instituto nacional de pesquisas da …...sobre anatomia das raízes. a todos os professores do...
TRANSCRIPT

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
ASPECTOS ETNOBOTÂNICOS E TAXONÔMICOS DE ARACEAE JUSS. NA
COMUNIDADE SANTA MARIA, BAIXO RIO NEGRO - AM
RINA FÁTIMA MARANHÃO DE OLIVEIRA
Manaus, Amazonas
Julho, 2011

ii
RINA FÁTIMA MARANHÃO DE OLIVEIRA
ASPECTOS ETNOBOTÂNICOS E TAXONÔMICOS DE ARACEAE JUSS. NA
COMUNIDADE SANTA MARIA, BAIXO RIO NEGRO - AM
Orientadora: Dra. Maria de Lourdes da Costa Soares Morais
Coorientador: Dr. Valdely Ferreira Kinupp
Dissertação apresentada ao Instituto
Nacional de Pesquisas da Amazônia
como parte dos requisitos para
obtenção do título de Mestre em
Botânica.
Manaus, Amazonas
Julho, 2011

iii
Banca examinadora do projeto de dissertação (aula de qualificação)
1. Dr. Ari de Freitas Hidalgo – UFAM
2. Dr. Danilo Filho – CPCA / INPA
3. Dr. Hiroshi Noda – CPCA / INPA
Banca examinadora do trabalho de conclusão do curso (defesa presencial)
1. Dr. Ari de Freitas Hidalgo – UFAM
2. Dr. Danilo Oliveira – UFRJ
3. Dra. Rita Mesquita – MUSA / INPA

iv
O48 Oliveira, Rina Fátima Maranhão de Aspectos etnobotânicos e taxonômicos de Araceae Juss. na comunidade Santa Maria, Baixo rio Negro, AM) / Rina Fátima Maranhão de Oliveira. --- Manaus : [s.n.], 2011. xvi, 119 f : il. color. Dissertação (mestrado) -- INPA, Manaus, 2011 Orientador : Maria de Lourdes da Costa Soares Coorientador : Valdely Ferreira Kinupp Área de concentração : Botânica
1. Araceae. 2. Etnobotânica. 3. Taxonomia. 4. Anatomia vegetal. 5. Heteropsis flexuosa. 6. Philodendron solimoesense. 7. Monstera obliqua. I. Título.
CDD 19. ed. 584.64
Sinopse: Foi realizado um levantamento etnobotânico com ênfase na família Araceae em uma comunidade ribeirinha, localizada no Baixo rio Negro, Amazonas, onde se registrou um alto Valor de Uso para espécies pertencentes a essa família, que fornecem, a partir de suas raízes aéreas, fibras usadas no artesanato. A partir deste levantamento, foram investigados os aspectos anatômicos das lâminas foliares e das raízes aéreas das espécies de Araceae mais utilizadas para este fim, Heteropsis flexuosa (Kunth) G.S.Bunting (cipó-titica), Monstera obliqua Miq. (ambé-sima) e Philodendron solimoesense A.C.Sm. (ambé-coroa) visando identificar as principais características que justificam o seu uso em artesanato e contribuir com o estudo taxonômico dessas espécies. Palavras chave: 1- Etnobotânica. 2- Taxonomia. 3- Anatomia vegetal. 4- Araceae.

v
Aos meus pais, Renan e Shirley, que sempre
estiveram ao meu lado me apoiando e me
orientando.
Aos moradores da comunidade Santa Maria, que
permitiram minha presença entre eles e
compartilharam comigo seus conhecimentos e
sua hospitalidade, especialmente à Dona Isabel e
seu irmão Alonso, que me acolheram em suas
casas e serão inesquecíveis para mim.
DEDICO

vi
AGRADECIMENTOS
A Deus, que me concedeu saúde e proteção durante esse curso.
Aos meus orientadores, Dra. Maria de Lourdes Soares e Dr. Valdely Kinupp, pelo
incentivo, credibilidade, paciência e amizade depositadas a mim, e principalmente por tudo o
que aprendi com eles.
À amiga Andréia Barroncas, que desde o início do meu mestrado se dispôs a me
ajudar nas questões que envolviam desde a área de estudo até a anatomia vegetal.
Aos colegas da Fundação Vitória Amazônica – FVA, Sérgio Borges e Euzilene da
Silva que me apoiaram em Novo Airão, durante minha busca pelas comunidades.
Ao casal Lukcelly Loureiro, que me indicou a comunidade Santa Maria, e seu marido
Antônio Craveiro, (Centro de Apoio ao Turista – CAT / Novo Airão), que me receberam
muito bem em sua cidade.
À Izanei, filha de Dona Isabel, que muito gentilmente, intercedeu junto a sua mãe,
possibilitando a minha estadia na comunidade com segurança e conforto.
À Geise Canalez e Francisco Oliveira do Centro Estadual de Unidades de Conservação
– CEUC, que prontamente me indicaram os passos para obter autorização de pesquisa em UC
estadual.
Ao Comitê de Ética em Pesquisas com Seres Humanos – CEP/INPA, que contribuiu
bastante com a melhora da qualidade do trabalho durante a análise dos documentos.
Às doutoras Veridiana Scudeller, Zilvanda Lourenço e ao Dr. Ulisses Albuquerque
pelas contribuições fornecidas durante a correção do meu plano de trabalho.
Aos doutores Ari Hidalgo (UFAM), Hiroshi Noda (CPCA/INPA) e Danilo Fernandes
Filho (CPCA/INPA), que aceitaram o convite para participar da banca julgadora na minha
aula de qualificação.
Ao Sr. Francisco Pinto, da Fundação Amazonas Sustentável - FAS, que contribuiu
com algumas informações sobre a comunidade Santa Maria.
À Dra. Maria Silvia Mendonça, que autorizou a utilização do Laboratório de Botânica
Agroflorestal – LABAF/UFAM e aos amigos Manoel Viana (Bacurau), Poliana Roversibe,
Mariana Cassino pela ajuda no trabalho e amizade.
Ao Eng. Jorge Alves de Freitas, da Coordenação de Pesquisa em Produtos Florestais -
CPPF/INPA, que, com muita paciência e dedicação me ajudou com os cortes e com as
medições das fibras das raízes aéreas das três espécies de Araceae.

vii
Ao Dr. Wilson Meireles e à Jackeline, do Laboratório Temático de Microscopia
Eletrônica – LTME/INPA, que nos deram toda a assistência durante a realização das
fotografias das lâminas histológicas.
Às sensacionais amigas de turma Nallaret, Luísa, Joana e irmãs postiças Juliana,
Mariana e Naia, pelo companheirismo e amizade.
Ao Dr. Luís Carlos Bonates (KK), com sua inestimável ajuda na compreensão da
anatomia vegetal e às amigas Catarina e Vanessa, do laboratório de anatomia vegetal do
INPA, pelos momentos de estudo e descontração.
Ao Dr. Antônio Carlos, chefe da Coordenação de Pesquisas em Botânica –
CPBO/INPA, e às amigas da secretaria Neide, Gisele e Jéssica.
Ao seu Sebastião Silvino (Sabá), que me ajudou na coleta de campo.
Ao amigo e agora irmão de consideração Gleison Viana, pela inestimável amizade e
contribuição nesse trabalho.
Ao Dr. Eduardo Gonçalves, pela identificação de Xanthossoma, e ao Dr. Charles
Zartman, do INPA, pela identificação de Humiriaceae.
Ao Sr. Felipe França, ilustrador botânico do INPA, pelo esforço em desenhar as
lindíssimas pranchas que ilustram o trabalho.
À professora Dra. Dalva Graciano (UnB) que me ajudou a esclarecer algumas dúvidas
sobre anatomia das raízes.
A todos os professores do programa do curso de botânica, que me enriqueceram com
seus conhecimentos e dedicação a essa disciplina tão nobre da biologia.
Aos doutores Ari Hidalgo (UFAM), Danilo Oliveira (UFRJ) e Rita Mesquita (INPA),
pelas contribuições finais na defesa pública.
A toda minha família, meus tios, tias, primos, primas, avós, que sempre me
incentivaram, principalmente aos meus pais, meu irmão e meu cachorro que estiveram sempre
aguentando minhas fases de nervosismo e me dando força e sabedoria para seguir em frente
superando a mais um desafio.

viii
“Um dia, o índio Pakalamoka foi à Mãe-do-Mato, e esta lhe ensinou como livrar os macuxis de seus
males. Ao nascer da lua, Pakalamoka dirigiu se a um lago que ela indicara e, ao chegar, viu um bando de
pássaros. Flechou a ave que voava mais alto e colocou em torno de onde ela caiu capim e gravetos, e fez uma fogueira. Deitou-se e dormiu. Ao acordar, viu ao redor
de si e do lago, tajás com folhas de vários feitios e cores deslumbrantes:
o tajá que defende a roça;
o tajá que o torna bom caçador e bom pescador; o tajá que o faz invisível aos inimigos e maus
espiritos; o tajá que o poupa das fadigas da guerra, da caça, etc.;
o tajá que o faz vencer as disputas tradicionais da tribo;
e o tajá que o faz amado.
Como lhe ensinara a Mãe-do-Mato, Pakalamoka arrancou os tajás necessários ao trabalho, à saúde, à paz, aos amores e à felicidade, e levou a seu povo. A
tribo tornou-se numerosa, valente; nunca mais lhe faltaram peixes nem caça; e cessaram as doenças”.
Nunes Pereira

ix
RESUMO
A família Araceae Juss. possui espécies bem conhecidas no âmbito econômico para o homem destacando-se por serem utilizadas como plantas medicinais, alimentícias, fibrosas, e principalmente ornamentais. As raízes aéreas das espécies dos gêneros Heteropsis Kunth e Philodendron Schott, pertencentes à família Araceae, tem sido usadas até hoje como fonte de fibras para confecção de cestos na região amazônica. Na região do Baixo Rio Negro, Heteropsis flexuosa (Kunth) G.S. Bunting, conhecida como “cipó-titica” é a espécie mais utilizada pelos extratores como fonte de fibras, quanto ao gênero Philodendron, existem várias espécies utilizadas pelos extratores para confecção de artesanato e outros utensílios. O objetivo geral deste trabalho foi realizar um levantamento das espécies da família Araceae com potencial econômico, utilizados pela comunidade Santa Maria, localizada na margem esquerda do rio Negro, na qual seus moradores possuem um modo de vida composto basicamente pela pesca e agricultura familiar, complementando estas atividades com a coleta de Produtos Florestais Não Madeireiros (PFNM`s). Para a coleta de informações etnobotânicas foram utilizadas ferramentas como o “rapport” e a “turnê guiada”. O registro das informações foi baseado em questionário semiestruturado e através de conversas informais. A análise das informações foi realizada através da totalização de usos. Foram encontradas 51 espécies de Araceae distribuídas em 15 gêneros. O gênero Philodendron foi o mais diverso com 22 espécies. No total, foram identificadas 23 espécies úteis, apresentando dentre estas, usos para o artesanato, alimentícias, medicinais, ornamentais e místicas. Dentre as que apresentaram maior valor de uso foram Philodendron solimoesense e Heteropsis flexuosa, espécies cujas raízes são utilizadas como fonte de fibra para a confecção de artigos artesanais. Foi detectado o uso em artesanato de outra espécie da família Araceae, Monstera obliqua Miq., conhecida pelos artesãos como “ambé-sima”. Foi realizado o tratamento taxonômico dessas três espécies onde foi levada em consideração a distribuição, dados fenológicos, afinidade e histórico dos espécimes e detalhes das estruturas vegetativas e reprodutivas da planta, os quais foram ilustrados em pranchas. Também foi elaborada uma chave de identificação contendo características da morfologia externa e interna das espécies. Quanto ao estudo anatômico da lâmina foliar e da raiz aérea das espécies, foram selecionados alguns caracteres como diagnóstico para a taxonomia, como tipo de estômatos, tipo de cristais, ocorrência de laticíferos, presença de hipoderme, ocorrência de bainha de fibras esclerenquimáticas, forma do cilindro central, dentre outros. Também foi feito o estudo das fibras das raízes, onde as três espécies apresentaram fibras longas do tipo libriforme com paredes espessas e lumens reduzidos.

x
ABSTRACT
The species of the Araceae family has in the economic well-known to the man standing out for being used as medicinal plants, food, fibers, and mainly ornamental. The aerial roots of the species of Philodendron Schott and Heteropsis Kunth belonging to the family Araceae, has been used until now as a source of fiber for making baskets in the Amazon region. In the Lower rio Negro, Heteropsis flexuosa (Kunth) G.S. Bunting, known as "cipó-titica" is the species most used by the extractors as a source of fiber, while the genus Philodendron, many species used by the extractors for making handicrafts and other utensils. The overall objective of this study was to survey the species of the family Araceae with economic potential, used by the community Santa Maria, located on the left bank of rio Negro, in which its residents have a way of life composed primarily by fishing and farming family, complementing these activities with the collection of Non Timber Forest Products (NTFP`s). In order to collect ethnobotanical information tools were used as the "rapport" and "guided tours". The recording of information was based on semi-structured questionnaire and through informal conversations. The analysis of information was realized through the aggregation of uses. We found 51 species of Araceae distributed in 15 genera. The genus Philodendron was the most diverse with 22 species. In total, 23 useful species were identified, presenting among these uses for crafts, food, medicinal, ornamental and mystical. Among those who had higher use value were Philodendron solimoesense and Heteropsis flexuosa, species whose roots are used as a fiber source for making arts and crafts. Was detected using in crafts of another species of the family Araceae, Monstera obliqua Miq., known for artisans such as "ambé-sima". The treatment of these three taxonomic species which was taken into account the distribution, phenological data, and historical affinity of the specimens and details of the vegetative and reproductive structures of plants, which were illustrated in boards. It was also prepared containing an identification key with characteristics of internal and external morphology of the species. As for the anatomical study of leaf and root shoots of species, some were selected as diagnostic characters for taxonomy, such as type of stomata, type of crystals, the occurrence of latex, presence of hypodermis, the occurrence of sclerenchyma sheath fibers, so the central cylinder, among others. Also the study was made of the fibers of the roots, where the three species had long fiber type libriforme with thick walls and small lumens.

xi
SUMÁRIO
INTRODUÇÃO.......................................................................................................... 1
OBJETIVOS
Geral....................................................................................................................... 4
Específicos.............................................................................................................. 4
REVISÃO BIBLIOGRÁFICA
A família Araceae Juss......................................................................................... 5
Anatomia................................................................................................................ 6
Usos da família no âmbito comercial e etnobotânico......................................... 8
Artesanato e construção................................................................................. 8
Medicinais........................................................................................................ 10
Místicos............................................................................................................ 12
Ornamentais.................................................................................................... 12
Alimentícios..................................................................................................... 13
REFERÊNCIAS......................................................................................................... 14
CAPÍTULO I - ETNOBOTÂNICA DE PLANTAS ÚTEIS, COM ÊNFASE EM ARACEAE JUSS. NA COMUNIDADE SANTA MARIA, BAIXO RIO NEGRO - AM
RESUMO.................................................................................................................... 23
ABSTRACT................................................................................................................ 23
INTRODUÇÃO.......................................................................................................... 24
OBJETIVOS............................................................................................................... 26
MATERIAL E MÉTODOS
Área de estudo....................................................................................................... 27
Aspectos ambientais.............................................................................................. 28
Aspectos sócio-econômicos................................................................................... 29
Coleta de informações etnobotânicas.................................................................. 30
Análise das informações....................................................................................... 31
Coleta de material botânico................................................................................. 31
Identificação e descrição das espécies................................................................. 32

xii
RESULTADOS E DISCUSSÃO............................................................................... 34
Perfil sócio-econômico dos informantes............................................................. 34
Diversidade de espécies úteis na comunidade Santa Maria ............................ 35
Artesanato............................................................................................................. 44
Coleta e beneficiamento do “ambé-coroa”........................................................ 46
Tecelagem............................................................................................................. 48
Alimentícias.......................................................................................................... 50
Medicinais............................................................................................................. 51
Ornamentais......................................................................................................... 52
Místicas................................................................................................................. 53
CONCLUSÃO............................................................................................................ 55
REFERÊNCIAS......................................................................................................... 56
CAPÍTULO II – TAXONOMIA E MORFOANATOMIA DE TRÊS ESPÉCIES DE ARACEAE JUSS. DA ÁREA DE PROTEÇÃO AMBIENTAL DA MARGEM ESQUERDA DO RIO NEGRO
RESUMO.................................................................................................................... 62
ABSTRACT................................................................................................................ 62
INTRODUÇÃO.......................................................................................................... 63
OBJETIVOS............................................................................................................... 65
MATERIAL E MÉTODOS
Área de estudo..................................................................................................... 66
Coleta de material botânico............................................................................... 66
Identificação das espécies................................................................................... 68
Morfologia interna da lâmina foliar.................................................................. 70
Morfologia interna das raízes aéreas e fibras................................................... 70
RESULTADOS
Tratamento taxonômico..................................................................................... 72
Descrição dos gêneros......................................................................................... 72
Heteropsis Kunth.................................................................................................. 72
Monstera Adanson............................................................................................... 72

xiii
Philodendron Schott……………………………………………………………. 73
Relações infragenéricas................................................................................. 74
Chave para identificação das espécies.............................................................. 75
Descrição das espécies........................................................................................ 76
Heteropsis flexuosa (Kunth) G.S. Bunting....................................................... 76
Morfologia interna............................................................................................. 81
Lâmina foliar................................................................................................. 81
Raiz aérea....................................................................................................... 82
Monstera obliqua Miq…………………………………………………………. 83
Morfologia interna............................................................................................. 88
Lâmina foliar................................................................................................. 88
Raiz aérea....................................................................................................... 89
Philodendron solimoesense A.C. Sm……………...…………………………... 90
Morfologia interna............................................................................................. 94
Lâmina foliar................................................................................................. 94
Raiz aérea....................................................................................................... 95
Principais características anatômicas distintivas da lâmina foliar e as
raízes aéreas entre as espécies de Araceae estudadas............................................. 97
Estudo das fibras................................................................................................ 98
DISCUSSÃO............................................................................................................... 100
CONCLUSÃO............................................................................................................ 105
REFERÊNCIAS........................................................................................................ 106
APÊNDICE A............................................................................................................ 111
APÊNDICE B .…………………………………………………………………… 112
APÊNDICE C............................................................................................................. 116
ANEXO A................................................................................................................... 118
ANEXO B.................................................................................................................... 119

xiv
LISTA DE FIGURAS
CAPÍTULO I - ETNOBOTÂNICA DE ESPÉCIES ÚTEIS, COM ÊNFASE EM ARACEAE JUSS. NA COMUNIDADE SANTA MARIA, BAIXO RIO NEGRO
Figura 1. Figura 1. Mosaico de Unidades de Proteção Ambiental do Baixo Rio Negro e as comunidades da APA da Margem Esquerda do Rio Negro – Setor Aturiá Apuauzinho.......................................................................................................................
27
Figura 2. Estrutura física da comunidade Santa Maria, Baixo rio Negro - AM.............. 28
Figura 3. Registro do trajeto mostrando as trilhas contemplando três ambientes visitados na APA da Margem Esquerda do Rio Negro – Setor Aturiá-Apuauzinho........ 30
Figura 4. Famílias com maior número de espécies úteis para a comunidade Santa Maria, Baixo rio Negro – AM......................................................................................... 39
Figura 5. Diversidade de gêneros encontrados no entorno da comunidade Santa Maria, Baixo rio Negro, AM............................................................................................ 40
Figura 6. Espécies com maior Valor de Uso na comunidade Santa Maria, Baixo rio Negro – AM...................................................................................................................... 43
Figura 7. Diversidade de gêneros cultivados nos quintais da comunidade Santa Maria, Baixo rio Negro............................................................................................................... 44
Figura 8. Utilização de fibras e artesanato da comunidade Santa Maria........................ 46
Figura 9. Etapas do beneficiamento do “ambé-coroa” (Philodendron solimoesense A.C. Sm)........................................................................................................................... 48
Figura 10. Etapas iniciais da confecção de um paneiro feito de “cipó-ambé” (Philodendron solimoesense A.C. Sm) por um artesão da comunidade Santa Maria................................................................................................................................. 49
Figura 11. Tecelagem do “paneiro” feito de “ambé-coroa” por artesão da comunidade Santa Maria....................................................................................................................... 49
Figura 12. Espécies de Araceae utilizadas para fins artesanais na comunidade Santa Maria, Baixo rio Negro – AM......................................................................................... 50
Figura 13. Espécies de Araceae cultivadas nos quintais da Comunidade Santa Maria, Baixo rio Negro – AM...................................................................................................... 54
CAPÍTULO II – TAXONOMIA E MORFOANATOMIA DE TRÊS ESPÉCIES DE ARACEAE JUSS. DA ÁREA DE PROTEÇÃO AMBIENTAL DA MARGEM ESQUERDA DO RIO NEGRO
Figura 1. Mosaico de Unidades de Proteção Ambiental do Baixo Rio Negro e as comunidades da APA da Margem Esquerda do Rio Negro – Setor Aturiá Apuauzinho. 66 Figura 2. Registro do trajeto mostrando as trilhas percorridas contemplando três ambientes visitados na APA da Margem Esquerda do Rio Negro – Setor Aturiá-Apuauzinho...................................................................................................................... 67 Figura 3. Desenho esquemático dos tipos de folhas e de inflorescências....................... 69
Figura 4. Aspecto geral e estruturas reprodutivas de Heteropsis flexuosa...................... 80
Figura 5. Estruturas da lâmina foliar de Heteropsis flexuosa.......................................... 81
Figura 6. Cortes transversais das raízes aéreas de Heteropsis flexuosa.......................... 82

xv
Figura 7. Aspecto geral e estruturas reprodutivas de Monstera obliqua......................... 87
Figura 8. Estruturas da lâmina foliar de Monstera obliqua............................................. 88
Figura 9. Cortes transversais das raízes aéreas de Monstera obliqua............................. 89
Figura 10. Aspecto geral e estruturas reprodutivas de Philodendron solimoesense....... 93
Figura 11. Estruturas da lâmina foliar de Philodendron solimoesense........................... 95
Figura 12. Cortes transversais da raiz aérea de Philodendron solimoesense.................. 96
Figura 13. Mapa de distribuição geográfica das espécies H. flexuosa, M. obliqua e P. solimoesense..................................................................................................................... 97

xvi
LISTA DE ABREVIAÇÕES E SIGLAS APA Área de Proteção Ambiental
CEUC Centro Estadual de Unidades de Conservação
FAO Food and Agriculture Organization of the United Nations
FVA Fundação Vitória Amazônica
INPA Instituto Nacional de Pesquisas da Amazônia
IPÊ Instituto de Pesquisas Ecológicas
ONG Organização Não-Governamental
RDS Reserva de Desenvolvimento Sustentável
SDS Secretaria de Desenvolvimento Sustentável
SEUC Sistema Estadual de Unidades de Conservação
UC Unidade de Conservação

1
INTRODUÇÃO
Apesar do grande conhecimento das espécies da família Araceae no âmbito
econômico, estudos voltados para a sua taxonomia ainda são considerados escassos, além
disso, muitas amostras encontradas nos herbários são mal coletadas, devido à especificidade
de coleta e herborização para a família. No entanto, muitos trabalhos tem sido desenvolvidos
nas últimas décadas, principalmente no que diz respeito à anatomia (e.g.: French, 1997;
Vianna et al., 2001; Keating, 2002; 2004; Ferreira et al., 2006; Mantovani et al., 2010),
evolução e filogenia (e.g.: Hay & Mabberley, 1991; French et al., 1995; Tam, 2004), tendo
como exemplo a ampliação da circunscrição de Araceae, com a inclusão dos gêneros da
família Lemnaceae (Souza & Lorenzi, 2008; APG III, 2009).
Araceae possui grande importância econômica, visto que diversas espécies podem ser
utilizadas na alimentação, no fornecimento de fibras para o artesanato, na medicina popular e,
principalmente, como ornamental, devido à beleza de suas folhagens e inflorescências (Soares
& Mayo, 1999). Neste trabalho, as espécies tratadas taxonomicamente apresentam-se como
hemiepífitas, que são plantas que se desenvolvem sobre um suporte e possuem raízes
conectadas com o solo, denominadas raízes alimentadoras ou aéreas (Croat, 1988).
As raízes aéreas das espécies dos gêneros Heteropsis e Philodendron, tem sido usadas
até hoje como fonte de fibras para confecção de cestos na região amazônica (Durigan &
Castilho, 2004; Soares, 2008). Na região do Baixo rio Negro, Heteropsis flexuosa (Kunth)
G.S. Bunting, conhecida como “cipó-titica” é a espécie mais utilizada pelos extratores como
fonte de fibras (Durigan & Castilho, 2004). Quanto ao gênero Philodendron, existem várias
espécies utilizadas pelos extratores para confecção de artesanato e outros utensílios em toda a
Amazônia brasileira e extrabrasileira (Leoni & Marques, 2008; Garcia, 2009; Souza, 2010).
A matéria-prima obtida através desses recursos é considerada um Produto Florestal
Não Madeireiro (PFNM), definido pela FAO (2002) como qualquer espécie de origem animal
ou vegetal que não seja madeira para fins comerciais, que além do serviço ecológico,
possibilitam incremento de renda familiar e emprego nas zonas rurais. Embora existam leis
para alguns produtos (palmito, seringueira, castanheira, copaíba, cipó-titica), o manejo dos
PFNMs não está sob regulamentação legal, o que, dificulta a fiscalização e a própria

2
exploração (Machado, 2008). O cipó-titica, por exemplo, foi abrangido pela legislação apenas
na década de 2000, no Amapá (Lei Estadual no. 0631/2001) e no Amazonas (IN 001/2008), a
fim de se normatizar o manejo adequado da espécie (Canalez, 2009).
Em virtude da ação predatória causada pela retirada das raízes dessas espécies, existe
hoje uma preocupação em controlar a extração desses recursos através do manejo sustentável.
Porém, para se alcançar esse objetivo é necessário investir no conhecimento científico e
entender como as populações tradicionais utilizam esses recursos visando obter estratégias
para a conservação do equilíbrio ecológico, bem como para a melhoria da qualidade de vida
dessas pessoas.
Muitos pesquisadores consideram a criação de Unidades de Conservação (UC’s) como
principal meio de se manter e conservar recursos genéticos existentes no país (Ming et al.
2003). A criação de UC´s propicia a proteção in situ tanto da biodiversidade como do
conhecimento tradicional. A conservação in situ é a conservação de ecossistemas e hábitats e
a manutenção e recuperação de populações viáveis de espécies em seus meios naturais e, no
caso de espécies domesticadas ou cultivadas, nos meios onde tenham se desenvolvido suas
propriedades características (BRASIL, 2000).
Uma das alternativas que tem se mostrado eficiente na preservação de recursos
florestais é a criação de Unidades de Conservação de Uso Sustentável, categoria na qual se
inclui as Áreas de Proteção Ambiental – APA e as Reservas Extrativistas – RESEX. Nessas
unidades, os moradores tradicionais são estimulados a tratar de forma sustentável o ambiente,
visto que estão ligados intimamente com a natureza e usufruindo diretamente de seus
recursos, o que lhes garante um vasto conhecimento sobre os mesmos. Nesse contexto, a
população tradicional pode indicar os usos de espécies vegetais potenciais e ensinar novos
modelos para uso e manejo dos mesmos (Posey, 1992).
As espécies apresentadas neste trabalho fazem parte de um amplo espectro de usos e
variedades que podem ser exploradas futuramente como incremento de renda familiar e como
alternativa de exploração racional da floresta. O artesanato, que já é uma prática associada aos
indígenas e às populações ribeirinhas do rio Negro, é um bom exemplo de geração de renda.
Além disso, segundo a FVA (2009), a venda de produtos com maior valor agregado em
substituição à venda da matéria-prima, reduz a pressão sobre o recurso, uma vez que é
necessária menor quantidade de cipós para se obter retorno financeiro semelhante,
melhorando a relação custo benefício desta atividade.
Na Amazônia, há muito ainda a ser pesquisado, tendo em vista a grande riqueza
biológica e cultural de seu vasto território. O grande número de populações indígenas e

3
ribeirinhas que vivem isoladas, associado às necessidades básicas de alimentação, construção
e remédio, faz com que a proporção de produtos florestais não madeireiros utilizados seja
muito alta em relação a outras regiões. No campo da investigação farmacológica, por
exemplo, Rodrigues (2006) ressalta que em muitos casos, o isolamento geográfico contribui
ao fortalecimento das práticas médicas locais e tradicionais.
Portanto, o desenvolvimento de uma pesquisa etnobotânica associada à taxonomia das
espécies de Araceae utilizadas na comunidade Santa Maria, no Baixo rio Negro, poderá
promover não só o aprofundamento do conhecimento científico dessas espécies, como
também subsídios para o manejo, visando à conservação deste, e de outros grupos de plantas,
possibilitando assim, a manutenção das tradições que sustentam sua cultura e economia.
No primeiro capítulo, serão apresentados aspectos sócioeconômicos de alguns
moradores e as espécies de Araceae ocorrentes na comunidade Santa Maria. Posteriormente,
serão apresentados tratamentos taxonômicos e morfoanatômicos das três espécies mais
utilizadas para artesanato na comunidade, Heteropsis flexuosa (Kunth) G.S. Bunting,
Monstera obliqua Miq. e Philodendron solimoesense A.C.Sm.

4
OBJETIVOS
Geral
Realizar levantamento etnobotânico das espécies com potencial econômico utilizados
pela comunidade, com ênfase na família Araceae Juss., complementando com estudos sobre a
morfologia interna e taxonomia das espécies com maior valor de uso para o artesanato.
Específicos
Identificar as espécies de Araceae e seus respectivos usos na comunidade Santa
Maria, na APA da Margem Esquerda do Rio Negro;
Descrever os aspectos da morfologia interna e taxonomia das espécies mais utilizadas
no artesanato pelos comunitários;
Classificar as espécies vegetais indicadas quanto à origem, formas de vida e áreas de
ocorrência;
Elaborar um guia básico de identificação com fotos coloridas das espécies ocorrentes
na comunidade e no seu entorno, visando o retorno dessas informações para a
população local e sociedade em geral no reconhecimento das espécies.

5
REVISÃO BIBLIOGRÁFICA
A família Araceae Juss.
Araceae é uma família subcosmopolita que possui atualmente 109 gêneros e cerca de 2.823
espécies concentradas predominantemente nos trópicos da América, no Sudeste da Ásia e no
Arquipélago da Malásia (Govaerts et al., 2002). Na América do Sul está representada por 38
gêneros e cerca de dois terços do total das espécies (Mayo et al., 1997; Soares & Mayo, 1999;
Soares & Jardim-Lima, 2005). No Brasil está distribuída em todos os domínios
fitogeográficos e dados preliminares indicam a existência de 35 gêneros perfazendo 458
espécies, das quais 286 são endêmicas (Coelho et al., 2010).
Suas espécies apresentam uma alta diversidade de hábito e uma grande variedade no
formato das folhas, demonstrando uma plasticidade fenotípica principalmente nas primeiras
fases de desenvolvimento, constituindo dessa forma um grupo complexo em termos de vida e
ecologia (Croat, 1990; Mayo et al., 1997; Soares & Mayo, 1999). O hábito varia desde ervas,
pequenas a gigantes, epífitas, hemiepífitas, geófitas, litófitas, helófitas, raramente flutuantes
ou aquáticas submersas; apresentam caule trepador, arborescente, ereto, reptante ou
subterrâneo. As folhas variam de simples a compostas; pecíolo geralmente conspícuo, exceto
no gênero Heteropsis, que apresenta pecíolos curtos. O formato da lâmina varia desde inteira,
ovada, cordada, sagitada, hastada, trífida ou trissecta, pedatífida, pinatífida, pedatissecta a
dracontioide. A grande maioria das espécies prefere ambientes úmidos como florestas pluviais
e brejos ou crescem próximas à água corrente (Mayo et al., 1997, Soares & Mayo, 1999).
Como características reprodutivas, Araceae apresenta inflorescência na forma de
espádice composta por inúmeras flores bissexuais ou unissexuais, associada a uma bráctea
chamada espata. A infrutescência é geralmente cilíndrica ou, às vezes, globosas. Os frutos são
tipicamente bagas suculentas, embora raramente secas, mais comumente vermelhas ou
laranjas, dependendo do gênero em questão (Mayo et al., 1997). Os mecanismos de
polinização são conhecidos em gêneros com flores unissexuais e o odor da inflorescência é o
fator mais importante para a atração dos polinizadores, que incluem abelhas da família
Trigoniidae e Euglossinidae, besouros da família Scarabaeidae e moscas da família
Drosophilidae (Mayo et al., 1997). A termogênese é um mecanismo de polinização, ocorrente
em Araceae, onde há elevação de calor volatilizando os odores das flores para atrair insetos
polinizadores. No caso de Philodendron solimoesense A.C. Sm., que ocorre na Amazônia e

6
em outras florestas tropicais úmidas, a temperatura aumenta em até 14°C acima da
temperatura ambiente da floresta ao redor (Exotic rain forest, 2010).
Historicamente, Jussieu (1789) estabeleceu Araceae como família natural, porém
reconheceu poucos gêneros, provavelmente devido à escassez de boas amostras de material de
taxa não europeu. Heinrich Wilhelm Schott foi o primeiro botânico a trabalhar somente com a
Araceae e fazer estudos comparativos minunciosos das inflorescências, flores e frutos de
Araceae, além disso, descreveu novos gêneros e espécies e criou a primeira classificação
natural principal para toda a família (Mayo et al., 1997). Posteriormente, o pesquisador que
mais contribuiu com a família foi Adolf Engler, fazendo a primeira modificação taxonômica
proposta por Schott na Flora brasiliensis em 1878. Em 1979, Bogner atualizou o sistema de
Engler, adicionando gêneros, na época, recentemente descritos e levando em conta novas
sinonímias.
Mais recentemente, trabalhos como o de Saguraki (1998) (“Taxonomia e filogenia das
espécies de Philodendron, seção Calostigma (Schott) Pfeiffer no Brasil”), Gonçalves &
Temponi (2004), (“A new Monstera (Araceae: Monsteroideae) from Brazil”), Temponi
(2007), (“Sistemática de Anthurium sect. Urospadix”), Soares et al., (2009), (“Two new
species and a new combination in Amazonian Heteropsis (Araceae)”) e APG II (2003) e III
(2003), por exemplo, vêm trazendo grandes contribuições para o entendimento da taxonomia
e filogenia da família no Brasil e no mundo.
Anatomia
Engler frequentemente usava características da anatomia para definir importantes
subdivisões na família Araceae. Desde o tratamento da família para a Flora Brasiliensis
(1878), o autor diferenciou do primeiro sistema de classificação proposto por Schott (1860),
com ênfase dada não só à morfologia vegetativa como também a anatomia. Solereder (1919)
revisou a anatomia de cerca de 23 gêneros de Aroideae e com a colaboração de Meyer (1928)
compilou observações sobre a anatomia geral de cerca da metade dos gêneros de Araceae.
Grayum (1984) revisou a literatura de anatomia vegetal em sua tese dando ênfase na anatomia
do pólen.
Trabalhos como os de Dahlgren & Cliford (1982), Dahlgren et al. (1985) foram
relevantes para o estudo da anatomia na família Araceae. No entanto, o pesquisador mais
prolífero para a anatomia sistemática da família Araceae foi James French, juntamente com
seus colaboradores. Entre suas contribuições destacam-se o estudo dos padrões de

7
espessamento na parede do endotécio nas subfamílias de Pothoideae e Monsteroideae
(French, 1985), observações nos padrões de vascularização de estames na família (French,
1986), reportando a ocorrência sistemática da hipoderme esclerótica em raízes de Araceae
(1987), na estrutura do óvulo e tricomas placentais (French, 1987) e registrando a ocorrência
de laticíferos anastomosados na família (French, 1988) onde, considerou a presença destes
laticíferos, como o caráter mais definitivo da subfamília Colocasioideae. French (1997)
também revisou a literatura sobre a anatomia interna e forneceu, apesar de não ilustrado, um
compreensivo sumário da anatomia de todos os órgãos e tecidos. Mais recentemente,
apresentam-se os estudos de Keating (2002), que publicou uma coletânea de trabalhos em
anatomia e investigação original de folha e pecíolo em vários gêneros de Araceae e os de
Keating (2004), citando a ocorrência de ráfides na família.
Estudos para o gênero Heteropsis Kunth ainda são muito escassos, conhecidos apenas
os de Potiguara & Nascimento (1994) que descreveram a anatomia dos órgãos vegetativos de
Heteropsis jenmanii Oliv., análises da folha de H. oblongifolia Kunth e H. spruceana Schott
por Keating (2002) e o estudo do colênquima peciolar de alguns gêneros de Araceae realizado
por Gonçalves et al. (2004), onde inclui H. oblongifolia. Pesquisas recentes para Heteropsis
foram realizadas por Soares (2008) em sua tese de doutorado, as quais abrangem além da
anatomia de quatro espécies, incluindo H. flexuosa, sistemática e ecologia de várias espécies
amazônicas ocorrentes na Reserva Florestal Adolpho Ducke.
A subfamília Monsteroideae, a qual pertence H. flexuosa e M. obliqua, foi circunscrita
por Engler em 1876 e baseada principalmente em estudos sobre a anatomia interna e externa.
Após muitos estudos e rearranjos cladísticos, as monsteroídeas têm sido analisadas utilizando
diferentes métodos. Quanto aos estudos sobre anatomia se destacam os trabalhos de Nicolson,
1960; Eyde et al., 1967; French & Tomlinson, 1981; French, 1985, 1986a, b, 1987; Ray,
1988; Carvell, 1992; e Dietmar-Benhke, 1995. Dentre os mais recentes para a espécie M.
obliqua em particular, cita-se o trabalho de Arunika et al. (2005), um interessante estudo
ontogenético, onde os autores afirmam que as perfurações na epiderme das folhas são
originadas por grupos de células vizinhas programadas para morrer simultaneamente.
Em relação ao gênero Philodendron, foram realizadas pesquisas como a de Pfitzer
(1872), que notou a ocorrência de aquíferos na epiderme. Ambronn (1879-81) observou o
colênquima em forma de anéis em Philodendron eximium Schott. French & Tomlinson (1981,
1984) publicaram estudos sobre os padrões de vascularização do caule no gênero, onde foi
encontrada uma anatomia vascular muito heterogênea. Bunting (1968), em seus trabalhos
voltados para a anatomia e taxonomia do complexo de Philodendron hederaceum (Jacq.)

8
Schott. Estudos de Schneider & Carlquist (1998), voltados para a presença de vasos do
xilema, em cinco gêneros da subfamília Philodendroideae como parte de uma série de estudos
sobre monocotiledôneas. Mais especificamente, para o subgênero Meconostigma, ao qual
pertence P. solimoesense, foi feita uma série de estudos por Mayo (1986), onde o autor
analisa e descreve a anatomia da inflorescência de várias espécies desse subgênero.
Usos da família no âmbito comercial e etnobotânico
A família Araceae possui espécies bem conhecidas no âmbito econômico para o
homem (Plowman, 1969), representando um grupo taxonômico de considerável importância
econômica, com espécies destacando-se por serem utilizadas como plantas medicinais,
alimentícias, fibrosas, e, principalmente, ornamentais (Croat, 1994; Mayo et al., 1997).
Artesanato e construção
Um aspecto importante, tanto para a cultura como para a economia, é o uso das raízes
aéreas das plantas de alguns gêneros da família como matéria-prima para artesanato. Van den
Berg (1984), em sua pesquisa etnobotânica no mercado do Ver-o-Peso em Belém, Estado do
Pará, registrou o “cipó-titica” (Heteropsis spruceana Schott), utilizada para fabricação de
cestos. Há muito tempo os indígenas e outros moradores que habitam a floresta amazônica
coletam fibras de Heteropsis para serem utilizadas em construções e trabalhos manuais
(Balée, 1994; Paz & Miño et al., 1995). No Parque Nacional do Jaú (PNJ), região do Baixo
rio Negro, a exploração destas fibras é a principal atividade da produção extrativista vegetal
(Durigan & Castilho, 2004).
No PNJ, além do “cipó-titica”, as raízes de espécies do gênero Philodendron,
conhecido como “cipó-ambé” ou simplesmente “ambé”, também são muito utilizadas como
matéria-prima para confecção de produtos como cestos, bolsas e paneiros. Crisci & Gancedo
(1971) relata que as raízes aéreas de P. bipinnatifidum Schott, além de medicinais, são muito
utilizadas pelos indígenas Guayaquí, do Paraguai, na confecção de arco e flecha.
De acordo com João Barbosa Rodrigues, os homens da tribo dos Crichaná,
supostamente os atuais Waimiri-Atroari, utilizavam cintos de raízes de aráceas como parte de
suas vestimentas (Milliken et al., 1992).

9
Na construção das malocas dos indígenas Waimiri-Atroari, os materiais são amarrados
com as raízes aéreas de Heteropsis spp., que também são amplamente utilizadas pelos
indígenas e colonos em cestarias. Os indígenas também usam o cipó-titica para ligar as seções
divididas de Iriartella setigera (Mart.) H.Wendl (Arecaceae) que compõem as portas da
maloca (Milliken et al., 1992).
Para os povos indígenas da bacia do Caura, no sul da Venezuela, cestas continuam a
desempenhar um papel central nas atividades de subsistência diária, mas elas também se
tornaram um dos principais itens do artesanato comercial. Cestas feitas de Heteropsis,
chamado Minñato na língua Ye’kwana, fornecem atualmente a principal fonte de renda em
dinheiro para as mulheres Ye’kwana (Knab-Vispo et al., 2003).
Na Amazônia brasileira e extrabrasileira, os povos tradicionais utilizam as raízes
aéreas das espécies dos gêneros Philodendron e Heteropsis na construção de cordas ou
amarras para confecção de cestos, chapéus e bolsas (Soares & Mayo, 1999; Plowden et al.,
2003; Durigan & Castilho, 2004).
Em decorrência do extrativismo intensivo dos cipós e a não reposição dessas plantas
nas Unidades de Conservação, o Estado do Amazonas, após alguns estudos científicos,
aprovou por meio da Secretaria de Desenvolvimento Sustentável – SDS, a Instrução
Normativa 001/2008 no intuito de ordenar e normatizar o manejo adequado da espécie a
extração de fibras vegetais como piaçava, cipó-titica e outros (SDS, 2008). Segundo Canalez
(2009), esta norma propõe promover suporte técnico e estrutura organizacional aos que
trabalham e vivem do extrativismo de fibras vegetais, a fim de agregar renda.
As espécies da família Araceae e seus subprodutos (fibras obtidas dos cipós), objetos
de estudo deste trabalho, são classificadas como produtos florestais não madeireiros (PFNM),
visto que estes, segundo definição da “Food and Agriculture Organization” (FAO), são todos
os materiais biológicos (diferentes de madeira e lenha) que podem ser extraídos de florestas
naturais, agroecossistemas e de árvores que crescem espontaneamente em áreas rurais e que
podem ter utilização doméstica, ser comercializados ou ter significado social, cultural ou
religioso (Vantomme, 2001).
Devido à fragilidade e importância na manutenção do equilíbrio das florestas
tropicais, considera-se boas práticas de manejo dos cipós de Araceae, segundo a SDS (2008):
deixar sem coletar todos os fios verdes na planta; não coletar os fios que estão enrolados ao
tronco da árvore onde está o cipó; não coletar quando a planta apresentar apenas um fio
maduro; realizar a coleta através da poda ou arrancando os fios mediante torção para evitar a
queda da planta; não coletar os fios maduros de acordo com o número que cada árvore suporte

10
apresenta adotando as seguintes medidas: a) metade dos fios maduros para as árvores que
tiverem menos de 20 fios; b) um terço dos fios maduros para as árvores que tiverem mais de
20 fios; as plantas de cipó-titica das áreas onde as coletas ocorrerem, deverão passar por um
período de descanso de no mínimo três anos para permitir a regeneração dos seus fios.
Medicinais
Schultes & Raffauf (1990) citam diversas espécies do gênero Anthurium Schott com
usos medicinais. Na região de Puerto Ospina, no alto Putumayo, Anthurium aureum Engler,
quando triturado e misturado com um óleo, tem sido eficaz como cataplasma no tratamento de
ferimentos nas pernas por ferroada de “arraia-lixa”. A decocção quente das folhas coriáceas
de Anthurium crassinervum (Jacq.) Schott var. caatingae R.E. Schultes é utilizada pelos
Kubeos, da região de Mitú, na lavagem de ouvido para aliviar infecção causada por fungos.
Os Andokes, indígenas da região do médio Caquetá, na Colômbia, aplicam a seiva de
Anthurium flexuosum Kunth em casos de cegueira e também é usado para problemas de
ameaça do baço. Os frutos maduros de Anthurium infectorium R.E. Schultes são usados pelos
indígenas Kubeos para pintar os dentes de roxo ou preto para as danças e cerimônias. Existe
uma crença entre os nativos que esta prática ajuda a preservar os dentes das cáries. Os
indígenas Kófans, do Equador Amazônico, esmagam todas as partes de Anthurium jenmanii
Engler, em água quente e aplicam externamente para aliviar rigidez no pescoço (Schultes &
Raffauf, 1990).
A decocção das folhas de Anthurium scandens (Aubl.) Engl. é considerado um bom
tratamento para picada de cobra, entre os indígenas Tikuna, do Alto Solimões. Esse povo
também emprega o sumo adstringente dos frutos carnosos para estancar o fluxo de pus nas
gengivas nos casos de piorreia (inflamação das estruturas em volta dos dentes), uma doença
comum na tribo (Schultes & Raffauf, 1990)
Os rizomas de Dracontium longipes Engl. (“milho-de-cobra”) são empregados na
medicina caseira na região amazônica contra mordida de cobra, crises de asma, amenorreia,
tosse-comprida, sarna, anemia, antiviral, tônica e antiinflamatória (Lorenzi & Matos, 2002).
Entre os indígenas Sionas, no Equador, as folhas de Monstera adansonii Schott são
trituradas, fervidas em água e a decocção é friccionada sobre furúnculos. A seiva de
Montrichardia arborescens Schott é cáustica e a raiz em pó é usada como diurético na
Amazônia (Schultes & Raffauf, 1990).

11
Na medicina popular e indígena vários autores têm relatado o uso de espécies do
gênero Philodendron Schott., como por exemplo Philodendron bipinnatifidum Schott,
espécies cujas folhas e caule produzem suco cáustico usado contra orquite (inflamação dos
testículos), reumatismo e úlceras. As sementes são usadas contra parasitas intestinais. O
decocto (fervura) das folhas frescas e cascas é indicado contra hidropsia (acúmulo de água nas
células) e externamente na forma de banhos contra erisipela e inflamações reumáticas e
orquite. O cataplasma das inflorescências frescas e amassadas é utilizado contra úlceras
(Lorenzi & Matos, 2002).
Plowman (1969) listou 26 espécies de Philodendron usadas em diversos tipos de
doença, dentre estas P. imbe Schott ex Endl., que ocorre na Amazônia, conhecida como “cipó-
de-imbé”, “cipó-guimbé”, “tracuá” e “curuba”, dos quais as folhas são usadas na cura de
doenças reumáticas, tumores e úlcera. As folhas de P. fragrantissimum (Hook) G. Don, P.
pedatum (Hook) Kunth e P. cuneatum Engl. são utilizadas pelos indígenas Taiwano do Rio
Kananarí (Colômbia) para tratar inflamação de pele. As folhas e pecíolos de P. hylaeae G.S.
Bunting, misturados à farinha de mandioca são utilizados no tratamento de combate a
parasitas intestinais.
Além das espécies de Philodendron, Plowman (1969) também comenta que as folhas
de Montrichardia linifera (Arruda) Schott, espécie que ocorre nos Estados de Pernambuco,
Piauí, Rio de Janeiro, norte do Brasil e no Suriname, são consideradas antirreumáticas e
eficazes contra úlceras. Além disso, suas raízes, apesar de tóxicas, são antidiuréticas.
O chá das folhas de Pistia stratiotes L. (“alface-d`água”, “mureru”) é considerado
diurético, também usado contra hematúria (sangue na urina), como expectorante,
antidisintérico, anti-hemorroidal, antidiabético e é empregado na medicina popular,
principalmente na região amazônica. Em forma de banho, o chá é indicado para aliviar a
inflamação causada por erisipela. Os Tikunas utilizam suas folhas moídas em mistura com sal
para aplicação localizada em cataplasma na eliminação de verrugas (Lorenzi & Matos, 2002).
A inflorescência perfumada de Spathiphyllum floribundum (Linden & André) N.E.
Britton é esfregada na testa para aliviar a dor de cabeça. A seiva do caule de Syngonium
yurimaguense Engl., é empregada pelos Tikunas para tratar da picada dolorosa da “formiga-
conga”. Urospatha caudata Schott, é utilizada na Amazônia brasileira, através do sumo de
seus rizomas, no tratamento para problemas de pele (Schultes & Raffauf, 1990).

12
Místicos
Van den Berg (1984), em levantamento etnobotânico, realizado em um mercado
popular na Amazônia, encontrou espécies utilizadas para uso religioso ou místico, como os
“tajás” (Caladium spp. e Philodendron spp.) que são utilizados em rituais afro-brasileiros.
Segundo Schultes & Raffauf apud Croat (1994) os indígenas Kubeo, da Colômbia, utilizam
Philodendron insigne Schott em rituais amorosos. Grande parte da população atribui às
plantas do gênero Dieffenbachia valor simbólico e poderes mágico, sendo comum no Brasil
seu cultivo em vasos que ornamentam ambientes e protegem de ações maléficas (Camargo,
1998).
Ornamentais
As Araceae são muito utilizadas para ornamentação, principalmente pela beleza e
exuberância de suas folhagens, Anthurium andraeanum Linden, Zantedeschia aethiopica (L.)
Spreng. e seus híbridos tem sido importantes fontes de flores de corte gerando lucros de
milhões de dólares em vários países (Bown, 1988).
Na América Tropical as principais espécies utilizadas no paisagismo são dos gêneros
Philodendron Schott, Monstera Adans., Epipremnum Schott, Spathiphyllum Schott,
Anthurium Schott, Dieffenbachia Schott, Syngonium Schott, Aglaonema Schott, Caladium
Vent. e Zantedeschia Spreng. (Bown, 1988). Destas, as mais importantes espécies de Araceae
provenientes da Amazônia são dos gêneros Caladium, Dieffenbachia e Philodendron (Graf,
1963; Souza & Lorenzi, 2008). Espécies de outros gêneros também são utilizadas como
ornamentais, por exemplo, Dracontium longipes (Lorenzi & Matos, 2002). Pistia stratiotes L.
destaca-se como espécie de uso ornamental sendo ocasionalmente cultivada em lagos
decorativos na Amazônia e em vários países e é também muito utilizada em aquários, lagos,
tanques e espelhos d’água (Lorenzi & Souza, 2008).
Na Amazônia, Van den Berg (1984) cita para uso ornamental o “antúrio” (Anthurium
bellum Schott), “brasileirinho” (Caladium humboldtianum Schott) e espécies de “tajás”
(Caladium spp. e Xanthosoma spp.). Muitas espécies do gênero Philodendron também são
utilizadas, dentre estas, Philodendron melinonii Brongn. ex Regel, conhecida popularmente
como “marta-rocha” ou filodendro-da-amazônia, é uma das mais consagradas como
ornamental na região amazônica (Lorenzi & Souza, 2008; Souza, 2010).

13
Alimentícios
Para uso alimentício, Posey (1984) relata que os indígenas Kayapós, cujo território
está situado na região do Planalto Central do Brasil, do sul do Pará até o norte do Mato
Grosso, cultivam variedades de plantas rizomatosas de Araceae e outras famílias em terrenos
bem drenados de encosta, reservados principalmente para fornecer alimentos em caso de
enchentes ou desastres agrícolas. Xanthosoma sagittifolium Schott, popularmente conhecida
como “taioba” e Colocasia esculenta (L.) Schott, mais conhecida por “inhame” ou “taro” são
dotadas de caule subterrâneo com reserva de amido (Lorenzi & Souza, 2008). Além disso,
seus rizomas podem ser usados na panificação, produção de álcool, colas, medicamentos e as
folhas também podem ser consumidas (Cardoso, 1997).
Amorphophallus paeoniifolius (Dennst.) Nicolson (“elephant yam”) é cultivada por
seus tubérculos ricos em amido na Ásia tropical, particularmente na Índia. Os rizomas de
Amorphophallus konjac K. Koch são muito utilizados no Japão, dos quais é extraído o
“konnyacu” um agente gelatinoso muito usado na indústria de alimentos. Cyrtosperma
merkusii (Hassk.) Schott, cresce desde o sudeste da Ásia até a Oceania e é usada como
comida para populações de baixa renda ou como ração para animais. Da mesma forma, na
América Tropical, utiliza-se Alocasia macrorrhizos (L.) Schott, que é exótica, invasora e
merece cautela no seu uso (Bown, 1988; Kinupp, 2007).
A mais importante planta cultivada do México é a Monstera deliciosa Liebm.,
popularmente conhecida como “banana-do-mato”, “fruta-pão-mexicana” ou “costela-de-
adão”, cuja infrutescência é comestível (Lorenzi & Souza, 2008). As infrutescências de
Philodendron bipinnatifidum Schott, quando maduras, são comestíveis, adocicadas e muito
saborosas. (Kinupp & Barros, 2007; Kinupp, 2007).

14
REFERÊNCIAS
APG II. 2003. An update of the Angiosperm Phylogeny Group classification for the orders
and families of flowering plants: APG II. Botanical Journal of the Linnean Society 141:
399-436.
APG III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders
and families of flowering plants: APG III. Botanical Journal of the Linnean Society.
Arunika H. L. A. N. Gunawardena; Sault K.; Donnelly, P.; Greenwood, J. S.; Dengler, N. G.
2005. Programmed cell death and leaf morphogenesis in Monstera obliqua (Araceae). Planta
(2005) 221: 607-618.
Balée, W. 1994. Footprints of the florest Ka`apor ethnobotany: the historical Ecology of
plant utilization by an Amazonian People. New York: Columbia Univ. Press. 396pp.
Bogner, J. 1979. A critical list of the Aroid Genera. Aroideana 1(3): 63-73.
Bown, D. 1988. Aroids. Plants of the Arum family. Timber Press: Portland, Oregon. 256pp.
BRASIL. 2000. Sistema Nacional de Unidades de Conservação da Natureza – SNUC: Lei
N° 9.985, de 18 de Julho de 2000. Brasília: MMA/SBF. 32pp.
Bunting, G.S. 1968. Vegetative anatomy and taxonomy of the Philodendron scandens
complex. Gentes Herbarium 10 (2): 136-168.
Camargo, M.T.L.A. 1998. Plantas medicinais e de rituais afro-brasileiros II: estudo
etnofarmacobotânico. Ícone, São Paulo, Brasil. 232 pp.
Canalez, G. G. 2009. Produtos florestais não madeireiros: aráceas epifíticas da Reserva
Extrativista Auatí-Paraná. Manaus : [s.n.].
Cardoso, M.O. (Org.) 1997. Hortaliças não-convencionais da Amazônia. Brasília: Embrapa
– CPAA. 150pp.
Carvell, W. N. 1992. Floral anatomy of the Pothoideae and Monsteroideae (Araceae).
American Journal of Botany 76 (6): 231.

15
Coelho, M.A.N., Soares, M.L., Sakuragui, C.M., Mayo, S., Andrade, I.M. de, Temponi, L.G.
2010. Anthurium in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de
Janeiro. (http://floradobrasil.jbrj.gov.br/2010/FB021967)
Crisci, J.V.; Gancedo, O.A. 1971. Sistemática y Etnobotanica del guembe (Philodendron
bipinnatifidum) uma importante arácea sudamericana. Rev. Mus. La Plata, Bot. 11 (65) :
285-302.
Croat, T.B. 1985. Collecting and preparing specimens of Araceae. Ann. Missouri Botanic
Garden. v. 17: p. 252-258.
Croat, T.B. 1990 “1988”. Ecology and life forms of Araceae. Aroideana, 11(3-4): 1-55.
Croat, T.B. 1994. The use of the New World Araceae as drug plants. J. Jap. Bot. 69: 185-
203.
Dahlgren, R.M.T. & Clifford, H. T. 1982. The Monocotyledons: A Comparative Study.
Academic Press, New York.
Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F. 1985. The Families of the Monocotyledons –
Structure, Evolution and Taxonomy. Springer-Verlag, Berlin. 520 pp.
Dietmar-Benhke, H. 1995. P-type sieve element and the systematics of the Arales (sensu
Cronquist 1988) – with S-type plastids in Pistia. Plant Systematics and Evolution 195: 87-
119.
Durigan, C.C.; Castilho, C.V. 2004. O extrativismo de cipós (Heteropsis spp., Araceae) no
Parque Nacional do Jaú. In: Janelas para a Biodiversidade no Parque Nacional do Jaú:
Uma estratégia para a Biodiversidade na Amazônia. Sérgio Henriques Borges; Simone
Iwanaga; Carlos César Durigan; Marcos Roberto Pinheiro (Eds.) Manaus: Fundação Vitória
Amazônica.
Engler, A. 1878. Araceae. In Martius, C.F.P. von, Flora brasiliensis 3(2): 56–88, t. 11-102.

16
Eyde, R. H.; Nicolson, D. H.; Sherwin, P. 1967. A survey of floral anatomy in Araceae.
American Journal of Botany 54: 478-497.
Exotic rain forest. 2010. Philodendron ‘Marijke’ A putative hybrid of Philodendron
goeldii G.M. Barroso and Philodendron solimoesense A.C. Sm. from the collection of
Joep Moonen in French Guiana. Disponível em:
<www.exoticrainforest.com/Philodendron%20Marijke.html> Acesso em: 12 mar. 2011.
FAO, Organización Mundial para la Alimentación y la Agricultura, 2002. Agricultura
Mundial: hacia los años 2015/2030. Informe resumido. Roma, Italia. 97pp.
Ferreira, L.S.; Marsola, F.J.; Teixeira, F.de P. 2006. Anatomia dos órgãos vegetativos de
Dieffenbachia picta Schott (Araceae) com ênfase na distribuição de cristais, laticíferos e grãos
de amido. Revista Brasileira de Farmacognosia. Brazilian Journal of Pharmacognosy,
16(Supl.): 664-670.
French, J.C. & Tomlinson, P.B. 1981. Vascular patterns in stems of Araceae: subfamily
Monsteroideae. American Journal of Botany 68:713-729.
French, J. C. & Tomlinson, P.B. 1984. Patterns of stem vaculature in Philodendron.
American Journal of Botany, Columbus, 71 (10): 1432-1433.
French, J.C. 1985a. Patterns of endothecial walls thickenings in Araceae: subfamilies
Pothoideae and Monsteroideae. American Journal of Botany 72 (3): 472-486.
French, J.C. 1985b. Patterns of endothecial walls thickenings in Araceae: subfamilies
Calloideae, Lasioideae and Philodendroideae. Botanical Gazette 146: 521-533.
French, J.C. 1986. Patterns of stamen vasculature in the Araceae. American Journal of
Botany 73: 434-449.
French, J.C. 1987a. Structure of ovular and placental trichomes of Araceae. Botanical
Gazette 148: 198-208.

17
French, J.C. 1987b. Systematic occurrence of a sclerotic hypodermis in roots of Araceae.
American Journal of Botany 74: 891-903.
French, J.C. 1988. Systematic occurrence of anastomosing laticifers in Araceae. Botanical
Gazette 149: 71-81.
French, J.C.; Chung, M.; Hur, Y. 1995. Chloroplast DNA phylogeny of the Ariflorae. In:
Rudall, P.J.; Cribb, P.J.; Cutler, D.F.; Humphries, C.J. (eds.), Monocotyledons: systematic
and evolution Academic Press, London, p.255-275.
French, J.C. 1997. Vegetative anatomy In: Mayo, S.J., Bogner, J. and Boyce, P. C. (Eds). The
genera of Araceae. Royal Botanic gardens, Kew 9-24 pp.
FVA – Fundação Vitória Amazônica. 2009. Monitoramento e educação ambiental:
ferramentas para a conservação. Relatório técnico final. Fundação Vitória Amazônica /
Projeto Corredores Ecológicos. Manaus – Amazonas.
Garcia, N. 2009. Efecto de la extracción de raíces aéreas sobre las poblaciones de “tripeperro”
(Philodendron longirrhizum, Araceae) en los Andes Centrales de Colombia. Caldasia 31 (1):
19-29.
Gonçalves, E.G.; Paiva, E.A.S.; Coelho, M.A.N. 2004. A preliminary survey of petiolar
collenchyma in the Araceae. Annals of the Missouri Botanical Garden. Vol 91, num. 3.
Gonçalves, E.G. & Temponi, L.G. 2004. A new Monstera (Araceae: Monsteroideae) from
Brazil. Brittonia, 56(1), 2004, pp. 72-74.
Govaerts, R.; Frodin, D.G.; Borgner, J., Boyce, P.; Cosgreff, B.; Croat, T.B.; Gonçalves,
E.G.; Gayum, M.; Hay, A.; Hetterscheid, W.; Landolt, E.; Mayo, S.J.; Murata, J.; Nguyen,
V.D.; Sakuragui, C.M.; Singh, Y.; Thompson, S.; Zhu, G. 2002. World checklist and
bibliography of Araceae (and Acoraceae). Royal Botanic Garden, Kew.
Graf, A.B. 1963. Exotica 3. Pictorial cyclopedia of exotic plants. Guide to care of plants
indoors. Roehrs Company, Rutherford, N.J., U.S.A. 1826pp.

18
Grayum, M.H. 1984. Palynology and phylogeny of the Araceae. (Ph. D. Thesis) University
of Michigan. 852pp.
Grenand, P.; C. Moretti; H. Jacquemin. 1987. Pharmacopées tradltionelles en Guyane.
ORSTOM, Paris.
Hay, A. & Mabberley, D.J. 1991. Transference of function and the origin of aroids : their
significance in early angiosperm evolution. Bot. Jahrb. 113 : 339-428.
Keating, R. C. 2002. Acoraceae and Araceae. In: Gregory, M. and Cutler, D. F. Anatomy of
the monocotiyledons. Oxyford University Press, New York, 322pp.
Keating, R. C. 2004. Systematic occurrence of raphide crystals in Araceae. Annals of the
Missouri Botanical Garden 91 (3): 495-504.
Kinupp , V. F. Plantas alimentícias não-convencionais da região metropolitana de Porto
Alegre, RS. 2007. 562 f. Tese (Doutorado em Fitotecnia) – Universidade Federal do Rio
Grande do Sul, Porto Alegre. Disponível em: <http://www.lume.ufrgs.br/handle/10183
/12870>. Acesso em: 10 mai. 2011.
Kinupp, V.F. & Barros, I.B.I. de. 2007. Riqueza de Plantas Alimentícias Não Convencionais
na Região Metropolitana de Porto Alegre, Rio Grande do Sul. Revista Brasileira de
Biociências, 5: 63-65.
Knab-Vispo, C., Hoffmann, B.; Moermond, T.; Conrad Vispo. 2003. Ecological Observations
on Heteropsis spp. (Araceae) in Southern Venezuela. Economic Botany, 57(3) 345-353.
Lorenzi, H. & Matos, F.J.A. 2002. Plantas medicinais no Brasil: nativas e exóticas. Nova
Odessa, SP: Instituto Plantarum. 512pp.
Lorenzi, H. & Souza, H. M. de. 2008. Plantas ornamentais no Brasil: arbustivas, herbáceas
e trepadeiras. 4ª edição. Ed. Nova Odessa, SP: Instituto Plantarum. 1088pp.

19
Machado, F.S. 2008. Manejo de Produtos Florestais Não Madeireiros: um manual com
sugestões para o manejo participativo em comunidades da Amazônia. PESACRE e
CIFOR: Rio Branco/Acre: 105pp.
Mantovani, A., Filartiga, A. Del P., Coelho, M. A. N. 2010. Anatomia comparada da folha e
espata de espécies de Anthurium (Araceae) ocorrentes na Mata Atlântica. Revista Brasil.
Bot., V.33, n.1, p.185-200.
Mayo, S.J.; Bogner, J.; Boyce, P. C. 1997. The Genera of Araceae. Royal Botanic Gardens,
Kew. 370pp.
Mayo, S.J. 1986. Systematics of Philodendron Schott (Araceae) with special reference to
inflorescence characters. PhD Thesis, University of Reading, Reading.
Milliken, W.; Miller, R.P.; Pollard, S.R.; Wandelli, E.V. 1997. The Ethnobotany of the
Waimiri Atroari Indians. Royal Botanic Gardens, Kew. 146pp.
Milliken, W. 1997. Traditional anti-malarial medicine in Roraima, Brasil. Economic Botany,
51 (3): 212-237.
Ming, L.C.; Silva, S.M.P. da; Silva, M.A.S. da; Hidalgo, A.F.; Marchese, J.A.; Chaves,
F.C.M. Manejo e cultivo de plantas medicinais: algumas reflexões sobre as perspectivas e
necessidades do Brasil. In: Coelho, M. de F.B; Junior, P.C.; Dombroski, J.L.D. (Org) 2003.
Diversos olhares em Etnobiologia, Etnoecologia e Plantas Medicinais. Anais do I
Seminário Mato-Grossense de Etnobiologia e Etnoecologia e II Seminário Centro-Oeste de
Plantas Medicinais. 1ª edição. Cuiabá: Unicen.
Nicolson, D. H. 1960. The occurrence of trichosclereids in the Monsteroideae (Araceae).
American Journal of Botany, 47: 598-602.
Paz y Miño, C.G.; Balslev, H.; Valença, R. 1995. Lianas útiles para los indígenas Siona-
Secoya de la Amazonia Ecuatoriana. Economic Botany, 49, 269-275.

20
Plowden, C.; Christopher Uhlb; Oliveira, F.A. 2003. The ecology and harvest potencial of
titica vine roots (Heteropsis flexuosa) in the easter Brasilian Amazon. Forest Ecology and
Management 182: 59-73.
Plowman, T. 1969. Folk uses of new world aroids. Economic botany, 23(2): 97-122.
Posey, D.A. 1992. Traditional knowledge, conservation, and the rain forest harvest. In: M.
Plotkin e L. Famolare (eds). Sustainable harvest and marketing of rain florest products.
Washington, Island Press/Conservation International. 325pp.
Ray, T. 1988. Survey of shoot organization in the Araceae. American Journal of Botany,
75(1): 56-84
Rodrigues, E. 2006. Plants and Animals Utilized as Medicines in the Jaú National Park (JNP),
Brazilian Amazon. In: Phytotherapy Research Phytother. Res. 20, 378-391
Sakuragui, C.M. 1998. Taxonomia e filogenia das espécies de Philodendron, seção
Calostigma (Schott) Pfeiffer no Brasil. Tese de doutorado, Universidade de São Paulo, São
Paulo.
Schott, H.W. 1860. Prodromus systematis aroideanum. Vienna: Typis congregationis
mechitharisticae, Vindobonae. 602pp.
Schultes, R.E. & Raffauf, R.F. 1990. The healing forest. Medicinal and toxic plants of the
Northwest Amazonia. Dioscorides Press. Portland, Oregon. 484pp.
SDS – Secretaria de Desenvolvimento Sustentável do Amazonas. 2008. Instrução
Normativa nº 001 de 11 de Fevereiro de 2008.
Soares, M.L. 1996. Levantamento florístico do gênero Philodendron Schott (Araceae) na
Reserva Florestal Adolfo Ducke – Manaus – AM. Recife: Dissertação mestrado, 1996.
179pp.

21
Soares, M. L. & Mayo, S. J. 1999. Araceae. p. 672-687. In: Ribeiro, J. E. L. S. et al. Flora da
Reserva Ducke. Guia de identificação das plantas vasculares de uma floresta de terra firme
na Amazônia Central. INPA, DFID, Manaus.
Soares, M.L. & Jardim-Lima. 2005. Amazonian Species of Araceae in the INPA Herbarium,
Manaus – Amazonas, Brasil. Aroideana 28: 134-152.
Soares, M.L. 2008. Sistemática e ecologia de Heteropsis Kunth (Araceae Juss.) com destaque
especial nas espécies ocorrentes na Reserva Florestal Adolpho Ducke, Manaus – Amazonas,
Brasil. Tese doutorado – INPA/UFAM. 207pp.
Soares, M.L.; Mayo, S.J.; Croat, T.B.; Gribel, R. 2009. Two new species and a new
combination in Amazonian Heteropsis (Araceae). Kew bulletin, vol. 64, n. 2, p. 263-270.
Solereder S. H. & Meyer, F. J. 1928. Systematische Anatomie der Monocotyledonen. Heft
III, ppt 100-169. Borntraeger, Berlin.
Souza, V.C. & Lorenzi, H. 2008. Botânica sistemática: guia ilustrado para identificação das
famílias de Fanerógamas nativas e exóticas no Brasil, baseado em APG II. 2ª edição: Nova
Odessa, SP: Instituto Plantarum. 703pp.
Tam, S.M.; Boyce, P.C.; Upson, T.M.; Barabé, D.; Bruneau, A.; Forest, F.; Parker, J.S. 2004.
Intergeneric and infrafamilial phylogeny of subfamily monsteroideae (araceae) revealed by
chloroplast trnl-f sequences. American Journal of Botany 91(3): 490-498.
Temponi, L.G. 2007. Sistemática de Anthurium sect. Urospadix (Araceae). Tese doutorado.
São Paulo.
Van den Berg, M.E. 1984. Ver-o-Peso. The ethnobotany of an Amazonian market. Advances
in economic botany, 140-149.
Vantomme, P. 2001. Production and trade opportunities for non-wood forest products,
particulary food products for niche markets. Geneva: Forest products Division (FAO).
http://www.fao.org/forestry/FOP/FOPW/NWFP/nwfp-e.stm

22
Vianna, W.O.; Soares, M.K.M.; Appezzato-da-Glória, B. 2001. Anatomia da raiz escora de
Philodendron bipinnatifidum Schott (Araceae). Acta bot. bras. 15(3): 313-320.

23
Etnobotânica de plantas úteis na comunidade Santa Maria, Baixo rio Negro
– AM, com ênfase nas espécies da família Araceae Juss.
RESUMO A família Araceae possui espécies bem conhecidas no âmbito econômico para o homem destacando-se por serem utilizadas como plantas medicinais, alimentícias, fibrosas e, principalmente, ornamentais. O objetivo geral deste trabalho foi realizar um levantamento das espécies da família Araceae utilizadas pelos moradores da comunidade Santa Maria, localizadas na Área de Proteção Ambiental da Margem Esquerda do rio Negro, Manaus, Amazonas. As informações etnobotânicas foram coletadas utilizando metodologias como o “rapport” e “turnê-guiada” e registradas através de questionário semiestruturado e conversas informais. A análise das informações foi realizada através do cálculo do índice de Valor de Uso. No total, foram encontradas 51 espécies de Araceae distribuídas em 15 gêneros, dos quais Philodendron foi o mais diverso com 22 espécies. Deste total foram identificadas 23 espécies úteis de Araceae, tanto na área de floresta como na comunidade, com indicações de usos para artesanato, alimentação, medicinal, ornamentação e misticismo. Dentre as espécies que apresentaram maior índice de Valor de Uso (VU) estavam Philodendron solimoesense A.C.Sm. (VU = 0,6) e Heteropsis flexuosa (Kunth) G.S. Bunting (VU = 0,5), cujas raízes são utilizadas como fonte de fibra para a confecção de artigos artesanais. Além dessas espécies, foi registrado também o uso de Monstera obliqua Miq., conhecida popularmente na região como “ambé-sima”, cujas raízes aéreas também são utilizadas como fonte de fibras para o artesanato.
ABSTRACT The species of the family Araceae has in the economic well-known to the man standing out for being used as medicinal plants, food, fibers and mainly ornamental. The overall objective of this study was to survey the species of the family Araceae used by residents of the community Santa Maria, located in the Environmental Protection Area of the Left Bank of the rio Negro, Manaus, Amazonas. The ethnobotanical data were collected using methodologies such as "rapport" and "guided-tour" and recorded semi-structured questionnaire and through informal conversations. The information analysis was done by calculating the index Use Value. In total, 51 species were found distributed in 15 genera Araceae, Philodendron of which was the most diverse with 22 species. Of the total 23 species were identified useful Araceae, both in the forest and the community, with indications of uses for crafts, food, medicinal, ornamental and mysticism. Among the species with the highest index Use Value (UV) were Philodendron solimoesense A.C.Sm. (UV = 0.6) and Heteropsis flexuosa (Kunth) G.S. Bunting (UV = 0.5), whose roots are used as a fiber source for making arts and crafts. Besides these species, was also recorded using Monstera obliqua Miq., popularly known locally as "ambe-sima," whose aerial roots are also used as a source of fiber crafts.

24
INTRODUÇÃO
Estudos etnobotânicos vêm ganhando destaque cada vez maior nos últimos anos, suas
implicações ideológicas, biológicas, ecológicas e filosóficas dão respaldo ao seu crescente
progresso metodológico e conceitual (Morais et al., 2003). Além disso, contribuem em
especial para o desenvolvimento planejado da região onde os dados foram coletados (Begossi
et al., 2002). O conhecimento obtido das populações tradicionais deve ser valorizado e
compartilhado com o objetivo de possibilitar e garantir o uso desses recursos por determinado
grupo de pessoas, no sentido de melhorar as condições de vida dessas populações.
No Brasil e em vários outros países, a intensificação dos trabalhos nessa área leva ao
conhecimento das espécies que são utilizadas, o que pode servir como instrumento para
delinear estratégias de utilização e conservação das espécies nativas e seus potenciais (Ming,
2000), uma vez que este é detentor de grande biodiversidade e também de povos com culturas
diversas que sabem a aplicabilidade de uma grande quantidade de espécies vegetais
(Elizabestsky, 2005).
Os primeiros levantamentos etnobotânicos realizados para a família Araceae foram
mais voltados para o âmbito da botânica econômica, visto que muitas de suas espécies são
utilizadas como ornamentais, alimentícias, medicinais, místicas e como matéria-prima para
confecção de artesanato.
As Araceae são muito utilizadas para ornamentação, principalmente pela beleza e
exuberância de suas folhagens, assim Anthurium andraeanum Linden, Zantedeschia
aethiopica (L.) Spreng. e seus híbridos têm sido importantes fontes de flores de corte gerando
lucros de milhões de dólares em vários países. Na América Tropical as principais espécies
utilizadas no paisagismo são dos gêneros Philodendron Schott, Monstera Adans.,
Epipremnum Schott, Spathiphyllum Schott, Anthurium Schott, Dieffenbachia Schott,
Syngonium Schott, Aglaonema Schott, Caladium Vent. e Zantedeschia Spreng. (Bown, 1988).
Vários gêneros geófitos que possuem caules subterrâneos ricos em amido, são
cultivados como alimento em países tropicais, como espécies de Alocasia Neck. ex Raf.,
Amorphophallus paeoniifolius (Dennst.) Nicolson e Cyrtosperma chamissonis (Schott) Merr.
Nos trópicos, as espécies alimentícias mais utilizadas são Colocasia esculenta (L.) Schott,
conhecidas como “inhame” ou “taro”, originários da Ásia, e Xanthosoma sagittifolium (L.)
Schott, conhecida como “taioba”, originária da América do Sul. Monstera deliciosa Liebm.,
também ornamental, é apreciada por seus frutos comestíveis, cujo sabor se parece com o de

25
“abacaxi”. As sementes de Typhonodorum, de Madagascar e Montrichardia, da América do
Sul, ambas utilizadas como alimento (Bogner, 1987; Bown, 1988).
Espécies medicinais de Araceae tem sido usadas para combater vermes parasitas,
febres, doenças de pele, desinterias, mordidas de cobra, ferimentos, tumores, asmas,
reumatismos e como laxativo. Schultes (1963), após convívio com algumas tribos da América
do Sul registrou o uso de partes de certas espécies pertencentes aos gêneros Anthurium,
Philodendron e Urospatha como contraceptivos femininos. Algumas espécies de Araceae são
consideradas tóxicas, como Epipremnum giganteum (Roxb.) Schott e Amorphophallus
prainiiHook. f., sendo utilizadas como constituintes de flechas venenosas usadas por
indígenas na Malásia.
Espécies vegetais fibrosas ocupam papel de destaque no cotidiano das comunidades
tradicionais amazônicas ao lado das espécies medicinais, alimentícias e madeireiras (Oliveira
et al., 2006). Esses povos tradicionais da região amazônica utilizam as raízes aéreas das
espécies do gênero Philodendron e Heteropsis na construção de cordas ou amarras para
embarcações, casas e em artesanatos na confecção de cestos, chapéus e bolsas (Milliken et al.,
1992; Soares & Mayo, 1999; Plowden et al., 2003; Durigan & Castilho, 2004). No entanto,
não se sabe realmente quantas e quais espécies pertencentes ao gênero Philodendron são
utilizadas para esse fim. As áreas de maior intensidade de exploração do “cipó-titica” no
Amazonas estão localizadas ao norte do Estado, ao longo do rio Negro, onde a tradição de
confeccionar utensílios como cestos e paneiros de cipós tem sido praticada principalmente
pelos homens e é passada de pai para filho.
Para garantir não só a preservação e conservação desses recursos, como também das
tradições locais, é necessário que haja um estudo detalhado e uma avaliação da importância
das plantas que estão sendo utilizadas por essas pessoas e de que forma elas vem sendo
utilizadas no cotidiano dessas comunidades. Portanto, foi de suma importância resgatar o
conhecimento dos usos das espécies mais utilizadas na Comunidade Santa Maria com intuito
de não só preservar o material genético como também sensibilizar a população local para a
valorização dos potenciais desses recursos que podem vir a fortificar sua economia e manter a
tradição local.

26
OBJETIVOS Geral
Realizar um levantamento etnobotânico das espécies utilizadas na comunidade Santa
Maria, Baixo rio Negro – AM, dando ênfase na família Araceae Juss. com potencial
econômico, visando o aproveitamento adequado e a conservação das mesmas.
Específicos
Identificar as espécies de Araceae e seus respectivos usos na comunidade Santa
Maria, na APA da Margem Esquerda do Rio Negro;
Registrar o conhecimento dos usos das espécies mais utilizadas nesta comunidade
com intuito de mostrar o valor das características etnoculturais locais para a
comunidade e população em geral;
Classificar as espécies vegetais indicadas quanto à origem, formas de vida e áreas de
ocorrência;
Elaborar um guia básico de identificação com fotos coloridas das espécies ocorrentes
na comunidade e no seu entorno, visando o retorno dessas informações para a
população local e sociedade em geral no reconhecimento das espécies.

27
MATERIAL E MÉTODOS
Área de estudo
Criada em abril de 1995, pelo decreto de lei N� 16.490, a APA do Rio Negro - Setor
Aturiá-Apuauzinho possui 586.422,00 hectares e está incluída na categoria de Uso
Sustentável e faz parte do mosaico de Unidades de Conservação do Baixo rio Negro, que tem
ao todo 1,8 milhões de hectares e integra o corredor central da Amazônia (Figura 1). Drenada
pelo rio Cuieiras, a 70 km de Manaus, a área representa uma zona de amortecimento para
UC’s de proteção integral, sendo habitada por mais de 600 famílias, totalizando 15
comunidades dispersas (FAS, 2009).
Figura 1. Mosaico de Unidades de Proteção Ambiental do Baixo Rio Negro e as comunidades da APA da Margem Esquerda do Rio Negro – Setor Aturiá Apuauzinho, seta mostrando comunidade Santa Maria. Fonte: Fundação Amazonas Sustentável – FAS, 2009.

28
A comunidade Santa Maria está localizada nas coordenadas S 2° 55’ 44,2” W 60° 27’
58,7”, na Área de Proteção Ambiental da Margem Esquerda do Rio Negro, Setor Aturiá –
Apuauzinho. Possui uma igreja católica e uma igreja evangélica; associação comunitária; uma
escola de Ensino Fundamental de 1ª a 9ª Série (Figura 2A); um centro comunitário utilizado
para os momentos de lazer e entretenimento dos comunitários, reuniões de planejamento e
demais ações coletivas (Figura 2C); um gerador comunitário, fornecendo energia
regularmente durante 10 horas por dia atendendo todos os moradores; um sistema de
abastecimento de água com rede de distribuição; um posto de saúde (Figura 2 B); sistemas de
transporte coletivos (recreio) até a sede do município. O deslocamento de Manaus a
comunidade leva em média três horas e meia para os barcos regionais e uma hora e quinze
minutos de lancha do tipo “voadeira” (FAS, 2009).
Figura 2. Estrutura física da comunidade Santa Maria, baixo rio Negro - AM. A) Escola Municipal Luiz Jorge Silva. B) Posto de saúde e C) Centro comunitário.
Aspectos ambientais
Segundo estudos do plano gestor, feitos para o Parque Estadual do Rio Negro – Setor
Sul, que faz divisa com a área de estudo, o clima é classificado como tropical chuvoso
apresentando uma média de temperatura superior a 18ºC no mês mais frio, segundo a
classificação de Koppen. Quanto à precipitação, existe uma variação sazonal, na qual as
chuvas se iniciam no período de dezembro a fevereiro e atingem o pico no período de março a
maio. Nos meses de junho a agosto ocorre a diminuição das chuvas e o período de setembro a
novembro é o mais seco na região (SDS, 2010).
A maior parte da terra-firme do Parque Estadual do Rio Negro Setor Sul apresenta
Floresta Ombrófila Densa. A Floresta de Terras Baixas ocorre nas proximidades do rio Apuaú
(SDS, 2010).
Quanto à formação geológica, grande parte do cráton, que é a porção sólida da crosta
terrestre constituída por rochas cristalinas, encontra-se recoberto pelas bacias Solimões e

29
Amazonas, que recebeu sucessivos pacotes sedimentares desde o paleozoico até o depósito
atual das planícies de inundação. Nas proximidades da área de estudo, os depósitos
sedimentares do Cretáceo (Formação Alter-do-Chão) predominam na geologia da superfície
(SDS, 2010).
O relevo acha-se talhado sobre os sedimentos consolidados da Formação Alter-do-
Chão, sob cobertura vegetal Floresta Densa e aberta com palmeiras (Radambrasil, 1978) e
Campinarana nas porções mais rebaixadas. Os solos são: Latossolo Vermelho Amarelo –
predominantemente, Podzólico Hidromórfico e Hidromórfico Gleizado (SDS, 2010).
Aspectos socio-econômicos
O Baixo rio Negro foi habitado principalmente pelos povos Tarumã, Manaós e Barés.
Estes povos indígenas passaram por processos históricos de transformação cultural e perda
populacional, chegando até a extinção absoluta de alguns deles, em decorrência dos
empreendimentos mercantis e religiosos, guerras e epidemias instaladas após chegada dos
colonizadores europeus. Os Barés localizam-se atualmente do Baixo ao Alto rio Negro,
enquanto que os Manaós e Tarumã foram praticamente extintos (Meira, 2006).
A reocupação do Baixo rio Negro nos últimos 60 anos foi muito influenciada pela
queda do preço da borracha e pelo início da urbanização e da industrialização da cidade de
Manaus. Atualmente, a economia na região do Baixo rio Negro depende quase totalmente de
produtos extrativistas como peixes ornamentais, fibras vegetais e produtos da agricultura
familiar (FVA, 2005), além do extrativismo florestal madeireiro e não madeireiro, e
atividades ligadas ao turismo e à venda de artesanato, com forte inserção do componente
“tradição cultural” (Illenseer, 2009).
No ano de 1964 chegaram os primeiros moradores da vila Santa Maria, a maioria
vinda do Alto rio Negro, principalmente dos municípios de São Gabriel da Cachoeira e Santa
Isabel, hoje a comunidade conta com cerca de 48 famílias (Vieira, 2010). Os moradores da
comunidade Santa Maria possuem um modo de vida composto basicamente pela pesca e
agricultura familiar, complementando estas atividades com a coleta de produtos florestais não
madeireiros (PFNM’s) da floresta para uso doméstico e comercial.

30
Coleta de informações etnobotânicas
Para a pesquisa etnobotânica foi obtida autorização da comunidade através de seus
representantes Arlem Francis Gomes (Presidente) e Ednaldo Loureiro (Vice-presidente), além
da autorização do Comitê de Ética em Pesquisa com Seres Humanos do Instituto Nacional de
Pesquisas da Amazônia – CEP/INPA (ANEXO A).
Foram realizadas cinco visitas entre os meses de dezembro de 2009 a setembro de
2010, o tempo de permanência na comunidade foi de três dias a uma semana. Portanto, foi
possível coletar amostras botânicas dos mesmos indivíduos em vários períodos do ano e
presenciar eventos como a colheita de produtos provenientes da roça e a tradicional festa de
Santo Antônio, realizada todos os anos na comunidade.
O auxílio de um “informante-chave” foi necessário devido ao seu amplo conhecimento
sobre a área, as espécies utilizadas como matéria-prima para confecção de artesanato e de
plantas de um modo geral. Cada informante indicava outra pessoa envolvida com artesanato
ou conhecedora sobre outras plantas e assim sucessivamente, obtendo-se então informações
através do método “bola de neve” (Bailey, 1982; Amorozo et al. 2002).
Foi utilizado o “rapport”, proposto por Albuquerque et al. (2004), onde o pesquisador
se apresenta na casa de cada informante evidenciando a importância do trabalho. A
metodologia de coleta nos quintais das casas e ambiente florestal do entorno da comunidade
Santa Maria, foi baseada no método de “turnê-guiada” por um coletor local de cipós, onde
foram realizados percursos em três trilhas nos ambientes de “igapó”, “terra-firme” e
“campinarana”, gravados pelo aparelho GPS (Global Positioning System), através da função
“registro de trajeto”, gerando um mapa com o trajeto percorrido de aproximadamente 12
quilômetros no total.
Figura 3. Registro do trajeto mostrando as trilhas contemplando três ambientes visitados na APA da Margem Esquerda do Rio Negro – Setor Aturiá-Apuauzinho.

31
O registro sobre as plantas utilizadas foi baseado em questionário semiestruturado
(APÊNDICE A), além de conversas informais realizadas durante a visita aos quintais e às
trilhas de coleta de cipós.
Análise das informações
A análise quantitativa das informações etnobotânicas foi baseada no cálculo do índice
de “Valor de Uso”, proposta por Phillips & Gentry (1993) e adaptada por Rossato et al.
(1999), onde o número de usos será simplesmente totalizado, pela categoria de uso da planta,
táxon da planta ou tipo de vegetação, entrevistando-se uma única vez cada informante.
O valor de uso de uma espécie pelo informante é representado pela fórmula:
VUs = ∑Uis / ns
Onde,
VUs = Valor de Uso da espécie;
Us = número de usos mencionados por cada informante para a espécie s;
ns = número total de informantes (n=13, total de informantes).
Coleta de material botânico
O trabalho foi realizado mediante autorização N°. 57/09 – CEUC/SDS, de acordo com
a Lei Complementar N°. 53, de 05 de junho de 2007, que instituiu o Sistema Estadual de
Unidades de Conservação do Amazonas (SEUC/AM), artigo 43 do Capítulo VI da Lei do
SEUC/AM, que trata das pesquisas científicas em Unidades de Conservação (ANEXO B).
Algumas espécies foram fotografadas quando os informantes autorizavam. Indivíduos
em estádio fértil ou estéril foram coletados e até mesmo em estádio juvenil, visando melhor
entendimento e para a circunscrição das espécies.
A metodologia da coleta e preparação de exsicatas das amostras seguiu as
recomendações de Croat (1985) e Soares & Mayo (1999). No campo, as amostras foram
acondicionadas em sacos plásticos de 50 litros ou prensadas provisoriamente somente entre
jornais e prensa de madeira e borrifadas com álcool de uso doméstico para manterem-se
conservadas. Posteriormente, estas foram transportadas até o Herbário INPA/Manaus (sala de

32
secagem), e desidratadas na estufa elétrica com temperatura de 45º a 50ºC, por um período de
quatro a seis dias até estarem totalmente desidratadas.
Após totalmente secas e devidamente montadas, as amostras foram identificadas,
catalogadas e incorporadas no Herbário INPA. Duplicatas foram doadas ao Herbário EAFM,
do Instituto Federal de Educação, Ciência e Tecnologia do Amazonas Campus Manaus-Zona
Leste.
Identificação e descrição das espécies
As identificações foram baseadas fundamentalmente nas obras de Schott (1860),
Engler (1878), Krause (1913), Mayo et al. (1997), Soares & Mayo (1999), Soares (2008) e
demais literaturas específicas, bem como por comparações com exsicatas depositadas no
Herbário INPA.
O trabalho de descrição foi baseado em observações de plantas vivas no campo e em
análises morfológicas comparativas com exsicatas incorporadas no Herbário INPA. As
análises foram realizadas em laboratório da Pós-graduação em Botânica do INPA e foram
baseadas na terminologia específica para a família, assim como em alguns casos por Radford
et al. (1974) e Lawrence (1977).
Foram tomadas as medidas dos entrenós, profilo, pecíolo, bainha, lâmina foliar,
pedúnculo, espata e suas divisões, espádice e suas divisões e das flores conforme
recomendação de Mayo (1991). Para as três espécies mais utilizadas no artesanato, foram
apresentadas pranchas, contendo ilustrações do hábito e detalhes de estruturas vegetativas e
florais da planta.
As descrições das espécies foram baseadas nos espécimes coletados na própria área de
estudo, observando-se os indivíduos em seu hábitat, principalmente às diferenças entre os
indivíduos da mesma população, características vegetativas e reprodutivas. Foram anotadas
características, tais como: hábito de crescimento; textura, cor e brilho da lâmina foliar;
presença, cor e brilho dos nectários extraflorais; forma geométrica da seção transversal do
pecíolo, brilho, cor e estrias longitudinais do pedúnculo; coloração das estruturas da
inflorescência e da infrutescência e outras informações relevantes.
Para cada espécie foi feita uma descrição morfológica acompanhada do nome popular,
distribuição, dados fenológicos quando possível, comentários sobre a afinidade e histórico dos
espécimes e uma relação do material examinado. Os dados fenológicos e distribuição do

33
material examinado foram baseados em anotações apostas em etiquetas de material
herborizado depositado no Herbário/INPA.

34
RESULTADOS E DISCUSSÃO
Perfil socio-econômico dos informantes
Neste estudo foram realizadas entrevistas com 13 moradores, dos quais oito pertencem
ao sexo feminino (61,5%) e cinco ao sexo masculino (38,5%). Sendo a pesquisa baseada no
método “bola de neve”, conclui-se que o conhecimento sobre plantas na comunidade está
mais associado às mulheres, no entanto, os conhecimentos ligados aos recursos vegetais
provenientes do interior da floresta como madeiras e fibras para o artesanato era de domínio
masculino. Esse resultado corrobora com Hanazaki et al. (2000), que afirmam que os homens
tendem a conhecer mais plantas nativas utilizadas para construções/tecnologias e as mulheres
a ter um conhecimento maior sobre plantas medicinais.
Em relação à origem dos informantes, 38,5% afirmaram ser descendentes de indígenas
das etnias Baré, Desana e Baniwa, o que contribui para a permanência de práticas como, por
exemplo, a utilização de plantas para o tratamento de doenças. Sete (54%) desses informantes
ainda possuem o conhecimento da “língua geral” (Nheengatú), porém poucos a utilizam para
se comunicar. A maior parte dos informantes (69,2%) é proveniente de outros municípios do
Amazonas, como Codajás, Manacapuru, Barcelos e principalmente São Gabriel da Cachoeira
e região do rio Içana. Apenas um (7,6%) informante era nascido na própria comunidade e três
(23,2%) da capital Manaus. Dentre os motivos da mudança para a comunidade estavam a
família, melhores condições de vida e dificuldade de adaptação à cidade. Esta realidade é
comum em comunidades ribeirinhas do Baixo rio Negro que, de acordo com trabalhos de
Cardoso (2008) e Scudeller et al. (2009), a maioria dos moradores são amazonenses
descendentes de indígenas do Alto rio Negro.
A agricultura e a pesca são as principais atividades, representadas por dez (77%) dos
informantes, enquanto que os outros dois (15,3%) eram comerciantes e um (7,6%) agente de
saúde. De acordo com Denevan (1992), a agricultura tradicional constitui-se como uma das
mais antigas e tradicionais formas de uso do solo desenvolvidas por povos indígenas e não-
indígenas da Amazônia. Apesar da agricultura e pesca serem as principais ocupações, sete
(54%) informantes também produzem artesanato, sendo que maior parte da matéria-prima é
retirada da própria floresta (madeira, fibras, sementes e argila) e serve como atrativo para o
turismo na região.
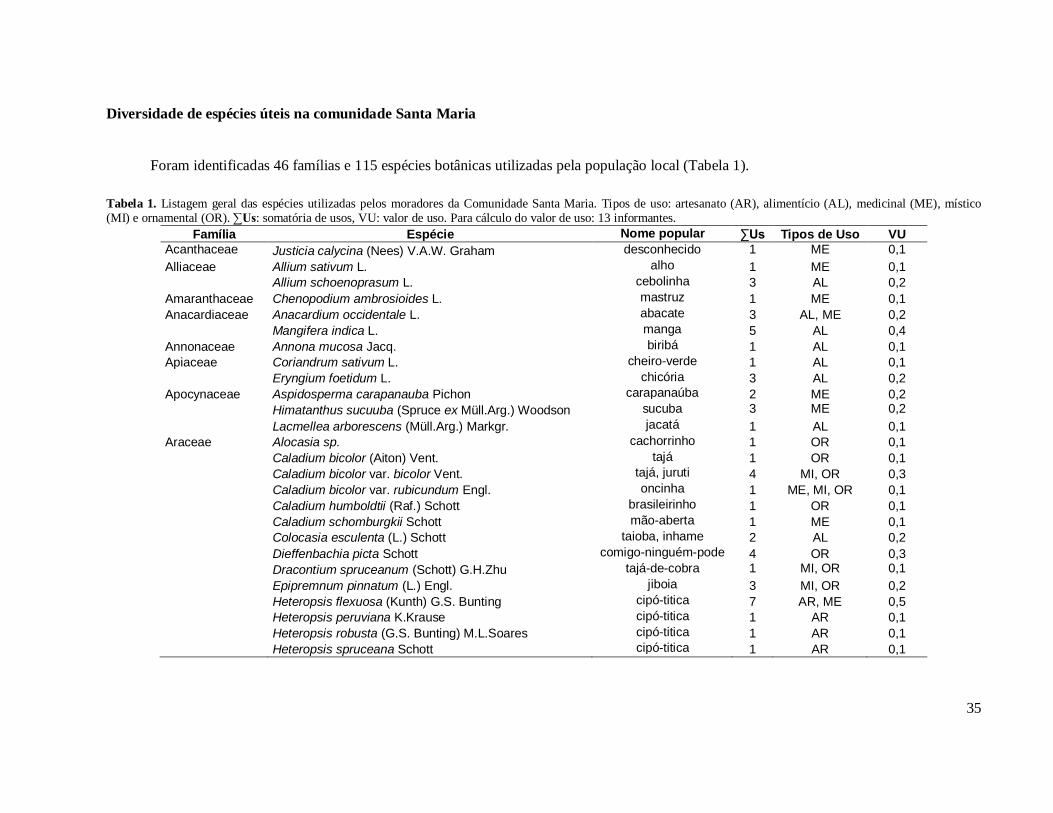
35
Diversidade de espécies úteis na comunidade Santa Maria
Foram identificadas 46 famílias e 115 espécies botânicas utilizadas pela população local (Tabela 1). Tabela 1. Listagem geral das espécies utilizadas pelos moradores da Comunidade Santa Maria. Tipos de uso: artesanato (AR), alimentício (AL), medicinal (ME), místico (MI) e ornamental (OR). ∑Us: somatória de usos, VU: valor de uso. Para cálculo do valor de uso: 13 informantes.
Família Espécie Nome popular ∑Us Tipos de Uso VU Acanthaceae Justicia calycina (Nees) V.A.W. Graham desconhecido 1 ME 0,1 Alliaceae Allium sativum L. alho 1 ME 0,1 Allium schoenoprasum L. cebolinha 3 AL 0,2 Amaranthaceae Chenopodium ambrosioides L. mastruz 1 ME 0,1 Anacardiaceae Anacardium occidentale L. abacate 3 AL, ME 0,2 Mangifera indica L. manga 5 AL 0,4 Annonaceae Annona mucosa Jacq. biribá 1 AL 0,1 Apiaceae Coriandrum sativum L. cheiro-verde 1 AL 0,1 Eryngium foetidum L. chicória 3 AL 0,2 Apocynaceae Aspidosperma carapanauba Pichon carapanaúba 2 ME 0,2 Himatanthus sucuuba (Spruce ex Müll.Arg.) Woodson sucuba 3 ME 0,2 Lacmellea arborescens (Müll.Arg.) Markgr. jacatá 1 AL 0,1 Araceae Alocasia sp. cachorrinho 1 OR 0,1 Caladium bicolor (Aiton) Vent. tajá 1 OR 0,1 Caladium bicolor var. bicolor Vent. tajá, juruti 4 MI, OR 0,3 Caladium bicolor var. rubicundum Engl. oncinha 1 ME, MI, OR 0,1 Caladium humboldtii (Raf.) Schott brasileirinho 1 OR 0,1 Caladium schomburgkii Schott mão-aberta 1 ME 0,1 Colocasia esculenta (L.) Schott taioba, inhame 2 AL 0,2 Dieffenbachia picta Schott comigo-ninguém-pode 4 OR 0,3 Dracontium spruceanum (Schott) G.H.Zhu tajá-de-cobra 1 MI, OR 0,1 Epipremnum pinnatum (L.) Engl. jiboia 3 MI, OR 0,2 Heteropsis flexuosa (Kunth) G.S. Bunting cipó-titica 7 AR, ME 0,5 Heteropsis peruviana K.Krause cipó-titica 1 AR 0,1 Heteropsis robusta (G.S. Bunting) M.L.Soares cipó-titica 1 AR 0,1 Heteropsis spruceana Schott cipó-titica 1 AR 0,1
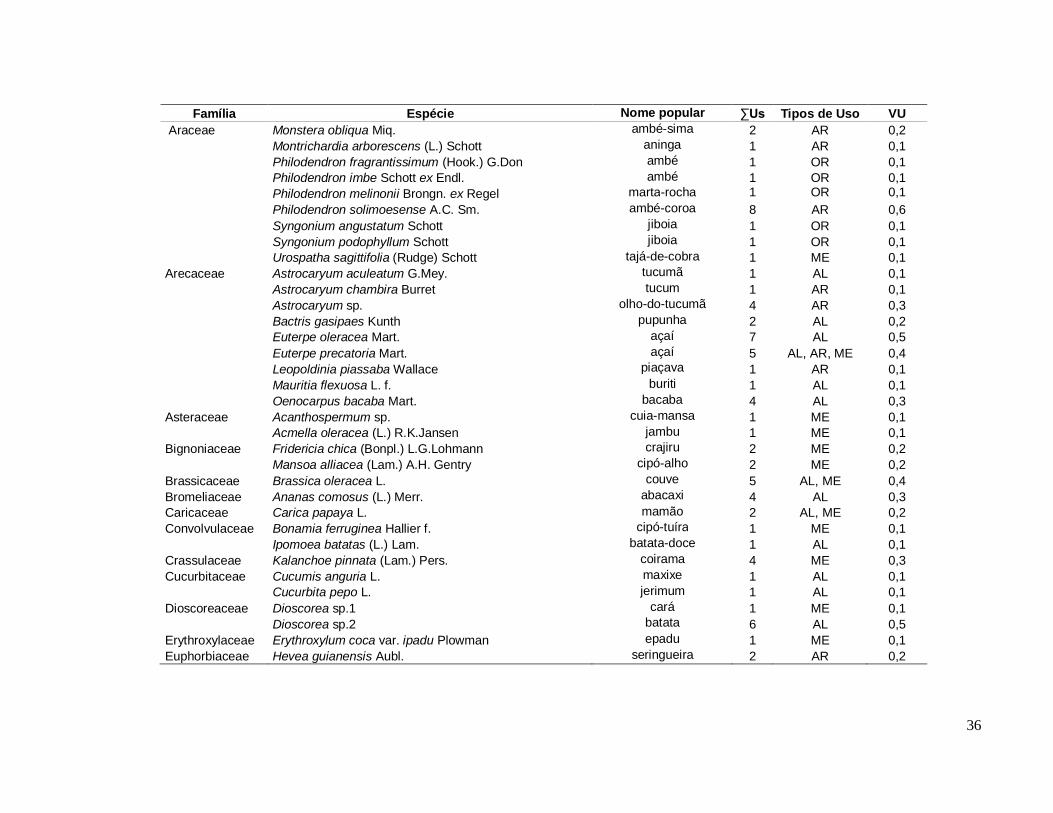
36
Família Espécie Nome popular ∑Us Tipos de Uso VU Araceae Monstera obliqua Miq. ambé-sima 2 AR 0,2 Montrichardia arborescens (L.) Schott aninga 1 AR 0,1 Philodendron fragrantissimum (Hook.) G.Don ambé 1 OR 0,1 Philodendron imbe Schott ex Endl. ambé 1 OR 0,1 Philodendron melinonii Brongn. ex Regel marta-rocha 1 OR 0,1 Philodendron solimoesense A.C. Sm. ambé-coroa 8 AR 0,6 Syngonium angustatum Schott jiboia 1 OR 0,1 Syngonium podophyllum Schott jiboia 1 OR 0,1 Urospatha sagittifolia (Rudge) Schott tajá-de-cobra 1 ME 0,1 Arecaceae Astrocaryum aculeatum G.Mey. tucumã 1 AL 0,1 Astrocaryum chambira Burret tucum 1 AR 0,1 Astrocaryum sp. olho-do-tucumã 4 AR 0,3 Bactris gasipaes Kunth pupunha 2 AL 0,2 Euterpe oleracea Mart. açaí 7 AL 0,5 Euterpe precatoria Mart. açaí 5 AL, AR, ME 0,4 Leopoldinia piassaba Wallace piaçava 1 AR 0,1 Mauritia flexuosa L. f. buriti 1 AL 0,1 Oenocarpus bacaba Mart. bacaba 4 AL 0,3 Asteraceae Acanthospermum sp. cuia-mansa 1 ME 0,1 Acmella oleracea (L.) R.K.Jansen jambu 1 ME 0,1 Bignoniaceae Fridericia chica (Bonpl.) L.G.Lohmann crajiru 2 ME 0,2 Mansoa alliacea (Lam.) A.H. Gentry cipó-alho 2 ME 0,2 Brassicaceae Brassica oleracea L. couve 5 AL, ME 0,4 Bromeliaceae Ananas comosus (L.) Merr. abacaxi 4 AL 0,3 Caricaceae Carica papaya L. mamão 2 AL, ME 0,2 Convolvulaceae Bonamia ferruginea Hallier f. cipó-tuíra 1 ME 0,1 Ipomoea batatas (L.) Lam. batata-doce 1 AL 0,1 Crassulaceae Kalanchoe pinnata (Lam.) Pers. coirama 4 ME 0,3 Cucurbitaceae Cucumis anguria L. maxixe 1 AL 0,1 Cucurbita pepo L. jerimum 1 AL 0,1 Dioscoreaceae Dioscorea sp.1 cará 1 ME 0,1 Dioscorea sp.2 batata 6 AL 0,5 Erythroxylaceae Erythroxylum coca var. ipadu Plowman epadu 1 ME 0,1 Euphorbiaceae Hevea guianensis Aubl. seringueira 2 AR 0,2

37
Família Espécie Nome popular ∑Us Tipos de Uso VU Euphorbiaceae Indeterminada batatão 2 AR, ME 0,2 Jatropha curcas L. pião-branco 1 MI 0,1 Jatropha gossypiifolia L. pião-roxo 8 ME, MI 0,6 Manihot esculenta Crantz macaxeira 11 AL 0,8 Fabaceae Bauhinia guianensis Aubl. escada-de-jabuti 1 ME 0,1 Inga edulis Mart. ingá 4 AL 0,3 Ormosia sp. tento 5 AR 0,4 Senna sp. desconhecido 1 OR 0,1 Swartzia sp. mirapiranga 1 AR 0,1 Humiriaceae Endopleura uchi (Huber) Cuatrec. uchi 2 ME 0,2 Icacinaceae Poraqueiba sericea Tul. mari 2 AL 0,2 Lamiaceae Indeterminada roxinha 1 OR 0,1 Mentha piperita L. hortelãzinho 1 ME 0,1 Ocimum campechianum Mill. alfavaca 2 AL, ME 0,2 Plectranthus barbatus Andrews boldo 2 ME 0,2 Lauraceae Cinnamomum zeylanicum Blume canela 1 ME 0,1 Persea americana Mill. abacate 4 AL, ME 0,3 Lecythidaceae Bertholletia excelsa Bonpl. castanha-do-brasil 2 AL, ME 0,2 Malvaceae Gossypium barbadense L. algodão 1 ME 0,1 Hibiscus rosa-sinensis L. papoula 1 OR 0,1 Theobroma grandiflorum (Willd. ex Spreng.) K.Schum. cupuaçu 6 AL 0,5 Marantaceae Calathea allouia (Aubl.) Lindl. ariá 5 AL 0,4 Indeterminada vai-vem 2 MI, OR 0,2 Ischnosiphon spp. arumã 4 AR 0,3 Melastomataceae Bellucia grossularioides (L.) Triana goiaba-de-anta 1 ME 0,1 Moraceae Artocarpus heterophyllus Lam. jaca 1 AL 0,1 Brosimum parinarioides Ducke leite-de-amapá 1 ME 0,1 Musaceae Musa paradisiaca L. banana 9 AL 0,7 Myrtaceae Psidium guajava L. goiaba 1 AL, ME 0,1 Syzygium jambolanum (Lam.) DC. azeitona-preta 4 AL, ME 0,3 Syzygium malaccense (L.) Merr. & L.M. Perry jambo 1 ME 0,1 Orchidaceae Cattleya violacea (Kunth) Rolfe parasita 1 OR 0,1 Indeterminada parasita 1 OR 0,1 Passifloraceae Passiflora edulis Sims maracujá 2 AL, ME 0,2
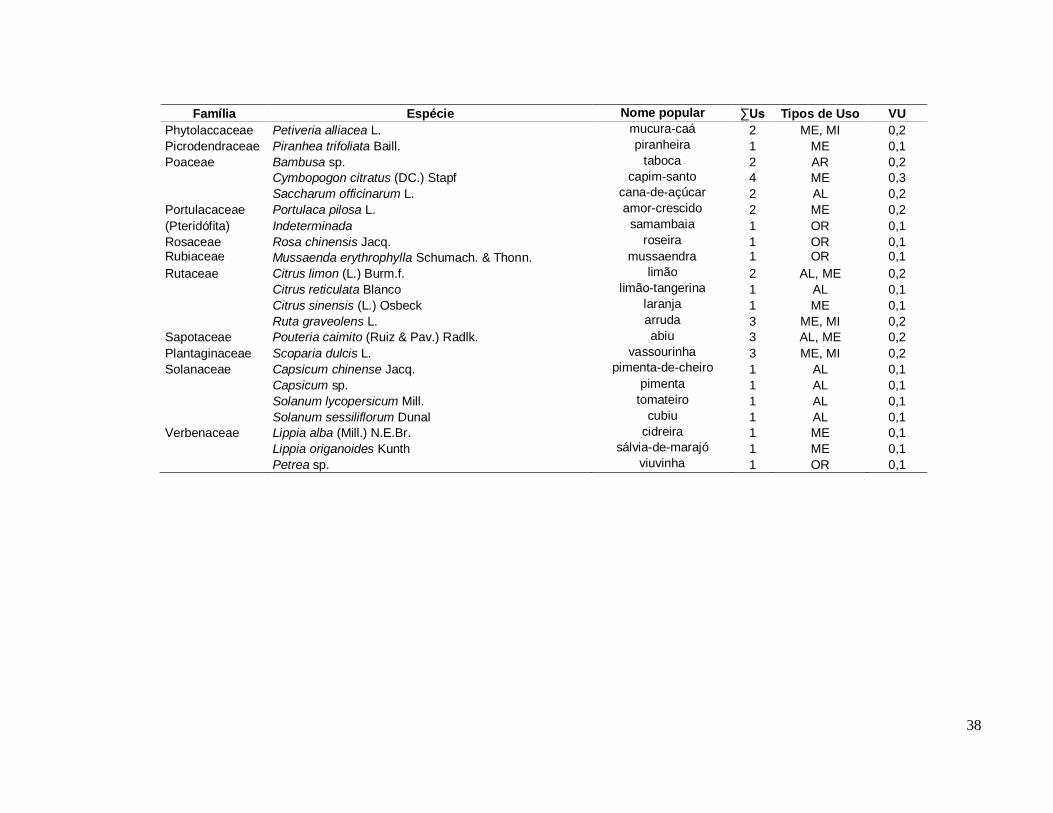
38
Família Espécie Nome popular ∑Us Tipos de Uso VU Phytolaccaceae Petiveria alliacea L. mucura-caá 2 ME, MI 0,2 Picrodendraceae Piranhea trifoliata Baill. piranheira 1 ME 0,1 Poaceae Bambusa sp. taboca 2 AR 0,2 Cymbopogon citratus (DC.) Stapf capim-santo 4 ME 0,3 Saccharum officinarum L. cana-de-açúcar 2 AL 0,2 Portulacaceae Portulaca pilosa L. amor-crescido 2 ME 0,2 (Pteridófita) Indeterminada samambaia 1 OR 0,1 Rosaceae Rosa chinensis Jacq. roseira 1 OR 0,1 Rubiaceae Mussaenda erythrophylla Schumach. & Thonn. mussaendra 1 OR 0,1 Rutaceae Citrus limon (L.) Burm.f. limão 2 AL, ME 0,2 Citrus reticulata Blanco limão-tangerina 1 AL 0,1 Citrus sinensis (L.) Osbeck laranja 1 ME 0,1 Ruta graveolens L. arruda 3 ME, MI 0,2 Sapotaceae Pouteria caimito (Ruiz & Pav.) Radlk. abiu 3 AL, ME 0,2 Plantaginaceae Scoparia dulcis L. vassourinha 3 ME, MI 0,2 Solanaceae Capsicum chinense Jacq. pimenta-de-cheiro 1 AL 0,1 Capsicum sp. pimenta 1 AL 0,1 Solanum lycopersicum Mill. tomateiro 1 AL 0,1 Solanum sessiliflorum Dunal cubiu 1 AL 0,1 Verbenaceae Lippia alba (Mill.) N.E.Br. cidreira 1 ME 0,1 Lippia origanoides Kunth sálvia-de-marajó 1 ME 0,1 Petrea sp. viuvinha 1 OR 0,1

39
Araceae foi a família que apresentou maior número de espécies úteis na comunidade
Santa Maria, com 23 espécies (18%), entre plantas cultivadas e nativas da floresta (Figura 4).
Também foi a família que apresentou maior número de tipos de usos em relação às outras
famílias, sendo utilizada para artesanato, alimento, medicinal, ornamental e uso místico
(Tabela 2). Em estudo realizado por Souza (2010), em comunidades da RDS Tupé,
Amazonas, Araceae também foi a família mais representativa, apresentando 14 espécies úteis
para a população local.
A família Arecaceae ficou em segundo lugar em relação a quantidade de espécies
úteis, as espécies mais citadas pelos informantes foram o “açaí” (Euterpe spp.), utilizada
como alimento (frutos), medicinal (raízes) e no artesanato (sementes), a “bacaba”
(Oenocarpus bacaba Mart.) e o “tucumã” (Astrocaryum aculeatum G.Mey.) utilizados na
alimentação. Em um levantamento etnobotânico, realizado por Oliveira et al. (2006), na
microrregião do Salgado, estado do Pará, estas duas famílias foram as mais importantes, no
entanto, Arecaceae ficou em primeiro lugar e Araceae em segundo com as espécies
Heteropsis flexuosa e Philodendron imbe, utilizadas pelos pescadores em trançados.
Figura 4. Famílias botânicas com maior número de espécies úteis para a comunidade Santa Maria, Baixo rio
Negro – AM.
A família Euphorbiaceae ficou em terceiro lugar pela predominância de grande
variedade de mandiocas cultivadas nos roçados, onde a maioria dos informantes as cultivam
para produzir farinha. Levando em consideração que a maior parte dos informantes é
descendente de indígenas, podemos afirmar que o cultivo da mandioca faz parte da cultura
alimentar desses povos. Segundo o IPÊ (2007), 70% das famílias indígenas da região do rio
Cuieiras, tarabalham com a agricultura de mandioca.

40
Neste estudo foram encontradas 51 espécies de Araceae distribuídas em 15 gêneros
entre plantas coletadas na floresta e na comunidade (Tabela 2). Deste total, 39 espécies (78%),
foram encontradas na floresta e 12 (22%) sendo cultivadas pelos moradores da comunidade
Santa Maria para vários tipos de uso. Dentre as espécies encontradas na área de floresta, o
gênero Philodendron apresentou 22 espécies, ou seja, 56% do total, sendo este, de acordo
com Madison (1979) e Soares (1996), o gênero de Araceae mais abundante e diverso na
Amazônia (Figura 5).
Figura 5. Diversidade de gêneros coletados na área florestal da comunidade Santa Maria, Baixo rio Negro, AM.

41
Tabela 2. Lista das espécies da família Araceae utilizadas pelos moradores da comunidade Santa Maria. Hábitos: epífita (E), hemiepífita (HE) e herbácea terrícola (HT). Ocorrência: área florestal (F), área da comunidade (C). Origem: Amazônia (Am); Mata Atlântica (M.A.); Pantanal (P), Caatinga (Ca); Cerrado (Ce). Uso: artesanato (AR), alimentício (AL), medicinal (ME), místico (MI) e ornamental (OR).
Táxon Registro herbário Nome popular Hábito Ocorrência Origem Uso Alocasia sp. cachorrinho HT C Ásia1 OR Anthurium atropurpureum R.E. Schult. & Maguire EAFM 4553 desconhecido E F Am2 _ Anthurium bonplandii Bunting EAFM 4563 desconhecido E F Am2 _ Anthurium eminens Schott EAFM 4559 desconhecido HE F Am, M.A.2 _ Anthurium gracile (Rudge) Schott desconhecido E F Am, Ca, Ce2 _ Anthurium pentaphyllum (Aubl.) G. Don EAFM 4567; INPA 239245 desconhecido HE F Am, M.A., P2 _ Caladium bicolor (Aiton) Vent. tajá HT C Am, Ca, Ce, M.A.2 OR Caladium bicolor var. bicolor tajá-pintado HT C Am, Ca, Ce, M.A.2 MI, OR Caladium bicolor var. rubicundum Engl. INPA 239225 oncinha HT C Am3 ME, MI, OR Caladium humboldtii Schott brasileirinho HT C Am2 OR Caladium schomburgkii Schott mão-aberta HT C Am2 ME, OR Colocasia esculenta (L.) Schott taioba HT C Ásia1 AL Dieffenbachia picta Schott comigo-ninguém-pode HT C América do Sul4 MI, OR Dieffenbachia parvifolia Engl. EAFM 4562 aninga HT F Am2 _ Dracontium spruceanum (Schott) G.H.Zhu INPA 239242 tajá-de-cobra HT F Am2 ME Epipremnum pinnatum (L.) Engl. jibóia HE C Ilhas Salomão1 MI, OR Heteropsis flexuosa (Kunth) G.S. Bunting EAFM 4564; INPA 239233;
INPA 239243 cipó-titica HE F Am, M.A.2 AR, ME
Heteropsis peruviana K.Krause EAFM 4578 cipó-titica HE F Am2 AR Heteropsis robusta (G.S. Bunting) M.L.Soares EAFM 4570 cipó-titica HE F Am2 AR Heteropsis spruceana Schott EAFM 4579 cipó-titica HE F Am2 AR Monstera obliqua Miq. EAFM 4550; INPA 239231 ambé-sima HE F Am, M.A.2 AR Montrichardia arborescens (L.) Schott EAFM 4581 aninga HT F Am, Ce, M.A.2 AR Philodendron asplundii Croat & M.L.Soares INPA 239235 ambé HE F Am2 _ Philodendron barrosoanum G.S.Bunting EAFM 4561; INPA 239230 ambé HE F Am2 _ Philodendron billietiae Croat EAFM 4574 ambé HE F Am2 _ Philodendron brevispathum Schott EAFM 4572 ambé HE F Am, Ce2 _ Philodendron distantilobum K.Krause EAFM 4575; INPA 239239 ambé HE F Am2 _

42
Táxon Registro herbário Nome popular Hábito Ocorrência Origem Uso Philodendron elaphoglossoides Schott EAFM 4556 ambé HE F Am2 _ Philodendron fragrantissimum (Hook.) G.Don EAFM 4569; INPA 239227 ambé HE F Am, M.A.2 OR Philodendron goeldii G.M.Barroso INPA 239237 ambé HE F Am2 _ Philodendron hastatum K.Koch & Sello EAFM 4568 ambé HE F M.A.2 _ Philodendron imbe Schott ex Endl. ambé HE C M.A.2 OR Philodendron linnaei Kunth INPA 239226; INPA
239228 ambé HE F Am, Ce2 _
Philodendron cf. maximum K. Krause EAFM 4573 ambé HE F Am2 _ Philodendron megalophyllum Schott EAFM 4552; INPA 239241 ambé HE F Am, Ce, M.A.2 _ Philodendron melinonii Brongn. ex Regel marta-rocha, tajá E F e C Am2 OR Philodendron ornatum Schott INPA 239232 ambé HE F Am, Ca, M.A.2 _ Philodendron pedatum (Hook.) Kunth EAFM 4560 ambé HE F Am, Ca, Ce, M.A.2 _ Philodendron platypodum Gleason EAFM 4557 ambé HE F Am2 _ Philodendron pulchrum G.M.Barroso EAFM 4555; INPA 239246 ambé HE F Am2 _ Philodendron rudgeanum Schott EAFM 4558 ambé HE F Am, M.A.2 _ Philodendron solimoesense A.C.Sm. EAFM 4551; INPA 239238 ambé-coroa HE F Am2 AR Philodendron surinamense (Miq.) Engl. EAFM 4565 ambé HE F Am, M.A.2 _ Philodendron tortum M.L.Soares & Mayo EAFM 4577 ambé HE F Am2 _ Philodendron wittianum Engl. EAFM 4576; INPA 239229 ambé HE F Am2 _ Rhodospatha oblongata Poepp. INPA 239236 desconhecido HE F Am, Ca, M.A.2 _ Rhodospatha venosa Gleason EAFM 4554 desconhecido HE F Am2 _ Syngonium angustatum Schott jiboia HE F Am, Ce2 OR Syngonium podophyllum Schott jiboia HE C Costa Rica e Panamá2 OR Urospatha sagittifolia (Rudge) Schott EAFM 4580 tajá HT F Am, Ca, Ce2 ME Xanthosoma striolatum Mart. ex Schott INPA 239234 tajá-bravo HT C Am2 _ 1. Lorenzi, H. e Souza, H.M. 2008. Plantas ornamentais do Brasil. Arbustivas, herbáceas e trepadeiras. 4a ed. Instituto Plantarum. 2. Coelho, M.A.N., Soares, M.L., Sakuragui, C.M., Mayo, S., Andrade, I.M. de, Temponi, L.G. 2010. Araceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. (http://floradobrasil.jbrj.gov.br/2010). 3. Engler, A. 1878. Araceae. In: de Martius, C.F.P. Flora Brasiliensis. 3(2): 182. (http://facultystaff.wwc.edu/~presslar/greenhouse/caladium/bicolor/var_rub.htm). 4. http://www.tropicos.org

43
O número de espécies de Araceae encontradas na região foi considerado elevado em
comparação a outros trabalhos realizados no Brasil. Em um levantamento realizado em nove
fragmentos de Mata Atlântica de Pernambuco, por exemplo, foram encontradas 18 espécies,
distribuídas em nove gêneros, sendo Philodendron também o mais diverso, com oito espécies
(Pontes et al., 2010). Segundo levantamento realizado por Canalez (2009), em Fonte Boa no
Amazonas, o gênero Philodendron foi o mais rico apresentando 16 espécies. Um dos motivos
do maior registro de espécies no presente estudo foi a coleta em mais de um ambiente
florestal (floresta de “floresta ombrófila”, “campinarana” e “igapó”).
Dentre as dez espécies com maior valor de uso, Araceae foi representada por
Philodendron solimoesense A.C.Sm. que se apresentou em terceiro lugar, com VU=0,6 e
Heteropsis flexuosa (Kunth) G.S.Bunting em quinto lugar com VU= 0,5 ambas para uso no
artesanato. A espécie que apresentou maior Valor de Uso foi Manihot esculenta Crantz
(Euphorbiaceae) com VU=0,8, seguida de Musa paradisiaca L. (Musaceae), com VU=0,7,
ambas para uso alimentício (Figura 6).
Figura 6. Espécies com maior Valor de Uso na comunidade Santa Maria, Baixo rio Negro – AM.
Foram encontradas 12 espécies de Araceae (22%) distribuídas em sete gêneros, sendo
cultivadas na área da comunidade para fins alimentícios, medicinais, ornamentais e místicos
(Figura 7). Destas, cinco pertencem ao gênero Caladium (Caladium bicolor Vent., C. bicolor
var. bicolor, C. bicolor var. rubicundum Engl., C. humboldtii Schott e C. schomburgkii

44
Schott) e duas espécies pertecem ao gênero Syngonium (Syngonium angustatum e S.
podophyllum), representando juntos 59% das espécies cultivadas. Os gêneros Alocasia,
Colocasia e Epipremnum são exóticos (Souza & Lorenzi, 2008). Muitas plantas, cultivadas
em quintais são obtidas em mercados municipais, ou trazidas de outras regiões pelos próprios
moradores, quando vindos de outras regiões.
Figura 7. Diversidade de gêneros cultivados na comunidade Santa Maria, Baixo rio Negro.
Artesanato
No entorno da comunidade Santa Maria foram identificadas quatro espécies de
Heteropsis (“cipó-titica”): H. flexuosa (Kunth) G.S. Bunting, H. peruviana K. Krause, H.
robusta (G.S. Bunting) M.L. Soares e H. spruceana Schott, utilizadas para o artesanato
(Figura 12 A-D). No entanto, H. flexuosa é a espécie de maior preferência para o artesanato e
construção, pois no momento da turnê-guiada, realizada com o informante-chave, este
apontou H. flexuosa, dentre as outras espécies coletadas do mesmo gênero, como a mais
procurada para o artesanato. Esta afirmação corrobora com Durigan & Castilho (2004), os
quais afirmam que na região do Baixo rio Negro, H. flexuosa é também a espécie mais
utilizada pelos extratores como fonte de fibras. Segundo um artesão da comunidade Santa
Maria, era comum misturar o “timbó-açu” (Thoracocarpus bissectus (Vell.) Harling –
Cyclanthaceae) à “piraíba” de “cipó-titica”, que é a forma de organização em feixes com
cerca de um metro de comprimento de raízes amarradas. Essa prática também foi relatada
pelos artesãos do povo Wai-Wai do Sudeste da Guiana, onde o “cipó-titica” é chamado de
“Nibbi”, também era substituído quando escasso ou ausente, pelas raízes de Cyclanthaceae ou

45
outras espécies de Araceae (Hoffman, 1997).
Philodendron solimoesense é a espécie mais utilizada do gênero Philodendron para o
artesanato na comunidade, conhecida como “cipó-ambé” ou “ambé-coroa”. Este último nome
se deve aos acúleos presentes em todo o comprimento da raiz aérea, lembrando o aspecto de
uma coroa (Figura 12 H). Outras espécies de Philodendron, também são utilizadas na região
amazônica, como Philodendron fragrantissimum (Hook.) G.Don, também conhecido como
“cipó-ambé” na RDS Amanã, que foi relatado pelos artesãos locais como matéria-prima para
confecção de “paneiros” e peneiras, os quais estão associados ao acondicionamento,
transporte da mandioca brava e à produção de farinha (Leoni & Marques, 2008). Em algumas
comunidades da RDS do Tupé, no Baixo rio Negro, foi registrado o uso de Philodendron
billietiae Croat como fonte de fibra para o artesanato (Souza, 2010).
Além dos gêneros Heteropsis e Philodendron, já considerados importantes para a
economia extrativista em várias regiões da Amazônia e do Brasil, também foi relatado o uso
das raízes de Monstera obliqua, conhecida na comunidade como “ambé-sima” (Figura 12 E).
Esta, por ter o diâmetro menor e ser menos resistente, é utilizada apenas para dar acabamento
nas bordas dos “paneiros”, que são utilizados principalmente para o transporte de produtos
extraídos da roça (Figura 8 B).
O caule de Montrichardia arborescens (L.) Schott, conhecida como “aninga”, foi
relatado para a confecção de pequenas canoas, que eram utilizadas como brinquedo, durante a
infância de alguns moradores da comunidade descendentes de indígenas do Alto rio Negro.
Atualmente, este recurso poderá servir para a produção de peças decorativas ou até mesmo
embalagens para algum tipo de produto como, por exemplo, bombons com recheios de geleias
de frutas regionais, o que já é visto na cidade de Manaus, e consequentemente contribuir para
a geração de renda na comunidade.
Apesar de ainda haver utilização dessas raízes como fonte de fibra para o artesanato
em municípios de todo o Baixo rio Negro, os mais jovens da comunidade não apresentam
interesse em aprender com os mais velhos o processo de fabricação dos cestos, paneiros e
outros utensílios, tornando o produto cada vez mais escasso e colocando em risco a tradição
cultural. Estes produtos exigem conhecimentos especializados nas etapas de coleta,
beneficiamento e confecção e não possuem substitutos no mercado devido às suas qualidades
funcionais (Leoni & Marques, 2008), uma vez que são recomendados para o
acondicionamento de produtos perecíveis como, por exemplo, o açaí, fruto amazônico
amplamente consumido atualmente em todo o Brasil e no mundo, pois em sacos de
polietileno o produto apodrece mais rapidamente (BRASÍLIA, 1998).

46
Segundo Pellin & Caiut (2006 apud. Valente, 2009), o artesanato é uma boa opção de
geração de emprego para as comunidades pela possibilidade de se conciliar a presença
humana com a conservação. Assim como em comunidades do Parque Nacional do Jaú,
reforça-se aos artesãos da comunidade Santa Maria, os benefícios de se trabalhar com
produtos de maior valor agregado em relação a produtos não beneficiados como matérias-
primas de outras atividades. A possibilidade de vendê-los, em substituição a venda da
matéria-prima, reduz o trabalho de coleta do recurso, uma vez que é necessária menor
quantidade de cipós para se obter retorno financeiro semelhante (FVA, 2009). Dessa forma, o
artesanato ajuda a diminuir a pressão sobre os recursos naturais e melhora a qualidade de vida
da comunidade.
Figura 8. Utilização de fibras e artesanato da comunidade Santa Maria. A) Cercado feito com estacas de madeira presas com raízes de “ambé-coroa”; B) Paneiro feito de “ambé-coroa” sendo utilizado na tradicional festa de Santo Antônio da comunidade; C) Cesta feita de “ambé-coroa”; D) Cesto feito de “cipó-titica”.
Coleta e beneficiamento do “ambé-coroa”
As raízes do “ambé-coroa” ou “cipó-ambé” (Philodendron solimoesense) são
encontradas tanto na floresta de terra-firme como no igapó e margens do rio. Na comunidade
Santa Maria a coleta é realizada em qualquer época do ano, porém os moradores retiram
menor quantidade de raízes, em comparação a outras comunidades do Baixo rio Negro, onde

47
a extração dos cipós para comercialização é retirada em escala maior, principalmente nos
períodos de cheia. Nas comunidades do Baixo rio Negro, tanto o “cipó-titica” como o “cipó-
ambé”, tem se consolidado após o declínio da exploração da borracha (Durigan & Castilho,
2004).
Os coletores escolhem as raízes maduras, com no mínimo 2 cm de diâmetro, e mais
escuras e com a entrecasca avermelhada. Para verificar se está verde por dentro, fazem um
corte utilizando uma faca ou até mesmo a própria unha. Outro critério que influencia na
escolha da raiz é a presença de nós, causada pela regeneração da raiz quando cortada ou
arrancada, portanto, são evitadas raízes com muitos nós a distâncias de menos de 1,0 – 1,5 m.
As raízes são transportadas até a comunidade a pé ou através de canoa na forma de
feixes amarrados nas extremidades, a “piraíba”, ou em forma de rolos de cipó. Na
comunidade, as raízes são guardadas ainda com casca em local protegido do sol e da chuva
podendo, dessa forma, durar até mais de um mês. O processo do beneficiamento do “ambé-
coroa” na comunidade Santa Maria consiste das seguintes fases:
I – Mondagem (retirada da casca):
Este procedimento consiste em descascar as raízes torcendo-as até ocasionar o seu
rompimento, depois, a casca é puxada ficando somente o cipó (Figura 9 A e B). O processo de
descascar seis fios dura cerca de 20 minutos. Ocasionalmente, a resina que se desprende da
raiz pode irritar bastante a pele, nesse caso, o artesão recomenda passar a raiz na brasa para
neutralizar sua causticidade.
II - Raspagem:
Tem como finalidade deixar somente a parte central do cipó, que será utilizada para o
artesanato. Nessa etapa usa-se uma faca não muito amolada para retirar a parte grossa com as
marcas dos espinhos para dar o aspecto liso à fibra (Figura 9 C).
III – Desfiamento:
Esse processo consiste em partir o cipó em vários fios no sentido vertical, na espessura
adequada para determinado uso (Figura 9 D). Geralmente o cipó é desfiado em até três fios.

48
IV - Acabamento:
O processo de acabamento tem a finalidade de deixar os fios todos da mesma espessura,
cerca de 0,7 cm de diâmetro para este produto e é feito com apoio dos pés e uma faca,
passando-a de uma extremidade a outra (Figura 9 E e 9 F). Os fios que saem mais finos
servem para fazer a borda do paneiro.
Figura 9. Etapas do beneficiamento do “ambé-coroa” (Philodendron solimoesense A.C. Sm.). A-B) retirada da casca; C) raspagem; D) desfiamento; E-F) acabamento. (Fotos: Rina Oliveira, Comunidade Santa Maria, junho/2010).
Tecelagem
Geralmente são necessários doze fios, de 4 a 8 m de comprimento para fazer até três
paneiros. Em um paneiro médio, com capacidade para 15 kg e cerca de 37-40 cm de largura,
são utilizados aproximadamente de 30 fios com 2-3 m de comprimento cada um. A tecelagem
começa pelo fundo ou base do paneiro, onde os fios devem ser separados três grupos de feixes
paralelos, um que fica na horizontal e dois que passam em sentidos transversais formando
uma cruz ou um X. Os fios deverão ser entrelaçados paralelamente aos fios iniciais nos três
sentidos diferentes (Figura 10).

49
Figura 10. Etapas iniciais da confecção de um paneiro feito de “cipó-ambé” (Philodendron solimoesense A.C. Sm.) por um artesão da comunidade Santa Maria. (Desenho esquemático: Rina Oliveira, fev/2011).
A tecelagem do fundo deve terminar com os trinta fios entrelaçados, três grupos com
cerca de 10 fios cada um. A partir daí, a tecelagem do “paneiro” começa-se escolhendo um fio
para formar o centro dos três grupos formando um hexágono (Figura 11-A). Após a confecção
do fundo do “paneiro”, novos fios vão sendo colocados nas laterais e trançados com os três
primeiros feixes de fios (Figura 11-B e 11-C). As fibras mais finas servem para fazer a borda
(Figura 11-D).
O tingimento das fibras é opcional e geralmente são utilizadas tinturas como o fruto de
Bixa orellana L. (“urucum”) e Fridericia chica (Bonpl.) L.G.Lohmann (“crajirú”). No
município de Novo Airão, a madeira conhecida por “mirapiranga” (Swartzia sp.) é utilizada
para confeccionar as alças.
Figura 11. Tecelagem do “paneiro” feito de “ambé-coroa” (Philodendron solimoesense A.C.Sm.) por artesão da comunidade Santa Maria, Manaus, AM. A) fundo do paneiro; B-C) artesão trançando as fibras de “ambé”; D) acabamento do “paneiro” com fibras mais finas de cipó nas bordas. (Fotos: Rina Oliveira, Comunidade Santa Maria, junho/2010)

50
Segundo artesãos do Parque Nacional do Jaú e Resex do rio Unini, em visita realizada
em setembro de 2009, o quilo do “cipó-ambé” (Philodendron sp.) bruto é vendido por R$
0.50 e se estiver beneficiado, ou seja, pronto para tecer, o preço sobe para R$ 2.50 ou mais.
Os cestos confeccionados na Comunidade Santa Maria custam de R$ 20,00 a R$ 30,00,
dependendo do tamanho. Em relação às fibras de “cipó-titica” (Heteropsis spp.), os preços
pagos nos rios Jaú e Unini eram de R$ 1,80/kg de “cipó titica” e R$ 1,20 a R$ 1,50/kg de
“timbó-açu” em levantamento realizado em 2004 (SDS, 2005).
Figura 12. Espécies de Araceae utilizadas para fins artesanais na comunidade Santa Maria, Baixo rio Negro – AM. (Fotos: Valdely Kinupp (A e E); Rina Oliveira (B, C, D e F); M.L. Soares (G e H), Comunidade Santa Maria, 2010).
Alimentícias
A única espécie alimentícia de Araceae registrada na comunidade Santa Maria foi
Colocasia esculenta, conhecida na comunidade como “taioba”, apesar de em outras regiões
do Brasil ser amplamente conhecida como “inhame” e fora do país como “taro”. Este último
nome comercial é mundialmente utilizado (Figura 13-G, 13-H). A parte da planta utilizada é o
rizoma, que deve ser cozido antes do consumo. A espécie conhecida como “taioba”,

51
Xanthosoma sagittifolium, é encontrada próximo ao Parque Estadual do Rio Negro Setor Sul,
sendo cultivada em roçados nas comunidades do rio Cuieiras (Cardoso, 2008).
Colocasia esculenta pertence à subfamília mais importante economicamente de
Araceae, Colocasioideae, com espécies amplamente cultivadas nos trópicos como cultura
alimentar. Tanto as folhas quanto os caules ricos em amido são utilizados como alimento por
humanos, assim como por animais domésticos (Madison, 1981). Esta planta já teve seu
consumo recomendado na região amazônica para combater a desnutrição (Barreto &
Cavalcanti, 1947), além disso, Paiva et al. (1981) apontam a variedade conhecida como
“curumim”, sendo a melhor para as condições de Manaus, cujos solos possuem baixa
fertilidade.
Medicinais
As espécies indicadas como medicinais pela população local foram Caladium bicolor
var. rubicundum Engl. (“oncinha”), Caladium schomburgkii Schott (“mão-aberta”),
Heteropsis spp. (“cipó-titica”) e Urospatha sagittifolia (Rudge) Schott (“tajá-de-cobra”)
(Tabela 3). No entanto, somente as duas espécies de Caladium são cultivadas nos quintais, as
outras duas são coletadas na mata. A aplicação destas plantas restringe-se ao uso externo, pois
sua ingestão não é aconselhável devido à irritação na mucosa causada por cristais de oxalato
de cálcio, presentes nas espécies da família Araceae (Keating, 2004).
No campo da investigação farmacológica, Rodrigues (2006) ressalta que em muitos
casos, o isolamento geográfico dessas populações, contribui ao fortalecimento das práticas
médicas locais e tradicionais. Apesar da presença do posto de saúde na Comunidade Santa
Maria e de relativa facilidade de acesso à capital Manaus, através dos barcos de recreios,
muitas pessoas da comunidade ainda recorrem ao conhecimento tradicional para obter
remédios a base de plantas medicinais.
Tabela 3. Espécies de Araceae utilizadas como plantas curativas na Comunidade Santa Maria, seus respectivos nomes populares, partes utilizadas e indicações de uso.
Nome científico Nome popular Parte usada Indicação de uso
Caladium bicolor var. rubicundum Engl. oncinha rizoma Infecções de pele em animais (“bicheiras”)
Caladium schomburgkii Schott mão-aberta folha Furúnculos e tumores Heteropsis spp. cipó-titica raiz aérea Ferrada de arraia Urospatha sagittifolia (Rudge) Schott tajá-de-cobra rizoma Mordida de cobra

52
Ornamentais
Foram encontradas 12 espécies, distribuídas em sete gêneros, utilizadas como
ornamentais na Comunidade Santa Maria (Figura 13). Essas espécies possuem fácil
propagação vegetativa e não são muito exigentes quanto ao uso de insumos de jardinagem
para sua manutenção. Concomitantemente ao uso ornamental, a espécie Caladium bicolor var.
rubicundum possui uso medicinal e místico, enquanto que Caladium schomburgkii também é
usada como planta medicinal (Figuras 13 D e F).
A maioria das espécies usada como ornamentais na comunidade são adquiridas em
mercados municipais procedentes de outras regiões do país. Além da beleza exuberante das
folhagens e inflorescências das espécies de Araceae, há uma grande facilidade de cultivo e
resistência de algumas espécies que contribuem para a sua propagação. Portanto, o cultivo de
espécies nativas é muito importante, principalmente das espécies que demonstram um
potencial ornamental e ainda são desconhecidas, no intuito de fornecer novas perspectivas de
mercado e fonte de renda para as comunidades amazônicas.
Dentro desse contexto, recomenda-se o uso ornamental de algumas espécies
encontradas na área da Comunidade Santa Maria, que apresentam potencial ornamental,
como: Anthurium atropurpureum R.E. Schult. & Maguire, A. gracile (Rudge) Schott, A.
eminens Schott, A. pentaphyllum (Aubl.) G. Don, Dracontium spruceanum (Schott) G.H. Zhu,
Monstera obliqua Miq. (esta bastante utilizada na cidade Manaus, e.g. em decoração interna
de shoppings e paisagismo em condomínios), Philodendron asplundii Croat & M.L.Soares, P.
barrosoanum G.S. Bunting, P. distantilobum K. Krause, P. hyleae G.S. Bunting, P. ornatum
Schott, P. pedatum (Hook.) Kunth, P. goeldii G.M. Barroso, P. linnaei Kunth, P. tortum
M.L.Soares & Mayo, Philodendron wittianum Engl. Algumas destas espécies já possuem uso
ornamental consagrado em várias regiões do Brasil, porém são pouco exploradas para esse
fim na comunidade. Este levantamento corrobora com Souza & Lorenzi (2008), os quais
afirmam que há uma quantidade muito maior de gêneros e espécies nativas de Araceae com
grande potencial ornamental ainda pouco explorado.
A Comunidade Santa Maria apresenta grande potencial eco-turístico, portanto
recomenda-se também a construção de um epifitário, não só para as espécies de Araceae,
apresentadas neste trabalho, como também para espécies epífitas e hemiepífitas de outras
famílias como Bromeliaceae, Cactaceae, Cyclanthaceae, Gesneriaceae e Orchidaceae, que
apresentam elevado valor ornamental, visando contemplar uma parte da beleza florística da
região na própria comunidade. Além de possibilitar uma exposição rápida das plantas no local

53
para os visitantes, o epifitário contribuirá na promoção do conhecimento e do interesse dos
jovens da comunidade em trabalhar na conservação da flora e de outros recursos da floresta.
Místicas
Epipremnum pinnatum Schott (“jiboia”), também utilizada como planta ornamental, é
considerada pelos moradores como “planta para atrair gente”, principalmente quando se trata
de estabelecimentos comerciais (Figura 13 J). No entanto, pela facilidade de propagação
vegetativa e hábito escandente, esta planta foi amplamente eliminada, pois segundo alguns
moradores, esta chegava a cobrir outras plantas quando adultas.
Dieffenbachia picta Schott (“comigo-ninguém-pode”), é outra planta utilizada em
frente às casas, tanto como ornamental, como para combater o “mau olhado” (Figura 13 I).
Algumas pessoas, apesar de relatarem não utilizar, conhecem através de seus pais e avós,
plantas consideradas místicas como o “pião-roxo” (Jatropha gossypiifolia L. –
Euphorbiaceae) citado pela maioria dos informantes.
Outra planta responsável por possuir um legado místico no imaginário de alguns
moradores é o Caladium bicolor, popularmente conhecida como “tajá” ou “uirapuru”, cujas
folhas variegadas são conhecidas em lendas como a do “tajá que pia” e por trazer “mau
agouro” para aqueles que a encontram no caminho (Figura 13 C). Existem variedades de
Caladium como o C. bicolor var. rubicundum Engl., “tajá-oncinha”, que deve seu nome
popular às pintas de suas folhas, parecidas com as da “onça-pintada”, possui uma lenda de
que precisa ser nutrida com sangue e restos de carne provenientes do preparo das refeições
durante as sextas-feiras de lua cheia (Figura 13 D). Segundo Cardoso (2008), em sua pesquisa
realizada no rio Cuieiras, a identificação e nomeação dos “tajás” foram baseadas na cor e
pigmentação de suas folhas, podendo estas serem “onças” (tajá-onça), “surucucus” (tajá-
surucucu) e “sussuaranas” (tajá-sussuarana).
Outro mito difundido foi constatado na comunidade através de conversas informais,
onde alguns extratores de cipó acreditam que o “cipó-titica” (Heteropsis spp.), nasce através
de uma formiga conhecida como “tucandeira” (Paraponera spp.). Este mito também é
conhecido por moradores de comunidades do Parque Nacional do Jaú, no Baixo rio Negro
(Durigan, 1998).

54
Figura 13. Espécies de Araceae cultivadas nos quintais da Comunidade Santa Maria, Baixo rio Negro – AM. (Fotos: Rina Oliveira, comunidade Santa Maria, jun/2010).

55
CONCLUSÃO
A comunidade Santa Maria demonstrou ser uma área rica na diversidade da família
Araceae. As espécies apresentaram uma grande importância na comunidade, sendo utilizadas
para fins artesanais, alimentícios, medicinais, ornamentais e místicos.
Deve ser incentivada a utilização de outras espécies com potencial fibroso, como
Monstera obliqua (“ambé-sima”), por exemplo, a qual foi registrada pela primeira vez neste
trabalho como fonte de fibras para o artesanato. Dessa forma, diminui-se o impacto sobre as
espécies de raízes com diâmetro maior como H. flexuosa e P. solimosense, que além de serem
mais procuradas, necessitam de mais tempo para regeneração.
A categoria de uso ornamental foi a que apresentou maior número de espécies de
Araceae, no entanto, constatou-se uma grande preferência por espécies exóticas, o que
significa pouca valorização das espécies locais com potencial ornamental.
Trabalhos como este, fornecem subsídios para conhecimento científico das espécies
locais da família Araceae, e, consequentemente, melhor aproveitamento das mesmas como
recurso a ser explorado de forma sustentável pelos moradores, através de seu cultivo na
comunidade.
Destaca-se a importância de popularizar o conhecimento sobre a família não só na
comunidade, enfatizando a necessidade de preservar os recursos vegetais como potenciais
elementos geradores de renda e também a importância ecológica desses recursos.

56
REFERÊNCIAS Albuquerque, U.P.; Lucena, R.F.P. 2004. Métodos e técnicas na pesquisa etnobotânica.
Recife: Livro rápido / NUPEEA. 189 p.
Amorozo, M.C.M.; Ming, L.C.; Silva, S.M.P. (orgs.). 2002. Métodos de coleta e análise de
dados em etnobiologia, etnoecologia e disciplinas correlatas. Rio Claro: UNESP/CNPq.
Barreto, J.B. & Cavalcanti, T.A. de A. 1947. Contribuição ao problema alimentar da
Amazônia. Memórias do Instituto Oswaldo Cruz. 45 (4) 853-875.
Bailey, K.D. 1982. Methods of social research. The Free Press, Macmillan Pubisher.
Bogner, J. 1987. Morphological variation in Aroids. Aroideana 10(2): 4-16.
Brasília, MMA / SUFRAMA / SEBRAE / GTA. 1998. Produtos Potenciais da Amazônia.
Opções de investimento na Amazônia Legal. Açaí. Obra em 19 v.; 51pp.
Canalez, G. de G. 2009. Produtos florestais não madeireiros: aráceas epifíticas da Reserva
Extrativista Auatí-Paraná. Manaus – Amazonas.
Cardoso, T.M. 2008. Etnoecologia, construção da diversidade agrícola e manejo da
dinâmica espaço-temporal dos roçados indígenas no rio Cuieiras, Baixo Rio Negro
(AM). Dissertação de mestrado. Manaus : [s.n.].
Croat, T.B. 1985. Collecting and preparing specimens of Araceae. Ann. Missouri Botanic
Garden. v. 17: p. 252-258.
Denevan, W.M. 2001. Cultivated landscapes of native Amazonia and the Andes. Oxford
University Press.
Durigan, C.C. 1998. Biologia e extrativismo do cipó-titica (Heteropsis spp. – Araceae)
Estudo para avaliação dos impactos da coleta sobre a vegetação de terra firme no

57
Parque Nacional do Jaú. Dissertação (mestrado) – Universidade do Amazonas, INPA,
Manaus.
Durigan, C.C.; Castilho, C.V. 2004. O extrativismo de cipós (Heteropsis spp., Araceae) no
Parque Nacional do Jaú. In: Janelas para a Biodiversidade no Parque Nacional do Jaú:
Uma estratégia para a Biodiversidade na Amazônia. Sérgio Henriques Borges; Simone
Iwanaga; Carlos César Durigan; Marcos Roberto Pinheiro (Eds.). Manaus: Fundação Vitória
Amazônica.
Engler, A. 1878. Araceae. In: Martins, C.E.P. von, Flora Brasiliensis 3 (2): 25-224, tt. 2-52.
FAS – FUNDAÇÃO AMAZONAS SUSTENTÁVEL – Programa bolsa floresta. 2009.
Relatório da precursora da APA do Rio Negro. Manaus – AM.
FVA – Fundação Vitória Amazônica. 2005. Uma análise crítica das unidades de
conservação do baixo rio Negro com propostas para as unidades estaduais. Encarte
Técnico n. 2. Documento técnico vinculado ao Relatório Técnico No. 1 do Projeto
Geopolítica da Conservação no Baixo Rio Negro, Amazônia Brasileira, implementado pela
Fundação Vitória Amazônica com recursos da Fundação Betty & Gordon Moore. Manaus,
AM.
FVA – Fundação Vitória Amazônica. 2009. Monitoramento e educação ambiental:
ferramentas para a conservação. Relatório técnico final. Fundação Vitória Amazônica/Projeto
Corredores Ecológicos. Manaus – Amazonas.
Hanazaki, N.; Tamashiro, J.Y.; Leitão Filho, H.F. & Begossi, A. 2000. Diversity of plant uses
in two Caiçara communities from the Atlantic Forest coast, Brazil. Biodiversity and
Conservation 9: 597-615.
Hoffman, B. 1997. The biology and use of “nibbi” Heteropsis flexuosa (Araceae): the
source of an aerial root fiber product in Guyana. Dissertation. Florida International
University, Miami.

58
IPÊ. 2007. Projeto Etnobotânica e Manejo Agroflorestal. Relatório Técnico Parcial, Novo
Airão.
Illenseer, R. 2009. Projeto Mosaico de Unidades de Conservação do Baixo Rio Negro.
Instituto de Pesquisas Ecológicas – IPÊ. Caderno do Grupo de Educação e Extensão
Socioambiental, Ano 1, n.1.
Keating, R. C. 2004. Systematic occurrence of raphide crystals in Araceae. Annals of the
Missouri Botanical Garden 91 (3): 495-504.
Krause, K. 1913. Araceae: Philondroideae – Philodendreae – Philodendrinae. Leipzig, In: A.
Engler, Das Pflanzenreich. 60 (IV 23 Db) p. 1-143.
Lawrence, G.H.M. 1977. Taxonomia das plantas das vasculares. Lisboa: Fundação
Calouste Gulbenkian, v.1, 854pp.
Leoni, J. M. e Marques, T. S. 2008. Conhecimento de artesãos sobre plantas utilizadas na
produção de artefatos - Reserva de Desenvolvimento Sustentável Amanã – AM. UAKARI,
v.4, n.2, p.67-77, dez.2008.
Madison, M. 1979. Notes on some aroids along the rio Negro, Brazil. Aroideana Vol. 2.
Madison, M. 1981. Notes on Caladium (Araceae) and its allies. Selbyana 5(3-4): 342-377.
Mayo, S.J. 1991. A revision of Philodendron subgenus Meconostigma (Araceae). Kew
Bulletin, v. 46, n. 4, p. 601 – 681.
Mayo, S.J.; Bogner, J.; Boyce, P. C. 1997. The Genera of Araceae. Royal Botanic Gardens,
Kew. 370pp.
Meira, M. 2006. Índios e brancos nas águas pretas: histórias do rio Negro. In: Forline, L.C.,
Murrieta, R.S.S. e Vieira, I.C. (orgs.). Amazônia além dos 500 anos. Belém: Museu
Paraense Emílio Goeldi, 183-226.

59
Morais, R.G. de; Jorge, S. da S.A.; Neto, G.G. 2003. Pesquisas regionais com informações
sobre plantas medicinais: a diversidade biológica e sócio-cultural de mato grosso em
foco. In: Diversos olhares em etnobiologia, etnoecologia e plantas medicinais. Cuiabá:
Unicen.
Oliveira, J.; Potiguara, R. C. V.; Lobato, L. C. B. 2006. Fibras vegetais utilizadas na pesca
artesanal na microrregião do Salgado, Pará. Boletim do Museu Paraense Emílio Goeldi.
Ciências Humanas, Belém, v. 1, n. 2, p. 113-127.
Paiva, W.O.; Weigel, P.; Filho, D.F.; Machado, F.M. 1981. Estudos preliminares em taioba
(Colocasia sp.) nas condições edafo-climáticas de Manaus. Acta Amazonica 11 (3): 417-421.
Phillips, G.; Gentry, A.H. 1993. The useful plants of Tambopata, Peru: I. Statistical
hypotheses tets with a new quantitative technique. Economic Botany 47(1): 15-32.
Phillips, O.L. 1996. Some quantitative methods for analyzing ethnobotanical knowledge. In:
Alexiades, M.N. Selected Guideline for ethnobotanical research. A field manual. The New
York Botanical Garden.
Pontes, T. A.; Andrade, I. M. de; Alves, M. 2010. Flora da Usina São José, Igarassu,
Pernambuco: Araceae. Rodriguésia 61(4): 2010.
Radford, A.E.; Dickison, W.C.; Massey, J.R. 1974. Vascular Plant Systematics. New York :
Harper e Row. 891pp.
Rodrigues, E. 2006. Plants and Animals Utilized as Medicines in the Jaú National Park (JNP),
Brazilian Amazon. In: Phytotherapy Research. Res. 20, 378–391.
Rossato, S.C.; Leitão-Filho, H.; Begossi, A. 1999. A ethnobotany of caiçaras of the Atlantic
Forest Coast (Brazil). Economic Botany, 53(4): 387-395.
Schott, H.W. 1860. Prodromus systematis aroideanum. Vienna: Typis congregationis
mechitharisticae, Vindobonae. 602pp.

60
Scudeller, V.V.; Veiga, J.B.; Araújo-Jorge, L.H. 2009. Etnoconhecimento de plantas de uso
medicinal nas comunidades São João do Tupé e Central (Reserva de Desenvolvimento
Sustentável do Tupé) In: Santos-Silva, E.N.; Scudeller V. V. (Orgs.). Biotupé: Meio Físico,
Diversidade Biológica e Sociocultural do Baixo Rio Negro, Amazônia Central, Vol. 2,
Editora UEA Ltda, Manaus.
SDS – Secretaria de Desenvolvimento Sustentável do Amazonas. 2008. Instrução
Normativa nº 001 de 11 de Fevereiro de 2008.
SDS - Secretaria de Desenvolvimento Sustentável do Amazonas. 2010. Plano de gestão
Parque Estadual do Rio Negro Setor Sul. (Consulta pública). Realização: Instituto de
Pesquisas Ecológicas (IPÊ).
Soares, M.L.C.; Mayo, S.J. 1999. Araceae. In: Ribeiro, J.E.L.S.; Vicentini, A.; Sothers, C.A.;
Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.; Martins, L.H.P.; Lohmann, L.G.; Assunção,
P.A.C.L.; Pereira, E.C.; Silva, C.F.; Mesquita, M.R.; Procópio, L. (Eds). Flora da Reserva
Ducke: Guia de identificação das plantas vasculares de uma floresta de terra – firme na
Amazônia Central. Instituto Nacional de Pesquisas da Amazônia, Manaus. p. 672-687.
Soares, M.L.; Jardim-Lima. 2005. Amazonian Species of Araceae in the INPA Herbarium,
Manaus – Amazonas, Brasil. Aroideana 28: 134-152.
Soares, M.L. 2008. Sistemática e ecologia de Heteropsis Kunth (Araceae Juss.) com
destaque especial nas espécies ocorrentes na Reserva Florestal Adolpho Ducke, Manaus
– Amazonas, Brasil. Tese doutorado – INPA/UFAM. 207pp.
Souza, C. C. V. 2010. Etnobotânica de quintais em três comunidades ribeirinhas na
Amazônia Central, Manaus – AM. Dissertação de Mestrado. Manaus – Amazonas.
Souza, V.C.; Lorenzi, H. 2008. Botânica sistemática: guia ilustrado para identificação das
famílias de Fanerógamas nativas e exóticas no Brasil, baseado em APG II. 2ª edição: Nova
Odessa, SP: Instituto Plantarum. 703pp.

61
Valente, T.P. 2009. Subsídios ao uso sustentável do cipó-preto – raízes de Philodendron
corcovadense Kunth (Araceae). Dissertação de mestrado. Curitiba : [s.n.]. 115pp.
Vieira, H. M. T. 2010. Histórico da comunidade Santa Maria. Documento pessoal.

62
Taxonomia e morfoanatomia de três espécies de Araceae Juss. utilizadas
para o artesanato na Área de Proteção Ambiental da Margem Esquerda do
Rio Negro.
RESUMO As raízes aéreas de espécies como o “cipó-titica”, pertencente ao gênero Heteropsis Kunth e “cipó-ambé” do gênero Philodendron Schott são muito utilizadas para fins artesanais na Amazônia, devido à resistência de suas fibras. Foram identificadas duas espécies de Araceae muito utilizadas para o artesanato na Comunidade Santa Maria, Heteropsis flexuosa (Kunth) G.S.Bunting, Philodendron solimoesense A.C.Sm. e outra espécie também utilizada como fonte de fibra, Monstera obliqua Miq., com baixo índice de uso, mas com potencial fibroso. Foi realizado o tratamento taxonômico dessas três espécies onde foi levada em consideração a distribuição, dados fenológicos, afinidade e histórico dos espécimes e detalhes das estruturas vegetativas e reprodutivas da planta, os quais foram ilustrados em pranchas. Foi também elaborada uma chave de identificação contendo características da morfologia externa e interna das espécies. Quanto ao estudo da morfologia interna da lâmina foliar e da raiz aérea das espécies, foram selecionados alguns caracteres como diagnóstico para a taxonomia, como tipo de estômatos, tipo de cristais, ocorrência de laticíferos, presença de hipoderme, ocorrência de bainha de fibras esclerenquimáticas, forma do cilindro central, dentre outros. Também foi feito o estudo das fibras das raízes, onde as três espécies apresentaram fibras longas do tipo libriforme com paredes espessas e lumens reduzidos. ABSTRACT The aerial roots of species such as the "cipó-titica", belonging to the genus Heteropsis Kunth and "cipó-ambe" of the genus Philodendron Schott are widely used for craft in the Amazon due to the strength of its fibers. We identified two species of Araceae widely used for crafts in Santa Maria Community, Heteropsis flexuosa (Kunth) G.S.Bunting, Philodendron solimoesense A.C.Sm. and other species also used as a source of fiber, Monstera obliqua Miq. with a low rate of use but with potential fibrous. The treatment of these three taxonomic species which was taken into account the distribution, phenological data, and historical affinity of the specimens and details of the vegetative and reproductive structures of plants, which were illustrated in boards. Was also prepared containing an identification key with characteristics of internal and external morphology of the species. Regarding the study of the internal morphology of leaf and root shoots of species, some characters have been selected as a diagnosis for the taxonomy as a type of stoma, type of crystals, the occurrence of latex, presence of hypodermis, the occurrence of sclerenchyma sheath fibers, shape of the central cylinder, among others. Also the study was made of the fibers of the roots, where the three species had long fiber type libriforme with thick walls and small lumens.

63
INTRODUÇÃO
Araceae é uma família subcosmopolita, que possui atualmente 109 gêneros e cerca de 2.823
espécies concentradas predominantemente nos trópicos da América, no Sudeste da Ásia e no
Arquipélago da Malásia (Govaerts et al., 2002). Na América do Sul está representada por 38
gêneros e cerca de dois terços do total das espécies (Mayo et al., 1997; Soares & Mayo, 1999;
Soares & Jardim-Lima, 2005). No Brasil possui 35 gêneros distribuídos em 458 espécies, das
quais 286 são endêmicas e encontram-se distribuídas em todos os domínios fitogeográficos do
país (Coelho et al., 2010).
As espécies dessa família representam um grupo taxonômico de considerável
importância econômica, com algumas espécies destacando-se por serem utilizadas como
medicinais, comestíveis, fibrosas, e, principalmente ornamentais, devido à beleza de suas
folhagens (Croat, 1994; Mayo et al., 1997). No entanto, na Amazônia, as raízes aéreas de
espécies como o “cipó-titica”, pertencente ao gênero Heteropsis Kunth (Potiguara &
Nascimento, 2004; Durigan & Castilho, 2004; Soares, 2008), e “cipó-ambé” do gênero
Philodendron Schott, são muito utilizadas para fins artesanais, devido à resistência de suas
fibras. Conforme detectado neste estudo, uma terceira espécie do gênero Monstera Adans.
também tem sido utilizada para a mesma finalidade. Estas espécies possuem hábito
hemiepifítico, segundo Croat (1990), algumas espécies de Araceae dividem-se em epífitas
verdadeiras; hemiepífitas primárias, as quais iniciam sua vida como epífitas verdadeiras, sem
contato com o solo e, quando mais tarde, emitem longas raízes em direção ao chão; e
hemiepífitas secundárias, que são aquelas que iniciam sua vida no solo e sobem em uma
árvore suporte quando adulta, podendo manter ou não ligação com o solo.
O gênero Heteropsis é composto de 17 espécies com uma grande concentração destas
na Amazônia e apenas duas espécies nativas da Mata Atlântica que são H. salicifolia Kunth e
H. rigidifolia Engl. (Soares et al., 2008). O gênero pertence à subfamília Monsteroideae Engl.
e está caracterizado pela ausência de laticíferos e tricosclereídeos. Pertecem também a esta
subfamília, as espécies do gênero Monstera, as quais são notáveis principalmente pelas
perfurações naturais em suas folhas, sendo muito valorizadas como ornamentais. O número e
o padrão das perfurações, quando presentes, ajudam na identificação das espécies. As plantas
jovens das espécies desse gênero podem ter folhas inteiras, as perfurações vão aparecendo
conforme as folhas vão se tornando maiores (Phillips, 1987). O gênero Philodendron é o
segundo maior gênero da família depois de Anthurium, possui cerca de 700 espécies (Coelho
& Mayo, 1998) e é bem diversificado no Estado do Amazonas (Mayo et al., 1995). Este

64
gênero pertence à subfamília Philodendroideae e está dividido em três subgêneros, o
subgênero Philodendron, Pteromischum e Meconostigma. Estes são bem distintos na
morfologia vegetativa, reprodutiva, e na distribuição geográfica (Mayo, 1997).
Estudos sobre a anatomia da família Araceae têm sido reportados principalmente por
French em vários trabalhos (French, 1987; French, 1988) nos quais foram relatados vários
tipos de estruturas secretoras, incluindo canais resiníferos, laticíferos, nectários extraflorais,
cavidades mucilaginosas e escâmulas intravaginais (Mayo et al., 1997).
Neste trabalho foram estudados aspectos morfoanatômicos de três espécies de
Araceae, Heteropsis flexuosa, Monstera obliqua e Philodendron solimoesense. Essas espécies
possuem hábito hemiepifítico e suas raízes aéreas são utilizadas como fonte de fibras para o
artesanato na Comunidade Santa Maria, fazendo parte da renda familiar. Características
anatômicas são importantes para a definição taxonômica de alguns grupos e são consideradas
tão valiosas quanto às da morfologia externa (Stuessy, 1990). Portanto, este trabalho teve
como objetivo investigar os caracteres consistentes da morfologia externa e interna das
lâminas foliares e raízes das três espécies para auxiliar na circunscrição das mesmas, visando
subsidiar plano de manejo sustentável.

65
OBJETIVOS
Geral
Realizar estudo da morfologia externa e interna de duas espécies (Heteropsis flexuosa
e Philodendron solimoesense) com maior valor de uso da família Araceae Juss. na
comunidade Santa Maria e de uma espécie citada por artesãos locais com índice de Valor de
Uso baixo, porém, diagnosticada com potencial para o artesanato (Monstera obliqua Miq.).
Específicos
Analisar a morfologia externa e interna dessas três espécies
Identificar caracteres anatômicos consistentes visando subsidiar o estudo taxonômico;
Elaborar um tratamento taxonômico das três espécies utilizadas para o artesanato na
comunidade;
Fazer estudo das fibras das raízes das espécies, visando à obtenção de características
que justifique seu uso em artesanato.

66
MATERIAL E MÉTODOS Área de estudo
A Comunidade Santa Maria está localizada nas coordenadas S 2° 55’ 44,2” W 60° 27’
58,7”, na Área de Proteção Ambiental da Margem Esquerda do Rio Negro, Setor Aturiá-
Apuauzinho, e faz parte do mosaico de Unidades de Conservação do Baixo rio Negro, que
tem ao todo 1,8 milhões de hectares e integra o corredor central da Amazônia (Figura 1).
Drenada pelo rio Cuieiras, a 70 km de Manaus, a área representa uma zona de amortecimento
para UC’s de proteção integral, sendo habitada por mais de 600 famílias, totalizando 15
comunidades dispersas (FAS, 2009).
Figura 1. Mosaico de Unidades de Proteção Ambiental do Baixo rio Negro e as comunidades da APA da Margem Esquerda do Rio Negro – Setor Aturiá-Apuauzinho, seta mostrando Comunidade Santa Maria. Fonte: SDS, 2009.
Coleta de material botânico
O trabalho foi realizado mediante autorização N°. 57/09 – CEUC/SDS, de acordo com
a Lei Complementar N°. 53, de 05 de junho de 2007, que institui o Sistema Estadual de
Unidades de Conservação do Amazonas (SEUC/AM), artigo 43 do Capítulo VI da Lei do
SEUC/AM, que trata das pesquisas científicas em Unidades de Conservação (ANEXO B).
Manaus
Santa Maria
APA da M.E. do Rio Negro,
setor Aturiá-Apuauzinho
Novo Airão

67
A metodologia de coleta na Comunidade Santa Maria, foi baseada no método de
“turnê guiada” por um coletor local de cipós, onde foram realizados percursos em três trilhas
de “igapó” e “terra-firme”, gravados pelo aparelho GPS (Global Positioning System), através
da função “registro de trajeto”, do qual foi gerado um mapa com o trajeto percorrido de
aproximadamente 12 quilômetros no total (Figura 2).
Figura 2. Registro do trajeto mostrando as trilhas percorridas contemplando três ambientes visitados na APA da Margem Esquerda do Rio Negro – Setor Aturiá-Apuauzinho.
As coletas botânicas e preparações das exsicatas das amostras seguiram as
recomendações de Croat (1985) e Soares & Mayo (1999). Para cada espécime, foram feitas
anotações em campo dos caracteres morfológicos impossíveis de serem vistos em material
herborizado e imprescindíveis na diferenciação das espécies, como o hábitat da planta,
coloração do caule, forma das gemas axilares, escâmulas, presença de ondulação nas margens
do pecíolo, coloração da lâmina foliar, visibilidade da venação, margem da lâmina foliar,
forma da lâmina, além da coloração dos elementos florais e dos frutos quando presentes. As
inflorescências dos espécimes foram colocadas em frascos de vidro contendo F.A.A. e /ou
álcool 70%, com a finalidade de preservar o material para posteriores análises morfológicas e
ilustrações dos detalhes florais. Quando possível, as plantas foram fotografadas.
As amostras foram acondicionadas em sacos plásticos de 50 litros ou prensadas
provisoriamente somente entre jornais e borrifadas com mistura de água e álcool de uso
doméstico para conservação das mesmas. Posteriormente, as amostras foram transportadas até

68
o Herbário INPA (sala de secagem) em Manaus, e desidratadas em estufa elétrica com
temperatura de 45º a 50ºC, por um período de quatro a seis dias até estarem totalmente
desidratadas.
Para o estudo anatômico foram retiradas cinco folhas por indivíduo, de três indivíduos
de cada espécie, sendo cada folha dividida em quatro regiões (meio-base, meio-meio, meio-
margem e meio-ápice), retirando-se do limbo foliar segmentos de 1 cm2, que foram
conservadas em álcool 70% glicerinado. As amostras das raízes aéreas maduras foram
coletadas no campo, selecionadas de três indivíduos, com comprimento e diâmetro variado.
Estas foram amostradas com 5 cm de comprimento das raízes adultas na porção apical. Após
a coleta o material foi acondicionado em frascos de vidro identificados e numerados contendo
solução de álcool a 70% glicerinado.
Identificação das espécies
As identificações das espécies, foram baseadas fundamentalmente nas obras de Schott
(1860), Engler (1878), Krause (1913), Mayo et al. (1997); Soares & Mayo (1999), Soares
(2008) e com auxílio de especialistas (Dra. Maria de Lourdes Soares, Dr. Valdely Kinupp e
Dr. Eduardo Gonçalves), bem como por comparações com exsicatas depositadas no Herbário
INPA.
Além da descrição baseada em observações de campo, foram também feitas análises
morfológicas comparativas com exsicatas incorporadas no herbário INPA. As análises foram
realizadas em laboratório da Pós-Graduação em Botânica do INPA e foram baseadas na
terminologia específica para a família, assim como em alguns casos por Radford et al. (1974)
e Lawrence (1977). Foram tomadas as medidas dos entrenós, profilo, pecíolo, bainha, lâmina
foliar, pedúnculo, espata e suas divisões, espádice e suas divisões e das flores conforme
recomendação de Mayo (1997) (Figura 3).

69
Figura 3. Desenho esquemático dos tipos de folhas e inflorescências. A) Lâmina foliar inteira. B) Lâmina foliar sagitada. C) Espádice com flores bissexuadas e espata em formato de bote. D) Espádice com flores unissexuadas e espata dividida em lâmina e tubo (Desenho esquemático: Rina Oliveira, mar/2011, adaptado de Mayo et al., 1997).
Para cada espécie foi feita uma descrição morfológica, seguida de sinonímias, nome
popular, distribuição, dados fenológicos quando possível, comentários sobre a afinidade e
histórico das espécies, uma relação do material examinado e pranchas contendo ilustrações do
hábito e detalhes de estruturas vegetativas e reprodutivas da planta com auxílio de
microscópio estereoscópico acoplado a câmara clara. Logo após a descrição taxonômica
segue a descrição da morfologia interna para cada espécie, acompanhada de pranchas com
fotos tiradas no Laboratório Temático de Microscopia eletrônica do INPA.
Os dados fenológicos e distribuição das espécies foram baseados nas informações das
etiquetas apostas nas amostras incorporadas no Herbário INPA. A apresentação das
descrições das espécies está em ordem alfabética.
As espécies férteis foram catalogadas e incorporadas ao Herbário INPA enquanto as
amostras estéreis foram doadas ao Herbário EAFM, do Instituto Federal de Educação, Ciência
e Tecnologia do Amazonas, Campus Manaus-Zona Leste.

70
Morfologia interna da lâmina foliar O trabalho de investigação da morfologia interna foi realizado no Laboratório de
Botânica Agro-Florestal (LABAF/UFAM) e no Laboratório de Anatomia do departamento de
Botânica – INPA.
A dissociação da epiderme foliar foi feita com mistura de Jeffrey (ácido crômico 1: 1
ácido nítrico), lavados em água, corados com Astrablau-Fucsina, desidratados em série
alcoólica e montados em gelatina glicerinada.
Os cortes histológicos foram feitos a mão livre, com lâminas de barbear, descorados
com hipoclorito de sódio e corados em Astrablau-Fucsina, desidratados em série alcoólica e
montados em gelatina glicerinada (Bonates, 1993).
Morfologia interna das raízes aéreas e fibras
No laboratório de Anatomia da Madeira da Coordenação de Pesquisas em Produtos
Florestais do INPA, as amostras das raízes aéreas das três espécies (cavacos) foram
seccionados em plano longitudinal com 5 mm de espessura, diafanizados em solução de
hipoclorito de sódio e água destilada (1:1), desidratadas em série alcoólica-etílica, coradas em
safranina por 15 min., colocadas em papel filtro para tirar excesso de corante, mergulhadas
em xilol para fixação e montadas em resina sintética “Permount”.
A análise das fibras das raízes foi realizada através da maceração ou dissociação dos
tecidos, sendo este o único método que revela as particularidades das células no seu todo. O
método empregado foi o do ácido acético e água oxigenada, onde para o preparo da solução
macerante utilizou-se: 50% de ácido acético glacial; 38% de água oxigenada 130 V; 12% de
água destilada.
Primeiramente foram colocadas em frascos de vidro, lascas pequenas e finas sem o
córtex na solução macerante de ácido acético. Após esse processo, os frascos foram vedados
com fita adesiva e colocadas na estufa por 24 horas a uma temperatura de 60°C.
Após a retirada do material dos frascos, estes permaneceram abertos por alguns
minutos para evaporar o cheiro de ácido acético. Posteriormente, foram tampados com uma
gaze fina de polielastano e lavados em água corrente até desaparecer o cheiro do ácido. Após
a lavagem, a gaze foi removida e o material aderido à gaze foi removido com o auxílio de

71
uma espátula ou agulha histológica fina e colocado dentro do frasco novamente. Este, com o
nível reduzido de água, foi fechado e agitado para a separação dos elementos constituintes do
lenho.
Após a maceração do material, este foi transferido, com o auxílio de uma pinça, para o
vidro de relógio. Depois de corado em safranina por cerca de 20 minutos, o material passou
pela série alcoólica (50%, 70%, 96% e 100%). Posteriormente, as amostras foram montadas
com glicerina entre lâminas e lamínulas para observação em microscópio óptico.

72
RESULTADOS Tratamento Taxonômico
Descrição dos gêneros
1. Heteropsis Kunth, Enum. PL. 3: 59 (1841). Tipo: Lectótipo H. salicifolia Kunth.
O gênero pertence à subfamília Monsteroideae, à tribo Heteropsideae, pela ausência
de laticíferos e tricosclereídeos. Plantas hemiepífitas com crescimento monopodial. Folhas:
dísticas, numerosas; pecíolo usualmente curto, canaliculado, embainhado, inteiramente
adnado ao internódio, parecendo somente um genículo apical livre, raramente pecíolo com
bainha livre (H. melinonii Brongn. ex Regel e H. steyermarkii G.S.Bunting); lâmina foliar
oblonga a lanceolada, coriácea a subcoriácea, ápice acuminado até atenuado, base cuneada a
arredondada; nervuras laterais primárias e interprimárias paralepinadas, formando uma
nervura submarginal coletora, uma elevada ordem de nervura reticulada. Inflorescência
solitária, espata em formato de bote, convoluta a semiconvoluta, alva-amarela, geralmente
decídua após antese; espádice cilíndrico a elipsoide, menor que a espata, estípite geralmente
curto ou ausente. Flores bissexuadas, perigônio ausente; estames 4 por flor, livres, filetes
achatados, anteras deiscentes, aberturas apicais; gineceu obpiramidal-prismático, ovário
bilocular, cada lóculo com 2 óvulos anátropos, placentação basal a sub-basal,
excepcionalmente axial; estigma oblongo ou arredondado. Fruto tipo baga. Semente ovoide-
elipsoide, endosperma ausente.
2. Monstera Adanson, Fam. Pl. 2: 470 (1763), nom. cons. TIPO: M. adansonii Schott
(Dracontium pertusum), typ. cons.
Tornelia Gutierrez ex Schott, Gen. Aroid. t. 74 (1858).
Serangium W. Wood ex R.A. Salisbury, Gen. Pl. Fragm. 5 (1866).
Plantas hemiepífitas, caules com crescimento simpodial, profilo decíduo. Folha
simples; pecíolo geniculado no ápice, canaliculado, bainha usualmente longa, persistente,
decompondo-se em massa membranosa a fibrosa ou inteiramente decídua; lâmina foliar

73
inteira, elíptica, oblíqua, oblonga a ovado-elíptica, geralmente fenestrada, raramente pinatífida
profundamente; nervuras laterais primárias pinadas, reunidas em uma nervura marginal
coletora próxima a margem, laterais secundárias mais ou menos paralelas às primárias,
frequentemente paralela-pinadas, às vezes reticuladas, laterais terciárias reticuladas, formando
uma alta ordem de nervuras reticuladas. Inflorescência uma a várias por simpódio floral;
pedúnculo mais curto que o pecíolo; espata caduca, não constricta, ereta aberta, ovada ou
oblongo-ovada, cuspidada, em forma de bote e um tanto convoluta na base, branca a cor de
rosa internamente, permanecendo aberta após antese, caduca; espádice séssil, subcilíndrico,
um tanto mais curto que a espata, homogêneo, sem regiões distintas. Flores bissexuais,
aclamídeas, poucas flores basais usualmente estéreis; estames 4, livres, filetes achatados,
conectivos delgados, teca oblongo-elipsoide, deiscência por um corte longitudinal; gineceu
obovoide a elipsoide, prismático, ovário bilocular, 2 óvulos por lóculo, anátropos, funículo
curto, placentação axial-basal, região estilar maciça, mais larga que o ovário, ápice truncado a
curtamente atenuado, estigma oblongo-elíptico a linear e longitudinal ou arredondado; flores
estéreis com 4 estaminoides, cônicos, pistilódio biloculares, prismáticos. Fruto 1-3,
desprendendo-se da região estilar prismática na maturidade, polposo internamente. Semente
ovoide a elipsoide, comprimida, testa lisa, embrião largo, endosperma ausente.
3. Philodendron Schott, Wiener Zeitschr Kunst, Lit., Theater und Mode 3: 780 (1829), nom.
et orth. cons. (‘Philodendrum’). LECTOTYPE: P. grandifolium (Jacq.) Schott (ver Britton e
Wilson, 1923).
Philodendrum Schott in Wiener Z. Kunst 1929 (3): 780 (1829), orth. rej. Arosma
Rafinesque, Fl. Tell. 3: 66 (1837, “1836”).
Telipodus Rafinesque, Fl. Tell. 3: 66 (1837, “1836”).
Thaumatophyllum Schott in Bonplandia 7: 31 (1859).
Elopium Schott in Oesterr. bot. Zeitschr. 15: 34 (1865).
Baursea Post & O. Kuntze, Lexicon Gen. Phanerog. 62 (1903).
Plantas hemiepífitas, epífitas rosuladas ou mais raramente ervas terrícolas; caule com
internós longos ou acaulescentes, muitas vezes produzindo ramos flageliformes. Folhas
numerosas, pequenas a gigantes, profilos de caules maduros marcescentes e decíduos ou
persistentes e membranáceos ou de componentes formando uma massa fibrosa; pecíolo longo,

74
conspícuo, algumas vezes verruculoso ou estriado, às vezes geniculado no ápice; bainha longa
ou inconspícua; lâmina foliar de forma extremamente variada, desde inteira, cordada, sagitada
ou hastada, trífida, trissecta, pinatilobada, pinatífida, bipinatífida, algumas vezes pedatissecta;
nervação peniparalelinérvea, as nervuras secundárias laterais mais finas e paralelas que as
nervuras laterais primárias, ocasionalmente as terciárias reticuladas transversalmente entre as
secundárias, às vezes sem a presença de nervuras laterais primárias. Inflorescência 1-11 por
simpódio floral; pedúnculo usualmente curto, cilíndrico-subcilíndrico, às vezes alargado no
ápice; espata comumente persistente até a completa maturação do fruto, raramente decídua
logo após a antese, levemente até fortemente constrita entre tubo e lâmina; tubo mais ou
menos ventricoso, frequentemente de coloração púrpura ou vermelha internamente; lâmina
abrindo completamente ou parcialmente na antese, fechando-se posteriormente, geralmente
esbranquiçada internamente; espádice séssil a estipitado, porção basal denominada zona
feminina formada por flores femininas ou pistiladas, geralmente mais curta que a zona
masculina fértil, constituída por estames férteis, situada no ápice do espádice e entre estas
uma zona masculina estéril intermediária, composta por estaminódios e geralmente mais curta
do que a zona masculina fértil, às vezes apresenta-se mais longa; raramente apresenta-se um
apêndice estaminodial no ápice do espádice. Flores unissexuais, nuas, estames 2-6 por flor
masculina, livres, prismáticos a obpiramidais, às vezes muito alongados e finos, anteras
sésseis, teca elipsóide a oblonga, alongada, deiscência por uma curta fenda lateral ou poro
subapical; flor feminina composta de um só gineceu ovoide, subcilíndrico ou obovóide,
ovário com (2-) 4-8 (-21) lóculos, óvulos 1-91 por lóculo, geralmente hemianátropos, funículo
com ou sem tricomas, estilete tão largo quanto o ovário, às vezes lobado, estigma discoide,
globoso, também às vezes lobado, frequentemente largo; fruto tipo baga, subcilíndrica a
obovoide, usualmente vermelha, amarela, branca ou vermelho-alaranjada; sementes
numerosas, variando muito de tamanho, testa frequentemente costelada.
Relações infragenéricas
O gênero Philodendron Schott está atualmente subdividido em três subgêneros
(Philodendron, Pteromischum e Meconostigma), dez seções e onze subseções. Estes
subgêneros são distintos tanto na morfologia vegetativa, floral e na distribuição geográfica. A
classificação do gênero foi elaborada a partir de Krause (1913), baseada no hábitat, forma da

75
folha, detalhes da inflorescência, das estruturas florais e a nomenclatura dos taxa
infragenéricos foi atualizada por Mayo (1990).
O subgênero Meconostigma é o menor dos três subgêneros e as principais
características são o hábito arborescente, caule duro e fibroso coberto por cicatrizes de folhas
velhas dispostas em espiral e estames finos e compridos. Além dessas características, Mayo
(1986) ampliou o número de caracteres que o diferencia dos dois outros subgêneros, sendo
estes: zona estéril do espádice longa; espata espessa; caule adulto composto
morfologicamente de unidades simpodiais de uma folha só e de um padrão caracterizado pela
supressão do entrenó hipodial; vascularização do gineceu mostrando um sistema axilar de
feixes, o qual é independente do sistema que fornece os óvulos através dos funículos; feixe
vascular do estame não bifurcado; dutos de resina na zona adaxial da espata e esta em seção
longitudinal possui um formato de “J”.
Chave para identificação das espécies 1. Planta hemiepífita primária ou raramente terrícola, escâmulas intravaginais
numerosas coroando os internós, raízes com acúleo em toda sua extensão, cilindro central da raiz aérea lobado, lâmina foliar sagitada, nervuras paralelinérvea sem nervura submarginal coletora. Estômatos paracíticos. Inflorescência com espata persistente......................................................................... Philodendron solimoesense
1’. Planta hemiepífita secundária, ausência de escâmulas nos internós, raízes aérea sem acúleo, cilindro central das raízes não lobado, lâmina foliar não sagitada, com nervura submarginal coletora. Inflorescência com espata caduca.
2. Lâmina foliar lanceolada - subobovada, ápice acuminado a atenuado, base aguda-acuneada, margem inteira, revoluta. Estômatos paracíticos. Inflorescência com espádice estipitado......................................................... Heteropsis flexuosa
2’. Lâmina foliar ovada-elíptica, fortemente fenestrada, ápice acuminado, margem geralmente rompida. Estômatos tetracíticos. Inflorescência com espádice séssil (sem estípite).......................................................................................... Monstera obliqua

76
Descrição das espécies
1. Heteropsis flexuosa (Kunth) G.S. Bunting, Revista Fac. Agric. Univ. Centr. Venezuela 10:
201 (1979). Tipo: Venezuela, Montana de Javita, Humboldt e Bonpland 978 (Holótipo: P
n.v.). (Figuras 4, 13)
Pothos flexuosus Kunth, Nov. Gen. Sp. 7: 151. 1825.
Heteropsis jenmanii Oliv., Hooker's Icon. Pl. 20: t. 1949 (1890). Tipo: Guyana,
Mazaruni River, Kalacoon, Maio 1889, G.S. Jenman 5000 (Holótipo: K n.v.)
Planta hemiepífita secundária, entrenós 2-4,5 cm de comp., subcilíndricos, verde-claro a
marrom escuro, estriado verticalmente quando seco, gema 0,5-1 mm de comp., geralmente
extrorsa, ápice agudo. Raiz aérea 3-8 mm diâm, contorno cilíndrico, marrom-castanho, estrias
longitudinais em toda sua extensão. Folha: pecíolo 0,5-1,3 cm comp., 1,3-2,3 mm diâm.,
fortemente canaliculado, margem lisa, geralmente retorcido, inteiramente geniculado; lâmina
foliar 14-24,5 cm comp., 3,5-7,3 cm larg., lanceolada a subobovada, subcoriácea, sublustrosa,
verde escuro na face adaxial, verde amarelado na face abaxial, margem revoluta, ápice
acuminado a atenuado, base aguda a cuneada, excepcionalmente obtusa, nervura central na
face adaxial achatada a entalhada, impressa e proeminente na face abaxial, amarelada,
nervuras laterais primárias e interprimárias obscuras em ambas as faces, nervura submarginal
coletora proeminente 1 – 3,5 mm distante da margem. Inflorescência: terminal e axilar,
entrenós do pedúnculo: 0,4 – 1,9 cm comp., 3,8 – 8,1 mm diâm., verde escuro; pedúnculo 4 –
7,5 cm comp., 4,3 – 6,2 mm diâm., verde escuro; espata 6,0 – 7 cm de comp., 21 – 31,3 mm
diâm., elipsoide, convoluta quando aberta em antese, amarela, mesmo tamanho do espádice,
ápice agudo; espádice 4 – 5,8 cm de comp., 10,6 – 15 mm diâm., elipsoide, subclavado, ápice
obtuso, amarelo claro, estipitado; estípite 0,4 – 1 cm comp., 5 – 7 mm diâm., verde,
subcilíndrico. Flores: estames 2,5 – 3,5 mm comp., 1 – 2 mm larg., antera 0,5 – 2 mm de
comp., 0,5 – 1,2 mm de diâm.; gineceu: 3,5 - 4 mm comp. 2 mm diâm., levemente
prismático, lóculo 2 por ovário; ovário 1 – 2 mm comp., 0,5 – 1 mm larg., oblongo; óvulos 1
– 2 por lóculo, ca. 0,5 mm comp., anátropos, subsésseis; placentação sub-basal; estigma
discoide-oblongo. Infrutescência: não vista.
Nome popular: cipó-titica (Brasil), Nibbi ou Mibi (Guiana), Tamshi (Peru).

77
Hábitat: planta exclusiva de Floresta Ombrófila Densa de Terra-firme.
Distribuição: Bolívia; Brasil (Acre, Amapá, Amazonas, Bahia, Maranhão, Pará, Pernambuco,
Rondônia e Roraima); Colômbia; Equador; Guianas; Peru; Suriname e Venezuela (Figura 13).
Dados fenológicos: coletada com flores todo o ano, com frutos nos meses outubro a
dezembro.
Material examinado: APA da Margem Esquerda do Rio Negro, Comunidade Santa Maria, 5
Mar 2010, R.F.M. Oliveira, G.P. Vianae e A.C. Padron s/n (EAFM, est.); 03 Jun 2010,
R.F.M. Oliveira, G.P. Viana e A.C. Padron 28 (INPA, fl); 11 Ago 2010 R.F.M. Oliveira, M.L.
Soares, S. Silvino e A.C. Padron 52 (INPA, fl.). BOLÍVIA: Pando: Abuna, 8 Jul. 1992, Al.
Gentry e Alan Perry 77987 (MO, fl.). BRASIL: Acre: Cruzeiro do Sul, 21 Ago. 1986, T.B.
Croat e Arito Rosas, Jr. 62435 (INPA, est.). Amapá: Porto Grande, 1 Mar. 2001, L.A.
Pereira, F. Cesarino, S.M. Alfaia e J. Reis 151 (INPA, fl.); 1 Mar. 2001, L.A. Pereira, F.
Cesarino, S.M. Alfaia e J.P. Reis, 162 (INPA, est.). Amazonas: Estirão do Equador, 8 Ago
1973, E. Lleras, W.C. Sterward, J.C. Ongley, D.F. Coelho, J.F. Ramos e J. F. Lima 17214
(INPA, fl.); Estrada Manaus – Itacoatiara, Nov. 1973, W. Steward e J. Ramos 17656 (INPA
fl.); Humaitá, 16 Jun. 1980, A. Janssen 474 (INPA, fl.); Manaus, 16 Ago. 1986, T.B. Croat
62208 (INPA, est.); São Gabriel da Cachoeira, 10 Fev. 2001, D. Hoffman, e M. Farias, 139
(INPA, fl.); 12 Out. 1976, Aldair s/n (INPA, est.); 20 Nov. 1968, D. Coelho e J. Lima s/n
(INPA, fl.). Pará: Itaituba, 20 Mai. 1983, I.L. Amaral, M.N. Silva, O.P. Monteiro, J. Lima, L.
Brako, W.D. Reese e M. Dibben 1352 (INPA, fr.); Oriximiná, 17 Ago. 2008, D.R. Oliveira e
C.A. Paulo Santos 124 (INPA, est.); Paragominas, 1 Mar. 1980, T. Plowman, G. Davidse,
N.A. Rosa, C.S. Rosário e M.R. dos Santos 9402 (INPA, est.); Tucuruí, 22 Out. 1983, E. Lima
e A. Silva 90 (INPA, fr.); Pernambuco: Recife, 14 Fev. 1990, M.L. Guedes 2267 (UFBA fl.);
Rondônia: Porto Velho, 26 Out. 1979, G. Vieira, J. Zarucchi, A.S.L. Silva, C.D.A. Mota e
J.F. Ramos 310 (INPA, fr.); 28 Jun. 1983, M.G. Silva 6506 (INPA, fl.). Roraima: Estrada
Manaus – Caracaraí, 16 Nov. 1977, W.C. Steward, I. Araújo, W.R. Buck, J.F. Ramos e J.
Ribamar 39 (INPA, fr.); 18 Fev. 1971, G.T. Prance, W. C. Steward, F.P. Harter, J.F. Ramos,
W.S. Pinheiro e O.P. Monteiro 10598 (INPA, fr.); 28 Fev. 1971, G.T. Prance, W.C. Sterward,
F.P. Harter, J.F. Ramos, W.S. Pinheiro e O.P. Monteiro 10762 (INPA, fl.). COLÔMBIA: 17
Set. 1991, H. Vester 569 (MO, est.). EQUADOR: Province Napo: 12 Nov. 1987, C.E. Ceron
2881 (MO, fl.); 19 Jun. 1991, W. Palacios e E. Freire 7396 (MO, fr.). 00º 52’S 76º 05’W, 9-

78
13 Jan. 1988, C.E. Ceron e F. Coelho 3295 (MO, fr.); 00º 57’S 76º 13’W, 9 Jan. 1988, D.
Neill et al. 8353 (MO, fr.). GUIANA: Edo. Bolívar: Set. 1986, Angel Fernandez 3322
(INPA fl.); 28 Mar. 1910, C.W. Anderson 432 (K est); 29 Jun. 1979, M.F. Prevost 670 (K
est.), 03º 37’ N 53º 12’ W, 27 Mar. 1983, S. Mori e J. Pipoly 15415 (P, MO, P fr.); 3º 37’N
53º 12’W, 19 Mar. 1996, B. Hoffman e D. Allicock 5060 (MO fr.); 15 Mai. 1986, S.A. Mori e
T.D. Pennington 18014 (MO fr.); 5º 30’ N 48º 22’ W, 6 Jul. 1982, T.B. Croat 53843 (MO
est.). PERU: Reserva Tambopata: 15 Mar. 1988, D. Bell et al. 88-178 (K fr.); 2 Jan. 1973,
B. Berlin 800 (MO fr.); 04º 55’ S 073º 45’ W, s.d., J. Baluarte s.n. (MO 5564058 est.); 14
Mar. 1980, V. Huashikat 2262 (MO fr). SURINAME: 15 Mar. 1949, A.M.E. Jonker et al.
2715 (IAN fr.); 4 Mar. 1918, B. Wallnofer 3803 (KEW fr.); 12 Fev. 1919, B.W. 4255 (P fl.);
30 Jun. 1924, B.W. 6555 (MO est.); 6 Fev. 1942, G. Stahel s.n. (K, MO fl.). VENEZUELA: 6
Fev. 1990, B. Stergios e J. Velazco 14706 (MO est.), 5º 37’ N 67º 22’ O, 29 Jan. 1991, G.A.
Romero et al. 2270 (MO fl.); 5º 37’ N 67º 22’ O, 6 Fev. 1992, G.A. Romero et al. 2317 (MO
est.); 5º 37’ N 67º 22’ O, 9 Fev. 1992, G.A. Romero et al. 2343 (MO fl.); 18 Abr. 1990, J.
Perez 962 (MO est.).
Comentários:
As espécies que compõem o gênero Heteropsis possuem folhas muito similares, no
entanto, alguns caracteres são peculiares a H. flexuosa como a gema com ápice acuminado e
extrorso, lâmina foliar discolor onde, se apresenta verde escuro na face adaxial e fortemente
amarelada na face abaxial. Estas características diferem de H. oblongifolia Kunth que é a
espécie mais semelhante de H. flexuosa entre as espécies do gênero.
Segundo Soares (2008), esta espécie tem a mais ampla distribuição entre os táxons
pertencentes ao gênero e que possivelmente seja a espécie mais explorada na Amazônia
brasileira e extrabrasileira, principalmente na região do Baixo rio Negro. As raízes
alimentadoras dessa espécie possuem características anatômicas e propriedades físicas e
mecânicas apropriadas para o uso em artesanato.
Na Comunidade de Santa Maria, o “cipó-titica” é utilizado para confecção de
artesanato como cestas, bolsas e vassouras, fazendo parte da economia familiar. Além disso, a
seiva de suas raízes aéreas foi citada por moradores como remédio para ferrada de “arraia”.

79
As raízes aéreas dessa espécie também são usadas como amarras em construções de casas e
embarcações rústicas pelos moradores da comunidade.
Em estudo anatômico, a espécie apresenta raízes com epiderme suberizada, dando
aspecto de velame, muito comum em orquídeas, além de apresentar uma bainha com várias
camadas de fibras esclerenquimáticas de comprimento longo e libriformes ao redor do
cilindro central. Os frutos de Heteropsis flexuosa apresentam cor verde a amarela,
dependendo do estádio de maturidade, conforme visto em informações apostas nas etiquetas
de amostras depositadas do Herbário INPA.
Apesar de não ter sido feito um delineamento amostral, foram identificadas 10
famílias de árvores suportes (forófitos) para H. flexuosa: Annonaceae (indeterminada),
Malvaceae (Scleronema micranthum (Ducke) Ducke), Boraginaceae (Cordia sp.),
Euphorbiaceae (Mabea sp.), Fabaceae-Faboideae (Aldina heterophyla Spruce ex Benth.),
Meliaceae (Guarea sp.), Moraceae (Brosimum sp. e Sorocea sp.), Myristicaceae (Iryanthera
sp., Virola calophylla Warb. e indeterminada), Sapotaceae (Pouteria sp.), Urticaceae
(Pourouma sp.). Mas, a família que se destacou foi Lecythidaceae com o gênero Eschweilera,
os “matamatás” (E. coriaceae (DC.) S.A.Mori e Eschweilera spp.).
Em trabalho realizado na Reserva Ducke, por Soares (2008), Lecythidaceae também
está entre as famílias com maior número de indivíduos de Heteropsis, incluindo também
Sapotaceae, Annonacae, Lauraceae, Moraceae, Fabaceae e Meliaceae, famílias também
identificadas como forófitos no presente estudo.

80
Figura 4. Heteropsis flexuosa (Kunth) G.S. Bunting. A) Hábito; B) Venação da lâmina foliar; C) Vista frontal do estame; D) Seção transversal do ovário; E) Seção longitudinal do gineceu (A – E preparados de Oliveira, R.F.M. et al., 28; ilustração de Felipe França, fev/2011).

81
Morfologia interna
Lâmina foliar
Epiderme: em vista frontal, a epiderme em ambas as faces, apresenta células
poligonais com paredes anticlinais retas ou ligeiramente onduladas e irregulares, recoberta por
cutícula lisa. Em vista transversal apresentam as paredes da face tanto adaxial quanto abaxial
externas lisas. Estômatos: são do tipo paracítico, raramente geminados e situam-se ao mesmo
nível da epiderme (Figura 5 A). As folhas são anfihipoestomáticas e apresentam um número
médio de 69,9 estômatos por mm2 na face abaxial e 5,5 na adaxial. Mesofilo: apresenta duas a
três camadas de parênquima paliçádico com células curtas e duas a seis camadas de células do
parênquima lacunoso com pequenos espaços intercelulares (Figura 5 E). Feixes
fibrovasculares: são colaterais e envolvido por duas a quatro camadas de fibras
esclerenquimáticas espessadas (Fgura 5 B). Idioblastos: foram observadas células solitárias,
aclorofiladas com uma parede espessa e estão dispersos por todo o mesofilo (Figura 5 D).
Laticíferos: não foi registrada a presença de laticíferos. Cristais: foram encontrados cristais
do tipo drusas dispersos no mesofilo (Figura 5 C) (Tabela 1).
Figura 5. Estruturas da lâmina foliar de Heteropsis flexuosa (Kunth) G.S. Bunting. A) Vista superficial da epiderme com estômatos paracíticos e geminados (seta); B-E) Cortes transversais; B) Feixe vascular colateral envolto por fibras esclerenquimáticas; C) Mesofilo contendo drusas (setas); D) Idioblasto; E) Feixes colaterais acessórios. Barras = 12,5 µm (A, B); 5 µm (C, D); 50 µm (E).

82
Raiz aérea
Periderme: na parte mais externa, encontra-se um súber formado a partir de várias
camadas de células, enfileiradas e originadas a partir do córtex (Figura 6 A-C). Lenticelas
também foram encontradas em algumas regiões da periderme (Figura 6 E). Córtex:
subjacente à periderme, encontra-se constituído por até 30 camadas de células isodiamétricas
de paredes delgadas relativamente esféricas, clorofiladas ou não, incolores em sua maioria,
vacuoladas com fibras esclerenquimáticas dispersas (Figura 6 B). No córtex ocorre ainda uma
bainha com cerca de seis a oito camadas de fibras esclerenquimáticas, com paredes espessas e
lúmens reduzidos (Figura 6 A e F). Endoderme: localizada logo abaixo da bainha de fibras
esclerenquimáticas, é formada por uma camada de células parenquimáticas achatadas e
aparentemente mais densas próximas ao periciclo (Figura 6 F e G). Cilindro central: há
alternância do xilema e floema, que nesse caso, também são circundados por fibras
esclerenquimáticas (Figura 6 F) (Tabela 2).
Figura 6. Cortes transversais das raízes aéreas de Heteropsis flexuosa (Kunth) G.S. Bunting. A) Aspecto geral mostrando a periderme, bainha de fibras esclerenquimáticas () e cilindro central; B) Súber e fibras esclerenquimáticas dispersas no córtex; C) Região de suberização da periderme; D) Células em divisão entre a região de suberização e córtex; E) Lenticela; F) Endoderme (seta) envolvendo o cilindro vascular e bainha de fibras esclerenquimáticas (); G) Endoderme e periciclo mais externamente ao cilindro central. PE= periderme; Co= córtex; Fi = fibra; CC= cilindro central; Su= súber; Sub = suberização; En= endoderme; Pc = periciclo. Barras = 50 µm (A); 12,5 µm (B); 5 µm (C,D,G); 25 µm (E, F).

83
2. Monstera obliqua Miq. Linnaea 18: 79. 1844. (Figuras 7, 13) Monstera boliviana Rusby. Bull. New York Bot. Gard. 6: 494. 1910.
Monstera expilata Schott. Prodr. Syst. Aroid.: 367. 1860.
Monstera falcifolia Engl. Bot. Jahrb. Syst. 37:117. 1905.
Monstera falcifolia var. latifolia K. Krause. Notizbl. Königl. Bot Gart. Berlin 11: 614.
1932.
Monstera fendleri Engl. Bot. Jahrb. Syst. 37: 116. 1905.
Monstera killipii K. Krause. Notizbl. Königl. Bot. Gart. Berlin 11: 614. 1932.
Monstera microstachys Schott. Oesterr. Bot. Wochenbl. 5: 274. 1855.
Monstera obliqua var. expilata (Schott) Engl. Martius, C.F.P. Von e auct. suc. (Eds.)
1878. Fl. Bras. 3(2):113.
Monstera sagotiana Engl. Bot. Jahrb. Syst. 37:117. 1905.
Monstera snethlagei K. Krause. Notizbl. Königl. Gart. Berlin 9:272. 1925.
Monstera unilatera Rusby. Bull. New York Bot. Gard. 6: 494. 1910.
Planta hemiepífita secundária. Caule ca. 0,6 cm diâm., crescimento simpodial,
profilo decíduo. Raiz aérea 2 – 4 mm diâm., contorno cilíndrico, cor castanho claro. Folha:
pecíolo 15 – 35 cm comp., bainha 5 – 24 cm de comp., 3 – 7 mm de diâm.; pecíolo livre 8,5 –
12 cm de comp., 2 – 6 mm diâm., geniculado no ápice, canaliculado na face adaxial,
arredondado na face abaxial; lâmina foliar 26 - 46 cm comp., 14 - 35 cm larg., inteira, elíptica
quando jovem a ovada-elíptica quando adulta, cartácea, fortemente fenestrada, ápice
acuminado, base arredondada a hastada, margem geralmente rompida, nervura central
levemente entalhada na face adaxial, proeminente e arredondada na face abaxial, nervuras
laterais primárias pinadas, laterais interprimárias paralelas às primárias, nervuras secundárias
formando uma nervura sub-marginal coletora bem próxima a margem, laterais terciárias
reticuladas. Inflorescência: uma por simpódio floral, pedúnculo: 11,5 – 17 cm comp., 7 mm
diâm.; espata 6 cm comp., 13,2 mm diâm., não constrita, ereta aberta, ovada ou oblongada,
decídua após antese; espádice 4,5 – 5 cm comp., 10 – 12 mm diâm., séssil. Flores
bissexuadas, aclamídeas; estames 4, 2,5 – 3 mm comp., livres, filetes 1 mm comp., 0,6 – 0,9
mm diâm., anteras 1,5 – 2 mm comp., 1 mm diâm., gineceu: estigma ca. 1 mm de comp.,
ovário 1,5 – 2,5 mm comp., 0,5 – 1 mm diâm., bilocular, 2 óvulos por lóculo, 0,6 – 1 mm
comp., placentação basal. Infrutescência não vista.

84
Nome popular: “costela-de-adão”, “ambé-sima” (Comunidade Santa Maria, Baixo rio Negro,
Amazonas, Brasil)
Hábitat: espécie encontrada em florestas de Terra-Firme.
Distribuição: Bolívia e Brasil (Figura 13).
Dados fenológicos: coletada com inflorescência nos meses de abril a dezembro e com frutos
nos meses fevereiro, março, junho, agosto, setembro, novembro.
Material examinado: APA da Margem Esquerda do Rio Negro Setor Aturiá-Apuauzinho, 6
Mar. 2010, R.F.M. Oliveira, G.P. Viana e A.C. Padron 24 (EAFM est.); 6 Mar. 2010, R.F.M.
Oliveira, G.P. Viana e A.C. Padron 26 (INPA fl.); 14 Jan 2011, R.F.M. Oliveira, Kinupp,
V.F., Viana, M.R. e Padron, A.C. 60 (EAFM, est.); BOLÍVIA: Pando: Rio Madeira, 19 Nov.
1968, G.T. Prance, W.A. Rodrigues, J.F. Ramos, L.G. Farias, 8640 (INPA fr.); 9 Jul. 1968,
G.T. Prance, W.A. Rodrigues, J.F. Ramos, L.G. Farias, 5696 (INPA fl.); 20 Jul. 1968, G.T.
Prance, E. Forero, B.L. Wrigley, J.F. Ramos, L.G. Farias 6211 (INPA fl.). BRASIL:
Amapá: RDS Iratapuru, 15 Nov. 2004, L.A. Pereira 767 (INPA fr.). Amazonas:
Anavilhanas, Ilha Três Bocas, 6 Jun. 1989, S. Mori, C. Gracie, W. Capraro, J. Mitchell, D.
Russell 20391 (INPA); Alto Rio Negro, Ilha Novo Retiro, 17 Abr. 1967, W. Rodrigues e L.
Coelho 8389 (INPA fl.); Barcelos, 04 Fev. 1984, I.L. Amaral, 1491 (INPA fr.); Boca do Acre,
03 Out. 1980, C.A. Cid e B.W. Nelson 2701 (INPA fr.); Lábrea, 31 Out. 1968, G.T. Prance,
J.F. Ramos e L.G. Farias 8119 (INPA fr.); Manacapuru, 2 Mai. 1957, W. Rodrigues 321
(INPA fl.); Manaus, 18 Abr. 1955, J. Chagas 973 (INPA est.); Manaus, 15 Fev. 1995, M.L.
Soares, C.F. da Silva, M.A.S. da. Costa 177 (INPA fr.); Manaus-Itacoatiara, Madeireira Mil,
11 Dez. 1999, M.L. Soares 476 (INPA fl.); Maraã, 1 Nov. 1982, I,L. Amaral, C.A. Cid, J.
Guedes, J. Lima 285 (INPA fr.); Paraná Autaz Mirim, 26 Mar. 1973, A. Loureiro et al. 37647
(INPA fr.); Pico Rondon, 6 Fev. 1984, J. Pipoly et al. 6687 (INPA fl.); Rio Javari, 20 Out.
1976, P.I.S. Braga e J.R. Nascimento 3247 (INPA fr.); Rio Purus, 14 Ago. 1956, J. Ruber
4716 (INPA est.); Rio Purus, 19 Jun. 1971, G.T. Prance et al. 13381 (INPA fl. e fr.); Sta.
Isabel do Rio Negro, 16 Ago. 1999, M.G.M. Roosmalen, I.L. Amaral, F.D. Matos, A. Lima, J.
Guedes 1071 (INPA fr.); Tabatinga, 14 Out. 1976, P.I.S. Braga, J. Ramos, J.P. Nascimento
3131 (INPA fr.); Tefé, 14 Dez. 1982, I.L. Amaral, C.A. Cid, T. Plowman, W. Davis, J. Lima,
J. Guedes, 757 (INPA fl.). Mato Grosso: Alta Floresta, 26 Set. 2006, D. Sasaki, I. Vieira,

85
J.H. Piva 425 (INPA fr.); Chapada dos Guimarães, 23 Jul. 1999, M. Macedo e I. Lopes s/n
(INPA est.); Cuiabá, 05 Jul. 2001, M. Macedo, E. Gonçalves, E. Silva 7559 (INPA fr.). Pará:
Itaituba, 24 Abr. 1983, I.L. Amaral, M.N. Silva, O.P. Monteiro, J. Lima, L. Brako, W.D.
Reese, M. Dibben 1049 (INPA fr.); 29 Abr. 1983, M.N. Silva 160 (INPA fl.); Oriximiná, 04
Jun. 1980, C. Davidson e G.Martinelli CD 10050 (INPA fr.). Rondônia: Ilha no rio Madeira,
28 Jul. 1968, G.T. Prance, E. Forero, B.L. Wrigley, J.F. Ramos, L.G. Farias 6546 (INPA fl. e
fr.); 29 Jun. 1968, G.T. Prance, D. Philcox, E. Forero, L.F. Coelho, J.F. Ramos, L.G. Farias
5339 (INPA fl. e fr.); 28 Jul. 1968, G.T. Prance, E. Forero, B.L. Wrigley, J.F. Ramos, L.G.
Farias 6546 (INPA fl e fr.); 29 Jun. 1968, G.T. Prance, D. Philcox, E. Forero, L.F. Coelho,
J.F. Ramos, L.G. Farias 5339 (INPA fl. e fr.); Rio Jaciparaná, 30 Jun. 1968, G.T. Prance, D.
Philcox, E. Forero, L.F. Coelho, J.F. Ramos, L.G. Farias 5379 (INPA fl.); 30 Jun. 1968, G.T.
Prance, D. Philcox, E. Forero, L.F. Coelho, J.F. Ramos, L.G. Farias 5379 (INPA fl.);
Rodovia Alvorada – Costa Marques, 02 Jul. 1983, M.G. Silva 6539 (INPA fr.); Sta. Bárbara,
16 Ago. 1968, G.T. Prance e J.F. Ramos 7010 (INPA fl.); Terra Indígena Uru-Eu-Wau-Wau,
29 Jan. 2002, L.R.U.C. Lopes, L.C. Maretto, J. Araújo 01 (INPA fl.).
Comentários: A grande variação de tamanho, forma e número de perfurações nas lâminas foliares de
M. obliqua, tem influenciado vários taxonomistas a descreverem nomes diferentes para essa
espécie. Isto pode ser confirmado pelo grande número de sinonímias existentes para a espécie.
A função das perfurações em Monstera é desconhecida, mas é provável que sirvam
para reduzir o efetivo do tamanho da folha e, portanto, propriedades de transferência de calor,
bem como os folíolos individuais de mais folhas compostas convencionais (Madison 1977).
Outra hipótese intrigante é a de que as perfurações servem como camuflagem, pela
interrupção do contorno da folha, ou como sinais para os herbívoros de que defesas químicas
ou físicas induzidas já podem estar presentes (Brown & Lawton, 1991). Segundo estudo
ontogenético de Arunika et al. (2005), as perfurações na epiderme das folhas são originadas
por grupos de células vizinhas programadas para morrer simultaneamente.
Os frutos variam de amarelo a alaranjado conforme a maturidade, informações estas,
retiradas das observações apostas em etiquetas do Herbário INPA.
Esta espécie possui raiz aérea com menor diâmetro em comparação às outras espécies
aqui estudadas. Porém, estas são utilizadas apenas para o acabamento dos artefatos

86
confeccionados pelos artesãos da Comunidade de Santa Maria, onde também registrou-se pela
primeira vez o uso de raízes dessa espécie para o artesanato.
Para esta espécie, foram identificadas oito famílias de árvores suportes (forófitos), a
saber: Apocynaceae (Aspidosperma sp.), Burseraceae (Protium sp.), Chrysobalanaceae
(Licania sp.), Lauraceae (Mezilaurus sp.), Melastomataceae (Bellucia glossularioides (L.)
Triana e Mouriri sp.), Myristicaceae (Virola sp., Iryanthera sp. e indeterminada), Rubiaceae
(indeterminada) e Sapindaceae (Cupania sp.). Constatou-se que as espécies do gênero
Protium spp. (“breu”), pertencentes à família Burseraceae, foram colonizados com mais
frequência por indivíduos de M. obliqua.

87
Figura 7. Monstera obliqua Miq. A) Hábito; B) Seção transversal da flor; C) Seção longitudinal da flor; D) Vista apical da flor; E) Vista frontal do estame (A – E preparados de Oliveira, R.F.M. et al., 26; ilustração de Felipe França, fev/2011).

88
Morfologia interna
Lâmina foliar
Epiderme: em vista frontal apresentam células retangulares e alongadas na face
adaxial e irregulares na face abaxial, recobertas por cutícula muito fina em ambas as faces
(Figura 8 A e B). Em corte transversal apresentam células grandes, paredes periclinais
externas planas e paredes periclinais internas arredondadas (Figura 8 F). Estômatos: são do
tipo tetracíticos e situam-se no mesmo nível da epiderme (Figuras 8 B e C). As folhas são
anfihipoestomáticas e apresentam um número médio de 63,6 estômatos por mm2 na face
abaxial e 7,95 na face adaxial, sendo possível observá-los apenas na região da nervura central.
Tricomas: não foram encontrados nessa espécie. Mesofilo: apresenta uma a duas camadas de
tecido paliçádico, irregular, repleto de cloroplastos e tecido lacunoso com células de braços
curtos. Feixes fibrovasculares: apresentam-se em forma de feixes colaterais, com feixes
acessórios na nervura central e rodeados por uma a três camadas de fibras esclerenquimáticas
(Figura 8 D e E). Idioblastos: foram encontradas células solitárias, aclorofiladas, com paredes
espessas dispersas pelo mesofilo (Figura 8 F). Laticíferos: não foi constatada a presença de
laticíferos. Cristais: foram vistos cristais do tipo ráfides (Figura 8 F) (Tabela 1).
Figura 8. Estruturas da lâmina foliar de Monstera obliqua Miq. A-C) Vista frontal. A) Epiderme adaxial; B) Epiderme abaxial; C) Estômato tetracícito; D-F) Cortes transversais; D e E) Feixe vascular colateral envolto por fibras esclerenquimáticas; F) Mesofilo contendo ráfides (seta). Barras = 50 µm (A); 25 µm (B); 5 µm (C); 50 µm (D); 12,5 µm (E, F).

89
Raízes aéreas
Epiderme: foi observada uma epiderme unisseriada com apêndices epidérmicos
(Figura 9 A e B). Exoderme: apresenta uma camada de células retangulares a subcilíndricas
(Figura 9 B). Córtex: é composto por aproximadamente 20 camadas de células
parenquimáticas com poucos espaços intercelulares e isodiamétricas (Figura 9 D). Observa-
se ainda, uma bainha (anel esclerenquimático) com cerca de duas a três camadas de fibras
esclerenquimáticas, com paredes espessas e lúmens reduzidos. Endoderme: formada por
uma camada de células parenquimáticas (Figura 9 A). Cilindro central: é constituído de
elementos de vaso rodeados por fibras esclerenquimáticas e raramente presença de
tilossomos (Figura 9 C e E) (Tabela 2).
Figura 9. Cortes transversais das raízes aéreas de Monstera obliqua Miq. A) Vista geral da epiderme, córtex e cilindro central; B) Epiderme estratificada evidenciando a exoderme (seta); C) Cilindro central mostrando elementos de vaso; D) Região do córtex; E) Elementos de vaso contendo tilossomos (seta) rodeados por fibras esclerenquimáticas. Barras = 25 µm (A); 12,5 µm (B,C,D); 5 µm (E).

90
3. Philodendron solimoesense A.C. Sm. in: Journal of the Arnold Arboretum 20: 289. 1939;
Bunting in Ver. Fac. Agron. (Maracay) 10: 246 (1980). Tipo: Brasil. Amazonas: rio Solimões,
Município: São Paulo de Olivença, Out-Dez. 1936, Krukoff 8861 (infr., Holótipo NY;
Isótipos VEN). (Figuras 10, 13)
P. ‘williamsii – speciosum complex’ Madison in Aroideana 2(3) (1979)
Planta hemiepífita primária, geralmente suspensa quando adulta, com caule ca. 7 cm diâm..
Raiz: aérea até 18 m de comp., 5 – 20 mm de diâmetro, cinza com acúleos; internós muito
curtos em planta adulta, escâmulas intravaginais numerosas, ca. 0,5 cm de comp., triangular a
acuminada, marrom claro na base e creme no ápice, coroando os internós; cicatriz foliar
elíptica a suborbicular 3,2 – 4 cm de comp., 2 – 2,8 cm de largura, cinza claro. Folha: profilo
45,5 cm de comp., 1,1 cm de largura, verde claro arredondado na face abaxial e canaliculado
na adaxial; pecíolo 63 – 76,5 cm de comp., 1 -1,5 cm diâm., fissurado verticalmente, verde
mais claro que a lâmina, aplainado na face adaxial, arredondado na abaxial; lâmina foliar
sagitada, margem frequentemente ondulada, verde escura na face adaxial e verde mais claro
na face abaxial; divisão anterior: 31 – 45,5 cm de comp., 25,5 – 33 cm de larg., ápice agudo a
acuminado, nervura central achatada e mais clara que a lâmina juntamente com as
secundárias, nervuras laterais primárias 4 - 6 por lado; divisões posteriores: 21 – 29 cm de
comprimento e 12 – 20 cm de largura, nervuras laterais primárias acroscópicas 2 – 3 pares e 2
laterais primárias basiscópicas, ausência de nervura submarginal coletora em ambas as
divisões. Inflorescência: uma por simpódio floral; pedúnculo 8 – 10,7 cm de comp., 1,5 – 2
cm de diâm., verde claro, estriado verticalmente; espata 29,5 – 32,5 cm de comp., 4 – 5 cm de
diâmetro, levemente constrita, verde claro na face externa, face interna creme com canais
resiníferos, com decorrência de 3,6 – 6,5 cm de comp.; espádice: zona feminina 6,5 – 10,5 cm
de comp., 21,5 – 29,4 mm de diâm. subcilíndrica; zona masculina estéril 9,6 – 10,5 cm de
comp., 22,5 – 26,2 mm de diâm. subcilíndrica; zona masculina fértil 8 – 10,7 cm de comp., 7
– 18,5 mm de diâm., elipsoide a subcilíndrica. Flores: estames 2 – 4mm de comp., 0,5 – 1
mm de larg.; estaminódios: 3,5 – 4 mm de comp., 1 – 2 mm de larg.; gineceu: 4 – 5,5 mm de
comp.; ovário 3 – 4 mm comp., 1 – 1.3 mm diâm., cilíndrico a delgado, multilocular,
recheados por substância mucilaginosa; óvulos 0,6 – 1,8 mm de comp., 4 – 5 por lóculo,
anátropos; placentação axial, bisseriada, óvulos inseridos da base até o ápice; estilete 4 – 5
mm de larg., estigma 0,5 - 1 mm larg. fortemente lobado. Infrutescência: não vista.

91
Nome popular: “cipó-ambé”, “ambé-coroa” (Comunidade de Santa Maria, Baixo rio Negro,
Amazonas, Brasil).
Hábitat: espécie encontrada em florestas de terra-firme, igapós, em margens de córregos e
baixio de solo arenoso.
Distribuição: Brasil (Figura 13).
Dados fenológicos: coletada com inflorescência durante os meses de junho a dezembro e com
frutos nos meses de março e novembro.
Material examinado: APA da Margem Esquerda do Rio Negro – Setor Aturiá-Apuauzinho,
6 Mar. 2010, R.F.M. Oliveira, G.P. Viana e A.C. Padron 25 (EAFM, est.) 10 Ago. 2010,
R.F.M. Oliveira, M.L. Soares, S. Silvino e A.C. Padron 31 (EAFM, fl.); 18 Set. 2010, R.F.M.
Oliveira, G.P. Viana e A.C. Padron 57 (INPA fl). BRASIL: Amapá: Mazagão, 15 Abr.
1986, M.J.P. Pires, N.T. Silva, P.B. Oliveira e M. Conceição 856 (INPA est.). Amazonas:
Estrada Manaus-Caracaraí, km 131, 22 Mar. 1974, A. Loureiro, O.P. Monteiro, A.P. de
Miranda, J. Lima e V.S. Albuquerque 48098 (INPA fr.); km 22, 11 Nov. 1966, G.T. Prance,
B.S. Pena e J.F. Ramos 3094 (INPA fr.); Manaus, 14 Jul. 1992, M. Nee 42972 (INPA fl. e
fr.); Maraã, Rio Ituxi, 12 Jul. 1971, G.T. Prance, P.J.M. Maas, A.A. Atchley, W.C. Steward,
D.B. Woolcott; D.F. Coelho; O.P. Monteiro, W.S. Pinheiro e J.F. Ramos 14156 (INPA fl.);
Rio Ixé, confluência com Rio Negro, 25 Out. 1987, D.W. Stevenson 830 (INPA fl.); Rio
Japurá: 5 Dez. 1982, T. Plowman, E.W. Davis, C.A. Cid, I.L. Amaral, J. Lima e J. Guedes
12248 (INPA fl.); Rio Negro, 40-50 km acima da foz do Rio Branco, 09 Out. 1978, Madison,
P.I.S. Braga e Kennedy P.F.E. 87 (INPA fl.); Entre Ilha Uabetuba e Ilha da Silva, 14 Out.
1971, G.T. Prance, P.J.M. Maas, D.B. Woolcott, D.F. Coelho, O.P. Monteiro e J.F. Ramos
15249 (INPA fl.); Mato Grosso: Paranatinga, 28 Nov. 1990, M. Macedo e S. Assumpção
2977 (INPA fr.) Rondônia: Porto Velho, 14 Jun. 1986, W. Thomas, M. Carvalho da Silva e
E.C. Moraes 5113 (INPA fl.).
Comentários:
P. solimoesense é uma das espécies pertencentes ao subgênero Meconostigma, além de
P. goeldii G.M. Barroso, P. venezuelense G.S. Bunting, que ocorrem também na Amazônia.

92
Estas espécies possuem gineceus mais derivados, com estiletes muito elaborados e geralmente
mais de 20 lóculos em cada ovário, o que contraria o fato desse subgênero ser menos bem
sucedido no ambiente florestal. A ocorrência de poucas espécies com hábito mais uniforme e
morfologia muito derivada do gineceu na bacia amazônica, também sugere que o subgênero
Meconostigma poderia ter colonizado essa região numa época relativamente recente em
contraste com a parte sudeste da área de distribuição (Mayo, 1988).
Mayo (1991) em uma revisão do subgênero Meconostigma cita para essa espécie 26-
34 (-47) lóculos por ovário. Infrutescência cerca de 12 cm comprimento, 4 – 4.5 cm diâmetro
com bagas variando de cor branca, laranja ou vermelha e cheiro adocicado, as sementes
ovoide a elíptica com funículo arilado.
As raízes aéreas desta espécie são muito utilizadas para artesanato na Comunidade de
Santa Maria no Baixo rio Negro. Estas apresentam as seguintes características morfológicas:
cor cinza, até 2 cm diâmetro, acúleos em toda sua extensão e em corte transversal, apresenta
um cilindro central em formato lobado. Ressalta-se que esta é, a primeira vez que se relata a
utilização das raízes desta espécie para uso em artefatos.
Apesar de não terem sido feitas coletas sistemáticas dos forófitos neste estudo,
observou-se que seis famílias de árvores suporte (forófitos) estavam colonizadas por P.
solimoesense. Estas são Arecaceae (Attalea sp.), Fabaceae (Dipteryx polyphylla (Huber)
Ducke, Swartzia polyphylla DC. e indeterminada), Humiriaceae (Vantanea celativenia
(Standl.) Cuatrec.), Lauraceae (Licaria chrysophylla (Meisn.) Kosterm.) e Lecythidaceae
(Eschweleira sp.). Dentre as espécies de forófitos, Swartzia polyphylla (“arabá”), apareceu
mais frequentemente com indivíduos de P. solimoesense.

93
Figura 10. Philodendron solimoesense A.C. Sm. A) Hábito; B) Vista lateral do óvulo C) Seção longitudinal do gineceu D) Vista apical da flor masculina estéril com quatro estaminódios; E) Visão apical da flor masculina estéril com três estaminódios; F) Vista frontal do estame; G) Seção transversal do ovário; H) Vista frontal do estaminódio (A – H preparados de Oliveira, R.F.M. et al., 31; ilustração de Felipe França, fev/2011).

94
Morfologia interna
Lâmina foliar
Epiderme: em vista frontal, as células apresentam paredes retas, cutícula levemente
rugosa, e mais espessa na face abaxial (Figura 11 A). Em corte transversal, as células
apresentam-se poligonais em ambas as faces (Figura 11 F). Estômatos: são do tipo paracítico,
ocorrendo em suaves depressões com câmaras subestomáticas de tamanhos variáveis (Figura
11 A, C e F). As folhas são anfihipoestomáticas, pois apresentam um número médio de 103,9
estômatos por mm2 na face abaxial e 39,1 na epiderme adaxial. Tricomas: em nosso estudo
não foi possível visualizar o tricoma inteiro, apenas a sua base, são solitários e raros, devido à
técnica utilizada para a dissociação das epidermes (Figura 11 B). Mesofilo: tecido paliçádico
apresenta-se compacto com duas camadas de células tecido lacunoso com 7 a 8 camadas
formado por parênquima lacunoso com grandes espaços intercelulares conferindo-lhe aspecto
aerenquimatoso (Figura 11F). Colênquima anelar: localizado logo abaixo da epiderme
inferior e situados contíguos aos feixes fibrovasculares, sendo constituído de três a quatro
camadas de células aclorofiladas (Figura 11 G). Feixes fibrovasculares: foram vistos feixes
colaterais envolvidos por 2 – 4 camadas de fibras esclerenquimáticas (Figura 11 D, 11 E).
Idioblastos: células apresentando substância ergástica de coloração escura (Figura 11 F).
Laticíferos: foram observados próximos aos feixes vasculares (Figura 11 F). Cristais: foram
evidenciados cristais do tipo drusa situados geralmente no tecido paliçádico (Figura 11 G)
(Tabela 1).

95
Figura 11. Esturturas da lâmina foliar de Philodendron solimoesense A.C. Sm. A e B) Vista frontal da epiderme. A) Estômato paracítico em visão superficial (seta); B) Bases de pelo (setas); C-F) Cortes transversais; C) Estômato submerso na epiderme evidenciando a câmara subestomática (); D) Feixe vascular central com feixes dispersos pelo mesofilo; E) Feixe vascular colateral envolto por fibras esclerenquimáticas; F) Cavidade resinífera (seta) e parênquima lacunoso aerenquimatoso com uma ampla câmara subestomática (); G) Mesofilo contendo drusas (seta). Barras = 12,5 µm (A, B); 5 µm (C); 50 µm (D); 12,5 µm (E); 25 µm (F); 5 µm (G).
Raízes aéreas
Periderme: apresenta-se espessa e é composta mais externamente por uma espessa
camada de súber (felema) (Figura 12 B, C e D). Córtex: apresenta-se mais internamente e é
composto por células dispostas de forma irregular apresentando espaços intercelulares
(Fgura 12 A). Na região fronteiriça com o cilindro vascular apresentam-se em arranjo radial,
onde também ocorrem dutos resiníferos e cristais na forma de ráfides (Figura 12 F).
Endoderme: constituída de uma camada pouco evidenciada de células com paredes
cilíndricas e espessamento secundário (Figura 12 F). Cilindro vascular: apresenta-se
lobado e possui um arranjo alterno e radial do xilema e floema (Figura 12 E) (Tabela 2).

96
Figura 12. Cortes transversais da raiz aérea de Philodendron solimoesense A.C. Sm. A) Regiões distintas: córtex e parte do cilindro central; B) Periderme evidenciando a região suberificada; C) Periderme (seta); D) Ráfides dispersas por todo o córtex (setas); E) Cilindro central lobado; F) Endoderme e cavidade resinífera (seta); G) Elementos de vaso rodeados por fibras esclerenquimáticas. Barras = 50 µm (A, B, C); 25 µm (D); 200 µm (E); 12,5 µm (F); 50 µm (G).

97
Figura 13. Mapa de distribuição geográfica das espécies Heteropsis flexuosa (Kunth) G.S. Bunting, Monstera obliqua Miq. e Philodendron solimoesense A.C. Sm.
Principais características anatômicas da lâmina foliar e das raízes aéreas das espécies estudadas de Araceae
Tabela 1. Características anatômicas distintivas da lâmina foliar das espécies estudadas de Araceae.
Caracteres da lâmina foliar Espécies 1 2 3
Epiderme diferenciada entre as faces adaxial e abaxial - + - Ornamentação cuticular em ambas as faces epidérmicas - - + Lâminas foliares anfihipostomáticas + + + Estômatos na epiderme adaxial somente na nervura central - + - Estômatos paracíticos + - + Estômatos paracíticos submersos - - + Estômatos tetracíticos - + - Resquícios de tricomas - - + Drusas no mesofilo + - + Ráfides no mesofilo - + - Idioblastos com substâncias ergásticas no mesofilo + + + Laticíferos - - + Colênquima anelar - - + Parênquima lacunoso aerenquimatoso - - + Feixes fibrovasculares colaterais + + +
1.Heteropsis flexuosa (Kunth) G.S.Bunting; 2. Monstera obliqua Miq.; 3. Philodendron solimoesense A.C.Sm. + = presente; - = ausente.

98
Tabela 2. Características anatômicas distintivas da raiz aérea das espécies estudadas de Araceae. Caracteres da raiz aérea Espécies
1 2 3 Periderme como camada de revestimento e proteção + - + Córtex com fibras esclerenquimáticas dispersas + - - Exoderme com uma camada de células retangulares a quadradas + - - Exoderme com células retangulares a subcilíndricas - + - Células suberizadas abaixo da exoderme + - + Drusas no córtex da raiz aérea + - + Canais resiníferos ou laticíferos - - + Córtex com células isodiamétricas de paredes delgadas relativamente esféricas, achatadas ou ovaladas
+ - -
Córtex com células parenquimáticas em forma esférica, relativamente isodiamétricas e justapostas - + - Fibras esclerenquimáticas dispersas no córtex + - - Endoderme com uma a duas camadas de células parenquimáticas ovais + + - Bainha de fibras esclerenquimáticas ao redor do cilindro central + - - Cilindro central arredondado + + - Cilindro central lobado - - + Braquiesclereídes nos elementos de vasos + - - Tilossomos em elementos de vaso - + - 1. Heteropsis flexuosa (Kunth) G.S.Bunting; 2. Monstera obliqua Miq.; 3. Philodendron solimoesense A.C.Sm. + = presente; - = ausente.
Estudo das fibras
As fibras das três espécies se apresentam libriformes, longas, de paredes celulares
secundárias grossas, lignificadas e lúmens reduzidos. Quanto ao comprimento, as fibras de
H. flexuosa variam de 2,050 a 3,600 µm e apresentam uma média de 2,765 µm, o que as
caracterizam como fibras longas. Quanto à espessura da parede, estas são classificadas
como delgadas a espessas que variam de 0,15 a 0,4 µm, com uma média de 0,3 µm, e
apresentam diâmetros maiores em relação ao lúmen que variam de 0,05 a 0,3 µm. O
diâmetro total das fibras varia de 0,23 a 0,69 µm e apresenta uma média de 0,5 µm.
M. obliqua apresenta um comprimento das fibras que varia de 1,560 a 3,010 µm e
apresentam uma média de 2,422 µm, caracterizadas como fibras longas. Quanto à espessura
da parede, estas são classificadas como delgadas a espessas que variam de 0,4 a 0,8 µm,
com uma média de 0,6 µm, e apresentam diâmetros maiores em relação ao lúmen que
variam de 0,4 a 1,1 µm. O diâmetro total das fibras varia de 1,03 a 1,95 µm e apresenta uma
média de 1,5 µm.
As fibras de P. solimoesense variam de 1,600 a 3,100 µm e apresentam uma média
de 2,224 µm, e são caracterizadas como fibras longas. Quanto à espessura da parede, estas
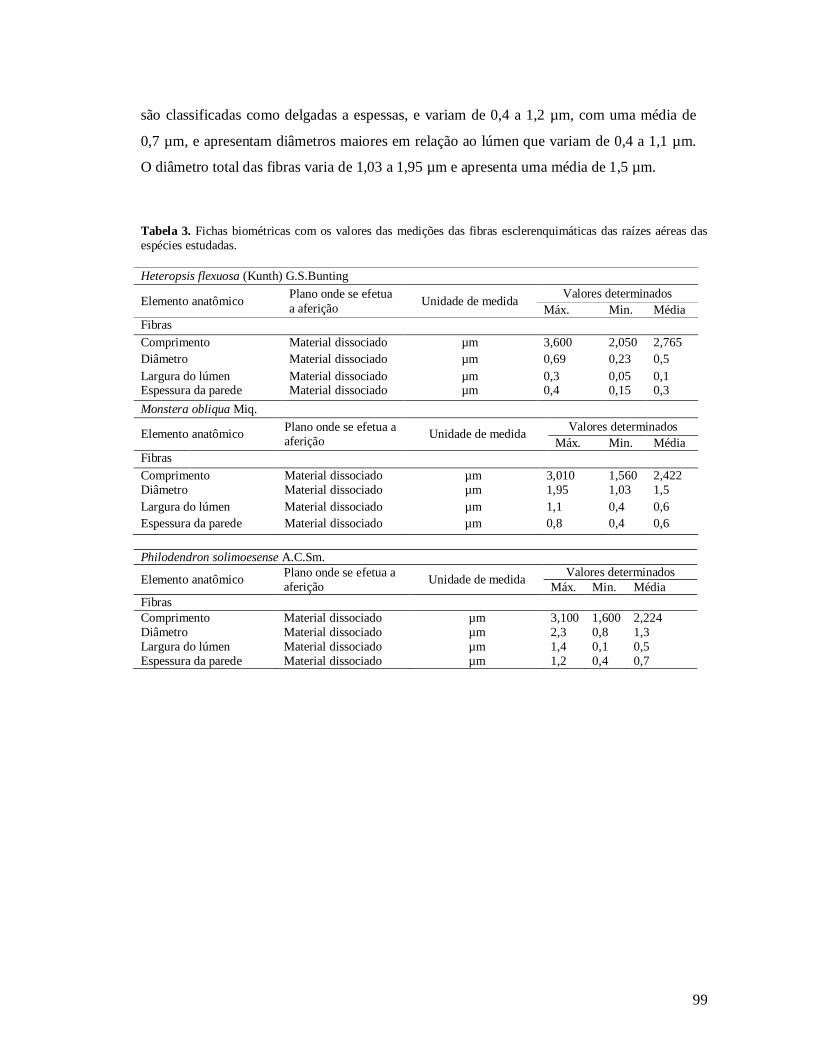
99
são classificadas como delgadas a espessas, e variam de 0,4 a 1,2 µm, com uma média de
0,7 µm, e apresentam diâmetros maiores em relação ao lúmen que variam de 0,4 a 1,1 µm.
O diâmetro total das fibras varia de 1,03 a 1,95 µm e apresenta uma média de 1,5 µm.
Tabela 3. Fichas biométricas com os valores das medições das fibras esclerenquimáticas das raízes aéreas das espécies estudadas. Heteropsis flexuosa (Kunth) G.S.Bunting
Elemento anatômico Plano onde se efetua a aferição Unidade de medida Valores determinados
Máx. Min. Média Fibras Comprimento Material dissociado µm 3,600 2,050 2,765 Diâmetro Material dissociado µm 0,69 0,23 0,5 Largura do lúmen Material dissociado µm 0,3 0,05 0,1 Espessura da parede Material dissociado µm 0,4 0,15 0,3 Monstera obliqua Miq.
Elemento anatômico Plano onde se efetua a aferição Unidade de medida Valores determinados
Máx. Min. Média Fibras Comprimento Material dissociado µm 3,010 1,560 2,422 Diâmetro Material dissociado µm 1,95 1,03 1,5 Largura do lúmen Material dissociado µm 1,1 0,4 0,6 Espessura da parede Material dissociado µm 0,8 0,4 0,6 Philodendron solimoesense A.C.Sm.
Elemento anatômico Plano onde se efetua a aferição Unidade de medida Valores determinados
Máx. Min. Média Fibras Comprimento Material dissociado µm 3,100 1,600 2,224 Diâmetro Material dissociado µm 2,3 0,8 1,3 Largura do lúmen Material dissociado µm 1,4 0,1 0,5 Espessura da parede Material dissociado µm 1,2 0,4 0,7

100
DISCUSSÃO Aspectos anatômicos das espécies
Lâmina foliar
Philodendron solimoesense foi a única espécie que apresentou uma cutícula rugosa em
ambas as faces da lâmina foliar. Enquanto que Heteropsis flexuosa e Monstera obliqua
apresentaram cutícula lisa. No entanto, Keating (2002), descreve cutícula lisa para as espécies
P. bipinnatifidum Schott, P. goeldii G.M. Barroso e P. leal-costae Mayo & Barroso, onde,
todas pertencentes também ao subgênero Meconostigma. O fato de a cutícula de P.
solimoesense ter se apresentado mais espessa, pode estar relacionado ao ambiente em que a
planta vive, ou seja, nas margens do rio ou em áreas de igapó, onde, em ambos os ambientes,
há alta incidência de luz refletida através da água. Segundo alguns autores (Withner et al.,
1974; Appezzato-da-Glória & Carmello-Guerreiro, 2006), a cutícula possui uma camada
brilhante e refletora que atua na proteção contra o excesso de luminosidade. Embora haja
diferença entre P. bipinnatifidum, P. goeldii, P. leal-costae e a espécie estudada do mesmo
subgênero, a cutícula não foi considerada um caráter de valor taxonômico para P.
solimoesense, por não ter sido feita uma avaliação mais aprofundada, visto que características
como a da epiderme variam conforme o ambiente em que o indivíduo se encontra.
Quanto à estrutura da epiderme em vista frontal, apenas em M. obliqua houve
diferenças significativas entre a face adaxial e abaxial. A primeira apresentou células
isodiamétricas, retangulares e alongadas, com estômatos somente na região da nervura,
enquanto que a abaxial apresentou células com paredes onduladas e estômatos distribuídos em
todas as regiões da lâmina. Enquanto que H. flexuosa e P. solimoesense não apresentaram
diferenças no formato das células.
A lâmina foliar das três espécies são anfi-hipoestomáticas. Os estômatos são
paracíticos em H. flexuosa e P. solimoesense, enquanto que em M. obliqua é tetracítico. No
entanto, os estômatos de P. solimoesense, diferem dos de H. flexuosa por apresentarem-se
submersos, ocorrendo em suaves depressões e possuem câmaras subestomáticas que variam
de tamanho, que chegam a formar grandes câmaras de ar. Esses resultados corroboram com
Grayum (1990), que cita que os estômatos paracíticos e folhas hipoestomáticas ou
anfiestomáticas, geralmente são comuns nos gêneros de Araceae.

101
Resquícios de tricomas raros e solitários foram observados na epiderme de P.
solimoesense, no entanto, em nosso estudo não foi possível classificá-los, devido a
possibilidade da técnica utilizada para a dissociação ter destruído os mesmos. De forma geral,
os tricomas são muito variáveis na sua estrutura e podem ter valor diagnóstico para a
taxonomia, nesse caso, Philodendron é um dos poucos gêneros de Araceae que apresentam
tricomas nas folhas (Bogner, 1987; Keating, 2002).
Constatou-se a presença de cristais nas folhas das três espécies. Foram vistas drusas
em todo o mesofilo de H. flexuosa e P. solimoesense. Drusas são comuns em dicotiledôneas,
mas relativamente raras em monocotiledôneas, onde são encontradas em alguns táxons como
Acorus, Araceae e Tofieldia (Prychid e Rudall, 1999). Em M. obliqua apresentou apenas
idioblastos com ráfides, no entanto, Keating (2002) cita drusas no mesofilo de Monstera
acuminata C. Koch, M. dilacerata C. Koch, M. membranacea Madison, M. oreophila
Madison, M. pittieri Engl., M. punctulata (Schott) e M. siltepecana Matuda.
Nas três espécies foram observados idioblastos no mesofilo. Em P. solimoesense, as
células clorenquimáticas situadas em torno dos feixes fibrovasculares contêm grandes
vacúolos possivelmente relacionados com reserva hídrica e/ou substâncias ergásticas que,
segundo Medeiros et al. (2003), podem atuar de forma isolada ou conjunta na proteção
alelopática a patógenos e insetos.
Laticíferos foram observados em P. solimoesense, próximo aos feixes vasculares.
Estes podem ser classificados de acordo com o tipo de substância encontrada em seu interior,
que podem ser óleos, resinas, ceras e borracha. No entanto, não foi realizado estudo
aprofundado com intuito de identificar a natureza destas substâncias na espécie. Porém,
podemos afirmar que se trata possivelmente de dutos resiníferos, frequentemente encontrados
em espécies do gênero Philodendron, inclusive no subgênero Meconostigma, com base em
estudos realizados por Keating (2002). A ausência de laticíferos e tricosclereídes caracterizam
taxonomicamente os gêneros Heteropsis e Anadendrum, pertencentes à subfamília
Monsteroideae (Mayo et al., 1997).
P. solimoesense foi a única espécie que apresentou um colênquima anelar, localizado
logo abaixo da epiderme inferior a qual é constituída de uma única camada contínua de
células aclorofiladas contendo substância densa, possivelmente mucilaginosa. Estas células
apresentam formato quadrangular a arredondado, parede pouco espessa e distribui-se de
forma compacta não se intercalando com o parênquima. Esta é interrompida por feixes de
fibras esclerenquimáticas de formato alongado com paredes grossas lignificadas e
extremidades afiladas. P. solimoesense também foi a única espécie que apresentou um

102
parênquima lacunoso aerenquimatoso, formado por espaços com amplas câmaras de ar
possivelmente relacionados com o período de submersão das raízes na época de inundação da
floresta.
As três espécies apresentam feixes fibrovasculares colaterais, nos quais as fibras
variam em diâmetro, apresentando-se mais espessas nas extremidades dos pólos. Segundo
Bonates (2007), esses espessamentos localizados nos pólos dos feixes colaboram para evitar o
colapso de células do mesofilo e do próprio sistema condutor quando há ocorrência de déficit
hídrico na planta. Fibras esclerenquimáticas em volta dos feixes vasculares encontrados nas
três espécies estudadas também foram relatadas para algumas espécies de Anthurium
(Mantovani et al., 2010).
Raízes aéreas
H. flexuosa e P. solimoesense foram as duas espécies que apresentaram um
espessamento secundário, com uma periderme espessa, com células dando origem ao súber
(felema) em direção à parte mais externa. Nas camadas de súber, H. flexuosa e P.
solimoesense apresentaram estruturas denominadas lenticelas, que são aberturas que ocorrem
na periderme, responsáveis pelas trocas gasosas. Castro (2009), afirma que é comum a
formação de lenticelas, para o provimento de aeração, em estruturas cobertas por camadas de
súber, o qual é impermeável ao oxigênio.
H. flexuosa e M. obliqua apresentaram uma bainha formando um anel com camadas de
fibras esclerenquimáticas, com paredes espessas e lúmens reduzidos, que de acordo com Van
Flett (1961), concede sustentação, resistência mecânica, elasticidade e impermeabilidade, o
que vem sendo confirmado por estudos como o de Soares (2008), para o gênero Heteropsis, e
pelo próprio histórico de uso das raízes dessa espécie como uma das mais procuradas pelos
extratores de cipós ao longo do rio Negro. No entanto, em P. solimoesense, as células
parenquimáticas do córtex apresentam-se dispostas de forma irregular e apresentam espaços
intercelulares esquizógenos. Também ocorrem cavidades resiníferas e cristais na forma de
ráfides, que segundo Keating (2004), são classificadas como forma de fuso “spindle-shaped”,
e alcançam paredes com maior espessura no subgênero Meconostigma, ao qual esta espécie
pertence. Segundo Dickison (2000) e Castro (2009), a camada de súber atua como proteção
mecânica e regula as variações de temperatura e umidade. O crescimento secundário radicular
em monocotiledôneas é extremamente raro, nesse caso, o meristema secundário de

103
espessamento é o tecido que produz raízes adventícias e forma ligações entre a vasculatura de
raiz, caule e folha (Michereff et al., 2005).
M. obliqua apresenta uma epiderme composta mais externamente por tricomas que
podem estar relacionados à função de aderência nas árvores suportes. A exoderme foi vista
apenas em M. obliqua com uma camada de células retangulares a subcilíndricas, apesar de
Soares et al., (2008) ter relatado a presença de uma exoderme também para a espécie H.
flexuosa. De acordo com Cutler (2007), a exoderme consiste de uma ou mais camadas, mais
externa ao córtex, caracterizada por células angulares com paredes geralmente lignificadas.
Essas camadas de células atuam na conservação da água no meio interno, através da
prevenção da transpiração e evaporação do tecido cortical (Bonates, 2007). No entanto,
estudos mais aprofundados em ontogenia são necessários para obtenção de uma afirmação
mais consistente a respeito dessa característica.
Em M. obliqua as células parenquimáticas apresentam-se de forma esférica e
relativamente isodiamétricas. Alguns autores têm relatado a ocorrência de cloroplastos no
córtex externo de raízes aéreas de algumas espécies da família (Mayo et al., 1997) e
ocorrência de colênquima em espécies de Monstera e Philodendron (Hinchee, 1981), o que
não foi constatado neste estudo, talvez por falta de um estudo mais acurado.
A endoderme das três espécies consistiu de uma camada de células achatadas
delimitando mais externamente o cilindro vascular. Menezes et al., (2005) comprovaram a
atividade meristemática da endoderme, através da proposta de uma nova interpretação do
meristema de espessamento primário para as monocotiledôneas.
O cilindro central das três espécies apresentou alternância dos feixes de xilema e
floema, os quais estão envolvidos por fibras esclerenquimáticas. Na região central observam-
se os anéis concêntricos formados por elementos de metaxilema de maiores diâmetros
rodeados de fibras esclerenquimáticas. Enquanto que em M. obliqua, o cilindro vascular é
constituído de elementos de vaso rodeados por fibras esclerenquimáticas e raramente
tilossomos, que segundo Cutler (2007), são células epiteliais que proliferam em cavidades
intercelulares. No entanto, a localização de tilossomos em elementos de vasos, como
apresentado aqui, pode indicar uma função “tampão”, proposta por Benzing et al. (1982) onde
estes atuariam como barreira para transpiração, uma vez que estes se apresentam compactos
quando secos e quando umedecidos aumentam sua superfície tornando-se frouxos. O cilindro
vascular das raízes aéreas de P. solimoesense apresentou-se lobado e diferenciou-se das
demais espécies, corroborando com estudos feitos por Vianna et al. (2001), que citam cilindro

104
lobado para P. bippinatifidum Schott, espécie pertencente ao subgênero Meconostigma,
característica peculiar e de importância taxonômica para espécies deste subgênero.
Fibras
As fibras foram classificadas, quanto à forma, como libriformes e quanto ao
comprimento, como curtas a longas. De acordo com as normas de procedimento em estudos
de anatomia de madeira, as fibras curtas são aquelas com comprimento que varia de 900 a
1600 µm e as longas são maiores ou iguais a 1600 µm (IBAMA, 1992). Quanto à espessura
da parede, estas são classificadas como delgadas a espessas, pois apresentam lúmen
distintamente perceptível. Este resultado corrobora com Soares (2007), que analisou quatro
espécies de Heteropsis ocorrentes na Reserva Ducke, Manaus, cujas fibras das raízes
também apresentaram paredes espessadas e lumens reduzidos. Em relação às
monocotiledôneas, as fibras de Bambusa vulgaris Schrad. ex J.C.Wendl.
(Poaceae) também foram classificadas como muito longas (comprimento médio: 2,299 mm)
(Guimarães Junior et al., 2010).
As espécies estudadas são caracterizadas pela resistência fornecida através de suas
fibras na confecção de artigos artesanais. A grande proporção de fibras é um caráter
constante em monocotiledôneas (Cutter 1986; Fahn 1990). Com relação ao comprimento
das fibras, H. flexuosa foi a que apresentou o maior tamanho atingindo até 3,600 µm. P.
solimoesense apresentou fibras com comprimento máximo de 3,100 µm, seguida de M.
obliqua, que apresentou fibras de até 3,010 µm. H. flexuosa, além de apresentar fibras com
maior comprimento, apresenta várias camadas de fibras esclerenquimáticas unidas em uma
bainha entre o tecido parenquimático que, de acordo com Van Flett (1961), concede
sustentação, resistência mecânica, elasticidade e impermeabilidade.
Apesar de M. obliqua apresentar menor diâmetro e menor concentração de fibras
entre as espécies estudadas, mas, esta espécie apresenta um grande potencial também para a
utilização em artefatos que necessitem de material resistente. No entanto, essas raízes
restringem-se somente para o uso em acabamento nas bordas de cestos e paneiros feitos
com fibras mais resistentes. Filho (1987), estudando espécies arbóreas, observou que há
uma relação entre a variação das dimensões das fibras com as propriedades físico-mecânicas
da madeira e da celulose. Este resultado pode ser um parâmetro que abrange as espécies
estudadas neste trabalho uma vez, que estas possuem fibras com potencial do ponto de vista
tecnológico.

105
CONCLUSÃO
Heteropsis flexuosa e Monstera obliqua apresentaram uma bainha de fibras
esclerenquimáticas (anel esclerenquimático), o que concerne maior resistência e
flexibilidade.
Philodendron solimoesense, apresentou um cilindro central lobado, que também
concerne flexibilidade, com elementos de vasos cercados de fibras esclerenquimáticas
longas reforçadas em ambas as espécies pelos tamanhos e espessuras de suas fibras.
Estas características internas das raízes apresentam grande utilidade para
identificação em campo através de um simples corte travsversal. Além disso, mesmo depois
de beneficiadas, é possível reconhecer a quais espécies pertencem essas raízes, através da
observação de sua anatomia, o que pode vir a ser bastante útil para reconhecimento da
origem de produtos produzidos com espécies de plantas amazônicas.
Apesar de as raízes de Monstera obliqua serem usadas apenas para o acabamento
dos artigos artesanais, este estudo mostra que suas raízes são possivelmente tão resistentes
quanto as raízes de H. flexuosa e P. solimoesense, no entanto, necessita de um estudo mais
acurado.
Esta é a primeira citação de raízes de P. solimoesense e M. obliqua para a utilização
em artesanato.
Estudos como este são de suma importância para identificar novas espécies com
potencial econômico e assim contribuir com o manejo adequado dessas espécies para que
esses recursos sejam conservados.

106
REFERÊNCIAS Appezzato-da-Glória, B. e Carmello-Guerreiro, S.M. 2006. Anatomia vegetal. 2ª. Ed. atual.
Viçosa : Ed. UFV, 2006. 438pp.
Benzing, D.H.; Ott, D.W. e Friedman, W.E. 1982. Roots of Sobralia macrantha
(Orchidaceae): structure and the velame – exodermis complex. American Journal of Botany
69 : 608-614.
Bogner, J. 1987. Morphological variation in aroids. Aroideana 10 (2): 4–16.
Bonates, L.C. de M. 1993. Estudos ecofisiológicos de Orchidaceae da Amazônia. II –
Anatomia ecológica foliar de espécies com metabolismo Cam de uma campina da Amazônia.
Acta Amazonica, 23(4): 315-348.
Bonates, L.C. de M. 2007. Anatomia ecológica da folha e da raiz e aspectos ecofisiológicos
de Orchidaceae epífitas de uma campina da Amazônia Central. Tese (doutorado). INPA /
UFAM.
Castro, E.M. 2009. Histologia vegetal: estrutura e função de órgãos vegetativos. Lavras:
UFLA, 234pp.
Coelho, M.A.N., Soares, M.L., Sakuragui, C.M., Mayo, S., Andrade, I.M. de, Temponi, L.G.
2010. Araceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de
Janeiro. (http://floradobrasil.jbrj.gov.br/2010/FB000051).
Croat, T.B. 1985. Collecting and preparing specimens of Araceae. Ann. Missouri Botanic
Garden. v. 17: p. 252-258.
Croat, T.B. 1990 “1988”. Ecology and life forms of Araceae. Aroideana 11(3-4): 1-55.
Croat, T.B. 1994. New species of Philodendron Subgenus Pteromischum (Araceae) from
Saül, French Guiana. Novon. St. Louis, v.4; p.211-219.

107
Cutler, D. F.; Botha, T.; Stevenson, D. W. 2001. Plant anatomy: an applied approach.
Blackwell Publishing. 302pp.
Cutter, E. G. 1986. Anatomia Vegetal. 2 ed., São Paulo, Roca, v. 2. 304pp.
Dickison, W.C. 2000. Integrative plant anatomy. Academic press. 533pp.
Durigan, C.C.; Castilho, C.V. 2004. O extrativismo de cipós (Heteropsis spp., Araceae) no
Parque Nacional do Jaú. In: Janelas para a Biodiversidade no Parque Nacional do Jaú:
Uma estratégia para a Biodiversidade na Amazônia. Sérgio Henriques Borges; Simone
Iwanaga; Carlos César Durigan; Marcos Roberto Pinheiro (Eds.) Manaus: Fundação Vitória
Amazônica. p. 231-244.
Eames, A. J. e MacDaniels, L. H. 1925. An introduction to plant anatomy. McGraw-Hill
Book Company, Inc, New York.
Engler, A. 1878. Araceae. In: Martius, C.F.P. von, Flora brasiliensis 3(2): 56–88, t. 11 - 102.
Fahn, A. 1990. Plant anatomy. 4 ed., Oxford, Pergamon Press, 588pp.
FAS – FUNDAÇÃO AMAZONAS SUSTENTÁVEL – Programa bolsa floresta. 2009.
Relatório da precursora da APA do Rio Negro. Manaus – AM.
French, J.C. 1997. Vegetative anatomy In: Mayo, S.J., Bogner, J. and Boyce, P. C. (Eds).
The genera of Araceae. Royal Botanic gardens, Kew 9 -24 p.
Govaerts, R.; Frodin, D.G.; Borgner, J., Boyce, P.; Cosgreff, B.; Croat, T.B.; Gonçalves,
E.G.; Gayum, M.; Hay, A.; Hetterscheid, W.; Landolt, E.; Mayo, S.J.; Murata, J.; Nguyen,
V.D.; Sakuragui, C.M.; Singh, Y.; Thompson, S.; Zhu, G. 2002. World checklist and
bibliography of Araceae (and Acoraceae). Royal Botanic Garden, Kew.
Grayum (1990) Grayum, M.H. 1990. Evolution and Phylogeny of the Araceae. Annals of the
Missouri Botanical Garden, 77: 628-697.

108
Guimarães Junior, M.; Novack, K.M.; Botaro, V.R. 2010. Caracterização anatômica da fibra
de bambu (Bambusa vulgaris) visando sua utilização em compósitos poliméricos. Revista
Iberoamericana de Polímeros, 11(7), 442-456.
Hinchee, M. A. W. 1981. Morphogenesis of aerial and subterranean roots of Monstera
deliciosa. Botanical Gazette 142: 347-359.
IBAMA – Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis.
Diretoria de Incentivo à Pesquisa e Divulgação. Laboratório de Produtos Florestais. Série
Técnica 15. 1992. Normas e procedimentos em estudos de anatomia da madeira: I –
Angiospermae II – Gimnospermae / Vera T. Rauber Coradin, Graciela I. Bolzon de Muniz.
Brasília. 17pp.
Keating, R. C. 2002. Acoraceae and Araceae. In: Gregory, M. and Cutler, D. F. Anatomy of
the monocotiyledons. Oxyford University Press, New York, 322pp.
Keating, R. C. 2004. Systematic occurrence of raphide crystals in Araceae. Annals of the
Missouri Botanical Garden 91 (3): 495-504.
Krause, K. 1913. Araceae: Philondroideae – Philodendreae – Philodendrinae. Leipzig, In:
Engler, Das Pflanzenreich. 60 (IV 23 Db) p. 1-143.
Lawrence, G.H.M. 1977. Taxonomia das plantas das vasculares. Lisboa: Fundação
Calouste Gulbenkian, v.1, 854pp.
Leoni, J. M. e Marques, T. S. 2008. Conhecimento de artesãos sobre plantas utilizadas na
produção de artefatos - Reserva de Desenvolvimento Sustentável Amanã – AM. UAKARI,
v.4, n.2, p.67-77.
Mantovani, A., Filartiga, A. Del P., Coelho, M. A. N. 2010. Anatomia comparada da folha e
espata de espécies de Anthurium (Araceae) ocorrentes na Mata Atlântica. Revista Brasil.
Bot., V.33, n.1, p.185-200.

109
Mayo, S.J. 1991. A revision of Philodendron subgenus Meconostigma (Araceae). Kew
Bulletin, v. 46, n. 4, p. 601 – 681.
Mayo, S.J.; Borgner, J. e Boyce, P.C. 1997. The genera of Araceae. Kew, Royal Botanic
Gardens.
Menezes, N.L. de; Silva, D.C.; Arruda, R.C.O.; Melo-de-Pina, G.F.; Cardoso, V.A.; Castro,
N.M.; Scatena, V.L.; Scremin-Dias, E. 2005. Meristematic activity of the endodermis and the
pericycle in the primary thicknening in monocotyledons. Considerations on the “PTM”.
Anais da Academia Brasileira de Ciências, jun., ano/vol. 77, num. 002. Rio de Janeiro,
Brasil. p. 259-274.
Michereff, S. J.; Andrade, D. E. G. T.; Menezes, M. 2005. Ecologia e manejo de patógenos
radiculares em solos tropicais. Imprensa Universitária, Recife : UFRPE 398pp.
Coelho, M.A.N.; Waechter, J.L.; Mayo, S. J. 2009. Revisão taxonômica das espécies de
Anthurium (Araceae) Seção Urospadix Subseção Flavescentiviridia. Rodriguésia 60 (4):
799-864.
Phillips, C. 1987. Monstera obliqua, featuring the “Laceleaf” form – an extreme variant
within a highly variable species. Aroideana v. 10(2) p. 20-22.
Potiguara, R.C.V.; Nascimento, M.E. 1994. Contribuição á anatomia dos órgãos vegetativos
de Heteropsis jenmanii Oliv. (ARACEAE). Boletim Museu Paraense Emílio Goeldi, 10(2):
237-247.
Prychid, C.J., Rudall, P.J. 1999. Calcium oxalate crystals in monocotyledons: a review of
their structure and systematics. Annals of Botany 84:725-739.
Radford, A.E.; Dickison, W.C.; Massey, J.R. et al. 1974. Vascular Plant Systematics. New
York : Harper e Row. 891pp.
Schott, H.W. 1860. Prodromus systematis aroideanum. Vienna: Typis congregationis
mechitharisticae, Vindobonae. 602pp.

110
Soares, M.L.C.; Mayo, S.J. 1999. Araceae. In: Ribeiro, J.E.L.S.; Vicentini, A.; Sothers, C.A.;
Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.; Martins, L.H.P.; Lohmann, L.G.; Assunção,
P.A.C.L.; Pereira, E.C.; Silva, C.F.; Mesquita, M.R.; Procópio, L. (Eds). Flora da Reserva
Ducke: Guia de identificação das plantas vasculares de uma floresta de terra-firme na
Amazônia Central. Instituto Nacional de Pesquisas da Amazônia, Manaus. p. 672-687.
Soares, M.L.; Jardim-Lima. 2005. Amazonian Species of Araceae in the INPA Herbarium,
Manaus – Amazonas, Brasil. Aroideana 28: 134-152.
Soares, M.L. 2008. Sistemática e ecologia de Heteropsis Kunth (Araceae Juss.) com destaque
especial nas espécies ocorrentes na Reserva Florestal Adolpho Ducke, Manaus – Amazonas,
Brasil. 207 p.: il. Tese doutorado – INPA/UFAM.
Souza, C. C. V. 2010. Etnobotânica de quintais em três comunidades ribeirinhas na
Amazônia Central, Manaus – AM. Dissertação de Mestrado. Manaus : [s.n.], 2010.
Stuessy, T.F. 1990. Plant taxonomy. The systematic evaluation of camparative data.
Columbia University Press, New York. 176pp.
Vianna, W.O.; Soares, M.K.M. & Appezzato-da-Glória, B. 2001. Anatomia da raiz escora de
Philodendron bipinnatifidum Schott (Araceae). Acta bot. bras. 15(3): 313-320.
Withner, C. L.; Nelson, P. K. ; Wejksnora, P. J. – 1974. The anatomy of orchids. In: The
orchids. Scientifics studies. Edited by C.L. Withner. Jonh Wiley, New York. 347 – 367.

111
APÊNDICE A – Questionário aos sujeitos da pesquisa – Comunidade Santa Maria, Baixo rio Negro – AM.
Pesquisadora responsável: Rina Fátima M. de Oliveira Nome/apelido:__________________________________________________________ Idade:___________ Tempo de moradia na comunidade:___________ 1) Quais as plantas utilizadas na sua casa e/ou terreno como ornamental (enfeite ou plantas de vaso):
Nome popular Onde ela foi
adquirida (área de ocorrência)?
Qual parte da planta é mais valorizada (flor, folhas, frutos,
etc.)?
Em que parte da casa ou terreno ela é cultivada? (roçado, frente/varanda, quintal, etc.). Por quê?
2) Quais as plantas são utilizadas na sua casa como medicinais (remédio):
Nome popular
Para que é utilizada?
Parte da planta utilizada
Forma de preparo
Modo de uso do remédio
Área de ocorrência
3) Quais as plantas utilizadas na sua propriedade como alimento/comestível?
Nome popular Parte da planta utilizada Forma de consumo Área de ocorrência 4) Quais as plantas utilizadas na sua propriedade como matéria-prima p/ o artesanato e construção (não-madeireiros)?
Nome popular
Parte da planta utilizada
Área de ocorrência
Em que época é coletada
Produtos obtidos da matéria-prima
5) Existem plantas utilizadas como místicas (simpatias) ou religiosas na sua casa?
Nome popular Para quê é utilizada Como é utilizada
Existe uma época específica para a
planta ser utilizada? Quando (se
houver)?
Área de ocorrência
6) Quais são as plantas, ou produtos obtidos destas, mais importantes para a economia da sua casa, ou seja, aquelas que você depende para ganhar dinheiro ou fazer alguma troca por outros produtos?
Nome popular
De que forma
Por quanto é vendida
Para quem é vendida
Em que época é vendida
Onde ela ocorre?

112
APÊNDICE B – Pranchas coloridas com fotos das espécies de Araceae encontradas no entorno da comunidade Santa Maria.

113

114

115

116
APÊNDICE C – Atas da aula de qualificação e defesa presencial.

117

118
ANEXO A – Autorização para a pesquisa obtida através do Comitê de Ética em Pesquisas com Seres Humanos – CEP/INPA.

119
ANEXO B – Autorização de coleta em Unidade de Conservação Estadual.