identificaÇÃo de marcadores moleculares … · as aves dos céus, e os peixes do mar, e tudo o...
TRANSCRIPT

TESE
IDENTIFICAÇÃO DE MARCADORES MOLECULARES ASSOCIADOS À RESISTÊNCIA
À FERRUGEM MARROM ( Puccinia melanocephala) EM CANA-DE-AÇÚCAR
FERNANDA RAQUEL CAMILO DOS SANTOS
Campinas, SP 2013

INSTITUTO AGRONÔMICO CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
IDENTIFICAÇÃO DE MARCADORES MOLECULARES ASSOCIADOS À RESISTÊNCIA À FERRUGEM MARROM ( Puccinia melanocephala) EM
CANA-DE-AÇÚCAR
FERNANDA RAQUEL CAMILO DOS SANTOS
Orientadora: Dra. Maria Imaculada Zucchi Co-Orientadora: Dra. Luciana Rossini Pinto
Tese submetida como requisito parcial para obtenção do grau de Doutor em Agricultura Tropical e Subtropical Área de Concentração em Genética, Melhoramento Vegetal e Biotecnologia.
Campinas, SP Julho, 2013



Aos meus pais, irmãs e aos meus
cunhados por sempre estarem ao
meu lado.
DEDICO
À memória da Paula, que mesmo
ausente se faz lembrar todos os dias.
OFEREÇO

AGRADECIMENTOS
Agradeço a Deus.
A FAPESP – Fundação de Amparo a Pesquisa do Estado de São Paulo, pela concessão da bolsa de estudos. A CAPES – Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, pela concessão da bolsa sanduíche.
Aos meus pais Luli e Rubius, as minhas irmãs Rúbia e Fabíola e aos meus cunhados Zé e Thiago pela base, apoio, incentivo e cuidados. E a todos os meus familiares, especialmente minhas tias Iris, Mércia e Tereza, família querida, além das crianças Fausto e Mocinha, pra eles tudo é festa.
A Dra. Maria Imaculada Zucchi, pela orientação, confiança e amizade. Pelo otimismo e boa vontade com seus orientados. Fui afortunada com esta orientadora, e como dizem que o aluno “pega” o jeito do orientador, ficarei feliz tendo ao menos parte do carisma, empolgação e otimismo da Maria. Com as mesmas palavras agradeço minha co-orientadora Dra. Luciana Rossini, com quem tudo começou.
Ao Dr. Marcos Landell e todos do Centro de Cana – IAC que me recepcionaram de braços abertos, e as amizades que lá fiz em especial Bruna, Cibele, Letícia, Natália e Thaís, e Dra. Silvana Creste.
Ao Dr. José Baldin Pinheiro, pela amizade e otimismo e todos os colegas do Laboratório de Diversidade Genética e Melhoramento da ESALQ que fazem o dia mais gostoso, em especial aos amigos Alessandro Alves, Carlos Batista, Jaqueline Campos, Mariza Monteiro, Nathália Spagnol, Carol Grando e Fatinha Bosseti.
Aos funcionários de campo do Centro de Cana em Ribeirão Preto que me levavam para o campo coletar colmos e palmito e pelo transporte do meu material para Piracicaba. Aos funcionários de campo da APTA de Piracicaba pela ajuda na instalação do experimento e ao Daniel Sarto pela ajuda nas avaliações de campo.
Ao Dr. Jorge da Silva por ter me recepcionado tão bem em seu laboratório e a todos da Texas A&M University pelos meses de tão harmoniosa e alegre convivência, em especial para Denise Rossi, Jong-Won Park, Maribel Leon, Nora Solis, Thiago Benatti e Victoria Mora.
A Dra. Luciana Carlini-Garcia e Rodrigo Gazaffi pela colaboração com a análise e interpretação dos dados, além da empolgação quanto aos resultados que me animaram na reta final. Aos membros da banca: Dra. Mariângela Cristofani, Dra. Silvana Creste, Dra. Luciana Carlini e Dra. Raffaella Rosseto pelas contribuições e correções, ao verem coisas que quem escreveu já não vê mais.
Ao Dr. Comstock e todos da USDA (Canal Point – Florida), por terem me recebido e me permitido participar da rotina do melhoramento de cana-de-açúcar em seu estabelecimento, e pelo caminhão em miniatura que me deram de presente. E especialmente aos amigos Dra. Lucimeres e Andrew Mignoault que lá fiz.
Aos amigos de longa data, pelos contatos virtuais que fazem tão bem, e pelos encontros esporádicos que renovam as forças, Ana Luiza, Giovana Matos, Joana, João Guilherme, Michelle dos Santos, Paula Lima, Rafael Gudar, Regina Priolli, Thiago Fonseca, Vanessa Silveira e Fábio Paiva.
Aos amigos das Repúblicas nas cidades em que morei nestes quatro anos de nômade que tive, pelos cafés da tarde, jantares e bons momentos compartilhados: Aditi, Ana Luiza, Carolina Parra, Diogo, Mateus, Rafael, Tales e Thiago, e a lista só aumenta.
Meus sinceros agradecimentos a todos os amigos que fizeram parte destes anos e a todos que com boa fé contribuíam comigo diretamente ou indiretamente.

“Quando vejo os teus céus, obra dos teus dedos, a lua e as estrelas que preparaste;
Que é o homem mortal para que te lembres dele? E o filho do homem, para que o visites? Pois pouco menor o fizeste do que os anjos, e
de glória e de honra o coroaste. Fazes com que ele tenha domínio
sobre as obras das tuas mãos; tudo puseste debaixo de seus pés:
Todas as ovelhas e bois, assim como os animais do campo,
As aves dos céus, e os peixes do mar, e tudo o que passa pelas veredas dos mares.
Ó SENHOR, Senhor nosso, quão admirável é o teu nome sobre toda a terra!”
Salmos 8:3-9

SUMÁRIO
RESUMO ......................................................................................................................................... VII ABSTRACT .................................................................................................................................... VIII 1INTRODUÇÃO ................................................................................................................................. 1 2REVISÃO BIBLIOGRÁFICA .......................................................................................................... 4 2.1Origem e evolução da cana-de-açúcar ............................................................................................ 4 2.2Ferrugem marrom – Puccinia melanocephala ................................................................................ 5 2.3Ferrugem marrom e o melhoramento genético de cana-de-açúcar no mundo ................................ 7 2.4Resistencia de plantas e variabilidade genética de P. melanocephala ............................................ 8 2.5Marcadores Moleculares ............................................................................................................... 10 2.6Mapas Genéticos ........................................................................................................................... 13 2.7Mapeamento de QTLs ................................................................................................................... 15 2.8Estudos da resistência da cana-de-açúcar à ferrugem marrom ..................................................... 18 3MATERIAL E MÉTODOS ............................................................................................................. 20 3.1População de mapeamento ............................................................................................................ 20 3.2Desenho experimental ................................................................................................................... 21 3.3Instalação do experimento ............................................................................................................ 22 3.4Avaliação fenotípica ..................................................................................................................... 24 3.5Avaliações realizadas .................................................................................................................... 24 3.6Avaliações com Marcadores Moleculares .................................................................................... 25 3.6.1Extração e quantificação do DNA genômico ............................................................................. 25 3.6.2Marcadores EST-SSR ................................................................................................................ 26 3.6.3Marcadores AFLP-RGA ............................................................................................................ 27 3.6.4Marcadores CISP ....................................................................................................................... 28 3.6.5Desenvolvimento de primers ..................................................................................................... 30 3.6.5.1Marcadores EST-SSR ............................................................................................................. 30 3.6.5.2 Marcadores TRAP.................................................................................................................. 31 3.7Construção do mapa genético ....................................................................................................... 34 3.7.1Análises de segregação dos locos .............................................................................................. 34 3.7.2Construção do mapa de ligação ................................................................................................. 34 3.8Mapeamento de QTLs ................................................................................................................... 35 3.9Análise de Marca Individual ......................................................................................................... 36 4RESULTADOS E DISCUSSÃO ..................................................................................................... 36 4.1Avaliação fenotípica da resistência da ferrugem marrom ............................................................. 36 4.2Obtenção de Polimorfismo ............................................................................................................ 40 4.2.1EST-SSR .................................................................................................................................... 40 4.2.2AFLP-RGA ................................................................................................................................ 44 4.2.3CISP 45 4.2.4TRAP ......................................................................................................................................... 46 4.3Análises de segregação e formação dos grupos de ligação ........................................................... 48 4.4Análise de Marca Individual ......................................................................................................... 62 5CONCLUSÃO ................................................................................................................................. 78 6REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................................. 79

vii
SANTOS, Fernanda Raquel Camilo dos. Identificação de marcadores moleculares associados à resistência à ferrugem marrom (Puccinia melanocephala) em cana-de-açúcar. 2013. 101f. Tese (Doutorado) – Instituto Agronômico – IAC, Campinas.
RESUMO O Brasil é, segundo o FAO (Food and Agricultural Organization of United States, 2010),o principal
produtor mundial de cana-de-açúcar. É responsável por mais da metade do açúcar comercializado
no mundo e o mercado do etanol produzido proveniente da cana-de-açúcar cresce a cada ano, uma
vez que a política nacional para a produção dessa cultura se orienta na sua expansão sustentável,
com base em critérios econômicos, ambientais e sociais. Programas de melhoramento genético
buscam cultivares resistentes a estresses bióticos, abióticos e adaptadas às regiões produtoras, como
melhor forma de aumentar a produtividade sustentável da cultura. O genoma de Saccharum spp. é
complexo e desafiante para estudos genéticos, e a geração de mapas genéticos e identificação de
QTLs envolvendo características de interesse agronômico contribuem para gerar conhecimento
genético das espécies e para o desenvolvimento de tecnologias que possibilitem auxiliar programas
de melhoramento genético. Este trabalho objetivou obter novas marcas polimórficas para
construção de mapa genético da população segregante derivada do cruzamento biparental entre o
clone IACSP95-3018 e a cultivar IACSP93-3046 do programa de melhoramento genético do
Instituto Agronômico (IAC), além de identificar QTLs associados à resistência à ferrugem marrom.
Quinhentos e trinta e oito novos alelos polimórficos foram gerados, e somados a outros 824 alelos
anteriormente obtidos. O mapa genético foi composto por 113 grupos de ligação, variando de duas a
vinte marcas por grupo. As análises de QTL identificaram 177 marcas diferentes em associação
com a ferrugem, e 346 associações considerando seis avaliações fenotípicas. Cinquenta e nove
alelos tiveram alta associação (p<0,01) com a ferrugem, e uma marca foi associada com todas as
seis avaliações.
Palavras–chave: cana-de-açúcar, Puccinia melanocephala, marcador molecular, mapeamento genético, QTL.

viii
SANTOS, Fernanda Raquel Camilo dos. Identification of molecular markers related with brown rust resistance (Puccinia melanocephala) in sugarcane. 2013. 101f. PhD thesis – Instituto Agronômico – IAC, Campinas.
ABSTRACT
According to the FAO (Food and Agricultural Organization of United States, 2010) Brazil is the
most important producer of sugarcane. It is responsible for more than half of commercialized sugar
in the world and the market for ethanol, derived from sugarcane, rises every year, and the national
politics for sugarcane production has guided itself to sustainable expansion, based on economics,
environmental and social issues. Sugarcane breeding programs are always looking for genotypes
biotic and abiotic resistant, and adapted to the productive areas, as the best way to increase its
sustainable productivity. The Saccharum spp. genome is complex and a challenge for genetics
studies. The linkage studies, such as genetics maps and QTL helps
mapping generation and QTL identification involving important agronomic traits helps to increase
genetic information about sugarcane. This present work aim to create new polymorphic markers to
construct a genetic map from IACSP95-3018 clone vs IACSP93-3046 cultivar from the IAC
breeding program, and to identify QTL associated with brown rust resistance. Five hundred and
thirty nine polymorphic alleles were generated from different molecular markers, and other
polymorphic alleles from this same cross were added to create a map. Together, they formed 113
linkage groups, ranging from two to twenty markers per group. The QTL analysis identified 177
different markers associated with brown rust, and 346 associations when considering the six
different phenotypic evaluations that were done. Fifty-nine alleles had high association (p<0,01)
with brown rust, and one of them was associated with all six evaluations.
Key words: sugarcane, Puccinia melanocephala, molecular markers, genetic map, QTL.

1
1. INTRODUÇÃO
A história do Brasil é intimamente relacionada à cultura da cana-de-açúcar, que
foi trazida ao país na época da colonização e serviu, graças ao seu potencial em
armazenar sacarose, de carro-chefe nas fazendas de monocultura da época. Mais de
quinhentos anos depois, continua a ser uma das principais culturas do país, sendo
utilizada na produção de açúcar, etanol, energia e cachaça, além do potencial para
extração de etanol de segunda geração a partir do seu bagaço e palhada ou de variedades
com alto teor de biomassa. Embora seja o maior produtor mundial da cana o país
continua a importar etanol, a demanda pelo produto é alta e o preço do combustível
oscila frequentemente em função das safras anuais. Além disso, o interesse na cultura é
também crescente por se tratar de energia renovável e menos poluente quando
comparada ao petróleo.
Dentre todas as espécies cultivadas atualmente a cana-de-açúcar tem
indiscutivelmente o genoma mais complexo (GRIVET &ARRUDA, 2001). Os híbridos
atuais possuem número cromossômico bastante variável, entre 100 a 130 cromossomos,
com 80% destes cromossomos derivados de S. officinarum, 10% de S. spontaneum e
10% dos cromossomos recombinantes entre os dois genomas (D'HONT et al., 1996), e
uma vez que a instabilidade meiótica e o alto grau de poliploidia resultam na
esterilidade de muitos genótipos, impossibilita o desenvolvimento e uso de linhagens de
cana (JACKSON, 2005), conferindo um desafio especial tanto para o melhoramento
quanto para estudos genéticos da espécie (BUTTERFIELD, 2006).
O objetivo principal dos programas de melhoramento de cana-de-açúcar é
promover novas cultivares que ampliem a produtividade de açúcar e álcool (LANDELL
&BRESSIANI, 2008) e mais recentemente a biomassa (LOUREIRO et al. 2010). Tais
programas são realizados através de seleções de genitores, hibridização e seleção na
progênie de indivíduos superiores durante vários estágios de propagação clonal
(LANDELL &BRESSIANI, 2008). Os cruzamentos biparentais são, segundo
LANDELL &BRESSIANI (2008) os mais populares nas estações de cruzamento do
Brasil, embora policruzamentos sejam realizados para a produção de grande quantidade
de sementes.

2
O sucesso de um programa de melhoramento, além da variabilidade genética
existente na população base a ser explorada depende também da sua herdabilidade. A
maioria dos caracteres agronômicos de cana-de-açúcar é de herança quantitativa
(HOGARTH et a., 1987) e as análises de QTLs para características como conteúdo de
açúcar e de fibra têm indicado muitos QTLs de pequeno efeito (AITKEN et al., 2006;
HOARAU et al., 2002; MING et al., 2002; PIPERIDIS et al.; 2008; PINTO et al.,
2010). Entretanto existem alguns exemplos de herança simples em cana-de-açúcar,
como para a resistência à mancha ocular, causada pelo fungo Bipolaris sacchari
(MUDGE et al., 1996) e resistência para mancha amarela, causada pelo fungo
Mycovellosiella koepkei (ALJANABI et al. 2007).
A herança da resistência da cana-de-açúcar à ferrugem marrom, causada pelo
fungo Puccinia melanocephala H. & P. Sydew, não é totalmente esclarecida na
literatura, sendo ora considerada de herança qualitativa (TAI et al.1981); ora
considerada governada por um ou poucos genes de efeito maior (DAUGROIS et al.,
1996; ASNAGHI et al., 2000; 2001; RABOIN et al., 2001; COSTET et al., 2012) e
ainda relatada a existência de cultivares que possuem genes de efeito maior, atuando
qualitativamente, podendo estar associados a outros genes de efeito menor, que atuam
quantitativamente na reação à ferrugem marrom e dessa forma a resistência para esta
doença em cana seria conferida pela somatória desses efeitos, sendo a parcela principal
determinada pelos genes de efeito maior (RAMDOYAL et al., 1996; RAMDOYAL et
al., 2000; McINTYRE et al., 2005).
Prejuízos causados pela ferrugem marrom da cana-de-açúcar são consideráveis
e ocorrem quando o ambiente apresenta as condições favoráveis de temperaturas
amenas (até 30ºC), folhas umedecidas e variedades suscetíveis à doença (RAID &
COMSTOCK, 2000). Além disso, o fungo causador da ferrugem marrom apresenta
curto ciclo de vida, permitindo o desenvolvimento rápido de epidemias, sendo o plantio
de cultivares resistentes o único método econômico para o controle da doença.
A ferrugem marrom foi durante décadas a principal doença causada por fungo
de importância econômica na cana-de-açúcar, porém em dezembro de 2009, foi
constatada na região de Araraquara – São Paulo, outra ferrugem, a alaranjada, causada
pelo fungo Puccinia kuehnii (W. Krüger) E.J. Butler. Em 2010 a ferrugem alaranjada já
estava disseminada em diversos municípios de São Paulo e também em alguns do
Paraná, Mato Grosso do Sul, Espírito Santo, Goiás e Minas Gerais (BARBASSO,

3
2010), sendo ambas as ferrugens, marrom e alaranjada, as de maior importância para a
agricultura canavieira (VIRTUDAZO et al. 2001a).
Os marcadores moleculares são uma ferramenta rápida e eficaz para estudos
genômicos (FERREIRA &GRATTAPAGLIA, 1998), e em teoria, desde que revele
polimorfismo entre indivíduos, todo fragmento de DNA pode ser utilizado como
marcador molecular, e quando comparados aos marcadores morfológicos, oferecem a
vantagem de não sofrem influências do ambiente e do estádio fisiológico de
desenvolvimento das plantas (SOUZA, 2001).
A partir do polimorfismo detectado no DNA é possível empregar estes dados
para caracterização de genótipos, estimativas de diversidade genética, mapeamento
genético e fazer inferências sobre as relações entre o genótipo e o fenótipo dos
indivíduos, mapeamento locos de caracteres quantitativos (QTLs) envolvidos com
fenótipos de interesse. Segundo BUTTERFIELD (2006) a importância de características
de resistência a doenças e pragas aliada ao fato de que podem ser influenciados pela
variação ambiental os tornam alvos apropriados para seleção assistida por marcadores.
A geração de mapas genéticos e identificação de QTLs envolvendo
características de interesse agronômico contribuem, portanto, para gerar conhecimento
genético das espécies e desenvolvimento de tecnologias que possibilitem auxiliar
programas de melhoramento genético.
Por se tratar de uma planta poliploide, o mapeamento genético em cana-de-
açúcar é diferente daquele tradicional realizado para plantas diplóides, pois o padrão de
bandas obtido não permite a identificação do genótipo (da SILVA et al., 1995). O
método utilizado para mapeamento genético em poliplóides é baseado na segregação de
marcadores de dose única proposto por WU et al. (1992). Neste método as bandas são
interpretadas como marcadores dominantes, com presença ou ausência do fragmento,
ignorando o caráter co-dominante dos marcadores microssatélites e RFLP (Restriction
Fragment Length Polymorphism). Apesar de a população de cana-de-açúcar segregante
mais bem estudada até o momento para mapeamento genético ser oriunda de
autofecundação (R570) (DAUGROIS et al., 1996; HOARAU et a., 2001; ASNAGHI et
al., 2004), as populações segregantes utilizadas para mapeamento em cana-de-açúcar
são geralmente cruzamentos biparentais ou irmãos completos, isso devido à dificuldade
de autofecundação e formação de linhas homozigóticas (da SILVA et al. 1995; MING et
al., 2001).

4
Este trabalho objetivou obter novas marcas polimórficas para construção de
mapa genético utilizando a população segregante derivada do cruzamento biparental
entre o clone IACSP95-3018 e a cultivar IACSP93-3046 do programa de melhoramento
genético do Instituto Agronômico (IAC), e identificar marcadores associados à
resistência à ferrugem marrom. Para tanto, foram utilizados quatro tipos diferentes de
marcadores moleculares: AFLP-RGA (Amplified Fragment Length Polymorphism-
Resistance Genes Analogues), TRAP (Target Region Amplification Polymorphism)
EST-SSR (Expressed Sequence Tag - Single Sequence Repeat) e CISP (Conserved
Intron Scanning), todos obtidos a partir de sequências expressas do genoma de cana-de-
açúcar.
2 REVISÃO BIBLIOGRÁFICA
2.1 Origem e evolução da cana-de-açúcar
Acredita-se que a cana-de-açúcar seja originária do sudeste da Ásia, sendo o
exato centro de origem do gênero Saccharum desconhecido. Diversos autores têm
reportado possíveis locais, estando a Índia, Indonésia, Papua Nova Guiné, China e ilhas
da Polinésia entre as regiões mais citadas (MUKHERJEE, 1957).
A cana pertence à Tribo Andropogoneae, família das Poaceaes. Esta tribo
inclui gramíneas tropicais e subtropicais, incluindo os gêneros de cereais sorgo e milho
Zea (D'HONT et al. 1998). O Gênero Saccharum compreende seis espécies,
S. spontaneum, S. officinarum, S. robustum, S. edule, S. barberi, e S. sinense (D'HONT
et al. 1998). Variedades atuais são complexos híbridos interespecíficos (Saccharum
spp.), provenientes da hibridação interespecífica de duas espécies dentro do gênero
Saccharum, Saccharum officinarum L. (2n = 80) e Saccharum spontaneum L. (2n=40 –
128) (PANGE & BABU 1960, PRICE 1963).
A espécie S. officinarum é octaploide, tem 80 cromossomos, com número
básico x=10, enquanto S. spontaneum apresenta número básico de cromossomo x=8,
podendo ser classificados de pentaplóides até hexadecaplóides (D'HONT et al. 1998).
Segundo PANGE & SRINIVASAN (1957) plantas de S. spontaneum
apresentam alta variação fenotípica e adaptabilidade, podendo ser encontradas em
diversos ambientes como África, Europa Oriental, grande parte da Ásia, incluindo as
regiões de ilhas. Já a espécie S. officinarum pode ser encontrada apenas em jardins

5
varietais, pois não há registro de sua ocorrência na natureza, e acredita-se ter sido
selecionada pelo homem de formas mutantes de S. robustum (x=10, 2n = 6x = 60),
espécie predominante na Nova Guiné (STEVENSON, 1965).
S. officinarum ou a "cana nobre" acumula níveis muito elevados de sacarose no
colmo, mas tem baixa resistência a doenças. Ao contrário, S. spontaneum, que acumula
pouca sacarose, é uma espécie altamente polimórfica, com níveis mais elevados de
resistência a doenças, adaptabilidade e tolerância a estresse (SREENIVASAN et al.
1987; MING et al. 2006).
Segundo D´HONT et al. (1996) a cultivar R570 (2n = 115) apresentou ter,
através de avaliação por GISH, 10% de dos cromossomos provenientes de S.
spontaneum e 80% por S. officinarum, e outros 10% eram claramente provenientes da
recombinação destas duas espécies, com poucos casos de dupla recombinação. Já na
cultivar NCo376 as proporções eram de 20:70:10, respectivamente.
O evento de hibridização que deu origem à nobilização das atuais cultivares de
Saccharum spp. envolveu S. spontaneum (evento mais comum de 2n=64, segundo
IRVINE 1999) como genitor masculino e S. officinarum (2n=80) como genitor
feminino. No cruzamento acredita-se que o gameta feminino não sofreu a redução
meiótica, gerando a progênie com 2n+n cromossomos (80+32 = 112). O
retrocruzamento da geração F1 voltado para S. officinarum resultou novamente na
transmissão 2n+n (80+56 = 136), mas nas próximas gerações a transferência de n foi
revertida ao normal (40+68 = 108), sendo que as canas nobilizadas apresentam o
número cromossômico variando de 100 a 130, com contribuição de 5 a 10% de S.
spontaneum (MING et al. 2006).
2.2 Ferrugem marrom – Puccinia melanocephala
A ferrugem marrom é causada pelo fungo basidiomiceto Puccinia
melanocephala H. & P. Sydew. Desde seu primeiro relato na Índia na década de 1950 a
ocorrência da ferrugem acompanha a das regiões canavieiras mundiais, estando presente
nos continentes Africano, Asiático, Americano e da Oceania; porém somente em 1978,
quando introduzida no Caribe foi considerada realmente uma doença de importância
econômica (RALOFF, 2001). Existem relatos de perdas de produção chegando a mais
de 50% no México em 1981-1982 (PURDY et al. 1983; COMSTOCK 1992), existe

6
ainda relatos de reduções na produtividade da ordem de sete e 10 toneladas por hectare
(SILVA et al., 2001).
No Brasil, a ferrugem marrom da cana-de-açúcar foi constatada pela primeira
vez no Município de Capivari (SP) em 1986 e se espalhou rapidamente por toda a
região Centro-Sul do país, chegando à região Nordeste, nos Estados de Pernambuco e
Alagoas já no ano seguinte (AMORIM 1987).
Os sintomas causados pela ferrugem marrom na cana-de-açúcar são
semelhantes aos sintomas de ferrugem em outras culturas, sendo limitados às partes
foliares da planta. Inicialmente são formadas manchas alongadas e amareladas, paralelas
às nervuras que vão aumentando de tamanho e adquirindo coloração vermelho-
castanho, devida à formação de urédias. Com o rompimento das urédias ocorre a
formação de pústulas, sintoma típico das ferrugens. Em variedades altamente suscetíveis
as pústulas agrupam-se e quando estas coalescem ocorre a necrose dos tecidos,
geralmente nas folhas mais baixas, reduzindo a vida da folha que perde sua função
fotossintética, prejudicando o crescimento e consequentemente, a produção de cana-de-
açúcar (SACILOTO, 2003). Para efeito de ilustração uma foto das ferrugens marrom e
alaranjada é apresentada na Figura 1.
Figura 1Folhas de clones de cana-de-açúcar com ferrugem marrom (folha superior) e ferrugem alaranjada (folha inferior). Canal Point, FL, EUA, 2013.
A ferrugem marrom é favorecida por temperaturas amenas a não muito altas
(17 a 30°C) e alta umidade relativa (>70%). O fungo tem curto ciclo de vida, permitindo
o aparecimento de epidemias dentro de 5 a 6 semanas, e a cultura, de aspecto
FRCSantos, 2013

7
inicialmente verde, se torna avermelhada devido à formação maciça de pústulas na
superfície abaxial das folhas (RYAN & EGAN, 1989).
Uma sequência de eventos ocorre para o estabelecimento da ferrugem marrom
na cana-de-açúcar, começando pela germinação dos urediniósporos que desenvolve um
tubo germinativo, este ao entrar em contato com um estômato forma um apressório,
formando uma alça de infecção que adentra a folha através do estômato e passando
pelas células-guardas irá formar uma vesícula subestomática, esta desenvolve hifas
infectivas que colonizam e se desenvolvem no mesófilo foliar.
A ponta da hifa infectiva se torna septada e origina uma estrutura chamada de
célula mãe do haustório que se posiciona ao lado da parede celular de uma célula do
mesófilo. A parede celular da célula é penetrada e ocorre a formação do haustório
(PURDY et al., 1983). Os urediniósporos, estruturas responsáveis pela disseminação e
infecção da doença, são dispersos pelo vento e respingos de chuva, e o transporte de
toletes contaminados é responsável pela disseminação a grandes distâncias.
2.3 Ferrugem marrom e o melhoramento genético de cana-de-açúcar no
mundo
A avaliação da ferrugem é geralmente realizada in loco por técnicos, que
percorrem o canavial observando as folhas e atribuindo uma nota segundo uma
determinada escala. Esta nota indica quão suscetível é a cultivar à ferrugem, de acordo
com a percentagem da concentração das pústulas (TAI et al., 1981; AMORIM et al.,
1987). TAI et al. (1981) descreveram a resistência à ferrugem marrom em cana-de-
açúcar de herança não aditiva, porém HOGARTH et al. (1983; 1993) mostram que a
maioria da variância genética é aditiva.
A herdabilidade da resistência à ferrugem marrom é alta (COMSTOCK et al.
1992; HOGARTH et al. 1993; RAMDOYAL et al. 2000), portanto a eliminação de
parentais suscetíveis é uma estratégia adequada para o desenvolvimento de variedades
resistentes. Diante disso a seleção de genótipos resistentes à ferrugem marrom ocorre
nos estágios iniciais de um programa de melhoramento genético.
Algumas cultivares que eram consideradas como resistentes, podem apresentar
um comportamento fora do padrão esperado dependendo da época ou local de cultivo.
Um exemplo é a cultivar RB83-5486, tida como resistente, mas que atualmente nos
estados de São Paulo, Espírito Santo e Minas Gerais vem sofrendo severos ataques de

8
ferrugem e sendo considerada de resistência intermediária à susceptível (SANTOS,
2008).
Também a cultivar NA5679, considerada como resistente em Pernambuco,
apresenta susceptibilidade no Espírito Santo e a cultivar SP79-1011 é considerada
susceptível no Centro-Sul e resistente no Nordeste. Assim testes de resistência devem
ser realizados em nível local, diretamente na região de plantio, objetivando identificar as
cultivares mais indicadas para as condições locais (SANTOS, 2008).
O controle da ferrugem é realizado através de cultivares resistentes,
encontrados nas áreas tradicionais de cultivo. A maioria das cultivares lançadas até o
momento são resistentes ao fungo, das 69 principais cultivares lançadas no Brasil,
apenas 14 são consideradas suscetíveis à P. melanochepala (SANTOS, 2008). O
crescimento da colheita mecânica (com ausência de queimadas), tende a favorecer o
aparecimento de novos patógenos além do aumento de doenças consideradas mantidas
sob controle.
2.4 Resistencia de plantas e variabilidade genética de P. melanocephala
Na natureza, a resistência de plantas a doenças é regra ao passo que a
suscetibilidade é exceção, pois há uma infinidade de microrganismos aos quais as
plantas estão diariamente expostas, e apenas uma pequena porção desses
microrganismos é capaz de causar doenças em plantas. Esse fato é garantido por
mecanismos pré-formados (espessura da cutícula, número e disposição de estômatos e
tricomas) e pós-formados (substâncias químicas como fungitóxicas e antiproteicas)
atuantes nessas plantas e que em conjunto é chamada de resistência de planta não
hospedeira (PASCHOLATI, 1995). Além destas barreiras físicas e químicas as plantas
possuem ainda um sistema de defesa que possibilita a identificação de microrganismos
patogênicos.
Quando determinado patógeno se torna adaptado a uma espécie de planta este
suprime, através de fatores de virulência, parte dos mecanismos de defesa da planta.
Porém quando a planta hospedeira possui genes específicos de resistência que a tornam
capazes de detectar a presença do patógeno e suprimir os seus fatores de virulência, o
estabelecimento deste na planta não ocorre. A evolução do patógeno ocorre então
quando ele se torna capaz de se estabelecer na planta sem que esta detecte sua presença
(BENT & MACKEY, 2007).

9
As interações plantas-patógenos são co-evolutivas, marcadas por relações
geneticamente controladas por patógenos e seus hospedeiros. Assim, quando ocorrem
modificações genéticas em um dos componentes, podem ocorrer modificações genéticas
na população do outro (CAMARGO, 1995). Uma metáfora popular na literatura vê a
co-evolução de genes R de resistência na planta e Avr de avirulência no hospedeiro
como uma corrida armamentista. Nesta, um gene R é derrotado como resultado de uma
mutação no gene de avirulência correspondente, que permite ao patógeno escapar ao
reconhecimento pelo hospedeiro (ELLIS et al., 2000), esta interação permite então que
novos níveis de relação entre esses organismos possam ser gerados.
Genes que conferem resistência a uma ampla variedade de doenças causadas
por vírus, bactérias, fungos e nematoides tem sido clonados e estudados (HULBERT et
al. 2001). Tais genes de resistência apresentam motivos conservados, que participam de
mecanismos de transdução de sinal, e as proteínas de resistência que eles codificam
apresentam diferentes domínios, sendo cinco principais :os que contêm regiões de
ligação de nucleotídeos (NBS – Nucleotide Binding Site) e regiões repetitivas ricas em
leucina (LRR – Leucine Rich Repeat) (NBS-LRR), domínios compostos por regiões
LRR e transmembranares (TM), domínios compostos por regiões LRR e proteína
quinase, domínio simples de proteína quinase (Pto) e o domínio serina-treonina quinase
(STK) (BAKER et al. 1997; HAMMOND & KANYUKA, 2007). É possível observar a
presença do domínio LRR atuando em vários dos domínios relacionados à resistência a
patógenos, e a maioria dos genes de resistência clonados até o momento codifica para
proteínas pertencentes aos domínios NBS e LRR (ELLIS et al. 2000; DANGL &
JONES 2001; MICHELMORE & MEYERS, 1998).
O domínio LRR consiste de uma sequência repetitiva de motivos, em média de
24 aminoácidos, de leucina e ocasionalmente com resíduos de asparagina e prolina em
regiões conservadas. Tal domínio tem sido proposto como participante em interações
proteína-proteína (KOBE &DEISENHOFER 1994) e é um candidato para funções de
reconhecimento patógeno-hospedeiro (TAKKEN &JOOSTEN 2000). Análises
genéticas têm demonstrado que genes análogos de resistência (RGAs) são amplamente
distribuídos no genoma, geralmente organizados em cluster e algumas vezes fortemente
ligados a locos de resistência (MEYERS et al. 1999).
A mutação é a mais importante fonte de variabilidade genética em fungos
fitopatogênicos (CAMARGO, 1995) e podem ser mutações gênicas ou cromossômicas.
As mutações gênicas podem ocorrer por substituição, deleção, e inversão de um ou mais

10
segmentos de DNA. As mutações cromossômicas correspondem à quebra de fragmentos
cromossômicos e podem ser deleções de segmentos, inserções e duplicações, inversões
e translocações, ocorrendo a troca de segmentos entre cromossomos não homólogos
(PASSAGLIA, 2003).
A existência de diferentes raças fisiológicas de P. melanocephala não é um
consenso, um estudo realizado na Flórida sugeriu a existência de quatro raças
patogênicas, levantando a hipótese de que variantes patogênicas surgem nas regiões
produtoras e são paralelas ao desenvolvimento de novas cultivares (SHINE; et al. 2005).
Porém ASNAGHI et al. (2001) demonstraram que o principal gene de resistência
relacionado à ferrugem marrom provoca resistência contra vários isolados coletados em
diferentes áreas geográficas, e ASNAGHI et al. (2004) relatam que a quebra de
resistência à ferrugem não foi observada nas Ilhas Reunião e em vários outros lugares
nas últimas décadas, apesar do intensivo cultivo da cultivar R570 nestes locais.
2.5 Marcadores Moleculares
Os marcadores moleculares passaram a ser utilizados em estudos genéticos de
plantas na década de 1980. Uma variedade de marcadores moleculares tem sido
utilizada na busca por polimorfismo, como os SSRs (single sequence repeat) ou
microssatélites, EST-SSR (Expressed Sequence Tag-SSR), AFLP (Amplified Fragment
Lenght Polymorphism), AFLP-RGA (AFLP-Resistence Gene Analogue), TRAP (Target
Region Amplification Polymorphism), e CISPs (Conserved Intron Scanning).
Marcadores microssatélites consistem de pequenas sequências com 1 a 6
nucleotídeos de comprimento repetidos em tandem, que ocorrem naturalmente no
genoma. São marcadores codominantes e sua frequência é relativamente alta
(FERREIRA & GRATTAPAGLIA, 1998). Acredita-se que durante a replicação de uma
região repetitiva, as fitas de DNA separam-se e unem-se novamente de forma incorreta,
o que geraria cópias de trechos de alelos com diferentes tamanhos ou números de
repetições, por meio da inserção ou deleção de uma unidade de repetição. O
polimorfismo também pode estar associado ao sistema de reparo pela DNA polimerase
ou ainda, ser consequência do processo de recombinação (FIELD & WILLS, 1996).
Outro fator que também pode ser responsável pela alta taxa de polimorfismo destes
marcadores é o crossing-over desigual, que por problemas no pareamento dessas

11
sequências durante o quiasma, aumenta a taxa de mutação das regiões microssatélites
(SCHLÖTTERER et al., 1998).
As regiões que contém as sequências simples repetidas são amplificadas por
PCR (Polimerase Chain Reaction) utilizando-se um par de primers específicos que
flanqueiam o microssatélite, e que geralmente são conservadas em indivíduos da mesma
espécie. Esta técnica revela polimorfismo em um loco devido a diferenças no número de
vezes (n) em que, por exemplo, um dinucleotídeo (AG)n se repete naquele loco. Cada
segmento amplificado de tamanho diferente representa um alelo diferente do mesmo
loco e, por este motivo, os marcadores microssatélites são multialélicos (FERREIRA &
GRATTAPAGLIA, 1998). Marcadores EST-SSRs são uma variação dos marcadores
SSR, onde os primers específicos são desenvolvidos para regiões microssatélites a partir
de sequências de etiquetas expressas e não de sequências genômicas como no SSR
tradicional (CORDEIRO et al. 2001). Tais marcadores podem facilitar a identificação
de regiões que controlam características agronômicas de interesse e são considerados
ideais para seleção assistida (CATO et al. 2001; MA et al. 2004). Em cana-de-açúcar,
marcadores do tipo microssatélites derivados das etiquetas de sequências expressas
(EST-SSRs) foram utilizados por OLIVEIRA et al. (2007), MACCHERONI et al.
(2009) e PINTO et. al (2010).
O AFLP baseia-se na amplificação do DNA via PCR para detectar diferenças
em um conjunto de fragmentos selecionados e digeridos com enzimas de restrição
(CAIXETA et al., 2006) e à praticidade da amplificação por PCR. A técnica baseia-se
na digestão do DNA genômico através de duas enzimas de restrição e na ligação de
adaptadores específicos complementares às regiões coesivas das enzimas utilizadas. Os
fragmentos gerados são então amplificados via PCR por primers pré-seletivos com
sequências complementares à dos adaptadores, objetivando aumentar a amplificação de
sequências; e por fim é realizada uma amplificação final através de primers que contém
nucleotídeos seletivos na sua extremidade 3´ (VOS et a. 1995).
Os marcadores AFLP-RGA são uma modificação na técnica de AFLP onde um
primer da amplificação seletiva do AFLP é substituído por um primer RGA,
degenerado ou não, podendo ser desenvolvido à partir de diversas regiões de genes de
resistência, incluindo regiões de domínios de ligação a nucleotídeo (NBS) ou de
domínio rico em repetições de leucina (LRR), e sua utilização pode ser útil na seleção
de marcas associadas a genes de resistência (SORIANO et al. 2005; ZHANG et al.
2007). Outra técnica de AFLP modificado foi proposta anteriormente, por HAYES

12
&MAROOF (2000), no qual a amplificação seletiva, diferente da técnica de AFLP
tradicional de VOS et al.(1995) utiliza um primer proveniente de região exclusivamente
NBS. Segundo ZHANG et al. (2007) uma vantagem do AFLP-RGA em relação à
técnicas como NBS-profiling (Van der LINDEN et al., 2004) e à técnica proposta por
HAYES & MAROOF (2000) é a da utilização de primers provenientes também de
regiões LRR, que são menos conservadas do que as regiões NBS e portanto mais
prováveis de haver polimorfismos. O AFLP-RGA, ainda segundo ZHANG e
colaboradores (2007) oferece grande flexibilidade para inúmeras combinações de
primers na varredura do genoma para RGAs, e, além disso, a distribuição dos RGAs ou
dos clusters de RGA no genoma das plantas amplia a cobertura da técnica AFLP-RGA,
representando um recurso de grande utilidade para o mapeamento de genes candidatos
para resistência a doenças em plantas. Marcadores RGA foram utilizados em cana por
ROSSI et al. (2003) e McINTYRE et al. (2005).
Há cerca de uma década atrás, HU & VICK (2003) desenvolveram a técnica
dos marcadores TRAPs, justificando que naquela época a tecnologia do sequenciamento
já estava gerando enormes quantidades de informação de sequências para várias
espécies, e utilizaram então ferramentas de bioinformática e ESTs para gerar
marcadores polimórficos de sequências de genes candidatos. O polimorfismo é gerado a
partir da combinação de um primer fixo e desenhado a partir de uma sequência EST de
interesse, e um primer reverso arbitrário, seguindo as especificações de LI & QUIROS
(2001) na técnica do marcador SRAP, onde um primer era constituído de CCGG para
anelar preferencialmente em regiões exônicas, e o outro primer, reverso, contém a
sequência AATT, para anelar preferencialmente em regiões com AT, frequentemente
encontradas em regiões promotoras e de íntrons, mais polimórficas que as regiões
exônicas. O polimorfismo gerado por marcadores TRAPs tem sido considerado
eficiente em estudos de diversidade genética de espécies componentes do complexo
Saccharum e cultivares de cana-de-açúcar (ALWALA et al. 2006; CRESTE et al. 2010;
SUMAN et al. 2012).
Conserved Intron Scaning Primers (CISPs) são marcadores moleculares que
exploram regiões genômicas de espécies que ainda são sub-caracterizadas, com a
finalidade de identificar polimorfismo em regiões intrônicas (FELTUS et al. 2006). Os
CISPs se utilizam da disponibilidade de genomas sequenciados de espécies modelo,
usando-as como base para espécies órfans de sequenciamento. Ele é baseado em
primers específicos obtidos de sequências EST do DNA de interesse. Os ESTs são

13
alinhados com um genoma modelo relacionado ao da espécie a ser estudada, e a partir
disso são desenhados primers em regiões exônicas altamente conservadas entre as duas
espécies e que flanqueiem regiões intrônicas observadas no genoma modelo. FELTUS
et al. (2006) relatam a técnica como eficiente tanto para explorar polimorfismo de DNA
quanto sequências conservadas não codificantes do genoma ou de genes candidatos,
além de servir como âncora para genômica comparativa entre diversas espécies. PARK
et al. (2012) desenvolveram cinco pares de primers CISPs específicos para o complexo
Saccharum utilizando o genoma do sorgo (Sorghum bicolor) como modelo para
identificar regiões intrônicas entre sequências ESTs conservadas. Os primers
denominados Ifs foram desenvolvidos para os seguintes genes envolvidos na biossíntese
da lignina: IF1 –Phenylalanine ammonialyase (PAL), IF3 - 4-coumarate coenzyme A
ligase (4-CL), IF5 - caffeoyl-CoA 3-O-methyltransferase (CCoAOMT), IF6 -
cinnamoyl-CoAreductase (CCR) e IF11 - peroxidase (POX). A lignina é um composto
secundário presente na parede celular de plantas, que além de ser importante para o
crescimento e desenvolvimento destas, participa também na resistência à estreses
biótico e abiótico da planta (BI et al., 2010).
2.6 Mapas Genéticos
A ordenação linear de marcadores genéticos em grupos de ligação constitui um
mapa de ligação. Para ordenar estas marcas STURTEVANTEM (1913), propôs a
utilização da frequência de recombinação como uma medida da distância entre dois
genes, permitindo a construção de mapas, nos quais a posição relativa dos marcadores
fenotípicos de um determinado organismo poderia ser representada. Apesar de o
mapeamento ter sido pensado há muitos anos, apenas a partir da década de 1980,
quando as técnicas de biologia molecular tornaram a utilização de marcadores
moleculares possíveis, que a criação de mapas genéticos tornou-se realidade
(BOTSTEIN et al., 1980). CARNEIRO &VIEIRA (2002) produziram uma revisão
bibliográfica sobre mapas genéticos em plantas, relatando desde os poucos primeiros
mapas produzidos a partir de marcadores morfológicos, passando pelas isoenzimas e
chegando aos variados mapas desenvolvidos utilizando-se os marcadores moleculares.
As populações de mapeamento são usualmente derivadas do cruzamento de
linhagens endogâmicas originando populações F2, de retrocruzamento, RIL (linhagens
puras recombinantes) e conjunto de linhagens duplo haploides. Nessas populações o
desequilíbrio de ligação é maximizado permitindo a análise de ligação. Uma vantagem

14
do uso de populações experimentais diploides é que existem somente dois alelos
segregando para cada loco e a fase de ligação desses alelos é conhecida.
Porém há espécies em que a obtenção de linhagens puras não é viável, devido à
forte depressão por endogamia e/ou longo período entre gerações. Dentro deste contexto
destaca-se a cana-de-açúcar, cuja depressão por endogamia é acentuada pelo elevado
nível de ploidia, e as populações segregantes são obtidas a partir de genitores não
endogâmicos. Nestes casos torna-se impossível identificar o genótipo a partir do padrão
de bandas obtido (da SILVA et al. 1995), pois os locos podem estar presentes em
dosagens variadas, desde nuliplex, que ocorre quando todos os alelos são nulos
(aaaaaaaa), simplex – quando apenas um alelo é informativo (Aaaaaaaa) até multiplex
(da SILVA & SORRELLS, 1996).
GRATTAPAGLIA & SEDEROFF (1994) propuseram a abordagem de
mapeamento denominada de duplo pseudo-testcross para gerar mapas genéticos de
espécies outcrossing (que não formam linhagens endogâmicas), eucalipto na ocasião do
estudo. O nome duplo do teste é devido ao fato de que após a genotipagem da
população segregante são selecionados para mapeamento apenas locos que segregam na
proporção 1:1, como nos cruzamentos entre linhagens. Neste mapeamento são gerados
dois conjuntos distintos de marcadores, sendo um para cada genitor do cruzamento,
resultando em dois mapas de ligação. Entretanto, tanto do ponto de vista biológico
como estatístico é mais interessante a obtenção de um mapa com as informações
integradas (GARCIA et al., 2006). Isso só pode ser feito com a presença de marcadores
em heterozigose em ambos parentais, que são utilizados para estabelecer relações de
ligação entre os marcadores segregando individualmente em cada parental (WU et al.,
2000).
WU et al. (2002) propuseram um método de mapeamento integrado dos
genitores, baseado em análises de máxima verossimilhança que permite estimar a fração
de recombinação e as fases de ligação entre locos em populações de mapeamento não
endogâmicas. Nesta abordagem são utilizadas para o mapeamento aquelas marcas que
apresentam dois tipos de segregação na progênie, na proporção de 1:1, que ocorre
quando uma única cópia do alelo está presente em um dos genitores e ausente no outro
(Aaaaaaaa x aaaaaaaa) e na proporção 3:1, que ocorre quando apenas uma única cópia
de um alelo está presente em ambos os genitores (Aaaaaaaa x Aaaaaaaa). A integração
dos mapas com segregação 1:1 só pode ser feito com a presença de alelos em comum
em ambos parentais, que são utilizados para estabelecer relações de ligação entre os

15
marcadores que segregam individualmente em cada parental. Esse tipo de mapa
integrado por marcadores que funcionam como pontes entre as marcas de segregação
1:1 apresentam ainda a vantagem do aproveitamento de mais marcas e consequente
maior saturação do mapa (WU et al., 2002; GARCIA et al., 2006).
GARCIA et al. (2006) comprovaram a melhora dos resultados ao utilizar esta
abordagem de WU et al. (2002) através do software OneMap (MARGARIDO et al.
2007) que tem implementado os algoritmos propostos por WU et al. (2002). A partir da
versão 2.0 (MARGARIDO et al., 2011) o software passou a utilizar a abordagem
multiponto (WU et a., 2002b) estimando simultaneamente a fração de recombinação e
as fases de ligação entre os marcadores por verossimilhança usando o modelo
markoviano oculto (HMM ou Hidden Markov Model).
Para o gênero Saccharum mapas de ligação vêm sendo construído ao longo das
últimas três décadas. Para S. spontaneum utilizando marcadores RFLP, RAPD (Al
JANABI et al., 1993; SOBRAL & HONNEYCUT1993; da SILVA et al., 1995), entre
cruzamentos de S. officinarum e S. robustum usando RAPD, RFLP e AFLP
(GUIMARÃES et al., 1999), e para Saccharum officinarum (MUDGE et al. 1996;
MING et al., 1998; MING et al., 2002). Alguns mapas genéticos foram feitos também
para cultivares de cana, como para a autofecundação da cultivar elite R570 de cana-de-
açúcar (GRIVET et al., 1996; HOARAU et al., 2001); para cultivares SP80-180 e SP80-
4966 (GARCIA et al. 2006; OLIVEIRA et al., 2007; PASTINA et al. 2012); e
recentemente um mapa para a cultivar LCP 85-383 popular em Lousiana foi construído
(SUMAN et al., 2011) e PALHARES et al., (2012) desenvolveram um mapa entre
clones IAC66-6 e TUC71-7.
A construção de mapa genético integrado foi utilizada por GARCIA et al
(2006), que comprovou a superioridade do método, após, OLIVEIRA et al. (2007) e
PALHARES et al. (2012) também utilizaram a técnica, todos através do software
OneMap (MARGARIDO et al., 2007; MARGARIDO et al., 2011).
2.7 Mapeamento de QTLs
Estudos de QTLs em plantas começam com a construção de uma população de
mapeamento através da escolha dos genitores, que difiram em uma ou mais
características de interesse. O mapeamento de QTLs consiste no procedimento de
estimar o número de regiões que controlam a variação dos caracteres em estudo, assim
como suas respectivas posições, efeitos e interações (ZENG et al., 1999). A grande

16
maioria das características herdáveis de importância econômica são quantitativas. Os
fenótipos resultantes apresentam uma variação contínua na população em vez de classes
discretas. Esse tipo de herança é determinado pela segregação de vários locos, cada um
com um pequeno efeito no fenótipo e sofrendo uma grande influência ambiental
(TANKSLEY, 1993).
A detecção e mapeamento de QTL são possíveis através de análise
concomitante de marcadores que apresentem polimorfismo e de características
fenotípicas contrastantes dos indivíduos. Assim como no mapeamento genético, o
princípio das análises de mapeamento de QTL é a existência de desequilíbrio gamético
de ligação decorrente da redução da frequência de recombinação entre genes situados
em regiões próximas entre si ao longo de determinado cromossomo, de outro modo os
alelos e regiões de QTL ocorrerão com distribuição independente, impossibilitando a
localização dos mesmos (COELHO, 2000).
As abordagens genético-estatísticas criadas para identificação de QTL foram
recebendo sucessivas melhorias ao longo dos anos, existindo atualmente de simples
abordagens até testes complexos. O método mais simples é o da análise de marca
individual, baseada em testes estatísticos como o teste t, análise de variância, regressão
linear simples (EDWARDS et al., 1987; STUBER et al., 1987) e teste da razão de
verossimilhanças (WELLER, 1986).
Tais abordagens testam a hipótese nula de que a média do caráter é
independente do genótipo do marcador. Quando esta hipótese é rejeitada considera-se
que pelo menos um QTL esteja ligado à marca em questão. Porém a Análise de Marca
Individual não permite a distinção entre um QTL de pequeno efeito, mas que esteja
situado muito próximo ao marcador, de um QTL que apresenta grande efeito, porém
que está situado distante do marcador, limitando a técnica mais à detecção do que
propriamente ao mapeamento de QTL (LYNCH &WALSH, 1998) já que se espera de
um mapeamento de QTL a posição no genoma e a estimação de seus efeitos. Além
disso, relacionar o QTL a uma marca específica tem a desvantagem da possibilidade da
ocorrência de recombinação entre a marca e o QTL (TANKSLEY, 1993).
Outra abordagem é a do mapeamento de intervalo simples (IM – Inverval
Mapping) proposto por LANDER & BOTSTEIN (1989), que utiliza o princípio da
máxima verossimilhança para verificar a possível presença de um QTL na região entre
um par de marcas adjacentes em um grupo de ligação. Assim, em um intervalo, várias
posições são testadas para a presença de um QTL. Porém o IM não permite eliminar de

17
forma satisfatória o surgimento dos chamados QTL fantasmas. Estes QTLs surgem
quando um QTL está localizado em intervalos adjacentes ao intervalo que está sendo
mapeado, fazendo com que os testes de hipótese apresentem falsos positivos. Isso
ocorre porque os métodos de IM não consideram todos os marcadores simultaneamente,
usando apenas dois deles ao mesmo tempo (marcas flanqueadoras), sendo difícil
diferenciar entre o efeito de um QTL verdadeiro e o efeito dos QTL fantasmas (LYNCH
&WALSH, 1998).
O mapeamento por intervalo composto ou CIM (Composite Interval Mapping),
proposto por ZENG (1993; 1994), combina mapeamento por intervalos simples e
regressão linear múltipla. Este método concentra os testes em uma região genômica por
vez, sendo que a estimativa da localização dos QTLs e seus efeitos ficam sem viés, pois
os marcadores podem ser utilizados como delimitadores para deixar mais clara a
posição dos QTLs, através de um adensamento, que pode ser feito por uma abordagem
de agrupamentos segregantes (LIU, 1998). Em CIM cofatores são usados para reduzir a
variação residual ao servirem de limite de cumprimento da análise, controlando assim o
background genético de outras marcas.
Finalmente o Mapeamento de Múltiplos intervalos (MIM – Multiple Inteval
Mapping, KAO &ZENG, 1997; KAO et al., 1999) utiliza o intervalo de múltiplos
marcadores simultaneamente para ajustar vários prováveis QTLs diretamente no modelo
para mapeamento de QTL. Essa abordagem aumentou a precisão e o poder da detecção
de QTLs, além de que a ocorrência de epistasia entre QTLs e valores de herdabilidade
dos caracteres quantitativos estudados pode ser analisada.
Todos estes métodos de mapeamento de QTLs apresentam em comum assumir
de que os fenótipos avaliados apresentam distribuição normal com variância igual em
ambos os pais. Sob estas condições o QTL pode ser testado por um simples teste
paramétrico (teste t no caso de análise de marca simples e LOD score no caso de
mapeamento por intervalo) (KRUGLYAK & LANDER, 1995), Em casos não
paramétricos considera-se que tais métodos não podem ser aplicados diretamente, e uma
abordagem de transformação deve ser utilizada para converter os dados fenotípicos em
uma distribuição mais próxima da normal (WRIGHT, 1968).
Porém algumas vezes não existem transformações que sirvam para a
transformação dos dados, ou mesmo que sim, os dados outliers podem ainda ser muito
dispersos. Existem ainda algumas alternativas, como a proposta por KRUGLYAK &
LANDER (1995) ou a de LI et al. (2006) que afirma que quando a característica

18
observada para identificação de QTLs apresenta cinco classes ou mais os dados podem
ser tratados como contínuos, sem a necessidade de transformação.
Assim como para a construção de mapas genéticos, a identificação de QTLs
em espécies não endogâmicas é complexa, e com agravantes para espécies poliploides,
esforços vêm sendo feitos por grupos de pesquisa genético-estatísticas para a
implementação de modelos que permitam a identificação de QTLs mais robustas que as
utilizadas atualmente. Em cana-de-açúcar trabalhos de mapeamento de QTLs têm sido
desenvolvidos, utilizando abordagens de Análise de Marcas Individuais ou Mapeamento
por Intervalo. MING et al. (2001, 2002), detectaram 36 QTLs relacionados à produção
de açúcar, número de colmo, conteúdo de fibra e cinzas. PINTO et al. (2010) detectaram
120 associações entre as características agronômicas de fibra, produtividade, POL, e
tonelada de cana-de-açúcar por hectare.
Marcas associadas à resistência a ferrugem marrom foram primeiramente
obtidas por GRIVET et al. (1996) que identificaram uma sonda RFLP ligada a um gene
de resistência a ferrugem com segregação mendeliana 3:1 na progênie, que foi
confirmado também por DAUGROIS et al. (1996).
Anos mais tarde três sondas RGA foram associadas à resistência a ferrugem
(McINTYRE et al. 2005a). McINTYRE et al. (2005b) trabalhando com marcadores do
tipo microssatélites genômicos e AFLP para a identificação de marcadores associados à
ferrugem marrom em cana-de-açúcar encontraram duas regiões genômicas, uma
proveniente do mapa referente ao genitor feminino e outra do genitor masculino, além
de um marcador não ligado de ambos os genitores associados com a resistência à
ferrugem marrom ao longo de dois anos de avaliações fenotípicas. Recentemente
COSTET et al. (2012) publicaram o desenvolvimento de primers específicos para o
gene Bru1, primeiramente observado na cultivar R570, que é considerado o gene maior
de resistência para ferrugem e está presente em muitas outras cultivares.
A identificação de QTLs pode ser particularmente útil para a identificação de
genótipos de interesse nos programas de melhoramento genético, além de contribuir
para o entendimento genético da característica, como número, posição, efeitos e
interações entre os QTLs.
2.8 Estudos da resistência da cana-de-açúcar à ferrugem marrom

19
O grupo de pesquisa do CIRAD, liderado pela pesquisadora Angelique D´Hont
estudou por muitos anos a resistência de cana-de-açúcar à ferrugem marrom. Usando
uma população de autofecundação da cultivar francesa R570, DAUGROIS et al. (1996)
identificaram um gene maior de resistência (Bru1) que segregava como um gene
simples dominante, esse gene estava ligado a uma sonda de RFLP (CDSR29), que
inicialmente não estava ligada a nenhum dos grupos de ligação formados para o mapa
daquela população. Um mapeamento adicional para a mesma população, realizado por
ASNAGHI et al. (2000) utilizando a sintenia entre Poaceaes determinou a localização
do gene de resistência no grupo de ligação VII do mapa da R570.
ASNAGUI et al. (2004) utilizaram marcadores AFLP em um bulk da progênie
de R570 de 695 indivíduos, e utilizando o mapa da mesma progênie desenvolvido por
HOARAU et al. (2001) que era o mais saturado na época; identificaram oito marcas
rodeando o gene Bru1, totalizando 10 cM entre a primeira e a última marca. As duas
marcas mais próximas estavam a 2,2 cM e 1,9 cM do gene. Considerando a relação
média entre distancias de mapa físico e genético para cana de 500Kb/cM os autores
comentaram a dificuldade por conta ainda da distância do alvo para se iniciar a técnica
de chromosome walking com a biblioteca BAC de cana-de-açúcar da R570 que já havia
sido construída (TOMKINS et al., 1999).
Segundo D´HONT et al. (2010) o grupo de pesquisa refinou então a busca no
cromossomo ortólogo de sorgo, clonando os dois marcadores AFLP mais próximos do
gene alvo (aaccac6 e attcag) e analisando com sondas RFLP na população de sorgo.
Ambos os locos foram mapeados no grupo de ligação quatro (GL4), próximos da marca
CDSR29. Estes resultados permitiram que a região do sorgo a ser explorada ficasse em
um intervalo de 8,8 cM. Alinharam então o GL4 do mapa de sorgo do grupo com o
mesmo homólogo do mapa saturado de sorgo de BOWERS et al. (2003), e nove marcas
RFLP foram alocadas na região alvo sobre os dois mapas e foram analisadas na
progênie da R570. Com isso conseguiram rodear o Bru1 com quatro novas marcas a 0,3
cM e 0,6 cM de cada lado do gene e reduzir o alvo em um intervalo de 0,9 cM. Naquele
momento havia disponível parte do mapa físico do sorgo, onde o grupo decidiu então
explorar este para refinar o mapa genético de cana, reduzindo a busca para 0,6 cM. Em
2005 a disponibilidade do sequenciamento genético do arroz possibilitou que maior
refinamento do mapa genético na região do Bru1 (Le CUNFF et al., 2008).
Após estes esforços uma nova população de 1600 indivíduos da
autofecundação de R570 foi construída para aumentar a resolução do mapa genético na

20
região de Bru1 e ao final o novo mapa saturado na região alvo incluía três marcas
mapeadas a 0,28 cM em ambos os lados de Bru1 e três marcas co-segregavam com o
gene. Foi então que o grupo decidiu que a resolução da área alvo do mapa genético
estava boa o suficiente para começar a construir o mapa físico para a região de interesse.
Sendo a cana poliploide, para diferenciar o haplótipo alvo determinaram qual
BAC continha a região de Bru1 utilizando cortes de enzimas e sondas de RFLP e
finalmente identificaram uma BAC correspondente à área alvo. Este BAC cobria apenas
parcialmente o alvo. Para buscar o restante do alvo construíram então uma nova
biblioteca BAC. Ao fim Le CUNFF et al. (2008) obtiveram uma região de grande
resolução próxima ao gene Bru1, com marcas beirando a 0,28 cM e 0,14 cM o alvo, e
desenvolveram um mapa físico do haplótipo, porém que ainda apresenta duas regiões de
gaps.
Recentemente, 16 anos após a descoberta do gene de maior efeito para
resistência à ferrugem, o grupo de pesquisa publicou um artigo (COSTET et al. 2012)
em que várias marcas ligadas ao Bru1 foram analisadas em um painel de 380 cultivares
e clones pré-comerciais onde dois marcadores moleculares PCR-específicos foram
ligados ao gene de efeito maior de resistência à ferrugem (Bru1), estando este gene
presente em 86% dos 194 acessos resistentes utilizados, revelando que constitui a
principal fonte de resistência das cultivares modernas.
Portanto neste estudo de COSTET et al. (2012) a ausência de marcas ligadas à
Bru1 foi observada também em cultivares resistentes, indicando a presença de outras
fontes de resistência à ferrugem marrom, o que corrobora com RABOIN et al. (2006),
que observou outro gene de resistência à ferrugem marrom utilizando uma população de
mapeamento entre o clone MQ76-53 e a cultivar R570, chamando-o de Bru2, que foi
considerado o segundo gene maior de resistência conhecido para resistência à ferrugem
marrom em cana-de-açúcar.
3 MATERIAL E MÉTODOS
3.1 População de mapeamento
A população de mapeamento é constituída por 219indivíduos, tomados ao
acaso, do cruzamento biparental entre o clone IACSP95-3018 como genitor feminino

21
esuscetível à ferrugem marrom; e a cultivar IACSP93-3046como genitor masculino e
resistente, considerada uma cultivar robusta e responsiva (LANDELL et al., 2005),
ambos do Programa de Melhoramento Cana do IAC.
3.2 Desenho experimental
O delineamento adotado foi uma reprodução do realizado na cidade de
Ribeirão Preto - SP, de blocos aumentados, considerando cinco blocos contendo 44
indivíduos da progênie cada um. Os tratamentos em comum para avaliação de efeito
ambiental foram os padrões SP81-3250 e RB835486, repetidos duas vezes por bloco,
além dos pais. A variedade suscetível IAC86-2480 foi adotada como linha infectora
(Figura 2).
Figura 2 Croqui de campo utilizado nos experimentos de Ribeirão Preto e em Piracicaba. PRG1 – Genitor feminino (IACSP95-3018). PRG2 – Genitor masculino (IACSP93-3046). P1 – Padrão 1 - SP81-3250. P2 - RB835486. Infectora - IAC86-2480. Bordadura - IACSP93-3046.

22
3.3 Instalação do experimento
Para a instalação do ensaio de avaliação da ferrugem marrom na Fazenda
Experimental de Piracicaba, primeiramente, foram preparadas as mudas para o plantio
gema a gema, onde gemas individuais são plantadas. Para isso, colmos dos 220
indivíduos da população foram coletados de um ensaio previamente instalado na
Fazenda Nova Aliança, no município Sales Oliveira – São Paulo (Figura 3). Os toletes
foram cortados na região das gemas, uma a uma, e estas foram plantadas em caixas
plásticas contendo substrato. As caixas foram mantidas em casa de vegetação até a
brotação. Após estabelecimento das plantas, seis plantas de cada genótipo da população
de mapeamento foram transplantadas para copos plásticos, devidamente identificados
(Figura 4).
Figura 3 Colmos coletados dos genótipos do experimento de Sales Oliveira – SP, que formam a população de mapeamento. Foram utilizados para a produção de mudas gema a gema.

23
Figura 4 - Plantas oriundas de gemas individualizadas, após transplante para copos
plásticos.
A área de plantio em Piracicaba foi preparada e o transporte do material foi
realizado de Ribeirão Preto para Piracicaba em um caminhão do Centro de Cana (Figura
5). Em Piracicaba foram plantadas, com auxilio de matracas, as mudas de cana-de-
açúcar. Foram plantados três clones de cada genótipo em um metro de parcela, com
espaçamento de um metro entre parcelas (Figura 5).
Figura 5 - Esquerda: Caminhão carregado com mudas dos clones da população para
serem transportados para Piracicaba. Direita - Área com experimento instalado em
Piracicaba.

24
3.4 Avaliação fenotípica
O nível de resistência das plantas à ferrugem foi obtido com base na infecção
natural utilizando a escala de notas descrita por AMORIM et al. (1987), a qual leva em
conta a concentração de manchas necróticas e pústulas na região foliar da planta
(porcentagem de área foliar atacada), variando de um (0%) a nove (>50%) (Figura 6).
Figura 6 - Escala de notas de agressividade da ferrugem marrom em cana-de-açúcar
(AMORIM, 1987).
3.5 Avaliações realizadas
As avaliações foram feitas em seis ocasiões, sendo as quatro primeiras
anteriores a esta tese: duas na cidade de Ribeirão Preto em cana planta (mesmo ensaio,
épocas diferentes) e duas em cana-soca (primeiro corte) (mesmo ensaio, épocas
diferentes) e outras duas avaliações em cana planta na cidade de Piracicaba, como é
demonstrado na Tabela 1. A primeira avaliação em cana planta tanto em Ribeirão Preto
quanto em Piracicaba foi realizada seis meses após o plantio, e a segunda avaliação foi
realizada aos oito meses após o plantio, também em ambas as cidades. Em cana-soca a
primeira avaliação foi realizada seis meses após o primeiro corte e a segunda seis meses
após a primeira avaliação.

25
Tabela 1 Avaliações de sintomas da ferrugem na população mapa em diferentes anos, locais e cortes. Avaliação Cidade Corte Mês e Ano
1º Ribeirão Preto Cana-planta Novembro 2005
2º Ribeirão Preto Cana-planta Janeiro 2006
3º Ribeirão Preto Cana-soca Janeiro 2007
4º Ribeirão Preto Cana-soca Março2007
5º Piracicaba Cana-planta Dezembro 2011
6º Piracicaba Cana-planta Fevereiro 2012
3.6 Avaliações com Marcadores Moleculares
3.6.1 Extração e quantificação do DNA genômico
A extração de DNA genômico foi realizada através do palmito fresco, coletado
em Sales Oliveira, SP, um dos campos experimentais da população de mapeamento. O
protocolo utilizado para a extração foi adaptado de AL JANABI et al. (1999) e permite
a extração de DNA de alta qualidade e quantidade, por utilizar a região meristemática
da planta. O tecido meristemático foi retirado individualmente de cada palmito e
macerado em cadinho de porcelana, juntamente com o tampão de extração.
Para cada amostra, foram utilizados 3,5 ml do macerado com tampão de
extração juntamente com 3,5 ml de mix de extração em um tubo tipo falcon de volume
total de 15 ml. As amostras foram agitadas para ressuspender o tecido no tampão e
levadas ao banho-maria à 65ºC durante o período 60 minutos, sendo agitados para nova
ressuspenção a cada 10 minutos. Depois de retiradas do banho-maria e já com as
amostras em temperatura ambiente, foram adicionados 7,0 ml de fenol-clorofórmio-
álcool isoamílico, na proporção de 25:24:1 partes. Os tubos foram agitados por 5
minutos por inversão obtendo-se uma emulsão homogênea. Em seguida, os tubos foram
centrifugados por 10 minutos a 3.000 rpm e o sobrenadante transferido para novos
tubos, contendo 600 µL de NaCl 5M; foram então adicionados cinco ml de isopropanol
gelado em cada amostra, e estas foram suavemente misturadas até a formação do
precipitado. As amostras foram levadas ao freezer (-20ºC) por uma hora. Os
precipitados foram então pescados de cada tubo com o auxílio de uma pipeta de vidro e

26
transferidos para tubos do tipo eppendorf de volume total 2,0ml contendo 500 µL de
álcool 70%, sendo invertidos suavemente para limpeza do precipitado, após isso foram
centrifugados por 5 minutos a 10.000 rpm e o álcool descartado; uma nova etapa de
limpeza foi realizada. Finalizada a limpeza o álcool foi descartado e as amostras ficaram
overnight sobre a bancada, com a tampa aberta para secagem do material. No dia
seguinte foi adicionado tampão TE (10 mM Tris-HCl, um mM de EDTA, pH 8,0)
acrescido de um µL de RNAse (10mg/mL) e deixado sobre bancada overnight. O DNA
foi então armazenado em geladeira.
Após a extração, os DNAs foram quantificados em gel de agarose 0,8% (p/v)
corado com brometo de etídeo na presença de um padrão de DNA do fago λ de
quantidades conhecidas. A avaliação do DNA em gel de agarose permitiu confirmar a
sua alta qualidade necessária para a realização da técnica de AFLP-RGA, a qual utiliza
a digestão do DNA com enzima de restrição.
3.6.2 Marcadores EST-SSR
Os marcadores microssatélites derivados de ESTs utilizados em testes de
amplificação e polimorfismo são oriundos de fontes diversas, sendo 17 pares de primers
EST-SSR desenvolvido junto com aqueles publicados por da SILVA (2001), porém
suas sequências não estão entre as que foram publicadas até o momento (comunicação
pessoal). Quarenta e seis pares de primers foram desenvolvidos por OLIVEIRA et al.
(2009) e PINTO et al. (2004; 2006), um par de primer desenvolvido por PARK et al.
(2012) a partir da sequência nucleotídica relacionada à lignina; além de dez pares de
primers que foram desenvolvidos para este estudo, totalizando 73 primers EST-SSR
avaliados em seis indivíduos e ambos os pais. Foram realizados dois tipos diferentes de
PCR para amplificação dos microssatélites, sendo uma para os géis que foram corados
com nitrato de prata, e outra para corrida eletroforética em sequenciador automático.
As reações de amplificação para géis corados em nitrato de prata foram
efetuadas em um volume final de 15 µl contendo 25 ng de DNA, 10 mM de Tampão
10x, 2 mM de MgCl2, 0,1 mM de dNTPs, 0,2 mM dos primers (forward e reverse) e 1U
de Taq polimerase. As amostras foram submetidas à desnaturação inicial de 94°C
durante três minutos, seguido de 30 ciclos compostos por etapas de 94°C por um
minuto, temperatura de anelamento específica para cada par de primers por um minuto e
a extensão a 72°C por 1 minuto, e um ciclo final de 72°C por dois minutos. Após a
amplificação, foram adicionados 7,5 µl de tampão formamida (formamida 98 %, EDTA

27
10 mM pH 8,0, azul de bromofenol 0,002 %, p/v e xileno cianol 0,002 %, p/v) ao
volume final de 15 µl da reação para a sua desnaturação à temperatura de 95 ºC por 3
minutos. Os produtos resultantes da amplificação foram submetidos à eletroforese em
gel desnaturante de poliacrilamida 6%, com tampão de corrida TBE 1X sob potência
constante de 55 W e duração de acordo com o tamanho esperado para cada EST-SSRs.
A visualização das bandas foi efetuada em solução de nitrato de prata conforme
protocolo descrito por CRESTE et al. (2001).
As reações de PCR para corrida elétroforética em sequenciador automático
(primers desenvolvidos por da Silva (Texas A&M University, comunicação pessoal) e
primers desenvolvidos neste trabalho) foram realizadas em volume final de 10 µL,
contendo 100ng de DNA genômico, 1U de Taq polimerase (NEB), 2 µL de tampão 10X
(NEB), 0,2 mM de cada dNTP e 0,02 µM do primer forward com cauda M13 na
extremidade 5´ (cacgacgttgtaaaacgaca), 0,2 µM de primer M13 IRDye 700 ou 800 e 0.2
µM do primer reverso. As reações de PCR foram incubadas por 5 minutos à 94ºC,
seguidos por 30 ciclos de desnaturação à 94ºC por 30 segundos, anelamento de 45
segundos à 56ºC, e elongação por 45 segundos à 72ºC. Outros oito ciclos de
desnaturação à 94ºC por 30 segundos, 53ºC por 45 segundos e 72ºC por 45 segundos; e
um ciclo final de 10 minutos à 72ºC. Os produtos de PCR foram visualizados em gel de
poliacrilamida através do Licor 4300 DNA analyzer (Licor) seguindo as recomendações
do fabricante.
3.6.3 Marcadores AFLP-RGA
Para a obtenção dos marcadores AFLP-RGAs, duzentos e cinquenta
nanogramas de DNA genômico foram digeridos com as enzimas de restrição EcoRI e
MseI (Invitrogen) em um volume final de reação de 20 uL contendo tampão 10X
OnePhorAll (GE). A reação de digestão foi conduzida a temperatura de 37ºC por 4
horas, seguida de 15 minutos a 65ºC para inativação da atividade enzimática, em
termociclador (BioRad). Em seguida, procedeu-se a ligação dos adaptadores em um
volume final de reação de 40 uL contendo 20 uL da reação de digestão em tampão 5
vezes (5X), 4 uM do adaptador para EcoRI (5 pmol) e 40 uM do adaptador para MseI
(50 pmol), 0,5 mM de ATP e 1 U de T4DNA ligase (Biolabs). A reação de ligação dos
adaptadores foi conduzida em termociclador à temperatura de 37ºC por 16 horas.
Após a ligação dos adaptadores, uma alíquota da reação foi diluída seis vezes
em água ultrapura e dois uL desta diluição foram utilizados na reação de pré-

28
amplificação de volume final de 15 uL contendo tampão 10X (200 mM Tris pH 8.4,
500 mMKCl), 0,5 mM de MgCl2, 0,16 mM de dNTPs e 3,3 uM de primer Eco+0 e
MseI+0 e 0,1U de Taq polimerase. As reações de amplificação foram conduzidas em
termociclador à 94ºC por 30 segundos, seguidas de temperatura de anelamento à 56ºC
por 60 segundos e extensão a 72ºC por 60 segundos, repetidas 29 vezes.
A reação de amplificação seletiva foi conduzida em um volume final de 10 ul
de reação contendo, dois UL (da reação de pré-amplificação diluída 10X em água
ultrapura), tampão 10X (200 mM Tris pH 8.4, 500 mMKCl), 0,25mM de dNTPs, 2,0
mM de MgCl2, 0,05 uM de primer Eco marcado com IR700 ou IR800 e 0,30 uM do
primer RGA. As condições de amplificação utilizadas constaram de 13 ciclos a 94ºC
por 30 segundos, 65ºC (-0,7 ºC/ ciclo) por 30 segundos e 72ºC por 1 minuto, seguido
por 23 ciclos de 94ºC por 30 segundos, 56ºC por 30 segundos e 72ºC por 1 minuto. Os
produtos de PCR foram visualizados em gel de poliacrilamida através do Licor 4300
DNA analyzer (Licor) seguindo as recomendações do fabricante.
3.6.4 Marcadores CISP
Foram utilizados os cinco primers CISPs, IF1, IF3, IF5, IF6 e IF11,
desenvolvidos por PARK et al. (2012) para amplificar os genótipos IACSP95-3018 e
IACSP93-3046, genitores da nossa população de mapeamento. As reações de
amplificação de PCR foram realizadas de acordo com o realizado pelos autores. Em
volume final de 30 µL de reação, contendo cerca de 200 ng de DNA genômico, 1,25 U
de Taq polimerase (NEB) 0,03 U de Pfu DNA polimerase (Stratagene), três µL tampão
de PCR Pfu 10x, 0,12 mM de Sulfato de Magnésio (MgSO4), 0,2 mM de cada dNTP e
0,33 µM de cada primer forward e reverse. A reação de PCR foi incubada por 4
minutos a 94 ºC, seguidos de 10 ciclos de desnaturação a 94 ºC por 30 segundos,
anelamento de 30 segundos a 65 ºC (-1 ºC por ciclo) e elongação a 72 ºC por 2 minutos,
e mais outros 25 ciclos de 94 ºC por 30 segundos, 30 segundos a 56 ºC, 72 ºC por 2
minutos, e seguidos por extensão final de 7 minutos a 72 ºC. Os produtos de PCR foram
visualizados em gel de agarose 2% corado com SYBR Safe (Invitrogen). Foi utilizado o
marcador de peso molecular 2-Log DNA ladder, da Biolabs.
Os amplificados foram visualizados em transiluminador ultravioleta, recortados
do gel cada um separadamente com lâminas estéreis e armazenados no freezer em tubos
do tipo eppendorf devidamente identificados. Foram escolhidos para sequenciamento os
amplificados dos CISPs IF3 (4-CL), e IF5(CCoAMMT).

29
Plasmídeos pGEM-T Easy Vector (Promega) foram utilizados segundo
especificações do fabricante para transformação e amplificação dos fragmentos
recortados do gel. A transformação foi realizada em células competentes DH5α, que
foram retiradas do freezer -80ºC e imediatamente mantidas no gelo. O volume total da
ligação foi adicionado às células competentes e mantido em gelo por dez minutos e
então transferido para banho-maria à 40ºC por 30 segundos e retornados ao gelo por
mais 5 minutos. Após o choque térmico foi adicionado um mL de meio LB e incubado a
37ºC durante uma hora. Cem microlitros desta solução foram espalhados em uma placa
de Petri contendo meio LB sólido com carbenicilina, IPTG e X-Gal, o restante foi
centrifugado durante um minuto a11GRPM, descartado o sobrenadante, e as células
foram ressuspendidas e recuperadas com o auxílio de uma pipeta e então espalhadas em
outra placa de petri com o mesmo meio LB. Tanto o meio LB sólido quanto líquido foi
realizado conforme protocolo pGEM-T Easy Vector da Promega. As placas foram então
mantidas a 37ºC overnight.
Dez colônias brancas e isoladas das demais foram escolhidas para
multiplicação. Com o auxílio de ponteiras as colônias foram retiradas e adicionadas em
tubo de ensaio contendo cinco mL de LB líquido e cinco uL (100µg/mL) de antibiótico
carbenicilina. Os tubos foram mantidos à 37ºC sob agitação de 250 rpm overnight.
Foi realizada extração do DNA plasmidial das 10 colônias de cada genitor
através do Kit Zyppy Plasmid Miniprep Kit (Zymo Research), segundo instruções do
fornecedor. A concentração do DNA foi estimada através de espectofotômetro
Nanodrop 1000 (Thermo Scientific). Para certificar que havia a presença do inserto no
plasmídeo foram utilizados 3µL do DNA em teste de digestão com a enzima EcoR1
(Biolabs), seguindo instrução do fabricante. Foram adicionados 5µL de tampão de
carregamento na digestão, e uma corrida eletroforética em gel de agarose 1% contendo
SYBR safe DNA gel stain (Invitrogen) foi realizada. Os amplificados foram visualizados
em transiluminador ultra-violeta Kodak Imagem System.
Os fragmentos de DNA foram então enviados para sequenciamento em
empresa especializada. As sequências obtidas foram então visualizadas no software
4Peaks (http://4peaks.en.softonic.com/mac), as bases da extremidade referentes ao
plasmídio foram retiradas e mantidas as provenientes de clonagem. Estas foram então
salvas em modo fasta, e alinhadas pelo método Clustal W utilizando o software MEGA
versão 5 (TAMURA et al., 2011), no modo default.

30
As sequências nucleotídicas foram analisadas no software CAPS designer
(MUELLER et al., 2005) com a finalidade de converter SNPs localizados entre os
genitores em marcadores CAPS. Foram selecionadas três enzimas de restrição, BsaHI,
BsiHKAI e HaeIII, para CISP 4-CL (IF3) e a enzima BstNI para CISP CCoAMMT
(IF5). Os genitores e seis indivíduos da progênie, sendo três resistentes (38, 39 e 141) e
três suscetíveis (32, 85 e 99) foram amplificados com os primers CISP IF3 e CISP IF5
para cada uma das enzimas utilizadas, e as respectivas enzimas de restrição foram
adicionadas aos 30 µL de volume final da reação de PCR. Para a digestão com HaeIII e
BsaHI foram utilizados 20U de enzima para cada amostra, incubadas por 3 horas a 37ºC
seguido de inativação por 20 minutos a 80ºC. Foram adicionados à reação de PCR
também 20U da enzima BsiHKAI, porém adicionados com 0,5 µL de BSA para cada
uma das oito amostras, a digestão foi realizada a 65ºC por três horas. A digestão com
BstNI foi realizada com 10U de enzima e 0,5 µL de BSA para cada amostra, a 60ºC
durante três horas. Todas as enzimas utilizadas são da marca Biolabs. A digestão dos
produtos de PCR foi analisada em gel de agarose 2%
3.6.5 Desenvolvimento de primers
Foi acessado o banco de dados SUCEST (Sugarcane Expressed Sequence Tag
Project) que contém dados de ESTs de cana-de-açúcar, e o download de todas as 43.143
sequências gênicas ali depositadas e suas respectivas anotações foi realizado. A partir
daí foram feitas buscas pelas palavras chaves rust, resistance, defense, LRR e R genes.
Ao todo foram separadas 868 acessos SAS (Sugarcane Assembled Sequences). Para
cada um destes foi realizada uma leitura mais detalhada da anotação e descartados
aqueles que estavam fora do propósito, restando então 594 SAS.
3.6.5.1 Marcadores EST-SSR
As 594 sequências nucleotídicas foram então utilizadas para busca de
sequências microssatélites utilizando-se o software Webtroll (MARTINS et al., 2009).
Os padrões de busca por microssatélites foram di, tri, tetra e pentanucleotídeos repetidos
em tandem, sendo o número mínimo de bases total igual a 18, ou seja, repetições
dinucleotídicas repetidas ao menos nove vezes, trinucleotídicas ao menos seis vezes,
tetranucleotídicas repetidas ao menos cinco vezes e penta com ao menos quatro
repetições. Doze SSRs atendiam estes pré-requisitos. Antes de desenhar os primers

31
destas doze sequências gênicas foi realizado BLASTn contra o genoma do sorgo
(Sorghum bicolor) no programa Phytozome (http://www.phytozome.net/sorghum)
usando os parâmetros default. Buscando evitar o desenvolvimento de primers que
pudessem englobar regiões intrônicas e ter como resultado fragmentos
consideravelmente maiores do que o esperado e também evitando que o primer perdesse
sua função de avaliar polimorfismo em regiões exônicas, foi observado então se havia
presença de regiões intrônicas entre as sequências, Dois dos doze genes apresentavam
interrupções intrônicas e foram descartados, sendo então desenvolvidos primers para as
dez sequências gênicas restantes.
Os parâmetros utilizados para desenvolvimento dos primers foram sequências
de 150 a 300 pares de bases, contendo de 40 a 60% de bases guanina e citosina, primers
de 18 a 22 bases e com temperatura de anelamento de 55 a 62ºC. A qualidade dos pares
de primers foi confirmada através do software NetPrimer
(http://www.premierbiosoft.com/netprimer/index.html).
3.6.5.2 Marcadores TRAP
As mesmas 594 sequências SAS selecionadas do banco de dados SUCEST e
que foram utilizadas para localização de microssatélites, excluindo-se aquelas dez
utilizadas para desenvolvimento dos microssatélites, foram utilizadas para desenhar
primers TRAPs fixos.
Com a finalidade de aprimorar a anotação destas e restringir a busca, as 584
sequências foram então analisadas no Blat2GO. Foram separadas 48 sequências que
apresentavam como função protein Kinase activity no item GO ID e outras 116
sequências com Processo Biológico relacionado à defense response. Estas 164
sequências foram então analisadas no Phytozome (http://www.phytozome.net/sorghum)
através de BLASTn contra o genoma do sorgo. Foram observadas, além de homologia
nas sequências, à qual cromossomo do sorgo estas sequências pertenciam. Diante disso
foram selecionadas todas as sete sequências que eram relatadas no cromossomo quatro
do sorgo, uma vez que se acredita que o gene principal de resistência à ferrugem esteja
relacionado a este cromossomo no sorgo (ASNAGHI et al., 2000). Foi desenvolvido
então um primer forward para cada uma das sete sequências através do Primer3
(ROZEN & SKALETSKY, 2000). Os parâmetros empregados foram os mesmos
utilizados por HU & VICK (2003) sendo o número requerido mínimo e máximo de

32
18nucleotídeos, e temperatura Tm máxima, mínima e ótima de 55ºC, 50ºC e 53ºC,
respectivamente. A qualidade destes primers foi checada utilizando-se o software
NetPrimer (http://www.premierbiosoft.com/netprimer/index.html).
Além dos sete primers relacionados ao cromossomo quatro do sorgo foram
desenvolvidos mais nove primers TRAPs selecionados aleatoriamente entre as
sequências previamente escolhidas, porém levando em consideração que estas fossem
relativamente grandes, e que tivessem homologia também com os cromossomos de
arroz e milho, usando também o BLASTn do Phytozome. As sequências dos primers
forward desenvolvidas, suas identidades no SUCEST e seus grupos de homologia no
sorgo estão demonstrados na Tabela 2.
Tabela 2 Marcadores TRAPs desenvolvidos, identificação das sequências no banco de dados SUCEST, cromossomo homólogo no sorgo e sequência dos primers forward.
TRAP Número SUCEST
Cromossomo(s)
referente no
Sorgo
Sequência dos Primers Forward
TRAP_Sucest_1 SCSGAD1005E03.g 4 TAACTGGGACACTCAACG
TRAP_Sucest_2 SCMCRT2088F10.g 4 TAGTAGCAAGCACGGAGA
TRAP_Sucest_3 SCCCCL4013C03.g 4 CTCACCGATCTCAACCTT
TRAP_Sucest_4 SCEZLB1013E06.g 4 GGGAGCATAGACAAGAGG
TRAP_Sucest_5 SCACLV1020F05.g 4 TAATGGTCAAGCAGGTGA
TRAP_Sucest_6 SCQSHR1021H04.g 4 GGTGGTGTAGTTGGTCCT
TRAP_Sucest_7 SCEPAM2014D05.g 4 TCACCCAGCACCTATACA
TRAP_Sucest_8 SCSGHR1067C04.g 2,1 AGCGGCAACTACCTCTAC
TRAP_Sucest_9 SCMCRT2087B07.g 2,1 GCCTCAATCTTCACCTTC
TRAP_Sucest_10 SCVPCL6041G02.g 2 CCTTGAAAGTTCCTCTGG
TRAP_Sucest_11 SCQSHR1023C04.g 9,5 AGTGAGGCACCGTAACTT
TRAP_Sucest_12 SCCCCL3080B03.g 8,3 CCGTTGGACCTTAGTTTC
TRAP_Sucest_13 SCSFSB1103A12.g 5 TAGGAGGCTGGAAAGAAG
TRAP_Sucest_14 SCCCCL4013C10.g 3 AGTGTCTCCAAGCACCTT
TRAP_Sucest_15 SCCCLR2C01A06.g 10 GAGTTTGGACAGGGCTAC
TRAP_Sucest_16 SCSGAD1006H08.g 8 ATTGTGAGGTTGGAGAGG
As reações de PCR foram realizadas seguindo o protocolo de SUMAN et al.
(2012) com algumas modificações. Em volume final de 15 µL, contendo 10 µM de cada
primer fixo, 3 µM de cada 2 primers arbitrários IRDye 700/800, 3µL de1,5 µL de
MgCl2 25 mM (Promega), 1,5 µL de tampão de reação 10× (Promega), 2 µL de 2,0

33
mMde dNTP mix (Biolabs), 1 U Taq DNA polimerase (Biolabs) e 100 ng de DNA
genômico. A reação foi realizada com os seguintes parâmetros: desnaturação a 94°C por
4 minutos seguidos de cinco ciclos de desnaturação a 94ºC por 1 minuto, anelamento a
35ºC por 1 minuto e elongação a 72ºC por um minuto. E mais 35 ciclos de 94ºC por 1
minuto, 56ºC por 1 minuto e 72ºC por 1 minuto. Além de um alongamento final de 72ºC
por 7 minutos.
Os produtos de PCR foram visualizados em gel de poliacrilamida através de
Licor 4300 DNA analyzer (Licor), na função AFLP durante 03h30min horas, seguindo
as recomendações do fabricante. O teste de amplificação e polimorfismo foi realizado
utilizando-se ambos os genitores e mais seis indivíduos da população de mapeamento,
além de em dois genótipos de Miscanthus (Tus05-02b e Tus05-08e) e um genótipo de
sorgo (ATX 3381). Cada um dos 16 primers fixos desenvolvidos foi testado com os
quatro primers arbitrários desenvolvidos por LI & QUIROS (2001), sendo estes:
arbitrário 1 – GACTGCGTACGAATTATT, arbitrário 2 –
GACTGCGTACGAATTTGC, arbitrário 3 – GACTGCGTACGAATTGAC e arbitrário
4 – GACTGCGTACGAATTTGA, todos marcados com fluorescência, contabilizando
um total de 64 combinações.
Os locos de todos os diferentes marcadores moleculares testados que
apresentaram polimorfismo foram então genotipados nos 220 indivíduos da população
de mapeamento, para isso em cada loco genotipados foram realizadas três PCRs e
quatro géis de poliacrilamida, incluindo os respectivos genitores em cada um dos géis
com pentes de 64 poços.
Os testes de polimorfismo dos primers desenvolvidos neste trabalho (EST-SSR
e TRAP) foram realizados entre os genitores IACSP95-3018 e IACSP93-3046, seis
genótipos da população, dois genótipos de Miscanthus (Tus05-02b e Tus05-08e) e um
genótipo de sorgo (ATX 3381), os três últimos cedidos gentilmente por Jong-Won Park.
Os demais testes de polimorfismo (AFLP-RGA e EST-SSR de literatura) foram
realizados entre os genitores da população e seis indivíduos da progênie.
Apenas bandas legíveis e polimórficas nos quatro géis da população foram
consideradas, sendo que fragmentos duvidosos não foram genotipados. Foram também
ignorados fragmentos menores de 100 pares de bases, evitando-se a genotipagem de
artefatos.

34
3.7 Construção do mapa genético
3.7.1 Análises de segregação dos locos
A codificação dos alelos foi a mesma para todos os marcadores utilizados,
sendo atribuído 1 para presença e 0 para ausência das bandas em todos os indivíduos da
população. Quando não foi possível determinar a presença ou não de banda foi
considerado dado perdido para o genótipo em questão.
Através do software R versão 2.13.0 foi realizado o teste de segregação através
do qui-quadrado (χ2), em relação às hipóteses de segregação com base na segregação de
marcadores de dose única de 1:1 e 3:1, que correspondem às configurações A0000000 x
00000000(ou vice-versa) e A0000000x A0000000, respectivamente, considerando Ao
alelo amplificado (banda) e 0como ausência de banda (alelo nulo).
A correção de Bonferroni foi realizada para minimizar os erros associados com
múltiplos testes, nesta correção o valor de χ2 crítico é obtido dividindo-se o valor de α
a 5% de probabilidade (0,05) pelo número total de marcadores. O nível de significância
usado para determinar a validade das razões de significância de 1:1 e 3:1 foi de 3,67e-05
(alfa = 0,05/1363). Após a correção as marcas com desvios significativos foram
desconsideradas.
3.7.2 Construção do mapa de ligação
Aos alelos polimórficos obtidos neste trabalho foram adicionados mais 824
alelos que foram gerados pelo grupo de pesquisa da Dra. Luciana Rossini Pinto, do
Centro de Cana do IAC, estes alelos adicionais são derivados de marcadores AFLP,
EST-SSR e SSR genômico.
As análises de ligação na população de mapeamento e a construção do mapa
genético foram realizadas usando-se o software OneMap (MARGARIDO et al. 2007
versão 2.0).
O LOD score é por definição a razão de chances (odds ratio) do logarítimo na
base dez do teste de razão de verossimilhança (LRT), que compara valores de
verossimilhança considerando diferentes valores de parâmetros r de fração de
recombinação (probabilidade de ocorrer um evento de recombinação entre os locos) até
que este atinja um valor estável. O valor de LRT é equivalente ao valor de qui quadrado
da soma das marcas utilizadas multiplicando por 0,2172. O valor mínimo do LOD foi
determinado então se utilizando o valor do qui quadrado das marcas genotipadas (após a

35
correção de Bonferroni), multiplicado por 0,2172. Diante disto os critérios para
estabelecer os grupos de ligação utilizados foram de LOD score ≥ 5,0 e fração de
recombinação para o teste de dois pontos entre todos os pares de marcas de ≤ 0,35. A
distância expressa em centiMorgans (cM) foi estimada usando a função de KOSAMBI
(1944).
Após o teste de ligação os marcadores que foram atribuídos a algum grupo de
ligação foram ordenados através de funções que variaram de acordo com o número de
marcas em cada grupo. Nos grupos com três a seis marcas ligadas foi utilizado o
comando compare, onde se testa todas as possíveis ordens escolhendo a de maior valor
de máxima verossimilhança, e grupos com mais de seis marcas ligadas foram ordenadas
usando o comando order, processo iniciado com subgrupo de marcas no qual compare é
feito e os demais marcadores são posteriormente adicionados utilizando o comando try.
Tais funções são idênticas aquelas utilizadas pelo programa MAPMAKER (LANDER
et al., 1987). Além disso, os heatmaps, que são as matrizes gráficas do LOD e das
frações de recombinação foram utilizadas para determinar a ordem final das marcas nos
respectivos grupos de ligação. Eles auxiliam na tomada de decisões durante o processo
de ordenação final das marcas, pois mostram ligações finais das marcas através de
cores, quanto mais quente maior a probabilidade de ligação destas, e quanto mais frias,
menor a chance de estarem ligadas.
3.8 Mapeamento de QTLs
O delineamento experimental utilizado nestes ensaios é o de blocos
aumentados de Federer (FEDERER 1956) que são, por construção, desbalanceados e
não ortogonais, e sua utilização é viável apenas nas fases iniciais de melhoramento, pois
geram estimativas de baixa precisão e seu uso conduz a média de tratamentos viciadas,
mesmo no caso de 100% de sobrevivência dos genótipos (RESENDE, 1999). Diante
disso é recomendado o tratamento dos dados associados com o BLUP (Best Linear
Unbiased Prediction), o qual ajusta as médias para as estimativas BLUE (Best Linear
Unbiased Estimative) dos efeitos fixos ou efeitos ambientais identificáveis.
Os dados não apresentavam normalidade e como são divididos em nove classes
nenhuma transformação ajustou os dados à normalidade. Segundo LI et al. (2006)
quando a característica observada para identificação de QTLs apresenta cinco classes ou
mais os dados são tratados como contínuos. Diante disso nenhuma transformação dos
dados foi realizada.

36
A análise para modelos mistos (proc mixed) foi realizada no SAS (Statistical
Analysis System), considerando o delineamento de blocos aumentados de Federer, em
que o efeito de testemunhas foi fixo e os efeitos blocos, genótipos dentro dos blocos e
erro foram aleatórios. Assim, temos estimativas de máxima verossimilhança das
variâncias residual, de progênies e de blocos e testes F para testemunhas. Obtiveram-se
então os melhores preditores lineares não viesados (BLUPs) para os efeitos de
progênies.
3.9 Análise de Marca Individual
As análises de QTLs foram realizadas no software R versão 2.13.02 utilizando
os valores de BLUPs obtidos para as notas de ferrugem marrom fenotipadas. A análise
de marcas individual baseia-se na comparação entre as médias dos genótipos
marcadores e é feita separadamente para cada loco. Se as médias são diferentes pode-se
inferir que pelo menos um QTL encontra-se próximo a este marcador. O modelo
utilizado é o de regressão linear conforme eq. (1), sendo “b0” a média do caráter, “b1” o
quanto o valor de um caráter aumenta ou diminui pela presença da banda, xj a variável
codificadora de presença ou ausência da banda e “e” o erro aleatório e j varia de 1 a 219
representando os indivíduos. A presença do QTL é testada determinando-se se b1 é
significativamente diferente de zero.
(1) Yj = b0 + b1 xj + e j
A estatística F testa a hipótese H0 (b1=0) vs H1(b1≠0). Assim, quanto menor o
p-valor menor a evidência de que H0 é verdadeiro, portanto, rejeita-se H0 e se aceita H1.
Caso o teste F seja significativo rejeita-se H0, acreditando que o marcador tenha
associação com o caráter quantitativo em questão.
4 RESULTADOS E DISCUSSÃO
4.1 Avaliação fenotípica da resistência da ferrugem marrom
As frequências das notas de ferrugem nas seis avaliações estão apresentadas na
Tabela 3. Observa-se que a frequência das notas diminui conforme aumenta a infestação
da doença, estando a grande maioria das notas entre um e três, consideradas como

37
resistentes (Figura 7). As figuras 8, 9e 10apresentam o histograma da frequência das
notas relacionadas às avaliações de Ribeirão Preto (cana planta), Ribeirão Preto (cana
soca) e Piracicaba (cana planta), respectivamente separadas para facilitar a visualização
dos dados.
As avaliações em cana planta (Ribeirão Preto e Piracicaba) que foram
realizadas aos seis meses após o plantio apresentaram maior quantidade de notas de 1 a
3, consideradas resistentes, do que nas avaliações realizadas aos oito meses após o
plantio. Nas duas segundas avaliações o número de notas referentes a plantas de
resistência intermediária e suscetível aumentaram. Esse resultado não confere com o
observado por TAI et al. (1984), onde o número de plantas de 5 a 7 meses que
apresentavam suscetibilidade era maior do que entre o número de plantas entre 7 e 9
meses após o plantio, porém os próprios autores comentam que a resistência pode ser
afetada pelas condições do ambiente.
Tabela 3.Frequência de notas de ferrugem da população de mapeamento, separadas pela cidade e corte. Ribeirão Preto Piracicaba
Cana planta Cana soca Cana planta
Nota Novembro 2005 Janeiro 2006 Janeiro 2007 Março 2007 Dezembro 2011 Fevereiro 2012
1 163 115 21 27 90 41
2 6 17 79 71 63 73
3 23 40 28 34 24 27
4 15 22 21 19 9 24
5 7 8 30 26 9 15
6 5 11 22 21 7 11
7 0 4 14 16 5 12
8 0 2 2 3 0 5
9 0 0 2 2 2 0
TOTAL 219 219 219 219 209 208
Podemos observar que a nota um aparece em maior frequência em todas as
quatro avaliações de cana planta, não ocorrendo o mesmo em cana soca, que teve maior
frequência de notas dois (Figura 7) e estabilidade na frequência das notas um, três,
quatro, cinco, seis e sete. Tal observação é esperada, uma vez que é sabido que a cana
soca apresenta maior suscetibilidade a várias doenças e patógenos em relação à cana
planta (ROTT et al., 2000).

38
Figura 7 - Histograma com a frequência das notas das seis avaliações de ferrugem na
população de estudo.
Comparando-se a frequência das notas de cana planta entre as cidades de
Ribeirão Preto e Piracicaba (Figuras 8e 10, respectivamente) é possível observar que na
cidade de Piracicaba houve uma maior distribuição das notas, não sendo a frequência da
nota um aparentemente superior, como acontece em Ribeirão Preto, onde a nota um é
responsável por 74e 52% da frequência das notas, para cada uma das avaliações. Em
Piracicaba a frequência da nota um atinge apenas 43 e 20% para cada uma das
avaliações. Com exceção da nota um (ambas as avaliações) e nota quatro (uma
avaliação), para todas as demais, a fenotipagem realizada na cidade de Piracicaba
apresentou maior amplitude na frequência de notas do que para a cidade de Ribeirão
Preto, aonde, como já comentado, a máxima nota de resistência é mais frequente.
Tal resultado é esperado, pois segundo LANDELL et al. 2005 o histórico da
região de Piracicaba demonstra que a região é mais propícia à ocorrência de ferrugem
marrom do que a região de Ribeirão Preto, tendo sido este um dos principais motivos de
realizar a avaliação também na cidade de Piracicaba.
0
50
100
150
1 2 3 4 5 6 7 8 9
Frequência das notas de ferrugem
Novembro de 2005 (RP, cana planta) Janeiro de 2006 (RP, cana planta)
Janeiro de 2007 (RP, cana soca) Março de 2007 (RP, cana soca)
Dezembro de 2011 (Piracicaba, cana planta) Fevereiro de 2012 (Piracicaba, cana planta)

39
Figura 8 Histograma com frequência das notas de ferrugem marrom na população de
mapeamento em duas avaliações realizadas em Ribeirão Preto, nos anos de 2005 e
2006, em cana planta.
Figura 10 Histograma com frequência das notas de ferrugem marrom na população de mapeamento em duas avaliações realizadas em Piracicaba, nos anos de 2011 e 2012, em cana planta.
020406080
100120140160
1 2 3 4 5 6 7 8 9
Ribeirão Preto - cana planta
Novembro de 2005 (RP, cana planta) Janeiro de 2006 (RP, cana planta)
020406080
100120140
1 2 3 4 5 6 7 8 9
Piracicaba - cana planta
Dezembro de 2011 (Piracicaba, cana planta) Fevereiro de 2012 (Piracicaba, cana planta)
Figura 9 Histograma com frequência das notas de ferrugem marrom na população de
mapeamento em duas avaliações realizadas em Ribeirão Preto, nos anos de 2006 e
2007, em cana soca.
020406080
100120140
1 2 3 4 5 6 7 8 9
Ribeirão Preto - cana soca
Janeiro de 2007 (RP, cana soca) Março de 2007 (RP, cana soca)

40
Considerando-se as avaliações independentemente e atribuindo resistência às
categorias de notas de um a três, e suscetibilidade às categorias de notas quatro a nove, a
frequência de resistentes é de três vezes para uma de suscetibilidade em três das seis
avaliações, e ao se considerar os dados somados de todas as avaliações, o teste de qui-
quadrado demonstra que a proporção é de exatas três notas resistentes para uma
suscetível. A segregação da resistência em 3:1 vai de encontro com características
oligogênicas de efeito maior pela qual a ferrugem marrom em cana-de-açúcar já foi
relatada (DAUGROIS et al. 1996).
4.2 Obtenção de Polimorfismo
4.2.1 EST-SSR Dentre os 17 primers desenvolvidos por Jorge da Silva (Texas A&M
University, comunicação pessoal) testados, 13 não amplificaram ou eram monomórficos
e quatro (SSR32, SSR37, SSR38-1, SSR52) foram polimórficos, sendo três deles com
um alelo e um com dois alelos polimórficos, totalizando cinco alelos genotipados. O par
de primers microssatélite desenvolvido por PARK et. al (2012) amplificou dez alelos,
sendo que todos foram polimórficos.
Dos dez pares de primers EST-SSR desenvolvidos a partir das sequências
selecionadas do SUCEST (Figura 11) cinco amplificaram os genótipos de cana-de-
açúcar, e todos apresentaram polimorfismo (SSR_sucest_rst4, SSR_sucest_rst5,
SSR_sucest_rst7, SSR_sucest_rst8 e SSR_sucest_rst9), tendo o número de alelos
variado de 1 à 6, totalizando 19 alelos genotipados na população mapa. Quatro dos
cinco locos amplificados em cana amplificaram também em Mischanthus e apenas um
no sorgo.

41
.
Figura 11.Gel de poliacrilamida do teste de polimorfismo de primers SSR desenvolvidos à partir de sequências obtidas no SUCEST. A identificação dos primers estão na parte superior do gel. Os dois primeiros indivíduos são os genitores da população, os próximos seis indivíduos são da progênie da população mapa; os dois próximos são a amplificação dos genótipos de Miscanthus, enquanto o último genótipo é a amplificação de Sorgo. Marcadores de pesos moleculares Concentrated 50-350 Size Standard (Licor) estão nas laterais do gel, sendo a marca 200 pb indicada na lateral direita.
Na Tabela 4 constam os resultados dos testes de polimorfismo, bem como a
identificação da sequência do SUCEST, a sequência dos primers desenvolvidos e os
motivos microssatélites.
200pb
EST-6 EST-7 EST-8 EST-9 EST-10
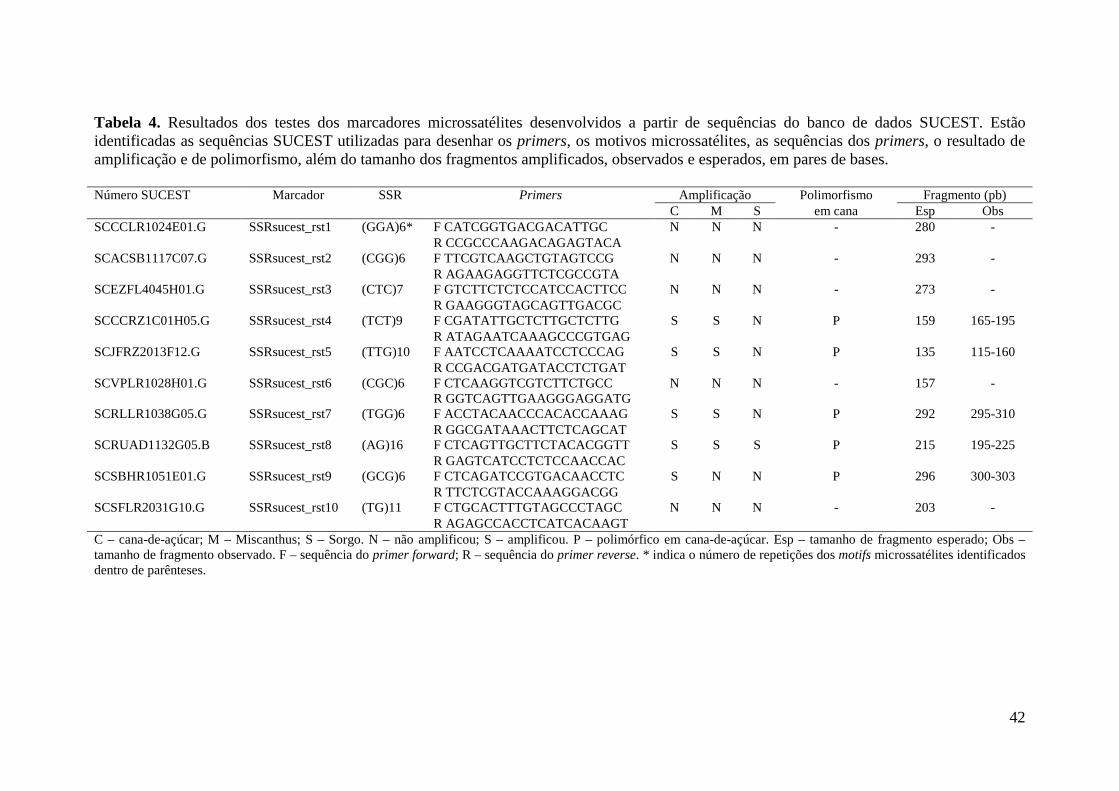
42
Tabela 4. Resultados dos testes dos marcadores microssatélites desenvolvidos a partir de sequências do banco de dados SUCEST. Estão identificadas as sequências SUCEST utilizadas para desenhar os primers, os motivos microssatélites, as sequências dos primers, o resultado de amplificação e de polimorfismo, além do tamanho dos fragmentos amplificados, observados e esperados, em pares de bases. Número SUCEST Marcador SSR Primers Amplificação Polimorfismo Fragmento (pb) C M S em cana Esp Obs SCCCLR1024E01.G SSRsucest_rst1 (GGA)6* F CATCGGTGACGACATTGC N N N - 280 - R CCGCCCAAGACAGAGTACA SCACSB1117C07.G SSRsucest_rst2 (CGG)6 F TTCGTCAAGCTGTAGTCCG N N N - 293 - R AGAAGAGGTTCTCGCCGTA SCEZFL4045H01.G SSRsucest_rst3 (CTC)7 F GTCTTCTCTCCATCCACTTCC N N N - 273 - R GAAGGGTAGCAGTTGACGC SCCCRZ1C01H05.G SSRsucest_rst4 (TCT)9 F CGATATTGCTCTTGCTCTTG S S N P 159 165-195 R ATAGAATCAAAGCCCGTGAG SCJFRZ2013F12.G SSRsucest_rst5 (TTG)10 F AATCCTCAAAATCCTCCCAG S S N P 135 115-160 R CCGACGATGATACCTCTGAT SCVPLR1028H01.G SSRsucest_rst6 (CGC)6 F CTCAAGGTCGTCTTCTGCC N N N - 157 - R GGTCAGTTGAAGGGAGGATG SCRLLR1038G05.G SSRsucest_rst7 (TGG)6 F ACCTACAACCCACACCAAAG S S N P 292 295-310 R GGCGATAAACTTCTCAGCAT SCRUAD1132G05.B SSRsucest_rst8 (AG)16 F CTCAGTTGCTTCTACACGGTT S S S P 215 195-225 R GAGTCATCCTCTCCAACCAC SCSBHR1051E01.G SSRsucest_rst9 (GCG)6 F CTCAGATCCGTGACAACCTC S N N P 296 300-303 R TTCTCGTACCAAAGGACGG SCSFLR2031G10.G SSRsucest_rst10 (TG)11 F CTGCACTTTGTAGCCCTAGC N N N - 203 - R AGAGCCACCTCATCACAAGT C – cana-de-açúcar; M – Miscanthus; S – Sorgo. N – não amplificou; S – amplificou. P – polimórfico em cana-de-açúcar. Esp – tamanho de fragmento esperado; Obs – tamanho de fragmento observado. F – sequência do primer forward; R – sequência do primer reverse. * indica o número de repetições dos motifs microssatélites identificados dentro de parênteses.

43
Pode-se ainda tentar ajustar as condições da reação de PCR para estes primers
que não amplificaram. Oito dos dez pares de primers SSR desenvolvidos apresentavam
sequências repetitivas trinucleotídicas, que tendem a prevalecer em regiões de proteínas,
comparando-se a repetições dinucleotídicas e tetranucleotídicas (VARSHNEY et al.
2002).
Outros 12 pares de primers EST-SSR desenvolvidos por PINTO et al. (2006);
OLIVEIRA et al. (2009) e MARCONI et al. (2011) foram utilizados. No total foram
genotipados na população mapa 22 primers provenientes de marcadores EST-SSRs, que
geraram um total de 86 marcas polimórficas. O número de alelos polimórficos variou de
1 a 10, com média de quatro marcas polimórficas por marcador EST-SSRs (Tabela 5).
Tabela 5. Marcadores microssatélites derivados de regiões expressas do genoma de cana-de-açúcar que foram genotipados na população de mapeamento, número de alelos polimórficos obtidos e fonte das sequências. EST-SSR NºA.P Fonte EST-SSR NºA.P Fonte do primer
SCB81 5 OLIVEIRA et al. 2009 SCC22 6 PINTO et al. 2006
SCB184 4 MARCONI et al. 2011 SSR32 1 Não publicados
SCB73 5 OLIVEIRA et al. 2009 SSR37 1 Não publicados
SCC17 7 PINTO et al. 2006 SSR38-1 2 Não publicados
SCC05 5 PINTO et al. 2006 SSR52 1 Não publicados
SCB64 2 OLIVEIRA et al. 2009 CISP-5 10 PARK et al. 2012
SCB52 1 OLIVEIRA et al. 2009 SSR_sucest_rst4 5 Desenvolvido
SCB67 2 OLIVEIRA et al. 2009 SSR_sucest_rst5 6 Desenvolvido
SCB221 7 MARCONI et al. 2011 SSR_sucest_rst7 3 Desenvolvido
SCC19 4 PINTO et al. 2006 SSR_sucest_rst8 4 Desenvolvido
SCB97 4 OLIVEIRA et al. 2009 SSR_sucest_rst9 1 Desenvolvido
NºA.P – número de alelos polimórficos obtidos para cada marca genotipada.
Os marcadores microssatélites são um dos mais utilizados, pois são
codominantes, específicos e de alta reprodutibilidade. Os SSRs derivados de ESTs
podem ser associados com genes de função conhecida e serem utilizados como
marcadores SSR funcionais (CATO et al. 2001; SILVA 2001; HACKAUF
&WEHLING, 2002).
O projeto SUCEST utilizou 26 bibliotecas de cDNA e produziu 237.954 ESTs
que foram reunidos em 43.000 clusters (VETTORE et al. 2001). Primers
microssatélites têm sido desenvolvidos de banco de dados ESTs, como o SUCEST por

44
exemplo, (CORDEIRO et al. 2001; PINTO et al. 2004; PINTO et al. 2006; OLIVEIRA
et al. 2009; MARCONI et al. 2011).
4.2.2 AFLP-RGA Considerando-se os critérios de qualidade visual e polimorfismo para escolha
das combinações de primers, foram utilizados 20 combinações para a genotipagem da
população mapa. As combinações selecionadas foram nomeadas com a junção dos
códigos dos primers seletivos AFLP-EcoR1 e RGAs, e estão descritos na Tabela 6, que
apresenta ainda o número de alelos totais observados e o número de alelos
polimórficos genotipados para cada combinação utilizada.
Tabela 6.Combinações de pares de primers AFLP-RGA genotipados na população mapa, número total de alelos observados e número de alelos polimórficos genotipados por combinação de primer.
Combinações de primers utilizadas Nº Alelos Observados Nº Alelos Genotipados
AFLP RGA Eco-AAA NLRR_F 33 06
Eco-AAA PTO2 53 08
Eco-AAA NLRR_R 29 06
Eco-AAC XLRR_F 56 12
Eco-AAC PTO1 28 11
Eco-AAG XLRR_F 20 03
Eco-AAG NLRR_F 42 01
Eco-AAG NLRR_R 35 01
Eco-ACA NLRR_R 45 11
Eco-ACA XLRR_F 16 02
Eco-ACC XLRR_R 42 17
Eco-ACC RLRR_F 43 08
Eco-ACG NLRR_R 31 05
Eco-ACG XLRR_F 36 07
Eco-ACG PTO2 43 15
Eco-ACT RLRR_F 28 10
Eco-ACT XLRR_F 41 07
Eco-AGA PTO1 33 06
Eco-AGA XLRR_F 62 12
Eco-AGG PTO2 46 11
Total 762 159
Um total de 762 alelos pôde ser observado, tendo variado de 16 a 62 por
combinação de primer, com média de 38 alelos por combinação de primer. O número

45
total de alelos polimórficos genotipados foi 159, com média de oito alelos polimórficos
por loco. A amplitude do número de alelos polimórficos genotipados por loco foi
grande, variando de um alelo (Eco-AAG_NLRR-F e Eco-AAG_NLRR-R) até 17 alelos
polimórficos (Eco-ACC_XLRR-F).
ZHANG et al. (2007) utilizaram vinte e duas combinações de primers AFLP-RGA em
duas espécies de algodão cultivado, Gossypium hirsutum L. e G. barbadense L.,
obtendo 446 bandas amplificadas, sendo 17% e 8,3% de bandas polimórficas dentro de
cada uma das espécies, respectivamente, e 57,4% de polimorfismo entre as espécies. Ao
se comparar os dados de AFLP-RGA com dados obtidos de marcadores AFLP e RGA
separadamente para obtenção de similaridade entre os genótipos avaliada os autores
consideram AFLP-RGA e AFLP apresentaram resultados semelhantes entre os oito
genótipos utilizados, já as quatro combinações de marcadores RGAs não puderam
discriminar os genótipos.
Os resultados obtidos no presente trabalho foram gerados a partir de 220 indivíduos
altamente poliploides, o que pode ter gerado um maior número de fragmentos
amplificados do que no trabalho de ZHANG et al. (2007). Em relação ao número de
bandas polimórficas estes autores identificaram mais polimorfismo entre os oito
indivíduos das duas diferentes espécies de algodão do que pudemos encontrar no
presente estudo, mesmo sendo a cana-de-açúcar cultivada proveniente de hibridação
interespecífica entre S. officinarum e S. spontaneum, as sucessivas gerações de
retrocruzamento voltadas para a cana nobre, S. officinarum, tornaram os híbridos atuais
formados por 80% do genoma desta espécie (D’HONT et al., 1996). Tal fato, aliado ao
fato de estarmos lidando com uma população de cruzamento biparental, pode ser
responsável pela menor observação de bandas polimórficas na população mapa em
estudo, mesmo o número de indivíduos sendo expressivamente maior do que o utilizado
por ZHANG et al. (2007). Além disso, no presente trabalho para a genotipagem da
população eram necessárias a realização de três PCRs e quatro géis de poliacrilamida,
fato que contribuía para a dificuldade de genotipagem.
4.2.3 CISP Nenhuma das quatro enzimas de restrição que poderiam identificar
polimorfismo entre os produtos de PCR dos genitores mostraram diferença entre os
tamanhos de fragmentos obtidos, e não puderam, portanto, ser utilizadas para
discriminar polimorfismo entre os genótipos.

46
4.2.4 TRAP Os resultado dos testes de amplificação e polimorfismo dos marcadores TRAPs
estão disponíveis na Tabela 7.
Tabela 7. Resultado de amplificação das 64 combinações de primers TRAP testadas em genótipos de Cana-de-açúcar, Miscanthus e Sorgo.
TRAPs
Arbitrário 1 Arbitrário 2 Arbitrário 3 Arbitrário 4 ca
na
mis
can
thu
s
sorg
o
can
a
mis
can
thu
s
sorg
o
can
a
mis
can
thu
s
sorg
o
can
a
mis
can
thu
s
sorg
o
TRAP_Sucest_1 N N N S S S N N N S S S
TRAP_Sucest_2 N N N S S S N N N S S S
TRAP_Sucest_3 N N N S S S S S S S S S
TRAP_Sucest_4 N N N S S S N N S S S S
TRAP_Sucest_5 N N N S S S S S S S S S
TRAP_Sucest_6 S S S S S S S S S S S S
TRAP_Sucest_7 S S S S S S S S S N N N
TRAP_Sucest_8 S S S N N N S S S N N N
TRAP_Sucest_9 S S S S S S S S S S S S
TRAP_Sucest_10 S S S S S S S S S S S S
TRAP_Sucest_11 S S S S S S S S S S S S
TRAP_Sucest_12 S S S S S S S S S S S S
TRAP_Sucest_13 S S S S S S S S S S S S
TRAP_Sucest_14 S S S S S S S S S S S S
TRAP_Sucest_15 S S S S S S S S S S S S
TRAP_Sucest_16 S S S S S S S S S S S S
N- não amplificou, S- amplificou.
Todos os marcadores TRAPs que amplificaram em cana também amplificaram
em Miscanthus e em sorgo, e uma combinação de primer amplificou apenas o sorgo.
Miscanthus é um gênero pertencente ao complexo Saccharum e estes primers poderão
ser utilizados em estudos de diversidade genética do complexo e gêneros relacionados,
como o sorgo.
Foram selecionados para genotipagem da população de mapeamento 25
combinações de primers mais polimórficas e com melhor qualidade de visualização dos
fragmentos, estando estas relacionadas na Tabela 8.
Foram genotipados 293 alelos polimórficos, sendo que o número de alelos
genotipados por combinação de primers TRAPs variou de um a 24, com média de

47
11,7alelos polimórficos por combinação. CRESTE et al. (2010) utilizaram 21
marcadores TRAPs para acessar a variabilidade genética entre genótipos de cana com
primers fixos desenvolvidos para metabolismo de sacarose e nove marcadores TRAPs
com primers fixos desenvolvidos para genes relacionados à tolerância a seca; obtiveram
a média de 16,2 e 17,2 alelos polimórficos por marcador, respectivamente, média maior
provavelmente pelo fato de terem utilizado genótipos de banco de germoplasma, sendo
que a utilização da população de mapeamento permite acessar o polimorfismo
proveniente de apenas dois genitores.
Tabela 8.Combinações de primers TRAPs genotipados nos 220 genótipos da população de mapeamento. As combinações genotipadas estão identificadas número de alelos polimórficos genotipados para a combinação em questão entre parênteses. TRAPs Primers arbitrários
Primer específico Arbitrário 1 Arbitrário 2 Arbitrário 3 Arbitrário 4
TRAP_Sucest_1 (10)
TRAP_Sucest_2 (11)
TRAP_Sucest_3 (13) (3)
TRAP_Sucest_4 (8) (7)
TRAP_Sucest_5 (14) (16)
TRAP_Sucest_6 (9)
TRAP_Sucest_7
TRAP_Sucest_8 (14) (24)
TRAP_Sucest_9 (7) (18)
TRAP_Sucest_10 (9) (3)
TRAP_Sucest_11
TRAP_Sucest_12 (5) (20)
TRAP_Sucest_13 (10) (16)
TRAP_Sucest_14 (15) (12)
TRAP_Sucest_15 (17) (12)
TRAP_Sucest_16 (13) (1)
SUMAN et al. (2012) utilizando 16 combinações de TRAPs com primers fixos
para lignina, obtiveram 99% das bandas amplificadas polimórficas. No presente
trabalho não contabilizamos o número de fragmentos amplificados, porém visivelmente
o número de bandas monomórficas é muito superior ao de bandas polimórficas. Assim
como relatado por HU & VICK (2003) os fragmentos chegaram até cerca de 900 pares
de bases, porém não foi possível realizar a leitura de bandas maiores, uma vez que estas
se tornam difusas no gel.

48
Os marcadores TRAPs têm sido utilizados principalmente para acessar a
diversidade genética do complexo Saccharum ou das espécies do gênero Saccharum e
variedades comerciais (ALWALA et al., 2006, CRESTE et al., 2010e SUMAN et al.,
2012). SUMAN et al. (2012) e ALWALA et al. (2006) sequenciaram alguns dos
fragmentos gerados pelos marcadores TRAPs utilizados, num esforço para verificar se o
TRAP corresponde à sequência do gene candidato para o qual o primer fixo foi
desenhado, e confirmaram que muitos dos fragmentos apresentaram relação positiva. A
técnica foi também utilizada em uma população recombinante de trigo (LIU et al.
2005).
Considerando a soma dos alelos obtidos dos 22 primers EST-SSR, das 20
combinações de AFLP-RGA e das 25 combinações de TRAP foram genotipados na
população de mapeamento 538 alelos polimórficos (Tabela 9).
Tabela 9. Marcadores moleculares utilizados, número de pares de primers ou combinações utilizadas na população de mapeamento genético e número de alelos polimórficos genotipados. Marcador Nº primers utilizados Nº de alelos genotipados
EST-SSR 22 86
AFLP-RGA 20 159
TRAP 25 293
Total 67 538
No total foram utilizados 1363 alelos para a construção do mapa genético,
sendo os 538 gerados neste trabalho e mais 824 alelos já genotipados para este mesmo
cruzamento.
4.3 Análises de segregação e formação dos grupos de ligação
Após as análises de segregação dos 1363 alelos, 784 apresentaram segregação
1:1 ou 3:1 e foram considerados para o mapeamento. A distorção de segregação de
marcas encontrada foi de 42%,maior do que as observadas por RABOIN (2006) e
GARCIA et al. (2006) que obtiveram distorção de segregação de 36% e 33%
respectivamente, usando marcadores AFLP, SSR e RFLP, maior também do que a
obtida por PALHARES et al. (2012), de 26% utilizando AFLP e EST-SSR, e
semelhante aos 41% apresentada por AITKEN et al. (2005) que utilizaram AFLP e

49
SSR. A proporção de polimorfismo em dose única detectada é relacionada à segregação
de cada loco e sua dosagem (da SILVA et al. 1993), portanto a alta distorção de
segregação em dose única observada neste cruzamento biparental pode indicar que a
população segregante utilizada apresenta variados níveis de ploidia e aneuploidia, fato
este comum em cana-de-açúcar. Inclusive, para esta mesma população de cruzamento
(IACSP95-3018 e IACSP93-3046), SERANG et al. (2012), utilizando SNPs e
algoritmos para predizer a ploidia da população, puderam observar grande variabilidade
de ploidia entre os indivíduos.
Para a análise de ligação dos locos as 784 marcas em dose única foram
classificadas de acordo com a origem do alelo, conforme a notação de WU et al. (2002)
e utilizadas no OneMap. Nesta notação os alelos provenientes do genitor IACSP95-
3018 foram denominados de D1.13 (configuração “ao x oo”); os alelos oriundos do
genitor IACSP93-3046 foram denominados D2.18 (configuração “oo x ao” ) e os alelos
provenientes de ambos os genitores foram denominados como C.8 (configuração “ao x
ao” ) sendo aa presença do alelo e o a ausência dele para todas as configurações.
Das 784 marcas, foram designadas 361, 215 e 208 marcas como C.8, D1.13 e
D2.18, respectivamente (Tabela 10). Foi então criada uma matriz contendo os locos e as
informações alélicas dos 220 indivíduos.
Tabela 10. Notação atribuída aos alelos de acordo com WU et al. (2002), configuração alélica das notações, número de alelos pertencentes à cada uma das notações e a porcentagem destes em relação ao total de 784 marcas em dose única. Notação Configuração Número de alelos %
D1.13 ao x oo 215 27,4%
D2.18 oo x ao 208 26,5%
C.8 ao x ao 361 46,1%
A partir dos critérios estabelecidos para a formação dos grupos 321 marcas não
foram ligadas a nenhuma outra (41%), valor semelhante aos 40% de marcas disponíveis
não ligadas no mapa construído por OLIVEIRA et al. (2007), as demais 463 marcas
foram distribuídas em 113 grupos de ligação, que variaram de duas marcas por grupo
(49 grupos) a 20 marcas (um grupo). A Tabela 11 apresenta a variação no número de
marcas obtida por grupo, sendo que oito grupos apresentaram ao menos dez marcas
ligadas.

50
O número de grupos de ligação obtido foi próximo ao número cromossômico
existente nas cultivares modernas de cana-de-açúcar, que apresentam entre 100 a 130
cromossomos (GRIVET & ARRUDA 2001; HOARAU et al. 2001) e corrobora com o
número de grupos de ligação encontrados em mapa genéticos de cana, AITKEN et al.
(2005) formaram 136 grupos de ligação, PALHARES et al. (2012) obtiveram 107
grupos e PASTINA et al. (2012) obtiveram 96 grupos.
Tabela 11. Número de grupos de ligação formados e número de marcas ligadas em cada grupo.
Número de GL Número de marcas ligadas no GL 49 2 21 3 12 4 10 5 7 6 3 8 3 9 2 11 1 12 2 13 1 14 1 17 1 20
GL – Grupo de Ligação
O número de marcas não ligadas, o tamanho reduzido da maioria dos grupos de
ligação e poucas marcas por grupo indicam que o mapa não está saturado e que
provavelmente a maioria dos grupos de ligação representam grupos não conectados
(PASTINA et al., 2012), este padrão vem sendo encontrado nos mapas genéticos de
cana-de-açúcar e demonstram a dificuldade de saturar os mapas e detectar portanto
regiões de QTLs com maior precisão. Segundo GARCIA et al. (2006), a presença de
regiões com gaps nos grupos de ligação é esperada, uma vez que apenas os marcadores
em dose única foram utilizados para o mapeamento. Isso resulta numa limitação do
mapa apenas em regiões onde esses marcadores são encontrados (AITKEN et al., 2007).
Os marcadores SNPs (single nucleotide polymorphism) são provavelmente
indicados para acessar mais regiões do genoma devido ao seu alto grau de
polimorfismo, contribuindo para uma maior saturação do mapa. Esta mesma população
está sendo utilizada para detecção de SNPs e nos próximos anos o número de marcas
ligadas crescerá (Luciana Rossini -IAC, comunicação pessoal).

51
Como exemplo, o grupo de ligação 46 teve oito marcas ligadas e seu heatmap,
que é a matriz gráfica do LOD e das frações de recombinações pode ser visualizado na
Figura 12. No exemplo é possível ver as cores quentes ao centro, e esfriando conforme
as marcas estão nas extremidades do grupo de ligação. Quando o heatmap apresenta
marcas ligadas com cores frias podemos tomar a decisão de dividir o grupo ou não.
Figura 12 Heatmap do grupo de ligação 46, composto por oito marcas que apresentam
uniformidade de cores, sugerindo que a ordenação obtida para os marcadores está
adequada.
De forma geral, o mapa genético não apresentou grandes problemas de ligação
e, não sendo necessário dividir grupos de ligação, o mapa final da população em estudo
(Figura 13) apresentou, portanto 113 grupos de ligação, com um comprimento total de
6.178,6 cM, praticamente o mesmo comprimento observado no mapa da população
SP80-180 x SP80-4966 desenvolvido por OLIVEIRA et al. (2007), de 6.261,1cM.O
cumprimento dos grupos variou de 0,0 (grupo 94) a 398,29 (grupo 38), com média de
54,67 cM por grupo de ligação. O grupo 94 é composto por apenas duas marcas, sendo
ambas derivadas de um mesmo marcador, que provavelmente remetem a um mesmo
alelo, explicando o valor de 0,0 cM.
A ligação entre marcas provenientes dos genitores IACSP95-3018 e IACSP93-
3046 por marcadores biparentais (codificação C.8) ocorreu em oito dos 113 grupos de
ligação. Essa integração de marcas foi próxima às dez observadas por PALHARES et
al. (2012) na integração entre marcas oriundas dos genitores IAC 66-6 e TUC 71.

52
Entretanto, OLIVEIRA et al. (2007) observaram apenas um grupo consistentemente
integrado por marcas dos dois genitores através de marca biparental. Os três mapas
genéticos aqui comparados são provenientes de diferentes cruzamentos, e conforme
PALHARES et al. (2012) a diferença no número de integração das marcas pode ser um
fato inerente à população estudada, como também ao incremento das análises
multiponto pelo software OneMap, uma vez que no trabalho de OLIVEIRA et al. (2007)
esta melhora não estava ainda disponível no software.

53
Figura 13 Mapa de ligação integrado para a população derivada do cruzamento entre os genitores IACSP93-3018 e IACSP95-3046. Os grupos
estão indicados com o número acima. À esquerda estão as posições estimadas e à direita o código atribuído aos marcadores. Em negrito as
marcas associadas negativamente (resistência) e sublinhado as marcas com associação positiva (suscetibilidade) com a ferrugem marrom.
1 2 3 4 5
6 7 8 9 10

54
Continuação.
11 12 13 14 15
16 17 18 19 20

55
Continuação.
21 22 23 24 25
26 27 28 29 30

56
Continuação.
31 32 33 34 35

57
Continuação.
36 37 38 39 40 41
42 43 44 45 46 47

58
Continuação.
48 49 50 51 52
53 54 55 56 57

59
Continuação
58 59 60 61 62
63 64 65 66 67 68
69 70 71 72 73 74

60
Continuação
75 76 77 78 79 80
81 82 83 84 85 86
87 88 89 90 91 92

61
Continuação
93 94 95 96 97 98
99 100 101 102 103 104
105 106 107 108 109 110
111 112 113

62
4.4 Análise de Marca Individual
A análise de marca individual para detecção de possíveis QTLs foi realizada
separadamente para cada uma das seis avaliações fenotípicas, sendo em Ribeirão Preto
duas vezes na mesma cana planta em tempos diferentes, também em Ribeirão Preto,
duas avaliações em tempos diferentes em cana-soca e em Piracicaba duas avaliações em
tempos diferentes em cana planta.
Foram utilizados todos os alelos com segregação, mesmo os não ligados ao
mapa, uma vez que a análise de marca simples utiliza cada marca isoladamente. Os
alelos que tiveram associação significativa com as notas de ferrugem, assim como o
efeito (R²) de cada um deles no aumento/diminuição das notas em % estão
demonstrados na Tabela 12.Cento e setenta e sete alelos foram associados com 5% de
nível de significância (p<0,05)considerando as seis avaliações fenotípicas, variando no
mínimo em34 até 63associações significativas por avaliação. O número de associações
com nível de significância a 1% (p<0,01) foi de 59 para todas as avaliações, variando de
6 a 16 por época avaliada. Todas as diferentes avaliações apresentaram marcas
associadas exclusivas. Dentre os alelos avaliados, 116 alelos associados estavam ligados
a algum dos 113 grupos de ligação do mapa gerado, representando 62% das marcas
associadas mapeadas. O grupo de ligação GL2 foi formado com 13 marcas, e foi o que
apresentou maior número de marcas associadas à ferrugem (6), e dez associações
considerando todas as avaliações.
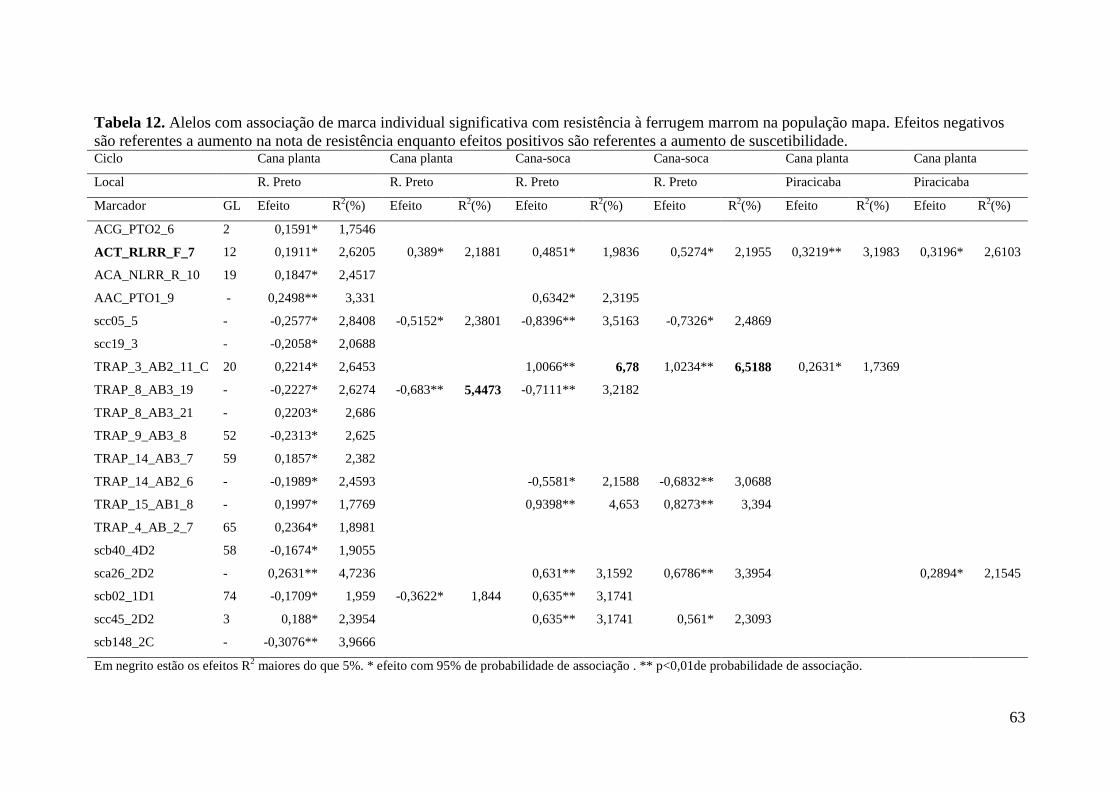
63
Tabela 12. Alelos com associação de marca individual significativa com resistência à ferrugem marrom na população mapa. Efeitos negativos são referentes a aumento na nota de resistência enquanto efeitos positivos são referentes a aumento de suscetibilidade. Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
ACG_PTO2_6 2 0,1591* 1,7546
ACT_RLRR_F_7 12 0,1911* 2,6205 0,389* 2,1881 0,4851* 1,9836 0,5274* 2,1955 0,3219** 3,1983 0,3196* 2,6103
ACA_NLRR_R_10 19 0,1847* 2,4517
AAC_PTO1_9 - 0,2498** 3,331 0,6342* 2,3195
scc05_5 - -0,2577* 2,8408 -0,5152* 2,3801 -0,8396** 3,5163 -0,7326* 2,4869
scc19_3 - -0,2058* 2,0688
TRAP_3_AB2_11_C 20 0,2214* 2,6453 1,0066** 6,78 1,0234** 6,5188 0,2631* 1,7369
TRAP_8_AB3_19 - -0,2227* 2,6274 -0,683** 5,4473 -0,7111** 3,2182
TRAP_8_AB3_21 - 0,2203* 2,686
TRAP_9_AB3_8 52 -0,2313* 2,625
TRAP_14_AB3_7 59 0,1857* 2,382
TRAP_14_AB2_6 - -0,1989* 2,4593 -0,5581* 2,1588 -0,6832** 3,0688
TRAP_15_AB1_8 - 0,1997* 1,7769 0,9398** 4,653 0,8273** 3,394
TRAP_4_AB_2_7 65 0,2364* 1,8981
scb40_4D2 58 -0,1674* 1,9055
sca26_2D2 - 0,2631** 4,7236 0,631** 3,1592 0,6786** 3,3954 0,2894* 2,1545
scb02_1D1 74 -0,1709* 1,959 -0,3622* 1,844 0,635** 3,1741
scc45_2D2 3 0,188* 2,3954 0,635** 3,1741 0,561* 2,3093
scb148_2C - -0,3076** 3,9666
Em negrito estão os efeitos R2 maiores do que 5%. * efeito com 95% de probabilidade de associação . ** p<0,01de probabilidade de associação.

64
Tabela 12.Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2 (%) Efeito R2 (%) Efeito R2 (%) Efeito R2 (%) Efeito R2 (%)
scb130_3C 83 -0,2012* 2,0598
scb130_10D1 - -0,1966* 2,6367 -0,2922* 2,1954
scb312_8D2 51 0,1717* 2,0146
scb292_6C 10 -0,2031* 2,372 -0,6987** 3,0318
scb268_4D1 - 0,1703* 1,9543 0,3546* 1,7778
scb208_4D2 67 0,1941* 2,5571 0,5644* 2,5166 0,5796* 2,4668
scb208_5D2 67 0,1916* 2,4876 0,4988* 1,8249
scb208_12D1 - -0,174* 1,8854
scb248_3C - 0,2395* 1,9035 0,5138* 1,8066
scb259_6C 2 0,1915* 1,7651 0,5166* 2,693 0,7387** 3,0525
Cir1_1C 101 -0,279* 2,4232
Cir12_2D1 2 -0,1616* 1,7582
Cir12_4C 101 -0,2638* 2,4399
Cir14_1D1 44 -0,1831* 2,2413
Cir35_2C - -0,2379* 2,7091
Cir35_4D1 19 0,1804* 2,3323 0,237* 1,9891
Cir36_7D2 51 0,2242** 3,5012
Cir50_1C - 0,2226* 2,2734 0,4267* 1,7518 0,6998* 2,6126 0,7575* 2,8455
Cir51_1D2 85 -0,1686* 1,9492 -0,5845* 2,6885 -0,5429* 2,1545 -0,2955* 2,7621

65
Tabela 12. Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
SMC415MS_7D2 57 -0,1623* 1,8345
SMC863CG_7D2 - -0,2569** 4,7691 -0,7782** 4,5649 -0,7491** 4,0212
aagcat26D2 - -0,1954* 2,5918 -0,5231* 2,1734 -0,5074* 1,8994
aagcat52C - -0,2154* 1,9078 -0,8889** 3,8015 -0,9294** 3,86
actctt11C - -0,2168* 2,647 -0,4928* 2,8695 -0,584* 2,2797
aagcag1D2 3 0,1883* 2,3338
aaccat1D1 19 0,1678* 1,8284
acgcag17C 80 0,2351** 2,9766
agcctc1D1 26 -0,1791* 2,0134
agcctc21D2 - -0,1756* 1,9309 -0,4818* 1,8186
accctc19C - 0,2118* 1,9335
agccag22C - 0,2088* 2,0703
TRAP_16_AB1_1 2 0,1603* 1,7872
TRAP_16_AB1_11 - 0,2099* 1,9243 0,5233* 2,4317 0,6533* 2,101
TRAP_8_AB1_6 97 0,1957* 2,4102 0,3655** 1,8964
TRAP_8_AB1_11 - 0,1848* 2,2123
ACA_NLRR_R_4 - -0,4428* 2,0486
ACT_XLRR_F_4 29 0,4227* 1,8075
scb221_2 41 0,4229* 1,8065

66
Tabela 12. Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
EST_Sucest_rst7_2_C - -0,9389** 6,4673
TRAP_5_AB3_6 - 0,4928* 1,8924
TRAP_16_AB_3_1 18 0.3818* 2,1220
TRAP_3_AB2_1 34 0,3835* 2,0781
TRAP_9_AB3_1 50 0,4186* 2,5728 0,2749* 1,9391
TRAP_14_AB2_10 60 0,5991* 2,7267
TRAP_14_AB2_11 - 0,5486* 1,9083 0,3783* 1,9488
scb100_2C 29 0,6432** 3,9252 0,5932* 1,8617 0,6079* 1,8159
scc36_3C - 0,553* 1,9991 0,8263* 2,469 0,889* 2,6565
sca31_2D1 31 0,3621* 1,8203 -0,3362* 2,8206
scb60_1D1 76 0,3565* 1,8106 0,5247* 2,1866
sca53_7D2 35 0,3899* 2,1634 0,2566* 2,0626 0,2733* 1,9111
scb01_3C 56 -0,5104* 2,5074 -0,6735* 2,436 -0,6719* 2,2512
scb292_12D1 2 0,3806* 2,0672 0,2684* 2,2628
scb292_13D1 2 0,3806* 2,0672 0,2684* 2,2628
scb288_2C - 0,7207** 4,1953
scb246_4D2 - 0,3788* 2,0342 0,2497* 1,9548 0,2931* 2,1926
scb226_5D2 95 0,3893* 2,1454
scb180_2C - -0,463* 2,049
scb194_1D1 38 -0,3553* 1,8032

67
Tabela 12Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
scb232_1C - -0,4635* 2,612
scb236_5D1 97 0,3892* 2,1232
Cir12_7D2 102 -0,4976** 3,7237 -0,3501** 3,2573
Cir21_1D2 103 0,4152* 2,5625 0,5358* 2,2939 0,5547* 2,3065
Cir31_2C - -0,5537** 3,5922
Cir36_2D1 51 0,3482* 1,7911
SMC843_1C 29 0,5151* 2,5527
SMC843_5D2 16 0,3686* 1,9454
SMC863CG_6C 35 0,6392** 3,2853 0,8178* 2,9031 0,8626** 3,0294
actctt8D1 - -0,4791* 1,9809 -0,5632* 2,5231 -0,2639* 2,2544 -0,3659** 3,3836
aagcag40C 102 -0,4416* 2,743 -0,5943* 2,0748 -0,5683* 1,7624
aaccat8C - -0,4938* 2,5702
aaccaa14D2 112 0,5783* 2,1628 -0,4938* 1,9236
agccag13D2 - -0,4533* 2,8999
TRAP_8_AB1_11 - 0,5089** 3,4668
ACC_XLRR_R_8 4 0,9938** 7,4777 1,1137** 8,7109 0,3817** 3,4256
ACT_RLRR_F_5 10 -0,5568* 2,4727 -0,6479** 3,1144
AAC_XLRR_F_7 - 0,5483* 2,2036 0,5462* 2,0354
TRAP_10_AB_3_8 20 0.7377* 2,3630 0.9681** 3,8260
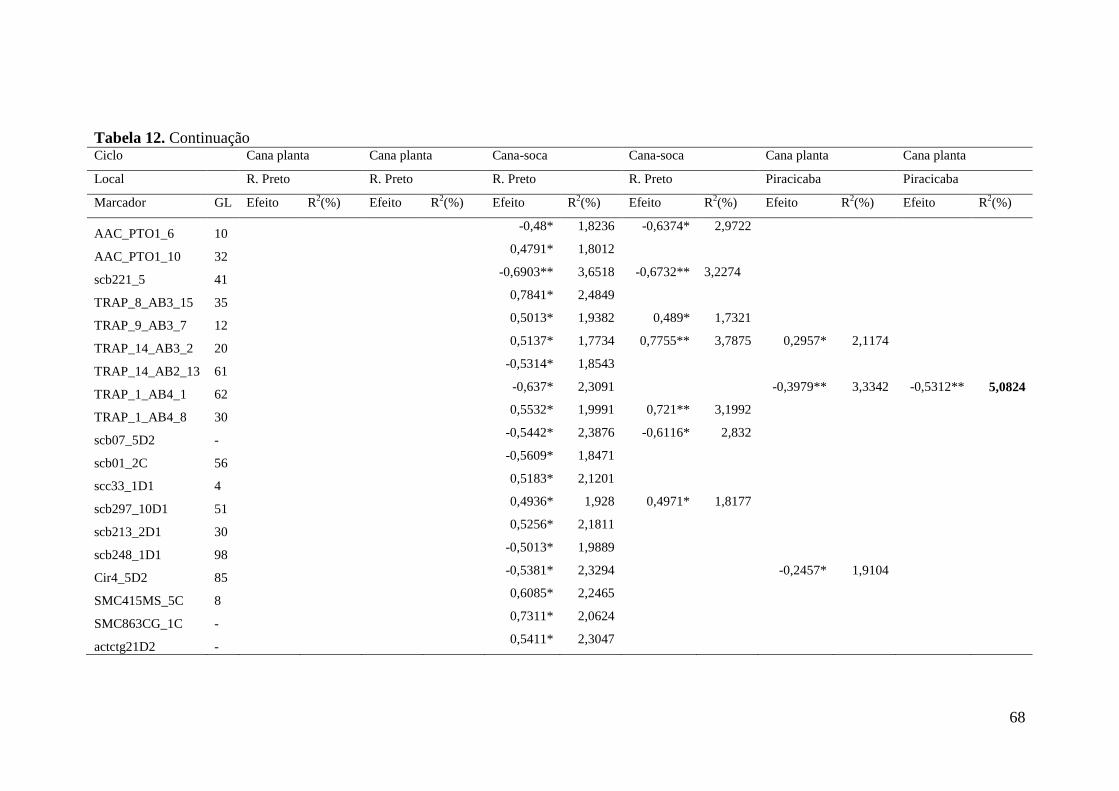
68
Tabela 12. Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
AAC_PTO1_6 10 -0,48* 1,8236 -0,6374* 2,9722
AAC_PTO1_10 32 0,4791* 1,8012
scb221_5 41 -0,6903** 3,6518 -0,6732** 3,2274
TRAP_8_AB3_15 35 0,7841* 2,4849
TRAP_9_AB3_7 12 0,5013* 1,9382 0,489* 1,7321
TRAP_14_AB3_2 20 0,5137* 1,7734 0,7755** 3,7875 0,2957* 2,1174
TRAP_14_AB2_13 61 -0,5314* 1,8543
TRAP_1_AB4_1 62 -0,637* 2,3091 -0,3979** 3,3342 -0,5312** 5,0824
TRAP_1_AB4_8 30 0,5532* 1,9991 0,721** 3,1992
scb07_5D2 - -0,5442* 2,3876 -0,6116* 2,832
scb01_2C 56 -0,5609* 1,8471
scc33_1D1 4 0,5183* 2,1201
scb297_10D1 51 0,4936* 1,928 0,4971* 1,8177
scb213_2D1 30 0,5256* 2,1811
scb248_1D1 98 -0,5013* 1,9889
Cir4_5D2 85 -0,5381* 2,3294 -0,2457* 1,9104
SMC415MS_5C 8 0,6085* 2,2465
SMC863CG_1C - 0,7311* 2,0624
actctg21D2 - 0,5411* 2,3047

69
Tabela 12. Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
aagcat2D2 - -0,648** 3,1208 -0,6426* 2,8506
aagcat13D1 - -0,5636* 2,3207 -0,6774** 3,1133
actctt13D1 - -0,6876** 3,7822 -0,582* 2,5173
aaccaa37C 20 0,5892* 2,3136 0,6176* 2,3606 0,3372** 3,1192
acgctt7D1 10 -0,5009* 1,8687 -0,6637** 3,0529
agactg13D1 4 0,5408* 2,3231
actcat35D2 - 0,5527* 2,4045 0,5567* 2,2389
TRAP_8_AB1_8 8 0,506* 1,8987 0,5415* 2,035 0,2505* 1,7454
TRAP_9_AB2_4 - -0,8692* 2,7141 -0,8764* 2,601
TRAP_15_AB1_15 60 0,6* 1,8189
scb07_7C 68 -0,5246* 1,7391
scb311_4C 86 0,5652* 1,819
scb285_3D2 22 -0,4999* 1,8303
Cir12_6C 46 -0,541* 1,7518
Cir36_6D2 - 0,5164* 1,9742
SMC1047HA6D2 36 0,5224* 2,0048
acgctt36D1 10 -0,6426* 2,9264
acgcag19C 107 0,7565* 1,7611
ACT_RLRR_F_9 - 0,2631* 2,1497

70
Tabela 12. Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
Cisp5_2 - -0,4536** 3,521
TRAP_12_AB3_6 - 0,2898* 2,4141
TRAP_12_AB3_8 57 0,255* 1,8236
TRAP_13_AB2_7 - 0,4327* 2,7071 0,4739* 2,7212
TRAP_14_AB3_6 55 -0,3638* 2,8261
TRAP_15_AB1_6 - -0,3581* 2,417 -0,4851* 3,7277
TRAP_15_AB1_7 62 -0,3008* 1,7763
scb07_10D1 68 0,2415* 1,7974
sca47_1C 73 -0,3055* 2,1925
scb55_2C - 0,3985** 3,0726
scb43_5C - -0,4882** 4,0048 -0,5161* 3,6949
scb300_4D1 89 0,3384** 3,5869 0,3* 2,3028
scb189_2D1 2 0,339** 3,5659
scb270_7D2 7 0,2793* 2,451
Cir32_8D2 56 0,253* 2,0016
Cir32_10D1 19 0,2614* 2,1425
Cir35_1D2 56 0,2814* 2,7258 0,4261** 4,6696
Cir51_2C 73 -0,3012* 2,0783
SMC36Buq_5D1 7 -0,2708* 2,2966
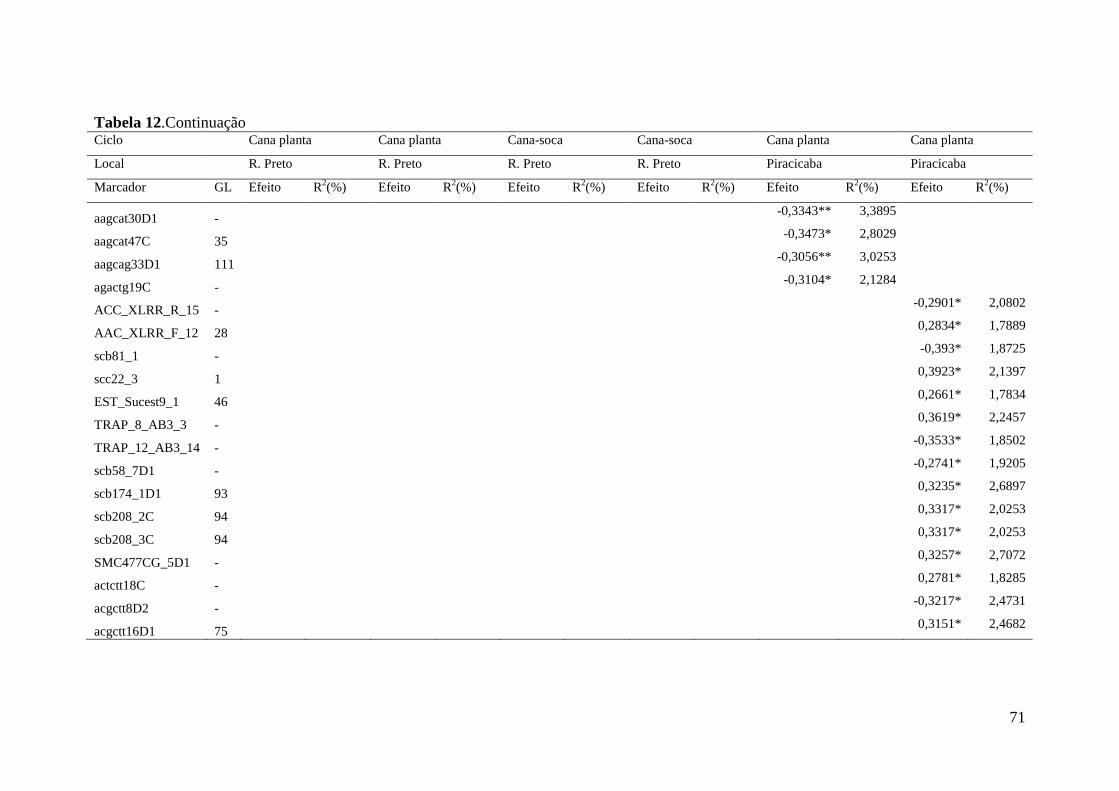
71
Tabela 12.Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
aagcat30D1 - -0,3343** 3,3895
aagcat47C 35 -0,3473* 2,8029
aagcag33D1 111 -0,3056** 3,0253
agactg19C - -0,3104* 2,1284
ACC_XLRR_R_15 - -0,2901* 2,0802
AAC_XLRR_F_12 28 0,2834* 1,7889
scb81_1 - -0,393* 1,8725
scc22_3 1 0,3923* 2,1397
EST_Sucest9_1 46 0,2661* 1,7834
TRAP_8_AB3_3 - 0,3619* 2,2457
TRAP_12_AB3_14 - -0,3533* 1,8502
scb58_7D1 - -0,2741* 1,9205
scb174_1D1 93 0,3235* 2,6897
scb208_2C 94 0,3317* 2,0253
scb208_3C 94 0,3317* 2,0253
SMC477CG_5D1 - 0,3257* 2,7072
actctt18C - 0,2781* 1,8285
acgctt8D2 - -0,3217* 2,4731
acgctt16D1 75 0,3151* 2,4682

72
Tabela 12. Continuação Ciclo Cana planta Cana planta Cana-soca Cana-soca Cana planta Cana planta
Local R. Preto R. Preto R. Preto R. Preto Piracicaba Piracicaba
Marcador GL Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%) Efeito R2(%)
acgcag21D1 110 0,2721* 1,8576
agccag24D2 - 0,2681* 1,8292
TRAP_9_AB_2_3 38 0,2777* 1,9819
TRAP_3_AB_3_2 6 0.4149** 4,2740
Total R2 99,9 (17,49) 99,5 (3,07) 100 (10,9) 98,8 (9,61) 99,9 (10,39) 98,8(10,77)
Número de
associações com P
< 0,05 (P < 0.01)
54(6) 49(8) 63(13) 49(16) 38(10) 34(6)
Número de
associações
comuns entre
avaliações da
mesma planta
7 36 10
Número de marcas
exclusivas
27(3) 23(5) 11(0) 10(0) 18(5) 15(1)

73
Foram detectadas associações positivas (57%), indicando que a marca está
relacionada ao aumento da severidade da doença, e negativas (43%), indicando que a
marca está associada à redução da nota da severidade da doença, ou seja, com plantas
mais resistentes.
Cinquenta e nove marcas foram associadas com nível significativo de 1%
(p<0,01), sendo 30 delas de associações negativas. Quarenta e nove por cento (29
alelos) destas associações ocorreram entre as duas avaliações de cana-soca e 16 (55%)
destas marcas em cana soca foram associações negativas.
A consistência de efeito positivo ou negativo dos locos que estiveram
associados em mais de uma avaliação foi alta, sendo que apenas três marcas tiveram
associações com efeitos contrários entre as avaliações. A Tabela 13 apresenta a relação
das 178 marcas associadas (as geradas neste trabalho e também aquelas que foram
adicionadas), separando o número de marcas por tipo de marcador e a porcentagem em
relação ao número de marcas geradas.
Tabela 13. Número de marcas associadas conforme o marcador molecular utilizado e porcentagem em relação ao número de marcas geradas por marcador. Marcas geradas neste trabalho Marcas previamente geradas Marcador
EST-SSR AFLP-RGA TRAP AFLP EST-SSR SSR Nº avaliação
associada
1 7 8 25 21 25 23
2 1 4 8 7 15 4
3 - 1 7 5 9 4
4 1 - 1 1 1 -
5 - - - - - -
6 - 1 - - - -
Total 9 14 41 34 50 30
% total marcas* 10,5 8,8 11,5 - - -
* Porcentagem de alelos associados à resistência à ferrugem em relação ao número total de marcas polimórficas de cada marcador utilizado neste trabalho.
Considerando-se os diferentes tipos de marcadores utilizados na análise de
associação à ferrugem que foram gerados neste trabalho, os marcadores TRAPs
apresentaram o maior número de associação (41 alelos), e maior porcentagem de
associação com a ferrugem em relação ao número de marcas polimórficas geradas
(11,5%). Utilizando marcadores TRAPs, LIU et al. (2005) identificaram QTLs para as
características altura e ZHANG et al. (2010) para a ferrugem em trigo.

74
A marca TRAP_3_AB2_11 foi associada positivamente em quatro épocas de
avaliação, sendo significativa a 1% nas duas avaliações de cana-soca, com variação
fenotípica de 6,7 e 6,5, (terceiro e quarto maiores valores observados), e seu primer fixo
foi desenhado em região que apresenta homologia no cromossomo quatro do sorgo,
onde o gene Bru1de resistência à ferrugem marrom é relacionado (ASNAGHI et al.,
2000). Este loco foi ligado no GL20, que, formado por mais outras cinco marcas, tem
duas delas (TRAP_14_AB3 e E-aac_M-caa37) também associadas positivamente à
ferrugem, ambas nas mesmas três épocas de avaliação (cana-soca avaliação 1 e 2 e cana
planta avaliação 1 Piracicaba). Estas três marcas associadas positivamente à ferrugem,
em pelo menos três épocas de avaliação, sugerem que esta seja uma região de QTL
relacionado à suscetibilidade da planta.
O marcador AFLP-RGA teve apenas 14 marcas associadas à ferrugem, porém
o único alelo que esteve associado (positivamente) em todas as seis épocas de avaliação
foi o AFLP-RGA (ACT_RLRR_F_7) e, os valores fenotípicos que foram explicados
nas seis associações variaram de 1,98 a 3,19, com média de 2,46.Considerando todos os
tipos de marcadores utilizados, os dois maiores valores fenotípicos (8,71 e 7,47)
explicados pela presença dos alelos são relacionados ao marcador AFLP-RGA
(ACC_XLRR_R_8). Ambos os valores fenotípicos estavam associados positivamente
em cana-soca; esse alelo ACC_XLRR_R_8 foi associado também a 1% em uma das
avaliações de cana planta em Piracicaba, com R2 de 3,4. Os primers RLRR e XLRR
foram desenvolvidos baseados em regiões repetitivas ricas em leucina do gene RPS2 do
patosistema arroz- Xathomonas compestris pv oryzae, entre as 14 associações com
primers AFLP-RGA cinco utilizam o primer XLRR e quatro utilizam o primer de
RLRR.
O par do primer RGA utilizado na combinação associada nas seis avaliações
(RLRR_R) foi utilizado, entre outros, por YAN et al. (2003) para identificar marcas
ligadas ao gene Yr5 relacionado à resistência a ferrugem em trigo (Puccinia striiformis
f. sp. tritici ), tendo esta marca co-segregado com o loco Yr5.Em cana-de-açúcar,
utilizando marcadores RFLP desenvolvidos a partir de regiões RGAs, ROSSI et al., em
2003, mapearam dois locos LRR-like próximos ao gene de resistência à ferrugem
localizado no mapa da cultivar R570 por ASNAGHI et al. (2000). Também em cana-de-
açúcar McINTYRE et al. (2005) desenvolveram e utilizaram 54 primers RGAs e
avaliaram a doença em uma população de mapeamento por dois anos, 3 marcas
mapeadas foram associadas à resistência a ferrugem nos dois anos.

75
ZHANG et al. (2007) ao implementar a técnica de AFLP-RGA em algodão
combinaram o primer AFLP Eco-ACG com o primer RGA Pto kin1, e obtiveram 19
fragmentos, sendo 15 deles polimórficos. Ao usarmos o primer RGA Pto kin2,
desenhado como par do Pto kin1(CHEN et al., 1998), foram contabilizadas 45 bandas,
provavelmente devido ao maior genoma da cana, e destas, também 15 alelos foram
polimórficos.
Entre as quatorze associações encontradas com alelos provenientes de AFLP-
RGA, quatro são provenientes de combinações como primer Pto kin, que desenvolvido
por CHEN et al. (1998), é baseado na sequência da proteína quinase do gene Pto de
tomate, que confere resistência à Pseudomonas syringae pv tomato, o gene Pto não
interage apenas com proteínas de avirulência de P. syringeae, mas também funciona
como um mediador de sinal de transdução. (MARTIN et al. 1993). A relação evolutiva
entre genes de resistência como o Pto kinase de tomate tem sido estudada (RIELY &
MARTIN, 2001; VLEESHOUWERS et al., 2001), porém como a maioria dos genes de
resistência codificam para NBS-LRR, os estudos evolucionários são primariamente
focados nesta classe (MEYERS et al., 2005).
Genes de variadas plantas relacionados à resistência a diversas doenças têm
sido intimamente relacionados ao gene Pto de resistência (XU & DENG 2010;
MARTINEZ et al. 2008; PERAZA-ECHEVERRIA et al. 2007; Di CASPRIO &
CIPRIANI 2003). Existem mais de 300 genes defensin-like em Arabdopsis, muitos
deles quinases que foram reportados estarem envolvidos em atividades de defesa
(SILVERSTEIN et al., 2005). Mas evidências genéticas indicam que outros genes
podem ser requeridos para a ativação da resposta de defesa, como é o caso do gene Prf,
necessário para a ativação do gene de avirulência Pto em tomate (SALMERON et al.,
1996; MACKEY et al., 2002; MUCYN et al., 2006) e uma das hipóteses da resistência à
Puccinia triticina em trigo entre os genes Lr21 e Lr40 (HUANG &GILL, 2001).
Comparado a bactérias, são poucos os exemplos estudados de genes de avirulência de
fungos, devido ao seu maior genoma e dificuldade de transformação.
Muitos possíveis QTLs foram relacionados à suscetibilidade a ferrugem
marrom, e ocorreu ainda de haverem efeitos positivo e negativos no mesmo grupo de
ligação formado, uma possível explicação para este fato seria a existência de epistasia, a
interação entre alelos de locos diferentes.
O número total de marcas associadas à ferrugem marrom foi bastante alto
quando comparado ao número de marcas associadas a outras características fenotípicas
descritas em cana de açúcar (MING et al., 2002; AITKEN et al., 2006; 2008; PINTO et

76
al., 2010) o alto valor pode ser atribuído à utilização do cruzamento biparental entre um
clone elite e uma variedade comercial, por acumularem alelos relacionados à
características de interesse no melhoramento genético.
O maior valor relativo ao aumento da resistência (efeito negativo) explicado
por marca associada (quinto maior efeito considerando ambos, positivo e negativo) foi
relacionado a um alelo EST-SSR (EST_Sucest_rst7_2), com R2 de 6,46. Este alelo foi
associado à somente uma época de avaliação (Ribeirão Preto, cana planta avaliação 2).
Também de efeito negativo, a marca Cir12_7 foi ligada a outra marca AFLP (aag-
cag_40) formando o grupo de ligação 102. Esta marca Cir12_7 está dentro da região de
saturação do gene Bru1 na cultivar R570 (COSTET et al.,2012).
O GL10 foi formado por oito marcas, cinco delas (SCB292_2;
ACT_RLRR_F_5; AAC_PTO1_6; E-ACG_M-CTT-7, e E-ACG_M_CTT_36) estão
associadas negativamente à ferrugem; sendo ligadas pelo alelo EST-SSR SCB292, cujo
alelo estava presente em ambos os genitores (configuração C), para as demais quatro
marcas apenas o genitor suscetível apresentava o alelo (configuração D1). Os cinco
alelos foram associados na segunda avaliação de cana-soca, sendo três delas a 1% e três
alelos foram associados também na primeira avaliação de cana-soca.
Os grupos de ligação foram atribuídos a grupos de homologia (GH) baseados
em marcadores microssatélites derivados de um mesmo loco (da SILVA et al. 1993).
Provenientes de 61 locos, as 261marcas microssatélites alocadas no mapa formaram 15
grupos de homologia. O número de grupos de homologia observados em mapas de
cana-de-açúcar variam entre oito (ROSSI et al., 2003; AITKEN et al., 2005;
PALHARES et al., 2012), 9 (SUMAN et al. 2011), 10 (GRIVET et al., 1996) e
11(PASTINA et al., 2012).
O número de GL em cada GH no presente trabalho variou de dois a 16 (Tabela
14), estando de acordo com o comumente observado, que variam de dois a mais de 20
(HOARAU et al. 2001; ROSSI et al. 2003; AITKEN et al. 2005; PASTINA et al.,
2012). D´HONT et al. (2010) comentam que não se sabe se a variação no número de GL
por GH reflete a real variação no número de cromossomos homólogos entre os GH de
cana-de-açúcar ou se reflete uma esparça cobertura de alguns cromossomos que são
representados por mais de um grupo de ligação. E para GH com grande número de GL
os autores comentam ainda que parte desta variação deve também refletir a diferença
básica no número de cromossomos nas espécies S. officinarum e S. spontaneum, que
deram origem aos atuais híbridos de cana.

77
Tabela 14. Grupos de ligação da população de cruzamento IACSP95-3018 e IACSP93-3046, e os grupos de homologia formados. Grupos de Homologia Grupos de Ligação Total I 1, 6, 7, 37, 43, 44, 61, 67, 68, 77, 81, 94, 98, 99,100 15
II 2, 10, 24, 30, 35, 46, 54, 71, 72, 76, 78, 79, 90, 101, 102, 107 16
III 4, 28, 55, 96, 104 5
IV 5, 23, 86, 87 4
V 8, 9, 19, 32, 56, 57, 63, 73, 74, 82, 83, 84, 85, 103, 105 15
VI 12, 56, 91 3
VII 14, 17, 58 3
VIII 16, 29, 69 3
IX 18, 41, 2
X 22, 51, 92 3
XI 31, 75 2
XII 34, 36, 38,50 4
XIII 39, 40 2
XIV 41, 66 2
XV 51, 88,92 3
Além da complexidade genética, a cana-de-açúcar é uma cultura perene em que
as plantas são usualmente colhidas em vários anos, assim as características são medidas
não apenas em diferentes locais, mas também ao longo de sucessivos cortes, e seus
QTLs são baseados em informações de séries de cortes e locais (PASTINA et al., 2012).
A avaliação de QTLs em cana-soca é extremamente importante, uma vez que,
segundo COLETI (2008) da área total de produção de cana, 80% são referentes à cana-
soca. O presente trabalho apresenta um mapa genético entre um cruzamento biparental
cuja progênie vem sendo estudada e em breve receberá novas marcas, além disso, traz
informações sobre novas marcas associadas à ferrugem em Ribeirão Preto e Piracicaba,
e em cana planta e cana soca.
Embora a presença do gene maior de resistência Bru1 pareça ser mantida ao
longo do tempo, o uso de fontes alternativas para a resistência em programas de
melhoramento é importante para ampliar a base genética da resistência (COSTET et al.,
2012; GLYNN et al., 2012). DAUGROIS et al. (1996) e RABOIN et al. (2006)
discutem que a presença destes genes de efeitos maiores já relatados não explicam
sozinhos a resistência, pois ainda assim há uma importante variação no nível de
suscetibilidade à ferrugem marrom. Casos em que a expressão fenotípica dos genes

78
representa pequenos valores podem ter na utilização de marcadores moleculares o
auxílio para a piramidação de genes (BRUNELLI et al. 2002). Com a saturação do
mapa genético apresentado será possível realizar uma avaliação mais robusta de QTLs
relacionados à ferrugem marrom, utilizando mapeamento por intervalo composto
(GAZAFFI, 2009), ampliando o conhecimento sobre esta população de cana-de-açúcar.
5 CONCLUSÃO
Existe variabilidade genética para a resistência à ferrugem marrom e para
construção de mapa genético de ligação para a população segregante derivada do
cruzamento biparental entre o clone IACSP95-3018 e a cultivar IACSP93-3046 do
programa de melhoramento genético do Instituto Agronômico (IAC).
Este trabalho evidencia as potencialidades do uso de marcas geradas a partir de
marcadores TRAPs, AFLP-RGA e EST-SSR, e genômicos AFLP e SSR para a
construção de mapa em cana-de-açúcar. O número de grupos de ligação gerado no mapa
está de acordo com o esperado para a cana-de-açúcar.
As marcas foram eficientes para a identificação de alelos associados à resistência
à ferrugem marrom, identificando 177 marcas diferentes em associação com a ferrugem,
e 346 associações considerando todas as seis diferentes avaliações fenotípicas.
Muitos alelos (59) tiveram alta associação (p<0,01) com a ferrugem marrom, e
uma marca foi associada com todas as seis avaliações, tendo influencia no aumento da
severidade da doença. As marcas desenvolvidas no presente trabalho podem ainda ser
testadas com novas fenotipagens relacionadas a doenças e outros caracteres de interesse
agronômico nesta população, ampliando a busca por QTLs. A validação destas marcas
em cultivares de resistência e suscetibilidade previamente conhecidas poderá auxiliar na
confirmação das associações de marca e fenótipo.

79
6 REFERÊNCIAS BIBLIOGRÁFICAS
AITKEN, K. S.; JACKSON, P. A.; MCINTYRE, C. L. A combination of AFLP and
SSR markers provides extensive map coverage and identification of homo(eo)logous
linkage groups in a sugarcane cultivar. Theoretical and Applied Genetics, New York,
v.110, p.789-801, 2005.
AITKEN, K. S.; JACKSON, P. A.; MCINTYRE, C. L. Quantitaive trait loci identified
for in a sugarcane (Saccharum spp.) cultivar x Saccharum officinarum population.
Theoretical and Applied Genetics, New York, v.112, p.1306-1317, 2006.
AITKEN, K. S.; JACKSON, P. A.; MCINTYRE, C. L. Construction of a genetic
linkage map for Saccharum officinarum incorporating both simplex and duplex markers
to increase genome coverage. Genome, Ottawa, v.50, p.742-756, 2007.
AITKEN, K. S., HERMANN, S.; KARNO, K.; BONNETT, G. D.; MCINTYRE, L. C.;
JACKSON, P. A. Genetic control of yield related stalk traits in sugarcane. Theoretical
and Applied Genetics, New York, v.117, p.1191-1203, 2008.
ALBINO, J. C.; CRESTE, S.; FIGUEIRA, A. Mapeamento genético no melhoramento
de cana-de-açúcar. Biotecnologia, online, v.36, p.82-91, 2006.
AL-JANABI, S.M.; HONEYCUTT, R.J.; McCLELLAND, M.; SOBRAL, B.W.S. A
genetic linkage map of Saccharum spontaneum L. “SES 208”. Genetics, Bethesda,
v.134, p. 1249-1260, 1993.
AL-JANABI, S. M.; FORGET, L.; DOOKUN, A. An improved and rapid protocol for
the isolation of polysaccharide and polyphenol-free sugarcane DNA. Plant Molecular
Biology Reporter, New York, n.17, p.1-8. 1999.
AL-JANABI, S.M.; PARMESSUR, Y.; KROSS, H.; DHAYAN, S.; SAUMTALLY, S.;
RAMDOYAL, K.; AUTREY, L.J.C.; DOOKUN-SAUMTALLY, A. Identification of a
major quantitative trait locus (QTL) for yellow spot (Mycovellosiella koepkei) disease
resistance in sugarcane. Molecular Breeding, Berlin, v. 19, p. 1-14, 2007.

80
ALWALA, S.; KIMBENG, C.A.; GRAVOIS, K.A.; BISCHOFF, K.P. Trap, a new tool
for sugarcane breeding: comparison with AFLP and coefficient of parentage. Journal
American Society Sugar Cane Technologists, Florida, v.26, p.62-86, 2006.
AMORIM, L. BERGAMIN FILHO, A. SANGUINO, A.; ARDOSO, C.O.N.;
MORAIS, V.A.; FERNANDES, C.R. Metodologia de Avaliação da Ferrugem da Cana-
de-Açúcar (Puccinia melanocephala). Boletim Técnico Copersucar, São Paulo, n.39,
p.13-16, 1987.
ASNAGUI, C.; PAULET, F.; KAYE, C.; GRIVET, L.; DEU, M.; GLASZMANN, J.C.;
D´HONT, A. Application of synteny across Poacea to determine the map location of
sugarcane rust resistant gene. Theoretical and Applied Genetics, New York, v.101(5-
6), p.962-969, 2000.
ASNAGHI, C.; D’HONT, A.; GLASZMANN, J.C.; ROTT, P. Resistance of sugarcane
cultivar R570 to Puccinia melanocephala isolates from different geographic locations.
Plant Disease, Saint Paul, v.85, n.3, p.282-286, 2001.
ASNAGHI C.; ROQUES, D.; RUFFEL, S.; KAYE, C.; HOARAU, J. Y.;
TÉLISMART, H.; GIRARD, J.C.; RABOIN, L.M.; RISTERUCCI, A.M.; GRIVET, L.;
D’HONT, A. Targeted mapping of a sugarcane rust resistance gene (Bru1) using bulked
segregant analysis and AFLP markers. Theoretical and Applied Genetics, New York,
v.108(4), p.759–764, 2004.
BAKER, B.; ZAMBRYSKI, P.; STASKEWICZ, B.; DINESH-KUMAR, S.P. Signaling
in plant-microbe interactions. Science, Washington, v.276, n.5313, p.726–733, 1997.
BARBASSO, D.; JORDÃO, H,; MACCHERONI, W.; BOLDINI, J.; SANGUINE, A.,
First report of Puccinia kuehnii causal agent of orange rust of sugarcane, in Brazil.
Plant Disease. Saint Paul, doi: 10.1094/PDIS-94-9-1170C, 2010.
BENT, A. F.; MACKEY, D. ELICITORS, EFFECTORS, AND R GENES: The new
paradigm and a lifetime supply of questions. Annual Review of Phytopathology, v. 45,
p: 399–436, 2007.

81
BI, C.; CHEN, F.; JACKSON, L.; GILL, B. S.; LI, W. Expression of Lignin
Biosynthetic Genes in Wheat during Development and upon Infection by Fungal
Pathogens. Plant Molecular Biology Reporter, Heidelberg, v.29, n.1, p.149–161,
2010.
BOTSTEIN, D.; WHITE, R.L.; SKOLNICK, M.; DAVIS, R.W. Construction of a
genetic linkage map in man using restriction fragment length polymorphisms.
American Journal of Human Genetics, Chicago, v.32, p.314-331, 1980.
BRUNELLI, K.R.; SILVA, H.P.; CAMARGO, L.E.A. Mapeamento de genes de
resistência quantitativa a Puccinia polysora em milho. Fitopatologia Brasileira v. 27,
p.134-140, 2002
BOWERS, J.E.; ABBEY, C.; ANDERSON, S.; CHANG, C.; DRAYE, X.; HOPPE,
A.H. A high density genetic recombination map of sequence-tagged sites for Sorghum,
as a framework for comparative structural and evolutionary genomics of tropical
grasses. Genetics, Bethesda, v.165, p. 367–386, 2003.
BUTTERFIELD, M. Marker assisted breeding in sugarcane: a complex polyploid .
2006. 91 f. PhD Thesis, University of Stellenbosch, South Africa.
CAIXETA, E.T.; OLIVEIRA, A.C.T.; DE BRITO, G.G.; SAKIYAMA, S.S. (2006)
Tipos de marcadores moleculares. Ed. BORÉM, A.; CAIXETA, E.T. In: Marcadores
Moleculares, Viçosa –MG, p. 09-78.
CAMARGO, L.E.A. Análise Genética da Resistência e da Patogenicidade. Ed.
BERGAMIN FILHO, A.; KIMATI, H.; AMORIN, L. In: Manual de Fitopatologia:
princípios e conceitos vol 1. Piracicaba: Editora Ceres, 1995, p.470-492.
CARNEIRO, M.S.; VIEIRA, M.L.C. Mapas genéticos em plantas. Bragantia,
Campinas, v.61, n.2, p.89-100, 2002.

82
CATO, S.A.; GARDNER, R.C.; KENT, J.; RICHARDSON, T.E. A rapid PCR-method
for genetically mapping ESTs. Theoretical and Applied Genetics, New York, v.102,
p.296-306, 2001.
CHEN, X.M.; LINE, R.F.; LEUNG, H. Genome scanning for resistance-gene analogs in
rice, barley, and wheat by high-resolution electrophoresis. Theoretical and Applied
Genetics, New York, v. 97, n. 3, p. 345–355, 1998.
COELHO, A.S.G. Considerações gerais sobre a análise de QTLs. In: Pinheiro, J.B.
(Ed.). Análise de QTL no melhoramento de plantas: segunda jornada em genética e
melhoramento de plantas. FUNAPE, Goiania, 2000, p. 1-20.
COLETI, J.T. Tratos culturais na cana soca. In: DINARDO-MIRANDA, L.L.;
LANDELL, M.G.A.; VASCONCELOS, A.C.M. (Ed.). Cana-de-açúcar. Campinas:
Instituto Agronômico, 2008. p. 613-630.
COMSTOCK, J.C; WU, K.K.; SCHNELL, R.J. Heritability of resistance to sugar cane
rust. Sugar Cane, Kent, v.6, p.7-10, 1992.
CORDEIRO, G.M., CASU, R.E., MCINTYRE, C.L., MANNERS, J.M., HENRY, R.J.
Microsatellite markers from sugarcane (Saccharum spp.) ESTs cross transferable to
erianthus and sorghum. Plant Science, Columbus, v.160, n.6, p.1115-1123, 2001.
COSTET, L.;LE CUNFF, L.;ROYAERT, S.;RABOIN, L.M.;HERVOUET, C.;TOUBI,
L.;TELISMART, H.; GARSMEUR,O.; ROUSSELLE, Y.; PAUQUET, J.;
NIBOUCHE, S.; GLASZMANN, J.C.; HOARAU, J.Y.; D’HONT, A. Haplotype
structure around Bru1 reveals a narrow genetic basis for brown rust resistance in
modern sugarcane cultivars. Theoretical and Applied Genetics, New York, v.125,
p.825-836, 2012.
CRESTE, S.; TULMANN NETO., A.; FIGUEIRA, A. Detection of single sequence
repeat polymorphisms in denaturing polyacrylamide sequencing gel by silver staining.
Plant Molecular Biology Reporter, Heidelberg, v.19, p.299-306, 2001.

83
CRESTE, S.; ACCORONI, K.A.C.; PINTO, L.R..; VENCKOVSKY, R.; GIMENES,
M.A.; XAVIER, M.A.; LANDELL, M.G.A. Genetic variability among sugarcane
genotypes based on polymorphisms in sucrose metabolism and drought tolerance genes.
Euphytica, Wageningen, v. 172, p. 435–446, 2010.
DANGL, J.L.; JONES, J.D.G. Plant pathogens and integrated defence responses to
infection. Nature, New York, v. 411, p. 826–833, 2001.
DA SILVA, J.A.G.; SORRELLS, M.E.; BURNQUIST, W.; TANKSLEY, S.D. RFLP
linkage map of Saccharum spontaneum. Genome, Ottawa, v.36, p.782-791, 1993.
DA SILVA, J.A.G.; HONNEYCUT, RJ.; BURNQUIST, W. L.; AL-JANABI, S. M.;
SORRELLS, M.E.; TANKSTEY, S.D.; SOBRAL, B.W.S. Saccharum spontaneum L.
'SES 208' genetic linkage map containing RFLP and PCR based markers. Molecular
Breeding, Dordrecht, v.1, p.165-179, 1995.
DA SILVA, J.A.; SORRELLS, M.E. Linkage analysis in polyploids using molecular
markers. In: JAUHAR, P. (Ed). Methods of Genome Analysis in Plants. ed. Florida:
CRC Press, 1996. p. 211-228.
DA SILVA, J. Preliminary analysis of microsatellite markers derived from sugarcane
expressed sequence tags (ESTs). Genetics and Molecular Biology, Ribeirão Preto,
v.24, n.1-4, p.155-159, 2001.
DAUGROIS J.H.; GRIVET L.; ROQUES D.; HOARAU J.Y.; LOMBARD H.;
GLASZMANN J.C.; D’HONT, A. A putative major gene for rust resistance linked with
an RFLP marker in sugarcane cultivar R570. Theoretical and Applied Genetics, New
York, v.92, p.1059-1064, 1996.
Di GASPERO, G.; CIPRIANI, G. Nucleotide binding site/leucine-rich repeats, Pto-like
and receptor-like kinases related to disease resistance in grapevine. Molecular Genetic
and Genomics, Berlin, v.269, p.612-623, 2003.
D’HONT, A.; GRIVET, L.; FELDMAN, P.; RAO, P.S.; BERDING, N.;
GLASZMANN, J.C. Characterization of the double genome structure of modern

84
sugarcane cultivars (Saccharum spp.) by molecular cytogenetics. Molecular and
General Genetics, New York, v.250, p.405-413. 1996.
D’HONT, A.; ISON, D.; ALIX, K.; ROUX, C.; GLASZMANN, J.C. Determination of
basic chromosome numbers in the genus Saccharum by physical mapping of ribosomal
RNA genes. Genome, Ottawa, v.41, p.221-225. 1998.
D´HONT, A.; GARSMEUR, O.; MCINTYRE, L. Mapping, Tagging, and Map-Based
Cloning of Simply Inherited Traits In: Genetics, Genomics and Breeding of Sugarcane.
Ed: Robert J. Henry; Chittaranjan Kole, Science Publishers 2010, 97-116
EDWARDS, M.D.; STUBER, C.W.; WENDEL, J.F. Molecular-marker-facilitated
investigations of quantitative-trait loci in maize. 1. Numbers, genomic distribution and
types of gene action. Genetics, Bethesda, v.116, p.113-125, 1987.
ELLIS, J.; DODDS, P.; PRYOR, T. Structure, function and evolution of plant disease
resistance genes. Current Opinion in Plant Biology , London, v.3, p. 278–84, 2000.
FEDERER, W. T. Augmented (or hoonuiaku) designs. Hawaiian Planters Record,
Honolulu, v.2, p.191-208, 1956.
FELTUS, F.; SINGH, H.; LOHITHASWA, H.; SCHULZE, S.R,; SILVA, T.D.;
PATERSON, A.H. A comparative genomics strategy for targeted discovery of single-
nucleotide polymorphisms and conserved-noncoding sequences in orphan crops. Plant
Physiology, Minneapolis, v.140(4), p.1183-1191, 2006.
FERREIRA, M.E.; GRATTAPAGLIA, D. Introdução ao uso de marcadores
moleculares em análises genéticas. Brasília: EMBRAPA, CENARGEN, 1998. 220p.
FIELD, D.; WILLS, C. Long, polymorphic microsatellites in simple organisms.
Proceedings of Biological Sciences. London, v 263, n.1367, p.209-215, 1996.
GARCIA, A.A.F.; KIDO, E.A.; MEZA, A.N.; SOUZA, H.M.B.; PINTO, L.R.;
PASTINA, M.M.; LEITE, C.S.; SILVA, J.A.G. DA; ULIAN, E.C.; FIGUEIRA,
A.V.O.; SOUZA, A. P. Development of an Integrated Genetic Map of a Sugarcane

85
(Saccharum spp.) Commercial Cross, based on a Maximum-Likelihood Approach for
Estimation of Linkage and Linkage Phases. Theoretical and Applied Genetics, New
York, v.112, p.298-314, 2006.
GAZAFFI, R. Desenvolvimento de modelo genético - estatístico para mapeamento
de QTLs em progênie de irmãos completos, com aplicação em cana-de-açúcar.
2009. 103f. Tese – ESALQ, Piracicaba.
GLYNN, N.C.; COMSTOCK, J.C.; McCorkle, K. Screening for Resistance to Brown
Rust of Sugarcane: Use of Bru1 resistance gene prospects and challenges. Journal
American Society of Sugar Cane Technologists, Belle Glade, v. 32, p. 82, 2012.
GRATTAPAGLIA, D.; SEDEROFF, R. Genetic linkage maps of Eucalyptus grandis
and Eucalyptus urophylla using a pseudo-testcross mapping strategy and RAPD
markers. Genetics, Bethesda, v.137, p.1121-1137, 1994.
GRIVET, L; D’HONT, A; ROQUES, D.; FELDMANN, P.; LANAUD, C.E.;
GLASZMANN, J.C. RFLP mapping in cultivated sugarcane (Saccharum spp.): Genome
Organization in a Highly Polyploid and Aneuploid interespecific Hybrid. Genetics,
Bethesda, v.142, p.987-1000, 1996.
GRIVET, L.; ARRUDA, P. Sugarcane genomics: depicting the complex genome of an
important tropical crop. Current Opinion in Plant Biology , Amsterdam, v.5, p.122-
127, 2001.
GUIMARÃES, C.T. Mapeamento comparativo e detecção de QTL’s em cana-de-açúcar
utilizando marcadores moleculares. Tese (Doutorado em Genética e Melhoramento) –
curso de Genética e Melhoramento, UFV, Viçosa, MG, 1999. 70p.
HACKAUF, B.,WEHLING, P. Identification of microsatellite polymorphisms in an
expressed portion of rye genome. Plant Breeding, v.121, p. 17–25, 2002.
HAMMOND-KOSACK, K.E.; KANYUKA, K.R. Genes in plants. Encyclopedia of
life sciences, Hertfordshire, 2007. DOI: 10.1002/9780470015902.a0020119

86
HAYES, A.J.; SAGHAI MAROOF, M.A. Targeted resistance gene mapping in soybean
using modified AFLPs. Theoretical and Applied Genetics, New York, v. 100, n. 8, p.
1279–1283, 2000.
HOARAU, J.Y.; OFFMANN, B.; D’HONT, A.; ROQUES, D.; RISTERUCCI, A.M.;
GLASZMANN, J.C.; GRIVET, L. Genetic dissection of a modern sugarcane cultivar
(Saccharum spp.) genome mapping with AFLP markers. Theoretical and Applied
Genetics, New York, v. 103, n. 1, p. 84–97, 2001.
HOARAU, J. Y.; GRIVET, L.; OFFMANN, B.; RABOIN, L. M.; DIORFLAR, J. P.;
PAYET, J.; HELLMANN, M.; D’HONT, A.; GLASZMANN, J. C. Genetic dissection
of a modern sugarcane cultivar (Saccharum spp.) II. Detection of QTLs for yield
components. Theoretical and Applied Genetics, v.105, p1027-1037, 2002.
HOGARTH, D.M.; RYAN, C.C.; TAYLOR, P.W.J. Quantitative inheritance of rust
resistance in sugar cane. Field Crops Research, Philidelphia, v.34, n.2, p.187-193,
1993.
HOGARTH, D. M.; CROSS, K. W. V. The inheritance of fiber content in sugar cane.
Proceeding Australian Society of Sugar Cane Technologie, p.93 98, 1987.
HOGARTH, D.M.; RYAN, C.C.; SKINNER, J.C. Inheritance of resistance to rust in
sugarcane - comments. Field Crops Research, Philidelphia, v.7, p. 313-316, 1983.
HU, J.; VICK, B. A. Target Region Amplification Polymorphism : A Novel Marker
Technique for Plant Genotyping. Plant Molecular Biology Reporter, Heidelberg, v.21,
p.289-294, 2003.
HUANG, L.; GILL, B.S. An RGA - like marker detects all known Lr21 leaf rust
resistance gene family members in Aegilops tauschii and wheat. Theoretical and
Applied Genetics, New York, v. 103, n. 6-7, p. 1007–1013, 2001.
HULBERT, S.H.; WEBB, C.A.; SMITH, S.M.; SUN, Q. Resistance gene complexes:
Evolution and utilization. Annual Review of Phytopathology, v.39, p. 285–312, 2001.

87
IRVINE, J.E. Saccharim species as horticultural classes. Theoretical and Applied
Genetics, New York, v. 98, p. 186-194, 1999.
JACKSON, P. Progress and prospects in genetic improvement in sucrose accumulation.
Field Crops Research, Philidelphia, v.92, p.277-290, 2005.
KAO, C.H.; ZENG, Z.B. General formulae for obtaining the MLEs and the asymptotic
variance- covariance matrix in mapping quantitative trait loci when using the EM
algorithm. Biometrics, Washington, v.53, p.653-665, 1997.
KAO, C.H.; ZENG, Z.B.; TEASDALE, R. Multiple interval mapping for quantitative
trait loci. Genetics, Bethesda, v.152, p.1203-1216, 1999.
KOBE, B., DEISENHOFER, J. The leucine-rich repeat: a versatile binding motif.
Trends Biochemical Science, Calgary, v.19, p.415-421, 1994.
KOSAMBI, D. D. The estimation of map distance from recombination values.
Annuaire Eugenetic, Washington, v.12, p.172-175, 1944.
KRUGLYAK, L.; LANDER, E.S. A nonparametric approach for mapping quantitative
trait loci. Genetics, Pittsburgh, v. 139, p. 1421-1428, 1995.
LANDELL, M.G.A.; PINTO, L. R.; CRESTE, S.; XAVIER, M.A.; ANJOS, I.A.;
VASCONCELOS, A.C.M.; BIDÓIA, M.A.P.; SILVA, D.N.; SILVA, M.A. Seleção de
novas variedades de cana-de-açúcar e seu manejo de produção. Informações
Agronômicas (Encarte Técnico), Piracicaba, SP , v. 110, p. 18-24, 12 set. 2005.
LANDELL, M.G.A.; BRESSIANI, J. Melhoramento genético, caracterização e manejo
varietal. In: DINARDO-MIRANDA, L.L.; LANDELL, M.G.A.; VASCONCELOS,
A.C.M. (Ed.). Cana-de-açúcar. Campinas: Instituto Agronômico, 2008. p.101-156.
LANDER, E.; GREEN, P.; ABRAHAMSON, J.; BARLON, A.; DALEY, M.;
LINCOLN, S.; NEWBURG, L. MAPMAKER: an interactive computer package for
constructing primary genetic linkages maps of experimental and natural populations.
Genomics, San Diego, v. 1, p. 174-181, 1987.

88
LANDER, E.S.; BOTSTEIN, D. Mapping Mendelian factors under lying quantitative
traits using RFLP linkage maps. Genetics, Bethesda, v.21, p.185-199, 1989.
Le CUNFF L, GARSMEUR O, RABOIN LM, PAUQUET J, TELISMART H, SELVI;
A, GRIVET L, PHILIPPE R, BEGUM D, DEU M, COSTET L, Wing R,
GLASZMANN JC, D’HONT A. Diploid/polyploid syntenic shuttle mapping and
haplotype-specific chromosome walking toward a rust resistance gene (Bru1) in highly
polyploidy sugarcane (2n ~ 12x ~ 115). Genetics. Bethesda, v.180, p.649–660, 2008.
LI, J.; WANG, S.; ZENG, Z.B. Multiple-Interval Mapping for Ordinal Traits. Genetics.
Bethesda, v. 176, p. 1649-1663, 2006.
LI, G.; QUIROS, C.F. Sequence-related amplified polymorphism (SRAP), a new
marker system based on a simple PCR reaction: its application to mapping and gene
tagging in Brassica. Theoretical and Applied Genetics, New York, v. 103, n. 2-3, p.
455–461, 2001.
LIU, B.H. Statistical genomics: linkage, mapping and QTL analysis. Boca Raton: CRC
Press, 1998. 611 p.
LIU, Z.H.; ANDERSON, J.A; HU, J.; FRIESEN, T.L.; RASMUSSEN, J.B.; FARIS,
J.D. A wheat intervarietal genetic linkage map based on microsatellite and target region
amplified polymorphism markers and its utility for detecting quantitative trait loci.
Theoretical and applied genetics, New York, v. 111, n. 4, p. 782–94, 2005.
LOUREIRO, M.E.; BARBOSA, M.H.P.; LOPES, F.J.F.; SILVÉRIO, F.O. Sugarcane
breeding and selection for more efficient biomass conversion in cellulosic ethanol. In:
Routes to Cellulosic Ethanol. Ed. BUCKERIDGE, M.S.; GOLDMAN, G.H., pp. 199–
240. Springer, New York, 2010.
LYNCH, M.; WALSH, B. Genetics and analysis of quantitative traits. Sunderland:
Sinauer, 1998. 980p.

89
MA, H.M.; SCHULZE, S.; LEE, S.; YANG, M.; MIRKOV, E.; IRVINE, J.; MOORE,
P.; PATERSON, A. An EST survey of the sugarcane transcriptome. Theoretical and
Applied Genetics, New York, v.108, p.851-863, 2004.
MACCHERONI, W.; JORDÃO, H.; DE GASPARI, R.; MOURA, G. L.;
MATSUOKA, S. Development of a dependable microsatellite-based fingerprinting
system for sugarcane. Sugar cane International [s.l.], v.27, n.2, p. 8-13, 2009.
MACKEY, D.; HOLT, B.F.; WIIG, A.; DANGL, J.L. RIN4 interacts with
Pseudomonas syringae type III effector molecules and is required for RPM1-mediated
resistance in Arabidopsis. Cell, Cambridge, v.108, p.743– 754, 2002.
MARCONI, T.G.; COSTA, E.A.; MIRANDA, H. MANCINI, M.C.; SILVA, C.B.C.;
OLIVEIRA, K.M.; PINTO, L.R.; MOLLINARI, M.; GARCIA, A.A.F.; SOUZA, A.P.
Functional markers for gene mapping and genetic diversity studies in sugarcane. BMC
Research Notes, v. 4, p.264, 2011.
MARGARIDO, G.R..; SOUZA, A.P.; GARCIA, A.A.F. OneMap: software for genetic
mapping in outcrossing species. Hereditas, Lund, v.144(3), p.78-79, 2007.
MARGARIDO; G.R.A.; MOLLINARI, M.; GARCIA, A.A.F. OneMap 2.0-1 -
Software for constructing genetic maps in experimental crosses: full-sib, RILs, F2 and
backcrosses. Cran-R, 2011.
MARTIN, G.B.; BROMMONSCHENKEL, S.H.; CHUNWONGSE, J.; FRARY, A.;
GANAL, M.W.; SPIVEY, R.; WU, T.; EARLE, E.D.; TANKSLEY, S.D. Map-based
cloning of a protein kinase gene conferring disease resistance in tomato. Science,
Washington, v. 262, p.1432-1436, 1993.
MARTINS, W.S.; LUCAS, D.C.S.; NEVES, K.F.S.; BERTIOLI, D.J. WebSat - A web
software for microsatellite marker development. Bioinformation. Biomedical
Informatics Publishing Group, Melbourne, v.3, n.6, p.282-283, 2009.

90
MARTINEZ, M.G.; CASTAGNARO, A.P.; DIAZ-RICCI, J.C. Genetic diversity of
Pto-like serine/threonine kinase disease resistance genes in cultivated and wild
strawberries. Journal of Molecular Evolution, New York, v.67, p.211-221, 2008.
McINTYRE, C.L.; CASU, R.E.; DRENTH, J.; KNIGHT, D.; WHAN, V.A.; CROFT,
B.J., MANNERS, J.M. Resistance genes analogues in sugarcane and their association
with quantitative trait loci for rust resistance. Genome, Ottawa, v.48, p.391-400, 2005a.
McINTYRE, C.L.; WHAN, V.A.; CROFT, B.; MAGAREY, R.; SMITH, G.R.
Identification and Validation of Molecular Markers Associated with Pachymetra Root
Rot and Brown Rust Resistance in Sugarcane Using Map-and Association-based
Approaches. Molecular Breeding, Dordrecht, v.16, p.151-161, 2005b.
MEYERS, B.C.; DICKERMAN, A.W.; MICHELMORE, R.W.;
SIVARAMAKRISHNAN, S.; SOBRAL, B.W.; YOUNG, N.D. Plant disease resistance
genes encode members of an ancient and diverse protein family within the nucleotide-
binding superfamily. Plant Journal, Malden, v.20, p.317-332, 1999.
MEYERS, B.C.; KAUSHIK, S.; NANDETY, R.S. Evolving disease resistance genes.
Current opinion in plant biology , London, v. 8, n. 2, p. 129–34, 2005.
MICHELMORE, R.W.; MEYERS, B.C. Clusters of resistance genes in plants evolve by
divergent selection and a birth-and-death process. Genome Research, Woodbury, v. 8,
p. 1113-1130, 1998.
MING, R.; LIU S.C.; LIN; Y.R.; da SILVA J.A.G.; WILSON, W.; BRAGA, D.
Detailed alignment of Saccharum and Sorghum chromosomes: comparative
organization of closely related diploid and polyploid genomes. Genetics, Bethseda,
v.150, p.1663–1682, 1998.
MING, R.; LIU, S.C.; MOORE, P.H.; IRVINE, J.E.; PATERSON, A.H. QTL Analysis
in a complex autopolyploid: genetic control of sugar content in sugarcane. Genome
Research, London, v.11, p.2075-2084, 2001.

91
MING, R.; WANG, Y.W.; DRAYE, X.; MOORE, P.H.; IRVINE, J.E.; PATERSON,
A.H. Molecular dissection of complex traits in autopolyploids: mapping QTL’s
affecting sugar yield and related traits in sugarcane. Theoretical and Applied
Genetics, New York, v.105, p.332-345, 2002.
MING, R.; MOORE, P.H.; WU, K.; D´HONT, A.; GLAZMANN, J.C.; TEW, T.L.;
MIRKOV, T.E.; DA SILVA, J.; JIFON, J.; RAI, M.; SCHNELL, R.J.; BRUMBLEY,
S.M.; LAKSHMANAN, P.; COMSTOCK, J.C.; PATERSON, A.H. Sugarcane
improvement through breeding and biotechnology. Plant Breeding Review, v.27, p.15-
118, 2006.
MUCYN, T. S.; CLEMENTE, A.; ANDRIOTIS, V. M. E. GILES E.D.;
STASKAWICZ, B.J.; RATHJEN, J.P. The tomato NBARC-LRR protein Prf interacts
with Pto kinase in vivo to regulate specific plant immunity. The Plant cell, Waterbury,
v. 18, n. 10, p. 2792–806, 2006.
MUDGE, J.; ANDERSEN, W.R.; KEHRER, R.; FAIRBANKS, D.J. A RAPD genetic
map of Saccharum officinarum. Crop Science, Madison, v. 36, p.1362–1366, 1996.
MUELLER, L.; SOLOW, T.; TAYLOR, N. The SOL Genomics Network. A
comparative resource for Solanaceae biology and beyond. Plant Physiology,
Minneapolis, v.138, p.1310-1317, 2005.
MUKHERJEE, S.K. Origin and distribution of Saccharum. Botanical Gazette,
Chicago, v. 119, p. 55–61, 1957.
OLIVEIRA, K. M.; PINTO, L. R.; MARCONI, T. G.; MARGARIDO, G.R.A.;
PASTINA, M.M.; TEIXEIRA, L.H.M.; FIGUEIRA, A.V.; ULIAN, E.C.; GARCIA,
A.A.F.; SOUZA, A.P. Functional integrated genetic linkage map based on EST-markers
for a sugarcane (Saccharum spp.) commercial cross. Molecular Breeding, Dordrecht,
v.20, n.3, p.189-208, 2007.
OLIVEIRA, K.M.; PINTO, L.R.; MARCONI, T.G.; MOLLINARI, M.; ULIAN, E.C.;
CHABREGAS, S.M.;FALCO, M.C.; BURNQUIST, W.;GARCIA, A.A.; SOUZA,

92
A.P. Characterization of new polymorphic functional markers for sugarcane. Genome,
Ottawa, v.52, n.2, p.191-209, 2009.
PALHARES, A. C.; RODRIGUES-MORAIS, T. B.; SLUYS, M.-A. VAN;
DOMINGUES, D.S.; MACCHERONI, W.; JORDÃO, H.; SOUZA, A.P.; MARCONI,
T.G.; MOLLINARI, M.; GAZAFFI, R.; GARCIA, A.A.F.; VIEIRA, M.L.C. A novel
linkage map of sugarcane with evidence for clustering of retrotransposon-based
markers. BMC genetics, London, v. 13, p. 51, 2012.
PANJE, R.R.; SRINIVASAN, K. Studies in Saccharum spontaneum. The geographical
distribution of spikelet lenght. Indian Journal of Sugarcane Research &
Development, Nova Dehli, v.1, p.1-8, 1957.
PANJE, R.R.; BABU, C. Studies in Saccharum spontaneum. Distribution and
geographical association of chromosome numbers. Cytologia, Tokyo, v.25, p.152-172,
1960.
PARK, J.W.; SOLIS-GRACIA, N.; TREVINO, C.; SILVA, J.A. Exploitation of
conserved intron scanning as a tool for molecular marker development in the
Saccharum complex. Molecular Breeding, Dordrecht, v.30, n.2, p.987-999, 2012.
PASTINA, M. M.; MALOSETTI, M.; GAZAFFI, R; MOLLINARI, M.;
MARGARIDO, G.R.A.; OLIVEIRA, K.M.; PINTO, L.R.; SOUZA, A.P.; van
EEUWIJK, F.A.; GARCIA, A.A.F. A mixed model QTL analysis for sugarcane
multiple-harvest-location trial data. Theoretical and applied genetics. New York, v.
124, n. 5, p. 835–49, 2012.
PASCHOLATI, S.F.; LEITE, B. Hospedeiro: mecanismos de resistência. In: Armando
Bergamin Filho; Hiroshi Kimati; Lilian Amorim. (Eds.). Manual de Fitopatologia -
Princípios e Conceitos. 3ed.São Paulo, SP: Editora Agronômica Ceres Ltda, 1995, v. 1,
p. 417-453.
PASSAGLIA, L.M.P. Mutação, mecanismos de reparo do DNA e recombinação. In:
ZAHA, A.; FERREIRA, H.B.; PASSAGLIA, L.M.P. (Eds.). Biologia molecular
básica. ed. Porto Alegre: Mercado Aberto, 2003. p. 116-158.

93
PERAZA-ECHEVERRIA, S.; JAMES-KAY, A.; CANTO-CANCHE, B.; CASTILLO-
CASTRO, E. Structural and phylogenetic analysis of Pto-type disease resistance gene
candidates in banana. Molecular Genetic and Genomics, Berlin, v.278, p.443-453,
2007.
PINTO, L.R.; OLIVEIRA, K.M.; ULIAN, E.C.; GARCIA, A.F.; SOUZA, A.P. Survey
in the sugarcane expressed sequence tag database (SUCEST) for simple sequence
repeats. Genome, Ottawa, v.47, p.795-804, 2004.
PINTO, L.R.; OLIVEIRA, K.M.; MARCONI, T.; GARCIA, A.A.F.; ULIAN, E.C.;
SOUZA, A.P.; Characterization of Novel Sugarcane Expressed Sequence Tag
Microsatellites And Their Comparison With Genomic SSRs. Plant Breeding, Berlin, v.
125, p. 378—384, 2006.
PINTO, L.R.; GARCIA A.A.F.; PASTINA, M.M.; TEIXEIRA, L.H.M. ; BRESSIANI,
J.A.; ULIAN, E.C.; BIDOIA, M.A.P.; SOUZA, A.P. Analysis of genomic and
functional RFLP derived markers associated with sucrose content, fiber and yield QTLs
in a sugarcane (Saccharum spp.) commercial cross. Euphytica, Wageningen, v.172,
p.313-327, 2010.
PIPERIDIS, N.; JACKSON, P.A.; D’HONT, A.; BESSE, P.; HOARAU, J.Y.;
COURTOIS, B.; AITKEN, K.S.; McINTYRE, C.L. Comparative genetics in sugarcane
enables structured map enhancement and validation of marker-trait associations.
Molecular Breeding, Berlin, v. 21, p. 233-247, 2008.
PRICE, S. Cytogenetics of modern sugar canes. Economic botany, v.17, p.97-105,
1963.
PURDY, L.H.; LIU, L.; DEAN, J.L. Sugarcane rust, a newly important disease. Plant
Disease, Saint Paul, v.69, p.1292-1296, 1983.
R Development Core Team (2011). R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-
07-0, Disponível em: <http://www.R-project.org/.>.

94
RABOIN L.,M.;OFFMANN, B.;HOARAU, J.Y.;NOTISE, J.;COSTET,
L.;TELISMART, H.;ROQUES, D.;ROTT, P.;D’HONT, A. Undertaking genetic
mapping of sugarcane smut resistance. Proceedings of Society African of Sugarcane
v. 75, p. 94–98, 2001.
RABOIN, L.M.; OLIVEIRA, K.M.; LECUNFF, L.; TELISMART, H.; ROQUES, D.;
BUTTERFIELD, M.; HOARAU, J.Y.; D‘HONT, A. Genetic mapping in sugarcane, a
high polyploid, using bi-parental progeny: identification of a gene controlling stalk
colour and a new rust resistance gene. Theoretical and Applied Genetics, New York,
v. 112: 1382–1391, 2006.
RAID, R.N., COMSTOCK, J.C., in: ROTT, P.; BAILEY, R.A.; COMSTOCK, J.C.;
CROFT, B.J.; SAUMTALLY, A.S. In: A guide to sugarcane disease. Ed. Repères.
2000. 343 p.
RALOFF, J. ILL Winds - drust storms ferry toxic agents between countries and even
continents: sugarcane rust. Science News, Washington, v.160, n.14, p.218, 2001.
RAMDOYAL, K.; SULLIVAN, S.; MEDAN, H.; BADALOO, G.; SAUMTALLY, S.;
DOMAINGUE, R. Trends in the inheritance of rust (Puccinia melanocephala H. and P.
Syd) in sugar cane. Sugar Cane, Kent, v.3, p.19-23, 1996.
RAMDOYAL, K.; SULLIVAN, S.; LIM SHIN CHONG, L.C.Y.; BADALOO, G.H.;
SAUMTALLY, S., DOMAINGUE, R. The genetics of rust resistance in sugar cane
seedling populations. Theoretical and Applied Genetics, New York, v.100, n3-4,
p.557-563, 2000.
RESENDE, M.D.V. Predição de valores genéticos, componentes de variância,
delineamentos de cruzamento e estrutura de populações no melhoramento florestal.
Tese de Doutorado, 1999. Universidade Federal do Paraná, Curitiba. 434 p.
RIELY, B.K.; MARTIN, G.B.: Ancient origin of pathogen recognition specificity
conferred by the tomato disease resistance gene Pto. Proceedings of the National
Academy of Sciences, Washington, v.98, p.2059-2064, 2001

95
RYAN, C.C.; EGAN, B.T. Rust. RICAUD, C.; EGAN, B.T.; GILLASPIE JUNIOR,
A.G.; HUGHES, C.G. In: Diseases of sugarcane. ed. Amsterdam: Elsevier, 1989.
p.189-210.
ROSSI, M.; ARAUJO, P. G.; PAULET, F.; GARSMEUR, O.; DIAS, V. M.; CHEN, H.;
VAN SLUYS, M. A.; D’HONT, A. D. Genomic distribution and characterization of
EST-derived resistance gene analogs (RGAs) in sugarcane. Molecular and Genetic
Genomics, Berlin, v.269, p.406-419, 2003.
ROTT, P.; BAILEY, R.A.; COMSTOCK, J.C.; CROFT, B.J.; SAUMTALLY, A.S. In:
A guide to sugarcane disease. Ed. Repères. 2000. 343 p.
ROZEN S.; SKALETSKY H. J. Primer3 on the WWW for general users and for
biologist programmers. In: KRAWETZ, S.; MISENER, S. (Ed.) Bioinformatics
Methods and Protocols: Methods in Molecular Biology. ed. Totowa: Humana Press,
2000. p. 365-386.
SAS Institute (1989) SAS/STAT user’s guide. Version 6, 4th ed. SAS Institute, Cary,
Carolina do Norte.
SACILOTO, R.F.Z. Inserção do gene PR5K em cana-de-açúcar visando induzir
resistência ao fungo da ferrugem Puccinia melanocephala. 2003. 86f. Dissertação
(Mestrado em Fitopatologia) - Escola Superior de Agricultura “Luiz de Queiroz”,
Universidade de São Paulo, Piracicaba.
SALMERON, J.; OLDROYD, G.; ROMMENS, C.; SCOFIELD, S.; KIM, H.;
LAVELLE, D.; DAHLBECK, D.; STASKAWICZ, B. Tomato Prf is a member of the
leucine-rich repeat class of plant disease resistance genes and lies embedded within the
Pto kinase gene cluster. Cell, Cambridge, v.86, p.123–133, 1996.
SANTOS, A.S. Doenças causadas por fungos. In: DINARDO-MIRANDA, L.L.;
LANDELL, M.G.A.; VASCONCELOS, A.C.M. (Ed.). Cana-de-açúcar. ed. Campinas:
Instituto Agronômico, 2008. p.423-435.

96
SCHLÖTTERER, C.; RITTER, R.; HARR, B.; BREM, G. High mutation rate of a long
microssatellite allele in Drosophila melanogaster provides evidence for allele-
specificmutation rates. Molecular Biology and Evolution, Chicago, v.15, n.10, p.1269-
1274, 1998.
SERANG, O.; MOLLINARI, M.; GARCIA, A.A.F. Efficient Exact Maximum a
Posteriori Computation for Bayesian SNP Genotyping in Polyploids. Plos One, online,
v. 7(2), p. e30906, 2012.
SHINE, J.M.; COMSTOCK, J.C.; DEAN, J.L. Comparison of five isolates of sugarcane
rust and differential reaction on six sugarcane clones. In: International Society Of
Sugar Cane Technologists Congress, 2005, Guatemala. Proceedings. Guatemala:
ISSCT, 2005. v.1, p.638-647.
SILVA, M.A.; DINARDO-MIRANDA, L.L.; ALMEIDA, A.M.; CAMPANA, M.P.;
COELHO, A.; LANDELL, M.G.A.; FIGUEIREDO, P.; PERECIN, D. Danos causados
pela ferrugem, Puccinia melanocephala, a variedades de cana-de-açúcar com diferentes
níveis de resistência. Summa phytopathologica, Botucatu, v.27, p.39-43, 2001.
SILVERSTEIN, K. A. T.; GRAHAM, M. A.; PAAPE, T. D.; VANDENBOSCH, K. A.
Genome Organization of More Than 300 Defensin-Like Genes in Arabidopsis. Plant
Physiology, Minneapolis, v. 138, n. June, p. 600–610, 2005.
SOBRAL, B.W.S.; HONEYCUTT, R.J. High output genetic mapping of polyploids
using PCR- generated markers. Theoretical and Applied Genetics, New York, v. 86,
p. 105-112, 1993.
SORIANO, J. M.; VILANOVA, S.; ROMERO, C.; LLÁCER, G.; BADENES, M. L.
Characterization and mapping of NBS-LRR resistance gene analogs in apricot (Prunus
armeniaca L.). Theoretical and applied genetics, New York, v.110, n.5, p.980-989,
2005.
SOUZA, A.P. Biologia molecular aplicada ao melhoramento. In: NASS, L.L.;
VALOIS, A.C.C.; MELO, I.S.; VALADARES-INGLIS, M.C. (Ed.). Recursos

97
genéticos e melhoramento de plantas. ed. Rondonóplis: Fundação M.T., 2001. p.939-
966.
SREENIVASAN, T.V.; AHLOOWALIA, B.S.; HEINZ, D.J. Cytogenetics. In:HEINZ,
D.J. (Ed.). Sugarcane improvement through breeding. ed. Amsterdam: Elsevier,
1987. p.211-253.
STEVENSON, G.C. Genetics and Breeding of Sugarcane. Tropical Science Series.
ed. London: Longmans, Green and Co Ltd., 1965. 284p.
STUBER, C.W.; EDWARDS, M.D.; WENDEL, J.F. Molecular maker-facilites
investigations of quantitative trait loci in maize. II. Factors influencing yield and its
components traits. Crop Science, Madison, v.27, p.639-648, 1987.
STURTEVANT, A. H. The linear arrangement of six sex-linked factors in Drosophila,
as shown by their mode of association. Journal of Experimental Zoology, Iowa,v.14,
p. 43-59. 1913.
SUMAN, A.; PAN, Y-B.; THONGTHAWEE, S.; BURNER, M.D.; KINBENG,
COLLINS, A.K. Genetic analysis of the sugarcane (Saccharum spp.) cultivar ‘LCP 85-
384’. I. Linkage mapping using AFLP, SSR, and TRAP markers. Theoretical and
applied genetics, New York, v. 123, p. 77-93, 2011.
SUMAN, A.; ALI, K.; ARRO, J.; ARNOLD, P.S.; KIMBENG A.C.; BAISAKH, N.
Molecular Diversity Among Members of the Saccharum Complex Assessed Using
TRAP Markers Based on Lignin-Related Genes. BioEnergy Research, New York, v.5,
n.1, p.197-205, 2012.
TAI, P.Y.P.; MILLER, J.D.; DEAN, J.L.; Inheritance of resistence to rust in sugarcane.
Field Crops Research, Philidelphia, v.4, p.261-268, 1981.
TAI, P.Y.P.; DEAN, J.L.; MILLER, J.D. Effect of selection for agronomic performance
on frequency of rust susceptibility in sugarcane. (1984) Ed. BEARDSLEY, D.L.;
CLAYTON, J.E.; MARTIN, F.A. In: American Society of Sugar Cane Technologists, p.
22-27.

98
TAKKEN, F.L.W.; JOOSTEN, M.A.H.J. Plant resistance genes: their structure,
function and evolution. European Journal of Plant Pathology, v.106, p.699-713,
2000.
TAMURA, K.; PETERSON, D.; PETERSON, N.; STECHER, G.; NEI, M.; KUMAR,
S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood,
evolutionary distance, and maximum parsimony methods. Molecular Biology and
Evolution, Oxford, v.28, n.10, p.2731-2739, 2011.
TANKSLEY, S.D. Mapping polygenes. Annual Review Genetics, Palo Alto, v.27,
p.205-233, 1993.
TOMKINS, J.P.; YU, Y.; MILLER-SMITH, H.; FRISCH D.A.; WOO, S.S.; WING, R.
A bacterial artificial chromosome library for sugarcane. Theoretical and Applied
Genetics, New York, v. 99, p. 419–424, 1999.
Van der LINDEN, C.G.; WOUTERS, D.C.; MIHALKA, V.; KOCHIEVA, E.Z,
SMULDERS, M.J.; VOSMAN, B. Efficient targeting of plant disease resistance loci
using NBS profiling. Theoretical and Applied Genetics, New York, v.109, p. 384–
393, 2004.
VARSHNEY, R.K., THIEL, T., STEIN, N., LANGRIDGE, P., GRANER, A. In silico
analysis on frequency and distribution of microsatellites in ESTs of some cereal species.
Cellular Molecular Biology, Lett. v. 7, p. 537–546, 2002.
VIRTUDAZO, E.V.; NAKAMURA H. KAKISHIMA, M. Phylogenetic analysis of
sugarcane rusts based on sequences of ITS, 5.8s rDNA and D1/D2 regions of LSU
rDNA. Journal of General Plant Pathology v.67, p.28-36, 2001a.
VETTORE, A.L.; SILVA, F.R.; KEMPER, E.L.; ARRUDA, P. The libraries that made
SUCEST. Genetics and Molecular Biology, Ribeirão Preto, v.24, p. 1-4, 2001.
VLEESHOUWERS V.G.; MARTENS A.; van DOOIJEWEERT, W.; COLON, L.T.;
GOVERS, F.; KAMOUN, S. Ancient diversification of the Pto kinase family preceded

99
speciation in Solanum. Molecular Plant-Microbe Interact , Saint Paul, v.14, p.996-
1005, 2001.
VOS, P.; HOGERS, R.; BLEEKER, M.; REIJANS, M.; VAN DE LEE, T.; HORNES,
M.; FRIJTERS, A.; POT, J.; PELEMAN, J.; KUIPER, M.; ZABEAU, M. AFLP: a new
technique for DNA fingerprinting. Nucleic Acids Research, Oxford, v.23, n.21,
p.4407-4414, 1995.
XU, Q.; DENG, X. Cloning and phylogenetic analyses of serine/threonine kinase class
defense-related genes in a wild fruit crop “chestnut rose”. BMC research notes,
London, v. 3, p. 202, 2010.
YAN, G. P.; CHEN, X M; LINE, R F; WELLINGS, C. R. Resistance gene-analog
polymorphism markers co-segregating with the YR5 gene for resistance to wheat stripe
rust. Theoretical and applied genetics. New York, v. 106, n. 4, p. 636–43, 2003.
WELLER, J.I. Maximum likelihood techniques for the mapping and analysis of
quantitative trait loci with the aid of genetic markers. Biometrics, Washington, v.42,
p.627-640, 1986.
WRIGHT, S. Evolution and the Genetics of Populations, Vol. I, Genetic and Biometric
Foundations. University of Chicago Press, Chicago, 1968.
WU, K.K.; BURNQUIST, W.; SORRELLS, M.E.; TEW, T.L.; MOORE, P.W.;
TANKSLEY, S.D. The detection and estimation of linkage of polyploids using single-
dose restriction fragments. Theoretical and Applied Genetics, New York, v.83, p.
294-300, 1992.
Wu, R.L.; Ma, C.X.; Wu, S.S.; Zeng, Z.B. Linkage mapping of sex-specific differences.
Genetic Research, London, v.79, p.85–96, 2002a.
WU, R.L; MA, C.X.; PAINTER, I.; ZENG, Z.B. Simultaneous maximum likelihood
estimation of linkage and linkage phases in outcrossing species. Theoretical
Population Biology New York, v.61, p.349-363, 2002b.

100
ZENG, Z.B. Theoretical basis of separation of multiple linked gene effects on mapping
quantitative trait loci. Proceedings of the National Academy of Science, Washington,
v.90, p.10972-10976, 1993.
ZENG, Z.B. Precision mapping of quantitative trait loci. Genetics, Bethesda, v.136,
p.1457-1468, 1994.
ZENG, Z.B., KAO, C.H.; BASTEN, C.J. Estimating the genetic architecture of
quantitative traits. Genetic Research, London, v. 74, p. 279-289, 1999.
ZHANG, J.; YUAN, Y.; NIU, C.; HINCHLIFFE, D.J.; LU, Y.; YU, S.; PERCY, R.G.;
ULLOA, M.; CANTRELL, R.G.AFLP-RGA Markers in Comparison with RGA and
AFLP in Cultivated Tetraploid Cotton. Crop Science, Madison, v.47, p.180-187, 2007.
ZHANG, N.; YUAN, S.; YANG, W.; LIU, D. Identification of Lr24 with targeted
region amplified polymorphism (TRAP) analysis in wheat. Frontiers of Agriculture in
China, Baodin, v. 4, n. 1, p. 18–23, 2010.