otimizaÇÃo de meio de cutivo para a produÇÃo de toxinas por bacillus thuringiensis subsp....
DESCRIPTION
OTIMIZAÇÃO DE MEIO DE CUTIVO PARA A PRODUÇÃO DE TOXINAS POR Bacillus thuringiensis SUBSP. israelensis EMPREGANDO RESÍDUOS AGROINDUSTRIAISTRANSCRIPT

ELISANGELA ANDRADE ANGELO
OTIMIZAÇÃO DE MEIO DE CUTIVO PARA A PRODUÇÃO
DE TOXINAS POR Bacillus thuringiensis SUBSP. israelensis
EMPREGANDO RESÍDUOS AGROINDUSTRIAIS
LONDRINA
2008

ELISANGELA ANDRADE ANGELO
OTIMIZAÇÃO DE MEIO DE CULTIVO PARA A PRODUÇÃO DE TOXINAS POR
Bacillus thuringiensis SUBSP. israelensis EMPREGANDO RESÍDUOS
AGROINDUSTRIAIS
Dissertação apresentado ao programa de Pós-graduação em Ciência de Alimentos da Universidade Estadual de Londrina, como requisito parcial para obtenção do título de mestre em Ciência de Alimentos.
Orientador: Profº Dr. Raúl Jorge Hernan Castro-Gómez
Co-Orientadora: Drª. Gislayne Fernandes Lemes Trindade Vilas-Bôas
Londrina
2008
1

ELISANGELA ANDRADE ANGELO
OTIMIZAÇÃO DE MEIO DE CULTIVO PARA A PRODUÇÃO DE TOXINAS POR
Bacillus thuringiensis SUBSP. israelensis EMPREGANDO RESÍDUOS
AGROINDUSTRIAIS
Dissertação de mestrado
COMISSÃO EXAMINADORA
___________________________________________________Profº Dr. Raúl Jorge Hernan Castro-Gómez (Orientador)
____________________________________________________(Membro)
_____________________________________________________(Membro)
Londrina, de Fevereiro de 2009
2

Dedicatória
3

AGRADECIMENTOS
4

ANGELO, Elisangela Andrade. Otimização de meio de cultivo para a produção de toxinas por Bacillus thuringiensis subsp. israelensis empregando resíduos agroindustriais. Dissertação de Mestrado (Mestrado em Ciência de Alimentos) - Universidade Estadual de Londrina. 2009.
RESUMO
O controle de insetos é realizado, em sua maioria, por produtos químicos, os quais ocasionam grandes prejuízos ambientais e à saúde humana, destacando-se ainda a rápida seleção de insetos resistentes. O controle biológico por entomopatógenos é uma alternativa eficiente, principalmente devido a sua alta especificidade, menor freqüência de resistência nos insetos alvos e baixo efeito residual no ambiente. Bacillus thuringiensis é uma bactéria Gram-positiva esporulante, produtora de cristais protéicos com atividade inseticida. Apesar do amplo uso B. thuringiensis no controle biológico, há poucos trabalhos publicados quanto a sua produção, visto que muitas informações são segredos industriais. Portando, o presente trabalho teve por objetivo otimizar um meio de cultura, empregando resíduos agroindústrias, para produção de toxinas por B. thuringiensis israelensis, utilizando a metodologia de superfície de resposta. Foi estudada também a cinética de crescimento e produção de toxina pela bactéria, bem como a toxicidade da cultura pós-fermentação, por um período de 28 dias. Como fontes de nitrogênio orgânico foram estudadas: farinha de soja, farinha de crisálida, peptona bacteriológica e uréia agroindustrial. Como fontes de carbono foram utilizadas sacarose, glicose e fécula de mandioca. Como fonte de nitrogênio inorgânico foi estudada sulfato de amônia. O parâmetro analisado para a otimização foi toxicidade, por meio de bioensaio contra Aedes aegypti. Foram estudadas também o percentual de esporulação e o crescimento (por meio da contagem de Unidades Formadoras de Colônia). A farinha de crisálida e a glicose foram as fontes mais adequada de nitrogênio orgânico e carbono, respectivamente, para a produção de toxinas. A superfície de resposta mostrou-se um método eficaz para a otimização, resultando em um meio com as seguintes faixas de concentrações das fontes de carbono e nitrogênio: glicose 0,3 a 0,55g/L; farinha de crisálida 55 a 58g/L e sulfato de amônia 0,6 a 1,1g/L. O meio otimizado resultou em uma toxicidade de 0,703 ppm (v/v), a qual é superior a de meios comumente utilizados para B. thuringiensis israelensis. A toxicidade da cultura aumentou com o tempo de cultivo, sendo máxima com 96 horas. A cultura manteve-se tóxica durante os 28 dias de análise, sendo que o armazenamento sem refrigeração mostrou-se mais eficaz (?)(Notexto não fica claro isto)). Esses resultados contribuem para o desenvolvimento de uma tecnologia local e para o aproveitamento de resíduos agroindustriais.
Palavras-chave: Bacillus thuringiensis, controle biológico, Aedes aegypti, superfície de
resposta, toxinas, produção, resíduos agroindustriais.
5

ANGELO, Elisangela Andrade. Medium optimization for toxins productions by Bacillus thuringiensis subsp. israelensis using agricultural byproducts. Dissertation (Marter’s degree dissertation – Food Science) – State University of Londrina. 2009.
ABSTRACT
Synthetic insecticides have been used in control of insects, but the use of these chemical insecticides has become problematic due to a lot of factors including physiological resistance in the vectors and environmental pollution. Hence, there has been an increased interest in recent years in the use of biological agents. This biological control is more specific, have lower frequency of resistance in insect targets and low residual effect on the environment. Bacillus thuringiensis is a Gram-positive spore-forming bacterium that produces a parasporal crystal protein which is toxic for many insects. This bacteria is the most widely used worldwide as a biological agent in controlling insects, but there is not very much information about its production. This fact is because many information are industrial secrecy. Therefore, the objective of this study was to optimize the production medium using agrindustrial byproducts, for toxins production by B. thuringiensi subsp. israelensis. This optimization was done with Response Surface Analyses (RSA) methodology. Also it was studied the kinetics of growth and production of the toxin by the bacteria, and it post-fermentation toxicity after 28 days store period. The organic nitrogen sources studied were soybean meal, Bombix mori pupae meal (BMP), bacterial peptone and agricultural urea. The carbon sources studied were glucose, sucrose and cassava starch. The inorganic nitrogen source studied was ammonium sulfate. The culture medium toxicity was evaluated by bioassays against Aedes aegypti and this parameter was the criteria used for the medium optimization. It was also studied the percentage of the bacteria sporulation and growth (Colony Forming Units, CFU,). The BMP and glucose were the most appropriate sources of nitrogen and carbon, respectively, for toxins production. The RSA, proved to be an effective method for medium optimization, resulting in a medium composition for maximal response with the following ranges of carbon and nitrogen concentrations: glucose 0,3 to 0,55gL-1; BMP 55 to 58gL-1 and ammonium sulfate 0,6 to 1,1gL-1. The optimized medium resulted in a toxicity of 0,703 ppm (v / v), which is above the mediums toxicity commonly obtained used B. thuringiensis israelensis. The culture medium toxicity increased with cultivation time until 96 hours. The culture medium toxicity remains stable under refrigeration temperatures during 28 days and when it was kept without refrigeration, the response was more effective. These results contribute to the development of a local technology and to promote the use of agroindustrial byproducts produced in the region.
Key-words: Bacillus thuringiensis, biological control, Aedes aegypti, response surface
methodology, toxins, production, agricultural byproduct.
6

ÍNDICE DE FIGURAS
Figura 1.1 Cultura lisada de B. thuringiensis subsp. israelensis, evidenciando os esporos elipsóides e os cristais protéicos arredondados.......................................................................15
Figura 1.2. Modos de ação das toxinas Cry.............................................................................18
Figura 1.3. Estrutura tridimensional das proteínas Cry, produzidas por B. thuringiensis... . .20
Figura 5.1. Cultura de B. thuringiensis israelensis em diferentes momentos de cultivo..........64
7

ÍNDICE DE TABELAS
Tabela 1.1 Principais ingredientes utilizados em meios de cultivo de bactérias entomopatogênicas e suas concentrações................................................................................26
Tabela 4.1 Níveis das fontes de nutrientes estudadas na etapa de seleção das fontes.............36
Tabela 4.2 Planejamento experimental utilizado na etapa de seleção das fontes de nutrientes...................................................................................................................................................36
Tabela 4.3 Planejamento fatorial fracionado 23-1.....................................................................37
Tabela 4.4 Níveis das variáveis no primeiro planejamento fatorial incompleto da etapa de otimização das fontes de nutrientes..........................................................................................38
Tabela 4.5 Níveis das variáveis no segundo planejamento fatorial incompleto da etapa de otimização das fontes de nutrientes..........................................................................................38
Tabela 4.6 Níveis das variáveis no planejamento fatorial completo (23) com repetição no ponto central e pontos estrelas.................................................................................................39
Tabela 4.7 Planejamento fatorial completo 23 com repetição no ponto central e pontos estrelas 39
Tabela 5.1 Efeito das fontes testadas sobre o bioensaio (etapa de seleção das fontes)...........45
Tabela 5.2 Efeito das fontes testadas sobre o bioensaio (primeiro planejamento fatorial incompleto – otimização das fontes).........................................................................................47
Tabela 5.3 Efeito das fontes testadas sobre o bioensaio (segundo planejamento fatorial incompleto – otimização das fontes).........................................................................................51
Tabela 5.4 Ensaios para confirmação do ponto central (g/L): 0,5 de glicose, 1,0 de sulfato de amônia e 50 de farinha de crisálida. Valor das variáveis e bioensaio.....................................53
Tabela 5.5 Respostas do Delineamento Composto Central Rotacional (planejamento fatorial completo, com repetição no ponto central e pontos estrelas)..................................................55
Tabela 5.6 Análise de variância (p<0,05) para a resposta bioensaio do delineamento composto central rotacional.....................................................................................................55
Tabela 5.7 Efeito das fontes de nutrientes sobre o bioensaio (delineamento composto central rotacional).................................................................................................................................58
Tabela 5.8 Valores previstos para o bionsaio em diferentes concentrações das fontes de nutrientes...................................................................................................................................59
Tabela 5.9 Custos dos meios de cultivo: Arcas adaptado e Farinha de crisálida otimizado.....61
8

9

ÍNDICE DE GRÁFICOS
Gráfico 5.1 Superfície de resposta para variáveis glicose e farinha de crisálida, tendo como resposta o bioensaio (primeiro planejamento fatorial incompleto).........................................48
Gráfico 5.2 Superfície de resposta para variáveis sulfato de amônia e farinha de crisálida, tendo como resposta o bioensaio (primeiro planejamento fatorial incompleto)......................48
Gráfico 5.3 Superfície de resposta (3 dimensões) para variáveis sulfato de amônia e glicose, tendo como resposta o bioensaio (primeiro planejamento fatorial incompleto)......................49
Gráfico 5.4 Superfície de resposta (plana) para variáveis sulfato de amônia e glicose, tendo como resposta o bioensaio (primeiro planejamento fatorial incompleto)................................50
Gráfico 5.5 Superfície de resposta para variáveis sulfato de amônia e glicose, tendo como resposta o bioensaio (segundo planejamento fatorial incompleto)..........................................52
Gráfico 5.6 Superfície de resposta para variáveis sulfato de amônia e farinha de crisálida, tendo como resposta o bioensaio (segundo planejamento fatorial incompleto)......................52
Gráfico 5.7 Superfície de resposta para variáveis farinha de crisálida e glicose, tendo como resposta o bioensaio (segundo planejamento fatorial incompleto)..........................................53
Gráfico 5.8 Superfície de resposta para variáveis sulfato de amônia e glicose, tendo como resposta o bioensaio (delineamento composto central rotacional)..........................................56
Gráfico 5.9 Superfície de resposta para variáveis farinha de crisálida e glicose, tendo como resposta o bioensaio (delineamento composto central rotacional)..........................................57
Gráfico 5.10 Superfície de resposta para variáveis farinha de crisálida e sulfato de amônia, tendo como resposta o bioensaio (delineamento composto central rotacional)......................57
Gráfico 5.11 Valores observados e previstos para o bioensaio (% de morte).........................59
Gráfico 5.12 Correlação entre a biomassa (mg/mL) mensuradas pelas metodologias de gravimetria e turbidimetria.......................................................................................................62
Gráfico 5.13 Crescimento microbiano (biomassa) e toxicidade (bioensaio) ao longo do cultivo (96 horas)......................................................................................................................63
Gráfico 5.14 Potencial hidrogeniônico (pH) ao longo do (96 horas)......................................65
Gráfico 5.15 Toxicidade da cultura durante o armazenamento................................................67
10

SUMÁRIO
1. INTRODUÇÃO....................................................................................................................12
2. REVISÃO BIBLIOGRÁFICA.............................................................................................142.1 Características gerais de Bacillus thuringiensis..............................................................142.2 Bacillus thuringiensis subsp. israelensis.........................................................................162.3 Proteínas Cry...................................................................................................................172.4 Outras toxinas produzidas por B. thuringiensis..............................................................212.5 Produção de B. thuringiensis..........................................................................................222.6 Meios de cultura..............................................................................................................242.7 Bioensaio.........................................................................................................................282.8 Planejamento de experimentos e uso do método de superfície de resposta....................29
3. OBJETIVOS.........................................................................................................................323.1 Objetivo Geral.................................................................................................................323.2 Objetivos Específicos......................................................................................................32
4. MATERIAIS E MÉTODOS.................................................................................................334.1 Microrganismo................................................................................................................334.2 Recuperação e manutenção da cultura............................................................................334.3 Preparo do pré-inóculo....................................................................................................334.4 Condições de cultivo.......................................................................................................344.5 Seleção das fontes de nutrientes.....................................................................................344.6 Otimização das concentrações das fontes de nutrientes selecionadas............................374.7 Estudo da cinética de crescimento e produção de toxinas..............................................404.8 Toxicidade da cultura de Bacillus thuringiensis subsp. israelensis duratne armazenamento.....................................................................................................................404.9 Métodos analíticos..........................................................................................................41
4.9.1 Bioensaio..................................................................................................................414.9.2 Unidades formadoras de colônia (UFC)..................................................................414.9.3 Quantificação de esporos.........................................................................................424.9.4 Biomassa seca (gravimetria)....................................................................................434.9.5 Turbidimetria...........................................................................................................434.9.6 Lâminas para análise microscópica.........................................................................44
4.10 Análise estatística..........................................................................................................44
5. RESULTADOS E DISCUSSÃO..........................................................................................455.1 Seleção das fontes de nutrientes.....................................................................................455.1 Otimização das concentrações das fontes de nutrientes selecionadas............................47
5.1.1 Planejamentos fatoriais incompletos........................................................................475.1.2 Planejamento fatorial completo...............................................................................54
5.3 Cinética de crescimento e produção de toxinas..............................................................625.4 Toxicidade da cultura de Bacillus thuringiensis subsp. israelensis durante armazenamento.....................................................................................................................67
6. CONSIDERAÇÕES FINAIS................................................................................................69
7. REFERÊNCIAS BIBLIOGRÁFICAS..................................................................................70
11

ANEXO 1:Processo de produção de farinha de crisálida pela indústria de fiação de seda Bratac s.a...................................................................................................................................75
ANEXO 2: Análise da constituição química de farinha de crisálida........................................76
12

1. INTRODUÇÃO
Estima-se que existam cerca de 2,5 milhões de espécies de insetos, das quais 10% são
consideradas pragas, isto é, causam danos ao ser humano ou perdas econômicas na pecuária
ou na agricultura. Muitas doenças de interesse da saúde pública, como malária, dengue, febre
amarela, filariose e doença de chagas são transmitidas por insetos vetores. Em geral, os países
mais atingidos por estas doenças são os tropicais e subtropicais, devido principalmente às suas
características climáticas favoráveis à proliferação de insetos. O crescente aquecimento global
demanda preocupação especial, estendendo as áreas propícias a esta proliferação também a
regiões atualmente com clima temperado.
O controle atual da população de insetos é realizado, principalmente, por inseticidas
químicos, cujo uso indiscriminado e massivo causa danos ambientais e à saúde humana.
Produtos químicos com alta toxicidade, porém com baixa especificidade e alto efeito residual,
prejudicam o meio ambiente, além de contribuírem para a existência de considerável número
de espécies de insetos com populações resistentes, inviabilizando a aplicação de tais produtos.
Uma alternativa aos inseticidas químicos é o controle biológico, o qual pode ser feito
com o uso racional de entomopatógenos, que constituem os componentes ativos dos
bioinseticidas ou inseticidas biológico.
Entre as vantagens dos bioinseticidas destacam-se: alta especificidade, menor risco
ambiental e à saúde humana, menor freqüência de resistência nos insetos alvo e a
possibilidade do entomopatógeno se multiplicar no ambiente e, com isso aumentar sua
permanência.
Como desvantagens dos bioinseticidas, destacam-se espectro de ação restrito e maior
suscetibilidade às condições ambientais. Estas desvantagens podem ser atenuadas com o uso
de boas formulações e estudos para aplicação do produto, a fim de torná-lo mais resistentes às
condições ambientais e com maior tempo de prateleira.
A comercialização de bioinseticidas corresponde a cerca de 5% do mercado de
pesticidas. Porém, o uso destes produtos vem crescendo dez vezes mais do que o uso de
inseticidas químicos. As bactérias destacam-se como promissoras no controle biológico. Entre
estes microrganismos, Bacillus thuringiensis (Bt) é o mais utilizado como bioinseticida.
Estima-se que os produtos a base dessa bactéria correspondam a cerca de 97% do mercado
mundial de bioinseticidas.
13

Entre as razões para o sucesso dos produtos à base de B. thuringiensis estão sua alta
toxicidade aos insetos alvos, capacidade de esporulação e o caráter monogênico de sua
principal toxina, o que facilita a manipulação genética da mesma.
A subespécie israelensis de B. thuringiensis mostrou-se eficaz no combate a dípteros,
ordem onde se encontram muitos gêneros com espécies vetores de doenças, como Aedes,
Culex e Anoplheles. Muitos produtos feitos a partir dessa subespécie vêm sendo utilizados há
mais de trinta anos em países africanos sob a recomendação da Organização Mundial da
Saúde (OMS).
Os maiores produtores de inseticidas a base de B. thuringiensis são Estados Unidos e
Canadá. O desenvolvimento de produção local, com linhagens mais adaptadas às condições
brasileiras e com meios de cultivo alternativos, poderia tornar o produto à base de B.
thuringiensis mais competitivo no Brasil.
Embora a produção de B. thuringiensis seja bem estudada, muitas informações
necessárias ao desenvolvimento de uma produção local não estão disponíveis na literatura.
Isto ocorre porque muitas informações são propriedades de indústrias que comercializam
produtos a base de B. thuringiensis.
Considerando a necessidade do Brasil em controlar os insetos vetores de doenças ou
pragas agrícolas, tendo em vista as vantagens do controle biológico e a dificuldade em se
obter informações sobre a produção de B. thuringiensis torna-se necessário o estudo e
desenvolvimento de tecnologia local para produção de bioinseticidas.
14

2. REVISÃO BIBLIOGRÁFICA
2.1 Características gerais de Bacillus thuringiensis
Bacillus. thuringiensis foi descrito em 1915 na Alemanha, isolado a partir de traça de
farinha (Anagasta kuehniella). Anteriormente, em 1902, no Japão, o pesquisador Ishiwata já
havia isolado uma bactéria a partir de Bombyx mori, que posteriormente se soube ser também
uma subspécie de B. thuringiensis. No entanto, a comercialização de produtos a base desta
bactéria só se iniciou em 1938 na França, com o bioinseticida comercializado sob o nome de
“Sporeine”. (BRAR et al., 2006; CAPALBO et al., 2004; VIDAURRE, 1996).
Pertencente à família Bacillaceae, a qual engloba a maioria das espécies formadoras
de esporos, B. thuringiensis é um bastonete gram positivo, com célula vegetativa de 1,0 a
1,2 µm de largura por 3,0 a 5,0 µm de comprimento, geralmente móveis. O esporo dessa
bactéria possui formato elipsoidal e localiza-se na região central ou paracentral quando no
interior da célula mãe. A espécie é aeróbia não estrita com faixa de temperatura de
crescimento entre 10 e 45°C. B. thuringiensis apresenta um amplo complexo enzimático, o
que lhe permite utilizar uma variedade de substratos. A principal característica que distingue a
espécie das outras do mesmo gênero é a presença intracelular de um cristal protéico (GLARE
& O’CALLAGHAN, 2000; HABIB & ANDRADE, 1998; MORAES et al., 2001; VILAS-
BÔAS et al., 2007).
Devido à suas características fisiológicas de esporulante e aeróbia não estrita, B.
thuringiensis apresenta resistência a condições adversas, o que contribui para seu alto
potencial no controle biológico (MORAES et al., 1998).
O cristal protéico típico de B. thuringiensis é, em geral, produzido durante a
esporulação e constitui o principal ingrediente ativo dos bioinseticidas a base dessa bactéria.
Este cristal é tóxico para várias espécies de insetos, destacando-se no controle de:
Lepidóptera, Díptera e Coleóptera. Porém, há subespécies de B. thuringiensis que apresentam
cristais tóxicos contra insetos dos gêneros Himenóptera, Hemíptera, Orthoptera, Phthraphera e
também para alguns nematóides, protozoários e ácaros (BRAR et al., 2006; GLARE &
O’CALLAGHAN, 2000; SCHNEPF et al., 1998).
15

Os cristais de B. thuringiensis são formados principalmente por proteínas
denominadas Cristal (Cry) e Citolíticas (Cyt), as quais são também conhecidas como δ-
endotoxinas (BRAVO et al., 2007). Ao final da esporulação, o cristal protéico corresponde a
cerca de 20% a 30% do peso seco da célula, sendo liberado no momento da lise celular
(ARANTES et al., 2002; GLARE & O’CALLAGHAN, 2000). A figura 1.1 apresenta esporos
e cristais de B. thuringiensis liberados com a lise celular, vistos em microscópio óptico com
aumento de 1000 vezes.
Figura 1.1 Cultura lisada de B. thuringiensis subsp. israelensis, evidenciando os esporos elipsóides (seta vermelha) e os cristais protéicos arredondados (seta azul). Aumento de 1000 vezes. Coloração: Amido-Black e Fucsina.Fonte: Cedido por Drª. Gislayne Fernandes Lemes Trindade Vilas-Bôas; Laboratório de Bioinseticida / Departamento de Biologia Geral/ Universidade Estadual de Londrina.
Bacillus thuringiensis já foi isolado de diversos ambientes, destacando-se solos,
insetos vivos ou mortos, grãos estocados, amostras de água entre outros. Embora muitos
sorotipos tenham sido isolados de amostras de solo, sabe-se que o esporo pode persistir por
muito tempo neste ambiente, porém essa bactéria não é capaz de se multiplicar naturalmente
no solo ou na água. Pesquisas recentes apontam o inseto como o nicho de B. thuringiensis,
pois é principalmente neste ambiente que ocorre sua multiplicação e interações genéticas
(RAYMOND et al., 2008; VILAS-BÔAS et al., 2000).
16
9 µm

2.2 Bacillus thuringiensis subsp. israelensis
A ordem de insetos Díptera possui muitos vetores de patógenos, destacando-se os
gêneros Aedes (Meigen, 1818), Culex (Linnaeus, 1758) e Anopheles (Meigen, 1818). Entre os
patógenos transmitidos por estes insetos destacam-se os causadores da dengue, malária,
filariose, encefalites e febre amarela. Muitos destes vetores já apresentam resistência a
inseticidas químicos. Durante muito tempo procurou-se uma subespécie de B. thuringiensis
capaz de controlar com eficiência as populações de Díptera. Em 1976/77, Goldberg e Margalit
isolaram e descreveram a variedade israelensis (Bti) a partir da água de uma lagoa de alta
salinidade. Esta subespécie apresentou-se com alto potencial no controle de larvas de algumas
espécies de Díptera. Desde então, Bti vem sendo bastante estudada e aplicada no controle
biológico (BECKER & MARGALIT, 1993; CAPALBO et al., 2004, GOLDEBERG &
MARGALIT, 1977; VIDAURRE, 1996).
Devido ao seu modo de ação complexo, é difícil encontrar mosquitos resistentes às
toxinas de B. thuringiensis israelensis. Esta subespécie tem sido utilizada em vários países da
África, na China, na Alemanha e na Austrália (BECKER & MARGALIT, 1993; ÖSKAN et
al., 2003; RUSSEL & KAY, 2008).
A segurança ambiental de B. thuringiensis israelensis tendo sido estudada e
numerosos testes com animais não-alvo tem sido realizados. Em alguns estudos com
mamíferos, por exemplo, fez-se a administração de doses altas de Bti (108 bactérias/animal)
via subcutânea, oral, intraperitonial, ocular e por inalação; estes testes revelaram que esta
subespécie é inócua para mamíferos. Além desses estudos, o fato de não haver registro de
malefícios a outros animais em locais onde Bti é utilizada, comprovam sua segurança
ambiental. Seu uso é permitido nos Estados Unidos desde 1981 (BECKER & MARGALIT,
1993).
O cristal protéico de Bti possui, em geral, a forma arredondada, como apresentado na
figura 1, sendo formado principalmente por 4 proteínas Cry: Cry4A (125KDa), Cry4B
(134KDa), Cry11A (67KDa) e Cry1Aa(27KDa).
17

2.3 Proteínas Cry
As proteínas Cry e Cyt são as principais constituintes dos cristais que caracterizam B.
thuringiensis. Estas proteínas são codificadas por genes que geralmente se localizam em
plasmídeos, e com menor freqüência localizam-se no cromossomo bacteriano. O cristal
protéico foi descoberto em 1953 por Hannay (BRAVO et al., 2007; HABIB & ANDRADE,
1986; LERECLUS, et al., 1993).
Atualmente estão descritas 436 proteínas Cry, sendo que a cada ano novas proteínas
são descobertas, só no ano de 2008 foram descritas 42 novas proteínas. De acordo com a
definição mais aceita, uma proteína é considera Cry quando forma uma inclusão paraesporal
(cristal) em B. thuringiensis, exibe algum grau de toxicidade a insetos alvos, ou alguma
proteína que tenha seqüência de aminoácidos similar a uma proteína Cry já descrita (BRAVO
et al., 2007; CRICKMORE et al., 2008, SCHNEPF, et al, 1998).
Uma mesma subespécie de B. thuringiensis pode conter várias cópias de um mesmo
gene cry ou de genes cry diferentes, sendo que as proteínas produzidas por estes genes
formarão um mesmo cristal. Dependendo das proteínas que constituem o cristal, este possui
um formato diferente, portanto há uma correlação entre a estrutura e composição protéica do
cristal. Como determinadas proteínas Cry são tóxicas para insetos específicos, o formato do
cristal pode fornecer indícios de qual inseto a subespécie em questão possui ação efetiva.
(LERECLUS et al., 1993; GLARE & O’CALLAGHAN, 2000).
As proteínas Cry são sintetizadas na forma de protoxinas, portanto, para poder agir é
necessária sua ativação, a qual ocorre no interior do inseto. Atualmente, há dois modelos
baseados em dados experimentais que explicam o modo de ação das toxinas Cry. As primeiras
etapas desses dois modelos são iguais: após a ingestão dos cristais, esses são solubilizados no
intestino do inseto, local com pH alcalino, liberando as protoxinas. Essas são clivadas por
proteases do próprio inseto, resultando em toxinas ativas, com cerca de 60kDa. A toxina ativa
é capaz de ligar-se a receptores específicos presentes nas células das microvilosidades
intestinais do inseto (BRAVO et al., 2007; SCHNEPF, et al., 1998).
O modelo de ação denominado “formação de poros” é o mais antigo e parece ser o
mais comum entre os diferentes gêneros de insetos (dípteros, lepidópteros e coleópteros).
Segundo esse modelo, a ligação da toxina com receptores específicos levaria a formação de
oligômeros de toxinas, os quais se ligariam a receptores secundários, que se encontram
firmemente ancorados na membrana da célula intestinal. Como resultado dessa ligação,
18

ocorreria a inserção da toxina oligomérica na membrana da célula intestinal,
resultando em poros nesse epitélio (BRAVO et al., 2007; BRAVO & SOBERÓN, 2008;
HABIB & ANDRADE, 1986).
O modelo mais atual que explica a ação de toxinas Cry é denominado “sinal de
transdução” e foi estudado em apenas poucos insetos-alvos. De acordo com esse modelo, a
ligação da proteína Cry com receptores específicos, induziria uma série de reações
intracelulares, que envolvem a proteína G e a adenilato ciclase, resultando no aumento da
AMP cíclico intracelular e ativação da proteína quinase A. Todas essas conseqüências levam
a um desequilíbrio da pressão interna celular, danificando-a (BRAVO & SOBERÓN, 2008).
A figura 1.2 apresenta um esquema que resume os modelos de ação por “formação de
poros” e por “sinal de transdução”.
Figura 1.2. Modos de ação das toxinas Cry. Os passos 1 a 3 são iguais nos dois modelos. O modelo “formação de poros” está representado na porção superior pelas números 4 a 6. A etapa 4 refere-se à oligomerização das toxinas, a 5 a ligação ao segundo receptor e a etapa 6 a inserção da toxina na membrana. O modelo “sinal de transdução” está representado na porção inferior pelo números 4a e 5a. A etapa 4a refere-se à ligação da toxina ao receptor e estimulação da proteína G, que estimularia a adenilato cilcase (etapa 5a) e após uma série de reações em cascata, comprometeriam o equilíbrio interno celular.Fonte: BRAVO & SOBERÓN, 2008.
19

A ação das toxinas, seja por qualquer um dos modelos, resulta na paralisia do aparelho
digestório, ocasionando morte por inanição. Em muitos insetos a causa da morte é septicemia
e paralisia geral dos movimentos, nesses casos a causa principal é a germinação de esporos
presentes no produto e não, necessariamente, devido á ação da toxina (BRAVO et al., 2007;
BRAVO & SOBERÓN, 2008; HABIB & ANDRADE, 1986; VALLETE-GELY et al, 2008).
Como grande parte dos genes cry localiza-se em plasmídeos conjugativos, os sorotipos
podem apresentar uma grande combinação de genes, o que resulta na ampla variedade de
perfis de toxicidade (CAPALBO et al., 2004; LERECLUS et al., 1993).
A transcrição da maioria dos genes cry é dependente do fator sigma, produzido
durante o processo de esporulação, por isso a maior parte das proteínas que compõem o cristal
é produzida durante essa etapa. Apenas, uma pequena parcela dos genes cry é regulada
independentemente da esporulação, sendo, portanto, expressos também durante o crescimento
vegetativo. Em ambos os casos, a proteína Cry é fruto do metabolismo secundário
(ARANTES et al., 2002; LERECLUS et al., 1993).
A massa molecular das proteínas Cry varia entre 40 e 140 KDa. A molécula destas
proteínas é globular, formada por três domínios estruturais ligados entre si por pequenas
pontes. Até o momento, seis proteínas Cry tiveram suas estruturas bem determinadas por
cristalografia por raios-X (figura1.3). Estas proteínas apresentaram um alto grau de
similaridade entre seus domínios, o que indica que apresentam um modo de ação semelhante.
O domínio 1 (N-terminal) é formado por cadeias polipeptídicas com formato de α-hélices,
sendo uma central (hidrofóbica), cercada por 6 cadeias marginais. O domínio 2 é composto
por três cadeias polipeptídicas antiparalelas com formato de β-folha pregueada. Já o domínio
3 apresenta o formato de β-sandwich (BRAVO et al., 2007).
Embora estudos atuais indiquem que várias porções dos diferentes domínios da
proteína se insiram na membrana no momento de formação dos poros, acredita-se que o
domínio 1 seja o principal responsável pela inserção (NAIR & DEAN, 2008). Os domínios 2
e 3 apresentam similaridades estruturais com cadeias polipeptídica que se ligam a
carboidratos e estariam envolvidos principalmente no reconhecimento de receptores presentes
na membrana das células dos insetos-alvo (BRAVO et al., 2007).
Durante muito tempo, a nomenclatura das proteínas Cry foi feita de acordo com o
inseto-alvo, desta forma, eram classificadas como Cry I as proteínas tóxicas a lepidópteras,
Cry II tóxicas a lepidópteras e dípteras, Cry III contra coleópteras, Cry IV contra dípteras.
Devido às constantes descobertas de novas proteínas Cry, tal sistema mostrou-se inconsistente
e, atualmente a classificação é feita baseando-se na seqüência de aminoácidos das proteínas.
20

Periodicamente o banco de dados on-line sobre proteínas de B. thuringiensis atualiza a
classificação e nomenclatura de tais proteínas (CRICKMORE et al., 1998; 2008).
Figura 1.3. Estrutura tridimensional das proteínas Cry, produzidas por B. thuringiensis. Nota-se que a estrutura geral das proteínas Cry é conservada.Fonte: BRAVO et al., 2007.
As proteínas Cyt são definidas como uma inclusão paraesporal de B. thuringiensis que
apresenta atividade hemolítica, ou alguma proteína que apresente seqüência de aminoácidos
similar a proteínas Cyt já descritas. Estas proteínas são formadas por apenas um domínio
composto por duas cadeias exteriores em forma de α-hélice, e uma cadeia interna em forma de
β-folha pregueada. Atualmente são conhecidas 27 proteínas Cyt (BRAVO et al., 2007;
CRICKMORE et al., 1998; 2008).
Assim como as proteínas Cry, as Cyt também são sintetizadas na forma de protoxinas.
No interior do inseto, as proteínas Cyt sofrem quebras, liberando a toxina ativa. Ao contrário
das proteínas Cry, Cyt não se liga a receptores específicos da membrana celular, e sim,
diretamente aos lipídios da membrana. Após sua ligação, as proteínas Cyt induzem a
formação de poros ou agem desestruturando a bicamada lipídica das membranas (BRAVO et
al., 2007).
Vários estudos têm demonstrado ocorrer um efeito sinergístico entre as proteínas Cry e
entre estas e as proteínas Cyt, ou seja, a maior toxicidade é devida à interação das proteínas,
onde uma potencializa o efeito da outra. (BRAVO et al., 2007; ÖSKAN, 2003).
21

2.4 Outras toxinas produzidas por B. thuringiensis
Além das proteínas Cry e Cyt, B. thuringiensis sintetiza outras toxinas que podem ou
não participar da ação entomopatogênica.
Em 1967 Heimpel sugeriu o nome de β-exotoxina para uma substância termoestável e
tóxica para alguns insetos. Devido a sua estrutura química o nome β-exotoxina vem sendo
substituído por thuringiensina. Estas proteínas possuem baixo peso molecular, a sua
toxicidade relaciona-se a inibição da RNA polimerase, pois é análoga a nucleotídeos. Embora
não possua especificidade, a ação de thuringiensina contribui para a toxidade global das
linhagens que a expressam (GLARE & O’CALLAGHAN, 2000; HABIB & ANDRADE,
1998; LERECLUS et al., 1993). Alguns trabalhos citam a importância dessa toxina no
controle de besouros e algumas espécies de ácaros (WU et al., 2002).
Muitos sorotipos de B. thuringiensis produzem proteínas denominadas Vip (vegetative
insecticidal proteins), as quais são sintetizadas na etapa vegetativa do crescimento. Apesar de
não integrarem o cristal protéico, estas proteínas contribuem para a toxicidade global das
linhagens e possuem forma de intoxicação similar às das proteínas Cry (CRICKMORE et al.,
2008; GLARE & O’CALLAGHAN, 2000).
Em 2000, Mizuki et al. descreveram uma nova toxina de B. thuringiensis a qual
passou a ser denominada Parasporina. Estas toxinas são definidas como proteínas paraesporal
de B. thuringiensis sem atividade hemolítica e que apresentam a capacidade de atacar células
cancerígenas (OHBA et al., 2008).
Bacillus thuringiensis sintetiza fosfolipases C, estas são produzidas ao final da fase
exponencial de crescimento e são excretadas para o meio de cultura (AGAISSE et al., 1995).
Essa bactéria sintetiza também quitinases e proteases que contribuem para romper a
membrana peritríquia presente no intestino dos insetos, contribuindo, dessa maneira, para a
toxicidade global das linhagens (GLARE & O’CALLAGHAN, 2000).
Além das toxinas, o esporo de B. thuringiensis também contribui parar sua toxicidade,
pois estes podem germinar no interior do inseto-alvo, ocasionando septicemia; ou
potencializarem o efeito da toxina em uma ação sinergística (GLARE & O’CALLAGHAN,
2000; RAYMOND et al., 2008).
22

2.5 Produção de B. thuringiensis
As primeiras indústrias a demonstrarem interesse no estudo e comercialização de B.
thuringiensis foram empresas de fermentação. Logo as indústrias de produtos químicos
perceberam a importância deste mercado de controle biológico e se lançaram em sua pesquisa
e produção (CABALBO & MORAES, 1988).
Nas décadas de 1970 e 1980, com o isolamento de novas linhagens e a possibilidade
da manipulação gênica, o interesse por B. thuringiensis aumentou bastante, principalmente no
setor privado (ARANTES et al., 2002; CAPALBO et al., 2004).
No Brasil, estudos realizados por diversos grupos em diferentes regiões têm tentado
implantar centros biotecnológicos onde ocorra a produção de produtos a base de B.
thuringiensis. Atualmente, tais centros estão em fase de estabelecimento dos métodos de
produção e execução dos testes preliminares. Como exemplos destes centros podem-se citar a
Probiom Tecnologia, situada na cidade de Campinas (São Paulo) e Bioticom, situada em
Recife (Pernambuco). A empresa Bthek Biotecnologia Ltda., com sede em Brasília, foi uma
das primeiras a formular e comercializar um produto nacional à base de B. thuringiensis.
Em geral, os produtos à base de B. thuringiensis são compostos por uma mistura de
cristais, esporos, células vegetativas e ingredientes secundários da formulação (CAPALBO et
al., 2004). Muitos destes produtos não são acessíveis aos consumidores, quer pelo preço, pela
falta de informação ou devido às condições legais estabelecidas pelo país. O desenvolvimento
de produções locais pode ser uma solução para tal situação. Neste caso, a produção pode ser
feita em um volume menor, escala piloto, usando-se materiais alternativos e abundantes na
região a ser atendida (SALAMA & MARGALIT, 1993; MORRIS et al., 1997).
Em geral, a produção de B. thuringiensis, assim como de outros produtos microbianos,
envolve as seguintes etapas: seleção da linhagem, estocagem, processo fermentativo,
recuperação do princípio ativo (esporos e cristais), formulação do produto e análise da
qualidade. Referencia!
Segundo Couch (2000) os principais critérios para seleção de uma subespécie
bacteriana para produção de bioinseticidas são: espectro de ação, potência por unidade de
volume da cultura, requerimentos nutricionais, facilidade de produção, estabilidade genética e
facilidade de estocagem.
A estocagem apropriada é de grande importância, pois a linhagem deve conservar seu
potencial tóxico e velocidade de crescimento. No caso de B. thuringiensis, as trocas de
23

plasmídeos ocorrem com certa freqüência e há relatos de perca de toxicidade após sucessivas
fermentações, sendo essencial a constante busca por novas linhagens e o monitoramento
durante o processo de fermentação (COUCH, 2000; BIZARRI et al, 2008).
A forma mais comum de produção de B. thuringiensis é por fermentação submersa
(líquida) descontínua, também conhecida como processo em batelada. Nesta fermentação um
recipiente é inoculado com o microrganismo e o meio de cultura líquido, não havendo
acréscimo ou retirada significativa do meio fermentado. Portanto, ocorre todo o
desenvolvimento da cultura, sendo retirado o produto apenas no final do processo. Em geral,
as proteínas Cry de B. thuringiensis são formadas no fim da fermentação, quando as
condições do meio se tornam desfavoráveis, sendo o processo em batelada satisfatório para tal
produção (MORAES et al., 2001).
Alguns pesquisadores têm estudando as fermentações sólida, contínua e em batelada
alimentada, porém até o momento, o mais viável para produção de proteínas Cry, ainda é o
processo em batelada (ADAMS et al., 2002; CHEN et al., 2003, MORAES et al., 2001,
VALLEJO et al., 1999).
Na fermentação industrial os passos a serem seguidos são: pré inóculo, geralmente
feito em frascos pequenos, pré fermentador, ?comumente com 1/5 do volume da fermentação
e o fermentador final. Durante todas as etapas deve-se analisar a cultura quanto à
contaminação, características morfológicas e potencial entomopatogênico. Em geral, os
reatores utilizados permitem o controle das principais condições de cultivo, as quais são:
temperatura, pH, aeração e agitação. O pré-inoculo, pré-fermentador e o fermentador em
volumes crescentes são feitos a fim de diminuir o tempo da fase lag, ou seja, o período de
adaptação do microrganismo às condições de cultivo. Em geral, pré fermentadores e
fermentador possuem os mesmos ingredientes no meio de cultivo. É importante limitar o
número de passos no processo de produção a fim de evitar contaminações e mudanças
indesejáveis no comportamento da bactéria (COUCH, 2000; MORAES et al., 2001).
Ao final da fermentação, a cultura de B. thuringiensis apresenta em média 6 a 8% de
sólidos, dos quais 1 a 3% são cristais e esporos. Há vários métodos que podem ser utilizados
para a recuperação destes cristais e esporos, sendo o mais comum a centrifugação e a micro-
filtração. É importante ressaltar que com tais processos, ocorre a recuperação principalmente
das proteínas Cry e muitas outras toxinas que podem contribuir para a toxicidade final do
produto são perdidas. Atualmente novas técnicas de recuperação e/ou concentração do
produto estão sendo desenvolvidas para complementar as mais utilizadas, destacando-se: a
liofilização e a flotação (BRAR et al., 2006; COUCH, 2000).
24

Após a recuperação, os produtos são formulados. No caso dos bioinseticidas, a
formulação tem três objetivos principais: conferir estabilidade ao produto durante a estocagem
e aplicação, facilitar a aplicação do produto e proteger o microrganismo das condições
adversas do ambiente. Há dois tipos básicos de formulação: sólida e liquida. Em geral, as
formulações sólidas são mais estáveis e preferíveis. As formulações são importantes, pois
podem permitir aos produtos a base de B. thuringiensis competir melhor no mercado de
produtos inseticidas (BRAR et al., 2006; COUCH, 2000).
Embora sejam segredos industriais, as formulações geralmente contam com uma
combinação de aditivos reconhecidos pela FDA (Food and drug Administration) ou pelo
órgão competente do país. É comum o uso de dispersantes, protetores e surfactantes (GLARE
& O’CALLAGHAN, 2000; BRAR et al., 2006).
Antes de ser comercializado, o produto formulado deve passar por testes de análise de
qualidade que atestem principalmente sua potência tóxica. Para B. thuringiensis, a toxicidade
é geralmente analisada por meio de bioensaio com o inseto-alvo (COUCH, 2000).
2.6 Meios de cultura
A escolha do meio de cultivo adequado é extremamente importante para o sucesso de
um produto. Esta escolha deve atender a dois requisitos principais; proporcionar uma boa
produção e não ser muito oneroso (COUCH, 2000).
Os meios de cultivo para B. thuringiensis geralmente possuem uma fonte de
nitrogênio, outra de carbono e sais minerais. Algumas vezes se adicionam ao meio alguns
tampões e anti-espumantes a fim de facilitar o processo (COUCH, 2000).
A fonte de carbono, além de fornecer matéria prima para muitos compostos celulares,
serve como fonte de energia. O nitrogênio é requerido principalmente para síntese de
proteínas e ácidos nucléicos. Os sais minerais atuam como co-fatores, sendo também
importantes no controle da osmolaridade celular.
Couch (2000) cita os principais componentes e suas concentrações utilizadas no
cultivo de B. thuringiensis na América do Norte (Tabela 1). Apesar das indústrias não
revelarem a composição de seus meios, sabe-se que estes geralmente são combinação dos
25

produtos listados. Muitos dos produtos utilizados são componentes indefinidos, porém não
devem apresentar muita variação de um lote para outro.
Meios alternativos vêm sendo estudados a fim de baratear o produto final,
principalmente nas produções regionais em escala piloto. Poopathi et al. (2002) obtiveram
resultados expressivos em cultivos em meio à base de batata. Vora e Shethna (1999)
estudaram um meio contendo extrato de soja suplementado com cistina, tendo bons resultados
para produção de d-endotoxina. Prabakaran et al. (2008) desenvolveram um meio de cultivo a
base de água de coco, obtendo resultados satisfatórios para produção de toxinas.
Vários estudos têm sido feitos utilizando-se como meio de cultivo para B.
thuringiensis águas residuais de indústrias e de estações de tratamento de água. Tais trabalhos
têm apresentado bons resultados, e muitas produções em escala piloto já utilizam estes
resíduos (LACHHAB et al., 2002; MONTIEL et al., 2001; VIDYARTHI et al., 2002;
YEZZA et al., 2005, 2006; CHANG et al, 2008).
A fonte de carbono para cultivo de B. thuringiensis varia muito conforme o objetivo
da fermentação, bem como qual linhagem está sendo cultivada. A maioria dos trabalhos
revela que a melhor fonte para crescimento vegetativo não corresponde a melhor fonte para
esporulação e formação dos cristais protéicos. Referencia!
Os estudos de Içgen et al. (2002b) indicam um efeito inibidor da glicose durante a
síntese da d-endotoxina. De acordo com este trabalho, as melhores fontes de carbono para
produção dos cristais tóxicos e esporulação são lactose, sacarose e inulina.
Segundo de Öskan et al. (2003), a glicose, amido e melaço inibem a produção de
toxina, em particular a proteína Cry4B (134KDa). Este estudo foi feito com B. thuringiensis
subsp. israelensis e ressalta o efeito repressor da glicose, a qual agiria em etapas de
fosforilação da proteína quinase Calfostina C, responsável pela produção das protoxinas Cry.
Segundo os autores, as melhores fontes de carbono para biossíntese da d-endotoxina seriam
dextrina, aveia, maltose, lactose, inulina, glicerol e sacarose, sendo a dextrina a mais
promissora de todas quanto à esporulação e produção de toxina.
26

Tabela 1.1 Principais ingredientes utilizados em meios de cultivo de bactérias
entomopatogênicas e suas concentrações.
Ingrediente Concentração (g/L)
Farinha de soja 20-40
Farinha da semente do algodão 14-30
Proteína de batata 15-40
Licor de milho ou sólidos derivados de
milho
15-30
Glicose 10-30
Peptona 2-5
Xarope de milho 20-45
Melaço 1.0-18.6
Glicerol 2.0-10
Amido de milho 10-15
Extrato de levedura 2.0
KH2PO4 1.0
K2HPO4 1.0
FeSO4 0.02
FeSO4.7H2O 0.0005-0.02
MgSO4. 7H2O 0.3
MnSO4. H2O 0.02
ZnSO4.7H2O 0.02
(NH4)2SO4 2
CuSO4.5H2O 0.005
CaCO3 1.0-1.5
PPG2000 2.0-5.0
Anti-espumante Hodag FD62 3.0
Anti-espumante SAG 5693 0.5-1.25
Anti-espumante Dow Corning 0.1
Fonte: Couch, 2000
27

Resultados diferentes dos pesquisadores Öskan et al. (2003) estão presentes nos
trabalhos de Vora e Shethna (1999) e Prabagaran et al. (2004), os quais indicam o melaço-de-
cana como uma boa fonte de carbono para produção de toxina. Estes estudos comprovam que
é necessário otimizar um meio para cada sorotipo, pois os resultados do estudo com um
determinado sorotipo muitas vezes não podem ser extrapolados para outros.
As fontes de nitrogênio podem ser tanto orgânicas quanto inorgânicas, sendo comum o
uso das duas fontes juntas. Os estudos de Arcas et al. (1984), Içgen et al. (2002b), Öskan et
al. (2003) e Zouari e Jaoua (1999) indicam que o uso de apenas nitrogênio inorgânico não é
aconselhável para o cultivo de B. thuringiensis, pois há diminuição de crescimento,
esporulação e biossíntese. Dentre as mais promissoras fontes de nitrogênio inorgânico
apontadas por estes autores, destacam-se os fosfatos [NH4H2PO4 e (NH4)2HPO4] e sulfatos por
otimizarem esporulação e muitas vezes a produção de toxinas. Entre as fontes orgânicas, o
extrato de levedura é bastante usado no cultivo de B. thuringiensis, porém peptona e soja
apresentam resultados promissores.
Como fontes alternativas de nitrogênio orgânico vêm se destacando: farinhas
derivadas de proteína animal (farinha de peixe e de crisálida), soja e alguns cereais (cevada e
trigo) (ALVES et al., 1997; DEVI, 2005; GHRIBI et al., 2005; MORRIS et al., 1997,
PRABAKARAN & BALAMARAN, 2006; ZOUARI & JAOUA, 1999).
A concentração de sais no meio de cultivo influencia diretamente a osmolaridade do
mesmo. Não há consenso sob qual porcentagem de sais é melhor, bem como quais os sais que
devem ser adicionados. Referencia!
Arcas et al. (1987) obtiveram uma boa produção de toxina com um meio contendo
osmolaridade de 808 miliosmol, sendo este um experimento clássico que resultou em um
meio bastante adotado para crescimento e produção de B. thuringiensis. É importante ressaltar
que embora os sais ajam como um todo na osmolaridade, cada íon fornecido pelos sais pode
ter um efeito diferente. Muitas vezes, produção e crescimento não se relacionam, sendo que
alguns íons melhoram um parâmetro e prejudicam ou são neutros para outro. Os principais
íons fornecidos pelos sais são: magnésio, cálcio, manganês, cobre e zinco.
Os estudos de Öskan et al. (2003) e Içgen et al. (2002a) revelaram que o manganês é
crítico para a diferenciação celular, sendo requerido para esporulação e formação do cristal de
toxina. Provavelmente este íon atue em processos de co-regulação da síntese da toxina. Os
melhores resultados aparecem quando a concentração de manganês varia entre 10-6 e 10-4M,
valores maiores que estes se tornam tóxicos para todos os processos celulares.
28

Assim como manganês, o magnésio influencia o metabolismo secundário, portanto,
com efeito na produção das proteínas Cry. Sua concentração ideal é em torno de 10 -3 M. O
cálcio é bastante importante no processo de esporulação, promovendo a estabilidade do cristal
protéico. A concentração de 10-3 M de cálcio estimula a esporulação, porém é inibidora para o
crescimento vegetativo e síntese protéica. Ao que parece, o cálcio não é essencial para a
síntese de toxina. Recomenda-se o uso de concentrações menores que 10 -3 M de cálcio, a fim
de não prejudicar a formação das proteínas Cry (IÇGEN et al., 2002a; ÖSKAN et al., 2003).
Metais como zinco, cobre e ferro apresentam resultados negativos para o crescimento,
esporulação e produção de toxina mesmo em concentrações pequenas como 10-7 M (IÇGEN et
al., 2002a; ÖSKAN et al., 2003).
2.7 Bioensaio
Um dos parâmetros mais importantes no estudo com B. thuringiensis é o seu potencial
tóxico ante o inseto-alvo. Embora muitos estudos utilizem a quantificação de proteínas Cry
para aferir a toxicidade de uma produção, sabe-se que o bioensaio é um método mais sensível,
pois fornece indícios não só quantitativos, mas também da eficácia da toxicidade da produção.
Liu e Tzeng (1998) indicam em seu trabalho a utilização do bioensaio na otimização de
produções, pois apenas contagem de esporos ou quantificação de proteínas não foram
suficientes.
O bioensaio pode ser entendido como um teste onde seres vivos são expostos a uma
determinada substância, sob condições controladas. No caso de B. thuringiensis, o bioensaio é
feito com larvas de insetos alvos segundo protocolos da Organização Mundial da Saúde
(OMS). (MORAES, et al., 2001; WORLD HEALTH ORGANIZATION, 2005).
Para B. thuringiensis israelensis, o bioensaio é feito com larvas de terceiro ou quarto
instar de dípteras dos gêneros Aedes, Culex, Anopheles, entre outros. Nesse ensaio, larvas
entre terceiro ou quarto instar são colocadas em diferentes recipientes, nos quais são aplicadas
diferentes concentrações do produto. As larvas são mantidas a 25-28°C por um período de 24
horas, quando então são mensuradas as quantidades de larvas mortas em cada uma das
concentrações. Faz-se então um cálculo da concentração letal para 50% (CL50) e 90% (CL90)
das larvas por meio de uma regressão linear log-probit, a qual pode ser feita com o uso de
29

programas de computador adequados. O bioensaio é feito em triplicata e há um controle
negativo, no qual não é aplicado o produto. Em alguns casos, como por exemplo, no estudo de
novas linhagens, padroniza-se uma concentração para as diferentes e linhagens e aplica-se a
cultura, só posteriormente realiza-se o bioensaio completo. Esse bioensaio tem como objetivo
fazer uma triagem inicial de novas linhagens (WORLD HEALTH ORGANIZATION, 2005).
O bioensaio pode ser expresso também em termos Unidades Internacionais (ITU),
nesse caso, a CL50 do produto em teste é comparada com o bioensaio de uma cepa de padrão
internacional a qual foi atribuída um valor arbitrário de potência. Para B. thuringiensis
israelensis, a cepa padrão é a IPS82, a qual tem um valor de potência contra A. aegypti de
15000 ITU/mg (WORLD HEALTH ORGANIZATION, 2005).
Muitos fatores podem influenciar em um bioensaio, destacando-se a espécie de inseto,
o instar, a densidade de larvas, qualidade da água e a temperatura, sendo crucial a
padronização do teste. Estudos revelam que A. aegypti é uma das espécies mais sensíveis à B.
thuringiensis israelensis, isto ocorre devido a fatores intrínsecos dessa espécie, tais como sua
alta velocidade de filtração (JARIC´-PERKUSIC et al, 2008; OTIENO-AYAYO et al., 2008).
2.8 Planejamento de experimentos e uso do método de superfície de resposta
Os processos fermentativos são influenciados por muitas variáveis, que podem
interagir entre si. O estudo da influência das variáveis bem como da suas interações sob um
determinado resultado é muito difícil de ser realizado sem um planejamento experimental
adequado. Desta maneira, o planejamento experimental tem por objetivo principal “projetar
um experimento de forma que ele seja capaz de fornecer exatamente o tipo de informação que
procuramos” (BARROS NETO et al., 1995).
O planejamento adequado permite executar o experimento de maneira mais organizada
e com um número de ensaios reduzidos ao mínimo necessário para se alcançar os objetivos de
interesse. Além disso, com o planejamento experimental é possível calcular o erro
experimental e até mesmo otimizar mais de uma resposta ao mesmo tampo (RODRIGUES &
IEMMA, 2005).
Até o início da década de 1970, os estudos sobre meios de cultivo para
microrganismos eram feitos variando-se uma variável a cada ensaio. Embora tenha sido
30

bastante comum, este método não permite o estudo das interações entre as variáveis, além de
muitas vezes exigir uma grande quantidade de ensaios, tornando-se dispendioso. No início
dos anos de 1970, os trabalhos dos pesquisadores Fisher e Box sobre desenho experimental se
popularizaram. De acordo com a proposta destes trabalhos, mais de uma variável muda
durante um ensaio, possibilitando estudar as interações entre uma variável e outra, além de
facilitar a modelagem matemática do processo (KENNEDY & KROUSE, 1999;
RODRIGUES & IEMMA, 2005).
Entre os métodos mais comuns de planejamento experimental destacam-se o
planejamento fatorial. Nessa metodologia, há combinação entre os valores de duas ou mais
variáveis. Esses valores são definidos a priori segundo os interesses do pesquisador, com base
em dados da literatura ou análises empíricas.
O planejamento fatorial pode ser completo ou incompleto, ele é dito completo quando
todas as combinações entre as variáveis nos seus diferentes valores são executadas em
ensaios. Já no planejamento fatorial incompleto, também chamado de fracionado, apenas um
número limitado das possíveis combinações das variáveis é realizado em ensaios (KENNEDY
& KROUSE, 1999; RODRIGUES & IEMMA, 2005).
Geralmente, o planejamento fracionado é utilizado quando a quantidade de variáveis e
valores é muito grande e a execução de todas as combinações se torna inviável, neste caso o
planejamento fracionado serve para fazer uma triagem de quais variáveis são importantes para
o processo e devem continuar no estudo. O planejamento fatorial completo geralmente é feito
em uma segunda etapa, quando as variáveis importantes já foram selecionadas e,
posteriormente se deseja estudar seus valores e interações com mais detalhes (KENNEDY &
KROUSE, 1999).
Denomina-se nível os valores que cada variável será estudada, no caso de se estudar 5
variáveis em 2 níveis cada, resultarão em 32 ensaios em um planejamento experimental
completo. Neste exemplo, o planejamento é dito 25, sendo o expoente a quantidade de
variáveis e o número elevado à quantidade de níveis de cada variável. Caso em uma etapa do
experimento realize-se apenas 16 ensaios dos 32 possíveis, o desenho passa a ser um fatorial
incompleto 25-1.
Tendo-se feito um planejamento experimental adequado, podem-se aplicar técnicas
que permitam o estudo da otimização do processo. Entre as técnicas estatísticas de otimização
mais utilizadas destaca-se a superfície de resposta. “A metodologia do planejamento fatorial,
associada à análise de superfície de respostas, é uma ferramenta fundamental na teoria
estatística, que fornece informações seguras sobre o processo, minimizando o empirismo que
31

envolve técnicas de tentativa e erro” (BOX et al, 1978 apud RODRIGUES & IEMMA, 2005).
Esta técnica foi desenvolvida por Box e Wilson no início da década de 1950 e consiste em
duas etapas principais que devem ser repetidas quantas vezes forem necessárias a fim de se
alcançar um ótimo. A primeira etapa é a de modelagem e geralmente é feita ajustando-se os
modelos matemáticos aos dados experimentais obtidos por meio de um planejamento fatorial
adequado. A segunda etapa consiste em um deslocamento geralmente no sentido de máxima
inclinação do modelo matemático (BARROS NETO et al., 1995). Em geral, nesta técnica a
resposta é representada em gráficos de superfícies que permitem visualizar melhor os
resultados contribuindo para sua interpretação (KENNEDY & KROUSE, 1999).
Alguns trabalhos já foram realizados com B. thuringiensis utilizando-se a metodologia
de superfície de resposta. Entre estes trabalhos destaca-se Prabagaran et al. (2004) os quais
estudaram o melhoramento de um meio a base de ingredientes alternativos para a produção de
toxina por uma subespécie de B. thuringiensis. Tokcaer et al. (2006) utilizaram a metodologia
de superfície de resposta para otimizar as condições de cultivo para produção de proteínas Cry
por B. thuringiensis. Ambos os trabalhos conseguiram melhorar a produção de toxinas pelas
subespécies estudadas.
32

3. OBJETIVOS
3.1 Objetivo Geral
Otimizar um meio de cultivo para a produção de toxinas de Bacillus thuringiensis
subsp. israelensis HD537 por fermentação submersa descontínua, utilizando resíduos
agroindustriais, com o intuito de controlar populações de Aedes aegypti.
3.2 Objetivos Específicos
Determinar a melhor fonte de nitrogênio entre: farinha de soja, farinha de crisálida,
peptona bacteriológica, uréia agroindustrial e sulfato de amônia para a produção de toxinas
por Bacillus thuringiensis subsp. israelensis HD537. Como? ASR?
Determinar a melhor fonte de carbono entre: sacarose, glicose e fécula de mandioca
para a produção de toxinas por Bacillus thuringiensis subsp. israelensis HD537. Como?
ASR?
Determinar as melhores concentrações de fontes de nutrientes selecionadas para
produção de toxinas por Bacillus thuringiensis subsp. israelensis HD537 utilizando a
metodologia de Analise de Superfície de Resposta.
Estudar a cinética de crescimento e produção de toxinas por Bacillus thuringiensis
subsp. israelensis HD537.
Verificar a estabilidade da toxicidade da cultura de Bacillus thuringiensis subsp.
israelensis HD537 por um período de 28 dias após sua fermentação.
33

4. MATERIAIS E MÉTODOS
4.1 Microrganismo.
O microrganismo utilizado neste estudo foi B. thuringiensis subsp. israelensis,
HD537, o qual foi gentilmente cedido pelo Laboratório de Bioinseticida do Departamento de
Biologia Geral da Universidade Estadual de Londrina.
Essa linhagem de B. thuringiensis foi isolada no Estado do Espírito Santo (Brasil) por
?????? (AUTOR, ANO) e caracterizado pelo Collaboration Centre for Entomopathogenic
Bacillus (Paris, França).
4.2 Recuperação e manutenção da cultura
A linhagem de B. thuringiensis israelensis HD537 foi inicialmente recuperada a partir
de cultura estoque em água e fitas de papel filtro. Após a recuperação, as culturas foram
mantidas sob refrigeração, em placas contendo agar nutriente, o qual continha (g/L): 5,0 de
peptona, 3,0 de extrato de carne e 15 de agar (BRASIL, 2004). Esta cultura foi replicada a
cada 15 dias.
4.3 Preparo do pré-inóculo
Antes do preparo do pré-inóculo, B. thuringiensis israelensis HD537 foi cultivado em
meio Arcas adaptado (ANDRADE, 2005), a fim de melhorar o crescimento do microrganismo
em meio líquido. Nessa etapa, a cultura foi mantida a 30ºC e 120 rpm (agitador rotativo)
durante 16 horas em frascos tipo Erlenmeyer de 500 mL de capacidade, contendo 50 ml de
34

meio, o qual continha (g/L): MgSO4 (1,0); KH2PO4, (1,0); K2HPO4 (1,0); MnSO4.H2O (0,3);
CaCl2 (0,4); (NH4)2SO4 (1,0), extrato de levedura (8,0) e glicose (4,5).
Após esse cultivo, o pré-inóculo foi feito transferindo-se 1,25 mL da cultura em meio
Arcas adaptado para um Erlenmeyer de 500 mL de capacidade, contendo 50 mL do meio a ser
testado. O pré-inóculo foi mantido a 30ºC, 120 rpm (agitador rotativo) por um período de 16
horas.
4.4 Condições de cultivo
O inoculo foi feito transferindo 1,25 mL do pré-inóculo para frascos Erlenmeyer de
500 ml de capacidade contendo 50 mL do meio estéril a ser estudado.
As culturas foram mantidas por um período de 72 ou 96 horas, a 30ºC e 120 rpm em
agitador rotativo. O tempo de cultivo foi baseado em testes preliminares e também em dados
da literatura (ANDRADE, 2005; COUCH, 2000).
4.5 Seleção das fontes de nutrientes
As fontes de nutriente estudadas bem como seus níveis iniciais foram escolhidos de
acordo com testes preliminares e dados da literatura. Como fontes de nitrogênio orgânico
foram estudadas:
Farinha de soja: escolhida por ser utilizada na produção do Laboratório de
Bioinseticida do Centro de Ciências Biológicas da Universidade Estadual de
Londrina;
Farinha de crisálida: esse ingrediente foi gentilmente cedido pela indústria de
fiação de seda Bratac s.a.. O processo de obtenção dessa farinha pela indústria
citada encontra-se no anexo 1. Esse ingrediente foi escolhido para o estudo com
base em testes preliminares e dados da literatura (ALVES et al., 1997);
35

Peptona bacteriológica: escolhido por ser ingrediente comum no cultivo industrial
de bactérias entomopatogênicas (COUCH, 2000);
Uréia agroindustrial: escolhido por ser comumente utilizada na agricultura como
fonte de nitrogênio e por apresentar um baixo custo.
Como fonte de nitrogênio inorgânico foi utilizado o sulfato de amônia ((NH4)2SO4), o
qual compõe o meio Arcas, comumente utilizado na fermentação de B. thuringiensis
(ARCAS, 1987).
As fontes de carbono estudadas foram:
Glicose: escolhida por ser comumente empregada no cultivo de B. thuringiensis
(ARCAS, 1987).
Sacarose: escolhida com base em estudos de Içgen et al. (2002b) e Öskan et al.
(2003).
Fécula de mandioca: escolhida por apresentar um baixo custo e como uma
alternativa ao uso de amido de milho, o qual é empregado em várias produções de
bactérias entomopatogênicas (COUCH, 2000).
A seleção das fontes foi feita por meio de um planejamento fatorial fracionado 28-4, no
qual cada variável foi estudada em dois níveis (-1 e +1) com variações extremas. Esses
experimentos foram feitos em dois blocos aleatórios O objetivo desta etapa foi criar um
contraste entre os níveis e, com isto realizar uma seleção qualitativa. Os níveis das diferentes
fontes de nutrientes bem como o planejamento fatorial encontra-se na tabela 4.1 e 4.2
respectivamente.
Além das fontes de nutrientes em estudo, o meio de cultivo continha uma mistura de
sais baseada no meio Arcas adaptado (ANDRADE, 2005), a qual não sofreu alteração e não
foi foco de estudo durante a pesquisa. Os sais presentes nessa mistura foram (g/L): MgSO4
(1,0); KH2PO4, (1,0); K2HPO4 (1,0); MnSO4.H2O (0,3) e CaCl2 (0,4).
As condições dos experimentos dessa etapa foram as mesmas descritas no item 4.3 por
um período de 72 horas.
A resposta analisada para a seleção das fontes foi bioensaio, conforme descrito no item
4.9.1. Além dessa resposta, foi analisado o efeito das fontes de nutrientes em estudo sobre o
crescimento e esporulação; para isso, foram feitas contagem de Unidades Formadoras de
Colônia e Contagem de esporos, como descritos no itens 4.9.2 e 4.9.3 respectivamente.
36

Tabela 4.1 Níveis das fontes de nutrientes estudadas na etapa de seleção das fontes
Fontes de nutrientes (Variáveis) Nível -1 Nível +1
Farinha de soja (g/L) 5,0 50
Farinha de crisálida (g/L) 2,0 20
Peptona de carne (g/L) 0,5 5
Uréia (g/L) 0,2 2,0
Sulfato de amônia (g/L) 0,2 2,0
Glicose (g/L) 1,0 10
Sacarose (g/L) 1,0 10
Fécula de mandioca (g/L) 1,5 15
Tabela 4.2 Planejamento experimental utilizado na etapa de seleção das fontes de nutrientes
Experimento Sacarose Glicose
Fécula de mandioca
Farinha de soja
Farinha de
crisálida
Peptona de
carne Uréia
Sulfato de
amônia1 -1 -1 -1 -1 -1 -1 -1 -12 1 -1 -1 -1 -1 1 1 13 -1 1 -1 -1 1 -1 1 14 1 1 -1 -1 1 1 -1 -15 -1 -1 1 -1 1 1 1 -16 1 -1 1 -1 1 -1 -1 17 -1 1 1 -1 -1 1 -1 18 1 1 1 -1 -1 -1 1 -19 -1 -1 -1 1 1 1 -1 110 1 -1 -1 1 1 -1 1 -111 -1 1 -1 1 -1 1 1 -112 1 1 -1 1 -1 -1 -1 113 -1 -1 1 1 -1 -1 1 114 1 -1 1 1 -1 1 -1 -115 -1 1 1 1 1 -1 -1 -116 1 1 1 1 1 1 1 1
37

4.6 Otimização das concentrações das fontes de nutrientes selecionadas
Essa etapa teve por objetivo otimizar a concentração das fontes de nutrientes
selecionadas como descrito no item 4.5. Inicialmente foram feitos dois planejamentos fatoriais
incompletos 23-1, onde os níveis das variáveis foram selecionados segundo dados da literatura.
Esses planejamentos fatoriais foram feitos em apenas um bloco com 9 experimentos, como
descrito na tabela 4.3..
Tabela 4.3 Planejamento fatorial fracionado 23-1
Experimentos Glicose
Sulfato de
amônia
Farinha de
crisálida1 -1 -1 -12 -1 0 13 -1 1 04 0 -1 15 0 0 06 0 1 -17 1 -1 08 1 0 -19 1 1 1
Os níveis das variáveis no primeiro e no segundo planejamento fatorial incompleto
encontram-se descritos nas tabelas 4.4 e 4.5, respectivamente. Esses níveis foram codificados
de acordo com a equação abaixo:
X codificado = Xi – X0
ΔX / 2
Onde:
X codificado: nível codificado da variável
Xi = valor real
X0 = valor no ponto central
ΔX = valor real máximo – valor real mínimo
38

Tabela 4.4 Níveis das variáveis no primeiro planejamento fatorial incompleto da etapa de
otimização das fontes de nutrientes
Variáveis Nível (-1) Nível (0) Nível (+1)Glicose (g/L) 2,00 6 10,00
Sulfato de amônia (g/L) 2,00 4 6,00Farinha de
Crisálida (g/L) 10,00 30 50,00
Tabela 4.5 Níveis das variáveis no segundo planejamento fatorial incompleto da etapa de
otimização das fontes de nutrientes
Variáveis Nível (-1) Nível (0) Nível (+1)Glicose (g/L) 0,5 1,25 2,0
Sulfato de amônia (g/L) 1,0 3,0 5,0Farinha de
Crisálida (g/L) 50,0 90,0 130,0
Após o segundo planejamento fatorial incompleto, foram feitos 4 experimentos com
valores para as variáveis próximos ao do experimento com melhor bioensaio obtido até então.
Com isto, determinou-se o ponto central para o próximo planejamento. Com base nesses
resultados, foi feito um fatorial completo central rotacional (23), com uma repetição no ponto
central mais os pontos axiais (“pontos estrelas”). Os níveis das variáveis utilizados nesse
planejamento fatorial completo encontram-se na tabela 4.6. Os 16 experimentos desse
planejamento fatorial completo central rotacional foram feitos em dois blocos, como descrito
na tabela 4.7.
As condições de cultivo dessa etapa de otimização foram as mesmas descritas no item
4.3 em todos os experimentos o cultivo durou 72 horas.
A resposta analisada para a otimização da concentração das fontes de carbono,
nitrogênio orgânico e inorgânico foi o bioensaio, como descrito no item 4.9.1. Além dessa
39

resposta, estudou-se também o crescimento (UFC) e esporulação, como descrito nos itens
4.9.2 e 4.9.3 respectivamente.
Tabela 4.6 Níveis das variáveis no planejamento fatorial completo (23) com repetição no
ponto central e pontos estrelas
Variáveis
Nível (-1,79)
Nível (-1) Nível (0) Nível (+1)
Nível (+1,79)
Glicose (g/L) 0,005 0,25 0,5 0,75 0,95Sulfato de
amônia (g/L) 0,10 0,5 1,0 1,5 1,90Farinha de
Crisálida (g/L) 32,11 40 50 60 67,89
Tabela 4.7 Planejamento fatorial completo 23 com repetição no ponto central (experimentos 9
e 10) e pontos estrelas (experimentos 11 a 16)
Experimentos
Blocos Glicose Sulfato de Amônia
Farinha de.
crisálida1 1 -1,00000 -1,00000 -1,000002 1 -1,00000 -1,00000 1,000003 1 -1,00000 1,00000 -1,000004 1 -1,00000 1,00000 1,000005 1 1,00000 -1,00000 -1,000006 1 1,00000 -1,00000 1,000007 1 1,00000 1,00000 -1,000008 1 1,00000 1,00000 1,00000
9 (C) 1 0,00000 0,00000 0,0000010 (C) 1 0,00000 0,00000 0,00000
11 2 -1,78885 0,00000 0,0000012 2 1,78885 0,00000 0,0000013 2 0,00000 -1,78885 0,0000014 2 0,00000 1,78885 0,0000015 2 0,00000 0,00000 -1,7888516 2 0,00000 0,00000 1,78885
40

4.7 Estudo da cinética de crescimento e produção de toxinas
Após a otimização das fontes nutrientes, foi feito um estudo a fim de se verificar a
curva de crescimento de B. thuringiensis israelensis HD537 no meio desenvolvido, bem como
em qual das etapas de crescimento ocorre a maior produção de toxinas: Para isso, foram
inoculadas culturas com o meio de cultivo com as fontes otimizadas e mantidas como descrito
no item 4.3. Essas culturas foram mantidas por um período de 96 horas.
Em intervalos predeterminados, foram retiradas alíquotas das culturas a fim de se
analisar seu crescimento, produção de toxinas e pH. Nessa etapa, o crescimento foi avaliado
por meio de gravimetria e turbidimetria, como descrito nos itens 4.9.4 e 4.9.5
respectivamente. A produção de toxinas foi avaliada por meio de bioensaio, como descrito no
item 4.9.1. O pH foi medido por meio de um aparelho potenciômetro modelo Foram feitas
ainda, lâminas para análise microscópica, como descrito no item 4.9.6.
Esse experimento foi repetido por 3 vezes, sendo que todos os métodos de análise de
crescimento e medição de pH foram feitos em duplicata e o bioensaio foi feito em triplicata.
4.8 Toxicidade da cultura de Bacillus thuringiensis subsp. israelensis durante
armazenamento
A toxicidade da cultura de Bacillus thuringiensis subsp. israelensis HD537 foi
estudada por um período de 28 dias. Para isso, foram feitas quatro culturas com o meio
otimizado, as quais foram fermentadas sob as condições do item 4.3 por um período de 72
horas.
Após o cultivo, duas culturas foram mantidas sob refrigeração em câmara fria a 5°C
(+/-2°C) e duas foram mantidas sob as condições ambientes, porém ao abrigo da luz. A cada 7
dias foram feitos bioensaio com cálculo da CL50 para essas culturas.
41

4.9 Métodos analíticos
4.9.1 Bioensaio
Os bioensaios foram realizados no Laboratório de Entomologia Médica, do professor
Dr. José Lopes, do Centro de Ciências Biológicas da Universidade Estadual de Londrina.
Estes testes foram feitos com base na metodologia da WHO 2005, com larvas entre terceiro e
quarto instar de A. aegypti (Linnaeus, 1762), as quais foram colocadas em recipientes
contendo 100 mL de água destilada. Após aplicação do produto, as larvas foram mantidas a
25°C por um período de 24 horas, quando então, procedeu-se a leitura do bioensaio quanto à
mortalidade das mesmas. Em cada recipiente utilizado no bioensaio foram colocadas 20
larvas.
Na etapa de seleção das fontes de nutrientes, as culturas dos diferentes experimentos
foram diluídas em água destilada. Procedeu-se, então, o bioensaio aplicando-se as culturas
diluídas, de maneira que a concentração final nos recipiente do bioensaio fosse de 10 ppm
(v/v). Essa concentração foi escolhida, após testes preliminares com diferentes concentrações.
Nas etapas de otimização das fontes e estudo da cinética, o bioensaio foi feito como
descrito para a etapa de seleção de fontes, porém a concentração final no recipiente do
bioensaio foi de 1 ppm (v/v).
Esses bioensaios foram expressos como porcentagem de morte das larvas e foram
feitos em triplicata.
Os bioensaios com o meio otimizado, bem como de pontos do estudo da cinética e da
etapa de verificação da toxicidade das culturas durante armazenamento foram expressos em
termos de CL50. Para isso, as culturas foram diluídas; e 5 ou 6 concentrações diferentes foram
aplicadas em recipientes dos bioensaios. Ao final do teste, foi feita a quantificação de larvas
mortas em cada uma das concentrações. A CL50 foi calculada por meio de análise de regressão
linear do log-Probit, utilizando-se o programa de computador microprobit (FINNEY, 1971).
Esse bioensaio foi feito em triplicata.
4.9.2 Unidades formadoras de colônia (UFC)
42

A quantificação das unidades formadoras de colônia foi feita diluindo-se 1 mL da
cultura em frascos contendo 9 mL de solução salina estéril de cloreto de sódio (NaCl) a
0.85%. Após diluições em série em frascos contendo salina, 0,5 mL das amostras diluídas
foram inoculadas por espalhamento superficial em placas contendo agar nutriente.
As placas foram mantidas a 30°C por um período de 24 horas, quando então se fez a
contagem das unidades formadoras de colônia. Foram desconsideradas as placas com mais de
300 UFC ou menos de 30 UFC. Considerando-se as diluições procedidas, fez-se o cálculo das
unidades formadoras de colônia na cultura inicial. Essa análise foi feita em triplicata e
expressa como logaritmo de UFC por mililitro de cultura.
4.9.3 Quantificação de esporos
Para a quantificação de esporos, os frascos de salina com cultura diluída, após
utilização na quantificação de UFC, foram mantidos a 80 °C por 20 minutos. Após esse
período, fez-se o espalhamento de 0,5 mL da cultura diluída como descrito para a
determinação de UFC. Essa análise foi feita em triplicata e expressa como porcentagem de
esporulação, a qual foi calculada com base nos valores de UFC.
4.9.4 Biomassa seca (gravimetria)
Na etapa de estudo da cinética, a biomassa seca foi utilizada para verificar o
crescimento do microrganismo. Para isso, 4 mL da cultura foram centrifugados a 4500g, 5°C
por 15 minutos. O sobrenadante foi utilizado para etapas posteriores e o precipitado foi
ressuspendido em 4mL solução salina de cloreto de sódio (0,85%) e centrifugado novamente
em iguais condições.
O sobrenadante da segunda centrifugação foi desprezado e o precipitado
ressuspendido novamente em 4mL da solução salina. Alíquotas de 1mL foram transferidas
para cadinhos previamente tarados, os quais foram submetidos a 105 °C até peso constante.
43

Essa análise foi feita em duplicata e expressa como miligramas de biomassa por
mililitro da cultura. A fim de não se aferir como biomassa algum componente do meio de
cultura, esse foi submetido a centrifugações, como descrito acima, e o valor de massa seca do
meio de cultura foi descontado do valor da biomassa seca dos experimentos.
4.9.5 Turbidimetria
Na etapa do estudo da cinética, o crescimento do microrganismo foi verificado
também por meio de teste de turbidimetria. Para isso, fez-se uma curva de calibração de uma
cultura em meio Arcas adaptado relacionando absorbância a 540 nm e biomassa seca
(mg/mL).
O segundo precipitado ressuspendido em salina do item 4.6.4, foi diluído em salina e
sua absorbância lida em espectrofotômetro a 540 nm. Por meio da curva de calibração,
calculou-se, então, a concentração da biomassa da cultura, expressa como miligramas de
biomassa por mililitros da cultura.
Essa análise foi feita em duplicata. Como o meio de cultura poderia conter alguns
pigmentos que interferissem na análise da absorbância, foi feita uma leitura do sobrenadante
do meio de cultura (sem crescimento microbiano) centrifugado, e essa absorbância foi
descontada do valor da absorbância dos experimentos.
4.9.6 Lâminas para análise microscópica
A fim de acompanhar a morfologia das células durante o estudo da cinética, bem como
para averiguar se havia contaminação nos demais experimentos, foram feitas lâminas para
análise em microscópio óptico.
Para confecção das lâminas, alíquotas das culturas foram retiradas com auxílio de uma
alça de platina. Após esfregaço e fixação por calor as lâminas foram coradas com amido black
(por 70 segundos), lavadas e coradas com fucsina (10 segundos). Essa coloração evidência os
esporos e cristais de B. thuringiensis.
44

4.10 Análise estatística
Os planejamentos experimentais, montagem dos gráficos de superfície de resposta,
bem como os testes de análise de variância (ANOVA) foram feitos utilizando-se o programa
Statistic versão 6.0. Os demais gráficos foram feitos com auxílio do programa Excel (versão
2003). A comparação entre as CL50 foi feita utilizando-se o teste de τ-Student.
45

5. RESULTADOS E DISCUSSÃO
5.1 Seleção das fontes de nutrientes
Embora tenha sido estudado o crescimento e esporulação, a otimização desse estudo
visou à produção de toxinas, por isso, o bioensaio foi a resposta utilizada na escolha das
fontes e na etapa posterior. A tabela 5.1 apresenta os efeitos das diferentes fontes de nutrientes
testadas sobre o bioensaio.
Tabela 5.1 Efeito das fontes testadas sobre o bioensaio (etapa de seleção das fontes). Em
vermelho, as variáveis com efeito significativo (ANOVA, p<0,05).
Efeito Probabilidade Coeficiente. Variáveis/Interações 0,517500 0,000233 0,517500(1) Sacarose -0,070000 0,047421 -0,035000(2) Glicose -0,035000 0,157299 -0,017500(3) Fécula de mandioca -0,440000 0,001289 -0,220000(4) Farinha de soja -0,535000 0,000872 -0,267500(5) Farinha de crisálida 0,060000 0,062957 0,030000(6)Peptona bacteriológica
0,045000 0,104467 0,022500
(7) Uréia agroindustrial -0,455000 0,001205 -0,227500(8) Sulfato de amônia 0,430000 0,001349 0,215000Interação 1 e 5 -0,045000 0,104467 -0,022500Interação 1 e 8 0,035000 0,157299 0,017500Interação 2 e 5 -0,010000 0,591752 -0,005000Interação 2 e 8 0,020000 0,333333 0,010000Interação 5 e 8 -0,045000 0,104467 -0,022500
Esse resultado apresentou um coeficiente de determinação (R) de 0,999 e um R
ajustado de 0,996. Isto indica que 99% dos resultados são explicados pelo efeito das variáveis,
sendo, portanto bastante confiável.
46

Com exceção da glicose (a tabela indica um efeito negativo), as demais fontes de
carbono apresentaram um efeito negativo sobre o bioensaio. Embora a glicose não tenha
apresentado efeito positivo, seu resultado não foi significativo, sendo esta a escolhida como
fonte de carbono para as etapas posteriores.
Diferente dos trabalhos de Içgen et al. (2002b) e Öskan et al. (2003), esses resultados
não apontam a sacarose como uma boa fonte para produção de toxinas, tendo em vista seu
efeito negativo para o bioensaio. O que ressalta a importância de um estudo direcionado para
cada linhagem específica, bem como para o resultado que se pretende otimizar.
Quanto ao crescimento, a análise das Unidades Formadoras de Colônia (log UFC/mL)
demonstrou que a sacarose e a glicose não tiveram efeito significativo sobre o crescimento, já
a fécula de mandioca apresentou um efeito negativo. Isso provavelmente ocorreu, devido a
maior complexidade estrutural da fécula de mandioca frente às outras fontes, o que dificultou
a utilização da mesma durante o crescimento microbiano. No trabalho de Öskan et al. (2003),
o amido, o qual possui estrutura semelhante à fécula, apresentou um efeito negativo sobre o
crescimento de B. thuringiensis israelensis HD500, justificado por sua estrutura química
complexa. Muitos meios que utilizam materiais complexos, como águas residuais do
processamento de amido, fazem um pré-tratamento antes de utilizá-la no meio de cultivo
(YEZZA et al., 2006; CHANG et al., 2008). (Verficar a presença de enzimas amiloliticas em
Bacillus thuringiensis)
Dentre as fontes de nitrogênio orgânico, apenas a uréia agroindustrial apresentou
efeito significativo, porém negativo sobre o bioensaio. Já a farinha de crisálida e peptona não
apresentaram efeito sobre o bioensaio, sendo portanto necessário escolher entre essas duas
fontes para as etapas de otimização. Com o intuito de aproveitar resíduos agroindustriais e
abaixar o custo da produção, foi escolhida para os testes posteriores a farinha de crisálida.
O sulfato de amônia apresentou um efeito significativo e positivo sobre o bioensaio, o
que, portanto, indica sua importância como fonte de nitrogênio na produção da toxina,
resultado semelhante aos estudos de Içgen et al. (2002a).
Em relação ao crescimento, dentre as fontes de nitrogênio, apenas a peptona
apresentou um efeito significativo e positivo. Porém, como o objetivo principal era a
otimização das toxinas, essa fonte não foi escolhida para as etapas posteriores.
Quanto à esporulação, nenhuma das fontes de carbono ou nitrogênio apresentou efeito
significativo sobre tal parâmetro. A taxa de esporulação foi alta em todos os experimentos
dessa etapa, em média de 74,45%.
47

5.1 Otimização das concentrações das fontes de nutrientes selecionadas
5.1.1 Planejamentos fatoriais incompletos
Após a seleção da glicose, farinha de crisálida e sulfato de amônio como fontes de
nutrientes, a concentração dessas fontes no meio de cultivo foi otimizada visando a melhora
no bioensaio.
Inicialmente foi feito um planejamento fatorial incompleto, cujos resultados dos
efeitos encontram-se na tabela 5.2 e nos gráfico 5.1 a 5.3.
Tabela 5.2 Efeito das fontes testadas sobre o bioensaio (primeiro planejamento fatorial
incompleto – otimização das fontes). Em vermelho, as variáveis com efeito significativo
(ANOVA, p<0,05). L representa o termo linear e Q o termo quadrático.
Efeito Probabilidade Coeficiente.Variáveis. 9,55556 0,006958 9,55556Glicose (L) 1,33333 0,566987 0,66667Glicose (Q) -3,33333 0,188893 -1,66667(2)Sulfato de amônia(L)
-0,66667 0,766450 -0,33333
Sulfato de amônia(Q)
3,66667 0,163685 1,83333
(3)Farinha de crisálida(L)
15,33333 0,015991 7,66667
Farinha de crisálida(Q)
-4,33333 0,125525 -2,16667
48

Gráfico 5.1 Superfície de resposta para variáveis glicose e farinha de crisálida, tendo como
resposta o bioensaio (primeiro planejamento fatorial incompleto). O sulfato de amônio está
fixado no valor codificado central.
Gráfico 5.2 Superfície de resposta para variáveis sulfato de amônia e farinha de crisálida,
tendo como resposta o bioensaio (primeiro planejamento fatorial incompleto). A glicose está
fixado no valor codificado central.
49

Os resultados desse primeiro planejamento fatorial apresentaram um coeficiente de
determinação de 0,97, já o coeficiente ajustado foi de 0,90; sendo satisfatório para estudos
biológicos.
Entre as variáveis testadas, a farinha de crisálida apresentou o termo linear altamente
significativo e positivo, o que indica que esta fonte influenciou muito o bioensaio (na
realidade influencio a toxicidade do meio de cultura utilizado no bioensaio), porém ainda não
se encontra no seu ponto ótimo (termo quadrático não significativo). Observando os gráficos
5.1 e 5.2, vê-se que a região de ótimo, representado em vermelho, está no sentido de aumento
da concentração da farinha de crisálida. Por isso, para o segundo planejamento incompleto, os
valores para a concentração de farinha de crisálida foram calculados a partir do ponto máximo
(+1) do primeiro planejamento, ou seja, houve um aumento na concentração dessa fonte.
Gráfico 5.3 Superfície de resposta (3 dimensões) para variáveis sulfato de amônia e glicose,
tendo como resposta o bioensaio (primeiro planejamento fatorial incompleto). A farinha de
crisálida está fixada no valor codificado +1.
50

Em relação à glicose e ao sulfato de amônio, nenhum dos dois apresentou o termo
quadrático ou linear significativo sobre o bioensaio. No entanto, os valores dessas variáveis
não foram fixados por esse experimento, por ele ser um fatorial incompleto, ou seja, poderia
haver interações e efeitos significativos futuros.
Observando-se os gráfico 5.3 e 5.4, vê-se que há dois pontos em direção ao ótimo para
essas duas variáveis, resultando em um gráfico com formato de “cela”. Como a produção visa
a melhora da resposta, conjuntamente com a diminuição dos custos, a otimização dessas duas
variáveis prosseguiu no sentido de menor valor, ou seja, a glicose em torno dos valor
codificados -1,37 a -1,0; já o sulfato de amônia em torno dos valores codificados -1,5 e a 0,5.
Gráfico 5.4 Superfície de resposta (plana) para variáveis sulfato de amônia e glicose, tendo
como resposta o bioensaio (primeiro planejamento fatorial incompleto). A farinha de crisálida
está fixada no valor codificado +1.
Portanto, para o segundo experimento os valores da concentração da farinha de
crisálida foram aumentados, já os concentrações de glicose e sulfato de amônia diminuíram,
como apresentado na tabela 4.5.
Os resultados com os efeitos das variáveis no segundo planejamento fatorial
incompleto encontram-se na tabela 5.3.
51

Tabela 5.3 Efeito das fontes testadas sobre o bioensaio (segundo planejamento fatorial
incompleto – otimização das fontes). Em vermelho, as variáveis com efeito significativo
(ANOVA, p<0,05). L representa o termo linear e Q o termo quadrático.
Efeitos Probabilidade Coeficiente.Variáveis. 19,7778 0,000012 19,7778Glicose (L) -19,3333 0,012189 -9,6667Glicose (Q) -13,6667 0,032552 -6,8333Sulfato de amônia(L) -14,6667 0,044148 -7,3333Sulfato de amônia(Q) -6,6667 0,258147 -3,3333Farinha de crisálida(L)
-32,0000 0,000431 -16,0000
Farinha de crisálida(Q)
3,3333 0,563070 1,6667
Os resultados desse segundo planejamento fatorial apresentaram um coeficiente de
determinação de 0,81, sendo satisfatório para estudos biológicos.
Observando-se os efeitos das variáveis percebe-se que o sulfato de amônia e a farinha
de crisálida apresentaram efeito quadrático não significativo, o que indica que essas fontes
ainda não se encontram em seu ponto ótimo. Essas variáveis apresentaram o termo linear
significativo e negativo, o que indica que para alcançar seu nível ótimo, a concentração dessas
variáveis deve diminuir.
Em relação à glicose, tanto o termo linear como o quadrático foram significativos, o
que pode indicar uma interação com outra variável ou dois destinos de otimização, o aumento
ou diminuição de sua concentração, o que levaria a formação do chamado “ponto de cela”.
Os gráficos 5.5 a 5.7 ilustram o efeito das variáveis sobre o bioensaio, no segundo
planejamento fatorial incompleto.
Embora nessa etapa as variáveis ainda não se encontrem no seu nível ótimo, já houve
uma melhoria na resposta; pois o maior valor do bioensaio melhorou de 24% no primeiro
planejamento incompleto para 60% de morte das larvas no segundo planejamento fatorial
incompleto. Esses valores referem-se à previsão da resposta segundo o modelo, os valores
reais para tal resposta foram de 22 e 68% de mortes no primeiro e segundo planejamento,
respectivamente.
52

Gráfico 5.5 Superfície de resposta para variáveis sulfato de amônia e glicose, tendo como
resposta o bioensaio (segundo planejamento fatorial incompleto). A farinha de crisálida está
fixada no valor codificado -1.
Gráfico 5.6 Superfície de resposta para variáveis sulfato de amônia e farinha de crisálida,
tendo como resposta o bioensaio (segundo planejamento fatorial incompleto). A glicose está
fixada no valor codificado -1.
53

Gráfico 5.7 Superfície de resposta para variáveis farinha de crisálida e glicose, tendo como
resposta o bioensaio (segundo planejamento fatorial incompleto). O sulfato de amônia está
fixado no valor codificado -1.
Tendo em vista os efeitos das variáveis sobre o bioensaio, a otimização prosseguiu
mapeando a região próxima ao ponto mínimo (-1) do segundo planejamento fatorial
incompleto. Nesse ponto, as variáveis apresentavam os seguintes valores (g/L): 0,5 de glicose,
1,0 de sulfato de amônia e 50 de farinha de crisálida.
Com o objetivo de verificar se o ponto mínimo (-1) do segundo experimento deveria
ser mesmo o ponto central em um planejamento fatorial completo, foram feitos três ensaios
(A, B e C) com valores próximos a esse ponto mínimo. Os valores das variáveis nesses três
ensaios bem como o resultado do bioensaio encontram-se na tabela 5.4.
Tabela 5.4 Ensaios para confirmação do ponto central (g/L): 0,5 de
glicose, 1,0 de sulfato de amônia e 50 de farinha de crisálida.
Valor das variáveis e bioensaio.
Ensaios Glicose
(g/L)
Sulfato de
amônia (g/L)
Farinha de
crisálida (g/L)
Bioensaio (% de
morte das larvas)
A 0,9 1,8 66 50
54

B 0,7 1,4 58 46
C 0,3 0,6 42 38
Tendo em vista que nenhum dos ensaios resultou em um bioensaio melhor do que do
ponto mínimo (-1) do segundo planejamento fatorial incompleto, esse ponto foi utilizado
como central no planejamento completo.
5.1.2 Planejamento fatorial completo
O planejamento fatorial completo apresenta melhor resolução, permitindo o estudo das
interações entre as variáveis, já que todos os experimentos possíveis são realizados. Nessa
etapa, esse planejamento “mapeou” a região próxima ao ponto central (g/L): 0,5 de glicose,
1,0 de sulfato de amônia e 50 de farinha de crisálida. Os níveis das variáveis no planejamento
fatorial completo encontram-se na tabela 4.7.
Inicialmente foram feitos os experimentos do bloco 1, porém analisando-se os
resultados verificou-se que o modelo linear não explicava adequadamente essa fermentação,
por isso, fez-se um segundo bloco incluindo os pontos estrelas. Esses pontos correspondem ao
planejamento inicial, porém com uma rotação de 45 ° em relação ao ponto de partida. Dessa
maneira, é possível estabelecer um modelo quadrático (equação de segundo grau) para o
processo em estudo.
Além dos pontos estrelas, o planejamento fatorial completo apresentou uma repetição
no ponto central, a qual permite a estimativa do erro puro, ou seja, o que é inerente ao
processo. Esse planejamento é também conhecido como Delineamento Composto Central
Rotacional (RODRIGUES & IEMMA, 2005).
Os resultados (bioensaio, UFC e porcentagem de esporulação) desse planejamento
fatorial completo, incluindo os pontos estrelas, encontram-se na tabela 5.4. Os resultados da
análise de variância e dos efeitos para a resposta bioensaio encontram-se nas tabelas 5.5 e 5.6
respectivamente.
55

Tabela 5.5 Respostas do Delineamento Composto Central Rotacional (planejamento fatorial
completo, com repetição no ponto central e pontos estrelas).
Ensaio
Bloco Glicose Sulfato de Amônia
Farinha de.
crisálida
Bioensaio (% morte)
log UFC % esporulação
1 1 -1,00000 -1,00000 -1,00000 33,33 12,602 92,392 1 -1,00000 -1,00000 1,00000 53,33 13,978 93,653 1 -1,00000 1,00000 -1,00000 6,67 14,544 78,124 1 -1,00000 1,00000 1,00000 20,00 14,431 81,955 1 1,00000 -1,00000 -1,00000 0,00 16,17 80,526 1 1,00000 -1,00000 1,00000 53,33 16,057 69,087 1 1,00000 1,00000 -1,00000 26,67 17,732 67,198 1 1,00000 1,00000 1,00000 33,33 15,903 74,78
9 (C) 1 0,00000 0,00000 0,00000 66,67 15,819 79,9810 (C) 1 0,00000 0,00000 0,00000 60,00 15,924 83,33
11 2 -1,78885 0,00000 0,00000 22,50 14,851 84,212 2 1,78885 0,00000 0,00000 5,00 14,505 85,3513 2 0,00000 -1,78885 0,00000 12,50 14,851 86,1814 2 0,00000 1,78885 0,00000 10,00 16,158 81,3415 2 0,00000 0,00000 -1,78885 7,50 15,556 83,50416 2 0,00000 0,00000 1,78885 10,00 15,982 79,34
Tabela 5.6 Análise de variância (p<0,05) para a resposta bioensaio do delineamento
composto central rotacional. Em vermelho, as variáveis com efeito significativo (ANOVA,
p<0,05). L representa o termo linear e Q o termo quadrático.
Soma dos quadrados
Graus de liberdade
Quadrado médio
F calculado
Probabilidade
Blocos 699,213 1 699,213 4,058296 0,100067Glicose (L) 68,056 1 68,056 0,395000 0,557271Glicose (Q) 1161,907 1 1161,907 6,743809 0,048432Sulfato de amônia(L) 231,940 1 231,940 1,346198 0,298329Sulfato de amônia(Q) 1334,659 1 1334,659 7,746478 0,038753Farinha de crisálida(L) 664,118 1 664,118 3,854601 0,106834Farinha de crisálida(Q) 1519,379 1 1519,379 8,818610 0,031171Interação glicose X sulfato (L)
555,444 1 555,444 3,223849 0,132522
Interação glicose X far. De crisálida (L)
88,844 1 88,844 0,515661 0,504843
Interação sulfato X farinha de crisálida (L)
355,644 1 355,644 2,064192 0,210292
Erro puro 861,462 5 172,292 Total 7540,666 15
56

O erro puro não foi significativo, bem como a variação referente aos blocos, portanto
os fatores que mais influenciaram a resposta foram a concentração das fontes de nutrientes.
Nessa etapa, todas as fontes apresentaram apenas o termo quadrático significativo, o que
indica que as mesmas já alcançaram seu ótimo de concentração para a resposta bioensaio.
Os gráficos 5.8 a 5.10 representam a superfície de resposta para o delineamento
composto central rotacional. Observa-se que a zona em vermelho escura, correspondente a
maior taxa de morte no bioensaio, encontra-se no centro do gráfico, ou seja, todo o seu
entorno foi mapeado. Isso significa que para alcançar os melhores resultados de bioensaio
pode-se trabalhar em intervalos de concentração das fontes que correspondem a tal área e, não
necessariamente com um único valor fixo de concentração; o que facilita o controle da
fermentação. Para o sulfato de amônia, a faixa de variação da região de ótimo está entre (g/L)
0,6 a 1,1; já para a glicose é de 0,3 a 0,55g/L, enquanto que para a farinha de crisálida tal
faixa encontra-se entre 55 a 58g/L.
Gráfico 5.8 Superfície de resposta para variáveis sulfato de amônia e glicose, tendo como
resposta o bioensaio (delineamento composto central rotacional). A farinha de crisálida está
fixada no valor ponto central.
57

Gráfico 5.9 Superfície de resposta para variáveis farinha de crisálida e glicose, tendo como
resposta o bioensaio (delineamento composto central rotacional). O sulfato de amônia
encontra-se fixado no valor ponto central.
Gráfico 5.10 Superfície de resposta para variáveis farinha de crisálida e sulfato de amônia,
tendo como resposta o bioensaio (delineamento composto central rotacional). A glicose
encontra-se fixada no valor ponto central.
58

Tabela 5.7 Efeito das fontes de nutrientes sobre o bioensaio (delineamento composto central
rotacional). Em vermelho, as variáveis com efeito significativo (ANOVA, p<0,05). L
representa o termo linear e Q o termo quadrático.
Efeito Probabilidade CoeficientePrincipal/interações 55,9605 0,002818 55,9605Blocos -14,7490 0,100067 -7,3745Glicose (L) -4,3479 0,557271 -2,1740Glicose (Q) -21,7725 0,048432 -10,8863Sulfato de amônia(L) -8,0267 0,298329 -4,0133Sulfato de amônia(Q) -23,3350 0,038753 -11,6675Farinha de crisálida(L) 13,5822 0,106834 6,7911Farinha de crisálida(Q) -24,8975 0,031171 -12,4488Interação glicose e sulf. de amônia (L)
16,6650 0,132522 8,3325
Interação glicose e far. de crisálida (L)L
6,6650 0,504843 3,3325
Interação sulf. de amônia e far. de crisálida (L)
-13,3350 0,210292 -6,6675
A tabela 5.6 apresenta o efeito das fontes de nutrientes sobre o bioensaio.
Considerando-se apenas os efeitos significativos e o coeficiente, pode-se construir a equação
quadrática a seguir que explique o efeito das fontes sobre o bioensaio:
Y= 55,96 – 10,89 A2 -11,67 B2 – 12,45 C2
Onde:
Y= valor do bioensaio em porcentagem de morte
A= concentração da glicose (valor codificado)
B= concentração do sulfato de amônia (valor codificado)
C= concentração da farinha de crisálida (valor codificado)
Com base na equação que representa o processo, foi possível prever os valores de
bioensaio para diferentes valores das fontes de nutrientes. O gráfico 5.10 relaciona os valores
previstos com os valores observados nos bioensaios; observa-se que houve uma correlação
considerável entre esses valores. O coeficiente de determinação desse delineamento (R) foi de
0,88; o que indica que 88% da variação é explicada pelo modelo.
59
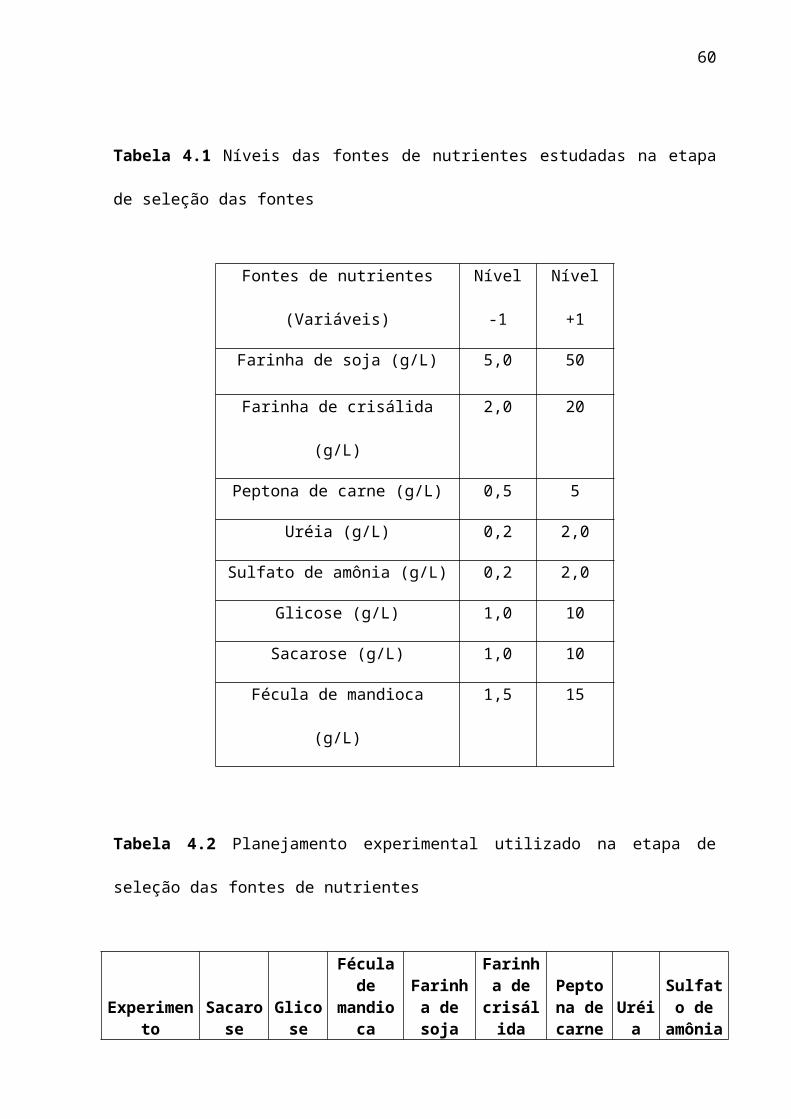
Gráfico 5.11 Valores observados e previstos para o bioensaio (% de morte).
A tabela 5.8 apresenta algumas combinações de valores das variáveis bem como o
bioensaio previsto. Observa-se que há uma ampla faixa onde a variação do bioensaio é
mínima. O valor máximo previsto pelo modelo foi de 56,82%, no entanto, o valor máximo de
morte das larvas observado nos ensaios desta etapa foi de 66,67%. Mesmo nos gráficos que
representam a superfície de resposta, a zona de maior morte no bioensaio corresponde a cerca
de 40%. Essas diferenças entre os valores máximos esperados e previstos justificam-se pelo
ajuste que o modelo matemático deve apresentar para que a maior parte dos dados seja
explicada por ele.
Tabela 5.8 Valores previstos para o bionsaio em diferentes concentrações das fontes de
nutrientes.
Previsão (bioensaio)
Glicose (codificado)
Glicose (real)
Sulfato (codificado)
Sulfato (real)
Farinha (codificado)
Farinha (real)
55,9605 0 0,5 g/L 0 1 g/L 0 50 g/L55,08828 -0,4 0,4 0 1 g/L 0 50 g/L50,73246 -0,8 0,3 0 1 g/L 0 50 g/L
56,2965 0 0,5 g/L -0,2 0,9 0 50 g/L54,16821 0 0,5 g/L -0,6 0,7 0 50 g/L
56,8208 0 0,5 g/L 0 1 g/L 0,2 5256,6851 0 0,5 g/L 0 1 g/L 0,4 54
55,55362 0 0,5 g/L 0 1 g/L 0,6 56
60
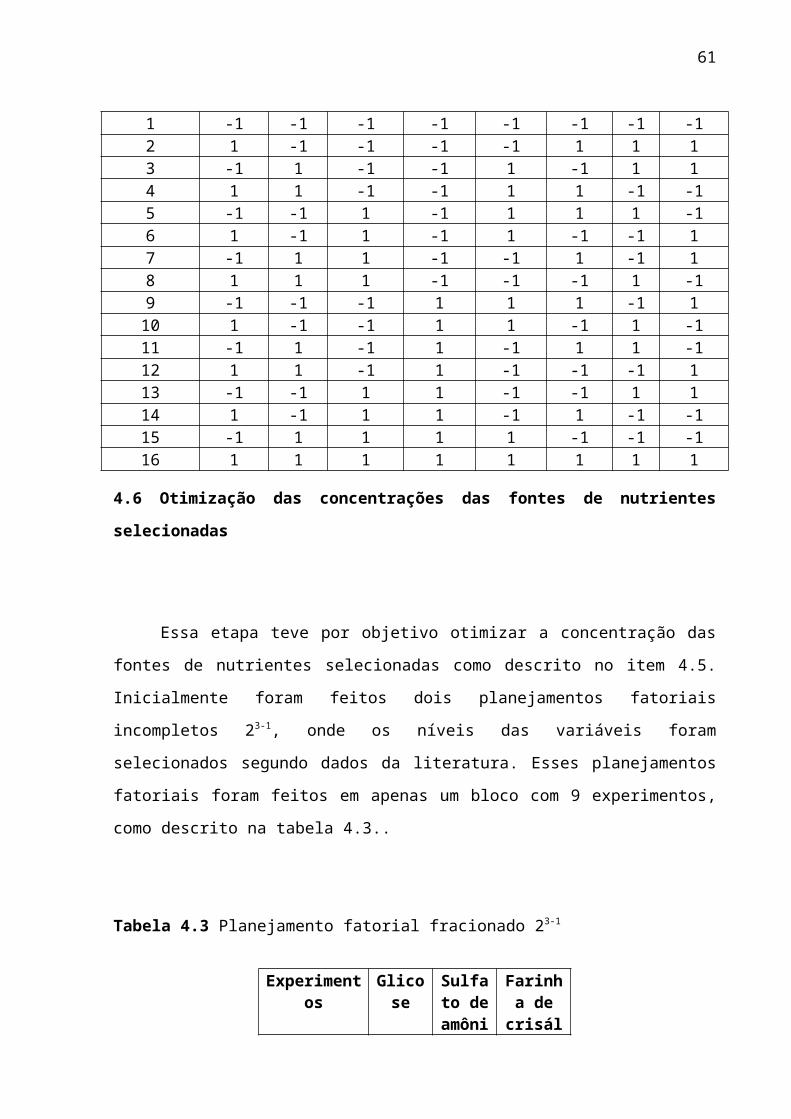
Em relação ao crescimento e esporulação, as concentrações testadas não apresentaram
efeitos significativos para tais parâmetros. No entanto, na região de ótimo para bioensaio o
log médio de UFC foi de 15,87 já a esporulação média foi de 81,65%, o que indica um bom
crescimento e esporulação.
Embora em um estudo de superfície de resposta o ótimo encontre-se em uma faixa e
não em um valor fixo das variáveis, para os experimentos seguintes foi adotado o valor das
variáveis no ponto máximo previsto de bioensaio. Esse ponto corresponde aos seguintes
valores das variáveis (g/L): 0,5 de glicose; 1,0 de sulfato de amônia e 52 de farinha de
crisálida. Além das fontes de carbono e de nitrogênio orgânico e inorgânico, o meio de cultura
otimizado continha a mistura de sais, citadas no item 4.5, a qual não foi objeto de estudo.
Muitos autores relatam o efeito repressor da glicose sobre a produção de toxinas por B.
thuringiensis; porém, esse fato não se observou nesse estudo, provavelmente porque a
concentração de glicose utilizada foi baixa (0,5g/L), quando comparada com a dos estudos
que observaram a repressão (em média 1,0g/L) (ÖSKAN et al.; 2003). Nas etapas iniciais da
otimização, quando a concentração de glicose era alta (entre 1,25 a 10g/L), os resultados do
bioensaio foram bem baixos, em torno de 12% de morte das larvas. Possivelmente, nessa
etapa a glicose estaria reprimindo a produção de toxina.
Assim como nos trabalhos de Içgen et al. (2002b), o sulfato de amônia mostrou-se
uma boa fonte de nitrogênio orgânico para a produção de toxinas por B. thuringiensis. A
concentração ótima dessa fonte (1,0g/L) foi metade do que geralmente se costuma utilizar na
indústria, o que leva a um barateamento da produção (COUCH, 2000).
Logo no início da descoberta de B. thuringiensis, em 1902, ele foi isolado a partir de
larvas infectadas de Bombix mori. Desde então, já se sabia que essa bactéria era capaz de se
desenvolver em tal meio (BRAR et al., 2006; CAPALBO et al., 2004; VIDAURRE, 1996).
Os bons resultados obtidos com o meio a base de farinha de crisálida confirmam esse fato.
Além disso, o fato desse meio ter alcançado resultados promissores reforça que o nicho de B.
thuringiensis é o próprio inseto, onde ocorrem suas interações genéticas e seu metabolismo
estaria mais adaptado (RAYMOND et al., 2008; VILAS-BÔAS et al., 2000).
A farinha de crisálida apresenta uma composição bastante rica, destacando
principalmente como fonte de proteínas e aminoácidos, portanto de nitrogênio orgânico. O
anexo 2 apresenta uma tabela com a constituição dessa farinha, essas análises foram feitas
pelo laboratório Labtec (Campinas) e, gentilmente cedida pelo professor Amarildo Passini
(Departamento de Agronomia, UEL). Embora apresente os minerais potássio, magnésio,
zinco, ferro e cobalto; a concentração desses é baixa. Portanto, ainda há necessidade de
61

suplementação mineral. Alves et al. (1997) estudou vários ingredientes alternativos para o
crescimento de B. thuringiensis kurstaki, a farinha de crisálida foi o mais promissor. Nesse
estudo, a concentração utilizada foi de 10g/L dessa farinha além de suplementação com sais e
fonte de carbono.
A CL50 média do meio otimizado foi de 0,703 ppm (v/v). Esse resultado foi
comparado com o bioensaio de cultura feita em meio Arcas adaptado (ANDRADE, 2005),
cuja CL50 média foi de 3,01 ppm (v/v). Esses valores diferiram estatisticamente, sendo que o
meio à base de crisálida apresenta uma toxicidade cerca de 4,3 vezes maior do que o meio
Arcas adaptado. (Não entendi)
A tabela 5.9 apresenta os custos referentes à matéria-prima para produção de 1 L de
meio a base de farinha de crisálida otimizado e do meio Arcas adaptado.
Tabela 5.9 Custos dos meios de cultivo: Arcas adaptado e Farinha de crisálidas otimizado.
Custos/Meios Meio Arcas
adaptado
Meio farinha de
crisálida otimizado
Mistura de sais
Extrato de levedura
Glicose
Farinha de crisálida
Sulfato de amônia
Custo total (1L)
Fonte: ??????????????? Cotação feita em ?????????????
Além do meio otimizado apresentar um menor custo, sua toxicidade é maior, o que
permite que ele seja diluído cerca de 4x mais do que o meio Arcas adaptado. Dessa maneira,
com 1L do meio crisálida otimizado é possível obter 4L de meio com igual toxicidade do
meio Arcas adaptado; o que resulta em um custo real do meio otimizado cerca de ???%
menor.
62

5.3 Cinética de crescimento e produção de toxinas
O estudo da cinética de crescimento de B. thuringiensis israelensis foi feito com o
objetivo de verificar em qual etapa a toxicidade da cultura era máxima e, com isso determinar
o tempo de fermentação. Segundo Couch (2000), as fermentações industriais de bactérias
entomopatogênicas duram de 62 a 92 horas. Nesse estudo, a cultura foi monitorada por um
período de 96 horas
O crescimento microbiano foi mensurado por meio de gravimetria e turbidimetria.
Esses dois métodos apresentaram uma correlação de 0,93; como apresentado no gráfico 5.12.
No entanto, a metodologia de gravimetria apresentou um desvio padrão menor (2,04) em
relação à turbidimetria (6,5) e, por isso, foi utilizada no estudo da cinética.
Gráfico 5.12 Correlação entre a biomassa (mg/mL) mensuradas pelas metodologias de
gravimetria e turbidimetria.
O gráfico 5.13 apresenta o crescimento microbiano e a toxicidade, mesurada por meio
de bioensaio, durante as 96 horas de cultivo.
63
R2 = 0,9333
0
2
4
6
8
10
12
14
16
18
0 5 10 15 20
Biomassa (gravimetria)
Bio
mas
sa (
turb
idim
etri
a)

Gráfico 5.13 Crescimento microbiano (biomassa) e toxicidade (bioensaio) ao longo do
cultivo (96 horas).
A fase de adaptação às condições de cultivo (lag) durou cerca de 6 horas; ou seja
apenas 6,25% do tempo de cultivo. A fase exponencial, a qual apresenta a maior taxa de
crescimento, correspondeu a cerca de 72,92% do tempo de cultivo; ao final da qual a
biomassa correspondia a 15,13mg/mL. Não é possível identificar a fase de declínio de uma
cultura microbiana por meio da metodologia de gravimetria, visto que essa mensura a
biomassa total, incluindo células vivas e mortas (REFERÊNCIA, ANO). Por isso, o tempo
restante da cultura foi considerado como fases estacionária e de declínio, a qual durou cerca
de 20,83% do tempo de cultivo.
A esporulação é uma forma de a bactéria sobreviver a situações adversas, visto que os
esporos são mais resistentes. Em geral, a esporulação ocorre na fase estacionária, quando a
bactéria já consumiu grande parte das fontes de nutrientes e esse fator passa a ser uma
limitante para o seu crescimento. Além disso, durante a fermentação, o microrganismo pode
produzir metabólitos que, com o acúmulo, acaba por inibir seu próprio crescimento
(REFERÊNCIA, ANO).
A maior parte das toxinas de B. thuringiensis é sintetizada durante a esporulação,
portanto, diferente de muitos cultivos, a fase estacionária é bastante importante na
fermentação dessa bactéria. (LERECLUS et al., 1993; GLARE & O’CALLAGHAN, 2000).
Observando-se o gráfico 5.13, percebe-se que a toxicidade da cultura foi aumentando
64
0
2
4
6
8
10
12
14
16
0 3 6 9 28 34 52 55 58 79 82 96
Tempo (horas)
Bio
mas
sa
0
20
40
60
80
100
120
Bio
ensa
io
Média biomassa gravimetria (mg/mL) Bioensaio (% de morte)

gradativamente, alcançando o melhor resultado por volta de 96 horas de cultivo, ou seja, já no
período estacionário.
A fim de se verificar com maior precisão qual o momento de maior toxicidade, foram
feitos bioensaios com cálculo de CL50 para a cultura com 72 horas e com 96 horas. O tempo
de 72 horas foi escolhido por ter sido o adotado nas etapas anteriores. A cultura com 96 horas
de cultivo apresentou uma CL50 média de 0,50; a qual foi significativamente menor do que a
cultura com 72 horas, a qual apresentou CL50 média de 0,70.
A figura 5.1 apresenta fotomicroscopia da cultura de B. thuringiensis israelensis em
diferentes momentos de cultivo.
Figura 5.1. Cultura de B. thuringiensis israelensis em diferentes momentos de cultivo. Aumento de 1000 vezes. Coloração: Amido-Black e Fucsina. Seta verde: células vegetativas. Seta azul clara: células em final de esporulação. Seta azul escura: cristais. Seta vermelha: esporos.
Analisando-se as fotomicroscopia, percebe-se que inicialmente predominavam células
vegetativas (6 horas). Já com 32 horas de cultivo, a maior parte das lâminas apresentava
células já em esporulação inicial, com esporos e cristais no seu interior. Às 56 horas foi
possível observar uma grande quantidade de esporos e cristais de toxina já no exterior da
65

célula, porém ainda era evidente uma grande quantidade de células em esporulação. A cultura
com 72 horas de cultivo apresentava praticamente apenas esporos e cristais fora das células,
com poucas células em final de esporulação. Às 96 horas de cultivo, foi possível observar
uma grande quantidade de cristais e esporos fora da célula, porém havia células vegetativas.
Provavelmente, às 96 horas de cultivo alguns esporos produzidos durante o cultivo
germinaram, resultando novamente em algumas células vegetativas.
Pela análise da toxicidade observa-se que o período de 96 horas seria o indicado para o
cultivo, porém, analisando-se as lâminas percebe-se que nesse momento alguns esporos já
estariam germinando. A maior toxicidade do período de 96 horas provavelmente ocorreu
porque a grande maioria das células da fase de crescimento já havia liberado seus cristais, ao
passo que com 72 horas algumas células, ainda que poucas, apresentavam-se no final de
esporulação. Uma alternativa para essa questão seria manter fermentar a cultura por 72 horas
e, posteriormente mantê-la em repouso por 24 horas, para que as células em final de
esporulação liberem suas toxinas. Dessa maneira, o tempo de cultivo não se estenderia muito,
não resultando em custo extra para a produção. MORAES et al.(2001) relatam essa estratégia
para alguns cultivos de B. thuringiensis.
Gráfico 5.14 Potencial hidrogeniônico (pH) ao longo do (96 horas).
66
6
6,5
7
7,5
8
8,5
0 20 40 60 80 100
Tempo (horas)
pH

O gráfico 5.14 apresenta os valores do potencial hidrogeniônico (pH) ao longo do cultivo
de 96 horas. Em geral, a cultura de B. thuringiensis, quando não há controle de pH, esse inicia
em torno de 7.0, diminui para cerca de 6,0, no início da esporulação. Ao final da esporulação
e com a lise celular o pH sobe para valores neutros e até básicos como 8.9 (ABDEL-
HAMEED, 2001; LUNA et al., 2004).
Nesse estudo, não se observou a queda inicial que, geralmente ocorre durante o início do
cultivo de B. thuringiensis. Provavelmente, isso ocorreu porque a concentração de glicose no
meio era baixa (0,5g/L), o que resultou em menor produção de ácidos orgânicos responsáveis
pela queda inicial do pH (CHANG, et al., 2008). Além disso, os sais dihidrogenofosfato de
potássio (KH2PO4) e fosfato de potássio dibásico anidro (K2HPO4) presentes no meio
provavelmente tamponaram essa leve diminuição inicial. Posteriormente, o pH aumentou para
cerca de 8,0 e manteve-se.
Os estudos de Öskan et al. (2003) sugerem o pH 7.0 com variação de ±0.3 como melhor
para o crescimento e a produção de toxinas por B. thuringiensis. Isso porque, algumas
proteínas Cry, principalmente Cry4Ba, mostram-se mais sensíveis a variações de pH, com
comprometimento pronunciado nos casos de alcalinização. Já os estudos de Içgen et al.
(2002a) indicam uma melhor produção com pH entre 5,5 e 6,5. Outros estudos indicam que
não há uma alteração significativa na produção de toxinas quando o pH do meio varia entre
6,5 a 8,0 (HOLMBERG, R, et al. apud. IÇGEN et al. 2002a).
No presente estudo, a produção de toxinas foi satisfatória, embora tenha ocorrido
variação de pH entre 6,4 e 8,2. Para uma precisão maior quanto a influência do pH na
produção de toxinas, esse deveria ter sido foco de estudo.
67

5.4 Toxicidade da cultura de Bacillus thuringiensis subsp. israelensis durante armazenamento
O gráfico 5.15 apresenta a toxicidade da cultura de B. thuringiensis israelensis,
avaliada por meio de bioensaio com cálculo CL50, por um período de 28 dias de
armazenamento.
Gráfico 5.15 Toxicidade da cultura durante o armazenamento.
Inicialmente o comportamento das duas culturas armazenadas foi igual, com
diminuição da toxicidade para cerca de 1,0 ppm (v/v). No entanto, com 21 e 28 dias de
armazenamento, a cultura não refrigerada apresentou quedas na CL50, chegando a 0,43 ppm
(v/v). Já a cultura refrigerada continuou a apresentar um aumento da CL50 até 21 dias de
armazenamento e, posteriormente, manteve-se em cerca de 1,20 ppm (v/v).
Geralmente, os estudos sobre armazenamento de B. thuringiensis apresentam dados
apenas de produtos formulados (ARAÚJO et al., 2008), no entanto, dados sobre o
armazenamento da cultura são importantes, pois servem para se comparar com o do produto
formulado e, assim, estabelecer a eficácia da formulação.
68
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0 5 10 15 20 25 30
Tempo (dias)
CL
50
(pp
m,
v/v)
Cultura com refrigeração Cultura sem refrigeração

Glare & O’Callaghan, 2000 relatam que os principais fatores que influenciam a
toxicidade de B. thuringiensis no meio ambiente são a radiação solar e temperatura. Couch
(2000) afirma que logo após a fermentação, é interessante manter a cultura a 4ºC por algumas
horas, para que ocorra a completa lise celular.
Nesse estudo, a melhor condição de armazenamento foi sem refrigeração.
Provavelmente, a baixa temperatura (5ºC +/-2) por um período prolongado de tempo foi
prejudicial para as toxinas, resultando em uma menor toxicidade. Já à temperatura ambiente,
pode ter possibilitado a lise de praticamente todas as células, e posterior germinação de
esporos tardios, resultando em uma melhor toxicidade. Esse resultado reforça a eficácia de se
deixar a cultura em repouso após sua fermentação, antes de formulá-la (MORAES et al.,
2001).
69

6. CONSIDERAÇÕES FINAIS (Conclusões?)
Entre as fontes de nitrogênio estudas (farinha de crisálida, peptona bacteriológica,
uréia agroindustrial e sulfato de amônia), a farinha de crisálida e o sulfato de amônia foram os
mais adequados para produção de toxinas por B. thuringiensis israelensis HD537.
Entre as fontes de carbono estudadas (sacarose, glicose e fécula de mandioca), a
glicose apresentou-se a mais adequada para produção de toxinas.
A metodologia de superfície de resposta mostrou-se eficaz na otimização do meio de
cultivo, resultando em um meio com as seguintes faixas de concentrações das fontes de
carbono e nitrogênio: glicose 0,3 a 0,55g/L; farinha de crisálida 55 a 58g/L e sulfato de
amônia 0,6 a 1,1g/L..
A otimização do meio alternativo à base de farinha de crisálida resultou em um meio
com CL50 de 0,703 ppm (v/v). Esse meio apresentou um custo ??? vezes menor do que o meio
de cultivo Arcas adaptado.
As maiores toxicidades da cultura de B. thuringiensis israelensis HD537 ocorreram
nas etapas finais de cultivo, sendo máxima em 96 horas.
O armazenamento da cultura de B. thuringiensis israelensis HD537 em condições não
refrigerada foi a mais adequada para manutenção de sua toxicidade por um período de 28
dias.
Indicam-se estudos posteriores com utilização da superfície de resposta para otimizar
as condições físicas de cultivo (pH, temperatura e aeração), visto que essas não foram objeto
de estudo.
Para uma melhoria maior da produção de B. thuringiensis israelensis HD537 indica-se
o estudo do aumento de escala e a de sua formulação.
70

7.REFERÊNCIAS BIBLIOGRÁFICAS
ABDEL-HAMEED, A. A stirred tank culture of Bacillus thuringiensis H-14 for production of the mosquitocidal d-endotoxin: matematical modelling and scalling-up strudies. World Journal of microbiology e Biotechnology, v.17, p.857-861; 2001.
ADAMS, T. T, et al. Solid state fermentation of broiler litter for production of biocontrol agents. Bioresearch Technology, v. 82, p. 33-41; 2002.
AGAISSE, H.; LERECLUS, D. How does Bacillus thuringiensis produce so much inseticidal crystal protein? Journal of Bacteriology, v. 177, p.6027-6032, 1995.
ALVES, L. F. A., et al., Production of Bacillus thuringiensis Berliner var. kurstaki grow in alternative media. Biocontrol Science and Technology, v. 7, p. 377-383, 1997.
ANDRADE, E. Efeito dos nutrientes e inoculo na biossíntese dos cristais protéicos de Bacillus thuringiensis var. israelensis. Trabalho de Conclusão de Curso. Universidade Estadual de Londrina, 2005, 39p.
ARANTES, O. M. N. et al Bacillus thuringiensis: estratégias no controle biológico. In: SERAFINE, L., A.et al (org) Biotecnologia: avanços na agricultura e na agroindústria. Caxias do Sul: Agropecuária, 2002. p.269-293.
ARCAS, J.; YANTORO, O.; ERTOLA, R. A new medium for growth delta-endotoxin production by Bacillus thuringiensis var. kurstaki. Biotechnology Letters, v.6, n. 8, p. 495-500, 1984.
ARCAS, J.; YANTORO, O.; ERTOLA, R. Effect of high concentration of nutrients on Bacillus thuringiensis cultures. Biotechnology Letters, v.9, n.2, p.105-110, 1987.
BARROS NETO, B. et al. Planejamento e otimização de experimentos. Campinas: Ed. da Unicamp, 1995, 300p.
BECKER, N.; MARGALIT, J. Use of Bacillus thuringiensis israelensis again mosquitoes and black flies. In: ENTWISTLE, P.F. et al. (Ed) Bacillus thuringiensis , an environmental biopesticide: theory and practice. Wiley, 1993. p.147-170.
BIZARRI, M. F. et al. Changes in the properties of Bacillus thuringiensis after prolonged culture in a rich medium. Journal of Applied Microbiology, v. 104, p.60-9, 2008.
BRAR, S. K. et al. Recent advances in downstream processing and formulations of Bacillus thuringiensis based biopesticides. Process Biochemistry, v.46, p. 323-342, 2006.
BRASIL, Agência Nacional da Vigilância Sanitária (Anvisa). Manual de Microbiologia Clínica para o Controle de Infecção em Serviços de Saúde. Brasília: Anvisa, 2004.
BRAVO, A.; SOBERÓN, M. How to cope with insect resistance to Bt toxins? Trends in Biotechnology v.26, n.10, p. 573-579, 2008.
71

BRAVO, A. et al. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon, v.49, p. 423-435, 2007.
CAPALBO, D. M. F.; MORAES, I.O. Aspectos da produção de Bacillus thuringiensis. In: Anais do congresso brasileiro de entomologia, Campinas. Fundação Cargill, 1988. p.70-80.
CAPALBO, D. M. F. et al. Bacillus thuringiensis: formulações e plantas transgênicas. In: BORÉM, A. (ed.). Biotecnologia e meio ambiente. Viçosa: Folha de Viçosa, 2004. p.309-350.
CHANG, M. et al., Starch processing wastewater as a new medium for production of Bacillus thuringiensis. World J Microbiol Biotechnol. v.24, p.441-7, 2008.
CHEN, S. et al. Fed-batch culture of Bacillus thuringiensis based on motile intensity. Journal of industrial Microbiology and Biotechnology, v. 30, p.677-681, 2003.
COUCH Terry L. Industrial fermentation and formulation of entomopathogenic bacteria. In: CHARLES, J. F. (org) Entomopathogenic bacteria: from laboratory to field applications. Kluwer: Academic Publishes, 2000. p. 297-316.
CRICKMORE, N. et al. "Bacillus thuringiensis toxin nomenclature" Disponível: <http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/>. Acesso em: 24 dez. 2008.
CRICKMORE, N. et al. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiology and molecular biology review, v. 62, n. 3, p. 807-813, 1998.
DEVI, P. S. V. et al. Barley-based medium for the cost-effective production of Bacillus thuringiensis. World Journal of Microbiology & Biotechnology, v. 21, p. 173-179, 2005.
FINNEY, D. J Probit Analysis, London: Cambridge Univ. Press, 1971, 3rd ed.
GHRIBI, D. et al. Improvement of bioinsecticides production through adaptation of Bacillus thuringiensis cells to heat treatment and NaCl addition. Journal of Applied Microbiology, v.98, p. 823-831, 2005.
GLARE, T. R.; O’CALLAGHAN. Bacillus thuringiensis Biology, Ecology and Safety. Chichester: John Wiley & Sons, 2000, 350p.
GOLDEBERG, L. J.; MARGALIT J. A bacterial spore demonstrating rapid larvicidal activity against Anopheles sergenii, Uranotaenia unguiculata, Culex uinivatus, Aedes aegypti and Culex pipiens. Mosquito News, v. 37, p. 355-358, 1977.
HABIB, M.E.M.; ANDRADE, C.F.S. Bactérias entomopatogênicas. In: ALVES, S. B. (coord) Controle microbiano de insetos. Piracicaba: Manole, 1986.p. 130-140.
HABIB, M.E.M.; ANDRADE, C.F.S. Bactérias entomopatogênicas. In: ALVES, S. B. (coord) Controle microbiano de insetos. Piracicaba: Manole, 1998. p. 383-446.
72

IÇGEN, Y.; IÇGEN, B.; ÖZCENGIZ, G. Regulation of crystal protein biosynthesis by Bacillus thuringiensis: I Effects of mineral elements and pH. Research in Microbiology, v. 153, p 599-604, 2002a.
IÇGEN, Y.; IÇGEN, B.; ÖZCENGIZ, G. Regulation of crystal protein biosynthesis by Bacillus thuringiensis: II Effects of carbon and nitrogen sources. Research in Microbiology, v. 153, p. 605-609, 2002b.
JARIC´-PERKUSIC et al. Influence of Molting on Efficacy of Two Functionally DifferentLarvicides: Bti and Temephos. Journal of Economic Entomology, v. 101, n. 10, p. 1204-1210, 2008.
KENNEDY, M.; KROUSE, D. Strategies for improving fermentation medium performance: a review. Journal of Industrial Microbiology & Biotechnology, v. 23, p. 456-475, 1999.
LACHHAB, K. et al. Production of Bacillus thuringiensis biopasticides using wastewaters sludge as raw material: effect of inoculum and sludge solids concentration. Process Biochemistry. v.37. p. 197-208, 2002.
LERECLUS, D. et al. Diversity of Bacillus thuringiensis toxins and genes. In ENTWISTLE, P.F. et al. (ed) Bacillus thuringiensis , an environmental biopesticide: theory and practice . Wiley, 1993. p.37-70.
LIU, B-L; TZENG, Y-M. Optimization of growth medium for the production of spores from Bacillus thuringiensis using response surface methodology. Bioprocess Engineering. v.18, p. 413-418, 1998.
LUNA, C. L. et al., Bacillus thuringiensis var. israelensis production involving re-use of theSupernatant. Biotechnology Letters v. 26, p. 143–145, 2004.
MONTIEL, M. L. T. et al. Wastewaters treatment sludge a raw material for the production of Bacillus thuringiensis based biopesticides. Water Ress., v. 35, n. 16, p. 3807-3816, 2001.
MORAES, I. O. et al. Produção de bactérias entomopatogênicas. In: ALVES, S. B. (ed.) Controle microbiano de insetos. Piracicaba: Manole, 1998. p. 815-844.
MORAES, I.O. et al.. Produção de bioinseticidas. In: LIMA. U.A. (coord) Biotecnologia industrial: processos fermentativos e enzimáticos. v 3. Porto Alegre: Edgar Blücher, 2001. p.245-265.
MORRIS, O. N. et al. Suitability of 30 agricultural products and by-products as nutrient sources for laboratory production of Bacillus thuringiensis subsp. Aizawai (HD133). Journal of invertebrate pathology, v. 70, p. 113-120, 1997.
NAIR, M. S.; DEAN, D. H., All Domains of Cry1A Toxins Insert into Insect Brush Border Membranes. The Journal of Biological Chemistry. v. 283, n. 39, p. 26324–26331, 2008.
OHBA, M. et al. "Parasporin nomenclature". Disponível em: <http://parasporin.fitc.pref.fukuoka.jp/>. Acesso em: 30 dez. 2008.
73

OTIENO-AYAYO et al., Variations in the mosquito larvicidal activities of toxins from Bacillus thuringiensis ssp. Israelensis. Environmental Microbiology v.10, n. 9, p. 2191-2199, 2008.
ÖSKAN, M. et al. Nutritional and cultural parameters influencing antidpteran delta-endotoxin production. Research in Microbiology, v.154, p.49-53, 2003
POOPATHI, S. et al. Development of low cost media for the culture of mosquito larvicides, Bacillus sphaericus and Bacillus thuringiensis sorovar. israelensis World journal of Microbiology & Biotechnology, v.18, p.209-216, 2002.
PRABAGARAN, S. R. et al. Media optimization of Bacillus thuringiensis PBT-372 using response surface methodology. Chemistry, Biochemistry and Eng. Q., v. 18, n. 2, p. 183-187, 2004.
PRABAKARAN, G.; BALARAMAN, K. Development of a cost-effective medium for the large scale production of Bacillus thuringiensis var. israelensis. Biological control, v. 36, p. 288-292, 2006.
PRABAKARAN, G. et al. Coconut water as a cheap source for the production of δ-endotoxin of Bacillus thuringiensis var. israelensis, a mosquito control agent. Acta Tropica v.105, p. 35–38, 2008.
RAYMOND, B. et al. Quantifying the reproduction of Bacillus thuringiensis HD1 in cadavers and live larvae of Plutella xylostella. Journal of Invertebrate Pathology v.98 p. 307-313, 2008.
RODRIGUES, M. I.; IEMMA, A. F., Planejamento de experimentos e otimização de processos: uma estratégia seqüencial de planejamentos. Campinas: Casa do pão, 2005, 326p.
RUSSEL, T. L.; KAY, B. H. Biologically based insecticides for the control of immature Australian mosquitoes: a review. Australian Journal of Entomology. v. 47, p. 232–242, 2008.
SALAMA, H. S.; MARGALIT, J. The use of Bacillus thuringiensis israelensis in development countries. In ENTWISTLE, P.F. et al. (ed) Bacillus thuringiensis , an environmental biopesticide: theory and practice. Wiley, 1993. p.237-254.
SCHNEPF, E. et al. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and Molecular Biology Review, v. 62, p. 775-806, 1998.
TOKCAER, Z. et al. Response surface optimization of antidipteran delta-endotoxin production by Bacillus thuringiensis subsp. israelensis HD 500. Process Biochemistry, v. 41, p. 350-355, 2006.
VALLEJO, F. et al. Production of Bacillus thuringiensis subsp. medellin by batch and fed-batch culture. Biotechnology techniques, v. 13, p.279-281, 1999.
VALLETE-GELLY, I. et al. Bacterial strategies to overcome insect defenses. Nature reviews: Microbiology. v. 6, p. 302-313, 2008.
74

VIDAURRE, T. J. C. Estudos de otimização da produção de bioinseticida bacteriano a partir do isolamento de nova linhagem de “Bacillus thuringiensis”. Tese de doutorado, Universidade Estadual de Campinas, 1996, 136p.
VIDYARTHI, A. S. et al. Studies on the production of B. thuringiensis based biopesticides using wastewaters sludge as a raw material. Water Research, v. 36, p. 4850-4860, 2002.
VILAS-BÔAS, L A. et al.. Survival and conjugation of Bacillus thuringiensis in soil microcosm. FEMS Microbiology Ecology, v.31, n.3, p.255-259, 2000.
VILAS-BÔAS, G. T.; et al. Biology and taxonomy of Bacillus cereus, Bacillus anthracis and Bacillus thuringiensis. Can. J. Microbiol. v. 53 p. 673-687, 2007.
VORA, D.; SHETNA, Y. I. Enhanced growth, sporulation and toxin production by Bacillus thuringiensis subsp kurstaki in oil seed meal extract media contain cystine. World Journal of Microbioly & Biotechnology, v. 15, p. 747-749. 1999
WORLD HEALTH ORGANIZATION (WHO). Guidelines for laboratory and field testing of mosquito larvicides. WHO/CDS/WHOPES/GCDPP/2005.13, 2005, 39p.
WU, W. et al. Effect of shear stress on cultivation of Bacillus thuringiensis for thuringiensin production. Applied microbiology biotechnology. v.58, p.175-177. 2002.
YEZZA, A. et al. Wastewaters sludge pre-treatment for enhancing entomotoxicity produced by Bacillus thuringiensis var. kurstaki. World Journal of Microbiology & Biotechnology, v. 21, p. 1165-1174, 2005.
YEZZA, A. et al. Bioconversion of industrial wastewaters and wastewaters sludge into Bacillus thuringiensis based biopesticides in pilot fermentor. Bioresource Technology, v. 97, p. 1850-1857, 2006.
ZOUARI, N.; JAOUA, S. The effect of complex carbon, and nitrogen, salt, Tween-80 and acetate on delta- endotoxin production by Bacillus thuringiensis subsp kurstaki. Journal of industrial microbiology e biotechnology. v.23, p.497-502. 1999.
ZOUARI, N.; JAOUA, S. The effect of complex carbon, and nitrogen, salt, Tween-80 and
acetate on delta- endotoxin production by Bacillus thuringiensis subsp kurstaki. Journal of
industrial microbiology e biotechnology. v.23, p.497-502. 1999.
75

ANEXO 1:Processo de produção de farinha de crisálida pela indústria de fiação de seda
Bratac s.a.
A matéria prima (casulos contendo as crisálidas do bicho da seda), após passarem pelo
processo industrial de fiação automático e fiação semi-automático, são transportados através
de canaletas para o setor de Sub-produtos, onde segue por meio de esteiras para a máquina
estopadeira. Essa máquina faz a separação das crisálidas dos resíduos de seda.
A máquina estopadeira é dividida em esteira de entrada, tanque de banho com soda
cáustica e vapor e dois tanques de enxágüe com água limpa.
Na esteira de entrada, um funcionário é responsável pela separação dos casulos que
ainda podem ser reutilizados no setor de fiação automático e semi-automático, o restante do
processo é todo automatizado.
As crisálidas seguem para os tanques de banho com solução de soda cáustica e vapor,
por meio de esteiras. Esse processo tem por objetivo retirar os resíduos de seda.
Após a passagem pela solução de soda cáustica, as crisálidas seguem em esteira direto
para o depósito da centrífuga, e em seguida, para a centrífuga. Após centrifugar, seguem em
esteira para o secador de crisálidas.
No secador, as crisálidas permanecem por 1 hora, sendo a temperatura de 130ºC no
primeiro compartimento e 118ºC no segundo compartimento. Após a secagem, as crisálidas
são encaminhadas para o resfriador, onde serão resfriadas pelo processo de exaustão.
Após a secagem e resfriamento, as crisálidas seguem para a mesa de classificação.
Nesse estágio de produção, é feito o exame da umidade das crisálidas. São realizadas duas
amostras de locais diferentes na peneira rotativa e duas amostras dos sacos embalados,
totalizando quatro amostras por turno, para monitoração da umidade para evitar possíveis
falhas,
As crisálidas secas seguem para o silo de armazenamento ou moinho.
No silo de armazenamento, as crisálidas secas permanecem até o momento da
embalagem como Crisálidas Integrais, ou seguem para o moinho, onde são trituradas e
embaladas como Farinha de crisálida, de acordo com o pedido do cliente.
Fonte: Manual de Boas Práticas de Fabricação da empresa Fiação de seda BRATAC s.a.
76

ANEXO 2: Análise da constituição química de farinha de crisálida.
RELATÓRIO DE ANÁLISESLABORATÓRIO CENTRAL – LABTEC
FONE: (19) 227-9994 ou 729-4441Rua das Magnólias, 2405
Jd Bandeirantes -CEP: 13050-070Campinas -SP Fax: (19)729-4443
e-mail: [email protected] da análise: 15/04/2003
Análise ResultadoProteína Bruta (%) 52.6600Extrato Etereo (%) 27.1700Cálcio (%) 0.1100Fósforo total (%) 0.6900Alanina (%) 2.8214Arginina (%) 2.7279Ácido Aspartico (%) 5.8778Glicina (%) 2.3481Isoleucina (%) 2.1805Leucina (%) 3.7528Ácido Glutâmico (%) 6.7585Lisina (%) 3.4737Cistina (%) 0.6224Metionina (%) 1.6891Fenilalanina (%) 2.6662Tirosina (%) 2.9083Treonina (%) 2.6158Triptofano (%) 0.6702Prolina (%) 2.6552Valina (%) 2.8380Histidina (%) 1.5121Serina (%) 2.6524Potássio (Mg/Kg) 7643.0900Magnésio (Mg/Kg) 3266.7500Manganês (Mg/Kg) não detectadoZinco (Mg/Kg) 162.290Ferro (Mg/Kg) 89.1100Cobre (Mg/Kg) 14.6500Cobalto (Mg/Kg) 8.5000
77

78