silvio pires gomes - usp€¦ · até tocar o céu se crer... acredite que nenhum de nós já...
TRANSCRIPT


SILVIO PIRES GOMES
Avaliação estereológica da
microestrutura do fígado em
animais desnutridos e submetidos à
renutrição protéica
São Paulo
2011

Silvio Pires Gomes
Avaliação estereológica da microestrutura do fígado em animais desnutridos e submetidos à renutrição protéica.
Tese apresentada ao Programa de Pós-Graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção
do título de Doutor em Ciências
Departamento:
Cirurgia
Área de Concentração:
Anatomia dos Animais Domésticos e Silvestres
Orientador:
Prof. Dr. Antonio Augusto Coppi
São Paulo
2011



FOLHA DE AVALIAÇÃO
Nome: GOMES, Silvio Pires
Título: Avaliação estereológica da microestrutura do fígado em animais desnutridos e submetidos à renutrição protéica.
Tese apresentada ao Programa de Pós-Graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Doutor em Ciências
Data: ______ /______/______
BANCA EXAMINADORA
Prof. Dr. ________________________________Instituição: ___________________
Assinatura: _____________________________Julgamento: ___________________
Prof. Dr.:________________________________Instituição: ___________________
Assinatura:_____________________________Julgamento: ___________________
Prof. Dr. ________________________________Instituição: ___________________
Assinatura:_____________________________Julgamento: ___________________
Prof. Dr. ________________________________Instituição: ___________________
Assinatura: _____________________________Julgamento: ___________________
Prof. Dr.:________________________________Instituição: ___________________
Assinatura:_____________________________Julgamento: ___________________

Testamento de um cão
Minhas posses materiais são poucas e eu deixo tudo para você...
Uma coleira mastigada em uma das extremidades, faltando dois botões, uma desajeitada
cama de cachorro e uma vasilha de água que se encontra rachada na borda.
Deixo para você a metade de uma bola de borracha, uma boneca rasgada que você vai
encontrar debaixo da geladeira, um ratinho de borracha sem apito que está debaixo do
fogão da cozinha e uma porção de ossos enterrados no canteiro de rosas e sob o assoalho
da minha casinha.
Além disso, eu deixo para você a memória, que, aliás, são muitas.
Deixo para você a memória de dois enormes e meigos olhos, marrons, de uma caudinha
curta e espetada, de um nariz molhado e de choradeira atrás da porta.
Deixo para você uma mancha no tapete da sala de estar junto à janela, quando nas tardes
de inverno eu me apropriava daquele lugar, como se fosse meu, e me enrolava feito uma
bolinha para pegar um pouco de sol.
Deixo para você um tapete esfarrapado em frente de sua cadeira preferida, o qual nunca foi
consertado com o tipo de linha certo e isso é verdade. Eu o mastiguei todinho, quando
ainda tinha cinco meses de idade, lembra-se?
Deixo para você um esconderijo que fiz no jardim debaixo dos arbustos perto da varanda da
frente, onde eu encontrava asilo durante aqueles dias de verão. Ele deve estar cheio de
folhas agora e por isso talvez você tenha dificuldades em encontrá-lo. Sinto muito!
Deixo também só para você, o barulho que eu fazia ao sair correndo sobre as folhas de
outono, quando passeávamos pelo bosque.
Deixo ainda, a lembrança de momentos pelas manhãs, quando saíamos junto pela margem
do riacho, e você me dava àqueles biscoitos de baunilha.
Recordo-me das suas risadas, porque eu não consegui alcançar aquele coelho impertinente.
Deixo-lhe como herança minha devoção, minha simpatia, meu apoio quando as coisas não
iam bem, meus latidos quando você levantava a voz aborrecida e a e minha frustração por
você ter ralhado comigo.
Eu nunca fui à igreja e nunca escutei um sermão. No entanto, mesmo sem haver falado
sequer uma palavra em toda a minha vida, deixo para você o exemplo de paciência, amor e
compreensão.
Sua vida tem sido mais alegre, porque eu estive ao seu lado!
(Autoria Desconhecida)

Indiferença
Não passe tão indiferente só porque eu não sou gente, só porque não sei falar. Também sou um ser vivente, sinto as dores que você sente mas não posso me expressar.
Sou um bicho abandonado, pela vida maltratado,
quase sempre escorraçado, até mesmo apedrejado! Vivo sedento e faminto,
ninguém quer saber o que sinto! Se ficar doente e triste
vejo logo um dedo em riste. E vem a sentença fatal:
-Melhor matar este animal! -Ele deve estar raivoso!
Para sua comodidade vive dizendo inverdade, fazendo muita maldade,
seu mentiroso. Mesmo que esteja raivoso, Você mata o próprio irmão, já foi descoberta a vacina. Mas para a sua raiva humana, ainda não existe remédio, nenhuma medicação, com toda sua evolução, na história da medicina!
Faz guerras, assalta, mata com ou sem razão,
às vezes por ambição! É bem pior que eu,
que chamam de vira-lata! Olhe bem para o meu semblante: -Estou triste, apavorado, pois, a qualquer instante, posso ser sacrificado! Mas você não se importa nem com o seu semelhante! -Você sim, está doente, egoísta, indiferente. Mas se algo ruim lhe acontece logo lembra que há Deus, chora, reza e faz prece...
Mas Deus só ajuda aquele que de todos se compadece. Lembre-se do que escreveu
São Francisco de Assis: - Quem maltrata um animal
jamais poderá ser feliz!
(Autoria Desconhecida)

Conquistando o Impossível
Acredite é hora de vencer
Essa força vem
De dentro de você
Você pode
Até tocar o céu se crer...
Acredite que nenhum de nós
Já nasceu com jeito
Pra super-herói
Nossos sonhos
A gente é quem constrói...
É vencendo os limites
Escalando as fortalezas
Conquistando o impossível
Pela fé...
Campeão, vencedor
Deus dá asas, faz teu voo
Campeão, vencedor
Essa fé que te faz imbatível
Te mostra o teu valor...
Acredite que nenhum de nós
Já nasceu com jeito Pra super-herói Nossos sonhos
A gente é quem constrói..
É vencendo os limites
Escalando as fortalezas
Conquistando o impossível
Pela fé...
Campeão, vencedor
Deus dá asas, faz teu voo
Campeão, vencedor
Essa fé que te faz imbatível Te mostra o teu valor...
Tantos recordes
Você pode quebrar
As barreiras
Você pode ultrapassar
E vencer...
Campeão, vencedor
Deus dá asas, faz teu voo
Campeão, vencedor
Essa fé que te faz imbatível
(Autora: Jamily)
Sabor de Mel
O agir de Deus é lindo
Na vida de quem é fiel
No começo tem provas amargas
Mas no fim tem o sabor do mel
Eu nunca vi um escolhido sem resposta
Porque em tudo Deus lhe mostra uma
solução
Até nas cinzas ele clama e Deus atende
Lhe protege
Lhe defende com as suas fortes mãos
Você é um escolhido
E a tua história não acaba aqui
Você pode estar chorando agora
Mas amanhã você irá sorrir.
Deus vai te levantar das cinzas e do pó
Deus vai cumprir tudo que tem te
prometido
Você vai ver a mão de Deus te exaltar
Quem te vê há de falar Ele é mesmo escolhido.
Vão dizer que você nasceu pra vencer
Que já sabiam porque você
Tinha mesmo cara de vencedor
E que se Deus quer agir ninguém pode
impedir
Então você verá cumprir cada palavra
Que o Senhor falou,
Quem te viu passar na prova
E não te ajudou
Quando ver você na benção
Vão se arrepender
Vai estar entre a platéia
E você no palco
Vai olhar e ver
Jesus brilhando em você
Quem sabe no teu pensamento
Você vai dizer
Meu Deus como vale a pena
A gente ser fiel
Na verdade a minha prova Tinha um gosto amargo
Mas minha vitória hoje
Tem sabor de mel.
A minha vitória hoje
Tem sabor de mel.
(Autor: Agailton Silva)

Ao que sempre me mostrou o quanto é importante correr atrás dos sonhos, mesmo que os caminhos sejam longos e que mesmo que coloquem pedras em seus
caminhos, mas com muita fé podemos fazer da dor um sorriso e saber que Deus estará sempre ao nosso lado, seja qual for o caminho escolhido por nós. Ele sabe
que é muito difícil ir atrás dos sonhos, mas nunca deixa ninguém estragar tudo aquilo que conseguimos construir, pois Deus nos mantém forte mesmo quando não conseguimos nos firmar em pé. Por isso agradeço a Deus por tudo que ele me deu
até hoje, pela ajuda incondicional quando eu mais precisei dele. Pois só ele consegue olhar o herói que temos dentro de nós, pois ele nos fortalece e nos
protege. Muito Obrigado Deus por abençoar todos os dias de minha vida.

Aos meus pais Joaquim Gomes e Zulmira Pires e a minha irmã Fátima Gomes e meu cunhado Evandro Bertoncin por estarem sempre ao meu lado em todas as
horas em que mais precisei e por saber que posso contar com todos em todos os momentos de minha vida. Pois todos sabem o quanto é importante o apoio da
família para que todos os nossos sonhos possam ser realizados, pois é através da família que vencemos os obstáculos que a vida faz enfrentarmos todos os dias. E
agradeço muito a minha mãe por sempre acreditar nos meus sonhos e da força que sempre me deu para continuar a seguir em frente e conseguir trilhar o meu futuro, a ela hoje dedico a minha tese de doutorado, pois sem ela não seria o que sou hoje e
sei que ela estará muito feliz hoje aqui do meu lado para realizar mais essa conquista.

Hoje quero agradecer ao meu cachorro Xuxo onde quer que esteja, estará sempre
em meus pensamentos e que nunca será esquecido ao longo de minha vida, como o “único” e fiel amigo que transformou toda a minha vida, pois foi por conta dele que fiz
a minha graduação em Medicina Veterinária. E hoje graças a ele estou aqui conquistando mais um título que será celebrado por mim em pensamento a ele, pela
eterna gratidão que teve comigo. Ainda que tenha passado quase 10 anos, nunca me esqueci de você meu eterno amigão.
Eu Te Amo Muito.

A minha cachorrinha Odara que faz dos dias tristes de minha vida muito mais alegre
e que sei que hoje ela é o anjo da minha vida e por toda essa fidelidade e amor que ela tem por mim eu tento ao máximo retribuir com muito amor, carinho e proteção
essa amizade fraternal que tenho por você. Quero que saiba que enquanto eu estiver por aqui nada de mal irá acontecer com você. Pois por você sempre farei qualquer coisa. Amo-Te minha cachorrinha mais carente do mundo. E agradeço
também ao Hércules por ser o fiel amigo de minha cachorra e por defendê-la
sempre que ela precisar.

AGRADECIMENTOS
São em momentos de mais tristezas que paramos para pensar no que
aconteceu em nossas vidas, nas pessoas que por elas passaram; as coisas
que fizemos e tudo que aprendemos. São nesses momentos que começamos
a lembrar de grandes amizades, que sempre estiveram presentes conosco
quando precisávamos e que nunca poderíamos esquecer.
(Autoria Desconhecida)
À Faculdade de Medicina Veterinária e Zootecnia da Universidade de São
Paulo, por proporcionar meu desenvolvimento científico, profissional e
pessoal.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico -
CNPQ (141901 / 2007 - 1) pelo apoio financeiro para a realização deste
trabalho.
Ao meu orientador, Prof. Dr. Antonio Augusto Coppi, pela amizade,
confiança durante todos esses anos que estivemos juntos e também pelo
aprendizado oferecido a mim e por me mostrar o quanto é importante uma
pesquisa científica séria e por estar sempre presente quando se precisa dele,
seja a qualquer hora do dia ou da noite e por ter me aceitado como seu
orientado durante todos os anos de meus estudos. Agradeço a Deus por hoje
poder ter feito e estar fazendo parte da sua equipe profissional o que me
honra muito.

Ao Laboratório de Estereologia e Anatomia Química – L.S.S.C.A., sob
orientação do Prof. Dr. Antonio Augusto Coppi pela oportunidade de fazer
parte do time de profissionais que nele estão comprometidos e pela
oportunidade do aprendizado durante a realização de minha tese.
Ao Programa de Pós-Graduação em Anatomia dos Animais Domésticos e
Silvestres da Universidade de São Paulo e pela oportunidade de crescer
profissionalmente me tornando um doutor em ciências.
Aos Professores Doutores do setor de Anatomia, pelo aprendizado durante
as aulas de Pós-Graduação.
Aos Professores Doutores Antonio Augusto Coppi, Francisco Blasquez,
Romeu de Souza, Márcia Machado, Laura Maifrino por ter aceitado o
convite para a minha banca de doutorado e por se deslocarem para que
pudessem me ajudar em mais essa etapa de minha vida. Meus mais sinceros
agradecimentos.
A Professora Doutora Patrícia Castelucci pela ajuda essencial durante todo o
meu mestrado e também pela ajuda oferecida durante o meu doutorado e

pela participação importantíssima durante a minha qualificação de doutorado
e pela amizade durante todos esses anos.
Ao Prof. Dr. Romeu Rodrigues de Souza pela amizade, confiança durante
todos esses anos e por aceitar o convite de participar da minha banca de
doutorado.
A minha grande amiga Tais Sasahara pela ajuda incondicional nesta minha
tese de doutorado e por mais de uma década de amizade, que fizeram com
que a nossa amizade ficasse ainda mais sincera ao longo dos anos.
Ao meu amigo Fernando Ladd por tudo que convivemos juntos durante estes
anos de L.S.S.C.A. e pela sua amizade que sempre que sempre tivemos e
que me faz acreditar que existem amigos para todas as ocasiões fora e dentro
do campo profissional. Conte comigo sempre.
A minha amiga Luciana Abrahão por ser essa amiga muito especial e que
possui um coração grandioso e saber que uma amizade sobrevive a todos os
momentos, seja qual for à circunstância e sei que hoje você se faz necessária
em minha vida. Espero poder ser seu amigo por muitos e muitos anos.
A minha amiga Andrea Almeida por tudo que vivemos juntos, por esses anos
de convivência, pelos momentos de luta, garra, força, dedicação e
descontração que passamos juntos durante todo esse tempo. Quero
agradecer pela amizade sincera que você sempre teve comigo e quero que
saiba que lhe considero muito. Obrigado pela ajuda em minha tese e pela
amizade que temos um pelo outro. Te adoro muito!!!
Ao meu amigo Demilto da Pureza pela verdadeira amizade que durante
esses anos pudemos manter sempre da maneira mais bonita que uma
amizade sincera poderia trilhar e caminhar. Pelo apoio, ajuda e por sempre

estender a mão para me ajudar em qualquer momento que eu precisar.
Amigão saiba que estarei sempre por aqui quando você precisar. Mesmo você
indo para tão longe espero que você continue a ser este meu amigo que me
honra muito.
Ao meu amigo Felipe da Roza pela amizade e pela ajuda que você sempre
me ofereceu durante todos esses anos e também pelos momentos de
descontração que é essencial em uma boa amizade.
A minha amiga Aliny Ladd pela amizade sincera e por tudo que passamos
durante essa longa caminhada. Quero lhe dizer que você é uma amiga muito
importante para mim e que tudo que você precisar pode contar comigo
sempre. Obrigado pela amizade minha amiga.
A minha fiel amiga Eliane Muniz por tudo que passamos juntos, por esses
anos de convivência, pelos momentos de luta, garra, força, dedicação e muita
descontração que passamos juntos por este pequeno período, mas que para
mim foi muito importante. Quero poder contar sempre com a sua amizade.
Pois ela é única. E tenho a certeza que ainda teremos mais pela frente nesta
nossa longa amizade.
A Ana Paula Frigo pelos momentos de descontração no L.S.S.C.A. e pela
amizade que foi construída durante este pequeno tempo de conhecimento.
Aos meus amigos e companheiros do L.S.S.C.A. (Laboratory of Stochastic
Stereology and Chemical Anatomy): pelo companheirismo e saiba que hoje
nos tornamos mais que companheiros de trabalhos, mas grandes amigos.
Obrigado também pelos momentos de descontração que vivemos durante
todos esses anos e por me aturarem arduamente durante anos.

A minha amiga Eunice Leite que durante esses 12 anos de amizade que
mostrou o quanto uma amizade pode ser valiosa e que a cada dia mais nossa
amizade se torna mais grandiosa e importante para mim.
Aos amigos de Pós-Graduação deste departamento agradeço por fazer parte
integrante dos momentos passados aqui na Anatomia, pois de uma forma
direta ou indireta fizeram parte da minha vida.
Aos funcionários da Biblioteca da FMVZ-USP.
Aos funcionários da FMVZ-USP, pela consideração, atenção e respeito dado
durante esta minha estada neste programa de Anatomia dos Animais
Domésticos e Silvestres, em especial: Maicon, Jackeline, Edinaldo (Índio),
Diogo, Ronaldo, João, Raimundo, Natália (Branca), Claudia.
.

RESUMO
GOMES, S. P. Avaliação estereológica da microestrutura do fígado em animais desnutridos e submetidos à renutrição protéica. [Stereological evaluation of
microstructure in the liver of animals denutrition of followed by proteic renutrition]. 2011. 83 f. Tese (Doutorado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2011.
A fome e a má nutrição permanecem entre os problemas mais devastadores que
enfrentam a maioria dos pobres e carentes do mundo. A desnutrição está
relacionada com o desequilíbrio do estado nutricional e ocorre sob diversas formas,
podendo afetar o fígado. A desnutrição no início da vida pode afetar profundamente
o corpo e o cérebro de ratos adultos. Alguns desses efeitos são permanentes
enquanto outros desaparecem quando os animais são submetidos ao processo de
renutrição. Sendo assim, outra maneira de interferir na regeneração hepática
fundamenta-se no fato de que os processos proliferativos do fígado podem ser
modulados por fatores alimentares. Experimentos de desnutrição protéico-energética
mostraram que o fígado de ratos desnutridos regenera-se mais lentamente após a
hepatectomia. O objetivo deste estudo foi avaliar os efeitos da desnutrição e
renutrição protéica na microestrutura dos fígados de camundongos. Foram utilizados
12 camundongos Swiss machos, subdividos nos seguintes grupos: Controle do
desnutrido (CD) (n=3), Desnutrido (D) (n=3), Renutrido (R) (n=3) e Controle do
renutrido (CR) (n=3). Os animais dos dois grupos controle e do grupo renutrido
receberam ração normoprotéica (12%) ao passo que o grupo desnutrido recebeu
ração hipoprotéica contendo (2%). Os seguintes parâmetros estereológicos do
fígado foram estimados: volume, número total e o volume menor (p= 0,001) em
relação ao CR (R= 8020 µm³ e CR= 10018 µm³). O número total de hepatócitos nos
animais desnutridos não apresentou diferença significativa (p= 0,055) em
comparação ao seu CR (D = 1.1 x 107
e CD=1.6 x 107) e o R não apresentou
diferença significativa (p= 0,055) em comparação ao seu CR (R= 2.8 x 107 e CR=
2.2 x 107). Portanto, a renutrição atenuou a atrofia do volume do hepatócito
ocasionada pelo processo de desnutrição (carência de proteína).
Palavras-chave: Estereologia. Desnutrição. Renutrição. Camundongo. Fígado.

ABSTRACT
GOMES, S. P. Stereological evaluation of microstructure in the liver of animals denutrition of followed by proteic renutrition. [Avaliação estereológica da
microestrutura do fígado em animais desnutridos e submetidos à renutrição protéica]. 2011. 83 f. Tese (Doutorado em Ciências) – Faculdade de Medicina
Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2011.
The hunger and the malnutrition remain the most devastating problems faced by the
majority of poor and needy people in the world. The malnutrition is related to an
unbalanced nutritional condition and it occurs in different ways may affecting the
liver. In the beginning of life, the malnutrition can affect profoundly the body and the
brain in adult rats. Most of these effects are permanent while others disappear when
the animal are submitted to a renutrition process. In this way, other manner to
interfere in the hepatic regeneration is based on the fact that the liver proliferative
processes might be modulating by food factors. The energetic protein malnutrition
experiments have showed that the undernourished rat livers regenerate slower after
hepatectomy. The aim of this study was to evaluate the effects of the caloric protein
malnutrition and renutrition in the liver microstructure. In this study were used 12
Swiss male mice divided in: control of the undernourished mice (UC) (n=3),
undernourished (U) (n=3), renourished (RN) (n=3) and control of the renourished
(RNC) (n=3). The animals from both control groups and the undernourished group
received normal protein food (12% protein) and the undernourished group received
hypoprotein food (2% protein). The following stereological parameters were
estimated: volume, total number and mean volume of the hepatocytes. The mean
volume of U was significantly lower (p=0.001) than the UC (U= 23.57 mm³ e UC=
46.5 2mm³) and RN was lower (p=0.001) than the RNC (RN= 38.89 mm³ e RNC=
52.03 mm³).The mean volume of hepatocytes in undernourished animals was lower
(p=0.001) in comparison with the respective control (U= 4505 µm³ e CD= 8170.13
µm³) and the RN was lower (p= 0.001) in relation to the RNC (RN= 8020 µm³ e
RNC= 10018 µm³). The total number of hepatocytes in undernourished animals did
not presented significant difference (p= 0.055) in comparison with the respective
control (U = 1.1 x 107
e UC= 1.6 x 107). So, the renutrition attenuated the hepatocyte
volume atrophy caused by the malnutrition process (protein deficiency).
Keywords: Stereology. Malnutrition. Renutrition. Mice. Liver.

SUMÁRIO
1 INTRODUÇÃO ............................................................................................................ 26 2 OBJETIVOS ................................................................................................................ 29
2.1 OBJETIVO PRINCIPAL ........................................................................................... 29 2.1.1 Estudo qualitatitvo .............................................................................................. 29 2.1.2 Estudo histoquantitativo (Estereológico) ....................................................... 29
2.2 OBJETIVOS SECUNDÁRIOS ................................................................................. 30 3 REVISÃO SISTEMÁTICA DA LITERATURA ........................................................... 32
3.1 ASPECTOS MORFOFUNCIONAIS DO FÍGADO .................................................. 32 3.2 ASPECTOS NUTRICIONAIS .................................................................................. 35 4 MATERIAIS E MÉTODOS ......................................................................................... 41
4.1 MATERIAIS .............................................................................................................. 41 4.1.1 Animais ................................................................................................................. 41
4.2 MÉTODOS................................................................................................................ 42 4.2.1 Rações .................................................................................................................. 42 4.2.2 Indução da desnutrição ..................................................................................... 43 4.2.3 Recuperação nutricional.................................................................................... 44 4.2.4 Obtenção de amostras de sangue ................................................................... 44
4.2.4.1 Sangue total ....................................................................................................... 44 4.2.4.2 Realização do hemograma ............................................................................... 44 4.2.4.3 Análises bioquímicas ......................................................................................... 45 4.2.5 Estudo histológico .............................................................................................. 45 4.2.6 Estudo estereológico ......................................................................................... 46
4.2.6.1 Estudo morfoquantitativo (estereológico) do fígado ........................................ 46 4.2.6.1.1 Volume do fígado (Vref) ................................................................................. 46 4.2.6.1.2 Coeficiente de erro ......................................................................................... 48 4.2.6.1.3 Número total de hepatócitos (N) .................................................................... 49
4.2.6.1.4 Volume médio dos hepatócitos ( Nv ) ............................................................. 50 5 ANÁLISE ESTATÍSTICA INFERENCIAL ................................................................. 52 6 RESULTADOS ............................................................................................................ 54 6.1 PESO CORPORAL .................................................................................................. 54 6.2 PESO DO FÍGADO .................................................................................................. 55 6.3 MICROESTRUTURA ............................................................................................... 56 6.4 VOLUME DO FÍGADO (Vref) .................................................................................. 57 6.4.1 Princípio de cavalieri .......................................................................................... 57
6.5 COEFICIENTE DE ERRO DO VOLUME DO FÍGADO (CE) ................................. 58 6.6 NÚMERO TOTAL DE HEPATÓCITOS (N) ............................................................ 60
6.7 VOLUME MÉDIO DOS HEPATÓCITOS ( Nv ) ...................................................... 60 6.8 AVALIAÇÕES BIOQUÍMICAS ................................................................................. 61 7 DISCUSSÃO ............................................................................................................... 65
7.1 MODELOS DE DESNUTRIÇÃO PROTÉICA ......................................................... 65 7.2 VOLUME DO FÍGADO (Vref) .................................................................................. 67 7.3 NÚMERO TOTAL DE HEPATÓCITOS (N) ............................................................ 68
7.4 VOLUME MÉDIO DOS HEPATÓCITOS ( Nv ) ....................................................... 70 7.5 AVALIAÇÕES BIOQUÍMICAS ................................................................................. 71 8 CONCLUSÕES ........................................................................................................... 74

REFERÊNCIAS .............................................................................................................. 76

LISTA DE TABELA
Tabela 1- Composição Percentual da Ração. Ração controle I e Ração
hipoproteíca II. ...........................................................................................43

LISTA DE FIGURAS
Figura 1 - Secção do fígado de animal controle do desnutrido. Sistema de pontos utilizado para a estimativa do volume do fígado a partir do Princípio de
Cavalieri. Escala de barra: 20 cm .................................................................48 Figura 2 - Fotomicrografia do fígado de um camundongo do grupo controle do
desnutrido mostrando a aplicação do disector óptico para estimativa do número total de hepatócitos. Técnica de coloração: Hematoxilina de
Mayer. Escala de barra: 20µm ......................................................................50 Figura 3 - Fotomicrografia do fígado de um camundongo do grupo desnutrido
mostrando a estimativa do volume médio dos hepatócitos pelo método rotator planar. Técnica de coloração: Hematoxilina de Mayer. Escala de
barra: 20µm ....................................................................................................51 Figura 4 - Foto macrografia mostrando comparativamente os fígados dos
camundongos dos grupos CD, D, R, CR. Escala de barra = 2 cm ............55 Figura 5 - Fotomicrografia mostrando a microestrutura do fígado de A: Controle do
desnutrido, B: Desnutrido, C: Renutrido e D: Controle do renutrido, exibindo perfis dos hepatócitos, onde: Núcleo (a) e Hepatócito (b).
Técnica de Coloração: Hematoxilina de Mayer. Escala de Barra: 20µm...57

LISTA DE GRÁFICOS
Gráfico 1 - Peso dos Animais (p= 0, 001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos .............................................................................. 54
Gráfico 2 - Peso dos Fígados (p= 0, 001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos .............................................................................. 55
Gráfico 3 - Volume dos Fígados estimados pelo Princípio de Cavalieri (p= 0, 001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos ............................... 58
Gráfico 4 - Coeficiente de erro dos fígados (à esquerda) e respectivos intervalos de confiança (à direta) estimados para os grupos Controle do Desnutrido (A e B) e Desnutrido (C e D) ................................................ 59
Gráfico 5 - Coeficiente de erro dos fígados (à esquerda) e respectivos intervalos de confiança (à direta) estimados para os grupos Renutridos (E e F) e Controle do Renutrido (G e H) ............................................................. 59
Gráfico 6 - Número total de hepatócitos (p= 0, 055*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos .............................................................. 60
Gráfico 7 - Volume médio dos hepatócitos (p= 0, 001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos .............................................................. 61
Gráfico 8 - Hemograma para os grupos CD, D, R e CR. Número de hemácias (A). A diferença entre os grupos para este parâmetro foi altamente significativa (p= 0, 001). Hematócrito (B). A diferença foi estatisticamente significativa (p= 0, 001). Hemoglobina (C). A diferença entre os grupos foi estatisticamente significativa (p= 0, 001). Leucócitos (D). Foi detectada diferença estatística entre os grupos (p= 0, 002). Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos ....................... 62
Gráfico 9 - Função hepática para os grupos CD, D, R e CR. Proteína total (A). Albumina (B). Globulina (C). ALT (D). Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos ....................................................................................................... 63
Gráfico 10 - Função hepática e glicose para os grupos CD, D, R e CR. FA (A), GGT (B). AST (C). Glicose (D). Foi detectada diferença estatística entre os grupos (p= 0, 038). Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos..... 63

Introdução

26
Gomes, SP Introdução
1 INTRODUÇÃO
De acordo com o Fundo das Nações Unidas para a Infância (UNICEF), a
desnutrição infantil é um problema de dimensões alarmantes em boa parte do
mundo. Associada à pobreza e à desigualdade, é um expressivo fator de
mortalidade de crianças nos países em desenvolvimento, apesar dos esforços
realizados nas últimas décadas para reduzir esse índice (UNICEF, 2006).
A desnutrição, segundo a Organização Mundial de Saúde, é um quadro de
carência protéico-calórica, que abrange formas graves de doenças, como o
marasmo (suprimento calórico insuficiente) e o Kwashiorkor (Privação protéica
grave), além de fases e formas moderadas (WHO, 1992).
A desnutrição infantil continua a ser um dos problemas mais importantes da
saúde pública do mundo atual, devido a sua magnitude e conseqüências
desastrosas para o crescimento (MONTE, 2000). A OMS estima que mais de 20
milhões de crianças nasça com baixo peso a cada ano, cerca de 150 milhões de
crianças com menos de 5 anos têm baixo peso para sua idade (ACC/SCN, 2000).
A desnutrição protéico-calórica é um dos maiores problemas de saúde pública
no mundo, representando nas nações subdesenvolvidas cerca de 40% das causas
dos pacientes hospitalizados. Considerando que o processo normal metabólico
precisa de proteínas, acredita-se que todos os tecidos serão afetados por um estado
precário de nutrição protéica embora não apresentem o mesmo grau e severidade
de modificação (SANT’ANA et al., 1997).
Em países industrializados também são descritos casos de desnutrição,
porém com maior acometimento de idosos (REDMOND et al., 1991b,c).
A desnutrição protéico-calórica acomete desde crianças provenientes de
comunidades carentes, até pacientes idosos, potencializando processos infecciosos
e aumentando o número de óbitos (CHANDRA, 1999).
A desnutrição protéico-energética é de longa data, considerada um problema
social, sendo inclusive apontada como um marcador de pobreza (ISSLER et al.,
1996), medida também através de habitações precárias, baixos salários,
analfabetismo, desemprego ou empregos mal remunerados (AZEREDO et al.,
1999).

27
Gomes, SP Introdução
A desnutrição no início da vida pode afetar profundamente o corpo e o
cérebro de ratos adultos. Alguns desses efeitos são permanentes enquanto outros
desaparecem quando os animais são submetidos ao processo de renutrição
(YUCEL; WARREN; GUMUSBURUN, 1994).
É sabido que o tamanho do fígado é influenciado pelo estado nutricional.
Sendo assim, outra maneira de interferir na regeneração hepática fundamenta-se no
fato de que os processos proliferativos do fígado podem ser modulados por fatores
alimentares. Experimentos de desnutrição protéico-energética mostraram que o
fígado de ratos desnutridos regenera-se mais lentamente após a hepatectomia
parcial (DOYLE; WILSON; HARTROFT, 1968).
Seu peso (órgão) em animais desnutridos chega a reduzir-se em 27%, por
hipoplasia e atrofia do mesmo (PARRA et al., 1995).
O fígado é um órgão-alvo na terapia gênica por ser uma região de muitas
alterações metabólicas. Ultimamente a transferência de genes estáveis aos
hepatócitos pode compensar deficiências que afetam suas funções (MACHADO et
al., 2002).
Analisando a literatura especializada verifica-se que embora os autores
mencionados acima, realizando experimentos em animais submetidos à desnutrição,
têm demonstrado algumas preocupações com a incidência desta condição durante o
período de gestação e lactação, mencionando seus efeitos na glândula hepática.
Constata-se uma carência principalmente de publicações enfocando os efeitos da
desnutrição, principalmente no que tange aos seus aspectos morfoquantitativos.
Desta forma o estudo da estrutura hepática de camundongos, sob os
aspectos quantitativos, estereológicos e morfométricos, durante estados de
desnutrição e renutrição protéica será fundamental apara aferir objetivamente (e não
empiricamente) os possíveis danos ocasionados pela mesma no fígado.
Ainda, o conhecimento destas informações quantitativas pode ser útil no
entendimento da patogênese hipoprotéica hepática e conseqüentemente no seu
tratamento, melhorando a qualidade de vida de pacientes animais e humanos.

Objetivos

29
Gomes, SP Objetivos
2 OBJETIVOS
Os objetivos foram divididos em: objetivo principal e objetivos secundários.
2.1 OBJETIVO PRINCIPAL
O objetivo deste estudo foi descrever os efeitos de uma dieta hipoprotéica
assim como da renutrição na microestrutura do fígado de camundongos.
2.1.1 Estudo qualitatitvo
Caracterização morfológica do fígado
2.1.2 Estudo histoquantitativo (Estereológico)
Volume do fígado (Vref)
Coeficiente de erro do volume do fígado (CE)
Intervalo de confiança para o volume do fígado
Número total de hepatócitos (N)
Volume médio dos hepatócitos ( Nv )

30
Gomes, SP Objetivos
2.2 OBJETIVOS SECUNDÁRIOS
Com o intuito de caracterizar o modelo experimental da desnutrição
hipoprotéica e da renutrição, os seguintes parâmetros foram avaliados:
Hemograma
Avaliações bioquímicas: (1) Proteína total; (2) Albumina; (3)
Globulina; (4) Alamina aminotransferase (ALT); (5) Fosfatase
alcalina (FA); (6) Gama glutamil transpeptidase (GGT); (7)
Aspartato aminotransferase (AST); (8) Glicose.

Revisão sistemática da literatura

32
Gomes, SP Revisão sistemática da literatura
3 REVISÃO SISTEMÁTICA DA LITERATURA
A literatura consultada para embasamento científico mais relevante será
discorrida a seguir:
3.1 ASPECTOS MORFOFUNCIONAIS DO FÍGADO
O fígado é o maior órgão sólido do corpo, constituído aproximadamente de
2% a 5% do peso de um homem adulto. Pode exercer até 100 funções diferentes, a
maioria das quais é exercida pelos hepatócitos (ROUILLER, 1964; GUYTON; HALL,
2002; GARTNER; HIAT, 2003; JUNQUEIRA; CARNEIRO, 2004).
O fígado é o maior tecido glandular do organismo, situado no quadrante
superior direito da cavidade abdominal, abaixo do diafragma. É um órgão
multifuncional e dentre suas principais funções destacam-se a síntese e a secreção
protéica, a secreção biliar, a regulação dos níveis glicêmicos e a metabolização de
drogas e carboidratos. Seu parênquima é constituído por hepatócitos, que perfazem
cerca de 80% da população celular (MACHADO et al., 2002).
São células altamente diferenciadas e estáveis, ou seja, apesar do baixo
índice mitótico, entram em divisão celular mediante estímulo tanto de natureza física,
quanto infecciosa, como tóxica. A lesão do parênquima hepático induzida por
quaisquer estímulos desencadeia o processo de reparação, que começou ser
elucidado com o advento da biologia molecular, cujas técnicas permitem maiores
esclarecimentos dos eventos que promovem e controlam a divisão celular, tão
importantes no prognóstico e patogênese das mais freqüentes doenças do órgão
como hepatite, cirrose e hepatomas (MACHADO et al., 2002).
A organização funcional do fígado reflete em extraordinárias funções. A maior
função do fígado envolve a degradação de aminoácidos, carboidratos, lipídios, e
vitaminas e seu subseqüente estoque, conversão metabólica, e liberação para o
sangue e bile. A bile desempenha um importante papel na digestão de lipídeos.
Todavia, muitas das funções do fígado estão inter-relacionadas, o que se torna
particularmente evidente quando surge anormalidade do fígado, visto que muitas de

33
Gomes, SP Revisão sistemática da literatura
suas funções são afetadas simultaneamente (ROUILLER, 1964; GUYTON; HALL,
2002; GARTNER; HIAT, 2003; JUNQUEIRA; CARNEIRO, 2004).
A anatomia macroscópica do fígado divide o órgão em lobos, delimitados por
sulcos ou fissuras, elementos anatômicos facilmente identificáveis na superfície da
glândula. (ROUILLER, 1964; GUYTON; HALL, 2002; GARTNER; HIAT, 2003;
JUNQUEIRA; CARNEIRO, 2004).
O fígado possui funções específicas, está interposto entre o trato digestivo (e
o baço) e o resto do corpo. Por essa interposição, o fígado é provido com um
suplemento duplo, pois ele recebe sangue da veia porta e da artéria hepática. A
principal função deste órgão é envolver eficientemente a captação de aminoácidos,
carboidratos, lipídeos e vitaminas, assim também fazendo seu armazenamento,
conversão metabólica e liberação no sangue e bile (BERK; POTHER; STREMME,
1987).
O fígado também faz a biotransformação hepática, que converte substâncias
hidrofóbicas insolúveis em água, que podem ser excretadas pela urina e bile (BERK;
POTHER; STREMME, 1987).
E por causa das funções metabólicas e secreção no sangue, o fígado é
considerado uma glândula endócrina. Ao mesmo tempo, a produção de bile e
secreção o faz também ser descrito como uma glândula exócrina (POPPER, 1988).
Histologicamente, o lóbulo hepático apresenta forma hexagonal, com uma
veia centro-lobular e com as tríades portais nas extremidades desse hexágono
imaginário. As colunas de hepatócitos dispõem-se lado a lado, a partir da veia
central e entre elas estão situados os sinusóides hepáticos, que transportam o
sangue que chega pelos ramos da veia porta e da artéria hepática e os chamados
espaços de Disse. Há ainda os canalículos biliares que transportam a bile até os
ductos biliares, nos espaços porta (GERBER; THUNG, 1983).
As células que compõe o fígado, hepatócitos, derivam do epitélio do intestino
primitivo (ALBERTS et al., 2004). São poliédricas, com seis ou mais superfícies,
variam de 12 a 25 μm de diâmetro. Contêm um e às vezes dois núcleos
arredondados que apresenta em seu interior uma rede e um ou dois nucléolos
refringentes. Alguns núcleos são poliplóides, contendo múltiplos do número haplóide
de cromossomos. Núcleos poliplóides são caracterizados pelo seu tamanho maior,
que é proporcional a ploidia (GOSS, 1988; JUNQUEIRA; CARNEIRO, 2004).

34
Gomes, SP Revisão sistemática da literatura
Aproximadamente 75% de sangue hepático vêm diretamente das vísceras
gastrointestinais e baço através da veia de porta. O sangue proveniente do sistema
porta traz drogas absorvidas diretamente pelo intestino para o fígado em forma
concentrada. Os hepatócitos são responsáveis pela síntese, pela degradação e pelo
armazenamento de um imenso número de substâncias, enzimas droga-
metabolizantes tem papel de desintoxicar muitos compostos, mas ativa a toxicidade
de outros (JAESCHKE; GORES; CDDERBAUM et al., 2002; ALBERTS; JOHNSON;
LEWIS et al., 2004).
Weibel et al. (1969) descreveram os dados morfométricos normais em fígado
de ratos. Segundo os autores, estudos pela microscopia de luz demonstraram que
96% do volume hepático dos ratos são constituídos pelo parênquima lobular, isto é,
pelos hepatócitos, sinusóides, espaços de Disse e capilares biliares, enquanto os
4% restantes constituem o parênquima não lobular, ou seja, espaços portais, veias
centro-lobulares e ramos da veia hepática.
O fígado possui outras funções não relacionadas com a digestão, como por
exemplo, a hematopoiese, a coagulação sanguínea, a fagocitose e a desintoxicação
(GUYTON; HALL, 2002).
Todas as células hepáticas (hepatócitos, células endoteliais, de Kupffer, de Ito
e ductais) proliferam para substituir a perda de tecido hepático. No entanto, os
hepatócitos são os primeiros a proliferar, sendo que as maiorias dos estudos
focalizam estas células por elas constituírem cerca de 90% da massa hepática e
60% do número total de células (RAMALHO et al., 1993; DIHEL; RAI, 1996;
MICHALOPOULUS; DEFRANCES, 1997).
Sendo o fígado o maior órgão sólido do corpo humano, o mesmo é constituído
por aproximadamente 2 a 5% do peso corpóreo em adultos e 5% nos recém-
nascidos. A anatomia do fígado em humanos constitui-se por dois lobos principais
(direito e esquerdo) e dois acessórios (quadrado e caudado). Já o fígado de ratos e
camundongos possui lobo lateral esquerdo, lateral direito, medial direito, medial
esquerdo, dois processos papilares e o processo caudado, e sendo assim em
camundongos há a presença da vesícula biliar. Os três últimos são considerados
lobos caudados. Entretanto, hoje os médicos gastroenterologistas dão mais enfoque
a anatomia vascular funcional do fígado (BISMUTH, 1988).
No histológico do fígado, podemos observar que o fígado é um órgão
multicelular, composto de hepatócitos e outras populações de células não

35
Gomes, SP Revisão sistemática da literatura
parênquimais. Além de três subpopulações de células sinusóides bem conhecidas
(células de Kupffer, células endoteliais e células estreladas hepáticas), existem
também células epiteliais dos ductos biliares e células mesoteliais da cápsula de
Glisson (GRISHAM, 1980, 1983).
No lóbulo, as células hepáticas dispõem-se em placas orientadas
radicalmente. Cada placa é constituída por células dispostas em uma só camada.
Essas placas são perfuradas e, freqüentemente, anastomosam-se, resultando num
labirinto complexo, que dá ao lóbulo um aspecto esponjoso. O espaço entre as
placas hepatocitárias é ocupado por capilares sinusóides, denominadas sinusóides
hepáticas (JUNQUEIRA; CARNEIRO, 1999).
3.2 ASPECTOS NUTRICIONAIS
A desnutrição no início da vida pode afetar profundamente o corpo e o
cérebro de ratos. Alguns desses efeitos são permanentes enquanto outros
desaparecem durante o período de reabilitação nutricional (YUCEL; WARREN;
GUMUSBURUN, 1994).
A subnutrição de ratos durante o início da vida é conhecida como causadora
de déficits permanentes no número total de células cerebrais determinados por
arranjos bioquímicos do conteúdo do DNA cerebral. Se isso é devido a uma perda
de neurônios ou uma falha de proliferação de células gliais permanece incerto a
partir dessas medidas bioquímicas (BEDI, 1991).
Pesquisas em animais revelaram que uma nutrição inadequada no período
pré-parto, como por exemplo, uma restrição severa de comida para a mãe, redução
severa do conteúdo em proteína da dieta da mãe, conduz a deficiências nutricionais
a um aumento de liberação de noradrenalina no cérebro de animais (ZANIN et al.,
2003).
O estado nutricional é determinado por um balanço entre o suprimento de
nutrientes e a utilização destes pelo organismo. A desnutrição está relacionada com
o desequilíbrio do estado nutricional e ocorre sob diversas formas, podendo ter
conseqüências reversíveis ou irreversíveis. Estudos específicos sobre os efeitos da
desnutrição nos neurônios evidenciaram uma redução de 27% no número de

36
Gomes, SP Revisão sistemática da literatura
neurônios no intestino delgado de ratos cujas mães sofreram restrição protéica
durante as duas últimas semanas de gestação (ZANIN et al., 2003).
Recentemente desenvolvidos, os métodos estereológicos têm sido usados em
experimentos para examinar os efeitos de dois níveis de renutrição durante o início
da vida pós-natal no número total de células granulares de ratos (BEDI, 1991).
Segundo Castelucci et al. (2002), a privação protéica da mãe durante a
gravidez, e dos recém-nascidos nos primeiros 21 dias reduziu o peso dos filhotes em
aproxidamente 50%.
Pode-se observar que durante o período pré-natal, após o nascimento, após o
desmame, os filhotes foram removidos de suas mães e após 21 dias sob dieta
controlada de proteínas, ou mudado de uma dieta hipoprotéica para uma dieta
normal (grupos renutridos). Os animais renutridos apresentaram um peso 20% a
baixo do controle (CASTELUCCI et al., 2002).
Segundo Castelucci et al.(2002), em uma análise quantitativa os animais do
grupo controle tiveram pesos corpóreos quase dobrados em relação àqueles
privados de proteínas aos 21 dias após o nascimento.
O peso do corpo dos animais controle foi de aproximadamente 60% maior do
que o grupo subnutrido aos 42 dias, contudo realimentando os ratos subnutridos
entre 21 dias e 42 dias reestabeleceu-se o peso em 20% do normal aos 42 dias
(CASTELUCCI et al., 2002).
De acordo com o protocolo de Castelucci et al. (2002); Brandão et al. (2003);
Gomes et al. (2006); Gomes et al. (2009) foram utilizados em seus experimentos
animais que recebiam uma dieta protéica de 20% de proteína-caseína (animais
nutridos) e um dieta protéica de 5% de proteína-caseína (animais desnutridos), a
partir do primeiro dia de gestação desses animais e após os mesmos seus filhotes
terem nascidos, os mesmo começaram a receber a mesma dieta utilizada pelas
fêmeas gestantes. Os animais por eles utilizados não possuíam os mesmos
parentescos e após seus 21 dias de vida, os filhotes foram escolhidos em cada
grupo sistematicamente aleatórios.
De acordo com Gomes et al. (2009) os animais privados de proteína-caseina
a 5% e os animais que receberam proteína-caseina a 20% tiveram seus pesos
corpóreos com significância e seus volumes altamente significativos em comparação
entre os grupos estudados.

37
Gomes, SP Revisão sistemática da literatura
As dietas dos ratos continham fibras, misturas de salina equilibrada e vitamina
na mesma quantidade. O grupo controle recebeu uma dieta com 20% de proteína e
o grupo desnutrido uma dieta hipoprotéica purificada de apenas 4% (REEVES;
NIELSEN; FAHEY, 1993). A fonte de proteína utilizada foi à caseína. Com exceção
do teor de proteínas, as duas dietas foram idênticas e isocalóricas. Ratos
alimentados com dieta hipoprotéica consumiram menos alimentos, em comparação
aos ratos do grupo controle, resultando em consumo reduzido de proteínas, perda
de peso corporal e uma diminuição da proteína plasmática, albumina e pré-albumina
(FOCK et al., 2008; VINOLO et al., 2008). O grupo controle recebeu uma dieta
adequada de proteínas e do grupo controle receberam uma dieta deficiente em
proteínas, mas o grupo de desnutridos teve uma ingestão alimentar
significativamente menor e experimentou uma perda de 20,2% em peso do corpo em
um período de 14 dias após a introdução da dieta hipoprotéica (FOCK et al., 2008).
Em uma dieta fazendo uso de suplementos glutamínicos para o grupo de
camundongos nutridos e outra dieta livre de suplementos glutamínicos para o grupo
de camundongos desnutridos, pode-se observar também uma diferença de peso
corpóreo entre os grupos. E em uma renutrição pode-se observar uma reversão do
quadro para o grupo dos animais nutridos (ROGERO et al., 2008).
Em outro experimento feito por Fock et al. (2007), os camundongos do grupo
controle utilizaram uma dieta do grupo controle de 20% de proteína e o grupo
desnutrido de 4% de proteína e pode-se averiguar que os camundongos do grupo
controle para o grupo desnutridos teve uma perda acentuada de 22% de massa
corporal quando comparados ao grupo nutrido, pela baixa porcentagem de proteína
em sua ingestão alimentar e observou-se uma anorexia aparente desses animais.
Walrand et al. (2000), submeteu ratos adultos de a um programa de
restrição alimentar, sendo um grupo de animais submetidos a uma desnutrição com
23% de proteína e 1,6 gramas de caseína e um grupo de animais renutrido com 27%
de proteína e 3,2 gramas de caseína, para poder investigar as alterações da
indução dessa dieta e também observar nos animais renutridos as alterações que
esta dieta causou no metabolismo desses animais. E pode ser observado que o
resultado obteve um P< 0, 005.
De acordo com Cruz-Orive (2005), tem que se haver uma variância geral
entre as fatias obtidas para o uso do Método de Cavalieri. As seções de Cavalieri

38
Gomes, SP Revisão sistemática da literatura
são amplamente utilizadas em estereologia para estimar as fatias do estudo a ser
realizado.
Considerando o uso do Cavalieri para estimar qualquer outra medida
geométrica, sejam células ou números de partículas. A variação de seus
estimadores corresponde dependendo do seu interesse de estudo. As observações
deste estudo são sistemáticas, portanto dependentes como um todo (CRUZ-ORIVE,
2005). E também foi publicado para contar as seções do Cavalieri por Cruz-Orive e
García-Fiñana (2005); e as fatias do Cavalieri por Cruz-Orive e Geiser (2004).
Segundo Marcos, Monteiro e Rocha (2007), o mesmo salienta que as
estimativas numéricas baseadas em métodos diferentes utilizados nas pesquisas,
devem sempre ser comparados com aqueles baseados em estereologia (quando
disponível). Considerando a natureza imparcial da última geração de métodos
estereológicos, estas estimativas devem ser sempre consideradas como excelência,
para fins de quantificação, ou seja, para apoiar, ou não, as estimativas derivadas de
métodos mais indiretos.
Com o número de hepatócitos obtidos na estereologia, é fundamentalmente
diferente das que são obtidas pela análise de imagem, porque os perfis dos
hepatócitos são simples (não corrigida) e são contadas as seções que aparecem,
enquanto na estereologia faz-se uma contagem de células tridimensional, em sua
proporção real - independentemente do seu tamanho, número, forma ou distribuição
espacial (HOWARD; REED, 2005).
Os dados de uma abordagem estereológica devem ser considerados como
padrão de excelência para os fins práticos, sendo que seus valores provenientes de
outros métodos de contagem ou potencialmente tendenciosos devem ser
comparados entre seus resultados (HOWARD; REED, 2005).
Para Halici et al. (2009), a estereologia utiliza-se dos princípios matemáticos e
é utilizado para avaliar as propriedades imparciais das estruturas existentes no
espaço 3D à partir de seções 2D do microscópio. O método estereológico da
dissecção óptica também pode ser efetivamente usado para estimar o número de
objetos conhecidos em lâminas histológicas. Também pode ser útil para examinar as
mudanças no número de quaisquer objetos de estudo e como um indicador de
eficácia terapêutica ou tratamentos com drogas tóxicas. A mesma teve como
objetivo de estudo estimar a densidade numérica dos hepatócitos para determinar se
esta é afetada pelo tratamento haloperidol em ratos. Neste estudo, foi utilizado o

39
Gomes, SP Revisão sistemática da literatura
método do “disector óptico” e também para analisar as amostras
histopatologicamente, tanto na microscopia de luz e da microscopia eletrônica.
Uma nova geração de métodos estereológicos vem sendo desenvolvidos
desde 1984, permitindo uma maior precisão: de uma precisa e eficiente estimativa
para a contagem total do número de células (HOWARD; REED, 1998). Por último,
duas abordagens imparciais estão disponíveis: o fracionamento através do
fractionator óptico (WEST; GUNDERSEN, 1991) e da precisão da técnica para o
volume (PAKKENBERG; GUNDERSEN, 1996). O último é uma análise em duas
etapas, envolvendo estimativas separadas e imparciais do volume referência e da
densidade numérica das partículas, e assim através dessas estimativas podemos
achar também o número total de partículas (MARCOS; MONTEIRO; ROCHA, 2004).
Garcia-Fiñana et al. (2002), utiliza-se para a contagem de partículas a técnica
da contagem de pontos baseada no Princípio de Cavalieri. Sendo que esta técnica
requer de seções histológicas planas onde sobre estas seções foi sobreposto um
sistema de pontos.
Em alguns estudos os coeficientes de erros do Princípio de Cavalieri são
estimados de acordo com Gundersen et al., (1999). Já outros coeficientes de erros
do princípio de Cavalieri são estimados de acordo com Garcia-Fiñana et al. (2002),
onde as fórmulas desse coeficiente de erro são baseados em modelos para uma
estrutura de covariância de dados sistemáticos, e eles são apenas aproximações -
em oposição ao projeto de estimadores imparciais. Dados este que indicam que os
coeficientes de erros foram utilizados baseando-se em um melhor parâmetro,
considerando sempre oportuno para verificá-los para uma reamostragem empírica
em um conjunto de dados completos.

Materiais e métodos

41
Gomes, SP Materiais e métodos
4 MATERIAIS E MÉTODOS
Os materiais utilizados e os métodos empregados neste trabalho serão
apresentados em detalhes abaixo.
4.1 MATERIAIS
O seguinte material foi empregado neste estudo:
4.1.1 Animais
O desenvolvimento da presente pesquisa foi realizado de acordo com os
princípios éticos de experimentação animal da Comissão de Ética da Faculdade de
Medicina Veterinária e Zootecnia da Universidade de São Paulo, sob o protocolo nº
1521/2008.
Foram utilizados até o momento 12 fígados provenientes de Camundongos
Swiss machos, com 90 dias de idade (grupos controle dos desnutridos (CD) e
desnutridos (D)) e 120 dias de idade (grupos renutridos (R) e controle dos renutridos
(CR)) machos. De acordo com o tratamento empregado e o tipo de dieta
disponibilizada, os animais foram divididos em 4 grupos:
a) Controle dos Desnutridos (CD) com 3 animais, tendo peso médio de 51,63
g (Desvio Padrão = 1,8)
b) Desnutridos (D) com 3 animais, tendo peso médio de 24,96 g (Desvio
Padrão = 1,7)
c) Renutridos (R) com 3 animais, tendo peso médio de 45,9 g (Desvio Padrão
= 3,4)
d) Controle dos Renutridos (CR) com 3 animais, tendo peso médio de 64,46 g
(Desvio Padrão = 1,9)

42
Gomes, SP Materiais e métodos
Os animais foram provenientes do Departamento de Análises Clínicas do
Biotério da Faculdade de Ciências Farmacêuticas da Universidade de São Paulo.
4.2 MÉTODOS
Os seguintes métodos serão empregados neste estudo:
4.2.1 Rações
Os animais receberam ração controle contendo 12% de proteína (Tabela 1,
Ração Controle I) e ração normoprotéica contendo 2% de proteína (Tabela 1, Ração
Hipoprotéica II), sendo que ambas serão preparadas no Laboratório Hematologia da
Faculdade de Ciências Farmacêuticas/USP na forma granulada e conservada a 4ºC
até o uso. Ambas as rações são isocalóricas e diferem apenas no conteúdo protéico,
sendo que foram preparadas de acordo com as recomendações de 1993 do Instituto
Americano de Nutrição para Camundongos adultos (REEVES; NIELSEN; FAHEY,
1993). A fonte de proteína que será utilizada é a caseína, sendo a concentração
protéica da caseína e das rações determinadas pelo método Micro-Kjeldahl
(INSTITUTO ADOLFO LUTZ, 1985). O teor de fosfato da mistura salínica que será
utilizada no preparo das rações será corrigido de acordo com a concentração
protéica de cada lote de caseína (controle e hipoprotéica) para se adequar aos
valores normais recomendados (REEVES; NIELSEN; FAHEY, 1993).

43
Gomes, SP Materiais e métodos
Tabela 1- Composição Percentual da Ração. Ração controle I e Ração hipoproteíca II.
COMPOSIÇÃO RAÇÃO CONTROLE I
(G/KG RAÇÃO)
RAÇÃO
HIPOPROTÉICA II
(G/KG RAÇÃO)
Proteína 120 20
Sacarose 100 100
Celulose 50 50
Óleo de Soja 40 40
Mistura Salínica1 35 35
Mistura Vitamínica1 10 10
L-Cistina 1,8 0, 257
Bitartarato de Colina 2,5 2,5
Tert-butilhidroquinona 0, 008 0, 008
Amido (q.s.p.) 1000 1000
1 As misturas salínica e vitamínica serão preparadas sob encomenda de acordo com as
recomendações de 1993 do Instituto Americano de Nutrição para camundongos adultos (REEVES, 1993).
4.2.2 Indução da desnutrição
Os animais foram alocados em gaiolas metabólicas individuais e foram
mantidos sob as mesmas condições ambientais, com ciclo de luz de 12h,
temperatura de 2210%, onde passaram por um período de adaptação aos
gaioleiros de 15 a 20 dias. Neste período, todos os animais receberam ração
normoprotéica e água ad libitum e tiveram seu peso avaliado. Após o período de
adaptação, tendo os animais atingido as estabilidades do peso, estes foram
separados em 2 grupos: controle do desnutrido e desnutrido. O grupo desnutrido
passou a receber ração hipoprotéica, enquanto o grupo controle do desnutrido
continuará a receber ração normoprotéica. Durante este período será realizada a
cada 48 horas a pesagem dos animais e o consumo de ração. Este tratamento
permaneceu durante 1 mês, quando então, os animais do grupo desnutrido foram
separados em 2 subgrupos: um foi eutanasiado, enquanto o outro foi submetido ao
processo de renutrição. O grupo controle também foi separado em dois subgrupos:
um deles foi eutanasiado simultaneamente ao grupo desnutrido; o outro continuou a
receber ração controle, acompanhando o grupo renutrido (BORELLI et al, 2007).

44
Gomes, SP Materiais e métodos
4.2.3 Recuperação nutricional
O processo de renutrição foi mantido por 30 dias, sendo que ao final deste
período, os animais dos grupos renutridos e controle do renutrido foram
eutanasiados. Durante o período da renutrição, os animais receberam água e ração
ad libitum (BORELLI et al, 2007).
4.2.4 Obtenção de amostras de sangue
As seguintes amostragens são empregadas neste estudo:
4.2.4.1 Sangue total
Amostras de sangue total foram obtidas por punção do plexo axilar de
camundongos previamente anestesiados com xilazina (16 mg/kg de peso corporal) e
cetamina (120 mg/kg de peso corporal), utilizando-se EDTA 10% (sal sódico) como
anticoagulante, na proporção de 50 µL de EDTA para cada 1,0 mL de sangue.
4.2.4.2 Realização do hemograma
Amostras de sangue total obtida de acordo com o descrito no item 4.2.4.1
foram utilizadas para a determinação do Número Total de Hemácias, Hematócrito,
Hemoglobina e Leucócitos. Estas análises foram realizadas no analisador
automático de células sangüíneas ABC Vet (ABX DIAGNOSTICS), o método e as
medições utilizadas foram: impedância, fotometria (BORELLI et al, 2007).

45
Gomes, SP Materiais e métodos
4.2.4.3 Análises bioquímicas
Amostras de sangue totais obtidas de acordo com o item 4.2.4.1 foram
utilizado os soros para a determinação dos seguintes parâmetros: proteína total,
albumina, globulina, alanina aminotransferase (ALT), fosfatase alcalina (FA), gama
glutamil transpeptifase (GGT), aspartato aminotransferase (AST) e glicose. Estas
análises foram realizadas no analisador clínico randômico Metrolab 2300 plus da
Wiener® Lab Group, o método utilizado foi através do colorímetro (SANTOS, 2009).
4.2.5 Estudo histológico
Aos 90 dias os animais do grupo controle do desnutrido e desnutrido foram
eutanasiados com uma sobredose de 100 mg/kg de Ketamina e 10 mg/kg de
Xilazina (DONOVAN; BROWN, 1991; FLECKNELL, 1996) ambas pela via
intramuscular e aos 120 dias os animais do grupo renutrido e controle do renutrido
também foram eutanasiados de acordo com o grupo de animais controle do
desnutrido e desnutrido.
Após a constatação do óbito dos animais, a cavidade torácica foi aberta e
uma cânula inserida dentro do ventrículo esquerdo do coração até atingir a aorta
ascendente. Primeiramente 100 ml de uma solução de lavagem composta de
tampão fosfato salino (PBS) (0.1M, pH7,4) e 2% de heparina foi perfundida.
Posteriormente, foram perfundidos 100 ml de solução fixadora composta por 4%
formoldeído em tampão fosfato de sódio (PSB) (0.1M, pH7,4). A perfusão sistêmica
(via aorta ascendente) foi conduzida utilizando-se uma bomba perfusora digital
(Masterflex) sob um fluxo pulsátil de 8 ml/min. Para promover uma melhor fixação
do fígado, uma perfusão retrógrada foi realizada a partir da aorta abdominal.
Para evidenciação do fígado foi promovida uma celiotomia pré-umbilical e
paracostal de ambos os lados para a retirada da musculatura abdominal (músculo
oblíquo externo, músculo oblíquo interno, músculo transverso abdominal e reto do
abdômen. Uma vez identificado o fígado, este foi retirado sob o microscópico
cirúrgico Leica®
M651 (6-40x).

46
Gomes, SP Materiais e métodos
Os fígados foram removidos dos animais e pesados antes que fossem
imersos na mesma solução fixadora e armazenados a temperatura de 4°C durante
uma semana.
Subseqüentemente, o fígado foi verticalizado no seu eixo longo, com a ajuda
do bar rotator “VUR section” (BADDELEY; GUNDERSEN; CRUZ-ORIVE, 1986),
embebidos em uma solução aquosa de ágar (10%) e seccionados exaustiva e
seriadamente em micrótomo de lâmina vibratória (vibrótomo) Leica® (VT 1000S) com
40μm de espessura. As secções foram coradas com Hematoxilina de Mayer por 1
minuto. As secções histológicas foram então desidratadas em concentrações
crescentes de etanol, diafanizadas em xilol e montadas com DPX sobre lâmina e
sob lamínula.
4.2.6 Estudo estereológico
As secções histológicas foram observadas usando o Microscópio Leica® DMR
equipado com câmera digital Olympus DP72 e analisadas usando o software NEW
CAST da Visiopharm Integrator System, versão 3.6.5.0.
4.2.6.1 Estudo morfoquantitativo (estereológico) do fígado
Os seguintes estudos através dos métodos estereológicos serão amostrados
neste estudo:
4.2.6.1.1 Volume do fígado (Vref)
O volume do fígado foi estimado usando-se o Princípio de Cavalieri
(MAYHEW e OLSEN, 1991; HOWARD e REED, 2005).

47
Gomes, SP Materiais e métodos
O fígado foi seccionado seriadamente em fatias de 40 µm de espessura.
Sobre estas fatias foi sobreposto um sistema de pontos (a/p= 0.378mm2), no
aumento de 1.0x, com a ajuda do estereomicroscópico Leica®
M165C (0.73 -12x)
(Figura 1).
Em cada fatia foi contado o número de pontos que tocavam o fígado
(parênquima e estroma). Para se estimar o volume do fígado, a partir do princípio de
Cavalieri, foi aplicada a seguinte fórmula (RIBEIRO et al., 2004; MAYHEW E
OLSEN,1991):
V = Σp x a/p x t
Onde:
V: Volume do fígado
∑p: Somatória dos pontos que tocam o fígado
a/p: Área associada a cada ponto do sistema teste
t: Espessura média das secções do fígado

48
Gomes, SP Materiais e métodos
Figura 1 - Secção do fígado de animal controle do desnutrido. Sistema de pontos utilizado
para a estimativa do volume do fígado a partir do Princípio de Cavalieri. Escala de barra: 20 cm
4.2.6.1.2 Coeficiente de erro
O coeficiente de erro do Princípio de Cavalieri foi estimado de acordo com a
recente literatura de Gundersen et al. (1999); García-Fiñana (2000); García-Fiñana e
Cruz-Orive (2000a) e García-Fiñana et al. (2002), aplicando a seguinte fórmula:
PnabV )(0724,0ˆ ;
2log2
1ˆ q ;
0ˆ q ;
121)( q ;
2
210
2 )()4)ˆ(3()()ˆ( iPccvcqVce ;

49
Gomes, SP Materiais e métodos
QCE
1
22 )(ˆ)
~( ipc PvVce ;
)~
()ˆ()~
( 222 VCEVCEVCE PC .
4.2.6.1.3 Número total de hepatócitos (N)
O número total de hepatócitos do (N) foi estimado multiplicando-se o número
total de partículas contadas pelo inverso das frações amostradas (MAYHEW, 1991)
(Figura 2).
ssfhsfasfQN
1.
1.
1.
Onde:
Q- é o número de partículas contadas pelo método do disector óptico que,
neste caso, são hepatócitos
ssf é a fração da amostragem das secções
asf é a fração das áreas amostradas para contagem
hsf é a fração da espessura do corte efetivamente utilizada para contagem
E para calcular o coeficiente de erro, utilizamos a seguinte fórmula
(PAKKENBERG; GUNDERSEN, 1988):
50
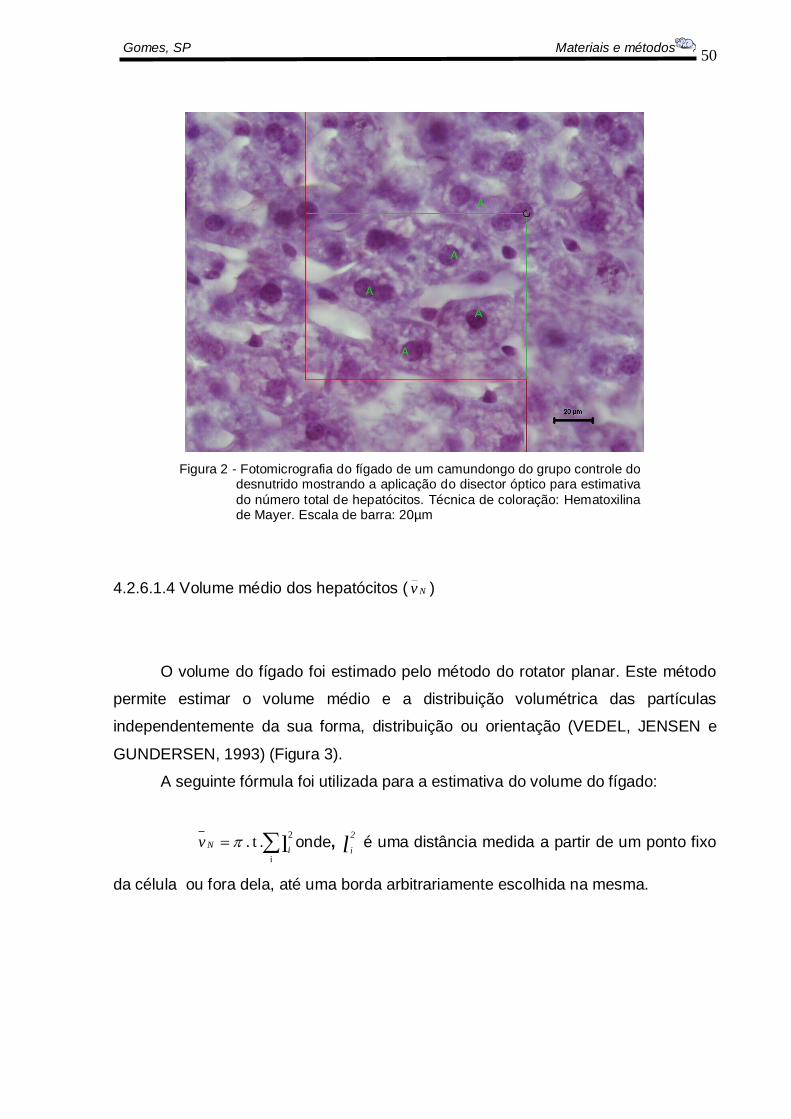
Gomes, SP Materiais e métodos
Figura 2 - Fotomicrografia do fígado de um camundongo do grupo controle do desnutrido mostrando a aplicação do disector óptico para estimativa
do número total de hepatócitos. Técnica de coloração: Hematoxilina de Mayer. Escala de barra: 20µm
4.2.6.1.4 Volume médio dos hepatócitos ( Nv )
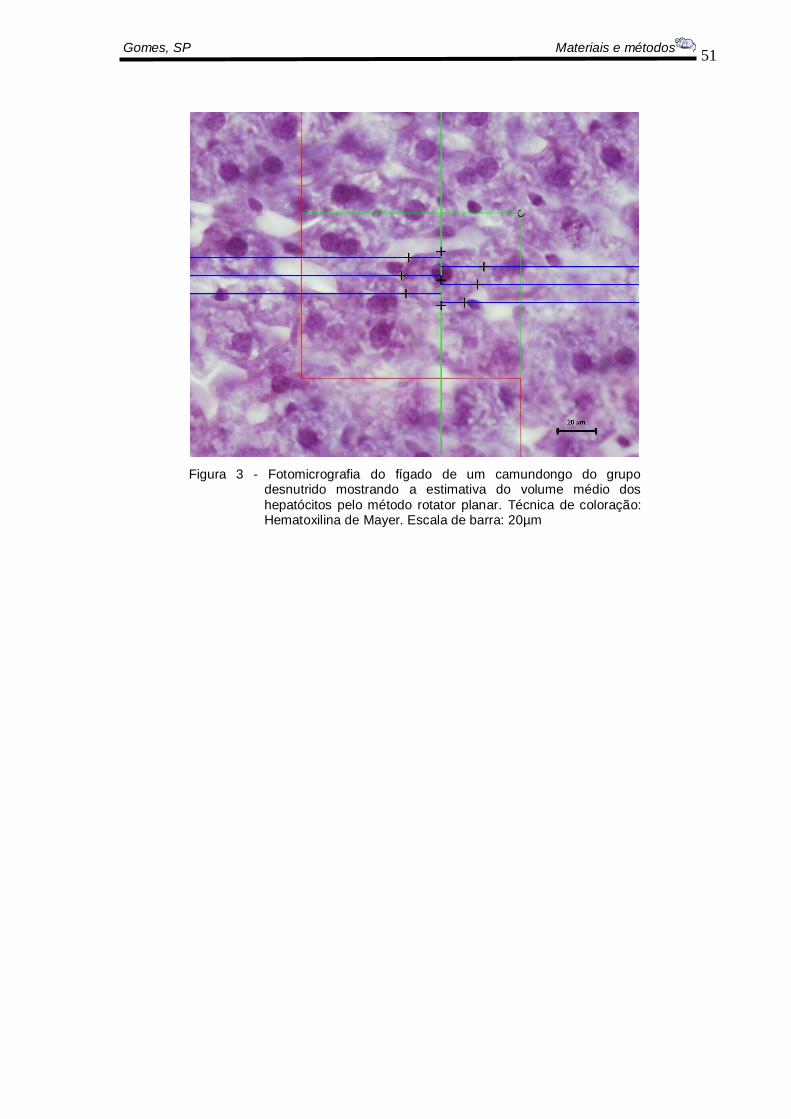
O volume do fígado foi estimado pelo método do rotator planar. Este método
permite estimar o volume médio e a distribuição volumétrica das partículas
independentemente da sua forma, distribuição ou orientação (VEDEL, JENSEN e
GUNDERSEN, 1993) (Figura 3).
A seguinte fórmula foi utilizada para a estimativa do volume do fígado:
i
2
l. t . i
Nv onde, l2
i é uma distância medida a partir de um ponto fixo
da célula ou fora dela, até uma borda arbitrariamente escolhida na mesma.
51
Gomes, SP Materiais e métodos
Figura 3 - Fotomicrografia do fígado de um camundongo do grupo desnutrido mostrando a estimativa do volume médio dos
hepatócitos pelo método rotator planar. Técnica de coloração: Hematoxilina de Mayer. Escala de barra: 20µm

52
Gomes, SP Análise estatística inferencial
5 ANÁLISE ESTATÍSTICA INFERENCIAL
A distribuição de freqüência dos parâmetros estereológicos e bioquímicos
foram analisadas pelo “one-way” ANOVA, realizada por meio do software MINITAB®
16 (2010). Quando diferenças significativas entre grupos foram detectadas (p˂ 0,05),
o teste post-hoc de Tukey foi utilizado para comparações múltiplas.
Os resultados são expressos pela média seguida pelo coeficiente de variação
(CV) calculado pela razão entre o desvio padrão e a média.

Resultados

54
Gomes, SP Resultados
6 RESULTADOS
Os resultados são oriundos de quatros grupos (animais controle do
desnutrido, desnutrido, renutrido e controle do renutrido). Desta forma os métodos
empregados serão relacionados neste capítulo.
6.1 PESO CORPORAL
Os pesos corporais dos animais foram: 51,63 gramas (Desvio Padrão= 1,8),
24,86 gramas (Desvio Padrão= 1,7), 45,90 gramas (Desvio Padrão= 3,4) e 64,46
gramas (Desvio Padrão= 1,9), respectivamente para os grupos CD, D, R e CR.
(Gráfico 1).
O peso corporal dos animais foi maior no CR em relação ao CD, D e R (p=
0,001).
Gráfico 1 - Peso dos animais (p= 0,001*) dos grupos controle do desnutrido,
desnutrido, renutrido e controle do renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos
55
Gomes, SP Resultados
6.2 PESO DO FÍGADO
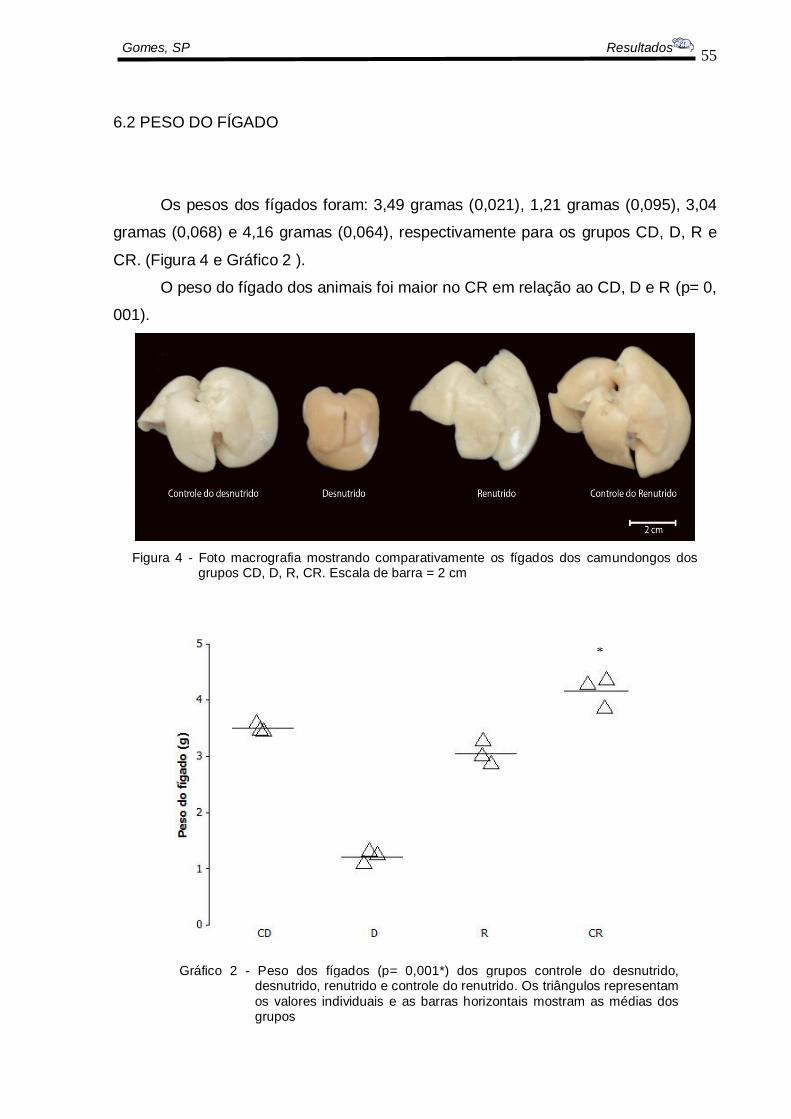
Os pesos dos fígados foram: 3,49 gramas (0,021), 1,21 gramas (0,095), 3,04
gramas (0,068) e 4,16 gramas (0,064), respectivamente para os grupos CD, D, R e
CR. (Figura 4 e Gráfico 2 ).
O peso do fígado dos animais foi maior no CR em relação ao CD, D e R (p= 0,
001).
Figura 4 - Foto macrografia mostrando comparativamente os fígados dos camundongos dos grupos CD, D, R, CR. Escala de barra = 2 cm
Gráfico 2 - Peso dos fígados (p= 0,001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos representam
os valores individuais e as barras horizontais mostram as médias dos grupos

56
Gomes, SP Resultados
6.3 MICROESTRUTURA
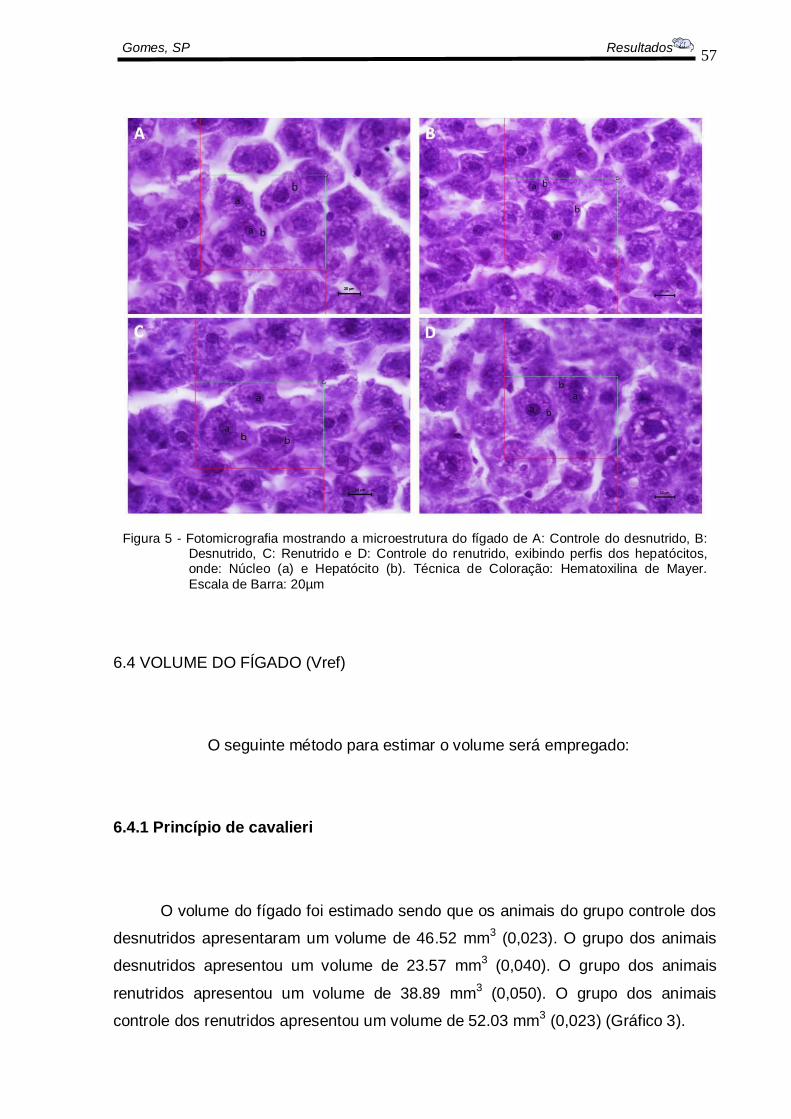
Histologicamente, o lóbulo hepático apresenta forma hexagonal, com uma
veia centro-lobular e com as tríades portais nas extremidades desse hexágono
imaginário. As colunas de hepatócitos (Figura 5) dispõem-se lado a lado, a partir da
veia central e entre elas estão situados os sinusóides hepáticos, que transportam o
sangue que chega pelos ramos da veia porta e da artéria hepática.
A estrutura do fígado (distribuição dos hepatócitos) não foi modificada nos
grupos avaliados, somente o volume que foi menor nos animais desnutridos e
renutridos.
Podemos observar que o fígado é um órgão multicelular, composto de
hepatócitos e outras populações de células não parênquimas. Além de três
subpopulações de células sinusóides bem conhecidas (células de Kupffer, células
endoteliais e células estreladas hepáticas), existem também células epiteliais dos
ductos biliares e células mesoteliais da cápsula de Glisson.
57
Gomes, SP Resultados
Figura 5 - Fotomicrografia mostrando a microestrutura do fígado de A: Controle do desnutrido, B: Desnutrido, C: Renutrido e D: Controle do renutrido, exibindo perfis dos hepatócitos, onde: Núcleo (a) e Hepatócito (b). Técnica de Coloração: Hematoxilina de Mayer.
Escala de Barra: 20µm
6.4 VOLUME DO FÍGADO (Vref)
O seguinte método para estimar o volume será empregado:
6.4.1 Princípio de cavalieri
O volume do fígado foi estimado sendo que os animais do grupo controle dos
desnutridos apresentaram um volume de 46.52 mm3 (0,023). O grupo dos animais
desnutridos apresentou um volume de 23.57 mm3 (0,040). O grupo dos animais
renutridos apresentou um volume de 38.89 mm3 (0,050). O grupo dos animais
controle dos renutridos apresentou um volume de 52.03 mm3 (0,023) (Gráfico 3).

58
Gomes, SP Resultados
O volume do fígado dos animais foi maior no CR em relação ao CD, D e R (p=
0,001).
Gráfico 3 - Volume dos fígados estimados pelo Princípio de Cavalieri (p= 0,001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do
renutrido. Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos
6.5 COEFICIENTE DE ERRO DO VOLUME DO FÍGADO (CE)
O coeficiente de erro do volume do fígado e os intervalos de confiança para o
volume do fígado foram estimados sendo que os animais do grupo controle dos
desnutridos apresentaram um coeficiente de erro do volume de 1,15%. O grupo dos
animais desnutridos apresentou um coeficiente de erro do volume de 1,74%. O
grupo dos animais renutridos apresentou um coeficiente de erro do volume de 1,2%.
E o grupo dos animais controle dos renutridos apresentou um coeficiente de erro do
volume de 1,02% (Gráficos 4 e 5).

59
Gomes, SP Resultados
Gráfico 4 - Coeficiente de erro dos fígados (à esquerda) e respectivos intervalos de
confiança (à direta) estimados para os grupos Controle do desnutrido (A e B) e Desnutrido (C e D)
Gráfico 5 - Coeficiente de erro dos fígados (à esquerda) e respectivos intervalos de confiança
(à direta) estimados para os grupos Renutridos (E e F) e Controle do renutrido (G e H)

60
Gomes, SP Resultados
6.6 NÚMERO TOTAL DE HEPATÓCITOS (N)
O número total de hepatócitos foi de 1.1 x 107
(0,086) para o grupo controle
do desnutrido. Tendo seu coeficiente de erro de 8,43%. Sendo que para o grupo
desnutrido o número total foi de 1.6 x 107 (0,094). Tendo seu coeficiente de erro de
6,93%. O número total para o grupo renutrido foi de 2.8 x 107 (0,176). Tendo seu
coeficiente de erro de 7,0%. Enquanto que para o grupo controle do renutrido o
número total foi de 2.2 x 107 (0,514). Tendo seu coeficiente de erro de 7,53%.
O grupo R foi maior em relação ao CD e o D (diferença significativa “border
line”, p= 0, 055) (Gráfico 6).
Gráfico 6 - Número total de hepatócitos (p= 0,055*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos
representam os valores individuais e as barras horizontais mostram as médias dos grupos
6.7 VOLUME MÉDIO DOS HEPATÓCITOS ( Nv )
O volume médio dos hepatócitos foi de 8.170,13 µm3
(0,094) para o grupo
controle do desnutrido. Sendo que para o grupo desnutrido o volume médio foi de
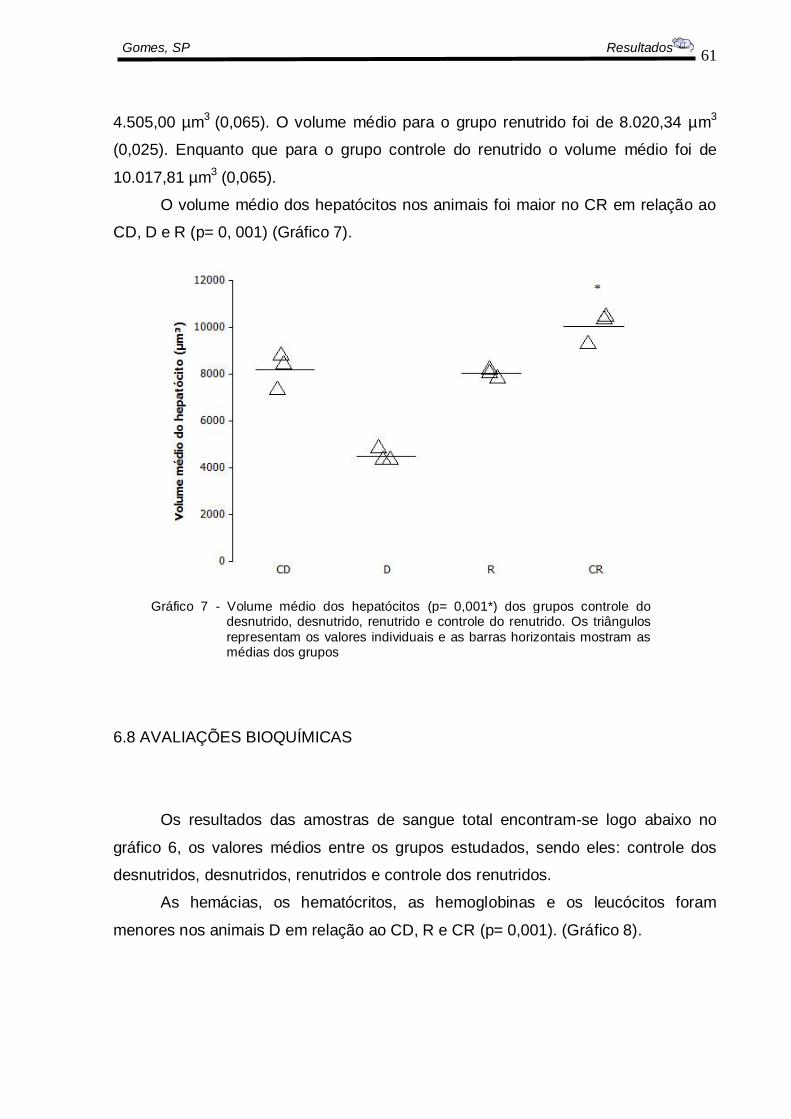
61
Gomes, SP Resultados
4.505,00 µm3
(0,065). O volume médio para o grupo renutrido foi de 8.020,34 µm3
(0,025). Enquanto que para o grupo controle do renutrido o volume médio foi de
10.017,81 µm3 (0,065).
O volume médio dos hepatócitos nos animais foi maior no CR em relação ao
CD, D e R (p= 0, 001) (Gráfico 7).
Gráfico 7 - Volume médio dos hepatócitos (p= 0,001*) dos grupos controle do desnutrido, desnutrido, renutrido e controle do renutrido. Os triângulos
representam os valores individuais e as barras horizontais mostram as médias dos grupos
6.8 AVALIAÇÕES BIOQUÍMICAS
Os resultados das amostras de sangue total encontram-se logo abaixo no
gráfico 6, os valores médios entre os grupos estudados, sendo eles: controle dos
desnutridos, desnutridos, renutridos e controle dos renutridos.
As hemácias, os hematócritos, as hemoglobinas e os leucócitos foram
menores nos animais D em relação ao CD, R e CR (p= 0,001). (Gráfico 8).
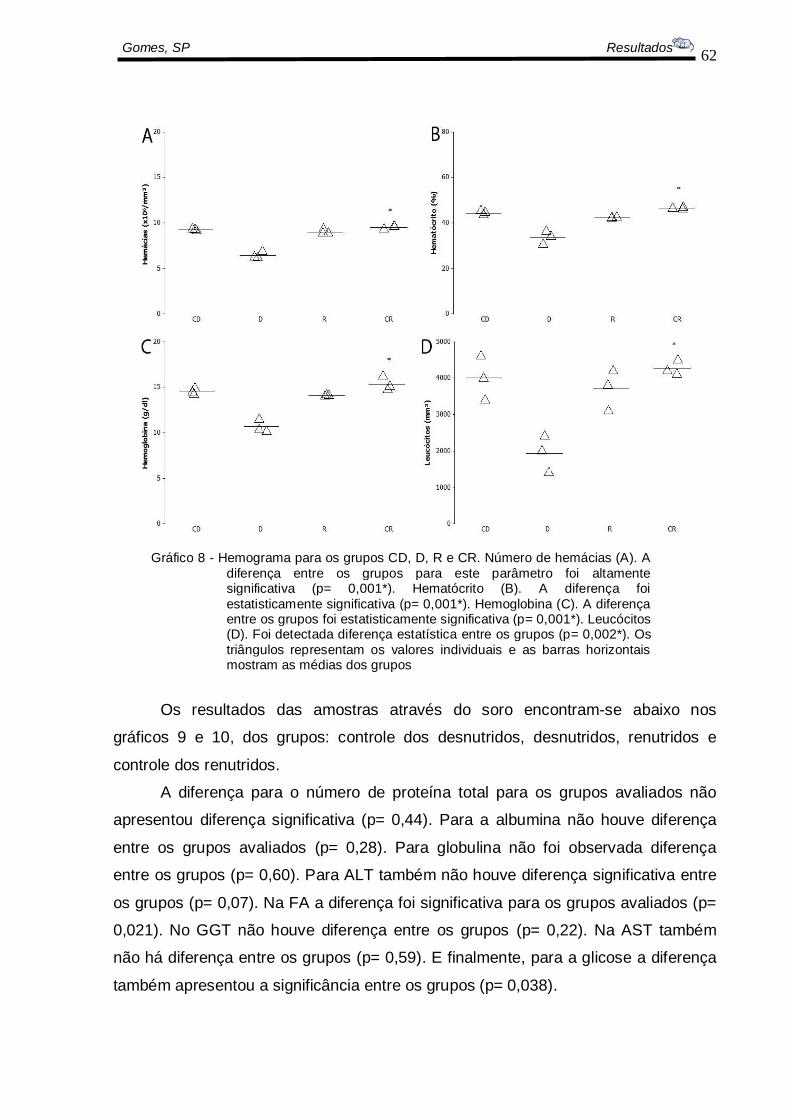
62
Gomes, SP Resultados
Gráfico 8 - Hemograma para os grupos CD, D, R e CR. Número de hemácias (A). A
diferença entre os grupos para este parâmetro foi altamente significativa (p= 0,001*). Hematócrito (B). A diferença foi
estatisticamente significativa (p= 0,001*). Hemoglobina (C). A diferença entre os grupos foi estatisticamente significativa (p= 0,001*). Leucócitos (D). Foi detectada diferença estatística entre os grupos (p= 0,002*). Os
triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos
Os resultados das amostras através do soro encontram-se abaixo nos
gráficos 9 e 10, dos grupos: controle dos desnutridos, desnutridos, renutridos e
controle dos renutridos.
A diferença para o número de proteína total para os grupos avaliados não
apresentou diferença significativa (p= 0,44). Para a albumina não houve diferença
entre os grupos avaliados (p= 0,28). Para globulina não foi observada diferença
entre os grupos (p= 0,60). Para ALT também não houve diferença significativa entre
os grupos (p= 0,07). Na FA a diferença foi significativa para os grupos avaliados (p=
0,021). No GGT não houve diferença entre os grupos (p= 0,22). Na AST também
não há diferença entre os grupos (p= 0,59). E finalmente, para a glicose a diferença
também apresentou a significância entre os grupos (p= 0,038).

63
Gomes, SP Resultados
Gráfico 9 - Função hepática para os grupos CD, D, R e CR. Proteína total (A).
Albumina (B). Globulina (C). ALT (D). Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos
Gráfico 10 - Função hepática e glicose para os grupos CD, D, R e CR. FA (A), GGT
(B). AST (C). Glicose (D). Os triângulos representam os valores individuais e as barras horizontais mostram as médias dos grupos

Discussão

65
Gomes, SP Discussão
7 DISCUSSÃO
A desnutrição é um problema de saúde pública de grande prevalência em
Países Subdesenvolvidos e tem sido foco de diversos estudos. Assim o objetivo do
presente estudo foi de estudar os efeitos da dieta hipoprotéica e da renutrição no
fígado de camundongos.
Sendo que o principal achado deste trabalho foi de que a renutrição consegue
atenuar às disfunções do fígado em decorrência da desnutrição.
7.1 MODELOS DE DESNUTRIÇÃO PROTÉICA
O que se pode encontrar na literatura, foram publicações relacionadas ao
sistema nervoso entérico em animais submetidos à desnutrição utilizando
quantificação morfométrica para calcular a densidade por área, e com diferentes
protocolos para induzir a desnutrição. E essa desnutrição fez com que houvesse
uma diminuição de 45% no intestino delgado do animal (BRANDÃO et al., 2003).
Muitos modelos usados eram eficientes e os animais no grupo nutridos
pesavam aproximadamente 60% a mais do que os ratos desnutridos, segundo
Castelucci et al. (2002).
Segundo Gomes et al. (2009), a dieta aplicada em seu estudo para a
verificação do pesos dos animais em comparação do grupo nutrido ao grupo
desnutrido chegou a ser de quase 4,63 vezes maior do grupo nutrido para o grupo
desnutrido, vale ressaltar que neste estudo o modelo de desnutrição foi diferente do
aplicado nesta pesquisa, onde Gomes et al. (2009), utilizou o protocolo de
desnutrição pré e pós natal, sendo que o animal nutrido recebia uma dieta protéica
de 20% proteína-caseína e para os desnutridos uma dieta hipoprotéica de 5% de
proteína-caseína.
De acordo com Gomes et al. (2009), através do uso de ferramentas com
design baseado na estereologia, pode-se encontrar resultados mais apurados
utilizando a mesma dieta utilizada por Castelucci et al. (2002).

66
Gomes, SP Discussão
Neste trabalho foi utilizada uma dieta com 2% de proteína oferecida aos
animais do grupo desnutridos e 12% de proteína para o grupo dos animais nutridos.
E pode-se notar que de acordo com as dietas utilizadas por Vinolo et al. (2008), o
estudo atual procurou compensar a ração menor da ingestão fornecida para os
animais desnutridos. E em conseqüência embora os animais ingerissem uma
quantidade mais baixa de alimentos, a dieta que foi preparada obteve os nutrientes
mínimos para os ratos adultos, com a exceção da porcentagem da proteína. E o
mesmo pode concluir que as diferenças observadas entre os grupos experimentais e
dos controles são predominantes por conta da redução da entrada de energia
ingerida pelos mesmos. Nesta dieta Vinolo et al. (2008), fez uso de uma dieta
contendo 20% de proteína para o grupo controle e 4% para o grupo desnutridos.
Segundo Nunes et al. (2002), os resultados obtidos em seu estudo
demonstraram que o paradigma de desnutrição aplicado é efetivo, pois
precocemente já é possível detectar diferença significativa de peso entre os animais
do grupo nutrido e desnutrido. O paradigma proposto, restrição de acesso ao
alimento, assemelha-se ao processo de desnutrição que ocorre em famílias
carentes, no qual a ingesta é inadequada, entre outras razões, pela não
disponibilidade de alimentos. Os possíveis efeitos do stress nos filhotes, pela
separação da mãe, foram minimizados pelo controle de condições ambientais
adequadas, entretanto esse efeito certamente pode ser extrapolável à condição da
desnutrição humana. Pode também ganho de peso constante durante a reabilitação
nutricional, entretanto os ratos do grupo desnutrido permaneciam com peso
significativamente inferior aos do grupo controle.
O uso de uma dieta para animais desnutridos com 23% de proteína e 27%
para o grupo de animais nutridos e o efeito da duração dessa dieta por 6 semanas
fez com que os animais neste estudo proposto pode ser observado que o resultado
obteve um P< 0,005 (WALRAND et al., 2000).
Em outro experimento feito por Fock et al. (2007), os camundongos do grupo
controle utilizaram uma dieta do grupo controle de 20% de proteína e o grupo
desnutrido de 4% de proteína e pode-se averiguar que os camundongos do grupo
controle para o grupo desnutridos teve uma perda de 22% de massa corporal
quando comparados ao grupo nutrido, pela baixa porcentagem de proteína em sua
ingestão alimentar e pode-se observar uma anorexia aparente desses animais.

67
Gomes, SP Discussão
E neste estudo proposto, podemos observar que tanto os pesos dos animais
e o peso dos fígados foram altamente significativos (vide resultados), dessa maneira
a dieta proposta de 2% de proteína para o grupo desnutrido e 12% para o grupo
controle está de acordo também com os trabalhos publicados por autores
supracitados a cima. Onde o peso corporal dos animais nutridos para os desnutridos
foram aproximadamente 2 vezes maior, e em comparação ao nutrido com o
renutrido foi aproximadamente de 1,14 vezes maior. E o peso do fígado dos animais
nutridos para os desnutridos foram aproximadamente 2,88 vezes maior, e em
comparação ao renutrido foi de aproximadamente 1,14 vezes maior.
O estado nutricional é determinado por um balanço entre o suprimento de
nutrientes e a utilização destes pelo organismo. A desnutrição está relacionada com
o desequilíbrio do estado nutricional e ocorre sob diversas formas, podendo ter
conseqüências reversíveis ou irreversíveis (ZANIN et al., 2003).
7.2 VOLUME DO FÍGADO (Vref)
O volume do fígado apresentou-se 49,3% menor nos animais desnutridos em
relação aos animais nutridos. E 16,4% menor em comparação aos animais
renutridos para os nutridos, atenuando a atrofia observada nos animais desnutridos.
Segundo Zanin et al. (2003), uma nutrição inadequada no período pré-parto,
com uma restrição severa de proteína para a mãe, conduz com a deficiências
nutricionais irreversíveis. Sendo assim, neste caso, o volume do fígado, é menos
tanto nos animais desnutridos e renutridos em relação ao que obteve uma dieta rica
em proteínas (animais controles).
Segundo Castelucci et al. (2002), em uma análise quantitativa os animais do
grupo normalmente alimentados tiveram pesos corpóreos quase dobrados em
relação àqueles privados de proteínas aos 21 dias após o nascimento.
Para avaliar o volume do fígado neste estudo foi usado o Princípio de
Cavalieri, que segundo Duran et al. (2007) também usou para poder avaliar o
volume do lobo direito do fígado para uma contagem baseada na técnica de pontos.
Mesmo que o Método de Cavalieri demande um maior tempo para a sua
execução, como foi realizado neste estudo, ele tem sido reportado como sendo um

68
Gomes, SP Discussão
método eficiente e acurado (GUNDERSEN; JENSEN, 1987; GUNDERSEN et al.,
1988; GRAVES; DOMMETT, 2000; VAN VRÉ et al., 2007) e neste trabalho em
particular obtivemos o volume do fígado feito no órgão todo. E podemos perceber
que houve uma diferença estatística entre os quatros grupos analisados e estudados
tamanha acurácia do método.
7.3 NÚMERO TOTAL DE HEPATÓCITOS (N)
Pudemos observar que a variabilidade entre os grupos foi pequena, o que nos
permitiu encontrar diferenças para os parâmetros estereológicos, e para o número
total de hepatócitos tivemos do que se chama de “border line”, onde a variabilidade
nos grupos R e CR estava um pouco mais alta e obtivemos p= 0,055.
E por estes motivos esperaremos que com a inclusão de mais dois animais
em cada grupo para o artigo científico possa possivelmente detectar um p< 0,05
para o número total de hepatócitos.
Segundo Odaci et al. (2009), para quantificar o número de hepatócitos foi
utilizado em seu estudo o método disector óptico para estimar tanto o número total
e densidade numérica dos hepatócitos nos fígados de ratos (GUNDERSEN et al.,
1988). Hepatócitos foram contados de acordo com as regras imparciais na apuração
dos disector óptico (HOWARD; REED, 1998). Foram utilizados frações para a
realização destes procedimentos para análise estereológica sendo que foram
aplicados vários estágios para uma subamostragem. Para cada efeito, uma
amostragem. O nosso experimento também baseou-se neste mesmo método para
que a acurácia do método utilizado nos números totais fossem mais próximo com o
número real do órgão.
Segundo Odaci et al. (2009), a densidade média numérica e o número total de
hepatócitos foram calculados para ambos os grupos experimenais e o grupo
controle, sendo que nos resultados encontrou uma diferença significativa apenas no
número total de hepatócitos.
Nos resultados obtidos por Odaci et al. (2009), encontrou-se um total de
hepatócitos de 1,57 x 106, e na nossa pesquisa encontramos para o número total

69
Gomes, SP Discussão
uma média de 1,1 x 107 para os animais controle do desnutrido de 90 Dias e para os
animais controle do renutrido um total médio de 2,2 x 107.
Sendo que neste experimento foi utlizado camundongos Swiss. E no
experimento do Odair et al., (2009) foram utilizados ratos Wistar.
No experimento feito Por Karbalay-Doust et al. (2008), foi utlizado o método
disector óptico (Howard et al., 1998) para estimar o número total de núcleos dos
hepatócitos, sendo que no estudo feito foi utilizado o rato controle quando
comparado ao animal tratado, obteve um aumento de 20%. Obtendo assim um
resultado para o número total no valor de 5.3 x 108. Ocasionado por uma hiperplasia.
E neste caso tivemos um aumento de 35,51% quando comparado o grupo controle
do grupo tratado.
Segundo o estudo desenvolvido por Marcos et al. (2006) foi utilizado para
estimar o número total de hepatócitos igualmente utilizado em nosso estudo da
amostragem sistemática e uniformemente aleatória, espera-se que a contagem
atinja todas as áeras dos cortes durante o experimento, para que não haja nenhum
desvio do material e não dê erro na estimativa final do número total.
O material, neste caso o fígado, utilizado no experimento de Marcos et al.
(2006), os materiais foram emblocados em parafina, já em nosso experimento o
fígado foi embebido em ágar, para que não houvesse uma retração final que
pudesse comprometer o resultado final do número total de hepatócitos.
Carthew et al. (1998) obteve um achado de 20% no animal controle e estimou
uma médio para o número total de 1,52 x 109. As diferenças em relação ao número
total que fora obtidas poderia ser atribuída à cepa de ratos diferentes e também pela
abordagem técnica e pelo plano de amostragem.
Todas as amostras por ele prevista para a obtenção de uma estimativa do
número total o seu coeficiênte de erro deu inferior a 10% (SCHMITZ et al., 2000). No
caso do nosso do estudo, obtivemos um coeficiênte de menos que 10%.
Recentemente desenvolvidos, os métodos estereológicos têm sido usados em
experimentos para examinar os efeitos de dois níveis de subnutrição durante o início
da vida pós-natal no nímero total de células de ratos (BEDI, 1991).
Os resultados obtidos no trabalho são provenientes de apenas três animais
por grupo. Embora não seja considerado um número suficiente para estatística
clássica, na estereologia a eficiência dos métodos de amostragem (SURS)
(GUNDERSEN et al., 1999) que são aplicados nos variados níveis hierárquicos do

70
Gomes, SP Discussão
volume do fígado, nos permite gerar dados com grau de confiabilidade e precisão
mesmo que ainda se tenha um número pequeno de indivíduos analisados por grupo
(CD, D,R e CR).
7.4 VOLUME MÉDIO DOS HEPATÓCITOS ( Nv )
Os hepatócitos dos animais desnutridos tiveram uma diminuição de 44,86%
em relação ao seu controle e no caso dos animais renutridos ao se controle teve
uma diminuição de 19,93%, atenuando a atrofia observada nos animais desnutridos.
No trabalho realizado por Karbalay-Doust (2008), também foi realizado o
método do nucleador, um estimador eficiente de volume no número de partículas
médio ponderadas, a seleção de células de acordo com sua densidade numérica.
Em nosso estudo também calculamos o volume médio só que usando o método do
rotatorA média do volume encontrada por Karbalay-Doust. (2008), foi de 5.30 µm
3,
sendo que nos camundongos de nosso experimento encontrou-se um volume médio
de 8.1 µm3, obtendo um aumento significante em relação ao grupo dos animais
acometidos.
Segundo Jack et al. (1990), o método nucleador, uma nova abordagem
estereológica, forneceu uma estimativa eficiente, imparcial do volume celular médio
de lesões focais e áreas extrafocal. Ele também forneceu uma amostra imparcial das
células para estimar o volume nuclear do hepatócito e da porcentagem de células
binucleadas. Os resultados mostraram um aumento no volume médio de células
mononucleares de 4700 µm3, demonstrando o efeito hipertrófico. Medições de
volume nuclear dos hepatócitos mononucleares e binucleadas foram utilizados para
avaliar a aploidia.
Já neste trabalho realizado usando o “one-way” ANOVA, usando o software
MINITAB® 16, obtivemos diferenças altamente significantes.
Uma vez que métodos estereológicos são utilizados para estimar volumes de
partículas sem assumir forma, tamanho, orientação ou distribuição das células
(GUNDERSEN et al., 1988). E o volume médio se revela muito mais acurado do que

71
Gomes, SP Discussão
a estimativa de tamanho baseado simplesmente em uma área seccional (TANDRUP,
2004).
7.5 AVALIAÇÕES BIOQUÍMICAS
Uma vez que a desnutrição afeta preferencialmente tecidos com alta taxa de
proliferação, o comprometimento da hemopoese nessas condições tem sido descrito
por diversos estudos (BORELLI; MARIANO; BOROJEVIC, 1995; BARON, 1997,
BORELLI et al., 2007). Como conseqüência, indivíduos desnutridos apresentam
tanto anemia (redução do número de hemácias, volume hematócrito e concentração
de hemoglobina), como leucopenia (IDOHOU-DOSSOU et al., 2003; BACHOU et al.,
2006), sendo o quadro hematológico um importante fator indicativo de desnutrição
(BACHOU et al., 2006). A presença de anemia e leucopenia nos animais desnutridos
reflete o comprometimento do tecido hemopoético frente à desnutrição. Uma análise
maior dos leucócitos presentes no sangue desses animais mostra que a leucopenia
encontrada é, principalmente, à custa de neutropenia e linfocitopenia.
Neste estudo, o que se pode notar foi que, todos os grupos, comparados ao
controle do desnutrido, apresentaram elevação da ALT, fato este que pode
evidenciar as alterações funcionais do fígado levada pela desnutrição utilizada neste
trabalho. Os índices dos animais do grupo desnutrido foram elevados, o que poderia
indicar um processo inflamatório mais intenso, esperado nos animais deste grupo
(COUTINHO, 2004).
Em relação à AST houve mais que o dobro da ALT, o que poderia exprimir
uma maior gravidade e profundidade da lesão. Essa elevação simultânea de AST e
FA no grupo controle desnutrido e renutrido leva-se a suposição de que há um
processo de colestase, sendo que o mesmo foi causado pela tensão contínua e
compressão focal nos ductos, que foram bem mais acentuadas (BARRETO, 1971).
Níveis altos de FA, como podem observar neste trabalho, mesmo no controle
do desnutrido e no controle do renutrido, tem sido encontradas nos resultados do
Centro de Bioterismo da Faculdade de Medicina da Universidade de São Paulo
(USP, 2004).

72
Gomes, SP Discussão
A concentração sérica das aminotransferases informa o grau de
comprometimento hepático, uma vez que a sua liberação no soro pode significar
morte celular.
As concentrações sérica de ALT é bem mais indicada para a avaliação da
integridade celular do fígado que a AST (AWADALLA et al, 1975). As alterações de
FA relaciona-se diretamente à obstrução biliar de causas intra e extra hepáticas, ou
uma lesão hepatocelular (LIMA et al., 2001).
Sabe-se que a desnutrição causa uma deterioração das funções hepáticas,
com alterações sorológicas relacionadas à ALT, AST e FA (ONYENEKE et al, 2003).
A hipoalbuminemia ocorreu apenas em função da desnutrição, uma vez que
essa alteração protéica não alterou a função hepática.
Enfim, podemos observar que a desnutrição pode interferir nos níveis e nos
fatores das funções hepáticas, porém, pode-se notar que essas alterações das
funções hepáticas foi muito mais importante quando mostrou-se que na renutrição
os índices continuam afetados.
De modo geral, os resultados deste trabalho indicam comprometimento dos
setores granulocítico, linfóide e eritróide, que pode decorrer não só da ausência de
proteínas, como também de alterações nos fatores que controlam a proliferação, a
diferenciação e a liberação das células do compartimento central para o
compartimento periférico, bem como dos fatores que regulam a saída destas células
do compartimento periférico para o compartimento extravascular (no caso dos
leucócitos) (BORELLI; MARIANO; BOROJEVIC, 1995; VITURI et al., 2000).
Com relação ao comportamento hematológico dos animais desnutridos,
observou-se uma diminuição significante do número de hemácias, da concentração
de hemoglobina e do volume hematócrito, bem como do número de leucócitos,
linfócitos e neutrófilos segmentados em relação aos animais desnutridos CD, R e
CR.
Esses resultados indicam reversão das alterações hematológicas encontradas
na desnutrição após retorno à ração normoprotéica. Os resultados obtidos para os
animais do grupo renutrido estão de acordo com achados na literatura e indicam que
o retorno à alimentação adequada tende a normalizar os parâmetros hematológicos
(WALRAND et al., 2000; VÁSQUEZ-GARIBAY et al., 2002; DEWAN et al., 2009),
sendo novamente o hemograma usado como indicativo do estado nutricional.

Conclusões

74
Gomes, SP Conclusões
8 CONCLUSÕES
Com base nos métodos empregados e nos resultados obtidos, podemos
concluir que os animais desnutridos apresentaram uma alteração no fígado com
uma redução em seu volume (atrofia), bem como uma perda do número total de
hepatócitos e uma atrofia dos mesmos. Nos renutridos houve atenuação da atrofia
do volume do fígado e no tamanho dos hepatócitos. Esta reversão foi de 83,6%.
Será que se esta renutrição perdurasse por mais um ou dois meses essa atenuação
seria de 100% ?

Referências

76
Gomes, SP Referências
REFERÊNCIAS
ACC/SCN. Nutrition throughout life. In:_____4th Report on the World Nutrition Situation. Geneva: ACC/SCN./World Health Organization, 2000.
ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WALTER, P. Histologia: vida e morte das células nos tecidos. In.:___________. Biologia Molecular da Célula. 4. ed. Porto Alegre: Artmed, 2004. p. 1277.
AWADALLA, H. N.; SHERIF, A. F.; SHAFEI, A. Z.; KHALIL, H. A.; GURGIS, F. K. Enzyme levels in homogenate of liver from mice infected with Schistosoma mansoni and from uninfected mice. International Journal of Parasitology, v. 5, p. 27-
31,1975. AZEREDO, V. B.; CARMO, M. G. T.; BOAVENTURA, G. T.; DIAS, M. M.; LOBO, F. P. Multimistura: uma alternativa alimentar. Rio de Janeiro: Independente, 1999. p.
79. BACHOU, H.; TUMWINE, J. K.; MWADIME, R. K. N.; TYLLESKÄR, T. Risk factors in hospital deaths in severely malnourished children in kampala, Uganda. BMC Pediatics, v. 6, n. 7, p. 1-9, 2006..
BADDELEY, A. J.; GUNDERSEN, H. J. G.; CRUZ-ORIVE, L. M. Estimation of surface área from Vertical Sections. Journal of Microscopy, v. 142, p. 259–276,
1986. BARON, R. B. Desnutrição protéico-calórica. In: BENNET, J. C.; PLUM, F, C. Tratado de medicina interna. 20. ed. Rio de Janeiro: Guanabara-koogan, 1997,
p.1273-1277. BARRETO, U. S. T. Alkaline phosphatase in schistosomiasis. Annals of Internal Medicine, v. 74, p. 450-451, 1971.
BEDI, K. S. Early-life undernutrition causes deficits in rat dentate gyrus granule cell number. Experientia, v. 4, p. 1073-1074,1991.
BERK, P. D.; POTHER, B. J.; STREMMED, W. The sinusoidal surface on the hepatocyte: a dynamic plasma membrane specialized for high volume molecular transit. In: REUTTER, W.; POPPER, H.; AREAS, I. M.; HEINRICH, P. C.; KEPPLER, D.; LANDMANN, L. (Ed.). Modulation of liver cell expression. Lancaster: MTP
Press Ltda, 1987. p.107-125. BISMUTH, H. Surgical anatomy and anatomical surgery of the liver. In: BLUMGARD, L. H. (Ed). Surgery of the liver and biliary Tract. Edinburgh: Churchill Livingstone,
1988, p.1-9.

77
Gomes, SP Referências
BORELLI, P.; MARIANO, M.; BOROJEVIC, R. Protein malnutrition: effect on myeloid cell production and cell mobilization into inflammatory reactions in mice. Nutrition Research, v. 15, n. 10, p. 1477-1485, 1995.
BORELLI, P.; FOCK, R. A.; BLATT, S.; PEREIRA, J.; BEUTLER, B.; TSUJITA, M.; SOUZA, A. C.; XAVIER, J. G. Effect of protein deprivation on erytropoiesis. British Journal of Nutrition, v. 97, n. 2, p. 307-314, 2007.
BRANDÃO M. C. S.; DE ANGELIS R. C.; DE SOUZA R. R.; LIBERTI E. A. Effects of pre-and postnatal protein deprivation on the myenteric plexus of the small intestine: a morphometric study in wealing rats. Nutrition Research, v. 23, p. 215-223, 2003.
CARTHEW, P.; EDWARDS, R. E.; NOLAN, B. M. New approaches to the quantitation of hypertrophy and hyperplasia in hepatomegaly. Toxicology Letters. v. 102/103, p. 411-415, 1998. CASTELUCCI, P.; SOUZA, R. R.; ANGELIS, R. C.; FURNESS, J. B.; LIBERTI, E. A. Effects of pré-and postnatal protein deprivation and postnatal refeeding on myenteric neurons of the rat large intestine: a quantitative morphological study. Cell and Tissue Research, v. 310, n. 1, p. 1-7, 2002.
CHANDRA R. R. Nutrition and immunology: from the clinic to cellular biology and back again. Proceedings of the Nutrition Society, v. 58, p. 681-683, 1999.
CRUZ-ORIVE, L. M.; GEISER, M. Estimation of particle number by stereology: an update. Journal of Aerosol Medicine, v. 17, p. 197-212, 2004.
CRUZ-ORIVE, L. M. A general variance predictor for cavalieri slices. Journal of Microscopy, v. 222, p. 158-165, 2005.
CRUZ-ORIVE, L. M.; GARCÍA-FIÑANA, M. A review of the article: comments on the shortcomings of predicting the precision of cavalieri volume estimates based upon assumed measurement functions, by Edmund Glaser. Journal of Microscopy ,v.
218, p. 6-8, 2005. COUTINHO, E. M. Malnutrition and hepatic fibrosis in murine schistosomiasis. Memórias do Instituto Oswaldo Cruz, v. 99,p. 85-92, 2004.
DEWAN, P.; FARIDI, M. M. A.; AGARWAL, K. N. Cytokine response to dietary rehabilitation with curd (Indian Dahi) & leaf protein concentrate in malnourished children. Indian Journal of Medical Research, v. 130, p. 31 – 36, 2009.
DIEHL, A. M.; RAI, R. Review: regulation of live regeneration by pro-inflamatory cytoknes. Journal of Gastroenterology and Hepatology, v. 11, n. 5, p. 466-470,
1996. DONOVAN, J.; BROWN P. Current Protocol in Immunology. New York: John
Wiley & Sons, 1991, p. 141–144.

78
Gomes, SP Referências
DOYLE, M.; WILSON,R. B.; HARTROFT,W. S. The effects of starvation on liver regeneration in rat after partial hepatectomy. Experimental and molecular Pathology, v. 9, p. 400-404, 1968.
DURAN, C.; AYDINLI, B.; TOKAT, Y.; YUZER, Y.; KANTARCI, M.; AKGUN, M.; PALAT, K. Y.; UNAL, B.; KILLI, R.; ATAMANALP, S. Stereological evaluation of liver volume in living donor liver transplantation using MDCT via the cavalieri method. Liver Transplantation, v.13, p. 693-698, 2007.
FLECKNELL, P. Laboratory animal anaesthesia. 2. ed. London: Academic Press,
London, 1996. p. 15–73. FOCK, R. A.; VINOLO, M. A.; ROCHA, V. M. S. ; SÁ ROCHA, L. C.; BORELLI, P. Protein-energy malnutrition decreases the expression of TLR-4/MD-2 and CD14 receptors in peritoneal macrophages and reduces the synthesis of TNF-alpha in response to lipopolysaccharide (LPS) in mice. Cytokine, v. 40, n. 2, p. 105-114,
2007. FOCK, R. A.; VINOLO, M. A. R.; CRISMA, A. R.; NAKAJIMA, K.; ROGERO, M. M.; BORELLI, P. Protein-energy malnutrition modifies the production of interleukin-10 in response to lipopolysaccharide (LPS) in a murine Model. Journal of Nutritional Science and Vitaminology, v.54, p. 371-377, 2008.
GARCÍA-FINÃNA, M. Muestro sistemático en R. aplicaciones a la estereologia.
Tesis (Doctoral)-Universidad de Cantabria, 2000. GARCÍA-FINÃNA, M.; CRUZ-ORIVE, L. M. New approximations for the efficiency of cavalieri sampling. Journal of Microscopy, v. 199, p. 224-238, 2000a.
GARCÍA-FINÃNA, M.; CRUZ-ORIVE, L. M.; MACKAY, C. E.; PAKKENBERG, B.; ROBERTS N. Comparison of MR imaging against physical sectioning to estimate the volume of human cerebral compartments. NeuroImage, v. 18, p. 505-516, 2002.
GARTNER, C. P.; HIAT, L. P. Tratado de histologia. 2. ed. Rio de Janeiro:
Guanabara koogan, 2003. p. 456. GERBER, M. A.; THUNG, S. N.; SHEN, S.; STROHMEYER, F. W.; ISHAK, K. G. Phenotypic characterization of hepatic proliferation. Antigenic expression by proliferation epithelial cells in fetal liver, massive hepatic necrosis and nodular transformations of the liver. American Journal Pathologic. v. 10, p. 70-74, 1983.
GOMES, O. A.; CASTELUCCI, P.; FONTES, R. B. V.; LIBERTI, E. A. Effects of preand postnatal protein and postnatal refeeding on myenteric neurons of the rat small intestine: a quantitative morphological study. Clinical Autonomic Neuroscience. v. 126, p. 277-284, 2006.
GOMES, S. P.; NYENGAARD, J. R.; MISAWA, R.; GIROTTI, P. A.; CASTELUCCI, P.; HERNANDEZ-BLAZQUEZ, F. J.; MELO, M. P.; RIBEIRO, A. A. C. Atrophy and neuron loss: effects of a protein-deficient diet on sympathetic neurons. Journal of Neuroscience Research, v. 87, p. 3568-3575, 2009.

79
Gomes, SP Referências
GOSS, C. M. Gray anatomia. Rio de Janeiro: Guanabara Koogan,1988. p. 1017-
1026. GRAVES, M. J; DOMMETT, D. M. F. Comparison of cardiac stroke volume measurement determined using stereological analysis of breath-holdcine MRI and phase contrast velocity mapping. The British Journal of Radiology, v. 73, p. 825-
832, 2000. GRISHAM, J. W. Cell types in long-term propagable cultures of rat liver. Animals of the New York Academy of Sciences, v. 349, p. 128-137, 1980.
GRISHAM, J. W. Cell types in rat liver cultures: their identification and isolation. Molecular and Cellular Biochemistry, v. 53-54, n. 1-2, p. 23-33, 1983. GUNDERSEN, H. J. G.; JENSEN, E. B. The efficiency of systematic sampling in stereology and its prediction. Journal of Microscopy, v. 147, p. 229-263, 1987.
GUNDERSEN, H. J.; BAGGER, P.; BENDTSEN, T.; EVANS, S. M.; KORBO, L.; MARCUSSEN, N.; MOLER, A.; NIELSEN, K.; NYENGAARD, J. R.; PAKKENBERG, B.; SORENSEN, F. B.; VESTERBY, A.; WEST, M.J. The new stereological tools: disector, fractionator, nucleator and point sampled intercepts and their use in pathological research and diagnosis. Acta Pathologica, Microbiologica et Immunologica Scandinavica, v. 96, n. 10, p. 858-881, 1988.
GUNDERSEN, H. J.; BENDTSEN, T. F; KORBO, L.; MARCUSSEN, N.; MOLLER, A.; NIELSEN, K.; NYENGAARD, J. R.; PAKKENBERG, B.; SORENSEN, F.B.; VESTERBY, A. Some new, simple and efficient stereological methods and their use in pathological research and diagnosis. Acta Pathologica, Microbiologica et Immunologica Scandinavica, v. 96, p. 379-394, 1988.
GUNDERSEN, H. J. G.; JENSEN, E. B. V.; KIEU, K.; JENSEN, J. The efficiency of systematic sampling in stereology - seconsidered. Journal of Microscopy, v.193, p.
199-211, 1999. GUYTON, A. C.; HALL, J. E. Tratado de fisiologia médica. 10. ed. Rio de Janeiro:
Guanabara Koogan, 2002, p. 973. HALICI, Z.; DURSUN, H.; KELES, O. N.; ODACI, E.; SULEYMAN, H.; AYDIN, N.; CADIRCI, E.; KALKAN, Y.; UNAL, B. Effect of chronic treatment of haloperidol on the rat liver: a stereological and histopathological study. Naunyn-Schmiedebergs Archives of Pharmacology, v. 379, p. 253-261, 2009.
HOWARD, C. V.; REED, M. G. Number quantification. Unbiased stereology. three-dimensional measurement in microscopy. Oxford: Bios Scientific Publishers. p. 69-
106, 1998. HOWARD, C. V.; REED, M. G. Unbiased Stereology. Three-Dimensional Measurement in Microscopy, 2. ed. Bios Scientific Publishers, 2005.

80
Gomes, SP Referências
INSTITUTO ADOLFO LUTZ. Normas Analíticas do Instituto Adolfo Lutz. 3 ed.
São Paulo: INSTITUTO ADOLFO LUTZ, 1985, p. 533. IDOHOU-DOSSOU, N.; WADE, S.; GUIRO, A. T.; SARR, C. S.; DIAHAM, B.; CISSÉ, D. Nutritional status of preschool senegalese children: long-term effects of early severe malnutrition. British Journal of Nutrition, v. 90, p. 1123-1132, 2003.
ISSLER, R. M. S.; GIUGLIANI, E. R. J.; KREUTZ, G. T.; MENESES, C. F.; JUSTO, E. B.; KREUTZ, V. M. Poverty levels and children’s health status: study of risk factors in an urban population of low socioeconomic level. Revista Saúde Pública, v. 30, n.
6, p. 506-511, 1996. JACK, E. M.; STAUBLI, W.; WAECHTER, F.; BENTLET, P.; SUTER, J.; BIERI, F. Ultrastructural changes in chemically induced preneoplastic focal lesions in the rat liver: a stereological study. Carcinogenesis. v. 11, p. 1531-1538, 1990.
JAESCHKE, H.; GORES, G. J.; CDDERBAUM, A. I.; HERNSON, J. A.; PESSAYRE, D.; LEMASTERS, J. J. Mechanisms of hepatotoxicity. Toxicological Sciences, v.
65, p. 166-176, 2002. JUNQUEIRA, L. C.; CARNEIRO, J. Histologia básica. 9. ed. Rio de Janeiro:
Guanabara Koogan, 1999. p. 273-282. JUNQUEIRA, L. C.; CARNEIRO, J. Histologia básica. 10 ed. Rio de Janeiro:
Guanabara Koogan, 2004. p. 488. KARBALY-DOUST, S.; NOORAFASHAN, A. Stereological study of the effects of nandrolone decanoate on the mouse liver. Micron, v. 40, p. 471-475, 2009.
LIMA, A. A. A. L.; RAMALHO, L. N. Z.; ZUCOLOTO, S.; BAGNATO, V. .; SILVA, JÚNIOR. O. C. Estudo da fosfatase alcalina em ratos cirróticos hepatectomizados após aplicação de laser. Acta Cirurgica Brasileira, v. 16, p. 16-17, 2001.
MACHADO, A. L. S.; ROCHA, R. F.; SANTOS, R. C. S. Hepatectomia parcial: estudo da resposta proliferativa. Revista Biociência de Taubaté, v. 8, n. 2, p. 27-35, 2002. MARCOS, R.; MONTEIRO, A. F.; ROCHA, E. Estimation of the number of stellate cells in a liver with the smooth fractionator. Journal of Microscopy, v. 215, p. 174-
182, 2004. MARCOS, R.; MONTEIRO, R. A. F.; ROCHA, E. Design-based stereological estimation of hepatocyte number, by combining the smooth optical fractionator and immunocytochemistry with anti-carcinoembryonic antigen polyclonal antibodies. Liver Internacional, v. 26, p. 116-124, 2006.
MARCOS, R.; MONTEIRO, R. A. F.; ROCHA, E. Determination of hepatocellularity number in the rat. Toxicology in Vitro, v. 21, p. 1692-1693, 2007.
MAYHEW, T. M.; OLSEN, D. R. Magnetic resonance imaging (MRI) and model-free estimates of brain volume determined using the cavalieri principle. Journal of

81
Gomes, SP Referências
Anatomy, v. 178, p. 133-144, 1991.
MICHALOPOULUS, G. K.; DEFRANCES, M. C. Liver Regeneration. Science, v. 276,
p. 60-66, 1997. MONTE, C. M. G. Desnutrição: um desafio secular à nutrição infantil. Jornal de Pediatria, v. 76, n. 3, p. 285-297, 2000.
NUNES, M. L.; BATISTA, B. B.; MICHELI. F.; BATISTELLA. V. Effects of early malnutrition and nutritional rehabilitation in rats. Journal of Pediatrics, v. 78, n. 1, p.
39-44, 2002. ODACI, E.; BILEN, H.; HACIMUFTUOGLU, A.; KELES, O. N.; CAN, I.; BILICI, M. Long-term treatments wit- and high-dose olanzapine change hepatocyte numbers in rats. A stereological and histopathological study. Archives of Medical Research, v.
40, p. 139-145, 2009. ONYENEKE, E.C.; NJOYA, H. K.; EBOT, M. N.; UCHUNGO, G. A.; ERIYAMREMU, G. E.; ALUMANAH, E. O. Liver function status in some nigerian children with protein energy malnutrition. Orient Journal of Medicine, v. 15, p. 58-63, 2003.
PAKKENBERG, B.; GUNDERSEN, H. J. G. Total number of neurons and glial cells in human brain nuclei estimated by the disector and fractionator. Journal of Microscopy, v. 150, p.1-20, 1988.
PAKKENBERG, B.; GUNDERSEN, H. J. G. The total number of neurons and glia cells in human brain nuclei estimated by disector and the fractionator. Journal of Microscopy, v. 215, p. 279-285, 1996.
PARRA,O. M.; SILVA, R. A. P. S.; SILVA, J. R. M. C.; HERNANDEZ-BLAZQUEZ, F. J.; PEDUTO, L.; SAAD, W. A.; SAAD JUNIOR, W. A. Reduction of liver mass due to malnutrition in rats. Correlation with emanciation of animals and size of organs not insert in the portal system, Revista Paulista de Medicina, v. 113, n. 3, p. 903-909,
1995. POPPER, H. Viral versus chemical hepatocarcinogenesis. Journal of Hepatology,
v. 6, p. 229-238, 1988. RAMALHO, F. S.; RAMALHO, L. N. Z.; ZUCOLOTO, S.; SILVA JR., O. C. Regeneração hepática. Algumas definições num universo de incertezas. Acta Cirúrgica Brasileira, v. 8, n. 4, p.177-189, 1993.
REDMOND, H. P. Immuno suppressive mechanisms in protein-calorie malnutrition. Surgery, v. 110, n. 2, p. 311-317, 1991c.
REDMOND, H. P. Protein-calorie malnutrition impairs host defense against candida albicans. Journal of Surgery Research, v. 50, n. 6, p. 552-559, 1991b.
REEVES, P. G.; NIELSEN, F. H.; FAHEY, G. C. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc Writing Committee

82
Gomes, SP Referências
on the Reformulation of the AIN-T6A Rodent Diet. American Institute of Nutrition,
n. 121, p. 1939-1951, 1993. RIBEIRO, A.A.; DAVIS, C.; GABELLA, G. Estimate of size and total number of neurons in superior cervical ganglion of rat, capybara and horse. Anatomy and Embryology (Berl), v. 208, n. 5, p. 367-380, 2004.
ROGERO, M. M.; BORELLI, P.; VINOLO, M. A. R.; FOCK, R. A.; PIRES, I. S. O.; TIRAPEGUI, J. Dietary glutamine supplementation affects macrophage function, hematopoiesis and nutritional status in early weaned mice. Clinical Nutrition, v. 27,
p. 386-397, 2008. ROUILLER, C. The liver. London: Academic Pres Inc.,1964. v. 2, p. 674.
SANT’ANA, D. M. G.; NETO, M. H.; SOUZA, R. E.; MOLINARI, S. L. Morphological and quantitative study of the myenteric plexus of the ascending colon of rats subjected to proteic desnutrition. Neuropsiquiatr, v. 4, n. 55, p. 687-695, 1997.
SANTOS, R. M. S. Avaliação do estado nutricional relativo à vitamina A e ferro em crianças HIV positivas ou com AIDS. Tese (Mestrado) – Universidade Federal
de Mato Grosso, 2009. SCHMITZ, C.; HOF, P. R. Recommendations for straightforward and rigorous methods os counting neurons based on a computer simulation approach. Journal of Chemical Neuroanatomy, v. 20, p. 93–114, 2000. TANDRUP, T. Unbiased estimates of number and size of rat dorsal root ganglion cells in studies of structure and cell survival. Journal of Neurocytology, v. 33, n. 5,
p. 173-192, 2004. UNICEF – Desnutrição: situação da infância brasileira 2006. Disponível em: <<http://www.unicef.org/brazil/Pags_040_051_Desnutricao.pdf>>, acesso em:
maio, 2010. USP - Universidade de São Paulo. Faculdade de Medicina. Parâmetros biológicos dos animais de experimentação no biotério da FMUSP, 2004. São Paulo. Disponível em: <http://www.tcirurgica.fm.usp.br/bioterio/asnell/bioquimica.doc>. Acesso
em: 27/12/2010. VAN VRÉ, E. A.; VAN BEUSEKOM; H. M.; VRINTS, C. J.; BOSMANS, J. M.; BULT, H.; VAN DER GIESSEN, W. J. Stereology: a simplified and more time-efficient method than planimetry for the quantitative analysis of vascular structures in different models of intimal thickening. Cardiovascular Pathology, v.16, p. 43-50, 2007.
VÁZQUEZ-GARIBAY, E.; CAMPOLLO-RIVAS, O.; ROMERO-VELARDE, E.; MÉNDEZ-ESTRADA, C.; GARCIA-IGLESIAS, T.; ALVIZO-MORA, J. G.; VIZMANOS-LAMOTTE, B. Effect of renutrition on natural and cell-mediated immune response in infants with severe malnutrition. Journal of Pediatric Gastroenterology and Nutrition, v. 34, n. 3, p. 296-301, 2002.

83
Gomes, SP Referências
VEDEL-JENSEN, E. B.; GUNDERSEN, H. J. G. The Rotator. Journal of Microscopy, v. 170, p. 35-44, 1993.
VINOLO, M. A. R.; CRISMA, A. R.; NAKAJIMA, K.; ROGERO, M. M.; FOCK, R. A.; BORELLI, P. Malnourished mice display an impaired hematologic response to granulocyte colony-stimultating factor administration. Nutrition Research, v. 28, p.
791-797, 2008. VITURI, C. L.; BORELLI, P.; ALVAREZ-SILVA, M.; TRENTIN, A. Z. Alterations of the bone marrow in extracelular matrix in undernourished Mice. Brazilian Journal of Medical and Biological Research, v. 33, p. 889-895, 2000.
WALRAND, S.; CHAMBON-SAVANOVITCH, C.; FELGINES, C.; JACQUES, C.; RAUL, F.; NORMAND, B.; FARGES, M. C.; BEAUFRÈRE, B.; VASSON, M. P.; CYNOBER, L. Aging: a barrier to renutrition? nutritional and immunologic evidence in rats. American Journal of Clinical Nutrition, v. 72, p. 816-824, 2000.
WALRAND, S.; MOREAU, K.; CALDEFIE, F.; TRIDON, A.; CHASSAGNE, J.; PORTEFAIX, G.; CYNOBER, L.; BEAUFRÈRE, B.; VASSON, M. P.; BOIRIE, Y. Specific and nonspecific immune responses to fasting and refeeding differ in healthy young adult and eldery persons. American Journal of Clinical Nutrition, v. 74, p.
670-678, 2000. WEIBEL, E. R.; STÄUBLI, W.; GNÄGI, H. R.; HESS, F. A. Correlated morphometric and biochemical studies on the liver cell. I. Morphometric model, stereologic methods, and normal morphometric data for rat liver. Journal Cell Biology, v. 42, p.
63-91, 1969. WEST, M. J. S.; GUNDERSEN, H. J. G. Unbiased stereological estimation of the number of neurons in the subdivisions of the rat hippocampus using the optical fractionator. Anatomical Record, v. 231, p. 482-497, 1991.
WHO. Expertt Committee on Nutrition. 6
th report. 1992. (Who Technical Report
Series, 45). YUCEL, F.; WARREN, A.; GUMUSBURUN, E. The effects of undernutrition on connectivity I the cerebellar cortex of adults rats. Journal of Anatomic, v. 184, p.59-
64, 1994. ZANIN, S. T.; MOLINARI, S. L.; SANT'ANA, D. M.; DE MIRANDA NETO, M. H. NADH-Diaphorase positive neurons of the jejunum of disnurtured adult rats (Rattus Norvegicus): quantitative aspects. Neuropsiquiatr, v. 61, n. 3, p. 650 -653, 2003. GARTNER, C. P.; HIAT, L.P. Tratado de Histologia. 2 ed. Rio de Janeiro: Ed.
Guanabara koogan, p. 456, 2003.