sanidade atualidades no circovírus suíno tipo 2 (pcv2) dra ... · esses agentes virais genética...
TRANSCRIPT

26
Sanidade
Suínos & Cia Ano VI - nº 35/2010
Atualidades no Circovírus Suíno Tipo 2 (PCV2)
Sanidade
Dra. Maria FortM. Veterinária e Doutora
Atualidades no Circovírus Suíno Tipo 2 (PCV2)
Introdução
Histórico do PCV2 e da circovirose
suína (PCVD)
Em 1974 Tisher e colabo-
radores descobriram um vírus que contaminava uma linhagem de célu-
las renais de suínos PK-15 (ATTC-CCL33), morfologicamente similar ao picornavírus(182). Mais tarde de-
monstrou-se que o agente contami-nante era um vírus do tipo filamento circular de DNA simples (ssDNA), o qual passou a ser denominado circo-
vírus suíno (PCV)(183). Embora pes-
quisas sorológicas tenham revelado a presença de anticorpos anti-PCV na
população de suínos, nenhuma infec-
ção pode ser atribuída ao vírus(184, 41, 2), o qual, consequentemente, foi consi-derado não patogênico.
Em 1991 uma nova doença
emergente, denominada síndrome do emagrecimento progressivo pós-
desmame (sigla em inglês, PMWS), foi relatada no oeste canadense(20, 97).
Essa nova síndrome caracterizava-se, clinicamente, por perda de peso progressiva, dificuldade respiratória e palidez da pele. Do ponto de vista da patologia, os suínos doentes apre-
sentavam um acometimento multis-
sistêmico, caracterizado por linfo-adenopatia generalizada, pneumonia intersticial, hepatite, esplenomega-
lia e úlceras gástricas(20, 97). Um ano
mais tarde um agente semelhante ao
PCV foi isolado de tecidos de suínos
afetados pela PMWS, nos EUA e na Europa(03, 43). A comparação entre o
PCV original não patogênico e o re-
cém descoberto circovírus, associado aos surtos de PMWS, revelou serem esses agentes virais genética e antige-
nicamente distintos(03, 120). Por isso, o PCV foi, subsequentemente, dividido em dois tipos: o vírus não patogênico derivado da célula PK-15 foi denomi-nado PCV do tipo 1 (PCV1), e o cir-covírus isolado de suínos acometidos
pela PMWS passou a ser denominado
PCV do tipo 2 (PCV2)(65, 120).
Além da PMWS, o PCV2 tem sido associado a outras doenças, como a síndrome de dermatite e nefropatia
suína (sigla em inglês, PDNS), falhas reprodutivas e o complexo da doen-
ça respiratória suína (sigla em inglês, PRDC). Em 2002, Allan e colabora-
dores propuseram o termo circovirose
suína (sigla em inglês, PCVD) para agrupar todas as doenças e síndromes
associadas ao PCV2. De modo simi-
lar, em março de 2006, a Associação Norte-americana de Veterinários Es-
pecialistas em Suínos (AASV) propôs uma nomenclatura semelhante: doen-
ças associadas ao circovírus suíno (si-gla em inglês, PCVAD).
Atualmente ainda não há um
consenso relativo a essa nomenclatura
e ambos os termos, PCVD e PCVAD, são usados na Europa e nos EUA, res-
pectivamente. Até os dias de hoje, da-
dos experimentais têm demonstrado
uma ligação efetiva entre o PCV2 e
o desenvolvimento de doenças apenas
com relação à PMWS(05, 16) e a falhas
reprodutivas(159, 115). A associação en-
tre o PCV2 e as demais condições tem
sido baseada em estudos retrospecti-vos e/ou casos clínicos, não havendo evidências experimentais dessas co-
nexões, até o momento(165).
Taxonomia
O PCV2 é membro de uma família de vírus ssDNA não enve-
lopados, pequenos e icosaédricos (família Circoviridae), que afeta os vertebrados(104). Em função da descri-
ção contínua de novos agentes seme-
lhantes ao circovírus nos últimos anos, a classificação da família Circoviri-dae tem sido e deverá ser submetida a uma reorganização(35). No momento, de acordo com o Comitê Internacional de Taxonomia Viral (sigla em inglês, ICTV) (www.ictvonline.org), essa fa-
mília está dividida – com base no ta-
manho de seus vírus e de sua dimen-
são gênica – nos gêneros Circovírus e Gyrovirus (Figura 1.1). Além do PCV, sete vírus aviários têm sido descritos com sendo do gênero Circovírus: o vírus da doença do bico e penas(150), o circovírus dos pombos(201), o circoví-rus dos canários(143), o circovírus dos gansos(187), o circovírus dos patos(69), o circovírus dos tentilhões(171) e o circo-
vírus das gaivotas(173). Embora ainda não incluídos na lista de taxonomia do

ICTV, casos recentes de circovirose acometendo corvos(177), estorninhos(74)
e cisnes(64) têm sido relatados. O vírus da anemia dos frangos é a única espé-
cie incluída no gênero Gyrovirus(186).
Morfologia e organização
molecular
O PCV2 é um dos menores ví-
rus conhecidos, com uma partícula vi-ral icosaédrica não envelopada de 17 ± 1,3 nm de diâmetro(183). A molécula
do genoma é do tipo ssDNA circular
covalentemente fechada, com aproxi-madamente 1,76 a 1,77 kilobases(120).
Após a infecção da célula pelo PCV2
o ssDNA converte-se em um DNA
de filamento duplo intermediário
(dsDNA), também conhecido como forma replicativa (sigla em inglês, RF). A RF é considerada um genoma codificado por dupla polaridade (posi-tivo para o DNA viral e negativo para
o complementar). Os genes do PCV2
estão organizados em 11 supostos quadros de leitura abertos (sigla em inglês, ORFs)(65). Entretanto, a ex-
pressão protéica é descrita por apenas
três deles. ORF1, também conhecido por gene Rep, localiza-se no filamen-
to positivo, orientado no sentido horá-
rio; ele codifica as proteínas replica-
ses não estruturais Rep e Rep’, de 314 e 178 aminoácidos (aa) de compri-mento, respectivamente(111, 24). ORF2 ou gene Cap, localizado no filamento complementar e orientado no sentido
anti-horário, codifica a única proteína estrutural (233-234 aa), o capsídeo (Cap)(112, 124). ORF3 está completa-
mente sobreposto ao gene ORF1 e lo-
caliza-se no filamento complementar, orientado no sentido anti-horário; ele codifica uma proteína não estrutural de 105 aa de comprimento. In vitro, a proteína ORF3 parece estar implicada na apoptose das células PK15, indu-
zida por vírus(99). Muito recentemente
foi demonstrado que leitões inocu-
lados com um PCV2 mutante ORF3 deficiente manifestavam viremia de-
crescente e menos lesões patológicas
associadas ao PCV2, em comparação a suínos inoculados com o PCV2 do
tipo selvagem. Por essa razão foi su-
gerido que a proteína ORF3 pode ter um papel essencial na replicação e na
patogenicidade viral(79).
Propriedades biológicas e físico-
químicas
O PCV2 é muito estável em
condições ambientais; entretanto, sua infectividade pode ser reduzida pela
exposição viral a altas temperaturas.
Deste modo, após pasteurização a 60º C por 10 minutos, seu poder de infecção decresce 1,6 log; após trata-
mento por calor seco (80º C por duas horas) decresce 0,75 log e 1,25 log após tratamento por calor extremo
(120º C por 30 minutos) (Welch et. al., 2006). A titulação viral pode ser reduzida também por desinfetantes comerciais à base de álcalis (ex.: hi-dróxido de sódio), agentes oxidantes (ex.: hipoclorito de sódio) ou amônia quaternária(157, 114).
Variação genética e antigênica
A homologia genética entre
os isolados de PCV2 é relativamente
alta(112, 121, 91), embora exista diversi-dade entre as populações do mesmo.
A análise filogenética mais recente mostrou que isolados de PCV2, de diferentes origens geográficas, varia-
vam em sua sequência genômica(65,
49, 112, 121). Mais tarde, com base na sequência de aa/genômica do PCV2 ou no polimorfismo de restrição ao comprimento dos fragmentos, foram identificados dois genogrupos diferenciados do PCV2(36,180, 18, 56, 129,
61). Dependendo do autor e da técni-
ca laboratorial empregada, diferentes nomenclaturas foram usadas, inicial-mente, para se referir aos genogrupos do PCV2 (Tabela 1.1).
Os dois grupos filogenéticos foram comumente referidos como
genótipos 1 e 2 na Europa e PCV2a
e PCV2b na América do Norte. Atu-
almente a nomenclatura norte-ame-
ricana tem sido adotada, consequen-
temente, os genótipos europeus 1 e 2 foram finalmente designados como PCV2a e PCV2b, respectivamente(167).
Recentemente, um novo genótipo foi identificado em amostras dinamar-quesas arquivadas(42) e identificado, após acordo, como PCV2c(167). Na Di-
namarca conclui-se que o PCV2c cir-
Figura 1.1. Classificação taxonômica do PCV2
Suínos & CiaAno VI - nº 35/2010
27
Sanidade

culou predominantemente nos anos
80, o PCV2a durante os anos 90 e o PCV2b entre 2001/2002 em diante, sugerindo uma mudança potencial
no genótipo dos isolados do PCV2
ao longo do tempo(42). Nesse mesmo
estudo, a análise das sequências de PCV2 disponíveis no Centro Nacio-
nal de Informação Biotecnológica
(sigla em inglês, NCBI), banco de dados de fevereiro/2007, evidenciou que o PCV2b tornou-se predominante ao longo do tempo em vários países, apontando para uma mudança no ge-
nótipo global, de PCV2a para PCV2b. Essa distribuição temporal dos genó-
tipos do PCV2 e a atual predominân-
cia do PCV2b a campo são amparadas por vários estudos epidemiológicos, realizados em nível global(10, 26, 56, 178,
181, 29). A detecção de cepas distintas
de PCV2 no mesmo suíno, seja per-tencendo ao mesmo genótipo ou a
genótipo diferente, tem sido também relatada(36,26, 56, 61,71).
Estudos recentes trouxeram
evidências do potencial homólogo de recombinação entre cepas do PCV2 que estejam coinfectando o mesmo animal(42) ou, simultaneamente, se replicando em cultivo celular(96). Os
autores desses estudos sugerem que o surgimento de novos isolados e a
mudança potencial de PCV2a para
PCV2b pode ser o resultado da re-
combinação entre cepas coexistentes no mesmo animal. Com base na ca-
pacidade de reação similar entre an-
ticorpos monoclonais e policlonais
às diferentes cepas de PCV2(06, 119), pensou-se inicialmente que não ha-
veria diferenças antigênicas entre as mesmas. Entretanto, estudos recen-
tes nos levaram a concluir o oposto.
Lefebvre et. al. (2008) descobriram que quatro entre 16 anticorpos mo-
noclonais (em inglês, mAbs) produ-
zidos contra a proteína Cap da cepa PCV2a não reagiram com cepas do
PCV2b, nem demonstraram uma afi-
nidade reduzida em comparação a cepas do PCV2a, quando submetidos a provas realizadas com uma imuno-
peroxidase monocamada (em inglês, IPMA) ou com neutralização viral. Adicionalmente, nenhum dos mAbs testados foi capaz de neutralizar to-
dos as sete cepas incluídas no estudo.
Caracterização adicional relativa à di-versidade antigênica dos isolados de PCV2 foi fornecida por Shang et. al.
(2009). Nesse estudo, mAbs específi-
cos contra a proteina Cap do PCV1 e
do PCV2 foram usados para mapear
epítopos comuns e tipo-específicos entre PCV1 e PCV2, além de identifi-
car diferenças antigênicas entre cepas de PCV2 de genótipos distintos. Dois
epítopos de célula linear B, específi-
cos para a proteína Cap do PCV2 e
dois epítopos compartilhados pelas
proteínas Cap do PCV1 e PCV2 fo-
ram identificados. A análise da di-versidade antigênica destes epítopos revelou três fenótipos antigênicos do PCV2 com diferentes comprimentos
de genoma, denominados PCV21766, PCV21767 e PCV21768. Apesar da exis-
tência de diferenças antigênicas, a imunidade induzida após a infecção de suínos com um genótipo conferiu
proteção contra o desafio subsequente com outro genótipo(136).
A infecção pelo PCV2 e a
síndrome do emagrecimento
progressivo pós-desmame
A infecção pelo PCV2 não é
igual à PMWS. No campo, a grande maioria das infecções por PCV2 são
subclínicas e apenas uma pequena proporção de suínos infectados pelo
PCV2 desenvolve a forma clínica da
doença. Atualmente a expressão ple-
na da PMWS é tida como dependente
de um co-fator (ou co-fatores) “ain-
da não claramente identificado(s)”, o qual desencadearia a progressão do PCV2 no sentido da doença(165).
Características clínicas e
epidemiológicas
Embora a PMWS não tenha sida relatada anteriormente a 1991(19), estudos retrospectivos realizados com amostras arquivadas evidenciaram a infecção pelo PCV2 na Alemanha, em 1962(73), na Bélgica, em 1969(158), no Reino Unido, em 1970(63), na Irlanda, em 1973(195), e no Canadá e na Espa-
nha em 1985(108, 153). Nos dias de hoje a infecção por PCV2 está tão espalha-
da na população de suínos domestica-
dos que quase nenhuma criação soro-
negativa poderia ser encontrada, no caso de um estudo epidemiológico(92,
103, 62). Em contraste, a prevalência da PMWS é geralmente baixa, variando entre 4% e 30%, embora uma morbi-dade de mais de 60% tenha sido re-
latada em algumas granjas(162). Desde
a sua primeira descrição, no Canadá, até os dias de hoje, a PMWS tem sido diagnosticada em países de todos os
cinco continentes, sendo considerada, atualmente, uma doença endêmica e de alto impacto econômico, na maioria dos países produtores de carne suína.
A infecção por PCV2 pode
ocorrer durante toda a vida produtiva
Tabela 1.1. Nomenclaturas publicadas referentes aos dois principais genogrupos do PCV2
Autorde Boisseson et
al., 2004Timmusk et al.,
2005Carman et al.,
2006Olvera et al., 2007 Grau-Roma et
al., 2008Gagnon et al.,
2007
GrupoI SG3 Modelo tipo 321 Grupo biológico 1 Genótipo 1 PCV2b
II SG1/SG2 Modelo tipo 422 Grupo biológico 2 Genótipo 2 PCV2a
Anuncio_Baycox_2010_210x297mm.indd 1
28
Suínos & Cia Ano VI - nº 35/2010
Sanidade

do suíno, mas a PMWS geralmente afeta animais entre oito e 16 semanas de idade(172, 62). A síndrome caracte-
riza-se, clinicamente, por retardo no crescimento, perda de peso e morte. Outros sintomas clínicos como di-
ficuldade respiratória, diarréia e pa-
lidez da pele também são descritos com frequência nos suínos acometi-dos. Setenta a 80% dos animais do-
entes morrem(162). A figura 1.2 mostra a condição de emagrecimento de um
suíno acometido pela PMWS, em comparação com um companheiro de
baia clinicamente sadio.
Patologia associada ao PCV2
Os achados patológicos mais
evidentes associados à PMWS, embo-
ra nem sempre presentes, são os linfo-
nodos intumescidos e os pulmões não
colapsados. Em menor extensão, a atrofia do timo, a presença de pontos esbranquiçados nos rins e a ulcera-
ção da porção esofágica do estomago
também podem ocorrer(155).
Ao contrário dos achados
macroscópicos, os quais são ines-
pecíficos e variáveis, as lesões mi-croscópicas associadas ao PCV2 são
únicas. São encontradas, tipicamente, lesões no tecido linfóide consisten-
tes com depleção linfocitária, com-
binadas com infiltração de células gigantes histiocíticas e/ou multinu-
cleadas. Corpos de inclusão basófi-
los intra-citoplasmáticos podem ser
encontrados em células de linhagem
monocíticas(155). A gravidade das le-
sões está diretamente relacionada ao
status da doença(164, 134). Além disso, suínos acometidos pela PMWS po-
dem ser diferenciados daqueles em condição subclínica, pela intensidade das lesões associadas ao PCV2.
Diagnóstico
A complexidade do diagnósti-
co da PMWS recai no fato de somente
a detecção sérica ou tecidual do PCV2
não ser conclusiva para o estabeleci-
mento do mesmo, ainda que na pre-
sença de sinais clínicos compatíveis
com a PMWS. Deste modo, a presen-
ça de infecções subclínicas é comum, e os sinais clínicos e macroscópicos
encontrados não são específicos, sen-
do compatíveis com muitas outras
doenças dos suínos, infecciosas ou não. Atualmente, três critérios devem ser observados para o estabelecimen-
to de um diagnóstico individual da
PMWS(165):
A. Sinais clínicos compatíveis
com a PMWS (retardo no crescimen-
to e refugagem);
B. Lesões histopatológicas, de moderadas a severas, caracterizadas por depleção linfocitária, juntamente com inflamação granulomatosa (Fi-gura 1.3-A);
C. Quantidade moderada a alta
de genoma de PCV2 ou antígeno anti-
PCV2 entre as lesões (Figura 1.3-B).
Um diagnóstico de infecção
subclínica por PCV2 é estabelecido quando, embora detectado no soro ou nos tecidos, a quantidade viral é pe-
quena e associada a um número míni-mo ou à ausência de lesões(134).
O desempenho das técnicas de
diagnóstico sorológico e virológico
tem sido comparado com a intenção
de se fazer uma diferenciação entre as infecções subclínicas e os casos de PMWS em suínos vivos. Pela análise
quantitativa de PCV2 no soro, Olve-
ra et. al. (2004) sugeriu 107 cópias de
Figura 1.2.: suíno acometido pela PMWS (A) e outro animal da mesma idade, clinicamente são (B).
Notar o retardo no crescimento e a coluna vertebral marcada, no animal doente.
Figura 1.3.: (A) Depleção de linfócitos moderada a severa e infiltração granulomatosa em tecido
linfóide, corado pelo método da hematoxilina/eosina. (B) DNA de PCV2 (quantidade de alta a
moderada) detectado em tecidos linfóide por hibridização in situ (em inglês, ISH), corado pelo
método fast green.
30
Suínos & Cia Ano VI - nº 35/2010
Sanidade

DNA/mL como o limiar potencial para
se distinguir entre PMWS e infecção
subclínica em suínos acometidos. O uso da mesma técnica, em combina-
ção com testes sorológicos (IPMA ou soro neutralização) permite a confir-mação ou a exclusão do diagnóstico
da PMWS(54, 62). Todavia, estes estudos concluíram que estas técnicas podem ser úteis para aprimorar o diagnóstico
da PMWS, mas não servem como al-ternativa à histopatologia e à detecção
do PCV2 nos tecidos.
Uma vez que casos individu-
ais de PMWS podem ser diagnos-
ticados em um plantel sem que haja aumento nos índices de mortalidade
e sem perdas econômicas associadas à doença(125), surgiu a necessidade do estabelecimento formal da definição de casos no plantel. Por essa razão o Consórcio da União Europeia para o
PCVD (www.pcvd.org, 2005) lançou uma proposta na qual dois critérios para o diagnóstico da PMWS no nível
da granja foram definidos:
Aumento significativo na 1.
mortalidade pós-desmame. Essa situa-
ção é considerada quando a mortalida-
de usual (em um período de um a dois meses) for igual ou maior que a média histórica da mortalidade (em um período de, pelo menos, três meses) mais 1,66 vezes o desvio padrão. De modo alternativo, o aumento poderá ser determinado estatisticamente por
meio do teste do qui-quadrado. Se os dados históricos não estiverem
disponíveis, o aumento na mortalida-
de poderá ser determinado quando a mortalidade pós-desmame do plantel
exceder o nível regional ou nacional
em 50% ou mais;
Cumprimento da definição 2.
de casos individuais de PMWS nos
animais da propriedade (é necessário haver um relato de, pelo menos, um a cada cinco suínos necropsiados).
Patogenia
A Figura 1.4 apresenta uma vi-são geral dos eventos que se sucedem no decurso da infecção pelo PCV2, além das diferenças – já discutidas – entre os suínos que desenvolvem PMWS e aqueles que permanecem infectados de modo subclínico. Pou-
co se sabe a respeito dos eventos que ocorrem nos estágios iniciais da infecção, e as células-alvo para a re-
plicação precoce do PCV2 ainda não
foram identificadas. Nos suínos infec-
tados pelo PCV2, a quantidade mais alta de vírus encontra-se no citoplas-
ma das células de linhagem monó-
citos/macrófagos(155, 160) e estudos in
vitro demonstram que o PCV2 é hábil para infectar esses tipos de células, permanecendo de modo persistente
por longos períodos de tempo sem
nenhuma replicação ativa aparente(58,
192). Já foi sugerido inclusive que, ape-
sar dos monócitos não serem o alvo
primário da replicação do PCV2, eles poderiam representar um mecanismo
de disseminação para o vírus por meio
do hospedeiro(192). Adicionalmente, o PCV2 é capaz de induzir uma di-minuição na capacidade funcional de
células dendríticas (sigla em inglês, DC) cultivadas in vitro(193). Esse fato
mais recente pode ser crítico, devido à função central desse tipo de célula
na mediação das respostas imune ina-
ta e vírus-específicas. Como a imuni-dade inata desenvolvida pelos suínos
na pós-infecção pelo PCV2 ainda não
foi muito estudada, a possibilidade do dano causado pelo PCV2 na fun-
cionalidade das DC in vitro ocorrer
também in vivo permanece desconhe-
cida. A confirmação ou não dessa teo-
ria traria informação no sentido de es-
clarecer quais eventos imunológicos estariam ocorrendo no estágio inicial
da infecção pelo PCV2.
A viremia causada pelo PCV2
é detectada, inicialmente, sete dias pós inoculação (PI) e os títulos virais aumentam, atingindo um pico entre os
dias 14 e 21 PI(156, 148, 136). Nesse período
o PCV2 pode estar presente em vários
órgãos, embora a carga viral mais alta seja detectada, tipicamente, no tecido linfóide. Além das células de linha-
gem monocíticas, o PCV2 tem sido detectado em células epiteliais dos
rins e do trato respiratório, em célu-
las endoteliais, linfócitos, enterócitos, hepatócitos, células da musculatura lisa e células pancreáticas (ácinais e dos dutos)(118, 155, 170, 160). Uma vez que o PCV2 não codifica o seu próprio DNA, polimerases e células na fase S são necessárias para que o vírus com-
plete o seu ciclo de infecção(185), o que nos faz considerar que as células com as mais altas taxas de mitose sejam as mais eficazes no seu processo de re-
plicação. Embora estudos anteriores sugiram que os linfócitos não sejam alvos para a replicação do PCV2(58,
192), estudos recentes sugerem o opos-
to. Desse modo, por meio da medida do Cap mRNA, a replicação do PCV2 foi revelada na população de linfó-
citos das células mononucleares do
sangue periférico (sigla em inglês, PBMC) e dos linfonodos brônquicos de suínos experimentalmente inocu-
lados com PCV2(202). Adicionalmente, estudos in vitro demonstraram que PBMC estimuladas pela Concana-
valina A (ConA)* são suscetíveis à replicação do PCV2(202, 95, 98). A carac-
terização adicional de subpopulações de leucócitos infectadas pelo PCV2
indicou que, principalmente, células T circulantes (CD4+ e CD8+) e, em menor extensão, linfócitos B (IgM+), podem apoiar a replicação do PCV2, enquanto monócitos derivados das PBMC aparentemente não(203, 95, 98).
Entre a segunda e a terceira semanas
PI desenvolveram-se respostas imu-
nes específicas ao PCV2(144, 148, 122). A
Habilidade do suíno em mon-
tar uma resposta imune adaptativa
adequada tem sido sugerida como determinante para evitar o progresso
* lecitina com efeito hemaglutinante, extraída de uma espécie de feijão.
Suínos & CiaAno VI - nº 35/2010
31
Sanidade

da infecção pelo PCV2, no sentido da PMWS. Deste modo, o início do desenvolvimento dos anticorpos anti-
PCV2 é tipicamente seguido pelo de-
créscimo nos títulos virais, os quais usualmente levam à resolução do pro-
cesso de viremia(148, 122, 54, 136). Naqueles suínos acometidos subclinicamente a quantidade de PCV2 detectada nos te-
cidos é baixa, resultando em menores ou em nenhuma alteração no sistema
imunológico, normalmente levando à recuperação. É importante notar, entretanto, que a viremia de longa duração causada pelo PCV2 e/ou a
detecção viral nos tecidos têm sido relatadas em suínos infectados expe-
rimentalmente de modo subclínico, a despeito da presença de altos títu-
los de anticorpos anti-PCV2(05, 144, 148).
Em termos de conversão, respostas humorais pobres(16) e, em particular, a ausência de anticorpos neutralizan-
tes (em inglês, NA) específicos anti-PCV2(123) estão relacionadas com um
aumento na replicação viral, o que re-
sulta em severas lesões no tecido lin-
fóide e em mudanças significativas no sistema imunológico. Esses eventos
levam os suínos ao estado de depres-
são característico da PMWS. Como já visto em condições experimentais, es-
tudos a campo realizados em granjas infectadas ou não pela PMWS tam-
bém revelam diferenças importantes entre os eventos que ocorrem durante episódios de infecções clínicas e sub-
clínicas devidas ao PCV2.
Da fase de lactação ao período
de engorda/terminação, a proporção de suínos positivos para o PCV2 e a
carga viral aumentam gradualmente
e coincidentemente com o desmame, fase de anticorpos antiPCV2 de ori-
gem materna (em inglês, MDA). A mais alta pressão de infecção ocorre
no momento em que surge a doença(17,
62). Suínos doentes mostram alta carga
viral no soro e em locais de excreção
em potencial, juntamente com uma resposta de anticorpos mais fraca, comparada a animais com infecção
subclínica(54, 62, 128). Naqueles suínos subclinicamente infectados, a dura-
ção da viremia é variável, entre cinco e 21 semanas. Uma alta proporção de
suínos pode se apresentar virêmica e, ao mesmo tempo, ter altos níveis de anticorpos antiPCV2(172).
Fatores que influenciam a
progressão da infecção por PCV2
para PMWS
Até os dias de hoje, os meca-
nismos pelos quais os suínos desen-
volvem a PMWS ou permanecem
subclinicamente infectados com o
Figura 1.4. Revisão dos eventos associados no decurso da infecção pelo PCV2, em um modelo experimental. Fatores potenciais envolvidos nessa
progressão, tanto em PMWS como em infecções subclínicas, estão indicados nas linhas destacadas. MDA = imunidade passiva; DC = células dendríticas;
MO = macrófagos.
32
Suínos & Cia Ano VI - nº 35/2010
Sanidade

PCV2 continuam desconhecidos. Inú-
meros fatores têm sido identificados como influenciadores no progresso da infecção pelo PCV2 em direção à
PMWS (Figura 1.4.) e, consequente-
mente, a patogenia da doença ainda não é totalmente compreendida. Sua
complexidade é adicionalmente in-
fluenciada pela falta de um modelo experimental universal para a doença
que possa ser reproduzido. Tentativas de reproduzir a PMWS somente com o PCV2 renderam resultados bem-sucedidos em um número limitado
de estudos(05, 83, 16, 67). Na maioria das
ocasiões a simples inoculação com
o PCV2 resulta em infecções sub-clínicas(13, 45, 107, 144, 01). As chances de
se chegar à doença clínica aumen-
tam quando o PCV2 é inoculado em combinação com outro patógeno que acometa o suíno, como o parvovírus suíno (PPV)(05, 45, 68), o vírus da síndro-
me respiratória e reprodutiva dos suí-
nos (PRRSv)(07, 156) ou o Mycoplasma
hyopneumoniae(131). Crescimento da
replicação do PCV2 e desenvolvi-
mento da PMWS têm sido induzidos também após a estimulação do siste-
ma imunológico, tanto pela injeção de produtos imunoestimulantes(87, 89,
60, 197), ou pela vacinação(130). Baseado
em todos esses dados, tem sido postu-
lado que o estímulo ao tecido linfói-de – seja por infecções concomitantes ou por qualquer outro fator imuno-estimulante – promova um ótimo am-
biente para a replicação do PCV2.
De modo interessante, a indu-
ção de um estado de imunossupressão
nos suínos, por meio da injeção de ciclosporina ou dexametasona, tam-
bém resultou em regulação posterior da replicação do PCV2 nos tecidos
dos animais infectados(88, 80, 122). Nesse
contexto, o aumento na carga viral foi atribuído ao enfraquecimento induzi-do à resposta imune dos suínos trata-
dos. Para aumentar o esclarecimento
sobre a patogenia da doença, vários
experimentos e estudos a campo têm sido realizados no sentido de identifi-
car os fatores que influenciam o efeito da PMWS. Desse modo, fatores ine-
rentes ao individuo, ao vírus e fatores influenciadores da interação vírus/hospedeiro têm sido relatados.
Com relação a fatores hospe-
deiro-dependentes, tem-se afirma-
do que a susceptibilidade à PMWS poderia ser determinada genetica-
mente, uma vez que alguns dados de campo ou experimentais indicam
certos cruzamentos ou linhagens ge-
néticas como mais frequentemente acometidos(102, 132, 133). Adicionalmen-
te, o fato dos suínos com antecedentes genéticos idênticos poderem expres-
sar diferentes sintomas clínicos após
a inoculação experimental do PCV2, também sugeriu a existência de va-
riabilidade genética interindividual. Muito recentemente foi relatado que o cromossomo suíno 13 poderia conter
genes que confeririam suscetibilidade à PMWS, e um gene candidato em potencial chegou a ser identificado(78).
Esse gene codifica para a proteína MyRIP, a qual está envolvida no trá-
fico citoplasmático de vesículas e na exocitose. Os autores especularam
que a persistência do PCV2 nas ve-
sículas celulares do sistema imune
inato poderia estar relacionada com
a falta de tráfico citosólico, media-
do por meio da MyRIP. Entretanto, a suscetibilidade à PMWS poderia não estar relacionada a nenhum poli-
morfismo na codificação de parte da proteína MyRIP, tampouco com a ex-
pressão diferencial desse gene entre
suínos sãos e doentes.
A respeito dos fatores vírus-
dependentes, demonstrou-se que su-
ínos inoculados experimentalmente
com um PCV2 submetido a um alto número de passagens em cultivo ce-
lular apresentaram tanto viremia, como lesões associadas ao PCV2 re-
duzidas, em comparação com suínos
inoculados com a cepa selvagem do
PCV2(52). Estas diferenças foram atri-
buídas a duas mutações aa na proteína Cap, sugerindo que mudanças míni-mas no genoma do PCV2 poderiam
contar como causadoras de diferenças
na virulência e na patologia associa-
das ao mesmo.
Tem sido afirmado também que o genoma do PCV2 poderia in-
fluenciar o progresso da infecção pelo vírus. Vários estudos epidemiológicos
demonstraram que isolados do PCV2 estavam associados à ocorrência de PMWS numa frequência maior, com-
parados com isolados pertencentes
ao genótipo PCV2a, o que sugere a existência de diferenças na virulência entre os genótipos do PCV2(18, 26, 56, 61,
199). Essa associação, entretanto, não pode ser reproduzida em um estudo controlado, delineado com esse pro-
pósito, usando isolados de PCV2 de cada genótipo(136).
A presença de anticorpos anti-
PCV2 passivos (adquiridos) no mo-
mento da exposição ao vírus é tam-
bém um importante fator a conside-
rar. Muitos estudos conduzidos com suínos, natural e experimentalmente infectados com o PCV2, revelaram que a replicação viral e a expressão da PMWS são altamente influen-
ciadas pela presença de anticorpos
anti-PCV2. Sob condições de campo, tem sido demonstrado que os suínos geralmente não desenvolvem PMWS
antes de quatro semanas de idade(152,
62) nem se infectam enquanto os anti-corpos anti-PCV2 de origem materna
(MDA) estão presentes(62). Os autores
desses estudos sugerem que a imuni-dade materna deveria conferir prote-
ção contra a infecção por PCV2 e a
doença. Sob condições experimentais, o efeito protetor dos MDA tem sido
provado e demonstrado como depen-
dente do título de anticorpos obtido por meio da ingestão do colostro(117,
139).
Suínos & CiaAno VI - nº 35/2010
33
Sanidade

Além dos fatores dependen-
tes do hospedeiro e do vírus, outros fatores principalmente relacionados
às rotinas de manejo implementadas no plantel, têm sido relatados como responsáveis pelo aumento do risco
da aquisição da PMWS(154, 103). Não
há dados científicos disponíveis, re-
lativos ao fato do manejo influenciar no surgimento da doença. Entretanto, sabe-se que certas rotinas, como a mistura de suínos com idades diferen-
tes ou baias muito extensas, podem aumentar o nível de estresse entre
os animais. Baixas no sistema imu-
ne relacionadas a fatores estressantes
podem explicar a alta ocorrência de PMWS, seguida de certas medidas de manejo.
Apesar de toda a pesquisa rea-
lizada, o exato mecanismo pelo qual todos os fatores anteriormente men-
cionados desencadearia o desenvolvi-
mento da PMWS não é conhecido. No
entanto, quando todos os dados acu-
mulados são considerados, torna-se evidente que suínos acometidos pela PMWS diferem daqueles com infec-
ção subclínica, tanto na carga viral do PCV2 quanto na extensão dos danos
causados ao sistema imune (Figura 1.4.). Ambos os fatos apontam para o equilíbrio entre a capacidade do suí-no em produzir uma resposta imune eficaz contra o PCV2 e a habilidade viral em evadir-se/ deprimir o contro-
le imunológico, como pontos chave na patogenia da PMWS.
Medidas de controle
Antes da disponibilidade de vacinas contra o PCV2, as medidas de prevenção contra a PMWS foca-
lizavam-se no controle dos fatores de risco envolvidos no progresso da
doença. Um plano de recomendações
contendo 20 pontos importantes foi
proposto, no sentido de ajudar os pro-
dutores a identificar e ajustar práticas de manejo que favoreciam o apareci-mento da mesma(105). Eram recomen-
dações focalizadas principalmente na redução da pressão de infecção
do PCV2 e de outros agentes infec-
ciosos e na minimização do estresse nos suínos. Os pontos principais do
plano de Madec incluem: 1) limitação
do contato entre suínos de leitegadas
distintas; 2) redução do estresse; 3)
otimização das condições de higiene, por meio de procedimentos de lim-
peza e desinfecção e 4) boa nutrição. A implementação do referido plano
reduziu significativamente as perdas associadas à PMWS, sendo o mesmo ainda considerado – nos dias de hoje – um modelo de estratégias de inter-
venção para o controle da referida
doença.
Atualmente, além de controlar os fatores desencadeantes da PMWS, a prevenção de surtos da doença pode
ser obtida de modo efetivo pelo con-
trole do PCV2 por meio da vacinação.
A informação sobre vacinas comer-ciais disponíveis contra o PCV2 está
revista na seção 3.2.2.1.
O PCV2 e o sistema imune
Imunopatogenia da infecção pelo PCV2
• Atividade imunomoduladora do
PCV2
A noção de que suínos aco-
metidos pela PMWS padecem de
uma imunodeficiência adquirida(33)
levou à especulação de que a infecção pelo PCV2 poderia modular a defesa
imune do hospedeiro. Recentemente, muitos estudos in vitro têm apoiado essa hipótese e esclarecido alguns
aspectos do complexo de interação
entre o PCV2 e as células do sistema
imune.
A infecção pelo PCV2 induz a uma diminuição na atividade funcional
das células dendríticas (DC), depen-
dendo da subpopulação envolvida(193).
Nas DC mielóides (mDC) a infecção pelo PCV2 não altera a habilidade das mesmas em processar e apresentar o
antígeno aos linfócitos T, não interfe-
rindo também em sua maturação(192,
193). Reciprocamente, a interação do PCV2 com as DC plasmocitóides
(pDC) – também conhecidas por cé-
A implementação do plano de Madec reduz significativamente as perdas associadas à PMWS.
34
Suínos & Cia Ano VI - nº 35/2010
Sanidade

lulas produtoras naturais de interferon
ou NIPC (sigla em inglês) – induziu a sinais inadequados de resposta ao perigo. Deste modo, a inibição indu-
zida pelo PCV2 ao interferon alpha (em inglês, IFN-α) e o fator de necro-
se tumoral alpha (em inglês, TNF-α), normalmente produzidos pelo NIPC a partir da interação com oligodeoxi-
nucleotídeos (em inglês, ODNs) com motivos centrais CpG (em inglês, CpG-ODN), consequentemente in-
terferem com a maturação das NIPC, do mesmo modo que com a matura-
ção parácrina das mDC(193). Devido à
importância das DC na mediação das
defesas inatas, a habilidade do PCV2 em interferir com o seu funcionamen-
to poderia representar uma grande
barreira ao desenvolvimento de uma resposta imune adequada, seja contra o próprio PCV2 como a qualquer ou-
tro patógeno(194).
Quando o PCV2 é adiciona-
do, in vitro, a uma cultura de macró-
fagos alveolares (em inglês, AM), observa-se uma produção alterada de certas citocinas e/ou quimiocinas. Desse modo, a infecção pelo PCV2 diminui a produção de O
2, radicais li-
vres e H2O
2 e aumenta a produção de
TNF-α, interleucina 8 (IL-8), fatores quimiotáticos derivados de macrófa-
gos alveolares-II (em inglês, AMCF-II), fatores estimuladores de colônias de granulócitos (G-CSF) e proteínas quimiotáticas de monócitos I (em inglês, MCP-I)(21). Especula-se que essa alteração na funcionalidade dos
AM infectados pelo PCV2 possa fa-
vorecer a sua disseminação, tornando também os suínos mais suscetíveis a infecções pulmonares oportunistas e
secundárias.
A adição do PCV2 a células
mononucleares do sangue periféri-
co (PBMC), obtidas tanto de suínos doentes quanto saudáveis, suprimiu a resposta da IL-4 e da IL-2 frente à fitohemaglutinina (em inglês, PHA) e promoveu a secreção das IL-10, IL-
1β e IL-8(31). De modo interessante, ao contrário do que foi observado para as pDC, o PCV2 induziu a produção de IFN-α nas PBMC(200). Adicionalmen-
te, o PCV2 parece modular a resposta imune específica desenvolvida pelos suínos frente a outros patógenos(82).
Desse modo, IL-12, IFN-α, IFN-γ e IL-2 revertem os efeitos das PBMC, após o estímulo do vírus da pseudo-
raiva ser dessensibilizado pelo PCV2. O efeito inibitório nas reações frente a IL-12, IFN-α e IFN-γ foi mediado pela liberação do IL-10 induzido pelo PCV2. O aumento do nível sérico
dessa citocina , in vivo, em suínos in-
fectados pelo PCV2 foi associado ao
desenvolvimento da PMWS(176).
A implicação de diferentes
componentes do PCV2 na modulação
da resposta imune tem sido também investigada. Vincent et. al. (2007) concluiu que a diminuição da capaci-dade funcional das DC induzida pelo PCV2 não requer a replicação viral, sendo mediada pelo DNA do vírus.
Os mesmos autores demonstraram
que uma concentração mínima de dsDNA (forma replicativa) foi neces-
sária para mediar a referida inibição. No caso de PBMC, a supressão da IL-2 e do IFN-γ induzida pelo PCV2, liberada mediante o retorno do antí-geno, está associada ao vírus total e a certos CpG-ODNs derivados de seu genoma. Em contrapartida, partículas virais PCV2 like (em inglês, VLP) não demonstram nenhum efeito su-
pressor e não modulam, tampouco, as respostas ao IFN-α(81). De fato, a mo-
dulação da produção de IFN-α pela PBMC poderia ser atribuída à pre-
sença do CpG-ODN no genoma do PCV2. Sequências com ambas as ati-vidades, IFN-α indutoras e inibidoras foram detectadas, mas os indutores do IFN-α predominaram(200, 81). A maio-
ria dos CpG-ODN inibitórios foram encontrados dentro do gene Rep(81).
Com relação à habilidade do PCV2 em induzir a IL-10, ela é mantida ape-
nas desde que o vírus total seja usado no estímulo às células. Nem VLPs, tampouco nenhum dos CpG-ODNs estudados foram identificados como indutores da IL-10(81).
Todos esses dados juntos sugerem que o PCV2 tem o potencial de evadir-se do controle imunológico
e de mediar a imunossupressão, por meio do enfraquecimento dos meca-
nismos de defesa do hospedeiro. Nos
dias de hoje ainda não se sabe por que apenas uma pequena proporção de suínos acometidos pelo PCV2 tem o
seu sistema imunológico comprome-
tido e incapaz de antagonizar o efeito imunomodulador do PCV2.
Imunossupressão em suínos acometidos pelo PCV2
A evidência mais contunden-
te da imunossupressão corresponde
à extensa lesão observada no tecido linfóide de suínos acometidos pela
PMWS. Ela inclui a depleção dos
linfócitos B e T combinada com um aumento no número de macrófagos e
a perda ou a redistribuição das célu-
las dendríticas interfoliculares(28). No
tecido linfóide a depleção dos linfóci-
tos T envolve, principalmente, células CD4+ e, em menor extensão, células CD8+(161). Outra característica da imu-
nossupressão em suínos acometidos
pela PMWS é a alteração dos sub-
conjuntos das PBMC. Em um estudo transversal, no qual casos naturais de PMWS foram comparados com
suínos sadios, foi relatado – no caso da doença – um decréscimo nas sub-
populações de IgM+, CD8+ e CD4+/
CD8+(30). A cinética dessa linfopenia, assim como o fenótipo das células
envolvidas, foi adicionalmente ca-
racterizada por Nielsen et. al. (2003), sob condições experimentais e usan-
do suínos livres de doenças específi-
cas (em inglês, SPF). Deste modo, a depleção dos linfócitos B (CD21+) e
T (CD3+) foi observada somente em
Suínos & CiaAno VI - nº 35/2010
35
Sanidade

suínos inoculados com o PCV2 que, mais tarde, desenvolveram PMWS a qual se iniciou no sétimo dia pós-infecção, tornando-se severa no mo-
mento da ocorrência dos sinais clíni-cos. Mudanças nos subconjuntos de células T envolveram, principalmen-
te, CD3+CD4+CD8+ células de me-
mória T. De modo oposto, naqueles suínos inoculados com o PCV2 que não manifestaram sinais clínicos, o número de linfócitos T citotóxi-
cos (CD3+CD4-CD8+) e linfócitos T
γδ (CD3+CD4-CD8-) aumentou, em comparação com o grupo controle de
suínos, sugerindo, em consequência, uma resposta ativa à infecção pelo
PCV2.
Depleção linfóide e leucope-
nia são características consistentes
de suínos acometidos pela PMWS.
Entretanto, ainda não se sabe se a perda de linfócitos é um efeito di-
reto da infecção pelo PCV2 ou uma
consequência indireta das respostas à mesma. Alguns autores afirmaram ser a depleção linfóide um resultado
da apoptose induzida pelo vírus(170,
84). No entanto, resultados contradi-tórios foram encontrados por outros
autores(110, 149). Em um estudo recente
observou-se que PBMC infectada por PCV2 apresentou mudanças morfoló-
gicas típicas de degeneração celular.
Estas mudanças foram correlatas com
um aumento no título viral, sugerindo que a infecção da PBMC pelo PCV2 poderia levar à morte celular(95). Foi demonstrado, in vivo, que os linfóci-tos B e T dão suporte à replicação do
PCV2(141, 202). Ainda que este último fato esteja implícito, a depleção se-
vera observada em suínos acometidos pela PMWS necessita ser elucidada.
Imunidade protetiva desenvolvida frente à infecção pelo PCV2
• Respostas humorais
A maioria dos inquéritos so-
rológicos publicados, relativos ao PCV2, baseiam-se na detecção do total de anticorpos anti-PCV2 (em inglês, TA), sem que haja determi-nação de sua atividade neutralizante. No campo, a soro conversão para o TA ocorre em ambos os casos: suí-nos acometidos pela PMWS e casos
de infecção sub-clínica(152, 92, 172, 62).
Enquanto alguns estudos não encon-
traram diferenças entre suínos aco-
metidos ou não pela PMWS, relativas a títulos de TA(92), outros trabalhos relataram respostas fracas em suínos
doentes(122, 62). Sob condições experi-mentais, respostas lentas ou baixos títulos de TA têm sido relatados, com relação à expressão da PMWS(16, 90,
156, 127, 123). Muitos estudos, a campo e experimentais, têm demonstrado que o PCV2 poderia persistir no sangue e
nos tecidos, na presença de altos títu-
los de TA(152, 92, 172, 116, 86, 107); entretanto, estes estudos não descriminam anti-
corpos neutralizados ou não.
Tem sido demonstrado que suí-nos infectados pelo PCV2 desenvol-
vem anticorpos neutralizantes (NAs) específicos, frente ao mesmo(144, 123,54).
Sob condições experimentais, NAs se desenvolveram entre dez e 28 dias pós-infecção(144, 123, 54) e baixos títulos têm sido relatados para um aumento da replicação do PCV2 e desenvol-
vimento da PMWS(123). Amostras de
soro longitudinais de dois estudos a
campo distintos, um deles concluído na Bélgica e o outro na Dinamarca, foram analisadas para a presença de
NA.
Demonstrou-se que NA de origem maternal foi transferido, de modo passivo, a todos os leitões. Os títulos de NA de origem maternal
desapareceram gradativamente, até aproximadamente dez semanas de idade nos suínos da Bélgica e três se-
manas pós-combinação dos lotes nos suínos dinamarqueses. Em ambos os casos, nenhum dos suínos que desen-
volveu PMWS soroconverteu para
NA. Adicionalmente, outro estudo demonstrou que os níveis de NA fo-
ram correlatos com o estado clínico-
patológico dos suínos naturalmente
infectados(54). Desse modo, suínos po-
sitivos para o PCV2, com títulos NA iguais ou acima de 1:512 foram mais
considerados como infectados sub-
clínicamente e aqueles com títulos ≤ 1:16 tiveram grande probabilidade de adquirir a PMWS.É importante notar que nem todos os suínos com baixos títulos de NA apresentaram baixos ní-veis de TA(123, 54). Este último fato su-
gere que alguns animais desenvolvem resposta humoral com baixos NAs ou que NA desenvolve-se mais tarde que a ausência de NA. Um atraso na resposta do NA tem sido relatado em
suínos infectados subclínicamente pelo PCV2(144, 54). Adicionalmente, há um estudo relatando a coexistência de altos títulos de NA e de uma alta
carga viral sérica e tecidual(54). Con-
siderados em conjunto, esses dados sugerem que a simples presença de anticorpos anti-PCV2 não garante to-
talmente a eliminação viral, quando a infecção já se instalou e aponta para um sistema composto por outros me-
canismos imunológicos distintos das
respostas humorais.
Respostas mediadas por células
Os preceitos das respostas
adaptativas mediadas por células
no controle da infecção e da doença
causadas pelo PCV2 ainda carecem
de um estudo mais profundo. Entre-
tanto, o fato dos suínos acometidos pelo PCV2 apresentarem uma dimi-
nuição em suas respostas relativas
às células T(30, 126), é sugestivo da sua contribuição no processo de proteção imunitária contra a infecção causada
pelo vírus. Adicionalmente, tem sido demonstrado que a imunossupressão artificialmente induzida pode poten-
cializar a replicação viral(88, 80, 122). Em
suínos gnotobióticos, o tratamento
36
Suínos & Cia Ano VI - nº 35/2010
Sanidade
com ciclosporina A (CyA), antes da inoculação do PCV2, resultou em aumento da replicação viral. Sugeriu-
se que esse efeito tenha sido parcial-mente mediado por uma diminuição
na capacidade de resposta celular, causada pela CyA(88, 122). Ademais, os níveis de expressão do mRNA do
IFN-γ, em PBMC de suínos inocula-
dos experimentalmente com o PCV2, foram considerados correlatos com a
replicação viral e o estado imunos-
supressor induzido pela CyA. Desse modo, uma expressão mais acentuada do mRNA do IFN-γ aparentemente tornou os suínos menos suscetíveis à
replicação do PCV2(22). Outro estudo, no qual os suínos foram imunodepri-midos com dexametasona, mostrou resultados semelhantes. Deste modo, a inoculação do PCV2 em suínos tra-
tados com dexametasona induziu a replicação viral e as lesões virais as-
sociadas ao processo, visto que nem o genoma do PCV2, tampouco as lesões associadas ao mesmo, foram
detectadas em suínos inoculados so-
mente com o PCV2(80).
Imunidade protetiva conferida pela vacinação contra o PCV2
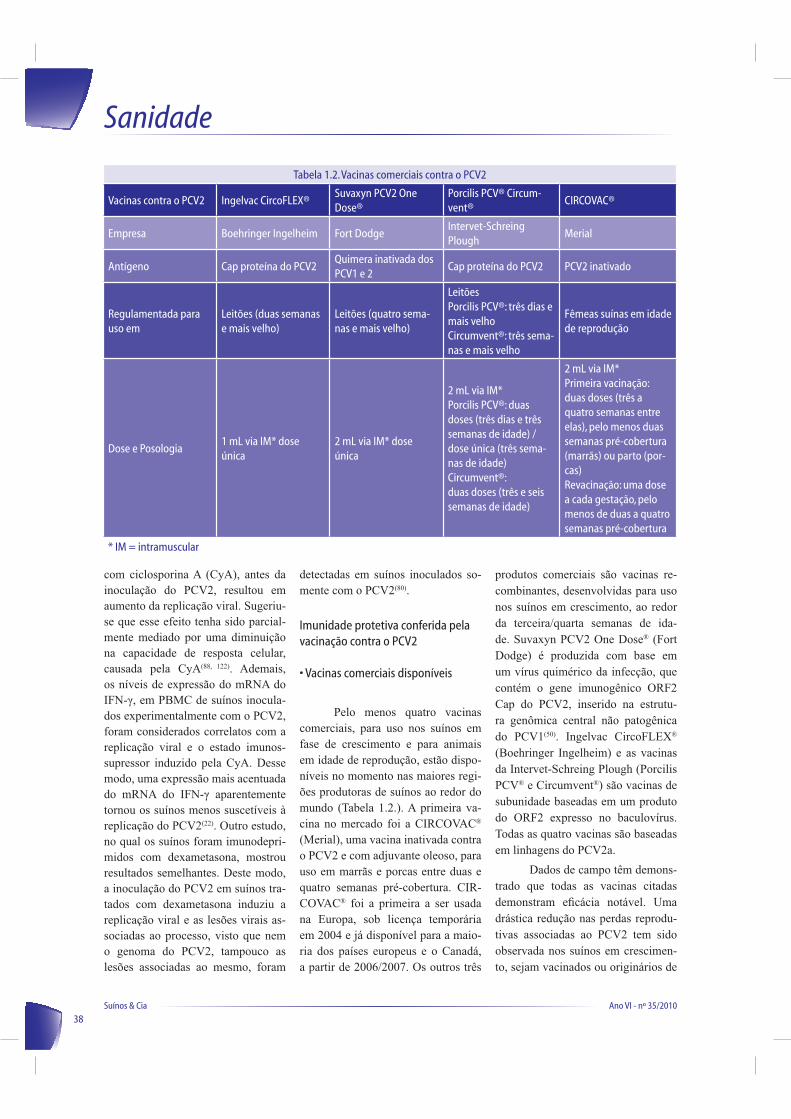
• Vacinas comerciais disponíveis
Pelo menos quatro vacinas comerciais, para uso nos suínos em fase de crescimento e para animais
em idade de reprodução, estão dispo-
níveis no momento nas maiores regi-
ões produtoras de suínos ao redor do
mundo (Tabela 1.2.). A primeira va-
cina no mercado foi a CIRCOVAC®
(Merial), uma vacina inativada contra o PCV2 e com adjuvante oleoso, para uso em marrãs e porcas entre duas e
quatro semanas pré-cobertura. CIR-
COVAC® foi a primeira a ser usada
na Europa, sob licença temporária em 2004 e já disponível para a maio-
ria dos países europeus e o Canadá, a partir de 2006/2007. Os outros três
produtos comerciais são vacinas re-
combinantes, desenvolvidas para uso nos suínos em crescimento, ao redor da terceira/quarta semanas de ida-
de. Suvaxyn PCV2 One Dose® (Fort Dodge) é produzida com base em um vírus quimérico da infecção, que contém o gene imunogênico ORF2 Cap do PCV2, inserido na estrutu-
ra genômica central não patogênica do PCV1(50). Ingelvac CircoFLEX®
(Boehringer Ingelheim) e as vacinas da Intervet-Schreing Plough (Porcilis PCV® e Circumvent®) são vacinas de
subunidade baseadas em um produto do ORF2 expresso no baculovírus. Todas as quatro vacinas são baseadas em linhagens do PCV2a.
Dados de campo têm demons-
trado que todas as vacinas citadas demonstram eficácia notável. Uma drástica redução nas perdas reprodu-
tivas associadas ao PCV2 tem sido
observada nos suínos em crescimen-
to, sejam vacinados ou originários de
Tabela 1.2. Vacinas comerciais contra o PCV2
Vacinas contra o PCV2 Ingelvac CircoFLEX®Suvaxyn PCV2 One Dose®
Porcilis PCV® Circum-vent®
CIRCOVAC®
Empresa Boehringer Ingelheim Fort DodgeIntervet-Schreing Plough
Merial
Antígeno Cap proteína do PCV2Quimera inativada dos PCV1 e 2
Cap proteína do PCV2 PCV2 inativado
Regulamentada para uso em
Leitões (duas semanas e mais velho)
Leitões (quatro sema-nas e mais velho)
LeitõesPorcilis PCV®: três dias e mais velhoCircumvent®: três sema-nas e mais velho
Fêmeas suínas em idade de reprodução
Dose e Posologia1 mL via IM* dose única
2 mL via IM* dose única
2 mL via IM*Porcilis PCV®: duas doses (três dias e três semanas de idade) / dose única (três sema-nas de idade)Circumvent®:duas doses (três e seis semanas de idade)
2 mL via IM*Primeira vacinação: duas doses (três a quatro semanas entre elas), pelo menos duas semanas pré-cobertura (marrãs) ou parto (por-cas)Revacinação: uma dose a cada gestação, pelo menos de duas a quatro semanas pré-cobertura
* IM = intramuscular
38
Suínos & Cia Ano VI - nº 35/2010
Sanidade

plantéis vacinados. Desse modo, me-
lhoras nos índices de ganho médio de
peso diário e na conversão alimentar, decréscimo nos níveis de mortalidade
e redução nos custos com medicação
são alguns dos benefícios observa-
dos em lotes vacinados(85, 46). A vaci-
nação das fêmeas tem demonstrado efeito benéfico no desempenho dos plantéis reprodutivos. Deste modo, a vacinação de marrãs e porcas tem
revelado um aumento no número de
leitões nascidos vivos e no número
de terminados/porca/ano, além de re-
duzir o número de mumificados por porca(179, 191). Recentemente, o efeito potencial protetor da vacinação de re-
produtoras na prevenção da infecção
fetal pelo PCV2 e das falhas reprodu-
tivas, foi investigado sob condições experimentais(160). Porcas-sentinela
para o PCV2 e prenhas, foram vaci-nadas ou receberam placebo no 28º dia de gestação, tendo sido inoculadas com um isolado de PCV2b ao redor do 56º dia.
Falhas reprodutivas não pude-
ram ser reproduzidas, mas a infecção das porcas-sentinela prenhas pelo
PCV2 resultou em infecção fetal, da qual a vacinação das porcas não foi suficiente para proteger. A vacinação de reprodutoras não preveniu a veicu-
lação do PCV2 por meio do colostro, sugerindo que a vacinação da porca pode não prevenir a transmissão ver-
tical.
Até os dias de hoje, pou-
cos estudos têm lidado com o(s) mecanismo(s) subjacente da proteção induzida pela vacina. É comum assu-
mir que o princípio básico da eficá-
cia da vacinação se baseia no efeito protetor dos anticorpos anti-PCV2, seja por aquisição passiva (vacinação das porcas) ou por indução ativa (va-
cinação dos leitões). Entretanto, bai-xa resposta de anticorpos ou falta de
desenvolvimento dos mesmos após a
vacinação nem sempre impede a pro-
teção. Fenaux et. al. (2004a) afirmam
que após a imunização com um vírus quimérico PCV1-2, mesmo nem todos os suínos soroconvertendo contra o
PCV2, ainda assim estarão protegidos de manifestar viremia pelo referido
agente e sinais clínicos após o desafio com o PCV2. Os autores desse estudo
sugeriram haver um sistema de con-
trole potencial da imunidade mediada
por células, na proteção induzida pela vacinação. Recentemente, foi relata-
do que o colostro de porcas SPF vaci-nadas continha células secretoras in-
terferon-gama (em inglês, IFN-γ-SC) PCV2 específicas(59). A transferência das mesmas à sua leitegada foi prova-
da, mas o efeito protetor das mesmas não pôde ser elucidado, uma vez que IFN-γ-SC foram apenas detectadas em leitões neonatos e durante um pe-
ríodo de tempo muito curto.
Protótipos de vacinas experimentais
Além das vacinas comerciais, outros protótipos de vacinas têm sido delineados e testados em diversos
modelos in vivo. Estes incluem va-
cinas inativadas(45), vacinas produ-
zidas a partir de DNA(14, 77, 11, 47, 169) e
vacinas de subunidade recombinantes com expressão de proteínas virais do
PCV2(101, 14, 76,197 , 47, 196, 12). Dados de es-
tudos realizados com as vacinas an-
teriormente mencionadas permitem a
compreensão de detalhes intrínsecos
relativos às respostas imune geradas
a partir de diferentes ORFs do PCV2 e suas correspondentes proteínas co-
dificadas. Blanchard et. al. (2003) descobriram que a proteína Cap ORF2-codificada era um potente imu-
nógeno, induzindo a proteção contra o desafio subsequente com o PCV2, ao passo que a proteína Rep, produto de ORF1 era fracamente imunogêni-ca. Proteção imunitária foi atribuída à forte resposta precoce de anticor-
pos induzida pela Cap, mas não pela Rep. No mesmo estudo, a proteção induzida pelo DNA e por vacinas de
subunidade também foi comparada.
Interessante notar que enquanto a res-
posta imune obtida pela vacina de su-
bunidade neutralizou a infecção pelo
PCV2, a vacinação com o DNA pare-
ceu promover a replicação do agente
nos suínos(15). Em contraste, utilizan-
do como modelo um trabalho realiza-
do com roedores, Shen et. al. (2008)
concluiu que a imunização com uma
vacina produzida a partir de um plas-
mídeo de ORF2 (ORF2p) resultou em
maior eficácia diante do desafio com
o PCV2, comparativamente à proteí-
na Cap.
Adicionalmente, ainda que
ambos os sistemas tenham eficácia
comparável na indução de respostas
linfo proliferativas e Cap específicas
para células T CD4+, a ORF2p foi
superior à proteína Cap no desenca-
deamento de células T CD8+ e restau-
ração das respostas dos anticorpos à
neutralização viral. Foi sugerido, por
essa razão, que as células T CD8+ e
as respostas NA teriam papel crucial
na indução da proteção vacinal contra
o PCV2. Outros estudos, realizados
com vetores virais vivos, mostraram
que viroses recombinantes que ex-
pressam Cap podem induzir repostas
humorais específicas e/ou respostas de
proliferação de linfócitos ao PCV2(76,
197, 196, 175, 47, 140, 197). Entretanto, a imuno-
genicidade desses produtos foi testada
na maioria das vezes em ratos, sendo
a eficácia em suínos demonstrada uma
única vez e em apenas um deles(197).
Confira na próxima edição a segunda
parte do trabalho de atualização sobre Circovírus Suino - PCV2.
Referências Bibliográficas estão
disponíveis no site:www.suinosecia.com.br
Suínos & CiaAno VI - nº 35/2010
39
Sanidade