metabolismo lipídico de ratos alimentados com diferentes...
TRANSCRIPT

Arq Bras Cardiol2002; 78: 25-31.
Aguila e colsMetabolismo lipídico de ratos
2 52 52 52 52 5
Instituto de Biologia – UERJ, Departamento de Nutrição Aplicada - Escola deNutrição - UNI-RIOCorrespondência: Márcia Barbosa Águila - Centro Biomédico/Instituto deBiologia UERJ - Av. 28 de Setembro, 87 (fds) - 20551-030 - Rio de Janeiro, RJE-mail: [email protected] para publicação em 22/2/01Aceito em 18/4/01
Objetivo - Avaliar o efeito de diferentes lipídios dieté-ticos sobre o metabolismo lipídico de ratos durante o en-velhecimento.
Métodos - Foram estudados 50 ratos machos (Wistar),desde o desmame até 12 e 18 meses de idade. Houve su-plementação dietética com óleo de soja (S), óleo de canola(CA), banha de porco e gema de ovo (BG) e óleo de ca-nola+banha de porco e gema de ovo (CA+BG). A pressãoarterial da cauda (PAC) foi aferida mensalmente, a relaçãocoração/corpo (RCC) foi determinada. Os ratos foram sa-crificados com 12 e com 18 meses e amostras de sangue fo-ram coletadas para análise lipídica: colesterol total (CT),LDL-C, VLDL-C, HDL-C, triglicerídios (TG) e glicose.
Resultados - O tipo de óleo ingerido pelos animaismodificou significativamente a PAC, RCC e os níveis de li-pídios sangüíneos em ratos aos 12 e 18 meses. Não houvediferença na curva de sobrevivência dos animais nos gru-pos. BG apresentou a maior PAC, CA foi o único que nãoalterou a PAC com o envelhecimento. Houve diminuiçãoda RCC nos animais BG e CA+BG. Aos 12 meses foram ob-servadas diferenças no CT, HDL-C, LDL-C, VLDL-C, TG eglicose. Aos 18 meses houve diferença significativa no CT,HDL-C e glicose. O maior valor de CT foi encontrado noCA e o menor no S.
Conclusão - Não ocorre aumento da PAC e sim melho-ra do perfil lipídico de ratos alimentados por longo tempocom dieta suplementada com CA, com elevação dos níveisde HDL-C em comparação com os outros tipos de dieta.
Palavras-chave: óleo de canola, lipoproteínas, ratos
Arq Bras Cardiol, volume 78 (nº 1), 25-31, 2002
Márcia Barbosa Águila, Carla Cota Loureiro, Alessandra da Rocha Pinheiro,Carlos Alberto Mandarim-de-Lacerda
Rio de Janeiro - RJ
Metabolismo Lipídico de Ratos Alimentados com DiferentesTipos de Lipídios
Artigo Original
Alguns trabalhos experimentais em animais relatamque o aumento da ingestão de colesterol resulta em sua ele-vação na corrente sangüínea 1-3, com o conseqüente aumen-to do risco para doenças cardiovasculares 4.
Os ácidos graxos n-3 são ácidos graxos poliinsatura-dos de cadeia longa, encontrados naturalmente em óleos depeixe e em algumas plantas. Os ácidos graxos da família n-3são os ácidos eicosapentaenóico (EPA) (C20: 5n-3),docosahexaenóico (DHA) (C22: 6n-3) e α -linolênico (ALN)(C18: 3n-3) 5,6. O uso do EPA e do DHA já está muito bemevidenciado, porém estão crescendo as evidências dos efei-tos metabólicos do ALN. O ALN é um ácido graxo derivadode vegetais, que pode ser encontrado na soja, canola, aça-frão e amendoim 7. Após a ingestão, o ALN pode serdessaturado e alongado para as outras formas de ácidosgraxos poliinsaturados de cadeia longa, o EPA e o DHA 8,9.O óleo de canola proporciona uma opção importante de fontedietética de ácido graxo da família n-3, principalmente para osvegetarianos e indivíduos que não consomem peixe 10.
Connor 11 listou os possíveis mecanismos da ingestão deácidos graxos n-3 sobre o metabolismo lipídico: 1) inibição dasíntese de VLDL; 2) diminuição da síntese de apolipoproteínaB; 3) aumento do catabolismo das VLDL; 4) diminuição da sín-tese de LDL e 5) redução da lipemia pós-prandial.
As lipoproteínas estão intimamente ligadas com o ris-co de doenças cardiovasculares, as lipoproteínas de baixadensidade (LDL) indicam risco aumentado, enquanto aslipoproteínas de alta densidade (HDL) são consideradas umfator de proteção 12,13. Quando o risco de doenças cardio-vasculares foi considerado em termos de HDL-C e LDL-C, aincidência de doenças cardiovasculares foi maior com o au-mento da concentração de LDL-C e diminuição de HDL-C 14.Tanto a LDL-C quanto a HDL-C são fatores de risco inde-pendentes para as doenças cardiovasculares 15. Isto é im-portante para estabelecer as condições que influenciam asmudanças nos níveis de LDL e HDL ao longo da vida.
Por causa da grande variabilidade genética e dos hábi-tos dietéticos das populações humanas, o modelo experi-mental com ratos mostra-se adequado para estudos expe-

2 62 62 62 62 6
Aguila e colsMetabolismo lipídico de ratos
Arq Bras Cardiol2002; 78: 25-31.
rimentais 16-19. O importante é que existe uma similaridadegeral entre o sistema cardiovascular de ratos com o de ou-tros mamíferos, incluindo o homem 20. Apesar da dificuldadede produzir hiperlipidemia e aterosclerose em ratos, dietasespeciais podem induzir o aumento do nível de colesterolsangüíneo e, também, induzir hipertensão arterial ou doen-ças renais 21, como tem sido observado em trabalhos préviosde nosso grupo 22-25.
O objetivo deste estudo é investigar a possível influ-ência de quatro dietas experimentais com diferentes tipos degordura dietética sobre indicadores cardiovasculares e ometabolismo lipídico de ratos durante o envelhecimento.
Métodos
Foram estudados 50 ratos machos (Wistar), obtidos decolônias mantidas pela Universidade do Estado do Rio deJaneiro. A pesquisa seguiu as normas estabelecidas noGuide for Care and Use of Laboratory Animals publicadaspelos US National Institutes of Health (NIH Publication No.85-23 revisados em 1985).
Os animais foram colocados em caixas de polipropile-no, em grupos de cinco, mantidos em temperatura (21±1°C)e umidade (60±10%) controladas. O ambiente foi submetidoa ciclos de 12h de claro e escuro (luz artificial, de 7h às 19h) eciclo de exaustão de ar (15min/h). Os grupos receberam águalivremente e a mesma dieta basal, composta de caseína, fari-nha de trigo, maisena, clara de ovo e mistura de vitaminas eminerais. As dietas tinham as seguintes características:47% de carboidratos, 29% de lipídios, 23% de proteínas,11,5% de fibras alimentares e teor de sódio de 0,05% (ou500mg/kg de dieta), a mesma quantidade para todas as die-tas. Os valores energéticos das dietas foram praticamenteiguais. As dietas eram preparadas semanalmente no labora-tório e armazenadas à temperatura de +4°C. Dependendo dogrupo de estudo houve suplementação da dieta com dife-rentes tipos de lipídios: com óleo de soja (grupo soja, S),com óleo de canola (grupo canola, CA), com banha de por-co e gema de ovo (grupo BG) e com óleo de canola+banhade porco e gema de ovo (grupo CA+BG).
Os animais receberam as dietas desde o desmame (21dias de vida). Cada grupo teve uma amostra de no mínimocinco animais sacrificados aos 12 e aos 18 meses. A pressãoarterial da cauda (verificada por pleistimografia, RTBP1007,Kent Scientific Co., Litchfield, CT, USA) e a massa corporaltotal foram verificadas mensalmente.
Na época prevista, os animais foram sacrificados,mantido o jejum desde às 19h do dia anterior. Foram aneste-siados com éter dietílico, o tórax foi aberto e coletada amos-tra de sangue do átrio direito. Em seguida, foi injetado 3mlde KCl a 10% no ventrículo esquerdo até parada cardíaca emdiástole. O coração foi retirado cortando os vasos da base,o mais curto possível, o volume e o peso cardíaco foram de-terminados por deslocamento de líquido (método deScherle) 26. A relação coração/corpo (RCC) foi determinadadividindo-se os respectivos pesos.
Foram analisados os parâmetros bioquímicos: coles-
terol total (CT), triglicerídeos (TG), colesterol da lipoproteínade baixa densidade (LDL-C), colesterol da lipoproteína de altadensidade (HDL-C), colesterol da lipoproteína de muito baixadensidade (VLDL-C) e glicose. O plasma foi separado dosangue total por centrifugação em temperatura ambiente du-rante 15min com 120g de gravidade e, em seguida, estocadosem uma temperatura de -80°C, até serem analisados no labora-tório 1,27. A glicose foi medida usando o método enzimáticocom kit Art. 07.3757.7 (U.S.#42954), o colestereol medido como kit colesterol 1.19738.0001. KgaA 64271, calibração SystemMulti-test, Merk Cat. nº 1.19720.0001 (Darmstadt, Alemanha),a HDL medida usando o kit Diasys Cat. nº 10.351.030, ostriglicerídios medidos usando o kit triglicerídios 1.19706.0001KgaA 64271, calibração System Multi-test, Merk Cat. nº1.19729.0001 (Darmstadt, Alemanha). A LDL, VLDL e fraçõesde quilomícrons foram precipitadas abundantemente pelaadição de ácido fosfotungstênico na presença de íons demagnésio. Após a centrifugação, o colesterol na fração HDL-C, mantido em suspensão, foi determinado. A LDL-C foi cal-culada de acordo com a fórmula de Friedwald 28.
O teste de Mantel-Haenezel (T) foi usado para compa-rar as curvas de sobrevivência dos animais dos diferentesgrupos. Se a hipótese nula é verdadeira, T poderia ser consi-derado com distribuição χ2 (qui-quadrado com um grau deliberdade) 29. A estatística descritiva foi calculada para to-dos os parâmetros. As diferenças entre os grupos foramtestadas, usando-se o teste não paramétrico de Mann-Whitney, dois a dois com nível de significância de 0,05 30.
Resultados
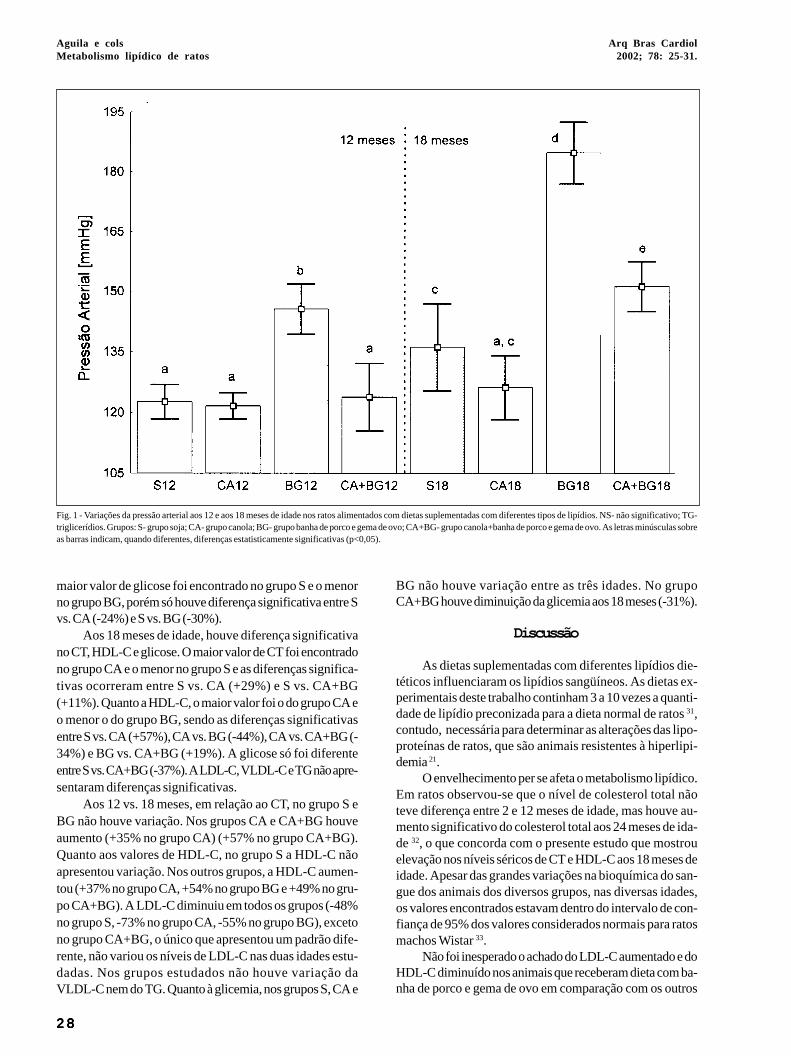
Os resultados estão resumidos na tabela I e nas figuras1 e 2. O tipo de óleo ingerido pelos animais modificou signifi-cativamente a pressão arterial da cauda, RCC e os níveis delipídios sangüíneos em ratos aos 12 e 18 meses de idade. Nãohouve diferença significativa na curva de sobrevivência dosanimais considerando os diferentes grupos. As variações dapressão arterial da cauda estão apresentadas na figura 1.
Aos 12 meses de idade, o grupo BG apresentou a maiorpressão arterial da cauda. As diferenças foram significati-vas entre as comparações: S vs. BG (+17%), CA vs. BG(+16%) e BG vs. CA+BG (-18%).
Aos 18 meses de idade, o aumento da pressão arterialda cauda no grupo BG foi ainda mais acentuado e o grupoCA+BG também apresentou aumento da pressão arterial dacauda. As diferenças foram significativas entre as compara-ções: S vs. BG (+29%), S vs. CA+BG (+12%), CA vs. BG(+32%), CA vs. CA+BG (+15%) e BG vs. CA+BG (-24%).
Aos 12 vs. 18 meses, o grupo CA foi o único que nãoalterou a pressão arterial da cauda com o envelhecimento.Os outros três grupos apresentaram elevação da pressãoarterial da cauda. O grupo S aumentou 10%, o grupo BG, 23%e o grupo CA+BG, 18%.
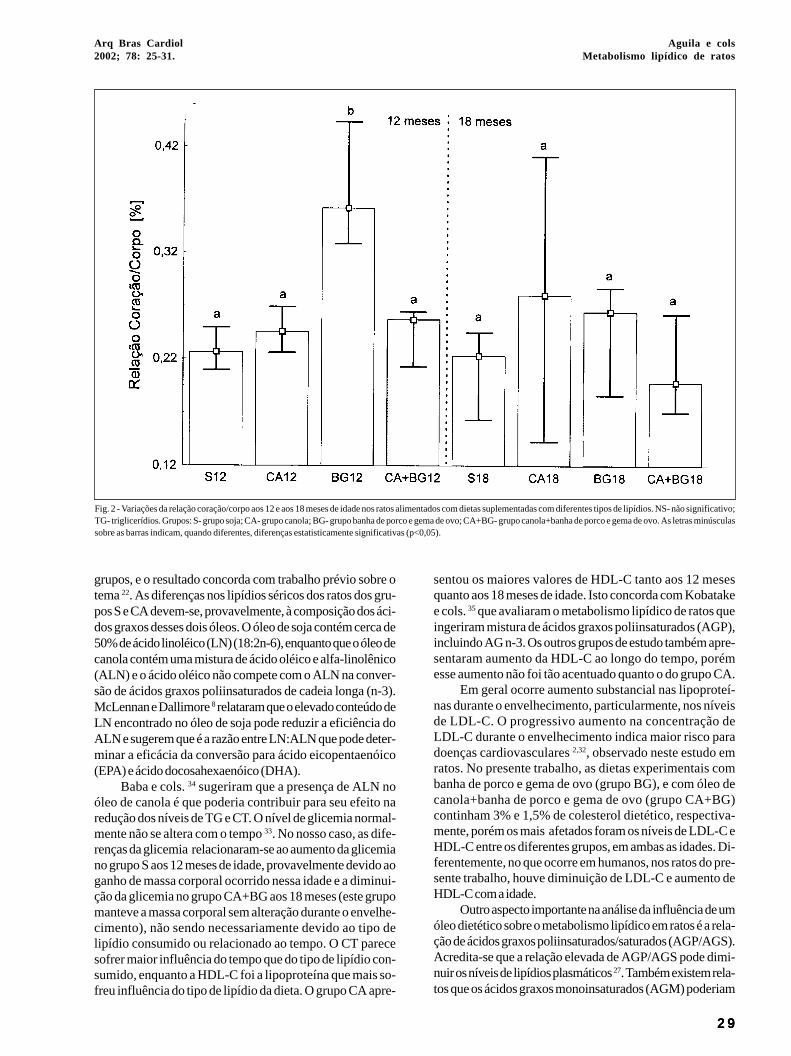
As variações da relação coração/corpo encontram-se nafigura 2. Aos 12 meses de idade, houve diferença significativaentre os grupos S vs. BG (+43%) e CA vs. BG (+31%); e aos 18meses de idade, não houve diferença entre os grupos estudados.
Arq Bras Cardiol2002; 78: 25-31.
Aguila e colsMetabolismo lipídico de ratos
2 72 72 72 72 7
Aos 12 vs. 18 meses, nos grupos S e CA não houve al-teração da RCC com o envelhecimento. Houve diminuiçãoda RCC no grupo BG e no grupo CA+BG. O grupo BG teveredução de 86% e o grupo CA+BG redução de 37% da RCC.Quanto à variação do peso corporal acha-se na tabela I. Aos12 meses de idade, houve diferença significativa entre osgrupos S vs. CA (-20%) e S vs. BG (-34%), entretanto aos 18meses de idade, não houve diferença entre os grupos estu-dados; e aos 12 vs.18 meses, não houve diferença entre asidades estudadas.
As variações da glicose e lipídios sangüíneos estãoapresentadas na tabela I.
Aos 12 meses de idade, observamos diferença no CT,HDL-C, LDL-C, VLDL-C, TG e Glicose. As diferenças foramsignificativas entre os seguintes grupos: em relação ao CT,entre os grupos S vs. CA+BG (-22%), CA vs. CA+BG (-27%)e BG vs. CA+BG (-37%); em relação a HDL-C, o maior valorfoi encontrado no grupo CA (+25% que no grupo S, +103%
que no grupo BG e +65% que no grupo CA+BG), o menorvalor de HDL-C foi encontrado no grupo BG, sendo que asdiferenças entre S vs. BG (-39%), S vs. CA+BG (-25%) tam-bém foram significativas. Aos 12 meses, todos os gruposaumentaram os níveis de LDL-C, menos o grupo CA+BG. ALDL-C foi maior no grupo BG e menor no grupo CA+BG,mas as diferenças, estatisticamente significativas ocorreramentre S vs. CA (+45%), S vs. BG (+46%), CA vs. CA+BG (-58%) e BG vs. CA+BG (-59%). A VLDL-C e TG também apre-sentaram o mesmo padrão de variação, os maiores valoresde VLDL-C e TG foram encontrados no grupo S e os meno-res no grupo BG. Aos 12 meses de idade, os animais maispesados eram os do grupo S. Houve diferenças significati-vas entre S vs. BG (-47% para VLDL-C e -46% para TG), CAvs. BG (-45% tanto para VLDL-C como para TG) e BG vs.CA+BG (+77% para VLDL-C e + 75% para TG). Aos 12 me-ses de idade, a glicemia teve alterações entre os grupos. O
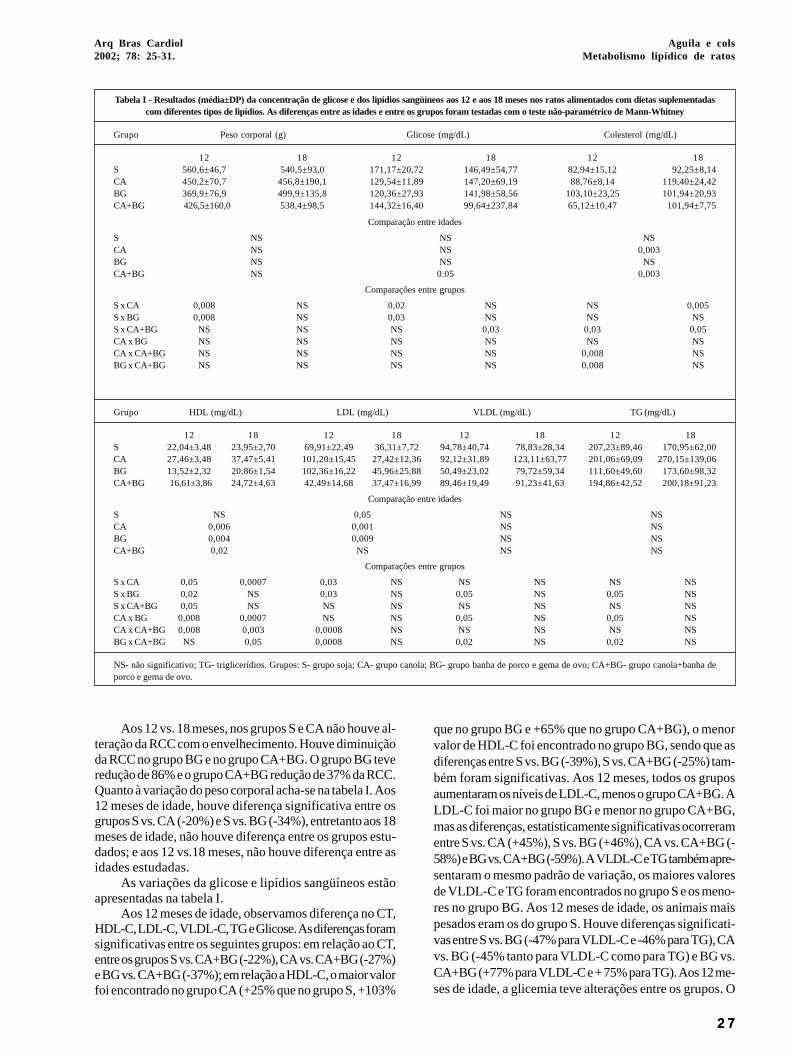
Tabela I - Resultados (média±DP) da concentração de glicose e dos lipídios sangüíneos aos 12 e aos 18 meses nos ratos alimentados com dietas suplementadascom diferentes tipos de lipídios. As diferenças entre as idades e entre os grupos foram testadas com o teste não-paramétrico de Mann-Whitney
Grupo Peso corporal (g) Glicose (mg/dL) Colesterol (mg/dL)
12 18 12 18 12 18S 560,6±46,7 540,5±93,0 171,17±20,72 146,49±54,77 82,94±15,12 92,25±8,14CA 450,2±70,7 456,8±190,1 129,54±11,89 147,20±69,19 88,76±8,14 119,40±24,42BG 369,9±76,9 499,9±135,8 120,36±27,93 141,98±58,56 103,10±23,25 101,94±20,93CA+BG 426,5±160,0 538,4±98,5 144,32±16,40 99,64±237,84 65,12±10,47 101,94±7,75
Comparação entre idades
S NS NS NSCA NS NS 0,003BG NS NS NSCA+BG NS 0.05 0,003
Comparações entre grupos
S x CA 0,008 NS 0,02 NS NS 0,005S x BG 0,008 NS 0,03 NS NS NSS x CA+BG NS NS NS 0,03 0,03 0,05CA x BG NS NS NS NS NS NSCA x CA+BG NS NS NS NS 0,008 NSBG x CA+BG NS NS NS NS 0,008 NS
Grupo HDL (mg/dL) LDL (mg/dL) VLDL (mg/dL) TG (mg/dL)
12 18 12 18 12 18 12 18S 22,04±3,48 23,95±2,70 69,91±22,49 36,31±7,72 94,78±40,74 78,83±28,34 207,23±89,46 170,95±62,00CA 27,46±3,48 37,47±5,41 101,20±15,45 27,42±12,36 92,12±31,89 123,11±63,77 201,06±69,09 270,15±139,06BG 13,52±2,32 20,86±1,54 102,36±16,22 45,96±25,88 50,49±23,02 79,72±59,34 111,60±49,60 173,60±98,32CA+BG 16,61±3,86 24,72±4,63 42,49±14,68 37,47±16,99 89,46±19,49 91,23±41,63 194,86±42,52 200,18±91,23
Comparação entre idades
S NS 0,05 NS NSCA 0,006 0,001 NS NSBG 0,004 0,009 NS NSCA+BG 0,02 NS NS NS
Comparações entre grupos
S x CA 0,05 0,0007 0,03 NS NS NS NS NSS x BG 0,02 NS 0,03 NS 0,05 NS 0,05 NSS x CA+BG 0,05 NS NS NS NS NS NS NSCA x BG 0,008 0,0007 NS NS 0,05 NS 0,05 NSCA x CA+BG 0,008 0,003 0,0008 NS NS NS NS NSBG x CA+BG NS 0,05 0,0008 NS 0,02 NS 0,02 NS
NS- não significativo; TG- triglicerídios. Grupos: S- grupo soja; CA- grupo canola; BG- grupo banha de porco e gema de ovo; CA+BG- grupo canola+banha deporco e gema de ovo.
2 82 82 82 82 8
Aguila e colsMetabolismo lipídico de ratos
Arq Bras Cardiol2002; 78: 25-31.
maior valor de glicose foi encontrado no grupo S e o menorno grupo BG, porém só houve diferença significativa entre Svs. CA (-24%) e S vs. BG (-30%).
Aos 18 meses de idade, houve diferença significativano CT, HDL-C e glicose. O maior valor de CT foi encontradono grupo CA e o menor no grupo S e as diferenças significa-tivas ocorreram entre S vs. CA (+29%) e S vs. CA+BG(+11%). Quanto a HDL-C, o maior valor foi o do grupo CA eo menor o do grupo BG, sendo as diferenças significativasentre S vs. CA (+57%), CA vs. BG (-44%), CA vs. CA+BG (-34%) e BG vs. CA+BG (+19%). A glicose só foi diferenteentre S vs. CA+BG (-37%). A LDL-C, VLDL-C e TG não apre-sentaram diferenças significativas.
Aos 12 vs. 18 meses, em relação ao CT, no grupo S eBG não houve variação. Nos grupos CA e CA+BG houveaumento (+35% no grupo CA) (+57% no grupo CA+BG).Quanto aos valores de HDL-C, no grupo S a HDL-C nãoapresentou variação. Nos outros grupos, a HDL-C aumen-tou (+37% no grupo CA, +54% no grupo BG e +49% no gru-po CA+BG). A LDL-C diminuiu em todos os grupos (-48%no grupo S, -73% no grupo CA, -55% no grupo BG), excetono grupo CA+BG, o único que apresentou um padrão dife-rente, não variou os níveis de LDL-C nas duas idades estu-dadas. Nos grupos estudados não houve variação daVLDL-C nem do TG. Quanto à glicemia, nos grupos S, CA e
BG não houve variação entre as três idades. No grupoCA+BG houve diminuição da glicemia aos 18 meses (-31%).
Discussão
As dietas suplementadas com diferentes lipídios die-téticos influenciaram os lipídios sangüíneos. As dietas ex-perimentais deste trabalho continham 3 a 10 vezes a quanti-dade de lipídio preconizada para a dieta normal de ratos 31,contudo, necessária para determinar as alterações das lipo-proteínas de ratos, que são animais resistentes à hiperlipi-demia 21.
O envelhecimento per se afeta o metabolismo lipídico.Em ratos observou-se que o nível de colesterol total nãoteve diferença entre 2 e 12 meses de idade, mas houve au-mento significativo do colesterol total aos 24 meses de ida-de 32, o que concorda com o presente estudo que mostrouelevação nos níveis séricos de CT e HDL-C aos 18 meses deidade. Apesar das grandes variações na bioquímica do san-gue dos animais dos diversos grupos, nas diversas idades,os valores encontrados estavam dentro do intervalo de con-fiança de 95% dos valores considerados normais para ratosmachos Wistar 33.
Não foi inesperado o achado do LDL-C aumentado e doHDL-C diminuído nos animais que receberam dieta com ba-nha de porco e gema de ovo em comparação com os outros
Fig. 1 - Variações da pressão arterial aos 12 e aos 18 meses de idade nos ratos alimentados com dietas suplementadas com diferentes tipos de lipídios. NS- não significativo; TG-triglicerídios. Grupos: S- grupo soja; CA- grupo canola; BG- grupo banha de porco e gema de ovo; CA+BG- grupo canola+banha de porco e gema de ovo. As letras minúsculas sobreas barras indicam, quando diferentes, diferenças estatisticamente significativas (p<0,05).
Arq Bras Cardiol2002; 78: 25-31.
Aguila e colsMetabolismo lipídico de ratos
2 92 92 92 92 9
Fig. 2 - Variações da relação coração/corpo aos 12 e aos 18 meses de idade nos ratos alimentados com dietas suplementadas com diferentes tipos de lipídios. NS- não significativo;TG- triglicerídios. Grupos: S- grupo soja; CA- grupo canola; BG- grupo banha de porco e gema de ovo; CA+BG- grupo canola+banha de porco e gema de ovo. As letras minúsculassobre as barras indicam, quando diferentes, diferenças estatisticamente significativas (p<0,05).
grupos, e o resultado concorda com trabalho prévio sobre otema 22. As diferenças nos lipídios séricos dos ratos dos gru-pos S e CA devem-se, provavelmente, à composição dos áci-dos graxos desses dois óleos. O óleo de soja contém cerca de50% de ácido linoléico (LN) (18:2n-6), enquanto que o óleo decanola contém uma mistura de ácido oléico e alfa-linolênico(ALN) e o ácido oléico não compete com o ALN na conver-são de ácidos graxos poliinsaturados de cadeia longa (n-3).McLennan e Dallimore 8 relataram que o elevado conteúdo deLN encontrado no óleo de soja pode reduzir a eficiência doALN e sugerem que é a razão entre LN:ALN que pode deter-minar a eficácia da conversão para ácido eicopentaenóico(EPA) e ácido docosahexaenóico (DHA).
Baba e cols. 34 sugeriram que a presença de ALN noóleo de canola é que poderia contribuir para seu efeito naredução dos níveis de TG e CT. O nível de glicemia normal-mente não se altera com o tempo 33. No nosso caso, as dife-renças da glicemia relacionaram-se ao aumento da glicemiano grupo S aos 12 meses de idade, provavelmente devido aoganho de massa corporal ocorrido nessa idade e a diminui-ção da glicemia no grupo CA+BG aos 18 meses (este grupomanteve a massa corporal sem alteração durante o envelhe-cimento), não sendo necessariamente devido ao tipo delipídio consumido ou relacionado ao tempo. O CT parecesofrer maior influência do tempo que do tipo de lipídio con-sumido, enquanto a HDL-C foi a lipoproteína que mais so-freu influência do tipo de lipídio da dieta. O grupo CA apre-
sentou os maiores valores de HDL-C tanto aos 12 mesesquanto aos 18 meses de idade. Isto concorda com Kobatakee cols. 35 que avaliaram o metabolismo lipídico de ratos queingeriram mistura de ácidos graxos poliinsaturados (AGP),incluindo AG n-3. Os outros grupos de estudo também apre-sentaram aumento da HDL-C ao longo do tempo, porémesse aumento não foi tão acentuado quanto o do grupo CA.
Em geral ocorre aumento substancial nas lipoproteí-nas durante o envelhecimento, particularmente, nos níveisde LDL-C. O progressivo aumento na concentração deLDL-C durante o envelhecimento indica maior risco paradoenças cardiovasculares 2,32, observado neste estudo emratos. No presente trabalho, as dietas experimentais combanha de porco e gema de ovo (grupo BG), e com óleo decanola+banha de porco e gema de ovo (grupo CA+BG)continham 3% e 1,5% de colesterol dietético, respectiva-mente, porém os mais afetados foram os níveis de LDL-C eHDL-C entre os diferentes grupos, em ambas as idades. Di-ferentemente, no que ocorre em humanos, nos ratos do pre-sente trabalho, houve diminuição de LDL-C e aumento deHDL-C com a idade.
Outro aspecto importante na análise da influência de umóleo dietético sobre o metabolismo lipídico em ratos é a rela-ção de ácidos graxos poliinsaturados/saturados (AGP/AGS).Acredita-se que a relação elevada de AGP/AGS pode dimi-nuir os níveis de lipídios plasmáticos 27. Também existem rela-tos que os ácidos graxos monoinsaturados (AGM) poderiam

3 03 03 03 03 0
Aguila e colsMetabolismo lipídico de ratos
Arq Bras Cardiol2002; 78: 25-31.
reduzir os lipídios plasmáticos tão eficientemente quanto osAGP 36,37. A relação AGP+AGM/AGS, entretanto, tem sidoconsiderada um indicador ainda melhor do efeito de um óleodietético sobre os lipídios plasmáticos 27. O óleo de canola, abanha de porco e a gema de ovo contêm maior quantidade deAGM do que o óleo de soja. Neste trabalho, as dietas dosgrupos CA, BG e CA+BG continham 58, 50 e 53% de AGM,respectivamente. De acordo com a hipótese AGP+AGM/AGS, o óleo de canola, que contém 92% de AGM+AGP, pode-ria favorecer o aumento significativo da HDL-C em compara-ção com os outros grupos. O óleo de soja contém pequenaquantidade de AGM (26%) e grande quantidade de AGP(61%) e um conteúdo relativamente alto de AGM+AGP (po-rém menor que o óleo de canola).
Huang e Chang 38 estudaram os efeitos de diferentes ti-pos de óleos dietéticos contendo relações fixas de ácidosgraxos (AGP/AGS = 1, AGP+AGM/AGS = 5,7 e 4) em ratos eobservaram que os AGM poderiam aumentar, ao invés de di-minuir, o CT, TG, LDL-C e o colesterol hepático. Relatou-seque uma relação maior de AGP/AGS poderia ser melhor 27. Nopresente trabalho, entretanto, os animais dos grupos CA e Stiveram diferenças nos lipídios sangüíneos, mesmo tendorelação similar de AGP/AGS das dietas. Uma discussão maisdetalhada sobre a composição das dietas foi apresentada emtrabalho anterior do grupo 25. Entre os lipídios utilizados nessetrabalho, a relação AGP+AGM/AGS foi maior no grupo CA(11%) e menor no grupo BG (2%), o que pode indicar que exis-te uma relação entre AGP+AGM/AGS e HDL-C. O conteúdode AGP foi menor nos grupos BG (15%) e CA+BG (21%), rela-tivamente elevado no grupo CA (33%) e mais alto no grupo S(61%). A relação de AGP/AGS no óleo de soja foi de 4,4 e noóleo de canola de 4,1, revelando que não há nenhuma relaçãoentre AGP/AGS e lipídios séricos de ratos. A relação AGP/AGS dos grupos S e CA foi cinco vezes maior do que a rela-ção AGP/AGS nos grupos CA+BG (0,8) e BG (0,4).
Em relação à biometria dos animais, as dietas suple-mentadas com diferentes lipídios dietéticos influenciaram aPA e a RCC dos ratos aos 12 e 18 meses de idade. Já é sabidoque o tipo de lipídio na dieta pode afetar a PA. Dietas comgrande quantidade de gordura saturada podem aumentar aPA, enquanto que dietas com maior teor de gordura po-liinsaturada e monoinsaturada diminuem a PA 39,40. A dietarica em colesterol causa, primariamente, sobrecarga de pres-são 40, o que poderia explicar o aumento observado na PAdos animais alimentados com dieta suplementada com ba-nha de porco e gema de ovo. Neste estudo, apesar do con-teúdo de sódio das quatro dietas experimentais ser igual, osanimais do grupo BG apresentaram maiores valores da PAaos 12 meses e o aumento foi mais acentuado aos 18 mesesde idade. Os animais do grupo CA+BG, que aos 12 mesesnão apresentaram alteração da PA, aos 18 meses de idade,mostraram elevação da PA.
A massa corporal dos animais variou muito ao longodo tempo e os animais do grupo BG aos 12 meses foram maisleves que os demais animais e os do grupo S, os mais pesa-dos. Porém, aos 18 meses, essa diferença no peso corporaldeixou de existir.
Finalmente, podemos sugerir que não ocorre aumentoda PA e sim melhora do perfil lipídico de ratos alimentadospor longo tempo com dieta suplementada com óleo decanola, com elevação dos níveis de HDL-C em comparaçãocom os outros tipos de dieta.
Agradecimentos
Ao Prof. José Firmino N. Neto (Chefe do LaboratórioCentral do Hospital Universitário Pedro Ernesto da UERJ)por seu auxílio na análise bioquímica do sangue dos ratos.Este trabalho foi financiado parcialmente pelo CNPq eFAPERJ.
1. Schaefer EJ, Levy RI, Ernest ND, Van Sant F, Brewew HB Jr. The effects of lowcholesterol high polyunsaturated fat, and low fat diets on plasma lipid andlipoprotein cholesterol levels in normal and hypercholesterolemic subjects. AmJ Clin Nutr 1981; 34: 1758-63.
2. Hopkins PN. Effects of dietary cholesterol on serum cholesterol: a meta-analysisand review. Am J Clin Nutr 1992; 32: 1281-9.
3. Mensink RP, Katan MB. Effect of fatty acids on serum lipids and lipoproteins: ameta-analysis of 27 trials. Arterioscl Thromb 1992; 12: 911-9.
4. Kromhout D, Bosschteter EB, Coulander CL. The inverse relation between fishconsumption and 20-year mortality from coronary heart disease. N Engl J Med1985; 312: 1205-9.
5. Goodnight SH, Cairns JA. Therapeutic use of n-3 fatty acids for vascular diseaseand thrombosis. Chest 1995; 108: 302S-4S.
6. Stone NJ. Fish consumption, fish oil, lipids, and coronary heart disease.Circulation 1996; 94: 2337-40.
7. O’Keefe JH, Harris W. From Inuit to implementation: omega-3 fatty acids come ofage. Mayo Clin Proc 2000; 75: 607-14.
8. McLennan PL, Dallimore JA. Dietary canola oil modifies myocardial fatty acidsand inhibits cardiac arrhythmias in rats. J Nutr 1995; 125: 1003-9.
9. Salem N Jr, Wegher B, Mena P, Uauy RD. Arachidonic and docosahexaenoic acidsare biosynthesized from their 18-carbon precursors in human infants. Proc NatlAcad Sci USA 1996; 93: 49-54.
10. McLennan PL. Relative effects of dietary saturated, monounsaturated, andpolyunsaturated fatty acids on cardiac arrhythmias in rats. Am J Clin Nutr 1993;57: 207-12.
11. Connor WE. The impact of dietary omega-3 fatty acids on the synthesis andclearance of apo b and chylomicrons. Proceedings of the Scientific Conference onOmega-3 fatty acids in Nutrition, Vascular Biology, and Medicine, Houston,Texas, 1994: 19-32.
12. Berliner JA, Navab M, Fogelman AM, et al. Atherosclerosis: Basic mechanisms,oxidation, inflammation, and genetics. Circulation 1995; 91: 2488-96.
13. Stein Y, Stein O, Dabach Y, Hollander G, Ben-Naim M, Halperin G. Anti-atherogenicity of high-density lipoprotein. Isr J Med Sci 1996; 32: 503-8.
14. Schaefer EJ, Lichtenstein AH, Lamon-Fava S, McNamara JR, Ordovas JM.Lipoproteins, nutrition, aging, and atherosclerosis. Am J Clin Nutr 1995;61(suppl): 726S-40S.
15. Assmann G, Schulte H, von Eckardstein A, Huang Y. High-density lipoproteincholesterol as a predictor of coronary heart disease risk. The PROCAM experien-ce and pathophysiological implications for reverse cholesterol transport.Atherosclerosis 1996; 124: S11-S20.
16. Batra S, Rakusan K. Capillary length, tortuosity, and spacing in rat myocardiumduring cardiac cycle. Am J Physiol 1992; 32 (Heart Circulation Physiol 32):H1369-H76.
Referências

Arq Bras Cardiol2002; 78: 25-31.
Aguila e colsMetabolismo lipídico de ratos
3 13 13 13 13 1
17. Cleutjens JPM, Kandala JC, Guarda E, Guntaka RV, Weber T. Regulation ofcollagen degradation in the rat myocardium after infarction. J Mol Cell Cardiol1995; 27: 1281-92.
18. Katstura J, Cheng W, Sarangarajan R, et al. Necrotic and apoptotic cardiomyocy-te cell death in the aging heart of Fisher 344 rats. Am J Physiol 1996; 271 (HeartCirculation Physiol 40): H1215-H28.
19. Kalisnik M, Porenta OV, Mattfeldt T. Stereological and morphometric studies ofmammalian myocardium: a review. Acta Stereol 1998; 17: 389-401.
20. Campbell SE, Gerdes AM, Smith TD. Comparison of regional differences incardiac cardiomyocyte dimensions in rats, hamsters, and guinea pigs. Anat Rec1986; 219: 53-9.
21. Gill III TJ, Smith GJ, Wissler RW, Kunz HW. The rat as an experimental animal.Science 1989; 21: 269-76.
22. Aguila MB, Apfel MIR, Mandarim-de-Lacerda CA. Stereology of the myocar-dium and blood biochemistry in aged rats fed with a cholesterol-rich and canolaoil diet (n-3 fatty acid rich). Basic Res Cardiol 1998; 93: 182-91.
23. Aguila MB, Mandarim-de-Lacerda CA. Numerical density of cardiac cardiomy-ocytes in aged rats fed a cholesterol-rich diet and a canola oil diet (n-3 fatty acidrich). Virchows Arch 1999; 434: 451-453.
24. Aguila MB, Mandarim-de-Lacerda CA. Effect of different high-fat diets on themyocardium stereology and blood pressure in rats. Pathol Res Pract 2000; 196:841-846.
25. Aguila MB, Mandarim-de-Lacerda CA. Blood pressure, ventricular volume andnumber of cardiomyocyte nuclei in rats fed for 12 months on diets differing in fatcomposition. Mech Ageing Dev 2001; 122: 77-88.
26. Scherle W. A simple method for volumetry of organs in quantitative stereology.Mikroskopie 1970; 26: 57-63.
27. Chen PR, Tsai CE. Various high monounsaturated edible oils might affect plasmalipids differently in man. Nut Res 1995; 15: 615-621.
28. Friedwald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-
density lipoprotein cholesterol in plasma, without use of preparativeultracentrifuge. Clin Chem 1972; 8: 99-502.
29. Matthews DE, Farewell V. Using and understanding medical statistics. Basel:Karger, 1996; 246.
30. Zar JH. Biostatistical analysis. Upper Saddle River: Prentice Hall, 1999; 663.31. Reeves PG, Nielsen FH, Fahey GC. AIN-93 purified diets for laboratory rodents:
final report of the American Institute of Nutrition Ad Hoc Writing Committee onthe Reformulation of the AIN-76A rodent diet. J Nutr 1993; 123: 1939-51.
32. Lacko AG, Davis JL. Age-related changes in rat and primate plasma cholesterolmetabolism. J Am Geriatr Soc 1979; 5: 212-7.
33. Tucker JT. Diseases of the Rat. Gunpower Square, London: Taylor & Francis,1997: 1-22.
34. Baba NH, Antoniades K, Habbal Z. Effects of dietary canola, olive, and linolenicacid enriched olive oils on plasma lipids, lipid peroxidation and lipoproteinlipase activity in rats. Nutr Res 1999; 19: 601-12.
35. Kobatake Y, Hirahara F, Innami S, Nishide E. Dietary effect of n-3 type po-lyunsaturated fatty acids on serum and liver lipid levels in rats. J Nut SciVitaminol 1983; 29:11-21.
36. Becker N, Illingworth DR, Alaupovic P, Connor W, Sundberg E. Effects ofsaturated, monounsaturated and w-6 polyunsaturated fatty acids on plasma lipids,lipoproteins and apoproteins in humans. Am J Clin Nutr 1983; 37: 355-360.
37. Mattson FH, Grundy SM. Comparison of the effects of dietary saturated,monounsaturated, and polyunsaturated fatty acids on plasma lipids andlipoproteins in man. J Lipid Res 1985; 26: 194-202.
38. Huang PC, Chang NW. Effects of dietary monounsaturated fatty acids on plasmalipids in humans (Abstract). 14th International Congress of NutritionalSciences, 1989.
39. Iacono JM, Dougherty RM. Blood pressure and fat intake. In: Laragh J, Brener BM(Eds). Hypertension, Physiology, Diagnosis and Treatment. New York: RavenPress, 1990: 257-76.
40. Pietinen P. Dietary fat and blood pressure. Ann Med 1994; 26: 465-8.
Editor da Seção de Fotografias Artísticas: Cícero Piva de AlbuquerqueCorrespondência: InCor - Av. Dr. Enéas C. Aguiar, 44 - 05403-000 - São Paulo, SP - E-mail: dclcicero@incor. usp.br
Igarapé do Rio Tapajós - PA Simone Augusta de Oliveira - SP