procesamiento del rna
TRANSCRIPT


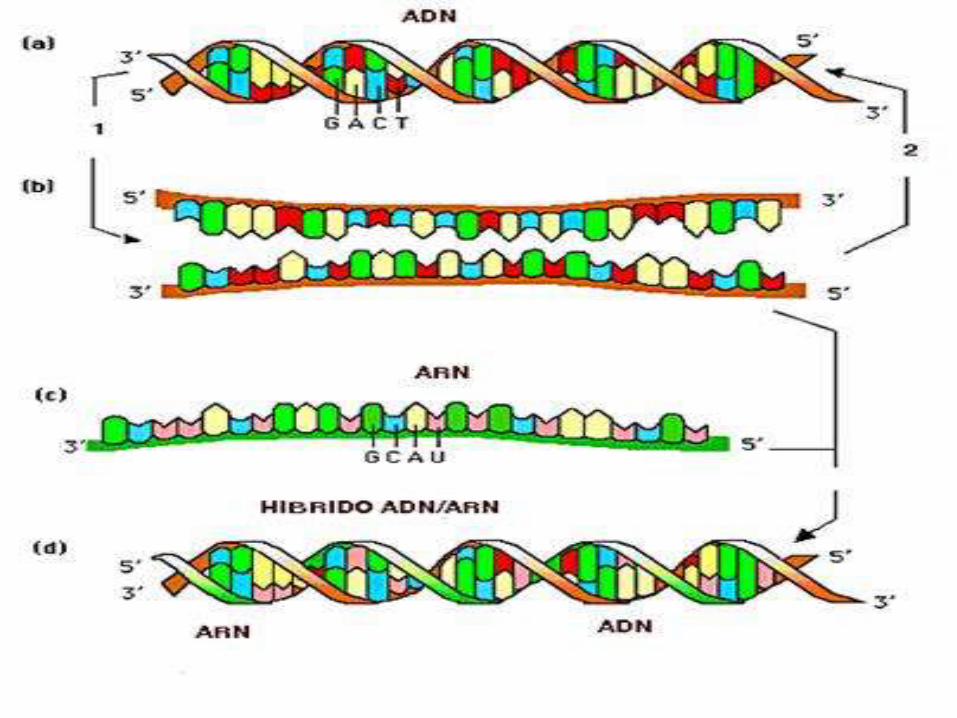

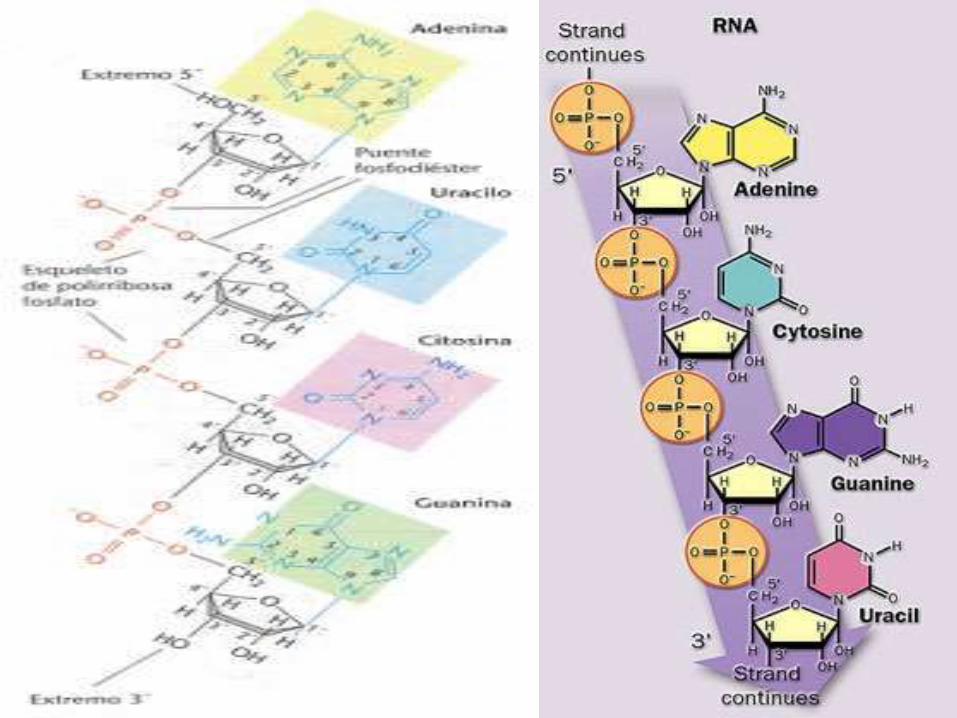
El ARN es un filamento de una sola cadena, es decir, no forma doble hélice.
La presencia de un oxígeno en la posición 2' de la ribosa impide que se forme la doble cadena de la manera en que se forma en el ADN.

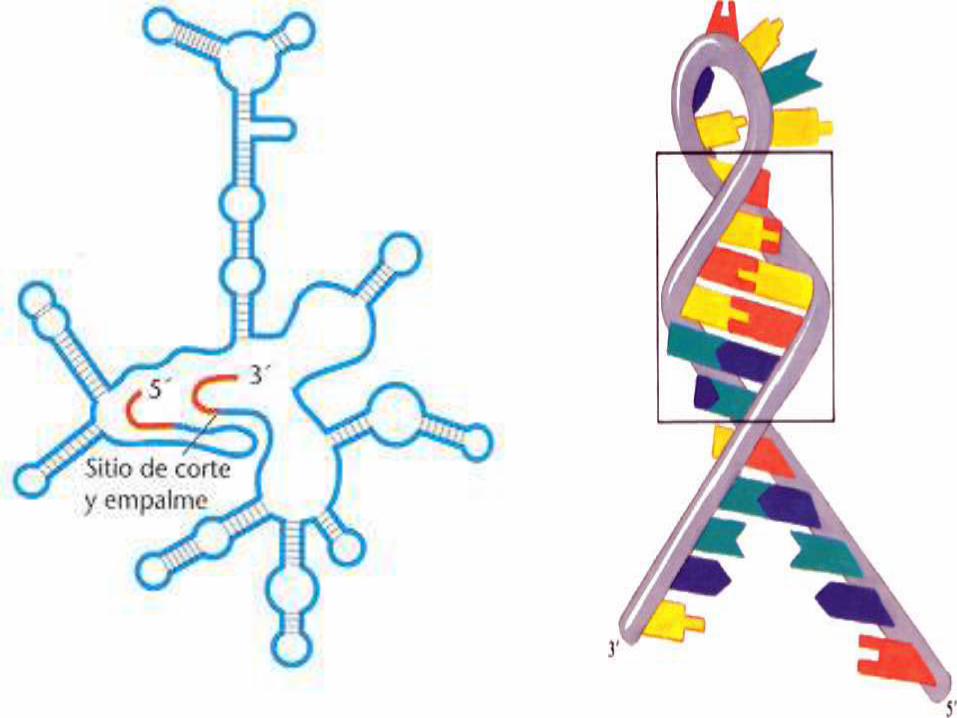
El filamento de ARN se puede enrollar sobre sí mismo mediante la formación de pares de bases en algunas secciones de la molécula, formando las denominadas estructura secundarias del ARN:

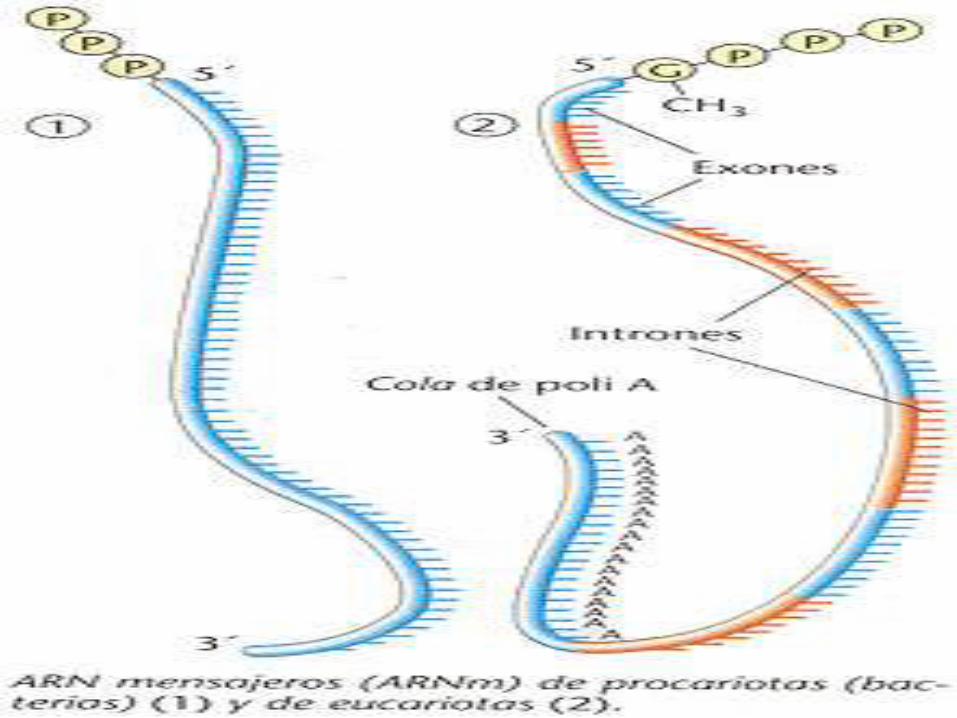
En procariotas el extremo 5' posee un grupo trifosfato y en eucariotas en el extremo 5' posee un grupo metil-guanosina unido al trifosfato, y el extremo 3' posee una cola de poli-A.

En la naturaleza existen diversos métodos de splicing del ARN.
El mecanismo de splicing depende de la estructura del fragmento de ARN que pasará por este proceso.

El Spliceosoma es un complejo formado por cinco ribonucleoproteínas nucleares pequeñas o snRNP (complejo formado por unas diez proteínas más una pequeña molécula de ARN).
El ARN de los snRNP es el encargado de reconocer el intrón.
Se han identificado dos tipos de spliceosomas, el mayor y el menor[cita requerida], cada uno de los cuales contiene diferentes tipos de snRNP.

5´ y 3´ Sitios de Splicing. Unión de manera covalente. Importante que no haya malos cortes. 5´AG/GU. 3´ AG/ G. Aumentadores del corte y empalme exónico
o ESE. Inclusión de un intrón o exclusión de un exón.


Esta formado por los snRNP U1, U2, U4, U5 y U6.
Reconoce la secuencia consenso GU (Guanina-Uracilo) del extremo 5’ del intrón así como la secuencia consenso AG del extremo 3’.
El 99% de los intrones lo hacen a través de este mecanismo.


Complejo E: U1 se une a la secuencia consenso GU del extremo 5’ del sitio de corte del intrón, junto con las proteínas accesorias ASF/SF2, U2AF, SF1/BBP.
Complejo A: U2 se une al sitio de ramificación e hidroliza ATP. El sitio de ramificación se sitúa a una distancia de 20-40 nucleótidos del extremo 3’ del intrón y en él se localiza la secuencia consenso CURAY.
Complejo B1: U5, U4 y U6 trimerizan, y U5 se une al exón 5’ y U6 a U2.
Complejo B2 – U1 es liberado, U5 pasa del exón al intrón y U6 se une al extremo 5’ del sitio de corte.
Complejo C1: U4 es liberado, U5 se une al sito de empalme del extremo 3’ del exón, U6 y U2 catalizan la reacción de transesterificación y el extremo 5’ del intrón es cortado; como resultado se forma una estructura en lazo característica denominada lariat.
Complejo C2: el extremo 3’ del intrón es cortado lo que provoca la liberación del lazo de ARN. A continuación los exones son ligados, lo que conlleva gasto de ATP. Por último, el complejo se disocia.

Es similar al Spliceosoma mayor aunque los introneseliminados mediante este mecanismo son escasos, y además presentan diferencias en los sitios de corte y empalme.
También se diferencian en las secuencias consenso reconocidas, que en este caso son AU y AC para los extremos 3’ y 5’, respectivamente.
Además, salvo la partícula snRNP U5, el resto son análogos funcionales denominadas U11 (análogo funcional de la U1), U12 (U2), U4atac (U4) y U6atac(U6).

También se puede denominar transempalme o empalme en trans.
Consiste en el empalme de exones de dos transcritos primarios distintos, con la consiguiente formación de un ARN híbrido.

Corte y empalme en el que el propio intrón actúa como catalizador en su eliminación, por lo que no se requiere de proteínas.
Cuando un fragmento de ARN tiene actividad catalítica se le denomina ribozima.
Para que el mecanismo de autosplicing sea preciso se requiere de la hidrólisis de ATP.
Existen dos tipos de intrones que actúan como ribozimas, los intrones del grupo I y los del grupo II.
La similitud en el mecanismo de corte y empalme de estos intrones y el spliceosoma sugiere que probablemente evolucionaron juntos aunque también se ha propuesto que el autosplicing surgió durante el mundo de ARN.

El grupo OH 3’ de un nucleósido libre de guanina o del propio intrón o un cofactor (GMP, GDP o GTP) ataca al fosfato del sitio de corte 5’. Lo que da lugar al corte del intrón por su extremo 5’ y a la formación del lariat(estructura en lazo).
El grupo OH 3’ del exón lleva a cabo un ataque nucleofílico contra el extremo 3’ del intrón, lo que origina su corte y la liberación de la estructura en lazo.
Los exones son unidos.

El grupo OH 2’ de una adenosina específica del intrónataca el sitio de corte 5’, originando la estructura en lazo (lariat).
El grupo OH 3’ del exón lleva a cabo un ataque nucleofílico contra el extremo 3’ del intrón, lo que origina su corte y la liberación de la estructura en lazo.
Los exones son unidos.
