nitrificação e desnitrificação em lagoas costeiras...
TRANSCRIPT

Universidade Federal do Rio de Janeiro
Instituto de Biologia
Programa de Pós-Graduação em Ecologia
Nitrificação e Desnitrificação em Lagoas
Costeiras Tropicais
Ana Lúcia Santoro
Abril de 2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2
Universidade Federal do Rio de Janeiro
Instituto de Biologia
Programa de Pós-Graduação em Ecologia
Banca: Prof. Dr. Alex Enrich Prast
Prof. Dr. Vera Lúcia M. Huszar
Prof. Dr. William Zamboni de Melo
Nitrificação e Desnitrificação em Lagoas
Costeiras Tropicais
Ana Lúcia Santoro
Orientador: Prof. Dr. Alex Enrich Prast
Dissertação apresentada ao Programa de Pós-
Graduação em Ecologia da Universidade Federal
do Rio de Janeiro como parte dos requisitos para a
obtenção do título de Mestre.
Abril de 2010

3
Agradecimentos
A Deus que, desde o início da minha jornada acadêmica, me capacitou e guiou os meus passos
em todos os momentos. Toda a honra seja dada a Ele.
À minha mãe, que tudo fez por mim. Mãe, foi por você que cheguei até aqui e é por você que
ainda irei muito além.
Ao meu maior presente, meu marido, que incondicionalmente esteve sempre comigo, ao meu
lado, de perto ou de longe, sempre torcendo e me apoiando com muito amor. Charlles, amo
muito você.
Ao meu primeiro e único orientador, desde o início muito mais que um orientador acadêmico,
um conselheiro de vida. Alex, muito obrigada pela confiança e carinho.
Ao Laboratório de Limnologia, onde tive a honra de aprender minhas primeiras lições
científicas. Amigos da limno, obrigada por tudo. Em especial, agradeço ao Rodrigo Coutinho e
ao Adriano Caliman pela coleta dos dados referentes ao capítulo III desta dissertação.
Ao Laboratório de Biogeoquímica, o qual tive o privilégio de ver nascer e a alegria de ajudá-lo a
crescer. Muito obrigada por todas as oportunidades.
Aos amigos ”acadêmicos”, de turma ou de lab, que muito me ajudaram direta ou indiretamente
nesta conquista. Queridos, a mensagem é genérica, mas o carinho é pessoal.
Aos professores da banca, Dr. Alex Enrich Prast, Dr. Vera Lúcia M. Huszar e Dr. William
Zamboni de Melo, por todas as contribuições que certamente enriqueceram esta dissertação.

4
ÍNDICE
RESUMO ................................................................................................................................ 5
ABSTRACT ............................................................................................................................ 6
INTRODUÇÃO GERAL ........................................................................................................ 7
CAPÍTULO I: Regulação Microbiológica da Disponibilidade de Nitrogênio em Ecossistemas
Aquáticos Continentais ............................................................................................................ 8
REFERÊNCIAS ......................................................................................................... 25
CAPÍTULO II: Controle da Nitrificação pela Salinidade em Lagoas Costeiras Salinas.......... 31
REFERÊNCIAS .......................................................................................................... 38
CAPÍTULO III: Taxas e Fatores Reguladores do Processo de Desnitrificação no Sedimento de
Duas Lagoas Costeiras Tropicais ............................................................................................. 40
REFERÊNCIAS .......................................................................................................... 49
CONCLUSÕES GERAIS ........................................................................................................ 51

5
RESUMO
Ecossistemas aquáticos continentais são de extrema importância para a ciclagem
global de nitrogênio. Nestes ambientes grandes quantidades de nitrogênio são fixadas,
tornando-se bio-disponíveis, e também emitidas para a atmosfera na forma de moléculas
reativas. O capítulo introdutório do presente manuscrito apresenta as principais vias de
transformação do nitrogênio em ecossistemas aquáticos continentais e a importância de cada
uma delas para a regulação da bio-disponibilidade deste macronutriente no sistema. O segundo
capítulo trata da influência da salinidade sobre a nitrificação, processo central do ciclo do
nitrogênio, e corroborando resultados obtidos anteriormente, a variação de salinidade interferiu
negativamente na atividade nitrificante principalmente sob baixas concentrações de sal. O
último capítulo evidencia formas diferenciadas de regulação do processo de desnitrificação em
duas lagoas costeiras de status tróficos distintos e, independente do tipo de regulação, as taxas
de desnitrificação foram extremamente baixas se comparadas às temperadas e os resultados
apontam para a escassez de nitrato como principal responsável.

6
ABSTRACT
Freshwater ecosystems are of extreme importance for the global cycling of
nitrogen. In these environments large amounts of nitrogen are fixed, making it bio-
available, and also emitted to the atmosphere as reactive molecules. The introductory
chapter of this manuscript presents the main ways of nitrogen transformation in
freshwater ecosystems and the importance of each to the regulation of bio-availability of
this macronutrient in the system. The second chapter deals with the influence of salinity
on nitrification, central process of the nitrogen cycle, and corroborating results obtained
previously, the change in salinity had a negative influence on nitrifying activity
especially under low salt concentrations. The final chapter highlights different forms of
regulation of denitrification in two coastal lagoons of different trophic status and
regardless of the type of regulation, denitrification rates were extremely low compared
to temperate ones and the results point to a shortage of nitrate as the main responsible.

7
INTRODUÇÃO GERAL
Ecossistemas aquáticos oceânicos e continentais são de extrema importância
para a ciclagem global de nitrogênio. Nestes ambientes grandes quantidades de
nitrogênio molecular são fixadas, ou seja, combinadas a carbono, hidrogênio ou
oxigênio, e se tornam disponíveis para a utilização da biota. Ecossistemas aquáticos
também são áreas de grande potencial de emissão de formas reativas de nitrogênio para
a atmosfera, como íon amônio (NH4+) e óxido nitroso (N2O).
O aumento da concentração de nutrientes, especialmente fósforo e nitrogênio, e
o conseqüente aumento da produtividade total nos ecossistemas aquáticos é chamado de
eutrofização. Com o crescente processo de eutrofização artificial (antropogênica) dos
ecossistemas aquáticos, por despejo de efluentes domésticos ou industriais, o papel
regulador dos microorganismos ligados ao ciclo do nitrogênio nestes ambientes tem
ganhado destaque. As formas reativas de nitrogênio produzidas em sistemas aquáticos e
liberadas para a atmosfera também tem recebido atenção crescente da comunidade
científica. Isto porque tais formas nitrogenadas contribuem para a formação de
nevoeiros fotoquímicos, chuvas ácidas e, no caso do óxido nitroso, possui poder estufa
até 300 vezes maior que o gás carbônico, além de contribuir para o consumo do O3
estratosférico.
No entanto, ainda se conhece muito pouco sobre o funcionamento do ciclo do
nitrogênio fora da região temperada. A grande maioria dos estudos relacionados aos
processos integrantes do ciclo do nitrogênio e seus fatores reguladores em ecossistemas
aquáticos contemplam ambientes temperados do hemisfério norte. Revisões e
estimativas globais acerca dos processos e produtos do ciclo do nitrogênio evidenciam a
escassez de dados tropicais.
A fim de contribuir para a compreensão global do funcionamento do ciclo do
nitrogênio, o presente estudo visou avaliar a ocorrência e os fatores reguladores dos
processos de nitrificação e desnitrificação, dois importantes processos do ciclo do
nitrogênio, em lagoas costeiras tropicais.

Capítulo I 8
REGULAÇÃO MICROBIOLÓGICA DA DISPONIBILIDADE DE NITR OGÊNIO EM
ECOSSISTEMAS AQUÁTICOS CONTINENTAIS
Ana Lúcia Santoro
ASPECTOS GERAIS
Os elementos químicos nitrogênio (N), carbono (C), fósforo (P), oxigênio (O) e enxofre
(S) são reconhecidamente indispensáveis à vida, e dentre estes, o nitrogênio é o que apresenta
maior abundância na atmosfera, hidrosfera e biosfera global (Galloway et al. 2003). O montante
total de nitrogênio na Terra, incluindo solo, água e atmosfera, é de aproximadamente 4 x 1021 g,
massa maior que a dos quatro outros elementos citados anteriormente juntos (Mackenzie 1998).
Contudo, mais de 99% desse montante não está disponível para mais de 99% dos organismos
vivos. A razão para isto está no fato do nitrogênio ser encontrado na natureza quase inteiramente
na forma de nitrogênio molecular (N2) (Galloway et al. 2003). Esta forma química não pode ser
assimilada pela maioria dos organismos, já que quebrar a ligação tripla que liga os dois átomos
de nitrogênio na molécula de N2 requer grande quantidade de energia que só é alcançada em
processos artificiais, envolvendo altas temperaturas, ou por um pequeno e especializado grupo
de microorganismos fixadores de N.
O nitrogênio é um macronutriente que constitui as células de todos os seres vivos, sendo
utilizado para a síntese de DNA, RNA e proteínas, e é um dos principais elementos limitantes à
produção primária em ecossistemas terrestres e aquáticos. Segundo Elser et al. (2007), fontes de
N inorgânico como amônia, nitrato e nitrito são severamente limitadas em ecossistemas
naturais. Apesar do entendimento comum de que a produção primária de águas continentais é
limitada por fósforo (P), muitos trabalhos mostram uma grande variedade destes sistemas
limitados por N (Elser et al. 1990; Francoeur 2001). Por outro lado, o excesso desse elemento
em ecossistemas aquáticos desencadeia o processo de eutrofização, um dos maiores problemas
dos últimos séculos (Smil 1997). Desta forma, a regulação da disponibilidade de nitrogênio, que
se dá fundamentalmente através dos processos microbiológicos relacionados à ciclagem deste
nutriente (Fig.1), é extremamente importante para a manutenção da dinâmica e da composição
dos ecossistemas.
A maioria dos processos do ciclo biogeoquímico global do nitrogênio é realizada por
bactérias, destacando os processos de fixação biológica de nitrogênio, nitrificação e
desnitrificação (Fig 1) (Klotz & Stein 2008). Os compostos nitrogenados presentes na natureza
são divididos em dois grupos: não-reativos e reativos. A molécula de N2 é considerada não-
reativa e as reativas (Nr) incluem todos os compostos biologicamente, fotoquimicamente ou
radiativamente ativos da atmosfera ou biosfera (Galloway et al. 2003). As formas nitrogenadas

Capítulo I 9
mais estudadas em ecossistemas aquáticos e terrestres são: N-orgânico particulado e dissolvido,
amônia (NH3) e íon amônio (NH4+), óxido nitroso (N2O), nitrito (NO2
–) e nitrato (NO3–).
Embora muitos rios e lagos submetidos à pressão antrópica possam apresentar elevadas
concentrações de nitrato na água, ecossistemas aquáticos em geral possuem potencial limitado
para acumulação de formas reativas de nitrogênio, uma vez que o tempo de residência destas
formas na água é bastante reduzido, sendo mais longo no sedimento destes ambientes
(Galloway et al. 2003). A zona de interface sedimento-água de ambientes aquáticos oferece
regiões de intensa deposição e degradação de matéria orgânica (Dean 1999; Heinen &
McManus 2004). Estas condições fomentam alta atividade biogeoquímica e a disponibilidade de
oxigênio pode diminuir e até se tornar nula já nos primeiros milímetros do sedimento (Sweerts
et al. 1989), fazendo deste o principal sítio de ocorrência dos processos relacionados ao ciclo do
nitrogênio em ecossistemas aquáticos.
Piña-Ochoa e Alvarez-Cobelas (2006) argumentam que a maioria dos estudos sobre
desnitrificação em ecossistemas aquáticos está restrita entre as latitudes 25 e 60º N, com
pouquíssimos dados nas regiões tropicais e polares. O mesmo pode ser dito em relação a todos
os outros processos do ciclo do nitrogênio, ainda muito pouco conhecidos fora da região
temperada.
FIXAÇÃO BIOLÓGICA DE NITROGÊNIO (FBN)
O nitrogênio é um elemento único entre os principais necessários à vida, já que seu ciclo
inclui um vasto reservatório atmosférico (N2) que precisa ser fixado (combinado a carbono,
hidrogênio ou oxigênio) antes de ser utilizado pela maioria dos organismos. A disponibilidade
FIG. 1: Esquema simplificado do ciclo do nitrogênio em ecossistemas aquáticos continentais. Em destaque os principais processos bacterianos envolvidos na regulação da biodisponibilidade de N nestes ambientes. FBN – Fixação biológica de nitrogênio; NOP – Nitrogênio orgânico particulado; NOD – Nitrogênio orgânico dissolvido; RDNA – Redução dissimilatória do nitrato a amônia; ANAMMOX – Oxidação anaeróbica da amônia.

Capítulo I 10
desta forma fixada controla, pelo menos em parte, a produtividade, a estocagem de carbono e a
composição de espécies de muitos ecossistemas.
Bactérias e, também, arqueias fixadoras de nitrogênio (diazotróficas) convertem
nitrogênio atmosférico (N2) em amônia (NH3) para própria assimilação. Esta fixação biológica
de nitrogênio neutraliza a remoção de nitrogênio biodisponível por processos microbianos como
desnitrificação e oxidação anaeróbica da amônia (ANAMMOX) e representa a principal fonte
de N para a maioria da biosfera, incapaz de assimilar N2 diretamente (Dekas et al. 2009).
O processo de Fixação Biológica de Nitrogênio (FBN) pode ser sucintamente
representado pela equação:
N2 + 8 H+ + 8 e– → 2 NH3 + H2
Este processo é catalisado pela enzima nitrogenase, que é restrita a alguns procariotas
autótrofos e heterótrofos. Estes organismos utilizam a nitrogenase em conjunto com outras
enzimas e co-fatores para facilitar a quebra altamente endergônica da ligação tripla da molécula
de N2 para produção de amônia (Postgate 1970). Em organismos heterótrofos, o N2 é reduzido a
partir do consumo de compostos orgânicos enquanto, em organismos autótrofos, os elétrons são
oriundos do processo de fotossíntese. Dentre os heterotróficos podem ser encontradas archaea e
bactérias aeróbias, anaeróbias, anaeróbias facultativas e microaerofílicas (Herbert, 1999).
Dentre os organismos autótrofos destaca-se o grupo das cianofíceas, principalmente as que
possuem heterocitos. As cianobactérias são responsáveis pela maior parte do N2 planctônico
fixado em ambientes aquáticos e esta habilidade às proporciona uma vantagem competitiva
durante períodos de escassez de nitrogênio na coluna d’água (Tonno & Noges 2003).
A ligação entre as duas moléculas de N2 é extremamente estável e a demanda energética
para sua quebra é elevada, sendo estimado um mínimo de 16 ATP para cada molécula fixada
(Postgate et al. 1988). Em decorrência do elevado custo metabólico, os organismos autótrofos
são responsáveis pela maior porcentagem do N fixado em ecossistemas aquáticos,
especialmente em ambientes rasos onde há grande penetração de luz (Herbert 1999). Segundo
Schindler (1977), nitrogênio não deveria ser um fator limitante em sistemas aquáticos já que os
organismos fixadores de nitrogênio presentes seriam capazes de converter nitrogênio
atmosférico em formas biodisponíveis. Deste modo, para que tal limitação ocorra, alguns fatores
devem limitar a fixação biológica de nitrogênio (Vitousek & Howarth 1991).
Em corpos aquáticos continentais tais fatores podem ser: disponibilidade de carbono
orgânico, oxigênio e metais traço, pH, salinidade (Herbert 1999), luz (Lewis & Levine 1984),
temperatura (Marcarelli & Wurtsbaugh 2006) e concentração de outros macronutrientes
(Howarth et al. 1988; Marcarelli & Wurtsbaugh 2006).

Capítulo I 11
Marcarelli & Wurtsbaugh (2006) mostram através de experimentos que em rios
oligotróficos dos Estados Unidos a FBN foi regulada pela disponibilidade de fósforo, e a
magnitude da resposta à disponibilidade deste nutriente foi modulada pela temperatura (Fig 2).
FIG. 2: Influência da temperatura e disponibilidades de nitrogênio e fósforo sobre as taxas de fixação biológica de nitrogênio (FBN) em rios oligotróficos. As barras representam ± erro padrão. Adaptado de Marcarelli & Wurtsbaugh (2006).
Os mesmos autores mostram em outro trabalho ( Marcarelli & Wurtsbaugh 2007 ) um
estímulo de até 2500% na fixação de N em resposta a adição de fósforo em ambientes aquáticos
continentais temperados. A adição de N, no entanto, com ou sem adição simultânea de fósforo,
diminuiu a atividade de fixação em até 73%. Enquanto a temperatura exerce influência direta
sobre a fisiologia dos organismos, sendo a FBN um processo de grande gasto energético, este é
interrompido quando há ilimitada disponibilidade de N para o crescimento dos organismos.
Diferente do que se pensava até então, Bryhn & Blenckner (2007) concluíram que é muito
pouco provável que a concentração de N2 limite a fixação biológica de nitrogênio em
ecossistemas aquáticos.
A figura 3 mostra que a resposta à luz da FBN foi modelada com sucesso utilizando
uma equação anteriormente desenvolvida para a resposta à luz da fotossíntese. Lewis & Levine
(1984) encubaram amostras de água de seis diferentes profundidades ao longo da coluna d’água
de um lago venezuelano e encontraram uma curva similar a esperada para o processo
fotossintético.
FIG. 3: Resposta da fixação biológica de nitrogênio (FBN) planctônica à um gradiente de intensidade luminosa em um sistema lacustre venezuelano. Adaptado de Lewis & Levine (1984).
F
BN
Tratamento frio Tratamento quente controle nitrogênio
fósforo
Luz (W.m-2)
Luz (Einst m-2 h-1)
FB
N (
nm
ol
N 2 h
-1)

Capítulo I 12
No Brasil, Enrich-Prast & Esteves (1998) mostraram um padrão de variação diária das
taxas de FBN estudando comunidades perifíticas associadas a uma espécie de macrófita
aquática em um lago amazônico. Tal variação foi atribuída a flutuações na disponibilidade de
carbono orgânico, gerando diferentes respostas à incidência de luz solar de comunidades
predominantemente compostas por organismos autótrofos e heterótrofos (Fig 4).
FIG. 4: Variação diária da luz solar ( ) e da fixação biológica de nitrogênio (FBN) ( ) em comunidades perifíticas dominadas por organismos autótrofos (A) e heterótrofos (B). Adaptado de Enrich-Prast & Esteves (1998).
Antes da extensiva alteração do ciclo do N provocada pela ação humana, de 90 a 130 Tg
de N eram fixados anualmente em ambientes terrestres (Galloway et al. 1995). Já a taxa
planctônica de fixação biológica de nitrogênio ainda é incerta sendo estimada entre 10 a 200 Tg
N por ano (Gruber & Sarmiento 1997).
AMONIFICAÇÃO
Ecossistemas aquáticos continentais, especialmente sistemas lacustres por sua posição
terminal na bacia de drenagem, recebem e acumulam grande quantidade de matéria orgânica
que é então estocada no sedimento (Heinen & McManus 2004). Esta condição torna o processo
de amonificação um importante provedor de Nr lábil nestes ambientes, embora este seja um
processo ainda pouco explorado.
Amonificação (ou mineralização orgânica de N) é um processo contínuo de
decomposição através do qual compostos orgânicos de N com alto peso molecular são
sequenciadamente hidrolisados em compostos mais simples, por meio da atividade de enzimas
extracelulares (Sinsabaugh et al. 1991). Em seguida, estes compostos, geralmente na forma de
aminoácidos, são quebrados resultando na liberação de NH4+ (Gardner et al. 1989). Dependendo
da complexidade estrutural da matéria orgânica, tanto uma simples reação de deaminação
quanto uma complexa série de etapas metabólicas podem ser denominadas amonificação.
Normalmente o processo é limitado pela taxa de hidrólise dos compostos de maior peso
molecular, mas limitações em qualquer etapa do contínuo de decomposição podem regular o
processo (Stanford & Smith 1972). A amonificação de macromoléculas de N pode ser
sumarizada da seguinte forma:
Tempo (h) Tempo (h)
F
BN
(nm
ol h
-1)
Luz (µE m
2 sec -1)
(B)

Capítulo I 13
Proteínas Peptídeos Aminoácidos Ácidos org. + NH4+
O processo de amonificação é realizado por uma série de gêneros bacterianos e de
fungos e corresponde à transformação de N orgânico particulado (NOP) e dissolvido (NOD) em
NH4+ como mostra a figura 1. A produção de amônia está acoplada à mineralização da matéria
orgânica uma vez que esta forma inorgânica de N é liberada paralelamente à oxidação ou a
fermentação do C orgânico à CO2 (Fig 5). No entanto, esta liberação de NH4+ durante a
mineralização da matéria orgânica depende da qualidade e especialmente da razão C:N do
substrato degradado (Lancelot & Billen 1985).
FIG. 5: Representação dos fluxos de carbono e nitrogênio associados à degradação bacteriana da matéria orgânica. Retirado de Lancelot & Billen (1985).
A qualidade da matéria orgânica disponível, ou seja, se esta é lábil ou refratária para
utilização pelos microorganismos decompositores, determina a rapidez com que o substrato é
mineralizado e por conseqüência a taxa de amonificação do ambiente. A qualidade da matéria
orgânica, por sua vez, depende da sua origem (Enriquez et al. 1993). O detrito de macrófitas
aquáticas pode consistir de 25 e 30% de fibra, com conteúdo de lignina de aproximadamente
8%, sendo sua taxa de mineralização muito baixa quando comparada a de detritos formados por
fitoplâncton que contém material nitrogenado mais lábil. Sumi & Koike (1990) mostram uma
correlação negativa significativa (-0,71; p < 0,05) entre a razão C:N do substrato e a taxa de
amonificação de sedimentos costeiros japoneses.
A temperatura ambiental também é capaz de interferir nas taxas locais de mineralização
da matéria orgânica e amonificação, principalmente por acelerar a taxa metabólica dos
organismos envolvidos. Zhang et al. (2008) encontraram correlação positiva significativa entre
o fluxo de amônia do sedimento para a coluna d’água e a temperatura da água em um lago
eutrófico chinês. Os autores atribuem a resposta dos fluxos de amônia à intensificação do
proteinases peptidases deaminação

Capítulo I 14
processo de amonificação com o aumento da temperatura. A figura 6 mostra o comportamento
observado do fluxo de amônia ao longo do gradiente de temperatura.
FIG. 6: Resposta do fluxo de amônia sedimento-água ao aumento de temperatura em um lago eutrófico. Adaptado de Zhang et al. (2008).
O processo de mineralização da matéria orgânica ocorre em condições aeróbias e
anaeróbias, observando-se em geral as maiores taxas em presença de oxigênio. Estudos como o
de Lillebo et al. (2007) no entanto, mostram que condições de depleção de oxigênio
favoreceram o processo de amonificação no sedimento de pequenos rios da região mediterrânea.
A taxa de amonificação no sedimento de sistemas aquáticos é então regulada
principalmente pelo aporte e qualidade da matéria orgânica local. Em ambientes rasos, o
processo de amonificação pode contribuir com até 80% do N requerido pelo fitoplâncton
(Hansen & Blackburn 1992). Tendo o íon NH4+ como principal produto final, este pode ser
transportado para a coluna d’água ou no sedimento, ser assimilado por organismos bentônicos
ou ainda oxidado a NO3- ou NO2
- via nitrificação (Sumi & Koike 1990).
NITRIFICAÇÃO
A atividade nitrificante é de fundamental importância para todos os ecossistemas uma
vez que este processo une as fases reduzidas e oxidadas do ciclo do nitrogênio (Fig. 1),
desempenhando papel central (Joye et al. 1999). Seus produtos (nitrito e nitrato) servem como
substrato para desnitrificação, promovendo o elo que permite a transferência de nitrogênio
biodisponível do ambiente para a atmosfera, sob a forma de N2 e N2O (Jenkins & Kemp 1984).
Em ambientes com baixa disponibilidade de nitrato o processo de nitrificação pode subsidiar até
75 % de todo o N2 produzido (Dong et al. 2006).
O processo de nitrificação é realizado por bactérias quimioautotróficas que obtém sua
energia através da oxidação de formas reduzidas de nitrogênio. Este é um processo que se dá em
duas etapas distintas, nas quais a amônia é oxidada a nitrito e posteriormente a nitrato como
ilustrado abaixo:
(I) NH 4+ + 1½ O2 → NO2
- + H2O + 2 H+
(II) NO 2- + ½ O2 → NO3
-
T
emp
erat
ura
(ºC
)
Fluxo de amônia (mg m-2 d-1)

Capítulo I 15
Embora baixos níveis de oxigênio (até 0,3 mg L-1) já sejam suficientes (Cavari 1977),
ambas as etapas ocorrem estritamente em condição aeróbica. Assim como os outros processos
do ciclo do nitrogênio, o sedimento é o principal sítio de ocorrência da nitrificação em
ambientes aquáticos. A distribuição vertical das bactérias nitrificantes neste substrato é
determinada principalmente pela penetração de O2, que normalmente é de cerca de 1,65 mm
dependendo de fatores como: tipo de sedimento, conteúdo de matéria orgânica, temperatura,
grau de mistura do sedimento e bioturbação (Revsbech et al. 1980). Nitrosospira e
Nitrosomonas são os gêneros mais conhecidos capazes de realizar a primeira etapa, a oxidação
da amônia a nitrito, enquanto a oxidação de nitrito a nitrato é realizada por vários gêneros,
sendo Nitrobacter o mais comumente encontrado em ambientes costeiros (Enrich-Prast 2005).
O carbono utilizado por bactérias nitrificantes para a construção de moléculas é obtido
por meio da fixação de CO2. Segundo Fenchel et al. (1998), bactérias nitrificantes precisam
oxidar 35 e 100 moles de amônia e nitrito respectivamente para fixação de 1 único mol de C,
sendo este um processo de baixíssimo ganho energético. Com isso, estes organismos possuem
crescimento muito lento. Enquanto o tempo de duplicação de bactérias heterotróficas é de cerca
de 1 hora, o tempo de duplicação de bactérias nitrificantes varia entre 7 e 10 horas em
laboratório, podendo chegar a semanas em ambientes naturais (Enrich-Prast et al. 2009).
Uma série de fatores é capaz de regular a atividade nitrificante. Alguns destes têm
impacto fisiológico direto sobre as bactérias, tais como temperatura, pH e disponibilidade de
NH4+, O2 e CO2 (Herbert 1999). A salinidade também pode desempenhar papel importante
(Risgaard-Petersen et al. 2004a), influenciando diretamente os organismos nitrificantes
(Rysgaard et al. 1999) e regulando a capacidade de absorção de NH4+ pelo sedimento (Boatman
& Murray 1982). Santoro & Enrich-Prast (2009) observaram um decaimento exponencial das
taxas de nitrificação no sedimento de lagoas costeiras do estado do Rio de Janeiro, Brasil, em
resposta ao aumento da salinidade na coluna d’água (Fig. 7).
FIG. 7: Modelo de decaimento exponencial entre as taxas de nitrificação no sedimento de lagoas costeiras e salinidade. Equação: y= 89,29*exp (-0,1285*x) + (-2,929); R2= 0,5345; n= 44. Adaptado de Santoro & Enrich-Prast (2009).
Salinidade (ppt)
N
itrifi
caçã
o (
nmol
NO 3
- cm
-3 h
-1)

Capítulo I 16
Outros fatores como incidência de luz e presença de organismos fotossintetizantes como
algas e macrófitas aquáticas modificam as condições químicas da interface sedimento-água,
principalmente através do estímulo à produção, no caso da luz, e produção propriamente dita de
O2 (Wilkie & Mulbry 2002; Munoz & Guieysse 2006). A atividade destes organismos aumenta
a penetração de oxigênio no sedimento, estimulando processos aeróbicos como a nitrificação
(Risgaardpedersen et al. 1994; Lorenzen et al. 1998; Travieso et al. 2006). Utilizando
microsensores de O2 e NO3-, Lorenzen et al. (1998) mostram o estímulo ao processo de
nitrificação realizado pela atividade fotossintética de microalgas bentônicas através de
microperfis no sedimento de um ecossistema lacustre dinamarquês (Fig 8).
FIG. 8: Concentrações de O2 (○, valores médios e desvio padrão, n= 6) e NO3- (□, valores médios e desvio padrão,
n=6), perfis calculados (linhas) e taxas de assimilação de NO3-, nitrificação e desnitrificação na ausência (A) e na
presença (B) de luz em sedimento lacustre colonizado por microalgas bentônicas. Adaptado de Lorenzen et al. (1998).
Por outro lado, Risgaard–Petersen et al. (2004b) mostram que algas microfitobentônicas
podem competir diretamente com bactérias nitrificantes por NH4+ (Fig 9). Estas algas
bentônicas possuem maior taxa de crescimento e assimilação de N, podendo limitar o processo
de nitrificação.
FIG. 9: Taxas de nitrificação no sedimento em três diferentes tratamentos: livre de algas bentônicas, com a presença de algas bentônicas e adição de NH4
+ e somente com a presença de algas. Adaptado de Risgaard-Petersen et al. (2004b).
assimilação
desnitrificação
nitrificação
assimilação
P
rofu
ndi
dade
Alga Alga + N Livre de Alga

Capítulo I 17
A excreção de parte dos produtos fotossintéticos por estes organismos representa uma
fonte adicional de C e pode também regular o processo de nitrificação no sedimento
(Middelburg et al. 2000). Elevadas concentrações de carbono orgânico reduzem as taxas de
nitrificação por estimularem o rápido crescimento de bactérias heterotróficas, que competem
com a comunidade nitrificante por oxigênio e amônia (Strauss & Lamberti 2000, 2002).
FIG. 9: Esquematização das hipóteses acerca dos efeitos da razão C:N e qualidade do carbono orgânico disponível sobre as taxas de nitrificação. (A) hipótese 1: sob elevada razão C:N bactérias heterotróficas são mais eficientes na competição por NH4
+, resultando em baixas taxas de nitrificação; sob baixa razão C:N a comunidade nitrificante não é exposta à forte competição por NH4
+, resultando em altas taxas de nitrificação. (B) hipótese 2: quando o carbono orgânico disponível no ambiente é lábil (facilmente assimilado por bactérias heterotróficas) o carbono orgânico presente terá um efeito negativo maior sobre as taxas de nitrificação do que quando este é mais refratário (assimilado com maior dificuldade por bactérias heterotróficas). Adaptado de Strauss & Lamberti (2000).
DESNITRIFICAÇÃO
A remoção biológica de nitrato de ecossistemas aquáticos é normalmente atribuída à
assimilação na biomassa de algas e microorganismos, produzindo N orgânico que será
remineralizado posteriormente, e à desnitrificação (Burgin & Hamilton 2007).
Desnitrificação é um processo de redução dissimilatória na qual o nitrato ou nitrito
(NO3- ou NO2
-) é reduzido anaerobicamente a N molecular (N2) por bactérias heterotróficas
anaeróbicas facultativas. Este processo requer simultaneamente um meio anóxico (ou subóxico,
< 0,2 mg O2 l-1) e disponibilidade de matéria orgânica e nitrato (Mosier et al. 2002). Na
ausência de oxigênio, bactérias desnitrificantes utilizam nitrato como aceptor e carbono
orgânico como doador de elétrons como mostra a reação a seguir:
NH4+
Alta razão C:N Baixa razão C:N
Bactéria nitrificante
Bactéria heterotrófica
Bactéria nitrificante
Bactéria heterotrófica
Bactéria nitrificante
Bactéria heterotrófica
Bactéria nitrificante
Bactéria heterotrófica
C orgânico lábil
C orgânico refratário
NH4+ NH4
+
NH4+ NH4
+
Baixas taxas de nitrificação Altas taxas de nitrificação
Baixas taxas de nitrificação Altas taxas de nitrificação

Capítulo I 18
4 NO3- + 5 (CH2O) + 5 H2O → 2 N2 + 5 CO2 + 8 H2O + 4 OH–
Uma vez que a maioria do N fixado disponível no sedimento de ambientes aquáticos,
principal sítio anóxico para ocorrência do processo, está na forma de amônio (NH4+), ao menos
que haja um fluxo de nitrato proveniente da coluna d’água para o sedimento, o processo de
desnitrificação depende das taxas locais de nitrificação. O nitrato produzido através da
nitrificação na zona aeróbica difunde no sedimento até a zona anóxica onde a desnitrificação
ocorre (Nielsen et al. 1990). Esta combinação de processos é conhecida na literatura como
nitrificação-desnitrificação acoplada, enquanto o processo subsidiado pelo nitrato proveniente
da coluna d’água pode ser referido como desnitrificação direta (Fennel et al. 2009).
Muitos microorganismos heterotróficos são capazes de utilizar nitrato e nitrito como
aceptores de elétrons em condições anaeróbias, sendo o gênero Pseudomonas o mais conhecido
e estudado em ambientes aquáticos (Enrich-Prast 2005). Na ausência de O2, a redução do nitrato
promove mais energia que a redução de outros compostos oxidados, como Mn+4, Fe+3 ou SO4–2
(Fenchel et al. 1998). Estudos recentes mostraram que foraminíferos bentônicos, eucariotos
unicelulares encontrados no sedimento de uma grande variedade de ecossistemas aquáticos,
acumulam estoques intracelulares de nitrato (Risgaard-Petersen et al. 2006) e também são
capazes de respirar este substrato via desnitrificação (Piña-Ochoa et al. 2010), explicando a
ocorrência destes organismos em condições anóxicas. Risgaard-Petersen et al. (2006) estimam
que o estoque de nitrato intracelular encontrado seja suficiente para subsidiar a respiração
anaeróbica destes organismos por mais um mês.
Por ter como produto formas gasosas de N (N2 e N2O como produto intermediário) que
não podem ser assimiladas pela maioria dos organismos e geralmente difundem para a
atmosfera, o processo de desnitrificação diminui a biodisponibilidade de N no ambiente
aquático. Ecossistemas aquáticos continentais são responsáveis por cerca de 20 % do total
global da atividade desnitrificante, sendo a desnitrificação realizada no sedimento a maior via de
escape de N fixado de sistemas aquáticos naturais ou alterados (Seitzinger et al. 2006). Se por
um lado, este processo regula a produção primária em ecossistemas limitados por nitrogênio,
por outro, promove controle do processo de eutrofização artificial em ecossistemas que recebem
efluentes com elevadas concentrações de compostos nitrogenados. Segundo Galloway et al.
(2003), áreas úmidas são tão eficientes na remoção de N reativo (Nr) através da desnitrificação
que ambientes deste tipo são freqüentemente construídos para remover o Nr de efluentes de uma
série de atividades antrópicas.
A maior limitação para ocorrência do processo de desnitrificação é o fato de que a
produção de nitrato (nitrificação) requer a presença de oxigênio (O2) enquanto a desnitrificação
requer condições subóxicas. Por isso a desnitrificação ocorre em interfaces óxico/anóxico,

Capítulo I 19
podendo esta interface ser uma separação no tempo ou no espaço (ou ambos) (Seitzinger et al.
2006). A profundidade da zona aeróbica no sedimento de sistemas aquáticos é diretamente
afetada pelo consumo de O2 no mesmo, o que se associa com as taxas de decomposição da
matéria orgânica e nitrificação (Seitzinger et al. 2006). Segundo Fennel et al. (2009), a taxa de
consumo de O2 no sedimento pode ser um preditor efetivo da atividade desnitrificante neste
substrato.
A taxa de consumo de oxigênio regula a profundidade da zona aeróbica e
conseqüentemente pode regular a desnitrificação de duas maneiras: (1) diretamente, através da
criação de condições anóxicas ou (2) indiretamente, controlando as taxas de nitrificação (Jensen
et al. 1994). Os resultados obtidos pelo estudo de Santoro et al. (dados ainda não publicados)
em duas lagoas costeiras brasileiras de status trófico distintos mostram a ocorrência dos dois
tipos de regulação supracitados (Fig 10). As taxas de desnitrificação no sedimento do ambiente
oligotrófico se correlacionaram principalmente com as taxas de consumo de oxigênio no
sedimento, indicando o tipo de regulação direta. Já no sistema eutrófico, com elevadas taxas de
consumos de oxigênio no sedimento, o processo de desnitrificação respondeu principalmente às
variações na disponibilidade de nitrato na coluna d’água, já que a anoxía do sedimento limitava
a produção de nitrato via nitrificação, indicando o tipo de regulação indireta.
FIG. 10: Análise de componentes principais (ACP). Estão posicionados no plano 1-2 os vetores: taxa de consumo de oxigênio no sedimento (SOC), concentração de nitrato ([NO3
-]) e amônia ([NH4+]) na coluna d’água e, como
variável complementar (*), a taxa de desnitrificação no sedimento (Denit) em duas lagoas costeiras, uma eutrófica (IMB) e outra oligotrófica (CAB). N=10. Quanto mais próximos os vetores, maior a correlação entre eles. Adaptado de Santoro et al. (em prep.).
Além da disponibilidade de O2, fatores como temperatura, salinidade, pH,
disponibilidade de C orgânico, nitrato e nitrito também podem regular a atividade desnitrificante
-1,0 -0,5 0,0 0,5 1,0
Factor 1 : 38,07%
-1,0
-0,5
0,0
0,5
1,0
Fac
tor
2 :
30,4
8%
[NH4+] CAB
SOC IMB
*Denit CAB
SOC CAB
[NH4+] IMB
[NO3-] CAB
*Denit IMB
[NO3-] IMB
Regulação direta
Regulação indireta

Capítulo I 20
(Cornwell et al. 1999; Pina-Ochoa & Alvarez-Cobelas 2006). A figura 11 mostra alguns dos
resultados obtidos na revisão sobre desnitrificação em ambientes aquáticos realizada por Pina-
Ochoa & Alvarez-Cobelas (2006). As taxas do processo foram significativamente maiores em
condições de maior disponibilidade de C orgânico e nitrato (Mann-Whitney, p< 0,05).
FIG. 11: Box-whisker plot para taxas de desnitrificação em diferentes ambientes aquáticos sob condições de baixa e alta disponibilidade de carbono orgânico (baixo < 10 mg g-1) e nitrato (baixo < 50 µM). Adaptado de Pina-Ochoa e Alvarez-Cobelas (2006).
O processo de bioturbação realizado por macroorganismos bentônicos que constroem
tubos e galerias, como poliquetas e quironomídeos, também pode ser um importante fator
regulador do processo de desnitrificação no sedimento. A atividade destes organismos não só
aumenta a extensão da zona de interface sedimento-água como também, ao bombear
intermitentemente água da camada d’água acima do sedimento para dentro dos tubos e galerias,
levam o nitrato disponível para as camadas mais profundas do sedimento, onde a desnitrificação
ocorre (Svensson & Leonardson 1996).
FIG. 12: Comportamento das taxas de desnitrificação direta (a; utilizando nitrato proveniente da coluna d’água) e via nitrificação (b; utilizando nitrato proveniente do processo de nitrificação no sedimento) ao longo do gradiente de colonização por macroorganismos bentônicos em sedimento lacustre temperado. As linhas baseiam-se na análise de regressão linear simples e os coeficientes de correlação de Spearman também podem ser observados (n=16). Adaptado de Svensson & Leonardson (1996).
D
esni
trifi
caçã
o (µ
mol
N m
-2 h
-1)
C ORGÂNICO NITRATO
baixo baixo alto alto
D
esni
trifi
caçã
o (µ
mol
N m
-2 h
-1)
Macrofauna (g PS m-2) Macrofauna (g PS m-2)
a b

Capítulo I 21
Embora o consumo de nitrato em sedimentos aquáticos seja normalmente atribuído em
grande parte ao processo de desnitrificação, as estimativas das taxas de desnitrificação através
de medições diretas normalmente não explicam o total de consumo deste substrato no sistema
(Seitzinger 1988; Burgin & Hamilton 2007). Uma possível explicação para essa discrepância é a
falta de métodos capazes de extrapolar taxas de desnitrificação locais para todo o ecossistema
(Cornwell et al. 1999). No entanto, uma explicação alternativa é que parte da remoção de nitrato
em ecossistemas aquáticos pode ser atribuída a outros processos como a redução dissimilatória
de nitrato à amônia (RDNA) (Tiedje et al. 1982) e a oxidação anaeróbica da amônia
(ANAMMOX) (Jetten 2001). O estudo de Laverman et al. (2007) sobre a influência da
salinização na remoção de nitrato em um lago costeiro mostra esta defasagem entre as taxas de
redução de nitrato e as taxas de desnitrificação, atribuindo esta diferença principalmente a
ocorrência do processo de DNRA (Fig 13).
FIG. 13: Taxas de redução dissimilatória de nitrato e desnitrificação no sedimento de um lago costeiro obtidas em experimentos com (barras pretas) e sem (barras brancas) a adição de NaCl. Em ambos os tratamentos as taxas de redução dissimilatória de nitrato foram maiores que as de desnitrificação. Estão representadas médias e desvio padrão. Adaptado de Laverman et al. (2007).
REDUÇÃO DISSIMILATÓRIA DO NITRATO A AMÔNIA
A existência da redução dissimilatória do nitrato a amônia (RDNA, ou em inglês
DNRA) é amplamente reconhecida há pelo menos 25 anos, no entanto, o interesse na potencial
importância deste processo como via de remoção do nitrato em escala ecossistêmica é mais
recente (Burgin & Hamilton 2007). Neste processo, realizado por alguns grupos de bactérias
fermentativas estritamente anaeróbicas (Keith & Herbert 1983), o nitrato é reduzido à amônia.
Ao contrário do processo de desnitrificação, a RDNA não elimina o nitrogênio do
ecossistema, mantendo-o disponível para os produtores primários e microorganismos, sendo a
amônia resultante uma forma inorgânica de N ainda mais lábil que o nitrato. Pouco ainda é
conhecido sobre o destino da amônia produzida pela RDNA, mas é possível que sob condições
apropriadas, esta seja novamente convertida a nitrato via nitrificação (Fig1). Por ser uma forma
T
axas
(nm
ol c
m-3 h
-1)
Redução de nitrato
Desnitrificação
Capítulo I 22
extremamente lábil de N inorgânico, a amônia produzida também pode ser assimilada na
biomassa de plantas e microorganismos (Burgin & Hamilton 2007).
Atualmente são reconhecias duas vias de ocorrência da RDNA: uma fermentativa e
outra ainda pouco estudada ligada a oxidação do enxofre. A via fermentativa possibilita o fluxo
de elétrons da matéria orgânica para a redução do nitrato via reações fermentativas (Megonigal
et al. 2004). Embora as condições que promovam a RDNA (fermentativa) e a desnitrificação
sejam similares (basicamente anoxía e disponibilidade de nitrato e carbono orgânico), a RDNA
parece ser favorecida em ambientes com pouca disponibilidade de nitrato e ricos em carbono
lábil (de fácil utilização por microorganismos), enquanto a desnitrificação seria favorecida em
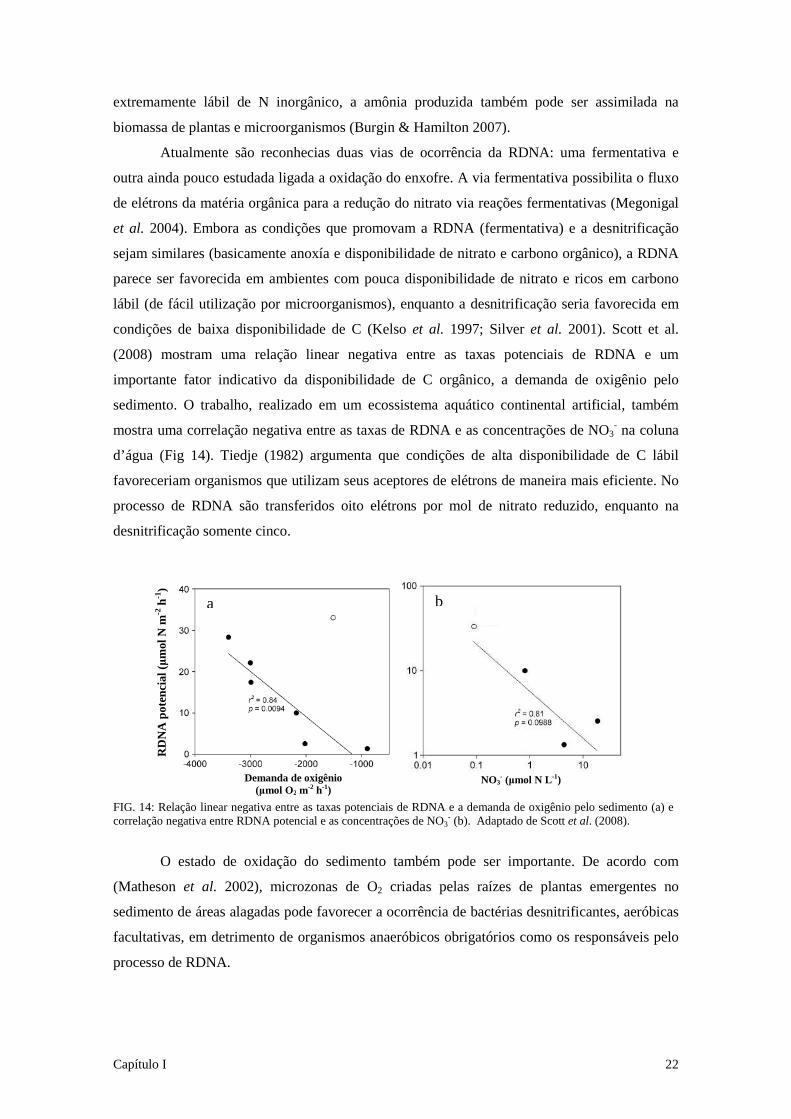
condições de baixa disponibilidade de C (Kelso et al. 1997; Silver et al. 2001). Scott et al.
(2008) mostram uma relação linear negativa entre as taxas potenciais de RDNA e um
importante fator indicativo da disponibilidade de C orgânico, a demanda de oxigênio pelo
sedimento. O trabalho, realizado em um ecossistema aquático continental artificial, também
mostra uma correlação negativa entre as taxas de RDNA e as concentrações de NO3- na coluna
d’água (Fig 14). Tiedje (1982) argumenta que condições de alta disponibilidade de C lábil
favoreceriam organismos que utilizam seus aceptores de elétrons de maneira mais eficiente. No
processo de RDNA são transferidos oito elétrons por mol de nitrato reduzido, enquanto na
desnitrificação somente cinco.
FIG. 14: Relação linear negativa entre as taxas potenciais de RDNA e a demanda de oxigênio pelo sedimento (a) e correlação negativa entre RDNA potencial e as concentrações de NO3
- (b). Adaptado de Scott et al. (2008).
O estado de oxidação do sedimento também pode ser importante. De acordo com
(Matheson et al. 2002), microzonas de O2 criadas pelas raízes de plantas emergentes no
sedimento de áreas alagadas pode favorecer a ocorrência de bactérias desnitrificantes, aeróbicas
facultativas, em detrimento de organismos anaeróbicos obrigatórios como os responsáveis pelo
processo de RDNA.
R
DN
A p
oten
cial
(µm
ol N
m-2
h-1
)
Demanda de oxigênio (µmol O2 m-2 h-1)
NO3- (µmol N L-1)
a b

Capítulo I 23
OXIDAÇÃO ANAERÓBICA DA AMÔNIA
A oxidação anaeróbica da amônia, processo quimiolitoautotrófico conhecido como
ANAMMOX (do inglês ‘anaerobic ammonium oxidation’), consiste na combinação de amônia
e nitrito sob condições anaeróbicas e foi descrito pela primeira vez por Mulder et al. (1995). A
amônia é utilizada como doadora e o nitrito como aceptor de elétrons, tendo como produto final
N2. O nitrito pode derivar-se da redução do nitrato realizada por bactérias desnitrificantes
(Kuypers et al. 2003) ou da oxidação da amônia através da atividade nitrificante. Damste et al.
(2002) mostram em biofilmes bacterianos que o aglomerado de bactérias que realizava o
processo de ANAMMOX era frequentemente encontrado próximo ao de bactérias nitrificantes.
O esquema de Dalsgaard et al. (2005) (Fig. 15) posiciona a zona de ocorrência do processo de
ANAMMOX ao longo de um perfil de profundidade no sedimento. O processo ocorreria então
no mesmo estrato da desnitrificação, já que as condições de ocorrência são praticamente as
mesmas para ambos os processos, e logo abaixo da zona óxica de ocorrência da nitrificação.
FIG. 15: Representação esquemática da distribuição de algumas formas dissolvidas de N na água intersticial e acima do sedimento (A) e indicação das zonas de profundidade de ocorrência dos processos indicados (B). Adaptado de Dalsgaard et al. (2005).
O processo foi descoberto em uma estação de tratamento de esgoto e desde então
estudos têm observado a ocorrência deste em zonas anóxicas de ecossistemas aquáticos
marinhos e continentais (Rysgaard et al. 2004; Schubert et al. 2006). Embora a importância
ecológica deste processo já tenha sido colocada em questão (Zehr & Ward 2002), ainda se sabe
relativamente pouco sobre o processo e sobre as bactérias envolvidas (Strous et al. 2006). O
tempo de duplicação de bactérias que realizam a ANAMMOX foi de 11 dias em condições
Águ
a P
rof.
dent
ro d
o se
dim
ento
Concentração Processos
Resp. aeróbica Nitrificação
Desnitrificação ANAMMOX
Mineralização anaeróbica

Capítulo I 24
laboratoriais ótimas e de 2 a 3 semanas em média segundo Strous & Jetten (2004). Culturas de
enriquecimento a partir de inóculos adequados geralmente necessitam de 100 a 200 dias, o que
dificulta muito o estudo destes organismos (van Dongen et al. 2001).
A ANAMMOX ocorre onde haja disponibilidade de amônia e nitrito sob condições
anóxicas, contribuindo para uma remoção permanente de nitrito do sistema (Burgin & Hamilton
2007). No entanto, o processo é inibido por alguns compostos orgânicos como piruvato, etanol e
glicose (Jetten et al. 1999), fazendo com que o processo de oxidação anaeróbica da amônia seja
mais expressivo em ecossistemas com limitada disponibilidade de C lábil ou com excesso de N
em relação a C (Dalsgaard et al. 2005). Considerando a coluna d’água de oceanos, o processo
de ANAMMOX pode contribuir com até 50% de todo o N2 produzido (Dalsgaard et al. 2003).
Pouco ainda é conhecido sobre o processo em sistemas aquáticos continentais, no entanto, é
esperado que este seja mais importante em lagos profundos e/ou oligotróficos (Burgin &
Hamilton 2007). De acordo com o estudo realizado por Schubert et al. (2006), de 7 a 13% do
total de N2 produzido no lago Tanganyika é derivado da oxidação anaeróbica da amônia.
Durante o processo de oxidação anaeróbica da amônia, também é possível observar a
fixação de CO2 pelos organismos, mas a via de ocorrência desta fixação ainda é desconhecida
(Schmid et al. 2003). Além da redução do nitrito para oxidação da amônia, as bactérias
responsáveis pela ANAMMOX também oxidam anaerobicamente parte do nitrito a nitrato e este
processo poderia suprir os elétrons necessários para a fixação de CO2 (deGraaf et al. 1996).
IMPACTOS ANTRÓPICOS
Antes da interferência humana, a síntese de formas reativas de N a partir de N2 ocorria
principalmente através de dois processos: relâmpagos e FBN. Tais formas reativas de N não se
acumulavam nos ecossistemas uma vez que os processos de FBN e desnitrificação se
compensavam (Ayres et al. 1994).
Segundo Galloway et al. (2003), ao longo das últimas décadas a produção de Nr por
atividades humanas tem sido maior que a produção natural de todos os sistemas terrestres
juntos. E de acordo com o mesmo autor, este aumento tem três causas principais: (1) o cultivo
extensivo de legumes, arroz e outras culturas que promovem a conversão de N2 em N orgânico
através da FBN; (2) a queima de combustíveis fósseis, que convertem tanto N2 como N fóssil
em formas reativas de N (NOx); e (3) o processo de Haber-Bosch que, converte N2 a NH3
artificialmente para sustentar a produção de alimentos e outras atividades industriais. A criação
de Nr apenas através do processo de Haber-Bosch aumentou de 0, antes de 1910, para mais de
100 Tg N por ano em 2000, com cerca de 85% desse montante sendo utilizado na produção de
fertilizantes (Galloway et al. 2002).
Alterações no ciclo do nitrogênio provocam múltiplas conseqüências e segundo
Galloway et al. (2008), as atividades humanas mais que dobraram o montante de N em

Capítulo I 25
circulação na biosfera. Na atmosfera, estas conseqüências incluem: aumento da concentração
global de oxido nitroso (N2O), um poderoso gás estufa com força radiativa 300 vezes maior que
a do CO2 e um dos responsáveis pelo consumo de O3 estratosférico; aumento substancial dos
fluxos de formas gasosas radioativas de nitrogênio (dois terços ou mais das emissões globais de
oxido nítrico e amônia são de origem antrópica); e contribuição substancial para a ocorrência de
chuvas ácidas e nevoeiro fotoquímico que afetam zonas urbanas e agrícolas em todo o mundo
(Vitousek et al. 1997).
Uma das principais vias de aporte de N antropogênico nos ecossistemas é a deposição
atmosférica (Elser et al. 2009), mas o aporte de esgoto doméstico e outros efluentes despejados
in natura em ambientes aquáticos também é um problema crescente e a concentração de nitrato
nos rios se correlaciona fortemente com a ocupação humana nas bacias (Howarth et al. 1996).
Em regiões de baixa fixação de nitrogênio a adição deste elemento geralmente aumenta a
produtividade e o estoque de carbono interno, desencadeando ou acelerando o processo de
eutrofização (Aber et al. 1991). A diversidade biológica também pode ser alterada e
normalmente diminui onde a adição de nitrogênio aumenta a produtividade dos ecossistemas
(Tilman 1987).
BIBLIOGRAFIA
Aber J.D., Melillo J.M., Nadelhoffer K.J., Pastor J. & Boone R.D. (1991) Factors Controlling Nitrogen Cycling and Nitrogen Saturation in Northern Temperate Forest Ecosystems. Ecological Applications, 1, 303-315
Ayres R.U., Schlesinger W.H. & Socolow R.H. (1994) Human impacts on the carbon and nitrogen cycles. In: Industrial Ecology and Global Change (eds. Socolow RH, Andrews C, Berkhout R & Thomas V), pp. 121 - 155. Cambridge University Press, Cambridge (MA)
Boatman C.D. & Murray J.W. (1982) Modeling Exchangeable Nh4+ Adsorption in Marine-Sediments - Process and Controls of Adsorption. Limnology and Oceanography, 27, 99-110
Bryhn A.C. & Blenckner T. (2007) Can nitrogen gas be deficient for nitrogen fixation in lakes? Ecological Modelling, 202, 362-372
Burgin A.J. & Hamilton S.K. (2007) Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Frontiers in Ecology and the Environment, 5, 89-96
Cavari B.Z. (1977) Nitrification Potential and Factors Governing Rate of Nitrification in Lake-Kinneret. Oikos, 28, 285-290
Cornwell J.C., Kemp W.M. & Kana T.M. (1999) Denitrification in coastal ecosystems: methods, environmental controls, and ecosystem level controls, a review. Aquatic Ecology, 33, 14
Dalsgaard T., Canfield D.E., Petersen J., Thamdrup B. & Acuna-Gonzalez J. (2003) N-2 production by the anammox reaction in the anoxic water column of Golfo Dulce, Costa Rica. Nature, 422, 606-608
Dalsgaard T., Thamdrup B. & Canfield D.E. (2005) Anaerobic ammonium oxidation (anammox) in the marine environment. Research in Microbiology, 156, 457-464

Capítulo I 26
Damste J.S.S., Strous M., Rijpstra W.I.C., Hopmans E.C., Geenevasen J.A.J., van Duin A.C.T., van Niftrik L.A. & Jetten M.S.M. (2002) Linearly concatenated cyclobutane lipids form a dense bacterial membrane. Nature, 419, 708-712
Dean W.E. (1999) The carbon cycle and biogeochemical dynamics in lake sediments. Journal of Paleolimnology, 21, 375-393
deGraaf A.A.V., deBruijn P., Robertson L.A., Jetten M.S.M. & Kuenen J.G. (1996) Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor. Microbiology-Uk, 142, 2187-2196
Dekas A.E., Poretsky R.S. & Orphan V.J. (2009) Deep-Sea Archaea Fix and Share Nitrogen in Methane-Consuming Microbial Consortia. Science, 326, 422-426
Dong L.F., Nedwell D.B. & Stott A. (2006) Sources of nitrogen used for denitrification and nitrous oxide formation in sediments of the hypernutrified Colne, the nutrified Humber, and the oligotrophic Conwy estuaries, United Kingdom. Limnology and Oceanography, 51, 545-557
Elser J.J., Andersen T., Baron J.S., Bergstrom A.K., Jansson M., Kyle M., Nydick K.R., Steger L. & Hessen D.O. (2009) Shifts in Lake N:P Stoichiometry and Nutrient Limitation Driven by Atmospheric Nitrogen Deposition. Science, 326, 835-837
Elser J.J., Bracken M.E.S., Cleland E.E., Gruner D.S., Harpole W.S., Hillebrand H., Ngai J.T., Seabloom E.W., Shurin J.B. & Smith J.E. (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 10, 1135-1142
Elser J.J., Marzolf E.R. & Goldman C.R. (1990) Phosphorus and Nitrogen Limitation of Phytoplankton Growth in the Fresh-Waters of North-America - a Review and Critique of Experimental Enrichments. Canadian Journal of Fisheries and Aquatic Sciences, 47, 1468-1477
Enrich-Prast A. (2005) Caminhos do nitrogênio em ecossistemas aquáticos continentais. In: Lições de Limnologia (eds. Roland F, Cesar D & Marinho M), pp. 209 - 227. Rima, São Carlos
Enrich-Prast A., Bastviken D. & Crill P.M. (2009) Chemosynthesis. In: Encyclopedia of Inland Waters, pp. 211 - 225
Enrich-Prast A. & Esteves F.A. (1998) Diurnal variation of rates of denitrification and nitrogen fixation of periphyton associated with Oryza glumaepatula Steud in an Amazonian Lake. Hydrobiologia, 368, 189-192
Enriquez S., Duarte C.M. & Sandjensen K. (1993) Patterns in Decomposition Rates among Photosynthetic Organisms - the Importance of Detritus C-N-P Content. Oecologia, 94, 457-471
Fenchel T., King G.M. & Blackburn T.H. (1998) Bacterial biogeochemistry: the ecophysiology of mineral cycling. 2 edn. Academic Press.
Fennel K., Brady D., DiToro D., Fulweiler R.W., Gardner W.S., Giblin A., McCarthy M.J., Rao A., Seitzinger S., Thouvenot-Korppoo M. & Tobias C. (2009) Modeling denitrification in aquatic sediments. Biogeochemistry, 93, 159-178
Francoeur S.N. (2001) Meta-analysis of lotic nutrient amendment experiments: detecting and quantifying subtle responses. Journal of the North American Benthological Society, 20, 358-368
Galloway J.N., Aber J.D., Erisman J.W., Seitzinger S.P., Howarth R.W., Cowling E.B. & Cosby B.J. (2003) The nitrogen cascade. Bioscience, 53, 341-356
Galloway J.N., Cowling E.B., Seitzinger S.P. & Socolow R.H. (2002) Reactive nitrogen: Too much of a good thing? Ambio, 31, 60-63
Galloway J.N., Schlesinger W.H., Levy H., Michaels A. & Schnoor J.L. (1995) Nitrogen-Fixation - Anthropogenic Enhancement-Environmental Response. Global Biogeochemical Cycles, 9, 235-252
Galloway J.N., Townsend A.R., Erisman J.W., Bekunda M., Cai Z.C., Freney J.R., Martinelli L.A., Seitzinger S.P. & Sutton M.A. (2008) Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science, 320, 889-892

Capítulo I 27
Gardner W.S., Chandler J.F. & Laird G.A. (1989) Organic Nitrogen Mineralization and Substrate Limitation of Bacteria in Lake-Michigan. Limnology and Oceanography, 34, 478-485
Gruber N. & Sarmiento J.L. (1997) Global patterns of marine nitrogen fixation and denitrification. Global Biogeochemical Cycles, 11, 235-266
Hansen L.S. & Blackburn T.H. (1992) Effect of Algal Bloom Deposition on Sediment Respiration and Fluxes. Marine Biology, 112, 147-152
Heinen E.A. & McManus J. (2004) Carbon and nutrient cycling at the sediment-water boundary in western Lake Superior. Journal of Great Lakes Research, 30, 113-132
Herbert R.A. (1999) Nitrogen cycling in coastal marine ecosystems. Fems Microbiology Reviews, 23, 563-590
Howarth R.W., Billen G., Swaney D., Townsend A., Jaworski N., Lajtha K., Downing J.A., Elmgren R., Caraco N., Jordan T., Berendse F., Freney J., Kudeyarov V., Murdoch P. & Zhu Z.L. (1996) Regional nitrogen budgets and riverine N&P fluxes for the drainages to the North Atlantic Ocean: Natural and human influences. Biogeochemistry, 35, 75-139
Howarth R.W., Marino R. & Cole J.J. (1988) Nitrogen-Fixation in Fresh-Water, Estuarine, and Marine Ecosystems .2. Biogeochemical Controls. Limnology and Oceanography, 33, 688-701
Jenkins M.C. & Kemp W.M. (1984) The Coupling of Nitrification and Denitrification in 2 Estuarine Sediments. Limnology and Oceanography, 29, 609-619
Jensen K., Sloth N.P., Risgaardpetersen N., Rysgaard S. & Revsbech N.P. (1994) Estimation of Nitrification and Denitrification from Microprofiles of Oxygen and Nitrate in Model Sediment Systems. Applied and Environmental Microbiology, 60, 2094-2100
Jetten M.S.M. (2001) New pathways for ammonia conversion in soil and aquatic systems. Plant and Soil, 230, 9-19
Jetten M.S.M., Strous M., van de Pas-Schoonen K.T., Schalk J., van Dongen U.G.J.M., van de Graaf A.A., Sogemann S., Muyzer G., van Loosdrecht M.C.M. & Kuenen J.G. (1999) The anaerobic oxidation of ammonium. Fems Microbiology Reviews, 22, 16
Joye S.B., Connell T.L., Miller L.G., Oremland R.S. & Jellison R.S. (1999) Oxidation of ammonia and methane in an alkaline, saline lake. Limnology and Oceanography, 44, 178-188
Keith S.M. & Herbert R.A. (1983) Dissimilatory Nitrate Reduction by a Strain of Desulfovibrio-Desulfuricans. Fems Microbiology Letters, 18, 55-59
Kelso B.H.L., Smith R.V., Laughlin R.J. & Lennox S.D. (1997) Dissimilatory nitrate reduction in anaerobic sediments leading to river nitrite accumulation. Applied and Environmental Microbiology, 63, 4679-4685
Klotz M.G. & Stein L.Y. (2008) Nitrifier genomics and evolution of the nitrogen cycle. Fems Microbiology Letters, 278, 146-156
Kuypers M.M.M., Sliekers A.O., Lavik G., Schmid M., Jorgensen B.B., Kuenen J.G., Damste J.S.S., Strous M. & Jetten M.S.M. (2003) Anaerobic ammonium oxidation by anammox bacteria in the Black Sea. Nature, 422, 608-611
Lancelot C. & Billen G. (1985) Carbon-Nitrogen relationship in nutrient metabolism of coastal marine ecosystem. In: Advances in Aquatic Microbiology (eds. Jannash HW & Leb Williams PJ), pp. 263 - 321. Academic Press London, London
Laverman A.M., Canavan R.W., Slomp C.P. & Van Cappellen P. (2007) Potential nitrate removal in a coastal freshwater sediment (Haringvliet Lake, The Netherlands) and response to salinization. Water Research, 41, 3061-3068
Lewis W.M. & Levine S.N. (1984) The Light Response of Nitrogen-Fixation in Lake Valencia, Venezuela. Limnology and Oceanography, 29, 894-900
Lillebo A.I., Morais M., Guilherme P., Fonseca R., Serafim A. & Neves R. (2007) Nutrient dynamics in Mediterranean temporary streams: A case study in Pardiela catchment (Degebe River, Portugal). Limnologica, 37, 337-348
Lorenzen J., Larsen L.H., Kjaer T. & Revsbech N.P. (1998) Biosensor determination of the microscale distribution of nitrate, nitrate assimilation, nitrification, and denitrification in

Capítulo I 28
a diatom-inhabited freshwater sediment. Applied and Environmental Microbiology, 64, 3264-3269
Mackenzie F.T. (1998) Our Changing Planet: An Introduction to Earth System Science and Global Environmental Change. 2 edn. Prentice-Hall, Upper Saddle River (NJ).
Marcarelli A.M. & Wurtsbaugh W.A. (2006) Temperature and nutrient supply interact to control nitrogen fixation in oligotrophic streams: An experimental examination. Limnology and Oceanography, 51, 2278-2289
Marcarelli A.M. & Wurtsbaugh W.A. (2007) Effects of upstream lakes and nutrient limitation on periphytic biomass and nitrogen fixation in oligotrophic, subalpine streams. Freshwater Biology, 52, 2211-2225
Matheson F.E., Nguyen M.L., Cooper A.B., Burt T.P. & Bull D.C. (2002) Fate of N-15-nitrate in unplanted, planted and harvested riparian wetland soil microcosms. Ecological Engineering, 19, 249-264
Megonigal J., Hines M. & Visscher P. (2004) Anaerobic metabolism: linkages to trace gases and aerobic processes. In: Biogeochemistry (ed. Schlesinger WH), pp. 317 - 424. Elsevier-Pergamon, Oxford, UK
Middelburg J.J., Barranguet C., Boschker H.T.S., Herman P.M.J., Moens T. & Heip C.H.R. (2000) The fate of intertidal microphytobenthos carbon: An in situ C-13-labeling study. Limnology and Oceanography, 45, 1224-1234
Mosier A.R., Doran J.W. & Freney J.R. (2002) Managing soil denitrification. Journal of Soil and Water Conservation, 57, 8
Mulder A., Vandegraaf A.A., Robertson L.A. & Kuenen J.G. (1995) Anaerobic Ammonium Oxidation Discovered in a Denitrifying Fluidized-Bed Reactor. Fems Microbiology Ecology, 16, 177-183
Munoz R. & Guieysse B. (2006) Algal-bacterial processes for the treatment of hazardous contaminants: A review. Water Research, 40, 2799-2815
Nielsen L.P., Christensen P.B., Revsbech N.P. & Sorensen J. (1990) Denitrification and Oxygen Respiration in Biofilms Studied with a Microsensor for Nitrous-Oxide and Oxygen. Microbial Ecology, 19, 63-72
Pina-Ochoa E. & Alvarez-Cobelas M. (2006) Denitrification in aquatic environments: A cross-system analysis. Biogeochemistry, 81, 111-130
Piña-Ochoa E., Høgslund S., Geslin E., Cedhagen T., Revsbech N.P., Nielsen L.P., Schweizer M., Jorissen F., Rysgaard S. & Risgaard-Petersen N. (2010) Widespread occurrence of nitrate storage and denitrification among Foraminifera and Gromiida. Proceedings of the National Academy of Sciences of the United States of America, 107, 6
Postgate J.R. (1970) Biological Nitrogen Fixation. Nature, 226, 25-& Postgate J.R., Kent H.M. & Robson R.L. (1988) Nitrogen fixation by Desulfovibrio. In: The
Nitrogen and Sulphur Cycles (eds. Cole JA & Ferguson JS), pp. 457 - 473. Cambridge University Press, Cambridge
Revsbech N.P., Jorgensen B.B. & Blackburn T.H. (1980) Oxygen in the Sea Bottom Measured with a Microelectrode. Science, 207, 1355-1356
Risgaard-Petersen N., Langezaal A.M., Ingvardsen S., Schmid M.C., Jetten M.S.M., Op den Camp H.J.M., Derksen J.W.M., Pina-Ochoa E., Eriksson S.P., Nielsen L.P., Revsbech N.P., Cedhagen T. & van der Zwaan G.J. (2006) Evidence for complete denitrification in a benthic foraminifer. Nature, 443, 93-96
Risgaard-Petersen N., Meyer R.L., Schmid M., Jetten M.S.M., Enrich-Prast A., Rysgaard S. & Revsbech N.P. (2004a) Anaerobic ammonium oxidation in an estuarine sediment. Aquatic Microbial Ecology, 36, 293-304
Risgaard-Petersen N., Nicolaisen M.H., Revsbech N.P. & Lomstein B.A. (2004b) Competition between ammonia-oxidizing bacteria and benthic microalgae. Applied and Environmental Microbiology, 70, 5528-5537
Risgaardpedersen N., Rysgaard S., Nielsen L.P. & Revsbech N.P. (1994) Diurnal-Variation of Denitrification and Nitrification in Sediments Colonized by Benthic Microphytes. Limnology and Oceanography, 39, 573-579

Capítulo I 29
Rysgaard S., Glud R.N., Risgaard-Petersen N. & Dalsgaard T. (2004) Denitrification and anammox activity in Arctic marine sediments. Limnology and Oceanography, 49, 1493-1502
Rysgaard S., Thastum P., Dalsgaard T., Christensen P.B. & Sloth N.P. (1999) Effects of salinity on NH4+ adsorption capacity, nitrification, and denitrification in Danish estuarine sediments. Estuaries, 22, 21-30
Santoro A.L. & Enrich-Prast A. (2009) Salinity control of nitrification in saline shallow coastal lagoons. Acta Limnologica Brasiliensia, 21, 6
Schindler D.W. (1977) Evolution of Phosphorus Limitation in Lakes. Science, 195, 260-262 Schmid M., Walsh K., Webb R., Rijpstra W.I.C., van de Pas-Schoonen K., Verbruggen M.J.,
Hill T., Moffett B., Fuerst J., Schouten S., Damste J.S.S., Harris J., Shaw P., Jetten M. & Strous M. (2003) Candidatus "Scalindua brodae", sp nov., Candidatus "Scalindua wagneri", sp nov., two new species of anaerobic ammonium oxidizing bacteria. Systematic and Applied Microbiology, 26, 529-538
Schubert C.J., Durisch-Kaiser E., Wehrli B., Thamdrup B., Lam P. & Kuypers M.M.M. (2006) Anaerobic ammonium oxidation in a tropical freshwater system (Lake Tanganyika). Environmental Microbiology, 8, 1857-1863
Scott J.T., McCarthy M.J., Gardner W.S. & Doyle R.D. (2008) Denitrification, dissimilatory nitrate reduction to ammonium, and nitrogen fixation along a nitrate concentration gradient in a created freshwater wetland. Biogeochemistry, 87, 99-111
Seitzinger S., Harrison J.A., Bohlke J.K., Bouwman A.F., Lowrance R., Peterson B., Tobias C. & Van Drecht G. (2006) Denitrification across landscapes and waterscapes: A synthesis. Ecological Applications, 16, 2064-2090
Seitzinger S.P. (1988) Denitrification in Fresh-Water and Coastal Marine Ecosystems - Ecological and Geochemical Significance. Limnology and Oceanography, 33, 702-724
Silver W.L., Herman D.J. & Firestone M.K. (2001) Dissimilatory nitrate reduction to ammonium in upland tropical forest soils. Ecology, 82, 2410-2416
Sinsabaugh R.L., Antibus R.K. & Linkins A.E. (1991) An Enzymatic Approach to the Analysis of Microbial Activity During Plant Litter Decomposition. Agriculture Ecosystems & Environment, 34, 43-54
Stanford G. & Smith S.J. (1972) Nitrogen Mineralization Potentials of Soils. Soil Science Society of America Proceedings, 36, 465-&
Strauss E.A. & Lamberti G.A. (2000) Regulation of nitrification in aquatic sediments by organic carbon. Limnology and Oceanography, 45, 1854-1859
Strauss E.A. & Lamberti G.A. (2002) Effect of dissolved organic carbon quality on microbial decomposition and nitrification rates in stream sediments. Freshwater Biology, 47, 65-74
Strous M. & Jetten M.S.M. (2004) Anaerobic oxidation of methane and ammonium. Annual Review of Microbiology, 58, 99-117
Strous M., Pelletier E., Mangenot S., Rattei T., Lehner A., Taylor M.W., Horn M., Daims H., Bartol-Mavel D., Wincker P., Barbe V., Fonknechten N., Vallenet D., Segurens B., Schenowitz-Truong C., Medigue C., Collingro A., Snel B., Dutilh B.E., Op den Camp H.J.M., van der Drift C., Cirpus I., van de Pas-Schoonen K.T., Harhangi H.R., van Niftrik L., Schmid M., Keltjens J., van de Vossenberg J., Kartal B., Meier H., Frishman D., Huynen M.A., Mewes H.W., Weissenbach J., Jetten M.S.M., Wagner M. & Le Paslier D. (2006) Deciphering the evolution and metabolism of an anammox bacterium from a community genome. Nature, 440, 790-794
Sumi T. & Koike I. (1990) Estimation of Ammonification and Ammonium Assimilation in Surficial Coastal and Estuarine Sediments. Limnology and Oceanography, 35, 270-286
Svensson J.M. & Leonardson L. (1996) Effects of bioturbation by tube-dwelling chironomid larvae on oxygen uptake and denitrification in eutrophic lake sediments. Freshwater Biology, 35, 289-300
Sweerts J., Stlouis V. & Cappenberg T.E. (1989) Oxygen Concentration Profiles and Exchange in Sediment Cores with Circulated Overlying Water. Freshwater Biology, 21, 401-409

Capítulo I 30
Tiedje J.M., Sexstone A.J., Myrold D.D. & Robinson J.A. (1982) Denitrification - Ecological Niches, Competition and Survival. Antonie Van Leeuwenhoek Journal of Microbiology, 48, 569-583
Tilman D. (1987) Secondary Succession and the Pattern of Plant Dominance Along Experimental Nitrogen Gradients. Ecological Monographs, 57, 189-214
Tonno I. & Noges T. (2003) Nitrogen fixation in a large shallow lake: rates and initiation conditions. Hydrobiologia, 490, 23-30
Travieso L., Benitez F., Sanchez E., Borja R. & Colmenarejo M.F. (2006) Production of biomass (algae-bacteria) by using a mixture of settled swine and sewage as substrate. Journal of Environmental Science and Health Part a-Toxic/Hazardous Substances & Environmental Engineering, 41, 415-429
van Dongen U., Jetten M.S.M. & van Loosdrecht M.C.M. (2001) The SHARON((R))-Anammox((R)) process for treatment of ammonium rich wastewater. Water Science and Technology, 44, 153-160
Vitousek P.M., Aber J.D., Howarth R.W., Likens G.E., Matson P.A., Schindler D.W., Schlesinger W.H. & Tilman D.G. (1997) Human alteration of the global nitrogen cycle: Sources and consequences. Ecological Applications, 7, 737-750
Vitousek P.M. & Howarth R.W. (1991) Nitrogen Limitation on Land and in the Sea - How Can It Occur. Biogeochemistry, 13, 87-115
Wilkie A.C. & Mulbry W.W. (2002) Recovery of dairy manure nutrients by benthic freshwater algae. Bioresource Technology, 84, 81-91
Zehr J.P. & Ward B.B. (2002) Nitrogen cycling in the ocean: New perspectives on processes and paradigms. Applied and Environmental Microbiology, 68, 1015-1024
Zhang M., Xu J. & Xie P. (2008) Nitrogen dynamics in large shallow eutrophic Lake Chaohu, China. Environmental Geology, 55, 1-8

Capítulo II 31
CONTROLE DA NITRIFICAÇÃO PELA SALINIDADE EM LAGOAS
COSTEIRAS SALINAS
Acta Limnol. Bras., 2009, vol. 21, no. 2, p. 263-267
Ana Lúcia Santoro & Alex Enrich Prast
Resumo: Objetivos: O objetivo do presente estudo foi mostrar a importância do controle da salinidade
sobre a atividade nitrificante no sedimento de lagoas costeiras rasas e salinas; Métodos: Foram
mensuradas as taxas potenciais de nitrificação no sedimento utilizando o método de produção de nitrato
ao longo de um curto período de tempo e a concentração de clorofila-a no sedimento foi utilizada como
indicativo da biomassa algal; Resultados: As lagoas estudadas apresentaram diferenças significativas nas
taxas potenciais de nitrificação, concentrações de clorofila-a no sedimento e salinidade da coluna d’água.
As taxas de nitrificação variaram de valores não detectáveis a 120 nmol NO3- cm-3 h-1 e se
correlacionaram negativamente com os valores de salinidade, apresentando um padrão de decaimento
exponencial com a variação de salinidade entre 0 e 30 ‰; Conclusões: A ausência de correlação entre
nitrificação potencial e clorofila-a pode ser atribuída à baixa densidade algal contemplada neste estudo.
Para lagoas com reduzida colonização algal no sedimento, como as deste estudo, a salinidade da água é
um importante fator de regulação do processo de nitrificação no sedimento.
Palavras-chave: nitrificação, salinidade, lagoas costeiras, lagos rasos, tropicos. INTRODUÇÃO
A nitrificação é definida como a oxidação de compostos reduzidos de nitrogênio e é um
processo microbiano utilizado para produção de energia. Este processo ocorre em duas etapas,
sendo a amônia (NH4+) oxidada a nitrito (NO2
-) e posteriormente a nitrato (NO3-) sob condições
óxicas (Fenchel et al. 1998). Uma vez que bactérias nitrificantes são aeróbicas obrigatórias, a
disponibilidade de oxigênio é um reconhecido fator controlador do processo de nitrificação no
sedimento.
Algas bentônicas ao colonizarem o sedimento modificam as condições químicas da
interface sedimento-água, especialmente graças à produção de O2 através da atividade
fotossintética e do seu potencial para assimilação de nutrientes (Wilkie & Mulbry 2002; Munoz
& Guieysse 2006). A atividade destes organismos aumenta a penetração de oxigênio no
sedimento, estimulando processos aeróbicos incluindo a nitrificação (Risgaardpedersen et al.
1994; Lorenzen et al. 1998; An & Joye 2001; Travieso et al. 2006). No entanto, parte dos
produtos fotossintéticos excretados representa uma fonte adicional de carbono para bactérias
heterotróficas, aumentando a atividade destes organismos no sedimento e podendo
conseqüentemente estimular a competição entre bactérias heterotróficas e nitrificantes
(Middelburg et al. 2000).

Capítulo II 32
Microalgas bentônicas são comumente encontradas em ambientes salinos. Porém, nestes
ambientes, o possível efeito positivo causado por algas bentônicas sobre o processo de
nitrificação pode ser tamponado por um efeito negativo causado pela salinidade. Segundo
Risgaard-Petersen et al. (2004), alterações na salinidade da água podem representar um
importante fator controlador da atividade nitrificante no sedimento de ecossistemas aquáticos.
A salinidade regula a capacidade de absorção de NH4+ do sedimento (Boatman &
Murray 1982) e causa um impacto fisiológico sobre as bactérias nitrificantes (Joye &
Hollibaugh 1995; Rysgaard et al. 1999), fazendo com que normalmente as taxas de nitrificação
sejam mais baixas em ambientes com elevada salinidade. Além disso, a reduzida solubilidade
do oxigênio em condições de alta salinidade diminui a penetração e a disponibilidade deste gás
no sedimento, reduzindo a espessura da zona de ocorrência da nitrificação (Revsbech et al.
1980).
Ainda assim, alguns estudos observam relações positivas entre salinidade e nitrificação.
Em sedimentos estuarinos, alguns trabalhos mostram maior atividade nitrificante sob salinidade
intermediária (Meyer et al. 2001; Magalhaes et al. 2005). Experimentos em laboratório
mostraram que algumas espécies de bactérias nitrificantes são capazes de se adaptar a variações
de salinidade e que, a tolerância a salinidade normalmente reflete o ambiente original (Finstein
& Bitzky 1972). Helder e DeVries (1983) observaram que bactérias oxidadoras de amônia
poderiam se adaptar e crescer sob um gradiente de salinidade entre 0 e 35 ppm, mas que sob
elevadas salinidades este crescimento ocorreria depois de uma fase ‘lag’ de até 20 dias. Esta
longa fase pré-crescimento pode ser uma desvantagem para comunidades nitrificantes de
ambientes instáveis com relação à salinidade, como lagoas costeiras. Estes ecossistemas são
submetidos a mudanças de salinidade relativamente rápidas como resposta a chuva, entrada de
água do mar e intensidade de evaporação.
O objetivo do presente estudo foi mostrar que a mudança da salinidade pode ser um
importante fator regulador da atividade nitrificante no sedimento de lagoas costeiras colonizadas
por algas bentônicas.
MATERIAL E MÉTODOS
Área de Estudo
O presente estudo foi realizado nas lagoas costeiras Pires, Preta, Catingosa e Visgueiro,
com áreas de 0,9, 2,2, 0,1 e 1,2 km2 respectivamente. As lagoas são localizadas no Parque
Nacional da Restinga de Jurubatiba (22º 22’ 30” S e 41º 15’ 42” O), uma das mais importantes
áreas costeiras de preservação no Brasil (nordeste do estado do Rio de Janeiro; Fig. 1). Esta
região apresenta temperatura média anual de 24ºC e período chuvoso mais intenso entre os
meses de Novembro e Abril (Carmouze et al. 1991). Os quatro ambientes estudados são rasos

Capítulo II 33
(profundidade média de 0,8 m) e paralelos à costa, sendo classificados como lagoas formadas
nas depressões arenosas que constituem o ecossistema de restinga (Martin & Domingues 1994).
Estes sistemas aquáticos são influenciados por aportes terrestres de compostos orgânicos ácidos
pigmentados e também por aportes salinos subterrâneos (Suzuki et al. 1998), fazendo com que a
água destas lagoas apresente uma ampla variação de coloração e salinidade em curtos intervalos
de tempo (Enrich-Prast et al. 2004). Com isso, é possível observar nestes ecossistemas uma
grande variação de salinidade e de colonização de algas bentônicas tanto temporalmente quanto
espacialmente, entre lagoas.
FIG 1: Ilustração das lagoas do Parque Nacional da Restinga de Jurubariba, com detalhe na posição das quatro lagoas estudadas.
Amostragem e análises
O sedimento de todas as lagoas foi amostrado em três diferentes meses (Outubro/05,
Dezembro/05 e Janeiro/06) no formato de testemunhos utilizando tubos de acrílico. Quatro
réplicas foram obtidas em cada um dos ambientes e somente o primeiro centímetro dos
testemunhos foi separado e homogeneizado para as análises de nitrificação potencial e
concentração de clorofila-a.
No momento das amostragens in situ foram mensurados os seguintes parâmetros
limnológicos da coluna d’água: pH (Analion PM 608 pH meter), salinidade e temperatura

Capítulo II 34
(Thermosalinometer YSI-30). A concentração de amônia foi mensurada de acordo com (Bower
& Holmhansen 1980).
As taxas potenciais de nitrificação no sedimento foram obtidas a partir da produção de
nitrato durante um período de incubação de 3 – 4 horas. Este curto período de incubação exclui
a possibilidade de crescimento da comunidade bacteriana nitrificante durante o experimento
(Kaplan 1983). A incubação foi realizada utilizando tubos de 50 ml posicionados sobre um
agitador a 100 rpm. Em cada um dos tubos foram adicionados 2 ml de sedimento previamente
homogeneizado, 30 ml de água da própria lagoa, coletada no momento da amostragem de
sedimento, além de 1 ml de NH4+Cl (20mM) e 1 ml de KH2PO4 (4mM) a fim de evitar qualquer
limitação por nutrientes (Belser 1979). A cada hora de incubação, 6 ml do conteúdo de cada um
dos tubos foi retirado e imediatamente centrifugado a 5000 rpm para posterior determinação das
concentrações de NO3- através de análise de redução utilizando cádmio em um sistema de
injeção de fluxo (ASIA/Ismatec).
As taxas potenciais de nitrificação no sedimento (TPN) foram então calculadas a partir
do ângulo de inclinação (α) da reta obtida com a regressão linear das concentrações de nitrato
obtidas ao longo do tempo de incubação, como mostra a seguinte equação:
TPN (nmol NO3- cm-3 h-1) = α (nmol NO3
- L-1 h-1) -----------------------------------
As concentrações de clorofila-a foram utilizadas como um índice de biomassa algal e
foi mensurada a partir de espectrofotometria após extração do pigmento utilizando acetona 90%
conforme (Dalsgaard et al. 2000).
Análise dos dados
Os dados obtidos no presente estudo não apresentaram distribuição normal
(Kolmogorov-Smirnov, p < 0,05) e mesmo após a transformação dos dados, estes não
apresentaram características paramétricas (Zar 1996). Sendo assim, o teste não paramétrico de
Kruskall-Wallis (p < 0,05) seguido do teste de comparação múltipla de Dunn (p < 0,05) foi
utilizado para identificação de possíveis diferenças entre os diferentes ambientes amostrados
com relação à concentração de clorofila-a, taxas potenciais de nitrificação e salinidade. O teste
de correlação não paramétrica de Spearman e um modelo de regressão linear (p < 0,05) também
foram utilizados para compreensão dos resultados.
Volume de água (L)
Volume de sedimento (cm-3)

Capítulo II 35
RESULTADOS
A produção de nitrato durante as incubações foi linear em todos os sedimentos
analisados (r2 = 0,75 – 0,99), indicando o início da atividade nitrificante já no início do período
de incubação. A produção linear de nitrato também caracteriza a ausência de crescimento
significativo da comunidade nitrificante durante o período de incubação.
As taxas potenciais de nitrificação foram significativamente diferentes entre algumas
das lagoas amostradas (Kruskall-Wallis; p < 0,05), variando entre valores abaixo do limite de
detecção do método até 120 nmol NO3- cm-3 h-1 (Fig. 2a). As maiores e as menores taxas deste
processo mensuradas neste estudo foram observadas nas lagoas Pires e Visgueiro,
respectivamente. Também foi possível observar diferenças significativas entre alguns dos
ambientes com relação às concentrações de clorofila-a no sedimento (Kruskall-Wallis; p <
0,05), no entanto, as lagoas Pires e Visgueiro não se diferenciaram estatisticamente neste
aspecto (Dunn test; p < 0,05). Essas duas lagoas apresentaram as maiores concentrações de
clorofila-a, mas a variação deste parâmetro durante o período de estudo foi muito restrita e as
maiores concentrações não passaram de 10,0 mg clo-a m-2 (Fig. 2b).
FIG 2: Taxas potenciais de nitrificação (a) e concentrações de clorofila-a (b) no sedimento das lagoas estudadas. Os símbolos representam medianas máximos e mínimos. Letras diferentes indicam diferença significativa (Kruskall-Wallis; p < 0,05).
Nitr
ifica
ção
(nm
ol N
O3- c
m-3 h
-1)
Clo
rofil
a-a
(mg
clo-
a m
-2)
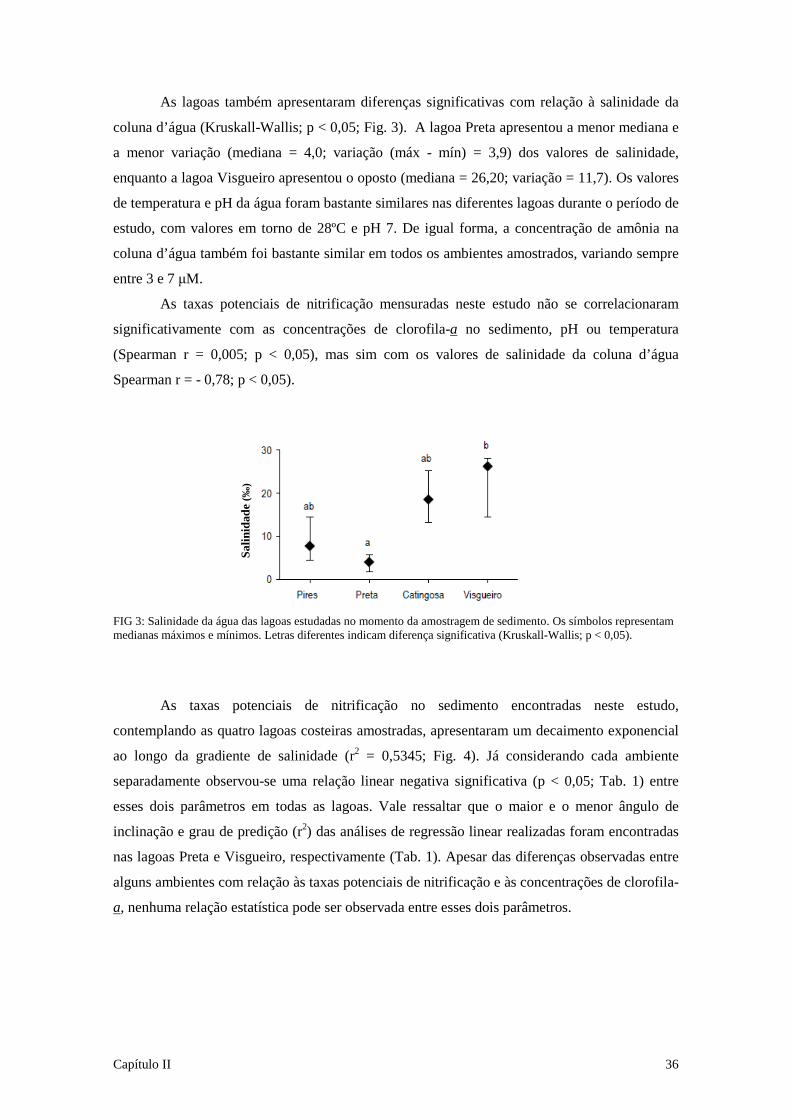
Capítulo II 36
As lagoas também apresentaram diferenças significativas com relação à salinidade da
coluna d’água (Kruskall-Wallis; p < 0,05; Fig. 3). A lagoa Preta apresentou a menor mediana e
a menor variação (mediana = 4,0; variação (máx - mín) = 3,9) dos valores de salinidade,
enquanto a lagoa Visgueiro apresentou o oposto (mediana = 26,20; variação = 11,7). Os valores
de temperatura e pH da água foram bastante similares nas diferentes lagoas durante o período de
estudo, com valores em torno de 28ºC e pH 7. De igual forma, a concentração de amônia na
coluna d’água também foi bastante similar em todos os ambientes amostrados, variando sempre
entre 3 e 7 µM.
As taxas potenciais de nitrificação mensuradas neste estudo não se correlacionaram
significativamente com as concentrações de clorofila-a no sedimento, pH ou temperatura
(Spearman r = 0,005; p < 0,05), mas sim com os valores de salinidade da coluna d’água
Spearman r = - 0,78; p < 0,05).
FIG 3: Salinidade da água das lagoas estudadas no momento da amostragem de sedimento. Os símbolos representam medianas máximos e mínimos. Letras diferentes indicam diferença significativa (Kruskall-Wallis; p < 0,05).
As taxas potenciais de nitrificação no sedimento encontradas neste estudo,
contemplando as quatro lagoas costeiras amostradas, apresentaram um decaimento exponencial
ao longo da gradiente de salinidade (r2 = 0,5345; Fig. 4). Já considerando cada ambiente
separadamente observou-se uma relação linear negativa significativa (p < 0,05; Tab. 1) entre
esses dois parâmetros em todas as lagoas. Vale ressaltar que o maior e o menor ângulo de
inclinação e grau de predição (r2) das análises de regressão linear realizadas foram encontradas
nas lagoas Preta e Visgueiro, respectivamente (Tab. 1). Apesar das diferenças observadas entre
alguns ambientes com relação às taxas potenciais de nitrificação e às concentrações de clorofila-
a, nenhuma relação estatística pode ser observada entre esses dois parâmetros.
Sal
inid
ade
(‰)

Capítulo II 37
FIG 4: Modelo de decaimento exponencial entre as taxas de nitrificação no sedimento e a salinidade da coluna d’água. Equation: y = 89.29 *exp (– 0.1285 * x) + (– 2.929); R2 = 0.5345; n = 44.
TAB. 1: Modelo de regressão linear aplicado entre as taxas de nitrificação no sedimento e a salinidade da coluna d’água para cada uma das lagoas estudadas.
DISCUSSÃO
As taxas potenciais de nitrificação no sedimento variaram em até duas ordens de
magnitude entre as diferentes lagoas estudadas. A ausência de uma relação positiva ou negativa
entre a biomassa de algas bentônicas, representada pela concentração de clorofila-a no
sedimento, e a atividade nitrificante pode ser atribuída à baixa densidade algal contemplada
durante o período de estudo, que foi de duas a três ordens de magnitude menor que a
normalmente encontrada em estudos deste tipo (Risgaard-Petersen 2003).
Apesar das concentrações de amônio no sedimento e o pH das lagoas estudadas não
apresentarem grande variação temporal ou ainda entre os diferentes ambientes durante o período
de estudo, pequenas variações destes parâmetros já podem ser importantes na regulação da
atividade nitrificante in situ. O efeito direto destes parâmetros, no entanto, não pôde ser avaliado
neste estudo por limitações laboratoriais.
A significativa relação observada entre as taxas de nitrificação no sedimento e a
salinidade indica que este parâmetro limnológico deve ser um dos principais fatores reguladores
da atividade nitrificante no sedimento de lagoas costeiras, especialmente em ambientes pouco
colonizados por algas bentônicas. Rysgaard et al. (1999) mostram um decaimento da atividade
nitrificante com o aumento da salinidade em estuários dinamarqueses, atribuindo esta queda
principalmente à redução da capacidade de absorção de NH4+ pelo sedimento. Segundo os
Salinidade (‰)
Nitr
ifica
ção
(nm
ol N
O3- c
m-3 h
-1)

Capítulo II 38
autores, a redução mais pronunciada na absorção de amônia pelo sedimento ocorreu no aumento
de salinidade entre 0 e 10 ‰. Esta faixa crítica de aumento da salinidade é comparável a faixa
de variação observada na lagoa Preta, onde foi encontrada a regressão linear mais acentuada (de
maior ângulo de inclinação) entre as taxas potenciais de nitrificação no sedimento e a salinidade
da água.
A influência da salinidade deve ser mais drástica em ambientes de baixa salinidade uma
vez que os organismos nitrificantes precisarão de mais tempo para se adaptar às variações entre
água doce e salobra. Os resultados obtidos por Helder & Devries (1983) mostraram que a
adaptação de bactérias nitrificantes de ambientes salinos à condição de água doce é
particularmente lenta. O modelo de regressão linear aplicado às diferentes lagoas estudadas
mostra uma relação negativa mais acentuada (de maior inclinação) nas lagoas Pires e Preta,
justamente as lagoas cuja mediana de salinidade foi menor que 10 ‰. Esta influencia negativa
mais acentuada sobre a atividade nitrificante em condições de baixa salinidade também explica
o decaimento exponencial observado entre nitrificação e salinidade.
Os resultados obtidos no presente estudo sugerem que um mesmo aumento de
salinidade deve ser mais crítico para a atividade nitrificante local em ambientes aquáticos de
água doce ou de baixa influência salina que em ecossistemas mais salinos. A salinidade deve ser
então um dos principais fatores reguladores da atividade nitrificante em ecossistemas costeiros,
especialmente nos de baixa salinidade.
REFERÊNCIAS
An S. & Joye S.B. (2001) Enhancement of coupled nitrification-denitrification by benthic photosynthesis in shallow estuarine sediments. Limnology and Oceanography, 46, 62-74
Belser L.W. (1979) Population Ecology of Nitrifying Bacteria. Annual Review of Microbiology, 33, 309-333
Boatman C.D. & Murray J.W. (1982) Modeling Exchangeable Nh4+ Adsorption in Marine-Sediments - Process and Controls of Adsorption. Limnology and Oceanography, 27, 99-110
Bower C.E. & Holmhansen T. (1980) A Salicylate-Hypochlorite Method for Determining Ammonia in Seawater. Canadian Journal of Fisheries and Aquatic Sciences, 37, 794-798
Carmouze J.P., Knoppers B. & Vasconcelos P. (1991) Metabolism of a Subtropical Brazilian Lagoon. Biogeochemistry, 14, 129-148
Dalsgaard T., Nielsen L.P., Brotas V., Viaroli P., Underwood G., Nedwell D.B., SUNDBACK K., RYSGAARD S., Miles A., Bartoli M., Dong L., Thornton D.C.O., Ottosen L.D.M., Castaldelli G. & Risgaard-Petersen N. (2000) Protocol handbook for NICE – Nitrogen Cycling in Estuaries: a project under the EU research programme: Marine
Science and Technology (MAST III). National Environmental Research Institute, Silkeborg. Enrich-Prast A., Bozelli R.L., Esteves F.D. & Meirelles-Pereira F. (2004) Lagoas Costeiras da
Restinga de Jurubatiba: Descrição de suas variáveis Limnológicas. In: Pesquisas de longa duração na restinga de Jurubatiba: Ecologia, história natural e conservação (eds. Rocha CFD, Esteves FA & Scarano FR), p. 376. RiMa Editora, São Carlos

Capítulo II 39
Fenchel T., King G.M. & Blackburn T.H. (1998) Bacterial biogeochemistry: the ecophysiology of mineral cycling. 2 edn. Academic Press.
Finstein M.S. & Bitzky M.R. (1972) Relationships of Autotrophic Ammonium-Oxidizing Bacteria to Marine Salts. Water Research, 6, 31-&
Helder W. & Devries R.T.P. (1983) Estuarine Nitrite Maxima and Nitrifying Bacteria (Ems-Dollard Estuary). Netherlands Journal of Sea Research, 17, 1-18
Joye S.B. & Hollibaugh J.T. (1995) Influence of Sulfide Inhibition of Nitrification on Nitrogen Regeneration in Sediments. Science, 270, 623-625
Kaplan W.A. (1983) Nitrification. In: Nitrogen in the marine environment (eds. CARPENTER EJ & CAPONE DJ), pp. 139 - 190. Academic Press, New York
Lorenzen J., Larsen L.H., Kjaer T. & Revsbech N.P. (1998) Biosensor determination of the microscale distribution of nitrate, nitrate assimilation, nitrification, and denitrification in a diatom-inhabited freshwater sediment. Applied and Environmental Microbiology, 64, 3264-3269
Magalhaes C.M., Joye S.B., Moreira R.M., Wiebe W.J. & Bordalo A.A. (2005) Effect of salinity and inorganic nitrogen concentrations on nitrification and denitrification rates in intertidal sediments and rocky biofilms of the Douro River estuary, Portugal. Water Research, 39, 1783-1794
Martin L. & Domingues J.M.L. (1994) Geological history as coastal lagoons. In: Coastal lagoon processes (ed. KJERVE B), pp. 41 - 68. Elsevier Oceanography Series, Netherlands
Meyer R.L., Kjaer T. & Revsbech N.P. (2001) Use of NOx- microsensors to estimate the activity of sediment nitrification and NOx- consumption along an estuarine salinity, nitrate, and light gradient. Aquatic Microbial Ecology, 26, 181-193
Middelburg J.J., Barranguet C., Boschker H.T.S., Herman P.M.J., Moens T. & Heip C.H.R. (2000) The fate of intertidal microphytobenthos carbon: An in situ C-13-labeling study. Limnology and Oceanography, 45, 1224-1234
Munoz R. & Guieysse B. (2006) Algal-bacterial processes for the treatment of hazardous contaminants: A review. Water Research, 40, 2799-2815
Revsbech N.P., Sorensen J., Blackburn T.H. & Lomholt J.P. (1980) Distribution of Oxygen in Marine-Sediments Measured with Microelectrodes. Limnology and Oceanography, 25, 403-411
Risgaard-Petersen N. (2003) Coupled nitrification-denitrification in autotrophic and heterotrophic estuarine sediments: On the influence of benthic microalgae. Limnology and Oceanography, 48, 93-105
Risgaard-Petersen N., Meyer R.L., Schmid M., Jetten M.S.M., Enrich-Prast A., Rysgaard S. & Revsbech N.P. (2004) Anaerobic ammonium oxidation in an estuarine sediment. Aquatic Microbial Ecology, 36, 293-304
Risgaardpedersen N., Rysgaard S., Nielsen L.P. & Revsbech N.P. (1994) Diurnal-Variation of Denitrification and Nitrification in Sediments Colonized by Benthic Microphytes. Limnology and Oceanography, 39, 573-579
Rysgaard S., Thastum P., Dalsgaard T., Christensen P.B. & Sloth N.P. (1999) Effects of salinity on NH4+ adsorption capacity, nitrification, and denitrification in Danish estuarine sediments. Estuaries, 22, 21-30
Suzuki M.S., Ovalle A.R.C. & Pereira E.A. (1998) Effects of sand bar openings on some limnological variables in a hypertrophic tropical coastal lagoon of Brazil. Hydrobiologia, 368, 111-122
Travieso L., Benitez F., Sanchez E., Borja R. & Colmenarejo M.F. (2006) Production of biomass (algae-bacteria) by using a mixture of settled swine and sewage as substrate. Journal of Environmental Science and Health Part a-Toxic/Hazardous Substances & Environmental Engineering, 41, 415-429
Wilkie A.C. & Mulbry W.W. (2002) Recovery of dairy manure nutrients by benthic freshwater algae. Bioresource Technology, 84, 81-91
Zar J.H. (1996) Biostatistical analysis. 3 edn. Upper Saddle River: Prentice Hall International Editions.

Capítulo III 40
TAXAS E FATORES REGULADORES DO PROCESSO DE DESNITRIFICAÇÃO NO
SEDIMENTO DE DUAS LAGOAS COSTEIRAS TROPICAIS
Ana Lúcia Santoro
Resumo: As taxas de desnitrificação no sedimento foram mensalmente mensuradas em duas lagoas
costeiras tropicais de diferentes status tróficos utilizando o método de pareamento isotópico de 15N.
Alguns parâmetros ambientais foram correlacionados às taxas de desnitrificação a fim de determinar os
principais fatores deste processo em cada um dos ambientes. As taxas de desnitrificação encontradas
foram extremamente baixas se comparadas às taxas de consumo de oxigênio pelo sedimento, com médias
mensais variando de 0,15 a 1,78 µmol N2 m-2 h-1 e de 426 a 4248 µmol O2 m
-2 h-1 respectivamente. O
consumo de oxigênio pelo sedimento foi em geral três vezes maior no ambiente eutrófico, enquanto a
média de desnitrificação não se diferenciou significativamente da encontrada no ambiente oligotrófico. A
baixa atividade desnitrificante foi atribuída tanto às baixas concentrações de NO3- na água (< 2,0 µM)
quanto ao reduzido fornecimento deste substrato via nitrificação. A presença de NH4+ na coluna d’água
em concentrações entre 2,7 e 46,8 µM também indicam a ausência de qualquer atividade nitrificante
expressiva tanto na lagoa oligotrófica quanto na eutrófica. Sedimentos de ambientes temperados com
taxas de consumo de oxigênio similares as encontradas aqui normalmente apresentam taxas de
nitrificação-desnitrificação de uma a duas ordens de magnitude maiores, indicando uma regulação
diferencial destes processos em ambientes tropicais.
INTRODUÇÃO
Desnitrificação é um processo de redução dissimilatória do nitrato no qual o nitrato
(NO3-) ou nitrito (NO2
-) é reduzido anaerobicamente a nitrogênio molecular (N2) por bactérias
heterotróficas anaeróbicas facultativas. A atividade desnitrificante requer um meio anóxico além
uma fonte de matéria orgânica e nitrato (Mosier et al. 2002). Uma vez que a maioria do
nitrogênio fixado disponível no sedimento está na forma de íon amônio (NH4+), ao menos que
haja um fluxo de nitrato da coluna d’água para dentro do sedimento, a desnitrificação no
sedimento depende das taxas locais de nitrificação (oxidação do amônio a nitrito ou nitrato). O
nitrato produzido na zona aeróbica do sedimento (ou proveniente da coluna d’água) difunde até
a zona subóxica onde o processo de desnitrificação ocorre (Nielsen et al. 1990). Esta
combinação de processos é comumente chamada de nitrificação-desnitrificação acoplada,
enquanto a desnitrificação subsidiada pelo fluxo físico de nitrato da coluna d’água para o
sedimento pode ser chamada de desnitrificação direta (Fennel et al. 2009).
Segundo Fennel et al. (2009), o consumo de oxigênio pelo sedimento (COS) deve ser
um preditor efetivo da atividade desnitrificante no sedimento. A profundidade da zona aeróbica

Capítulo III 41
no sedimento de ecossistemas aquáticos é diretamente afetada pelo COS, que por sua vez é
associado às taxas de decomposição da matéria orgânica e nitrificação (Seitzinger et al. 2006).
Assume-se que sistemas aquáticos continentais contribuam com cerca de 20 % do total
global de desnitrificação (Seitzinger et al. 2006), sendo a desnitrificação no sedimento a maior
via de perda de nitrogênio fixado de sistemas aquáticos naturais ou alterados. No entanto, a
grande maioria dos estudos sobre desnitrificação em ecossistemas aquáticos contempla zonas
temperadas do hemisfério norte, com pouquíssimos dados em regiões tropicais (Pina-Ochoa &
Alvarez-Cobelas 2006).
O objetivo do presente estudo foi avaliar possíveis mudanças temporais nas taxas de
desnitrificação no sedimento e os fatores reguladores deste processo em duas lagoas costeiras
tropicais com diferentes status tróficos.
ÁREA DE ESTUDO
As lagoas Imboassica (22º 25’ S e 42º 56’ O) e Cabiúnas (22º 00’ S e 42º 00’ O) estão
situadas na costa norte do estado do Rio de Janeiro, sudeste do Brasil (Fig. 1). Esta região
possui temperatura média mensal máxima e mínima entre 20,7 ºC em Julho e 26,2 ºC em
fevereiro (INMET 1992). Os dois ambientes são alongados perpendicularmente à linha da costa,
rasos, relativamente próximos, distando um do outro cerca de 30 km, e possuem área de bacia
hidrográfica similar, 50 e 45 km2 respectivamente.
FIG. 1: Área de estudo com destaque para as duas lagoas amostradas.
Rio de Janeiro State
Macaé City
Imboassica
41o 45’ W
22o 15’ S
Macaé City
5 0 5 10
Kilometers
Cabiúnas

Capítulo III 42
A lagoa Imboassica, com área de 3,26 km2, localiza-se em uma planície fluvio-marinha
quaternária (CPRM 2000). A área de bacia deste sistema caracteriza-se por uso predominante de
pastagem extensiva; no entanto, esta lagoa recebe aportes expressivos de nutrientes de fontes
urbanas. Descargas permanentes de esgoto doméstico contribuíram para a eutrofização da lagoa
Imboassica principalmente após 1995 (Esteves 1998). Durante o período de amostragem (2001)
a lagoa Imboassica foi classificada como mesoeutrófica a eutrófica, com concentração média de
fósforo total de 52 mg m-3 (Salas & Martino 2001).
A lagoa Cabiúnas é menor (0,34 km2 de área) e menos impactada por atividade
humanas já que esta está localizada dentro de uma área de conservação (Parque Nacional da
Restinga de Jurubatiba). Este ecossistema está situado em uma planície marinha arenosa
quaternária, onde a vegetação terrestre original, chamada de restinga, está preservada (CPRM
2000). A lagoa Cabiúnas foi classificada como oligotrófica durante o período de estudo, com
concentração média de fósforo total de 11 mg m-3 (Salas & Martino 2001).
MATERIAL E MÉTODOS
As taxas de desnitrificação e consumo de oxigênio no sedimento foram mesuradas
mensalmente de fevereiro a outubro de 2001 de acordo com Dalsgaard et al. (2000). A cada
mês, água (utilizando garrafa de van D’orn) e testemunhos de sedimento (utilizando tubos de
acrílico; 15,8 x 3,6 cm) foram coletados na área central de cada lagoa e imediatamente
transportados para um laboratório de campo.
No laboratório, as amostras de sedimento de cada uma das lagoas foram dispostas em
pequenos tanques contendo água da própria lagoa e foram mantidas aeradas sob temperatura
local durante duas horas para estabilização. As taxas de consumo de oxigênio e desnitrificação
no sedimento foram então determinadas através de incubações de 2 a 3 horas de duração.
O consumo de oxigênio no sedimento foi o primeiro processo mensurado e depois de
um novo período de estabilização de 2 horas foram então obtidas as taxas de desnitrificação.
Este último processo foi mensurado através do método de pareamento isotópico proposto por
Nielsen (1992), com adição de 1 ml 15NO3- (5 mM) em cada um dos testemunhos alcançando
concentração de aproximadamente 100 µM na coluna d’água. Assumindo uma mistura
randômica do 15NO3- adicionado com o 14NO3
- presente na coluna d’água e produzido no
sedimento pela nitrificação, a formação de 29N2 (14N15N) e 30N2 (
15N15N) ao final da incubação é
utilizada para calcular as taxas reais e potenciais de desnitrificação, assim como as frações Dn
(nitrificação-desnitrificação) e Dw (desnitrificação utilizando nitrato proveniente da coluna
d’água).
As concentrações de oxigênio foram mensuradas com o auxílio de um oxímetro (YSI –
55), já as concentrações de NO3- + NO2
- (Golterman et al. 1978) e amônio (Bower &

Capítulo III 43
Holmhansen 1980) por espectrofotometria. As espécies isotópicas de nitrogênio foram
analisadas utilizando um espectômetro de massas (Europa Scientific).
Uma análise de componentes principais (ACP) foi realizada utilizando a matriz de
correlação entre os reconhecidamente principais fatores reguladores diretos ou indiretos do
processo de desnitrificação: consumo de oxigênio no sedimento (um indicativo das condições
aeróbicas locais) e disponibilidades de nitrato e amônia. Tal matriz foi utilizada para detectar
relações dominantes entre as citadas variáveis ambientais e a desnitrificação no sedimento, a fim
de compreender os principais processos reguladores em cada sistema estudado. Os valores de
COS das diferentes lagoas apresentaram distribuição Gaussiana (Normality test, p < 0,05) e
foram comparados estatisticamente através de uma análise de teste t não pareado ao nível de
significância de 5 %. Já as taxas totais de desnitrificação, com distribuição não paramétrica
(Normality test, p < 0,05), foram comparadas estatisticamente utilizando o teste de Mann-
Whitney (p < 0,05). As frações Dn e Dw foram comparadas através do teste não paramétrico de
Kruskal-Wallis seguido do pós teste de Dunn, ambos ao nível de significância de 5 %.
RESULTADOS
As concentrações de amônia na coluna d’água ficaram abaixo de 2,5 µM em todos os
meses amostrados na lagoa Cabiúnas, enquanto os mesmos valores variaram entre < 2,0 µM e
46,78 µM na lagoa Imboassica. Os dois ambientes apresentaram concentrações de nitrato na
coluna d’água abaixo de 2,0 µM durante todo o período de estudo (Fig. 2).
FIG. 2: Concentrações de NH4+ (A) e NO3
- (B) na coluna d’água das lagoas Imboassica ( • ) e Cabiúnas ( ◦ ) durante os meses de estudo. Notar a diferença de escala dos gráficos.
As taxas de COS foram significativamente maiores na lagoa Imboassica (t test, p <
0,05), com valores médios entre 1838 µmol O2 m-2 h-1 no mês de maio e 5258 µmol O2 m
-2 h-1
no mês de Abril. Já a lagoa Cabiúnas apresentou valores médios de COS entre 426 µmol O2 m-2
h-1 em Outubro e 1294 µmol O2 m-2 h-1 no mês de junho (Fig. 3C e D).
As taxas reais de desnitrificação variaram de 0,013 a 4,347 µmol N2 m-2 h-1 na lagoa
Cabiúnas e de 0,214 a 1,671 µmol N2 m-2 h-1 na lagoa Imboassica (Fig. 3A e B). A lagoa
Feb Mar Apr May Jun Aug Oct0
5
10
30
50 A
µµ µµM
Feb Mar Apr May Jun Aug Oct0
1
2 B
Fev Abr Mai Ago Out Fev Abr Mai Ago Out
NH
4+
µM
NO
3-

Capítulo III 44
Cabiúnas apresentou taxas um pouco maiores nos primeiros três meses de estudo, mas de
maneira geral não houve diferença significativa (Mann-Whitney, p < 0,05) entre as lagoas com
relação a este processo.
FIG. 3: Taxas reais de desnitrificação (Dn-nitrificação-desnitrificação acoplada, barras claras e Dw-desnitrificação direta, barras escuras) e taxas de consumo de oxigênio no sedimento (COS) nas lagoas Imboassica e Cabiúnas em cada um dos meses de estudo. Os símbolos representam media e desvio padrão.
As taxas reais de desnitrificação foram divididas em nitrificação-desnitrificação (Dn) e
desnitrificação direta (Dw). A importância relativa da fração Dn nas duas lagoas estudadas é
facilmente observada nas figuras 3A e 3B. Esta fração variou de 11,4 a 88,9 % da taxa total de
desnitrificação na lagoa Imboassica e de 44,5 a 98,7 % na lagoa Cabiúnas. As taxas Dn e Dw
não se diferenciaram significativamente entre elas na lagoa Imboassica, mas na lagoa Cabiúnas
as taxas Dn prevaleceram de maneira significativa (Dunn, p < 0,05).
Uma relação linear significativa (F test, p < 0,05) entre as taxas Dn e o COS pôde ser
observada em ambos os sistemas amostrados (Fig. 4). No entanto, a lagoa Cabiúnas apresentou
os maiores valores de ângulo de inclinação e coeficiente de determinação (R).
Feb Mar Apr May Jun Aug Oct0
1
2
3 Imboassica
Den
itrifi
catio
nµµ µµ
mol
N2
m-2
h-1
Feb Mar Apr May Jun Aug Out0
1000
2000
3000
4000
5000
SO
Cµµ µµ
mol
O2
m-2
h-1
Feb Mar Apr May Jun Aug Oct0
1000
2000
3000
4000
5000
Feb Mar Apr May Jun Aug Oct0
1
2
3 Cabiúnas
Des
nitr
ifica
ção
(µm
ol N
2 m
-2 h
-1)
CO
S (
µmol
O2
m-2
h-1
)
Fev Abr Mai Ago Out
Fev Abr Mai Ago Out Fev Abr Mai Ago Out
Fev Abr Mai Ago Out

Capítulo III 45
FIG. 4: Regressão linear entre as taxas de nitrificação-desnitrificação (Dn) e consumo de oxigênio no sedimento (COS) nas lagoas Imboassica (•, N = 47) e Cabiúnas (◦, N = 31). Ambas as relações são significativas ao nível de 5 % (F test). Os três primeiros eixos gerados pela análise de componentes principais (ACP) podem
ser considerados significativos segundo Legendre & Legendre (1998), contemplando 90,46 %
da variância dos dados (Tab.1) e fornecendo uma excelente visualização dos dados (Fig. 5).
TAB. 1: Contribuição das variáveis para cada um dos fatores da ACP baseado nas relações de correlação.
Variable PCA1 (38.07 %) PCA2 (30.48 %) PCA3 (21.91 %)
SOC IMB 0.0022 0.1784 0.3862
SOC CAB 0.2135 0.1616 0.1338
[NH4+] IMB 0.0754 0.0075 0.4663
[NH4+] CAB 0.2682 0.1914 0.0131
[NO3-] IMB 0.1348 0.3360 0.0000
[NO3-] CAB 0.3059 0.1252 0.0005
Uma maior ou menor aproximação entre duas ponto-variáveis no plano gerado pela
ACP representa uma correlação mais forte ou mais fraca entre as variáveis, especialmente
quando estas estão afastadas do ponto central (0.0; 0.0) (Valentin 2000). Deste modo é possível
observar que as taxas de desnitrificação no sedimento das lagoas Cabiúnas e Imboassica se
correlacionaram fortemente com o consumo de oxigênio no sedimento (COS) e com a
disponibilidade de nitrato in situ, respectivamente.
0 1000 2000 3000 4000 5000 60000.0
0.5
1.0
1.5 y= 0.0006049x - 0.0007579; R= 0.192y= 0.00007302x + 0.2153; R= 0.08
SOC (µµµµmol O 2 m-2 h-1)
Dn
( µµ µµm
ol N
2 m
-2 h
-1)

Capítulo III 46
FIG. 5: Análise de Componentes Principais (ACP). Estão dispostos nos planos 1-2 (A) e 2-3 (B) os vetores: taxa de consumo de oxigênio no sedimento (SOC) e concentrações de nitrato ([NO3
-]) e amônia ([NH4+]) na coluna d`água
das lagoas Imboassica (IMB) e Cabiúnas (CAB). O vetor taxa de denitrificação no sediment (Desnit.) está disposto como variável complementar (*). N=10. Os círculos mostram os fatores mais correlacionados com as taxas de desnitrificação em cada uma das lagoas.
Uma atividade potencial de desnitrificação crescente poderia ser esperada com o
aumento do COS, já que este último deve criar um ambiente favorável ao processo de
desnitrificação sem limitação de substrato. Os resultados do presente estudo mostram uma
correlação positiva significativa entre o COS e as taxas de desnitrificação potencial (D15) na
lagoa Cabiúnas (Spearman, p < 0,05; n = 35) mas não na lagoa Imboassica (Spearman, p < 0,05;
n = 47). Também somente na lagoa Cabiúnas as taxas D15 e Dn apresentaram entre elas uma
relação linear significativa (F test, p < 0,0001) (Fig. 6A e B).
FIG. 6: Regressão linear entre as taxas potenciais de desnitrificação (D15) e de nitrificação-desnitrificação (Dn) no sediment das lagoas Imboassica (N = 50) e Cabiúnas (N = 38). A relação é significativa ao nível de 1% (F test) somente na lagoa Cabiúnas (B).
A B
-1,0 -0,5 0,0 0,5 1,0
Factor 2 : 30,48%
-1,0
-0,5
0,0
0,5
1,0
Fac
tor
3 :
21,9
1%
SOC CAB
*Denit CAB[NO3
-] IMB [NO3-] CAB
[NH4+] CAB
*Denit IMB
[NH4+] IMB
SOC IMB
-1,0 -0,5 0,0 0,5 1,0
Factor 1 : 38,07%
-1,0
-0,5
0,0
0,5
1,0F
acto
r 2
: 30
,48%
[NH4+] CAB
SOC IMB
*Denit CAB
SOC CAB
[NH4+] IMB
[NO3-] CAB
*Denit IMB
[NO3-] IMB
0.00 0.25 0.50 0.75 1.00 1.25 1.500
50
100
150
200Imboassica
Dn (µµµµmol m -2 h-1)
D15
( µµ µµm
ol15
NO
3- m-2
h-1
)
0 1 2 3 4 5 6 70
50
100
150
200
y = 31.355x + 6.1644; R = 0.8153
Cabiúnas
Dn (µµµµmol m -2 h-1)

Capítulo III 47
DISCUSSÃO
O presente estudo mostra taxas de desnitrificação no sedimento até duas ordens de
magnitude abaixo da média estimada para lagoas costeiras e concentrações de nitrato quase 50
vezes menores que a média de sistemas aquáticos temperados (Pina-Ochoa & Alvarez-Cobelas
2006). A compilação realizada por Fennel et al. (2009), com maioria dos dados em sistemas
temperados, utilizou uma concentração de 40 µM como média de nitrato na água e observou,
para ambientes abaixo desta média, taxas de desnitrificação em torno de 75 µmol N m-2 h-1,
valores tanto de concentração de nitrato quanto de desnitrificação muito acima dos apresentados
aqui.
As taxas de COS observadas na lagoa Cabiúnas são comparáveis as taxas encontradas
em um lago temperado meso-eutrófico (Sweerts et al. 1991), enquanto as taxas da lagoa
Imboassica foram maiores que as mensuradas em uma lagoa eutrófica rasa temperada (Dedieu
et al. 2007a). Essas diferenças podem ser atribuídas principalmente à alta produtividade
existente nos trópicos, que promove maior entrada de matéria orgânica que, por sua vez,
fomenta maiores taxas de respiração no sedimento.
Os ângulos de inclinação da regressão linear entre as taxas de nitrificação-
desnitrificação e de consumo de oxigênio no sedimento do presente estudo (0,0006 para
Cabiúnas e 0,00007 para Imboassica) são muito menores que os ângulos de 0,12 e 0,09 obtidos
para ambientes temperados por Seitzinger et al. (2006) e Fennel et al. (2009) respectivamente.
As taxas de nitrificação-desnitrificação, em especial, são muito mais baixas que as apresentadas
nos dois estudos supracitados, evidenciando a reduzida atividade nitrificante nos sistemas
tropicais estudados. Já entre as duas lagoas tropicais, o ângulo de inclinação da regressão gerada
para o sistema oligotrófico foi uma ordem de magnitude maior que o observado para o sistema
meso-eutrófico, cujas taxas de nitrificação-desnitrificação praticamente não responderam as
variações de COS.
A taxa de COS regula a espessura da camada óxica do sedimento e consequentemente
pode regular o processo de desnitrificação de duas maneiras diferentes: diretamente, através da
criação de condições anóxicas ou, indiretamente, influenciando a atividade nitrificante (Jensen
et al. 1994). A matriz de correlação entre as variáveis ambientais e as taxas de desnitrificação
expressa através dos planos da ACP sugere que a atividade desnitrificante é regulada de maneira
diferente em cada um dos ambientes amostrados.
O tipo de regulação direta do COS sobre a desnitrificação parece ocorrer na lagoa
Cabiúnas. Este ambiente apresenta menores taxas de COS, sendo este o fator que se
correlacionou mais fortemente com as taxas de desnitrificação de acordo também com a ACP.
Por outro lado, o processo de desnitrificação é normalmente limitado pela disponibilidade de
nitrato ou nitrito em sedimentos ricos em matéria orgânica lábil (Fennel et al. 2009). As taxas
extremamente altas de COS da lagoa Imboassica parecem estar regulando indiretamente o

Capítulo III 48
processo de desnitrificação através da limitação da atividade nitrificante, já que como pode ser
visto através da análise de ACP, a disponibilidade de nitrato na água foi o fator que mais se
correlacionou com as taxas de desnitrificação durante o período de estudo. O intenso
metabolismo e consumo de oxigênio pode eventualmente até extinguir a camada óxica do
sedimento, limitando a nitrificação e mantendo as baixas concentrações de nitrato. Dedieu et al.
(2007b) já observou uma relação sazonal negativa entre COS e as taxas de Dn no sedimento de
uma lagoa eutrófica temperada.
Em ecossistemas tropicais como a lagoa Imboassica, com baixas concentrações de
nitrato, altas temperaturas e intenso consumo de oxigênio no sedimento durante todo o ano, os
processos de nitrificação e nitrificação-desnitrificação (responsável por até 98 % da
desnitrificação total da lagoa Imboassica) podem ser permanentemente limitados. Além disso, a
lagoa Imboassica caracteriza-se por esporádicos grandes aportes salinos através do rompimento
da barra de areia que a separa do mar (Esteves 1998). Esta característica faz com que a região
central da lagoa, onde os testemunhos de sedimento foram coletados, apresente grande variação
esporádica de salinidade. Tal variação de salinidade já se mostrou um importante fator limitante
da atividade nitrificante em lagoas costeiras (Santoro & Enrich-Prast 2009).
A relação significativa entre a atividade potencial de desnitrificação e as taxas Dn na
lagoa Cabiúnas aponta para a importância da nitrificação como o processo provedor de substrato
também neste sistema. A atividade potencial de desnitrificação só foi observada quando ou onde
havia suporte da atividade nitrificante, gerando uma relação linear significativa. É possível que
a escassez de substrato no sedimento da lagoa Imboassica, com atividade nitrificante quase
inexistente, subsidie uma reduzida comunidade desnitrificante que não responderia às adições
de nitrato. Além disso, o processo de RDNA é favorecido em detrimento da desnitrificação em
condições de alta disponibilidade de matéria orgânica (Tiedje et al. 1982; Christensen et al.
2000). Deste modo, o pouco nitrato disponível na lagoa Imboassica estaria subsidiando
principalmente uma comunidade RDNA, capaz de excluir competitivamente a comunidade
desnitrificante.
As taxas de desnitrificação e principalmente de nitrificação-desnitrificação obtidas neste
estudo foram extremamente baixas, tanto na lagoa oligotrófica quanto na meso-eutrófica,
evidenciando a reduzida atividade nitrificante no sedimento destes sistemas. Esta atividade
parece ser inexpressiva não só no sedimento, mas nos sistemas de maneira geral, já que as
concentrações de nitrato na coluna d’água não acompanharam a disponibilidade de NH4+
durante os meses de estudo.
A ocorrência limitada do processo de nitrificação pode ser a principal responsável pela
baixa atividade desnitrificante observada particularmente em ambientes aquáticos tropicais ricos
em matéria orgânica. Estudos adicionais são necessários para complementar os resultados

Capítulo III 49
obtidos aqui e melhor compreender o papel da nitrificação como processo chave em sistemas
aquáticos tropicais e seus fatores reguladores.
REFERÊNCIAS
Bower C.E. & Holmhansen T. (1980) A Salicylate-Hypochlorite Method for Determining
Ammonia in Seawater. Canadian Journal of Fisheries and Aquatic Sciences, 37, 794-798
Christensen P.B., Rysgaard S., Sloth N.P., Dalsgaard T. & Schwaerter S. (2000) Sediment mineralization, nutrient fluxes, denitrification and dissimilatory nitrate reduction to ammonium in an estuarine fjord with sea cage trout farms. Aquatic Microbial Ecology, 21, 73-84
CPRM (2000) Unidades morfoesculturais e geomorfológicas de Macaé. In. CPRM, Rio de Janeiro
Dalsgaard T., Nielsen L.P., Brotas V., Viaroli P., Underwood G., Nedwell D.B., Sundback K., Rysgaard S., Miles A., Bartoli M., Dong L., Thornton D.C.O., Ottosen L.D.M., Castaldelli G. & Risgaard-Petersen N. (2000) Protocol handbook for NICE – Nitrogen Cycling in Estuaries: a project under the EU research programme: Marine Science and Technology (MAST III). National Environmental Research Institute, Silkeborg.
Dedieu K., Rabouille C., Gilbert F., Soetaert K., Metzger E., Simonucci C., Jezequel D., Prevot F., Anschutz P., Hulth S., Ogier S. & Mesnage V. (2007a) Coupling of carbon, nitrogen and oxygen cycles in sediments from a Mediterranean lagoon: a seasonal perspective. Marine Ecology-Progress Series, 346, 45-59
Dedieu K., Rabouille C., Thouzeau G., Jean F., Chauvaud L., Clavier J., Mesnage V. & Ogier S. (2007b) Benthic O-2 distribution and dynamics in a Mediterranean lagoon (Thau, France): An in situ microelectrode study. Estuarine Coastal and Shelf Science, 72, 393-405
Esteves F.A. (1998) Lagoas Costeiras: Origem, funcionamento e possibilidades de manejo. In: Ecologia de Lagoas Costeiras (ed. Esteves FA), pp. 63-87. NUPEM, Macaé
Fennel K., Brady D., DiToro D., Fulweiler R.W., Gardner W.S., Giblin A., McCarthy M.J., Rao A., Seitzinger S., Thouvenot-Korppoo M. & Tobias C. (2009) Modeling denitrification in aquatic sediments. Biogeochemistry, 93, 159-178
Golterman H.L., Clymo R.S. & Ohnstad M.A.M. (1978) Methods for physical and chemical analisys of freshwater. BlackwellScientific Publications, Oxford.
INMET (1992) Relatório Anual do Instituto Nacional de Meteorologia - 1991. INMET, Rio de Janeiro.
Jensen K., Sloth N.P., Risgaardpetersen N., Rysgaard S. & Revsbech N.P. (1994) Estimation of Nitrification and Denitrification from Microprofiles of Oxygen and Nitrate in Model Sediment Systems. Applied and Environmental Microbiology, 60, 2094-2100
Legendre P. & Legendre L. (1998) Numerical Ecology. 2 edn. Elsevier Science BV, Amsterdam.
Mosier A.R., Doran J.W. & Freney J.R. (2002) Managing soil denitrification. Journal of Soil and Water Conservation, 57, 8
Nielsen L.P. (1992) Denitrification in Sediment Determined from Nitrogen Isotope Pairing. Fems Microbiology Ecology, 86, 357-362
Nielsen L.P., Christensen P.B., Revsbech N.P. & Sorensen J. (1990) Denitrification and Oxygen Respiration in Biofilms Studied with a Microsensor for Nitrous-Oxide and Oxygen. Microbial Ecology, 19, 63-72
Pina-Ochoa E. & Alvarez-Cobelas M. (2006) Denitrification in aquatic environments: A cross-system analysis. Biogeochemistry, 81, 111-130
Salas H. & Martino P. (2001) Metodologías simplificadas para la evaluacíon de eutroficacion en lagos cálidos tropicales. OPS/OMS, Lima.

Capítulo III 50
Santoro A.L. & Enrich-Prast A. (2009) Salinity control of nitrification in saline shallow coastal lagoons. Acta Limnologica Brasiliensia, 21, 6
Seitzinger S., Harrison J.A., Bohlke J.K., Bouwman A.F., Lowrance R., Peterson B., Tobias C. & Van Drecht G. (2006) Denitrification across landscapes and waterscapes: A synthesis. Ecological Applications, 16, 2064-2090
Sweerts J., Bargilissen M.J., Cornelese A.A. & Cappenberg T.E. (1991) Oxygen-Consuming Processes at the Profundal and Littoral Sediment Water Interface of a Small Meso-Eutrophic Lake (Lake Vechten, the Netherlands). Limnology and Oceanography, 36, 1124-1133
Tiedje J.M., Sexstone A.J., Myrold D.D. & Robinson J.A. (1982) Denitrification - Ecological Niches, Competition and Survival. Antonie Van Leeuwenhoek Journal of Microbiology, 48, 569-583
Valentin J.L. (2000) Ecologia Numérica: uma introdução a analise multivariada de dados ecológicos. Editora Interciência, Rio de Janeiro.

Conclusões Gerais 51
CONCLUSÕES GERAIS
O capítulo introdutório do presente manuscrito apresentou as principais vias de
transformação do nitrogênio em ecossistemas aquáticos continentais e a importância de cada
uma delas para a regulação da bio-disponibilidade deste macronutriente no sistema. Esta
regulação é extremamente importante uma vez que o montante de nitrogênio bio-disponível no
ambiente é capaz de limitar ou superabundar a produtividade primária e o estoque de carbono
do mesmo, influenciando toda a estrutura do ecossistema.
Neste contexto destacam-se os processos de desnitrificação e nitrificação, por
representarem a principal via de eliminação de nitrogênio bio-disponível de ambientes
aquáticos. O primeiro processo tem influência direta na bio-disponibilidade de nitrogênio,
enquanto o segundo interfere indiretamente uma vez que subsidia o primeiro através do
fornecimento de substrato.
Os dois últimos capítulos apresentados aqui fornecem, através da avaliação destes dois
processos em lagoas costeiras tropicais, informações acerca do funcionamento do ciclo do
nitrogênio em sistemas tropicais e muito particulares, já que se caracterizam como ambientes de
água doce com influência marinha.
O segundo capítulo mostrou então a influência da salinidade sobre o processo de
nitrificação. Corroborando resultados obtidos anteriormente, a variação de salinidade interferiu
negativamente na atividade nitrificante principalmente sob baixas concentrações de sal. Foi
possível concluir através dos resultados obtidos, entre eles o decaimento exponencial das taxas
de nitrificação ao longo do gradiente de salinidade, que uma variação de salinidade deve ser
mais crítica para a atividade nitrificante local em ambientes aquáticos de água doce ou de baixa
influência salina. Este deve ser então um dos principais fatores reguladores do processo de
nitrificação em ambientes tropicais costeiros com influência salina reduzida ou esporádica,
justificando as reduzidas taxas de nitrificação mostradas aqui se comparadas às obtidas em
sistemas temperados.
O processo de nitrificação também foi muito importante para a interpretação dos dados
do último capítulo, que evidenciou formas diferenciadas de regulação do processo de
desnitrificação em duas lagoas costeiras de status tróficos distintos. No entanto, independente
do tipo de regulação, as taxas de desnitrificação obtidas também foram muito baixas se
comparadas às temperadas e os resultados apontam para a escassez de nitrato como principal
responsável.
Com a ausência de fontes externas de nitrato a atividade nitrificante se torna ainda mais
importante para a ocorrência da desnitrificação. Entretanto, os resultados do último capítulo
mostram a deficiência do processo de nitrificação tanto no sedimento quanto nas lagoas como
um todo. As elevadas taxas tropicais de consumo de oxigênio somadas às maiores temperaturas,

Conclusões Gerais 52
e a reduzida e/ou esporádica influência salina no caso dos ambientes costeiros, não favorecem a
ocorrência da nitrificação. Este que é o processo central do ciclo do nitrogênio, se fortemente
limitado em sistemas tropicais, se torna um processo chave capaz de alterar o funcionamento do
ciclo do nitrogênio como um todo.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo