tese corrigida depois da defesa - universidade estadual do ... · espécie do bicho da seda; a...
TRANSCRIPT

Felix, S.P.
1
1.1 - INTRODUÇÃO
1.1 - Mamona: Aspectos gerais
A mamona é uma euphorbiacea, cuja origem é dada ora sendo asiática, ora
africana e, até mesmo, como planta nativa da América. Esta planta foi introduzida
em quase todo o mundo, principalmente nas regiões tropicais e subtropicais. No
Brasil a mamona é conhecida desde a era colonial quando dela se extraía o óleo
para lubrificar as engrenagens dos inúmeros engenhos de cana. Ela está largamente
difundida por todo o Brasil, não havendo praticamente terreno baldio, mata ou
lavoura abandonada onde não cresça (FORNAZIERI, 1986).
A mamona (Figura 1) se classifica da seguinte forma (disponível em: < http:
//www.ncbi.nih.gov/ > Acesso em: 25/03/07).
Figura 1 - Mamona. (www.paraíba.pb.gov.br/)
Super Reino: Eucaryota
Reino: Viridiplantae
Filo: Streptophyta
Superdivisão: Spermatophyta
Divisão: Magnoliophyta
Classe: Magnoliopside
Subclasse: Rosidae
Ordem: Malpighiales
Família: Euphorbiacea
Subfamília: Acalyphoideae
Tribo: Acalypheae
Gênero: Ricinus
Espécie: Ricinus communis

Felix, S.P.
2
1.2 - Importância Econômica
Da mamona aproveita-se tudo: as folhas servem de alimento para uma
espécie do bicho da seda; a haste, além de celulose própria para a fabricação de
papel, fornece matéria-prima para a produção de tecidos grosseiros; além dos
principais produtos derivados da semente da mamona que são o óleo e a torta
residual da extração do óleo. As fábricas de óleo de mamona atualmente existentes
industrializam toda a produção, obtendo-se como subproduto, a torta de mamona
(FORNAZIERI, 1986).
1.2.1 - Óleo de mamona
O óleo extraído das sementes de mamona possui um mercado internacional
crescente, garantido por mais de 700 diferentes aplicações que incluem desde uso
medicinal em cosméticos até a substituição ao petróleo na fabricação de plásticos e
lubrificantes. O produto também é utilizado na fabricação de tintas e isolantes, na
produção de fibra ótica, vidro à prova de balas e próteses ósseas. Além disso, é
indispensável para impedir o congelamento de combustíveis e lubrificantes de
aviões e foguetes espaciais quando atingem baixíssimas temperaturas. Além destas
aplicações, o óleo de mamona é empregado para produção de corantes, anilinas,
desinfetantes, germicidas, colas e aderentes, base para fungicidas e inseticidas,
tintas de impressão e vernizes. O óleo de mamona transformado em plástico, sob a
ação de reatores nucleares, adquire a resistência do aço, mantendo a leveza da
matéria plástica. Uma das aplicações de grande valor econômico do óleo de
mamona é na fabricação do nylon e da matéria plástica empregada na fabricação de
espumas plásticas, onde o óleo de mamona confere ao material texturas variáveis,
desde a macia e esponjosa até a dura e rígida (FORNAZIERI, 1986). Atualmente, no
Brasil, as sementes de mamona estão sendo cogitadas como uma das principais
fontes para a produção de Biodiesel.
1.2.1.1 - Biodiesel
O Biodiesel é um combustível renovável, biodegradável, não corrosivo e
ambientalmente correto, sucedâneo ao óleo diesel mineral. Possui um grande apelo
ambiental, por ser obtido de fontes naturais renováveis, tais como óleos vegetais
(por exemplo, girassol, nabo forrageiro, algodão, mamona, soja e canola) e gordura

Felix, S.P.
3
animal, mas mais especialmente por diminuir as emissões de gases poluentes
durante a combustão quando comparado aos combustíveis fósseis (PIRES et al.,
2004).
Alguns pesquisadores admitem que o óleo de mamona é o melhor para
produzir biodiesel, por ser o único solúvel em álcool (necessário para o processo de
produção), e não necessitar de calor, reduzindo dessa maneira o gasto de energia
que requerem outros óleos vegetais para sua transformação em combustível
(PARENTE, 2004).
A utilização do biodiesel já havia sido proposta em 1986 por Fornazieri. O
biodiesel substitui total ou parcialmente o óleo diesel de petróleo em motores
ciclodiesel automotivos (de caminhões, tratores, caminhonetes, automóveis, etc) ou
estacionários (geradores de eletricidade, ou calor, entre outros), podendo ser usado
puro ou misturado ao diesel em diversas proporções. No Brasil, a Lei nº 11.097, de
13 de janeiro de 2005, introduz o biodiesel na matriz energética brasileira,
estabelecendo a obrigatoriedade da adição de um percentual mínimo de biodiesel ao
óleo diesel comercializado ao consumidor, em qualquer parte do território nacional.
Esse percentual obrigatório será de 5% em oito anos após a publicação da referida
lei, havendo um percentual obrigatório intermediário de 2% em três anos após a
publicação da mesma (disponível em: < http: //www.biodiesel.org.br/ > Acesso em:
15/01/07).
O Brasil, país que possui excelentes condições edafo-climáticas, tem
potencial de abastecer com biodiesel o consumo mundial, podendo substituir em
60% o óleo diesel de petróleo, levando em consideração que 1.000 kg de óleo
vegetal produz 1000 litros de biodiesel, sendo que, 1000 kg de mamona produzem
470 kg de óleo vegetal (PARENTE, 2004). Em conseqüência disso, o Governo
Federal lançou em 2004 o Programa Nacional de Produção e Uso de Biodiesel
(PNPB) que é um programa interministerial que objetiva a implementação de forma
sustentável, tanto técnica, como economicamente, a produção e uso do Biodiesel no
Brasil.
A área plantada necessária para atender ao percentual de mistura de 2%, de
biodiesel ao diesel de petróleo é estimada em 1,5 milhões de hectares, o que
equivale a 1% dos 150 milhões de hectares plantados e disponíveis para agricultura
no Brasil. As regras permitem a produção a partir de diferentes oleaginosas e rotas
tecnológicas, possibilitando a participação do agronegócio e da agricultura familiar

Felix, S.P.
4
(disponível em: < http: //www.biodiesel.org.br/ > Acesso em: 15/01/07). Contudo,
discussões a respeito do biodiesel têm procurado priorizar oleaginosas que
propiciem maior emprego de mão-de-obra, e inserir regiões que estejam à margem
do processo de desenvolvimento econômico. Nesse contexto, a cultura da mamona
vem despertando maior interesse, tanto por parte do programa nacional como do
programa baiano à medida que a região do semi-árido nordestino apresenta-se com
maior potencial para a implementação dos programas (PIRES et al., 2004).
Então, as vantagens econômicas e ambientais, o cultivo da mamona e a
produção industrial de biodiesel a partir desta, ou seja, as cadeias produtivas do
biodiesel, têm grande potencial de geração de empregos, promovendo, dessa forma,
a inclusão social, especialmente quando se considera o amplo potencial produtivo
da agricultura familiar predominante nestas regiões (disponível em: < http:
//www.biodiesel.org.br/ > Acesso em: 15/01/07).
1.2.2 - Torta de mamona
A torta residual da extração do óleo de mamoneira tem sido utilizada como
um co-produto também amparado pelo Programa Nacional de Produção e Uso de
Biodiesel, pois é produzido na proporção aproximada de 1,2 toneladas para cada
tonelada de óleo extraído (AZEVEDO & LIMA, 2001).
O seu principal uso é como adubo orgânico, apresentando potencial
fertilizante. Nestas condições a torta de mamona apresenta elevadíssima
porcentagem de matéria orgânica e riqueza dos macroelementos. A adição de torta
de mamona ao solo, com dosagens variando de acordo com a cultura e o tipo de
solo, e rico ou não em nutrientes, além de suprir as necessidades nutricionais das
plantas aumenta o pH , reduzindo a acidez total, elevando o conteúdo de carbono e
promovendo melhoria geral na parte física do mesmo. A utilização da torta no solo,
além de reduzir os nematóides e elevar o poder tampão e a capacidade de troca de
cátions do solo, tem propriedade de reduzir a densidade aparente do ambiente em
todos os tipos de solos, o que interfere positivamente no crescimento e no
desenvolvimento radicular, com renovação mais rápida e adequada do oxigênio.
Apesar de apresentar alto teor de proteínas, não se recomenda o uso da torta
para ração, pois é tóxica devido à presença da proteína ricina, e do complexo
alergênico, denominado nas décadas passadas de CB-1A (Castor Bean Allergen)
que é uma mistura de proteínas de baixo peso molecular (YOULE & HUANG, 1978).

Felix, S.P.
5
Atualmente sabe-se que o complexo alergênico CB-1A representa cerca de 12,5%
do peso da torta, como determinado pelo teste de precipitação de antígenos
diluídos. Este complexo é formado por cerca de 20 isoformas de proteínas com
massa molecular entre 10 e 14 kDa, sendo pertencentes à classe das albuminas 2S
(MACHADO et al., 2003). Duas isoformas alergênicas, Ric c 1 e Ric c 3 já se
encontram seqüenciadas com características biológicas bem determinadas
(MACHADO & SILVA, 1992; SILVA JR. et al., 1996).
Para o aproveitamento da torta deve se ter cuidado com o método usado no
processo industrial de detoxificação e de desalerginização, para não afetar a
qualidade final da torta ou farelo. Existem diversos métodos para alcançar a
detoxificação, como por exemplo: o cozimento por uma ou duas horas, ou ferver por
curtos períodos de tempo uma mistura de torta moída e água, com mudança da
água após cada fervura. Este é um processo caro e as usinas de óleo preferem
vender a torta apenas como fertilizante.
A presença de proteínas alergênicas em sementes de mamona já é
conhecida há muitos anos, no entanto, os tratamentos que são aplicados hoje para a
torta de mamona se referem à detoxificação, mas são ineficientes para a
desalerginização da torta (ANANDAN et al., 2005). Não há estudos que avaliem os
problemas provocados pela manipulação inadequada da torta.
1.3 - Alérgenos de Plantas
O termo alérgeno é utilizado para descrever duas ou três propriedades
moleculares distintas: i) a propriedade para sensibilizar (isto é, induzir a produção de
anticorpos de alta afinidade, particularmente da classe IgE, pelo sistema imune); ii) a
propriedade de se ligar aos anticorpos IgE; iii) e ainda a propriedade para ativar uma
reação alérgica (isto é, desencadear sintomas alergênicos em uma pessoa
sensibilizada) (AALBERSE, 2000). Alérgenos de plantas em geral, são proteínas de
defesa da planta, que permitem a ela resistir aos estresses bióticos e abióticos.
Muitos tecidos de plantas, que são consumidos por humanos, contêm milhares
destas proteínas alergênicas. Aproximadamente 0,5% da população dos Estados
Unidos é afetada por vários estágios da alergia alimentar mediada por IgE
específicas para alérgenos de plantas (BREITENEDER & RADAUER, 2004).

Felix, S.P.
6
Os alérgenos de plantas são classificados dentro de famílias e superfamílias,
baseados em sua estrutura e função. Elas são agrupadas dentro de uma mesma
família se possuirem 30% (ou mais) de resíduos de aminoácidos idênticos ou ainda,
se tiverem baixa homologia, mas apresentarem função e estrutura tridimensional
similares sugerindo uma origem evolucionária comum (BREITENEDER &
RADAUER, 2004). Adicionalmente tornou-se evidente que o nível de exposição e as
propriedades do alérgeno em si são importantes para a determinação do potencial
alergênico (BREITENEDER & MILLS, 2005).
Os alérgenos de origem vegetal, mais abundantes, pertencem às
superfamílias Cupin e Prolamina. As albuminas 2S pertencem à família das
prolaminas. Existem também outros alérgenos pertencentes aos grupos das
“proteínas relacionadas à patogênese” e Profilinas (BREITENEDER & RADAUER,
2004).
1.3.1 - Superfamília Cupin:
As proteínas da superfamília Cupin são funcionalmente diversas.
Compartilham um domínio estrutural beta-barril, responsável pelo nome da família
(do latin cupa, barril). Domínios simples de cupin apresentam um domínio
conservado, já bicupins apresentam dois domínios. Bicupins incluem as globulinas,
que são proteínas de reserva de sementes, além de serem o componente majoritário
da nossa dieta. Com base no coeficiente de sedimentação, as globulinas são
divididas em duas famílias: Vicilinas 7S e Legumininas 11S (DUNWELL et al., 1998).
Essas proteínas são alérgenos potenciais encontrados em plantas, como
amendoim, soja, lentilha, noz, avelã e algodão (BREITENEDER & RADAUER, 2004).
1.3.2 - Superfamília Prolamina:
A existência dessa família é baseada na presença de um esqueleto
conservado de oito resíduos de cisteína. Todas as proteínas dessa superfamília são
de baixo peso molecular, além de serem ricas em cisteínas, apresentarem estrutura
tridimensional semelhante rica em α-hélice, e serem estáveis ao tratamento térmico
e a proteólise (BREITENEDER & MILLS, 2005). Compreende três grupos principais
de alérgenos de plantas: proteínas transportadoras de lipídeos não específicos
(nsLTPs) , inibidores de alfa amilase/tripsina de sementes de cereais, prolamina de
cereais e albuminas 2S (BREITENEDER & RADAUER, 2004).

Felix, S.P.
7
1.3.2.1 - Albuminas 2S
É o principal grupo de proteínas de reserva presente nas dicotiledôneas, além
de serem os principais alérgenos da mamona. Apresentam massa molecular de
10.000 - 14.000 Da, altos teores de arginina, serina e glutamina. Sabe-se que
algumas delas são inibidores de proteases, outras são inibidores de α-amilases, e
algumas podem ainda apresentar propriedades alergênicas (MACHADO & SILVA,
1992). Apresentam uma distribuição característica de oito cisteínas em um padrão
conservado de pontes de enxofre e geralmente são compostas de 2 cadeias
polipeptídicas diferentes de 3 a 5 e de 8 a 10 kDa ligadas por duas pontes dissulfeto.
Elas ainda apresentam duas ligações intracadeias, o que as tornam proteínas muito
estáveis e compactas (PANTOJA-UCEDA et al., 2004).
As albuminas 2S são sintetizadas em tempos específicos durante o
desenvolvimento da semente e depositadas dentro dos vacúolos (corpúsculos
protéicos) durante o desenvolvimento da semente, para então serem degradadas
durante a germinação, dando suporte ao crescimento da semente (REGENTE & LA
CANAL, 2001). Elas são sintetizadas no retículo endoplasmático rugoso, como um
precursor protéico de alto peso molecular. Posteriormente este precursor é clivado
proteoliticamente, gerando a perda de um peptídeo ligante e de outros pequenos
peptídeos (SHEWRY et al., 1995). A glicosilação dessas proteínas pode ocorrer
durante a síntese protéica e os carboidratos incorporados são, em sua maioria,
manose e glicosamina (BEWLEY & BLACK, 1994).
Acreditava-se que as albuminas 2S fossem inativas metabolicamente, mas
atualmente, devido à sua capacidade inibidora de proteinases, às propriedades
alergênicas (MACHADO & SILVA JR, 1992), e à ação antifúngica (AGGIZIO et al.,
2003), acredita-se que elas estejam envolvidas em funções de defesa constitutivas
da planta (REGENTE & LA CANAL, 2001).
As propriedades alergênicas das albuminas 2S são resistentes à
desnaturação térmica e química, podendo, mesmo, após os tratamentos de
desnaturação, redução e alquilação, desencadear alergia por contato, bem como por
inalação (VIEIRA, 2002).
Historicamente, sempre houve tentativas de isolar e identificar os alérgenos
de Ricinus communis. Em 1943, Spies e Coulson isolaram da semente de mamona
uma fração proteica de baixo peso molecular, estável ao calor, que foi denominada

Felix, S.P.
8
CB-1A. Em 1977, Li e colaboradores isolaram e caracterizaram uma proteína das
sementes de Ricinus communis de baixo peso molecular com alto “teor” de
glutamina e um espectro não comum de UV que mostrou propriedades similares
àquelas do alérgeno de Ricinus communis. Em 1978, Youle e Huang concluíram que
CB-1A era a albumina 2S de reserva caracterizada por Li e colaboradores em 1997.
Em 1982, Sharief e Li seqüenciaram uma proteína das sementes de Ricinus
communis (Ric c 1), de coeficiente de sedimentação 2S, constituída de duas
subunidades unidas por pontes de enxofre. A menor contendo 34 aminoácidos (Ric c
1 leve) com massa molecular aparente de 4 kDa e a subunidade maior composta de
61 aminoácidos (Ric c 1 pesada) com massa molecular de 7 kDa.
Machado e Silva em 1992 isolaram e seqüenciaram uma segunda albumina
2S diferente da apresentada por Sharief e Li, denominada de Ric c 3, tendo peso
molecular em torno de 11 kDa, presente no mesmo precursor (29 kDa) de Ric c 1
(Figura 2). A estrutura primária documentada corresponde aos resíduos de 18 - 49
(cadeia leve) e 66 - 130 (cadeia pesada) do precursor protéico. De forma similar, a
albumina 2S descrita por Sharief e Li (1982) corresponde aos resíduos 136 - 169
(cadeia leve) e 173 - 237 (cadeia pesada). A segunda albumina 2S apresenta
resíduos conservados de cisteína nas duas cadeias o que é uma característica típica
das albuminas 2S (SILVA JR. et al., 1996).

Felix, S.P.
9
Erro!
Erro!
Figura 2 - Esquema do processamento do precursor das isoformas Ric c 3 e Ric c 1. Em A)
Precursor intacto com Peptídeo sinal em bege, pontes de enxofre em amarelo, Ric c 3 e Ric
c 1 respectivamente em vermelho (cadeias leves) e em marrom (cadeias pesadas),
peptídeos de ligação em azul; em B) Perda do peptídeo sinal; em C) Perda dos peptídeos
de ligação com conseqüente separação das duas isoformas.
Machado e colaboradores em 2003 forneceram dados bioquímicos e
imunológicos para a presença de pelo menos nove frações diferentes de albumina
2S. Essas nove frações apresentam uma composição similar de aminoácidos, com
alta quantidade de glutamina, além da similaridade na sequência N-terminal, sendo
que sete dessas proteínas apresentaram potencial alergênico.
Em 2004, Mayerhoffer caracterizou um epitopo, cuja seqüência peptídica está
presente também em outros alérgenos. Felix em 2006 caracterizou cinco epitopos
alergênicos nas isoformas de albumina 2S, Ric c 1 e Ric c 3, de mamona, capazes
de promover a desgranulação de mastócitos. Esses epitopos foram caracterizados
utilizando peptídeos sintéticos, os quais foram produzidos com base nas seqüências
das cadeias leves e pesadas de Ric c 1 e de Ric c 3, as principais isoformas de
albumina 2S de Ricinus communis. Estes peptídeos foram sintetizados pelo método
de fase sólida pela pesquisadora Maria Aparecida Juliano da Universidade Federal
A B A B
C

Felix, S.P.
10
de São Paulo - Departamento de Biofísica. As seqüências dos peptídeos, bem como
suas localizações nas isoformas Ric c 1 e Ric c 3 são apresentadas na figura 3.
Já foi observado também que muitos alérgenos de nozes e sementes são
albuminas 2S, por exemplo, Ber e 1 da castanha do Maranhão (Bertholletia excelsa),
Jur r 1 de noz (Juglans regia), Ses i 2 de algodão (BREITENEDER & RADAUER,
2004) e SFA-8 de girassol (Helianthus annuus) sendo que este último possui
estrutura tridimensional muito semelhante a Ric c 3 de Ricinus communis
(PANTOJA-UCEDA et al., 2004).
RIC c 3 cadeia leve: 35 resíduos; P1- Resíduos 1 a 15 e P2- Resíduos 16 a 31.
ESKGEREGSSSQQCRQEVQRKDLSSCERYLRQSSS
RIC c 3 cadeia pesada: 65 resíduos; P4- Resíduos 20 a 39 e P5- Resíduos 40 a 54.
QQQESQQLQQCCNQVKQVRDECQCEAIKYIAEDQIQQGQLHGEESERVAQRAGE
IVSSCGVRCMR
RIC c 1 cadeia leve: 34 resíduos; P3- Resíduos 11 a 24.
PSQQGCRGQIQEQQNLRQCQEYIKQQVSGQGPRR
RIC c 1 cadeia pesada: 64 resíduos; P0-Resíduos 22 a 40
QERSLRGCCDHLKQMQSQCRCEGLRQAIEQQQSQGQLQGQDVFEAFRTAANLPS
MCGVSPTSRF
Figura 3 - Seqüências dos peptídeos sintéticos e suas localizações nas isoformas Ric c 1 e Ric c 3.
P2 P1
P5 P4
P3
P0

Felix, S.P.
11
1.4 - Aeroalérgenos
A asma é uma manifestação de hipersensibilidade imediata e de reações de
fase tardia no pulmão que afeta 5-30 % das crianças e 2-30 % de adultos. Já a rinite,
uma manifestação alérgica mais comum do trato respiratório superior afeta 40 % da
população. Muitos estudos têm mostrado que essas doenças coexistem em 98,9%
dos casos (GIOULEKAS et al., 2004). Uma das causas da rinite e da asma é a
inalação de aeroalérgenos (proteínas alergênicas presentes no ar).
O pólen de diversas oleaginosas amplamente utilizadas na agricultura, como
girassol, soja, milho e mamona, é rico em proteínas alergênicas. Assim, a alergia
desencadeada por pólens, é um problema que pode afetar grande parte das
populações que residem próximo ao plantio destas oleaginosas, bem como do
trabalhador que manipula tais sementes (THORPE et al., 1988).
1.5 - Hipersensibilidade
O Brasil, apesar de ser um dos maiores produtores de mamona da atualidade,
não dispõe de estudos detalhados sobre casos de hipersensibilidade causados por
albumina 2S desta semente. Com o intuito de gerar conhecimentos nesta área,
estudos comparativos das propriedades químicas e biológicas relacionadas ao
processo de hipersensibilidade desencadeado por albuminas 2S, vêm sendo
realizados em nosso laboratório (VIEIRA, 2002; MAYERHOFFER, 2002;
MAYERHOFFER, 2004; FELIX, 2006).
Os distúrbios causados pelas respostas imunes anormais são chamados de
reações de hipersensibilidade. Quatro tipos são descritos (tipos I, II, III, IV). (ROITT,
1998).
A hipersensibilidade imediata é causada pelos anticorpos IgE e pelos
mastócitos, também chamada de hipersensibilidade do tipo I. Os anticorpos não-IgE
poderão causar lesões recrutando e ativando as células inflamatórias e interferindo
nas funções celulares normais. Alguns desses anticorpos são específicos para os
antígenos de células particulares ou da matriz extracelular e são encontrados
ligados a estas células ou tecidos, ou livres na circulação; as doenças induzidas por
esses anticorpos são classificadas como distúrbios de hipersensibilidade do tipo II.
Outros anticorpos podem formar complexos imunes na circulação, e os complexos

Felix, S.P.
12
são subseqüentemente depositados, principalmente nos vasos sanguíneos, e
causam lesão. As doenças causadas por estes complexos imunes são classificadas
como hipersensibilidade do tipo III. Finalmente, a lesão tecidual pode ser devida aos
linfócitos T que ativam mecanismos efetores da reação de hipersensibilidade tardia
(DTH) ou lisam diretamente células-alvo; esses estados são classificados como
distúrbios de hipersensibilidade do tipo IV (ROITT et al., 1998). O processo de
alergia envolve reações imunes humorais (do tipo I) e mediadas por células (do tipo
IV).
1.5.1 - Hipersensibilidade imediata (Tipo I)
A indução de uma resposta imune aos antígenos estranhos requer a
cooperação entre as células apresentadoras de antígenos (APC), as populações de
linfócitos T e linfócitos B. Inicialmente as APCs (macrófagos ou células dendríticas)
internalizam os antígenos. Estes sofrem clivagem proteolítica e os fragmentos
peptídicos gerados são expostos nas membranas externas da APC em associação
com moléculas do complexo de histocompatibilidade principal (MHC II). Os linfócitos
T auxiliares (Th1 e/ou Th2) reconhecem esses epitopos expostos e uma vez
ativados promovem uma mudança de classe de imunoglobulinas nas células B
através de citocinas, iniciando a resposta imunológica. A ativação de clones de
células Th2, específicos para alérgenos, é essencial para o desenvolvimento de
doenças atópicas (ABBAS, 2003). Diferente de células Th1, as células Th2 ativadas
por contato com APCs, produzem quantidades relativamente grandes de
interleucinas 4 (IL-4) e 5 (IL-5) que induz o “switching” dos anticorpos IgM para
produção de anticorpos IgE. Os anticorpos IgE, liberados pelos plasmócitos, se
associam aos receptores específicos (FcεRI) na superfície dos mastócitos
encontrados no tecido, e basófilos circulantes no sangue (LICHTENSTEIN, 1993).
Essa é a primeira sensibilização. Com uma subseqüente exposição ao mesmo
antígeno (segunda sensibilização), ocorrerá uma ligação cruzada entre segmentos
específicos do antígeno (epitopos que ligam IgE) com as moléculas de IgE pré-
ligadas nos mastócitos e basófilos, ativando os mensageiros intracelulares (ABBAS,
2003). Cascatas envolvendo enzimas tirosina quinase, fosfolipase C, proteína
quinase C e influxo de íons Ca2+ induzem a desgranulação dos mastócitos. Dentre
as substâncias liberadas, responsáveis por muitos sintomas alérgicos da

Felix, S.P.
13
hipersensibilidade imediata (Figura 4), encontram-se agentes quimiotáticos,
ativadores inflamatórios e espasmógenos (LICHTENSTEIN, 1993).
Como conseqüência tem-se rápida exsudação vascular de plasma,
vasodilatação, broncoconstrição e mais tarde inflamação. Os infiltrados inflamatórios
dessa reação de fase tardia são enriquecidos em eosinófilos, basófilos e células
Th2. Casos extremos de hipersensibilidade imediata podem resultar da anafilaxia e
da asma brônquica, podendo levar ao óbito (ABBAS, 2003).
A síntese de IgE é regulada pela herança, exposição ao antígeno e citocinas
secretadas por células T. Sendo assim, os indivíduos atópicos (predispostos a
hipersensibilidade imediata) têm mais IgE no sangue do que os não atópicos.
A hipersensibilidade imediata se apresenta de diversas formas nos vários
órgãos envolvendo diferentes mediadores e células-alvo (ROITT et al., 1998). Em
vários casos pode ocorrer reação cruzada, que acontece quando dois ou mais
antígenos possuem partes de sua estrutura primária (epitopos contínuos) e/ou
terciária (epitopos comformacionais) com características similares.
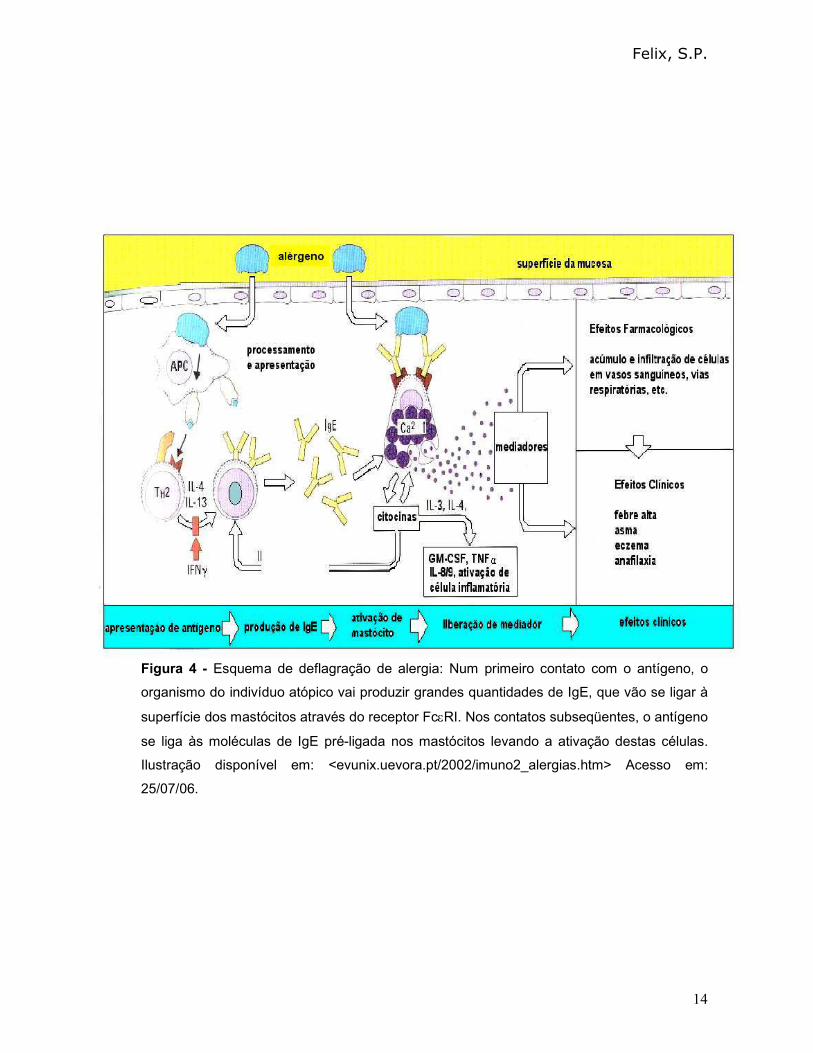
Felix, S.P.
14
Figura 4 - Esquema de deflagração de alergia: Num primeiro contato com o antígeno, o
organismo do indivíduo atópico vai produzir grandes quantidades de IgE, que vão se ligar à
superfície dos mastócitos através do receptor FcεRI. Nos contatos subseqüentes, o antígeno
se liga às moléculas de IgE pré-ligada nos mastócitos levando a ativação destas células.
Ilustração disponível em: <evunix.uevora.pt/2002/imuno2_alergias.htm> Acesso em:
25/07/06.

Felix, S.P.
15
1.5.1.1 - Epitopos
A seqüência de aminoácidos reconhecida pela molécula de anticorpo é muito
menor que a macromolécula imunogênica. Por isso, a ligação do anticorpo ocorre
somente numa porção específica do antígeno. Esta região é chamada de epitopo ou
determinante antigênico. Os antígenos podem ter múltiplos epitopos e cada um pode
se ligar a uma molécula de anticorpo (ABBAS, 2003).
Em proteínas, os epitopos contínuos são formados por aminoácidos
adjacentes. Já os epitopos conformacionais são formados pelos resíduos de
aminoácidos que não estão em seqüência, porém tornam-se espacialmente
justapostos na proteína enovelada. Os epitopos contínuos são mantidos após uma
desnaturação, contudo, os epitopos conformacionais são perdidos (ABBAS, 2003).
Vieira em 2002 observou que ambas as isoformas isoladas do “pool” de
albuminas 2S, Ric c 1 e Ric c 3, mesmo após serem submetidas à desnaturação,
são capazes de desencadear a desgranulação de mastócitos, indicando a presença
de epitopos contínuos nas duas isoformas. Mayerhoffer em 2004 verificou que após
a clivagem enzimática dessas isoformas, dois peptídeos presentes na cadeia
pesada de Ric c 3, induziram uma resposta significativa quanto a desgranulação dos
mastócitos. Neste mesmo trabalho, Mayerhoffer caracterizou um epitopo linear, cuja
seqüência peptídica está presente também em outros alérgenos. Felix em 2006
caracterizou cinco epitopos lineares alergênicos nas isoformas de albumina 2S, Ric
c 1 e Ric c 3, de mamona. Na seqüência de todos os peptídeos identificados por
Felix em 2006 e Mayerhoffer em 2004 era possível observar a presença de pelo
menos dois resíduos de aminoácidos dicarboxílicos. Esta característica nos fez
acreditar que os grupamentos carboxílicos laterais destas cadeias fossem
importantes na interação com as IgE.
1.6 - Mastócitos
Os mastócitos são células derivadas da medula óssea, que residem no tecido
conjuntivo, em todo corpo. Porém, são localizados predominantemente nas
proximidades dos vasos sanguíneos e nervos, abaixo dos epitélios e mucosas,
estando presentes também em órgãos linfóides. São caracterizados pela extrema
abundância de grânulos, os quais preenchem todo o citoplasma a ponto de,
freqüentemente, impedir a visibilidade do núcleo (DA SILVA & MOTA, 2003). Esses
grânulos são constituídos principalmente por glicoproteínas e coram-se

Felix, S.P.
16
metacromaticamente devido ao seu conteúdo em proteoglicanos, destacando-se
facilmente em cortes corados por azul de toluidina (JUNQUEIRA & CARNEIRO,
1995). Na maioria das espécies, os mastócitos são extremamente ricos em
histamina e heparina, e, no rato e no camundongo, são, além disso, ricos em
serotonina (DA SILVA & MOTA, 2003).
Os mastócitos expressam em sua superfície grande número de receptores Fc
de alta afinidade para IgE (FcεRI). Em conseqüência, a superfície de cada célula
mostra-se recoberta por moléculas de IgE, derivadas de linfócitos que foram
absorvidas da circulação, que atuam como receptores para antígenos específicos. A
ativação dos mastócitos se dá pela ligação cruzada das moléculas do receptor
FcεRI, causada pela ligação de antígenos multivalentes às moléculas IgE ligada ao
receptor (ABBAS, 2003). Essa ativação resulta em três respostas biológicas:
secreção dos conteúdos pré-formados dos grânulos; síntese e secreção de
mediadores lipídicos; e secreção de citocinas. As funções efetoras dos mastócitos
são mediadas por estas moléculas solúveis liberadas das células sob ativação.
Essas moléculas podem ser divididas em mediadores pré-formados, que incluem as
aminas biogênicas, ou vasoativas e as macromoléculas dos grânulos, e os
mediadores recém-sintetizados, que incluem mediadores derivados de lipídeos e
citocinas. Nos mastócitos humanos o único mediador da classe das aminas
biogênicas presentes em quantidades significantes é a histamina (ABBAS, 2003). Os
mais importantes mediadores lipídicos são produtos do metabolismo do ácido
araquidônico, tais como prostaglandina D2; leucotrienos, especialmente o
leucotrieno C4 (LTC4); e o fator ativador de plaquetas (PAF) (BOYCE, 2003).
Histamina, prostaglandina D2 e leucotrieno C4 contribuem para a modulação
da mucosa pela indução de edema e da secreção de muco e, no caso de asma,
broncoconstrição. Mastócitos também secretam citocinas pró-inflamatórias incluindo,
IL-4, IL-5 e IL-13, que regulam a síntese de IgE e o desenvolvimento de inflamação
eosinofílica (BRADDING,1999).
Os mastócitos são historicamente conhecidos pelos seus envolvimentos na
hipersensibilidade do tipo 1, mas possuem funções protetoras e homeostáticas. Eles
reconhecem diretamente os produtos de infecções bacterianas através de várias
proteínas receptoras na superfície, liberando proteases, citocinas e mediadores que
recrutam neutrófilos, limitando o alcance da infecção bacteriana e facilitando o
reparo do tecido (BOYCE, 2003).

Felix, S.P.
17
1.7 - Imunoterapia
Para amenizar os sintomas da hipersensibilidade imediata, os pesquisadores
têm procurado investir na imunoterapia. Stephen e colaboradores em 2004
exploraram o fato de que a resposta a um determinado antígeno é dirigida por Th1
ou Th2, dependendo da rota e dose do antígeno, além da natureza de apresentação
deste. Sendo assim, tem sido empregado o uso de adjuvantes (MPL, nucleotídeos
derivados de bactérias), que induzem resposta Th1 ao invés de resposta Th2 que
favorecem o “switching” de classe de IgE para IgG. Esta estratégia é apresentada na
patente americana de número US 6.610.297 B1. Uma outra abordagem similar foi
apresentada na patente internacional PCT WO 2004/019978 A1. Neste caso, ácido
nucléico recombinante induz também imunidade tipo Th1 e inibe a produção de IgE.
Essas estratégias apresentam uma vantagem, pelo fato de o efeito persistir por
aproximadamente três anos após o término do tratamento (Stephen et al., 2004).
Outra terapia é a hipossensibilização, onde extratos preparados diretamente
das fontes de alérgenos são injetados por via subcutânea nos pacientes em
quantidades pequenas, porém crescentes, em intervalos determinados de tempo.
Como resultado, os níveis de IgE específica diminuem e os de IgG aumentam
muitas vezes. Entretanto, este tipo de tratamento apresenta um ponto fraco. O
extrato injetado contém, além do alérgeno conhecido, outros potenciais alérgenos e
componentes não alergênicos. Assim, tem se observado que indivíduos atópicos são
freqüentemente co-sensibilizados contra diferentes alérgenos durante a
hipossensibilização (STEPHEN et al., 2004). Extratos de alérgenos hidrolisados
também têm sido utilizados para induzir um estado de anergia de célula T específica
e tolerância imunológica em seres humanos alérgicos, terapia esta apresentada na
patente PI 9714899-7 A.
Anticorpos anti-IgE omalizumab, que são anticorpos monoclonais humanizados
estão sendo utilizados como bloqueadores da resposta aos antígenos. O bloqueio
consiste na ligação do omalizumab com a região constante de IgE (Cε3), que
normalmente se ligaria no receptor FcεRI do mastócito. Com isso, não ocorrerá a
hipersensibilização tipo I (SOLÈR, 2002). Com esse efeito terapêutico, o nível de IgE
tem sido reduzido no soro, além de diminuir a chance de uma nova síntese de IgE.
No entanto, os sintomas reaparecem em poucas semanas depois do término do

Felix, S.P.
18
tratamento (SOLÈR, 2002).
Outra abordagem imunoterápica é mediada pela coagregação inibitória do
receptor FcγRIIb com alta afinidade ao receptor de IgE, FcεRI. Com o aumento do
nível de IgG, este se liga no seu receptor, induzindo a fosforilação do receptor de
IgE, inibindo assim o sinal que seria desencadeado por ele (DAERON et al., 1995).
Considerando que o conhecimento das estruturas dos alérgenos concomitante
com a elucidação dos epitopos alergênicos é um passo fundamental para o
tratamento imunoterápico, diversos grupos têm se envolvido neste desafio utilizando
várias estratégias metodológicas. Muitas delas envolvem ensaios que avaliam
unicamente a capacidade das IgE de se ligarem a um determinado peptídeo. A
estratégia empregada por Mayerhoffer em 2004 e por Felix em 2006 foi baseada em
um procedimento que avaliava a interação IgE-epitopo de uma forma mais funcional,
uma vez que estava centrada na visualização dos mastócitos desgranulados após a
primeira sensibilização com soro total anti-albuminas 2S, produzidos em ratos, e
após a segunda sensibilização com alérgenos de Ricinus communis. Os trabalhos
produzidos por Mayerhoffer e Felix foram criticados por terem sido realizados em
ratos uma vez que, nestes animais, a resposta alérgica poderia ser tanto mediada
por IgE como por IgG2a. Neste sentido, neste projeto procuramos qualificar esta
resposta, desenvolver novas estratégias para quantificar o processo de
desgranulação dos mastócitos, e realizar alguns experimentos para avaliar se estes
epitopos também são reconhecidos por IgE humana.

Felix, S.P.
19
2 - OBJETIVOS
• Otimizar procedimento para quantificar a desgranulação de mastócitos por
dosagem da histamina liberada dos grânulos;
• Investigar se a desgranulação de mastócitos de ratos provocada por albumina
2S de Ricinus communis é mediada por IgE ou por uma subclasse de IgG;
• Investigar se IgE presente em soro de indivíduos atópicos reconhecem os
epitopos de albumina 2S de Ricinus communis, previamente identificados em
ratos;
• Avaliar quais aminoácidos dos epitopos alergênicos poderiam estar
envolvidos na ligação com a imunoglobulina E;
• Testar a eficiência de aminoácidos livres no bloqueio da ligação entre
imunoglobulina E e epitopos alergênicos;
• Avaliar a existência de reações cruzadas entre alérgenos de fontes
alimentares e albuminas 2S de Ricinus communis;
• Avaliar a existência de reações cruzadas entre aeroalérgenos e albuminas 2S
de Ricinus communis;
• Avaliar a eficiência dos bloqueadores químicos como protetores de reações
cruzadas entre alérgenos de Ricinus communis e de alérgenos de fontes
alimentares e inalantes.

Felix, S.P.
20
3 - METODOLOGIA
3.1 - Obtenção de albumina 2S:
Utilizamos as albuminas 2S isoladas das sementes de Ricinus communis,
cultivar IAC226, já disponíveis no laboratório, as quais foram extraídas pela
metodologia descrita por Thorpe e colaboradores em 1988, segundo as adaptações
propostas por Marcondes de Souza em 1997.
3.2 - Obtenção de soro anti-albumina 2S de rato:
O “pool” de soro anti-albuminas 2S foi produzido na Universidade Federal
Fluminense, em colaboração com o Dr. Maurício Verícimo. Para tanto, 10 ratos R/A
tor, bons produtores de IgE, foram imunizados por injeção intraperitoneal de 0,5 mL
de salina contendo 0,01 mg de CB-1A (conjunto de albumina 2S obtidos da
variedade de mamona Amarelo de Irecê) e 5,0 mg de hidróxido de alumínio. Um
mês após a 1ª imunização, os animais receberam uma dose reforço de antígeno.
Nesta etapa, a mesma quantidade do antígeno foi misturada com 2,5 mg de
hidróxido de alumínio. Os animais foram sangrados, por punção cardíaca, 7 dias
após o reforço, sendo que volumes de soro iguais de cada animal foram recolhidos,
reunidos e guardados em alíquotas de 100 µL.
3.3 - Avaliação da alergenicidade:
Para analisar as propriedades alergênicas empregamos ensaio de
desgranulação de mastócito obtido de lavado peritoneal de rato.
3.3.1 - Obtenção dos mastócitos de rato:
Ratos Wistar (não imunizados), de aproximadamente 250 g cada foram
empregados como fonte de mastócitos. Os ratos foram sacrificados por asfixia em
CO2 e a cavidade peritoneal foi lavada com 20 mL de DMEM contendo 12 U/mL de
heparina. O lavado foi retirado da cavidade peritoneal com auxílio de pipeta Pasteur
após incisão no peritônio do animal.

Felix, S.P.
21
O lavado peritoneal do rato (cerca de 15 mL) foi colocado em placa de Petri
por 30 minutos a 37oC para separar mastócitos de macrófagos. Após esse tempo,
2/3 do meio de cultura foi retirado cuidadosamente da superfície com auxílio de
pipeta Pasteur e descartado. O líquido remanescente (cerca de 4-5 mL) contendo os
mastócitos foi transferido para um tubo Falcon de 50 mL. A suspensão final de
células foi dividida em alíquotas de 100 µL.
3.3.2 - Ensaios de desgranulação:
Avaliamos a ativação dos mastócitos mediada ou não por imunoglobulinas. As
alíquotas de 100 µL da preparação enriquecida em mastócitos foram submetidas ao
tratamento com 1 µL de soro total anti-albumina 2S e com 10 µL da amostra a ser
testada (10 µg/mL). A mistura era incubada por 1 hora a 37o C. Nos controles de
ativação inespecífica, o soro era omitido do ensaio.
Para a avaliação da desgranulação, uma alíquota de 10 µL era utilizada para
a contagem de mastócitos por microscopia ótica, o remanescente era reservado
para a dosagem de histamina.
3.3.2.1 - Avaliação do percentual de desgranulação por microscopia ótica:
A contagem diferencial das células íntegras e coradas foi feita em câmara de
Neubauer, nos quatro quadrantes, através da observação em microscópio óptico
Zeiss Axioplan. Para tanto, 5 µL da suspensão de células, após os diversos tipos de
incubação foram misturadas durante 15 min com 5 µL de solução aquosa contendo
0,1% de azul de toluidina, 10% de formaldeído e 1% de ácido acético, pH 2,8 para
evidenciar a desgranulação.
Como controle negativo de sensibilização induzida, mastócitos sem
tratamento prévio foram também incubados com o corante nas condições citadas e
foram observados ao microscópio ótico. A contagem destas células, íntegras e
desgranuladas, permitiu uma avaliação do procedimento de obtenção.
Foram utilizados três ratos para cada análise. Cada experimento foi feito em
duplicata, onde eram empregados mastócitos do mesmo animal.
3.3.2.2 - Dosagem de histamina:
Para esta dosagem empregamos um processo de cromatografia de troca
catiônica, seguido por derivatização pós-coluna, onde a histamina reagiu com OPA,

Felix, S.P.
22
produzindo um composto fluorescente. Vários gradientes de eluição foram testados
e o processo utilizado baseou-se na eluição isocrática empregando-se NaOH 0,2M
como eluente. Uma curva padrão foi feita empregando-se de 1pmol a 1 nmol de
histamina fornecida pela r-Biopharm.
Os 90 µL remanescentes da suspensão de células foram centrifugados à
4000 g por 10 minutos e 20 µL do sobrenadante foram utilizados para a dosagem da
histamina liberada, por processos cromatográficos. O restante da amostra (70 µL) foi
sonicado por 30 segundos para rompimento da membrana dos mastócitos. Uma
alíquota de 20 µL da amostra sonicada foi também analisada por cromatografia para
dosagem da histamina. O processo de sonicação provocava o rompimento de
praticamente todas as células e os valores da quantificação de histamina eram
tomados como 100%.
Uma planilha de valores foi montada no programa excel. Nesta os valores de
histamina liberada pelos alérgenos, pelo processo de sonicação e o número de
células eram considerados. A partir destas informações a porcentagem de histamina
liberada era calculada.
A histamina foi dosada após separação dos componentes do meio reacional
(DMEM) por cromatografia de troca catiônica, empregando um sistema HPLC.
A eluição da histamina retida foi feita com NaOH 0,2 M empregando um fluxo
de 0,6 mL/min. A detecção foi feita por derivatização pós coluna, utilizando o
reagente o-phthaldialdeido *. Inicialmente o eluato era neutralizado com a solução A
e depois reagia com a solução B. Utilizamos um detector de fluorescência da
Shimatzu, modelo RF 535, onde a excitação era feita a 375 nm e a emissão lida a
460 nm.
* preparo de reagente o-phthaldialdeído (OPA):
- Solução A:
Preparo de 1L do tampão com a seguinte composição química:
40,7 g de Na2CO3 (0,384M); 13,6 g de H3BO3 (0,216M); 18,8 g de K2SO4
(0,108M)
Os reagentes foram dissolvidos em água até um volume final de 1L. Ficando
o pH da solução próximo a 10, sem ajuste necessário.

Felix, S.P.
23
- Solução B – OPA 0,08%:
400 mg de OPA foi dissolvido em 7 mL de etanol e em seguida foi adicionado
1 mL de 2-mercaptoetanol. O volume foi ajustado a 500 mL com a solução A.
3.4 - Separação das imunoglobulinas E das imunoglobulinas G de rato por
cromatografia de afinidade em batelada:
Para investigar se a desgranulação de mastócitos de ratos provocada por
albumina 2S de Ricinus communis é mediada por IgE ou por uma subclasse de IgG
foi feito cromatografia de afinidade em batelada, a fim de separar essas duas
classes de imunoglobulinas para analisá-las posteriormente, por ensaios de
desgranulação.
Para isso, aproximadamente 100 µL de proteína G acoplada a Sepharose
(Gibco BRL EUA) foram colocados em um eppendorf. A resina (proteína G acoplada
a Sepharose) foi lavada cinco vezes com 100µL de tampão NaHCO3 0,2M com NaCl
0,5M pH 8,3 (tampão de equilíbrio). Um volume de 30µL de soro total anti-albumina
2S de rato R/A tor foi misturado com a resina já lavada. A essa mistura foram
adicionados 50µL de tampão de equilíbrio e, em seguida, a mistura foi incubada por
16 horas sob agitação a 4ºC.
Após a incubação, o sobrenadante (teoricamente com IgE purificadas) foi
retirado e estocado a 4ºC para análises posteriores (ensaios de “dot blotting” e
desgranulação de mastócitos). Depois da retirada do sobrenadante, a resina foi
lavada dez vezes com 100µL do tampão de equilíbrio, para retirar o resto do material
não ligado.
A IgG, associada a proteína G acoplada a Sepharose, foi eluída usando
100µL de uma solução 0,5M de ácido acético pH 3,0-2,5. Como a solução de ácido
acético é muito agressiva, foi necessário transferir a fração de IgG eluída
imediatamente para uma solução neutralizante de 1M TRIS-HCl pH 9,0. Após a
neutralização, a solução foi estocada a 4ºC para análises posteriores (“dot blotting” e
desgranulação de mastócitos).
Para estocar a resina, esta foi lavada três vezes com 100µL do ácido acético
1 M pH 2,5, seguido de três lavagens com 100µL de tampão de equilíbrio, uma vez
com 100µL de Guanidina 4-6M por 30-60 minutos a 4ºC e, finalmente colocada em
100µL de etanol 20%.

Felix, S.P.
24
3.4.1 - Comprovação da ausência de IgG no sobrenadante:
Para avaliar a eficiência da precipitação em retirar as subclasses de
imunoglobulina G, o sobrenadante foi analisado por “dot blotting”, empregando como
anticorpo secundário o anti-IgG2a de rato produzido em cabra (Serotec).
A membrana de nitrocelulose foi dividida em quadrados de 2 cm e colocada
em uma placa de 24 poços. Posteriormente, cada membrana foi umedecida em
tampão fosfato de sódio 0,1 M contendo NaCl 0,5 M pH 7,6 (PBS) e deixada secar a
temperatura ambiente. Cerca de 10 µL da solução estoque (10 µg/mL) de albumina
2S e 10µL de Tampão NaHCO3 0,2M pH 8,3 contendo NaCl 0,5M pH 8,3 (tampão
utilizado durante a precipitação, presente na Fração IgE) foram aplicados sobre as
membranas. Após a secagem das amostras nas membranas, estas foram
mergulhadas em tampão bloqueador (PBS + Leite em pó Molico 2%) e mantidas
nesta solução por uma hora. Posteriormente, cada membrana foi incubada com 1µL
do sobrenadante da cromatografia de afinidade em batelada diluído em 200 µL do
tampão de bloqueio por um período de duas horas a temperatura ambiente. Em
seguida, as membranas foram lavadas cinco vezes com PBS por uma hora, e
incubadas com anticorpo anti-IgG2a de rato complexado a peroxidase (1:2000 em
tampão bloqueador) por uma hora. A revelação foi feita utilizando uma mistura
contendo 5 mg de DAB dissolvidos em 4,9 mL de água ultrapura, 300µL de imidazol
0,1M, 100µL de tampão Tris-HCl 2M ( pH 7,5) e 5µL de peróxido de hidrogênio 30%.
Como controles, foram utilizados 10µL de água destilada (controle negativo) e
10 µL da solução estoque (10 µg/mL) de albumina 2S (controle positivo), só que
nessas amostras o anticorpo primário foi o soro total anti-albumina 2S (1:800),
produzido em rato R/A tor (antes da separação por cromatografia de afinidade em
batelada).
3.4.2 - Comprovação da presença de IgG na fração ligada:
Para comprovação da presença de IgG na fração associada à resina de
Sepharose, a qual foi eluída com ácido acético, foi também feito um ensaio de “dot
blotting” conforme descrito no item 3.4.1. Neste caso, 1 µL da fração eluída foi
dissolvida em 100µL do tampão bloqueador como anticorpo primário. Controles
adicionais utilizando a solução de eluição e o tampão de neutralização foram
incluídos.

Felix, S.P.
25
3.4.3 - Análise da desgranulação dos mastócitos de rato, sensibilizados pelas
frações enriquecidas em IgE ou enriquecidas em IgG, utilizando albumina 2S e
seus peptídeos:
Para analisar a desgranulação de mastócitos de rato, sensibilizados pelas
frações enriquecidas em IgE (FE) e fração enriquecida em IgG (FG), utilizando
albumina 2S e seus peptídeos (P0, P1, P2, P3, P4, P5), os mastócitos de rato,
obtidos por sedimentação em placa de Petri, foram tratados com:
a) 5 µL de FE e 10 µL da solução estoque de albumina 2S (10 µg/mL);
b) 10 µL de FG e 10 µL da solução estoque de albumina 2S (10 µg/mL);
c) 10 µL da solução estoque (10 µg/mL) de cada um dos peptídeos sintéticos
(avaliação da desgranulação não mediada por imunoglobulinas);
d) 1 µL soro total anti-albumina 2S e 10 µL da solução estoque (10 µg/mL) de cada
um dos peptídeos sintéticos;
e) 5 µL de FE e 10 µL da solução estoque (10 µg/mL) de cada um dos peptídeos
sintéticos;
f) 10 µL de FG e 10 µL da solução estoque (10 µg/mL) de cada um dos peptídeos
sintéticos.
Como controle positivo, a fração enriquecida em mastócitos (100 µL) foi
tratada com 1 µL de soro total anti-albuminas 2S e com 10 µL da solução estoque de
albuminas 2S (10 µg/mL).
A determinação do percentual de desgranulação foi feita por microscopia
ótica, como descrito no item 3.3.2.
3.5 - Obtenção de IgE humana:
Soro de indivíduos atópicos (com níveis de IgE elevados) foi utilizado como
fonte de IgE humano.
Estes soros foram coletados no Laboratório de Côrreas, localizado na cidade
de Petrópolis-RJ, para controle de saúde de indivíduos atópicos. O soro excedente,
após análises, foi cedido pelo Laboratório de Côrreas. A dosagem de IgE nestes
soros foi feita pelo método de quimiluminescência e aqueles que apresentavam

Felix, S.P.
26
valores de IgE total acima de 140 KU/L, valor máximo de referência para adultos
não atópicos, foram utilizados. Para efeito de inviolabilidade à intimidade, os nomes
dos indivíduos foram omitidos. O Laboratório de Corrêas participa do controle de
qualidade da Sociedade Brasileira de Análises Clínicas.
Os soros destes indivíduos foram investigados quanto à sua capacidade de
reconhecimento de albuminas 2S de Ricinus communis pela técnica de “dot blotting”
descrita no item 3.4.1. Neste caso membranas de nitrocelulose foram incubadas
com 10 µL da solução estoque (10 µg/mL) de albumina 2S, e após o bloqueio, com 1
µL de soro de cada paciente diluído em 50 µL de tampão de bloqueio. O anticorpo
secundário utilizado foi o anti-IgE humano complexado a biotina (1:2000). Para
revelação empregamos o sistema peroxidase complexada a streptavidina (1:1000).
3.5.1 - Identificação de epitopos alergênicos de albumina 2S de R. communis,
por “dot blotting”, utilizando soro do indivíduo atópico:
Para identificação de epitopos alergênicos de albumina 2S de R. communis
foi feito “dot blotting” conforme descrito no item 3.4.1. Os antígenos utilizados foram
10 µL da solução estoque (10 µg/mL) de albumina 2S, de Ric c 1 e de Ric c 3 e de
peptídeos sintéticos. Como anticorpo primário usamos o soro do indivíduo atópico
que reconheceu, com maior intensidade, as albuminas 2S de R. communis na
diluição (1:50) e como secundário utilizamos o anticorpo anti-IgE humano
complexado a biotina (1:2000). Para revelação empregamos o sistema peroxidase
complexada a streptavidina (1:1000).
3.5.2 - Separação das imunoglobulinas E e das imunoglobulinas G de humano,
por cromatografia de afinidade em batelada:
A separação das imunoglobulinas E das imunoglobulinas G humanas por
cromatografia de afinidade em batelada, foi feita conforme descrito no item 3.4,
porém o soro utilizado foi o soro contendo IgE do indivíduo atópico, reativo à
albumina 2S de R. communis.
3.5.2.1 - Comprovação da presença de IgE no sobrenadante:
Para comprovação da presença de IgE no sobrenadante foi feito “dot blotting”
conforme descrito no item 3.4.1, porém além dos antígenos descritos, foi também
utilizado 10µL de tampão contendo ácido acético 1M pH 2,5, ácido acético 0,5M pH

Felix, S.P.
27
2,5 e TRIS-HCl pH 9,0 (tampão utilizado durante a precipitação, presente na fração
IgG); como anticorpo primário foi utilizado 1µL da fração eluída, dissolvida em 18 µL
do tampão bloqueador ou 1µL do sobrenadante dissolvido em 25µL do tampão
bloqueador, cada um em uma membrana contento albumina 2S de R. communis. Já
o anticorpo secundário utilizado foi o anticorpo anti-IgE humano complexado a
biotina (1:2000). Para revelação empregamos o sistema peroxidase complexada a
streptavidina (1:1000).
Como controles, foram utilizados 10µL de água destilada (controle negativo) e
10 µL da solução estoque (10 µg/mL) de albumina 2S (controle positivo), sendo que
nessas amostras o anticorpo primário foi o soro total anti-albumina 2S do indivíduo
atópico mais reativo à albumina 2S de R. communis (antes da separação por
cromatografia de afinidade em batelada) (1:50).
3.6 - Determinação dos aminoácidos envolvidos na interação IgE/epitopo:
3.6.1 – Empregando IgE de ratos
Como descrito anteriormente, na seqüência de todos os peptídeos
identificados por Felix em 2006 e Mayerhoffer em 2004, era possível observar a
presença de pelo menos dois resíduos de aminoácidos dicarboxílicos. Esta
característica nos fez acreditar que os grupamentos carboxílicos laterais destas
cadeias fossem importantes na interação com as IgE. Assim, como estratégia para
investigar quais aminoácidos dos epitopos alergênicos poderiam se ligar às IgE,
mastócitos sensibilizados com soro total anti-albuminas 2S foram incubados com
uma mistura dos aminoácidos livres, presentes em cada peptídeo sintético, mas
sempre omitindo o ácido aspártico e o ácido glutâmico. O princípio básico desta
estratégia seria que os aminoácidos livres, importantes para o reconhecimento pelas
IgE, se ligariam a estas imunoglobulinas, mas não de forma cruzada e, desta
maneira, impediriam o reconhecimento dos alérgenos pelas IgE.
Tanto para albumina 2S como para os peptídeos sintéticos, a mistura de
aminoácidos foi preparada da seguinte forma: cada um dos aminoácidos livres (10
µL da solução estoque de 10 µg/mL), correspondentes à seqüência de aminoácidos
de cada peptídeo, exceto ácido glutâmico (Glu) e ácido aspártico (Asp). O ácido
glutâmico e o ácido aspártico, nas mesmas concentrações, foram utilizados
individualmente e separadamente da mistura de aminoácidos.

Felix, S.P.
28
Inicialmente alíquotas de 100 µL dos mastócitos de rato obtidos através de
sedimentação em placa de Petri, foram incubados com 1 µL de soro total anti-
albumina 2S.
Após a incubação com o soro anti-albumina 2S, os mastócitos, teoricamente
contendo IgEs ligadas, foram tratados com 10 µL da mistura de aminoácidos, ou
ácido glutâmico, ou ácido aspártico, por 5 minutos. Este meio foi então incubado
com cada uma das frações alergênicas.
- No ensaio para albumina 2S: 10 µL da solução estoque (10 µg/mL) de albumina
2S foram adicionados em cada solução de mastócitos (com IgEs ligadas) contendo
cada mistura específica de aminoácidos baseada na seqüência da albumina 2S, Asp
e Glu, separadamente.
- No ensaio para peptídeos sintéticos: 10 µL da solução estoque (10 µg/mL) de
cada um dos seis peptídeos sintéticos foram adicionados em cada solução de
mastócitos (com IgEs ligadas) contendo cada mistura específica de aminoácidos
baseada na seqüência de cada peptídeo sintético, Asp e Glu, separadamente;
Em todos os ensaios, os controles positivos (mastócitos tratados com soro total
anti-albuminas 2S na presença de albuminas 2S), os controles negativos (mastócitos
sem tratamento) e, a detecção do percentual de desgranulação por microscopia
ótica, foram feitos como descrito no item 3.3.2.
A suspensão de células remanescente (a que não foi corada - 90 µL) foi
utilizada para dosagem de histamina, conforme descrito a seguir no item 3.3.2.2.
3.6.2 - Investigação da capacidade ligante de aminoácidos livres na porção Fab
da IgE humana:
A investigação da capacidade ligante de aminoácidos livres (considerados
agentes bloqueadores de IgE ligadas em células de ratos), na porção Fab da IgE
humana foi feita através dos experimentos de ELISA. Inicialmente o soro contendo
IgE, do indivíduo atópico mais reativo à albumina 2S de R. communis, foi utilizado
como anticorpo primário na diluição (1:50). Este, foi tratado, na proporção de 1:10,
com a solução estoque (10 µg/mL) de ácido aspártico ou ácido glutâmico, por 5
minutos.
A placa de ELISA foi sensibilizada com 20 µg de cada uma das amostras
(albumina 2S, cadeia leve de Ric c 1, cadeia pesada de Ric c 1, cadeia leve de Ric c
3, cadeia pesada de Ric c 3 e cada um dos seis peptídeos sintéticos) diluídas em

Felix, S.P.
29
100 µL de tampão carbonato/bicarbonato 0,05M, pH 9,6, durante 18 horas, a 4ºC.
Após esse tempo, a placa foi lavada duas vezes com PBS contendo 0,05% de
Tween 20 (300 µL/poço durante 1 hora). Em seguida, foram adicionados 300 µL de
tampão bloqueador (gelatina 1% em PBS contendo Tween), permanecendo na placa
por 1 hora à temperatura ambiente. Após esse tempo de bloqueio, a placa foi
novamente lavada com PBS contendo Tween 0,05% (300 µL/poço durante 1 hora), e
foram adicionados 50 µL/poço do soro do indivíduo atópico reativo à albumina 2S,
previamente tratado (diluído 1:50 em tampão bloqueador), durante 1 hora à
temperatura ambiente. Posteriormente, a placa foi novamente lavada com PBS
contendo Tween, por três vezes de 5 minutos e, em seguida, foi incubada com o
anticorpo anti-IgE de humano complexado a biotina (diluído 1:2000 em tampão
bloqueador), por 45 minutos à temperatura ambiente. A placa foi lavada com PBS
contendo Tween, por três vezes de 5 minutos. Para revelação empregamos o
sistema peroxidase complexada a streptavidina (diluído 1:1000 em tampão
bloqueador) por 30 minutos. Posteriormente, a placa foi lavada 3 vezes com PBS
contendo Tween 0,05%. Em seguida, a revelação foi feita com 50 µL da solução de
revelação (10 mg de OPD; 10 µL de H2O2 30%; 6,5 mL de ácido cítrico 0,1 M; 7,0 mL
de fosfato de sódio 0,2 M e 9,0 mL de H2O destilada). A reação foi interrompida
adicionando 50 µL de H2SO4 por poço. A leitura da placa foi realizada a 492 nm.
Como controle positivo foram utilizadas cada uma das amostras com o soro
do indivíduo atópico reativo à albumina 2S sem o tratamento descrito anteriormente
(sem os aminoácidos bloqueadores), e como controles negativos foram utilizados
albumina 2S sem o anticorpo primário e água destilada.
3.7 - Avaliação da resposta cruzada entre albumina 2S e alérgenos alimentares
e inalantes (aeroalérgenos) padronizados pelo método descrito por Bradford,
através da desgranulação de mastócitos de rato:
A resposta dos mastócitos de rato foi avaliada com os diversos tipos de
substâncias alergênicas (alérgenos alimentares e alérgenos inalantes). A
determinação foi realizada por contagem dos mastócitos íntegros e desgranulados.
Nos diversos ensaios utilizamos como fonte de IgE o soro de rato (RA/tor) imunizado
contra o “pool” de albumina 2S.

Felix, S.P.
30
3.7.1 - Resposta a partir da desgranulação mediada por alérgenos alimentares
do Kit de teste da FDA Allergenic LTDA:
A padronização das concentrações das proteínas presentes nos extratos
alergênicos (produzidos pela FDA Allergenic LTDA) foi feita pelo método descrito por
Bradford em 1976, onde a curva-padrão foi feita utilizando ovalbumina (OVA).
Os alérgenos presentes no kit de alimentos da FDA Allergenic LTDA são das
seguintes fontes: aves, camarão, carne bovina, carne suína, clara, tomate, gema,
leite, peixes, arroz, glúten, morango, milho, trigo, soja, pimenta, abacaxi, cacau, caju,
amendoim, laranja, limão.
Assim, os mastócitos obtidos através de sedimentação em placa de Petri,
foram tratados: a) Com 10 µl da solução estoque (10 µg/mL) de cada um dos
extratos de alérgenos alimentares; b) Com 1µL soro anti-albumina 2S e com 10 µL
da solução estoque (10 µg/mL) de cada um dos extratos dos alérgenos alimentares.
Controles positivos (mastócitos tratados com soro anti-albuminas 2S na
presença de albuminas 2S), negativos (mastócitos sem tratamento) e a detecção do
percentual de desgranulação, por microscopia ótica, foram feitos como descrito no
item 3.3.2.
3.7.2 - Resposta a partir da desgranulação mediada por alérgenos inalantes da
bateria de alérgenos do Kit de teste da FDA Allergenic LTDA:
Os alérgenos que compõe este kit de inalantes da FDA Allergenic LTDA são
extratos das fontes listadas a seguir: poeira domiciliar, ácaros, fungos do ar, lã,
gramíneas, flores, tabaco, epitélio de gato, epitélio de cão, epitélio de eqüino, epitélio
bovino, epitélio mix, penas mix, algodão, crina, capim, linho, macela, barata, pireto,
seda, taboa.
Assim, os mastócitos obtidos através de sedimentação em placa de Petri,
foram tratados: a) Com 10 µl da solução estoque (10 µg/mL) de cada um dos
extratos de aeroalérgenos que o teor de proteínas também foi padronizado por
Bradford; b) Com 1µL soro anti-albumina 2S e com 10 µL da solução estoque (10
µg/mL) de cada um dos extratos dos alérgenos inalantes.
Controles positivos (mastócitos tratados com soro anti-albuminas 2S na
presença de albuminas 2S), negativos (mastócitos sem tratamento) e a detecção do
percentual de desgranulação, por microscopia ótica, foram feitos como descrito no
item 3.3.2.

Felix, S.P.
31
3.8 - Avaliação da possibilidade de uso de aminoácidos dicarboxilícos como
bloqueadores para a resposta cruzada de alérgenos alimentares e inalantes:
Nestes ensaios empregamos um procedimento similar ao descrito no item 3.6.1.
Para tanto, os mastócitos foram sensibilizados com soro total anti-albuminas 2S, e
incubados com Asp e Glu, separadamente. Posteriormente a esta suspensão, 10 µL
da solução estoque (10 µg/mL) de cada alérgeno, que apresentou resposta cruzada
com albumina 2S de R. communis , foram adicionados.
Em todos os ensaios, os controles positivos (mastócitos tratados com soro total
anti-albuminas 2S na presença de albuminas 2S), os controles negativos (mastócitos
sem tratamento) e, a detecção do percentual de desgranulação por microscopia
ótica, foram feitos como descrito no item 3.3.2.

Felix, S.P.
32
4 - RESULTADOS
4.1 - Avaliação da atividade alergênica.
A atividade alergênica foi investigada por ensaios de ativação dos mastócitos,
onde a desgranulação foi desencadeada por albumina 2S e foi avaliada por
visualização e contagem das células íntegras e desgranuladas no microscópio ótico
e/ou por quantificação da histamina liberada. As figuras 5A e 5B mostram mastócitos
sem qualquer tratamento para ativação e a figura 5C e 5D mastócitos sensibilizados
por soro e albuminas 2S de R. communis.
Figura 5 - Mastócitos íntegros (A/B) e mastócitos desgranulados (C/D), ambos corados por
azul de toluidina. Aumento de 400X.
A B
C D

Felix, S.P.
33
Após contagem dos mastócitos íntegros e desgranulados, observamos que a
fração enriquecida com os mastócitos (controle negativo) apresentou cerca de 30%
de desgranulação, tanto na ausência, como na presença de soro total, indicando que
este percentual de desgranulação é inerente ao processo empregado para o
isolamento dos mastócitos ou reflete as condições fisiológicas do animal.
Valores similares foram encontrados quando albumina 2S foi incubada com
os mastócitos na ausência de soro total. No entanto, quando os mastócitos foram
sensibilizados com o soro total e albumina 2S foi adicionada, observamos uma
desgranulação de cerca de 70% das células (Tabela I). Substâncias que induziam
desgranulação acima de 50% foram consideradas potencialmente alergênicas.
Tabela I - Avaliação do percentual de desgranulação de mastócitos de
rato, mediada por albumina 2S.
Amostras
% de desgranulação
na ausência de soro
total (±±±± D.P)
% de desgranulação
na presença de soro
total (±±±± D.P)
Controle
negativo
27,6 (± 1,2)
30,0 (± 1,0)
Albumina 2S
(ct+)
34,6 (± 1,3)
70,0 (± 1,0)
Em todos os ensaios, após a visualização e contagem no microscópio ótico,
uma alíquota era retirada para quantificação de histamina.
Para dosagem de histamina, empregamos inicialmente um processo de
cromatografia de troca iônica, onde o material era fixado em uma coluna Shim-pack
– amino –Li da Shimadzu, normalmente utilizada para a separação de componentes
de fluídos fisiológicos. O material ligado era eluído por dois tampões (Tampão citrato
- ácido perclórico pH 3,2 e citrato - ácido bórico, pH 10,0) e posteriormente por
NaOH 0,2M. A histamina era eluída com cerca de 5 minutos de lavagem com este
último eluente. Este processo durava cerca de 150 minutos. Baseado nesta
observação, otimizamos processos cromatográficos, onde ajustamos um método
isocrático, de 10 minutos, onde somente NaOH 0,2 M, fluxo 0,6 mL por minuto foi
Felix, S.P.
34
empregada como um eluente. Nesta nova condição, os vários componentes do meio
DMEM utilizado nos ensaios de desgranulação eram eluídos logo no início do
processo, separando-se da histamina. O método mostrou-se linear para
concentrações entre 1pmol e 125 pmols de histamina.
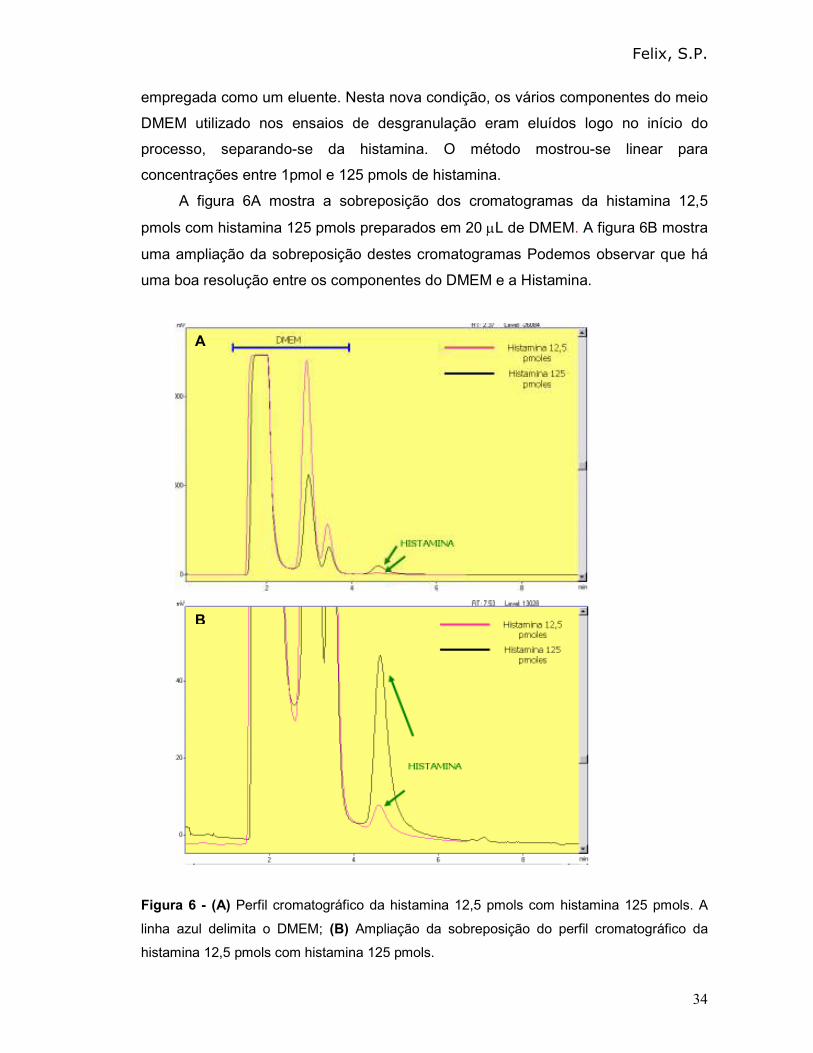
A figura 6A mostra a sobreposição dos cromatogramas da histamina 12,5
pmols com histamina 125 pmols preparados em 20 µL de DMEM. A figura 6B mostra
uma ampliação da sobreposição destes cromatogramas Podemos observar que há
uma boa resolução entre os componentes do DMEM e a Histamina.
Figura 6 - (A) Perfil cromatográfico da histamina 12,5 pmols com histamina 125 pmols. A
linha azul delimita o DMEM; (B) Ampliação da sobreposição do perfil cromatográfico da
histamina 12,5 pmols com histamina 125 pmols.
A
B
Felix, S.P.
35
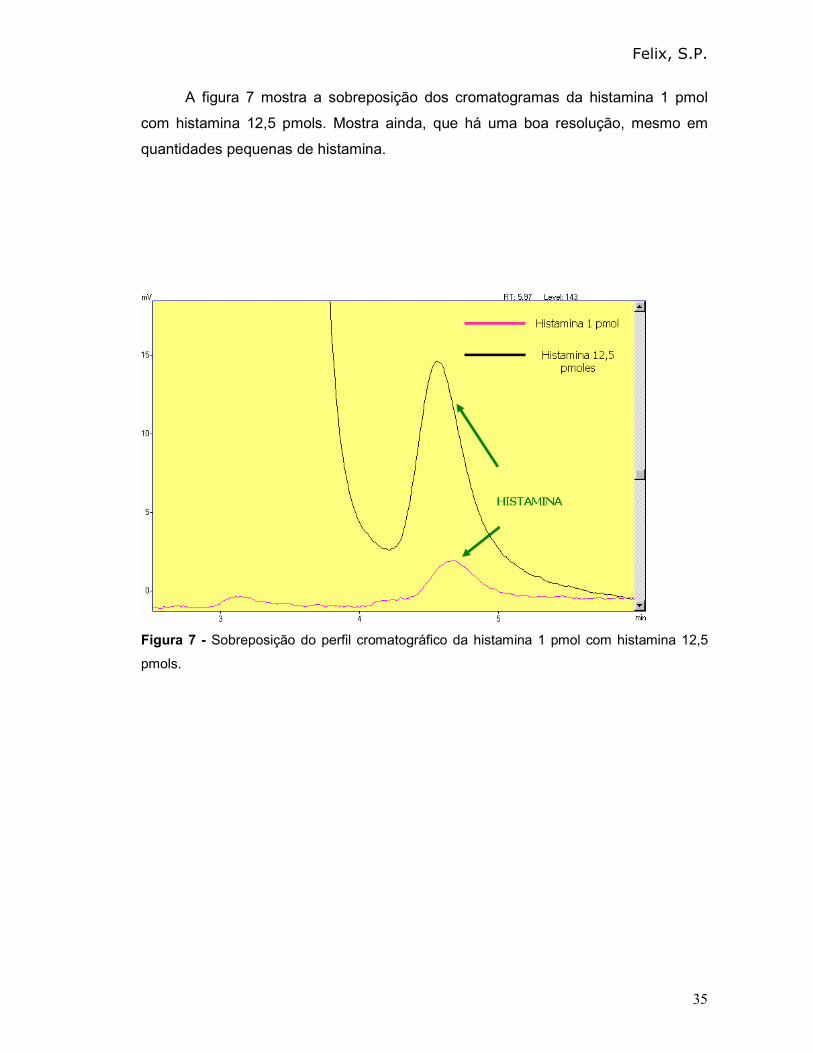
A figura 7 mostra a sobreposição dos cromatogramas da histamina 1 pmol
com histamina 12,5 pmols. Mostra ainda, que há uma boa resolução, mesmo em
quantidades pequenas de histamina.
Figura 7 - Sobreposição do perfil cromatográfico da histamina 1 pmol com histamina 12,5
pmols.
Felix, S.P.
36
Para avaliar o percentual de liberação de histamina, foi necessário,
inicialmente, ajustarmos uma metodologia para romper os mastócitos a fim de liberar
completamente o conteúdo de histamina contido nos grânulos. Tempos gradativos
de exposição ao ultrassom (10 a 60 segundos) foram empregados, sendo observado
que 30 segundos era a condição mais adequada para liberação total de histamina.
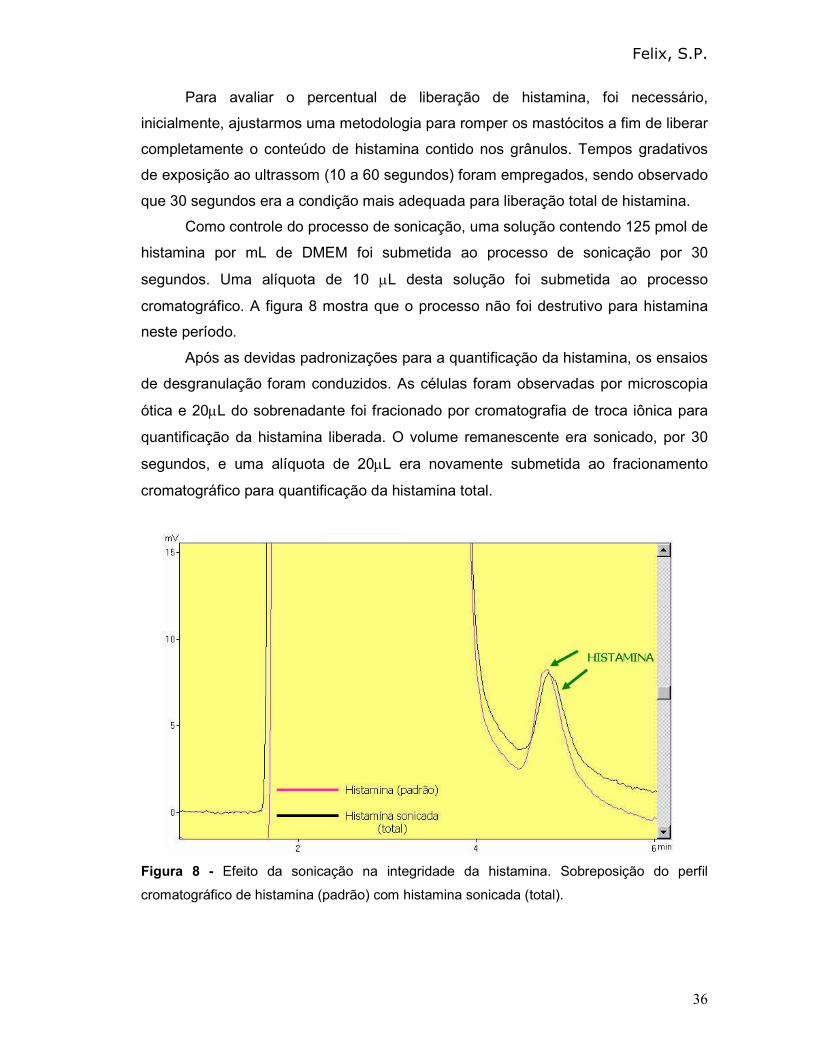
Como controle do processo de sonicação, uma solução contendo 125 pmol de
histamina por mL de DMEM foi submetida ao processo de sonicação por 30
segundos. Uma alíquota de 10 µL desta solução foi submetida ao processo
cromatográfico. A figura 8 mostra que o processo não foi destrutivo para histamina
neste período.
Após as devidas padronizações para a quantificação da histamina, os ensaios
de desgranulação foram conduzidos. As células foram observadas por microscopia
ótica e 20µL do sobrenadante foi fracionado por cromatografia de troca iônica para
quantificação da histamina liberada. O volume remanescente era sonicado, por 30
segundos, e uma alíquota de 20µL era novamente submetida ao fracionamento
cromatográfico para quantificação da histamina total.
Figura 8 - Efeito da sonicação na integridade da histamina. Sobreposição do perfil
cromatográfico de histamina (padrão) com histamina sonicada (total).
Felix, S.P.
37
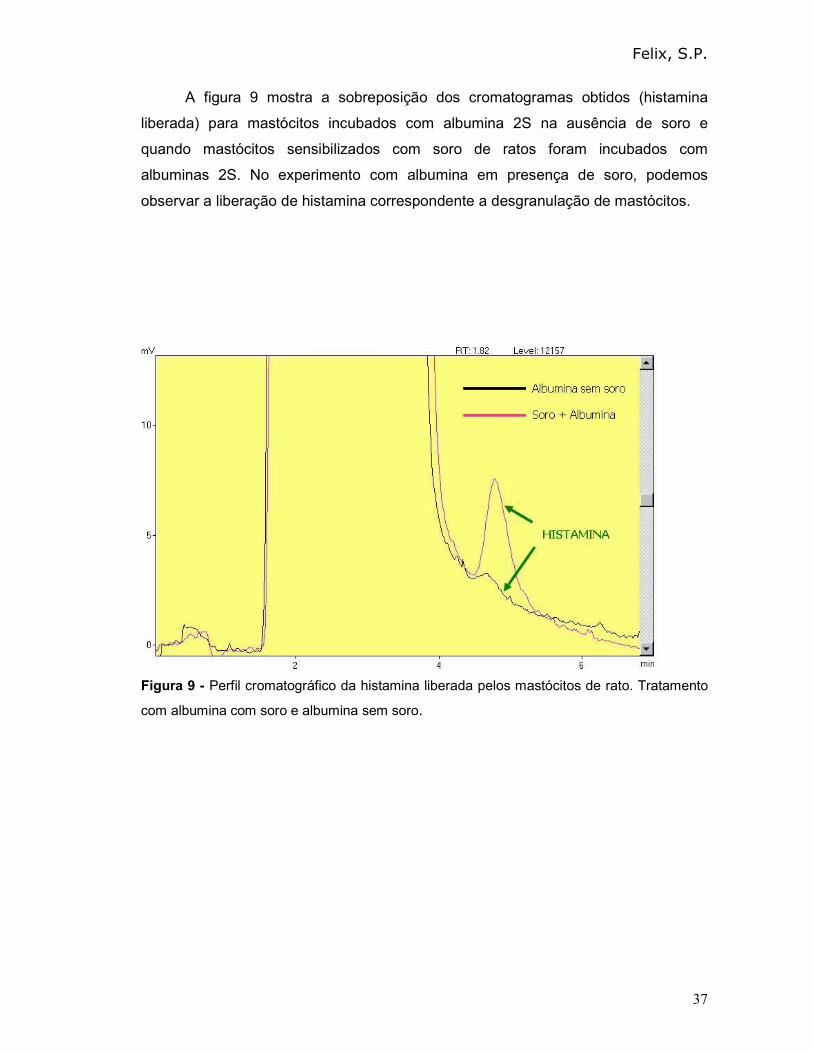
A figura 9 mostra a sobreposição dos cromatogramas obtidos (histamina
liberada) para mastócitos incubados com albumina 2S na ausência de soro e
quando mastócitos sensibilizados com soro de ratos foram incubados com
albuminas 2S. No experimento com albumina em presença de soro, podemos
observar a liberação de histamina correspondente a desgranulação de mastócitos.
Figura 9 - Perfil cromatográfico da histamina liberada pelos mastócitos de rato. Tratamento
com albumina com soro e albumina sem soro.
Felix, S.P.
38
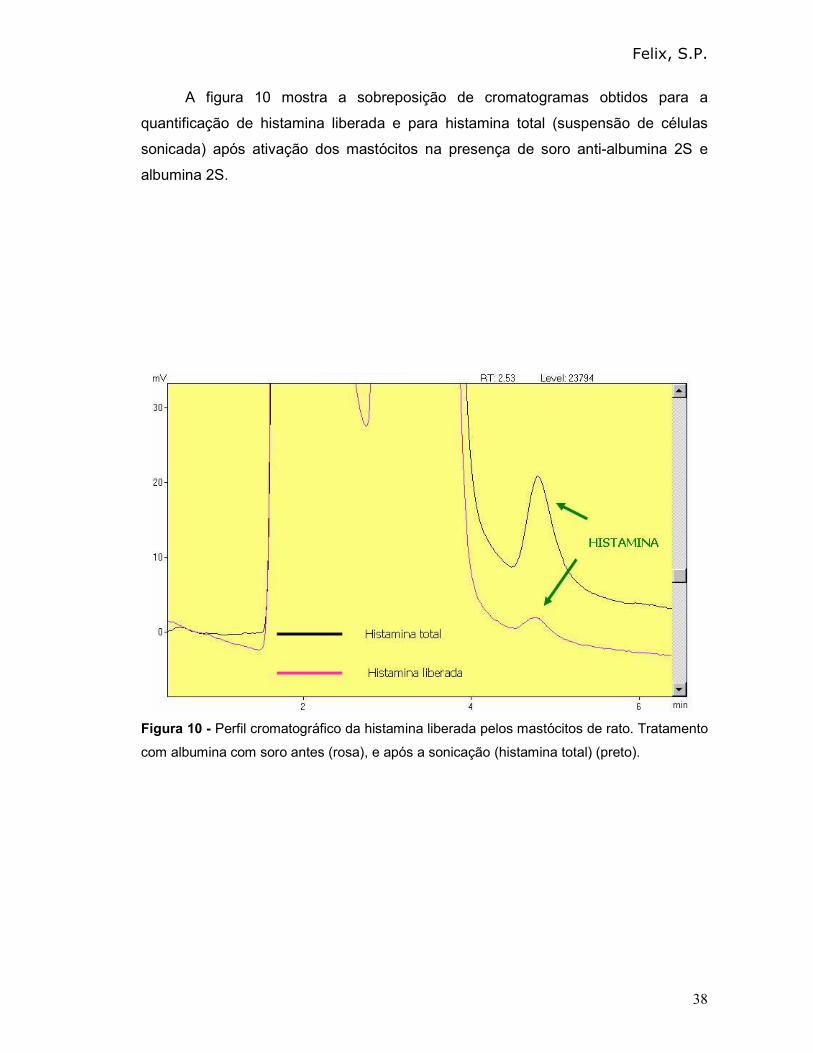
A figura 10 mostra a sobreposição de cromatogramas obtidos para a
quantificação de histamina liberada e para histamina total (suspensão de células
sonicada) após ativação dos mastócitos na presença de soro anti-albumina 2S e
albumina 2S.
Figura 10 - Perfil cromatográfico da histamina liberada pelos mastócitos de rato. Tratamento
com albumina com soro antes (rosa), e após a sonicação (histamina total) (preto).
Felix, S.P.
39
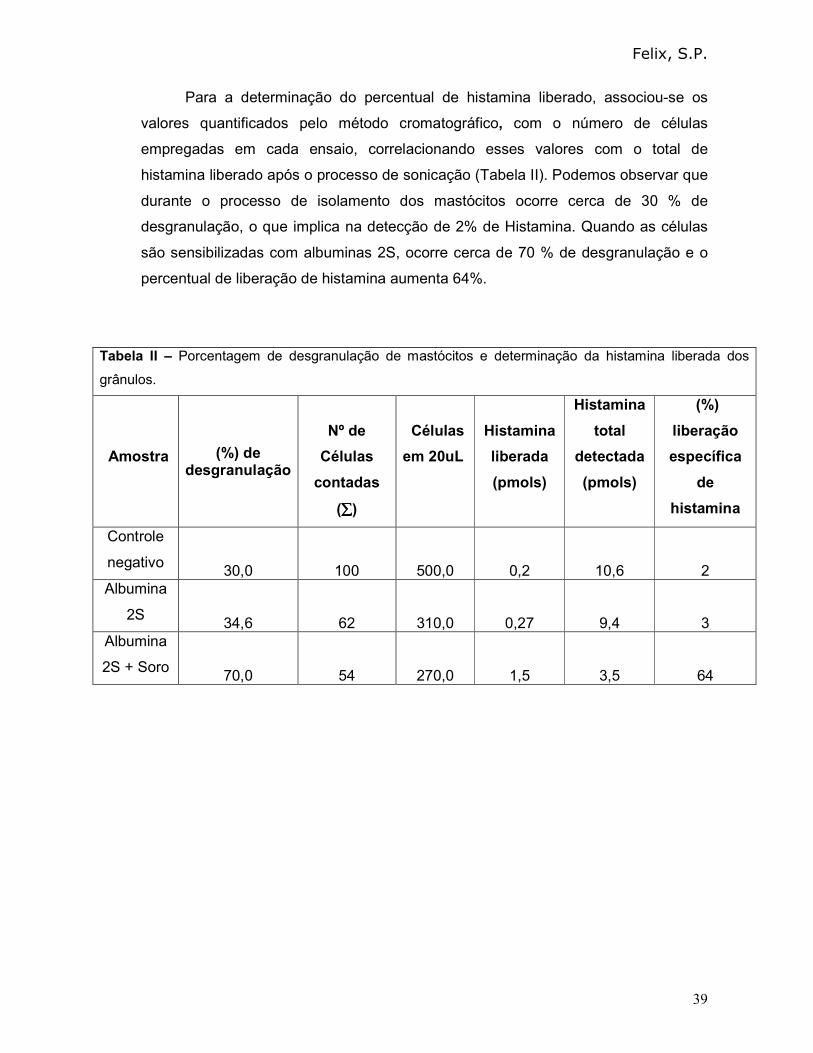
Para a determinação do percentual de histamina liberado, associou-se os
valores quantificados pelo método cromatográfico, com o número de células
empregadas em cada ensaio, correlacionando esses valores com o total de
histamina liberado após o processo de sonicação (Tabela II). Podemos observar que
durante o processo de isolamento dos mastócitos ocorre cerca de 30 % de
desgranulação, o que implica na detecção de 2% de Histamina. Quando as células
são sensibilizadas com albuminas 2S, ocorre cerca de 70 % de desgranulação e o
percentual de liberação de histamina aumenta 64%.
Tabela II – Porcentagem de desgranulação de mastócitos e determinação da histamina liberada dos
grânulos.
Amostra (%) de desgranulação
Nº de
Células
contadas
(∑∑∑∑)
Células
em 20uL
Histamina
liberada
(pmols)
Histamina
total
detectada
(pmols)
(%)
liberação
específica
de
histamina
Controle
negativo 30,0 100 500,0 0,2 10,6 2
Albumina
2S 34,6 62 310,0 0,27 9,4 3
Albumina
2S + Soro 70,0 54 270,0 1,5 3,5
64
Felix, S.P.
40
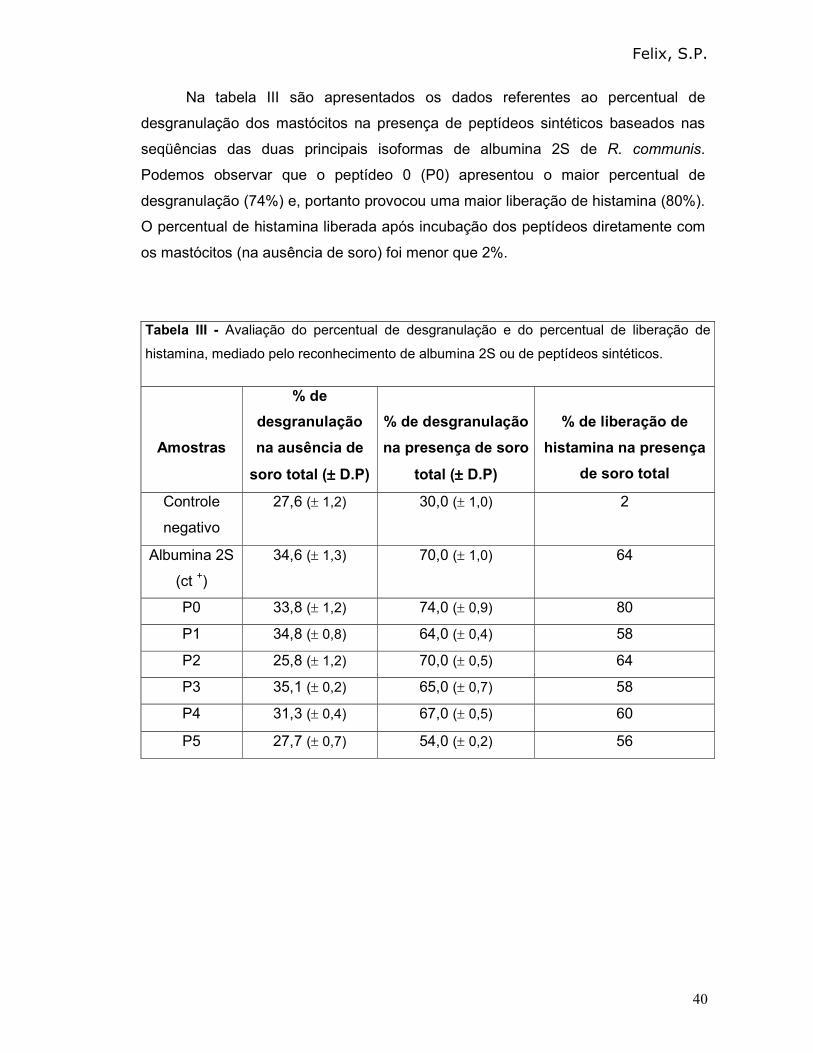
Na tabela III são apresentados os dados referentes ao percentual de
desgranulação dos mastócitos na presença de peptídeos sintéticos baseados nas
seqüências das duas principais isoformas de albumina 2S de R. communis.
Podemos observar que o peptídeo 0 (P0) apresentou o maior percentual de
desgranulação (74%) e, portanto provocou uma maior liberação de histamina (80%).
O percentual de histamina liberada após incubação dos peptídeos diretamente com
os mastócitos (na ausência de soro) foi menor que 2%.
Tabela III - Avaliação do percentual de desgranulação e do percentual de liberação de
histamina, mediado pelo reconhecimento de albumina 2S ou de peptídeos sintéticos.
Amostras
% de
desgranulação
na ausência de
soro total (±±±± D.P)
% de desgranulação
na presença de soro
total (±±±± D.P)
% de liberação de
histamina na presença
de soro total
Controle
negativo
27,6 (± 1,2) 30,0 (± 1,0) 2
Albumina 2S
(ct +)
34,6 (± 1,3) 70,0 (± 1,0) 64
P0 33,8 (± 1,2) 74,0 (± 0,9) 80
P1 34,8 (± 0,8) 64,0 (± 0,4) 58
P2 25,8 (± 1,2) 70,0 (± 0,5) 64
P3 35,1 (± 0,2) 65,0 (± 0,7) 58
P4 31,3 (± 0,4) 67,0 (± 0,5) 60
P5 27,7 (± 0,7) 54,0 (± 0,2) 56

Felix, S.P.
41
4.2 - Avaliação da separação das imunoglobulinas IgE e IgG, por cromatografia
de afinidade em batelada.
• Análise do sobrenadante (depletado de IgG)
A comprovação da ausência de IgG, no sobrenadante obtido após
cromatografia de afinidade em batelada foi feita por imunodetecção (”dot blotting”),
utilizando como anticorpo secundário anti-IgG2a de rato complexado a peroxidase
(1:2000) (Figura 11). Esta figura mostra que o “pool” de albumina 2S foi reconhecido
pela IgG presente no soro antes da precipitação (dot A2). Porém, quando o
sobrenadante obtido após a precipitação foi utilizado como fonte de anticorpo
primário, tanto o tampão NaHCO3 0,2M pH 8.3 com NaCl 0,5M pH 8.3) (dot A3)
como a albumina 2S (dot A4) reagiram negativamente, indicando que as IgG foram
retiradas pelo processo de precipitação.
Figura 11 - “dot blotting” comprovando a ausência de IgG, no sobrenadante obtido após
cromatografia de afinidade em batelada, utilizando soro de rato: (A1) controle negativo
(água destilada) com anticorpo total anti-albumina 2S rato; (A2) Controle positivo (albumina
2S) com anticorpo total anti-albumina 2S rato; (A3) Tampão NaHCO3 0,2M pH 8.3 com NaCl
0,5M pH 8.3; (A4) albumina 2S com o sobrenadante como anticorpo primário.
1 2 3 4
A

Felix, S.P.
42
• Análise da fração ligada (fração IgG)
Inicialmente, o material ligado a proteína G acoplada a Sepharose, foi eluído
como descrito no item 3.4. A comprovação da presença de IgG, na fração eluída foi
feita por imunodetecção (“dot blotting”), utilizando o anticorpo secundário anti-IgG2a
de rato complexado a peroxidase (1:2000) (Figura 12).
Neste caso, as membranas foram previamente incubadas com tampão ou
com o “pool” de albuminas 2S.
Como mostrado na figura 12, o soro total (dot 2) e a fração eluída (fontes do
anticorpo primário, IgG) (dot 4) reconheceram as albuminas 2S, indicando que as
IgG ficaram ligadas na resina durante o processo de precipitação.
Figura 12 - “dot blotting” comprovando a presença de IgG, na fração eluída, obtida após
cromatografia de afinidade em batelada, utilizando soro de rato: (A1) controle negativo
(água destilada) com anticorpo total anti-albumina 2S rato; (A2) Controle positivo (albumina
2S) com anticorpo total anti-albumina 2S rato; (A3) Tampão contendo ácido acético 1M pH
2.5 com ácido acético 0,5M pH 2.5 com TRIS-HCl pH 9.0; (A4) albumina 2S com a fração
eluída como anticorpo primário.
O soro utilizado nos experimentos descritos, foi coletado de ratos R/A tor
reconhecidos como bons produtores de IgE. Assim, após os ensaios descritos
acima, denominamos o sobrenadante livre de IgG, como fração enriquecida em IgE
(FE) e o material eluído como fração enriquecida em IgG (FG).
1 2 3 4
A
Felix, S.P.
43
4.3 - Quantificação da desgranulação dos mastócitos de rato, sensibilizados
por FE e por FG.
Como já descrito na literatura, ratos podem produzir IgG2a e IgE que
poderiam se associar aos mastócitos e assim poderiam ser mediadores na interação
com os epitopos de albuminas 2S. Para verificarmos que classe de imunoglobulina
estaria envolvida neste processo, os mastócitos foram sensibilizados ou com soro
total, ou com FE ou com FG.
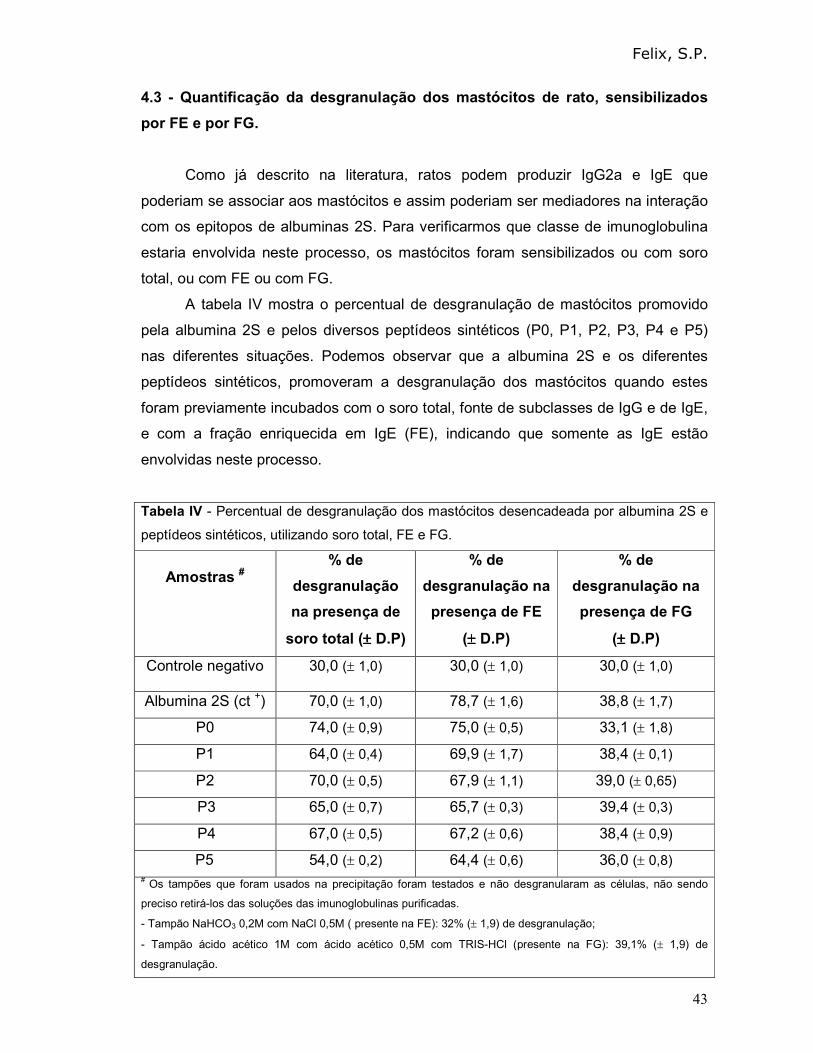
A tabela IV mostra o percentual de desgranulação de mastócitos promovido
pela albumina 2S e pelos diversos peptídeos sintéticos (P0, P1, P2, P3, P4 e P5)
nas diferentes situações. Podemos observar que a albumina 2S e os diferentes
peptídeos sintéticos, promoveram a desgranulação dos mastócitos quando estes
foram previamente incubados com o soro total, fonte de subclasses de IgG e de IgE,
e com a fração enriquecida em IgE (FE), indicando que somente as IgE estão
envolvidas neste processo.
Tabela IV - Percentual de desgranulação dos mastócitos desencadeada por albumina 2S e
peptídeos sintéticos, utilizando soro total, FE e FG.
Amostras #
% de
desgranulação
na presença de
soro total (±±±± D.P)
% de
desgranulação na
presença de FE
(±±±± D.P)
% de
desgranulação na
presença de FG
(±±±± D.P)
Controle negativo 30,0 (± 1,0) 30,0 (± 1,0) 30,0 (± 1,0)
Albumina 2S (ct +) 70,0 (± 1,0) 78,7 (± 1,6) 38,8 (± 1,7)
P0 74,0 (± 0,9) 75,0 (± 0,5) 33,1 (± 1,8)
P1 64,0 (± 0,4) 69,9 (± 1,7) 38,4 (± 0,1)
P2 70,0 (± 0,5) 67,9 (± 1,1) 39,0 (± 0,65)
P3 65,0 (± 0,7) 65,7 (± 0,3) 39,4 (± 0,3)
P4 67,0 (± 0,5) 67,2 (± 0,6) 38,4 (± 0,9)
P5 54,0 (± 0,2) 64,4 (± 0,6) 36,0 (± 0,8) # Os tampões que foram usados na precipitação foram testados e não desgranularam as células, não sendo
preciso retirá-los das soluções das imunoglobulinas purificadas.
- Tampão NaHCO3 0,2M com NaCl 0,5M ( presente na FE): 32% (± 1,9) de desgranulação;
- Tampão ácido acético 1M com ácido acético 0,5M com TRIS-HCl (presente na FG): 39,1% (± 1,9) de
desgranulação.
Felix, S.P.
44
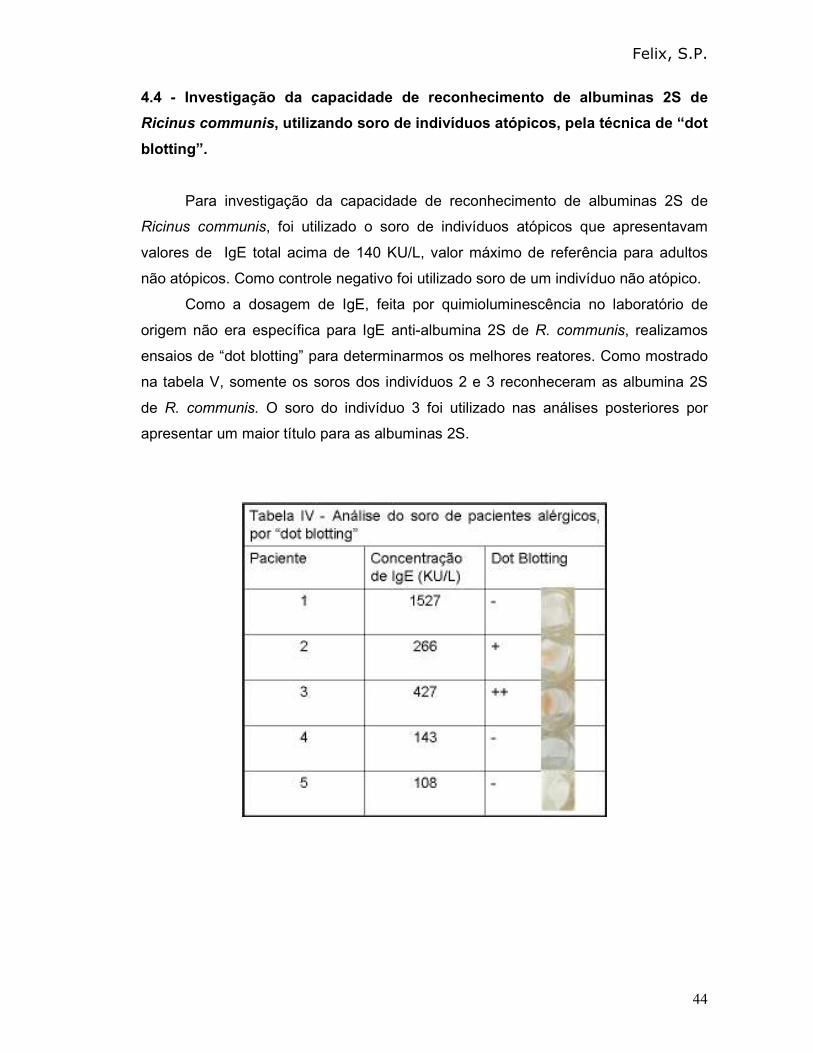
4.4 - Investigação da capacidade de reconhecimento de albuminas 2S de
Ricinus communis, utilizando soro de indivíduos atópicos, pela técnica de “dot
blotting”.
Para investigação da capacidade de reconhecimento de albuminas 2S de
Ricinus communis, foi utilizado o soro de indivíduos atópicos que apresentavam
valores de IgE total acima de 140 KU/L, valor máximo de referência para adultos
não atópicos. Como controle negativo foi utilizado soro de um indivíduo não atópico.
Como a dosagem de IgE, feita por quimioluminescência no laboratório de
origem não era específica para IgE anti-albumina 2S de R. communis, realizamos
ensaios de “dot blotting” para determinarmos os melhores reatores. Como mostrado
na tabela V, somente os soros dos indivíduos 2 e 3 reconheceram as albumina 2S
de R. communis. O soro do indivíduo 3 foi utilizado nas análises posteriores por
apresentar um maior título para as albuminas 2S.

Felix, S.P.
45
4.5 - Identificação de epitopos alergênicos de albumina 2S de R. communis,
por “dot blotting”, utilizando soro do indivíduo atópico selecionado.
Como mencionado anteriormente, as duas principais isoformas de albuminas
2S de R. communis são Ric c 1 e Ric c 3. Cada uma destas proteínas é constituída
de duas cadeias polipeptídicas. As isoformas separadas das cadeias, já estavam
disponíveis em nosso laboratório, as quais foram empregadas nos ensaios de
reconhecimento pelo soro do indivíduo atópico selecionado. Como podemos verificar
na figura 13, ambas as isoformas foram reconhecidas (linha A) com intensidade
equivalente àquela do reconhecimento do “pool” de albuminas 2S. Na linha B,
observamos que as cadeias desnaturadas e alquiladas também foram reconhecidas
por este soro, indicando a presença de epitopos contínuos. Observamos ainda, na
figura 13, linha C que os epitopos reconhecidos por soro de ratos imunizados foram
também reconhecidos pelo soro do indivíduo atópico.

Felix, S.P.
46
Figura13 - “dot blotting” utilizando peptídeos sintéticos, Ric c 1 e Ric c 3 (os principais
alérgenos de R. communis): (A1) controle negativo (água destilada) com o soro do indíviduo
selecionado; (A2) albumina 2S sem soro; (A3) Quimiotripsina + soro; (A4) albumina 2S
(controle positivo) + soro; (A5) Ric c 1 + soro; (A6) Ric c 3 + soro; (B1) Cadeia leve de Ric c
1 + soro; (B2) Cadeia pesada de Ric c 1 + soro; (B3) Cadeia leve de Ric c 3 + soro; (B4)
Cadeia pesada de Ric c 3 + soro; (C1) P0 + soro; (C2) P1 + soro; (C3) P2 + soro; (C4) P3 +
soro; (C5) P4 + soro ; (C6) P5 + soro.
1 2 3 4 5 6
A B C

Felix, S.P.
47
4.6 - Avaliação da presença das imunoglobulinas E no sobrenadante, obtido
após a cromatografia de afinidade em batelada, utilizando soro do indivíduo
atópico.
A literatura não apresenta relatos de que a ativação dos mastócitos, em
humanos, possa ser mediada por IgG. Mesmo assim, adotamos um procedimento
de separação de IgG de IgE, similar ao que aplicamos para o soro de ratos.
A avaliação do reconhecimento das albuminas 2S foi então testada na
presença das frações isoladas, FE ou FG. A figura 14 mostra que o “pool” de
albumina 2S foi reconhecido tanto pelo soro total do índividuo atópico selecionado
(dot A2), quanto pela FE (dot A3), mas não pela FG (dot A4).
Figura 14 - “dot blotting” comprovando a presença de IgE, no sobrenadante obtido após
cromatografia de afinidade em batelada, utilizando soro do indivíduo atópico selecionado:
(A1) controle negativo (água destilada) com soro total; (A2) Controle positivo (albumina 2S)
com soro total; (A3) albumina 2S com FE como anticorpo primário; (A4) albumina 2S com
FG como anticorpo primário; (A5) Tampão contendo ácido acético 1M pH 2.5, ácido acético
0,5M, pH 2,5 e TRIS-HCl pH 9,0; (A6) Tampão NaHCO3 0,2M pH 8,3 contendo NaCl 0,5M
pH 8,3.

Felix, S.P.
48
4.7 - Identificação da natureza química dos aminoácidos, presentes nos
diferentes epitopos, envolvidos na interação com a IgE.
Nestes ensaios, os mastócitos sensibilizados com soro total de ratos foram
incubados com aminoácidos livres, a fim de avaliar a ocupação dos sítios das IgE
que poderiam ser responsáveis pelo reconhecimento de epitopos alergênicos.
Como foi proposto por Felix em 2006 que o grupamento γ carboxílico do ácido
glutâmico e/ou do ácido aspártico pudesse estar envolvido na ligação com as IgEs
ligadas nos mastócitos, esses dois aminoácidos foram omitidos do preparo da
mistura de aminoácidos.
Na tabela VI é mostrado o percentual de desgranulação de mastócitos
promovido pela Albumina 2S e pelos diversos peptídeos sintéticos (P0, P1, P2, P3,
P4 e P5), antes e depois da incubação prévia com a mistura de aminoácidos, livres
de aminoácidos dicarboxílicos. Como podemos observar, não houve mudança
significativa nos valores de desgranulação dos mastócitos quando estes foram
previamente incubados com estes aminoácidos. Ou seja, estes aminoácidos não
ocuparam os sítios de reconhecimento das IgE destinados ao reconhecimento dos
epitopos de albumina 2S.
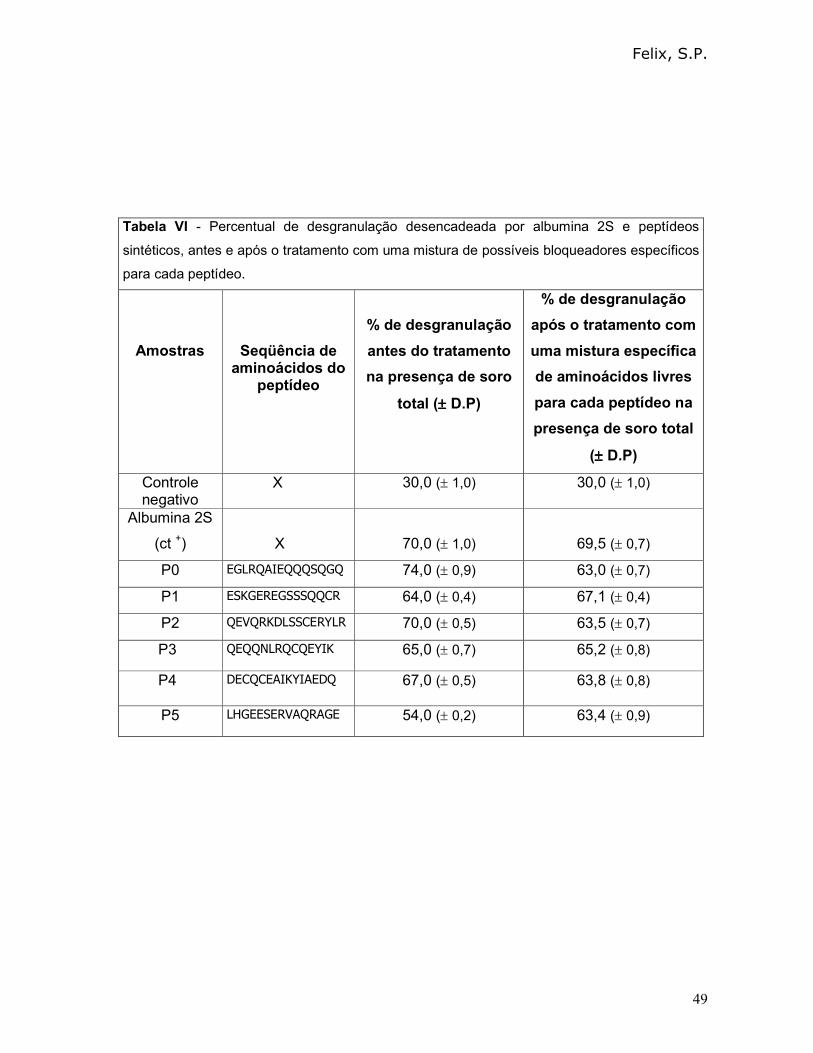
Felix, S.P.
49
Tabela VI - Percentual de desgranulação desencadeada por albumina 2S e peptídeos
sintéticos, antes e após o tratamento com uma mistura de possíveis bloqueadores específicos
para cada peptídeo.
Amostras
Seqüência de
aminoácidos do peptídeo
% de desgranulação
antes do tratamento
na presença de soro
total (±±±± D.P)
% de desgranulação
após o tratamento com
uma mistura específica
de aminoácidos livres
para cada peptídeo na
presença de soro total
(±±±± D.P)
Controle negativo
X 30,0 (± 1,0) 30,0 (± 1,0)
Albumina 2S
(ct +)
X
70,0 (± 1,0)
69,5 (± 0,7)
P0 EGLRQAIEQQQSQGQ 74,0 (± 0,9) 63,0 (± 0,7)
P1 ESKGEREGSSSQQCR 64,0 (± 0,4) 67,1 (± 0,4)
P2 QEVQRKDLSSCERYLR 70,0 (± 0,5) 63,5 (± 0,7)
P3 QEQQNLRQCQEYIK
65,0 (± 0,7) 65,2 (± 0,8)
P4 DECQCEAIKYIAEDQ
67,0 (± 0,5) 63,8 (± 0,8)
P5 LHGEESERVAQRAGE
54,0 (± 0,2) 63,4 (± 0,9)
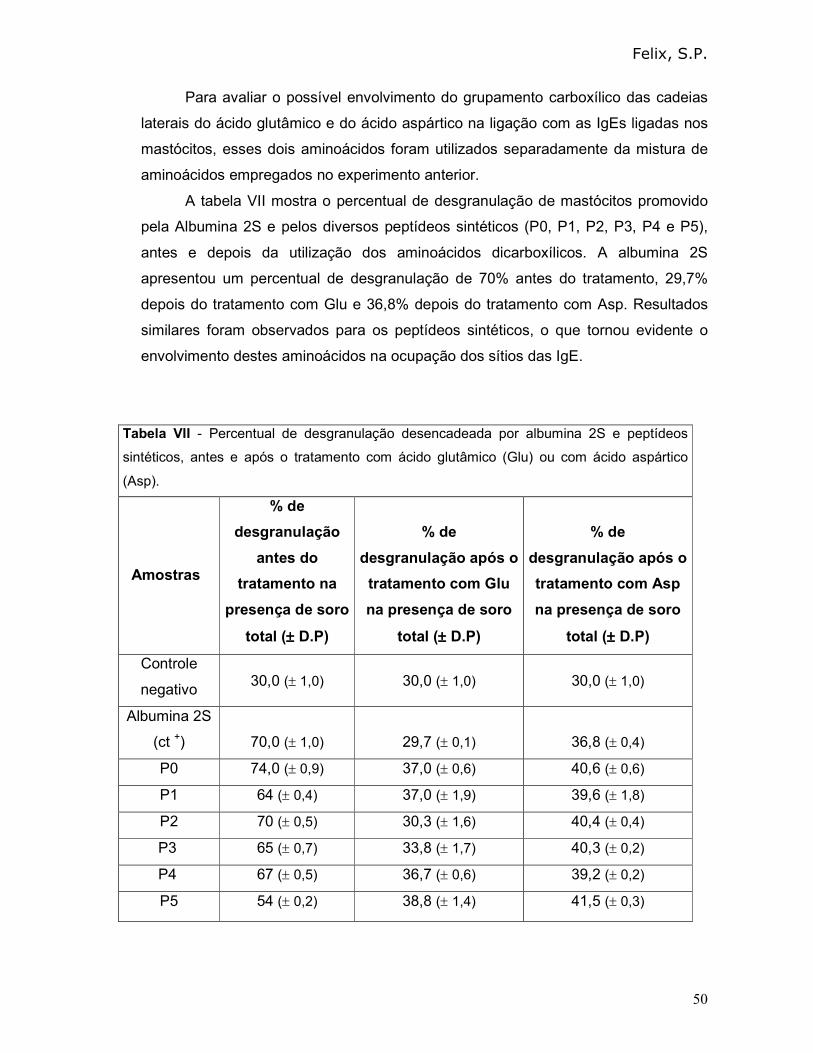
Felix, S.P.
50
Para avaliar o possível envolvimento do grupamento carboxílico das cadeias
laterais do ácido glutâmico e do ácido aspártico na ligação com as IgEs ligadas nos
mastócitos, esses dois aminoácidos foram utilizados separadamente da mistura de
aminoácidos empregados no experimento anterior.
A tabela VII mostra o percentual de desgranulação de mastócitos promovido
pela Albumina 2S e pelos diversos peptídeos sintéticos (P0, P1, P2, P3, P4 e P5),
antes e depois da utilização dos aminoácidos dicarboxílicos. A albumina 2S
apresentou um percentual de desgranulação de 70% antes do tratamento, 29,7%
depois do tratamento com Glu e 36,8% depois do tratamento com Asp. Resultados
similares foram observados para os peptídeos sintéticos, o que tornou evidente o
envolvimento destes aminoácidos na ocupação dos sítios das IgE.
Tabela VII - Percentual de desgranulação desencadeada por albumina 2S e peptídeos
sintéticos, antes e após o tratamento com ácido glutâmico (Glu) ou com ácido aspártico
(Asp).
Amostras
% de
desgranulação
antes do
tratamento na
presença de soro
total (±±±± D.P)
% de
desgranulação após o
tratamento com Glu
na presença de soro
total (±±±± D.P)
% de
desgranulação após o
tratamento com Asp
na presença de soro
total (±±±± D.P)
Controle
negativo
30,0 (± 1,0)
30,0 (± 1,0)
30,0 (± 1,0)
Albumina 2S
(ct +)
70,0 (± 1,0)
29,7 (± 0,1)
36,8 (± 0,4)
P0 74,0 (± 0,9) 37,0 (± 0,6) 40,6 (± 0,6)
P1 64 (± 0,4) 37,0 (± 1,9) 39,6 (± 1,8)
P2 70 (± 0,5) 30,3 (± 1,6) 40,4 (± 0,4)
P3 65 (± 0,7) 33,8 (± 1,7) 40,3 (± 0,2)
P4 67 (± 0,5) 36,7 (± 0,6) 39,2 (± 0,2)
P5 54 (± 0,2) 38,8 (± 1,4) 41,5 (± 0,3)

Felix, S.P.
51
4.8 - Avaliação dos efeitos dos aminoácidos dicarboxílicos, no bloqueio da
desgranulação de mastócitos de rato causado por albumina 2S e peptídeos
sintéticos, por dosagem de histamina:
A figura 15 mostra o cromatograma obtido após fracionamento por
cromatografia de troca iônica do sobrenadante do ensaio de desgranulação dos
mastócitos causada pelo conjunto de Albuminas 2S. Em preto podemos observar o
teor de histamina liberado quando os mastócitos foram pré-sensibilizados com soro
total de rato. Em rosa podemos observar a liberação de histamina quando os
mastócitos, pré-sensibilizados com soro total eram pré-incubados com ácido
glutâmico. Nesta situação verificamos que o ácido glutâmico impediu a liberação de
histamina.
Figura 15 - Perfil cromatográfico da histamina liberada pelos mastócitos de rato. Tratamento
com albumina 2S com soro e albumina 2S com soro incubado previamente com o bloqueador
ácido glutâmico (Glu).

Felix, S.P.
52
A figura 16 mostra os cromatogramas obtidos após fracionamento por
cromatografia de troca iônica dos sobrenadantes dos ensaios de desgranulação dos
mastócitos causada pelos peptídeos sintéticos. Na figura 16 e tabela VIII, podemos
observar que o ácido glutâmico reduziu a liberação de histamina, de 80% para 52%,
quando os mastócitos foram sensibilizados pelo peptídeo P0 (16A). Por outro lado, o
ácido glutâmico reduziu significativamente a liberação de histamina quando os
outros peptídeos sintéticos foram empregados (figuras 16B a 16F), sendo que o
bloqueio foi completo para os peptídeos P3 e P5.
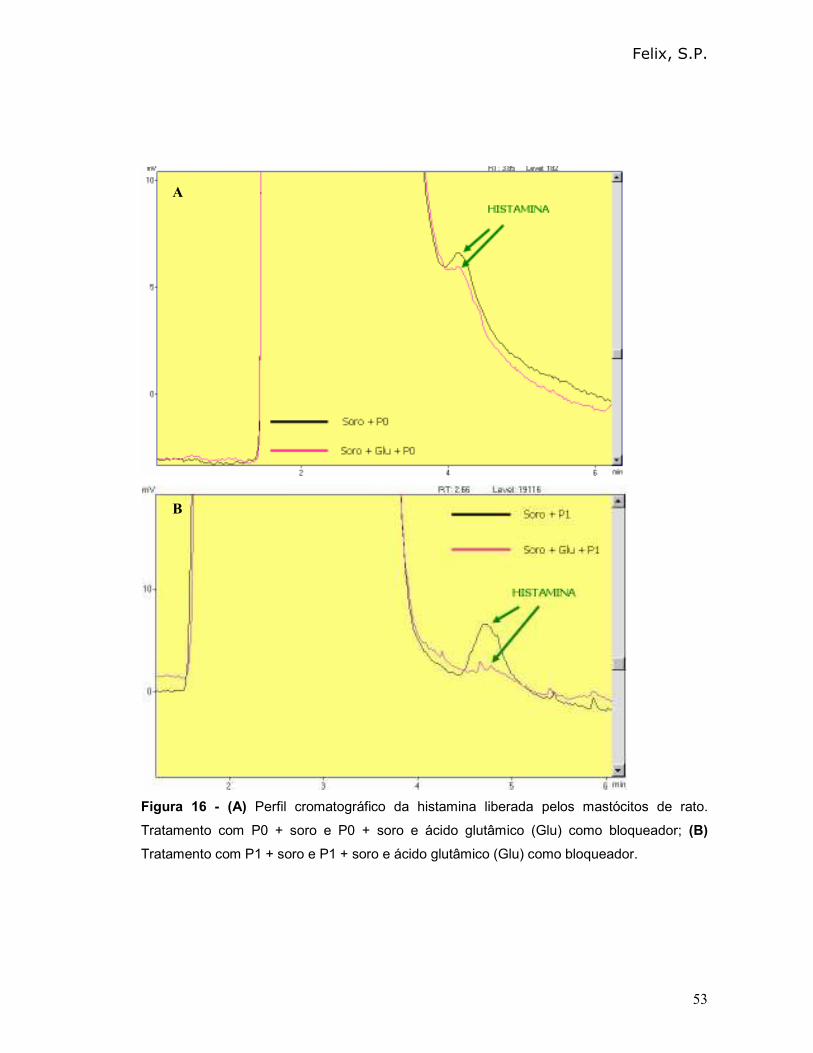
Felix, S.P.
53
Figura 16 - (A) Perfil cromatográfico da histamina liberada pelos mastócitos de rato.
Tratamento com P0 + soro e P0 + soro e ácido glutâmico (Glu) como bloqueador; (B)
Tratamento com P1 + soro e P1 + soro e ácido glutâmico (Glu) como bloqueador.
B
A
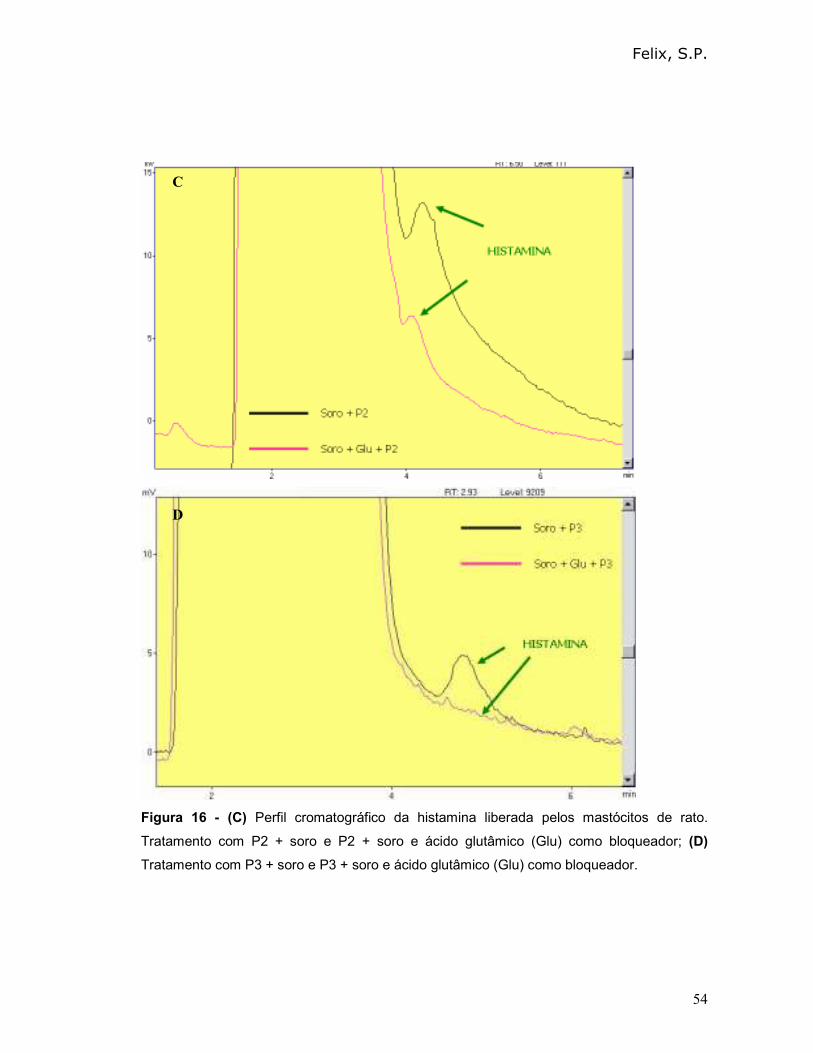
Felix, S.P.
54
Figura 16 - (C) Perfil cromatográfico da histamina liberada pelos mastócitos de rato.
Tratamento com P2 + soro e P2 + soro e ácido glutâmico (Glu) como bloqueador; (D)
Tratamento com P3 + soro e P3 + soro e ácido glutâmico (Glu) como bloqueador.
D
C
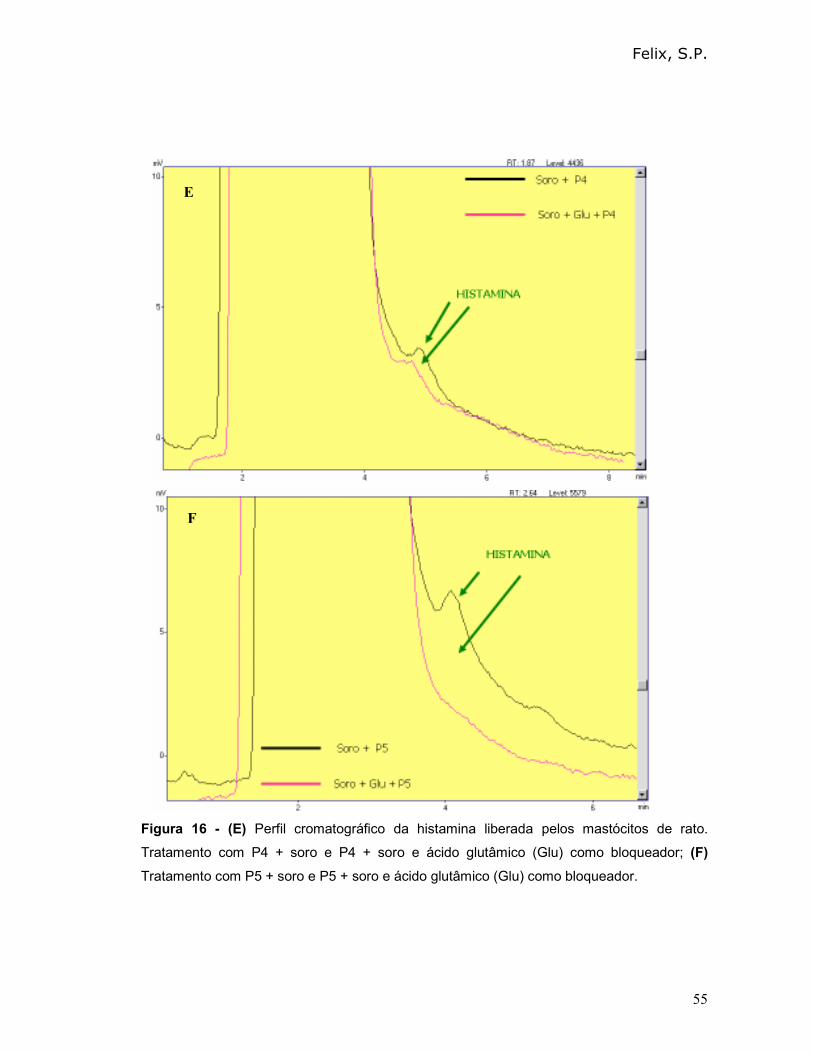
Felix, S.P.
55
Figura 16 - (E) Perfil cromatográfico da histamina liberada pelos mastócitos de rato.
Tratamento com P4 + soro e P4 + soro e ácido glutâmico (Glu) como bloqueador; (F)
Tratamento com P5 + soro e P5 + soro e ácido glutâmico (Glu) como bloqueador.
F
E
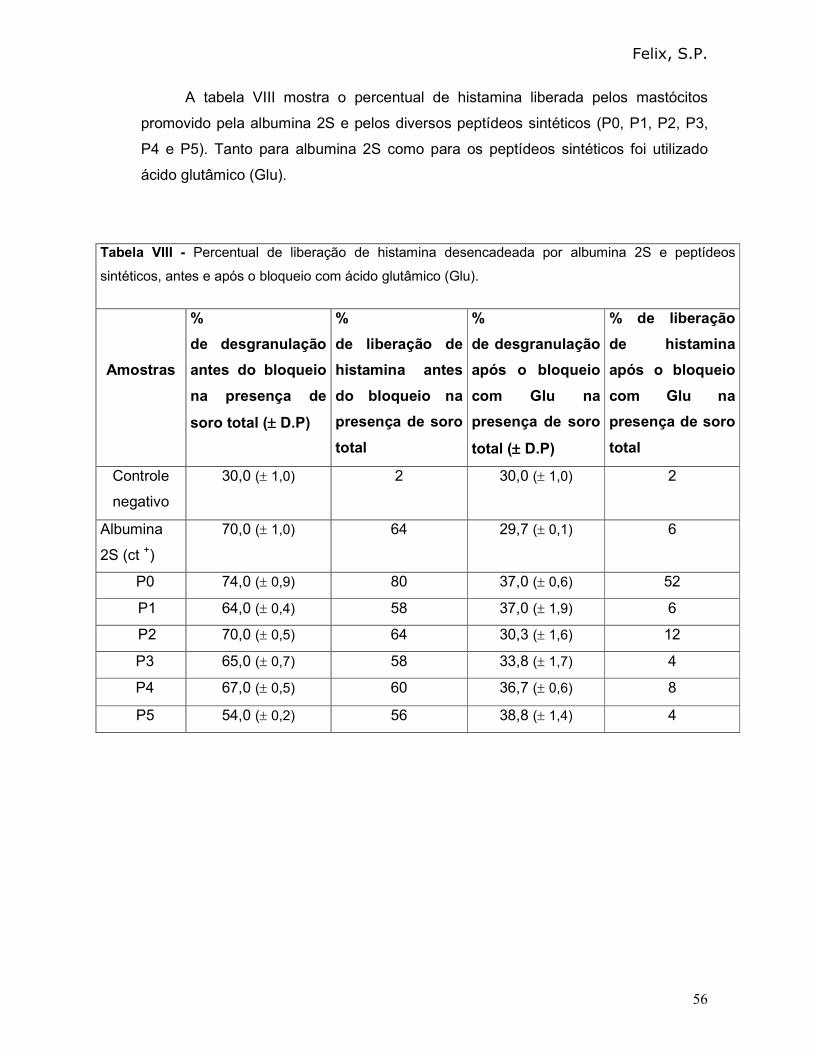
Felix, S.P.
56
A tabela VIII mostra o percentual de histamina liberada pelos mastócitos
promovido pela albumina 2S e pelos diversos peptídeos sintéticos (P0, P1, P2, P3,
P4 e P5). Tanto para albumina 2S como para os peptídeos sintéticos foi utilizado
ácido glutâmico (Glu).
Tabela VIII - Percentual de liberação de histamina desencadeada por albumina 2S e peptídeos
sintéticos, antes e após o bloqueio com ácido glutâmico (Glu).
Amostras
%
de desgranulação
antes do bloqueio
na presença de
soro total (±±±± D.P)
%
de liberação de
histamina antes
do bloqueio na
presença de soro
total
%
de desgranulação
após o bloqueio
com Glu na
presença de soro
total (±±±± D.P)
% de liberação
de histamina
após o bloqueio
com Glu na
presença de soro
total
Controle
negativo
30,0 (± 1,0) 2 30,0 (± 1,0) 2
Albumina
2S (ct +)
70,0 (± 1,0) 64 29,7 (± 0,1) 6
P0 74,0 (± 0,9) 80 37,0 (± 0,6) 52
P1 64,0 (± 0,4) 58 37,0 (± 1,9) 6
P2 70,0 (± 0,5) 64 30,3 (± 1,6) 12
P3 65,0 (± 0,7) 58 33,8 (± 1,7) 4
P4 67,0 (± 0,5) 60 36,7 (± 0,6) 8
P5 54,0 (± 0,2) 56 38,8 (± 1,4) 4

Felix, S.P.
57
4.9 - Investigação da capacidade ligante de aminoácidos livres na porção Fab
da IgE humana, por ELISA.
Para avaliar se a ação bloqueadora do grupamento γ carboxílico do ácido
glutâmico estendia-se à IgE humana, foi feita a investigação da capacidade ligante
de aminoácidos livres na porção Fab da IgE humana por ELISA.
Na tabela IX são mostrados os valores obtidos quando albumina 2S, Ric c 1,
Ric c 3 e os diversos peptídeos sintéticos (P0, P1, P2, P3, P4 e P5), na presença ou
ausência do ácido glutâmico como bloqueador.
Na coluna 2 da tabela, podemos observar que tanto o “pool” de proteínas
nativas (albumina 2S) como as isoformas isoladas, Ric c 1 e Ric c 3, foram
reconhecidas pela IgE presente no soro do indivíduo atópico selecionado. Sendo
que Ric c 1 comportou-se como o mais alergênico. Entre os peptídeos, o P2
apresentou uma maior leitura. Ao analisarmos a coluna 3, podemos observar que o
ácido glutâmico provavelmente se ligou à IgE limitando a ligação dos diversos
epitopos. A eficiência de proteção foi máxima para os peptídeos P2, P3, P4, P5.
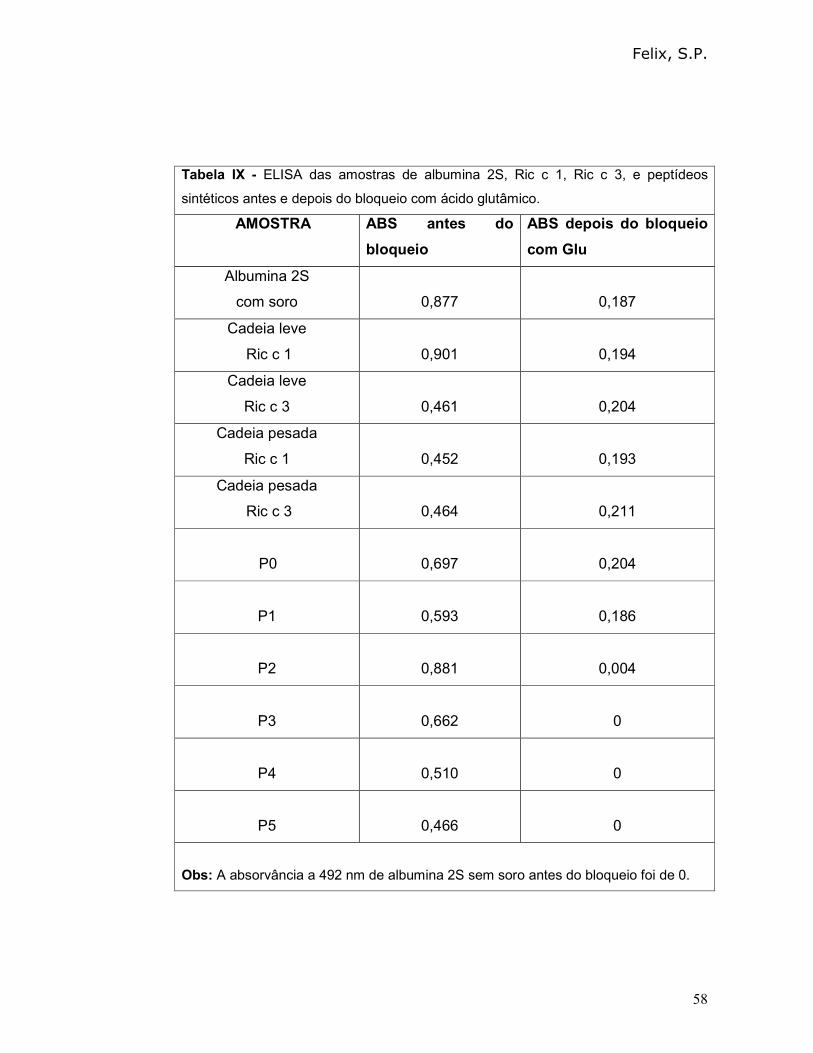
Felix, S.P.
58
Tabela IX - ELISA das amostras de albumina 2S, Ric c 1, Ric c 3, e peptídeos
sintéticos antes e depois do bloqueio com ácido glutâmico.
AMOSTRA ABS antes do
bloqueio
ABS depois do bloqueio
com Glu
Albumina 2S
com soro
0,877
0,187
Cadeia leve
Ric c 1
0,901
0,194
Cadeia leve
Ric c 3
0,461
0,204
Cadeia pesada
Ric c 1
0,452
0,193
Cadeia pesada
Ric c 3
0,464
0,211
P0
0,697
0,204
P1
0,593
0,186
P2
0,881
0,004
P3
0,662
0
P4
0,510
0
P5
0,466
0
Obs: A absorvância a 492 nm de albumina 2S sem soro antes do bloqueio foi de 0.

Felix, S.P.
59
4.10 - Avaliação da existência de reações cruzadas entre albumina 2S de
Ricinus communis e alérgenos alimentares, através da desgranulação de
mastócitos de rato.
A tabela X mostra o percentual de desgranulação de mastócitos promovidos
por alérgenos alimentares. Foram obtidos antígenos derivados de alimentos de
diversas naturezas como vegetais, frutos-do-mar, aves, bovinos e suínos. O teor
protéico das diferentes preparações comerciais de alérgenos alimentares foi
inicialmente quantificado pelo método descrito por Bradford. Após esta dosagem
foram feitas diluições correspondentes para que, em cada ensaio de desgranulação,
fossem utilizados 10 µL da solução estoque (10 µg/mL) de extrato alergênico para
100 µL de células, e 1 µL de soro anti-albumina 2S.
Admitimos a existência de reação cruzada nas situações onde observou-se
um potencial de desgranulação de mastócitos acima de 50,0 %. Dados obtidos para
camarão (53,8 %), peixes (57,6 %), glúten (56,3 %), trigo (56,0 %), soja (62,5 %),
amendoim (51,4 %) e milho (54.4 %) mostram reação cruzada com albumina 2S de
R. communis.
Quando pimenta foi utilizada como fonte, os resultados não foram conclusivos
devido à presença de pigmentos nestes extratos.
Apesar de ter havido desgranulação para o alérgeno de morango, esta não é
desencadeada por IgE, já que mesmo sem o soro anti-IgE houve a desgranulação
dos mastócitos quando este alérgeno foi incubado diretamente com as células.
A porcentagem de desgranulação sem o soro, apresentada na tabela X, prova
que a desgranulação de mastócitos foi mediada por anticorpo e não por outros
fatores.
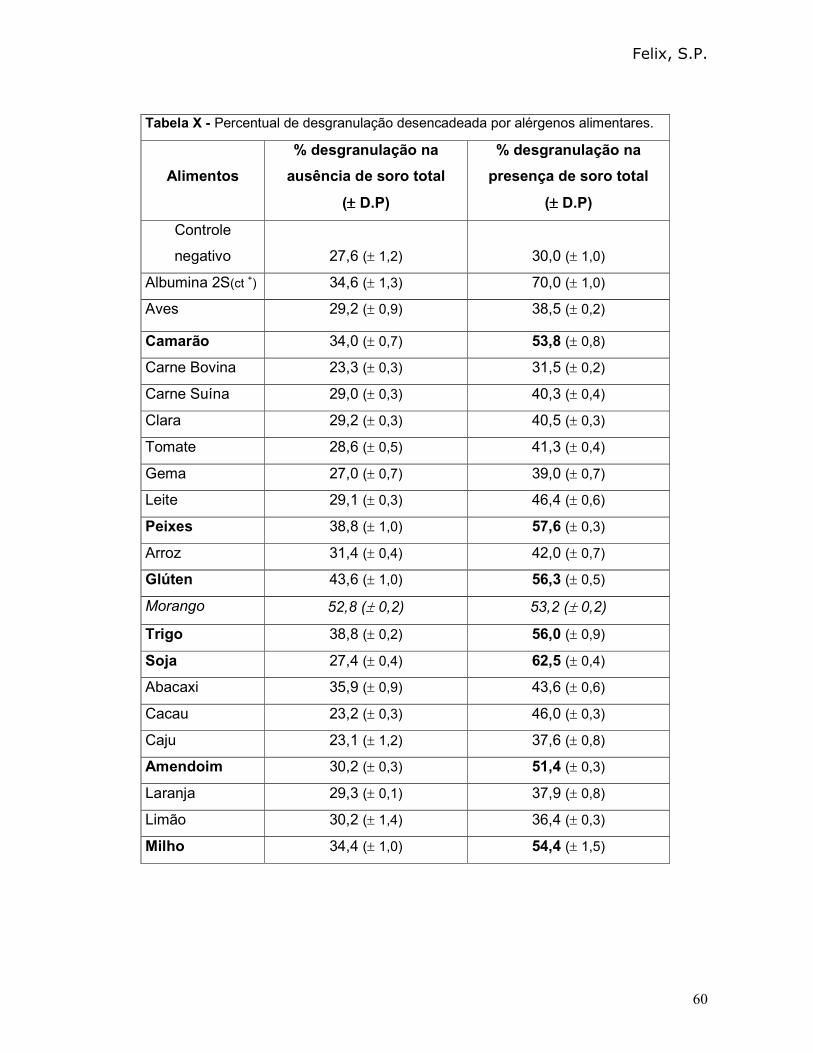
Felix, S.P.
60
Tabela X - Percentual de desgranulação desencadeada por alérgenos alimentares.
Alimentos
% desgranulação na
ausência de soro total
(±±±± D.P)
% desgranulação na
presença de soro total
(±±±± D.P)
Controle
negativo
27,6 (± 1,2)
30,0 (± 1,0)
Albumina 2S(ct +) 34,6 (± 1,3) 70,0 (± 1,0)
Aves 29,2 (± 0,9) 38,5 (± 0,2)
Camarão 34,0 (± 0,7) 53,8 (± 0,8)
Carne Bovina 23,3 (± 0,3) 31,5 (± 0,2)
Carne Suína 29,0 (± 0,3) 40,3 (± 0,4)
Clara 29,2 (± 0,3) 40,5 (± 0,3)
Tomate 28,6 (± 0,5) 41,3 (± 0,4)
Gema 27,0 (± 0,7) 39,0 (± 0,7)
Leite 29,1 (± 0,3) 46,4 (± 0,6)
Peixes 38,8 (± 1,0) 57,6 (± 0,3)
Arroz 31,4 (± 0,4) 42,0 (± 0,7)
Glúten 43,6 (± 1,0) 56,3 (± 0,5)
Morango 52,8 (± 0,2) 53,2 (± 0,2)
Trigo 38,8 (± 0,2) 56,0 (± 0,9)
Soja 27,4 (± 0,4) 62,5 (± 0,4)
Abacaxi 35,9 (± 0,9) 43,6 (± 0,6)
Cacau 23,2 (± 0,3) 46,0 (± 0,3)
Caju 23,1 (± 1,2) 37,6 (± 0,8)
Amendoim 30,2 (± 0,3) 51,4 (± 0,3)
Laranja 29,3 (± 0,1) 37,9 (± 0,8)
Limão 30,2 (± 1,4) 36,4 (± 0,3)
Milho 34,4 (± 1,0) 54,4 (± 1,5)

Felix, S.P.
61
4.11 - Avaliação da existência de reações cruzadas entre albumina 2S de
Ricinus communis e alérgenos inalantes, através da desgranulação de
mastócitos de rato.
A tabela XI mostra o percentual de desgranulação de mastócitos promovidos
por alérgenos inalantes, cuja concentração também foi padronizada pelo método
descrito por Bradford. Os maiores níveis de desgranulação de mastócitos foram
observados para poeira domiciliar com 51,4%; fungos do ar com 51,9% e tabaco
55,0%.
Apesar de ter havido desgranulação para o alérgeno de macela, esta não é
desencadeada por IgE, já que mesmo sem o soro anti IgE houve a desgranulação
dos mastócitos quando este alérgeno foi incubado diretamente com as células.
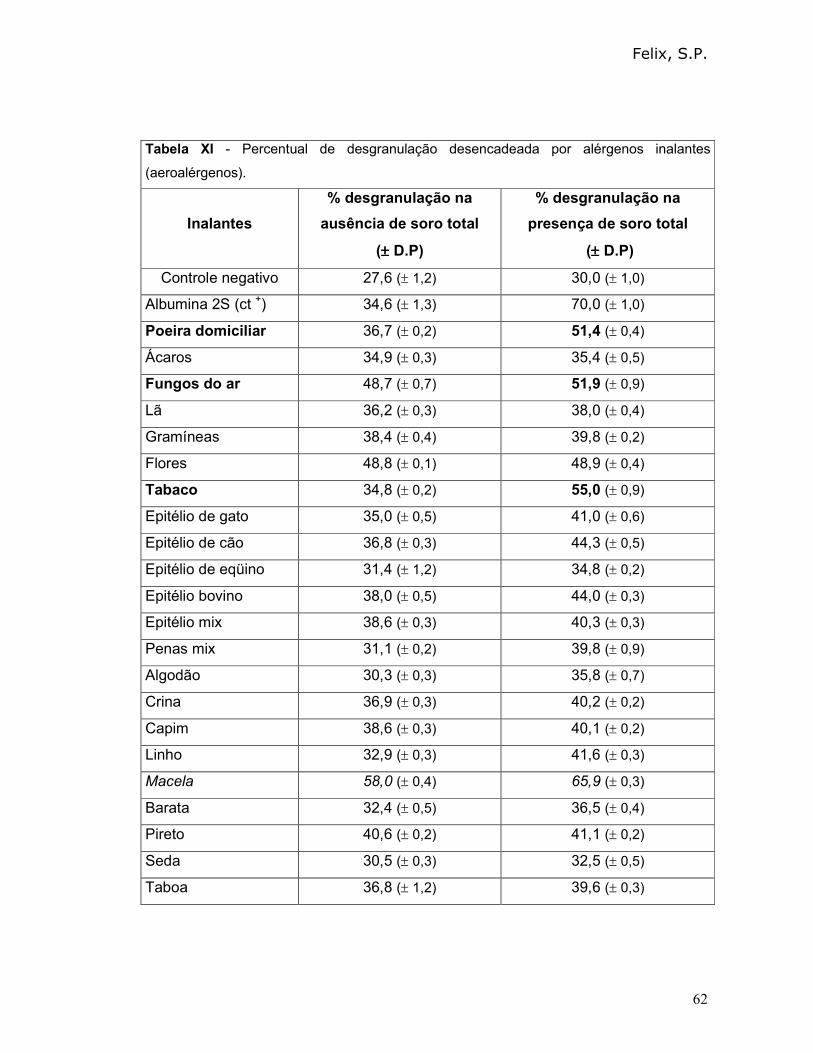
Felix, S.P.
62
Tabela XI - Percentual de desgranulação desencadeada por alérgenos inalantes
(aeroalérgenos).
Inalantes
% desgranulação na
ausência de soro total
(±±±± D.P)
% desgranulação na
presença de soro total
(±±±± D.P)
Controle negativo 27,6 (± 1,2) 30,0 (± 1,0)
Albumina 2S (ct +) 34,6 (± 1,3) 70,0 (± 1,0)
Poeira domiciliar 36,7 (± 0,2) 51,4 (± 0,4)
Ácaros 34,9 (± 0,3) 35,4 (± 0,5)
Fungos do ar 48,7 (± 0,7) 51,9 (± 0,9)
Lã 36,2 (± 0,3) 38,0 (± 0,4)
Gramíneas 38,4 (± 0,4) 39,8 (± 0,2)
Flores 48,8 (± 0,1) 48,9 (± 0,4)
Tabaco 34,8 (± 0,2) 55,0 (± 0,9)
Epitélio de gato 35,0 (± 0,5) 41,0 (± 0,6)
Epitélio de cão 36,8 (± 0,3) 44,3 (± 0,5)
Epitélio de eqüino 31,4 (± 1,2) 34,8 (± 0,2)
Epitélio bovino 38,0 (± 0,5) 44,0 (± 0,3)
Epitélio mix 38,6 (± 0,3) 40,3 (± 0,3)
Penas mix 31,1 (± 0,2) 39,8 (± 0,9)
Algodão 30,3 (± 0,3) 35,8 (± 0,7)
Crina 36,9 (± 0,3) 40,2 (± 0,2)
Capim 38,6 (± 0,3) 40,1 (± 0,2)
Linho 32,9 (± 0,3) 41,6 (± 0,3)
Macela 58,0 (± 0,4) 65,9 (± 0,3)
Barata 32,4 (± 0,5) 36,5 (± 0,4)
Pireto 40,6 (± 0,2) 41,1 (± 0,2)
Seda 30,5 (± 0,3) 32,5 (± 0,5)
Taboa 36,8 (± 1,2) 39,6 (± 0,3)

Felix, S.P.
63
4.12 - Análise do efeito de aminoácidos dicarboxílicos como bloqueadores
utilizando diversos extratos alergênicos que reagiram cruzado com albumina
2S de Ricinus communis.
Como observamos que os ácidos dicarboxilícos estavam envolvidos na
interação entre IgE e os epitopos de albuminas 2S de R. communis, utilizamos
esses dois aminoácidos dicarboxílicos como possíveis agentes para bloquear a
resposta cruzada.
A tabela XII mostra o percentual de desgranulação de mastócitos promovido
por alérgenos alimentares e inalantes, que apresentaram reação cruzada com
albumina 2S, cujos mastócitos sensibilizados por soro anti-albumina 2S foram
previamente incubados com ácido aspártico ou glutâmico separadamente. Podemos
observar que ocorreu uma redução da resposta cruzada quando Asp e Glu foram
utilizados, sendo que a redução do percentual de desgranulação para todos os
alérgenos utilizados foi mais efetivo quando o ácido glutâmico foi empregado.
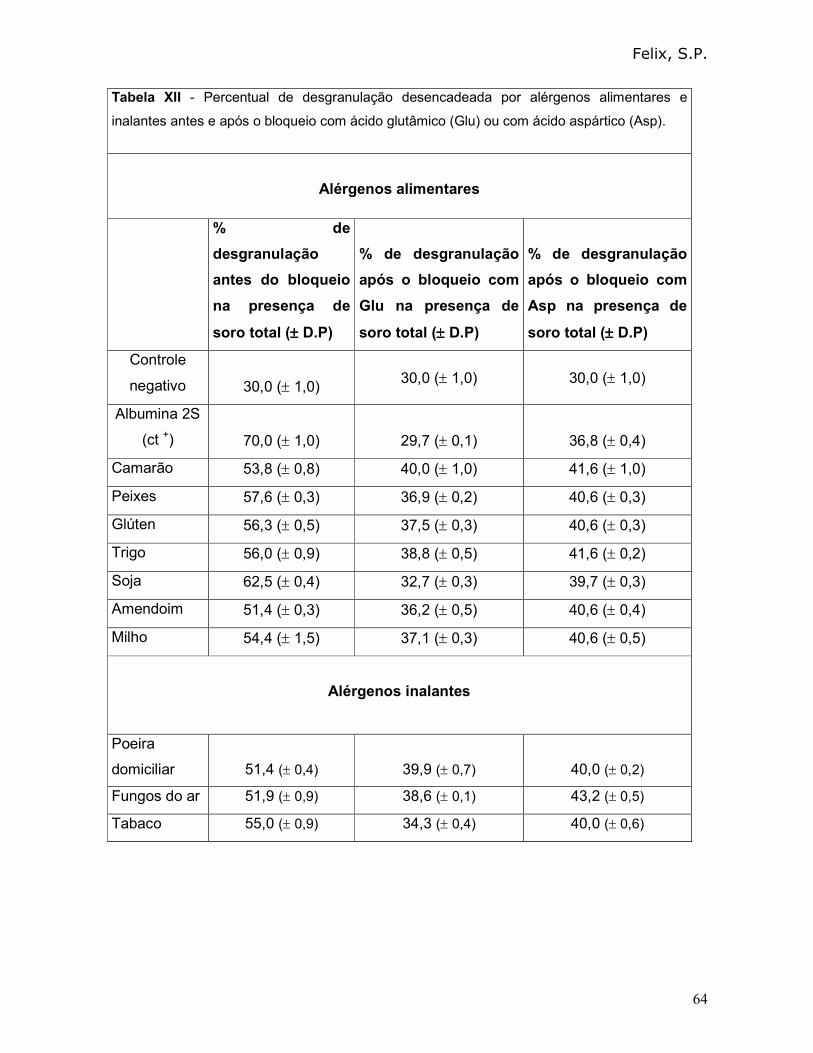
Felix, S.P.
64
Tabela XII - Percentual de desgranulação desencadeada por alérgenos alimentares e
inalantes antes e após o bloqueio com ácido glutâmico (Glu) ou com ácido aspártico (Asp).
Alérgenos alimentares
% de
desgranulação
antes do bloqueio
na presença de
soro total (±±±± D.P)
% de desgranulação
após o bloqueio com
Glu na presença de
soro total (±±±± D.P)
% de desgranulação
após o bloqueio com
Asp na presença de
soro total (±±±± D.P)
Controle
negativo
30,0 (± 1,0)
30,0 (± 1,0)
30,0 (± 1,0)
Albumina 2S
(ct +)
70,0 (± 1,0)
29,7 (± 0,1)
36,8 (± 0,4)
Camarão 53,8 (± 0,8) 40,0 (± 1,0) 41,6 (± 1,0)
Peixes 57,6 (± 0,3) 36,9 (± 0,2) 40,6 (± 0,3)
Glúten 56,3 (± 0,5) 37,5 (± 0,3) 40,6 (± 0,3)
Trigo 56,0 (± 0,9) 38,8 (± 0,5) 41,6 (± 0,2)
Soja 62,5 (± 0,4) 32,7 (± 0,3) 39,7 (± 0,3)
Amendoim 51,4 (± 0,3) 36,2 (± 0,5) 40,6 (± 0,4)
Milho 54,4 (± 1,5) 37,1 (± 0,3) 40,6 (± 0,5)
Alérgenos inalantes
Poeira
domiciliar
51,4 (± 0,4)
39,9 (± 0,7)
40,0 (± 0,2)
Fungos do ar 51,9 (± 0,9) 38,6 (± 0,1) 43,2 (± 0,5)
Tabaco 55,0 (± 0,9) 34,3 (± 0,4) 40,0 (± 0,6)

Felix, S.P.
65
5 - DISCUSSÃO
As alergias constituem um mal que afeta cerca de 25% da população mundial.
Este número está em crescimento diretamente relacionado com a toxicidade do
meio ambiente, exposição à poeira doméstica e pólen de diversas plantas
(GIOULEKAS et al., 2004). Além disto, indivíduos sensibilizados por determinada
substância alergênica podem desencadear processos alérgicos quando em contato
com outros alérgenos. O pólen e as sementes de diversas oleaginosas, como
girassol, mamona, soja e milho são ricos nestas proteínas alergênicas. Assim, a
alergia desencadeada por pólens, é um problema que pode afetar grande parte das
populações que residem próximo ao plantio destas oleaginosas, bem como do
trabalhador que manipula tais sementes (THORPE et al., 1988). Atualmente, a única
forma utilizada para selecionar os trabalhadores que manipulam a mamona é expô-
los ao contato com a planta por um curto período de tempo. Os trabalhadores que
apresentarem sintomas de alergia são considerados atópicos e são afastados do
trabalho.
No atual contexto mundial no qual se procura substituir combustíveis derivados
de petróleo pelos derivados de matéria orgânica (biodiesel, álcool) com fins de
obtenção de créditos de carbono, é de se esperar que o plantio de oleaginosas
como mamona cresça de forma exponencial, aumentando enormemente o risco de
sensibilização da população a estas substâncias alergênicas. Esta situação implica
que novas estratégias de tratamento para a alergia, visando principalmente alergias
provocadas por alérgenos de oleaginosas, sejam desenvolvidas.
A proposta desse trabalho resume-se na investigação de qual classe ou
subclasse de imunoglobulina é responsável pela alergia causada por albumina 2S
de Ricinus communis, na avaliação de se os epitopos reconhecidos por

Felix, S.P.
66
imunoglobulina de rato são também reconhecidos por IgE humana, e na confirmação
da existência de respostas cruzadas entre alérgenos de mamona e alérgenos de
outras fontes; além de investigar a ação de agentes bloqueadores de
imunoglobulina.
As albuminas 2S são encontradas em quase todos os tipos de sementes
(PASTORELLO et al., 1998). Na maioria das espécies, a albumina 2S consiste em
duas cadeias de aproximadamente 9 e 4 kDa, que são sintetizadas como um único
precursor ligado por pontes de enxofre antes do processamento vacuolar (clivagem
proteolítica) (PATRICK et al., 1996). As albuminas 2S de Ricinus communis já foram
descritas como proteínas alergênicas, quando foram relatados casos de
hipersensibilidade em populações que residiam próximo aos moinhos de
processamento de óleo de mamona (THORPE et al., 1988).
Estudos bioquímicos anteriores do nosso grupo, realizados por Mayerhoffer em
2004, confirmaram o potencial alergênico do “pool” de albuminas 2S, mais
especificamente as cadeias Ric c 1 e Ric c 3 que induziram a desgranulação dos
mastócitos de rato mesmo quando desnaturadas, além de identificar dois epitopos
presentes na cadeia pesada de Ric c 3 que induziram respostas significativas. No
entanto, a hidrólise tríptica impediu a identificação de epitopos lineares nas demais
subunidades das isoformas das albuminas 2S.
A fim de identificar quais os epitopos de albumina 2S de Ricinus communis são
alergênicos, Mayerhoffer em 2004 e Felix em 2006 analisaram a alergenicidade de
seis peptídeos sintéticos os quais foram produzidos com base nas seqüências das
cadeias leves e pesadas de Ric c 1 e de Ric c 3, as principais isoformas de albumina
2S de Ricinus communis. Quatro dos epitopos de ligação com IgE foram
identificados na proteína Ric c 1 e dois na proteína Ric c 3. A partir dessa análise foi

Felix, S.P.
67
observado que todos os seis peptídeos induziram positivamente a desgranulação
dos mastócitos de rato (MAYERHOFFER, 2004 & FELIX, 2006).
Sabe-se que a alergia causada pela albumina 2S de R. communis e outros
alérgenos se dá pela ligação do antígeno com a IgE pré-ligada nos mastócitos e
basófilos. No entanto, em ratos, nosso modelo experimental, alguns alérgenos
deflagram a alergia se ligando com a IgG2a, como mostram estudos realizados por
Dybing e colaboradores em 2004 quando ratos são sensibilizados com alguns
aeroalérgenos presentes em partículas do ar. Além disso, Jonge e colaboradores em
2007 sensibilizaram ratos da linhagem “Brown Norway” com alguns alérgenos
alimentares, e observaram expressiva resposta mediada por IgG2a.
Logo, a fim de identificar qual imunoglobulina é responsável pela alergia
causada por albumina 2S de R. communis e alérgenos de outras fontes, fizemos
uma cromatografia de afinidade em batelada com soro total de rato, para separar as
imunoglobulinas E das imunoglobulinas G, processo que foi certificado por ensaios
de “dot blotting”. Posteriormente, testes de desgranulação de mastócitos foram feitos
utilizando as imunoglobulinas separadas e, a partir dessa análise, ficou comprovado
que para as albuminas 2S de R. communis, foi a IgE a responsável por tal evento da
resposta alérgica (desgranulação de mastócitos) (TABELA IV).
Como já descrito anteriormente, Felix em 2006 identificou os epitopos de
albumina 2S de Ricinus communis que são responsáveis pela desgranulação de
mastócitos de ratos. A fim de avaliar se esses epitopos reconhecidos por IgE de rato
são também reconhecidos por IgE humana foi feito “dot bloting” (Figura 13) e ELISA
(TABELA IX). A partir dessas análises comprovamos que a IgE humana reconhece
os mesmos epitopos que a IgE de rato. Assim, acreditamos que ao aprofundarmos
os estudos nesse assunto, poderemos desenvolver novas estratégias para
prevenção e/ou tratamento de indivíduos atópicos, já que esses epitopos, que foram

Felix, S.P.
68
reconhecidos pela IgE humana, podem ser bons candidatos para o desenho de
bloqueadores de IgE que visam a redução de efeitos anafiláticos, ou para o
desenvolvimento de variantes hipoalergênicas de albumina 2S; semelhante ao que
foi proposto em 2004 na patente americana (US 2004/0115220 A1). Esta propõe um
novo tratamento baseado na utilização de epitopos de IgE, no qual, as novas
regiões apresentadas podem ser alvo para a imunoprofilaxia ou imunoterapia
passiva ou ativa. Estes peptídeos são usados no tratamento da alergia ou podem
ser usados como vacina para induzir anticorpos próprios durante a imunoprofilaxia
ou imunoterapia.
Pons e colaboradores em 2004 mostraram que devido à homologia entre
aminoácidos das proteínas alergênicas de reserva das sementes de soja com
proteínas alergênicas de amendoim, é possível utilizar a imunoterapia de indivíduos
alérgicos à soja para indivíduos alérgicos ao amendoim. Sendo assim, acredita-se
que o mesmo possa ser feito com a albumina 2S de Ricinus communis e os
alérgenos de diversas fontes alimentares e inalantes que reagiriam positivamente ao
teste de reação cruzada. Para confirmar então se a IgE anti-albumina 2S reagiria
aos alérgenos de diversas fontes alimentares e inalantes do kit FDA Allergenic LTDA
padronizados pelo método descrito por Bradford, avaliamos as respostas cruzadas
entre esses alérgenos e foi observada reação cruzada para os alérgenos: camarão,
peixe, glúten, trigo, soja, amendoim, milho, poeira domiciliar, fungos do ar e tabaco.
Não foi possível verificar se a pimenta causava reação cruzada, pois esta
apresentava aspecto turvo ao se observar em microscópio ótico impossibilitando
assim a avaliação da desgranulação dos mastócitos. No caso do alérgeno de
morango e do alérgeno de macela, apesar de ter havido desgranulação, esta não foi
desencadeada por IgE, já que mesmo sem o anti-soro houve a desgranulação dos

Felix, S.P.
69
mastócitos quando estes alérgenos foram incubados diretamente com as células
(TABELA X e XI).
Diversas abordagens imunoprofiláticas e imunoterapêuticas passivas e ativas
que interferem com o evento clássico das reações alérgicas do tipo I, o mecanismo
de liberação de histamina mediado por IgEs, têm sido investigadas. O uso de
adjuvantes que induzem resposta Th1 ao invés de resposta Th2, além do switching
de classe de IgE para IgG tem sido uma dessas abordagens. A IgG atua bloqueando
o efeito do IgE, pois quando o IgG se liga no seu receptor, ele induz fosforilação no
receptor de IgE, inibindo assim a cascata de sinalização que seria desencadeada
por ele (STEPHEN et al., 2004; DAERON et al., 1995).
Além disso, a tecnologia do DNA recombinante causou uma tremenda explosão
de conhecimento sobre doenças causadas por alérgenos. Usando esta tecnologia,
quase um repertório completo de alérgenos tem sido produzido como moléculas
recombinantes e então utilizadas em uso clínico para diagnóstico e monitoramento
de doenças alérgicas (LINHART & VALENTA, 2005).
Outras estratégias visam a prevenção da ligação da região Fc da IgE aos
receptores FcεRI dos mastócitos, ou a ligação competitiva de IgE aos receptores por
peptídeos derivados da porção constante de IgE. A patente americana US
2002/0141989 A1 propõe esta estratégia de tratamento. Contudo, nenhuma
estratégia visa o bloqueio da fração Fab das imunoglobulinas E, evitando com que o
antígeno específico se ligue e deflagre a alergia. Visto que epitopos nada mais são
do que pequenas seqüências de aminoácidos reconhecidas pela molécula de
anticorpo, com o intuito de identificar bloqueadores específicos para tal região da
IgE, utilizamos os 20 aminoácidos livres, como possíveis agentes que poderiam se
associar às IgE ligadas nos mastócitos de rato.

Felix, S.P.
70
Para isso, utilizamos soro de ratos imunizados com albumina 2S de R.
communis, onde foram empregados os 20 aminoácidos livres em combinações
diferentes, previamente à adição de albumina 2S de R. communis e peptídeos
sintéticos separadamente. A efetividade do bloqueio foi então analisada por testes
de desgranulação de mastócitos. Os ácidos dicarboxílicos foram omitidos da mistura,
já que Felix em 2006 observou que nos seis peptídeos sintéticos, produzidos a partir
da seqüência da albumina 2S de R. communis, existiam pelo menos dois resíduos
de ácidos glutâmicos alternados. Essa característica nos fez acreditar na hipótese
de que os grupamentos γ carboxílicos deste aminoácido estivessem envolvidos na
ligação com as IgEs ligadas nos mastócitos. A partir do resultado de desgranulação
de mastócitos de rato, observamos que o bloqueio da ligação entre epitopo e IgE
não foi efetivo quando a mistura de aminoácidos livres, que omitia os aminoácidos
dicarboxílicos, foi empregado. Mas quando foi utilizado os ácidos glutâmico e
aspártico separadamente, observamos o bloqueio dessa ligação entre epitopo e IgE,
com níveis de desgranulação próximos ao controle negativo.
A histamina, que é um dos principais mediadores liberados pela desgranulação
dos mastócitos durante a deflagração da alergia, foi dosada e os valores obtidos
foram correlacionados com os resultados dos ensaios de desgranulação de
mastócitos, quando o ácido glutâmico foi utilizado como bloqueador.
Nossos experimentos evidenciaram que os aminoácidos dicarboxílicos agiram
como agentes bloqueadores de IgE, ou seja, esses aminoácidos impediram que a
albumina 2S de R. communis e os peptídeos sintéticos, produzidos a partir desse
alérgeno, se ligassem as IgE pré-ligadas nos mastócitos, evitando assim a
desgranulação destes e consequentemente a liberação de histamina e uma possível
deflagração da alergia. Isso mostra que esses bloqueadores são bons candidatos

Felix, S.P.
71
para o desenvolvimento de drogas para tratamento de alergias causadas por
mamona.
Os resultados obtidos a partir do teste de desgranulação de mastócitos
mostram que esses bloqueadores também foram efetivos para os alérgenos que
reagiram cruzado com o alérgeno de mamona (TABELA XII). Em virtude desses
resultados, o tratamento baseado neste princípio, que venha a ser desenvolvido
para alérgenos de mamona será polivalente, porque poderá ser utilizado também
contra alérgenos de outras fontes.
No entanto, muitos estudos ainda precisam ser feitos até que seja desenvolvido
um medicamento a partir desses bloqueadores, pois embora o ácido glutâmico e o
ácido aspártico sejam produzidos pelo organismo naturalmente, e serem
precursores da síntese de aminoácidos e muitas outras moléculas biologicamente
importantes, são quase totalmente absorvidos, o que facilitaria sua administração via
oral (REEDS et al., 1996), eles podem apresentar possíveis limitações quanto a seu
uso. Além de serem rapidamente metabolizados, o que diminuiria sua permanência
no organismo e consequentemente sua eficácia, existem alguns dados que mostram
que excesso desses aminoácidos no organismo pode comprometer a visão (SMITH,
1990).
Sendo assim, espera-se que os dados obtidos por este trabalho contribuam,
não só para a formação do conhecimento científico relevante nesta área, mas
também para o desenvolvimento de novas abordagens quanto ao tratamento e/ou
prevenção da alergia causada por Ricinus communis e conseqüentemente das
demais fontes alergênicas.

Felix, S.P.
72
6 - CONCLUSÃO
⇒ Uma metodologia para determinação do potencial alergênico, baseada em
ativação de mastócitos, quantificada por visualização por microscopia ótica e
dosagem de histamina foi desenvolvida
⇒ A reação de hipersensibilidade provocada pelas albuminas 2S de R.
communis, em ratos, é mediada por IgE
⇒ Os epitopos alergênicos das albuminas 2S mapeados em roedores, foram
também reconhecidos por IgE humana
⇒ Indivíduos atópicos, apresentando sintomas de alergia não associadas
diretamente à mamona, apresentaram IgE que reconheceram as albuminas 2S de
R.communis
⇒ Os resíduos de aminoácidos dicarboxílicos estão envolvidos na ligação da
IgE aos epitopos alergênicos
⇒ Existe reação cruzada entre albuminas 2S de R. communis com alérgenos
de diversas fontes alimentares e inalantes
⇒ Ácido glutâmico e ácido aspártico são agentes bloqueadores de IgE, e
portanto são candidatos potenciais para o tratamento da hipersensibilidade do tipo I,
podendo ser efetivo para alergia desencadeada não só por mamona, mas também
por camarão, peixe, glúten, trigo, soja, amendoim, milho, poeira domiciliar, fungos do
ar e tabaco

Felix, S.P.
73
7 - REFERÊNCIAS BIBLIOGRÁFICAS
AALBERSE, R.C. Structural biology of allergens. Journal Allergy and Clinical Immunology, 106 (nº 2): 228-238, 2000. ABBAS, A. K.; LICHTMAN, A.H.; POBER, J.S. Imunologia Celular e Molecular. 4ª edição. Editora Revinter, 2003. Cap. 19 p.424-444. AGGIZIO, A.P.; CARVALHO, A.O.; RIBEIRO,S.F.F.;MACHADO,O.L.T.; AVES,E.W.; BLOCH JR, C.; OKOROKOV, L.A.; SAMARAO, S.S.; PRATES, M.V.; GOMES, V.M. A 2S albumin homologous protein from passion fruit seeds inhibits the fungal growth and acidification of the médium by Fusarium oxisporum. Archives of Biochemistry and Biophics, 46: 188-195, 2003. ANANDAN S, ANIL KUMAR GK, GHOSH J AND RAMACHANDRA KS. Effect of different physical and chemical treatments on detoxification of ricin in castor cake. Animal Feed Science And Technology, 120: 159-168, 2005. AZEVEDO, D.M.P.; LIMA, E.F. O agronegócio da mamona no Brasil. Campina Grande: Embrapa Algodão, 2001, p. 350. Berrens, L.; Romano, M.L.G.; Câmara, M.T.G.; inventores. Processo para produzir um extrato provendo tolerância para aplicação a um indivíduo a fim de prover tolerância contra material alergênico, extrato de material de fonte alergênica, preparação e uso da mesma. Patente brasileira PI 9714899-7 A, 2000. BEWLEY, D.J. & BLACK, M. Physiology of development and germination. Seeds. 2. New York and London: Plenum Press ed. Cap 1 e 4, 1996. BRADDING, P. Allergen immunotherapy, mast cell. Clinical and experimental Allergy. 29:1445-1448, 1999 BRADFORD, M.M. A rapid and sensitive method for the quantification of proteins utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248-254, 1976. BREITENEDER, H. & MILLS, C., E.N. Plant food allergens – structural and functional aspects of allergenicity. Biotechnology Advances. 23: 395-399, 2005. BREITENEDER H. & RADAUER C. A classification of plant food allergens. J. Allergy Clin Immunol, 113:821-30, 2004. BOYCE, J.A. Mast cells: beyond IgE. J. Allergy and Clinical Immunology. 11: 25-32, 2003. Chua, K.Y.; Liew, L.N.; inventores. Recombinat nucleic acid useful for inducing protective immune response against allergens. Inventores: Patente internacional PCT número WO 2004/019978 A1, 2003.

Felix, S.P.
74
DAERON, M.; MALBEC, O.; LATOUR, S.; AROCK, M.; FRIDMAN, W.H. Regulation of high-affinity IgE receptor-mediated mast cell activation by murine low-affinity IgG receptors. J Clin Invest . 95:577-85, 1995. DA SILVA, D.; MOTA, I. Imunologia Básica e aplicada. 5ª edição. Editora Guanabara Koogan S.A. p. 156-164, 2003. De Bassols, V., inventor. Vaccine. United States patent US 2004/0115220 A1, 2004 Jul. DYBING, E.; LØVDAL, T.; HETLAND, R. B. Respiratory allergy adjuvant and inflammatory effects of urban ambient particles. Toxicology, 198: 307–314, 2004. DUNWELL J.M. Cupins: a new superfamily of functionally diverse proteins that include germins and plant storage proteins. Biotechnol Genet Eng Rev, 15:1-32, 1998. FELIX, S.P. Caracterização de epitopos ligantes de IgE em alérgenos de Ricinus communis e investigação de respostas cruzadas entre alérgenos. Monografia – Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, Rio de Janeiro, Brasil, 2006. FORNAZIERI, A. Mamona, Uma rica fonte de óleo e divisas. Coleção Brasil Agrícola, Editora Ícone, São Paulo. 1986. GIOULEKAS D. PAPAKOSTA D., DAMIALIS A., SPIEKSMA F., GIULEKA P., PATAKAS D. Allergenic pollen records (15 years) and sensitization in patients with respiratory allergy in Thessaloniki, Greece. Allergy, 59:174-184, 2004. JONGE, J. D.; KNIPPELS, L.M.J.; EZENDAMB; ODINK, J.; PENNINKS, A.H.; LOVEREN, H. V. The importance of dietary control in the development of a peanut allergy model in Brown Norway rats. Methods, 41: 99–111, 2007. JUNQUEIRA, L.C.; CARNEIRO, J. Histologia Básica. 8ª edição. Editora Guanabara Koogan, p. 69-93, 1995. Kricek, F., Stadler, B.; Vogel, M., inventors. Anti-Idiotypic antibodies against antibodies which inhibit the binding of immunoglobulins E to its high affinity receptors. United States patent US 2002/0141989 A1, 2002 Oct. Kricek, F.; Stadler, B.; inventores. Peptide Immunogenens for vaccination against and treatment of allergy. Patente Americana número US 6,610,297 B1, 2003. LI, S.S.L.; LIN, T.T.S.; FORDE, M.D. Isolation and characterization of a low-molecular weigth seed protein fron Ricinus communis. Biochim. Biophys. Acta, 492: 364-369, 1977. LICHTENSTEIN, L.M. Allergy and the immune Systen. Scientific American, 85-93, 1993.

Felix, S.P.
75
LINHART, B.; VALENTA, R. Molecular design of allergy vaccines. Current Opinion in Immunology, 17: 646-655, 2005. MACHADO, O.L.T.; MARCONDES, J.A.; DE SOUZA-SILVA, F.; HANSEN, E.; RIBEIRO, P.D.; VERÍSSIMO, M.; KANASHIRO, M.; KIPNIS, T.L.; DA SILVA JR., J.G.; DOS SANTOS, M.F. AND COSTA E SILVA, M.C. Characterization of allergenic 2S albumin isoforms from Ricinus communis seeds. Allergologie, 26:45-51, 2003. MACHADO, O.L.T.; SILVA, J.G. An allergenic 2S storage protein from Ricinus communis seeds which is part of the albumin precursor predict by c-DNA data. Brazilian Journal of Medical and Biological research, 25: 567-582, 1992. MARCONDES-DE-SOUZA, J. A. Estudo comparativo das proteínas alergênicas presentes em sementes de Ricinus communis. Monografia - Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, Rio de Janeiro Brasil, 1997. MAYERHOFFER, R.D.O. Peptídeos isolados de moléculas de MHC de classe II de macrófagos murinos pulsados com albumina 2S de sementes de Ricinus communis. Monografia - Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, Rio de Janeiro Brasil, 2002. MAYERHOFFER, R.D.O. Identificação de peptídeos alergênicos de Albumina 2S de Ricinus communis (Mamona). Tese (mestrado em Ciências Biológicas) - Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, Rio de Janeiro Brasil, 2004. PANTOJA-UCEDA, D.; SHEWRY, P.R.; BRUIX, M.; TATHAN, A.S.; SANTORO, J.; RICO, M. Solution structure of a methionine-rich 2S albumin from sunflower seeds: relationship to its allergenic and emulsifying properties. Biochemistry, 43: 6976-6986, 2004. PARENTE, E.J.S: Minicurso do Congresso Brasileiro de Mamona. Campina Grande.2004. CD-ROM PASTORELLO, E.A.; FARIOLLI, L.; PRAVETTONI, V.; ISPANO, M.; CONTI, A., ANSALONI, R. Sensitization to the major allergen of Brazil nut is correlated with a clinical expression of allergy. Journal Allergy Clinical Immunology, 102; 1021-1027, 1998. PATRICK, J.E.; THOYS; NAPIER, J.; MILLICHIP, M.; STOBART, K.; GRIFFITHS, W. T.; TATHAM, A. S.; SHEWRY, P. R. Characterization of sunflower seed albumin which oil bodies. Plant Science, 118: 119-125, 1996. PIRES, M.M.; ALVES, J.M.; NETO, J.A.A.; ALMEIDA, C.M.; SOUZA, G.S.; CRUZ, R.S.; MONTEIRO, R.; LOPES, B.S.; ROBRA, S. Biodiesel de mamona: uma avaliação econômica. I Congresso Brasileiro de Mamona. Campina grande PB, 2004. CD-ROM PONS, L.; PONNAPAN, U.; HALL, R. A; SIPSON, P.; COCKRELL, G.; WEST, M.; SAMPSON, H. A.; HELM, R. M.; BURKS, A. W. Soy immunotherapy for peanut-

Felix, S.P.
76
allergic mice:Modulation of the peanut-allergy response. J. Allergy clinical Immunology, 114: 915-921, 2004. REEDS, P.J.; BURRIN, D.G.; JAHOOR F.; WYKES L.; HENRY, J.; FRAZER, M.E. Enteral glutamate is almost completely metabolized in first pass by the gastrointestinal tract of infant pigs. Am.J.Physiol, 270 : 413-418, 1996. REGENTE, M. & LA CANAL, L. Are storage 2S albumins also defensive proteins? Physiological and Molecular Plant Pathology, 59: 275-276, 2001. ROITT, I.; BROSTOPFF, J.; MALE, D. Hypersensitivity – Type I and Hypersensitivity tipe IV. Immunology. 4. Barcelona: Grafos S. A., 22.2-22.17 e 25.1- 25.8, 1998. SHARIEF, F.S.; Li, S.S. Aminoacid sequence of small and large subunits of seed storage protein from Ricinus communis. Journal of Biological Chemistry, 257:14753-14759, 1982. SHEWRY, P.R.; NAPIER, J.A.; TATHAM, A.S. Seed storage proteins: structures and biosynthesis. Plant Cell, 7:945-56, 1995. SILVA JR.; J.G., M.A.T.; MACHADO, O.L.T., IZUMI, C.; PANDOVAN, J.C.; CHAIT, B.T., MIRZA, U.A. &GREENE, L.J. Aminoacid sequence of New 2S Albumin from Ricinus communis with is Part of 29-kDa Precursor Protein. Archives of Biochemistry and Biophysics, 336: 10-18, 1996. SOLÈR M. Anti-IgE-antibodies in the treatment of allergic diseases. Rev Fr Allergol Immunol Clin, 42: 45-9, 2002. SMITH, R.J. Glutamine metabolism and its physiologic importance.. J. Parent. Enter. Nutr. 14: 40-44, 1990. SPIES, J. R., COULSON, E. J. The chemistry of allergens VIII. Isolation and properties of an active protein-polysaccharidic fraction, CB-1A, from castor bean. Journal of American Chemical Society, 65: 1720-1725,1943. STEPHEN J. T.; JAMES N. F.; KAYHAN N.; STEPHEN R. D. Mechanisms of immunotherapy. J. Allergy Clin Immunol, 113:1025-34, 2004. THORPE, S.C.; KEMEDY, D.M.; PANZANI, R.C.; MC GULR, B.; LORD, M. Allergy to castor bean II - Identification of the major allergens in castor bean seeds. Journal of Allergy and Clinical Immunology, 82: 67-72, 1988. VIEIRA, M.R. Desgranulação de mastócitos por isoformas de albumina 2S de sementes de R. communis. Monografia - Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, Rio de Janeiro Brasil, 2002. YOULE, R.J., HUANG, A.H.C. Evidence that Castor Bean allergens are Albumin storage proteins in protein bodies of castor bean. Plant Physiol, 61: 1040-1042, 1978.