nitrogen. Twenty-four from 24 isolates from different hosts were incubated in medium containing the following treatments (carbon x nitrogen sources): i) asparagine x dextrose, ii) dextrose x tryptone, iii) dextrose x potassium nitrate, iv) sucrose x asparagine, v) sucrose x tryptone, vi) x sucrose x potassium nitrate vii) sorbitol x asparagine, viii) sorbitol x tryptone, ix) sorbitol x potassium nitrate, x) control [PDA] grown in two replications. The plates were incubated under 12 hours of light at 25 °C, in a completely randomized design, with the factor consisting C/N sources of 10 treatments, isolated factor 24 treatments, two replicates, totaling 480 experimental unit. Mycelial growth was evaluated during the seven-day period, calculated by regression mycelial growth rate (mm.dia-1) and the area under the mycelial growth progress curve (AACPCM). The combination sorbitol x potassium nitrate promoted the highest growth rates for the árvore-da-fortuna isolated, mulberry and banana. Regarding the C/N combinations, the statistical analysis revealed significant differences among the isolates, under the effect of C/N interaction, as well as independent factors, mycelial growth, sporulation and dry weight of the mycelium. Isolates of yucca and cassava were the resulting higher mycelial growth. Through this work we can see that isolated as yucca and cassava have higher metabolic activity than the other, and the source of C originated dextrose is promoting increased production of mycelium. No N source was highlighted.
Palavras-chave: micelial raise, carbon, nitrogên, dry weight, nutrition.
8

INTRODUÇÃO
O gênero Colletotrichum gloeosporioides, é um dos mais importantes fungos
fitopatogênicos do mundo, principalmente nas regiões tropicais e subtropicais. Suas espécies
ocasionam doenças de grande expressão econômica em leguminosas, cereais, hortaliças e
culturas perenes, incluindo diversas frutíferas (SERRA et al., 2008; SUTTON, 1992).
Existem registros por volta de 688 táxons, representados por variedades e formeae
speciales pertencentes ao gênero Colletotrichum gloeosporioides. Este fungo pertence ao
Reino Fungi, grupo dos Fungos Mitospóricos e sub-grupo dos Coelomicetos, e sua fase
teleomórfica pertencente ao gênero Glomerella sp. incluindo-se no Reino Fungi, Divisão
Ascomicota (KIRK et al.,2001).
Espécies de Colletotrichum gloeosporioides, são tradicionalmente diferenciados com
base em caracteres morfológicos e culturais, que devem ser considerados simultaneamente e
nunca isoladamente. Morfologicamente o fungo apresenta acérvulos circulares, conidióforos
curtos e simples. Conídios hialinos, gutulados, ovais, oblongos ou falcados. A massa de
conídios possui coloração rósea à salmão, e pode estar presente no acérvulo. Suas setas
podem ser longas, septadas e pigmentadas (GALLI, 1978). A antracnose é o nome da doença
causada por este patógeno, afeta principalmente frutíferas, hortaliças e as cultivares atacando
ramos novos, folhas, inflorescências e frutos. Nas folhas há o aparecimento de manchas
escuras, enegrecimento de nervuras e de contornos irregulares. As inflorescências afetadas
apresentam flores escuras, ou manchas petalares esparsas, apresentando aspecto de requeima,
crestamento ou morte do limbo em sua totalidade. As lesões na ráquis podem levar a queda
dos frutos, antes da maturação fisiológica, ou mumificação quando ainda novos. No período
de maturação, há o aparecimento de lesões escuras e deprimidas na superfície da planta, que
podem atingir também a polpa. A doença pode ocasionar prejuízos que variam em função do
grau de suscetibilidade da planta hospedeira e das condições ambientais (BERGER, 1998;
SERRA et al., 2008).
As antracnoses podem provocar perdas de até 100% na produção quando os fatores
cultivar suscetível, ambiente favorável ao patógeno e sementes infectadas estiverem
simultaneamente presentes durante o período de cultivo (SILVA, 2004). A disseminação
planta ocorre principalmente através dos respingos de chuva, pois os conídios estão
aglutinados por uma substância gelatinosa hidrossolúvel, e não são facilmente carregados pelo
vento. Os conídios uma vez em contato com o hospedeiro, sob condições de umidade,
9

germinaram e o micélio resultante, com prévia formação de apressórios, penetra diretamente
através da cutícula pela emissão do tubo de infecção (GALLI et al.,1978).
O carbono (C) e nitrogênio (N) são os mais importantes e essenciais elementos para
nutrição dos fungos, sendo utilizados no processo infectivo, crescimento e reprodução
(SANGEETHA & RAWAL, 2008). A nutrição de C/N exerce efeito nos processos
fisiológicos de Colletotrichum gloeosporioides, principalmente aqueles relacionados ao
crescimento, produção de conídios, germinação e peso seco. Podem ocasionar diferenças
bioquímicas entre Colletotrichum spp., pela sua habilidade em usar determinada fonte de
carbono e nitrogênio (COUTO & MENEZES, 2004). As populações de C. gloeosporioides
podem responder de forma diferenciada aos diferentes tipos de fontes de C ou N ou podem
preferir um tipo de elemento químico específico.
O objetivo desta monografia é caracterizar quanto ao uso de fontes de carbono e
nitrogênio dos isolados de Colletotrichum gloesporioides, que foram coletados de diferentes
hospedeiros.
10

REVISÃO LITERATURA
Esta espécie bastante polífaga e presente e muitas hospedeiras é considerada uma
espécie complexa devido apresentar imensa variabilidade genética dentre os isolados
analisados, contudo apresenta imensa variabilidade biológica e molecular (BROWN et al.,
1996; OHNSTON e JONES, 1997).Este táxon pertencente ao subgrupo dos Coelomicetos
apresenta como teleomorfo o ascomiceto Glomerella cingulata (Stonem). Spauld. e Von
Schrenk (1903) (INDEX FUNGORUM, 2014).
Em meio de cultura possui colônias variáveis, de coloração cinza claro a cinza escuro,
seu micélio é aéreo, formando aglomerações associadas ao acérvulo, no lado reverso da placa
também possui coloração branca acinzentada com a idade. As setas podem ou não estarem
presentes. Escleródios são ausentes. Apressórios clavados, sua forma pode apresentar-se
como ovados, obovados, algumas vezes lobados, de coloração marrom escura, apresenta
dimensões de 6-20 x 4-12 μm, seus conídios formados em massas de colorações salmão,
possuem formato reto, cilíndrico, ápice obtuso, base truncada, 12-17 x 3,5-6 μm. A produção
de conídios é bastante heterogênea em meio de cultura (SUTTON, 1992).
A variabilidade fisiológica de C. gloeosporioides é representada por 21 formas
especiales e variedades [C.gloeosporioides (C.g.) f.sp. alatae, C.g.f.sp. gloeosporioides,
C.g.f.sp. heveae, C.g.f.sp. melongenae, C.g.f.sp. nectrioides, C.g.f.sp. aeschynomenes, C.g.
f.sp. clidemiae, C.g.f.sp. cucurbitae, C.g.f.sp. cuscutae, C.g.f.sp. manihotis, C.g.f.sp. pilosae,
C.g. f.sp. uredinicola, C.g. var. aleritidis, C.g. var. cephalosporioides, C.g. var.
gloeosporioides, C.g. var. gomphrenae, C.g. var. hederae, C.g. var. minus, C.g. var. minus,
C.g. var. nectrioidea] registradas em literatura (INDEX FUNGORUM, 2014).
Os conídios germinam de seis a nove horas após o contato inicial com o hospedeiro, se
as condições lhes forem favoráveis. Há formação do tubo germinativo, seguido do apressório,
que penetra mecanicamente pela cutícula e pela epiderme da plantas. O aparecimento de
sintomas pode ser observado a partir de sexto dia após o início da infecção (SUTTON, 1992;
FREEMAN et al., 1998; LOPES, 2001).
As dificuldades encontradas na identificação das espécies de Colletotrichum sp. estão
relacionadas a grande diversidade fenotípica, influência de fatores ambientais na estabilidade
de critérios de identificação. O círculo de hospedeiros ou hospedeiro de origem não é um
critério confiável para identificação de espécies, por exemplo, como C. gloeosporioides e
C.acutatum que infectam as mesmas hospedeiras em complexos (FREEMAN et al., 1998;
11

PERES et al. 2005). É frequente a ocorrência de mais de uma espécie de Colletotrichum
associada a uma mesma hospedeira e uma mesma espécie pode atacar múltiplas hospedeiras
(FREEMAN et al., 1998). As três espécies causam antracnose, mas podridão do pedúnculo,
inflorescências e frutos conhecida como “flor preta” e atribuída a C.acutatum outras culturas
como a amêndoa (Prunus amygdalus L.) (FOSTER &ADASKAVEG, 1999), maçã (Malus
domestica Borkh.) (BERNSTEIN et al., 1995), abacate (Persea americana Mill.)
(JOHNSTON & JONES, 1997), pêssego (Prunus persica (L.) Batsch) (ADASKAVEG &
HARTIN, 997); citros (BROWN et al., 1996; GOES & KIMATI, 1997) e algumas solanáceas
(TOZZE Jr. et al., 2006) são infectadas por C. acutatum e C gloeosporioides.
A nutrição de C/N exerce efeito nos processos fisiológicos de fungos, principalmente
aqueles relacionados ao crescimento, produção de conídios, germinação e peso seco
(COCHRANE, 1958) e permite também, estabelecer diferenças entre os isolados e
Colletotrichum spp. pela habilidade em usar determinada fonte de carbono e nitrogênio.
A caracterização de populações de patógenos permite-nos identificar fontes de
variabilidade fisiológica e morfológica, indicando surgimento de subpopulações ou táxons
infra-supra-específicos que podem ser responsáveis por quebra de resistência ao hospedeiro.
Esta identificação implica na descoberta do processo de co-evolução do patógeno sobre o
hospedeiro sendo esta descoberta uma importante ferramenta em programas de
melhoramento.
Os complexos de espécies atuando sobre os hospedeiros podem resultar em
mecanismos de ação diferenciados e expressando fenotipicamente o que se conhece por
resistência a fungicidas quando que na verdade são nichos de parasitismo modificados e que
não são afetados pela atividade química de fungicidas.
A divergência existente em espécies ou populações de Colletotrichum spp. existentes
na natureza é bastante ampla, tornando aspectos da identificação utilizando critérios
morfológicos um procedimento complexo e impreciso. A própria espécie de Colletotrichum
gloeosporioides que apresenta ampla gama de hospedeiras (inespecificidade), não apresenta
diferenças morfológicas para pode apresentar diferenças fisiológicas de interação planta-
patógeno.
Agregar outros métodos de diferenciação bioquímica permite separar, e reconhecer
estas diferenças fisiológicas e/ou bioquímicas que está população desenvolve permitir maior
robustez na caracterização e reconhecimento das populações presentes no ambiente.
12

O consumo de C e N representa um dos principais componentes inorgânicos
metabolizados pelas hifas fúngicas, desta forma o consumo diferencial permitirá separar
grupos de fungos com similaridades fisiológicas e bioquímicas, e por meio de analogias estas
similaridades apresentarão confluências com outros critérios de caracterização da população
estudada.
13

MATERIAIS E MÉTODOS
O experimento foi realizado no Laboratório de Microbiologia e Fitopatologia do
Instituto Federal Goiano Campus-Urutaí, sendo representado pelos fatores tratamento (nove
tipos de combinações C/N), fator hospedeiro de origem 24 tipos (mangaba, negramina, conde,
abacate, banana, manga, goiaba folha, falso-massambará, caqui, antúrio, árvore-da-fortuna,
uva, cana, mandioca, goiaba fruto, amora, banana fruto, iuca, chuchu, soja, tomate, coco,
mamão e dracena), sendo assim um experimento inteiramente casualizado em fatorial (fator
tratamento e fator hospedeiro de origem) com duas repetições e duas testemunhas.
As fontes de carbono utilizadas neste estudo (dextrose, sacarose e sorbitol) foram
combinadas com três fontes de nitrogênio (asparagina, peptona e nitrato de potássio), na
proporção de 10:1 (10 g de C para 1 g de N). O meio basal para adição das combinações
carbono/nitrogênio foi composto de: 0,5 g de MgSO4.7H2O; 1,0 g de KH2PO4; 17 g de ágar,
1.000 mL de água destilada (LILLY & BARNETT, 1951), sendo o pH ajustado para 5,5.
Após a autoclavagem, os meios foram vertidos em placas de Petri, num volume aproximado
de 20 mL/placa. Discos de micélio (6,0 mm de diâmetro) dos 24 isolados (Tab. 1), oriundos
de colônias matrizes jovens (cinco dias de crescimento), foram transferidos para o centro das
placas de Petri contendo os tratamentos (CxN), e as culturas foram incubadas em condições
de claro contínuo e temperatura de 25 °C, durante sete dias. A avaliação do crescimento
micelial consistiu na determinação do diâmetro das colônias (mm) de cada isolado e
combinações de C/N (Tab. 4), usando-se a média de duas leituras efetuadas em dois sentidos
diametralmente opostos.
Para a determinação do peso seco, os isolados foram cultivados individualmente
durante cinco dias em frascos de Erlenmeyer, sem agitação das culturas, contendo 50 mL de
cada combinação C/N, adicionados ao meio líquido basal. As condições de incubação foram
as mesmas anteriormente citadas. Ao final do período de incubação, as culturas foram
filtradas em gaze dupla, e as massas micelial coletadas foram depositadas em caixas de papel
alumínio, com peso previamente determinado e isentas de umidade. As caixas foram
colocadas em estufa a 50 °C, durante quatro dias e, ao final deste período, foram determinadas
por diferença o peso seco da massa micelial de cada isolado, o qual foram expressos em mg.
14

Nome científico Família botânica Local de coleta1 Abacate Persea americana Lauraceae Urutaí/GO2 Amora Morus nigra Moraceae Urutaí/GO3 Antúrio Anthurium andraeanum Araceae Ipameri/GO4 Árvore da Fortuna Polycias frusticosa Araliaceae Urutaí/GO5 Banana Musa paradisiaca Musaceae Brasília/DF6 Banana (folha) Musa paradisiaca Musaceae Brasília/DF7 Cana Saccharum officinalis Poaceae Urutaí/GO8 Caqui Diospyros kaki Ebenaceae Urutaí/GO9 Chuchu Sechium edule Cucurbitaceae Brasília/DF
10 Coco Cocos nucifera Arecaceae Urutaí/GO11 Conde Annona coriacea Annonaceae Urutaí/GO12 Dracena Dracena fragans Liliaceae Urutaí/GO13 Falso Massambará Sorghum arundinaceum Poaceae Urutaí/GO14 Goiaba Psidium guajava Myrtaceae Brasília/DF15 Goiaba (folha) Psidium guajava Myrtaceae Brasília/DF16 Iuca Yucca elephantipes Agavaceae Brasília/DF17 Mamão Carica papaia Caricaceae Brasília/DF18 Mandioca Manihot esculenta Euphorbiaceae Urutaí/GO19 Manga Anacardium mangiferae Anacardiaceae Brasília/DF20 Mangaba Hancornia speciosa Apocynaceae Goiânia/Go21 Negramina Sipurana guianensis Siparunaceae Urutaí/GO22 Soja Glycine max Fabacaeae Urutaí/GO23 Tomate Solanum lycopersicum Solanaceae Urutaí/GO
24 Uva Vitis vinifera Vitaceae Brasília/DF
Hospedeiro
Tabela 1. Informações dos hospedeiros por onde os isolados foram obtidos considerando o nome científico, família botânica e local de coleta.
O diâmetro micelial foi avaliado diariamente durante um período de 7 dias,
resultando no progresso temporal. A integralização dos valores de dias e diâmetro da colônia
matematicamente resultou no valor de AACPCM. Com os dados do diâmetro da colônia
coletados em diferentes avaliações, nos tratamentos, calculou-se a taxa de crescimento (TC)
e a área abaixo da curva de progresso do crescimento micelial (AACPCM) pelo método da
integralização trapezoidal (BERGER, 1988). Para o cálculo da AACPCM, utilizou-se a
seguinte fórmula: , em que: n = número
de avaliações, y = diâmetro da colônia (mm), t = tempo quando da avaliação do diâmetro da
colônia, (yi + yi + 1) = altura média do retângulo entre os pontos yi e yi + 1, ti + 1 =
diferença da base do retângulo entre os pontos ti + 1 e ti. A curva de progresso do
crescimento micelial pode ser expressa pela proporção de diâmetro micelial x tempo.
Os valores do peso seco micelial e de AACPCM foram utilizados para a realização
da análise de variância (ANOVA), os fatores individuais e as interações do experimento em
15

fatorial. Além da ANOVA foi realizado o teste Tukey, para identificar a significância das
medidas dos fatores individuais utilizando o programa “SAS for windows”.
16

RESULTADOS E DISCUSSÃO
Todos os isolados estudados foram identificados como pertencentes a espécie
Colletotrichum gloeosporioides. A principais fonte de carbono para os isolados testados
continha em suas combinações dextrose e sorbitol, pois os maiores valores de AACPCM e
taxa de Crescimento (TC), foram para tratamentos que continham essas fontes de C. A
sacarose não demonstrou uma fonte de C eficiente para a morfofisiologia dos isolados (Fig
1). Cinquenta % dos isolados (árvore-da-fortuna, banana fruto, caqui e conde) apresentaram
maiores valores de AACPCM para a combinação dextrose e nitrato de potássio. Trinta e oito
% dos isolados (banana folha, banana fruto e caqui (folha), tiveram TC maiores para
isolados também submetidos a combinação de dextrose e nitrato de postássio (Fig 1). Os
isolados de conde, chuchu e árvore da fortuna apresentaram os menores valores médios de
AACPCM e TC.
Em geral, as médias do crescimento micelial dos isolados nas combinações das fontes
de carbono (dextrose, sacarose, sorbitol) e como fontes de nitrogênio (asparagina, peptona e
nitrato de potássio), demonstram a diferenças na habilidade em utilizar diferentes fontes de
carbono e nitrogênio, entre isolados de uma mesma espécie, e esta informação também foi
constatada por, Tandon & Chandra (1962), para C. gloeosporioides.
As combinações dextrose/ peptona e sacarose/peptona foram as que induziram
maiores médias de peso seco da massa micelial, 5,3 mg e 5,4 mg, respectivamente,
destacando-se significativamente das médias das demais fontes C/N. Na combinação
sacarose/peptona, não ocorreram diferenças significativas entre os isolados, o mesmo
acontecendo nas combinações de dextrose/asparagina (T1) e sorbitol/peptona(T8). (Tabela 2,
3 e 4). Mata Neto et al. (2011) registraram o efeito de C/N no crescimento de isolados de
Colletotrichum gloesporioides. E verificaram esses resultados.
Analisando-se os dados do crescimento micelial e peso seco do micélio, constatou-se
uma correlação positiva entre o crescimento micelial e o peso seco do micélio, r(Pearson) =
0,5121, ou seja, os isolados que demonstraram maior habilidade de crescimento no meio
sólido, tiveram também maior peso seco da massa micelial no meio líquido.
Segundo Tandon & Chandra (1962), um bom crescimento micelial está associado a
uma boa esporulação. No presente estudo foi verificado que alguns isolados apresentaram
boa produção de conídios, aliada a elevado crescimento em meio sólido e líquido, 17

confirmando os dados dos autores mencionados. Às vezes, um meio ótimo para o
crescimento rápido resulta na exaustão dos nutrientes e os metabólitos secundários liberados
pelo fungo nesse meio, inibem a produção de conídios (GRIFFIN,1994). Por outro lado, o
crescimento micelial reduzido pode estimular a esporulação naquele substrato. As principais
fontes de carbono para os isolados testados continham em suas combinações dextrose e
sorbitol, pois os maiores valores de AACPCM e TC, foram para tratamentos que continham
essas fontes de C. A sacarose não demonstrou uma fonte de C eficiente para a
morfofisiologia dos isolados.
Tabela 2. Valores de F da análise de variância dos fatores hospedeiros de origem e tratamentos e os valores dos parâmetros das variáveis dependentes peso seco de micélio e área abaixo da curva de progresso do crescimento micelial (AACPCM).
NS não significativo
**significativo a 1%.
A análise de variância para o arranjo fatorial para os fatores (variáveis independentes)
hospedeiros de origem, tratamentos com carbono e nitrogênio e a interação das variáveis
independentes analisou-se as medias e os desvios padrões das variáveis dependentes peso
seco de micélio e dos valores de área abaixo da curva de progresso do crescimento micelial
(AACPCM), sendo que rejeitou-se a hipótese de nulidade (~0,001) para a variável
independentes apenas para o fator tratamento C/N (F=7,52**), não rejeitando para diferenças
entre hospedeiros de origem (F= 1,48ns) e interação (F=0,000002ns). Para a variável
dependente AACPCM rejeitou-se a hipótese de nulidade para as variáveis hospedeiros de
18
FatoresValor F
Peso Seco do micélio AACPCM
Hospedeiro de origem 1,48ns 8,35**
Tratamentos (C/N) 7,52** 3,23**
Interação 0,02ns 3,27 ns

origem (F=8,35**) e tratamentos C/N (F=3,23**), não rejeitando-se a hipótese de nulidade
para interação entre os dois fatores (Tab. 2)
19

Tabela 3. Médias dos pesos secos de micélio (PS) e dos Valores de área abaixo da curva de progresso do crescimento micelial (AACPCM) dos isolados submetidos a diferentes concentrações de carbono e nitrogênio (C/N).Or. Hospedeiros de origem PS (g)* Hospedeiros de origem AACPCM*
1 Mangaba 3,16 a Iuca 16,4 a
2 Negramina 3,16 ab Mandioca 15,89 a
3 Conde 3,16 ab Antúrio 15,7 ab
4 Abacate 3,16 ab Tomate 15,6 bc
5 Banana 3,16 ab Uva 15,6 bc
6 Manga 3,16 ab Chuchu 15,5 bc
7 Goiaba (folha) 3,16 ab Soja 15,3 bc
8 Falso massambará 3,16 ab Árvore da fortuna 16,3 cd
9 Caqui 3,16 ab Abacate 15,1 de
10 Antúrio 3,16 ab Coco 14,8 ef
11 Árvore da Fortuna 3,16 ab Manga 14,6 fg
12 Uva 3,16 ab Cana 14,3 fg
13 Cana 3,16 ab Dracena 14,3 gh
14 Mandioca 3,16 ab Negramina 14,1 gh
15 Goiaba (Fruto) 3,16 ab Conde 13,4 gh
16 Amora 3,16 ab Mangaba 13,4 gh
17 Banana (fruto) 3,16 ab Amora 13,1 gh
18 Iuca 3,16 ab Goiaba (fruto) 12,8 gh
19 Chuchu 3,16 ab Falso massambará 12,8 gh
20 Soja 3,16 ab Goiaba 12,7 gh
21 Tomate 3,16 ab Mamão 12,5 gh
22 Coco 3,16 ab Banana (fruta) 12,2 gh
23 Mamão 3,16 ab Banana 12,2 gh
24 Dracena 3,16 ab Caqui 11,9 h
*Médias seguidas de mesma letra na coluna não diferem entre si ao teste Tukey (P~0,05)
20
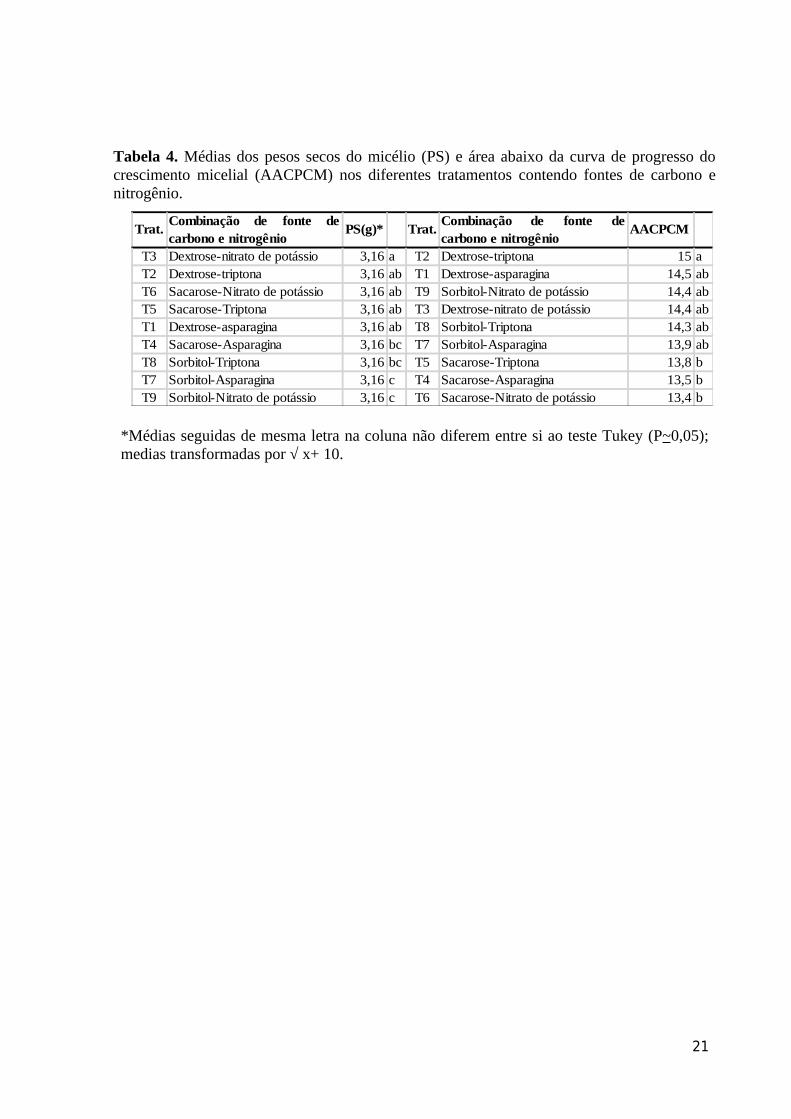
Tabela 4. Médias dos pesos secos do micélio (PS) e área abaixo da curva de progresso do crescimento micelial (AACPCM) nos diferentes tratamentos contendo fontes de carbono e nitrogênio.
Trat.Combinação de fonte decarbono e nitrogênio
PS(g)* Trat.Combinação de fonte decarbono e nitrogênio
AACPCM
T3 Dextrose-nitrato de potássio 3,16 a T2 Dextrose-triptona 15 aT2 Dextrose-triptona 3,16 ab T1 Dextrose-asparagina 14,5 abT6 Sacarose-Nitrato de potássio 3,16 ab T9 Sorbitol-Nitrato de potássio 14,4 abT5 Sacarose-Triptona 3,16 ab T3 Dextrose-nitrato de potássio 14,4 abT1 Dextrose-asparagina 3,16 ab T8 Sorbitol-Triptona 14,3 abT4 Sacarose-Asparagina 3,16 bc T7 Sorbitol-Asparagina 13,9 abT8 Sorbitol-Triptona 3,16 bc T5 Sacarose-Triptona 13,8 bT7 Sorbitol-Asparagina 3,16 c T4 Sacarose-Asparagina 13,5 bT9 Sorbitol-Nitrato de potássio 3,16 c T6 Sacarose-Nitrato de potássio 13,4 b
*Médias seguidas de mesma letra na coluna não diferem entre si ao teste Tukey (P~0,05); medias transformadas por √ x+ 10.
21
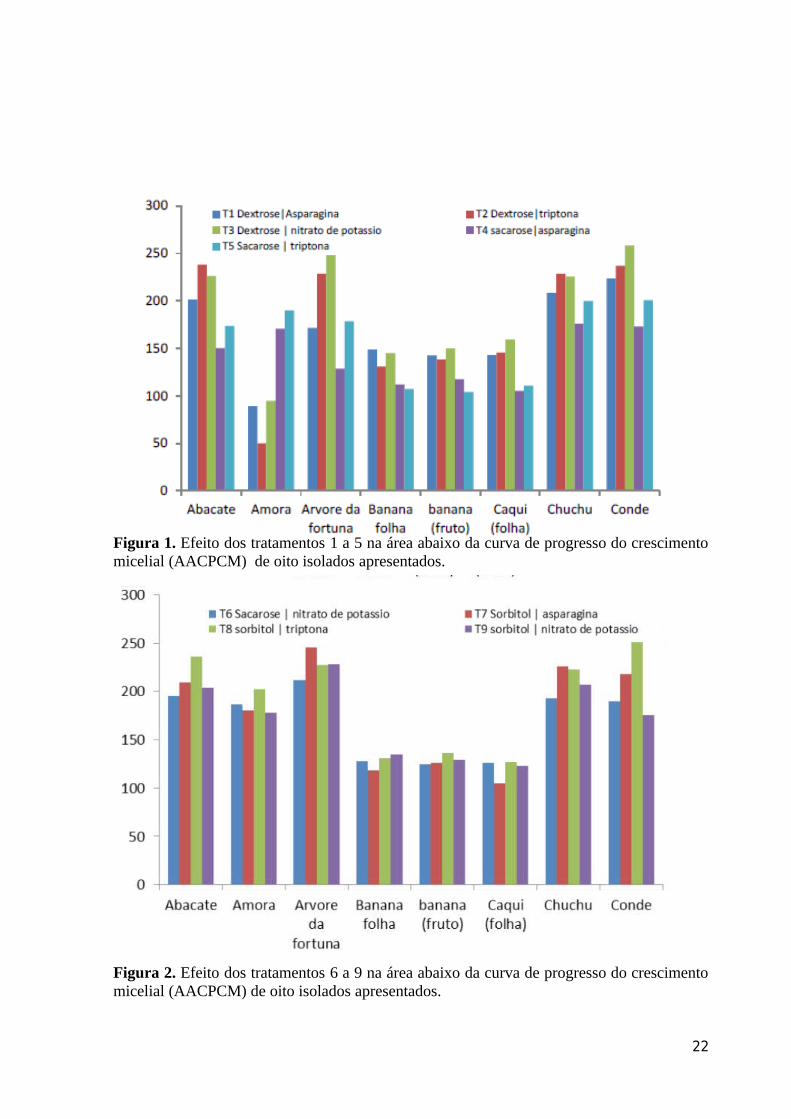
Figura 1. Efeito dos tratamentos 1 a 5 na área abaixo da curva de progresso do crescimento micelial (AACPCM) de oito isolados apresentados.
Figura 2. Efeito dos tratamentos 6 a 9 na área abaixo da curva de progresso do crescimento micelial (AACPCM) de oito isolados apresentados.
22
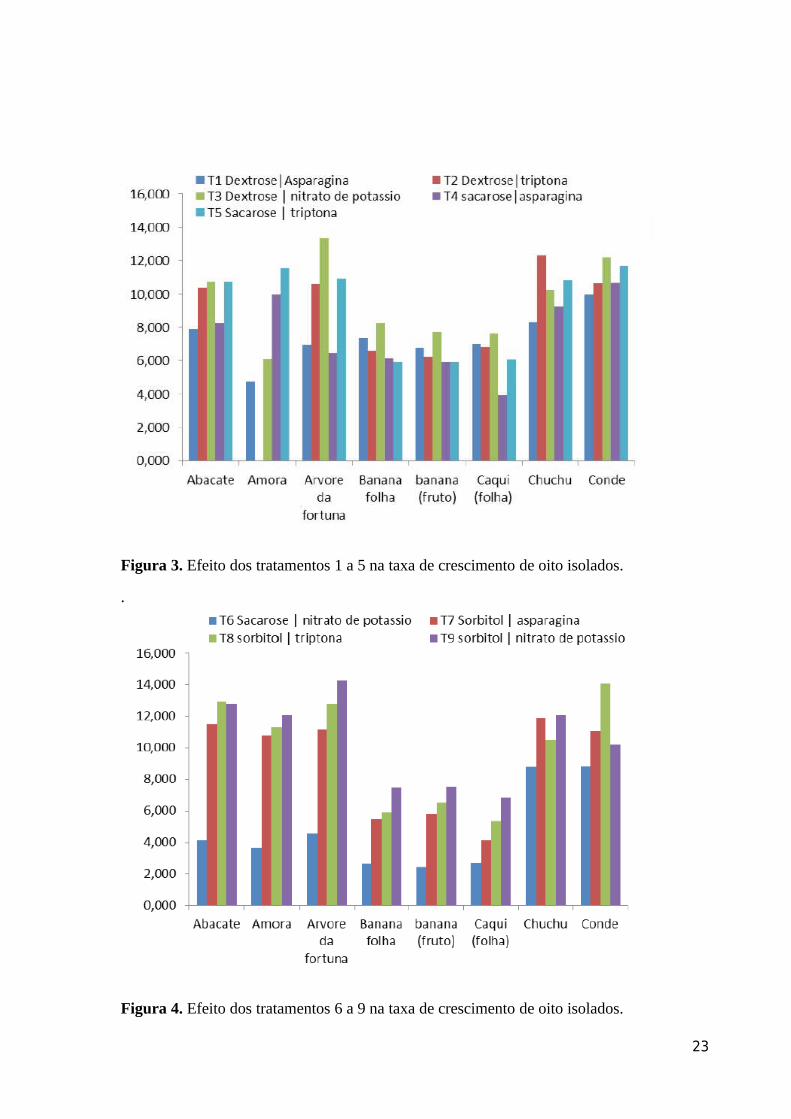
Figura 3. Efeito dos tratamentos 1 a 5 na taxa de crescimento de oito isolados.
.
Figura 4. Efeito dos tratamentos 6 a 9 na taxa de crescimento de oito isolados.
23

CONCLUSÕES
Através desse trabalho foi verificado uma variabilidade quanto ao uso de fontes de C e
N, evidenciando a combinação dextrose e nitrato de potássio, excluindo o uso de sacarose.
Foi verificado uma imensa variabilidade não tendenciosa por isolados e não
padronizada para o gênero C.gloeosporioides. quanto ao uso de fontes de C e N, evidenciando
a combinação dextrose e nitrato de potássio como mais promissora para crescimento e
esporulação de Colletotrichum sp. A ferramenta de crescimento de C/N mostrou-se um
promissor marcador de variabilidade para identificação de características distintivas para
trabalhos de caracterização de população de isolados.
Através desse trabalho podemos verificar que isolados como de iuca e mandioca possuem
atividade metabólica mais elevada em relação aos demais, e a fonte de C originada de
dextrose é a que promove maior produção de micélio. Nenhuma fonte de N mereceu destaque
dextrose x triptona.
24

BIBLIOGRAFIA
BERGER, R.D. The analysis of the effects of control measures on the development of
epidemics. In: KRANZ, J.; ROTEM, J. (Ed.). Experimental techniques in plant disease
epidemiology. Heidelberg: Springer-Verlang, p.137-151.1988.
BROWN, A.E., SREENIVASAPRASAD, S. & TIMMER, W.L. Molecular characterization
of slow-orange key lime anthracnose strains of Colletotrichum from citrus as C. acutatum.
Phytopathology 86:523-527. 1996.
COCHRANE, V.W. Physiology of fungi. New York. John Wiley & Sons Inc. 1958.
COUTO, E.F. MENEZES, M. Caracterização fisiomorfológica de isolados deColletotrichummusae. FitopatologiaBrasileira, v.29, p.406-412, 2004.FORSTER, H. & ADASKAVEG, J.E. Identification of subpopulations of
Colletotrichumacutatum and epidemiology of almond anthracnose in California.
Phytopathology 89:1056-1065. 1999.
FREEMAN, S., KATAN, T. & SHABI, E. Characterization of Colletotrichum species
responsible for anthracnose diseases of various fruits. Plant Disease 82:596-605. 1998.
GALLI, F., TOKESHI, H., CARVALHO, P.C.T., BALMER, E., KIMATI, H., CARDOSO,
C.O.N.SALGADO, C.L. Manual de Fitopatologia: doenças das plantas e seu controle. 2ª ed.
São Paulo: Agronômica Ceres, 1978.
GOES, A. & KIMATI, H. Caracterização morfológica de isolados de Colletotrichum
acutatum e C.gloeosporioides associados à queda prematura dos frutos cítricos.
SummaPhytopatologica 23:5-13. 1997.
GUNNEL, P & GUBLER, WD Taxonomy and morphology of Colletotrichum species
pathogenic to strawberry. Mycologia 84:157-165. 1992.
INDEX FUNGORUM, Disponível em http://www.indexfungorum.org/Names/Names.aps,
acessado em abril de 2010.
25

JOHNSTON, PR & JONES, D Relationships among Colletotrichum isolates from fruit-rots
assessed using rDNA sequences. Mycologia 89:420-430, 1997.
KIRK, P.M., CANNON, P.F., DAVID, J.C., STALPERS, J.A. Dictionary of the Fungi. 9th Ed.
CABI Publishing, Wallingford, UK, 2001.
LILLY, VG & BARNETT, HC Physiology of fungi. New York. McGraw-Hill, 1951.
LOPEZ, A.M.Q. Taxonomia, patogênese e controle de espécies do gênero Colletotrichum.
Revisão Anual de Patologia de Plantas 9:291-337. 2001.
PERES, NA, TIMMER, LW, ADASKAVEG, JE & CORRELL, JC. Lifestyles of
Colletotrichumacutatum. Plant Disease 89:784-796. 2005.
SANGEETHA, C.G., RAWAL, R. D. Nutritional Studies of
Colletotrichumgloeosporioides(Penz.) Penz. andSacc. The incitant of Mango Anthracnose.
World Journal of Agricultural Sciences,vol. 4 n.6,p.717-720, 2008.
SUTTON, B. C The genus Glomerella and its anamorph Colletotrichum.In BAILEY, J.A,
JEGER, M. J. Colletotrichum: Biology pathology and control. British Society for Plant
Pathology, CAB International, 1992.
TANDON, R.N. & CHANDRA, S. The nutrition of Colletotrichum gloeosporioides PENZ.
Mycopathology et. Mycology Applied 18:213-224. 1962.
TOZZE JR., H.J., MELLO, M.B.A. & MASSOLA JR., N.S. Morphological and physiological
characterization of Colletotrichum sp. isolates from solanaceous crops. Summa
Phytopathologica 32:71-79. 2006.
26