morfoanatomia de espécies lacustres de monocotiledôneas do
TRANSCRIPT

Universidade Estadual de Feira de Santana Departamento de Ciências Biológicas
Programa de Pós-Graduação em Botânica
Morfoanatomia de Espécies Lacustres de
Monocotiledôneas do Semi-árido da Bahia, Brasil
Kelly Regina Batista Leite
FEIRA DE SANTANA – BA 2008

ii
Universidade Estadual de Feira de Santana Departamento de Ciências Biológicas
Programa de Pós-Graduação em Botânica
Morfoanatomia de Espécies Lacustres de
Monocotiledôneas do Semi-árido da Bahia, Brasil
Kelly Regina Batista Leite
Tese apresentada ao Programa de Pós-Graduação em Botânica da Universidade Estadual de Feira de Santana, como parte dos requisitos necessários para obtenção do título de Doutor em Botânica.
Profa. Dra. Vera Lúcia Scatena (UNESP – Rio Claro) Orientadora
Prof. Dr. Flávio França (UEFS)
Co-Orientador
FEIRA DE SANTANA – BA 2008

iii
BANCA EXAMINADORA
Profa. Dra. Vera Lúcia Scatena (UNESP-Rio Claro) Orientadora e Presidente da Banca
Alessandra Ike Coan (UFPR)
Delmira Costa da Silva (UESC)
Cláudia Elena Carneiro (UEFS)
Francisco de Assis Ribeiro dos Santos (UEFS)

iv
...
e aprendi que se depende sempre de tanta muita diferente gente
toda pessoa sempre é as marcas das lições diárias de outras tantas pessoas
e é tão bonito quando a gente entende que a gente é tanta gente onde quer que a gente vá
é tão bonito quando a gente sente que nunca está sozinho
por mais que pense estar
e é tão bonito quando a gente pisa firme nessas linhas que estão
nas palmas de nossas mãos
é tão bonito quando a gente vai à vida nos caminhos onde bate
bem mais forte o coração
...
(Gonzaguinha)

v
À Deus, pois só a Ti devemos toda a honra e toda a glória.
Aos meus pais, João e Airma, aos meus irmãos, Mônica e Lincoln, e por último, mas muito importante, ao meu marido Vlamir, eu dedico.

vi
AGRADECIMENTOS
Ao Senhor Deus, pela coragem que me deu para enfrentar mais este desafio e por
permitir que eu chegasse até o final dele. Obrigada meu Pai por ter me dado uma família e
amigos que me ajudaram em todos os momentos desta jornada.
Ao Programa de Pós-Graduação em Botânica da Universidade Estadual de Feira de
Santana (PPGBot/UEFS) pela oportunidade de realização do curso.
À Profa. Dra. Vera Lúcia Scatena, minha orientadora, pela coragem de ter assumido a
orientação à distância, pela confiança, paciência e estímulo, pelas conversas científicas e
sobre a vida pessoal e profissional, pela segurança e tranqüilidade que sempre me passou
quando eu ligava desesperada e, principalmente, por ter entendido e apoiado todas as
mudanças que aconteceram na minha vida ao longo desses quatro anos (que não foram
poucas). Obrigada não apenas pela orientação, mas pelo seu carinho e amizade.
Ao Prof. Dr. Flávio França por ter me apresentado às plantas aquáticas e por ter
aceitado ser meu co-orientador; pelo incentivo, confiança e apoio em todos os momentos.
Ao Prof. Dr. Francisco de Assis Ribeiro dos Santos. Você foi o responsável pelos
primeiros vôos alçados, por me mostrar como era importante sair e conhecer outras
realidades. Obrigada pela amizade e incentivo desde a graduação.
Ao Prof. Dr. Luciano Paganucci de Queiroz, curador do Herbário HUEFS, onde
trabalhei alguns anos e durante os dois anos e meio do início da tese. Obrigada pelo incentivo
e pelo apoio no curso de doutorado, mesmo quando me ausentava do trabalho.
Ao Prof. Dr. Cássio van den Berg pela GRANDE ajuda na análise de similaridade.
Ao Prof. Dr. Marccus Alves (UFPE) pela atenção dispensada quando da minha estada
em Recife e por ter me ensinado a diferenciar as espécies de Cyperaceae com as quais eu
trabalhava.
Ao estagiários do Laboratório de Morfo-Taxonomia Vegetal da UFPE que me
receberam tão bem, em especial, à Msc. Shirley Martins, por ter cedido sua casa quando

vii
estive em Recife para aprender mais sobre Cyperaceae. Obrigada pelas conversas e pela
amizade que daí surgiu.
À Profa. Dra. Joana Angélica Guimarães da Luz, diretora do Instituto de Ciências
Ambientais e Desenvolvimento Sustentável (ICADS) da Universidade Federal da Bahia, pela
amizade e por ter concedido minhas licenças para poder concluir a tese.
Aos meus colegas de trabalho da UFBA-Barreiras, Prof. Jorge Costa, Profa. Loislene
Brito, Profa. Luci Ribeiro, Profa. Renata Canalle (que agora está na UFPI) e Profa. Agda
Rocha (que está na Infraero), por terem entendido minhas ausências por conta da tese e pela
amizade nesta nova etapa da vida que começamos juntos em Barreiras. Agradeço
principalmente à Luci por ter ficado com minha turma para que eu pudesse me ausentar e a
Jorge Costa, por compreender, aceitar e confiar numa vice-coordenadora de Colegiado que se
ausentava tanto, OBRIGADA PELO INCENTIVO!!!
À Adriana e Gardênia, secretárias do PPGBot/UEFS, pela atenção, paciência e
disponibilidade em atender a todos nós, sempre precisando de muitas coisas.
Aos amigos do Herbário HUEFS, Elaine, Renata, Silvinha, Téo e Zezé. Quero incluir
aqui, Úlvia (Élvia), Andréa Karla, Paty, Dani, Milene, Paula Dib e Cosme. Obrigada pelos
muitos anos de convivência, pelas conversas e alegrias proporcionadas, sinto muita falta de
vocês.
Aos meus GRANDES amigos do Laboratório de Micromorfologia Vegetal – Bárbara,
Cláudia, Hilder, Jaílson, Lázaro, Luciene, Marcos, Mari, Paulino, Ricardo, Rita e Teresa e aos
estagiários com quem convivi trocando informações, idéias, técnicas e é claro, agradáveis
momentos de descontração durante a hora do café ou nos almoços do laboratório. Sinto muita
falta dessa família!!!
Ao meu amigo Paulino, companheiro de longas datas, de conversas e de dúvidas... só
nós dois sabemos como é bom entendermos um ao outro.
À Dione e Adriano, funcionários do Laboratório de Águas e Saneamento do
Departamento de Tecnologia da Universidade Estadual de Feira de Santana, pelas análises de

viii
água proveniente das lagoas. À Alê, Adriana, Aline e Rita, da UNESP de Rio Claro, pela paciência em me ensinar
algumas técnicas e pelas conversas em dias difíceis e de saudade. Agradeço principalmente a
Aline e Alê pelas dicas valiosas sobre a tese.
Ao meu amigo estatístico Marcelo de Paula, colega da UFBA-Barreiras, que tanto me
ajudou. Obrigada Lindinho.
Aos meus queridos alunos da UFBA pelo incentivo e torcida
À Aline, Agda, Maria Antonia e Andria, as meninas do pensionato de Dona Maria (in
memorian) e seu Sebastião. Quando estava em Rio Claro, vocês muito me ajudaram a
diminuir a saudade que sentia de casa.
À Dani pela ajuda na confecção dos mapas e Gisele Rocha pela atenção e paciência na
obtenção das imagens ao MEV.
Agradeço imensamente à Profa. Néa Andrade de Macêdo pela dedicação e paciência
com que me ensinou a gostar da anatomia vegetal.
Aos meus pais João e Airma, aos meus irmãos Mônica e Lincoln e a Aninha. O final
desta jornada significa uma conquista, que sei, também é de vocês. Obrigada por sempre
estarem ao meu lado, mesmo eu estando sempre longe e por terem entendido a minha
ausência e humor algumas vezes instável.
Ao meu querido marido Vlamir, meu amor, meu porto seguro e meu companheiro de
todas as horas. Você não tem noção do quanto me ajudou, me deu coragem, apoio, força,
incentivo, carinho, tranqüilidade e, principalmente, compreendeu a minha ausência, até
mesmo quando eu estava ao seu lado. Sem você tudo teria sido muito mais difícil. Essa
conquista é nossa e para a nossa família, que já está crescendo...
O final desta tese significa apenas que mais uma etapa foi cumprida, não a última;
significa que sentirei saudades de muitas pessoas, coisas, lugares e situações; e se os últimos
quatro anos foram os mais difíceis saibam também, que eles foram os mais felizes.
OBRIGADA A TODOS!!!!

ix
SUMÁRIO
SUMÁRIO ........................................................................................................... ix
LISTA DE FIGURAS ......................................................................................... x
RESUMO .......................................................................................................... xiv
ABSTRACT ....................................................................................................... xv
INTRODUÇÃO GERAL .................................................................................... 1
REFERÊNCIAS BIBLIOGRÁFICAS .............................................................. 6
CAPÍTULO I ....................................................................................................... 8
Anatomia de espécies anfíbias de Cyperaceae de lagoas do semi-árido, Bahia, Brasil ..................................................................................................................... 8
CAPÍTULO II .................................................................................................... 42
Anatomia de espécies emergentes de lagoas temporárias da Bahia, Brasil ........ 42
CAPÍTULO III .................................................................................................. 74
Convergência adaptativa em espécies de monocotiledôneas lacustres do semi-árido da Bahia, Brasil .......................................................................................... 74
CONSIDERAÇÕES FINAIS ........................................................................... 93

x
LISTA DE FIGURAS INTRODUÇÃO GERAL
Figura 1. Lagoa temporária do semi-árido baiano no período seco ................................................ 5
Figura 2. Lagoa temporária do semi-árido baiano no período alagado ........................................... 5
CAPÍTULO I
Figuras 1-12. Aspectos morfológicos de espécies de Cyperaceae: 1-3: Cyperus odoratus L. 1.
Hábito, 2. Detalhe do sistema subterrâneo. 3. Secção transversal da região mediana do
escapo. 4-6: Oxycaryum cubense (Poep. & Kunth) Lye 4. Hábito. 5. Detalhe do
sistema subterrâneo. 6. Secção transversal da região mediana do escapo. 7-9: Pycreus
macrostachyos (Lam.) Raynal 7. Hábito. 8. Detalhe do sistema subterrâneo. 9. Secção
transversal da região mediana do escapo. .......................................................................... 29
Figuras 10-19. Fotomicrografias de secções transversais da região mediana de raízes de
espécies de Cyperaceae: 10-12. Cyperus odoratus L. 10. Aspecto geral. 11. Detalhe da
epiderme e exoderme. 12. Detalhe do cilindro vascular. 13-16. Oxycaryum cubense
(Poep. & Kunth) Lye. 13. Aspecto geral - raiz adulta. 14. Detalhe da epiderme e
exoderme. 15. Detalhe do cilindro vascular. 16. Aspecto geral - raiz jovem. 17-19.
Pycreus macrostachyos (Lam.) Raynal. 17. Aspecto geral. 18. Detalhe da epiderme e
exoderme. 19. Detalhe do cilindro vascular.. .................................................................... 30
Figuras 20-26. Fotomicrografias de secções transversais da região mediana de rizomas de
espécies de Cyperaceae: 20-21. Cyperus odoratus L. 20. Aspecto geral. 21. Detalhe do
cilindro vascular. 22-24: Oxycaryum cubense (Poep. & Kunth) Lye. 22. Aspecto geral
do rizoma. 23. Região do nó do estolão. 24. Região do entrenó do estolão. 25-26.
Pycreus macrostachyos (Lam.) Raynal. 25. Aspecto geral. 26. Detalhe do cilindro
vascular .............................................................................................................................. 32
Figuras 27-35. Fotomicrografias de secções paradérmicas e transversais de folhas de espécies
de Cyperaceae: 27-32. Vista frontal da epiderme. 27-28. Cyperus odoratus L. 27. Face
adaxial. 285. Face abaxial com estômatos. 29-30. Oxycaryum cubense (Poep. &
Kunth) Lye. 29. Face adaxial. 30. Face abaxial com estômatos. 31-32. Pycreus

xi
macrostachyos (Lam.) Raynal. 31. Face adaxial. 32. Face abaxial com estômatos. 33-
35. Bordo foliar. 33. Cyperus odoratus L. 34. Oxycarium cubense (Poep. & Kunth)
Lye. 35. Pycreus macrostachyos (Lam.) Raynal ............................................................... 34
Figuras 36-45. Fotomicrografias de secções transversais de folhas de espécies de Cyperaceae:
36-37. Cyperus odoratus L. - nervura central e mesofilo, respectivamente. 38-40.
Oxycarium cubense (Poep. & Kunth) Lye - nervura central na região mediana, na
região apical, e mesofilo na região mediana, respectivamente. 41-44. Pycreus
macrostachyos (Lam.) Raynal – 41-42. nervura central na região mediana e basal,
respectivamente. 43. Detalhe do diafragma. 44. Detalhe do mesofilo. 45. Cyperus
odoratus - detalhe do cristal cônico.. ................................................................................. 36
Figuras 46-56. Fotomicrografias de secções transversais da região mediana de escapos de
espécies de Cyperaceae: 46-48. Cyperus odoratus L. - aspecto geral, detalhe do
córtex e da epiderme, respectivamente. 49-51. Oxycarium cubense (Poep. & Kunth)
Lye - aspecto geral, detalhe do feixe vascular composto e simples, respectivamente.
52-54. Pycreus macrostachyos (Lam.) Raynal - aspecto geral, detalhe dos feixes
vasculares com estrutura Kranz e do feixe vascular colateral, respectivamente. 55-56.
Detalhe da epiderme. 55. Oxycaryum cubense (Poep. & Kunth) Lye. 56. Pycreus
macrostachyos (Lam.) Raynal. Legenda: Cl = clorênquima; Ff = feixe de fibras; Hi
=hipoderme; La = lacuna de ar. ..... ...........................................................................39
CAPÍTULO II
Figuras 1-6. Aspectos morfológicos de espécies de monocotiledôneas: 1-2. Echinodorus
grandiflorus (Alismataceae). 1.Hábito. 2. Detalhe do sistema subterrâneo. 3-4.
Eichhornia paniculata (Pontederiaceae). 3. Hábito. 4. Detalhe do sistema subterrâneo.
5-6: Habenaria repens (Orchidaceae). 5. Hábito. 6. Detalhe do sistema subterrâneo. ..... 62
Figuras 7-18. Fotomicrografias de aspectos anatômicos de secções transversais de raízes de
espécies de monocotiledôneas: 7-12. Echinodorus grandiflorus (Alismataceae). 7-8.
Aspecto geral da raiz jovem. 9. Raiz adulta. 10. Detalhe da epiderme e exoderme. 11.
Detalhe do cilindro vascular. 12. Detalhe das células braciformes que se rompem e
formam as lacunas de ar - setas. 13-15. Eichhornia paniculata (Pontederiaceae) – raiz
adulta. 13. Aspecto geral. 14. Detalhe da epiderme e exoderme. 15. Detalhe do
cilindro vascular. 16-18. Habenaria repens (Orchidaceae) – raiz adulta. 16. Aspecto

xii
geral. 17. Detalhe da epiderme e córtex. 18. Detalhe da endoderme com estrias de
Caspary e cilindro vascular. ............................................................................................... 63
Figuras 19-25. Fotomicrografias de aspectos anatômicos de secções transversais da região
mediana do rizoma de espécies de Monocotiledôneas: 19-20. Echinodorus
grandiflorus (Alismataceae): 19. Aspecto geral. 20. Detalhe da região do cilindro
vascular. 21-23: Eichhornia paniculata (Pontederiaceae): 21. Aspecto geral do córtex
com lacunas de ar e raízes intracorticais. 22. Detalhe do córtex e cilindro vascular. 23.
Detalhe da saída de raiz. 24-25: Habenaria repens (Orchidaceae): 24. Aspecto do
córtex com lacunas de ar. 25. Detalhe da endoderme e cilindro vascular. ........................ 66
Figuras 26-31. Fotomicrografias de aspectos anatômicos da região mediana da epiderme foliar
em vista frontal, de espécies de Monocotiledôneas: 26-27. Echinodorus grandiflorus
(Alismataceae) - Face adaxial e abaxial, respectivamente. 28-29: Eichhornia
paniculata (Pontederiaceae) - Face adaxial e abaxial, respectivamente. 30-31:
Habenaria repens (Orchidaceae) - Face adaxial e abaxial, respectivamente. ................... 67
Figuras 32-42. Fotomicrografias de aspectos anatômicos da região mediana de secções
transversais de folhas de espécies de Monocotiledôneas: 32-34. Echinodorus
grandiflorus (Alismataceae). 32. Aspecto geral da nervura central. 33. Detalhe do
mesofilo com nervuras menores. 34. Detalhe do diafragma que limita as lacunas de ar.
35-37. Eichhornia paniculata (Pontederiaceae). 35. Aspecto geral da nervura central.
36. Detalhe do mesofilo. 37. Detalhe do mesofilo contendo ráfides. 38-39. Habenaria
repens (Orchidaceae). 38. Aspecto geral da nervura central. 39. Detalhe do mesofilo.
40-43. Bordo foliar. 40. E. grandiflorus. 41. E. paniculata. 42. H. repens. ...................... 69
Figuras 43-48. Fotomicrografias de aspectos anatômicos da região mediana de secções
transversais de escapos de espécies de Monocotiledôneas: 43-44. Echinodorus
grandiflorus (Alismataceae) - Aspecto geral e detalhe do córtex, respectivamente. 45-
46. Eichhornia paniculata (Pontederiaceae) - Aspecto geral e detalhe do córtex,
respectivamente. 47-48. Habenaria repens Orchidaceae) - Aspecto geral e detalhe do
córtex, respectivamente. .................................................................................................... 71
CAPÍTULO III
Figuras 1-8. Aspectos anatômicos da exoderme, endoderme e lacunas de ar de raízes e
rizomas de espécies de monocotiledôneas, em secções transversais. 1. Pycreus

xiii
macrostachyos (Cyperaceae) - aspecto geral da raiz. 2. Oxycarium cubense
(Cyperaceae) - detalhe da epiderme e exoderme. 3. Eichhornia paniculata
(Pontederiaceae) - detalhe da epiderme e exoderme. 4. Pycreus macrostachyos
(Cyperaceae) - detalhe da endoderme e cilindro vascular. 5. Eichhornia paniculata
(Pontederiaceae) - detalhe da endoderme. 6. Oxycarium cubense (Cyperaceae)-detalhe
do rizoma com lacunas de ar no córtex. 7-8. Rizoma de Eichhornia paniculata
(Pontederiaceae): 7. Detalhe da endoderme e cilindro vascular. 8. Detalhe do córtex. .... 89
Figuras 9-19. Aspectos anatômicos de folhas e escapos de espécies de monocotiledôneas da
região mediana, em secções transversais. 9-14. Detalhe do mesofilo. 9. Cyperus
odoratus (Cyperaceae). 10. Oxycaryum cubense (Cyperaceae). 11. Pycreus
macrostachyos (Cyperaceae). 12. Echinodorus grandiflorus (Alismataceae). 13.
Eichhornia paniculata (Pontederiaceae). 14. Habenaria repens (Orchidaceae). 15-19.
Escapo: aspecto geral, detalhe do córtex e cilindro vascular. 15. Cyperus odoratus
(Cyperaceae). 16. Oxycarium cubense (Cyperaceae). 17. Echinodorus grandiflorus
(Alismataceae). 18. Eichhornia paniculata (Pontederiaceae). 19. Habenaria repens
(Orchidaceae). .................................................................................................................... 90

xiv
RESUMO
Cyperus odoratus L., Oxycaryum cubense (Poepp. & Kunth) Lye and Pycreus
macrostachyos Lam. Raynal (Cyperaceae) apresentam forma de vida anfíbia e Echinodorus
grandiflorus (Cham. & Schltdl.) Micheli (Alismataceae), Eichhornia paniculata (Spreng.)
Solms Laubach (Pontederiacae) e Habenaria repens Nutt. (Orchidaceae) têm forma de vida
emergente e todas as espécies pertencentes a ambas as formas de vida ocorrem nas lagoas
temporárias do semi-árido baiano. Essas lagoas apresentam-se secas no período de estiagem e
inundadas no período de chuvas. Apresentam diversidade de espécies como conseqüência da
variação espacial e temporal do período chuvoso na região. Estudou-se a morfoanatomia dos
órgãos vegetativos dessas macrófitas aquáticas coletadas em diferentes lagoas da região,
considerando-se as formas de vida anfíbia e emergente, para levantar características
anatômicas adaptativas. Também verificou-se quais desses caracteres anatômicos indicam
convergência adaptativa das espécies ao ambiente lacustre temporário. A forma de vida
anfíbia das espécies de Cyperaceae foi caracterizada pela presença de lacunas de ar nos
diferentes órgãos; presença de células buliformes e hipoderme nas folhas; presença; redução
de elementos xilemáticos; e pouca lignificação nas paredes das células dos diferentes órgãos.
As espécies emergentes apresentam maior semelhança anatômica nos órgãos subterrâneos do
que nos órgãos aéreos, e se caracterizam por apresentar células epidérmicas com cutícula fina;
presença de lacunas de ar; idioblastos contendo compostos fenólicos e cristais; redução do
tecido vascular e de sustentação em todos os órgãos. Caracteres anatômicos relacionados ao
sistema dérmico, como epiderme unisseriada, com cutícula fina; fundamental, como presença
de lacunas de ar, diminuição dos tecidos de sustentação, exoderme e endoderme com várias
camadas de células; e vascular, como redução no número e no espessamento de elementos
xilemáticos indicam a convergência adaptativa dessas espécies às lagoas temporárias da
Bahia.

xv
ABSTRACT
Cyperus odoratus L., Oxycaryum cubense (Poepp. & Kunth) Lye and Pycreus
macrostachyos Lam. Raynal (Cyperaceae) present amphibious life form and Echinodorus
grandiflorus (Cham. & Schltdl.) Micheli (Alismataceae), Eichhornia paniculata (Spreng.)
Solms Laubach (Pontederiacae) and Habenaria repens Nutt. (Orchidaceae) are emergent
species that occur in temporary lagoons from semi-arid regions. These lagoons are dry during
the drought season and full of water in wet one. Because of the variation in the rain season
there are a great diversity of species in these lagoons. It was studied the morphoanatomy of
vegetative organs from the aquatic plants collected at different lagoons of semi-arid regions,
from state of Bahia. It was considered the life form to study the anatomic adaptative features,
and which of these features indicate the adaptative convergence of species from the temporary
lagoons environment. The amphibious life form of the species of Cyperaceae had been
characterized by the presence of air canals in the different organs; presence bulliforms cells
and hypoderm on the leaves; reduction of xylem elements; and little lignification in the cell
walls from different organs. The emergent species present more similarity in the
subterraneous organs than in the aerial one and are characterized by presence of epidermal
cells with thin cuticles; presence of air canals; idioblasts conteining phenolic compounds and
crystals; and smaller amounts of vascular and sustentation tissues. Anatomic features related
with dermic system, like uniseriate epidermal, with thin cuticle; fundamental system shows
the presence of air canals, reduction of the amounts sustentation tissue, exoderm and
endoderm with many cell layers; and vascular system show a reduction in the number and
thickening of xylem elements indicated the adaptative convergence of these species of the
temporary lagoons from Bahia state.

1
INTRODUÇÃO GERAL
O semi-árido envolve a região Nordeste do Brasil e norte de Minas Gerais ocupando
cerca de 900.000 km2 incluindo parte dos Estados do Piauí, Ceará, Rio Grande do Norte,
Paraíba, Pernambuco, Alagoas, Sergipe, Bahia e Minas Gerais (Queiroz et al. 2006).
A região é caracterizada por clima quente, baixa pluviosidade (entre 250 e 800 mm
anuais) e duas estações bem definidas durante o ano: estação chuvosa (chamada de inverno)
que dura de três a cinco meses, com chuvas irregulares, torrenciais, locais, de pouca duração,
e estação seca (chamada de verão), que dura de sete a nove meses, quase sem chuvas (Maia
2004).
O clima da região é extremo, exigindo que animais e plantas sejam adaptados a longos
períodos de seca e/ou enchentes, apesar da semi-aridez dominante na região (Sandes et al.
2006). Segundo esses autores, ocorre predomínio de rios e de corpos de água temporários,
com biota aquática diversa, e muitas espécies endêmicas, que aponta a caatinga como
portadora de grande heterogeneidade de fauna e flora.
Os limites da caatinga são considerados coincidentes com os da região semi-árida e
ocorre variação na vegetação, tanto do ponto de vista fisionômico, quanto florístico e
morfofuncional, e essa variação se dá em função das condições de relevo, solo, déficit hídrico
e topografia, com reconhecidos tipos vegetacionais: campos rupestres, florestas, cerrados,
brejos e ambientes aquáticos (Queiroz et al. 2006).
Numa região onde as temperaturas são sempre elevadas, as zonas úmidas ou lagoas
temporárias naturais são consideradas essenciais para a população local, constituindo-se
verdadeiras manchas de biodiversidade de fauna e flora aquática. Estas lagoas estão entre os
ambientes mais produtivos no acúmulo de energia, pois o armazenamento de água e a
retenção de nutrientes têm um papel de destaque na biodiversidade e na produtividade
econômica da região (Maltchik 2000).

2
A vegetação aquática do semi-árido nordestino pode ser encontrada nos espelhos
d’água naturais formados por depressão de relevo. Aí acumula a água superficial, resultante
dos poucos períodos chuvosos da região, originando as lagoas temporárias da caatinga
(Maltchik 2000).
Pesquisas científicas sobre áreas alagáveis no Brasil são necessárias e de extrema
urgência, visto que estas estão submetidas a todo tipo de degradação, principalmente pela sua
utilização inadequada através de atividades agropastoris (Esteves 1988).
No caso da flora aquática, muitas espécies apresentam diferenças morfológicas e
adaptações especiais devido à incidência de luz, sazonalidade do regime pluviométrico e
amplitude e duração de cada fase do ciclo hidrológico, cheia e seca (Maltchik et al. 1999).
Dentre as famílias com maior número de representes no ambiente aquático da região semi-
árida, pode-se citar Alismataceae, Araceae, Asteraceae, Cyperaceae, Leguminosae,
Lentibulariaceae, Melastomataceae, Plantaginaceae, Poaceae, Pontederiaceae, Rubiaceae e
Typhaceae(Queiroz et al. 2006).
A readaptação das plantas ao modo de vida aquático ocorreu em função de
modificações anatômicas, como redução do sistema de sustentação, redução do número ou
ausência de estômatos, cloroplastos localizados preferencialmente na parte superior das folhas
e redução do número e grau de lignificação dos elementos condutores do xilema (Pompêo &
Moschini-Carlos 2003).
A partir de estudos com plantas que crescem desde em brejos até ambientes
verdadeiramente aquáticos, surgiu o termo macrófitas aquáticas, aceito e adotado pelo
International Program of Biology (Esteves 1988). As macrófitas aquáticas têm a capacidade
de ocupar vários ambientes em função de apresentarem diferentes graus de plasticidade
morfoanatômica e fisiológica, o que lhes garante vantagem adaptativa, permitindo sua
colonização e sobrevivência sob condições que seriam consideradas adversas para outras
espécies (Bradshaw 1965; Redbo-Torstensoon 1994).

3
Segundo Irgang et al. (1984), a distribuição das macrófitas é variável, ocupando
diversos gradientes de profundidade num perfil aquático e apresentando as seguintes formas
de vida: Anfíbia - vive bem tanto em área alagada como fora da água; Emergente - enraizada
no fundo, parcialmente submersa; Flutuante fixa - enraizada no fundo, com caule e/ou ramos
e/ou folhas flutuantes; Flutuante livre - não enraizada no fundo, podendo ser levada pela
correnteza; Submersa fixa - enraizada no fundo, com caule e folhas submersas, geralmente
com a flor para fora d’água; Submersa livre - não enraizada no fundo, totalmente submersa,
emergindo somente a flor; Epífita - se instala sobre outras plantas aquáticas.
Na literatura, de forma geral, estudos com macrófitas aquáticas de lagoas temporárias
da região semi-árida brasileira apresentam levantamento florístico de determinada área e os
estudos anatômicos são relativamente escassos (Matias et al. 2007, 2008). Na Bahia, mais
especificamente, é pouco conhecida a flora aquática lêntica em se tratando de plantas
vasculares. Para o Estado existem levantamentos florísticos de lagoas (Bezerra & França
1999; França et al. 2003) e de lagoa costeira (Neves et al. 2006).
Buscou-se neste trabalho, dados sobre a anatomia de espécies de macrófitas aquáticas,
especialmente algumas monocotiledôneas, através de sua caracterização morfoanatômica, a
fim de contribuir para o conhecimento de suas formas de vida e da convergência adaptativa
entre elas. Para tanto, escolhemos: a) espécies de diferentes gêneros da mesma família
(Cyperaceae) com a mesma forma de vida (anfíbia) – capítulo 1 da tese; b) espécies de
diferentes famílias com a mesma forma de vida (emergente) – capítulo 2; e c) levantar
características anatômicas que indicassem convergência adaptativa – capítulo 3.
Área de estudo
O Estado da Bahia caracteriza-se por apresentar um balanço hídrico negativo (inferior
a 800 mm), forte insolação, temperaturas relativamente altas e regime de chuvas marcado pela

4
escassez, irregularidade e concentração de precipitações num período de três meses (Eiten &
Goodland 1979). Este Estado faz parte da região semi-árida do Brasil e apresenta rios, riachos
e lagoas temporários com ocorrência de cheia e seca de diferentes freqüências, intensidades e
duração, sendo que o período de cheia é um fator determinante na ocorrência e na composição
da estrutura de macrófitas aquáticas (Fig 1 -2).
As espécies estudadas neste trabalho foram coletadas em períodos sucessivos de cheia,
de março a junho, em algumas lagoas do semi-árido da Bahia, localizadas no município de
Angüera, ao longo da rodovia BA 052 (Estrada do Feijão).
Viagens de coleta
As viagens de coleta foram realizadas mensalmente ao longo dos anos 2005 e 2006.
As coletas foram realizadas no período de cheias. Essas coletas mensais foram feitas
procurando conhecer melhor a área de estudo e acompanhar o desenvolvimento das espécies
escolhidas.
Parte do material coletado, após identificação prévia, foi enviada para identificação
pelos especialistas Dr. Marccus Alves (Cyperaceae), Dra. Lígia Queiroz Matias
(Alismataceae), Dr. Cassio van den Berg (Orchidaceae) e Dr. Flávio França (Pontederiaceae)
e a seguir incorporados ao acervo do Herbário da Universidade Estadual de Feira de Santana
(HUEFS). Outra parte do material foi utilizada para o estudo morfoanatômico.

5
Figura 1. Lagoa temporária do semi-árido baiano no período seco
Figura 2. Lagoa temporária do semi-árido baiano no período alagado

6
REFERÊNCIAS BIBLIOGRÁFICAS
Bradshaw, A.D. 1965. Evolutionary significance of phenotypic plasticity in plants. Advances
Genetics. 13: 115-155.
Bezerra, M.G. & França, F. 1999. Arales de Açudes em uma área do semi-árido baiano.
Sitientibus 20: 45-54.
Eiten, G. & Goodland, R. 1979. Ecology and management of semi-arid ecosystems in Brazil.
Pp. 277-300. In: Management of Semi-Arid Ecosystems, B.H.Walker (ed.). Elsevier
Scientific Publishing Company, Amsterdam.
Esteves, F.A. 1988. Fundamentos de Limnologia. 2a. Ed. Interciência. Rio de Janeiro. 602p.
França, F.; Melo, E. de; Góes Neto, A.; Araújo, D.; Bezerra, M.G.; Ramos, H.M.; Castro, I. &
Gomes, D. 2003. Flora vascular de açudes de uma região do semi-árido da Bahia, Brasil.
Acta Botanica Brasilica 17(4): 549-559.
Irgang, B. E., Pedralli, G. & Waechter, J. L. 1984. Macrófitas aquáticas da Estação Ecológica
do Taim, Rio Grande do Sul, Brasil. Rossléria 6: 395-404.
Maia, G.N. 2004. Caatinga: árvores e arbustos e suas utilidades. 1ª. ed. São Paulo, D&Z
Computação Gráfica e Editora.
Maltchik, L., Costa, M.A.J. & Duarte, M.D.C 1999. Inventory of brazilian semiarid shallow
lakes. Anais da Academia Brasileira de Ciências 71: 801-808.
Maltchik, L. 2000. As lagoas temporárias do semi-árido. Ciência Hoje 28(167): 67-70
Matias, L.Q., Soares, A. & Scatena, V.L. 2007. Systematic consideration of petiole anatomy
of species of Echinodorus Richard (Alismataceae) from north-eastern Brazil. Flora 202:
395-402.
Matias, L.Q., Soares, A. & Scatena, V.L. 2008. Anatomy of Echinodorus (Alismataceae)
scapes from northeastern Brazil as applied to taxonomy. Edinburgh Journal of Botany
65(1): 11-21.

7
Neves, E.L., Leite, K.R.B.; França, F. & Melo, E. 2006. Plantas aquáticas vasculares em uma
lagoa de planície costeira no município de Candeias, Bahia, Brasil. Sitientibus - Série
Ciências Biológicas 6 (1): 24-29.
Pompêo, M.L.M. & Moschini-Carlos, V. 2003. Macrófitas aquáticas e perifíton: Aspectos
ecológicos e metodológicos. São Carlos, Rima, 124p.
Queiroz, L.P.; Conceição, A.A. & Giulietti, A.M. 2006. Nordeste semi-árido: caracterização
geral e lista das fanerógamas. In: Diversidade e caracterização das fanerógamas do semi-
árido brasileiro. Recife, Associação Plantas do Nordeste. Vol I.
Redbo-Torstensoon, P. 1994. Variation in plastic response to a salinity gradient within a
population the halophytic plant Spergularia marina. Oikos 70:349358.
Sandes, M.A.L., Meyer, M., Souza Junior, N.N. & Moreira, M.C.B. 2006. Comunidade
Fitoplanctônica e qualidade da água do Rio de Contas (Alto e Médio Contas – Semi-
árido/BA). In. Modelos de Gestão das Águas Superficiais e Subterrâneas. 6:41-53.

8
CAPÍTULO I
Anatomia de espécies anfíbias de Cyperaceae de lagoas do
semi-árido, Bahia, Brasil

9
RESUMO
Foi estudada a anatomia de raízes, rizomas, folhas e escapos de Cyperus odoratus L.,
Oxycaryum cubense (Poep. & Kunth) Lye e Pycreus macrostachyos (Lam.) Raynal
(Cyperaceae-Poales) de lagoas do semi-árido da Bahia visando caracterizar anatomicamente
as espécies e levantar estruturas adaptativas à forma de vida anfíbia. As espécies apresentam
raízes com epiderme unisseriada, córtex com exoderme e lacunas de ar. Os rizomas
apresentam feixes vasculares anfivasais ou colaterais no cilindro vascular. As folhas
apresentam epiderme unisseriada, com células de parede periclinal externa mais espessada
que a interna, estômatos somente na face abaxial, parênquima clorofiliano alternado com
lacunas de ar e feixes vasculares colaterais. Os escapos apresentam epiderme unisseriada,
feixes de fibras distribuídos perifericamente alternados com parênquima clorofiliano, com
lacunas de ar e com feixes vasculares colaterais. Oxycaryum cubense difere de Cyperus
odoratus e Pycreus macrostachyos por apresentar raízes com pelos longos, rizomas e estolões
com feixes vasculares colaterais, folhas com bordo agudo, e ausência de estrutura Kranz nas
folhas e escapos. Características anatômicas como presença de lacunas de ar observadas
principalmente nas raízes, presença de células buliformes na face adaxial da epiderme foliar,
de hipoderme, de redução de elementos xilemáticos e pouca lignificação nas paredes das
células dos tecidos dos diferentes órgãos são consideradas importantes na adaptação dessas
plantas anfíbias às lagoas temporárias do semi-árido da Bahia.
Palavras-chaves: Cyperaceae, Poales, anatomia, semi-árido, lagoas temporárias

10
ABSTRACT
We examined the anatomy of roots, rhizomes, leaves, and scapes of Cyperus odoratus
L., Oxycaryum cubense (Poep. & Kunth) Lye, and Pycreus macrostachyos (Lam.) Raynal
(Cyperaceae-Poales) from temporary lakes in the semi-arid region of Bahia in order to
characterize the anatomy of these species and examine their structural adaptations to
amphibious lifestyle. All species demonstrated roots with uniseriate epidermal cells, cortex
with exoderm and air canals. The rhizomes demonstrated amphivasal or collateral vascular
bundles in the vascular cylinder. The leaves demonstrated uniseriate epidermal cells with
external periclinal cell walls thicker that the inner walls, stomata on abaxial leaf surface,
chlorenchyma alternating with air canals, and collateral vascular bundles. The scapes
demonstrated uniseriate epidermal cells, fibrous groups distributed peripherally and alternated
with chlorenchyma, with air canals, and collateral vascular bundles. Oxycaryum cubense
differs from Cyperus odoratus and Pycreus macrostachyos by having root with long hairs;
rhizomes and stolons with collateral vascular bundles; foliar blades with acute margins, and
lacking Kranz structures in leaves and scapes. Anatomic characters with presence of large air
canals were observed principally in the roots, the presence of bulliform cells on the adaxial
face of the leaf epidermis, and hypodermis, reduction of the numbers of xylem vascular
elements and little lignification of the cell walls of the different plant organs are considered
important features in the adaptation of these amphibious plants that inhabit temporary lakes in
the semi-arid region of Bahia State.
Key-words: Cyperaceae, anatomy, semi-arid, Bahia, temporary lakes

11
INTRODUÇÃO
A família Cyperaceae está incluída na ordem Poales (APG II 2003), no mesmo clado
que agrupa as famílias Bromeliaceae, Eriocaulaceae, Juncaceae, Mayacaceae, Poaceae,
Rapateaceae, Typhaceae e Xyridaceae entre outras. Cyperus odoratus L., Pycreus
macrostachyos (Lam.) Raynal e Oxycaryum cubense (Poep. & Kunth) Lye, selecionadas para
este trabalho pertencem à diferentes gêneros de Cyperoideae e à tribo Cypereae (Goetghebeur
1998), sendo os gêneros Cyperus L. e Pycreus P. Beauv considerados os mais próximos
filogeneticamente.
Cyperaceae é uma família cosmopolita, constituindo a terceira maior família em
diversidade de espécies entre as monocotiledôneas, apresentando cerca de 104 gêneros e mais
de 5000 espécies, de hábito terrestre ou aquático, formando grandes populações em diversos
habitats, principalmente em ambientes abertos e úmidos das regiões tropicais e subtropicais
(Goetghebeur 1998). Cerca de 31 gêneros de Cyperaceae contêm espécies aquáticas que
apresentam variadas formas de vida (Cook 1996) e colonizam principalmente ambientes
lênticos, merecendo especial atenção as espécies encontradas nas lagoas temporárias (França
et al. 2003; Matias et al. 2003).
Os ecossistemas aquáticos temporários, conhecidos como alagados, brejos ou áreas
inundáveis, são típicos da região Nordeste do Brasil, apresentando-se secos nos períodos de
estiagem e retornando ao estado alagado na época das chuvas. São caracterizados por
apresentar grande diversidade vegetacional, com espécies tolerantes e adaptadas a solos que
sazonalmente sofrem inundações (Nielsen & Orcutt 1996).
Estudos com espécies de Cyperaceae de ambiente aquático se restringem à listas
taxonômicas ou aspectos ecológicos, apresentando alta freqüência de espécies da família nas
floras de lagoas temporárias. A alta representatividade dessa família em ambientes aquáticos é
devida à presença de rizomas, tubérculos e estolões, que permite maior eficiência na

12
propagação vegetativa (Pott et al. 1989; Bove et al. 2003; França et al. 2003; Matias et al.
2003).
Estudos anatômicos com espécies de Cyperaceae têm se mostrado úteis na taxonomia
de vários grupos, principalmente com espécies brasileiras de Hypolytrum Rich. (Alves et al.
2002) e Bulbostylis DC. (Prata et al. 2007). Com espécies aquáticas brasileiras foram
desenvolvidos estudos de estruturas anatômicas das brácteas de Cyperus giganteus Vahl
(Rodrigues & Estelita 2003) e estudos sobre a atividade meristemática da endoderme de
Cyperus papyrus L. (Menezes et al. 2005).
Dentre as várias espécies de Cyperaceae aquáticas, ocorrentes nas lagoas temporárias
do semi-árido da Bahia, buscou-se estudar aquelas que apresentam forma de vida anfíbia para
tentar entender quais características podem ser consideradas adaptativas e que permitem o
sucesso das espécies.
Dessa forma, este trabalho objetivou caracterizar anatomicamente os órgãos
vegetativos de Cyperus odoratus L., Pycreus macrostachyos (Lam.) Raynal e Oxycaryum
cubense (Poep. & Kunth) Lye (Cyperaceae) ocorrentes em lagoas do semi-árido da Bahia,
buscando levantar características adaptativas à forma de vida anfíbia.

13
MATERIAL E MÉTODOS
Espécies de Cyperus odoratus L., Oxycaryum cubense (Poep. & Kunth) Lye e Pycreus
macrostachyos (Lam.) Raynal, foram coletadas mensalmente em quatro lagoas diferentes do
semi-árido da Bahia, localizadas entre as coordenadas 12°16’32”S/39°03’22”W e
12°10’00”S/39°12’36”W ao longo da rodovia BA 052 (Estrada do Feijão) no município de
Angüera, em períodos sucessivos de cheia.
As lagoas onde os espécimes foram coletados não apresentam comunicação, são
formadas por depressão do relevo e alimentadas por água da chuva, permanecendo cheias por
um tempo maior que o período das chuvas, sendo este tempo suficiente para o
estabelecimento de uma flora rica e com alta diversidade de espécies vegetais (Maltchik 1999;
França et al. 2003). O critério utilizado para a escolha das espécies de diferentes gêneros de
Cyperaceae foi devida a maior representatividade das espécies nas lagoas temporárias do
semi-árido da Bahia e por apresentarem a mesma forma de vida, anfíbia.
Para a análise anatômica foram utilizadas plantas adultas fixadas em FAA 70℅ e
posteriormente preservadas em álcool etílico a 70℅ (Johansen 1940). O material estudado
encontra-se depositado no Herbário da Universidade Estadual de Feira de Santana (HUEFS):
Cyperus odoratus: Leite et al. 477; Leite 486; Leite & França 495; Leite & França 519; Leite
& França 520. Oxycaryum cubense: Leite et al. 475; Leite & França 522; Leite & França
523; Leite & França 524; Leite & França 525. Pycreus macrostachyos: Leite 486A; Leite
487A; Leite & França 496; Leite & França 500; Leite & Lessa 507.
A partir do material fixado foram feitas secções nas regiões apical, mediana e basal de
raízes, rizomas, estolões, folhas e escapos. Foram utilizados pelo menos três indivíduos de
cada espécie provenientes de diferentes lagoas. As secções transversais e longitudinais dos
diferentes órgãos foram feitos à mão livre, com o auxílio de lâmina de barbear, clarificadas

14
com hipoclorito de sódio a 50% e submetidas ao processo de dupla coloração com azul de
astra e safranina (modificado de Bukatsch 1972 apud Kraus & Arduim 1997).
Para dissociação da epiderme foliar, foi utilizado o método de Jeffrey (Johansen 1940)
cujas porções foram coradas com Safranina aquosa a 1%.
Algumas peças dos diferentes órgãos foram submetidas à desidratação em série etílica
e incluídas em historresina Leica [2-hidroxietil-metacrilato] (Feder & O´Brien 1968). O
material incluído foi seccionado em micrótomo rotativo, com 10 µm de espessura. As secções
anatômicas foram submetidas à coloração com ácido-periódico-reativo de Schiff (PAS) e azul
de toluidina (Feder & O´Brien 1968) e montadas em lâminas permanentes com Entellan.
Todo o material foi fotomicrografado em microscópio Olympus BX40 e as lâminas
depositadas no acervo do Laboratório de Micromorfologia vegetal da Universidade Estadual
de Feira de Santana.

15
RESULTADOS
Os caracteres morfológicos observados nas espécies estudadas podem ser observados
nas figuras 1-9 e os anatômicos nas figuras 10-56.
Cyperus odoratus, Oxycaryum cubense e Pycreus macrostachyos (Cyperaceae) são
plantas herbáceas, anfíbias, tolerantes à seca, com rizoma vertical, folhas lineares, filotaxia
espiralada, formando touceiras (Fig. 1, 4 e 7). O rizoma na porção basal é envolvido por
bainhas de folhas velhas, de onde crescem as raízes adventícias (Fig. 2, 5 e 8), e na porção
apical, por folhas jovens e escapos. Oxycaryum cubense apresenta estolão que cresce a partir
do rizoma (Fig. 4).
Os esquemas das secções transversais da região mediana dos escapos das espécies
estudadas apresentam contorno triangular (Fig. 3, 6 e 9). Internamente à epiderme ocorrem
feixes de fibras de esclerênquima. Em Oxycaryum cubense (Fig. 6), esses feixes estão
localizados próximos entre si e o escapo é fistuloso. Ocorre maior proporção de aerênquima
em Cyperus odoratus e Pycreus macrostachyos (Fig. 3 e 9).
As raízes das espécies estudadas, em secção transversal, apresentam epiderme, córtex
heterogêneo e cilindro vascular (Fig. 10, 13, 17). A epiderme é unisseriada, com células de
paredes finas (Fig. 11, 17, 18). Os pêlos absorventes ocorrem isolados, sendo mais abundantes
e alongados em Oxycaryum cubense (Fig. 14).
O córtex é heterogêneo e apresenta-se dividido em externo, mediano e interno. O
córtex externo é constituído pela exoderme, que apresenta células hexagonais, com paredes
levemente espessadas. (Fig10-11, 13-14, 17-18).
O córtex mediano ocupa a maior porção das raízes. É formado por dois tipos distintos
de células: aquelas arredondadas, pequenas, formando de uma a três fileiras longitudinais,
intercaladas por células maiores, braciformes (Fig. 10, 13, 17).

16
As raízes de todas as espécies apresentam células corticais jovens com redução de
espaços intercelulares e são dispostas em fileiras radiais como evidenciado no estágio inicial
do desenvolvimento de Oxycaryum cubense (Fig. 16). À medida que a raiz se desenvolve as
células do córtex mediano tornam-se braciformes e, na maturidade, suas paredes se rompem
formando lacunas de ar. São observados restos das paredes tangenciais das células rompidas
(Fig. 10, 13, 17). As células arredondadas do córtex mediano permanecem com as paredes
íntegras formando fileiras longitudinais (Fig. 10 e 17).
O córtex interno é formado por células parenquimáticas arredondadas, menores do que
as demais, sem espaços intercelulares (Fig. 10, 12-13, 15, 17, 19). Essas células apresentam
paredes levemente espessadas na maturidade. Observa-se pouca lignificação nas paredes das
células do córtex mediano (Fig. 10, 13, 17). A endoderme é unisseriada, com células de
paredes finas (Fig. 10, 12, 15, 19).
O periciclo é unisseriado e apresenta células de paredes finas (Fig. 10, 12-13, 15, 19).
O cilindro vascular tem estrutura poliarca e pode apresentar vários elementos de metaxilema
(Fig. 12). Observa-se redução no número de elementos xilemáticos em todas as raízes das
espécies estudadas (Fig. 12, 15, 19).
Os rizomas das espécies estudadas apresentam epiderme, córtex e cilindro vascular
(Fig. 20-26). A epiderme é constituída por células de paredes finas (Fig. 20, 25). O córtex
apresenta células parenquimáticas de paredes finas e arredondadas (Fig. 20, 22, 25), podendo
apresentar raízes adventícias intracorticais (Fig. 20, 25). A endoderme é unisseriada, com
células de paredes finas (Fig. 26). O córtex de Oxycaryum cubense (Fig. 22) apresenta amplas
lacunas de ar.
O cilindro vascular é delimitado pelo periciclo (Fig. 20-21, 22, 25-26) e apresenta
distribuição atactostélica (Fig. 20, 22, 25). É constituído por feixes vasculares anfivasais em
Cyperus odoratus (Fig. 21) e Pycreus macrostachyos (Fig. 26) e por feixes vasculares
colaterais em Oxycaryum cubense (Fig. 22). Em todos esses feixes vasculares, é observada a

17
redução do número de elementos xilemáticos quando comparado com os elementos de floema
(Fig. 21-24, 26).
Oxycaryum cubense apresenta estolão além de rizoma. Na região do nó (Fig. 23),
observam-se plexos vasculares, que irão vascularizar folhas e raízes. Na região do entrenó
(Fig. 24), a epiderme é unisseriada, com células de paredes espessadas. O córtex é formado
por células arredondadas, onde estão distribuídos os feixes vasculares colaterais e as lacunas
de ar (Fig. 24). Apenas as células dos feixes vasculares apresentam paredes lignificadas (Fig.
20-26).
As folhas de todas as espécies de Cyperaceae estudadas, em vista frontal, apresentam
células epidérmicas alongadas no sentido longitudinal da lâmina, com paredes levemente
sinuosas (Fig. 27-32). As folhas são hipoestomáticas, com estômatos do tipo paracítico,
distribuídos em fileiras longitudinais (Fig. 28, 30, 32).
Em secção transversal da folha, a epiderme é uniestratificada com células epidérmicas
da face adaxial maiores em relação às da face abaxial (Fig. 33-42, 44) e com paredes
periclinais externas mais espessadas que as internas (Fig. 33-42). O bordo foliar é
arredondado em Cyperus odoratus e Pycreus macrostachyos (Fig. 33, 35) e agudo em
Oxycaryum cubense (Fig. 34) com esclerênquima adjacente à epiderme.
Nas três espécies estudadas, na face adaxial da epiderme, principalmente na altura da
nervura central, ocorrem células buliformes que são maiores do que as demais células
epidérmicas (Fig. 36, 38-39, 41), estando ausentes apenas na região basal das lâminas (Fig.
42). Internamente à epiderme, ocorrem feixes de fibras de esclerênquima distribuídos ao
longo de toda a extensão da lâmina foliar (Fig. 33-42).
O mesofilo apresenta hipoderme voltada à face adaxial (Fig. 33-42), com parênquima
clorofiliano paliçádico voltado à face abaxial em Oxycaryum cubense (Fig. 38-40) e disposto
de forma radiada em torno dos feixes vasculares em Cyperus odoratus (Fig. 36-37) e Pycreus
macrostachyos (Fig. 41-42, 44). Cyperus odoratus e Pycreus macrostachyos apresentam

18
feixes vasculares com anatomia Kranz do tipo “clorociperóide”. No mesofilo de todas as
espécies ocorre parênquima clorofiliano braciforme (Fig. 37, 40-42), que constitui diafragmas
que limitam lacunas de ar (Fig. 43).
Os feixes vasculares das folhas são colaterais (Fig. 33-42, 44), grandes e pequenos,
envolvidos por bainha dupla. A bainha externa, parenquimática, constitui a endoderme e a
interna, de células de paredes espessadas constitui o periciclo. Este último pode envolver
completamente os feixes vasculares e formar calotas de fibras junto ao xilema (Fig. 38-39). A
redução dos elementos xilemáticos pode ser facilmente observada (Fig. 36-42, 44). As lacunas
de ar ocorrem em toda a extensão do limbo foliar, sempre intercaladas com feixes vasculares
ou logo abaixo deles (Fig. 36-42, 44). As células da epiderme e do mesofilo não apresentam
lignificação, com exceção dos feixes de fibras e dos feixes vasculares (Fig. 36-42, 44-45).
Os escapos das espécies de Cyperaceae estudadas, em secção transversal, apresentam
epiderme uniestratificada, com células de paredes finas (Fig. 48, 50, 53, 55-56). Os estômatos
estão situados levemente acima das demais células epidérmicas (Fig. 53, 56). Logo abaixo da
epiderme ocorrem feixes de fibras de esclerênquima distribuídos em toda a extensão do
escapo (Fig. 46-50, 52-53, 55-56).
O parênquima clorofiliano é paliçádico e está situado perifericamente, com três a
quatro camadas de células. Em Cyperus odoratus (Fig. 48) e Pycreus macrostachyos (Fig.
53), o parênquima paliçádico apresenta-se radiado em torno dos feixes vasculares,
constituindo anatomia Kranz.
Os feixes vasculares são colaterais e distribuídos por todo o parênquima, com maior
concentração na periferia. São envolvidos de forma parcial (Fig. 47-48, 52) ou total (Fig. 49-
51) pelo periciclo espessado. Observa-se redução do número de elementos xilemáticos em
comparação com os elementos floemáticos (Fig. 46-54). Logo abaixo do parênquima
clorofiliano ocorrem lacunas de ar (Fig. 46, 49, 52). Apenas as células dos feixes de fibras e
dos feixes vasculares apresentam paredes lignificadas (Fig. 47-56).

19
Corpos silicosos do tipo cônico foram observados na epiderme das folhas (Fig. 45) e
dos escapos (Fig. 55), além de cristais prismáticos e ráfides, formando idioblastos na região
que delimita a lacuna de ar.

20
DISCUSSÃO
As espécies estudadas neste trabalho são macrófitas aquáticas, anfíbias, que ocorrem
em lagoas temporárias, constituindo um grupo interessante de plantas devido à sua facilidade
de crescimento.
Cyperus odoratus, Oxycaryum cubense e Pycreus macrostachyos (Cyperaceae),
mesmo pertencendo a gêneros diferentes, apresentam alguns aspectos morfológicos e
anatômicos semelhantes. São plantas afetadas por contínua condição de estresse, caracterizada
por períodos de chuva e de seca na região. O estresse pode ser induzido por fatores bióticos e
abióticos, traduzindo-se no desenvolvimento de características morfológicas e anatômicas
adaptativas, principalmente nos órgãos de absorção e na presença de aerênquima, conforme
referido por Nielsen & Orcutt (1996).
As diferenças anatômicas observadas entre as espécies estudadas, como raízes com
pêlos longos, presença de estolão com feixes vasculares colaterais, folhas com bordo agudo e
ausência de anatomia Kranz em Oxycaryum cubense podem estar relacionadas com a herança
genética. Essas diferenças podem ser entendidas por ser este um gênero considerado mais
distante filogeneticamente de Cyperus e Pycreus, que já pertenceram ao mesmo gênero,
Cyperus (Goetghebeur 1998).
Acredita-se que o sucesso ecológico das espécies estudadas deva-se ao fato delas
apresentarem sistema subterrâneo com rizoma, que segundo Font Quer (1989), além de
facilitar a propagação vegetativa, protege a planta de rigores ambientais.
Nas raízes de Oxycaryum cubense, ocorrem pêlos absorventes longos e em
abundância. Essa característica provavelmente está relacionada à forma de vida, que no início
do seu desenvolvimento é epífita de outras plantas flutuantes como Salvinia sp., Azzola sp.,
Pistia stratiotes L. e Eichhornia crassipes (Mart.) Solms (Tur 1971; Neves et al. 2006). Na
fase adulta e na época de seca, essa espécie se fixa ao substrato em decomposição que está na

21
margem da lagoa, comportando-se como anfíbia (Pott & Pott 2000). A presença desses pêlos
radiculares longos e abundantes em sua raiz, além de exercer função de absorção, deve
também atuar na estabilidade da planta conforme acontece em outras espécies aquáticas (Tur
1971).
Em todos os órgãos das espécies estudadas observou-se a presença de lacunas de ar,
principalmente nas raízes, que apresentam grande tamanho quando comparadas com a
dimensão do córtex. A sobrevivência de plantas que ocorrem em solos alagados, em geral,
está relacionada à presença de raiz cujo córtex contém grande quantidade de espaços de ar
(Nielsen & Orcutt 1996). Adaptações ao ambiente aquático podem ser consideradas
características genéticas (Davy et al. 1990; Nielsen & Orcutt 1996) ou fisiológicas. Neste
último caso, as raízes das plantas que crescem em ambientes que são inundadas
periodicamente respondem ao estresse com a formação de grande quantidade de raízes
adventícias e de lacunas de ar, como observado nas espécies aqui estudadas. Lacunas de ar
podem ser interpretadas como aerênquima que pode ser lisígeno ou esquizógeno segundo
Nielsen & Orcutt (1996), ou esquizolisígeno segundo Smirnoff & Crawford (1983). Esses
autores relatam que esses aerênquimas irão estocar oxigênio.
Para Luxmoore et al. (1972) e Nielsen & Orcutt (1996), a formação de lacunas de ar
em órgãos vegetativos, principalmente raízes e rizomas, pode estar relacionada à temperatura
e à intensa luminosidade. Na área de coleta das espécies aqui estudadas, a temperatura e a
luminosidade são elevadas. Além desses fatores, acredita-se que essas lacunas de ar também
estão relacionadas à baixa aeração da água da lagoa. A baixa aeração faz com que o etileno se
acumule nas células corticais estimulando a atividade da celulase, que leva à plasmólise e em
seguida à morte celular, formando as lacunas de ar, conforme o que foi apresentado para
outras plantas de ambientes alagados (Kawase & Whitmoyer 1980; Coan et al. 2002).
Observou-se também em todas as espécies a relação entre a endoderme meristemática
e as células que vão formar o aerênquima, pois as células que arredondadas que se mantém

22
íntegras na raiz são provenientes da endoderme enquanto as células braciformes que formam
o aerênquima provavelmente têm origem do meristema fundamental.
Lacunas de ar encontradas também nas folhas e escapos das espécies de Cyperaceae
aqui estudadas, também foram observadas em outras macrófitas aquáticas por Sculthorpe
(1967), Nielsen & Orcutt (1996) e Kawase & Whitmoyer (1980). Essas lacunas promovem a
oxigenação através de difusão entre órgãos aéreos e submersos. São capazes de transportar o
O2 formado durante a fotossíntese nas folhas para outros órgãos da planta, indicando a
continuidade dessas lacunas de ar ao longo de todo o vegetal, corroborando o que foi relatado
para outras espécies aquáticas por Boeger (1994).
A presença de células braciformes limitando as lacunas de ar nos diferentes órgãos das
espécies estudadas, que forma o diafragma, é responsável pela condução de gases além de
atuar de forma mecânica em órgãos submersos conforme o que foi demonstrado em outras
plantas aquáticas por Sculthorpe (1967), Kaul (1971) e Fahn (1990).
Nas folhas, a maior sinuosidade observada nas paredes das células epidérmicas de C.
odoratus e P. macrostachyos em relação à O. cubense possivelmente está relacionada ao local
de coleta e desenvolvimento das espécies, pois O.cubense além de ter seu desenvolvimento
inicial como epífita, foi coletada em local mais próximo à margem da lagoa e as outras duas
espécies em ambiente mais seco.
Nas folhas das espécies estudadas, ocorrem células buliformes que podem estar
relacionadas à abertura e fechamento laterais do limbo durante o dia em função da variação da
intensidade de luz e temperatura, comum na região semi-árida da Bahia. Esta característica de
abertura e fechamento do limbo através de células buliformes foi observada também em
espécies de Poaceae (Beal 1886) e outras Cyperaceae (Mani 1964). Nas espécies anfíbias de
Cyperaceae aqui estudadas, esse movimento do limbo foliar se restringe às regiões mediana e
apical da lâmina, uma vez que a porção basal está em contato direto com a água. Uma
possível interpretação para esta característica é que, como esse movimento foliar, ocorre nas

23
horas mais quentes do dia e de maior intensidade luminosa, certamente leva a uma diminuição
da área da superfície foliar exposta à luz e, assim, reduz a transpiração e a temperatura da
folha, facilitando a fotossíntese.
Nas folhas, logo abaixo da epiderme adaxial, ocorre hipoderme parenquimática,
constituída por células translúcidas, comum a todas as espécies e mais evidente em
Oxycaryum cubense. A hipoderme, conforme já descrita por Koyama (1965), para
Cyperaceae, pode estar relacionada ao acúmulo de água, constituindo-se, portanto, aqüífera,
ajudando na manutenção das espécies em períodos de seca.
Em todas as espécies de Cyperaceae estudadas, foram observados corpos silicosos na
forma de cristais cônicos na epiderme e ráfides nas células que formam as lacunas de ar. Os
cristais cônicos ocorrem sempre na parede periclinal interna das células epidérmicas de folhas
e escapos e juntos aos feixes de fibras que estão associados ou não aos feixes vasculares,
corroborando o que foi apresentado por Metcalfe (1971) para outras espécies da família.
A presença de cristais nas plantas é considerada por Metcalfe (1971), Prychid &
Rudall (1999) e Lucas et al. (2000) como suporte mecânico e atua na proteção contra a
herbivoria, podendo ainda promover a formação de lacunas de ar em plantas aquáticas, além
de se projetar para dentro delas.
Para Prychid & Rudall (1999) os cristais de oxalato de cálcio são o produto final do
metabolismo celular e representam uma forma de estocar cálcio e ácido oxálico, evitando que,
em alta concentração, sejam tóxicos para a célula ou tecido, promovendo o equilíbrio iônico.
Essas autoras relataram que pode haver reabsorção do oxalato de cálcio pelas plantas em
períodos de baixa concentração deste composto no solo.
Para algumas espécies anfíbias, a passagem da forma terrestre para aquática e vice-
versa, envolve ajustes estruturais e fisiológicos, conforme pode ser observado em Cyperus
odoratus e Pycreus macrostachyos, que apresentam feixes vasculares com estrutura Kranz.
Segundo Nielsen & Orcutt (1996), a maioria das plantas terrestres e aquáticas, apresenta

24
fotossíntese C3, sendo encontrados metabolismo C4 ou CAM em espécies sujeitas à altas
temperaturas e baixa disponibilidade de água, constituindo um mecanismo fisiológico de
adaptação ao processo fotossintético.
Nas espécies estudadas, esses feixes estão dispostos ao longo de toda a lâmina foliar e
na região periférica do escapo, sempre intercalados por amplas lacunas de ar, o que pode
indicar a adaptação delas à condição de alagamento. Além disso, elas já estão prontas para
assumir a condição de plantas terrestres quando o ambiente secar. Como apenas Oxycaryum
cubense não apresentou anatomia Kranz, acredita-se que essa característica esteja relacionada
também à sua forma de crescimento, que no início do desenvolvimento é epífita e quando
adulta torna-se anfíbia.
As espécies de Cyperaceae aqui estudadas, quando comparadas às espécies terrestres
de Cyperaceae estudadas por Metcalfe (1971) apresentaram reduzida lignificação,
característica comum de plantas aquáticas, pois a água exerce a função de sustentação em
alguns órgãos. No entanto, por serem anfíbias essas espécies apresentam maior lignificação
nas paredes de suas células do que outras macrófitas aquáticas (Coan et al. 2002; Matias et al.
2007; Rodrigues et al. 2007; Fabbri et al. 2005).
Assim, a forma de vida anfíbia das espécies de Cyperaceae aqui estudadas pode ser
caracterizada anatomicamente pela presença de lacunas de ar nos diferentes órgãos,
principalmente no sistema subterrâneo; pela presença de células buliformes, e hipoderme nas
folhas; além da presença de corpos silicosos; da redução de elementos xilemáticos e da pouca
lignificação nas paredes das células nos diferentes órgãos. Essas características são
consideradas importantes na adaptação dessas espécies ao ambiente aquático.

25
REFERÊNCIAS BIBLIOGRÁFICAS
Alves, M.V.; Estelita, M.E.M.; Wanderley, M.G.L. & Thomas, W.W. 2002. Aplicações
taxonômicas da anatomia foliar das espécies brasileiras de Hypolytrum Rich. (Cyperaceae).
Revista Brasileira de Botânica 25(1): 1-9.
Angiosperm Philogeneyic Group (APG). 2003. An update of the Angiosperm Phylogeny
Group classification for the orders and families of flowering plants. APG II. Botanical
Journal of Linnean Society 141(4): 399-346.
Beal, W.J. 1886. The bulliform or hygroscopic cells of grasses and sedges compared.
Botanical Gazette 11(12): 321-326.
Boeger, M.R.T. 1994. Adaptações morfológicas e anatômicas de Polygonum spectabile Mart.
(Polygonaceae) às inundações na várzea Amazônica. Amazoniana 13(1/2): 3-11.
Bove, C.P.; Gil, A.S.B.; Moreira, C.B. & Anjos, R.F.B. 2003. Hidrófitas fanerogâmicas de
ecossistemas aquáticos temporários da planície costeira do estado do Rio de Janeiro,
Brasil. Acta Botanica Brasilica 17(1): 119-135.
Coan, A.I.; Scatena, V.L. & Giulietti, A.M. 2002. Anatomia de algumas espécies aquáticas de
Eriocaulaceae brasileiras. Acta Botanica Brasilica 16(4): 371-384.
Cook, C.D.K. 1996. Water plants of world. Amsterdan. SPB Academic Publishing.
Davy, A.J.; Noble, S.M. & Oliver, R.P. 1990. Genetic variation and adaptation to flooding in
plants. Aquatic Botany 38(1): 91-108.
Fabbri, L.T., Rua, G.H. & Bartoloni, N. 2005. Different patterns of aerênquima formation in
two hygrophytic species of Paspalum (Poaceae) as response to flooding. Flora 200: 354-
360.
Fahn, A. 1990. Plant anatomy. Oxford. Pergamon Press.
Feder, N. & O Brian, T.P. 1968. Plant microthecnique: some principles and new methods.
American Journal of Botany 55(1): 123-142.

26
França, F.; Melo, E. de; Góes Neto, A.; Araújo, D.; Bezerra, M.G.; Ramos, H.M.; Castro, I. &
Gomes, D. 2003. Flora vascular de lagoas de uma região do semi-árido da Bahia, Brasil.
Acta Botanica Brasilica 17(4): 549-559.
Font Quer, P. 1989. Diccionario de Botánica. Barcelona. Editorial Labor.
Goetghebeur, P. 1998. Cyperaceae. Pp. 141-190. In: K. Kubitzki, H. Huber, P. Rudall, P.
Stevens & T. Stützel (eds.) The families and genera of vascular plants. Berlin. Spreng-
Verlag. v. IV.
Johansen, D.A. 1940. Plant microtechnique. New York. McGraw Hill Book Co.
Kaul, R.B. 1971. Diafragms and aerenchyma in Scirpus validus. American Journal of
Botany 58(9): 808.
Kawase, M. & Whitmoyer, R.E. 1980. Aerenchyma development in waterlogged plants.
American Journal of Botany 67(1): 18-22.
Koyama, 1965. Interrelationships between the tribes Lagenocarpeae and Sclerieae
(Cyperaceae). Bulletim Torrey Botanical Club 92(4): 250-265.
Kraus, J.E. & Arduin, M. 1997. Manual básico de metodos em morfologia vegetal. Rio de
Janeiro. Seropédica.
Lucas, P.W.; Turner, I.M.; Dominy, N.J. & Yamashita, N. 2000. Mechanical defences to
herbivory. Annals of Botany 86(5): 913-920.
Luxmoore, R.J.; Sojka, R.E. & Stolzy, L.H. 1972. Root porosity and growth responses of
wheat to aeration and light intensity. Soil and Science 113(5): 354-357.
Maltchik, L. 1999. Inventory of brazilian semiarid shallow lakes. Anais da Academia
Brasileira de Ciências 71(4-I): 801-808.
Mani, A.P. 1964. Bulliform cells in Cyperus. Science and Culture 30(5): 241-242.
Matias, L.Q., E.R. Amado & E.P. Nunes. 2003. Macrófitas aquáticas da lagoa Jijoca de
Jericoacoara, Ceará, Brasil. Acta Botanica Brasilica 17(4): 623-623.

27
Matias, L.Q., Soares, A. & Scatena, V.L. 2007. Systematic consideration of petiole anatomy
of species of Echinodorus Richard (Alismataceae) from north-eastern Brazil. Flora 202:
395-402.
Menezes, N.L.; Silva, D.C.; Arruda, R.C.O.; Mello-de-Pinna, G.F.; Cardoso, V.A.; Castro,
N.M.; Scatena, V.L. & Scremin-Dias, E. 2005. Meristematic activity of the endodermis
and the pericycle in the primary thickening in monocotyledons. Considerations on the
“PTM”. Anais da Academia Brasileira de Ciências 77(2): 259-274.
Metcalfe, C.R. 1971. Anatomy of the monocotyledons. Cyperaceae. Oxford. Claredon Press.
v.V.
Neves, E.L., Leite, K.R.B.; França, F. & Melo, E. 2006. Plantas aquáticas vasculares em uma
lagoa de planície costeira no município de Candeias, Bahia, Brasil. Sitientibus - Série
Ciências Biológicas 6 (1): 24-29.
Nielsen, E.T. & Orcutt, D.M. 1996. Physiology of plants under stress abiotic factors. John
Wiley & Sons, Inc. United States of America.
Pott, V.J.; Bueno, N.C.; Pereira, R.A.C.; Sallis, S.M. &Vieira, N.L. 1989. Distribuição de
macrófitas aquáticas numa lagoa na Fazenda Nhumirim, Nhecolândia, Pantanal, MS.
Acta Botanica Brasilica 3(2): 153-168.
Pott, V.J. & Pott, A. 2000. Plantas aquáticas do pantanal. Brasília. Embrapa.
Prata, A.P., Menezes, N.L., Mazzoni-Viveiros, S.C., Wanderley, M.G.L. & Thomas, W.W.
2007. Anatomia do escape e rizoma de espécies brasileiras de Bulbostylis Kunth
(Cyperacae). Revista Brasil. Bot. 30(2): 245-256.
Prychid, C.J. & Rudall, P.J. 1999. Calcium oxalate crystals in Monocotyledons: a review of
their structure and systematic. Annals of Botany 84(6): 725-739.
Rodrigues, A.C. & Estelita, M.E.M. 2003. Origin and structure of the Kranz tissue in bracts of
Cyperus giganteus Vahl (Cyperaceae). Revista Brasileira de Botânica 26(4): 445-452.

28
Rodrigues, S., Scremin-Dias, E., Medeiros, S.C.H. & Souza, M.C. 2007. Alterações
estruturais do caule e da folha de Ludwigia inclinata (L.f.) M. Gómez, desenvolvidos
emersos e submersos. Revista Brasileira de Biociências 5(1): 174-176.
Sculthorpe, C.D. 1967. The biology of aquatic vascular plants. London, Edward Arnold.
Smirnoff, C., N. & Crawford, R.M.M. 1983. Variation in the structure and responses to
flooding of root aerenchyma in some wetland plants. Annals of Botany 51(2): 237-249.
Tur, N.M. 1971. Nuevos casos de epifitismo acuático. Bolletim Sociedade Argentina de
Botánica 13(4): 243-249.

29
Figuras 1-9. Aspectos morfológicos de espécies de Cyperaceae: 1-3: Cyperus odoratus L. 1.
Hábito, 2. Detalhe do sistema subterrâneo. 3. Secção transversal da região mediana
do escapo. 4-6: Oxycaryum cubense (Poep. & Kunth) Lye 4. Hábito. 5. Detalhe do
sistema subterrâneo. 6. Secção transversal da região mediana do escapo. 7-9: Pycreus
macrostachyos (Lam.) Raynal 7. Hábito. 8. Detalhe do sistema subterrâneo. 9.
Secção transversal da região mediana do escapo. Legenda: ▬ = epiderme, ● = feixe
de fibras, ▓ = feixe vascular, ▒ = lacuna de ar, □ = parênquima.

30
2
3 c m
3 c m
3 cm
5 m
m
400
mµ
400
mµ
400
mµ
1 ,5
cm
1,5
c m
1
4
5
6
7
3
8
9

31
Figuras 10-19. Fotomicrografias de secções transversais da região mediana de raízes de
espécies de Cyperaceae: 10-12. Cyperus odoratus L. 10. Aspecto geral. 11. Detalhe
da epiderme e exoderme. 12. Detalhe do cilindro vascular. 13-16. Oxycaryum
cubense (Poep. & Kunth) Lye. 13. Aspecto geral - raiz adulta. 14. Detalhe da
epiderme e exoderme. 15. Detalhe do cilindro vascular. 16. Aspecto geral - raiz
jovem. 17-19. Pycreus macrostachyos (Lam.) Raynal. 17. Aspecto geral. 18. Detalhe
da epiderme e exoderme. 19. Detalhe do cilindro vascular. Legenda: En =
endoderme; Ex = exoderme; La = lacuna de ar; Mx = metaxilema; Pe = periciclo.
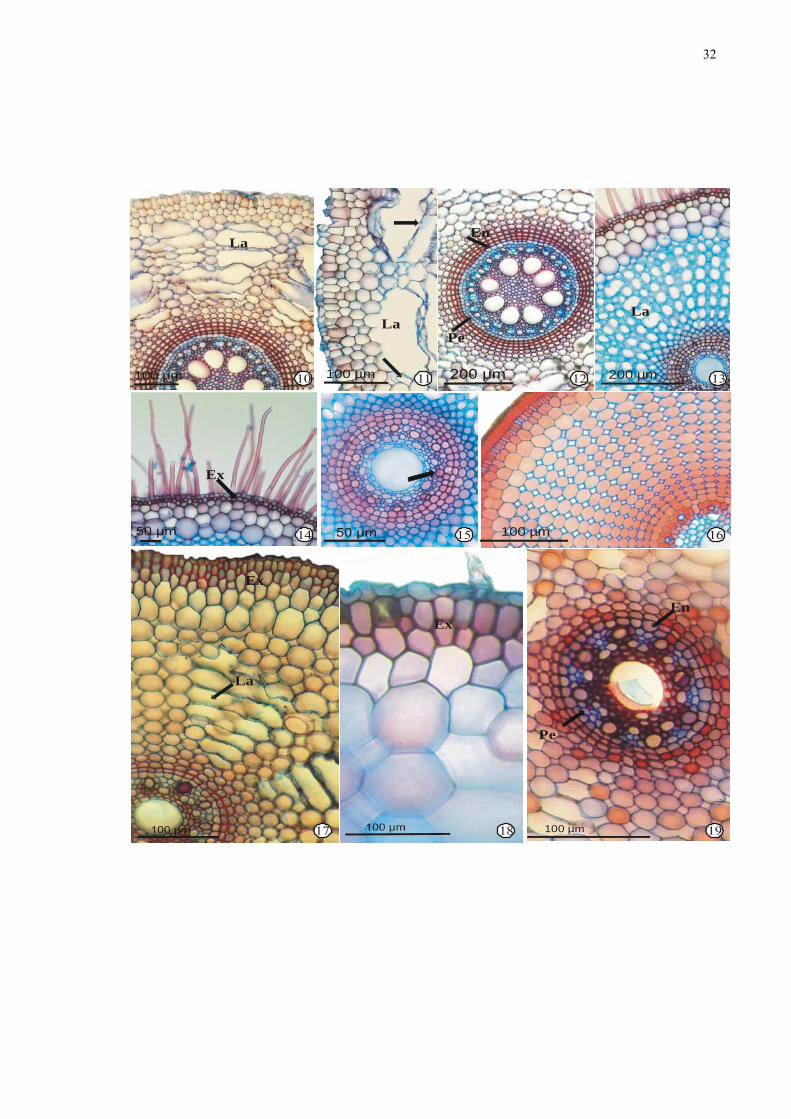
32
100 µm
100 µm
100 µm
100 µm 200 µm
La
LaLa
En
Pe
En
Pe
Ex
La
Ex
Ex
1210
14 15 16
17
13
18 19
11

33
Figuras 20-26. Fotomicrografias de secções transversais da região mediana de rizomas de
espécies de Cyperaceae: 20-21. Cyperus odoratus L. 20. Aspecto geral. 21. Detalhe
do cilindro vascular. 22-24: Oxycaryum cubense (Poep. & Kunth) Lye. 22. Aspecto
geral do rizoma. 23. Região do nó do estolão. 24. Região do entrenó do estolão. 25-
26. Pycreus macrostachyos (Lam.) Raynal. 25. Aspecto geral. 26. Detalhe do
cilindro vascular. Legenda: En = endoderme; Pe = periciclo.
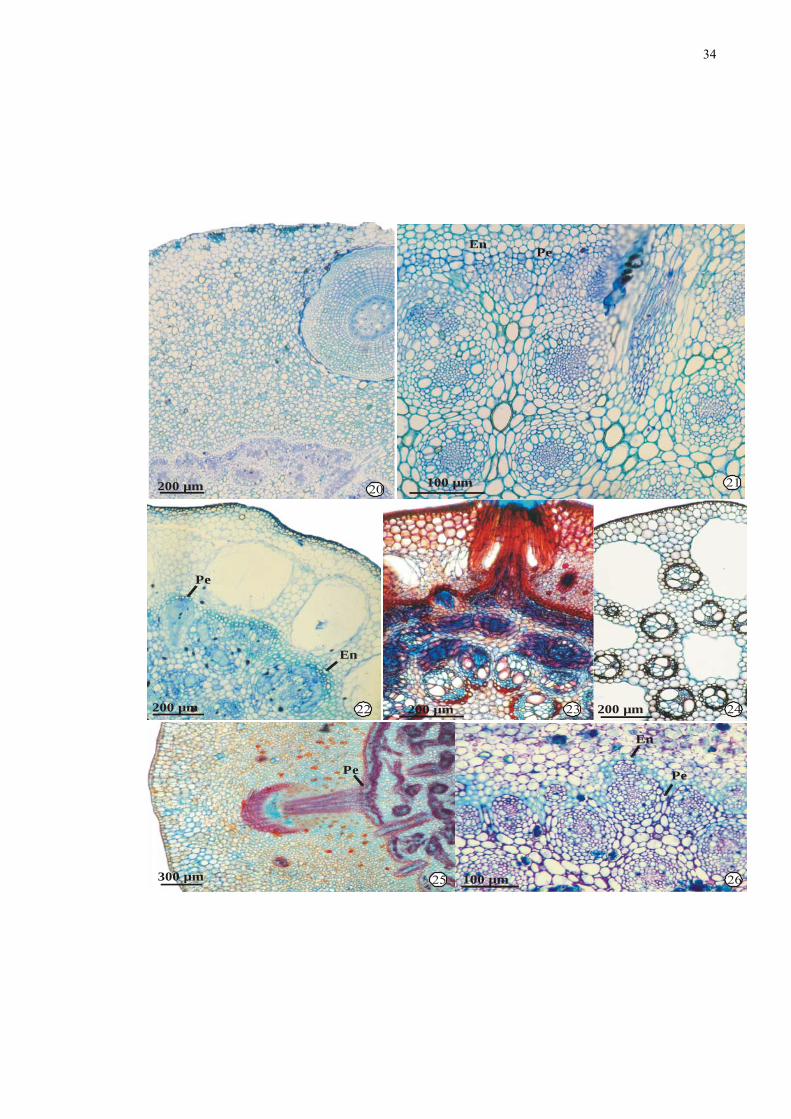
34
200 µm 100 µm
200 µm200 µm 200 µm
300 µm 100 µm
EnPe
En
Pe
Pe
En
Pe
20 21
22 23 24
25 26

35
Figuras 27-35. Fotomicrografias de secções paradérmicas e transversais de folhas de espécies
de Cyperaceae: 27-32. Vista frontal da epiderme. 27-28. Cyperus odoratus L. 27. Face
adaxial. 285. Face abaxial com estômatos. 29-30. Oxycaryum cubense (Poep. & Kunth) Lye.
29. Face adaxial. 30. Face abaxial com estômatos. 31-32. Pycreus macrostachyos (Lam.)
Raynal. 31. Face adaxial. 32. Face abaxial com estômatos. 33-35. Bordo foliar. 33. Cyperus
odoratus L. 34. Oxycarium cubense (Poep. & Kunth) Lye. 35. Pycreus macrostachyos (Lam.)
Raynal. Legenda: Cl = clorênquima; Ff = feixe de fibras; Hi = hipoderme; La= lacuna de ar.
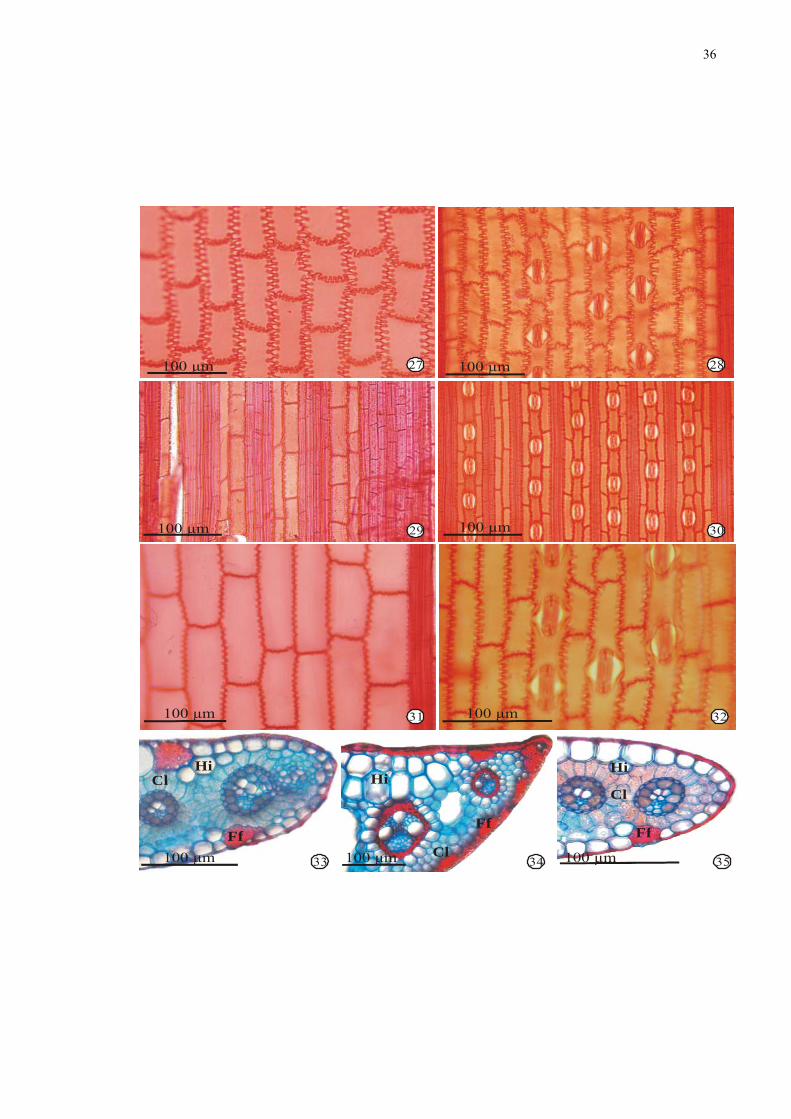
36
HiHi
HiCl
Cl
Cl
FfFfFf
100 µm 100 µm 2827
100 µm 100 µm 3029
100 µm 100 µm31 32
100 µm 100 µm 100 µm3433 35

37
Figuras 36-45. Fotomicrografias de secções transversais de folhas de espécies de Cyperaceae:
36-37. Cyperus odoratus L. - nervura central e mesofilo, respectivamente. 38-40. Oxycarium
cubense (Poep. & Kunth) Lye - nervura central na região mediana, na região apical, e
mesofilo na região mediana, respectivamente. 41-44. Pycreus macrostachyos (Lam.) Raynal –
41-42. nervura central na região mediana e basal, respectivamente. 43. Detalhe do diafragma.
44. Detalhe do mesofilo. 45. Cyperus odoratus - detalhe do cristal cônico. Legenda: Cb =
célula buliforme; Cl = clorênquima; Cs = Corpo silicoso; Ff = feixe de fibras; Hi =
hipoderme; La = lacuna de ar.
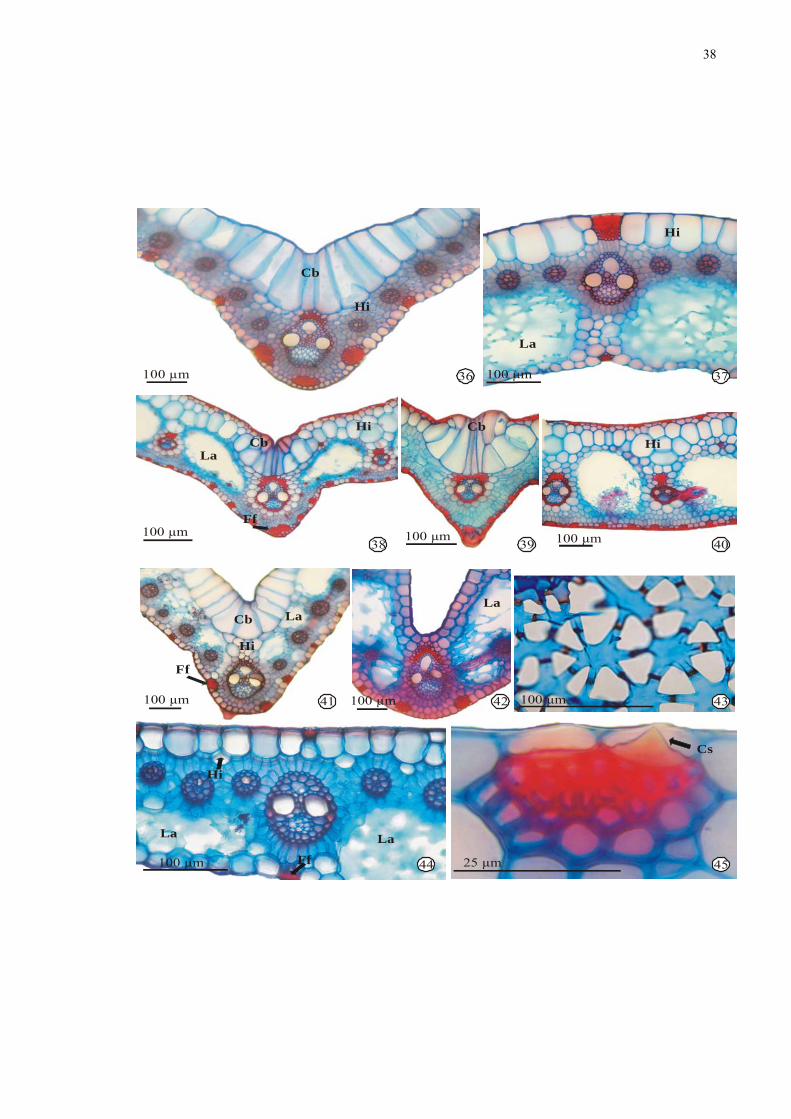
38
100 µm 100 µm
100 µm 100 µm 100 µm
100 µm 100 µm
25 µm
Cs
Cb
CbCb
Cb
Hi
Hi
Hi
Hi
Hi
La
LaLa
Ff
Ff
100 µm
Hi
Ff
La La
100 µm
36 37
38 39 40
41 42 43
44 45
La

39
Figuras 46-56. Fotomicrografias de secções transversais da região mediana de escapos de
espécies de Cyperaceae: 46-48. Cyperus odoratus L. - aspecto geral, detalhe do córtex e da
epiderme, respectivamente. 49-51. Oxycarium cubense (Poep. & Kunth) Lye - aspecto geral,
detalhe do feixe vascular composto e simples, respectivamente. 52-54. Pycreus
macrostachyos (Lam.) Raynal - aspecto geral, detalhe dos feixes vasculares com estrutura
Kranz e do feixe vascular colateral, respectivamente. 55-56. Detalhe da epiderme. 55.
Oxycaryum cubense (Poep. & Kunth) Lye. 56. Pycreus macrostachyos (Lam.) Raynal.
Legenda: Cl = clorênquima; Ff = feixe de fibras; Hi =hipoderme; La = lacuna de ar.
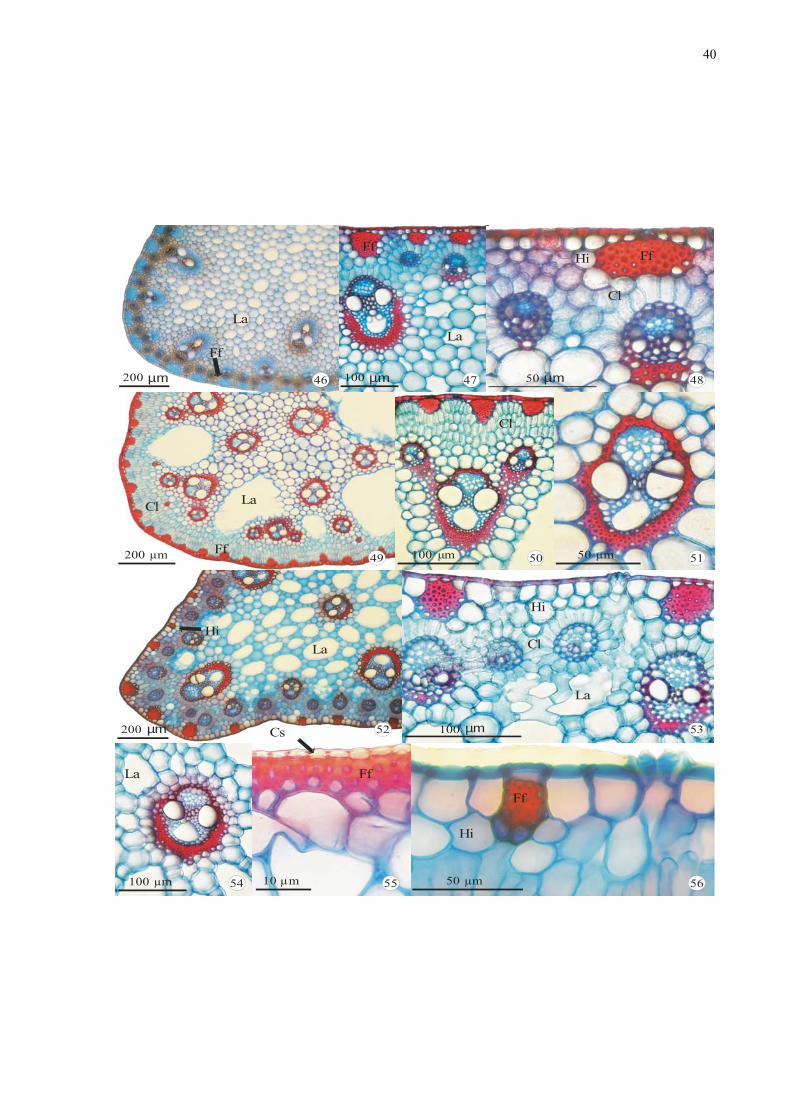
40
100 mµ 50 mµ
50 mµ100 mµ
100 mµ200 mµ
100 mµ
200 mµ
200 mµ
50 mµ10 mµ
La
LaLa
La
La
La
Cs
Ff
Ff
FfFf
Ff
Ff
Hi
Cl
Cl
Cl
Cl
Hi
Hi
Hi
46 47 48
49 50 51
52 53
54 55 56

41

42
CAPÍTULO II
Anatomia de espécies emergentes de lagoas temporárias da
Bahia, Brasil

43
RESUMO
Estudou-se anatomia de raízes, rizomas, folhas e escapos de Echinodorus grandiflorus
(Alismataceae), Eichhornia paniculata (Pontederiaceae) e Habenaria repens (Orchidaceae) de
lagoas temporárias, procurando levantar características anatômicas adaptativas à forma de vida
emergente. Todas as espécies apresentam células epidérmicas com cutícula fina em todos os órgãos;
raízes com exoderme e lacunas de ar no córtex; rizomas com lacunas de ar no córtex; epiderme
foliar com células de paredes finas, anfiestomáticas, com mesofilo dorsiventral, e com
esclerênquima associado aos feixes vasculares; exceto Habenaria repens que é hipoestomática, com
mesofilo regular. Todas as espécies apresentam lacunas de ar no mesofilo e feixes vasculares
colaterais; escapos com epiderme unisseriada, córtex com lacunas de ar e feixes vasculares
distribuídos de forma atactostélica. Células epidérmicas com cutícula fina, presença de lacunas de
ar, idioblastos contendo compostos fenólicos e cristais e redução do tecido de sustentação, são
características presentes nos diferentes órgãos das espécies estudadas que permitiram agrupá-las
como emergentes. Como elas pertencem a grupos taxonômicos distintos, acredita-se que essas
estruturas anatômicas se traduzem em convergências adaptativas à forma de vida emergente.
Palavras-chaves: Alismataceae, Pontederiaceae, Orchidaceae, anatomia, lagoas temporárias

44
ABSTRACT
It was studied anatomy of roots, rhizomes, leaves and scapes of Echinodorus grandiflorus
(Alismataceae), Eichhornia paniculata (Pontederiaceae) and Habenaria repens (Orchidaceae) of
temporary lagoons, to achieve the adaptative anatomical characteristics to the emergent form of life.
All the species demonstrated epidermal cells with thin cuticles on all the organs; roots with an
exoderm and air canals in the cortex; rhizomes with air canals in the cortex; foliar epidermis with
cells of thin walls, anphiestomatic, with a dorsiventral mesophyll, and with sclerenchym associated
to the vascular bundle; except in Habenaria repens that is hypoestomatic, with regular mesophyll.
All the species demonstrsted air canals in the mesophyll and collateral vascular bundles; scapes
with uniseriate epiderms, cortex with air canals and vascular bundles distributed of atactostelic
form. Epidermal cells with thin cuticles, presence of air canals, idioblasts containing phenolic
compounds and crystals and reduced amount of sustentation tissue are characteristic present in the
different organs of the species studied here, allowing them to be grouped emergent plants.
Considering that the species are from different taxonomic group, these anatomical similarities
represent adaptative convergence to an emergent plants.
Key words - Alismataceae, Pontederiaceae, Orchidaceae, anatomy, lagoons temporaries

45
INTRODUÇÃO
A presença de rios e lagoas intermitentes é predominante na caatinga durante a estação
chuvosa, sendo catalogadas 7.358 lagoas no Estado da Bahia. Por apresentarem duração da lâmina
d’água efêmera, favorecem o estabelecimento de uma flora diferenciada, com espécies bem
adaptadas a esse ecossistema (Maltchik et al. 1999, Kercher & Zedler 2004).
Nessas áreas alagáveis ocorrem espécies de monocotiledôneas como Cyperaceae, Poaceae,
Araceae, Alismataceae, Pontederiaceae, Nymphaeaceae e Orchidaceae entre outras (França et al.
2003, Matias et al. 2003). Representantes dessas famílias podem apresentar uma ou mais formas de
vida. Neste trabalho estudou-se apenas algumas espécies emergentes, como Echinodorus
grandiflorus (Alismataceae), Eichhornia paniculata (Pontederiaceae) e Habenaria repens
(Orchidaceae), por serem comuns em estudos fitossociológicos de ambientes aquáticos (França et
al. 2003, Matias et al. 2003).
Alismataceae apresenta espécies de hábito aquático, nativas de regiões tropicais e
temperadas, com forma de vida emergente ou flutuante (Matias et al. 2007, 2008). Pontederiaceae
têm representantes amplamente distribuídos nas regiões tropicais e subtropicais, sendo
exclusivamente aquáticos e com forma de vida flutuante ou emergente (Barrett 2004). Orchidaceae
é uma família com distribuição pantropical, com raros representantes aquáticos (Stern 1997).
No Brasil, sob o ponto de vista anatômico, foram estudadas macrófitas aquáticas de
Scrophulariaceae por Bona & Morretes (2003), de Cyperaceae por Rodrigues & Estelita (2003) e de
Alismataceae do semi-árido nordestino por Matias et al. (2007, 2008). Nestes estudos, as estruturas
anatômicas são relacionadas com possíveis adaptações ao ambiente.
O conhecimento da biologia das macrófitas aquáticas de lagoas temporárias do semi-árido
pode fornecer dados importantes sobre a ecologia das formas de vida apresentadas e do
ecossistema, visando sua conservação, uma vez que essas áreas estão sujeitas à utilização

46
inadequada por atividades agropastoris. A maioria das espécies que ocorrem em lagoas temporárias
apresenta ciclo de vida curto em função da pluviometria, facilitando estudos de sucessão ecológica
e de estratégias de resistência morfológica, anatômica e fisiológica (Pimenta et al. 1998, Kercher &
Zedler 2004).
Baseado no exposto, este trabalho visou caracterizar anatomicamente os órgãos vegetativos
de Echinodorus grandiflorus (Alismataceae), Eichhornia paniculata (Pontederiacae) e Habenaria
repens (Orchidaceae). Como essas espécies são freqüentes em lagoas temporárias do semi-árido da
Bahia, procurou-se verificar a existência de adaptações anatômicas à forma de vida emergente, que
indiquem convergências entre elas.

47
MATERIAL E MÉTODOS
Os espécimes de Echinodorus grandiflorus (Cham. & Schltdl.) Micheli (Alismataceae),
Eichhornia paniculata (Spreng.) Solms Laubach (Pontederiaceae) e Habenaria repens Nutt.
(Orchidaceae) foram selecionados para este estudo por apresentarem, nas lagoas temporárias, a
mesma forma de vida, emergente, de acordo com levantamento feito por França et al. (2003). As
coletas foram feitas em quatro diferentes lagoas do semi-árido baiano, no município de Angüera -
BA, e em período chuvoso, sob as coordenadas 12°16’32”S e 39°03’22”W (Lagoa 1), 12°13’24”S e
39°06’59”W (Lagoa 2), 12°11’38”S e 39°09’07”W (Lagoa 3), 12°10’00”S e 39°12’36”W (Lagoa
4).
Esses corpos d’água temporários são comuns no nordeste brasileiro, em função da
sazonalidade de chuvas e depressões de relevo, favorecendo acúmulo de água superficial e não
existindo nenhum tipo de comunicação entre elas. A vegetação aquática desses ambientes é
dinâmica, tem rápido estabelecimento e a sucessão de espécies ocorre em função do regime de
chuvas e do período de estabilidade da água nas lagoas, que se mantêm vivas enquanto houver
umidade no solo (Maltchik et al. 1999).
Para a análise anatômica, utilizou-se plantas adultas fixadas em FAA a 70% e
posteriormente preservadas em álcool etílico a 70% (Johansen 1940). O material estudado encontra-
se depositado no Herbário da Universidade Estadual de Feira de Santana (HUEFS): Echinodorus
grandiflorus: Leite et al. 480; Leite & França 497; Leite & Lessa 508; Leite & França 515; Leite &
França 516; Leite & França 517. Eichhornia paniculata: Leite et al. 482; Leite & França 485; Leite
& França 498; Leite & Lessa 509; Leite & França 518. Habenaria repens: Leite et al. 476; Leite &
França 483; Leite 487; Leite & Lessa 507.
A partir do material fixado foram selecionadas peças de regiões apical, mediana e basal de
raízes, rizomas, folhas e escapos. Nas regiões apical e basal das raízes, as secções foram feitas à

48
uma altura de até cinco milímetros à partir do ápice e da base. Foram utilizados pelo menos três
indivíduos de cada espécie. Secções transversais dos diferentes órgãos foram feitos à mão livre,
com o auxílio de lâmina de barbear e submetidas ao processo de dupla coloração com azul de astra
e safranina (modificado de Bukatsch 1972 apud Kraus & Arduim 1997). Para dissociação da
epiderme foliar, foi utilizado o método de Jeffrey (Johansen 1940) cujas porções foram coradas com
safranina a 1%.
Algumas peças dos diferentes órgãos foram incluídas em historresina (Feder & O´Brien
1968) e posteriormente seccionadas a 5µm de espessura em micrótomo rotativo. O processo de
coloração foi feito com ácido-periódico-reativo de Schiff (PAS) e azul de toluidina (Feder &
O´Brien 1968) e as secções foram montadas em lâminas permanentes com Entellan.
Testes microquímicos foram realizados em material fresco, em secções obtidas à mão livre
de todos os órgãos vegetativos. Para tanto, algumas secções foram submetidas ao tratamento com
cloreto férrico para verificação de compostos fenólicos, fenol para sílica (Johansen 1940) e ácido
clorídrico a 10% para oxalato de cálcio (Kraus & Arduim 1997).

49
RESULTADOS
Echinodorus grandiflorus (Alismataceae), Eichhornia paniculata (Pontederiaceae) e
Habenaria repens (Orchidaceae) têm seus aspectos morfológicos e anatômicos representados
respectivamente através das figuras 1-6 e 7-48.
As espécies estudadas Echinodorus grandiflorus (Alismataceae) (Fig. 1), Eichhornia
paniculata (Pontederiaceae) (Fig. 3) e Habenaria repens (Orchidaceae) (Fig. 5) são macrófitas
aquáticas, emergentes, pois o nível de água chega até a metade do tamanho das plantas. Elas
crescem logo após o período de chuvas e permanecem vivas enquanto houver disponibilidade de
água no solo, desaparecendo no período seco. Apresentam rizoma vertical, envolvido por bainhas
de folhas senescentes, de onde partem as raízes adventícias (Fig. 2, 4, 6).
São herbáceas e apresentam folhas simples. Echinodorus grandiflorus apresenta folhas
ovaladas, filotaxia rosulada e escapo triangular sustentando inflorescência tipo verticilastro (Fig. 1).
Eichhornia paniculata tem folhas cordiformes e filotaxia rosulada. Nesta espécie, algumas folhas
são dissimilares e da região lateral superior do pecíolo cresce o escapo que sustenta inflorescência
em panícula (Fig. 3). Habenaria repens apresenta folhas lanceoladas e filotaxia alterna. O escapo é
cilíndrico e sustenta flores em racemo (Fig. 5).
Levando-se em consideração que as espécies estudadas pertencem a três diferentes famílias
de macrófitas aquáticas, suas descrições anatômicas se referem principalmente ao sucesso delas no
ambiente aquático.
As raízes das espécies estudadas, em secção transversal, apresentam córtex diferenciado
(Fig. 8-9, 13, 16). Na raiz jovem das espécies estudadas, em secção transversal, próximo à região
meristemática, inicia-se a diferenciação dos tecidos, como pode ser observado em Echinodorus
grandiflorus (Fig. 7). Nesta fase de desenvolvimento das raízes jovens, observam-se células
epidérmicas de paredes delgadas, com cutícula fina, alongadas transversalmente ao eixo de

50
crescimento da raiz (Fig. 7-8). Observa-se também o início do desenvolvimento de lacunas de ar no
córtex mediano.
As raízes apresentam córtex externo constituído pela exoderme (hipoderme) formada por
duas a três camadas de células hexagonais e com parede periclinal externa mais espessada que as
demais em Echinodorus grandiflorus (Fig. 10) e Eichhornia paniculata (Fig. 13-14). Em
Habenaria repens essas células são arredondadas e sem espessamento de parede (Fig. 16-17).
O córtex mediano ocupa a maior porção das raízes, e é formado por tipos distintos de células
parenquimáticas: arredondadas em Habenaria repens (Fig. 16-17), alongadas em Eichhornia
paniculata (Fig. 13) e braciformes em Echinodorus grandiflorus (Fig. 9, 12). As paredes das células
do córtex mediano se afastam formando lacunas de ar esquizógenas (Fig. 16-17) e/ou se rompem,
formando lacunas de ar lisígenas (Fig. 8-9, 12, 13).
O córtex interno em Echinodorus grandiflorus (Fig. 7-11) e Eichhornia paniculata (Fig. 13,
15) apresentam várias camadas de células arredondadas de tamanho menor que as demais e com
redução de espaço intercelular. A camada mais interna, a endoderme, é unisseriada, com células de
paredes delgadas, onde se evidenciam estrias de Caspary (Fig. 11, 18 - setas). Em Habenaria
repens, o córtex interno apresenta uma única camada de células, a endoderme (Fig. 16-18). O
cilindro vascular de todas as espécies estudadas apresentam vários elementos de metaxilema
ocupando a região central (Fig. 8, 13, 15-16).
Os rizomas das espécies estudadas apresentaram epiderme unisseriada, com cutícula fina, e
córtex diferenciado, com células parenquimáticas de paredes finas e arredondadas (Fig. 21, 24). A
presença de amplas lacunas de ar no córtex dos rizomas é marcante nas três espécies (Fig. 19, 21-
22, 24). Observam-se raízes intracorticais (Fig. 19, 21, 23). Em Eichhornia paniculata (Fig. 23),
observa-se a saída de uma raiz adventícia e a conexão dos tecidos dessa nova raiz com aqueles do
rizoma. Todas as espécies apresentam rizoma com córtex interno constituído de várias camadas de

51
células de paredes levemente espessadas (Fig. 20, 21, 24) e endoderme e periciclo unisseriados (Fig.
20, 22, 25).
As folhas de Echinodorus grandiflorus e Eichhornia paniculata são anfiestomáticas e as de
Habenaria repens são hipoestomáticas. Em vista frontal, Echinodorus grandiflorus (Fig. 26-27)
apresenta células epidérmicas com paredes sinuosas e, em Eichhornia paniculata (Fig. 28-29) e
Habenaria repens (Fig. 30-31), as células epidérmicas apresentam paredes retas em ambas as faces.
Echinodorus grandiflorus e Eichhornia paniculata apresentam estômatos paracíticos (Fig. 26-29) e
Habenaria repens estômatos anomocíticos (Fig. 31).
Em secção transversal da folha, a epiderme de todas as espécies é uniestratificada, com a
parede periclinal externa mais espessada que a interna e com cutícula fina (Fig. 32-33, 35-42).
Habenaria repens apresenta células epidérmicas da face adaxial maiores do que as da face abaxial
(Fig. 38-39).
O mesofilo é do tipo dorsiventral em Echinodorus grandiflorus e Eichhornia paniculata
(Fig. 33, 36 respectivamente) e homogêneo em Habenaria repens com redução dos espaços
intercelulares (Fig. 39). Nas espécies com mesofilo dorsiventral, o parênquima clorofiliano
paliçádico apresenta de duas a três camadas de células voltadas à face adaxial da lâmina foliar (Fig.
33, 36-37). Em Echinodorus grandiflorus, o parênquima lacunoso apresenta cerca de cinco camadas
de células, com redução dos espaços intercelulares (Fig. 33) e em Eichhornia paniculata logo
abaixo do parênquima paliçádico podem ser observadas amplas lacunas de ar (Fig. 35-36).
O mesofilo das espécies estudadas apresenta feixes vasculares colaterais dispostos em uma
única fileira central (Fig. 33) em Echinodorus grandiflorus e Habenaria repens, e em duas fileiras,
alternando os menores com os maiores, em Eichhornia paniculata (Fig. 35-36). Os feixes
vasculares apresentam fibras pericíclicas, exceto em Habenaria repens (Orchidaceae), que é
destituída de tecido de sustentação.

52
A nervura central da folha de Habenaria repens (Fig. 38) apresenta apenas um feixe
vascular central enquanto que Echinodorus grandiflorus (Fig. 32) e Eichhornia paniculata (Fig. 35)
apresentam um feixe vascular maior nas posições adaxial e abaxial respectivamente, e de dois a seis
feixes vasculares menores. Logo abaixo da epiderme ocorre colênquima lacunar. Os feixes
vasculares apresentam periciclo espessado (Fig. 32, 35).
O bordo foliar é obtuso em Echinodorus grandiflorus e Eichhornia paniculata (Fig. 40-41,
respectivamente) e agudo em Habenaria repens (Fig. 42). Nessa região, logo abaixo da epiderme,
ocorrem células de parênquima em Echinodorus grandiflorus e Habenaria repens (Fig. 40, 42,
respectivamente) e parênquima clorofiliano em Eichhornia paniculata (Fig. 41).
As lacunas de ar presentes no mesofilo das espécies estudadas são maiores em Eichhornia
paniculata (Fig. 35-36). Elas são delimitadas por diafragmas constituídos de células braciformes
(Fig. 34-seta).
Os escapos das espécies estudadas, em secção transversal, apresentam epiderme com
cutícula fina, córtex e cilindro vascular distintos (Fig. 43-48, respectivamente). A epiderme é
uniestratificada, com células de paredes finas em Echinodorus grandiflorus e Eichhornia paniculata
(Fig. 44, 46) e parede periclinal externa levemente mais espessada que a interna em Habenaria
repens (Fig. 48).
O córtex é constituído por células parenquimáticas arredondadas, intercaladas por lacunas de
ar e feixes vasculares em Echinodorus grandiflorus (Fig. 43-44) e Eichhonia paniculata (Fig. 45-
46) e intercaladas apenas por lacunas de ar em Habenaria repens (Fig. 47-48). A endoderme, que
limita o cilindro vascular, é cilíndrica e constitui-se de células de paredes finas (fig. 43-44, 46-48).
O periciclo é constituído por uma a duas camadas de células de paredes levemente espessadas nas
três espécies estudadas (Fig. 43-44, 46-48). Os feixes vasculares do cilindro vascular são colaterais
e apresentam distribuição atactostélica (Fig. 43-48). Nesta região também ocorrem lacunas de ar
com dimensões variadas (Fig. 43-48).

53
Nas raízes, rizomas, folhas e escapos das espécies estudadas foram observados idioblastos
contendo compostos fenólicos (Fig. 8-seta, 21-24, 32, 40-41) e cristais na forma de ráfides e drusas
(Fig. 22, 37-setas).

54
DISCUSSÃO
Os órgãos vegetativos de Echinodorus grandiflorus (Alismataceae), Eichhornia paniculata
(Pontederiaceae) e Habenaria repens (Orchidaceae), espécies de monocotiledôneas distintas
filogeneticamente, apresentam em comum: cutícula fina, idioblastos contendo cristais e compostos
fenólicos, lacunas de ar e redução do tecido de sustentação. Essas características representam
convergências adaptativas dessas espécies ao ambiente aquático e à forma de vida emergente.
A cutícula que recobre a parede periclinal externa dos órgãos emersos e submersos das
espécies estudadas neste trabalho apresentou-se fina, assim como em outras espécies de
Angiospermas que crescem em locais com disponibilidade de água. Para Sculthorpe (1967), a
dificuldade de obter oxigênio é um dos fatores responsáveis pela presença de cutícula fina,
principalmente, nos órgãos submersos de macrófitas aquáticas e não apenas à disponibilidade de
água. Esse caráter pode favorecer a passagem de gases dissolvidos na água para o interior do órgão.
Nos órgãos emersos das espécies estudadas, a presença de cutícula fina pode estar relacionada à
diminuição da intensidade luminosa durante o período de chuvas, época em que as espécies foram
coletadas, além da constante umidade do solo.
A presença de cutícula mesmo fina nas espécies estudadas além de favorecer a aeração, nos
órgãos emersos e submersos, lhes confere proteção. Nas raízes, pode atuar juntamente com a
exoderme como uma barreira a mais contra ataque de patógenos, conforme o que foi relatado por
Peterson (1992) para outras espécies aquáticas.
A maior densidade estomática nas folhas de E. paniculata seguido de E. grandiflorus em
relação à H. repens pode ser associada ao tipo de mesofilo dorsiventral e homogêneo observado,
respectivamente, nas espécies além de estar associada à mecanismos fisiológicos de trocas gasosas
uma vez que em E. paniculata os estômatos se localizam em ambas as faces epidérmicas estando
mais em contato com as lacunas de ar em E. paniculata.

55
Lacunas de ar estão presentes nos órgãos vegetativos das três espécies estudadas. Nessas
espécies que têm forma de vida emergente, as lacunas ar presentes nas raízes se desenvolvem na
diferenciação dos tecidos, sendo mais freqüentes no córtex das raízes, rizomas e escapos do que nas
folhas. São menores nas folhas de Echinodorus grandiflorus (Alismataceae) e Habenaria repens
(Orchidaceae) que se encontram sempre emersas, mesmo no período chuvoso, ficando submersas
apenas as raízes e rizomas.
Para Peterson (1992), a sobrevivência de plantas que vivem em solos alagados está
diretamente relacionada à presença de espaços de ar. Para Armstrong et al. (1991), estes espaços de
ar favorecem a sobrevivência da raiz em ambientes mais profundos. No caso das plantas emergentes
estudadas, que já crescem sob condição de alagamento, elas apresentam lacunas de ar nas raízes
desde o início de seu desenvolvimento. O mesmo foi observado em raízes adventícias de outras
plantas emergentes, que crescem em solos inundados, por Seago & Marsh (1989) em Typha glauca
Godr. (Typhaceae), por Schussler & Longstreth (1996) em Saggitaria lancifolia L. (Alismataceae) e
por Seago Jr. et al. (2000) em Pontederia cordata L. (Pontederiaceae).
Tanto nas espécies emergentes estudadas neste trabalho como naquelas estudadas por Seago
& Marsh (1989), Schussler & Longstreth (1996) e por Seago Jr. et al. (2000), as espécies
apresentam a característica comum de desenvolvimento de lacunas de ar no ápice da raiz, como
uma convergência adaptativa ao ambiente aquático e à forma de vida.
Provavelmente as lacunas de ar das plantas estudadas neste trabalho são de origem
esquizógena na raiz jovem, rizoma, folha e escapo, e lisígena na região cortical da raiz adulta.
Mesmo não tendo sido realizados estudos ontogenéticos, a partir de análise anatômica das espécies
e por comparação ao que foi apresentado por Pimenta et al. (1998) e por Schussler & Longstreth
(1996), que afirmam que as plantas desenvolvem aerênquima esquizógeno e apenas
secundariamente pode ocorrer formação de aerênquima lisígeno. Esses autores atribuem a formação
de aerênquima, em órgãos subterrâneos, à baixa oxigenação do solo úmido. Segundo Peterson

56
(1992), a hipoxia promove a concentração de etileno em células corticais estimulando a atividade da
celulase e formando as lacunas de ar.
Para Sculthorpe (1967) e Armstrong et al. (1991), a presença de lacunas de ar, tanto de
origem lisígena como esquizógena, é um caráter comum em plantas aquáticas e é uma forma de
assegurar o suprimento de O2 às células ou órgãos com pouco oxigênio.
Acredita-se que o desenvolvimento das lacunas de ar seja anterior ao espessamento da
exoderme e endoderme nas espécies emergentes assim como observado Seago et al. (2000) em
Pontederia cordata. Para estes autores, a relação entre esses tecidos possibilita a ocupação dessas
plantas em áreas específicas no ambiente aquático de água doce e mesmo que haja redução no
espessamento da exoderme e/ou endoderme, a estrutura e função desses tecidos na raiz está
relacionada ao controle da permeabilidade de íons e gases, ao movimento da água no corpo da
planta, ao suporte estrutural oferecido pelo aerênquima, à flexibilidade ou rigidez à flutuação, à
sustentação da raiz no substrato e ao bloqueio da entrada de patógenos.
Delimitando as lacunas de ar presentes nos diferentes órgãos vegetativos das espécies
estudadas, foram observados diafragmas constituídos por células braciformes cujos braços
apresentam comprimentos variados. Concorda-se com Peterson (1992) em que os diafragmas
aumentam a resistência mecânica dos órgãos e impedem a passagem de água sem impedir a
passagem de O2 entre uma lacuna de ar e outra. Segundo Sculthorpe (1967), as lacunas de ar
presentes no mesofilo de folhas e escapos têm a função de facilitar o acúmulo de O2, transportando-
o lacuna a lacuna até os órgãos submersos ou subterrâneos.
A presença de idioblastos contendo compostos fenólicos e cristais na forma de ráfides ou
drusas presentes nos órgãos vegetativos das espécies emergentes estudadas, também ocorre em
outros grupos de Angiospermas, principalmente entre as monocotiledôneas. Acredita-se que a
presença destas estruturas esteja relacionada a mecanismos de defesa das plantas, conferindo-lhes
proteção contra o ataque de patógenos e herbívoros (Metcalfe 1985, Mauseth 1988).

57
Essas substâncias ergásticas podem aparecer isoladas ou combinadas. Apresentam
importância no ambiente aquático, pois servem como proteção para a planta que pode ser
consumida por diversos organismos (Sculthorpe 1967). Além disso, apresenta-se como uma
evidência filogenética, como já observado por Mauseth (1988) e Prychid & Rudall (1999, 2000),
através da presença constante de cristais em espécies de Monocotiledôneas.
Nos diferentes órgãos das espécies estudadas, ocorre redução do tecido de sustentação,
aqueles que ocorrem estão associados aos sistemas vasculares. A redução acentuada de elementos
de sustentação provenientes do meristema fundamental deve estar relacionada ao hábito aquático, à
pouca luminosidade e ao curto tempo de vida dessas plantas, além de não necessitarem de força
mecânica para se sustentarem uma vez que crescem em ambiente lêntico temporário. Esses órgãos
vegetativos têm como elementos de sustentação mecânica os tecidos que formam o sistema vascular
e os que formam as lacunas de ar e diafragmas conforme relatado para outras espécies aquáticas por
Sculthorpe (1967) e Stern (1997). Concorda-se com Sculthorpe (1967) em que os tecidos de
sustentação teriam sofrido redução em algumas formas biológicas em função de a água assumir o
principal papel de sustentação através do aerênquima bastante desenvolvido.
É importante ressaltar a semelhança do parênquima cortical dos órgãos subterrâneos (raiz e
rizoma) de Echinodorus grandiflorus (Alismataceae) e Eichhornia paniculata (Pontederiaceae),
cujos representantes são exclusivamente aquáticos, com Habenaria repens (Orchidaceae). Segundo
Stern (1997), esta última é a única espécie aquática de Habenaria, gênero constituído de plantas
terrestres de locais úmidos e sombreados. Certamente esta é uma característica indicativa de
convergência adaptativa das espécies à forma de vida.
As semelhanças anatômicas nos órgãos subterrâneos e a análise de similaridade realizada
nos leva a acreditar que estes mecanismos estruturais adaptativos foram desenvolvidos devido à sua
forma de vida emergente. A aeração dos órgãos subterrâneos é um mecanismo de sobrevivência que
favorece a reprodução das espécies em função do curto tempo de vida que apresentam. Também,

58
independente do ambiente, lótico ou lêntico, as características observadas nos órgãos submersos das
diferentes espécies representam convergências adaptativas mais evidentes do que aquelas
observadas nos órgãos aéreos.

59
REFERÊNCIAS BIBLIOGRÁFICAS
ARMSTRONG, W., JUSTIN, S.H.F.W., BECKETT, P.M. & LYTHE, S. 1991. Root adaptation to
soil waterlogging. Aquatic Botany 39(1): 57-73.
BARRETT, S.C. 2004. Pontederiaceae. In Flowering plants of the neotropics. (N.Smith, S. Mori, A.
Henderson, D.Stevenson & S.Heald, eds.). Princeton University Press, Princeton, p.474-476.
BONA, C. & MORRETES, B.L. 2003. Anatomia das raízes de Bacopa salzmanii (Benth.) Wettst.
Ex edwall e Bacopa monnierioides (Cham.) Robinson (Scrophullariaceae) em ambientes
aquático e terrestre. Acta Botanica Brasilica 17(1): 155-170.
FEDER, N. & O’BRIAN, T.P. 1968. Plant microthecnique: some principles and new methods.
American Journal of Botany 55(1): 123-142.
FRANÇA, F., MELO, E., GÓES-NETO, A., ARAÚJO, D., BEZERRA, M.G., RAMOS, H.M.;
CASTRO, I. & GOMES, D. 2003. Flora vascular de açudes de uma região do semi-árido da
Bahia, Brasil. Acta Botanica Brasilica 17(4): 549-559.
JOHANSEN, D.A. 1940. Plant microtechnique. McGraw Hill Book Co., New York.
KERCHER, S.M. & ZEDLER, J.B. 2004. Flood tolerance in wetland angiosperms: a comparison of
invasive and noninvasive species. Aquatic Botany 80(2): 89-102.
KRAUS, J.E. & ARDUIM, M. 1997. Manual básico de metodos em morfologia vegetal. EDUSP,
São Paulo.
MALTCHIK, L., COSTA, M.A.J. & DUARTE, M.D.C 1999. Inventory of brazilian semiarid
shallow lakes. Anais da Academia Brasileira de Ciências 71: 801-808.
MATIAS, L.Q., AMADO, E.R. & NUNES, E.P. 2003. Macrófitas aquáticas da lagoa Jijoca de
Jericoacoara, Ceará, Brasil. Acta Botanica Brasilica 17(4): 623-623.

60
MATIAS, L.Q., SOARES, A. & SCATENA, V.L. 2007. Systematic consideration of petiole
anatomy of species of Echinodorus Richard (Alismataceae) from north-eastern Brazil. Flora
202: 395-402.
MATIAS, L.Q., SOARES, A. & SCATENA, V.L. 2008. Anatomy of Echinodorus (Alismataceae)
scapes from northeastern Brazil as applied to taxonomy. Edinburgh Journal of Botany 65(1):
11-21.
MAUSETH, J.D. 1988. Plant anatomy. The Benjamin Cumming Publishing Company, California.
METCALFE, C.R. 1985. Secreted mineral substances - Crystals. In Anatomy of the dicotyledons:
Wood structure and conclusion of the general introduction (C.R. Metcalfe, & L. Chalk, eds.).
Claredon Press, Oxford. v.2, p.82-91.
PETERSON, R.L. 1992. Adaptations of root structure in relation to biotic and abiotic factors.
Cannadian Journal of Botany 70(4): 661-675.
PIMENTA, J.A., BIANCHINI, E. & MEDRI, M.E. 1998. Adaptations to flooding by tropical trees:
morphological and anatomical modifications. In Ecophysiological strategies of xerophytic and
amphibious plants in the neotropics (F.R. Scarano & A.C. Franco, eds). Series Oecologia
Brasiliensis, PPGE-UFRJ, Rio de Janeiro, v.4, p.157-176.
PRYCHID, C.J. & RUDALL, P.J. 1999. Calcium oxalate crystals in Monocotyledons: a review of
their structure and systematic. Annals of Botany 84(6): 725-739.
PRYCHID, C.J. & RUDALL, P.J. 2000. Distribution of calcium oxalate crystals in
monocotyledons. In Monocots: Systematic and evolution (K.L. Wilson & D.A.Morrison, eds.).
Csiro, Melbourne. p.159-162.
RODRIGUES, A.C. & ESTELITA, M.E.M. 2003. Origin and structure of the Krans tissue in bracts
of Cyperus giganteus Vahl (Cyperaceae). Revista Brasileira de Botanica 26(4): 445-452.

61
SCHUSSLER, E.E. & LONGSTRETH, D.J. 1996. Aerenchyma develops by cell lysis in roots and
cell separation in leaf petioles in Sagittaria lancifolia (Alismataceae). American Journal of
Botany 83(10): 1266-1273.
SCULTHORPE, C.D. 1967. The biology of aquatic vascular plants. Edward Arnold, London.
SEAGO Jr., J.L. & MARSH, L. 1989. Adventicious root development in Typha glauca, with
emphasis on the cortex. American Journal of Botany 76(6): 909-923.
SEAGO Jr., J.L., PETERSON, C.A. & ENSTONE, D.E. 2000. Cortical development in roots of the
aquatic plant Pontederia cordata (Pontederiaceae). American Journal of Botany 87(8): 1116-
1127.
SNEATH, P.H.A. & SOKAL, R.R. 1973. Numerical Taxonomy. San Francisco, W. H. Freeman.
STERN, W.L. 1997. Vegetative anatomy of subtribe Habenariinae (Orchidaceae). Botanical Journal
of the Linnean Society 125(3): 211-227.

62
Figuras 1-6. Aspectos morfológicos de espécies de monocotiledôneas: 1-2. Echinodorus
grandiflorus (Alismataceae). 1.Hábito. 2. Detalhe do sistema subterrâneo. 3-4. Eichhornia
paniculata (Pontederiaceae). 3. Hábito. 4. Detalhe do sistema subterrâneo. 5-6: Habenaria
repens (Orchidaceae). 5. Hábito. 6. Detalhe do sistema subterrâneo.

63
1
5
2
6
3 4
4 cm
2 cm
2,7 cm
2 cm
4 cm
2 cm

64
Figuras 7-18. Fotomicrografias de aspectos anatômicos de secções transversais de raízes de espécies
de monocotiledôneas: 7-12. Echinodorus grandiflorus (Alismataceae). 7-8. Aspecto geral da raiz
jovem. 9. Raiz adulta. 10. Detalhe da epiderme e exoderme. 11. Detalhe do cilindro vascular. 12.
Detalhe das células braciformes que se rompem e formam as lacunas de ar - setas. 13-15.
Eichhornia paniculata (Pontederiaceae) – raiz adulta. 13. Aspecto geral. 14. Detalhe da epiderme e
exoderme. 15. Detalhe do cilindro vascular. 16-18. Habenaria repens (Orchidaceae) – raiz adulta.
16. Aspecto geral. 17. Detalhe da epiderme e córtex. 18. Detalhe da endoderme com estrias de
Caspary e cilindro vascular. Legenda: Co-coifa, En-endoderme, Ex-exoderme, Fl-floema, La-lacuna
de ar, Mx-metaxilema, Pe-priciclo, Xi-xilema.

65
9
11
12
151413
16 17 18
8200 µm
200 µm
100 µm
200 µm
100 µm 100 µm 100 µm
200 µm 200 µm 100 µm
La
La
La
LaLa
La
La
Ex
Ex
Ex
Ex Co
En Pe
PeEn
Pe
EnMx
Mx
XiFl
Ex
Cf
Pe
En
Pe En
Ex
100 µm 10
7100 µm
63

66
Figuras 19-25. Fotomicrografias de aspectos anatômicos de secções transversais da região mediana
do rizoma de espécies de Monocotiledôneas: 19-20. Echinodorus grandiflorus
(Alismataceae): 19. Aspecto geral. 20. Detalhe da região do cilindro vascular. 21-23:
Eichhornia paniculata (Pontederiaceae): 21. Aspecto geral do córtex com lacunas de ar e
raízes intracorticais. 22. Detalhe do córtex e cilindro vascular. 23. Detalhe da saída de raiz.
24-25: Habenaria repens (Orchidaceae): 24. Aspecto do córtex com lacunas de ar. 25.
Detalhe da endoderme e cilindro vascular. Legenda: Cf-composto fenólico, Cr-cristal, En-
endoderme, La-lacuna de ar, Mx-metaxilema, Pe-priciclo.
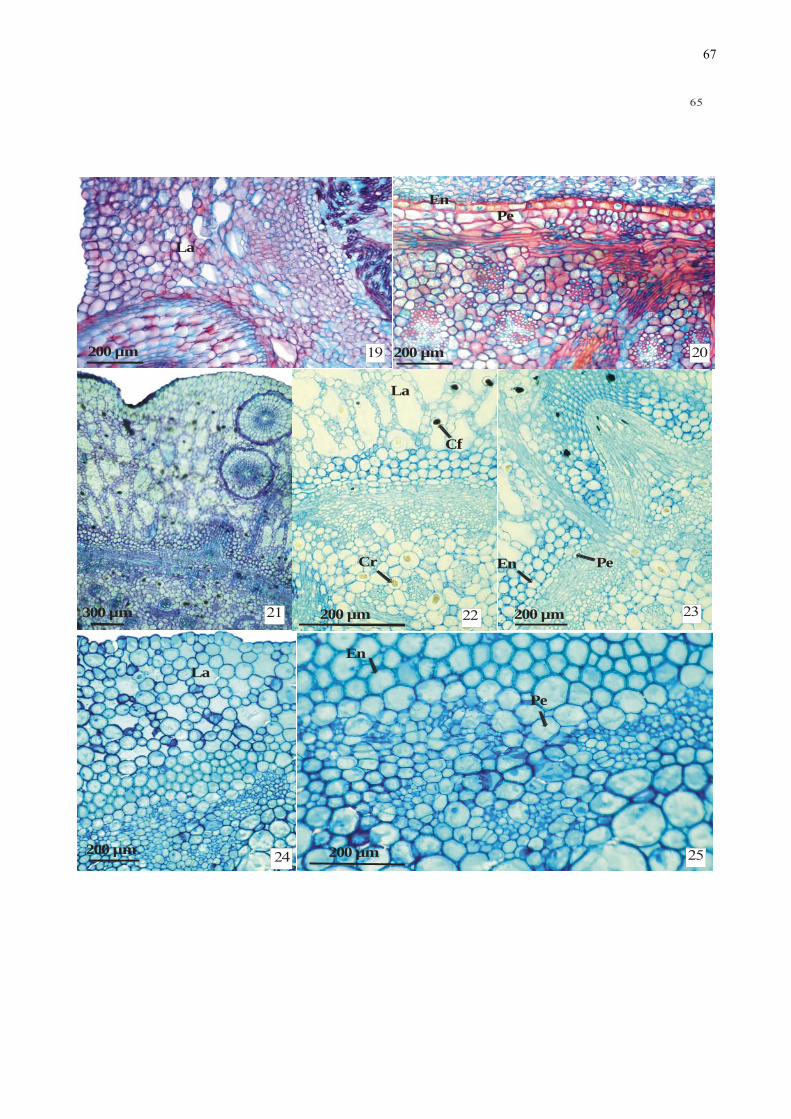
67
25
21 22 23
24
200 µm 19 200 µm 20
300 µm 200 µm 200 µm
200 µm 200 µm
EnPe
En Pe
En
Pe
La
La
La
Cf
Cr
65

68
Figuras 26-31. Fotomicrografias de aspectos anatômicos da região mediana da epiderme foliar em
vista frontal, de espécies de Monocotiledôneas: 26-27. Echinodorus grandiflorus
(Alismataceae) - Face adaxial e abaxial, respectivamente. 28-29: Eichhornia paniculata
(Pontederiaceae) - Face adaxial e abaxial, respectivamente. 30-31: Habenaria repens
(Orchidaceae) - Face adaxial e abaxial, respectivamente.

69
26 27
2928
3130
200 µm
200 µm200 µm
200 µm200 µm
200 µm
67

70
Figuras 32-42. Fotomicrografias de aspectos anatômicos da região mediana de secções transversais
de folhas de espécies de Monocotiledôneas: 32-34. Echinodorus grandiflorus (Alismataceae). 32.
Aspecto geral da nervura central. 33. Detalhe do mesofilo com nervuras menores. 34. Detalhe do
diafragma que limita as lacunas de ar. 35-37. Eichhornia paniculata (Pontederiaceae). 35. Aspecto
geral da nervura central. 36. Detalhe do mesofilo. 37. Detalhe do mesofilo contendo ráfides. 38-39.
Habenaria repens (Orchidaceae). 38. Aspecto geral da nervura central. 39. Detalhe do mesofilo. 40-
43. Bordo foliar. 40. E. grandiflorus. 41. E. paniculata. 42. H. repens. Legenda: Cf–composto
fenólico, Cr-cristal, Df-diafragma, Es-estômato, La- lacuna de ar.

71
32
33
34
36
35
38
42
39
4140
200 µm
200 µm
200 µm
200 µm
200 µm
La
Es
Es
Es Es
La
LaDf
Df
Es37200 µm
Cr
Cr
La
Es
200 µmEs
200 µm 200 µm 200 µm
400 µm
69

72
Figuras 43-48. Fotomicrografias de aspectos anatômicos da região mediana de secções transversais
de escapos de espécies de Monocotiledôneas: 43-44. Echinodorus grandiflorus (Alismataceae) -
Aspecto geral e detalhe do córtex, respectivamente. 45-46. Eichhornia paniculata (Pontederiaceae)
- Aspecto geral e detalhe do córtex, respectivamente. 47-48. Habenaria repens Orchidaceae) -
Aspecto geral e detalhe do córtex, respectivamente. Legenda: Cf – composto fenólico, Cr-cristal,
Df-diafragma, En-Endoderme, Es-estômato, Fp-fibras pericíclicas, La- lacuna de ar.

73
48
4443
4645 4646464646
200 µm 200 µm
200 µm 200 µm
200 µm400 µm 47
La
La
LaLa
La
La
Df
Pe
Fp
Fp
FpCf
Cf
Cf
Cf
En
La
En
71

74
CAPÍTULO III
Convergência adaptativa em espécies de monocotiledôneas
lacustres do semi-árido da Bahia, Brasil

75
RESUMO
Macrófitas aquáticas podem apresentar formas de vida variadas, o que lhes permite
colonizar diferentes áreas ao longo de um perfil ambiental, inclusive lagoas temporárias do
semi-árido nordestino. A plasticidade apresentada nos tecidos de proteção, fundamental e
vascular dos órgãos vegetativos das macrófitas garante sua sobrevivência sob condição de
alagamento. Com isso, este trabalho buscou levantar caracteres anatômicos que indicassem a
convergência adaptativa, de espécies anfíbias e emergentes de lagoas temporárias da Bahia.
Cyperus odoratus L., Oxycaryum cubense (Poep. & Kunth) Lye e Pycreus macrostachyos
(Lam.) Raynal (Cyperaceae) – anfíbias, e Echinodorus grandiflorus (Cham. & Schltdl.)
Micheli (Alismataceae), Eichhornia paniculata (Spreng.) Solms Laubach (Pontederiacae),
Habenaria repens Nutt. (Orchidaceae) – emergentes, apresentam características anatômicas
que as convergem ao tipo de ambiente aquático no qual são encontradas, considerando que
essas espécies pertencem à grupos taxonômicos distintos. São consideradas características
anatômicas convergentes: presença de lacunas de ar e epiderme unisseriada em todos os
órgãos vegetativos; exoderme e endoderme constituídas de várias camadas de células de
paredes com espessamentos variados nas raízes; redução do tecido de sustentação e de
elementos de transporte do xilema em todos os órgãos, quando comparadas às demais
espécies do gênero que são terrestres. Essas características são adaptativas às espécies de
monocotiledôneas anfíbias e emergentes que ocorrem nas lagoas temporárias da Bahia.
Palavras-chave: Anatomia ecológica, macrófitas aquáticas, convergência adaptativa, lagoas do
semi-árido

76
ABSTRACT
Aquatic plants can present different life forms, that allow the colonization them
different areas along an environmental gradient, including temporary lagoons of the semi-arid
Northeastern Brazil. The plasticity demonstrated in the tissues of the vegetative organs of the
aquatic plants ensure their survival under wetting condition. The anatomic adaptative features
which indicate the adaptative convergence of amphibious and emergent species of temporary
lagoons from State of Bahia were assessed. Cyperus odoratus L., Oxycaryum cubense (Poep.
& Kunth) Lye and Pycreus macrostachyos (Lam.) Raynal (Cyperaceae) - amphibious and
Echinodorus grandiflorus (Cham. & Schltdl.) Micheli (Alismataceae), Eichhornia paniculata
(Spreng.) Solms Laubach (Pontederiacae), Habenaria repens Nutt. (Orchidaceae) – emergent,
demonstrated anatomical characteristics that converge them to the type of aquatic
environment in which they are found, considering that those species belong to different
taxonomic groups. Convergent anatomical characteristics are considered: presence of air
canals and uniseriate epidermis in all the vegetative organs; exoderm and endoderm are
composed of several cells layers with varied cell walls thickness; reduced amounts of tissue of
sustentation and xylem transport elements in all of their organs when compared to terrestrial
species of the same genus. These are the adaptative characteristics of species of amphibious
and emergent monocotyledons inhabiting temporary lagoons in Bahia State.
Key-words: ecologic anatomy, aquatic plants, adaptative convergence, lagoons of semi-arid

77
INTRODUÇÃO
Os processos adaptativos que ocorrem nas diferentes formas de vida de macrófitas
aquáticas são resultantes da informação genética e de adaptações (Davy et al. 1990, Futuyma
1993). Segundo os autores, as adaptações ocorrem em resposta a grande número de fatores
bióticos e abióticos, que individualmente, podem afetar a capacidade de uma espécie
sobreviver e se reproduzir em um determinado local.
As macrófitas aquáticas, das lagoas temporárias do semi-árido brasileiro, variam tanto
na sua estrutura externa quanto interna, o que permite agrupá-las em formas de vida
específicas (Irgang et al. 1984), de acordo com a zonação ecológica.
No semi-árido, a sobrevivência das macrófitas aquáticas depende de sua tolerância aos
extremos ambientais, períodos secos e chuvosos. Essa tolerância adaptativa ao ambiente
aquático temporário ocorre através de um conjunto de respostas plásticas, manifestadas por
características fenológicas, morfológicas, anatômicas e fisiológicas (Wells & Pigliucci 2000).
A estas respostas plásticas dá-se o nome de plasticidade fenotípica (Wells & Pigliucci
2000), que é a propriedade que o genótipo tem de alterar a sua expressão fenotípica em
resposta às mudanças ambientais. A plasticidade fenotípica pode ser entendida também como
a habilidade de um organismo em reagir ao ambiente alterando sua forma, estado, movimento
ou velocidade de reação à determinada condição ambiental (West-Eberhard 2003).
O baixo nível de oxigênio encontrado no ambiente lêntico é um dos maiores
problemas das macrófitas aquáticas (Matthews & Seymour 2006), pois a hipoxia pode levar
ao aumento da produção de íons Fe2+ e Mn2+, tóxicos para as plantas, produzindo danos
irreversíveis aos órgãos subterrâneos. Como resposta, para evitar esses danos essas plantas
apresentam convergência anatômica que pode ser traduzida, principalmente, pela presença de
lacunas de ar, encontradas nos diferentes órgãos vegetativos, formadas pelo espaçamento
entre as células ou pela desintegração delas (Sculthorpe 1967; Keddy 2000).

78
Plantas aquáticas de diferentes grupos taxonômicos podem adaptar-se a uma área com
igual variação temporal e espacial do nível de água do ecossistema, podendo apresentar
características ecológicas, e às vezes morfológicas, semelhantes. Através dessas semelhanças
ecológicas podemos enquadrá-las em diferentes formas de vida sem, no entanto, saber se
ocorres semelhanças também nas estruturas internas.
Desta forma, este trabalho teve como objetivo levantar características anatômicas de
algumas espécies de monocotiledôneas anfíbias e emergentes, de lagoas temporárias da Bahia,
que indicassem convergência adaptativa ao ambiente.

79
MATERIAL E MÉTODOS
Representantes de Cyperus odoratus L., Oxycaryum cubense (Poep. & Kunth) Lye,
Pycreus macrostachyos (Lam.) Raynal (Cyperaceae) - anfíbias, e de Echinodorus
grandiflorus (Cham. & Schltdl.) Micheli (Alismataceae), Eichhornia paniculata (Spreng.)
Solms Laubach (Pontederiaceae) e Habenaria repens Nutt. (Orchidaceae) – emergentes,
foram coletados durante o período chuvoso, em lagoas temporárias do semi-árido nordestino
da Bahia. Essas lagoas foram formadas pelo acúmulo de água superficial, não apresentavam
comunicação entre si, e estavam localizadas ao longo da rodovia BA 052 (Estrada do Feijão).
Todos os indivíduos foram coletados em áreas com semelhantes condições ambientais,
como altitude, condição geológica e temperatura atmosférica, havendo variação de tamanho e
profundidade das lagoas.
Para a análise anatômica foram utilizadas peças dos órgãos vegetativos de plantas
adultas, fixadas em FAA a 70% e posteriormente preservadas em álcool etílico a 70%
(Johansen 1940). As espécies estudadas foram coletadas e incorporadas ao acervo do Herbário
da Universidade Estadual de Feira de Santana (HUEFS): Cyperus odoratus: Leite et al. 477;
Leite 486; Leite & França 495; Leite & França 519; Leite & França 520; Leite & França 521.
Oxycaryum cubense: Leite et al. 475; Leite & França 522; Leite & França 523; Leite &
França 524; Leite & França 525. Pycreus macrostachyos: Leite 486A; Leite 487A; Leite &
França 496; Leite & França 500; Leite & Lessa 507. Echinodorus grandiflorus: Leite et al.
480; Leite 528; Leite & França 495; Leite & Lessa 508; Leite & França 515; Leite & França
516; Leite & França 517. Eichhornia paniculata: Leite et al. 482; Leite & França 485; Leite
& França 498; Leite & Lessa 509; Leite & França 518. Habenaria repens: Leite et al. 476;
Leite & França 483; Leite 487; Leite & Lessa 507.
Foram seccionadas regiões medianas de raízes, rizomas, folhas e escapos, de pelo
menos três indivíduos diferentes de cada espécie. Secções transversais foram feitas à mão
livre, com auxílio de lâmina de barbear, coradas com azul de astra e safranina (modificado de
Bukatsch 1972 apud Kraus & Arduim 1997) e montadas em lâminas semi-permantes com
gelatina glicerinada.

80
Algumas peças dos órgãos vegetativos foram incluídas em historresina (Feder &
O´Brien 1968) e seccionadas em micrótomo rotativo. O processo de coloração foi feito com
ácido-periódico-reativo de Schiff (PAS) e azul de toluidina (Feder & O´Brien 1968) e as
secções foram montadas em lâminas permanentes com Entellan.

81
RESULTADOS
Presença de lacunas de ar, exoderme e endoderme com uma ou várias camadas de
células de espessamento variado e redução de elementos xilemáticos e de sustentação podem
ser observados através das figuras 1-19.
Nas macrófitas aquáticas aqui estudadas a aeração ao longo do corpo vegetal, é uma
evidência observada através da presença de lacunas de ar presentes em todos os órgãos. Nas
raízes e rizomas, as lacunas de ar são amplas em todas as espécies e ocupam a região mediana
do córtex (Fig. 1, 6, 8). O córtex mediano é maior em extensão quando comparado com o
córtex externo e interno (Fig. 1, 6, 8).
Nas folhas e escapos as lacunas de ar são amplamente desenvolvidas na maioria das
espécies (Fig 9-11, 16-17), com exceção de Eichhornia paniculata (Fig. 13) e Habenaria
repens (Fig. 14), que são pequenas.
Todas as espécies estudadas apresentam epiderme revestindo todos os órgãos
vegetativos. Esta se apresenta unisseriada, constituída de células isodiamétricas, na sua
maioria, sem espessamento de parede (Fig. 1-3, 6, 8-19). Apenas os órgãos aéreos apresentam
variação no espessamento da cutícula, que é maior especialmente na folha e escapo de
Cyperus odoratus, Oxycaryum cubense e Pycreus macrostachyos (Fig. 9-11, 15-16).
Nas raízes e rizomas de todas as espécies estudadas ocorre córtex diferenciado, com
exoderme formada por várias camadas de células com paredes espessadas (Fig. 1-3, 6, 8). O
espessamento das células da exoderme pode apresentar-se regular (Fig. 1), mais intenso na
primeira camada (Fig. 2), ou na parede periclinal externa que é mais espessada que a interna
(Fig. 3). Nas raízes e rizomas as células da endoderme também apresentam paredes com
espessamentos variados, conforme a espécie (Fig. 1, 4-7), com exceção de Habenaria repens
que não apresenta endoderme com células de paredes espessadas.

82
As espécies estudadas neste trabalho, apresentam redução nos tecidos de sustentação
em todos os órgãos vegetativos. Quando presente, os tecidos de sustentação ocorrem apenas
nos órgãos aéreos como folhas e escapos, na forma de feixes de fibras esclerenquimáticas
(Fig.9-11, 15, 16) e na forma de fibras pericíclicas que envolvem os feixes vasculares (Fig. 9-
19).
Nas folhas de algumas espécies os feixes de fibras esclerenquimáticas ocupam posição
logo abaixo da epiderme (Fig. 9-11). As células que formam o periciclo são espessadas e
envolvem total (Fig. 10, 15) ou parcialmente (Fig. 12) os feixes vasculares de folhas e
escapos, além do cordão pericíclico dos escapos (Fig. 17-19). Apenas Habenaria repens não
apresentou tecido de sustentação nas folhas.
Em todas as espécies estudadas foi observada redução no número de elementos
xilemáticos tanto nos órgãos subterrâneos quanto nos aéreos, quando comparadas essas
espécies com as demais espécies terrestres do mesmo gênero, disponíveis na literatura. Essa
redução pode ser traduzida na menor quantidade de xilema quando comparada com floema
(Fig. 4, 6, 9-19).

83
DISCUSSÃO
As lacunas de ar presentes em todos os órgãos vegetativos das espécies anfíbias e
emergentes estão ligadas ao sistema de condução de oxigênio por todo o corpo da planta e são
consideradas de extrema importância no sucesso adaptativo dessas macrófitas aquáticas.
Segundo Matthews & Seymour (2006), nas plantas aquáticas, a condução de oxigênio ocorre
250.000 vezes mais rápido do que a da água graças ao eficiente sistema de distribuição de ar
pelas lacunas.
Apesar de não termos realizado estudos ontogenéticos, as lacunas de ar das raízes e
rizomas das espécies estudadas parecem ser do tipo lisígenas, e aquelas das folhas e escapos,
do tipo esquizógenas, de acordo com a estrutura anatômica observada nos órgãos adultos. De
acordo com Sculthorpe (1967) a incidência de luz nos órgãos aéreos está diretamente
relacionada à formação de lacunas de ar esquizógenas.
As espécies anfíbias e emergentes estudadas se enquadram na definição de Levitt
(1980), no que diz respeito à resistência ao estresse hídrico, sendo classificadas como plantas
que evitam o estresse produzindo aerênquima. Segundo o autor, a falta de oxigênio nas raízes
é o principal fator que induz a formação de lacunas de ar.
Por outro lado, nos órgãos aéreos das espécies estudadas, a formação de lacunas de ar
pode ser devida à produção do oxigênio produzido pela fotossíntese, conforme observado por
Sculthorpe (1967). Segundo o autor, todo o gás produzido na fotossíntese que não é
transportado para os órgãos subterrâneos se acumula nos espaços intercelulares dos órgãos
aéreos favorecendo sua distensão.
A presença de apenas uma camada de células epidérmicas, com fina camada de
cutícula nos órgãos subterrâneos e de espessura variável nos órgãos aéreos das espécies
estudadas, corrobora o que foi observado por Sculthorpe (1967) e Matias et al. (2008) para
outras macrófitas aquáticas. De acordo com os autores, a presença de uma camada de células

84
epidérmicas com cutícula fina ou espessa, vai depender da condição ambiental a que essas
plantas estão submetidas.
O número de camadas de células da exoderme das raízes e rizomas das espécies
estudadas foi maior quando comparado com outras espécies anfíbias de Cyperaceae como
Cyperus giganteus Vahl. (Rodrigues & Estelita 2003) e de Scrophulariaceae como Bacopa
salzmanii (Benth.) Wettst. ex Edwall e Bacopa monnierioides B.L.Rob. (Bona e Morretes
2003). Concorda-se com Enstone & Peterson (2005) que afirmam que a hipoxia induz ao
maior desenvolvimento de exoderme e do espessamento de suas paredes. Também concorda-
se com Hose et al. (2001) que afirmam que o espessamento e o aumento do número de
camadas de células da exoderme é comum em plantas aquáticas.
A grande maioria das espécies de macrófitas aquáticas estudadas apresenta endoderme
na raiz e rizoma, constituídas de várias camadas de células com paredes espessadas. Segundo
Sculthorpe (1967), Mauseth (1988) e Seago et al. (2000), raízes e rizomas de macrófitas
aquáticas que vivem em locais sujeitos ao estresse hídrico apresentam endoderme reforçada
primeiramente com suberina e em menor quantidade com lignina, além de mais de uma
camada de células. Assim como a endoderme, a exoderme, também constituída por células de
paredes espessadas e com várias camadas de células, atua seletivamente na passagem de
diferentes substâncias para o interior do órgão.
A redução de tecidos de sustentação nos diferentes órgãos das espécies estudadas é
comum, pois essas espécies ocorrem em ambiente de água parada, lêntico. De acordo com
Sculthorpe (1967), o tecido de sustentação nessas plantas, quando comparadas às espécies
terrestres, é extremamente reduzido, pois essas espécies não necessitam de força mecânica
para se sustentar.
Nas espécies estudadas, acredita-se que a maior proporção de tecidos de sustentação
nos representantes de Cyperaceae deve-se ao grupo taxonômico ao qual as espécies pertencem
e talvez esse caráter seja resultante da informação genética do grupo. Além disso, apresentam

85
forma de vida anfíbia, capaz de suportar períodos de cheia e seca das lagoas temporárias do
semi-árido. Por outro lado, Echinodorus grandiflorus (Alismataceae), Eichhornia paniculata
(Pontederiaceae) e Habenaria repens (Orchidaceae) mesmo pertencendo a grupos
taxonômicos distintos, apresentam redução nos tecidos de sustentação, como resposta
adaptativa.
A redução do tecido vascular nas espécies estudadas, relacionada principalmente à
diminuição do número e lignificação dos elementos xilemáticos, é uma característica de
plantas aquáticas reconhecida por vários autores (Sculthorpe 1967, Bristow 1975, Mauseth
1988, Bona & Morretes 2003). Segundo Bristow (1975), principalmente em
monocotiledôneas, o sistema vascular é reduzido e pobremente lignificado, o que leva a
degeneração de alguns elementos condutores do xilema, mas não afeta a função de transporte.
Em Habenaria repens a reduzida quantidade de tecido protetor, fundamental e
vascular não lhe permite suportar a falta de água, o que foi observado também por Stern
(1997) para esta espécie e para outras Habenariinae (Orchidaceae). Esta característica não foi
observada nas demais espécies estudadas e, ao menor sinal de aumento de temperatura do
local, Habenaria repens desapareceu completamente. Essa característica, de acordo com
Lewitt (1980), indica que esta planta apresenta resistência à adaptação, diferindo das demais
espécies que apresentam capacidade de adaptação.
Acredita-se que características anatômicas relacionadas ao sistema dérmico como:
apenas uma camada de células epidérmicas e a presença de espessura reduzida de cutícula; ao
sistema fundamental como: presença de lacunas de ar em todos os órgãos vegetativos e
diminuição nos tecidos de sustentação, exoderme e endoderme com várias camadas de células
com espessamento de parede variado; e vascular como: diminuição no número e no
espessamento dos elementos xilemáticos confirma a convergência adaptativa das plantas
estudadas ao ambiente lacustre temporário. Essas características indicam convergência ao

86
ambiente aquático e apresentam variação plástica na forma de vida anfíbia e emergente das
macrófitas lacustres estudadas.

87
REFERÊNCIAS BIBLIOGRÁFICAS
Bona, C. & Morretes, B.L. 2003. Anatomia das raízes de Bacopa salzmanii (Benth.) Wettst.
Ex Edwall e Bacopa monnierioides (Cham.) Robinson (Scrophullariaceae) em ambientes
aquático e terrestre. Acta Botanica Brasilica 17(1): 155-170.
Bristow, J.M.1975. The structure and function of roots in aquatic vascular plants. Pp. 221-
236. In The development and function of roots. J.G. Torrey & D.T.Clark, eds. Academic
Press. 618p
Davy, A.J., Noble, S.M. & Oliver, R.P. 1990. Genetic variation and adaptation to flooding in
plants. Aquatic botany 38(1): 91-108.
Enstone, D.E. & Peterson, C.A. 2005. Suberin lamella development in maize seedling roots
grown in aerated and stagnant conditions. Plant, Cell and Environment 28: 444-455.
Feder, N. & O’Brian, T.P. 1968. Plant microthecnique: some principles and new methods.
American Journal of Botany 55(1): 123-142.
Futuyma, D.J. 1993. Biologia evolutiva. 2ª Ed., Ribeirão Preto, FUNPEC
Hose, E., Clarkson, D.T., Steudle, E. Schreiber & Hartung, W. 2001. The exodermis: a variable
apoplastic barrier. Journal of Experimental Botany 52(365): 2245-2264.
Irgang, B. E., Pedralli, G. & Waechter, J. L. 1984. Macrófitas aquáticas da Estação Ecológica
do Taim, Rio Grande do Sul, Brasil. Rossléria 6: 395-404.
Johansen, D.A. 1940. Plant microtechnique. McGraw Hill Book Co., New York.
Keddy, P.A. 2000. Wetland ecology: principles and conservation. Cambridge University Press.
United Kingdom. 614p.
Kraus, J.E. & Arduim, M. 1997. Manual básico de métodos em morfologia vegetal. EDUSP,
São Paulo.
Lewitt, J. 1980. Responses of plants to environmental stresses: water, radiation, salt and other
stresses. Vol II, pp. 213-228. Academic Press, Inc. London.

88
Matias, L.Q., Soares, A. & Scatena, V.L. 2008. Anatomy of Echinodorus (Alismataceae)
scapes from northeastern Brazil as applied to taxonomy. Edinburgh Journal of Botany
65(1): 11-21.
Matthews, P.G.D. & Seymour, R.S. 2006. Anatomy of the gas canal system of Nelumbo
nucifera. Aquatic Botany 85(2): 147-154.
Mauseth, J.D. 1988. Plant anatomy. The Benjamin Cumming Publishing Company,
California.
Rodrigues, A.C. & Estelita, M.E.M. 2003. Origin and structure of the Krans tissue in bracts of
Cyperus giganteus Vahl (Cyperaceae). Revista Brasileira de Botanica 26(4): 445-452.
Sculthorpe, C.D. 1967. The biology of aquatic vascular plants. Edward Arnold, London.
Seago, J.L., Peterson, C.A. & Enstone, D.E. 2000. Cortical development in roots of the
aquatic plant Pontederia cordata (Pontederiaceae). American Journal of Botany 87(8):
1116-1127.
Stern, W.L. 1997. Vegetative anatomy of subtribe Habenariinae (Orchidaceae). Botanical
Journal of the Linnean Society 125(3): 211-227.
Wells, C.L. & Pigliucci, M. 2000. Adaptative phenotypic plasticity: the case of heterophylly
in aquatic plants. Perspectives in plant ecology, evolution ans systematic 3(1): 1-18.
West-Eberhard, M.J. 2003. Developmental plasticity and evolution. Oxford, University Press.
794p.

89
Figuras 1-8. Aspectos anatômicos da exoderme, endoderme e lacunas de ar de raízes e
rizomas de espécies de monocotiledôneas, em secções transversais. 1. Pycreus
macrostachyos (Cyperaceae) - aspecto geral da raiz. 2. Oxycarium cubense
(Cyperaceae) - detalhe da epiderme e exoderme. 3. Eichhornia paniculata
(Pontederiaceae) - detalhe da epiderme e exoderme. 4. Pycreus macrostachyos
(Cyperaceae) - detalhe da endoderme e cilindro vascular. 5. Eichhornia paniculata
(Pontederiaceae) - detalhe da endoderme. 6. Oxycarium cubense (Cyperaceae)-
detalhe do rizoma com lacunas de ar no córtex. 7-8. Rizoma de Eichhornia
paniculata (Pontederiaceae): 7. Detalhe da endoderme e cilindro vascular. 8. Detalhe
do córtex. (Barras - Fig. 1-5, 8 = 50µm; Fig. 6-7 = 100 µm). Legenda: En-
endoderme; Ep–epiderme; Ex–Exoderme; La–lacuna de ar.
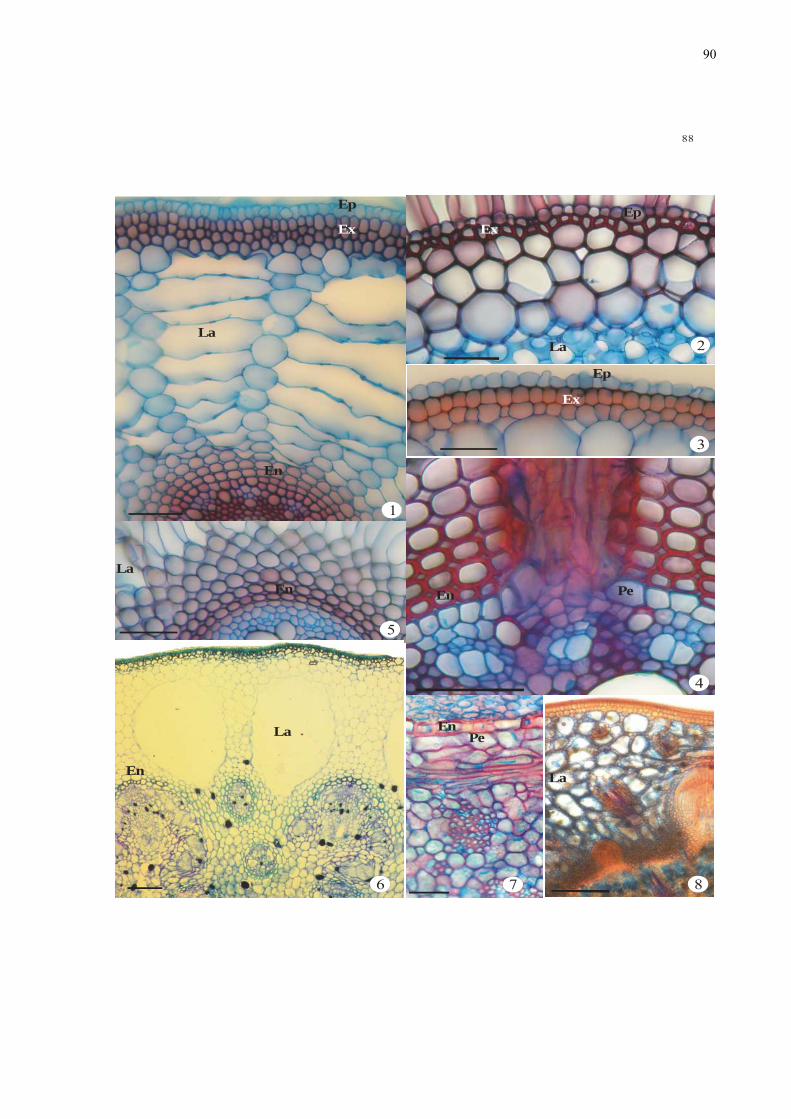
90
1
3
2
4
5
7 8
La
La
La
La
En
En En
En
Pe
Pe
Ex Ex
Ex
6
En
La
Ep Ep
Ep
88

91
Figuras 9-19. Aspectos anatômicos de folhas e escapos de espécies de monocotiledôneas da
região mediana, em secções transversais. 9-14. Detalhe do mesofilo. 9. Cyperus odoratus
(Cyperaceae). 10. Oxycaryum cubense (Cyperaceae). 11. Pycreus macrostachyos
(Cyperaceae). 12. Echinodorus grandiflorus (Alismataceae). 13. Eichhornia paniculata
(Pontederiaceae). 14. Habenaria repens (Orchidaceae). 15-19. Escapo: aspecto geral, detalhe
do córtex e cilindro vascular. 15. Cyperus odoratus (Cyperaceae). 16. Oxycarium cubense
(Cyperaceae). 17. Echinodorus grandiflorus (Alismataceae). 18. Eichhornia paniculata
(Pontederiaceae). 19. Habenaria repens (Orchidaceae). (Barras - Fig. 9-14 = 100 µm; Fig. 15-
19 = 200 µm). Legenda: Ep–epiderme; Ff-feixe de fibras esclerenquimáticas; Fl–floema; Fp-
fibra pericíclica; La–lacuna de ar; Pe-periciclo.
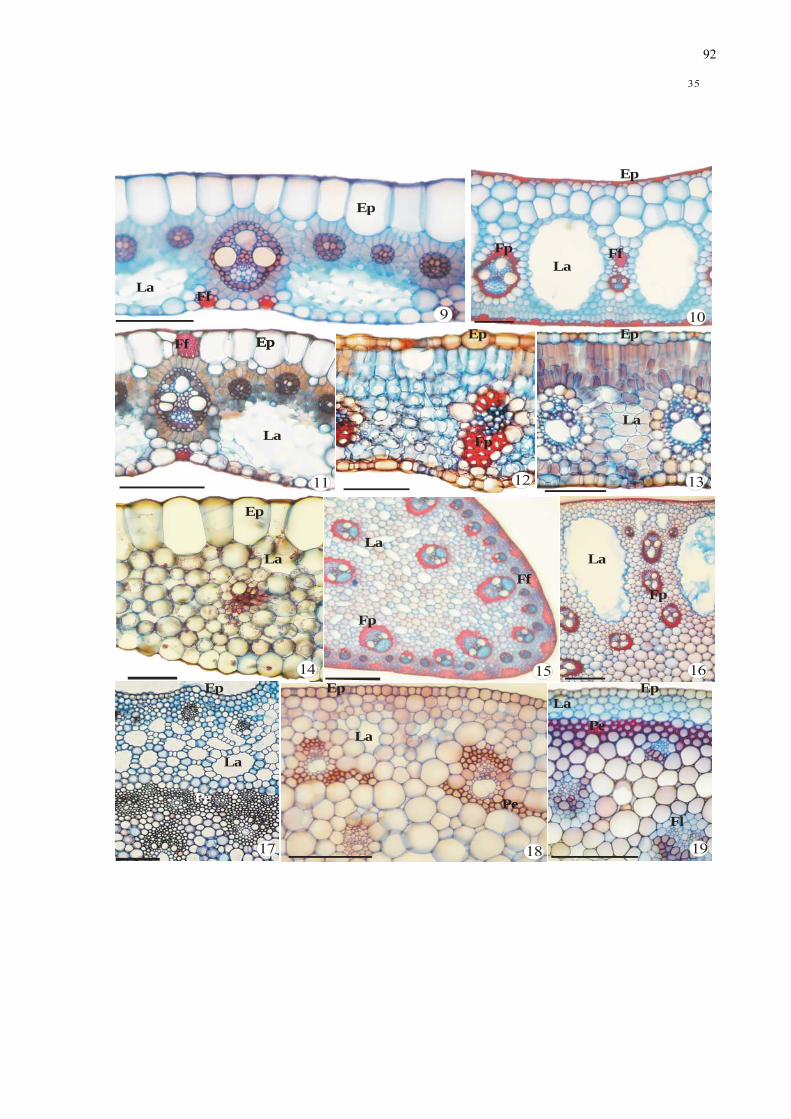
92
9 10
11 12 13
14 15 16
191817
LaLa
LaLa
LaLa
La
La
La
LaPe
PePe
Ff
Ff
Ff
Fp
Ff
Fl
Fp
Ep
EpEp
Ep
EpEp
Ep
Ep Ep Ep
Fp
Fp
35

93
CONSIDERAÇÕES FINAIS
- Cyperus odoratus, Oxycaryum cubense, Pycreus macrostachyos (Cyperaceae) –
anfíbias; Echinodorus grandiflorus (Alismataceae), Eichhornia paniculata (Pontederiaceae) e
Habenaria repens (Orchidaceae) – emergentes, estão bem adaptadas aos solos inundáveis das
lagoas temporárias da Bahia. As variações plásticas observadas nos diferentes tecidos dos
diferentes órgãos vegetativos dessas espécies ocorrem em função das formas de vida por elas
apresentadas.
- O estudo morfoanatômico dos órgãos vegetativos dessas espécies de
monocotiledôneas, pertencentes a diferentes famílias e gêneros, apresentaram características
comuns às demais plantas aquáticas. Sobretudo, este estudo indica que características como
presença de epiderme com apenas uma camada de células com cutícula fina e lacunas de ar
em todos os órgãos vegetativos, exoderme e endoderme multisseriadas com células de paredes
espessadas nas raízes e rizomas, redução no número de tecidos vasculares e de tecidos de
sustentação nos diferentes órgãos são comuns e convergentes ao ambiente aquático.
- Verificou-se em todos os órgãos vegetativos de Cyperus odoratus, Oxycaryum
cubense e Pycreus macrostachyos, espécies anfíbias de Cyperaceae, que embora pertencentes
a diferentes gêneros, apresentam características anatômicas comuns. Cutícula com
espessamento variado nas células epidérmicas; espessamento das paredes das células da
exoderme e endoderme de raízes e rizomas; presença de folhas anfiestomáticas, com células
buliformes e hipoderme; lacunas de ar, redução do número de elementos xilemáticos e de
tecidos de sustentação em todos os órgãos são características que convergem estas plantas ao
ambiente aquático e à forma de vida anfíbia.
- As espécies Echinodorus grandiflorus (Alismataceae), Eichhornia paniculata
(Pontederiaceae) e Habenaria repens (Orchidaceae) embora pertençam a grupos taxonômicos
diferentes (famílias distintas), apresentaram características comuns à forma de vida

94
emergente. Todos os órgãos vegetativos apresentam epiderme com apenas uma camada de
células, recoberta por cutícula fina, exoderme e endoderme multisseriada com células de
paredes espessadas nas raízes e rizomas; presença de lacunas de ar, idioblastos com
compostos fenólicos e cristais e ausência de esclerênquima.
- Com os dados gerados a partir deste trabalho com macrófitas aquáticas anfíbias e
emergentes e unindo-os futuramente com os de outras formas de vida que se pretende estudar,
procura-se contribuir para o conhecimento de suas adaptações ao ambiente aquático, mesmo
que temporário. Esses dados poderão contribuir para a compreensão dos processos ecológicos,
além de subsidiar estudos taxonômicos, filogenéticos e fisiológicos.