introduÇÃo staphylococcus aureus - livros grátislivros01.livrosgratis.com.br/cp060283.pdf ·...
TRANSCRIPT

1
1. INTRODUÇÃO
1.1. Staphylococcus aureus As bactérias classificadas como Staphylococcus aureus subespécie
aureus (freqüentemente referidas como Staphylococcus aureus) estão
alocadas na família Staphylococcaceae. Estes microrganismos são
encontrados na microbiota anfibiôntica presente na pele e nas membranas
mucosas de humanos, assim como de outros mamíferos e pássaros. No
homem, elevadas concentrações desses cocos Gram-positivos podem estar
presentes nas axilas, na parte anterior das narinas e no períneo e, menores
quantidades, na orofaringe, boca, vagina, trato intestinal e glândulas mamárias
(CHILLER, SELKIN & MURAKAWA, 2001; MURRAY et al., 2003).
A patogênese de infecções causadas pelos S. aureus pode ser dividida
em cinco estágios principais: (1) colonização, (2) infecção localizada,
(3) disseminação sistêmica e/ou sepse, (4) infecção disseminada, (5) toxemias
(ARCHER, 1998). A taxa de colonização por S. aureus em indivíduos sadios
pode chegar a aproximadamente 30% (SOARES et al., 1997). Acredita-se que
a colonização preceda à infecção. Assim, por exemplo, um abscesso localizado
na pele poderia surgir após o microrganismo ter sido introduzido, no local da
infecção, a partir de um sítio inicial de colonização. As infecções localizadas na
pele e nos tecidos moles podem se apresentar sob a forma de celulite,
ulcerações infectadas e impetigo bolhoso, e nos folículos pilosos como
foliculite, furunculose ou carbunculose, sendo esta última caracterizada pela
presença de vários furúnculos pequenos e coalescentes, localizados,
principalmente, na região da cabeça e pescoço. A partir da infecção na pele,
principalmente as mais profundas, o microrganismo pode ganhar a corrente
sangüínea e se disseminar para órgãos distantes, podendo levar, como
conseqüência, ao choque séptico, endocardite, osteomielite, infecção urinária,
artrite séptica, meningite, pneumonia e/ou carbúnculo renal. Este último quadro
é caracterizado por lesões sob a forma de abscesso no córtex renal, acomete
pacientes sem sintomas urológicos, e está sempre associado a bacteremia e a
presença de S. aureus na urina (JEMNI et al., 1992; PROJAN & NOVICK,
1997). Os S. aureus podem ainda causar doenças relacionadas à produção de
toxinas como, a síndrome da pele escaldada, a síndrome do choque tóxico e a

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2
intoxicação estafilocócica; sendo essas doenças resultantes da produção de
uma variedade de exotoxinas específicas, pelo microrganismo, as quais
funcionam como superantígenos (SAg; ARCHER, 1998; TENOVER &
GAYNES, 2000).
1.2. Staphylococcus aureus Resistentes à Meticilina (MRSA) No começo da década de 1960 a meticilina foi lançada no comércio
como o primeiro derivado semi-sintético da penicilina, contendo uma
modificação química que tornou a molécula capaz de resistir à ação da
penicilinase estafilocócica. Porém, logo após a introdução desse antibiótico na
clínica médica surgiram os S. aureus resistentes à meticilina (MRSA, do inglês:
Methicillin-Resistant Staphylococcus aureus; BARBER, 1961).
O marco inicial (tempo zero) na escala evolutiva dos MRSA foi a
aquisição do gene mecA (codificador da PBP2a), o qual confere resistência à
meticilina ou às outras penicilinas semi-sintéticas, como a oxacilina. O gene
mecA é um segmento de DNA de 2,1 kb, o qual não é nativo da espécie
S. aureus, e que se encontra inserido em um grande bloco de DNA exógeno,
conhecido como elemento mec ou cassete cromossômico estafilocócico mec
(SCCmec; do inglês: staphylococcal cassette chromosome mec; KATAYAMA,
ITO & HIRAMATSU, 2000). O SCCmec é um elemento genético móvel,
integrado ao cromossomo dos MRSA, flanqueado por seqüências diretas e
repetidas. Este segmento de DNA, variando em torno de 21-67 kb, carreia
genes reguladores para a expressão do gene mecA (mecI: codificador de uma
proteína repressora do mecA e mecRI: codificador de uma proteína
transmembranosa sensora de β-lactâmicos e transdutora de sinal). Além disso,
o SCCmec também carreia o chamado complexo gênico ccr, o qual codifica
recombinases responsáveis pela mobilidade (inserção/excisão) do SCCmec no
cromossomo bacteriano (KATAYAMA, ITO & HIRAMATSU, 2000; HIRAMATSU
et al., 2002; ITO, et al., 2004). O SCCmec encontra-se integrado em um sítio
específico (attBscc), localizado na orfX (ORF, do inglês: open reading frame)
no terminal 3`, próximo à origem de replicação dos S. aureus. Ito e
colaboradores (ITO et al., 2001) identificaram, inicialmente, três diferentes
elementos SCCmec. Esses elementos são diferentes entre si e cada SCCmec
carreia uma combinação característica dos complexos gênicos ccr e mec e,

3
ainda, podem possuir ou não outros determinantes de resistência. O SCCmec
tipo I é encontrado nas cepas de MRSA que predominavam na década de
1960, anos relativamente recentes da quimioterapia antimicrobiana. Esta ilha
não contém gene de resistência a antibióticos, exceto o mecA. Ao contrário,
tanto o SCCmec tipo II, como o tipo III, detectados em amostras que se
tornaram dominantes a partir da década de 1980, carreiam múltiplos genes de
resistência. Alguns desses genes encontram-se no transposon (Tn) Tn554, que
codifica resistência à eritromicina e espectinomicina e no ΨTn554, que codifica
resistência ao cádmio. Foi sugerido que uma cópia do elemento de inserção
(IS; do inglês: insertion element) IS431, proximamente localizada ao gene
mecA, funcionaria como hot spot (região de alta freqüência de recombinação),
facilitando assim a aquisição de outros genes de resistência a antimicrobianos.
Os três tipos de SCCmec, citados acima, têm sido identificados em amostras
de MRSA associadas às infecções hospitalares.
Posteriormente, foi descrito o elemento SCCmec tipo IV, tanto em
amostras de MRSA hospitalares quanto nas adquiridas na comunidade (CA-
MRSA). Este elemento não carreia nenhum gene de resistência a
antimicrobianos, com exceção do gene mecA, e apresenta uma nova
combinação dos complexos ccr e mec (ITO et al., 2001). Em 2004, Ito e
colaboradores, descobriram um novo elemento mec em amostras de CA-
MRSA, o SCCmec tipo V. Este elemento possui 28 kb de tamanho e não
carreia nenhum outro gene de resistência, além do mecA (ITO et al., 2004).
Portanto, o complexo mec foi dividido em cinco principais classes A-E, como
descrito na tabela 1 (DEURENBERG et al., 2007).
Tabela 1. Principais classes de complexos mec.
Classe Estrutura SCCmec Espécies
A mecI–mecRI–mecA–IS431 II, III Staphylococcus aureus
B ΨIS1272–∆mecRI–mecA–IS431a I, IV Staphylococcus aureus
C IS431–∆mecRI–mecA–IS431 V Staphylococcus aureus
D ∆mecRI–mecA–IS431 b – Staphylococcus caprae
E ∆mecRI–mecA–IS431 c – Staphylococcus aureus
aΔ representa uma deleção no regulador mecRI; bclasse D: mecRI apresenta uma deleção de 847pb; cclasse E: mecRI apresenta uma deleção de 976pb. Adaptado de DEURENBERG et al., 2007.

4
A classe A contém o complexo mec completo (mecI-mecRI-mecA) e as
classes B e C contêm os genes reguladores do mecA não-funcionais, devido à
presença das seqüências de inserção ΨIS1272 e/ou IS431 (KATAYAMA, ITO &
HIRAMATSU, 2001; HIRAMATSU et al., 2004). Por sua vez, as classes D e E,
apesar de apresentarem estruturas do complexo mec semelhantes, divergem
entre si quanto ao número de pares de bases (pb) deletadas na região do DNA
englobando os genes mecI e mecRI. A classe D apresenta uma deleção de
847 pb, enquanto a classe E de 976 pb (HIRAMATSU et al., 1992; KATAYAMA,
ITO & HIRAMATSU, 2001). Além disso, outros elementos genéticos também
podem estar presentes nos diferentes SCCmec, dentre eles os plasmídios
pUB110 e pT181, e o transposon Tn554. Desta forma, devido às características
acima citadas, o SCCmec pode ser definido como uma ilha de resistência a
antimicrobianos (HIRAMATSU et al., 2002). Variantes dos tipos principais de
SCCmec também têm sido descritas, com base nas diferenças de seqüências
nucleotídicas observadas em regiões conhecidas como regiões J (do inglês:
junkyard regions; ITO, KUWAHARA & HIRAMATSU, 2007). Oliveira e
colaboradores, mais recentemente, propuseram um novo tipo de elemento
SCCmec, o tipo VI, em substituição ao tipo IV alótipo 4 (um subtipo ccrAB),
uma vez que esse elemento encontra-se em diversas amostras pertencentes a
uma mesma linhagem estafilocócica (OLIVEIRA, MILHEIRIÇO & DE
LENCASTRE, 2006).
Desde a emergência do MRSA tem sido observado um aumento
crescente de infecções por esses microrganismos. Acredita-se que dois fatores
possam estar envolvidos neste cenário: a expansão clonal de linhagens de
MRSA já existentes e a conversão de algumas linhagens de S. aureus
susceptíveis à meticilina (MSSA) em MRSA, através da transferência horizontal
do elemento mec (NOTO et al., 2008). No final da década de 1980, surtos
hospitalares causados pelos MRSA aumentaram consideravelmente. Cepas
resistentes a diversos outros antimicrobianos, exceto à vancomicina, estavam
freqüentemente envolvidas nesses surtos. Atualmente a sigla MRSA implica
em resistência cruzada a todos os antibióticos β-lactâmicos (AL-MASSAUDI,
DAY & RUSSEL, 1991; TOMASZ, 1994; TEIXEIRA et al., 1995).

5
A emergência e a ampla disseminação dos MRSA é resultante, em
parte, da forte pressão seletiva exercida pela imensa quantidade de agentes
antimicrobianos lançados no ambiente hospitalar. Além disso, a convergência
de diversos processos evolucionários raros, relacionados com a sobrevivência
e adaptação da bactéria neste cenário, certamente contribuiu para
disseminação global de clones epidêmicos de MRSA que estamos assistindo
na atualidade. Os determinantes genéticos que definem a superior
epidemicidade desses clones ainda não são conhecidos, mas parecem estar
associados a genes que são responsáveis pela colonização efetiva do
hospedeiro (AMARAL et al., 2005; DE LENCASTRE, OLIVEIRA & TOMASZ,
2007).
Outra característica evolucionária importante, também observada nas
populações de MRSA, é a distribuição de sua população em uma estrutura
altamente clonal (FENG et al., 2008). Com base nessa informação, para melhor
se estudar a epidemiologia local ou global dos MRSA, diversos métodos de
tipagem molecular vêm sendo utilizados. Uma das metodologias preconizadas,
e considerada mais discriminativa, portanto indicada em análises locais, é a
técnica de eletroforese sob campos elétricos alternados, conhecida pela sigla
PFGE (PFGE, do inglês: pulsed-field gel electrophoresis). Contudo, esta
técnica apresenta como sua principal desvantagem a dificuldade de
correlacionar, de forma precisa, os diferentes perfis de fragmentação genômica
obtidos a partir de clones de EMRSA (MRSA, do inglês: Epidemic Methicillin-
Resistant Staphylococcus aureus) em diferentes laboratórios. Entretanto, tal
correlação é possível quando se acrescentam aos géis de PFGE padrões com
tamanho de fragmentos conhecidos ou representantes de clones internacionais
circulantes na atualidade. Outros métodos moleculares, baseados em técnicas
de seqüenciamento de DNA, foram também desenvolvidos. Tais métodos
apresentam como principal vantagem a possibilidade de se gerar perfis
genéticos padronizados, permitindo a comparação acurada entre as diferentes
linhagens bacterianas. Uma dessas metodologias foi a tipagem através de
seqüenciamentos de multilócus enzimáticos (MLST, do inglês: multi locus
sequence typing; ENRIGHT et al., 2000). O MLST é atualmente um método de
tipagem molecular muito utilizado e fundamenta-se no seqüenciamento de sete
genes metabólicos constitutivamente expressos pela bactéria (para os

6
S. aureus: arcC, aroE, glpF, gmk, pta, tpi e yqiL). Para cada um desses genes,
seqüências diferentes (com pequenas variações de nucleotídeos) são referidas
como alelos. O conjunto formado pelos diferentes alelos, de cada um dos sete
loci, fornece o perfil alélico, que define, por sua vez, a seqüência-tipo (ST) da
amostra analisada (ENRIGHT et al., 2000). A partir dessa nova abordagem,
clones epidêmicos de MRSA, anteriormente nomeados de acordo com a região
onde foram primeiramente detectados (como exemplo, o clone epidêmico
brasileiro) ou de acordo com o grupo de pacientes dos quais foram isolados
(como exemplo, o clone pediátrico), passam agora a ser designados por sua
ST e também pelo seu tipo de SCCmec (tabela 2).
Amostras clínicas de MRSA de uma mesma linhagem, ou de linhagens
geneticamente relacionadas, encontram-se hoje disseminadas por todo o
mundo, sendo capazes de causar infecções nosocomiais pandêmicas,
atingindo grandes extensões geográficas intercontinentais (TEIXEIRA et al.,
1995; VAN BELKUM et al., 1997a,b; DE SOUZA et al., 1998; SÁ-LEÃO et al.,
1999; COIMBRA et al., 2000; SOARES et al., 2001; COIMBRA et al., 2003).
Dessa forma, os MRSA tornaram-se os mais importantes patógenos
hospitalares, em termos de incidência e de gravidade das infecções, sendo
reconhecidos como um sério problema global de Saúde Pública (COIMBRA et
al., 2003). Um fato que chama atenção, com relação aos MRSA epidêmicos, é
a velocidade com que a diseminação desses clones ocorre (GRUNDMANN et
al., 2006). Nosso grupo foi o primeiro a demonstrar a ampla disseminação de
um clone de MRSA através de áreas geográficas de dimensões continentais
(TEIXEIRA et al., 1995). Detectamos amostras do clone epidêmico brasileiro
(ST239-SCCmecIIIA) em hospitais localizados de Manaus a Porto Alegre.
Estudos mais recentes, publicados também pelo nosso grupo (COIMBRA et al.,
2003), demonstraram que a linhagem ST5-SCCmecII (CNI/J; clone Nova
Iorque/Japão), descrita inicialmente na cidade de Nova Iorque (EUA) e em
Tóquio (Japão; AIRES DE SOUSA et al., 2000), estava, na realidade,
disseminada em hospitais de diversos estados do leste dos EUA (COIMBRA et
al., 2003). Desse modo, a cada ano, um grande número de novos estudos é
publicado na literatura científica, apontando linhagens específicas de MRSA
como sendo responsáveis por grandes surtos em hospitais distribuídos
mundialmente (DEURENBERG et al., 2007; KENNEDY et al., 2008). A

7
disseminação geográfica das principais linhagens de MRSA envolvidas nessas
verdadeiras “pandemias hospitalares” está descrita na tabela 2.
Tabela 2. Distribuição geográfica das principais linhagens de MRSA envolvidas em surtos hospitalares.
Clonea Linhagem (ST-SCCmec)b Disseminação geográficac
Arcaico 250-I Alm, Ast, Din, EUA, RU, Sui, Uga
Berlin 45-IV Alm, Arm, Ast, Bel, Esp, EUA, Fin, Hol, Hun, Nor, Sue, Sui
CEB/Húngaro 239-III Aga, Alm, Arg, Ast, Aus, Bra, Chi, Chn, Cin, Cor, Esl, Esp, EUA, Fin, Gre, Hol, Ids, Ind, Ita, Mon, Pol, Por, RG, RT, RU, Sri, Sue, Tal, Tun, Uru, Vie
CI 247-I Alm, Bel, Cro, Din, Esl, Esp, EUA, Fin, Fra, Hol, Ita, Pol, Por, RT, RU, Sue, Sui
CNI/J 5-II Alm, Ast, Bel, Can, Cin, Cor, Din, EUA, Fin, Fra, Irl, Jap, Mex, RU, Sue, Uru
CP 5-IV ou VI Aga, Arg, Ast, Bra, Col, Cor, Din, Esp, EUA, Fra, Nor, Pol, Por, RU, Sue, Uru
Irlandês 1 8-II Ast, EUA, Irl, RU
Sudeste alemão 228-I Alm, Bel, Din, Esl, Esp, Ita, Sui
EMRSA 2, 6 8-IV Alm, Ast, Bel, EUA, Fin, Fra, Hol, Irl, Nor, RU,Tai
EMRSA 3 5-I Arg, Esl, Nor, Pol, RU
EMRSA 15 22-IV Alm, Ast, Bel, Cin, Din, Esp, Irl, Kuw, Nor, NZL, Por, RT, RU, Sue
EMRSA 16 36-II Ast, Bel, Can, Din, Esp, EUA, Fin, Gre, Irl, Mex, Nor, RU, Sue, Sui
aCEB, clone epidêmico brasileiro; CI, clone ibérico; CNI/J, clone Nova Iorque/Japão; CP, clone pediátrico; EMRSA, MRSA epidêmico. bST-SCCmec, classificação da linhagem baseada na seqüência-tipo, obtida a partir da tipagem por MLST, e do tipo de SCCmec encontrado. cAga, Argélia; Alm, Alemanha; Arg, Argentina; Arm, Armênia; Ast, Austrália; Aus, Áustria; Bel, Bélgica; Bra, Brasil; Can, Canadá; Chi, Chile; Chn, China; Cin, Cingapura; Col, Colômbia; Cor, Coréia; Cro, Croácia; Din, Dinamarca; Esl, Eslovênia; Esp, Espanha; EUA, Estados Unidos da América; Fin, Finlândia; Fra, França; Gre, Grécia; Hol, Holanda; Hun, Hungria; Ids, Indonésia; Ind, Índia; Irl, Irlanda; Ita, Itália; Jap, Japão; Kuw, Kuwait; Mex, México; Mon, Mongólia; Nor, Noruega; NZl, Nova Zelândia; Pol, Polônia; Por, Portugal; RG, República da Georgia; RT, República Tcheca; RU, Reino Unido; Sri, Sri Lanka; Sue, Suécia; Sui, Suíça; Tai, Taiwan; Tal, Tailândia; Tun, Tunísia; Uga, Uganda; Uru, Uruguai; Vie, Vietnam. Adaptado de CAMPANILE et al., 2001; JEMILI-BEN, BOUTIBA-BEN & BEN, 2006; DEURENBERG et al., 2007; BARTELS et al., 2008.
Devido ao fato da maioria desses clones apresentar resistência a uma
gama de agentes antimicrobianos, a vancomicina tem sido amplamente
utilizada, em todo o mundo, como a droga de escolha no tratamento empírico
das infecções nosocomiais causadas pelos MRSA (TEIXEIRA et al., 1995;

8
COIMBRA et al, 2003; ELIOPOULOS, 2005). Entretanto, a utilização desse
quimioterápico (ministrado exclusivamente por via endovenosa) aumenta o
tempo de internação do paciente e também os riscos com os efeitos colaterais,
promovidos por sua toxicidade. Tais fatores, aliados ao valor comercial
relativamente elevado do tratamento do paciente infectado por MRSA, acabam
contribuindo para um aumento da morbidade e da mortalidade dos pacientes, o
qual é geralmente mais acentuado nos países menos favorecidos, e ainda para
um aumento significativo dos custos hospitalares.
Do mesmo modo, como observado para os diversos antimicrobianos
utilizados no tratamento de infecções associadas aos MRSA e, como
conseqüência da pressão seletiva exercida pelo intenso uso de vancomicina,
foi relatado em 1997, em um hospital japonês, o primeiro caso de falha
terapêutica da vancomicina (CDC, 1997; HIRAMATSU et al., 1997). Foi
observado que a amostra de S. aureus, isolada da ferida cirúrgica de um
recém-nascido, submetido a uma cirurgia cardíaca, apresentava resistência
intermediária à vancomicina (VISA, do inglês: vancomycin-intermediate
S. aureus; concentração mínima inibitória, CMI, entre 8-16 μg/mL). Nenhuma
das amostras VISA identificadas naquele, e em outros estudos, carreavam o
gene vanA ou qualquer outro determinante genético de resistência à
vancomicina (HIRAMATSU et al., 1997; SMITH et al., 1999; OLIVEIRA et al.,
2001; DENIS et al., 2002; KRZYSZTON-RUSSJAN et al., 2002). Por outro lado,
o espessamento e a compactação da matriz da parede celular, observados nas
amostras VISA, têm sido implicados como um possível mecanismo responsável
por esse fenótipo de resistência (SIERADZKI et al., 1997; HANAKI et al., 1998;
HIRAMATSU, 2001; WALSH & HOW, 2002; CUI et al., 2003). Posteriormente,
um novo caso envolvendo uma cepa resistente à vancomicina foi relatado em
junho de 2002, em Michigan, EUA (CHANG et al., 2003). Entretanto, nesse
relato a cepa de S. aureus isolada apresentava resistência plena à vancomicina
(VRSA, do inglês: vancomycin-resistant S. aureus, CMI, ≥32 μg/mL; NCCLS,
2003). Os testes genotípicos realizados com esta cepa indicaram a presença
de vanA, sugerindo que sua aquisição pelo VRSA tenha, provavelmente,
ocorrido através da transferência de material genético a partir de uma cepa de
Enterococcus faecalis resistente à vancomicina (VRE; do inglês: vancomycin-

9
resistant Enterococcus) carreadora de vanA, também isolada do mesmo sítio
infectado pelo MRSA (CHANG et al., 2003). Apenas dois meses após esse
último relato, mais um caso de VRSA foi detectado nos EUA, desta vez na
Pensilvânia (TENOVER et al., 2004; WHITENER et al., 2004). Felizmente, a
publicação de relatos sobre a emergência de VRSA em outros países tem sido
rara (TIWARI & SEN, 2006; WEIGEL et al., 2007; SAHA, et al., 2008), e até o
momento, não houve registro de isolamento de amostras de VRSA no Brasil.
Entretanto, um estudo realizado no ano de 2000, em nosso país, pelo nosso
grupo, demonstrou a presença de amostras VISA em um hospital localizado no
Piauí. Neste episódio, um dos pacientes envolvidos foi a óbito, provavelmente,
devido à falha no tratamento com a vancomicina (DOS SANTOS SOARES et
al., 2000). Posteriormente, outros pesquisadores também detectaram amostras
VISA em nosso meio (OLIVEIRA et al., 2001). Em conjunto, tais estudos
indicam a emergência de amostras de S. aureus apresentando resistência à
vancomicina, em diversos países, e alertam as autoridades da saúde para a
necessidade de uma vigilância rigorosa e constante nos hospitais, com relação
a prevenção e ao controle das infecções causadas pelos MRSA.
No final da década de 1990, dois novos agentes antimicrobianos, a
linezolida (da classe das oxazolidinonas) e a quinupristina-dalfopristina, este
último também conhecido como sinercide (um antibiótico semi-sintético da
classe das estreptograminas; MALBRUNY et al., 2002), foram desenvolvidos
por indústrias farmacêuticas. Tais drogas foram introduzidas no comércio com
indicações para pacientes portadores de quadros infecciosos severos
causados por patógenos Gram-positivos resistentes à vancomicina (DIEKEMA
& JONES, 2001; TVERDEK, CRANK & SEGRETI, 2008). Porém, a emergência
de amostras resistentes a esses agentes antimicrobianos já vem sendo
relatada em nosocômios de diversos países (TSIODRAS et al., 2001;
MALBRUNY et al., 2002; FARRELL et al., 2004; MEKA et al., 2004; PICAZO et
al., 2006; MENDES et al., 2008). Além disso, a quinupristina-dalfopristina
parece não ser a escolha mais indicada para o tratamento de pacientes com
pneumonia causada por MRSA. Essa observação baseia-se no fato da
utilização dessa droga ter sido associada a uma alta taxa de mortalidade
(25,3%) entre pacientes apresentando esse quadro infeccioso. E ainda, o alto
custo desse antimicrobiano, os riscos potenciais de seus efeitos colaterais e as

10
múltiplas interações com outras drogas restringem sua utilização no tratamento
de infecções severas por MRSA (TVERDEK, CRANK & SEGRETI, 2008). Uma
amostra clínica de MRSA resistente à linezolida também já foi isolada em 2006,
em nosso país (GALES et al., 2006).
A disseminação global de um pequeno número de linhagens específicas
e epidêmicas de MRSA representa elevados custos sociais e econômicos em
todo o mundo (COIMBRA et al., 2003). Um estudo realizado em 2002 avaliou o
impacto financeiro gerado nos gastos públicos pela política de vigilância
epidemiológica contra os MRSA, entre os anos de 1991 e 2000, no Centro
Médico Universitário Utrecht, Holanda. Também foram avaliadas as despesas
com o pagamento de médicos e outros profissionais dos departamentos de
cirurgia, microbiologia e farmácia e de funcionários da subdivisão de higiene e
prevenção de infecção bem como de outros serviços daquele hospital. A essas
despesas também foram associados os gastos com materiais, meios de
culturas, medicações específicas, descontaminação, descarte de materiais e
indumentárias. Além disso, durante o período de 10 anos desse estudo, foram
também estimados os prejuízos gerados com as 78.000 culturas adicionais
realizadas, assim como os 2.265 dias de trabalho perdidos com hospitalizações
e licenças temporárias devido a infecções e descolonização de MRSA em 29
profissionais de saúde daquele hospital. Os autores concluíram que a rigorosa
política de vigilância contra os MRSA adotada pelo hospital holandês, durante o
período avaliado no estudo, custou aos cofres públicos um total de 2.800.000
euros. Os mesmos autores projetaram ainda as conseqüências logísticas e
financeiras que seriam geradas caso essa política rígida de vigilância não fosse
adotada naquele hospital. Baseados nessas análises, os autores especularam
que os custos associados à utilização de antibióticos alternativos e de última
geração, que seriam requeridos no cenário de alta endemicidade de MRSA,
deveriam ser, pelo menos, 2 vezes maior que o montante gasto nos 10 anos
com as medidas de controle (VRIENS et al., 2002). No Brasil, foi realizada
recentemente (2005) uma análise do custo-benefício com a utilização da
linezolida e da vancomicina em pacientes com quadros de pneumonias
nosocomiais causadas por MRSA e associadas à ventilação mecânica
(MACHADO et al., 2005). Baseados nos valores unitários de cada
medicamento e de todos os dispositivos e equipamentos médicos necessários

11
à sua administração (dados do INCOR – Instituto do Coração do HC/FMUSP
obtidos em maio de 2004), foi demonstrado que, em 11 dias de internação, o
custo diário por paciente com a linezolida chegou a R$ 439,06 (total de
R$ 4.829,66) enquanto que com a vancomicina foi de R$ 255,01 (total de
R$ 2.805,11). Os mesmos autores concluíram, entretanto, que mesmo a
linezolida apresentando um custo direto 60% mais elevado que a vancomicina,
sua utilização ao final do tratamento representaria uma redução de custo de
41,3% por paciente em comparação à utilização da vancomicina (levando-se
em consideração o custo-benefício, uma vez que a taxa de cura dos pacientes
tratados com a primeira foi de 62,2% enquanto que com a segunda foi de
apenas 21,2%; MACHADO et al., 2005). Tais dados corroboram a problemática
causada pelos MRSA em quadros de infecções nosocomiais, impactando
sócio-economicamente a política de saúde pública tanto de países com uma
vigilância epidemiológica mais rigorosa como de países com um sistema de
vigilância deficiente.
Contudo, as infecções causadas pelos MRSA não estão associadas
somente às infecções hospitalares (HA-MRSA, do inglês: hospital-associated
methicillin-resistant Staphylococcus aureus). Clones de MRSA podem estar
também associados a infecções que ocorrem na comunidade (CA-MRSA, do
inglês: community-associated methicillin-resistant Staphylococcus aureus).
Apesar dos surtos de infecções severas por MRSA em ambiente comunitário
serem raros, estes também têm sido motivo de grande preocupação por parte
de pesquisadores e profissionais da área da saúde de todo o mundo, uma vez
que podem atingir crianças e adultos sadios da comunidade. Tal preocupação
é ainda sustentada pelo fato dos antibióticos β-lactâmicos serem, amplamente,
utilizados no tratamento empírico dos diversos tipos de infecções causadas
pelos S. aureus na comunidade (DIEP et al., 2004). Além disso, tem sido
descrita a emergência de algumas linhagens de CA-MRSA apresentando uma
combinação “bem equilibrada” (em termos de vantagens adaptativas) de alguns
genes de resistência a drogas, elementos genéticos móveis (que favorecem de
alguma forma sua disseminação) e fatores de virulência, não apenas na
comunidade, mas também no ambiente hospitalar. Como conseqüência, não
são raros os eventos de substituição de algumas linhagens endêmicas de
MRSA hospitalar por algumas cepas de CA-MRSA bem adaptadas

12
(GRUNDMANN et al., 2006). Assim, é provável que diversos fatores
relacionados não somente ao perfil de resistência, mas também ao potencial de
virulência (poder de disseminação e manutenção das diferentes populações de
S. aureus) nos mais variados e hostis ambientes, sejam responsáveis pelo
sucesso observado desse patógeno bacteriano.
Como apontamos anteriormente, o primeiro estudo que ilustra bem a
última das características mencionadas (amplo poder de disseminação e
manutenção no hospedeiro) foi realizado no Brasil, em 1995, pelo nosso grupo
(TEIXEIRA et al., 1995). Naquele estudo foi demonstrado que a disseminação
hospitalar da linhagem ST239-SCCmecIIIA abrangia uma área de cerca de
5.300 Km, sendo esta detectada em diversos nosocômios localizados entre
Manaus e Porto Alegre (TEIXEIRA et al., 1995). Assim, o amplo poder de
disseminação de um clone de MRSA (o ST239-SCCmecIIIA) ficou evidente
(TEIXEIRA et al., 1995). A extraordinaria capacidade de expansão dessa
linhagem é demonstrada ainda pelo fato do ST239 ser hoje o MRSA mais
disseminado em hospitais de todo o mundo (tabela 2; GRUNDMANN et al.,
2006; BARTELS et al., 2008). Dados recentes demonstram que a incidência
das infecções nosocomiais causadas pelo ST239 são alarmantes, podendo
chegar a taxas tão elevadas quanto 90%, como foi verificado em hospitais
distribuídos pelo continente asiático (FEIL et al., 2008).
Além dos fatores associados diretamente com o microrganismo, como
os relacionados a sua virulência e patogenicidade, existem ainda diversos
fatores de risco associados ao hospedeiro e que contribuem para o aumento da
incidência das infecções nosocomiais causadas pelos MRSA. Dentre essas
condições estão incluídos os extremos de idades, representados pelos recém-
nascidos pré-maturos (que não apresentam o sistema imunológico
completamente formado) e pelos idosos (apresentando
imunocomprometimentos), o uso prolongado de antimicrobianos (afetando o
funcionamento de certos órgãos, devido aos seus efeitos colaterais e
toxicidade, e também exercendo uma pressão seletiva para a emergência de
multirresistência aos antimicrobianos), as doenças de base (comprometendo
parte da defesa imune), as deficiências imunológicas, o uso de terapias
imunossupressoras e a pré-exposição a procedimentos médicos invasivos (tais
como cirurgias, uso de cateteres endovenosos, ventilação mecânica, próteses

13
ou de outros dispositivos médicos). Com relação a esses últimos, os S. aureus
pertencem a um grupo de microrganismos capazes de se depositar sobre
superfícies lisas, como as superfícies plásticas dos biopolímeros, formando um
verdadeiro filme de bactérias, conhecido como biofilme. Por este motivo, o
biofilme tem sido reconhecido como um importante fator de virulência
associado, principalmente, às infecções nosocomiais.
1.3. Produção de Biofilme em Staphylococcus aureus
Nas últimas duas décadas diversas publicações têm apontado o gênero
Staphylococcus como um gênero bacteriano capaz de formar biofilme,
causando um novo tipo de infecção, as quais poderiam ser agrupadas e melhor
definidas como: “infecções crônicas associadas a biopolímeros” (GÖTZ, 2002).
Através de microscopia eletrônica de varredura, já havia sido observado que
esses microrganismos eram capazes de formar múltiplas camadas de células,
agregadas por uma matriz amorfa, na superfície do biopolímero (PETERS,
LOCCI, & PULVERER, 1981; CHRISTENSEN et al.,1982; MARRIE, NELLIGAN
& COSTERTON, 1982). O biofilme costuma ser definido como uma
comunidade microbiana (caracterizada por células de diferentes gêneros e
espécies que se aderem a um substrato, ou interface, e também a outras
células) inserida em uma matriz constituída de uma substância polimérica, de
natureza extracelular. Essa comunidade apresenta um fenótipo diferente no
que diz respeito ao crescimento celular, expressão gênica e síntese protéica
Na natureza, tal comunidade microbiana é formada por múltiplas espécies,
enquanto que os biofilmes associados às infecções nosocomiais são, de modo
geral, formados por uma única espécie (DONLAN & COSTERTON, 2002).
A composição química do principal polissacarídeo presente no biofilme
produzido pelos estafilococos é formada por uma homoglicana linear composta
de glicosaminoglicana, β-1,6-ligada (PNAG, do inglês: polymeric N-
acetylglucosamine), apresentando pelo menos 130 resíduos de 2-deoxi-2-
amino-D-glicopiranosil, dos quais 80-85% são N-acetilados, sendo o restante
não N-acetilados e contendo uma pequena quantidade de fosfato e resíduos
succinil éster-ligados (15-20%), tornando-o aniônico. Assim, de acordo com a
sua função de agregação intercelular, esse polissacarídeo foi denominado
adesina polissacarídica intercelular (PIA; do inglês: polysaccharide intercellular

14
adhesin; MACK, et al., 1996). A síntese de PIA in vitro ocorre a partir de
produtos codificados pelo operon ica (intercellular adhesion; CRAMTON et al.,
1999), mas até o momento pouco se sabe sobre as funções das proteínas
IcaA, D, B e C, codificadas por esse operon. Entretanto, diversas evidências
científicas indicam que a composição do biofilme sintetizado pelas espécies do
gênero Staphylococcus seja de natureza distinta. Estudos parecem apontar
importantes diferenças na composição da matriz do biofilme formado por
S. epidermidis e pelo S. aureus, sugerindo que proteínas, ou mesmo DNA,
possam ter um papel mais importante na formação do biofilme do S. aureus do
que o PIA/PNAG (O’NEILL et al. 2008; IZANO et al., 2008).
Uma importante característica dos filmes biológicos é que seu
microambiente fornece uma série de vantagens para as células que se
encontram ali inseridas (sésseis) em relação às demais células, que se
encontram na fase planctônica (dispersas fora dessa estrutura). Uma das
vantagens é a retenção e concentração, na matriz polimérica, de diversos
requerimentos nutricionais, tais como, carbono, nitrogênio e fosfato
(BEVERIDGE et al., 1997). Outra vantagem do crescimento bacteriano no
ambiente do biofilme é a resistência que essa estrutura fornece às células
contra uma gama de estratégias de remoção bacteriana intrínsecas ou
extrínsecas ao hospedeiro. Exemplos dos efeitos provocados pela barreira
física do biofilme seriam: a dificuldade de penetração de antibióticos até as
camadas mais internas dessa estrutura, a inibição da fagocitose, da ação de
radicais livres de oxigênio e proteases, da ação do complemento e de
anticorpos, dentre outros (BROWN, ALLISON, & GILBERT, 1988; IWATSUKI et
al., 2006; ULRICH et al., 2007).
Apesar da atuação do biofilme como uma barreira de difusão, diminuindo
a penetração dos agentes antimicrobianos, a resistência à drogas pode
também estar relacionada aos baixos níveis metabólicos e de crescimento
bacteriano, no ambiente do biofilme (XU, MCFETERS & STEWART, 2000). No
caso da vancomicina, ao contrário do observado para os demais grupos de
antibióticos, foi demonstrado em um modelo in vivo, utilizando cateter de
hemodiálise, que altas concentrações dessa droga foram encontradas nas
camadas mais profundas do biofilme estafilocócico após sua administração
(WILCOX et al., 2001). Os autores sugeriram que esta droga não seria efetiva

15
para a erradicação dos estafilococos, no filme biológico, pelo fato de possuir
grande afinidade de ligação aos componentes da matriz do biofilme, tornando-a
indisponível para atuar de forma eficaz no alvo bacteriano. Ao contrário da
vancomicina, baixas concentrações de linezolida foram encontradas nas
camadas mais internas do biofilme, apesar deste antimicrobiano ter diminuído a
carga bacteriana do filme biológico acumulado no cateter. Os autores
sugeriram que pelo fato da linezolida não se ligar aos componentes da matriz
polimérica do biofilme, esta droga poderia estar sendo removida durante a
etapa de lavagem para a retirada das células planctônicas do cateter, o que
explicaria a detecção de baixos níveis da mesma (CHIBA et al., 1998 apud
WILCOX et al., 2001). Finalmente, os autores concluíram que, apesar da
utilização desses antimicrobianos levar a uma diminuição da massa
estafilocócica na estrutura do biofilme, nenhum deles foi capaz de eliminar
100% das bactérias (WILCOX et al., 2001). Outro fator, muito importante,
relacionado ao filme biológico é o aumento da oportunidade de troca de
material genético entre os microrganismos localizados neste ambiente, uma
vez que estes se encontram muito próximos fisicamente devido à alta
densidade populacional e pela ligação célula-célula. Assim, trocas genéticas
podem levar a um fenótipo de múltipla resistência aos antimicrobianos, e ainda,
provavelmente, a um aumento da patogenicidade, devido a aquisição de genes
relacionados aos fatores de virulência, dentre outras possíveis aquisições
(PARSEK & SINGH, 2003; DE ARAÚJO et al., 2006). E ainda, o acúmulo de
ácidos oriundos do metabolismo bacteriano pode levar a alterações de pH
nesse microambiente, antagonizando a ação de agentes antibióticos que
estejam sendo administrados ao paciente e que estejam conseguindo penetrar
nessa barreira formada pelo filme biológico (STEWART & CONSTERTON,
2001). Por outro lado, o aumento populacional bacteriano no biofilme
representa um potencial para a dispersão de microcolônias, através da ação
mecânica do fluxo de fluídos ou de processos bacterianos (regulados de
acordo com sinais ambientais), resultando na sua migração e adesão a novos
sítios de colonização no hospedeiro (BOYD & CHAKRABARTY, 1994;
CONLON, HUMPHREYS & O’GARA, 2004). Por outro lado, acredita-se que o
ambiente de biofilme possa facilitar a sobrevivência bacteriana em superfícies
inanimadas. Em um estudo recente, foi demonstrado que a maioria dos

16
patógenos nosocomiais, mais incidentes, pode sobreviver sobre superfícies
secas e inanimadas por longos períodos, como no caso dos MRSA (cuja
persistência durou 7 meses; KRAMER et al., 2006).
Através da observação experimental in vitro, por microscopia, do biofilme
bacteriano, foram sugeridas duas etapas para que ocorra sua formação: a
primeira etapa estaria relacionada à adesão bacteriana inicial à superfície do
polímero. Já a segunda etapa estaria associada à deposição (acúmulo) das
células de acordo com o crescimento bacteriano, formando múltiplas camadas
de um agregado de bactérias somadas ao acúmulo de material amorfo,
constituindo, deste modo, o biofilme (GÖTZ, 2002). Na primeira etapa a
bactéria pode se ligar diretamente, de forma não específica, ao polímero.
Acredita-se que este tipo de ligação deva ocorrer, principalmente, devido às
interações eletrostáticas e hidrofóbicas que ocorrem entre as proteínas
ancoradas na superfície bacteriana e a matriz polimérica do biomaterial
(FLEER & VERHOEF, 1989; MAIRA-LITRÁN et al., 2002).
Várias proteínas foram envolvidas nessa etapa inicial de formação do
biofilme (HEILMANN et al., 1997). Em S. aureus, foi observado que todas as
cepas carreadoras da proteína associada a biofilme, conhecida como Bap
(Bap, do inglês: biofilm-associated protein), são altamente aderentes e
fortemente produtoras de biofilme. Entretanto, até o momento, a proteína Bap
(codificada por um gene localizado em uma ilha de patogenicidade, SaPIbov2)
encontra-se apenas em amostras isoladas de mastite bovina e parece estar
associada tanto a primeira como a segunda fase de formação de biofilme
(CUCARELLA et al., 2004; TORMO et al., 2005; VAUTOR et al., 2008). Outros
exemplos de proteínas que têm sido citadas como envolvidas na adesão
celular primária às superfícies inertes seriam as proteínas ClpC e ClpX,
pertencentes ao complexo proteolítico Clp (Clp, do inglês: caseinolytic
protease; FREES et al., 2004) e a proteína A (O'SEAGHDHA et al., 2006). Um
outro componente dos S. aureus que também parece envolvido na produção de
biofilme é o ácido teicóico. Segundo Gross e colaboradores as interações entre
as cargas elétricas dessa estrutura e das superfícies de vidro ou de poliestireno
seria mais um fator facilitador da aderência do S. aureus nas etapas iniciais de
formação de biofilme (GROSS et al., 2001).

17
Ainda na primeira etapa de formação do biofilme, uma forma de
interação indireta da bactéria com o polímero também pode ocorrer. Esta via de
interação alternativa é promovida pela ligação da bactéria às proteínas da
matriz extracelular, presentes no plasma do hospedeiro, tais como a
fibronectina, o fibrinogênio, o colágeno, a laminina, a trombospondina, a
elastina, etc. (HEILMANN et al., 2003), que por sua vez se depositam sobre
superfícies plásticas. Neste tipo de interação indireta, através da ligação do
S. aureus ao filme condicionante (formado pela deposição das proteínas
plasmáticas ao biomaterial) estariam envolvidos, por exemplo, o fator
clumping A (ClfA) implicado na aderência a tubos de polietileno e de cloreto de
polivinil recobertos por proteínas plasmáticas obtidas de cão (VAUDAUX et al.,
1995) e a autolisina, que dentre outras funções também apresenta propriedade
de ligação à fibronectina, fibrinogênio e vitronectina (HEILMANN et al., 2005).
O envolvimento da proteína A na aderência dos S. aureus à matriz extracelular
do hospedeiro foi demonstrado em estudos utilizando tecidos epiteliais das vias
aéreas de humanos (MONGODIN et al., 2000).
A segunda etapa de formação de biofilme tem início quando a bactéria já
se encontra aderida, direta ou indiretamente, à superfície do polímero. Em
S. epidermidis, tal evento começa com o acúmulo de PIA/PNAG. Porém, em
S. aureus, o(s) principal(is) componente(s) desta matriz não está(ão)
claramente elucidado(s). De qualquer forma, acredita-se que através dessa
matriz as células bacterianas são unidas umas às outras, formando um
agregado celular (GÖTZ, 2002).
As infecções estafilocócicas associadas aos biopolímeros podem ocorrer
de forma precoce ou tardia após a implantação do biomaterial. A forma precoce
ocorre alguns dias ou semanas após a cirurgia ou implantação do dispositivo
no paciente, e em muitos casos, a introdução do agente etiológico se dá
durante a realização desses procedimentos médicos. Por outro lado, as
infecções tardias começam após um longo período (várias semanas, meses ou
anos) entre a implantação de uma prótese e o aparecimento da infecção.
Assim, devido à capacidade de se aderir a tais superfícies inertes, os S. aureus
têm se tornado os principais agentes de infecções hospitalares associados a
biopolímeros em todo o mundo (JAMES & JAYAKRISHNAN, 2003; IZANO et
al., 2008). Dentre as diversas formas de manifestações clínicas das doenças

18
estafilocócicas, as bacteremias causadas pelos S. aureus, são sem dúvida, um
dos quadros infecciosos de maior gravidade. Um dos principais fatores que
contribui para o desenvolvimento deste quadro é a utilização de cateteres
endovenosos centrais, que aumentam a predisposição de pacientes
hospitalizados para desenvolver infecções associadas à formação de biofilme.
Diversos autores já relataram que mais da metade dos casos de bacteremias e
a maioria dos casos de endocardite causados por MRSA, em ambiente
hospitalar, são originados a partir de procedimentos cirúrgicos ou uso deste
tipo de dispositivo implantável (PETTI & FOWLER, 2002; TAK et al. 2002;
GIACOMETTI et al., 2005). Um estudo recente que confirma essa observação
foi publicado por Edgeworth e colaboradores. Neste estudo, envolvendo um
surto, durante um período de 2 anos, causado por uma variante (TW) do MRSA
ST239, na unidade de tratamento intensivo do Hospital-Escola Guy’s and St.
Thomas’ (Londres, Inglaterra), foi observada uma forte correlação entre os
altos indices de bacteremias causadas por essa linhagem epidêmica de MRSA
(47% dos casos) e a utilização de acessos vasculares em 59% dos pacientes
internados naquela unidade de tratamento (EDGEWORTH et al., 2007).
Uma importante dificuldade enfrentada por profissionais da saúde no
tratamento de infecção associadas a biomateriais é o fato de que, mesmo com
o início de uma terapia antimicrobiana empírica, a probabilidade de se obter
uma resposta satisfatória com a terapia antibiótica é muito baixa e, em alguns
casos, nula. Sendo assim, não raramente, a retirada do dispositivo médico é a
única alternativa para o reestabelecimento da saúde do paciente (WILCOX et
al., 2001; GIACOMETTI et al., 2005; STOODLEY et al., 2005). Porém, com
freqüência, a substituição do implante infectado acaba contribuindo para uma
maior debilidade do paciente, além de gerar prejuízos econômicos, e de não
garantir a ausência de complicações futuras, promovidas pelo risco continuado
de recorrência ou, até mesmo, de nova infecção (FUX et al., 2003).
1.4. Principais Sistemas Reguladores de Fatores de Virulência em
Staphylococcus aureus
Por serem patógenos extremamente versáteis, os S. aureus apresentam
outras notáveis características além da sobrevivência em sítios inanimados.
Como dito anteriormente, essa espécie bacteriana é capaz de colonizar e

19
infectar vários nichos no hospedeiro animal, como pele, membranas mucosas e
tecidos. Por isso, esses microrganismos são responsáveis por uma série de
doenças, desde infecções de pele e toxemias até doenças sistêmicas severas
(ARCHER, 1998; TENOVER & GAYNES, 2000; NOVICK, 2003).
Essa versatilidade depende de um amplo sistema de genes acessórios,
muitas vezes envolvidos na patogênese, que são responsáveis pela adaptação
da bactéria ao ambiente hostil do hospedeiro. A maior parte dos genes
envolvidos na virulência bacteriana codifica para proteínas de superfície ou
para proteínas secretadas para o ambiente extracelular. São exemplos de
proteínas associadas à parede bacteriana, a proteína A (SpA), a coagulase
(Coa), proteínas que se ligam à fibronectina A e B (FnBPA e B), que se ligam
ao fibrinogênio (FbPs) ou ao colágeno (CoBPs), e como exoproteínas, as α-, β-,
δ-, γ-hemolisinas, as leucocidinas, proteases, lipases, superantígenos
enterotoxinas, TSST-1, toxinas esfoliativas (ETA-D), dentre outras agressinas
que causam danos na célula hospedeira, degradando componentes da matriz
extracelular, danificando membranas ou as que estão envolvidas na evasão da
bactéria ao sistema de defesa específico do hospedeiro (PROJAN & NOVICK,
1997; WESSON et al., 1998; JARRAUD et al., 2000; NOVICK, 2000; AMARAL
et al., 2005). Além disso, a invasão de células epiteliais potencializa a
habilidade bacteriana de persistir no organismo do hospedeiro, aumentando,
conseqüentemente, a morbidade e o estado de portador (NOVICK, 2003;
AMARAL et al., 2005).
Esses fatores de virulência dos S. aureus encontram-se finamente
regulados por uma complexa e intrincada rede de sistemas reguladores da
virulência dos S. aureus, tais como: o agr, sar, sigB, arl, dentre outros (NOVICK
et al., 2000; SAID-SALIM et al., 2003; BRONNER, MONTEIL & PRÉVOST,
2004). O principal regulador de genes envolvidos na virulência dos
estafilococos é o agr (agr, do inglês: accessory gene regulator). Este sistema
funciona como um sensor de densidade populacional (quorum-sensing) que
promove a regulação negativa de proteínas de superfície, envolvidas na
colonização do hospedeiro, e a regulação positiva de proteínas extracelulares,
mais envolvidas nos processos de agressão ao hospedeiro (JANZON &
ARVIDSON, 1990; NOVICK et al., 1993, 1995; MORFELDT, TEGMARK &

20
ARVIDSON, 1996; JI, BEAVIS & NOVICK, 1997; LINA et al., 1998; NOVICK &
MUIR, 1999; LYON et al, 2002; ZHANG et al, 2002).
A estrutura do lócus agr é constituída de duas regiões divergentes de
transcrição que codificam os RNA conhecidos como RNAII e RNAIII,
coordenadas pelos promotores agrP2 e agrP3, respectivamente (figura 1;
JARRAUD et al., 2000). O RNAII corresponde à molécula transcrita pelo
operon P2 que é composto de quatro genes: agrB, D, C e A. O gene agrD
codifica para uma proteína que dá origem ao peptídeo autoindutor do sistema
Agr, conhecido como AIP (AIP, do inglês: autoinducing peptide) ou peptídeo
Agr (JI, BEAVIS & NOVICK, 1997; JARRAUD et al., 2000). Os peptídeos
indutores, produzidos a partir do lócus agr, possuem algumas variações na
seqüência de aminoácidos e nos domínios histidina-quinase de seus
receptores. A partir dessas diferenças foram determinados quatro tipos
polimórficos de agr, até o momento: AgrI, II, III e IV (JI, BEAVIS & NOVICK,
1997; JARRAUD et al., 2000; 2002). A biossíntese do AIP ocorre em algumas
etapas, como veremos a seguir. Primeiramente, a proteína AgrD (produto do
agrD) é sintetizada em nível ribossomal e ancorada a parte interna da
bicamada de fosfolipídios da membrana celular bacteriana, através de uma
hélice com domínios anfipáticos na sua região N-terminal, que a dirige ao
mesmo compartimento onde se encontra a proteína AgrB (produto do gene
agrB; ZHANG, LIN & JI 2004). Chegando até a AgrB, começa o processamento
da proteína AgrD, que dará origem ao peptídeo Agr. A AgrB efetua a digestão
proteolítica da AgrD e, posteriormente, uma ligação tioéster é formada, sendo
seu produto, já em forma de AIP, secretado pela AgrB. Já as proteínas AgrC e
AgrA participam de um sistema de tradução de sinais via dois componentes
que é responsável pela ativação do lócus agr. Com a secreção de AIP no meio
extracelular, tal peptídeo se liga hidrofobicamente à proteína AgrC, que
encontra-se exposta na membrana da célula bacteriana e que apresenta,
portanto, um domínio transmembranoso e um outro domínio citoplasmático
histidina-quinase (WRIGHT III et al., 2004). Assim, quando o peptídeo AIP se
liga à AgrC, essa se autofosforila em seu domínio histidina-quinase,
transferindo posteriormente o grupamento fosfato para o domínio aspartato da
proteína AgrA. Uma vez fosforilada, a AgrA torna-se ativada e, como apresenta
o papel de reguladora de resposta do sistema de dois componentes, se liga

21
(através de seu segundo domínio) à região promotora agrP2 e agrP3, ativando
a transcrição dos RNAII e RNAIII, respectivamente (figura 1). Cabe ressaltar
que a ativação e expressão do lócus agr pode ser pontencializada quando a
proteína AgrA atua em sinergismo com a proteína SarA (codificada em trans,
por outro operon), sendo capazes de se ligar a regiões distintas dos
promotores agrP2P3, ativando assim a transcrição de todo o operon agr
(JANZON & ARVIDSON, 1990; NOVICK et al., 1993, 1995; MORFELDT,
TEGMARK & ARVIDSON, 1996; JI, BEAVIS & NOVICK, 1997; LINA et al.,
1998; NOVICK & MUIR, 1999; LYON et al., 2002; ZHANG et al., 2002;
NOVICK, 2003). Figura 1. Diagrama esquemático do sistema Agr, mostrando os dois promotores agr divergentes (P2 e P3) e suas regiões de transcrição. O operon P2 é responsável pela transcrição do RNAII codificando os genes agrBDCA. Dois desses, agrC e agrA codificam proteínas do sistema de tradução de sinais, via dois componentes, e os outros dois, agrB e agrD, codificam para proteínas que irão gerar um peptídeo autoindutor (AIP), ligante da proteína receptora de sinal (AgrC). Quando o AIP se liga a AgrC leva à fosforilação da histidina-quinase no domínio carboxi-terminal citoplasmático. A seguir o grupamento fosfato é transferido para a AgrA, que uma vez fosforilada, ativa a transcrição dos promotores agrP2P3. A ativação do promotor P3 é responsável pela transcrição do RNAIII (molécula efetora do agr).
O RNAIII, por sua vez, é a molécula efetora do lócus agr dos
estafilococos, que contém uma pequena ORF que codifica para a δ-hemolisina

22
em seu terminal 5` e uma região não-codificadora, esta última responsável pela
regulação em trans (à distância), de diversos fatores de virulência, em nível de
tradução direta ou indireta. Porém a δ-hemolisina não apresenta funções nesta
regulação (NOVICK, 2003). Desta forma, quando um processo infeccioso
estafilocócico se inicia, como a densidade populacional bacteriana é ainda
baixa (correspondendo ao ínicio/metade da fase logarítimica do crescimento),
pouco peptídeo Agr é produzido, levando, conseqüentemente, a expressão de
RNAIII em níveis basais. Como o RNAIII é responsável pela regulação direta e
negativa de genes codificadores de proteínas de superfície, a menor expressão
desse RNA regulador determina uma maior biossíntese de tais proteínas,
essenciais para a aderência/invasão do patógeno na fase de colonização do
hospedeiro. Foi demonstrado que, a regulação direta e negativa acontece
através do pareamento de domínios de seqüências ricas em citosinas, na
região conservada da molécula de RNAIII, com seqüências RBS (RBS, do
inglês: ribosome-binding site) de seus mRNA-alvos (mecanismo antisensing;
BENITO et al., 2000; NOVICK, 2003; GEISINGER et al., 2006; BOISSET et al.,
2007). Um exemplo seria a repressão da expressão da proteína A (codificada
pelo gene spa). Neste caso, um dos domínios localizados no terminal 3’ do
RNAIII se liga ao RBS do mRNA spa, formando uma dupla fita de RNA
(RNAdf), impedindo o acoplamento do ribossomo para a posterior tradução do
RNAspa e induzindo uma rápida e específica degradação deste RNAdf pela
ação da endoribonuclease III (RNase III; HUNTZINGER et al., 2005).
Por outro lado, nas etapas mais avançadas do processo infeccioso, nas
quais a bactéria já se encontra melhor adaptada ao nicho em que se localiza e
em alta densidade populacional (equivalendo ao final da fase logarítimica e
início da fase estacionária de crescimento), uma maior quantidade de peptídeo
Agr é excretado, promovendo uma maior ativação do sistema Agr e,
conseqüentemente, uma maior transcrição de RNAIII. Assim, além da
regulação direta e negativa, esse RNA regulador também é responsável pela
regulação indireta e positiva de toxinas secretadas (agressinas) envolvidas na
agressão ao hospedeiro. Na regulação indireta e positiva, o RNAIII interage
com o mRNA de um gene que codifica para a proteína repressora Rot (Rot, do
inglês: repressor of toxins), envolvida na inibição da expressão dos genes

23
codificadores de exoproteínas (BENITO et al., 2000; NOVICK, 2003). Rot é um
regulador negativo, constitutivamente expresso nos S. aureus, que atua na
inibição da transcrição de uma série de agressinas estafilocócicas (SAID-
SALIM et al., 2003). Assim, como para o RNAspa, a complementariedade de
bases entre seqüências do RNAIII e do RNArot promovem a degradação do
RNAdf através da RNase III, diminuindo a síntese de Rot, liberando,
conseqüentemente, a síntese de diversas toxinas estafilocócicas que estavam
reprimidas pelo Rot (NOVICK et al., 1995; MORFELDT, TEGMARK &
ARVIDSON, 1996; GEISINGER et al., 2006; BOISSET et al., 2007).
Outro importante integrante da rede reguladora da virulência dos
S. aureus é o regulador de genes acessórios sarA (sar, do inglês:
staphylococcal accessory regulator). Este lócus regulador codifica uma proteína
de 14,5 kDa (SarA) com propriedades de ligação a moléculas de DNA, e que é
requerida, pelo menos sob algumas condições de crescimento bacteriano, para
a máxima ativação em trans dos promotores do lócus agr e,
conseqüentemente, da expressão da molécula de RNAIII (regulação agr-
dependente; HEINRICHS, BAYER & CHEUNG, 1996; PRAGMAN e
SCHILIEVERT, 2004). A expressão de sarA é promovida por três promotores,
sarP1, sarP2 e sarP3, responsáveis pela síntese de três transcritos
sobrepostos (figura 2).
sarAP1P3 P2
sarB
sarC
sarA
Figura 2. Esquema do operon sarA. Os retângulos P1, P2 e P3 representam as posições dos promotores P1, P2 e P3. As setas representam os transcritos de sar. Pontilhados cinza representam a ORF sarA e a sobreposição da mesma nos transcritos sarC e sarB. Adaptado de BAYER, HEINRICHS & CHEUNG, 1996.
Desta forma, o operon sarA pode ser expresso durante todas as fases
de crescimento bacteriano, sendo sarP1 e sarP2 responsáveis por sua
expressão no início/metade da fase exponencial de crescimento e, sarP3,
responsável por sua expresão no final da fase exponencial/início da fase

24
estacionária ou ainda durante condições de estresse ambiental (MANNA,
BAYER & CHEUNG, 1998). Nesta última, a ativação de sarA, promovida por
sarP3, é dependente do fator σB (DEORA, TSENG & MISRA, 1997). Além
disso, SarA também participa da regulação positiva e negativa de vários fatores
de virulência dos S. aureus, podendo esta regulação ocorrer de forma
independente da ativação do lócus agr (agr-independente). Dados obtidos a
partir da análise da estrutura secundária de SarA indicam que seu papel como
ativador na expressão gênica seria, possivelmente, promovido pelo seu
domínio hélice-volta-hélice (helix-turn-helix) que permite sua ligação aos sulcos
maiores (formados pela estrutura em alfa-hélice) das moléculas de DNA-alvo,
enquanto as regiões flanqueadoras a este domínio seriam responsáveis pela
interação com os sulcos menores do mesmo DNA. Desta forma, o complexo
SarA-DNA facilitaria a transcrição dos genes-alvo (LIU et al., 2006). Por outro
lado, tal propriedade de ligação a regiões promotoras pode, para certos genes,
funcionar como um mecanismo de repressão, caso a conformação produzida
pela interação SarA-DNA dificulte a transcrição gênica, como sugerido, por
exemplo, para o gene spa. Tem sido proposto que a regulação agr-
independente, promovida por SarA, possa ainda ocorrer através da ligação
desta proteína aos trancritos de genes que codificam para certos fatores de
virulência, aumentando assim a estabilidade do mRNA, durante a fase
logarítmica do crescimento bacteriano (CHEUNG et al., 2004; ROBERTS et al.,
2006). Dentre os fatores de virulência regulados positivamente pelo SarA,
incluem-se as proteínas de ligação à fibronectina (FnBPs) e ao fibrinogênio
(FbBP), a TSST-1 e a enterotoxina estafilocócica B (SEB). Dentre os fatores
regulados negativamente podemos citar as diversas proteases estafilocócicas
(CHAN & FOSTER, 1998; CHEUNG et al., 2004). Por outro lado, com o avanço
das pesquisas em torno de SarA, novas proteínas homólogas Sar foram
descobertas em S. aureus (tabela 3).

25
Tabela 3. Membros pertencentes à família de proteínas homólogas Sar.
Proteínas* Principais funções sugeridas Referência
MgrAa Regulador de autólise e do agr TRUONG-BOLDUC, ZHANG, & HOOPER, 2003
Rot Repressor da síntese de toxinas, com função oposta ao agr
SAID-SALIM et al., 2003
SarA Ativador de genes por vias dependentes ou independentes do agr
CHAN & FOSTER, 1998; CHEUNG et al., 2004
SarR Regulador negativo de sarA e positivo do agr CHEUNG et al., 2004
SarSb Ativador da síntese da proteína A TEGMARK et al., 2000
SarT Ativador de sarS e repressor da síntese de α-hemolisina
CHEUNG et al., 2004
SarU Regulador positivo do agr CHEUNG et al., 2004
SarV Regulador de autólise reprimido por SarA e MgrA
MANNA et al., 2004
SarX Regulador negativo do agr e ativado por MgrA MANNA & CHEUNG, 2006
SarY Função desconhecida KURODA et al., 2001
SarZ Regulador positivo de hlac KAITO et al., 2006
*Proteínas homólogas Sar codificadas no genoma da cepa de S. aureus N315. aMgrA, regulador também conhecido como Rat ou NorR. bSarS, ativador também conhecido como SarH1. chla, gene que codifica para a α-hemolisina. Adaptado de CHEUNG et al., 2008.
Uma dessas proteínas é a MgrA, pertencente a subfamília de
reguladores MarR (TRUONG-BOLDUC, ZHANG, & HOOPER, 2003). A função
deste regulador foi primeiramente evidenciada em um estudo que demonstrou
que uma mutação no gene mgrA resultava em um fenótipo de autólise
aumentado e em uma diminuição da transcrição dos genes agr, sarS e sarX
(INGAVALE et al., 2005). Posteriormente, estudos de perfis transcricionais
(LUONG et al., 2006) revelaram que MgrA estava envolvida na regulação de
355 genes, promovendo a regulação positiva de 175 genes (dentre estes,
genes codificadores de exoproteínas) e a negativa dos 180 genes restantes
(dentre os quais encontram-se genes codificadores de proteínas de superfície;
INGAVALE et al., 2005; LUONG et al., 2006). Adicionalmente, o efeito de MgrA
na autólise bacteriana parece ser mediado por sarV, que é um regulador
positivo de diversas enzimas autolíticas, como veremos a seguir.
Outro membro da família Sar, é a proteína Rot, uma repressora da
biossíntese de toxinas estafilocócicas (SAID-SALIM et al., 2003). A função de
Rot foi investigada a partir de estudos utilizando a tecnologia de RNA-
microarrays, que revelaram que tal proteína estava envolvida na repressão da

26
transcrição de 168 genes, incluindo, os genes codificadores das α- e β-
hemolisinas, de proteases, de uma lípase e urease estafilocócicas. Rot parece
também atuar na regulação positiva da proteína A e de SarS, influenciando a
expressão desses genes de forma antagônica ao agr (SAID-SALIM et al.,
2003).
Além destas proteínas homólogas Sar também é membro desta famíla o
repressor SarR. Esta proteína, da mesma forma como SarA, é capaz de se
ligar a regiões promotoras, promovendo, através da mudança conformacional
gerada pelo híbrido SarR-DNA, a repressão de genes-alvo. Foi observado que
SarR atua como um modulador negativo da expressão do operon sar (MANNA
& CHEUNG, 2001; CHEUNG et al., 2004).
A proteína SarS, também conhecida como SarH1, teve sua identificação
baseada em sua habilidade de ligação aos promotores agrP3, hla (codificador
da α-hemolisina) e ssp (codificador de uma serina protease; TEGMARK,
KARLSSON, & ARVIDSON, 2000). SarS, diferentemente das proteínas SarA e
SarR (com estruturas diméricas), apresenta-se em uma estrutura monomérica,
podendo atuar na ativação da transcrição de spa ou na repressão da
transcrição de hla (TEGMARK et al., 2000; CHEUNG et al., 2004). Análises
transcricionais indicaram que sarS é reprimido pelo agr e mgrA e ativado pelas
proteases ClpX e ClpP (FREES, SORENSEN, & INGMER, 2005). Em um
estudo recente (2006) foi sugerido que SarA pode atuar tanto como um
importante repressor quanto como um modesto ativador de hla. No primeiro
caso, a repressão de hla ocorreria devido a regulação positiva promovida por
SarA sobre sarS (OSCARSSON, et al., 2006). Contudo, este resultado foi
obtido a partir de experimentos realizados com a cepa de S. aureus, SH1000
(uma cepa laboratorial que apresenta baixa produção de α-toxina), sendo,
portanto, necessárias informações adicionais sobre este dado em amostras
clínicas de S. aureus (CHEUNG et al., 2008).
Mais um integrante da família de proteínas homólogas Sar, é a proteína
SarT, que é normalmente reprimida pelos reguladores agr e sarA. A ativação
de sarT resulta na regulação positiva de sarS (CHEUNG et al., 2004), levando
a repressão de hla e a ativação de spa (SCHMIDT et al., 2001, SCHMIDT,
MANNA & CHEUNG, 2003).

27
Adjacentemente ao gene sarT localiza-se o gene de mais um homólogo
Sar, o sarU. Este gene codifica para uma proteína dimérica que é transcrita de
forma divergente a sarT. sarU é reprimido por SarT, que por sua vez, é
regulado negativamente pelo agr. Como sarU é em ativador da expressão do
agr, quando este último encontra-se mais expresso, menor a síntese de SarT e
maior a de SarU, o que, conseqüentemente, amplifica o sinal do agr (MANNA &
CHEUNG, 2003, CHEUNG et al., 2004, 2008).
O gene sarV (codificador de SarV, outro homólogo Sar) é muito pouco
expresso (sendo muitas vezes não detectado) sob condições de crescimento
bacteriano in vitro. Entretanto, estudos utilizando cepas de S. aureus mutantes
em mgrA e sarA apresentam um aumento significativo na expressão desse
gene, indicando que a repressão de sarV seja promovida por mgrA e sarA
(CHEUNG et al., 2004). Ao contrário do observado nos mutantes mgrA e sarA,
o mutante sarV foi mais resistente a lise da parede celular mediada pela ação
de antibiótico ou detergente, quando comparado com sua cepa isogênica
selvagem (hiper-expressando sarV e apresentando uma maior susceptibilidade
à lise). De acordo com esses resultados, sarV parece participar de uma via de
modulação de autólise comum nos S. aureus (CHEUNG et al., 2004).
Assim como SarV, SarX é mais um homólogo Sar formado por um único
domínio protéico. SarX tem sua síntese ativada pela MgrA (MANNA &
CHEUNG, 2006) e apresenta a função de repressor do agr. Desta forma,
quando MgrA é expresso, ativa a expressão do agr e de sarX. Este último, por
sua vez, promove a repressão do agr de forma dependente de mgrA,
demonstrando a complexidade e o intrincamento de toda a rede reguladora da
família de homólogos Sar.
De acordo com o genoma da cepa de S. aureus N315, sarY é mais um
gene putativo codificador de uma proteína homóloga Sar. Entretanto, sua
função não foi ainda elucidada (KURODA et al., 2001; CHEUNG et al. 2008).
Finalmente, SarZ é o último dos membros da família de reguladores
homólogos Sar descrito até o momento. SarZ também apresenta homologia
com a subfamília MarR, (CHEUNG et al., 2004) e foi identificado recentemente,
em 2006. Kaito e colaboradores (KAITO et al., 2006) identificaram este novo
homólogo Sar a partir de experimentos que evidenciaram a habilidade desta
proteína de restaurar a atividade hemolítica em um mutante de S. aureus em

28
cvfA (um regulador de virulência com atividade de fosfodiesterase), o qual era
defectivo para a expressão de hla. Através de modelos de infecção em
camundongos e bicho-da-seda foi observado que o mutante sarZ derivado da
cepa de S. aureus RN4220 apresentou uma virulência atenuada. Contudo
estes resultados devem ser interpretados com cautela, uma vez que a cepa
RN4220 foi geneticamente manipulada através de mutagêneses com
nitrosoguanidina. E ainda, quando as mutações em sarZ foram transduzidas na
cepa 8325-4 (defectiva para os genes rsbU e sigB) foi observada apenas uma
sutil diminuição da atividade hemolítica, sendo este fenótipo parcialmente
restaurado com a complementação de sarZ em trans. Isto sugere que outros
genes, downstream a sarZ, possam contribuir para a expressão do fenótipo
hemolítico (CHEUNG et al., 2008).
Como dito anteriormente, além dos reguladores agr e sar, outros
reguladores também participam da modulação dos diversos fatores de
virulência dos S. aureus. O sigB, por sua vez, codifica um fator sigma
alternativo (σB) para a transcrição de genes mais envolvidos na resposta dos
S. aureus às condições ambientais adversas, geralmente observadas na fase
estacionária do crescimento bacteriano. Algumas dessas variações ambientais
podem ocorrer, por exemplo, no interior do biofilme maduro, levando a um
estado de estresse bacteriano, gerado pelo esgotamento de nutrientes e de
íons essenciais para o metabolismo, pelos extremos de pH (devido ao acúmulo
de produtos metabólicos bacterianos naquele microambiente) e também pelo
desequilíbrio osmótico. Outro exemplo seria a variação de temperatura
observada no local da infecção e, ainda as condições promovidas pela
resposta inflamatória do hospedeiro, em resposta a ação de agressinas
secretadas pelas bactérias (LORENZ et al., 2008). Por outro lado, quando os
S. aureus encontram-se em condições ideais de crescimento, sigB torna-se
inativo, devido à repressão promovida pela proteína RsbW (um fator anti-σ;
PANÉ-FARRÉ et al., 2006).
Similarmente ao sistema Agr, o lócus arl também codifica um sistema
regulador de virulência via dois componentes, o ArlR-ArlS. Este sistema está
diretamente envolvido na regulação de alguns genes de virulência, tais como o
hld (codificador da δ-hemolisina), hlc (codificador da hemolisina Hlc), lukD e

29
lukE (codificadores de leucocidinas) e genes relacionados ao metabolismo
bacteriano. Entretanto, sua função parece estar mais relacionada ao processo
de autólise bacteriana e a atividade proteolítica extracelular, apresentando
também um papel na regulação negativa da aderência dos S. aureus a
polímeros de poliestireno (FOURNIER & HOOPER, 2000). Apesar do agr e de
sarA serem requeridos para regulação de spa, através da ativação de ArlRS,
este último pode promover a diminuição da transcrição do agr e aumentar a
transcrição de sarA (FOURNIER, KLIER, & RAPOPORT, 2001); demonstrando
assim a complexidade e intrincamento desses sistemas de regulação.
1.5. Regulação de genes envolvidos na formação de biofilme glicose-induzido ica-independente em Staphylococcus aureus
Muitos estudos vêm indicando a existência de um ou mais mecanismos
de formação de biofilme PIA/PNAG-independente, tanto em S. epidermidis
como em S. aureus (BEENKEN et al., 2004; FITZPATRICK, HUMPHREYS &
O’GARA, 2005; ROHDE et al., 2005, 2007, KOGAN et al., 2006). Beenken e
colaboradores, através de experimentos de DNA-microarrays, utilizando uma
amostra clínica de S. aureus, demonstraram que mutações deletérias no lócus
ica, levando à completa inibição de PIA/PNAG, causavam apenas um pequeno
impacto na formação de biofilme, que continuava a ser produzido e acumulado
normalmente, tanto em ensaios realizados in vitro como in vivo (BEENKEN et
al., 2004).
Um gene indicado como aparentemente envolvido na regulação da
produção de biofilme ica-independente de S. aureus é o rbf (rbf, do inglês:
regulator of biofilm formation). Esse gene codifica para a proteína Rbf que
pertence à família AraC/XylS (cujas proteínas apresentam domínios de ligação
a moléculas de DNA) e que, portanto, não apresenta características de
proteínas de superfície e nem de secretadas. Segundo Lim e colaboradores, a
Rbf parece regular positivamente uma proteína de 190 kDa, envolvida na
produção de biofilme em resposta a certas alterações ambientais, tais como as
promovidas por variações nas concentrações de glicose e NaCl. Ainda de
acordo com os autores deste trabalho, a Rbf atuaria, provavelmente, através de
uma via independente de ica, uma vez que a transcrição deste operon não foi
afetada em um mutante isogênico para o rbf (LIM et al., 2004). Posteriormente,

30
em outro trabalho, foi observado que a indução da síntese de biofilme por NaCl
foi mais freqüente em amostras de MSSA do que em MRSA. Ao contrário do
que foi demonstrado para as amostras de MSSA, foi ainda observado que a
indução por NaCl não levou à produção de PIA/PNAG pelos MRSA, apesar do
aumento da transcrição do icaADBC detectado, (O’NEILL et al., 2007). Em um
estudo anterior, já havia sido demonstrado que o desenvolvimento de biofilme
em amostras de MRSA seria, principalmente, induzido por glicose, além de ser
ica-independente, estando aparentemente relacionado à produção de uma ou
mais proteínas com características de adesinas (FITZPATRICK, HUMPHREYS
& O'GARA, 2005).
Mais recentemente (2008), utilizando amostras clínicas de S. aureus,
O’Neill e colaboradores, aprofundando as investigações anteriores, avaliaram
quais seriam as possíveis proteínas que estariam envolvidas na estrutura do
biofilme. Foi demonstrado que as FnBPAB estavam associadas ao biofilme
glicose-induzido, ica-independente, tanto em MSSA como MRSA, em
condições experimentais de formação de biofilme sob fluxo ou estáticas
(O’NEILL et al., 2008).
A SasG é uma proteína de S. aureus, descrita recentemente, que foi
também associada à produção de biofilme ica-independente (CORRIGAN et
al., 2007). Esta proteína é homóloga à proteína Aap, sendo também, esta
última, relacionada à acumulação do biofilme em amostras clínicas de
S. epidermidis (DE ARAÚJO et al., 2006). A SasG foi ainda associada à
aderência bacteriana à superfície de células epiteliais nasais, demonstrando
seu envolvimento tanto na formação de biofilme como na colonização do
hospedeiro (CORRIGAN et al., 2007).
O envolvimento do ácido teicóico, outra estrutura também relacionada ao
processo inicial de aderência bacteriana a superfícies, através de mecanismo
ica-independente, foi descrito por Gross e colaboradores em 2001. Os autores
deste trabalho demonstraram que amostras de S. aureus dltA-nocautes e,
portanto, deficientes do resíduo de D-alanina em seu ácido teicóico,
apresentaram um fenótipo de biofilme negativo (GROSS et al., 2001). Um outro
estudo que corrobora o envolvimento do ácido teicóico na fase inicial de
formação de biofilme dos S. aureus foi recentemente publicado. Neste estudo
Vergara-Irigaray e colaboradores (2008), utilizando construções nocautes no

31
gene tagO (envolvido na síntese de ácido teicóico da parede celular),
demonstraram que os mutantes apresentaram menor habilidade para formar
biofilme. Porém, essa diminuição do biofilme não pôde ser relacionada a um
possível efeito dessa mutação na diminuição de PIA/PNAG ou em seu
ancoramento à superfície da célula (VERGARA-IRIGARAY et al., 2008).
Apesar dos estudos citados acima, pouco se conhece sobre a exata
composição do biofilme ica-independente dos S. aureus. Portanto, mais
estudos são necessários para a elucidação de tais mecanismos e das vias
envolvidas na regulação do filme biológico desenvolvido pelos S. aureus.
Entretanto, alguns estudos têm demonstrado que sistemas reguladores
da virulência dos S. aureus parecem estar também envolvidos na regulação do
biofilme, um desses sistemas é o ArlR-ArlS, um sistema de tradução de sinais
via dois componentes. Mutações neste lócus resultaram em um aumento da
aderência inicial e na produção de PIA/PNAG pela cepa de S. aureus 15981.
Entretanto, a ativação da síntese de biofilme neste mutante não foi afetada pela
deleção do operon ica, indicando que PIA/PNAG não é um componente
essencial da matriz do biofilme produzido pela amostra 15981 (TOLEDO-
ARANA et al., 2005). Similarmente, a deleção do gene da principal autolisina
dos S. aureus, atl (regulada pelo sistema ArlRS), não exerceu efeito no fenótipo
do biofilme do mutante arlRS (TOLEDO-ARANA et al., 2005). Por outro lado,
estudos prévios já haviam demonstrado uma dramática diminuição na atividade
proteolítica extracelular em mutações em arlS (FOURNIER & HOOPER, 2000).
Desta forma, é interessante citar que a ativação da síntese de biofilme no
mutante arlRS, descrita por Toledo-Arana e colaboradores (2005), foi sarA-
dependente (sabendo-se que SarA regula negativamente a produção de
proteases extracelulares). Assim, é provável que a ativação do biofilme nos
mutantes arlRS possa ter ocorrido pela inibição da atividade proteolítica, a qual,
quando ativada, degradaria uma proteína importante para a formação deste
fenótipo de biofilme. Em contraste ao observado com as mutações em sarA, a
deleção do agr ativou o biofilme no mutante arlRS (TOLEDO-ARANA et al.,
2005). Acredita-se que a regulação negativa do agr sobre este biofilme tenha
ocorrido de forma indireta. Tal conclusão se deve ao fato desse sistema arlRS
atuar regulando positivamente o lócus agr (LIANG et al., 2005).

32
Não apenas o estudo citado acima, mas também outros estudos têm
demonstrado que mutações no lócus sarA resultam na reduzida capacidade de
amostras de S. aureus em formar biofilme (BEEKEN, BLEVINS & SMELTZER,
2003; VALLE et al., 2003; CONLON, HUMPHREYS & O’GARA, 2004). Tal fato,
como dito anteriormente, parece ser devido à função reguladora negativa de
SarA sobre as proteases, uma vez que proteínas parecem ter um papel
importante na formação do biofilme estafilocócico (KARLSSON et al., 2001;
O´NEILL et al., 2007). Ao contrário de sarA, foi observado que mutações no
lócus agr de 21 amostras clínicas de S. aureus não acarretaram nenhuma
alteração significativa (efeito neutro) no fenótipo de acumulação de biofilme de
16 destas e induziram um aumento de sua síntese (regulação negativa) em
apenas 5 (23%) das demais amostras analisadas (O’NEILL et al., 2007). Em
oposição a este estudo, no ano de 2000, Vuong e colaboradores, haviam
relatado que de um total de 105 amostras de S. aureus estudadas, 78% das
amostras agr negativas foram capazes de produzir biofilme, mas que apenas
5% das agr positivas expressaram o filme biológico. Deste modo, este estudo
parece sugerir que o agr deva ter um profundo impacto no fenótipo de
formação de biofilme. E ainda, foi concluido, pelos autores, que o efeito do agr
no biofilme não estaria relacionado à expressão diferenciada da proteína Atl
(principal autolisina de parede celular) ou do PIA/PNAG, mas parecia estar
associado, pelo menos em parte, à propriedade surfactante da δ-hemolisina
(codificada na região do RNAIIIagr). Foi sugerido que tal propriedade, quando
presente, poderia impedir interações hidrofóbicas entre a célula e a superfície
do polímero (VUONG et al., 2000).
Em conjunto, esses dados parecem indicar a participação de diversas
proteínas estafilocócicas na intermediação da aderência bacteriana inicial a
polímeros e na acumulação do biofilme glicose-induzido e ica-independente em
amostras de S. aureus, apontando para um mecanismo multifatorial. Em
adição, tais dados evidenciam a complexidade da rede genética reguladora
envolvida e sugerem que os loci agr e sarA parecem estar (direta ou
indiretamente) comprometidos na modulação deste fenótipo de produção de
biofilme.

33
2. OBJETIVOS
Nesta Tese nós partimos do pressuposto que a variante epidêmica do
CEB, predominante em hospitais brasileiros, deveria apresentar alguma
vantagem adaptativa que explicasse sua ampla colonização hospitalar, em
relação às demais variantes. Estudos anteriores de nosso laboratório
demonstraram que a variante predominante do CEB apresentava maior
capacidade para se aderir e invadir células epiteliais brônquicas humanas
(AMARAL et al. 2005). Como a formação de biofilme é considerado um
importante mecanismo de virulência nas infecções associadas a biopolímeros
biomateriais, neste trabalho de tese nós nos propomos a estudar
comparativamente a produção de biofilme em um total de 199 amostras de
S. aureus pertencentes ou não ao CEB e estudar os mecanismos moleculares
envolvidos na virulência.
Objetivos específicos:
(i) Analisar comparativamente a produção de biofilme em amostras
pertencentes ao CEB, a clones de MRSA raros nos hospitais
brasileiros e em amostras de MSSA, através de ensaios de
formação de biofilme em microplacas de poliestireno inerte.
(ii) Analisar a expressão do lócus agr nas amostras acima, através da
análise da atividade da δ-hemolisina e de ensaios de Northern-
blotting.
(iii) Analisar os mecanismos moleculares envolvidos na atenuação do
lócus agr, detectada neste estudo em algumas representantes de
amostras CEB, envolvendo: Northern-blotting para se estudar a
expressão do mRNA sarA e seqüenciamento de DNA para os loci
agr e sarA.

34
3. MATERIAIS E MÉTODOS
3.1. Amostras bacterianas Para os experimentos de biofilme foi utilizado um total de 199 amostras
clínicas de S. aureus (149 MRSA e 50 MSSA), isoladas de humanos, em
diversos hospitais localizados no país, e cuja caracterização molecular foi
realizada previamente (AMARAL et al., 2005). Destas, 100 eram amostras
relacionadas ao CEB (ST239-SCCmecIIIA), sendo 50 classificadas como
variante epidêmica do CEB, pois apresentavam um perfil predominante e
idêntico de bandas no PFGE, entre as amostras CEB estudadas. Tal perfil foi
designado por nós como “perfil A1”. As outras 50 amostras CEB possuiam um
perfil no PFGE que variava de 1 a 6 bandas do perfil apresentado pela amostra
BMB9393 (protótipo do perfil A1) e foram aqui agrupadas como “perfil não-A1”.
Por sua vez, as 49 amostras restantes de MRSA possuiam perfis de PFGE que
diferiam em mais de 6 bandas do perfil da amostra BMB9393 e foram
classificadas como clones esporádicos, uma vez que eram raramente
observados em hospitais brasileiros. Finalmente, as 50 amostras de S. aureus
remanescentes eram MSSA. (AMARAL et al., 2005). A amostra de
Streptococcus pyogenes 75194 foi utilizada neste estudo como controle
negativo do biofilme, uma vez que não adere às superfícies lisas, e a amostra
de Staphylococcus epidermidis 70D como controle positivo, por se tratar de
uma espécie altamente produtora de biofilme.
As amostras de S. aureus RN6390B e RN4220 (ambas apresentando
loci agr e sarA funcionais) e a contrução agr-nula RN6911 (Δagr::tetM) foram
utilizadas como amostras-padrão ou controle em vários experimentos aqui
realizados. Estas últimas amostras nos foram gentilmente cedidas pelo Dr.
Richard P. Novick (New York University School of Medicine, EUA).
As amostras deste estudo, BMB9393 (agr-funcional) e GV69 (agr-
atenuado) foram utilizadas como protótipos.
Todas as amostras encontravam-se estocadas a –70ºC, em TSB (Tryptic
Soy Broth) adicionado de glicerol, para uma concentração final de 10% (p/v).

35
3.2. Ensaio de biofilme O ensaio de biofilme foi realizado em microplaca de poliestireno inerte
(96 poços; Nunc; Nunclon). A partir do estoque, a amostra bacteriana foi
semeada, com o auxílio de uma alça bacteriológica, em placas de Petri
contendo TSA (Tryptic Soy Agar) e incubada por 24 h a 37ºC.
Posteriormente, 3 colônias foram repicadas para 2 mL de TSB e a
cultura incubada sob agitação (200 rpm) por 18 h. A cultura foi diluída (1:100)
em TSB suplementado com glicose, para uma concentração final de 1%
(SAKOULAS et al., 2002). A partir de então, 200 µL foram aplicados em cada
um dos poços da placa de microtitulação e esta incubada por 20 h a 37ºC.
Foram utilizados 4 poços da placa de microtitulação para cada amostra. Após
incubação, a microplaca foi lida em leitor de ELISA (microplate reader model
550; Bio-Rad), no qual foi feita uma primeira leitura para medir a densidade
óptica (DO) do crescimento bacteriano a 570 nm. A seguir, o sobrenadante da
cultura foi removido e os poços foram gentilmente lavados por 4 vezes com
solução salina estéril (NaCl a 0,85% p/v). Na etapa seguinte, as células
aderidas foram fixadas a 65ºC por 1 h. Posteriormente, foram aplicados 200 μL
de cristal violeta para coloração de Gram em cada poço e, após 2 min, cada
poço foi lavado gentilmente (por pelo menos 4 vezes) com água destilada
estéril para a retirada de excesso de resíduos do corante. A placa foi seca em
forno a 45ºC (SAKOULAS et al., 2002) e a DO do biofilme foi medida a 570 nm.
A fórmula abaixo foi utilizada para se calcular a unidade de biofilme (UB).
DO do Biofilme = Unidade de Biofilme (UB) DO do Crescimento
A maior leitura da DO do biofilme da amostra de S. pyogenes 75194 foi
utilizada como base para a comparação e interpretação da unidade de biofilme
das amostras de S. aureus estudadas. Assim, se a UB fosse menor ou igual a
2 vezes (2X) o valor da maior leitura da UB do biofilme do S. pyogenes, a
amostra era considerada como não produtora. Se o valor da UB estivesse entre
2-4X o valor da maior leitura da UB do biofilme do S. pyogenes, a amostra era
considerada fracamente produtora. Entre 4-8X, a amostra era considerada
moderadamente produtora, e acima de 8X, fortemente produtora.

36
A técnica de triagem em microplaca de poliestireno inerte tem sido
amplamente utilizada para se testar a produção de biofilme em S. aureus
(BEENKEN et al. 2003; TOLEDO-ARANA et al. 2005; O’NEILL et al. 2008) e
tem sido considerada um excelente método de triagem quando se deseja testar
múltiplos microrganismos (CASSAT, LEE & SMELTZER, 2006).
Adicionalmente, recentes estudos têm obtido resultados semelhantes ao comparar esta técnica com outros métodos, nos quais o desenvolvimento do
biofilme ocorre sob condições de fluxo ou in vivo (BEENKEN et al., 2004).
3.2.1. Cálculos estatísticos Os valores das UB obtidos foram utilizados nos cálculos estatísticos
(teste t de Students’) para averiguação da hipótese nula, em populações não
pareadas, com variâncias desconhecidas, estipulando-se α = 0,05 (DUNM,
1964a).
3.3. Triagem de atividade da δ-hemolisina
Inicialmente, para avaliarmos a atividade do lócus agr, entre as amostras
CEB (perfil A1 e não-A1) estudadas, realizamos uma triagem que se baseia na
detecção da atividade da δ-hemolisina em hemácias de carneiro. Tal estratégia
tem sido utilizada para esta finalidade (NOVICK, et al., 1995; SAKOULAS et al.,
2002; ADHIKARI, ARVIDSON & NOVICK, 2007) e se fundamenta no fato de
que o gene que codifica para a δ-hemolisina (hld) encontra-se sobreposto ao
gene que codifica para o RNAIII. Desta forma, a tradução da δ-hemolisina é
uma indicação de que o RNAIII está sendo transcrito. Para a realização do
teste, as amostras pertencentes ao CEB e as amostras-controle RN6390B (agr-
funcional), RN4220 (produtora de β-hemolisina) e RN6911 (Δagr-nula) foram
semeadas por esgotamento em placas de TSA e incubadas a 37ºC por 24 h.
Colônias isoladas foram repicadas em TSB e a cultura incubada a 37ºC por
18 h, sob agitação (250 rpm). Após incubação, a cepa RN4220 foi estriada
verticalmente no centro da placa contendo ágar sangue desfibrinado de
carneiro (AS) a 5% (v/v; PlastLabor). A seguir, na mesma placa, foram também
estriadas, perpendicularmente à RN4220 e proximamente (sem tocar) à estria
desta, as amostras CEB (perfil A1 e não-A1) e amostras-controle.

37
A caracterização da δ-hemolisina, através deste teste, baseia-se na
ocorrência de um sinergismo entre a δ-hemolisina e a β-hemolisina. A
interpretação deste teste foi realizada de acordo com os padrões hemolíticos
descritos anteriormente por Adhikari e colaboradores (ADHIKARI, ARVIDSON
& NOVICK, 2007).
3.3.1. Cálculos estatísticos A proporção de indivíduos dentro dos diferentes grupos de amostras
(CEB A1 e não-A1) que produziram δ-hemólise forte, fraca ou não detectada foi
utilizada nos cálculos estatísticos, utilizando-se o teste do Chi-quadrado para
averiguação da hipótese nula, com variância igual a 2 e α = 0,05 (DUNM,
1964b).
O mesmo teste também foi utilizado nos cálculos estatísticos para a
análise da proporção de indivíduos dentro dos diferentes grupos de amostras
produtoras de biofilme (fenótipos forte/moderado e fraco/não produtor) que
apresentaram δ-hemólise forte, fraca ou não detectada.
3.4. Experimentos de Northern-blotting Os experimentos de Northern-blotting foram realizados para
representantes de amostras CEB que apresentaram atividade de δ-hemolisina
visivelmente diminuida no screening em AS. Também foram utilizadas para o
controle destes experimentos 2 amostras clínicas de MSSA, além da amostra
BMB9393 (protótipo agr-funcional) e a amostra padrão agr-funcional RN6390B. O RNA total foi obtido utilizando-se o kit RNeasy Mini Kit (Qiagen). O RNA foi
quantificado no GeneQuant RNA/DNA calculator da Pharmacia Biotech. Um
volume contendo 10 μg da preparação de RNA foi aplicado em um gel de
agarose a 1%, em presença de tampão MOPS e formaldeído, conforme
recomendado no manual do fabricante do RNeasy Mini Kit. A transferência do
RNA do gel de agarose para a membrana de náilon (Hybond-N+, Amersham
Pharmacia) foi realizada através de vácuo (Vacum geneXL; Pharmacia) por 2 h
a 55 mm Hg, conforme recomendações do fabricante. Após transferência, a
membrana foi colocada sobre papel de filtro e deixada por 30 min à

38
temperatura ambiente. Após secagem, a membrana foi colocada em um forno
a 80°C por 2 h para fixação do RNA.
3.4.1. Obtenção da sonda para o rnaIII, sarA e 16S rrna
3.4.1.1. Extração do DNA
A técnica utilizada foi a descrita por Sambrook, Fritsch & Maniatis (1989)
com algumas modificações, conforme relatamos a seguir. Para a obtenção do
DNA genômico da amostra RN6390B, esta foi cultivada em TSB (37°C/18 h, a
200 rpm), centrifugada a 2500 x g e o sedimento lavado 2X com TE (Tris 1 M,
pH 7,5; Sigma, EDTA 0,1 mM, pH 8,0). O sedimento foi homogeneizado e
acrescido de lisostafina, para uma concentração final de 5.000 U/mL (Sigma),
para degradação da parede bacteriana. Após incubação por 1 h a 37°C. Os
protoplastos obtidos foram lisados em SDS para uma concentração final de 1%
(p/v). A digestão protéica foi realizada por 1 h a 37°C com proteinase K, para
uma concentração final de 455 U/mL (Sigma). Posteriormente, igual volume de
uma mistura de fenol-clorofórmio (1:1; v/v), previamente saturado com tampão,
foi adicionado. O tubo foi agitado, manual e vigorosamente, por cerca de 10
min e centrifugado a 2500 x g por 30 min. A fase aquosa superior foi retirada,
transferida para novo tubo descartável cônico e adicionada de RNAse para
uma concentração final de 600 UK/mL (Sigma). O lisado bacteriano foi
incubado por 1 h a 37°C. Após esse período, igual volume de fenol-clorofórmio
foi acrescentado e o tubo novamente agitado por 10 min. Posteriormente, foi
realizada uma centrifugação a 2500 x g por 30 min. Novamente, a fase aquosa
superior foi transferida para um novo tubo descartável, onde foi acrescentado o
dobro do volume de etanol absoluto gelado. O lisado foi gentilmente misturado
com o etanol através de movimentos circulares, manuais, por
aproximadamente 5 min, de forma a enrolar o DNA. O DNA assim precipitado
foi coletado com auxílio de uma alça bacteriológica descartável e transferido
para um tubo estéril. Posteriormente, duas lavagens com álcool etílico a 70%
(v/v) foram realizadas, sendo o material centrifugado em microcentífuga por
10 min, sob máxima rotação. Finalmente, o DNA foi deixado por 15 min na
estufa (37°C) para a evaporação do etanol. O DNA foi diluído em água

39
bidestilada estéril e estocado a -20°C até o momento do uso (SAMBROOK,
FRITSCH & MANIATIS, 1989).
3.4.1.2. Amplificação das sondas para o rnaIII, sarA e 16S rrna As sondas específicas foram obtidas através do método da reação em
cadeia da polimerase (PCR; do inglês: polymerase chain reaction), utilizando-
se DNA total obtido da cepa RN6390B. Primers específicos foram utilizados
para amplificar fragmentos internos de DNA correspondentes a 212 pb para o
rnaIII, 460 pb para o sarA e de 368 pb para o 16S rrna. Os seguintes primers
foram utilizados: rnaIII (primer 1: 5´ catagcactgagtccaagga 3´ e primer 3:
5´ caatcggtgacttagtaaaatg 3´; desenhados a partir da seqüência com número
de acesso no GenBank X52543), sarA (primer SARAF1:
5´ gtatcatctatcaaacttcacc 3´ e primer SARAF2: 5´ ggcaaatgtatcgagcaagatg 3´;
desenhados a partir da seqüência com número de acesso no GenBank
NC_009782) e 16S rrna (primer F16S: 5´ aacgcattaagcactccgc 3´ e primer
R16S: 5´ gtgtgtagcccaaatcataa 3´; desenhados a partir da seqüência com
número de acesso no GenBank NC_009782). Para as respectivas reações de
amplificação os seguintes reagentes foram utilizados: mistura de dNTPs (dNTP
mix), para uma concentração final de 200 μM para cada dNTP; 10 pmol de
cada primer; 0,125 U TaqDNA polimerase (Gibco); 200 ng do DNA molde
(dosado espectrofotometricamente); 1,5 μM de MgCl2 e 5 μL do tampão da
enzima 10 vezes (10X) concentrado, em um volume final de 50 μL de reação.
Todas as amplificações acima foram realizadas no termociclador
GeneAmp PCR System 2.400 (PE Applied Biosystems), programado de acordo
com as seguintes etapas: pré-desnaturação a 94ºC/4 min (sem a adição da
Taq), posteriormente, adicionou-se a Taq e realizou-se 30 ciclos: desnaturação
(94ºC/30 s); anelamento (55ºC/30 s) e extensão (72ºC/1 min). Finalmente, foi
realizada uma extensão final (72ºC/4 min; INNIS et al., 1990).
3.4.1.3. Eletroforese do produto da amplificação Após amplificação, uma alíquota de cada amplicon, e do marcador de
corrida “low DNA mass ladder” (Invitrogen), foram adicionados,
separadamente, ao corante de corrida contendo glicerol (azul de bromofenol a

40
0,25% (p/v); xyleno-cianol FF a 0,25% (p/v); glicerol a 30% (p/v) em água) e os
DNA aplicados nas canaletas de um mini-gel de agarose preparado a 1,5% em
tampão TAE 1X (tris-acetato, 40 mM; EDTA 1 mM, pH 8,0). O gel foi,
posteriormente, tratado com brometo de etídio a 0,5 μg/mL e o DNA visualizado
com auxílio de um transiluminador de luz ultravioleta (UV) e fotografado com
filme Polaroid tipo 667 (SAMBROOK, FRITSCH & MANIATIS, 1989).
3.4.1.4. Purificação e marcação das sondas para o rnaIII, sarA e 16S rrna
A purificação do amplicon, diretamente a partir da reação de PCR, foi
realizada utilizando-se o kit “QIAquick PCR purification” (Qiagen), de acordo
com as recomendações do fabricante. O produto purificado foi marcado com o
kit “alkphos direct nucleic acid labelling and detection system” (Amersham
Biosciences do Brasil Ltda) de acordo com a recomendação do fabricante. Para
hibridização do RNA com as sondas marcadas foi utilizado o sistema acima,
conforme recomendação do fabricante (Amersham Biosciences do Brasil Ltda).
Finalmente, para detecção do sinal, a membrana foi exposta a um filme de
raios-X (X-OMAT-XK1; Kodak, São Paulo, Brasil), por pelo menos 15 min, e o
filme revelado automaticamente (X-OMAT/M20-BR; Kodak). Em alguns
experimentos de Northern-blotting, as sondas foram retiradas da membrana
através incubação em SDS (duodecil sulfato de sódio) 0,5% a 60°C, durante
60 min e de lavagem com Tris 100 mM (pH 8.0) por 5 min em temperatura
ambiente, e a membrana re-utilizada para novas hibridizações com outras
sondas.
3.5. Seqüenciamento dos loci agr e sarA Para o seqüenciamento completo do operon agr foram utilizadas 2
amostras pertencentes ao CEB (GV24 e GV69). O seqüenciamento parcial
(correspondente a região trailer do RNAIIagr) do lócus agr da amostra protótipo
CEB (agr-funcional) BMB9393 também foi realizado. Além disso, por
constatarmos erros na seqüência da amostra RN6390B, depositada no
GenBank, o seqüenciamento completo do operon agr desta amostra foi
também realizado. Para o sequenciamento das regiões contíguas upstream e
downstream aos promotores de sarA (sarP1, sarP2 e sarP3) foram utilizadas

41
as três amostras CEB acima. Para as reações de sequenciamento, o DNA
cromossômico das amostras foi extraído conforme descrito anteriormente no
item 3.4.1.1.
3.5.1. Desenho dos primers para a amplificação de fragmentos de DNA do operon agr e da região promotora de sarA
Inicialmente, os desenhos dos primers 1 (forward) e 2 (reverse) para a
amplificação do operon agr foram baseados nas seqüências do lócus agr das
cepas agr+ tipo I designadas RN6390B e RN4220, disponíveis no GenBank sob
os números de acesso X52543 e AF230358, respectivamente. Posteriormente,
foi feita uma análise visual das regiões que flanqueiam os fragmentos de DNA
a serem amplificados, visando-se escolher seqüências de nucleotídeos
variando entre 18 e 22 bases, com conteúdo de G e C em torno de 50% e
evitando-se seqüências repetitivas ou que pudessem vir a formar estruturas
secundárias. Uma vez realizada essa primeira análise da seqüência, os primers
desenhados foram analisados com o auxílio do programa Primer Analysis, da
Cybergene (http://www.cybergene.se/primer.html), visando-se à detecção de
palíndromos e hairpins.
Por se tratar de um operon com mais de 3.000 pb, foram necessários
desenhos de vários sets de primers para que todo o operon agr fosse
seqüenciado. Assim sendo, após o seqüenciamento da região dos promotores
P2 e P3 do operon agr (primeira região seqüenciada da fita 5’-3’), os primers
forwards para a continuação do seqüenciamento foram desenhados com base
nos eletrosferogramas resultantes do seqüenciamento de cada fragmento de
DNA amplificado. Os desenhos dos primers reverses para o seqüenciamento
da fita 3’-5’ foram, por sua vez, baseados nas seqüências do operon agr de
S. aureus tipo I, disponíveis no GenBank (X52543/RN6390B;
AF230358/RN4220 e AF210055/CMRSA-1), conforme especificado acima.
Os desenhos dos primers sarP1F (forward) e sarP1R (reverse), sarP2F
e sarP2R e sarP3F e sarP3R, que foram utilizados para a amplificação das
regiões contíguas upstream e downstream aos três promotores de sarA, sarP1,
sarP2 e sarP3, respectivamente, foram baseados na seqüência da cepa de
S. aureus RN6390B, disponível no GenBank sob o número de acesso U46541.

42
Todos os primers utilizados neste estudo e os seus respectivos números
de acesso no GenBank encontram-se descritos na tabela 4.
3.5.2. Desenho do programa de PCR (polymerase chain reaction) As amplificações das seqüências genômicas do operon agr e das
regiões promotoras de sarA foram realizadas como descrito anteriormente no
item 3.4.1.2. com uma única modificação: a temperatura programada de
anelamento variou de acordo com a Tm°C calculada para cada primer (tabela
4). A eletroforese dos produtos das amplificações e a purificação dos mesmos
foram realizadas como descrito anteriormente nos itens 3.4.1.3. e 3.4.1.4.,
respectivamente.
3.5.3. Seqüenciamento dos produtos de PCR purificados Os seqüenciamentos foram realizados através da técnica de primer
walking utilizando-se um seqüenciador automático MegaBACE 1000
(Amersham Biosciences). As reações de seqüenciamento foram realizadas de
acordo com o protocolo para o MegaBACE 1000, utilizando o DYEnamic ET
Dye Terminator Cycle Sequencing Kit (com Thermo Sequenase™ II DNA
Polimerase). As seqüências foram analisadas pelo software Sequence
Analyser utilizando o Base Caller Cimarron 3.12.
3.5.4. Edição das seqüências As seqüências oriundas dos seqüenciamentos realizados neste estudo
foram analisadas visualmente, comparando-se o nucleotídeo da seqüência
linear com o pico do gráfico do eletrosferograma correspondente, obtido
através do software Chromas (http://www.technelysium.com.au/chromas.html).
3.6.5. Alinhamento múltiplo das seqüências O alinhamento das seqüências correspondentes às regiões amplificadas
das amostras CEB, RN6390B e das seqüências de S. aureus não pertencente
ao CEB (RN9107 e D22, ambas agr tipo I, disponíveis no GenBank sob os
números de acesso DQ229854 e DQ157983, respectivamente) foi realizado
através do software CLUSTALW (THOMPSON, HIGGINS & GIBSON, 1994).

43
Posteriormente, foi realizada uma análise visual do alinhamento das
seqüências obtidas, visando-se o mapeamento de possíveis mutações nas
regiões amplificadas das amostras CEB, em relação às seqüências de
S. aureus não pertencente ao CEB previamente publicadas e a da amostra
RN6390B.
3.6. Modelagem da estrutura secundária da molécula de RNAm agr Para a predição da estrutura secundária da molécula de RNAII (RNA
mensageiro do lócus agr), através de modelagem computacional, foi utilizado o
software RNAstructure versão 4.2 © 1996-2004 (MATHEWS et al., 2004).

44
4. RESULTADOS
4.1. Produção de biofilme em microplaca de poliestireno
Uma vez que a produção de biofilme tem sido associada ao aumento da
persistência bacteriana no hospedeiro e à patogênese de diversas infecções
nosocomiais; e pelo fato de amostras CEB (ST239) estarem ampla e
mundialmente disseminadas em diversos hospitais, formulamos a hipótese de
que tais amostras poderiam apresentar uma superior produção de biofilme.
Para averiguarmos essa hipótese, foi realizada uma análise da formação
de biofilme sobre poliestireno inerte, sob condições estáticas, em 4 diferentes
populações de S. aureus compostas por 50 amostras pertencentes a variante
epidêmica do CEB (CEB A1), 50 variantes não-epidêmicas do CEB (CEB não-
A1), 49 amostras de MRSA pertencentes a clones não relacionados ao CEB
(clones esporádicos, CE) e 50 amostras de S. aureus sensíveis à meticilina
(MSSA). Todas as amostras utilizadas neste estudo encontram-se relacionadas
na tabela 4.
Após o cálculo das unidades de biofilme (UB; tabela 5), observamos que
a média dos valores das UB (UBmédia) para as amostras CEB A1, foi de 2,09,
sendo significativamente maior que as médias calculadas para as amostras de
MSSA (UBmédia de 0,5; p<0,001), clones esporádicos de MRSA (UB Bmédia de 0,6;
p<0,001) e outras amostras pertencentes a subclones CEB não-A1, (UBmédia de
0,6; p<0,001). Portanto, o valor da UBmédia obtido para a população de
amostras pertencentes ao CEB A1 foi de aproximadamente 3 a 4 vezes maior
que os observados para os três outros grupos de S. aureus analisados. A figura
3 apresenta uma fotografia do biofilme produzido sobre a superfície de
poliestireno, por amostras representantes dos 4 diferentes fenótipos: forte,
moderado, fraco e não produtor. Nesta podemos observar o grande acúmulo
de biofilme (fenótipo forte produtor) desenvolvido pela variante CEB A1,
comparável somente, em quantidade, ao biofilme formado pelos S. epidermidis.
Além disso, nesta mesma figura, encontra-se representada a distribuição das
proporções de cada população estudada versus o fenótipo de produção de
biofilme apresentado.
Os dados deste estudo demonstraram que somente 2% das amostras
CEB A1 foram classificadas como não produtoras de biofilme, enquanto que um

45
número significativamente maior de amostras não produtoras de biofilme (16%,
18,3% e 26%) foi observado dentre as variantes do CEB não-A1, as amostras
pertencentes aos clones esporádicos de MRSA e as amostras de MSSA,
respectivamente. Um dos fatores marcantes, que também contribuiu para o
incremento nos valores da UB para a população formada pelas amostras CEB
A1, foi que a maioria dessas amostras (74%) apresentou um fenótipo forte
produtor de biofilme (figura 3). Porém, foi observado que nas outras
populações de S. aureus analisadas, as amostras classificadas como
produtoras de biofilme apresentaram, predominantemente, um fenótipo de
biofilme fraco, com taxas de 54%, 35% e 50%, para CEB não-A1, clones de
MRSA esporádicos e MSSA, respectivamente (figura 3).
4.2. Screening da expressão de δ-hemolisina
Uma vez que o agr tem sido implicado na modulação de diversos fatores
de virulência de S. aureus, e que dados da literatura científica indicam que este
quorum-sensing regula tanto positivamente como negativamente diversos
fatores de virulência dos S. aureus, testamos a hipótese de que a atividade do
lócus agr na população de amostras CEB estudadas poderia se encontrar de
alguma forma alterada, uma vez que dentre os membros dessa população se
destacava uma variante com habilidade superior para produzir e acumular
biofilme (variante A1). Inicialmente, devido ao grande número de amostras a
serem analisadas, optamos por uma triagem rápida da produção de δ-
hemolisina (codificada pelo gene hld, cujo mRNA é co-transcrito na molécula
de RNAIII), utilizando ágar sangue de carneiro. Assim, a presença da δ-
hemolisina, significa que o agr está sendo expresso (ativo) e a diminuição ou
ausência de sua detecção poderia significar que o agr encontra-se inibido
(expressão do RNAIII atenuada ou ausente).
A hemólise causada pela δ-hemolisina pode ser claramente observada,
na placa de ágar sangue de carneiro, pela formação de uma zona hemolítica
que lembra uma ponta de seta, na interseção das estrias de semeaduras da
amostra teste (produtora de δ-hemolisina) com a RN4220 (produtora de β-
hemolisina). A forma em ponta de seta é proveniente do sinergismo entre a β-
hemolisina da amostra RN4220 e a δ-hemolisina da amostra teste (figura 4B).

46
Nas amostras que produzem α-hemolisina (regulada positivamente pelo
RNAIII), a formação de ponta de seta (devido ao sinergismo entre β- e δ-
hemolisinas) não é tão clara ou não se forma. Adicionalmente, a expressão de
α-hemolisina é positivamente regulada pelo agr e na ausência da atividade
deste a α-hemolisina não é expressa (figura 4B). Desta forma, a presença de
uma zona clara e estreita de hemólise ao redor da estria bacteriana é
considerada positiva para δ-hemolisina (mesmo quando não há formação de
ponta de seta). A β-hemólise se caracteriza por uma zona de hemólise mais
escura (turva) e, freqüentemente mais ampla, ao redor da estria bacteriana. Do
total das amostras CEB testadas para a produção de δ-hemolisina em placas
de AS, em 31% a δ- e α-hemolisinas não foram detectadas (figura 4A), ou seja,
não houve formação de ponta de seta ou de zona hemolítica estreita ao redor
da estria bacteriana teste, indicando que o sistema Agr deveria estar atenuado
ou totalmente inibido nessas amostras. É importante ressaltar que dentre as 31
amostras não δ-hemolíticas, apenas 3 (10%) eram representantes CEB tipo A1
e 28 (90%) CEB não-A1. E ainda, do total de 58 amostras CEB (A1 e não-A1),
apresentando um fenótipo de acumulação de biofilme forte ou moderado, 43
amostras (74%) expressaram normalmente a δ-hemolisina (tendo como
parâmetro de comparação as amostras protótipo BMB9393, apresentando uma
atividade de δ-hemólise em ponta de seta; e GV69, apresentando atividade de
δ-hemólise muito fraca ou em algumas vezes imperceptível). Tais dados
sugerem uma forte associação entre o caráter de biofilme forte/moderado e a
regulação positiva do agr (p<0,0001). Por outro lado, uma relação inversa foi
observada para muitas das amostras que possuíam o lócus agr inibido. Ou
seja, 62% das amostras nas quais a δ-hemólise estava inibida apresentaram
fenótipo de biofilme fraco/não produtor (p<0,0001; figura 5).
4.3. Experimentos de Northern-blotting utilizando sonda específica para o RNAIII Os resultados obtidos na triagem fenotípica para a avaliação da
atividade do lócus agr indicavam fortemente que certas representantes CEB
(majoritariamente as classificada no grupo CEB não-A1) apresentavam o lócus
agr inibido. Desta forma, de modo a confirmar os dados acima, 7

47
representantes das amostras estudadas (5 apresentado atividade de δ-
hemólise inibida e 2 apresentando atividade normal de δ-hemólise,
representada pela formação de ponta de seta na placa de AS) foram
escolhidas, aleatoriamente, para confirmação do funcionamento do lócus agr,
através dos experimentos de Northern-blotting. Além disso, utilizou-se a
amostra RN6390B (apresentando agr-funcional) como controle do experimento.
Todas as 5 amostras analisadas, cujo lócus agr parecia inibido,
apresentaram uma nítida diminuição da expressão do RNAIII, quando o sinal
quimioluminescente foi comparado com o das 2 outras amostras clínicas de
S. aureus (que apresentavam atividade normal de δ-hemólise) e com a amostra
RN6390B (figura 6A). Na figura 6B, a amostra BMB9393 (CEB A1),
apresentando uma atividade de δ-hemólise em ponta de seta, e a amostra
GV69 (CEB A1), apresentando atividade de δ-hemólise extremamente fraca ou
algumas vezes imperceptível, tiveram a expressão do lócus agr
genotipicamente comparadas, confirmando a atenuação da expressão do
RNAIII na GV69 em relação à expressão detectada para a BMB9393, que
transcreveu normalmente essa molécula reguladora (tendo como parâmetro de
comparação a expressão da sonda controle para o 16S rRNA, hibridizado na
mesma membrana). Desta forma, as amostras BMB9393 (CEB A1) e GV69
(CEB A1) foram escolhidas como protótipos neste estudo e colocadas lado a
lado nos experimentos de Northern-blotting e em outros experimentos deste
trabalho.
Os resultados dos experimentos de Northern-blotting claramente
demonstraram que a transcrição do RNAIII em algumas das amostras
estudadas estava atenuada, confirmando os dados obtidos através da triagem
em placas de ágar sangue, que indicavam que várias amostras CEB
(principalmente as amostras não-A1) apresentavam uma atenuação
significativa no lócus agr.
4.4. Seqüenciamento do lócus agr
A partir deste ponto, concentramos nossos estudos na tentativa de
compreendermos os mecanismos envolvidos na atenuação do lócus agr nas
amostras CEB. Inicialmente, partimos da hipótese de que tal atenuação poderia

48
ser devido a uma mutação no operon agr, levando a uma menor transcrição do
RNAIII. Para testarmos tal suposição, realizamos o seqüenciamento completo
dos loci agr de 2 representantes CEB A1 (GV24 e GV69), com o objetivo de
averiguarmos se a atenuação natural do agr observada nessas amostras, tanto
a partir do teste de triagem, quanto a partir dos experimentos de Northern-
blotting, seria devido a uma possível mutação pontual nesse lócus regulador
global. O agr da cepa RN6390B também foi completamente seqüenciado neste
estudo, uma vez que detectamos erros importantes na seqüência original
depositada no GenBank (número de acesso X52543). Inicialmente, verificamos
que as duas amostras CEB apresentavam um polimorfismo agr do tipo I.
Comparando essas seqüências, verificamos que as 2 amostras CEB
primeiramente seqüenciadas apresentavam uma mutação pontual (substituição
de guanina por timina) na região do DNA corresponde ao 3’ trailer do RNAII
policistrônico do agr, quando o alinhamento múltiplo dessas seqüências foi
realizado em conjunto com a seqüência da amostra RN6390B (apresentando
polimorfismo agr tipo I) e com seqüências completas do lócus agr das amostras
de MSSA RN9107 e D22 (agr tipo I) que encontram-se depositadas no
GenBank (números de acesso: DQ229854 e DQ157983, respectivamente). O
resultado desse alinhamento pode ser observado na figura 7.
Para averiguarmos se a mutação pontual encontrada no lócus agr seria
responsável por uma alteração conformacional na molécula de RNAIIagr, o que
poderia explicar a atenuação do lócus agr nas representantes CEB, realizamos
uma modelagem computacional da estrutura secundária da seqüência do
RNAII, através do software RNAstructure versão 4.2. Como resultado, a
estrutura secundária predita do RNAII para as amostras CEB apresentou uma
alteração significativa nos hairpins na região 3’ trailer do RNAII quando
comparada com a estrutura predita obtida a partir da seqüência da cepa
RN6390B (figura 8).
Posteriormente, a região 3’ do RNAII de outra representante CEB A1,
porém apresentando o agr funcional, a amostra protótipo BMB9393, também
teve o seu lócus agr parcialmente seqüenciado, com o intuito de averiguarmos
se essa mutação era, realmente, uma característica das variantes agr
atenuadas ou uma característica comum dessa linhagem. Após

49
seqüenciamento, edição e alinhamento, verificamos que a amostra BMB9393,
assim como as demais amostras CEB seqüenciadas, também apresentava a
mesma mutação pontual na região do DNA correspondente ao 3’ trailer do
RNAII (figura 7). Portanto, este resultado parecia indicar que tal mutação era
uma característica comum às amostras pertencentes a esta linhagem e, desta
forma, não poderia ser diretamente responsabilizada pela atenuação do lócus
agr observada nas amostras CEB GV24 e GV69.
4.5. Determinação do papel do sarA na atenuação do agr detectada nas amostras CEB
Como SarA é uma proteína reguladora transcricional que atua
positivamente na expressão do agr, se ligando nas regiões dos promotores P2
e P3 deste lócus, nossa próxima etapa foi testar a hipótese de que o sarA
poderia estar atenuado e, conseqüentemente, influenciando à expressão do
agr nessas amostras. Decidimos assim, verificar tal suposição através de
experimentos de Northern-blotting com uma sonda específica para sarA, e
utilizando RNA total das amostras CEB A1 protótipos: GV69 (agr-atenuada) e
BMB9393 (agr-funcional). Os resultados destes experimentos demonstraram,
claramente, uma reduzida expressão do mRNAsarA para a amostra GV69
quando comparados aos resultados da amostra BMB9393 (figura 6B). Nesta
mesma figura, pode-se observar ainda o ensaio controle com a sonda
específica 16S rrna, gene constitutivamente expresso nos S. aureus,
confirmando que o sinal mais fraco observado na GV69 não foi resultado de
uma aplicação de menor quantidade de RNA total na canaleta correspondente,
mas sim de uma diminuição na expressão do mRNAsarA, uma vez que
utilizamos a mesma membrana (após retirada da sonda) para realizar os
experimentos com a sonda para o 16S rrna. Assim, a figura 6B representa
fotografias de uma mesma membrana hibridizada, separadamente, com as
sondas específicas para o agr, sarA e 16S rrna, após retirada de cada sonda
através de fervura em SDS. Conseqüentemente, nossa suspeita de que uma
atenuação na expressão do sarA poderia estar envolvida no mecanismo de
atenuação do lócus agr foi confirmada (figura 6B).
Para averiguarmos se essa atenuação na transcrição do mRNAsarA
estava sendo causada por uma mutação no operon sar, ou se era causada por

50
um defeito upstream sar, realizamos o seqüenciamento das regiões contíguas
(upstream e downstream) aos 3 promotores sar nas três representantes CEB,
para as quais o agr havia sido seqüenciado: GV24, GV69 e BMB9393. Após o
alinhamento múltiplo das seqüências sar obtidas para as amostras CEB com as
seqüências das amostras de MSSA, RN9107 e D22, disponíveis no GenBank
(números de acesso DQ229854 e DQ157983, respectivamente), não foi
evidenciada nenhuma alteração na região abrangendo os promotores de sar
nas seqüências CEB das amostras analisadas.

51
5. DISCUSSÃO
Os Staphylococcus aureus são patógenos que apresentam um potencial
de variabilidade genética, relativamente, elevado, o que contribuiu para sua
rápida adaptação ao hospedeiro e capacidade para causar doenças
(KENNEDY et al., 2008). Esta característica marcante pôde ser, repetidamente,
verificada através da emergência de cepas que adquiriram, de forma
excepcionalmente rápida, mecanismos de resistência a virtualmente quase
todos os agentes antimicrobianos, logo após a introdução dos mesmos na
prática médica (OLIVEIRA, TOMASZ & DE LENCASTRE, 2002; NOTO et al.,
2008). O MRSA ST239-SCCmecIIIA, denominado clone epidêmico brasileiro
(CEB), teve seu amplo poder de disseminação (através de distâncias de
dimensões continentais) reconhecido pela primeira vez, na literatura científica,
em um estudo de nosso grupo que relatou seu espalhamento em hospitais
localizados de norte a sul do Brasil (TEIXEIRA et al. 1995). Posteriormente, foi
relatado por Edgeworth e colaboradores (EDGEWORTH et al., 2007) que este
clone era geneticamente semelhante a um clone anteriormente detectado na
Inglaterra, na década de 1980, o qual naquela época era denominado de
EMRSA-1 (MARPLES, RICHARDSON & DE SAXE, 1986). Atualmente
amostras de MRSA ST239 vêm sendo detectadas em hospitais de todo o
mundo (JEMILI-BEN, BOUTIBA-BEN & BEN, 2006; DEURENBERG et al.,
2007). Este clone, que parece descender da recombinação de duas linhagens
dominantes, ST8 e ST30 (ROBINSON & ENRIGHT, 2004), faz parte de um
grupo seleto de clones específicos de MRSA pandêmicos que se encontram
amplamente disseminados, em diversos hospitais dos 5 continentes
(DEURENBERG et al., 2007), sendo este, na atualidade, o clone de MRSA
mais disseminado no mundo.
Foi sugerido, anteriormente, que o sucesso de clones pandêmicos de
MRSA, como patógenos hospitalares, seria reflexo de sua rápida evolução,
frente à pressão exercida pela utilização intensa e indiscriminada de
antimicrobianos em todo o mundo (DAY et al., 2001). Porém, além do fenótipo
de multirresistência, os S. aureus apresentam outras características notáveis,
como a capacidade de sobreviver por longos períodos em superfícies

52
inanimadas (KRAMER, SCHWEBKE & KAMPF, 2006) e de colonizar e/ou
infectar vários nichos no hospedeiro, incluindo pele, membranas mucosas,
sangue e vários outros tecidos e órgãos, inclusive os vitais. Devido a essas e
outras propriedades, esses microrganismos são hoje responsáveis por uma
série de doenças, desde infecções de pele, toxemias, e até doenças sistêmicas
extremamente severas (ARCHER, 1998; TENOVER & GAYNES, 2000;
NOVICK, 2003). Porém, o papel dos diferentes fatores de virulência no
desenvolvimento das infecções estafilocócicas permanece até o momento por
ser totalmente esclarecido.
Pelo fato do ST239 ser, no momento, o clone pandêmico de MRSA mais
disseminado, e o biofilme estar sendo apontado como um importante fator de
virulência em infecções hospitalares, principalmente nas associadas aos
biopolímeros, decidimos testar a hipótese de que as amostras CEB poderiam
estar acumulando mais biofilme do que outras amostras de MRSA. Neste
estudo desenvolvemos então um modelo experimental para avaliar a habilidade
desses estafilococos para desenvolver biofilme. O modelo escolhido foi o da
formação do filme bacteriano sobre poliestireno inerte, em condições estáticas
de crescimento bacteriano, em presença de glicose. Este foi selecionado por
ser o teste de triagem indicado para se avaliar um número elevado de amostras
(CASSAT, LEE & SMELTZER, 2007). Além disso, a maioria dos estudos
anteriores, publicados, sobre desenvolvimento do biofilme em S. aureus, não
detectou diferenças significativas no biofilme formado, sob condições de
crescimento bacteriano estáticas (sem troca do meio de cultura) ou de fluxo
(com fluxo contínuo de meio de cultura novo), in vivo ou in vitro (BEENKEN et
al. 2004; TOLEDO-ARANA et al. 2005; O’NEILL et al. 2008). Os resultados
aqui apresentados demonstraram que a maioria das amostras CEB testadas foi
capaz de produzir biofilme (91%). Por outro lado, biofilme foi também uma
característica comum entre as outras amostras de MRSA (82%) e de MSSA
(74%) analisadas. Essas elevadas taxas de produção de biofilme observadas
em S. aureus devem, certamente, contribuir não somente na patogênese das
infecções associadas à biomateriais, como também para uma maior
sobrevivência desses microrganismos sobre superfícies inanimadas, como
mobiliário e utilitários hospitalares, equipamentos e materias de uso médicos,
como por exemplo, um simples estetoscópio (HILL, KING & DAY, 2006). Em

53
estudos anteriores de nosso grupo foram detectadas taxas de produção de
biofilme de 76% para uma coleção de amostras hospitalares de S. epidermidis,
enquanto que amostras isoladas da comunidade apresentavam uma
percentagem significativamente menor, de cerca de 60% (DE ARAÚJO et al.,
2006). Assim, a partir do presente estudo, podemos concluir que a produção de
biofilme (em amostras hospitalares) de S. aureus é uma característica tão (ou
mais) comum quanto a observada em amostras de S. epidermidis.
É importante ressaltar aqui que a variante CEB A1 apresentou um
significante aumento na acumulação do biofilme (3-4 vezes maior) em relação
às outras amostras analisadas (p<0.001). Assim, é razoável supor que tal
propriedade poderia ser, pelo menos em parte, responsável pela ampla
propagação dessa linhagem pandêmica em hospitais de todo o mundo. Chama
atenção o fato de amostras de MRSA relacionadas ao ST239 terem sido
relatadas como agentes de infecções nosocomiais na Alemanha, Argélia,
Argentina, Austrália, Áustria, Brasil, Chile, China, Cingapura, Coréia, Eslovênia,
Espanha, EUA, Finlândia, Grécia, Holanda, Índia, Indonésia, Itália, Mongólia,
Polônia, Portugal, Reino Unido, República da Geórgia, República Tcheca, Sri
Lanka, Suécia, Tailândia, Tunísia, Uruguai e Vietnam (CAMPANILE et al.,
2001; JEMILI-BEN, BOUTIBA-BEN & BEN, 2006; DEURENBERG et al., 2007;
BARTELS et al., 2008). Um estudo recente de Edgeworth e colaboradores
(2007) demonstrou que pacientes que adquiriram infecções causadas por uma
variante (TW) do ST239, em um hospital do Reino Unido, apresentaram maior
risco de desenvolver bacteremia (47% vs. 13%; p<0.001) e de apresentar
cultura positiva para MRSA, a partir de ponta de cateter (59% vs. 26%;
p<0.001). Porém, foi observada entre tais pacientes menor chance de
colonização nasal por TW (p<0.001). Não sabemos se TW corresponde a
variante A1 de nosso estudo (a qual apresentou elevada capacidade para
acumular biofilme), mas de qualquer modo, nos parece lógico supor que uma
maior acumulação do filme biológico desta variante ST239 possa ter sido o
mecanismo responsável pela sua superior capacidade para causar bacteremia
(EDGEWORTH et al., 2007). Portanto, um experimento interessante seria
testar a hipótese de que essas duas variantes (A1 e TW) se tratam de um
mesmo indivíduo. A maioria das amostras utilizadas em nosso estudo não

54
estava envolvida em infecções na corrente sangüínea e, portanto, a associação
entre produção de biofilme e bacteremias não pôde ser aqui avaliada.
A versatilidade bacteriana depende de um amplo conjunto de genes
acessórios, muitos dos quais envolvidos na patogênese, e que são
responsáveis pela adaptação da bactéria ao ambiente hostil do hospedeiro. Os
S. aureus produzem uma gama de fatores de virulência, tantos associados a
sua superfície como liberados para o meio extracelular. Apesar de sabermos
que a patogênese das infecções estafilocócicas é multifatorial, como dissemos
anteriormente, pouco se conhece com relação aos principais mecanismos
associados a essas doenças (PROJAN & NOVICK, 1997; WESSON et al, 1998; JARRAUD et al., 2000; NOVICK, 2000; GRUNDMANN et al., 2006). Em
estudo anterior, evidenciamos que amostras CEB A1 apresentavam maior
habilidade para aderir e colonizar a superfície de células epiteliais brônquicas
humanas. Nossos dados comprovaram que a invasão maciça promovida pelas
amostras CEB A1, nas células epiteliais respiratórias 16HBEo–, foi paralela a
maior capacidade dessas amostras de se ligar à fibronectina humana
imobilizada, através de um mecanismo integrina RGD-dependente. Assim, é
possível que a capacidade do MRSA de se ligar eficientemente à fibronectina e
invadir, maciçamente, células epiteliais respiratórias humanas aumente a sua
habilidade para persistir no organismo do hospedeiro, contribuindo,
conseqüentemente, para o estado de portador e aumento da morbidade
(AMARAL et al., 2005). Dados recentes de O'Neill e colaboradores
demonstraram que as FnBPAB parecem também estar envolvidas na produção
de biofilme ica-independente, glicose-induzido, tanto em amostras de MRSA
como em amostras de MSSA (O'NEILL et al., 2008). Assim, nos parece
provável que uma maior expressão de FnBPAB pelas amostras CEB A1
poderia ser responsável, pelo menos em parte, pela elevada acumulação de
biofilme observada neste estudo. Seria interessante se estudos fossem
realizados para se comprovar tal hipótese. Apesar desses dados recentes, a
composição do biofilme ica-independente dos S. aureus não se encontra ainda
totalmente elucidada, uma vez que outras moléculas protéicas têm sido
também apontadas como mediadoras deste biofilme (O’NEILL et al., 2008). Resumindo, nossos resultados (AMARAL et al. 2005) representam os
primeiros dados experimentais, registrados na literatura científica, demostrando

55
que, dentro de uma mesma linhagem de MRSA, variantes epidêmicas,
apresentando características específicas associadas à virulência do
microrganismo, podem prevalescer, expressando, de modo diferencial, fatores
que conferem propriedades vantajosas para sua sobrevivência, persistência e
disseminação global.
Estudos envolvendo genômica comparativa, entre as amostras
protótipos GV69 (biofilme moderado; agr atenuado) e BMB9393 (biofilme forte;
agr funcional) realizados pelo nosso grupo (dados não publicados), através da
metodologia de microarrays da Affymetrix, demonstraram que essas duas
variantes apresentavam, basicamente, o mesmo core genômico e que as
principais diferenças entre essas amostras estavam relacionadas a elementos
móveis (principalmente bacteriófagos), à presença de cls1 (gene que codifica
para cardiolipina sintetase) somente na amostra BMB9393 e, ainda, em relação
a diferenças relacionadas à presença de alguns genes hipotéticos. É possível
que esses elementos móveis possam criar regiões de instabilidade no
cromossomo desses microrganismos gerando mutações que afetariam a
expressão de determinados genes, ou ainda, poderiam ocorrer mutações
pontuais em genes de virulência ou em seus reguladores que resultariam na
virulência diferenciada observada entre essas duas variantes.
De modo similar, em um amplo estudo sobre genômica comparativa dos
MRSA, através de microarrays, foi também demonstrado que a principal
diferença entre as amostras estudadas encontrava-se relacionada à presença
ou ausência de determinados bacteriófagos, inclusive em amostras dentro de
uma mesma linhagem (LINDSAY et al., 2006). Da mesma forma, ao realizar um
estudo de genômica comparativa, através de microarrays, com 16 amostras
ST239 (variante TW), aqueles autores, como nós, não detectaram genes que
poderiam estar especificamente ligados a maior patogenicidade da variante
epidêmica (EDGEWORTH et al., 2007). Recentemente, Kennedy e
colaboradores (2008) realizaram estudos de genômica comparativa, através do
seqüenciamento completo de 10 amostras representantes do clone de CA-
MRSA, conhecido como USA300, amplamente disseminado nos EUA. Esses
autores demonstraram que 8 das 10 amostras analisadas apresentaram
poliformismo de apenas um nucleotídeo (SNP; do inglês: single nucleotide
polymorphism), o que indica divergência recente. Inexplicavelmente, duas

56
dessas amostras apresentaram reduzida mortalidade em um modelo animal de
sepse, comparada com a cepa de referência (p = 0,0002). Tais dados em
conjunto corroboram a idéia de que diferenças genéticas mínimas no genoma
dos MRSA poderiam exercer profunda influência sobre a expressão da
virulência (KENNEDY et al., 2008).
No Brasil, e em outros países, incluindo os Estados Unidos,
aproximadamente 60% dos S. aureus envolvidos em casos de infecção
hospitalar são MRSA (GALES et al., 2000; NNIS system, 2004). Paralelamente
ao aumento dos procedimentos invasivos, na medicina moderna, tem sido
observado um aumento significativo no número de infecções nosocomiais
associadas ao biofilme, pois parte significativa dessas doenças está associada
ao uso de cateteres e proteses médicas. Do mesmo modo, mais recentemente,
detectou-se um aumento significativo nos índices de mortalidade em
bacteremias causadas pelos MRSA (ALLARD et al., 2008; LIBERT et al.,
2008). Quadros de endocardites associadas a bacteremias causadas por
MRSA (com taxas variando entre 12% e 31% dos casos analisados) também
foram relatados em uma recente revisão publicada sobre o assunto
(BAMBERGER, 2007). A capacidade dos S. aureus de produzirem biofilme
sobre tais dispositivos médicos implantáveis é, sem dúvida, um grave problema
de saúde pública global, e resolver tal questão nos parece um dos maiores
desafios a serem enfrentados pela humanidade, para os próximos anos, com
relação às infecções nosocomiais.
Segundo Cramton e colaboradores, o lócus ica, responsável pela
biossíntese de PIA/PNAG (adesina intercelular polissacarídica/poli-N-acetil-
glicosamina), presente no genoma dos S. aureus e dos S. epidermidis, seria
essencial para a produção de biofilme, uma vez que a deleção do ica na cepa
de S. aureus, ATCC 35556, inibiu o desenvolvimento dessa estrutura em
superfícies de poliestireno, sob condições estáticas (CRAMTON et al., 1999).
Entretanto, estudos posteriores demonstraram que a biossíntese do biofilme
induzido por glicose em S. aureus pode ocorrer de forma independente de ica.
Mutantes obtidos por inserção de transposon, isogênicos à cepa de S. aureus
parental MT23142 (uma derivada da cepa NCTC 8325-4), desenvolveram um
mecanismo alternativo, ica-independente, para a produção de biofilme sob
condições estáticas, na ausência do sistema de dois componentes ArlRS

57
(FOURNIER & HOOPER, 2000). Desenvolvimento de biofilme ica-
independente também pôde ser observado, mais recentemente, quando O’Neill
e colaboradores, em 2007, utilizando mutantes icaADBC-deletados não
detectaram efeito algum dessa deleção sobre a produção de biofilme induzido
pela glicose em 6 amostras de MRSA. Entretanto, os mesmos autores
verificaram a completa interrupção na biosíntese de biofilme para 4 amostras
de MSSA (O’NEILL et al., 2007). Neste mesmo estudo, o tratamento do
biofilme ica-independente com proteases foi capaz de eliminar completamente
o biofilme formado, sugerindo uma composição protéica, o que também foi
confirmado por estudo recente realizado por nós com amostras CEB A1
(COELHO et al., 2008).
Boles e Horswill, em 2008, em estudos utilizando um mutante Δica::tet,
derivado do S. aureus SH1000, verificaram que a inibição do agr aumentava o
biofilme ica-independente nesta amostra e que sua ativação, no biofilme pré-
formado, levava à dispersão (destacamento) do biofilme. Como os autores
detectaram, no sobrenadante do biofilme, um paralelo aumento da serina
protease (regulada positivamente pelo agr), sugeriram que este mecanismo
poderia estar envolvido na dispersão do biofilme mediada pelo agr. Além disso,
observaram que o duplo mutante nos genes agr-regulados aur e splABCDEF
(codificando para metalo e serina proteases, respectivamente) apresentou uma
diminuição da atividade proteásica, aumento da acumulação e diminuição do
destacamento do biofilme (BOLES & HORSWILL, 2008). Diversas proteínas,
tanto reguladas positivamente pelo agr, tais como as α- (Hla) e δ-hemolisinas
(Hld; VUONG et al., 2000; CAIAZZA & O’TOOLE, 2003; ADHIKARI et al.,
2007), como as reguladas negativamente, incluindo as proteínas de ligação à
fibronectina A e B (FnBPAB; O'NEILL et al., 2008), têm sido implicadas na
formação/acumulação do biofilme glicose-induzido, ica-independente. Foi
demonstrado, recentemente, que na realidade é o pH moderadamente ácido,
induzido pelo crescimento na presença de glicose, o fator que induz este tipo
de biofilme dos S. aureus (O'NEILL et al., 2008). Porém, esses estudos são
ainda incipientes e mais dados são necessários para a elucidação da(s)
mólecula(s)-chave envolvida(s) no desenvolvimento do biofilme ica-

58
independente pelos S. aureus e suas implicações na patogênese das infecções
causadas pelos MRSA.
Foi observado, previamente, que um mutante nulo para o gene hla,
derivado da cepa NCTC8325-4, foi incapaz de colonizar eficientemente
superfícies plásticas, tanto sob condições estáticas como de fluxo (CAIAZZA &
O'TOOLE, 2003). Recentemente, Yarwood e colaboradores demonstraram o
surgimento de colônias variantes agr-inibidas, no ambiente do biofilme,
apresentando a expressão de vários fatores de virulência alterados, inclusive
uma diminuição da expressão de hla (YARWOOD, et al., 2007). Sabe-se que a
transcrição do hla, assim como a de outros genes codificadores de toxinas
estafilocócicas, é positivamente regulada pelo RNAIII (molécula efetora do
lócus agr). Em um outro estudo interessante, realizado com um modelo animal
de abscesso, foi verificado que a expressão do quorum-sensing Agr podia
variar dependendo da localização da infecção, nos diferentes nichos do animal
(VAN WAMEL et al., 2002).
Outra hemolisina estafilocócica, a δ-hemolisina, foi associada ao
biofilme. Foi sugerido que as propriedades surfactantes desta hemolisina,
codificada pelo hld, o qual se encontra sobreposto na região do transcrito
RNAIII, poderiam estar envolvidas no aumento do biofilme formado por um
mutante designado mut6, obtido por inserção de transposon no lócus agr,
(VUONG et al., 2000). Mais recentemente, estudos realizados por O’Neill e
colaboradores (2008) demonstraram que a perda da sortase (enzima
responsável pelo ancoramento ao peptídeoglicana das proteínas apresentando
o sinal LPXTG) reduziu o fenótipo de biofilme na construção srtA nocaute de
S. aureus (SRTA1), uma derivada da amostra SH1000, a qual, por sua vez, é
uma derivada da NTCT8325-4 apresentando um sigB funcional. Tais dados
corroboram com uma natureza protéica do biofilme de S. aureus. Além disso,
os autores demonstraram que a introdução de dupla mutação em fnbAB
(codificando para as FnBPA e B) reduziu severamente a formação de biofilme
na construção fnbA::tetM, fnbB::ermA, designada DU5883, também derivada
da NTCT8325-4. Adicionalmente, observaram que a promoção do filme
bacteriano pelas FnBP ocorreu em nível de acumulação e não de adesão
primária ao polímero. Mutação em fnbA ou fnbB, isoladamente, não afetou,

59
substancialmente, a expressão do biofilme e, a complementação do duplo
mutante fnbAB, através de plasmídio carreando fnbA ou fnbB, restaurou a
expressão do biofilme. Os autores ainda demonstraram que a formação do
biofilme promovida pelas FnBP era dependente de SarA, não dependia dos
domínios de ligação à fibronectina ou ao fibrinogênio, e que mutações em fnBP
não tiveram efeito sobre as amostras de MSSA analisadas (O’NEILL
et al., 2008). Porém, como as FnBPs são reguladas negativamente pelo Agr,
seria de se esperar que a atenuação do agr levasse ao aumento da
acumulação do biofilme e não à diminuição. Em nossos estudos, na amostra
agr-atenuada GV69 o sarA também encontra-se atenuado, o que poderia
também estar contribuindo para a diminuição do biofilme nesta amostra.
Estudos recentes de Boles e Horswill (2008) demonstraram que SarA inibe a
ação de certas proteases bacterianas, inibindo a dispersão do filme biológico e
aumentando assim sua acumulação. Ao contrário, a inibição de sarA levaria a
um aumento da dispersão, levando, conseqüentemente, à diminuição do
biofilme. Outras proteínas de superfície, como a proteína Bap (presente em 5%
dos S. aureus bovinos; LASA & PANEDÉS, 2006) e a proteína SasG de
S. aureus (Aap – homóloga de S. epidermidis; CORRIGAN et al., 2007), têm
sido implicadas no desenvolvimento de biofilme dos S. aureus.
A proteína SasG está presente na superfície dos S. aureus sob a forma
de fibrilas peritríqueas (CORRIGAN et al., 2007). Através de estudos de
aderência, utilizando microscopia de transmissão, verificamos que fibrilas
peritríqueas estão claramente envolvidas na aderência da variante CEB A1,
através de um delicado contato às extremidades de não mais que três
microvilosidades da célula brônquica humana (AMARAL et al., 2005). A
presença do gene sasG foi também detectada em amostras CEB A1, através
de experimentos de microarrays realizados, previamente, pelo nosso grupo
(Lócus Affymetrix: SA2505; dados não publicados). Assim é possível que tal
proteína possa estar também implicada, tanto no aumento da adesividade,
como da acumulação de biofilme observado em amostras CEB A1. Não existe
até o momento na literatura científica, que seja do nosso conhecimento,
nenhum estudo sobre a regulação gênica da proteína SasG.
Acredita-se que o sucesso dos S. aureus como patógenos deve-se,
dentre outros fatores, à fina regulação, coordenada de forma harmônica e

60
global, de um vasto repertório de sistemas genéticos, responsáveis pela
modulação de um conjunto de fatores de virulência. Dentre esses reguladores
podemos destacar o agr, sar, sigB, arl e sae (NOVICK et al., 2000; SAID-
SALIM et al., 2003; BRONNER, MONTEIL & PRÉVOST, 2004). Assim, parece
lógica a suposição de que a atenuação, ou maior expressão, desses sistemas
reguladores globais, em amostras clínicas, possa representar um importante
mecanismo para a adaptação bacteriana ao hospedeiro e assim contribuir para
o sucesso de clones específicos de MRSA (ou mesmo de variantes de um
mesmo clone), apresentando um maior potencial patogênico.
Um dos primeiros estudos sobre a formação de biofilme induzido por
glicose, sob superfície de poliestireno, foi publicado em 2000, na Alemanha.
Nessa investigação ficou confirmada, estatisticamente, a forte correlação entre
o genótipo agr-nulo das cepas RN6911(Δagr::tetM) e mut6 (mutante por
inserção do transposon no gene agrC), com o aumento da formação de
biofilme por estas amostras, em comparação às cepas selvagens isogênicas
(VUONG et al., 2000). Contudo, em um estudo publicado por Beenken e
colaboradores, em 2003, nos EUA, utilizando 8 construções agr-nulas, o agr
não apresentou efeito relevante sobre a formação do biofilme, exceto em uma,
a amostra RN6911 (BEENKEN et al., 2003). Similarmente, no mesmo ano,
também foi demonstrado, por outro grupo na Espanha, que deleções no agr,
em duas cepas de S. aureus, 15981 e v329, não resultaram em alteração
importante na formação de biofilme glicose-induzido, em condições de fluxo ou
estáticas, e ainda indicaram a participação da proteína SarA na ativação da
síntese de biofilme em S. aureus (VALLE et al., 2003).
Como pode ser observado, pesquisadores têm estudado os mecanismos
moleculares e genéticos envolvidos no desenvolvimento do biofilme pelos
S. aureus, mas, apesar do envolvimento, pelo menos em parte, do sistema de
quorum-sensing Agr, descrito por alguns, os dados relatados são
extremamente contraditórios. Uma das hipóteses para explicar tais
controvérsias foi levantada por Yarwood e colaboradores que demonstraram
que a modulação da formação do biofilme de S. aureus, promovida pelo agr,
poderia divergir sob diferentes condições ambientais (YARWOOD et al., 2004).
É importante ressaltar, que na maioria desses estudos, envolvendo agr e
biofilme, poucas cepas foram utilizadas, sendo uma grande parte destas

61
oriunda de construções genéticas derivadas da cepa NCTC8325-4, incluindo a
RN6911, que possui uma mutação natural devido a uma deleção de 11pb no
gene rsbU (codificador de um regulador positivo do σB; PALMA & CHEUNG,
2001), além de uma mutação nonsensing no agrA (ADHIKARI et al., 2007).
Mais recentemente, O’Neill e colaboradores, ainda na tentativa de
elucidar o papel do agr na formação no filme biológico de S. aureus,
observaram um pequeno impacto do quorum-sensing Agr na formação de
biofilme, uma vez que mutações nesse lócus apresentaram aumento
(>2 vezes) na produção de biofilme em apenas 5 dos 21 mutantes agr-nocaute
(23%) analisados (O’NEILL et al., 2007). Esses mesmos autores sugeriram que
SarA possuia um papel mais relevante no desenvolvimento do biofilme em
S. aureus (O’NEILL et al., 2007). A importância do sarA na formação de
biofilme também já havia sido relatada nos estudos de Beenken e
colaboradores, citados acima. Esses autores demonstraram que se sarA fosse
deletado, isoladamente ou simultaneamente ao agr, nas amostras de S. aureus
analisadas observava-se uma redução na capacidade de formação de biofilme.
Desta forma foi concluído que mutações no sarA seriam responsáveis pela
inibição do fenótipo de síntese de biofilme, tanto em condições estáticas,
quanto de fluxo (BEENKEN et al., 2003). Como SarA é um inibidor de
proteases, é possível que a deleção de sarA leve a um destacamento do
biofilme e, conseqüentemente, a diminuição de sua acumulação (BOLES &
HORSWILL, 2008). Estudos recentes de nosso laboratório (COELHO et al., 2008), utilizando
14 construções nocaute Δagr::tetM derivadas de amostras clínicas circulantes
na atualidade, e construções complementadas com plasmídio contendo rnaIII,
indicaram que o agr afetou, significativamente, a acumulação do biofilme de
S. aureus (MRSA ou MSSA), podendo este efeito ser tanto positivo, como
negativo, ou ainda neutro. Demonstramos ainda que tal divergência foi
independente da linhagem estudada, mas que podia ser predita pela
quantidade de biofilme acumulado pela amostra selvagem isogênica;
indicando, assim, a natureza multifatorial do biofilme; ou seja, que mecanismos
diversos devem estar envolvidos na sua formação/acumulação.

62
Os estudos de Boles & Horswill (2008), com um mutante agr nulo
derivado da amostra SH1000, demonstraram que a construção apresentando o
agr deletado possuia uma maior propensão para acumulação de biofilme,
enquanto a amostra selvagem apresentava maior tendência para dispersão,
sugerindo que o agr teria um papel importante no destacamento do biofilme,
por ativar certas proteases bacterianas, degradando a matriz proteinácea do
filme biológico (BOLES & HORSWILL, 2008). Ao contrário, no estudo aqui
apresentado verificamos que a maior acumulação de biofilme, na variante CEB
A1, estava associada, com extrema significância, à funcionalidade do lócus agr
(p<0.0001). Paralelamente, as amostras CEB não-A1, as quais acumulavam
menos biofilme, apresentaram o agr inibido (p<0.0001). Nossos resultados
sugerem que 31% das amostras CEB analisadas apresentaram atenuação
natural do lócus agr e que esta inibição parece ser determinada geneticamente,
uma vez que a maioria das amostras CEB, agr atenuadas, foi classificada
como CEB-não A1. Os resultados divergentes entre nossos dados e os obtidos
pelos autores acima poderiam ser explicados pelo fato daqueles autores terem
utilizado somente um mutante agr, derivado do S. aureus SH1000, ou ainda,
pela ativação e maior expressão de determinadas proteases nas amostras CEB
não-A1 pela concomitante atenuação de sarA, como observado na amostra
GV69. Esta última proposição nos parece instigante e interessante e, portanto,
deveria também ser testada. Cabe ressaltar que diferenças importantes na
expressão de proteases foram detectadas, através de estudos de microarrays,
realizados com as amostras GV69 e BMB9393 (dados não publicados). Como o agr regula negativamente FnBP, nos parece, em uma primeira
análise, que a regulação de FnBP pelo agr não poderia ser o mecanismo chave
na acumulação do biofilme observado nas amostras CEB A1; pois se assim
fosse, as amostras agr atenuadas deveriam apresentar uma capacidade de
biofilme aumentada em relação ao CEB A1, e não diminuida como foi aqui
demonstrado. Corroborando com esta hipótese, dados recentes de nosso
laboratório (COELHO et al., 2008) indicaram que a inibição ou deleção do agr
em amostras que acumulam fortemente o biofilme, incluindo o CEB A1,
diminuiram significativamente a expressão do filme biológico; apesar de não ter
eliminado totalmente o mesmo. Como não existem estudos sobre o papel do

63
agr na expressão da proteína SasG não podemos avaliar uma possível
influência do RNAIII (na modulação de SasG) no fenômeno aqui observado.
A circulação de amostras clínicas agr-inibidas foi verificada também por
Fowler e colaboradores (2004) que descreveram uma associação significativa
entre amostras de S. aureus isoladas de bacteremias persistentes e inibição da
atividade da δ-hemolisina, sugerindo uma disfunção no lócus agr. Os autores
observaram que tais amostras compartilhavam o mesmo perfil de PFGE e
maior resistência às atividades microbicidas de proteínas plaquetárias
induzidas pela trombina (FOWLER et al., 2004). Esses resultados reforçam os
dados obtidos em nossos estudos e parecem demonstrar que a detecção de
amostras clínicas apresentando agr atenuado não parece ser um fenômeno
isolado e que poderia, portanto, possuir uma significativa importância clínica.
Os ensaios de triagem para a detecção da atividade da δ-hemolisina e
Northern-blotting para a análise da transcrição do RNAIII aqui realizados,
confirmaram que indivíduos dentro de uma mesma população clonal (clone
epidêmico brasileiro), apresentando backgrounds genéticos bastante
semelhantes, podem apresentar alterações importantes na expressão do agr
(principal quorum-sensing dos S. aureus). Estudos anteriores realizados por
Papakyriacou e colaboradores (PAPAKYRIACOU et al., 2000) demonstraram
que um clone de MRSA (CMRSA-3) envolvido em infecções hospitalares no
Canadá, no final da década de 1990 e início da de 2000, parecia estar mais
associado à infecção do que à colonização, quando comparado a outras cepas
de MRSA. Semelhantemente aos resultados por nós obtidos, estudando uma
única representante deste clone, aqueles autores verificaram que esta possuia
o agr naturalmente atenuado e um aumento na expressão da atividade de
ligação à fibronectina humana. Traber e Novick, demonstraram, em 2006, que
a ausência da produção de α- e δ-hemolisinas na cepa RN4220 (uma cepa
laboratorial derivada da cepa clínica RN450) ocorreu devido a uma mutação na
extremidade 3’ do gene agrA, que levou a uma alteração do tipo frameshift,
ocasionando a adição de 3 resíduos aminoacil à região C-terminal da proteína
AgrA. Foi relatado ainda, que essa mutação não inativaria o lócus agr, mas
atrasaria sua ativação em 2-3h, resultando em falhas das traduções das α- e δ-
hemolisinas. Esses autores concluíram então, que a ocorrência dessa mutação

64
pode não se tratar apenas de um fenômeno laboratorial, mais do que isso,
poderia representar um mecanismo natural de modulação da atividade do agr
(TRABER & NOVICK, 2006). Porém, em nosso estudo, demonstramos que a
menor expressão do RNAIII foi paralela a uma expressão diminuida de SarA
(proteína reguladora global da transcrição dos S. aureus). Entretanto, o(s)
defeito(s) responsável(is) por tais alterações não estavam localizados no agr e
nem nas regiões promotoras do operon sar, mas sim em uma região upstream ao sar. Estudos sobre a expressão gênica global através de microarrays
(Affymetrix), utilizando cDNA das protótipos BMB9393 e GV69, realizados
anteriormente pelo nosso grupo (dados não publicados), revelaram que não
somente as expressões de sarA e do agr estavam afetadas na amostra CEB A1
agr-atenuada GV69, como também a expressão de um inibidor de sarA,
designado sarR. Esses dados sugerem que o aumento da expressão de sarR
na fase estacionária de crescimento, observado na GV69, foi o mecanismo
regulador responsável pela atenuação do sarA e, conseqüentemente, do agr,
observada nas amostras CEB. SarR parece atuar se ligando à região
promotora sarA, através de seu promotor P1, para regular negativamente sua
transcrição, e assim reduzir a expressão da proteína SarA (HSIEH, TSENG &
STEWART, 2008).
De modo semelhante ao fenômeno aqui descrito, a seleção experimental
no ambiente de biofilme, de variantes de uma cepa de S. aureus apresentando
inibição no agr, foi relatada por Yarwood e colaboradores (YARWOOD et al.,
2007). Esses autores também verificaram que membros da família gênica sar
(sar homólogos) estavam envolvidos na atenuação observada no lócus agr
nessas variantes. Foi observado naquele estudo que a transcrição de sarU
(regulador positivo indireto do agr, por se ligar à região promotora de sarT,
regulador negativo do agr) estava também diminuida nas amostras agr-
atenuadas. Esses dados, em conjunto com os nossos resultados, sugerem que
a atenuação natural do agr, em amostras clínicas, ocorre de forma indireta,
devido à intervenção de outros fatores upstream a este quorum-sensing via sar
homólogos. De qualquer modo, uma vez que tanto o agr quanto o sarA são loci
reguladores globais da virulência dos S. aureus, o padrão de acumulação de
biofilme, observado na variante CEB-A1, deve estar relacionado, pelo menos
em parte, com as diferenças observadas entre essa variante e as variantes

65
CEB não-A1, quanto ao perfil de expressão desses e de outros genes
reguladores, como o sarA e o sarR.
Day e colaboradores, em um interessante estudo sobre evolução clonal
em S. aureus, demonstraram que a variante predominante de um determinado
complexo clonal (CC), definido por tipagem de seqüenciamento de multilócus
enzimáticos (MLST), era sempre apontada (em um modelo de algoritmos
matemáticos) como o ancestral predito, quando diversos CC de S. aureus
foram estudados. Além disso, seus estudos indicaram que amostras
relacionadas aos clones patogênicos de S. aureus apresentavam maior
capacidade de colonização nasal do que as amostras relacionadas a clones
isolados da microbiota anfibiôntica da narina. Esses autores sugeriram que as
variantes predominantes dos clones patogênicos eram, dentre os membros da
população clonal, as que retinham maior potencial de aderência às células
nasais (DAY et al., 2001). Desta forma, nossos resultados estão de acordo com
os estudos de Day e colaboradores e levantam a possibilidade que a variante
CEB A1 possa ter sido a ancestral da população CEB estudada. Em conjunto,
esses resultados sustentam a hipótese que o domínio de clones de S. aureus
resistentes a uma grande variedade de drogas, em um cenário epidemiológico
particular, parece resultar não somente da pressão seletiva exercida pelo uso
continuado e intenso de antimicrobianos, mas também, provavelmente, das
influências promovidas pelas associações dinâmicas dos diferentes fatores
envolvidos na co-evolução bactéria-hospedeiro, o que poderia culminar na
seleção de patógenos melhor adaptados (condicionados) para causar
infecções nosocomiais. Assim, parece provável que, de tempos em tempos, ao
longo da história da evolução dos S. aureus, cepas com maior capacidade de
virulência possam emergir.
Concluindo, o sucesso que temos observado de alguns clones
específicos, hospitalares e pandêmicos de MRSA, como no caso do ST239,
poderia ser creditado a uma superior capacidade de colonização; além da
aquisição da resistência aos β-lactâmicos conferir uma vantagem óbvia no
ambiente nosocomial, em relação aos demais competidores sensíveis. Em
adição a maior acumulação de biofilme aqui observada, dados anteriores,
obtidos por nós, demonstraram que a variante CEB predominante (A1)

66
apresenta maior adesividade e invasibilidade para células epiteliais
respiratórias humanas (AMARAL et al., 2005).
Assim, é possível que diferentes combinações de características
genotípicas e fenotípicas poderiam produzir a variabilidade necessária para a
expressão de determinados conjuntos de fatores bacterianos (apropriados para
a colonização e infecção), nos diferentes nichos do hospedeiro.
Adicionalmente, tais características poderiam contribuir para a epidemicidade
de determinados clones pandêmicos, ou mesmo de variantes dentro de um
mesmo clone. Dados obtidos por Edgeworth e colaboradores (2007),
demonstrando uma nítida associação entre o desenvolvimento de bacteremia e
a detecção de uma variante do ST239, designada TW (EDGEWORTH et al.,
2007), corroboram com esta hipótese e, juntamente com os dados obtidos a
partir de nossos estudos sobre a virulência experimental diferenciada do CEB
(AMARAL et al., 2005), representam os primeiros dados da literatura científica
indicando que certos clones de MRSA podem ter evoluído de forma a se tornar
patógenos nosocomiais de sucesso. Outras implicações seriam as de que
estes resultados parecem demonstrar que, pelo menos em certos casos, o
MLST pode não possuir amplo significado epidemiológico (TURNER & FEIL,
2007), uma vez que variantes de um mesmo clone podem se tornar
epidêmicas, sendo inclusive responsáveis por surtos de dimensões
continentais.
O ônus para esta acelerada evolução dos MRSA, que estamos
testemunhando, na atualidade, se volta para os governos e autoridades de
saúde, que necessitam criar sistemas eficazes de monitoramento desses
clones multirresistentes e responsáveis por surtos pandêmicos; visando
organizar e promover dados para o pronto controle de sua disseminação, bem
como para o estabelecimento de protocolos de tratamento eficazes das
doenças a eles associadas. Por outro lado, a grande responsabilidade e
desafio dos profissionais da saúde estaria em exercer um controle rígido e
eficaz da disseminação desses clones no ambiente hospitalar, de forma que os
hospitais possam continuar sendo locais onde as expectativas sejam de cura e
não de aquisição de mais doenças.

67
Tabela 4. Primers utilizados nos seqüenciamentos parciais do operon agr e das regiões contíguas upstream e downstream aos promotores de sarA de cepas pertencentes ao CEB e da RN6390.
Primers Forward (5’–3’) Reverse (5’–3’) TaºC* N°. acesso GenBank
Promotores agrP2P3
catagcactgagtccaagga agacatgcacatggtgcacat 56 X52543; AF230358
Gene agrB taaatcgtataatgacagtgag ggtgttatgtagaaagtattat 52 X52543; AF230358; AF210055
Gene agrD ttgcaagtacgattagggatg gagagtgtgatagtaggtgg 55 X52543; AF230358
Gene agrC (1º set)
aacattggtaacatcgcagc cgactatcttactgcttac 51 X52543; AF210055
Gene agrC (2º set)
agcttttttgtagtttatgtgac gtcacagtattggtattattctt 55 X52543; AF210055
Gene agrC (3º set)
tcgagaagatgacatgcctg tggaaattgccctcgcaact 55 X52543
Gene agrA (1º set)
caactagccataaggatgtga gtggcagtaattcagtgtatg 55 X52543
Gene agrA (2º set)
agttgcagcgatggattttatt gcactgtatagctggcttttt 55 X52543
Promotores sarP2
tgggtagtatgctttgacaca ctttaacgtgctatagattggt 55 U46541
Promotores sarP3
ctttaacgtgctatagattggt gcgctaaatcgtttcattaaata 55 U46541
Promotores sarP1
gcgctaaatcgtttcattaaata gggcaaatgtatcgagcaag 55 U46541
*Ta°C, temperatura de anelamento dos primers forward e reverse para a amplificação através de PCR dos fragmentos de DNA que foram seqüenciados.
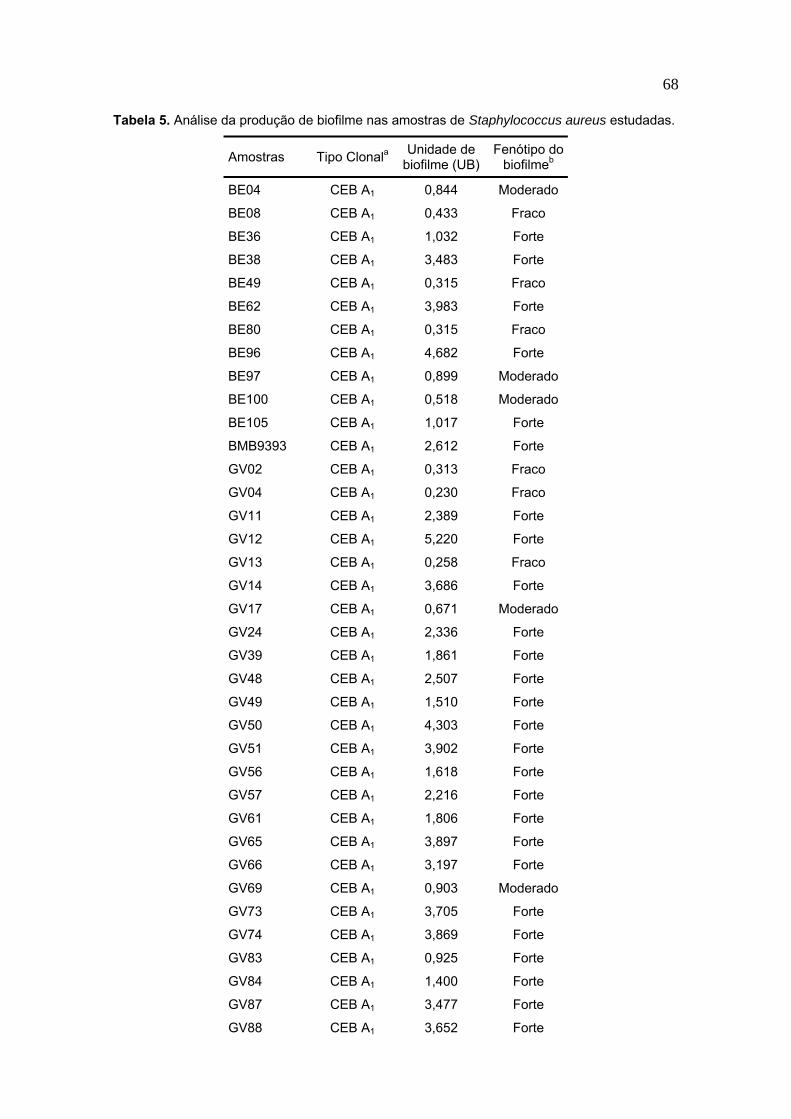
68
Tabela 5. Análise da produção de biofilme nas amostras de Staphylococcus aureus estudadas.
Amostras Tipo Clonala Unidade de biofilme (UB)
Fenótipo do biofilmeb
BE04 CEB A1 0,844 Moderado
BE08 CEB A1 0,433 Fraco
BE36 CEB A1 1,032 Forte
BE38 CEB A1 3,483 Forte
BE49 CEB A1 0,315 Fraco
BE62 CEB A1 3,983 Forte
BE80 CEB A1 0,315 Fraco
BE96 CEB A1 4,682 Forte
BE97 CEB A1 0,899 Moderado
BE100 CEB A1 0,518 Moderado
BE105 CEB A1 1,017 Forte
BMB9393 CEB A1 2,612 Forte
GV02 CEB A1 0,313 Fraco
GV04 CEB A1 0,230 Fraco
GV11 CEB A1 2,389 Forte
GV12 CEB A1 5,220 Forte
GV13 CEB A1 0,258 Fraco
GV14 CEB A1 3,686 Forte
GV17 CEB A1 0,671 Moderado
GV24 CEB A1 2,336 Forte
GV39 CEB A1 1,861 Forte
GV48 CEB A1 2,507 Forte
GV49 CEB A1 1,510 Forte
GV50 CEB A1 4,303 Forte
GV51 CEB A1 3,902 Forte
GV56 CEB A1 1,618 Forte
GV57 CEB A1 2,216 Forte
GV61 CEB A1 1,806 Forte
GV65 CEB A1 3,897 Forte
GV66 CEB A1 3,197 Forte
GV69 CEB A1 0,903 Moderado
GV73 CEB A1 3,705 Forte
GV74 CEB A1 3,869 Forte
GV83 CEB A1 0,925 Forte
GV84 CEB A1 1,400 Forte
GV87 CEB A1 3,477 Forte
GV88 CEB A1 3,652 Forte
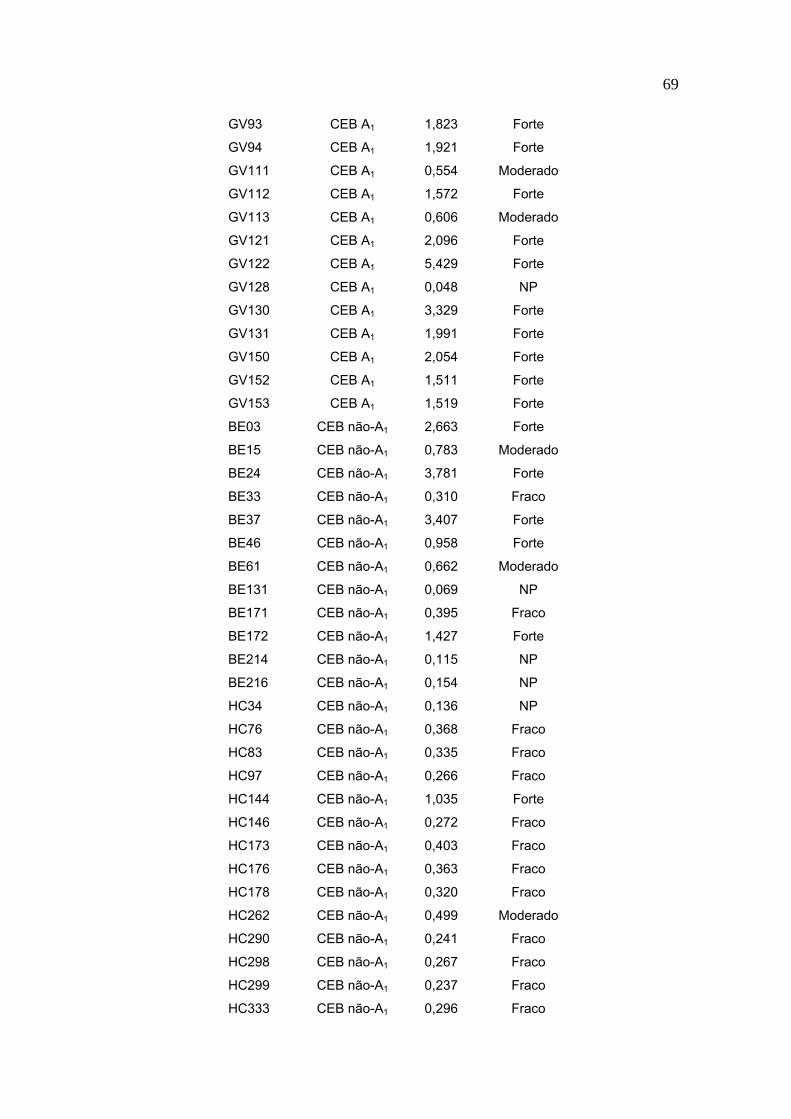
69
GV93 CEB A1 1,823 Forte
GV94 CEB A1 1,921 Forte
GV111 CEB A1 0,554 Moderado
GV112 CEB A1 1,572 Forte
GV113 CEB A1 0,606 Moderado
GV121 CEB A1 2,096 Forte
GV122 CEB A1 5,429 Forte
GV128 CEB A1 0,048 NP
GV130 CEB A1 3,329 Forte
GV131 CEB A1 1,991 Forte
GV150 CEB A1 2,054 Forte
GV152 CEB A1 1,511 Forte
GV153 CEB A1 1,519 Forte
BE03 CEB não-A1 2,663 Forte
BE15 CEB não-A1 0,783 Moderado
BE24 CEB não-A1 3,781 Forte
BE33 CEB não-A1 0,310 Fraco
BE37 CEB não-A1 3,407 Forte
BE46 CEB não-A1 0,958 Forte
BE61 CEB não-A1 0,662 Moderado
BE131 CEB não-A1 0,069 NP
BE171 CEB não-A1 0,395 Fraco
BE172 CEB não-A1 1,427 Forte
BE214 CEB não-A1 0,115 NP
BE216 CEB não-A1 0,154 NP
HC34 CEB não-A1 0,136 NP
HC76 CEB não-A1 0,368 Fraco
HC83 CEB não-A1 0,335 Fraco
HC97 CEB não-A1 0,266 Fraco
HC144 CEB não-A1 1,035 Forte
HC146 CEB não-A1 0,272 Fraco
HC173 CEB não-A1 0,403 Fraco
HC176 CEB não-A1 0,363 Fraco
HC178 CEB não-A1 0,320 Fraco
HC262 CEB não-A1 0,499 Moderado
HC290 CEB não-A1 0,241 Fraco
HC298 CEB não-A1 0,267 Fraco
HC299 CEB não-A1 0,237 Fraco
HC333 CEB não-A1 0,296 Fraco
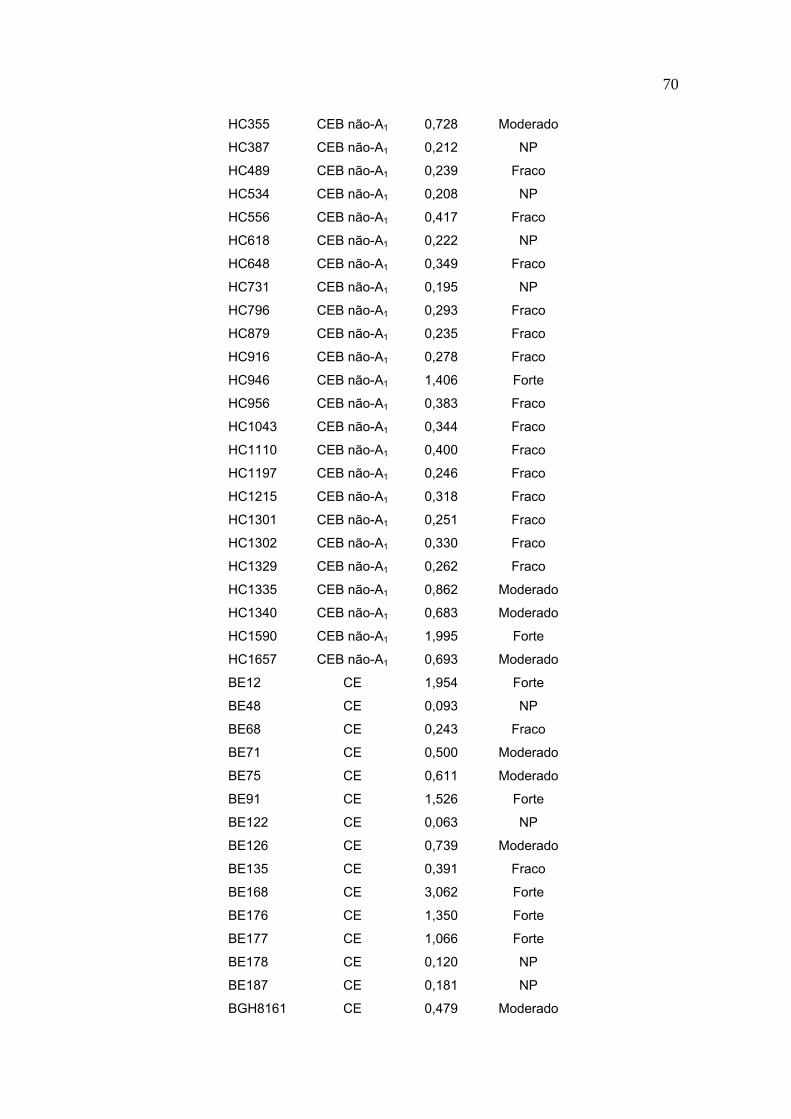
70
HC355 CEB não-A1 0,728 Moderado
HC387 CEB não-A1 0,212 NP
HC489 CEB não-A1 0,239 Fraco
HC534 CEB não-A1 0,208 NP
HC556 CEB não-A1 0,417 Fraco
HC618 CEB não-A1 0,222 NP
HC648 CEB não-A1 0,349 Fraco
HC731 CEB não-A1 0,195 NP
HC796 CEB não-A1 0,293 Fraco
HC879 CEB não-A1 0,235 Fraco
HC916 CEB não-A1 0,278 Fraco
HC946 CEB não-A1 1,406 Forte
HC956 CEB não-A1 0,383 Fraco
HC1043 CEB não-A1 0,344 Fraco
HC1110 CEB não-A1 0,400 Fraco
HC1197 CEB não-A1 0,246 Fraco
HC1215 CEB não-A1 0,318 Fraco
HC1301 CEB não-A1 0,251 Fraco
HC1302 CEB não-A1 0,330 Fraco
HC1329 CEB não-A1 0,262 Fraco
HC1335 CEB não-A1 0,862 Moderado
HC1340 CEB não-A1 0,683 Moderado
HC1590 CEB não-A1 1,995 Forte
HC1657 CEB não-A1 0,693 Moderado
BE12 CE 1,954 Forte
BE48 CE 0,093 NP
BE68 CE 0,243 Fraco
BE71 CE 0,500 Moderado
BE75 CE 0,611 Moderado
BE91 CE 1,526 Forte
BE122 CE 0,063 NP
BE126 CE 0,739 Moderado
BE135 CE 0,391 Fraco
BE168 CE 3,062 Forte
BE176 CE 1,350 Forte
BE177 CE 1,066 Forte
BE178 CE 0,120 NP
BE187 CE 0,181 NP
BGH8161 CE 0,479 Moderado
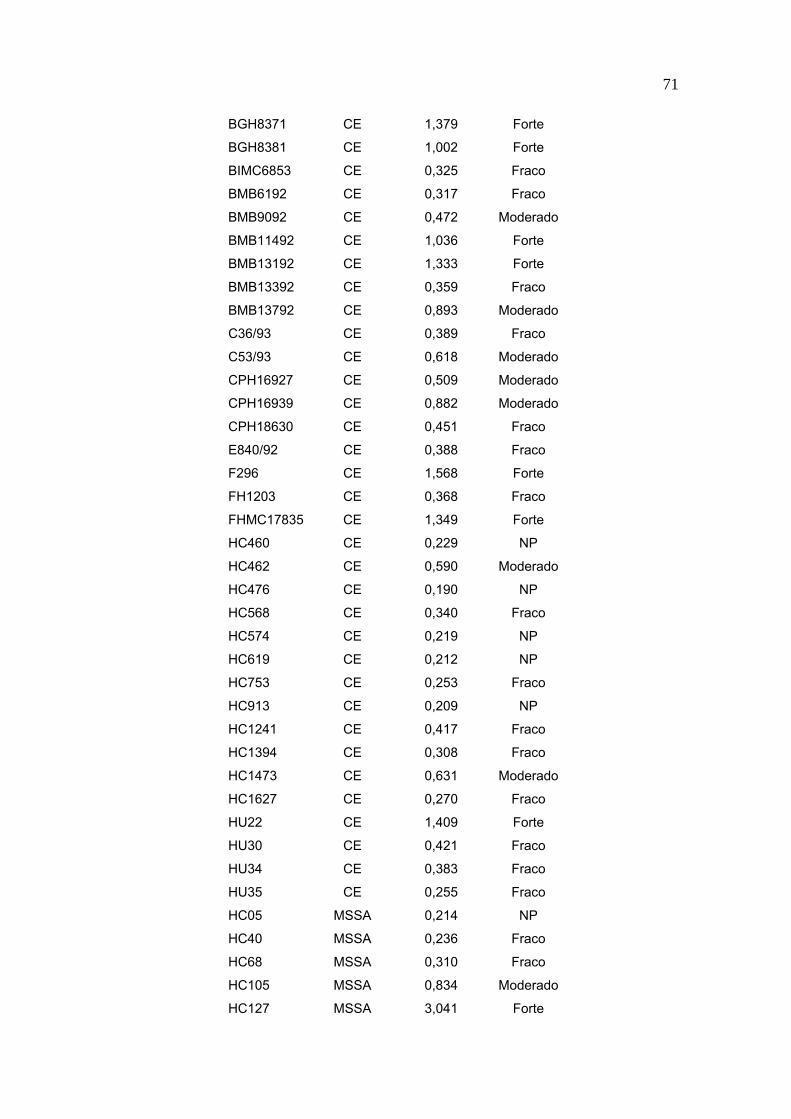
71
BGH8371 CE 1,379 Forte
BGH8381 CE 1,002 Forte
BIMC6853 CE 0,325 Fraco
BMB6192 CE 0,317 Fraco
BMB9092 CE 0,472 Moderado
BMB11492 CE 1,036 Forte
BMB13192 CE 1,333 Forte
BMB13392 CE 0,359 Fraco
BMB13792 CE 0,893 Moderado
C36/93 CE 0,389 Fraco
C53/93 CE 0,618 Moderado
CPH16927 CE 0,509 Moderado
CPH16939 CE 0,882 Moderado
CPH18630 CE 0,451 Fraco
E840/92 CE 0,388 Fraco
F296 CE 1,568 Forte
FH1203 CE 0,368 Fraco
FHMC17835 CE 1,349 Forte
HC460 CE 0,229 NP
HC462 CE 0,590 Moderado
HC476 CE 0,190 NP
HC568 CE 0,340 Fraco
HC574 CE 0,219 NP
HC619 CE 0,212 NP
HC753 CE 0,253 Fraco
HC913 CE 0,209 NP
HC1241 CE 0,417 Fraco
HC1394 CE 0,308 Fraco
HC1473 CE 0,631 Moderado
HC1627 CE 0,270 Fraco
HU22 CE 1,409 Forte
HU30 CE 0,421 Fraco
HU34 CE 0,383 Fraco
HU35 CE 0,255 Fraco
HC05 MSSA 0,214 NP
HC40 MSSA 0,236 Fraco
HC68 MSSA 0,310 Fraco
HC105 MSSA 0,834 Moderado
HC127 MSSA 3,041 Forte
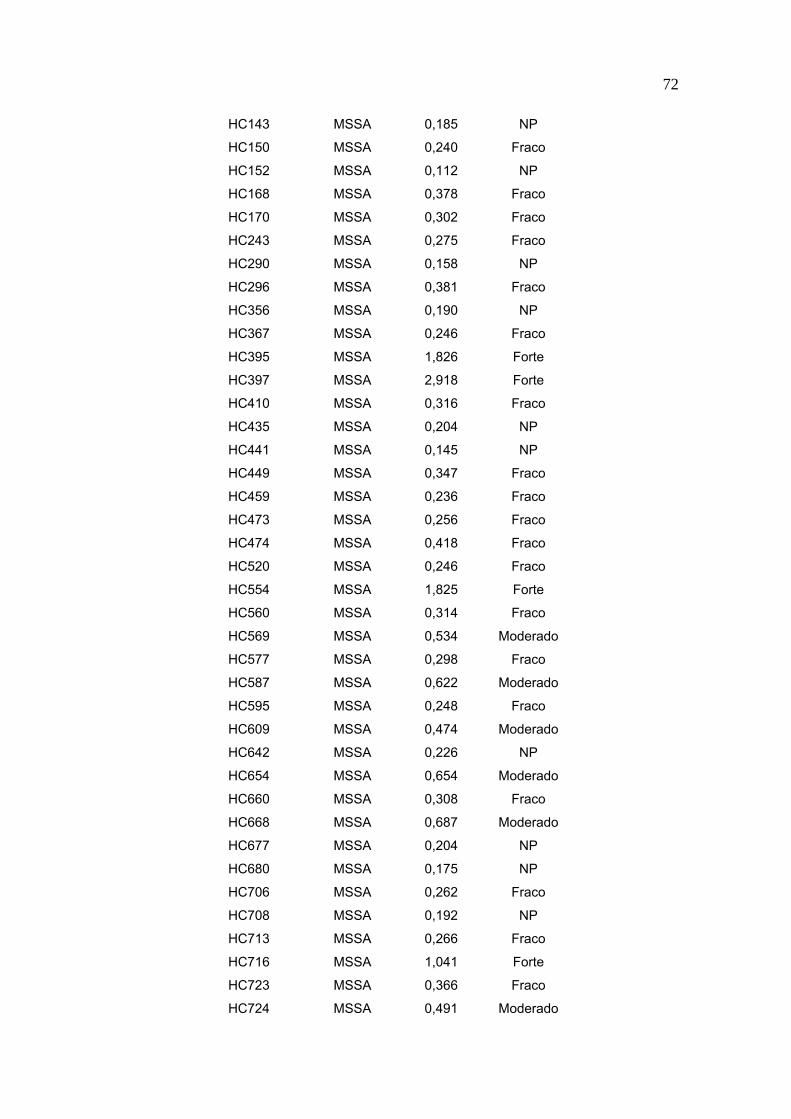
72
HC143 MSSA 0,185 NP
HC150 MSSA 0,240 Fraco
HC152 MSSA 0,112 NP
HC168 MSSA 0,378 Fraco
HC170 MSSA 0,302 Fraco
HC243 MSSA 0,275 Fraco
HC290 MSSA 0,158 NP
HC296 MSSA 0,381 Fraco
HC356 MSSA 0,190 NP
HC367 MSSA 0,246 Fraco
HC395 MSSA 1,826 Forte
HC397 MSSA 2,918 Forte
HC410 MSSA 0,316 Fraco
HC435 MSSA 0,204 NP
HC441 MSSA 0,145 NP
HC449 MSSA 0,347 Fraco
HC459 MSSA 0,236 Fraco
HC473 MSSA 0,256 Fraco
HC474 MSSA 0,418 Fraco
HC520 MSSA 0,246 Fraco
HC554 MSSA 1,825 Forte
HC560 MSSA 0,314 Fraco
HC569 MSSA 0,534 Moderado
HC577 MSSA 0,298 Fraco
HC587 MSSA 0,622 Moderado
HC595 MSSA 0,248 Fraco
HC609 MSSA 0,474 Moderado
HC642 MSSA 0,226 NP
HC654 MSSA 0,654 Moderado
HC660 MSSA 0,308 Fraco
HC668 MSSA 0,687 Moderado
HC677 MSSA 0,204 NP
HC680 MSSA 0,175 NP
HC706 MSSA 0,262 Fraco
HC708 MSSA 0,192 NP
HC713 MSSA 0,266 Fraco
HC716 MSSA 1,041 Forte
HC723 MSSA 0,366 Fraco
HC724 MSSA 0,491 Moderado

73
HC742 MSSA 0,133 NP
HC752 MSSA 0,387 Fraco
HC764 MSSA 0,312 Fraco
HC921 MSSA 0,270 Fraco
HC1017 MSSA 0,262 Fraco
HC1080 MSSA 0,065 NP
aCEB A1: amostras de MRSA do CEB apresentando o perfil de PFGE predominante (A1); CEB não-A1: amostras de MRSA pertencentes a subclones do CEB (perfil de PFGE diferindo de 1 a 6 bandas do perfil do CEB A1); CE: amostras de MRSA pertencentes a clones esporádicos (raramente detectados em hospitais e com perfil de PFGE diferindo em mais de 6 bandas do perfil do CEB A1); MSSA: amostras de S. aureus susceptíveis à meticilina. bFenótipo do biofilme: classificação do fenótipo de produção de biofilme com base na comparação da maior leitura da UB do biofilme (0,115) da amostra de S. pyogenes 75194. NP: amostras de S. aureus não produtoras de biofilme (UB ≤ 0,230); Fraco: amostras de S. aureus fracamente produtoras de biofilme (UB ≥ 0,231 e ≤ 0,459); Moderado: amostras de S. aureus moderadamente produtoras de biofilme (UB ≥ 0,460 e ≤ 0,919); Forte: amostras de S. aureus fortemente produtoras de biofilme (UB ≥ 0,920).

74
Figura 3. Distribuição das amostras de acordo com o fenótipo de biofilme detectado e fotografia dos diferentes fenótipos produzidos sobre superfície de poliestireno inerte.
0%
10%
20%
30%
40%
50%
60%
70%
80%
CE CEB não-A1
Per
cent
agem
de
amos
tras
CEB A1MSSA
Fraco
Forte Moderado
S. epidermidis Branco
Não produtor
MSSA: amostras de S. aureus susceptíveis à meticilina; CE: amostras de MRSA pertencentes a clones esporádicos (raramente detectados em hospitais e com perfil de PFGE diferindo em mais de 6 bandas do perfil do CEB A1); CEB não-A1: amostras de MRSA pertencentes a subclones do CEB (perfil de PFGE diferindo de 1 a 6 bandas do perfil do CEB A1); CEB A1: amostras de MRSA do CEB apresentando um perfil de PFGE predominante (A1). Branco: TSB suplementado com glicose a 1%; S. epidermidis cepa 70D: controle positivo. Fenótipos do biofilme descritos como forte, moderado, fraco e não produtor foram atribuídos de acordo com o cálculo das Unidades de Biofilme (UB). A ilustração representativa dos fenótipos de biofilme detectados (no canto direito da figura) corresponde à acumulação de biofilme das amostras CEB A1 (GV88, forte; GV17, moderado; GV02, fraco e GV128, não produtor).
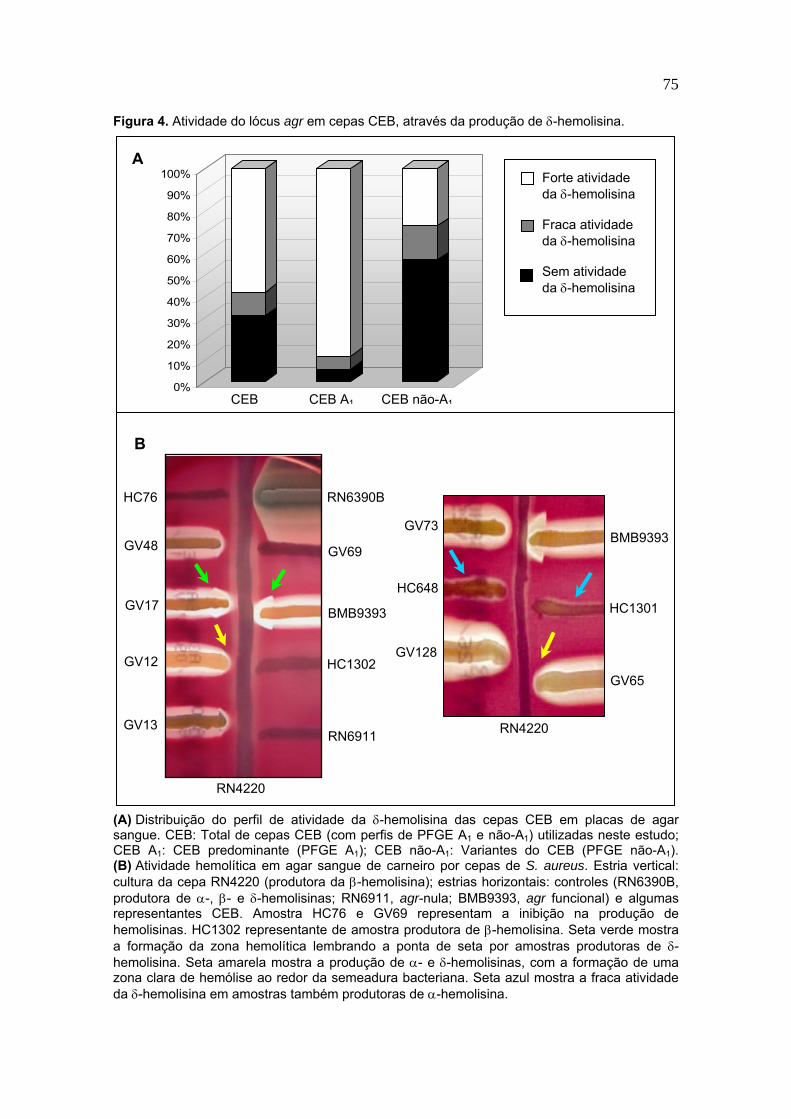
75
Figura 4. Atividade do lócus agr em cepas CEB, através da produção de δ-hemolisina.
57,2%
5,8% 5,8%
RN6390B
GV69
BMB9393
HC1302
RN6911GV13
GV12
GV17
GV48
HC76
RN4220
GV73
HC648
GV128
BMB9393
HC1301
GV65
RN4220
B
AForte atividade da δ-hemolisina Fraca atividade da δ-hemolisina Sem atividade da δ-hemolisina
CEB A1 CEB não-A1
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
CEB
(A) Distribuição do perfil de atividade da δ-hemolisina das cepas CEB em placas de agar sangue. CEB: Total de cepas CEB (com perfis de PFGE A1 e não-A1) utilizadas neste estudo; CEB A1: CEB predominante (PFGE A1); CEB não-A1: Variantes do CEB (PFGE não-A1). (B) Atividade hemolítica em agar sangue de carneiro por cepas de S. aureus. Estria vertical: cultura da cepa RN4220 (produtora da β-hemolisina); estrias horizontais: controles (RN6390B, produtora de α-, β- e δ-hemolisinas; RN6911, agr-nula; BMB9393, agr funcional) e algumas representantes CEB. Amostra HC76 e GV69 representam a inibição na produção de hemolisinas. HC1302 representante de amostra produtora de β-hemolisina. Seta verde mostra a formação da zona hemolítica lembrando a ponta de seta por amostras produtoras de δ-hemolisina. Seta amarela mostra a produção de α- e δ-hemolisinas, com a formação de uma zona clara de hemólise ao redor da semeadura bacteriana. Seta azul mostra a fraca atividade da δ-hemolisina em amostras também produtoras de α-hemolisina.

76
Figura 5. Associação entre a atividade do lócus agr, através da produção de δ-hemolisina, e o fenótipo de acumulação de biofilme em cepas CEB (PFGE A1 e não-A1).
Produtor de biofilme forte ou moderado
Produtor de biofilme fraco ou não produtor
agr funcional agr inibido
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
agr funcional: o nível de expressão do lócus agr foi avaliado através da detecção da atividade da δ-hemolisina nas amostras CEB A1 e não-A1 estudadas. agr inibido: representa a expressão atenuada, ou completamente inibida do lócus agr, através da detecção de pouca ou de nenhuma δ-hemolisina produzida pelas amostras CEB estudadas.

77
Figura 6. Experimentos de Northern-blotting utilizando RNA total de amostras de S. aureus e sondas específicas para os loci agr, sarA e 16S rrna.
GV69 BMB9393
rnaIII
sarA
16Srrna
B
rnaIII
A
1 2 3 4 5 6 7 8
(A) Northern-blotting utilizando uma sonda específica para uma região interna do rnaIIIagr. Números de 1 a 5: amostras representativas do CEB apresentando uma atenuação do agr. Números 6 e 7: amostras clínicas de MSSA apresentando o agr funcional. Número 8: amostra controle RN6390B com agr-funcional. (B) Northern-blotting utilizando sondas específicas para uma região interna do rnaIIIagr, sarA e 16S rrna. GV69 e BMB9393 (protótipos das variantes CEB A1 apresentando agr-atenuado e -funcional; respectivamente).

78
Figura 7. Alinhamento múltiplo das seqüências da região 3’ trailer do RNAIIagr demonstrando uma mutação pontual nas amostras CEB (GV69 e BMB9393) em comparação com outras amostras de S. aureus (RN6390B, RN9107 e D22).
A seqüência das amostras de S. aureus RN6390B, RN9107 e D22 estão disponíveis no GenBank sob os números de acesso X52543, DQ229854 e DQ157983, respectivamente. Destaque em cinza: mutação pontual com a substituição de uma guanina (G) por uma timina (T). Os asterísticos e os pontos representam o consenso e a continuidade das seqüências, respectivamente.

79
Figura 8. Estrutura secundária da molécula de RNAIIagr predita por análise computacional.
Representação das estruturas secundárias da molécula de RNAIIagr predita para as cepas CEB GV24, GV69 e BMB9393 (A) e para a cepa RN6390B (B). Representação, em maior aumento, das áreas alteradas das estruturas secundárias da molécula de RNAIIagr predita paras as cepas CEB (C) e para a cepa RN6390B (D).

80
6. BIBLIOGRAFIA
ADHIKARI, R.P.; ARVIDSON, S. & NOVICK, R.P. A nonsense mutation in agrA accounts for the defect in agr expression and the avirulence of Staphylococcus aureus 8325-4 traP::kan. Infect Immun., 4534–4540, 2007.
AIRES DE SOUSA, M.; DE LENCASTRE, H.; SANCHES, I.S.; KIKUCHI, K.; TOTSUKA, K. & TOMASZ, A. Similarity of antibiotic resistance patterns and molecular typing properties of methicillin-resistant Staphylococcus aureus isolates widely spread in hospitals in New York City and in a hospital in Tokyo, Japan. Microb. Drug. Resist., 6, 253-258, 2000.
ALLARD, C.; CARIGNAN, A.; BERGEVIN, M.; BOULAIS, I.; TREMBLAY, V.; ROBICHAUD, P.; DUPERVAL, R. & PEPIN, J. Secular changes in incidence and mortality associated with Staphylococcus aureus bacteraemia in Quebec, Canada, 1991-2005. Clin. Microbiol. Infect., 14, 421-428, 2008.
AL-MASSAUDI, S.B.; DAY, M.J. & RUSSEL, AD. Antimicrobial resistance and gene transfer in Staphylococcus aureus. J. Appl. Bacteriol; 70, 279- 290, 1991.
AMARAL, M.M.; COELHO, L.R.; FLORES, R.P.; SOUZA, R.R.; SILVA-CARVALHO, M.C.; TEIXEIRA, L.A.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M. The predominant variant of the Brazilian epidemic clonal complex of methicillin-resistant Staphylococcus aureus has an enhanced ability to produce biofilm and to adhere to and invade airway epithelial cells. J. Infect. Dis.,192, 801-810, 2005.
ARCHER, G.L. Staphylococcus aureus: a well-armed pathogen. Clin. Infect. Dis., 26, 1179-1181, 1998.
BAMBERGER, D.M. Bacteremia and endocarditis due to methicillin-resistant Staphylococcus aureus: the potential role of daptomycin. Ther. Clin. Risk Manag., 3, 675-684, 2007.
BARBER, M. Methicillin-resistant staphylococci. J. Clin. Pathol.,14, 385-393, 1961.
BARTELS, M.D.; NANUASHVILI, A.; BOYE, K.; ROHDE, S.M.; JASHIASHVILI, N.; FARIA, N.A.; KERESELIDZE, M.; KHAREBAVA, S. & WESTH, H. Methicillin-resistant Staphylococcus aureus in hospitals in Tbilisi, the Republic of Georgia, are variants of the Brazilian clone. Eur. J. Clin. Microbiol. Infect. Dis., 27, 757-760, 2008.
BAYER, M.G.; HEINRICHS, J.H. & CHEUNG, A.L. The molecular architecture of the sar locus in Staphylococcus aureus. J. Bacteriol.,178, 4563-4570, 1996.
BEENKEN, K.E.; BLEVINS, J.S. & SMELTZER, M.S. Mutation of sarA in Staphylococcus aureus limits biofilm formation. Infect. Immun., 71,4206-4211, 2003.
BEENKEN, K.E.; DUNMAN, P.M.; MCALEESE, F.; MACAPAGAL, D.; MURPHY, E.; PROJAN, S.J.; BLEVINS, J.S. & SMELTZER, M.S. Global gene expression in Staphylococcus aureus biofilms. J. Bacteriol., 186, 4665-4684, 2004.
BENITO, Y.; KOLB, F.A.; ROMBY, P.; LINA, G.; ETIENNE, J. & VANDENESCH, F. Probing the structure of RNAIII, the Staphylococcus aureus agr regulatory RNA, and identification of the RNA domain involved in repression of protein A expression. RNA, 6, 668-679, 2000.
BEVERIDGE, T.J.; MAKIN, S.A.; KADURUGAMUWA, J.L. & LI, Z. Interactions between biofilms and the environment. FEMS Microbiol. Rev., 20, 291-303, 1997.

81
BOISSET, S.; GEISSMANN, T.; HUNTZINGER, E.; FECHTER, P.; BENDRIDI, N.; POSSEDKO, M.; CHEVALIER, C.; HELFER, A.C.; BENITO, Y.; JACQUIER, A.; GASPIN, C.; VANDENESCH, F. & ROMBY, P. Staphylococcus aureus RNAIII coordinately represses the synthesis of virulence factors and the transcription regulator Rot by an antisense mechanism. Genes Dev., 21, 1353-1366, 2007.
BOLES, B.R. & HORSWILL, A.R. agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog., 4, 1-13, 2008.
BOYD, A. & CHAKRABARTY, A.M. Role of alginate lyase in cell detachment of Pseudomonas aeruginosa. Appl. Environ. Microbiol., 60, 2355-2359, 1994.
BRONNER, S.; MONTEIL, H. & PRÉVOST, G. Regulation of virulence determinants in Staphylococcus aureus: complexity and applications. FEMS Microbiol. Rev., 8, 183-200, 2004.
BROWN, M.R.; ALLISON, D.G. & GILBERT, P. Resistance of bacterial biofilms to antibiotics, a growth-rate related effect? J. Antimicrob. Chemother., 22, 777-780, 1988.
CAIAZZA, N.C. & O'TOOLE, G.A. Alpha-toxin is required for biofilm formation by Staphylococcus aureus. J. Bacteriol.,185, 3214-3217, 2003.
CAMPANILE, F., CAFISO, V., CASCONE, C., GIANNINO, V., DI MARCO, O. & STEFANI, S. Clonal diffusion and evolution of mecA and Tn554 polymorfhisms in methicillin-resistant Staphylococccus aureus in Italy. Infect. Med., 9, 30-38, 2001.
CASSAT, J.; DUNMAN, P.M.; MURPHY, E.; PROJAN, S.J.; BEENKEN, K.E.; PALM, K.J.; YANG, S.J.; RICE, K.C.; BAYLES, K.W. & SMELTZER, M.S. Transcriptional profiling of a Staphylococcus aureus clinical isolate and its isogenic agr and sarA mutants reveals global differences in comparison to the laboratory strain RN6390. Microbiol., 152, 3075-3090, 2006.
CASSAT, J.E.; LEE, C.Y. & SMELTZER, M.S. Investigation of biofilm formation in clinical isolates of Staphylococcus aureus. Methods Mol. Biol., 391, 127-144, 2007.
CDC (Centers for Disease Control and Prevention). Reduced susceptibility of Staphylococcus aureus to vancomycin - Japan, 1996. MMWR. Morb. Mortal. Wkly. Rep., 46, 624–626, 1997.
CHAN, P.F. & FOSTER, S.J. Role of SarA in virulence determinant production and environmental signal transduction in Staphylococcus aureus. J. Bacteriol., 6232–6241, 1998.
CHANG, S.; SIEVERT, D.M.; HAGEMAN, J.C.; BOULTON, M.L.; TENOVER, F.C.; DOWNES, F.P.; SHAH, S.; RUDRIK, J.T.; PUPP, G.R.; BROWN, W.J.; CARDO, D.; FRIDKIN, S.K & Vancomycin-Resistant Staphylococcus aureus Investigative Team. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N. Engl. J. Med., 348, 1342-1347, 2003.
CHEUNG, A.L.; BAYER, A.S.; ZHANG, G.; GRESHAM, H. & XIONG, Y.Q. Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. FEMS Immunol. Med. Microbiol., 40, 1-9, 2004.
CHEUNG, A.L.; NISHINA, K.A.; TROTONDA, M.P. & TAMBER, S. The SarA protein family of Staphylococcus aureus. Int. J. Biochem. Cell. Biol., 40, 355-361, 2008.

82
CHILLER, K.; SELKIN, B.A. & MURAKAWA, G.J. Skin microflora and bacterial infection of the skin. J. Investig. Dermatol. Simp. Proc., 6, 170-174, 2001.
CHRISTENSEN, G.D.; SIMPSON, W.A.; BISNO, A.L. & BEACHEY, E.H. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun., 37, 318-326, 1982.
COELHO, L.R.; SOUZA, R.R.; FERREIRA, F.A.; GUIMARÃES, M.A.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M.S. agr-RNAIII molecule divergently regulates glucose-induced biofilm formation in Staphylococcus aureus clinical isolates in a lineage-independent fashion. Microbiol., submitted, 2008.
COIMBRA, M.V.; TEIXEIRA, L.A.; RAMOS, R.L.; PREDARI, S.C.; CASTELLO, L.; FAMIGLIETTI, A.; VAY, C.; KLAN, L. & FIGUEIREDO, A.M.S. Spread of the Brazilian epidemic clone of a multirresistant MRSA in two cities in Argentina. J. Med. Microbiol., 49, 187-192, 2000.
COIMBRA, M.V.S.; SILVA-CARVALHO, M.C.; WISPLINGHOFF, H.; HALL, G.O.; TALLENT, S.; WALLACE, S.; EDMOND, M.B.; FIGUEIREDO, A.M.S. & WENZEL, R.P. Clonal spread of methicillin-resistant Staphylococcus aureus in large geographic area of the United States. J. Hosp. Infect., 53, 103-110, 2003.
CONLON, K.M.; HUMPHREYS, H. & O'GARA, J.P. Inactivations of rsbU and sarA by IS256 represent novel mechanisms of biofilm phenotypic variation in Staphylococcus epidermidis. J. Bacteriol., 186, 6208-6219, 2004.
CORRIGAN, R.M.; RIGBY, D.; HANDLEY, P. & FOSTER, T.J. The role of Staphylococcus aureus surface protein SasG in adherence and biofilm formation. Microbiology, 153, 2435-2446, 2007.
CRAMTON, S.E.; GERKE, C.; SCHNELL, N.F.; NICHOLS, W.W. & GOTZ, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun., 67, 5427-5433, 1999.
CUCARELLA, C.; TORMO, M.A.; UBEDA, C.; TROTONDA, M.P.; MONZON, M.; PERIS, C.; AMORENA, B.; LASA, I. & PENADES, J.R. Role of biofilm-associated protein bap in the pathogenesis of bovine Staphylococcus aureus. Infect. Immun., 72, 2177-2185, 2004.
CUI, L.; MA, X.; SATO, K.; OKUMA, K.; TENOVER, F.C.; MAMIZUKA, E.M.; GEMMELL, C.G.; KIM, M.N.; PLOY, M.C.; EL-SOLH, N.; FERRAZ, V. & HIRAMATSU, K. Cell wall thickening is a common feature of vancomycin resistance in Staphylococcus aureus. J. Clin. Microbiol., 41, 5–14, 2003.
DAY, N.P.; MOORE, C.E.; ENRIGHT, M.C.; BERENDT, A.R.; SMITH, J.M.; MURPHY, M.F.; PEACOCK, S.J.; SPRATT, B.G. & FEIL, E.J. A link between virulence and ecological abundance in natural populations of Staphylococcus aureus. Science. 292, 114-116, 2001.
DE ARAÚJO, G.L.; COELHO, L.R.; DE CARVALHO, C.B.; MACIEL, R.M.; CORONADO, A.Z.; ROZENBAUM, R.; FERREIRA-CARVALHO, B.T.; FIGUEIREDO, A.M. & TEIXEIRA, L.A. Commensal isolates of methicillin-resistant Staphylococcus epidermidis are also well equipped to produce biofilm on polystyrene surfaces. J. Antimicrob. Chemother., 57, 855-864, 2006.
DE LENCASTRE, H.; OLIVEIRA, D. & TOMASZ, A. Antibiotic resistant Staphylococcus aureus: a paradigm of adaptive power. Curr. Opin. Microbiol., 10, 428-435, 2007.

83
DE SOUZA, M.A.; SANCHES, I.S.; FERRO, M.L.; VAZ, M.J.; SARAIVA, Z.; TENDEIRO, T.; SERRA, J. & DE LENCASTRE, H. Intercontinental spread of a multidrug-resistant methicillin resistant Staphylococcus aureus clone. J. Clin. Microbiol., 36: 2590-2596, 1998.
DENIS, O.; NONHOFF, C.; BYL, B.; KNOOP, C.; BOBIN-DUBREUX, S. & STRUELENS, M.J. Emergence of vancomycin-intermediate Staphylococcus aureus in a Belgian hospital: microbiologic and clinical features. J. Antimicrob. Chemother., 50, 383–391, 2002.
DEORA, R.; TSENG, T. & MISRA, T.K. Alternative transcription factor sigmaSB of Staphylococcus aureus: characterization and role in transcription of the global regulatory locus sar. J. Bacteriol., 6355–6359, 1997.
DEURENBERG, R.H.; VINK, C.; KALENIC, S.; FRIEDRICH, A.W.; BRUGGEMAN, C.A. & STOBBERINGH, E.E. The molecular evolution of methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect., 13, 222-235, 2007.
DIEKEMA, D.J. & JONES, R.N. Oxazolidinone antibiotics. Lancet., 358, 1975-1982, 2001.
DIEP, B.; GEORGE, F.; SENSABAUGH, S.; NARAPORN, S.; SOMBOONA, D.; HEATHER, A.; CARLETON, & PERDREAU-REMING, F. Widespread skin and soft-tissue infections due to two methicillin-resistant Staphylococcus aureus Strains Harboring the Genes for Panton-Valentine Leucocidin. J. Clin. Microbiol., 42, 2080-2084, 2004.
DONLAN, R.M. & COSTERTON, J.W. Biofilms, survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev., 15, 167-193, 2002.
DOS SANTOS SOARES, M.J.; DA SILVA-CARVALHO, M.C.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M. Spread of methicillin-resistant Staphylococcus aureus belonging to the Brazilian epidemic clone in a general hospital and emergence of heterogenous resistance to glycopeptide antibiotics among these isolates. J. Hosp. Infect., 44, 301-308, 2000.
DUNM, O.J. Enumeration data: the chi-square test. Basic statistics: a primer for the biomedical sciences. 112-115, John Wiley & Sons, Inc. USA, 1964b.
DUNM, O.J. Tests of hypotheses on population means: Unknow variances, paired data. Basic statistics: a primer for the biomedical sciences. 78-95, John Wiley & Sons, Inc. USA. 1964a.
EDGEWORTH, J.D.; YADEGARFAR, G.; PATHAK, S.; BATRA, R.; COCKFIELD, J.D.; WYNCOLL, D.; BEALE, R. & LINDSAY, J.A. An outbreak in an intensive care unit of a strain of methicillin-resistant Staphylococcus aureus sequence type 239 associated with an increased rate of vascular access device-related bacteremia. Clin. Infect. Dis., 15, 493-501, 2007.
ELIOPOULOS, G.M. Antimicrobial agents for treatment of serious infections caused by resistant Staphylococcus aureus and enterococci. Eur. J. Clin. Microbiol. Infect. Dis., 24, 826-831, 2005.
ENRIGHT, M.C.; DAY, N.P.J.; DAVIES, C.E.; PEACOCK, S.J. & SPRATTB. G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol., 1008–1015, 2000.
FARRELL, D.J.; MORRISSEY, I.; BAKKER, S.; BUCKRIDGE, S. & FELMINGHAM, D. In vitro activities of telithromycin, linezolid, and quinupristin-dalfopristin against Streptococcus pneumoniae with macrolide resistance due to ribosomal mutations. Antimicrob. Agents Chemother., 48, 3169-3171, 2004.

84
FEIL, E.J.; NICKERSON, E.K.; CHANTRATITA, N.; WUTHIEKANUN, V.; SRISOMANG, P.; COUSINS, R.; PAN, W.; ZHANG, G.; XU, B.; DAY, N.P. & PEACOCK, S.J. Rapid detection of the pandemic methicillin-resistant Staphylococcus aureus clone ST 239, a dominant strain in Asian hospitals. J. Clin. Microbiol., 46, 1520-1522, 2008.
FITZPATRICK, F.; HUMPHREYS, H. & O'GARA, J.P. Evidence for icaADBC-independent biofilm development mechanism in methicillin-resistant Staphylococcus aureus clinical isolates. J. Clin. Microbiol., 43,1973-1976, 2005.
FLEER, A. & VERHOEF, J. An evaluation of the role of surface hydrophobicity and extracellular slime in the pathogenesis of foreign-body-related infections due to coagulase-negative staphylococci. J. Invest. Surg., 2, 391-396, 1989.
FOURNIER, B. & HOOPER, D.C. A new two-component regulatory system involved in adhesion, autolysis, and extracellular proteolytic activity of Staphylococcus aureus. J. Bacteriol.; 182,3955-3964, 2000.
FOURNIER, B.; KLIER, A. & RAPOPORT, G. The two component system ArlS–ArlR is a regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol., 247–261, 2001.
FOWLER, V.G.J.R. ; SAKOULAS, G. ; MCINTYRE, L.M. ; MEKA, V.G. ; ARBEIT, R.D. ; CABELL, C.H. ; STRYJEWSKI, M.E. ; ELIOPOLUS, G.M. ; RELLER, L.B. ; COREY, G.R. ; JONES, T. ; LUCINDO, N. ; YEAMAN, M.R. & BAYER, A.S. Persistent bacteremia due to methicillin-resistant Staphylococcus aureus infection is associated with agr dysfunction and low-level in vitro resistance to thrombin-induced platelet microbicidal protein. J. Infect. Dis., 90, 1140-1149, 2004.
FREES, D.; CHASTANET, A.; QAZI, S.; SØRENSEN, K.; HILL, P.; MSADEK, T. & INGMER, H. Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus. Mol. Microbiol., 54,1445-1462, 2004.
FREES, D.; SØRENSEN, K. & INGMER, H. Global virulence regulation in Staphylococcus aureus, pinpointing the roles of ClpP and ClpX in the sar/agr regulatory network. Infect. Immun., 73, 8100-8108, 2005.
FUX, C.A.; STOODLEY, P.; HALL-STOODLEY, L. & CONSTERTON, J.W. Bacterial biofilmes: a diagnostic and therapeutic challenge. Expert Rev. Anti Infect. Ther., 1, 667-683, 2003.
GALES, A.C.; JONES, R.N.; PFALLER, M.A.; GORDON, K.A. & SADER, H.S. Two-year assessment of the pathogen frequency and antimicrobial resistance patterns among organisms isolated from skin and soft tissue infections in Latin American hospitals: results from the SENTRY antimicrobial surveillance program, 1997-98. SENTRY Study Group. Int J. Infect. Dis., 4, 75-84, 2000.
GALES, A.C.; SADER, H.S.; ANDRADE, S.S.; LUTZ, L.; MACHADO, A. & BARTH, A.L. Emergence of linezolid-resistant Staphylococcus aureus during treatment of pulmonary infection in a patient with cystic fibrosis. Intern. J. Antimicrob. Agents, 27, 300–302, 2006.
GEISINGER, E.; ADHIKARI, R.P.; JIN, R.; ROSS, H.F. & NOVICK, R.P. Inhibition of rot translation by RNAIII, a key feature of agr function. Mol. Microbiol. 61, 1038–1048, 2006.
GIACOMETTI, A.; CIRIONI, O.; GHISELLI, R.; ORLANDO, F.; MOCCHEGIANI, F.; SILVESTRI, C.; LICCI, A.; DE FUSCO, M.; PROVINCIALI, M.; SABA, V. & SCALISE, G. Comparative efficacies of quinupristin-dalfopristin, linezolid, vancomycin, and ciprofloxacin in treatment, using the antibiotic-lock technique, of experimental catheter-related infection due to Staphylococcus aureus. Antimicrob. Agents Chemother., 49, 4042-4045, 2005.

85
GÖTZ, F. Staphylococcus and biofilms. Mol. Microbiol., 43, 1367-1378, 2002.
GROSS, M.; CRAMTON, S.E.; GOTZ, F. & PESCHEL, A. Key role of teichoic acid net charge in Staphylococcus aureus colonization of artificial surfaces. Infect. Immun., 69, 3423-3426, 2001.
GRUNDMANN, H.; AIRES-DE-SOUSA, M.; BOYCE, J. & TIEMERSMA, E. Emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet, 368, 874–885, 2006.
HANAKI, H.; KUWAHARA-ARAI, K.; BOYLE-VAVRA, S.; DAUM, R.S.; LABISCHINSKI, H. & HIRAMATSU, K. Activated cell-wall synthesis is associated with vancomycin resistance in methicillin-resistant Staphylococcus aureus clinical strains Mu3 and Mu50. J. Antimicrob. Chemother., 42, 199–209, 1998.
HEILMANN, C.; HARTLEIB, J.; HUSSAIN, M.S. & PETERS, G. The multifunctional Staphylococcus aureus autolysin aaa mediates adherence to immobilized fibrinogen and fibronectin. Infect Immun., 73, 4793-4802, 2005.
HEILMANN, C.; HUSSAIN, M.; PETERS, G. & GOTZ, F. Evidence for autolysin-mediated primary attachment of Staphylococcus epidermidis to a polystyrene surface. Mol. Microbiol., 24,1013-1024, 1997.
HEILMANN, C.; THUMM, G.; CHHATWAL, G.S.; HARTLEIB, J.; UEKOTTER, A. & PETERS, G. Identification and characterization of a novel autolysin (Aae) with adhesive properties from Staphylococcus epidermidis. Microbiol., 149, 2769-2778, 2003.
HEINRICHS, J.H.; BAYER, M.G. & CHEUNG, A.L. Characterization of the sar locus and its interaction with agr in Staphylococcus aureus. J. Bacteriol., 178, 418–423, 1996.
HILL, C.; KING, T. & DAY, R. A strategy to reduce MRSA colonization of stethoscopes. J. Hosp. Infect., 62, 122-123, 2006.
HIRAMATSU, K. Vancomycin-resistant Staphylococcus aureus: a new model of antibiotic resistance. Lancet Infect. Dis., 1, 147–155, 2001.
HIRAMATSU, K.; ASADA, K.; SUZUKI, E.; OKONOGI, K. & YOKOTA, T. Molecular cloning and nucleotide sequence determination of the regulator region of mecA gene in methicillin-resistant Staphylococcus aureus (MRSA). FEBS Lett. 298, 133–136, 1992.
HIRAMATSU, K.; HANAKI, H.; INO, T.; YABUTA, K.; OGURI, T. & TENOVER, F.C. Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycinsusceptibility. J. Antimicrob. Chemother., 40, 135–136, 1997.
HIRAMATSU, K.; WATANABE, S.; TAKEUCHI, F.; ITO, T. & BABA, T. Genetic characterization of methicillin-resistant Staphylococcus aureus. Vaccine, 22S, S5–S8, 2004.
HIRAMATSU, K.; KATAYAMA, Y.; YAZAWA, H. & ITO, T. Molecular genetics of methicillin-resistant Staphylococcus aureus. Int. J. Med. Microbiol., 292, 67-74, 2002.
HSIEH, H.Y.; TSENG, C.W. & STEWART, G.C. Regulation of Rot expression in Staphylococcus aureus. J. Bacteriol., 190, 546-554, 2008.

86
HUNTZINGER, E.; BOISSET, S.; SAVEANU, C.; BENITO, Y.; GEISSMANN, T.; NAMANE, A.; LINA, G.; ETIENNE, J.; EHRESMANN, B.; EHRESMANN, C.; JACQUIER, A.; VANDENESCH, F. & ROMBY, P. Staphylococcus aureus RNAIII and the endoribonuclease III coordinately regulate spa gene expression. EMBO J., 24, 824-835, 2005.
INGAVALE, S.; VAN WAMEL, W.; LUONG, T.T.; LEE, C.Y. & CHEUNG, A.L. Rat/MgrA, a regulator of autolysis, is a regulator of virulence genes in Staphylococcus aureus. Infect. Immun., 73, 1423-1431, 2005.
INNIS, M.A., GELFAND, D.H., SNINSKY, J.J. & WHITE, T.J. PCR protocols a guide to methods and applications. Academic Press, Inc. San Diego, California, USA, 1990.
ITO, T., KUWAHARA, K. & HIRAMATSU, K. Staphylococcal cassette chromosome mec (SCCmec) analysis of MRSA. Methods Mol. Biol., 391, 87-102, 2007.
ITO, T.; KATAYAMA, Y.; ASADA, K. & MORI, N. Structural comparison of three types of staphylococcal cassette chromosome mec integrated in the chromosome in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother., 45, 1323-1336, 2001.
ITO, T.; MA, X.X.; TAKEUCHI, F.; OKUMA, K.; YUZAWA, H. & HIRAMATSU, K. Novel type V staphylococcal cassette chromosome mec driven by a novel cassette chromosome recombinase, ccrC. Antimicrob. Agents Chemother., 48, 2637-2651, 2004.
IWATSUKI, K.; YAMASAKI, O.; MORIZANE, S. & OONO, T. Staphylococcal cutaneous infections: invasion, evasion and aggression. J. Dermatol. Sci., 42, 203-214, 2006.
IZANO, E.A.; AMARANTE, M.A.; KHER, W.B. & KAPLAN, J.B. Differential roles of poly-N-acetylglucosamine surface polysaccharide and extracellular DNA in Staphylococcus aureus and Staphylococcus epidermidis biofilms. Appl. Environ. Microbiol., 74, 470-476, 2008.
JAMES, N.R. & JAYAKRISHNAN, A. Surface thiocyanation of plasticized poly(vinyl chloride) and its effect on bacterial adhesion. Biomaterials., 24, 2205-2212, 2003.
JANZON, L. & ARVIDSON, S. The role of the delta-lysin gene (hld) in the regulation of virulence genes by the accessory gene regulator (agr) in Staphylococcus aureus. EMBO J., 9, 1391-1399, 1990.JARRAUD et al., 2000
JARRAUD, S.; LYON, G.J.; FIGUEIREDO, A.M.; GERARD, L.; VANDENESCH, F.; ETIENNE, J., MUIR, T.W. & NOVICK, R.P. Exfoliatin-producing strains define a fourth agr specificity group in Staphylococcus aureus. J. Bacteriol., 82, 6517-6522, 2000.
JEMILI-BEN, J.M.; BOUTIBA-BEN, B.I. & BEN, R.S. Identification of staphylococcal cassette chromosome mec encoding methicillin resistance in Staphylococcus aureus isolates at Charles Nicolle Hospital of Tunis. Pathol. Biol., 54, 453-455, 2006.
JEMNI, L.; MDIMAGH, L.; JEMNI-GHARBI, H.; JEMNI, M.; KRAIEM, C. & ALLEGUE, M. Kidney carbuncle: diagnostic, bacteriological and therapeutic considerations. Apropos of 11 cases. J. Urol. (Paris)., 98, 228-231, 1992.
JI, G.; BEAVIS, R. & NOVICK, R.P. Bacterial interference caused by autoinducing peptide variants. Science. 276, 2027-2030, 1997.
KAITO, C.; MORISHITA, D.; MATSUMOTO, Y.; KUROKAWA, K. & SEKIMIZU, K. Novel DNA binding protein SarZ contributes to virulence in Staphylococcus aureus. Mol. Microbiol., 62, 1601-1617, 2006.

87
KARLSSON, A.; SARAVIA-OTTEN, P.; TEGMARK, K.; MORFELDT, E. & ARVIDSON, S. Decreased amounts of cell wall-associated protein A and fibronectin-binding proteins in Staphylococcus aureus sarA mutants due to up-regulation of extracellular proteases. Infect. Immun., 69,4742-4748, 2001.
KATAYAMA, Y.; ITO. T. & HIRAMATSU, K. Genetic organization of the chromosome region surrounding mecA in clinical staphylococcal strains: role of IS431-mediated mecI deletion in expression of resistance in mecA-carrying, low-level methicillin-resistant Staphylococcus haemolyticus. Antimicrob. Agents Chemother., 45, 1955-1963, 2001.
KATAYAMA, Y.; ITO, T. & HIRAMATSU, K. A new class of genetic element, Staphylococcus aureus chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother., 44, 1549-1555, 2000.
KENNEDY, A.D.; OTTO, M.; BRAUGHTON, K.R.; WHITNEY, A.R.; CHEN, L.; MATHEMA, B.; MEDIAVILLA, J.R.; BYRNE, K.A.; PARKINS, L.D.; TENOVER, F.C.; KREISWIRTH, B.N.; MUSSER, J.M. & DELEO, F.R. Epidemic community-associated methicillin-resistant Staphylococcus aureus: recent clonal expansion and diversification. Proc. Natl. Acad. Sci., USA., 105, 1327-1332, 2008.
KOGAN, G.; SADOVSKAYA, I.; CHAIGNON, P.; CHOKR, A. & JABBOURI, S. Biofilms of clinical strains of Staphylococcus that do not contain polysaccharide intercellular adhesin. FEMS Microbiol. Lett., 255,11-16, 2006.
KRAMER, A.; SCHWEBKE, I. & KAMPF, G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect. Dis., 6, 130, 2006.
KRZYSZTON-RUSSJAN, J.; GNIADKOWSKI, M.; POLOWNIAK-PRACKA, H.; HAGMAJER, E. & HRYNIEWICZ, W. The first Staphylococcus aureus isolates with reduced susceptibility to vancomycin in Poland. J. Antimicrob. Chemother., 50, 1065–1069, 2002.
KURODA, M.; OHTA, T.; UCHIYAMA, I.; BABA, T.; YUZAWA, H.; KOBAYASHI, I.; CUI, L.; OGUCHI, A.; AOKI, K.; NAGAI, Y.; LIAN, J.; ITO, T.; KANAMORI, M.; MATSUMARU, H.; MARUYAMA, A.; MURAKAMI, H.; HOSOYAMA, A.; MIZUTANI-UI, Y.; TAKAHASHI, N.K.; SAWANO, T.; INOUE, R.; KAITO, C.; SEKIMIZU, K.; HIRAKAWA, H.; KUHARA, S.; GOTO, S.; YABUZAKI, J.; KANEHISA, M.; YAMASHITA, A.; OSHIMA, K.; FURUYA, K.; YOSHINO, C.; SHIBA, T.; HATTORI, M.; OGASAWARA, N.; HAYASHI, H. & HIRAMATSU, K. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet., 357, 1225-1240, 2001.
LASA, I. & PENADÉS, J.R. Bap: a family of surface proteins involved in biofilm formation. Res. Microbiol., 157, 99-107, 2006.
LIANG, X.; ZHENG, L.; LANDWEHR, C.; LUNSFORD, D.; HOLMES, D. & JI, Y. Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J. Bacteriol., 187, 5486-5492, 2005.
LIBERT, M.; ELKHOLTI, M.; MASSAUT, J.; KARMALI, R.; MASCART, G. & CHERIFI, S. Risk factors for meticillin resistance and outcome of Staphylococcus aureus bloodstream infection in a Belgian university hospital. J. Hosp. Infect., 68, 17-24, 2008.
LIM, Y.; JANA M.; LUONG, T.T. & LEE, C.Y. Control of glucose- and NaCl-induced biofilm formation by rbf in Staphylococcus aureus. J. Bacteriol., 186,722-729, 2004.
LINA, G.; JARRAUD, S.; JI, G.; GREENLAND, T.; PEDRAZA, A.; ETIENNE, J.; NOVICK, R.P. & VANDENESCH, F. Transmembrane topology and histidine protein kinase activity of AgrC, the agr signal receptor in Staphylococcus aureus. Mol. Microbiol., 28, 655-662, 1998.

88
LINDSAY, J.A.; MOORE, C.E.; DAY, N.P.; PEACOCK, S.J.; WITNEY, A.A.; STABLER, R.A.; HUSAIN, S.E.; BUTCHER, P.D. & HINDS, J. Microarrays reveal that each of the ten dominant lineages of Staphylococcus aureus has a unique combination of surface-associated and regulatory genes. J. Bacteriol., 188, 669-676, 2006.
LIU, Y.; MANNA, A.C.; PAN, C.H.; KRIKSUNOV, I.A.; THIEL, D.J.; CHEUNG, A.L. & ZHANG, G. Structural and function analyses of the global regulatory protein SarA from Staphylococcus aureus. Proc. Natl. Acad. Sci. U S A., 103, 2392-2397, 2006.
LORENZ, U.; HÜTTINGER, C.; SCHÄFER, T.; ZIEBUHR, W.; THIEDE, A.; HACKER, J.; ENGELMANN, S.; HECKER, M. & OHLSEN, K. The alternative sigma factor sigma B of Staphylococcus aureus modulates virulence in experimental central venous catheter-related infections. Microb. Infect.,10, 217-223, 2008.
LUONG, T.T.; DUNMAN, P.M.; MURPHY, E.; PROJAN, S.J. & LEE, C.Y. Transcription Profiling of the mgrA Regulon in Staphylococcus aureus. J. Bacteriol., 188, 1899-1910, 2006.
LYON, G.J.; WRIGHT, J.S.; MUIR, T.W. & NOVICK, R.P. Key determinants of receptor activation in the agr autoinducing peptides of Staphylococcus aureus. Biochemistry., 41,10095-10104, 2002.
MACHADO, A.R.L.; ARNS, C.C.; FOLLADOR, W. & GUERRA, A. Cost-effectiveness of linezolid versus vancomycin in mechanical ventilation-associated nosocomial pneumonia caused by methicillin-resistant Staphylococcus aureus. Brazilian J. Infect. Dis., 9, 191-200, 2005.
MACK, D.; FISCHER, W.; KROKOTSCH, A.; LEOPOLD, K.; HARTMANN, R.; EGGE, H. & LAUFS, R. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear beta-1,6-linked glucosaminoglycan, purification and structural analysis. J. Bacteriol., 178, 175-183, 1996.
MAIRA-LITRÁN, T.; KROPEC, A.; ABEYGUNAWARDANA, C.; JOYCE, J.; MARK, G. III, GOLDMANN, D.A. & PIER, G.B. Immunochemical properties of the staphylococcal poly-N-acetylglucosamine surface polysaccharide. Infect. Immun., 70, 4433-4440, 2002.
MALBRUNY, B.; CANU, A.; BOZDOGAN, B.; FANTIN, B.; ZARROUK, V.; DUTKA-MALEN, S.; FEGER, C. & LECLERCQ, R. Resistance to quinupristin-dalfopristin due to mutation of L22 ribosomal protein in Staphylococcus aureus. Antimicrob. Agents Chemother., 46, 2200-2207, 2002.
MANNA, A. & CHEUNG, A.L. Characterization of sarR, a modulator of sar expression in Staphylococcus aureus. Infect. Immun., 69, 885-896, 2001.
MANNA, A.C. & CHEUNG, A.L. Expression of SarX, a negative regulator of agr and exoprotein synthesis, is activated by MgrA in Staphylococcus aureus. J. Bacteriol., 188, 4288-4299, 2006.
MANNA, A.C. & CHEUNG, A.L. sarU, a sarA homolog, is repressed by SarT and regulates virulence genes in Staphylococcus aureus. Infect Immun., 71, 343-353, 2003.
MANNA, A.C.; BAYER, M.G. & CHEUNG, A.L. Transcriptional analysis of different promoters in the sar locus in Staphylococcus aureus. J. Bacteriol., 3828–3836, 1998.

89
MANNA, A.C.; INGAVALE, S.S.; MALONEY, M.; VAN WAMEL, W. & CHEUNG, A.L. Identification of sarV (SA2062), a new transcriptional regulator, is repressed by SarA and MgrA (SA0641) and involved in the regulation of autolysis in Staphylococcus aureus. J. Bacteriol., 186, 5267-5280, 2004.
MARPLES, R.R.; RICHARDSON, J.F. & DE SAXE, M.J. Bacteriological characters of strains of Staphylococcus aureus submitted to a reference laboratory related to methicillin resistance. J. Hyg; 96, 217–223, 1986.
MARRIE, T.J.; NELLIGAN, J. & COSTERTON, J.W. A scanning and transmission electron microscopic study of an infected endocardial pacemaker lead. Circulation., 66 1339-1341, 1982.
MATHEWS, D.H.; DISNEY, M.D.; CHILDS, J.L.; SCHROEDER, S.J.; ZUKER, M. & TURNER, D.H. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proc. Natl. Acad. Sci. USA., 101, 7287-7292, 2004.
MEKA, V.G.; PILLAI, S.K.; SAKOULAS, G.; WENNERSTEN, C.; VENKATARAMAN, L.; DEGIROLAMI, P.C.; ELIOPOULOS, G.M.; MOELLERING, R.C. JR. & GOLD, H.S. Linezolid resistance in sequential Staphylococcus aureus isolates associated with a T2500A mutation in the 23S rRNA gene and loss of a single copy of rRNA. J. Infect. Dis., 190, 311-317, 2004.
MENDES, R.E.; DESHPANDE, L.M.; CASTANHEIRA, M.; DIPERSIO, J.; SAUBOLLE, M.A. & JONES, R.N. First report of cfr-mediated resistance to linezolid in human staphylococcal clinical isolates recovered in the United States. Antimicrob. Agents Chemother., 52, 2244-2246, 2008.
MONGODIN, E.; BAJOLET, O.; HINNRASKY, J.; PUCHELLE, E. & DE BENTZMANN, S. Cell wall-associated protein A as a tool for immunolocalization of Staphylococcus aureus in infected human airway epithelium. J. Histochem. Cytochem., 48, 523-534, 2000.
MORFELDT, E.; TEGMARK, K., & ARVIDSON, S. Transcriptional control of the agr-dependent virulence gene regulator, RNAIII, in Staphylococcus aureus. Mol. Microbiol., 21, 1227-1237, 1996.
MURRAY, P.R.; BARON, E.J.; JORGENSEN, J.H.; PFALLER, M.A. & YOLKEN, R.H. Staphylococcus, Micrococcus and other catalase- positive cocci that grow aerobically. In, Manual of clinical Microbiology, 8th ed. ASM Press, Washington, D.C., USA, 1, 384-404, 2003.
NCCLS (National Committee for Clinical Laboratory Standards). Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically, 6th ed. Approved standard, M7-A6. Wayne, Pennsylvania: National Committee for Laboratory Standards, 2003.
NNIS SYSTEM. NATIONAL NOSOCOMIAL INFECTIONS SURVEILLANCE. (NNIS) System report, data summary from January 1992 through June 2004, issue October 2004. Am. J. Infec., 32, 470-485, 2004.
NOTO, M.J.; KREISWIRTH, B.N.; MONK, A.B. & ARCHER, G.L. Gene acquisition at the insertion site for SCCmec, the genomic island conferring methicillin resistance in Staphylococcus aureus. J. Bacteriol., 1276-1283, 2008.
NOVICK, R.P. & MUIR, T.W. Virulence gene regulation by peptides in staphylococci and other Gram-positive bacteria. Curr. Opin. Microbiol., 2, 40-45, 1999.

90
NOVICK, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol.. 48, 1429-1449, 2003.
NOVICK, R.P. Pathogenicity factors and their regulation. In: Gram-positive pathogens. 392-407. Edited by V.A. Fischetti, R.P. Novick, J.J. Ferretti, D.A Portnoy, and J.I. Rood. ASM Press, Washington, D.C. USA, 2000.
NOVICK, R.P.; PROJAN, S.J.; KORNBLUM, J.; ROSS, H.F.; JI, G.; KREISWIRTH, B.; VANDENESCH, F. & MOGHAZEH, S. The agr P2 operon: an autocatalitic sensory transduction system in Staphylococcus aureus. Mol. Gen. Genet., 248, 446-458, 1995.
NOVICK, R.P.; ROSS, H.F.; PROJAN, S.J.; KORNBLUM, J.; KREISWIRTH, B. & MOGHAZEH, S. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J., 12, 3967-3975, 1993.
OLIVEIRA, D.C.; TOMASZ, A. & DE LENCASTRE, H. Secrets of success of a human pathogen: molecular evolution of pandemic clones of meticillin-resistant Staphylococcus aureus. Lancet Infect. Dis., 2, 180-189, 2002.
OLIVEIRA, D.C.; MILHEIRIÇO, C. & DE LENCASTRE, H. Redefining a structural variant of staphylococcal cassette chromosome mec, SCCmec type VI. Antimicrob. Agents Chemother., 50, 3457-3459, 2006.
OLIVEIRA, G.A.; DELL’AQUILA, A.M.; MASIERO, R.L.; LEVY, C.E.; GOMES, M.S.; CUI, L.; HIRAMATSU, K. & MAMIZUKA, E.M. Isolation in Brazil of nosocomial Staphylococcus aureus with reduced susceptibility to vancomycin. Infect. Control. Hosp. Epidemiol., 22, 443–448, 2001.
O'NEILL, E.; POZZI, C.; HOUSTON, P.; HUMPHREYS, H.; ROBINSON, D.A.; LOUGHMAN, A.; FOSTER, T.J. & O'GARA, J.P. A novel Staphylococcus aureus biofilm phenotype mediated by the fibronectin-binding proteins, FnBPA and FnBPB. J. Bacteriol., 190, 3835-3850, 2008.
O'NEILL, E.; POZZI, C.; HOUSTON, P.; SMYTH, D.; HUMPHREYS, H.; ROBINSON, D.A., & O’GARA, J.P. Association between methicillin susceptibility and biofilm regulation in Staphylococcus aureus isolates from device-related infections. J. Clin. Microbiol., 1379–1388, 2007.
O'SEAGHDHA, M.; VAN SCHOOTEN, C.J.; KERRIGAN, S.W.; EMSLEY, J.; SILVERMAN GJ, COX D, LENTING PJ, FOSTER TJ. Staphylococcus aureus protein A binding to von Willebrand factor A1 domain is mediated by conserved IgG binding regions. FEBS J., 273, 4831-4841, 2006.
OSCARSSON, J.; KANTH, A.; TEGMARK-WISELL, K. & ARVIDSON, S. SarA is a repressor of hla (alpha-hemolysin) transcription in Staphylococcus aureus: its apparent role as an activator of hla in the prototype strain NCTC 8325 depends on reduced expression of sarS. J. Bacteriol., 188, 8526-8533, 2006.
PALMA, M., & CHEUNG, A.L. σB activity in Staphylococcus aureus is controlled by RsbU and an additional factor(s) during bacterial growth. Infect. Immun., 69, 7858–7865, 2001.
PANÉ-FARRÉ, J.; JONAS, B.; FÖRSTNER, K.; ENGELMANN, S. & HECKER, M. The σB regulon in Staphylococcus aureus and its regulation. Int. J. Med. Microbiol., 296, 237–258, 2006.

91
PAPAKYRIACOU, H.; VAZ, D.; SIMOR, A.; LOUIE, M. & MCGAVIN, M.J. Molecular analysis of the accessory gene regulator (agr) locus and balance of virulence factor expression in epidemic methicillin-resistant Staphylococcus aureus. J. Infect. Dis., 181, 990-1000, 2000.
PARSEK, M.R. & SINGH, P.K. Bacterial biofilms, an emerging link to disease pathogenesis. Annu. Rev. Microbiol., 57, 677-701, 2003.
PETERS, G.; LOCCI, R. & PULVERER, G. Microbial colonization of prosthetic devices. II. Scanning electron microscopy of naturally infected intravenous catheters. Zentralbl Bakteriol Mikrobiol Hyg [B]., 173, 293-299, 1981.
PETTI, C.A. & FOWLER, V.G. JR. Staphylococcus aureus bacteremia and endocarditis. Infect. Dis. Clin. North Am.,16, 413-435. 2002.
PICAZO, J.J.; BETRIU, C.; RODRÍGUEZ-AVIAL, I.; CULEBRAS, E.; GÓMEZ, M. & LÓPEZ, F.; GRUPO VIRA. Antimicrobial resistance surveillance: VIRA STUDY 2006. Enferm. Infect. Microbiol. Clin., 24, 617-628, 2006.
PRAGMAN, A.A. & SCHLIEVERT, P.M. Virulence regulation in Staphylococcus aureus: the need for in vivo analysis of virulence factor regulation. FEMS Immunol. Med. Microbiol., 42, 147-154, 2004.
PROJAN, S.J. & NOVICK, R.P. 1997. The molecular basis of pathogenecity. In: The staphylococci in human disease. 55-82. Edited by Crossley, K.B., Archer, G.L. New York: Churchill Livingstone. USA, 1997.
ROBERTS, C.; ANDERSON, K.L.; MURPHY, E.; PROJAN, S.J.; MOUNTS, W.; HURLBURT, B.; SMELTZER, M.; OVERBEEK, R.; DISZ, T. & DUNMAN, P.M. Characterizing the effect of the Staphylococcus aureus virulence factor regulator, SarA, on log-phase mRNA half-lives. J. Bacteriol., 188, 2593-2603, 2006.
ROBINSON, D.A. & ENRIGHT, M.C. Evolution of Staphylococcus aureus by large chromosomal replacements. J. Bacteriol., 186, 1060–1064, 2004.
ROHDE, H.; BURANDT, E.C.; SIEMSSEN, N.; FROMMELT, L.; BURDELSKI, C.; WURSTER, S.; SCHERPE, S.; DAVIES, A.P.; HARRIS, L.G.; HORSTKOTTE, M.A.; KNOBLOCH, J.K.; RAGUNATH, C.; KAPLAN, J.B. & MACK, D. Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prosthetic hip and knee joint infections. Biomaterials, 28, 1711-1720, 2007.
ROHDE, H.; BURDELSKI, C.; BARTSCHT, K.; HUSSAIN, M.; BUCK, F.; HORSTKOTTE, M.A.; KNOBLOCH, J.K.; HEILMANN, C.; HERRMANN, M. & MACK, D. Induction of Staphylococcus epidermidis biofilm formation via proteolytic processing of the accumulation-associated protein by staphylococcal and host proteases. Mol. Microbiol., 55,1883-1895, 2005.
SAHA, B.; SINGH, A.K.; GHOSH, A. & BAL, M. Identification and characterization of a vancomycin-resistant Staphylococcus aureus isolated from Kolkata (South Asia). J. Med. Microbiol., 57, 72-79, 2008.
SAID-SALIM, B.; DUNMAN, P.M.; MCALEESE, F.M.; MACAPAGAL, D.; MURPHY, E.; MCNAMARA, P.J.; ARVIDSON, S.; FOSTER, T.J.; PROJAN, S.J. & KREISWIRTH, B.N. Global regulation of Staphylococcus aureus genes by Rot. J. Bacteriol., 185, 610-619, 2003.

92
SAKOULAS, G.; ELIOPOULOS, G.M.; MOELLERING R.C.JR.; WENNERSTEN, C.; VENKATARAMAN, L.; NOVICK, R.P. & GOLD, H.S. Accessory gene regulator (agr) locus in geographically diverse Staphylococcus aureus isolates with reduced susceptibility to vancomycin. Antimicrob. Agents Chemother., 46, 1492-1502, 2002.
SA-LEAO, R.; SANTOS-SANCHES, I.; DIAS, D.; PERES, I.; BARROS, R.M. & DE LENCASTRE, H. Detection of an archaic clone of Staphylococcus aureus with low-level resistance to methicillin in a pediatric hospital in Portugal and in international samples, relics of a formerly widely disseminated strain? J. Clin. Microbiol., 37, 1913-1920, 1999.
SAMBROOK, J.; FRITSCH, E.F. & MANIATIS, T. Molecular cloning: a Laboratory Manual. Edited by Cold. Spring Harbor. NY. USA, 1989.
SCHMIDT, K.A.; MANNA, A.C. & CHEUNG, A.L. SarT influences sarS expression in Staphylococcus aureus. Infect Immun., 71, 5139-5148, 2003.
SCHMIDT, K.A.; MANNA, A.C., GILL, S. & CHEUNG, A.L. SarT, a repressor of alpha-hemolysin in Staphylococcus aureus. Infect. Immun., 4749-4758, 2001.
SIERADZKI, K. & TOMASZ, A. Inhibition of cell wall turnover and autolysis by vancomycin in a highly vancomycin-resistant mutant of Staphylococcus aureus. J. Bacteriol., 179, 2557-2566, 1997.
SMITH, T.L.; PEARSON, M.L.; WILCOX, K.R.; CRUZ, C.; LANCASTER, M.V.; ROBINSON-DUNN, B.; TENOVER, F.C.; ZERVOS, M.J.; BAND, J.D.; WHITE, E. & JARVIS, W.R. Emergence of vancomycin resistance in Staphylococcus aureus. Glycopeptide-intermediate Staphylococcus aureus Working Group. N. Engl. J. Med., 340, 493–501, 1999.
SOARES, M.J.S.; TEIXEIRA, L.A.; NUNES, M.R.; CARVALHO, M.C.S.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M.S. Analysis of different molecular methods for typing methicillin-resistant Staphylococcus aureus isolates belonging to the Brazilian epidemic clone. J. Med. Microbiol., 50, 732-742, 2001.
SOARES, M.J.S.; TOKUMARU-MIYAZAKI, N.H.; NOLETO, A.L.S. & FIGUEIREDO, A.M.S. Enterotoxin production by Staphylococcus aureus clones and detection of Brazilian epidemic MRSA clone (III::B:A) among isolates from food handler workers. J. Med. Microbiol., 48, 1-8, 1997.
STEWART, P.S. & COSTERTON, J.W. Antibiotic resistance of bacteria in biofilms. Lancet., 358, 135-138, 2001.
STOODLEY, P.; KATHJU, S.; HU, F.Z.; ERDOS, G.; LEVENSON, J.E.; MEHTA, N. DICE, B.; JOHNSON, S.; HALL-STOODLEY, L.; NISTICO, L.; SOTEREANOS, N.; SEWECKE, J.; POST, J.C. & EHRLICH, G.D. Molecular and imaging techniques for bacterial biofilms in joint arthroplasty infections. Clin. Orthop. Rel. Res., 437:31–40, 2005.
TAK, T.; REED, K.D.; HASELBY, R.C.; MCCAULEY, C.S.J.R. & SHUKLA, S.K. An update on the epidemiology, pathogenesis and management of infective endocarditis with emphasis on Staphylococcus aureus. W.M.J., 101, 24-33, 2002.
TEGMARK, K.; KARLSSON, A. & ARVIDSON, S. Identification and characterization of SarH1, a new global regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol., 398- 409, 2000.
TEIXEIRA, L.A.; RESENDE, C.A.; ORMOND, L.R.; ROSEMBAUM, R.; FIGUEIREDO, A.M.S.; DE LENCASTRE, H. & TOMASZ, A. Geografic spread of epidemic multirresistant Staphylococcus aureus clone in Brazil. J. Clin. Microbiol.; 33, 2400-2404, 1995.

93
TENOVER, F.C. & GAYNES, R.P. The epidemiology of Staphylococus aureus infections. In: Gram-positive pathogens. 414-421. Edited by Fischetti, V.A., Novick, R.P., Ferretti, J.J., Portnoy, D.A. & Rood, J.I. Washington, D.C.: ASM Press, USA, 2000.
TENOVER, F.C.; WEIGEL, L.M.; APPELBAUM, P.C.; MCDOUGAL, L.K.; CHAITRAM, J.; MCALLISTER, S.; CLARK, N.; KILLGORE, J.; O'HARA, C.M.; JEVITT, L.;PATEL, J.B. & BOZDOGAN,
B. Vancomycin-Resistant Staphylococcus aureus Isolate from a Patient in Pennsylvania. Antimicrob. Agents Chemother.; 48, 275-280, 2004.
THOMPSON, J.D.; HIGGINS, D.G. & GIBSON, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22, 4673-4680, 1994.
TIWARI, H.K. & SEN, M.R. Emergence of vancomycin resistant Staphylococcus aureus (VRSA) from a tertiary care hospital from northern part of India. BMC Infect. Dis., 6,156, 2006.
TOLEDO-ARANA, A.; MERINO, N.; VERGARA-IRIGARA,Y. M.; DEBARBOUILLE, M.; PENADES, J.R. & LASA, I. Staphylococcus aureus develops an alternative, ica-independent biofilm in the absence of the arlRS two-component system. J. Bacteriol., 187, 5318-5329, 2005.
TOMASZ, A. Multiple-antibiotic resistant pathogenic bacteria. A report on the Rockefeller University workshop. New Engl. J. Med., 330, 1247-1251, 1994.
TORMO, M.A.; KNECHT, E.; GOTZ, F.; LASA, I. & PENADES, J.R. Bap-dependent biofilm formation by pathogenic species of Staphylococcus, evidence of horizontal gene transfer? Microbiology., 151, 2465-2475, 2005.
TRABER, K. & NOVICK, R. A slipped-mispairing mutation in AgrA of laboratory strains and clinical isolates results in delayed activation of agr and failure to translate δ- and α-haemolysins. Mol. Microbiol., 59, 1519–1530, 2006.
TRUONG-BOLDUC, Q.C.; ZHANG, X. & HOOPER, D.C. Characterization of NorR protein, a multifunctional regulator of norA expression in Staphylococcus aureus. J. Bacteriol., 185, 3127-3138, 2003.
TSIODRAS, S.; GOLD, H.S.; SAKOULAS, G.; ELIOPOULOS, G.M.; WENNERSTEN, C.; VENKATARAMAN, L.; MOELLERING, R.C. & FERRARO, M.J. Linezolid resistance in a clinical isolate of Staphylococcus aureus. Lancet., 358, 207-208, 2001.
TURNER, K.M. & FEIL, E.J. The secret life of the multilocus sequence type. Int. J. Antimicrob. Agents., 29, 129-135, 2007.
TVERDEK, F.P.; CRANK, C.W. & SEGRETI, J. Antibiotic therapy of methicillin-resistant Staphylococcus aureus in critical care. Crit. Care Clin., 24, 249-260, 2008.
ULRICH, M.; BASTIAN, M.; CRAMTON, S.E.; ZIEGLER, K.; PRAGMAN, A.A.; BRAGONZI, A.; MEMMI, G.; WOLZ, C.; SCHLIEVERT, P.M.; CHEUNG, A. & DÖRING, G. The staphylococcal respiratory response regulator SrrAB induces ica gene transcription and polysaccharide intercellular adhesin expression, protecting Staphylococcus aureus from neutrophil killing under anaerobic growth conditions. Mol. Microbiol., 65, 1276-1287, 2007. Erratum in: Mol. Microbiol., 66, 278, 2007.
VALLE, J.; TOLEDO-ARANA, A.; BERASAIN, C.; GHIGO, J.; AMORENA, B.; PENADES, J.R. & LASA, I. SarA and not σB is essential for biofilm development by Staphylococcus aureus. Mol. Microbiol., 48, 1075-1087, 2003.

94
VAN BELKUM, A.; VAN LEEUWEN, W.; VERKOOYEN, R.; SACILIK, S.C.; COKMUS, C. & VERBRUGH, H. Dissemination of a single clone of methicillin-resistant Staphylococcus aureus among Turkish hospitals. J. Clin. Microbiol., 35, 978-981, 1997a.
VAN BELKUM, A.; VANDENBERGH, M.; KESSIE, G.; QADRI, S.M.; LEE, G.; VAN DEN BRAAK, N.; VERBRUGH, H. & AL-AHDAL, M.N.. Genetic homogeneity among methicillin-resistant Staphylococcus aureus strains from Saudi Arabia. Microb. Drug Resist., 3, 365-369, 1997b.
VAN WAMEL, W.; XIONG, Y.Q.; BAYER, A.S.; YEAMAN, M.R.; NAST, C.C. & CHEUNG, A.L. Regulation of Staphylococcus aureus type 5 capsular polysaccharides by agr and sarA in vitro and in an experimental endocarditis model. Microb. Pathog., 33, 73-79, 2002.
VAUDAUX, P.E.; FRANÇOIS, P.; PROCTOR, R.A.; MCDEVITT, D.; FOSTER, T.J.; ALBRECHT, R.M.; LEW, D.P.; WABERS, H. & COOPER, S.L. Use of adhesion-defective mutants of Staphylococcus aureus to define the role of specific plasma proteins in promoting bacterial adhesion to canine arteriovenous shunts. Infect. Immun., 63, 585-590, 1995. Erratum in: Infect. Immun., 63, 3239, 1995.
VAUTOR, E.; ABADIE, G.; PONT, A. & THIERY, R. Evaluation of the presence of the bap gene in Staphylococcus aureus isolates recovered from human and animals species. Vet. Microbiol., 127, 407-411, 2008.
VERGARA-IRIGARAY, M.; MAIRA-LITRÁN, T.; MERINO, N.; PIER, G.B.; PENADÉS, J.R. & LASA, I. Wall teichoic acids are dispensable for anchoring the PNAG exopolysaccharide to the Staphylococcus aureus cell surface. Microbiol., 154, 865-877, 2008.
VRIENS, M.; BLOK, H.; FLUIT, A.; TROELSTRA, A.; VAN DER WERKEN, C. & VERHOEF, J. Costs associated with a strict policy to eradicate methicillin-resistant Staphylococcus aureus in a Dutch University Medical Center: a 10-year survey. Eur. J. Clin. Microbiol. Infect. Dis., 21, 782-786, 2002.
VUONG, C.; SAENZ, H.L.; GOTZ, F. & OTTO, M. Impact of the agr quorum-sensing system on adherence to polystyrene in Staphylococcus aureus. J. Infect. Dis., 182,1688-1693, 2000.
WALSH, T.R. & HOWE, R.A. The prevalence and mechanisms of vancomycin resistance in Staphylococcus aureus. Annu. Rev. Microbiol., 56, 657–675, 2002.
WEIGEL, L.M.; DONLAN, R.M.; SHIN, D.H.; JENSEN, B.; CLARK, N.C.; MCDOUGAL, L.K.; ZHU, W.; MUSSER, K.A.; THOMPSON, J.; KOHLERSCHMIDT, D.; DUMAS, N.; LIMBERGER, R.J. & PATEL, J.B. High-level vancomycin-resistant Staphylococcus aureus isolates associated with a polymicrobial biofilm. Antimicrob. Agents Chemother., 51, 231-238, 2007.
WESSON, C.A.; LIOU, L.E.; TODD, K.M.; BOHACH, G.A.; TRUMBLE, W.R., & BAYLES, K.W. Staphylococcus aureus Agr and Sar global regulators influence internalization and induction of apoptosis. Infect. Immun., 66, 5238-5243, 1998.
WHITENER, C.J.; PARK, S.Y.; BROWNE, F.A, PARENT, L.J.; JULIAN, K.; BOZDOGAN, B.; APPELBAUM, P.C.; CHAITRAM, J.; WEIGEL, L.M.; JERNIGAN, J.; MCDOUGAL, L.K.; TENOVER, F.C. & FRIDKIN, S.K. Vancomycin-resistant Staphylococcus aureus in the absence of vancomycin exposure. Clin. Infect. Dis., 38, 1049-1055, 2004.
WILCOX, M.H., KITE, P., MILLS, K. & SUGDEN, S. In situ measurement of linezolid and vancomycin concentrations in intravascular catheter-associated biofilm. J. Antimcrob. Chemother., 47, 171-175, 2001.

95
WRIGHT, J.S. III; LYON, G.J.; GEORGE, E.A.; MUIR, T.W. & NOVICK, R.P. Hydrophobic interactions drive ligand-receptor recognition for activation and inhibition of staphylococcal quorum sensing. Proc. Natl. Acad. Sci., USA., 101, 16168-16173, 2004.
XU, K.D.; MCFETERS, G.A. & STEWART, P.S. Biofilm resistance to antimicrobial agents. Microbiology., 146, 547-549, 2000.
YARWOOD, J.M.; BARTELS, D.J.; VOLPER, E.M. & GREENBERG, E.P. Quorum sensing in Staphylococcus aureus biofilms. J. Bacteriol.,186, 1838-1850, 2004.
YARWOOD, J.M.; PAQUETTE, K.M.; TIKH, I.B.; VOLPER, E.M. & GREENBERG, E.P. Generation of virulence factor variants in Staphylococcus aureus biofilms. J. Bacteriol., 189, 7961-7967, 2007.
ZHANG, L.; GRAY, L.; NOVICK, R.P. & JI, G. Transmembrane topology of AgrB, the protein involved in the post-translational modification of AgrD in Staphylococcus aureus. J. Biol. Chem., 277, 34736-34742, 2002.

96
7. ARTIGOS PUBLICADOS RELACIONADOS A ESTA TESE
AMARAL, M.M.; COELHO, L.R.; FLORES, R.P.; SOUZA, R.R.; SILVA-CARVALHO, M.C.; TEIXEIRA, L.A.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M. The predominant variant of the Brazilian epidemic clonal complex of methicillin-resistant Staphylococcus aureus has an enhanced ability to produce biofilm and to adhere to and invade airway epithelial cells. J. Infect. Dis.,192, 801-810, 2005.
COELHO, L.R.; SOUZA, R.R.; FERREIRA, F.A.; GUIMARÃES, M.A.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M.S. agr-RNAIII molecule divergently regulates glucose-induced biofilm formation in Staphylococcus aureus clinical isolates in a lineage-independent fashion. Microbiol., aceite condicionado à revisão, 2008.
SOUZA, R.R.; COELHO, L.R.; BOTELHO, A.M.N.; RIBEIRO, A.; RITO, P.N.; VIANA, V.V.; TEIXEIRA, L.A.; FERREIRA-CARVALHO, B.T. & FIGUEIREDO, A.M.S. Biofilm formation and prevalence of lukF-pv, seb, sec, and tst genes among hospital- and community-acquired isolates of some international MRSA linages. Clin. Microbiol. Infect., em revisão, 2008.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo