tagusensis wells 1982 e a carcinofauna … · 2 ficha catalográfica comissão julgadora: profa....
TRANSCRIPT

0
Natália Matos de Menezes
RELAÇÃO ESPÉCIE-ÁREA ENTRE O
CORAL BIOINVASOR TUBASTRAEA
TAGUSENSIS WELLS 1982 E A
CARCINOFAUNA ASSOCIADA
Salvador
2012

1
Natália Matos de Menezes
RELAÇÃO ESPÉCIE-ÁREA ENTRE O
CORAL BIOINVASOR TUBASTRAEA
TAGUSENSIS WELLS 1982 E A
CARCINOFAUNA ASSOCIADA
Salvador
2012
Dissertação apresentada ao Instituto de
Biologia da Universidade Federal da Bahia,
para a obtenção de Título de Mestre em
Ecologia e Biomonitoramento.
Orientador: Dr. Rodrigo Johnsson
Co-orientadora: Dra. Elizabeth Neves

2
Ficha Catalográfica
Comissão Julgadora:
Profa. Dra. Vera Solferini Prof. Dr. Ruy K. P. Kikuchi
Prof. Dr. Rodrigo Johnsson
Menezes, Natalia Matos de
Relação espécie-área entre o coral bioinvasor
Tubastraea tagusensis Wells 1982 e a carcinofauna associada,
62 páginas.
Orientador: Rodrigo Johnsson
Co-orientadora: Elizabeth Neves
Dissertação (Mestrado) - Instituto de Biologia da
Universidade Federal da Bahia, 2012.
Palavras-Chaves: 1.Tubastraea. 2.Crustacea. 3. Facilitação. 4.
Relação espécie-área. Universidade Federal da Bahia.
Instituto de Biologia.

3
Dedico este trabalho aos meus pais
e meu irmão com muito carinho

4
“Se queremos progredir, não devemos repetir a história,
mas fazer uma nova história”
(Mahatma Gandhi)

5
AGRADECIMENTOS
A Deus.
Aos meus pais Iraldo Barbosa de Menezes e Leta Lucia Brandão Matos
de Menezes que são para mim exemplos de caráter, de força e
determinação. Agradeço pelo amor e carinho dedicado e por sempre
acreditarem em mim. Ao meu irmão Gustavo Matos de Menezes pelos
incentivos, momentos de alegria e crescimento pessoal que me
proporcionou no período do mestrado.
Aos meus orientadores Rodrigo Johnsson e Elizabeth Neves que me
deram a oportunidade de desenvolver o projeto de Mestrado. A eles
dedico toda a minha gratidão pelo trabalho de me iniciar na vida
acadêmica, pelos conhecimentos científicos, pela educação na vida
profissional, e pelos momentos de descontração.
A equipe do LABIMAR (Laboratório de Crustacea, cnidaria e Fauna
associada) pelo auxílio em diversas partes do trabalho, pelas opiniões
informais, pelas ricas e provocantes discussões relacionadas ao tema
“Zoologia vrs. Ecologia”, pelo companheirismo, por tornarem o meu
período de mestrado muito mais divertido do que eu esperava.
Superaram minhas expectativas!
Um agradecimento especial para Roberta Canário, Cristiano Bahia,
Barbara Conceição e Marcos Nogueira pela paciência na ajuda com a
identificação dos grupos de Crustacea.
Ao Prof. Dr. Cláudio Sampaio (UFAL) pelas informações sobre
localização da T. tagusensis, e pelas fotografias cedidas.
A Marcos De Paula “Marquinhos Conspirador” que contribuiu para a
realização das atividades de campo.
A Gabriel Barros pelo auxílio na coleta.

6
A Igor Cruz que me incentivou na realização deste trabalho, me dando
suporte principalmente no trabalho de campo e na padronização das
fotografias.
A Gilane Couto e Diogo Brito, por me proporcionarem momentos de
troca e aprofundamento de conhecimentos sobre o tema Bioinvasão,
principalmente durante o curso INTRODUÇÃO A BIOINVASÃO que
ministramos juntos.
A Pós-graduação em Ecologia e Biomonitorament da UFBA. A
secretária Jussara pela competência e pela imensa paciência em resolver
os problemas infindáveis dos alunos. Aos colegas de turma e aos
professores Pedro Rocha, Francisco Barros, Eduardo Mariano, Charbel
Niño El-Hani, Iara Sordi, Mauro Ramalho, Blandina Viana e Eduardo
Mendes pela dedicação e pela contribuição na minha formação de
Ecóloga.
Aos funcionários do Instituto de Biologia que auxiliaram no dia-a-dia
do meu trabalho, seja na limpeza do ambiente, seja na segurança, seja
pela simpatia de um simples cumprimento de “bom dia” rotineiro.
Aos meus amigos por serem essas pessoas maravilhosas que deixam a
minha vida mais FELIZ! Agradeço a Marina Bonfim, André Mendonça,
Wilton Fahning, Rafael Abreu, Rosara Lopes, Aiucha Almeida, Luciana
Rebouças pelos momentos de “reciclagem de idéias”, tão importantes
para o bom andamento da dissertação.
A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
(CAPES) pela concessão da bolsa de mestrado e a Pós-graduação em
Ecologia e Biomonitoramento pelo auxílio financeiro para a realização
das atividades de campo.

7
ÍNDICE
Introdução Geral 01
Capa de rosto do artigo 26
Abstract/Resumo 27
1.1. Introdução 28
1.2. Material e Métodos 33
1.3. Resultados 40
1.4. Discussão 52
1.5. Agradecimentos 62
1.6. Referências 62
Conclusões Gerais 72
Apêndice 1, 2
Anexo

8
ÍNDICE DE FIGURAS
Figura 1. Mapa indicando a área de estudo e a localização geográfica do Naufrágio
Cavo Artemidi, Salvador, Bahia......................................................................................34
Figura 2. Local de coleta e procedimentos de campo. A. Croqui do Naufrágio Cavo
Artemidi retirado do site http://www.naufragiosdobrasil.com.br B. Disposição das
colônias no local de coleta. C. Procedimento de redirada das colônias. D.
Acondicionamento das colônias e respectiva fauna associada em saco plástico. E.
Bombona onde as colônias foram depositadas e levadas para laboratório......................37
Figura 3. Tratamento das colônias de Tubastraea tagusensis. A. Branqueamento das
colônias com hipoclorito de sódio 2%. B. Secagem das colônias. C. Mesa de fotografia.
D. Fotografia da colônia com escala e identificação. E. Medição da área das colônias
com o programa Nis-Elements D....................................................................................38
Figura 4. Procedimento de análise dos crustáceos associados. A. Filtragem da fauna
associada desprendida das colônias. B. Fixação do filtrado em potes de vidro com álcool
70%. C. Triagem dos indivíduos com auxílio de lupa, placa de petri e pinça. D.
Separação dos crustáceos em grandes grupos. E. Separação em morfotipos de cada
grande grupo por colônia. F. Armazenamento das amostras em potes maiores com
álcool 70%.......................................................................................................................39
Figura 5. Número de grupos de Crustacea e de cada grande grupo de Crustacea em
relação ao volume de cada colônia de T.
tagusensis...............................................................46
Figura 6. Média móvel do número de espécies de Crustacea e de cada grande grupo de
Crustacea em relação a média móvel do volume de cada colônia de T.
tagusensis..............47
Figura 7. Regressão linear simples entre o volume das colônias de T. tagusensis e os
táxons de Crustacea.........................................................................................................48
Figura 8. Curvas de rarefação (acúmulo de espécies) com base nos valores de Sobs
(Mao Tau) gerados através de 999 aleatorizações para as colônias de T. tagusensis com
diferentes tamanhos. Em vermelho e laranja os dados da soma das colônias. Em azul, os
dados de uma única colônia.............................................................................................49
Figura 9. Abundâncias de cada táxon ao longo do aumento da área das colônias. Área
das colônias (m) pelo número de indivíduos...................................................................50

9
ÍNDICE DE TABELAS
Tabela 1. Identificação dos taxa encontrados, frequência de ocorrênica no total de
colônias, número total de indivíduos e abundância relativa de cada grupo. A
classificação seguiu Martin e Davis (2001).....................................................................42
Tabela 2. Modelos e resultados da regressão linear simples entre a área das colônias de
T. tagusensis e os taxa de Crustacea................................................................................44

0

1
INTRODUÇÃO GERAL
Bioinvasão é fundamentalmente um processo natural que compreende a chegada,
estabelecimento e expansão de uma espécie fora da sua distribuição geográfica original,
de forma a causar impactos ambientais (Carlton 1996). Entretanto, a frequência de
ocorrência deste processo atingiu taxas sem precedentes causadas, principalmente, pela
ação das atividades humanas que levam a quebra de barreiras geográficas de disperssão
(Lodge 1993; Elton 1980). Em ambientes marinhos, as principais vias de introdução são
a navegação, através do transporte não intencional de espécies exóticas por água de
lastro e/ou bioincrustação, e das atividades de maricultura de organismos exóticos e
suas espécies associadas (Carlton 1987). Dentre os impactos, destacam-se problemas
relacionados a pragas (patógenos e parasitas) em atividades de aquicultura, prejuízos em
sistemas de produção de energia devido a organismos incrustantes e, principalmente, a
perda da biodiversidade através da competição ou predação a organismos nativos,
formação de híbridos ou mesmo pela modificação de estrutura física dos ambientes
levando, consequentemente, a alteração das comunidades e ecossistemas (Crooks 2002;
Whitfield et al. 2002; Chandrasekaran et al. 2008; Lesser e Slattery 2011). De fato,
invasões biológicas têm sido consideradas o segundo maior motivo de perda de
biodiversidade no mundo, perdendo apenas para processos que envolvem perda de
habitat (Vitousek et al. 1996; Rosenzweig 2001)
Estudos recentes têm destacado as bioinvasões causadas por espécies capazes de
modificar o ambiente, alterando as comunidades diretamente ou facilitando invasão de
outras espécies (Crooks 2002; Simberloff 2006; Wallentinus e Nyberg 2007). Dentre
estas, plantas e invertebrados marinhos sésseis são os mais citados por proporcionarem
uma grande heterogeneidade espacial que aumenta o nicho fundamental de diversas
espécies através de interações, tais como fornecimento de abrigo contra
predação/estresse ambiental e disponibilidade para recrutamento (Bertness e Leonard
1997; Hacker e Gaines 1997; Bruno et al. 2003; Sousa et al. 2009; Heiman e Micheli
2010). Conforme já exposto, quando introduzidos em sua forma juvenil e/ou adulta,
estes organismos podem trazer outras espécies exóticas associadas (processo
comumente chamado de “invasão cruzada”), modificar o ambiente de forma a favorecer
ou não grupos de espécies nativas, ou permitir que espécies exóticas detectadas que

2
antes não conseguiam se estabelecer, passem a aumentar a sua população (Simberloff
2006; Sousa et al. 2009).
Dentre os diversos táxons considerados modificadores de ambiente, os corais
escleractíneos possuem uma baixa representatividade como bioinvasores. Estes são
organismos que adquiriram evolutivamente a habilidade de secretar um esqueleto
calcário que auxilia na sua proteção (Stanley 2002), mas que, simultaneamente, constrói
estruturas espaciais complexas capazes de abrigar uma fauna associada altamente
diversa em relações simbióticas, facultativas e/ou obrigatórias (Vitopil e Willis 2001).
Por outro lado, os corais são extremamente sensíveis à variação ambiental e,
consequentemente, tendem a não resistir ao transporte por água de lastro ou
bioincrustação (Veron 1995). Na literatura, os registros citam apenas cinco espécies de
corais exóticos: Oculina patagonica (D‟Angelis D‟Ossat 1908), nativa do Oceâno
Atlântico Sul, descoberta no Mediterrâneo em 1960 (Sartoretto et al. 2008); Fungia
scutaria Lamarck, 1801 , originário do Indo-Pacífico e estabelecida em recifes da costa
da Jamaica (Bush et al. 2004); e as congeneres do Indo-Pacífico Tubastraea tagunsensis
Wells, 1982, Tubastraea coccinea Lesson, 1829 e Tubastraea micranthus (Ehrenberg
1834) que estão ocorrendo no Atlântico ocidental (De Paula e Creed 2004; Sammarco et
al. 2010). Outros grupos de cnidários também tem sido encontrados, como a Carijoa
(Telesto) riisei (Duchassaing & Michelotti 1860) do Atlântico considerada invasora no
Havaí (Grigg 2003) e a Stereonephthya aff. curvata (Kükental 1911), de origem
desconhecida, que vem se estabelecendo no Brasil (Ferreira 2003). Entretanto, são
espécies ainda pouco estudadas.
Embora seja incomum a ocorrência de corais bioinvasores, um grupo tem se
destacado pela grande capacidade de adaptação ao ambiente. As espécies do gênero
Tubastraea Lesson 1829, conhecidas como „coral sol‟ devido aos tentáculos geralmente
em tons de amarelo intenso, têm ampliado rapidamente sua distribuição ao longo do
Oceano Atlântico, acreditando-se que a via de introdução tenha sido por bioincrustação
em cascos de navios ou plataformas de petróleo, onde são freqüentemente encontradas
(Sammarco et al. 2004). Ao longo do Atlântico Norte, Tubastraea coccinea foi
inicialmente registrada em Porto Rico (1943), alastrando-se por diversas regiões no
Golfo do México, Florida, Caribe, porém, principalmente em ambientes artificiais

3
(Cairns 2000; Fenner 2001; Fenner e Banks 2004). Tubastraea micranthus foi também
observada em substratos artificiais na região do Golfo do México, entretanto, ainda em
baixas densidades (Sammarco et al. 2010). No Brasil, o genêro foi citado pela primeira
vez em Castro e Pires (2001). Posteriormente, De Paula e Creed (2004) procederam a
identificação de T. coccinea e T. tagusensis para comunidades coralíneas de costões
rochosos na região da Baía de Ilha Grande, no Rio de Janeiro, onde atualmente são bem
distribuidas (De Paula e Creed 2004, 2005). Recentemente, a ocorrência de T. coccinea
e T. tagusensis foi ampliada, sendo registradas nas Ilhas de Búzios e Ilha de Vitória, no
estado de São Paulo (Mantelatto et al. 2011), e observada em 2008 sobre substratos
artificiais e recifais no Estado da Bahia, setor nordeste do Brasil (Sampaio C. – pers.
commun.; e obs. pess. in situ).
Pertencente a família Dendrophyilliidae (Gray, 1847), Tubastraea compreende
formas coloniais azooxanteladas com notável facilidade de ocupação do ambiente
(Glynn et al. 2008; Paz-García 2007). O monitoramento realizado na região da Baía de
Ilha Grande tem revelado que Tubastraea adaptou-se facilmente às condições locais,
aumentando sua abundância e distribuição, e causando modificações na estrutura das
comunidades de algumas localidades (Lages et al. 2011). De fato, diversos estudos
evidenciam o alto potencial reprodutivo e a grande capacidade competitiva destas
espécies.
Aspectos da biologia reprodutiva do gênero baseiam-se principalmente nos
estudos histológicos de Glynn et al. (2008) para colônias de T. coccinea do Pacífico
Oriental. De acordo com o autor, trata-se de uma espécie hermafrodita e incubadora
com rápido amadurecimento sexual (jovens colônias com 2-10 pólipos foram
observadas com gônadas férteis). Plânulas possuem entre 0,5-1,5 mm de comprimento –
ou 3,0-5,0 mm e fixam-se em fios de muco que se desprendem da abertura oral do
pólipo até o substrato estabelecendo-se próximo às colônias parentais em padrão
tipocamente filopátrico (Paz-Garcia et al. 2007).
Além do grande potencial reprodutivo, outros traços caracterizam o gênero como
potencial invasor, como uma ampla distribuição batimétrica, chegando a ocupar
profundidades entre 0 a 108 m (Glynn et al. 2008); preferências pouco seletivas por
substrato – o assentamento pode ocorrer sobre madeira, granito, concreto, aço e

4
cerâmica; a presença de defesas químicas contra predadores generalistas (como peixes
pertencentes as famílias Pomacentridae, Ostracidae e Haemulidae) e contra algumas
algas competidoras como Cladophora sp., and Lithophyllum sp.. Adicionalmente, foram
também encontradas nas adjacências de outros cnidários conhecidos como fortes
competidores, incluindo zoantídeos (Zoanthus sp., Palythoa caribaeorum (Duchassaing
e Michelotti 1860)), corais (Phyllangia americana Edwards & Haime, 1849 , Astrangia
rathbuni Vaughan, 1906) e octocorais (Carijoa (Telesto) riisei (Duchassaing &
Michelotti 1860)) (Paula e Creed 2005). De fato, Creed (2006) verificou que a
proximidade entre Tubastraea e o coral endêmico Mussismilia hispida (Verrill
1901) poderia levar à redução, ou mesmo, exclusão da espécie nativa. Até o momento,
apenas a esponja Desmapsamma anchorata (Carter 1882) foi relatada recobrindo
Tubastraea, tratando-se de um competidor superior (Meurer 2010).
Embora diversos estudos tenham comprovado o potencial invasor do gênero ao
longo do litoral brasileiro, aspectos como a possibilidade das colônias terem trazido
espécies associadas e a capacidade de abrigar organismos nativos de hábito críptico,
endolítico ou até parasita, papel de extrema importância exercido por corais
escleractínios, são ainda pouco explorados. Diversos estudos têm relatado associações
de corais escleractíneos com inúmeros poliquetos, moluscos e crustáceos (Young 1986;
Preston e Doherty 1990/1994; Sin 1999; Nogueira et al. 2003; Oigman-Pszczol e Creed
2004; Johnsson et al. 2006). Para Tubastraea em especial, podem ser citados a
associação com copépodes sifonostomatóideos em exemplares das ilhas Molucas
(Indonésia) e Taiwan (China) (Humes 1997; Cheng et al. 2011), o registro de
Latopilumnus tubicolus, um caranguejo que habita pequenos orifícios („pits‟),
construídos no esqueledo de T. michranthus nas Filipinas e Japão (Türkay e
Schuhmacher 1985), e a ocorrência de pequenos peixes recifais dormindo entre os
pólipos de T. coccinea no Caribe (Wirtz 2006). Assim, existe uma lacuna no
conhecimento no que se refere ao potencial deste gênero como facilitador (vetor) da
introdução de outras espécies exóticas, ou como facilitador (nicho espacial) para
espécies nativas.
Dentre os organismos que podem se beneficiar dessa introdução, destacam-se os
crustáceos crípticos que dependem de substratos fornecidos por organismos bentônicos

5
para proteção, captação de alimento e além disso, representam uma biomassa
substancial nestes ambientes (Gerlach 1978). Crustáceos ocupam uma importante
posição na base da cadeia trófica uma vez que reciclam detritos, exercem importante
papel como herbívoros e transferem energia para níveis tróficos mais altos (Preston e
Doherty 1994). Na literatura, vários autores relatam a interação entre corais e
crustáceos, incluindo casos de parasitismo, comensalismo e associações facultativas
(Goh et al. 1989). Copépodes endógenos ou epibiontes foram encontrados habitando
espécies de coral como Tubastraea (Cheng et al. 2011), Psammocora (Humes 1997),
Parahalomitra (Humes 1997), Galaxea (Humes 1996). Caranguejos Brachyura e
Porcellanidae também são comuns em corais podendo apresentar associações
facultativas ou obrigatórias como as que acontecentem com os gêneros Trapezia,
Tetralia e com a família Cryptochiridae (Abele 1976; Gotelli et al. 1985; Goh et al.
1989; Sin 1999; Vitopill e Wills 2001; Carricart-Ganivet et al. 2004; Johnsson et al.
2006; Gheerardyn et al. 2008); por fim, cirripédios também têm sido encontradas como
epibiontes de corais (Nogueira 2003; Tsang et al. 2009).
Relações como as mencionadas acima, comumente chamadas interações
“hóspede-hospedeiro”, tendem a assumir padrões espaciais diretamente correlacionados
à diversidade de espécies (Kuris et al. 1980). Dentre eles, destaca-se a relação espécie-
área, que prevê que o número de espécies é uma função simples ou direta da extensão
geográfica ocupada pela biota, sendo usualmente expressa pelas seguintes funções:
onde S = número de espécies, A = área do habitat, k e z = constantes (intercepto e
coeficiente angular, respectivamente) (Scheiner 2003; Dengler 2009). Trata-se de um
dos modelos mais estudados na ecologia desde o início da sua formulação no início do
séc. XX (Gleason 1922; Arrhenius 1923; Preston 1962), principalmente pela sua
aplicabilidade em trabalhos de distribuição da diversidade e conservação dos
organismos em diferentes escalas espaciais e temporais (Simberloff e Abele 1976;
Fahrig 1998; Lomolino e Weiser 2001; Scheiner 2003; Turner e Tjorve 2005).
Para compreender biologicamente esta relação, três modelos foram
desenvolvidos, podendo ter explicabilidade distinta a depender da escala espacial

6
(Turner e Tjorve 2005), a saber: 1) Hipótese de área „per se’, derivada da Teoria do
Equilíbrio da Biogeografia de Ilhas que foi originalmente descrita para descrever
padrões de riqueza de espécie em ilhas oceânicas (MacArthur e Wilson 1963/67). Esta
postula que o número de espécies em uma determinada ilha é uma função do tamanho
da área, da sua distancia em relação a fonte de propágulos e do balanço entre as taxas de
imigração e extinção de espécies. Nesse modelo, a taxa de imigração depende da
distância da área em questão em relação à área fonte, porém é independente do tamanho
da ilha. Já a taxa de extinção é inversamente proporcional ao tamanho da área. Esta
hipótese é aplicada para regiões isoladas e tem sido utilizada em escalas intermediárias
de paisagens e regionais nas quais dominam processos evolutivos (Turner e Tjorve
2005). 2) Hipótese da heterogeneidade do habitat proposta por Williams (1964)
considera que áreas maiores possuem maior riqueza por possuir maior diversidade de
habitats que possibilitam a abertura de mais nichos vagos. A partição de nicho entre
espécies em paisagens heterogêneas diminui a competição interespecífica e mantém a
coexistência e a riqueza de espécies, sendo relevante em diferentes escalas (Turner e
Tjorve 2005). 3) Hipótese de amostragem passiva (Connor e McCoy 1979) propõe que
áreas maiores possuem uma riqueza maior de espécies simplesmente por possuir uma
abundância maior de indivíduos, sendo considerada como uma hipótese nula e adequada
a escalas locais (Turner e Tjorve 2005).
Entretanto, de acordo com Kuris (1980), a aplicabilidade dessas hipóteses para a
interação entre organismos, ou seja, para a relação hóspede-hospedeiro, deve considerar
algumas modificações em função do ciclo de vida do hospedeiro. Dentre elas, destaca-
se o fato das (1) distâncias entre as fontes flutuarem ao longo do tempo devido a
mobilidade dos hospedeiros - rapidamente para organismos vágeis e lentamente para
organismos sedentários ou sésseis; (2) diferenças sazonais no comportamento ou nas
condições fisiológicas podem afetar a presença dos hóspedes; (3) a qualidade do
hospedeiro varia através de modificações ecológicas no tempo como resultado do
crescimento e envelhecimento; por fim, (4) como a vida dos hospedeiros é efêmera,
assembléias de espécies parasitas raramente alcançam um equilíbrio mínimo ao longo
da vida do hospedeiro.
Adicionalmente, interações interespecíficas também têm sido utilizadas para
explicar padrões de relação espécie-área. De acordo com Simberloff (1978), a

7
distribuição e abundância dos organismos em diferentes áreas pode estar diretamente
relacionada a fenômenos como competição e predação, em que cada espécie interage
simultaneamente com diversas outras em intensidade e modo distinto. O autor (op. cit.)
comenta, ainda, que os resultados dessas interações são mais evidentes em estágios
avançados de sucessão, ou seja, após um longo período de colonização das espécies alí
presentes, e que os efeitos podem ser mais ou menos proeminentes a depender do grupo
observado. Alguns trabalhos mostraram que a presença de determinadas espécies exclui
a presença de outras ao longo do tempo (Connell 1961; Paine 1974, 1980). Por outro
lado, trabalhos abordando o mesmo tema, porém restritos a um determinado táxon,
mostraram ausência da interferência de interações, havendo influência apenas da área no
aumento do tamanho das populações (Gotelli e Abele 1983).
Diversos autores tentaram aplicar as teorias acimas mencionadas em situações
envolvendo hospedeiros como “ilhas”. Por exemplo, pode-se citar estudos com plantas
hospedeiras de insetos herbívoros (Lawton 1986; Grez 1992; Athen e Tscharntke,
1999), peixes em “patches” de recifes de coral (Chittaro 2002; Belmaker et al. 2007;
Hattori e Shibuno 2010), diversidade de briófitas e micro invertebrados associados a
rochedos de rios (Jacobsen 2005; Heino e Kai Korsu 2008) e, por fim, decápodes
associados a corais escleractíneos (Barry 1865; Abele 1976; Abele e Patton 1976;
Gotelli e Abele 1983; Vytopill e Wills 2001). Entretanto, os trabalhos de Grez (1992),
Belmaker et al. (2007) e Abele e Patton (1976) chamam a atenção para o fato de que,
apesar de ser frequentemente ignorada, a amostragem passiva é a mais propensa a
influenciar a relação espécie-área em escalas menores, devendo ser primariamente
considerada neste caso.
Embora existam estudos realizados com a fauna associada a organismos
invasores, estes são ainda restritos (Torchin et al. 2003). Além disso, os padrões de
distribuição de diversidade como os acima mencionados (relação espécie-área) têm sido
negligenciados (Castilla et al. 2004; Sousa et al. 2009; Heiman and Micheli 2010).
Desta forma, considerando a ampla capacidade de expansão de Tubastraea, a escassez
de estudos que consideram corais bioinvasores como prováveis vetores de espécies
exóticas e hospedeiros para espécies nativas e a complexidade intrínseca a esse tipo de
associação, relacionada principalmete a padrões espaciais de diversidade e à interação
entre os organismos hóspedes, o objetivo deste trabalho é avaliar a relação espécie –

8
área entre o coral bioinvasor Tubastraea tagusensis e a carcinofauna associada em
colônias provenientes do Naufrágio Cavo Artemidi, localizado na entrada da Baía
de Todos-os-Santos – BA, primeiro provável sítio de ocorrência da espécie no
Estado da Bahia. Trata-se do primeiro ensaio utilizando um coral escleractíneo e uma
espécie com potencial bioinvasor elevado na projeção de uma análise de relação
espécie-área. Estudos pretéritos são escassos e restritos a riqueza de Decapoda
associados a corais do Indo-Pacífico (Abele e Patton 1976; Gotelli e Abele 1983). Os
objetivos específicos do trabalho sequem discriminados: (1) Registrar a carcinofauna
que está se associando a T. tagusensis para utilização da mesma como nicho, seja ela
exótica ou nativa, discutindo suas implicações. (2) Descrever a relação espécie-área
entre T. tagusensis e a carcinofauna associada. (3) Testar primariamente se os padrões
de relação espécie-área observados estão de acordo com a hipótese de amostragem
passiva, determinada por Connor e McCoy (1979). (4) Testar se os padrões de relação
espécie-área observados podem ser explicados com base na relação entre táxons de
Crustacea encontrados.
REFERÊNCIAS
Arrhenius O (1923) Statistical investigations in the constitution of plant
associations. Ecology 4:68-73.
Athen O, Tscharntke T (1999) Insect communities of phragmites habitats used for
sewage purification: effects of age and area of habitats on species richness and
herbivore-parasitoid interactions. Limnologica 29:71-74.
Abele LG (1976) Comparative species richness in fluctuating and constant
environments: coral-associated decapod crustacea. Science 192:461-463.
Abele LG, Patton WK (1976) The size of coral heads and the community biology of
associated decapod crustaceans. Journ Biogeogr 3:35-47.
Barry CK (1965) Ecological study of the decapods crustaceans commensal with the
branching coral Pocillopora meandrina var. nobilis Verrill. Dissertation, University
of Hawaii.
Belmaker J, Ben-Moshe N, Ziv Y, Shashar N (2007) Determinants of the steep
species–area relationship of coral reef fishes. Coral Reefs 26:103-112.
Bertness MD, Leonard GH (1997) The role of positive interactions in communities:
lessons from intertidal habitats. Ecology 78(7):1976-1989.

9
Bruno JF, Stachowicz JJ, Bertness MD (2003) Inclusion of facilitation into
ecological theory. Trends Ecol Evol 18(3):119-125.
Bush SL, Precht WF, Woodley JD, Bruno JF (2004) Indo-Pacific mushroom corals
found on Jamaican reefs. Coral Reefs 23:234.
Cairns SD (2000) Revision of the shallow- water azooxanthellate Scleractinia of the
Western Atlantic. Stud Nat Hist Carib 75:1–240.
Carlton JT (1987) Patterns of transoceanic marine biological invasions in the Pacific
Ocean. Bull Mar Sci 41(2):452-465.
Carlton JT (1996) Biological invasions and cryptogenic species ecology 77(6):
1653-1655.
Carricart-Ganivet JP, Carrera -Parra LF, Quan-Young LI, García-Madrigal MS
(2004) Ecological note on Troglocarcinus corallicola (Brachyura: Cryptochiridae)
living in symbiosis with Manicina areolata (Cnidaria: Scleractinia) in the Mexican
Caribbean. Coral Reefs 23:215–217.
Castilla JC, Lagos NA, Cerda M (2004) Marine ecosystem engineering by the alien
ascidian Pyura praeputialis on a mid-intertidal rocky shore. Mar Ecol Prog Ser 268:
119–130.
Castro CB, Pires DO (2001) Brazilian Coral Reefs: what we already know and what
is still missing. Bull Mar Sci 69(2):357-371.
Chandrasekaran S, Nagendran NA, Pandiaraja D, Krishnankutty N, Kamalakannan
B (2008) Bioinvasion of Kappaphycus alvarezii on corals in the Gulf of Mannar,
India. Science 94(9):1167-1172.
Cheng Y, Dai C, Chang W (2011) A New Siphonostomatoid copepod associated
with the ahermatypic coral Tubastraea aurea from Taiwan. Zool Stud 50(5):605-
610.
Chittaro PM (2002) Species-area relationships for coral reef fish assemblages of St.
Croix, US Virgin Islands. Mar Ecol Progr Ser 233:253–261.
Connor EF, McCoy ED (1979) The statistics and biology of the species-area
relationship. Am Nat 113:791-833.
Connell JH (1961) The influence of interspecific competition and other factors on
the distribution of the barnacle Chthamalus Stellatus. Ecology 42(4):710-723.
Creed JC (2006) Two invasive alien azooxanthellate corals, Tubastraea coccinea
and Tubastraea tagusensis, dominate the native zooxanthellate Mussismilia hispida
in Brazil. Nota in Coral Reefs 25:350.

10
Crooks JA (2002) Characterizing ecosystem-level consequences of biological
invasions: the role of ecosystem engineers. Oikos 97:153–166.
Dengler J (2009) Which function describes the species–area relationship best? A
review and empirical evaluation. J Biogeogr 36:728–744.
Elton CS (1980) The ecology of invasions: by animals and plants. Chapman and
Hall, London
Fahrig L (1998) When does fragmentation of breeding habitat affect population
survival? Ecol Mod 105:273–292.
Fenner D (2001) Biogeography of three caribbean corals (Scleractinia) and the
invasion of Tubastraea coccinea into the Gulf of Mexico. Bull Mar Sci 69(3):1175-
1189.
Fenner D, Banks K (2004) Orange cup coral Tubastraea coccinea invades Florida
and the Flower Garden Banks, Northwestern Gulf of Mexico. Coral Reefs, 23:505-
507.
Ferreira CEL (2003) Non-indigenous corals at marginal sites. Coral Reefs 22:498.
Gerlach SA (1978) Food-chain relationships in subtidal silty sand marine sediments
and the role of meiofauna in stimulating bacterial productivity. Oecologia 33: 55–
69.
Gheerardyn H, Troch M, Ndaro SGM, Raes M, Vincx M, Vanreusel A. (2008)
Community structure and microhabitat preferences of harpacticoid copepods in a
tropical reef lagoon (Zanzibar Island, Tanzania). Jour Mar Biol Assoc UK 88(4):
747–758.
Gleason HA (1922) On the relation between species and area. Ecology 3(2): 158-
162.
Glynn PW, Colley SB, Maté JL, Cortés J, Guzman HM, Bailey RL, Feingold JS,
Enochs IC (2008) Reproductive ecology of the azooxanthellate coral Tubastraea
coccinea in the Equatorial Eastern Pacific: Part V. Dendrophylliidae. Mar Biol 153:
529–544.
Goh BPL, Chou LM, Ng PKL (1989) Anomuran and Brachyuran crab symbionts of
Singapure hard coral of the families Acroporidae, Agaricidae and Pocilloporidae.
Indo-Malayan Zool 6:25-44.
Gotelli NJ, Gilchrist SL, Abele LG (1985) Population biology of Trapezia spp. and
other coral-associated decapods. Mar Ecol Prog Ser 21:89-98.
Gotelli NJ, Abele LG (1983) Community patterns of coral-associated decapods. Mar
Ecol 13:131-139.

11
Grez AA (1992) Riqueza de species de insectos herbívoros y tamanho de parche de
vegetacion huesped: uma contrastacion experimental. Rev Chil Hist Nat 60:115-
120.
Grigg RW (2003) Invasion of a deep water coral bed by an alien species, Carijoa
riisei. Coral Reefs 22: 121−122.
Hacker SD, Gaines SD (1997) Some implications of the direct positive interactions
for community species diversity. Ecology 78(7):1997-2003.
Hattori A, Shibuno T (2010) The effect of patch reef size on fish species richness in
a shallow coral reef shore zone where territorial herbivores are abundant. Ecol Res
25:457–468.
Heiman KW, Micheli F (2010) Non-native ecosystem engineer alters estuarine
communities. Integr Comp Biol 1–11.
Heino J, Korsu K (2008) Testing species–stone area and species–bryophyte cover
relationships in riverine macroinvertebrates at small scales. Freshw Biol 53:558–
568.
Humes AG (1996) New genera of Copepoda (Poecilostomatoida) from the
scleractinian coral Psammocora in New Caledonia. Zool Journ Linn Soc 118:50-82.
Humes AG (1997) Two siphonostomatoid copepods (Coralliomyzontidae)
associated with the ahermatypic coral Tubastraea in the Moluccas. Hydrobiologia
344: 195–203.
Humes AG (1997) Two new copepod genera (Poecilostomatoida) associated with
the scleractinian coral Psammocora in New Caledonia. Zool Scrip 26:51-60.
Jacobsen D (2005) Temporally variable macroinvertebrate–stone relationships in
streams. Hydrobiologia 544:201–214.
Johnsson R, Neves EG, Franco MO, Lang FS (2006) The association of two gall
crabs (Brachyura: Cryptochiridae) with the reef-building coral Siderastrea stellata
Verrill, 1868. Hydrobiologia 559:379-384.
Kuris AM, Blaustein AR, Alió JJ (1980) Host as islands. Amer Nat. 116(4):570-
586.
Lages BG, Fleury BG, Ferreira CEL, Pereira RC (2006) Chemical defense of an
exotic coral as invasion strategy. J Exp Mar Biol Ecol 328: 127−135.
Lages BG, Fleury BG, Menegola C, Creed JC (2011) Change in tropical rocky shore
communities due to an alien coral invasion. Mar Ecol Prog Ser 438:85–96.
Lawton JH (1983) Plant architecture and the diversity of phytophagous insects. Rev
Entol 28:23-39.

12
Lesser MP, Slattery M (2011) Phase shift to algal dominated communities at
mesophotic depths associated with lionfish (Pterois volitans) invasion on a
Bahamian coral reef. Biol Invasions 13:1855–1868.
Lodge DM (1993) Biological invasions: lessons for ecology. Trends Ecol Evol 8:
133-137.
Lomolino MV, Weiser MD (2001) Towards a more general species-area
relationship: diversity on all islands, great and small. Journ Biogeograp 28: 431-
445.
MacArthur R, Wilson EO (1967) The theory of island biogeography. Princeton
University Press, Princeton
Martin JW e Davis GE (2001) An Updated Classification of the Recent Crustacea.
Nat Hist Mus Los Angeles Sci Ser 39:1-124.
Mantelatto MC, Creed JC, Mourão GG, Migotto AE, Lindner A (2011) Range
expansion of the invasive corals Tubastraea coccinea and Tubastraea tagusensis in
the Southwest Atlantic. Nota in Coral Reefs 1:1
Meurer BC, Lages NS, Pereira O, Palhano S, Magalhães GM (2010) First record of
native species of sponge overgrowing invasive corals Tubastraea coccinea and
Tubastraea tagusensis in Brazil. Mar Biod Rec 3:1-3.
Nogueira JMM (2003) Fauna living in colonies of Mussismilia hispida (Verrill)
(Cnidaria: Scleractinia) in four South Eastern Brasil Islands. Braz Arch Biol Tech
46(3):421-432.
Oigman–Pszczol SS, Creed JC (2004). Distribution and abundance of fauna on
living tissues of two Brazilian hermatipic corals Mussismilia hispida (Verrill 1902)
and Siderastrea stellata Verrill, (1868). Hidrobiologia 563: 143-154.
Paine T (1974) Intertidal community structure experimental studies on the
relationship between a dominant competitor and its principal predator. Oecologia
15:93-120.
Paine T (1980) Food Webs: Linkage, Interaction strength and community
infrastructure. Journ Anim Ecol 49(3):666-685.
Paula AF, Creed JC (2004) Two species of the coral Tubastraea (Cnidaria,
Scleractinia) in Brazil: a case of accidental introduction. Bull Mar Sci 74(1): 175–
183.
Paula AF, Creed JC (2005) Spatial distribution and abundance of nonindigenous
coral genus Tubastraea (Cnidaria, Scleractinia) around Ilha Grande, Brazil. Braz J
Biol 65(4): 661-673.
Paz-García DA (2007). Larval release from Tubastraea coccinea in the Gulf of
California, Mexico. Nota in Coral Reefs, 26:433.

13
Preston FW (1962) The canonical distribution of commonness and rarity: Part I
Ecology 43(2):185-215.
Preston EM (1971) Niche overlap and competition among five sympatric congeneric
species of Xanthid crabs. Dissertation University of Hawaii.
Preston NP, Doherty PJ (1990) Cross-shelf patterns in the community structure of
coral-dwelling Crustacea in the central region of the Great Barrier Reef. I. Agile
shrimps. Mar Ecol Prog Ser 66:47-61.
Preston NP, Doherty PJ (1994) Cross-shelf patterns in the community structure of
coral-dwelling Crustacea in the central region of the Great Barrier Reef. 11.
Cryptofauna. Mar Ecol Prog Ser 104:27-38.
Rosenzweig ML (2001) The four questions: What does the introduction of exotic
species do to diversity? Evolut Ecol Res 3:361–367.
Sammarco PW, Atchison AD, Boland GS (2004) Expansion of coral communities
within the Northern Gulf of Mexico via offshore oil and gas platforms. Mar Ecol
Progr Ser 280:129–143.
Sammarco PW, Porter SA, Cairns SD (2010) A new coral species introduced into
the Atlantic Ocean - Tubastraea micranthus (Ehrenberg 1834) (Cnidaria, Anthozoa,
Scleractinia): An invasive threat? Aquat Inv 5(2):131-140.
Sartoretto S, Harmelin J, Bachet F, Bejaoui N, Lebrun O, and Zibrowius H. (2008)
The alien coral Oculina patagonica De Angelis, 1908 (Cnidaria, Scleractinia) in
Algeria and Tunisia. Aquat Inv 3(2):173-180.
Scheiner SM (2003) Six types of species-area curves. Glob Ecol Biogeograp 12:
441-447.
Simberloff DS, Abele LG (1976) Island biogeography theory and conservation
practice. Science 191(4224):285-286.
Simberloff D (1978) using island biogeographic distributions to determine if
colonization is stochastic. Amer Nat 112(986):713-726.
Simberloff D (2006) Invasional meltdown 6 years later: important phenomenon,
unfortunate metaphor, or both? Ecology 9:912–919.
Sin T (1999) Distribution and host specialization in Tetralia crabs (Crustacea:
Brachyura) symbiotic with corals in the Great Barrier Reef, Australia. Coral Reef
65(3):839–850.
Sousa R, Gutiérrez JL, Aldridge DC (2009) Non-indigenous invasive bivalves as
ecosystem engineers. Biol Inv 11:2367-2385.

14
Stanley GD (2002). The evolution of modern corals and their early history. Earth
Sci Rev 60:195-225.
Torchin ME, Lafferty KD, Dobson AP, McKenzie VJ, Kuris AM (2003) Introduced
species and their missing parasites. Nature 421:628-630.
Tsang LM, Chan BKK, Shih F, Chu KH, Chen CA (2009) Host-associated
speciation in the coral barnacle Wanella milleporae (Cirripedia: Pyrgomatidae)
inhabiting the Millepora coral. Mol Ecol 18:1463–1475.
Türkay M, Schuhmacher HT (1985) Latopilumnus tubicolus n. gen. n. sp., eineneue
korallenassoziierte Krabbe, die die Bildungeiner Wohnhöhle induziert. (Crustacea:
Decapoda: Pilumnidae). Senckenbergiana mark 17:55-63.
Turner WR, Tjørve E (2005) Scale-dependence in species-area relationships.
Ecography, 28: 721-730.
Veron JEN (1995) Corals in space and time: The biogeography and evolution of the
Scleractinia. UNSW Press, Sydney
Vitousek PM, D' Antonio CM, Loope L, Westbrooks R (1996) Biological invasions
as global environmental changes. Amer Sci 84:468-478.
Vitopil E, Willis BL (2001) Epifaunal community in Acropora spp. (Scleractinia) on
the Great Barrier Reef: implications of coral morphology and habitat complexity.
Coral Reefs 20:281-288.
Wallentinus I, Nyberg CD (2007) Introduced marine organisms as habitat modifiers.
Mar Poll Bull 55:323–332.
Whitfield PE, Gardner T, Vives SP, Gilligan MR, Courtenay WR, Ray GC, Hare JA
(2002) Biological invasion of the Indo-Pacific lionfish Pterois volitans along the
Atlantic coast of North America. Mar Ecol Prog Ser 235:289–297.
Williams CB (1964) Patterns in the balance of nature. Academic Press, London.
Wirtz P (2006) Let sleeping fish lie. G Mar Env 3:27.
Young PS (1986) Análise qualitativa e quantitativa da fauna associada a corais
hermatípicos (Coelenterata: Scleractinia) nos recifes de João Pessoa, PB. Rev Braz
Zool 3:99-126.

15
Natalia Matos de Menezes, Elizabeth Neves, Rodrigo Johnsson
RELAÇÃO ESPÉCIE-ÁREA EM COLÔNIAS DO CORAL BIOINVASOR
TUBASTRAEA TAGUSENSIS WELLS 1982 E A CARCINOFAUNA ASSOCIADA
Endereço dos autores:
Universidade Federal da Bahia, Instituto de Biologia, Depto. de Zoologia. Av. Adhemar
de Barros s/n, Campus Ondina - CEP: 40170-290, Salvador, BA – Brasil.
E-mail: [email protected], [email protected],
Autor para correspondência: Natalia Matos de Menezes –
[email protected], phone: + 55 xx 0713283-6561

16
RESUMO
Estudos recentes têm voltado a atenção para bioinvasão causada por organismos
modificadores do ambiente. Dentre eles, plantas e invertebrados marinhos sésseis são os
mais citados por criarem estruturas espaciais complexas que aumentam o nicho
fundamental de diversas espécies através de “facilitação”. Corais escleractínios exóticos
não são comuns. Entretanto, as espécies Tubastraea tagusensis e T. coccinea têm se
destacado como bioinvasores de ambientes de águas rasas no Caribe e no Sudoeste
Atlântico. No Brasil, ambas foram registradas inicialmente no sudeste (23°S).
Recentemente, ambas as espécies foram observadas no nordeste, entre 12 e 15 m em um
naufrágio na Baía de Todos-os-Santos (13°S). Embora alguns estudos tenham
proporcionado informações sobre o seu potencial de reprodução e de competição,
poucos estão relacionados às “facilitação”. Neste trabalho, além de registrar a interação
de T. tagusensis com crustáceos no Naufrágio Cavo Artemidi, foram analisados padrões
de relação espécie-área entre T. tagusensis e os crustáceos associados. Trinta amostras
foram removidas e examinadas. O material foi fixado e os organismos associados foram
triados para identificação. As colônias foram branqueadas para a medição do volume.
Foram encontrados indivíduos pertencentes aos grupos Copepoda, Ostracoda,
Amphipoda, Isopoda, Tanaidacea e Decapoda. Relação significativamente positiva entre
o volume e o número de taxa de crustáceos foi encontrada apenas para Copepoda e
Ostracoda. Tal resultado não pode ser atribuído a amostragem passiva nem a relação
entre os crustáceos associados a T. tagusensis, sustentando a necessidade de mais
estudos para entender os processos biológicos que explicam os padrões encontrados.
PALAVRAS CHAVES: Tubastraea, Crustacea, “facilitação”, fauna associada, relação
espécie-área, Atlântico Sul Ocidental

17
ABSTRACT
Recent studies have highlighted bioinvasions caused by engineering organisms.
Among them, plants and sessile marine invertebrates are known for creating complex
spatial structures, increasing the fundamental niche of many species by “facilitative
interactions”. Records of exotic corals are not common. However, Tubastraea coccinea
and T. tagusensis have been pointed out as bioinvaders to shallow-water environments
along the Caribbean and Southwestern Atlântic. In Brazil, T. coccinea and T. tagusensis
were firstly recorded to Southwestern (23°S). Nowadays, both species have been
reported to Northwestern at 12-15m depth on an 80‟s shipwreck (13ºS). Data of
geographic expansion and potential of competition are partially available; however,
facilitative interactions have not been approached. In this work, besides recording
associated crustaceans to T. tagusensis from Cavo Artemidi shipwreck, we analyzed
typical diversity patterns related to specie-area relationship for these groups. A total of
thirty colonies was sampled and examined. The material was fixed, and associated
organisms were sorted out for identification. The colonies were bleached for volume
measurements. The inventory of crustacean fauna was diverse, revealing the occurrence
of Copepoda, Ostracoda, Amphipoda, Isopoda, Tanaidacea and Decapoda. Therefore,
the results support an interesting scenario: the invasive T. tagusensis has been used as a
new niche for native species. Positive specie-area relationship was observed just for
Copepoda and Ostracoda. This result cannot be attributed to passive sampling event or
to correlation among associated crustaceans, showing the need for more studies to
understand the biological process that explain the founded patterns.
KEY WORDS: Tubastraea, Crustacea, facilitation, associated fauna, specie-area
relationschip, Southwestern Atlântic

18
INTRODUÇÃO
Estudos recentes têm destacado as bioinvasões causadas por espécies capazes de
modificar o ambiente, alterando as comunidades diretamente ou facilitando invasão de
outras espécies (Crooks 2002; Simberloff 2006; Wallentinus e Nyberg 2007). Dentre
estas, plantas e invertebrados marinhos sésseis são os mais citados por proporcionarem
uma grande heterogeneidade espacial que aumenta o nicho fundamental de diversas
espécies através de facilitação, tais como fornecimento de abrigo contra
predação/estresse ambiental e disponibilidade para recrutamento (Bertness e Leonard
1997; Hacker e Gaines 1997; Bruno et al. 2003; Sousa et al. 2009; Heiman e Micheli
2010). Conforme já exposto, quando introduzidos, estes organismos podem trazer outras
espécies exóticas associadas (“invasão cruzada”), modificar o ambiente de forma a
favorecer ou não grupos de espécies nativas, ou permitir que espécies exóticas
“detectadas” que antes não conseguiam se estabelecer, passem a aumentar a sua
população (Simberloff 2006; Sousa et al. 2009).
Dentre os diversos grupos considerados modificadores de ambiente, corais
escleractíneos, conhecidos por construirem um esqueleto calcário que auxilia na sua
proteção (Stanley 2002), mas que ao mesmo tempo constrói estruturas espaciais
complexas capazes de abrigar uma fauna associada altamente diversa em relações
simbióticas, possuem uma baixa representatividade como bioinvasores. Na literatura, os
registros citam apenas cinco espécies de corais exóticos: Oculina patagonica, nativa do
Oceâno Atlântico Sul descoberta no Mediterrâneo em 1960 (Sartoretto et al. 2008);
Fungia scutaria, originário do Indo-Pacífico e é considerado estabelecido na costa da
Jamaica (Bush et al. 2004); e as congeneres originárias do Indo-Pacífico Tubastraea
tagunsensis Wells, 1982, Tubastraea coccinea Lesson, 1829 e Tubastraea micranthus
(Ehrenberg 1834), que estão ocorrendo no Atlântico Ocidental (Fenner 2000; De Paula
e Creed 2004, Sammarco et al. 2010; Fenner e Banks 2004). Devido a elevada
incidência em navios e plataforma de petróleo, acredita-se que estas foram introduzidas
através de bioincrustação (Sammarco et al. 2004).
Embora seja incomum a ocorrência de corais invasores, o gênero Tubastraea
Lesson 1829 tem se destacado pela grande capacidade de adaptação no ambiente, o alto
potencial reprodutivo e de competição com espécies nativas (Lages et al. 2006; Creed

19
2006; Glynn et al. 2008; Creed e Paula 2007; Paz-García 2007; Lages 2010). No Brasil,
o genêro foi citado pela primeira vez por Castro e Pires (2001). Posteriormente, De
Paula e Creed (2004) procederam a identificação de T. coccinea e T. tagusensis para
comunidades coralíneas de costões rochosos na região da Baía de Ilha Grande, no Rio
de Janeiro, onde atualmente são bem distribuídas (De Paula e Creed 2004, 2005). A
ocorrência de T. coccinea e T. tagusensis foi ampliada, sendo registradas em São Paulo,
Santa Catarina (Mantelatto et al. 2011), e mais recentemente, observada em substratos
artificiais e recifais no Estado da Bahia, setor nordeste do Brasil (Sampaio C. – pers.
commun.; e obs. pess. in situ).
Apesar de diversos estudos terem comprovado o potencial de invasão de
Tubastraea ao longo do litoral brasileiro, aspectos como a possibilidade das colônias
terem trazido espécies associadas e sua capacidade de prover habitat para diversos
organismos, papel de extrema importância exercido por corais escleractíneos, são ainda
pouco explorados (Meurer 2010). Diversos estudos têm relatado associações de corais
escleractíneos com inúmeros invertebrados marinhos (Young 1986; Preston e Doherty
1990/1994; Sin 1999; Nogueira et al. 2003; Oigman-Pszczol e Creed 2004; Johnsson et
al. 2006). Para Tubastraea em especial, podem ser citados a associação com copépodes
sifonostomatóides em exemplares provenientes das ilhas Molucas (Indonesia) e Taiwan
(China) (Humes 1997, Cheng et al. 2011), o registro de Latopilumnus tubicolus, um
caranguejo que habita pequenos orifícios („pits‟), construídos no esqueledo de T.
michranthus nas Filipinas e Japão (Türkay e Schuhmacher 1985), e a ocorrência de
pequenos peixes recifais dormindo entre os pólipos de T. coccinea no Caribe (Wirtz
2006). Assim, existe uma lacuna no conhecimento relativo ao potencial deste gênero
como facilitador (vetor) da introdução de outras espécies exóticas, ou como facilitador
(nicho espacial) para espécies nativas.
Dentre os organismos que podem se beneficiar dessa introdução, destacam-se os
crustáceos crípticos que dependem de substratos fornecidos por organismos bentônicos
para proteção e captação de alimento além de representarem uma biomassa substancial
nestes ambientes (Gerlach, 1978; Preston e Doherty 1994). Diversis artigos registram a
interação entre corais e crustaceos associados, tais como copépodes, caranguejos e
cirripédios (Abele 1976; Gotelli et al. 1985; Goh et al. 1989; Humes 1996; Humes
1996; Humes 1997 a, b; Sin 1999; Vitopil e Wills 2001; Nogueira 2003; Carricart-

20
Ganivet et al. 2004; Johnsson et al. 2006; Gheerardyn et al. 2008; Tsang et al. 2009;
Cheng et al. 2011; Badaró et al. no prelo).
Relações como as mencionadas acima, comumente chamadas interações
“hospede-hospedeiro”, tendem a assumir padrões espaciais diretamente atrelados a
diversidade das espécies (Kuris et al. 1980). Dentre eles, destaca-se a relação espécie-
área, que prevê que o número de espécies é uma função simples ou direta da extensão
geográfica ocupada pela biota, sendo mais usualmente expressada pelas seguintes
funções:
onde S = número de espécies, A = área do habitat, k e z = constantes (intercepto e
coeficiente angular, respectivamente) (Scheiner 2003; Dengler 2009). Trata-se de um
dos modelos mais estudados na ecologia desde o início da sua formulação no início do
séc. XX (Gleason 1922; Arrhenius 1923; Preston 1962), principalmente pela sua
aplicabilidade em trabalhos de distribuição da diversidade e conservação dos
organismos em diferentes escalas espaciais e temporais (Simberloff e Abele 1976;
Fahrig 1998; Lomolino e Weiser 2001; Scheiner 2003; Turner e Tjorve 2005).
Para compreender biologicamente esta relação, três modelos foram
desenvolvidos, podendo ter diferentes explicabilidades a depender da escala (Turner e
Tjorve 2005), a saber: 1) Hipótese de área „per se’, derivada da Teoria do Equilíbrio da
Biogeografia de Ilhas que foi originalmente descrita para descrever padrões de riqueza
de espécie em ilhas oceânicas (MacArthur e Wilson 1963/67). Este modelo, que tem
sido utilizado em grandes escalas (Turner e Tjorve 2005), postula que o número de
espécies em uma determinada ilha é uma função do tamanho da área, da sua distancia
em relação a fonte de propágulos e do balanço entre as taxas de imigração e extinção de
espécies. 2) Hipótese da heterogeneidade do habitat proposta por Williams (1964).
Neste caso, a hipótese considera que áreas maiores possuem maior riqueza por possuir
maior diversidade de habitats que possibilita a abertura de mais nichos disponíveis,
sendo relevante em diferentes escalas (Turner e Tjorve 2005). 3) Hipótese de
amostragem passiva (Connor e McCoy 1979). Esta propõe que áreas maiores possuem
uma riqueza maior de espécies espécies simplesmente por possuirem uma abundância

21
maior de indivíduos, sendo considerada como uma hipótese nula e adequada a escalas
locais (Turner e Tjorve 2005).
Atrelado ao efeito da área, interações interespecíficas têm sido mencionadas
como determinantes nos padrões de abundância e co-ocorrência entre espécies
associadas (Connel 1961; Preston 1971). De acordo com Simberloff (1978), a
distribuição dos organismos em diferentes áreas pode estar diretamente relacionada a
fenômenos como competição e predação, em que cada espécie interage
simultaneamente com diversas outras em intensidades e modos diferentes. O autor
comenta ainda que, os resultados dessas interações são mais evidenciados em estágios
avançados de sucessão, ou em áreas maiores. Alguns trabalhos mostraram que a
presença de determinadas espécies exclui a presença de outras ao longo do tempo,
influenciando na formação do padrão de relação espécie-área (Connell 1961; Paine
1974, 1980).
Embora existam estudos realizados com a fauna associada a organismos
invasores, estes são ainda escassos e restritos (Torchin et al. 2003). Além disso, os
padrões de distribuição de diversidade como os acima mencionados tem sido
negligenciados (Castilla et al. 2004; Sousa et al. 2009; Heiman and Micheli 2010).
Desta forma, consideranto a ampla capacidade de expansão do gênero Tubastraea, a
inexistência de estudos que investigam a atuação do gênero como provável hospedeiro
para espécies nativas e a complexidade intrínseca a esse tipo de associação, relacionada
principalmente a padrões espaciais de diversidade e à interação entre os organismos
hóspedes, o objetivo deste trabalho é avaliar a relação espécie – área entre o coral
bioinvasor Tubastraea tagusensis Wells 1982 e a carcinofauna associada em
colônias provenientes do Naufrágio Cavo Artemidi, localizado na entrada da Baía
de Todos os Santos, BA. Objetivos específicos seguem discriminados: (1) registrar a
carcinofauna que está se associando a T. tagusensis, seja ela exótica ou nativa,
discutindo suas implicações; (2) descrever a relação espécie-área entre a espécie T.
tagusensis e a carcinofauna associada; (3) testar primariamente se os padrões de relação
espécie-área observados estão de acordo com a hipótese de amostragem passiva,
determinada por Connor e McCoy (1979); (4) testar se os padrões de relação espécie-
área observados podem ser explicados com base na relação entre os táxons de Crustacea
encontrados.

22
MATERIAL E MÉTODOS
Área de estudo
A amostragem foi realizada no Naufrágio Cavo Artemidi (13°13‟.310‟‟S,
38°31‟.551‟‟O), um antigo cargueiro grego carregado com 16.800 toneladas de ferro
gusa provenientes de Minas Gerais que veio a pique em 1980 no Banco de Santo
Antônio, aproximadamente a 4 km de Salvador (Figura 1). Trata-se de uma região
caracterizada pela cobertura essencialmente arenosa e pela presença de “dunas
hidráulicas” devido as correntes nas marés mais intensas. Atualmente, o navio encontra-
se entre 9 a 29 m de profundidade onde desenvolveu-se ao longo de 30 anos um recife
artificial formado principalmente por colônias de Siderastrea, Missismilia hispida e pela
presença massiça de esponjas e cirripédios. Devido a grande dimensão (cerca de 180m
de comprimento) e beleza natural, o naufrágio passou a ser conhecido como “o Gigante
do Litoral Brasileiro”. É um ponto turístico amplamente visado pelas operadoras
brasileiras de mergulho. De acordo com a Lei Nº 10.166, de 27 de dezembro de 2000,
naufrágio é um ambiente que deve ser protegido devido à importância histórica e
cultural (patrimônio histórico) e devido à função que exerce como recife artificial capaz
de auxiliar na manutenção da diversidade da biota marinha.
Geograficamente, o Cavo Artemidi está localizado na entrada da Baía de Todos-
os-Santos (BTS), a segunda maior baía costeira do Brasil (1200 Km2), sendo
diretamente influenciado pelas suas características ambientais. A BTS possui uma alta
diversidade de ecossistemas marinhos, incluindo cerca de 91 ilhas internas, recifes de
corais, estuários, manguezais e recifes artificiais (mais de 50 naufrágios) (Cirano e
Lessa 2007; Cruz et al. 2009). É uma região altamente suscetível a invasões biológicas
por causa da presença de três importantes portos comerciais, intenso tráfego de navios e
embarcações particulares capazes de introduzir organismos exóticos através de
bioincrustação ou água de lastro, os principais mecanismos de introdução de espécies
marinhas. Além disso, a região tem sido impactada pela poluição industrial e doméstica,
sedimentação e superexploração dos recursos naturais, fatores que afetam as
comunidades alí existentes e consequentemente, podem diminuir sua resistência à
bioinvasão (Celino e Queiroz 2006; Amado-Filho et al. 2008; Felizzola et al. 2008).

23
Figura 1. Mapa indicando a área de estudo e a localização geográfica do Naufrágio
Cavo Artemidi, Salvador, Bahia.
Procedimentos de campo
As coletas foram realizadas nos dias 28/11/10 e 29/12/2010 e o acesso ao local
foi através de barco apropriado para mergulho. A amostragem procedeu durante 2
mergulhos autônomos de 30 min de duração, realizada por uma dupla de mergulho entre
uma profundidade de 12 e 15 m na proa do navio, onde as colônias estão localizadas.
Foram removidas 30 colônias com diferentes tamanhos de modo aleatório. Indivíduos
com sinais de doença ou morte parcial não foram coletados no intuito de evitar a
influência destes fatores nos organismos associados, fato previamente previsto por
Kuris (1980). Os individuos foram cuidadosamente retirados com auxílio de martelo e
ponteira e imediatamente inseridos em sacos plásticos para evitar a perda da
carcinofauna associada (Figura 2-C,D). No barco, o material foi devidamente
etiquetado, de forma que cada colônia recebeu um código de identificação, e por fim
depositado em uma bombona de 50 litros para transporte ao laboratório (Figura 2-E).
Procedimentos em laboratório
Em laboratório, o tratamento do material e coleta de dados envolveu dois
protocolos distintos de atuação: a triagem e identificação da carcinofauna associada e a
análise do volume de cada colônia coletada.
Triagem e identificação da carcinofauna associada

24
Inicialmente, a água de cada saco plástico, contendo colônias de Tubastraea e os
crustáceos associados desprendidos durante os procedimentos de campo, foi filtrada em
peneira com malha de 200 micrômetros e acondicionada em vidros contendo álcool
70%. Cada colônia foi re-examinada cuidadosamente para a retirada da fauna críptica e
outros organismos que por ventura tenham ficado aderidos. Cada pote de vidro de
alcool 70% contendo a carcinofauna associada foi etiquetada com o mesmo código da
colônia correspondente. A fauna associada a cada colônia foi posteriormente
identificada ao menor nível taxonômico possível, e foi realizada uma contagem do
número de indivíduos coletados de cada grupo. A observação dos indivíduos foi feita
com o auxílio de lupa (OLIMPUS SZ51) e microscópio (OLYMPUS CX31), placas de
petri, laminas, lamínulas e pinças (Figura 4).
Branqueamento e medição da área da colônia
Após a retirada da fauna associada, as colônias de Tubastraea foram tratadas
com hipoclorito de sódio à 2% para remoção do tecido e mantidas ao ar livre para a
secagem. Testemunhos foram preservados secos. Em seguida os espécimes foram
analisados com auxílio de bibliografia adequada para confirmação da identidade
taxonômica. Por fim, para estimar a área, cada colônia foi fotografada com máquina
digital (NIKON, modelo COOLPIX 995) em uma mesa fotográfica, com distância fixa.
As imagens foram analisadas no software NIS-ELEMENTS D (www.nis-
elements.com/d.html), onde a área em cm2 que cada colônia preenchia na fotografia foi
mensurada (Figura 3 ). A altura das colônias foram medidas com auxílio de um
paquímetro. Para estimar o volume, foi feita uma multiplicação entre a área e o volume.
Ao término das duas etapas anteriores, foram elaboradas tabelas no Microsoft
Excel 2010 incluindo área de cada colônia, os grupos contidos em cada uma delas e o
número de indivíduos de cada grupo. Por fim, o material examinado foi depositado nas
coleções de Crustacea e Cnidaria do Museu de Zoologia / UFBA.
Tratamento estatístico
Primariamente, foi realizada uma análise descritiva, onde foram compilados
dados de riqueza e abundância dos taxa encontrados, e a sua frequência de ocorrência
nas colônias (Tabela 1, Figura 9).
Para analisar a relação espécie-área, no intuito de observar a distribuição dos
dados, foram feitos gráficos de dispersão e gráficos de média móvel, onde a variável

25
independente (eixo x) era a área das colônias e as variáveis dependentes (eixo y) foram
número de taxa de Crustacea em geral, e dos grupos Copepoda, Ostracoda, Amphipoda,
Isopoda, Tanaidacea e Decapoda (Figuras 5 e 6). A média móvel representou a média de
cinco amostras, sendo uma central, duas anteriores e duas posteriores, e foi utilizada
para facilitar a observação dos padrões encontrados, uma vez que esta atenua a variação.
Posteriormente, os dados foram transformados para Log (x+1) para realização de uma
regressão linear simples (Log-log) (Figura 7). As premissas analisadas foram a
normalidade dos resíduos e a ausência de relação entre o eixo x e os resíduos. O nível de
significância utilizado foi 0,05.
No intuito de testar se a amostragem passiva foi o principal processo que
determinou as relações espécie-área encontradas, foram realizadas curvas de rarefação
(curva de acúmulo de espécies Sobs (Mao Tau) (Colwell et al. 2004)) para comparar se
o somatório do número de espécies em colônias pequenas era o mesmo que o
encontrado em apenas uma colônia grande (Figura 8). Para isso, foram feitas
combinações de forma que a soma da área das primeiras colônias dessem um valor
equivalente (aproximado) a uma de tamanho maior, até chegar ao valor máximo
encontrado. Tais combinações seguem discriminadas:
3 colônias = 6,4 cm²; 1 colônia = 7 cm²
4 colônias = 14 cm²; 1 colônia = 14 cm²
5 colônias = 22,3 cm²; 1 colônia = 23 cm²
6 colônias = 31 cm²; 1 colônia = 30 cm²
7 colônias = 41 cm²; 1 colônia = 42 cm²
As curvas de rarefação foram geradas através de 999 aleatorizações, cada uma
com uma disposição aleatória da sequência das espécies encontradas. A partir deste
número, foram calculados a média do número de espécies para cada número de
indivíduos amostrados e os seus respectivos intervalos de confiança 95%.
Por fim, foram aplicadas correlações de Spearman entre os taxóns de crustáceos
no sentido de entender a interferência da interação entre eles na relação espécie-área
(Figura 10). O nível de significância utilizado foi 0,05, porém corrigido pela correção de
Bonferroni, passou para 0,0009. Os programas utilizados foram o Statistica v.7, o
EstimateS v.8.2.0 e o Microsoft Excel 2010.

26
Foto:
Leia Figueiredo
Figura 2. Local de coleta e procedimentos de campo. A. Croqui do Naufrágio Cavo
Artemidi retirado do site http://www.naufragiosdobrasil.com.br B. Disposição das
colônias no local de coleta. C. Procedimento de redirada das colônias. D.
Acondicionamento das colônias e respectiva fauna associada em saco plástico. E.
Bombona onde as colônias foram depositadas e levadas para laboratório.
E D
C B
A

27
Figura 3. Tratamento das colônias de Tubastraea tagusensis. A. Branqueamento das
colônias com hipoclorito de sódio 2%. B. Secagem das colônias. C. Mesa de fotografia.
D. Fotografia da colônia com escala e identificação. E. Medição da área das colônias
com o programa Nis-Elements D.
A
E
D C
B B A
D

28
E
Figura 4. Procedimento de análise dos crustáceos associados. A. Filtragem da fauna
associada desprendida das colônias. B. Fixação do filtrado em potes de vidro com álcool
70%. C. Triagem dos indivíduos com auxílio de lupa, placa de petri e pinça. D.
Separação dos crustáceos em grandes grupos. E. Separação em morfotipos de cada
grande grupo por colônia. F. Armazenamento das amostras em potes maiores com
álcool 70%.
C
F
D
A B

29
RESULTADOS
O volume das 30 colônias de Tubastraea tagusensis fotografadas variou entre
0,53cm² a 246,4cm². Em relação aos crustáceos associados, foram encontrados seis
grandes grupos, sendo eles Copepoda, Amphipoda, Ostracoda, Decapoda, Isopoda e
Tanaidacea, que representaram respectivamente 27,27%, 27,27%, 18,18%, 12,73%,
10,91%, e 3,64% da riqueza encontrada. Em termos de abundância, Copepoda se
destacou como o mais representativo, com 68,79%, seguidos por Amphipoda 12,99%,
Ostracoda 10,0%, Isopoda 6,97%, Decapoda 0,77% e Tanaidacea 0,42%. Por fim,
Copepoda foi também o mais frequente ocorrendo em todas as colônias, seguido por
Ostracoda 86,7%, Isopoda 86,70%, Amphipoda 80%, Decapoda 73,30% e Tanaidacea
30,00% (Tabela 1).
Copepoda foi representado por duas ordens, sete famílias e quinze morfotipos
(Harpaticoida e Sophonostomatoida). Em Harpaticoida, estiveram presentes cinco
famílias e dez morfotipos, sendo elas cinco morfotipos de Thalestridae, dois de
Peltidiidae, um de cada uma das seguintes famílias: Harpaticidae, Laophontidae e
Tegastidae. Em Siphonostomatoida, foram observadas duas famílias e cinco morfotipos,
sendo quatro morfotipos de Asterocheres e um de Artotrogidae. O morfotipo
Thalestridae 1 foi o grupo mais abundante entre os Crustacea (40,56%) além de ocorrer
em 100% das colônias. Os táxons Talestridae 2, 3, 4, 5, Harpaticidae e Laophontidae
foram menos abundantes (entre 1,94 e 6,59%), embora a sua frequência de ocorrência
nas colônias tenha se mantido elevada (entre 43,33 e 76,67%). Os outros grupos,
Peltidiidae 1, 2, Tegastidae, Asterocheridae 1, 2, 3, 4 e Artotrogidae foram muito menos
abundantes (0,04 a 0,5 %) e muito menos frequentes (3,33 a 13,33%) (Tabela 1).
Amphipoda foi representado por duas ordens, seis famílias e também quize
morfotipos. Apenas um morfotipo foi observado na subordem Caprellidea (2,71% da
abundância e 46,67% de frequência nas colônias). A subordem Gammaridea foi mais
diversa, com cinco famílias e quatorze morfotipos. Eurisidae foi a mais comum, com
nove morfotipos, Isaeidae com dois morfotipos e Leuchotoidae, Talitridae,
Ischyroceridae com apenas um morfotipo cada. Entre os Gammaridea, os táxons mais
representativos foram Eurisidae 1 (2,44% da abundância e 43,33% de frequência nas
colônias) e Isaeidae 1 (3,68% da abundância e 40,00% de frequência nas colônias). Os

30
outros grupos variaram a abundância entre 0,08 e 0,97% e a frequência entre 3,33 e 30%
(Tabela 1).
Ostracoda apresentou duas subclasses e dez morfotipos. Destes, dois morfotipos
pertenceram a subclasse Myodocopa e oito a subclasse Podocopa. Os taxa mais
abundantes foram Podocopa 4 e 5 e representaram 4,15 e 2,64%. Os táxons Podocopa 1,
2, 3, 4 e 5 foram os mais frequentes, ocorrendo entre 43,33 e 46,67% das colônias. Os
outros grupos, Podocopa 6, 7, 8 e Miodocopa 1 e 2 foram menos abundantes (0,04 à
0,19%) e frequentes (3,33 à 13,33%) (Tabela 1).
Isopoda apresentou duas subordens, três famílias e seis morfotipos. A subordem
Asellota foi composta pela família Munnidae com dois morfotipos e pela família
Janiridae com três morfotipos do gênero Ianiropsis. Os táxons Ianiropsis 1 e 3 e
Munnidae 1 foram os mais abundantes (variando entre 1,43 e 3,10%) e frequentes
(variando entre 23,33 e 66,67%). Os outros grupos foram menos representativos
variando entre 0,04 e 0,54% de abundância e 3,33 16,37% de frequência, sendo que a
família Anthuridae apareceu somente uma vez (Tabela 1).
Por fim, os táxons Decapoda e Tanaidacea foram os grupos mais raros. Para o
primeiro foram registradas seis espécies, sendo elas Pilumnus quoyi, Pilumnus
spinosissimus, Micropanope nuttingi, Mithrax caribbaeus, Hexapanopeus angustifrons,
Pitho lherminieri pertencentes a família Xanthidae (infraordem Brachyura), e
Megalobrachium soriatum, pertencente a família Porcellanidae (infraordem Anomura),
que variaram a sua abundância entre 0,04 e 0,23% e sua frequência de ocorrência nas
colônias entre 3,33 e 13,33%. O segundo, Tanaidacea, apresentou apenas dois
morfotipos pertencentes as famílias Nototanaidae (0,39% da abundância e 26,67% de
frequencia nas colônias) e Colleteidae (0,04% da abundância e 3,33% de frequência nas
colônias), sendo o primeiro mais frequente (Tabela 1).

31
Tabela 1. Identificação dos taxa encontrados, frequência de ocorrênica no total de
colônias, número total de indivíduos e abundância relativa de cada grupo. A
classificação seguiu Martin e Davis (2001).
Taxa Nº de
ocorrência
nas colonias
Frequência nas
colônias
(%)
Número
de
indivíduos
Abundância
relativa
(%)
Classe Maxillopoda
SubClasse Copepoda 100 68,79
Ordem Harpaticoida
Thalestridae 1 30 100,00 1046 40,56
Thalestridae 2 22 73,33 141 5,47
Thalestridae 3 22 73,33 97 3,76
Thalestridae 4 13 43,33 50 1,94
Thalestridae 5 15 50,00 158 6,13
Harpaticidae 21 70,00 81 3,14
Laophontidae 23 76,67 170 6,59
Peltidiidae 1 4 13,33 13 0,50
Peltidiidae 2 4 13,33 4 0,16
Tegastidae 4 13,33 4 0,16
Ordem Siphonostomatoida
Asterocheridae 1 1 3,33 1 0,16
Asterocheridae 2 3 10,00 4 0,04
Asterocheridae 3 2 6,67 2 0,16
Asterocheridae 4 1 3,33 1 0,08
Artotrogidae 2 6,67 2 0,04
Classe Ostracoda 86,7 10,04
Subclasse Myodocopa
Myodocopa 1 4 13,33 5 0,19
Myodocopa 2 3 10,00 4 0,16
Subclasse Podocopa
Podocopa 1 14 46,67 28 1,09
Podocopa 2 14 46,67 21 0,81
Podocopa 3 9 30,00 22 0,85
Podocopa 4 19 63,33 107 4,15
Podocopa 5 15 50,00 68 2,64
Podocopa 6 1 3,33 1 0,04
Podocopa 7 2 6,67 2 0,08
Podocopa 8 1 3,33 1 0,04

32
Taxa Nº de
ocorrência
nas colonias
Frequência.
nas colônias
(%)
Número
de
indivíduos
Abundância
relativa
(%)
Classe Malacostraca
Subclasse Eeumalacostraca
Superordem Peracarida
Ordem Amphipoda 80,00 12,99
Subordem Caprellidea
Caprellidae 14 46,67 70 2,71
Subordem Gammaridea
Eurisidae 1 13 43,33 63 2,44
Eurisidae 2 3 10,00 5 0,19
Eurisidae 3 3 10,00 3 0,12
Eurisidae 4 6 20,00 13 0,50
Eurisidae 5 9 30,00 25 0,97
Eurisidae 6 8 26,67 20 0,78
Eurisidae 7 4 13,33 4 0,16
Eurisidae 8 2 6,67 4 0,16
Eurisidae 9 2 6,67 15 0,58
Leocothoidae 1 3,33 2 0,08
Talitridae 4 13,33 5 0,19
Isaeidae 1 12 40,00 95 3,68
Isaeidae 2 1 3,33 3 0,12
Ischyroceridae 2 6,67 8 0,31
Ordem Isopoda 86,7 6,97
Ianiropsis 1 20 66,67 80 3,10
Ianiropsis 2 7 16,67 14 0,54
Ianiropsis 3 12 40,00 43 1,67
Munnidae 1 7 23,33 37 1,43
Munnidae 2 5 16,67 5 0,19
Anthuridae 1 3,33 1 0,04
Ordem Tanaidacea 30,00 0,42
Nototannaidae 8 26,67 10 0,39
Colleteidae 1 3,33 1 0,04
Superordem Eucarida
Ordem Decapoda 73,30 0,77
Infraordem Anomura
Megalobrachium soriatum 2 6,67 4 0,16
Infraordem Brachyura
Pilumnus quoyi 3 10,00 3 0,12
Pilumnus spinosissimus 1 3,33 1 0,04
Micropanope nuttingi 4 13,33 6 0,23
Mithrax caribbaeus 1 3,33 1 0,04
Hexapanopeus angustifrons 1 3,33 1 0,04
Pitho lherminieri 4 13,33 4 0,16
TOTAL 2579 100,00
Relação espécie-área

33
Observando os gráficos da Figura 5 e 6, foi possível notar que Crustacea como
um todo apresentou um crescimento inicial do número de espécies em relação á área de
T. tagusensis seguido de uma grande oscilação nesses valores, porém tendendo a uma
estabilização no final. Entretanto, quando os gráficos de dispersão dos grandes grupos
dentro de Crustacea são observados isoladamente, percebe-se que apenas Copepoda e
Ostracoda possuiram uma distribuição semelhante à encontrada para Crustacea (Figura
5, 6). Os outros grupos apresentaram um padrão diferente do esperado para a relação
espécie-área, mostrando uma grande aleatorização dos dados (Figura 5 e 6).
Em concordância com os gráficos, a regressão entre a área das colônias e o
número de taxa mostrou relação somente para Crustacea em geral (R²=0,33; F=14,35;
p<0,05), Copepoda (R²=0,52; F=31,19; p<0,05) e Ostracoda (R²=0,27; F=10,56;
p<0,05), sendo que o grupo Copepoda apresentou a maior relação (R²=0,52). Para os
outros grupos, as relações não foram significativas (Figura 7). Os valores de “z”
variaram entre 0,02 e 0,28, com os valores mais altos obtidos para os grupos que
mostraram uma relação significativa entre o número de táxons e a área das colônias
(Crustacea: z = 0,27; Copepoda: z = 0,24; Ostracoda: z = 0,28) (Figura 7).
Tabela 2. Modelos e resultados da regressão linear simples entre o volume das colônias
de T. tagusensis e os taxa de Crustacea. * = os dados não atendenram as premissas do
teste
Taxa Modelo R² F P-value
Crustacea y = 0,63 + 0,27.x 0,33 14,35 <0,05
Copepoda y = 0,37 + 0,24.x 0,52 31,19 <0,05
Ostracoda y = -0,01 + 0,28.x 0,27 10,56 <0,05
Amphipoda y = 0,31 + 0,09.x 0,02 0,65 0,42
Isopoda y = 0,11 + 0,11.x 0,08 2,29 0,14
Tanaidacea *
Decapoda y = -0,05 + 0,11.x 0,07 2,34 0,13

34
Teste de hipótese
As análises de rarefação mostraram que a comparação entre colônias com área
até 14,9 cm², não apresentou diferença caso várias colônias de tamanho reduzido sejam
comparadas com uma maior de tamanho equivalente (Figura 8, Gráficos 1 e 2).
Entretanto, a partir desse tamanho, observou-se que o somatório do número de espécies
de colônias menores tende a ser maior do que o número de espécies de uma única
colônia com tamanho equivalente (Figura 8, Gráficos 3, 4 e 5). Ou seja, a hipótese da
amostragem passiva é aceita quando colônias com até 14,9 cm² são comparadas. A
partir desse tamanho, os resultados aceitam a hipótese de que outros processos
biológicos influenciam a riqueza de taxa de Crustacea em colônias de T. tagusensis.
Correlação entre grupos de Crustacea associados
A observação da análise de correlação de Spearman entre todos os taxa
encontrados mostrou ausência de correlações negativas significativas e a presença de
cinco correlações positivas significativas (Thalestridae3/Leuchotoidae,
Thalestridae3/Colleteidae, Peltidiidae1/Munnidae1, Asterocheridae2/Podocopa4 e
Artotrogidae/Isaeidae1) (Apêndice 2). A despeito dos resultados da correlação, através
dos gráficos de abundância dos grupos em cada colônia, é posível fazer algumas
considerações (Figura 9). Alguns grupos de ocorrencia mais rara restringiram a colônias
pequenas, como Peltidiidae 1 e 2, Tegastidae, Eurisidae 1, 2, 4, 5, 8 e Leucothoidae. Já,
outros grupos foram mais abundantes ou se restringiram a colônias grandes (maiores
que 8,9 cm²), como Asterocheridae 1, 2, 3, 4, Artotrogidae, Eurisidae 3, Munnidae 1,
Anthuridae 1, Colleteidae, Podocopa 6, 8, Myodocopa 2, Megalobrachyum soriatum,
Pilumnus quoyi, Pilumnus spinosissimus, Micropanope nittingi, Mithrax caribbaeus,
Hexapanopeus angustifrons, Pitho lherminieri.
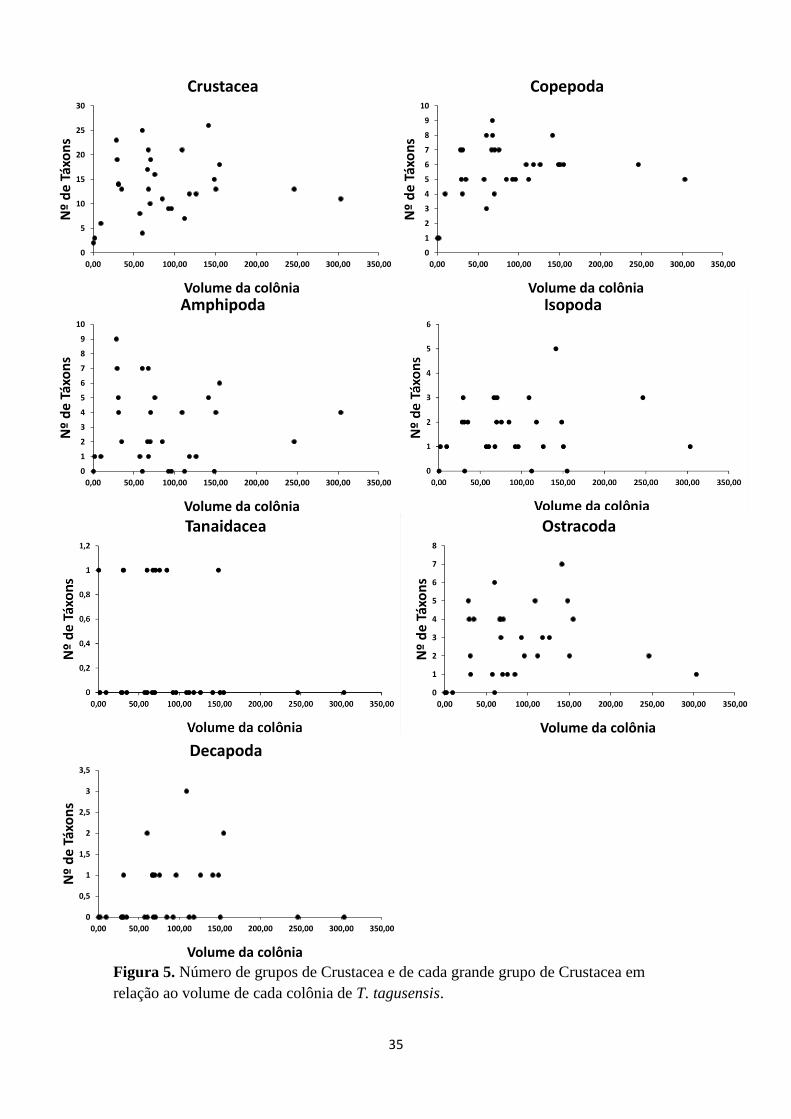
35
Figura 5. Número de grupos de Crustacea e de cada grande grupo de Crustacea em
relação ao volume de cada colônia de T. tagusensis.
0
1
2
3
4
5
6
7
8
9
10
0,00 50,00 100,00 150,00 200,00 250,00 300,00 350,00
Nº
de
Táx
on
s
Volume da colônia
Copepoda
0
1
2
3
4
5
6
7
8
9
10
0,00 50,00 100,00 150,00 200,00 250,00 300,00 350,00
Nº
de
Táx
on
s
Volume da colônia
Amphipoda
0
1
2
3
4
5
6
7
8
0,00 50,00 100,00 150,00 200,00 250,00 300,00 350,00
Nº
de
Táx
on
s
Volume da colônia
Ostracoda
0
0,5
1
1,5
2
2,5
3
3,5
0,00 50,00 100,00 150,00 200,00 250,00 300,00 350,00
Nº
de
Táx
on
s
Volume da colônia
Decapoda
0
5
10
15
20
25
30
0,00 50,00 100,00 150,00 200,00 250,00 300,00 350,00
Nº
de
Táx
on
s
Volume da colônia
Crustacea
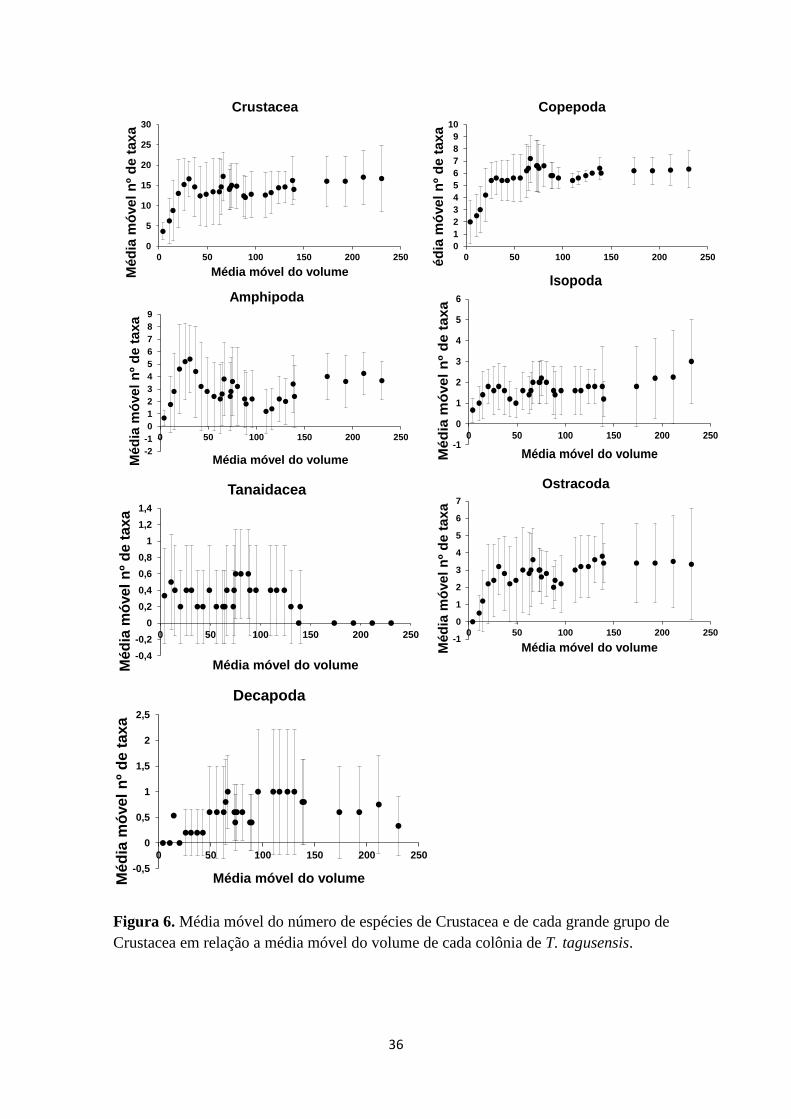
36
Figura 6. Média móvel do número de espécies de Crustacea e de cada grande grupo de
Crustacea em relação a média móvel do volume de cada colônia de T. tagusensis.
0
5
10
15
20
25
30
0 50 100 150 200 250
Mé
dia
mó
ve
l n
º d
e t
axa
Média móvel do volume
Crustacea
0
1
2
3
4
5
6
7
8
9
10
0 50 100 150 200 250
Mé
dia
mó
ve
l n
º d
e t
axa
Média móvel do volume
Copepoda
-2
-1
0
1
2
3
4
5
6
7
8
9
0 50 100 150 200 250
Mé
dia
mó
ve
l n
º d
e t
axa
Média móvel do volume
Amphipoda
-1
0
1
2
3
4
5
6
0 50 100 150 200 250M
éd
ia m
óvel
nº
de t
axa
Média móvel do volume
Isopoda
-0,4
-0,2
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0 50 100 150 200 250
Méd
ia m
óvel
nº
de t
axa
Média móvel do volume
Tanaidacea
-0,5
0
0,5
1
1,5
2
2,5
0 50 100 150 200 250
Méd
ia m
óvel
nº
de t
axa
Média móvel do volume
Decapoda
-1
0
1
2
3
4
5
6
7
0 50 100 150 200 250
Méd
ia m
óvel
nº
de t
axa
Média móvel do volume
Ostracoda

37
Scatterplot (Spreadsheet5 8v*30c)
Ostracoda = -0,0025+0,2799*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
Ostr
aco
da
Scatterplot (Spreadsheet5 8v*30c)
Amphipoda = 0,3107+0,0984*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Am
ph
ipo
da
Scatterplot (Spreadsheet5 8v*30c)
Tanaidacea = 0,192-0,0571*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
-0,05
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
Ta
na
ida
ce
a
Scatterplot (Spreadsheet5 8v*30c)
Isopoda = 0,1846+0,112*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
Iso
po
da
Scatterplot (Spreadsheet5 8v*30c)
Crustacea = 0,6294+0,2704*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
0,4
0,6
0,8
1,0
1,2
1,4
1,6
Cru
sta
ce
a
Scatterplot (Spreadsheet5 8v*30c)
Copepoda = 0,3743+0,2358*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
1,1
Co
pe
po
da
Scatterplot (Spreadsheet5 8v*30c)
Decapoda = -0,0459+0,1057*x
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6
log (x+1) do volume das colônias
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
De
ca
po
da
Figura 7. Regressão linear simples entre o volume das colônias de T. tagusensis e os
táxons de Crustacea.

38
0
5
10
15
20
25
0 20 40
N
de
táxon
s
Amostras
3 cols. = 6,4 cm²; 1 col. = 7 cm²
Sobs (Mao Tau) único
Sobs 95% CI LowerBound único
Sobs 95% CI UpperBound único
Sobs (Mao Tau)
Sobs 95% CI LowerBound
Sobs 95% CI UpperBound
0
5
10
15
20
25
30
35
40
0 200 400
N
de
táxon
s
Amostras
6 cols. = 31 cm²; 1 col. = 30 cm²
Sobs (Mao Tau) único
Sobs 95% CI LowerBound único
Sobs 95% CI UpperBound único
Sobs (Mao Tau) soma
Sobs 95% CI LowerBound soma
Sobs 95% CI UpperBound soma
0
5
10
15
20
25
30
35
40
45
0 200 400 600
N
de T
áxons
Amostras
7 cols. = 41 cm²; 1 col. = 42 cm²
Sobs (Mao Tau) soma
Sobs 95% CI LowerBound soma
Sobs 95% CI UpperBound soma
Sobs (Mao Tau)
Sobs 95% CI LowerBound
Sobs 95% CI UpperBound
0
2
4
6
8
10
12
14
16
18
0 50 100 150
N
de táxons
Amostras
4 cols. = 14 cm²; 1 col. = 14 cm²
Sobs (Mao Tau) único
Sobs 95% CI LowerBound único
Sobs 95% CI UpperBound único
Sobs (Mao Tau) soma
Sobs 95% CI LowerBound soma
Sobs 95% CI UpperBound soma
-5
0
5
10
15
20
25
30
35
0 100 200
N
de táxons
Amostras
5 cols. = 22,3 cm²; 1 col. = 23 cm²
Sobs (Mao Tau) único
Sobs 95% CI LowerBound único
Sobs 95% CI UpperBound único
Sobs (Mao Tau) soma
Sobs 95% CI LowerBound soma
Sobs 95% CI UpperBound soma
Figura 8. Curvas de rarefação (acúmulo de espécies) com base nos valores de Sobs
(Mao Tau) gerados através de 999 aleatorizações para as colônias de T. tagusensis com
diferentes tamanhos. Em vermelho e laranja os dados da soma das colônias. Em azul, os
dados de uma única colônia.
1 2
3 4
5

39
Thalestridae 5
Harpaticidae
Peltidiidae 1
Peltidiidae 2
Laophontidae
Tegastidae
Asterocheridae 1
Asterocheridae 2
Asterocheridae 3
Asterocheridae 4
Artotrogidae
Eurisidae 1
Eurisidae 2
Eurisidae 3
Eurisidae 4
Eurisidae 5
Eurisidae 6
Eurisidae 7
Eurisidae 8
Leucothoidae
Talitridae
Isaeidae 1
1 5
51 0
2 04 06 0
1 5
51 0
8
246
0 . 40 . 81 . 2
1 02 03 0
1 . 2
0 . 40 . 8
1 . 2
0 . 40 . 8
1 . 0
2 . 0
0 . 40 . 8
1 . 2
1 . 2
0 . 40 . 8
0 . 4
0 . 8
3 0
1 02 0
3 . 5
0 . 51 . 52 . 5
0 . 20 . 61 . 0
135
1357
1357
0 . 2
0 . 61 . 0
0 . 51 . 52 . 53 . 5
0 . 5
1 . 5
2 . 5
0 . 5
1 . 5
2 . 5
1 02 03 0
Eurisidae 9
1 5
5
1 0
0,4 1 4 7 8 1110 12 12 13 13 14 14 15 15 20 21 252423 28 30 42 42158 17 19 2415
Thalestridae 1
Thalestridae 2
Thalestridae 3
Thalestridae 4
1 5
51 0
5 01 0 01 5 0
2 5
5
1 5
Figura 9. Abundâncias de cada táxon ao longo do aumento da área das colônias. Área
das colônias (m) pelo número de indivíduos.

40
Isaeidae 2
Ischyroceridae
Caprellidae
Ianiropsis 1
Ianiropsis 2
Ianiropsis 3
Munnidae 1
Munnidae 2
Anthuridae
Nototanaidae
Colleteidae
Podocopa 1
Podocopa 2
Podocopa 3
Podocopa 4
Podocopa 5
Podocopa 6
Myodocopa 1
Myodocopa 2
Podocopa 7
Podocopa 8
Megalobrachiumsoriatum
Pilumnus quoyi 0 . 4
0 . 81 . 2
1 . 02 . 03 . 0
0 . 4
0 . 8
1 . 2
1 02 03 04 0
1 0
3 0
5 0
1 0
3 0
5 0
51 01 5
26
1 0
135
51 0
1 5
0 . 20 . 61 . 0
0 . 20 . 61 . 0
0 . 51 . 52 . 53 . 5
0 . 20 . 61 . 0
1
35
1
3
5
13
5
51 01 52 0
0 . 5
1 . 5
2 . 5
5
1 01 5
0 . 20 . 61 . 0
0 . 20 . 61 . 0
1 . 0
2 . 0
0 . 81 . 2
Pilumnus spinosissimus 0 . 4
Micropanope nuttingi
1 . 5
2 . 5
Mithrax caribbaeus
Hexapanopeus angustifrons
Pitho lherminieri
0 . 5
0 . 4
0 . 81 . 2
0 . 40 . 8
1 . 2
0 . 4
0 . 81 . 2
0,4 1 4 7 8 1110 12 12 13 13 14 14 15 15 20 21 252423 28 30 42 42158 17 19 2415
Figura 9. Abundâncias de cada táxon ao longo do aumento da área das colônias. Área
das colônias (m) pelo número de indivíduos.

41
DISCUSSÃO
Grupos de Crustacea associados a T. Tagusensis
Os grandes grupos de Crustacea registrados em Tubastraea tagusensis são
comuns em ambientes recifais, podendo ser encontrados em diversos organismos
marinhos sésseis como esponjas, ascídias, cirripédios, corais, algas e gramas marinhas.
A proporção da abundância e da riqueza observada, entretanto, varia a depender do
hospedeiro. Nogueira et al. (2003), analisando a fauna asscoada a endêmica Mussismilia
hispida, encontrou os mesmos grupos registrados neste trabalho, sendo que Copepoda
foi o mais abundante (mais de 50% de todos os indivíduos). Preston e Doherty (1994),
fazendo o mesmo para Pocillopora verrucosa em diversas regiões da Grande Barreira
da Austrália, encontraram que em geral, Copepoda foi o grupo dominante, seguido por
Ostracoda, Amphipoda, Cumacea, Tanaidacea e Isopoda, sendo que Decapoda não
entrou no estudo. Por sua vez, analisando a criptofauna de crustacea em colônias de
Madracis mirabilis, Lewis e Snelgrove (1990), revelaram que Amphipoda foi o grupo
mais abundante, seguido por Isopoda, Copepoda e Decapoda. Em termos de riqueza,
entretanto, Copepoda teve os maiores valores, seguido por Decapoda, Amphipoda e
Isopoda.
Considerando caracteristicas biológicas espécificas de cada grupo, alguns
aspectos sobre habito de vida podem ser discutidos. Copepoda apresentou uma grande
variedade de hábitos entre os grupos encontrados. As famílias Asterocheridae e
Artotrogidae pertencem a Ordem Siphonostomatoida, conhecida principalmente por
possuir organismos com o labro e o lábio fundidos formando um sifão que utilizam para
alimentação, sendo tipicamente parasitas. Diversos trabalhos relataram a associação de
espécies dessas famílias com corais (Humes 1996, 1997b; Johnsson e Neves 2005),
inclusive com o gênero Tubastraea (Humes 1997a). Além deles, foram encontradas
famílias da ordem Harpacticoida, que apresenta grupos com hábitos de vida bentônicos,
porém, diversificados. Thalestridae, Peltidiidae, Tegastidae e Harpaticidae são
conhecidos por possuírem a forma do corpo comprimida dorsoventralmente ou
lateralmente, cefalossomo arredondado ou alargado e a primeira perna (P1) fortemente
preênsil, o que facilita a sua aderência na superfície áspera de esqueletos de corais, mas
principalmente de algas (Hicks 1977; Bell et al. 1987; Klumpp et al. 1988; Arroyo
2004, 2006; Shimono et al. 2004). Preston e Doherty (1994) discutem que o fato de

42
corais abrigarem algas filamentosas epilíticas fornece um microhabitat para
microcrustáceos herbívoros como os mencionados acima. Isso pode justificar a
ocorrência dessas famílias associadas ao coral invasor. Por fim, outra família
encontrada, Laophontidae, possui um corpo caracteristicamente fusiforme e é típica de
sedimento. De acordo com Jacoby e Greenwood (1988) e Preston e Doherty (1994),
grupos demersais que aparecem associados a corais podem emergir do sedimento ou da
base das colônias. Por outro lado, Gheerardyn et al. (2008) discutem que o sedimento
retido pelos corais também forma microhabitats que permitem a ocorrência de táxon
característicos de sedimento, fato possível devido a morfologia de T. tagusensis que
possibilita a deposição de sedimento no espaço entre pólipos (Figura 3).
Os outros grupos encontrados possuem predominantemente um hábito de vida
transitório, podendo ocupar diversos substratos. Começando com os Ostracoda, embora
existam estudos mostrando que alguns grupos são bastante abundantes em ambientes
recifais (Nogueira 2003), informações específicas sobre a biologia e o tipo de interação
desses organismos com seus hospedeiros são escassas. De acordo com Whatley e Wall
(1975) e Athersuch (1979), muitas espécies são pastadoras e se alimentam de bactérias e
microalgas, especialmente diatomáceas que crescem sobre macroalgas. Em um trabalho
sobre os Ostracoda da costa da Ilha de Trindade, Ghilardi (2004) encontrou apenas
indivíduos da Ordem Podocopa, sendo o gênero Paranecidae predominante de
ambientes recifais.
Em relação aos grupos de Peracarida, a grande maioria é amplamente
distribuída, podendo ocupar diversos substratos. Dentre os Amphipoda, caprelídeos e
gamarídeos já foram registrados em bancos de alga e de grama marinha recifais
(Karaçuha et al. 2009), recifes de ostras e mexilhões, manguezais, esponjas, tunicados,
briozoários, hidróides (Jones 1948; Guerra-Garcia e Garcia-Gomes 2001; Ribeiro et al.
2003; Diaz et al. 2005; Gonzales et al. 2008) e corais (Preston e Doherty 1994; Guerra-
Garcia 2002). Caprellidea utiliza o organismo hospedeiro para se fixar, ou se camuflar,
enquanto captura o seu alimento da coluna d‟água e por isso tendem a ocorrer em locais
com hidrodinâmica moderada, o que provávelmente é proporcionado por microhabitats
fornecidos por corais (Guerra-Garcia 2003; Diaz et al. 2005; Gonzales et al. 2008). Os
Gammaridea podem viver em tubos construídos por eles próprios, ou ser ebipiontes,
podendo se locomover entre substratos, sendo comumente onívoros, comedores de

43
partícula (Ischyroceridae, Leucothoidae), herbívoros, predadores ou parasitas (Garcia e
Leite 2010). De acordo com Chenelot et al. (2008), Eurisidae e Isaeidae são
característicos de substratos duros como recifes de coral. Em Isopoda, os Asellota,
como os Janiridae (Ianiropsis sp.) e Munnidae são em geral onívoros (Macdonald 2010)
e comuns em ambientes recifais (Achultz 1969). Já Anthuridae, são pricipalmente
carnívoros e predadores e habitam tubos de areia, ora construidos por eles mesmos, ora
por outros animais, ou são também encontrados em animais incrustantes ou algas
(Achultz 1969). Por fim, em relação à Tanaidacea, o pouco que se conhece sobre sua
história de vida sugere que são consumidores raptoriais e se alimentam de detritos e dos
próprios organismos aos quais se associam (Holdich and Jones 1983), o que sugere que
estes organismos possam estar se alimentando de T. tagusensis.
Os Decapoda possuem relações de especificidade coral-decapoda conhecidas,
como as realizadas entre os gêneros Tetralia e Trapezia e corais acroporídeos e,
supstamente, os realizados entre a família Cryptochiridae e corais do gênero Siderastrea
(Sin 1999, Sin e Lee 2000; Johnsson et al. 2006; Badaró et al. no prelo). Entretanto, as
espécies encontradas neste trabalho apresentam interação facultativa com corais,
podendo ocupar outros substratos como areia, conchas quebradas cascalho fundos de
rochas esponjas e algas calcárias (Melo 1996). Em geral, utilizam esses habitats
principalmente para proteção contra predador e estresse físico.
Poucos dos grupos encontrados foram anteriormente registrados para a costa da
Bahia, e se restringem aos asterocherideos (Bispo et al., 2006), aos decapodes (Melo,
1996, Almeida et al., 2010), e aos caprelideos(Young, 1998). Os outros grupos,
copépodes Thalestridae, Peltidiidae, Tegastidae, Harpaticidae, Laophontidae e
Artotrogidae, os anfípodes Ischyroceridae, Leucothoidae, Eurisidae, Isaeidae, Talitridae,
os isópodes Anthuridae, Munnidae e Janniridae, e os tanaidáceos Tanaidomorpha foram
relatados para o Brasil, mas não para a Bahia (Kornicker and Sohn, 1976; Young, 1998;
Machado and Drozinski, 2002; Coimbra and Bergue, 2003; Ramos et al., 2004). No
Nordeste, Neves and Johnsson (2008) relataram Artotrogidae para Pernambuco, Myers
and McGrath (1984) relataram Ischyroceridae em Pernambuco, Schellenberg (1938)
registrou Leucothoidae para Pernambuco, Soares 1979 observou Talitridae para
Pernambuco e Silva 2010 observou tanaidomorpha como resultado do programa
Revizee-NE. Os outros grupos foram registrados apenas para a costa sudeste do Brasil

44
Figura 10. Incorporação da
facilitação no coceito de nicho. (a-i)
Conceito de nicho sem a considerar
a facilitação. (a-ii) Conceito de
nicho incorporando a facilitação –
neste caso, processos podem
aumetar o recurso fornecido pelo
nicho fundamental e a mitigar o
efeito de fatores que encolhem o
nicho realizado. Figura modificada
de Bruno et al. (2003).
(Young, 1998). Assim, este trabalho expande a ocorrência desses grupos na costa
brasileira, além de descrever T. tagusensis como novo substrato para eles. Espécies
exóticas não foram encontradas, mas esta possibilidade não pode ser descartada. Futuras
identificações a nível específico poderão revelar ou não a ocorrencia de espécies
exóticas.
Por fim, o registro inédito da associação entre a espécie invasora T. tagusensis e
crustáceos na costa brasileira remete a uma nova abordagem que tem ganhado espaço
nos estudos de ecologia: a incorporação da “facilitação” como um fator determinante
nos processos ecológicos. Até pouco tempo atrás, as relações negativas de competição,
predação e a influência dos fatores abióticos sobre os organismos foram a “âncora” das
explicações da organização das comunidades naturais dando origem a modelos
fundamentais como as equações de Lotka e Volterra e o Princípio de Exclusão
Competitiva de Gause (Bruno et al. 2003). A partir do final do século XX, a
“facilitação” passou a ganhar importância e ser reconhecida não apenas como simples
registros, mas sim, como “peça chave” para o entendimento de diversos padrões como
os de crescimento das populações, distribuição, composição e diversidade das
comunidades (Jones 1994; Stachowicz 2001; Bruno e Bertness 2001). Basicamente,
facilitação influencia um princípio clássico da ecologia, o de nicho. Enquanto interações
negativas afetam o nicho realizado das espécies, tendendo a reduzi-lo, a facilitação
tende a aumentá-lo, de forma que estas podem alcançar populações de tamanhos
maiores que os previstos pelo nicho fundamental (Figura 10).

45
Rodriguez (2006) chama a atenção para a influência dessa mudança de
paradigmas na literatura relacionada a bioinvasão. Ou seja, embora trabalhos sobre
competição, predação e fatores abióticos sejam mais comuns na literatura corrente sobre
o tema, a quantidade de estudos sobre “facilitação” tem aumentado, o que mostra a sua
relevância. De acordo com a revisão realizada pelo autor, entre os anos de 1993 e 2004,
cerca de 61 trabalhos sobre o impacto de facilitadores exóticos foram encontrados,
sendo 39% relacionados a habitats terrestres, 36% a ambientes de água doce a apenas
25% a ambientes marinhos. Em relação á T. tagusensis, até o momento, sabe-se que a
espécie, juntamente com a congênere T. coccinea está modificando comunidades
bentônicas da Baía de Ilha Grande (Lages et al 2011). Os estudos, entretanto, fornecem
informações específicas apenas para modificação na comunidade de grupos como
esponjas, cnidários, e urocordados,deixando a desejar em relação a grupos que vivem
associados como os crustáceos, poliquetos e “peixes”.
Embora este trabalho não tenha realizado nenhuma comparação da fauna
associada a T. tagusensis com a fauna associada a espécies nativas, não podendo
indicar, portanto, modificações na comunidade de crustáceos crípticos associados a
coral, os novos registros indicam que T. tagusensis está servindo de substrato para
determinados táxons de Crustacea locais, corroborando as expectativas de que um coral
escleractíneo, mesmo se tratando de uma espécie invasora, pode abrigar organismos
nativos. Considerando que T. tagusensis está expandindo rapidamente sua população e
modificando comunidades macrobentônicas (Lages et al. 2011), este trabalho dá
subsídios para a realização de estudos que visem conhecer a influencia dessas
modificações nas comunidades microbentonicas.
Relação espécie-área
Analisando a relação espécie-área encontrada para Crustacea como um todo,
percebe-se que o resultado significativo encontrado na regressão (R²=0,33; p<0,05),
juntamente com o alto valor de z (0,27) foi principalmente determinado pelos taxa
Copepoda e Ostracoda, que apresentaram valores semelhantes (R²=0,52; p<0,05; z=0,24
e R²=0,27; p<0,05; z=0,28 respectivamente). Os outros grupos, entretanto, não
apresentaram relação significativa, possuindo valores de z baixos e aparentemente, não
influenciando os resultados gerais. De acordo com Preston (1962), Azovsky (2002) e

46
Forero-Medina e Vieira (2007), tal variação também tem sido encontrada na literatura e
está relacionada principalmente a influência da escala nos fatores que determinam a
relação espécie-área. Ou seja, a ocorrência ou não dos padrões típicos de relação
espécie-área depende de fatores como a escala de variação temporal, a escala de
variação no gradiente ambiental e a escala do organismo que se está estudando (e.g.
escala da área de habitação, escala de tamanho).
A interferência de gradientes temporais e ambientais na relação espécie-área tem
sido mostrada em alguns estudos. Nour et al. (1999) e Murgui (2007), estudando a
diversidade de aves em fragmentos de mata, encontraram que a riqueza de espécies em
fragmentos de floresta é maior em determinadas estações do ano, mas que a relação
espécie-área encontrada é muito semelhante. Ou seja, a sazonalidade influência na
variação dos valores do intercepto, mas não nos valores de z das regressões. Da mesma
forma, Abele (1976) observou resultados semelhantes quando analisaram a influência
da estabilidade ambiental na relação entre o número de espécies de Decapoda e colônias
de Pocillopora damicornis. Entretanto, Francesco et al. (2007) encontraram, além de
variação no intercepto, variações também nos valores de z quando analisou a influencia
da variação de umidade (regiões úmidas e secas) sobre o efeito da pastagem na relação
entre número de plantas e a área. No presente trabalho, o fato das únicas duas coletas
terem sido realizadas na mesma época e em apenas um ambiente anulou o efeito desses
dois fatores.
Em relação a escala do organismo em estudo, duas características são
comumente citadas: a área de habitação e o tamanho do corpo. Área de habitação tem
como definição “a expressão espacial do comportamento animal para prover a sua
sobrevivência e reprodução” (Burt 1943), e em geral, não atinge o potencial de
mobilidade dos organismos, que tende a ser restrita a uma única região ou hospedeiro.
De acordo com Borger et al. (2008), quanto maior a área de habitação de um organismo,
maior deve ser a escala espacial de estudo para que seja possível perceber a influência
deste fator sobre processos ecológicos. Assim, é possível que a T. tagusensis tenha
servido como área de habitação para Copepoda e Ostracoda, mas que para os outros
grupos, seja necessário uma escala diferenciada de estudo para se obter uma relação
espécie-área.

47
Outro fator importante na determinação da relação espécie-área é o tamanho dos
indivíduos. Interrelações entre a diversidade de espécies e tamanho do corpo tem sido
amplamente discutida desde as publicações de Robert May (May 1988). Em geral,
organismos menores dividem o ambiente mais finamente e possuem também uma área
de habitação menor. Já organismos maiores possuem uma área de habitação maior,
sendo necessário uma área mínima para se detectar padrões de diversidade. Mac Arthur
e Wilson (1967) observou que antes desse tamanho mínimo, não é possível observar
uma relação espécie-área em ilhas. Assim, neste trabalho a escala espcacial de estudo
parece ter sido ideal para organismos pequenos de baixa mobilidade como os Copepoda
e Ostracoda, enquanto que para os outros grupos seria necessária uma escala maior para
detectar os padrões.
Em contraponto à ausência de relação espécie-área encontrada para a maioria
dos táxons de Crustacea estudados, os resultados de Copepoda e Ostracoda apresentam
uma típica relação encontrada na literatura, já tendo sido encontrados valores
semelhantes em diversos trabalhos (Mac Arthur e Wilson 1967, Connor e McCoy 1979;
Grez 1992; Belmaker et al. 2007; Báldi 2008). Entretanto, as explicações biológicas
tentem a variar (Turner e Tjorve 2005, Figura 12), sendo necessários testes empíricos
para esclarecer os processos
Figura 12. Escala de fatores que influenciam a relação espécie-área. Tons mais escuros
representam áreas de maior efeito. Os valores da abscissa são aproximados e vão
depender do táxons e locais estudados. Essa figura deve ser considerada como hipótese
de trabalho baseada em conhecimentos correntes e deve ser ser refinada através de

48
fatores adicionais para ser melhor compreendida. Figura modificada de Turner and
Tjørve (2005).
Para estudos em pequena escala espacial, a aceitção da hipótese de amostragem
passiva é bastante comum. Diversos estudos teóricos e evidências empíricas tem
encontrado resultados semelhantes ao encontrado neste trabalho (Chittaro 2002;
Williams 1995; He and Legendre 1996; Condit et al. 1996). Grez et al. (1992),
analisando a taxa de imigração/extição de insetos herbívoros em “patches” com uma
única planta observou que embora as taxas de imigração aumentassem com o aumento
da área, a taxa de extinção permanencia a mesma, concluindo asssim, que a amostragem
passiva explicava melhor os resultados encontrados do que a hipótese de área per se.
Belmaker et al. (2007), analisando a relação espécie-área entre peixes e “patches”
através de um modelo de randomização dos seus dados, também observou que a
amostragem passiva foi o que mais importou, embora isso não tenha significado que a
comunidade tenha se mostrou totalmente randômica. Abele e Patton (1976), entretanto,
comparando diversas réplicas de número de Dacapoda associados a duas colônias de
Pocillopora versos uma colônia observou que na maioria dos casos, duas colônias
pequenas tinham mais espécies que apenas uma colônia de tamanho grande equivalente
e atribuiu isso ao alcançe do equilíbrio no número de espécies em colônias maiores.
Apesar disso, neste trabalho a hipótese foi rejeitada quando colônias a partir de 14cm²
foram analisadas (Gráficos C, D e E da Figura 8).
Para entender a diferença encontrada nos gráficos C, D e E da Figura 8, em que
o número de espécies em colônias maiores é menor do que o somatório do número de
espécies de colônias menores, pode-se lançar mão de algumas explicações. Dentre elas,
destacam-se a “Hipótese de Heterogeneidade do Habitat”, importante nas mais variadas
escalas, “Teoria de Biogeografia de Ilhas”, que embora seja mais eficiente em explicar
processos na escala da paisagem, tem sido utilizada em estudos de micro escala, como
nas diversas relações hóspede-hospedeiros citadas por Kuris (1980), e as interações
negativas entre os organismos.
A “Hipótese de Heterogeneidade do Habitat” é citada em diversos trabalhos, que
ora corroboram a idéia de que áreas maiores possuem uma maior riqueza devido a maior
heterogeneidade de habitats, ora mostram que nem sempre isso acontece. De fato, em

49
casos mais raros, áreas menores podem ser mais heterogêneas suportando uma maior
riqueza que áreas maiores (Báldi 2008). Apesar disso, tal hipótese não se aplica ao
presente trabalho uma vez que a área em questão, colônias de T. tagusensis, não varia os
tipos de estrutura com o aumento da área. Embora características como número de
pólipos, espaço entre os coralitos e diâmetro dos coralitos possam ter variado com o
tamanho das colônias, fala-se em (1) aumento do número e do tamanho das estruturas,
mas não em (2) adição de uma nova estrutura. Beck (2000) distingue estes dois
processos como “complexidade do habitat” e “heterogeneidade do habitat”,
respectivamente, e chama a atenção para a confusão que é feita na literatura em relação.
A heterogeneidade do habitat, tal qual observada por Connor e McCoy (1979), se refere
apenas ao segundo exemplo. Assim, visto que as colônias estudadas não apresentam
variação da heterogeneidade, mas sim um incremento da complexidade, concordamos
que a hipótese da heterogeneidade não poderia ser uma possível explicação para o
resultado encontrado.
Em relação a “Teoria de Biogeografia de Ilhas”, uma vez que premissas como a
taxa de imigração e extinção e a distância entre “patches” não foram analisadas,
discussões sobre a explicabilidade desta teoria para os resultados encontrados não é
possível.
Já, as análises de correlação entre os crustáceos permitiram fazer algumas
inferências sobre as interações entre os crustáceos (Tabela 2). O fato dos dados não
evidenciarem um padrão de interações negativas como competição ou predação entre os
táxons foram também encontrados na literatura. Simberloff (1978), em seus estudos
sobre sucessão de insetos em ilhas de mangue, observou que plantas e invertebrados não
são tão influenciados por interações competitivas como são os vertebrados. O autor
argumenta que a comunidade de Arthropoda é determinada principalmente pela
capacidade de dispersão e manutenção da sua população (nascimento e morte) no
ambiente, e não pela competição, uma vez que os recursos raramente são limitados para
estes organismos. O mesmo foi encontrado por Gotelli e Abele (1983) para decápodes
em colônias de Pocillopora damicornis. O mesmo pode estar justificando a falta de
correlação negativa encontrada neste trabalho.

50
Em relação as correlações significativamente positivas encontradas (n=5)
(Apêndice 2), estas podem se tratar simplesmente de um reflexo da relação espécie-área,
umas vez que o número de indivíduos tende a aumentar com o tamanho da área. Ou,
podem também corresponder à interações de facilitação em que a presença de uma
espécie favoreceria a presença de outra, auxiliando na formação da relação espécie-área.
Entretanto os mecanismos que explicariam tal influência não são claros. De fato, a
incorporação dos processos de facilitação na ecologia é recente e ainda não se sabe até
onde e em quais condições este pode influenciar na estruturação da comunidade e,
consequentemente, na relação espécie-área.
Embora os testes estatísticos não tenham explicitado padrões de interação
negativa, foi possível fazer algumas observações com base nos gráficos de abundância
de cada grupo (Figura 9). Alguns táxons menos frequentes ocorreram apenas em
colônias menores, enquanto outros ocorreram apenas em colônias maiores. De acordo
com Bruno (2002), certas espécies só conseguem ocorrer no início da colonização,
quando competidores superiores ainda não estão dominando o ambiente. Já outros, só
ocorrem em áreas maiores por terem mais facilidade de chegar atravéz da dispersão. E
este é um dos fatores que causa a mudança na composição e a estabilização da curva de
relação espécie-área. Assim, a argumentação deste autor pode explicar a ocorrência de
grupos como Peltidiidae 1 e 2, Tegastidae, Eurisidae 1, 2, 4, 5, 8 e Leucothoidae,
principalente em colônias pequenas, e a ocorrência de grupos Asterocheridae 1, 2, 3, 4,
Artotrogidae, Eurisidae 3, Munnidae 1, Anthuridae 1, Colleteidae, Podocopa 6, 8,
Myodocopa 2, Megalobrachyum soriatum, Pilumnus quoyi, Pilumnus spinosissimus,
Micripanope nittingi, Mithrax caribbaeus, Hexapanopeus angustifrons, Pitho
lherminieri, principalmente em colônias maiores.
Por fim, é grande o número de possibilidades que podem explicar a rejeição da
hipótese de amostragem passiva. Entre elas encontram-se a relação das taxas de
imigração e extinção e as interações entre os entre os Crustacea e outros grupos como
poliquetas, moluscos, algas e fungos, que precisam ser estudados para fornecer
respostas mais concretas. Embora exista a necessidade de mais estudos para elucidar
outros aspéctos da relação espécie-área entre a carcinofauna associada a T. tagusensis,
este trabalho sustenta que ela ocorre para os grupos Copepoda e Ostracoda e que esta
não é resultado da amostragem passiva.

51
AGRADECIMENTOS
A equipe do LABIMAR (Laboratório de Crustacea, Cnidaria e fauna associada)
pelo suporte em diversas partes do trabalho. A Roberta Canário, Cristiano Bahia,
Barbara Conceição, Marcos Nogueira pelo auxílio na identificação dos grupos de
Crustacea, ao Prof. Dr. Cláudio Sampaio (UFAL) pelo auxílio com informações sobre
localização da T. tagusensis, a Marcos De Paula por contribuir na realização das
atividades de campo, a Gabriel Barros pelo auxílio na coleta, a Igor Cruz que me
incentivou na realização deste trabalho, auxiliando nas atividades de campo e na
padronização das fotografias. A Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (CAPES) pela concessão da bolsa de mestrado, à Pós-graduação em Ecologia e
Biomonitoramento que contribuiu de diversas formas para a realização deste trabalho.
REFERÊNCIAS
Almeida, A.O., Souza, G.B.G., Boehs, G. and Bezerra, L.E.A. 2010. Shallow-water
anomuran and brachyuran crabs (Crustacea: Decapoda) from southern Bahia, Brazil.
Latin American Journal of Aquatic Research 38(3): 329-376.
Abele LG (1976) Comparative species richness in fluctuating and constant
environments: Coral-Associated Decapod Crustacea. Science 192:461-463.
Abele LG, Patton WK (1976) The size of coral heads and the community biology of
associated decapod crustaceans. Journ Biogeogr 3:35-47.
Achults GA (1969) How to know the marine Isopod crustaceans. V.C. Brown, New
Jersey.
Amado-Filho GM, Salgado LT, Rebelo MF, Rezende CE, Karez CS, Pfeiffer WC
(2008). Heavy metals in benthic organisms from Todos os Santos Bay, Brazil. Braz
Journ Biol 68:95-100.
Arroyo NL, Maldonado M, Pérez- Portela R, Benito J (2004) Distribution patterns of
meiofauna associated with a sublittoral Laminaria bed in the Cantabrian Sea (north-
eastern Atlantic). Mar Biol 144:231–242.
Arroyo NL, Maldonado M, Walter K (2 0 0 6 ) Within- and between- plant distribut ion
of harpacticoid copepods in a North Atlantic bed of Laminaria ochroleuca J. Mar Biol
86:309-316.
Arrhenius O (1923) Statistical investigations in the constitution of plant associations.
Ecology 4:68-73.

52
Athersuch J (1979) The ecology and distribution of the littoral ostracods of Cyprus.
Journ Natl Hist 13:135–160.
Azovsky AI (2000) Concept of scale in marine ecology: linking the words or the
worlds? Ecology 1:28-34.
Azovsky AI (2002) Size-dependent species-area relationships in benthos: is the world
more diverse for microbes? Ecography 25:273 – 282.
Badaró MFS, Neves EG, Castro P, Johnsson, R no prelo. Description of a new genus of
Cryptochiridae (Decapoda, Brachyura) associated with Siderastrea (Anthozoa,
Scleractinia), with notes on feeding habits. Sci Mar
Báldi A (2008) Habitat heterogeneity overrides the species–area relationship. J
Biogeogr 35:675–681.
Beck MW (2000) Separating the elements of habitat structure: independent effects of
habitat complexity and structural components on rocky intertidal gastropods. J Exp Mar
Biol Ecol 249:29–49.
Bell SS, Walters K, Hall MO (1987) Habitat utilization by harpacticoid copepods: a
morphometric approach Mar. Ecol. Prog. Ser. 35:59-64.
Belmaker J, Ben-Moshe N, Ziv Y, Shashar N (2007) Determinants of the steep species–
area relationship of coral reef fishes. Coral Reefs 26:103–112.
Bertness MD, Leonard GH (1997) The role of positive interactions in communities:
lessons from intertidal habitats. Ecology 78(7):1976-1989.
Bispo, R., R. Johnsson, E.G. Neves. 2006. A new species of Asterocheres (Copepoda,
Siphonostomatoida, Asterocheridae) associated to Placospongia cristata Boury-Esnault
(Porifera) in Bahia State, Brazil. Zootaxa 1351: 23-34.
Borger L, Dalziel BD, Fryxell JM (2008) Are there general mechanisms of animal home
range behaviour? A review and prospects for future research. Ecology 11: 637–650.
Bruno JF, Bertness MD (2001) Habitat modification and facilitation in benthic marine
communities. Mar Commun Ecol 201-218.
Bruno JF, Stachowicz JJ, Bertness MD (2003) Inclusion of facilitation into ecological
theory. Trends Ecol Evol 18(3):119-125.
Bruno JF (2002) Causes of landscape-scale rarity in cobble beach plant communities.
Ecology 83(8):2304–2314.
Burt WH (1943). Territoriality and home range concepts as applied to mammals. J.
Mammal 24:346–352.
Bush SL, Precht WF, Woodley JD, Bruno JF (2004) Indo-Pacific mushroom corals
found on Jamaican reefs. Coral Reefs 23:234.

53
Carricart-Ganivet JP, Carrera -Parra LF, Quan-Young LI, García-Madrigal MS (2004)
Ecological note on Troglocarcinus corallicola (Brachyura: Cryptochiridae) living in
symbiosis with Manicina areolata (Cnidaria: Scleractinia) in the Mexican Caribbean.
Coral Reefs 23:215–217.
Castilla JC, Lagos NA, Cerda M (2004) Marine ecosystem engineering by the alien
ascidian Pyura praeputialis on a mid-intertidal rocky shore. Mar Ecol Prog Ser 268:
119–130.
Castro CB, Pires DO (2001) Brazilian Coral Reefs: what we already know and what is
still missing. Bull Mar Sci 69(2):357-371.
Celino JJ, Queiroz AFS (2006). Fonte e grau de contaminação por hidrocarbonetos
policíclicos aromáticos (HPAs) de baixa massa molecular em sedimentos da Baía de
Todos os Santos. Rev Esc Minas 59:265-270.
Chandrasekaran S, Nagendran NA, Pandiaraja D, Krishnankutty N, Kamalakannan B
(2008) Bioinvasion of Kappaphycus alvarezii on corals in the Gulf of Mannar, India.
Science 94(9):1167-1172.
Chenelot H, Stephen J, Hoberg M (2008) Invertebrate communities associated with
various substrates in the Nearshore Eastern Aleutian Islands, with emphasis on thick
crustose coralline algae. Proc Amer Acad Under Sci.
Cheng Y, Dai1 Chang-Feng, Chang Wen-Been (2011) A New Siphonostomatoid
copepod associated with the ahermatypic coral Tubastraea aurea from Taiwan. Zool
Stud 50(5):605-610.
Chittaro PM (2002) Species-area relationships for coral reef fish assemblages of St.
Croix, US Virgin Islands. Mar Ecol Progr Ser 233:253–261.
Cirano M, Lessa GC (2007). Oceanographic characteristics of Baía de Todos-os-Santos,
Brazil. Rev Bras Geof 25:363-387.
Colwell RK, Mao CX, Chang J (2004) Interpolating, extrapolating, and comparing
incidence-based species accumulation curves. Ecology 85(10):2717–2727.
Condit R, Hubbell SP, Lafrankie JV, Sukumar R, Manokaran N, Foster RB., Ashton PS
(1996) Species-area and species-individual relationships for tropical trees: A
Comparison of Three 50-ha Plots. Journal of Ecology 84(4):549-562.
Connell JH (1961) The influence of interspecific competition and other factors on the
distribution of the barnacle Chthamalus Stellatus. Ecology 42(4):710-723.
Connor EF, McCoy ED (1979) The statistics and biology of the species-area
relationship. Am Nat 113:791-833.
Coimbra, J.C., C.T. Bergue. 2003. A new recent marine Ostracoda species
(Hemicytheridae) from Brazil. Iheringia 93(3): 243-247.

54
Creed JC (2006) Two invasive alien azooxanthellate corals, Tubastraea coccinea and
Tubastraea tagusensis, dominate the native zooxanthellate Mussismilia hispida in
Brazil. Nota in Coral Reefs 25:350.
Creed JC, Paula AF (2007) Substratum preference during recruitment of two invasive
alien corals onto shallow-subtidal tropical rocky shores. Mar Ecol Prog Ser 330:101–
111.
Crooks JA (2002) Characterizing ecosystem-level consequences of biological invasions:
the role of ecosystem engineers. Oikos 97:153–166.
Cruz ICS, Kikuchi RKP, Leão ZMAN (2008). Use of the video transect method for
characterizing the Itacolomis reefs, Eastern Brazil. Braz Journ Ocean 56:271-280.
Dengler J (2009) Which function describes the species–area relationship best? A review
and empirical evaluation. J Biogeogr 36:728–744.
Díaz YJ, Guerra-Garcíab JM, Martín A (2005) Caprellids (Crustacea: Amphipoda:
Caprellidae) from shallow waters of the Caribbean coast of Venezuela. Org. Divers.
Evol. 10:1-25.
Dial KP, Marzluff JM (1988) Are the smallest organisms the most diverse? Ecology
69:1620-1624.
Fahrig L (1998) When does fragmentation of breeding habitat affect population
survival? Ecol Mod 105:273–292.
Felizzola JF, Wagener ALR, Almeida AC, Lin WO (2008). Butyltin speciation in
sediments from Todos os Santos Bay (Bahia, Brazil) by GC-PFPD. Química Nova
31:89-93.
Fenchel T (1993) There are more small than large species? Oikos 68(2): 375-378.
Fenner D (2001) Biogeography of three caribbean corals (Scleractinia) and the invasion
of Tubastraea coccinea into the Gulf of Mexico. Bull Mar Sci 69(3):1175-1189.
Fenner D, Banks K (2004) Orange cup coral Tubastraea coccinea invades Florida and
the Flower Garden Banks, Northwestern Gulf of Mexico. Coral Reefs, 23:505-507.
Forero-Medina G, Vieira MV (2007) Conectividade funcional e a importância da
interação organismo-paisagem. Oecol Bras 11(4):493-502.
Francesco B, Jan L, Maria-Teresa S (2007) Grazing effects on the species-area
relationship: Variation along a climatic gradient in NE Spain. Veg Sci 18:25-34.
García AF, Leite FPP (2010) Distribuição de anfípodes, de acordo com suas estratégias
de forrageamento, em praias do Litoral Norte de São Paulo. XVIII Congresso interno de
iniciação científica da Unicamp, Campinas

55
Gerlach SA (1978) Food-chain relationships in subtidal silty sand marine sediments and
the role of meiofauna in stimulating bacterial productivity. Oecologia 33: 55–69.
Ghilardi VG (2004) Origem e taxonomia dos Ostracodes (Crustacea) da Ilha de
trindade, Brasil. Dissertação, Universidade Federal do Rio Grande do Sul.
Gheerardyn H, Troch M, Ndaro SGM, Raes M, Vincx M, Vanreusel A. (2008)
Community structure and microhabitat preferences of harpacticoid copepods in a
tropical reef lagoon (Zanzibar Island, Tanzania). Jour Mar Biol Assoc UK 88(4): 747–
758.
Gleason HA (1922) On the relation between species and area. Ecology 3(2): 158-162.
Glynn PW, Colley SB, Maté JL, Cortés J, Guzman HM, Bailey RL, Feingold JS,
Enochs IC (2008) Reproductive ecology of the azooxanthellate coral Tubastraea
coccinea in the Equatorial Eastern Pacific: Part V. Dendrophylliidae. Mar Biol 153:
529–544.
Goh BPL, Chou LM, Ng PKL (1989) Anomuran and Brachyuran crab symbionts of
Singapure hard coral of the families Acroporidae, Agaricidae and Pocilloporidae. Indo-
Malayan Zool 6:25-44.
González AR, Guerra-García JM, Maestre MJ, Ruiz-Tabares A, Espinosa F, Gordillo I,
Sánchez-Moyano JE, García-Gómez· JC (2008) Community structure of caprellids
(Crustacea: Amphipoda: Caprellidae) on seagrasses from southern Spain. Mar Res
62:189–199.
Gotelli NJ, Gilchrist SL, Abele LG (1985) Population biology of Trapezia spp. and
other coral-associated decapods. Mar Ecol Prog Ser 21:89-98.
Gotelli NJ, Abele LG (1983) Community patterns of coral-associated decapods. Mar
Ecol 13:131-139.
Grez AA (1992) Riqueza de species de insectos herbívoros y tamanho de parche de
vegetacion huesped: uma contrastacion experimental. Rev Chil Hist Nat 60:115-120.
Guerra-García JM (2002) Redescription of five rare caprellids (Crustacea: Amphipoda:
Caprellidea) collected from Tanzanian coasts. Mar Res 55:221–231.
Guerra-García JM, Garcia-Gomes JC (2001) the spacial distribution of caprelidea
(Crustacea: amphipoda) a stress bioindicator in Ceuta (North Africa: Gilbraltar Area)
Mar Ecol 22(4):357-367.
Hacker SD, Gaines SD (1997) Some implications of the direct positive interactions for
community species diversity. Ecology 78(7):1997-2003.
He F, Legendre P (1996) On specie area relation. Amer Nat 148(4):719-737.
Heiman KW, Micheli F (2010) Non-native ecosystem engineer alters estuarine
communities. Integr Comp Biol 1–11.

56
Hicks GRF (1977) Species associations and seasonal population densities of marine
phytal harpacticoid copepods from Cook Strait. J Mar Fresh Res 11(4): 621-43.
Holdich DH, Jones JA (1993) The distribution and ecology of British shallow-water
tanaid crustaceans (Peracarida, Tanaidacea). Journ Nat Hist 17(2):157-183.
Hopkins B (1957) The concept of minimal area. J Ecol 45:441-449.
Humes AG (1996) New genera of Copepoda (Poecilostomatoida) from the scleractinian
coral Psammocora in New Caledonia. Zool Journ Linn Soc 118:50-82.
Humes AG (1997) Two siphonostomatoid copepods (Coralliomyzontidae) associated
with the ahermatypic coral Tubastraea in the Moluccas. Hydrobiologia 344: 195–203 a.
Humes AG (1997) Two new copepod genera (Poecilostomatoida) associated with the
scleractinian coral Psammocora in New Caledonia. Zool Scrip 26:51-60 b.
Jacoby CA, Greenwood JG (1988). Spatial, temporal, and behavioral patterns in
emergence of zooplankton in the Lagoon of Heron Reef. Great Barrier Reef,
Australia. Mar Biol 97:309-328.
Johnsson RJTS (1992) A relação de espécie-area em colônias de Polydorella sp.
(Polichaeta, Spionidae) e a fauna de poliquetos acompanhantes em Ponta de Itaipú, RJ,
Brasil. Monografia, Universidade Federal Rio de Janeiro
Johnsson R, Neves EG (2005) A revision of Metapontius (Siphonostomatoida:
Artotrogidae) with the description of a new species associated with an octocoral from
Eniwetok Atoll, Marshall Islands (USA). Zootaxa 1035:51-59.
Johnsson R, Neves EG, Franco MO, Lang FS (2006) The association of two gall crabs
(Brachyura: Cryptochiridae) with the reef-building coral Siderastrea stellata Verrill,
1868. Hydrobiologia 559:379-384.
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos
69:373-386.
Jones SN (1948) The Ecology of the amphipoda of the south of the isle of Man. J Mar
Biol Assoc UK 27(2):400-439.
Kornicker, L.S. and I. G. Sohn. 1976. Phylogeny, ontogeny, and morphology of living
and fossil Thaumatocypridacea (Myodocopa: Ostracoda). Smithsonian Institution Press
219: 1-124.
Karaçuha ME, Sezgin M, Dağli E (2009) Temporal and spatial changes of crustaceans
in mixed eelgrass beds, Zostera marina L. and Z. noltii Hornem., at the Sinop peninsula
coast (the southern Black Sea, Turkey). Turk J Zool 33:375-386.
Klumpp DW, McKinnon AD, Mundy CN (1988) Motile cryptofauna of a coral reef:
abundance, distribution and trophic potential. Mar Ecol Prog Ser 45: 95-108.

57
Kuris AM, Blaustein AR, Alió JJ (1980) Host as islands. Amer Nat 116(4):570-586.
Lages BG, Fleury BG, Ferreira CEL, Pereira RC (2006) Chemical defense of an exotic
coral as invasion strategy. J Exp Mar Biol Ecol 328: 127−135.
Lages BG, Fleury BG, Pinto AC, Creed JC (2010) Chemical defenses against generalist
fish predators and fouling organisms in two invasive ahermatypic corals in the genus
Tubastraea. Marine Ecology 31:473–482.
Lages BG, Fleury BG, Menegola C, Creed JC (2011) Change in tropical rocky shore
communities due to an alien coral invasion. Mar Ecol Prog Ser 438:85–96.
Lewis JB, Snelgrove PVR (1990) Corallum morphology and composition of
crustacean cryptofauna of the hermatypic coral Madracis mirabilis. Mar Biol 106:
267-272.
Lomolino MV, Weiser MD (2001) Towards a more general species-area relationship:
diversity on all islands, great and small. Journ Biogeograp 28: 431-445.
MacArthur R, Wilson EO (1967) The theory of island biogeography. Princeton
University Press, Princeton
Machado, C.P. and N.G.S. Drozinski. 2002. Taxonomia e distribuição de
Actinocythereis brasiliensis sp. nov. (Podocopida, Trachyleberididae) na plataforma
continental brasileira. Iheringia 92(3): 5-12.
Macdonald TA, Burd BJ, Macdonald VI, Roodselar AV (2010) Taxonomic and feeding
guild classification for the marine benthic macroinvertebrates of the Strait of Georgia,
British Columbia. Can Tech Rep Fish Aquat Sci 1-63.
Martin JW e Davis GE (2001) An Updated Classification of the Recent Crustacea. Nat
Hist Mus Los Angeles Sci Ser 39:1-124.
Mantelatto MC, Creed JC, Mourão GG, Migotto AE, Lindner A (2011) Range
expansion of the invasive corals Tubastraea coccinea and Tubastraea tagusensis in the
Southwest Atlantic. Nota in Coral Reefs 1:1
May RM (1988), How Many Species are there on earth? Science 241(4872): 1441-
1449.
Melo GAS (1996) Manual de identificação dos Brachyura (Caranguejos e Siris) do
Litoral Brasileiro. Plêiade, São Paulo
Meurer BC, Lages NS, Pereira O, Palhano S, Magalhães GM (2010) First record of
native species of sponge overgrowing invasive corals Tubastraea coccinea and
Tubastraea tagusensis in Brazil. Mar Biod Rec 3:1-3.

58
Moreira J, Gestoso L, Troncoso JS (2008). Diversity and temporal variation of peracarid
fauna (Crustacea: Peracarida) in the shallow subtidal of a sandy beach: Playa América
(Galicia, NW Spain). Mar Ecol 29:12–18.
Murgui E (2007) Effects of seasonality on the species–area relationship: a case study
with birds in urban parks. Global Ecol Biogeogr 16:319-329.
Myers, A.A. and D. McGrath. 1984. A revision of the north-east Atlantic species of
Ericthonius (Crustacea:Amphipoda). Journal of the Marine Biological Association of the
United Kingdom 64: 379-400.
Neves, E.G., R. Johnsson. 2008. Three new species of Artotrogidae (Copepoda,
Siphonostomatoida) from the southeastern coast of Pernambuco State, Brazil. Zootaxa,
1932: 47-60.
Nogueira JMM (2003) Fauna living in colonies of Mussismilia hispida (Verrill)
(Cnidaria: Scleractinia) in four South Eastern Brasil Islands. Braz Arch Biol Tech
46(3):421-432.
Nohara T, Tabuki R (1990) Seasonal distribution of Ostracoda on two species of marine
plants and two holothurians in Okinawa, Japan. In Whatley R, Maybury C. Ostracoda
and Global Events. Chap-man & Hall, London.
Nour N, Damme R, Matthysen E, Dhondt AA (1999) Forest birds in forest fragments:
are fragmentation effects independent of season? Bird Study 46(3):279-288.
Oigman–Pszczol SS, Creed JC (2004). Distribution and abundance of fauna on living
tissues of two Brazilian hermatipic corals Mussismilia hispida (Verrill 1902) and
Siderastrea stellata Verrill, (1868). Hidrobiologia 563: 143-154.
Paine T (1974) Intertidal community structure experimental studies on the
relationship between a dominant competitor and its principal predator. Oecologia
15:93-120.
Paine T (1980) Food Webs: Linkage, interaction strength and community infrastructure.
Journ Anim Ecol 49(3):666-685.
Paula AF, Creed JC (2004) Two species of the coral Tubastraea (Cnidaria, Scleractinia)
in Brazil: a case of accidental introduction. Bull Mar Sci 74(1): 175–183.
Paula AF, Creed JC (2005) Spatial distribution and abundance of nonindigenous coral
genus Tubastraea (Cnidaria, Scleractinia) around Ilha Grande, Brazil. Braz J Biol 65(4):
661-673.
Paz-García DA (2007). Larval release from Tubastraea coccinea in the Gulf of
California, Mexico. Nota in Coral Reefs, 26:433.
Preston FW (1962) The canonical distribution of commonness and rarity: Part I Ecology
43(2):185-215.

59
Preston EM (1971) niche overlap and competition among five sympatric congeneric
species of Xanthid crabs. Dissertation, University of Hawaii.
Preston NP, Doherty PJ (1990) Cross-shelf patterns in the community structure of coral-
dwelling Crustacea in the central region of the Great Barrier Reef. I. Agile shrimps. Mar
Ecol Prog Ser 66:47-61.
Preston NP, Doherty PJ (1994) Cross-shelf patterns in the community structure of coral-
dwelling Crustacea in the central region of the Great Barrier Reef. 11. Cryptofauna. Mar
Ecol Prog Ser 104:27-38.
Ramos, M.I.F., R.C. Whatley, J.C. Coimbra (2004) Sub-recent marine Ostracoda
(Pontocyprididae and Bairdiidae) from the southern Brazilian continental shelf. Revista
Brasileira de Paleontologia 7(3): 311-318.
Ribeiro SM, Omena EP, Muricy G (2003) Macrofauna associated to Mycale
microsigmatosa (Porifera, Demospongiae) in Rio de Janeiro State, SE Brazil. Estuar
Coast Shelf Sci 57:951–959.
Rodriguez LF (2006) Can invasive species facilitate native species? Evidence of how,
when, and why these impacts occur. Biol Invas 8: 927–939.
Sammarco PW, Atchison AD, Boland GS (2004) Expansion of coral communities
within the Northern Gulf of Mexico via offshore oil and gas platforms. Mar Ecol Progr
Ser 280:129–143.
Sammarco PW, Porter SA, Cairns SD (2010) A new coral species introduced into the
Atlantic Ocean - Tubastraea micranthus (Ehrenberg 1834) (Cnidaria, Anthozoa,
Scleractinia): An invasive threat? Aquat Inv 5(2):131-140.
Sartoretto S, Harmelin J, Bachet F, Bejaoui N, Lebrun O, and Zibrowius H. (2008) The
alien coral Oculina patagonica De Angelis, 1908 (Cnidaria, Scleractinia) in Algeria and
Tunisia. Aquat Inv 3(2):173-180.
Schellenberg, A. 1938. Brazilianische Amphipodevmit biologischen Bemerkungen.
Zoologische Jahrbücher, Systematik, Ökologie und Geographie der Tiere 71: 203-218.
Scheiner SM (2003) Six types of species-area curves. Glob Ecol Biogeograp 12: 441-
447.
Stachowicz JJ (2001) mutualism, facilitation, and the structure of ecological
communities. BioScience 51(3):235-246.
Shimono T, Iwasaki N, Kawai H (2004) A new species of Dactylopusioides (Copepoda:
Harpacticoida: Thalestridae) infesting a brown alga, Dictyota dichotoma in Japan.
Hydrobiologia 523:9–15.
Simberloff DS, Abele LG (1976) Island biogeography theory and conservation practice.
Science 191(4224): 285-286.

60
Simberloff D (1978) using island biogeographic distributions to determine if
colonization is stochastic. Amer Nat 112(986):713-726.
Simberloff D (2006) Invasional meltdown 6 years later: important phenomenon,
unfortunate metaphor, or both? Ecology 9:912–919.
Sin T (1999) Distribution and host specialization in Tetralia crabs (Crustacea:
Brachyura) symbiotic with corals in the Great Barrier Reef, Australia. Coral Reefs,
65(3):839-850.
Sin TM, Lee AC (2000) Host specialisation in trapeziid crabs: consequences for rarity
at local scales. Proceedings 9th International Coral Reef Symposium 23-27.
Silva, C.L.A. 2010. Taxonomia e aspectos ecológicos de Tanaidacea (Crustacea:
peracarida) da plataforma continental e bancos oceânicos do nordeste do Brasil,
coletados durante o programa Revizee-NE. Dissertação, Universidade Federal de
Pernambuco, 262p.
Soares, C.M.A. 1979. Estudo ecológico da região de Itamaracá, Pernambuco, Brasil.
Anfípodos das famílias Talitridae e Amphitoidae. Trabalhos Oceanográficos da
Universidade Federal de Pernambuco 14: 93-104.
Sousa WP (1979) Experimental investigations of disturbance and ecological succession
in a rocky intertidal algal community. Ecol. Monogr. 49:227-254.
Sousa R, Gutiérrez JL, Aldridge DC (2009) Non-indigenous invasive bivalves as
ecosystem engineers. Biol Inv 11:2367-2385.
Stanley GD (2002). The evolution of modern corals and their early history. Earth Sci
Rev 60:195-225.
Stransky B, Brandt A (2010) Occurrence, diversity and community structures of
peracarid crustaceans (Crustacea, Malacostraca) along the southern shelf of Greenland.
Polar Biol 33:851–867.
Torchin ME, Lafferty KD, Dobson AP, McKenzie VJ, Kuris AM (2003) Introduced
species and their missing parasites. Nature 421:628-630.
Tsang LM, Chan BKK, Shih F, CHU KH, Chen CA (2009) Host-associated speciation
in the coral barnacle Wanella milleporae (Cirripedia: Pyrgomatidae) inhabiting the
Millepora coral. Mol Ecol 18:1463–1475.
Türkay M, Schuhmacher HT (1985) Latopilumnus tubicolus n. gen. n. sp., eine neue
korallenassoziierte Krabbe, die die Bildung einer Wohnhöhle induziert. (Crustacea:
Decapoda: Pilumnidae). Senckenbergiana mark 17:55-63.
Turner WR, Tjørve E (2005) Scale-dependence in species-area relationships.
Ecography, 28: 721-730.

61
Vitopil E, Willis BL (2001) Epifaunal community in Acropora spp. (Scleractinia) on the
Great Barrier Reef: implications of coral morphology and habitat complexity. Coral
Reefs 20:281-288.
Wallentinus I, Nyberg CD (2007) Introduced marine organisms as habitat modifiers.
Mar Poll Bull 55:323–332.
Whatley RC, Wall DR (1975) The relationship be-tween Ostracoda and algae in littoral
and sublittoral marine environments. Bull Amer Paleon Soc 65: 173–203.
Williams CB (1964) Patterns in the balance of nature. Academic Press, London.
Wirtz P (2006) Let sleeping fish lie. G Mar Env 3: 27.
Young PS (1986) Análise qualitativa e quantitativa da fauna associada a corais
hermatípicos (Coelenterata: Scleractinia) nos recifes de João Pessoa, PB. Rev Braz Zool
3:99-126.
Zavaleta ES, Hobbs RJ, Mooney HA (2001) Viewing invasive species removal in a
whole-ecosystem context. Trend Ecol Evol 16(8):454-459.
Young, P.S. 1998. Catalogue of Crustacea of Brazil. Rio de Janeiro: Museu Nacional
720p.

62
CONCLUSÕES GERAIS
Os resultados mostram que o coral invasor Tubastraea tagusensis está servindo
de substrato para crustáceos dos grupos Copepoda, Ostracoda, Amphipoda,
Isopoda, Tanaidacea e Decapoda, corroborando as expectativas.
Dentre os crustáceos encontrados, há a possibilidade de parasitas estarem
utilizando Tubastraea como recurso alimentar.
Representantes de Decapoda exóticos não foram encontrados. Assim, os
registros faunísticos sustentam que T. tagusensis está sendo ocupada por uma
fauna nativa.
Em relação aos outros grupos, devido a dificuldade de identificação em nível
específico, não foi possível avaliar se estes eram exóticos ou não (Nem em nível
de gênero e família).
Relação espécie-área foi encontrada somente para os grupos Copepoda e
Ostracoda, fato que foi atribuido no trabalho a influência da escala em
parâmetros do hábito de vida (área de habitação e tamanho do corpo) dos táxons
encontrados.
A hipótese de amostragem passiva foi aceita para colônias de até 14 cm² e
rejeitada para colônias acima de 14 cm², indicando que em escalas maiores
outros processos biológicos que não a amostragem passiva podem estar
influencindo os padrões de relação espécie-área encontrados.
As correlações entre os táxons de Crustacea encontrados não apresentaram um
padrão de interações negativas que poderiam indicar os processos ecológicos
responsáveis pela estruturação da comunidade.
Interações positivas e interações negativas com outros táxons não pertencentes à
Crustacea necessitam ser verificados para explicar a relação espécie-área
encontrada.
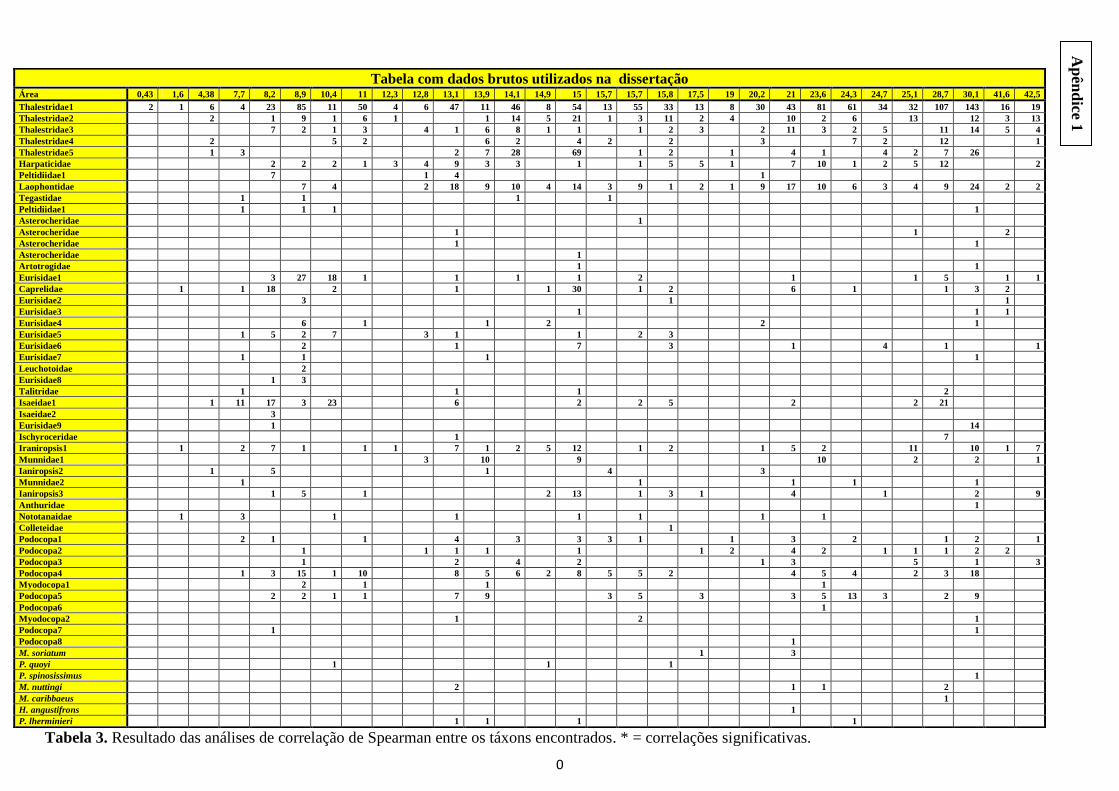
0
Tabela 3. Resultado das análises de correlação de Spearman entre os táxons encontrados. * = correlações significativas.
Tabela com dados brutos utilizados na dissertação Área 0,43 1,6 4,38 7,7 8,2 8,9 10,4 11 12,3 12,8 13,1 13,9 14,1 14,9 15 15,7 15,7 15,8 17,5 19 20,2 21 23,6 24,3 24,7 25,1 28,7 30,1 41,6 42,5
Thalestridae1 2 1 6 4 23 85 11 50 4 6 47 11 46 8 54 13 55 33 13 8 30 43 81 61 34 32 107 143 16 19
Thalestridae2 2 1 9 1 6 1 1 14 5 21 1 3 11 2 4 10 2 6 13 12 3 13
Thalestridae3 7 2 1 3 4 1 6 8 1 1 1 2 3 2 11 3 2 5 11 14 5 4
Thalestridae4 2 5 2 6 2 4 2 2 3 7 2 12 1
Thalestridae5 1 3 2 7 28 69 1 2 1 4 1 4 2 7 26
Harpaticidae 2 2 2 1 3 4 9 3 3 1 1 5 5 1 7 10 1 2 5 12 2
Peltidiidae1 7 1 4 1
Laophontidae 7 4 2 18 9 10 4 14 3 9 1 2 1 9 17 10 6 3 4 9 24 2 2
Tegastidae 1 1 1 1
Peltidiidae1 1 1 1 1
Asterocheridae 1
Asterocheridae 1 1 2
Asterocheridae 1 1
Asterocheridae 1
Artotrogidae 1 1
Eurisidae1 3 27 18 1 1 1 1 2 1 1 5 1 1
Caprelidae 1 1 18 2 1 1 30 1 2 6 1 1 3 2
Eurisidae2 3 1 1
Eurisidae3 1 1 1
Eurisidae4 6 1 1 2 2 1
Eurisidae5 1 5 2 7 3 1 1 2 3
Eurisidae6 2 1 7 3 1 4 1 1
Eurisidae7 1 1 1 1
Leuchotoidae 2
Eurisidae8 1 3
Talitridae 1 1 1 2
Isaeidae1 1 11 17 3 23 6 2 2 5 2 2 21
Isaeidae2 3
Eurisidae9 1 14
Ischyroceridae 1 7
Iraniropsis1 1 2 7 1 1 1 7 1 2 5 12 1 2 1 5 2 11 10 1 7
Munnidae1 3 10 9 10 2 2 1
Ianiropsis2 1 5 1 4 3
Munnidae2 1 1 1 1 1
Ianiropsis3 1 5 1 2 13 1 3 1 4 1 2 9
Anthuridae 1
Nototanaidae 1 3 1 1 1 1 1 1
Colleteidae 1
Podocopa1 2 1 1 4 3 3 3 1 1 3 2 1 2 1
Podocopa2 1 1 1 1 1 1 2 4 2 1 1 1 2 2
Podocopa3 1 2 4 2 1 3 5 1 3
Podocopa4 1 3 15 1 10 8 5 6 2 8 5 5 2 4 5 4 2 3 18
Myodocopa1 2 1 1 1
Podocopa5 2 2 1 1 7 9 3 5 3 3 5 13 3 2 9
Podocopa6 1
Myodocopa2 1 2 1
Podocopa7 1 1
Podocopa8 1
M. soriatum 1 3
P. quoyi 1 1 1
P. spinosissimus 1
M. nuttingi 2 1 1 2
M. caribbaeus 1
H. angustifrons 1
P. lherminieri 1 1 1 1
Ap
ênd
ice 1
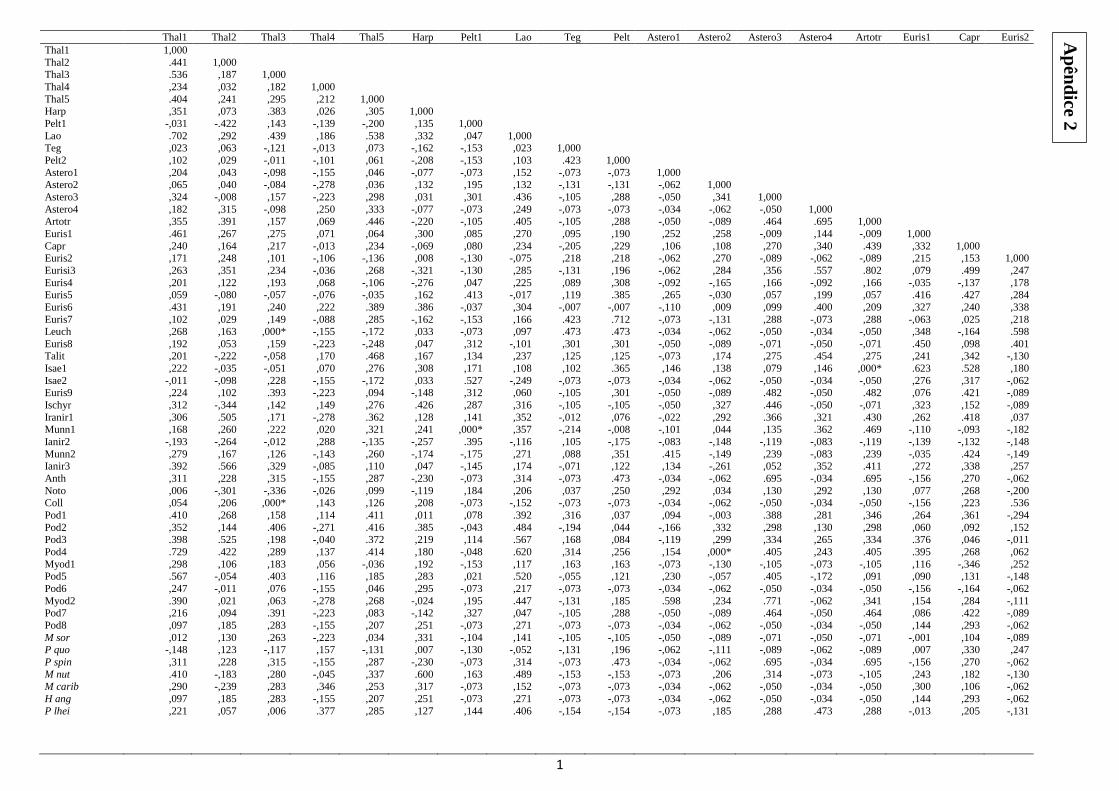
1
Thal1 Thal2 Thal3 Thal4 Thal5 Harp Pelt1 Lao Teg Pelt Astero1 Astero2 Astero3 Astero4 Artotr Euris1 Capr Euris2
Thal1 1,000
Thal2 .441 1,000
Thal3 .536 ,187 1,000
Thal4 ,234 ,032 ,182 1,000
Thal5 .404 ,241 ,295 ,212 1,000 Harp ,351 ,073 .383 ,026 ,305 1,000
Pelt1 -,031 -.422 ,143 -,139 -,200 ,135 1,000
Lao .702 ,292 .439 ,186 .538 ,332 ,047 1,000 Teg ,023 ,063 -,121 -,013 ,073 -,162 -,153 ,023 1,000
Pelt2 ,102 ,029 -,011 -,101 ,061 -,208 -,153 ,103 .423 1,000
Astero1 ,204 ,043 -,098 -,155 ,046 -,077 -,073 ,152 -,073 -,073 1,000 Astero2 ,065 ,040 -,084 -,278 ,036 ,132 ,195 ,132 -,131 -,131 -,062 1,000
Astero3 ,324 -,008 ,157 -,223 ,298 ,031 ,301 .436 -,105 ,288 -,050 ,341 1,000
Astero4 ,182 ,315 -,098 ,250 ,333 -,077 -,073 ,249 -,073 -,073 -,034 -,062 -,050 1,000
Artotr ,355 .391 ,157 ,069 .446 -,220 -,105 .405 -,105 ,288 -,050 -,089 .464 .695 1,000
Euris1 .461 ,267 ,275 ,071 ,064 ,300 ,085 ,270 ,095 ,190 ,252 ,258 -,009 ,144 -,009 1,000
Capr ,240 ,164 ,217 -,013 ,234 -,069 ,080 ,234 -,205 ,229 ,106 ,108 ,270 ,340 .439 ,332 1,000 Euris2 ,171 ,248 ,101 -,106 -,136 ,008 -,130 -,075 ,218 ,218 -,062 ,270 -,089 -,062 -,089 ,215 ,153 1,000
Eurisi3 ,263 ,351 ,234 -,036 ,268 -,321 -,130 ,285 -,131 ,196 -,062 ,284 ,356 .557 .802 ,079 .499 ,247
Euris4 ,201 ,122 ,193 ,068 -,106 -,276 ,047 ,225 ,089 ,308 -,092 -,165 ,166 -,092 ,166 -,035 -,137 ,178 Euris5 ,059 -,080 -,057 -,076 -,035 ,162 .413 -,017 ,119 .385 ,265 -,030 ,057 ,199 ,057 .416 .427 ,284
Euris6 .431 ,191 ,240 ,222 .389 .386 -,037 ,304 -,007 -,007 -,110 ,009 ,099 .400 ,209 ,327 ,240 ,338
Euris7 ,102 ,029 ,149 -,088 ,285 -,162 -,153 ,166 .423 .712 -,073 -,131 ,288 -,073 ,288 -,063 ,025 ,218 Leuch ,268 ,163 ,000* -,155 -,172 ,033 -,073 ,097 .473 .473 -,034 -,062 -,050 -,034 -,050 ,348 -,164 .598
Euris8 ,192 ,053 ,159 -,223 -,248 ,047 ,312 -,101 ,301 ,301 -,050 -,089 -,071 -,050 -,071 .450 ,098 .401 Talit ,201 -,222 -,058 ,170 .468 ,167 ,134 ,237 ,125 ,125 -,073 ,174 ,275 .454 ,275 ,241 ,342 -,130
Isae1 ,222 -,035 -,051 ,070 ,276 ,308 ,171 ,108 ,102 .365 ,146 ,138 ,079 ,146 ,000* .623 .528 ,180
Isae2 -,011 -,098 ,228 -,155 -,172 ,033 .527 -,249 -,073 -,073 -,034 -,062 -,050 -,034 -,050 ,276 ,317 -,062 Euris9 ,224 ,102 .393 -,223 ,094 -,148 ,312 ,060 -,105 ,301 -,050 -,089 .482 -,050 .482 ,076 .421 -,089
Ischyr ,312 -,344 ,142 ,149 ,276 .426 ,287 ,316 -,105 -,105 -,050 ,327 .446 -,050 -,071 ,323 ,152 -,089
Iranir1 ,306 .505 ,171 -,278 .362 ,128 ,141 ,352 -,012 ,076 -,022 ,292 .366 ,321 .430 ,262 .418 ,037 Munn1 ,168 ,260 ,222 ,020 ,321 ,241 ,000* ,357 -,214 -,008 -,101 ,044 ,135 .362 .469 -,110 -,093 -,182
Ianir2 -,193 -,264 -,012 ,288 -,135 -,257 .395 -,116 ,105 -,175 -,083 -,148 -,119 -,083 -,119 -,139 -,132 -,148
Munn2 ,279 ,167 ,126 -,143 ,260 -,174 -,175 ,271 ,088 ,351 .415 -,149 ,239 -,083 ,239 -,035 .424 -,149 Ianir3 .392 .566 ,329 -,085 ,110 ,047 -,145 ,174 -,071 ,122 ,134 -,261 ,052 ,352 .411 ,272 ,338 ,257
Anth ,311 ,228 ,315 -,155 ,287 -,230 -,073 ,314 -,073 .473 -,034 -,062 .695 -,034 .695 -,156 ,270 -,062
Noto ,006 -,301 -,336 -,026 ,099 -,119 ,184 ,206 ,037 ,250 ,292 ,034 ,130 ,292 ,130 ,077 ,268 -,200 Coll ,054 ,206 ,000* ,143 ,126 ,208 -,073 -,152 -,073 -,073 -,034 -,062 -,050 -,034 -,050 -,156 ,223 .536
Pod1 .410 ,268 ,158 ,114 .411 ,011 ,078 .392 ,316 ,037 ,094 -,003 .388 ,281 ,346 ,264 ,361 -,294
Pod2 ,352 ,144 .406 -,271 .416 .385 -,043 .484 -,194 ,044 -,166 ,332 ,298 ,130 ,298 ,060 ,092 ,152 Pod3 .398 .525 ,198 -,040 .372 ,219 ,114 .567 ,168 ,084 -,119 ,299 ,334 ,265 ,334 .376 ,046 -,011
Pod4 .729 .422 ,289 ,137 .414 ,180 -,048 .620 ,314 ,256 ,154 ,000* .405 ,243 .405 .395 ,268 ,062
Myod1 ,298 ,106 ,183 ,056 -,036 ,192 -,153 ,117 ,163 ,163 -,073 -,130 -,105 -,073 -,105 ,116 -,346 ,252 Pod5 .567 -,054 .403 ,116 ,185 ,283 ,021 .520 -,055 ,121 ,230 -,057 .405 -,172 ,091 ,090 ,131 -,148
Pod6 ,247 -,011 ,076 -,155 ,046 ,295 -,073 ,217 -,073 -,073 -,034 -,062 -,050 -,034 -,050 -,156 -,164 -,062
Myod2 .390 ,021 ,063 -,278 ,268 -,024 ,195 .447 -,131 ,185 .598 ,234 .771 -,062 ,341 ,154 ,284 -,111 Pod7 ,216 ,094 .391 -,223 ,083 -,142 ,327 ,047 -,105 ,288 -,050 -,089 .464 -,050 .464 ,086 .422 -,089
Pod8 ,097 ,185 ,283 -,155 ,207 ,251 -,073 ,271 -,073 -,073 -,034 -,062 -,050 -,034 -,050 ,144 ,293 -,062
M sor ,012 ,130 ,263 -,223 ,034 ,331 -,104 ,141 -,105 -,105 -,050 -,089 -,071 -,050 -,071 -,001 ,104 -,089 P quo -,148 ,123 -,117 ,157 -,131 ,007 -,130 -,052 -,131 ,196 -,062 -,111 -,089 -,062 -,089 ,007 ,330 ,247
P spin ,311 ,228 ,315 -,155 ,287 -,230 -,073 ,314 -,073 .473 -,034 -,062 .695 -,034 .695 -,156 ,270 -,062
M nut .410 -,183 ,280 -,045 ,337 .600 ,163 .489 -,153 -,153 -,073 ,206 ,314 -,073 -,105 ,243 ,182 -,130 M carib ,290 -,239 ,283 ,346 ,253 ,317 -,073 ,152 -,073 -,073 -,034 -,062 -,050 -,034 -,050 ,300 ,106 -,062
H ang ,097 ,185 ,283 -,155 ,207 ,251 -,073 ,271 -,073 -,073 -,034 -,062 -,050 -,034 -,050 ,144 ,293 -,062
P lhei
,221 ,057 ,006 .377 ,285 ,127 ,144 .406 -,154 -,154 -,073 ,185 ,288 .473 ,288 -,013 ,205 -,131
Ap
ênd
ice 2

2
Eurisi3 Euris4 Euris5 Euris6 Euris7 Leuch Euris8 Talit Isae1 Isae2 Euris9 Ischyr Iranir1 Munn1 Ianir2 Munn2 Ianir3 Anth
Thal1
Thal2
Thal3
Thal4
Thal5 Harp
Pelt1
Lao Teg
Pelt2
Astero1 Astero2
Astero3
Astero4
Artotr
Euris1
Capr Euris2
Eurisi3
Euris4 1,000 Euris5 -,119 1,000
Euris6 -,073 ,237 1,000
Euris7 .527 ,119 -,007 1,000 Leuch .446 ,265 ,318 .473 1,000
Euris8 ,267 .445 ,159 ,301 .719 1,000 Talit -,194 ,235 .418 ,125 -,073 -,105 1,000
Isae1 -,206 .686 .393 ,102 ,206 .364 .536 1,000
Isae2 -,092 ,358 -,110 -,073 -,034 .670 -,073 ,303 1,000 Euris9 ,176 ,160 -,159 ,301 -,050 .429 -,105 ,103 .670 1,000
Ischyr -,133 ,050 ,357 -,105 -,050 -,071 .708 .420 -,050 -,071 1,000
Iranir1 ,155 ,140 ,240 ,181 -,022 ,145 ,210 ,230 ,232 .367 -,003 1,000 Munn1 ,089 -,024 ,025 ,260 -,101 -,146 ,022 -,187 -,101 ,145 -,146 .389 1,000
Ianir2 ,194 -,027 -,265 ,061 -,083 ,272 -,174 -,006 .479 ,272 -,119 -,102 -,024 1,000
Munn2 -,022 ,051 -,093 ,351 -,083 -,119 ,079 ,134 -,083 ,251 -,119 ,154 -,056 -,199 1,000 Ianir3 ,275 ,218 .584 ,122 ,304 ,319 -,054 ,143 ,134 ,255 -,210 .458 ,091 -,198 ,164 1,000
Anth ,323 -,119 -,110 .473 -,034 -,050 -,073 -,146 -,034 .719 -,050 ,276 ,289 -,083 .415 ,219 1,000
Noto -,106 .368 -,017 ,037 -,111 -,160 .439 ,319 -,111 -,160 ,120 ,202 ,051 -,073 ,174 -,167 -,111 1,000 Coll -,092 ,318 ,345 -,073 -,034 -,050 -,073 ,230 -,034 -,050 -,050 ,111 -,101 -,083 -,083 ,255 -,034 -,111
Pod1 -,190 ,051 ,203 ,037 -,164 -,057 .472 ,204 ,094 ,213 ,306 ,332 -,059 -,025 .469 ,159 ,199 ,122
Pod2 ,010 -,156 ,258 ,200 ,130 -,018 ,124 -,039 -,166 ,096 ,187 ,128 .417 -,270 ,063 ,098 ,284 -,129 Pod3 ,125 -,058 .376 ,084 ,199 ,065 ,140 ,126 -,119 ,065 ,096 .646 ,279 -,133 ,077 ,311 ,199 ,039
Pod4 ,351 ,191 ,198 .373 ,298 ,252 ,239 ,258 ,044 ,268 ,201 .439 ,218 -,021 ,281 ,306 ,320 ,088
Myod1 .540 -,029 ,015 .441 .527 ,341 -,153 -,103 -,073 -,105 -,105 ,021 ,303 ,053 -,175 ,096 -,073 -,029 Pod5 ,127 -,007 ,063 ,267 ,080 ,116 ,006 ,053 ,080 ,278 ,243 -,065 ,099 ,098 .410 ,094 ,299 ,004
Pod6 -,092 -,119 -,110 -,073 -,034 -,050 -,073 -,146 -,034 -,050 -,050 ,111 .405 -,083 -,083 -,146 -,034 ,292
Myod2 ,074 ,215 ,009 ,185 -,062 -,089 ,174 ,156 -,062 ,355 ,327 ,280 ,044 -,148 .457 ,127 .536 ,291 Pod7 ,166 ,172 -,159 ,288 -,050 .446 -,105 ,113 .695 .999 -,071 .366 ,135 ,285 ,239 ,254 .695 -,160
Pod8 -,092 -,119 ,248 -,073 -,034 -,050 -,073 ,146 -,034 -,050 -,050 ,177 -,101 -,083 .415 ,279 -,034 -,111
M sor -,133 -,171 ,108 -,105 -,050 -,071 -,105 ,007 -,050 -,071 -,071 -,022 -,146 -,119 ,251 ,301 -,050 -,160 P quo ,129 ,349 ,074 -,131 -,062 -,089 -,130 ,261 -,062 -,089 -,089 ,040 -,182 -,148 -,149 ,196 -,062 ,042
P spin ,323 -,119 -,110 .473 -,034 -,050 -,073 -,146 -,034 .719 -,050 ,276 ,289 -,083 .415 ,219 1.000 -,111
M nut M carib
H ang
P lhei
-,194 -,092
-,092
,024
-,073 -,119
-,119
,084
,347 ,248
,248
,226
-,153 -,073
-,073
,135
-,073 -,034
-,034
-,073
-,104 -,050
-,050
-,105
.480
.527
-,073
.403
,327 ,327
,146
,058
-,073 -,034
-,034
-,073
-,104 -,050
-,050
-,105
.731
.719
-,050
,275
,148 -,221
,177
,163
,036 -,101
-,101
,298
-,174 -,083
-,083
,061
,070 -,083
.415
,088
-,098 -,146
,279
-,045
-,073 -,034
-,034
-,073
,191 -,111
-,111
,191

3
Coll Pod1 Pod2 Pod3 Pod4 Myod1 Pod5 Pod6 Myod2 Pod7 Pod8 M sor P quo P spin M nut M carib H ang P lhei
Thal1 Thal2
Thal3
Thal4 Thal5
Harp
Pelt1 Lao
Teg
Pelt2
Astero1
Astero2
Astero3 Astero4
Artotr
Euris1 Capr
Euris2
Eurisi3 Euris4
Euris5 Euris6
Euris7
Leuch Euris8
Talit
Isae1 Isae2
Euris9
Ischyr Iranir1
Munn1
Ianir2 Munn2
Ianir3
Anth Noto 1,000
Coll -,164 1,000
Pod1 -,166 ,046 1,000 Pod2 -,119 .399 ,235 1,000
Pod3 -,011 .562 ,165 ,345 1,000
Pod4 -,073 -,215 ,202 -,067 .470 1,000 Myod1 -,172 ,297 ,311 -,070 .558 ,328 1,000
Pod5 -,034 -,164 ,284 -,119 ,154 .454 ,230 1,000
Pod6 -,062 .371 ,134 ,192 .423 -,130 .472 -,062 1,000 Myod2 -,050 ,211 ,085 ,057 ,262 -,105 ,273 -,050 ,341 1,000
Pod7 -,034 ,281 ,344 ,318 ,088 -,073 ,161 -,034 -,062 -,050 1,000
Pod8 -,050 ,095 ,346 ,153 -,080 -,105 ,231 -,050 -,089 -,071 .719 1,000 M sor .557 -,295 -,298 -,214 -,053 -,130 -,192 -,062 -,111 -,089 -,062 -,089 1,000
P quo -,034 ,199 ,284 ,199 ,320 -,073 ,299 -,034 .536 .695 -,034 -,050 -,062 1,000
P spin -,073 ,301 .455 ,180 ,280 ,106 .386 .436 ,206 -,105 .436 ,273 -,130 -,073 1,000 M nut -,034 ,094 ,130 -,119 ,044 -,073 ,080 -,034 -,062 -,050 -,034 -,050 -,062 -,034 .509 1,000
M carib -,034 ,281 ,344 ,318 ,088 -,073 ,161 -,034 -,062 -,050 1.000 .719 -,062 -,034 .436 -,034 1,000
H ang -,073 ,347 ,119 ,154 .384 ,125 .382 -,073 ,185 -,105 -,073 -,105 -,131 -,073 ,153 -,073 -,073 1,000

4