interaÇÃo de macrÓfagos peritoneais com subpopulaÇÕes de … · 2013-10-30 · interaÇÃo de...
TRANSCRIPT

CURSO SUPERIOR DE TECNOLOGIA EM BIOTECNOLOGIA
INTERAÇÃO DE MACRÓFAGOS PERITONEAIS COM
SUBPOPULAÇÕES DE TOXOPLASMA GONDII QUE EXPÕEM OU
NÃO FOSFATIDILSERINA
THIAGO ALVES TEIXEIRA DOS SANTOS
RIO DE JANEIRO 2009

THIAGO ALVES TEIXEIRA DOS SANTOS
Aluno do Curso de Biotecnologia
INTERAÇÃO DE MACRÓFAGOS PERITONEAIS COM
SUBPOPULAÇÕES DE TOXOPLASMA GONDII QUE EXPÕEM OU
NÃO FOSFATIDILSERINA
Monografia apresentada como
requisito parcial para a obtenção
do grau de Tecnólogo em
Biotecnologia pelo Centro
Universitário Estadual da Zona
Oeste.
Rio de Janeiro
2009

ii
INTERAÇÃO DE MACRÓFAGOS PERITONEAIS COM
SUBPOPULAÇÕES DE TOXOPLASMA GONDII QUE EXPÕEM OU
NÃO FOSFATIDILSERINA
Elaborado por Thiago Alves Teixeira dos Santos
Aluno do curso de Biotecnologia da UEZO
Este trabalho de Graduação foi analisado e aprovado com
Grau: 10,0
Rio de Janeiro, 24 de Julho de 2009
Jéssica Manya Bittencourt Dias Vieira, Doutora em Ciências (Microbiologia)
Maria de Fátima Sarro da Silva, Doutora em Ciências (Microbiologia)
Sergio Henrique Seabra, Doutor em Ciências (Biofísica)
Orientador e Presidente da Banca
RIO DE JANEIRO, RJ - BRASIL
JULHO DE 2009

iii
Resumo
A exposição de Fosfatidilserina (PS) na superfície membranar é o principal evento para
indicar apoptose. Esta exposição é fundamental para que o Fator de Transformação de
Crescimento beta1 (TGF-1) sinalize e induza uma resposta anti-inflamatória durante a
fagocitose de células apoptóticas. Quando o parasita protozoário expõe PS, inibe a
atividade inflamatória do macrófago por reproduzir a sinalização das células apoptóticas. A
Toxoplasmose é uma doença mundial causada pelo Toxoplasma gondii. No entanto, a
invasão ativa do T.gondii inibe a produção de NO, propiciando a persistência da infecção.
Nosso grupo tem mostrado que o mecanismo utilizado pelo T.gondii para inibir a produção
de NO é similar ao apresentado na Leishmania quanto a dependência da exposição de PS.
Em experimentos realizados, observou-se que, após a separação das populações PS+ e PS-,
o índice de infectividade das interações com parasitas PS- foi bastante reduzido em relação
à infecção gerada pela população PS+ e população total. Resultados obtidos por
microscopia eletrônica de varredura mostram que a subpopulação de parasitos PS+ penetra
no macrófago principalmente por penetração ativa, enquanto a subpopulação PS- invade
por um mecanismo fagocítico. Análise do nitrito contido no sobrenadante das interações
demonstram a inibição da produção de NO pelos macrófagos na infecção gerada pelos
parasitos PS+. O Resultado do teste in vivo e análise do baço e do fígado das cobaias
indicam que a subpopulação PS- causa um grande processo inflamatório no hospedeiro,
enquanto a subpopulação PS+ mata por hiperparasitemia. Estes resultados sugerem que as
subpopulações de T.gondii PS+ e PS- podem invadir macrófagos de diferentes formas e
que o desenvolvimento do parasita depende da população total.
Palavras-chave: Toxoplasma gondii; Macrófago; Óxido nítrico; Fosfatidilserina

iv
Abstract
The exposure of phosphatidylserine (PS) on the surface membrane is the main event to
indicate apoptosis. This exposure is essential for the growth factor beta1 (TGF-1) signalis
and induces a anti-inflammatory response during phagocytosis of apoptotic cells. When the
protozoan parasite exposes PS, inhibits the macrophage activity inflammatory signaling by
mimicking the apoptotic cells. Toxoplasmosis is a worldwide disease caused by
Toxoplasma gondii. However, active invasion of T.gondii inhibits production of NO,
leading to persistence of infection. Our group has shown that the mechanism used by
T.gondii to inhibit NO is similar to that presented by Leishmania related to the dependence
of the exposure of PS. In experiments, it was observed that after the separation PS- and
PS+ populations, the index of infectivity of interactions with PS- parasites was rather small
in relation to infection caused by PS+ population and total population. Results obtained by
scanning electron microscopy show that the subpopulation of parasites PS+ penetrates the
macrophages mainly by active penetration, while the PS- subpopulation invades by a
phagocytic mechanism. Analysis of nitrite in the supernatant of the interactions
demonstrate the inhibition of NO production by macrophages in parasite infection
generated by PS+. The result of the in vivo test in mice and spleen and liver analysis
indicate that PS- subpopulations cause a large inflammatory process in the host, while the
subpopulation PS+ kills per hyperparasitemy. These results suggest that subpopulations of
T.gondii PS + and PS- can invade macrophages in various ways and the development of
the parasite depends on the total population.
Keywords: Toxoplasma gondii, macrophages, nitric oxide; phosphatidylserine Keywords:
Toxoplasma gondii, macrophages, nitric oxide; phosphatidylserine

Sumário
Resumo.................................................................................................................................iii
Abstract................................................................................................................................iv
1. Introdução ......................................................................................................................... 7
1.1. O Toxoplasma gondii .................................................................................................. 7
1.2. O macrófago ................................................................................................................ 9
1.3. Parasito X Célula ....................................................................................................... 10
2. Objetivo ........................................................................................................................... 12
3. Materiais e métodos........................................................................................................ 13
3.1. Macrófagos peritoneais ............................................................................................. 13
3.2. Obtenção de taquizoítas de Toxoplasma gondii ........................................................ 13
3.3. Isolamento de subpopulações PS+ e PS-................................................................... 13
As subpopulações PS+ e PS- foram isoladas da seguinte forma:..................................... 13
3.4. Análise por Citometria de fluxo do método de separação das populações ............... 14
3.5. Ativação de macrófagos peritoneais e interação com parasitas ................................ 14
3.6. Preparo de amostras para análise em microscópio óptico e índice de Infecção ........ 14
3.7. Preparo de amostras para microscopia eletrônica de varredura ................................ 15
3.8. Análise da produção de óxido nítrico ........................................................................ 15
3.9. Teste in vivo............................................................................................................... 15
3.10. Preparo de amostras para microscopia eletrônica de transmissão e microscopia óptica ................................................................................................................................ 15
4. Resultados ....................................................................................................................... 17
4.1. Análise da eficácia do método de isolamento de subpopulações de T.gondii. .......... 17
4.2. Índice de infecção da interação realizada entre as subpopulações isoladas e a população total de T.gondii com macrófagos................................................................... 18
4.3. Microscopia eletrônica de varredura das interações entre a subpopulação de T.gondii PS- e PS+ com macrófagos. ............................................................................................. 19
4.4. Dosagem do nitrito contido no sobrenadante da interação das subpopulações e população total (controle) de T.gondii com macrófagos .................................................. 20
4.5. Índice de mortalidade de camundongos C57/BL-6 durante as infecções com as subpopulações isoladas de T.gondii e a população total (controle).................................. 21

6
4.6. Microscopia óptica de tecido do baço e do fígado de camundongo C57/BL-6 após a interação com subpopulação de T. gondii PS + e PS- e população total.......................... 22
4.7. Microscopia Eletrônica de Transmissão do baço e do fígado de camundongos C57/BL-6 após a interação com as subpopulações e população total de T. gondii. ......... 23
5. Discussão ......................................................................................................................... 24
6. Conclusão ........................................................................................................................ 27
7. Referências ...................................................................................................................... 28

7
1. Introdução
1.1. O Toxoplasma gondii
A Toxoplasmose é uma doença mundial causada pelo Toxoplasma gondii e os
países que apresentam maiores taxas de infecção localizam-se em áreas de clima quente e
úmido e em altitudes menores. Em vários lugares do mundo, cerca de 95% da população
encontra-se infectada pelo T. gondii.
O T. gondii foi descrito primeiramente no Brasil por Splendore (1908) em um
coelho e, por Nicolle & Manceaux (1908) no Norte da África em um roedor
(Ctenodactylus gondi). Trata-se de um protozoário parasítico intracelular obrigatório capaz
de infectar uma grande variedade de vertebrados, inclusive seres humanos (LYONS &
JOHNSON, 1995; HOWE et al., 1996). Pertencente ao Filo Apicomplexa, à Classe
Sporozoea, à Sub-classe Coccidia, à Ordem Eucoccidiida, à Família Sarcocystidae e à
Sub-família Toxoplasmatinae (LEVINE et al., 1980).
O ciclo de vida do T.gondii (figura 1) é heterogêneo, tendo como hospedeiros
definitivos membros da família felidae e como hospedeiros intermediários outros
mamíferos e algumas aves. As principais formas infectivas do parasita são os bradizoítos,
que formam cistos teciduais, e os esporozoítos, que formam os oocistos.

8
Figura 1: Ciclo de vida do T. gondii (DUBEY, 1998)
A infecção origina-se principalmente por via oral, através da ingestão de água e
vegetais contendo oocistos ou carne contaminada por cistos teciduais. (WONG &
REMINGTON, 1993). Após a ingestão, através da degradação enzimática, a parede
externa dos cistos teciduais e dos oocistos é rompida, liberando, assim, as formas
infectantes esporozoítos e bradizoítos, respectivamente, no intestino do hospedeiro, onde
as células são invadidas rapidamente pelos parasitos, que se diferencia em uma forma de
multiplicação rápida, a taquizoíta, que se multiplica assexuadamente. Os taquizoítas são
liberados na corrente sanguínea após o rompimento da parede da célula hospedeira,
possibilitando, assim, a infecção de outras células, o que possibilita sua propagação por
todo o organismo. Esta infecção inicial é dada como a fase aguda da doença. Quando o
sistema imunológico do hospedeiro começa a atuar sobre os taquizoítas, o parasito sofre
uma nova diferenciação em bradizoítos, que é uma forma de desenvolvimento lenta,
formando os cistos teciduais, o que caracteriza a fase crônica da infecção (REY, 2002).
Em membros da família felidae, após a ingestão de alimentos contendo cistos, os
bradizoítos liberados dos enterócitos, multiplicam-se por uma série de esquizogonias, e se
diferenciam em gametas masculinos e femininos. A fusão desses gametas resulta na
formação de um zigoto diplóide (oocisto) que é liberado no ambiente através das fezes do

9
animal, onde se divide em 8 progênies haplóides denominadas esporozoítos (WONG &
REMINGTON, 1993). O T. gondii também pode ser transmitido pela via transplacentária
ao feto durante a infecção aguda de fêmeas no período de gestação (FRENKEL, 1990).
1.2. O macrófago
O macrófago é a célula mais diferenciada do sistema mononuclear fagocítico (VAN
FURTH, 1972) com medida típica entre 25 a 50 m de diâmetro. Sua superfície apresenta
grande número de projeções, pseudópodes e invaginações, e o citoplasma grande com
número de vesículas e vacúolos, que podem ser de secreção ou de reserva (SANTOS & DE
SOUZA, 1983). Filamentos de actina e microtúbulos estão espalhados pelo citoplasma
com forte concentração próximo à superfície celular e estão envolvidos na mobilidade dos
pseudópodos (DE SOUZA & BENCHIMOL, 1985), bem como no tráfico vesicular e de
organelas (ALLISON et al., 1971). O núcleo do macrófago apresenta-se com formato
irregular e em cortes finos mostra alguns lóbulos. Na microscopia óptica, o macrófago
possui um núcleo mais regular, com formato riniforme. A cromatina encontra-se dispersa.
Lisossomos estão presentes espalhados pelo citoplasma. A região do Golgi é bem
desenvolvida, posicionando-se ao redor do centríolo. O retículo endoplasmático rugoso
também é bem desenvolvido, estando geralmente perto do núcleo e do Complexo de Golgi
(Ian, 1973). As mitocôndrias encontram-se em grande número irradiando da região
perinuclear para a periferia celular (CARVALHO & DE SOUZA, 1989).
A ativação de macrófagos pode ser definida como sendo a aquisição da capacidade
de realizar funções complexas (ADANS & HAMILTON, 1984), funções essas que o
macrófago residente não realiza. Sabe-se que a ativação dos macrófagos está relacionada à
resposta imunológica inata ou específica na qual o antígeno (patógeno) inoculado é
apresentado a linfócitos T que secretam citocinas, como o interferon- (IFN-), que irão
ativar os macrófagos podendo, estes, serem mais ativados se entrarem em contato com
lipopolissacarídeo ou fator de necrose tumoral (TNF-) (ADANS & HAMILTON, 1984).
O IFN- é o principal sinal para ativação de macrófagos e está relacionado com
ativação anti-parasitária, que é baseada na produção de óxido nítrico (NO) (MUNÕZ-

10
FERNANDES et al., 1992). Macrófagos de camundongo tratados com IFN- em
combinação com lipopolissacarídeo bacteriano (LPS) vão expressar a enzima óxido
sintetase induzível (iNOS) (STUEHR & MARLETTA, 1987). Esta enzima, em
macrófagos de camundongo, possui duas variantes: uma encontrada no citosol (130 kDa) e
a outra associada a pequenas vesículas citoplasmáticas (135 Kda). Estas pequenas
vesículas não correspondem a lisossomos ou peroxissomos (VODOVOTZ et al., 1995). A
reação que leva a produção de NO pode ser descrita como se segue:
L - arginina iNOS NO + L – citrulina
Quadro 1: reação de produção de NO
Através da quantificação dos íons nitrito e nitrato, produtos finais gerados pela
estabilização do NO, é possível avaliar a produção de NO pelos macrófagos em cultura
(MOSHAGE et al., 1995)
1.3. Parasito X Célula
A fagocitose é a primeira barreira que protozoários invasores precisam burlar para
sobreviver. O processo de entrada se inicia com a adesão do microorganismo na superfície
celular do macrófago. Após a adesão à superfície e ingestão do protozoário, ocorre a
formação do vacúolo fagocítico, que se funde com lisossomas. As enzimas lisossomais
degradam o que se encontra no interior do fagolisossoma. Certos parasitos protozoários
como o T. cruzi escapam desses vacúolos sem danos expressivos, outros como o T. gondii
não permitem a fusão dos lisossomas com o vacúolo, enquanto que parasitas como
Leishmania sp. conseguem sobreviver dentro de vacúolos acidificados que contêm enzimas
lisossomais (revisto em DE SOUZA, 1994).
Já foi descrito que amastigotas (BALANCO et al., 2001) e promastigotas
metacíclicas (TRIPATHI & GUPTA, 2003) de L. amazonensis expressam fosfatidilserina
(PS) do lado externo da membrana plasmática (BARCINSKI et al., 2003). A exposição de
Fosfatidilserina (PS) na superfície da membrana plasmática é o principal evento para

11
indicar apoptose. Esta exposição é fundamental para que o Fator de Transformação de
Crescimento beta1 (TGF-1) sinalize e induza uma resposta antiinflamatória durante a
fagocitose de células apoptóticas. Quando o parasita protozoário expõe PS, inibe a
atividade inflamatória do macrófago por reproduzir a sinalização das células apoptóticas.
Nosso grupo tem mostrado que o mecanismo utilizado pelo T.gondii para inibir o
NO é similar ao apresentado na Leishmania quanto à dependência da exposição de PS.
Estudos da interação de macrófagos com T. gondii vem demonstrando que o parasito
também possui capacidade de inibir a ativação de macrófagos. Foi demonstrado que
macrófagos de camundongo ativados com IFN- e LPS infectados com taquizoítas de T.
gondii tem a produção de NO inibida devido à redução da expressão da iNOS em
macrófagos infectados (SEABRA et al., 2002). Taquizoítas de T. gondii apresentam baixa
infectividade quando colocados para interagir com células mutantes (sem receptores para
TGF-), comparados a interação com células parentais (SEABRA et al., 2002). Sugere-se
que o T.gondii utiliza estes receptores, podendo ativá-los durante a entrada. Tem-se
descrito a exposição de PS nas formas infectivas de protozoários parasitas como T. cruzi
(DAMATTA, et al., 2007).
Estes protozoários parasitos como o Toxoplasma gondii possuem mecanismos de
invasão às células hospedeiras muito investigados, tais como: fagocitose (JOINER &
DUBREMETZ, 1993) na qual o parasito não tem participação ativa na entrada e também
por penetração ativa, onde o parasito entra em contato com a membrana da célula
hospedeira, força-a, formando vacúolo rente ao seu corpo.
Estudos têm sido realizados com a finalidade de conhecer processos relacionados à
infecção com estes parasitos como mecanismos de invasão e infecção gerados em células
fagocíticas como o macrófago.

12
2. Objetivo
Avaliar o comportamento da infecção realizada por subpopulações PS+ e PS- isoladas.

13
3. Materiais e métodos
3.1. Macrófagos peritoneais
Macrófagos peritoneais foram obtidos por lavagem peritoneal de camundongos
suiços (CF1) com 5 ml de solução de Hank. Os macrófagos foram plaqueados em
lamínulas de vidro em placas de 24 poços. Após 1 hora de aderência a 37°C, as células
eram lavadas com solução de Hank a 37°C e cultivadas com meio Eagle modificado por
Dulbeccus (DMEM) contendo 5% de soro fetal bovino (SFB) a 37°C, em atmosfera de 5%
de CO2.
3.2. Obtenção de taquizoítas de Toxoplasma gondii
Taquizoítas de T. gondii (cepa RH) foram mantidos por inoculação na cavidade
peritoneal de camundongos a cada 2 ou 3 dias. Após esse período, era feito lavado
peritoneal injetando 3 ml de solução de Hank. O lavado foi centrifugado a 100g por 5
minutos, o sobrenadante coletado e centrifugado a 1000g por 10 minutos. Os parasitos
eram ressuspensos em DMEM e contados.
3.3. Isolamento de subpopulações PS+ e PS-
As subpopulações PS+ e PS- foram isoladas da seguinte forma:
Os parasitos foram coletados do camundongo infectado, purificados e em seguida foi
adicionado Anexina V incorporada à microesferas magnéticas, que reconhece e se liga
à Fosfatidilserina. Após a ligação das microesferas aos parasitos, foi feita a passagem
destes pela coluna magnética, onde somente os parasitos PS+ ficam aderidos à coluna,
que depois foi eluída para coleta destes.

14
3.4. Análise por Citometria de fluxo do método de separação das populações
Após a separação realizada na coluna magnética, as subpopulações de T.gondii
isoladas foram incubadas com iodeto de propidium por 1h à temperatura ambiente. O
controle foi feito com a população total dos parasitos, também incubada com iodeto de
propidium. Após incubação, os parasitos foram analisados em um citômetro de fluxo BD
Xcalibur. As análises em histograma e gráfico de pontos foram realizadas no programa
WinMDI 2.8 para PC.
3.5. Ativação de macrófagos peritoneais e interação com parasitas
Interações foram feitas com macrófagos: a) residentes, b) ativados com 50 U/ml de
interferon- recombinante de camundongo (IFN-; Sigma) e 100 ng/ml de
lipopolissacarídeo bacteriano (LPS; Sigma). As células eram lavadas e os protozoários PS-
, PS+ isolados e população total colocados na proporção de 10 parasitas por macrófago por
1 e 24 horas
3.6. Preparo de amostras para análise em microscópio óptico e índice de Infecção
Após o processo de fixação, as células foram lavadas, coradas com solução de
Giemsa (diluído em água destilada na proporção de 1:10), desidratadas numa série de
soluções acetona-xilol, montadas sobre Entellan e observadas em microscópio óptico
Axioplan – ZEISS. O índice de infecção (I.I.) foi obtido da seguinte forma:
I.I.= % células infectadas (nº parasitos intracelulares/ nº total de células infectadas).
Quadro 2: cálculo realizado para obtenção do índice de infecção

15
3.7. Preparo de amostras para microscopia eletrônica de varredura
As lamínulas foram fixadas em fixador de Karnovsky (paraformaldeído 4%,
glutaraldeído 2,5%, tampão cacodilato de sódio 0,1M, pH 7.4), desidratadas nas seguintes
concentrações de acetona: 30%, 40%, 50%, 70% e 100%, secas em ponto crítico e
recobertas com ouro através de metalização com 25 nm de espessura.
3.8. Análise da produção de óxido nítrico
A produção de NO foi avaliada indiretamente pela leitura colorimétrica do nitrito
nos diferentes sobrenadantes das culturas pelo reagente de Griess. Os sobrenadantes eram
misturados na proporção de 1:1 com o reagente de Griess (1 volume de 1 % de
Sulfanilamida em 5 % de ácido ortofosfórico em água deionizada com volume igual de 0,1
% de N-[1-Naphthyl] Ethylenediamina em água deionizada). Após 10 minutos, a mistura
era lida em leitor de ELISA (540 nm) e a quantificação da produção de NO baseada na
curva padrão feita com quantidades de nitrito de Sódio definidas, diluídas no meio de
cultura (GREEN et al., 1982).
3.9. Teste in vivo
Oito camundongos C57/BL-6 foram inoculados com a subpopulação de parasitos
PS-, oito com a subpopulação PS+ e oito com a população total (controle), na concentração
de 105 parasitos por mL. Baços e fígados desses camundongos foram coletados e
processados para microscopia óptica e eletrônica de transmissão após a morte dos mesmos.
3.10. Preparo de amostras para microscopia eletrônica de transmissão e microscopia
óptica
A etapa de fixação foi feita com 2.5% de glutaraldeído e 4% de paraformaldeído
em tampão cacodilato de sódio 0,1M (pH 7.4). Antes da pós-fixação, as amostras foram
lavadas com tampão cacodilato de sódio e então pós-fixadas com solução contendo 1% de
tetróxido de ósmio. Após este processo, as amostras foram desidratadas com acetona em

16
diferentes concentrações: 30%, 40%, 50%, 70% e 100%. A inclusão foi feita com resina
epóxi. Seções ultrafinas de 80 nm de espessura foram coradas com acetato de uranila e
citrato de chumbo (amostras para microscopia de transmissão) e com coradas com solução
de Giemsa (amostras para microscopia óptica).
17
4. Resultados
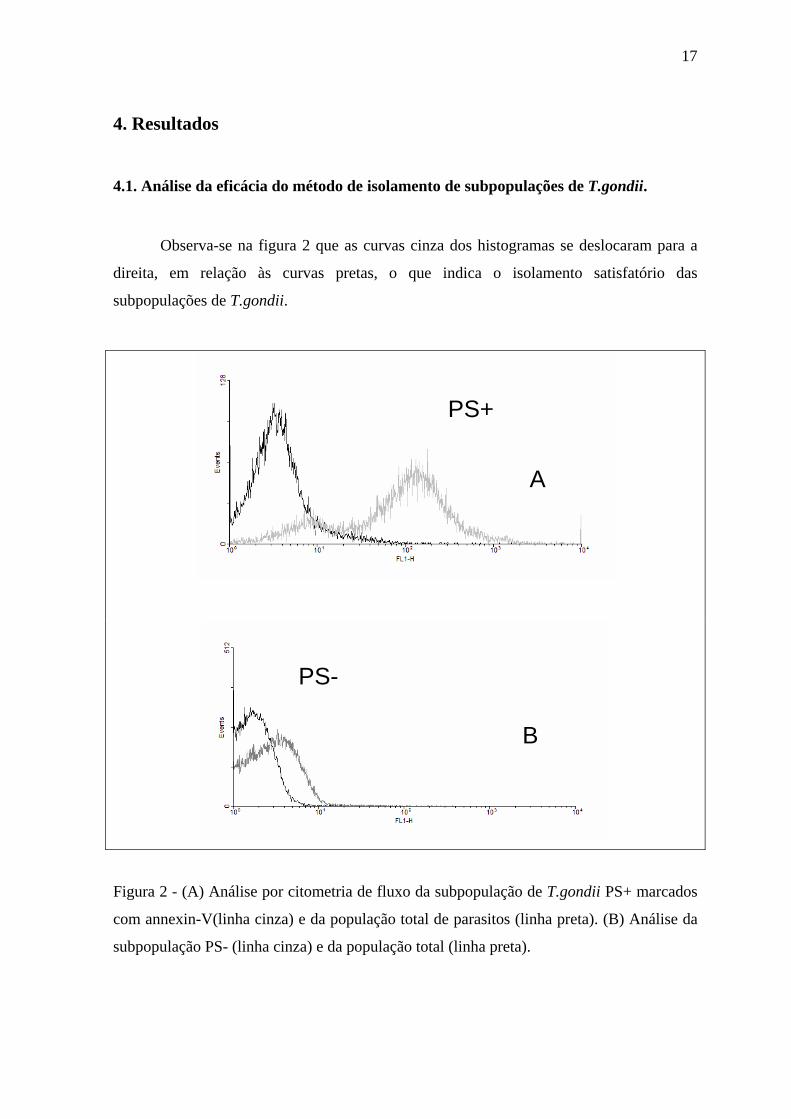
4.1. Análise da eficácia do método de isolamento de subpopulações de T.gondii.
Observa-se na figura 2 que as curvas cinza dos histogramas se deslocaram para a
direita, em relação às curvas pretas, o que indica o isolamento satisfatório das
subpopulações de T.gondii.
A
PS+
B
PS-
Figura 2 - (A) Análise por citometria de fluxo da subpopulação de T.gondii PS+ marcados
com annexin-V(linha cinza) e da população total de parasitos (linha preta). (B) Análise da
subpopulação PS- (linha cinza) e da população total (linha preta).
18
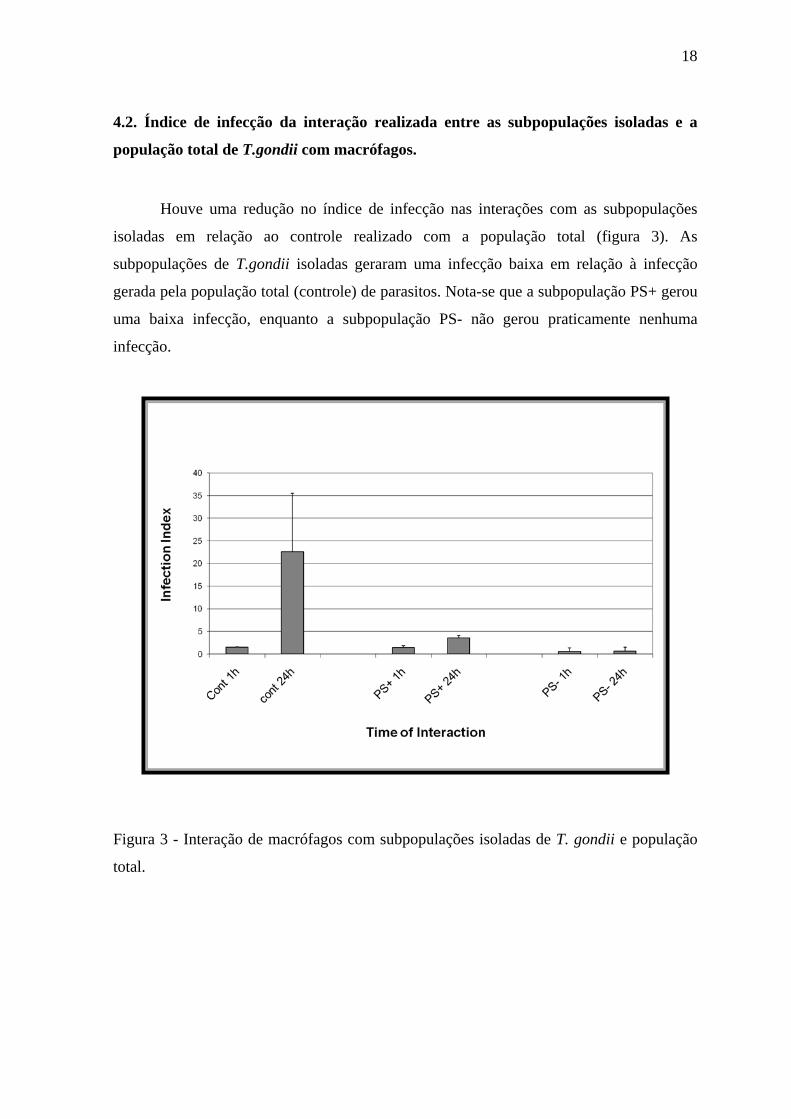
4.2. Índice de infecção da interação realizada entre as subpopulações isoladas e a
população total de T.gondii com macrófagos.
Houve uma redução no índice de infecção nas interações com as subpopulações
isoladas em relação ao controle realizado com a população total (figura 3). As
subpopulações de T.gondii isoladas geraram uma infecção baixa em relação à infecção
gerada pela população total (controle) de parasitos. Nota-se que a subpopulação PS+ gerou
uma baixa infecção, enquanto a subpopulação PS- não gerou praticamente nenhuma
infecção.
Figura 3 - Interação de macrófagos com subpopulações isoladas de T. gondii e população
total.
19
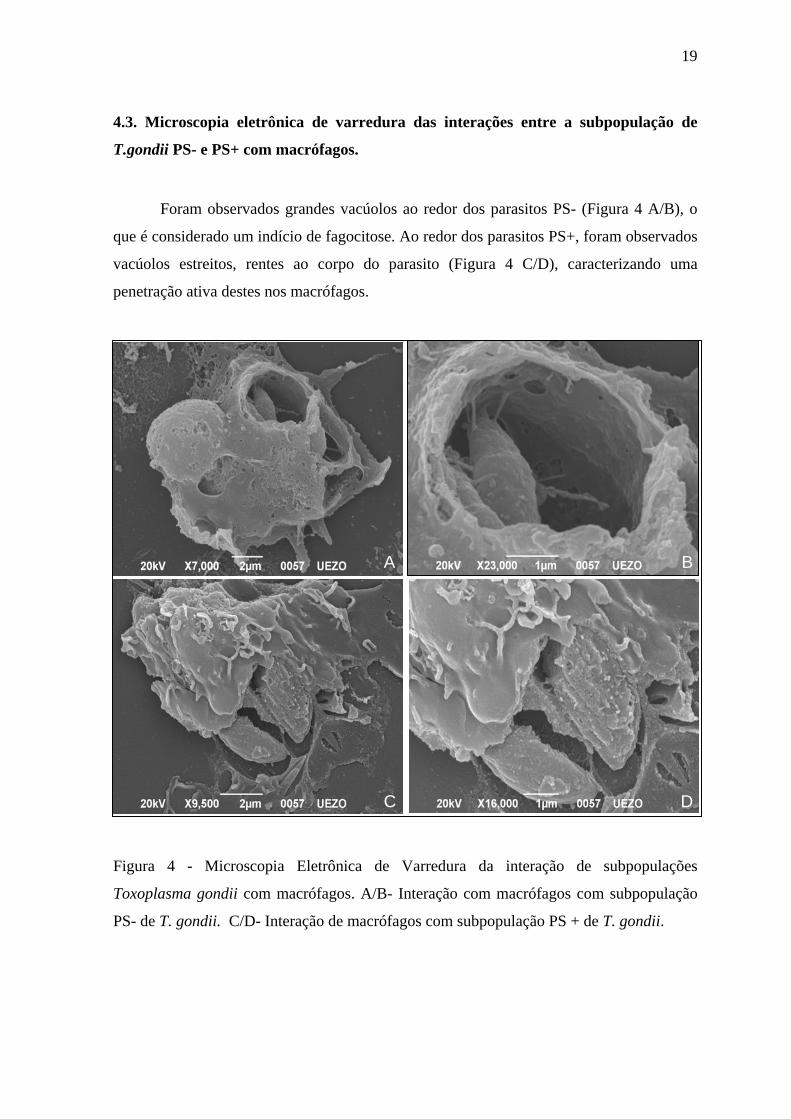
4.3. Microscopia eletrônica de varredura das interações entre a subpopulação de
T.gondii PS- e PS+ com macrófagos.
Foram observados grandes vacúolos ao redor dos parasitos PS- (Figura 4 A/B), o
que é considerado um indício de fagocitose. Ao redor dos parasitos PS+, foram observados
vacúolos estreitos, rentes ao corpo do parasito (Figura 4 C/D), caracterizando uma
penetração ativa destes nos macrófagos.
A B
C D
Figura 4 - Microscopia Eletrônica de Varredura da interação de subpopulações
Toxoplasma gondii com macrófagos. A/B- Interação com macrófagos com subpopulação
PS- de T. gondii. C/D- Interação de macrófagos com subpopulação PS + de T. gondii.
20
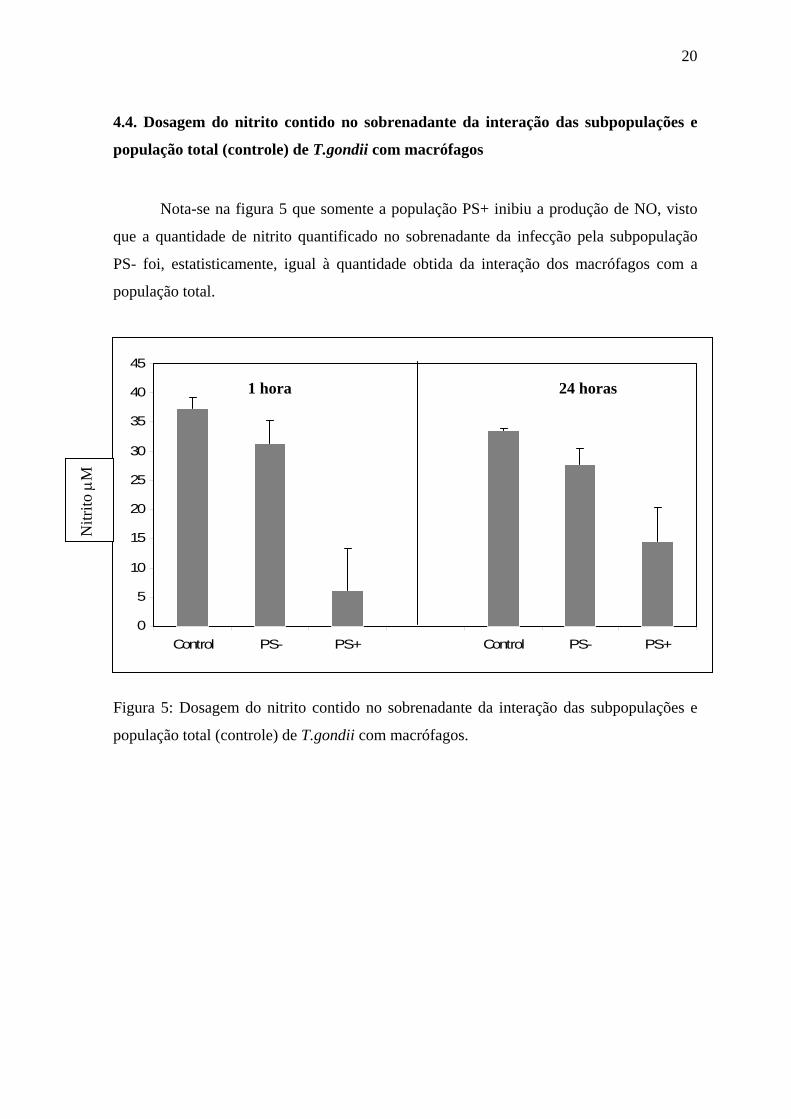
4.4. Dosagem do nitrito contido no sobrenadante da interação das subpopulações e
população total (controle) de T.gondii com macrófagos
Nota-se na figura 5 que somente a população PS+ inibiu a produção de NO, visto
que a quantidade de nitrito quantificado no sobrenadante da infecção pela subpopulação
PS- foi, estatisticamente, igual à quantidade obtida da interação dos macrófagos com a
população total.
0
5
10
15
20
25
30
35
40
45
Control PS- PS+ Control PS- PS+
24 horas 1 hora
Nit
rito
M
Figura 5: Dosagem do nitrito contido no sobrenadante da interação das subpopulações e
população total (controle) de T.gondii com macrófagos.
21
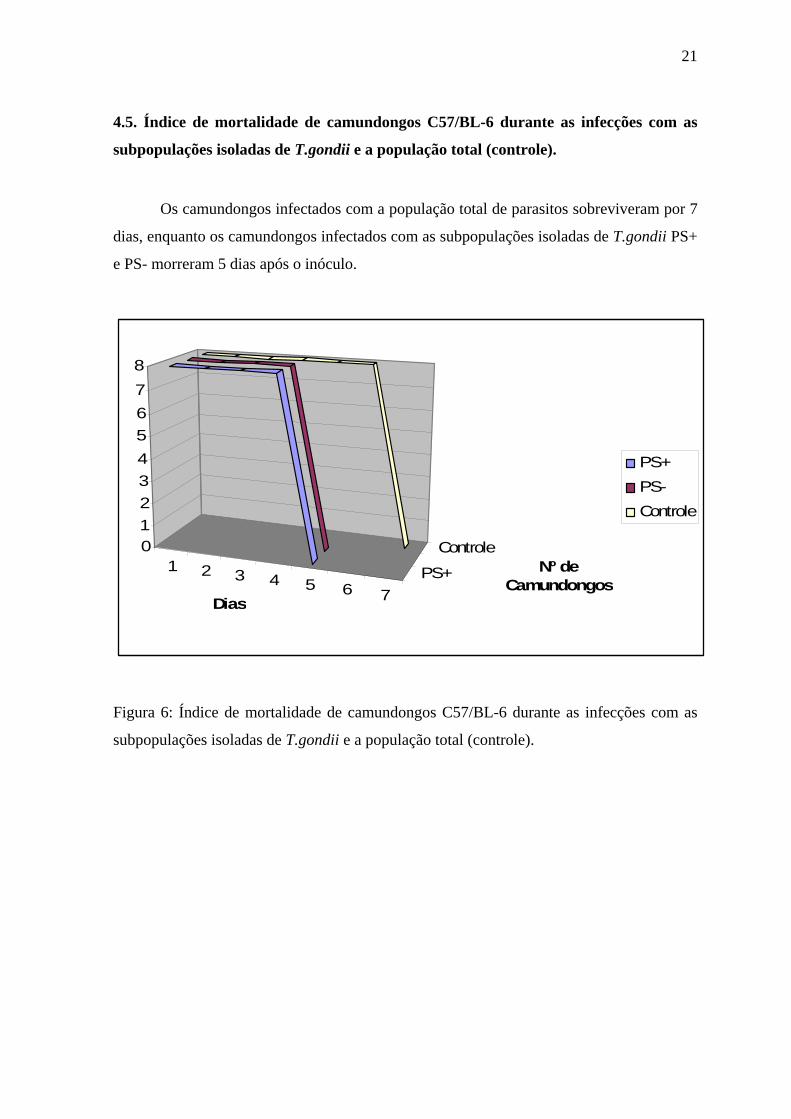
4.5. Índice de mortalidade de camundongos C57/BL-6 durante as infecções com as
subpopulações isoladas de T.gondii e a população total (controle).
Os camundongos infectados com a população total de parasitos sobreviveram por 7
dias, enquanto os camundongos infectados com as subpopulações isoladas de T.gondii PS+
e PS- morreram 5 dias após o inóculo.
1 2 3 4 5 6 7
PS+
Controle01
2
3
4
5
6
7
8
Dias
Nº de Camundongos
PS+
PS-
Controle
Figura 6: Índice de mortalidade de camundongos C57/BL-6 durante as infecções com as
subpopulações isoladas de T.gondii e a população total (controle).
22
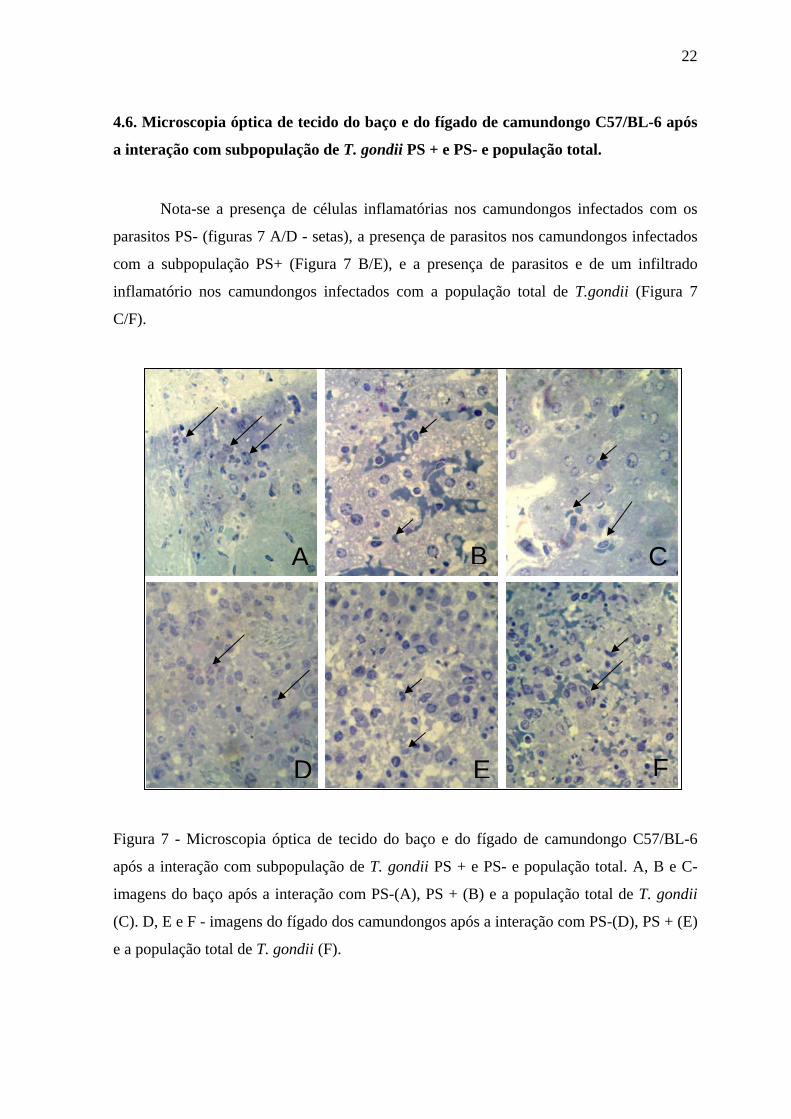
4.6. Microscopia óptica de tecido do baço e do fígado de camundongo C57/BL-6 após
a interação com subpopulação de T. gondii PS + e PS- e população total.
Nota-se a presença de células inflamatórias nos camundongos infectados com os
parasitos PS- (figuras 7 A/D - setas), a presença de parasitos nos camundongos infectados
com a subpopulação PS+ (Figura 7 B/E), e a presença de parasitos e de um infiltrado
inflamatório nos camundongos infectados com a população total de T.gondii (Figura 7
C/F).
B CA
FD E
Figura 7 - Microscopia óptica de tecido do baço e do fígado de camundongo C57/BL-6
após a interação com subpopulação de T. gondii PS + e PS- e população total. A, B e C-
imagens do baço após a interação com PS-(A), PS + (B) e a população total de T. gondii
(C). D, E e F - imagens do fígado dos camundongos após a interação com PS-(D), PS + (E)
e a população total de T. gondii (F).
23
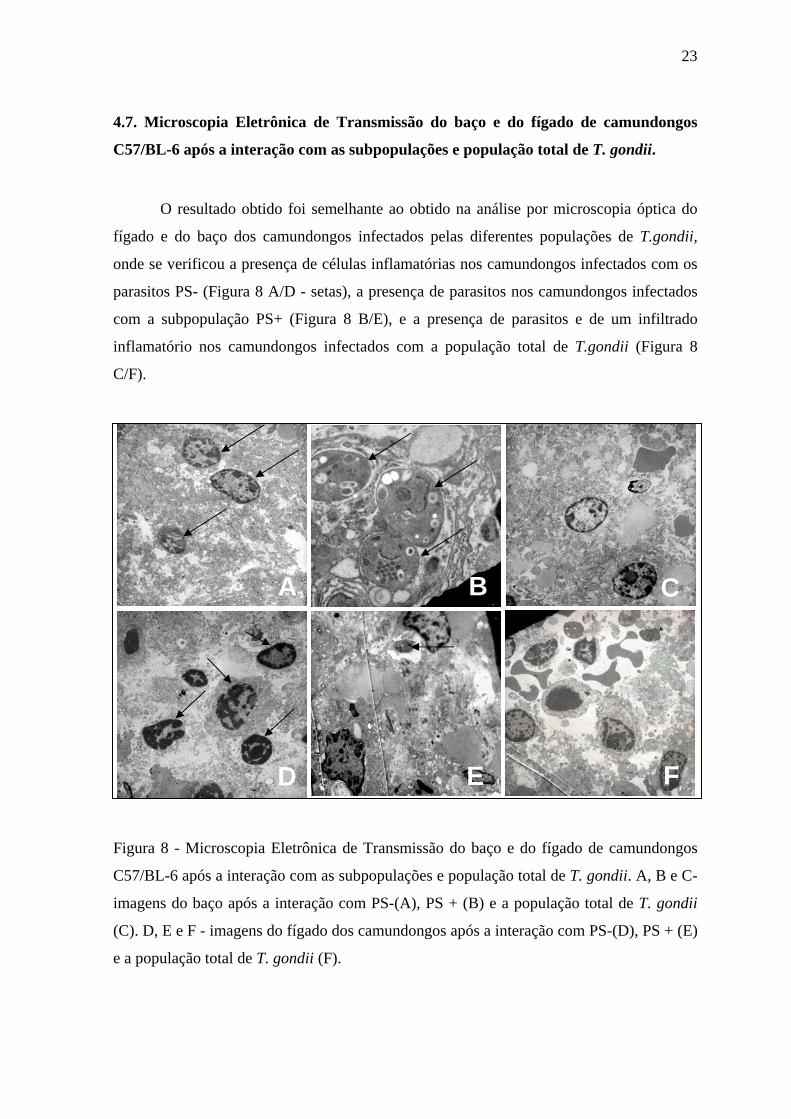
4.7. Microscopia Eletrônica de Transmissão do baço e do fígado de camundongos
C57/BL-6 após a interação com as subpopulações e população total de T. gondii.
O resultado obtido foi semelhante ao obtido na análise por microscopia óptica do
fígado e do baço dos camundongos infectados pelas diferentes populações de T.gondii,
onde se verificou a presença de células inflamatórias nos camundongos infectados com os
parasitos PS- (Figura 8 A/D - setas), a presença de parasitos nos camundongos infectados
com a subpopulação PS+ (Figura 8 B/E), e a presença de parasitos e de um infiltrado
inflamatório nos camundongos infectados com a população total de T.gondii (Figura 8
C/F).
A B
ED
C
F
Figura 8 - Microscopia Eletrônica de Transmissão do baço e do fígado de camundongos
C57/BL-6 após a interação com as subpopulações e população total de T. gondii. A, B e C-
imagens do baço após a interação com PS-(A), PS + (B) e a população total de T. gondii
(C). D, E e F - imagens do fígado dos camundongos após a interação com PS-(D), PS + (E)
e a população total de T. gondii (F).

24
5. Discussão
O Toxoplasma gondii é um protozoário parasítico intracelular obrigatório e
causador da Toxoplasmose, uma doença mundial que atinge principalmente países de
clima quente e úmido, localizados em altitudes menores, infectando diversos tipos de
vertebrados, inclusive humanos (LYONS & JOHNSON, 1995; HOWE et al., 1996). O
parasito penetra no organismo do hospedeiro através da ingestão de vegetais e água
infectados por oocistos (esporozoítos) ou carne contaminada por cistos teciduais
(bradizoítos), podendo ainda ocorrer a infecção via transplacentária, quando a
contaminação da mãe ocorre durante o período gestacional. Após penetrar no organismo
hospedeiro por via oral, a parede dos cistos é degradada pela ação de enzimas estomacais e
os parasitos são liberados, diferenciando-se em taquizoítas, que é uma forma de rápida
reprodução. Os taquizoítas invadem as células do hospedeiro e multiplicam-se
assexuadamente, (endodiogenia) nos hospedeiros intermediários, ou sexuadamente, em
membros da família felidae, o que caracteriza a fase aguda da doença. Quando o sistema
imunológico do hospedeiro passa a desempenhar uma grande atividade contra os parasitos,
estes invadem as células e se diferenciam em bradizoítos, que é uma forma de proliferação
lenta do T.gondii, formando os cistos teciduais, caracterizando a fase crônica da doença.
O sucesso da infecção do organismo pelo parasito depende dos mecanismos de
evasão deste da resposta imunológica do hospedeiro. Têm-se descrito em estudos com
Leishmania amazonensis que a exposição de fosfatidilserina na superfície dos parasitos é
um dos principais mecanismos de inibição da resposta imunológica. A fosfatidilserina é
um fosfolipídio que compõe a membrana celular que, em situações normais, fica voltado
para o meio intracelular. Quando uma célula entra em processo de apoptose, a
fosfatidilserina é exposta na parte extracelular da membrana, funcionando como
sinalizador inibindo o desencadeamento de uma resposta inflamatória influenciada pelos
macrófagos. Este trabalho foi baseado nesta capacidade que certa população de T.gondii
possui de expor fosfatidilserina em sua superfície, o que mimetiza o estado de apoptose
celular, inibindo uma resposta anti-parasitária do organismo hospedeiro. Em busca da
compreensão do comportamento da infecção originada por estas subpopulações de
parasitos, a que expõe fosfatidilserina (PS+) e a que não expõe fosfatidilserina (PS-), foram

25
realizadas interações in vitro e testes in vivo após o processo de isolamento dessas
subpopulações, através de separação magnética.
Com o objetivo de comprovar que o mecanismo de isolamento de subpopulações
empregado separava satisfatoriamente as populações PS+ e PS-, após o processo de
separação, os parasitos foram processados e submetidos à citometria de fluxo. De acordo
com a plotagem dos histogramas obtidos por citometria de fluxo após a análise da
população total de parasitos e das subpopulações isoladas (figura 2) foi possível verificar
que o sistema de separação empregado fornece um isolamento satisfatório das
subpopulações, o que é primordial para que os resultados obtidos nos experimentos
posteriores sejam validados.
Através da interação entre macrófagos peritoneais de camundongos com as
subpopulações isoladas de T. gondii e população total nos períodos de 1 hora e 24 horas
(figura 3), foi observado que as subpopulações isoladas de T.gondii estabeleceram uma
infecção menor que a população total (controle). Para melhor compreensão deste resultado,
foi realizada a análise por microscopia eletrônica de varredura dessas interações, o que
possibilitou a verificação de que os parasitos PS- invadem os macrófagos através de
mecanismos fagocíticos, através da observação do grande vacúolo ao redor do parasito
(figuras 4A e 4B), enquanto os parasitos PS+ invadem os macrófagos por penetração ativa,
visto o vacúolo rente ao corpo do parasito (figuras 4C e 4D). Com este resultado, conclui-
se que o índice de infecção na interação com a subpopulação PS+ foi maior que o índice
gerado pela infecção por parasitos PS- devido aos diferentes métodos de invasão celular
utilizados e ao estabelecimento do mecanismo de evasão pelos parasitos, onde os que
expõem fosfatidilserina penetram nos macrófagos de maneira ativa e, além disso, é capaz
de mimetizar a sinalização de uma célula em apoptose, inibindo, assim, a produção de NO
no interior do vacúolo. Já os parasitos PS- negativos, além de penetrarem nos macrófagos
por fagocitose, não são capazes de estabelecer o mecanismo de evasão, não inibindo,
assim, a atividade anti-parasitária dos macrófagos, o que explica a praticamente ausência
de infecção na interação por estes parasitos.
A dosagem do nitrito presente no sobrenadante nas interações por colorimetria
torna possível a elucidação da taxa de produção de óxido nítrico (NO) pelos macrófagos,

26
que, normalmente, produzem o NO no interior do vacúolo durante sua atividade anti-
parasitária, afim de destruir os parasitos que se encontram no vacúolo. A quantificação do
nitrito contido no sobrenadante das interações com as subpopulações isoladas, assim como
com a população total de T.gondii (figura 5) evidenciam que somente a subpopulação PS+
inibiu a produção de NO, visto que a quantidade de nitrito quantificado no sobrenadante da
infecção pela subpopulação PS- foi, estatisticamente, igual à quantidade obtida da
interação dos macrófagos com a população total. Este resultado confirma a idéia de que
somente os parasitos que expõem fosfatidilserina são capazes de inibir a produção de NO
pelos macrófagos.
Com a avaliação in vivo do comportamento das infecções com as subpopulações
PS- e PS+ total de T.gondii, verificou-se que os camundongos infectados pelas
subpopulações isoladas de parasitos tiveram uma morte precoce em relação aos
camundongos infectados pela população total de parasitos (figura 6). Através da avaliação
do fígado e do baço dos camundongos infectados por microscopia óptica (figura 7) e
microscopia eletrônica de transmissão (figura 8), sugere-se que os camundongos infectados
pela subpopulação PS+ morreram por hiperparazitemia (nota-se a presença de muitos
parasitos nas figuras 7B, 7E, 8B e 8E), visto que o sistema imunológico destes não foi
ativado de modo a conter e controlar a proliferação do parasito no organismo, e que os
camundongos infectados pela subpopulação PS- morreram devido o grande processo
inflamatório gerado por seus sistemas imunológicos (nota-se a presença de células
inflamatórias nas figuras 7A, 7D, 8A e 8D) já que os parasitos que não expõem
fosfatidilserina não são capazes de estabelecer a inibição da atividade antiparasitária. No
fígado e baço de camundongos infectados com a população total (controle) observa-se a
presença de parasitos e de infiltrado inflamatório (figuras 7C, 7F, 8C e 8F,
respectivamente), sugerindo que a sobrevida desses camundongos em relação aos outros
infectados com as subpopulações isoladas deve-se ao estado de equilíbrio entre a resposta
imunológica do hospedeiro e o desenvolvimento do parasito.

27
6. Conclusão
Com os resultados obtidos, conclui-se que para a infecção pelo parasito
Toxoplasma gondii ser estabelecida satisfatoriamente, possibilitando o desenvolvimento do
parasito, é necessária a presença da população total destes. Este fato pode se dar através da
necessidade do controle da resposta imunológica, de modo que seja estabelecido um limite
na proliferação dos parasitos no organismo, garantindo, assim, a sobrevivência do
hospedeiro e, consequentemente, do T.gondii.

28
7. Referências
Adams, D.O. & Hamilton, T.A. - The cell biology of macrophage activation. Ann.
Rev. Immunol. (1984), 2: 283-318.
Adams, D.O. and Hamilton, T.A. (1988). Phagocytic cells. Cytotoxic activities of
macrophages. In Inflamation. Basic principles and clinical correlates (ed. J.I. Galin, I.M.
Goldstein, and R. Snyderman), pp. 471-92. Raven, New York
Adams, L.B., Hibbs, J.B., Jr., Taintor, R.R. & Krahenbuhl, J.L. Microbiostatic
effect of murine-activated macrophages for Toxoplasma gondii role for synthesis of
inorganic nitrogen oxides from L-arginine. J. Immunol. (1990), 144: 2725-2729.
Albina, J. E., Cui, S., Mateo, R. B., & Reichnner, J. S. Nitric oxide mediated
apoptosis in murine peritoneal macrophages. J. Immunol. (1993), 150: 5080-5085.
Araújo-Jorge, T.C., Waghabi, M.C., Hasslocher-Moreno, A.M., Xavier, S.S.,
Higuchi, M.L., Keramidas, M., Bailly, S. & Feige, J.-J.Implication of transforming growth
factor-1 in Chagas disease Myocardiopathy. J. Infect. Dis. (2002), 186: 1823-1828.
Auger, M.J. & Ross, J.A. 1992. The biology of the macrophage. In The
Macrophage. (C.E. Lewis and J.O'D. McGee, eds) IRL Press, New York, NY. 1-74.
Balanco, J.M.F., Moreira, M.E.C., Bonomo, A.C., Bozza, P.T., Amarante-Mendes,
G., Pitmez, C., & Barcinski, M.A. Apoptotic mimicry by an obligate intracellular parasite
downregulates macrophage microbicidal activity. Curr. Biol. (2001), 11: 1870-1873.
Balestiere, F.M., Queiroz, A.R., Scavone, C., Costa, V.M., Barral-Netto, M.,
Abrahamsohn, I.A. Leishmania (L.) amazonensis-induced inhibition of nitric oxide synthesis in host macrophages. Microbes Infect. (2002), 4: 23-29.
Barcinski, M.A., Moreira, M.E.C., Balanco, J.M.F., Wanderley, J.L.M., &
Bonomo, A.C. The role of apoptotic mimicry in host-parasite interplay: is death the only
alternative for altruistic behavior? Kinetop. Biol. Dis. (2003), 2: 6-7.

29
Barral-Neto, M. Barral, A., Brownell, C. E., Skeiky, Y. A. W., Ellingsworth, L. R.,
Twardzik, D. R., and Reed, S. G. Transforming growth factor- in leishmanial infection: a
parasite escape mechanism. Science (1992), 257: 545-548.
Barral, A., Barral-Neto, M., Yong, E. C., Brownell, C. E., Twardik, D. R., & Reed,
S. G. Transforming growth factor as a virulence mechanism for Leishmania brasiliensis.
Proc. Natl. Acad. (1993), 90: 3442-3446.
Bermudez, L. E., Covaro, G., & Remington, J. Infection of murine macrophage
with Toxoplasma gondii is associated with released of transforming grown factor- and
down regulation of expression of tumor necrosis factor receptors. Infec. Immun. (1993),
61: 4126-4130.
Bhone, W., Heesemann, J., & Gross, U. Reduced replication of Toxoplasma gondii
is necessary for induction of bradyzoite-specific antigens: a possible role for nitric oxide in
triggering stage conversion. Infec. Immun. (1994), 62: 1761-1767.
Bogdan, C. & Röllinghoff, M. How do protozoan parasites survive inside
macrophages? Parasitol. Today (1999), 15: 22-28.
Bratton, D.L., Fadok, V.A., Richter, D.A., Kailey, J.M., Frasch, S.C., Nakamura,
T., & Henson, P.M. Polyamine regulation of plasm membrane phospholipid flip-flop
during apoptosis. J. Biol. Chem. (1999), 274: 28113-28120.
Butcher, B. A. & Denkers, E. Y. Mechanism of entry determines the ability of
Toxoplasma gondii to inhibit macrophage proinflammatory cytokine production. Infect.
Immun. (2002), 70: 5216-5224.
Cohn, Z.A. - The activation of mononuclear phagocytes: fact, fancy and future. J.
Immunol. (1978), 121: 813-816.

30
DaMatta, R.A., Seabra, S.H., Manhaes DS, L. & de Souza, W. Nitric oxide is not
involved in the killing of Trypanosoma cruzi by chicken macrophages. Parasitol. Res.
(2000), 86: 239-243.
De Souza, W. Estrategias parasitarias de los protozoos intracelulares. Invest.
Ciencia. (1994)212: 56-63.
Dubey, J.P., Lindsay, D.S., Speer, C.A. Structures of Toxoplasma gondii
tachyzoites, bradyzoites and sporozoites and biology and development of tissue cysts.
Clin. Microbiol. Rev. (1998), 11: 267-299.
Furtado, G. C., Slowik, M., Kleimman, H. K. & Joiner, K. A. Laminin enhances
biding of Toxoplasma gondii tachyzoites to J774 murine macrophage cells. Immunity.
(1992), 60: 2337-2342.
Greenberg, S. S., Jie, O., Zhao, X., & Wang, J. Role of PKC and tyrosine kinase in
ethanol-mediated inhibition of LPS-inducible nitric oxide synthase. Alcohol (1998), 16:
167-175.
Liew, F. Y. & Cox, F. E. G. Nonspecific defence mechanism: the role of nitric
oxide. Immunol. Today. (1991), 12: A17-A21.
Liew, F. Y., Millot, S., Parkinson, C., Palmer, R. M. J., & Moncada, S. Macrophage
killing of Leishmania parasite in vivo is mediated by nitric oxide from L-arginine. J.
Immunol. (1990), 144: 4794-4797.
Lüder, C. G. K., Algner, N., Lang, C., Bleicher, N., & Gross, U. Reduced
expression of inducible nitric oxide synthase after infection with Toxoplasma gondii
facilitates parasite replication in activated murine macrophages. Int. J. Parasitol. (2003),
33: 833-844.
MacMicking, J., Xie, Q.-W., & Nathan, C. Nitric oxide and macrophage function.
Annu. Rev. Immunol. (1997), 15: 323-350.

31
Malkovsky, M., & Sondel, P.M. Interleukin 2 and its receptor: structure, function
and therapeutic potential. Blood Rev. (1987), 1: 254-266.
Ming, M., Ewen, M. E., & Pereira, M. E. A. Trypanosome invasion of mammalian
cells requires activation of the TGF signaling pathway. Cell. (1995), 82: 287-296.
Norris, K. A., Schrimpf, J. E., Flynn, J. L., & Morris Jr., S. M. Enhancement of
macrophage microbicidal activity: supplemental arginine and citrulline augment nitric
oxide production in murine paritoneal macrophage and promote intracellular killing of
Trypanosoma cruzi. Infect. Immun. (1995), 63: 2793-2796.
Pakianathan, D. R., & Kuhn, E. Trypanosoma cruzi affects nitric oxide production
by murine peritoneal macrophages. J. Parasitol. (1994), 80: 432-437.
REY, L. Bases da parasitologia Médica. 2.ed. LOCAL: Guanabara Koogan,2002.
P.102-105.
Roberts, A.B. TGF- signalling from receptors to the nucleus. Mic. Infect. (1999),
1: 1265-1273.
Robinson, J. P. Oxygen & Nitrogen reactive metabolites and phagocytic cells.
Phagocyte Function: A guide for Research and Clinical Evaluation. Ed. J. P. Robinson
and George F. Babcock, pp.217, 1998.
Rosen, G.M., Pou, S., Ramos, C.L., Cohen, M.S. & Britigan, B.E. Free radicals
and phagocytic cells. FASEB J. (1995), 9: 200-209.
Scharton-Kersten, T. M., Yap, G., Magran, J., & Sher, A. Inducible nitric oxide is
essential for host control of persistent but not acute infection with the intracellular
pathogen Toxoplasma gondii. J. Exp. Med. (1997), 185: 1261-1273.

32
Seabra, S.H., de Souza, W., & DaMatta, R.A. Toxoplasma gondii partially inhibits
nitric oxide production of activated murine macrophages. Exp. Parasitol. (2002), 100: 62-
70.
Sheffler, L. A., Wink, D. A., Mellilo, G., & Cox, G. W. Exogenous nitric oxide
regulates INF- plus lipopolyssacharide-induced nitric oxide synthase expression in mouse
macrophages. J. Immunol. (1995), 155: 886-894.
Sibley, L. D., Lawson, R., & Weidner, E. Superoxide dismutase & catalase in
Toxoplasma gondii. Mol. Biochem. Parasitol. (1986), 19: 83.
Sibley, L.D. Invasion of vertebrate cells by Toxoplasma gondii. Trends Cell Biol.
(1995), 5: 129-132.
Sibley, L. D. Toxoplasma gondii: perfecting an intracellular life style. Traffic.
(2003), 4: 581-586.
Silva, J. S., Vespa, G. N. R., Cardoso, M. A. G., Aliberti, J. C. S., & Cunha, F. Q.
Tumor necrosis factor alpha mediates resistance to Trypanosoma cruzi infection in mice by
inducing nitric oxide production in infected gamma interferon-activated macrophages.
Infect. Immun. (1995), 63: 4862-4867.
Stenger, S., Thüring, H., Röllinghoff, M., & Bogdam, C. Tissue expression of
inducible nitric oxide synthase is closely associated with resistance to Leishmania major.
J. Exp. Med. (1994), 180: 793-793.
Stuehr, D.J. & Marletta, M.A. Induction of nitrite/nitrate synthesis in murine
macrophages by BCG infection, lymphokines, or Interferon-. J. Immunol. (1987), 139:
518-525.
Stuehr, D.J. & Marletta, M.A. Mamalian nitrate biosynthesis: mouse macrophages
produce nitrite and nitrate in response to Escherichia coli lipopolysacharide. Proc. Natl.
Acad. Sci. USA (1985), 82: 7738-7742.

33
Subauste, C. S., & Remington, J. S. Immunity to Toxoplasma gondii. Curr. Opn.
Immunol. (1993), 5: 532-537.
Thardin, J. F., M`Rini, C., Beraud, M., Vandaele, J., Frisach, M. F., Bessieres, M.
H., Seguela, J. P., & Pipy, B. Eicosanoid production by mouse peritoneal macrophages
during Txoplasma gondii penetration: role of parasite and host cell phospholipases. Infect.
Immun. (1993), 61: 1432-1441.
Triphati, A., & Gupta, C.M. Transbilayer translocation of membrane
phosphatidylserine and its role in macrophage invasion in Leishmania promastigotes. Mol.
Biochem. Parasitol. (2003), 128: 1-9.
Vieira, M., Dutra, J.M.F., Carvalho, T.M.U., Cunha-e-Silva, N., Souto-Padrón, T.,
& de Souza, W. Cellular signalling during the macrophage invasion by Trypanosoma
cruzi. Histochem. Cell Biol. (2002), 118: 491-499.
Vodovotz, Y., Russell, D., Xie, Q-W., Bogdan, C. & Nathan, C. Vesicle membrane
association of nitric oxide synthase in primary mouse macrophages. J. Immunol. (1995),
154: 2914-2925.
Wong, S.Y., Remington, J.S. Biology of Toxoplasma gondii. AIDS, v.7, n.3, p.299-
316, 1993.