nematoide metodologia
TRANSCRIPT

Resumo — Neste trabalho foram utilizadas duas espécies de ocafezal.asp?
SE=4&ID=27nematóides de galhas provenientes de cafeeiro, Meloidogyne
paranaensis e Meloidogyne incógnita raça 2, com o objetivo de estudar a seletividade
fisiológica ou preferência de hospedeiros dessas duas espécies às plantas de café e
tomate. As duas espécies de Meloidogyne foram multiplicadas nessas plantas durante
dois anos. Plântulas de tomate cv. Santa Cruz e café cvs. Mundo Novo e Catuaí, com
aproximadamente 15 cm de comprimento, foram inoculadas com 10.000 ovos/planta,
sendo o delineamento inteiramente ao acaso, com seis repetições. Após três meses
foram avaliadas as plantas de tomate e, após oito meses, as plantas de café. A
multiplicação dos nematóides foi avaliada pelo índice de galhas ou ootecas e pelo fator
de reprodução. Sobrevivência diferencial de indivíduos em populações de M. incognita
e M. Paranaensis foi observada nos dois cultivares de cafeeiros, quando essas plantas
foram inoculadas com nematóides multiplicados em tomateiros. Essas duas
populações de nematóides perderam significativamente a virulência ao cafeeiro após,
aproximadamente, oito gerações em tomateiro. Tais populações são aparentemente
polimórficas nos locus de virulência, de maneira que os alelos de virulência só podem
ser detectados quando as populações multiplicadas em tomateiro são inoculadas em
cafeeiro. As populações de nematóides das galhas multiplicadas em cafeeiro
mostraram virulência intermediária quando inoculadas em tomateiro ‘Santa Cruz’.
Introdução — Sobrevivência diferencial ou seletividade fisiológica de indivíduos de
uma determinada espécie ou população são o resultado da variabilidade genética
dentro dessa população. Pesquisas realizadas nos últimos 15 anos demonstraram que
populações de organismos contêm níveis nítidos de diversidade genética. Essa
variação genética existe em todas as populações de nematóides, em muitos locus,
tendo múltiplos alelos. A identificação dessa variação tem sido demonstrada de várias
formas: preferência do hospedeiro ou raças fisiológicas do parasito, perfis de
isoenzimas e proteínas e análises do DNA. Devido à praticidade da seleção de
hospedeiros, uma das primeiras formas de variação a serem reconhecidas é a
diferença inter ou intra-específica na preferência do hospedeiro Nas espécies
partenogenéticas, como é o caso de Meloidogyne spp., a ocorrência de mutações
deverá ser mantida nas gerações subseqüentes, se os hospedeiros e as condições

climáticas forem favoráveis, sobretudo em climas amenos, que permitem várias
gerações do parasito por ano. Mesmo densidades populacionais mínimas podem ser
mantidas, pois para essas espécies não há necessidade do encontro de machos e
fêmeas (Caswell & Roberts, 1987).
Os nematóides de galhas (Meloidogyne spp.) estão amplamente distribuídos nas
plantações de café do Brasil, onde causam grandes perdas para os produtores e para a
economia do País. Meloidogyne incógnita ( Kofoid & White) Chitwood, M. paranaensis
(Carneiro et al., 1996), M. exígua Goeldi e M. coffeicola (Lordello & Zamith) têm sido
reportados em plantações de café nos Estados do ocafezal.asp?SE=6&ID=40Paraná,
São Paulo e Minas Gerais por muitos anos, havendo flutuações na predominância de
uma espécie em relação a outras. Acredita-se que M. coffeicola tenha sido erradicada
de várias plantações, embora ainda venha ocorrendo esporadicamente (Carneiro &
Almeida, 2000). A utilização de populações de Meloidogyne spp. multiplicadas em
tomateiros é normalmente empregada nos ensaios para seleção de resistência
genética em várias culturas, pois o tomateiro propicia alta reprodução da maior parte
das espécies de Meloidogyne. A perda de virulência de M. paranaensis e M. incógnita
ao cafeeiro, quando multiplicados, sucessivamente, em plantas de tomateiros cv. Santa
Cruz, tem sido relatada por nematologistas (Carneiro, R. e Gonçalves, W., informação
pessoal).
O objetivo deste trabalho foi comprovar a seletividade fisiológica ou preferência de
hospedeiros de duas espécies de Meloidogyne, em cafeeiro e tomateiro suscetíveis,
quando inoculados com populações multiplicadas, durante dois anos consecutivos,
nessas duas plantas.
Material e métodos — Neste trabalho foram utilizadas duas espécies de nematóides
de galhas provenientes do cafeeiro, M. paranaensis e M. incógnita raça 2, tendo sido
ambas purificadas previamente pelo método descrito por Carneiro et al. (1996) e
multiplicadas, durante dois anos, em tomateiros (Lycopersicon esculentum) cv. Santa
Cruz e cafeeiros (Coffea arábica) cv. Mundo Novo.
Suspensões de ovos dessas duas espécies de nematóides foram extraídas pelo
método de Hussey & Barker (1973), utilizando-se uma solução a 0,5% de NaOCl.
Plântulas de tomate cv. Santa Cruz e cafeeiros cvs. Mundo Novo e Catuaí, com

aproximadamente 15 cm de comprimento, foram inoculadas com 10.000 ovos/planta,
sendo o delineamento inteiramente ao acaso, com seis repetições. Após três meses
foram avaliadas as plantas de tomate e, após oito meses, as plantas de café. A
reprodução dos nematóides foi avaliada pelo índice de galhas ou ootecas, segundo
Hartman & Sasser (1985) e pelo número total de ovos e juvenis de segundo estádio
(J2) no sistema radicular e no solo (NOJ), sendo os ovos extraídos pelo método acima
mencionado, utilizando-se uma solução de 1,0% de NaOCl. Os J2 do solo foram
extraídos através do método de Jenkins (1964). A partir dos valores de NOJ,
calcularam-se os fatores de reprodução (FR), que são os valores de NOJ/10000.
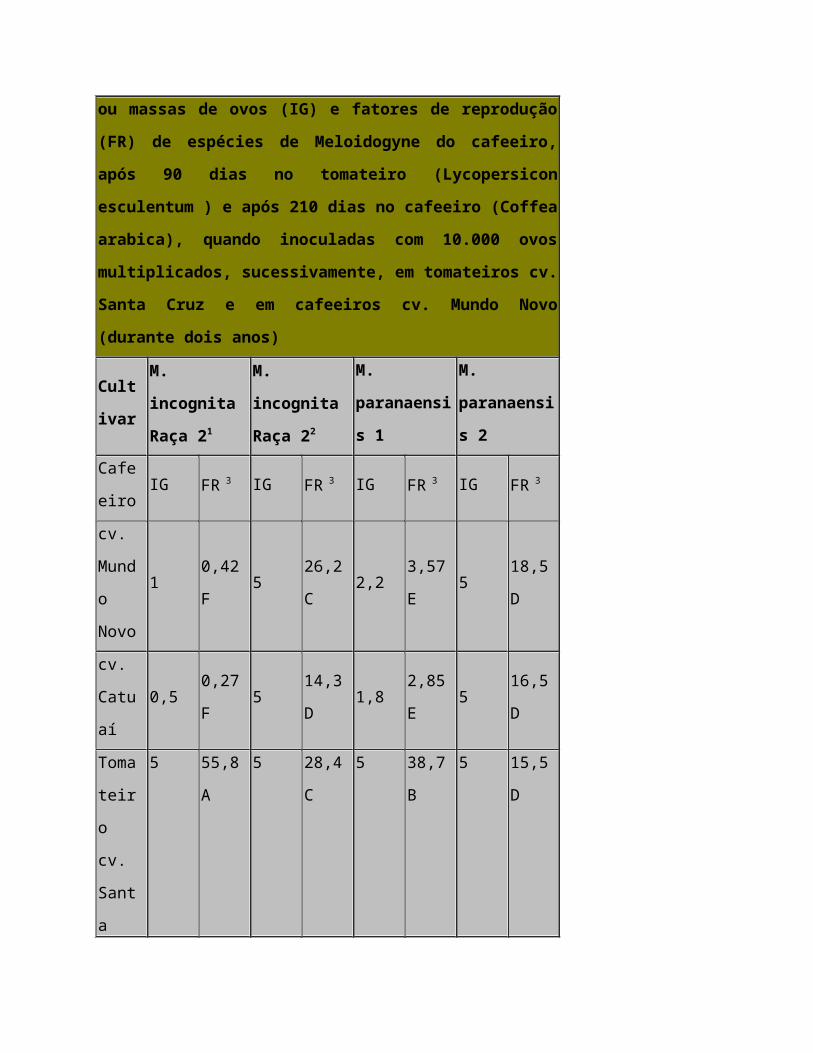
Resultados e discussão — Através da análise dos resultados (Tabela 1), pode-se
observar que as plantas de cafeeiro infestadas com inóculo de Meloidogyne spp.,
proveniente do tomateiro, apresentaram um índice de galhas (IG) e um fator de
reprodução (FR) significativamente inferiores aos das plantas com inóculo proveniente
do cafeeiro. Embora tenha ocorrido adaptação seletiva de M. paranaensis e M.
incógnita raça 2 do cafeeiro ao tomateiro, foi no caso desta última espécie que a
adaptação foi ainda mais intensa (Tabela 1). Embora o cv. Mundo Novo tenha
apresentado fatores de reprodução superiores aos do cv. Catuaí, ocorreram poucas
diferenças estatísticas entre os dois cultivares de cafeeiros testados, mostrando que
ambos são suscetíveis quando inoculados com populações provenientes do cafeeiro e
praticamente resistentes quando inoculados com populações provenientes do tomateiro
(Tabela 1). Ocorreu também redução significativa do FR em tomateiros cv. Santa Cruz,
tanto para M. incógnita como para M. paranaensis, quando se utilizou inóculo
proveniente do cafeeiro, quando comparado com o inóculo proveniente do tomateiro
(Tabela 1). Dessa maneira, pode-se comprovar neste trabalho que, embora a
adaptação seletiva tenha ocorrido no cafeeiro e tomateiro, ela foi superior no tomateiro.
Um exemplo interessante de um fenômeno similar ao descrito é o comportamento de
populações de M. incognita, M. arenaria e M. javanica, que se tornam virulentas ao
longo do tempo aos cultivares de tomate resistentes, quebrando a resistência ao gene
Mi. Tais populações são aparentemente polimórficas nos locus de virulência, de
maneira que os alelos virulentos só podem ser detectados quando expostos aos
cultivares de tomateiro resistentes (Prot, 1984). Isolados desses Meloidogyne spp.

virulentos ao gene Mi podem ser selecionados ao longo de seis a doze gerações, com
reinoculações sucessivas em plantas contendo o gene Mi (Netscher, 1977).
Populações de M. incógnita e M. paranaensis do cafeeiro devem ser também
polimórficas nos locus de virulência, de maneira que reinoculações sucessivas no
tomateiro selecionaram, ao longo do tempo, indivíduos altamente virulentos a essa
planta, restando poucos indivíduos virulentos ao cafeeiro, e para M. incógnita a pressão
de seleção no tomateiro foi mais acentuada do que para M. paranaensis (Tabela 1).
Mais estudos são necessários no sentido de precisar o número de gerações
necessárias para que as populações de Meloidogyne spp. se adaptem ao tomateiro,
perdendo, significativamente, a sua virulência ao cafeeiro.
Esses resultados devem servir de alerta aos nematologistas e melhoristas quanto ao
uso de inóculos provenientes de tomateiros para seleção de genótipos de cafeeiro com
resistência a Meloidogyne spp.
Conclusões — Populações de M. incógnita raça 2 e M. paranaensis provenientes do
cafeeiro, quando inoculadas em tomateiros cv. Santa Cruz por dois anos consecutivos,
perderam significativamente a virulência a cafeeiros altamente suscetíveis. Essas duas
espécies de nematóides das galhas multiplicadas em plantas de café mostraram uma
virulência intermediária quando inoculadas em tomateiro ‘Santa Cruz’.
Referências bibliográficas
CARNEIRO, R.M.D.G.; ALMEIDA, M.R.A & CARNEIRO, R.G. Enzyme phenotypes of
Brazilian populations of Meloidogyne spp. Fundamental and Applied Nematology,
v.19, n.6, p.555-560. 1996.
CARNEIRO, R.M.D.G. & ALMEIDA, M.R.A. Distribution of Meloidogyne spp. on
coffee in Brazil: identification, characterization and intraspecific variability.
Mejoramiento sostenible del café Arabica por los recursos genéticos, asistido pôr
los marcadores moleculares, com enfasis en la resistência a los nematodos.
Publication Especial CATIE/IRD: 43-48. 2000.
CASWELL, E.P. & ROBERTS, P.A. Nematode population genetics. Pp.390-396 in J.
A. Veech & D. W. Dickson eds. Vistas on Nematology. De Leon Springs, Florida,
USA:E. O. Painter Printing Co. 1987.

HARTMAN, K.M. & SASSER, J.N. Identification of Meloidogyne species on the basis of
differential host test and perineal pattern morphology. In: Carter, C. C. & Sasser, J. N.
eds. An advanced treatise on Meloidogyne, vol. II, Methodology. Raleigh: North
Carolina State University Graphics: p.69-77. 1985.
HUSSEY, R. S. & BARKER, K.R. A comparison of methods of collecting inocula for
Meloidogyne spp., including a new technique. Plant Disease Reporter, v.57, p.1025-
1028. 1973.
JENKINS, W.R. A rapid centrifugal-flotation technique for separating nematodes
from soil. Plant Disease Reporter, v.48, 692p. 1964.
NETSCHER, C. Observations and preliminary studies on the occurrence of
resistance breaking biotypes of Meloidogyne spp. on tomato. Cah. ORSTOM, Ser.
Biol., v.11, p.173-178. 1977.
PROT, J.C. A naturally occurring resistance breaking biotype of Meloidogyne arenaria
on tomato: Reproduction and pathogenicity on tomato cultivars Roma and Rossol.
Revue de Nématologie, v.7,: p.23-28. 1984.
Regina M. Gomes Carneiro e Camila.L. Jorge, são pesquisadoras da Embrapa
(Empresa Brasileira de Pesquisa Agropecuária).
Fonte: II Simpósio de Pesquisa dos Cafés do Brasil - Setembro de 2001.
TABELA 1 — Valores médios dos índices de galhas ou
massas de ovos (IG) e fatores de reprodução (FR) de
espécies de Meloidogyne do cafeeiro, após 90 dias no
tomateiro (Lycopersicon esculentum ) e após 210 dias no
cafeeiro (Coffea arabica), quando inoculadas com 10.000
ovos multiplicados, sucessivamente, em tomateiros cv.
Santa Cruz e em cafeeiros cv. Mundo Novo (durante dois
anos)
Cultiv
ar
M. incognita
Raça 21
M. incognita
Raça 22
M.
paranaensis
1
M.
paranaensis
2
Cafeei
roIG FR 3 IG FR 3 IG FR 3 IG FR 3

cv.
Mund
o
Novo
1 0,42 F 5 26,2 C 2,2 3,57 E 5 18,5 D
cv.
Catuaí0,5 0,27 F 5 14,3 D 1,8 2,85 E 5 16,5 D
Tomat
eiro
cv.
Santa
Cruz
5 55,8 A 5 28,4 C 5 38,7 B 5 15,5 D
1 Inóculo de nematóides provenientes de plantas de
tomateiro.2 Inóculo de nematóides provenientes de plantas de
cafeeiro.3 Fatores de reprodução médios seguidos das mesmas
letras não diferem ente si pelo teste de Tukey a 5% de
probabilidade

Secretaria de Agricultura, Irrigação e Reforma Agrária
Revista Bahia Agrícola
V.2, n.3, novembro de 1998
indice.htm -
indice indice.htm -
indice
Pesquisa Agrícola
Flutuação populacional de Meloidogyne exigua
(Goeldi, 1887) em cafeeiros no município
da Barra do Choça - BA
Sandra Elizabeth Souza*
Luiz Humberto Souza**
Florisvalda da Silva Santos***
Rivaldo Vicente Silva***
INTRODUÇÃO
O planalto de Vitória da Conquista tem no café um importante pólo agrícola do
estado, capaz de gerar emprego, renda e desenvolvimento na região. Nos anos
80, a produção atingiu um milhão de sacas, entretanto, a produção média atual é
de seiscentos mil sacas. Verifica-se que, caso houvesse manejo adequado das
lavouras e aplicação de tecnologias conhecidas, a capacidade de produção
poderia ser dobrada.
Representando um importante ponto de estrangulamento da lavoura cafeeira no
planalto de Vitória da Conquista, destaca-se a ocorrência do nematóide das
galhas, do gênero Meloidogyne, constatado, por Sharma & Sher (1973); Lordello
(1978) e Meloidogyne exigua por Souza et al., 1997. Conforme relatos históricos,
a infestação de Meloidogyne exigua Goeldi, 1887, dizimou os cafezais da antiga
província do Rio de Janeiro no final do século passado (Jobert,1892).
Em áreas infestadas com as espécies do gênero Meloidogyne, o crescimento,
translocação de água e nutrientes e a produção do cafeeiro é seriamente
comprometida (Arruda, 1960). Uma resposta do cafeeiro à presença de
Meloidogyne na raiz é, a formação de células gigantes (galhas) no sistema
radicular, resultado da hipertrofia de células do cilindro central ao redor do corpo
do nematóide que ali se desenvolve comprimindo os vasos do xilema, reduzindo
a absorção e transporte de água e nutrientes. Em conseqüência deste
parasitismo a parte aérea de cafeeiro exibe sintomas de deficiências
nutricionais, queda de folhas, ocorrência do bicho mineiro e cercosporiose com
altas infestações. O sistema radicular é reduzido e observa-se uma queda na
produção.
A fêmea de Meloidogyne instalada no interior de uma raiz parasitada deposita
seus ovos ali mesmo ou no exterior, rompendo o córtex radicular e emergindo
até a superfície. Após a eclosão as larvas, denominadas infestantes ou juvenis,
migram no solo até as raízes novas do cafeeiro, penetrando-as e tornando-se
sedentárias. Durante seu ciclo de vida passam por metamorfose originando
fêmeas, caracterizadas pelo formato de pêra ou machos de aspecto vermiforme
e alongado (Lordello, 1978; Taylor & Sasser, 1978).
Uma fêmea de Meloidogyne coloca em média 400 a 500 ovos podendo chegar
até 2000 ovos (Taylor & Sasser, 1978) e da habilidade da eclosão de suas
larvas e da migração até o hospedeiro depende o seu ciclo de vida. Em média
temperaturas de 20° - 45°C são as mais favoráveis à oviposição de Meloidogyne
exigua, alcançando o ótimo a 22°C (Tronconi et al., 1986; Lima, 1984). Jahen
(1989) verificou que não existe um nascimento uniforme de larvas, podendo
alguns ovos entrarem ou não em diapausa e alguns não mais se
desenvolverem, se as condições não forem favoráveis. Os ovos de Meloidogyne,
sobrevivem a uma ampla variação de umidade do solo, mas em solos muito
secos ocorre pouca ou nenhuma eclosão. O mesmo pode ocorrer com relação
ao volume de oxigênio, decorrente da estrutura do solo (Laughlin & Lordello,
1977).

O sistema ecológico no qual vive os fitonematóides é uma complexa inter-
relação entre nematóides, plantas hospedeiras, microclima, propriedades físicas,
biológicas e químicas do solo. As larvas juvenis de Meloidogyne, após eclosão
enfrentam barreiras físicas do solo durante sua migração até as raízes do
hospedeiro. Esta movimentação mínima, mas de supra importância para a sua
sobrevivência ocorre entre as partículas de solo, no filme de água, podendo ser
nulo o seu deslocamento se a umidade for insuficiente ou excessiva. Ocorrem
circunstâncias nas quais os nematóides podem sobreviver em condições de
seca por muitos anos (Norton, 1978). Solos saturados geralmente são
desfavoráveis pois induzem a produção de toxinas por microrganismos
anaeróbios (Laughlin & Lordello, 1977).
Freqüentemente, as precipitações são mostradas como sendo fatores relevantes
para o crescimento das plantas, bem como responsáveis pelas flutuações
estacionais dos fitonematóides. Conforme Ferris et al. (1971) citado por Carneiro
et al. (1980), plantas e nematóides apresentam semelhança na necessidade de
água. Wallace (1971) sugere que a pluviosidade, embora seja um fator de fácil
obtenção, não apresenta as vantagens dos componentes diretos tal como, a
umidade do solo.
O complexo cafeeiro tem no solo um importante fator de estudo da dinâmica de
vida dos nematóides das galhas do café. O conhecimento da influência dos
fatores edáficos que interagem com a flutuação estacional devem ser a base dos
estudos para planos de manejo das lavouras infestadas e controle de altos
níveis populacionais, ano após ano, acompanhando sua migração.
O presente trabalho estuda a relação da dinâmica populacional, através do
número de galhas na raiz e do número de juvenis de Meloidogyne exigua no
solo, com a umidade do solo e a pluviosidade, em cafeeiros infestados no
campo, com o objetivo de identificar a melhor época para iniciar o controle e o
manejo da população destes fitonematóides.

MATERIAL E MÉTODOS
O experimento foi instalado na Fazenda Palmeira, no município da Barra do
Choça-BA, região do planalto de Vitória da Conquista, localizado no Sudoeste
do Estado da Bahia, com altitude média de 800m, temperatura média em torno
de 20°C e precipitação anual entre 500 e 1200mm (média mínima e máxima). O
ensaio foi montado num delineamento experimental de blocos ao acaso com 4
repetições em plantações de cafeeiros com quinze anos da variedade Catuaí
vermelho e Catuaí amarelo cultivados em espaçamento de 4,0 x 1,5m, sem
irrigação e em Latossolo Vermelho Amarelo, de textura média, altamente
infestados com Meloidogyne exigua. Esta espécie foi identificada mediante o
estudo da configuração perineal das fêmeas, conforme a técnica de Taylor &
Sasser (1978). A parcela experimental foi composta de dez plantas na linha de
plantio, utilizando-se 6 plantas centrais como parcela útil e as demais como
bordadura.
As coletas de raízes e solo na rizosfera do cafeeiro foram feitas durante um ano
a cada 40 dias, processadas e analisadas no Laboratório de Fitopatologia e
Nematologia da UESB/Vitória da Conquista, sendo monitorada também as
variações de umidade do solo na data da coleta, determinada pelo método da
estufa a 105°C (Fontes, 1992).
A flutuação populacional dos nematóides foi avaliada quanto ao número de
galhas em 2g de raízes e número de juvenis livres em 300cm3 de solo, extraídas
pelo método da flutuação centrífuga em solução de sacarose (Jenkis, 1964). As
estimativas populacionais foram efetuadas a partir de contagens realizadas em
lâmina de Peters, através de microscopia óptica. Os dados de pluviosidade
mensal foram obtidos através da leitura dos pluviômetros instalados no local do
experimento.

RESULTADOS E DISCUSSÕES
A umidade do solo na rizosfera do cafeeiro pode ser um dos fatores que
interagem com a população de Meloidogyne exigua no solo. Na Figura 1,
observa-se que a umidade de solo acima de 10%, condicionou um melhor
crescimento de juvenis. No período compreendido pelos meses de junho, julho,
agosto e setembro, a maior umidade do solo proporcionou maiores índices de
juvenis de Meloidogyne. Estes resultados concordam com Wallace, 1971;
Carneiro et al., 1980; Pinochet et al., 1986; Huang, 1992 e 1997.
O registro das precipitações pluviométricas demonstra ocorrência de dois picos
de chuvas no local do experimento, sendo um em novembro com total acima de
150mm e outro em março em torno de 450mm (Figura 2). Verifica-se na Figura 1
que, os maiores índices de juvenis de Meloidogyne exigua no solo ocorreram
nos meses de julho a início de setembro/96, coincidindo com o período de
baixas precipitações pluviométricas (Figura 2). Nos maiores picos de
pluviosidade, detecta-se menores índices de larvas no solo, exceto ao que se
refere ao mês de março, significando ocorrência da eclosão de ovos e migração
dos juvenis para o solo neste período de alta precipitação.
Com relação a formação de células gigantes, verifica-se uma tendência de
aumento no número de galhas nos meses de agosto e novembro seguido de
pequeno decréscimo, e um outro período de elevação deste índice,
principalmente, nos meses de março a maio, mostrando maior atividade e
concentração de juvenis de Meloidogyne exigua parasitando as células das
raízes novas do cafeeiro, conforme indica a Figura 2. De modo geral observa-se
durante a estação seca, um menor número de galhas, aumentando rapidamente
no início da estação chuvosa, permanecendo alto durante essa estação.
O maior número de juvenis no solo e menor número de galhas na raiz do
cafeeiro no período de julho a início de setembro, indica que nem todas os
juvenis penetraram nas raízes, talvez pela escassez de raízes novas emitidas
pelo cafeeiro devido as baixas precipitações . No período das maiores

precipitações, ocorreu rápida infecção, constatada pelo aumento número de
galhas, reduzindo o número de juvenis no solo, determinada pela abundância de
sítios de infecção (raízes novas), emitidas pelo cafeeiro neste período. De fato,
Mendes et al.(1977); Nakasono et al. (1980) e Jaehn et al. (1983) observaram
que a penetração de juvenis de Meloidogyne exigua ocorre somente em raízes
novas na região do meristema apical. Souza em 1990, estudando a variação
sazonal de Meloidogyne exigua em cafeeiros, observou rápida infecção,
constatada pelo aumento do número de galhas no início da estação chuvosa no
Sul de Minas Gerais.
CONCLUSÕES
Neste primeiro ano de avaliação foi possível encontrar duas épocas para
iniciar o controle e o manejo da população de Meloidogyne exigua, a primeira
ocorrendo entre os meses de julho a início de setembro e a segunda entre os
meses de março a maio.
Modificações ocorridas com a umidade do solo e a pluviosidade
provavelmente produziram interferências na flutuação de juvenis de
Meloidogyne exigua no solo e no número de galhas do meristema radicular
em cafeeiros no campo.
Com a continuação dos estudos, será possível inferir com maior precisão
sobre os efeitos do clima, solo e planta sobre os nematóides no cafeeiro e
desta forma ajudar o agricultor no aumento da eficiência dos métodos
escolhidos para controlar e manejar a população de Meloidogyne exigua.

REFERÊNCIAS BIBLIOGRÁFICAS
ARRUDA, H.V. de. Redução no crescimento de cafeeiro com um ano de campo,
devido ao parasitismo de nematóides. Bragantia, v.19, p.179-82, 1960.
CARNEIRO, R. M. G. et al. Flutuação populacional de quatro espécies de
nematóides parasitos da cana-de-açúcar (Saccharum Officinarum L.) em relação
a pluviosidade e umidade do Solo. Revista Brasileira de Nematologia, v.4, p.127-
35, 1980.
FONTES, L. E. F. Caracterização física do solo: Uma nova abordagem. Viçosa:
UFV, 1992. 44 p.
HUANG, S. P. Nematóides que atacam olerícolas e seu controle. Informe
agropecuário, v.16, n.172, p.31-6, 1992.
HUANG, S. P. Ecologia de fitonematóides. In: VARGAS, M. A. T.; HUNGRIA, M.
Biologia dos solos dos cerrados. Planaltina: EMBRAPA-CPAC/Nobel, 1997.
p.447-3.
JAEHN, A . Efeito da temperatura na biologia de três raças de Meloidogyne
incógnita (Tylenchida e Meloidogynidae) em cafeeiro (Coffea arabica) e
estimativa do número de gerações para o Estado de São Paulo. Piracicaba/SP:
ESALQ, 1989. 101p. (Tese D.S.).
JAEHN, A. et al. Efeito de nitrogênio e de potássio em Meloidogyne incognita
( Kofoid e White, 1919). Chtiwood, 1949, como parasito do cafeeiro. Sociedade
Brasileira de Nematologia, Piracicaba, v.7, p.189-208, 1983.
JENKIS, W. R. A rapid centrifugal flotation technique for separating nematodes
from soil. Plant Disease Reporter, Washington, v.48, n.9,p.692, 1964.
JOBERT, M. C. Sur une maladie du cafier observee au Bresil. Archives do
Museu Nacional, Rio de Janeiro, v.8, p.103-5, 1892.

LAUGHLIN, C.W. & LORDELLO, L.G.E. Sistemas de manejo de nematóides:
relações entre a densidade de populações e os dados da planta. Nematologia
Brasileira, Piracicaba, v.2, p.15-24, 1977.
LIMA, R. D’ARC de. Embriogênese, desenvolvimento pós-embriogênico e
caracterização morfométrica de Meloidogyne exigua (Goeldi, 1887). Viçosa:
UFV, 1984. 59p. (Tese M.S.).
LORDELLO, L.G.E. Nematóides das plantas cultivadas. São Paulo: Nobel, 1978,
314 p.
MENDES, B.V. et al. Observações histopatológicas de raízes de cafeeiro
parasitadas por Meloidogyne exigua Goeldi, 1887. Nematologia Brasileira,
Piracicaba, v.2, p.207-9, 1977.
NAKASONO, K. et al. Desenvolvimento de raízes de cafeeiros novos
transplantados e penetração por Meloidogyne exigua. Nematologia Brasileira,
Piracicaba, v.2, p.257-65, 1980.
NORTON, H. V. Ecology of plant-parasitic nematodes. New York: John Wiley &
Sons, 1978. 268 p.
PINOCHET, J. et al. Flutuacion estacional de poblaciones de nematodes en dos
cafetales en Panamá. Turrialba, v.36, n.2, p.149-56, 1986.
SOUZA, S.E. Dinâmica populacional de Meloidogyne exigua (GOELDI, 1887) em
cafeeiros novos (Coffea arabica L.). Lavras, MG: Universidade Federal de
Lavras, 1990. 89 p. (Tese M.S.).
SOUZA, S. E. et al. Ocorrência de nematóide das galhas em cafeeiros no
município da Barra do Choça-BA. In: CONGRESSO BRASILEIRO DE
NEMATOLOGIA,20., Gramado, 1997. Anais...
SHARMA, R. D. & SHER, S. A. Nematodes associated with coffee in Bahia,
Brazil. Arquivo do Instituto Biológico, São Paulo, v.40, n.2, p.131-5, 1973.

TAYLOR, A.L. & SASSER, J.N. Biology, identification and control of root-knot
nematodes. Internacional Meloidogyne Project. North Carolina: State University,
1978. 111p.
TRONCONI, M.N. et al. Influência da temperatura na patogenicidade e
reprodução de Meloidogyne exigua em mudas de cafeeiro. Nematologia
Brasileira, Piracicaba, v.10, p.69-83, 1986.
WALLACE, H.R. Abiotic influences in the soil environment. In: ZUCKERMAN,
R.M. et al. (eds.). Plants Parasitic Nematodes. New York: Academic Press, 1971.
v. 1. p. 257-80.

Page 1
DISTRIBUIÇÃO QUALI-QUANTITATIVA DE NEMATÓIDES FITOPARASITOS
EM ÁREAS DE PRODUÇÃO DE ALGODÃO EM MATO GROSSO DO
SUL*Guilherme Lafourcade Asmus(1). (1) Embrapa Agropecuária Oeste, Caixa
Postal 661, 79804-970, Dourados, MS, [email protected]. RESUMO
Nos anos agrícolas de 2001/02 e 2002/03, foram amostrados em Mato Grosso
do Sul 184 talhões de lavouras nas regiões de Aral Moreira, Chapadão do Sul,
Costa Rica, Deodápolis, Dois Irmãos do Buriti, Eldorado, Guia Lopes da Laguna,
Itaquiraí, Maracaju, Mundo Novo, Naviraí, Nioaque, Ponta Porã, São Gabriel do
Oeste, Sidrolândia e Sonora. As amostras foram compostas de 10 a 15
subamostras de solo tomadas à profundidade de 0,0 – 0,20 m, na rizosfera do
algodoeiro, embaladas em sacos plásticos, armazenadas em caixas de isopor e
levadas ao laboratório, onde foi realizada a extração, identificação e
quantificação dos nematóides presentes. Os resultados obtidos evidenciaram
que existe grande variabilidade na distribuição dos nematóides fitoparasitos
entre as diferentes regiões amostradas. A freqüência média de ocorrência de M.
incognita, R. reniformis e P. brachyurus foi de, respectivamente, 27,7; 16,8 e
65,2%. As maiores densidades populacionais e freqüências de ocorrência de M.
incognita e R. reniformis foram observadas onde a cotonicultura é praticada
sucessivamente por vários anos em monocultura. Na região mais nova de
produção de algodão no estado (Chapadão do Sul, Costa Rica e São Gabriel do
Oeste), caracterizadas como áreas de cerrado, foram observadas altas
freqüências de ocorrência de P. brachyurus (respectivamente, 81,8; 78,9 e
86,9%), porém em baixas populações. INTRODUÇÃO O conhecimento da
distribuição quali-quantitativa de espécies de nematóides fitopatogênicos
configura-se como uma das informações mais importantes para o manejo de
áreas infestadas. Há três principais espécies de nematóides que atacam o
algodoeiro no Brasil: o nematóide de galhas (Meloidogyne incognita), o
nematóide reniforme (Rotylenchulus reniformis) e o nematóide das lesões
radiculares (Pratylenchus brachyurus). A recente introdução do nematóide de

cisto na região central do País, desencadeou uma mudança do perfil dos
sistemas de produção de grãos, proporcionando o avanço da cultura do algodão
para uso, em rotação, em áreas tradicionalmente cultivadas com soja. Dessa
forma, a cultura do algodoeiro tem logrado uma expressiva expansão de área no
Estado de Mato Grosso do Sul. Tal expansão da região tradicional no centro-sul
do estado para a região de cerrados, trouxe consigo o que há de mais moderno
em termos de tecnologia e, a despeito dos altos investimentos necessários,
possibilitou a diversificação da produção – condição indispensável para a
estabilidade do negócio agrícola. Se por um lado os benefícios obtidos com a
cultura do algodão tornam esta planta atrativa para cultivos em larga escala, é
bem verdade que o uso continuado do solo pela mesma pode acabar por
inviabilizar a produção. Entre outros *Trabalho parcialmente financiado pela
FUNDECT/SEPROD/PLUMA, MS.
--------------------------------------------------------------------------------
Page 2
problemas advindos do uso continuado do algodão em monocultivo, revestem-se
de importância os nematóides fitoparasitos. Este trabalho teve por objetivo
conhecer a distribuição de nematóides fitoparasitos em lavouras de algodoeiro
do Estado de Mato Grosso do Sul. MATERIAL E MÉTODOS Nos anos agrícolas
de 2001/02 e 2002/03, foram amostrados 184 talhões de lavouras de algodoeiro
nos municípios de Aral Moreira, Chapadão do Sul, Costa Rica, Deodápolis, Dois
Irmãos do Buriti, Eldorado, Guia Lopes da Laguna, Itaquiraí, Maracaju, Mundo
Novo, Naviraí, Nioaque, Ponta Porã, São Gabriel do Oeste, Sidrolândia e
Sonora. Cada amostra foi composta de 10 a 15 subamostras de solo tomadas à
profundidade de 0,0 - 0,20 m, na rizosfera do algodoeiro. Após homogeneizadas,
as amostras foram embaladas em sacos plásticos, armazenadas em caixas de
isopor e levadas ao laboratório, onde ficaram sob refrigeração até que fossem
processadas. Cada amostra composta foi representativa de áreas (talhões) de
aproximadamente 100 hectares, à exceção daquelas obtidas em pequenas
propriedades. A extração dos nematóides do solo foi realizada pelo método de

peneiramento e centrifugação (Jenkins, 1964). As suspensões de nematóides
obtidas foram submetidas a 55ºC por 5 minutos e armazenadas em formalina a
2%. As contagens do número de espécimes por amostra foi realizada em duas
alíquotas de 0,5 mL em lâmina de Peters, sob microscópio óptico. As espécies
Rotylenchulus reniformis e Pratylenchus brachyurus foram confirmadas através
da análise de características morfométricas (Handoo & Golden, 1989; Robinson
et al., 1997). Toda amostra com presença de formas jovens de nematóide de
galhas (Meloidogyne) foi colocada em vaso de argila para o qual foi
transplantada uma muda de tomateiro “Rutgers”. Após 35 a 40 dias, as fêmeas
obtidas de galhas formadas no tomateiro foram utilizadas para a confirmação da
espécie através da análise da configuração perineal (Taylor et al., 1955).
RESULTADOS E DISCUSSÃO Os resultados obtidos evidenciaram uma grande
variabilidade na distribuição dos nematóides fitoparasitos nas diferentes regiões
de produção de algodão de Mato Grosso do Sul (Tabela 1). A freqüência média
de ocorrência de M. incognita, R. reniformis e P. brachyurus nas áreas
amostradas foi de, respectivamente, 27,7%; 16,8% e 65,2%. A população média
observada das três espécies foi de, respectivamente, 260, 997 e 42
nematóides/200 mL de solo. É interessante observar que as maiores densidades
populacionais e freqüências de ocorrência de M. incognita e R. reniformis foram
observadas onde a cotonicultura é praticada sucessivamente por vários anos em
monocultivo. Nestas áreas predominam pequenas propriedades, que não
utilizam rotação de culturas. Na região mais nova de produção de algodão no
estado (Chapadão do Sul, Costa Rica e São Gabriel do Oeste), áreas
caracterizadas como de cerrado, foram observadas altas freqüências de
ocorrência de P. brachyurus (respectivamente, 81,8%; 78,9% e 86,9%), porém
em baixas populações. A relativamente alta freqüência de P. brachyurus
configura-se numa novidade e também motivo de preocupação em áreas de
produção de algodão devido a

Page 3
escassez de informações sobre as relações entre esse nematóide e a cultura .
Além disso outras culturas importantes para os sistemas de produção de grãos
em Mato Grosso, como o milho e a soja, também são suscetíveis a este
fitonematóide. Até o momento, não foram estabelecidos os limites populacionais
de danos causados por fitonematóides em algodoeiro no País. No entanto,
tomando como base dados da literatura internacional (Starr, 1998), com exceção
de Chapadão do Sul e Costa Rica, em todos os municípios onde M. incognita
estava presente, foram encontradas populações superiores ao limite de dano (>
200 formas infecticvas (J2)/200 mL). No caso de R. reniformis, observou-se
populações acima do limiar de danos (entre 600 e 2000 nematóides/200 mL) em
Deodápolis, Naviraí, Nioaque e Aral Moreira; nesta última ocorrendo em
populações variando entre 2680 e 9230 nematóides/200 mL. De uma maneira
geral, as área novas de cultivo de algodoeiro no Estado, especialmente
Chapadão do Sul e Costa Rica, ainda encontram-se com baixa freqüência dos
principais fitonematóides, e especial atenção deve ser dada ao manejo dessas
áreas para que possam continuar livres de seus danos. REFERÊNCIAS
BIBLIOGRÁFICAS HANDOO, Z. A.; GOLDEN, A. M. A key and diagnostic
compendium to the species of the genus Pratylenchus Filipjev, 1936 (lesion
nematodes). Journal of Nematology, St. Paul, v. 21, n. 2, p. 202-218, 1989.
JENKINS, W. R. A rapid centrifugal-flotation technique for separating nematodes
from soil. Plant Disease Reporter, Beltsville, v. 48, n. 9, p. 692, 1964.
ROBINSON, A. F.; INSERRA, R. N.; CASWELL-CHEN, E. P.; VOVLAS, N.;
TROCCOLI, A. Rotylenchulus species: identification, distribution, host ranges,
and crop plant resistance. Nematropica, Auburn, v. 27, n. 2, p. 127-180, 1997.
STARR, J. L. Cotton. In: BARKER, K. R.; PEDERSON, G. A.; WINDHAM, G. L.
(Ed.). Plant and nematode interactions. Madison: American Society of Agronomy,
1998. cap. 17, p. 359-380. TAYLOR, A. L.; DROPKIN, V. H.; MARTIN, G. C.
Perineal patterns of root-knot nematodes. Phytopathology, St. Paul, v. 45, n. 1, p.
26-45, 1955. Tabela 1. Freqüência de ocorrência e variação populacional de
Meloidogyne incognita, Rotylenchulus reniformis e Pratylenchus brachyurus

associados ao algodoeiro em municípios produtores de Mato Grosso do Sul, nas
safras 2001/02 e 2002/03. Embrapa Agropecuária Oeste, Dourados, MS, 2003.
Meloidogyne incognita Rotylenchulus reniformis Pratylenchus brachyurus
Município N1Freqüência (%) Pop. Média2Freqüência (%) Pop. Média2
Freqüência (%) Pop. Média2
--------------------------------------------------------------------------------
Page 4
Aral Moreira 3 0,0 - 100,0 5167 (2680-9230) 0,0 - Chapadão do Sul 33 9,1 53,4
(10-120) 0,0 - 81,8 52,2 (10-220) Costa Rica 19 5,3 20,0 (20) 0,0 - 78,9 30,0 (10-
110) Deodápolis 12 75,0 199,4 (20-850) 58,3 1159,3 (40-3930) 16,7 15,0 (10-20)
Dois Irmãos do Buriti 3 66,7 1080 (370-1790) 33,3 130,0 (130) 66,7 80,0 (50-
110) Eldorado 7 28,6 795,0 (590-1000) 0,0 - 85,7 36,7 (10-60) Guia Lopes da
Laguna 4 25,0 1440,0 (1440) 0,0 - 100,0 37,7 (10-70) Itaquiraí 11 45,4 94,0 (10-
250) 0,0 - 72,7 38,7 (20-90) Maracaju 15 0,0 - 6,7 150,0 (150) 33,3 24,0 (20-30)
Mundo Novo 6 66,7 155,0 (10-350) 0,0 - 66,7 20,0 (10-40) Naviraí 18 66,7 140,0
(10-530) 55,5 176,0 (30-920) 50,0 30,0 (10-70) Nioaque 15 46,7 318,6 (20-770)
40,0 718,3 (10-1930) 86,7 54,6 (10-230) Ponta Porã 8 0,0 - 0,0 - 12,5 10,0 (10)
São Gabriel d’Oeste 23 21,7 222,0 (10-1050) 8,7 375,0 (210-540) 86,9 47,0 (10-
120) Sidrolândia 3 0,0 - 0,0 - 66,7 55,0 (30-80) Sonora 4 0,0 - 25,0 190,0 (190)
25,0 60,0 (60) TOTAL 184 27,7 260,3 (10-1790) 16,8 996,9 (30-9230) 65,2 41,6
(10-220) 1Número de amostras. 2Médias calculadas a partir das amostras em
que o nematóide ocorreu. Os dados entre parênteses indicam a variação da
população, em número de nematóides/200 mL de solo.

FUNGOS MICORRÍZICOS ARBUSCULARES EM POMARES IRRIGADOS DE CITROS Nas últimas décadas têm se multiplicado as evidências do efeito benéfico das associações micorrízicas com diversas plantas superiores de importância econômica. Em algumas espécies vegetais é tão acentuada a dependência à presença desses fungos que, na ausência total da simbiose, não respondem satisfatoriamente à adubação fosfatada. As fruteiras destacam-se como o grupo de plantas onde as micorrizas merecem especial atenção no que se refere aos efeitos benéficos da simbiose. Muitos avanços científicos foram obtidos com essas plantas, principalmente devido à condição de dependência micorrízica, importância econômica da fruticultura e especialmente porque nesse grupo a infecção micorrízica foi inicialmente explorada. As plantas cítricas, por possuírem um sistema radicular com pêlos absorventes pouco desenvolvidos, exibem elevada dependência à micorrização e as vantagens mútuas dessa associação são expressas em maior disponibilidade de nutrientes, especialmente o fósforo, para o porta-enxerto cítrico e no fornecimento de fotossintatos para o endófito. A magnitude das respostas varia muito com o local, tipo de organismo, condições do solo (pH, nutrientes disponíveis, aspectos físicos), entre outros fatores. O presente estudo objetivou avaliar a intensidade de colonização natural de raízes dos citros por fungos MA em pomares irrigados da região semi-árida do Estado da Bahia, além de estabelecer relações com as combinações copa/porta-enxerto. O trabalho foi conduzido no município de Iaçu, BA, em pomar de citros com quatro anos de idade, cultivado sob condições irrigadas. O sistema de irrigação utilizado foi microaspersão, com uma vazão média de 30 l/hora, sendo que o raio de alcance do jato d'água dos microaspersores, nas diversas parcelas, em torno de 4m. O delineamento experimental foi inteiramente casualizado, com 4 tratamentos e 4 repetições, onde cada planta constituía uma unidade experimental. Os tratamentos consistiram em diferentes combinações copa/porta-enxerto: 1) lima ácida Tahiti (Citrus� � latifolia Tanaka)/citrumelo Swingle (C. paradisi Macf. x P. trifoliata (L.) Raf.), 2) lima� � ácida Tahiti/limão Cravo (C. limonia Osbeck), 3) lima da Pérsia (C. limettioides� � � � Tanaka)/limão Cravo e 4) laranja Pera (C. sinensis L.)/limão Cravo. � � � � � �Foram coletadas amostras de solo e raízes finas para a determinação de esporos e colonização micorrízica nas plantas cítricas. A amostragem foi repetida 6 vezes, em intervalo bimestral, e as amostras foram retiradas em quatro diferentes pontos, na projeção das copas das plantas, a uma profundidade de 0 a 20 cm da superfície. A colonização micorrízica foi estimada após clareamento das raízes em 10% KOH e coloração em azul de trypan a 0,05% em glicerol. A densidade populacional de fungos MA foi avaliada após extração dos esporos em flutuação centrífuga em solução de sacarose. A contagem desses esporos foi processada em câmara de Peter com auxílio de um microscópio ótico. A lima ácida Tahiti enxertada em citrumelo Swingle apresentou a maior colonização� � � � micorrízica, particularmente no período de avaliação final do experimento. Não se constatou diferença significativa na infecção micorrízica dos demais tratamentos, exceto na combinação lima da Pérsia/limão Cravo que apresentou o� � menor índice de colonização radicular.

Glomus intraradices e G. etunicatum foram as espécies com maior registro de ocorrência em todos os tratamentos. Essa frequência de distribuição dos FMA sugere maior adaptabilidade dessas espécies às condições edafoclimáticas e, principalmente, maior compatibilidade com a planta cítrica hospedeira. Referências Bibliográficas BALOTA, E. L.; KANASHIRO, M. A microbiologia do solo na cultura dos citros. Laranja, Cordeirópolis, v.19, n.1, 167-183, 1998.GIOVANNETTI, M.; MOSSE, B. An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytologist, Oxford, v.84, p.489-500, 1980.JENKINS, W. R. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant Disease Reporter, Beltsville, v.48, p.692, 1964.PHILLIPS, J. M.; HAYMAN, D. S. Improved procedures for clearing roots and staining parasitic and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society, Cambridge, v.55, p.158-161, 1970.REINHARDT, D. H. Potencial da fruticultura irrigada nos Tabuleiros Costeiros. Cruz das Almas, BA: EMBRAPA/CNPMF, 1996. 23p. (EMBRAPA CNPMF. Documentos, 69).�SCHENCK, N.; PÉREZ, Y. Manual for the identification of VA mycorrhizal fungi. INVAM, Gainesville, FL, 1988. 241p.ZAMBOLIM, L.; SIQUEIRA, J. O. Importância e potencial das associações micorrízicas para a agricultura. Belo Horizonte:EPAMIG, 1985. 36p. (Documentos, 26).
Antonio Alberto Rocha Oliveira e Ygor da Silva Coelho
Pesquisadores, Embrapa Mandioca e Fruticultura ([email protected])
Claúdia Melo da Paixão e Robélia Dias Tosta Amorim
Alunas do Curso de Mestrado da Escola de Agronomia da UFBA

NEMATÓIDES ASSOCIADOS A PLANTAS FRUTÍFERAS EM ALGUNSESTADOS
BRASILEIROSJORGE TEODORO DE SOUZA 1CLÉBER MAXIMINIANO 2VICENTE
PAULO CAMPOS3RESUMO - Com o objetivo de estudar a ocorrência e adistribuição
de nematóides em plantas frutíferas,analisaram-se 172 amostras de solo e raízes,
retiradasde abacaxizeiro [Ananas comosus (L.) Merril], acerola(Malpighia glabra L.),
bananeira (Musa sp), citros(Citrus spp.), figueira (Ficus carica L.), fruta-pão(Artocarpus
incisa L.) e pessegueiro (Prunus persicaL.), provenientes de 31 municípios dos Estados
daBahia, Ceará, Minas Gerais, Pernambuco e São Paulo.Os gêneros de fitonematóides
encontrados, com aspercentagens de ocorrência em relação ao número totalde
amostras coletadas, foram: Helicotylenchus Steiner,1945 (84,4 %), Meloidogyne
Goeldi, 1887 (50,9%),Criconemella De Grisse & Loof, 1965 e TylenchulusCobb, 1913
(24,9%), Pratylenchus Filipjev, 1936(15,6%), Aphelenchoides Fischer, 1894
(8,1%),Radopholus Thorne, 1949 (6,9%), Xiphinema Cobb,1913 (6,4 %), Rotylenchulus
Linford & Oliveira, 1940,Discocriconemella De Grisse & Loof, 1965,Paratylenchus
Micoletzky, 1922, RotylenchusFilipjev, 1936 e Scutellonema Andrássy, 1958(0,6%). As
espécies mais importantes e freqüênciasde ocorrência em relação ao número total
deamostras foram: Helicotylenchus dihystera (Cobb,1893) Sher, 1961 (54,3%), H.
multicinctus (Cobb,1893) Golden, 1956 (46,8%), Meloidogyne incognita(Kofoid & White,
1919) Chitwood, 1949 (26%),Tylenchulus semipenetrans Cobb, 1913 (24,9%),
M.javanica (Treub, 1885) Chitwood, 1949 (24,3%),Pratylenchus brachyurus (Godfrey,
1929) Filipjev &Schuurmans Steckhoven, 1941 (18,8%) eRadopholus similis Cobb,
1913 (6,9%).TERMOS PARA INDEXAÇÃO: Plantas frutíferas, fitonematóides,
ocorrência, distribuição.NEMATODES ASSOCIATED WITH FRUIT CROPS IN
SOMEBRAZILIAN STATESABSTRACT - The occurrence and distribution ofnematodes
parasites of fruit crops were studied in 172soil and root samples of pineapple [Ananas
comosus L.Merril], Barbados cherry (Malpighia glabra L.),banana (Musa sp), citrus
(Citrus spp), fig (Ficus caricaL.), bread fruit (Artocarpus incisa L.) and peach(Prunus
persica L.), from 31 counties of Bahia, Ceará,Minas Gerais, Pernambuco and São
Paulo states,Brazil. Plant parasitic nematode genera with theirpercentage occurrence in
relation to the total numberof samples were: Helicotylenchus Steiner, 1945(84.4%),
Meloidogyne Goeldi, 1887 (50.9%),Criconemella De Grisse & Loof, 1965 and

TylenchulusCobb, 1913 (24.9%), Pratylenchus Filipjev, 1936 (15.6%),Aphelenchoides
Fischer, 1894 (8%), Radopholus Thorne,1949 (6.9%), Xiphinema Cobb, 1913
(6.4%),Rotylenchulus Linford & Oliveira, 1940,Discocriconemella De Grisse & Loof,
1965, ParatylenchusMicoletzky, 1922, Rotylenchus Filipjev, 1936 andScutellonema
Andrássy, 1958 (0.6%). The mostimportant species and their frequency of occurrence
inrelation to the total number of samples were:Helicotylenchus dihystera (Cobb, 1893)
Sher, 1961(54,3%), H. multicinctus (Cobb, 1893) Golden, 1956(46.8%), Meloidogyne
incognita (Kofoid & White, 1919)Chitwood, 1949 (26%), Tylenchulus semipenetrans
Cobb,1913 (24.9%), M. javanica (Treub, 1885) Chitwood, 1949(24.3%), Pratylenchus
brachyurus (Godfrey, 1929)Filipjev & Schuurmans Steckhoven, 1941 (18.8%)
andRadopholus similis Cobb, 1913 (6.9 %).INDEX TERMS: Fruit crops, plant
nematodes, occurrence, distribution.1. Eng. Agr., M.Sc., Departamento de
Fitopatologia, UNIVERSIDADE FEDERAL DE LAVRAS(UFLA).2. Eng. Agr.,
Departamento de Fitopatologia/ UFLA, Caixa. Postal 37, 37.200 - 000, Lavras - MG.3.
Eng. Agr., MSc., Ph.D., Prof. Titular, Departamento de Fitopatologia/UFLA
--------------------------------------------------------------------------------
Page 2
Ciênc. e Agrotec., Lavras, v.23, n.2, p.353-357, abr./jun., 1999354INTRODUÇÃOVárias
espécies de plantas frutíferas cultivadasno Brasil possuem importantes problemas
decorrentesdo ataque de fitonematóides. O abacaxizeiro, por exem-plo, [Ananas
comosus (L.) Merril] pode sofrer prejuízosde até 42 %, devido ao ataque de nematóides
(Raski eKrusberg, 1984). No Brasil, nematóides têm sido asso-ciados a essa cultura
nos Estados da Bahia, Maranhão,Espírito Santo, São Paulo, Pernambuco, Rio de
Janeiro,Minas Gerais, Goiás e Sergipe (Manso et al., 1994). Asperdas causadas por
Radopholus similis Cobb, 1913 nabananeira, variam de 15 a 50% (Tarté e
Pinochet,1981). Nas plantas cítricas (Citrus spp) existem muitasespécies de
nematóides associadas (Manso et al., 1994).Tylenchulus semipenetrans Cobb, 1913 é
considerado onematóide mais importante economicamente para a ci-tricultura brasileira
(Lordello e Lordello, 1991), com am-pla disseminação no país (Ferraz, 1980). Essa
espé-cie pode causar prejuízos de 10 a 30 %, dependendodas condições ambientais e
demais condições locais(Duncan, 1995). Heterodera fici Kirjanova, 1954 é onematóide

mais prejudicial à figueira (Franco ePenteado, 1986), contudo, Meloidogyne
incognita(Kofoid & White, 1919) Chitwood, 1949 encontra-se mais disseminado no
Brasil (Campos, 1997).O presente trabalho foi realizado com o objetivode atualizar
informações acerca da ocorrência e distri-buição de nematóides associados a plantas
de acerola,abacaxizeiro, bananeira, plantas cítricas, plantas defruta-pão, figueira e
pessegueiro nos Estados da Bahia,São Paulo, Minas Gerais, Ceará e
Pernambuco.MATERIAL E MÉTODOSAmostras de 500 cm3de solo e 80 g de raízes
fo-ram coletadas em 31 municípios dos Estados de MinasGerais, São Paulo, Ceará,
Pernambuco e Bahia, numtotal de 172 amostras, obtidas de acerola, bananeira,citros,
figueira, fruta-pão e pessegueiro. As raízes foramlavadas, dissecadas ao microscópio
estereoscópico einvestigadas quanto à presença de fêmeas adultaspertencentes ao
gênero Meloidogyne, mesmo nãoapresentando galhas. Nas infestadas, foram
obtidas10 fêmeas, que foram dispostas lado a lado em gotasde água destilada sobre
lâmina de vidro, recobertascom lamínula 24x50mm e esmagadas para a obser-vação
da configuração perineal. A análise da confi-guração perineal foi feita conforme os
padrões contidosem Taylor e Sasser (1978). Após a análise sob micros-cópio
estereoscópio, as raízes foram trituradas em li-quidificador por 45 segundos, passadas
em peneiras de20 e 325 malhas, sendo que o material coletado na úl-tima peneira foi
centrifugado em sacarose, adaptando-se à técnica de Jenkins (1964). Os nematóides
do soloforam extraídos usando-se o método de Jenkins(1964), a partir de alíquotas de
100 cm3de solo. Assuspensões obtidas tanto do solo quanto das raízesforam
observadas sob microscópio ótico a 250X paraa identificação do gênero. A seguir, os
nematóidesforam fixados em TAF (4,5 ml de trietanolamina,15 ml de formaldeído a 38
% e 80,5 ml de águadestilada) e montados em lâminas semipermanentes,que foram
utilizadas em estudos morfológicos emorfométricos para a identificação das
espécies.Das amostras coletadas, 72,2% provieram doEstado de Minas Gerais, 12,7 %
de Pernambuco, 6,9%da Bahia, 5,8% do Ceará e 2,3% de São Paulo. O maiornúmero
de amostras foi coletado de bananeira (62,4%),seguido por citros (31,2%),
abacaxizeiro, acerola e fi-gueira (1,2 %) e fruta-pão (0,6 %).RESULTADOS E
DISCUSSÕESForam encontrados 14 gêneros de fitonematói-des, com as seguintes
percentagens de ocorrência emrelação ao número total de amostras coletadas: Helico-

tylenchus Steiner, 1945 (84,4%); Meloidogyne sp. (50,9%); Criconemella De Grisse e
Loof, 1965 e TylenchulusCobb, 1913 (24,9%); Pratylenchus Filipjev, 1936 (15,6%);
Aphelenchoides Fisher, 1894 (8,1%); RadopholusThorne, 1949 (6,9%); Xiphinema
Cobb, 1913 (6,4%);Rotylenchulus Linford e Oliveira, 1940; Discocricone-mella De
Grisse e Loof, 1965; Paratylenchus Mico-letzky, 1925; Rotylenchus Filipjev, 1936 e
ScutellonemaAndrássy, 1958 (0,6%) (Tabela 1).As espécies mais importantes e
respectivas fre-qüências de ocorrência em relação ao número total deamostras foram:
Helicotylenchus dihystera Cobb, 1893(54,3%), H. multicinctus (46,8%), M. incognita
(26%),T. semipenetrans (24,9%), M. javanica Treub, 1885(24,3%), P. brachyurus
Godfrey, 1929 (18,8%) e R.similis (6,9%).Plantas de acerola foram encontradas parasi-
tadas por nematóides das galhas pela primeira vezem Minas Gerais (Tabela 1). A
suscetibilidade daplanta de acerola à M. javanica e M. incognita foiconfirmada pelos
testes de hospedabilidade realiza-dos por Ferraz, Monteiro e Inomoto (1989). Essefato
representa um risco para os plantios da regiãomineira dessa frutífera, uma vez que o
ataque denematóides das galhas pode resultar em severos da-nos para essa planta
(Franco e Ponte, 1989; Sharmae Junqueira, 1993).
--------------------------------------------------------------------------------
Page 3
Ciênc. e Agrotec., Lavras, v.23, n.2, p.353-357, abr./jun., 1999355TABELA 1 -
Nematóides parasitos de plantas frutíferas identificados em amostras coletadas em
municípios de al-guns Estados brasileiros.FrutíferaNematóides IdentificadosMunicípio/
EstadoNodeAmostrasAbacaxizeiro M. incognita, H. dihystera, P.
brachyurusGuaraçaí/SP2Acerola M. incognitaM. javanica, Criconemella sp.,
Helicotylenchus sp.Belo Horizonte/MGParaisópolis/ MG11Bananeira M. javanica, M.
incognita, Meloidogyne sp., H. dihystera, H.multicinctus, Helicotylenchus sp.,
Criconemella sp.Capitólio/MG4M. incognita, Helicotylenchus sp.Carmo da
cachoeia/MG1M. incognita, H. multicinctusCarmo de Minas/MG2M. javanica,
Helicotylenchus sp., Xiphinema sp.Coqueiral/MG2Meloidogyne sp., M. incognita, M.
javanica, H. dihystera, H.multicinctus, Criconemella sp., P. brachyurus, R.
similis,Aphelenchoides sp.Janaúba/MG21M. javanica, M. incognita, H. dihystera,
Criconemella sp.Lavras/MG8M. javanica, H. dihystera, Aphelenchoides

sp.Machado/MG4M. incognita, H. multicinctus, P. brachyurus, R.
similisMauriti/CE10BananeiraM. javanica, H. multicinctusMontes Claros/MG1H.
dihystera, Xiphinema sp.Patos de Minas/MG1M. javanica, M. incognita, H. dihystera, H.
multicinctus,Aphelenchoides sp., P. brachyurusPerdões/MG6M. incognita, Meloidogyne
sp. Helicotylenchus sp., Aphelenchoidessp., P. brachyurusPorteirinha/MG4M. incognita,
M. javanica, H. multicinctus, R. similisRecife/PE4R. similis, Xiphinema sp.Ribeirão
Pires/SP1M. javanica, H. multicinctus, Aphelenchoides sp.Salvador/BA2M. javanica, H.
dihysteraSanta Rita do Sapu-caí/MG2M. incognita, M. javanica, H. dihystera, H.
multicinctus, Helicotylenchus sp., P. brachyurus, Aphelenchoides sp., Criconemella
sp.Santo Antônio do Ampa-ro/MG8M. incognita, M. javanica, Meloidogyne sp., H.
multicinctus, P.brachyurus, R. similis, Scutellonema sp., Paratylenchus sp.São Vicente
Ferrer/PE18M. incognita, H. multicinctus, Aphelenchoides sp.Wagner/BA9Continua...
--------------------------------------------------------------------------------
Page 4
Ciênc. e Agrotec., Lavras, v.23, n.2, p.353-357, abr./jun., 1999356TABELA 1-
Continuação....FrutíferaNematóides IdentificadosMunicípio/
EstadoNodeAmostrasCitrosHelicotylenchus sp., P. brachyurus, Criconemella sp.,
TylenchulussemipenetransAlfenas/MG14H. dihystera, Helicotylenchus sp., T.
semipenetransCampo Belo/MG1H. dihystera, Pratylenchus sp., Aphelenchoides sp.,
Criconemella sp.T. semipenetrans, R. reniformisLavras/MG7Helicotylenchus sp., P.
brachyurus, Criconemella sp.Pará de Minas/MG3H. dihystera, Helicotylenchus sp., P.
brachyurus, Aphelenchoidessp.,Criconemella sp., T. semipenetransPerdões/MG26P.
coffeae, Criconemella sp., T. semipenetransSão Carlos/SP1H. dihystera,
Helicotylenchus sp., T. semipenetrans, Xiphinema sp.São João Del Rei/MG2FigueiraH.
dihystera, Helicotylenchus sp., Discocriconemella sp.Caldas/MG1M. incognita,
Helicotylenchus sp.Salinas/MG1Fruta-PãoHelicotylenchus sp., Rotylenchus sp.,
Xiphinema sp., Criconemella sp.Nova Viçosa/MG1Pessegueiro M. arenaria,
Aphelenchoides sp.Pouso Alegre/MG2H. dihystera, Xiphinema sp., Aphelenchoides
sp.Santana da Vargem/MG1De um total de 107 amostras retiradas de bana-nais, 12
coletadas de municípios nos Estados do Ceará,Minas Gerais, Pernambuco e São
Paulo (Tabela 1), en-contravam-se infestadas por R. similis, que é o nema-tóide mais

danoso à cultura. Esse nematóide provocalesões nas raízes de sustentação, com o
conseqüentetombamento das plantas. Cuidados especiais deverãoser tomados pelos
produtores para evitar a dissemi-nação de R. similis, que é feita, principalmente,
pormeio de mudas infestadas. Helicotylenchus spp. eMeloidogyne spp. ocorreram,
respectivamente, em90,7 % e 75,9 % das amostras coletadas dos bana-nais.De 54
amostras coletadas da rizosfera de plan-tas cítricas, constatou-se a presença de T.
semipenetransem 43, evidenciando a ampla disseminação desse ne-matóide no Brasil.
R. reniformis foi constatado em La-vras - MG, parasitando Citrus sp. Essa espécie foi
de-tectada anteriormente parasitando plantas cítricas emMinas Gerais (Ferraz, 1980);
entretanto, a extensão dosdanos causados às plantações citrícolas
permanecedesconhecida.M. arenaria foi constatada pela primeira vez pa-rasitando
pessegueiro em Minas Gerais, em amostrasprovenientes do município de Pouso Alegre
(Tabela 1).Essa espécie já havia sido encontrada em mudas de pes-segueiro
provenientes do Estado do Rio Grande do Sul(Santos e Lozano, 1988).Em conclusão,
foram encontrados 14 gêneros defitonematóides associados ao abacaxizeiro, plantas
deacerola, bananeira, plantas cítricas, figueira, plantas defruta-pão e pessegueiro nos
Estados da Bahia, Ceará,Minas Gerais, São Paulo e Pernambuco. As espécies
demaior ocorrência nos pomares amostrados foram: H.dihystera (54,3 %), H.
multicinctus (46,8 %), M. in-cognita (26 %), T. semipenetrans (24,9 %), M.
javanica(24,3 %), P. brachyurus (18,8 %) e R. similis (6,9 %).REFERÊNCIAS
BIBLIOGRÁFICASCAMPOS, V.P. Nematóides na cultura da figueira.Informe
Agropecuário, Belo Horizonte, v.18,n.188, p.33-38, 1997.
--------------------------------------------------------------------------------
Page 5
Ciênc. e Agrotec.,