mauro sérgio ywazaki efeito da loperamida e do farelo de ...§ão mauro.pdf · influência dos...
TRANSCRIPT

0
Programa de Pós-graduação em Ciência Animal
Mestrado
Mauro Sérgio Ywazaki
Efeito da loperamida e do farelo de trigo na dieta de leitões
recém desmamados sobre o controle da diarréia e a
maturação intestinal
LONDRINA - PR
2009

1
Mauro Sérgio Ywazaki
Efeito da loperamida e do farelo de trigo na dieta de leitões recém
desmamados sobre o controle da diarréia e a maturação intestinal
Dissertação apresentada ao curso de Pós-Graduação,
em Ciência Animal, da Universidade Estadual de
Londrina, como requisito parcial à obtenção do título de
Mestre.
ORIENTADOR: Prof. Dr. Caio Abércio da Silva
LONDRINA - PR
2009

2
Candidato: Mauro Sérgio Ywazaki
Título da dissertação:
Efeito da loperamida e do farelo de trigo na dieta de leitões recém
desmamados sobre o controle da diarréia e a maturação intestinal
Comissão examinadora:
Prof. Dr. Caio Abércio da Silva (Orientador) – Universidade Estadual de Londrina
Profa. Dra. Nilva Aparecida Nicolao Fonseca – Universidade Estadual de Londrina
Profa. Dra. Maria Marta Loddi – Universidade Estadual de Ponta Grossa
LONDRINA - PR
2009

3
Dedico este trabalho aos meus pais por abdicarem da minha companhia pelos estudos, além do mais concedendo todo apoio e as condições necessárias para a minha melhor formação.

4
AGRADECIMENTOS
Agradeço a Deus pelos desafios pelos quais passei em minha vida, que
ajudaram no meu amadurecimento, e pelas alegrias que vivi permitindo ter
esperanças e sonhos.
Agradeço aos meus pais pelo apoio constante e eterno.
Agradeço aos meus irmãos, Valter e Cristina, pelo carinho sempre presente.
Agradeço à Sthefânia pelo companheirismo e dedicação ao nosso amor e
toda sua familia que me deram conselhos em todos os sentidos.
Agradeço a todos da minha família que fazem parte da minha vida.
Agradeço à todos mestres (professores) que passaram por mim, mediando o
conhecimento, abrindo oportunidades e mostrando os caminhos.
Agradeço em especial ao Caio, não somente pela sua qualidade de professor
(transmitindo seus conhecimentos e experiências), mas também pela sua amizade
que compreendeu e incentivou o meu caminho, e com certeza meus maiores
agradecimentos e respeito serão poucos do que muito que foi oferecido.
Agradeço a todos os meus amigos (do colégio, da faculdade, da pós
graduação, da Espanha...) que, mesmo distantes, a amizade nunca morrerá.
Agradeço também aos animais, que me levaram a escolher esta profissão
que amo e respeito.
Agradeço a Universidade Estadual de Londrina e a Universidade Autonoma
de Barcelona pelo curso de pós graduação.
Muito Obrigado !!!

5
“Se um dia tiver que escolher entre o mundo e o
amor... lembre-se: se escolher o mundo ficará
sem o amor, mas se escolher o amor, com ele
conquistará o mundo.”
(Albert Einstein)

6
Sumário
ABREVIAÇÕES UTILIZADAS ............................ ....................................................... 9
LISTA DE TABELAS .................................. ............................................................. 10
LISTA DE FIGURAS .................................. .............................................................. 11
RESUMO ................................................................................................................. 12
ABSTRACT .......................................... ................................................................... 13
1- INTRODUÇÃO ..................................................................................................... 14
2. REVISÃO DE LITERATURA .......................... ...................................................... 15
2.1 - A característica do desmame precoce ...............................................................15
2.2 - Influência da nutrição na microbiologia intestinal .............................................16
2.3 - Influência da nutrição no desenvolvimento do trato digestivo ........................17
2.4 - Importância dos ácidos graxos de cadeia curta ................................................18
2.5 - DIETA COM SUPLEMENTAÇÃO DE FIBRA ....................................................20
2.5.1 - Definição ..............................................................................................................20
2.5.2 - Classificação .......................................................................................................20
2.5.3 - Digestibilidade ....................................................................................................21
2.5.4 - Tempo de trânsito do conteúdo digestivo ......................................................22
2.5.5 - Substrato de fermentação.................................................................................23
2.5.6 - Mudanças nas propriedades físico químicas do conteúdo digestivo .........24
2.6 - LOPERAMIDA ........................................................................................................24
2.6.1 - Indicação .............................................................................................................24
2.6.2 - Mecanismo de ação ...........................................................................................25
2.6.3 - Patofisiologia do pós desmame .......................................................................26
3 - REFERÊNCIAS .................................................................................................. 28
4 - OBJETIVOS ..................................... ................................................................... 35
4.1 - Objetivo geral .........................................................................................................35
4.2 - Objetivo específico ................................................................................................35
5 - ARTIGO PARA PUBLICAÇÃO ........................ ................................................... 36

7
RESUMO ................................................................................................................. 37
ABSTRACT .......................................... ................................................................... 38
1. INTRODUÇÃO ..................................................................................................... 39
2. MATERIAL E MÉTODOS ............................. ........................................................ 41
2.1 Animais e estabelecimento .....................................................................................41
2.2 Procedimentos experimentais e de amostragem ................................................41
2.3 Procedimentos analíticos ........................................................................................42
2.4 Análises estatísticas ................................................................................................43
3. RESULTADOS ..................................... ................................................................ 45
3.1 Experimento I ............................................................................................................45
3.1.1 Desempenho animal, padrão sanitário e digestibilidade dos nutrientes ......45
3.1.2 Fermentação dos produtos finais e mudanças quantitativas das populações
microbianas nas fezes ................................................................................................................46
3.2 Experimento 2 ...........................................................................................................46
3.2.1 Digestão e morfometria da mucosa intestinal ..................................................46
3.2.2 Características físico-químicas, parâmetros fermentativos e população
microbiana do conteúdo digestivo no cólon ............................................................................47
4. DISCUSSÃO ........................................................................................................ 48
4.1 A influência do farelo de trigo na adaptação dos leitões após o desmame ....48
4.2 A influência da loperamida na adaptação dos leitões frente a dieta ................50
5. CONCLUSÕES .................................................................................................... 53
6. RECONHECIMENTOS ......................................................................................... 54
7. REFERÊNCIAS .................................................................................................... 63

8
Esta dissertação corresponde à parte de um projeto de pesquisa que resultará numa
tese de doutorado do Departamento de Alimentos e Produção Animal da
Universidade Autônoma de Barcelona (UAB), Espanha.
Na qualidade de membro colaborador do grupo, por ocasião de nossa estada na
supracitada instituição, assumimos a responsabilidade do presente trabalho com a
orientação do Prof. Dr. Caio Abércio da Silva e participação dos alunos de pós-
graduação e professores do Departamento de Alimentos da UAB.

9
ABREVIAÇÕES UTILIZADAS
AGCC: ácidos graxos de cadeia curta
AGV: ácidos graxos voláteis
CAD: capacidade de água disponível
CRF: fator de liberação de corticotrofina
E. coli: Escherichia coli
FAO: organização das nações unidas para a agricultura e alimentação
FDA: fibra em detergente ácido
FDN: fibra em detergente neutro
FNT-α: fator-alfa de necrose tumoral
FOS: frutoligossacarídeos
FT: farelo de trigo
GPD: ganho de peso diário
IL-1: interleucinas-1
IL-6: interleucinas-6
LDA: lignina em detergente ácido
LOP: loperamida
MO: matéria orgânica
MOR: receptores µ-opióide
MOS: mananoligosacarídeos
MS: matéria seca
PB: proteína bruta
PNA: polissacarídeos não amiláceos
SID: digestibilidade ileal aparente
SNC: sistema nervoso central
TGI: trato gastrointestinal
TMT: tempo mínimo de trânsito
WHO: organização mundial da saúde

10
LISTA DE TABELAS
Tabela 1. Composição e análises química da dieta…….……...…………………….. 55
Tabela 2. Desempenho zootécnico dos leitões recém desmamados....................... 56
Tabela 3. Mortalidade, leitões com diarréia e coeficientes de digestibilidade aparente
da matéria orgânica e proteína bruta, em leitões recém desmamados…………….. 57
Tabela 4. Concentração dos ácidos graxos de cadeia curta (AGCC) e população
bacteriana (enterobacteria e lactobacilli) nas fezes dos leitões 13 dias após
desmame.……………………………………..…………………………………………… 59
Tabela 5. Concentração de ácidos graxos de cadeia curta (AGCC) e ácido láctico e
população bacteriana (enterobactéria e lactobacilli) do conteúdo digestivo de cólon
em leitões após 15 dias de desmame.......……………………………………………... 60
Tabela 6. Coeficiente de digestibilidade aparente total da matéria orgânica e amido,
e análise histológica do jejuno e cólon dos leitões após 15 dias do desmame.….... 61

11
LISTA DE FIGURAS
Figura 1. Representação esquemática da composição dos carboidratos, fibra PNA
(Polissacarídeos não amiláceos), de acordo com o método de Van Soest (FDN: fibra
em detergente neutro; FDA: fibra em detergente ácido; LDA: Lignina em detergente
ácido) (Adaptado por Gerrits and Verstengen, 2006).................................................21
Figura 2. Tempo mínimo de trânsito (h) dos leitões…….…………….............………58
Figura 3. Capacidade de retenção de água (CRA) e capacidade de água disponível
(CAD) do conteúdo digestivo no cólon dos leitões.....................................................62

12
Efeito da loperamida e do farelo de trigo na dieta de leitões recém
desmamados sobre o controle da diarréia e a maturaç ão intestinal
RESUMO
O pós desmame na produção intensiva dos suínos é considerado um momento
crítico, sendo as infecções intestinais uma das maiores preocupações. Um dos
objetivos da nutrição dos leitões recém desmamados seria estimular o consumo,
propiciar o equilíbrio das populações microbianas do trato digestivo e promover o
desenvolvimento da morfologia intestinal e, para que tais resultados possam ser
encontrados é fundamental o conhecimento da patofisiologia do pós desmame e a
influência dos compostos da dieta na morfofisiologia animal. Portanto no presente
trabalho foram conduzidos dois experimentos com leitões recém desmamados
utilizando o farelo de trigo (FT) como fonte de fibra insolúvel com a possível hipótese
de acelerar o ritmo de trânsito intestinal e o medicamento loperamida (LOP) para
retardar o ritmo de trânsito intestinal, sendo avaliados o desempenho zootécnico,
padrão sanitário, atividade microbiana, as populações bacterianas, as características
do conteúdo digestivo e a morfologia intestinal. No experimento I, 128 leitões recém
desmamados foram distribuídos aleatoriamente em um arranjo fatorial 2x2, inclusão
de FT (0 vs 40 g/kg) e administração da LOP (0 vs 0,07 mg/kg PV). No experimento
II, 24 leitões foram distribuídos em três tratamentos: uma dieta controle; dieta com
80 g/kg de FT e dieta com 80 g/kg de FT combinada com o tratamento LOP (FT-
LOP). No Experimento I, a LOP aumentou o consumo médio diário e o ganho de
peso médio diário dos animais (P = 0,001 e P = 0,007, respectivamente). O mesmo
resultado foi obtido quando o FT foi associado com a LOP. O grupo FT-LOP também
apresentou uma maior concentração de ácidos graxos de cadeia curta (P = 0,013),
ácido acético (P = 0,004), e ácido propiônico (P = 0,093). No entanto a inclusão do
FT reduziu a digestibilidade da matéria orgânica e da proteína bruta (P = 0,001) e
demonstrou uma tendência em diminuir a população de enterobactérias (P = 0,089).
No Experimento II o FT aumentou a concentração de ácido butírico (P = 0,086). As
considerações finais indicam que a inclusão de níveis moderados de FT pode
modificar a microbiota intestinal e associado com a LOP pode beneficiar a saúde
animal e o desempenho.
Palavras Chave: Fibra; Microbiota; Trânsito intesti nal; Suínos

13
Administration of loperamide and addition of wheat bran to the
diets of weaner pigs decreases the incidence of diarrhea and
enhances their gut maturation
ABSTRACT
Post weaning intensive pig production is considered a critical period, being the
intestinal infections one of the major problems. One of the goals of nutrition of newly
weaned piglets are stimulate consumption, providing a balance of the digestive tract
microbial populations and improving the development of intestinal morphology, and
to ensure that such results can be found is fundamental knowledge of the post
weaning pathophysiology and the influence of compound animal diet in
morphophysiology. Therefore in the present work were conducted two experiments
with newly weaned piglets using wheat bran (WB) as insoluble fiber source with
possible chance to accelerate the intestinal transit and loperamide (LOP) to slow the
intestinal transit, being evaluated the performance, health status, microbial activity
and population, physicochemical characteristics of the hindgut digesta and intestinal
morphology. In experiment I, a total of 128 early weaned pigs were randomly
distributed in a 2x2 factorial combination of WB inclusion (0 vs 40 g/kg) and LOP
administration (0 vs 0,07 mg/kg BW). For experiment II a total of 24 piglets were
allotted to three dietary treatments: a diet control (CT); a diet with 80 g/kg of WB and
the combination of 80 g/kg WB with treatment LOP (WB-LOP). In Experiment I, LOP
improved the ADFI and ADG of the animals (P = 0,001 and P = 0,007, respectively).
The same result was obtained when WB was combined with LOP. The WB-LOP
group also showed a higher concentration of SCFA (P = 0,013), acetic acid (P =
0,004), and propionic acid (P = 0,093). On the other hand WB inclusion reduced the
OM and CP digestibility (P = 0,001) and tended to decrease the enterobacteria
population (P = 0,089). In Experiment II WB increased the butyric acid concentration
(P = 0,086). The conclusion indicated that the inclusion of a moderate level of WB
can modify the intestinal microflora and activity combined with LOP may be beneficial
to animal health and performance.
Key words: Fibre; Intestinal transit; Microbiota; S wine

14
1- INTRODUÇÃO
Os primeiros dias após o desmame, que ocorre em torno de 21 a 28 dias de
idade, são críticos na produção intensiva de suínos, devido aos diversos fatores que
comprometem à saúde do leitão, dentre elas as mudanças sociais, ambientais e
nutricionais que ocorrem (Weary et al., 2008). Este conjunto de fatores pode alterar
o comportamento animal, determinando um consumo insuficiente de ração até um
estado de anorexia, que pode comprometer a integridade intestinal resultando nos
transtornos digestivos (Montagne et al., 2003).
Com o objetivo de minimizar os impactos negativos após o desmame, o uso
de antibióticos na ração animal tem sido a principal estratégia de controle. Entretanto
existe uma preocupação crescente quanto a este procedimento pelos riscos da
geração de resistência bacteriana com destaque para a Salmonella (Close, 2000),
um sério problema de saúde pública.
Muitos países tem adotado uma postura rigorosa com relação aos antibióticos
na produção animal. A comunidade européia baniu totalmente esta classe de
produto como promotor de crescimento. Outros países como o Brasil, proibiram e/ou
restringiram o uso de alguns antibióticos (Porfírio, 1998), baseados em normas
estabelecidas pelo Ministério da Agricultura e pelo Codex Alimentarius da
FAO/WHO.
A ausência ou restrição na administração de antibióticos na ração pode
permitir com mais facilidade o aumento na incidência de diversas desordens
intestinais nos animais, sendo constante a busca por produtos alternativos e novas
práticas de manejo (Hampson et al., 2001), especialmente de ordem alimentar,
criando condições para definição de um equilíbrio intestinal, capaz de manter a
estrutura e funcionalidade do sistema digestivo (Van Dijk et al., 1999).
Neste aspecto, dentre muitas propostas avaliadas, a incorporação na ração
para leitões recém desmamados, de fibra predominantemente insolúvel, demonstrou
efeitos positivos no desenvolvimento intestinal e nas funções digestivas (Wellock,
2007) com redução na proliferação de patógenos no trato (Bertschinger et al., 1983).
Portanto a adição de fibra insolúvel, em quantidades ponderáveis, na dieta de
leitões recém desmamados, pode melhorar a saúde animal e o desempenho
zootécnico

15
2. REVISÃO DE LITERATURA
2.1 - A característica do desmame precoce
O período pós desmame na suinocultura intensiva ocorre de forma precoce,
sendo considerada a etapa mais crítica na vida do leitão (Hampson et al., 2001).
Neste momento ocorrem diversas mudanças repentinas para o animal, tanto social,
como a separação da mãe e o contato com animais de outras leitegadas, quanto
ambiental e nutricional, pela mudança de uma dieta líquida ideal baseada em
lactose, lipídios e proteínas do leite, para uma dieta sólida baseada em diferentes
proteínas, lipídios e carboidratos de origem vegetal e animal. Neste particular, o leite
materno oferece também componentes “não nutritivos” que facilitam a digestão e
previnem as enfermidades, pois aportam proteção imunológica pelas
imunoglobulinas e modulam a população microbiana intestinal (Weary et al., 2008).
Este conjunto de alterações define-se como um momento de estresse com aumento
dos níveis do fator de liberação de corticotrofina (CRF) e cortisol séricos (Moeser et
al., 2007), que influenciam no comportamento animal, determinando um consumo
insuficiente ou até mesmo um estado de anorexia (Lallès et al., 2004) que repercute
no desequilíbrio intestinal.
Somado ao estresse do período, a imaturidade fisiológica do leitão, que pode
comprometer a funcionalidade gastrointestinal, com redução das atividades
enzimáticas, baixa capacidade de diminuição do pH gástrico, resultam no deficiente
aproveitamento dos nutrientes (Lallès et al., 2007) e no aparecimento de
enfermidades.
Estudos de prevalência de enteropatógenos em leitões recém desmamados
com diarréia, demonstraram que a população de Escherichia coli, foi o agente
bacteriano mais isolado nos animais em fase de creche no Brasil (Menin et al., 2008)
e demais países.
A E.coli se estabelece no trato gastrointestinal, logo após o nascimento,
entretanto, devido a uma imunossupressão ou injúria na barreira gastrointestinal do
hospedeiro, as cepas de E. coli podem causar um processo infeccioso limitado ou
até disseminado.

16
2.2 - Influência da nutrição na microbiologia intestinal
A colonização do trato gastrointestinal pelas populações bacterianas é
influenciada por diversos fatores tanto do hospedeiro quanto das próprias
populações microbianas. Com relação aos principais fatores do hospedeiro que
afetam no processo da colonização microbiana do trato digestivo estão as reações
imunológicas, a presença de receptores intestinais, a taxa de passagem do
conteúdo digestivo e as secreções gástricas (Stewart et al., 1993). O mecanismo de
ação da microbiota benéfica na prevenção da colonização de patógenos
oportunistas pode ocorrer através da competição dos receptores do epitélio, de
nutrientes, pela produção de substâncias antimicrobianas (bacteriocinas e ácidos
orgânicos), pela estimulação do sistema imune e através de mudanças no meio
ambiente intestinal decorrente da metabolização de alguns compostos (Mandalari et
al., 2008). A dieta é considerada a principal responsável por criar as condições
favoráveis para a obtenção do equilíbrio intestinal entre o hospedeiro e a microbiota
benéfica (Van Dijk et al., 1999).
Predominantemente nos suínos, a microbiota do trato digestivo difere
quantitativamente e qualitativamente ao longo do trato gastrointestinal, com maiores
concentrações de microrganismos no ceco e cólon (Simpson et al., 1999).
Na literatura é amplamente citada a importância da microbiota, devido a sua
participação no desenvolvimento da morfologia intestinal, na defesa contra bactérias
oportunistas, na modulação do sistema imune e na digestão de nutrientes
(Wostmann, 1996).
O surgimento de novas técnicas de biologia molecular, como os métodos
baseados na codificação do gene 16S rDNA, vem sendo utilizadas para identificar as
espécies bacterianas do trato digestivo de diversos hospedeiros, dentre eles o suíno
(Leser et al., 2002).
Durante o período de aleitamento os grupos bacterianos que predominam no
trato gastrointestinal dos leitões são dos gêneros Lactobacillus, Bifidobacterium e
Streptococcus, os quais podem utilizar os substratos do leite (Swords et al., 1993) e
geralmente colaboram para a saúde animal. Após o desmame e início do consumo
animal por uma dieta sólida, composta por ingredientes de origem vegetal, torna-se
complexa a compreensão das populações microbianas na promoção da saúde

17
intestinal, entretanto, pode-se dizer que o aumento da biodiversidade do
ecossistema intestinal previne a colonização de patógenos oportunistas
(Konstantinov et al., 2004). Alguns estudos com a adição de cepas de Lactobacillus
na dieta obtiveram efeitos positivos no desempenho animal (Konstantinov et al.,
2008) e diminuíram a população de Clostridium e Enterobactérias (Tortuero et al.,
1995).
Segundo Barrow et al. (1977), a colonização dos Lactobacillus no trato
gastrointestinal controla a população de Escherichia coli devido ao aumento na
produção de ácido láctico, que diminui o pH do estômago e do duodeno, embora
segundo Konstantinov et al. (2008) não tenha sido evidenciada uma diferença
significativa do valor do pH entre uma dieta contendo o probiótico e uma dieta
controle.
2.3 - Influência da nutrição no desenvolvimento do trato digestivo
Para a melhor compreensão da problemática do desmame precoce no nível
fisiológico do trato digestivo, é necessário reconhecer primariamente o importante
papel deste sistema na digestão e absorção.
No intestino grosso há uma intensa atividade microbiana, que atuam sobre os
componentes que não foram digeridos ou completamente absorvidos, como as
fibras, alguns lipídios, proteínas insolúveis e as secreções endógenas, sendo assim
aproveitados sob a forma de ácidos graxos voláteis e de vitaminas (Lallès et al.,
2007).
O hospedeiro contribui para a prevenção da colonização de bactérias
oportunistas mediante as barreiras físicas (estrutura das criptas e vilosidades) e as
barreiras químicas (secreção de mucinas, citoquinas e peptídeos antimicrobianos)
(Moal, 2006), porém após o desmame há uma deficiência destas barreiras
decorrente do baixo consumo de alimento. O resultado deste déficit refletirá na
deterioração da estrutura e funcionalidade digestiva com predisposição à má
absorção, desidratação, diarréia e infecções intestinais (Hampson et al., 2001),
levando a estase do crescimento. Neste período, através de análises histológicas,

18
verificou-se uma redução no tamanho da vilosidade e um aumento na profundidade
de cripta (Pluske et al., 1996).
As vilosidades intestinais, além de contribuírem na defesa contra uma
infecção, têm também como função aumentar a superfície de absorção dos
nutrientes previamente digeridos, sendo assim associadas como um importante
parâmetro morfológico de avaliação do desenvolvimento animal, como foi
demonstrado por Pluske et al. (1997), que observaram uma relação direta entre o
ganho de peso diário com o tamanho da vilosidade.
O aumento da profundidade da cripta está associado com o incremento do
número de células secretórias, que podem influenciar na patogênese da diarréia
(Pierce, 2005). No entanto, outros estudos observaram que o aumento da
profundidade de cripta pode ser devido à maior produção de ácidos graxos de
cadeia curta (AGCC) no intestino grosso, estimulando a multiplicação celular no
intestino delgado (Montagne et al., 2003).
2.4 - Importância dos ácidos graxos de cadeia curta
A fermentação do carboidrato no trato digestivo do suíno resulta na produção
de AGCC, ácido láctico e gases (hidrogênio, carbono, dióxido). Pesquisas
demonstraram que os produtos advindos da fermentação dos carboidratos em
monogástricos possuem diversas funções importantes para o desenvolvimento
animal.
Os principais AGCC que se destacam são o acetato, o propionato e o butirato,
os quais apresentam maiores concentrações no lúmen do cólon proximal devido à
intensa atividade microbiana que há neste local, sendo as bactérias dos gêneros
Lactobacillus e Bifidobacterium as principais responsáveis pelo metabolismo destes
componentes (Topping et al., 2001).
Pesquisas verificaram que os AGCC, principalmente o butirato, considerado
uma importante fonte de energia para as células epiteliais do intestino, é
responsável por funções como imunomoduladoras e antiinflamatórias (Luhrs et al.,
2002), e podem acelerar a proliferação das células epiteliais do intestino, aumentar a
resistência do hospedeiro contra os microrganismos oportunistas, inibir a

19
proliferação das bactérias patogênicas e estimular a absorção de água e sódio
(Krishnan et al. 1999), podendo assim, serem considerados também como agentes
anti-diarréicos.
Sabe-se que o crescimento dos microrganismos oportunistas, incluindo a E.
coli patogênica e a Salmonella, é favorecida pelo pH próximo da neutralidade,
enquanto o pH ácido é mais favorável para o desenvolvimento das bactérias
residentes, como os Lactobacillus (Drasar & Barrow, 1985). Esta correlação pode ser
observada através de um estudo in vitro em condições anaeróbicas que verificou
que altas concentrações de ácidos graxos voláteis (AGV) inibiram a presença da
Salmonella typhi (Kunte et al., 1998).
É importante salientar que em leitões lactentes o pH é baixo devido à
produção de ácido láctico. Assim as bactérias resistentes ao ambiente ácido
persistem e proliferam no intestino proximal, ocasionando maior proteção contra a
penetração de patógenos sensíveis ao meio ácido (Radecki & Yokoyama, 1991).
Entretanto, após o desmame, que está correlacionado com um baixo consumo, pode
ocorrer desestabilização desta proteção adquirida. Estudos em ratos demonstraram
que a restrição de alimento resultou em um aumento do pH do conteúdo cecal, o
qual foi correlacionado com as baixas concentrações de AGV (Illman et al., 1986)
Pesquisas com suínos, com cânulas no intestino, observaram que as
concentrações de AGV diminuem significativamente no período pós desmame,
correspondendo com um aumento de E. coli ileal e concomitantemente diminuição
das concentrações de Lactobacillus (Franklin, 2002).
Portanto, no período pós-desmame uma das estratégias de controle da
proliferação das bactérias patogênicas seria manter o ambiente gastrointestinal nas
condições ácidas através da dieta e da adição de agentes acidificantes ou por meio
da adição de substratos que ao serem metabolizados pela microbiota produzam os
AGV, principalmente os AGCC (Campbell et al., 1997).
Os acidificantes raramente afetam o pH abaixo da região proximal do intestino
delgado, entretanto, componentes da dieta que promovem a produção de AGV
podem afetar o pH nas regiões distais do trato intestinal, incluindo o ceco e o
intestino grosso (Campbell et al., 1997).
A determinação da concentração dos AGCC nas fezes não é
necessariamente representativa na região proximal do cólon, pois pode ser afetada
pelo tempo de trânsito e absorção intestinal. Segundo Buraczewska et al. (1988), o

20
grau de fermentação da fibra em detergente neutro (FDN) no intestino delgado de
suínos varia entre 10 a 32 %, dependendo da fonte de FDN, e a digestão da matéria
seca ocorre em torno de 14-39% no intestino grosso (Metz et al., 1985), sendo
rapidamente metabolizada e absorvida na forma de AGCC (Rechkemmer et al.,
1988), que pode representar uma fonte de energia para as atividades metabólicas.
2.5 - DIETA COM SUPLEMENTAÇÃO DE FIBRA
2.5.1 - Definição
O conceito da fibra dietética é definido como componentes da dieta de origem
vegetal que são resistentes às enzimas digestivas, não sendo absorvidos pelo
organismo animal (Kritchevsky, 1988).
Embora exista uma grande variedade de componentes não convencionais
associados com a fibra dietética, como as ceras, taninos, glicoproteínas, ácido fítico,
amido resistente, quitina, quitosano e até minerais, devido à baixa digestibilidade
destes componentes não fazem parte oficial da fibra dietética (Dreher, 1987).
2.5.2 - Classificação
As fibras podem ser classificadas em seu grau de polimerização:
polissacarídeos estruturais (celulose, hemicelulose, pectina e amido resistente),
polissacarídeos não estruturais (gomas e mucilagens) e compostos não
polissacarídeos como a lignina, amido e outros açúcares. E podem ser também
classificadas em relação ao seu grau de solubilidade em água como solúveis e
insolúveis, como demonstrado na Figura 1.

21
Figura 1 Representação esquemática da composição dos carboidratos, fibra PNA
(Polissacarídeos não amiláceos), de acordo com o método de Van Soest (FDN: fibra
em detergente neutro; FDA: fibra em detergente ácido; LDA: Lignina em detergente
ácido) (Adaptado por Gerrits and Verstengen, 2006)
2.5.3 - Digestibilidade
Os impactos dos diferentes tipos de fibra vão depender da sua solubilidade,
estrutura química e da composição dos açúcares.
O aumento nos níveis de fibra na dieta dos suínos reduz linearmente a
digestibilidade aparente ileal da matéria seca, dos minerais e da proteína.
A fibra influencia a digestibilidade dos outros constituintes da dieta, o que
pode ser parcialmente explicado por acelerar a taxa de passagem, diminuir a
superfície de contato das enzimas com o conteúdo digestivo (Knudsen et al., 1993),
e unir-se com os aminoácidos e peptídeos limitando a absorção destes compostos
(Sauer et al., 1991). Estes fatores podem explicar parcialmente o aumento da
excreção de nitrogênio, pois na literatura é citado que em uma dieta com fibras há
um aumento de perdas endógenas do elemento, incluindo mucos, descamação
epitelial, suco pancreático (Ikegami et al., 1990) e da própria população microbiana,
também responsável pela produção de nitrogênio. Apesar da diminuição da

22
digestibilidade dos nutrientes devido à adição de fibra na dieta, esta perda pode ser
compensada com um aumento no consumo de ração (Molist et al., 2009)
2.5.4 - Tempo de trânsito do conteúdo digestivo
O trato digestivo dos suínos pode ser dividido em três compartimentos: o
estômago, o intestino delgado e o intestino grosso. O estômago representa o maior
volume de todo o TGI e tem como função misturar, estocar, digerir o alimento e
controlar o esvaziamento para o duodeno, que é influenciado pelo tamanho da
partícula, viscosidade, osmolaridade e os constituintes da dieta como os lipídios,
proteínas, amido e os polissacarídeos não amiláceos (Anguita et al., 2007).
No período de desmame a taxa de esvaziamento gástrico é lenta devido ao
baixo consumo de alimentos associado com o estresse pós desmame (Lallès et al.,
2007).
O conceito do tempo de trânsito está correlacionado com a digestão,
absorção e fermentação microbiana no trato gastrointestinal (Van Soest et al., 1983),
embora seja possível que a característica física da dieta, como um alto teor de
lignina, participe no aumento da motilidade por estímulo mecânico. (Kass et al.,
1980).
As fibras predominantemente insolúveis, como o farelo de trigo, não têm uma
capacidade de retenção de água significativa como às fibras solúveis, porém ambas
aumentam o ritmo de trânsito e a massa fecal (Spiller et al., 1993). Sugere-se que o
farelo de trigo estimula a motilidade intestinal mecanicamente, independente da
capacidade de retenção de água (Tomlin & Read, 1988).
Em suínos, a fração de polissacarídeos não amiláceos (PNA) do trigo
aumentou a secreção pancreática em comparação com a fração amilácea
(Zebrowska & Low, 1987), embora não tenha sido claramente evidenciada uma
aceleração no tempo de trânsito. No trabalho de Bardon e Fioramonti (1983),
realizado com suínos alimentados com rações suplementadas com PNA, houve uma
aceleração no trânsito. Den Hartog et al. (1985) observaram que dietas com altos
níveis de PNA para suínos retardaram o trânsito no estômago e intestino delgado,
porém aceleraram no intestino grosso.

23
Malbert et al. (1994) sugerem que as concentrações de AGV ileal podem
afetar os padrões de esvaziamento gástrico e as contrações ileais nos suínos. Os
autores observaram que a adição de AGV no íleo diminuiu as contrações gástricas,
enquanto que aumentou as contrações ileais. É possível que a diminuição das
concentrações de AGV, observada em leitões recém desmamados possa provocar
um retardamento do alimento na região intestinal, contribuindo para a proliferação
dos patógenos (Mathew et al., 1996).
2.5.5 - Substrato de fermentação
A fermentação das fontes de carboidratos é a principal responsável na
produção do butirato, e devido à sua importante função para a manutenção da
saúde intestinal (Hamer et al., 2007), as indústrias alimentares se interessaram em
adicionar fontes de fibra na alimentação, principalmente para aumentar a
concentração de butirato no cólon. Porém, a quantidade produzida neste segmento
é dependente da estrutura química, como a solubilidade e o grau de polimerização
da dieta fibrosa.
As fibras insolúveis, como a celulose e a lignina, possuem baixo grau de
fermentação, mas estão associadas com um aumento da massa fecal e na
diminuição do tempo de trânsito no cólon. O farelo de trigo, em uma análise in vitro,
apresentou uma grande produção de ácido butírico (Edwards e Eastwood, 1992) e
de acordo com McIntyre et al. (1991), o produto é uma fonte de fibra de fermentação
lenta, podendo ter maior influência na região distal do cólon, ao contrário das fibras
de fermentação rápida. A composição química do farelo de trigo é constituída
principalmente por celulose insolúvel e arabinoxilanos (Selvendran, 1984).
As fibras solúveis se caracterizam por serem rapidamente degradadas no
cólon, são altamente fermentáveis e geram grandes quantidades de AGCC no
intestino (Rose et al., 2007) e seus efeitos metabólicos no trato gastrointestinal,
retardam o esvaziamento gástrico e o trânsito intestinal. Porém foram associados
com aumento na incidência das desordens entéricas (Hopwood et al., 2004). Alguns
exemplos de fontes de fibra solúvel são a oligofrutose, a inulina, o farelo de aveia e o
amido de milho.

24
2.5.6 - Mudanças nas propriedades físico químicas do conteúdo digestivo
As implicações nutricionais e fisiológicas dos complexos carboidratos são
dependentes das propriedades físico-químicas dos polissacarídeos. Solubilidade,
tamanho da partícula, capacidade de retenção de água são alguns fatores
relacionados com os efeitos da dieta fibrosa no trato gastrointestinal (Institute of
Food Technologists, 1989)
As propriedades físico-químicas do PNA são importantes para determinar
seus efeitos metabólicos (Low et al., 1986). O aumento da capacidade de retenção
de água pode estar correlacionado com o aumento na produção de AGCC (Molist et
al., 2009) e na difusão dos produtos da digestão até a superfície da mucosa (Dierick
et al., 1989).
2.6 - LOPERAMIDA
2.6.1 - Indicação
A loperamida é um derivado sintético da piperidina (C5H11N) (Einarson et al.,
2000) e um agonista opióide, sendo um medicamento de uso humano.
A loperamida conceitualmente é um medicamento utilizado para o tratamento
sintomático da diarréia, devido aos seus efeitos na diminuição da motilidade e
favorecimento do aumento na absorção dos solutos e água pelo epitélio. Além
destas propriedades clássicas, a propriedade do peptídeo opióide e a expressão dos
receptores µ-opióide periférico (MOR) pelas células envolvidas nas respostas
inflamatórias (Blalock, 1998) conduzem para novas pesquisas, demonstrando as
funções dos moduladores MOR, na regulação do sistema imune e reações
inflamatórias.

25
2.6.2 - Mecanismo de ação
A loperamida é um agonista dos receptores µ-opióide periféricos (MOR)
(DeHaven-Hudkins et al., 1999).
Estudos anatomicos, moleculares e eletrofisiológicos demonstraram que
existem três receptores opióides (µ, δ, κ) expressados dentro dos neurônios
sensoriais encontrados no sistema nervoso central (Mansour, 1995) e periférico
(Sternini, 2001) em animais e humanos, presentes em diversos tecidos, incluindo o
intestino (particularmente nos linfócitos plexos mioentéricos e submucosa) (Mccarthy
et al., 2001).
A loperamida atua na diminuição das atividades dos plexos mioentéricos
reduzindo a motilidade e a secreção dos músculos lisos circulares e longitudinais da
parede intestinal (Turnberg, 1983).
Recentes estudos demonstraram que a ativação destes receptores pelos
agonistas pode provocar diversas conseqüências, resultando em uma atenuação da
excitabilidade dos terminais periféricos da nocicepção e uma diminuição da
propagação das ações potenciais dos neurônios sensoriais. Tais mecanismos
incluem o aumento do nível de potássio e inibição da liberação do cálcio dependente
pelos precursores da pró-nocicepeção, e compostos pró-inflamatórios provenientes
das terminações dos nervos sensoriais periféricos. Portanto, podem ser
considerados não somente como anti-nociceptivo, mas também como
antiinflamatórios (Stein, 1995).
Diversas evidências reiteram a potencial atividade antiinflamatória pelos
agonistas periféricos µ e κ (Pierre, 2004).
Estudos in vivo evidenciaram que a ativação do MOR influenciou as funções
regulatórias do sistema imune relatados em diversos modelos animais com doenças
autoimunes (Sacerdote et al., 1999) e inflamatórias (Mousa et al., 2001).
Uma expressiva analgesia periférica pode ser obtida pela interação dos
receptores opióides periféricos com os peptídeos opióides liberados pelas células
imunes, o mecanismo da migração do conteúdo opióide das células para o tecido
inflamado está sob investigação. A finalidade do mecanismo de migração das
células utilizadas pelo sistema imune não é somente para combater os patógenos,
mas também para o controle da dor dentro dos tecidos. Portanto a dor pode ser

26
exacerbada pela inibição da imigração dos opióides produzidos pelas células (Stein,
1999).
Trabalhos recentes relataram que os efeitos analgésicos dos opióides não
ocorrem exclusivamente dentro do sistema nervoso central (SNC), como foi
preconizado há décadas atrás. Foi observado que os receptores opióides periféricos
dos nervos sensoriais são controlados durante o desenvolvimento inflamatório. A
ligação endógena dos peptídeos opióides são expressos pelas células residentes
imunes dentro do tecido periférico inflamado (Janson & Stein, 2003 ).
2.6.3 - Patofisiologia do pós desmame
É amplamente estabelecido que o consumo é um importante fator para o
desenvolvimento do sistema digestivo dos suínos (Lallès et al, 2004), mas após o
desmame, que é caracterizado por um período de estresse, este consumo pode ser
insuficiente (Montagne et al.,2003), contribuindo para o desenvolvimento de um
processo inflamatório intestinal (McCracken et al., 1999).
Sabe-se que estes fatores de estresse são frequentemente associados com o
início do desenvolvimento das enfermidades entéricas (Melin et al., 2004), mas a
fisiopatologia deste processo é desconhecida.
Moeser et al. (2007) demonstraram que após o desmame há um aumento nos
níveis dos hormônios CRF e cortisol na circulação sanguínea, sendo estes
associados com o estresse. Estudos in vitro demonstraram que o CRF aplicado
diretamente nos tecidos colônicos aumentaram a motilidade, a permeabilidade e a
secreção de íons, indicando uma ação direta do CRF nas células intestinais
(Saunders et al., 2002).
O comprometimento da mucosa intestinal e o aumento da permeabilidade,
típicos após o desmame, podem permitir a passagem dos antígenos (bactérias,
toxinas, e nutrientes alérgenos) do lúmen para a lâmina própria, podendo cruzar o
epitélio até os tecidos sub-epiteliais, induzindo a uma resposta imune frente a uma
infecção que desencadeará uma resposta inflamatória local (Berkes et al., 2003),
caracterizada pela liberação de citoquinas pró-inflamatórias, como as interleucinas-1
(IL-1), interleucinas-6 (IL-6), fator-alfa de necrose tumoral (FNT-α), os quais são

27
responsáveis pelo comportamento convalescente (Kent, 1996). De acordo com estes
resultados o pós desmame dos leitões está associado com um aumento das
citoquinas mRNA pró-inflamatórias presentes no seu intestino (Pié et al., 2004).
As citoquinas possuem efeitos diretos na motilidade e na secreção
gastrointestinal (Bueno, 2000), além de influenciar o hipotálamo gerando uma
desconexão da demanda metabólica pela regulação do consumo. É provável que a
anorexia ou o consumo insuficiente possa ser um sinal de defesa dada pela resposta
imune do hospedeiro, devido a maior seletividade das dietas, ou seja, selecionar o
alimento para minimizar o risco de infecção ou em busca de compostos anti-
infecciosos (Kyriazakis, 2003).
Estímulos de estresse e liberação dos fatores de corticotrofina e citoquinas
podem liberar os peptídeos opióides, provocando a analgesia local, enquanto que
uma supressão do sistema imune exclui estes efeitos como também inibe a síntese
ou a liberação das citoquinas (Stein et al., 1990). Estes resultados estão de acordo
com o conceito de que os peptídeos opióides endógenos podem ser secretados
pelas células imunes, a ocupação dos receptores opióides dos nervos sensoriais
e/ou a liberação dos neuropéptidos pró-inflamatório (Stein 1995). Os efeitos
analgésicos dos opióides periféricos podem ser observados principalmente sob
condições patológicas como a inflamação, neuropatia, distensão do cólon. (Gebhart
et al., 2006).

28
3 - REFERÊNCIAS
ANGUITA M., et. al. Effect of coarse ground corn, sugar beet pulp and w heat bran on the voluntary intake and physicochemical ch aracteristics of digesta of growing pigs. Livestock Science. v. 107, n. 2-3, p. 182-191, 2007.
BARDON, T.; FIORAMONTI, J. Nature of the effects of bran on digestive transit time in pigs. British Journal of Nutrition; v. 50, p. 685-690, 1983.
BARROW, P.A.; FULLER, A.; NEWPORT, M. J. Changes in the microflora and physiology of the anterior intestinal tract of pigs weaned at 2 days, with special reference to the pathogenesis of diarrhoea. Infection and Immunity, v. 18, p. 586-595, 1977.
BERKES, J., et. al. Intestinal epitelial responses to enteric pathogens : effects on the tight junction barrier, ion transport, and infl ammation. Gut, v. 52 439–451, 2003.
BERTSCHINGER, H. U., et. al. Role of nutrition in the pathogenesis of porcine Escherichia coli enterotoxaemia. Annals of Veterinary Research. v.14, n. 4, p. 469-472, 1983.
BLALOCK, J.E. Beta-endorphin in immune cells. Immunology Today. v.19, p. 191-192, 1998.
BURACZEWSKA, L., et al. Ileal and total digestibility of NDF and ADF of dif ferent feedstuffs in the pig. In: BURACZEWSKA L.; BURACZEWSKI S.; PASTUCZEWSKA B.; ZEBROWSKA T. (ed.) Digestive Physiology in the Pig. Jablonna, Poland. Proc. 4th Int. Seminar. Institute of Animal Physiology and Nutrition, 1988. p. 224-228.
CAMPBELL, J. M.; FAHEY G. C.; WOLF B. W. Selected indigestible oligosaccharides affect large bowel mass, cecal and fecal short chain fatty acids, pH and microflora in rats. Journal of Nutrition. v. 127, p. 130-136, 1997.
CLOSE, W. H. Producing Pigs without Antibiotic Growth Promoters. In______. Proceedings of the 2000 Banff Pork Seminar. University of Alberta, Faculty of Extension, Edmonton, Alberta. 2000. p. 47-56.
COELHO, A.M.; FIORAMONTI, J.; BUÉNO, L. Systemic lipopolysaccharide influences rectal sensitivity in rats: role of mast cells, cytokines, and vagus nerve. American Journal of Physiology - Gastrointestinal and Liver Physiology. v. 279, p. 781-790, 2000.
DEHAVEN-HUDKINS, D.L., et. al. Loperamide (ADL 2-1294), an opioid antihyperalgesic agent with peripheral selectivity. Journal of Pharmacology and Experimental Therapeutics. v. 289, p. 494-502, 1999.

29
DEN HARTOG, L. A., et al. Digestive Physiology in the Pig. In: Just A.; Jorgensen H.; Fernandez J.A. National Institute of Animal Science Publication, Copenhagen: National Institute of Animal Science n. 580, 1985. p. 199-202.
DIERICK, N., et al. A. Approach to the energetic importance of fibre diges tion in pigs. I. Importance of fermentation in the overall energy supply. Animal Feed Science Technology v. 23, p. 141, 1989.
DRASAR, B.S.; BARROW, P.A. Aspects of microbiology 10 In: Schlessinger, D. Intestinal Microbiology. American Society of Microbiology. Washington,D.C., 1985. p. 28-38.
DREHER, M. L. Dietary fiber and its link to disease. In: DREHER M.L. Handbook Dietary Fiber. An Applied Approach, New York and Basel: Marcel Dekker, Inc. 1987. p. 281-322.
EDWARDS, C. A.; EASTWOOD, M. A. Comparison of the effects of ispaghula and wheat bran on rat caecal and colonic fermentati on. Gut. v. 33, p. 1229-1233, 1992.
EINARSON A., et al. Prospective, controlled, multicentre study of loper amide in pregnancy. Canadian Journal of Gastroenterology. v. 14, n.3, 2000.
FRANKLIN, M. A, et al. Characterization of microbial populations and volat ile fatty acid concentrations in the jejunum, ileum, an d cecum of pigs weaned at 17 vs 24 days of age. Journal of Animal Science. v. 80, n.11, p. 2904-2910, 2002.
GEBHART, G.F. et. al. Peripheral opioid modulation of visceral pain. Issue New Medications For Drug Abuse, v. 909, p. 41 – 50, 2006.
GERRITS, W. J. J.; VERSTEGEN, M. W. El Papel De La Fibra Dietética En Alimentación Animal. In: FEDNA, 22. 2006 Barcelona, Spain. Proceedings of the XXII Curso De Especialización. Federación Española para el Desarrollo de la Nutrición Animal, 2006. p.19-35.
HAMER, H. M., et. al. Review article: the role of butyrate on colonic fun ction. Alimentary Pharmacology & Therapeutics. v. 27, p. 104-119, 2007.
HAMPSON, D. J.; PLUSKE, J. R.; PETHICK, D. W. Dietary manipulation of enteric disease. Pig News and Information. v.22, p. 21-28, 2001.
HOPWOOD, D.E. et al. Addition of pearl barley to a rice-based diet for n ewly weaned piglets increases the viscosity of the intes tinal contents, reduces starch digestibility and exacerbates post-weaning c olibacillosis. British Journal of Nutrition v.92, p.419-427, 2004. IKEGAMI, S., et. al. Effect of viscous indigestible polysaccharides on pancreatic-biliary secretion and digestive organs i n rats. Journal of Nutrition. v. 120, p. 353, 1990.

30
ILLMAN, B. J.; TOPPING D. L.; TRIMBLE R. P. Effects of food restriction and starvation refeeding on volatile fatty acid concent rations in the rat. Journal of Nutrition. v. 116, p. 1694-1700, 1986.
INSTITUTE OF FOOD TECHNOLOGISTS. Dietary fiber: A scientific status summary by the Institute of Food. Technologists' Ex pert Panel on Food Safety & Nutrition. Food Technology. p. 133-139, 1989.
JANSON, W., STEIN, C. Peripheral opioid analgesia. Current Pharmaceutical Biotechnology. v.4, p. 270–274, 2003.
KASS, M. L., et al. Utilization of dietary fiber from alfalfa by growin g swine. I. Apparent digestibility of diet components in specif ic segment of the gastrointestinal tract. Journal of Animal Science. v. 50, p. 175-191, 1980.
KENT, S., et al. Mechanisms of sickness-induced decreases in food-mo tivated behavior. Neuroscience Biobehavioral Reviews. v. 20, n.1, p. 171–175, 1996.
KNUDSEN B.; JENSEN K. E.; B. B; HANSEN, I. Digestion of polysaccharides and other major components in the small and large intes tine of pigs fed on diets consisting of oat fractions rich in b-D-glucan. British Journal of Nutrition, v. 70, p. 531-556, 1993.
KONSTANTINOV, S. R., et al. Microbial diversity studies of the porcine gastrointestinal ecosystem during weaning transitio n. Animal Research. v. 53, p. 317-324, 2004.
KONSTANTINOV, S.R., et. al. Feeding of Lactobacillus sobrius reduces Escherichia coli F4 levels in the gut and promotes growth of infected piglets. FEMS Microbiology Ecology Published. v. 66, n.3, p.599-607, 2008.
KRISHNAN, S.; RAMAKRISHNA, B. S; BINDER, H. J. Stimulation of sodium chloride absorption from secreting rat colon byshor t-chain fatty acids. Digestive Diseases Sciences. v. 44, p. 1924–1930, 1999.
KRITCHEVSKY, D. Dietary Fiber. Annual Review of Nutrition. v. 8, p. 30-328, 1988.
KUNTE, D. P., et. al. Inactivation of Salmonella typhi by high levels of volatile fatty acids during anaerobic digestión. Journal of Applied Microbiology. v. 84, p. 138-142, 1998.
KYRIAZAKIS, I. The control and prediction of food intake in sickne ss and in health. In: WISEMAN J.; M. A. VARLEY M.A.; KEMP B. Perspectives in Pig Science., U.K.: Nottingham University Press, 2003 . p. 381–403.
LALLÈS, J. P., et al. Gut function and dysfunction in young pigs. Physiology. Animal Research. v. 53, p. 301-316, 2004.
LALLÈS, J. P.; BOSI, P.; SMIDT, H.; STOKES, C. R. Weaning - A challenge to gut physiologists. Livestock Science. v. 108, n. 1-3, p. 82-93, 2007.

31
LESER, T. D., et. al. Culture-independent analysis of gut bacteria: the p ig gastrointestinal tract microbiota revisited. Applied and Environmental Microbiology. v. 68, p. 673-690, 2002.
LOW, A. G., et al. Influence of wheat bran, cellulose, pectin and low or high viscosity guar gum on glucose and water absorption from pig jejunum. Proceedings of the Nutrition Society, v. 45, p. 55, 1986.
LUHRS, H., et. al. Butyrate inhibits NF-kappaB activation in Lamina Pr opria Macrophages of Patients with Ulcerative Colitis. Scandinavian Journal of Gastroenterology, v. 37, p. 458-466, 2002.
MALBERT, C. H., et. al. Remote effects of ileo-colic SCFA levels on gastric motility and emptying. Proc. 6th Int. Symp. Digestive Physiology of Pigs. Bad Doberan, Germany, p. 283-286, 1994.
MANDALARI, G., et. al., Potential Prebiotic Properties of Almond (Amygdalus communis L.). Seeds Applied and Environmental Microbiology, v. 74, n. 14, p. 4264-4270, 2008.
MANSOUR, A., et al. Immunohistochemical localization of the cloned µ op ioid receptor in the rat CNS. Journal of Chemical Neuroanatomy. v.8, p. 283-305, 1995.
MATHEW, A. G., et. al. Effect of weaning on ileal short chain fatty acid concentrations in pigs. Nutrition Research. v. 16, p. 1689-1698, 1996.
MCCARTHY, L., et al. Expression of functional µ-opioid receptors during T cell development. Journal of Neuroimmunology. v. 114, p. 173-180, 2001.
MCCRACKEN, B.A., et al. Weaning anorexia may contribute to local inflammation in the piglet small intestine. Journal of Nutrition. v. 129, p. 613-619, 1999.
MCINTYRE, A., et. al. Differents fibers have different regional effects o n luminal contents of rat colon. Gastroenterology, v. 101 p. 1274–1281, 1991.
MENIN, A., et. al. Enteropathogenic bacterial agents in pigs of differ ent age groups and profile of resistance in strains of Esch erichia coli and Salmonella spp. to antimicrobial agents. Ciência Rural v. 38, n. 6, Santa Maria, Sept. 2008.
METZ, S. H. M.; DEKKER, R. A.; EVERTS, H. Digestive Physiology in the Pig. In JUST, A.; JORGENSEN, H.; FERNANDEZ, J.A. National Institute of Animal Science Publication Copenhagen: National Institute of Animal Science, n. 580, 1985., p. 227-230.
MOAL, V. L. L.; SERVIN, A. L. The front line of enteric host defense against unwelcome intrusion of harmful microorganisms: Muci ns, antimicrobial peptides, and microbiota. Clinical Microbiology Reviews, v. 19, p. 315-337, 2006.
MOESER, A.J. et. al. Stress signaling pathways activated by weaning medi ate intestinal dysfunction in the pig. American Journal of Physiology - Gastrointestinal and Liver Physiology. v.292, p. 173–181, 2007.

32
MOLIST, F., et. al. Effects of the insoluble and soluble dietary fibre on the physicochemical properties of digesta and the micro bial activity in early weaned piglets . Animal Feed Science and Technology, v. 149, p. 346–353, 2009.
MONTAGNE, L.; PLUSKE, J. R.; HAMPSON, D. J. A review of interactions between dietary fibre and the intestinal mucosa, an d their consequences on digestive health in young non-ruminant animals. Animal Feed Science and Technology, v.108, n. 1-4, p. 95-117, 2003.
MOUSA, S.A., et al. Beta-Endorphin-containing memory-cells and mu-opioi d receptors undergo transport to peripheral inflamed tissue. Journal of Neuroimmunology. v.115, p.71-78, 2001.
PIÉ, S., et al. Weaning is associated with an upregulation of expre ssion of inflammatory cytokines in the intestine of piglets. Journal of Nutrition. v. 134, p. 641–647, 2004.
PIERCE, K. M., et al. Dietary manipulation post weaning to improve piglet performance and gastro-intestinal health. Animal Science, v. 81, p. 347-356, 2005.
PIERRE J-M RIVIÈRE. Peripheral kappa-opioid agonists for visceral pain. British Journal of Pharmacology. v.141, ed. 8, p. 1331-1334, April, 2004.
PLUSKE, J. R.; WILLIAMS, I. H.; AHERNE, F. X. Maintenance of villous height and crypt depth in piglets by providing continuous nutrition after weaning. Animal Science, v. 62, p. 131-144, 1996.
PLUSKE, J.R.; HAMPSON, D.J.; WILLIAMS, I.H. Factors influencing the structure and function of the small intestine in the weaned p ig: a review. Livestock Production Science, v. 51, p. 215-236, 1997.
PORFÍRIO, T. A. Aspectos regulatórios dos resíduos de drogas de uso veterinário em carcaças. In______. Aditivos na produção de ruminantes. Aditivos na Produção de não-ruminantes. Fronteiras do Melhoramento Genético Animal. 35ª Reunião Anual da SBZ. Botucatu, 1998. p. 269-287.
RADECKI, S. V.; YOKOYAMA, M. T. Intestinal bacteria and their influence on swine nutrition. In: MILLER, E. R.; ULLREY, D.E.; LEWIS, A. Swine Nutrition. Stoneham: Butterworth-Heinemann, 1991. p. 439-447.
RECHKEMMER, G.; RÖNNAU, K.; VON ENGELHARDT, W. Fermentation of polysaccharides and absorption of short chain fatty acids in the mammalian hindgut. Comparative Biochemistry and Physiology 90A, p. 563-568, 1988.
ROSE, D. J., et. al. Influence of dietary fiber on inflammatory bowel di sease and colon cancer: importance of fermentation pattern. Nutrition Review, v. 65: p. 51–62, 2007.
SACERDOTE, P. et. al. Hypothalamic beta-endorphin concentrations are decreased in animal models of autoimmune disease. Annals of the New York Academy of Sciences. v. 876, p. 305-308, 1999.

33
SAUER, W. C., et al.The effect of source of fiber on the ileal and feca l amino acid digestibility and bacterial nitrogen excretion in growing pigs. Journal of Animal Science. v. 69, p. 4070, 1991.
SAUNDERS, P.R., et.al. Peripheral corticotropin-releasing factor induces diarrhea in rats: role of CRF1 receptor in fecal wa tery excretion. European Journal of Pharmacology v. 435, p. 231–235, 2002.
SELVENDRAN, R. R. The plant cell wall as a source of dietary fiber: c hemistry and structure. American Journal of Clinical Nutrition, v. 39, p. 320-337, 1984.
SIMPSON, J. M., et. al. Application of denaturant gradient gel electrophore sis for the analysis of the porcine gastrointestinal mi crobiota. Journal of Microbiological Methods. v. 36, n.3, p. 167-179, 1999.
SPILLER, G. A. ed. Handbook of Dietary Fiber in Human Nutrition. 2nd ed.n. Boca Raton, FL: CRC Press Inc., p. 419-438, 1993.
STEIN C., et. al. Peripheral opioid receptors mediating antinocicepti on in inflammation: Evidence for involvement of mu, delta , and kappa receptors. Journal Pharmacology and Experimental Therapeutics. v. 248, p. 1269–1275, 1989.
STEIN, C. The control of pain in peripheral tissue by opioids . New England Journal of Medicine. v. 332, p. 1685-1690, 1995.
STEIN, C.et al. Opioids from immunocytes interact with receptors on sensory nerves to inhibit nociception in inflammation. Proceedings of the National Academy Sciences. USA, v. 87, p. 5935-5939, 1990.
STERNINI, C. Receptors and transmission in the brain-gut axis: p otential for novel therapies. III. µ-opioid receptors in the ent eric nervous system. American Journal of Physiology - Gastrointestinal and Liver Physiology. v. 281, ed.1, p. 8-15, 2001.
STEWART, C.S., et al. Recent advances in probiosis in pigs: observations on the microbiology of the pig gut. In: GARNSWORTHY, P.C.; COLE D.J.A. Recent Advances in Animal Nutrition, Nottingham University Press, 1993. p. 197–220.
SWORDS, W. E., et. al. Postnatal changes in selected bacterial groups of the pig colonic microflora. Biology of the Neonate. v. 63, n. 3, p. 191-200, 1993.
TOMLIN J.; READ N. Laxative properties of indigestible particles. British Medical Journal, v. 297, p. 1175-1176, 1988.
TOPPING, D. L.; CLIFTON P. M. Short-chainfatty acids and human colonic function:roles of resistant starch and nonstarch po lysaccharides. Physiological Reviews. v. 81, p. 1031–64, 2001.
TORTUERO, F, et al. Response of piglets to oral administration of lacti c acid bacteria. Journal of Food Protection. v. 58, n. 12, p. 1369-1374, 1995.

34
TURNBERG, L.A. Antisecretory activity of opiates in vitro and in v ivo in man. Scandinavian Journal Gastroenterology Supplement. v. 84, p. 79-83, 1983.
VAN DIJK, J. E., MOUWEN, J. M.; KONINKX, J.F. Review on Histology and Absorptive Capacity of the Gastrointestinal Epithel ium. In: Nutrition and Gastrointestinal Physiology - Today and Tomorrow. Jansman, A. J. M.; J. Huisman, eds. TNO, Wageningen, Netherlands. p. 1-8. 1999.
VAN SOEST, P. J., UDEN, P.; WRICK, K. L. Nutrition Reports International. v.27, p. 17-28, 1983.
WEARY, D. M.; JASPER, J.; HÖTZEL, M.J. Understanding weaning distress. Applied Animal Behaviour Science, v.110, n. 1-2, p. 24-41, 2008.
WELLOCK I. J.; HOUDIJK J. G. M., KYRIAZAKIS I. Effect of dietary non-starch polysaccharide solubility and inclusion level on gu t health and the risk of post weaning enteric disorders in newly weaned piglets. Livestock Science, v. 108, n. 1-3, p. 186-189, 2007.
WOSTMANN, B.S. Immunology, including radiobiology and transplantat ion. VII. In: WOSTMANN, B.S., ed. Germfree and gnotobiotic animal models. Background and applications. Boca Raton, FL: CRC Press, 1996. p.101–25.
ZEBROWSKA, T.; LOW, A. G. The Influence of Diets Based on Whole Wheat, Wheat Hour and Wheat Bran on Exocrine Pancreatic Se cretion in Pigs. Journal of Nutrition. v. 117, p. 1212-1216, 1987.

35
4 - OBJETIVOS
4.1 - Objetivo geral
• Avaliar a influência do farelo de trigo e da loperamida aos leitões recém
desmamados.
4.2 - Objetivo específico
• Avaliar o desempenho zootécnico, padrão sanitário e a digestibilidade
aparente da matéria orgânica e da proteína ao longo trato digestivo do leitão.
• Avaliar o tempo mínimo de trânsito digestivo orofecal
• Quantificar as populações microbianas e a concentração dos ácidos graxos
voláteis no conteúdo digestivo do cólon proximal e nas fezes
• Avaliar as características histológicas e morfométricas da mucosa intestinal
• Avaliar as propriedades físico-químicas do conteúdo digestivo do cólon
proximal

36
5 - ARTIGO PARA PUBLICAÇÃO
Segundo as normas da revista British Journal of Nutrition
Efeito da loperamida e do farelo de trigo na dieta de leitões recém
desmamados sobre o controle da diarréia e a maturaç ão intestinal

37
Efeito da loperamida e do farelo de trigo na dieta de leitões recém desmamados
sobre o controle da diarréia e a maturação intestinal
RESUMO
Em dois experimentos com leitões recém desmamados foram avaliadas a influência
da inclusão do farelo de trigo e da loperamida no desempenho, padrão sanitário,
atividade microbiana e as populações, as características do conteúdo digestivo e a
morfologia intestinal. Nestes experimentos o farelo de trigo (FT) foi utilizado como
fonte de fibra insolúvel com a possível hipótese de acelerar o ritmo de trânsito
intestinal e o medicamento loperamida (LOP) para retardar o ritmo de trânsito
intestinal. No experimento I, 128 leitões recém desmamados foram distribuídos
aleatoriamente em um arranjo fatorial 2x2, inclusão de FT (0 vs 40 g/kg) e
administração da LOP (0 vs 0.07 mg/kg PV). No experimento II, 24 leitões foram
distribuídos em três tratamentos de dietas: uma dieta controle (CT); dieta com 80
g/kg de FT e a combinação de 80 g/kg de FT com o tratamento LOP (FT-LOP). No
Experimento I, LOP aumentou o consumo médio diário e o ganho de peso médio
diário dos animais (P = 0,001 e P = 0,007, respectivamente). O mesmo resultado foi
obtido quando o FT foi associado com a LOP. O grupo FT-LOP também apresentou
uma maior concentração de ácidos graxos de cadeia curta (P = 0,013), ácido acético
(P = 0,004), e ácido propiônico (P = 0,093). No entanto a inclusão do FT reduziu a
digestibilidade da matéria orgânica e da proteína bruta (P = 0,001) e demonstrou
uma tendência em diminuir a população de enterobactérias (P = 0,089). No
Experimento II, o FT aumentou a concentração de ácido butírico (P = 0,086). As
considerações finais indicam que a inclusão de níveis moderados de FT pode
modificar a microbiota intestinal e associado com a LOP pode beneficiar a saúde
animal e o desempenho.
Palavras-chave: Fibra; Microbiota; Suínos; Trânsito intestinal

38
Administration of loperamide and addition of wheat bran to the diets of weaner pigs
decreases the incidence of diarrhea and enhances their gut maturation
ABSTRACT
In two experiments with newly weaned piglets were evaluated the effect of the
inclusion of wheat bran and loperamide in animal performance, health, microbial
activity pattern and populations, the characteristics of content intestinal tract and
morphology. These experiments the wheat bran (WB) was used as a source of
insoluble fiber with possible chance to accelerate the intestinal transit and loperamide
(LOP) to slow the intestinal transit. In experiment I, a total of 128 newly weaned
piglets were randomly distributed in a 2 x 2 factorial arrangement, inclusion of WB (0
vs 40 g/kg) and administration of LOP (0 vs 0.07 mg/kg PV). In experiment II, a total
of 24 piglets were distributed on three treatments diets: a diet control (CT); diet with
80 g/kg WB and combination of 80 g/kg WB with treatment LOP (WB-LOP). In
experiment I, LOP improved the average daily feed intake and average daily gain of
the animals (P = 0.01 and p = 0,007, respectively). The same result was achieved
when the WB was associated with the LOP. WB-LOP group also submitted a greater
concentration of short-chain fatty acids (P = 0,013), acetic acid (P = 0,004), and
propionic acid (P = 0,093). However, the inclusion of WB reduced organic matter
digestibility and crude protein (P = 0,001) and showed a tendency to reduce the
population of enterobacteriaceae (P = 0,089). In experiment II, WB increased
concentration of butyric acid (P = 0,016). The final considerations indicate that the
inclusion of moderate levels of WB can modify the intestinal microbiota and
associated with the LOP can benefit animal health and performance.
Keywords: Fibre; Intestinal transit; Microbiota; Swine

39
1. INTRODUÇÃO
O desmame é uma fase crítica para os leitões. Este período de transição está
associado com a anorexia, a deterioração da funcionalidade digestiva e a
acumulação de alimentos não digeridos resultantes de uma ineficiente digestão nos
primeiros dias após desmame(1). Durante esta fase, os leitões são mais suscetíveis
aos quadros diarréicos, devido à proliferação e fixação de cepas de Escherichia coli
B-hemoliticas(2) na mucosa intestinal. Este transtorno digestivo, entretanto, pode ser
melhorado pela adição de fontes de fibra na dieta de leitões(3).
Uma explicação fisiológica sustenta que as fibras para os animais jovens constituem
a principal fonte de energia dietética para a microbiota intestinal e, portanto, podem
promover uma interação entre o hospedeiro e a microbiota entérica comensal(4). A
adição de fibras na dieta é capaz de reduzir a fermentação protéica do conteúdo
digestivo(5), normalizar a funcionalidade do cólon, do intestino delgado e da
morfologia da mucosa intestinal, embora ainda haja contradições nos efeitos dos
polissacarídeos não amiláceos (PNA) na saúde do leitão.
Sendo assim, alguns estudos demonstraram que a adição de fontes de PNA
predominantemente insolúvel ou de fermentação lenta(6) ou PNA solúvel que não
incrementa a viscosidade(7) reduz os sinais clínicos associados com as infecções e
aumenta a estrutura e a funcionalidade intestinal. Em contrapartida, dietas contendo
fontes de PNA solúveis promoveram aumento na viscosidade do conteúdo digestivo,
como por exemplo, a cevada sem casca ou a goma guar, que foram associadas com
o aumento na incidência das desordens entéricas(8).
Em estudos anteriores foi observado que a adição de farelo de trigo na dieta de
leitões recém desmamados promoveu uma mudança benéfica na colonização
microbiana do trato digestivo, com aumento na produção de butirato no intestino
grosso e menores contagens de enterobactérias no conteúdo digestivo no cólon(3) e
na mucosa intestinal(9). O farelo de trigo é uma fonte de PNA predominantemente
insolúvel, parcialmente resistente à degradação no trato gastrointestinal dos animais
monogástricos, com efeitos na redução do tempo de trânsito no intestino delgado e
intestino grosso(10;11). O butirato é considerado a principal fonte de energia para os

40
colonócitos e tem sua produção aumentada com a diminuição do tempo de trânsito
gastrointestinal(12) ou pela taxa de diluição in vitro(13).
A loperamida (LOP) é conhecida como medicamento capaz de aumentar o tempo de
trânsito intestinal. O princípio da LOP consiste basicamente em diminuir o
peristaltismo e a secreção de fluídos, resultando em maior tempo de trânsito e um
aumento na absorção dos fluídos e eletrólitos pelo trato TGI(14). Estes efeitos foram
densamente empregados para retardar o tempo de trânsito oro-cecal em
humanos(15), ratos (16) e também suínos (17). Baseado nestes estudos o objetivo do
trabalho foi confirmar os prováveis efeitos antagônicos no tempo de trânsito
digestivo pelo tratamento dos animais com loperamida e pela inclusão do farelo de
trigo na dieta para leitões recém desmamados, e avaliar os seus efeitos nos
aspectos produtivos, digestivos e microbiológicos com relação à mudança do tempo
de trânsito do conteúdo.

41
2. MATERIAL E MÉTODOS
2.1 Animais e estabelecimento
Dois experimentos foram realizados nas instalações de pesquisa animal da
Universidade Autônoma de Barcelona, sendo antes aprovados pelo Comitê de
Revisão dos Protocolos desta instituição. As condições de alimentação, supervisão,
instalação, desenvolvimento e sacrifício foram conformes as normas de
procedimentos da União Européia. No Experimento I, foram utilizados no total 128
leitões de linhagens comerciais ([Large White x Landrace] x Pietrain), os quais não
receberam o manejo de creep feed e foram desmamados aos 24 dias de idade com
peso médio de 6.4 ± 1.17 kg. Os leitões foram conduzidos de uma propriedade
comercial para a unidade experimental e alojados em 32 baias (4 animais/baia).
Cada baia possuía comedouros e bebedouros do tipo nipple para assegurar o
acesso à alimentação e água ad libitum. As baias foram divididas em quatro
tratamentos com 8 repetições por tratamento, em um arranjo fatorial 2×2 que
incluíram dois níveis de farelo de trigo (FT) na dieta (0 vs. 40 g/kg, Controle vs. FT,
respectivamente) e dois níveis na administração de loperamida (LOP) (0 ou 0.07
mg/Kg PV, logo 0 vs. LOP, respectivamente). Para o Experimento II, um total de 24
leitões com peso médio de 7.4 ± 1.17 kg de mesma origem genética, sexo e idade
como o experimento anterior, foram distribuídos aleatoriamente em 12 baias (2
animais/baia). As baias foram distribuídas em três tratamentos que incluíram uma
dieta controle (CT), dieta com adição de 8% FT e a adição de 8% FT associado à
administração de LOP (0.07 mg/Kg PV).
2.2 Procedimentos experimentais e de amostragem
No Experimento I os animais receberam as dietas desde o primeiro dia do
experimento até o 13º dia. A loperamida (Fortasec®, Esteve, Barcelona) foi
administrada via oral, diariamente pela manhã, para o grupo LOP e para o grupo

42
controle foi administrado água via oral, para simular a mesma intervenção entre os
tratamentos. Foram divididos em dois períodos experimentais (1ª semana e 2ª
semana) para registrar o peso vivo (PV), consumo de ração e o padrão sanitário dos
leitões. No 10º dia do experimento, 0.15% de óxido de cromo foram adicionados na
dieta para determinar a digestibilidade aparente total. No 12º dia do experimento,
amostras de fezes foram coletadas para determinar o cromo e a concentração de
AGCC, e contagem de lactobacilli e enterobacterias. No 12º dia do experimento,
0.25% de óxido de ferro foi incluído na dieta para a determinação do tempo mínimo
de trânsito (TMT), que corresponde ao tempo entre a administração e o
aparecimento do marcador vermelho nas fezes por baia(18). Para o Exp. II todos os
animais receberam as dietas experimentais durante 15 dias. No 15º dia do
experimento, os animais foram sacrificados com injeção intravenosa de pentobarbital
sódico (200 mg/kg PV). Os animais foram sangrados, e o abdômen foi
imediatamente aberto e todo o TGI foi atado e removido. Amostras do cólon
consistiam no conjunto de todos os conteúdos colônicos. Metade das amostras
coletadas foi congelada a nitrogênio líquido, e posteriormente secada a 103ºC para a
remoção completa de água. A outra metade foi dividida em quatro alíquotas: 3 g
foram coletadas, em tubo screw cap de 10 ml sendo anteriormente registrados os
seus pesos, para a análise da capacidade de retenção da água; o restante foi
recolhido em tubos para a realização de testes de capacidade de água disponível,
AGCC e populações microbianas. Segmentos de 4 cm da região mediana do jejuno
e de 4 cm da região mediana do cólon foram removidos, abertos longitudinalmente e
fixados por imersão em 10% (v/v) da solução tampão de formalina para estudo
histológico.
2.3 Procedimentos analíticos
As análises químicas e energéticas das dietas (Tabela 1) foram realizadas de acordo
com os procedimentos padrões da Associação Oficial das Análises Químicas(19). A
concentração do óxido crômico na ração e no conteúdo digestivo foi determinada
pela espectofotometria de absorção atômica conforme o método de Williams et
al.(20). A capacidade de retenção da água do conteúdo de cólon fresco foi

43
determinada pela centrifugação (2500 x g / 25 min), segundo Anguita et al.(21); a
capacidade de água disponível foi determinada com a razão da fase líquida para a
fase sólida obtida após o procedimento padrão de permanência de três horas em
temperatura ambiente do conteúdo digestivo fresco em tubos testes. O DNA das
fezes e do conteúdo do cólon foi extraído e purificado usando o Kit comercial
QIAamp DNA Stool Mini Kit (Qiagen, West Sussex, UK) com algumas modificações,
como descrito por Castillo et al.(22). As enterobacterias e lactobacilli foram
quantificados por PCR em tempo real usando SyBR Green dye segundo Castillo et
al.(22). A razão lactobacilli:enterobacteria foi calculada pela subtração do log 16S
rDNA gene lactobacilli copies/g Matéria Fresca (MF) menos log 16S rDNA gene
enterobacteria copies/g MF. As concentrações de ácidos graxos de cadeia curta e
ácido láctico foram determinadas por cromatografia gasosa após submeter as
amostras a um tratamento ácido-base, seguido pela extração em éter e derivação,
como descrito por Jensen et al.(23). Amostras de tecidos para estudo histológico
foram desidratadas e fixadas na cera da parafina, seccionadas a 4 µm e coradas
com hematoxilina e eosina. Parâmetros morfométricos foram realizados por
microscopia óptica (BHS, Olympus, Spain). Mensurou-se a altura das vilosidades e a
profundidade das criptas, e o número de células caliciformes nas criptas. As
medidas foram feitas em 10 criptas e 10 vilosidades íntegras de cada secção
intestinal de cada amostra. A altura da vilosidade e profundidade de cripta foram
medidas usando o micrometro ocular linear (Olympus, Ref 209-35040, Microplanet,
Barcelona, Spain). Com base na morfologia celular, diferenças entre as células
caliciformes e linfócitos foram claramente distinguíveis no aumento de 400x. A
densidade celular foi expressa pelo número de linfócitos por 1,000 µm2. Todas
análises morfométricas foram feitas pela mesma pessoa, a qual não tinha
conhecimento dos tratamentos.
2.4 Análises estatísticas
No Experimento I resultados do desempenho produtivo, contagem microbiana,
digestibilidade da matéria orgânica e da proteína bruta, TMT e AGCC nas fezes
foram submetidos à análise de variância utilizando o procedimento GLM(24). Os

44
dados foram analisados como um arranjo fatorial de 2 x 2 de tratamentos, com a
dieta e o tratamento LOP como os fatores, em quatro blocos casualizados. O
resultado do desempenho produtivo foi ajustado pelo peso vivo inicial através da
análise de covariância. No Experimento II resultados da digestibilidade da matéria
orgânica (MO) e amido, características físico-químicas, AGCC e ácido láctico e
população microbiana do conteúdo de cólon e a morfometria da mucosa intestinal
foram submetidos à análise de variância, considerando a dieta como fator de
classificação utilizando o procedimento GLM(24). Diferenças significativas foram
consideradas P<0,05. Tendências para 0,05<P<0,15 foram também consideradas.

45
3. RESULTADOS
3.1 Experimento I
3.1.1 Desempenho animal, padrão sanitário e digestibilidade dos nutrientes
Resultados do consumo e desempenho estão demonstrados na Tabela 2. Os leitões
que receberam o tratamento LOP apresentaram maior consumo diário (P = 0.001)
que o tratamento sem LOP. Diferenças foram mais evidentes nos animais
alimentados com a dieta com FT, demonstrando uma tendência da interação do FT
e a LOP durante a segunda semana (P = 0.070) e no período total (P = 0.069).
Efeito significativo dos tratamentos experimentais foi observado para o ganho de
peso médio diário dos animais durante a 1ª e 2ª semana após desmame. Os
tratamentos com LOP e FT determinaram aumento para o ganho de peso animal.
Para o grupo tratado com LOP e FT foi demonstrado o maior aumento no GPD
(interações entre FT e LOP, P = 0.047). Devido aos diferentes resultados do
consumo e GDP, os animais tratados com FT demonstraram melhor eficiência
alimentar (P = 0.013) durante as duas primeiras semanas, enquanto o grupo LOP
determinou melhor eficiência alimentar durante a 1ª semana (P = 0.005).
A Tabela 3 demonstra o número de leitões com diarréia e a taxa de mortalidade,
como também a digestibilidade da matéria orgânica e da proteína bruta (PB). O
tratamento LOP reduziu o número de animais que apresentaram diarréia (P = 0.029),
embora, não tenha sido encontrada diferença significativa na mortalidade entre os
tratamentos. A incorporação de FT na dieta reduziu a digestibilidade no trato
digestivo da MO (P = 0.001) e PB (P = 0.001). Por outro lado, o tratamento LOP
tendenciou a incrementar o coeficiente de digestibilidade da MO (P = 0.061) e
aumentou a digestibilidade da PB (P = 0.026), especialmente com a dieta FT (P
interação = 0.074 e 0.116 para a digestibilidade da PB e MO, respectivamente). Na
Fig. 3, está apresentada o tempo mínimo de trânsito (TMT) registrado no 13º dia
após o desmame. Não foram encontradas diferenças significativas entre os

46
tratamentos, entretanto, o tratamento LOP mostrou tendência (P = 0.070) em
aumentar o TMT, comparado com os demais tratamentos.
3.1.2 Fermentação dos produtos finais e mudanças quantitativas das populações
microbianas nas fezes
A concentração total de ácidos graxos de cadeia curta (AGCC) e a contagem
microbiana nas fezes estão demonstradas na Tabela 4. Uma interação significativa
foi observada entre os grupos FT e LOP (P = 0.013) na concentração total de AGCC.
A administração de LOP aumentou a concentração de AGCC nas fezes dos leitões
alimentados com FT, enquanto nenhuma diferença foi observada no grupo CT. O
tratamento LOP aumentou a concentração de ácido acético (P = 0.004) e indicou
aumentar o ácido propiônico (P = 0.093) nos leitões suplementados com FT.
Também promoveu incremento (P = 0.009) do ácido butírico em ambos os grupos de
tratamento (CT e FT). No entanto, leitões suplementados com FT na dieta
mostraram tendência em reduzir (P = 0.062) as concentrações de isoácidos. Os
leitões suplementados com FT na dieta também apresentaram capacidade (P =
0.089) de diminuir a contagem de enterobactéria nas fezes comparados com a dieta
CT. Não foram observadas diferenças significativas na contagem de lactobacilli ou
associação com o tratamento LOP.
3.2 Experimento 2
3.2.1 Digestão e morfometria da mucosa intestinal
A digestibilidade dos nutrientes e os resultados histológicos estão demonstrados na
Tabela 6. O tratamento com LOP aumentou (P = 0.010) a digestibilidade da MO e
também promoveu mudanças significativas na morfometria do jejuno, com aumento
(P = 0.031) na razão altura vilosidade – profundidade de cripta, comparado com a

47
dieta FT. Nenhuma outra diferença significativa foi observada em qualquer das
variáveis estudadas no jejuno. No cólon não foram observadas diferenças
significativas entre os tratamentos na morfometria e nos parâmetros celulares.
3.2.2 Características físico-químicas, parâmetros fermentativos e população
microbiana do conteúdo digestivo no cólon
Resultados relacionados com as características físico-químicas do conteúdo
digestivo (capacidade de retenção de água e capacidade de água disponível) estão
apresentados na Fig. 4. Animais alimentados com FT e LOP aumentaram (P =
0.001) a capacidade de retenção de água comparada com a dieta controle. No
entanto, animais alimentados com FT demonstraram as menores concentrações (P
= 0.018) da capacidade de água disponível comparada com a dieta controle. A
concentração dos AGCC e a contagem microbiológica do conteúdo digestivo estão
apresentadas na Tabela 5. Não foram encontradas diferenças significativas entre os
tratamentos nas concentrações do ácido acético, propiônico, isoácidos e láctico,
embora a concentração do ácido butírico indicou tendência de aumento (P = 0.086)
nos animais tratados com FT comparado com a dieta controle. Não foram
encontradas diferenças significativas na população de enterobactérias. Nenhum
outro resultado significativo foi observado na contagem dos lactobacilli e na razão
lactobacilli:enterobacteria.

48
4. DISCUSSÃO
4.1 A influência do farelo de trigo na adaptação dos leitões após o desmame
A fibra tornou-se um dos principais ingredientes da dieta para a associação com a
nutrição de animais jovens. Estudos demonstraram que a adição de fontes de PNA,
predominantemente insolúvel de baixa fermentação (como o farelo de aveia), na
dieta de leitões recém desmamados pode diminuir a incidência de diarréias e
melhorar o desempenho animal (6;25). A razão dos possíveis efeitos, contudo, é ainda
polêmico, mas os pesquisadores sugerem que possa haver mudanças na
quantidade e na atividade metabólica através da seleção dos componentes pela
microbiota intestinal.
O FT (a membrana externa do grão de trigo) foi escolhido para este estudo devido
às altas proporções do PNA como a celulose insolúvel e arabinoxilanos(26) e pelo
tamanho maior da partícula. Os resultados demonstraram que os animais
alimentados com o tratamento FT apresentaram redução na digestibilidade aparente
total da MO e PB, contudo melhoraram a eficiência alimentar. Diversos trabalhos
relataram que as dietas com fibra reduziram a digestibilidade total das proteínas e
energia. Na prática, os componentes da fibra diluem os nutrientes da ração devido à
fração PNA ser digerida em menor tempo que outras frações, como os amidos, PB
ou lipídios (27;28). Além disso, mudanças nas características físicas do conteúdo
intestinal decorrentes da presença dos componentes específicos da fibra podem
aumentar a viscosidade, influenciar no esvaziamento gástrico ou diminuir a difusão
ou mobilidade das enzimas, substratos e nutrientes e menor superfície de absorção.
A consequência é que a fibra pode reduzir a digestibilidade dos lipídios(29) ou
aumentar a excreção de N endógeno(30). Este efeito foi quantitativamente descrito
por Le Goff et al.(31). Os resultados encontrados verificaram que a fração FDN é
significativa no coeficiente de energia, com aproximadamente 0.1% redução por 1 g
FDN/kg MO.

49
Na literatura, é conceitualmente aceito que o efeito da fibra na dieta dos animais em
crescimento pode também reduzir o consumo voluntário e o GPD. No presente
estudo os leitões alimentados com FT não diminuíram o consumo e mostraram
melhor eficiência alimentar. Alguns autores (3;6;32) relataram que níveis moderados de
FT ou casca de aveia na dieta pós desmame aumentaram o consumo animal. Os
autores sugerem que os animais jovens necessitam de um mínimo de fibra para uma
correta funcionalidade do trato digestivo. Porém, não se deve excluir a possibilidade
que o aumento da eficiência alimentar, observado no presente estudo, poderia ser
devido, parcialmente, ao aumento da massa dos órgãos internos, possivelmente
pelo maior peso dos conteúdos intestinais(33).
A inclusão de FT na dieta reduziu a cadeia de ácidos graxos ramificados e apontou
tendência a diminuir a população de enterobactérias nas fezes. Resultados
semelhantes indicaram que a incorporação de FT na dieta diminuiu a população de
enterobactérias no conteúdo cecal(3) e a ligação das E. Coli K88 na mucosa ileal
após uma infecção experimental(9). Embora a enterobacteria esteja constituída por
diversas espécies bacterianas, a diminuição pode indicar uma mudança benéfica na
composição das populações microbianas. Neste sentido, diferentes autores
demonstraram que a inclusão de carboidratos fermentáveis na dieta dos leitões
recém desmamados pode reduzir a fermentação protéica ao longo do TGI (4;34)
sendo observada neste trabalho a redução dos ácidos ramificados. A fermentação
protéica no trato digestivo é considerada como potencial risco a disbiose e
proliferação das bactérias patogênicas(35). Leitões alimentados com FT na dieta
também indicam aumentar a concentração de ácido butírico no cólon. Neste
contexto, o butirato é considerado um importante metabólito, porque é a principal
fonte de energia para os colonócitos e pode apresentar efeitos benéficos na mucosa
ceco-cólon inflamada(13). É aceitável que o amido e os farelos oriundos do trigo ou
aveia estimulem a formação de butirato(36), enquanto as frações ricas em xilanos e
pectinas são todas associadas com uma baixa formação deste ácido graxo(37).
Entretanto, a fibra é também capaz de modificar as propriedades físico-químicas do
conteúdo digestivo. O FT, por possuir alto conteúdo de fibra insolúvel, está
associado à melhora dos quadros de constipação em humanos (38) e relacionados à
redução do tempo de trânsito no intestino delgado e grosso dos suínos(11). No

50
presente trabalho não foi evidenciada uma diferença considerável no tempo de
trânsito com a suplementação do FT, mas sim um significativo aumento na
capacidade de retenção de água e uma redução na porcentagem da capacidade de
água disponível do conteúdo digestivo colônico. Os resultados demonstraram uma
maior capacidade de ligação da água ao longo da cadeia PNA insolúvel comparado
com outros componentes, como o amido ou proteína(21). Além do mais, estas
mudanças podem apontar que as propriedades físico-químicas do conteúdo
digestivo podem ter importância em alguns processos do TGI, como o esvaziamento
gástrico, a motilidade do intestino delgado ou a fermentação no lúmen intestinal.
Alguns relatos têm sugerido que uma dieta mais fibrosa e com maior granulometria
pode modificar as propriedades físico-químicas e microbiológicas do conteúdo
digestivo, com diminuição no nível de sobrevivência de algumas enterobactérias,
como a Salmonella(39). Sugere-se que os processos anterointestinais, como a
distribuição do ácido clorídrico dentro do conteúdo estomacal, são favorecidos
quando a dieta tem estrutura mais grossa e uma maior capacidade de retenção de
água, os quais podem diminuir a contagem de Salmonella e a sua chance de
alcançar o intestino delgado.
4.2 A influência da loperamida na adaptação dos leitões frente à dieta
Loperamida é um derivado opióide sintético frequentemente utilizado como um
medicamento antidiarreico em humanos. Este diminui a motilidade dos músculos
lisos circulares e longitudinais da parede intestinal, retarda o fluxo entérico e
estimula a absorção da água colônica(40). Enquanto LOP é amplamente utilizada em
adultos, há uma preocupação sobre a segurança deste medicamento para crianças
e durante o curso de uma diarréia infecciosa. A contra-indicação nestes casos vem
quando diante de uma infecção bacteriana invasiva, pelo risco de agravamento da
sintomatologia devido a estase do conteúdo digestivo, permitindo a translocação
bacteriana. Os resultados do estudo indicaram que a LOP mostrou tendência para o
aumento do TMT ao longo do TGI e aumentou a digestibilidade aparente total da MO
e PB, especialmente com a associação ao FT. Estes resultados estão pertinentes
com um incremento significativo da altura da vilosidade e a razão entre altura da

51
vilosidade/profundidade de cripta no jejuno (Exp. 2), a qual é considerada critério útil
para a estimação da capacidade digestiva do intestino delgado(41). Em contraste,
outros autores relataram que a LOP inibiu a secreção pancreaticobiliar (bilirubina e
amilase), atuando no nervo que supre o pâncreas e a vesícula biliar(42;43).
Animais tratados com LOP também aumentaram o consumo diário de ração e o
GPD durante as primeiras duas semanas após desmame. A LOP é conhecida por
afetar não somente os receptores opiódes relacionados com a motilidade intestinal,
mas também aqueles relacionados à analgesia que estão presentes nos nervos
sensoriais periféricos e que estão regulados durante o desenvolvimento da
inflamação(44). Após o desmame, os estresses que os animais sofrem conduzem a
um período de anorexia que pode contribuir a uma inflamação local no intestino
delgado(45). Além disso, a redução do consumo durante os primeiros dias após
desmame é considerada a causa do dano da barreira intestinal dos leitões e de
encurtamento das vilosidades. O tratamento dos animais com LOP após desmame
parece ter reduzido a inflamação intestinal e melhorado o comportamento animal;
resultando em um maior consumo, ganho de peso, e melhoria da integridade da
mucosa. Esta hipótese está em acordo com Bowden(46), que relata os efeitos
positivos dos medicamentos analgésicos, antiinflamatórios e agentes bloqueadores
dos receptores muscarínicos no apetite dos suínos.
O aumento da digestibilidade do FT com LOP está associado com um aumento
significativo das concentrações de AGCC, ácido acético e propiônico nas fezes, o
qual provavelmente resulta na maturação avançada do trato colônico digestivo.
Graham et al.(47) e Le Goff et al.(31) sugerem que a capacidade da degradação da
fibra no intestino dos suínos aumenta com a idade, principalmente devido ao
aumento do tempo de trânsito digestivo e da atividade metabólica microbiana. No
presente estudo, o efeito LOP poderia estar associado com o tempo mais longo da
fermentação colônica. Entretanto, não se podem excluir as mudanças das atividades
metabólicas da microbiota enquanto se adaptam aos substratos. A adaptação dos
microorganismos pode ser devido à microbiota intestinal responsável pela eficiência
na fermentação do amido resistente(48) ou PNA(49). Entretanto, no presente estudo, a
LOP não promoveu mudanças na contagem microbiana nas fezes e no conteúdo
colônico digestivo, mas aumentou as concentrações do ácido butírico nas fezes de

52
leitões alimentados com ambas dietas. Alguns estudos relataram que a mudança do
tempo de trânsito pode alterar a atividade bacteriana e modificar os tipos
bacterianos, afetando a proporção individual de AGCC(29;50). O lactato produzido
pelas bactérias ácido lácticas tem sido recentemente sugerido como a principal
precursora da síntese do butirato(29), com a formação de butirato aumentado pela
diminuição do tempo de trânsito do TGI(41), ou a maior taxa de diluição in vitro(51). Os
resultados obtidos aparentam ser contraditórios com as referências acima.
Entretanto, as mudanças observadas na digestão dos tratamentos LOP podem ter
determinados mudanças relevantes na fermentação dos substratos na região distal
do TGI. Deve ser ressaltado que quantidades significativas de lactato podem ser
encontradas no intestino grosso dos leitões, mas não nos animais adultos(37), que
pode representar a maturação precoce do lúmen intestinal nos animais tratados com
LOP.

53
5. CONCLUSÕES
A inclusão moderada de níveis de fibra insolúvel como ingrediente através do FT
pode modificar a atividade microbiota intestinal e, quando associada à loperamida na
dieta após desmame, os efeitos benéficos podem ser ampliados, melhorando a
saúde animal e o desempenho produtivo.

54
6. RECONHECIMENTOS
Esta pesquisa foi sustentada pela Espanha CICYT (projeto AGL2005-07438-C02-
01). Agradecimentos ao Ministério da Educação, Cultura e Esporte, Bolsas de
Estudos para a pesquisa da Espanha.

55
Tabela 1 .Composição e análises química da dieta
aDietas experimentais: CT, dieta controle; FT, Dieta com farelo de trigo. bFarinha de peixe: produto desidratado, e desengordurado pelo processo de baixa temperatura (<70ºC) e pressão. cSuplemento por kilograma na ração: Vitamina A, 5000 UI; Vitamina D3, 1000 UI; Vitamin E 15.0 mg; Vitamina B, 11.3 mg; Vitamina B2, 3.5 mg; Vitamin B6, 1.5 mg; vitamina B12, 0.025 mg; Pantotenato de cálcio, 10.0 mg; Niacina, 15.0 mg; Biotina, 15.0 mg; Ácido fólico, 0.1 mg; Vitamina K3, 2.0 mg; Fe, 80.0 mg; Cu, 6.0 mg; Co, 0.7 mg; Zn, 60.0 mg; Mn, 30.0 mg; I, 0.7 mg; Se, 0.1 mg e Etoxiquin, 0.15 mg.
Dietaª Experimento 1 Experimento 2 CT FT CT FT
Ingredientes (g/kg) Milho 332.1 290.3 331.1 280.0 Cevada 211.6 210.0 238.2 210.0 Soro de leite em pó 130.0 130.0 130.0 130.0 Soro de leite gorduroso 100.0 100.0 100.0 100.0 Proteína de soja 90.0 90.0 92.5 87.1 Glúten de trigo 58.1 55.7 30.0 30.0 Farinha de peixeb 40.0 40.0 40.0 40.0 Farelo de trigo - 40.0 - 80.0 Óleo de girassol - 6.5 - 6.5 Fosfato bicálcico 10.3 9.6 9.1 8.4 Carbonato de cálcio 9.3 9.7 11.7 10.6 L-lisina HCL 6.8 6.7 5.2 5.2 DL-Metionina 1.5 1.5 4.1 4.1 L-Treonina 2.1 2.1 2.3 2.3 L-Triptofano 0.7 0.7 0.6 0.6 Premix Vitaminico e mineralc 4.0 4.0 5.0 5.0 Sal 3.4 3.3 - - Óxido de Cromo - - 0.15 0.15 Valores calculados (g/kg na ração) EM (MJ/kg) 14.6 14.6 14.4 14.3 Proteína bruta 208.8 209.1 189.6 190.9 SID lisina 13.75 13.71 12.14 12.12 SID metionina 4.84 4.79 6.94 6.91 SID treonina 8.72 8.68 8.28 8.23 SID triptofano 2.67 2.67 2.27 2.29 SID isoleucina 7.77 7.73 7.02 6.96 SID valina 9.07 9.05 8.3 8.26 Análises químicas e energéticas (g/kg na ração) Matéria seca 908.2 907.6 903.0 903.0 GE (MJ/Kg) 17.1 17.5 17.8 17.7 Proteína bruta (CP; N x 6.25) 200.8 205.6 208.0 210.8 Fibra em Detergente Neutro 74.0 87.0 85.3 106.3 Fibra em Detergente Ácido 25.0 29.0 18.9 22.9 Extrato Etéreo 67.4 80.5 71.0 75.0 Cinzas 55.0 57.0 64.0 66.0
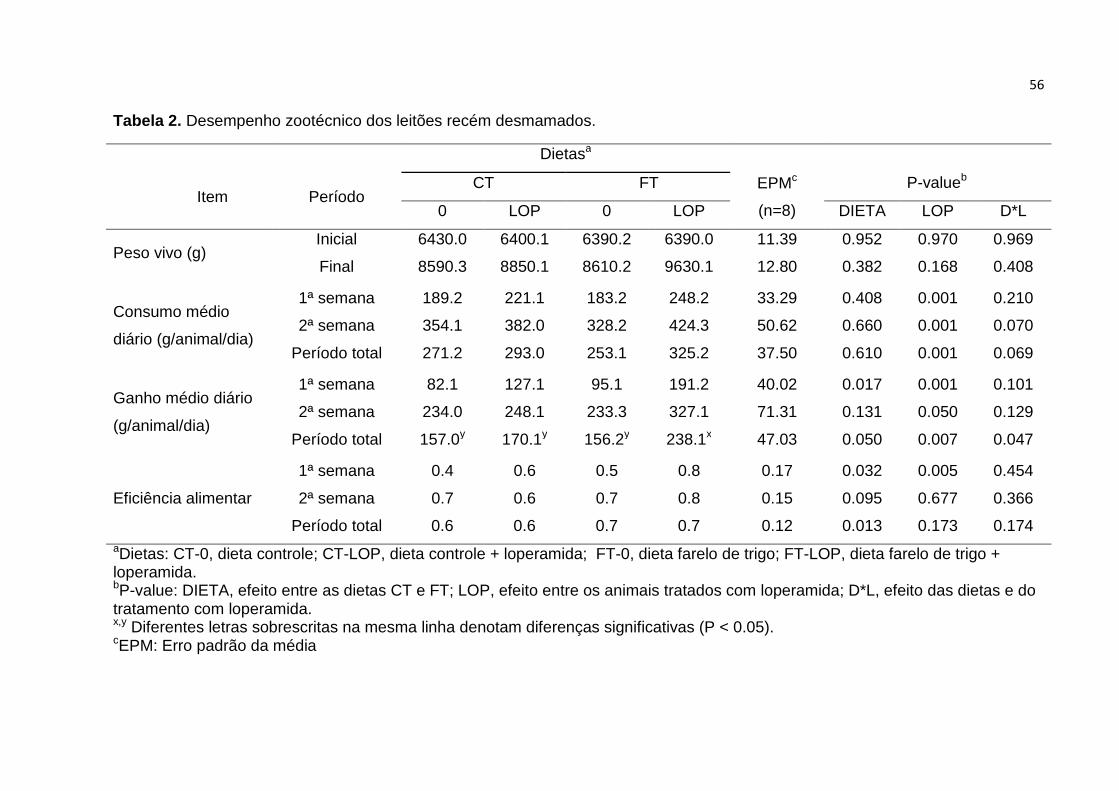
56
Tabela 2. Desempenho zootécnico dos leitões recém desmamados.
aDietas: CT-0, dieta controle; CT-LOP, dieta controle + loperamida; FT-0, dieta farelo de trigo; FT-LOP, dieta farelo de trigo + loperamida. bP-value: DIETA, efeito entre as dietas CT e FT; LOP, efeito entre os animais tratados com loperamida; D*L, efeito das dietas e do tratamento com loperamida. x,y Diferentes letras sobrescritas na mesma linha denotam diferenças significativas (P < 0.05). cEPM: Erro padrão da média
Dietasa
Item Período CT FT EPMc
(n=8)
P-valueb
0 LOP 0 LOP DIETA LOP D*L
Peso vivo (g) Inicial 6430.0 6400.1 6390.2 6390.0 11.39 0.952 0.970 0.969
Final 8590.3 8850.1 8610.2 9630.1 12.80 0.382 0.168 0.408
Consumo médio
diário (g/animal/dia)
1ª semana 189.2 221.1 183.2 248.2 33.29 0.408 0.001 0.210
2ª semana 354.1 382.0 328.2 424.3 50.62 0.660 0.001 0.070
Período total 271.2 293.0 253.1 325.2 37.50 0.610 0.001 0.069
Ganho médio diário
(g/animal/dia)
1ª semana 82.1 127.1 95.1 191.2 40.02 0.017 0.001 0.101
2ª semana 234.0 248.1 233.3 327.1 71.31 0.131 0.050 0.129
Período total 157.0y 170.1y 156.2y 238.1x 47.03 0.050 0.007 0.047
Eficiência alimentar
1ª semana 0.4 0.6 0.5 0.8 0.17 0.032 0.005 0.454
2ª semana 0.7 0.6 0.7 0.8 0.15 0.095 0.677 0.366
Período total 0.6 0.6 0.7 0.7 0.12 0.013 0.173 0.174
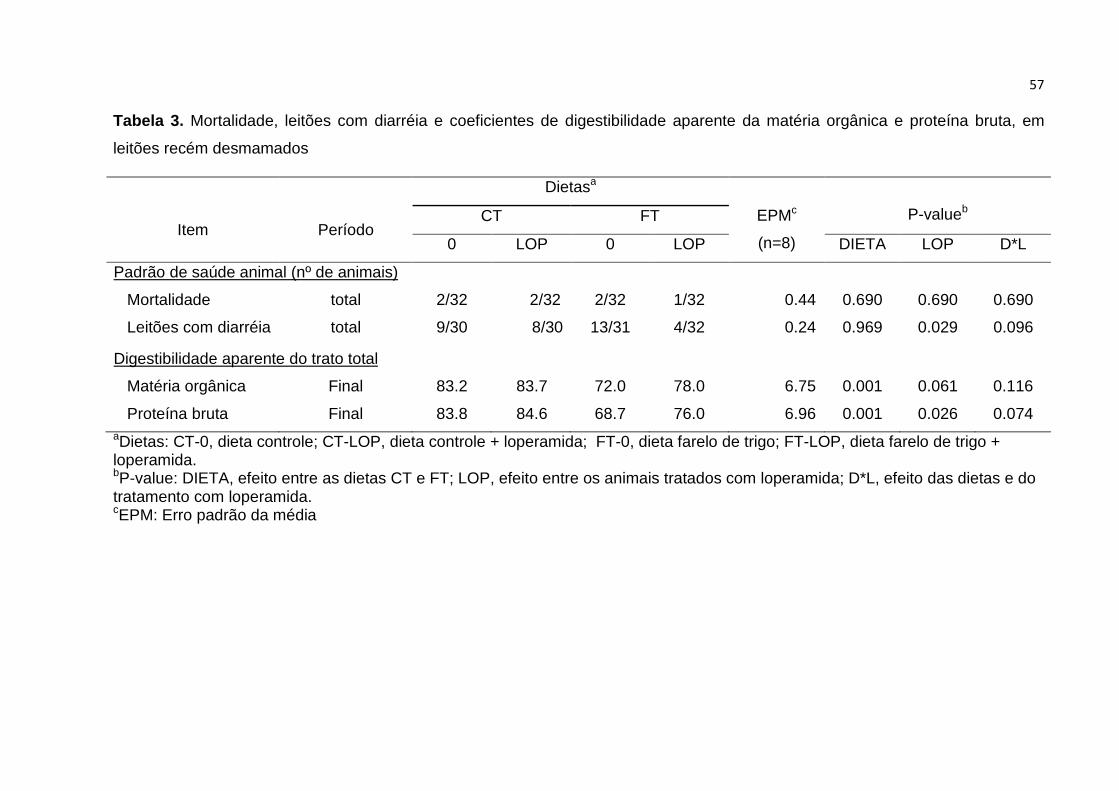
57
Tabela 3. Mortalidade, leitões com diarréia e coeficientes de digestibilidade aparente da matéria orgânica e proteína bruta, em
leitões recém desmamados
aDietas: CT-0, dieta controle; CT-LOP, dieta controle + loperamida; FT-0, dieta farelo de trigo; FT-LOP, dieta farelo de trigo + loperamida. bP-value: DIETA, efeito entre as dietas CT e FT; LOP, efeito entre os animais tratados com loperamida; D*L, efeito das dietas e do tratamento com loperamida. cEPM: Erro padrão da média
Dietasa
EPMc
(n=8)
P-valueb Item Período
CT FT
0 LOP 0 LOP DIETA LOP D*L
Padrão de saúde animal (nº de animais)
Mortalidade total 2/32 2/32 2/32 1/32 0.44 0.690 0.690 0.690
Leitões com diarréia total 9/30 8/30 13/31 4/32 0.24 0.969 0.029 0.096
Digestibilidade aparente do trato total
Matéria orgânica Final 83.2 83.7 72.0 78.0 6.75 0.001 0.061 0.116
Proteína bruta Final 83.8 84.6 68.7 76.0 6.96 0.001 0.026 0.074
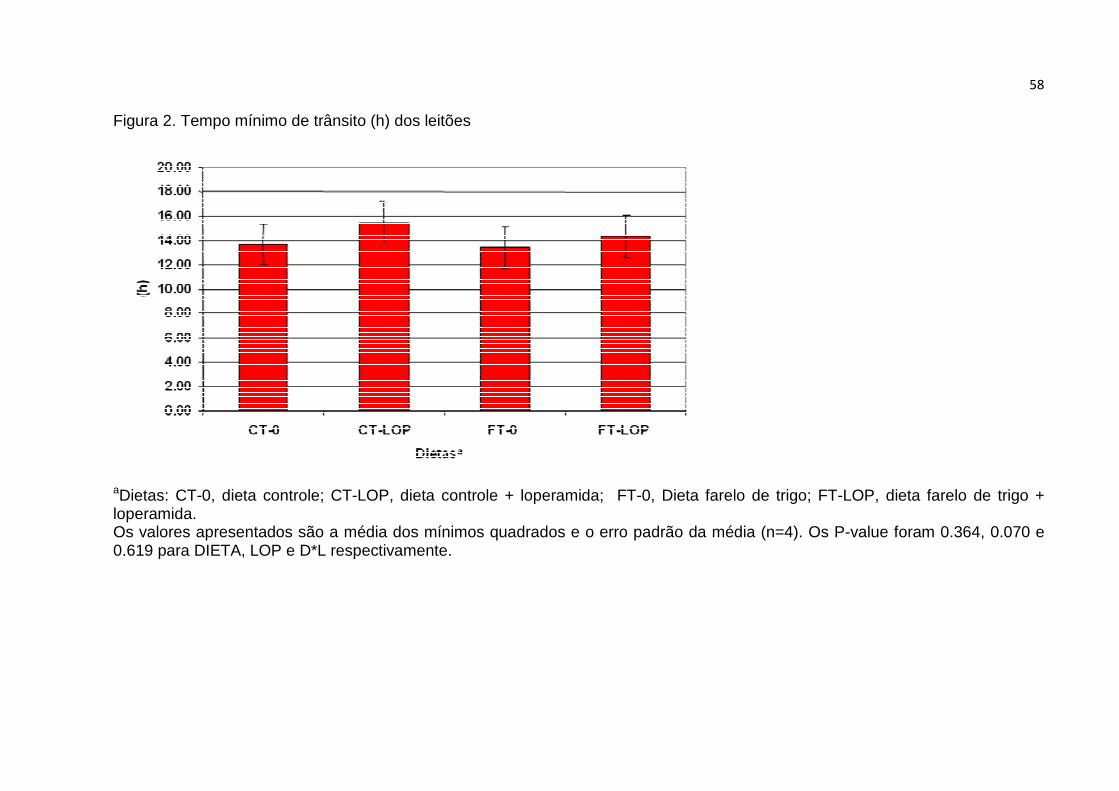
58
Figura 2. Tempo mínimo de trânsito (h) dos leitões
aDietas: CT-0, dieta controle; CT-LOP, dieta controle + loperamida; FT-0, Dieta farelo de trigo; FT-LOP, dieta farelo de trigo + loperamida. Os valores apresentados são a média dos mínimos quadrados e o erro padrão da média (n=4). Os P-value foram 0.364, 0.070 e 0.619 para DIETA, LOP e D*L respectivamente.
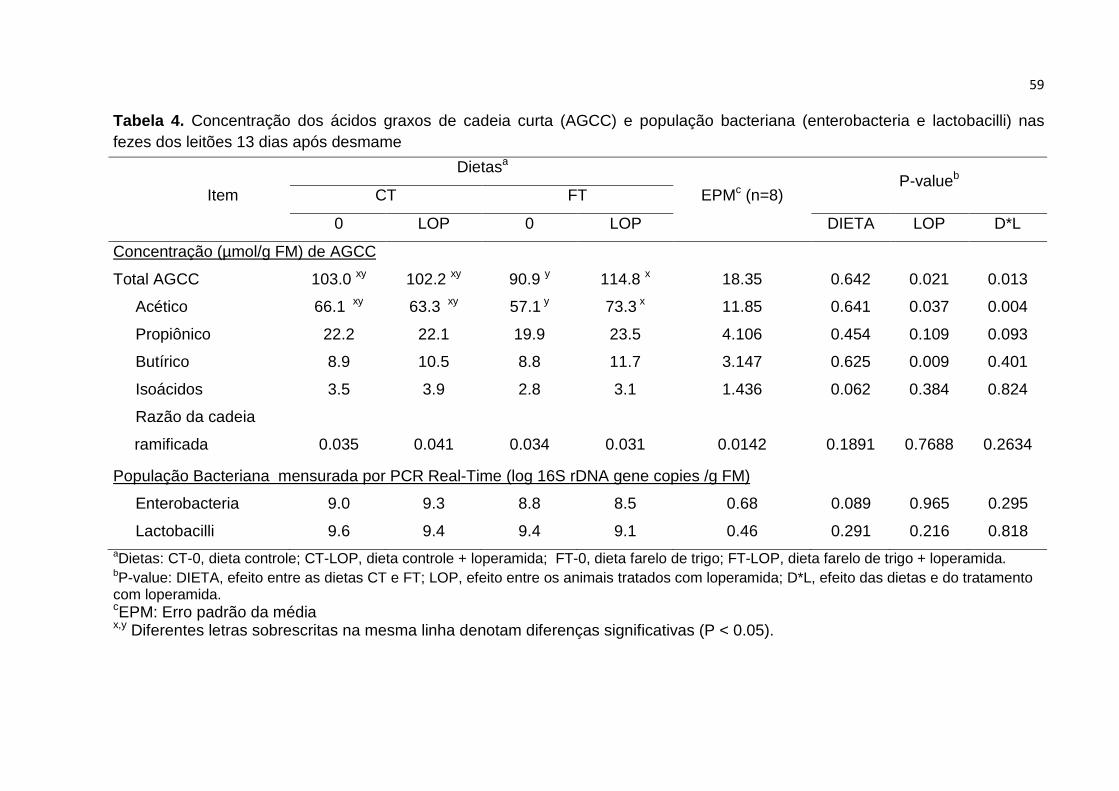
59
Tabela 4. Concentração dos ácidos graxos de cadeia curta (AGCC) e população bacteriana (enterobacteria e lactobacilli) nas fezes dos leitões 13 dias após desmame
aDietas: CT-0, dieta controle; CT-LOP, dieta controle + loperamida; FT-0, dieta farelo de trigo; FT-LOP, dieta farelo de trigo + loperamida. bP-value: DIETA, efeito entre as dietas CT e FT; LOP, efeito entre os animais tratados com loperamida; D*L, efeito das dietas e do tratamento com loperamida. cEPM: Erro padrão da média x,y Diferentes letras sobrescritas na mesma linha denotam diferenças significativas (P < 0.05).
Item
Dietasa
EPMc (n=8) P-valueb
CT FT
0 LOP 0 LOP DIETA LOP D*L
Concentração (µmol/g FM) de AGCC
Total AGCC 103.0 xy 102.2 xy 90.9 y 114.8 x 18.35 0.642 0.021 0.013
Acético 66.1 xy 63.3 xy 57.1 y 73.3 x 11.85 0.641 0.037 0.004
Propiônico 22.2 22.1 19.9 23.5 4.106 0.454 0.109 0.093
Butírico 8.9 10.5 8.8 11.7 3.147 0.625 0.009 0.401
Isoácidos 3.5 3.9 2.8 3.1 1.436 0.062 0.384 0.824
Razão da cadeia
ramificada 0.035 0.041 0.034 0.031 0.0142 0.1891 0.7688 0.2634
População Bacteriana mensurada por PCR Real-Time (log 16S rDNA gene copies /g FM)
Enterobacteria 9.0 9.3 8.8 8.5 0.68 0.089 0.965 0.295
Lactobacilli 9.6 9.4 9.4 9.1 0.46 0.291 0.216 0.818
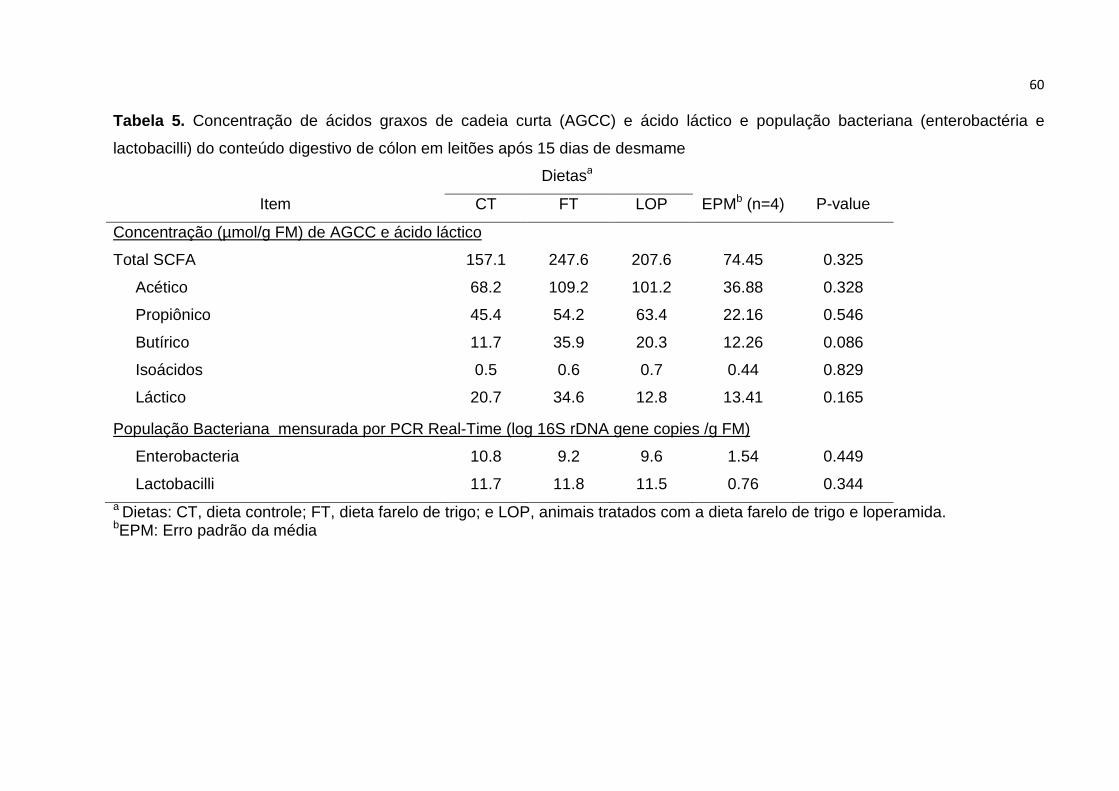
60
Tabela 5. Concentração de ácidos graxos de cadeia curta (AGCC) e ácido láctico e população bacteriana (enterobactéria e
lactobacilli) do conteúdo digestivo de cólon em leitões após 15 dias de desmame
Dietasa
Item CT FT LOP EPMb (n=4) P-value
Concentração (µmol/g FM) de AGCC e ácido láctico
Total SCFA 157.1 247.6 207.6 74.45 0.325
Acético 68.2 109.2 101.2 36.88 0.328
Propiônico 45.4 54.2 63.4 22.16 0.546
Butírico 11.7 35.9 20.3 12.26 0.086
Isoácidos 0.5 0.6 0.7 0.44 0.829
Láctico 20.7 34.6 12.8 13.41 0.165
População Bacteriana mensurada por PCR Real-Time (log 16S rDNA gene copies /g FM)
Enterobacteria 10.8 9.2 9.6 1.54 0.449
Lactobacilli 11.7 11.8 11.5 0.76 0.344 a Dietas: CT, dieta controle; FT, dieta farelo de trigo; e LOP, animais tratados com a dieta farelo de trigo e loperamida. bEPM: Erro padrão da média
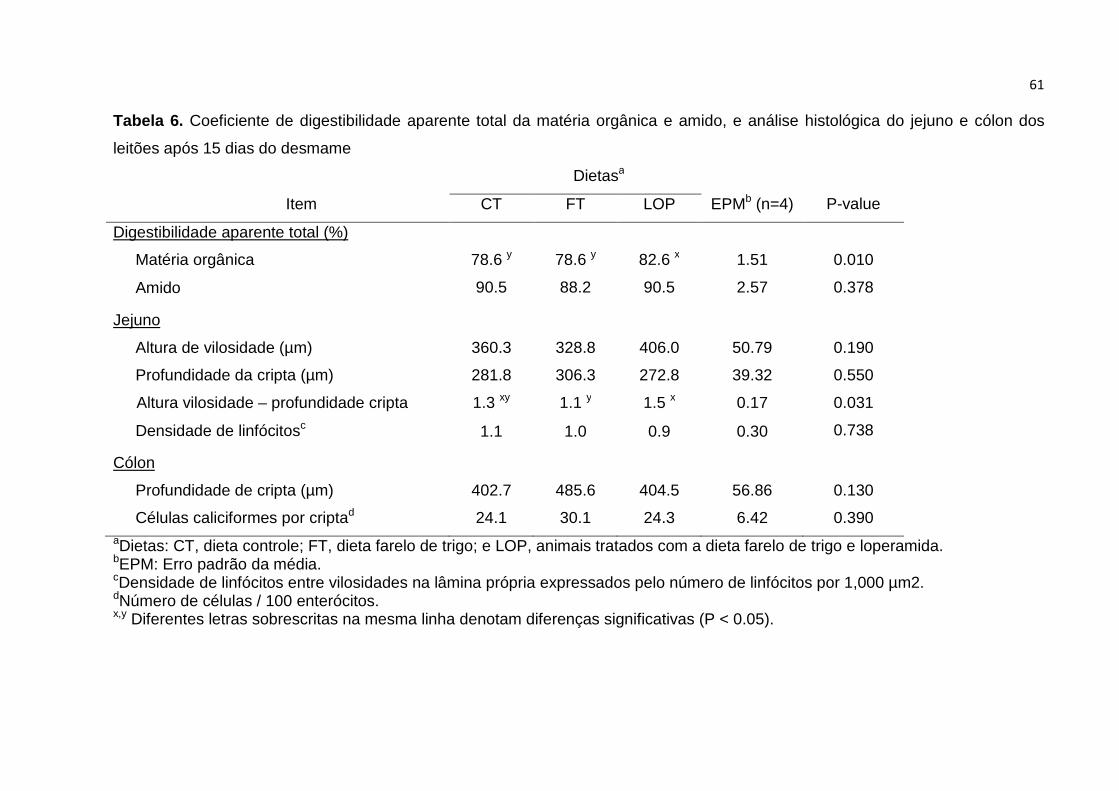
61
Tabela 6. Coeficiente de digestibilidade aparente total da matéria orgânica e amido, e análise histológica do jejuno e cólon dos
leitões após 15 dias do desmame
Dietasa
Item CT FT LOP EPMb (n=4) P-value
Digestibilidade aparente total (%)
Matéria orgânica 78.6 y 78.6 y 82.6 x 1.51 0.010
Amido 90.5 88.2 90.5 2.57 0.378
Jejuno
Altura de vilosidade (µm) 360.3 328.8 406.0 50.79 0.190
Profundidade da cripta (µm) 281.8 306.3 272.8 39.32 0.550
Altura vilosidade – profundidade cripta 1.3 xy 1.1 y 1.5 x 0.17 0.031
Densidade de linfócitosc 1.1 1.0 0.9 0.30 0.738
Cólon
Profundidade de cripta (µm) 402.7 485.6 404.5 56.86 0.130
Células caliciformes por criptad 24.1 30.1 24.3 6.42 0.390 aDietas: CT, dieta controle; FT, dieta farelo de trigo; e LOP, animais tratados com a dieta farelo de trigo e loperamida. bEPM: Erro padrão da média. cDensidade de linfócitos entre vilosidades na lâmina própria expressados pelo número de linfócitos por 1,000 µm2. dNúmero de células / 100 enterócitos. x,y Diferentes letras sobrescritas na mesma linha denotam diferenças significativas (P < 0.05).
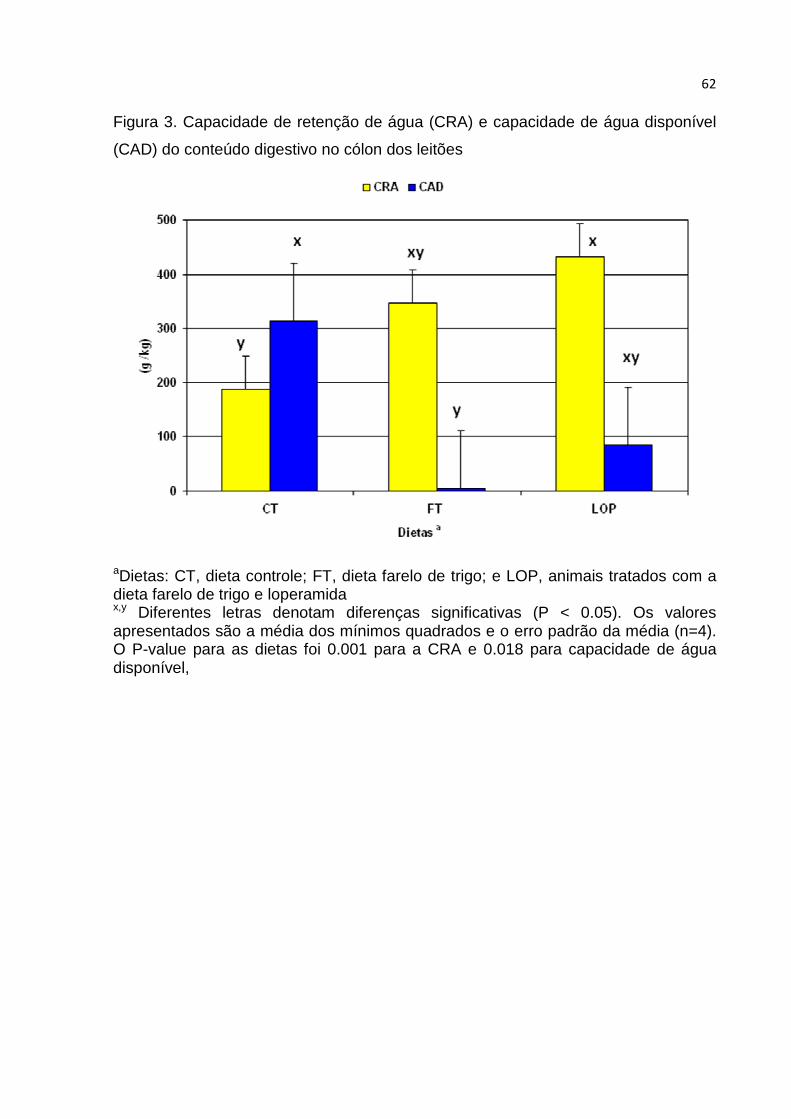
62
Figura 3. Capacidade de retenção de água (CRA) e capacidade de água disponível
(CAD) do conteúdo digestivo no cólon dos leitões
aDietas: CT, dieta controle; FT, dieta farelo de trigo; e LOP, animais tratados com a dieta farelo de trigo e loperamida x,y Diferentes letras denotam diferenças significativas (P < 0.05). Os valores apresentados são a média dos mínimos quadrados e o erro padrão da média (n=4). O P-value para as dietas foi 0.001 para a CRA e 0.018 para capacidade de água disponível,

63
7. REFERÊNCIAS
(1) LALLES, J.P. et al. Weaning - A challenge to gut physiologists.
Livestock Sci, v.108, p.82-93, 2007.
(2) FAIRBROTHER, J.M.; NADEAU, E.; GYLES, C.L. Escherichia coli in
postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and
prevention strategies. Anim. Health Res. Ver. v.6, p.17-39, 2005.
(3) MOLIST, F. et al. Effects of the insoluble and soluble dietary fibre
on the physicochemical properties of digesta and th e microbial activity in early
weaned piglets. Anim Feed Sci. Technol. v.149, p.346-353, 2009.
(4) AWATI, A. et al. Effect of inclusion of fermentable carbohydrates in
the diet on fermentation end-product profile in fec es of weanling piglets. J.
Anim. Sci. v.84, p.2133-2140, 2006.
(5) HERMES, R.G. et al. Effect of dietary level of protein and fiber on
the productive performance and health status of pig lets. Under revision, 2009.
(6) MATEOS, G.G. et al. Inclusion of oat hulls in diets for young pigs
based on cooked maize or cooked rice. Anim. Sci. v.82, p.57-63, 2007.
(7) WELLOCK, I.J. et al. The consequences of non-starch
polysaccharide solubility and inclusion level on th e health and performance of
weaned pigs challenged with enterotoxigenic Escheri chia coli. Br. J. Nutr. v.99,
p.520-530, 2007.
(8) HOPWOOD, D.E. et al. Addition of pearl barley to a rice-based diet
for newly weaned piglets increases the viscosity of the intestinal contents,
reduces starch digestibility and exacerbates post-w eaning colibacillosis. Br. J.
Nutr. v.92, p.419-427, 2004.
(9) MOLIST, F. et al. Effect of wheat bran on the health and
performance of weaned pigs challenged with Escheric hia coli K88 +. Livestock
Sci. Under revision, 2009.
(10) CUMMINGS, J.H.; STEPHEN, A.M. The Role of Dietary Fiber in the
Human-Colon. Can. Med. Assoc. J. v.123, p.1109-1114, 1980.

64
(11) WILFART, A. et al. Digesta transit in different segments of the
gastrointestinal tract of pigs as affected by insol uble fibre supplied by wheat
bran. Br. J. Nutr. v.98, p.54-62, 2007.
(12) LEWIS, S.J.; HEATON, K.W. Increasing butyrate concentration in
the distal colon by accelerating intestinal transit . Gut. v.41, p.245-251, 1997.
(13) OUFIR, L.E. et al. Relationships between transit time in man and in
vitro fermentation of dietary fiber by fecal bacter ia. Eur. J. Clin. Nutr. v.54, p.603-
609, 2000.
(14) BAKER, D.E. Loperamide: A pharmacological review. Rev. Gastr. Dis.
v.7, p.S11-S18, 2007.
(15) STEPHEN, A.M.; WIGGINS, H.S.; CUMMINGS, J.H. Effect of
Changing Transit-Time on Colonic Microbial-Metaboli sm in Man. Gut. v.28,
p.601-609, 1987.
(16) MITTELSTADT, S.; HEMENWAY, C.; SPRUELL, R. Effects of fasting
on evaluation of gastrointestinal transit with char coal meal. J. Pharmacol.
Toxicol. Meth. v.52, p.154-158, 2005.
(17) AWOUTERS, F, et al. Loperamide - Survey of Studies on
Mechanism of Its Antidiarrheal Activity. Dig. Dis. Sci. v.38, p.977-995, 1993.
(18) SCHURCH, A.F. et al. The Use of Chromic Oxide in Digestibility
Studies with Pigs Fed Adlibitum in the Barn. J. Anim. Sci. v.11, p.261-265, 1952.
(19) AOAC. In: Associ.Offic.Anal.Chem. (Ed.). Official methods of analysis
Arlington, VA (USA), 1995.
(20) WILLIAMS, C.H.; IISMAA, O.; DAVID, D.J. Determination of Chromic
Oxide in Faeces Samples by Atomic Absorption Spectr ophotometry. J. Agric.
Sci. v.59, p.381-385, 1962.
(21) ANGUITA, M., et al. Effect of coarse ground corn, sugar beet pulp
and wheat bran on the voluntary intake and physicoc hemical characteristics of
digesta of growing pigs. Livestock Sci. v.107, p.182-191, 2007.
(22) CASTILLO, M., et al. The response of gastrointestinal microbiota to
avilamycin, butyrate, and plant extracts in early-w eaned pigs. J. Anim. Sci. v.84,
p.2725-2734, 2006.
(23) JENSEN, B.B.; JORGENSEN, H. Effect of Dietary Fiber on Microbial
Activity and Microbial Gas-Production in Various Re gions of the
Gastrointestinal-Tract of Pigs. Appl. Environ. Microbiol. v.60,p.1897-1904, 1994.

65
(24) SAS Inc., SAS® User's Guide: Statistics, Version 8 Edition. SAS Inc.,
Cary, NC, USA, 1999.
(25) KIM, J.C.,et al. Addition of oat hulls to an extruded rice-based die t
for weaner pigs ameliorates the incidence of diarrh oea and reduces indices of
protein fermentation in the gastrointestinal tract. Br. J. Nutr. v.99, p.1217-1225,
2008.
(26) KNUDSEN, K.E.B. Carbohydrate and lignin contents of plant
materials used in animal feeding. Anim. Feed Sci. Technol. v.67, p.319-338, 1997.
(27) MORALES, J., et al. Comparative digestibility and lipogenic activity
in Landrace and Iberian finishing pigs fed ad libit um corn- and corn-sorghum-
acorn-based diets. Livestock Prod. Sci. v.77, p.2-3, 2002.
(28) KNUDSEN, K.E.B., et al. Rye bread enhances the production and
plasma concentration of butyrate but not the plasma concentrations of glucose
and insulin in pigs. J. Nutr. v.135, p.1696-1704, 2005.
(29) FREIRE, J.P.B., et al. Effect of dietary fibre source on total tract
digestibility, caecum volatile fatty acids and dige stive transit time in the
weaned piglet. Anim. Feed Sci. Technol. v.87, p.71-83, 2000.
(30) SCHULZE, H., et al. Dietary Level and Source of Neutral Detergent
Fiber and Ileal Endogenous Nitrogen Flow in Pigs. J. Ani. Sci. v.73, p.441-448,
1995.
(31) LE GOFF, G.; VAN MILGEN, J.; NOBLET, J. Influence of dietary fibre
on digestive utilization and rate of passage in gro wing pigs, finishing pigs and
adult sows. Anim. Sci. v.74, p.503-515, 2002.
(32) KYRIAZAKIS, I.; EMMANS, G.C.; ANDERSON, D.H. Do Breeds of
Pig Differ in the Efficiency with Which They Use A Limiting Protein Supply. Br.
J. Nutr. v.74, p.183-195, 1995.
(33) POND, W.G., et al. Markers for Estimating Digesta Flow in Pigs and
the Effects of Dietary Fiber. J. Anim. Sci. v.63, p.1140-1149, 1986.
(34) BIKKER, P, et al.The effect of dietary protein and fermentable
carbohydrates levels on growth performance and inte stinal characteristics in
newly weaned piglets. J. Anim. Sci. v.84, p.3337-3345, 2006.
(35) PROHASZKA, L.; BARON, F. The predisposing role of high dietary
protein supplies in enteropathogenic Escherichia co li infections of weaned
pigs. Zentralbl Veterinarmed B. v.27, p.222-232, 1980.

66
(36) BUGAUT, M. Occurrence, absorption and metabolism of short
chain fatty acids in the digestive tract of mammals . Comp. Bioch. Physiol. v.86,
p.439-472, 1987.
(37) HUGHES, A.S., et al. In vitro fermentation by human fecal
microflora of wheat arabinoxylans. J. Agric. Food Chem. v.55, p.4589-4595, 2007.
(38) CANN, P.A.; READ, N.W.; HOLDSWORTH, C.D. What Is the Benefit
of Coarse Wheat Bran in Patients with Irritable Bow el Syndrome. Gut. v.25,
p.168-173, 1984.
(39) MIKKELSEN, L.L., et al. Effects of physical properties of feed on
microbial ecology and survival of Salmonella enteri ca serovar Typhimurium in
the pig gastrointestinal tract. Appl. Environ. Microbiol. v.70, p.3485-3492, 2004.
(40) SCHILLER, L.R., et al. Mechanism of the antidiarrheal effect of
loperamide. Gastroenterol. v.86, p.1475-1480, 1984.
(41) MONTAGNE, L.; PLUSKE, J.R.; HAMPSON, D.J. A review of
interactions between dietary fibre and the intestin al mucosa, and their
consequences on digestive health in young non-rumin ant animals. Anim. Feed
Sci. Technol. v.108, p.95-117, 2003.
(42) APPIA, F., et al. Loperamide induced inhibition of pancreatic
secretion in rats. Eur. J. Pharmacol. v.103, p.71-79, 1984.
(43) THIMISTER, P.W.L., et al. Inhibition of pancreaticobiliary secretion
by loperamide in humans. Hepatology v.26, p.256-261, 1997.
(44) STEIN, C.; MACHELSKA, H.; SCHAFER, A. Peripheral analgesic and
antiinflammatory effects of opioids. Zeitschrift Rheumatol. v.60, p.416, 2001.
(45) MCCRACKEN, B.A., et al. Weaning anorexia may contribute to local
inflammation in the piglet small intestine. J. Nutr. v.129, p.613-619, 1999.
(46) BOWDEN, H.C.; SHARMAN, D.F.; STEPHENS, D.B. Effects of
antiinflammatory analgesic drugs and muscarinic rec eptor blocking agents on
appetite in the pig. J. Vet. Pharmacol. Ther. v.10, p.269-272, 1987.
(47) GRAHAM, H.; HESSELMAN, K.; AMAN, P. The influence of wheat
bran and sugar-beet pulp on the digestibility of di etary components in a cereal
based pig diet. J. Nutr. v.116, p.242-251, 1986.
(48) LE BLAY, G., et al. Enhancement of butyrate production in the rat
caecocolonic tract by long term ingestion of resist ant potato starch. Br. J. Nutr.
v.82, p.419-426, 1999.

67
(49) LOW, A.G. Role of dietary fibre in pig diets. In Recent Advances in
Animal Nutrition, [DJ Cole and W Haresign, editors]. London: Butterworth-
Heinemann, p. 87-112, 1985.
(50) LEWIS, S.; COCHRANE, S. Alteration of sulfate and hydrogen
metabolism in the human colon by changing intestina l transit rate. Am. J.
Gastroenterol. v.102, p.624-633, 2007.
(51) LETERME, P.; SEVE, B.; THEWIS, A. The current 15N-leucine
infusion technique is not suitable for quantitative measurements of ileal
endogenous amino acid flows in pigs. J. Nutr. v.128, p.1961-1968, 1998.