teorias evolutivas
TRANSCRIPT

9
TTTTTeorias Evolutivaseorias Evolutivaseorias Evolutivaseorias Evolutivaseorias Evolutivas
Roberto IannuzziMarina Bento Soares
A palavra evolução tem sua origem no termolatino evolutio, que significa desenrolar (Salzano, 1993).Erroneamente associado à ideia de progresso, este ter-mo envolve apenas o sentido de mudança. Portanto,subentende-se que evoluir implica em mudar. Todas asteorias evolutivas que estejam relacionadas a fenôme-nos biológicos, físicos ou sociais são teorias de mudan-ça. Deste modo, o conceito de evolução pode ser apli-cado tanto às mudanças que operam no mundo biológi-co quanto às mudanças culturais e tecnológicas.
O tema abordado neste capítulo refere-se à evo-lução que ocorre no mundo orgânico, relativo aos seresvivos, denominada evolução biológica.
Evolução biológica compreende a modificaçãosofrida por populações de organismos através do tem-po; tempo este que ultrapassa o período de vida de umaúnica geração (Futuyma, 1993). As mudanças conside-radas evolutivas são aquelas herdadas via material ge-nético. Contidas no conceito de evolução biológica es-tão as ideias de:
• mudança contínua dos organismos através do tempo;• irreversibilidade das mudanças (verdadeira para a
maioria dos acontecimentos evolutivos);• divergência de características entre os organismos,
refletida pela diversidade encontrada no mundo bio-lógico (Salzano, 1993).
Histórico do Pensamento Evolutivo
Até o século XVIII acreditava-se que o planetaTerra era muito jovem, tendo sido criado por Deus hápoucos milhares de anos e que os seres vivos, igual-mente, eram criações divinas. O naturalista e filósofoCharles Bonnet (1720-1793) ilustrava bem esta concep-ção de mundo. Para ele, os seres vivos e o universoorganizavam-se numa escala graduada, do menos parao mais perfeito. Os seres vivos dispunham-se hierarqui-camente em uma cadeia, formando uma “escada davida”, com o homem ocupando o topo. A cadeia eraestática, sem mudanças desde o ato da criação (Barbe-rena, 1984).
Ao final do século XVIII, os geólogos reconhece-ram que as rochas sedimentares haviam sido depositadasem diferentes épocas, propondo que a Terra poderia sermuito mais antiga do que se imaginava. A grande contri-buição da geologia foi a descoberta do “tempo profun-do”, o que limitou temporalmente a existência e, con-sequentemente, a importância do ser humano na históriado planeta. A ideia, até então vigente, de uma Terra jo-vem governada pela espécie humana foi substituída pelanoção de uma vastidão quase incomensurável de tempo,com a presença do Homem restrita aos instantes finaisdo tempo geológico (Gould, 1991).
3ª Prova

138 Paleontologia
3ª Prova
A noção do “tempo profundo” deve-se a JamesHutton (1726-1797). Em 1788, ele publicou um ensaiointitulado Theory of Earth (Teoria da Terra), criando oprincípio do uniformitarismo, o qual sustentava queos mesmos processos físicos que atuam no presente fo-ram também os responsáveis por eventos passados. Ba-seando-se em observações de campo, reconheceu queas inconformidades, as quais representavam hiatos detempo entre dois episódios na formação das rochas, eramas evidências mais consistentes da quase inconcebívelvastidão do tempo. Desta forma, observou que a histó-ria da Terra incluía múltiplos ciclos de deposição esoerguimento. Segundo Hutton, a Terra era muito anti-ga “sem vestígio de um começo – sem perspectiva de um fim”(Gould, 1991).
Além disso, a descoberta cada vez mais frequen-te de fósseis nos séculos XVIII e XIX mostrou que ou-tras formas de vida tinham aparecido e desaparecidoem várias épocas ao longo do tempo. Os fósseis, quecaracterizavam os diferentes estratos de rocha, foramgeralmente considerados como reflexo de uma suces-são de catástrofes. Georges Cuvier (1769-1832), o fun-dador da anatomia comparada, reconstituindo esquele-tos de animais fósseis, percebeu as interrupções no re-gistro sedimentar e fossilífero, e começou a elaborar aideia sobre a sucessão dos diversos tipos de animaisatravés do tempo. Criou a teoria do catastrofismo, pos-tulando que as descontinuidades que separavam os di-ferentes animais fósseis eram consequência de catás-trofes que dizimavam faunas inteiras. Acreditava queapós cada evento de extinção em determinada área,faunas provenientes de outros locais ocupavam os am-bientes agora vagos (um ir e vir de faunas).
A teoria do uniformitarismo de Hutton foi re-tomada e vigorosamente defendida por Charles Lyell(1797-1875) na sua obra chamada Principles of Geology(Princípios da Geologia/1830-1833). Ele argumentavaque todos os eventos passados, sem exceção, poderiamser explicados pela ação de fenômenos hoje atuantes.Deste modo, a ideia central de seu trabalho era de que“o presente é a chave para o passado”. Lyell não era adep-to da sucessão temporal de fósseis, para ele uma espé-cie extinta poderia reaparecer, desde que houvesse con-dições ambientais propícias para tal. Também não acre-ditava em episódios de extinção e/ou origem em massa.Para ele, o surgimento das diferentes formas de vida es-tava distribuído de modo uniforme ao longo do tempogeológico.
Nem o uniformitarismo de Lyell, nem tampoucoo catastrofismo de Cuvier admitiam a ideia de evolução
biológica. Mas pensamentos sobre evolução dos seresvivos já floresciam no final do século XVIII. Em 1766,Georges-Louis Leclerc, “Conde de Buffon” (1707-1788)foi o primeiro a sugerir que as diferentes espécies teriamsurgido por variação a partir de ancestrais comuns. Noentanto, foram Jean-Baptiste Lamarck (1744-1829) eCharles Robert Darwin (1809-1882) os primeiros cien-tistas que procuraram explicar a diversidade dos seresvivos através da ação de um mecanismo evolutivo, comoserá visto a seguir.
Lamarckismo
Em sua obra intitulada Philosophie Zoologique (Fi-losofia Zoológica/1809), Lamarck propôs que as for-mas de vida progridem, por transformação gradual, emdireção a uma maior complexidade e perfeição. Sus-tentou que o caminho da progressão é determinado poruma “força interna” influenciada por “circunstânciasexternas”, ou seja, mudanças no ambiente trazem anecessidade de alterações no organismo. Desta forma,o organismo responde usando alguns órgãos mais queoutros. Segundo ele, “o uso e o desuso dos órgãos” alterama morfologia destes, a qual é transmitida para as gera-ções subsequentes. Um exemplo clássico, segundo opróprio Lamarck, seria a evolução do pescoço da girafa.De acordo com sua teoria, os ancestrais da girafa pode-riam ter sido animais de pescoço curto, que na tentativade alcançarem as folhas mais altas das árvores para ali-mentarem-se, esticavam seus pescoços. Este compor-tamento intencional e repetitivo fez com que, geraçãoapós geração, os pescoços fossem se tornando gradati-vamente mais compridos, devido ao esforço. Assim foiatingido o estágio de desenvolvimento do pescoço dasatuais girafas. Lamarck, deste modo, enfatizou a adap-tabilidade e a transmissão de caracteres adquiridos.
Lamarck foi duramente criticado por Cuvier, poiseste argumentava que o registro fóssil não revelava sé-ries graduais intermediárias de ancestrais e descenden-tes. Também Lyell se mostrou contrário às ideias deLamarck, já que num mundo uniformitarista não havialugar para a evolução. Vários experimentos realizadosposteriormente demonstraram que características adqui-ridas ao longo da vida não são hereditárias. O biólogoalemão August Weismann (1834-1914), por exemplo,ao cortar a cauda de camundongos por várias geraçõessucessivas, constatou que os camundongos desprovi-dos de cauda sempre davam origem a descendentescom cauda.

139Capítulo 9 – Teorias Evolutivas
3ª Prova
Atualmente, a teoria de Lamarck não é aceita,pois foi comprovado que as características adquiridasdurante a vida não são hereditárias, apenas mudançasao nível genético podem ser herdadas pelos descen-dentes.
Lamarckismo
Princípios fundamentais:
– lei do uso e do desuso dos órgãos;
– lei da transmissão de características adquiridas degeração para geração.
Conceitos equivocados:
– as modificações adquiridas durante a vida em fun-ção do uso ou desuso dos órgãos não são transmi-tidas hereditariamente.
Contribuição para a teoria evolutiva:
– ênfase na capacidade de adaptação dos organis-mos ao meio e consequente modificação destesao longo das sucessivas gerações.
Darwinismo
Após a teoria de Lamarck, nenhum mecanismoevolutivo satisfatório foi reconhecido, até que a teoriade Charles Darwin e Alfred Wallace (1823-1913) fosseapresentada.
A história sobre a concepção do Darwinismoinicia-se com a viagem ao redor do mundo empreen-dida pelo jovem Charles Darwin, entre 1831 e 1836,a bordo do navio do governo inglês H.M.S. Beagle, naqualidade de naturalista. A partir de suas observaçõesacerca da diversidade de fauna e flora, constatadasdurante essa viagem, Darwin voltou à Inglaterra coma ideia de que os seres vivos se modificavam com otempo. Mas um mecanismo que explicasse como es-tes evoluíam era ainda desconhecido. Já em sua casa,em Downe (Inglaterra), Darwin interessou-se pelacriação de animais domésticos. Ele percebeu que oscriadores sempre escolhiam determinadas caracterís-ticas nos animais e as selecionavam através das vári-as gerações, até obterem uma nova raça. As novasraças eram mantidas somente se os seus membrosfossem acasalados entre si. Este método era denomi-nado de “seleção artificial”. Esta “seleção artificial”forneceu a Darwin pistas de como ocorria a seleção
das características nos animais selvagens e a conse-quente modificação destes ao longo do tempo. En-tão, em 1838, Darwin leu o ensaio de Thomas RobertMalthus (1765-1834) intitulado An essay on the princi-ples of population (Um ensaio sobre os princípios dapopulação, 1803), o qual proferia que enquanto aspopulações humanas cresciam em progressão geo-métrica, a oferta de alimentos crescia em progressãoaritmética. Darwin percebeu que isto não era obser-vado na natureza, já que as populações de seres vivosse mantêm num nível mais ou menos constante. Poroutro lado, ele observou que os indivíduos dessaspopulações apresentavam variabilidade, inclusive emcaracterísticas essenciais à sua adaptação ao ambien-te. Constatou também que, devido a esta variabilida-de, muitos indivíduos morriam precocemente, outrosnão eram bem-sucedidos reprodutivamente, e algunseram longevos e capazes de produzir descendentesférteis em boa quantidade. Desta forma, concluiu quedeveria existir uma “luta pela sobrevivência” (i.e.competição), onde apenas os mais aptos seriam sele-cionados ao longo do tempo, transmitindo suas ca-racterísticas vantajosas aos seus descendentes. A esteprocesso denominou seleção natural, em oposiçãoà “seleção artificial”. As diferenças entre as popula-ções iriam se acumulando com a sucessão das váriasgerações, a ponto destas se diferenciarem dos tiposoriginais, constituindo-se em novas formas ou “es-pécies”.
Segundo vários autores, as ideias de Darwin es-tão marcadas pelas concepções políticas dominantes nasociedade inglesa da época; o progresso era de ocorrên-cia lenta, gradual e constante dentro da estabilidademonárquica (Freire-Maia, 1988). Dessa forma, as ideiasde Lyell foram de grande importância na teoriaDarwinista. Para que ocorresse evolução, Darwin acre-ditava que deveria haver uma certa estabilidade noambiente. Sustentou que as falhas no registro geológi-co foram as responsáveis pela ausência de formas inter-mediárias entre as “espécies”. Acreditava que a modi-ficação era direcional, lenta e gradual, sempre enfati-zando a seleção natural como a principal causa da evo-lução. Assim, a seleção natural ocorreria em um ambi-ente calmo, sem catástrofes, e num intervalo de tempolongo.
Darwin passou 20 anos testando experimental-mente suas convicções, analisando as evidências acu-muladas durante sua viagem ao redor do mundo e es-crevendo um longo tratado sobre o assunto. No ano de1858, Darwin recebeu uma carta de um jovem natura-

140 Paleontologia
3ª Prova
lista, Alfred Russel Wallace, na qual este propunha omesmo mecanismo de evolução dos seres vivos conce-bido por ele. Darwin, reconhecendo o valor do trabalhode Wallace, escreveu um breve resumo de suas teorias.Os dois trabalhos foram então apresentados concomi-tantemente em uma reunião da Sociedade Linneana,em Londres: Sobre a tendência das espécies em formar vari-edades e Sobre a perpetuação das variedades e espécies pormeio da seleção natural. Cabe ressaltar que a teoria daseleção natural de Wallace foi elaborada com o mesmorigor científico, mas sem o mesmo embasamento expe-rimental da teoria darwiniana. Enquanto Wallace escre-veu apenas um ensaio com os resultados de suas pes-quisas, Darwin escreveu vários livros relatando seusexperimentos e observações. Apesar da obra de Darwinser muito mais difundida, Wallace merece ser reconhe-cido como codescobridor deste mecanismo evolutivo,ou seja, a seleção natural.
Em 1859, foi publicada a 1ª. edição da principalobra de Darwin – The Origin of Species (A Origem dasEspécies) – onde ele apresentava uma síntese de am-plo alcance sobre a teoria da evolução, recorrendo atodas as fontes relevantes de informações que pudes-sem corroborar suas ideias (i. e. o registro fóssil, a bioge-ografia, estudos sobre anatomia e embriologia compa-radas, modificações em animais domesticados, entre ou-tras) .
Apesar dos evolucionistas atuais aceitarem so-mente parte do pensamento de Darwin, sua teoria temum reconhecimento quase universal nos meios cientí-ficos. Desde a publicação da Origem das Espécies, oconhecimento sobre as causas e caminhos das mudan-ças evolutivas tem aumentado enormemente, tendosido amplamente reformulado sob a óptica da modernabiologia evolutiva.
Darwinismo
Princípios fundamentais:
– devido à desproporção entre o crescimento popu-lacional e a quantidade de espaço e alimentodisponíveis, deve existir uma luta pela sobrevi-vência entre os indivíduos;
– como resultado desta luta, apenas os mais aptospermaneceriam vivos, transmitindo suas caracte-rísticas aos seus descendentes (seleção natural).
Destes princípios depreendem-se duas te-ses principais:
– os organismos são produtos de uma história dedescendência com modificação lenta e gradual, apartir de ancestrais comuns;
– o principal mecanismo de evolução é o da sele-ção natural das características hereditárias dos or-ganismos.
Importância do Darwinismo:
– ter sido a base da moderna teoria evolutiva.
Teoria Sintética da Evolução
O primeiro quarto do século XX foi inauguradopelo redescobrimento das leis de Mendel e marcadopelo desenvolvimento explosivo da Genética.
Em sua teoria, Darwin não havia conseguidoexplicar a natureza da herança das características nosorganismos. De acordo com o pensamento da época,ele acreditava que a herança ocorria através de umafusão de elementos paternos e maternos, como umamistura de tintas. Mas uma hereditariedade desse tipolevaria a uma rápida redução da variabilidade das carac-terísticas nos organismos. Outro problema a ser resolvi-do era como se dava a conservação e o ressurgimentode características, apesar da reprodução sexual. Nenhu-ma solução satisfatória foi encontrada até 1900, quandohouve a divulgação das experiências do monge GregorJohann Mendel (1822-1884), apresentadas em 1865. Aessência da hereditariedade mendeliana é que os fato-res hereditários, os genes, reunidos em um indivíduo,devido à fertilização de um ovo, separar-se-ão novamen-te nas células reprodutivas (gametas) do indivíduo.Mendel, através de experiências com o cruzamento deplantas, constatou que a variação genética podia sermantida indefinidamente numa população com repro-dução sexuada. Isto porque este tipo de reprodução per-mite a separação (segregação) e a reunião dos genes deuma população ao longo das gerações, fato este mate-maticamente demonstrado por ele.
Desde 1901, quando alguns pesquisadores che-garam à conclusão da existência dos genes (fatores he-reditários), confirmando todas as proposições de Mendel(o pai da Genética), houve um acentuado impulso noconhecimento científico sobre os genes e suas proprie-dades.

141Capítulo 9 – Teorias Evolutivas
3ª Prova
Genes, cromossomos e mutações
Os genes são formados por DNA (ácidodesoxirribonucleico) e localizam-se nas células dosorganismos. Eles se encontram em organismossexuados, aos pares, reunidos em filamentos que seorganizam em cromossomos (macromoléculasnucleoproteicas portadoras dos genes). Qualquertipo de alteração nos genes ou em sua ordenaçãonos cromossomos, ocorrida nas células reprodutivas(gametas), pode causar o aparecimento de novascaracterísticas hereditárias. Alterações desta natu-reza recebem o nome de mutação.
A partir de 1940, os trabalhos de Ronald Fisher,John Haldane, Sewall Wright, Theodosius Dobzansky,Ernest Mayr, Julian Huxley, George Simpson e G.Ledyard Stebbins reinterpretaram, à luz de novos da-dos provindos da genética e outras áreas do conheci-mento biológico (botânica, zoologia, paleontologia,ecologia), os conceitos obtidos a partir do darwinismo.Assim nasceu uma nova teoria evolutiva denominadateoria sintética da evolução ou teoria neodarwinista.A teoria darwiniana, quanto à seleção natural, não so-freu alteração em nenhum de seus aspectos essenciaisem virtude da teoria genética, mas sim foi enriquecidapor uma teoria da hereditariedade que permitiu dar umamaior precisão ao conhecimento da evolução, resolven-do alguns problemas até então sem solução (Futuyma,1993).
A genética veio mostrar que as característicasdos organismos são determinadas por fatores herdáveis,os genes. As modificações (= mutações) ocorridas aoacaso nos genes, quando não letais aos organismos, sãoa fonte de origem das novas características nos seresvivos e, por conseguinte, responsáveis pela diversida-de morfológica observada nas populações. A manuten-ção da variabilidade nas características dos organismosé garantida inicialmente pela reprodução, onde ocorrea recombinação dos genes dentro da população e, pos-teriormente, pelo mecanismo da seleção natural.
A estrutura básica da Teoria Sintética clássica éque a evolução é um fenômeno de duas faces: a produ-ção da variabilidade gênica e a sua manutenção. Os fa-tores que determinam alterações no conjunto gênico(conjunto de todos os genes presentes em determinadapopulação) são denominados “fatores evolutivos”. Ateoria sintética reconhece como causas da evolução os
seguintes fatores evolutivos: mutação, recombinaçãogênica, deriva genética, seleção natural e migra-ção. Mutação é um fator de produção e aumento davariabilidade gênica. Recombinação e migração sãofatores de aumento da variabilidade gênica. Derivagenética, seleção natural e também a migração são fa-tores de redução da variabilidade gênica (figura 9.1).
Genótipo versus Fenótipo
O genótipo é o conjunto dos genes de umúnico indivíduo. O conjunto dos diferentes genó-tipos (= tipos gênicos) de uma população forma oseu conjunto gênico, patrimônio gênico ougenoma.
O conjunto de todas as características (mor-fológicas, fisiológicas, comportamentais e outras)de um indivíduo, manifestadas ao longo da vida eque se desenvolvem pela interação dos genes como meio ambiente, é denominado fenótipo.
a. Mutação
É a fonte básica de toda a variação hereditária; osdemais mecanismos evolutivos utilizam a variabilida-de que é produzida pelas mutações. Por isso, as muta-ções têm sido chamadas de “matéria-prima da evolu-ção” (vide também quadro sobre genes, cromossomose mutações). A mutação pode ser gênica (alteração deum gene) ou cromossômica (alteração do número ouestrutura dos cromossomos).
b. Recombinação gênica
É um processo que reorganiza os genes já exis-tentes nos cromossomos. O mecanismo primário derecombinação gênica é a reprodução sexuada, onde ospares de cromossomos permutam (trocam) pedaços en-tre si durante a formação das células reprodutivas(gametas). Para alguns geneticistas, a recombinação en-volve também a segregação (separação) independentedesses pares de cromossomos nas células reprodutivas,durante a meiose (redução do número de cromossomos).Ambos mecanismos possibilitam um grande númerode combinações gênicas, dando origem a vários tiposde gametas que irão se unir na fecundação. Através darecombinação gênica, uma população pode ter sua vari-abilidade gênica aumentada, sem adição de novosgenes. A recombinação gênica leva à formação de no-vos genótipos, expondo-os à seleção natural e a outrosagentes (Araújo, 1984).
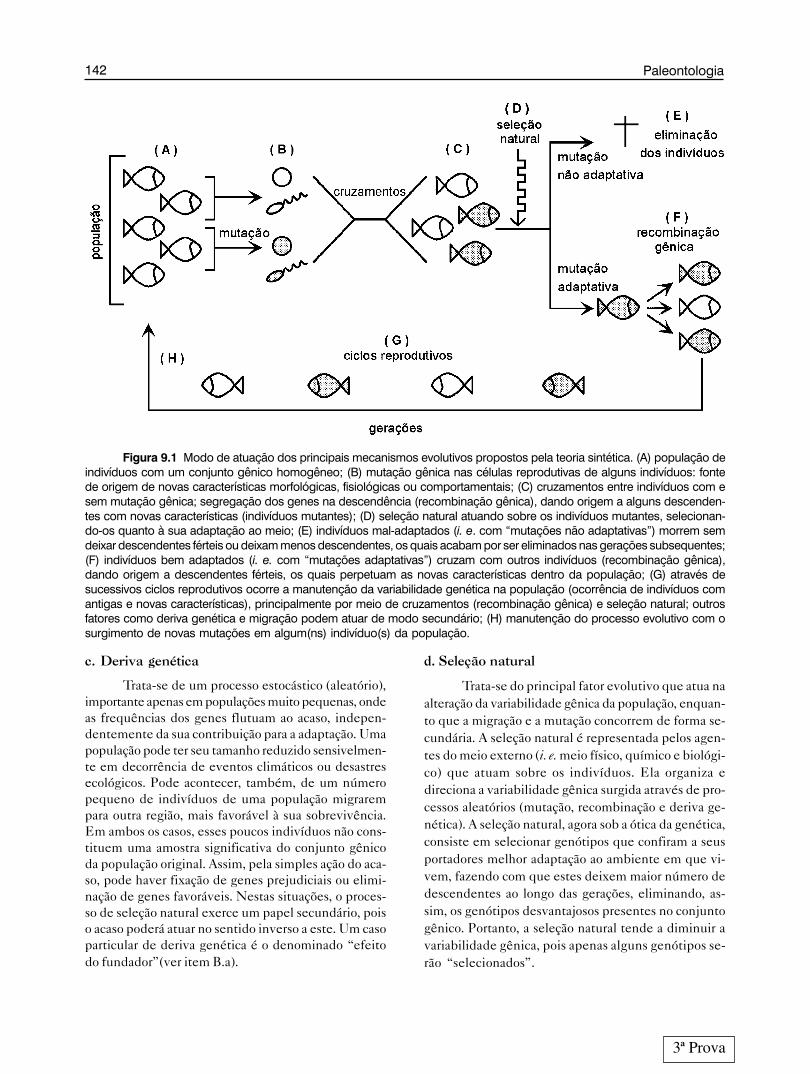
142 Paleontologia
3ª Prova
c. Deriva genética
Trata-se de um processo estocástico (aleatório),importante apenas em populações muito pequenas, ondeas frequências dos genes flutuam ao acaso, indepen-dentemente da sua contribuição para a adaptação. Umapopulação pode ter seu tamanho reduzido sensivelmen-te em decorrência de eventos climáticos ou desastresecológicos. Pode acontecer, também, de um númeropequeno de indivíduos de uma população migrarempara outra região, mais favorável à sua sobrevivência.Em ambos os casos, esses poucos indivíduos não cons-tituem uma amostra significativa do conjunto gênicoda população original. Assim, pela simples ação do aca-so, pode haver fixação de genes prejudiciais ou elimi-nação de genes favoráveis. Nestas situações, o proces-so de seleção natural exerce um papel secundário, poiso acaso poderá atuar no sentido inverso a este. Um casoparticular de deriva genética é o denominado “efeitodo fundador”(ver item B.a).
d. Seleção natural
Trata-se do principal fator evolutivo que atua naalteração da variabilidade gênica da população, enquan-to que a migração e a mutação concorrem de forma se-cundária. A seleção natural é representada pelos agen-tes do meio externo (i. e. meio físico, químico e biológi-co) que atuam sobre os indivíduos. Ela organiza edireciona a variabilidade gênica surgida através de pro-cessos aleatórios (mutação, recombinação e deriva ge-nética). A seleção natural, agora sob a ótica da genética,consiste em selecionar genótipos que confiram a seusportadores melhor adaptação ao ambiente em que vi-vem, fazendo com que estes deixem maior número dedescendentes ao longo das gerações, eliminando, as-sim, os genótipos desvantajosos presentes no conjuntogênico. Portanto, a seleção natural tende a diminuir avariabilidade gênica, pois apenas alguns genótipos se-rão “selecionados”.
Figura 9.1 Modo de atuação dos principais mecanismos evolutivos propostos pela teoria sintética. (A) população deindivíduos com um conjunto gênico homogêneo; (B) mutação gênica nas células reprodutivas de alguns indivíduos: fontede origem de novas características morfológicas, fisiológicas ou comportamentais; (C) cruzamentos entre indivíduos com esem mutação gênica; segregação dos genes na descendência (recombinação gênica), dando origem a alguns descenden-tes com novas características (indivíduos mutantes); (D) seleção natural atuando sobre os indivíduos mutantes, selecionan-do-os quanto à sua adaptação ao meio; (E) indivíduos mal-adaptados (i. e. com “mutações não adaptativas”) morrem semdeixar descendentes férteis ou deixam menos descendentes, os quais acabam por ser eliminados nas gerações subsequentes;(F) indivíduos bem adaptados (i. e. com “mutações adaptativas”) cruzam com outros indivíduos (recombinação gênica),dando origem a descendentes férteis, os quais perpetuam as novas características dentro da população; (G) através desucessivos ciclos reprodutivos ocorre a manutenção da variabilidade genética na população (ocorrência de indivíduos comantigas e novas características), principalmente por meio de cruzamentos (recombinação gênica) e seleção natural; outrosfatores como deriva genética e migração podem atuar de modo secundário; (H) manutenção do processo evolutivo com osurgimento de novas mutações em algum(ns) indivíduo(s) da população.

143Capítulo 9 – Teorias Evolutivas
3ª Prova
e) Migração
Constitui outro fator capaz de alterar a variabili-dade em uma população. A mudança na variabilidadepode ser compreendida ao se imaginar uma populaçãocom uma dada constituição genética, recebendomigrantes de outra população com diferente constitui-ção genética, que passam a cruzar com a população re-sidente. Os imigrantes introduzirão genes novos e con-tribuirão para o aumento da variabilidade gênica da po-pulação. Porém, através da migração é estabelecido um“fluxo gênico” (intercâmbio gênico) que tende a dimi-nuir as diferenças genéticas entre as populações. Logo,a migração é um fator que promove a uniformidadegenética entre as populações, podendo, deste modo,contribuir para a redução da variabilidade gênica do con-junto gênico total das populações envolvidas.
Teoria sintética
Princípios Fundamentais
– as populações apresentam variação genética quesurge através de mutação ao acaso (i.e. nãodirigida adaptativamente) e recombinação gênica(novas combinações genotípicas);
– as populações evoluem por alterações nas fre-quências dos genes geradas pelos seguintes fato-res evolutivos: mutação, recombinação gênica,deriva genética, migração e, principalmente, pelaseleção natural;
Méritos da Teoria Sintética:
– reuniu, sob uma teoria consistente, a essência dateoria de Darwin (a seleção natural) e os conheci-mentos do campo da genética, extraídos inicial-mente dos trabalhos de Mendel;
– conseguiu fornecer explicações consistentes so-bre a origem e a manutenção da variabilidade dascaracterísticas nas populações de cada espécie esobre as diferenças entre as espécies, fazendo agrande síntese de todos os dados biológicos e pa-leontológicos;
– definiu as populações, e não os indivíduos, comoas “unidades evolutivas”.
A elucidação dos princípios básicos da evoluçãoatravés da teoria sintética retirou do indivíduo o papelde promotor da evolução. Uma vez que boa parte dateoria pôde ser formalizada em termos de modelos es-tatísticos (frequências gênicas e genotípicas, por exem-
plo), as populações de indivíduos semelhantes torna-ram-se as “unidades evolutivas”. Mas o que são indiví-duos semelhantes? São os indivíduos de uma mesmaespécie! E o que é uma espécie? A resposta a esta ques-tão era crucial para se estudar a evolução. Sem umaclara idéia do que é uma espécie, não se poderia levaradiante uma série de experimentos necessários à com-provação, em termos empíricos, da existência da evo-lução biológica como postulada pela teoria sintética.Devido a isso, a partir de 1940, vários pesquisadoresesforçaram-se em definir corretamente o que são as“espécies biológicas” para poder explicar como elassurgem com base nos mecanismos propostos pela teo-ria sintética. É o que será abordado a seguir.
Espécie e Especiação
A. Conceito Biológico de Espécie
A teoria sintética postula que a evolução ocorreao nível da espécie e a mudança evolutiva provém,principalmente, da seleção natural agindo sobre espé-cies existentes, de forma a transformá-las ou diversificá-las, originando novas espécies. Para melhor compreen-der esta definição é importante conhecer o mais difun-dido conceito biológico de espécie, originalmenteconcebido por Mayr em 1942 e aprimorado por ele em1977.
Espécies são agrupamentos de populações naturaisintercruzantes, reprodutivamente isoladas de outros grupossemelhantes (Mayr, 1977).
Este conceito realça o fato de que:
• uma espécie é constituída por populações, ou seja, agru-pamentos de indivíduos que ocorrem em uma mesmaárea geográfica, em um mesmo intervalo de tempo;
• as espécies são reais e possuem um conjunto gênicoque é compartilhado por todos os seus membros.
Assim sendo, os membros de uma espécie cons-tituem-se em:
– uma comunidade reprodutiva: na qual os indivíduosprocuram-se uns aos outros com a finalidadereprodutiva;
– uma unidade ecológica: que interage com outras es-pécies com as quais compartilha o meio ambientecomo uma unidade;
– uma unidade genética: que consiste num grande con-junto gênico em intercomunicação, enquanto o in-divíduo é meramente um componente com uma pe-quena porção do conteúdo genético total deste con-junto.

144 Paleontologia
3ª Prova
Além disso, este conceito salienta o isolamentoreprodutivo como principal fator determinante de umaespécie. Daí advém o fato de que entre duas popula-ções de espécies distintas não deve haver intercâmbiogênico.
Novos conceitos de espécie biológica têm sur-gido nos últimos anos, uma vez que o conceito de Mayrestá restrito, basicamente, aos organismos sexuados. Omais inovador deles é o chamado “conceito de coe-são”, formulado e proposto por Templeton (1981). Esteconceito procura ressaltar os mecanismos que mantêmas espécies “coesas”, ao contrário do conceito de Mayrque valoriza os mecanismos de isolamento reprodutivo.A partir destes novos conceitos uma espécie biológicatem sido definida em três distintos níveis: o genético-molecular, o morfológico e o ecológico.
Com relação à definição de espécie ao nívelgenético-molecular, os fósseis dos organismos têm, sal-vo raríssimas exceções (p. e.: insetos conservados emâmbar, mamutes congelados), as suas característicasgenético-moleculares totalmente destruídas durante osprocessos de fossilização. Portanto, na paleontologianão há normalmente como definir uma espécie em ter-mos genético-moleculares.
A definição de espécie ao nível morfológico éa mais clássica e usual em paleontologia, tendo emvista que as características morfológicas são as quemais se preservam nos fósseis. Desde o século XVIII,quando a paleontologia presenciou um grande incre-mento dentro dos meios científicos, vindo assim atornar-se um ramo independente da ciência, o con-ceito de espécie em termos morfológicos é pratica-mente o único que tem sido utilizado. Uma espéciedefinida com base nestes critérios denomina-se demorfoespécie ou espécie paleontológica . Amorfoespécie é, portanto, uma espécie estabelecidaem termos das diferenças morfológicas. A espéciepaleontológica expressa meramente o grau de dife-rença morfológica entre os organismos fossilizados.Ela pode, indiretamente, refletir a existência de es-pécies distintas, uma vez que uma significativa dife-renciação morfológica entre as formas, normalmen-te, conduz ao isolamento reprodutivo.
Já a definição de espécie ao nível ecológico éhabitualmente utilizada de maneira indireta pelos pa-leontólogos. As adaptações de um organismo ao meio(relação forma/função em paleontologia) e o hábitatonde ele vive sempre foram considerados pelos paleon-tólogos em suas discussões a respeito dos fósseis (taxa).
Entretanto, este tipo de informação nunca é formalmen-te incorporado à definição de uma espécie em paleon-tologia. Isto pode ser explicado pelo fato de que: a) osfósseis representam evidências bastante incompletasdos organismos do passado; b) esses organismos, emgeral, não foram fossilizados exatamente em seus am-bientes de vida; c) os antigos ambientes de vida dessesorganismos, quando preservados, demonstram, atravésdas rochas sedimentares, um registro incompleto de to-dos os seus parâmetros (bióticos ou abióticos). Destaforma, torna-se bastante difícil e impreciso ao paleon-tólogo a incorporação de informações paleoecológicasàs definições de espécie. No entanto, se tais informa-ções forem suficientemente bem conhecidas, elas po-dem vir a ser anexadas às definições das espécies pa-leontológicas, tendo em vista os conceitos adotados nabiologia moderna.
Especiação
Entende-se por especiação os processos que es-tão envolvidos no surgimento de novas espécies.Especiação consiste, em última análise, no desenvolvi-mento de barreiras ao intercâmbio gênico entre as popu-lações. Isto porque o processo de especiação ocorre apartir do acúmulo de diferenças entre os conjuntos gênicos(diferenciação genética) em populações de uma mesmaespécie. Estas diferenças gênicas, no entanto, podem serperdidas a qualquer momento, caso não haja um isola-mento reprodutivo (interrupção do fluxo gênico), poisum intercruzamento entre os indivíduos de diferentespopulações de uma mesma espécie promove a homoge-neização dos conjuntos gênicos destas (vide item Migra-ção). Apenas com o isolamento reprodutivo as diferen-ças genéticas obtidas entre as populações têm possibili-dades reais de se preservarem ao longo do tempo.
Não esquecer que são os genes que codifi-cam e determinam todas as características dos seresvivos. Logo, quanto mais diferenças gênicas exis-tam entre duas populações, mais acentuadas serãoas diferenças em termos morfológicos, fisiológicose comportamentais entre elas, pois diferentes genescodificam diferentes produtos gênicos, que por suavez determinam distintas características.
A diferenciação genética (gênica e cromossô-mica) das populações em espécies distintas deve-seà ação, conjunta ou não, dos fatores evolutivos sele-ção natural e deriva genética.

145Capítulo 9 – Teorias Evolutivas
3ª Prova
Portanto, o desenvolvimento de mecanismosque propiciem o isolamento reprodutivo das popula-ções de organismos é fundamental para a origem dasnovas espécies. Isolamento geográfico e isolamentoecológico têm sido apontados como fatores que auxili-am na manutenção das diferenças obtidas entre as po-pulações. O isolamento geográfico (= separação espaci-al) pode propiciar a diferenciação genética, a qual podelevar a um isolamento reprodutivo. Já o isolamentoecológico tem base genética e constitui, em si, umabarreira de isolamento reprodutivo. Ele é muito impor-tante em plantas e em animais ecologicamente especia-lizados.
O isolamento reprodutivo é gerado por meca-nismos genéticos (intrínsecos). Templeton (1981) divi-de os tipos de especiação com base nos mecanismosgenéticos em duas categorias. A primeira – Transiliên-cia – apresenta mecanismos que envolvem alteraçõesgenéticas (gênicas ou cromossômicas) ou a formaçãode híbridos férteis, com posterior diferenciação denova(s) espécie(s). Essa categoria, por não ser testávelno âmbito da paleontologia, é de pouco interesse aqui.Apenas o mecanismo “efeito do fundador”, que envol-ve deriva genética e está incluído nesta categoria, temreal interesse paleontológico, como verificar-se-á adi-ante. Já os mecanismos da segunda categoria – Diver-gência –, por envolverem aspectos geográficos e eco-lógicos, são os únicos passíveis de verificação pa-leontológica.
Existem três tipos básicos de especiação a partirdos mecanismos de Divergência, os quais, do ponto devista estritamente geográfico, podem ser classificadosem: especiação alopátrica, especiação simpátrica eespeciação parapátrica. A classificação geográfica des-tes tipos de especiação é útil em termos paleontológi-cos, tendo sido adotada por Mayr (1942, apud Grant, 1991).Porém, não indica os mecanismos básicos envolvidos noprocesso de especiação. Por isso, Templeton (1981) pro-curou substituir estes nomes por outros que refletissemtais mecanismos: especiação adaptativa, especiaçãopor hábitat e especiação clinal.
a. Especiação alopátrica (“especiação adaptativa”)
Duas populações de uma mesma espécie diver-gem para duas áreas distintas como consequência de umaseparação espacial (alopatria), ocasionada pelosurgimento de uma barreira geográfica (montanha, mar,rio, floresta, deserto, etc.). Se as duas áreas diferirem emsuas características ecológicas e/ou as duas populações
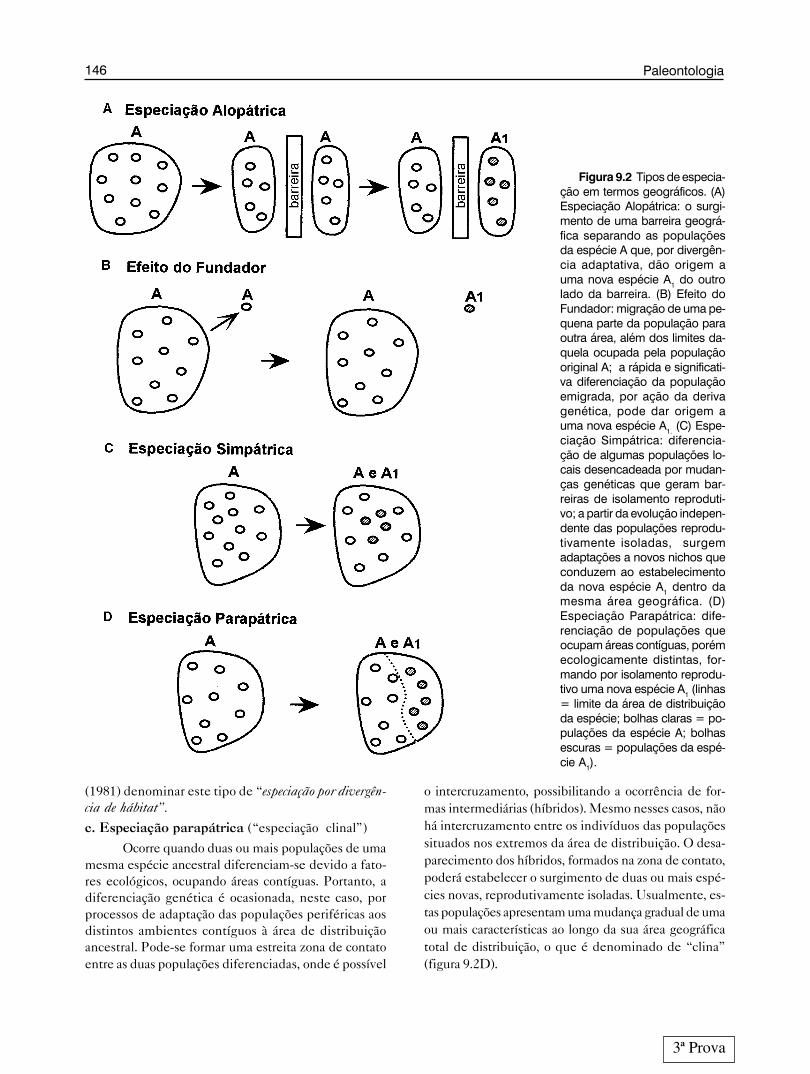
ficarem separadas por tempo suficiente para haver diver-gência genética, mecanismos de isolamento reprodutivopoderão surgir. Em consequência disso, haverá especi-ação. Segundo Templeton (1981) a diferenciação gené-tica entre as populações separadas é, principalmente,desencadeada por processos de adaptação aos distintosambientes (i. e. fatores ecológicos bióticos e abióticos)gerados pelo estabelecimento da barreira (figura 9.2A).
Um tipo especial de especiação alopátrica, o efei-to do fundador, proposto por Mayr (1963, apud Mayr,1977), merece destaque. A partir de uma população gran-de, um pequeno número de indivíduos migra para alémdos limites de distribuição da espécie, levando consigouma pequena amostra do conjunto gênico da popula-ção original. A situação de população pequena e perifé-rica propicia a ação da deriva genética, a qual leva àprodução, nesta população, de genótipos diferentes dosnormalmente encontrados na população inicial (Araú-jo, 1984). Como este fenômeno é verificado apenas empopulações muito pequenas, o que ocorre com maiorfrequência é a extinção destas, devido à fixação de ca-racterísticas não adaptativas ou deletérias. No entanto,o efeito do fundador, nos casos bem-sucedidos, podeconduzir a uma especiação mais rápida e de maior mag-nitude, graças a uma ação conjunta da deriva genética eda seleção natural, propiciando diferenças mais acentu-adas entre as novas espécies (figura 9.2B). Por suas ca-racterísticas este tipo de especiação foi fundamental naelaboração da “teoria do equilíbrio pontuado” deEldredge & Gould, 1972, como será visto mais adiante.
b. Especiação simpátrica (“especiação por hábitat”)
Especiação sem separação geográfica. Ocorrequando duas ou mais populações, derivadas de umapopulação original, coexistem em um mesmo territó-rio, sem intercruzarem-se. Normalmente, neste tipode especiação acontece, logo de início, uma modifi-cação genética que impede o cruzamento entre algunsdos indivíduos da mesma população, criando assimuma nova população reprodutivamente isolada, den-tro do mesmo território. A partir disso, as duas popula-ções geradas evoluem separadamente, acumulando di-ferenças genéticas (= mudanças morfológicas, fisioló-gicas, comportamentais) que conduzirão à formaçãode novas espécies ecologicamente diferenciadas, a fimde possibilitar sua coexistência dentro da mesma área(figura 9.2C). Portanto, em um processo simpátrico oisolamento reprodutivo inicial é fundamental para queocorra a especiação. A diferenciação ecológica (= com-patibilização ecológica) é consequência do processoe não desencadeadora deste, apesar de Templeton
146 Paleontologia
3ª Prova
(1981) denominar este tipo de “especiação por divergên-cia de hábitat”.
c. Especiação parapátrica (“especiação clinal”)
Ocorre quando duas ou mais populações de umamesma espécie ancestral diferenciam-se devido a fato-res ecológicos, ocupando áreas contíguas. Portanto, adiferenciação genética é ocasionada, neste caso, porprocessos de adaptação das populações periféricas aosdistintos ambientes contíguos à área de distribuiçãoancestral. Pode-se formar uma estreita zona de contatoentre as duas populações diferenciadas, onde é possível
o intercruzamento, possibilitando a ocorrência de for-mas intermediárias (híbridos). Mesmo nesses casos, nãohá intercruzamento entre os indivíduos das populaçõessituados nos extremos da área de distribuição. O desa-parecimento dos híbridos, formados na zona de contato,poderá estabelecer o surgimento de duas ou mais espé-cies novas, reprodutivamente isoladas. Usualmente, es-tas populações apresentam uma mudança gradual de umaou mais características ao longo da sua área geográficatotal de distribuição, o que é denominado de “clina”(figura 9.2D).
Figura 9.2 Tipos de especia-ção em termos geográficos. (A)Especiação Alopátrica: o surgi-mento de uma barreira geográ-fica separando as populaçõesda espécie A que, por divergên-cia adaptativa, dão origem auma nova espécie A1 do outrolado da barreira. (B) Efeito doFundador: migração de uma pe-quena parte da população paraoutra área, além dos limites da-quela ocupada pela populaçãooriginal A; a rápida e significati-va diferenciação da populaçãoemigrada, por ação da derivagenética, pode dar origem auma nova espécie A1. (C) Espe-ciação Simpátrica: diferencia-ção de algumas populações lo-cais desencadeada por mudan-ças genéticas que geram bar-reiras de isolamento reproduti-vo; a partir da evolução indepen-dente das populações reprodu-tivamente isoladas, surgemadaptações a novos nichos queconduzem ao estabelecimentoda nova espécie A1 dentro damesma área geográfica. (D)Especiação Parapátrica: dife-renciação de populações queocupam áreas contíguas, porémecologicamente distintas, for-mando por isolamento reprodu-tivo uma nova espécie A1 (linhas= limite da área de distribuiçãoda espécie; bolhas claras = po-pulações da espécie A; bolhasescuras = populações da espé-cie A1).

147Capítulo 9 – Teorias Evolutivas
3ª Prova
Padrões de Especiaçãoem Paleontologia
Uma maneira clássica de visualizar a especiaçãoem paleontologia refere-se ao número de espécies quesão geradas ao longo do tempo por uma linhagem deorganismos (linhagem evolutiva). Como os fósseis sãoencontrados em estratos de rochas que se sucedem domais antigo para o mais jovem, este tipo de classifica-ção é bastante útil na paleontologia.
O mais clássico padrão de especiação concebidotem como princípio a modificação lenta e gradual dasespécies ao longo do tempo e é denominado especiaçãofilética ou gradual. Neste padrão uma única espécie,pelo lento acúmulo de novas características genéticas,vai se transformando em outra espécie (figura 9.3A). Aespeciação filética ocorre através de anagênese. Estetipo de especiação fornece aos paleontólogos as chama-das cronoespécies, que são espécies paleontológicas(i. e. morfologicamente distintas) que se sucedem ao lon-go de uma mesma linhagem evolutiva (Futuyma, 1993).Cronoespécies, portanto, são geradas por anagênese enão são equivalentes às espécies biológicas, pois neces-sariamente não implicam na ocorrência de processos deespeciação biológica.
Anagênese versus Cladogênese
Anagênese é um processo evolutivo queleva a mudanças direcionais através do tempo den-tro de uma única linhagem. Ocorre nos organismos,independente da ocorrência do processo de espe-ciação (figura 9.4A e B).
Cladogênese corresponde a um processoevolutivo que gera ramificações nas linhagens deorganismos ao longo de sua história evolutiva. Im-plica obrigatoriamente em especiação biológica (fi-gura 9.4C).
Portanto, a ação da anagênese (ou dos pro-cessos anagenéticos) pode ou não conduzir à espe-ciação, já a cladogênese (ou os processos cladoge-néticos) sempre resulta em um evento de especia-ção.
O padrão de especiação gradual foi concebido apartir da análise do registro fóssil à luz das ideiasdarwinistas. Apesar de atualmente este padrão ser criti-cado por alguns evolucionistas, ele constitui-se em um
dos pressupostos básicos do gradualismo, como será vistoa seguir. Além da evolução filética (gradual), são tam-bém apresentadas por vários autores a especiaçãoredutiva (fusão de espécies através da formação de hí-bridos) e a especiação aditiva (multiplicação do nú-mero de espécies) (figura 9.3B e C). A especiação aditivaé gerada por cladogênese. Deve-se salientar que a es-peciação aditiva corresponde aos modelos de especia-ção biológica apresentados no item anterior.
Microevolução versus Macroevolução
Os termos microevolução e macroevoluçãoforam criados por Richard Goldschmidt (1940) com oobjetivo de separar os mecanismos neodarwinistas, res-ponsáveis pela diferenciação entre as espécies, daque-les que originam grupos taxonômicos hierarquicamentesuperiores (gêneros, famílias, ordens, classes, filos) (Araú-jo, 1984). Pode-se definir microevolução como o con-junto de mudanças que ocorrem dentro das populações eque podem levar até sua diferenciação em espécies(Freire-Maia, 1988). Já a macroevolução, também co-nhecida como “evolução acima do nível das espécies”,abrange as mudanças suficientemente grandes para dis-tinguir os taxa superiores (Futuyma, 1993).
Desde a primeira metade do século XX, algunscientistas já questionavam a evolução lenta e gradual,tipicamente darwiniana. A partir da observação de queo registro fóssil revela descontinuidades marcantes, ouseja, não existem formas intermediárias indicando umatransformação lenta e gradual entre os taxa superiores,foram apresentados modelos macroevolutivos que seconfrontavam com a teoria neodarwinista. Os macro-evolucionistas, destacando-se Richard Goldschmidt eGeorge Simpson, argumentavam que não se podia com-preender a macroevolução baseando-se apenas nos pro-cessos neodarwinistas de acúmulo lento e gradual demicromutações. Goldschmidt (1940) propôs o termo“mutação sistêmica” ou “macromutação” para desig-nar uma grande mutação capaz de fazer, por exemplo,surgir uma ave a partir de um réptil. Em 1944, Simpsonlevou a paleontologia aos domínios da teoria sintéticaatravés de sua obra intitulada Tempo and mode in evolution(Tempo e modo em evolução), a qual causou um gran-de impacto na época (Barberena, 1984). A partir dasdescontinuidades do registro fóssil, Simpson criou aexpressão “evolução quântica” para denominar a ocor-rência de uma mudança significativa, sem meio-termo.Para ele, a origem dos grandes grupos sistemáticos se-

148 Paleontologia
3ª Prova
ria sempre produzida por uma evolução deste tipo.Simpson não acreditava nas macromutações de Golsch-midt (1940), mas apontava a deriva genética como oprocesso mais consistente para fazer a ligação entre amicro e a macroevolução.
A ênfase nos processos macroevolutivos foi de-saparecendo do cenário científico com a morte dos seusmaiores expoentes, sendo que nunca se chegou a umacordo sobre qual seria o mecanismo ou fator capaz degerar as grandes novidades evolutivas (Freire-Maia,1988). Deste modo, a visão gradualista manteve-secomo uma das principais diretrizes dos processosevolutivos.
Segundo a teoria sintética, a evolução em todosos níveis deve ser explicada pelos fatores microevo-lutivos (mutação, recombinação, migração, seleção na-tural, deriva genética), ou seja, as mudanças macro-evolutivas são compostas por eventos microevolutivos,não havendo justificativa para se supor a ocorrência defenômenos diferentes dos atualmente propostos (Araú-jo, 1984).
Apesar disso, dentro da biologia evolutiva aindanão existe um ponto de vista consensual que expliquea origem dos taxa superiores. Até o momento não fo-ram encontradas respostas completamente satisfatóriasdentro da Teoria Sintética para questões como (Freire-
Maia, 1988): Por quais mecanismos as diferenças entrefamílias, ordens, classes e filos evoluíram? Como surgi-ram os vertebrados a partir de invertebrados? Como sedeu a origem dos novos órgãos (p. ex.: bexiga natatóriados peixes, mamas dos mamíferos, etc.)? Segundo osdefensores da macroevolução, os neodarwinistas escla-recem em parte os fenômenos capazes de gerar as gran-des novidades evolutivas, ou seja, descrevem os fenô-menos, mas, no entanto, não fornecem explicaçõessatisfatórias sobre como e por que estes ocorreram. Naopinião de Freire-Maia (1988), uma vez que o registrofóssil é o único meio através do qual a macromutaçãopode ser estudada, não há justificativas para se explicara evolução nos seus níveis mais altos a partir somentede inferências microevolutivas. Segundo Futuyma(1993), a macroevolução constitui-se ainda hoje em umdos tópicos mais desafiadores e difíceis de serem resol-vidos dentro da biologia evolutiva.
Gradualismo versus Pontuísmo
Como visto anteriormente, para os darwinistas eneodarwinistas, a evolução, mesmo ao nível dos taxasuperiores, é relativamente lenta e gradual. Por isso, aespeciação realizada segundo o modelo neodarwinistafoi denominada de gradualismo filético.
Figura 9.3 Modos de especiação em paleontologia. (A) Gradual ou filética: quando há a transformação gradual deuma espécie em outra. (B) Redutiva (fusão de espécies): quando há a formação de híbridos a partir de duas espéciesancestrais distintas. (C) Aditiva (multiplicação de espécies): quando há o surgimento de novas espécies a partir de umaespécie ancestral.

149Capítulo 9 – Teorias Evolutivas
3ª Prova
Figura 9.4 Diagramas representativos dos proces-sos de anagênese e cladogênese. As ordenadas repre-sentam o tempo e as abscissas representam as mudançasmorfológicas que ocorrem ao longo do tempo. (A) Proces-so de anagênese onde não ocorreu especiação (=cronoclina). (B) Processo de anagênese resultando emespeciação filética, onde a linhagem da espécie ancestralA deu origem às espécies A1 e A2, sucessivamente. (C)Processo de cladogênese com nítida especiação da li-nhagem da espécie ancestral A, dando origem às espéci-es A1 e A2.
Segundo Eldredge & Gould (1972), os princi-pais postulados do gradualismo filético são:
• novas espécies surgem pela transformação de umapopulação ancestral em descendentes modificados;
• a transformação envolve grande número de indivídu-os, usualmente toda a população ancestral;
• a transformação ocorre em toda a área de distribuiçãogeográfica da espécie ancestral ou numa grande por-ção da mesma;
• idealmente, o registro fóssil da origem de uma novaespécie deveria estar constituído por uma longasequência de formas intermediárias, quase impercep-tivelmente gradacionais, ligando, de forma contínua,o ancestral ao descendente;
• os hiatos morfológicos encontrados no registro fóssilsão devidos às falhas do registro geológico (que ébastante incompleto).
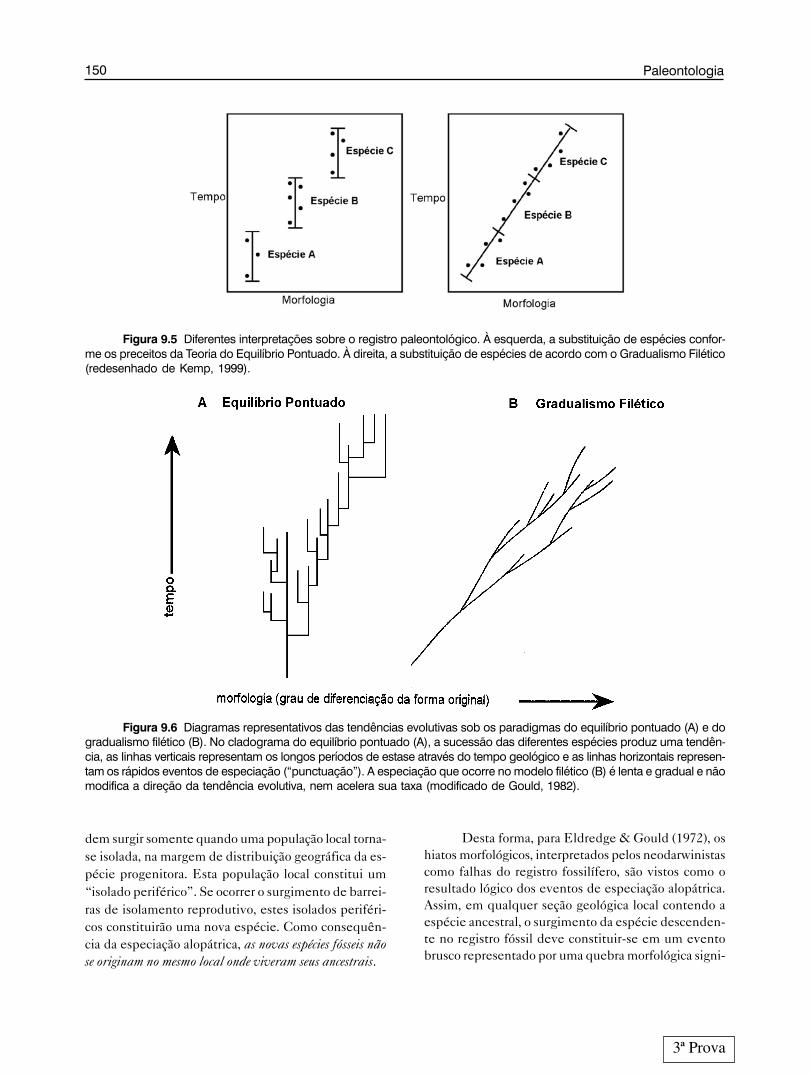
Até meados de 1970, era esta a visão dominadapelo paradigma do gradualismo filético, que se tinhasobre especiação. Não concordando com este tipo deinterpretação, os paleontólogos Eldredge & Gould(1972) apresentaram uma nova teoria evolutiva deno-minada Teoria do Equilíbrio Pontuado, conhecidatambém como pontuísmo ou pontualismo.
A expressão “equilíbrio pontuado” foi empre-gada por esses autores para ilustrar a situação onde umaespécie era substituída instantaneamente no registrofóssil por uma espécie descendente, mas sem a presen-ça de formas intermediárias entre elas (fig. 9.5), comoera preconizado pelo gradualismo filético.
Para Eldredge & Gould (1972), a evolução nãodeve ser vista como um desdobramento gradual, massim como um processo onde ocorre um equilíbriohomeos-tático – estase evolutiva – pontuado, apenasraramente, por rápidos e episódicos eventos de especi-ação (Barberena, 1984) (figura 9.6). É importante sali-entar que a estase compreende um longo período detempo em que não acontecem mudanças morfológicassignificativas, o que não quer dizer total estagnação.Ocorrem, neste período, apenas pequenas mudanças nãodirecionais (oscilações genéticas decorrentes dos pro-cessos microevolutivos) em torno da média das carac-terísticas das populações. Apesar da especiação poderdurar alguns milhares de anos, ela ocorre várias vezesem termos de tempo geológico. Os autores estimamque a duração das fases de “rápida especiação” devecompreender cerca de 1% ou menos do período de estase,dentro do tempo total de existência das espécies.
Para explicar os eventos de especiação em suateoria, Eldredge & Gould (1972) basearam-se no mo-delo de especiação alopátrica dos isolados periféricos(efeito do fundador) de Mayr (1963, 1970 apud Mayr,1977). Os autores consideram que novas espécies po-
150 Paleontologia
3ª Prova
dem surgir somente quando uma população local torna-se isolada, na margem de distribuição geográfica da es-pécie progenitora. Esta população local constitui um“isolado periférico”. Se ocorrer o surgimento de barrei-ras de isolamento reprodutivo, estes isolados periféri-cos constituirão uma nova espécie. Como consequên-cia da especiação alopátrica, as novas espécies fósseis nãose originam no mesmo local onde viveram seus ancestrais.
Desta forma, para Eldredge & Gould (1972), oshiatos morfológicos, interpretados pelos neodarwinistascomo falhas do registro fossilífero, são vistos como oresultado lógico dos eventos de especiação alopátrica.Assim, em qualquer seção geológica local contendo aespécie ancestral, o surgimento da espécie descenden-te no registro fóssil deve constituir-se em um eventobrusco representado por uma quebra morfológica signi-
Figura 9.5 Diferentes interpretações sobre o registro paleontológico. À esquerda, a substituição de espécies confor-me os preceitos da Teoria do Equilíbrio Pontuado. À direita, a substituição de espécies de acordo com o Gradualismo Filético(redesenhado de Kemp, 1999).
Figura 9.6 Diagramas representativos das tendências evolutivas sob os paradigmas do equilíbrio pontuado (A) e dogradualismo filético (B). No cladograma do equilíbrio pontuado (A), a sucessão das diferentes espécies produz uma tendên-cia, as linhas verticais representam os longos períodos de estase através do tempo geológico e as linhas horizontais represen-tam os rápidos eventos de especiação (“punctuação”). A especiação que ocorre no modelo filético (B) é lenta e gradual e nãomodifica a direção da tendência evolutiva, nem acelera sua taxa (modificado de Gould, 1982).

151Capítulo 9 – Teorias Evolutivas
3ª Prova
ficativa entre as duas (punctuação). Este evento marcaa migração da espécie descendente, vinda da área peri-fericamente isolada onde se desenvolveu, para a áreada espécie ancestral. A quebra morfológica reflete afase de “rápida especiação”ocorrida na população localinicial isolada (= isolado periférico). Com a chegada daespécie descendente, a espécie ancestral pode conti-nuar existindo normalmente, retrair-se geograficamen-te ou extinguir-se (figura 9.7). Portanto, se a especiaçãoocorrer rapidamente em pequenas populações, distan-tes da área em que a espécie ancestral é mais abundan-te, muito raramente encontrar-se-á uma evidência realdesse evento no registro fossilífero (Barberena, 1984),tendo em vista que os organismos raros têm poucachance de serem fossilizados.
Muita polêmica criou-se após a publicação dateoria do equilíbrio pontuado, tendo os seus autores re-cebido várias críticas por parte dos neodarwinistas. Em1977, Gould & Eldredge publicaram um novo trabalhorediscutindo sua teoria e manifestando-se sobre os tra-balhos publicados pelos autores que criticaram opontuísmo. Gould & Eldredge (1977) salientam quesua teoria não invalida a teoria sintética, apenas contra-ria o ponto de vista de que a evolução lenta, gradual econtínua seja o modo predominante dentro das mudan-ças evolutivas. Para eles, o equilíbrio pontuado é o pro-cesso evolutivo mais comum na natureza.
Os neodarwinistas afirmam que quando há osurgimento de novas espécies em isolados periféricos, o
número de indivíduos inicialmente é baixo e, portanto,a probabilidade de se encontrar algum fóssil desses in-divíduos é muito pequena ou nula. A nova espécie sóterá maior probabilidade de deixar fósseis quando esti-ver dominando o ambiente, o que significa um aumen-to do número de indivíduos. Neste momento, ela pos-sivelmente já será uma espécie totalmente diferencia-da. Os neodarwinistas admitem também rápidas mu-danças nas taxas evolutivas, e alegam que eventos derápida especiação são constatados hoje em dia e podemser explicados pelos mecanismos microevolutivos. Se-gundo Salzano (1993), a polêmica entre neodarwinistase pontuístas acontece devido a problemas ao nível deescala temporal. Uma mudança que opera por milharesde gerações é classificada como lenta pelos geneticistas,mas pode ser considerada instantânea pelos paleontólo-gos, devido à baixa resolução do registro geológico.Em geologia, milhares de anos podem estar materiali-zados e condensados em uma fina camada (de espessu-ra centimétrica) de rocha sedimentar.
Uma questão ainda em aberto é sobre o fenômenoda estase. Os neodarwinistas acreditam que a estase ocorraapenas em frequências muito baixas. O exemplo clássicoseria o dos “fósseis vivos”, que possuem baixas taxasevolutivas e são raros na natureza. Eles são também afavor de que, na maior parte das vezes, quando ocorrembaixas taxas evolutivas, como no exemplo citado, ostaxa continuam a evoluir por anagênese (mudanças
Figura 9.7 Esquema mostrando como ocorre o processo de especiação do ponto de vista geográfico, segundo aTeoria do Equilíbrio Pontuado de Eldredge & Gould (1972). Observa-se que a especiação ocorrida nas pequenas popula-ções dos isolados periféricos (= especiação peripátrica) é o principal mecanismo responsável pelo estabelecimento dopadrão do tipo equilíbrio pontuado (redesenhado de Kemp, 1999).

152 Paleontologia
3ª Prova
direcionais), veja figura 9.4A. Já os pontuístas insistemna ocorrência da estase (mudanças não direcionais) nagrande maioria das espécies. Desta forma, eles não acre-ditam que ocorra evolução por anagênese (i. e. através debaixas taxas evolutivas), veja figura 9.4B, mas apenaspor cladogênese (i. e. através de altas taxas evolutivas),veja figura 9.4C. Segundo Gould (1982), em termos deestudos paleontológicos, os fenômenos de estase são pra-ticamente ignorados. Os paleontólogos preocupam-seapenas com eventos de rápida especiação, quando de-veriam na verdade investigar os períodos de estase, poisestes constituem-se em dados reais bem documenta-dos no registro geológico.
Debatendo o Registro Fóssil
Desde a proposição da Teoria do Equilíbrio Pon-tuado, uma sorte de pesquisadores tratou de avaliar, oumesmo, reavaliar o registro fóssil com o intuito de cons-tatar ou não a ocorrência do fenômeno de estase e even-tos de especiação do tipo “pontuado”, como sugeridopor Eldredge & Gould (1972) e Gould & Eldredge(1977). Como resultado, um número cada vez maior deestudos bastante detalhados foi realizado com base nosregistros fósseis considerados os mais completos possí-veis, utilizando-se para isso de análises estatísticas apro-priadas.
Um dos primeiros estudos utilizados com a fina-lidade de constatar a existência do fenômeno da estaseentre as espécies fósseis, provém do trabalho de Coope(1979) que estudou detalhadamente besourosquaternários. Ele analisou o registro de espécies debesouros nos últimos 1,7 milhão de anos e não encon-trou nenhuma mudança evolutiva nestas; todas as espé-cies conhecidas ao longo do Quaternário são idênticas,em termos morfológicos, às espécies modernas. Segun-do Kemp (1999), este se configura em um dos melhoresexemplos de estase conhecidos, uma vez que as espé-cies não se modificaram mesmo durante um período deintensas flutuações climáticas, como foi o Quaternário.
Ainda em relação à constatação da ocorrência daestase no registro, Cheetham (1987) estudou 46 carac-terísticas mensuráveis em um gênero de briozoário doCaribe, através de um período de cerca de 4,5 Ma, paradepósitos do Mioceno superior e Plioceno inferior. Eleverificou que praticamente não ocorreu nenhuma mu-dança nessas características dentro das espécies anali-sadas, exceto por umas poucas que apresentaram algu-
ma alteração ao longo do tempo. Cheetham (1987) con-cluiu que apesar das espécies não terem exibido umacompleta estase, a análise demonstrou que a maioriadas suas características manteve-se estabilizada por lon-gos intervalos de tempo, como preconizado por Eldred-ge & Gould (1972) e Gould & Eldredge (1977).
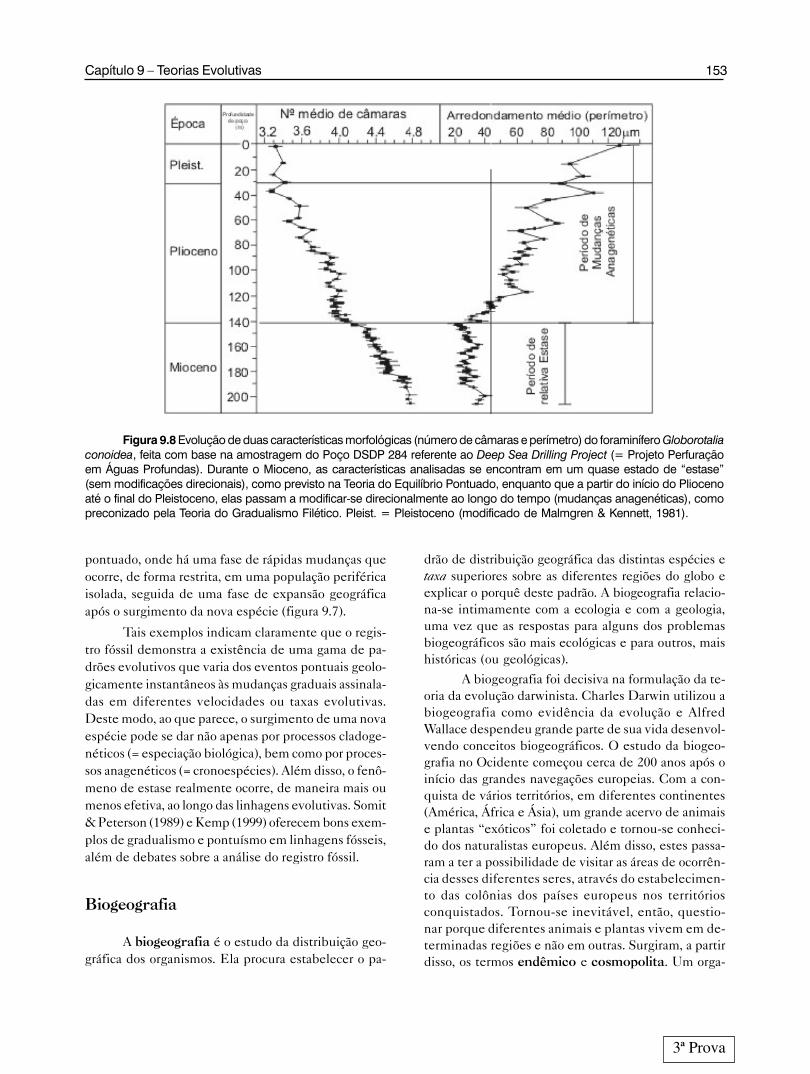
No entanto, outro estudo (Malmgren & Kennet,1981) envolvendo foraminíferos planctônicos e bentô-nicos, recuperados de testemunhos de sondagem per-furados em depósitos marinhos do Pacífico Sul, mos-trou claramente a ocorrência de mudanças graduais dascaracterísticas dentro das espécies, o que se configuraem um processo evolutivo anagenético. As amostragensforam feitas em intervalos de cerca de 100 000 anos,distribuídos ao longo dos últimos 8 Ma, o que corres-ponde a um período que vai Mioceno tardio ao Pleisto-ceno tardio. Porém, esta análise demonstrou efetiva-mente que além de processos anagenéticos, as caracte-rísticas podem também apresentar momentos de “es-tagnação” ou mudanças não direcionais, típicas do fe-nômeno da estase (figura 9.8).
Há na literatura um bom número de exemplosda ocorrência de processos anagenéticos e, portanto, deeventos de especiação graduais. No entanto, há igual-mente, bons exemplos sobre mudanças pontuais naslinhagens evolutivas, ou seja, caracterizadas por rápi-dos eventos de especiação. Raros são os casos nos quaisambos os processos são constatados ao longo de uma mes-ma linhagem filética, como o citado acima (Malmgren &Kennet, 1981) e o apresentado por Geary (1990) queestudou cuidadosamente gastrópodes do Mioceno daEurásia. Essa autora verificou inicialmente um eventode estase na espécie Melanopsis impressa por um inter-valo de 7 Ma. Então, nos 2 Ma subsequentes, esta espé-cie começou a apresentar mudanças graduais em algu-mas de suas características, o que resultou, ao final des-te período, em uma nova espécie denominada deMelanopsis fossilis. Este representa um claro exemplodo surgimento de uma nova espécie através de um pro-cesso lento e gradual, com preservação das formas in-termediárias, como preconizado pelo modelo do gra-dualismo filético. Entretanto, Geary (1990) assinalou,também para este mesmo período, o surgimento de umanova espécie, Melanopsis vindobonensis, que aparentemen-te evolui a partir de Melanopsis fossilis de maneira bastan-te rápida, surgindo de modo instantâneo no registro (me-nos de 500 mil anos), exceto por uma única localidadena qual foram encontradas formas intermediárias. Estecaso, por outro lado, encaixa-se perfeitamente no pa-drão de especiação descrito pelo modelo do equilíbrio
153Capítulo 9 – Teorias Evolutivas
3ª Prova
pontuado, onde há uma fase de rápidas mudanças queocorre, de forma restrita, em uma população periféricaisolada, seguida de uma fase de expansão geográficaapós o surgimento da nova espécie (figura 9.7).
Tais exemplos indicam claramente que o regis-tro fóssil demonstra a existência de uma gama de pa-drões evolutivos que varia dos eventos pontuais geolo-gicamente instantâneos às mudanças graduais assinala-das em diferentes velocidades ou taxas evolutivas.Deste modo, ao que parece, o surgimento de uma novaespécie pode se dar não apenas por processos cladoge-néticos (= especiação biológica), bem como por proces-sos anagenéticos (= cronoespécies). Além disso, o fenô-meno de estase realmente ocorre, de maneira mais oumenos efetiva, ao longo das linhagens evolutivas. Somit& Peterson (1989) e Kemp (1999) oferecem bons exem-plos de gradualismo e pontuísmo em linhagens fósseis,além de debates sobre a análise do registro fóssil.
Biogeografia
A biogeografia é o estudo da distribuição geo-gráfica dos organismos. Ela procura estabelecer o pa-
drão de distribuição geográfica das distintas espécies etaxa superiores sobre as diferentes regiões do globo eexplicar o porquê deste padrão. A biogeografia relacio-na-se intimamente com a ecologia e com a geologia,uma vez que as respostas para alguns dos problemasbiogeográficos são mais ecológicas e para outros, maishistóricas (ou geológicas).
A biogeografia foi decisiva na formulação da te-oria da evolução darwinista. Charles Darwin utilizou abiogeografia como evidência da evolução e AlfredWallace despendeu grande parte de sua vida desenvol-vendo conceitos biogeográficos. O estudo da biogeo-grafia no Ocidente começou cerca de 200 anos após oinício das grandes navegações europeias. Com a con-quista de vários territórios, em diferentes continentes(América, África e Ásia), um grande acervo de animaise plantas “exóticos” foi coletado e tornou-se conheci-do dos naturalistas europeus. Além disso, estes passa-ram a ter a possibilidade de visitar as áreas de ocorrên-cia desses diferentes seres, através do estabelecimen-to das colônias dos países europeus nos territóriosconquistados. Tornou-se inevitável, então, questio-nar porque diferentes animais e plantas vivem em de-terminadas regiões e não em outras. Surgiram, a partirdisso, os termos endêmico e cosmopolita. Um orga-
Figura 9.8 Evolução de duas características morfológicas (número de câmaras e perímetro) do foraminífero Globorotaliaconoidea, feita com base na amostragem do Poço DSDP 284 referente ao Deep Sea Drilling Project (= Projeto Perfuraçãoem Águas Profundas). Durante o Mioceno, as características analisadas se encontram em um quase estado de “estase”(sem modificações direcionais), como previsto na Teoria do Equilíbrio Pontuado, enquanto que a partir do início do Pliocenoaté o final do Pleistoceno, elas passam a modificar-se direcionalmente ao longo do tempo (mudanças anagenéticas), comopreconizado pela Teoria do Gradualismo Filético. Pleist. = Pleistoceno (modificado de Malmgren & Kennett, 1981).

154 Paleontologia
3ª Prova
nismo cosmopolita é aquele dotado de uma grandedistribuição geográfica (em escala continental). Já umorganismo endêmico (indígena ou nativo) é aquele quepossui uma distribuição restrita a uma determinada re-gião geográfica (Mendes, 1988). O grau de endemismodepende do nível taxonômico abordado. Assim, umafamília pode ser cosmopolita (ocorrer em quase todosos continentes, p. ex.), enquanto um gênero ou umaespécie desta mesma família pode ser endêmico(a) deuma região (em um único continente).
A origem da biogeografia é atribuída a Buffonque, em 1761, declarou que o “Velho” e o “Novo” mun-dos não tinham espécies de mamíferos em comum. Estaobservação levou à formulação da “Lei de Buffon”, aqual postulava que diferentes regiões do globo, emboraàs vezes compartilhando as mesmas condições ambien-tais, eram habitadas por diferentes espécies de animaise plantas (Myers & Giller, 1988). Alfred Wallace e ou-tros biogeógrafos pioneiros perceberam que muitos taxaendêmicos apresentavam distribuições que se sobrepu-nham parcialmente dentro da mesma área geográfica –distribuições congruentes (Futuyma, 1993). A partir dis-so, estabeleceram os chamados reinos biogeográficos,os quais representam extensas áreas geográficas (emescala continental) caracterizadas por um grande con-junto de taxa endêmicos. Um exemplo clássico é a faunade marsupiais da Austrália, onde cangurus, coalas e ou-tros mamíferos são endêmicos daquele continente. Nointerior e nas bordas dos reinos, as barreiras à dispersãodefinem, muitas vezes, os limites entre associações deespécies. Qualquer obstáculo à dispersão, seja de natu-reza abiótica (parâmetros físico-químicos) ou biótica(parâmetros biológicos), denomina-se barreira bioge-ográfica. Há vários tipos de barreiras que influem nadistribuição dos organismos:
• no meio terrestre: mares, grandes rios, cadei-as de montanhas, desertos, tipos de clima,descontinuidades na vegetação;
• no meio aquático: variações de salinidade,temperatura, profundidade, tipos de substratoe luminosidade.
A parte da biogeografia que estuda a distribuiçãogeográfica dos organismos do passado recebe o nome depaleobiogeografia. Através da aplicação de seus con-ceitos, é possível detectar a influência das “paleobarrei-ras” na distribuição dos organismos pretéritos (Mendes,1988). Daí a importância de se entender, em paleontolo-gia, os princípios básicos da biogeografia.
O estudo das distribuições geográficas dos orga-nismos pode ser dividido em biogeografia histórica e
biogeografia ecológica. O botânico Augustin Pyramede Candolle (1778-1841), ao definir estes termos, reco-nheceu que as explicações “ecológicas” dependiam de“causas físicas operando no tempo presente”, enquanto queas “históricas” estavam relacionadas a “causas que nãomais existem hoje” (Nelson & Platnick, 1984). Os bioge-ógrafos históricos tenderiam, portanto, a ver as distri-buições como consequência de acontecimentos passa-dos, como, por exemplo, a deriva dos continentes, aopasso que os biogeógrafos ecológicos tenderiam a rela-cionar as distribuições com fatores contemporâneos,como as interações interespecíficas ou a distribuição doshábitats (Futuyma, 1993). Com base nisso, Wiley (1981)define:
• biogeografia histórica, como o estudo dasdistribuições espacial e temporal dos organis-mos (ao nível taxonômico), sendo as explica-ções para estas distribuições baseadas em even-tos históricos passados;
• biogeografia ecológica, como o estudo dadispersão dos organismos (ao nível do indiví-duo ou de populações locais) e dos mecanis-mos que a mantêm ou a modificam.
Esses dois enfoques sobrepõem-se e, na maioriados casos, tanto processos históricos como ecológicosdevem ser levados em conta na compreensão dos pa-drões de distribuição dos organismos abordados. Mui-tos padrões históricos têm, em última análise, bases eco-lógicas, e a dispersão tem um componente histórico,bem como um ecológico. Entretanto, a meta e o inte-resse do biogeógrafo ecológico relacionam-se mais coma ecologia do que com a sistemática, enquanto que ooposto é verdadeiro para o biogeógrafo histórico. O pri-meiro está interessado na dinâmica das biotas como umaunidade ecológica e o segundo está interessado na ori-gem e relação de parentesco entre as biotas (Wiley,1981).
Para os paleontólogos, a biogeografia histórica éo que normalmente interessa, uma vez que a biogeo-grafia ecológica, por definição, tem pouca aplicabilidadeem paleontologia, seja porque trata da distribuição dosorganismos atuais, seja porque trabalha apenas ao níveldas evidências ecológicas, difíceis de serem totalmen-te elucidadas em estudos paleontológicos. Além disso,o registro fóssil é um importante elemento dentro docampo da biogeografia histórica, auxiliando inclusivena compreensão da distribuição dos grupos atuais, comoserá visto adiante.

155Capítulo 9 – Teorias Evolutivas
3ª Prova
A. Biogeografia Histórica: Dispersãoversus Vicariância
Inicialmente, deve-se analisar quais são as cau-sas da distribuição dos organismos segundo a biogeo-grafia histórica. A atual distribuição de um táxon emuma determinada área hoje pode ser devido a duasrazões históricas:
• seus ancestrais originalmente já ocorriam lá e seusdescendentes sobreviveram até os dias presentes;
• seus ancestrais originalmente ocorriam em algum ou-tro lugar e depois eles (ou seus descendentes) disper-saram-se para esta nova área, onde seus descendentessobrevivem até hoje (Nelson & Platnick, 1984).
Estas duas razões caracterizam dois tipos de ex-plicação histórica: dispersão e vicariância. Disper-
são ocorre quando parte de uma população ancestral,de distribuição limitada por uma barreira biogeográfi-ca, consegue atravessá-la, colonizando uma nova áreaalém da barreira. Se os membros da população coloni-zadora permanecerem isolados dos membros da área deorigem, eles eventualmente evoluirão para um táxondistinto (figura 9.9A). Vicariância ocorre quando umapopulação ancestral é dividida em pelo menos duassubpopulações devido ao surgimento de uma barreirageográfica intransponível; com o tempo as subpopu-lações evoluem para diferentes taxa (figura 9.9B). Por-tanto, no fenômeno da vicariância o aparecimento dabarreira é visto como “a causa da disjunção” (separaçãoda população em subpopulações). Logo, a barreira tema idade da disjunção. No fenômeno de dispersão a bar-reira é anterior à disjunção e não pode ser apontadacomo a causa desta (Nelson & Platnick, 1984). Deve-
Figura 9.9 Processos de distribui-ção geográfica versus especiação. (A) Dis-persão: (1) população ancestral da espé-cie A distribuída em uma determinada árealimitada por uma barreira geográfica; (2)dispersão de parte da população atravésda barreira; (3) formação de duas popula-ções da espécie A; (4) subsequente dife-renciação das populações com osurgimento da espécie A1. (B) Vicariância:(1) população ancestral da espécie A dis-tribuída em uma determinada área; (2)aparecimento de uma barreira biogeográ-fica dentro da área de distribuição da es-pécie A; (3) formação de duas populaçõesda espécie A de cada lado da barreira; (4)subsequente diferenciação das popula-ções com o surgimento da espécie A1 (mo-dificado de Nelson & Platnick, 1984).

156 Paleontologia
3ª Prova
se ressaltar que a distribuição disjunta de um táxon oude um grupo taxonômico qualquer recebe o nome de“padrão vicariante”. A constatação de um padrãovicariante de distribuição não explica em si o fenôme-no (dispersão x vicariância) que o originou, pois o pa-drão vicariante (distribuição disjunta) pode surgir nãosó através das barreiras ocasionadas pelas mudanças nadistribuição das terras e continentes (ação da deriva con-tinental), mas também pela extinção de populações in-termediárias relacionada a alterações no ambiente(Futuyma, 1993).
Como exemplo de dispersão temos a atual distri-buição de certos mamíferos placentários (gatos, antas,veados) na América do Sul, uma vez que estes emigra-ram da América do Norte, no final do Plioceno, atravésdo Istmo do Panamá (Mendes, 1988). Em termos devicariância, a distribuição das aves ratitas (emas, avestru-zes, casuares, emus) na América do Sul, África eAustralásia, pode ser explicada pela fragmentação doantigo supercontinente do Gondwana durante oMesozoico. Este evento provocou o isolamento destegrupo de aves em subgrupos, os quais sobreviveram in-dependentemente em cada um dos atuais continentesresultantes da fragmentação do Gondwana (Futuyma,1993).
Por último, deve-se salientar que as explicaçõesrelativas à dispersão e à vicariância diferem entre si, tam-bém, quanto ao seu grau de generalização: a dispersãotende a explicar a distribuição de um grupo particular deorganismos e a vicariância, a distribuição de muitos gru-pos ao mesmo tempo. Para a dispersão é fator importan-te o “modo de dispersão” de um grupo particular, sen-do que este varia de grupo para grupo. Portanto, umaexplicação possível para um grupo de aranhas, por exem-plo, pode não ser aceitável para um grupo de aves. Asexplicações relativas à dispersão têm sua generaliza-ção limitada a grupos que tenham modos de dispersãosemelhantes. Por outro lado, para a vicariância o modode dispersão em si não é importante. Os fatores causaisda vicariância são mudanças geológicas e ecológicasque afetam simultaneamente muitos grupos de orga-nismos com diferentes modos de dispersão (Nelson &Platnick, 1984). Logo, as explicações vicariantes ten-dem a explicar padrões de distribuição que requeremuma explanação mais geral (Wiley, 1981).
B. Métodos em Biogeografia Histórica
Uma das melhores ferramentas para explicar a dis-tribuição de um grupo é um bom registro fóssil. O registro
fóssil pode ser empregado como um fator decisivo paradeterminar-se quando e onde um grupo surgiu e, conse-quentemente, se ele poderia ou não ter sido posterior-mente fragmentado pela deriva continental (Futuyma,1993).
A distribuição pretérita de um grupo muitas ve-zes esclarece se a sua distribuição atual deve-se à dis-persão ou à vicariância. Por exemplo, atualmente, asantas (Tapiridae) distribuem-se pela América Tropical eMalásia. O grupo surgiu no Terciário, quando os atuaiscontinentes já se achavam separados. Fósseis detapirídeos são encontrados na América do Norte, Ásia eEuropa desde o início do Terciário superior até o Pleis-toceno, mostrando que este grupo possuía uma ampladistribuição no passado. Na América do Sul, os tapirídessó chegaram no Pleistoceno, vindos da América do Norte,através do Istmo do Panamá. A atual distribuição disjuntadas antas deve-se à extinção de populações intermediári-as na Europa, Ásia e América do Norte e representa vestí-gio de um grupo que se dispersou através da Eurásia eAméricas. Já grupos mais antigos indicam, através do re-gistro fóssil, que a deriva dos continentes pode explicarmelhor suas distribuições atuais. Os sapos pipídeos daAmérica do Sul e África, por exemplo, são conhecidos emambos os continentes, desde o final do Mesozoico, quan-do estes ainda estavam parcialmente unidos. Portanto, aposterior separação da América do Sul e África explica aatual distribuição disjunta deste grupo.
Em termos de grupos extintos, a mesma meto-dologia pode ser aplicada. As glossopterídeas foramplantas muito comuns no antigo supercontinente doGondwana no final do Paleozoico. A atual distribuiçãodisjunta de seu registro fóssil na Austrália, África, Amé-rica do Sul, Índia e Antártica deve-se à fragmentaçãoposterior do Gondwana, pois essas plantas exclusiva-mente terrestres jamais poderiam ter cruzado os exten-sos mares oceânicos que hoje separam aqueles conti-nentes. Portanto, a distribuição pretérita das glossopterí-deas por todo o Gondwana deve-se, possivelmente, afenômenos de dispersão a partir de um centro de origemsituado em alguma região do antigo supercontinente.Já a distribuição dos marsupiais fósseis (grupo dos cangu-rus, coalas, gambás) pode ser explicada pela deriva con-tinental. Esses animais originaram-se no Gondwana, ten-do ficado isolados na Austrália, América do Sul e Antár-tica desde o final do Mesozoico, devido inicialmente àfragmentação do Pangea, que isolou o Gondwana dosdemais continentes e, posteriormente, devido à com-pleta fragmentação do Gondwana. Assim sendo, os mar-supiais formaram três grandes faunas distintas sobre cadaum dos três continentes austrais mencionados. A fauna

157Capítulo 9 – Teorias Evolutivas
3ª Prova
antártica extinguiu-se durante o Terciário devido aoresfriamento e congelamento deste continente. Asfaunas da América do Sul e Austrália evoluíram inde-pendentemente, dando origem a duas faunas endêmicasdistintas. Portanto, a distribuição disjunta dos marsupi-ais fósseis (e dos atuais também!) pode ser explicadapor isolamento devido à deriva continental, seguida daextinção de populações intermediárias.
Na ausência de um registro fóssil adequado, ahistória da distribuição de um grupo pode ser inferida,muitas vezes, a partir da análise filogenética. Nestecaso, a biogeografia histórica depende inteiramente deuma taxonomia filogeneticamente correta. Não se podetentar explicar a distribuição de um táxon sem que seusmembros possuam realmente um ancestral comum, ouem outras palavras, constituam um grupo monofilético.Muitas vezes é possível inferir que um grupo se origi-nou numa área e se dispersou para outra, se a filogeniado grupo for determinada pela cladística ou por méto-dos semelhantes (Futuyma, 1993).
Filogenia, monofilia e polifilia
Filogenia é a genealogia de um grupo detaxa, tais como espécies, gêneros, etc. As análisesfilogenéticas, através da sistemática filogenéti-ca, procuram estabelecer a relação de parentesco
entre os taxa de um grupo particular. Este tipo deanálise resulta na construção de árvores filogené-ticas (p. ex.: os cladogramas) que procuram repre-sentar as diversas linhas de descendência dos taxaanalisados.
Grupos de taxa podem ser mono ou poli-filéticos. Os grupos monofiléticos são aqueles emque os taxa derivam de um único ancestral comum.Grupos polifiléticos são aqueles em que os taxa ana-lisados derivam de dois ou mais ancestrais.
O estudo da biogeografia associado à sistemáti-ca filogenética é denominado de biogeografia filoge-nética. Este método baseia-se em conceber uma hipó-tese de filogenia detalhada para um grupo de organis-mos e inferir a sua história biogeográfica a partir dafilogenia estabelecida. Os pontos básicos da biogeo-grafia filogenética são, segundo Wiley (1981):
• as espécies estreitamente relacionadas tendem a subs-tituir umas às outras no espaço geográfico, apresen-tando padrões disjuntos (= vicariantes) de distribui-ção; taxa superiores (gêneros, famílias, ordens) po-dem também ser vicariantes, mas frequentemente de-monstram graus de superposição geográfica;
• se diferentes grupos de organismos monofiléticosdemonstram o mesmo padrão geográfico, eles prova-velmente dividem a mesma história biogeográfica.
Figura 9.10 Cladogramas como indicadores de relações filogenéticas. No cladograma apresentado pode-se verifi-car que: as características unicamente divididas entre os taxa 3 e 4 definem este grupo e não são encontradas nos taxa 1 e2; as características unicamente divididas entre os taxa 2, 3 e 4 definem este grupo e não são encontradas no táxon 1. Se ascaracterísticas únicas de cada grupo indicam a existência de um ancestral comum, então os cladogramas representamrelações evolutivas de parentesco. No caso deste cladograma pode-se admitir que os taxa 3 e 4 têm um ancestral comum C, oqual não é ancestral dos taxa 1 ou 2; que os taxa 2, 3 e 4 têm um ancestral comum B, o qual não é ancestral do táxon 1; e quetodos os taxa possuem um único ancestral comum A. Com isso, o cladograma representa a filogenia (= história evolutiva)deste grupo de taxa e mostra a sequência de divergência (ou diferenciação) dos taxa a partir de seus ancestrais comuns. Ogrupo em questão pode ser considerado monofilético, pois todos os seus taxa (1, 2, 3 e 4) possuem um ancestral comum A
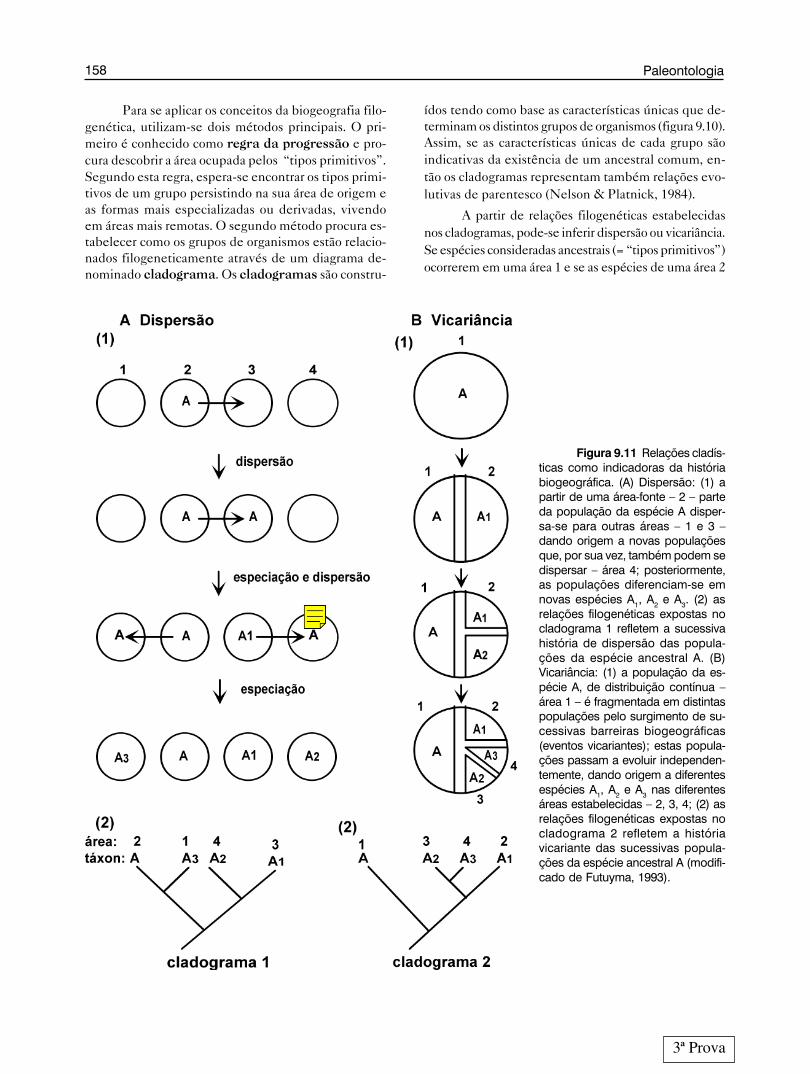
158 Paleontologia
3ª Prova
Para se aplicar os conceitos da biogeografia filo-genética, utilizam-se dois métodos principais. O pri-meiro é conhecido como regra da progressão e pro-cura descobrir a área ocupada pelos “tipos primitivos”.Segundo esta regra, espera-se encontrar os tipos primi-tivos de um grupo persistindo na sua área de origem eas formas mais especializadas ou derivadas, vivendoem áreas mais remotas. O segundo método procura es-tabelecer como os grupos de organismos estão relacio-nados filogeneticamente através de um diagrama de-nominado cladograma. Os cladogramas são constru-
ídos tendo como base as características únicas que de-terminam os distintos grupos de organismos (figura 9.10).Assim, se as características únicas de cada grupo sãoindicativas da existência de um ancestral comum, en-tão os cladogramas representam também relações evo-lutivas de parentesco (Nelson & Platnick, 1984).
A partir de relações filogenéticas estabelecidasnos cladogramas, pode-se inferir dispersão ou vicariância.Se espécies consideradas ancestrais (= “tipos primitivos”)ocorrerem em uma área 1 e se as espécies de uma área 2
Figura 9.11 Relações cladís-ticas como indicadoras da históriabiogeográfica. (A) Dispersão: (1) apartir de uma área-fonte – 2 – parteda população da espécie A disper-sa-se para outras áreas – 1 e 3 –dando origem a novas populaçõesque, por sua vez, também podem sedispersar – área 4; posteriormente,as populações diferenciam-se emnovas espécies A1, A2 e A3. (2) asrelações filogenéticas expostas nocladograma 1 refletem a sucessivahistória de dispersão das popula-ções da espécie ancestral A. (B)Vicariância: (1) a população da es-pécie A, de distribuição contínua –área 1 – é fragmentada em distintaspopulações pelo surgimento de su-cessivas barreiras biogeográficas(eventos vicariantes); estas popula-ções passam a evoluir independen-temente, dando origem a diferentesespécies A1, A2 e A3 nas diferentesáreas estabelecidas – 2, 3, 4; (2) asrelações filogenéticas expostas nocladograma 2 refletem a históriavicariante das sucessivas popula-ções da espécie ancestral A (modifi-cado de Futuyma, 1993).

159Capítulo 9 – Teorias Evolutivas
3ª Prova
forem “grupo irmão” (ou seja, intimamente relaciona-das) das espécies da área 1, provavelmente a área 1 é aárea de origem do grupo todo. Neste caso, temos umahistória de dispersão (figura 9.11A). Por outro lado, seuma ampla região habitada por um táxon sofrer uma su-cessiva fragmentação em áreas menores e isoladas, asrelações entre os taxa derivados do táxon ancestral de-vem refletir a história da fragmentação. Esta é uma histó-ria de vicariância (figura 9.11B).
Pode-se ainda, em vista a compreender umprocesso vicariante, elaborar o chamado cladograma ge-ográfico. Este é um cladograma que especifica apenasa relação entre as várias áreas em estudo, sem referên-cia alguma aos organismos que as habitam. Tais clado-gramas são obtidos a partir tanto de informações bioló-gicas (análise dos cladogramas dos organismos), quantode informações geológicas (análise dos eventos geoló-gicos). Eles são muito fáceis de ser construídos ao nívelcontinental, mas são muito difíceis de ser elaboradospara pequenas áreas geográficas, uma vez que são ne-cessários dados geológicos e geográficos bastante pre-cisos. A técnica consiste em comparar um cladogramageográfico das áreas estudadas com os vários cladogra-mas dos distintos organismos (cladogramas biológicos)que as habitam. Se houver uma correspondência entreo cladograma geográfico e os cladogramas biológicos,então pode-se inferir que as causas associadas com ocladograma geográfico possam, também, ter causado assequências filogenéticas de diferenciação dos taxa (ouespeciação, no caso de espécies) observadas nos clado-gramas biológicos (Wiley, 1981).
Referências
ARAÚJO, A. M. 1984. Microevolução e Macroevolução:a perspectiva da genética. In: CONGRESSO BRA-SILEIRO DE GEOLOGIA, 33. Rio de Janeiro, SBG,Anais, v. 1, p. 451-460.
BARBERENA, M. C. 1984. Microevolução e Macro-evolução: a perspectiva da paleontologia. In: CON-GRESSO BRASILEIRO DE GEOLOGIA, 33. Riode Janeiro, SBG. Anais, v. 1, p. 465-476.
CHEETHAM, A. H. 1987. Time of evolution in aNeogene bryozoan: are trends in single morpholo-gical characters misleading? Paleobiology, 13: 286-296.
COOPE, G. R. 1979. Late Cenozoic fossil Coleoptera:evolution, biogeography, and ecology. Annual Reviewof Ecology and Systematics, 10: 247-267.
DARWIN, C. 1859. The Origin of Species. John Murray,
London, 490 p.
ELDREDGE, N. & GOULD, S. J. 1972. Punctuatedequilibria: an alternative to phyletic gradualism, In:SCHOPF, T.J.M. (ed.). Models in paleobiology.Freeman, Cooper e Co, San Francisco, p. 82-115.
FREIRE-MAIA, N. 1988. Teoria da Evolução: DeDarwin à Teoria Sintética. São Paulo, Ed. Itatiaia,Ed. Univ. de São Paulo, 415 p.
FUTUYMA, D.J. 1993. Biologia Evolutiva. SociedadeBrasileira de Genética/CNPq, Ribeirão Preto, 631 p.
GEARY, D. H. 1990. Patterns of evolutionary tempoand mode in the radiation of Melanopsis (Gastropoda;Melanopsidae). Paleobiology, 16: 492-511.
GOLDSCHMIDT, R. 1940. The material basis of evolu-tion. Yale University Press, New Haven, Con-necticut.
GOULD, S. J. 1982. The meaning of ponctuatedequilibrium and its role in validating a hierarchicalapproach to macroevolution. In: MILKMAN, R.(ed.). Perspectives on Evolution, Sinauer Associates,Inc. publishers Sunderland, Massachusets, p. 5-104.
GOULD, S. J. 1991. Seta do Tempo Ciclo do Tempo, mito emetáfora na descoberta do tempo geológico. Companhiadas Letras, 221 p.
GOULD, S. J. & ELDREDGE, N. 1977. PunctuatedEquilibria: The Tempo and Mode of EvolutionReconsidered. Paleobiology, 3: 115-151.
GRANT, V. 1991. The Evolutionary Process, a CriticalStudy of Evolutionary Theory. Columbia UniversityPress, New York, 487 p.
HUTTON, J. 1788. Theory of Earth. Transactions of theRoyal Society of Edimburg, 1: 209-305.
KEMP, T. S. 1999. Fossils & Evolution. Oxford UniversityPress, Oxford, 284 p.
LAMARCK, J. B. 1809. Philosophie zoologique. Paris,Denter.
LYELL, C. (1830-1833). Principles of geology, being anattempt to explain the former changes of the earth’s surfaceby reference to causes now in operation, 3 v. London,John Murray.
MALMGREN, B. A. & KENNET, J. P. 1981. Phyleticgradualism in a Late Cenozoic planktonic foramini-feral lineage; DSDP site 284, southwest Pacific.Paleobiology, 7: 230-240.
MALTHUS, T. R. 1798. An essay on the principles ofpopulation. London, J. Johnson.

160 Paleontologia
3ª Prova
MAYR, E. 1977, Populações, espécies e evolução. Editorada Universidade de São Paulo, Série 3ª (CiênciaPuras), v. 5, 485 p.
MENDES, J. C. 1988. Paleontologia Básica. T. A. Queiroz(ed.). Editora da Universidade de São Paulo, 347 p.
MYERS, A. A. & GILLER, P. S. 1988. Analyticalbiogeography and integrated approach to the study ofanimal and plant distributions. London, Chapman &Hall, 578 p.
NELSON, G. & PLATNICK, N. 1984. Introduction.In: HEAD, J. J. (ed.). Biogeography. CarolinaBiological Supply Company, p. 3-16.
SALZANO, F. M. 1993. Biologia, Cultura e Evolução.Porto Alegre, Editora da Universidade/UFRGS, Sín-tese Universitária/8, Porto Alegre, 111 p.
SIMPSON, G. G. 1944. Tempo and Mode in Evolution.Columbia University Press, New York, 237 p.
SOMIT, A. & PETERSON, S. A. 1989. The dynamics ofevolution: the punctuated equilibrium debate in the na-tural and social sciences. Cornell University Press,Ithaca, New York.
TEMPLETON, A. R.1981. Mechanisms of speciation– a population genetic approach. Annual Review ofEcology and Systematics, 12: 23-48.
WILEY, E. O.1981. Phylogenetics, the Theory and Practiceof Phylogenetic Systematics. John Wyley & Sons, NewYork, 439 p.