jonato prestes - unimep.br · 3 efeito do exercÍcio fÍsico sobre a funcionalidade de linfÓcitos...
TRANSCRIPT

1
UNIVERSIDADE METODISTA DE PIRACICABA FACULDADE DE CIÊNCIAS DA SAÚDE
MESTRADO EM EDUCAÇÃO FÍSICA
DISSERTAÇÃO DE MESTRADO
EFEITO DO EXERCÍCIO FÍSICO SOBRE A FUNCIONALIDADE DE LINFÓCITOS E CITOCINAS CIRCULANTES DE RATOS
JONATO PRESTES
PIRACICABA-SP Fevereiro de 2006

2
UNIVERSIDADE METODISTA DE PIRACICABA
FACULDADE DE CIÊNCIAS DA SAÚDE MESTRADO EM EDUCAÇÃO FÍSICA
EFEITO DO EXERCÍCIO FÍSICO SOBRE A FUNCIONALIDADE DE LINFÓCITOS E CITOCINAS CIRCULANTES DE RATOS
JONATO PRESTES
PIRACICABA-SP Fevereiro de 2006
Dissertação apresentada ao Curso de Mestrado em Educação Física da Universidade Metodista de Piracicaba, para obtenção do título de Mestre em Educação Física na área de concentração “Performance Humana”, sob orientação da Profa. Dra. Cláudia Regina Cavaglieri.

3
EFEITO DO EXERCÍCIO FÍSICO SOBRE A FUNCIONALIDADE DE LINFÓCITOS E CITOCINAS CIRCULANTES DE RATOS
BANCA EXAMINADORA:
PROFa. DRa. CLÁUDIA REGINA CAVAGLIERI (ORIENTADORA)
PROF. DR. CARLOS ALBERTO DA SILVA
PROF. DR. SÉRGIO EDUARDO DE ANDRADE PEREZ
PROFa. DRa. ADRIANNE CHRISTINE PALANCH (SUPLENTE)
UNIVERSIDADE METODISTA DE PIRACICABA FACULDADE DE CIÊNCIAS DA SAÚDE
PIRACICABA-SP 2006

4
O mundo é grande e cabe
nesta janela sobre o mar.
O mar é grande e cabe
na cama e no colchão de amar.
O amor é grande e cabe
no breve espaço de beijar
(Carlos Drummond de Andrade in “Amar se Aprende Amando”)

5
DEDICATÓRIA
A DEUS que me deu luz, indicou o caminho e me acompanhou em todos os
passos da minha vida até hoje, e que com certeza continuará me guiando para
caminhos ainda mais gloriosos e abençoados.
Aos meus Pais, Jauri de Oliveira Prestes e Hedvirges Prestes, por terem me
dado tanto apoio e carinho até hoje. Este é mais um dos sonhos realizados,
que estarei sempre dedicando a vocês. Sonho que se realizou pela existência
destes dois maravilhosos seres humanos.
Neste momento só posso agradecer a Anelena Bueno Frollini, por toda a
compreensão, ajuda, cumplicidade, amor e por ter sempre estado ao meu lado.
Uma pessoa que mudou minha vida.

6
AGRADECIMENTO ESPECIAL
A Professora DRa. Cláudia Regina Cavaglieri, uma pessoa formidável, por ter
acreditado no meu trabalho, na minha pessoa. Obrigado pela oportunidade de
me tornar este Homem que sou hoje, é com muita honra e dedicação que eu
tive o prazer de ter recebido sua orientação e mais do que tudo, sua amizade.
Muito obrigado Cláudia você é inesquecível e possue uma inteligência
inestimável.
Ao grupo de Imunologia do exercício, em especial aos meus amigos Rodrigo
Dias, Clílton Krauss de Oliveira Ferreira e Felipe Donatto que foram pessoas
que sempre me ajudaram e me fizeram crescer muito na pesquisa e na vida.
As pessoas que moraram comigo nestes dois anos e me suportaram, dando
amizade e carinho, grandes homens com muito conhecimento científico,
grandes futuros nomes da Educação Física no Brasil, Christiano Bertoldo
Urtado, Gerson do Santos Leite, Vinicius Guzzoni, Henrique Del Bem.
Ao Professor Denis Foschini pelas oportunidades oferecidas e pela amizade.

7
AGRADECIMENTOS
A Patrícia Carla Paulino pelos dias em que nos auxiliou brilhantemente no
laboratório de Fisiologia, sempre demonstrando paciência, carinho e amizade.
A Melissa Victo, Márcia Guerreschi, Sandra Brambila pelo apoio nos
experimentos realizados para a concretização deste trabalho.
As Professoras DRa. Rozangela Verlengia e DRa. Adrianne Christine Palanch
pelos conselhos, amizade e ajuda nos tempos difíceis.
A Professora DRa. Slivia Crepaldi Alves pelo o incentivo e oportunidades
abertas neste período, uma grande mestre.
A todos os professores da Pós-graduação pelo conhecimento transmitido e
pela amizade e ao programa de Mestrado em Educação Física da Unimep
como um todo.
A todos os colegas da Pós-graduação, pela convivência agradável.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pelo apoio financeiro na forma de bolsa de estudos, por meio do programa
CAPES/PROSUP, ao PIBIC/CNPQ pelas bolsas cedidas aos alunos de
iniciação científica que participaram deste trabalho e ao FAP/UNIMEP pela
apoio financeiro ao projeto.

8
SUMÁRIO
1. INTRODUÇÃO ........................................................................................................ 1 2. REVISÃO DE LITERATURA .................................................................................. 4 2.1 Sistema Imunológico ......................................................................................... 4 2.2 Efeitos do Exercício sobre o Sistema Imunitário ............................................... 8 3. OBJETIVOS DA PESQUISA ................................................................................ 24 3.1 Objetivo Geral ................................................................................................ 24 3.2 Objetivos Específicos ..................................................................................... 24 4. MATERIAIS E MÉTODOS .................................................................................... 25 4.1 Animais ............................................................................................................ 25 4.2 Grupos Experimentais ..................................................................................... 25 4.3 Grupo Controle ................................................................................................ 25 4.4 Grupos Submetidos a uma (1) sessão de Exercício Físico ............................. 26 4.5 Grupo Exercitado cinco (5) sessões ................................................................ 26 4.6 Exercício Físico ............................................................................................... 27 4.7 Descrição Metodológica dos Parâmetros Analisados ..................................... 27 4.7.1 Leucometria ............................................................................................ 27 4.7.2 Leucograma Diferencial - % Linfócitos Circulantes ................................ 28 4.7.3 Obtenção dos Linfócitos dos Linfonodos Mesentéricos.......................... 28 4.7.4 Protocolo IL-2 ......................................................................................... 30 4.7.5 Protocolo IL-6 ......................................................................................... 32 4.7.6 Protocolo TNF-α ..................................................................................... 34 4.7.7 Potencial Transmembrânico de Mitocôndria .......................................... 36 4.7.8 Viabilidade de Linfócitos Circulantes ...................................................... 37 4.7.9 Fragmentação de DNA ........................................................................... 37 4.8 Análise Estatística ........................................................................................... 38 5. RESULTADOS ...................................................................................................... 39 5.1 Leucometria ..................................................................................................... 39 5.2 Leucograma Diferencial - Número total de Linfócitos Circulantes .................. 41 5.3 Linfócitos do Linfonodo Mesentérico ............................................................... 43 5.4 Viabilidade de Linfócitos .................................................................................. 45 5.5 Potencial Transmembrânico Mitocondrial de Linfócitos Circulantes ............... 46 5.6 Fragmentação de DNA .................................................................................... 47 5.7 Concentração Sérica de IL-2 ........................................................................... 48 5.8 Concentração Sérica de IL-6 .......................................................................... 50 5.9 Concentração Sérica de TNF-α ....................................................................... 52 5.10 Representação esquemática geral dos resultados.........................................54 6. DISCUSSÃO...........................................................................................................55 7. CONCLUSÕES.......................................................................................................70 8. REFERÊNCIAS BIBLIOGRÁFICAS.......................................................................72 9. ANEXO: Publicações referentes ao mestrado....................................................88

9
LISTA DE ABREVIATURAS
ACTH
Hormônio adrenocorticotrópico
AMPc Adenosina monofosfato cíclico
Bax Gene específico de apoptose
Bcl-2 Proteína anti-apoptótica
Bcl-xL Gene específico de apoptose
Bcl-xs Gene específico de apoptose
Células NK Células destruidoras naturais
Células Th Linfócitos T auxiliares
CD Grupo de diferenciação
CK Creatina cinase
CRF Fator de liberação de corticotropina
CRP Proteína-C reativa
DNA Ácido desoxirribonucléico
EDTA Etilenodiaminotetracetato
ELISA Ensaio imunoenzimático em fase sólida
EPM Erro padrão da média
Fas Receptor de morte celular
HSP72 Proteínas de choque térmico
ICAM-1 Molécula de adesão intercelular-1
IgA Imunoglobulina A
IgG Imunoglobulina G
IL-1 Interleucina-1

10
IL-1β Interleucina-1 beta
IL-1ra Interleucina1 receptor antagonista
IL-2 Interleucina-2
IL-4 Interleucina-4
IL-5 Interleucina-5
IL-6 Interleucina-6
IL-8 Interleucina-8
IL-10 Interleucina-10
Integrinas-β2 Integrinas-beta 2
Interferon-γ Interferon-gama
ITRS Infecção do trato respiratório superior
ITRSs Infecções do trato respiratório superior
LTCs Linfócitos T citolíticos
NO Óxido Nítrico
PBS Solução fosfato - salina
PHA Fitoemaglutinina
rpm Rotações por minuto
SOCS Supressores da sinalização de citocinas
STATs Proteínas sinalizadoras de transcrição
gênica
sTNF-R Receptor solúvel de TNF-α
TNF-α Fator de Necrose Tumoral-alfa
TGF- β Fator de Crescimento de
Transformação-beta

11
VCAM-1 Molécula vascular de adesão
intercelular-1
VIP Peptídeo intestinal vasoativo
VO2max Consumo máximo de oxigênio
VO2pico Pico de consumo máximo de oxigênio
durante o exercício físico

12
LISTA DE FIGURAS
Figura 1. Ilustração do modelo da curva em “J” da relação entre carga de
exercício e surgimento de infecções..................................................................15
Figura 2. Ilustração da proposta da curva em “S” que apresenta a relação entre
carga de exercício e surgimento de infecções...................................................16
Figura 3. Ilustração do procedimento de diluição dos calibradores para IL-2...31
Figura 4. Ilustração do procedimento de diluição dos calibradores para IL-6...33
Figura 5. Ilustração do procedimento de diluição dos calibradores para TNF-α
...........................................................................................................................35
Figura 6. Leucometria para o grupo controle e grupos 5, 15 minutos, exaustão e
exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................40
Figura 7. Leucograma diferencial – Número total de linfócitos circulantes para o
grupo controle e grupos 5, 15 minutos, exaustão e exercitado 5 sessões em
intensidade leve e moderada.............................................................................42
Figura 8. Número total de linfócitos provenientes do linfonodo mesentérico para
o grupo controle e grupos 5, 15 minutos, exaustão e exercitado 5 sessões em
intensidade leve e moderada.............................................................................44
Figura 9. Viabilidade de linfócitos circulantes para o grupo controle e grupos 5,
15 minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................45

13
Figura 10. Potencial transmembrânico mitocondrial de linfócitos circulantes
para o grupo controle e grupos 5, 15 minutos, exaustão e exercitado 5 sessões
em intensidade leve e moderada.......................................................................46
Figura 11. Fragmentação de linfócitos circulantes para o grupo controle e
grupos 5, 15 minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................47
Figura 12. Concentração sérica de IL-2 para o grupo controle e grupos 5, 15
minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................49
Figura 13. Concentração sérica de IL-6 para o grupo controle e grupos 5, 15
minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................51
Figura 14. Concentração sérica de TNF-α para o grupo controle e grupos 5, 15
minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................53

14
LISTA DE TABELAS
Tabela 1. Leucometria para o grupo controle e grupos 5, 15 minutos, exaustão
e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................39
Tabela 2. Leucograma diferencial – Número total de linfócitos circulantes para o
grupo controle e grupos 5, 15 minutos, exaustão e exercitado 5 sessões em
intensidade leve e moderada.............................................................................41
Tabela 3. Número total de linfócitos provenientes do linfonodo mesentérico
para o grupo controle e grupos 5, 15 minutos, exaustão e exercitado 5 sessões
em intensidade leve e moderada.......................................................................43
Tabela 4. Concentração sérica de IL-2 para o grupo controle e grupos 5, 15
minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................48
Tabela 5. Concentração sérica de IL-6 para o grupo controle e grupos 5, 15
minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................50
Tabela 6. Concentração sérica de TNF-α para o grupo controle e grupos 5, 15
minutos, exaustão e exercitado 5 sessões em intensidade leve e
moderada...........................................................................................................52
Tabela 7. Representação esquemática geral dos resultados............................54

15
RESUMO
O objetivo deste estudo foi verificar os efeitos do exercício físico (natação)
sobre leucócitos totais, contagem e função de linfócitos e citocinas circulantes.
Foram utilizados ratos da linhagem Wistar, + 2 meses, peso + 200g, divididos
em 4 grupos (n=6 ou n=3 por grupo), sendo: 1) grupo controle sedentário, 2)
grupos exercitados em intensidade leve durante 5, 15 minutos e exaustão (5L
15L, e EXL), 3) grupos exercitados em intensidade moderada (sobrecarga de
5% do peso corporal acoplado na região dorsal) na mesma duração (5M, 15M
e EXM) e 4) grupos exercitados durante 5 sessões de volume progressivo na
mesma semana (5, 15, 30, 45 e 60 minutos por dia) em ambas intensidades,
(5SL e 5SM). As análises realizadas foram: leucometria, leucograma
diferencial, contagem de linfócitos dos linfonodos mesentéricos, concentração
de citocinas circulantes (IL-2, IL-6 e TNF-α); potencial transmembrânico
mitocondrial, viabilidade e fragmentação de DNA de linfócitos determinadas por
citometria de fluxo. Aplicou-se o teste estatístico ANOVA, seguido de Post Hoc
Tukey com p*≤0.05, para comparação entre todos os grupos. Observamos
aumento no número de leucócitos totais e linfócitos circulantes, para todos os
grupos exercitados em relação ao controle. Houve aumento no número de
linfócitos mesentéricos nos grupos 5L, 5M e 15M; redução nos grupos EXL,
5SL e 5SM quando comparados com o controle. Na fragmentação de DNA de
linfócitos, foi detectada redução nos grupos 15L, 5M e 15M em relação ao
controle; redução na viabilidade de linfócitos apenas no grupo 15M. Foram
observadas reduções nas concentrações de TNF-α em todos os grupos
exercitados em intensidade moderada e no 5SL; aumento na IL-6 no EXL.
Concluímos que, o exercício realizado durante 5-15 minutos, até a exaustão ou
após 5 sessões de volume progressivo, pode modular os leucócitos circulantes,
promovendo leucocitose; alterar contagem e função de linfócitos, induzindo a
linfocitose e reduzindo a fragmentação de DNA; o exercício prolongado (10
horas em média) eleva a IL-6 e exercícios com sobrecarga promovem queda
na TNF-α circulante. O exercício de curta duração mesmo nas intensidades
leve e moderada, não é inócuo com relação à resposta imune.
Palavras-chave: exercício físico, função imune, linfócitos, citocinas

16
ABSTRACT
The purpose of this issue was to verify the effects of physical exercise
(swimming) on leukocytes count, lymphocytes count and function and
circulating levels of cytokines. Wistar rats were used (2 months old), with an
average weight of 200g, divided into four groups (n=6 or n=3 each group): 1) a
sedentary control group, 2) groups exercised at low intensity for 5, 15 minutes
and exhaustion (5L 15L, and EXL), 3) groups exercised at moderate intensity
(additional load of 5% of their body weight adapted on their backs) with the
same duration (5M, 15M and EXM) and 4) groups exercised for 5 progressive
volume sessions (5, 15, 30, 45 and 60 minutes each day) in the same week at
low and moderate intensities, (5SL and 5SM). The analyses performed were:
leukometry, leukogram differential, lymphocytes from lymph nodes count, seric
cytokines (IL-2, IL-6 and TNF-α) determined by ELISA method, mitochondrial
transmembrane potential, viability and DNA fragmentation of circulating
lymphocytes were measured by flow cytometry. The statistical analyses was
done applying the ANOVA test, followed by Tukey’s post hoc test (p*≤0.05), for
comparison between all the groups. A significant increase in total leukocytes
and in blood lymphocytes was observed for all exercised groups, as compared
to control. There was an increase in the number of lymphocytes from lymph
nodes in 5L, 5M and 15M; decrease in EXL, 5SL and 5SM groups related to
control. In lymphocytes DNA fragmentation, it was observed decrease in 15L,
5M and 15M as compared to control; decrease in lymphocytes viability, only in
15M group. It was observed decrease in TNF-α levels in all moderate intensity
exercised groups and 5SL; increase in IL-6 for the EXL group. We concluded
that, the exercise performed during 5-15 minutes, to exhaustion or after 5
sessions of progressive volume, can modulate circulating leukocytes, inducing
leukocytosis, modulate lymphocytes count and function, promoting
lymphocytosis and decrease in DNA fragmentation; prolonged exercise (10
hours in average) increase IL-6; exercise with additional loads decrease TNF- α
plasma levels. The exercise at short duration, even in low and moderate
intensities groups, was efficient to elicit an immune response.
KEY WORDS: Physical excercise, immune function, lymphocytes, cytokines

17
1. INTRODUÇÃO
A intensidade, duração e a freqüência do exercício exercem papel chave
na determinação das respostas imunológicas, podendo aumentar ou reduzir tal
função (MATTHEWS et al., 2002; MEYER et al., 2001; NIEMAN, 1994b;
ROWBOTTOM e GREEN, 2000). A prática de atividade física regular realizada
em intensidade moderada pode levar a redução na ocorrência de infecções,
especialmente do trato respiratório superior (ITRSs) (MATTHEWS et al., 2002).
Por outro lado, treinamentos de alto volume e intensidade, realizados por
atletas têm sido relacionados com aumentos da susceptibilidade a ITRSs
(NIEMAN e PEDERSEN, 1999; NIEMAN, 1998). O período de “janela aberta”
no qual atletas estão mais susceptíveis a infecções ocorre depois de sessões
exaustivas de exercício (PEDERSEN e ULLUM, 1994). Treinamentos intensos
podem ainda, reduzir a função de linfócitos ou acelerar o processo de apoptose
nestas células (PHANEUF e LEEUWENBURGH, 2001; QUADRILATERO e
HOFFMAN-GOETZ, 2005). Contudo, o exercício pode, paradoxalmente, tanto
promover melhora como debilitar a resposta imune; sendo esta dependente do
tipo de exercício e do nível de aptidão física de cada indivíduo (McCARTHY e
DALE, 1988; NEHLSEN-CANNARELLA et al., 1991; MINETTO et al., 2005).
Assim, para uma preparação física voltada a se chegar ao “desempenho
ótimo”, tanto do ponto de vista físico, nutricional ou psicológico, é necessário
ter conhecimento dos riscos que a atividade física pode trazer às funções
orgânicas, minimizando-os e enaltecendo apenas os benefícios, tanto em
atletas como para a população em geral.
O exercício físico regular é geralmente considerado como fator modulador
do estado de saúde, incluindo redução na pressão arterial, redução de peso
corporal, melhoria na tolerância à glicose e possivelmente contribui para uma
redução nas ITRSs (MACKINNON, 1997). Entre atletas e treinadores, existe
um senso comum que esforços intensos reduzem a resistência imunológica
constituindo um fator predispositor para as ITRSs, sendo que, evidências
epidemiológicas confirmam este fato (MACKINNON, 1997; NIEMAN e
PEDERSEN, 1999; NIEMAN, 1998).

18
O risco de ITRS parece estar especialmente elevado durante a primeira
ou segunda semana após eventos como maratona e após 1-9h de exercícios
de endurance prolongados (NIEMAN et al., 1990). Nessas situções, os
mecanismos de defesa contra agentes invasores estão reduzidos,
caracterizando o período como “janela aberta”, ou seja, aumento na
susceptibilidade a doenças e infecções, principalmente oportunistas (NIEMAN
et al., 1990).
Os mecanismos envolvidos neste processo não foram ainda
completamente elucidados. Estes efeitos podem ocorrer devido à ação de
hormônios do estresse, interações neuro-endócrinas, liberação de citocinas, de
fatores hematológicos, nutricionais ou diminuição dos níveis circulantes de
glutamina (SHEPARD, 1998).
Exercícios físicos intensos e de curta duração podem elevar o número
total de leucócitos no sangue numa relação direta à sua intensidade, sendo
que, este aumento ocorre principalmente na série granulocítica e em especial
nos poliformonucleares (BENONI et al., 1995; HOST et al., 1995; SAXTON et
al., 2003). O número de monócitos e de linfócitos também aumenta, mas em
menor escala (HOST et al., 1995; MOOREN et al., 2004; NIEMAN, 1994b)
sendo que, as células “Destruidoras Naturais” (NK) são as que demonstram
maiores alterações em relação à subpopulação dos linfócitos (MILES et al.,
2002; SHEPHARD e SHEK, 1995; TIMMONS et al., 2006).
Um dos mecanismos propostos para explicar esta linfocitose passageira,
poderia ser a liberação de adrenalina provocada pelo exercício (AZENABOR e
HOFFMAN-GOETZ, 2000). Após cinco minutos do término do exercício, a
contagem de linfócitos começa a diminuir provavelmente pelos efeitos
posteriores do cortisol liberado durante o mesmo (PEDERSEN et al., 1997),
sendo que, em geral de quatro a seis horas depois de encerrado o exercício
físico e, provavelmente após 24 horas de repouso, a contagem dos linfócitos se
normaliza (HOST et al., 1995; MEYER et al., 2004).
Aumento no tráfico de linfócitos é tipicamente controlado pelas
concentrações de catecolaminas, especialmente em atividades de curta
duração (BENSCHOP et al., 1996; KAPPEL et al., 1991; LANDMANN, 1992).
Reduções tardias observadas no número de linfócitos circulantes sofrem
influências do cortisol, particularmente em exercícios de duração mais

19
prolongada. Diversas substâncias podem modular o número e a função de
linfócitos, incluindo os hormônios do estresse e seus respectivos ciclos
circadianos, prostaglandinas, β-endorfinas e citocinas (JOZSA et al., 2005;
KAPPEL et al., 1991; PEDERSEN et al., 1997).
O exercício físico afeta a produção sistêmica de citocinas,
principalmente o Fator de Necrose Tumoral-alfa (TNF-α) (MOLDOVEANU et
al., 2000; RIVIER et al., 1994), além das interleucinas-1 beta (IL-1β)
(MOLDOVEANU et al., 2000), IL-6 (JANKORD e JEMIOLO, 2004;
MOLDOVEANU et al., 2000; OSTROWSKI et al, 2000), interferons e outras
citocinas (NEMET et al., 2004; RIVIER et al., 1994). A IL-6 é uma citocina pró-
inflamatória chave na fase aguda da resposta inflamatória (OSTROWSKI et al.,
2000). A Concentração plasmática de interleucina-2 (IL-2) reduziu 50%
imediatamente após uma corrida de 5Km, retornando aos valores basais em
torno de 2 horas depois do exercício, e aumentando em cerca de 50% 24 horas
depois da prova (ESPERSEN et al., 1990). A demanda imposta pelo exercício
e a resultante redução na atividade de células NK durante a recuperação estão
relacionadas a uma redução na expressão dos receptores de membrana de IL-
2 em células NK (McFARLIN et al., 2004). Em adultos, foram demonstrados
aumentos significativos na concentração de TNF-α circulante em resposta ao
exercício extenuante (OSTROWSKI et al., 1999), porém, outro estudo não
registrou nenhuma mudança na concentração da mesma citocina (SUZUKI et
al., 2000).
A morte celular programada ou apoptose é um importante mecanismo
para regulação da resposta imune e pode ser induzida pelo exercício físico
(GRANVILLE et al., 1998; OPFERMAN e KORSMEYER, 2003). A corrida de
alta intensidade em esteira foi associada com a expressão de proteínas pró-
apoptóticas (caspase-3 e citocromo citosólica-c) e a redução na expressão da
proteína anti-apoptótica Bcl-2 em linfócitos intestinais c, quando comparado a
um grupo controle não exercitado (QUADRILATERO e HOFFMAN-GOETZ,
2005).
Considerando as alterações imunes apresentadas, o objetivo geral deste
trabalho foi analisar o efeito do exercício físico (natação) em diferentes
intensidades e volumes sobre o sistema imune, mais especificamente sobre

20
número e funcionalidade de linfócitos, bem como a concentração de citocinas
circulantes em ratos.
2. REVISÃO DE LITERATURA
2.1 - Sistema Imunológico
O sistema imunológico é extremamente complexo, pois envolve vários
órgãos, tecidos, tipos de células e moléculas. As células do sistema imune
sintetizam e reorganizam uma variedade de moléculas, incluindo anticorpos,
proteínas de complemento, fatores de crescimento, citocinas e receptores para
as próprias moléculas. A eficiência das respostas imunológicas depende da
interação entre estas células e as moléculas (KELLEY, 2001).
A proteção do organismo é monitorada por dois tipos de imunidade: a
inata e a adquirida, que agem de maneira cooperativa. A inata (natural) possui
o mesmo mecanismo de defesa contra a maioria dos agentes infecciosos, por
isto é considerada um sistema de baixa especificidade. Epitélios, proteínas
sangüíneas, fagócitos, como os neutrófilos e macrófagos, participam deste
mecanismo de defesa. A imunidade adquirida ou específica, responde de
maneira particular aos vários tipos de antígenos, sendo, portanto especializada.
Este sistema é eficiente principalmente por sua memória, que lhe confere
respostas mais vigorosas aos mesmos agentes. Os linfócitos e os anticorpos
por eles produzidos são responsáveis pela ativação deste mecanismo de
defesa (ABBAS et al., 2003).
Estas respostas imunes são altamente dependentes da habilidade dos
leucócitos em migrarem do sangue para os tecidos periféricos, em locais de
inflamação. A migração, rolamento, ativação e forte adesão dos leucócitos
compreendem o clássico paradigma do recrutamento inflamatório celular
(CAVAGLIERI et al., 2000; PATEL et al., 2000).
Famílias específicas de moléculas de adesão mediam cada passo desta
cascata. A migração e o rolamento são as primeiras reações deste processo, e
são predominantemente mediadas por moléculas de adesão, chamadas
selectinas. Três tipos de selectinas foram identificadas selectina-P, E e L. A
selectina-P (CD62P) é primariamente utilizada para armazenar grânulos nas
plaquetas sangüíneas e células endoteliais, sendo rapidamente translocada

21
(dentro de minutos) para a superfície da membrana celular, através da ativação
celular (ADAMS e SHAW, 1994). A selectina-E (CD62E) também é produzida
nas células endoteliais e é gradualmente expressa (dentro de horas) na
superfície da membrana celular, seguida de estimulação por citocinas. A
selectina-L (CD62L) é constantemente expressa em todos os leucócitos, sendo
importante componente no processo inicial de diapedese da adesão celular
endotelial leucocitária, estando associada com o trânsito e inflamação dos
leucócitos (ADAMS e SHAW, 1994).
Fatores indutores, principalmente quimiotáticos derivados do endotélio,
como a interleucina-8 (IL-8) presente na parede dos vasos, podem ativar
moléculas de adesão leucocitárias, como as integrinas. Estas moléculas são
encontradas em vários tipos de leucócitos e mediam a adesão endotelial das
mesmas (BUNTING et al., 2002). As integrinas-β2 (complexo CD11/CD18) ou
integrinas de leucócitos elevam-se rapidamente depois de ativadas,
promovendo forte junção do leucócito ao endotélio vascular e subseqüente
migração transendotelial (NIELSEN e LYBERG, 2004).
NIESEN e LYBERG (2004) propuseram que a molécula de adesão
intercelular-1 (ICAM-1) e a molécula vascular de adesão celular-1 (VCAM-1),
conhecidos ligantes para as integrinas e exercem papel essencial na adesão
de leucócitos ao endotélio vascular.
A inflamação é um processo de defesa que envolve células sangüíneas e
proteínas de um tecido em resposta a uma injúria, infecção, trauma ou reação
imunológica (KELLEY, 2001). O recrutamento de leucócitos circulantes, para o
início da resposta inflamatória, ocorre devido à influência de citocinas
(KATSUHIKO et al., 2003).
As citocinas são hormônios protéicos mediadores e reguladores de
respostas imunes e inflamatórias, produzidas pelas próprias células de defesa,
durante as fases de ativação da imunidade inata e específica. Citocinas pró-
inflamatórias favorecem a produção de reações inflamatórias, sendo estas as
interleucinas: IL-1, IL-6, IL-8, Fator de Necrose Tumoral-α (TNFα) e aquelas
produzidas por células Th1 (IL-2 e interferon-γ) (KELLY, 2001; PLAYFAIR e
LYDYARD, 1999). As citocinas com ação antiinflamatória favorecem a
produção de imunoglobulina E, e ativação e/ou produção de eosinófilos, neste
grupo temos o receptor antagonista de IL-1 (IL-1ra), o Fator de Crescimento de

22
Transformação-β (TGF-β) e as citocinas produzidas por células Th2 (IL-4, IL-5
e IL-10). Um desequilíbrio entre as citocinas pró e anti-inflamatórias pode
induzir respostas inflamatórias ou de hipersensibilidade (alergias) (KELLY,
2001; PLAYFAIR e LYDYARD, 1999).
Referente à inflamação, foi demonstrado que exposições a agentes
externos (bactérias, traumas e exercício) aumentam as proteínas de choque
térmico extracelulares (eHSP72), que agem como “sinalizadores de perigo”
para o sistema imune, potencializando a resposta do óxido nítrico (NO) contra
bactérias e facilitando a recuperação da inflamação bacteriana (FLESHNER e
CAMPISI, 2003).
As células envolvidas com a inflamação aguda são as polimorfonucleares,
já na inflamação crônica observa-se um aumento de células mononucleares,
como, macrófagos, linfócitos T e plasmócitos. Os primeiros mediadores da
resposta inflamatória são a histamina e a serotonina, provenientes
principalmente, de basófilos, que são responsáveis pela vasodilatação e
aumento da permeabilidade vascular. Os mediadores secundários podem ser,
o ácido araquidônico, as enzimas de neutrófilos, as linfocitocinas (citocinas da
imunidade específica) e as monocitocinas da imunidade inata (ABBAS et al.,
2003).
A capacidade do sistema imunológico de não reagir contra seus próprios
antígenos é conhecida como tolerância. O organismo deve ser auto tolerante,
reconhecendo e respondendo apenas aos antígenos estranhos ao corpo. A
resposta do sistema imunológico à estimulação antigênica é conhecida como
auto-imunidade, que pode desencadear as doenças auto-imunes (ABBAS et
al., 2003).
Linfócitos, macrófagos e neutrófilos desempenham um papel central na
resposta imunitária e inflamatória. Linfócitos são células circulantes, têm sua
origem nos tecidos linfóides primários (timo e medula óssea), podendo migrar
para os órgãos linfóides secundários (baço, linfonodos e placas de Peyer).
Estas células encontram-se em estado quiescente até serem estimuladas a
proliferar, por exemplo, durante uma infecção por vírus ou bactérias (STITES et
al., 2000).
Os linfócitos podem ser divididos em duas classes, linfócitos T e linfócitos
B. Estas células se originam na medula óssea, porém as células T

23
amadurecem no timo, enquanto que os linfócitos B amadurecem no mesmo
local de origem. Após o período de maturação, estas células, junto com células
acessórias do sistema imune, se dirigem para tecidos linfóides periféricos
espalhados por todo o organismo. Além de diferentes locais de
amadurecimento, estas células também diferem em suas funções. Os linfócitos
B são responsáveis pela imunidade humoral, mediada por anticorpos por eles
produzidos, que atingem antígenos extracelulares e suas toxinas. Os linfócitos
T conferem a imunidade celular (microorganismos intracelulares), não
produzem anticorpos, mas secretam citocinas e apresentam capacidade de
lisar células que apresentam antígenos estranhos (ABBAS et al., 2003; STITES
et al., 2000).
As células T expressam diferentes proteínas de membrana, as quais são
responsáveis por sua caracterização. Estas células são classificadas a partir de
procedimentos experimentais, onde em cultura, observa-se a formação de
grupos celulares que apresentam a mesma origem e, portanto, as mesmas
características morfofuncionais. Desta forma, as células T são denominadas
pela sigla CD (Grupo de diferenciação), termo que corresponde aos
agrupamentos observados em cultura. Esta sigla é acrescida de um número
que corresponde ao tipo de proteína. Assim, os linfócitos T são divididos em
duas classes, que expressam em suas membranas diferentes proteínas. As
células T auxiliares apresentam a proteína CD4 expressa, estas células
estimulam o crescimento e a diferenciação de células B e ainda ativam
macrófagos através das citocinas secretadas. A outra classe de linfócitos é
conhecida como linfócitos T citotóxicos (LTCs), estes participam da lise de
células infectadas por vírus e células tumorais, e também ativam macrófagos
pela liberação de citocinas. Os LTCs expressam em suas membranas a
proteína CD8. A maioria dos tecidos possui a proporção de CD4 e CD8 de 2:1
(ABBAS et al., 2003; STITES et al., 2000).
Na década de 80, o grupo do Prof. Eric Newsholme determinou que
macrófagos e linfócitos utilizam glutamina em altas taxas como substrato
energético (CURI et al.,1986) e, posteriormente PITHON-CURI et al. (1997)
verificaram que os neutrófilos também utilizam glutamina. A partir destes
estudos, postulou-se que fatores nutricionais poderiam modular a
funcionalidade destas células imunes.

24
2.2 - Efeitos do Exercício Físico sobre o Sistema Imunológico
Estudos realizados in vivo demonstram que a resposta imunitária é
totalmente autônoma. Porém, existe uma nítida interação funcional deste
sistema com o sistema neuroendócrino, que modula a resposta imunitária
frente ao estresse fisiológico, psicológico ou patológico (FLESHNER, 2000;
JONSDOTTIR, 2000). Foi verificado que à prática regular de exercício físico
está associada à redução de alguns tipos de câncer, principalmente o de cólon
e o de mama (DE CARO et al., 2006; GALVÃO e NEWTON, 2005; HARDMAN,
2001; HAYDON et al., 2006; McTIERNAN et al., 1998). Um dos possíveis
mecanismos para esta relação é o fato de que, o exercício físico altera a
concentração e a sensibilidade de alguns hormônios, como por exemplo: a
insulina e o estradiol (YU et al., 2002). Outro possível mecanismo seria a
modulação na funcionalidade do sistema imunitário. HOFFMAN-GOETZ (1999)
observou que o estradiol induziu a diminuição da proliferação de linfócitos T e B
em camundongos fêmeas, sendo que este efeito foi mascarado quando os
animais foram submetidos também ao exercício físico. No entanto, HAYES et
al. (2003) indicaram que apesar das pesquisas demonstrarem que um
programa adequado de exercícios pode induzir efeitos imunomodulatórios
positivos em indivíduos saudáveis, até o momento, não está totalmente
esclarecido o papel do exercício na recuperação da resposta imune, depois de
tratamento quimioterápico contra o câncer.
O exercício físico é caracterizado pelo nosso organismo como estímulo
estressante, e este produz uma descarga simpática e de corticosteróides por
meio do eixo hipotálamo-hipófise-adrenal. Os estímulos estressantes atuam por
intermédio de neurônios aferentes ou diretamente sobre o hipotálamo,
promovendo a secreção do Fator de Liberação de Corticotropina (CRF) e
conseqüentemente a liberação de corticotropina (ACTH) pela hipófise.
SHEPARD et al. (2000) observaram que a expressão das moléculas de adesão
de vários subtipos de leucócitos, incluindo as seletinas, integrinas e membros
das imunoglobulinas, foi alterada pelo exercício agudo e crônico,
provavelmente devido à ação da adrenalina liberada no exercício.

25
A liberação de catecolaminas predomina principalmente, nos primeiros
minutos de exercício, conseqüentemente contribuindo para a elevação dos
linfócitos circulantes (ARLT e HEWISON, 2004; RONSEN et al., 2004). As
concentrações plasmáticas de adrenalina e noradrenalina aumentam quase
que linearmente com a duração do exercício dinâmico e exponencialmente com
a intensidade (SUGIURA et al., 2002). A expressão de receptores beta
adrenérgicos (β-adrenérgicos) nas células T, B, NK, macrófagos e neutrófilos
fornecem base molecular para que estas sejam células-alvo para sinalização
das catecolaminas (NAGAO et al., 2000). Existe uma relação entre número de
receptores adrenérgicos nas subpopulações de linfócitos e sua resposta ao
exercício (WEISE et al, 2004). Os receptores β-adrenérgicos presentes nos
linfócitos estão ligados intracelularmente ao sistema adelinato ciclase, para
geração de adenosina monofosfato cíclico (AMPc), como segundo mensageiro,
atuando no processo de ativação e diferenciação desta célula imune (WIGAL et
al., 2003).
O número de receptores β-adrenérgicos nas células NK aumenta durante
o exercício, sendo que, quando administrado β-bloqueadores adrenérgicos,
este efeito é inibido (PEDERSEN e TOFT, 2000). A adrenalina também
contribui para o recrutamento de células NK, a partir das paredes endoteliais
para a circulação geral, linfonodos, baço e intestino (PEDERSEN e
STEENSBERG, 2002). Dentre os subtipos de linfócitos, as células NK possuem
o maior número de receptores β-adrenérgicos, os linfócitos B e citotóxicos
número intermediário e os linfócitos T auxiliares, o menor número.
(PEDERSEN e STEENSBERG, 2002).
Os corticosteróides podem suprimir várias reações inflamatórias e
imunitárias. Em camundongos, ratos e coelhos, os glicocorticóides provocam
extensa destruição linfóide. Por outro lado, os linfócitos de cobaias, macacos e
seres humanos mostram-se altamente resistentes a lise induzida por esteróides
(JONSDOTTIR, 2000). Os efeitos antiinflamatórios e imunossupressores dos
corticosteróides podem ser decorrentes de suas ações sobre o trânsito celular
e funcionalidade dos leucócitos (STITES et al., 2000). De fato, a infusão de
corticosteróides intravenosa em humanos, causa redução no número de
linfócitos, monócitos e aumento no número de neutrófilos que alcançam seus
valores máximos 4 horas após a administração (PEDERSEN e HOFFMAN-

26
GOETZ, 2000). Em doses suprafisiológicas, os corticosteróides induzem a
morte celular de linfócitos T e B imaturos (SAPOLSKY et al., 2000).
A incubação de timócitos e esplenócitos com corticosteróides, na
presença de concentrações próximas aquelas observadas em exercícios
máximos, induz a apoptose destas células (HOFFMAN-GOETZ e
ZAJCHOWSKI, 1999). A apoptose induzida pelo exercício pode contribuir para
o quadro de linfopenia e redução da resposta imune depois do exercício
intenso (RONSEN et al., 2001a; 2004). A concentração de cortisol aumenta em
resposta ao exercício intenso e principalmente no de longa duração
(BUTCHER et al., 2005; DAVIS et al., 2000; DIMITRIOU et al., 2002; WEISE et
al., 2004). Esta elevação pode promover a entrada de neutrófilos provenientes
da medula óssea para a circulação, enquanto inibe a entrada de linfócitos,
facilitando seu regresso para outros tecidos periféricos (BUTCHER et al.,
2005). Desta forma, o cortisol que poderá permanecer elevado até 1,5 hora
após os exercícios de endurance, promove redução no número de linfócitos
circulantes (KANALEY et al., 2001).
Adicionalmente, o cortisol reduz os receptores de IL-1 e IL-2 nas células
T. A conseqüência imediata destas ações é a redução da atividade e
capacidade proliferativa das células NK e B (RONSEN et al., 2001a; 2004). A
longo prazo, elevações crônicas de cortisol podem aumentar a razão
catabólica, modificando as reservas de aminoácidos disponíveis para o
crescimento e proliferação de linfócitos (ARLT e HEWISON, 2004; RONSEN et
al., 2004).
Estudos observaram que, exercícios físicos intensos e de curta duração
elevaram o número total de leucócitos no sangue numa relação diretamente
proporcional à intensidade do exercício, sendo que, este aumento ocorre
principalmente na série granulocítica e em especial nos poliformonucleares
(BENONI et al., 1995; HOST et al., 1995). O número de monócitos e de
linfócitos também aumenta, mas em menor escala (HOST et al., 1995;
MOOREN et al., 2004; NIEMAN, 1994b). Dentre a subpopulação de linfócitos,
as células “Destruidoras Naturais” (NK) são as que mais aumentam (TIMMONS
et al., 2006). Um dos mecanismos propostos para explicar esta linfocitose
passageira, pode ser em decorrência dos efeitos da adrenalina induzida pelo
exercício (ORTEGA et al., 2003). O número de linfócitos começa a diminuir

27
cinco minutos após o término do exercício, provavelmente devido ao efeito
persistente do cortisol liberado durante o mesmo, diferentemente da adrenalina
que decresce logo após o fim do exercício físico (HOST et al., 1995). Em geral,
quatro a seis horas depois de encerrado o exercício físico e, com certeza após
24 horas de repouso, o número de linfócitos circulantes retorna aos valores
basais (MEYER et al., 2004). Com relação à função dos demais linfócitos, foi
observado também que o cortisol pode reduzir a capacidade mitogênica, nas
primeiras horas após o término do exercício físico (HOST et al., 1995; MALM,
2004).
Foi verificado um aumento médio de 3.5 vezes no número de monócitos
CD14+ CD16+, mesmo num curtíssimo período de exercício (1 minuto). Em
condições de repouso, a maioria (>75%) dos monócitos CD14+ CD16+ ficam
aderidos à parede endotelial dos vasos sangüíneos, e não no sangue
periférico. A rápida mobilização destas células a partir das paredes endoteliais,
em condições de estresse, deve servir para fornecer uma grande população
ativa de células para defesa em locais de lesão e infecção (STEPPICH et al.,
2000).
Em outro estudo, com maior volume de exercício, 10 homens fisicamente
ativos de idade entre 18-25 anos realizaram duas sessões de endurance de 20
minutos cada (5 min a 50% do VO2max e 15 min a 70% VO2max) no mesmo
dia, separadas por quatro horas de descanso, observou-se aumento no número
total de leucócitos circulantes, neutrófilos e contagem de linfócitos, com pouco
efeito sobre a atividade da célula NK. As alterações no sistema imune frente a
duas sessões de endurance foram mais pronunciadas, quando comparadas a
uma única sessão (MACFARLIN et al., 2003).
Recentemente, NIEMAN et al. (2005) analisaram o efeito da caminhada
de 30 minutos sobre a resposta imune. Esta atividade é executada em grande
escala pela população, com objetivos de melhorar a aptidão física e saúde.
Neste estudo, foram avaliadas 17 mulheres com idades entre 25 e 55 anos,
saudáveis que realizavam caminhadas a pelo menos 3 meses anteriores ao
experimento, com uma freqüência de 2-7 dias por semana. Verificou-se que
esta atividade realizada na intensidade de 60%-65% do VO2max foi associada
com modestas e temporárias alterações na contagem de leucócitos
(especialmente neutrófilos e células NK), proliferação de linfócitos induzida pela

28
fitoemaglutinina (PHA), e concentração plasmática de IL-6. Nenhuma mudança
na concentração plasmática de cortisol, produção de IgA salivar, ou
concentração plasmática de IL-1ra foi observada (NIEMAN et al., 2005).
Os efeitos citados acima contrastam com a maioria das alterações
registradas após exercício intenso e prolongado, como uma maratona
(NIEMAN et al., 2001). Estes dados foram observados em praticantes regulares
de exercício físico, sendo assim, devemos ter cautela ao extrapolar estes
efeitos e aplicá-los a indivíduos sedentários.
Em animais experimentais, DAVIS et al. (2004) verificaram que 1 hora de
exercício moderado, durante 6 dias, melhorou a competência da resposta de
macrófagos ao vírus da herpes tipo 1. Adicionalmente, observaram redução na
susceptibilidade à ITRSs. No entanto, mais estudos devem ser realizados para
estabelecer uma relação entre mudanças imunes agudas e melhora da
resistência imunológica.
Nesta linha de pensamento, outro estudo com ratos, demonstrou que a
corrida de alta intensidade em esteira está associada com aumento na
expressão de proteínas pró-apoptóticas (caspase-3 e citocromo citosólica-c) e
redução na expressão da proteína anti-apoptótica Bcl-2 em linfócitos intestinais
c, quando comparados ao grupo controle não exercitado. Adicionalmente,
devido ao fato de a Bcl-2 e a citocromo c serem proteínas específicas de
mitocôndria, foi sugerido que a apoptose de linfócitos intestinais durante o
exercício, pode ser desencadeada por um mecanismo oxidativo-dependente
através de uma via mitocondrial (apoptótica intrínseca) (QUADRILATERO e
HOFFMAN-GOETZ, 2005).
No caso do treinamento de força, característico de ações excêntricas, a
proteólise muscular pode estar aumentada. Neste sentido, WILLOUGHBY et al.
(2003) concluíram que uma única sessão deste tipo de exercício em indivíduos
destreinados, é efetiva no aumento da atividade da protease caspase-3
apoptótica e de suas vias proteolíticas relacionadas, associadas com lesão
muscular e força muscular reduzida. Isso indica que o exercício excêntrico
pode induzir apoptose no músculo esquelético através do aumento da atividade
da caspase-3.
Muitos dos eventos fisiológicos atribuídos a apoptose celular podem
ocorrer após exercícios prolongados e/ou intensos (GRANVILLE et al., 1998;

29
OPFERMAN e KORSMEYER, 2003). Mudanças nas características
bioquímicas e moleculares da apoptose como: níveis aumentados de caspase
3 (PATEL e HOFFMAN-GOETZ, 2002), aumento do receptor Fas (receptor de
morte celular) (MOOREN et al., 2002), liberação de Ca2+ (AZENABOR e
HOFFMAN-GOETZ, 2000), externalização de fosfatidilserina (HOFFMAN-
GOETZ e QUADRILATERO, 2003), despolarização da membrana mitocondrial
(HSU et al., 2002), expressão dos genes específicos de apoptose como Bax,
Bcl-xs, e Bcl-xL, condensação de cromatina (KUMAZAKI et al., 2002), e
fragmentação de DNA (LIN et al., 1999) foram observadas em leucócitos de
humanos e ratos depois de exercício extenuante. A apoptose e necrose foi
observada em linfócitos depois de exercício intenso agudo aeróbio
(HOFFMAN-GOETZ e DUERRSTEIN, 2004).
Contudo, admite-se que exercícios muito intensos são capazes de
danificar uma quantidade de tecido muscular suficiente para desencadear uma
resposta inflamatória aguda, que segundo ORTEGA et al. (2003) envolve
reações complexas moduladas pelo sistema imunitário através da liberação de
citocinas.
As citocinas são glicoproteínas, produzidas por diferentes tipos de células
do sistema imunitário que tem como função principal mediar a comunicação
entre as células do sistema imunitário e as de outros tecidos (MOLDOVEANU
et al., 2001). As citocinas inflamatórias são moduladas por vários estímulos,
incluindo o exercício físico, trauma e infecção. Este tipo de atividade afeta a
produção sistêmica de citocinas, principalmente o Fator de Necrose Tumoral
alfa (TNF-α) (MOLDOVEANU et al., 2000; RIVIER et al., 1994), além das
interleucinas-1 beta (IL-1β) (MOLDOVEANU et al., 2000), IL-6 (JANKORD e
JEMIOLO, 2004; MOLDOVEANU et al., 2000; OSTROWSKI et al, 2000),
interferons e outras citocinas (GARCIA et al., 1999; NEMET et al., 2004;
RIVIER et al., 1994).
Vários mecanismos foram propostos na tentativa de explicar a
suscetibilidade de atletas de endurance à infecções respiratórias (NIEMAN,
1994b). Foi observada uma relação direta entre o aumento da concentração
plasmática de IL-6, exercícios extenuantes e aumento de sepsis e infecções
respiratórias (OSTROWSKI et al., 2000; TOTH et al., 2006; XING et al., 1998;
YENDE et al., 2006). A IL-6 é uma citocina pró-inflamatória chave na fase

30
aguda da resposta inflamatória. É produzida por diferentes tipos de células, as
quais são originalmente estimuladas pelos monócitos. Foi verificada uma
ligação entre sua concentração plasmática e a intensidade da corrida
(OSTROWSKI et al., 2000), o que poderíamos hipoteticamente relacionar com
o aumento proporcional de lesão muscular.
A redução na concentração plasmática de glutamina pode também estar
associada como um possível fator causal da supressão imunológica (CURI,
2000; KEAST et al., 1995; NEWSHOLME et al., 1987; 1988; 1989; 1997;
PARRY-BILLINGS et al., 1990; ROWBOTTOM et al., 1996). No caso de
ciclistas, a relação glutamina/glutamato provou ser um importante indicativo de
treinamento excessivo, que se caracteriza por um período de queda na
capacidade de performance e a recuperação pode levar de alguns dias a várias
semanas. Apesar de não terem sido detectadas mudanças estatisticamente
significativas nas células do sistema imune e nas concentrações basais de
citocinas circulantes (IL-6 e TNF-α), os autores não descartaram que este
mecanismo pode estar envolvido no processo de supertreinamento (HALSON
et al., 2003). Ainda, RIETJENS et al. (2005) também registraram que em
ciclistas bem treinados, as mudanças na distribuição dos leucócitos não foram
consistentes, nem sensíveis o bastante para detectar a síndrome do excesso
de treinamento em seus estados iniciais.
Em condições normais, as concentrações plasmáticas de glutamina são
mantidas por um balanço entre a produção e utilização de glutamina por vários
órgãos. Dentre estes, o cérebro, os pulmões, fígado, músculo esquelético e
possivelmente o tecido adiposo liberam glutamina. No entanto, células do
sistema imune, fígado, rins e trato gastrintestinal são utilizadores primários da
mesma. Para determinar se uma célula produz ou consume glutamina analisa-
se a direção de uma única reação reversível, na qual a glutamina é sintetizada
da amônia e glutamato pela glutamina sintetase, e a glutaminase catalisa a
reação reversa para formar amônia e glutamato através da glutamina
(CASTELL, 2003; NEWSHOLME e CALDER, 1997).
Usualmente, sessões de treinamento e competição no ciclismo de rua
demoram várias horas. Está bem estabelecido que o exercício prolongado
pode induzir imunossupressão temporária chamada de “janela aberta”
(NIEMAN, 1999; PEDERSEN e ULLUM, 1994). NIEMAN (1994a) propôs a

31
curva em “J”, que descreve a relação entre intensidade do exercício e
possibilidade de infecção. De acordo com esta hipótese, o exercício moderado
protegerá o indivíduo de infecções, enquanto que, o exercício intenso
aumentará o número de episódios infecciosos num determinado período
(Figura 1).
Porém, nos atletas avaliados para formulação desta curva em “J”, não foi
distinguido a diferença entre carga “alta” e de “elite” para o exercício. Sendo
que, quando atletas de elite são inclusos no modelo, a relação entre carga de
exercício e risco de infecções sugerida deveria ter a forma de “S”, onde as
intensidades consideradas de “elite”, teriam menor possibilidade de induzir a
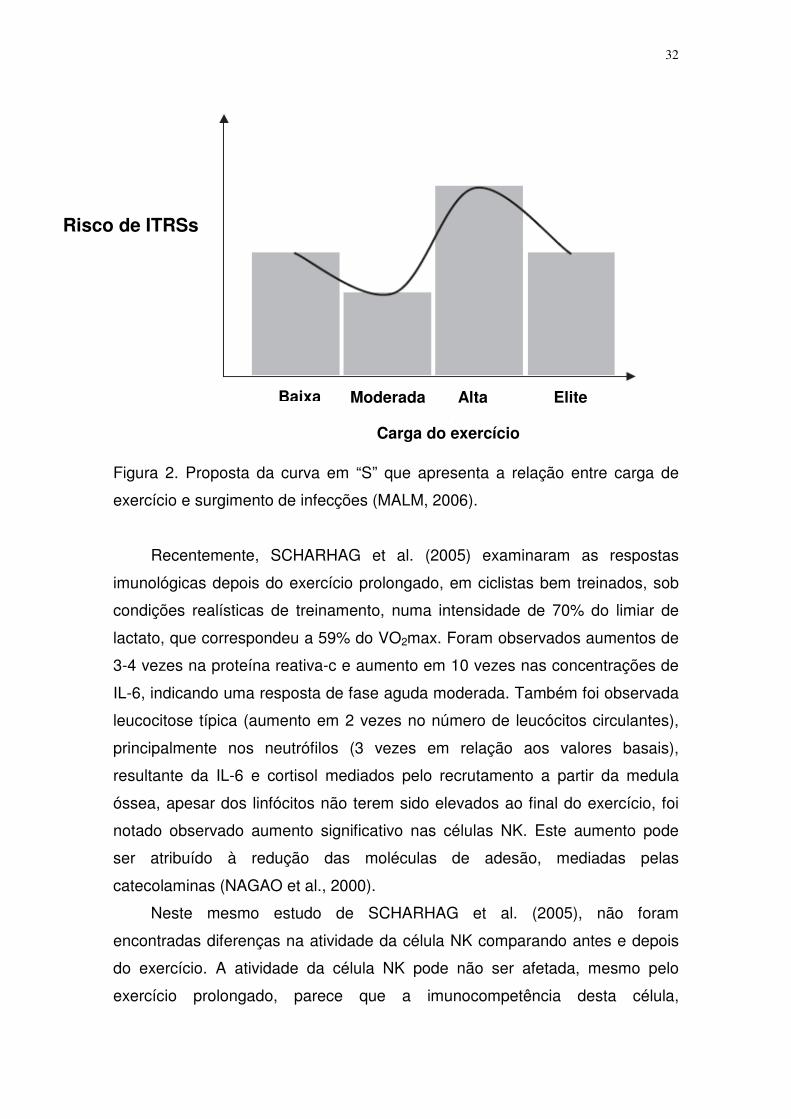
infecções em relação as intensidades “altas” (Figura 2) (MALM, 2006).
Para melhor visualização e entendimento destas duas propostas,
apresentamos abaixo os dois modelos de relação da carga de exercício e
possibilidade de infecções.
Figura 1. Modelo da curva em “J” da relação entre carga de exercício e
surgimento de infecções (NIEMAN, 1994a).
Sedentário Moderado Alto
Média
Abaixo da Média
Acima da Média
Risco de ITRSs
32
Figura 2. Proposta da curva em “S” que apresenta a relação entre carga de
exercício e surgimento de infecções (MALM, 2006).
Recentemente, SCHARHAG et al. (2005) examinaram as respostas
imunológicas depois do exercício prolongado, em ciclistas bem treinados, sob
condições realísticas de treinamento, numa intensidade de 70% do limiar de
lactato, que correspondeu a 59% do VO2max. Foram observados aumentos de
3-4 vezes na proteína reativa-c e aumento em 10 vezes nas concentrações de
IL-6, indicando uma resposta de fase aguda moderada. Também foi observada
leucocitose típica (aumento em 2 vezes no número de leucócitos circulantes),
principalmente nos neutrófilos (3 vezes em relação aos valores basais),
resultante da IL-6 e cortisol mediados pelo recrutamento a partir da medula
óssea, apesar dos linfócitos não terem sido elevados ao final do exercício, foi
notado observado aumento significativo nas células NK. Este aumento pode
ser atribuído à redução das moléculas de adesão, mediadas pelas
catecolaminas (NAGAO et al., 2000).
Neste mesmo estudo de SCHARHAG et al. (2005), não foram
encontradas diferenças na atividade da célula NK comparando antes e depois
do exercício. A atividade da célula NK pode não ser afetada, mesmo pelo
exercício prolongado, parece que a imunocompetência desta célula,
Alta Elite Moderada Baixa
Carga do exercício
Risco de ITRSs

33
aumentada ou atenuada é mais uma questão de redistribuição numérica. A
capacidade fagocitária de neutrófilos e monócitos não foi afetada pelo
exercício, tendo sido observada apenas pequena redução na capacidade
oxidativa de neutrófilos. Sendo assim, o ciclismo prolongado em intensidade
moderada parece não afetar substancialmente a função de células na primeira
linha de defesa, parecendo ser seguro do ponto de vista imunológico.
Foi sugerido que indivíduos com alta aptidão cardiorrespiratória tem
respostas celulares imunes atenuadas frente ao exercício agudo (20 minutos
de corrida em esteira a 65-70% do VO2pico estimado. Devido ao fato de
indivíduos fisicamente ativos terem apresentado uma menor resposta imune
relacionado a sua aptidão cardiovascular, estes exibiriam respostas imunes
ainda menores a mesma quantidade absoluta de exercício realizada por
indivíduos menos aptos (HONG et al., 2005). Um papel protetor do exercício
contra a imunossupressão induzida pelo estresse foi encontrada em estudos
com animais (FLESHNER et al., 2002; MORASKA e FLESHNER, 2001). O
exercício regular inibiu as reduções no número de células T CD4+ normalmente
observadas depois de exercício exaustivo em ratos (FU et al., 2003).
No entanto, a imunoproteção contra vários estressores em indivíduos
fisicamente ativos permanecem inconclusivos, a demarginação atenuada de
linfócitos em resposta ao stress físico e fisiológico claramente demonstra que a
aptidão física altera a resposta imune em humanos (HONG et al., 2004).
Com a intenção de analisar as associações entre longos períodos de
treinamento, com diferentes intensidades e durações por mais de 10 anos, os
parâmetros imunes celulares e humorais basais em atletas mulheres de meia
idade foram avaliados e comparados com um grupo controle. Neutrófilos,
eosinófilos, basófilos, linfócitos, monócitos e subpopulações de linfócitos, como
total de células T, B, células T CD4+, células T CD8+, e células NK do grupo
com treinamento de alta intensidade e longa duração (atletas de competição) e
do grupo de treinamento moderado (atletas recreacionais) não foram
estatisticamente diferentes do que o grupo controle não exercitado.
Desta maneira, neste estudo não foram obtidos dados para dar suporte a
imunossupressão induzida pelo treinamento de alta intensidade e longa
duração, a não ser pela correlação negativa com células T auxiliares CD4+ e
razão CD4/CD8, sendo considerado um marcador indireto de supressão da

34
imunidade celular. No entanto, foi demonstrada uma correlação positiva entre o
VO2max do grupo exercício moderado com IgG (imunoglobulina G), sendo esta
intensidade considerada benéfica para sistema imune humoral (BUYUKYAZI,
et al., 2004).
Através da análise do impacto de seis semanas de treinamento intenso de
natação em ratos a atividade proliferativa das células T e B foram reduzidas,
sendo que estes são indicativos para imunidade celular e humoral. Concluindo
que o treinamento pesado de longa duração resulta em maior suscetibilidade a
doenças e infecção, podendo ser, em parte, responsável pela síndrome do
sobretreinamento (PEIJIE et al., 2003).
No aspecto das concentrações de imunoglobulina-A salivar (IgA), esta
mostrou correlação inversa com o número de infecções do trato respiratório
superior, o que indica que a mensuração de IgA durante um período de
treinamento ou exercícios, pode predizer imunossupressão (GLEESON e PINE,
2000). PUTLUR et al. (2004), registraram concentrações reduzidas de IgA
salivar em jogadoras de futebol universitárias, no curso de 9 semanas de uma
temporada competitiva quando comparadas ao grupo controle, sendo que, a
freqüência de lesões e doenças também foi superior neste grupo.
Citocinas e Exercício Físico
Interleucina-2 (IL-2)
A IL-2 exerce efeitos regulatórios na maioria das células corporais,
especialmente células imunes, sendo primariamente produzidas pelos linfócitos
T e linfócitos NK. Esta citocina exerce um importante papel nas respostas
celulares humorais e inflamatórias (JANEWAY e TRAVERS, 1996). O exercício
demonstrou suprimir a função dos linfócitos T e NK, possivelmente assim,
comprometendo a produção de citocinas a partir destas células (STARKIE et
al., 2001c). As ações da IL-2 incluem a estimulação da proliferação e
diferenciação de células B e T, elevação da citotoxidade de linfócitos tanto
pelas células NK como pelas células citotóxicas, ativação de

35
monócitos/macrófagos, e a liberação de outras citocinas como TNF-α e IFNΎ
(STEENSBERG et al., 2001b).
A concentração plasmática de IL-2 pode exibir padrão complexo de
mudança em resposta ao exercício. Por exemplo, em corredores treinados, a
concentração plasmática de IL-2, mensurada por radioimunoensaio, reduziu
50% imediatamente depois uma prova de 5 Km, retornando aos valores basais
em torno de 2 horas depois do exercício, e então aumentou cerca de 50% 24
horas depois da prova (ESPERSEN et al., 1990). A demanda imposta pelo
exercício e a resultante redução na atividade de células NK durante a
recuperação estão relacionadas a uma redução na expressão dos receptores
de membrana de IL-2 em células NK (McFARLIN et al., 2004). Desta maneira,
o exercício de endurance de alta intensidade demonstrou causar uma
significativa redução no número de células T (IBFELT et al., 2002).
Foi sugerido que a redução transitória na concentração de IL-2
imediatamente depois do exercício pode refletir um concomitante aumento no
número de linfócitos ativados que expressam o receptor de IL-2, sendo que,
mais células com mais receptores removeriam rapidamente a IL-2 da
circulação. Este conceito é sustentado por duas observações: 1) alta elevação
no número de células NK CD16+ imediatamente depois do exercício, estas
células exibem maior densidade de receptores de IL-2 entre todos os linfócitos
(SHEK et al., 1995); e 2) rápido aumento na presença do receptor de IL-2 na
urina 1 hora depois de corrida de distância (SPRENGER et al., 1992).
Em contraste, a IL-2, mensurada por ELISA, não foi detectável no
plasma 24 horas depois de 20 Km de corrida, ou até 2 dias depois de 2 horas
de ciclismo a 70-75% do VO2max em ciclistas treinados (MACKINNON, 1999),
nem 5 dias depois contrações excêntricas máximas dos extensores do cotovelo
(NOSAKA e CLARKSON, 1996).
A produção de IL-2 foi reduzida em células do baço quando comparados
ratos exercitados (corrida extenuante aguda em esteira em velocidades
gradualmente maiores até fadiga voluntária) com grupo controle 2 dias depois
do exercício. As células do baço dos animais controle, em cultura, produziram
significativamente mais IL-2 do que as células dos animais exercitados. Em
períodos posteriores da cultura celular, não houve mais nenhuma diferença na
produção de IL-2. Estas alterações na cinética da IL-2 e outras citocinas (IL-6,

36
IL-12 e TNF-α) podem estar associadas ao aparecimento de infecções
oportunistas devido a imunossupressão transitória observada, no entanto, mais
estudos são necessários para esclarecer e confirmar os mecanismos
fisiológicos destes fatos (KOHUT et al., 2001).
Interleucina-6 (IL-6)
A interleucina-6 (IL-6) é uma molécula de sinalização intercelular
tradicionalmente associada com o controle e coordenação de respostas
imunes, esta é primeiramente secretada pelos macrófagos e linfócitos em
resposta a lesão ou infecção (PEDERSEN e TOFT, 2000). Com relação ao
músculo esquelético, a elevação de citocinas proinflamatórias como a IL-6 está
inserida no contexto de impactos deletérios, geralmente associada como
conseqüência de lesão no tecido muscular induzida por atividades de alta
intensidade ou ações excêntricas (KELLER, et al., 2001). No entanto, a
produção da IL-6 durante o exercício pode estar presente mesmo na ausência
de lesão muscular induzida pelo exercício (STARKIE et al., 2001b).
Quando os conteúdos de glicogênio estiverem comprometidos, pode
ocorrer crise energética no músculo em contração, afetando tanto o
metabolismo dos carboidratos como o lipídico, se o período de recuperação
entre as sessões de exercício for pequeno e a intensidade do trabalho elevada
(STICH et al., 2000). Por exemplo, existem observações de que esta citocina
direta ou indiretamente media efeitos catabólicos sobre o músculo esquelético
(GRIMBLE, 2003).
HADDAD et al., (2005) realizaram a infusão de IL-6 no músculo
esquelético in vivo em ratas. A dose foi determinada a partir de uma recente
observação que analisou a concentração de IL-6 encontrada depois de
exercício extenuante em humanos, que foi de 22 ng/l (STEENSBERG et al.,
2001a). Neste caso, devido à ausência dos efeitos sistêmicos (infusão direta no
músculo) foi detectada significativa atrofia muscular em ratas saudáveis. As
mudanças moleculares e celulares observadas nos músculos com infusão de
IL-6 sugerem que, mecanismos de retro alimentação negativa, via supressores
da sinalização de citocinas (SOCS) foram ativados, alterando o balanço da
fosforilação de proteínas sinalizadoras da via de transdução para ativação de

37
transcrição gênica (STATs), em favor de um perfil mais catabólico (HADDAD et
al., 2005).
Similarmente o seu papel no sistema imune, a liberação de IL-6 induzida
pelo exercício assumidamente regula componentes da fase de resposta aguda,
incluindo fase aguda de síntese proteica pelo fígado e liberação de
glicocorticóides via estimulação do eixo hipotalâmico-hipofisário-adrenal
(PEDERSEN et al, 2001b). Em outro contexto, a IL-6 parece estar relacionada
ao crescimento deficiente em crianças sob vários estados de doença
(BENEDETTI, et al., 1997) e ao processo de sarcopenia (redução do número
de sarcômeros em série) em idosos (BARBIERI et al., 2003).
Concentrações elevadas de IL-6 foram registradas imediatamente após
uma maratona e também em resposta ao exercício físico (PEDERSEN e
HOFFMAN-GOETZ, 2000; STEENSBERG et al., 2002). Em relação a
exercícios de características concêntricas e excêntricas, a IL-6 é produzida em
maiores quantidades do que qualquer outra citocina (PEDERSEN et al.,
2001a).
Em conseqüência de duas sessões prolongadas de ciclismo realizadas
por 9 atletas de endurance bem treinados, pode-se observar que uma segunda
sessão de alta intensidade no mesmo dia foi associada com aumentos mais
pronunciados na IL-6 e IL-1ra comparado com uma única sessão de exercício
similar. No entanto, uma tendência de atenuação na resposta aumentada
destas citocinas foi observada quando o período de intervalo entre as duas
sessões de exercício foi entendido de 3 para 6h e uma refeição adicional foi
fornecida (RONSEN et al., 2002). Os autores argumentaram que estas
respostas aumentadas frente ao exercício podem estar ligadas à depleção de
glicogênio muscular e aumentada mobilização de substrato por outros tecidos.
SUZUKI et al. (2003), demonstraram que uma maratona de 42.195-Km
induziu a liberação sistêmica de IL-6 e IL-8, sendo que estas citocinas mediam
o recrutamento e à ativação de neutrófilos e monócitos. No caso de garotas de
14-16 anos de idade, a pratica de uma sessão de pólo aquático resultou em
significativos aumentos nas concentrações circulantes de IL-6, mas o efeito
destas mudanças em resposta ao exercício sobre o processo de crescimento e
desenvolvimento em adolescentes ainda precisa ser mais bem estudado
(NEMET et al., 2003).

38
Foi sugerido que as elevações na IL-6 em resposta ao exercício podem
exercer um papel antiinflamatório, principalmente pela inibição na produção de
TNF-α, uma citocina tipicamente proinflamatória (PEDERSEN et al., 2003a).
Este mecanismo ocorre à medida que a de IL-6 estimula a produção de IL-1ra,
que se liga e bloqueia o receptor da IL-1β, exercendo assim, potentes efeitos
antiinflamatórios (HORN et al., 2000). Com o exercício, o pico de IL-1ra foi
encontrado 1-2h depois do pico de IL-6, sendo assumido que as concentrações
de IL-1ra refletem a produção de IL-6 (OSTROWSKI et al., 1999).
Adicionalmente, a IL-6 mostrou ter participação no controle de vias metabólicas
durante o exercício (PEDERSEN et al., 2003b).
Apesar de inconclusivo, existem evidências que a IL-6 pode afetar o
metabolismo lipídico, sendo que, a infusão de IL-6 em ratos aumentou as
concentrações de ácidos graxos e triacilgliceróis de uma forma dose
dependente. A hipertrialcilglicerídemia foi causada pela secreção renal
aumentada da mesma e liberação não reduzida (NONOGAKI et al., 1995).
Contudo, ratos deficientes em IL-6 desenvolveram obesidade antecipadamente
e quando foram tratados com IL-6 durante 18 dias, houve uma redução
significativa no peso corporal destes animais (WALLENIUS et al., 2002).
Neste aspecto, a eficácia da IL-6 foi testada como fator lipolítico em
mulheres ativas saudáveis. Os resultados encontrados foram que a infusão de
IL-6 aumentou a lipólise na ausência de hipertrialcilglicerídemia, mudanças nas
catecolaminas, glucagon, ou insulina, sem causar efeitos colaterais. Assim,
esta citocina é uma importante moduladora do metabolismo lipídico em
humanos, devido ao aumento na oxidação de gorduras e reesterificação de
ácidos graxos (GERRIT et al., 2003).
Fator de Necrose Tumoral-α (TNF-αααα)
O Fator de Necrose Tumoral (TNF-α) está inserido no grupo de citocinas
proinflamatórias atuando de maneira significativa na coordenação das
respostas inflamatórias do organismo. Tendo em vista que o exercício físico
pode romper a homeostasia celular, não é surpreendente que este possa
elevar as concentrações plasmáticas de várias citocinas. O TNF-α induz

39
efetivamente respostas inflamatórias locais e auxilia no controle de infecções.
No entanto, a produção sistêmica de TNF-α pode levar a sepsi, choque, e
morte (GOEBEL et al., 2000).
Sessões de exercício que induzem lesão de células musculares e altas
sobrecargas metabólicas causam uma liberação seqüencial de TNF-α e outras
citocinas tipicamente proinflamatórias (IL-1β e IL-6), o TNF-α por sua vez
estimula a produção de IL-6, que induz a fase de resposta aguda e a produção
de IL-1ra. (HENSON et al., 2000). As citocinas inflamatórias auxiliam na rápida
regulação da migração de neutrófilos e, posteriormente monócitos para áreas
de células musculares lesadas a outros tecidos metabolicamente ativos para
iniciar reparo (NIEMAN et al., 2001).
O exercício de endurance associado com microlesão muscular induz a
maiores respostas inflamatórias destas citocinas do que ciclismo, tênis,
patinação, que são concêntricos ou intermitentes em intensidade (NIEMAN et
al., 2000). Em adultos, foram demonstrados aumentos significativos no TNF-α
circulante em resposta ao exercício extenuante (OSTROWSKI et al., 1999),
porém, outro estudo não registrou nenhuma mudança na mesma citocina
(SUZUKI et al., 2000). No entanto, RHIND et al. (2001), mostraram que a
exposição prolongada ao frio (mesmo com a manutenção da temperatura
corporal normal) reduz a expressão intracelular de TNF-α e induz a redução
das concentrações séricas desta citocina depois do exercício.
Além disso, a grande quantidade de pesquisas examinando os efeitos
imunológicos das elevações no TNF-α, recentes estudos tem centrado sobre os
feitos metabólicos desta citocina. Esta mostrou ser expressa no músculo
esquelético humano (SAGHIZADEH et al., 1996; STARKIE et al., 2001a),
estando associado também com a resistência à insulina e a diabetes tipo 2
(KERN et al., 2001). Possivelmente, o músculo esquelético pode estar
associado aos aumentos do TNF-α (STEENSBERG et al., 2002). Apesar de
altas concentrações do RNAm de TNF-α terem sido demonstradas no tecido
muscular em pacientes com resistência a insulina, está menos claro se existe
uma relação causal entre insulina e TNF-α (KROGH-MADSEN et al., 2004).

40
3 - Objetivos da pesquisa
3.1 - Objetivo Geral
Investigar o efeito de exercícios físicos nas intensidades leve e moderada,
sobre o número e funcionalidade de linfócitos e, concentração de citocinas
circulantes em ratos, usando como modelo de exercício a natação.
3.2 - Objetivos Específicos
Avaliar os efeitos do exercício físico leve e moderado sobre:
1) Número total de leucócitos circulantes;
2) Leucograma diferencial (percentual de monócitos, neutrófilos e linfócitos
circulantes);
3) O número de linfócitos teciduais originários dos linfonodos mesentéricos;
4) O número de linfócitos circulantes;
5) A concentração sérica de Interleucina-2 (IL-2), Interleucina-6 (IL-6) e Fator
de Necrose Tumoral-α (TNF-α);
6) Funcionalidade de linfócitos circulantes (viabilidade, potencial
transmembrânico mitocondrial e fragmentação de DNA).

41
4. MATERIAIS E MÉTODOS
4.1 - Animais
Procedência e manutenção
Ratos da linhagem Wistar (Rathus novergicus var, albinus, Rodentia,
Mamalia), com 2 meses de idade, que foram obtidos do biotério Central da
Universidade Metodista de Piracicaba. Todo o experimento foi conduzido de
acordo com a política para pesquisas com animais experimentais do American
College of Sports Medicine.
Os animais receberam água e alimentação ad libitum e foram mantidos
em ambiente com temperatura constante de 23°C ± 2°C e ciclo claro/escuro de
12/12 horas, com luz acesa a partir das 6 horas, e mantidos em gaiolas
coletivas (5 animais por gaiola). Antes de iniciar o período experimental, os
animais permaneceram por 48 horas em adaptação às condições do biotério de
pesquisa.
4.2 - Grupos experimentais
Os animais foram divididos em 3 grupos: a) grupo controle – animal
sedentário; b) grupo com atividade física aguda - animal submetido à natação
durante 5, 15 minutos ou até a exaustão; c) grupo exercitado por 5 sessões.
4.3 - Grupo Controle
Composto por animais sedentários, não tendo nenhuma participação nos
protocolos de exercício físico (n-6), sendo identificados com o código (C).

42
4.4 - Grupos Submetidos a uma (1) sessão de Exercício Físico
Composto por animais sedentários, que realizaram uma única sessão de
exercício físico, com duração de 5, 15 minutos ou exaustão (n-6), tanto na
intensidade leve como na intensidade moderada.
Os animais que se exercitaram na intensidade leve foram
respectivamente identificados com os códigos (5L), (15L) e (EXL), e os animais
que se exercitaram na intensidade moderada foram identificados
respectivamente como (5M), (15M) e (EXM).
4.5 - Grupo Exercitado durante cinco (5) sessões
Composto por animais sedentários, que foram submetidos a 5 sessões
de natação, tanto na intensidade leve, como na intensidade moderada. Os
animais realizaram 5 sessões (1 por dia / durante 5 dias), sendo que as
sessões seguiram uma ordem crescente no aumento do volume (5, 15, 30, 45
e 60 minutos).
Os animais que realizaram 5 sessões de exercício em intensidade leve
foram identificados com o código (5SL) sendo que, os animais que realizaram a
5 sessões em intensidade moderada foram identificados com o código (5SM).
Com exceção da dosagem sérica de citocinas circulantes onde utilizamos n=3
animais, em todas as outras dosagens descritas foi utilizado n=6 animais para
cada grupo experimental.

43
4.6 - Exercício Físico
Para realização do exercício na intensidade moderada, cargas adicionais
com 5% do peso corporal dos animais foram acopladas em suas regiões
dorsais, o que corresponde a uma intensidade abaixo do limiar anaeróbio
(GOBATTO et al., 2001; VOLTARELLI et al., 2002), sendo que, no exercício de
intensidade leve não se fez uso de cargas adicionais.
Todos os protocolos foram realizados no período da tarde, entre 14 e 17
horas. Os animais dos grupos experimentais foram submetidos separadamente
ao exercício físico e sacrificados de 3 a 4 minutos após o final das sessões de
exercício estipuladas, para análise das variáveis sanguíneas e teciduais. O
sangue foi previamente armazenado em tubo de vidro com EDTA (100 µl para
3,5 ml de sangue), e mantido em gelo para análise da leucometria e
leucograma diferencial. Todas as coletas sangüíneas e teciduais foram
realizadas no mesmo período do dia (entre 14 e 17 horas), incluindo o grupo
controle, submetendo todos os animais à mesma influência do ritmo circadiano
hormonal, minimizando desta maneira, possíveis alterações fisiológicas
decorrentes de coletas realizadas em momentos diferentes do dia, que
poderiam alterar nossos resultados.
4.7 - Descrição Metodológica dos Parâmetros Analisados
4.7.1 - Leucometria
O protocolo para a determinação da leucometria foi realizado através
das seguintes etapas: Os animais foram sacrificados após o exercício físico de
natação ou sem exercício (grupo controle); O sangue foi colhido em tubo de
vidro que continha EDTA - anticoagulante para hematologia (100 µl para 3,5 ml
de sangue); Uma alíquota de 10 µL do sangue foi retirada, colocada em um
tubo de plástico e acrescentado 190 µL do corante TURKEY (Sigma, St. Louis,
MO, USA); Com pipeta o tubo foi homogeneizado; Foi preenchida a câmara de
Neubauer, e feita à contagem total dos leucócitos no microscópio (Cálculo:
número encontrado na soma dos 4 quadrantes ÷ 4 (número de quadrantes) x

44
20 (diluição) = número x 104); Observação: Os resultados foram expressos x
106. Esta metodologia foi realizada acompanhando as especificações propostas
por DORNFEST et al. (1986) e GADEBERG et al. (1979).
4.7.2 - Leucograma Diferencial - % de Linfócitos Circulantes
Os animais foram sacrificados após o exercício físico de natação ou sem
exercício (grupo controle); O sangue foi colhido em tubo de vidro que continha
EDTA - anticoagulante para hematologia (100 µl para 3,5 ml de sangue); O
tubo foi homogeneizado e preparado para fazer o esfregaço; Foi pego uma
lâmina bem limpa e seca (preparada 24h antes), colocado 7,5 µl de sangue
sobre a lâmina e com a lâmina extensora a gota de sangue foi pressionada
com ângulo de 45º em relação à extremidade da lâmina, no qual foi feito o
esfregaço; A arrasto foi procedido em direção à outra ponta da lâmina com
velocidade constante; Foi seca a temperatura ambiente (2 a 3 minutos) sendo
precedida a coloração (Coloração: Foi colocado sobre a lâmina de esfregaço
3ml MAY GRUNWALD e GIEMSA (corantes) (Sigma, St. Louis, MO, USA);
Após 4 minutos, foi colocado 5 ml de água destilada em cima da lâmina e 2
minutos foram esperados; A lâmina foi lavada com água corrente, deixando-a
inclinada para secar em temperatura ambiente); Após a secagem a leitura foi
procedida, em objetiva de imersão com óleo de imersão; A leitura do
leucograma diferencial foi feita no aparelho LEUCOTRON TP; O resultado
expressa o número diferencial dos tipos de leucócitos (neutrófilos, basófilos,
monócitos, eosinófilos e linfócitos); Foi enfatizado o percentual de linfócitos
circulantes seguindo as descrições de DORNFEST et al. (1986) e GADEBERG
et al. (1979).
4.7.3 - Obtenção dos linfócitos dos linfonodos mesentéricos
O protocolo para a determinação do número total dos linfócitos dos
linfonodos mesentéricos foi realizado através das seguintes etapas: Os animais
foram sacrificados após o exercício físico de natação ou sem exercício (grupo
controle); Após a colocação do animal na placa de sacrifício, realizou-se um
corte sagital, na região da parede abdominal, e posteriormente a musculatura

45
do abdômen foi aberta; Foram localizados e retirados os linfonodos
mesentéricos, sendo em seguida separado o tecido ganglionar linfóide do
tecido gorduroso; Para a obtenção dos linfócitos mesentéricos foi utilizado um
sistema de dois cilindros de aço inox de diferentes diâmetros de modo que um
se encaixe no outro, contendo cada cilindro um sistema de malhas na sua
extremidade. Os linfócitos foram colocados entre os dois cilindros, e através de
movimentos normais circulares os mesmos foram comprimidos (maceração),
obtendo-se linfócitos íntegros e isolados. Acrescentou-se 10 ml de PBS gelado
com o auxílio de uma seringa; Com o auxilio de um papel de filtro, o líquido foi
filtrado no tubo de ensaio 1; O líquido já no tubo de ensaio 1 de plástico foi
Centrifugado por 1 minuto a 2000 rpm ou até o Pelet ser formado; O
sobrenadante foi descartado e ressuspendido o Pelet no agitador em 10 ml de
PBS gelado (diluição 10 X) tubo 1; Foi pego 100 µL da amostra do tubo 1,
colocado em outro tubo e acrescentado 900 µL de PBS gelado, tubo 2 (diluição
10X); 100 µL da amostra do tubo 2 foi pego e colocado em um tubo de plástico;
Neste tubo de plástico foi acrescido 100 µL de Triplan Blue (Sigma, St. Louis,
MO, USA) e o conteúdo foi homogeneizado com pipeta; Foi preenchida a
câmara de Neubauer e feita a contagem do número total de linfócitos dos
linfonodos mesentéricos no microscópio (Cálculo: número encontrado na soma
dos 4 quadrantes ÷ 4 (número de quadrantes) x 200 (diluição) = número x 104);
Observação: Os resultados foram expressos x 106. A Obtenção dos linfócitos
dos linfonodos mesentéricos foi realizada seguindo a metodologia descrita por
SERRANO et al. (1993).
Determinação sérica de IL-2, IL-6 e TNF-αααα
As dosagens de IL-2, IL-6 e TNF-α foram realizadas a partir da coleta do
soro do sangue dos animais e determinadas pelo método ELISA (Ensaio
imunoenzimático em fase sólida), seguindo as especificações correspondentes
ao Kit (R&D Systems, Minneapolis, MN) para cada uma das citocinas em ratos
de acordo com as descrições de CAVAGLIERI et al. (2003). Todas as amostras
para determinação das concentrações das citocinas circulantes foram
realizadas em duplicata, para garantir a precisão dos resultados obtidos.

46
4.7.4 - Protocolo para dosagem de IL-2
Preparação do reagente
Colocar todos os reagentes em temperatura ambiente antes do uso.
Kit controle de IL-2 de rato – reconstituir o kit controle com 1mL de água
destilada.
Conjugado de IL-2 de rato – para preparar o conjugado suficiente para uma
placa, acrescentar 0.5mL do conjugado concentrado a 11mL do diluente do
conjugado. Usar um recipiente estéril, e proteja da luz.
Tampão de lavagem – adicionar 625mL de água destilada + o frasco de 25mL
do tampão de lavagem.
Solução substrato – o reagente A e B de cor devem ser misturados juntos em
volumes iguais para uso dentro de 15 minutos (deste volume será usado 100µL
por orifício). Proteger da luz.
Padrão de IL-2 de rato – reconstituir a solução padrão de IL-2 com 2mL do
calibrador diluente RD5-4. Não substituir outros diluentes. Essa reconstituição
produz uma solução estoque de 2.000pg por mL. Misturar a solução por no
mínimo 5 minutos com agitação leve, antes de fazer as diluições. Pipetar 200µL
do calibrador diluente RD5-4 dentro de cada tubo utilizado. Usar a solução
estoque para produzir uma diluição de 2x em série (como na figura abaixo).
Misturar em cada tubo antes da próxima transferência. A solução padrão e IL-2
do rato não diluída servem como um maior padrão (2.000pg por mL). O
calibrador diluente RD5-4 serve como padrão zero (0pg por mL).
47
Figura 3. Ilustração do procedimento de diluição dos calibradores para IL-2.
Procedimento do ensaio
Todos os reagentes e amostras foram colocados em temperatura
ambiente antes de usar. É recomendado que todas as amostras padrões e
controles sejam analisados em duplicata.
1. Preparar reagentes, diluições da curva padrão, controle e amostras como
direcionado na sessão anterior.
2. Utilizar somente as colunas necessárias.
3. Acrescentar 50µL do diluente de ensaio RD1-21 em cada pequeno orifício.
4. Acrescentar 50µL do padrão, controle ou amostra por orifício. Misturar
girando a placa gentilmente por 1 minuto. Cobrir com um adesivo fornecido.
Encubar por 2 horas em temperatura ambiente.
5. Aspirar cada orifício e lavar, repetindo o processo 4x para um total de 5
lavagens. Lavar preenchendo cada orifício com 400µL do tampão de
lavagem, usando uma pipeta multicanal e um becker grande. Completar a
remoção do líquido em cada passo é essencial para boa performance.
Depois da última lavagem, remover qualquer tampão de lavagem
remanescente por aspiração. Inverter a placa e bater contra papéis toalha
limpos.

48
6. Acrescentar 100µL do conjugado de IL-2 de rato em cada orifício. Cobrir
com uma nova fita adesiva. Encubar por 2 horas em temperatura ambiente.
7. Repetir a aspiração/lavagem como no passo 5.
8. Acrescentar 100µL da solução substrato em cada orifício. Encubar por 30
minutos em temperatura ambiente. Proteger da luz.
9. Acrescentar 100µL da solução de parada em cada orifício. Mover a placa
gentilmente para garantir mixagem.
Preparação da amostra
75µL da amostra + 75µL do calibrador diluente RD5-4 e misturar bem.
4.7.5 – Protocolo para dosagem de IL-6
Preparação do reagente
Colocar todos os reagentes em temperatura ambiente antes do uso.
Kit controle de IL-6 de rato – reconstituir o kit controle com 1mL de água
destilada.
Tampão de lavagem – adicionar 25mL do tampão de lavagem para 625mL de
água destilada.
Solução substrato – o reagente A e B de cor devem ser misturados juntos em
volumes iguais para uso dentro de 15 minutos (deste volume será usado 100µL
por orifício). Proteger da luz.
Padrão de IL-6 de rato – reconstituir a solução padrão de IL-6 com 1mL do
calibrador diluente RD5-16. Não substituir outros diluentes. Essa reconstituição
produz uma solução estoque de 4.000pg por mL. Misturar a solução por no
mínimo 5 minutos com agitação leve, antes de fazer as diluições. Utilizar tubos
de polipropileno - Pipetar 200µL do calibrador diluente RD5-16 dentro de cada
tubo utilizado. Usar a solução estoque para produzir uma diluição de 2x em
série (como na figura abaixo). Misturar em cada tubo antes da próxima
transferência. A solução padrão e IL-6 do rato não diluída serve como um maior

49
padrão (4.000pg por mL). O calibrador diluente RD5-16 serve como padrão
zero (0pg por mL).
Figura 4. Ilustração do procedimento de diluição dos calibradores para IL-6.
Procedimento do ensaio
Todos os reagentes e amostras foram colocados em temperatura
ambiente antes de usar. É recomendado que todas as amostras padrões e
controles sejam analisados em duplicata.
1. Preparar reagentes, diluições da curva padrão, controle e amostras como
direcionado na sessão anterior.
2. Utilizar somente as colunas necessárias.
3. Acrescentar 50µL do diluente de ensaio RD1-54 em cada pequeno orifício.
4. Acrescentar 50µL do padrão, controle ou amostra por orifício. Misturar
girando a placa gentilmente por 1 minuto. Cobrir com um adesivo fornecido.
Encubar por 2 horas em temperatura ambiente.
5. Aspirar cada orifício e lavar, repetindo o processo 4x para um total de 5
lavagens. Lavar preenchendo cada orifício com 400µL do tampão de lavagem,

50
usando uma pipeta multicanal e um becker grande. Completar a remoção do
líquido em cada passo é essencial para boa performance. Depois da última
lavagem, remova qualquer tampão de lavagem remanescente por aspiração.
Inverter a placa e bater contra papéis toalha limpos.
6. Acrescentar 100µL do conjugado de IL-6 de rato em cada orifício. Cobrir com
uma nova fita adesiva. Encubar por 2 horas em temperatura ambiente.
7. Repetir a aspiração/lavagem como no passo 5.
8. Acrescentar 100µL da solução substrato em cada orifício. Encubar por 30
minutos em temperatura ambiente. Proteger da luz.
9. Acrescentar 100µL da solução de parada em cada orifício. Mover a placa
gentilmente para garantir mixagem.
Preparação da amostra
Utilizar tubos de polipropileno. 75µL da amostra + 75µL do calibrador
diluente RD5-16 e misturar bem.
4.7.6 - Protocolo para dosagem de TNF-αααα
Preparação do reagente
Colocar todos os reagentes em temperatura ambiente antes do uso.
Kit controle de TNF-α de rato – reconstituir o kit controle com 1mL de água
destilada.
Tampão de lavagem – adicionar 625mL de água destilada + o frasco de 25mL
do tampão de lavagem.
Solução substrato – o reagente A e B de cor devem ser misturados juntos em
volumes iguais para uso dentro de 15 minutos (deste volume será usado 100µL
por orifício). Proteger da luz.
Padrão de TNF-α de rato – reconstituir a solução padrão de TNF-α com 2mL
do calibrador diluente RD5-17. Não substituir outros diluentes. Essa
reconstituição produz uma solução estoque de 800pg por mL. Misturar a
solução por no mínimo 5 minutos com agitação leve, antes de fazer as

51
diluições. Utilizar tubos de polipropileno -Pipetar 200µL do calibrador diluente
RD5-17 dentro de cada tubo. Utilizado a solução estoque para produzir uma
diluição em série (como na figura abaixo). Misturar em cada tubo antes da
próxima transferência. A solução padrão e TNF-α do rato não diluída servem
como um maior padrão (800pg por mL). O calibrador diluente RD5-17 serve
como padrão zero (0pg por mL).
Figura 5. Ilustração do procedimento de diluição dos calibradores para TNF-α.
Procedimento do ensaio
Todos os reagentes e amostras foram colocados em temperatura
ambiente antes de usar. É recomendado que todas as amostras padrões e
controles sejam analisados em duplicata.
1. Preparar reagentes, diluições da curva padrão, controle e amostras como
direcionado na sessão anterior.
2. Utilizar somente as colunas necessárias.
3. Acrescentar 50µL do diluente de ensaio RD1-41 em cada pequeno orifício.
4. Acrescentar 50µL do padrão, controle ou amostra por orifício. Misturar
girando a placa gentilmente por 1 minuto. Cobrir com um adesivo fornecido.
Encubar por 2 horas em temperatura ambiente.

52
5. Aspirar cada orifício e lavar, repetindo o processo 4x para um total de 5
lavagens. Lavar preenchendo cada orifício com 400µL do tampão de lavagem,
usando uma pipeta multicanal e um becker grande. Completar a remoção do
líquido em cada passo é essencial para boa performance. Depois da última
lavagem, remover qualquer tampão de lavagem remanescente por aspiração.
Inverter a placa e bater contra papéis toalha limpos.
6. Acrescentar 100µL do conjugado de TNF-α de rato em cada orifício. Cobrir
com uma nova fita adesiva. Encubar por 2 horas em temperatura ambiente.
7. Repetir a aspiração/lavagem como no passo 5.
8. Acrescentar 100µL da solução substrato em cada orifício. Encubar por 30
minutos em temperatura ambiente. Proteger da luz.
9. Acrescentar 100µL da solução de parada em cada orifício. Mover a placa
gentilmente para garantir mixagem.
Preparação da amostra
75µL da amostra + 75µL do calibrador diluente RD5-17 e misturar bem.
4.7.7 - Potencial transmembrânico da mitocôndria
As células foram centrifugadas a 1000g durante 15 minutos a 4°C, e o
Pelet foi ressuspendido em 1000 µL de PBS. A rodamina 123 é um corante
catiônico permeável à membrana, excitável por laser argônio que emite
fluorescência sendo rapidamente seqüestrado pela mitocôndria sem a indução
de efeitos citotóxicos (PITHON-CURI et al., 2003). A rodamina 123 (Molecular
Probes, Eugene, OR, USA). (5 µM) foi acrescentada e as células foram então
incubadas durante 15 minutos a 37°C no escuro. As células foram lavadas
duas vezes em PBS e incubadas durante 30 minutos a 30°C no escuro. A
fluorescência de 10000 eventos foi determinada por citometria de fluxo usando
o canal FL1 (fluorescência verde_530/30 nm). Esta técnica foi aplicada somente
nos grupos exercitados de curta duração (controle, 5 minutos leve e moderado
e 15 minutos leve e moderado nos linfócitos circulantes). Os valores foram

53
expressos em percentual de linfócitos circulantes com alteração do potencial
transmembrânico mitocondrial.
4.7.8 - Viabilidade de Linfócitos circulantes
Os linfócitos foram centrifugados a 1000g durante 15 minutos a 4°C, e o
Pelet obtido foi ressuspendido em 500 µL de solução salina tampão (PBS).
Posteriormente, foi acrescentado 50 µL de solução com iodeto de propídeo
(ICN Biomedicals, Costa Mesa, CA, USA) (50 mg/mL in PBS), e as células
foram analisadas com citometro de fluxo FACSCalibur (Becton Dickinson, San
Jose, CA, USA). O iodeto de propídeo é um componente fluorescente
altamente solúvel em água que é excluído das células viáveis. Nas células que
perderam a integridade da membrana, este se liga ao DNA intercalando-se
entre as bases, com pequena ou nenhuma preferência de seqüência. A
fluorescência foi mensurada utilizando o canal FL2 (fluorescência laranja-
vermelho 585/42 nm). 10000 eventos foram analisados por experimento. As
células com a fluorescência de iodeto de propídeo foram avaliadas pelo
software CellQuest (Becton Dickinson). Esta técnica foi aplicada somente nos
grupos exercitados de curta duração (controle, 5 minutos leve e moderado e 15
minutos leve e moderado nos linfócitos circulantes) (CURY-BOAVENTURA et
al., 2005). Os valores foram expressos em percentual de linfócitos viáveis.
4.7.9 - Fragmentação de DNA
A fragmentação de DNA foi analisada por citometria de fluxo depois que
as células foram misturadas ao iodeto de propídeo (ICN Biomedicals, Costa
Mesa, CA, USA), de acordo com o método descrito por NICOLETTI et al.
(1991). As células foram ressuspendidas numa solução que continha
detergentes permeabilizantes, que incorporaram a coloração no DNA. Em
seguida, as células foram centrifugadas a 1000g durante 15 minutos a 4°C. O
Pelet foi gentilmente ressuspendido em 300 µL de solução hipotônica que
continha 50 µg/mL de iodeto de propídeo, 0.1% citrato de sódio (Merck,
Darmstadt, Germany), e 0.1% Triton X-100 (Sigma, St. Louis, MO, USA). As
células foram então incubadas durante 2 h a 4°C. A fluorescência foi

54
mensurada e analisada por citometria de fluxo como descrito acima. Esta
técnica foi aplicada somente nos grupos exercitados de curta duração
(controle, 5 minutos leve e moderado e 15 minutos leve e moderado nos
linfócitos circulantes). Os valores foram expressos em percentual de linfócitos
circulantes com fragmentação de DNA.
4.8 - Análise Estatística
Todos os dados foram expressos como média ± Erro Padrão da Média
(EPM). A análise estatística foi realizada inicialmente pelo teste de normalidade
Kolmorogorov-Smirnov e pelo teste de homocedasticidade (critério de Bartlett).
Para as variáveis analisadas, que apresentaram distribuição normal e
homocedasticidade, foi utilizado a Anova e teste F sendo que, quando a
diferença apresentada era significante, aplicou-se o teste de Tukey HSD para
as comparações múltiplas. Em todos os cálculos foi fixado um nível crítico de
5% (p<0,05).
O software utilizado em todos os testes estatísticos foi o Statistica® 6.1.

55
5. Resultados
5.1 - Tabela 1. Leucometria
Todos os grupos exercitados apresentaram leucocitose quando
comparados ao controle (p* ≤ 0,05), na intensidade leve, 5L aumentou
110,44%, 15L 130,1%, EXL 84,5% e no grupo 5SL aumento de 295%.Na
intensidade moderada, 5M aumentou 207%, 15M 181,8%, EXL 90,53% e 5SM
64,32%. Na comparação entre os grupos, foi detectado maior número de
leucócitos circulantes (45,9%) no grupo 5M quando comparado ao 5L;
leucocitose 22,5% maior no 15M em relação ao 15L e menor leucocitose
(140,32%) no grupo 5SM em relação ao grupo 5SL (Tabela 1, Figura 6).
Leucometria x 106 Média + EPM
C 4,12 0,14
5L 8,67 0,87*
15L 9,48 0,75*
EXL 7,6 0,33*
5SL 16,27 0,46*
5M 12,65 0,74*#
15M 11,61 0,5*#
EXM 7,85 0,35*
5SM 6,77 0,23*#
Valores = média + Erro padrão da média. C= Grupo controle, 5L= Exercício
leve 5 minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão
em intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve
5M= Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos,
EXM= Exercício até a exaustão em intensidade moderada, 5SM= Grupo
exercitado 5 sessões em intensidade moderada, (n=6 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle. #Diferença significativa entre grupos de mesma duração, porém com
intensidades diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com
5SM) (p < 0,05).

56
C 5L 15L EXL 5SL 5M 15M EXM 5SM0
2
4
6
8
10
12
14
16
18
#
##
**
**
*
*
**
Leu
com
etri
a -
Nº
de
Leu
cóci
tos
x 10
6
Grupos Experimentais
Figura 6 – Leucometria. Valores = média + Erro padrão da média. C= Grupo
controle, 5L= Exercício leve 5 minutos, 15L= Exercício leve 15 minutos, EXL=
Exercício até a exaustão em intensidade leve, 5SL= Grupo exercitado 5
sessões em intensidade leve 5M= Exercício moderado 5 minutos, 15M=
Exercício moderado 15 minutos, EXM= Exercício até a exaustão em
intensidade moderada, 5SM= Grupo exercitado 5 sessões em intensidade
moderada, (n=6 para todos os grupos experimentais). *Diferença significativa
quando comparado ao grupo controle. #Diferença significativa entre grupos de
mesma duração, porém com intensidades diferentes (5L com 5M, 15L com
15M, EXL com EXM e 5SL com 5SM) (p < 0,05).

57
5.2 - Tabela 2. Número total de Linfócitos Circulantes.
Foi observada linfocitose estatisticamente significante em todos os grupos
exercitados em relação ao controle, (p* ≤ 0,05). Sendo que, o grupo 5L exibiu
aumento de 97%, 15L aumentou 165%, EXL 87,55% e o grupo 5SL aumentou
3 vezes mais em comparação ao controle, apresentando a maior elevação nos
linfócitos circulantes (Tabela 2, Gráfico 7). Na intensidade moderada, o grupo
5M aumentou 2,5 vezes, 15M aumentou 190,57%, EXM 94,34% e 5SM
100,4%, todos em relação ao controle (Tabela 2, Gráfico 7). Na comparação
entre os grupos, foi observado maior número de linfócitos circulantes no grupo
5M (78%) quando comparado ao grupo 5L; 5SL apresentou linfocitose 116,38%
maior do que o grupo 5SM (Tabela 2, Gráfico 7).
N0. Linfócitos Circulantes x 106 Média + EPM
C 2,65 0,0008
5L 5,24 0,0166*
15L 7,03 0,008*
EXL 4,97 0,007*
5SL 11,49 0,0207*
5M 9,33 0,0072*#
15M 7,7 0,0061*
EXM 5,15 0,0061*
5SM 5,31 0,0061*#
Valores = média + Erro padrão da média. C= Grupo controle, 5L= Exercício
leve 5 minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão
em intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve
5M= Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos,
EXM= Exercício até a exaustão em intensidade moderada, 5SM= Grupo
exercitado 5 sessões em intensidade moderada, (n=6 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle. #Diferença significativa entre grupos de mesma duração, porém com
intensidades diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com
5SM) (p < 0,05).

58
C 5L 15L EXL 5SL 5M 15M EXM 5SM0
2
4
6
8
10
12
#
#
**
*
*
*
*
*
*
N0 . T
ota
l de
Lin
fóci
tos
Cir
cula
nte
s x
106
Grupos Experimentais
Figura 7 - Leucograma Diferencial. Valores = média + Erro padrão da média.
C= Grupo controle, 5L= Exercício leve 5 minutos, 15L= Exercício leve 15
minutos, EXL= Exercício até a exaustão em intensidade leve, 5SL= Grupo
exercitado 5 sessões em intensidade leve 5M= Exercício moderado 5 minutos,
15M= Exercício moderado 15 minutos, EXM= Exercício até a exaustão em
intensidade moderada, 5SM= Grupo exercitado 5 sessões em intensidade
moderada, (n=6 para todos os grupos experimentais). *Diferença significativa
quando comparado ao grupo controle. #Diferença significativa entre grupos de
mesma duração, porém com intensidades diferentes (5L com 5M, 15L com
15M, EXL com EXM e 5SL com 5SM) (p < 0,05).

59
5.3 - Tabela 3. Número total de Linfócitos do linfonodo mesentérico.
Foram observados aumentos estatisticamente significantes nos grupos 5L
(31,8%), 5M (28,13%) e 15M (45,43%) no número total de linfócitos
mesentéricos quando comparados ao controle (p* ≤ 0,05). Houve diminuição de
18,26% no grupo EXL, 24,84% no 5SL e 53,93% no 5SM em relação ao
controle (Tabela 3, Figura 8). Na comparação entre grupos, o grupo 15M
demonstrou maior número total de linfócitos mesentéricos (44,74%) em
comparação ao 15L; similarmente, o grupo EXM apresentou maior número de
linfócitos teciduais (31,7%) quando comparado ao EXL; no caso dos grupos
exercitados durante 5 sessões, observamos valor 63,14% menor nos linfócitos
teciduais no grupo 5SM em relação ao 5SL (Tabela 3, Figura 8).
Linfócitos Mesentéricos x 106 Média + EPM
C 157 5,28
5L 206,92 13,32*
15L 157,75 9,57
EXL 128,33 4,56*
5SL 118 7,30*
5M 201,17 2,71*
15M 228,33 6,09*#
EXM 169 5,43#
5SM 72,33 3,84*#
Valores = média + Erro padrão da média. C= Grupo controle, 5L= Exercício
leve 5 minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão
em intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve
5M= Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos,
EXM= Exercício até a exaustão em intensidade moderada, 5SM= Grupo
exercitado 5 sessões em intensidade moderada, (n=6 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle. #Diferença significativa entre grupos de mesma duração, porém com
intensidades diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com
5SM) (p < 0,05).

60
C 5L 15L EXL 5SL 5M 15M EXM 5SM0
50
100
150
200
250
#
#
#
*
*
*
**
*
Nº
Tota
l de
Lin
fóci
tos
Mes
enté
rico
s x
106
Grupos Experimentais
Figura 8 – Número total de Linfócitos do linfonodo mesentérico. Valores =
média + Erro padrão da média. C= Grupo controle, 5L= Exercício leve 5
minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão em
intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve 5M=
Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos, EXM=
Exercício até a exaustão em intensidade moderada, 5SM= Grupo exercitado 5
sessões em intensidade moderada, (n=6 para todos os grupos experimentais).
*Diferença significativa quando comparado ao grupo controle. #Diferença
significativa entre grupos de mesma duração, porém com intensidades
diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com 5SM) (p <
0,05).

61
5.4 - Viabilidade de linfócitos circulantes
Quando realizamos a comparação dos grupos exercitados nas
intensidades leve e moderada com o grupo controle, observamos diminuição
significativa de 10% na viabilidade de linfócitos circulantes apenas no grupo
15M (p* ≤ 0,05). Na comparação entre grupos, não constatamos alterações
estatisticamente significantes (Figura 9).
C 5L 15L 5M 15M0
20
40
60
80
100
*
% li
nfó
cito
s vi
ávei
s
Grupos Experimentais
Figura 9 - Viabilidade de linfócitos circulantes. Valores = média + Erro
padrão da média. C= Grupo controle, 5L= Exercício leve 5 minutos, 15L=
Exercício leve 15 minutos, 5M= Exercício moderado 5 minutos, 15M= Exercício
moderado 15 minutos, (n=6 para todos os grupos experimentais). *Diferença
significativa quando comparado ao grupo controle (p < 0,05).
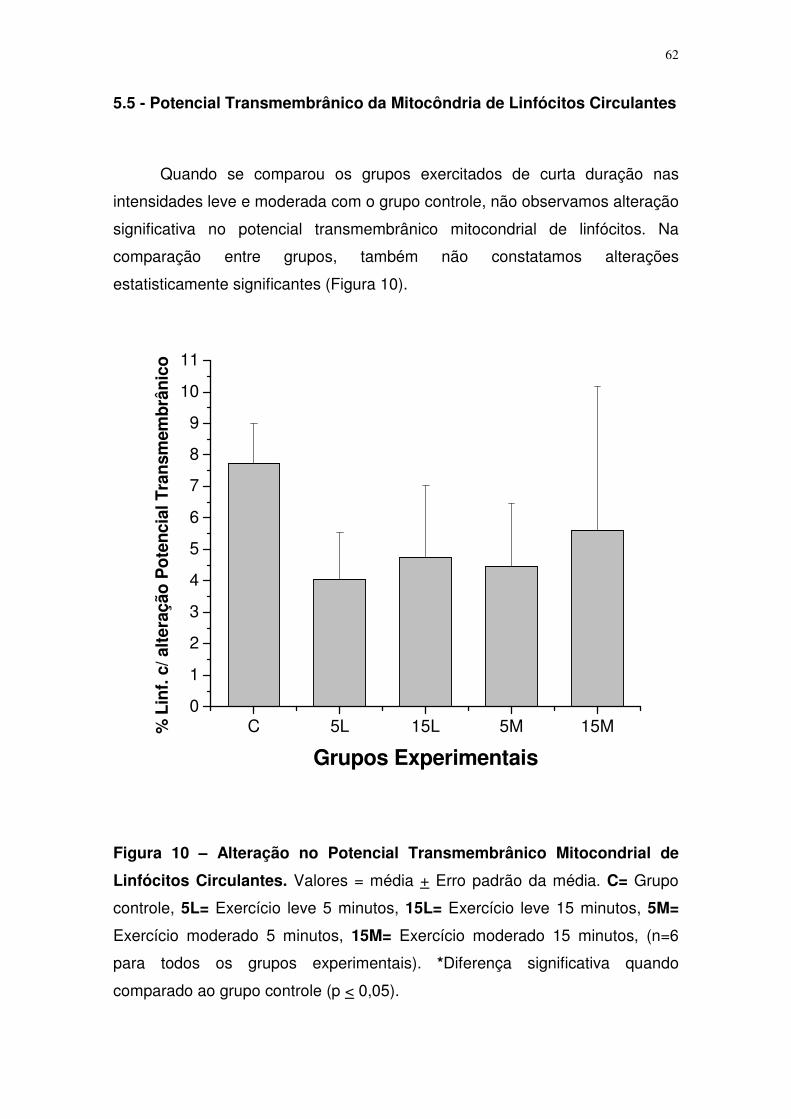
62
5.5 - Potencial Transmembrânico da Mitocôndria de Linfócitos Circulantes
Quando se comparou os grupos exercitados de curta duração nas
intensidades leve e moderada com o grupo controle, não observamos alteração
significativa no potencial transmembrânico mitocondrial de linfócitos. Na
comparação entre grupos, também não constatamos alterações
estatisticamente significantes (Figura 10).
C 5L 15L 5M 15M0
1
2
3
4
5
6
7
8
9
10
11
% L
inf.
c/ a
lter
ação
Po
ten
cial
Tra
nsm
emb
rân
ico
Grupos Experimentais
Figura 10 – Alteração no Potencial Transmembrânico Mitocondrial de
Linfócitos Circulantes. Valores = média + Erro padrão da média. C= Grupo
controle, 5L= Exercício leve 5 minutos, 15L= Exercício leve 15 minutos, 5M=
Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos, (n=6
para todos os grupos experimentais). *Diferença significativa quando
comparado ao grupo controle (p < 0,05).

63
5.6 - Fragmentação de DNA de linfócitos circulantes
Pudemos observar redução estatisticamente significante de 35,84% no
grupo 15L quando comparado com o controle, similarmente, os grupos 5M e
15M também apresentaram queda na fragmentação de DNA de linfócitos
circulantes em relação ao grupo controle (50% e 57%, respectivamente, p* ≤
0,05). Quando realizamos a comparação entre os grupos exercitados,
observamos diminuição significativa de 32,8% na fragmentação do DNA de
linfócitos no grupo 15M em relação ao 15L (Figura 11).
C 5L 15L 5M 15M0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
#**
*
% d
e lin
fóci
tos
com
fra
gm
enta
ção
do
DN
A
Grupos Experimentais
Figura 11 - Fragmentação de DNA de linfócitos circulantes. Valores =
média + Erro padrão da média. C= Grupo controle, 5L= Exercício leve 5
minutos, 15L= Exercício leve 15 minutos, 5M= Exercício moderado 5 minutos,
15M= Exercício moderado 15 minutos, (n=6 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle (p
< 0,05).

64
5.7 - Tabela 4. Concentração sérica de IL-2.
Não foram encontradas diferenças estatisticamente significativas em
nenhum dos grupos exercitados quando comparados com o grupo controle,
tanto nas intensidades leve como moderada. Na comparação entre os grupos
exercitados, também não foram observadas alterações estatisticamente
significativas na concentração sérica de IL-2 (p < 0,05) (Tabela 4 e Figura 12).
IL-2 (pg/ml) Média + EPM
C 56,2 7,01
5L 53,14 2,54
15L 48,97 2,68
EXL 52,31 3,36
5SL 54,53 5,3
5M 48,4 2,88
15M 49,46 1,63
EXM 59,8 4,4
5SM 52,58 4,09
Valores = média + Erro padrão da média. C= Grupo controle, 5L= Exercício
leve 5 minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão
em intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve
5M= Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos,
EXM= Exercício até a exaustão em intensidade moderada, 5SM= Grupo
exercitado 5 sessões em intensidade moderada, (n=3 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle. #Diferença significativa entre grupos de mesma duração, porém com
intensidades diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com
5SM) (p < 0,05).
.

65
C 5L 15L EXL 5SL 5M 15M EXM 5SM0
10
20
30
40
50
60
70IL
-2 (
pg
/ml)
Grupos Experimentais
Figura 12 - Concentração sérica de IL-2. Valores = média + Erro padrão da
média. C= Grupo controle, 5L= Exercício leve 5 minutos, 15L= Exercício leve
15 minutos, EXL= Exercício até a exaustão em intensidade leve, 5SL= Grupo
exercitado 5 sessões em intensidade leve 5M= Exercício moderado 5 minutos,
15M= Exercício moderado 15 minutos, EXM= Exercício até a exaustão em
intensidade moderada, 5SM= Grupo exercitado 5 sessões em intensidade
moderada, (n=3 para todos os grupos experimentais). *Diferença significativa
quando comparado ao grupo controle. #Diferença significativa entre grupos de
mesma duração, porém com intensidades diferentes (5L com 5M, 15L com
15M, EXL com EXM e 5SL com 5SM) (p < 0,05).
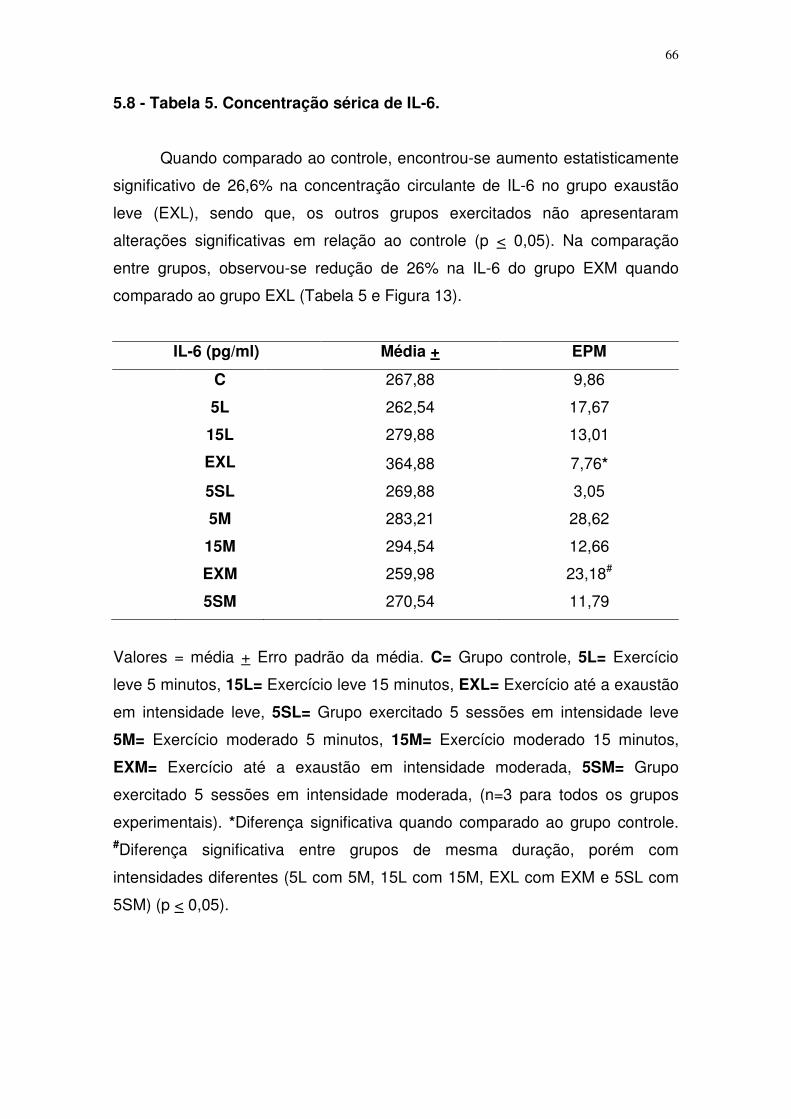
66
5.8 - Tabela 5. Concentração sérica de IL-6.
Quando comparado ao controle, encontrou-se aumento estatisticamente
significativo de 26,6% na concentração circulante de IL-6 no grupo exaustão
leve (EXL), sendo que, os outros grupos exercitados não apresentaram
alterações significativas em relação ao controle (p < 0,05). Na comparação
entre grupos, observou-se redução de 26% na IL-6 do grupo EXM quando
comparado ao grupo EXL (Tabela 5 e Figura 13).
IL-6 (pg/ml) Média + EPM
C 267,88 9,86
5L 262,54 17,67
15L 279,88 13,01
EXL 364,88 7,76*
5SL 269,88 3,05
5M 283,21 28,62
15M 294,54 12,66
EXM 259,98 23,18#
5SM 270,54 11,79
Valores = média + Erro padrão da média. C= Grupo controle, 5L= Exercício
leve 5 minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão
em intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve
5M= Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos,
EXM= Exercício até a exaustão em intensidade moderada, 5SM= Grupo
exercitado 5 sessões em intensidade moderada, (n=3 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle. #Diferença significativa entre grupos de mesma duração, porém com
intensidades diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com
5SM) (p < 0,05).

67
C 5L 15L EXL 5SL 5M 15M EXM 5SM0
50
100
150
200
250
300
350
400
#
*IL
-6 (
pg
/ml)
Grupos Experimentais
Figura 13 - Concentração sérica de IL-6. Valores = média + Erro padrão da
média. C= Grupo controle, 5L= Exercício leve 5 minutos, 15L= Exercício leve
15 minutos, EXL= Exercício até a exaustão em intensidade leve, 5SL= Grupo
exercitado 5 sessões em intensidade leve 5M= Exercício moderado 5 minutos,
15M= Exercício moderado 15 minutos, EXM= Exercício até a exaustão em
intensidade moderada, 5SM= Grupo exercitado 5 sessões em intensidade
moderada, (n=3 para todos os grupos experimentais). *Diferença significativa
quando comparado ao grupo controle. #Diferença significativa entre grupos de
mesma duração, porém com intensidades diferentes (5L com 5M, 15L com
15M, EXL com EXM e 5SL com 5SM) (p < 0,05).

68
5.9 - Tabela 6. Concentração sérica de TNF-α.
Foram detectadas reduções significativas na concentração circulante de
TNF-α nos grupos exercitados 5SL (24,72%), 5M (23,44%), 15M (23,07%),
EXM (26,19%, maior redução) e 5SM (23,81%) quando comparados ao
controle. Na comparação entre os grupos exercitados, observamos redução
estatisticamente significativa de 20% no grupo 15M em relação ao grupo 15L,
na TNF-α circulante (p < 0,05) (Tabela 6 e Figura 14).
TNF-α (pg/ml) Média + EPM
C 5,46 0,2
5L 4,84 0,27
15L 5,25 0,2
EXL 4,62 0,24
5SL 4,11 0,06*
5M 4,18 0,16*
15M 4,2 0,08*#
EXM 4,03 0,03*
5SM 4,16 0,19*
Valores = média + Erro padrão da média. C= Grupo controle, 5L= Exercício
leve 5 minutos, 15L= Exercício leve 15 minutos, EXL= Exercício até a exaustão
em intensidade leve, 5SL= Grupo exercitado 5 sessões em intensidade leve
5M= Exercício moderado 5 minutos, 15M= Exercício moderado 15 minutos,
EXM= Exercício até a exaustão em intensidade moderada, 5SM= Grupo
exercitado 5 sessões em intensidade moderada, (n=3 para todos os grupos
experimentais). *Diferença significativa quando comparado ao grupo controle. #Diferença significativa entre grupos de mesma duração, porém com
intensidades diferentes (5L com 5M, 15L com 15M, EXL com EXM e 5SL com
5SM) (p < 0,05).

69
C 5L 15L EXL 5SL 5M 15M EXM 5SM0
1
2
3
4
5
6
# *****
TN
F-a
lph
a (p
g/m
l)
Grupos Experimentais
Figura 14 - Concentração sérica de TNF-α. Valores = média + Erro padrão da
média. C= Grupo controle, 5L= Exercício leve 5 minutos, 15L= Exercício leve
15 minutos, EXL= Exercício até a exaustão em intensidade leve, 5SL= Grupo
exercitado 5 sessões em intensidade leve 5M= Exercício moderado 5 minutos,
15M= Exercício moderado 15 minutos, EXM= Exercício até a exaustão em
intensidade moderada, 5SM= Grupo exercitado 5 sessões em intensidade
moderada, (n=3 para todos os grupos experimentais). *Diferença significativa
quando comparado ao grupo controle. #Diferença significativa entre grupos de
mesma duração, porém com intensidades diferentes (5L com 5M, 15L com
15M, EXL com EXM e 5SL com 5SM) (p < 0,05).
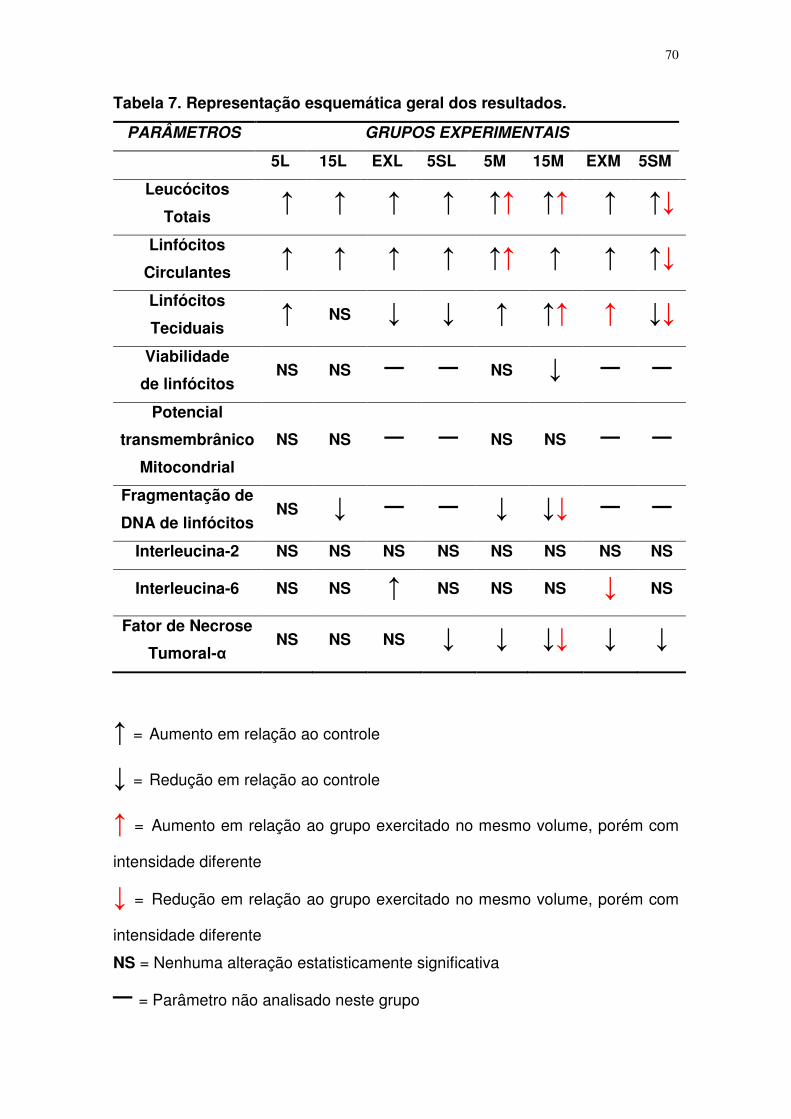
70
Tabela 7. Representação esquemática geral dos resultados.
PARÂMETROS GRUPOS EXPERIMENTAIS
5L 15L EXL 5SL 5M 15M EXM 5SM
Leucócitos
Totais ↑ ↑ ↑ ↑ ↑↑ ↑↑ ↑ ↑↓
Linfócitos
Circulantes ↑ ↑ ↑ ↑ ↑↑ ↑ ↑ ↑↓
Linfócitos
Teciduais ↑ NS ↓ ↓ ↑ ↑↑ ↑ ↓↓
Viabilidade
de linfócitos NS NS ─ ─ NS ↓ ─ ─
Potencial
transmembrânico
Mitocondrial
NS NS ─ ─ NS NS ─ ─
Fragmentação de
DNA de linfócitos NS ↓ ─ ─ ↓ ↓↓ ─ ─
Interleucina-2 NS NS NS NS NS NS NS NS
Interleucina-6 NS NS ↑ NS NS NS ↓ NS
Fator de Necrose
Tumoral-α NS NS NS ↓ ↓ ↓↓ ↓ ↓
↑ = Aumento em relação ao controle
↓ = Redução em relação ao controle
↑ = Aumento em relação ao grupo exercitado no mesmo volume, porém com
intensidade diferente
↓ = Redução em relação ao grupo exercitado no mesmo volume, porém com
intensidade diferente
NS = Nenhuma alteração estatisticamente significativa
─ = Parâmetro não analisado neste grupo

71
6. Discussão
A leucocitose ocorre em resposta ao estresse físico agudo, assim como
em exercícios físicos intensos e de curta duração, sendo um fenômeno bem
documentado na literatura (GOEBEL e MILLS, 2000). Os efeitos deste tipo de
exercício físico sobre o aumento no número de leucócitos circulantes são
mediados, pelo menos em parte, pela ativação do sistema nervoso simpático
(MILLS et al., 1999) e aumento agudo dos níveis plasmáticos de catecolaminas
durante o exercício (DISHMAN, 1997; MAZZEO, 1991). A leucocitose pode
aumentar linearmente de acordo com a elevação da intensidade do exercício,
que poderá também promover maior resposta das catecolaminas (BAIN et al.,
2000; PEDERSEN e HOFFMAN-GOETZ, 2000).
Em estudo realizado por HONG et al. (2005), foram utilizados indivíduos
saudáveis divididos em 2 grupos, grupo fisicamente ativo e grupo não ativo
fisicamente, que realizaram 20 minutos de exercício físico em esteira a uma
intensidade de 65-70% VO2max. Foi observada leucocitose similar ao aumento
das concentrações de catecolaminas, que não diferiu entre os dois grupos. Os
valores retornaram próximo aos valores de repouso após 10-15 minutos do
final do teste de esteira.
Sabe-se que o treinamento é capaz de alterar a freqüência cardíaca de
repouso e durante o exercício, ou seja, a promoção da bradicardia
(SIGVARDSSON et al., 1977). Além de promover uma regulação negativa nos
receptores β-adrenérgicos dos linfócitos em situação de repouso (OHMAN et
al., 1987). Assim, o destreinamento ou a baixa aptidão física direciona o
organismo a uma maior concentração plasmática de noradrenalina
(SOTHMANN et al., 1991). As catecolaminas liberadas durante o exercício
físico provocam efeitos característicos como aumento da freqüência cardíaca e
da força de contração do miocárdio proporcionando maior circulação sanguínea
pelo corpo. Dessa forma mais células são liberadas para a corrente sanguínea,
aumentando a quantidade de células circulantes, caracterizando a leucocitose.
Esta leucocitose induzida pelo exercício físico também foi encontrada em
nosso trabalho, através dos aumentos significantes detectados no número de
leucócitos totais circulantes em todos os grupos exercitados quando
comparados ao grupo controle não exercitado, sendo que, na intensidade leve,

72
o grupo 5L aumentou 110,44%, 15L (130,1%), EXL (84,5%) e 5SL (295%)
(Tabela 1 e Figura 4). Já os grupos exercitados em intensidade moderada
apresentaram maiores aumentos com relação à leucocitose, 5M (207%), 15M
(181,8%), EXM (90,53%), com exceção do grupo exercitado 5 sessões em
intensidade moderada (5SM, que apresentou aumento de 64,32%) (Tabela 1 e
Figura 6). Corroborando com os trabalhos comentados acima, no presente
estudo a intensidade do exercício promoveu maior leucocitose nos grupos
exercitados durante 5 e 15 minutos com sobrecarga adicional (45,9%, 5M>5L e
22,5% 15M>15L, respectivamente) quando comparados aos grupos
exercitados em intensidade leve na mesma duração (Tabela 1 e Figura 6).
A leucocitose menos pronunciada no grupo exercitado 5 sessões em
intensidade moderada (140,32% menor) quando comparado ao grupo
exercitado 5 sessões em intensidade leve pode estar relacionada a carga
adicional utilizada neste grupo. Em virtude da maior intensidade, os animais
podem não ter tido tempo suficiente para adaptação, uma vez que os
resultados encontrados nesse grupo são muito semelhantes aos grupos
exercitados até a exaustão (Tabela 1 e Figura 6). De fato, a intensidade
moderada pode ter imposto maior estresse ao grupo 5SM, induzindo a menor
contagem de leucócitos totais observada no mesmo. No entanto, acreditamos
ser necessária a realização de mais estudos que elucidem os mecanismos
moduladores da contagem total de leucócitos frente à realização de 5 sessões
de exercício com volumes crescentes numa mesma semana, em diferentes
intensidades.
Considerando estes aspectos relacionados ao aumento no número de
leucócitos circulantes, os linfócitos liberados da medula para a circulação geral,
durante o exercício, contribuíram para um aumento do número de células
marginadas nos vasos (HOGG, 1987). Disponibilizando maior número de
células para a circulação em decorrência do exercício. Os leitos
microvasculares pulmonar e esplênico são importantes reservatórios de
leucócitos marginados (HOGG, 1987). Neste sentido, VAN EEDEN et al. (1997)
e LAWRENCE et al. (1996) comentaram que os polimorfonucleares imaturos
liberados da medula óssea têm se mostrado preferencialmente marginados nos
capilares pulmonares.

73
O aumento no percentual de linfócitos circulantes (linfocitose) frente ao
exercício é passageiro, e, provavelmente se deve a liberação dos hormônios do
estresse, principalmente a adrenalina (SAXTON et al., 2003), aumentando a
liberação de células de seus compartimentos para a circulação sanguínea.
O exercício físico regular em intensidade submáxima pode não alterar as
subpopulações de linfócitos (PEDERSEN e HOFFMAN-GOETZ, 2000). Ainda,
em estudo realizado por HONG et al. (2005), onde foram realizados 20 minutos
de exercício físico de esteira a uma intensidade de 65-70% VO2max foram
observados aumentos no percentual de linfócitos circulantes. Em outro estudo,
McFARLIN et al. (2003) utilizaram 10 homens fisicamente ativos de idade entre
18-25 anos, que realizaram duas sessões de endurance de curta duração (20
minutos cada, 5 min a 50% do VO2max e 15 min a 70% VO2max) no mesmo
dia. Foram observados aumentos na contagem de linfócitos, mas mínimo efeito
sobre a atividade da célula NK.
Recentemente, NIEMAN et al. (2005) analisaram o efeito da caminhada
de 30 minutos na resposta imune. Neste estudo foram avaliadas 17 mulheres
com idades entre 25 e 55 anos, que já realizavam caminhadas pelo menos a
três meses, com uma freqüência de 2-7 dias por semana. Sumariamente,
verificou-se que a caminhada numa intensidade de 60%-65% do VO2max, foi
associada com aumentos padrões, porém temporários na contagem de células,
sendo que os linfócitos circulantes contribuíram com 40% na elevação total.
Os resultados dos estudos comentados acima são similares aos
observados no presente estudo. Pois verificamos aumento no percentual de
linfócitos circulantes em todos os grupos exercitados: 5 minutos leve (5L, 97%
de aumento), 15 minutos leve (15L, 165% de aumento), exercitado até a
exaustão leve (EXL, 87,55% de aumento) e exercitado 5 sessões em
intensidade leve (5SL, 333,6% de aumento). No exercício moderado,
observamos também aumento em todos os grupos: 5 minutos (5M, 252% de
aumento), 15 minutos (15M, 190,57% de aumento), exaustão (EXM, 94,34% de
aumento) e exercitado 5 sessões (5SM, 100% de aumento) quando
comparados ao controle (Tabela 2 e Figura 7).
No entanto, quando foi realizada a comparação entre as intensidades,
observamos maior número de linfócitos estatisticamente significativo, apenas
no grupo 5M (78% maior) quando comparado a 5L (Tabela 2 e Figura 7).

74
Adicionalmente, foi detectado menor número de linfócitos circulantes no grupo
5SM (116,38% menor) em relação ao grupo 5SL (Tabela 2 e Figura 7).
Similarmente a contagem total de leucócitos, a maior intensidade imposta aos
animais do grupo 5SM exerceu influência na contagem de linfócitos circulantes,
reduzindo o número deste subtipo celular. O período de exercício realizado (1
semana) parece não ter sido suficiente para adaptação a sobrecarga adicional.
LAPERRIERE et al. (1991; 1994) mostraram que 10 semanas de
treinamento em aerobiose promoveu um aumento do número de células CD4+,
CD8+ e linfócitos B, coletados no estado de repouso em homens sedentários
saudáveis. A intensidade utilizada foi de 70-80% da freqüência cardíaca
máxima predita para a respectiva idade / volume de 45 minutos, 3 vezes por
semana, utilizando-se como modelo de exercício o ciclismo. Porém,
PEDERSEN et al. (2000) observaram que em 7 semanas de treinamento,
somente o número de células NK estava aumentado sob condições de
repouso. Em estudo realizado por NIEMAN et al. (2000) que utilizaram 40
mulheres jovens (20 corredoras de elite e 19 mulheres não atletas), a atividade
das células NK estava significantemente aumentada nas corredoras de elite em
comparação com o grupo controle não atleta.
NIELSEN et al. (1997) demonstraram que aproximadamente 2/3 do total
de linfócitos liberados para a circulação geral, durante o exercício, são
provenientes do baço, órgão este que provavelmente teria contribuído para a
linfocitose nos grupos exercitados. Estando de acordo com GABRIEL e
KINDERMANN (1998), que propuseram os órgãos linfóides secundários como
responsáveis pelo aumento de células circulantes. Se o exercício for realizado
por longos períodos ou em alta intensidade, a concentração total de linfócitos
nos órgão linfóides secundários pode diminuir, levando ao quadro de linfopenia,
o que se deve à falta de células maduras que podem ser recrutadas, e à
redistribuição dos linfócitos para a circulação geral (PEDERSEN e HOFFMAN-
GOETZ, 2000; PEDERSEN e NIEMAM, 1998; RONSEN et al., 2001b).
Neste sentido, estes resultados estão de acordo com as da nossa
pesquisa, pois para curta duração foram observados aumentos nos linfócitos
provenientes dos linfonodos mesentéricos em relação ao controle, o grupo 5L
(aumentou 31,8%), 5M (aumentou 28,13%) e 15M (aumentou 45,43%),
possivelmente pelos mesmos mecanismos fisiológicos propostos para a

75
linfocitose circulante (Tabela 3 e Figura 8). No caso de exercício com maior
volume, foram observadas reduções significativas na contagem de linfócitos
mesentéricos para os grupos exaustão leve (EXL, redução de 18,26%),
exercitado 5 sessões em intensidade leve (5SL, redução de 24,84%) e
exercitado 5 sessões em intensidade moderada (5SM, redução de 54%),
quando comparados ao controle sedentário (Tabela 3 e Figura 8).
Estes linfócitos que possivelmente saíram do compartimento linfóide
(linfonodo mesentérico), podem ter contribuído para linfocitose circulante
observada nestes mesmos grupos. Novamente, em dois grupos exercitados a
intensidade promoveu maiores aumentos no número de células (neste caso
dos linfócitos teciduais), já que, observamos linfocitose tecidual mais
pronunciada no grupo 15M (44,74% maior) em relação ao 15L e também no
grupo EXM (31,7% maior) quando comparado ao grupo EXL. Finalmente, nos
grupos que realizaram 5 sessões encontramos menor número de linfócitos
teciduais (63,14%) no grupo 5SM em relação ao 5SL (Tabela 3 e Figura 8).
Este padrão foi evidenciado neste mesmo grupo, para as variáveis leucometria
e linfócitos circulantes, provavelmente, em decorrência do pequeno período
disponível para a adaptação a intensidade moderada (1 semana) e pelo maior
estresse imposto ao grupo 5SM.
Vale destacar que provavelmente nossos animais podem ter sido
submetidos a um estresse maior, em virtude do ciclo de luz ao qual foram
expostos e o horário a qual foram submetidos ao exercício. Sabe-se que no
ciclo de luz normal, os ratos, animais noturnos, apresentam um pico de
corticosteróides próximo da inversão da luz (final da tarde) (KALSBEEK et al.,
2003). Neste sentido, estes foram exercitados no período normalmente de
repouso (durante o dia, período da tarde, entre 14:00 e 17:00 horas).
Entretanto, esta condição foi amenizada, uma vez que, todos os grupos foram
submetidos à mesma condição de ciclo de luz no biotério, horário do exercício
e sacrifício (inclusive o grupo controle, porém não exercitado).
A apoptose e necrose foram observadas em linfócitos depois de exercício
aeróbio intenso e agudo (HOFFMAN-GOETZ e DUERRSTEIN, 2004).
Mudanças no potencial transmembrânico mitocondrial, força de direcionamento
para formação do ATP, constitui um passo obrigatório na indução da morte
apoptótica (KROEMER et al., 1998). A despolarização mitocondrial precede

76
sinais nucleares de apoptose à ativação de endonucleases endógenas e
caspases, que resultam em fragmentação de DNA irreversível da célula
(KROEMER et al., 1998).
Depois de exercícios extenuantes, a despolarização da membrana
mitocondrial pode ocorrer em neutrófilos de ratos (LAGRANHA et al., 2004),
monócitos e linfócitos de humanos, e linfócitos intestinais de camundongos
(QUADRILATERO e HOFFMAN-GOETZ, 2004). Durante a disfunção
mitocondrial, a citocromo-c é liberada no citosol ativando caspases e
eventualmente promovendo apoptose (GREEN e REED, 1998). Resultados
similares também foram observados por CONCORDET e FEERRY (1993), que
verificaram apoptose em timócitos de ratos que realizaram duas corridas em
esteira até a exaustão, evidenciada pela fragmentação de DNA. Nesta linha de
pensamento, em ratos, foi demonstrado que a corrida de alta intensidade em
esteira foi associada com a expressão de proteínas pró-apoptóticas (caspase-3
e citocromo citosólica-c) e a redução na expressão da proteína anti-apoptótica
Bcl-2 em linfócitos intestinais c, quando comparados ao grupo controle não
exercitado (QUADRILATERO e HOFFMAN-GOETZ, 2005).
MOOREN et al. (2002) relataram que o exercício exaustivo (corrida em
esteira e 80% do VO2max) induz a apoptose em linfócitos periféricos enquanto
que o exercício moderado (corrida em esteira e 60% do VO2max) não induz em
indivíduos saudáveis. HOFFMAN-GOETZ et al., (1999) não observaram
apoptose de timócitos em camundongos submetidos à corrida submáxima em
esteira durante 90 minutos.
Como mencionado acima, o exercício moderado não induziu a apoptose
de linfócitos circulantes, o que também não foi observado no presente estudo,
nas intensidades leve e moderada. De fato, nosso trabalho verificou que o
exercício de 15 minutos em intensidade leve promoveu redução de 35,84% na
porcentagem de linfócitos com fragmentação de DNA. Nos grupos 5 e 15
minutos em intensidade moderada observamos reduções de 50% e 57%,
respectivamente, em relação ao controle (Figura 11). Apesar de obtermos
redução de 10% na viabilidade no grupo 15 minutos moderado (Figura 9), uma
porcentagem menor de linfócitos estaria entrando em apoptose devido à
fragmentação de DNA (Figura 11). Portanto, a redução na viabilidade dos
linfócitos ocorreu por outro mecanismo, não investigado neste estudo. Ainda na

77
intensidade moderada, o grupo 15M apresentou um percentual menor de
linfócitos com fragmentação de DNA (32,8%) na comparação com o grupo
exercitado durante 15 minutos em intensidade leve (15L) (Figura 11). São raros
os trabalhos que avaliam a apoptose de linfócitos em exercícios de curta
duração, neste caso observamos diminuição na apoptose de linfócitos
circulantes pela fragmentação de DNA. Sendo assim, estes resultados devem
ser analisados cuidadosamente e os mecanismos envolvidos devem ser
totalmente esclarecidos, antes de serem relacionados possíveis benefícios do
exercício em seres humanos.
No caso do potencial transmembrânico, não encontramos diferenças
estatisticamente significantes em nenhum dos grupos exercitados (Figura 10).
Estes resultados são diferentes dos apresentados por LAGRANHA et al (2004)
e por QUADRILATERO e HOFFMAN-GOETZ, 2004, que utilizaram duração
mais prolongada e tipo de exercício diferente do nosso estudo. Desta forma,
são necessários mais trabalhos que avaliem os efeitos do exercício de curta
duração sobre a o potencial transmembrânico mitocondrial.
Outro fator imunológico importante que pode ser modulado pelo
exercício/treinamento físico é a concentração das citocinas circulantes. Estas
glicoproteínas atuam como uma rede de ligação para auxiliar na regulação da
sinalização das respostas imunes inatas e específicas. Existem diversas
explicações possíveis para a variabilidade dos resultados na responsividade
das citocinas pró e antiinflamatórias em relação ao exercício (PEDERSEN et
al., 1998a). Primeiro, a aptidão física, o tipo de atividade física, a intensidade e
duração do exercício, podem afetar o perfil das citocinas. Têm sido relatado
aumento na concentração de citocinas, depois do exercício envolvendo
componente excêntrico, e também no exercício concêntrico. (PEDERSEN,
2000). A especificidade e a sensibilidade dos kits utilizados em estudos com
exercício físico podem justificar as variações nos resultados (PEDERSEN,
2000).
Sabe-se que os inibidores de citocinas e as citocinas antiinflamatórias
restringem a magnitude e duração da resposta inflamatória ao exercício. A
presença de múltiplas citocinas (TNF-α, IL-1β, IL-6, IL-2 receptores e
interferon-γ) na urina depois do exercício demonstra que a expressão das

78
citocinas em resposta ao exercício é plenamente possível (SPRENGER et al,
1992).
Neste sentido MCFARLIN et al. (2003), realizaram um estudo com treze
homens de dezoito a vinte e seis anos, os quais se exercitaram em uma única
sessão por uma hora na bicicleta ergométrica com 75% a 80% do VO2max. As
amostras de sangue para mensurar as concentrações de citocinas circulantes
foram coletadas antes, imediatamente após e, duas horas e quatro horas após
o término do exercício. Os resultados demonstraram que não houve alteração
estatisticamente significativa na IL-2 quando não utilizada a ingestão de
carboidratos. Os estudos citados acima corroboram com nossos resultados,
pois quando foram comparados todos os grupos exercitados com o grupo
controle e entre os grupos, não observamos nenhuma alteração na
concentração sérica de IL-2 (Tabela 4 e Figura 12).
NIEMAN et al. (2001) demonstraram que este padrão de mudança nas
concentrações plasmáticas de citocinas não foi diferente entre homens e
mulheres ou entre indivíduos jovens e idosos que competiram uma maratona.
Sendo que, as concentrações de IL-2 permaneceram próximos aos valores pré-
prova ou em concentrações indetectaveis. Esses resultados são praticamente
idênticos aqueles de SUZUKI et al. (2000), que também mensuraram esta
citocina num grupo de 16 homens corredores de maratona antes e depois de
uma prova.
CASTELL et al. (1997) registraram que as concentrações plasmáticas de
IL-2 foram aumentadas 16 horas depois de uma corrida de maratona. BAUM et
al. (1997) apesar de terem observado que 30 minutos depois do exercício
exaustivo, a produção de interferon-γ induzida com fitoemaglutinina foi
suprimida, resultados inconsistentes foram encontrados para IL-2. Após
exercício concêntrico, realizado em bicicleta ergométrica durante 1 hora a 75%
do VO2max, não se detectou a produção in vitro de IL-2 (HAAHR et al., 1991).
A concentração plasmática de IL-2 pode exibir respostas complexas
depois do exercício. Por exemplo, ESPERSEN et al. (1990) encontraram
redução de 50% na concentração plasmática de IL-2 imediatamente depois de
uma corrida de 5Km, em corredores de elite. No entanto, os valores retornaram
as concentrações basais depois de duas horas após o exercício, e aumentando
50% 24h depois da corrida.

79
TOFT et al. (2002) registraram que depois de 60 minutos de exercício
em bicicleta ergométrica realizando a seguinte seqüência: 0-6 minutos a 50%,
6-12 minutos a 75%, 12-20 minutos a 100%, 20-25 minutos a 130%, 25-40
minutos a 100%, e 40-60 minutos a 75% do VO2max, a IL-6 aumentou
progressivamente depois desta seqüência, consistentemente com a hipótese
que este aumento está relacionado com lesão muscular.
Estudos prévios demonstraram que o exercício concêntrico, como
pedalar, resulta em aumentos menos pronunciados na IL-6 comparado com o
exercício excêntrico, como a corrida (NIEMAN et al., 1990; STARKIE et al.,
2001a). Contudo, durante a corrida intensa, como a maratona, aumentos de
100 vezes na concentração de IL-6 tem sido consistentemente demonstrados
(OSTROWSKI et al., 1999; OSTROWSKI et al., 1998a; STARKIE et al., 2001b;
TOFT et al., 2000). Num destes estudos, os autores especularam que o
aumento na IL-6 no plasma pode ter sido causado pela lesão muscular porque
estes registraram concentrações marcadamente elevadas de creatina kinase
(CK), depois do exercício comparado com o repouso (STARKIE et al., 2001a).
No entanto, estes pesquisadores também hipotetizaram que as elevações na
IL-6 plasmática durante uma maratona pode ser causada pela endotoxemia,
secundariamente a redução no fluxo sangüíneo visceral, suficiente para induzir
isquemia (BAGBY et al., 1996; CAMUS et al., 1997). Os maiores valores na
concentração de IL-6 registrados para corredores podem ser devido ao maior
estresse mecânico muscular imposto por esta atividade (SUZUKI et al., 2003).
As concentrações plasmáticas de IL-6 aumentadas depois do exercício
foram então associadas com lesão muscular (BRUUNSGAARD et al., 1997).
No entanto, estudos posteriores estabeleceram que a IL-6 pode aumentar
mesmo na ausência de lesão no tecido muscular (CROISIER et al., 1999;
KELLER et al., 2001; OSTROWSKI et al., 1998b; OSTROWSKI et al., 1998a;
STEENSBERG et al., 2001c; STEENSBERG et al., 2000). A IL-6 plasmática
durante o exercício aumenta com a intensidade e duração da atividade
(OSTROWSKI et al., 1998a).
Em estudo realizado com 12 atletas competidores homens (9 ciclistas, 3
triatletas; idade média 26 anos, altura média 179 cm e peso corporal de 71Kg)
que realizaram uma sessão de ciclismo de carga constante com duração de 4
horas na intensidade de 70% do limiar anaeróbio individual, numa pista de

80
400m, foi observado aumento na IL-6 de 10 vezes em relação aos valores
basais (SCHARHAG et al., 2005). O aumento na concentração plasmática de
IL-6 foi comparável ao aumento depois de 2-2.5 horas de teste em
cicloergômetro a 75% do VO2max (HISCOCK et al., 2003; NIEMAN et al.,
1998). Sendo menos pronunciado do que depois do ciclismo competitivo
(250Km de corrida de bicicleta, duração de 6.5 horas) (GANNON, et al., 1997),
e, foi maior em relação a um único teste ciclo ergométrico máximo ou repetitivo
anaeróbio (MEYER et al., 2001).
Especificamente em atletas de alto nível, neste caso, 9 homens (4
triatletas e 5 patinadores de velocidade; 21 a 27 anos de idade, 74,7Kg em
média) que realizaram sessões de exercício em cicloergômetro a 75% do
consumo máximo de oxigênio que duraram 75 minutos, foram detectados
aumentos significativos na concentração de IL-6 (RONSEN et al., 2002). De
maneira esperada, neste estudo, quando os indivíduos realizaram mais do que
uma sessão no mesmo dia, as concentrações de IL-6 foram significativamente
superiores em relação a uma única sessão (RONSEN et al., 2002).
Nesta linha de trabalhos, NEMET et al. (2003) também observaram
aumentos substanciais nas concentrações plasmáticas de IL-6 em 10
indivíduos do sexo feminino saudáveis, de 14-16 anos de idade que realizaram
uma única e típica sessão de 1 hora e 30 minutos de pólo aquático, sendo que
as amostras sangüíneas foram coletadas antes e logo após o exercício.
Caracterizando esta sessão com uma profunda resposta proinflamatória e
catabólica. A IL-6 sabidamente aumenta com o exercício em adultos, e mais
recentemente, isto foi demonstrado também em crianças submetidas à corrida
em esteira a 75% do VO2max durante 60 minutos (NEMET et al., 2002).
Apesar da interleucina-6 ser tradicionalmente considerada como um
mediador catabólico, estudos recentes sugerem que esta também possui
efeitos positivos sobre o crescimento em determinadas situações, incluindo o
exercício físico. Outros membros da família das citocinas IL-6 (fator de inibição
leucêmico) regulam o crescimento muscular (REARDON et al., 2001). A IL-6
sabidamente estimula a angiogênese mediada por fatores de crescimento
vasculares (fator de crescimento fibroblasto-2) (BLOTNICK et al., 1994) e fator
de crescimento vascular endotelial (FREEMAN et al., 1995).

81
A interleucina-6 é uma substância biologicamente ativa que não é
secretada apenas pelas células do sistema imunológico durante condições
inflamatórias, mas é também liberada do tecido adiposo (FRUBECK et al.,
2001) e pela contração muscular na ausência de inflamação (STEENSBERG et
al., 2000).
Na perspectiva metabólica, a IL-6 pode possuir efeitos lipolíticos em
associação com o exercício físico. HALL et al. (2003) propuseram que esta
citocina é um potente modulador do metabolismo de gorduras, através do
aumento da oxidação de ácidos graxos e a reesterificação dos mesmos, sem
causar hipertrialcilglicerídemia, em 18 homens ativos que receberam a infusão
de IL-6. WALLENIUS et al. (2002), acrescentaram que ratos deficientes de IL-6
desenvolveram obesidade prematuramente. Em soma, quando os ratos foram
tratados com infusão de IL-6 durante 18 dias, houve redução significativa no
peso corporal destes animais. Neste sentido, nos nossos resultados pudemos
observar aumentos significativos de 26,6% na IL-6 no grupo exaustão leve em
comparação ao controle e redução de 26% no grupo exercitado até a exaustão
em intensidade moderada (duração do tempo de nado inferior a 50 minutos em
média) em relação ao grupo exercitado até exaustão em intensidade leve
(Tabela 5 e Figura 13). Vale destacar, que no grupo exaustão leve os animais
nadaram em média 10 horas, sendo este exercício de característica aeróbia e
de longa duração, priorizando o metabolismo lipídico e a produção desta
citocina (Tabela 5 e Figura 13).
No músculo esquelético, KELLER et al. (2001) analisaram 6 homens,
fisicamente ativos, porém destreinados (idade média 26 anos, peso corporal
médio 78.1Kg e altura média de 1.87m) que realizaram uma sessão de 60
minutos em cicloergômetro a 70% do VO2max. Observaram que o exercício
prolongado ativa a transcrição do gene de IL-6 no músculo esquelético. Esta
resposta foi particularmente aumentada quando o conteúdo de glicogênio
muscular era baixo. A partir destes achados, os mesmos autores
estabeleceram que o tecido muscular é uma fonte de produção de IL-6 durante
o exercício prolongado e que a concentração de glicogênio muscular pode ser
um determinante crítico regulando a resposta desta molécula glicoproteica ao
exercício.

82
Quando realizada a infusão de concentrações relativamente modestas
de IL-6 (como aquelas que podem estar presentes depois do exercício ou em
baixos níveis inflamatórios crônicos) diretamente num músculo alvo in vivo, foi
observada significativa atrofia muscular em idosos (HADDAD et al., 2005).
Finalmente, PEDERSEN et al. (2001a) demonstraram que a interleucina-
6 é produzida localizadamente pelo músculo esquelético e liberada na
circulação em grandes quantidades. Foi hipotetizado esta citocina pode exercer
um papel importante na manutenção da homeostasia da glicose durante o
exercício prolongado, com ações similares a de um hormônio, otimizando a
resposta metabólica durante a atividade muscular.
Nesta linha de pensamento, durante o exercício pode ocorrer uma crise
energética (depleção de glicogênio) ou lesão tecidual que promovem a
sinalização para produção e liberação aumentadas de IL-6 (GLEESON, 2000).
Suas concentrações plasmáticas estimulam então a glicogenólise hepática e
liberação de glicose, ajudando a manter a glicemia sangüínea. A IL-6 também
promove síntese de proteínas de fase aguda e fatores de crescimento
insulínicos, que exercem importante papel na adaptação muscular ao exercício
(GLEESON, 2000). Estes efeitos metabólicos de regulação poderiam estar
provavelmente presentes em nossos resultados, devido às mudanças nesta
citocina terem sido detectadas apenas no grupo exaustão leve (média de 10
horas de natação), evidenciando a necessidade de manutenção da glicemia e
grande utilização de ácidos graxos (Tabela 5 e Figura 13).
Foi sugerido que a adrenalina pode ser um gatilho para resposta da IL-6
durante o exercício (PAPANICOLAOU et al., 1996). Pois, a infusão de
adrenalina induziu aumento de 6 vezes nesta citocina. No entanto, o exercício
promoveu aumentos de 30 vezes nas concentrações circulantes de IL-6
(STEENSBERG et al., 2001c). Adicionalmente, em outro estudo, ambas as
pernas, exercitada e não exercitada foram expostas às mesmas concentrações
hormonais, porém apenas a perna exercitada liberou IL-6 (STEENSBERG et
al., 2000). A noradrenalina foi sugerida como um potencial estímulo para a
elevação de IL-6 induzida pelo exercício (MAZZEO et al., 2001). Numerosos
trabalhos têm demonstrado que a adrenalina e a noradrenalina podem induzir a
produção de IL-6 em repouso (DERIJK et al., 1994; STEENSBERG et al.,
2001c; SONDERGAARD et al., 2000). No entanto, as catecolaminas não

83
exercem papéis fundamentais neste processo (PEDERSEN e STEENSBERG,
2002).
Diferentes células e tecidos produzem IL-6, e as principais fontes
durante infecções são provenientes de monócitos e macrófagos estimulados.
Um estudo demonstrou que estas células não eram as principais fontes de
elevação da IL-6 durante o exercício (STARKIE et al., 2001b). De fato, foi
recentemente demonstrado que o músculo posterior da coxa liberou IL-6 e que
esta liberação pode contribuir para as elevadas concentrações plasmáticas de
desta mesma citocina (STEENSBERG et al., 2000). No músculo esquelético, a
expressão de IL-6 aumenta após o exercício e precede as microrupturas nas
miofibrilas induzidas por este tipo de atividade. O controle da produção de
citocinas como: IL-6, IL-1β e TNF-α podem regular o processo de dor muscular
tardia decorrente de atividades esportivas (TOMIYA et al., 2004). De fato, a IL-
6 induz a proliferação de células satélites, que são células quiescentes
precursoras da formação de novos miotubos e auxiliam na regeneração
muscular (JONSDOTTIR et al., 2000).
Concentrações de glicocorticóides acima das variações fisiológicas,
entre as concentrações circadianas basais e as concentrações induzidas pelo
estresse do exercício, afetam diferentemente a produção de citocinas
específicas. A IL-6 é resistente à supressão dos glicocorticóides, no entanto, a
TNF-α é muito sensitiva aos glicocorticóides, indicando que as variações
fisiológicas nesta classe de hormônios podem exercer importante papel na
regulação da produção de TNF-α e também de outras citocinas como a IL-1β e
IL-6 (DeRIJK et al, 1997).
Interessantemente, a TNF-α pode aumentar a esterificação de ácidos
graxos e a formação de diacilglicerol, que é um importante ativador de algumas
isoformas de proteínas cinases C, podendo interferir com sinalização da
insulina via fosforilação de serina do IRS-1 (receptor de insulina-1) (YU et al.,
2002). Este pode ser um potencial mecanismo pelo qual a TNF-α causa
resistência à insulina (DYCK et al., 2006).
GOEBEL et al. (2000) observaram que o exercício aumentou a produção
in vitro de TNF-α em 32 indivíduos saudáveis (13 mulheres e 19 homens), com
idades entre 21 e 49 anos, que realizaram um exercício físico durante 15 a 18

84
minutos em cicloergômetro, pedalando a 70rpm (rotações por minuto) numa
intensidade de 75% do VO2max estimado.
O exercício extenuante induz aumentos na concentração de citocinas no
sangue. As concentrações de TNF-α apresentam aumento de duas a três
vezes depois de exercícios extenuantes (OSTROWSKI et al., 1999). É
relevante destacar que as citocinas têm meia vida curta, os tempos nos quais
as amostras são coletadas influenciam nos resultados (PEDERSEN et al,
1998b). Outros fatores que também podem alterar os dados obtidos estão
relacionados à duração, intensidade e tipo de exercício físico executado.
O aumento do TNF-α com relação ao metabolismo das células do
sistema imunitário, também pode estar envolvido em aumentar a captação de
glicose nestas células, com o intuito de melhorar sua resposta. Sabe-se que a
glicose e a glutamina são os principais substratos energéticos das células do
sistema imunitário, inclusive os neutrófilos (PITHON-CURI et al., 1997).
Em meninas saudáveis, de 14 a 16 anos, que realizaram uma única
sessão de polo aquático, com duração de 1 hora e 30 minutos, sendo que as
amostras de sangue foram coletadas antes e depois do exercício físico.
Encontraram uma pequena, porém significante redução de 20% sobre TNF-α.
(NEMET et al., 2003). No entanto, outros não registraram nenhuma mudança
(SUZUKI et al., 2000).
RHIND et al. (2001) demonstraram que a exposição prolongada ao frio
(apesar da manutenção da temperatura corporal) provoca redução na
expressão intracelular de TNF-α e leva a uma redução nas concentrações
plasmáticas de TNF-α depois do exercício.
SPRENGER et al. (1992) observaram elevadas concentrações de TNF-α
no estado de repouso, em 22 corredores quando comparados com indivíduos
controle, que não exibiram concentrações circulantes mensuráveis dessa
citocina. A correlação significante foi encontrada entre as concentrações
plasmáticas de TNF-α e IL-1β em indivíduos treinados e controle, sugerindo
que essas citocinas podem ser secretadas pelo mesmo tipo de estresse físico,
como a corrida. Com relação a outras citocinas, as concentrações plasmáticas
de TNF-α podem não mudar durante o exercício. Isso foi observado por
ULLUM et al. (1994) que não encontraram mudanças durante e depois de 6h

85
de ciclismo em indivíduos treinados, corroborando com outros estudos
(DRENTH et al., 1995; SMITH et al., 1992; SPRENGER et al., 1992).
TIMMONS et al. (2004) realizaram um estudo com doze meninos (9,8
anos de idade em média) e dez homens (22,1 anos de idade em média)
saudáveis, os quais pedalaram por 60 minutos a 70% do VO2max. As amostras
de sangue foram coletadas antes, imediatamente e 60 minutos após o
exercício. Concluíram que não houve aumento nas concentrações de TNF-α.
Os resultados destes estudos citados apresentaram estabilidade ou
diminuição nas concentrações plasmáticas de TNF-α, podem ser relacionados
de maneira similar com os dados encontrados em nossa pesquisa, pois foi
observada redução média de 23% nesta citocina nos grupos exercitados com
sobrecarga (Tabela 6 e Figura 14). A intensidade do exercício parece exercer
efeito sobre a concentração sérica de TNF-α, já que foi observada maior
redução de 20% no grupo exercitado durante 15 minutos em intensidade
moderada quando comparado ao grupo exercitado em intensidade leve no
mesmo volume (Tabela 6 e Figura 14).
Interessantemente, a resposta das citocinas ao exercício físico difere
daquela pronunciada nas diferentes infecções (FEBBRAIO e PEDERSEN,
2002; PEDERSEN e HOFFMAN-GOETZ, 2000; PEDERSEN et al., 2001a;
SUZUKI et al., 2002). O fato de que citocinas proinflamatórias, TNF-α e IL1-β,
em geral não aumentam com o exercício indica que a cascata de citocinas
induzida pelo exercício difere substancialmente da cascata de citocinas
induzida pelas infecções (OSTROWSKI et al., 2000). Recentemente,
PETERSEN e PEDERSEN (2005), propuseram que o exercício físico provoca
primariamente a liberação de IL-6, seguido de aumentos na IL-1ra, IL-10
(citocinas antiinflamatórias), proteína-C reativa (CRP) e receptor solúvel de
TNF-α (sTNF-R), inibindo ainda a produção de TNF-α. Em nosso trabalho,
pudemos registrar reduções na TNF-α nos grupos exercitados com sobrecarga
de 5%, além do grupo exercitado 5 sessões em intensidade leve (Tabela 6 e
Figura 14).
Outros estudos serão necessários para esclarecer se o aumento,
manutenção ou ainda redução destas citocinas poderia estar envolvido com a
melhora da resposta das células do sistema imunitário frente ao exercício.

86
7. Conclusões
Concluímos neste estudo que, o exercício físico tanto de curta duração,
como realizado até a exaustão, ou mesmo durante 5 sessões de volume
progressivo nas intensidades leve e moderada, pode modular o número de
leucócitos totais, promovendo aumento na contagem destas células. O
aumento na intensidade do exercício (de leve para moderada) pode induzir a
leucocitose mais pronunciada. Pudemos observar também que, quando uma
maior intensidade é exposta durante 5 sessões, pode-se alterar a resposta
leucocitária total, reduzindo a leucometria, o que nos leva a crer que os animais
ainda não estavam adaptados à esta carga.
No caso da contagem de linfócitos circulantes, o exercício promoveu
linfocitose. No exercício de natação com duração curta (5 a 15 minutos), o
aumento na contagem de linfócitos circulantes é acompanhado pelo aumento
no número de linfócitos teciduais, neste sentido, a duração do exercício pode
alterar a resposta dos linfócitos teciduais. De fato, após o exercício mais
prolongado (60 minutos a 10 horas de duração em média), os linfócitos
provenientes dos linfonodos mesentéricos foram reduzidos, migrando para a
corrente sangüínea e contribuindo na linfocitose induzida por este tipo de
atividade.
Com relação à funcionalidade de linfócitos em exercícios físicos
realizados por breves períodos (5 e 15 minutos, nas intensidades leve e
moderada), estes modularam agudamente o sistema imune promovendo
redução na viabilidade de linfócitos circulantes e redução na fragmentação de
DNA nesta célula. O exercício, mesmo quando realizado em curta duração e
intensidade moderada, não é inócuo frente à resposta imune.
A concentração sérica de TNF-α foi reduzida apenas no exercício
realizado com intensidade moderada e durante 5 sessões de volume
crescente. No entanto, para IL-6, o exercício mais prolongado (10 horas em
média) promoveu maiores modificações, induzindo aumentos expressivos.
A imunologia do exercício necessita prontamente de futuras
investigações devido à redução funcional de células imunes, que podem
ocorrer em indivíduos sedentários que se submetem a sessões agudas de
exercício, em alguns momentos, sem orientação profissional e/ou com

87
treinamentos inadequados, com relação à intensidade e duração que podem
extrapolar suas limitações físicas. Finalmente os efeitos positivos e negativos,
crônicos e agudos do exercício, em diferentes intensidades sobre o sistema
imune precisam ser melhor elucidados. Proporcionar a prática de treinamento
físico com benefícios e promoção da saúde, tanto em atletas como em
indivíduos sedentários, é de primordial importância para os profissionais da
área da fisiologia do exercício.
Para profissão de educadores físicos, fica a mensagem de que, as
alterações provocadas por qualquer tipo de exercício físico, não são apenas
estéticas ou musculares. Estas modificações induzidas no organismo devem
ser visualizadas de acordo com todo sistema biológico que compõem o ser
humano. Desta maneira, a compreensão das respostas imunológicas
resultantes de uma única sessão de exercício ou do treinamento físico
realizado em longo prazo, podem auxiliar na ampliação do conhecimento
científico e na adequação da prescrição individualizada do esforço.

88
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABBAS, A.K.; LICHTMAN, A.H.; PODER, J.S. Citocinas. In: _____. Imunologia celular e molecular. 4 ed. Rio de Janeiro: Revinter, cap.11, p.235-240,
2003. ADAMS, D.H.; SHAW, S. Leukocyte-endothelial interations and regulation of
leukocyte migration. Lancet., 343: 831-836, 1994. ARLT, W.; HEWISON, M. Hormones and immune function: implications of
aging. Aging Cell., 4: 209–216, 2004. AZENABOR, A.A.; HOFFMAN-GOETZ, L. Effect of exhaustive exercise on
membrane estradiol concentration, intracellular calcium, and oxidative damage in mouse thymic lymphocytes. Free Radic Biol Med., 28: 84-90, 2000.
BAGBY, G.J.; CROUCH, L.D.; SHEPHERD, R.E.; HOOFFMAN-GOETZ. Exercise and cytokines: spontaneous and elicited response. Exerc Immune Function., 55-78, 1996.
BAIN, B.J.; PHILLIPS, D.; THOMSON, K.; RICHARDSON, D.; GABRIEL, I. Investigation of the effect of marathon running on leucocyte counts of subjects of different ethnic origins: relevance to the aetiology of ethnic neutropenia. British J Haematol., 108: 483-487, 2000.
BARBIERI, M.; FERRUCCI, L.; RAGNO, E.; CORSI, A.; BANDINELLI, S.; BONAFE, M.; OLIVIERI, F.; GIOVAGNETTI, S.; FRANCESCHI, C.; GURALNIK, J.M.; PAOLISSO, G. Chronic inflamation and the effect of IGF-I on muscle strength and power in older persons. Am J Physiol Endocrinol Metab., 284: E481-E487, 2003.
BENEDETTI, F.; ALONZI, T.; MORETTA, A.; LAZZARO, D.; COSTA.; POLI, V.; MARTINI, A.; CILIBERTO, G.; FATTORI, E. Interleukin 6 causes growth impairment in transgenic mice through a decrease in insulin-like growth factor-1. J Clin Invest., 99: 643-650, 1997.
BENONI, G.; BELLAVITE, P.; ADAMI, A.; CHIRUMBOLO, S.; LIPPI, G.; BROCCO, G.; GIULINI, G. M.; CUZZOLIN. L. Changes in several neutrophil functions in basketball players before, during and after the sports season. Int J Sports Med., 16:34-37, 1995.
BENSCHOP, R.J.; RODRIGUEZ-FUERHAHN, M.; SCHEDLOWSKI, M. Cathecolamine-induced leukocytosis: early observations, current findings and future directions. Brain Behav Immun., 10: 77-91, 1996.
BLOTNICK, S.; PEOPLES, G.E.; FREEMAN, M.R.; EBERLEIN, T.J.; KLAGSBRUN, M. T lymphocytes synthesize and export heparin-binding epidermal growth factor-like growth factor and basic fibroblast growth factor, mitogens for vascular cells and fibroblasts: differential production and release by CD4+ and CD8+ T cells. Proc Natl Acad Sci USA., 91: 2890-2894, 1994.
BRUUNSGAARD, H.; GALBO, H.; HALKJAER-KRISTENSEN, J.; JOHANSEN, T.L.; MACLEAN, D.A.; PEDERSEN, B.K. Exercise-induce increase in serum interleukin-6 in humans is related to muscle damage. J Physiol Lond., 499: 833-841, 1997.
BUNTING, M.; HARRIS, E.S.; McINTYRE, T.M.; PRESCOTT, S.M.; ZIMMERMAN, G.A. Leukocyte adhesion deficiency syndromes: adhesion and tethering defects involving beta 2 integrins and selectin ligands. Curr Opin Hematol., 9: 30-35, 2002.

89
BUTCHER, S.K.; KILLAMPALLI, V.; LASCELLES, D.; WANG, K.; ALPAR, E. K.; LOR, J.M. Raised cortisol:DHEAS ratios in the elderly after injury: potential impact upon neutrophil function and immunity. Aging Cell., 4: 319–324, 2005.
BUYUKYAZI, G.; KUTUKCULER, N.; KUTLU, N.; GENEL, F.; KARADENIZ, G.; OZKUTUK, N. Differences in the cellular and humoral immune system between middle-aged men with different intensity and duration of physycally training. J Sports Med Phys Fitness., 44: 207-214, 2004.
CAMUS, G.; POORTMANS, J.; NYS, M.; DEBY-DUPONT, G.; DUCHATEAU, J.; DEBY, C.; LAMY, M. Mild endotoxaemia and the inflammatory response induced by a marathon race. Clin Sci (Colch)., 92:415-422, 1997.
CASTELL, L. Glutamine supplementation in vitro and in vivo, in exercise and immunodepression. Sports Med., 33(5): 323-345, 2003.
CASTELL, L.M.; POORTMANS, J.R.; LECLERCQ, R.; BRASSEUR, M.; DUCHATEAU, J.; NEWSHOLME, E.A. Some aspects of the acute phase response after a marathon race, and the effects of glutamine supplementation. Eur J Appl Physiol., 75:47-53, 1997.
CAVAGLIERI, C.R.; MARTINS, E.F.; COLLEONE, V.V.; RODRIGUES, C.; VECCHIA, M.G.; CURI, R. Fiber-rich diets alter rat intestinal leukocytes metabolism. J Nutrition Biochem., 11:555-561, 2000.
CAVAGLIERI, C.R.; NISHIYAMA, A.; FERNANDES, L.C.; CURI, R.; MILES, E.A.; CALDER, P.C. Differential effects of short-chain fatty acids on proliferation and production of pro- and anti-inflammatory cytokines by cultured lymphocytes. Life Sci., 73(13): 1683-90, 2003.
CONCORDET, J.P.; FERRY, A. Physiological programmed cell death in thymocytes is induced by physical stress (exercise). Am J Physiol., 265: C626-C629, 1993.
CROISIER, J.L.; CAMUS, G.; VENNEMAN, I.; DEBY-DUPONT, G.; JUCHMES-FERIR A.; LAMY, M.; CRIELAARD, J.M.; DEBY, C.; DUCHATEAU, J. Effects of training on exercise-induced muscle damage and interleukin 6 production. Muscle Nerve.,22:208-212, 1999.
CURI, R. Glutamina: metabolismo e aplicações clínicas e no esporte. Ed. Sprint, São Paulo, 2000.
CURI, R.; NESWHOLME, P.; NEWSHOLME, E.A. Intracellular distribution of some enzymes of the glutamine utilization pathway in rat lymphocytes. Biochem Biophys Res Commun., 138: 318-332, 1986.
CURY-BOAVENTURA, M.F.; POMPÉIA, C.; CURI, R. Comparative toxicity of oleic acid and linoleic acid on Raji cells. Nutrition., 21: 395-405, 2005.
DAVIS, S.N.; GALASSETTI, P.;. WASSERMAN, D.H.; TATE, D. Effects of Gender on Neuroendocrine and Metabolic Counterregulatory Responses to Exercise in Normal Man. J Clin Endocrinol Metab., 85: 224–230, 2000.
DAVIS, J.M.; MURPHY, E.A.; BROWN, A.S.; CARMICHAEL, M.D.; GHAFFAR, A.; MAYER, E.P. Effects of moderate exercise and oat beta-glucan on innate immune function and susceptibility to respiratory infection. Am J Physiol Regul Integr Comp Physiol., 286: R366-R372, 2004.
DE CARO, E.; FIOREDDA, F.; CALEVO, M.G.; SMERALDI, A.; SAITTA, M.; HANAU, G.; FARACI, M.; GRISOLIA, F.; DINI, G.; PONGIGLIONE, G.; HAUPT, R. Exercise capacity in apparently healthy survivors of cancer. Arch Dis Child.,; 91: 47–51, 2006.

90
DERIJK, R.H.; BOELEN, A.; TILDERS, F.J.; BERKENBOSCH, F.Induction of plasma interleukin-6 by circulating adrenaline in the rat. Psychneuroendocrinol., 19:155-163, 1994.
DERIJK, R.; MICHELSON, D.; KARP, B.; PETRIDES, J.; GALLIVEN, E.; DEUSTER, P.; PACIOTTI, G.; GOLD, P.W. Exercise and circadian rhythm-induced variations in plasma cortisol differentially regulate interleukin-1β (IL-1β), IL-6, and tumor necrosis fator-α (TNFα) production in humans: high sensitivity of TNFα and resistance of IL-6. J Clin Endocrinol Metab., 82(7): 2182-2191, 1997.
DIMITRIOU, L.; SHARP N.C.C.; DOHERTY, M. Circadian effects on the acute responses of salivary cortisol and IgA in well trained swimmers. Br J Sports Med., 36: 260-264, 2002.
DISHMAN, R.K. Brain monoamines, exercise, and behavioral stress: animal models. Med Sci Sports Exerc., 29: 63-74, 1997.
DORNFEST, B.S.; LAPIN, D.M.; NAUGHTON, B.A.; ADU, S.; KORN, L.; GORDON, A.S. Phenylhydrazine-induced leukocytosis in the rat. J Leuk Biol., 39: 37-48, 1986.
DRENTH, J. P.; VAN UUM, S. H.; VAN DEUREN, M.; PESMAN, G. J.; VAN DER VEN-JONGEKRIJG, J.; VAN DER MEER J. W. Endurance run increases circulating IL-6 and IL-1ra but downregulates ex vivo TNF-alpha and IL-1 beta production. J Appl Physiol., 79: 1497, 1995.
DYCK, D.J.; HEIGENHAUSER, G.J.F; BRUCE, C.R. The role of adipokines as regulators of skeletal muscle fatty acid metabolism and insulin sensitivity. Acta Physiol., 186: 5-16, 2006.
ESPERSEN, G.T.; ELBAEK, A.; ERNST, E.; TOFT, E.; KAALUND, S.; JERSILD, C.; GRUNNET, N. Effect of physycal exercise on cytokines and lymphocyte subpopulations in human peripheral blood. APMIS., 98: 395-400, 1990.
FEBBRAIO, M.A.; PEDERSEN, B.K. Muscle-derived interleukin-6: mechanisms for activation and possible biological roles. FASEB J., 16: 1335–1347, 2002.
FLESHNER, M. Exercise and neuroendocrine regulation of antibody product protective effect of physical activity on stress-induced suppression of the specific antibody response. Int J Sports Med., 21(suppl 1):S31-S32, 2000.
FLESHNER, M.; CAMPISI, J. Stress-Induced Extracellular HSP72 is a Functionally Significant Danger Signal to the Immune System. Med Sci Sports Exerc., 35(suppl 1): S9-S10, 2003.
FLESHNER, M.; CAMPISI, J.; DEAK, T.; GREENWOOD, B.N.; KINTZEL, J.A.; LEEM, T.H.; SMITH, T.P.; SORENSEN, B. Acute stressor exposure facilitates innate immunity more in physically active than in sedentary rats. Physiol Regul Integr Comp Physiol., 282: R1680-R1686, 2002.
FREEMAN, M.R.;SCHNECK, F.X.; GAGNON M.L.; CORLESS, C.; SOKER, S.; NIKNEJAD, K.; PEOPLES, G.E.; KLAGSBRUN, M. Peripheral blood T lymphocytes and infiltrating human cancers express vascular endothelial growth factor: a potential role for T cells in angiogenesis. Cancer Res., 55:4140-4145, 1995.
FRUBECK, G.; GÓMEZ-AMBROSE, J.; MURUZÁBAL, F.J.; BURELL, M.A.; The adipocyte: a model for integration of endocrine and metabolic signalling in energy metabolism regulation. Am J Physiol., 280: E827-E847, 2001.

91
FU, S.C.; QIN, L.; LEUNG, C.K.; CHAN, B.P.; CHAN, K.M. Regular moderate exercise training prevents decrease of CD4+ T-limphocytes induced by a single bout of strenuous exercise in mice. Can J Appl Physiol., 28: 370-381, 2003.
GABRIEL, H.H.; KINDERMANN, W. Adhesion molecules during immune response to exercise. Can J Physiol Pharmacol., 76: 512-23, 1998.
GADEBERG, O.V.; RHODES, J.M.; LARSEN, S.O. Isolation of human peripheral bloodhit2hit2 monocytes: A comparative methodological study. J Immunol Methods., 31 : 1-10, 1979.
GALVÃO, D.A.; NEWTON, R.U. Review of Exercise Intervention Studies in Cancer Patients. J CLIN ONCOL., 23: 899-909, 2005.
GANNON, G.; RHIND, S.; SUZUI, M.; SHEK, P.N.; SHEPHARD, R.J. Circulating levels of peripheral blood leucocytes and cytokines following competitive cycling. Can J Appl Physiol., 22:133-147, 1997.
GARCIA, C.; PITHON-CURI, T.C.; FIRMANO, M.L.; PIRES-DE-MELO, M.; NEWSHOLME, P.; CURI, R. Effect of adrenaline on glucose and glutamine metabolism and superoxide production by rat neutrophils. Clin Sci., 96: 549-55, 1999.
GERRIT, V.; STEENSBERG, A.; SACCHETTI, M.; FISCHER, C.; KELLER, C.; SCHJERLING, P.; HISCOCK, N.; MOLLER, K.; SALTIN, B.; FEBBRAIO, M.A.; PEDERSEN, B.K. Interleukin-6 Stimulates Lipolysis and Fat Oxidation in Humans. J Clin Endocrinol Metab., 88(7): 3005-3010, 2003.
GLEESON, M. Interleukins and exercise (perspectives). J Physiol., 529: 1, 2000.
GLEESON, M.; PYNE, D.B. Special features for the Olympics: effects of exercise on the immune system exercise effects on mucosal immunity. Immunol Cell Biol., 78: 536-544, 2000.
GOBATTO, C.A.; MELLO, M.A.R.; SIBUYA, C.Y.; AZEVEDO, J.R.M.; SANTOS, L.A.; KOKUBUN, E. Maximal lactate steady state in rats submitted to swimming exercise. Comp Bioch Physiol, 130: 21-27, 2001.
GOEBEL, M.U.; MILLS, P.J. Acute psychological stress and exercise and changes in peripheral leukocyte adhesion molecule expression and density. Psych Med., 62: 664-670, 2000.
GOEBEL, M.U.; MILLS, P.J.; IRWIN, M.R.; ZIEGLER, M.G. Interleukin-6 and Tumor Necrosis Factor-α Production After Acute Phychological Stress, Exercise, and Infused Isoproterenol: Differential Effects and Pathways. Psych Med., 62: 591-598, 2000.
GRANVILLE, D.J.; CARTHY, C.M.; HUNT, D.W.; McMANUS, B.M. Apoptosis: molecular aspects to cell death and disease. Lab Invest., 78: 893-913, 1998.
GREEN, D.R.; REED, J.C. Mitochondria and apoptosis. Science., 281: 1309 –1312, 1998.
GRIMBLE, R.F.; Inflammatory response in the eldery. Curr Opin Clin Nutr Metab Care., 6: 21-29, 2003.
HAAHR, P.M.; PEDERSEN, B.K.; FOMSGAARD, A.; TVEDE, N.; DIAMANT, M.; KLARLUND, K.; HALKJAER-KRISTENSEN, J.; BENDTZEN, K. Effect of physical exercise on in vitro production of interleukin 1, interleukin 6, tumor necrosis factor-alpha, interleukin 2 and interferon-gamma. Int J Sports Med., 12:223-237, 1991.

92
HADDAD, F.; ZALDIVAR, F.; COOPER, D.M.; ADAMS, G.R. IL-6-induced skeletal muscle atrophy. J Appl Physiol., 98: 911-917, 2005.
HALL, G.V.; STEENSBERG, A.; MASSIMO, S.; FISCHER, C.; KELLER, C.; SCHJERLING, P.; HISCOCK, N.; MOLLER, K.; SALTIN, B.; FEBBRAIO, M.A.; PEDERSEN, B.K. Interleukin-6 stimulates Lipolysis and Fat Oxidation in Humans. J Clin Endocrinol Metabolism., 88(7): 305-310, 2003.
HALSON, S.L.; LANCASTER, G.I.; JEUKENDRUP, A.E.; GLEESON, M. Immunological Responses to Overreaching in Cyclists. Med Sci Sports Exerc., 35(5): 854-861, 2003.
HARDMAN, A.E. Physical activity and cancer risk. Proc Nutr Soc., 60(1):107-13, 2001.
HAYDON, A.M.M.; MACINNIS, R.J.; ENGLISH, D.R.; GILES, G.G. Effect of physical activity and body size on survival after diagnosis with colorectal cancer. Gut., 55: 62–67, 2006.
HAYES, S.C.; ROWBOTTON, D.; DAVIES, P.S.W.; PARKER, T.W.; BASHFORD, J. Immunological Changes after Cancer Treatment and Participation in an Exercise Program. Med Sci Sports Exerc., 35(1): 2-9, 2003.
HENSON, D.A.; NIEMAN, D.C.; NEHLSEN-CANNARELLA, S.L.; FAGOAGA, O.R.; SHANNON, M.; BOLTON, M.R.; DAVIS, J.M.; GAFFNEY, C.T.; KELLN, W.J.; AUSTIN, M.D.; HJERTMAN, J.M.E.; SCHILLING, B.K. Influence of carbohydrate on cytokine and phagocytic responses to 2h of rowing. Med Sci Sports Exerc., 32: 1384-1389, 2000.
HISCOCK, N.; PETERSEN, E.; KRZYWKOWSKI, K.; BOZA, J.; HALKJAER-KRISTENSEN, J.; PEDERSEN, B.K. Glutamine supplementation further enhances exercise-induced plasma IL-6. J Appl Physiol., 95: 145-8, 2003.
HOFFMAN-GOETZ, L. Effect of estradiol and exercise on lymphocyte proliferation responses in female mice. Physiol Behav., Dec 1-15; 68 (1-2): 169-74, 1999.
HOFFMAN-GOETZ, L.; DUERRSTEIN, L. The effect of chronic and acute exercise on thymocyte apoptosis and necrosis in ovariectomized mice given dietary genistein. J Sports Med Phys Fitness., 44: 281-287, 2004.
HOFFMAN-GOETZ, L.; QUADRILATERO, J. Treadmill exercise in mice increases intestinal lymphocyte loss via apoptosis. Acta Physiol. Scand., 179: 289-297, 2003.
HOFFMAN-GOETZ, L.; ZAJCHOWSKI, S. In vitro apoptosis of lymphocytes after exposure to levels of corticosterone observed following submaximal exercise. J Sports Med Phys Fitness., 39: 269-274, 1999.
HOGG J. C., Neutrophil kinetics and lung injury. Physiol Rev., 67: 1249-1295, 1987.
HONG, S.; FARAG, N.H.; NELESEN, R.A.; ZIEGLER, M.G.; MILLS, P.J. Effects of regular exercise on lymphocytes subsets and CD62L after psychological vs. physical stress. J Psychosom Res., 56: 363-370, 2004. HONG, S.; JOHNSON, T.A.; FARAG, N.H.; GUY, H.J.; MATTHEWS, S.C.;
ZIEGLER, M.G.; MILLS, P.J. Attenuation of T-limphocyte demargination and adhesion molecule expression in response to moderate exercise in physically fit individuals. J Appl Physiol., 98: 1057-1063, 2005.

93
HORN, F.; HENZE, C.; HEIDRICH, K. Interleukin-6 signal transduction and lymphocyte function. Immunobiol., 202: 151-167, 2000.
HOST, C.R.; NORTON, K.I.; OLDS, T.S.; LOWE, E.L.A. The effects of altered exercise distribution on lymphocyte subpopulations. Eur J Appl Physiol., 72:157-164, 1995.
HSU, T.G.; HSU, K.M.; KONG, C.W.; LU, F.J.; CHENG, H.; TSAI, K. Leukocyte mitonchondria alterations after aerobic exercise in trained human subjects. Med Sci Sports Exerc., 34: 438-442, 2002.
IBFELT, T.; PETERSEN, E.W.; BRUUNSGAARD, H.; SANDMAND, M.; PEDERSEN, B.K. Exercise-induced change in type 1 cytokine-producing CD8+ T cells is related to a decrease in memory T cells. J Appl Physiol., 93: 645-648, 2002.
JANEWAY, C.A.; TRAVERS, P. Immunobiology: The Immune System in Health and Disease. 2nd ed. London: Current Biology Ltd., 1996.
JANKORD, R.; JEMIOLO, B. Influence of Physical Activity on Serum IL-6and IL-10 Levels in Healthy Older Men. Med Sci Sports Exerc., 36: 960-964, 2004.
JONSDOTTIR, I.H. Special feature for the Olympics: effects of exercise on the immune system: neuropeptides and their interaction with immune function. Immunol Cell Biol., 78: 562-70, 2000. JONSDOTTIR, I.H.; SCHJERLING, P.; OSTROWSKI, K. Muscle contractions
induce interleukin-6 mRNA production in rat muscle. J Physiol., 528: 157-163, 2000.
JOZSA, R.; OLAH, A.; CORNÉLISSEN, G.; CSERNUS, V.; OTSUKA, K.; ZEMAN, M.; NAGY, G.; KASZAKI, J.; STEBELOVA, K.; CSOKAS, N.; PAN, W.; HEROLD, M.; BAKLEN, E.E.; HALBERG, F. Circadian and extracircadian exploration during daytime hours of circulating corticosterone and other endocrine chronomes. Biomed Pharmacoth., 59: S109-S116, 2005.
KALSBEEK, A.; RUITER, M.; LA FLEUR, S.E.; VAN HEIJNINGEN, C.; BUIJS, R.M. The diurnal modulation of hormonal responses in the rat varies with different stimuli. J Neuroendocrinol., 15: 1144-1155, 2003.
KANALEY, J.A.; WELTMAN, J.Y.; PIEPER, K.S.; WELTMAN, A.; HARTMAN, M.L. Cortisol and Growth Hormone Responses to Exercise at Different Times of Day. J Clin Endocrinol Metab., 86: 2881–2889, 2001.
KAPPEL, M., TVEDE, N., GALBO, H., HAAHR, P.M., KJAER, M., LINSTOW, M., KLARLUND, K. AND PEDERSEN B.K. Evidence that the effect of physical exercise on NK cell activity is mediated by epinephrine. J Appl Physiol., 70: 2530-2534, 1991.
KATSUHIKO, S.; SHIGEYUKI, N.; MUTSUO, Y.; QIANG, L.; SHYGEYOSHI, K.; NORIYOSHI, O.; TAKASHI, K.; TAKASHI, U.; KAZUO, S. Impact of a Competitive Marathon Race on Systemic Cytokine and Neutrophil Responses. Med Sci Sports Exerc., 35(2): 348-355, 2003.
KEAST, D.; ARNSTEIN, D.; HARPER, W.; FRY, R.; MORTON, A. Depression of plasma glutamine concentration after exercise stress and its possible influence on the immune system. Med J Austr., 162: 15-18, 1995.
KERN, P.A.; RANGANATHAN, S.; LI, C.; WOOD, L.; RANGANATHAN, G. Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. Am J Physiol Endocrinol Metab., 280: E745-E751, 2001.

94
KELLER, C.; STEENSBERG, A.; PILEGAARD, H.; OSADA, T.; SALTIN, B.;
PEDERSEN, B.K.; NEUFER, P.D. Transcriptional activation of the IL-6 gene in human contracting skeletal muscle: influence of muscle glycogen content. Faseb J., 15:2748-2750, 2001.
KELLEY, D.S. Modulation of human immune and inflammatory responses by dietary fatty acids. Nutrition.,; 17(7-8): 669-73, 2001.
KELLY, D.A. The use of anti-interleukin-2 receptor antibodies in pediatric liver transplantation. Pediatr Transplant., 5(6): 386-9, 2001.
KOHUT, M.L.; BOEHM, G.W.; MOYNIHAN, J.A. Prolonged exercise suppresses antigen-specific cytokine response to upper respiratory infection. J Appl Physiol., 90: 678-684, 2001.
KROGH-MADSEN, R.; PLOMGAARD, P.; KELLER, P.; KELLER, C.; PEDERSEN, B.K. Insulin stimulates interleukin-6 and tumor necrosis factor-α gene expression in human subcutaneous adipose tissue. Am J Physiol Endocrinol Metab., 286: E234-E238, 2004.
KROEMER, G.; DALLAPORTA, B.; RESCHE-RIGON, M. The mitochondrial death/life regulator in apoptosis and necrosis. Ann Rev Physiol., 60: 619-642, 1998.
KUMAZAKI, T.M.; SASAKI, M.; NISHIYAMA, M.; TERANISHI, Y.; SUMIDA, H.; MITSUI, Y. Effect of BCL-2 down-regulation on cellular life span. Biogerontol., 3(5): 291-300, 2002.
LANDMANN, R. Beta-adrenergic receptors I human leucocyte subpopulations. Eur J Clin Invest., 22 (Suppl. 1): 30-36, 1992.
LAGRANHA, C.J.; SENNA, S.M.; DE LIMA, T.M.; DOI, S.Q.; CURI, R.; PITHON-CURI, T.C. Beneficial effect of glutamine on exercise-induced apoptosis of rat neutrophils. Med Sci Sports Exerc., 36(2): 210-217, 2004.
LaPERRIERE, A.; ANTONI, M.H.; IRONSON, G.; PERRY, A.; McCABE, P.; KLIMAS, N.; HELDER, L.; SCHNEIDERMAN, N.; FLETCHER M.A. Effects of aerobic exercise training on lymphocyte subpopulations. Int J Sports Med., 15 (Suppl 3): S127-S130, 1994.
LaPERRIERE, A.; FLETCHER, M.A.; ANTONI, M.H.; KLIMAS, N.G.; IRONSON, G.; SCHNEIDERMAN, N. Aerobic exercise training in ADIS risk group. Int J Sports Med., 12 (Suppl 1): S53-S57, 1991.
LAWRENCE, E.; VAN EEDEN, S.F.; ENGLISH, D.; HOGG, J.C. Polymorphonuclear leukocyte migration in streptococcal pneumonia: comparison and older PMN with those recently released from the marrow. Am J Respir Cell Mol Biol., 14: 217-224, 1996.
LIN, Y.S.; KUO, H.L.; KUO, C.F.; WANG, S.T.; YANG, B.C.; CHEN, H.I. Antioxidant administration inhibits exercise-induced thymocyte apoptosis in rats. Med Sci Sports Exerc., 31:1594-1598, 1999.
MALM, C. Susceptibility to infections in elite athletes: the S-curve. Scand J Med Sci Sports., 16: 4-6, 2006.
MALM, C. Exercise immunology: The current state of man and mouse. Sports Med., 34: 555-566, 2004.
McCARTHY D. A.; DALE M. M.; The leucocytosis of exercise. A review and model. Sports Med., 6(6): 333-63, 1988.

95
MACFARLIN, B.K.; MITCHELL, J.B.; MACFARLIN, M.A.; STEINHOFF, G.M. Repeated Endurance Exercise Affects Leukocyte Number but Not NK Cell Activity. Med Sci Sports Exerc., 35: 1130-1138, 2003.
MACKINNON, L.T. Advances in Execise Immunology. United States: Human Kinectics, 1999.
MACKINNON, L.T. Immunity in athletes. Int J Sports Med., 18: 562-568, 1997. MATTHEWS, C.E., OCKENE, I.S., FREEDSON, P.S., ROSAL, M.C.,
MERRIAM, P.A.; HEBERT, J.R. Moderate to vigorous physical activity and risk of upper-respiratory tract infection. Med Sci Sports Exerc., 34: 1242-1248, 2002.
MAZZEO R. S., Catecholamine responses to acute and chronic exercise. Med Sci Sports Exerc., 23: 839-845, 1991.
MAZZEO, R.S.; DONOVAN, D.; FLESHNER, M. et al. Interleukin-6 response to exercise and high-altitude exposure: influence of alpha-adrenergic blockade. J Appl Physiol., 91:2143-2149, 2001.
McFARLIN, B.K.; FLYNN, M.G.; STEWART, L.K.; TIMMERMAN, K.L. Carbohydrate intake during endurance exercise increases natural Killer cell responsiveness to IL-2. J Appl Physiol., 96: 271-275, 2004.
MCFARLIN, B.K.; MITCHELL, J.B.; MCFARLIN, M.A.; STEINHOFF, G.M. Repeated endurance exercise affects leukocyte number but not NK cell activity. Med Sci Sports Exerc., 35(7): 1130-8, 2003.
McTIERNAN, A.; ULRICH, C.; SLATE, S.; POTTER, J. Physical activity and cancer etiology:associations and mechanisms. Cancer Causes Control., 9(5):487-509, 1998.
MEYER, T.; FAUDE, O.; URHAUSEN, A.; SCHARHAG, J.; KINDERMANN, W. Different Effects of Two Regeneration Regimens on Immunological Parameters in Cyclists. Med Sci Sports Exerc., 36(10):1743-1749, 2004.
MEYER, T.; GABRIEL, H.; RÄTZ, M.; MULLER, H.J.; KINDERMANN, W. Anaerobic exercise induces moderate acute phase response. Med Sci Sports Exerc., 33: 549-555, 2001.
MILES M. P.; MACKINNON L. T.; GROVE D. S.; WILLIAMS N. I.; BUSH J. A.; MARX J. O.; KRAEMER W. J.; MASTRO A. M., The relationship of natural killer cell counts, perforin mRNA and CD2 expression to post-exercise natural killer cell activity in humans. Acta Physiol Scand., 174: 317-325, 2002.
MILLS P. J.; REHMAN J.; ZIEGLER M. G.; CARTER S. M.; DIMSDALE J. E.; MAISEL A. S., Nonselective beta blockade attenuates the recruitment of CD62L(-)T lymphocytes following exercise. Eur J Appl Physiol., 79: 531-534, 1999.
MINETTO, M.; RAINOLDI, A.; GAZZONI, M.; TERZOLO, M.; BORRIONE, P.; TERMINE, A.; SABA, L.; DOVIO, A.; ANGELI, A.; PACCOTI, P. Differential responses of serum and salivary interleukin-6 to acute strenous exercise. Eur J Appl Physiol., 93: 679-686, 2005.
MOLDOVEANU, A.I.; SHEPARD, R.J.; SHEK, P.N. Exercise elevates plasma levels but not gene expression of IL1 beta, IL-6, and TNF-alpha in blood mononuclear cells. J Appl Physiol., 89:1499-504, 2000.
MOLDOVEANU, A. SHEPHARD, R.J.; SHEK, P.N. The cytokine response to physical activity and training. Sports Med., 31: 115-44, 2001.

96
MOOREN, F. C.; BLOMING, D.; LECHTERMANN, A.; LERCH, M.M.; VOLKER, K. Lymphocyte apoptosis after exhaustive and moderate exercise. J Appl Physiol., 93:147-153, 2002.
MOOREN, F. C.; LECHTERMANN, A.; VOLKER, K. Exercise-Induced Apoptosis of Lymphocytes Depends on Training Status. Med Sci Sports Exerc., 36:1476-1483, 2004.
MORASKA, A.; FLESHNER, M. Voluntary physical activity prevents stress-induced behavioral depresión and anti-KLH antibody supression. Am J Physiol Regul Integr Comp Physiol., 281: R484-R489, 2001.
NAGAO, F.; SUZUI, M.; TAKEDA, K.; YAGITA, H.; OKUMURA, K. Mobilization of NK cells by exercise: downmodulation of adhesion molecules on NK cells by catecholamines. Am J Physiol Regul Integr Comp Physiol., 279: R1251-R1256, 2000.
NEHLSEN-CANNARELLA S. L.; NEIMAN D. C.; BASLK-LAMBERTON A. J.; A.J.; MARKOFF, P.A.; CHRITTON, D.B.; GUSEWITCH, G.; LEE, J.W. The effect of moderate exercise training an immune response. Med Sci Sports Exerc, 23: 64-70, 1991.
NEMET, D.; OH, Y.; KIM, H.S.; HIIL, M.A.; COOPER, D.M. The effect of intense exercise on inflammatory cyotkines and growth mediators in adolescent boys. Pediatrics., 110: 681-698, 2002.
NEMET, D.; PONTELLO, A.M.; ROSE-GOTTRON, C.; COOPER, D.M. Cytokines and Growth Factors during and after a Wrestling Season in Adolescent Boys. Med Sci Sports Exerc., 36:794-800, 2004.
NEMET, D.; ROSE-GOTTRON, C.M.; MILLS, P.J.; COOPER, D.M. Effect of Water Polo Practice on Cytokines, Growth Mediators, and Leukocytes in Girls. Med Sci Sports Exerc., 35: 356-363, 2003.
NEWSHOLME, E.A.; CALDER, P.C. The proposed role of glutamine in some cells of the immune system and speculative consequence for the whole animal. Nutrition., 13:728-30, 1997.
NEWSHOLME, E.A.; NEWSHOLME, P.; CURI, R.; CRABTREE, B.; ARDAWI, M.S.M. Glutamine metabolism in different tissues its physiological and pathological importance. In: Perspectives Clin Nutrition., Ed. J. M. KINNEY & P.R. BORUM. Urban and Schwarzenberg, Baltimore-Munich, 71-98, 1989.
NEWSHOME, E.A.; NEWSHOLME, P.; CURI, R.; CHALLONER, E.; ARDAWI, M.S.M. A role for muscle in the immune system and its importance in surgery, trauma, sepsis and burns. Nutrition., 4:261-8, 1988.
NEWSHOLME, E.A.; NEWSHOLME, P.; CURI, R. The role of the citric acid cycle in cells of the immune system and its importance in sepsis, trauma and burns. Biochem Soc Symp., 54:145-61, 1987.
NICOLETTI, I.; MIGLIORATI, G.; PAGLIACCI, M.C.; GRIGNANI, F.;RICCARDI, C. Arapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J Immunol Methods., 139:271–9, 1991.
NIELSEN, H.G.; LYBERG, T. Long-Distance Running Modulates the Expression of Leukocyte and Endothelial Adhesion Molecules. Scand J Immunol., 60: 356-362, 2004.

97
NIELSEN, H.B.; SECHER, N.H.; KRISTENSEN, J.H.; CHRISTENSEN, N.J.; ESPERSEN, K.; PEDERSEN, B.K. Splenectomy impairs lymphocytosis during maximal exercise. Am J Physiol Regul Integr Comp Physiol., 272: R1847-R1852, 1997.
NIEMAN, D.C. Exercise, infection, and immunity. Int J Sports Med., 15: S131-S141,1994a.
NIEMAN, D.C. Exercise, upper respiratory tract infection, and the immune system. Med Sci Sports Exerc., 26:128-139,1994b.
NIEMAN, D. C. Exercise and resistance to infection. Can Physiol Pharmacol., 76: 573-80, 1998.
NIEMAN, D.C. Nutrition, exercise and immune system function. Clin Sports Med., 18: 537-548, 1999.
NIEMAN, D.C.; HENSON, D.A.; AUSTIN, M.D.; BROWN, V.A. Imune Response to a 30-Minute Walk. Med Sci Sports Exerc., 37:57-62, 2005.
NIEMAN, D.C.; HENSON, D.A.; SMITH, L.L.; UTTER, A.C.; VINCI, D.M.; DAVIS, J.M.; KAMINSKY, D.E.; SHUTE, M. Cytokine changes after a marathon race. J Appl Physiol., 91:109-114, 2001.
NIEMAN, D.C.; JOHANSSEN, L.M.; LEE, J.W.; ARABATZIS, K. Infections episodes in runners before and after the Los Angeles Marathon. J Sports Med Phys Fitness., 30: 321-328, 1990.
NIEMAN, D.C.; KERNODLE, M.W.; HENSON, D.A.; SONNENFELD, G.; DAVIS, J.M. The acute response of the immune system to tennis drills in adolescent athletes. Res Q Exerc Sport., 71: 403-408, 2000.
NIEMAN, D. C.; PEDERSEN B. K. Exercise and immune function: recent developments. Sports Med., 27(2): 73-80, 1999.
NONOGAKI, K.; FULLER, G.M.; FUENTES, N.L.; MOSER, A.H.; STAPRANS, I.; GRUNFELD, C.; FEINGOLD, K.R. Interleukin-6 stimulates hepatic triglyceride secretion in rats. Endocrinol., 136: 2143-2149, 1995.
NOSAKA, K.; CLARKSON, P.M. Changes in indicators of inflammation after after eccentric exercise of the elbow flexors. Med Sci Sports Exerc., 28: 953-961, 1996.
OHMAN E. M.; BUTLER J.; KELLY J.; HORGAN J.; O’MALLEY K., Beta-adrenoreceptor adaptation to endurance training. J Cardiovasc Pharmacol., 10: 728-731, 1987.
OPFERMAN, J.T.; KORSMEYER, S.J. Apoptosis in the development and maintenance of the immune system. Nat Immunol., 4(5): 410-415, 2003.
OSTROWSKI, K.; HERMANN, C.; BANGASH, A.; SCHJERLING, P.; NIELSEN, J.N.; PEDERSEN, B.K. A trauma-like elevation in plasma cytokines in humans in response to treadmill running. J Physiol (Lond.)., 508:949-953, 1998a.
OSTROWSKI, K.; ROHDE, T.; ASP, S.; SCHJERLING, P.; PEDERSEN, B.K. Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans. J Physiol., 515: 287-291, 1999.
OSTROWSKI, K.; ROHDE, T.; ZACHO, M.; ASP, S.; PEDERSEN, B.K. Evidence that IL-6 is produced in human skeletal muscle during prolonged running. J Physiol (Lond.)., 508:949-953, 1998b.
OSTROWSKI, K.; SCHJERLING, P.; PEDERSEN, B.K. Physical activity and plasma interleukin-6 in humans – effect of intensity of exercise. Eur J Appl Physiol., 83:512-515, 2000.

98
ORTEGA, E. Neuroendocrine mediators in the modulation of phagocytosis by exercise: Physiological implications. Exerc Immunol Rev., 9:70-93, 2003.
PAPANICOLAOU, D.A.; PETRIDES, J.S.; TSIGOS, C. BINA, S.; KALOGERAS, K.T.; WILDER, R.; GOLD, P.W.; DEUSTER, P.A.; CHROUSOS, G.P. Exercise stimulates interleukin-6 secretion: inhibition by glucocorticoids and correlation with catecholamines. Am J Physiol., 271:E601-E605, 1996.
PARRY-BILLINGS, M.; EVANS, J.; CALDER, P.C.; NEWSHOLME, E.A. Does glutamine contribute to immunosupression after major burns? Lancet., 336:523-525, 1990.
PATEL, H.; HOFFMAN-GOETZ, L. Effects of oestrogen and exercise on caspase-3 activity in primary and secondary lymphoid compartments in ovariectomized mice. Acta Physiol Scand., 176: 177-184, 2002.
PATEL, K.D.; CUVELIER, S.L.; WIEHLER. Selectins: critical mediators of leucocyte recruitment. Semin Immunol., 14: 73-81, 2000.
PEDERSEN, B.K. Exercise and cytokines. Immunol Cell Biol., 78:532-535, 2000.
PEDERSEN, M.; BRUUNSGAARD, H.; WEIS, N.; HENDEL, H.W.; ANDREASSEN, B.U.; ELDRUP, E.; DELA, F.; PEDERSEN, B.K. Circulating levels of TNF-α and IL-6-relation to truncal fat mass and muscle mass in healthy eldery individuals and in patients with Type-2 diabetes. Mech Ageing Dev., 124: 495-502, 2003b.
PEDERSEN, B.K.; BRUUNSGAARD, H.; KLOKKER, M.; KAPPEL, M.; MACLEAN, D.A.; NIELSEN, H.B.; ROHDE, T.; ULLUM, H.; ZACHO, M. Exercise-induced immunomodulation: possible roles of neuroendocrino factors and metabolic factors. Int J Sports Med., 18(suppl 1): S2-7, 1997.
PEDERSEN, B.K.; HELGE, J.W.; RICHTER, E.A.; ROHDE, T.; KIENS, B. Training and natural immunity: effects of diets rich in fat or carbohydrate. Eur J Appl Physiol., 82: 98-102, 2000.
PEDERSEN, B.K.; HOFFMAN-GOETZ, L. Exercise and the Immune System: Regulation, Integration and Adaptation. Physiol Rev., 80: 1055-1081, 2000.
PEDERSEN, B.K.; NIEMAN, D.C. Exercise immunology: integration and regulation. Trends Immunol Today. 19(5):204-206, 1998a.
PEDERSEN, B.K.; ROHDE, T.; OSTROWSKI K. Recovery of the immune system after exercise. Acta Physiol Scand, 162: 325-332, 1998b.
PEDERSEN, B.K.; STEENSBERG, A. Exercise and hypoxia: effects on leukocytes and interleukin-6 – shared mechanisms?. Med Sci Sports Exerc., 34(12): 2004-2012, 2002.
PEDERSEN, B.K.; STEENSBERG, A.; FISCHER, C.; KELLER, C.; PLOMGAARD, P.; FEBBRAIO, M.; SALTIN, B. Searching for the exercise factor: is IL-6 a candidate? J Muscle Res Cell Motil., 24: 113-119, 2003a.
PEDERSEN, B.K.; STEENSBERG, A.; SCHJERLING, P. Muscle-derived interleukin-6: possible biological effects. J Physiol., 536: 329-337, 2001a.
PEDERSEN, B.K.; TOFT, A.D. Effects of exercise on lymphocytes and cytokines. Br J Sports Med., 34: 246-251, 2000.
PEDERSEN, B.K.; ULLUM, H. NK cell response to physycal activity: mechanisms of action. Med Sci Sports Exerc., 26: 140-146, 1994.
PEDERSEN, B.K.; WOODS, J.A.; NIEMAN, D.C. Exercise-induced immune changes- an influence on metabolism? Trends Immunol., 22: 473-475, 2001b.

99
PEIJIE, C.; HONGWU, L.; FENGPENG, X.; JIE, R.; ZHUANG, J. Heavy load exercise induced dysfunction of immunity and neuroendocrine responses in rats. Life Sci., 72: 2255-2262, 2003.
PETERSEN, A.M.W.; PEDERSEN, B.K. The anti-inflammatory effect of exercise. J Appl Physiol., 98: 1154-1162, 2005.
PHANEUF, S.; LEEUWENBURGH, C. Apoptosis and exercise. Med Sci Sports Exerc., 33: 393-396, 2001.
PITHON-CURI, T.C.; PIRES DE MELO, M.; DE AZEVEDO, R.; ZORN, T.M.T.; CURI, R. Glutamine utilization by rat neutrophils. Presence of phosphate-dependent glutaminase. Am J Physiol., 273: C1124-29, 1997.
PITHON-CURI, T.C.; SCHUMACHER, R.I.; FREITAS, J.J.; LAGRANHA, C.; NEWSHOLME, P.; PALANCH, A.C.; DOI, S.Q.; CURI, R. Glutamine delays spontaneous apoptosis in neutrophils. Am J Physiol - Cell Physiol., 284: C1355–C1361, 2003.
PLAYFAIR, J. H. L.; LYDYARD, P.M. Imunologia Médica. Ed. Revinter, Rio de Janeiro, 1999.
PUTLUR, P.; FOSTER, C.; MISKOWSKI, J.A.; KANE, M.K.; BURTON, S.E.; CHEETT, T.P.; McGUIGAN, M.R. Alteration of Immune Function in women collegiate soccer players and college students. J Sports Sci Med., 3: 234-243, 2004.
QUADRILATERO, J.; HOFFMAN-GOETZ, L. N-Acetyl-L-cysteine prevents exercise-induced intestinal lymphocyte apoptosis by maintaining intracellular glutathione levels and reducing mitochondrial membrane depolarization. Bioch Bioph Research Commun., 319: 894-901, 2004.
QUADRILATERO, J.; HOFFMAN-GOETZ, L. N-Acetyl-L-Cysteine Inhibits Exercise-Induced Lymphocyte Apoptotic Protein Alterations. Med Sci Sports Exerc., 37(1): 53-56, 2005.
REARDON, K.A.; DAVIS, J.; KAPSA, R.M.; CHOONG, P.; BYRNE, E. Myostatin, insulin-like growth factor-1, and leukemia inhibitory factor mRNAs are upregulated in chronic human disuse muscle atrophy. Muscle Nerve., 24: 893-899, 2001.
RIETJENS, G.J.W.M.; KUIPERS, H.; ADAM, J.J.; SARIS, W.H.M.; VAN BREDA, E.; VAN HAMONT, D.; KEIZER, H.A. Physiological, Biochemical and Psychological Markers of Strenuous Training-Induced Fatigue. Int J Sports Med., 26: 16-26, 2005.
RHIND, S.G.; CASTELLANI, J.W.; BRENNER, I.K.M.; SHEPHARD, R.J.; ZAMECNIK, J.; MONTAIN, S.J.; YOUNG, A.J.; SHEK, P.N. Intracellular monocyte and serum cytokine expression is modulated by exhausting exercise and cold exposure. Am J Physiol Regul Integr Comp Physiol., 281: R66-R75, 2001.
RIVIER, A.; PÈNE, J.; CHANEZ, P.; ANSELME, F.; CAILAUD, C.; PREFÁUT, C.; GODARD, PH.; BOUSQUET, J. Release of cytokines by Blood Monocytes During Strenuous Exercise. Int J Sports Med., 15:192-198, 1994.
RONSEN, O.; LEA, T.; BAHR, R.; PEDERSEN, B.K. Enhanced plasma IL-6 and IL-1ra responses to repeated vs. single bouts of prolonged cycling in elite athletes. J Appl Physiol., 92: 2547-2553, 2002.

100
RONSEN, O.; BORSHEIM, E.; BAHR, ROALD.; PEDERSEN, B.K.; HAUG, E.; KJELDSEN-KRAGH, J.; HOSTMARK, A.T. Immuno-endocrine and metabolic responses to long distance ski racing in world-class male and female cross-country skiers. Scand J Med Sci Sports., 14: 39–48, 2004.
RONSEN, O.; HOLM, K.; STAFF, H.; OPSTAD, P.K.; PEDERSEN, B. K.; BAHR, R. No effect of seasonal variation in training load on immuno-endocrine responses to acute exhaustive exercise. Scand J Med Sci Sports., 11: 141–148, 2001a.
RONSEN, O.; PEDERSEN, B.K.; ORITSLAND, B.K.; BAHR, R.; KJEDSEN-KRAGH, J. Leukocyte counts and lymphocyte responsiveness associated with repeat bouts of strenuous endurance exercise. J Appl Physiol., 91:245-435, 2001b.
ROWBOTTON, D.; KEAST, D.; MORTON, A. R. The merging role of glutamine as an indicator of exercise stress and overtraining. Sports Med., 21:80-97, 1996.
ROWBOTTOM, D.G.; GREEN, K.J. Acute exercise effects on the immune system. Med Sci Sports Exerc., 32(Suppl 7): S396-405, 2000.
SAGHIZADEH, M.; ONG, J.M.; GARVEY, W.T.; HENRY, R.R.; KERN, P.A. The expression of TNF alpha by human muscle. Relationship to insulin resistance. J Clin Invest., 97: 1111-1116, 1996.
SAPOLSKY, R.M.; ROMERO, L.M.; MUNCK, A.U. How Do Glucocorticoids Influence Stress Responses?Integrating Permissive, Suppressive, Stimulatory, and Preparative Actions. Endocrine Rev., 21: 55–89, 2000.
SAXTON J. M.; CLAXTON D.; WINTER E.; POCKLEY A. G. Peripheral blood leukocyte functional responses to acute eccentric exercise in humans are influenced by systemic stress, but not by exerxise-induced muscle damage. Clinical Sci (Lond)., 104: 69-77, 2003.
SCHARHAG, J.; MEYER, T.; GABRIEL, H.H.W.; SCHLICK, B.; FAUDE, O.; KINDERMANN, W. Does prolonged cycling of moderate intensity affect immune cell function. Br J Sports Med., 39: 171-177, 2005.
SERRANO, M.A.R.; CURI, R.; PARRY-BILLINGS, M.; WILLIANS, J.F.; NEWSHOLME, E.A. Effects of glucocorticoids on lymphocytes metabolism. Am J Physiol., 264: E24-E28, 1993.
SHEK, P.N.; SABISTON, B.H.; BUGUET, A., RADOMSKI, M.W. Strenuous exercise and immunological changes: A multiple-time-point analysis of leukocyte subsets, CD4/CD8 ratio, immunoglobulin production and NK cell response. Int J Sports Med., 16: 466-474, 1995.
SHEPARD, R.J. Exercise, immune function and HIV infection. J Sports Med Phys Fitness., 38(2): 101-10, 1998.
SHEPARD, R.J.; GANNON, G.; HAY, J.B.; SHEK, P.N. Adhesion molecule expression in acute and chronic exercise. Crit Ver Immunol., 20(3): 245-66, 2000.
SHEPHARD, R.J.; SHEK, P.N. Exercise, aging and immune function. Int J Sports Med, 16: 1-6, 1995.
SIGVARDSSON, K.; SVANFELDT, E.; KILBOM, A. Role of adrenergic nervous system in development of training-induced bradycardia. Acta Physiol Scand., 101: 481-488, 1977.
SMITH, J. A.; TELFORD, R. D.; BAKER, M. S.; HAPEL, A. J.; WEIDEMANN, M. J. Cytokine immunoreactivity in plasma does not change after moderate endurance exercise. J Appl Physiol., 73: 1396, 1992.

101
SONDERGAARD, S.R.; OSTROWSKI, K.; ULLUM, H.; PEDERSEN, B.K. Changes in plasma concentrations of interleukin-6 and interleukin-1 receptor antagonists in response to adrenaline infusion in humans. Eur J Appl Physiol., 83:95-98, 2000.
SPRENGER, H.; JACOBS, C.; NAIN, M.; GRESSNER, A.M.; PRINZ, H.; WESEMANN, W.; GEMSA, D. Enhanced release of cytokines, interleukin- 2 receptors, and neopterin after long-distance running. Clin Immunol Immunopathol., 53: 188-195, 1992.
STARKIE, R.L.; ROLLAND, J.; ANGUS, D.J.; ANDERSON, M.J.; FEBBRAIO, M.A. Circulating monocytes are not the source of elevations in plasma IL-6 and TNF-alpha levels after prolonged running. Am J Physiol., 280: C769-C774, 2001a.
STARKIE, R.L.; ROLLAND, J.; FEBBRAIO, M.A. Effect of adrenergic blockade on lymphocyte cytokine production at rest and during exercise. Am J Physiol Cell Physiol., 281: C1233-C1240, 2001b.
STEENSBERG, A.; FEBBRAIO, M.A.; OSADA, T.; SCHJERLING, P.; VAN HALL, G.; SALTIN, B.; PEDERSEN, B.K. Interleukin-6 production in contracting human skeletal muscle in influenced by pre-exercise muscle glycogen content. J Physiol., 537: 633-639, 2001a.
STEENSBERG, A.; KELLER, C.; STARKIE, R.L.; OSADA, T.; FEBBRAIO, M.A.; PEDERSEN, B.K. IL-6 and TNF-α expression in, and release from, contracting human skeletal muscle. Am J Physiol Endocrinol Metab., 283: E1272-E1278, 2002.
STEENSBERG, A.; TOFT, A.D.; BRUUNSGAARD, H.; SANDMAND, M.; HALKJAER-KRISTENSEN, J.; PEDERSEN, B.K. Strenuous exercise decreases the percentage of type 1 T cells in the circulation. J Appl Physiol., 91: 1708-1712, 2001b.
STEENSBERG, A.; TOFT, S.P.; SCHJERLING, A.D.; HALKJAER-KRISTENSEN, J.; PEDERSEN, B.K. Plasma interleukin-6 during strenous exercise: role of adrenaline. Am J Physiol., 281:1001-1004, 2001c.
STEENSBERG, A.; VAN HALL, G.; OSADA, T.; SACCHETTI, M.; SALTIN, B.; PEDERSEN, B.K. Production of IL-6 in contracting human skeletal muscles can account for the exercise-induced increase in plasma IL-6. J. Physiol. (Lond.)., 529:237-242, 2000.
STEPPICH, B.; DAYYANI, F.; GRUBER, R.; LORENZ, R.; MACK, M.; ZIEGLER-HEITBRO, H.W.L. Selective Mobilization of CD14+ CD16+ monocytes by exercise. Am J Cell Physiol., 279: C578-C586, 2000.
STICH, V.; DE GLISEZINSKI, I.; BERLAN, M.; BULOW, J.; GALITZKY, J.; HARANT, I.; SULJKOVICOVA, H.; LAFONTAN, M.; RIVIERI, D.; CRAMPES, F. Adipose tissue lipolysis is increased during a repeated bout of aerobic exercise. J Appl Physiol., 88: 1277-1283, 2000.
STITES, D.P.; TERR, A. I.; PARSLOW, T.G. Imunologia Médica. Ed. Guanabara Koogan, 9ª edição, Rio de Janeiro, 2000.
SOTHMANN, M.S.; HART, B.A.; HORN, T.S. Plasma catecholamine response to acute psychological stress in humans: relation to aerobic fitness and exercise training. Med Sci Sports Exerc., 23: 860-867, 1991.
SUGIURA, H.; NISHIDA, H.; SUGIURA, H.; MIRBOD, S.M. Immunomodulatory action of chronic exercise on macrophage and lymphocyte cytokine production in mice. Acta Physiol Scand., 174: 247-256, 2002.

102
SUZUKI, K.; NAKAJI, S.; YAMADA, M.; LIU, Q.; KURAKAKE, S.; OKAMURA, N.; KUMAE, T.; UMEDA, T.; SUGAWARA, K. Impact of a Competitive Marathon Race on Systemic Cytokine and Neutrophil Responses. Med Sci Sports Exerc., 35(2): 348-355, 2003.
SUZUKI, K.; YAMADA, M.; KURAKAKE, S.; OKAMURA, N.; YAMAYA, K.; LIU, Q.; KUDOH, S.; KOWATARI, K.; NAKAJI, S.; SUGAWARA, K. Circulating cytokines and hormones with immunosuppressive but neutrophil-priming potentials rise after endurance exercise in humans. Eur J Appl Physiol., 81: 281-287, 2000.
TIMMONS, B.W.; TARNOPOLSKY, M.A.; BAR-OR, O. Immune responses to strenuous exercise and carbohydrate intake in boys and men. Pediatric Research., 56: 227-234, 2004.
TIMMONS, B.W.; TARNOPOLSKY, M.A.; BAR-OR, O. Sex-based effects on the distribution of NK cells subsets in response to exercise and carbohydrate intake in adolescents. J Appl Physiol., 12 [article in press], 2006.
TOFT, A. D.; JENSEN, L. B.; BRUUNSGAARD, H.; IBFELT, T.; HALKEJAER-KRISTENSES, J.; FEBBRAIO, M.; PEDERSEN, B. K. Cytokine response to eccentric exercise in young and elderly humans. Am J Physiol Cell Physiol., 283, C289-C295, 2002.
TOFT, A.D.; THORN, M.; OSTROWSKI, K.; ASP, S.; MOLLER, K.; IVERSEN, S.; HERMANN, C.; SONDERGAARD, S.R.; PEDERSEN, B.K. N-3 polyunsaturated fatty acids do not affect cytokines response to strenous exercise. J Appl Physiol., 89: 2401-2406, 2000.
TOMIYA, A,; AIZAWA, T.; NAGATOMI, R.; SENSUI, H.; KOKUBUN, S. Myofibers Express IL-6 After Eccentric Exercise. Am J Sports Med., 32: 503-508, 2004.
TOTH, M.J.; ADES, P.A.; LEWINTER, M.M.; TRACY, R.P.; TCHERNOF, A. Skeletal muscle myofibrillar mRNA expression in heart failure: relationship to local and circulating hormones. J Appl Physiol., 100: 35 – 41, 2006.
ULLUM, H.; HAAHR, P. M.; DIAMANT, M.; PALMO, J.; HALKJAER-KRISTENSEN, J.; PEDERSEN, B. K. Bicycle exercise enhances plasma IL-6 but does not change IL-1 alpha, IL-1 beta, IL-6, or TNF-alpha pre-mRNA in BMNC. J Appl Physiol., 77: 93, 1994.
VAN EEDEN, S.; KITAGAWA, Y.; KLUT, M.E.; LAWRENCE, E.; HOGG, J.C. Polymorphonuclear leukocytes released from of bone marrow preferentially sequester in lung microvessels. Microcirculation., 4: 369-380, 1997.
VOLTARELLI, F.A.; GOBATTO, C.A.; MELLO, M.A.R. Determination of anaerobic threshold in rats using the lactate minimum test. Braz J Med Biol Res., 35: 1389-1394, 2002.
WALLENIUS, V.; WALLENIUS, K.; AHRÉN, B.; RUDLING, M.; CARLSTEN, H.; DICKSON, S.L.; OHLSSON, C.; JANSSON, J-O. Interleukin-6-deficient mice develop mature-onset obesity. Nat Med., 8: 75-79, 2002.
WEISE, M.; DRINKARD, B.; MEHLINGER, S.L.; HOLZER, S.M.; EISENHOFER, G.; CHARMANDARI, E.; CHROUSOS, G.P.; MERKE, D.P. Stress Dose of Hydrocortisone Is Not Beneficial in Patients with Classic Congenital Adrenal Hyperplasia Undergoing Short-Term, High-Intensity Exercise. J Clin Endocrinol Metab., 89: 3679–3684, 2004.

103
WIGAL, S.B.; NEMET, D.; SWANSON, J.M.; REGINO, R.; TRAMPUSH, J.; ZIEGLER, M.G.; COOPER, D.M. Catecholamine Response to Exercise in Children with Attention Deficit Hyperactivity Disorder. Pediatric Res., 53: 756-761, 2003.
WILLOUGHBY, D.S.; TAYLOR, M.; TAYLOR, L. Glucocorticoid receptor and ubiquitin expression after repeated eccentric exercise. Med Sci Sports Exerc., 35(12): 2023-31, 2003.
YENDE, S.; WATERER, G.W.; TOLLEY, E.A.; NEWMAN, A.B.; BAUER, D.C.; TAAFFE, D.R.; JENSEN, R.; CRAPO, R.; RUBIN, S.; NEVITT, M.; SIMONSICK, E.M.; SATTERFIELD, S.; HARRIS, T.; KRITCHEVSKY, S.B. Inflammatory markers are associated with ventilatory limitation and muscle dysfunction in obstructive lung disease in well functioning elderly subjects. Thorax., 61: 10–16, 2006.
YU, C.; CHEN, Y.; CLINE, G.W.; ZHANG, D.; ZONG, H.; WANG, Y.; BERGERON, R.; KIM, J.K.; CUSHMAN, S.W.; COONEY, G.J.; ATCHESON, B.; WHITE, M.F.; KRAEGEN, E.W.; SHULMAN, G.I. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-Kinase activity in muscle. J Biol Chem., 277: 50230-50236, 2002.
XING, Z.; GAUDIE, J.; COX, G.; BAUMANN, H.; JORDANA, M.; LEI, X. F.; ACHONG, M.K. J Clin Invest., 101: 311-320, 1998.

104
9. ANEXO: PUBLICAÇÕES REFERENTES AO MESTRADO
TRABALHOS RESUMIDOS PUBLICADOS EM ANAIS DE EVENTO
PRESTES, J.; GUERESCHI, M.G.; BRANBILLA, S.B.; DIAS, R.; FERREIRA, C. K.O.; CAVAGLIERI, C.R.; PALANCH, A.C. Alterações da ultra-estrutura de linfócitos dos linfonodos mesentéricos de ratos submetidos a 5 minutos de exercício físico agudo de intensidade leve e moderada In: 13° Simpósio Internacional de Iniciação Científica, 13º SIICUSP. Ribeirão Preto: USP Ribeirão Preto, 4807-4807, 2005.
PRESTES, J.; PALANCH, A.C.; FROLLINI, A.B.; FERREIRA, C.K.O.; DIAS, R.; DONATTO, F.; GUERESCHI, M.G.; URTADO, C.B.; LEITE, G.S.; CAVAGLIERI, C.R. Efeito do exercício físico realizado até exaustão, nas intensidades leve e moderada sobre a concentração sérica de IL-6 In: I Congresso Paulista da Sociedade Brasileira de Fisiologia do Exercício. IV Workshop em Fisiologia do exercício da UFSCAr. SãoCarlos: UFSCAr, t43-t43, 2005.
RAMALLO, B.T.; PRESTES, J.; FOSCHINI, D.; CAVAGLIERI, C.R. Efeitos agudos do método de múltiplas séries sobre as células do sistema imune In: 13° Simpósio Internacional de Iniciação Científica, 13º SIICUSP. Ribeirão Preto: USP Ribeirão Preto, 3110–3110, 2005.
VEIGA, A.; PRESTES, J.; RAMALLO, B.T.; FOSCHINI, D.; FERREIRA, C. K.O.; DONATTO, F.; URTADO, C.B.; CAVAGLIERI, C.R.; et. al. Efeitos agudos do treinamento de força sobre as células do sistema imunológico em praticantes de musculação em academias In: 8º Congresso de Iniciação e Produção Científica & 7º Seminário de extensão Metodista, 8º Congresso de Iniciação e Produção Científica e 7º Seminário de extensão Metodista. São Paulo: UMESP, 2971–2971, 2005.
PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício em intensidade leve sobre o número de leucócitos circulantes In: XX Reunião Anual da Federação de Sociedades de Biologia Experimental, FESBE. São Paulo: FESBE, 81–81, 2005.

105
PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício físico em intensidade leve sobre a concentração de linfócitos mesentéricos e circulantes de ratos sedentários In: XX Reunião Anual da Federação de Sociedades de Biologia Experimental, FESBE 2005. São Paulo: FESBE, 81-81, 2005.
DIAS, R.; PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício físico em intensidade leve sobre a glicemia, lactemia e hematócrito de ratos sedentários In: XX Reunião Anual da Federação de Sociedades de Biologia Experimental, FESBE 2005. São Paulo: FESBE, 81-81, 2005.
PRESTES, J.; CAVAGLIERI, C.R.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B.; PALANCH, A.C. Efeitos da adaptação ao exercício físico em intensidade leve sobre a glicemia, lacticemia e hematócrito de ratos sedentários In: IV Congresso Iternacional de Educação Física e Motricidade Humana, Motriz. Rio Claro: Divisa, 11:S69-S69, 2005.
PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; DIAS, R.; BRANBILLA, S.B.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício físico nas intensidades leve e moderada sobre a concentração de linfócitos mesentéricos In: XVIII Simpósio Internacional de Ciências do Esporte. Atividade Física e Esporte no Ciclo da vida. São Caetano do Sul: Celafiscs,13:163-163,2005.
PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício físico em intensidade leve na concentração de linfócitos teciduais e circulantes de ratos sedentários In: IV Congresso Internacional de Educação Física e Motricidade Humana, Motriz. Rio Claro: Divisa, 11:S145-S146, 2005.
DIAS, R.; PRESTES, J.; FERREIRA, C.K.; DONATTO, F.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação do exercício físico em intensidade leve sobre a glicemia, lactemia e hematócrito de ratos sedentários In: Congresso Internacional de Pegagogia do Esporte. Educação Física e Esportes: formação e intervenção profissional. Maringá-PR: STHAMPA, 2:120-121, 2005.

106
PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de duas sessões de exercício físico sobre a concentração de linfócitos mesentéricos de ratos sedentários In: XXVIII Simpósio Internacional de Ciências do Esporte. Atividade Física e Esporte no ciclo da vida. São Caetano do Sul: Celafiscs, 13:164-164, 2005.
DONATTO, F.; PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de duas sessões de exercício físico sobre a glicemia, lactacidemia de ratos sedentário In: XVIII Simpósio Internacional de Ciências do Esporte. Atividade Física e Esporte no ciclo da vida. São Caetano do Sul: Celafiscs, 13:S164-S164, 2005.
FROLLINI, A.B.; PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de duas sessões de exercício sobre o número de leucócitos circulantes de ratos In: XXVIII Simpósio Internacional de Ciências do Esporte. Atividade Física e Esporte no ciclo da vida. São Caetano do Sul: Celafiscs, 13:164–164, 2005.
PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; FOSCHINI, D.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de duas sessões de exercício sobre o número de linfócitos In: Semana de Reunião da Biologia. IV SERBIO. Piracicaba: SERBIO, 27–27, 2005.
PRESTES, J.; FERREIRA, C. K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de um minuto de exercício físico em intensidade leve sobre a concentração de linfócitos mesentéricos e circulantes de ratos sedentários In: Congresso Internacional de Pedagogia do Esporte. Educação Física e Esportes: formação e intervenção profissional. Maringá-PR: STHAMPA, 2:120-120, 2005.
FERREIRA, C. K.O.; PRESTES, J.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de um minuto de exercício físico em intensidade leve sobre a glicemia, lactemia e hematócrito em ratos sedentários In: Congresso Internacional de Pedagogia do Esporte. Educação Física e Esportes: formação e intervenção profissonal. Maringá-PR: STHAMPA, 2:132-133, 2005.

107
PRESTES, J.; FERREIRA, C. K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de um minuto de exercício físico em intensidade leve sobre o número de leucócitos circulantes de ratos sedentários In: Congresso Internacional de Pedagogia do Esporte. Educação Física e Esporte: Formação e intervenção profissional. Maringá-PR: STHAMPA, 2:132-132, 2005.
PRESTES, J.; VEIGA, A.; RAMALLO, B.T.; URTADO, C.B.; FERREIRA, C.K.O.; FOSCHINI, D.; DONATTO, F.; LEITE, G.S.; CURI, T.P.; CAVAGLIERI, C.R.; et. al. Efeitos de uma partida de basquetebol sobre as células do sistema imunológico de atletas profissionais In: 8º Congresso de Iniciação e Produção Científica & 7º Seminário de extensão Metodista. 8º Congresso de Iniciação e Produção Científica & 7º Seminário de extensão Metodista. São Paulo: UMESP, 2972-2972, 2005.
GUERESCHI, M.G.; PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; BRANBILLA, S.B.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Exercício físico agudo de intensidade leve e moderada altera a morfofisiologia de linfócitos circulantes de ratos In: XVIII Simpósio Internacional de Ciências do Esporte. Atividade Física e Esporte no ciclo da vida. São Caetano do Sul: Celafiscs, 13: 288-288, 2005.
FROLLINI, A.B.; PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; DIAS, R.; CAVAGLIERI, C.R. Influência do exercício físico realizado até exaustão, nas intensidades leve e moderada, sobre a concentração sérica de citocinas In: 13° Simpósio Internacional de Iniciação Científica, 13º SIICUSP. Ribeirão Preto: USP RIBEIRÃO PRETO, 2450-2450, 2005.
FERREIRA, C.K.O.; PRESTES, J.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeito do Exercício agudo com intensidades leve e moderada sobre a concentração plasmática de TNF-alpha In: XIX Reunião Anual Federação de Sociedades de Biologia Experimental. São Paulo: FESBE, 146-146, 2004.
PRESTES, J.; FERREIRA, C.K.O.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da Atividade Física aguda sobre as variáveis metabólicas de ratos In: XIX Reunião Anual Federação de Sociedades de Biologia Experimental. São Paulo: FESBE, 146-146, 2004.

108
TRABALHOS COMPLETOS PUBLICADOS EM ANAIS DE EVENTOS
PRESTES, J.; CAVAGLIERI, C.R. Determinação do limiar anaeróbio em ratos submetidos a um teste incremental na natação In: 3º Congresso de Pós Graduação, 3º Mostra Acadêmica Unimep. Piracicaba: Unimep, 2005.
PRESTES, J.; CAVAGLIERI, C.R.; PALANCH, A.C; GUERESCHI, M.G.; FERREIRA, C.K.O. The physical exercise effects on the immune system of rats In: Workshop sobre Atividade Física e Saúde. Piracicaba: UNIMEP, 2004.
ARTIGOS RESUMIDOS PUBLICADOS EM PERIÓDICOS
PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de duas sessões de exercício físico sobre a concentração de linfócitos mesentéricos de ratos. Revista brasileira de ciência e movimento. 13:S164-S164, 2005.
PRESTES, J.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício físico em intensidade leve sobre a glicemia, lacticemia e hematócrito de ratos sedentários. Motriz. 11:S69- S69, 2005.
PRESTES, J.; GUERESCHI, M.G.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; DIAS, R., BRANBILLA, S.B. CAVAGLIERI, C.R.; PALANCH, A.C. Exercício físico agudo de intensidade leve e moderada altera a morfofisiologia de linfócitos circulantes de ratos. Revista brasileira de ciência e movimento. 13:S288 - S288, 2005.
GUERESCHI, M.G.; PRESTES, J.; FERREIRA, C. K.O.; DONATTO, F.; CAVAGLIERI, C.R.; PALANCH, A.C. Alterações Morfofisiológicas em monócitos e macrófagos de ratos submetidos a exercício agudo de intensidade leve e moderada. Revista brasileira de ciência e movimento. 13:S82 - S82, 2005.
FOSCHINI, D.; PRESTES, J.; et. al. Dano muscular induzido por uma sessão de treinamento de força com múltiplas séries. Revista brasileira de ciência e movimento. 13:S174- S174, 2005.

109
FERREIRA, C.K.O.; PRESTES, J.; PALANCH, A.C.; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício em intensidade leve no número, viabilidade e funcionalidade de nacrófagos teciduais de ratos sedentários. Motriz. 11:S81 - S81, 2005.
DIAS, R.; PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DONATTO, F.; GUERESCHI, M.G.; BRANBILLA, S.B.; PALANCH, A.C.; CAVAGLIERI, C. R. Efeitos da adaptação ao exercício físico nas intesnidades leve e moderada sobre o percentual de monócitos e neutrófilos. Revista brasileira de ciência e movimento. 13:S166-S166, 2005.
DONATTO, F.; PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos de duas sessões de exercício físico sobre a glicemia, lactacidemia de ratos sedentário. Revista brasileira de ciência e movimento. 13:S164-S164, 2005.
FROLLINI, A.B.; PRESTES, J.; PALANCH, A.C.; CAVAGLIERI, C.R.; FERREIRA, C.K.O.; DONATTO, F..; GUERESCHI, M.G.; DIAS, R.; BRANBILLA, S.B. Efeitos de duas sessões de exercício sobre o número de leucócitos circulantes de ratos. Revista brasileira de ciência e movimento. 13:S164-S164, 2005.
GUERESCHI, M.G.; PRESTES, J.; FROLLINI, A.B.; FERREIRA, C.K.O.; DIAS, R.; BRANBILLA, S.B.; DONATTO, F.; CAVAGLIERI, C.R.; PALANCH, A.C. Alterações Morfológicas em monócitos e macrófagos de ratos submetidos a exercício físico agudo de intensidade leve e moderada. Revista brasileira de ciência e movimento. 10:S82-S82, 2004.
FERREIRA, C.K.O.; PRESTES, J.; DONATTO, F.; GUERESCHI, M.G.; PALANCH, A.C.; CAVAGLIERI, C.R. Respostas agudas ao exercício físico de natação nas intensidades leve e moderada sobre o número, viabilidade e funcionalidade de neutrófilos de ratos. Revista brasileira de ciência e movimento. 10:S95-S95, 2004.
ARTIGOS COMPLETOS PUBLICADOS EM PERIÓDICOS
PRESTES, J.; CAVAGLIERI, C.R.; FERREIRA, C.K.O.; DONATTO, F.; et al. Efeitos do fator de crescimento insulínico-I (IGF-I) sobre o músculo esquelético e suas relações com o exercício físico. Revista Brasileira de Ciência e Movimento. No prelo, 1, 2006.

110
PRESTES, J.; BUCCI, M.; CAVAGLIERI, C.R.; et. al. Metabolismo Lipídico: Suplementação e Performance Humana. Saúde em Revista. No prelo, 18, 2006.
PRESTES, J.; DIAS, R.; DONATTO, F.; FERREIRA, C.K.O.; FROLLINI, A.B.; GUERRESCHI, M.; PALANCH, A.; CAVAGLIERI, C.R. Efeito de duas sessões de exercício sobre o número de leucócitos totais e linfócitos circulantes. Revista Mackenzie de Educação Física. Artigo submetido, 2006.
PRESTES, J.; DIAS, R.; DONATTO, F.; FERREIRA, C.K.O.; FROLLINI, A.B.;
GUERRESCHI, M.; PALANCH, A.; VERLENGIA, R.; PITHON-CURI, T.; CURI, R.; CAVAGLIERI, C.R. Lymphocytes function and cytokines levels can be modulated by short duration physical exercise in different intensities? Eur J Sports Sci. Artigo submetido, 2006.
PRÊMIOS E TÍTULOS
MENSÃO HONROSA PARA PAINÉIS da XX Reunião Anual da Federação de Sociedades de Biologia Experimental, FESBE 2005. FERREIRA, C. KO.; PRESTES, J.; DONATTO, F.; GUERESCHI, M.G.; DIAS, R.; PALANCH, A.C.; CAVAGLIERI, C.R. Efeitos da adaptação ao exercício físico em intensidade leve sobre o número, viabilidade e funcionalidade de macrófagos peritoneais de ratos In: FESBE 2005. 81 – 81, 2005.
OUTRAS INFORMAÇÕES RELEVANTES
Participação no projeto de pesquisa intitulado: “Efeitos da suplementação de glutamina sobre a performance de jovens nadadores”. Projeto cadastrado no CNPQ.
Participação no grupo de pesquisa intitulado: “Endocrinologia e Performance Humana”.